
宿主細胞およびその使用方法
本発明は、前記ピキア株において機能的遺伝子産物をコードする少なくとも1つの核酸配列および/または少なくとも1つの機能的遺伝子産物を発現するのに必要な少なくとも1つの核酸配列が遺伝子改変された遺伝子改変ピキア株を提供するものであり、ここで前記遺伝子産物は、遺伝的修飾ピキア株においてタンパク質分解および/または糖鎖形成を担う。特に、機能的遺伝子産物または前記遺伝子産物の発現をコードする核酸配列であるPEP4、PRB1、YPS1、YPS2、YMP1、YMP2、YMP3およびPMT4が遺伝的に修飾されるピキア株が提供される。前記遺伝子改変宿主細胞の野生型親が、配列番号4、配列番号6および配列番号8のアミノ酸1−865と少なくとも60%の配列同一性を有するポリペプチドをコードする遺伝子を含み、前記野生型宿主細胞と比べると、前記遺伝子改変宿主細胞内で、前記遺伝子産物が低減するか排除されるように、前記遺伝子が前記宿主細胞のゲノム内で遺伝的に修飾される遺伝子改変宿主細胞も本明細書では提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本明細書は、米国仮特許出願第61/155,706(出願日2009年2月26日)の権利を主張し、参照によりその全体を本明細書に援用する。
【0002】
本発明は、生化学エンジニアリングの分野に関する。特に、本発明は遺伝子改変ピキア株およびその中にポリペプチドを産生する方法に関する。
【背景技術】
【0003】
治療用ポリペプチドおよびタンパク質は、バクテリア細胞、大腸菌細胞、真菌または酵母細胞微生物細胞、昆虫細胞、および哺乳類細胞を含む種々の宿主細胞内で発現させることができる。メチロトローフ酵母ピキア・パストリスなどの真菌宿主は、治療用タンパク質発現に顕著な特長を有する。例えば、真菌宿主は、多量の内因性タンパク質を分泌しないし、強い誘導性プロモータを有し、限定的な化学培地で増殖でき、高力価の組換えタンパク質を産生する(Cregg et al., Mol. Biotech. 16:23−52 (2000))。酵母および糸状菌は両方とも、細胞内組換えタンパク質のおよび分泌された組換えタンパク質の産生に使用され、成功を収めている(Cereghino, J. L. and J. M. Cregg 2000 FEMS Microbiology Reviews 24(1): 45 66; Harkki, A., et al. 1989 Bio−Technology 7(6): 596; Berka, R. M., et al. 1992 Abstr. Papers Amer. Chem.Soc.203: 121−BIOT; Svetina, M., et al. 2000 J. Biotechnol. 76(23): 245−251)。ピキアは、組換え型ヒト血清アルブミン(HSA)の発現に関して注目に値する宿主細胞である。しかしながら、HASと遺伝的に融合したポリペプチドを含む他の治療用ポリペプチドの発現は、好ましくないタンパク質分解および糖鎖形成という技術的障壁に直面している。
【0004】
ピキア・パストリスに発現する異種タンパク質は付加的マンノース糖を含んでいて、「高マンノース」グリカン並びにタンパク質上に負電荷を与えるマンノシルリン酸基を生み出す。高マンノースグリカンまたは荷電したマンナンを有するグリコシル化タンパク質はヒトに免疫応答を誘発する危険性が高い(Takeuchi, Trends in Glycosci & Glycotech., 9:S29−S35 (1997); Rosenfeld et al., J.Biol.Chem., 249:2319−2321 (1974))。したがって、糖鎖形成のパターンがヒトのパターンと同一であるか少なくとも類似しているような治療用のペプチド類、ポリペプチドおよび/またはタンパク質を真菌宿主組織において産生することが望ましい。
【0005】
従って、野生型系統と比較すると減少したタンパク質分解および/または糖鎖形成を有する異種ペプチド類、ポリペプチドおよび/またはタンパク質を産生することができる酵母系統、特にピキアの必要とされている。さらに加えて、酵母系統内で遺伝子を同定する必要があり、特にタンパク分解性と糖鎖形成経路に関わるタンパク質の産生を担うピキア株内で同定する必要がある。
【発明の概要】
【0006】
本発明の1つの態様において、遺伝子改変ピキア株が提供され、機能的遺伝子産物をコードする少なくとも1つの核酸配列、および/またはピキア株の少なくとも1つの遺伝子産物の発現に必要な少なくとも1つの核酸が遺伝子改変され、前記遺伝子産物は、前記遺伝子改変ピキア株においてタンパク質分解および/または糖鎖形成を担う。
【0007】
少なくとも1つの異種ポリペプチドを産生する方法も本発明で提供され、その方法には、本発明の遺伝子改変ピキア株内で前記異種ポリペプチドを発現させるステップも含まれる。別の態様において、本発明は、本発明の宿主細胞で異種的に発現したペプチド、ポリペプチド、およびタンパク質を提供する。
【0008】
本発明の別の態様において、配列番号4、配列番号6および配列番号8と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%配列同一性を有する単離したポリペプチドが提供される。前記宿主細胞の前記野生型の親が配列番号4、6および8と少なくとも60%の配列同一性を持つポリペプチドをコード化する遺伝子を含み、前記野生型宿主細胞と比較して、前記遺伝子改変宿主細胞内で遺伝子産物または活性が低下するか除去されるように、前記遺伝子が前記宿主細胞のゲノム内で改変される、遺伝子改変宿主細胞系統をもさらに提供する。
【図面の簡単な説明】
【0009】
【図1】親和性精製およびLC/MS分析を使用して同定した新規ピキアプロテアーゼYmp1を示す図である。
【図2】アミノ酸1−857を配列番号15として示すYMP1(配列番号4)の予測構造である。
【図3】ymp1突然変異系統のプロテアーゼ活性アッセイを示す図である。
【図4】ymp1突然変異系統のザイモグラムアッセイを示す図である。
【図5】異種タンパク質(配列番号1)のタンパク質分解に関するymp1ノックアウト効果を示す図である。
【図6】異種タンパク質(配列番号1)のタンパク質分解に関するysp2ノックアウト効果を示す図である。
【図7】異種タンパク質(配列番号1)のタンパク質分解に関するysp1ノックアウト効果を示す図である。
【図8】異種タンパク質(配列番号1)のタンパク質分解に関するpep4ノックアウト効果を示す図である。
【図9】ymp1、ymp3突然変異系統の緩衝液安定性試験を示す図である。
【図10】野生型(WT)、pmt1およびpmt4ピキア変異体の細胞壁溶解酵素アッセイ結果を示す図である。
【図11】pmt4突然変異系統由来の異種タンパク質(配列番号1)の糖鎖形成分析を示す図である。
【発明を実施するための形態】
【0010】
本明細書で使用する「宿主細胞(1つまたは複数)」は、単離したポリヌクレオチド配列を導入(例えば、形質転換、感染、もしくは形質移入)した細胞または導入(例えば、形質転換、感染、もしくは形質移入)可能な細胞を意味する。本発明の宿主細胞は、以下に制限されるものではないが、バクテリア細胞、真菌細胞、酵母細胞、微生物細胞、昆虫細胞、および哺乳類細胞を含んで良い。酵母および/または糸状真菌起源の本発明の宿主細胞は、これに制限されるものではないが、以下の科、属、および種を含んで良い:ピキア・パストリス(Pichia pastoris)、ピキア・フィンランディカ(Pichia finlandica)、ピキア・トレハロフィア(Pichia trehalophila)、ピキア・コクラメ(Pichia koclamae)、ピキア・メンブラナエファシエンス(Pichia membranaefaciens)、ピキア・メタノリカ(Pichia methanolica)、ピキア・ミヌタ(Pichia minuta)(オガタ・エミヌタ(Ogataea minuta)、ピキア・リンドネリ(Pichia lindneri))、ピキア・オプティアエ(Pichia opuntiae)、ピキア・サーモトレランス(Pichia thermotolerans)、ピキア・サリクタリア(Pichia salictaria)、ピキア・ゲルカム(Pichia guercum)、ピキア・ピジュペリ(Pichia pijperi)、ピキア・スティプティス(Pichia stiptis)、ピキア菌類(Pichia sp.)、サッカロミセス・カステリ(Saccharomyces castelii、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス・クルイベリ(Saccharomyces kluyveri)、サッカロミセス菌類(Saccharomyces sp.)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、クリベロマイセス菌類(Kluyveromyces sp.)、クリベロマイセス・ラクティス(Kluyveromyces lactis)、カンジダ・アルビカンス(Candida albicans)、カンジダ菌類(Candida sp.)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、アスペルギルス・ニデュランス(Aspergillus nidulans)、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・オリザエ(Aspergillus oryzae)、トリコデルマ・リーセイ(Trichoderma reesei)、クリソスポリウム・ラクノウェンス(Chrysosporium lucknowense)、フザリウム菌類(Fusarium sp.)、フザリウム・グラミネアラム(Fusariumu gramineum)、フザリウム・ベネナタム(Fusariumu venenatum)、ニセツリガネゴケ(Physcomitrella patens)およびアカパンカビ(Neurospora crassa)。
【0011】
当業者に周知である「形質転換した」とは、外部DNAまたはRNAの導入による
有機体のゲノムもしくはエピソームの特異的修飾、または他の安定的な外部DNAまたはRNAの導入を意味する。
【0012】
当業者に周知である「形質移入した」とは、以下に限定されないが、組換えDNAもしくはRNAを含む外部DNAまたはRNAを微生物に導入することである。
【0013】
当業者に周知である「同一性」とは、2個以上のポリペプチド配列、場合によっては2個以上のポリヌクレオチド配列を比較することで判定される関係のことである。当業界では、「同一性」は、ポリペプチド配列間、場合によってはポリヌクレオチド配列間の文字列の整合性によって判定される配列関連度をも意味する。「同一性」は、限定されないが、以下の文献に記述されている既存の方法で直ちに計算できる:(Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; and Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991; and Carillo, H., and Lipman, D., SIAM J. Applied Math.,48:1073(1988)。同一性を決定する方法は、テストされる配列間で最大の整合性が得られるように設計される。さらに、同一性を決定する方法は、市場で入手可能なコンピュータープログラムにコード化されている。2個の配列間の同一性を決定するコンピュータープログラム方法は、以下に限定されないが、GCGプログラムパッケージ(Devereux, J., et al., Nucleic Acids Research 12(1): 387 (1984)), BLASTP, BLASTN, and FASTA (Altschul, S.F. et al., J. Molec. Biol. 215: 403−410 (1990))を含む。ブラストXプログラムは、NBCIとその他のソースから公的に入手出来る。(BLAST Manual, Altschul, S., et al., NCBI NLM NIH Bethesda, MD 20894; Altschul, S., et al., J. Mol. Biol. 215: 403−410 (1990))。よく知られたSmith Watermanアルゴリズムも、同一性を決定するのに使って良い。
【0014】
ポリペプチド配列比較のためのパラメータは、以下を含む:
Algorithm: Needleman and Wunsch, J. Mol Biol. 48: 443−453 (1970)
Comparison matrix: BLOSSUM62 from Hentikoff and Hentikoff, Proc. Natl. Acad. Sci. USA. 89:10915−10919 (1992)
ギャップペナルティー: 12
ギャップ長ペナルティー: 4
これらのパラメータを使う有用なプログラムは、「ギャップ」プログラムとして遺伝学コンピュータグループ、Madison WI.から公的に入手可能である。上述のパラメータは、ペプチド比較(終端ギャップについてはペナルティなしである)のためのデフォルトパラメータである。
【0015】
ポリヌクレオチド比較のためのパラメータは以下を含む:Algorithm: Needleman and Wunsch, J. Mol Biol. 48: 443−453 (1970)
比較マトリックス:整合=+10、非整合=0
ギャップペナルティー:50
ギャップ長ペナルティー:3
入手可能先:「ギャップ」プログラム(Genetics Computer Group, Madison WI.)
これらは、核酸比較のためのデフォルトパラメータである。
【0016】
ポリヌクレオチドおよび場合によってはポリペプチドの「同一性」は、以下の(1)と(2)に説明される。
【0017】
(1)ポリヌクレオチドの実施形態は、参照配列、例えば配列番号3と少なくとも50、60、70、80、85、90、95、97ならびに100%同一性を有するポリヌクレオチド配列を備えた単離したポリヌクレオチドをさらに含む。ここで上記ポリヌクレオチド配列は、配列番号3の参照配列と同一であってもよく、またはその参照配列と比較してある特定の整数の数だけヌクレオチドに変化があって良く、ここで上記変化は、遷移および塩基転換、または挿入を含む少なくとも1つのヌクレオチドの欠失、置換からなる群から選択されるものであり、さらに、上記変化は、参照ヌクレオチド配列の5’または3’末端位置で生じてもよいし、参照配列のヌクレオチド間で個々にまたは参照配列内の1つ以上の連続するグループで分散されることにより、これらの末端位置の間のいずれかの位置で生じても良く、上記ヌクレオチドの変化数は、配列番号3のヌクレオチドの全数に、同一性の%を表す整数を掛け、それを100で割り、次に、その結果を上記配列IDNO:3内のヌクレオチドの全数から差し引くことによって決定される。言い換えれば、
【数1】

式中、nnはヌクレオチドの変化数、xnは配列番号3のヌクレオチドの全数、yは95%では0.95、97%では0.97または100%では1.00であり、・は乗算演算子を表す記号であり、かつxnとyの非整数の積はもっとも近い整数に切り捨て、それをxnから差し引く。ポリペプチドをコードするポリヌクレオチド配列の変化は、そのコード配列にナンセンス、ミスセンスまたはフレームシフトの変異をもたらし、それによって、かかる変化後ポリヌクレオチドによってコードされたポリペプチドを変化させる。
【0018】
(2)ポリペプチド実施形態は、参照配列、例えば配列番号4と少なくとも50、60、70、80、85、90、95、97ならびに100%同一性を有するポリペプチドを備えた単離したポリペプチドをさらに含み、ここで上記ポリペプチド配列は、参照配列と同一であってもよく、またはその参照配列と比較してある特定の整数の数だけアミノ酸変化があって良く、ここで上記変化は、保存的または非保存的置換または挿入を含む少なくとも1つのアミノ酸の欠失、置換からなる群から選択されるものであり、さらに、上記変化は、参照ポリペプチド配列のアミノ基またはカルボキシ基末端位置で生じてもよいし、または参照配列のアミノ酸間で個々に、または参照配列内の1つ以上の連続するグループで分散されることにより、これらの末端位置の間のいずれかの位置で生じても良く、さらに、上記アミノ酸の変化数は、アミノ酸の全数に同一性の%を表す整数を掛け、それを100で割り、次に、その結果を上記アミノ酸の全数から差し引くことによって決定される。言い換えれば、
【数2】
式中、naはアミノ酸の変化数、xaは配列のアミノ酸の全数、yは95%では0.95、97%では0.97または100%では1.00であり、・は乗算演算子を表す記号であり、またxaとyの非整数の積はもっとも近い整数へ切り捨て、それをxaから差し引く。
【0019】
「単離する」とは、自然な状態から、人の手を加えて変えることを意味し、すなわち、自然に起こることであれば、元の環境から変化したか元の環境から取り除かれるか、または両方を意味する。例えば、生体内に自然に存在するポリヌクレオチドまたはポリペプチドは単離していない。しかし、限定するものではないが、そのようなポリヌクレオチドまたはポリペプチドを細胞に再び導入する場合を含め、自然な状態において共存している物質から分離されたポリヌクレオチドまたはポリペプチドは、「単離されている」と呼ぶ。
【0020】
「単離している」または「実質的に純粋な」核酸またはポリヌクレオチド(例えば、RNA、DNAまたは混合ポリマー)とは、自然の宿主細胞内に未変性のポリヌクレオチドを必然的に伴っている他の細胞成分(例えば、リボソーム、ポリメラーゼおよびそれに自然に付随するゲノム配列)から実質的に分離されたものである。この用語は、(1)自然に発生する環境から取り除かれた、(2)「単離されたポリヌクレオチド」を自然の中で含んでいるポリヌクレオチドの全てまたは一部を伴わない、(3)自然界では連結していないポリヌクレオチドと動作可能に連結する、または(4)自然界では生じない、核酸またはポリヌクレオチドを包含する。用語「単離されている」または「実質的に純粋」とは、組換えまたはクローン化DNA分離株、化学的に合成されたポリヌクレオチド類似体、または異種システムによって生物学的に合成されたポリヌクレオチド類似体との関連で使用できる。
【0021】
しかしながら、「単離される」ことは、これまで記述してきた核酸またはポリヌクレオチドそれ自体を自然の環境から物理的に除去することを必ずしも要求しない。例えば、生体内ゲノムの内因性核酸配列は、この内因性核酸配列の発現が変化するように、例えば、増加、減少、または除去されるように、異種配列が内因性核酸配列に隣接して置かれている場合に、「単離されている」と本明細書では見なされる。この文脈において、異種配列は、異種配列それ自体が内因性(宿主細胞またはその子孫由来)であるか外来性(異なる宿主細胞またはその子孫由来)であるかを問わず、内因性核酸配列に自然界で隣接している配列ではない。例えば、プロモータ配列は、遺伝子が発現パターンを変化させるように、(例えば、相同組換えによって)宿主細胞のゲノム内の遺伝子の元来のプロモータで置き換えてもよい。この遺伝子は、自然の状態で隣接している配列から少なくとも一部は分離されているので、「単離されている」ことになる。
【0022】
核酸はまた、ゲノム内の対応する核酸に自然界では生じない任意の修飾を含むならば、「単離された」と見なされる。例えば、内因性コード配列は、人工的に、例えば、人間の介入により導入された挿入、欠失、または点変異を含むならば、「単離された」と見なされる。「単離された核酸」はまた、宿主細胞染色体の異種部位に組み込まれた核酸とエピソームとして存在する核酸コンストラクトも含む。さらに、「単離された核酸」は、実質的にその他の細胞の材料を使用せず、または組換え技術で産生する場合に実質的に培地を必要とせず、または化学的に合成する場合に実質的に化学前駆物質もしくはその他の化学物質も必要としない。
【0023】
本明細書で使用する場合、「機能的遺伝子産物をコードする核酸配列」とは、遺伝子のコード部分のあらゆる部分を意味する。機能的遺伝子産物をコードする核酸配列は、少なくとも1つの活性を行うことのできる酵素の一部であっても良いし、酵素全体であっても良い。
【0024】
本明細書で使用する場合、「少なくとも1つの遺伝子産物の発現に必要な核酸」とは、遺伝子の一部をコードし、および/または遺伝子産物をコードする核酸に動作可能に連結するが必ずしもコード配列を含まない核酸配列を意味する。例えば、少なくとも1つの遺伝子産物の発現に必要な核酸配列は、限定されないが、エンハンサー、プロモータ、制御配列、開始コドン、終止コドン、ポリアデニル化配列、および/またはコーディング配列を含む。
【0025】
本明細書で使用する「タンパク質分解」または「細胞内でタンパク質分解を担う遺伝子産物」とは、少なくとも1つのペプチド、ポリペプチドおよび/またはタンパク質の切断を引き起こすことができる任意のペプチド、ポリペプチド、タンパク質および/または酵素またはその部分を意味する。タンパク質分解を担う遺伝子産物は、切断(即ち、ペプチダーゼ)に直接関与しても良く、ペプチダーゼ合成経路の役割として間接的に関与してもよい。細胞内のタンパク質分解を担う遺伝子産物の例として、限定されないが、アスパルチルプロテアーゼ、セリンプロテアーゼ、分泌アスパルチルプロテアーゼ、分泌セリンプロテアーゼ、酵母メチロトローフプロテアーゼ、DDP IV様エンドペプチダーゼ、金属エンドペプチダーゼ、Prb1様セリンプロテアーゼ、Prb1 セリンプロテアーゼ、およびCPY様カルボキシペプチダーゼが挙げられる。分泌セリンプロテアーゼのように、細胞から分泌されてもタンパク質分解活性の一部または全部を維持しているプロテアーゼも、この定義に当てはまる。分泌プロテアーゼは、細胞内部および/または外部のタンパク質分解に関与する場合がある。
【0026】
本明細書で使用する場合、「糖鎖形成」または「細胞内の糖鎖形成に関与する遺伝子産生物」とは、細胞内において少なくとも1つの糖類部分をポリペプチドに付加する機能、または少なくとも1本の糖類鎖を伸長する機能に関わるペプチド、ポリペプチド、タンパク質および/または酵素もしくはその部分のことを意味する。細胞内で糖鎖形成を担う遺伝子産物は、糖類を細胞内で直接ポリペプチドに付加する作用に関与し、例えば、限定されないが、マンノシルトランスフェラーゼである。マンノシルトランスフェラーゼは、Dol−P−Manからの残基をペプチド、ポリペプチド、および/またはタンパク質上のセリンおよび/またはスレオニン残基に伝達でき、あるいは、GPD−Manからのマンノース残基を糖類に伝達するように作用し、このようにして、糖類鎖を伸長する。あるいは、糖鎖形成の機能を担う遺伝子産物は、糖鎖形成経路の部分であるかもしれないし、また細胞内で多糖類をポリペプチドに付加する機能に間接的に関与しているかもしれない。細胞内の糖鎖形成に関与する遺伝子産物の例には、限定されないが、マンノシルトランスフェラーゼ類が含まれる。
【0027】
本明細書で使用する場合、「酵母メチロトローフプロテアーゼ1(Ymp1)活性」とは、配列番号4として本明細書で同定されたタンパク質が遂行できる任意の活性を意味する。例えば、酵母メチロトローフプロテアーゼ1活性またはYmp1活性は、限定されないが、ペプチド、ポリペプチド、またはタンパク質をタンパク分解的に切断する酵素能力を含む。特に、Ymp1活性は、限定されないが、ヒトGLP−1および/またはヒトアルブミンなどのGLP−1アゴニストの断片および/または変異体を含むポリペプチドを切断する酵素能力を意味する。少なくとも1つのYmp1活性は、限定されないが、少なくとも1つのセリンプロテアーゼ活性を含む。
【0028】
「ポリヌクレオチド(1つまたは複数)」とは、一般に、非修飾RNAもしくはDNAまたは修飾RNAもしくはDNAであり得る、任意のポリリボヌクレオチドまたはポリデオキシリボヌクレオチドを意味する。「ポリヌクレオチド(1つまたは複数)」としては、単鎖および二重鎖DNA、単鎖および二重鎖領域、または単鎖、二重鎖および三重鎖領域の混合物であるDNA、単鎖および二重鎖RNA、単鎖および二重鎖領域の混合物であるRNA、単鎖または、より典型的には、二重鎖、または三重鎖領域のDNAおよびRNAを含むハイブリッド分子、または単鎖および二重鎖領域の混合物が挙げられるが、これらに限定されない。加えて、本明細書で使用する場合、「ポリヌクレオチド」とは、RNAもしくはDNA、またはRNAおよびDNAの両方を含む三重鎖領域を意味する。かかる領域の鎖は、同じ分子に由来してもよいし、異なる分子に由来してもよい。この領域は1つ以上の分子をすべて含み得るが、より典型的には、一部の分子の領域のみを含む。三重らせん状領域の1つの分子は、しばしばオリゴヌクレオチドである。本明細書で使用する場合、「ポリヌクレオチド(1つまたは複数)」という用語は、1つ以上の修飾塩基を含む上記のDNAまたはRNAも含む。したがって、安定性または他の理由のために修飾された骨格のDNAまたはRNAは、本明細書で意図する「ポリヌクレオチド(1つまたは複数)」である。さらに2例のみ挙げると、異常な塩基(イノシンなど)、または修飾塩基(トリチル化塩基など)を含むDNAまたはRNAも、本明細書において使用されるポリヌクレオチドである。当業者に知られている多くの有用な目的に適う多種多様の修飾がDNAおよびRNAで行われていることは理解されるであろう。本明細書で使用する場合、「ポリヌクレオチド(1つまたは複数)」という用語は、ポリヌクレオチドのかかる化学的、酵素的もしくは代謝的に修飾された形態、ならびにウイルスおよび細胞(例えば、単純細胞および複雑細胞が挙げられる)に特徴的なDNAおよびRNAの化学的形態を包含する。「ポリヌクレオチド(1つまたは複数)」は、しばしばオリゴヌクレオチド(1つまたは複数)と呼ばれる短いポリヌクレオチドも包含する。
【0029】
「ポリペプチド(1つまたは複数)」とは、ペプチド結合または修飾ペプチド結合により互いに結合した2つ以上のアミノ酸を含む任意のペプチドまたはタンパク質を意味する。「ポリペプチド(1つまたは複数)」とは、一般的にペプチド、オリゴペプチドまたはオリゴマーと呼ばれる短鎖と、一般にタンパク質と呼ばれるより長い鎖との両方を意味する。ポリペプチドは、20個の遺伝子コードアミノ酸以外のアミノ酸を含んでよい。「ポリペプチド(1つまたは複数)」とは、天然プロセス(プロセシングおよび他の翻訳後の修飾など)、または化学的修飾技術のいずれかにより修飾されたアミノ酸配列を含む。かかる修飾は、基礎テキストおよびより詳細な研究論文、ならびに膨大な研究文献に詳述されており、当業者にとって公知である。このような種類の修飾は、所定のポリペプチドのいくつかの部位に同じ種類の修飾が同程度または異なる程度で存在し得ることが理解されるであろう。所定のポリペプチドは、多くの種類の修飾を含んでよい。修飾は、ポリペプチドのどの場所(ペプチド骨格、アミノ酸側鎖およびアミノ末端もしくはカルボキシ末端を含む)でも生じ得る。修飾としては、アセチル化、アシル化、ADPリボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトールの共有結合、架橋、環化、ジスルフィド結合形態、脱メチル化、共有架橋形態、システイン形態、ピログルタミン酸形態、ホルミル化、γ−カルボキシル化、GPIアンカー形態、水酸化、ヨウ素化、メチル化、ミリストイル化、酸化、タンパク質分解プロセシング、リン酸化反応、プレニル化、ラセミ化、グリコシル化、脂質結合、硫酸化、グルタミン酸残基のγ−カルボキシル化、ヒドロキシル化、およびADP−リボシル化、セレノイル化、硫酸化、転移RNA媒介性のアミノ酸のタンパク質への付加(アルギニル化、およびユビキチン化など)が挙げられる。例えば、PROTEINS − STRUCTURE AND MOLECULAR PROPERTIES, 2nd Ed., T. E. Creighton, W. H. Freeman and Company, New York, 1993 and Wold, F., Posttranslational Protein Modifications: Perspectives and Prospects, pgs. 1−12 in POSTTRANSLATIONAL COVALENT MODIFICATION OF PROTEINS, B. C. Johnson, Ed., Academic Press, New York,(1983); Seifter, et al., Meth. Enzymol. 182:626−646(1990)およびRattan, et al., “Protein Synthesis: Posttranslational Modifications and Aging”, Ann NY Acad Sci 663:48−62(1992)を参照されたい。ポリペプチドは、分枝または環状であり、分枝があってもなくても良い。環状、分枝および分枝した環状ポリペプチドは、翻訳後の自然なプロセスから生み出されたものであっても、完全に合成的な方法によるものであっても良い。
【0030】
「組換え発現系(1つまたは複数)」とは、本発明の発現系またはその部分またはポリヌクレオチドであって、本発明のポリヌクレオチドおよびポリペプチドの産生のために宿主細胞または宿主細胞可溶化液へ導入、形質移入、または形質転換されたものである。
【0031】
本明細書において使用される場合、「変異体(1つまたは複数)」という用語は、参照ポリヌクレオチドまたは参照ポリペプチドとは異なるが、本質的な特性を保持するポリヌクレオチドまたはポリペプチドである。ポリヌクレオチドの典型的な変異体は、別の参照ポリヌクレオチドとはヌクレオチド配列が異なる。変異体のヌクレオチド配列の変化により、参照ポリヌクレオチドによりコードされたポリペプチドのアミノ酸配列が変化する場合もあるし、しない場合もある。以下に論じるように、ヌクレオチド変化により、参照配列によりコードされたポリペプチドのアミノ酸が置換、付加、欠失、融合および切断されてよい。ポリペプチドの典型的な変異体は、別の参照ポリペプチドとはアミノ酸配列が異なる。一般に、参照ポリペプチド配列とその変異体は、全体的に近似し同じ領域が多いので、その相違は限定されている。変異体と参照ポリペプチドのアミノ酸配列は、1つ以上の置換、付加、欠失の任意の組み合わせにより異なってよい。置換または挿入したアミノ酸残基は、遺伝子コードでコードされたものであってもよいし、そうでなくてもよい。
【0032】
本発明は、保存アミノ酸置換によって参照対象とは異なり、残基が同様な特徴を有する別のもので置換されている本発明の各ポリペプチドの変異体も含む。典型的なそのような置換は、アラニン、バリン、ロイシンおよびイソロイシンとの間、セリンとトレオニンとの間、酸性残基のアスパラギン酸とグルタミン酸との間、アスパラギンとグルタミンとの間、塩基性残基であるリシンとアルギニンとの間、芳香族残基のフェニルアラニンとチロシンとの間で生じる。特に、いくつか、つまり5−10、1−5、1−3、1−2または1つのアミノ酸が、任意の組み合わせで置換、消去、または付加された変異体が存在する。ポリヌクレオチドまたはポリペプチドの変異体は、天然(対立遺伝子変異体など)であってもよく、天然では知られていない変異体であってもよい。ポリヌクレオチドおよびポリペプチドの非天然変異体は、当業者に周知の、変異誘発技術により生成してもよいし、直接合成によって生成しても良いし、他の組換え技術によって生成しても良い。
【0033】
「微生物(1つまたは複数)」とは、(i)原核生物を意味し、限定されないが、以下の属のメンバーを含む:連鎖球菌、ブドウ球菌、百日咳菌、コリネバクテリウム、マイコバクテリウム、ナイセリア、ヘモフィルス、放線菌、ストレプトミセス、ノカルジア、エンテロバクター、エルシニア、フランシセラ、パスツレラ、モラクセラ、アシネトバクター、エリシペロスリクス、ブランハメラ、アクチノバチルス、ストレプトバチルス、リステリア、カリマトバクテリウム、ブルセラ菌、桿菌、クロストリジウム、トレポネーマ、大腸菌類、サルモネラ菌、クレブシエラ、ビブリオ、プロテウス、エルウィニア、ボレリア、レプトスピラ、スピリルム、カンピロバクター、赤痢菌、レジオネラ、シュードモナス、エロモナス、リケッチア、クラミジア、ボレリアおよびマイコプラズマ。
【0034】
さらに、限定されないが、以下の種またはグループのメンバーを含む:グループ A連鎖球菌、グループ B連鎖球菌、グループ C連鎖球菌、グループ DS連鎖球菌、グループ G連鎖球菌、肺炎球菌、化膿性連鎖球菌、ストレプトコッカス・アガラクチア、連鎖球菌 faecalis、フェシウム菌、連鎖球菌 durans、淋菌、髄膜炎菌、黄色ブドウ球菌、表皮ブドウ球菌、ジフテリア菌、ガードネレラ菌、結核菌、マイコバクテリウム・ボビス、マイコバクテリウム・ウルセランス、ライ菌、Actinomyctes israelii、リステリア菌、百日咳菌、パラ百日咳菌、気管支敗血症菌、大腸菌、志賀赤痢菌、インフルエンザ菌、ヘモフィルス・エジプチウス、パラインフルエンザ菌、軟性下疳菌、ボルデテラ、チフス菌、シトロバクター・フロインデイ、プロテウス・ミラビリス、プロテウス・ブルガリス、ペスト菌、肺炎かん菌、セラチア‐マルセッセンス、セラチア・リクファシエンス、コレラ菌、Shigella dysenterii、シゲラ・フレックスネリ、緑膿菌、野兎病菌、ウシ流産菌、炭疽菌、セレウス菌、ウェルシュ菌、破傷風菌、ボツリヌス菌、梅毒トレポネーマ、リケッチア・リケッチイおよびトラコーマ病原体。(ii)限定されないが、Archaebacterを含む古細菌、および(iii)単細胞または糸状真核生物であり、限定されないが以下を含む:原虫、菌類、酵母類属のメンバー、クリベロマイセス、またはカンジダ、および酵母類ceriviseae種のメンバー、クリベロマイセス・ラクティス、またはカンジダ・アルビカンス。
【0035】
「細菌(1つまたは複数)(l)」とは、(i)原核生物を意味し、限定されないが以下の属のメンバーを含む:連鎖球菌、ブドウ球菌、ボルデテラ、コリネバクテリウム、マイコバクテリウム、ナイセリア、ヘモフィルス、放線菌、ストレプトミセス、ノカルジア、エンテロバクター、エルシニア、フランシセラ、パスツレラ、モラクセラ、アシネトバクター、エリシペロスリクス、ブランハメラ、アクチノバチルス、ストレプトバチルス、リステリア、カリマトバクテリウム、ブルセラ菌、桿菌、クロストリジウム、トレポネーマ、大腸菌類、サルモネラ菌、クレブシエラ、ビブリオ、プロテウス、エルウィニア、ボレリア、レプトスピラ、スピリルム、カンピロバクター、赤痢菌、レジオネラ、シュードモナス、エロモナス、リケッチア、クラミジア、ボレリアおよびマイコプラズ。さらに、限定されないが以下の種またはグループのメンバーを含む:グループ A連鎖球菌、グループ B連鎖球菌、グループ C連鎖球菌、グループ DS連鎖球菌、グループ G連鎖球菌、肺炎球菌、化膿性連鎖球菌、ストレプトコッカス・アガラクチア、連鎖球菌 faecalis、フェシウム菌、連鎖球菌 durans、淋菌、髄膜炎菌、黄色ブドウ球菌、表皮ブドウ球菌、ジフテリア菌、ガードネレラ菌、結核菌、マイコバクテリウム・ボビス、マイコバクテリウム・ウルセランス、ライ菌、Actinomyctes israelii、リステリア菌、百日咳菌、パラ百日咳菌、気管支敗血症菌、大腸菌、志賀赤痢菌、インフルエンザ菌、ヘモフィルス・エジプチウス、パラインフルエンザ菌、軟性下疳菌、ボルデテラ、チフス菌、シトロバクター・フロインデイ、プロテウス・ミラビリス、プロテウス・ブルガリス、ペスト菌、肺炎かん菌、セラチア‐マルセッセンス、セラチア・リクファシエンス、コレラ菌、Shigella dysenterii、シゲラ・フレックスネリ、緑膿菌、野兎病菌、ウシ流産菌、炭疽菌、セレウス菌、ウェルシュ菌、破傷風菌、ボツリヌス菌、梅毒トレポネーマ、リケッチア・リケッチイおよびトラコーマ病原体、ならびに(ii)古細菌(限定されないがArchaebacterを含む)。
【0036】
本明細書で使用する場合、「異種ポリペプチド(1つまたは複数)」とは、形質転換した宿主細胞または対象となる微生物によって人工的に合成され、組換えDNAによってその宿主細胞または微生物に導入されたポリペプチドを意味する。例えば、ピキアは、ヒト血清アルブミンの発現用の宿主細胞として作用するが、非形質転換または形質導入されなかったピキアでは起こらない。異種ポリペプチドは、単離を促進するように修飾されたポリペプチドを含んでも良い。
【0037】
本明細書で使用する場合、「親和性タグ」とは、他の物質または分子への選択的親和性を得ることのできる分子に関係する任意の部分を意味する。例えば、親和性タグは、
分子にカラム内のパッキング物質との選択的親和性を提供することにより分子の精製を促進するのに使って良い。親和性タグの非限定的例はhis−タグである。
【0038】
核酸は、別の核酸配列と機能的な関係をもつように置かれると「動作可能に連結」する。例えば、プレ配列または分泌性リーダーは、ポリペプチドの分泌に関与するタンパク質前駆体として発現するならば、動作可能にポリペプチドのDNAに連結する。プロモータまたはエンハンサーは、それが配列の転写に影響するならば、動作可能にコード配列に連結する。または、翻訳を促進するように置かれるならば、リボソーム結合部位は動作可能にコード配列に連結する。一般に、「動作可能に連結する」とは、連結しているDNA配列が連続していることを意味し、分泌性リーダーの場合には、連続しかつリーディングフェーズにあることを意味する。しかしながら、エンハンサーは連続している必要はない。連鎖は、適切な制限酵素部位で連結して完結する。そのような部位が存在しない場合は、合成オリゴヌクレオチドアダプタまたはリンカーが通常のやり方に従って使われる。
【0039】
本明細書で使用される場合、細胞を「収集する」とは、細胞培養から細胞を採取することを意味する。
細胞は、収集する間濃縮され、例えば、遠心分離または濾過によって培養ブロスから分離される。細胞の収集は、細胞を溶解して、限定されないが、ポリペプチドおよびポリヌクレオチドなどの細胞内物質を得る段階を含んで良い。限定されないが、異種的に発現するポリペプチドを含む特定の細胞の物質は、培養中に細胞から遊離することがあるということを当業者は理解されたい。従って、対象の産物(例えば、異種的に発現したポリペプチド)は、細胞を収集した後も培養ブロスにとどまり続ける場合がある。
【0040】
さらに、組換えDNAコンストラクトが選択可能なマーカーをコードする方法も提供される。そのような選択可能なマーカーは陽性または陰性の選択を提供する。上記選択可能なマーカーを発現するステップ、選択段階の少なくとも1つの第1の形質転換した細胞によって産生した選択可能なマーカーの量を、選択段階の少なくとも1つの第2の形質転換した細胞によって産生した選択可能なマーカーの量と比較することを含む方法(第1と第2の形質転換した細胞は同じ選択可能なマーカーを産生する)も提供される。当業界で知られているように、選択可能なマーカーは、限定されないが、ジヒドロ葉酸還元酵素(dhfr)、β−ガラクトシダーゼ、蛍光タンパク質、分泌型のヒト胎盤のアルカリホスファターゼ、β−グルクロニダーゼ、酵母選択可能マーカーLEU2およびURA3、アポトーシス耐性遺伝子、およびアンチセンスオリゴヌクレオチド、並びにネオマイシン(neo)、カナマイシン、ジェネテシン、ハイグロマイシンB、ピューロマイシン、ゼオシン、ブラストサイジン、ストレプトスライシン、ビアラホス、フレオマイシン、およびアンピシリンを含む抗生物質存在下で成長する能力を与える抗生物質耐性遺伝子を含む。同様に当業界で知られているように、細胞は、限定されないが、視覚的点検または選択可能マーカーの発現を検出することができるBD蛍光標示式細胞分取器などの細胞選別機を含む種々の手段で選別ができる。
【0041】
当業界で理解されている用語「野生型」とは、遺伝的修飾なしに自然の集団で起こりうる宿主細胞またはポリペプチドまたはポリヌクレオチド配列を意味する。例えば、「宿主細胞の野生型親」とは、宿主細胞のゲノム内で遺伝的修飾がなされるまたは生じる前の無修飾の宿主細胞系統を意味する。
【0042】
本明細書で使われる場合、「力価収率」とは、溶液(例えば、培養ブロスまたは細胞−溶解混合物または緩衝液)中の産物(例えば、異種性に発現したポリペプチド)の濃度を意味し、通常mg/Lまたはg/Lで表現される。収率の増加とは、2個の規定された条件下で産生した産物の濃度の絶対的または相対的増加を意味する。
【0043】
本明細書で使われる場合、「インクレチンホルモン」とは、インスリン分泌を増強する、あるいは分泌レベルまたはインスリンを増加させるあらゆるホルモンを意味する。インクレチンホルモンの1つの例は、GLP−1である。GLP−1は、食物摂取に応答して腸のL細胞よって分泌されたインクレチンである。健常人の場合、GLP−1は、膵臓のグルコース−依存インスリン分泌を刺激することによって食後血糖値を調整する重要な働きをし、その結果末梢内のグルコース吸収を増加させる。GLP−1はまた、グルカゴン分泌を抑制し、肝臓のグルコース出力を低下させる。さらに、GLP−1は胃排出時間を遅延させ、小腸の運動性を緩慢にし食物吸収を遅らせる。GLP−1は、グルコース依存インスリン分泌に関わる遺伝子の転写を刺激し、β細胞新生を促進することによって、連続したβ細胞応答能を促進する(Meier, et al.Biodrugs 2003; 17 (2): 93−102)。
【0044】
本明細書に使用する場合、「GLP−1活性」とは、自然に起こるヒトGLP−1の1以上の活性を意味する。これらの活性は、限定されないが、血液および/または血漿グルコースの低下、グルコース依存インスリン分泌の刺激あるいは分泌レベルまたはインスリンの上昇、グルカゴン分泌の抑制、フルクトサミンの低下、脳へのグルコース送達および代謝の増加、胃内容排出の遅延、β細胞応答能の促進、および/または組織の新生を含む。GLP−1活性に付随したこれらの活性とその他の活性は、GLP−1活性またはGLP−1アゴニストを有する組成によって直接または間接的に引き起こされる。例えば、GLP−1活性を有する組成は、直接または間接的にグルコース−依存性を刺激するが、一方インスリン産生を刺激すると、哺乳類の血漿グルコースレベルを間接的に減少させる。
【0045】
本明細書で使用する場合、「インクレチンミメティック」とは、インスリン分泌を増強させる、あるいは分泌レベルまたはインスリンを増加できるできる化合物を意味する。インクレチンミメティックは、哺乳類において、インスリン分泌を刺激し、β細胞新生を増加させ、β細胞アポトーシスを抑制し、グルカゴン分泌を抑制し、胃内容排出を遅延させて満腹感を誘発させる能力がある。インクレチンミメティックは、GLP−1活性を有する任意のポリペプチドを含み、限定されないが、エキセンディン3およびエキセンディン4、その任意の断片および/または変異体および/または抱合体が含まれる。
【0046】
本明細書で使用する場合、「断片」とは、ポリペプチドに関して使用される場合、完全な天然ポリペプチドのアミノ酸配列とすべてではないが部分的に同じであるアミノ酸配列を有するポリペプチドである。断片は、「独立」していてもよいし、より大きなポリペプチドに含まれ、単一のより大きなポリペプチド内の単一の連続領域として部分または領域を形成してもよい。例として、天然GLP−1の断片は、天然アミノ酸1〜36位中のアミノ酸7〜36位を含む。さらに、ポリペプチドの断片は、天然配列の一部の変異体であってもよい。例えば、天然GLP−1のアミノ酸7〜30位を含むGLP−1の断片は、その一部の配列内にアミノ酸置換を有する変異体であってもよい。
【0047】
本明細書で使用する場合、「抱合体」または「抱合した」とは、互いに結合している2つの分子を意味する。例えば、第一のポリペプチドは、第二のポリペプチドに共有結合していてもよいし非共有結合していてもよい。第一のポリペプチドは、化学的リンカーにより共有結合していてもよいし、第二のポリペプチドに遺伝的に融合していてもよいが、第一ポリペプチドおよび第二ポリペプチドは共通のポリペプチド骨格を共有する。本発明の抱合体は、少なくとも1つのヒト血清アルブミンに抱合した治療ポリペプチドを含んで良い。その他の抱合体は、限定されないが、トランスフェリン、単鎖可変領域および/または少なくとも1つの抗体のFc領域に抱合した少なくとも1つの治療用ポリペプチドも含む。抱合体は、リンカーを含んでもよく、含まなくても良い。
【0048】
本明細書で使用する場合、「直列方向の」とは、同じ分子の一部として互いに隣接する2つ以上のポリペプチドを意味する。それらは共有結合か非共有結合のいずれかで結合していてよい。2つ以上の直列方向のポリペプチドは、同じポリペプチド骨格部分を形成していてよい。直列方向のポリペプチドは、順方向であっても逆方向であってもよく、および/または他のアミノ酸配列により分離されていてもよい。
【0049】
「ドメイン抗体」または「dAb」は、抗原と結合できる「単一可変ドメイン」と同義であると見なすことができる。単一可変ドメインは、ヒト抗体可変ドメインであることもあるが、さらにげっ歯類(例えば、WO 00/29004に開示されたもの)、テンジクサメおよびラクダVHH dAbsなどの他の種由来の単一抗体可変ドメインも含む。ラクダVHHは、自然界では軽鎖を持たない重鎖抗体を産生するラクダ、ラマ、アルパカ、ヒトコブラクダ、およびグアナコを含む種由来の免疫グロブリン単一可変ドメインポリペプチドである。そのようなVHHドメインは、当業者で入手可能な標準技術に従ってヒト化でき、そのようなドメインは「ドメイン抗体」と見なされる。本明細書で使用するVHは、ラクダVHH領域を含む。
【0050】
「単一可変ドメイン」という句は、異なる可変領域またはドメインとは独立に特異的に抗原またはエピトープに結合する抗原結合タンパク質可変ドメイン(例えば、VH、VHH、VL)を意味する。
【0051】
本明細書で使用する場合、「抗原結合タンパク質」とは、限定されないが、抗原と結合することができる可変ドメインとドメイン抗体などのドメインの抗体、抗体断片、ならびにその他のタンパク質コンストラクトを意味する。
本明細書で使用する場合、遺伝子改変宿主細胞内で比較される酵素またはその断片または酵素活性の「減少量」とは、非遺伝子改変宿主細胞と比較した時、少なくとも1つの酵素をより少なく産生、または少なくとも1種類の酵素活性をより少なく示す遺伝子改変宿主細胞を意味する。典型的に、遺伝子改変宿主細胞で産生した酵素活性は、遺伝的修飾前の同種の野生型系統と比較される。しかしながら、遺伝子改変宿主と、同属であるが異なる種もしくは系統からの野生型宿主または別の遺伝子改変系統を有する野生型宿主との比較を行うこともできる。少なくとも1つの酵素または酵素活性の減少には、少なくとも1つの酵素または酵素活性完全消滅が含まれ、その場合、少なくとも1つの酵素のうちのいずれも遺伝子改変宿主細胞内で産生されない、および/または少なくとも1つの酵素のうちのいずれも機能的でなく、活性を示さない。さらに、少なくとも1つの酵素活性の減少もこの定義の範囲内に含まれる。即ち、1つ以上の活性を有する酵素は、第1の活性量を維持し、一方同じ酵素の第2の活性は減少させることができる。
【0052】
本明細書で使用する場合、「厳密な条件」あるいは「厳密なハイブリッド形成法条件」とは、ハイブリッド形成が、配列間で少なくとも70%、あるいは少なくとも80%、あるいは少なくとも90%の同一性がある場合にのみ起こることを意味する。厳密なハイブリッド形成条件の例には、50%ホルムアミド、5xSSC(150mM NaCl、 15mMクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5xデンハート液、10%デキストラン硫酸、および20μg/mlの変性させ剪断したサケ精子DNAを含む溶液中で42℃で一晩インキュベーションし、続いてフィルタを約65℃で0.1xSSCを使って洗浄する工程が含まれる。ハイブリッド形成と洗浄条件は良く知られており、Sambrook, et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor, N.Y.,(1989), 特に Chapter 11に例証されており、参照によりその開示の全体をここに組み入れる。
【0053】
本明細書に使用する場合、「遺伝的修飾」または「遺伝的に修飾された」とは、1つ以上の塩基または細胞DNA配列(1つまたは複数)の断片の抑制、置換、欠失および/または挿入を意味する。そのような遺伝的修飾は、in vitroで(直接的に単離したDNA上で)またはin situで、例えば遺伝子工学技術あるいは細胞を変異原性薬剤に曝露することによって得ることができる。変異原性薬剤は、例えば、エネルギー性光線(エックス線、γ光線、紫外線など)などの物理的薬剤、またはアルキル化薬剤(EMS、NQOなど)、バイアルキレーティング薬剤、挿入剤などのDNAの異なる官能基と反応することのできる化学薬剤を含む。遺伝的修飾はまた、例えばRothsteinらによって明らかになった方法に従う遺伝的破壊によって得られる(Meth.Enzymol. 194:281−301(1991))。この方法に従うと、遺伝子の一部分または全てが相同組換えを通してin vitro修飾したバージョンによって置換される。遺伝的修飾はまた、DNA配列へ任意の変異挿入(トランスポゾン、ファージなど)することにより得ることができる。本明細書に使用する場合、「遺伝的に修飾した」とは、ポリペプチド、または核酸もしくはアミノ酸のそれぞれを少なくとも1つ欠失、置換もしくは抑制したポリペプチドをコードする遺伝子を意味することができる。例えば、野生型品種の少なくとも1つのアミノ酸を置換したポリペプチドは遺伝的修飾と見なされるだろう。
遺伝的修飾は、細胞の機序によって逆転するか、または減弱する。あるいは、変異は非逆転および/または非漏出性でありうる。「漏出変異」は、野生型機能の全部ではないが一部の不活性化を生じる変異を含む。
本発明の宿主細胞によって遂行される遺伝的修飾は、その細胞のDNA配列のコーディング領域および/または遺伝子の発現に影響を及ぼす領域に位置する。従って、本発明の修飾は一般的に、遺伝子産生またはタンパク質分解および/または糖鎖形成に関わるタンパク質および/または酵素の遺伝子産生の制御または促進に影響を及ぼす。本発明の細胞において、異種的に発現したポリペプチドをタンパク分解的に切断および/またはグリコシル化する能力が低下するのは、改変された生物学的特性を有する1つ以上の酵素の産生、かかる1つ以上の酵素の産生欠如、または1つ以上の酵素の低レベルの産生による構造上および/または立体構造上の変化が原因である。
【0054】
本発明1つの態様において、機能的遺伝子産物をコードする少なくとも1つの核酸配列および/または少なくとも1つの遺伝子産物が発現するのに必要な少なくとも1つの核酸が遺伝子改変された、遺伝的修飾ピキア株が提供される。ここで上記遺伝子産物は、遺伝的修飾ピキア株においてタンパク質分解および/または糖鎖形成を担う。本発明の遺伝子改変ピキア株は、限定されないが、次の機能的遺伝子産物少なくとも1つまたはかかる遺伝子産物の発現をコードする少なくとも1つの核酸配列が遺伝的修飾を受けているピキアを含む:PEP4、PRB1、YPS1、YPS2、YMP1、DAP2、GRH1、PRD1、YSP3、PRB3、YMP2、および/またはYMP3。さらに本発明に含まれるのは、遺伝子改変ピキアであって、このピキアは、野生型系統と比較して下記の酵素および/または酵素タイプの少なくとも1つにおいて、活性量が低下したか、全くないか、および/または少なくとも1つの活性が減少している:アスパルチルプロテアーゼ、セリンプロテアーゼ、分泌アスパルチルプロテアーゼ、分泌セリンプロテアーゼ、酵母メチロトローフのプロテアーゼ、DPP IV様エンドペプチダーゼ、金属エンドペプチダーゼ、Prb1様セリンプロテアーゼ、Prb1セリンプロテアーゼ、CPY様カルボキシペプチダーゼおよび/またはマンノシルトランスフェラーゼ。さらに、遺伝子修飾系統は下記から選択された酵素を産生することができる:アスパルチルプロテアーゼ、セリンプロテアーゼ、分泌アスパルチルプロテアーゼ、分泌するセリンプロテアーゼ、酵母メチロトローフプロテアーゼ、DPP IV様エンドペプチダーゼ、金属エンドペプチダーゼ、Prb1様セリンプロテアーゼ、Prb1セリンプロテアーゼ、CPY様カルボキシペプチダーゼおよび/またはマンノシルトランスフェラーゼ。ここで上記酵素は、野生型系統および/または酵素と比較すると、上記酵素の少なくとも1つの活性を低下させている。
【0055】
本発明の遺伝子改変ピキア株はまた、機能的遺伝子産物または上記遺伝子産物の発現をコードする以下の核酸配列のうち少なくとも1つが遺伝子改変されたピキア株を含む:OCH1、PMT1、PMT2、および/またはPMT4。本発明の遺伝子改変ピキアは、野生型系統と比較して、少なくとも1つのマンノシルトランスフェラーゼの量を減少させるか、全くなしにするかまたは活性を低下させる。さらに、遺伝子改変系統は、野生型酵素と比較して活性を低下させた少なくとも1つのマンノシルトランスフェラーゼなど、糖鎖形成に付随する酵素を産生してもよい。
【0056】
本発明の遺伝子改変ピキアは、限定されないが、野生型X−33またはSMD1163ピキア由来の遺伝子改変型を含む。本発明の遺伝子改変ピキアは、限定されないが、以下の野生型由来の遺伝子改変型を含む:ピキア・パストリス、ピキア・フィンランディカ、ピキア・トレハロヒィア、ピキア・コクラメ、ピキア・メンブラナエファシエンス、ピキア・メタノリカ、ピキア・ミヌタ(オガタ・エミヌタ、ピキア・リンドネリ)、ピキア・オプティアエ、ピキア・サーモトレランス、ピキア・サリクタリア、ピキア・ゲルカム、ピキア・ピジュペリ、ピキア・スティプティス、ピキア菌種。
【0057】
本発明の別の態様において、機能的遺伝子産物または上記遺伝子産物の発現をコードする次の核酸配列が遺伝的に改変された、改変ピキア株が提供される:PEP4、PRB1、YPS1、YPS2、YMP1、YMP2、YMP3およびPMT4。本発明の遺伝子改変ピキアは、野生型系統と比較して、以下の遺伝子産物のうち少なくとも1つについてその量を減少させるか、全くなしにするかまたは活性を低下させる:アスパルチルプロテアーゼ、セリンプロテアーゼ、セリン分泌プロテアーゼおよびマンノシルトランスフェラーゼ。本発明の遺伝子改変ピキアはまた、野生型系統と比較して、以下の遺伝子産物のうち少なくとも1つについてその活性を低下させる:少なくとも1つのアスパルチルプロテアーゼ、少なくとも1つのセリンプロテアーゼ、少なくとも1つのセリン分泌プロテアーゼおよび/または少なくとも1つのマンノシルトランスフェラーゼ。
【0058】
本発明の遺伝子改変ピキアは更に、少なくとも1つの異種ポリペプチドを発現できるポリヌクレオチドを含む。少なくとも1つの異種ポリペプチドを発現できるポリヌクレオチドは、限定されないが、ベクター、宿主細胞のゲノムに形質転換されたDNA、ウイルスまたはウイルスの一部分、および/またはプラスミドを含む。異種ポリペプチドを発現できるポリヌクレオチドはピキアのゲノムに形質転換されても良く、および/または発現ベクターの一部分であっても良く、および/またはエピソーム発現系であっても良い。
【0059】
当業者に理解されているように、DNAはいくつかの異なる方法によって宿主細胞に形質転換される。酵母の場合、エレクトロポレーション法、塩化リチウム法、またはスフェロプラスト法など、任意の簡便なDNA伝達方法を使用して良い。高密度の発酵に適した安定な系統を産生するするために、DNAを宿主染色体に組み込むことが望ましい。組込みは、当業者に周知の技術を使って相同組換え経由で行われる。例えば、少なくとも1つの異種タンパク質を発現可能なDNAには、宿主生物体の配列に相同的な隣接する配列を提供してもよい。このような方法により、所望または必須の遺伝子を破壊することなしに、宿主ゲノムの限定された部位で組込みが行われる。あるいは、少なくとも1つの異種タンパク質を発現可能なDNAは、、宿主染色体の望まれない遺伝子の部位に組み込まれ、遺伝子または遺伝子産物の発現の破壊または欠失を引き起こす。例えば、YMP1、YMP2、YMP3、PEP4、PRB1、YPS1、YPS2、DAP2、GRH1、PRD1、YSP3、PRB3、配列番号10、配列番号12および/または配列番号14の遺伝子の部位への組み込みは、異種タンパク質の発現を可能にするが、一方酵母タンパク質分解に関わる酵素の発現を妨げる。その他の実施形態において、DNAは、染色体、プラスミド、レトロウイルスベクター経由で、または宿主ゲノムにランダムに組込むことによって、宿主に導入することができる。
【0060】
本発明の別の態様において、本発明の宿主細胞内で発現した少なくとも1つの異種ポリペプチドは少なくとも1つのGLP−1アゴニストを含む。ある態様では、GLP−1アゴニストは、以下の群から選択される:インクレチンホルモンならびに/またはその断片、変異体および/もしくは抱合体、インクレチンミメティックならびに/またはその断片、変異体および/もしくは抱合体。いくつかの態様において、少なくとも1つの異種ポリペプチドは少なくとも1つのGLP−1活性を有する。
【0061】
GLP−1活性を有するポリペプチドは、ヒトGLP−1の少なくとも1つの断片および/または変異体を含み得る。ヒトGLP−1の天然に生じた2つの断片を配列番号2に示す。
7 8 9 10 11 12 13 14 15 16 17
His-Ala-Glu-Gly-Thr-Phe-Thr-Ser-Asp-Val-Ser-
18 19 20 21 22 23 24 25 26 27 28
Ser-Tyr-Leu-Glu-Gly-Gln-Ala-Ala-Lys-Glu-Phe-
29 30 31 32 33 34 35 36 37
Ile-Ala-Trp-Leu-Val-Lys-Gly-Arg-Xaa (配列番号2)
ここで37位のXaaはGly(以下、「GLP−1(7−37)」として示される)、または−NH2(以下、「GLP−1(7−36)」として示される)である。GLP−1断片としては、ヒトGLP−1のアミノ酸7〜36位(GLP−1(7−36))を含む、あるいはそれで構成されるGLP−1分子を挙げ得るが、これらに限定されない。GLP−1の変異体またはその断片としては、野生型GLP−1または配列番号2に示されるGLP−1の天然に生じた断片における、1つ、2つ、3つ、4つ、5つまたはそれ以上のアミノ酸置換を挙げ得るが、これらに限定されない。変異体GLP−1またはGLP−1の断片としては、野生型GLP−1のアラニン8に類似のアラニン残基の置換であり、かかるアラニンがグリシンに変異している(以下、「A8G」として示される)(例えば、全体を参照として本明細書に援用する米国特許第5,545,618号に開示の突然変異体を参照されたい)場合を挙げ得るが、これらに限定されない。
【0062】
別の態様において、GLP−1活性を有する少なくとも1つのポリペプチドは、ヒト血清アルブミンと融合したヒトGLP−1の少なくとも1つの断片および/または変異体を含む。別の態様では、GLP−1の少なくとも1つの断片および変異体はGLP−1(7−36(A8G))を含む。GLP−1の少なくとも1つの断片および変異体は、ヒト血清アルブミンと遺伝的に融合する。別の態様において、本発明の異種ポリペプチドは、少なくとも2個のGLP−1(7−36(A8G))を直列に含み、かつ、遺伝的にヒト血清アルブミンと融合する。2個のGLP−1(7−36(A8G))は、ヒト血清アルブミンのN末端で遺伝的に融合する。いくつかの場合には、異種ポリペプチドは配列番号1を含む。
HGEGTFTSDVSSYLEGQAAKEFIAWLVKGRHGEGTFTSDVSSYLEGQAAKEFIAWLVKGR 60
DAHKSEVAHRFKDLGEENFKALVLIAFAQYLQQCPFEDHVKLVNEVTEFAKTCVADESAE 120
NCDKSLHTL FGDKLCTVATLRETYGEMADCCAKQEPERNECFLQHKDDNPNLPRLVRPE 180
VDVMCTAFHDNEETFLKKYLYEIARRHPYFYAPELLFFAKRYKAAFTECCQAADKAACLL 240
PKLDELRDEGKASSAKQRLKCASLQKFGERAFKAWAVARLSQRFPKAEFAEVSKLVTDLT 300
KVHTECCHGDLLECADDRADLAKYICENQDSISSKLKECCEKPLLEKSHCIAEVENDEMP 360
ADLPSLAADFVESKDVCKNYAEAKDVFLGMFLYEYARRHPDYSVVLLLRLAKTYETTLEK 420
CCAAADPHECYAKVFDEFKPLVEEPQNLIKQNCELFEQLGEYKFQNALLVRYTKKVPQVS 480
TPTLVEVSRNLGKVGSKCCKHPEAKRMPCAEDYLSVVLNQLCVLHEKTPVSDRVTKCCTE 540
SLVNRRPCFSALEVDETYVPKEFNAETFTFHADICTLSEKERQIKKQTALVELVKHKPKA 600
TKEQLKAVMDDFAAFVEKCCKADDKETCFAEEGKKLVAASQAALGL 674
(配列番号1)
【0063】
さらに別の実施形態では、本発明の宿主細胞で発現した少なくとも1つの異種ポリペプチドは、少なくとも1つの抗原結合タンパク質、少なくとも1つの単一可変ドメイン、および/または少なくとも1つのドメイン抗体のうち1つ以上を含む。少なくとも1つの抗原結合ドメインを含むポリペプチドはまた、少なくとも1つのポリペプチドおよび/またはペプチド受容体アゴニストおよび/または拮抗物質を含む。いくつかの場合には、ポリペプチドアゴニストは、GLP―1受容体アゴニストであっても良い。当業者に理解されているように、2個以上の異種ポリペプチドが、同じ細胞内で発現する。例えば、GLP−1活性を有する異種ポリペプチドは、抗原結合タンパク質と同じ細胞内に発現させることができる。GLP−1活性を有する異種ポリペプチドは、発現に必要な核酸配列に動作可能に連結した抗原結合タンパク質と同じポリヌクレオチドから発現させても良い。あるいは、例えば、GLP−1活性を有するポリペプチドは、発現に必要な異なるポリヌクレオチド配列と動作可能に連結した同じエピソームのDNAまたはゲノムから、または別々のベクター上に位置するDNA配列から、抗原結合タンパク質などの第2の異種ポリペプチドとは独立に、発現させても良い。
【0064】
さらに、次のような遺伝子改変ピキア株も提供される:野生型ピキアと比較すると上記系統の上記少なくとも1つの異種ポリペプチドがタンパク質分解を低下させている。さらに、本発明の遺伝子改変ピキアは、野生型ピキアと比較すると、上記系統の上記少なくとも1つの異種ポリペプチドが糖鎖形成を低下させているか糖鎖形成をしないことを示す。
【0065】
別の態様では、本発明の遺伝子改変ピキアにおいて、上記異種ポリペプチドを発現させるステップを含む異種ポリペプチドの産生方法が提供される。さらに、本発明の遺伝子改変ピキアの任意の1つで産生する異種ポリペプチドも提供される。
【0066】
さらに本発明の別の態様では、配列番号3、5、および7と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%配列同一性を有する単離したポリヌクレオチドが提供される。さらに本発明の別の態様では、配列番号4、6、および8と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%配列同一性を有する単離したポリヌクレオチドが提供される。さらに実施形態は、遺伝子改変宿主細胞の野生型親が、配列番号4、6、8と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%の配列同一性を有するポリペプチドをコードする遺伝子を含む、遺伝子改変宿主細胞を含む。ここで上記遺伝子は、野生型宿主細胞と比較すると、上記遺伝子改変宿主細胞内で、上記遺伝子産物が低下するか除去されるように、上記宿主細胞のゲノム内で遺伝子改変される。さらに、野生型親が配列番号3、5、および7と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%の配列同一性を有する遺伝子を含み、上記遺伝子が、上記遺伝子改変宿主細胞のゲノム内で遺伝子改変されている、遺伝子改変宿主細胞も提供される。本発明の遺伝子改変宿主細胞は、上記野生型親宿主細胞と比較すると、プロテアーゼ活性が低下しているか除去されている。本発明の遺伝子改変宿主細胞は、メチロトローフ酵母プロテアーゼ(Ymp1)活性が低下しているか除去されている。いくつかの場合、宿主細胞はピキアである。
【0067】
さらに本発明の別の態様では、配列番号9、11および13と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%配列同一性を有する単離したポリヌクレオチドが提供される。さらに本発明の別の態様では、配列番号10、12および14と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%配列同一性を有する単離したポリヌクレオチドが提供される。
【0068】
当業者に理解されているように、培養下の宿主細胞によって産生された1以上の酵素の酵素活性は、培養の増殖条件によって影響されることがある。例えば、培養液中のの宿主細胞により産生されるプロテアーゼのタンパク分解活性は、下記の1以上の条件を変化させることによって低下させることができるだろう。条件:pH、溶存酸素、温度、モル浸透圧濃度、1以上の培地成分、特異的プロテアーゼ阻害薬、増殖時間および/または割合、細胞濃度、培養持続時間、および/またはグルコース摂食割合(例えば、流加培養)。タンパク水解物複合体を培養に加えると、タンパク質分解の抑制が極めて効率的である。さらに、最大の効果を得るために、条件を培養中1以上の特定時間に変化させてもよい。同様に、培養で産生されるタンパク質の糖鎖形成も、同様な因子によって影響されることがある。したがって、培養中、タンパク分解活性や糖鎖形成活性などの宿主細胞の酵素活性を低下させる増殖条件は、上記にリストされた1以上の非限定要因を調整することにより最適化することができる。
【0069】
さらに、当業界で理解されているように、宿主細胞の異種タンパク質の産生は、上記の同じ因子の多くを制御することによって増加させることができる。さらに加えて、限定されないが、ラパマイシンの増殖培地への付加を含め、ベクターのコピー数を増加させる因子を加えることによって、産生を増加させることもできる。産生を増加させることのできる他の因子には、限定されないが、タンパク質ジスルフィド異性化酵素(PDI)などの1以上のシャペロンタンパク質の同時発現も含まれる。さらに、ヘモグロビン(HB)は、宿主細胞の少なくとも1つの異種ポリペプチドと同時発現させることができ、酸化的代謝に必要な酸素の利用性を高め、従ってポリペプチド産生を増加させる。
【0070】
産生中に宿主細胞から分泌された異種タンパク質は、分泌を促進するリーダー配列を含んでも良い。リーダー配列は、分泌を改善させることにより異種的に発現したタンパク質の全体的な産生と回収を向上させるように、修飾してもよい。例えば、多様な分泌タンパク質由来の異なるリーダー配列を異種タンパク質に動作可能に連結し、発現改善について評価してもよい。あるいは、所定のリーダー配列は、改善されたリーダー配列変異体の同定のため、部位特異的変異誘発、または、コンビナトリアルライブラリのアプローチ手段によって修飾してもよい。異種タンパク質発現レベルを改善するため、2個以上のリーダーペプチドの領域を含むキメラのリーダー配列をを見つけてもよい。
【0071】
実施例
以下の実施例はこの発明の様々な非限定的態様を説明する。
【0072】
実施例1−Ymp1新規なプロテアーゼ
新規なプロテアーゼを親和性精製およびLC/MS分析を使ってピキア培養上清から同定した。図1は、ゲル上に同定された新規なピキアメチロトローフ酵母プロテアーゼ(Ymp1)を表す図である。B378(ピキアヌル系統)からの浄化ろ過された上清を、ビオチン標識したアプロチニンで一晩インキュベートした。上清を2mLアビジンカラム(ピアス)に供し、リン酸緩衝液で洗浄した。カラムを、0.1Mトリス、2M塩化ナトリウム、pH7.5、次に0.1M酢酸ナトリウム、2M塩化ナトリウム、pH4.5そして最終的にビオチン溶出緩衝液(ピアス)で経時的に溶出させた。溶出液を濃縮して、SDS−PAGEを行った。もっとも顕著なバンドであるバンド1と2をゲルから切り出し、還元し、アルキル化し、in situでトリプシンを使用して消化した。バンドからのトリプシンペプチドを液体クロマトグラフィー・タンデム質量分析で分析した。未解釈の配列データを、Mascotタンパク質同定ソフトウェアを使ってピキアコンティグ−ペプチドデータベース(Integrated Genetics)内で検索した。
【0073】
バンド1と2の両方から産生したペプチド類は、ピキアゲノムデータベース内のある翻訳領域(ORF)と整合した。遺伝子を単離して配列を求めると、1704アミノ酸の新規なタンパク質(配列IDNO:4)をコードする5115bp核酸(配列番号3)からなる翻訳領域を含んでいた。タンパク質全体の予想した分子量は、185.8KDである。ピキア(配列番号3に示される)からのこの遺伝子のポリヌクレオチド配列は、GenBankのどの配列とも相同性を有意には示さない。コード化したアミノ酸配列は、2個の構造領域含む:N末端領域(AA1−865)は、ボタンタケ属lixiiセリンエンドペプチダーゼと30.2%の同一性を共有し、C末端領域(AA1305−1449)は、乳酸桿菌ロイテリからのLPXTG−モチーフ細胞壁アンカー領域タンパク質と48%の同一性を共有する。しかしながら、配列番号4とヒポクレアlixiiセリンエンドペプチダーゼとの全長比較は、14.8%の同一性を示したに過ぎない。ここで2つの配列のグローバルアラインメントを計算するWilliam Pearson’s LALIGNプログラムバージョン2.2u(Myers and Miller、CABIOS(1989)4:11−17)を使用した。このプロテアーゼの予想された構造を図2に示す。そのC末端構造と配列は、全ての種の全てのセリンプロテアーゼの中でユニークである。遺伝子は、メチロトローフピキア培養に出現するので、酵母メチロトローフプロテアーゼ(YMP1)と名付けられた。これは、そのユニークな構造にもとづくプロテアーゼの新規なクラスである。Ymp1は、LC/質量分析(データを示さず)を使ったピキア培養上清のグローバル解析に基づくピキアメチロトローフ培地でもっとも大量に分泌するタンパク質の一つである。
【0074】
ATGTTCCTCA AAAGTCTCCT TAGTTTTGCG TCTATCCTAA CGCTTTGCAA GGCCTGGGAT
CTGGAAGATG TACAAGATGC ACCAAAGATC AAAGGTAATG AAGTACCCGG TCGCTATATC
ATTGAGTATG AAGAAGCTTC CACTTCAGCA TTTGCTACCC AACTGAGAGC TGGGGGATAT
GACTTTAACA TCCAATACGA CTACTCAACT GGTTCCCTTT TCAACGGAGC ATCTGTTCAA
ATCAGCAACG ATAACAAAAC CACTTTCCAG GATTTGCAAA GTTTGCGTGC AGTCAAAAAT
GTTTACCCAG CTACTCTCAT TACATTAGAT GAAACATTTG AGCTTGCTGA CACGAAGCCA
TGGAACCCTC ATGGAATTAC CGGTGTCGAT TCTTTGCATG AGCAAGGATA TACTGGTAGT
GGTGTTGTTA TTGCAGTTAT CGATACTGGT GTTGACTATA CACACCCTGC TCTGGGTGGT
GGTATCGGAG ATAATTTCCC TATCAAAGCT GGTTATGATT TGTCTTCCGG TGATGGTGTC
ATCACGAATG ATCCTATGGA TTGTGACGGT CATGGTACCT TTGTATCCTC CATCATTGTT
GCAAATAACA AAGATATGGT TGGTGTTGCA CCAGATGCTC AGATTGTCAT GTACAAAGTG
TTCCCCTGTT CTGATAGTAC TTCGACTGAC ATAGTTATGG CGGGTATGCA AAAGGCCTAT
GATGATGGTC ACAAGATTAT TTCGCTATCA CTGGGATCTG ACTCGGGGTT TTCCAGTACT
CCAGCTTCCT TAATGGCCAG CAGGATTGCT CAAGACAGAG TTGTTTTGGT GGCTGCTGGT
AACTCTGGAG AACTTGGTCC ATTCTATGCC TCCTCCCCTG CTTCTGGGAA ACAAGTCATT
TCAGTTGGAT CTGTTCAAAA CGAACAATGG ACAACCTTTC CAGTAACCTT TACCTCTTCA
AACGGTGAAT CAAGGGTTTT TCCTTACCTC GCTTACAATG GTGCACAGAT TGGATTTGAT
GCCGAGCTTG AGGTTGATTT TACCGAAGAA AGAGGATGCG TCTATGAACC AGAGATCTCC
GCAGATAATG CGAATAAAGC TATTTTGTTA AGAAGGGGCG TCGGCTGTGT TGAAAACTTG
GAATTCAATT TATTGTCTGT GGCTGGTTAC AAGGCTTACT TCTTGTACAA CTCATTTTCA
AGACCATGGA GTCTCTTGAA TATTTCTCCA CTGATTGAGC TAGACAACGC TTACTCTCTT
GTTGAAGAGG AAGTTGGAAT ATGGGTGAAA ACCCAAATCG ACGCCGGTAA CACCGTCAAG
TTAAAGGTGA GCACGAGTGA CCAAATGTTG CCATCTGATA AAGAGTATTT GGGAGTTGGA
AAGATGGATT ATTACTCCTC TCAAGGACCT GCTTATGAGC TTGAATTTTT CCCAACGATA
TCCGCTCCAG GTGGAGACAG TTGGGGCGCT TGGCCCGGTG GGCAATACGG TGTTGCCTCA
GGAACAAGTT TTGCTTGCCC CTATGTTGCA GGTCTTACAG CTCTTTATGA ATCGCAGTTT
GGAATTCAAG ATCCCCAGGA CTATGTGAGA AAATTAGTCT CCACAGCTAC CGATCTTCAA
TTATTTGACT GGAACGCAGT GAAACTTGAG ACCTCTATGA ATGCTCCACT TATTCAACAG
GGAGCTGGTC TAGTGAACGC TCTTGGTTTG TTTGAGACTA AGACTGTGAT CGTGTCTGCT
CCTTATTTGG AGCTCAATGA CACCATCAAT AGAGCCAGTG AGTATACCAT TCAAATTAAG
AATGAGAACT CTGAGACTAT TACCTATCAA GTTGTTCACG TTCCGGGAAC TACTGTCTAC
TCTAGATCAG CTTCTGGGAA CATCCCATAC CTGGTCAATC AAGATTTTGC ACCTTACGGT
GATAGTGATG CTGCGACAGT TGCTCTATCC ACAGAAGAGT TGGTTTTGGG ACCAGGAGAA
GTTGGTGAAG TCACTGTGAT CTTCTCTACA GAAGAAATTG ATCAAGAAAC TGCTCCAATT
ATTCAGGGTA AGATTACATT TTATGGTGAT GTCATACCGA TTGCTGTTCC TTATATGGGA
GTTGAAGTTG ATATTCATTC CTGGGAGCCT CTCATTGAGA GGCCTTTATC AGTGAGAATG
TATTTGGATG ATGGTTCCTT AGCATATGTT GATGATGATC CTGATTATGA GTTCAATGTG
TATGACTGGG ATTCTCCTAG ATTTTATTTT AACCTGAGAT ATGCAACCAA AGAAGTATCG
ATTGACTTGG TGCACCCTGA TTATAGCATT GAGAACGACT ACGAATGGCC TTTAGTTTCC
GGACACAACA ACTATTATGG TCCCGTGGGA TACGACTACG ATTATACCTC GGGTCAAGCC
TTTTTGCCTC GTTACTTTCA ACAACGTATT AACGAACTTG GATATCTTTC TTTTTCCAGA
TTTGCTAACT TTTCTGTAGT TCCTGCTGGT GAATACAAAG CTCTATTTAG AGTTTTGCTA
CCATATGGAG ACTTTTGGAA CAAAGAAGAC TGGCAATTGT TTGAATCCCC AGTGTTTAAC
GTCCTCGCTC CACCGAATGA AGAAAACACT ACTGAAGAGC CAACTGAGGA ATCCAGCGAG
GAGCCTACCG AAGAGTCAAC GTCTGAGTCA ACTGAAGAGC CCTCTTCTGA GTCAACTGAG
AAATCTAGCG AGGTGCCAAC TGAAGAAATT ACTGAAGATG CAACATCCAC AATTGATGAT
GATGAAGCAT CCACCGAAAG CTCTACTGAA GAACCAAGTG CTCAGCCCAC CGGTCCTTAC
TCTGATTTGA CTGTCGGTGA GGCCATTACC GACGTTAGTG TCACCAGTTT GAGGACAACT
GAAGCATTTG GATACACTTC CGACTGGTTG GTTGTGTCTT TCACTTTCAA CACTACTGAC
AGAGATATTA CTCTCCCACC TTACGCTGTT GTACAAGTAA CTATCCCAAA TGAACTTCAA
TTCATTGCTC ATCCAGAATA CGCCCCATAC CTTGAGCCCT CATTGCAAGT TTTCTACACT
AAGAATGAAA GATTAATTAT GACTAGTCAG TTCAACTACG ACACCAGAGT CATCGACTTC
AAGTTTGACA ATCGAGACCA AGTAATAACT CAAGTGGAGG GAGTTGTTTA TTTCACGATG
AAACTAGAAC AAGATTTCAT TTCTGCATTG GCCCCAGGTG AATACGATTT TGAATTTCAT
ACATCCGTTG ATTCTTATGC TTCGACCTTT GACTTTATTC CATTGATTAG ATCCGAGCCA
ATCAAATTGA TAGCAGGTGC ACCAGACGAA GTTGAATGGT TTATTGATAT TCCAAGTGCA
TACAGCGATT TGGCAACGAT AGATATTAGT TCTGATATCG ATACTAATGA TAATTTGCAG
CAGTACTTCT ATGATTGCTC AAAGCTCAAG TACACTATTG GAAAAGAGTT TGATCAGTGG
GGTAATTTTA CAGCTGGATC AGATGGTAAC CAATACAGCA ATACCACCGA TGGGTATGTT
CCAATTACTG ATTCTACCGG CTCTCCAGTA GCTGAAGTTC AATGTTTAAT GGAAAGTATC
TCATTGAGTT TCACAAATAC TCTTGCTGAG GATGAAGTAT TGAGAGTTGT TCTTCACTCT
TCTGCGTTTA GACGTGGTTC ATTCACCATG GCCAACGTGG TAAACGTTGA CATTACAGCT
GGTGGATTGG CAAAAAGAGA ACTCTTCTCT TATATATTGG ATGAAAATTA CTATGCTAGT
ACTGGATCTG AGGGGTTGGC ATTTGACGTA TTTGAAGTTG CTGATCAGGT CGAGGAGCCA
ACTGAGGAGT CAACCTCAGA GGAATCTACT GAACAGGAAA CTTCCACCGA GGAACCTACC
GAGGAATCAA CTGAACCTAC TGAGGAATCT ACCCAGGAAC CTACTGAAGA GCCCACCGAC
GAGCCTACTT CTGAGTCAAC TGAGGAACCT TCTGAGGAGC CAACTTCTGA CGATCTCTCA
ATTGACCCAA CTGCTGTACC TACCGATGAA CCTACTGAAG AGCCAACTGA GGAGCCTACT
TCTGAGTCAA CTGAGGAACC TTCTGAGGAG CCAACTTCTG ACGATCTCTC AATTGACCCA
ACTGCTGTAC CTACCGATGA ACCTACTGAA GAGCCAACTG AGGAGCCTAC TTCTGAGTCA
ACTGAGGAAC CTTCTGAGGA GCCAACTTCT GACGATCTCT CAATTGACCC AACTGCTGTA
CCTACCGATG AACCTACTGA AGAGCCAACT GAGGAGCCGA CCTCTGAGAC TACCGATGAT
CCATCGATAG CACCTACTGC TGTGCCAACT TCCGACACAT CTTCTGGACA ATCGGTGGTT
ACTCAAAACA CTACAGTCAC TCAGACTACC ATCACTTCAG TCTGTAATGT TTGTGCTGAG
ACCCCTGTAA CAATCACTTA CACTGCACCA GTTGTGACTA AGCCAGTTTC TTACACCACC
GTTACTTCAG TTTGCCATGT ATGTGCAGAG ACACCAATCA CAGTTACCTT GACGTTGCCA
TGTGAAACCG AAGACGTGAC AAAGACTGCC GGCCCTAAGA CTGTCACTTA CACCGAAGTT
TGCAACTCCT GTGCTGACAA GCCTATCACT TACACCTACA TCGCTCCAGA GTACACTCAA
GGTGCCGAAC GTACAACAGT TACATCGGTT TGCAACGTTT GTGCTGAGAC ACCTGTAACG
CTAACATACA CTGCGCCGAA AGCCAGTCGT CATACAGTTC CTTCACAATA TTCAAGTGCC
GGAGAGCTCA TTTCATCCAA GGGGATCACG ATTCCTACTG TTCCTGCCCG TCCAACTGGT
ACTTATAGTA AGTCTGTTGA CACTAGCCAA CGTACACTCG CTACCATTAC AAAATCTTCA
GATGAGTCTA ACACTGTTAC CACTACTCAA GCCACACAAG TTTTGAGCGG TGAATCCAGT
GGAATTCAAG CTGCTTCAAA CAGCACGAGC ATCTCAGCTC CAACTGTCAC TACAGCTGGG
AACGAGAACT CTGGATCTAG ATTTTCGTTT GCTGGACTAT TCACAGTTCT GCCTCTTATC
TTGTTCGTTA TATAA(配列番号3)
【0075】
配列番号3の遺伝子配列によってコードしたポリペプチド配列は、配列番号4として以下に示される。
MFLKSLLSFA SILTLCKAWD LEDVQDAPKI KGNEVPGRYI IEYEEASTSA FATQLRAGGY
DFNIQYDYST GSLFNGASVQ ISNDNKTTFQ DLQSLRAVKN VYPATLITLD ETFELADTKP
WNPHGITGVD SLHEQGYTGS GVVIAVIDTG VDYTHPALGG GIGDNFPIKA GYDLSSGDGV
ITNDPMDCDG HGTFVSSIIV ANNKDMVGVA PDAQIVMYKV FPCSDSTSTD IVMAGMQKAY
DDGHKIISLS LGSDSGFSST PASLMASRIA QDRVVLVAAG NSGELGPFYA SSPASGKQVI
SVGSVQNEQW TTFPVTFTSS NGESRVFPYL AYNGAQIGFD AELEVDFTEE RGCVYEPEIS
ADNANKAILL RRGVGCVENL EFNLLSVAGY KAYFLYNSFS RPWSLLNISP LIELDNAYSL
VEEEVGIWVK TQIDAGNTVK LKVSTSDQML PSDKEYLGVG KMDYYSSQGP AYELEFFPTI
SAPGGDSWGA WPGGQYGVAS GTSFACPYVA GLTALYESQF GIQDPQDYVR KLVSTATDLQ
LFDWNAVKLE TSMNAPLIQQ GAGLVNALGL FETKTVIVSA PYLELNDTIN RASEYTIQIK
NENSETITYQ VVHVPGTTVY SRSASGNIPY LVNQDFAPYG DSDAATVALS TEELVLGPGE
VGEVTVIFST EEIDQETAPI IQGKITFYGD VIPIAVPYMG VEVDIHSWEP LIERPLSVRM
YLDDGSLAYV DDDPDYEFNV YDWDSPRFYF NLRYATKEVS IDLVHPDYSI ENDYEWPLVS
GHNNYYGPVG YDYDYTSGQA FLPRYFQQRI NELGYLSFSR FANFSVVPAG EYKALFRVLL
PYGDFWNKED WQLFESPVFN VLAPPNEENT TEEPTEESSE EPTEESTSES TEEPSSESTE
KSSEVPTEEI TEDATSTIDD DEASTESSTE EPSAQPTGPY SDLTVGEAIT DVSVTSLRTT
EAFGYTSDWL VVSFTFNTTD RDITLPPYAV VQVTIPNELQ FIAHPEYAPY LEPSLQVFYT
KNERLIMTSQ FNYDTRVIDF KFDNRDQVIT QVEGVVYFTM KLEQDFISAL APGEYDFEFH
TSVDSYASTF DFIPLIRSEP IKLIAGAPDE VEWFIDIPSA YSDLATIDIS SDIDTNDNLQ
QYFYDCSKLK YTIGKEFDQW GNFTAGSDGN QYSNTTDGYV PITDSTGSPV AEVQCLMESI
SLSFTNTLAE DEVLRVVLHS SAFRRGSFTM ANVVNVDITA GGLAKRELFS YILDENYYAS
TGSEGLAFDV FEVADQVEEP TEESTSEEST EQETSTEEPT EESTEPTEES TQEPTEEPTD
EPTSESTEEP SEEPTSDDLS IDPTAVPTDE PTEEPTEEPT SESTEEPSEE TSDDLSIDPT
AVPTDEPTEE PTEEPTSEST EEPSEEPTSD DLSIDPTAVP TDEPTEEPTE EPTSETTDDP
SIAPTAVPTS DTSSGQSVVT QNTTVTQTTI TSVCNVCAET PVTITYTAPV VTKPVSYTTV
TSVCHVCAET PITVTLTLPC ETEDVTKTAG PKTVTYTEVC NSCADKPITY TYIAPEYTQG
AERTTVTSVC NVCAETPVTL TYTAPKASRH TVPSQYSSAG ELISSKGITI PTVPARPTGT
YSKSVDTSQR TLATITKSSD ESNTVTTTQA TQVLSGESSG IQAASNSTSI SAPTVTTAGN
ENSGSRFSFA GLFTVLPLIL FVI.(配列番号4)
【0076】
実施例2−新規なプロテアーゼ
ノックアウト系統(実施例3を参照)のピキアymp1の上澄みから、実施例1に記述したのと同様に、親和性精製とLC/MS分析を使ってさらに2つの新規なメチロトローフ酵母プロテアーゼ(Ymp2およびYmp3)を同定した。B580(ピキアymp1欠失系統)からの浄化ろ過された上清を、ビオチン標識したAEBSF(4−(2−アミノエチル)ベンゼンスルホニルフッ化物塩酸塩で一晩インキュベートした。上清を、10mLアビジンカラムに充填し(ピアス)、そしてリン酸緩衝液で洗浄した。カラムを、0.1Mトリス、2M塩化ナトリウム、pH7.5、次に0.1M酢酸ナトリウム、2M塩化ナトリウム、pH4.5、そして最終的にビオチン溶出緩衝液(ピアス)で経時的に溶出した。溶出物を8M尿素を加えて変性させ、20mMジチオスレイトール(DTT)と反応させてシステイン残基を還元し、次に、20mMヨードアセトアミドと反応させてアルキル化した。試料を100mMアンモニウム炭酸水素塩(pH8.5)で希釈して2M尿素とした後、タンパク質はトリプシンで消化した(Sequencing grade modified trypsin, Promega, Madison, WI)。消化の結果生じたペプチド類混合物をPepCleanスピンカラム(Pierce, Rockford, IL)を使って脱塩し、LC−MS/MS systemを使って分析した。このシステムでは、75μm内径逆相C18カラムを持つ高圧液体クロマトグラフィー(HPLC)が、オンラインでイオントラップ質量分析計(Thermo, Palo Alto, CA)と連結されていた。LC−MS/MS分析から得た質量分析データを使って、GenBankからの最近の非重複性タンパク質データベースとピキアデータベース(Integrated Genetics)内を、Mascotタンパク質同定ソフトウェアを使用して検索した。
【0077】
ペプチドの整合により、YMP2およびYMP3と名付けられた2つの遺伝子を同定し周知のプロテアーゼとの相同性を比べた。YMP2は、205アミノ酸の新規なタンパク質(配列番号6)をコードする618bp核酸(配列番号5)のオープンリーディングフレームを含む。タンパク質全体の予想される分子量は22.9KDである。ピキア(配列番号5に示す)からのこの遺伝子のポリヌクレオチド配列は、GenBankのいかなる配列とも相同性を有意に共有しない。しかしながら、コード化したアミノ酸配列は、ピキア・パストリスからのアルギニン/アラニンアミノペプチダーゼと29%の同一性を共有する。
ATGGCTCCCA GAACACTACC AGAAGACTTA ATTCCCTCCC TATACGACTT GCACATCTAC
AACTTCCAAC CCGAAAAAAA GACTTATGAT GGAGACATTG TCATCCACTT GGAGGTGAAG
GAGCCCACTG ATGAAGTGGT CTTCAATGCC AAGGATTTGG AATTGAAAGA CGTACATGTC
TTCCACAATG TCAACAAGTC TGAAAACGAA ATCCCCGTTA AGGAGATTGT TGATAACGAG
CTCATCACAA TTAAGCTCAA AGAGAAGGTT ACTTCCGGAA CGTTGCTGGT GAATATTTCC
TTCACCGGTA ACATTCAATC TGATAAAATT GGATTTTACA AGGGAGACAC AGATGTGGAA
GGAAGAGTCA CATACACTAC AAACCTTACC ACTCCAAATG CCAGGTTGGC ATTCCCATGT
CTTGATAACA TATTGTTGAA AGCTCCATTC AAGTTCGGAG TAACTGCCAA TCCAGGACAA
TTAGTGAGTT CCATTTTGGA TCTAAGCTCT GAGGCTGACG TCTTGAATGA CAATGACGAT
GTGATTGGTA CGAGATACCA ATACCAAGTG AGTGAGCCAA TAGCCCCAGC TTTACTGGAG
TGGACCATTC ATATTTAA (配列番号5)
【0078】
配列番号5の遺伝子配列によってコードしたポリペプチド配列は配列番号6として以下に示される。
MAPRTLPEDL IPSLYDLHIY NFQPEKKTYD GDIVIHLEVK EPTDEVVFNA KDLELKDVHV
FHNVNKSENE IPVKEIVDNE LITIKLKEKV TSGTLLVNIS FTGNIQSDKI GFYKGDTDVE
GRVTYTTNLT TPNARLAFPC LDNILLKAPF KFGVTANPGQ LVSSILDLSS EADVLNDNDD
VIGTRYQYQV SEPIAPALLE WTIHI (配列番号6)
【0079】
YMP3は、410のアミノ酸(配列番号8)の新規なタンパク質をコードする1233bp核酸(配列番号7)のオープンリーディングフレームを含む。タンパク質全体の予想される分子量は46.0KDである。このアミノ酸配列は、ピキアstipitisからのロイシンアミノペプチダーゼと47%の同一性を共有する。
ATGGTCAAAC TCATATCAAT TATAGCCCTA GTTCAACTTG TCTCTGCGAC AATTGTACCT
TGGAATCTCC AGAACGTCTT ATCTGACGTC CATCACCCTT CTCTCCATCT CTTGGATTAT
ATTCAATCCT TGAAGAACGA GGTAATGTTC GATGGCGACG ATCGCAGAAT AATCAAGTTA
GGCCCCCAAG AATACCGTAT TATCACTGAA AAAGAGAAAT ACCAGTTGAA AACAGAGGGG
ATATCATTTA TCGATGTCAC CTATCAGCAT GGAGACAATG TAGAGCTGCT CTATTCCAGT
GCGCCAGTTA CCGTTCCAGA CTATCTTTAT CCGTCCAATG ATACTTTCCA TTTCAAACAA
GTAAATTCTT TGATAGGTGA GATTGACATT GGCAGAATGC AGGCGTTTTT GGGAAGGTTC
TCTAGCTTCT TTACAAGATT TTACAAATCT GACAAGGGGT TGCAGAGTTC TATCTGGTTA
CAAGGTGAAT TGGTTCAATT GGCCTTGAAA GATCCATCGA GGTTCAATGT TACTACTGTG
GAACACCCTT GGAAGCAGAA TTCTGCCATC TTTACGATAT ACGGTGAAAA TGTTGATCCT
TCGAAAGGAA AAGGGGACAT TGTAGTAGTG GGATGCCATC AAGATTCCAT AAACTTGCTT
TTCCCCAACA TTCTCCGTGC TCCAGGGGCT GATGATGATG GATCTGGTGT AACTTCCAAC
CTTGAAGCGC TCAGAATCAT AGTTGAAAGT GGCCTCAAGT TTCACAATAC AGTAGAGTTT
CACTTTTATT CTGCCGAAGA AGGAGGACTA CTTGGCTCCC AGCAAATTTT CAGCTCGTAT
AGAGCTGCAG AAGAGACTGT TGTTGCTATG CTACAACAGG ACATGACTGG ATACATCCAA
AAAGCTTTAG ACCACGGGGA ATCCGACCAC TTCGGGCTAA TCACTGACCA TACAAACGCA
AATCTGAATA GCTTCCTTGC ACTTTTAATC GATGCATACA CTTCAATTCC CTACAAAGAA
ACCGAATGTG GGTATGCCTG CTCAGATCAT AGTTCTGCCT TGGAACATGG TTATCCATCT
GCCATGGTCT TTGAAAGTAG TTTTGCCTAC ACAAATCCCT TCATCCATAG CACCCAAGAC
ACAATTGACA AGATCAATTT TCCACATATG GCAGAGCAT GTCAAGTTGG TCCTGGGTTA
CGTTGTAGAG TTGGGATTAG AACATTTTAG GTGA (配列番号7)
【0080】
配列番号7の遺伝子配列によってコードしたポリペプチド配列は配列番号8として以下に示される。
MVKLISIIAL VQLVSATIVP WNLQNVLSDV HHPSLHLLDY IQSLKNEVMF DGDDRRIIKL
GPQEYRIITE KEKYQLKTEG ISFIDVTYQH GDNVELLYSS APVTVPDYLY PSNDTFHFKQ
VNSLIGEIDI GRMQAFLGRF SSFFTRFYKS DKGLQSSIWL QGELVQLALK DPSRFNVTTV
EHPWKQNSAI FTIYGENVDP SKGKGDIVVV GCHQDSINLL FPNILRAPGA DDDGSGVTSN
LEALRIIVES GLKFHNTVEF HFYSAEEGGL LGSQQIFSSY RAAEETVVAM LQQDMTGYIQ
KALDHGESDH FGLITDHTNA NLNSFLALLI DAYTSIPYKE TECGYACSDH SSALEHGYPS
AMVFESSFAY TNPFIHSTQD TIDKINFPHM AEHVKLVLGY VVELGLEHFR (配列番号8)
【0081】
さらに、3個の他の潜在的プロテアーゼが、全て周知の酵母プロテアーゼを使って、ピキアデータベース内で同定された。これらのプロテアーゼおよび対応するそれぞれのポリペプチド配列をコードする同定されたポリヌクレオチド配列は、配列番号9〜14として以下に示す。
ATGAAATCGG TTATTTGGAG CCTTCTATCT TTGCTAGCAT TGTCGCAGGC ATTGACTATT
CCATTGCTGG AAGAGCTTCA ACAGCAAACA TTTTTTAGCA AGAAAACCGT TCCTCAACAA
GTTGCTGAAT TGGTGGGCAC CCATTACTCT AAGGATGAGA TAATCAGTCT ATGGAAGGAC
ATTGAGCTGG ATGTACCCAG GGAAAAGATC CAAGAGGCCT TCGATAAGTT CGTAAAACAA
TCAACTGCCA CTTCCCCCGT TAGAAATGAA TTTCCCTTGT CTCAGCAAGA TTGGGTGACA
GTGACCAACA CCAAGTTTGA TAATTATCAA TTGAGGGTTA AAAAATCCCA CCCTGAAAAG
CTAAACATTG ATAAGGTAAA GCAATCTTCG GGATACCTGG ATATCATTGA TCAAGATAAG
CATCTTTTCT ATTGGTTTTT TGAATCCCGA AATGATCCGT CCACAGACCC AATCATCCTA
TGGTTGAATG GTGGACCCGG CTGCTCTTCT ATTACAGGGT TGCTATTCGA AAAGATTGGC
CCCAGTTACA TCACCAAAGA GATTAAGCCG GAACATAATC CTTATTCATG GAACAACAAT
GCTAGTGTTA TCTTCCTTGA GCAACCGGTT GGAGTAGGAT TTTCTTACTC TTCTAAGAAA
GTCGGTGATA CTGCAACTGC TGCCAAAGAT ACATATGTGT TTTTGGAGCT TTTCTTCCAA
AAGTTTCCTC AGTTCCTGAC CTCTAATCTG CACATTGCTG GGGAATCGTA TGCTGGCCAT
TATTTGCCCA AGATTGCTTC TGAGATTGTG TCTCACGCAG ACAAGACGTT TGACCTTTCA
GGAGTCATGA TCGGTAATGG TCTTACTGAT CCTCTAATTC AGTATAAGTA CTATCAGCCA
ATGGCCTGTG GAAAAGGTGG CTACAAGCAG GTCATTTCGG ACGAGGAATG TGATGAATTG
GATAGGGTCT ATCCAAGATG TGAACGTTTA ACGCGGGCAT GTTATGAGTT CCAAAATTCA
GTTACTTGTG TTCCGGCAAC ACTTTATTGC GACCAAAAGC TACTGAAGCC GTACACTGAC
ACTGGCTTGA ATGTCTATGA TATTCGTACA ATGTGCGATG AAGGGACTGA TTTGTGTTAC
AAAGAACTGG AATACGTGGA GAAGTACATG AACCAGCCTG AAGTGCAGGA AGCCGTGGGC
TCTGAAGTCA GTTCTTACAA AGGTTGTGAC GATGATGTCT TCTTAAGATT TTTGTACTCT
GGCGATGGAT CTAAGCCTTT CCACCAGTAT ATCACGGATG TTCTCAATGC AAGTATTCCG
GTTCTGATTT ACGCAGGTGA TAAAGATTAT ATCTGTAATT GGCTAGGAAA CCAAGCTTGG
GTCAATGAGC TAGAATGGAA CTTGTCTGAG GAATTCCAGG CAACTCCGAT TCGACCGTGG
TTCACTTTGG ACAATAACGA TTATGCAGGA AACGTACAAA CTTATGGAAA CTTTTCCTTT
CTAAGAGTAT TTGATGCTGG TCACATGGTT CCTTACAATC AACCAGTCAA CGCACTTGAC
ATGGTTGTCA GATGGACACA CGGTGATTTC TCATTTGGTT ATTAA (配列番号:9)
MKSVIWSLLS LLALSQALTI PLLEELQQQT FFSKKTVPQQ VAELVGTHYS KDEIISLWKD
IELDVPREKI QEAFDKFVKQ STATSPVRNE FPLSQQDWVT VTNTKFDNYQ LRVKKSHPEK
LNIDKVKQSS GYLDIIDQDK HLFYWFFESR NDPSTDPIIL WLNGGPGCSS ITGLLFEKIG
PSYITKEIKP EHNPYSWNNN ASVIFLEQPV GVGFSYSSKK VGDTATAAKD TYVFLELFFQ
KFPQFLTSNL HIAGESYAGH YLPKIASEIV SHADKTFDLS GVMIGNGLTD PLIQYKYYQP
MACGKGGYKQ VISDEECDEL DRVYPRCERL TRACYEFQNS VTCVPATLYC DQKLLKPYTD
TGLNVYDIRT MCDEGTDLCY KELEYVEKYM NQPEVQEAVG SEVSSYKGCD DDVFLRFLYS
GDGSKPFHQY ITDVLNASIP VLIYAGDKDY ICNWLGNQAW VNELEWNLSE EFQATPIRPW
FTLDNNDYAG NVQTYGNFSF LRVFDAGHMV PYNQPVNALD MVVRWTHGDF SFGY (配列番号10)
ATGATATTAC ACACCTATAT TATTCTCTCG TTATTGACTA TATTTCCTAA AGCTATTGGT
CTGTCCTTGC AGATGCCAAT GGCCTTGGAA GCTAGTTATG CCTCATTAGT GGAGAAAGCA
ACCCTCGCTG TTGGACAAGA AATTGATGCC ATACAAAAGG GTATTCAGCA AGGTTGGTTG
GAAGTAGAGA CAAGATTTCC AACTATAGTG TCACAGTTAT CCTATAGTAC TGGCCCAAAA
TTTGCGATCA AGAAGAAAGA TGCAACTTTT TGGGATTTCT ATGTTGAAAG TCAAGAGTTG
CCAAACTACC GAATTAATGA AAATCTGAAA CCAATTTTCA ACCCCTATTC GTGGAATGGT
AATGCTTCAA TCATCTACTT AGATCAACCG GTCAATGTTG GGTTTTCTTA TTCTTCATCA
TCGGTGAGTA ACACTGTTGT TGCGGGAGAA GATGTGTATG CATTTCTTCA GCTTTTTTTT
CAACACTTCC CGGAATATCA AACTAATGAC TTTCATATTG CCGGTGAATC TTATGCAGGA
CATTACATTC CGGTGTTTGC AGACGAAATT TTGAGTCAAA AGAACAGAAA TTTCAATCTT
ACTTCAGTCT TGATCGGAAA TGGATTAACT GACCCTTTGA CTCAATACCG ATATTACGAG
CCAATGGCTT GTGGTGAAGG TGGTGCCCCG TCAGTACTGC CTGCCGATGA GTGCGAAAAT
ATGCTAGTTA CCCAAGATAA ATGTTTGTCT TTAATTCAAG CATGCTACGA CTCACAGTCG
GCATTCACAT GCGCACCGGC TGCCATTTAT TGTAATAACG CTCAGATGGG ACCCTATCAG
AGAACTGGGA AGAATGTGTA TGATATTCGT AAGGAATGTG ATGGTGGATC CTTGTGCTAT
AAGGACCTTG AATTCATCGA TACCTACTTA AATCAAAAGT TTGTTCAAGA TGCTTTGGGC
GCCGAGGTCG ATACCTATGA ATCTTGCAAT TTTGAAATCA ACAGAAACTT TTTATTTGCT
GGAGATTGGA TGAAACCTTA TCATGAACAT GTCAGCAGTC TCTTGAACAA AGGTTTGCCC
GTTTTGATTT ACGCAGGGGA CAAAGATTTC ATTTGCAACT GGTTGGGTAA TCGAGCATGG
ACTGATGTCT TGCCGTGGGT TGATGCTGAT GGTTTTGAAA AAGCCGAAGT CCAAGATTGG
TTGGTTAATG GAAGGAAGGC TGGTGAATTT AAGAACTATA GCAACTTCAC CTACCTAAGG
GTTTATGATG CTGGTCATAT GGCCCCATAT GATCAGCCAG AGAATTCTCA TGAAATGGTC
AATAGATGGA TATCCGGAGA CTTTAGCTTT CACTAG (配列番号11)
MILHTYIILS LLTIFPKAIG LSLQMPMALE ASYASLVEKA TLAVGQEIDA IQKGIQQGWL
EVETRFPTIV SQLSYSTGPK FAIKKKDATF WDFYVESQEL PNYRINENLK PIFNPYSWNG
NASIIYLDQP VNVGFSYSSS SVSNTVVAGE DVYAFLQLFF QHFPEYQTND FHIAGESYAG
HYIPVFADEI LSQKNRNFNL TSVLIGNGLT DPLTQYRYYE PMACGEGGAP SVLPADECEN
MLVTQDKCLS LIQACYDSQS AFTCAPAAIY CNNAQMGPYQ RTGKNVYDIR KECDGGSLCY
KDLEFIDTYL NQKFVQDALG AEVDTYESCN FEINRNFLFA GDWMKPYHEH VSSLLNKGLP
VLIYAGDKDF ICNWLGNRAW TDVLPWVDAD GFEKAEVQDW LVNGRKAGEF KNYSNFTYLR
VYDAGHMAPY DQPENSHEMV NRWISGDFSF H (配列番号12)
ATGCAATTGC GTCATTCCGT TGGATTGGCT ATCTTATCTG CCATAGCAGT CCAAGGATTG
CTAATTCCTA ACATTGAGTC ATTACCCAGC CAGTTTGGTG CTAATGGTGA CAGTGAACAA
GGTGTATTAG CCCACCATGG TAAACATCCT AAAGTTGATA TGGCTCACCA TGGAAAGCAT
CCTAAAATCG CTAAGGATTC CAAGGGACAC CCTAAGCTTT GCCCTGAAGC TTTGAAGAAG
ATGAAAGAAG GCCACCCTTC GGCTCCAGTC ATTACTACCC ATTCCGCTTC TAAAAACTTA
ATCCCTTACT CTTATATTAT AGTCTTCAAG AAGGGTGTCA CTTCAGAGGA TATCGACTTC
CACCGTGACC TTATCTCCAC TCTTCATGAA GAGTCTGTGA GCAAATTAAG AGAGTCAGAT
CCAAATCACT CATTTTTCGT TTCTAATGAG AATGGCGAAA CAGGTTACAC CGGTGACTTC
TCCGTTGGTG ACTTGCTCAA GGGTTACACC GGATACTTCA CGGATGACAC TTTAGAGCTT
ATCAGTAAGC ATCCAGCAGT TGCTTTCATT GAAAGGGATT CGAGAGTATT TGCCACCGAT
TTTGAAACTC AAAACGGTGC TCCTTGGGGT TTGGCCAGAG TCTCTCACAG AAAGCCTCTT
TCCCTAGGCA GCTTCAACAA GTACTTATAT GATGGAGCTG GTGGTGAAGG TGTTACTTCC
TATGTTATCG ATACAGGTAT CCACGTCACT CACAAAGAAT TCCAGGGTAG AGCATCTTGG
GGTAAGACCA TTCCAGCTGG AGACGTTGAT GACGATGGAA ACGGTCACGG AACTCACTGT
GCTGGTACCA TTGCTTCTGA AAGCTACGGT GTTGCCAAGA AGGCTAATGT TGTTGCCATC
AAGGTCTTGA GATCTAATGG TTCTGGTTCG ATGTCAGATG TTCTGAAGGG TGTTGAGTAT
GCCACCCAAT CCCACTTGGA TGCTGTTAAA AAGGGCAACA AGAAATTTAA GGGCTCTACC
GCTAACATGT CACTGGGTGG TGGTAAATCT CCTGCTTTGG ACCTTGCAGT CAATGCTGCT
GTTAAGAATG GTATTCACTT TGCCGTTGCA GCAGGTAACG AAAACCAAGA TGCTTGTAAC
ACCTCGCCAG CAGCTGCTGA GAATGCCATC ACCGTCGGTG CATCAACCTT ATCAGACGCT
AGAGCTTACT TTTCTAACTA CGGTAAATGT GTTGACATTT TCGCTCCAGG TTTAAACATT
CTTTCTACCT ACACTGGTTC GGATGACGCA ACTGCTACCT TGTCTGGTAC TTCAATGGCC
TCTCCTCACA TTGCTGGTCT GTTGACTTAC TTCCTATCAT TGCAGCCTGC TGCTGGATCT
CTGTACTCTA ACGGAGGATC TGAGGGTGTC ACACCTGCTC AATTGAAAAA GAACCTCCTC
AAGTATGCAT CTGTCGGAGT ATTAGAGGAT GTTCCAGAAG ACACTCCAAA CCTCTTGGTT
TACAATGGTG GTGGACAAAA CCTTTCTTCT TTCTGGGGAA AGGAGACAGA AGACAATGTT
GCTTCCTCCG ACGATACTGG TGAGTTTCAC TCTTTTGTGA ACAAGCTTGA ATCAGCTGTT
GAAAACTTGG CCCAAGAGTT TGCACATTCA GTGAAGGAGC TGGCTTCTGA ACTTATTTAG
(配列番号13)
MQLRHSVGLA ILSAIAVQGL LIPNIESLPS QFGANGDSEQ GVLAHHGKHP KVDMAHHGKH
PKIAKDSKGH PKLCPEALKK MKEGHPSAPV ITTHSASKNL IPYSYIIVFK KGVTSEDIDF
HRDLISTLHE ESVSKLRESD PNHSFFVSNE NGETGYTGDF SVGDLLKGYT GYFTDDTLEL
ISKHPAVAFI ERDSRVFATD FETQNGAPWG LARVSHRKPL SLGSFNKYLY DGAGGEGVTS
YVIDTGIHVT HKEFQGRASW GKTIPAGDVD DDGNGHGTHC AGTIASESYG VAKKANVVAI
KVLRSNGSGS MSDVLKGVEY ATQSHLDAVK KGNKKFKGST ANMSLGGGKS PALDLAVNAA
VKNGIHFAVA AGNENQDACN TSPAAAENAI TVGASTLSDA RAYFSNYGKC VDIFAPGLNI
LSTYTGSDDA TATLSGTSMA SPHIAGLLTY FLSLQPAAGS LYSNGGSEGV TPAQLKKNLL
KYASVGVLED VPEDTPNLLV YNGGGQNLSS FWGKETEDNV ASSDDTGEFH SFVNKLESAV
ENLAQEFAHS VKELASELI (SEQ 番号14)
【0082】
実施例3−プロテアーゼノックアウト
いくつかのピキアノックアウト系統を、以下の方法で産生させた。ノックアウトされる5’末端遺伝子(200−500塩基対)を、5’遺伝子特異的プライマー、および選択マーカー発現カセット(例えば、KanMX)の5’末端と相補的な20bp配列を持つ3’遺伝子特異的プライマーを使って、PCR法でピキアゲノムDNAから増幅させた。ノックアウトされる3’末端遺伝子(200−500塩基対)を、KanMXの発現カセットの3’末端、および3’遺伝子特異的プライマーと相補的な20bp配列の5’遺伝子特異的プライマーを使って、PCR法でピキアゲノムDNAから増幅させた。別々のPCR法反応において、適当量の5’産物と3’産物、KanMX発現カセットを含むDNA、前回使用した5’と3’遺伝子特異的プライマーを加えて、ノックアウト遺伝子の5’と3’の相同領域に隣接するKanMX発現カセットを有するノックアウトカセットを作った。DNA断片を精製し、標準電気穿孔法方法を使ってピキアに形質転換した。組換え型細胞を、抗菌(例えば、KAN)を含むYPDプレート上で選択し、特異的プライマーを使用してPCR法で選別した。結果として得られたノックアウト系統をゲノム配列決定とサザンブロット法で確認した。
【0083】
次に、単一および複数ノックアウトを、異種ポリペプチドを発現することができるDNAを使って形質転換した。各ノックアウト系統を、配列番号1に記述されている治療用タンパク質をコードする少なくとも1つのベクターを使って形質転換した。形質転換したノックアウトの培養物を生長させ、上清を収集した。分泌した産物を、タンパク質分解による産物分解を検出するために、NuPAGE MOPS ゲルで泳動させた。複数バンドの半精製した異種タンパク質は、プロテアーゼ活性を示すと解釈し、一方、適度の分子量の異種タンパク質の単一バンドは、プロテアーゼ活性がないと解釈した。図5−8を参照されたい。
【0084】
発酵上清試料のプロテアーゼ活性を、蛍光ラベルおよび励起と発光がそれぞれ波長340と405nmのクエンチャーを備える合成ペプチドを使って測定した。ペプチド基質を、培養上清試料でインキュベートし、基質の分解割合は、蛍光性HPLCを使ってモニターした。結果は、ピーク領域/時間が増加する方向に表される。図3において、プロテアーゼ活性は、ymp1突然変異系統の発酵上清で約68〜285であり、一方コントロール系統のプロテアーゼ活性は約1541〜1909であった。したがって、ペプチドに関して突然変異系統はプロテアーゼ活性の劇的な低下を立証した。
【0085】
ザイモグラムゲルは、カゼインまたはゼラチンを基質として使ってプロテアーゼ活性を測定する別の方法である。この実験において、カゼインまたゼラチンを還元、非変性の条件下でマトリックスに取り込んだアクリルアミドゲル上で発酵試料を分離した。分離に引き続いて、タンパク質バンドを再生し、熟成中の緩衝液をプロテアーゼ活性に必要な補助因子を提供するのに適用した。ゲルは、37℃で長期間インキュベートしクマシー・ブルーで染色した。タンパク分解活性領域を、青色を背景としたゲル上の明瞭な領域として同定した。活性なプロテアーゼは基質を分解し、ゲル上の領域をクマシー・ブルーに結合させなかった。活性なプロテアーゼを欠乏するゲル上の領域は染色剤が基質に結合するのを可能にし、よって背景を青色にした。図4は、ymp1突然変異系統および2つのコントロール系統からの発酵上清を使ったプロテアーゼ活動領域を表す図である。コントロール系統は、高分子量と低分子量の両領域にわたって活性なプロテアーゼを有した。しかしながら、高分子量領域のシグナルは、ymp1突然変異系統では大幅に減少したか、失われた。これらのデータは、高分子量領域での活性が、ymp1の作用に依るものであろうことを示す。ノックアウトに関する結果の概要を表5に示す。
【0086】
緩衝液安定性アッセイを使ったさらなる試験によって、YMP1およびYPS1をピキア宿主系統から欠失させた場合、切断効率および部位変化が実証された。このアッセイでは、配列番号1で記述されているタンパク質標準を培養上清でインキュベートし、切断されたタンパク質産物を質量分光光度法で分析し、切断部位および切断産物の割合を示した(表1−4)。
【0087】
発酵ブロスB804(pmt4を欠失した親系統SMD1163、実施例4を参照)の上清を使って、5個の主要な切断部位、リジン28/グリシン29、フェニルアラニン36/スレオニン37、セリン38/アスパラギン酸39、チロシン43/ロイシン44およびリジン50/グルタミン酸51を検出した(表1)。
【表1】
【0088】
SMD1163_pmp4(B688)からYMP1を欠失させると、フェニルアラニン36/スレオニン37、セリン38/アスパラギン酸39およびチロシン43/ロイシン44の切断部位が減少するが、しかしリジン50/グルタミン酸51とリジン28/グリシン29の切断を維持、または増加させた(表2)。
【表2】
【0089】
SMD1163_pmt4(B805)からyps1プロテアーゼを欠失させても任意の切断部位は除去されなかったが、リジン50/グルタミン酸51部位において切断量は減少した(表3)。
【表3】
【0090】
Ymp1およびYps1プロテアーゼを両方ともSMD1163_pmt4(B803)から欠失させると、切断は有意に減少し、かつ、リジン28/グリシン29のみで起こった。
【表4】
【0091】
SMD1163内で、ymp2、ymp3を個々にノックアウトにした。これらのノックアウト系統とSMD1163コントロールついて振盪フラスコ誘導を実施し、培養上清試料を、誘導後24時間で取り出した。配列番号1に記述されているタンパク質標準を30℃で24時間これらの試料と一緒にインキュベートし、SDS PAGEゲルによって分析した。
【0092】
PAGE分析から、ymp3の欠失は、タンパク質標準の切断を有意に減らすが、ymp2の欠失は、タンパク質分解には効果が少なかった(図9を参照)。
【表5】
【0093】
ピキア内において本明細書で同定した6個のポリペプチドは、Pep4、Prb1、Yps1、Yps2および新しく発見されたYmp1、Ymp3であり、これらは異種ポリペプチド(配列番号1)分解に関与する。これらの遺伝子の遺伝的ノックアウトによって異種タンパク質発現の生産性が有意に改善できる。
【0094】
実施例4 マンノシルトランスフェラーゼノックアウト
ピキア野生型株GS115から産生した異種ポリペプチド(配列番号1)をLC/MSで分析すると、複数の162Da質量増加アイソフォームが観察された。多重酵素消化とLC/MSを使ってさらなる分析を行うと実証するHSA部分で糖鎖形成が発生することが示され、エドマン分解法および炭水化物組成分析(データ示さず)を使ってその結果を再確認した。炭水化物組成分析により、これらの修飾はO−マンノース糖鎖形成によるものであることが確認された、この糖鎖形成はピキア・パストリス内で産生される一般的なタンパク質翻訳後修飾である。
【0095】
ピキア株のマンノシルトランスフェラーゼノックアウトを実施例2に記述したプロテアーゼノックアウトと同様な方法を使って作製した。すなわち、KanMX発現カセットを酵母:OCH1、PMT1、PMT2、およびPMT4内で糖鎖形成を担うこれらの遺伝子のうち1つに挿入した。ノックアウト系統を、配列番号1を発現することのできるベクターを使って形質転換した。これらのノックアウト系統の表現型の変化をザイモリアーゼ感受性アッセイ(図10)を使って実証した。この実験において、指数関数的に成長する酵母培養をザイモリアーゼ5U/mlを追加してYPDでOD600=0.3に希釈した。培養液を、室温で穏やかに振盪しながらインキュベートした。試料を、2時間の間15分置きに収集し、細胞死の指標としてOD600測定を行った。
【0096】
マンノシルトランスフェラーゼ遺伝子は、細胞壁完全性のために必須である。pmt1およびpmt4ノックアウト系統は、部分的または不完全な細胞壁を持つことがあり、ザイモリアーゼ処置への感受性を増大させる場合がある。このデータは、pmt1およびpmt4変異体の両方がザイモリアーゼ処置により急激に死滅したことを示す。
【0097】
異種タンパク質を、遺伝子改変された系統内での異種産生と野生型系統とを比較しながら、糖鎖形成について評価した。ノックアウトについての結果概要を以下の表6に示す。さらに図11を参照されたい。Pmt4が単独でピキア内のHSA糖鎖形成を担っている結果が示された。pmt4単一ノックアウト系統が糖鎖形成のないヒトHASを産生することができる。
【表6】
【0098】
実施例5−シャペロンタンパク質との同時発現
いくつかのシャペロンタンパク質は、宿主細胞内の異種タンパク質と同時発現でき、異種タンパク質産生を増加させることができるだろう。酵母内の異種タンパク質と同時発現して異種タンパク質産生を増加させるシャペロンタンパク質の例を以下の表7に示す。
【表7】
【技術分野】
【0001】
本明細書は、米国仮特許出願第61/155,706(出願日2009年2月26日)の権利を主張し、参照によりその全体を本明細書に援用する。
【0002】
本発明は、生化学エンジニアリングの分野に関する。特に、本発明は遺伝子改変ピキア株およびその中にポリペプチドを産生する方法に関する。
【背景技術】
【0003】
治療用ポリペプチドおよびタンパク質は、バクテリア細胞、大腸菌細胞、真菌または酵母細胞微生物細胞、昆虫細胞、および哺乳類細胞を含む種々の宿主細胞内で発現させることができる。メチロトローフ酵母ピキア・パストリスなどの真菌宿主は、治療用タンパク質発現に顕著な特長を有する。例えば、真菌宿主は、多量の内因性タンパク質を分泌しないし、強い誘導性プロモータを有し、限定的な化学培地で増殖でき、高力価の組換えタンパク質を産生する(Cregg et al., Mol. Biotech. 16:23−52 (2000))。酵母および糸状菌は両方とも、細胞内組換えタンパク質のおよび分泌された組換えタンパク質の産生に使用され、成功を収めている(Cereghino, J. L. and J. M. Cregg 2000 FEMS Microbiology Reviews 24(1): 45 66; Harkki, A., et al. 1989 Bio−Technology 7(6): 596; Berka, R. M., et al. 1992 Abstr. Papers Amer. Chem.Soc.203: 121−BIOT; Svetina, M., et al. 2000 J. Biotechnol. 76(23): 245−251)。ピキアは、組換え型ヒト血清アルブミン(HSA)の発現に関して注目に値する宿主細胞である。しかしながら、HASと遺伝的に融合したポリペプチドを含む他の治療用ポリペプチドの発現は、好ましくないタンパク質分解および糖鎖形成という技術的障壁に直面している。
【0004】
ピキア・パストリスに発現する異種タンパク質は付加的マンノース糖を含んでいて、「高マンノース」グリカン並びにタンパク質上に負電荷を与えるマンノシルリン酸基を生み出す。高マンノースグリカンまたは荷電したマンナンを有するグリコシル化タンパク質はヒトに免疫応答を誘発する危険性が高い(Takeuchi, Trends in Glycosci & Glycotech., 9:S29−S35 (1997); Rosenfeld et al., J.Biol.Chem., 249:2319−2321 (1974))。したがって、糖鎖形成のパターンがヒトのパターンと同一であるか少なくとも類似しているような治療用のペプチド類、ポリペプチドおよび/またはタンパク質を真菌宿主組織において産生することが望ましい。
【0005】
従って、野生型系統と比較すると減少したタンパク質分解および/または糖鎖形成を有する異種ペプチド類、ポリペプチドおよび/またはタンパク質を産生することができる酵母系統、特にピキアの必要とされている。さらに加えて、酵母系統内で遺伝子を同定する必要があり、特にタンパク分解性と糖鎖形成経路に関わるタンパク質の産生を担うピキア株内で同定する必要がある。
【発明の概要】
【0006】
本発明の1つの態様において、遺伝子改変ピキア株が提供され、機能的遺伝子産物をコードする少なくとも1つの核酸配列、および/またはピキア株の少なくとも1つの遺伝子産物の発現に必要な少なくとも1つの核酸が遺伝子改変され、前記遺伝子産物は、前記遺伝子改変ピキア株においてタンパク質分解および/または糖鎖形成を担う。
【0007】
少なくとも1つの異種ポリペプチドを産生する方法も本発明で提供され、その方法には、本発明の遺伝子改変ピキア株内で前記異種ポリペプチドを発現させるステップも含まれる。別の態様において、本発明は、本発明の宿主細胞で異種的に発現したペプチド、ポリペプチド、およびタンパク質を提供する。
【0008】
本発明の別の態様において、配列番号4、配列番号6および配列番号8と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%配列同一性を有する単離したポリペプチドが提供される。前記宿主細胞の前記野生型の親が配列番号4、6および8と少なくとも60%の配列同一性を持つポリペプチドをコード化する遺伝子を含み、前記野生型宿主細胞と比較して、前記遺伝子改変宿主細胞内で遺伝子産物または活性が低下するか除去されるように、前記遺伝子が前記宿主細胞のゲノム内で改変される、遺伝子改変宿主細胞系統をもさらに提供する。
【図面の簡単な説明】
【0009】
【図1】親和性精製およびLC/MS分析を使用して同定した新規ピキアプロテアーゼYmp1を示す図である。
【図2】アミノ酸1−857を配列番号15として示すYMP1(配列番号4)の予測構造である。
【図3】ymp1突然変異系統のプロテアーゼ活性アッセイを示す図である。
【図4】ymp1突然変異系統のザイモグラムアッセイを示す図である。
【図5】異種タンパク質(配列番号1)のタンパク質分解に関するymp1ノックアウト効果を示す図である。
【図6】異種タンパク質(配列番号1)のタンパク質分解に関するysp2ノックアウト効果を示す図である。
【図7】異種タンパク質(配列番号1)のタンパク質分解に関するysp1ノックアウト効果を示す図である。
【図8】異種タンパク質(配列番号1)のタンパク質分解に関するpep4ノックアウト効果を示す図である。
【図9】ymp1、ymp3突然変異系統の緩衝液安定性試験を示す図である。
【図10】野生型(WT)、pmt1およびpmt4ピキア変異体の細胞壁溶解酵素アッセイ結果を示す図である。
【図11】pmt4突然変異系統由来の異種タンパク質(配列番号1)の糖鎖形成分析を示す図である。
【発明を実施するための形態】
【0010】
本明細書で使用する「宿主細胞(1つまたは複数)」は、単離したポリヌクレオチド配列を導入(例えば、形質転換、感染、もしくは形質移入)した細胞または導入(例えば、形質転換、感染、もしくは形質移入)可能な細胞を意味する。本発明の宿主細胞は、以下に制限されるものではないが、バクテリア細胞、真菌細胞、酵母細胞、微生物細胞、昆虫細胞、および哺乳類細胞を含んで良い。酵母および/または糸状真菌起源の本発明の宿主細胞は、これに制限されるものではないが、以下の科、属、および種を含んで良い:ピキア・パストリス(Pichia pastoris)、ピキア・フィンランディカ(Pichia finlandica)、ピキア・トレハロフィア(Pichia trehalophila)、ピキア・コクラメ(Pichia koclamae)、ピキア・メンブラナエファシエンス(Pichia membranaefaciens)、ピキア・メタノリカ(Pichia methanolica)、ピキア・ミヌタ(Pichia minuta)(オガタ・エミヌタ(Ogataea minuta)、ピキア・リンドネリ(Pichia lindneri))、ピキア・オプティアエ(Pichia opuntiae)、ピキア・サーモトレランス(Pichia thermotolerans)、ピキア・サリクタリア(Pichia salictaria)、ピキア・ゲルカム(Pichia guercum)、ピキア・ピジュペリ(Pichia pijperi)、ピキア・スティプティス(Pichia stiptis)、ピキア菌類(Pichia sp.)、サッカロミセス・カステリ(Saccharomyces castelii、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス・クルイベリ(Saccharomyces kluyveri)、サッカロミセス菌類(Saccharomyces sp.)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、クリベロマイセス菌類(Kluyveromyces sp.)、クリベロマイセス・ラクティス(Kluyveromyces lactis)、カンジダ・アルビカンス(Candida albicans)、カンジダ菌類(Candida sp.)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、アスペルギルス・ニデュランス(Aspergillus nidulans)、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・オリザエ(Aspergillus oryzae)、トリコデルマ・リーセイ(Trichoderma reesei)、クリソスポリウム・ラクノウェンス(Chrysosporium lucknowense)、フザリウム菌類(Fusarium sp.)、フザリウム・グラミネアラム(Fusariumu gramineum)、フザリウム・ベネナタム(Fusariumu venenatum)、ニセツリガネゴケ(Physcomitrella patens)およびアカパンカビ(Neurospora crassa)。
【0011】
当業者に周知である「形質転換した」とは、外部DNAまたはRNAの導入による
有機体のゲノムもしくはエピソームの特異的修飾、または他の安定的な外部DNAまたはRNAの導入を意味する。
【0012】
当業者に周知である「形質移入した」とは、以下に限定されないが、組換えDNAもしくはRNAを含む外部DNAまたはRNAを微生物に導入することである。
【0013】
当業者に周知である「同一性」とは、2個以上のポリペプチド配列、場合によっては2個以上のポリヌクレオチド配列を比較することで判定される関係のことである。当業界では、「同一性」は、ポリペプチド配列間、場合によってはポリヌクレオチド配列間の文字列の整合性によって判定される配列関連度をも意味する。「同一性」は、限定されないが、以下の文献に記述されている既存の方法で直ちに計算できる:(Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; and Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991; and Carillo, H., and Lipman, D., SIAM J. Applied Math.,48:1073(1988)。同一性を決定する方法は、テストされる配列間で最大の整合性が得られるように設計される。さらに、同一性を決定する方法は、市場で入手可能なコンピュータープログラムにコード化されている。2個の配列間の同一性を決定するコンピュータープログラム方法は、以下に限定されないが、GCGプログラムパッケージ(Devereux, J., et al., Nucleic Acids Research 12(1): 387 (1984)), BLASTP, BLASTN, and FASTA (Altschul, S.F. et al., J. Molec. Biol. 215: 403−410 (1990))を含む。ブラストXプログラムは、NBCIとその他のソースから公的に入手出来る。(BLAST Manual, Altschul, S., et al., NCBI NLM NIH Bethesda, MD 20894; Altschul, S., et al., J. Mol. Biol. 215: 403−410 (1990))。よく知られたSmith Watermanアルゴリズムも、同一性を決定するのに使って良い。
【0014】
ポリペプチド配列比較のためのパラメータは、以下を含む:
Algorithm: Needleman and Wunsch, J. Mol Biol. 48: 443−453 (1970)
Comparison matrix: BLOSSUM62 from Hentikoff and Hentikoff, Proc. Natl. Acad. Sci. USA. 89:10915−10919 (1992)
ギャップペナルティー: 12
ギャップ長ペナルティー: 4
これらのパラメータを使う有用なプログラムは、「ギャップ」プログラムとして遺伝学コンピュータグループ、Madison WI.から公的に入手可能である。上述のパラメータは、ペプチド比較(終端ギャップについてはペナルティなしである)のためのデフォルトパラメータである。
【0015】
ポリヌクレオチド比較のためのパラメータは以下を含む:Algorithm: Needleman and Wunsch, J. Mol Biol. 48: 443−453 (1970)
比較マトリックス:整合=+10、非整合=0
ギャップペナルティー:50
ギャップ長ペナルティー:3
入手可能先:「ギャップ」プログラム(Genetics Computer Group, Madison WI.)
これらは、核酸比較のためのデフォルトパラメータである。
【0016】
ポリヌクレオチドおよび場合によってはポリペプチドの「同一性」は、以下の(1)と(2)に説明される。
【0017】
(1)ポリヌクレオチドの実施形態は、参照配列、例えば配列番号3と少なくとも50、60、70、80、85、90、95、97ならびに100%同一性を有するポリヌクレオチド配列を備えた単離したポリヌクレオチドをさらに含む。ここで上記ポリヌクレオチド配列は、配列番号3の参照配列と同一であってもよく、またはその参照配列と比較してある特定の整数の数だけヌクレオチドに変化があって良く、ここで上記変化は、遷移および塩基転換、または挿入を含む少なくとも1つのヌクレオチドの欠失、置換からなる群から選択されるものであり、さらに、上記変化は、参照ヌクレオチド配列の5’または3’末端位置で生じてもよいし、参照配列のヌクレオチド間で個々にまたは参照配列内の1つ以上の連続するグループで分散されることにより、これらの末端位置の間のいずれかの位置で生じても良く、上記ヌクレオチドの変化数は、配列番号3のヌクレオチドの全数に、同一性の%を表す整数を掛け、それを100で割り、次に、その結果を上記配列IDNO:3内のヌクレオチドの全数から差し引くことによって決定される。言い換えれば、
【数1】
式中、nnはヌクレオチドの変化数、xnは配列番号3のヌクレオチドの全数、yは95%では0.95、97%では0.97または100%では1.00であり、・は乗算演算子を表す記号であり、かつxnとyの非整数の積はもっとも近い整数に切り捨て、それをxnから差し引く。ポリペプチドをコードするポリヌクレオチド配列の変化は、そのコード配列にナンセンス、ミスセンスまたはフレームシフトの変異をもたらし、それによって、かかる変化後ポリヌクレオチドによってコードされたポリペプチドを変化させる。
【0018】
(2)ポリペプチド実施形態は、参照配列、例えば配列番号4と少なくとも50、60、70、80、85、90、95、97ならびに100%同一性を有するポリペプチドを備えた単離したポリペプチドをさらに含み、ここで上記ポリペプチド配列は、参照配列と同一であってもよく、またはその参照配列と比較してある特定の整数の数だけアミノ酸変化があって良く、ここで上記変化は、保存的または非保存的置換または挿入を含む少なくとも1つのアミノ酸の欠失、置換からなる群から選択されるものであり、さらに、上記変化は、参照ポリペプチド配列のアミノ基またはカルボキシ基末端位置で生じてもよいし、または参照配列のアミノ酸間で個々に、または参照配列内の1つ以上の連続するグループで分散されることにより、これらの末端位置の間のいずれかの位置で生じても良く、さらに、上記アミノ酸の変化数は、アミノ酸の全数に同一性の%を表す整数を掛け、それを100で割り、次に、その結果を上記アミノ酸の全数から差し引くことによって決定される。言い換えれば、
【数2】
式中、naはアミノ酸の変化数、xaは配列のアミノ酸の全数、yは95%では0.95、97%では0.97または100%では1.00であり、・は乗算演算子を表す記号であり、またxaとyの非整数の積はもっとも近い整数へ切り捨て、それをxaから差し引く。
【0019】
「単離する」とは、自然な状態から、人の手を加えて変えることを意味し、すなわち、自然に起こることであれば、元の環境から変化したか元の環境から取り除かれるか、または両方を意味する。例えば、生体内に自然に存在するポリヌクレオチドまたはポリペプチドは単離していない。しかし、限定するものではないが、そのようなポリヌクレオチドまたはポリペプチドを細胞に再び導入する場合を含め、自然な状態において共存している物質から分離されたポリヌクレオチドまたはポリペプチドは、「単離されている」と呼ぶ。
【0020】
「単離している」または「実質的に純粋な」核酸またはポリヌクレオチド(例えば、RNA、DNAまたは混合ポリマー)とは、自然の宿主細胞内に未変性のポリヌクレオチドを必然的に伴っている他の細胞成分(例えば、リボソーム、ポリメラーゼおよびそれに自然に付随するゲノム配列)から実質的に分離されたものである。この用語は、(1)自然に発生する環境から取り除かれた、(2)「単離されたポリヌクレオチド」を自然の中で含んでいるポリヌクレオチドの全てまたは一部を伴わない、(3)自然界では連結していないポリヌクレオチドと動作可能に連結する、または(4)自然界では生じない、核酸またはポリヌクレオチドを包含する。用語「単離されている」または「実質的に純粋」とは、組換えまたはクローン化DNA分離株、化学的に合成されたポリヌクレオチド類似体、または異種システムによって生物学的に合成されたポリヌクレオチド類似体との関連で使用できる。
【0021】
しかしながら、「単離される」ことは、これまで記述してきた核酸またはポリヌクレオチドそれ自体を自然の環境から物理的に除去することを必ずしも要求しない。例えば、生体内ゲノムの内因性核酸配列は、この内因性核酸配列の発現が変化するように、例えば、増加、減少、または除去されるように、異種配列が内因性核酸配列に隣接して置かれている場合に、「単離されている」と本明細書では見なされる。この文脈において、異種配列は、異種配列それ自体が内因性(宿主細胞またはその子孫由来)であるか外来性(異なる宿主細胞またはその子孫由来)であるかを問わず、内因性核酸配列に自然界で隣接している配列ではない。例えば、プロモータ配列は、遺伝子が発現パターンを変化させるように、(例えば、相同組換えによって)宿主細胞のゲノム内の遺伝子の元来のプロモータで置き換えてもよい。この遺伝子は、自然の状態で隣接している配列から少なくとも一部は分離されているので、「単離されている」ことになる。
【0022】
核酸はまた、ゲノム内の対応する核酸に自然界では生じない任意の修飾を含むならば、「単離された」と見なされる。例えば、内因性コード配列は、人工的に、例えば、人間の介入により導入された挿入、欠失、または点変異を含むならば、「単離された」と見なされる。「単離された核酸」はまた、宿主細胞染色体の異種部位に組み込まれた核酸とエピソームとして存在する核酸コンストラクトも含む。さらに、「単離された核酸」は、実質的にその他の細胞の材料を使用せず、または組換え技術で産生する場合に実質的に培地を必要とせず、または化学的に合成する場合に実質的に化学前駆物質もしくはその他の化学物質も必要としない。
【0023】
本明細書で使用する場合、「機能的遺伝子産物をコードする核酸配列」とは、遺伝子のコード部分のあらゆる部分を意味する。機能的遺伝子産物をコードする核酸配列は、少なくとも1つの活性を行うことのできる酵素の一部であっても良いし、酵素全体であっても良い。
【0024】
本明細書で使用する場合、「少なくとも1つの遺伝子産物の発現に必要な核酸」とは、遺伝子の一部をコードし、および/または遺伝子産物をコードする核酸に動作可能に連結するが必ずしもコード配列を含まない核酸配列を意味する。例えば、少なくとも1つの遺伝子産物の発現に必要な核酸配列は、限定されないが、エンハンサー、プロモータ、制御配列、開始コドン、終止コドン、ポリアデニル化配列、および/またはコーディング配列を含む。
【0025】
本明細書で使用する「タンパク質分解」または「細胞内でタンパク質分解を担う遺伝子産物」とは、少なくとも1つのペプチド、ポリペプチドおよび/またはタンパク質の切断を引き起こすことができる任意のペプチド、ポリペプチド、タンパク質および/または酵素またはその部分を意味する。タンパク質分解を担う遺伝子産物は、切断(即ち、ペプチダーゼ)に直接関与しても良く、ペプチダーゼ合成経路の役割として間接的に関与してもよい。細胞内のタンパク質分解を担う遺伝子産物の例として、限定されないが、アスパルチルプロテアーゼ、セリンプロテアーゼ、分泌アスパルチルプロテアーゼ、分泌セリンプロテアーゼ、酵母メチロトローフプロテアーゼ、DDP IV様エンドペプチダーゼ、金属エンドペプチダーゼ、Prb1様セリンプロテアーゼ、Prb1 セリンプロテアーゼ、およびCPY様カルボキシペプチダーゼが挙げられる。分泌セリンプロテアーゼのように、細胞から分泌されてもタンパク質分解活性の一部または全部を維持しているプロテアーゼも、この定義に当てはまる。分泌プロテアーゼは、細胞内部および/または外部のタンパク質分解に関与する場合がある。
【0026】
本明細書で使用する場合、「糖鎖形成」または「細胞内の糖鎖形成に関与する遺伝子産生物」とは、細胞内において少なくとも1つの糖類部分をポリペプチドに付加する機能、または少なくとも1本の糖類鎖を伸長する機能に関わるペプチド、ポリペプチド、タンパク質および/または酵素もしくはその部分のことを意味する。細胞内で糖鎖形成を担う遺伝子産物は、糖類を細胞内で直接ポリペプチドに付加する作用に関与し、例えば、限定されないが、マンノシルトランスフェラーゼである。マンノシルトランスフェラーゼは、Dol−P−Manからの残基をペプチド、ポリペプチド、および/またはタンパク質上のセリンおよび/またはスレオニン残基に伝達でき、あるいは、GPD−Manからのマンノース残基を糖類に伝達するように作用し、このようにして、糖類鎖を伸長する。あるいは、糖鎖形成の機能を担う遺伝子産物は、糖鎖形成経路の部分であるかもしれないし、また細胞内で多糖類をポリペプチドに付加する機能に間接的に関与しているかもしれない。細胞内の糖鎖形成に関与する遺伝子産物の例には、限定されないが、マンノシルトランスフェラーゼ類が含まれる。
【0027】
本明細書で使用する場合、「酵母メチロトローフプロテアーゼ1(Ymp1)活性」とは、配列番号4として本明細書で同定されたタンパク質が遂行できる任意の活性を意味する。例えば、酵母メチロトローフプロテアーゼ1活性またはYmp1活性は、限定されないが、ペプチド、ポリペプチド、またはタンパク質をタンパク分解的に切断する酵素能力を含む。特に、Ymp1活性は、限定されないが、ヒトGLP−1および/またはヒトアルブミンなどのGLP−1アゴニストの断片および/または変異体を含むポリペプチドを切断する酵素能力を意味する。少なくとも1つのYmp1活性は、限定されないが、少なくとも1つのセリンプロテアーゼ活性を含む。
【0028】
「ポリヌクレオチド(1つまたは複数)」とは、一般に、非修飾RNAもしくはDNAまたは修飾RNAもしくはDNAであり得る、任意のポリリボヌクレオチドまたはポリデオキシリボヌクレオチドを意味する。「ポリヌクレオチド(1つまたは複数)」としては、単鎖および二重鎖DNA、単鎖および二重鎖領域、または単鎖、二重鎖および三重鎖領域の混合物であるDNA、単鎖および二重鎖RNA、単鎖および二重鎖領域の混合物であるRNA、単鎖または、より典型的には、二重鎖、または三重鎖領域のDNAおよびRNAを含むハイブリッド分子、または単鎖および二重鎖領域の混合物が挙げられるが、これらに限定されない。加えて、本明細書で使用する場合、「ポリヌクレオチド」とは、RNAもしくはDNA、またはRNAおよびDNAの両方を含む三重鎖領域を意味する。かかる領域の鎖は、同じ分子に由来してもよいし、異なる分子に由来してもよい。この領域は1つ以上の分子をすべて含み得るが、より典型的には、一部の分子の領域のみを含む。三重らせん状領域の1つの分子は、しばしばオリゴヌクレオチドである。本明細書で使用する場合、「ポリヌクレオチド(1つまたは複数)」という用語は、1つ以上の修飾塩基を含む上記のDNAまたはRNAも含む。したがって、安定性または他の理由のために修飾された骨格のDNAまたはRNAは、本明細書で意図する「ポリヌクレオチド(1つまたは複数)」である。さらに2例のみ挙げると、異常な塩基(イノシンなど)、または修飾塩基(トリチル化塩基など)を含むDNAまたはRNAも、本明細書において使用されるポリヌクレオチドである。当業者に知られている多くの有用な目的に適う多種多様の修飾がDNAおよびRNAで行われていることは理解されるであろう。本明細書で使用する場合、「ポリヌクレオチド(1つまたは複数)」という用語は、ポリヌクレオチドのかかる化学的、酵素的もしくは代謝的に修飾された形態、ならびにウイルスおよび細胞(例えば、単純細胞および複雑細胞が挙げられる)に特徴的なDNAおよびRNAの化学的形態を包含する。「ポリヌクレオチド(1つまたは複数)」は、しばしばオリゴヌクレオチド(1つまたは複数)と呼ばれる短いポリヌクレオチドも包含する。
【0029】
「ポリペプチド(1つまたは複数)」とは、ペプチド結合または修飾ペプチド結合により互いに結合した2つ以上のアミノ酸を含む任意のペプチドまたはタンパク質を意味する。「ポリペプチド(1つまたは複数)」とは、一般的にペプチド、オリゴペプチドまたはオリゴマーと呼ばれる短鎖と、一般にタンパク質と呼ばれるより長い鎖との両方を意味する。ポリペプチドは、20個の遺伝子コードアミノ酸以外のアミノ酸を含んでよい。「ポリペプチド(1つまたは複数)」とは、天然プロセス(プロセシングおよび他の翻訳後の修飾など)、または化学的修飾技術のいずれかにより修飾されたアミノ酸配列を含む。かかる修飾は、基礎テキストおよびより詳細な研究論文、ならびに膨大な研究文献に詳述されており、当業者にとって公知である。このような種類の修飾は、所定のポリペプチドのいくつかの部位に同じ種類の修飾が同程度または異なる程度で存在し得ることが理解されるであろう。所定のポリペプチドは、多くの種類の修飾を含んでよい。修飾は、ポリペプチドのどの場所(ペプチド骨格、アミノ酸側鎖およびアミノ末端もしくはカルボキシ末端を含む)でも生じ得る。修飾としては、アセチル化、アシル化、ADPリボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスファチジルイノシトールの共有結合、架橋、環化、ジスルフィド結合形態、脱メチル化、共有架橋形態、システイン形態、ピログルタミン酸形態、ホルミル化、γ−カルボキシル化、GPIアンカー形態、水酸化、ヨウ素化、メチル化、ミリストイル化、酸化、タンパク質分解プロセシング、リン酸化反応、プレニル化、ラセミ化、グリコシル化、脂質結合、硫酸化、グルタミン酸残基のγ−カルボキシル化、ヒドロキシル化、およびADP−リボシル化、セレノイル化、硫酸化、転移RNA媒介性のアミノ酸のタンパク質への付加(アルギニル化、およびユビキチン化など)が挙げられる。例えば、PROTEINS − STRUCTURE AND MOLECULAR PROPERTIES, 2nd Ed., T. E. Creighton, W. H. Freeman and Company, New York, 1993 and Wold, F., Posttranslational Protein Modifications: Perspectives and Prospects, pgs. 1−12 in POSTTRANSLATIONAL COVALENT MODIFICATION OF PROTEINS, B. C. Johnson, Ed., Academic Press, New York,(1983); Seifter, et al., Meth. Enzymol. 182:626−646(1990)およびRattan, et al., “Protein Synthesis: Posttranslational Modifications and Aging”, Ann NY Acad Sci 663:48−62(1992)を参照されたい。ポリペプチドは、分枝または環状であり、分枝があってもなくても良い。環状、分枝および分枝した環状ポリペプチドは、翻訳後の自然なプロセスから生み出されたものであっても、完全に合成的な方法によるものであっても良い。
【0030】
「組換え発現系(1つまたは複数)」とは、本発明の発現系またはその部分またはポリヌクレオチドであって、本発明のポリヌクレオチドおよびポリペプチドの産生のために宿主細胞または宿主細胞可溶化液へ導入、形質移入、または形質転換されたものである。
【0031】
本明細書において使用される場合、「変異体(1つまたは複数)」という用語は、参照ポリヌクレオチドまたは参照ポリペプチドとは異なるが、本質的な特性を保持するポリヌクレオチドまたはポリペプチドである。ポリヌクレオチドの典型的な変異体は、別の参照ポリヌクレオチドとはヌクレオチド配列が異なる。変異体のヌクレオチド配列の変化により、参照ポリヌクレオチドによりコードされたポリペプチドのアミノ酸配列が変化する場合もあるし、しない場合もある。以下に論じるように、ヌクレオチド変化により、参照配列によりコードされたポリペプチドのアミノ酸が置換、付加、欠失、融合および切断されてよい。ポリペプチドの典型的な変異体は、別の参照ポリペプチドとはアミノ酸配列が異なる。一般に、参照ポリペプチド配列とその変異体は、全体的に近似し同じ領域が多いので、その相違は限定されている。変異体と参照ポリペプチドのアミノ酸配列は、1つ以上の置換、付加、欠失の任意の組み合わせにより異なってよい。置換または挿入したアミノ酸残基は、遺伝子コードでコードされたものであってもよいし、そうでなくてもよい。
【0032】
本発明は、保存アミノ酸置換によって参照対象とは異なり、残基が同様な特徴を有する別のもので置換されている本発明の各ポリペプチドの変異体も含む。典型的なそのような置換は、アラニン、バリン、ロイシンおよびイソロイシンとの間、セリンとトレオニンとの間、酸性残基のアスパラギン酸とグルタミン酸との間、アスパラギンとグルタミンとの間、塩基性残基であるリシンとアルギニンとの間、芳香族残基のフェニルアラニンとチロシンとの間で生じる。特に、いくつか、つまり5−10、1−5、1−3、1−2または1つのアミノ酸が、任意の組み合わせで置換、消去、または付加された変異体が存在する。ポリヌクレオチドまたはポリペプチドの変異体は、天然(対立遺伝子変異体など)であってもよく、天然では知られていない変異体であってもよい。ポリヌクレオチドおよびポリペプチドの非天然変異体は、当業者に周知の、変異誘発技術により生成してもよいし、直接合成によって生成しても良いし、他の組換え技術によって生成しても良い。
【0033】
「微生物(1つまたは複数)」とは、(i)原核生物を意味し、限定されないが、以下の属のメンバーを含む:連鎖球菌、ブドウ球菌、百日咳菌、コリネバクテリウム、マイコバクテリウム、ナイセリア、ヘモフィルス、放線菌、ストレプトミセス、ノカルジア、エンテロバクター、エルシニア、フランシセラ、パスツレラ、モラクセラ、アシネトバクター、エリシペロスリクス、ブランハメラ、アクチノバチルス、ストレプトバチルス、リステリア、カリマトバクテリウム、ブルセラ菌、桿菌、クロストリジウム、トレポネーマ、大腸菌類、サルモネラ菌、クレブシエラ、ビブリオ、プロテウス、エルウィニア、ボレリア、レプトスピラ、スピリルム、カンピロバクター、赤痢菌、レジオネラ、シュードモナス、エロモナス、リケッチア、クラミジア、ボレリアおよびマイコプラズマ。
【0034】
さらに、限定されないが、以下の種またはグループのメンバーを含む:グループ A連鎖球菌、グループ B連鎖球菌、グループ C連鎖球菌、グループ DS連鎖球菌、グループ G連鎖球菌、肺炎球菌、化膿性連鎖球菌、ストレプトコッカス・アガラクチア、連鎖球菌 faecalis、フェシウム菌、連鎖球菌 durans、淋菌、髄膜炎菌、黄色ブドウ球菌、表皮ブドウ球菌、ジフテリア菌、ガードネレラ菌、結核菌、マイコバクテリウム・ボビス、マイコバクテリウム・ウルセランス、ライ菌、Actinomyctes israelii、リステリア菌、百日咳菌、パラ百日咳菌、気管支敗血症菌、大腸菌、志賀赤痢菌、インフルエンザ菌、ヘモフィルス・エジプチウス、パラインフルエンザ菌、軟性下疳菌、ボルデテラ、チフス菌、シトロバクター・フロインデイ、プロテウス・ミラビリス、プロテウス・ブルガリス、ペスト菌、肺炎かん菌、セラチア‐マルセッセンス、セラチア・リクファシエンス、コレラ菌、Shigella dysenterii、シゲラ・フレックスネリ、緑膿菌、野兎病菌、ウシ流産菌、炭疽菌、セレウス菌、ウェルシュ菌、破傷風菌、ボツリヌス菌、梅毒トレポネーマ、リケッチア・リケッチイおよびトラコーマ病原体。(ii)限定されないが、Archaebacterを含む古細菌、および(iii)単細胞または糸状真核生物であり、限定されないが以下を含む:原虫、菌類、酵母類属のメンバー、クリベロマイセス、またはカンジダ、および酵母類ceriviseae種のメンバー、クリベロマイセス・ラクティス、またはカンジダ・アルビカンス。
【0035】
「細菌(1つまたは複数)(l)」とは、(i)原核生物を意味し、限定されないが以下の属のメンバーを含む:連鎖球菌、ブドウ球菌、ボルデテラ、コリネバクテリウム、マイコバクテリウム、ナイセリア、ヘモフィルス、放線菌、ストレプトミセス、ノカルジア、エンテロバクター、エルシニア、フランシセラ、パスツレラ、モラクセラ、アシネトバクター、エリシペロスリクス、ブランハメラ、アクチノバチルス、ストレプトバチルス、リステリア、カリマトバクテリウム、ブルセラ菌、桿菌、クロストリジウム、トレポネーマ、大腸菌類、サルモネラ菌、クレブシエラ、ビブリオ、プロテウス、エルウィニア、ボレリア、レプトスピラ、スピリルム、カンピロバクター、赤痢菌、レジオネラ、シュードモナス、エロモナス、リケッチア、クラミジア、ボレリアおよびマイコプラズ。さらに、限定されないが以下の種またはグループのメンバーを含む:グループ A連鎖球菌、グループ B連鎖球菌、グループ C連鎖球菌、グループ DS連鎖球菌、グループ G連鎖球菌、肺炎球菌、化膿性連鎖球菌、ストレプトコッカス・アガラクチア、連鎖球菌 faecalis、フェシウム菌、連鎖球菌 durans、淋菌、髄膜炎菌、黄色ブドウ球菌、表皮ブドウ球菌、ジフテリア菌、ガードネレラ菌、結核菌、マイコバクテリウム・ボビス、マイコバクテリウム・ウルセランス、ライ菌、Actinomyctes israelii、リステリア菌、百日咳菌、パラ百日咳菌、気管支敗血症菌、大腸菌、志賀赤痢菌、インフルエンザ菌、ヘモフィルス・エジプチウス、パラインフルエンザ菌、軟性下疳菌、ボルデテラ、チフス菌、シトロバクター・フロインデイ、プロテウス・ミラビリス、プロテウス・ブルガリス、ペスト菌、肺炎かん菌、セラチア‐マルセッセンス、セラチア・リクファシエンス、コレラ菌、Shigella dysenterii、シゲラ・フレックスネリ、緑膿菌、野兎病菌、ウシ流産菌、炭疽菌、セレウス菌、ウェルシュ菌、破傷風菌、ボツリヌス菌、梅毒トレポネーマ、リケッチア・リケッチイおよびトラコーマ病原体、ならびに(ii)古細菌(限定されないがArchaebacterを含む)。
【0036】
本明細書で使用する場合、「異種ポリペプチド(1つまたは複数)」とは、形質転換した宿主細胞または対象となる微生物によって人工的に合成され、組換えDNAによってその宿主細胞または微生物に導入されたポリペプチドを意味する。例えば、ピキアは、ヒト血清アルブミンの発現用の宿主細胞として作用するが、非形質転換または形質導入されなかったピキアでは起こらない。異種ポリペプチドは、単離を促進するように修飾されたポリペプチドを含んでも良い。
【0037】
本明細書で使用する場合、「親和性タグ」とは、他の物質または分子への選択的親和性を得ることのできる分子に関係する任意の部分を意味する。例えば、親和性タグは、
分子にカラム内のパッキング物質との選択的親和性を提供することにより分子の精製を促進するのに使って良い。親和性タグの非限定的例はhis−タグである。
【0038】
核酸は、別の核酸配列と機能的な関係をもつように置かれると「動作可能に連結」する。例えば、プレ配列または分泌性リーダーは、ポリペプチドの分泌に関与するタンパク質前駆体として発現するならば、動作可能にポリペプチドのDNAに連結する。プロモータまたはエンハンサーは、それが配列の転写に影響するならば、動作可能にコード配列に連結する。または、翻訳を促進するように置かれるならば、リボソーム結合部位は動作可能にコード配列に連結する。一般に、「動作可能に連結する」とは、連結しているDNA配列が連続していることを意味し、分泌性リーダーの場合には、連続しかつリーディングフェーズにあることを意味する。しかしながら、エンハンサーは連続している必要はない。連鎖は、適切な制限酵素部位で連結して完結する。そのような部位が存在しない場合は、合成オリゴヌクレオチドアダプタまたはリンカーが通常のやり方に従って使われる。
【0039】
本明細書で使用される場合、細胞を「収集する」とは、細胞培養から細胞を採取することを意味する。
細胞は、収集する間濃縮され、例えば、遠心分離または濾過によって培養ブロスから分離される。細胞の収集は、細胞を溶解して、限定されないが、ポリペプチドおよびポリヌクレオチドなどの細胞内物質を得る段階を含んで良い。限定されないが、異種的に発現するポリペプチドを含む特定の細胞の物質は、培養中に細胞から遊離することがあるということを当業者は理解されたい。従って、対象の産物(例えば、異種的に発現したポリペプチド)は、細胞を収集した後も培養ブロスにとどまり続ける場合がある。
【0040】
さらに、組換えDNAコンストラクトが選択可能なマーカーをコードする方法も提供される。そのような選択可能なマーカーは陽性または陰性の選択を提供する。上記選択可能なマーカーを発現するステップ、選択段階の少なくとも1つの第1の形質転換した細胞によって産生した選択可能なマーカーの量を、選択段階の少なくとも1つの第2の形質転換した細胞によって産生した選択可能なマーカーの量と比較することを含む方法(第1と第2の形質転換した細胞は同じ選択可能なマーカーを産生する)も提供される。当業界で知られているように、選択可能なマーカーは、限定されないが、ジヒドロ葉酸還元酵素(dhfr)、β−ガラクトシダーゼ、蛍光タンパク質、分泌型のヒト胎盤のアルカリホスファターゼ、β−グルクロニダーゼ、酵母選択可能マーカーLEU2およびURA3、アポトーシス耐性遺伝子、およびアンチセンスオリゴヌクレオチド、並びにネオマイシン(neo)、カナマイシン、ジェネテシン、ハイグロマイシンB、ピューロマイシン、ゼオシン、ブラストサイジン、ストレプトスライシン、ビアラホス、フレオマイシン、およびアンピシリンを含む抗生物質存在下で成長する能力を与える抗生物質耐性遺伝子を含む。同様に当業界で知られているように、細胞は、限定されないが、視覚的点検または選択可能マーカーの発現を検出することができるBD蛍光標示式細胞分取器などの細胞選別機を含む種々の手段で選別ができる。
【0041】
当業界で理解されている用語「野生型」とは、遺伝的修飾なしに自然の集団で起こりうる宿主細胞またはポリペプチドまたはポリヌクレオチド配列を意味する。例えば、「宿主細胞の野生型親」とは、宿主細胞のゲノム内で遺伝的修飾がなされるまたは生じる前の無修飾の宿主細胞系統を意味する。
【0042】
本明細書で使われる場合、「力価収率」とは、溶液(例えば、培養ブロスまたは細胞−溶解混合物または緩衝液)中の産物(例えば、異種性に発現したポリペプチド)の濃度を意味し、通常mg/Lまたはg/Lで表現される。収率の増加とは、2個の規定された条件下で産生した産物の濃度の絶対的または相対的増加を意味する。
【0043】
本明細書で使われる場合、「インクレチンホルモン」とは、インスリン分泌を増強する、あるいは分泌レベルまたはインスリンを増加させるあらゆるホルモンを意味する。インクレチンホルモンの1つの例は、GLP−1である。GLP−1は、食物摂取に応答して腸のL細胞よって分泌されたインクレチンである。健常人の場合、GLP−1は、膵臓のグルコース−依存インスリン分泌を刺激することによって食後血糖値を調整する重要な働きをし、その結果末梢内のグルコース吸収を増加させる。GLP−1はまた、グルカゴン分泌を抑制し、肝臓のグルコース出力を低下させる。さらに、GLP−1は胃排出時間を遅延させ、小腸の運動性を緩慢にし食物吸収を遅らせる。GLP−1は、グルコース依存インスリン分泌に関わる遺伝子の転写を刺激し、β細胞新生を促進することによって、連続したβ細胞応答能を促進する(Meier, et al.Biodrugs 2003; 17 (2): 93−102)。
【0044】
本明細書に使用する場合、「GLP−1活性」とは、自然に起こるヒトGLP−1の1以上の活性を意味する。これらの活性は、限定されないが、血液および/または血漿グルコースの低下、グルコース依存インスリン分泌の刺激あるいは分泌レベルまたはインスリンの上昇、グルカゴン分泌の抑制、フルクトサミンの低下、脳へのグルコース送達および代謝の増加、胃内容排出の遅延、β細胞応答能の促進、および/または組織の新生を含む。GLP−1活性に付随したこれらの活性とその他の活性は、GLP−1活性またはGLP−1アゴニストを有する組成によって直接または間接的に引き起こされる。例えば、GLP−1活性を有する組成は、直接または間接的にグルコース−依存性を刺激するが、一方インスリン産生を刺激すると、哺乳類の血漿グルコースレベルを間接的に減少させる。
【0045】
本明細書で使用する場合、「インクレチンミメティック」とは、インスリン分泌を増強させる、あるいは分泌レベルまたはインスリンを増加できるできる化合物を意味する。インクレチンミメティックは、哺乳類において、インスリン分泌を刺激し、β細胞新生を増加させ、β細胞アポトーシスを抑制し、グルカゴン分泌を抑制し、胃内容排出を遅延させて満腹感を誘発させる能力がある。インクレチンミメティックは、GLP−1活性を有する任意のポリペプチドを含み、限定されないが、エキセンディン3およびエキセンディン4、その任意の断片および/または変異体および/または抱合体が含まれる。
【0046】
本明細書で使用する場合、「断片」とは、ポリペプチドに関して使用される場合、完全な天然ポリペプチドのアミノ酸配列とすべてではないが部分的に同じであるアミノ酸配列を有するポリペプチドである。断片は、「独立」していてもよいし、より大きなポリペプチドに含まれ、単一のより大きなポリペプチド内の単一の連続領域として部分または領域を形成してもよい。例として、天然GLP−1の断片は、天然アミノ酸1〜36位中のアミノ酸7〜36位を含む。さらに、ポリペプチドの断片は、天然配列の一部の変異体であってもよい。例えば、天然GLP−1のアミノ酸7〜30位を含むGLP−1の断片は、その一部の配列内にアミノ酸置換を有する変異体であってもよい。
【0047】
本明細書で使用する場合、「抱合体」または「抱合した」とは、互いに結合している2つの分子を意味する。例えば、第一のポリペプチドは、第二のポリペプチドに共有結合していてもよいし非共有結合していてもよい。第一のポリペプチドは、化学的リンカーにより共有結合していてもよいし、第二のポリペプチドに遺伝的に融合していてもよいが、第一ポリペプチドおよび第二ポリペプチドは共通のポリペプチド骨格を共有する。本発明の抱合体は、少なくとも1つのヒト血清アルブミンに抱合した治療ポリペプチドを含んで良い。その他の抱合体は、限定されないが、トランスフェリン、単鎖可変領域および/または少なくとも1つの抗体のFc領域に抱合した少なくとも1つの治療用ポリペプチドも含む。抱合体は、リンカーを含んでもよく、含まなくても良い。
【0048】
本明細書で使用する場合、「直列方向の」とは、同じ分子の一部として互いに隣接する2つ以上のポリペプチドを意味する。それらは共有結合か非共有結合のいずれかで結合していてよい。2つ以上の直列方向のポリペプチドは、同じポリペプチド骨格部分を形成していてよい。直列方向のポリペプチドは、順方向であっても逆方向であってもよく、および/または他のアミノ酸配列により分離されていてもよい。
【0049】
「ドメイン抗体」または「dAb」は、抗原と結合できる「単一可変ドメイン」と同義であると見なすことができる。単一可変ドメインは、ヒト抗体可変ドメインであることもあるが、さらにげっ歯類(例えば、WO 00/29004に開示されたもの)、テンジクサメおよびラクダVHH dAbsなどの他の種由来の単一抗体可変ドメインも含む。ラクダVHHは、自然界では軽鎖を持たない重鎖抗体を産生するラクダ、ラマ、アルパカ、ヒトコブラクダ、およびグアナコを含む種由来の免疫グロブリン単一可変ドメインポリペプチドである。そのようなVHHドメインは、当業者で入手可能な標準技術に従ってヒト化でき、そのようなドメインは「ドメイン抗体」と見なされる。本明細書で使用するVHは、ラクダVHH領域を含む。
【0050】
「単一可変ドメイン」という句は、異なる可変領域またはドメインとは独立に特異的に抗原またはエピトープに結合する抗原結合タンパク質可変ドメイン(例えば、VH、VHH、VL)を意味する。
【0051】
本明細書で使用する場合、「抗原結合タンパク質」とは、限定されないが、抗原と結合することができる可変ドメインとドメイン抗体などのドメインの抗体、抗体断片、ならびにその他のタンパク質コンストラクトを意味する。
本明細書で使用する場合、遺伝子改変宿主細胞内で比較される酵素またはその断片または酵素活性の「減少量」とは、非遺伝子改変宿主細胞と比較した時、少なくとも1つの酵素をより少なく産生、または少なくとも1種類の酵素活性をより少なく示す遺伝子改変宿主細胞を意味する。典型的に、遺伝子改変宿主細胞で産生した酵素活性は、遺伝的修飾前の同種の野生型系統と比較される。しかしながら、遺伝子改変宿主と、同属であるが異なる種もしくは系統からの野生型宿主または別の遺伝子改変系統を有する野生型宿主との比較を行うこともできる。少なくとも1つの酵素または酵素活性の減少には、少なくとも1つの酵素または酵素活性完全消滅が含まれ、その場合、少なくとも1つの酵素のうちのいずれも遺伝子改変宿主細胞内で産生されない、および/または少なくとも1つの酵素のうちのいずれも機能的でなく、活性を示さない。さらに、少なくとも1つの酵素活性の減少もこの定義の範囲内に含まれる。即ち、1つ以上の活性を有する酵素は、第1の活性量を維持し、一方同じ酵素の第2の活性は減少させることができる。
【0052】
本明細書で使用する場合、「厳密な条件」あるいは「厳密なハイブリッド形成法条件」とは、ハイブリッド形成が、配列間で少なくとも70%、あるいは少なくとも80%、あるいは少なくとも90%の同一性がある場合にのみ起こることを意味する。厳密なハイブリッド形成条件の例には、50%ホルムアミド、5xSSC(150mM NaCl、 15mMクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5xデンハート液、10%デキストラン硫酸、および20μg/mlの変性させ剪断したサケ精子DNAを含む溶液中で42℃で一晩インキュベーションし、続いてフィルタを約65℃で0.1xSSCを使って洗浄する工程が含まれる。ハイブリッド形成と洗浄条件は良く知られており、Sambrook, et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor, N.Y.,(1989), 特に Chapter 11に例証されており、参照によりその開示の全体をここに組み入れる。
【0053】
本明細書に使用する場合、「遺伝的修飾」または「遺伝的に修飾された」とは、1つ以上の塩基または細胞DNA配列(1つまたは複数)の断片の抑制、置換、欠失および/または挿入を意味する。そのような遺伝的修飾は、in vitroで(直接的に単離したDNA上で)またはin situで、例えば遺伝子工学技術あるいは細胞を変異原性薬剤に曝露することによって得ることができる。変異原性薬剤は、例えば、エネルギー性光線(エックス線、γ光線、紫外線など)などの物理的薬剤、またはアルキル化薬剤(EMS、NQOなど)、バイアルキレーティング薬剤、挿入剤などのDNAの異なる官能基と反応することのできる化学薬剤を含む。遺伝的修飾はまた、例えばRothsteinらによって明らかになった方法に従う遺伝的破壊によって得られる(Meth.Enzymol. 194:281−301(1991))。この方法に従うと、遺伝子の一部分または全てが相同組換えを通してin vitro修飾したバージョンによって置換される。遺伝的修飾はまた、DNA配列へ任意の変異挿入(トランスポゾン、ファージなど)することにより得ることができる。本明細書に使用する場合、「遺伝的に修飾した」とは、ポリペプチド、または核酸もしくはアミノ酸のそれぞれを少なくとも1つ欠失、置換もしくは抑制したポリペプチドをコードする遺伝子を意味することができる。例えば、野生型品種の少なくとも1つのアミノ酸を置換したポリペプチドは遺伝的修飾と見なされるだろう。
遺伝的修飾は、細胞の機序によって逆転するか、または減弱する。あるいは、変異は非逆転および/または非漏出性でありうる。「漏出変異」は、野生型機能の全部ではないが一部の不活性化を生じる変異を含む。
本発明の宿主細胞によって遂行される遺伝的修飾は、その細胞のDNA配列のコーディング領域および/または遺伝子の発現に影響を及ぼす領域に位置する。従って、本発明の修飾は一般的に、遺伝子産生またはタンパク質分解および/または糖鎖形成に関わるタンパク質および/または酵素の遺伝子産生の制御または促進に影響を及ぼす。本発明の細胞において、異種的に発現したポリペプチドをタンパク分解的に切断および/またはグリコシル化する能力が低下するのは、改変された生物学的特性を有する1つ以上の酵素の産生、かかる1つ以上の酵素の産生欠如、または1つ以上の酵素の低レベルの産生による構造上および/または立体構造上の変化が原因である。
【0054】
本発明1つの態様において、機能的遺伝子産物をコードする少なくとも1つの核酸配列および/または少なくとも1つの遺伝子産物が発現するのに必要な少なくとも1つの核酸が遺伝子改変された、遺伝的修飾ピキア株が提供される。ここで上記遺伝子産物は、遺伝的修飾ピキア株においてタンパク質分解および/または糖鎖形成を担う。本発明の遺伝子改変ピキア株は、限定されないが、次の機能的遺伝子産物少なくとも1つまたはかかる遺伝子産物の発現をコードする少なくとも1つの核酸配列が遺伝的修飾を受けているピキアを含む:PEP4、PRB1、YPS1、YPS2、YMP1、DAP2、GRH1、PRD1、YSP3、PRB3、YMP2、および/またはYMP3。さらに本発明に含まれるのは、遺伝子改変ピキアであって、このピキアは、野生型系統と比較して下記の酵素および/または酵素タイプの少なくとも1つにおいて、活性量が低下したか、全くないか、および/または少なくとも1つの活性が減少している:アスパルチルプロテアーゼ、セリンプロテアーゼ、分泌アスパルチルプロテアーゼ、分泌セリンプロテアーゼ、酵母メチロトローフのプロテアーゼ、DPP IV様エンドペプチダーゼ、金属エンドペプチダーゼ、Prb1様セリンプロテアーゼ、Prb1セリンプロテアーゼ、CPY様カルボキシペプチダーゼおよび/またはマンノシルトランスフェラーゼ。さらに、遺伝子修飾系統は下記から選択された酵素を産生することができる:アスパルチルプロテアーゼ、セリンプロテアーゼ、分泌アスパルチルプロテアーゼ、分泌するセリンプロテアーゼ、酵母メチロトローフプロテアーゼ、DPP IV様エンドペプチダーゼ、金属エンドペプチダーゼ、Prb1様セリンプロテアーゼ、Prb1セリンプロテアーゼ、CPY様カルボキシペプチダーゼおよび/またはマンノシルトランスフェラーゼ。ここで上記酵素は、野生型系統および/または酵素と比較すると、上記酵素の少なくとも1つの活性を低下させている。
【0055】
本発明の遺伝子改変ピキア株はまた、機能的遺伝子産物または上記遺伝子産物の発現をコードする以下の核酸配列のうち少なくとも1つが遺伝子改変されたピキア株を含む:OCH1、PMT1、PMT2、および/またはPMT4。本発明の遺伝子改変ピキアは、野生型系統と比較して、少なくとも1つのマンノシルトランスフェラーゼの量を減少させるか、全くなしにするかまたは活性を低下させる。さらに、遺伝子改変系統は、野生型酵素と比較して活性を低下させた少なくとも1つのマンノシルトランスフェラーゼなど、糖鎖形成に付随する酵素を産生してもよい。
【0056】
本発明の遺伝子改変ピキアは、限定されないが、野生型X−33またはSMD1163ピキア由来の遺伝子改変型を含む。本発明の遺伝子改変ピキアは、限定されないが、以下の野生型由来の遺伝子改変型を含む:ピキア・パストリス、ピキア・フィンランディカ、ピキア・トレハロヒィア、ピキア・コクラメ、ピキア・メンブラナエファシエンス、ピキア・メタノリカ、ピキア・ミヌタ(オガタ・エミヌタ、ピキア・リンドネリ)、ピキア・オプティアエ、ピキア・サーモトレランス、ピキア・サリクタリア、ピキア・ゲルカム、ピキア・ピジュペリ、ピキア・スティプティス、ピキア菌種。
【0057】
本発明の別の態様において、機能的遺伝子産物または上記遺伝子産物の発現をコードする次の核酸配列が遺伝的に改変された、改変ピキア株が提供される:PEP4、PRB1、YPS1、YPS2、YMP1、YMP2、YMP3およびPMT4。本発明の遺伝子改変ピキアは、野生型系統と比較して、以下の遺伝子産物のうち少なくとも1つについてその量を減少させるか、全くなしにするかまたは活性を低下させる:アスパルチルプロテアーゼ、セリンプロテアーゼ、セリン分泌プロテアーゼおよびマンノシルトランスフェラーゼ。本発明の遺伝子改変ピキアはまた、野生型系統と比較して、以下の遺伝子産物のうち少なくとも1つについてその活性を低下させる:少なくとも1つのアスパルチルプロテアーゼ、少なくとも1つのセリンプロテアーゼ、少なくとも1つのセリン分泌プロテアーゼおよび/または少なくとも1つのマンノシルトランスフェラーゼ。
【0058】
本発明の遺伝子改変ピキアは更に、少なくとも1つの異種ポリペプチドを発現できるポリヌクレオチドを含む。少なくとも1つの異種ポリペプチドを発現できるポリヌクレオチドは、限定されないが、ベクター、宿主細胞のゲノムに形質転換されたDNA、ウイルスまたはウイルスの一部分、および/またはプラスミドを含む。異種ポリペプチドを発現できるポリヌクレオチドはピキアのゲノムに形質転換されても良く、および/または発現ベクターの一部分であっても良く、および/またはエピソーム発現系であっても良い。
【0059】
当業者に理解されているように、DNAはいくつかの異なる方法によって宿主細胞に形質転換される。酵母の場合、エレクトロポレーション法、塩化リチウム法、またはスフェロプラスト法など、任意の簡便なDNA伝達方法を使用して良い。高密度の発酵に適した安定な系統を産生するするために、DNAを宿主染色体に組み込むことが望ましい。組込みは、当業者に周知の技術を使って相同組換え経由で行われる。例えば、少なくとも1つの異種タンパク質を発現可能なDNAには、宿主生物体の配列に相同的な隣接する配列を提供してもよい。このような方法により、所望または必須の遺伝子を破壊することなしに、宿主ゲノムの限定された部位で組込みが行われる。あるいは、少なくとも1つの異種タンパク質を発現可能なDNAは、、宿主染色体の望まれない遺伝子の部位に組み込まれ、遺伝子または遺伝子産物の発現の破壊または欠失を引き起こす。例えば、YMP1、YMP2、YMP3、PEP4、PRB1、YPS1、YPS2、DAP2、GRH1、PRD1、YSP3、PRB3、配列番号10、配列番号12および/または配列番号14の遺伝子の部位への組み込みは、異種タンパク質の発現を可能にするが、一方酵母タンパク質分解に関わる酵素の発現を妨げる。その他の実施形態において、DNAは、染色体、プラスミド、レトロウイルスベクター経由で、または宿主ゲノムにランダムに組込むことによって、宿主に導入することができる。
【0060】
本発明の別の態様において、本発明の宿主細胞内で発現した少なくとも1つの異種ポリペプチドは少なくとも1つのGLP−1アゴニストを含む。ある態様では、GLP−1アゴニストは、以下の群から選択される:インクレチンホルモンならびに/またはその断片、変異体および/もしくは抱合体、インクレチンミメティックならびに/またはその断片、変異体および/もしくは抱合体。いくつかの態様において、少なくとも1つの異種ポリペプチドは少なくとも1つのGLP−1活性を有する。
【0061】
GLP−1活性を有するポリペプチドは、ヒトGLP−1の少なくとも1つの断片および/または変異体を含み得る。ヒトGLP−1の天然に生じた2つの断片を配列番号2に示す。
7 8 9 10 11 12 13 14 15 16 17
His-Ala-Glu-Gly-Thr-Phe-Thr-Ser-Asp-Val-Ser-
18 19 20 21 22 23 24 25 26 27 28
Ser-Tyr-Leu-Glu-Gly-Gln-Ala-Ala-Lys-Glu-Phe-
29 30 31 32 33 34 35 36 37
Ile-Ala-Trp-Leu-Val-Lys-Gly-Arg-Xaa (配列番号2)
ここで37位のXaaはGly(以下、「GLP−1(7−37)」として示される)、または−NH2(以下、「GLP−1(7−36)」として示される)である。GLP−1断片としては、ヒトGLP−1のアミノ酸7〜36位(GLP−1(7−36))を含む、あるいはそれで構成されるGLP−1分子を挙げ得るが、これらに限定されない。GLP−1の変異体またはその断片としては、野生型GLP−1または配列番号2に示されるGLP−1の天然に生じた断片における、1つ、2つ、3つ、4つ、5つまたはそれ以上のアミノ酸置換を挙げ得るが、これらに限定されない。変異体GLP−1またはGLP−1の断片としては、野生型GLP−1のアラニン8に類似のアラニン残基の置換であり、かかるアラニンがグリシンに変異している(以下、「A8G」として示される)(例えば、全体を参照として本明細書に援用する米国特許第5,545,618号に開示の突然変異体を参照されたい)場合を挙げ得るが、これらに限定されない。
【0062】
別の態様において、GLP−1活性を有する少なくとも1つのポリペプチドは、ヒト血清アルブミンと融合したヒトGLP−1の少なくとも1つの断片および/または変異体を含む。別の態様では、GLP−1の少なくとも1つの断片および変異体はGLP−1(7−36(A8G))を含む。GLP−1の少なくとも1つの断片および変異体は、ヒト血清アルブミンと遺伝的に融合する。別の態様において、本発明の異種ポリペプチドは、少なくとも2個のGLP−1(7−36(A8G))を直列に含み、かつ、遺伝的にヒト血清アルブミンと融合する。2個のGLP−1(7−36(A8G))は、ヒト血清アルブミンのN末端で遺伝的に融合する。いくつかの場合には、異種ポリペプチドは配列番号1を含む。
HGEGTFTSDVSSYLEGQAAKEFIAWLVKGRHGEGTFTSDVSSYLEGQAAKEFIAWLVKGR 60
DAHKSEVAHRFKDLGEENFKALVLIAFAQYLQQCPFEDHVKLVNEVTEFAKTCVADESAE 120
NCDKSLHTL FGDKLCTVATLRETYGEMADCCAKQEPERNECFLQHKDDNPNLPRLVRPE 180
VDVMCTAFHDNEETFLKKYLYEIARRHPYFYAPELLFFAKRYKAAFTECCQAADKAACLL 240
PKLDELRDEGKASSAKQRLKCASLQKFGERAFKAWAVARLSQRFPKAEFAEVSKLVTDLT 300
KVHTECCHGDLLECADDRADLAKYICENQDSISSKLKECCEKPLLEKSHCIAEVENDEMP 360
ADLPSLAADFVESKDVCKNYAEAKDVFLGMFLYEYARRHPDYSVVLLLRLAKTYETTLEK 420
CCAAADPHECYAKVFDEFKPLVEEPQNLIKQNCELFEQLGEYKFQNALLVRYTKKVPQVS 480
TPTLVEVSRNLGKVGSKCCKHPEAKRMPCAEDYLSVVLNQLCVLHEKTPVSDRVTKCCTE 540
SLVNRRPCFSALEVDETYVPKEFNAETFTFHADICTLSEKERQIKKQTALVELVKHKPKA 600
TKEQLKAVMDDFAAFVEKCCKADDKETCFAEEGKKLVAASQAALGL 674
(配列番号1)
【0063】
さらに別の実施形態では、本発明の宿主細胞で発現した少なくとも1つの異種ポリペプチドは、少なくとも1つの抗原結合タンパク質、少なくとも1つの単一可変ドメイン、および/または少なくとも1つのドメイン抗体のうち1つ以上を含む。少なくとも1つの抗原結合ドメインを含むポリペプチドはまた、少なくとも1つのポリペプチドおよび/またはペプチド受容体アゴニストおよび/または拮抗物質を含む。いくつかの場合には、ポリペプチドアゴニストは、GLP―1受容体アゴニストであっても良い。当業者に理解されているように、2個以上の異種ポリペプチドが、同じ細胞内で発現する。例えば、GLP−1活性を有する異種ポリペプチドは、抗原結合タンパク質と同じ細胞内に発現させることができる。GLP−1活性を有する異種ポリペプチドは、発現に必要な核酸配列に動作可能に連結した抗原結合タンパク質と同じポリヌクレオチドから発現させても良い。あるいは、例えば、GLP−1活性を有するポリペプチドは、発現に必要な異なるポリヌクレオチド配列と動作可能に連結した同じエピソームのDNAまたはゲノムから、または別々のベクター上に位置するDNA配列から、抗原結合タンパク質などの第2の異種ポリペプチドとは独立に、発現させても良い。
【0064】
さらに、次のような遺伝子改変ピキア株も提供される:野生型ピキアと比較すると上記系統の上記少なくとも1つの異種ポリペプチドがタンパク質分解を低下させている。さらに、本発明の遺伝子改変ピキアは、野生型ピキアと比較すると、上記系統の上記少なくとも1つの異種ポリペプチドが糖鎖形成を低下させているか糖鎖形成をしないことを示す。
【0065】
別の態様では、本発明の遺伝子改変ピキアにおいて、上記異種ポリペプチドを発現させるステップを含む異種ポリペプチドの産生方法が提供される。さらに、本発明の遺伝子改変ピキアの任意の1つで産生する異種ポリペプチドも提供される。
【0066】
さらに本発明の別の態様では、配列番号3、5、および7と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%配列同一性を有する単離したポリヌクレオチドが提供される。さらに本発明の別の態様では、配列番号4、6、および8と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%配列同一性を有する単離したポリヌクレオチドが提供される。さらに実施形態は、遺伝子改変宿主細胞の野生型親が、配列番号4、6、8と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%の配列同一性を有するポリペプチドをコードする遺伝子を含む、遺伝子改変宿主細胞を含む。ここで上記遺伝子は、野生型宿主細胞と比較すると、上記遺伝子改変宿主細胞内で、上記遺伝子産物が低下するか除去されるように、上記宿主細胞のゲノム内で遺伝子改変される。さらに、野生型親が配列番号3、5、および7と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%の配列同一性を有する遺伝子を含み、上記遺伝子が、上記遺伝子改変宿主細胞のゲノム内で遺伝子改変されている、遺伝子改変宿主細胞も提供される。本発明の遺伝子改変宿主細胞は、上記野生型親宿主細胞と比較すると、プロテアーゼ活性が低下しているか除去されている。本発明の遺伝子改変宿主細胞は、メチロトローフ酵母プロテアーゼ(Ymp1)活性が低下しているか除去されている。いくつかの場合、宿主細胞はピキアである。
【0067】
さらに本発明の別の態様では、配列番号9、11および13と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%配列同一性を有する単離したポリヌクレオチドが提供される。さらに本発明の別の態様では、配列番号10、12および14と少なくとも50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、99%、および/または100%配列同一性を有する単離したポリヌクレオチドが提供される。
【0068】
当業者に理解されているように、培養下の宿主細胞によって産生された1以上の酵素の酵素活性は、培養の増殖条件によって影響されることがある。例えば、培養液中のの宿主細胞により産生されるプロテアーゼのタンパク分解活性は、下記の1以上の条件を変化させることによって低下させることができるだろう。条件:pH、溶存酸素、温度、モル浸透圧濃度、1以上の培地成分、特異的プロテアーゼ阻害薬、増殖時間および/または割合、細胞濃度、培養持続時間、および/またはグルコース摂食割合(例えば、流加培養)。タンパク水解物複合体を培養に加えると、タンパク質分解の抑制が極めて効率的である。さらに、最大の効果を得るために、条件を培養中1以上の特定時間に変化させてもよい。同様に、培養で産生されるタンパク質の糖鎖形成も、同様な因子によって影響されることがある。したがって、培養中、タンパク分解活性や糖鎖形成活性などの宿主細胞の酵素活性を低下させる増殖条件は、上記にリストされた1以上の非限定要因を調整することにより最適化することができる。
【0069】
さらに、当業界で理解されているように、宿主細胞の異種タンパク質の産生は、上記の同じ因子の多くを制御することによって増加させることができる。さらに加えて、限定されないが、ラパマイシンの増殖培地への付加を含め、ベクターのコピー数を増加させる因子を加えることによって、産生を増加させることもできる。産生を増加させることのできる他の因子には、限定されないが、タンパク質ジスルフィド異性化酵素(PDI)などの1以上のシャペロンタンパク質の同時発現も含まれる。さらに、ヘモグロビン(HB)は、宿主細胞の少なくとも1つの異種ポリペプチドと同時発現させることができ、酸化的代謝に必要な酸素の利用性を高め、従ってポリペプチド産生を増加させる。
【0070】
産生中に宿主細胞から分泌された異種タンパク質は、分泌を促進するリーダー配列を含んでも良い。リーダー配列は、分泌を改善させることにより異種的に発現したタンパク質の全体的な産生と回収を向上させるように、修飾してもよい。例えば、多様な分泌タンパク質由来の異なるリーダー配列を異種タンパク質に動作可能に連結し、発現改善について評価してもよい。あるいは、所定のリーダー配列は、改善されたリーダー配列変異体の同定のため、部位特異的変異誘発、または、コンビナトリアルライブラリのアプローチ手段によって修飾してもよい。異種タンパク質発現レベルを改善するため、2個以上のリーダーペプチドの領域を含むキメラのリーダー配列をを見つけてもよい。
【0071】
実施例
以下の実施例はこの発明の様々な非限定的態様を説明する。
【0072】
実施例1−Ymp1新規なプロテアーゼ
新規なプロテアーゼを親和性精製およびLC/MS分析を使ってピキア培養上清から同定した。図1は、ゲル上に同定された新規なピキアメチロトローフ酵母プロテアーゼ(Ymp1)を表す図である。B378(ピキアヌル系統)からの浄化ろ過された上清を、ビオチン標識したアプロチニンで一晩インキュベートした。上清を2mLアビジンカラム(ピアス)に供し、リン酸緩衝液で洗浄した。カラムを、0.1Mトリス、2M塩化ナトリウム、pH7.5、次に0.1M酢酸ナトリウム、2M塩化ナトリウム、pH4.5そして最終的にビオチン溶出緩衝液(ピアス)で経時的に溶出させた。溶出液を濃縮して、SDS−PAGEを行った。もっとも顕著なバンドであるバンド1と2をゲルから切り出し、還元し、アルキル化し、in situでトリプシンを使用して消化した。バンドからのトリプシンペプチドを液体クロマトグラフィー・タンデム質量分析で分析した。未解釈の配列データを、Mascotタンパク質同定ソフトウェアを使ってピキアコンティグ−ペプチドデータベース(Integrated Genetics)内で検索した。
【0073】
バンド1と2の両方から産生したペプチド類は、ピキアゲノムデータベース内のある翻訳領域(ORF)と整合した。遺伝子を単離して配列を求めると、1704アミノ酸の新規なタンパク質(配列IDNO:4)をコードする5115bp核酸(配列番号3)からなる翻訳領域を含んでいた。タンパク質全体の予想した分子量は、185.8KDである。ピキア(配列番号3に示される)からのこの遺伝子のポリヌクレオチド配列は、GenBankのどの配列とも相同性を有意には示さない。コード化したアミノ酸配列は、2個の構造領域含む:N末端領域(AA1−865)は、ボタンタケ属lixiiセリンエンドペプチダーゼと30.2%の同一性を共有し、C末端領域(AA1305−1449)は、乳酸桿菌ロイテリからのLPXTG−モチーフ細胞壁アンカー領域タンパク質と48%の同一性を共有する。しかしながら、配列番号4とヒポクレアlixiiセリンエンドペプチダーゼとの全長比較は、14.8%の同一性を示したに過ぎない。ここで2つの配列のグローバルアラインメントを計算するWilliam Pearson’s LALIGNプログラムバージョン2.2u(Myers and Miller、CABIOS(1989)4:11−17)を使用した。このプロテアーゼの予想された構造を図2に示す。そのC末端構造と配列は、全ての種の全てのセリンプロテアーゼの中でユニークである。遺伝子は、メチロトローフピキア培養に出現するので、酵母メチロトローフプロテアーゼ(YMP1)と名付けられた。これは、そのユニークな構造にもとづくプロテアーゼの新規なクラスである。Ymp1は、LC/質量分析(データを示さず)を使ったピキア培養上清のグローバル解析に基づくピキアメチロトローフ培地でもっとも大量に分泌するタンパク質の一つである。
【0074】
ATGTTCCTCA AAAGTCTCCT TAGTTTTGCG TCTATCCTAA CGCTTTGCAA GGCCTGGGAT
CTGGAAGATG TACAAGATGC ACCAAAGATC AAAGGTAATG AAGTACCCGG TCGCTATATC
ATTGAGTATG AAGAAGCTTC CACTTCAGCA TTTGCTACCC AACTGAGAGC TGGGGGATAT
GACTTTAACA TCCAATACGA CTACTCAACT GGTTCCCTTT TCAACGGAGC ATCTGTTCAA
ATCAGCAACG ATAACAAAAC CACTTTCCAG GATTTGCAAA GTTTGCGTGC AGTCAAAAAT
GTTTACCCAG CTACTCTCAT TACATTAGAT GAAACATTTG AGCTTGCTGA CACGAAGCCA
TGGAACCCTC ATGGAATTAC CGGTGTCGAT TCTTTGCATG AGCAAGGATA TACTGGTAGT
GGTGTTGTTA TTGCAGTTAT CGATACTGGT GTTGACTATA CACACCCTGC TCTGGGTGGT
GGTATCGGAG ATAATTTCCC TATCAAAGCT GGTTATGATT TGTCTTCCGG TGATGGTGTC
ATCACGAATG ATCCTATGGA TTGTGACGGT CATGGTACCT TTGTATCCTC CATCATTGTT
GCAAATAACA AAGATATGGT TGGTGTTGCA CCAGATGCTC AGATTGTCAT GTACAAAGTG
TTCCCCTGTT CTGATAGTAC TTCGACTGAC ATAGTTATGG CGGGTATGCA AAAGGCCTAT
GATGATGGTC ACAAGATTAT TTCGCTATCA CTGGGATCTG ACTCGGGGTT TTCCAGTACT
CCAGCTTCCT TAATGGCCAG CAGGATTGCT CAAGACAGAG TTGTTTTGGT GGCTGCTGGT
AACTCTGGAG AACTTGGTCC ATTCTATGCC TCCTCCCCTG CTTCTGGGAA ACAAGTCATT
TCAGTTGGAT CTGTTCAAAA CGAACAATGG ACAACCTTTC CAGTAACCTT TACCTCTTCA
AACGGTGAAT CAAGGGTTTT TCCTTACCTC GCTTACAATG GTGCACAGAT TGGATTTGAT
GCCGAGCTTG AGGTTGATTT TACCGAAGAA AGAGGATGCG TCTATGAACC AGAGATCTCC
GCAGATAATG CGAATAAAGC TATTTTGTTA AGAAGGGGCG TCGGCTGTGT TGAAAACTTG
GAATTCAATT TATTGTCTGT GGCTGGTTAC AAGGCTTACT TCTTGTACAA CTCATTTTCA
AGACCATGGA GTCTCTTGAA TATTTCTCCA CTGATTGAGC TAGACAACGC TTACTCTCTT
GTTGAAGAGG AAGTTGGAAT ATGGGTGAAA ACCCAAATCG ACGCCGGTAA CACCGTCAAG
TTAAAGGTGA GCACGAGTGA CCAAATGTTG CCATCTGATA AAGAGTATTT GGGAGTTGGA
AAGATGGATT ATTACTCCTC TCAAGGACCT GCTTATGAGC TTGAATTTTT CCCAACGATA
TCCGCTCCAG GTGGAGACAG TTGGGGCGCT TGGCCCGGTG GGCAATACGG TGTTGCCTCA
GGAACAAGTT TTGCTTGCCC CTATGTTGCA GGTCTTACAG CTCTTTATGA ATCGCAGTTT
GGAATTCAAG ATCCCCAGGA CTATGTGAGA AAATTAGTCT CCACAGCTAC CGATCTTCAA
TTATTTGACT GGAACGCAGT GAAACTTGAG ACCTCTATGA ATGCTCCACT TATTCAACAG
GGAGCTGGTC TAGTGAACGC TCTTGGTTTG TTTGAGACTA AGACTGTGAT CGTGTCTGCT
CCTTATTTGG AGCTCAATGA CACCATCAAT AGAGCCAGTG AGTATACCAT TCAAATTAAG
AATGAGAACT CTGAGACTAT TACCTATCAA GTTGTTCACG TTCCGGGAAC TACTGTCTAC
TCTAGATCAG CTTCTGGGAA CATCCCATAC CTGGTCAATC AAGATTTTGC ACCTTACGGT
GATAGTGATG CTGCGACAGT TGCTCTATCC ACAGAAGAGT TGGTTTTGGG ACCAGGAGAA
GTTGGTGAAG TCACTGTGAT CTTCTCTACA GAAGAAATTG ATCAAGAAAC TGCTCCAATT
ATTCAGGGTA AGATTACATT TTATGGTGAT GTCATACCGA TTGCTGTTCC TTATATGGGA
GTTGAAGTTG ATATTCATTC CTGGGAGCCT CTCATTGAGA GGCCTTTATC AGTGAGAATG
TATTTGGATG ATGGTTCCTT AGCATATGTT GATGATGATC CTGATTATGA GTTCAATGTG
TATGACTGGG ATTCTCCTAG ATTTTATTTT AACCTGAGAT ATGCAACCAA AGAAGTATCG
ATTGACTTGG TGCACCCTGA TTATAGCATT GAGAACGACT ACGAATGGCC TTTAGTTTCC
GGACACAACA ACTATTATGG TCCCGTGGGA TACGACTACG ATTATACCTC GGGTCAAGCC
TTTTTGCCTC GTTACTTTCA ACAACGTATT AACGAACTTG GATATCTTTC TTTTTCCAGA
TTTGCTAACT TTTCTGTAGT TCCTGCTGGT GAATACAAAG CTCTATTTAG AGTTTTGCTA
CCATATGGAG ACTTTTGGAA CAAAGAAGAC TGGCAATTGT TTGAATCCCC AGTGTTTAAC
GTCCTCGCTC CACCGAATGA AGAAAACACT ACTGAAGAGC CAACTGAGGA ATCCAGCGAG
GAGCCTACCG AAGAGTCAAC GTCTGAGTCA ACTGAAGAGC CCTCTTCTGA GTCAACTGAG
AAATCTAGCG AGGTGCCAAC TGAAGAAATT ACTGAAGATG CAACATCCAC AATTGATGAT
GATGAAGCAT CCACCGAAAG CTCTACTGAA GAACCAAGTG CTCAGCCCAC CGGTCCTTAC
TCTGATTTGA CTGTCGGTGA GGCCATTACC GACGTTAGTG TCACCAGTTT GAGGACAACT
GAAGCATTTG GATACACTTC CGACTGGTTG GTTGTGTCTT TCACTTTCAA CACTACTGAC
AGAGATATTA CTCTCCCACC TTACGCTGTT GTACAAGTAA CTATCCCAAA TGAACTTCAA
TTCATTGCTC ATCCAGAATA CGCCCCATAC CTTGAGCCCT CATTGCAAGT TTTCTACACT
AAGAATGAAA GATTAATTAT GACTAGTCAG TTCAACTACG ACACCAGAGT CATCGACTTC
AAGTTTGACA ATCGAGACCA AGTAATAACT CAAGTGGAGG GAGTTGTTTA TTTCACGATG
AAACTAGAAC AAGATTTCAT TTCTGCATTG GCCCCAGGTG AATACGATTT TGAATTTCAT
ACATCCGTTG ATTCTTATGC TTCGACCTTT GACTTTATTC CATTGATTAG ATCCGAGCCA
ATCAAATTGA TAGCAGGTGC ACCAGACGAA GTTGAATGGT TTATTGATAT TCCAAGTGCA
TACAGCGATT TGGCAACGAT AGATATTAGT TCTGATATCG ATACTAATGA TAATTTGCAG
CAGTACTTCT ATGATTGCTC AAAGCTCAAG TACACTATTG GAAAAGAGTT TGATCAGTGG
GGTAATTTTA CAGCTGGATC AGATGGTAAC CAATACAGCA ATACCACCGA TGGGTATGTT
CCAATTACTG ATTCTACCGG CTCTCCAGTA GCTGAAGTTC AATGTTTAAT GGAAAGTATC
TCATTGAGTT TCACAAATAC TCTTGCTGAG GATGAAGTAT TGAGAGTTGT TCTTCACTCT
TCTGCGTTTA GACGTGGTTC ATTCACCATG GCCAACGTGG TAAACGTTGA CATTACAGCT
GGTGGATTGG CAAAAAGAGA ACTCTTCTCT TATATATTGG ATGAAAATTA CTATGCTAGT
ACTGGATCTG AGGGGTTGGC ATTTGACGTA TTTGAAGTTG CTGATCAGGT CGAGGAGCCA
ACTGAGGAGT CAACCTCAGA GGAATCTACT GAACAGGAAA CTTCCACCGA GGAACCTACC
GAGGAATCAA CTGAACCTAC TGAGGAATCT ACCCAGGAAC CTACTGAAGA GCCCACCGAC
GAGCCTACTT CTGAGTCAAC TGAGGAACCT TCTGAGGAGC CAACTTCTGA CGATCTCTCA
ATTGACCCAA CTGCTGTACC TACCGATGAA CCTACTGAAG AGCCAACTGA GGAGCCTACT
TCTGAGTCAA CTGAGGAACC TTCTGAGGAG CCAACTTCTG ACGATCTCTC AATTGACCCA
ACTGCTGTAC CTACCGATGA ACCTACTGAA GAGCCAACTG AGGAGCCTAC TTCTGAGTCA
ACTGAGGAAC CTTCTGAGGA GCCAACTTCT GACGATCTCT CAATTGACCC AACTGCTGTA
CCTACCGATG AACCTACTGA AGAGCCAACT GAGGAGCCGA CCTCTGAGAC TACCGATGAT
CCATCGATAG CACCTACTGC TGTGCCAACT TCCGACACAT CTTCTGGACA ATCGGTGGTT
ACTCAAAACA CTACAGTCAC TCAGACTACC ATCACTTCAG TCTGTAATGT TTGTGCTGAG
ACCCCTGTAA CAATCACTTA CACTGCACCA GTTGTGACTA AGCCAGTTTC TTACACCACC
GTTACTTCAG TTTGCCATGT ATGTGCAGAG ACACCAATCA CAGTTACCTT GACGTTGCCA
TGTGAAACCG AAGACGTGAC AAAGACTGCC GGCCCTAAGA CTGTCACTTA CACCGAAGTT
TGCAACTCCT GTGCTGACAA GCCTATCACT TACACCTACA TCGCTCCAGA GTACACTCAA
GGTGCCGAAC GTACAACAGT TACATCGGTT TGCAACGTTT GTGCTGAGAC ACCTGTAACG
CTAACATACA CTGCGCCGAA AGCCAGTCGT CATACAGTTC CTTCACAATA TTCAAGTGCC
GGAGAGCTCA TTTCATCCAA GGGGATCACG ATTCCTACTG TTCCTGCCCG TCCAACTGGT
ACTTATAGTA AGTCTGTTGA CACTAGCCAA CGTACACTCG CTACCATTAC AAAATCTTCA
GATGAGTCTA ACACTGTTAC CACTACTCAA GCCACACAAG TTTTGAGCGG TGAATCCAGT
GGAATTCAAG CTGCTTCAAA CAGCACGAGC ATCTCAGCTC CAACTGTCAC TACAGCTGGG
AACGAGAACT CTGGATCTAG ATTTTCGTTT GCTGGACTAT TCACAGTTCT GCCTCTTATC
TTGTTCGTTA TATAA(配列番号3)
【0075】
配列番号3の遺伝子配列によってコードしたポリペプチド配列は、配列番号4として以下に示される。
MFLKSLLSFA SILTLCKAWD LEDVQDAPKI KGNEVPGRYI IEYEEASTSA FATQLRAGGY
DFNIQYDYST GSLFNGASVQ ISNDNKTTFQ DLQSLRAVKN VYPATLITLD ETFELADTKP
WNPHGITGVD SLHEQGYTGS GVVIAVIDTG VDYTHPALGG GIGDNFPIKA GYDLSSGDGV
ITNDPMDCDG HGTFVSSIIV ANNKDMVGVA PDAQIVMYKV FPCSDSTSTD IVMAGMQKAY
DDGHKIISLS LGSDSGFSST PASLMASRIA QDRVVLVAAG NSGELGPFYA SSPASGKQVI
SVGSVQNEQW TTFPVTFTSS NGESRVFPYL AYNGAQIGFD AELEVDFTEE RGCVYEPEIS
ADNANKAILL RRGVGCVENL EFNLLSVAGY KAYFLYNSFS RPWSLLNISP LIELDNAYSL
VEEEVGIWVK TQIDAGNTVK LKVSTSDQML PSDKEYLGVG KMDYYSSQGP AYELEFFPTI
SAPGGDSWGA WPGGQYGVAS GTSFACPYVA GLTALYESQF GIQDPQDYVR KLVSTATDLQ
LFDWNAVKLE TSMNAPLIQQ GAGLVNALGL FETKTVIVSA PYLELNDTIN RASEYTIQIK
NENSETITYQ VVHVPGTTVY SRSASGNIPY LVNQDFAPYG DSDAATVALS TEELVLGPGE
VGEVTVIFST EEIDQETAPI IQGKITFYGD VIPIAVPYMG VEVDIHSWEP LIERPLSVRM
YLDDGSLAYV DDDPDYEFNV YDWDSPRFYF NLRYATKEVS IDLVHPDYSI ENDYEWPLVS
GHNNYYGPVG YDYDYTSGQA FLPRYFQQRI NELGYLSFSR FANFSVVPAG EYKALFRVLL
PYGDFWNKED WQLFESPVFN VLAPPNEENT TEEPTEESSE EPTEESTSES TEEPSSESTE
KSSEVPTEEI TEDATSTIDD DEASTESSTE EPSAQPTGPY SDLTVGEAIT DVSVTSLRTT
EAFGYTSDWL VVSFTFNTTD RDITLPPYAV VQVTIPNELQ FIAHPEYAPY LEPSLQVFYT
KNERLIMTSQ FNYDTRVIDF KFDNRDQVIT QVEGVVYFTM KLEQDFISAL APGEYDFEFH
TSVDSYASTF DFIPLIRSEP IKLIAGAPDE VEWFIDIPSA YSDLATIDIS SDIDTNDNLQ
QYFYDCSKLK YTIGKEFDQW GNFTAGSDGN QYSNTTDGYV PITDSTGSPV AEVQCLMESI
SLSFTNTLAE DEVLRVVLHS SAFRRGSFTM ANVVNVDITA GGLAKRELFS YILDENYYAS
TGSEGLAFDV FEVADQVEEP TEESTSEEST EQETSTEEPT EESTEPTEES TQEPTEEPTD
EPTSESTEEP SEEPTSDDLS IDPTAVPTDE PTEEPTEEPT SESTEEPSEE TSDDLSIDPT
AVPTDEPTEE PTEEPTSEST EEPSEEPTSD DLSIDPTAVP TDEPTEEPTE EPTSETTDDP
SIAPTAVPTS DTSSGQSVVT QNTTVTQTTI TSVCNVCAET PVTITYTAPV VTKPVSYTTV
TSVCHVCAET PITVTLTLPC ETEDVTKTAG PKTVTYTEVC NSCADKPITY TYIAPEYTQG
AERTTVTSVC NVCAETPVTL TYTAPKASRH TVPSQYSSAG ELISSKGITI PTVPARPTGT
YSKSVDTSQR TLATITKSSD ESNTVTTTQA TQVLSGESSG IQAASNSTSI SAPTVTTAGN
ENSGSRFSFA GLFTVLPLIL FVI.(配列番号4)
【0076】
実施例2−新規なプロテアーゼ
ノックアウト系統(実施例3を参照)のピキアymp1の上澄みから、実施例1に記述したのと同様に、親和性精製とLC/MS分析を使ってさらに2つの新規なメチロトローフ酵母プロテアーゼ(Ymp2およびYmp3)を同定した。B580(ピキアymp1欠失系統)からの浄化ろ過された上清を、ビオチン標識したAEBSF(4−(2−アミノエチル)ベンゼンスルホニルフッ化物塩酸塩で一晩インキュベートした。上清を、10mLアビジンカラムに充填し(ピアス)、そしてリン酸緩衝液で洗浄した。カラムを、0.1Mトリス、2M塩化ナトリウム、pH7.5、次に0.1M酢酸ナトリウム、2M塩化ナトリウム、pH4.5、そして最終的にビオチン溶出緩衝液(ピアス)で経時的に溶出した。溶出物を8M尿素を加えて変性させ、20mMジチオスレイトール(DTT)と反応させてシステイン残基を還元し、次に、20mMヨードアセトアミドと反応させてアルキル化した。試料を100mMアンモニウム炭酸水素塩(pH8.5)で希釈して2M尿素とした後、タンパク質はトリプシンで消化した(Sequencing grade modified trypsin, Promega, Madison, WI)。消化の結果生じたペプチド類混合物をPepCleanスピンカラム(Pierce, Rockford, IL)を使って脱塩し、LC−MS/MS systemを使って分析した。このシステムでは、75μm内径逆相C18カラムを持つ高圧液体クロマトグラフィー(HPLC)が、オンラインでイオントラップ質量分析計(Thermo, Palo Alto, CA)と連結されていた。LC−MS/MS分析から得た質量分析データを使って、GenBankからの最近の非重複性タンパク質データベースとピキアデータベース(Integrated Genetics)内を、Mascotタンパク質同定ソフトウェアを使用して検索した。
【0077】
ペプチドの整合により、YMP2およびYMP3と名付けられた2つの遺伝子を同定し周知のプロテアーゼとの相同性を比べた。YMP2は、205アミノ酸の新規なタンパク質(配列番号6)をコードする618bp核酸(配列番号5)のオープンリーディングフレームを含む。タンパク質全体の予想される分子量は22.9KDである。ピキア(配列番号5に示す)からのこの遺伝子のポリヌクレオチド配列は、GenBankのいかなる配列とも相同性を有意に共有しない。しかしながら、コード化したアミノ酸配列は、ピキア・パストリスからのアルギニン/アラニンアミノペプチダーゼと29%の同一性を共有する。
ATGGCTCCCA GAACACTACC AGAAGACTTA ATTCCCTCCC TATACGACTT GCACATCTAC
AACTTCCAAC CCGAAAAAAA GACTTATGAT GGAGACATTG TCATCCACTT GGAGGTGAAG
GAGCCCACTG ATGAAGTGGT CTTCAATGCC AAGGATTTGG AATTGAAAGA CGTACATGTC
TTCCACAATG TCAACAAGTC TGAAAACGAA ATCCCCGTTA AGGAGATTGT TGATAACGAG
CTCATCACAA TTAAGCTCAA AGAGAAGGTT ACTTCCGGAA CGTTGCTGGT GAATATTTCC
TTCACCGGTA ACATTCAATC TGATAAAATT GGATTTTACA AGGGAGACAC AGATGTGGAA
GGAAGAGTCA CATACACTAC AAACCTTACC ACTCCAAATG CCAGGTTGGC ATTCCCATGT
CTTGATAACA TATTGTTGAA AGCTCCATTC AAGTTCGGAG TAACTGCCAA TCCAGGACAA
TTAGTGAGTT CCATTTTGGA TCTAAGCTCT GAGGCTGACG TCTTGAATGA CAATGACGAT
GTGATTGGTA CGAGATACCA ATACCAAGTG AGTGAGCCAA TAGCCCCAGC TTTACTGGAG
TGGACCATTC ATATTTAA (配列番号5)
【0078】
配列番号5の遺伝子配列によってコードしたポリペプチド配列は配列番号6として以下に示される。
MAPRTLPEDL IPSLYDLHIY NFQPEKKTYD GDIVIHLEVK EPTDEVVFNA KDLELKDVHV
FHNVNKSENE IPVKEIVDNE LITIKLKEKV TSGTLLVNIS FTGNIQSDKI GFYKGDTDVE
GRVTYTTNLT TPNARLAFPC LDNILLKAPF KFGVTANPGQ LVSSILDLSS EADVLNDNDD
VIGTRYQYQV SEPIAPALLE WTIHI (配列番号6)
【0079】
YMP3は、410のアミノ酸(配列番号8)の新規なタンパク質をコードする1233bp核酸(配列番号7)のオープンリーディングフレームを含む。タンパク質全体の予想される分子量は46.0KDである。このアミノ酸配列は、ピキアstipitisからのロイシンアミノペプチダーゼと47%の同一性を共有する。
ATGGTCAAAC TCATATCAAT TATAGCCCTA GTTCAACTTG TCTCTGCGAC AATTGTACCT
TGGAATCTCC AGAACGTCTT ATCTGACGTC CATCACCCTT CTCTCCATCT CTTGGATTAT
ATTCAATCCT TGAAGAACGA GGTAATGTTC GATGGCGACG ATCGCAGAAT AATCAAGTTA
GGCCCCCAAG AATACCGTAT TATCACTGAA AAAGAGAAAT ACCAGTTGAA AACAGAGGGG
ATATCATTTA TCGATGTCAC CTATCAGCAT GGAGACAATG TAGAGCTGCT CTATTCCAGT
GCGCCAGTTA CCGTTCCAGA CTATCTTTAT CCGTCCAATG ATACTTTCCA TTTCAAACAA
GTAAATTCTT TGATAGGTGA GATTGACATT GGCAGAATGC AGGCGTTTTT GGGAAGGTTC
TCTAGCTTCT TTACAAGATT TTACAAATCT GACAAGGGGT TGCAGAGTTC TATCTGGTTA
CAAGGTGAAT TGGTTCAATT GGCCTTGAAA GATCCATCGA GGTTCAATGT TACTACTGTG
GAACACCCTT GGAAGCAGAA TTCTGCCATC TTTACGATAT ACGGTGAAAA TGTTGATCCT
TCGAAAGGAA AAGGGGACAT TGTAGTAGTG GGATGCCATC AAGATTCCAT AAACTTGCTT
TTCCCCAACA TTCTCCGTGC TCCAGGGGCT GATGATGATG GATCTGGTGT AACTTCCAAC
CTTGAAGCGC TCAGAATCAT AGTTGAAAGT GGCCTCAAGT TTCACAATAC AGTAGAGTTT
CACTTTTATT CTGCCGAAGA AGGAGGACTA CTTGGCTCCC AGCAAATTTT CAGCTCGTAT
AGAGCTGCAG AAGAGACTGT TGTTGCTATG CTACAACAGG ACATGACTGG ATACATCCAA
AAAGCTTTAG ACCACGGGGA ATCCGACCAC TTCGGGCTAA TCACTGACCA TACAAACGCA
AATCTGAATA GCTTCCTTGC ACTTTTAATC GATGCATACA CTTCAATTCC CTACAAAGAA
ACCGAATGTG GGTATGCCTG CTCAGATCAT AGTTCTGCCT TGGAACATGG TTATCCATCT
GCCATGGTCT TTGAAAGTAG TTTTGCCTAC ACAAATCCCT TCATCCATAG CACCCAAGAC
ACAATTGACA AGATCAATTT TCCACATATG GCAGAGCAT GTCAAGTTGG TCCTGGGTTA
CGTTGTAGAG TTGGGATTAG AACATTTTAG GTGA (配列番号7)
【0080】
配列番号7の遺伝子配列によってコードしたポリペプチド配列は配列番号8として以下に示される。
MVKLISIIAL VQLVSATIVP WNLQNVLSDV HHPSLHLLDY IQSLKNEVMF DGDDRRIIKL
GPQEYRIITE KEKYQLKTEG ISFIDVTYQH GDNVELLYSS APVTVPDYLY PSNDTFHFKQ
VNSLIGEIDI GRMQAFLGRF SSFFTRFYKS DKGLQSSIWL QGELVQLALK DPSRFNVTTV
EHPWKQNSAI FTIYGENVDP SKGKGDIVVV GCHQDSINLL FPNILRAPGA DDDGSGVTSN
LEALRIIVES GLKFHNTVEF HFYSAEEGGL LGSQQIFSSY RAAEETVVAM LQQDMTGYIQ
KALDHGESDH FGLITDHTNA NLNSFLALLI DAYTSIPYKE TECGYACSDH SSALEHGYPS
AMVFESSFAY TNPFIHSTQD TIDKINFPHM AEHVKLVLGY VVELGLEHFR (配列番号8)
【0081】
さらに、3個の他の潜在的プロテアーゼが、全て周知の酵母プロテアーゼを使って、ピキアデータベース内で同定された。これらのプロテアーゼおよび対応するそれぞれのポリペプチド配列をコードする同定されたポリヌクレオチド配列は、配列番号9〜14として以下に示す。
ATGAAATCGG TTATTTGGAG CCTTCTATCT TTGCTAGCAT TGTCGCAGGC ATTGACTATT
CCATTGCTGG AAGAGCTTCA ACAGCAAACA TTTTTTAGCA AGAAAACCGT TCCTCAACAA
GTTGCTGAAT TGGTGGGCAC CCATTACTCT AAGGATGAGA TAATCAGTCT ATGGAAGGAC
ATTGAGCTGG ATGTACCCAG GGAAAAGATC CAAGAGGCCT TCGATAAGTT CGTAAAACAA
TCAACTGCCA CTTCCCCCGT TAGAAATGAA TTTCCCTTGT CTCAGCAAGA TTGGGTGACA
GTGACCAACA CCAAGTTTGA TAATTATCAA TTGAGGGTTA AAAAATCCCA CCCTGAAAAG
CTAAACATTG ATAAGGTAAA GCAATCTTCG GGATACCTGG ATATCATTGA TCAAGATAAG
CATCTTTTCT ATTGGTTTTT TGAATCCCGA AATGATCCGT CCACAGACCC AATCATCCTA
TGGTTGAATG GTGGACCCGG CTGCTCTTCT ATTACAGGGT TGCTATTCGA AAAGATTGGC
CCCAGTTACA TCACCAAAGA GATTAAGCCG GAACATAATC CTTATTCATG GAACAACAAT
GCTAGTGTTA TCTTCCTTGA GCAACCGGTT GGAGTAGGAT TTTCTTACTC TTCTAAGAAA
GTCGGTGATA CTGCAACTGC TGCCAAAGAT ACATATGTGT TTTTGGAGCT TTTCTTCCAA
AAGTTTCCTC AGTTCCTGAC CTCTAATCTG CACATTGCTG GGGAATCGTA TGCTGGCCAT
TATTTGCCCA AGATTGCTTC TGAGATTGTG TCTCACGCAG ACAAGACGTT TGACCTTTCA
GGAGTCATGA TCGGTAATGG TCTTACTGAT CCTCTAATTC AGTATAAGTA CTATCAGCCA
ATGGCCTGTG GAAAAGGTGG CTACAAGCAG GTCATTTCGG ACGAGGAATG TGATGAATTG
GATAGGGTCT ATCCAAGATG TGAACGTTTA ACGCGGGCAT GTTATGAGTT CCAAAATTCA
GTTACTTGTG TTCCGGCAAC ACTTTATTGC GACCAAAAGC TACTGAAGCC GTACACTGAC
ACTGGCTTGA ATGTCTATGA TATTCGTACA ATGTGCGATG AAGGGACTGA TTTGTGTTAC
AAAGAACTGG AATACGTGGA GAAGTACATG AACCAGCCTG AAGTGCAGGA AGCCGTGGGC
TCTGAAGTCA GTTCTTACAA AGGTTGTGAC GATGATGTCT TCTTAAGATT TTTGTACTCT
GGCGATGGAT CTAAGCCTTT CCACCAGTAT ATCACGGATG TTCTCAATGC AAGTATTCCG
GTTCTGATTT ACGCAGGTGA TAAAGATTAT ATCTGTAATT GGCTAGGAAA CCAAGCTTGG
GTCAATGAGC TAGAATGGAA CTTGTCTGAG GAATTCCAGG CAACTCCGAT TCGACCGTGG
TTCACTTTGG ACAATAACGA TTATGCAGGA AACGTACAAA CTTATGGAAA CTTTTCCTTT
CTAAGAGTAT TTGATGCTGG TCACATGGTT CCTTACAATC AACCAGTCAA CGCACTTGAC
ATGGTTGTCA GATGGACACA CGGTGATTTC TCATTTGGTT ATTAA (配列番号:9)
MKSVIWSLLS LLALSQALTI PLLEELQQQT FFSKKTVPQQ VAELVGTHYS KDEIISLWKD
IELDVPREKI QEAFDKFVKQ STATSPVRNE FPLSQQDWVT VTNTKFDNYQ LRVKKSHPEK
LNIDKVKQSS GYLDIIDQDK HLFYWFFESR NDPSTDPIIL WLNGGPGCSS ITGLLFEKIG
PSYITKEIKP EHNPYSWNNN ASVIFLEQPV GVGFSYSSKK VGDTATAAKD TYVFLELFFQ
KFPQFLTSNL HIAGESYAGH YLPKIASEIV SHADKTFDLS GVMIGNGLTD PLIQYKYYQP
MACGKGGYKQ VISDEECDEL DRVYPRCERL TRACYEFQNS VTCVPATLYC DQKLLKPYTD
TGLNVYDIRT MCDEGTDLCY KELEYVEKYM NQPEVQEAVG SEVSSYKGCD DDVFLRFLYS
GDGSKPFHQY ITDVLNASIP VLIYAGDKDY ICNWLGNQAW VNELEWNLSE EFQATPIRPW
FTLDNNDYAG NVQTYGNFSF LRVFDAGHMV PYNQPVNALD MVVRWTHGDF SFGY (配列番号10)
ATGATATTAC ACACCTATAT TATTCTCTCG TTATTGACTA TATTTCCTAA AGCTATTGGT
CTGTCCTTGC AGATGCCAAT GGCCTTGGAA GCTAGTTATG CCTCATTAGT GGAGAAAGCA
ACCCTCGCTG TTGGACAAGA AATTGATGCC ATACAAAAGG GTATTCAGCA AGGTTGGTTG
GAAGTAGAGA CAAGATTTCC AACTATAGTG TCACAGTTAT CCTATAGTAC TGGCCCAAAA
TTTGCGATCA AGAAGAAAGA TGCAACTTTT TGGGATTTCT ATGTTGAAAG TCAAGAGTTG
CCAAACTACC GAATTAATGA AAATCTGAAA CCAATTTTCA ACCCCTATTC GTGGAATGGT
AATGCTTCAA TCATCTACTT AGATCAACCG GTCAATGTTG GGTTTTCTTA TTCTTCATCA
TCGGTGAGTA ACACTGTTGT TGCGGGAGAA GATGTGTATG CATTTCTTCA GCTTTTTTTT
CAACACTTCC CGGAATATCA AACTAATGAC TTTCATATTG CCGGTGAATC TTATGCAGGA
CATTACATTC CGGTGTTTGC AGACGAAATT TTGAGTCAAA AGAACAGAAA TTTCAATCTT
ACTTCAGTCT TGATCGGAAA TGGATTAACT GACCCTTTGA CTCAATACCG ATATTACGAG
CCAATGGCTT GTGGTGAAGG TGGTGCCCCG TCAGTACTGC CTGCCGATGA GTGCGAAAAT
ATGCTAGTTA CCCAAGATAA ATGTTTGTCT TTAATTCAAG CATGCTACGA CTCACAGTCG
GCATTCACAT GCGCACCGGC TGCCATTTAT TGTAATAACG CTCAGATGGG ACCCTATCAG
AGAACTGGGA AGAATGTGTA TGATATTCGT AAGGAATGTG ATGGTGGATC CTTGTGCTAT
AAGGACCTTG AATTCATCGA TACCTACTTA AATCAAAAGT TTGTTCAAGA TGCTTTGGGC
GCCGAGGTCG ATACCTATGA ATCTTGCAAT TTTGAAATCA ACAGAAACTT TTTATTTGCT
GGAGATTGGA TGAAACCTTA TCATGAACAT GTCAGCAGTC TCTTGAACAA AGGTTTGCCC
GTTTTGATTT ACGCAGGGGA CAAAGATTTC ATTTGCAACT GGTTGGGTAA TCGAGCATGG
ACTGATGTCT TGCCGTGGGT TGATGCTGAT GGTTTTGAAA AAGCCGAAGT CCAAGATTGG
TTGGTTAATG GAAGGAAGGC TGGTGAATTT AAGAACTATA GCAACTTCAC CTACCTAAGG
GTTTATGATG CTGGTCATAT GGCCCCATAT GATCAGCCAG AGAATTCTCA TGAAATGGTC
AATAGATGGA TATCCGGAGA CTTTAGCTTT CACTAG (配列番号11)
MILHTYIILS LLTIFPKAIG LSLQMPMALE ASYASLVEKA TLAVGQEIDA IQKGIQQGWL
EVETRFPTIV SQLSYSTGPK FAIKKKDATF WDFYVESQEL PNYRINENLK PIFNPYSWNG
NASIIYLDQP VNVGFSYSSS SVSNTVVAGE DVYAFLQLFF QHFPEYQTND FHIAGESYAG
HYIPVFADEI LSQKNRNFNL TSVLIGNGLT DPLTQYRYYE PMACGEGGAP SVLPADECEN
MLVTQDKCLS LIQACYDSQS AFTCAPAAIY CNNAQMGPYQ RTGKNVYDIR KECDGGSLCY
KDLEFIDTYL NQKFVQDALG AEVDTYESCN FEINRNFLFA GDWMKPYHEH VSSLLNKGLP
VLIYAGDKDF ICNWLGNRAW TDVLPWVDAD GFEKAEVQDW LVNGRKAGEF KNYSNFTYLR
VYDAGHMAPY DQPENSHEMV NRWISGDFSF H (配列番号12)
ATGCAATTGC GTCATTCCGT TGGATTGGCT ATCTTATCTG CCATAGCAGT CCAAGGATTG
CTAATTCCTA ACATTGAGTC ATTACCCAGC CAGTTTGGTG CTAATGGTGA CAGTGAACAA
GGTGTATTAG CCCACCATGG TAAACATCCT AAAGTTGATA TGGCTCACCA TGGAAAGCAT
CCTAAAATCG CTAAGGATTC CAAGGGACAC CCTAAGCTTT GCCCTGAAGC TTTGAAGAAG
ATGAAAGAAG GCCACCCTTC GGCTCCAGTC ATTACTACCC ATTCCGCTTC TAAAAACTTA
ATCCCTTACT CTTATATTAT AGTCTTCAAG AAGGGTGTCA CTTCAGAGGA TATCGACTTC
CACCGTGACC TTATCTCCAC TCTTCATGAA GAGTCTGTGA GCAAATTAAG AGAGTCAGAT
CCAAATCACT CATTTTTCGT TTCTAATGAG AATGGCGAAA CAGGTTACAC CGGTGACTTC
TCCGTTGGTG ACTTGCTCAA GGGTTACACC GGATACTTCA CGGATGACAC TTTAGAGCTT
ATCAGTAAGC ATCCAGCAGT TGCTTTCATT GAAAGGGATT CGAGAGTATT TGCCACCGAT
TTTGAAACTC AAAACGGTGC TCCTTGGGGT TTGGCCAGAG TCTCTCACAG AAAGCCTCTT
TCCCTAGGCA GCTTCAACAA GTACTTATAT GATGGAGCTG GTGGTGAAGG TGTTACTTCC
TATGTTATCG ATACAGGTAT CCACGTCACT CACAAAGAAT TCCAGGGTAG AGCATCTTGG
GGTAAGACCA TTCCAGCTGG AGACGTTGAT GACGATGGAA ACGGTCACGG AACTCACTGT
GCTGGTACCA TTGCTTCTGA AAGCTACGGT GTTGCCAAGA AGGCTAATGT TGTTGCCATC
AAGGTCTTGA GATCTAATGG TTCTGGTTCG ATGTCAGATG TTCTGAAGGG TGTTGAGTAT
GCCACCCAAT CCCACTTGGA TGCTGTTAAA AAGGGCAACA AGAAATTTAA GGGCTCTACC
GCTAACATGT CACTGGGTGG TGGTAAATCT CCTGCTTTGG ACCTTGCAGT CAATGCTGCT
GTTAAGAATG GTATTCACTT TGCCGTTGCA GCAGGTAACG AAAACCAAGA TGCTTGTAAC
ACCTCGCCAG CAGCTGCTGA GAATGCCATC ACCGTCGGTG CATCAACCTT ATCAGACGCT
AGAGCTTACT TTTCTAACTA CGGTAAATGT GTTGACATTT TCGCTCCAGG TTTAAACATT
CTTTCTACCT ACACTGGTTC GGATGACGCA ACTGCTACCT TGTCTGGTAC TTCAATGGCC
TCTCCTCACA TTGCTGGTCT GTTGACTTAC TTCCTATCAT TGCAGCCTGC TGCTGGATCT
CTGTACTCTA ACGGAGGATC TGAGGGTGTC ACACCTGCTC AATTGAAAAA GAACCTCCTC
AAGTATGCAT CTGTCGGAGT ATTAGAGGAT GTTCCAGAAG ACACTCCAAA CCTCTTGGTT
TACAATGGTG GTGGACAAAA CCTTTCTTCT TTCTGGGGAA AGGAGACAGA AGACAATGTT
GCTTCCTCCG ACGATACTGG TGAGTTTCAC TCTTTTGTGA ACAAGCTTGA ATCAGCTGTT
GAAAACTTGG CCCAAGAGTT TGCACATTCA GTGAAGGAGC TGGCTTCTGA ACTTATTTAG
(配列番号13)
MQLRHSVGLA ILSAIAVQGL LIPNIESLPS QFGANGDSEQ GVLAHHGKHP KVDMAHHGKH
PKIAKDSKGH PKLCPEALKK MKEGHPSAPV ITTHSASKNL IPYSYIIVFK KGVTSEDIDF
HRDLISTLHE ESVSKLRESD PNHSFFVSNE NGETGYTGDF SVGDLLKGYT GYFTDDTLEL
ISKHPAVAFI ERDSRVFATD FETQNGAPWG LARVSHRKPL SLGSFNKYLY DGAGGEGVTS
YVIDTGIHVT HKEFQGRASW GKTIPAGDVD DDGNGHGTHC AGTIASESYG VAKKANVVAI
KVLRSNGSGS MSDVLKGVEY ATQSHLDAVK KGNKKFKGST ANMSLGGGKS PALDLAVNAA
VKNGIHFAVA AGNENQDACN TSPAAAENAI TVGASTLSDA RAYFSNYGKC VDIFAPGLNI
LSTYTGSDDA TATLSGTSMA SPHIAGLLTY FLSLQPAAGS LYSNGGSEGV TPAQLKKNLL
KYASVGVLED VPEDTPNLLV YNGGGQNLSS FWGKETEDNV ASSDDTGEFH SFVNKLESAV
ENLAQEFAHS VKELASELI (SEQ 番号14)
【0082】
実施例3−プロテアーゼノックアウト
いくつかのピキアノックアウト系統を、以下の方法で産生させた。ノックアウトされる5’末端遺伝子(200−500塩基対)を、5’遺伝子特異的プライマー、および選択マーカー発現カセット(例えば、KanMX)の5’末端と相補的な20bp配列を持つ3’遺伝子特異的プライマーを使って、PCR法でピキアゲノムDNAから増幅させた。ノックアウトされる3’末端遺伝子(200−500塩基対)を、KanMXの発現カセットの3’末端、および3’遺伝子特異的プライマーと相補的な20bp配列の5’遺伝子特異的プライマーを使って、PCR法でピキアゲノムDNAから増幅させた。別々のPCR法反応において、適当量の5’産物と3’産物、KanMX発現カセットを含むDNA、前回使用した5’と3’遺伝子特異的プライマーを加えて、ノックアウト遺伝子の5’と3’の相同領域に隣接するKanMX発現カセットを有するノックアウトカセットを作った。DNA断片を精製し、標準電気穿孔法方法を使ってピキアに形質転換した。組換え型細胞を、抗菌(例えば、KAN)を含むYPDプレート上で選択し、特異的プライマーを使用してPCR法で選別した。結果として得られたノックアウト系統をゲノム配列決定とサザンブロット法で確認した。
【0083】
次に、単一および複数ノックアウトを、異種ポリペプチドを発現することができるDNAを使って形質転換した。各ノックアウト系統を、配列番号1に記述されている治療用タンパク質をコードする少なくとも1つのベクターを使って形質転換した。形質転換したノックアウトの培養物を生長させ、上清を収集した。分泌した産物を、タンパク質分解による産物分解を検出するために、NuPAGE MOPS ゲルで泳動させた。複数バンドの半精製した異種タンパク質は、プロテアーゼ活性を示すと解釈し、一方、適度の分子量の異種タンパク質の単一バンドは、プロテアーゼ活性がないと解釈した。図5−8を参照されたい。
【0084】
発酵上清試料のプロテアーゼ活性を、蛍光ラベルおよび励起と発光がそれぞれ波長340と405nmのクエンチャーを備える合成ペプチドを使って測定した。ペプチド基質を、培養上清試料でインキュベートし、基質の分解割合は、蛍光性HPLCを使ってモニターした。結果は、ピーク領域/時間が増加する方向に表される。図3において、プロテアーゼ活性は、ymp1突然変異系統の発酵上清で約68〜285であり、一方コントロール系統のプロテアーゼ活性は約1541〜1909であった。したがって、ペプチドに関して突然変異系統はプロテアーゼ活性の劇的な低下を立証した。
【0085】
ザイモグラムゲルは、カゼインまたはゼラチンを基質として使ってプロテアーゼ活性を測定する別の方法である。この実験において、カゼインまたゼラチンを還元、非変性の条件下でマトリックスに取り込んだアクリルアミドゲル上で発酵試料を分離した。分離に引き続いて、タンパク質バンドを再生し、熟成中の緩衝液をプロテアーゼ活性に必要な補助因子を提供するのに適用した。ゲルは、37℃で長期間インキュベートしクマシー・ブルーで染色した。タンパク分解活性領域を、青色を背景としたゲル上の明瞭な領域として同定した。活性なプロテアーゼは基質を分解し、ゲル上の領域をクマシー・ブルーに結合させなかった。活性なプロテアーゼを欠乏するゲル上の領域は染色剤が基質に結合するのを可能にし、よって背景を青色にした。図4は、ymp1突然変異系統および2つのコントロール系統からの発酵上清を使ったプロテアーゼ活動領域を表す図である。コントロール系統は、高分子量と低分子量の両領域にわたって活性なプロテアーゼを有した。しかしながら、高分子量領域のシグナルは、ymp1突然変異系統では大幅に減少したか、失われた。これらのデータは、高分子量領域での活性が、ymp1の作用に依るものであろうことを示す。ノックアウトに関する結果の概要を表5に示す。
【0086】
緩衝液安定性アッセイを使ったさらなる試験によって、YMP1およびYPS1をピキア宿主系統から欠失させた場合、切断効率および部位変化が実証された。このアッセイでは、配列番号1で記述されているタンパク質標準を培養上清でインキュベートし、切断されたタンパク質産物を質量分光光度法で分析し、切断部位および切断産物の割合を示した(表1−4)。
【0087】
発酵ブロスB804(pmt4を欠失した親系統SMD1163、実施例4を参照)の上清を使って、5個の主要な切断部位、リジン28/グリシン29、フェニルアラニン36/スレオニン37、セリン38/アスパラギン酸39、チロシン43/ロイシン44およびリジン50/グルタミン酸51を検出した(表1)。
【表1】
【0088】
SMD1163_pmp4(B688)からYMP1を欠失させると、フェニルアラニン36/スレオニン37、セリン38/アスパラギン酸39およびチロシン43/ロイシン44の切断部位が減少するが、しかしリジン50/グルタミン酸51とリジン28/グリシン29の切断を維持、または増加させた(表2)。
【表2】
【0089】
SMD1163_pmt4(B805)からyps1プロテアーゼを欠失させても任意の切断部位は除去されなかったが、リジン50/グルタミン酸51部位において切断量は減少した(表3)。
【表3】
【0090】
Ymp1およびYps1プロテアーゼを両方ともSMD1163_pmt4(B803)から欠失させると、切断は有意に減少し、かつ、リジン28/グリシン29のみで起こった。
【表4】
【0091】
SMD1163内で、ymp2、ymp3を個々にノックアウトにした。これらのノックアウト系統とSMD1163コントロールついて振盪フラスコ誘導を実施し、培養上清試料を、誘導後24時間で取り出した。配列番号1に記述されているタンパク質標準を30℃で24時間これらの試料と一緒にインキュベートし、SDS PAGEゲルによって分析した。
【0092】
PAGE分析から、ymp3の欠失は、タンパク質標準の切断を有意に減らすが、ymp2の欠失は、タンパク質分解には効果が少なかった(図9を参照)。
【表5】
【0093】
ピキア内において本明細書で同定した6個のポリペプチドは、Pep4、Prb1、Yps1、Yps2および新しく発見されたYmp1、Ymp3であり、これらは異種ポリペプチド(配列番号1)分解に関与する。これらの遺伝子の遺伝的ノックアウトによって異種タンパク質発現の生産性が有意に改善できる。
【0094】
実施例4 マンノシルトランスフェラーゼノックアウト
ピキア野生型株GS115から産生した異種ポリペプチド(配列番号1)をLC/MSで分析すると、複数の162Da質量増加アイソフォームが観察された。多重酵素消化とLC/MSを使ってさらなる分析を行うと実証するHSA部分で糖鎖形成が発生することが示され、エドマン分解法および炭水化物組成分析(データ示さず)を使ってその結果を再確認した。炭水化物組成分析により、これらの修飾はO−マンノース糖鎖形成によるものであることが確認された、この糖鎖形成はピキア・パストリス内で産生される一般的なタンパク質翻訳後修飾である。
【0095】
ピキア株のマンノシルトランスフェラーゼノックアウトを実施例2に記述したプロテアーゼノックアウトと同様な方法を使って作製した。すなわち、KanMX発現カセットを酵母:OCH1、PMT1、PMT2、およびPMT4内で糖鎖形成を担うこれらの遺伝子のうち1つに挿入した。ノックアウト系統を、配列番号1を発現することのできるベクターを使って形質転換した。これらのノックアウト系統の表現型の変化をザイモリアーゼ感受性アッセイ(図10)を使って実証した。この実験において、指数関数的に成長する酵母培養をザイモリアーゼ5U/mlを追加してYPDでOD600=0.3に希釈した。培養液を、室温で穏やかに振盪しながらインキュベートした。試料を、2時間の間15分置きに収集し、細胞死の指標としてOD600測定を行った。
【0096】
マンノシルトランスフェラーゼ遺伝子は、細胞壁完全性のために必須である。pmt1およびpmt4ノックアウト系統は、部分的または不完全な細胞壁を持つことがあり、ザイモリアーゼ処置への感受性を増大させる場合がある。このデータは、pmt1およびpmt4変異体の両方がザイモリアーゼ処置により急激に死滅したことを示す。
【0097】
異種タンパク質を、遺伝子改変された系統内での異種産生と野生型系統とを比較しながら、糖鎖形成について評価した。ノックアウトについての結果概要を以下の表6に示す。さらに図11を参照されたい。Pmt4が単独でピキア内のHSA糖鎖形成を担っている結果が示された。pmt4単一ノックアウト系統が糖鎖形成のないヒトHASを産生することができる。
【表6】
【0098】
実施例5−シャペロンタンパク質との同時発現
いくつかのシャペロンタンパク質は、宿主細胞内の異種タンパク質と同時発現でき、異種タンパク質産生を増加させることができるだろう。酵母内の異種タンパク質と同時発現して異種タンパク質産生を増加させるシャペロンタンパク質の例を以下の表7に示す。
【表7】
【特許請求の範囲】
【請求項1】
ピキア株において機能的遺伝子産物をコードする少なくとも1つの核酸配列および/または少なくとも1つの機能的遺伝子産物の発現に必要な少なくとも1つの核酸配列が遺伝子改変される遺伝子改変ピキア株であって、前記遺伝子産物が、前記遺伝子改変ピキア株においてタンパク質分解および/または糖鎖形成に関与するものである、遺伝子改変ピキア株。
【請求項2】
以下の機能的遺伝子産物:PEP4、PRB1、YPS1、YPS2、YMP1、YMP2、YMP3、DAP2、GRH1、PRD1、YSP3、およびPRB3のうち少なくとも1つ、または前記遺伝子産物の発現をコードする少なくと1つの核酸配列が遺伝子改変される、請求項1に記載の遺伝子改変ピキア株。
【請求項3】
以下の酵素および/または酵素タイプ:アルギニン/アラニン様アミノペプチダーゼ、アスパルチルプロテアーゼ、セリンプロテアーゼ、分泌アスパルチルプロテアーゼ、分泌セリンプロテアーゼ、メチロトローフ酵母プロテアーゼ、DPP IV様エンドペプチダーゼ、金属エンドペプチダーゼ、Prb 1様セリンプロテアーゼ、Prb 1セリンプロテアーゼ、空胞のプロテアーゼB様、ロイシン様アミノペプチダーゼ、CPY様カルボキシペプチダーゼおよび/またはマンノシルトランスフェラーゼのうち少なくとも1つを、野生株と比べて、その量を減少、除去および/または活性を低下させて産生する、請求項1または2に記載の遺伝子改変ピキア株。
【請求項4】
機能的遺伝子産物または前記遺伝子産物の発現をコードする以下の核酸配列:OCH1、PMT1、PMT2、および/またはPMT4のうち少なくとも1つが遺伝子改変される、請求項1〜3のいずれか1項に記載の遺伝子改変ピキア株。
【請求項5】
野生型X−33またはSMD1163の遺伝子改変形態である、請求項1〜4のいずれか1項に記載の遺伝子改変ピキア株。
【請求項6】
野生型ピキア・パストリス、ピキア・フィンランディカ、ピキア・トレハロフィア、ピキア・コクラメ、ピキア・メンブラナエファシエンス、ピキア・メタノリカ、ピキア・ミヌタ(オガタ・エミヌタ、ピキア・リンドネリ)、ピキア・オプティアエ、ピキア・サーモトレランス、ピキア・サリクタリア、ピキア・ゲルカム、ピキア・ピジュペリ、ピキア・スティプティス、ピキア菌種の遺伝子改変形態である、請求項1〜5のいずれか1項に記載の遺伝子改変ピキア株。
【請求項7】
機能的遺伝子産物または前記遺伝子産物の発現をコードする以下の核酸配列:PEP4、PRB1、YPS1、YPS2、YMP1、YMP2、YMP3、およびPMT4が遺伝子改変される、請求項1〜6のいずれか1項に記載の遺伝子改変ピキア株。
【請求項8】
野生型株と比べて、量を減少、除去および/または活性を低下させた少なくとも1つのマンノシルトランスフェラーゼを産生する、請求項1〜7のいずれか1項に記載の遺伝子改変ピキア株。
【請求項9】
以下の機能的遺伝子産物:配列番号4、配列番号6、配列番号8、配列番号10、および/または配列番号12のうち少なくとも1つ、または前記遺伝子産物の発現をコードする少なくとも1つの核酸配列が遺伝子改変される、請求項1から請求項8のいずれか1項に記載の遺伝子改変ピキア株。
【請求項10】
少なくとも1つの異種ポリペプチドを発現することのできる少なくとも1つのポリヌクレオチドをさらに含んでなる、請求項1〜9のいずれか1項に記載の遺伝子改変ピキア株。
【請求項11】
少なくとも1つの異種ポリペプチドを発現することのできる前記ポリヌクレオチドが、ベクターおよび/またはプラスミドからのエピソーム発現である、請求項10に記載の遺伝子改変ピキア株。
【請求項12】
少なくとも1つの異種ポリペプチドを発現することのできる前記ポリヌクレオチドが、ピキアのゲノムに形質転換される、請求項11に記載の遺伝子改変ピキア株。
【請求項13】
前記少なくとも1つの異種ポリペプチドが、少なくとも1つのGLP−1アゴニストを含んでなる、請求項10〜12のいずれか1項に記載の遺伝子改変ピキア株。
【請求項14】
前記少なくとも1つのGLP−1アゴニストが、インクレチンのホルモン並びに/またはその断片、変異体および/もしくは抱合体、インクレチンのミメティック並びに/またはその断片、変異体および/もしくは抱合体からなる群から選択される、請求項13に記載の遺伝子改変ピキア株。
【請求項15】
少なくとも1つの異種ポリペプチドが、GLP−1活性を有する、請求項10〜14のいずれか1項に記載の遺伝子改変ピキア株。
【請求項16】
前記GLP−1活性を有する少なくとも1つのポリペプチドが、ヒト血清アルブミンと融合したヒトGLP−1の少なくとも1つの断片および/または変異体を含んでなる、請求項15に記載の遺伝子改変ピキア株。
【請求項17】
前記GLP−1の少なくとも1つの断片および変異体が、GLP−1(7−36(A8G))を含んでなる、請求項16に記載の遺伝子改変ピキア株。
【請求項18】
前記GLP−1の少なくとも1つの断片および変異体が、ヒト血清アルブミンと遺伝的に融合する、請求項17に記載の遺伝子改変ピキア株。
【請求項19】
GLP−1の少なくとも1つの断片および変異体が、ヒト血清アルブミンと直列に、かつ、遺伝的に融合する少なくとも2つのGLP−1(7−36(A8G))を含んでなる、請求項18に記載の遺伝子改変ピキア株。
【請求項20】
前記少なくとも2つのGLP−1(7−36(A8G))が、ヒト血清アルブミンのN末端で遺伝的に融合する、請求項19に記載の遺伝子改変ピキア株。
【請求項21】
前記異種ポリペプチドが、配列番号1を含んでなる、請求項10〜20のいずれか1項に記載の遺伝子改変ピキア株。
【請求項22】
前記少なくとも1つの異種ポリペプチドが、少なくとも1つの抗原結合タンパク質、少なくとも1つの単一可変ドメイン、および/または少なくとも1つのドメイン抗体のうち1つ以上を含んでなる、請求項10〜21のいずれか1項に記載の遺伝子改変ピキア。
【請求項23】
野生型ピキアと比べて、前記株が、前記株中で前記少なくとも1つの異種ポリペプチドのタンパク質分解の減少を示す、請求項10〜22のいずれか1項に記載の遺伝子改変ピキア株。
【請求項24】
野生型ピキアと比べて、前記株が、前記株中で前記少なくとも1つの異種ポリペプチドの糖鎖形成の減少を示すか、糖鎖形成を示さない、請求項10〜23のいずれか1項に記載の遺伝子改変ピキア株。
【請求項25】
異種ポリペプチドを産生する方法であって、請求項1〜24のいずれか1項に記載の遺伝子改変ピキアにおいて前記異種ポリペプチドを発現させることを含んでなる、方法。
【請求項26】
請求項1〜25のいずれか1項に記載の遺伝子改変ピキアで産生される異種ポリペプチド。
【請求項27】
遺伝子改変宿主細胞であって、前記遺伝子改変宿主細胞の野生型親は、配列番号4のアミノ酸1−865(配列番号15)と少なくとも60%の配列同一性を有するポリペプチドをコードする遺伝子を含み、前記遺伝子は、野生型宿主細胞と比べると、前記遺伝子産物が前記遺伝子改変の宿主細胞内で減少するか除去されるように前記宿主細胞のゲノム内で遺伝子改変される、遺伝子改変宿主細胞。
【請求項28】
前記野生型親が、配列番号4の全体の長さに渡って少なくとも60%の配列同一性を有するポリペプチドをコードする遺伝子を含んでなる、請求項27に記載の遺伝子改変宿主細胞。
【請求項29】
遺伝子改変宿主細胞であって、前記遺伝子改変宿主細胞の野生型親が、配列番号3と少なくとも60%の配列同一性を有する遺伝子を含み、前記遺伝子が、前記遺伝子改変宿主細胞のゲノム内で遺伝子改変される、遺伝子改変宿主細胞。
【請求項30】
前記野生型親宿主細胞と比べると、プロテアーゼ活性が減少しているか除去されている、請求項27または29に記載の遺伝子改変宿主細胞。
【請求項31】
酵母メチロトローフプロテアーゼ1活性が減少しているか除去されている、請求項27〜30に記載の遺伝子改変宿主細胞。
【請求項32】
ピキアである、請求項27〜31に記載の遺伝子改変宿主細胞。
【請求項33】
配列番号3と少なくとも60%の配列同一性を有するポリヌクレオチドを含んでなる、単離されたポリヌクレオチド。
【請求項34】
配列番号4と少なくとも60%の配列同一性を有する、単離されたポリペプチド。
【請求項35】
配列番号5と少なくとも60%の配列同一性を有するポリヌクレオチドを含んでなる、単離されたポリヌクレオチド。
【請求項36】
配列番号6と少なくとも60%の配列同一性を有する、単離されたポリペプチド。
【請求項37】
配列番号7と少なくとも60%の配列同一性を有するポリヌクレオチドを含んでなる、単離されたポリヌクレオチド。
【請求項38】
配列番号8と少なくとも60%の配列同一性を有する、単離されたポリペプチド。
【請求項39】
配列番号9と少なくとも60%の配列同一性を有するポリヌクレオチドを含んでなる、単離されたポリヌクレオチド。
【請求項40】
配列番号10と少なくとも60%の配列同一性を有する、単離されたポリペプチド。
【請求項41】
配列番号11と少なくとも60%の配列同一性を有するポリヌクレオチドを含んでなる、単離されたポリヌクレオチド。
【請求項42】
配列番号12と少なくとも60%の配列同一性を有する、単離されたポリペプチド。
【請求項43】
遺伝子改変ピキア株であって、前記ピキアの野生型株のゲノムが、配列番号3、5、7から選択される遺伝子と少なくとも85%の配列同一性を有する遺伝子を含み、配列番号3、5、および7と少なくとも85%の配列同一性を有する前記遺伝子が、欠失、変異、または破壊された時、前記遺伝子改変ピキア株が、前記野生型株よりも前記破壊された遺伝子の遺伝子産生物をより少なく産生する、遺伝子改変ピキア株。
【請求項44】
前記ピキアの野生型株のゲノムが、配列番号3と少なくとも85%の配列同一性を有する遺伝子を含み、配列番号3が、欠失、変異、または破壊された時、前記遺伝子改変ピキア株が、前記野生型株よりも配列番号4をより少なく産生する、請求項43に記載の遺伝子改変ピキア株。
【請求項45】
前記野生型株よりも分泌セリンプロテアーゼ活性をより少なく産生する、請求項44に記載の遺伝子改変ピキア株。
【請求項46】
前記ピキアの野生型株のゲノムが、配列番号5と少なくとも85%の配列同一性を有する遺伝子を含み、配列番号5が、欠失、変異、または破壊された時、前記遺伝子改変ピキア株が、前記野生型株よりも配列番号6をより少なく産生する、請求項43に記載の遺伝子改変ピキア株。
【請求項47】
前記野生型株よりもアルギニン/アラニンアミノペプチダーゼ様活性をより少なく産生する、請求項46に記載の遺伝子改変ピキア株。
【請求項48】
前記ピキアの野生型株のゲノムが、配列番号7と少なくとも85%の配列同一性を有する遺伝子を含み、配列番号7が、欠失、変異、または破壊された時、前記遺伝子改変ピキア株が、前記野生型株よりも配列番号8をより少なく産生する、請求項43に記載の遺伝子改変ピキア株。
【請求項49】
前記野生型株よりもロイシン/アミノペプチダーゼ様活性をより少なく産生する、請求項48に記載の遺伝子改変ピキア株。
【請求項1】
ピキア株において機能的遺伝子産物をコードする少なくとも1つの核酸配列および/または少なくとも1つの機能的遺伝子産物の発現に必要な少なくとも1つの核酸配列が遺伝子改変される遺伝子改変ピキア株であって、前記遺伝子産物が、前記遺伝子改変ピキア株においてタンパク質分解および/または糖鎖形成に関与するものである、遺伝子改変ピキア株。
【請求項2】
以下の機能的遺伝子産物:PEP4、PRB1、YPS1、YPS2、YMP1、YMP2、YMP3、DAP2、GRH1、PRD1、YSP3、およびPRB3のうち少なくとも1つ、または前記遺伝子産物の発現をコードする少なくと1つの核酸配列が遺伝子改変される、請求項1に記載の遺伝子改変ピキア株。
【請求項3】
以下の酵素および/または酵素タイプ:アルギニン/アラニン様アミノペプチダーゼ、アスパルチルプロテアーゼ、セリンプロテアーゼ、分泌アスパルチルプロテアーゼ、分泌セリンプロテアーゼ、メチロトローフ酵母プロテアーゼ、DPP IV様エンドペプチダーゼ、金属エンドペプチダーゼ、Prb 1様セリンプロテアーゼ、Prb 1セリンプロテアーゼ、空胞のプロテアーゼB様、ロイシン様アミノペプチダーゼ、CPY様カルボキシペプチダーゼおよび/またはマンノシルトランスフェラーゼのうち少なくとも1つを、野生株と比べて、その量を減少、除去および/または活性を低下させて産生する、請求項1または2に記載の遺伝子改変ピキア株。
【請求項4】
機能的遺伝子産物または前記遺伝子産物の発現をコードする以下の核酸配列:OCH1、PMT1、PMT2、および/またはPMT4のうち少なくとも1つが遺伝子改変される、請求項1〜3のいずれか1項に記載の遺伝子改変ピキア株。
【請求項5】
野生型X−33またはSMD1163の遺伝子改変形態である、請求項1〜4のいずれか1項に記載の遺伝子改変ピキア株。
【請求項6】
野生型ピキア・パストリス、ピキア・フィンランディカ、ピキア・トレハロフィア、ピキア・コクラメ、ピキア・メンブラナエファシエンス、ピキア・メタノリカ、ピキア・ミヌタ(オガタ・エミヌタ、ピキア・リンドネリ)、ピキア・オプティアエ、ピキア・サーモトレランス、ピキア・サリクタリア、ピキア・ゲルカム、ピキア・ピジュペリ、ピキア・スティプティス、ピキア菌種の遺伝子改変形態である、請求項1〜5のいずれか1項に記載の遺伝子改変ピキア株。
【請求項7】
機能的遺伝子産物または前記遺伝子産物の発現をコードする以下の核酸配列:PEP4、PRB1、YPS1、YPS2、YMP1、YMP2、YMP3、およびPMT4が遺伝子改変される、請求項1〜6のいずれか1項に記載の遺伝子改変ピキア株。
【請求項8】
野生型株と比べて、量を減少、除去および/または活性を低下させた少なくとも1つのマンノシルトランスフェラーゼを産生する、請求項1〜7のいずれか1項に記載の遺伝子改変ピキア株。
【請求項9】
以下の機能的遺伝子産物:配列番号4、配列番号6、配列番号8、配列番号10、および/または配列番号12のうち少なくとも1つ、または前記遺伝子産物の発現をコードする少なくとも1つの核酸配列が遺伝子改変される、請求項1から請求項8のいずれか1項に記載の遺伝子改変ピキア株。
【請求項10】
少なくとも1つの異種ポリペプチドを発現することのできる少なくとも1つのポリヌクレオチドをさらに含んでなる、請求項1〜9のいずれか1項に記載の遺伝子改変ピキア株。
【請求項11】
少なくとも1つの異種ポリペプチドを発現することのできる前記ポリヌクレオチドが、ベクターおよび/またはプラスミドからのエピソーム発現である、請求項10に記載の遺伝子改変ピキア株。
【請求項12】
少なくとも1つの異種ポリペプチドを発現することのできる前記ポリヌクレオチドが、ピキアのゲノムに形質転換される、請求項11に記載の遺伝子改変ピキア株。
【請求項13】
前記少なくとも1つの異種ポリペプチドが、少なくとも1つのGLP−1アゴニストを含んでなる、請求項10〜12のいずれか1項に記載の遺伝子改変ピキア株。
【請求項14】
前記少なくとも1つのGLP−1アゴニストが、インクレチンのホルモン並びに/またはその断片、変異体および/もしくは抱合体、インクレチンのミメティック並びに/またはその断片、変異体および/もしくは抱合体からなる群から選択される、請求項13に記載の遺伝子改変ピキア株。
【請求項15】
少なくとも1つの異種ポリペプチドが、GLP−1活性を有する、請求項10〜14のいずれか1項に記載の遺伝子改変ピキア株。
【請求項16】
前記GLP−1活性を有する少なくとも1つのポリペプチドが、ヒト血清アルブミンと融合したヒトGLP−1の少なくとも1つの断片および/または変異体を含んでなる、請求項15に記載の遺伝子改変ピキア株。
【請求項17】
前記GLP−1の少なくとも1つの断片および変異体が、GLP−1(7−36(A8G))を含んでなる、請求項16に記載の遺伝子改変ピキア株。
【請求項18】
前記GLP−1の少なくとも1つの断片および変異体が、ヒト血清アルブミンと遺伝的に融合する、請求項17に記載の遺伝子改変ピキア株。
【請求項19】
GLP−1の少なくとも1つの断片および変異体が、ヒト血清アルブミンと直列に、かつ、遺伝的に融合する少なくとも2つのGLP−1(7−36(A8G))を含んでなる、請求項18に記載の遺伝子改変ピキア株。
【請求項20】
前記少なくとも2つのGLP−1(7−36(A8G))が、ヒト血清アルブミンのN末端で遺伝的に融合する、請求項19に記載の遺伝子改変ピキア株。
【請求項21】
前記異種ポリペプチドが、配列番号1を含んでなる、請求項10〜20のいずれか1項に記載の遺伝子改変ピキア株。
【請求項22】
前記少なくとも1つの異種ポリペプチドが、少なくとも1つの抗原結合タンパク質、少なくとも1つの単一可変ドメイン、および/または少なくとも1つのドメイン抗体のうち1つ以上を含んでなる、請求項10〜21のいずれか1項に記載の遺伝子改変ピキア。
【請求項23】
野生型ピキアと比べて、前記株が、前記株中で前記少なくとも1つの異種ポリペプチドのタンパク質分解の減少を示す、請求項10〜22のいずれか1項に記載の遺伝子改変ピキア株。
【請求項24】
野生型ピキアと比べて、前記株が、前記株中で前記少なくとも1つの異種ポリペプチドの糖鎖形成の減少を示すか、糖鎖形成を示さない、請求項10〜23のいずれか1項に記載の遺伝子改変ピキア株。
【請求項25】
異種ポリペプチドを産生する方法であって、請求項1〜24のいずれか1項に記載の遺伝子改変ピキアにおいて前記異種ポリペプチドを発現させることを含んでなる、方法。
【請求項26】
請求項1〜25のいずれか1項に記載の遺伝子改変ピキアで産生される異種ポリペプチド。
【請求項27】
遺伝子改変宿主細胞であって、前記遺伝子改変宿主細胞の野生型親は、配列番号4のアミノ酸1−865(配列番号15)と少なくとも60%の配列同一性を有するポリペプチドをコードする遺伝子を含み、前記遺伝子は、野生型宿主細胞と比べると、前記遺伝子産物が前記遺伝子改変の宿主細胞内で減少するか除去されるように前記宿主細胞のゲノム内で遺伝子改変される、遺伝子改変宿主細胞。
【請求項28】
前記野生型親が、配列番号4の全体の長さに渡って少なくとも60%の配列同一性を有するポリペプチドをコードする遺伝子を含んでなる、請求項27に記載の遺伝子改変宿主細胞。
【請求項29】
遺伝子改変宿主細胞であって、前記遺伝子改変宿主細胞の野生型親が、配列番号3と少なくとも60%の配列同一性を有する遺伝子を含み、前記遺伝子が、前記遺伝子改変宿主細胞のゲノム内で遺伝子改変される、遺伝子改変宿主細胞。
【請求項30】
前記野生型親宿主細胞と比べると、プロテアーゼ活性が減少しているか除去されている、請求項27または29に記載の遺伝子改変宿主細胞。
【請求項31】
酵母メチロトローフプロテアーゼ1活性が減少しているか除去されている、請求項27〜30に記載の遺伝子改変宿主細胞。
【請求項32】
ピキアである、請求項27〜31に記載の遺伝子改変宿主細胞。
【請求項33】
配列番号3と少なくとも60%の配列同一性を有するポリヌクレオチドを含んでなる、単離されたポリヌクレオチド。
【請求項34】
配列番号4と少なくとも60%の配列同一性を有する、単離されたポリペプチド。
【請求項35】
配列番号5と少なくとも60%の配列同一性を有するポリヌクレオチドを含んでなる、単離されたポリヌクレオチド。
【請求項36】
配列番号6と少なくとも60%の配列同一性を有する、単離されたポリペプチド。
【請求項37】
配列番号7と少なくとも60%の配列同一性を有するポリヌクレオチドを含んでなる、単離されたポリヌクレオチド。
【請求項38】
配列番号8と少なくとも60%の配列同一性を有する、単離されたポリペプチド。
【請求項39】
配列番号9と少なくとも60%の配列同一性を有するポリヌクレオチドを含んでなる、単離されたポリヌクレオチド。
【請求項40】
配列番号10と少なくとも60%の配列同一性を有する、単離されたポリペプチド。
【請求項41】
配列番号11と少なくとも60%の配列同一性を有するポリヌクレオチドを含んでなる、単離されたポリヌクレオチド。
【請求項42】
配列番号12と少なくとも60%の配列同一性を有する、単離されたポリペプチド。
【請求項43】
遺伝子改変ピキア株であって、前記ピキアの野生型株のゲノムが、配列番号3、5、7から選択される遺伝子と少なくとも85%の配列同一性を有する遺伝子を含み、配列番号3、5、および7と少なくとも85%の配列同一性を有する前記遺伝子が、欠失、変異、または破壊された時、前記遺伝子改変ピキア株が、前記野生型株よりも前記破壊された遺伝子の遺伝子産生物をより少なく産生する、遺伝子改変ピキア株。
【請求項44】
前記ピキアの野生型株のゲノムが、配列番号3と少なくとも85%の配列同一性を有する遺伝子を含み、配列番号3が、欠失、変異、または破壊された時、前記遺伝子改変ピキア株が、前記野生型株よりも配列番号4をより少なく産生する、請求項43に記載の遺伝子改変ピキア株。
【請求項45】
前記野生型株よりも分泌セリンプロテアーゼ活性をより少なく産生する、請求項44に記載の遺伝子改変ピキア株。
【請求項46】
前記ピキアの野生型株のゲノムが、配列番号5と少なくとも85%の配列同一性を有する遺伝子を含み、配列番号5が、欠失、変異、または破壊された時、前記遺伝子改変ピキア株が、前記野生型株よりも配列番号6をより少なく産生する、請求項43に記載の遺伝子改変ピキア株。
【請求項47】
前記野生型株よりもアルギニン/アラニンアミノペプチダーゼ様活性をより少なく産生する、請求項46に記載の遺伝子改変ピキア株。
【請求項48】
前記ピキアの野生型株のゲノムが、配列番号7と少なくとも85%の配列同一性を有する遺伝子を含み、配列番号7が、欠失、変異、または破壊された時、前記遺伝子改変ピキア株が、前記野生型株よりも配列番号8をより少なく産生する、請求項43に記載の遺伝子改変ピキア株。
【請求項49】
前記野生型株よりもロイシン/アミノペプチダーゼ様活性をより少なく産生する、請求項48に記載の遺伝子改変ピキア株。
【図1】


【図2】

【図3】
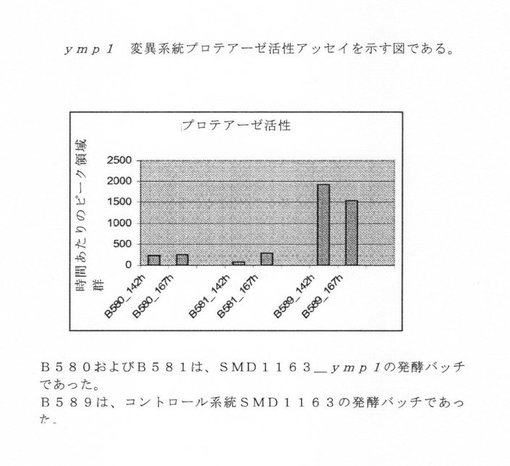
【図4】
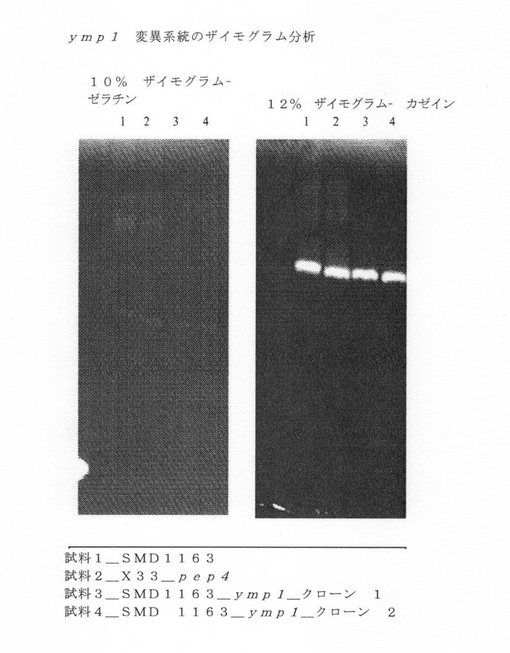
【図5】
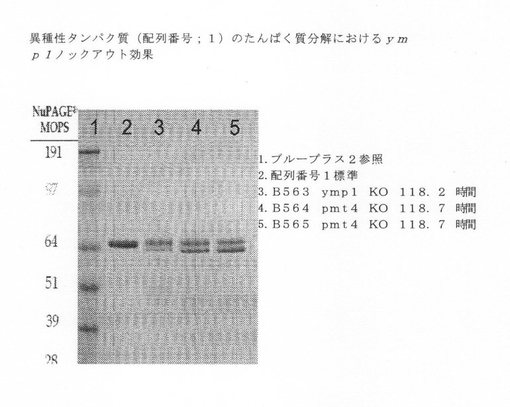
【図6】
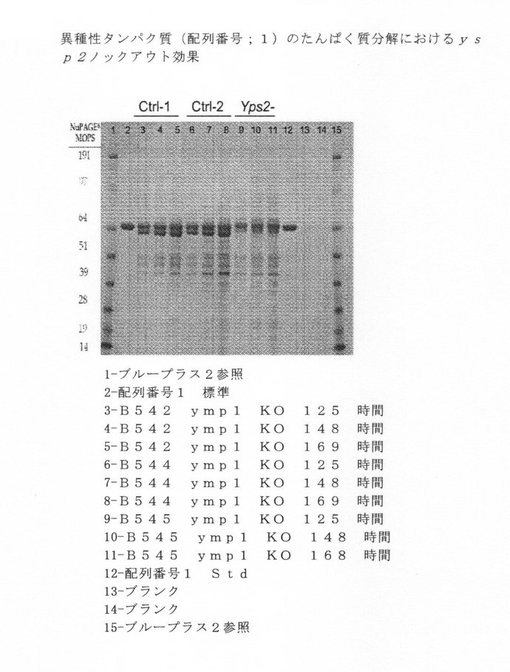
【図7】
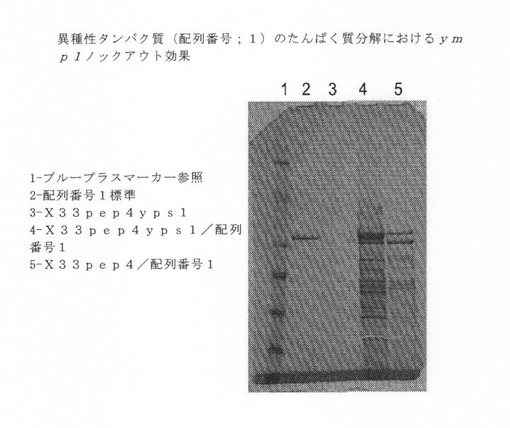
【図8】
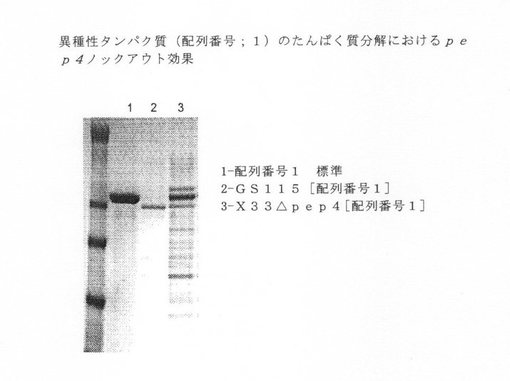
【図9】
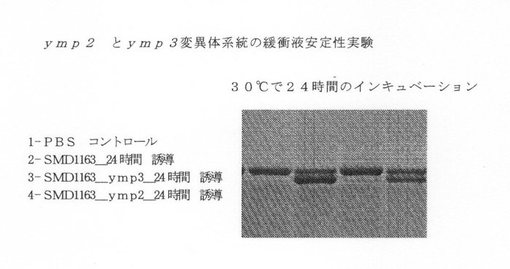
【図10】

【図11】
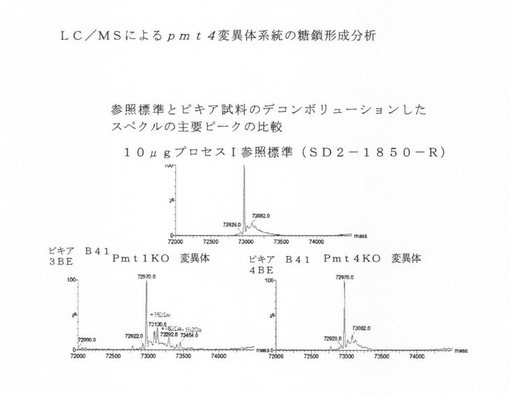
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2012−518999(P2012−518999A)
【公表日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願番号】特願2011−552128(P2011−552128)
【出願日】平成22年2月24日(2010.2.24)
【国際出願番号】PCT/US2010/025223
【国際公開番号】WO2010/099195
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(591002957)グラクソスミスクライン・リミテッド・ライアビリティ・カンパニー (341)
【氏名又は名称原語表記】GlaxoSmithKline LLC
【Fターム(参考)】
【公表日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願日】平成22年2月24日(2010.2.24)
【国際出願番号】PCT/US2010/025223
【国際公開番号】WO2010/099195
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(591002957)グラクソスミスクライン・リミテッド・ライアビリティ・カンパニー (341)
【氏名又は名称原語表記】GlaxoSmithKline LLC
【Fターム(参考)】
[ Back to top ]