
官能化生分解性ポリマー粒子の製造方法と、そのポリマー粒子を応用した医薬担体

【課題】ポリマー粒子の新規製造方法の提供。
【解決手段】ポリマー粒子の製造方法であって、A)融合タンパク質をコードする核酸を含んで成る細胞を用意するステップ、ここで当該の融合タンパク質はポリマーシンターゼと下記(a)〜(c)を含んで成る:(a)物質もしくはカップリング試薬もしくはそれらの組み合わせと結合することのできる結合ドメイン、又は(b)タンパク質、又は(c)それらの組み合わせ、そしてB)前記細胞を培養培地の中で培養して、当該細胞に前記核酸由来の融合タンパク質を産生させかつポリマー粒子を産生させるステップ;そしてC)前記培養細胞から前記ポリマー粒子を分離し、ポリマー粒子を含んで成る組成物を生成するステップ、を含んで成る方法。
【解決手段】ポリマー粒子の製造方法であって、A)融合タンパク質をコードする核酸を含んで成る細胞を用意するステップ、ここで当該の融合タンパク質はポリマーシンターゼと下記(a)〜(c)を含んで成る:(a)物質もしくはカップリング試薬もしくはそれらの組み合わせと結合することのできる結合ドメイン、又は(b)タンパク質、又は(c)それらの組み合わせ、そしてB)前記細胞を培養培地の中で培養して、当該細胞に前記核酸由来の融合タンパク質を産生させかつポリマー粒子を産生させるステップ;そしてC)前記培養細胞から前記ポリマー粒子を分離し、ポリマー粒子を含んで成る組成物を生成するステップ、を含んで成る方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、官能化生分解性ポリマー粒子の製造方法と、そのポリマー粒子を応用した医薬担体に関する。
【背景技術】
【0002】
多数ある活性成分の有効性は、その活性成分が体内の望む標的部位に正確に輸送されるかどうかにかかっている。というのも、使用される活性成分は特定の場所においてだけ十分な作用を及ぼすことができ、他の場所では有効でなかったりマイナスの効果さえ有することがしばしばあるからである。他の場所では有効でなかったりマイナスの効果さえ有するというのは、がん疾患と闘う場合、中でも中枢神経系(CNS)に発生するがん疾患と闘う場合に特に当てはまる。
【0003】
活性成分を動物の体内で輸送する際に起こる可能性のある問題の好例は、血液-脳関門(BBB)である。CNSを目指す活性成分は、まず最初にこの関門を通過する必要がある。血管と細胞からなるこのネットワークはCNSを保護しており、水に溶けない物質がCNSに入るのを阻止する。脂溶性物質は、単純な拡散によってこの関門を簡単に通過することができる。しかし極性物質やイオンの輸送には能動輸送システムと受動輸送システムが必要である。しかし例えばCNS用に新たに発見されたすべての薬のうちの98%を超えるものは水溶性でないため、BBBを通過できず、したがって標的部位に到達することができない。
【0004】
この例は、動物の体内で有効成分を輸送する間に起こる可能性のある問題を示すためのものである。例えばBBBの場合には、膜の透過性を変化させることによって活性成分の透過性をより大きくすることが試みられてきた。活性成分に対するBBBの透過性は、例えば人工的に浸透圧を大きくすることによって、あるいはブラジキニン類似体を投与することによって実現できる。Sanovichら(Sanovich, E.他、Brain Res.、第705巻(1〜2)、125〜135ページ、1995年)は、例えばブラジキニン類似体であるRMP-7をランタンと同時に投与することによってBBBの透過性が増大することを明らかにした。上記のメカニズムのうちの1つによってBBBを開かせることの基本的な1つの欠点は、あらゆる物質に対する透過性が大きくなるため、CNS内の標的細胞にダメージを与える可能性のある毒性物質に対する透過性も大きくなることである。
【0005】
可能な別の方法は、活性成分を化学的に修飾して親油性を大きくし、その結果としてBBBをより容易に通過できるようにするというものである。例えばクロランブシル誘導体についてそのことが示されている(Greig, N.H.他、Cancer Chemother. Pharmacol.、第25巻(5)、320〜325ページ、1990年)。
【0006】
上記の2つの系とは別の方法は、ナノ粒子またはミクロン粒子を使用するというものである。この場合には、膜の透過性も活性成分も変化させない。Kreuter(Kreuter, J.、J. Anat.、第189巻(3)、503〜505ページ、1996年)は、この目的で使用するポリマー粒子を化学的な方法(例えば乳化重合、界面重合、脱溶媒和、蒸発、溶媒沈降)で実質的に製造できることを示している。
【0007】
ドイツ国特許第197 45 950 A1号によると、極めて多彩な物質(ポリマー(この場合にはポリシアノアクリル酸ブチル)、固体または液体の脂質、o/wエマルジョン、w/o/wエマルジョン、リン脂質小胞)から調製したポリマー粒子に医薬物質を付着させ、CNSにその医薬物質を輸送する。
【0008】
膜(特にBBBの膜)を通じてナノ粒子またはミクロン粒子を輸送する際の別の極めて重要な因子は、ポリマー粒子のサイズである。以前の研究結果によると、270nmまでのサイズのポリマー粒子がBBBを通過できる(Lockman, P.R.他、Drug Develop. Indust. Pharmacy、第28巻(1)、1〜12ページ、2002年)。
【0009】
したがって使用するポリマー粒子が特定のサイズになっていることが重要である。Lockmanによると(Lockman, P.R.他、Drug Develop. Indust. Pharmacy、第28巻(1)、1〜12ページ、2002年)、上記の方法で製造したポリマー粒子のサイズは、一般に、フォトン相関分光法によって測定される。ブラウン運動に基づくこの方法では、レーザー・ビームを用いてポリマー粒子の測定を行ない、光の変化の時間依存性から粒子のサイズを決定する。
しかし得られるポリマー粒子のサイズを決定するこの分析法は、時間とコストが非常にかかる。
【発明の概要】
【発明が解決しようとする課題】
【0010】
したがって本発明の目的は、生物学的に活性な物質を輸送するためのすぐに使用できて低コストの系であって、動物の体内で活性成分の輸送を効果的かつ信頼性よく行なうことのできるものを提供することである。
【課題を解決するための手段】
【0011】
本発明で目的とする“生物学的に活性な物質”は、生物の側で生物学的な応答を開始させることのできるあらゆる物質である。このような物質には、生体内で特定の反応を触媒する酵素およびアブザイムや、生体内に存在することによってその生物に間接的な応答を起こさせるタンパク質(例えば抗体)だけでなく、非生物起源の無機分子や有機分子、すなわち自然に存在する生物によっては形成されず、人工的に製造される無機分子や有機分子などが含まれる。たいていの医薬活性成分も後者のグループに属する。生物学的に活性な物質は、その性質に応じ、さらに別の生物学的に活性な物質に結合させるのに適している場合もある。
【0012】
上記の目的を実現するため、生分解性ポリマー粒子の製造方法を提供する。この方法は、ポリマー粒子のサイズを制御するタンパク質をコードしている少なくとも1つの誘導遺伝子を微生物に導入するステップと、少なくとも1つの誘導遺伝子を誘導したこの微生物を、この微生物が生分解性ポリマー粒子を産生するのに適した培地の中で培養するステップを含んでいる。この方法により、生体適合性のある生分解性ポリマー粒子であって生物学的に活性な物質を輸送するのに適したものを製造することができる。そのとき、形成されるポリマー粒子のサイズを制御することにより、必要とされる望むサイズの粒子を製造することができる。特定のサイズのポリマー粒子を製造できるため、望むサイズのポリマー粒子の収率が増大する。そのためこの方法の効率が大きくなってコストの削減に役立つ。さらに、本発明の方法により、BBBを通過して輸送するための粒子サイズに関して上記の条件を満たすポリマー粒子を製造することができる。この方法のおかげで、微生物が天然に作り出すこの種のポリマー粒子よりも小さいポリマー粒子を製造することが特に可能になる。微生物が天然に作り出すポリマー粒子の平均サイズは300〜500nmである(Wieczorek, R.他、J. Bacteriol.、第177巻(9)、2425〜2435ページ、1997年)のに対し、本発明の方法ではポリマー粒子の産生を制御することが可能であり、ポリマー粒子はこの平均値よりもはるかに小さくなる。
【0013】
粒子のサイズを決定する遺伝子を上流の誘導プロモータ(例えばアラビノースによって誘導されるBADプロモータ)で誘導する。この目的で利用する微生物は、ポリマー粒子のサイズを制御する遺伝子を持っていないか、この遺伝子が不活化され、本発明の方法で説明した少なくとも1つの誘導遺伝子(この誘導遺伝子は、ポリマー粒子のサイズを制御するタンパク質をコードしている)によって置換されている。ここでは、ベクターを用いてこの遺伝子を細胞に導入する。ベクターについては、この明細書の実験の項でさらに詳しく説明する。その結果、初めて、生体適合性がある決まったサイズの生分解性ポリマー粒子を微生物プロセスにおいて製造することが可能になる。
【0014】
ポリマー粒子は、細胞内の細胞質含有物として堆積される。ポリマー粒子のコアは、ポリヒドロキシアルキルカルボン酸(特にポリヒドロキシアルカン酸)からなり、タンパク質とリン脂質でできた外殻膜によって囲まれている。外殻膜は、脂質と、その中に埋め込まれたタンパク質からなる。ポリマー粒子のコアを形成するポリヒドロキシアルキルカルボン酸は、融点が50℃〜176℃、結晶度が30%〜70%であり、破断値における伸長度が5%〜300%である。
【0015】
バイオテクノロジーによる培養に一層適しているが、遺伝子構成のために上記のポリマー粒子を形成することができない微生物(例えば、GRAS生物に分類されるある種の形態の大腸菌)でもこの有利な方法を利用できるようにするため、ポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子と、ポリマー粒子のサイズを制御する少なくとも1つの誘導遺伝子を導入する。代謝に影響を与えてポリマー粒子を形成させ、したがって形成されるポリマー粒子の組成に影響を与えることのできるあらゆるタンパク質を考慮することができる。ポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子としては、ここでは、チオラーゼ、レダクターゼ、ポリマーシンターゼのいずれかをコードしているものを選択する。ポリマーシンターゼとしては、ポリマーが形成される最終ステップの触媒となることができるあらゆるタンパク質が使用される。ポリマーの形成は、本発明で説明するポリマーシンターゼ以外にも、例えばリパーゼによっても起こる。ポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子としては、ラルストニア・ユートロファからのphaAチオラーゼ、phaBケトアシルレダクターゼ、phaCシンターゼのいずれかをコードしているものを選択することが好ましい。これらの追加遺伝子を導入すると、細胞は、ポリマー粒子の形成を可能にするタンパク質を産生できるようになる。ポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子を意図的に選択すると、得られるポリマー粒子の組成に影響を及ぼすこともできる。ポリマー粒子の形成に通じる代謝経路に関係するタンパク質をコードしている個々の遺伝子は、それぞれ異なる基質特異性を持っている可能性、あるいはそれぞれ異なる反応生成物を形成する可能性、あるいは代謝経路における分岐経路を阻止する可能性がある。そのためポリマー粒子の形成に関係する基質と分子に意図的に影響を及ぼすことができる。
【0016】
脂肪酸の代謝が変化した微生物(例えば実施例4に記載した大腸菌の突然変異体)の内部でポリマー粒子が産生されるようにするために必要なのは、ポリマー粒子のサイズを制御する少なくとも1つの誘導遺伝子を別にすると、ポリマーシンターゼだけである。どの生物を利用するかに応じ、さらに別の遺伝子を細胞内に導入して上記の条件下でポリマー粒子を産生できるようにする。細胞がポリマー粒子の形成に必要なすべての遺伝子を含んでいない場合でも、欠けているタンパク質によって産生される中間体を栄養媒体を通じて細胞に供給すると、ポリマー粒子の産生を進行させることができる。しかしポリマー粒子を形成するのに少なくとも1つのポリマーシンターゼは常に必要である。
【0017】
ポリマー粒子の性質は、その組成を制御すると影響を受ける可能性がある。性質に影響を与えることにより、例えばポリマー粒子の生分解速度に影響を及ぼすことが可能になる。生体内で形成されるポリマー粒子の組成に影響を与えるためには、ポリマー粒子の形成に関係するタンパク質をコードしている別の遺伝子を意図的に選択して細胞に導入するという上記の可能性は別にすると、チオラーゼおよび/またはポリマーシンターゼをコードしている少なくとも1つの追加遺伝子を細胞内に導入することが特に好ましい。チオラーゼとポリマーシンターゼの基質特異性が異なるため、中間体と最終生成物が異なり、したがって形成されるポリマー粒子のコアの組成が異なることになる。
【0018】
ポリマー粒子の産生原理をの一例を図2に示してある。ポリマー生合成のための活性化前駆体は、原則として、中心的な代謝物であるアセチルCoAに由来するもの、あるいはクエン酸サイクル、脂肪酸β酸化、新規な脂肪酸の生合成という主要な代謝経路の中間体に由来するもの、あるいはアミノ酸代謝に由来するものが可能である。脂肪酸を炭素源として使用する場合には、PHA生合成のための活性化前駆体として機能する中間体(アシルCoA、特に3-ヒドロキシアシルCoA)を脂肪酸β酸化によって製造する。
【0019】
粒子のサイズを制御する際には、ポリマー粒子のサイズを制御するタンパク質をコードしている少なくとも1つの誘導遺伝子は、フェイジン様タンパク質のファミリーに由来するものであり、ラルストニア・ユートロファからのフェイジン遺伝子phaPと、シュードモナス・オレオボランスからのフェイジン遺伝子phaFを含むグループの中から選択することが好ましい。フェイジンは分子量が14〜28kDaの両親媒性タンパク質であり、ポリマー粒子の疎水性表面にしっかりと結合する。
【0020】
ポリマーの組成が異なるポリマー粒子は、物理的特性が異なり、生物学的に活性な物質(特に薬理学的に活性な成分)を異なる速度で放出する。例えばC6〜C14の3-ヒドロキシ脂肪酸からなるポリマー粒子は、ポリマーの結晶度が低いためにポリマーの分解速度がより大きい。ポリマー骨格上に比較的大きな側鎖を有するポリマー成分のモル比が大きくなると、通常は結晶度が小さくなり、弾力特性がより顕著になる。本発明で説明した方法に従ってポリマーの組成を制御することにより、ポリマー粒子の生分解性に影響を及ぼすことが可能になり、したがって生物学的に活性な物質(特に薬理学的に活性な成分)の放出速度にも影響を及ぼすことが可能になる。
【0021】
ポリマー粒子を形成するための基質として、側部官能基を有する少なくとも1つの脂肪酸を培地に導入することが好ましい。導入することが特に好ましいのは、少なくとも1つのヒドロキシ脂肪酸および/または少なくとも1つのメルカプト脂肪酸および/または少なくとも1つのβ-アミノ脂肪酸である。“側部官能基を有する脂肪酸”は、飽和脂肪酸または不飽和脂肪酸を意味するものとする。その中には、メチル基、アルキル基、ヒドロキシル基、フェニル基、スルフヒドリル基、第一級アミノ基、第二級アミノ基、第三級アミノ基、アルデヒド基、ケト基、エーテル基、カルボキシル基、O-エステル基、チオエステル基、カルボン酸アミド基、ヘミアセタール基、アセタール基、リン酸モノエステル基、リン酸ジエステル基を含むグループの中から選択した側部官能基を有する脂肪酸も含まれる。
【0022】
ポリマー粒子は、基質特異性が異なるタンパク質をコードしているさまざまな遺伝子を使用することによる影響と、使用する培地に存在している添加物、基質、反応条件の影響の両方を遺伝子レベルで受けるため、使用する基質は、ポリマー粒子に望む組成と性質がどのようなものであるかによって決まる。
【0023】
形成されるポリマー粒子のサイズをより正確に制御するため、ポリマー粒子のサイズを制御するのに十分な量の基質を培地に添加する。すると粒子のサイズをより効果的に制御することが可能になる。
【0024】
本発明の方法でポリマー粒子を産生させるのに用いる微生物は、ラルストニア、アルカリゲネス、シュードモナス、ハロビフォルマを含む属の中から選択する。使用する微生物の選択は、ラルストニア・ユートロファ、アルカリゲネス・ラトゥス、大腸菌、シュードモナス・フラギ、シュードモナス・プティダ、シュードモナス・オレオボランス、緑膿菌、シュードモナス・フルオレセンス、ハロビフォルマ・ハロテレストリスを含むグループの中から行なうことが好ましい。このグループには、生体適合性のある生分解性ポリマー粒子を自然に産生することのできる微生物と、遺伝子の構成上それが不可能な微生物(例えば大腸菌)の両方が含まれる。後者の微生物に対し、ポリマー粒子の産生を可能にするのに必要な遺伝子を、本発明の方法を利用して導入する。たとえ微生物が、代謝が異なるためにポリマー粒子を形成するのに必要な基質を産生することができなくても、原則として、培養可能なあらゆる微生物を利用して上記の方法でポリマー粒子を産生させることができる。微生物が必要な基質を産生できない場合には、必要な基質を培地に添加した後、細胞内に導入した遺伝子が発現しているタンパク質を利用してポリマー粒子に変換する。
【0025】
細胞からポリマー粒子を得るためには、培養した微生物を公知の方法で破壊した後、ポリマー粒子を細胞残留物から分離する。この方法で得られるポリマー粒子のサイズの範囲は、標準的な方法(例えば排除クロマトグラフィまたは密度勾配遠心分離)を利用して望むサイズのポリマー粒子を選択することにより、さらに狭めることができる。
【0026】
本発明の方法で製造したポリマー粒子のタンパク質と脂質からなる外殻膜を修飾し、動物の体内で活性成分を輸送するのにより好ましい粒子特性を持たせることができる。そのためには、ポリマー粒子の表面に位置する脂質層を本発明の方法で得られたポリマー粒子から分離し、異なる組成の脂質層で置換する。
【0027】
脂質層を異なる組成の脂質層で置換するとき、新しい脂質層の重要な性質は、生物の膜を通過するポリマー粒子の輸送に影響を与えるという性質である。外殻膜の脂質層が標的とする膜の脂質層と適合している場合には、粒子の取り込み増大が観察される可能性がある(Fernart, L.他、J. Pharmacol. Exp. Ther.、第291巻(3)、1017〜1022ページ、1999年)。したがってポリマー粒子の脂質層を取り除き、異なる組成の脂質層で置き換えるとよい。これは、アセトン抽出によって、あるいはホスホリパーゼまたは非変性性洗浄剤を用いることによって実現することが好ましい。ここで、今や実質的に脂質を含まなくなったポリマー粒子に適切な両親媒性分子の混合物を添加し、望む組成の脂質層を得る。新しい脂質層は、リン脂質とエーテル脂質を含むグループの中から選択した両親媒性分子の混合物からなることが好ましい。
【0028】
上記の洗浄剤を用いて脂質だけでなくポリマー粒子の表面に位置するタンパク質(ただしポリマーシンターゼは除く)も除去し、そのタンパク質を、官能化したタンパク質で置換することもできる。裸のポリマー粒子をこのようにして製造し、そのポリマー粒子に対して数多くの修飾を行なうことができる。これについてはあとで詳しく説明する。微生物ハロビフォルマ・ハロテレストリスから得られるポリマー粒子は、このタイプの修飾に特に適している。
【0029】
ポリマー粒子をその後どのように利用するかに応じ、粒子サイズを少なくとも1つの誘導遺伝子で制御し、形成されるポリマー粒子の直径を10nm〜3μmにする。ポリマー粒子の直径は10nm〜900nmであることが好ましく、10nm〜100nmであることが特に好ましい。本発明の特別な一実施態様では、サイズの制御を、培地内における基質の利用可能性を制御することによって、あるいは2つのメカニズムを合わせることによって実現することもできる。
【0030】
本発明の方法で製造したポリマー粒子を官能化するには、ポリマー粒子のサイズを制御するタンパク質をコードしている少なくとも1つの誘導遺伝子が、ポリマー粒子結合ドメインと、少なくとも1つの結合ドメインとを含んでいて、この少なくとも1つの結合ドメインが、生物学的に活性な物質および/またはカップリング試薬と結合可能である必要がある。同様に、ポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子が、ポリマー結合ドメインと、少なくとも1つの結合ドメインとを含んでいて、この少なくとも1つの結合ドメインが、生物学的に活性な物質および/またはカップリング試薬と結合可能であることが特に好ましい。
【0031】
“結合ドメイン”は、細胞内に以前に導入した少なくとも1つの誘導遺伝子を修飾することによって、および/またはポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子を修飾することによって得られるタンパク質の一部を意味する。生物学的に活性な物質および/またはカップリング試薬と結合できる結合ドメインは、オリゴペプチド、酵素、アブザイム、非触媒性タンパク質を含むグループの中から選択する。このグループは、フラグ・エピトープまたは少なくとも1つのシステインを含んでいることが特に好ましい。このグループは、実験の項で機能性タンパク質という呼び方もする。このグループのメンバーに関するコード配列を、細胞内に以前に導入した少なくとも1つの誘導遺伝子のコード配列、および/またはポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子のコード配列に導入する。このようにすると、ポリマー粒子の形成、またはポリマー粒子の形成制御に関係するだけでなく、遺伝子の修飾によって得られる結合ドメインのおかげで生物学的に活性な物質および/またはカップリング試薬と結合することもできるタンパク質が産生される。この結合ドメインは、ポリマー粒子を産生させた後、カップリング試薬を用いて表面に位置するタンパク質を化学的に修飾することによっても得られる(実施例8を参照のこと)。
【0032】
この目的で使用する“カップリング試薬”は、一方の側には生物学的に活性な物質または別のカップリング試薬を結合させ、他方の側には結合ドメインを結合させるのに適した無機化合物または有機化合物である。
【0033】
この構造により、生物学的に活性な物質を輸送するのに適した多機能ポリマー粒子を産生させることができる。“ポリマー粒子結合ドメイン”は、ポリマー粒子の疎水性表面に結合できるタンパク質の一部であることが好ましい。ポリマー粒子の表面に結合するタンパク質の一部を含むポリマー粒子のドメインの選択は、ここでは、ポリマーデポリメラーゼ、ポリマー調節因子、粒子サイズ制御タンパク質を含む一群のタンパク質の中から行なう。これらのタンパク質は、ポリマー粒子を形成することのできる微生物(特にラルストニア、アルカリゲネス、シュードモナスという属からの微生物)に由来することが好ましい。粒子サイズ制御タンパク質は、ここでは、フェイジン様タンパク質のファミリーに由来することが好ましく、ラルストニア・ユートロファとシュードモナス・オレオボランスからのフェイジンを使用することがさらに好ましい。
【0034】
本発明の目的に合った“ポリマー調節因子”は、ポリマー粒子の形成に関係する遺伝子phaA、phaB、phaCの転写を調節するタンパク質である。ポリマー調節因子は、粒子の表面に結合することによって転写調節がなされなくする。このような調節因子の一例は、ラルストニア・ユートロファからのフェイジン・リプレッサ(phaR)である。このphaRは、フェイジン様遺伝子のプロモータに結合する。phaRの発現産物は、形成されるポリマー粒子のサイズを調節し、その遺伝子が読まれるのを阻止する。フェイジン・リプレッサは形成されるポリマー粒子の表面に結合するため、プロモータのこの部位が放出され、遺伝子の転写開始が可能になる。
【0035】
カップリング試薬および/または生物学的に活性な物質を結合させるのにポリマーシンターゼの結合ドメインを利用するというアイディアは、このタンパク質のポリマー粒子結合ドメインとポリマー粒子のコアの間の結合の安定性が増大していることから生まれた。
発明者は、驚くべきことに、変性剤(例えばドデシル硫酸ナトリウム(SDS)、尿素、グアニジウムヒドロクロリド、ジチオトレイトール)を用いても、酸性条件を利用しても、この結合を生分解性ポリマーのコアから切り離せないことを発見した。ここでは、ラルストニア・ユートロファ、シュードモナス・オレオボランス、シュードモナス・プティダ、緑膿菌に由来するポリマーシンターゼを使用することが好ましい。
【0036】
ポリマー粒子の表面に結合するタンパク質を遺伝子工学によって修飾してもポリマー粒子の形成に関係するタンパク質の機能には影響がないというのが、本発明による方法の特別な利点である。例えばあるタンパク質をポリマーシンターゼのN末端に融合して生物学的に活性な物質および/またはカップリング試薬のための結合ドメインを作っても、ポリマーシンターゼの機能は保持される。しかしタンパク質の機能が融合によって損なわれても、この欠点は、同じ機能を実行するタンパク質をコードしている活性状態の別の遺伝子の存在によって消すことができる。
【0037】
このようにして、タンパク質(特にポリマーシンターゼ)の結合ドメインを通じたポリマー粒子と生物学的に活性な物質および/またはカップリング試薬の間の安定な結合を保証することができる。
【0038】
発現すると粒子の表面に結合するタンパク質をコードする遺伝子を遺伝子工学によって修飾している間に、異なる修飾状態の遺伝子を細胞の中に導入することもできる。結合ドメインが異なるいろいろなタンパク質が発現した状態でポリマー粒子が形成されると、異なる結合ドメインを利用して粒子の表面を多官能化することができる。この方法により、官能化したポリマー粒子を直接的かつ効率的に大量生産することができる。
【0039】
次に、カップリング試薬を使用し、ポリマー粒子の表面に結合したタンパク質を官能化する。カップリング試薬の選択は、ビス(2-オキソ-3-オキサゾリジニル)ホスホニッククロリド(BOP-Cl)、ブロモトリスピロリジノホスホニウムヘキサフルオロリン酸(PyBroP)、ベンゾトリアゾール-1-イル-オキシ-トリスピロリジノホスホニウムヘキサフルオロリン酸(PyBOP)、n-ヒドロキシスクシンイミドビオチン、2-(1H-ベンゾトリアゾール-1-イル)-1,1,3,3-テトラメチルウロニウムヘキサフルオロリン酸(HBTU)、ジシクロヘキシルカルボジイミド、ジスクシンイミジル炭酸、1-(3-ジメチルアミノプロピル)-3-エチルカルボジイミド(EDC)、ビス(2-オキソ-3-オキサゾリジニル)ホスフィン、ジイソプロピルカルボジイミド(DIPC)、2-(1H-ベンゾトリオキサゾリル)-1,1,3,3-テトラメチルウロニウムテトラフルオロホウ酸(TBTU)、2-(5-ノルボルネン-2,3-ジカルボキシイミド)-1,1,3,3-テトラメチルウロニウムテトラフルオロホウ酸(TNTU)、パラニトロフェニルクロロギ酸、O-(n-スクシンイミジル)-1,1,3,3-テトラメチルウロニウムテトラフルオロホウ酸(TSTU)からなるグループの中から行なうことが好ましい。
【0040】
使用する生物学的に活性な物質は、殺虫剤、除草剤、薬理学的に活性な物質、タンパク質であることが好ましい。
【0041】
薬理学的に活性な物質の選択は、ジデオキシイノシン、フロクスウリジン、6-メルカプトプリン、ドキソルビシン、ダウノルビシン、1-ダルビシン、シスプラチン、メトトレキセート、タキソール、抗生物質、抗血液凝固薬、殺菌薬、抗不整脈薬、活性成分の前駆体、例示した活性成分の誘導体からなるグループの中から行なう。
【0042】
タンパク質の選択は、インスリン、カルシトニン、ACTH、グルカゴン、ソマトスタチン、ソマトトロピン、ソマトメジン、副甲状腺ホルモン、エリスロポエチン、視床下部放出因子、プロラクチン、甲状腺刺激ホルモン、エンドルフィン、エンケファリン、バソプレッシン、天然には存在しないアヘン製剤、スーパーオキシドディスムターゼ、抗体、インターフェロン、アスパラギナーゼ、アルギナーゼ、アルギニンデアミナーゼ、アデノシンデアミナーゼ、リボヌクレアーゼ、トリプシン、キモトリプシン、ペプシンからなるグループの中から行なうことが好ましい。
【0043】
本発明の別の側面は、生分解性ポリマー粒子を試験管内で製造する方法であって、ポリマー粒子の形成に適した溶液を少なくとも1つの基質とともに提供するステップと、この溶液に、ポリマー粒子のサイズを制御するのに適したタンパク質を導入するステップと、ポリマー粒子の形成に関係する別の少なくとも1つのタンパク質を導入するステップを含む方法である。この試験管内での方法には、ポリマー粒子のサイズを早くもポリマー粒子の製造中に制御できるため、形成されたポリマー粒子のサイズを決定し、個々のサイズのクラスに分類するというコストと時間がかかる作業を回避できるという利点もある。
【0044】
この明細書で使用する別の少なくとも1つのタンパク質は、ポリマーシンターゼである。このポリマーシンターゼの選択は、ラルストニア・ユートロファ、シュードモナス・オレオボランス、シュードモナス・プティダ、緑膿菌からのポリマーシンターゼを含むグループの中から行なうことが好ましい。
【0045】
微生物から単離したタンパク質と酵素を実験室で使用する試験管内での合成は、通常は、酵素と(場合によっては)酵素の基質をあらかじめ単離して精製する必要があるため、生体内での合成とは異なって非常にコストがかかる。生体適合性のある生分解性ポリマー粒子を試験管内で合成するための本発明による特別な一実施態様では、ポリマー粒子を形成するため、ポリマー粒子の形成に適した溶液に、少なくとも1つの脂肪酸(特に好ましいのは、β-メルカプト脂肪酸および/またはβ-アミノ脂肪酸)と、アシルCoAオキシダーゼまたはそれ以外の酸化・活性化酵素を添加する。R/S-3-ヒドロキシ脂肪酸とアシルCoAシンテターゼの代わりにこれらの基質を使用すると、CoAのリサイクル・システムが得られる。このリサイクル・システムにおいて、アシルCoAオキシダーゼは、脂肪酸を活性化して酸化させる一方で、CoAを消費し、ATPを加水分解する。重合の間、ポリマーシンターゼはCoAを除去するが、そのCoAは、アシルCoAオキシダーゼが使用できる。その結果、この試験管内での方法において顕著なコスト削減を実現することが可能になる。
【0046】
試験管内でポリマー粒子を製造する方法の別の利点は、少なくとも1つの基質を、ポリマー粒子のサイズを制御するのに十分な量、溶液に添加することにある。
【0047】
それに加え、形成されるポリマー粒子のサイズは、形成されるポリマー粒子のサイズを制御するのに十分な量のポリマーシンターゼを溶液に添加することによってさらに制御することもできる。生体内での方法と同様、試験管内での方法は、ポリマー粒子のサイズをより正確に制御することもできるため、望むサイズのポリマー粒子の収率が増大し、この方法の効率とコスト効率が向上する。
【0048】
しかし試験管内での方法では、粒子のサイズは、実際には、ポリマー粒子のサイズを制御するタンパク質を溶液の中に導入することによって制御する。このタンパク質は、フェイジン様タンパク質のファミリーに由来するものであり、ラルストニア・ユートロファからのフェイジンと、シュードモナス・オレオボランスからのフェイジンを含むグループの中から選択することが好ましい。
【0049】
少なくとも1つの基質と使用する酵素を選択することにより、生体内での方法と同様、ポリマーの組成を制御すること、そしてその結果として異なる性質を持ったポリマーのコアを得ることも可能になる。2つ以上の基質を用いる場合には、酵素から形成されるポリマーのタイプに応じ、例えばポリマーのコアの組成が異なるポリマー粒子を得ることが可能である。すでに説明したように、この方法で製造されるポリマーは、ポリマー粒子に極めて多彩な性質を付与する。
【0050】
試験管内でポリマー粒子を製造する方法では、ポリマー粒子の表面にある脂質層の組成は、リン脂質とエーテル脂質からなるグループの中から選択した少なくとも1つの両親媒性分子を溶液に添加することによって制御する。(添加する両親媒性分子がないときには、試験管内で得られる粒子は、タンパク質によってだけ取り囲まれる。そのため別のタイプの粒子になる。)このようにして、例えば動物の体内にある天然の膜に特別に合わせた特定の表面電荷を有するポリマー粒子を製造することができる。その後、そのポリマー粒子がこの膜を通過することになる。試験管内での方法で外殻膜の脂質層を修飾することの利点は、生体内での方法のように、その脂質層をその後修飾しなくても済むことである。
両親媒性分子(例えばリン脂質またはエーテル脂質)がすでに最初の溶液に添加されているため、望む組成の脂質膜が最初から得られる。
【0051】
試験管内での方法の別の特徴は、少なくとも1つの薬理学的に活性な物質を、ポリマー粒子の形成に適した溶液に添加するというものであり、生体内での方法でもこの特徴が利用されている。この物質は、ポリマー粒子を合成している間にポリマー粒子に組み込まれ、その後、粒子マトリックスを通じた拡散によって、あるいはポリマー粒子の分解によって動物の体内に放出させることができる。後者の場合には、本発明の方法によって製造したポリマー粒子において、ポリマーのコアの組成を上記のように制御することによってポリマー粒子の生分解速度を調節できるというさらに別の利点も有する。このようにして、活性成分を特定の期間にわたって連続的に放出させることが可能になる。
【0052】
官能化は、活性成分を成長中のポリマー粒子に組み込むこと以外に、生体内での方法のように、ポリマー粒子の形成に適した溶液に導入する少なくとも1つのタンパク質を選択するとき、導入した少なくとも1つのタンパク質が、ポリマー粒子結合ドメインと、少なくとも1つの結合ドメインとを備えていて、この少なくとも1つの結合ドメインが、生物学的に活性な物質および/またはカップリング試薬と結合可能になっているようにすることによって実現される。この場合に試験管内での方法でポリマー粒子を形成するのに用いるタンパク質は、すでに説明した生体内での方法から得ることができ、そのタンパク質は、上記のどのタイプの製造方法であるかに応じ、対応する性質を示す。次に、ポリマー粒子の形成に関係するそのタンパク質を、ポリマー粒子の形成に適した溶液に添加する。生体内での方法と同様、この方法で製造したポリマー粒子は、製造後に、すでに説明したカップリング試薬を用いた修飾によって、あるいはポリマー粒子の表面に結合しているタンパク質の結合ドメインに結合する生物学的に活性な物質を添加することによって、さらに修飾することもできる。
【0053】
本発明にはさらに、サイズが決まっていて両親媒性分子からなる表面層を有するポリマー粒子であって、その内部には、ポリマーデポリメラーゼ、ポリマー調節因子、ポリマーシンターゼ、粒子サイズ制御タンパク質を含むグループの中から選択した少なくとも1つのタンパク質が導入されており、少なくとも1つのこのタンパク質が、ポリマー粒子結合ドメインと、生物学的に活性な物質および/またはカップリング試薬と結合可能な結合ドメインとを備えているものが含まれる。好ましい一実施態様では、上記の少なくとも1つのタンパク質は、すでに説明した方法で製造する。
【0054】
本発明のポリマー粒子は有利な性質を持っているため、医薬調製物、殺虫剤、除草剤の製造に特に適している。この医薬調製物は、中枢神経系の疾患を治療するのに適していることが好ましい。本発明によるポリマー粒子は修飾できるため、BBBを通過する条件を満たすことができる。
【0055】
粒子サイズの制御、外殻膜の組成の制御、粒子表面の官能化、その中でも特に粒子表面の官能化が可能であることは、生分解性ポリマー粒子が生物学的に活性なあらゆる種類の物質を輸送するための担体として適しており、ポリマー粒子をその作用部位に狙いを定めて輸送できることを意味する。多価により、例えば少なくとも1つの薬理学的に活性な物質だけでなく抗体も粒子の表面に結合させることが可能になる。抗体には結合の特異性があるため、標的部位に正確に導くことができる。上記の利点ならびにその他の利点を、以下の実施例においてさらに詳しく説明する。
【図面の簡単な説明】
【0056】
【図1】生体内で産生される生分解性ポリマー粒子と、その表面に結合したタンパク質および脂質の概略図である。略号は、以下のタンパク質に関係する。A:ポリマーデポリメラーゼB:フェイジン(ラルストニア・ユートロファにおけるコード遺伝子の名称;phaP、ラルストニア・ユートロファにおけるコード遺伝子の名称;phaF)C:ポリマーシンターゼD:リン脂質E:ポリマー調節因子(例えばフェイジン・リプレッサ(ラルストニア・ユートロファからのPhaR))
【図2】ラルストニア・ユートロファにおいて可能な生分解性ポリマー合成法の一例である。基質であるアセチルCoAから出発して3段階のプロセスで単純なポリヒドロキシアルカン酸ポリヒドロキシブチル酸(PHB)を製造する。PHBのC4繰り返し単位は、β-ヒドロキシブチル酸である。合成の最終ステップにより、表面にポリマーシンターゼが結合したポリマー粒子が形成される。
【図3】生体内での実験結果であり、フェイジンの発現を増やした(溶液中の誘導因子の量を増やした)ときのポリマー粒子の直径の挙動を示している。
【図4】生体内での実験結果であり、フェイジンの発現を増やした(溶液中の誘導因子の量を増やした)ときの細胞内のポリマー粒子の数を示している。
【図5】試験管内の実験結果であり、ポリマーシンターゼに対するフェイジンの量の比がポリマー粒子の直径に及ぼす影響を示している。
【図6】試験管内の実験結果であり、ポリマーシンターゼに対する基質の比がポリマー粒子の直径に及ぼす影響を示している。
【図7】(ラルストニア・ユートロファからの遺伝子PhaPを含む)プラスミドpBBad-Pと(シュードモナス・オレオボランスからの遺伝子PhaFを含む)プラスミドpBBad-Fを構成する際の出発プラスミドとして使用するベクターpBBad22K(Sukchawalit, R.他、FEMS Microbiol Lett.、第181巻(2)、217〜223ページ、1999年)を示している。
【図8】ラルストニア・ユートロファからの粒子サイズ制御タンパク質フェイジンを発現させるPhaP遺伝子を含むベクターpBBad-Pを示している。
【図9】ポリマー(ポリヒドロキシブチル酸)を貯蔵させるプラスミドpBHR68(Spiekermann, P.他、Arch. Microbiol.、第171巻、73〜80ページ、1999年)を示している。このプラスミドは、phaAチオラーゼ、phaBケトアシルレダクターゼ、phaCシンターゼを発現させるための生合成オペロンを形成するラルストニア・ユートロファからの遺伝子phBRe、phbARe、phbCReを含んでいる。
【図10】ポリマー(ポリヒドロキシアルカン酸)を貯蔵させるプラスミドpBHR71(Langenbach, S.他、FEMS Microbiol. Lett.、第150巻、303〜309ページ、1997年)を示している。このプラスミドは、ポリマーシンターゼを発現させる遺伝子phaC1を含んでいる。
【図11】シュードモナス・オレオボランスからの粒子サイズ制御フェイジン様タンパク質を発現させる遺伝子PhaFを含むベクターpBBad-Fを示している。
【図12】図10のpBHR71プラスミドから出発して構成したベクターpBHR71-システインを示している。この構造では、ポリマーシンターゼをコードしている遺伝子phaC1がN末端に2個のシステイン残基を備えているため、生物学的に活性な物質の直接的な結合、またはカップリング試薬を通じた生物学的に活性な物質の結合が可能になる。
【図13】図10のpBHR71プラスミドから出発して同様に構成したベクターpBHR71-FLAGを示している。この構造では、ポリマーシンターゼをコードしている遺伝子phaC1がN末端に2個のフラグ・エピトープを備えている。このようにすると、フラグ遺伝子配列のSpeI制限部位を通じて機能性タンパク質の遺伝子配列を生体内で組み込むことができるだけでなく、すでに発現しているポリマーシンターゼにカップリング試薬および/または生物学的に活性な物質を試験管内で付着させることもできる。使用する生物学的に活性な物質は、直接に、あるいはカップリング試薬を通じて結合されるが、この物質により、標的部位(例えば細胞表面)への結合が可能になったり、生物学的活性(特に酵素活性)が与えられたりする。
【発明を実施するための形態】
【0057】
実施例1
生体内で産生される生分解性ポリマー粒子のサイズ制御
実施例1.1
【0058】
ラルストニア・ユートロファにおけるポリマー粒子の産生
【0059】
ラルストニア・ユートロファ(以前はアルカリゲネス・ユートロファス)の“ノックアウト”突然変異体(York, G.M.他、J. Bacteriol.、第183巻、4217〜4226ページ、2001年)を用いて生分解性ポリマー粒子を産生させた。この突然変異体は、ポリマー粒子の表面タンパク質フェイジン(phaP(-))の発現をコードしている遺伝子に欠陥があるため、phaP遺伝子によってコードされているフェイジンをもはや発現しない。また、この遺伝子を含まず、ポリマー粒子の生合成に必要な別の遺伝子も含まない微生物を用いて生分解性ポリマー粒子を産生させることもできる。可能な微生物の具体例としては、大腸菌、ハロビフォルマ・ハロテレストリスが挙げられよう。一実施態様では、すでに説明した生分解性ポリマー粒子を自然状態では産生できない大腸菌(実施例2を参照のこと)を用いる。
【0060】
次に、ベクターを用いて後者の微生物を形質転換する。ラルストニア・ユートロファの場合にはフェイジンをコードしているphaP遺伝子とプロモータ配列を含んでいる。この遺伝子は、誘導プロモータ(アラビノースによって誘導されるBADプロモータであることが好ましい)によって制御される。
【0061】
クローニング・ステップ
【0062】
phaPをコードしているラルストニア・ユートロファからのDNA配列を用いてクローニングを行なう。この配列は、“GenBank”にAF079155として登録されている(Hanley, S.Z.他、FEBS Letters、第447巻、99〜105ページ、1999年)。この配列を、誘導プロモータ(PBAD)を含むベクターpBBad22K(Sukchawalit, R.他、FEMS Microbiol. Lett.、第181巻(2)、217〜223ページ、1999年、図7を参照のこと)のNcoI/BamHI制限部位に挿入する。クローニングを目的として、対応するNcoI制限部位とBamHI制限部位を、フェイジンをコードしている配列に、プライマー5'-aaaggccccatggtcctcaccccggaaca-3'(配列ID番号1、NcoI制限部位に下線を付した)と5'-aaaggccggatcctcagggcactaccttcatcg-3'(配列ID番号2、BamHI制限部位に下線を付した)を用いたPCR突然変異誘発によって挿入する。得られるDNA断片(配列ID番号3)を制限酵素NcoIとBamHIを用いて加水分解し、同様にして加水分解したベクターpBBad22Kと連結する(図7)。このプラスミドは、今やタンパク質フェイジンのヌクレオチド配列を含んでいるため、今後はpBBad-Pと呼ぶことにする(図8)。このプラスミドを微生物に導入し、この微生物に関して従来技術で知られている形質転換技術を用いて形質転換する。pBBad-PのBADプロモータの誘導因子はアラビノースである。アラビノースは、使用している微生物の体内でこの遺伝子の発現を定量的に制御することができる。この制御メカニズムを利用してフェイジン遺伝子の発現を制御し、最終的には細胞内に存在するフェイジンの量によってポリマー粒子のサイズを制御する。
【0063】
実施例1.2
もともとポリマー粒子を形成することができなかった微生物によるポリマー粒子の産生
【0064】
もともとこのタイプのポリマー粒子を形成することができなかった微生物では、別のベクターまたは同じベクターが、ポリマー粒子の形成に関係する別の遺伝子を含んでいる。
ポリマー粒子を産生する微生物に導入する必要がある遺伝子の数と種類は、使用する微生物の基本的な構成によって決まる。最も簡単な場合、例えば大腸菌では、図2に示した方法でポリマー粒子を製造するのに少なくとも1つのチオラーゼ、1つのレダクターゼ、1つのポリマーシンターゼが必要である。特別な突然変異を含む生物(例えば実施例4からの大腸菌株)を利用する場合には、ポリマー粒子を製造するのに上記の遺伝子よりも少数で十分である。ポリマー合成の代謝経路から所定の基質前駆体が生物にすでに供給されている場合にもこのことが当てはまる。
【0065】
別のプラスミドを用いてポリマー粒子の形成に必要な遺伝子を細胞内に導入する場合には、ラルストニア・ユートロファの染色体DNAに由来する5.2kbのSmaI/EcoRI断片を含むプラスミドpBHR68(図9)が特に適している。このプラスミドは、ラルストニア・ユートロファに生分解性ポリマー粒子を産生させるための生合成オペロンを含んでいる(Spiekermann, P.他、Arch. Microbiol.、第171巻、73〜80ページ、1999年)。
【0066】
実施例1.3
pBBad-Pベクターを用いた粒子サイズの制御
【0067】
このようにして改変した微生物を、ルリア肉汁培地の中で30℃にて培養する。アラビノースを用いてプロモータを誘導すると、後期対数増殖期へと進む。培養を開始して24時間後、細胞内の粒子のサイズを測定する。そのために遠心分離で細胞を栄養培地から分離して破壊する。次に、トンネル電子顕微鏡に分析用ゲル濾過クロマトグラフィを組み合わせてポリマー粒子のサイズを測定する。ポリマー粒子のカウント数は、光学的顕微鏡のもとで無傷の細胞を調べることによって決定し、細胞1個当たりのポリマー粒子の数を数える。
【0068】
結果からわかるように、発現を制御することにより、個々の細胞内で形成されるポリマー粒子の平均サイズ(図3を参照のこと)とその平均数(図4を参照のこと)の両方を制御することができる。制御されたやり方でフェイジンの数を増やすと、ポリマー粒子の平均直径が小さくなり、それと同時にポリマー粒子の平均数が増える。このようにして、望む直径を有するポリマー粒子の収率を明らかに大きくすることができる。このようにすると、ポリマー粒子が、より早く、しかもよりコスト効率よく産生される。
【0069】
実施例1.4
基質利用可能性による粒子サイズの制御
【0070】
ポリマー粒子のサイズを制御する別のメカニズムは、ポリマー粒子の合成に必要な基質またはポリマーシンターゼの利用可能性を調節するというものである。ポリマーシンターゼの利用可能性は、アンチセンス法によって、あるいは遺伝子調節法によって、あるいは栄養培地においてポリマーシンターゼの形成に必要な基質がどれだけ利用できるかによって制御することができる。
【0071】
栄養培地における基質の利用可能性による粒子サイズ制御の一例は、存在する炭素源の濃度を減らすことにより、形成されるポリマー粒子の直径を制御する実験である。上記のプラスミドpBHR68(図9)と、生分解性ポリマー粒子を産生させるためのラルストニア・ユートロファ由来の生合成オペロン(図8)とを含む大腸菌株を、1.5%(w/v)のグルコースを含むM9培地の中で30℃にて増殖させる。定常的増殖期の初期には、グルコースなしのM9培地を添加することによって炭素源の濃度が元の値の1/50になる。それ以外は一定の増殖条件下でさらに20時間にわたって微生物を培養する。テストが終了したとき、微生物は、平均直径が130nmのポリマー粒子を含んでいる。
【0072】
実施例2
生体内で産生されるポリマー粒子の組成に影響を与える方法
【0073】
生分解性ポリマー粒子の組成は、細胞内に、phaPをコードしている遺伝子に加え、シンターゼ(特にチオラーゼ、または別のポリマーシンターゼ)の基質を提供する酵素をコードしている別の遺伝子を導入することによって変化させることができる。その結果、細胞は、ポリマー粒子が形成されている間に一連のさまざまなモノマーを伸長しているポリマー鎖に組み込むことができるため、異なる組成のポリマーからなるコアを有するポリマー粒子を作り出す。
【0074】
このことに関して挙げられる具体例は、生体内と試験管内の両方の方法で、基質特異性があるために伸長しているポリマー鎖に3-ヒドロキシ脂肪酸(C4〜C16)を異なった形で組み込むさまざまなポリマーシンターゼである。例えば、C4脂肪酸(C4)から生分解性ポリマー鎖を産生するラルストニア・ユートロファ由来のポリマーシンターゼや、アエロモナス・プンクタータ(C4とC6)、チオカプサ・フェンニギイ(C4とC8)、緑膿菌(C6〜C14)に由来するポリマーシンターゼを用いることができる。同時に2つ以上のポリマーシンターゼを細胞内に導入すると、非常に多彩な性質を有するポリマー粒子を産生させることができる。連続バッチでは、ランダムな突然変異誘発によって基質特異性が変化した新しいポリマーシンターゼも得ることができる(試験管内での進化)。
【0075】
形成されるポリマー粒子の組成に別の方法で影響を与えることもできる。さまざまな炭素源、さまざまな炭素源の前駆体、ポリマー粒子の形成につながる代謝経路の中間体などを提供することは、形成されるポリマーの性質に影響が及ぶ。さらに、ポリマー粒子の形成に関係する代謝経路は、阻害剤や、問題になっている代謝経路をノックアウトした突然変異体や、酵素の発現によって制御することができる。すると代謝経路が変化して別の中間体または最終生成物が形成される。
【0076】
脂肪酸の代謝に影響を与えるため、以下の酵素を用いる。すなわち、(R)-特異的エノイル-CoAヒドラターゼ、トランスアシラーゼ、ケトアシル-CoA/ACPレダクターゼである。これらの酵素は、ポリマー粒子が形成される際に、脂肪酸が代謝されるときの中間体を変化させ、異なる生成物(例えば側鎖が異なる脂肪酸)を提供するという性質を持っている。
これらの酵素は、ポリマーシンターゼのために供給する(R)-ヒドロキシアシル-CoA基質の鎖の長さに関して異なる特異性を有する。その結果、側鎖の長さが異なる結合ブロックがポリマーの内部に得られるため、異なる組成のポリマーになる。
【0077】
しかし細胞内に別の酵素を導入することは、代謝経路に影響を与える唯一の方法ではない。代謝経路の阻害剤を培地に添加することもできる。そのような阻害剤の具体例をいくつか挙げると、アクリル酸とトリクロサン(同意語:TCCまたは5-クロロ-2-(2,4-ジクロロフェノキシ)フェノール)である。
【0078】
実施例3
高度に結晶化したコアを有する生分解性ポリマー粒子を生体内において(R)-3-ヒドロキシブチル酸から産生させる方法
【0079】
この実験は、実施例1ですでに紹介したプラスミドpBBad-PとpBHR68を含む大腸菌株を用いて実施した。培養は、グルコースを炭素源として、実施例1に示した条件下で行なった。形成されるポリマー粒子は(R)-3-ヒドロキシブチル酸からなり、直径は、培地への誘導因子(アラビノース)の添加量に応じ、50〜500nmである。このようにして形成されるポリマー粒子の平均サイズは、ほんの20〜50nmほどしか違いがない。プラスミドpBBad-Pを用いて導入したフェイジンによって実現される制御特性を明らかにするため、上記の大腸菌株を用い、プラスミドpBBad-Pがない状態で実験を繰り返す。この場合には、直径が150〜250nmのポリマー粒子が得られる。この対照実験と比較すると、フェイジンを用いて粒子サイズを制御することで、より大きなポリマー粒子だけでなく、より小さなポリマー粒子も製造することができる。
【0080】
ポリマー粒子をサイズを基準にしてさらに分類することは、公知の方法で実現できる。
例えば、排除クロマトグラフィ、密度勾配遠心分離、5mMのリン酸塩バッファ(pH7.5)中での限外濾過などの方法がある。
【0081】
実施例4
あまり結晶化していないコアを有する生分解性ポリマー粒子を生体内において(R)-3-ヒドロキシ脂肪酸から産生させる方法
【0082】
この実験では、(R)-3-ヒドロキシ脂肪酸からなるコアを有するエラストマー性の生分解性ポリマー粒子を大腸菌の中で産生させる。ポリマー鎖は平均で6〜14個の炭素原子からなる。これは、シュードモナス・オレオボランスからのphaF遺伝子の発現を大腸菌の中で調節することによって実現される。phaF遺伝子は、フェイジンをコードしているラルストニア・ユートロファ由来の遺伝子phaPおよびポリマーシンターゼのための緑膿菌由来の遺伝子(phaC)に非常に似ている。この実施例では、phaF遺伝子の発現産物を用いて粒子サイズを制御する。ポリマー粒子は、シュードモナスのポリマーシンターゼ(phaC)だけを用いて脂肪酸の代謝を変化させた大腸菌の中で産生させることができる(Langenbach, S.他、FEMS Microbiol. Lett.、第150巻、303〜309ページ、1997年)。脂肪酸の代謝を変化させ、脂肪酸を炭素源として用いた場合には脂肪酸β酸化のCoA活性化中間体(例えばエノイル-CoA)が蓄積されてそれがポリマー合成の前駆体となるようにする。そのためには、大腸菌のfadB突然変異体を用いる(Langenbach, S.他、FEMS Microbiol. Lett.、第150巻、303〜309ページ、1997年)か、対応する脂肪酸の代謝を阻害する阻害剤を用いる(例えばアクリル酸;Qi, Q.他、FEMS Microbiol. Lett.、第167巻、89〜94ページ、1998年)。突然変異体または阻害された微生物を用いる場合には、そうでない場合に必要なケトアシルレダクターゼとチオラーゼはもはや必要ない。次に、大腸菌に固有のエノイル-CoAヒドラターゼを触媒としてエノイル-CoAからR-3-ヒドロキシアシル-CoAが形成される。次に、このR-3-ヒドロキシアシル-CoAがポリマーシンターゼによって変換されてポリ-(R)-3-ヒドロキシ脂肪酸になり、それが、形成されるポリマー粒子のポリマー・コアを形成する。
【0083】
クローニング
【0084】
Prietoら(Prieto, M.A.他、J. Bacteriol.、第181巻(3)、858〜868ページ、1999年)によってすでに報告されており、“GenBank”データベースにAJ010393という番号で登録されているシュードモナス・オレオボランスのphaF遺伝子を、ベクターpBBad-F(図11)を用いて大腸菌に導入して形質転換させる。このベクターは、ベクターpBBad22K(図7)を元にして得られる。ここでの個々のクローニング・ステップは、実施例1に記載したステップに対応している。phaF遺伝子をクローニングしてベクターpBBad22KのNcoI/BamHI制限部位に入れる。phaF遺伝子のPCR突然変異誘発に用いるプライマーは、以下の配列を有する:5'-aaagggccatggctggcaagaagaattccgagaa-3'(配列ID番号4、NcoI制限部位には下線を付した)と5'-aaagggggatcctcagatcagggtaccggtgcctgtctg-3'(配列ID番号5、BamHI制限部位には下線を付した)。次に、得られるDNA断片(配列ID番号6)をクローニングして上記のプラスミドpBBad22Kに導入する。phaFの配列を含むようになったこのプラスミドを、今後はpBBad-F(図11)と呼ぶ。このプラスミドに加え、ポリマーシンターゼのためのヌクレオチド配列を含むプラスミドpBHR71(図10;Langenbach, S.他、FEMS Microbiol. Lett.、第150巻、303〜309ページ、1997年)も用いて大腸菌を形質転換する。
【0085】
その後に発現させるための炭素源は、脂肪酸デカン酸である。フェイジンphaF遺伝子の発現は、ここでも、誘導因子であるアラビノースによって制御する。テストは、実施例1にすでに記載したようにして行なう。あらかじめ使用した誘導因子の量に応じ、この方法で形成されるポリマー粒子は直径が100〜500nmになる。
【0086】
実施例4
試験管内で産生させる生分解性ポリマー粒子のサイズ制御
【0087】
試験管内での産生において形成される生分解性ポリマー粒子のサイズは、ポリマーシンターゼ、フェイジン、フェイジン様タンパク質いずれかの利用可能性と、基質および代謝中間体の利用可能性によっても制御される。
【0088】
必要な酵素と基質は、生分解性ポリマー粒子を産生させるための試験管内のバッチでまず最初に利用できる必要がある。ラルストニア・ユートロファまたは緑膿菌からの組み換えポリマーシンターゼをこのテストで用いる(Gerngross, T.U.とMartin, D.P.、Proc. Natl. Acad. Sci. USA、第92巻、6279〜6283ページ、1995年;Qi, Q.他、Appl. Microbiol. Biotechnol.、第54巻、37〜43ページ、2000年)。このポリマーシンターゼは、(ヒスチジン-タグ融合体またはNi2+-NTAアガロースを用いた)アフィニティ・クロマトグラフィによって精製する。粒子サイズを制御するため、ラルストニア・ユートロファと緑膿菌からのポリマー粒子サイズ制御タンパク質を同様にして精製する。このタンパク質をラルストニア・ユートロファの中で発現させるため、実施例1ですでに用いたのと同じベクターを用いてフェイジン遺伝子を発現させる。生分解性ポリマー粒子を試験管内で産生させるための反応バッチは、ポリマーシンターゼとポリマー粒子サイズ制御タンパク質フェイジン以外に、ポリマー粒子を合成するための基質としてR-3-ヒドロキシブチリル-CoAまたはR-3-ヒドロキシデカノイル-CoAと、50mMのリン酸塩バッファ(pH7.5)と、1mMのMgCl2と、安定化のための5%グリセロール(v/v)も含んでいる。R-3-ヒドロキシブチリル-CoAまたはR-3-ヒドロキシデカノイル-CoAをポリマーシンターゼのための前駆体として使用しているため、チオラーゼとレダクターゼは使用する必要がない(図2を参照のこと)。
【0089】
図5の結果は、ポリマーシンターゼに対するフェイジンの量の比がポリマー粒子の直径に及ぼす影響を示している。それに対して図6は、ポリマーシンターゼに対する基質の比がポリマー粒子の直径に及ぼす影響を示している。これらの結果は、存在しているポリマーシンターゼの量によってポリマー粒子のサイズを調節できることも示している。したがって粒子サイズの効果的な調節は、本発明の方法によって実現される。
【0090】
実施例5
試験管内で産生されるポリマー粒子の組成に影響を及ぼす方法
【0091】
実施例5.1
ポリマーの組成に影響を及ぼす方法
【0092】
生分解性ポリマー粒子の組成は、さまざまな基質を反応バッチに添加することによって、および/または基質スペクトルが異なるさまざまなポリマーシンターゼ(図2を参照のこと)をそれぞれの場合に使用することによって、変えることができる。このようにして、ポリマー粒子の形成中に伸長しているポリマー鎖に一連の異なるモノマーが組み込まれ、ポリマーの組成が異なるポリマー粒子が産生される。緑膿菌からのポリマーシンターゼを用いているため、炭素原子を6〜14個有するR-3-ヒドロキシ脂肪酸構成ブロックが、伸長しているポリマー鎖に組み込まれる。例えば基質が1つだけ供給される場合には、緑膿菌からのポリマーシンターゼを用いると単独重合ポリマー粒子が得られる。
【0093】
ポリマー粒子の組成をいかにして変えるかのさらに別の具体例を実施例2に示す。
【0094】
実施例5.2
ポリマー粒子の膜組成に影響を及ぼす方法
【0095】
その後に例えば活性成分を導入する細胞の膜に融合させるためには、生分解性ポリマー粒子の表面にあるリン脂質層の組成を制御できる必要がある。この制御は、ポリマー粒子の形成に適した溶液を用意して試験管内で生分解性ポリマー粒子を産生させる(実施例4)ことによって実現される。膜は、さまざまな両親媒性分子の混合物を添加することにより、対応する条件に個別に合わせることができる。
【0096】
リン脂質を試験管内の反応バッチに添加する。所定数のポリマーシンターゼ分子がポリマー粒子の形成に関与する。重合が進行するにつれ、本発明のポリマー粒子はより大きくなり、表面にあるポリマーシンターゼはもはやポリマー粒子の表面を完全に覆うことができなくなる。その結果、(ポリマーの)疎水性領域が露出し、そこに両親媒性分子が自発的に付着する。
【0097】
生体内で産生された粒子の場合には、すでに存在しているリン脂質層をまず最初に除去する。そのためには、アセトン抽出を行なうか、非変性性洗浄剤を用いてリン脂質層を破壊する。次に、すでに説明したように、今や実質的に脂質を含まないポリマー粒子に、適切な両親媒性分子の混合物を添加する。ここでは、負に帯電したリン脂質またはホスファチジルコリンを使用することが好ましい。膜の組成を意図的に制御する一実施態様では、オクチルグリコシド(PH7.5)を1%含むPBSの中にポリマー粒子を懸濁させ、撹拌しながら過剰なホスファチジルコリンに対する透析を行なう。得られる粒子表面は、脳毛細管内皮細胞(BCEC)との融合に特に適している。
【0098】
実施例5.3
成長しているポリマー粒子にさまざまな物質を組み込む方法
【0099】
生分解性ポリマー粒子のコアへの物質の取り込みは、親油性蛍光染料であるナイル・レッド(シグマ社、セントルイス、ミズーリ州、アメリカ合衆国)またはローダミン123(Spiekermann, P.他、Arch. Microbiol.、第171巻、73〜80ページ、1999年)の助けを借りてすでに定量的に調べられてきた。すでに説明したように、生体内または試験管内での産生の間に蛍光染料ナイル・レッドを培地または反応バッチに添加すると、産生されるポリマー粒子が染色されるのを観察することができる。すなわちポリマー粒子の染色が、ポリマー粒子の合成中という早い時期から始まる。この染色は、細胞から単離する前にさえ始まる。
【0100】
薬理作用を有する活性成分を添加すると、ポリマー粒子のコアに組み込まれる可能性がある。例えば、ポリマー粒子を産生させるための試験管内での実験において、非極性抗がん剤であるパクリタキセルを反応バッチに添加する。水に溶けないパクリタキセルをポリマー粒子の疎水性ポリマー・コアに溶かした後に濃縮する。ここではパクリタキセルは、単にポリマー粒子を形成するために溶液に添加する。すると活性成分が粒子内に濃縮される。このことを確認するため、形成されたポリマー粒子を反応バッチから取り出し、溶液中にパクリタキセルが存在しているかどうかをHPLCによって調べる。溶液中のパクリタキセルの濃度が低下しているというのは、ここでは、パクリタキセルがポリマー粒子に組み込まれたことを示している。生体内でポリマー粒子が分解していくにつれ、活性成分が放出される。ポリマー粒子を含まない反応バッチを対照として用いる。この場合には、ポリマー粒子の形成に適した溶液中でパクリタキセルの濃度が低下することはない。
【0101】
実施例6
粒子表面の官能化
【0102】
生物学的に活性な物質は、ポリマー粒子の表面にすでに存在しているタンパク質と結合することができる。図1に示したすべてのタンパク質を考えることができる。その結果、複数の“架橋”戦略が得られる。これらの戦略により、粒子表面に結合するタンパク質を通じて生物学的に活性な物質を共有結合させることが可能になる。
【0103】
実施例6.1
薬理学的に活性な成分を付着させるために表面タンパク質を意図的に修飾する方法
【0104】
ポリマー粒子を官能化する方法の一例は、ヒドラゾンが結合したドキソルビシンを緑膿菌からのポリマーシンターゼphaC1に付着させるというものである(ヒドラゾンが結合したドキソルビシンは、King, H.D.他、Biconjugate Chem.、第10巻、279〜288ページ、1999年に記載されている)。ポリマーシンターゼphaC1は、ポリマー粒子の表面に結合しており、N末端に挿入された2つのシステイン残基を含んでいる。このポリマーシンターゼのシステイン残基が結合ドメインを形成しており、この結合ドメインを通じて生物学的に活性な物質を結合させることができる。
【0105】
クローニング
【0106】
システイン残基のトリプレット・コードをPCR突然変異誘発によってクローニングし、phaC1をコードしている遺伝子に組み込む。次に、この遺伝子をクローニングして図10に示したプラスミドpBHR71のXbaI制限部位とBamHI制限部位に組み込む。XbaI制限部位とBamHI制限部位も、このPCR突然変異誘発においてこの遺伝子に挿入する。phaC1をコードしている遺伝子のPCR突然変異誘発に使用するプライマーは、以下の配列を有する:N末端のプライマー5'-gggcTCTAGAaataaggagatatacatatgtgttgtaagaacaataacgagctt-3'(配列ID番号7、XbaI制限部位は大文字、システイン・トリプレットには下線)と、C末端のプライマー5'-aaacgcGGATCCttttcatcgttcatgca-3'(配列ID番号8、BamHI制限部位は大文字)。次に、このようにして得られたDNA断片(配列ID番号9)をXbaIとBamHIを用いて加水分解し、同様にして加水分解したプラスミドpBHR71と連結させる。得られるプラスミド(pBHR71-Cys(図12)と呼ぶ)を用いて大腸菌を公知の方法で形質転換する。この大腸菌は、ポリマーシンターゼが表面に存在するポリマー粒子を形成することができる。このポリマーシンターゼは、さまざまな物質(特に医薬物質)を付着させるための2つのシステイン残基を備えている。
【0107】
実施例6.1.1
ドキソルビシン(同意語:ヒドロキシダウノルビシン)を付着させる方法
【0108】
今や、ヒドラゾンを媒介とした“架橋”を、表面に露出したシステイン残基を通じて実現することができる。そのためには以下の方法を利用する。すなわち、ポリマーシンターゼが結合している単離したポリマー粒子100mg(合計1ml)を、ヘリウムを灌流させたPBS(pH7.5)と5mMのジチオトレイトール(DTT)の中で37℃にて3時間にわたって培養する。
この処理により、ポリマーシンターゼに含まれるジスルフィド架橋が減少する。次に、4℃にて40,000×gで30分間にわたって遠心分離を行なうことにより、低分子量の化合物を除去する。次に、減少したポリマー粒子を、ヒドラゾンが結合したドキソルビシンを10μモル含む1mlのPBSバッファを用いて懸濁させ、4℃にて30分間にわたって培養する。この期間が過ぎた後、上記の条件下で再び遠心分離を行ない、処理したポリマー粒子を洗浄する。その後、結合しなかったドキソルビシンをHPLCで検出する。ドキソルビシンがポリマーシンターゼにうまく結合したことは、濃度の低下によって確認される。
【0109】
実施例6.2
生物学的に活性な物質、特に薬理学的に活性な物質をポリマー粒子の結合ドメインに付着させる方法
【0110】
活性成分もポリマー粒子に結合させることができる。そのときには、ポリマー粒子の表面に結合したタンパク質の結合ドメインを通じて結合させる。そのため、結合ドメインをまず最初に作り出す必要がある。生物学的に活性な物質の結合は、ポリマー粒子の表面結合タンパク質(例えばポリマーデポリメラーゼ、フェイジン、フェイジン様タンパク質、ポリマーシンターゼ、ポリマー調節因子)を遺伝子改変することによって実現する。その結果、そのタンパク質が外に向いた結合ドメインを形成し、その結合ドメインにカップリング試薬または生物学的に活性な物質を結合させることができる。ポリマー粒子の上記表面タンパク質が、カップリング試薬または生物学的に活性な物質を直接付着させることのできるタンパク質と融合するとき、ポリマー粒子の表面タンパク質との融合後、表面タンパク質と融合タンパク質の両方の機能が完全に維持されていることが重要である。
【0111】
一実施態様では、2つのフラグ・エピトープを有するポリマー粒子を、緑膿菌からのポリマーシンターゼphaC1のN末端に直接融合させる。これらのフラグ・エピトープにより、市販されている抗フラグmAbs(モノクローナル抗体)(抗フラグM2、シグマ-オールドリッチ社)を結合させることができ、さらに、この方法がうまくいったことを確認するための酵素マーカーを結合させることができる。この実施例では、二次抗体/アルカリホスファターゼ共役体(抗マウス・アルカリホスファターゼ、シグマ-オールドリッチ社)を酵素マーカーとして使用する。次に、アルカリホスファターゼの活性がポリマー粒子の表面に及ぼす効果を光度計で測定する。
【0112】
実施例6.2.1
フラグ-phaC1融合タンパク質を有するポリマー粒子の産生
【0113】
以下のオリゴヌクレオチドを用いてフラグ-ポリマーシンターゼ融合タンパク質を産生させる:5'-tatgACTAGTgattataaagatgatgatgataaaca-3'と5'-tatgtttatcatcatcatctttataatcACTAGTca-3'(配列ID番号10と配列ID番号11、SpeI制限部位は大文字、フラグ・エピトープには下線)。ハイブリダイゼーションによって二本鎖DNAを得るため、これら2つのオリゴヌクレオチドを同じモル数(それぞれ10μM)混合し、室温(RT)にて30分間にわたって培養する。このようにして形成される二本鎖DNAは、フラグ・エピトープ(DYKDDDDK)をコードしており、NdeI制限部位(CA∇TATG/GTAT?AC)の張り出した末端部と相補的な張り出し末端部(TA)を有する。このDNA断片を制限酵素NdeIを用いて加水分解してクローニングし、同様に加水分解したベクターpHBR71(図10)に組み込む。このようにして得られるプラスミドpBHR71-FLAG(図13)は配列ID番号12の遺伝子を含んでおり、N末端にフラグが融合した(タンパク質のこの融合部分が結合ドメインを形成する)ポリマーシンターゼを発現させる。今やこの結合ドメインにより、生物学的に活性な物質および/またはカップリング試薬を結合させることができる。クローニングの間に同様にして導入する単一のSpeI制限部位は、機能性タンパク質をコードしている望む任意の別のDNA断片を挿入するのに利用できる。
【0114】
上記構造体を官能化する一例として以下の方法を利用する。すでにプラスミドpBBad-Fを含んでいて脂肪酸の代謝が変化した大腸菌株(実施例3を参照のこと)を、pBHR71-FLAGプラスミドを用いて形質転換すると、ポリマー粒子が発現する。ポリマー粒子は、細胞を破壊することによってその細胞から単離し、PBSバッファ(pH7.5)を用いて3回洗浄する。次に、ポリマー粒子を、フラグ・エピトープに結合する抗フラグ抗体とともに室温にて30分間にわたって培養する。次に、ポリマー粒子をすでに説明したようにして再び洗浄した後、二次抗体/アルカリホスファターゼ共役体とともにPBSバッファ中で室温にて30分間にわたって培養する。この30分間の培養が終わった後、粒子を0.1MのトリスHCl(pH8.5)の中で洗浄した後、アルカリホスファターゼの基質として2mg/mlのリン酸p-ニトロフェニルを粒子懸濁液に添加する。アルカリホスファターゼの活性を分光法によって410nmで測定する。結合ドメインを持たない(すなわちフラグ・エピトープのない)ポリマーシンターゼを含むポリマー粒子を負の対照として用いる。添加したリン酸p-ニトロフェニルは対照の中では変換されないため、410nmでの分光測定結果はネガティブである。ポリマー粒子が実際に形成されたという事実だけが、フラグ・エピトープの組み込みがポリマーシンターゼの機能に影響を与えなかったことの証拠である。
【0115】
この方法は、ポリマー粒子の上記表面タンパク質のうちの別の1つを用いて実施することもできる。2つ以上の表面タンパク質を修飾する場合には、非常に多彩な複数の物質をポリマー粒子に結合させることができる。そのためポリマー粒子が多価になって多くの異なった用途に適したものになる。
【0116】
前の実施例では、その後、形成されたポリマー粒子のすでに発現している表面タンパク質に生物学的に活性な物質を結合させたが、表面タンパク質にその物質を直接融合させた後にその物質を発現させることももちろん可能である。そのため、タンパク質(例えば酵素)のコード配列をpBHR71-FLAGプラスミドのphaC1遺伝子のC末端に融合させる。
【0117】
phaC1遺伝子と融合できるようにするため、挿入するタンパク質をコードしているDNA断片と、ポリマーシンターゼのC末端断片をコードしているDNA断片を、それぞれPCRによって得る。これら2つの断片を、張り出しているプライマーを用いて挿入した制限部位によって連結させる。このハイブリッド遺伝子の5'末端では、開始コドンから7ntの距離のところに、張り出したプライマーによってリボソーム結合部位(GAGGAG)と制限部位が挿入されている。使用するベクター(ここでは例えばpHBR71-FLAG・ベクター)がすでにリボソーム結合部位を備えている場合には、リボソーム結合部位をさらに挿入する必要はもはやない。このハイブリッド遺伝子の3'末端に位置する挿入された制限部位も、発現ベクターpHBR71-FLAGに意図的に挿入することが今や可能である。ハイブリッド遺伝子の5'末端と3'末端の制限部位は、ハイブリッド遺伝子の内部にもう一度現われることがないように選択し、しかも発現ベクターへの組み込みが現在のプロモータと並行して実施できるように選択する必要がある。この実施例では、SpeI制限部位を含むプライマーを用いて大腸菌からのlacZ遺伝子をPCRによって増幅する:5'-ggACTAGTatgaccatgattacggattcactggc-3'(配列ID番号13、SpeI制限部位は大文字)と5'-ccACTAGTtttttgacaccagaccaactggtaatggtagcg-3'(配列ID番号14、SpeI制限部位は大文字)。さらに、これらプライマーを用いて停止コドンをlacZ遺伝子の配列から除去し、連続的なリーディングフレームを得る。得られるDNA断片(配列ID番号15)をクローニングしてpHBR71-FLAG・プラスミドのSpeI制限部位に直接組み込む。得られる融合タンパク質は、β-ガラクトシダーゼ活性を有するポリマー粒子を形成する。対応するポリマー粒子を単離し、β-ガラクトシダーゼ活性を持つことを、基質としてのo-ニトロフェニル-β-D-ガラクトピラノシド(カルバイオケム社)を用いて還元条件下で明らかにする。
【0118】
実施例7
ポリマー粒子の表面タンパク質とポリマー・コアの間の結合の安定性
【0119】
本発明のために行なった研究により、変性剤(例えばドデシル硫酸ナトリウム(SDS)、尿素、グアニジウムヒドロクロリド、ジチオトレイトール)を用いた処理を行なっても、酸性条件を利用しても、ポリマーシンターゼを生分解性ポリマーのコアから引き離せないことが明らかになった。これは、ポリマー粒子と、ポリマーシンターゼのポリマー粒子結合ドメインとの間に共有結合が存在していることを示している。結合の安定性が増大しているため、ポリマー粒子に結合した物質やポリマー粒子に組み込まれた物質を標的部位に輸送することが可能になる。表面に結合したポリマーシンターゼのN末端断片(N末端から保存されたα/β-ヒドロラーゼ・ドメインの最初の部分まで)は極めて変化しやすいため、遺伝子工学の方法を利用して機能性タンパク質で置換することができる。このようにして、ポリマーシンターゼの活性と、ポリマー粒子の合成を維持する(Rehm, B.H.A.他、Biochem. Biophys. Acta、第1594巻、178〜190ページ、2002年)。その結果、安定性が増大した表面の官能化が実現する。生物学的に活性な物質および/またはカップリング試薬が結合する結合ドメインが異なっているさまざまなタンパク質の混合物は、必要ならば同時に使用する。するとポリマー粒子の表面が多価になる。これらのタンパク質を利用するには、試験管内であれば、異なる結合ドメインを有する精製したタンパク質を合成バッチに添加し、生体内であれば、結合ドメインを有する1つのタンパク質をそれぞれがコードしている遺伝子を対応する微生物の内部で発現させる。
【0120】
実施例7.1
ポリマー粒子の表面タンパク質をさらに修飾する可能性
【0121】
ラルストニア・ユートロファに由来する表面タンパク質フェイジン(phaP)のC末端断片(アラニン141から先のアミノ酸残基)は親水性であるため、機能性タンパク質と置換することができる。そのときフェイジンがポリマー粒子の結合ドメインを通じてポリマー粒子の表面に固着するのが妨げられることはない。
【0122】
この固着は疎水性相互作用に基づいており、可逆的である(Hanley, S.Z.他、FEBS Letters、第447巻、99〜105ページ、1999年)。細胞内ポリマーデポリメラーゼのこのC末端断片は、遺伝子工学の方法によって機能性タンパク質と融合させる。そのため、その後生物学的に活性な物質および/またはカップリング試薬を付着させることによってポリマー粒子の表面を多価にすることができる。
【0123】
ラルストニア・ユートロファの細胞内ポリマーデポリメラーゼのC末端(180番目から先のアミノ酸残基)は、この酵素をポリマー粒子のコアと結合させる(Saegusa, H.、J. Bacteriol.、第183巻(1)、94〜100ページ、2001年)。細胞内ポリマーデポリメラーゼのこのC末端断片は、遺伝子工学の方法によって機能性タンパク質と融合させる。そのためポリマー粒子の表面を多価にすることができる。
【0124】
シュードモナス・オレオボランスに由来する遺伝子phaIとphaFが発現してポリマー粒子の表面に結合しているタンパク質のN末端(140番目までのアミノ酸残基)は、このタンパク質をポリマー粒子のポリエステル・コアと結合させる(Prieto, M.A.他、J. Bacteriol.、第181巻(3)、858〜868ページ、1999年)。遺伝子phaIとphaFの発現産物のこのN末端は、遺伝子工学の方法によって機能性タンパク質と融合させるため、得られる結合ドメインは、生物学的に活性な物質および/またはカップリング試薬を付着させることによってポリマー粒子の表面を多価にすることができる。
【0125】
実施例8
カップリング試薬を用いてポリマー粒子の表面タンパク質の共有結合を修飾する方法
【0126】
生分解性ポリマー粒子の表面にある図1に示したタンパク質は、所定のアミノ酸に特異的に結合する特異的標識物質(例えばリシンに対して特異的に結合するn-ヒドロキシスクシンイミドビオチン)で処理することができる。するとヨードアセトアミドを媒介として生物学的に活性な物質(例えばビオチン)をシステインと連結させることができるようになる。ビオチンなどの分子は、ポリマー粒子の表面タンパク質に生物学的に活性な物質をさらに連結させる。生物学的に活性な物質としては、酵素に結合して生分解性ポリマー粒子の表面タンパク質を徐々に官能化することのできる例えばアビジン、ストレプトアビジンなどが挙げられる(Rehm, B.H.A.他、J. Bacteriol.、第176巻、5639〜5647ページ、1994年)。この官能化は、抗体または薬理学的に活性な成分を付着させることによって進行する可能性がある。所定の膜を通過する輸送および/または所定の膜との融合にとって有利な特定の表面電荷をポリマー粒子に与えるため、いろいろな表面電荷を有する分子も付着させることができる。
【0127】
ポリマー粒子の表面タンパク質に含まれるリシン残基をビオチンで標識するには、n-ヒドロキシスクシンイミドビオチンを用いる。この実験では、緑膿菌からのポリマーシンターゼphaC1を含むポリマー粒子を、プラスミドpHBR71を含む組み換え大腸菌から単離する。単離後、ポリマー粒子をPBS(pH8.0)の中で3回洗浄し、次いでn-ヒドロキシスクシンイミドビオチン(シグマ-オールドリッチ社)を、最終濃度が5mMになるまで溶液に添加する。4℃にて5分間にわたって培養した後に再び洗浄することによって反応を終了させる。
粒子の表面に結合したビオチンの検出には、基質としてストレプトアビジン-アルカリホスファターゼ共役体(シグマ-オールドリッチ社)とリン酸o-ニトロフェニル(カルバイオケム社)を用いる。n-ヒドロキシスクシンイミドビオチンで処理していない粒子が含まれている対照バッチでは、粒子はアルカリホスファターゼ活性を示さない。
【0128】
産生させたポリマー粒子の表面タンパク質を活性化させることのできる生物学的に活性な物質を連結させるための他の多数のカップリング試薬が、上記の具体例以外にも存在している。
【技術分野】
【0001】
本発明は、官能化生分解性ポリマー粒子の製造方法と、そのポリマー粒子を応用した医薬担体に関する。
【背景技術】
【0002】
多数ある活性成分の有効性は、その活性成分が体内の望む標的部位に正確に輸送されるかどうかにかかっている。というのも、使用される活性成分は特定の場所においてだけ十分な作用を及ぼすことができ、他の場所では有効でなかったりマイナスの効果さえ有することがしばしばあるからである。他の場所では有効でなかったりマイナスの効果さえ有するというのは、がん疾患と闘う場合、中でも中枢神経系(CNS)に発生するがん疾患と闘う場合に特に当てはまる。
【0003】
活性成分を動物の体内で輸送する際に起こる可能性のある問題の好例は、血液-脳関門(BBB)である。CNSを目指す活性成分は、まず最初にこの関門を通過する必要がある。血管と細胞からなるこのネットワークはCNSを保護しており、水に溶けない物質がCNSに入るのを阻止する。脂溶性物質は、単純な拡散によってこの関門を簡単に通過することができる。しかし極性物質やイオンの輸送には能動輸送システムと受動輸送システムが必要である。しかし例えばCNS用に新たに発見されたすべての薬のうちの98%を超えるものは水溶性でないため、BBBを通過できず、したがって標的部位に到達することができない。
【0004】
この例は、動物の体内で有効成分を輸送する間に起こる可能性のある問題を示すためのものである。例えばBBBの場合には、膜の透過性を変化させることによって活性成分の透過性をより大きくすることが試みられてきた。活性成分に対するBBBの透過性は、例えば人工的に浸透圧を大きくすることによって、あるいはブラジキニン類似体を投与することによって実現できる。Sanovichら(Sanovich, E.他、Brain Res.、第705巻(1〜2)、125〜135ページ、1995年)は、例えばブラジキニン類似体であるRMP-7をランタンと同時に投与することによってBBBの透過性が増大することを明らかにした。上記のメカニズムのうちの1つによってBBBを開かせることの基本的な1つの欠点は、あらゆる物質に対する透過性が大きくなるため、CNS内の標的細胞にダメージを与える可能性のある毒性物質に対する透過性も大きくなることである。
【0005】
可能な別の方法は、活性成分を化学的に修飾して親油性を大きくし、その結果としてBBBをより容易に通過できるようにするというものである。例えばクロランブシル誘導体についてそのことが示されている(Greig, N.H.他、Cancer Chemother. Pharmacol.、第25巻(5)、320〜325ページ、1990年)。
【0006】
上記の2つの系とは別の方法は、ナノ粒子またはミクロン粒子を使用するというものである。この場合には、膜の透過性も活性成分も変化させない。Kreuter(Kreuter, J.、J. Anat.、第189巻(3)、503〜505ページ、1996年)は、この目的で使用するポリマー粒子を化学的な方法(例えば乳化重合、界面重合、脱溶媒和、蒸発、溶媒沈降)で実質的に製造できることを示している。
【0007】
ドイツ国特許第197 45 950 A1号によると、極めて多彩な物質(ポリマー(この場合にはポリシアノアクリル酸ブチル)、固体または液体の脂質、o/wエマルジョン、w/o/wエマルジョン、リン脂質小胞)から調製したポリマー粒子に医薬物質を付着させ、CNSにその医薬物質を輸送する。
【0008】
膜(特にBBBの膜)を通じてナノ粒子またはミクロン粒子を輸送する際の別の極めて重要な因子は、ポリマー粒子のサイズである。以前の研究結果によると、270nmまでのサイズのポリマー粒子がBBBを通過できる(Lockman, P.R.他、Drug Develop. Indust. Pharmacy、第28巻(1)、1〜12ページ、2002年)。
【0009】
したがって使用するポリマー粒子が特定のサイズになっていることが重要である。Lockmanによると(Lockman, P.R.他、Drug Develop. Indust. Pharmacy、第28巻(1)、1〜12ページ、2002年)、上記の方法で製造したポリマー粒子のサイズは、一般に、フォトン相関分光法によって測定される。ブラウン運動に基づくこの方法では、レーザー・ビームを用いてポリマー粒子の測定を行ない、光の変化の時間依存性から粒子のサイズを決定する。
しかし得られるポリマー粒子のサイズを決定するこの分析法は、時間とコストが非常にかかる。
【発明の概要】
【発明が解決しようとする課題】
【0010】
したがって本発明の目的は、生物学的に活性な物質を輸送するためのすぐに使用できて低コストの系であって、動物の体内で活性成分の輸送を効果的かつ信頼性よく行なうことのできるものを提供することである。
【課題を解決するための手段】
【0011】
本発明で目的とする“生物学的に活性な物質”は、生物の側で生物学的な応答を開始させることのできるあらゆる物質である。このような物質には、生体内で特定の反応を触媒する酵素およびアブザイムや、生体内に存在することによってその生物に間接的な応答を起こさせるタンパク質(例えば抗体)だけでなく、非生物起源の無機分子や有機分子、すなわち自然に存在する生物によっては形成されず、人工的に製造される無機分子や有機分子などが含まれる。たいていの医薬活性成分も後者のグループに属する。生物学的に活性な物質は、その性質に応じ、さらに別の生物学的に活性な物質に結合させるのに適している場合もある。
【0012】
上記の目的を実現するため、生分解性ポリマー粒子の製造方法を提供する。この方法は、ポリマー粒子のサイズを制御するタンパク質をコードしている少なくとも1つの誘導遺伝子を微生物に導入するステップと、少なくとも1つの誘導遺伝子を誘導したこの微生物を、この微生物が生分解性ポリマー粒子を産生するのに適した培地の中で培養するステップを含んでいる。この方法により、生体適合性のある生分解性ポリマー粒子であって生物学的に活性な物質を輸送するのに適したものを製造することができる。そのとき、形成されるポリマー粒子のサイズを制御することにより、必要とされる望むサイズの粒子を製造することができる。特定のサイズのポリマー粒子を製造できるため、望むサイズのポリマー粒子の収率が増大する。そのためこの方法の効率が大きくなってコストの削減に役立つ。さらに、本発明の方法により、BBBを通過して輸送するための粒子サイズに関して上記の条件を満たすポリマー粒子を製造することができる。この方法のおかげで、微生物が天然に作り出すこの種のポリマー粒子よりも小さいポリマー粒子を製造することが特に可能になる。微生物が天然に作り出すポリマー粒子の平均サイズは300〜500nmである(Wieczorek, R.他、J. Bacteriol.、第177巻(9)、2425〜2435ページ、1997年)のに対し、本発明の方法ではポリマー粒子の産生を制御することが可能であり、ポリマー粒子はこの平均値よりもはるかに小さくなる。
【0013】
粒子のサイズを決定する遺伝子を上流の誘導プロモータ(例えばアラビノースによって誘導されるBADプロモータ)で誘導する。この目的で利用する微生物は、ポリマー粒子のサイズを制御する遺伝子を持っていないか、この遺伝子が不活化され、本発明の方法で説明した少なくとも1つの誘導遺伝子(この誘導遺伝子は、ポリマー粒子のサイズを制御するタンパク質をコードしている)によって置換されている。ここでは、ベクターを用いてこの遺伝子を細胞に導入する。ベクターについては、この明細書の実験の項でさらに詳しく説明する。その結果、初めて、生体適合性がある決まったサイズの生分解性ポリマー粒子を微生物プロセスにおいて製造することが可能になる。
【0014】
ポリマー粒子は、細胞内の細胞質含有物として堆積される。ポリマー粒子のコアは、ポリヒドロキシアルキルカルボン酸(特にポリヒドロキシアルカン酸)からなり、タンパク質とリン脂質でできた外殻膜によって囲まれている。外殻膜は、脂質と、その中に埋め込まれたタンパク質からなる。ポリマー粒子のコアを形成するポリヒドロキシアルキルカルボン酸は、融点が50℃〜176℃、結晶度が30%〜70%であり、破断値における伸長度が5%〜300%である。
【0015】
バイオテクノロジーによる培養に一層適しているが、遺伝子構成のために上記のポリマー粒子を形成することができない微生物(例えば、GRAS生物に分類されるある種の形態の大腸菌)でもこの有利な方法を利用できるようにするため、ポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子と、ポリマー粒子のサイズを制御する少なくとも1つの誘導遺伝子を導入する。代謝に影響を与えてポリマー粒子を形成させ、したがって形成されるポリマー粒子の組成に影響を与えることのできるあらゆるタンパク質を考慮することができる。ポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子としては、ここでは、チオラーゼ、レダクターゼ、ポリマーシンターゼのいずれかをコードしているものを選択する。ポリマーシンターゼとしては、ポリマーが形成される最終ステップの触媒となることができるあらゆるタンパク質が使用される。ポリマーの形成は、本発明で説明するポリマーシンターゼ以外にも、例えばリパーゼによっても起こる。ポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子としては、ラルストニア・ユートロファからのphaAチオラーゼ、phaBケトアシルレダクターゼ、phaCシンターゼのいずれかをコードしているものを選択することが好ましい。これらの追加遺伝子を導入すると、細胞は、ポリマー粒子の形成を可能にするタンパク質を産生できるようになる。ポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子を意図的に選択すると、得られるポリマー粒子の組成に影響を及ぼすこともできる。ポリマー粒子の形成に通じる代謝経路に関係するタンパク質をコードしている個々の遺伝子は、それぞれ異なる基質特異性を持っている可能性、あるいはそれぞれ異なる反応生成物を形成する可能性、あるいは代謝経路における分岐経路を阻止する可能性がある。そのためポリマー粒子の形成に関係する基質と分子に意図的に影響を及ぼすことができる。
【0016】
脂肪酸の代謝が変化した微生物(例えば実施例4に記載した大腸菌の突然変異体)の内部でポリマー粒子が産生されるようにするために必要なのは、ポリマー粒子のサイズを制御する少なくとも1つの誘導遺伝子を別にすると、ポリマーシンターゼだけである。どの生物を利用するかに応じ、さらに別の遺伝子を細胞内に導入して上記の条件下でポリマー粒子を産生できるようにする。細胞がポリマー粒子の形成に必要なすべての遺伝子を含んでいない場合でも、欠けているタンパク質によって産生される中間体を栄養媒体を通じて細胞に供給すると、ポリマー粒子の産生を進行させることができる。しかしポリマー粒子を形成するのに少なくとも1つのポリマーシンターゼは常に必要である。
【0017】
ポリマー粒子の性質は、その組成を制御すると影響を受ける可能性がある。性質に影響を与えることにより、例えばポリマー粒子の生分解速度に影響を及ぼすことが可能になる。生体内で形成されるポリマー粒子の組成に影響を与えるためには、ポリマー粒子の形成に関係するタンパク質をコードしている別の遺伝子を意図的に選択して細胞に導入するという上記の可能性は別にすると、チオラーゼおよび/またはポリマーシンターゼをコードしている少なくとも1つの追加遺伝子を細胞内に導入することが特に好ましい。チオラーゼとポリマーシンターゼの基質特異性が異なるため、中間体と最終生成物が異なり、したがって形成されるポリマー粒子のコアの組成が異なることになる。
【0018】
ポリマー粒子の産生原理をの一例を図2に示してある。ポリマー生合成のための活性化前駆体は、原則として、中心的な代謝物であるアセチルCoAに由来するもの、あるいはクエン酸サイクル、脂肪酸β酸化、新規な脂肪酸の生合成という主要な代謝経路の中間体に由来するもの、あるいはアミノ酸代謝に由来するものが可能である。脂肪酸を炭素源として使用する場合には、PHA生合成のための活性化前駆体として機能する中間体(アシルCoA、特に3-ヒドロキシアシルCoA)を脂肪酸β酸化によって製造する。
【0019】
粒子のサイズを制御する際には、ポリマー粒子のサイズを制御するタンパク質をコードしている少なくとも1つの誘導遺伝子は、フェイジン様タンパク質のファミリーに由来するものであり、ラルストニア・ユートロファからのフェイジン遺伝子phaPと、シュードモナス・オレオボランスからのフェイジン遺伝子phaFを含むグループの中から選択することが好ましい。フェイジンは分子量が14〜28kDaの両親媒性タンパク質であり、ポリマー粒子の疎水性表面にしっかりと結合する。
【0020】
ポリマーの組成が異なるポリマー粒子は、物理的特性が異なり、生物学的に活性な物質(特に薬理学的に活性な成分)を異なる速度で放出する。例えばC6〜C14の3-ヒドロキシ脂肪酸からなるポリマー粒子は、ポリマーの結晶度が低いためにポリマーの分解速度がより大きい。ポリマー骨格上に比較的大きな側鎖を有するポリマー成分のモル比が大きくなると、通常は結晶度が小さくなり、弾力特性がより顕著になる。本発明で説明した方法に従ってポリマーの組成を制御することにより、ポリマー粒子の生分解性に影響を及ぼすことが可能になり、したがって生物学的に活性な物質(特に薬理学的に活性な成分)の放出速度にも影響を及ぼすことが可能になる。
【0021】
ポリマー粒子を形成するための基質として、側部官能基を有する少なくとも1つの脂肪酸を培地に導入することが好ましい。導入することが特に好ましいのは、少なくとも1つのヒドロキシ脂肪酸および/または少なくとも1つのメルカプト脂肪酸および/または少なくとも1つのβ-アミノ脂肪酸である。“側部官能基を有する脂肪酸”は、飽和脂肪酸または不飽和脂肪酸を意味するものとする。その中には、メチル基、アルキル基、ヒドロキシル基、フェニル基、スルフヒドリル基、第一級アミノ基、第二級アミノ基、第三級アミノ基、アルデヒド基、ケト基、エーテル基、カルボキシル基、O-エステル基、チオエステル基、カルボン酸アミド基、ヘミアセタール基、アセタール基、リン酸モノエステル基、リン酸ジエステル基を含むグループの中から選択した側部官能基を有する脂肪酸も含まれる。
【0022】
ポリマー粒子は、基質特異性が異なるタンパク質をコードしているさまざまな遺伝子を使用することによる影響と、使用する培地に存在している添加物、基質、反応条件の影響の両方を遺伝子レベルで受けるため、使用する基質は、ポリマー粒子に望む組成と性質がどのようなものであるかによって決まる。
【0023】
形成されるポリマー粒子のサイズをより正確に制御するため、ポリマー粒子のサイズを制御するのに十分な量の基質を培地に添加する。すると粒子のサイズをより効果的に制御することが可能になる。
【0024】
本発明の方法でポリマー粒子を産生させるのに用いる微生物は、ラルストニア、アルカリゲネス、シュードモナス、ハロビフォルマを含む属の中から選択する。使用する微生物の選択は、ラルストニア・ユートロファ、アルカリゲネス・ラトゥス、大腸菌、シュードモナス・フラギ、シュードモナス・プティダ、シュードモナス・オレオボランス、緑膿菌、シュードモナス・フルオレセンス、ハロビフォルマ・ハロテレストリスを含むグループの中から行なうことが好ましい。このグループには、生体適合性のある生分解性ポリマー粒子を自然に産生することのできる微生物と、遺伝子の構成上それが不可能な微生物(例えば大腸菌)の両方が含まれる。後者の微生物に対し、ポリマー粒子の産生を可能にするのに必要な遺伝子を、本発明の方法を利用して導入する。たとえ微生物が、代謝が異なるためにポリマー粒子を形成するのに必要な基質を産生することができなくても、原則として、培養可能なあらゆる微生物を利用して上記の方法でポリマー粒子を産生させることができる。微生物が必要な基質を産生できない場合には、必要な基質を培地に添加した後、細胞内に導入した遺伝子が発現しているタンパク質を利用してポリマー粒子に変換する。
【0025】
細胞からポリマー粒子を得るためには、培養した微生物を公知の方法で破壊した後、ポリマー粒子を細胞残留物から分離する。この方法で得られるポリマー粒子のサイズの範囲は、標準的な方法(例えば排除クロマトグラフィまたは密度勾配遠心分離)を利用して望むサイズのポリマー粒子を選択することにより、さらに狭めることができる。
【0026】
本発明の方法で製造したポリマー粒子のタンパク質と脂質からなる外殻膜を修飾し、動物の体内で活性成分を輸送するのにより好ましい粒子特性を持たせることができる。そのためには、ポリマー粒子の表面に位置する脂質層を本発明の方法で得られたポリマー粒子から分離し、異なる組成の脂質層で置換する。
【0027】
脂質層を異なる組成の脂質層で置換するとき、新しい脂質層の重要な性質は、生物の膜を通過するポリマー粒子の輸送に影響を与えるという性質である。外殻膜の脂質層が標的とする膜の脂質層と適合している場合には、粒子の取り込み増大が観察される可能性がある(Fernart, L.他、J. Pharmacol. Exp. Ther.、第291巻(3)、1017〜1022ページ、1999年)。したがってポリマー粒子の脂質層を取り除き、異なる組成の脂質層で置き換えるとよい。これは、アセトン抽出によって、あるいはホスホリパーゼまたは非変性性洗浄剤を用いることによって実現することが好ましい。ここで、今や実質的に脂質を含まなくなったポリマー粒子に適切な両親媒性分子の混合物を添加し、望む組成の脂質層を得る。新しい脂質層は、リン脂質とエーテル脂質を含むグループの中から選択した両親媒性分子の混合物からなることが好ましい。
【0028】
上記の洗浄剤を用いて脂質だけでなくポリマー粒子の表面に位置するタンパク質(ただしポリマーシンターゼは除く)も除去し、そのタンパク質を、官能化したタンパク質で置換することもできる。裸のポリマー粒子をこのようにして製造し、そのポリマー粒子に対して数多くの修飾を行なうことができる。これについてはあとで詳しく説明する。微生物ハロビフォルマ・ハロテレストリスから得られるポリマー粒子は、このタイプの修飾に特に適している。
【0029】
ポリマー粒子をその後どのように利用するかに応じ、粒子サイズを少なくとも1つの誘導遺伝子で制御し、形成されるポリマー粒子の直径を10nm〜3μmにする。ポリマー粒子の直径は10nm〜900nmであることが好ましく、10nm〜100nmであることが特に好ましい。本発明の特別な一実施態様では、サイズの制御を、培地内における基質の利用可能性を制御することによって、あるいは2つのメカニズムを合わせることによって実現することもできる。
【0030】
本発明の方法で製造したポリマー粒子を官能化するには、ポリマー粒子のサイズを制御するタンパク質をコードしている少なくとも1つの誘導遺伝子が、ポリマー粒子結合ドメインと、少なくとも1つの結合ドメインとを含んでいて、この少なくとも1つの結合ドメインが、生物学的に活性な物質および/またはカップリング試薬と結合可能である必要がある。同様に、ポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子が、ポリマー結合ドメインと、少なくとも1つの結合ドメインとを含んでいて、この少なくとも1つの結合ドメインが、生物学的に活性な物質および/またはカップリング試薬と結合可能であることが特に好ましい。
【0031】
“結合ドメイン”は、細胞内に以前に導入した少なくとも1つの誘導遺伝子を修飾することによって、および/またはポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子を修飾することによって得られるタンパク質の一部を意味する。生物学的に活性な物質および/またはカップリング試薬と結合できる結合ドメインは、オリゴペプチド、酵素、アブザイム、非触媒性タンパク質を含むグループの中から選択する。このグループは、フラグ・エピトープまたは少なくとも1つのシステインを含んでいることが特に好ましい。このグループは、実験の項で機能性タンパク質という呼び方もする。このグループのメンバーに関するコード配列を、細胞内に以前に導入した少なくとも1つの誘導遺伝子のコード配列、および/またはポリマー粒子の形成に関係するタンパク質をコードしている別の少なくとも1つの遺伝子のコード配列に導入する。このようにすると、ポリマー粒子の形成、またはポリマー粒子の形成制御に関係するだけでなく、遺伝子の修飾によって得られる結合ドメインのおかげで生物学的に活性な物質および/またはカップリング試薬と結合することもできるタンパク質が産生される。この結合ドメインは、ポリマー粒子を産生させた後、カップリング試薬を用いて表面に位置するタンパク質を化学的に修飾することによっても得られる(実施例8を参照のこと)。
【0032】
この目的で使用する“カップリング試薬”は、一方の側には生物学的に活性な物質または別のカップリング試薬を結合させ、他方の側には結合ドメインを結合させるのに適した無機化合物または有機化合物である。
【0033】
この構造により、生物学的に活性な物質を輸送するのに適した多機能ポリマー粒子を産生させることができる。“ポリマー粒子結合ドメイン”は、ポリマー粒子の疎水性表面に結合できるタンパク質の一部であることが好ましい。ポリマー粒子の表面に結合するタンパク質の一部を含むポリマー粒子のドメインの選択は、ここでは、ポリマーデポリメラーゼ、ポリマー調節因子、粒子サイズ制御タンパク質を含む一群のタンパク質の中から行なう。これらのタンパク質は、ポリマー粒子を形成することのできる微生物(特にラルストニア、アルカリゲネス、シュードモナスという属からの微生物)に由来することが好ましい。粒子サイズ制御タンパク質は、ここでは、フェイジン様タンパク質のファミリーに由来することが好ましく、ラルストニア・ユートロファとシュードモナス・オレオボランスからのフェイジンを使用することがさらに好ましい。
【0034】
本発明の目的に合った“ポリマー調節因子”は、ポリマー粒子の形成に関係する遺伝子phaA、phaB、phaCの転写を調節するタンパク質である。ポリマー調節因子は、粒子の表面に結合することによって転写調節がなされなくする。このような調節因子の一例は、ラルストニア・ユートロファからのフェイジン・リプレッサ(phaR)である。このphaRは、フェイジン様遺伝子のプロモータに結合する。phaRの発現産物は、形成されるポリマー粒子のサイズを調節し、その遺伝子が読まれるのを阻止する。フェイジン・リプレッサは形成されるポリマー粒子の表面に結合するため、プロモータのこの部位が放出され、遺伝子の転写開始が可能になる。
【0035】
カップリング試薬および/または生物学的に活性な物質を結合させるのにポリマーシンターゼの結合ドメインを利用するというアイディアは、このタンパク質のポリマー粒子結合ドメインとポリマー粒子のコアの間の結合の安定性が増大していることから生まれた。
発明者は、驚くべきことに、変性剤(例えばドデシル硫酸ナトリウム(SDS)、尿素、グアニジウムヒドロクロリド、ジチオトレイトール)を用いても、酸性条件を利用しても、この結合を生分解性ポリマーのコアから切り離せないことを発見した。ここでは、ラルストニア・ユートロファ、シュードモナス・オレオボランス、シュードモナス・プティダ、緑膿菌に由来するポリマーシンターゼを使用することが好ましい。
【0036】
ポリマー粒子の表面に結合するタンパク質を遺伝子工学によって修飾してもポリマー粒子の形成に関係するタンパク質の機能には影響がないというのが、本発明による方法の特別な利点である。例えばあるタンパク質をポリマーシンターゼのN末端に融合して生物学的に活性な物質および/またはカップリング試薬のための結合ドメインを作っても、ポリマーシンターゼの機能は保持される。しかしタンパク質の機能が融合によって損なわれても、この欠点は、同じ機能を実行するタンパク質をコードしている活性状態の別の遺伝子の存在によって消すことができる。
【0037】
このようにして、タンパク質(特にポリマーシンターゼ)の結合ドメインを通じたポリマー粒子と生物学的に活性な物質および/またはカップリング試薬の間の安定な結合を保証することができる。
【0038】
発現すると粒子の表面に結合するタンパク質をコードする遺伝子を遺伝子工学によって修飾している間に、異なる修飾状態の遺伝子を細胞の中に導入することもできる。結合ドメインが異なるいろいろなタンパク質が発現した状態でポリマー粒子が形成されると、異なる結合ドメインを利用して粒子の表面を多官能化することができる。この方法により、官能化したポリマー粒子を直接的かつ効率的に大量生産することができる。
【0039】
次に、カップリング試薬を使用し、ポリマー粒子の表面に結合したタンパク質を官能化する。カップリング試薬の選択は、ビス(2-オキソ-3-オキサゾリジニル)ホスホニッククロリド(BOP-Cl)、ブロモトリスピロリジノホスホニウムヘキサフルオロリン酸(PyBroP)、ベンゾトリアゾール-1-イル-オキシ-トリスピロリジノホスホニウムヘキサフルオロリン酸(PyBOP)、n-ヒドロキシスクシンイミドビオチン、2-(1H-ベンゾトリアゾール-1-イル)-1,1,3,3-テトラメチルウロニウムヘキサフルオロリン酸(HBTU)、ジシクロヘキシルカルボジイミド、ジスクシンイミジル炭酸、1-(3-ジメチルアミノプロピル)-3-エチルカルボジイミド(EDC)、ビス(2-オキソ-3-オキサゾリジニル)ホスフィン、ジイソプロピルカルボジイミド(DIPC)、2-(1H-ベンゾトリオキサゾリル)-1,1,3,3-テトラメチルウロニウムテトラフルオロホウ酸(TBTU)、2-(5-ノルボルネン-2,3-ジカルボキシイミド)-1,1,3,3-テトラメチルウロニウムテトラフルオロホウ酸(TNTU)、パラニトロフェニルクロロギ酸、O-(n-スクシンイミジル)-1,1,3,3-テトラメチルウロニウムテトラフルオロホウ酸(TSTU)からなるグループの中から行なうことが好ましい。
【0040】
使用する生物学的に活性な物質は、殺虫剤、除草剤、薬理学的に活性な物質、タンパク質であることが好ましい。
【0041】
薬理学的に活性な物質の選択は、ジデオキシイノシン、フロクスウリジン、6-メルカプトプリン、ドキソルビシン、ダウノルビシン、1-ダルビシン、シスプラチン、メトトレキセート、タキソール、抗生物質、抗血液凝固薬、殺菌薬、抗不整脈薬、活性成分の前駆体、例示した活性成分の誘導体からなるグループの中から行なう。
【0042】
タンパク質の選択は、インスリン、カルシトニン、ACTH、グルカゴン、ソマトスタチン、ソマトトロピン、ソマトメジン、副甲状腺ホルモン、エリスロポエチン、視床下部放出因子、プロラクチン、甲状腺刺激ホルモン、エンドルフィン、エンケファリン、バソプレッシン、天然には存在しないアヘン製剤、スーパーオキシドディスムターゼ、抗体、インターフェロン、アスパラギナーゼ、アルギナーゼ、アルギニンデアミナーゼ、アデノシンデアミナーゼ、リボヌクレアーゼ、トリプシン、キモトリプシン、ペプシンからなるグループの中から行なうことが好ましい。
【0043】
本発明の別の側面は、生分解性ポリマー粒子を試験管内で製造する方法であって、ポリマー粒子の形成に適した溶液を少なくとも1つの基質とともに提供するステップと、この溶液に、ポリマー粒子のサイズを制御するのに適したタンパク質を導入するステップと、ポリマー粒子の形成に関係する別の少なくとも1つのタンパク質を導入するステップを含む方法である。この試験管内での方法には、ポリマー粒子のサイズを早くもポリマー粒子の製造中に制御できるため、形成されたポリマー粒子のサイズを決定し、個々のサイズのクラスに分類するというコストと時間がかかる作業を回避できるという利点もある。
【0044】
この明細書で使用する別の少なくとも1つのタンパク質は、ポリマーシンターゼである。このポリマーシンターゼの選択は、ラルストニア・ユートロファ、シュードモナス・オレオボランス、シュードモナス・プティダ、緑膿菌からのポリマーシンターゼを含むグループの中から行なうことが好ましい。
【0045】
微生物から単離したタンパク質と酵素を実験室で使用する試験管内での合成は、通常は、酵素と(場合によっては)酵素の基質をあらかじめ単離して精製する必要があるため、生体内での合成とは異なって非常にコストがかかる。生体適合性のある生分解性ポリマー粒子を試験管内で合成するための本発明による特別な一実施態様では、ポリマー粒子を形成するため、ポリマー粒子の形成に適した溶液に、少なくとも1つの脂肪酸(特に好ましいのは、β-メルカプト脂肪酸および/またはβ-アミノ脂肪酸)と、アシルCoAオキシダーゼまたはそれ以外の酸化・活性化酵素を添加する。R/S-3-ヒドロキシ脂肪酸とアシルCoAシンテターゼの代わりにこれらの基質を使用すると、CoAのリサイクル・システムが得られる。このリサイクル・システムにおいて、アシルCoAオキシダーゼは、脂肪酸を活性化して酸化させる一方で、CoAを消費し、ATPを加水分解する。重合の間、ポリマーシンターゼはCoAを除去するが、そのCoAは、アシルCoAオキシダーゼが使用できる。その結果、この試験管内での方法において顕著なコスト削減を実現することが可能になる。
【0046】
試験管内でポリマー粒子を製造する方法の別の利点は、少なくとも1つの基質を、ポリマー粒子のサイズを制御するのに十分な量、溶液に添加することにある。
【0047】
それに加え、形成されるポリマー粒子のサイズは、形成されるポリマー粒子のサイズを制御するのに十分な量のポリマーシンターゼを溶液に添加することによってさらに制御することもできる。生体内での方法と同様、試験管内での方法は、ポリマー粒子のサイズをより正確に制御することもできるため、望むサイズのポリマー粒子の収率が増大し、この方法の効率とコスト効率が向上する。
【0048】
しかし試験管内での方法では、粒子のサイズは、実際には、ポリマー粒子のサイズを制御するタンパク質を溶液の中に導入することによって制御する。このタンパク質は、フェイジン様タンパク質のファミリーに由来するものであり、ラルストニア・ユートロファからのフェイジンと、シュードモナス・オレオボランスからのフェイジンを含むグループの中から選択することが好ましい。
【0049】
少なくとも1つの基質と使用する酵素を選択することにより、生体内での方法と同様、ポリマーの組成を制御すること、そしてその結果として異なる性質を持ったポリマーのコアを得ることも可能になる。2つ以上の基質を用いる場合には、酵素から形成されるポリマーのタイプに応じ、例えばポリマーのコアの組成が異なるポリマー粒子を得ることが可能である。すでに説明したように、この方法で製造されるポリマーは、ポリマー粒子に極めて多彩な性質を付与する。
【0050】
試験管内でポリマー粒子を製造する方法では、ポリマー粒子の表面にある脂質層の組成は、リン脂質とエーテル脂質からなるグループの中から選択した少なくとも1つの両親媒性分子を溶液に添加することによって制御する。(添加する両親媒性分子がないときには、試験管内で得られる粒子は、タンパク質によってだけ取り囲まれる。そのため別のタイプの粒子になる。)このようにして、例えば動物の体内にある天然の膜に特別に合わせた特定の表面電荷を有するポリマー粒子を製造することができる。その後、そのポリマー粒子がこの膜を通過することになる。試験管内での方法で外殻膜の脂質層を修飾することの利点は、生体内での方法のように、その脂質層をその後修飾しなくても済むことである。
両親媒性分子(例えばリン脂質またはエーテル脂質)がすでに最初の溶液に添加されているため、望む組成の脂質膜が最初から得られる。
【0051】
試験管内での方法の別の特徴は、少なくとも1つの薬理学的に活性な物質を、ポリマー粒子の形成に適した溶液に添加するというものであり、生体内での方法でもこの特徴が利用されている。この物質は、ポリマー粒子を合成している間にポリマー粒子に組み込まれ、その後、粒子マトリックスを通じた拡散によって、あるいはポリマー粒子の分解によって動物の体内に放出させることができる。後者の場合には、本発明の方法によって製造したポリマー粒子において、ポリマーのコアの組成を上記のように制御することによってポリマー粒子の生分解速度を調節できるというさらに別の利点も有する。このようにして、活性成分を特定の期間にわたって連続的に放出させることが可能になる。
【0052】
官能化は、活性成分を成長中のポリマー粒子に組み込むこと以外に、生体内での方法のように、ポリマー粒子の形成に適した溶液に導入する少なくとも1つのタンパク質を選択するとき、導入した少なくとも1つのタンパク質が、ポリマー粒子結合ドメインと、少なくとも1つの結合ドメインとを備えていて、この少なくとも1つの結合ドメインが、生物学的に活性な物質および/またはカップリング試薬と結合可能になっているようにすることによって実現される。この場合に試験管内での方法でポリマー粒子を形成するのに用いるタンパク質は、すでに説明した生体内での方法から得ることができ、そのタンパク質は、上記のどのタイプの製造方法であるかに応じ、対応する性質を示す。次に、ポリマー粒子の形成に関係するそのタンパク質を、ポリマー粒子の形成に適した溶液に添加する。生体内での方法と同様、この方法で製造したポリマー粒子は、製造後に、すでに説明したカップリング試薬を用いた修飾によって、あるいはポリマー粒子の表面に結合しているタンパク質の結合ドメインに結合する生物学的に活性な物質を添加することによって、さらに修飾することもできる。
【0053】
本発明にはさらに、サイズが決まっていて両親媒性分子からなる表面層を有するポリマー粒子であって、その内部には、ポリマーデポリメラーゼ、ポリマー調節因子、ポリマーシンターゼ、粒子サイズ制御タンパク質を含むグループの中から選択した少なくとも1つのタンパク質が導入されており、少なくとも1つのこのタンパク質が、ポリマー粒子結合ドメインと、生物学的に活性な物質および/またはカップリング試薬と結合可能な結合ドメインとを備えているものが含まれる。好ましい一実施態様では、上記の少なくとも1つのタンパク質は、すでに説明した方法で製造する。
【0054】
本発明のポリマー粒子は有利な性質を持っているため、医薬調製物、殺虫剤、除草剤の製造に特に適している。この医薬調製物は、中枢神経系の疾患を治療するのに適していることが好ましい。本発明によるポリマー粒子は修飾できるため、BBBを通過する条件を満たすことができる。
【0055】
粒子サイズの制御、外殻膜の組成の制御、粒子表面の官能化、その中でも特に粒子表面の官能化が可能であることは、生分解性ポリマー粒子が生物学的に活性なあらゆる種類の物質を輸送するための担体として適しており、ポリマー粒子をその作用部位に狙いを定めて輸送できることを意味する。多価により、例えば少なくとも1つの薬理学的に活性な物質だけでなく抗体も粒子の表面に結合させることが可能になる。抗体には結合の特異性があるため、標的部位に正確に導くことができる。上記の利点ならびにその他の利点を、以下の実施例においてさらに詳しく説明する。
【図面の簡単な説明】
【0056】
【図1】生体内で産生される生分解性ポリマー粒子と、その表面に結合したタンパク質および脂質の概略図である。略号は、以下のタンパク質に関係する。A:ポリマーデポリメラーゼB:フェイジン(ラルストニア・ユートロファにおけるコード遺伝子の名称;phaP、ラルストニア・ユートロファにおけるコード遺伝子の名称;phaF)C:ポリマーシンターゼD:リン脂質E:ポリマー調節因子(例えばフェイジン・リプレッサ(ラルストニア・ユートロファからのPhaR))
【図2】ラルストニア・ユートロファにおいて可能な生分解性ポリマー合成法の一例である。基質であるアセチルCoAから出発して3段階のプロセスで単純なポリヒドロキシアルカン酸ポリヒドロキシブチル酸(PHB)を製造する。PHBのC4繰り返し単位は、β-ヒドロキシブチル酸である。合成の最終ステップにより、表面にポリマーシンターゼが結合したポリマー粒子が形成される。
【図3】生体内での実験結果であり、フェイジンの発現を増やした(溶液中の誘導因子の量を増やした)ときのポリマー粒子の直径の挙動を示している。
【図4】生体内での実験結果であり、フェイジンの発現を増やした(溶液中の誘導因子の量を増やした)ときの細胞内のポリマー粒子の数を示している。
【図5】試験管内の実験結果であり、ポリマーシンターゼに対するフェイジンの量の比がポリマー粒子の直径に及ぼす影響を示している。
【図6】試験管内の実験結果であり、ポリマーシンターゼに対する基質の比がポリマー粒子の直径に及ぼす影響を示している。
【図7】(ラルストニア・ユートロファからの遺伝子PhaPを含む)プラスミドpBBad-Pと(シュードモナス・オレオボランスからの遺伝子PhaFを含む)プラスミドpBBad-Fを構成する際の出発プラスミドとして使用するベクターpBBad22K(Sukchawalit, R.他、FEMS Microbiol Lett.、第181巻(2)、217〜223ページ、1999年)を示している。
【図8】ラルストニア・ユートロファからの粒子サイズ制御タンパク質フェイジンを発現させるPhaP遺伝子を含むベクターpBBad-Pを示している。
【図9】ポリマー(ポリヒドロキシブチル酸)を貯蔵させるプラスミドpBHR68(Spiekermann, P.他、Arch. Microbiol.、第171巻、73〜80ページ、1999年)を示している。このプラスミドは、phaAチオラーゼ、phaBケトアシルレダクターゼ、phaCシンターゼを発現させるための生合成オペロンを形成するラルストニア・ユートロファからの遺伝子phBRe、phbARe、phbCReを含んでいる。
【図10】ポリマー(ポリヒドロキシアルカン酸)を貯蔵させるプラスミドpBHR71(Langenbach, S.他、FEMS Microbiol. Lett.、第150巻、303〜309ページ、1997年)を示している。このプラスミドは、ポリマーシンターゼを発現させる遺伝子phaC1を含んでいる。
【図11】シュードモナス・オレオボランスからの粒子サイズ制御フェイジン様タンパク質を発現させる遺伝子PhaFを含むベクターpBBad-Fを示している。
【図12】図10のpBHR71プラスミドから出発して構成したベクターpBHR71-システインを示している。この構造では、ポリマーシンターゼをコードしている遺伝子phaC1がN末端に2個のシステイン残基を備えているため、生物学的に活性な物質の直接的な結合、またはカップリング試薬を通じた生物学的に活性な物質の結合が可能になる。
【図13】図10のpBHR71プラスミドから出発して同様に構成したベクターpBHR71-FLAGを示している。この構造では、ポリマーシンターゼをコードしている遺伝子phaC1がN末端に2個のフラグ・エピトープを備えている。このようにすると、フラグ遺伝子配列のSpeI制限部位を通じて機能性タンパク質の遺伝子配列を生体内で組み込むことができるだけでなく、すでに発現しているポリマーシンターゼにカップリング試薬および/または生物学的に活性な物質を試験管内で付着させることもできる。使用する生物学的に活性な物質は、直接に、あるいはカップリング試薬を通じて結合されるが、この物質により、標的部位(例えば細胞表面)への結合が可能になったり、生物学的活性(特に酵素活性)が与えられたりする。
【発明を実施するための形態】
【0057】
実施例1
生体内で産生される生分解性ポリマー粒子のサイズ制御
実施例1.1
【0058】
ラルストニア・ユートロファにおけるポリマー粒子の産生
【0059】
ラルストニア・ユートロファ(以前はアルカリゲネス・ユートロファス)の“ノックアウト”突然変異体(York, G.M.他、J. Bacteriol.、第183巻、4217〜4226ページ、2001年)を用いて生分解性ポリマー粒子を産生させた。この突然変異体は、ポリマー粒子の表面タンパク質フェイジン(phaP(-))の発現をコードしている遺伝子に欠陥があるため、phaP遺伝子によってコードされているフェイジンをもはや発現しない。また、この遺伝子を含まず、ポリマー粒子の生合成に必要な別の遺伝子も含まない微生物を用いて生分解性ポリマー粒子を産生させることもできる。可能な微生物の具体例としては、大腸菌、ハロビフォルマ・ハロテレストリスが挙げられよう。一実施態様では、すでに説明した生分解性ポリマー粒子を自然状態では産生できない大腸菌(実施例2を参照のこと)を用いる。
【0060】
次に、ベクターを用いて後者の微生物を形質転換する。ラルストニア・ユートロファの場合にはフェイジンをコードしているphaP遺伝子とプロモータ配列を含んでいる。この遺伝子は、誘導プロモータ(アラビノースによって誘導されるBADプロモータであることが好ましい)によって制御される。
【0061】
クローニング・ステップ
【0062】
phaPをコードしているラルストニア・ユートロファからのDNA配列を用いてクローニングを行なう。この配列は、“GenBank”にAF079155として登録されている(Hanley, S.Z.他、FEBS Letters、第447巻、99〜105ページ、1999年)。この配列を、誘導プロモータ(PBAD)を含むベクターpBBad22K(Sukchawalit, R.他、FEMS Microbiol. Lett.、第181巻(2)、217〜223ページ、1999年、図7を参照のこと)のNcoI/BamHI制限部位に挿入する。クローニングを目的として、対応するNcoI制限部位とBamHI制限部位を、フェイジンをコードしている配列に、プライマー5'-aaaggccccatggtcctcaccccggaaca-3'(配列ID番号1、NcoI制限部位に下線を付した)と5'-aaaggccggatcctcagggcactaccttcatcg-3'(配列ID番号2、BamHI制限部位に下線を付した)を用いたPCR突然変異誘発によって挿入する。得られるDNA断片(配列ID番号3)を制限酵素NcoIとBamHIを用いて加水分解し、同様にして加水分解したベクターpBBad22Kと連結する(図7)。このプラスミドは、今やタンパク質フェイジンのヌクレオチド配列を含んでいるため、今後はpBBad-Pと呼ぶことにする(図8)。このプラスミドを微生物に導入し、この微生物に関して従来技術で知られている形質転換技術を用いて形質転換する。pBBad-PのBADプロモータの誘導因子はアラビノースである。アラビノースは、使用している微生物の体内でこの遺伝子の発現を定量的に制御することができる。この制御メカニズムを利用してフェイジン遺伝子の発現を制御し、最終的には細胞内に存在するフェイジンの量によってポリマー粒子のサイズを制御する。
【0063】
実施例1.2
もともとポリマー粒子を形成することができなかった微生物によるポリマー粒子の産生
【0064】
もともとこのタイプのポリマー粒子を形成することができなかった微生物では、別のベクターまたは同じベクターが、ポリマー粒子の形成に関係する別の遺伝子を含んでいる。
ポリマー粒子を産生する微生物に導入する必要がある遺伝子の数と種類は、使用する微生物の基本的な構成によって決まる。最も簡単な場合、例えば大腸菌では、図2に示した方法でポリマー粒子を製造するのに少なくとも1つのチオラーゼ、1つのレダクターゼ、1つのポリマーシンターゼが必要である。特別な突然変異を含む生物(例えば実施例4からの大腸菌株)を利用する場合には、ポリマー粒子を製造するのに上記の遺伝子よりも少数で十分である。ポリマー合成の代謝経路から所定の基質前駆体が生物にすでに供給されている場合にもこのことが当てはまる。
【0065】
別のプラスミドを用いてポリマー粒子の形成に必要な遺伝子を細胞内に導入する場合には、ラルストニア・ユートロファの染色体DNAに由来する5.2kbのSmaI/EcoRI断片を含むプラスミドpBHR68(図9)が特に適している。このプラスミドは、ラルストニア・ユートロファに生分解性ポリマー粒子を産生させるための生合成オペロンを含んでいる(Spiekermann, P.他、Arch. Microbiol.、第171巻、73〜80ページ、1999年)。
【0066】
実施例1.3
pBBad-Pベクターを用いた粒子サイズの制御
【0067】
このようにして改変した微生物を、ルリア肉汁培地の中で30℃にて培養する。アラビノースを用いてプロモータを誘導すると、後期対数増殖期へと進む。培養を開始して24時間後、細胞内の粒子のサイズを測定する。そのために遠心分離で細胞を栄養培地から分離して破壊する。次に、トンネル電子顕微鏡に分析用ゲル濾過クロマトグラフィを組み合わせてポリマー粒子のサイズを測定する。ポリマー粒子のカウント数は、光学的顕微鏡のもとで無傷の細胞を調べることによって決定し、細胞1個当たりのポリマー粒子の数を数える。
【0068】
結果からわかるように、発現を制御することにより、個々の細胞内で形成されるポリマー粒子の平均サイズ(図3を参照のこと)とその平均数(図4を参照のこと)の両方を制御することができる。制御されたやり方でフェイジンの数を増やすと、ポリマー粒子の平均直径が小さくなり、それと同時にポリマー粒子の平均数が増える。このようにして、望む直径を有するポリマー粒子の収率を明らかに大きくすることができる。このようにすると、ポリマー粒子が、より早く、しかもよりコスト効率よく産生される。
【0069】
実施例1.4
基質利用可能性による粒子サイズの制御
【0070】
ポリマー粒子のサイズを制御する別のメカニズムは、ポリマー粒子の合成に必要な基質またはポリマーシンターゼの利用可能性を調節するというものである。ポリマーシンターゼの利用可能性は、アンチセンス法によって、あるいは遺伝子調節法によって、あるいは栄養培地においてポリマーシンターゼの形成に必要な基質がどれだけ利用できるかによって制御することができる。
【0071】
栄養培地における基質の利用可能性による粒子サイズ制御の一例は、存在する炭素源の濃度を減らすことにより、形成されるポリマー粒子の直径を制御する実験である。上記のプラスミドpBHR68(図9)と、生分解性ポリマー粒子を産生させるためのラルストニア・ユートロファ由来の生合成オペロン(図8)とを含む大腸菌株を、1.5%(w/v)のグルコースを含むM9培地の中で30℃にて増殖させる。定常的増殖期の初期には、グルコースなしのM9培地を添加することによって炭素源の濃度が元の値の1/50になる。それ以外は一定の増殖条件下でさらに20時間にわたって微生物を培養する。テストが終了したとき、微生物は、平均直径が130nmのポリマー粒子を含んでいる。
【0072】
実施例2
生体内で産生されるポリマー粒子の組成に影響を与える方法
【0073】
生分解性ポリマー粒子の組成は、細胞内に、phaPをコードしている遺伝子に加え、シンターゼ(特にチオラーゼ、または別のポリマーシンターゼ)の基質を提供する酵素をコードしている別の遺伝子を導入することによって変化させることができる。その結果、細胞は、ポリマー粒子が形成されている間に一連のさまざまなモノマーを伸長しているポリマー鎖に組み込むことができるため、異なる組成のポリマーからなるコアを有するポリマー粒子を作り出す。
【0074】
このことに関して挙げられる具体例は、生体内と試験管内の両方の方法で、基質特異性があるために伸長しているポリマー鎖に3-ヒドロキシ脂肪酸(C4〜C16)を異なった形で組み込むさまざまなポリマーシンターゼである。例えば、C4脂肪酸(C4)から生分解性ポリマー鎖を産生するラルストニア・ユートロファ由来のポリマーシンターゼや、アエロモナス・プンクタータ(C4とC6)、チオカプサ・フェンニギイ(C4とC8)、緑膿菌(C6〜C14)に由来するポリマーシンターゼを用いることができる。同時に2つ以上のポリマーシンターゼを細胞内に導入すると、非常に多彩な性質を有するポリマー粒子を産生させることができる。連続バッチでは、ランダムな突然変異誘発によって基質特異性が変化した新しいポリマーシンターゼも得ることができる(試験管内での進化)。
【0075】
形成されるポリマー粒子の組成に別の方法で影響を与えることもできる。さまざまな炭素源、さまざまな炭素源の前駆体、ポリマー粒子の形成につながる代謝経路の中間体などを提供することは、形成されるポリマーの性質に影響が及ぶ。さらに、ポリマー粒子の形成に関係する代謝経路は、阻害剤や、問題になっている代謝経路をノックアウトした突然変異体や、酵素の発現によって制御することができる。すると代謝経路が変化して別の中間体または最終生成物が形成される。
【0076】
脂肪酸の代謝に影響を与えるため、以下の酵素を用いる。すなわち、(R)-特異的エノイル-CoAヒドラターゼ、トランスアシラーゼ、ケトアシル-CoA/ACPレダクターゼである。これらの酵素は、ポリマー粒子が形成される際に、脂肪酸が代謝されるときの中間体を変化させ、異なる生成物(例えば側鎖が異なる脂肪酸)を提供するという性質を持っている。
これらの酵素は、ポリマーシンターゼのために供給する(R)-ヒドロキシアシル-CoA基質の鎖の長さに関して異なる特異性を有する。その結果、側鎖の長さが異なる結合ブロックがポリマーの内部に得られるため、異なる組成のポリマーになる。
【0077】
しかし細胞内に別の酵素を導入することは、代謝経路に影響を与える唯一の方法ではない。代謝経路の阻害剤を培地に添加することもできる。そのような阻害剤の具体例をいくつか挙げると、アクリル酸とトリクロサン(同意語:TCCまたは5-クロロ-2-(2,4-ジクロロフェノキシ)フェノール)である。
【0078】
実施例3
高度に結晶化したコアを有する生分解性ポリマー粒子を生体内において(R)-3-ヒドロキシブチル酸から産生させる方法
【0079】
この実験は、実施例1ですでに紹介したプラスミドpBBad-PとpBHR68を含む大腸菌株を用いて実施した。培養は、グルコースを炭素源として、実施例1に示した条件下で行なった。形成されるポリマー粒子は(R)-3-ヒドロキシブチル酸からなり、直径は、培地への誘導因子(アラビノース)の添加量に応じ、50〜500nmである。このようにして形成されるポリマー粒子の平均サイズは、ほんの20〜50nmほどしか違いがない。プラスミドpBBad-Pを用いて導入したフェイジンによって実現される制御特性を明らかにするため、上記の大腸菌株を用い、プラスミドpBBad-Pがない状態で実験を繰り返す。この場合には、直径が150〜250nmのポリマー粒子が得られる。この対照実験と比較すると、フェイジンを用いて粒子サイズを制御することで、より大きなポリマー粒子だけでなく、より小さなポリマー粒子も製造することができる。
【0080】
ポリマー粒子をサイズを基準にしてさらに分類することは、公知の方法で実現できる。
例えば、排除クロマトグラフィ、密度勾配遠心分離、5mMのリン酸塩バッファ(pH7.5)中での限外濾過などの方法がある。
【0081】
実施例4
あまり結晶化していないコアを有する生分解性ポリマー粒子を生体内において(R)-3-ヒドロキシ脂肪酸から産生させる方法
【0082】
この実験では、(R)-3-ヒドロキシ脂肪酸からなるコアを有するエラストマー性の生分解性ポリマー粒子を大腸菌の中で産生させる。ポリマー鎖は平均で6〜14個の炭素原子からなる。これは、シュードモナス・オレオボランスからのphaF遺伝子の発現を大腸菌の中で調節することによって実現される。phaF遺伝子は、フェイジンをコードしているラルストニア・ユートロファ由来の遺伝子phaPおよびポリマーシンターゼのための緑膿菌由来の遺伝子(phaC)に非常に似ている。この実施例では、phaF遺伝子の発現産物を用いて粒子サイズを制御する。ポリマー粒子は、シュードモナスのポリマーシンターゼ(phaC)だけを用いて脂肪酸の代謝を変化させた大腸菌の中で産生させることができる(Langenbach, S.他、FEMS Microbiol. Lett.、第150巻、303〜309ページ、1997年)。脂肪酸の代謝を変化させ、脂肪酸を炭素源として用いた場合には脂肪酸β酸化のCoA活性化中間体(例えばエノイル-CoA)が蓄積されてそれがポリマー合成の前駆体となるようにする。そのためには、大腸菌のfadB突然変異体を用いる(Langenbach, S.他、FEMS Microbiol. Lett.、第150巻、303〜309ページ、1997年)か、対応する脂肪酸の代謝を阻害する阻害剤を用いる(例えばアクリル酸;Qi, Q.他、FEMS Microbiol. Lett.、第167巻、89〜94ページ、1998年)。突然変異体または阻害された微生物を用いる場合には、そうでない場合に必要なケトアシルレダクターゼとチオラーゼはもはや必要ない。次に、大腸菌に固有のエノイル-CoAヒドラターゼを触媒としてエノイル-CoAからR-3-ヒドロキシアシル-CoAが形成される。次に、このR-3-ヒドロキシアシル-CoAがポリマーシンターゼによって変換されてポリ-(R)-3-ヒドロキシ脂肪酸になり、それが、形成されるポリマー粒子のポリマー・コアを形成する。
【0083】
クローニング
【0084】
Prietoら(Prieto, M.A.他、J. Bacteriol.、第181巻(3)、858〜868ページ、1999年)によってすでに報告されており、“GenBank”データベースにAJ010393という番号で登録されているシュードモナス・オレオボランスのphaF遺伝子を、ベクターpBBad-F(図11)を用いて大腸菌に導入して形質転換させる。このベクターは、ベクターpBBad22K(図7)を元にして得られる。ここでの個々のクローニング・ステップは、実施例1に記載したステップに対応している。phaF遺伝子をクローニングしてベクターpBBad22KのNcoI/BamHI制限部位に入れる。phaF遺伝子のPCR突然変異誘発に用いるプライマーは、以下の配列を有する:5'-aaagggccatggctggcaagaagaattccgagaa-3'(配列ID番号4、NcoI制限部位には下線を付した)と5'-aaagggggatcctcagatcagggtaccggtgcctgtctg-3'(配列ID番号5、BamHI制限部位には下線を付した)。次に、得られるDNA断片(配列ID番号6)をクローニングして上記のプラスミドpBBad22Kに導入する。phaFの配列を含むようになったこのプラスミドを、今後はpBBad-F(図11)と呼ぶ。このプラスミドに加え、ポリマーシンターゼのためのヌクレオチド配列を含むプラスミドpBHR71(図10;Langenbach, S.他、FEMS Microbiol. Lett.、第150巻、303〜309ページ、1997年)も用いて大腸菌を形質転換する。
【0085】
その後に発現させるための炭素源は、脂肪酸デカン酸である。フェイジンphaF遺伝子の発現は、ここでも、誘導因子であるアラビノースによって制御する。テストは、実施例1にすでに記載したようにして行なう。あらかじめ使用した誘導因子の量に応じ、この方法で形成されるポリマー粒子は直径が100〜500nmになる。
【0086】
実施例4
試験管内で産生させる生分解性ポリマー粒子のサイズ制御
【0087】
試験管内での産生において形成される生分解性ポリマー粒子のサイズは、ポリマーシンターゼ、フェイジン、フェイジン様タンパク質いずれかの利用可能性と、基質および代謝中間体の利用可能性によっても制御される。
【0088】
必要な酵素と基質は、生分解性ポリマー粒子を産生させるための試験管内のバッチでまず最初に利用できる必要がある。ラルストニア・ユートロファまたは緑膿菌からの組み換えポリマーシンターゼをこのテストで用いる(Gerngross, T.U.とMartin, D.P.、Proc. Natl. Acad. Sci. USA、第92巻、6279〜6283ページ、1995年;Qi, Q.他、Appl. Microbiol. Biotechnol.、第54巻、37〜43ページ、2000年)。このポリマーシンターゼは、(ヒスチジン-タグ融合体またはNi2+-NTAアガロースを用いた)アフィニティ・クロマトグラフィによって精製する。粒子サイズを制御するため、ラルストニア・ユートロファと緑膿菌からのポリマー粒子サイズ制御タンパク質を同様にして精製する。このタンパク質をラルストニア・ユートロファの中で発現させるため、実施例1ですでに用いたのと同じベクターを用いてフェイジン遺伝子を発現させる。生分解性ポリマー粒子を試験管内で産生させるための反応バッチは、ポリマーシンターゼとポリマー粒子サイズ制御タンパク質フェイジン以外に、ポリマー粒子を合成するための基質としてR-3-ヒドロキシブチリル-CoAまたはR-3-ヒドロキシデカノイル-CoAと、50mMのリン酸塩バッファ(pH7.5)と、1mMのMgCl2と、安定化のための5%グリセロール(v/v)も含んでいる。R-3-ヒドロキシブチリル-CoAまたはR-3-ヒドロキシデカノイル-CoAをポリマーシンターゼのための前駆体として使用しているため、チオラーゼとレダクターゼは使用する必要がない(図2を参照のこと)。
【0089】
図5の結果は、ポリマーシンターゼに対するフェイジンの量の比がポリマー粒子の直径に及ぼす影響を示している。それに対して図6は、ポリマーシンターゼに対する基質の比がポリマー粒子の直径に及ぼす影響を示している。これらの結果は、存在しているポリマーシンターゼの量によってポリマー粒子のサイズを調節できることも示している。したがって粒子サイズの効果的な調節は、本発明の方法によって実現される。
【0090】
実施例5
試験管内で産生されるポリマー粒子の組成に影響を及ぼす方法
【0091】
実施例5.1
ポリマーの組成に影響を及ぼす方法
【0092】
生分解性ポリマー粒子の組成は、さまざまな基質を反応バッチに添加することによって、および/または基質スペクトルが異なるさまざまなポリマーシンターゼ(図2を参照のこと)をそれぞれの場合に使用することによって、変えることができる。このようにして、ポリマー粒子の形成中に伸長しているポリマー鎖に一連の異なるモノマーが組み込まれ、ポリマーの組成が異なるポリマー粒子が産生される。緑膿菌からのポリマーシンターゼを用いているため、炭素原子を6〜14個有するR-3-ヒドロキシ脂肪酸構成ブロックが、伸長しているポリマー鎖に組み込まれる。例えば基質が1つだけ供給される場合には、緑膿菌からのポリマーシンターゼを用いると単独重合ポリマー粒子が得られる。
【0093】
ポリマー粒子の組成をいかにして変えるかのさらに別の具体例を実施例2に示す。
【0094】
実施例5.2
ポリマー粒子の膜組成に影響を及ぼす方法
【0095】
その後に例えば活性成分を導入する細胞の膜に融合させるためには、生分解性ポリマー粒子の表面にあるリン脂質層の組成を制御できる必要がある。この制御は、ポリマー粒子の形成に適した溶液を用意して試験管内で生分解性ポリマー粒子を産生させる(実施例4)ことによって実現される。膜は、さまざまな両親媒性分子の混合物を添加することにより、対応する条件に個別に合わせることができる。
【0096】
リン脂質を試験管内の反応バッチに添加する。所定数のポリマーシンターゼ分子がポリマー粒子の形成に関与する。重合が進行するにつれ、本発明のポリマー粒子はより大きくなり、表面にあるポリマーシンターゼはもはやポリマー粒子の表面を完全に覆うことができなくなる。その結果、(ポリマーの)疎水性領域が露出し、そこに両親媒性分子が自発的に付着する。
【0097】
生体内で産生された粒子の場合には、すでに存在しているリン脂質層をまず最初に除去する。そのためには、アセトン抽出を行なうか、非変性性洗浄剤を用いてリン脂質層を破壊する。次に、すでに説明したように、今や実質的に脂質を含まないポリマー粒子に、適切な両親媒性分子の混合物を添加する。ここでは、負に帯電したリン脂質またはホスファチジルコリンを使用することが好ましい。膜の組成を意図的に制御する一実施態様では、オクチルグリコシド(PH7.5)を1%含むPBSの中にポリマー粒子を懸濁させ、撹拌しながら過剰なホスファチジルコリンに対する透析を行なう。得られる粒子表面は、脳毛細管内皮細胞(BCEC)との融合に特に適している。
【0098】
実施例5.3
成長しているポリマー粒子にさまざまな物質を組み込む方法
【0099】
生分解性ポリマー粒子のコアへの物質の取り込みは、親油性蛍光染料であるナイル・レッド(シグマ社、セントルイス、ミズーリ州、アメリカ合衆国)またはローダミン123(Spiekermann, P.他、Arch. Microbiol.、第171巻、73〜80ページ、1999年)の助けを借りてすでに定量的に調べられてきた。すでに説明したように、生体内または試験管内での産生の間に蛍光染料ナイル・レッドを培地または反応バッチに添加すると、産生されるポリマー粒子が染色されるのを観察することができる。すなわちポリマー粒子の染色が、ポリマー粒子の合成中という早い時期から始まる。この染色は、細胞から単離する前にさえ始まる。
【0100】
薬理作用を有する活性成分を添加すると、ポリマー粒子のコアに組み込まれる可能性がある。例えば、ポリマー粒子を産生させるための試験管内での実験において、非極性抗がん剤であるパクリタキセルを反応バッチに添加する。水に溶けないパクリタキセルをポリマー粒子の疎水性ポリマー・コアに溶かした後に濃縮する。ここではパクリタキセルは、単にポリマー粒子を形成するために溶液に添加する。すると活性成分が粒子内に濃縮される。このことを確認するため、形成されたポリマー粒子を反応バッチから取り出し、溶液中にパクリタキセルが存在しているかどうかをHPLCによって調べる。溶液中のパクリタキセルの濃度が低下しているというのは、ここでは、パクリタキセルがポリマー粒子に組み込まれたことを示している。生体内でポリマー粒子が分解していくにつれ、活性成分が放出される。ポリマー粒子を含まない反応バッチを対照として用いる。この場合には、ポリマー粒子の形成に適した溶液中でパクリタキセルの濃度が低下することはない。
【0101】
実施例6
粒子表面の官能化
【0102】
生物学的に活性な物質は、ポリマー粒子の表面にすでに存在しているタンパク質と結合することができる。図1に示したすべてのタンパク質を考えることができる。その結果、複数の“架橋”戦略が得られる。これらの戦略により、粒子表面に結合するタンパク質を通じて生物学的に活性な物質を共有結合させることが可能になる。
【0103】
実施例6.1
薬理学的に活性な成分を付着させるために表面タンパク質を意図的に修飾する方法
【0104】
ポリマー粒子を官能化する方法の一例は、ヒドラゾンが結合したドキソルビシンを緑膿菌からのポリマーシンターゼphaC1に付着させるというものである(ヒドラゾンが結合したドキソルビシンは、King, H.D.他、Biconjugate Chem.、第10巻、279〜288ページ、1999年に記載されている)。ポリマーシンターゼphaC1は、ポリマー粒子の表面に結合しており、N末端に挿入された2つのシステイン残基を含んでいる。このポリマーシンターゼのシステイン残基が結合ドメインを形成しており、この結合ドメインを通じて生物学的に活性な物質を結合させることができる。
【0105】
クローニング
【0106】
システイン残基のトリプレット・コードをPCR突然変異誘発によってクローニングし、phaC1をコードしている遺伝子に組み込む。次に、この遺伝子をクローニングして図10に示したプラスミドpBHR71のXbaI制限部位とBamHI制限部位に組み込む。XbaI制限部位とBamHI制限部位も、このPCR突然変異誘発においてこの遺伝子に挿入する。phaC1をコードしている遺伝子のPCR突然変異誘発に使用するプライマーは、以下の配列を有する:N末端のプライマー5'-gggcTCTAGAaataaggagatatacatatgtgttgtaagaacaataacgagctt-3'(配列ID番号7、XbaI制限部位は大文字、システイン・トリプレットには下線)と、C末端のプライマー5'-aaacgcGGATCCttttcatcgttcatgca-3'(配列ID番号8、BamHI制限部位は大文字)。次に、このようにして得られたDNA断片(配列ID番号9)をXbaIとBamHIを用いて加水分解し、同様にして加水分解したプラスミドpBHR71と連結させる。得られるプラスミド(pBHR71-Cys(図12)と呼ぶ)を用いて大腸菌を公知の方法で形質転換する。この大腸菌は、ポリマーシンターゼが表面に存在するポリマー粒子を形成することができる。このポリマーシンターゼは、さまざまな物質(特に医薬物質)を付着させるための2つのシステイン残基を備えている。
【0107】
実施例6.1.1
ドキソルビシン(同意語:ヒドロキシダウノルビシン)を付着させる方法
【0108】
今や、ヒドラゾンを媒介とした“架橋”を、表面に露出したシステイン残基を通じて実現することができる。そのためには以下の方法を利用する。すなわち、ポリマーシンターゼが結合している単離したポリマー粒子100mg(合計1ml)を、ヘリウムを灌流させたPBS(pH7.5)と5mMのジチオトレイトール(DTT)の中で37℃にて3時間にわたって培養する。
この処理により、ポリマーシンターゼに含まれるジスルフィド架橋が減少する。次に、4℃にて40,000×gで30分間にわたって遠心分離を行なうことにより、低分子量の化合物を除去する。次に、減少したポリマー粒子を、ヒドラゾンが結合したドキソルビシンを10μモル含む1mlのPBSバッファを用いて懸濁させ、4℃にて30分間にわたって培養する。この期間が過ぎた後、上記の条件下で再び遠心分離を行ない、処理したポリマー粒子を洗浄する。その後、結合しなかったドキソルビシンをHPLCで検出する。ドキソルビシンがポリマーシンターゼにうまく結合したことは、濃度の低下によって確認される。
【0109】
実施例6.2
生物学的に活性な物質、特に薬理学的に活性な物質をポリマー粒子の結合ドメインに付着させる方法
【0110】
活性成分もポリマー粒子に結合させることができる。そのときには、ポリマー粒子の表面に結合したタンパク質の結合ドメインを通じて結合させる。そのため、結合ドメインをまず最初に作り出す必要がある。生物学的に活性な物質の結合は、ポリマー粒子の表面結合タンパク質(例えばポリマーデポリメラーゼ、フェイジン、フェイジン様タンパク質、ポリマーシンターゼ、ポリマー調節因子)を遺伝子改変することによって実現する。その結果、そのタンパク質が外に向いた結合ドメインを形成し、その結合ドメインにカップリング試薬または生物学的に活性な物質を結合させることができる。ポリマー粒子の上記表面タンパク質が、カップリング試薬または生物学的に活性な物質を直接付着させることのできるタンパク質と融合するとき、ポリマー粒子の表面タンパク質との融合後、表面タンパク質と融合タンパク質の両方の機能が完全に維持されていることが重要である。
【0111】
一実施態様では、2つのフラグ・エピトープを有するポリマー粒子を、緑膿菌からのポリマーシンターゼphaC1のN末端に直接融合させる。これらのフラグ・エピトープにより、市販されている抗フラグmAbs(モノクローナル抗体)(抗フラグM2、シグマ-オールドリッチ社)を結合させることができ、さらに、この方法がうまくいったことを確認するための酵素マーカーを結合させることができる。この実施例では、二次抗体/アルカリホスファターゼ共役体(抗マウス・アルカリホスファターゼ、シグマ-オールドリッチ社)を酵素マーカーとして使用する。次に、アルカリホスファターゼの活性がポリマー粒子の表面に及ぼす効果を光度計で測定する。
【0112】
実施例6.2.1
フラグ-phaC1融合タンパク質を有するポリマー粒子の産生
【0113】
以下のオリゴヌクレオチドを用いてフラグ-ポリマーシンターゼ融合タンパク質を産生させる:5'-tatgACTAGTgattataaagatgatgatgataaaca-3'と5'-tatgtttatcatcatcatctttataatcACTAGTca-3'(配列ID番号10と配列ID番号11、SpeI制限部位は大文字、フラグ・エピトープには下線)。ハイブリダイゼーションによって二本鎖DNAを得るため、これら2つのオリゴヌクレオチドを同じモル数(それぞれ10μM)混合し、室温(RT)にて30分間にわたって培養する。このようにして形成される二本鎖DNAは、フラグ・エピトープ(DYKDDDDK)をコードしており、NdeI制限部位(CA∇TATG/GTAT?AC)の張り出した末端部と相補的な張り出し末端部(TA)を有する。このDNA断片を制限酵素NdeIを用いて加水分解してクローニングし、同様に加水分解したベクターpHBR71(図10)に組み込む。このようにして得られるプラスミドpBHR71-FLAG(図13)は配列ID番号12の遺伝子を含んでおり、N末端にフラグが融合した(タンパク質のこの融合部分が結合ドメインを形成する)ポリマーシンターゼを発現させる。今やこの結合ドメインにより、生物学的に活性な物質および/またはカップリング試薬を結合させることができる。クローニングの間に同様にして導入する単一のSpeI制限部位は、機能性タンパク質をコードしている望む任意の別のDNA断片を挿入するのに利用できる。
【0114】
上記構造体を官能化する一例として以下の方法を利用する。すでにプラスミドpBBad-Fを含んでいて脂肪酸の代謝が変化した大腸菌株(実施例3を参照のこと)を、pBHR71-FLAGプラスミドを用いて形質転換すると、ポリマー粒子が発現する。ポリマー粒子は、細胞を破壊することによってその細胞から単離し、PBSバッファ(pH7.5)を用いて3回洗浄する。次に、ポリマー粒子を、フラグ・エピトープに結合する抗フラグ抗体とともに室温にて30分間にわたって培養する。次に、ポリマー粒子をすでに説明したようにして再び洗浄した後、二次抗体/アルカリホスファターゼ共役体とともにPBSバッファ中で室温にて30分間にわたって培養する。この30分間の培養が終わった後、粒子を0.1MのトリスHCl(pH8.5)の中で洗浄した後、アルカリホスファターゼの基質として2mg/mlのリン酸p-ニトロフェニルを粒子懸濁液に添加する。アルカリホスファターゼの活性を分光法によって410nmで測定する。結合ドメインを持たない(すなわちフラグ・エピトープのない)ポリマーシンターゼを含むポリマー粒子を負の対照として用いる。添加したリン酸p-ニトロフェニルは対照の中では変換されないため、410nmでの分光測定結果はネガティブである。ポリマー粒子が実際に形成されたという事実だけが、フラグ・エピトープの組み込みがポリマーシンターゼの機能に影響を与えなかったことの証拠である。
【0115】
この方法は、ポリマー粒子の上記表面タンパク質のうちの別の1つを用いて実施することもできる。2つ以上の表面タンパク質を修飾する場合には、非常に多彩な複数の物質をポリマー粒子に結合させることができる。そのためポリマー粒子が多価になって多くの異なった用途に適したものになる。
【0116】
前の実施例では、その後、形成されたポリマー粒子のすでに発現している表面タンパク質に生物学的に活性な物質を結合させたが、表面タンパク質にその物質を直接融合させた後にその物質を発現させることももちろん可能である。そのため、タンパク質(例えば酵素)のコード配列をpBHR71-FLAGプラスミドのphaC1遺伝子のC末端に融合させる。
【0117】
phaC1遺伝子と融合できるようにするため、挿入するタンパク質をコードしているDNA断片と、ポリマーシンターゼのC末端断片をコードしているDNA断片を、それぞれPCRによって得る。これら2つの断片を、張り出しているプライマーを用いて挿入した制限部位によって連結させる。このハイブリッド遺伝子の5'末端では、開始コドンから7ntの距離のところに、張り出したプライマーによってリボソーム結合部位(GAGGAG)と制限部位が挿入されている。使用するベクター(ここでは例えばpHBR71-FLAG・ベクター)がすでにリボソーム結合部位を備えている場合には、リボソーム結合部位をさらに挿入する必要はもはやない。このハイブリッド遺伝子の3'末端に位置する挿入された制限部位も、発現ベクターpHBR71-FLAGに意図的に挿入することが今や可能である。ハイブリッド遺伝子の5'末端と3'末端の制限部位は、ハイブリッド遺伝子の内部にもう一度現われることがないように選択し、しかも発現ベクターへの組み込みが現在のプロモータと並行して実施できるように選択する必要がある。この実施例では、SpeI制限部位を含むプライマーを用いて大腸菌からのlacZ遺伝子をPCRによって増幅する:5'-ggACTAGTatgaccatgattacggattcactggc-3'(配列ID番号13、SpeI制限部位は大文字)と5'-ccACTAGTtttttgacaccagaccaactggtaatggtagcg-3'(配列ID番号14、SpeI制限部位は大文字)。さらに、これらプライマーを用いて停止コドンをlacZ遺伝子の配列から除去し、連続的なリーディングフレームを得る。得られるDNA断片(配列ID番号15)をクローニングしてpHBR71-FLAG・プラスミドのSpeI制限部位に直接組み込む。得られる融合タンパク質は、β-ガラクトシダーゼ活性を有するポリマー粒子を形成する。対応するポリマー粒子を単離し、β-ガラクトシダーゼ活性を持つことを、基質としてのo-ニトロフェニル-β-D-ガラクトピラノシド(カルバイオケム社)を用いて還元条件下で明らかにする。
【0118】
実施例7
ポリマー粒子の表面タンパク質とポリマー・コアの間の結合の安定性
【0119】
本発明のために行なった研究により、変性剤(例えばドデシル硫酸ナトリウム(SDS)、尿素、グアニジウムヒドロクロリド、ジチオトレイトール)を用いた処理を行なっても、酸性条件を利用しても、ポリマーシンターゼを生分解性ポリマーのコアから引き離せないことが明らかになった。これは、ポリマー粒子と、ポリマーシンターゼのポリマー粒子結合ドメインとの間に共有結合が存在していることを示している。結合の安定性が増大しているため、ポリマー粒子に結合した物質やポリマー粒子に組み込まれた物質を標的部位に輸送することが可能になる。表面に結合したポリマーシンターゼのN末端断片(N末端から保存されたα/β-ヒドロラーゼ・ドメインの最初の部分まで)は極めて変化しやすいため、遺伝子工学の方法を利用して機能性タンパク質で置換することができる。このようにして、ポリマーシンターゼの活性と、ポリマー粒子の合成を維持する(Rehm, B.H.A.他、Biochem. Biophys. Acta、第1594巻、178〜190ページ、2002年)。その結果、安定性が増大した表面の官能化が実現する。生物学的に活性な物質および/またはカップリング試薬が結合する結合ドメインが異なっているさまざまなタンパク質の混合物は、必要ならば同時に使用する。するとポリマー粒子の表面が多価になる。これらのタンパク質を利用するには、試験管内であれば、異なる結合ドメインを有する精製したタンパク質を合成バッチに添加し、生体内であれば、結合ドメインを有する1つのタンパク質をそれぞれがコードしている遺伝子を対応する微生物の内部で発現させる。
【0120】
実施例7.1
ポリマー粒子の表面タンパク質をさらに修飾する可能性
【0121】
ラルストニア・ユートロファに由来する表面タンパク質フェイジン(phaP)のC末端断片(アラニン141から先のアミノ酸残基)は親水性であるため、機能性タンパク質と置換することができる。そのときフェイジンがポリマー粒子の結合ドメインを通じてポリマー粒子の表面に固着するのが妨げられることはない。
【0122】
この固着は疎水性相互作用に基づいており、可逆的である(Hanley, S.Z.他、FEBS Letters、第447巻、99〜105ページ、1999年)。細胞内ポリマーデポリメラーゼのこのC末端断片は、遺伝子工学の方法によって機能性タンパク質と融合させる。そのため、その後生物学的に活性な物質および/またはカップリング試薬を付着させることによってポリマー粒子の表面を多価にすることができる。
【0123】
ラルストニア・ユートロファの細胞内ポリマーデポリメラーゼのC末端(180番目から先のアミノ酸残基)は、この酵素をポリマー粒子のコアと結合させる(Saegusa, H.、J. Bacteriol.、第183巻(1)、94〜100ページ、2001年)。細胞内ポリマーデポリメラーゼのこのC末端断片は、遺伝子工学の方法によって機能性タンパク質と融合させる。そのためポリマー粒子の表面を多価にすることができる。
【0124】
シュードモナス・オレオボランスに由来する遺伝子phaIとphaFが発現してポリマー粒子の表面に結合しているタンパク質のN末端(140番目までのアミノ酸残基)は、このタンパク質をポリマー粒子のポリエステル・コアと結合させる(Prieto, M.A.他、J. Bacteriol.、第181巻(3)、858〜868ページ、1999年)。遺伝子phaIとphaFの発現産物のこのN末端は、遺伝子工学の方法によって機能性タンパク質と融合させるため、得られる結合ドメインは、生物学的に活性な物質および/またはカップリング試薬を付着させることによってポリマー粒子の表面を多価にすることができる。
【0125】
実施例8
カップリング試薬を用いてポリマー粒子の表面タンパク質の共有結合を修飾する方法
【0126】
生分解性ポリマー粒子の表面にある図1に示したタンパク質は、所定のアミノ酸に特異的に結合する特異的標識物質(例えばリシンに対して特異的に結合するn-ヒドロキシスクシンイミドビオチン)で処理することができる。するとヨードアセトアミドを媒介として生物学的に活性な物質(例えばビオチン)をシステインと連結させることができるようになる。ビオチンなどの分子は、ポリマー粒子の表面タンパク質に生物学的に活性な物質をさらに連結させる。生物学的に活性な物質としては、酵素に結合して生分解性ポリマー粒子の表面タンパク質を徐々に官能化することのできる例えばアビジン、ストレプトアビジンなどが挙げられる(Rehm, B.H.A.他、J. Bacteriol.、第176巻、5639〜5647ページ、1994年)。この官能化は、抗体または薬理学的に活性な成分を付着させることによって進行する可能性がある。所定の膜を通過する輸送および/または所定の膜との融合にとって有利な特定の表面電荷をポリマー粒子に与えるため、いろいろな表面電荷を有する分子も付着させることができる。
【0127】
ポリマー粒子の表面タンパク質に含まれるリシン残基をビオチンで標識するには、n-ヒドロキシスクシンイミドビオチンを用いる。この実験では、緑膿菌からのポリマーシンターゼphaC1を含むポリマー粒子を、プラスミドpHBR71を含む組み換え大腸菌から単離する。単離後、ポリマー粒子をPBS(pH8.0)の中で3回洗浄し、次いでn-ヒドロキシスクシンイミドビオチン(シグマ-オールドリッチ社)を、最終濃度が5mMになるまで溶液に添加する。4℃にて5分間にわたって培養した後に再び洗浄することによって反応を終了させる。
粒子の表面に結合したビオチンの検出には、基質としてストレプトアビジン-アルカリホスファターゼ共役体(シグマ-オールドリッチ社)とリン酸o-ニトロフェニル(カルバイオケム社)を用いる。n-ヒドロキシスクシンイミドビオチンで処理していない粒子が含まれている対照バッチでは、粒子はアルカリホスファターゼ活性を示さない。
【0128】
産生させたポリマー粒子の表面タンパク質を活性化させることのできる生物学的に活性な物質を連結させるための他の多数のカップリング試薬が、上記の具体例以外にも存在している。
【特許請求の範囲】
【請求項1】
生分解性ポリマー粒子の製造方法であって、
a)少なくとも1つの誘導遺伝子を微生物に導入するステップ、ここで当該遺伝子はポリマー粒子のサイズを制御するタンパク質をコードし、そしてラルストニア・ユートロファ(Ralstonia eutropha)からのフェイジン遺伝子phaAと、シュードモナス・オレオボランス(Pseudomonas oloeovorans)からのフェイジン遺伝子phaFを含むグループの中から選択され;
b)前記ポリマー粒子の形成に関係するタンパク質をコードしているさらに別の少なくとも1つの遺伝子を導入するステップ;
ここで、ステップa)及びb)において前記微生物に導入した少なくとも1つの上記誘導遺伝子が、ポリマー粒子結合ドメインと、少なくとも1つの結合ドメインとを備え、この少なくとも1つの結合ドメインが、生物学的に活性な物質および/またはカップリング試薬と結合可能であり、そして
c) a)に記載した少なくとも1つの誘導遺伝子を誘導した微生物を、その微生物が生分解性ポリマー粒子を産生するのに適した培地の中で培養するステップを含む方法。
【請求項2】
ポリマー粒子の形成に関係するタンパク質をコードしているさらに別の少なくとも1つの上記遺伝子が、チオラーゼ、レダクターゼ、ポリマーシンターゼのいずれかをコードしている、請求項1に記載の方法。
【請求項3】
ポリマー粒子の形成に関係するタンパク質をコードしているさらに別の少なくとも1つの上記遺伝子が、ラルストニア・ユートロファからのphaAチオラーゼ、phaBケトアシルレダクターゼ、phaCシンターゼのいずれかをコードしている、請求項1に記載の方法。
【請求項4】
チオラーゼおよび/またはポリマーシンターゼをコードしている少なくとも1つの別の遺伝子を細胞内で誘導させる、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
側部官能基を有する少なくとも1つの脂肪酸、その中でも特に好ましくは、少なくとも1つのヒドロキシ脂肪酸および/または少なくとも1つのメルカプト脂肪酸および/または少なくとも1つのβ-アミノ脂肪酸を、ポリマー粒子を形成するための基質として培地に導入する、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
上記基質を、ポリマー粒子のサイズを制御するのに十分な量、培地に添加する、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
使用する上記微生物の選択を、ラルストニア、アルカリゲネス、シュードモナス、ハロビフォルマを含む属の中から行なう、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
使用する上記微生物の選択を、ラルストニア・ユートロファ、アルカリゲネス・ラトゥス、大腸菌、シュードモナス・フラギ、シュードモナス・プティダ、シュードモナス・オレオボランス、緑膿菌、シュードモナス・フルオレセンス、ハロビフォルマ・ハロテレストリスを含むグループの中から行なう、請求項7に記載の方法。
【請求項9】
培養した微生物を公知の方法で破壊した後、ポリマー粒子を細胞残留物から分離する、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
ポリマー粒子の表面に位置する脂質層を請求項9に記載の方法によって得られたポリマー粒子から分離し、別の組成の脂質層で置換する、請求項9に記載の方法。
【請求項11】
粒子サイズを少なくとも1つの上記誘導遺伝子によって制御し、形成されるポリマー粒子の直径を10nm〜3μm、好ましくは10nm〜900nm、さらに好ましくは10nm〜100nmにする、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
上記ポリマー粒子結合ドメインが、ポリマー粒子の表面に結合するタンパク質の一部を含んでおり、そのタンパク質の選択を、ポリマーデポリメラーゼ、ポリマー調節因子、ポリマーシンターゼ、粒子サイズ制御タンパク質を含むグループの中から行なう、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
生物学的に活性な物質および/またはカップリング試薬と結合可能である少なくとも1つの上記結合ドメインの選択を、オリゴペプチド、酵素、アブザイム、非触媒性タンパク質を含むグループの中から行なう、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
生分解性ポリマー粒子を試験管内で製造する方法であって、
a)少なくとも1つの基質を含んでいてポリマー粒子の形成に適した溶液を用意するステップと;
b)ポリマー粒子のサイズを制御するのに適したタンパク質をこの溶液に導入するステップと;
c)ポリマー粒子の形成に関係する別の少なくとも1つのタンパク質を導入するステップと含み、
ここで、ステップa)及びb)において導入した少なくとも1つタンパク質が、ポリマー粒子結合ドメインと、少なくとも1つの結合ドメインとを備え、この少なくとも1つの結合ドメインが、生物学的に活性な物質および/またはカップリング試薬と結合可能であるものであるように選択される、方法。
【請求項15】
ステップa)において、ポリマー粒子の形成に適した溶液に、少なくとも1つの脂肪酸と、アシルCoAオキシダーゼとを添加してポリマー粒子を形成する、請求項14に記載の方法。
【請求項16】
ステップa)において、ポリマー粒子の形成に適した溶液に、少なくとも1つの基質を、ポリマー粒子のサイズを制御するのに十分な量添加する、請求項14または15に記載の方法。
【請求項17】
ステップb)において、フェイジン様タンパク質のファミリーに由来するタンパク質を導入することによりポリマー粒子のサイズを制御する、請求項14〜16のいずれか1項に記載の方法。
【請求項18】
ステップb)において、ラルストニア・ユートロファからのフェイジンと、シュードモナス・オレオボランスからのフェイジンを含むグループの中から選択したタンパク質を導入することによりポリマー粒子のサイズを制御する、請求項17に記載の方法。
【請求項19】
ポリマー粒子の形成に関係する、ステップc)で用いる別の少なくとも1つの上記タンパク質が、ポリマーシンターゼである、請求項14〜18のいずれか1項に記載の方法。
【請求項20】
ポリマー粒子の形成に関係する、ステップc)で用いる別の少なくとも1つの上記タンパク質が、ラルストニア・ユートロファ、シュードモナス・オレオボランス、シュードモナス・プティダ、緑膿菌からのポリマーシンターゼを含むグループの中から選択したポリマーシンターゼである、請求項19に記載の方法。
【請求項21】
上記溶液に上記ポリマーシンターゼを、ポリマー粒子のサイズを制御するのに十分な量添加する、請求項19または20に記載の方法。
【請求項22】
ステップb)において、少なくとも1つの薬理学的に活性な物質を上記溶液に添加する、請求項14〜21のいずれか1項に記載の方法。
【請求項23】
リン脂質とエーテル脂質からなるグループの中から選択した少なくとも1つの両親媒性分子をステップa)からの溶液に添加することにより、ポリマー粒子の表面にある脂質層の組成を制御する、請求項14〜22のいずれか1項に記載の方法。
【請求項24】
上記ポリマー粒子結合ドメインが、ポリマー粒子の表面に結合するタンパク質の一部であり、そのタンパク質の選択を、ポリマーデポリメラーゼ、ポリマー調節因子、ポリマーシンターゼ、粒子サイズ制御タンパク質を含むグループの中から行なう、請求項14〜23のいずれか1項に記載の方法。
【請求項25】
生物学的に活性な物質および/またはカップリング試薬と結合可能である少なくとも1つの上記結合ドメインの選択を、オリゴペプチド、酵素、アブザイム、非触媒性タンパク質を含むグループの中から行なう、請求項14〜24のいずれか1項に記載の方法。
【請求項26】
サイズが決まっていて両親媒性分子からなる表面層を有するポリヒドロキシルカルボキシレートのポリマー粒子であって、その内部には、ポリマーデポリメラーゼ、ポリマー調節因子、ポリマーシンターゼ、粒子サイズ制御タンパク質を含むグループの中から選択した少なくとも1つのタンパク質が導入されているポリマー粒子において、少なくとも1つのこのタンパク質が、ポリマー粒子結合ドメインと、生物学的に活性な物質および/またはカップリング試薬と結合可能な結合ドメインとを含んでいる、ポリマー粒子。
【請求項27】
請求項1〜25のいずれか1項に記載の方法で製造した、請求項26に記載のポリマー粒子。
【請求項28】
請求項26または27に記載のポリマー粒子を利用して製造した医薬調製物、殺虫剤、除草剤。
【請求項29】
中枢神経系疾患の治療に適している、請求項28に記載の医薬調製物。
【請求項1】
生分解性ポリマー粒子の製造方法であって、
a)少なくとも1つの誘導遺伝子を微生物に導入するステップ、ここで当該遺伝子はポリマー粒子のサイズを制御するタンパク質をコードし、そしてラルストニア・ユートロファ(Ralstonia eutropha)からのフェイジン遺伝子phaAと、シュードモナス・オレオボランス(Pseudomonas oloeovorans)からのフェイジン遺伝子phaFを含むグループの中から選択され;
b)前記ポリマー粒子の形成に関係するタンパク質をコードしているさらに別の少なくとも1つの遺伝子を導入するステップ;
ここで、ステップa)及びb)において前記微生物に導入した少なくとも1つの上記誘導遺伝子が、ポリマー粒子結合ドメインと、少なくとも1つの結合ドメインとを備え、この少なくとも1つの結合ドメインが、生物学的に活性な物質および/またはカップリング試薬と結合可能であり、そして
c) a)に記載した少なくとも1つの誘導遺伝子を誘導した微生物を、その微生物が生分解性ポリマー粒子を産生するのに適した培地の中で培養するステップを含む方法。
【請求項2】
ポリマー粒子の形成に関係するタンパク質をコードしているさらに別の少なくとも1つの上記遺伝子が、チオラーゼ、レダクターゼ、ポリマーシンターゼのいずれかをコードしている、請求項1に記載の方法。
【請求項3】
ポリマー粒子の形成に関係するタンパク質をコードしているさらに別の少なくとも1つの上記遺伝子が、ラルストニア・ユートロファからのphaAチオラーゼ、phaBケトアシルレダクターゼ、phaCシンターゼのいずれかをコードしている、請求項1に記載の方法。
【請求項4】
チオラーゼおよび/またはポリマーシンターゼをコードしている少なくとも1つの別の遺伝子を細胞内で誘導させる、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
側部官能基を有する少なくとも1つの脂肪酸、その中でも特に好ましくは、少なくとも1つのヒドロキシ脂肪酸および/または少なくとも1つのメルカプト脂肪酸および/または少なくとも1つのβ-アミノ脂肪酸を、ポリマー粒子を形成するための基質として培地に導入する、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
上記基質を、ポリマー粒子のサイズを制御するのに十分な量、培地に添加する、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
使用する上記微生物の選択を、ラルストニア、アルカリゲネス、シュードモナス、ハロビフォルマを含む属の中から行なう、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
使用する上記微生物の選択を、ラルストニア・ユートロファ、アルカリゲネス・ラトゥス、大腸菌、シュードモナス・フラギ、シュードモナス・プティダ、シュードモナス・オレオボランス、緑膿菌、シュードモナス・フルオレセンス、ハロビフォルマ・ハロテレストリスを含むグループの中から行なう、請求項7に記載の方法。
【請求項9】
培養した微生物を公知の方法で破壊した後、ポリマー粒子を細胞残留物から分離する、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
ポリマー粒子の表面に位置する脂質層を請求項9に記載の方法によって得られたポリマー粒子から分離し、別の組成の脂質層で置換する、請求項9に記載の方法。
【請求項11】
粒子サイズを少なくとも1つの上記誘導遺伝子によって制御し、形成されるポリマー粒子の直径を10nm〜3μm、好ましくは10nm〜900nm、さらに好ましくは10nm〜100nmにする、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
上記ポリマー粒子結合ドメインが、ポリマー粒子の表面に結合するタンパク質の一部を含んでおり、そのタンパク質の選択を、ポリマーデポリメラーゼ、ポリマー調節因子、ポリマーシンターゼ、粒子サイズ制御タンパク質を含むグループの中から行なう、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
生物学的に活性な物質および/またはカップリング試薬と結合可能である少なくとも1つの上記結合ドメインの選択を、オリゴペプチド、酵素、アブザイム、非触媒性タンパク質を含むグループの中から行なう、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
生分解性ポリマー粒子を試験管内で製造する方法であって、
a)少なくとも1つの基質を含んでいてポリマー粒子の形成に適した溶液を用意するステップと;
b)ポリマー粒子のサイズを制御するのに適したタンパク質をこの溶液に導入するステップと;
c)ポリマー粒子の形成に関係する別の少なくとも1つのタンパク質を導入するステップと含み、
ここで、ステップa)及びb)において導入した少なくとも1つタンパク質が、ポリマー粒子結合ドメインと、少なくとも1つの結合ドメインとを備え、この少なくとも1つの結合ドメインが、生物学的に活性な物質および/またはカップリング試薬と結合可能であるものであるように選択される、方法。
【請求項15】
ステップa)において、ポリマー粒子の形成に適した溶液に、少なくとも1つの脂肪酸と、アシルCoAオキシダーゼとを添加してポリマー粒子を形成する、請求項14に記載の方法。
【請求項16】
ステップa)において、ポリマー粒子の形成に適した溶液に、少なくとも1つの基質を、ポリマー粒子のサイズを制御するのに十分な量添加する、請求項14または15に記載の方法。
【請求項17】
ステップb)において、フェイジン様タンパク質のファミリーに由来するタンパク質を導入することによりポリマー粒子のサイズを制御する、請求項14〜16のいずれか1項に記載の方法。
【請求項18】
ステップb)において、ラルストニア・ユートロファからのフェイジンと、シュードモナス・オレオボランスからのフェイジンを含むグループの中から選択したタンパク質を導入することによりポリマー粒子のサイズを制御する、請求項17に記載の方法。
【請求項19】
ポリマー粒子の形成に関係する、ステップc)で用いる別の少なくとも1つの上記タンパク質が、ポリマーシンターゼである、請求項14〜18のいずれか1項に記載の方法。
【請求項20】
ポリマー粒子の形成に関係する、ステップc)で用いる別の少なくとも1つの上記タンパク質が、ラルストニア・ユートロファ、シュードモナス・オレオボランス、シュードモナス・プティダ、緑膿菌からのポリマーシンターゼを含むグループの中から選択したポリマーシンターゼである、請求項19に記載の方法。
【請求項21】
上記溶液に上記ポリマーシンターゼを、ポリマー粒子のサイズを制御するのに十分な量添加する、請求項19または20に記載の方法。
【請求項22】
ステップb)において、少なくとも1つの薬理学的に活性な物質を上記溶液に添加する、請求項14〜21のいずれか1項に記載の方法。
【請求項23】
リン脂質とエーテル脂質からなるグループの中から選択した少なくとも1つの両親媒性分子をステップa)からの溶液に添加することにより、ポリマー粒子の表面にある脂質層の組成を制御する、請求項14〜22のいずれか1項に記載の方法。
【請求項24】
上記ポリマー粒子結合ドメインが、ポリマー粒子の表面に結合するタンパク質の一部であり、そのタンパク質の選択を、ポリマーデポリメラーゼ、ポリマー調節因子、ポリマーシンターゼ、粒子サイズ制御タンパク質を含むグループの中から行なう、請求項14〜23のいずれか1項に記載の方法。
【請求項25】
生物学的に活性な物質および/またはカップリング試薬と結合可能である少なくとも1つの上記結合ドメインの選択を、オリゴペプチド、酵素、アブザイム、非触媒性タンパク質を含むグループの中から行なう、請求項14〜24のいずれか1項に記載の方法。
【請求項26】
サイズが決まっていて両親媒性分子からなる表面層を有するポリヒドロキシルカルボキシレートのポリマー粒子であって、その内部には、ポリマーデポリメラーゼ、ポリマー調節因子、ポリマーシンターゼ、粒子サイズ制御タンパク質を含むグループの中から選択した少なくとも1つのタンパク質が導入されているポリマー粒子において、少なくとも1つのこのタンパク質が、ポリマー粒子結合ドメインと、生物学的に活性な物質および/またはカップリング試薬と結合可能な結合ドメインとを含んでいる、ポリマー粒子。
【請求項27】
請求項1〜25のいずれか1項に記載の方法で製造した、請求項26に記載のポリマー粒子。
【請求項28】
請求項26または27に記載のポリマー粒子を利用して製造した医薬調製物、殺虫剤、除草剤。
【請求項29】
中枢神経系疾患の治療に適している、請求項28に記載の医薬調製物。
【図1】



【図2】
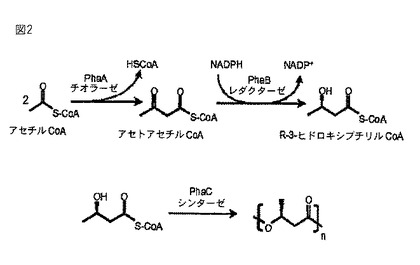
【図3】
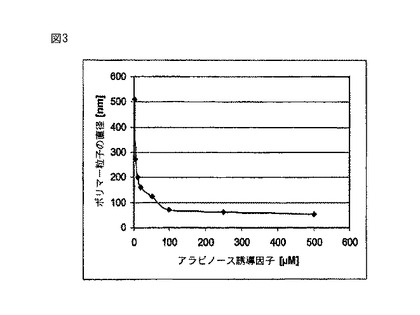
【図4】
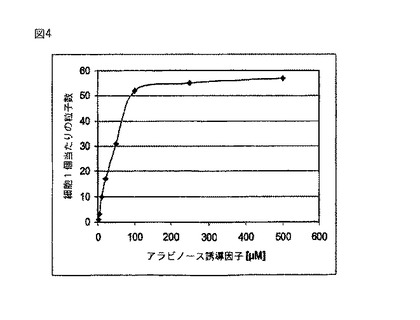
【図5】
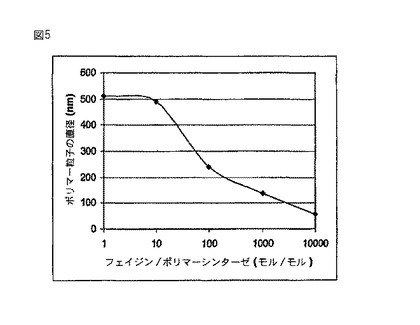
【図6】
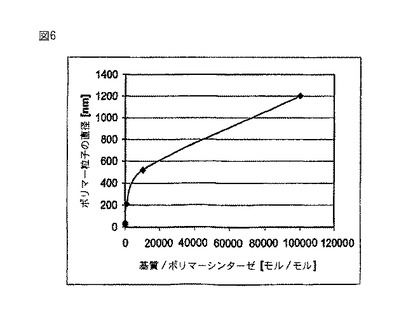
【図7】
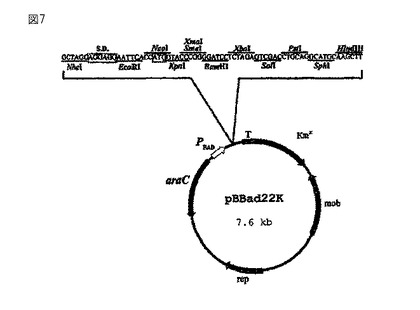
【図8】
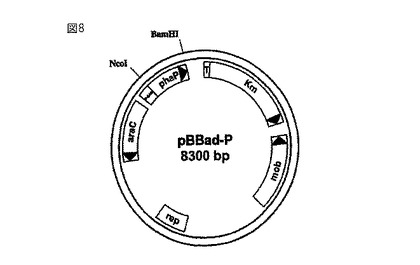
【図9】
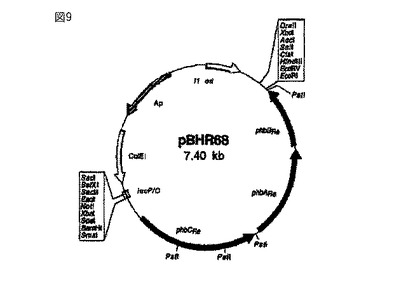
【図10】
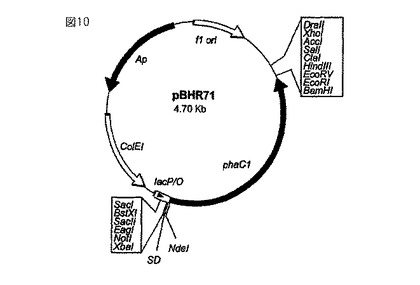
【図11】
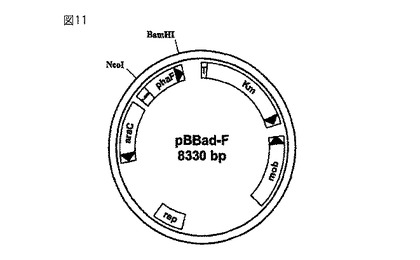
【図12】
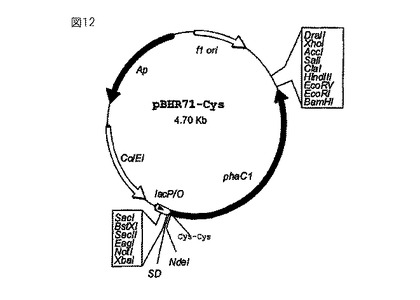
【図13】
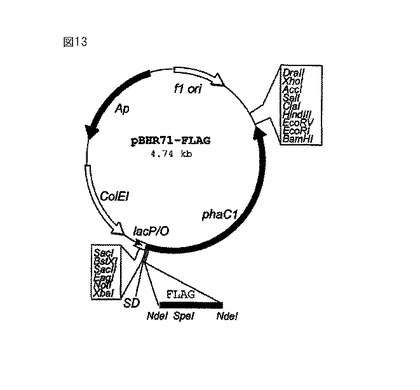
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公開番号】特開2012−249634(P2012−249634A)
【公開日】平成24年12月20日(2012.12.20)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−152732(P2012−152732)
【出願日】平成24年7月6日(2012.7.6)
【分割の表示】特願2004−531694(P2004−531694)の分割
【原出願日】平成15年8月22日(2003.8.22)
【出願人】(512179005)ポリバティックス リミティド (1)
【Fターム(参考)】
【公開日】平成24年12月20日(2012.12.20)
【国際特許分類】
【出願番号】特願2012−152732(P2012−152732)
【出願日】平成24年7月6日(2012.7.6)
【分割の表示】特願2004−531694(P2004−531694)の分割
【原出願日】平成15年8月22日(2003.8.22)
【出願人】(512179005)ポリバティックス リミティド (1)
【Fターム(参考)】
[ Back to top ]