
安定化されたインスリン様増殖因子ポリペプチド
本発明は、IGFからのE−ペプチドの天然の生理的切断が妨げられる、IGF−1又はIGF−2配列及びE−ペプチド配列を有する安定化されたポリペプチドに関する。
【発明の詳細な説明】
【技術分野】
【0001】
(背景技術)
インスリン様増殖因子(IGF)は、細胞がこれら細胞の生理的環境と情報伝達をするために使用する複合系の一部である。この複合系(インスリン様増殖因子軸と呼ばれることが多い。)は、2つの細胞表面受容体(IGF−1R及びIGF−2R)、2つのリガンド(IGF−1及びIGF−2)、6個の高親和性IGF結合タンパク質のファミリー(IGFBP1−6)及び関連するIGFBP分解酵素(プロテアーゼ)からなる。この系は、正常な生理の制御だけでなく、多くの病理的状態に対しても重要である(Glass、Nat Cell Biol 5:87−90、2003)。
【0002】
IGF軸は、細胞増殖の促進及び細胞死(アポトーシス)の抑制に関与することが示されている。IGF−1は、ヒト成長ホルモン(hGH)による刺激の結果として、主に、肝臓により分泌される。ヒトの身体における殆ど全ての細胞、特に筋肉、軟骨、骨、肝臓、腎臓、神経、皮膚及び肺における細胞は、IGF−1により影響を受ける。インスリン様の効果に加えて、IGF−1はまた細胞増殖を制御することもできる。IGF−1及びIGF−2は、IGF結合タンパク質として知られる遺伝子産物のファミリーにより制御される。これらのタンパク質は、IGF受容体への結合を妨害することによるIGF作用の阻害ならびに受容体への送達促進及び血流中のIGF半減期の延長を通じたIGF作用の促進の両方に関与する複合的な方法でIGF作用の調節に寄与する。特性が分かっている少なくとも6種類の結合タンパク質(IGFBP1−6)がある。
【0003】
その成熟型において、ソマトメジンとも呼ばれるヒトIGF−1(gpetlcgaelvdalqfvcgdrgfyfnkptgygsssrrapqtgivdeccfrscdlrrlemycaplkpaksa;配列番号1)は、培養において幅広い範囲の細胞の増殖を刺激することが示されている70アミノ酸の小タンパク質である。成熟タンパク質は、最初に、3つの既知のスプライシング変異mRNAによりコードされる。各mRNAのオープンリーディングフレームは、特定のIGF−1mRNAに依存して、70アミノ酸IGF−1及びC末端の特定のE−ペプチドを含有する前駆タンパク質をコードする。これらのE−ペプチドは、Ea(rsvraqrhtdmpktqkevhlknasrgsagnknyrm;配列番号2)、Eb(rsvraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk;配列番号3)及びEc(rsvraqrhtdmpktqkyqppstnkntksqrrkgstfeerk;配列番号4)ペプチドと呼ばれ、35から87アミノ酸長の範囲であり、N末端において共通配列領域及びC末端において可変配列領域を含む。例えば、IGF−1−Eaに対する野生型オープンリーディングフレームは、105アミノ酸(gpetlcgaelvdalqfvcgdrgfyfnkptgygsssrrapqtgivdeccfrscdlrrlemycaplkpaksa rsvraqrhtdmpktqkevhlknasrgsagnknyrm;配列番号5)のポリペプチドをコードする。生理的発現において、E−ペプチドは、内在性プロテアーゼにより前駆体から切断され、生物活性があることが知られている成熟70アミノ酸IGF−1が生じる。ある種の関連において、IGF−1のN末端アミノ酸の1から3個は生理的条件下で切断され、これにより67−70の間のアミノ酸を有する活性型IGF−1が生じることが知られている。ヒトIGF−2に対する1つのE−ペプチド
【0004】
【化1】

のみが156アミノ酸前駆体
【0005】
【化2】
に対して確認されていることを除き、IGF−2遺伝子発現及びプロセシングは同様の特性を特徴とする。IGF−1及びIGF−2の両方は、患者の血清中で内在性プロテアーゼにより急速に分解されるので、優れた薬物候補とはならないと思われる。意図されているあるストラテジーは、IGF−1の結合タンパク質の1つと複合体を形成することにより、薬物としてIGF−1を安定化することである。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Glass、Nat Cell Biol 5:87−90、2003
【発明の概要】
【0007】
発明の要旨
本発明は、このE−ペプチドを実質的に含有する前駆IGF−1又はIGF−2タンパク質が生物活性を有し、血清の存在下で安定化され、その結果、医薬品として有用であるIGF−1又はIGF−2ポリペプチドが得られるという事実に基づく。本発明の組成物において、例えば、E−ペプチドの位置1のアルギニン又は位置2のセリン(野生型前駆IGF−1における位置71及び72に対応する。)の何れかの変異又は欠失によって、IGF−1からのE−ペプチドの通常の切断が回避される。IGF−2において、例えば、E−ペプチドの位置1のアルギニン又は位置2のアスパラギン酸(野生型前駆IGF−2における位置68及び69に対応する。)の何れかの変異又は欠失によって、切断が回避される。IGF前駆タンパク質の他の修飾によって、この切断を回避又は減少させることができる。
【0008】
さらに、IGF−1前駆アミノ酸配列のさらなる修飾は、さらなる医薬的有益性を与え得る。例えば、本発明のポリペプチドは、IGF−1受容体に対する親和性の向上又は抑制性のIGF−1もしくはIGF−2結合タンパク質に対する結合能の低下を示し得る。
【0009】
明確性及び一貫性のために、この明細書を通じて、及び特許請求の範囲において、IGF−1もしくはIGF−2前駆体又は成熟タンパク質におけるアミノ酸残基の付番は、シグナルペプチドなしの野生型前駆タンパク質配列の付番に基づく。
【0010】
従って、本発明は、プロテアーゼによるIGF−1からのE−ペプチドの切断がIGF−1前駆タンパク質の修飾により減少する、IGF−1前駆タンパク質を含有するポリペプチドを含む。E−ペプチドは、Ea、Eb又はEcペプチドであり得る。前駆体のN末端において、前駆タンパク質のアミノ酸G1、P2又はE3が、R36(例えばR36A)及びR37(例えばR37A)など、欠失又は変異させられ得る。
【0011】
前駆タンパク質は、例えば、Ebのアミノ酸N95とT96との間へのEaのアミノ酸93−102の挿入によって、N−結合型グリコシル化コンセンサス配列 NXS/Tをさらに含み得る。一般に、前駆タンパク質は、前駆タンパク質のアルギニン側鎖など、前駆タンパク質のアミノ酸側鎖に共有結合されたオリゴ糖を含み得る。
【0012】
さらに、非天然アミノ酸(例えば、アセチレン又はアジド基を含むもの)により前駆タンパク質の残基を置換することができる。このような非天然アミノ酸は、前駆タンパク質の側鎖へのポリ(エチレングリコール)部分の連結を促進することができる。通常のタンパク質PEG化ストラテジーは当技術分野で周知である。
【0013】
前駆タンパク質は、前駆タンパク質のC末端に連結された1以上のさらなるE−ペプチドをさらに含み得る。例えば、ポリペプチドは、N末端からC末端まで、(1)第一のEbペプチドを有するIGF−1前駆タンパク質(このタンパク質中、G1、P1及びE1が欠失しており、R36もしくはR37の何れか又は両方が変異しており、R71及びS72が欠失しており、第一のEbペプチドの最後の7個のC末端アミノ酸が欠失している。);(2)第二のEbペプチド(このペプチド中、R71、S72及び第二のEbペプチドの最後の7個のC末端アミノ酸が欠失している。);(3)第三のEbペプチド(このペプチド中、R71、S72及び第三のEbペプチドの最後の7個のC末端アミノ酸が欠失している。);及び(4)第四のEbペプチド(このペプチド中、R71及びS72が欠失している。)を含み得る。
【0014】
IGF−1からのE−ペプチドの切断を妨害する効率的な手段は、R71又はS72の欠失又は変異である。
【0015】
同様に、本発明は、プロテアーゼによるIGF−2からのE−ペプチドの切断が、IGF−2前駆タンパク質の修飾により減少する、IGF−2前駆タンパク質を含む。特に、R68又はD69の欠失又は変異は、IGF−2前駆タンパク質のプロテアーゼ消化を回避する有効な手段であり得る。
【0016】
さらに、本明細書中に記載の有益性を与えるために、IGF−1の何れかのE−ペプチドをIGF−2と組み合わせることができ、IGF−2の何れかのE−ペプチドをIGF−1と組み合わせることができる。
【0017】
本発明は、さらに、本発明のポリペプチドの治療的有効量を投与することにより、筋骨格疾患、糖尿病、神経細胞死を治療する方法を含む。同様に、本発明は、筋骨格疾患、糖尿病、神経細胞死又は貧血の治療用の医薬品の製造のための本発明のポリペプチドの使用を含む。
【0018】
別の実施形態において、本発明は、E−ペプチドがないが、PEG化の部位として非天然アミノ酸がそこに導入されている、PEG化IGF−1を含む。E−ペプチドがない、本明細書中に記載のような、非天然アミノ酸を含有する何れの修飾PEG化IGF−1もまた本発明に含まれる。
【0019】
本発明はまた、所望の効果を得るために本発明のポリペプチドの有効量を投与する、獣医学の方法及び使用も含む。
【0020】
獣医学での使用は、(i)動物における増殖の速度及び/又は程度を促進すること、(ii)食物の身体組織への変換の効率を促進すること、(iii)泌乳動物において乳汁産生を促進すること、(iv)悪液質、外傷性傷害又は他の消耗疾患に付随する消耗性の症状に対して動物を治療すること及び(v)新生仔の健康を増進するために泌乳非ヒト動物を処置することを含む。
【0021】
全ての引用参考文献又は書類は、参照により本明細書中に組み込まれる。
【図面の簡単な説明】
【0022】
【図1】図1A−1Cは、10%ヒト血清の存在下又は非存在下で37℃にて0又は16時間温置した後の、本発明のポリペプチド及び野生型IGF−1前駆体のウエスタンブロットである。様々なIGF−1コンストラクトをコードする発現ベクターをCos7細胞へと遺伝子移入し、条件培養液を得た。「3mut」は、次の3セットの修飾:G1、G2及びE3の欠失;AlaへのArg37の変異(R37A);及びR71及びS72の欠失を有する、hIGF−1−E−ペプチド前駆体を指す。図1Aは、Eaを含有する、野生型及び3mut前駆体に対する(IGF−1に対する抗体を用いた)ウエスタンブロットの結果を示す。図1Bは、Ebを含有する、野生型及び3mut前駆体に対する(hIGF−1に対する抗体を用いた)ウエスタンブロットの結果を示す。図1Cは、Ecを含有する、野生型及び3mut前駆体に対する(hIGF−1に対する抗体を用いた)ウエスタンブロットの結果を示す。
【図2A】図2A−2Dは、様々なIGF−1ポリペプチド(「リガンド」))の生物学的活性を示す線グラフである。Cos7により発現されたポリペプチドによるC2C12筋芽細胞の刺激により、生物学的活性を測定した。次に、総AKT及びリン酸化AKTの相対量について、刺激したC2C12細胞をアッセイした。Long−R3−IGF−1は、E3R変異及びさらなる13アミノ酸のN末端延長ペプチドを有する成熟ヒトIGF−1アミノ酸配列からなる市販の試薬(Sigma製品番号1−1271)である。図2AはIGF−1−Ea3mutの活性を示す。図2BはIGF−1−Eb3mutの活性を示す。図2Cは、IGF−1−Eab3mut(Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されている、3mutコンストラクト)の活性を示す。図2Dは、IGF−1−Ec3mutの活性を示す。
【図2B】図2A−2Dは、様々なIGF−1ポリペプチド(「リガンド」))の生物学的活性を示す線グラフである。Cos7により発現されたポリペプチドによるC2C12筋芽細胞の刺激により、生物学的活性を測定した。次に、総AKT及びリン酸化AKTの相対量について、刺激したC2C12細胞をアッセイした。Long−R3−IGF−1は、E3R変異及びさらなる13アミノ酸のN末端延長ペプチドを有する成熟ヒトIGF−1アミノ酸配列からなる市販の試薬(Sigma製品番号1−1271)である。図2AはIGF−1−Ea3mutの活性を示す。図2BはIGF−1−Eb3mutの活性を示す。図2Cは、IGF−1−Eab3mut(Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されている、3mutコンストラクト)の活性を示す。図2Dは、IGF−1−Ec3mutの活性を示す。
【図2C】図2A−2Dは、様々なIGF−1ポリペプチド(「リガンド」))の生物学的活性を示す線グラフである。Cos7により発現されたポリペプチドによるC2C12筋芽細胞の刺激により、生物学的活性を測定した。次に、総AKT及びリン酸化AKTの相対量について、刺激したC2C12細胞をアッセイした。Long−R3−IGF−1は、E3R変異及びさらなる13アミノ酸のN末端延長ペプチドを有する成熟ヒトIGF−1アミノ酸配列からなる市販の試薬(Sigma製品番号1−1271)である。図2AはIGF−1−Ea3mutの活性を示す。図2BはIGF−1−Eb3mutの活性を示す。図2Cは、IGF−1−Eab3mut(Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されている、3mutコンストラクト)の活性を示す。図2Dは、IGF−1−Ec3mutの活性を示す。
【図2D】図2A−2Dは、様々なIGF−1ポリペプチド(「リガンド」))の生物学的活性を示す線グラフである。Cos7により発現されたポリペプチドによるC2C12筋芽細胞の刺激により、生物学的活性を測定した。次に、総AKT及びリン酸化AKTの相対量について、刺激したC2C12細胞をアッセイした。Long−R3−IGF−1は、E3R変異及びさらなる13アミノ酸のN末端延長ペプチドを有する成熟ヒトIGF−1アミノ酸配列からなる市販の試薬(Sigma製品番号1−1271)である。図2AはIGF−1−Ea3mutの活性を示す。図2BはIGF−1−Eb3mutの活性を示す。図2Cは、IGF−1−Eab3mut(Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されている、3mutコンストラクト)の活性を示す。図2Dは、IGF−1−Ec3mutの活性を示す。
【図3A】図3A−3D及び図4A−4Dは、リガンド結合に反応する受容体リン酸化についてアッセイすることによって、本発明のIGF−1前駆ポリペプチドが適切な受容体に対する選択性を維持するか否かを示す線グラフである。図3A及び図3Bは、IGF−1受容体(図3A)及びインスリン受容体(図3B)に対する、IGF−1−Ea3mutの受容体選択性を試験する。図3C及び3Dは、IGF−1受容体(図3C)及びインスリン受容体(図3D)に対する、IGF−1−Eb3mutの受容体選択性を試験する。
【図3B】図3A−3D及び図4A−4Dは、リガンド結合に反応する受容体リン酸化についてアッセイすることによって、本発明のIGF−1前駆ポリペプチドが適切な受容体に対する選択性を維持するか否かを示す線グラフである。図3A及び図3Bは、IGF−1受容体(図3A)及びインスリン受容体(図3B)に対する、IGF−1−Ea3mutの受容体選択性を試験する。図3C及び3Dは、IGF−1受容体(図3C)及びインスリン受容体(図3D)に対する、IGF−1−Eb3mutの受容体選択性を試験する。
【図3C】図3A−3D及び図4A−4Dは、リガンド結合に反応する受容体リン酸化についてアッセイすることによって、本発明のIGF−1前駆ポリペプチドが適切な受容体に対する選択性を維持するか否かを示す線グラフである。図3A及び図3Bは、IGF−1受容体(図3A)及びインスリン受容体(図3B)に対する、IGF−1−Ea3mutの受容体選択性を試験する。図3C及び3Dは、IGF−1受容体(図3C)及びインスリン受容体(図3D)に対する、IGF−1−Eb3mutの受容体選択性を試験する。
【図3D】図3A−3D及び図4A−4Dは、リガンド結合に反応する受容体リン酸化についてアッセイすることによって、本発明のIGF−1前駆ポリペプチドが適切な受容体に対する選択性を維持するか否かを示す線グラフである。図3A及び図3Bは、IGF−1受容体(図3A)及びインスリン受容体(図3B)に対する、IGF−1−Ea3mutの受容体選択性を試験する。図3C及び3Dは、IGF−1受容体(図3C)及びインスリン受容体(図3D)に対する、IGF−1−Eb3mutの受容体選択性を試験する。
【図4A】図4A及び4Bは、IGF−1受容体(図4A)及びインスリン受容体(図4B)に対する、IGF−1−Ec3mutの受容体選択性を試験する。図4C及び4Dは、IGF−1受容体(図4C)及びインスリン受容体(図4D)に対する、IGF−1−Eab3mutの受容体選択性を試験する。「IGF1−R3」は、上述のLong−R3−IGF−1を指す。「IGF1Eab」として挙げられたポリペプチドは、Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されたコンストラクトを指す。
【図4B】図4A及び4Bは、IGF−1受容体(図4A)及びインスリン受容体(図4B)に対する、IGF−1−Ec3mutの受容体選択性を試験する。図4C及び4Dは、IGF−1受容体(図4C)及びインスリン受容体(図4D)に対する、IGF−1−Eab3mutの受容体選択性を試験する。「IGF1−R3」は、上述のLong−R3−IGF−1を指す。「IGF1Eab」として挙げられたポリペプチドは、Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されたコンストラクトを指す。
【図4C】図4A及び4Bは、IGF−1受容体(図4A)及びインスリン受容体(図4B)に対する、IGF−1−Ec3mutの受容体選択性を試験する。図4C及び4Dは、IGF−1受容体(図4C)及びインスリン受容体(図4D)に対する、IGF−1−Eab3mutの受容体選択性を試験する。「IGF1−R3」は、上述のLong−R3−IGF−1を指す。「IGF1Eab」として挙げられたポリペプチドは、Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されたコンストラクトを指す。
【図4D】図4A及び4Bは、IGF−1受容体(図4A)及びインスリン受容体(図4B)に対する、IGF−1−Ec3mutの受容体選択性を試験する。図4C及び4Dは、IGF−1受容体(図4C)及びインスリン受容体(図4D)に対する、IGF−1−Eab3mutの受容体選択性を試験する。「IGF1−R3」は、上述のLong−R3−IGF−1を指す。「IGF1Eab」として挙げられたポリペプチドは、Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されたコンストラクトを指す。
【図5】図5は、様々なリガンドによる(C2C12筋芽細胞の3から4日の分化の結果としての)C2C12筋管の刺激における相対的AKTリン酸化を示すウエスタンブロットである。IGF−1Eb−マルチマーは、図6Aで図式的に示されるコンストラクトを指す。
【図6】図6A及び6Bは、本発明の2つのポリペプチドの概略図である。図6Aは、4セットの修飾:G1、G2及びE3の欠失;R37のAへの変異;R71及びS72の欠失;及び最後の7個のC末端アミノ酸の欠失を有するIGF−1−Eb前駆ポリペプチドを示す。さらに、本ポリペプチドのC末端での、2個のさらなるEbペプチドの付加(しかしR71及びS72はなく、最後の7個のC末端アミノ酸がない。)及び最後のEbペプチドの付加(しかしR71及びS72がない。)により、本ポリペプチドを長くする。このコンストラクトはIGF−1−Ebマルチマーと呼ぶことが多い。図6Bは、4セットの修飾:G1、P2及びE3の欠失;R37のAへの変異;R71及びS72の欠失;及びEbのアミノ酸95と96との間へのEaアミノ酸93から102の挿入があるIGF−1−Eab前駆ポリペプチドを示す。
【図7A】図7Aは、対応する動物IGF−1とのヒトIGF−1(配列番号1)の配列アラインメントである。分析した全ての動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。コチョウザメ(S2はP2に取って代わる。)を除く分析した全ての種においてG1、P2、E3は保存されている。R36及びR37は分析した全ての種において保存されている。
【図7B】図7Bは、ヒトIGF−1(配列番号1)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。系統樹の下は、タンパク質配列に対する100残基あたりの「アミノ酸置換」の数を示すスケールである。非ギャップミスマッチの数からの、サイレント置換について補正された距離値を計算するために、Kimuraの距離式を使用する。計算した値は、部位あたりの相違数の平均数であり、0から1の間となる。0は完全に同一であることを表し、1は同一性がないことを表す。系統樹のスケールは、これらの値に100を掛けたものを使用する。
【図8A】図8Aは、様々な動物EaペプチドとのヒトEaペプチド(配列番号2)の配列アラインメントである。分析した全動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。R71及びS72は分析した全ての種で保存されている。
【図8B】図8Bは、ヒトIGF−1 Eaペプチド(配列番号2)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。
【図9A】図9Aは、様々な動物EbペプチドとのヒトEbペプチド(配列番号3)の配列アラインメントである。分析した全動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。R71及びS72は分析した全ての種で保存されている。
【図9B】図9Bは、ヒトIGF−1 Ebペプチド(配列番号3)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。
【図10A】図10Aは、様々な動物EcペプチドとのヒトEcペプチド(配列番号4)の配列アラインメントである。分析した全動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。R71及びS72は分析した全ての種で保存されている。
【図10B】図10Bは、ヒトIGF−1 Ecペプチド(配列番号4)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。
【図11A】図11Aは、対応する動物IGF−2とのヒトIGF−2(配列番号7)の配列アラインメントである。分析した全動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。R68は分析した全ての種で保存されており;D69はチンパンジー(その位置はヒスチジン残基になっている。)を除き保存されている。
【図11B】図11Bは、ヒトIGF−2(配列番号7)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。
【図12A】図12Aは、様々な動物IGF−2E−ペプチドとのヒトIGF−2E−ペプチド(配列番号6)の配列アラインメントである。分析した全動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。R68は分析した全ての種で保存されており;D69はチンパンジー(その位置はヒスチジン残基になっている。)を除き保存されている。
【図12B】図12Bは、ヒトIGF−2Eペプチド(配列番号6)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。
【0023】
発明の詳細な説明
本発明は、活性型IGF−1又はIGF−2をそのE−ペプチドから放出することに関与する典型的なプロテアーゼ切断を妨害、減少又は回避するために修飾されているE−ペプチドを実質的に含有する新規のIGF−1及びIGF−2前駆ポリペプチドに関する。本発明のポリペプチドの有用性は、このような前駆ポリペプチドが生物学的に活性であり、安定であり、医薬品として有益であるという驚くべき発見に基づく。
【0024】
活性IGF前駆ポリペプチドについてのスクリーニング
次のアッセイを用いて本発明の何れかのポリペプチドの有用性を評価することができる。
【0025】
安定性
本発明のポリペプチドは、有効な薬物であるために、ヒト血清中など、内在性プロテアーゼ存在下で十分な安定性を有しなければならない。安定性を評価するために、10%ウシ胎仔血清、100U/mLペニシリン及び100μg/mLストレプトマイシンを含有するDMEM培地中で本ポリペプチドをコードする発現ベクターをCos7細胞(ATCC)に遺伝子移入することができる。分泌されたポリペプチドを含有する培地をさらなる分析に供する、又は、代って、この発現ベクターは、Cos7培養において発現されたポリペプチドの効率的な精製を促進するために、ポリペプチド中でヘキサ−ヒスチジンタグなどの容易に利用可能なタグをコードすることができる。いかに調製しても、ポリペプチド試料を正常ヒト血清(Sigma)又はPBS中で様々な時間(例えば、0、1、5、10及び16時間)温置し、ポリアクリルアミドゲル電気泳動に供し、ニトロセルロース上にブロットし、ヒトIGF−1又はIGF−2に対する一次抗体及び二次抗体(例えばホースラディッシュペルオキシダーゼ標識されたもの)を用いて関連のあるタンパク質を可視化する。いくつもの同様のブロッティング及び検出技術(蛍光色素又は放射性核種を使用するものものある。)を使用することができる。IGF−1又はIGF−2バンドの強度に対する前駆体バンドの強度から、様々な条件下で前駆ポリペプチドが切断される度合いが示されるはずである。本発明のポリペプチドが37℃で16時間ヒト血清に曝露されると、切断された成熟IGFに対する非切断前駆体の比が約1:2から1:0.1、例えば約1:1から1:0.5、特に約1:1の比又は約1:0.5の比となり得る。
【0026】
AKTリン酸化
本発明のポリペプチドは、IGF−1受容体を通じて信号を送る能力を維持しなければならない。(IGF−1及びIGF−2の両者ともIGF−1受容体を通じて信号を送る。)。このシグナリング能を調べるために、下流の細胞内標的であるAKTが細胞表面でのリガンド結合に反応してリン酸化されるか否かを評価することができる。AKTリン酸化の分析のために、血清不含培地中でC2C12筋芽細胞を飢餓状態にし、次いで、様々なリガンドで刺激する。細胞を溶解し、遠心により透明化する。PathScan phospho AKT(Ser473)サンドイッチELISAキット及びPathScan AKTサンドイッチELISAキット(Cell Signaling)をそれぞれ用いて、ELISAによって、AKTリン酸化及び総AKTレベルを分析する。
【0027】
IGF−1受容体特異性
本発明のポリペプチドは、好ましくは、IGF−1受容体に対して特異性を維持し、関連するインスリン受容体への結合は低親和性でなければならない。受容体特異性を評価するために、IGF−1受容体又はインスリン受容体を過剰発現する血清飢餓状態のNIH3T3細胞にポリペプチド試料を添加し、細胞を溶解し、DuoSet ICヒト蛍光体−IGF−1受容体及びインスリン受容体ELISAキット(R&D Systems)を用いたELISAに溶解液を供することによって、IGF−1受容体リン酸化又はインスリン受容体リン酸化のレベルを調べる。
【0028】
肥大のマウスモデルにおけるインビボ試験
既に筋肉肥大が誘発されている状況下で本発明のポリペプチドが骨格筋量を増加させるように作用し得るか否かを調べるために、処置及び未処置動物を運動させ、本ポリペプチドを投与された動物の筋肉が未処置動物よりも大きくなるか否かを調べることができる。
【0029】
運動モデル
当技術分野で公知のあるモデルは、使用者が荷重を変化させることができる自発運動用の回し車の使用に基づく(例えば、Konhilasら、Am J Physiol Heart Circ Physiol 289:H455−H465、2005を参照。)。自発運動用かご型車輪は、強制運動モデルに共通する心身の損傷を排除し、従って、筋肉量の増加が所望される比較的健康な個体において使用される候補薬物を評価するのにより適している。
【0030】
何れかの適切なマウス系統を使用することができる。例えば、雄C57B1/6Jマウスを無作為に実験(例えばIGF前駆ポリペプチドを投与される。)及び対照群に振り分けることができる。運動用回し車を含むケージに動物を1匹ずつ収容し;あまり運動しない対照動物を回し車のない同一ケージに収容する。運動用回し車は、Allenら、J Appl Physiol 90:1900−1908、2001に記載されている。簡潔に述べると、このシステムは、車輪の回転により起動されるデジタル電磁カウンタ(モデルBC600、Sigma Sport、Olney、IL)を備えた5.0cmの幅のランニング面のある11.5cmの直径の車輪(モデル6208、Petsmart、Phoenix、AZ)からなる。さらに、各車輪は、荷重の調整ができる抵抗機構により作製されている。これは、ステンレススチールの釣り糸をケージの上に連結し、車輪荷重に関与しないように、回転軸でケージ車輪に固定された固定滑車の周りにワイヤを巻きつけることにより行われる。このワイヤを再びスプリングと螺子でケージの上に固定する。この設計によって、車輪荷重を細かく調整することが可能となり、車輪荷重が車輪の回転全体に均一に分布される。運動期間の持続時間を通じて、各運動動物に対して、走った時間及び距離について毎日の運動値を記録する。全動物に水及び標準的な硬いげっ歯類用の餌を随意に給餌した。自発運動(かご型車輪を与えるもの)は、全群に対して約12週齢の平均週齢で開始することができる。各群は、50日間、動物が約19週齢になるまで、実験群に依存して、様々な抵抗下で走行を続ける。車輪が僅かに動くまで車輪に既知の重量を吊り下げることにより、車輪の荷重を決定する。全ての運動群は、最初の週、かご型車輪に荷重をかけずに開始する。しかし、「荷重なし」状態は実際には2gであり、これは、車輪内部及び摩擦荷重を維持するのに必要な荷重として測定される。1週間の車輪順化期間を考慮し、より大きい荷重の場合(この場合は2週間後に変化させることができる。)を除き1週間間隔で車輪荷重を変化させることができる。荷重の範囲は、2gから12gの何れかであり得る。特定の運動期間の終了直後に吸入麻酔下での頸椎脱臼によって運動及び静止対照動物を安楽死させる。体重を測定し、特定の筋肉を迅速に切り出し、洗浄し、後日の組織学的又は生化学的アッセイのために凍結する。
【0031】
別の運動肥大モデルも当業者にとって利用可能である。例えば、Lermanら、J Appl Physiol 92:2245−2255、2002に記載のトレッドミル運動モデルを参照のこと。
【0032】
クレンブテロール注入モデル
クレンブテロールは、筋肉量が増加することが確認されている増殖促進特性を有するβ2−アドレナリン作動性アゴニストである。クレンブテロール作用の正確な作用機構は不明であるが、筋肉タンパク質分解の低下が提案されている。病院において、クレンブテロールは抗喘息薬として使用されているが、殆ど、ヒト及びショー動物の両方において筋肉量を増加させるための筋肉増強剤として誤用されていると思われる。
【0033】
筋肉肥大を誘発するために、3、7又は14日間、5匹のマウスに毎日、クレンブテロール(3mg/kg、皮下(s.c.))を注射する。PBSを注射したマウスを陰性対照とする。治療に対する何らかの有害反応(即ち、毛並みの悪さ、嗜眠性)について動物を毎日監視する(目視)。クレンブテロール処置は、マウスがより恐怖を感じるか又は攻撃的になる可能性があるので、群で飼育する場合、特に闘争についてマウスを監視すべきである。マウスは動くことができ、普通に飲食可能である。第3、7又は14日に安楽死させるまでマウスを毎日監視し、さらなる分析のために組織を回収する。
【0034】
筋肉萎縮モデルにおけるインビボ試験
様々な骨格筋萎縮モデルにおいて、通常筋肉量が低下する条件下で筋肉量を維持する能力について本発明のIGF前駆ポリペプチドを試験することができる。IGF前駆ポリペプチドが筋肉量を増加させることができるか否かを調べるために、下記に記載の実施例モデルにを用いて、当業者は、IGF前駆ポリペプチドの投与及び使用を含む制御実験を容易に設計し、実施することができる。
【0035】
例えば、C57B16/2雄マウスをThe Jackson Laboratoriesから購入する。各実験の開始時にマウスが約9週齢となるようにマウスを購入する。一般に、通常のげっ歯類用飼料付きのマイクロアイソレーターケージにマウスを収容する。各実験の開始時にマウスの体重を測定する。各実験の終了時に、通常、CO2吸入の後に頸椎脱臼を行い、マウスを安楽死させ、さらなる処理のために筋肉組織を回収する。「最後の体重」を得るためにマウスの体重を測定する。回収することができる骨格筋は、前脛骨筋、長趾伸筋、ヒラメ筋及び腓腹筋である。場合によって回収する他の組織は、心臓、肝臓、脾臓、腎臓、精巣及び脳である。全ての筋肉及び組織を完全に切り出し、0.0001gまで測定可能な秤で重量を測定する。次に、後のRNA及びタンパク質抽出のために組織を液体窒素中で瞬間凍結するか又はコルクディスク上でOCT中で瞬間凍結物を包埋する。後の凍結切片作製のためにコルクディスク上で凍結させた筋肉を、液体窒素により濃厚なスラッシュ状になるまで冷却したイソペンタンに浸漬する。全試料を−80℃で保存する。
【0036】
デキサメタゾン処置
マウスにおいて筋肉萎縮を誘導する薬理学的方法は、毎日デキサメタゾン20mg/kgを腹腔内投与することである。デキサメタゾンは、ホルモンの糖質コルチコイドクラスの合成メンバーである。これは、抗炎症剤及び免疫抑制剤として作用し、ヒドロコルチゾンの約40倍の効力を有する。例えば、関節リウマチなど、多くの炎症及び自己免疫状態を治療するためにデキサメタゾンが使用される。これはまた、抗腫瘍治療のある種の副作用を軽減するために、化学治療を行っている癌患者にも投与される。デキサメタゾンは、マウス及びヒト患者の両方において筋肉萎縮を引き起こす。
【0037】
3、7又は14日間、マウスにデキサメタゾンを腹腔内(ip)注入する。最終日に、CO2を用いて被験動物を安楽死させ、足の筋肉を回収する。治療に対する何らかの有害反応(即ち、毛並みの悪さ、嗜眠性)について動物を毎日監視する(目視)。マウスは通常動くことができ、普通に飲食可能である。PBSを注入したマウスが陰性対照である。
【0038】
ギブス固定
様々な筋肉群を物理的に非活動性にした結果、これらの筋肉の萎縮が起こる。足首関節固定(「ピンヒール」又はギブス)は、ラット及びマウスの後肢筋肉組織の身体拘束を誘発するのに非常に有用で再現性のある方法であることが分かっている。
【0039】
固定化のためにイソフルオランでマウスに麻酔をかける。関節の周囲に軽量ギブス材料(VET−LITE)を用いて足首及び膝関節を90°に固定する。この材料を温水に浸し、次いで肢周囲に巻き付け、踵及び股関節は自由にする。ギブス材料が乾燥するまで関節を90°の位置に維持する。反対側の肢を対照とする。次に、マウスを麻酔から回復させ、普通のマイクロアイソレーターケージに収容する。観察によるとギブスは過剰ストレスを引き起こさず、動物はケージ中を自由に動き回り、飲食する。しかし、体重、活動性及び過敏性に影響を与える何らかの有害反応についてマウスを毎日監視する。
【0040】
マウスにギブスを嵌めたら、咀嚼する可能性があるので、ギブスがずれていないことを確認するためにその動物を毎日監視する。麻酔の回復後、動物は、動き、飲食することができ、特別なベッディング、ケージ収容又は他の支援は必要ない。
【0041】
除神経
一般に、除神経のためにイソフルオランガスでマウスに麻酔をかける。無菌の手術手順を用いて(ベタジン洗浄を3回行い、最後の洗浄はエタノール)、右坐骨神経を大腿の中ほどで切り離し、2から5mmの小片を切り出す。反対側の肢を対照とする。
【0042】
より具体的には、クリップ縫合で皮膚切開を閉じ、麻酔から回復させる前に動物にブプレノルフィンを1回投与する。手術から3、7又は14日後、CO2吸入後に頸椎脱臼を行い動物を安楽死させ、組織学的及び生化学的分析のために筋肉(腓腹筋複合体、前脛骨筋、長趾伸筋、ヒラメ筋)を取り出す。
【0043】
坐骨神経を横に切断する場合、関与する筋肉の骨格筋萎縮を誘発するために、手術を行った肢を固定化する。他は、麻酔の回復後、動物は動き、飲み、食べることができ、特別なベッディング、ケージ収容又は他の支援は必要ない。
【0044】
しかし、手術直後及び回復期(1−2時間)を通じて、動物を監視する。さらに、切開部位及び一般的な動物の健康状態を手術後3日間監視する。手術後7−10日後にクリップ縫合を除去する。
【0045】
遺伝学的モデル
筋肉萎縮のモデルとして、遺伝子操作したトランスジェニックマウスを使用することもできる。例えば、いわゆるMini Mice(The Jackson Laboratory、ストック番号003258)は、IGF−1遺伝子においてノックアウト変異があり、その結果、出生後の成長が異常に低下し、体重及び身長が低い。さらなる情報に関しては、Powell−Braxton、Genes Dev 7:2609−2617、1993を参照のこと。さらに、いわゆるMidi Mice(The Jackson Laboratory、ストック番号003259)は、IGF−1遺伝子において異なる変異があり、その結果、成獣で低体重となり、他の心血管表現型を示す矮小体型となる。さらなる情報に関しては、Lemboら、J Clin Invest 98:2648−2655、1996を参照のこと。
【0046】
IGF前駆体における重要及び最適な変異又は修飾
重要な変異
本発明は、一部分において、そのE−ペプチドを実質的に含有するIGF前駆ポリペプチドが生物活性を有し、血清の存在下で安定であるという観察に基づく。二塩基性のプロテアーゼ部位を標的とする内在性プロテアーゼによりE−ペプチドが切断されないことを確認するために、一般に、前駆体のE−ペプチドの2個のN末端二塩基性アミノ酸の何れかを欠失させるか、変異させる又は隠蔽する。IGF−1のある例において、これらの2個のアミノ酸はR71及びS72であり、一方、IGF−2の別の例において、これらの第一の2個のアミノ酸はR68及びD69である。
【0047】
様々な修飾によってこの切断の妨害が可能となる:
(1)1又は両方の二塩基性残基の欠失
(2)1又は両方の二塩基性残基の、アラニンなどの非塩基性アミノ酸への変異
(3)二塩基性残基の間への1以上の非塩基性アミノ酸の挿入
(4)プロテアーゼ部位を隠蔽するのに二塩基性残基に十分近い位置にグリコシル化部位を置くこと
(5)下記に記載のような、非天然アミノ酸による、何れかの二塩基性残基の置換又は二塩基性残基付近もしくはその間への挿入を用いた部位特異的PEG化。
【0048】
さらに、残基K68及びK65は、IGF−1/E−ペプチド切断に関与すると考えられ、従って、上述のような二塩基性アミノ酸に対する何らかの方策にこれらの残基の変異又は欠失を組み込むことができる。
【0049】
成熟IGFのN末端での変異
本発明のある実施形態において、IGF前駆ポリペプチドは、最初の数個のN末端アミノ酸の欠失又は変異を有する。IGF−1の場合、最初の3個のN末端アミノ酸の何れかを欠失させる又は変異させることができ、一方、IGF−2の場合、最初の6個のN末端アミノ酸の何れかを欠失させる又は変異させることができる。ある種のN末端アミノ酸がインビボで天然に切断されることが観察されており、これらの変異又は欠失を導入することによって、IGF結合タンパク質(IGFBP)との本発明のポリペプチドのインビボでの会合が最小限に抑えられる。IGF−1及びIGF−2のIGF−1受容体との相互作用はIGFBPにより制御される。全ての6個のIGFBPが、IGF作用(特にIGFBP5)を阻害することが示されているが、いくつかの例においては促進作用が観察されている。循環中のIGFの少なくとも99%が通常はIGFBPに結合している。新生仔期間後、循環中で最も豊富なIGFBPはIGFBP3であり、これは同様の親和性でIGF−1及びIGF−2の両方に結合できる。天然の短縮型IGF−1(G1、G2及びE3が欠失している。)は、IGFBP3に結合するが、天然のIGF−1よりも数倍親和性が低い。さらに、G3はIGFBP結合に重要であり、G6はIGF−2ペプチドにおいて同様の役割を果たす。
【0050】
従って、IGF−1前駆体のある実施例において、G1、P2又はE3の何れかを欠失させる又は変異させる(何れかのみ又は組み合わせて)ことができる。変異が所望される場合、アラニンへの変異を導入することができる。別の実施例において、IGF−2前駆体の別の例で、P4、S5及びE6の何れかを欠失させる又は変異させる(何れかのみ又は組み合わせて)ことができる。変異が所望される場合、アラニンへの変異を導入することができる。
【0051】
残基36及び37での変異
IGF−1をセリンプロテアーゼで切断することができる。R36又はR37の何れかのAへの変異によって、R36とR37との間のこの予想される切断部位におけるIGF−1の切断を妨げることができる。IGF−2のある例において、この有害な切断を妨げるために、R38を変異させる又は欠失させることができる。
【0052】
グリコシル化の使用
哺乳動物又はN−結合型グリコシル化が可能な他の真核細胞で発現される場合、IGF又は前駆体のE−ペプチド部分の何れかにN−結合型グリコシル化部位を付加することによって、本発明のポリペプチドのインビボ半減期を延長させることができる。Eaのこれらの部分は、N−X−S/T(式中、Xは何れかのアミノ酸であり得、この3残基の第三のアミノ酸はS又はTの何れかである。)のコンセンサスN−結合型グリコシル化配列に適合するので、インビトロで、IGF−1EaがN92及びN100でグリコシル化されることが分かっている。また、コンセンサスの隣接アミノ酸の状況が、アスパラギンがどの程度強くグリコシル化されるかに影響することも分かっている。従って、Eb又はEcにグリコシル化部位を導入するためのあるストラテジーは、Eb又はEcのほぼ同じ部分にコンセンサス配列周囲のEaアミノ酸を挿入することである。このストラテジーの特定の実施を下記実施例で例示する。何らかの事象において、当業者にとって公知の周囲環境のアミノ酸を含む何れかの他のコンセンサスN−結合型グリコシル化部位を本発明の前駆ポリペプチドに挿入することができる。さらに、本発明のポリペプチドの産生のために使用される特定の宿主を選択することによって、本発明のポリペプチドのO−結合型グリコシル化を行うことができる。例えば、IGF−1発現のためにある種の酵母株を使用すると、セリン又はスレオニンにおいてオリゴ糖が付加される。例えば、米国特許第5,273,966号を参照のこと。
【0053】
ポリ(エチレングリコール)の付加
ポリ(エチレングリコール)への連結(PEG;PEG化)は、治療用タンパク質薬の半減期を延長させるのに有益であることが証明されている。本発明のIGF前駆ポリペプチドのPEG化の結果、同様の医薬的長所が得られ得ると予想される。IGF−1のPEG化の方法は当技術分野で周知である。例えば、米国特許出願公開2006/0154865(これは、リジン−モノPEG化IGF−1の有用な特性について述べている。)を参照のこと。このようなリジン−モノPEG化を本発明の前駆IGFポリペプチドに適用することができる。さらに、非天然アミノ酸を導入することによって、本発明のポリペプチドの何れかの部分においてPEG化を行うことができる。Deitersら、J Am Chem Soc 125:11782−11783、2003;Wang及びSchultz、Science 301:964−967、2003;Wangら、Science 292:498−500、2001;Zhangら、Science 303:371−373、2004又は米国特許第7,083,970号に記載の技術によって、ある種の非天然アミノ酸を導入することができる。簡潔に述べるとこれらの発現系のいくつかは、本発明のポリペプチドをコードするオープンリーディングフレームに、アンバーTAGなど、ナンセンスコドンを導入するための部位特異的変異誘発を含む。次に、導入されたナンセンスコドンに特異的なtRNAを使用することができる宿主にこのような発現ベクターを導入し、最適な非天然アミノ酸を入れる。本発明のポリペプチドに対して部分を連結するために有益な特定の非天然アミノ酸には、アセチレン及びアジド側鎖を有するものが含まれる。次に、タンパク質のこれらの選択部位において、これらの新規アミノ酸を含有するIGF前駆ポリペプチドのPEG化を行うことができる。さらに、E−ペプチドがないこのようなPEG化IGF分子もまた治療薬として有用である。
【0054】
E−ペプチドのマルチマー
ある種の薬理学的関連において、薬物が血液脳関門又の一方又は他方に残ることを確実にするためにペプチド又はタンパク質のサイズを大きくすることは有益である。E−ペプチドが結合されたままである場合でも成熟IGF分子は比較的短いペプチドなので、本発明のポリペプチドのサイズを大きくすることは有益であり得る。これを行うある手段は、下記のある実施例で例示されるように、IGF前駆ポリペプチドのC末端においてE−ペプチドのマルチマーを提供することである。
【0055】
E−ペプチドのC末端の欠失
Ebの位置81のフリーのシステインによって、ホモ二量体形成又は、本発明のポリペプチドに存在する場合、薬物の活性が低下し得る他の影響が起こり得ることが疑われる。このように、EbのC81の欠失又は変異によって、薬物活性を最適化することができる。特定の例において、Ebの最後の7個のアミノ酸(即ちアミノ酸81−87)の欠失は有益である。
【0056】
他の変異又は修飾
本発明のIGF前駆ポリペプチドに組み込まれ得るIGFのさらなる変異又は修飾は米国特許第5,077,216号;及び米国特許出願公開2005/0287151、2006/0211606及び2006/0166328に記載されている。
【0057】
本発明は、実質的にそのE−ペプチドを含有する全ての既知及び未知の非ヒト動物前駆IGF−1又はIGF−2配列を含むことが意図され、本発明の修飾により、E−ペプチドの通常の切断が回避される又は減少する。
【0058】
使用されるべきIGFの好ましい型は、治療を受ける対象の種に依存する。
【0059】
IGFは種が適合することが好ましく、例えば、ウシを治療する場合、IGFの好ましい型はウシIGFである。
【0060】
配列相同性が高いためIGFの全ての型が様々な対象において有効であると思われるが、種が合致することにより、異種由来のIGFに対する免疫反応の誘導から引き起こされる有害な免疫学的合併症の可能性が回避される。
【0061】
本発明のある実施形態において、修飾非ヒト動物前駆IGF−1配列が提供される。
【0062】
E−ペプチドの通常の切断が脊椎動物由来の本発明の修飾により回避又は減少される、E−ペプチドを実質的に含有する前駆IGF−1配列が好ましい。
【0063】
例えば、このような配列には、以下に限定されないが、天然、合成又は組み換えの何れかの何らかのソースからの、マウス、ラット、ウシ、ブタ、ウマ、ヒツジ、ヤギ、鳥類、イヌ、ネコ、魚類など由来の配列が含まれる。
【0064】
本発明の別の実施形態において、修飾非ヒト動物前駆IGF−2配列が提供される。
【0065】
そのE−ペプチドの正常な切断が脊椎動物由来の本発明の修飾により回避されるか又は減少する、E−ペプチドを実質的に含有する前駆IGF−2配列が好ましい。
【0066】
例えば、このような配列には、以下に限定されないが、天然、合成又は組み換えの何れかの何らかのソースからの、マウス、ラット、ウシ、ブタ、ウマ、ヒツジ、ヤギ、鳥類、イヌ、ネコ、魚類など由来の配列が含まれる。
【0067】
IGF前駆ポリペプチドの治療用途
適応症例
本発明はまた、筋骨格疾患の治療又は予防用の医薬品の製造における本発明のIGF前駆ポリペプチドの使用も含む。さらに、本発明は、このような個体が筋骨格系疾患のリスクを有する又はその疾患に罹患しているにせよそうでないにせよ、個体において筋肉又は骨量を増加させるためのIGF前駆ポリペプチドの使用を含む。
【0068】
特に、筋骨格系疾患は筋萎縮であり得る。コルチゾール、デキサメタゾン、ベタメタゾン、プレドニゾン、メチルプレドニゾロン又はプレドニゾロンなどの糖質コルチコイドでの治療によるものを含め、筋萎縮には多くの原因がある。筋萎縮はまた、神経外傷性傷害による除神経の結果又は変性、代謝性もしくは炎症性ニューロパシーの結果(例えば、ギリアン−バレー症候群、末梢性ニューロパシー又は環境有害物質もしくは薬物への曝露)でもあり得る。さらに、筋萎縮は、成人の運動ニューロン疾患、小児脊髄性筋萎縮症、若年性脊髄性筋萎縮症、多発性導体ブロックを伴う自己免疫性運動神経障害、発作又は脊髄損傷による麻痺、外傷性傷害による骨格の不動化、長期にわたる床上安静、故意の不活発、不随意の不活発、代謝性ストレス又は栄養失調、癌、AIDS、飢餓、横紋筋融解症、甲状腺疾患、糖尿病、良性先天性低血圧、セントラルコア病、ネマリンミオパシー、筋管(中心核)ミオパシー、熱傷、慢性閉塞性肺疾患、肝臓疾患、敗血症、腎不全、うっ血性心不全又は加齢の結果であり得る。
【0069】
筋骨格系疾患はまた、デュシェンヌ型、ベッカー型、筋強直性、顔面肩甲上腕筋、エメリー・ドレフュス型、眼咽頭筋、肩甲上腕、肢帯、先天型筋ジストロフィー又は遺伝性末梢性ミオパシーなどの筋ジストロフィー症候群でもあり得る。筋骨格系疾患はまた、骨粗しょう症、骨折、低身長又は小人症でもあり得る。
【0070】
IGF−1はまたIGF−1受容体及びインスリン受容体のヘテロ二量体と結合することもできるので、IGF−1は、インスリン非依存性糖尿病の治療薬として示唆されている。従って、糖尿病を治療するために本発明のポリペプチドを使用することができる。
【0071】
IGF−1は、神経栄養性であり、ニューロンの生存を促進する。筋萎縮性側索硬化症(ALS)、脳萎縮、加齢及び認知症でみられるような、運動ニューロン死の症例を治療するためにIGF−1を使用することができることが示唆されている。従って、ALS、脳萎縮又は認知症など、ニューロン死と関連する状態を治療するために本発明のポリペプチドを使用することができる。
【0072】
IGF−1は、白血球及び赤血球細胞集団両方を増加させ、エリスロポエチンの投与に対して付加的効果がある。従って、貧血を治療するために、本発明のポリペプチドを使用することができる。
【0073】
IGF−1及びIGF−2はユビキタスであり、細胞分裂及び脊椎動物の成長の必須の制御因子なので、動物における成長を外部から促進又は維持するために様々な獣医学的方法においてこれらを有利に使用し得る。ある一部の例には、以下に限定されないが、
(i)動物における成長の速度及び/又は程度を促進すること(例えば、ブタ、ウシ、家禽及び魚類において筋肉成長を促進すること)、
(ii)例えばブタ、ウシ、家禽及び魚類において、食物の身体組織への変換の効率を促進すること(脂肪に対するやせの割合)及び
(iii)泌乳動物、例えば、乳牛、ヒツジ、ヤギなどにおいて乳汁産生を促進することが含まれる。他の獣医学的治療適用には、以下に限定されないが、
(iv)例えばイヌ、ネコ及びウマなどのペット動物において、悪液質、外傷性傷害又は他の消耗疾患に付随する消耗性の症状に対して動物を治療すること及び
(v)新生仔の状態を向上させるために例えば泌乳雌ブタにおいて、新生仔の健康を増進するために泌乳動物を処置することが含まれる。
【0074】
投与方法
遺伝子送達ビヒクルの使用を含む様々な方法において本発明のポリペプチドを送達することができる。本発明のポリペプチドの治療のための送達、例えば、細胞の遺伝子移入、遺伝子治療、送達ビヒクル又は医薬的に許容可能な担体による直接投与、本ポリペプチドをコードする核酸を含有する組み換え細胞を提供することによる間接的送達のために、タンパク質又は核酸などの物質の治療的送達のための当技術分野で公知の方法を使用することができる。
【0075】
様々な送達系が公知であり、本発明のポリペプチドを投与するために、例えばリポソーム中でのカプセル封入、微小粒子、マイクロカプセル、タンパク質を発現させることができる組み換え細胞、受容体介在エンドサイトーシス(例えば、Wu及びWu、J Biol Chem 262:4429−4432、1987)、レトロウイルス、アデノ関連ウイルス、アデノウイルス、ポックスウイルス(例えば、アビポックスウイルス、特に鶏痘ウイルス)又は他のベクターなどの一部としての核酸のコンストラクトなどを使用することができる。導入方法は、腸内又は非経口であり得、以下に限定されないが、皮内、筋肉内、腹腔内、静脈内、皮下、肺、鼻内、眼内、硬膜外及び経口経路が含まれる。何れかの従来の経路により、例えば点滴又はボーラス注射により、上皮又は皮膚粘膜層(例えば、口腔粘膜、直腸及び腸粘膜など)を通じた吸収により本ポリペプチドを投与することができ、他の生物学的に活性のある物質とともに投与し得る。投与は全身的又は局所であり得る。さらに、脳室内及び鞘内注入を含む何らかの適切な経路により、中枢神経系に本発明の医薬組成物を導入することが所望され得;例えば、Ommayaリザーバーなどのリザーバーに連結した脳室内カテーテルにより脳室内注射を促進し得る。例えば、吸入器又はネブライザー及びエアロゾル化剤による処方物の使用により、肺投与を使用することもできる。
【0076】
具体的な実施形態において、治療を必要とする部位に局所的に本発明の医薬組成物を投与することが所望され得;例えば、限定されないが、術中の局所点滴、局所適用により、例えば注射によって、カテーテルによって、又はインプラントによって、多孔、非多孔又はゼラチン性材料であるインプラント(シアラスティック(sialastic)膜などの膜を含む。)、繊維又は市販の皮膚代替物によって、これを遂行し得る。
【0077】
別の実施形態において、小胞、特にリポソームにおいて、活性物質を送達することができる(Langer、Science 249:1527−1533、1990参照)。さらに別の実施形態において、制御放出系において活性物質を送達することができる。ある実施形態において、ポンプを使用し得る。別の実施形態において、ポリマー材料を使用することができる(Howardら、J Neurosurg 71:105、1989)。別の実施形態において、本発明の活性物質が本発明のポリペプチドをコードする核酸である場合、適切な核酸発現ベクターの一部としてそれを構築すること及び、例えばレトロウイルスベクターを使用することによって又は直接注入によって又は微小粒子ボンバーメント(例えば遺伝子銃;Biolistic、Dupont)によって又は脂質もしくは細胞表面受容体での被覆又は遺伝子移入剤によって又は核に入ることが知られているホメオボックス様ペプチドへ連結してそれを投与することによって(例えば、Joliotら、Proc.Natl.Acad.Sci.USA 88:1864−1868、1991)など、細胞内に入るようにそれを投与することによって、そのコードされるタンパク質の発現を促進するためにインビボで核酸を投与することができる。あるいは、細胞内に核酸を導入し、相同組み換えによって、発現のために宿主細胞DNA内に組み込むことができる。
【0078】
細胞の遺伝子移入及び遺伝子治療
本発明は、インビトロ及びインビボでの細胞の遺伝子移入のための本発明のポリペプチドをコードする核酸の使用を包含する。標的細胞及び生物の遺伝子移入のために、多くの周知のベクターの何れかにこれらの核酸を挿入することができる。ベクター及び標的細胞の相互作用を通じて、エクスビボ及びインビボで細胞に核酸を遺伝子移入する。治療反応を誘発するのに十分な量で対象に本組成物を(例えば筋肉への注射により)投与する。
【0079】
別の態様において、本発明は、標的部位、即ち標的細胞又は組織、動物を処置する方法(本発明のポリペプチドをコードする核酸を細胞に遺伝子移入することを含む。)を提供するが、この核酸は、標的融合ポリペプチドをコードする核酸に操作可能に連結された誘導可能なプロモーターを含む。
【0080】
併用療法
多くの実施形態において、1以上のさらなる化合物又は治療と組み合わせて本発明のポリペプチドを投与することができる。例えば、1以上の治療用化合物とともに複数のポリペプチドを同時投与することができる。併用療法は、同時に行うか又は交互に行う投与を包含し得る。さらに、この組み合わせは、急性投与は長期投与を包含し得る。テストステロン又は特異的アンドロゲン受容体調節因子(SARM)などの同化剤と組み合わせて本発明のポリペプチドを投与することができる。さらなる同化剤には、成長ホルモン(GH)又は、GH放出を誘導する分子が含まれる。グレリンは食欲を増進させ得るので、悪液質に対する併用療法においてグレリンは特に有用である。同様に、同化作用を向上させるために本発明のポリペプチドをタンパク質サプリメントと組み合わせることができるか又は体重を増加させるために理学療法又は運動と組み合わせることができる。ミオスタチンを阻害する何らかの分子も併用療法に対する候補である。
【0081】
医薬組成物
本発明はまた、本発明のIGF前駆タンパク質及び医薬的に許容可能な担体を含む医薬組成物も提供する。「医薬的に許容可能な」という用語は、連邦もしくは州政府の監督官庁により承認されている又は米国薬局方又は動物もしくはヒトでの使用に関する他の一般に認められている薬局方に列挙されていることを意味する。「担体」という用語は、治療薬と一緒に投与される、希釈剤、アジュバント、賦形剤又はビヒクルを指す。このような医薬担体は、水及び油(石油、動物、植物又は合成起源の油を含む。ピーナツ油、ダイズ油、鉱物油、ゴマ油など)などの滅菌液であり得る。適切な医薬賦形剤には、デンプン、グルコース、ラクトース、スクロース、ゼラチン、麦芽、米、小麦粉、チョーク、シリカゲル、ステアリン酸ナトリウム、モノステアリン酸グリセロール、タルク、塩化ナトリウム、脱脂粉乳、グリセロール、プロピレン、グリコール、水、エタノールなどが含まれる。必要に応じて、本組成物はまた、少量の湿潤もしくは乳化剤又はpH緩衝剤も含有し得る。これらの組成物は、溶液、懸濁液、乳剤、錠剤、丸剤、カプセル、粉末、徐放製剤などの形態を取り得る。伝統的な結合剤及びトリグリセリドなどの担体とともに坐薬として本組成物を処方することができる。経口製剤は、医薬グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなどの標準的担体を含み得る。適切な医薬担体の例は、E.W.Martinによる「Remington’s Pharmaceutical Sciences」に記載されている。
【0082】
いくつかの実施形態において、静脈内投与に適した医薬組成物として常法に従い本組成物を処方する。必要な場合、本組成物は、可溶化剤及び注射部位の痛みを緩和するためのリドカインなどの局所麻酔も含み得る。本組成物を点滴により投与すべき場合、医薬グレードの滅菌水又は食塩水を含有する点滴ボトルを用いて投与することができる。本組成物を注射により投与すべき場合、投与前に成分を混合することができるように、注射用滅菌水又は食塩水のアンプルを提供し得る。
【0083】
中性又は塩の形態で本発明のポリペプチドを処方することができる。医薬的に許容可能な塩には、例えば塩酸、リン酸、酢酸、シュウ酸、酒石酸など由来ものなどの遊離アミノ基とともに形成されるもの、ナトリウム、カリウム、アンモニウム、カルシウム、水酸化鉄、イソプロピルアミン、トリエチルアミン、2−エチルアミノエタノール、ヒスチジン、プロカインなど由来のものなどの遊離カルボキシル基とともに形成されるものなどが含まれる。
【0084】
現在の記述に基づく標準的な臨床技術により、状態又は疾病の治療に有効である本発明のポリペプチドの量を決定することができる。さらに、最適投与量範囲の決定を促進するためにインビトロアッセイを場合によっては使用し得る。処方において使用すべき正確な用量もまた、投与経路及び状態の重症度に依存し、医師の判断及び各対象の事情に従い決定すべきである。しかし、静脈内投与のための適切な投与量範囲は通常、体重1kgあたり活性化合物約20−5000mgである。鼻内投与のための適切な投与量範囲は通常、約0.01pg/kg体重から1mg/kg体重である。有効用量は、インビトロ又は動物モデル試験系から得られた用量反応曲線から推定し得る。特に、可能性がある用法用量は、約60から120μg/kg体重で、皮下注射で1日2回であり得る。
【0085】
動物への使用
健康な動物に投与する場合、疾患に罹患している動物と投与量が異なり得る。例えば下記のような筋芽細胞増殖アッセイ(実施例79)又は哺乳動物上皮組織アッセイ(実施例80)など、当技術分野で公知のアッセイを用いて、適切な投与量の評価が当業者により容易に行われ得る。IGFを測定するための一般的なアッセイも実施例81におけるものなど、当技術分野で公知である。
【0086】
当業者にとって当然のことながら、ある種の動物は、光周期の長さにより影響を受ける季節性の妊孕性を示す。獣医学における方法又は使用の何らかの実施形態は、場合によっては、所望の効果を達成するために、その動物の生殖サイクル内の特定の時間に治療法を開始することを含み得る。当業者は、生殖状態及びサイクルを容易に決定することができること、必要に応じて、適切な方法の使用により同調させることができることを知っている。
【0087】
動物への適応のために使用する場合、本発明のIGF−1又はIGF−2ペプチドを経口用ドレンチ又は動物に対する経口もしくは固形飼料に対するサプリメントとして使用することもできる。
【0088】
本発明はさらに、次の実施例を記載するが、これに限定されない。
【実施例】
【0089】
(実施例1)
次の修飾を含有するhIGF−1−Ea前駆ポリペプチドをコードするDNA発現ベクターを構築した:G1の欠失、P2の欠失及びE3の欠失;R37のAへの変異;及びR71の欠失及びS72の欠失。これらの変異を、本開示を通じて「3mut」と呼ぶことがある。この結果、次の分泌タンパク質配列が得られる:
【0090】
【化3】
【0091】
10%ウシ胎仔血清、100U/mLペニシリン及び100μg/mLストレプトマイシンを含有するDMEM中でCos7細胞(ATCCより入手可能)を維持し、10cmプレートあたり1x106個の細胞の密度で播種した。これらの細胞培養物に対して、製造者の説明書に従い、Fugene(Roche)を用いて発現プラスミド8μgを遺伝子移入した。遺伝子移入から24時間後、細胞を1度洗浄し、血清不含培地中で48時間培養した。上清を回収し、−80℃で保存した。
【0092】
ヒト血清中でのポリペプチドの安定性を評価するために、野生型(wt)hIGF−1Ea及びhIGF−1Ea3mutで遺伝子移入したCos7細胞から回収した上清を10%ヒト血清(Sigma)非存在下又は存在下の何れかで37℃で16時間温置した。18%SDS−PAGEにより試料を分離し、ヒトIGF−1に対するヤギポリクローナル抗体を用いて免疫ブロッティングを行った。図1Aの結果から、16時間、血清とともに温置後、hIGF−1Eaが実質的に分解された一方、hIGF−1Ea3mutは安定であったことが示された。密度測定から、切断されたIGF−1に対する非切断IGF−1の比が約1:6.2であり、一方hIGF−1Ea3mutの場合の比は約1:0.68であることが示され、このことから、これらの変異の結果、ポリペプチドが安定化されることが分かる。
【0093】
hIGF−1Ea3mutがIGF−1Rを通じてシグナルを送ることができることを確認するために、本ポリペプチドと接触させた細胞のAKTリン酸化を測定した。C2C12をATCCから購入し、10%ウシ胎仔血清(AMIMED)、100U/mLペニシリン(Invitrogen)、100μg/mLストレプトマイシン(Invitrogen)及び2mMグルタミン(Invitrogen)を含有する高グルコース入りのダルベッコの改変イーグル培地(DMEM)(Invitrogen)中で維持した。AKTリン酸化の分析のために、6ウェルプレートのウェルあたり0.15x106個の細胞密度でC2C12細胞を播種し、増殖培地中で72時間培養した。血清不含培地中で細胞を4時間飢餓状態にし、次いで37℃で30分間、様々なリガンドで刺激した。様々なプロテアーゼ阻害剤を含有するPhosphoSafe緩衝液(Cell Signaling)で細胞を溶解し、4℃で14,000xgで15分間遠心することにより透明化した。PathScan phospho AKT(Ser473)サンドイッチELISAキット及びPathScan AKTサンドイッチELISAキット(Cell Signaling)をそれぞれ用いてELISAによって、AKTリン酸化及び総AKTレベルを分析した。AKTリン酸化の結果を図2Aでまとめるが、これから、hIGF−1Ea3mutが、long−R3−IGF1陽性対照試薬及び組み換えIGF−1と同程度にIGF−1R細胞経路を活性化できることが示される。さらに、図5のデータから、hIGF−1Ea3mutがAKTリン酸化へと導いたことが直接示される。
【0094】
次に、hIGF−1Ea3mutの受容体特異性がIGF−1Rで残っていることを確認するために、IGF−1R又はインスリン受容体(InsR)の何れかを過剰発現するNIH3T3の培養物に様々なリガンドを添加した。Cos7細胞に対して上記で述べたものと同じ条件下でこれらの細胞を培養した。IGF−1R及びInsRリン酸化の分析のために、6ウェルプレートのウェルあたり0.2x106個の細胞密度でNIH3T3−IGF1R及びNIH3T3−InsR細胞を播種し、増殖培地中で24時間培養した。血清不含培地中で18時間細胞を飢餓状態にし、次いで37℃で10分間、様々なリガンドで刺激した。AKT実験のために上記で述べたように細胞を溶解し、DuoSet ICヒト蛍光体−IGF1R及び−InsR ELISAキット(R&D Systems)を用いてELISAによってIGF−1R及びInsRリン酸化レベルを分析した。図3A及び3Bでまとめた結果から、このIGF−1前駆ポリペプチドがIGF−1受容体に対して特異性を保持し、関連するインスリン受容体と低親和性で結合するはずであることが示される。
【0095】
(実施例2)
次の変異:G1の欠失、P2の欠失及びE3の欠失;R37のAへの変異;及びR71の欠失及びS72の欠失(即ち「3mut」)を含有するhIGF−1−Eb前駆ポリペプチドをコードするDNA発現ベクターを構築した。この結果、次の分泌タンパク質配列が得られる:
【0096】
【化4】
【0097】
上記実施例1に記載の手順に従い、このポリペプチドをアッセイした。図1B及び密度測定の使用から、切断されたIGF−1に対する非切断IGF−1の比が約1:9であり、hIGF−1Eb3mutの場合の比が約1:1であることが示され、このことから、これらの修飾の結果、ポリペプチドが安定化されることが分かった。図2Bから、hIGF−1Eb3mutがlong−R3−IGF1陽性対照試薬及び組み換えIGF−1と同程度にIGF−1R細胞経路を活性化できたことが示される。さらに、図5のデータから、hIGF−1Eb3mutがAKTリン酸化へと導いたことが直接示される。図3C及び3Dでまとめた結果から、このIGF−1前駆ポリペプチドがIGF−1受容体に対して特異性を保持しており、関連するインスリン受容体と低親和性で結合するはずであることが示される。
【0098】
(実施例3)
次の変異:G1の欠失、P2の欠失及びE3の欠失;R37のAへの変異;及びR71の欠失及びS72の欠失(即ち「3mut」)を含有するhIGF−1−Ec前駆ポリペプチドをコードするDNA発現ベクターを構築した。この結果、次の分泌タンパク質配列が得られる:
tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnkntksqrrkgstfeerk(配列番号10)
【0099】
上記実施例1に記載の手順に従い、このポリペプチドをアッセイした。図1C及び密度測定の使用から、切断されたIGF−1に対する非切断IGF−1の比が約1:5であり、hIGF−1Ec3mutの場合の比が約1:0.96であることが示され、このことから、これらの修飾の結果、ポリペプチドが安定化されることが分かった。図2Dから、hIGF−1Ec3mutがlong−R3−IGF1陽性対照試薬及び組み換えIGF−1と同程度にIGF−1R細胞経路を活性化できたことが示される。さらに、図5のデータから、hIGF−1Ec3mutがAKTリン酸化へと導いたことが直接示される。図4A及び4Bでまとめられた結果から、このIGF−1前駆ポリペプチドがIGF−1受容体に対して特異性を保持しており、関連するインスリン受容体と低親和性で結合するはずであることが示される。
【0100】
(実施例4)
hIGF−1−Ebペプチドに対して次の修飾:G1の欠失、P2の欠失及びE3の欠失;R37のAへの変異;及びR71の欠失及びS72の欠失(即ち「3mut」);及びEbのアミノ酸95と96との間のEaアミノ酸93から102の挿入を含有するhIGF−1−Eabキメラ前駆ポリペプチドをコードするDNA発現ベクターを構築した。この挿入によって、N92に推定N−結合型グリコシル化シグナルが生成する。この結果、次の分泌タンパク質配列が得られる。
【0101】
【化5】
【0102】
上記実施例1に記載の手順のいくつかに従い、このポリペプチドをアッセイした。図2Cから、hIGF−1Eab3mutがlong−R3−IGF1陽性対照試薬及び組み換えIGF−1と同程度にIGF−1R細胞経路を活性化できたことが示される。図4C及び4Dでまとめられた結果から、このIGF−1前駆ポリペプチドがIGF−1受容体に対して特異性を保持しており、インスリン受容体を活性化しないことが示される。
【0103】
(実施例5)
次の変異:G1の欠失、P2の欠失及びE3の欠失、R36の欠失、R37の欠失、R71の欠失、S72の欠失、Ebの最後の7個のC末端アミノ酸の欠失;及び、2つのさらなるEbペプチド(両方とも、R71及びS72及び最後の7個のC末端のアミノ酸がない。)及び4番目及び最後のEbペプチド(R71及びS72がない。)のこの前駆体のC末端への挿入を含有するhIGF−1−Ebマルチマー前駆ポリペプチドをコードするDNA発現ベクターを構築した。図6Aは、このコンストラクトの概略図を示す。この結果、次の分泌タンパク質配列が得られる。
【0104】
【化6】
【0105】
実施例1に記載のようにこのポリペプチドをAKTリン酸化アッセイに供した。図5から、このhIGF−1−EbマルチマーがIGF−1R経路を通じてシグナルを送ることができたことが示される。
【0106】
(実施例6)
図6Bで概略が示されるような本発明のhIGF−1−Eb前駆ポリペプチドを発現させることができる。このコンストラクトは、次の修飾:G1の欠失、P2の欠失及びE3の欠失、R36の欠失、R37の欠失、R71の欠失、S72の欠失;及びEbのアミノ酸95と96との間のEaアミノ酸93−102の挿入(これにより、位置N92及びN100でN結合型グリコシル化部位が生じる。)を含有する。この結果、次の分泌タンパク質配列が得られる。
【0107】
【化7】
【0108】
(実施例7)
次の修飾:P4の欠失、S5の欠失及びE6の欠失;R38のAへの変異;及びR68の欠失及びD69の欠失を有する本発明のhIGF−2−E前駆ポリペプチドを発現させることができる。この結果、次の分泌タンパク質配列が得られる。
【0109】
【化8】
【0110】
(実施例8)
次の変異:G1の欠失及びP2の欠失;E3のXへの変異(Xは、PEG化された非天然アミノ酸である。);R37のAへの変異;及びR71の欠失及びS72の欠失を有する本発明のhIGF−1−Ea前駆ポリペプチドを発現させることができる。この結果、次の分泌タンパク質配列が得られる。
【0111】
【化9】
【0112】
(実施例9−78)
(△=欠失)
9)hIGF−1−Ea:△G1、△P2、△E3;R36A;△R71
【0113】
【化10】
【0114】
10)hIGF−1−Ea:△G1、△P2、△E3;R36A;△S72
【0115】
【化11】
【0116】
10)hIGF−1−Ea:△G1、△P2、△E3;R36A;△R71、△S72
【0117】
【化12】
【0118】
11)hIGF−1−Ea:△G1、△P2、△E3;R37A;△R71
【0119】
【化13】
【0120】
12)hIGF−1−Ea:△G1、△P2、△E3;R37A;△S72
【0121】
【化14】
【0122】
13)hIGF−1−Ea:△G1、△P2、△E3、△R37;△R71
【0123】
【化15】
【0124】
14)hIGF−1−Ea:△G1、△P2、△E3、△R37;△S72
【0125】
【化16】
【0126】
15)hIGF−1−Ea:△G1、△P2、△E3;△R37;△R71、△S72
【0127】
【化17】
【0128】
16)hIGF−1−Eb:△G1、△P2、△E3;R36A;△R71
【0129】
【化18】
【0130】
17)hIGF−1−Eb:△G1、△P2、△E3;R36A;△S72
【0131】
【化19】
【0132】
18)hIGF−1−Eb:△G1、△P2、△E3;R37A;△R71
【0133】
【化20】
【0134】
19)hIGF−1−Eb:△G1、△P2、△E3;R37A;△S72
【0135】
【化21】
【0136】
20)hIGF−1−Eb:△G1、△P2、△E3;R37A;△R71、△S72
【0137】
【化22】
【0138】
21)hIGF−1−Eb:△G1、△P2、△E3、△R37;△R71
【0139】
【化23】
【0140】
22)hIGF−1−Eb:△G1、△P2、△E3、△R37;△S72
【0141】
【化24】
【0142】
23)hIGF−1−Eb:△G1、△P2、△E3、△R37;△R71、△S72
【0143】
【化25】
【0144】
24)hIGF−1−Ec:△G1、△P2、△E3;R36A;△R71
【0145】
【化26】
【0146】
25)hIGF−1−Ec:△G1、△P2、△E3;R36A;△S72
【0147】
【化27】
【0148】
26)hIGF−1−Ec:△G1、△P2、△E3;R36A;△R71、△S72
【0149】
【化28】
【0150】
27)hIGF−1−Ec:△G1、△P2、△E2;R37A;△R71
【0151】
【化29】
【0152】
28)hIGF−1−Ec:△G1、△P2、△E3;R37A;△S72
【0153】
【化30】
【0154】
29)hIGF−1−Ec:△G1、△P2、△E3;R37A;△R71、△S72
【0155】
【化31】
【0156】
30)hIGF−1−Ec:△G1、△P2、△E3、△R37、△R71
【0157】
【化32】
【0158】
31)hIGF−1−Ec:△G1、△P2、△E3、△R37、△S72
【0159】
【化33】
【0160】
32)hIGF−1−Eab:△G1、△P2、△E3;R36A;△R71;Ebのaa95と96との間のEa aa93−102の挿入(即ち「Eab」)
【0161】
【化34】
【0162】
33)hIGF−1−Eab:△G1、△P2、△E3;R37A;△R71
【0163】
【化35】
【0164】
34)hIGF−1−Eab:△G1、△P2、△E3、△R37、△R71
【0165】
【化36】
【0166】
35)hIGF−1−Eab:△G1、△P2、△E3;R36A;△S72
【0167】
【化37】
【0168】
36)hIGF−1−Eab:△G1、△P2、△E3;R37A;△S72
【0169】
【化38】
37)hIGF−1−Eab:△G1、△P2、△E3、△R37、△S72
【0170】
【化39】
【0171】
38)hIGF−1−Eab:△G1、△P2、△E3;R36A;△R71、△S72
【0172】
【化40】
【0173】
39)hIGF−1−Eab:△G1、△P2、△E3、△R37、△R71、△S72
【0174】
【化41】
【0175】
40)hIGF−1−Ea:△P2、△E3;R37A;△R71、△S72
【0176】
【化42】
【0177】
41)hIGF−1−Eb:△P2、△E3;R37A;△R71、△S72
【0178】
【化43】
【0179】
42)hIGF−1−Ebマルチマー:(△G1、△P2、△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0180】
【化44】
【0181】
43)hIGF−1−Ebマルチマー:(△P2、△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0182】
【化45】
【0183】
44)hIGF−1−Ec:△P2、△E3;R37A;△R71、△S72
【0184】
【化46】
【0185】
45)hIGF−1−Ea:△E3;R37A;△R71、△S72
【0186】
【化47】
【0187】
46)hIGF−1−Eb:△E3;R37A;△R71、△S72
【0188】
【化48】
【0189】
47)hIGF−1−Ebマルチマー:(△G1、△P2、△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0190】
【化49】
【0191】
48)hIGF−1−Ebマルチマー:(△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0192】
【化50】
【0193】
49)hIGF−1−Ec:△E3;R37A;△R71、△S72
【0194】
【化51】
【0195】
50)hIGF−1−Ea:△E3;R37A;△R71、△S72
【0196】
【化52】
【0197】
51)hIGF−1−Eb:△E3;R37A;△R71、△S72
【0198】
【化53】
【0199】
52)hIGF−1−Ec:△E3;R37A;△R71、△S72
【0200】
【化54】
【0201】
53)hIGF−1−Eab:△E3;R37A;△R71、△S72
【0202】
【化55】
【0203】
54)hIGF−1−Ebマルチマー:(△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0204】
【化56】
【0205】
55)hIGF−1−Ea:E3A;R37A;△R71、△S72
【0206】
【化57】
【0207】
56)hIGF−1−Eb:E3A;R37A;△R71、△S72
【0208】
【化58】
【0209】
57)hIGF−1−Ec:E3A;R37A;△R71、△S72
【0210】
【化59】
【0211】
58)hIGF−1−Eab:E3A;R37A;△R71、△S72
【0212】
【化60】
【0213】
59)hIGF−1−Ebマルチマー:(E3A;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0214】
【化61】
【0215】
60)hIGF−1−Ea:△P2、△E3;R37A;△R71、△S72
【0216】
【化62】
【0217】
61)hIGF−1−Eb:△P2、△E3;R37A;△R71、△S72
【0218】
【化63】
【0219】
62)hIGF−1−Ec:△P2、△E3;R37A;△R71、△S72
【0220】
【化64】
【0221】
63)hIGF−1−Eab:△P2、△E3;R37A;△R71、△S72
【0222】
【化65】
【0223】
64)hIGF−1−Ebマルチマー:(△P2、△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0224】
【化66】
【0225】
65)MGF−1−Eb:△G1、△P2;E3X;R37A;△R71、△S72
【0226】
【化67】
【0227】
66)hIGF−1−Ec:△G1、△P2;E3X;R37A;△R71、△S72
【0228】
【化68】
【0229】
67)hIGF−1−Eab:△G1、△P2;E3X;R37A;△R71、△S72
【0230】
【化69】
【0231】
68)hIGF−1−Ebマルチマー:(△G1、△P2;E3X;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0232】
【化70】
【0233】
69)hIGF−1−Ea:△G1、△P2、△E3;R37A;△R71;S72X
【0234】
【化71】
【0235】
70)hIGF−1−Eb:△G1、△P2、△E3;R37A;△R71;S72X
【0236】
【化72】
【0237】
71)hIGF−1−Ec:△G1、△P2、△E3;R37A;△R71;S72X
【0238】
【化73】
【0239】
72)hIGF−1−Eab:△G1、△P2、△E3;R37A;△R71;S72X
【0240】
【化74】
【0241】
73)hIGF−1−Ebマルチマー:(△G1、△P2、△E3;R37A)−Eb(△R71;S72X;△C末端7aa)−2xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0242】
【化75】
【0243】
74)hIGF−1−Ea:△G1、△P2、△E3;R37A;△R71、△S72;N92X
【0244】
【化76】
【0245】
75)hIGF−1−Eb:△G1、△P2、△E3;R37A;△R71、△S72;C142X
【0246】
【化77】
【0247】
76)hIGF−1−Eab:△G1、△P2、△E3;R37A;△R71、△S72;C151X
【0248】
【化78】
【0249】
78)hIGF−1−Ebマルチマー(△G1、△P2、△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72;C71X)
【0250】
【化79】
【0251】
(実施例79)
筋芽細胞増殖アッセイ
筋芽細胞増殖アッセイは、信頼できるIGF活性のインビトロ指標を与え、胚性筋芽細胞及び成体衛星細胞に影響を与える因子に対するモデルとして使用される。この系において活性のある因子は、筋芽細胞の初代培養において同様に作用する。本発明のペプチドによるインビトロでの筋芽細胞増殖の促進から、筋芽細胞増殖を向上させること及び、従って子宮内での最終的な筋繊維数を向上させることにおいて、その活性が示される。さらに、筋芽細胞増殖の同様の促進から、(例えば衛星筋肉細胞増殖の刺激を介して)成体の筋肉肥大を促進するために本発明のペプチドを使用することができることが示される。
【0252】
(実施例80)
哺乳動物上皮組織アッセイ
泌乳動物において、乳房上皮組織は乳汁を産生し分泌する細胞なので、この組織の量は、乳汁産生の制限要因である。インビトロ系を用いて、増殖させ乳成分の産生量を向上させるために、本発明の修飾IGF−1又はIGF−2により、動物の乳線から得られる上皮細胞を刺激することができる。さらに、あるこのようなインビトロ細胞系で増殖させるために刺激した乳房上皮細胞を、除去した乳房脂肪体に再移植し、雌泌乳動物において増殖及び/又は乳汁を産生させるために刺激することができる。
【0253】
(実施例81)
血液又は他の体液中のIGF−1又はIGF−2の測定
用量反応曲線によって投与につき非経口投与されるペプチドの有効量を測定することができる。例えば、投薬を決定するために、処置すべき対象の血液又は体液中の本発明の修飾IGFペプチドを測定することができる。あるいは、対象にペプチドの漸増量を投与し、修飾IGF−1及びIGF−2に対する対象の血清レベルを調べることができる。修飾IGF−1又はIGF−2のこれらの血清レベルに基づくモル基準で、使用すべきペプチドの量を計算することができる。
【0254】
ペプチドの適切な投薬を決定するためのある方法は、体液又は血液などの生体液中の本発明のIGFペプチドを測定することを伴う。RIA及びELISAを含む何らかの手段によって、このようなレベルの測定を行うことができる。IGFレベルを測定した後、単回投与又は複数回投与を用いて本ペプチドと生体液を接触させる。この接触段階の後、生体液中のIGFレベルを再測定する。生体液IGFレベルが、所望の効率(そのためにこの分子が投与されるべきである。)を得るのに十分な量低下した場合、最大効率が得られるようにその分子の用量を調整することができる。インビトロ又はインビボでこの方法を行い得る。好ましくは、インビボで、即ち、生体液を対象から採取した後にこの方法を行い、IGFレベルを測定し、単回又は複数回投与により本明細書中のペプチドを哺乳動物に投与し(即ち、動物に投与することによりこの接触段階を行う。)、次いで動物から採取した生体液からIGFレベルを再測定する。
【0255】
投薬を決定するための別の方法は、本ペプチドに対する抗体又はLIFA形式でのペプチドに対する別の検出方法を使用することである。
【0256】
(実施例82)
hIGF−1−Ec 3mutのインビボ薬物動態
1mg/kgのrhIGF−1及び1.55mg/kgのhIGF−1−Ec 3mut(実施例3に記載)を、成熟雄マウス(n=3/群)に静脈内(i.v.)ボーラス注射した。試験物質の投与から5、15、30及び60分後、連続血液検体を回収した。rhIGF−1及びhIGF−1−Ec 3mutの血清濃度をELISAにより測定した。このアッセイはhIGF−1に特異的である。
【0257】
rhIGF−1及びhIGF−1−Ec 3mutの等モル用量をマウスにi.v.投与した。この結果から、試験した全ての時間点で、rhIGF−1と比較して、hIGF−1−Ec 3mutタンパク質のレベルが有意に高いことが分かり、このことから、hIGF−1−Ec 3mutは70アミノ酸−long IGF−1よりも代謝的に安定であることが示される。
【0258】
【表1】
【技術分野】
【0001】
(背景技術)
インスリン様増殖因子(IGF)は、細胞がこれら細胞の生理的環境と情報伝達をするために使用する複合系の一部である。この複合系(インスリン様増殖因子軸と呼ばれることが多い。)は、2つの細胞表面受容体(IGF−1R及びIGF−2R)、2つのリガンド(IGF−1及びIGF−2)、6個の高親和性IGF結合タンパク質のファミリー(IGFBP1−6)及び関連するIGFBP分解酵素(プロテアーゼ)からなる。この系は、正常な生理の制御だけでなく、多くの病理的状態に対しても重要である(Glass、Nat Cell Biol 5:87−90、2003)。
【0002】
IGF軸は、細胞増殖の促進及び細胞死(アポトーシス)の抑制に関与することが示されている。IGF−1は、ヒト成長ホルモン(hGH)による刺激の結果として、主に、肝臓により分泌される。ヒトの身体における殆ど全ての細胞、特に筋肉、軟骨、骨、肝臓、腎臓、神経、皮膚及び肺における細胞は、IGF−1により影響を受ける。インスリン様の効果に加えて、IGF−1はまた細胞増殖を制御することもできる。IGF−1及びIGF−2は、IGF結合タンパク質として知られる遺伝子産物のファミリーにより制御される。これらのタンパク質は、IGF受容体への結合を妨害することによるIGF作用の阻害ならびに受容体への送達促進及び血流中のIGF半減期の延長を通じたIGF作用の促進の両方に関与する複合的な方法でIGF作用の調節に寄与する。特性が分かっている少なくとも6種類の結合タンパク質(IGFBP1−6)がある。
【0003】
その成熟型において、ソマトメジンとも呼ばれるヒトIGF−1(gpetlcgaelvdalqfvcgdrgfyfnkptgygsssrrapqtgivdeccfrscdlrrlemycaplkpaksa;配列番号1)は、培養において幅広い範囲の細胞の増殖を刺激することが示されている70アミノ酸の小タンパク質である。成熟タンパク質は、最初に、3つの既知のスプライシング変異mRNAによりコードされる。各mRNAのオープンリーディングフレームは、特定のIGF−1mRNAに依存して、70アミノ酸IGF−1及びC末端の特定のE−ペプチドを含有する前駆タンパク質をコードする。これらのE−ペプチドは、Ea(rsvraqrhtdmpktqkevhlknasrgsagnknyrm;配列番号2)、Eb(rsvraqrhtdmpktqkyqppstnkntksqrrkgwpkthpggeqkegteaslqirgkkkeqrreigsrnaecrgkkgk;配列番号3)及びEc(rsvraqrhtdmpktqkyqppstnkntksqrrkgstfeerk;配列番号4)ペプチドと呼ばれ、35から87アミノ酸長の範囲であり、N末端において共通配列領域及びC末端において可変配列領域を含む。例えば、IGF−1−Eaに対する野生型オープンリーディングフレームは、105アミノ酸(gpetlcgaelvdalqfvcgdrgfyfnkptgygsssrrapqtgivdeccfrscdlrrlemycaplkpaksa rsvraqrhtdmpktqkevhlknasrgsagnknyrm;配列番号5)のポリペプチドをコードする。生理的発現において、E−ペプチドは、内在性プロテアーゼにより前駆体から切断され、生物活性があることが知られている成熟70アミノ酸IGF−1が生じる。ある種の関連において、IGF−1のN末端アミノ酸の1から3個は生理的条件下で切断され、これにより67−70の間のアミノ酸を有する活性型IGF−1が生じることが知られている。ヒトIGF−2に対する1つのE−ペプチド
【0004】
【化1】
のみが156アミノ酸前駆体
【0005】
【化2】
に対して確認されていることを除き、IGF−2遺伝子発現及びプロセシングは同様の特性を特徴とする。IGF−1及びIGF−2の両方は、患者の血清中で内在性プロテアーゼにより急速に分解されるので、優れた薬物候補とはならないと思われる。意図されているあるストラテジーは、IGF−1の結合タンパク質の1つと複合体を形成することにより、薬物としてIGF−1を安定化することである。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Glass、Nat Cell Biol 5:87−90、2003
【発明の概要】
【0007】
発明の要旨
本発明は、このE−ペプチドを実質的に含有する前駆IGF−1又はIGF−2タンパク質が生物活性を有し、血清の存在下で安定化され、その結果、医薬品として有用であるIGF−1又はIGF−2ポリペプチドが得られるという事実に基づく。本発明の組成物において、例えば、E−ペプチドの位置1のアルギニン又は位置2のセリン(野生型前駆IGF−1における位置71及び72に対応する。)の何れかの変異又は欠失によって、IGF−1からのE−ペプチドの通常の切断が回避される。IGF−2において、例えば、E−ペプチドの位置1のアルギニン又は位置2のアスパラギン酸(野生型前駆IGF−2における位置68及び69に対応する。)の何れかの変異又は欠失によって、切断が回避される。IGF前駆タンパク質の他の修飾によって、この切断を回避又は減少させることができる。
【0008】
さらに、IGF−1前駆アミノ酸配列のさらなる修飾は、さらなる医薬的有益性を与え得る。例えば、本発明のポリペプチドは、IGF−1受容体に対する親和性の向上又は抑制性のIGF−1もしくはIGF−2結合タンパク質に対する結合能の低下を示し得る。
【0009】
明確性及び一貫性のために、この明細書を通じて、及び特許請求の範囲において、IGF−1もしくはIGF−2前駆体又は成熟タンパク質におけるアミノ酸残基の付番は、シグナルペプチドなしの野生型前駆タンパク質配列の付番に基づく。
【0010】
従って、本発明は、プロテアーゼによるIGF−1からのE−ペプチドの切断がIGF−1前駆タンパク質の修飾により減少する、IGF−1前駆タンパク質を含有するポリペプチドを含む。E−ペプチドは、Ea、Eb又はEcペプチドであり得る。前駆体のN末端において、前駆タンパク質のアミノ酸G1、P2又はE3が、R36(例えばR36A)及びR37(例えばR37A)など、欠失又は変異させられ得る。
【0011】
前駆タンパク質は、例えば、Ebのアミノ酸N95とT96との間へのEaのアミノ酸93−102の挿入によって、N−結合型グリコシル化コンセンサス配列 NXS/Tをさらに含み得る。一般に、前駆タンパク質は、前駆タンパク質のアルギニン側鎖など、前駆タンパク質のアミノ酸側鎖に共有結合されたオリゴ糖を含み得る。
【0012】
さらに、非天然アミノ酸(例えば、アセチレン又はアジド基を含むもの)により前駆タンパク質の残基を置換することができる。このような非天然アミノ酸は、前駆タンパク質の側鎖へのポリ(エチレングリコール)部分の連結を促進することができる。通常のタンパク質PEG化ストラテジーは当技術分野で周知である。
【0013】
前駆タンパク質は、前駆タンパク質のC末端に連結された1以上のさらなるE−ペプチドをさらに含み得る。例えば、ポリペプチドは、N末端からC末端まで、(1)第一のEbペプチドを有するIGF−1前駆タンパク質(このタンパク質中、G1、P1及びE1が欠失しており、R36もしくはR37の何れか又は両方が変異しており、R71及びS72が欠失しており、第一のEbペプチドの最後の7個のC末端アミノ酸が欠失している。);(2)第二のEbペプチド(このペプチド中、R71、S72及び第二のEbペプチドの最後の7個のC末端アミノ酸が欠失している。);(3)第三のEbペプチド(このペプチド中、R71、S72及び第三のEbペプチドの最後の7個のC末端アミノ酸が欠失している。);及び(4)第四のEbペプチド(このペプチド中、R71及びS72が欠失している。)を含み得る。
【0014】
IGF−1からのE−ペプチドの切断を妨害する効率的な手段は、R71又はS72の欠失又は変異である。
【0015】
同様に、本発明は、プロテアーゼによるIGF−2からのE−ペプチドの切断が、IGF−2前駆タンパク質の修飾により減少する、IGF−2前駆タンパク質を含む。特に、R68又はD69の欠失又は変異は、IGF−2前駆タンパク質のプロテアーゼ消化を回避する有効な手段であり得る。
【0016】
さらに、本明細書中に記載の有益性を与えるために、IGF−1の何れかのE−ペプチドをIGF−2と組み合わせることができ、IGF−2の何れかのE−ペプチドをIGF−1と組み合わせることができる。
【0017】
本発明は、さらに、本発明のポリペプチドの治療的有効量を投与することにより、筋骨格疾患、糖尿病、神経細胞死を治療する方法を含む。同様に、本発明は、筋骨格疾患、糖尿病、神経細胞死又は貧血の治療用の医薬品の製造のための本発明のポリペプチドの使用を含む。
【0018】
別の実施形態において、本発明は、E−ペプチドがないが、PEG化の部位として非天然アミノ酸がそこに導入されている、PEG化IGF−1を含む。E−ペプチドがない、本明細書中に記載のような、非天然アミノ酸を含有する何れの修飾PEG化IGF−1もまた本発明に含まれる。
【0019】
本発明はまた、所望の効果を得るために本発明のポリペプチドの有効量を投与する、獣医学の方法及び使用も含む。
【0020】
獣医学での使用は、(i)動物における増殖の速度及び/又は程度を促進すること、(ii)食物の身体組織への変換の効率を促進すること、(iii)泌乳動物において乳汁産生を促進すること、(iv)悪液質、外傷性傷害又は他の消耗疾患に付随する消耗性の症状に対して動物を治療すること及び(v)新生仔の健康を増進するために泌乳非ヒト動物を処置することを含む。
【0021】
全ての引用参考文献又は書類は、参照により本明細書中に組み込まれる。
【図面の簡単な説明】
【0022】
【図1】図1A−1Cは、10%ヒト血清の存在下又は非存在下で37℃にて0又は16時間温置した後の、本発明のポリペプチド及び野生型IGF−1前駆体のウエスタンブロットである。様々なIGF−1コンストラクトをコードする発現ベクターをCos7細胞へと遺伝子移入し、条件培養液を得た。「3mut」は、次の3セットの修飾:G1、G2及びE3の欠失;AlaへのArg37の変異(R37A);及びR71及びS72の欠失を有する、hIGF−1−E−ペプチド前駆体を指す。図1Aは、Eaを含有する、野生型及び3mut前駆体に対する(IGF−1に対する抗体を用いた)ウエスタンブロットの結果を示す。図1Bは、Ebを含有する、野生型及び3mut前駆体に対する(hIGF−1に対する抗体を用いた)ウエスタンブロットの結果を示す。図1Cは、Ecを含有する、野生型及び3mut前駆体に対する(hIGF−1に対する抗体を用いた)ウエスタンブロットの結果を示す。
【図2A】図2A−2Dは、様々なIGF−1ポリペプチド(「リガンド」))の生物学的活性を示す線グラフである。Cos7により発現されたポリペプチドによるC2C12筋芽細胞の刺激により、生物学的活性を測定した。次に、総AKT及びリン酸化AKTの相対量について、刺激したC2C12細胞をアッセイした。Long−R3−IGF−1は、E3R変異及びさらなる13アミノ酸のN末端延長ペプチドを有する成熟ヒトIGF−1アミノ酸配列からなる市販の試薬(Sigma製品番号1−1271)である。図2AはIGF−1−Ea3mutの活性を示す。図2BはIGF−1−Eb3mutの活性を示す。図2Cは、IGF−1−Eab3mut(Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されている、3mutコンストラクト)の活性を示す。図2Dは、IGF−1−Ec3mutの活性を示す。
【図2B】図2A−2Dは、様々なIGF−1ポリペプチド(「リガンド」))の生物学的活性を示す線グラフである。Cos7により発現されたポリペプチドによるC2C12筋芽細胞の刺激により、生物学的活性を測定した。次に、総AKT及びリン酸化AKTの相対量について、刺激したC2C12細胞をアッセイした。Long−R3−IGF−1は、E3R変異及びさらなる13アミノ酸のN末端延長ペプチドを有する成熟ヒトIGF−1アミノ酸配列からなる市販の試薬(Sigma製品番号1−1271)である。図2AはIGF−1−Ea3mutの活性を示す。図2BはIGF−1−Eb3mutの活性を示す。図2Cは、IGF−1−Eab3mut(Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されている、3mutコンストラクト)の活性を示す。図2Dは、IGF−1−Ec3mutの活性を示す。
【図2C】図2A−2Dは、様々なIGF−1ポリペプチド(「リガンド」))の生物学的活性を示す線グラフである。Cos7により発現されたポリペプチドによるC2C12筋芽細胞の刺激により、生物学的活性を測定した。次に、総AKT及びリン酸化AKTの相対量について、刺激したC2C12細胞をアッセイした。Long−R3−IGF−1は、E3R変異及びさらなる13アミノ酸のN末端延長ペプチドを有する成熟ヒトIGF−1アミノ酸配列からなる市販の試薬(Sigma製品番号1−1271)である。図2AはIGF−1−Ea3mutの活性を示す。図2BはIGF−1−Eb3mutの活性を示す。図2Cは、IGF−1−Eab3mut(Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されている、3mutコンストラクト)の活性を示す。図2Dは、IGF−1−Ec3mutの活性を示す。
【図2D】図2A−2Dは、様々なIGF−1ポリペプチド(「リガンド」))の生物学的活性を示す線グラフである。Cos7により発現されたポリペプチドによるC2C12筋芽細胞の刺激により、生物学的活性を測定した。次に、総AKT及びリン酸化AKTの相対量について、刺激したC2C12細胞をアッセイした。Long−R3−IGF−1は、E3R変異及びさらなる13アミノ酸のN末端延長ペプチドを有する成熟ヒトIGF−1アミノ酸配列からなる市販の試薬(Sigma製品番号1−1271)である。図2AはIGF−1−Ea3mutの活性を示す。図2BはIGF−1−Eb3mutの活性を示す。図2Cは、IGF−1−Eab3mut(Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されている、3mutコンストラクト)の活性を示す。図2Dは、IGF−1−Ec3mutの活性を示す。
【図3A】図3A−3D及び図4A−4Dは、リガンド結合に反応する受容体リン酸化についてアッセイすることによって、本発明のIGF−1前駆ポリペプチドが適切な受容体に対する選択性を維持するか否かを示す線グラフである。図3A及び図3Bは、IGF−1受容体(図3A)及びインスリン受容体(図3B)に対する、IGF−1−Ea3mutの受容体選択性を試験する。図3C及び3Dは、IGF−1受容体(図3C)及びインスリン受容体(図3D)に対する、IGF−1−Eb3mutの受容体選択性を試験する。
【図3B】図3A−3D及び図4A−4Dは、リガンド結合に反応する受容体リン酸化についてアッセイすることによって、本発明のIGF−1前駆ポリペプチドが適切な受容体に対する選択性を維持するか否かを示す線グラフである。図3A及び図3Bは、IGF−1受容体(図3A)及びインスリン受容体(図3B)に対する、IGF−1−Ea3mutの受容体選択性を試験する。図3C及び3Dは、IGF−1受容体(図3C)及びインスリン受容体(図3D)に対する、IGF−1−Eb3mutの受容体選択性を試験する。
【図3C】図3A−3D及び図4A−4Dは、リガンド結合に反応する受容体リン酸化についてアッセイすることによって、本発明のIGF−1前駆ポリペプチドが適切な受容体に対する選択性を維持するか否かを示す線グラフである。図3A及び図3Bは、IGF−1受容体(図3A)及びインスリン受容体(図3B)に対する、IGF−1−Ea3mutの受容体選択性を試験する。図3C及び3Dは、IGF−1受容体(図3C)及びインスリン受容体(図3D)に対する、IGF−1−Eb3mutの受容体選択性を試験する。
【図3D】図3A−3D及び図4A−4Dは、リガンド結合に反応する受容体リン酸化についてアッセイすることによって、本発明のIGF−1前駆ポリペプチドが適切な受容体に対する選択性を維持するか否かを示す線グラフである。図3A及び図3Bは、IGF−1受容体(図3A)及びインスリン受容体(図3B)に対する、IGF−1−Ea3mutの受容体選択性を試験する。図3C及び3Dは、IGF−1受容体(図3C)及びインスリン受容体(図3D)に対する、IGF−1−Eb3mutの受容体選択性を試験する。
【図4A】図4A及び4Bは、IGF−1受容体(図4A)及びインスリン受容体(図4B)に対する、IGF−1−Ec3mutの受容体選択性を試験する。図4C及び4Dは、IGF−1受容体(図4C)及びインスリン受容体(図4D)に対する、IGF−1−Eab3mutの受容体選択性を試験する。「IGF1−R3」は、上述のLong−R3−IGF−1を指す。「IGF1Eab」として挙げられたポリペプチドは、Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されたコンストラクトを指す。
【図4B】図4A及び4Bは、IGF−1受容体(図4A)及びインスリン受容体(図4B)に対する、IGF−1−Ec3mutの受容体選択性を試験する。図4C及び4Dは、IGF−1受容体(図4C)及びインスリン受容体(図4D)に対する、IGF−1−Eab3mutの受容体選択性を試験する。「IGF1−R3」は、上述のLong−R3−IGF−1を指す。「IGF1Eab」として挙げられたポリペプチドは、Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されたコンストラクトを指す。
【図4C】図4A及び4Bは、IGF−1受容体(図4A)及びインスリン受容体(図4B)に対する、IGF−1−Ec3mutの受容体選択性を試験する。図4C及び4Dは、IGF−1受容体(図4C)及びインスリン受容体(図4D)に対する、IGF−1−Eab3mutの受容体選択性を試験する。「IGF1−R3」は、上述のLong−R3−IGF−1を指す。「IGF1Eab」として挙げられたポリペプチドは、Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されたコンストラクトを指す。
【図4D】図4A及び4Bは、IGF−1受容体(図4A)及びインスリン受容体(図4B)に対する、IGF−1−Ec3mutの受容体選択性を試験する。図4C及び4Dは、IGF−1受容体(図4C)及びインスリン受容体(図4D)に対する、IGF−1−Eab3mutの受容体選択性を試験する。「IGF1−R3」は、上述のLong−R3−IGF−1を指す。「IGF1Eab」として挙げられたポリペプチドは、Eaアミノ酸93から102がEbのアミノ酸95と96との間に挿入されたコンストラクトを指す。
【図5】図5は、様々なリガンドによる(C2C12筋芽細胞の3から4日の分化の結果としての)C2C12筋管の刺激における相対的AKTリン酸化を示すウエスタンブロットである。IGF−1Eb−マルチマーは、図6Aで図式的に示されるコンストラクトを指す。
【図6】図6A及び6Bは、本発明の2つのポリペプチドの概略図である。図6Aは、4セットの修飾:G1、G2及びE3の欠失;R37のAへの変異;R71及びS72の欠失;及び最後の7個のC末端アミノ酸の欠失を有するIGF−1−Eb前駆ポリペプチドを示す。さらに、本ポリペプチドのC末端での、2個のさらなるEbペプチドの付加(しかしR71及びS72はなく、最後の7個のC末端アミノ酸がない。)及び最後のEbペプチドの付加(しかしR71及びS72がない。)により、本ポリペプチドを長くする。このコンストラクトはIGF−1−Ebマルチマーと呼ぶことが多い。図6Bは、4セットの修飾:G1、P2及びE3の欠失;R37のAへの変異;R71及びS72の欠失;及びEbのアミノ酸95と96との間へのEaアミノ酸93から102の挿入があるIGF−1−Eab前駆ポリペプチドを示す。
【図7A】図7Aは、対応する動物IGF−1とのヒトIGF−1(配列番号1)の配列アラインメントである。分析した全ての動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。コチョウザメ(S2はP2に取って代わる。)を除く分析した全ての種においてG1、P2、E3は保存されている。R36及びR37は分析した全ての種において保存されている。
【図7B】図7Bは、ヒトIGF−1(配列番号1)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。系統樹の下は、タンパク質配列に対する100残基あたりの「アミノ酸置換」の数を示すスケールである。非ギャップミスマッチの数からの、サイレント置換について補正された距離値を計算するために、Kimuraの距離式を使用する。計算した値は、部位あたりの相違数の平均数であり、0から1の間となる。0は完全に同一であることを表し、1は同一性がないことを表す。系統樹のスケールは、これらの値に100を掛けたものを使用する。
【図8A】図8Aは、様々な動物EaペプチドとのヒトEaペプチド(配列番号2)の配列アラインメントである。分析した全動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。R71及びS72は分析した全ての種で保存されている。
【図8B】図8Bは、ヒトIGF−1 Eaペプチド(配列番号2)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。
【図9A】図9Aは、様々な動物EbペプチドとのヒトEbペプチド(配列番号3)の配列アラインメントである。分析した全動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。R71及びS72は分析した全ての種で保存されている。
【図9B】図9Bは、ヒトIGF−1 Ebペプチド(配列番号3)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。
【図10A】図10Aは、様々な動物EcペプチドとのヒトEcペプチド(配列番号4)の配列アラインメントである。分析した全動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。R71及びS72は分析した全ての種で保存されている。
【図10B】図10Bは、ヒトIGF−1 Ecペプチド(配列番号4)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。
【図11A】図11Aは、対応する動物IGF−2とのヒトIGF−2(配列番号7)の配列アラインメントである。分析した全動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。R68は分析した全ての種で保存されており;D69はチンパンジー(その位置はヒスチジン残基になっている。)を除き保存されている。
【図11B】図11Bは、ヒトIGF−2(配列番号7)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。
【図12A】図12Aは、様々な動物IGF−2E−ペプチドとのヒトIGF−2E−ペプチド(配列番号6)の配列アラインメントである。分析した全動物種及びその配列に対するそれらの対応するGenBank登録番号を与える。R68は分析した全ての種で保存されており;D69はチンパンジー(その位置はヒスチジン残基になっている。)を除き保存されている。
【図12B】図12Bは、ヒトIGF−2Eペプチド(配列番号6)と比較した、分析したアミノ酸配列の系統発生を示すグラフである。
【0023】
発明の詳細な説明
本発明は、活性型IGF−1又はIGF−2をそのE−ペプチドから放出することに関与する典型的なプロテアーゼ切断を妨害、減少又は回避するために修飾されているE−ペプチドを実質的に含有する新規のIGF−1及びIGF−2前駆ポリペプチドに関する。本発明のポリペプチドの有用性は、このような前駆ポリペプチドが生物学的に活性であり、安定であり、医薬品として有益であるという驚くべき発見に基づく。
【0024】
活性IGF前駆ポリペプチドについてのスクリーニング
次のアッセイを用いて本発明の何れかのポリペプチドの有用性を評価することができる。
【0025】
安定性
本発明のポリペプチドは、有効な薬物であるために、ヒト血清中など、内在性プロテアーゼ存在下で十分な安定性を有しなければならない。安定性を評価するために、10%ウシ胎仔血清、100U/mLペニシリン及び100μg/mLストレプトマイシンを含有するDMEM培地中で本ポリペプチドをコードする発現ベクターをCos7細胞(ATCC)に遺伝子移入することができる。分泌されたポリペプチドを含有する培地をさらなる分析に供する、又は、代って、この発現ベクターは、Cos7培養において発現されたポリペプチドの効率的な精製を促進するために、ポリペプチド中でヘキサ−ヒスチジンタグなどの容易に利用可能なタグをコードすることができる。いかに調製しても、ポリペプチド試料を正常ヒト血清(Sigma)又はPBS中で様々な時間(例えば、0、1、5、10及び16時間)温置し、ポリアクリルアミドゲル電気泳動に供し、ニトロセルロース上にブロットし、ヒトIGF−1又はIGF−2に対する一次抗体及び二次抗体(例えばホースラディッシュペルオキシダーゼ標識されたもの)を用いて関連のあるタンパク質を可視化する。いくつもの同様のブロッティング及び検出技術(蛍光色素又は放射性核種を使用するものものある。)を使用することができる。IGF−1又はIGF−2バンドの強度に対する前駆体バンドの強度から、様々な条件下で前駆ポリペプチドが切断される度合いが示されるはずである。本発明のポリペプチドが37℃で16時間ヒト血清に曝露されると、切断された成熟IGFに対する非切断前駆体の比が約1:2から1:0.1、例えば約1:1から1:0.5、特に約1:1の比又は約1:0.5の比となり得る。
【0026】
AKTリン酸化
本発明のポリペプチドは、IGF−1受容体を通じて信号を送る能力を維持しなければならない。(IGF−1及びIGF−2の両者ともIGF−1受容体を通じて信号を送る。)。このシグナリング能を調べるために、下流の細胞内標的であるAKTが細胞表面でのリガンド結合に反応してリン酸化されるか否かを評価することができる。AKTリン酸化の分析のために、血清不含培地中でC2C12筋芽細胞を飢餓状態にし、次いで、様々なリガンドで刺激する。細胞を溶解し、遠心により透明化する。PathScan phospho AKT(Ser473)サンドイッチELISAキット及びPathScan AKTサンドイッチELISAキット(Cell Signaling)をそれぞれ用いて、ELISAによって、AKTリン酸化及び総AKTレベルを分析する。
【0027】
IGF−1受容体特異性
本発明のポリペプチドは、好ましくは、IGF−1受容体に対して特異性を維持し、関連するインスリン受容体への結合は低親和性でなければならない。受容体特異性を評価するために、IGF−1受容体又はインスリン受容体を過剰発現する血清飢餓状態のNIH3T3細胞にポリペプチド試料を添加し、細胞を溶解し、DuoSet ICヒト蛍光体−IGF−1受容体及びインスリン受容体ELISAキット(R&D Systems)を用いたELISAに溶解液を供することによって、IGF−1受容体リン酸化又はインスリン受容体リン酸化のレベルを調べる。
【0028】
肥大のマウスモデルにおけるインビボ試験
既に筋肉肥大が誘発されている状況下で本発明のポリペプチドが骨格筋量を増加させるように作用し得るか否かを調べるために、処置及び未処置動物を運動させ、本ポリペプチドを投与された動物の筋肉が未処置動物よりも大きくなるか否かを調べることができる。
【0029】
運動モデル
当技術分野で公知のあるモデルは、使用者が荷重を変化させることができる自発運動用の回し車の使用に基づく(例えば、Konhilasら、Am J Physiol Heart Circ Physiol 289:H455−H465、2005を参照。)。自発運動用かご型車輪は、強制運動モデルに共通する心身の損傷を排除し、従って、筋肉量の増加が所望される比較的健康な個体において使用される候補薬物を評価するのにより適している。
【0030】
何れかの適切なマウス系統を使用することができる。例えば、雄C57B1/6Jマウスを無作為に実験(例えばIGF前駆ポリペプチドを投与される。)及び対照群に振り分けることができる。運動用回し車を含むケージに動物を1匹ずつ収容し;あまり運動しない対照動物を回し車のない同一ケージに収容する。運動用回し車は、Allenら、J Appl Physiol 90:1900−1908、2001に記載されている。簡潔に述べると、このシステムは、車輪の回転により起動されるデジタル電磁カウンタ(モデルBC600、Sigma Sport、Olney、IL)を備えた5.0cmの幅のランニング面のある11.5cmの直径の車輪(モデル6208、Petsmart、Phoenix、AZ)からなる。さらに、各車輪は、荷重の調整ができる抵抗機構により作製されている。これは、ステンレススチールの釣り糸をケージの上に連結し、車輪荷重に関与しないように、回転軸でケージ車輪に固定された固定滑車の周りにワイヤを巻きつけることにより行われる。このワイヤを再びスプリングと螺子でケージの上に固定する。この設計によって、車輪荷重を細かく調整することが可能となり、車輪荷重が車輪の回転全体に均一に分布される。運動期間の持続時間を通じて、各運動動物に対して、走った時間及び距離について毎日の運動値を記録する。全動物に水及び標準的な硬いげっ歯類用の餌を随意に給餌した。自発運動(かご型車輪を与えるもの)は、全群に対して約12週齢の平均週齢で開始することができる。各群は、50日間、動物が約19週齢になるまで、実験群に依存して、様々な抵抗下で走行を続ける。車輪が僅かに動くまで車輪に既知の重量を吊り下げることにより、車輪の荷重を決定する。全ての運動群は、最初の週、かご型車輪に荷重をかけずに開始する。しかし、「荷重なし」状態は実際には2gであり、これは、車輪内部及び摩擦荷重を維持するのに必要な荷重として測定される。1週間の車輪順化期間を考慮し、より大きい荷重の場合(この場合は2週間後に変化させることができる。)を除き1週間間隔で車輪荷重を変化させることができる。荷重の範囲は、2gから12gの何れかであり得る。特定の運動期間の終了直後に吸入麻酔下での頸椎脱臼によって運動及び静止対照動物を安楽死させる。体重を測定し、特定の筋肉を迅速に切り出し、洗浄し、後日の組織学的又は生化学的アッセイのために凍結する。
【0031】
別の運動肥大モデルも当業者にとって利用可能である。例えば、Lermanら、J Appl Physiol 92:2245−2255、2002に記載のトレッドミル運動モデルを参照のこと。
【0032】
クレンブテロール注入モデル
クレンブテロールは、筋肉量が増加することが確認されている増殖促進特性を有するβ2−アドレナリン作動性アゴニストである。クレンブテロール作用の正確な作用機構は不明であるが、筋肉タンパク質分解の低下が提案されている。病院において、クレンブテロールは抗喘息薬として使用されているが、殆ど、ヒト及びショー動物の両方において筋肉量を増加させるための筋肉増強剤として誤用されていると思われる。
【0033】
筋肉肥大を誘発するために、3、7又は14日間、5匹のマウスに毎日、クレンブテロール(3mg/kg、皮下(s.c.))を注射する。PBSを注射したマウスを陰性対照とする。治療に対する何らかの有害反応(即ち、毛並みの悪さ、嗜眠性)について動物を毎日監視する(目視)。クレンブテロール処置は、マウスがより恐怖を感じるか又は攻撃的になる可能性があるので、群で飼育する場合、特に闘争についてマウスを監視すべきである。マウスは動くことができ、普通に飲食可能である。第3、7又は14日に安楽死させるまでマウスを毎日監視し、さらなる分析のために組織を回収する。
【0034】
筋肉萎縮モデルにおけるインビボ試験
様々な骨格筋萎縮モデルにおいて、通常筋肉量が低下する条件下で筋肉量を維持する能力について本発明のIGF前駆ポリペプチドを試験することができる。IGF前駆ポリペプチドが筋肉量を増加させることができるか否かを調べるために、下記に記載の実施例モデルにを用いて、当業者は、IGF前駆ポリペプチドの投与及び使用を含む制御実験を容易に設計し、実施することができる。
【0035】
例えば、C57B16/2雄マウスをThe Jackson Laboratoriesから購入する。各実験の開始時にマウスが約9週齢となるようにマウスを購入する。一般に、通常のげっ歯類用飼料付きのマイクロアイソレーターケージにマウスを収容する。各実験の開始時にマウスの体重を測定する。各実験の終了時に、通常、CO2吸入の後に頸椎脱臼を行い、マウスを安楽死させ、さらなる処理のために筋肉組織を回収する。「最後の体重」を得るためにマウスの体重を測定する。回収することができる骨格筋は、前脛骨筋、長趾伸筋、ヒラメ筋及び腓腹筋である。場合によって回収する他の組織は、心臓、肝臓、脾臓、腎臓、精巣及び脳である。全ての筋肉及び組織を完全に切り出し、0.0001gまで測定可能な秤で重量を測定する。次に、後のRNA及びタンパク質抽出のために組織を液体窒素中で瞬間凍結するか又はコルクディスク上でOCT中で瞬間凍結物を包埋する。後の凍結切片作製のためにコルクディスク上で凍結させた筋肉を、液体窒素により濃厚なスラッシュ状になるまで冷却したイソペンタンに浸漬する。全試料を−80℃で保存する。
【0036】
デキサメタゾン処置
マウスにおいて筋肉萎縮を誘導する薬理学的方法は、毎日デキサメタゾン20mg/kgを腹腔内投与することである。デキサメタゾンは、ホルモンの糖質コルチコイドクラスの合成メンバーである。これは、抗炎症剤及び免疫抑制剤として作用し、ヒドロコルチゾンの約40倍の効力を有する。例えば、関節リウマチなど、多くの炎症及び自己免疫状態を治療するためにデキサメタゾンが使用される。これはまた、抗腫瘍治療のある種の副作用を軽減するために、化学治療を行っている癌患者にも投与される。デキサメタゾンは、マウス及びヒト患者の両方において筋肉萎縮を引き起こす。
【0037】
3、7又は14日間、マウスにデキサメタゾンを腹腔内(ip)注入する。最終日に、CO2を用いて被験動物を安楽死させ、足の筋肉を回収する。治療に対する何らかの有害反応(即ち、毛並みの悪さ、嗜眠性)について動物を毎日監視する(目視)。マウスは通常動くことができ、普通に飲食可能である。PBSを注入したマウスが陰性対照である。
【0038】
ギブス固定
様々な筋肉群を物理的に非活動性にした結果、これらの筋肉の萎縮が起こる。足首関節固定(「ピンヒール」又はギブス)は、ラット及びマウスの後肢筋肉組織の身体拘束を誘発するのに非常に有用で再現性のある方法であることが分かっている。
【0039】
固定化のためにイソフルオランでマウスに麻酔をかける。関節の周囲に軽量ギブス材料(VET−LITE)を用いて足首及び膝関節を90°に固定する。この材料を温水に浸し、次いで肢周囲に巻き付け、踵及び股関節は自由にする。ギブス材料が乾燥するまで関節を90°の位置に維持する。反対側の肢を対照とする。次に、マウスを麻酔から回復させ、普通のマイクロアイソレーターケージに収容する。観察によるとギブスは過剰ストレスを引き起こさず、動物はケージ中を自由に動き回り、飲食する。しかし、体重、活動性及び過敏性に影響を与える何らかの有害反応についてマウスを毎日監視する。
【0040】
マウスにギブスを嵌めたら、咀嚼する可能性があるので、ギブスがずれていないことを確認するためにその動物を毎日監視する。麻酔の回復後、動物は、動き、飲食することができ、特別なベッディング、ケージ収容又は他の支援は必要ない。
【0041】
除神経
一般に、除神経のためにイソフルオランガスでマウスに麻酔をかける。無菌の手術手順を用いて(ベタジン洗浄を3回行い、最後の洗浄はエタノール)、右坐骨神経を大腿の中ほどで切り離し、2から5mmの小片を切り出す。反対側の肢を対照とする。
【0042】
より具体的には、クリップ縫合で皮膚切開を閉じ、麻酔から回復させる前に動物にブプレノルフィンを1回投与する。手術から3、7又は14日後、CO2吸入後に頸椎脱臼を行い動物を安楽死させ、組織学的及び生化学的分析のために筋肉(腓腹筋複合体、前脛骨筋、長趾伸筋、ヒラメ筋)を取り出す。
【0043】
坐骨神経を横に切断する場合、関与する筋肉の骨格筋萎縮を誘発するために、手術を行った肢を固定化する。他は、麻酔の回復後、動物は動き、飲み、食べることができ、特別なベッディング、ケージ収容又は他の支援は必要ない。
【0044】
しかし、手術直後及び回復期(1−2時間)を通じて、動物を監視する。さらに、切開部位及び一般的な動物の健康状態を手術後3日間監視する。手術後7−10日後にクリップ縫合を除去する。
【0045】
遺伝学的モデル
筋肉萎縮のモデルとして、遺伝子操作したトランスジェニックマウスを使用することもできる。例えば、いわゆるMini Mice(The Jackson Laboratory、ストック番号003258)は、IGF−1遺伝子においてノックアウト変異があり、その結果、出生後の成長が異常に低下し、体重及び身長が低い。さらなる情報に関しては、Powell−Braxton、Genes Dev 7:2609−2617、1993を参照のこと。さらに、いわゆるMidi Mice(The Jackson Laboratory、ストック番号003259)は、IGF−1遺伝子において異なる変異があり、その結果、成獣で低体重となり、他の心血管表現型を示す矮小体型となる。さらなる情報に関しては、Lemboら、J Clin Invest 98:2648−2655、1996を参照のこと。
【0046】
IGF前駆体における重要及び最適な変異又は修飾
重要な変異
本発明は、一部分において、そのE−ペプチドを実質的に含有するIGF前駆ポリペプチドが生物活性を有し、血清の存在下で安定であるという観察に基づく。二塩基性のプロテアーゼ部位を標的とする内在性プロテアーゼによりE−ペプチドが切断されないことを確認するために、一般に、前駆体のE−ペプチドの2個のN末端二塩基性アミノ酸の何れかを欠失させるか、変異させる又は隠蔽する。IGF−1のある例において、これらの2個のアミノ酸はR71及びS72であり、一方、IGF−2の別の例において、これらの第一の2個のアミノ酸はR68及びD69である。
【0047】
様々な修飾によってこの切断の妨害が可能となる:
(1)1又は両方の二塩基性残基の欠失
(2)1又は両方の二塩基性残基の、アラニンなどの非塩基性アミノ酸への変異
(3)二塩基性残基の間への1以上の非塩基性アミノ酸の挿入
(4)プロテアーゼ部位を隠蔽するのに二塩基性残基に十分近い位置にグリコシル化部位を置くこと
(5)下記に記載のような、非天然アミノ酸による、何れかの二塩基性残基の置換又は二塩基性残基付近もしくはその間への挿入を用いた部位特異的PEG化。
【0048】
さらに、残基K68及びK65は、IGF−1/E−ペプチド切断に関与すると考えられ、従って、上述のような二塩基性アミノ酸に対する何らかの方策にこれらの残基の変異又は欠失を組み込むことができる。
【0049】
成熟IGFのN末端での変異
本発明のある実施形態において、IGF前駆ポリペプチドは、最初の数個のN末端アミノ酸の欠失又は変異を有する。IGF−1の場合、最初の3個のN末端アミノ酸の何れかを欠失させる又は変異させることができ、一方、IGF−2の場合、最初の6個のN末端アミノ酸の何れかを欠失させる又は変異させることができる。ある種のN末端アミノ酸がインビボで天然に切断されることが観察されており、これらの変異又は欠失を導入することによって、IGF結合タンパク質(IGFBP)との本発明のポリペプチドのインビボでの会合が最小限に抑えられる。IGF−1及びIGF−2のIGF−1受容体との相互作用はIGFBPにより制御される。全ての6個のIGFBPが、IGF作用(特にIGFBP5)を阻害することが示されているが、いくつかの例においては促進作用が観察されている。循環中のIGFの少なくとも99%が通常はIGFBPに結合している。新生仔期間後、循環中で最も豊富なIGFBPはIGFBP3であり、これは同様の親和性でIGF−1及びIGF−2の両方に結合できる。天然の短縮型IGF−1(G1、G2及びE3が欠失している。)は、IGFBP3に結合するが、天然のIGF−1よりも数倍親和性が低い。さらに、G3はIGFBP結合に重要であり、G6はIGF−2ペプチドにおいて同様の役割を果たす。
【0050】
従って、IGF−1前駆体のある実施例において、G1、P2又はE3の何れかを欠失させる又は変異させる(何れかのみ又は組み合わせて)ことができる。変異が所望される場合、アラニンへの変異を導入することができる。別の実施例において、IGF−2前駆体の別の例で、P4、S5及びE6の何れかを欠失させる又は変異させる(何れかのみ又は組み合わせて)ことができる。変異が所望される場合、アラニンへの変異を導入することができる。
【0051】
残基36及び37での変異
IGF−1をセリンプロテアーゼで切断することができる。R36又はR37の何れかのAへの変異によって、R36とR37との間のこの予想される切断部位におけるIGF−1の切断を妨げることができる。IGF−2のある例において、この有害な切断を妨げるために、R38を変異させる又は欠失させることができる。
【0052】
グリコシル化の使用
哺乳動物又はN−結合型グリコシル化が可能な他の真核細胞で発現される場合、IGF又は前駆体のE−ペプチド部分の何れかにN−結合型グリコシル化部位を付加することによって、本発明のポリペプチドのインビボ半減期を延長させることができる。Eaのこれらの部分は、N−X−S/T(式中、Xは何れかのアミノ酸であり得、この3残基の第三のアミノ酸はS又はTの何れかである。)のコンセンサスN−結合型グリコシル化配列に適合するので、インビトロで、IGF−1EaがN92及びN100でグリコシル化されることが分かっている。また、コンセンサスの隣接アミノ酸の状況が、アスパラギンがどの程度強くグリコシル化されるかに影響することも分かっている。従って、Eb又はEcにグリコシル化部位を導入するためのあるストラテジーは、Eb又はEcのほぼ同じ部分にコンセンサス配列周囲のEaアミノ酸を挿入することである。このストラテジーの特定の実施を下記実施例で例示する。何らかの事象において、当業者にとって公知の周囲環境のアミノ酸を含む何れかの他のコンセンサスN−結合型グリコシル化部位を本発明の前駆ポリペプチドに挿入することができる。さらに、本発明のポリペプチドの産生のために使用される特定の宿主を選択することによって、本発明のポリペプチドのO−結合型グリコシル化を行うことができる。例えば、IGF−1発現のためにある種の酵母株を使用すると、セリン又はスレオニンにおいてオリゴ糖が付加される。例えば、米国特許第5,273,966号を参照のこと。
【0053】
ポリ(エチレングリコール)の付加
ポリ(エチレングリコール)への連結(PEG;PEG化)は、治療用タンパク質薬の半減期を延長させるのに有益であることが証明されている。本発明のIGF前駆ポリペプチドのPEG化の結果、同様の医薬的長所が得られ得ると予想される。IGF−1のPEG化の方法は当技術分野で周知である。例えば、米国特許出願公開2006/0154865(これは、リジン−モノPEG化IGF−1の有用な特性について述べている。)を参照のこと。このようなリジン−モノPEG化を本発明の前駆IGFポリペプチドに適用することができる。さらに、非天然アミノ酸を導入することによって、本発明のポリペプチドの何れかの部分においてPEG化を行うことができる。Deitersら、J Am Chem Soc 125:11782−11783、2003;Wang及びSchultz、Science 301:964−967、2003;Wangら、Science 292:498−500、2001;Zhangら、Science 303:371−373、2004又は米国特許第7,083,970号に記載の技術によって、ある種の非天然アミノ酸を導入することができる。簡潔に述べるとこれらの発現系のいくつかは、本発明のポリペプチドをコードするオープンリーディングフレームに、アンバーTAGなど、ナンセンスコドンを導入するための部位特異的変異誘発を含む。次に、導入されたナンセンスコドンに特異的なtRNAを使用することができる宿主にこのような発現ベクターを導入し、最適な非天然アミノ酸を入れる。本発明のポリペプチドに対して部分を連結するために有益な特定の非天然アミノ酸には、アセチレン及びアジド側鎖を有するものが含まれる。次に、タンパク質のこれらの選択部位において、これらの新規アミノ酸を含有するIGF前駆ポリペプチドのPEG化を行うことができる。さらに、E−ペプチドがないこのようなPEG化IGF分子もまた治療薬として有用である。
【0054】
E−ペプチドのマルチマー
ある種の薬理学的関連において、薬物が血液脳関門又の一方又は他方に残ることを確実にするためにペプチド又はタンパク質のサイズを大きくすることは有益である。E−ペプチドが結合されたままである場合でも成熟IGF分子は比較的短いペプチドなので、本発明のポリペプチドのサイズを大きくすることは有益であり得る。これを行うある手段は、下記のある実施例で例示されるように、IGF前駆ポリペプチドのC末端においてE−ペプチドのマルチマーを提供することである。
【0055】
E−ペプチドのC末端の欠失
Ebの位置81のフリーのシステインによって、ホモ二量体形成又は、本発明のポリペプチドに存在する場合、薬物の活性が低下し得る他の影響が起こり得ることが疑われる。このように、EbのC81の欠失又は変異によって、薬物活性を最適化することができる。特定の例において、Ebの最後の7個のアミノ酸(即ちアミノ酸81−87)の欠失は有益である。
【0056】
他の変異又は修飾
本発明のIGF前駆ポリペプチドに組み込まれ得るIGFのさらなる変異又は修飾は米国特許第5,077,216号;及び米国特許出願公開2005/0287151、2006/0211606及び2006/0166328に記載されている。
【0057】
本発明は、実質的にそのE−ペプチドを含有する全ての既知及び未知の非ヒト動物前駆IGF−1又はIGF−2配列を含むことが意図され、本発明の修飾により、E−ペプチドの通常の切断が回避される又は減少する。
【0058】
使用されるべきIGFの好ましい型は、治療を受ける対象の種に依存する。
【0059】
IGFは種が適合することが好ましく、例えば、ウシを治療する場合、IGFの好ましい型はウシIGFである。
【0060】
配列相同性が高いためIGFの全ての型が様々な対象において有効であると思われるが、種が合致することにより、異種由来のIGFに対する免疫反応の誘導から引き起こされる有害な免疫学的合併症の可能性が回避される。
【0061】
本発明のある実施形態において、修飾非ヒト動物前駆IGF−1配列が提供される。
【0062】
E−ペプチドの通常の切断が脊椎動物由来の本発明の修飾により回避又は減少される、E−ペプチドを実質的に含有する前駆IGF−1配列が好ましい。
【0063】
例えば、このような配列には、以下に限定されないが、天然、合成又は組み換えの何れかの何らかのソースからの、マウス、ラット、ウシ、ブタ、ウマ、ヒツジ、ヤギ、鳥類、イヌ、ネコ、魚類など由来の配列が含まれる。
【0064】
本発明の別の実施形態において、修飾非ヒト動物前駆IGF−2配列が提供される。
【0065】
そのE−ペプチドの正常な切断が脊椎動物由来の本発明の修飾により回避されるか又は減少する、E−ペプチドを実質的に含有する前駆IGF−2配列が好ましい。
【0066】
例えば、このような配列には、以下に限定されないが、天然、合成又は組み換えの何れかの何らかのソースからの、マウス、ラット、ウシ、ブタ、ウマ、ヒツジ、ヤギ、鳥類、イヌ、ネコ、魚類など由来の配列が含まれる。
【0067】
IGF前駆ポリペプチドの治療用途
適応症例
本発明はまた、筋骨格疾患の治療又は予防用の医薬品の製造における本発明のIGF前駆ポリペプチドの使用も含む。さらに、本発明は、このような個体が筋骨格系疾患のリスクを有する又はその疾患に罹患しているにせよそうでないにせよ、個体において筋肉又は骨量を増加させるためのIGF前駆ポリペプチドの使用を含む。
【0068】
特に、筋骨格系疾患は筋萎縮であり得る。コルチゾール、デキサメタゾン、ベタメタゾン、プレドニゾン、メチルプレドニゾロン又はプレドニゾロンなどの糖質コルチコイドでの治療によるものを含め、筋萎縮には多くの原因がある。筋萎縮はまた、神経外傷性傷害による除神経の結果又は変性、代謝性もしくは炎症性ニューロパシーの結果(例えば、ギリアン−バレー症候群、末梢性ニューロパシー又は環境有害物質もしくは薬物への曝露)でもあり得る。さらに、筋萎縮は、成人の運動ニューロン疾患、小児脊髄性筋萎縮症、若年性脊髄性筋萎縮症、多発性導体ブロックを伴う自己免疫性運動神経障害、発作又は脊髄損傷による麻痺、外傷性傷害による骨格の不動化、長期にわたる床上安静、故意の不活発、不随意の不活発、代謝性ストレス又は栄養失調、癌、AIDS、飢餓、横紋筋融解症、甲状腺疾患、糖尿病、良性先天性低血圧、セントラルコア病、ネマリンミオパシー、筋管(中心核)ミオパシー、熱傷、慢性閉塞性肺疾患、肝臓疾患、敗血症、腎不全、うっ血性心不全又は加齢の結果であり得る。
【0069】
筋骨格系疾患はまた、デュシェンヌ型、ベッカー型、筋強直性、顔面肩甲上腕筋、エメリー・ドレフュス型、眼咽頭筋、肩甲上腕、肢帯、先天型筋ジストロフィー又は遺伝性末梢性ミオパシーなどの筋ジストロフィー症候群でもあり得る。筋骨格系疾患はまた、骨粗しょう症、骨折、低身長又は小人症でもあり得る。
【0070】
IGF−1はまたIGF−1受容体及びインスリン受容体のヘテロ二量体と結合することもできるので、IGF−1は、インスリン非依存性糖尿病の治療薬として示唆されている。従って、糖尿病を治療するために本発明のポリペプチドを使用することができる。
【0071】
IGF−1は、神経栄養性であり、ニューロンの生存を促進する。筋萎縮性側索硬化症(ALS)、脳萎縮、加齢及び認知症でみられるような、運動ニューロン死の症例を治療するためにIGF−1を使用することができることが示唆されている。従って、ALS、脳萎縮又は認知症など、ニューロン死と関連する状態を治療するために本発明のポリペプチドを使用することができる。
【0072】
IGF−1は、白血球及び赤血球細胞集団両方を増加させ、エリスロポエチンの投与に対して付加的効果がある。従って、貧血を治療するために、本発明のポリペプチドを使用することができる。
【0073】
IGF−1及びIGF−2はユビキタスであり、細胞分裂及び脊椎動物の成長の必須の制御因子なので、動物における成長を外部から促進又は維持するために様々な獣医学的方法においてこれらを有利に使用し得る。ある一部の例には、以下に限定されないが、
(i)動物における成長の速度及び/又は程度を促進すること(例えば、ブタ、ウシ、家禽及び魚類において筋肉成長を促進すること)、
(ii)例えばブタ、ウシ、家禽及び魚類において、食物の身体組織への変換の効率を促進すること(脂肪に対するやせの割合)及び
(iii)泌乳動物、例えば、乳牛、ヒツジ、ヤギなどにおいて乳汁産生を促進することが含まれる。他の獣医学的治療適用には、以下に限定されないが、
(iv)例えばイヌ、ネコ及びウマなどのペット動物において、悪液質、外傷性傷害又は他の消耗疾患に付随する消耗性の症状に対して動物を治療すること及び
(v)新生仔の状態を向上させるために例えば泌乳雌ブタにおいて、新生仔の健康を増進するために泌乳動物を処置することが含まれる。
【0074】
投与方法
遺伝子送達ビヒクルの使用を含む様々な方法において本発明のポリペプチドを送達することができる。本発明のポリペプチドの治療のための送達、例えば、細胞の遺伝子移入、遺伝子治療、送達ビヒクル又は医薬的に許容可能な担体による直接投与、本ポリペプチドをコードする核酸を含有する組み換え細胞を提供することによる間接的送達のために、タンパク質又は核酸などの物質の治療的送達のための当技術分野で公知の方法を使用することができる。
【0075】
様々な送達系が公知であり、本発明のポリペプチドを投与するために、例えばリポソーム中でのカプセル封入、微小粒子、マイクロカプセル、タンパク質を発現させることができる組み換え細胞、受容体介在エンドサイトーシス(例えば、Wu及びWu、J Biol Chem 262:4429−4432、1987)、レトロウイルス、アデノ関連ウイルス、アデノウイルス、ポックスウイルス(例えば、アビポックスウイルス、特に鶏痘ウイルス)又は他のベクターなどの一部としての核酸のコンストラクトなどを使用することができる。導入方法は、腸内又は非経口であり得、以下に限定されないが、皮内、筋肉内、腹腔内、静脈内、皮下、肺、鼻内、眼内、硬膜外及び経口経路が含まれる。何れかの従来の経路により、例えば点滴又はボーラス注射により、上皮又は皮膚粘膜層(例えば、口腔粘膜、直腸及び腸粘膜など)を通じた吸収により本ポリペプチドを投与することができ、他の生物学的に活性のある物質とともに投与し得る。投与は全身的又は局所であり得る。さらに、脳室内及び鞘内注入を含む何らかの適切な経路により、中枢神経系に本発明の医薬組成物を導入することが所望され得;例えば、Ommayaリザーバーなどのリザーバーに連結した脳室内カテーテルにより脳室内注射を促進し得る。例えば、吸入器又はネブライザー及びエアロゾル化剤による処方物の使用により、肺投与を使用することもできる。
【0076】
具体的な実施形態において、治療を必要とする部位に局所的に本発明の医薬組成物を投与することが所望され得;例えば、限定されないが、術中の局所点滴、局所適用により、例えば注射によって、カテーテルによって、又はインプラントによって、多孔、非多孔又はゼラチン性材料であるインプラント(シアラスティック(sialastic)膜などの膜を含む。)、繊維又は市販の皮膚代替物によって、これを遂行し得る。
【0077】
別の実施形態において、小胞、特にリポソームにおいて、活性物質を送達することができる(Langer、Science 249:1527−1533、1990参照)。さらに別の実施形態において、制御放出系において活性物質を送達することができる。ある実施形態において、ポンプを使用し得る。別の実施形態において、ポリマー材料を使用することができる(Howardら、J Neurosurg 71:105、1989)。別の実施形態において、本発明の活性物質が本発明のポリペプチドをコードする核酸である場合、適切な核酸発現ベクターの一部としてそれを構築すること及び、例えばレトロウイルスベクターを使用することによって又は直接注入によって又は微小粒子ボンバーメント(例えば遺伝子銃;Biolistic、Dupont)によって又は脂質もしくは細胞表面受容体での被覆又は遺伝子移入剤によって又は核に入ることが知られているホメオボックス様ペプチドへ連結してそれを投与することによって(例えば、Joliotら、Proc.Natl.Acad.Sci.USA 88:1864−1868、1991)など、細胞内に入るようにそれを投与することによって、そのコードされるタンパク質の発現を促進するためにインビボで核酸を投与することができる。あるいは、細胞内に核酸を導入し、相同組み換えによって、発現のために宿主細胞DNA内に組み込むことができる。
【0078】
細胞の遺伝子移入及び遺伝子治療
本発明は、インビトロ及びインビボでの細胞の遺伝子移入のための本発明のポリペプチドをコードする核酸の使用を包含する。標的細胞及び生物の遺伝子移入のために、多くの周知のベクターの何れかにこれらの核酸を挿入することができる。ベクター及び標的細胞の相互作用を通じて、エクスビボ及びインビボで細胞に核酸を遺伝子移入する。治療反応を誘発するのに十分な量で対象に本組成物を(例えば筋肉への注射により)投与する。
【0079】
別の態様において、本発明は、標的部位、即ち標的細胞又は組織、動物を処置する方法(本発明のポリペプチドをコードする核酸を細胞に遺伝子移入することを含む。)を提供するが、この核酸は、標的融合ポリペプチドをコードする核酸に操作可能に連結された誘導可能なプロモーターを含む。
【0080】
併用療法
多くの実施形態において、1以上のさらなる化合物又は治療と組み合わせて本発明のポリペプチドを投与することができる。例えば、1以上の治療用化合物とともに複数のポリペプチドを同時投与することができる。併用療法は、同時に行うか又は交互に行う投与を包含し得る。さらに、この組み合わせは、急性投与は長期投与を包含し得る。テストステロン又は特異的アンドロゲン受容体調節因子(SARM)などの同化剤と組み合わせて本発明のポリペプチドを投与することができる。さらなる同化剤には、成長ホルモン(GH)又は、GH放出を誘導する分子が含まれる。グレリンは食欲を増進させ得るので、悪液質に対する併用療法においてグレリンは特に有用である。同様に、同化作用を向上させるために本発明のポリペプチドをタンパク質サプリメントと組み合わせることができるか又は体重を増加させるために理学療法又は運動と組み合わせることができる。ミオスタチンを阻害する何らかの分子も併用療法に対する候補である。
【0081】
医薬組成物
本発明はまた、本発明のIGF前駆タンパク質及び医薬的に許容可能な担体を含む医薬組成物も提供する。「医薬的に許容可能な」という用語は、連邦もしくは州政府の監督官庁により承認されている又は米国薬局方又は動物もしくはヒトでの使用に関する他の一般に認められている薬局方に列挙されていることを意味する。「担体」という用語は、治療薬と一緒に投与される、希釈剤、アジュバント、賦形剤又はビヒクルを指す。このような医薬担体は、水及び油(石油、動物、植物又は合成起源の油を含む。ピーナツ油、ダイズ油、鉱物油、ゴマ油など)などの滅菌液であり得る。適切な医薬賦形剤には、デンプン、グルコース、ラクトース、スクロース、ゼラチン、麦芽、米、小麦粉、チョーク、シリカゲル、ステアリン酸ナトリウム、モノステアリン酸グリセロール、タルク、塩化ナトリウム、脱脂粉乳、グリセロール、プロピレン、グリコール、水、エタノールなどが含まれる。必要に応じて、本組成物はまた、少量の湿潤もしくは乳化剤又はpH緩衝剤も含有し得る。これらの組成物は、溶液、懸濁液、乳剤、錠剤、丸剤、カプセル、粉末、徐放製剤などの形態を取り得る。伝統的な結合剤及びトリグリセリドなどの担体とともに坐薬として本組成物を処方することができる。経口製剤は、医薬グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなどの標準的担体を含み得る。適切な医薬担体の例は、E.W.Martinによる「Remington’s Pharmaceutical Sciences」に記載されている。
【0082】
いくつかの実施形態において、静脈内投与に適した医薬組成物として常法に従い本組成物を処方する。必要な場合、本組成物は、可溶化剤及び注射部位の痛みを緩和するためのリドカインなどの局所麻酔も含み得る。本組成物を点滴により投与すべき場合、医薬グレードの滅菌水又は食塩水を含有する点滴ボトルを用いて投与することができる。本組成物を注射により投与すべき場合、投与前に成分を混合することができるように、注射用滅菌水又は食塩水のアンプルを提供し得る。
【0083】
中性又は塩の形態で本発明のポリペプチドを処方することができる。医薬的に許容可能な塩には、例えば塩酸、リン酸、酢酸、シュウ酸、酒石酸など由来ものなどの遊離アミノ基とともに形成されるもの、ナトリウム、カリウム、アンモニウム、カルシウム、水酸化鉄、イソプロピルアミン、トリエチルアミン、2−エチルアミノエタノール、ヒスチジン、プロカインなど由来のものなどの遊離カルボキシル基とともに形成されるものなどが含まれる。
【0084】
現在の記述に基づく標準的な臨床技術により、状態又は疾病の治療に有効である本発明のポリペプチドの量を決定することができる。さらに、最適投与量範囲の決定を促進するためにインビトロアッセイを場合によっては使用し得る。処方において使用すべき正確な用量もまた、投与経路及び状態の重症度に依存し、医師の判断及び各対象の事情に従い決定すべきである。しかし、静脈内投与のための適切な投与量範囲は通常、体重1kgあたり活性化合物約20−5000mgである。鼻内投与のための適切な投与量範囲は通常、約0.01pg/kg体重から1mg/kg体重である。有効用量は、インビトロ又は動物モデル試験系から得られた用量反応曲線から推定し得る。特に、可能性がある用法用量は、約60から120μg/kg体重で、皮下注射で1日2回であり得る。
【0085】
動物への使用
健康な動物に投与する場合、疾患に罹患している動物と投与量が異なり得る。例えば下記のような筋芽細胞増殖アッセイ(実施例79)又は哺乳動物上皮組織アッセイ(実施例80)など、当技術分野で公知のアッセイを用いて、適切な投与量の評価が当業者により容易に行われ得る。IGFを測定するための一般的なアッセイも実施例81におけるものなど、当技術分野で公知である。
【0086】
当業者にとって当然のことながら、ある種の動物は、光周期の長さにより影響を受ける季節性の妊孕性を示す。獣医学における方法又は使用の何らかの実施形態は、場合によっては、所望の効果を達成するために、その動物の生殖サイクル内の特定の時間に治療法を開始することを含み得る。当業者は、生殖状態及びサイクルを容易に決定することができること、必要に応じて、適切な方法の使用により同調させることができることを知っている。
【0087】
動物への適応のために使用する場合、本発明のIGF−1又はIGF−2ペプチドを経口用ドレンチ又は動物に対する経口もしくは固形飼料に対するサプリメントとして使用することもできる。
【0088】
本発明はさらに、次の実施例を記載するが、これに限定されない。
【実施例】
【0089】
(実施例1)
次の修飾を含有するhIGF−1−Ea前駆ポリペプチドをコードするDNA発現ベクターを構築した:G1の欠失、P2の欠失及びE3の欠失;R37のAへの変異;及びR71の欠失及びS72の欠失。これらの変異を、本開示を通じて「3mut」と呼ぶことがある。この結果、次の分泌タンパク質配列が得られる:
【0090】
【化3】
【0091】
10%ウシ胎仔血清、100U/mLペニシリン及び100μg/mLストレプトマイシンを含有するDMEM中でCos7細胞(ATCCより入手可能)を維持し、10cmプレートあたり1x106個の細胞の密度で播種した。これらの細胞培養物に対して、製造者の説明書に従い、Fugene(Roche)を用いて発現プラスミド8μgを遺伝子移入した。遺伝子移入から24時間後、細胞を1度洗浄し、血清不含培地中で48時間培養した。上清を回収し、−80℃で保存した。
【0092】
ヒト血清中でのポリペプチドの安定性を評価するために、野生型(wt)hIGF−1Ea及びhIGF−1Ea3mutで遺伝子移入したCos7細胞から回収した上清を10%ヒト血清(Sigma)非存在下又は存在下の何れかで37℃で16時間温置した。18%SDS−PAGEにより試料を分離し、ヒトIGF−1に対するヤギポリクローナル抗体を用いて免疫ブロッティングを行った。図1Aの結果から、16時間、血清とともに温置後、hIGF−1Eaが実質的に分解された一方、hIGF−1Ea3mutは安定であったことが示された。密度測定から、切断されたIGF−1に対する非切断IGF−1の比が約1:6.2であり、一方hIGF−1Ea3mutの場合の比は約1:0.68であることが示され、このことから、これらの変異の結果、ポリペプチドが安定化されることが分かる。
【0093】
hIGF−1Ea3mutがIGF−1Rを通じてシグナルを送ることができることを確認するために、本ポリペプチドと接触させた細胞のAKTリン酸化を測定した。C2C12をATCCから購入し、10%ウシ胎仔血清(AMIMED)、100U/mLペニシリン(Invitrogen)、100μg/mLストレプトマイシン(Invitrogen)及び2mMグルタミン(Invitrogen)を含有する高グルコース入りのダルベッコの改変イーグル培地(DMEM)(Invitrogen)中で維持した。AKTリン酸化の分析のために、6ウェルプレートのウェルあたり0.15x106個の細胞密度でC2C12細胞を播種し、増殖培地中で72時間培養した。血清不含培地中で細胞を4時間飢餓状態にし、次いで37℃で30分間、様々なリガンドで刺激した。様々なプロテアーゼ阻害剤を含有するPhosphoSafe緩衝液(Cell Signaling)で細胞を溶解し、4℃で14,000xgで15分間遠心することにより透明化した。PathScan phospho AKT(Ser473)サンドイッチELISAキット及びPathScan AKTサンドイッチELISAキット(Cell Signaling)をそれぞれ用いてELISAによって、AKTリン酸化及び総AKTレベルを分析した。AKTリン酸化の結果を図2Aでまとめるが、これから、hIGF−1Ea3mutが、long−R3−IGF1陽性対照試薬及び組み換えIGF−1と同程度にIGF−1R細胞経路を活性化できることが示される。さらに、図5のデータから、hIGF−1Ea3mutがAKTリン酸化へと導いたことが直接示される。
【0094】
次に、hIGF−1Ea3mutの受容体特異性がIGF−1Rで残っていることを確認するために、IGF−1R又はインスリン受容体(InsR)の何れかを過剰発現するNIH3T3の培養物に様々なリガンドを添加した。Cos7細胞に対して上記で述べたものと同じ条件下でこれらの細胞を培養した。IGF−1R及びInsRリン酸化の分析のために、6ウェルプレートのウェルあたり0.2x106個の細胞密度でNIH3T3−IGF1R及びNIH3T3−InsR細胞を播種し、増殖培地中で24時間培養した。血清不含培地中で18時間細胞を飢餓状態にし、次いで37℃で10分間、様々なリガンドで刺激した。AKT実験のために上記で述べたように細胞を溶解し、DuoSet ICヒト蛍光体−IGF1R及び−InsR ELISAキット(R&D Systems)を用いてELISAによってIGF−1R及びInsRリン酸化レベルを分析した。図3A及び3Bでまとめた結果から、このIGF−1前駆ポリペプチドがIGF−1受容体に対して特異性を保持し、関連するインスリン受容体と低親和性で結合するはずであることが示される。
【0095】
(実施例2)
次の変異:G1の欠失、P2の欠失及びE3の欠失;R37のAへの変異;及びR71の欠失及びS72の欠失(即ち「3mut」)を含有するhIGF−1−Eb前駆ポリペプチドをコードするDNA発現ベクターを構築した。この結果、次の分泌タンパク質配列が得られる:
【0096】
【化4】
【0097】
上記実施例1に記載の手順に従い、このポリペプチドをアッセイした。図1B及び密度測定の使用から、切断されたIGF−1に対する非切断IGF−1の比が約1:9であり、hIGF−1Eb3mutの場合の比が約1:1であることが示され、このことから、これらの修飾の結果、ポリペプチドが安定化されることが分かった。図2Bから、hIGF−1Eb3mutがlong−R3−IGF1陽性対照試薬及び組み換えIGF−1と同程度にIGF−1R細胞経路を活性化できたことが示される。さらに、図5のデータから、hIGF−1Eb3mutがAKTリン酸化へと導いたことが直接示される。図3C及び3Dでまとめた結果から、このIGF−1前駆ポリペプチドがIGF−1受容体に対して特異性を保持しており、関連するインスリン受容体と低親和性で結合するはずであることが示される。
【0098】
(実施例3)
次の変異:G1の欠失、P2の欠失及びE3の欠失;R37のAへの変異;及びR71の欠失及びS72の欠失(即ち「3mut」)を含有するhIGF−1−Ec前駆ポリペプチドをコードするDNA発現ベクターを構築した。この結果、次の分泌タンパク質配列が得られる:
tlcgaelvdalqfvcgdrgfyfnkptgygsssrapqtgivdeccfrscdlrrlemycaplkpaksavraqrhtdmpktqkyqppstnkntksqrrkgstfeerk(配列番号10)
【0099】
上記実施例1に記載の手順に従い、このポリペプチドをアッセイした。図1C及び密度測定の使用から、切断されたIGF−1に対する非切断IGF−1の比が約1:5であり、hIGF−1Ec3mutの場合の比が約1:0.96であることが示され、このことから、これらの修飾の結果、ポリペプチドが安定化されることが分かった。図2Dから、hIGF−1Ec3mutがlong−R3−IGF1陽性対照試薬及び組み換えIGF−1と同程度にIGF−1R細胞経路を活性化できたことが示される。さらに、図5のデータから、hIGF−1Ec3mutがAKTリン酸化へと導いたことが直接示される。図4A及び4Bでまとめられた結果から、このIGF−1前駆ポリペプチドがIGF−1受容体に対して特異性を保持しており、関連するインスリン受容体と低親和性で結合するはずであることが示される。
【0100】
(実施例4)
hIGF−1−Ebペプチドに対して次の修飾:G1の欠失、P2の欠失及びE3の欠失;R37のAへの変異;及びR71の欠失及びS72の欠失(即ち「3mut」);及びEbのアミノ酸95と96との間のEaアミノ酸93から102の挿入を含有するhIGF−1−Eabキメラ前駆ポリペプチドをコードするDNA発現ベクターを構築した。この挿入によって、N92に推定N−結合型グリコシル化シグナルが生成する。この結果、次の分泌タンパク質配列が得られる。
【0101】
【化5】
【0102】
上記実施例1に記載の手順のいくつかに従い、このポリペプチドをアッセイした。図2Cから、hIGF−1Eab3mutがlong−R3−IGF1陽性対照試薬及び組み換えIGF−1と同程度にIGF−1R細胞経路を活性化できたことが示される。図4C及び4Dでまとめられた結果から、このIGF−1前駆ポリペプチドがIGF−1受容体に対して特異性を保持しており、インスリン受容体を活性化しないことが示される。
【0103】
(実施例5)
次の変異:G1の欠失、P2の欠失及びE3の欠失、R36の欠失、R37の欠失、R71の欠失、S72の欠失、Ebの最後の7個のC末端アミノ酸の欠失;及び、2つのさらなるEbペプチド(両方とも、R71及びS72及び最後の7個のC末端のアミノ酸がない。)及び4番目及び最後のEbペプチド(R71及びS72がない。)のこの前駆体のC末端への挿入を含有するhIGF−1−Ebマルチマー前駆ポリペプチドをコードするDNA発現ベクターを構築した。図6Aは、このコンストラクトの概略図を示す。この結果、次の分泌タンパク質配列が得られる。
【0104】
【化6】
【0105】
実施例1に記載のようにこのポリペプチドをAKTリン酸化アッセイに供した。図5から、このhIGF−1−EbマルチマーがIGF−1R経路を通じてシグナルを送ることができたことが示される。
【0106】
(実施例6)
図6Bで概略が示されるような本発明のhIGF−1−Eb前駆ポリペプチドを発現させることができる。このコンストラクトは、次の修飾:G1の欠失、P2の欠失及びE3の欠失、R36の欠失、R37の欠失、R71の欠失、S72の欠失;及びEbのアミノ酸95と96との間のEaアミノ酸93−102の挿入(これにより、位置N92及びN100でN結合型グリコシル化部位が生じる。)を含有する。この結果、次の分泌タンパク質配列が得られる。
【0107】
【化7】
【0108】
(実施例7)
次の修飾:P4の欠失、S5の欠失及びE6の欠失;R38のAへの変異;及びR68の欠失及びD69の欠失を有する本発明のhIGF−2−E前駆ポリペプチドを発現させることができる。この結果、次の分泌タンパク質配列が得られる。
【0109】
【化8】
【0110】
(実施例8)
次の変異:G1の欠失及びP2の欠失;E3のXへの変異(Xは、PEG化された非天然アミノ酸である。);R37のAへの変異;及びR71の欠失及びS72の欠失を有する本発明のhIGF−1−Ea前駆ポリペプチドを発現させることができる。この結果、次の分泌タンパク質配列が得られる。
【0111】
【化9】
【0112】
(実施例9−78)
(△=欠失)
9)hIGF−1−Ea:△G1、△P2、△E3;R36A;△R71
【0113】
【化10】
【0114】
10)hIGF−1−Ea:△G1、△P2、△E3;R36A;△S72
【0115】
【化11】
【0116】
10)hIGF−1−Ea:△G1、△P2、△E3;R36A;△R71、△S72
【0117】
【化12】
【0118】
11)hIGF−1−Ea:△G1、△P2、△E3;R37A;△R71
【0119】
【化13】
【0120】
12)hIGF−1−Ea:△G1、△P2、△E3;R37A;△S72
【0121】
【化14】
【0122】
13)hIGF−1−Ea:△G1、△P2、△E3、△R37;△R71
【0123】
【化15】
【0124】
14)hIGF−1−Ea:△G1、△P2、△E3、△R37;△S72
【0125】
【化16】
【0126】
15)hIGF−1−Ea:△G1、△P2、△E3;△R37;△R71、△S72
【0127】
【化17】
【0128】
16)hIGF−1−Eb:△G1、△P2、△E3;R36A;△R71
【0129】
【化18】
【0130】
17)hIGF−1−Eb:△G1、△P2、△E3;R36A;△S72
【0131】
【化19】
【0132】
18)hIGF−1−Eb:△G1、△P2、△E3;R37A;△R71
【0133】
【化20】
【0134】
19)hIGF−1−Eb:△G1、△P2、△E3;R37A;△S72
【0135】
【化21】
【0136】
20)hIGF−1−Eb:△G1、△P2、△E3;R37A;△R71、△S72
【0137】
【化22】
【0138】
21)hIGF−1−Eb:△G1、△P2、△E3、△R37;△R71
【0139】
【化23】
【0140】
22)hIGF−1−Eb:△G1、△P2、△E3、△R37;△S72
【0141】
【化24】
【0142】
23)hIGF−1−Eb:△G1、△P2、△E3、△R37;△R71、△S72
【0143】
【化25】
【0144】
24)hIGF−1−Ec:△G1、△P2、△E3;R36A;△R71
【0145】
【化26】
【0146】
25)hIGF−1−Ec:△G1、△P2、△E3;R36A;△S72
【0147】
【化27】
【0148】
26)hIGF−1−Ec:△G1、△P2、△E3;R36A;△R71、△S72
【0149】
【化28】
【0150】
27)hIGF−1−Ec:△G1、△P2、△E2;R37A;△R71
【0151】
【化29】
【0152】
28)hIGF−1−Ec:△G1、△P2、△E3;R37A;△S72
【0153】
【化30】
【0154】
29)hIGF−1−Ec:△G1、△P2、△E3;R37A;△R71、△S72
【0155】
【化31】
【0156】
30)hIGF−1−Ec:△G1、△P2、△E3、△R37、△R71
【0157】
【化32】
【0158】
31)hIGF−1−Ec:△G1、△P2、△E3、△R37、△S72
【0159】
【化33】
【0160】
32)hIGF−1−Eab:△G1、△P2、△E3;R36A;△R71;Ebのaa95と96との間のEa aa93−102の挿入(即ち「Eab」)
【0161】
【化34】
【0162】
33)hIGF−1−Eab:△G1、△P2、△E3;R37A;△R71
【0163】
【化35】
【0164】
34)hIGF−1−Eab:△G1、△P2、△E3、△R37、△R71
【0165】
【化36】
【0166】
35)hIGF−1−Eab:△G1、△P2、△E3;R36A;△S72
【0167】
【化37】
【0168】
36)hIGF−1−Eab:△G1、△P2、△E3;R37A;△S72
【0169】
【化38】
37)hIGF−1−Eab:△G1、△P2、△E3、△R37、△S72
【0170】
【化39】
【0171】
38)hIGF−1−Eab:△G1、△P2、△E3;R36A;△R71、△S72
【0172】
【化40】
【0173】
39)hIGF−1−Eab:△G1、△P2、△E3、△R37、△R71、△S72
【0174】
【化41】
【0175】
40)hIGF−1−Ea:△P2、△E3;R37A;△R71、△S72
【0176】
【化42】
【0177】
41)hIGF−1−Eb:△P2、△E3;R37A;△R71、△S72
【0178】
【化43】
【0179】
42)hIGF−1−Ebマルチマー:(△G1、△P2、△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0180】
【化44】
【0181】
43)hIGF−1−Ebマルチマー:(△P2、△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0182】
【化45】
【0183】
44)hIGF−1−Ec:△P2、△E3;R37A;△R71、△S72
【0184】
【化46】
【0185】
45)hIGF−1−Ea:△E3;R37A;△R71、△S72
【0186】
【化47】
【0187】
46)hIGF−1−Eb:△E3;R37A;△R71、△S72
【0188】
【化48】
【0189】
47)hIGF−1−Ebマルチマー:(△G1、△P2、△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0190】
【化49】
【0191】
48)hIGF−1−Ebマルチマー:(△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0192】
【化50】
【0193】
49)hIGF−1−Ec:△E3;R37A;△R71、△S72
【0194】
【化51】
【0195】
50)hIGF−1−Ea:△E3;R37A;△R71、△S72
【0196】
【化52】
【0197】
51)hIGF−1−Eb:△E3;R37A;△R71、△S72
【0198】
【化53】
【0199】
52)hIGF−1−Ec:△E3;R37A;△R71、△S72
【0200】
【化54】
【0201】
53)hIGF−1−Eab:△E3;R37A;△R71、△S72
【0202】
【化55】
【0203】
54)hIGF−1−Ebマルチマー:(△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0204】
【化56】
【0205】
55)hIGF−1−Ea:E3A;R37A;△R71、△S72
【0206】
【化57】
【0207】
56)hIGF−1−Eb:E3A;R37A;△R71、△S72
【0208】
【化58】
【0209】
57)hIGF−1−Ec:E3A;R37A;△R71、△S72
【0210】
【化59】
【0211】
58)hIGF−1−Eab:E3A;R37A;△R71、△S72
【0212】
【化60】
【0213】
59)hIGF−1−Ebマルチマー:(E3A;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0214】
【化61】
【0215】
60)hIGF−1−Ea:△P2、△E3;R37A;△R71、△S72
【0216】
【化62】
【0217】
61)hIGF−1−Eb:△P2、△E3;R37A;△R71、△S72
【0218】
【化63】
【0219】
62)hIGF−1−Ec:△P2、△E3;R37A;△R71、△S72
【0220】
【化64】
【0221】
63)hIGF−1−Eab:△P2、△E3;R37A;△R71、△S72
【0222】
【化65】
【0223】
64)hIGF−1−Ebマルチマー:(△P2、△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0224】
【化66】
【0225】
65)MGF−1−Eb:△G1、△P2;E3X;R37A;△R71、△S72
【0226】
【化67】
【0227】
66)hIGF−1−Ec:△G1、△P2;E3X;R37A;△R71、△S72
【0228】
【化68】
【0229】
67)hIGF−1−Eab:△G1、△P2;E3X;R37A;△R71、△S72
【0230】
【化69】
【0231】
68)hIGF−1−Ebマルチマー:(△G1、△P2;E3X;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0232】
【化70】
【0233】
69)hIGF−1−Ea:△G1、△P2、△E3;R37A;△R71;S72X
【0234】
【化71】
【0235】
70)hIGF−1−Eb:△G1、△P2、△E3;R37A;△R71;S72X
【0236】
【化72】
【0237】
71)hIGF−1−Ec:△G1、△P2、△E3;R37A;△R71;S72X
【0238】
【化73】
【0239】
72)hIGF−1−Eab:△G1、△P2、△E3;R37A;△R71;S72X
【0240】
【化74】
【0241】
73)hIGF−1−Ebマルチマー:(△G1、△P2、△E3;R37A)−Eb(△R71;S72X;△C末端7aa)−2xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72)
【0242】
【化75】
【0243】
74)hIGF−1−Ea:△G1、△P2、△E3;R37A;△R71、△S72;N92X
【0244】
【化76】
【0245】
75)hIGF−1−Eb:△G1、△P2、△E3;R37A;△R71、△S72;C142X
【0246】
【化77】
【0247】
76)hIGF−1−Eab:△G1、△P2、△E3;R37A;△R71、△S72;C151X
【0248】
【化78】
【0249】
78)hIGF−1−Ebマルチマー(△G1、△P2、△E3;R37A)−3xEb(△R71、△S72、△C末端7aa)−Eb(△R71、△S72;C71X)
【0250】
【化79】
【0251】
(実施例79)
筋芽細胞増殖アッセイ
筋芽細胞増殖アッセイは、信頼できるIGF活性のインビトロ指標を与え、胚性筋芽細胞及び成体衛星細胞に影響を与える因子に対するモデルとして使用される。この系において活性のある因子は、筋芽細胞の初代培養において同様に作用する。本発明のペプチドによるインビトロでの筋芽細胞増殖の促進から、筋芽細胞増殖を向上させること及び、従って子宮内での最終的な筋繊維数を向上させることにおいて、その活性が示される。さらに、筋芽細胞増殖の同様の促進から、(例えば衛星筋肉細胞増殖の刺激を介して)成体の筋肉肥大を促進するために本発明のペプチドを使用することができることが示される。
【0252】
(実施例80)
哺乳動物上皮組織アッセイ
泌乳動物において、乳房上皮組織は乳汁を産生し分泌する細胞なので、この組織の量は、乳汁産生の制限要因である。インビトロ系を用いて、増殖させ乳成分の産生量を向上させるために、本発明の修飾IGF−1又はIGF−2により、動物の乳線から得られる上皮細胞を刺激することができる。さらに、あるこのようなインビトロ細胞系で増殖させるために刺激した乳房上皮細胞を、除去した乳房脂肪体に再移植し、雌泌乳動物において増殖及び/又は乳汁を産生させるために刺激することができる。
【0253】
(実施例81)
血液又は他の体液中のIGF−1又はIGF−2の測定
用量反応曲線によって投与につき非経口投与されるペプチドの有効量を測定することができる。例えば、投薬を決定するために、処置すべき対象の血液又は体液中の本発明の修飾IGFペプチドを測定することができる。あるいは、対象にペプチドの漸増量を投与し、修飾IGF−1及びIGF−2に対する対象の血清レベルを調べることができる。修飾IGF−1又はIGF−2のこれらの血清レベルに基づくモル基準で、使用すべきペプチドの量を計算することができる。
【0254】
ペプチドの適切な投薬を決定するためのある方法は、体液又は血液などの生体液中の本発明のIGFペプチドを測定することを伴う。RIA及びELISAを含む何らかの手段によって、このようなレベルの測定を行うことができる。IGFレベルを測定した後、単回投与又は複数回投与を用いて本ペプチドと生体液を接触させる。この接触段階の後、生体液中のIGFレベルを再測定する。生体液IGFレベルが、所望の効率(そのためにこの分子が投与されるべきである。)を得るのに十分な量低下した場合、最大効率が得られるようにその分子の用量を調整することができる。インビトロ又はインビボでこの方法を行い得る。好ましくは、インビボで、即ち、生体液を対象から採取した後にこの方法を行い、IGFレベルを測定し、単回又は複数回投与により本明細書中のペプチドを哺乳動物に投与し(即ち、動物に投与することによりこの接触段階を行う。)、次いで動物から採取した生体液からIGFレベルを再測定する。
【0255】
投薬を決定するための別の方法は、本ペプチドに対する抗体又はLIFA形式でのペプチドに対する別の検出方法を使用することである。
【0256】
(実施例82)
hIGF−1−Ec 3mutのインビボ薬物動態
1mg/kgのrhIGF−1及び1.55mg/kgのhIGF−1−Ec 3mut(実施例3に記載)を、成熟雄マウス(n=3/群)に静脈内(i.v.)ボーラス注射した。試験物質の投与から5、15、30及び60分後、連続血液検体を回収した。rhIGF−1及びhIGF−1−Ec 3mutの血清濃度をELISAにより測定した。このアッセイはhIGF−1に特異的である。
【0257】
rhIGF−1及びhIGF−1−Ec 3mutの等モル用量をマウスにi.v.投与した。この結果から、試験した全ての時間点で、rhIGF−1と比較して、hIGF−1−Ec 3mutタンパク質のレベルが有意に高いことが分かり、このことから、hIGF−1−Ec 3mutは70アミノ酸−long IGF−1よりも代謝的に安定であることが示される。
【0258】
【表1】
【特許請求の範囲】
【請求項1】
プロテアーゼによるIGF−1からのE−ペプチドの切断が、非ヒト動物IGF−1前駆タンパク質の修飾により減少する、非ヒト動物IGF−1前駆タンパク質を含むポリペプチド。
【請求項2】
非ヒト動物IGF−1前駆タンパク質が非ヒト脊椎動物種由来である、請求項1に記載のポリペプチド。
【請求項3】
非ヒト脊椎動物種が、マウス、ラット、ウシ、ブタ、ウマ、ヒツジ、ヤギ、鳥類、イヌ、ネコ又は魚類から選択される、請求項2に記載のポリペプチド。
【請求項4】
前駆タンパク質がEaペプチドを含む、請求項1から請求項3の何れか1項に記載のポリペプチド。
【請求項5】
Eaペプチドが図8Aの非ヒト動物配列から選択される、請求項4に記載のポリペプチド。
【請求項6】
前駆タンパク質がEbペプチドを含む、請求項1から請求項3の何れか1項に記載のポリペプチド。
【請求項7】
Ebペプチドが図9Aの非ヒト動物配列から選択される、請求項6に記載のポリペプチド。
【請求項8】
Ebの最後の7個のC末端アミノ酸が欠失している、請求項6又は請求項7に記載のポリペプチド。
【請求項9】
前駆タンパク質がEcペプチドを含む、請求項1から請求項3の何れか1項に記載のポリペプチド。
【請求項10】
Ecが図10Aの非ヒト動物配列から選択される、請求項9に記載のポリペプチド。
【請求項11】
前駆タンパク質のG1が欠失している又は変異させられている、請求項1から請求項10の何れかに記載のポリペプチド。
【請求項12】
前駆タンパク質のP2が欠失している又は変異させられている、請求項1から請求項11の何れかに記載のポリペプチド。
【請求項13】
前駆タンパク質のE3が欠失している又は変異させられている、請求項1から請求項12の何れかに記載のポリペプチド。
【請求項14】
前駆タンパク質のR36が欠失している又は変異させられている、請求項1から請求項13の何れかに記載のポリペプチド。
【請求項15】
R36がアラニンに変異させられている、請求項1から請求項14の何れかに記載のポリペプチド。
【請求項16】
前駆タンパク質のR37が欠失している又は変異させられている、請求項1から請求項15の何れかに記載のポリペプチド。
【請求項17】
R37がアラニンに変異させられている、請求項1から請求項16の何れかに記載のポリペプチド。
【請求項18】
N−結合型グリコシル化コンセンサス配列NXS/Tをさらに含む、請求項1から請求項17の何れかに記載のポリペプチド。
【請求項19】
Ebのアミノ酸N95とT96との間に挿入されたEaのアミノ酸93−102をさらに含む、請求項6に記載のEbポリペプチド。
【請求項20】
前駆タンパク質のアミノ酸側鎖に共有結合されたオリゴ糖をさらに含む、請求項1から請求項19の何れかに記載のポリペプチド。
【請求項21】
前駆タンパク質のアルギニン側鎖にオリゴ糖が共有結合されている、請求項20に記載のポリペプチド。
【請求項22】
前駆タンパク質の残基が非天然アミノ酸により置換されている、請求項1から請求項21の何れかに記載のポリペプチド。
【請求項23】
非天然アミノ酸がアセチレン又はアジド基を含む、請求項22に記載のポリペプチド。
【請求項24】
前駆タンパク質の側鎖に共有結合されているポリ(エチレングリコール)部分をさらに含む、請求項1から請求項23の何れかに記載のポリペプチド。
【請求項25】
前駆タンパク質のC末端に連結されているさらなるE−ペプチドをさらに含む、請求項1から請求項24の何れかに記載のポリペプチド。
【請求項26】
N末端からC末端までにて、
第一のEbペプチドを含むIGF−1前駆タンパク質(このタンパク質中、G1、P1及びE1が欠失しており、R36及びR37が欠失しており、R71及びS72が欠失しており、第一のEbペプチドの最後の7個のC末端アミノ酸が欠失している。)と;
第二のEbペプチド(このペプチド中、R71、S72及び第二のEbペプチドの最後の7個のC末端アミノ酸が欠失している。)と;
第三のEbペプチド(このペプチド中、R71、S72及び第三のEbペプチドの最後の7個のC末端アミノ酸が欠失している。)と;及び
第四のEbペプチド(このペプチド中、R71及びS72が欠失している。)と、
を含むポリペプチド。
【請求項27】
前駆タンパク質のR71又はS72が欠失している又は変異させられている、請求項1から請求項26の何れかに記載のポリペプチド。
【請求項28】
プロテアーゼによるIGF−2からのE−ペプチドの切断が、非ヒト動物IGF−2前駆タンパク質の修飾により減少する、非ヒト動物IGF−2前駆タンパク質を含むポリペプチド。
【請求項29】
非ヒト脊椎動物種が、マウス、ラット、ウシ、ブタ、ウマ、ヒツジ、ヤギ、鳥類、イヌ、ネコ又は魚類から選択される、請求項28に記載のポリペプチド。
【請求項30】
前駆タンパク質がIGF−2のEペプチドを含む、請求項28から請求項39の1項に記載のポリペプチド。
【請求項31】
Eペプチドが図12Aからの配列から選択される、請求項30に記載のポリペプチド。
【請求項32】
前駆タンパク質のR68又はD69が欠失している又は変異させられている、請求項28から請求項32の何れか1項に記載のポリペプチド。
【請求項33】
請求項1から請求項32の何れか1項に記載の本発明のポリペプチドの有効量を投与することを含む、非ヒト動物における成長の速度及び/又は程度を促進する方法。
【請求項34】
非ヒト動物が、ブタ、ウシ、ヒツジ、家禽及び魚類を含む食肉生産動物である、請求項33に記載の方法。
【請求項35】
請求項1から請求項32の何れか1項に記載の本発明のポリペプチドの有効量を投与することを含む、非ヒト動物における食物の身体組織への変換の効率を促進する方法。
【請求項36】
非ヒト動物が、ブタ、ウシ、ヒツジ、家禽及び魚類を含む食肉生産動物である、請求項35に記載の方法。
【請求項37】
請求項1から請求項32の何れか1項に記載の本発明のポリペプチドの有効量を投与することを含む、非ヒト泌乳動物において乳汁産生を促進する方法。
【請求項38】
非ヒト動物が乳牛、ブタ、ヒツジ及びヤギを含む泌乳動物である、請求項37に記載の方法。
【請求項39】
請求項1から請求項32の何れか1項に記載の本発明のポリペプチドの有効量を投与することを含む、悪液質、外傷性傷害又は他の消耗疾患に付随する消耗性の症状に対して非ヒト動物を治療する方法。
【請求項40】
非ヒトが、イヌ、ネコ及びウマを含むペット動物である、請求項39に記載の方法。
【請求項41】
請求項1から請求項32の何れか1項に記載の本発明のポリペプチドの有効量を投与することを含む、新生仔の健康を増進するために非ヒト泌乳動物を処置する方法。
【請求項42】
非ヒト動物が、乳牛、ブタ、ヒツジ及びヤギを含む泌乳動物である、請求項41に記載の方法。
【請求項43】
請求項33から請求項42の何れか1項に記載の方法における使用のための動物用組成物の製造のための、請求項1から請求項32の何れか1項に記載のポリペプチドの使用。
【請求項44】
前記動物用組成物が、経口用ドレンチ、経口飼料に対するサプリメント又は固形飼料に対するサプリメントである、請求項43に記載のポリペプチドの使用。
【請求項1】
プロテアーゼによるIGF−1からのE−ペプチドの切断が、非ヒト動物IGF−1前駆タンパク質の修飾により減少する、非ヒト動物IGF−1前駆タンパク質を含むポリペプチド。
【請求項2】
非ヒト動物IGF−1前駆タンパク質が非ヒト脊椎動物種由来である、請求項1に記載のポリペプチド。
【請求項3】
非ヒト脊椎動物種が、マウス、ラット、ウシ、ブタ、ウマ、ヒツジ、ヤギ、鳥類、イヌ、ネコ又は魚類から選択される、請求項2に記載のポリペプチド。
【請求項4】
前駆タンパク質がEaペプチドを含む、請求項1から請求項3の何れか1項に記載のポリペプチド。
【請求項5】
Eaペプチドが図8Aの非ヒト動物配列から選択される、請求項4に記載のポリペプチド。
【請求項6】
前駆タンパク質がEbペプチドを含む、請求項1から請求項3の何れか1項に記載のポリペプチド。
【請求項7】
Ebペプチドが図9Aの非ヒト動物配列から選択される、請求項6に記載のポリペプチド。
【請求項8】
Ebの最後の7個のC末端アミノ酸が欠失している、請求項6又は請求項7に記載のポリペプチド。
【請求項9】
前駆タンパク質がEcペプチドを含む、請求項1から請求項3の何れか1項に記載のポリペプチド。
【請求項10】
Ecが図10Aの非ヒト動物配列から選択される、請求項9に記載のポリペプチド。
【請求項11】
前駆タンパク質のG1が欠失している又は変異させられている、請求項1から請求項10の何れかに記載のポリペプチド。
【請求項12】
前駆タンパク質のP2が欠失している又は変異させられている、請求項1から請求項11の何れかに記載のポリペプチド。
【請求項13】
前駆タンパク質のE3が欠失している又は変異させられている、請求項1から請求項12の何れかに記載のポリペプチド。
【請求項14】
前駆タンパク質のR36が欠失している又は変異させられている、請求項1から請求項13の何れかに記載のポリペプチド。
【請求項15】
R36がアラニンに変異させられている、請求項1から請求項14の何れかに記載のポリペプチド。
【請求項16】
前駆タンパク質のR37が欠失している又は変異させられている、請求項1から請求項15の何れかに記載のポリペプチド。
【請求項17】
R37がアラニンに変異させられている、請求項1から請求項16の何れかに記載のポリペプチド。
【請求項18】
N−結合型グリコシル化コンセンサス配列NXS/Tをさらに含む、請求項1から請求項17の何れかに記載のポリペプチド。
【請求項19】
Ebのアミノ酸N95とT96との間に挿入されたEaのアミノ酸93−102をさらに含む、請求項6に記載のEbポリペプチド。
【請求項20】
前駆タンパク質のアミノ酸側鎖に共有結合されたオリゴ糖をさらに含む、請求項1から請求項19の何れかに記載のポリペプチド。
【請求項21】
前駆タンパク質のアルギニン側鎖にオリゴ糖が共有結合されている、請求項20に記載のポリペプチド。
【請求項22】
前駆タンパク質の残基が非天然アミノ酸により置換されている、請求項1から請求項21の何れかに記載のポリペプチド。
【請求項23】
非天然アミノ酸がアセチレン又はアジド基を含む、請求項22に記載のポリペプチド。
【請求項24】
前駆タンパク質の側鎖に共有結合されているポリ(エチレングリコール)部分をさらに含む、請求項1から請求項23の何れかに記載のポリペプチド。
【請求項25】
前駆タンパク質のC末端に連結されているさらなるE−ペプチドをさらに含む、請求項1から請求項24の何れかに記載のポリペプチド。
【請求項26】
N末端からC末端までにて、
第一のEbペプチドを含むIGF−1前駆タンパク質(このタンパク質中、G1、P1及びE1が欠失しており、R36及びR37が欠失しており、R71及びS72が欠失しており、第一のEbペプチドの最後の7個のC末端アミノ酸が欠失している。)と;
第二のEbペプチド(このペプチド中、R71、S72及び第二のEbペプチドの最後の7個のC末端アミノ酸が欠失している。)と;
第三のEbペプチド(このペプチド中、R71、S72及び第三のEbペプチドの最後の7個のC末端アミノ酸が欠失している。)と;及び
第四のEbペプチド(このペプチド中、R71及びS72が欠失している。)と、
を含むポリペプチド。
【請求項27】
前駆タンパク質のR71又はS72が欠失している又は変異させられている、請求項1から請求項26の何れかに記載のポリペプチド。
【請求項28】
プロテアーゼによるIGF−2からのE−ペプチドの切断が、非ヒト動物IGF−2前駆タンパク質の修飾により減少する、非ヒト動物IGF−2前駆タンパク質を含むポリペプチド。
【請求項29】
非ヒト脊椎動物種が、マウス、ラット、ウシ、ブタ、ウマ、ヒツジ、ヤギ、鳥類、イヌ、ネコ又は魚類から選択される、請求項28に記載のポリペプチド。
【請求項30】
前駆タンパク質がIGF−2のEペプチドを含む、請求項28から請求項39の1項に記載のポリペプチド。
【請求項31】
Eペプチドが図12Aからの配列から選択される、請求項30に記載のポリペプチド。
【請求項32】
前駆タンパク質のR68又はD69が欠失している又は変異させられている、請求項28から請求項32の何れか1項に記載のポリペプチド。
【請求項33】
請求項1から請求項32の何れか1項に記載の本発明のポリペプチドの有効量を投与することを含む、非ヒト動物における成長の速度及び/又は程度を促進する方法。
【請求項34】
非ヒト動物が、ブタ、ウシ、ヒツジ、家禽及び魚類を含む食肉生産動物である、請求項33に記載の方法。
【請求項35】
請求項1から請求項32の何れか1項に記載の本発明のポリペプチドの有効量を投与することを含む、非ヒト動物における食物の身体組織への変換の効率を促進する方法。
【請求項36】
非ヒト動物が、ブタ、ウシ、ヒツジ、家禽及び魚類を含む食肉生産動物である、請求項35に記載の方法。
【請求項37】
請求項1から請求項32の何れか1項に記載の本発明のポリペプチドの有効量を投与することを含む、非ヒト泌乳動物において乳汁産生を促進する方法。
【請求項38】
非ヒト動物が乳牛、ブタ、ヒツジ及びヤギを含む泌乳動物である、請求項37に記載の方法。
【請求項39】
請求項1から請求項32の何れか1項に記載の本発明のポリペプチドの有効量を投与することを含む、悪液質、外傷性傷害又は他の消耗疾患に付随する消耗性の症状に対して非ヒト動物を治療する方法。
【請求項40】
非ヒトが、イヌ、ネコ及びウマを含むペット動物である、請求項39に記載の方法。
【請求項41】
請求項1から請求項32の何れか1項に記載の本発明のポリペプチドの有効量を投与することを含む、新生仔の健康を増進するために非ヒト泌乳動物を処置する方法。
【請求項42】
非ヒト動物が、乳牛、ブタ、ヒツジ及びヤギを含む泌乳動物である、請求項41に記載の方法。
【請求項43】
請求項33から請求項42の何れか1項に記載の方法における使用のための動物用組成物の製造のための、請求項1から請求項32の何れか1項に記載のポリペプチドの使用。
【請求項44】
前記動物用組成物が、経口用ドレンチ、経口飼料に対するサプリメント又は固形飼料に対するサプリメントである、請求項43に記載のポリペプチドの使用。
【図1】

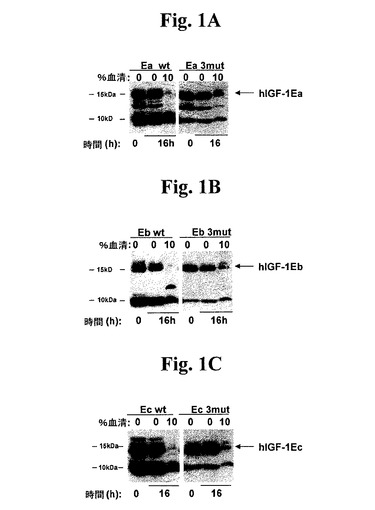
【図2A】
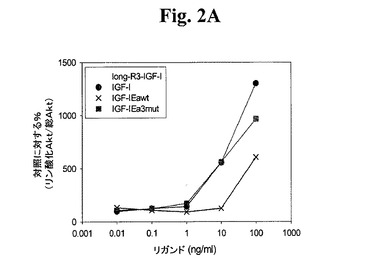
【図2B】
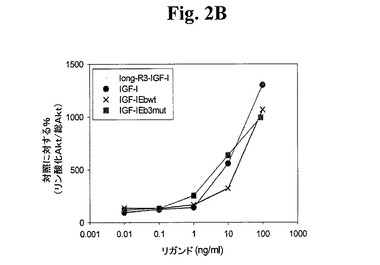
【図2C】
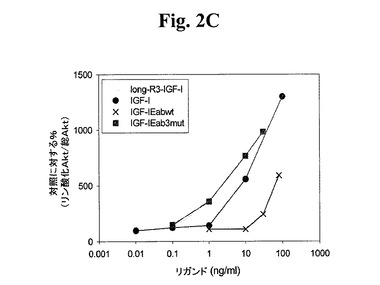
【図2D】
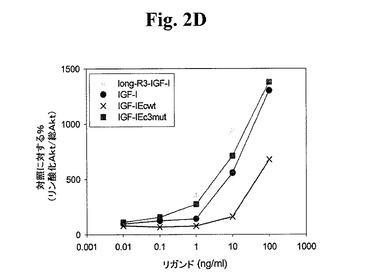
【図3A】
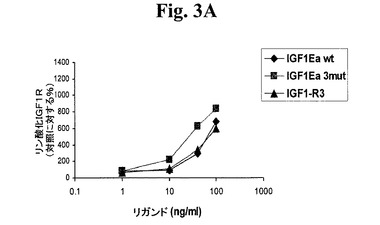
【図3B】
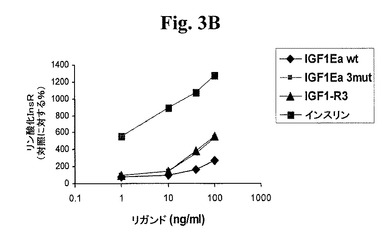
【図3C】
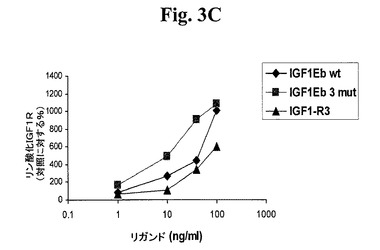
【図3D】
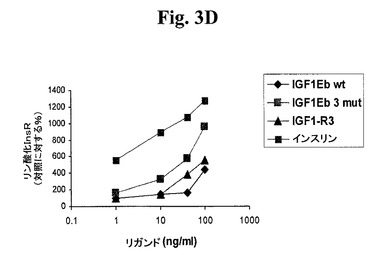
【図4A】
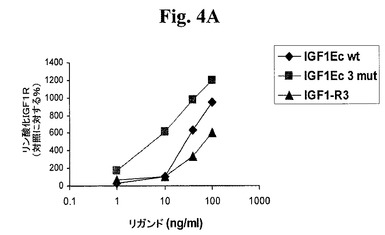
【図4B】
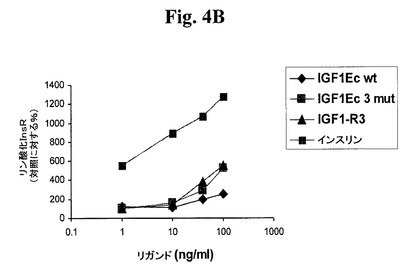
【図4C】
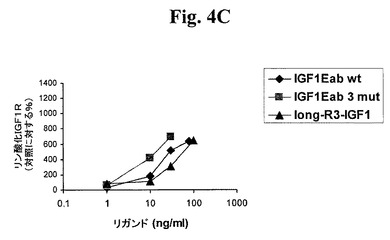
【図4D】
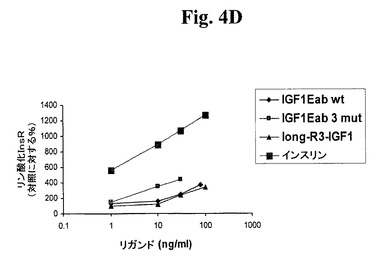
【図5】
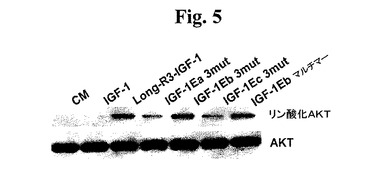
【図6】
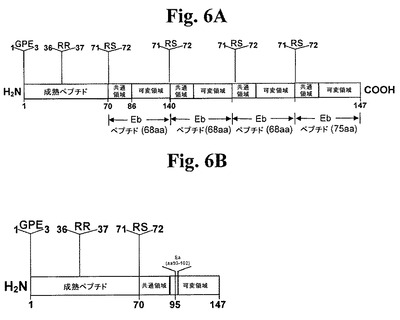
【図7A】

【図7B】

【図8A】

【図8B】

【図9A】

【図9B】
【図10A】

【図10B】
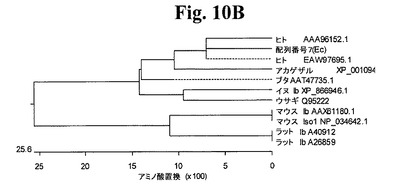
【図11A】

【図11B】
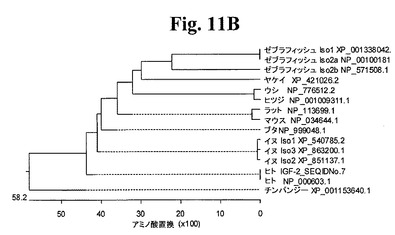
【図12A】

【図12B】
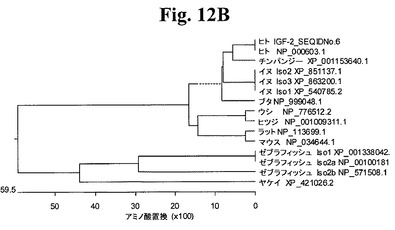
【図2A】
【図2B】
【図2C】
【図2D】
【図3A】
【図3B】
【図3C】
【図3D】
【図4A】
【図4B】
【図4C】
【図4D】
【図5】
【図6】
【図7A】
【図7B】
【図8A】
【図8B】
【図9A】
【図9B】
【図10A】
【図10B】
【図11A】
【図11B】
【図12A】
【図12B】
【公表番号】特表2009−539805(P2009−539805A)
【公表日】平成21年11月19日(2009.11.19)
【国際特許分類】
【出願番号】特願2009−513697(P2009−513697)
【出願日】平成19年6月6日(2007.6.6)
【国際出願番号】PCT/EP2007/055608
【国際公開番号】WO2007/141309
【国際公開日】平成19年12月13日(2007.12.13)
【出願人】(597011463)ノバルティス アクチエンゲゼルシャフト (942)
【Fターム(参考)】
【公表日】平成21年11月19日(2009.11.19)
【国際特許分類】
【出願日】平成19年6月6日(2007.6.6)
【国際出願番号】PCT/EP2007/055608
【国際公開番号】WO2007/141309
【国際公開日】平成19年12月13日(2007.12.13)
【出願人】(597011463)ノバルティス アクチエンゲゼルシャフト (942)
【Fターム(参考)】
[ Back to top ]