
対立遺伝子排除
本発明は、ヒトを除くトランスジェニック哺乳類におけるVH重鎖のみ抗体の生成方法に関する。特に、本発明は、複数の異種VH重鎖遺伝子座を哺乳類において発現させる段階を含む、ヒトを除くトランスジェニック哺乳類におけるVH重鎖のみ抗体の産生方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒトを除くトランスジェニック哺乳類における、抗原曝露に応答する親和性成熟した機能性重鎖のみ抗体の多様なレパートリーの改良された製造方法およびその使用に関する。本発明はまた、複数の遺伝子座からのクラス特異的重鎖のみ抗体の多様なレパートリーの製造に関する。
【0002】
特に、本発明は、任意のクラスまたは複数のクラスの混合物のヒト抗原特異的な高親和性の重鎖のみ抗体の生成、ならびに十分に機能性のVH抗原結合ドメインの単離および発現のための方法に関する。
【0003】
本発明の方法を用いて生成された重鎖のみ抗体も記載される。
【0004】
以下の説明において、全てのアミノ酸残基位置番号は、Kabat et al.により考案された番号付け方式に従って示される[1]。
【背景技術】
【0005】
<抗体>
抗体の構造は当技術分野で周知である。大部分の天然抗体は四量体であり、2本の重鎖と2本の軽鎖を含む。重鎖はそれぞれの重鎖に沿ってほぼ中間に位置するヒンジドメイン間のジスルフィド結合を介して相互に連結されている。軽鎖はそれぞれの重鎖とヒンジドメインのN末端側で会合している。それぞれの軽鎖は、通常、各重鎖とヒンジドメインに近いジスルフィド結合により結合している。
【0006】
抗体分子が正常に折り畳まれると、それぞれの鎖は、より線状のポリペプチド配列により連結された多数の別個の球状のドメインに折り畳まれる。例えば、軽鎖は、可変(VL)および定常(CL)ドメインに折り畳まれる。重鎖は、単一の可変ドメインVH、第1の定常ドメイン(CH1)、ヒンジドメインおよび2または3個のさらなる定常ドメインを有する。重鎖定常ドメインおよびヒンジドメインは一体となって抗体重鎖の定常領域として一般に公知な構造を形成する。重(VH)および軽(VL)鎖可変ドメインの相互作用は、抗原結合領域(Fv)の形成をもたらす。重鎖および軽鎖の相互作用は、重鎖のCH1ドメインと軽鎖のCκまたはCλドメインにより促進される。一般に、VHおよびVLの両方は抗原結合に必要とされるが、重鎖二量体およびアミノ末端断片は軽鎖の不在下で活性を保持することが示されている[2]。
【0007】
重(VH)および軽(VL)鎖の両方の可変ドメインの中で、一部の短いポリペプチドセグメントは例外的な可変性を示す。これらのセグメントは超可変領域または相補性決定領域(CDR)と称される。間にあるセグメントはフレームワーク領域(FR)と呼ばれる。VHおよびVLドメインのそれぞれの中に、3つのCDR(CDR1〜CDR3)がある。
【0008】
哺乳類において、5つのクラスの抗体(IgA、IgD、IgE、IgGおよびIgM)が存在し、ヒトには4つのIgGサブタイプと2つのIgAサブタイプが存在する。
【0009】
【表1】

抗体クラスはその生理学的機能において異なる。例えば、IgGは、成熟した免疫応答において主要な役割を果たす。IgMは、補体結合および凝集に関与する。IgAは、分泌物(涙、唾液、初乳、粘液)中の抗体の主要なクラスであり、従って局所免疫に役割を果たす。天然抗体のエフェクター機能は、重鎖定常領域によってもたらされる。
【0010】
IgAは、粘液を含有する部位(例えば、腸内、気道内または泌尿生殖器路内)に見出すことができ、病原体による粘膜部位のコロニー形成を防ぐ。IgDは主にB細胞上の抗原受容体として機能する。IgEはアレルゲンと結合して肥満細胞からのヒスタミン放出(アレルギーの基礎をなす機構)を誘発し、また、蠕虫(虫)に対する保護をもたらす。IgGは(その4つのアイソタイプにおいて)、侵入してくる病原体に対して抗体に基づく免疫性の大部分をもたらす。IgMは、B細胞の表面に発現され、B細胞の媒介する免疫性初期段階において(すなわち、病原体を排除するために十分なIgGが存在する前に)病原体を排除するための非常に高い親和性を有する分泌型としても存在する。
【0011】
正常なヒトB細胞は、単一の重鎖遺伝子座を14番染色体上に含有し、その遺伝子座から重鎖をコードする遺伝子が再構成により形成される。マウスにおいて、重鎖遺伝子座は12番染色体に位置する。正常な重鎖遺伝子座は、複数のV遺伝子セグメント、多数のD遺伝子セグメントおよび多数のJ遺伝子セグメントを含む。VHドメインの大部分はV遺伝子セグメントによってコードされるが、それぞれのVHドメインのC末端はD遺伝子セグメントとJ遺伝子セグメントによってコードされる。B細胞におけるVDJの再構成と、その後の親和性成熟は、それぞれのVHドメインにその抗原結合特異性をもたらす。正常なH2L2四量体の配列解析は、多様性が主にVDJ再構成と体細胞超変異の組合せに起因することを実証する[3]。ヒトゲノムには50個を超えるヒトV遺伝子セグメントが存在し、そのうちの39個のみが機能性である。
【0012】
完全なヒト抗体(H2L2)は、現在、トランスジェニックマウスから抗原曝露に応答して導くことができる。そのようなトランスジェニックマウスは単一のヒト重鎖遺伝子座および別個の軽鎖遺伝子座を含む。対応するマウス重鎖および軽鎖遺伝子座は、マウス抗体の不在下でヒト抗体のみが産生されるように欠失または抑制される([4〜10])。
【0013】
新しい分子生物学技法の出現で、重鎖のみ抗体(軽鎖を欠く)の存在が、人におけるB細胞増殖性障害において同定され(重鎖疾患)、さらにマウスモデル系においても同定された。分子レベルでの重鎖疾患の分析により、ゲノムレベルでの突然変異および欠失が重鎖CH1ドメインの不適切な発現をもたらし、軽鎖と結合する能力を欠く重鎖のみ抗体の発現を引き起こしたことが示された[11、12]。
【0014】
ラクダ科動物が、遺伝子の自然突然変異の結果として、軽鎖との結合を媒介するCH1ドメインが存在しないために軽鎖と結合することのできない機能性IgG2およびIgG3重鎖のみ二量体を生成することが示された[13]。ラクダ科動物重鎖のみ抗体の特性を特徴付けるのは、ヒトおよび正常なラクダ科動物VHドメインに対して改良された溶解度をもたらすラクダ科動物VHドメインの特定のサブセットである。この特定のサブセット中のラクダ科動物VHドメインは通常VHHドメインと称される。
【0015】
サメなどの種が、おそらく哺乳類T細胞受容体または抗体軽鎖に類縁であり、重鎖のみ様結合タンパク質ファミリーを生成することも示されている[14]。
【0016】
ラクダ科動物重鎖のみ抗体の産生のために、ラクダ科動物生殖細胞系列の重鎖遺伝子座は、可能性のある重鎖定常領域の一部または全部をコードする遺伝子セグメントを含む。成熟の間、再構成されたVHHDJ結合ドメインは、ヒンジドメインをコードする遺伝子セグメントの5’末端上にスプライスされて、CH1ドメインを欠き、従って軽鎖と会合することのできない重鎖をコードする、再構成された遺伝子がもたらされる。
【0017】
ラクダ科動物VHHドメインは、特徴的な多数のアミノ酸を37、44、45および47の位置に含有する[49]。これらの保存されたアミノ酸は、重鎖のみ抗体に溶解性を付与するために重要であると考えられる。特定のラクダ科動物VHドメインだけが、改良された溶解度特性を有するVHHドメインである。それらはVHサブファミリーに限定され、そのため限定された範囲の抗原だけに増殖的に応答する。
【0018】
重鎖のみモノクローナル抗体は、標準的なクローニング技術によりラクダ科動物脾臓のB細胞から、またはファージもしくはその他のディスプレイ技術によりB細胞mRNAから回復することができる[18]。ラクダ科動物に由来する重鎖のみ抗体は高親和性である。重鎖のみ抗体をコードするmRNAの配列解析は、正常な四量体抗体の産生においても観察されるように、多様性が主にVHHDJ再構成と体細胞超変異の組合せに起因することを実証する[49]。
【0019】
天然のラクダ科動物およびヒトVHドメインの重要かつ共通する特徴は、それぞれのドメインが、最適な溶解度および結合親和性のためにVLドメインでの二量化に依存することなく、モノマーとして結合することである。
【0020】
近年、ヒトを除くトランスジェニック哺乳類における重鎖のみ抗体の産生のための方法が開発された(WO02/085945号およびWO02/085944号参照)。潜在的に任意のクラス(IgM、IgG、IgD、IgAまたはIgE)の、任意の哺乳類に由来する機能性重鎖のみ抗体は、抗原曝露の結果としてヒトを除くトランスジェニック哺乳類(好ましくはマウス)から生成することができる。これらの初期研究は2つのラマV遺伝子セグメントの使用に頼り、抗体応答の限定されたレパートリーに悩まされた。
【0021】
Janssens et al.[15]は、トランスジェニックマウスにおける重鎖のみ抗体の誘導のための方法を開発した。B細胞での抗体成熟の結果として高い結合親和性を有するこれらの重鎖のみ抗体は、抗原曝露の結果として誘導され、確立されたハイブリドーマ技術を用いて選択され、軽鎖(例えばIgG、IgA、IgM)の不在下で任意のクラスの抗体として、またはVH結合ドメイン単独として生成され得る。これらの重鎖のみ抗体は、全てのヒトDおよびJ遺伝子セグメントと結合した2つのラマVHH(クラス3)遺伝子セグメントならびに全てのヒト定常領域をコードする遺伝子セグメントを含有する生殖細胞系列(すなわち非再構成)配置中の抗体重鎖遺伝子座から誘導された。それぞれの定常領域をコードする遺伝子セグメントは、CH1ドメインの欠失を有し、軽鎖の結合を妨げた。さらに、この遺伝子座は、3’末端に抗体LCRを含有し、B系列の細胞における高いレベルの発現を確保する。この遺伝子座を受精卵のマイクロインジェクションによりマウスに導入した。
<抗体を原料とした産物の製造>
抗体を原料とした産物の遺伝子工学による製造、特にヒトまたはヒト化抗体を原料とした産物の製造は、新規なクラスの医薬、診断法および試薬の生成をもたらすと同時に、新規な産業、雇用および富の創造の機会の創出をもたらした(www.drugresearcher.com、www.leaddiscovery.co.uk参照)。抗体を原料とした産物は通常、天然の四量体抗体から導かれる。抗体を原料とした産物の製造に関する特許および特許出願は多くある。これらの特許および特許出願は、(例えばトランスジェニックマウスからの)誘導の経路、製造の経路および産物に特異的な物質の事柄に関する。そのような抗体を原料とした産物は、抗体断片を介して完全な四量体抗体から一本鎖Fv(scFv)分子までさまざまである。
【0022】
抗体を原料とした産物は、21世紀に着手される新規な医薬品の高い割合を占める。モノクローナル抗体療法は、関節リウマチおよびクローン病の治療のための好ましい経路として既に容認され、癌の治療において見事な進展がある。また、抗体を原料とした産物は、心血管疾患および感染症の治療のために開発中である。大部分の販売されている抗体を原料とした産物は、標的リガンド(例えばTNFα)上の単一の明確なエピトープを認識し、結合する。
【0023】
近年、高親和性VHドメインは、ディスプレイライブラリー中の無作為化されたヒトVHドメインから選択されるか、またはラクダ科動物の抗原曝露によって自然に産生された重鎖のみ抗体から誘導されるか、またはラクダ科動物で作られたVHドメインライブラリーから誘導されている。これらの高親和性VHドメインは抗体を原料とした産物に組み込まれている。これらのVHドメインは、VHHドメインとも呼ばれ、多数の古典的なVHドメインとは違い、特に、軽鎖の不在下で重鎖の改良された溶解度および安定性を確保する多数の突然変異を提示する。これらの変化の中で最も顕著なものは、荷電アミノ酸が45位に存在することである[16]。
【0024】
多数のグループが、天然の四量体抗体から誘導される重鎖のみ抗体の作製に取り組んできた。Jaton et al.は(同文中に2およびその他の参照文献が引用される)、親和性精製された、特性が十分明らかであるウサギ抗体の縮小された重鎖成分の分離、それに続いてその後の個々の重鎖の再生を記載している。再生された重鎖の免疫学的特性解析により、軽鎖を含まない重鎖ホモ二量体が、単独で、能力のある結合抗原であることが実証された。
【0025】
その後、Ward et al.[18]は、クローン化されたマウスVH領域が、大腸菌発現系で可溶性タンパク質モノマーとして発現されると、高い親和性で抗原と結合する能力を保持することを明白に実証した。Ward et al.[18]は、VHドメインの単離および特性解析を記載し、古典的なモノクローナル抗体産生(最後の段落参照)と比較した場合のこのアプローチの潜在的な商業上の利益を示した。彼らはまた、通常は軽鎖と会合する重鎖から単離されたVHドメインが天然の四量体抗体の溶解性に欠けていることを認めている。従って、Ward et al.[18]は、これらの分子を表現するために「粘着性のある」という用語を用い、この「粘着性」は改良された溶解度特性を有するVHドメインの設計によって対処することができると提案した。
【0026】
VH溶解度の改良は、その後、ファージディスプレイを用いる無作為化された部位特異的なアプローチの組合せを用いて対処されてきた。例えば、Davies and Riechmann[17]は(WO92/01047号参照)、溶解度を改良すると同時に結合特異性を維持するためにラクダ科動物重鎖のみ抗体のVHドメインの特徴の一部をファージディスプレイと組み合わせて組み込んだ。
【0027】
ファージライブラリーに由来する単離されたヒトVHドメインに関する複数の独立した研究により、VHドメインの抗原特異的な結合が実証されたが、これらのVHドメインは溶解度が低いことが分かった。さらに、ファージアレイに提示される特異的結合特性を有するヒトVHドメインの選択が操作された抗体のためのビルディングブロックを形成することが示唆された[18]。
【0028】
ヒトVHドメインは改良された溶解度特性のために操作されてよく[19、20]、または溶解度はインビボで自然選択により獲得されてもよい[21]。しかし、VH結合ドメインがファージライブラリーから誘導されている場合、例えば親和性ホットスポットのランダム化を含む親和性改善策の適用にもかかわらず、抗原に対する固有の親和性は低マイクロモルから高ナノモルの範囲のままである[22]。
【0029】
ファージまたは代替ディスプレイ技術により生成されたヒトVHまたはラクダ科動物VHドメインは、体細胞突然変異の結果としての改良された特性の利点および正常な抗体結合部位のCDR3領域におけるDおよびJ遺伝子セグメントの組換えにより提供されるさらなる多様性を欠く。一部のラクダ科動物VH(VHH)ドメインは、ヒトVHに対して溶解度の点で利点を示すものの、ヒトにおいて抗原性であることが判明し、さらに、ラクダ科動物VHがラクダ科動物の免疫化またはファージディスプレイ技術により生成されねばならないという不利点を被る可能性がある。
【0030】
ファージ由来ヒトVH領域は、高い親和性結合特性を達成するためにそれらが多くの回のパニングおよびその後の変異誘発を必要とするので、使用することが困難である。ラクダ科動物VHHドメインは、ファージもしくは類似のディスプレイライブラリーから単離された場合に同じ困難な手順を必要とするか、または、古典的なハイブリドーマ技術に従わない大型動物(同様に古典的な抗体を作成するラマまたはラクダ)の免疫化を必要とする。さらに、ラクダ科動物の結合ドメインは抗原性であることが分かり、ヒト化を必要とし得る。
【0031】
抗原曝露の結果としてのヒトを除くトランスジェニック哺乳類における重鎖のみ抗体の産生(WO02/085945号およびWO02/085944号参照)は、これらの多くの問題を克服する。
【0032】
しかし、当技術分野において、重鎖のみ抗体の多様性およびインビボでB細胞の応答を最大化する、特に、クラス特異的ヒト重鎖のみ抗体の機能性レパートリーおよび多様な臨床、工業および研究用途で使用するための最大の抗原結合可能性を保持する機能性VH重鎖のみ結合ドメインを生成する必要性が残っている。
【発明の開示】
【0033】
本発明者らは、驚くべきことに、先行技術の制限を克服し、クラス特異的な重鎖のみ抗体を生成するために用いられるヒトを除くトランスジェニック哺乳類に存在する重鎖のみの遺伝子座の数を増加させることにより、抗体応答のレパートリーを大いに増大させることができることを示した。
【0034】
本発明は、ヒトを除くトランスジェニック哺乳類が複数の重鎖のみの遺伝子座を有する場合、それらの遺伝子座は対立遺伝子排除の対象となるという発見に依存する。その結果、ただ1つの遺伝子座が確率的に選択され、首尾よく組換えられ、結果として重鎖のみ抗体の産生がもたらされる。従って、複数のVH重鎖遺伝子座を同じヒトを除くトランスジェニック哺乳類に用いて、哺乳類から入手可能な抗体レパートリーおよび多様性を最大化することができる。抗原に曝露された場合、ヒトを除くトランスジェニック哺乳類は、残りの遺伝子座を除いて特異的抗原曝露に応答するために最も適しているV遺伝子セグメントを含む遺伝子座を「選択する」。
【0035】
本発明の方法により生成され得る重鎖のみ抗体は、ヒトを除くトランスジェニック哺乳類が一連の遺伝子座から「選択する」ことが可能であることの結果として高い結合親和性を示し、それから一般に伸張されたCDR3ループの不在下でV、DおよびJ遺伝子セグメントの再構成ならびに体細胞突然変異が起こり得る。本質的に正常なB細胞の成熟は、高いレベルの重鎖のみ抗体が(CH1ドメインが組換え遺伝子座に存在する全ての抗体クラスから排除されているという条件で)単離された血漿中に存在する状態で観察される。B細胞の成熟および構築された二量体(例えばIgG)または多量体(例えばIgM)の分泌は、軽鎖遺伝子の存在または発現に依存しない。
【0036】
従って、本発明の第1の態様では、その哺乳類に複数の異種のVH重鎖遺伝子座を供与する段階を含む、ヒトを除くトランスジェニック哺乳類におけるVH重鎖のみ抗体の産生のための方法が提供され、各VH重鎖遺伝子座は、1個以上のV遺伝子セグメント、1個以上のD遺伝子セグメント、1個以上のJ遺伝子セグメント、および発現した場合、CH1ドメインを含まず、前記遺伝子座の少なくとも1つからVH重鎖のみ抗体を発現する重鎖定常領域をコードする遺伝子セグメントを含む。
【0037】
好ましくは、それぞれのVH重鎖遺伝子座は、1つ以上のV遺伝子セグメント、例えば1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、45、50または60個のV遺伝子セグメントを含み、それは任意の脊椎動物種に由来してもよい。
【0038】
一実施形態では、各遺伝子座は、ただ1つのV遺伝子セグメントを含んでよい。本実施形態の一つの代替形態では、各V遺伝子セグメントは他の全てのV遺伝子セグメントとは異なっている。第2の代替形態では、各V遺伝子セグメントは、他の全てのV遺伝子セグメントと同一である。この第2の代替形態では、各遺伝子座中の残りの遺伝子セグメントは、他の全ての遺伝子座中の遺伝子セグメントと同じであってもよいし、異なっていてもよい。
【0039】
従って、非ヒト哺乳類は単一のVH重鎖遺伝子座の複数のコピーを含有し得ることが想定される。これは、B細胞において生産的な再構成が起こり、従って有用な重鎖のみ抗体の産生を可能にする機会を最適化するという利点を有する。
【0040】
非ヒト哺乳類が、多数の異なるVH重鎖遺伝子座を含有するならば、これは、所望の特異性を有する重鎖のみ抗体を得る機会をさらに最適化する。
【0041】
もう1つの実施形態では、それぞれの遺伝子座は複数のV遺伝子セグメントを含む。本実施形態では、任意の1つの遺伝子座中のV遺伝子セグメントは、全て同じ種の生物に由来してよい、例えば、全てのV遺伝子セグメントがヒト起源であってよい。あるいは、任意の1つの遺伝子座中のV遺伝子セグメントは異なる種の生物に由来してよい。例えば、一部のV遺伝子セグメントがヒトに由来し、他のものはラクダ科動物またはサメに由来してよい。
【0042】
好ましくは、V遺伝子セグメントはヒト起源である。
【0043】
用語「V遺伝子セグメント」は、ラクダ科動物およびヒトを含む脊椎動物に由来する任意の天然のV遺伝子セグメントを包含する。V遺伝子セグメントは、D遺伝子セグメント、J遺伝子セグメント、および重鎖定常(エフェクター)領域(CH1エキソン以外の数個のエキソンを含み得る)をコードする遺伝子セグメントと組み換えて、核酸が発現されるとVH重鎖のみ抗体を生成できなければならない。
【0044】
V遺伝子セグメントはまた、D遺伝子セグメント、J遺伝子セグメント、および重鎖定常領域(1個以上のエキソンを含むがCH1エキソンは含まない)をコードする遺伝子セグメントと組み換えて、本明細書に定義される重鎖のみ抗体を生成できる相同体、誘導体またはタンパク質断片をコードする任意の遺伝子配列をその範囲内に含む。V遺伝子セグメントは、例えばT細胞受容体遺伝子座または免疫グロブリン軽鎖遺伝子座に由来してよい。
【0045】
好ましくは、本発明の複数の重鎖遺伝子座は、複数の遺伝子座に分散された、改良された溶解度特性を有する39個の機能性ヒトV遺伝子セグメントおよび操作された前記ヒトV遺伝子セグメントの変異体のうちの、任意の数または任意の組合せを含む。これらは、任意の数の遺伝子座、例えば8つのV遺伝子セグメントを含む4つの遺伝子座に加えて7つのV遺伝子セグメントを含む1つの遺伝子座;4つのV遺伝子セグメントを含む7つの遺伝子座に加えて3つのV遺伝子セグメントを含む1つの遺伝子座;またはそれぞれ1つのV遺伝子セグメントを含む39の遺伝子座上であってよい。
【0046】
ヒトV遺伝子は7つのファミリー、VH1〜VH7に分類され、各ファミリー内の個々の遺伝子に番号が付けられている。それぞれの遺伝子が使用される頻度は、特定の免疫応答の変動する要件に依存する。例えば、ファミリーVH3の遺伝子は、細菌性抗原に応答する際にファミリーVH5の遺伝子と比較して選択的に使用され得る。従って、本発明のさらに好ましい実施形態では、特定の抗原に対する抗体応答を生成するために有用であることが示されたV遺伝子セグメントのグループを別々のヒトを除くトランスジェニック哺乳類の系へ分類する。V遺伝子セグメントはファミリーに従って分類されるか、またはそれらは個々の機能に従って分類される。例えば、ファミリーVH3のV遺伝子が細菌性抗原に対する抗体応答を生成するために有用であることが示されるならば、細菌性抗原に対する重鎖のみ抗体を生成するために特に有用であるヒトを除くトランスジェニック哺乳類を生成するためにファミリーVH3のV遺伝子を用いることができる。あるいは、ファミリーVH3およびVH5の数個の個別の遺伝子が細菌性抗原に対する免疫応答を生成するために有用であることが示されるならば、それらは一緒に分類されて、細菌性抗原に対する重鎖のみ抗体を生成するために特に有用であるヒトを除くトランスジェニック哺乳類を生成するために用いることができる。
【0047】
本発明の状況において、用語「異種の」とは、本明細書に記載される核酸配列または遺伝子座が、それが位置する哺乳類に対して内在性でないことを意味する。
【0048】
「VH重鎖遺伝子座」は、本発明の状況において、重鎖エフェクター領域をコードする1つ以上の遺伝子セグメント(それぞれCH1ドメインを欠く)と操作によって連結されている、1つ以上のV遺伝子セグメント、1つ以上のD遺伝子セグメント、および1つ以上のJ遺伝子セグメントを含む、VHドメインをコードする最小のマイクロ遺伝子座に関する。好ましくは、抗体レパートリーの可変性の主な原因は、V、DおよびJ遺伝子セグメントの選択により、さらにV−DおよびD−J接合により形成されるCDR3領域である。
【0049】
本発明の利点は、再構成されたVH遺伝子配列において得られた抗体レパートリーおよび多様性が、同じヒトを除くトランスジェニック哺乳類における複数のVH重鎖遺伝子座の使用によって最大化することができることである。Janssens et al.、2006[15]は、上に記載されるトランスジェニック遺伝子座が、再構成および対立遺伝子排除に関して正常な免疫グロブリン遺伝子座のような挙動をすることを示した。このことは、同じ動物において(異なる染色体上に)複数の遺伝子座を有して、対立遺伝子排除を利用することにより起こり得るVH組換えの数を最大化する可能性を開いた。各トランスジェニック遺伝子座は、1から40を超えるVH領域を含有する。遺伝子座の1つを無作為に選択して組換えを開始し、最初の組換えが非生産的などであった場合に生産的な組換えが遺伝子座の1つから産出されるまで次の遺伝子座がその後に続く、対立遺伝子排除のプロセスは、組み合わせた遺伝子座中に存在する全てのVH領域が実際に全体的な組換えプロセスの一部となることを確保するものである。
【0050】
Janssens et al.、[15]はまた、驚くべきことに、同じヒトを除くトランスジェニック哺乳類において複数の重鎖遺伝子座が同じ染色体上で直列になっている場合、その結果、生産的な再構成および抗体の発現が複数の重鎖遺伝子座から起こり得ることを見出した。そのような場合、その後のハイブリドーマクローンはポリクローナルである。当業者であれば、これが抗体の多様性を増加させるためのもう一つの機構であることを理解する。
【0051】
従って、本発明の別の態様では、本明細書に記載されるように、ヒトを除くトランスジェニック哺乳類において少なくとも2つ以上のVH重鎖遺伝子座が同じ染色体上で直列になって存在し、その結果、生産的な再構成および抗体の発現が2つ以上の遺伝子座から同時に起こり得る方法が提供される。
【0052】
好ましくは、多数の重鎖遺伝子座は最初に別々にヒトを除くトランスジェニック哺乳類に導入され、単一の重鎖遺伝子座を有するヒトを除くトランスジェニック哺乳類を作り出す。次に、VH領域の数を最大化するためにこれらの動物を交配させて複数の重鎖遺伝子座を有する後代を生成し、最大の多様性をもたらす。遺伝子座はまた、新しい1ラウンドの遺伝子組換えによって加えてもよい。新しい遺伝子座は、既に1以上の異種のVH重鎖遺伝子座を含むヒトを除く哺乳類由来の卵に注入される。異種のVH重鎖遺伝子座の安定な融合により、利用可能なVH領域の増加および、従って多様性がもたらされる。異種のVH重鎖遺伝子座を、さらなる異種のVH重鎖遺伝子座とともに含む、ヒトを除くトランスジェニック哺乳類に由来するES細胞の安定なトランスフェクションは、ES細胞技術を胚融合および胚盤胞注入による遺伝子組換えに使用することのできるヒトを除く哺乳類(例えばマウス)において多様性を増加させる代替経路を提供する。
【0053】
好ましくは、それぞれの異なる重鎖遺伝子座は、ヒトを除くトランスジェニック哺乳類のゲノム中に単一コピーとして存在する。
【0054】
さらに、複数のV、DおよびJ遺伝子セグメントを使用することにより、得られる抗体レパートリーおよび多様性のさらなる増加がもたらされる。その後の体細胞突然変異は、VLおよびLc(軽鎖)抗体遺伝子座の必要なく最小の遺伝子座(マイクロ遺伝子座)を使用しながら達成される。
【0055】
本発明の状況において、用語「D遺伝子セグメント」および「J遺伝子セグメント」には、DおよびJ遺伝子セグメントの天然の配列が含まれる。好ましくは、DおよびJ遺伝子セグメントは、V遺伝子セグメントと同じ脊椎動物に由来する。例えば、V遺伝子セグメントがヒトに由来する場合、必要に応じて可溶化されたまたは操作されたDおよびJ遺伝子セグメントも、ヒトに由来することが好ましい。あるいは、V遺伝子セグメントは、例えばラクダ科動物に由来し、DおよびJ遺伝子セグメントはヒトに由来してもよく、逆も同様である。
【0056】
D遺伝子セグメントおよびJ遺伝子セグメントという用語はまた、得られるセグメントが本明細書に記載される重鎖抗体遺伝子座の残りの成分と組み換えられて本明細書に記載の重鎖のみ抗体を生成することができる限り、誘導体、相同体およびその断片をその範囲内に含む。DおよびJ遺伝子セグメントは天然の供給源から誘導されてもよいし、それらは当業者が熟知ある、本明細書に記載される方法を用いて合成されてもよい。V、DおよびJ遺伝子セグメントは組換えが可能であり、好ましくは、体細胞突然変異を受ける。
【0057】
DおよびJ遺伝子セグメントは、単一の脊椎動物種に由来することが好ましい。これはいずれの脊椎動物種であってもよいが、好ましくはヒトである。
【0058】
好ましくは、各VH重鎖遺伝子座は、1〜40(2、3、4、5、6、7、8、9、10、12、14、16、18、20、30または40)以上のD遺伝子セグメントを含む。D遺伝子セグメントはいずれの脊椎動物種に由来してもよいが、最も好ましくは、D遺伝子セグメントはヒトD遺伝子セグメント(通常25個の機能性D遺伝子セグメント)である。
【0059】
好ましくは、それぞれのVH重鎖遺伝子座は、1〜20(2、3、4、5、6、7、8、9、10、12、14、16、18または20)以上のJ遺伝子セグメントを含む。J遺伝子セグメントはいずれの脊椎動物種に由来してもよいが、最も好ましくは、J遺伝子セグメントはヒトJ遺伝子セグメント(通常6つのJ遺伝子セグメント)である。
【0060】
各VH重鎖遺伝子座は、同一のDおよびJ遺伝子セグメントを含有してよい。あるいは、各VH重鎖遺伝子座は、DおよびJ遺伝子セグメントの異なる組合せを含有してよい。例えば、各VH重鎖遺伝子座がただ1つのV遺伝子セグメントを含有し、このセグメントが各遺伝子座において同一である場合、生産的な再構成を得るための機会をさらに最適化するために、各遺伝子座においてDおよびJ遺伝子セグメントの異なる組合せを使用することが有利である。しかし、各VH重鎖遺伝子座が1つ以上の異なるV遺伝子セグメントを含有する場合、各遺伝子座においてDおよびJ遺伝子セグメントの同一の組合せを使用することが有利であろう。
【0061】
好ましくは、VH重鎖遺伝子座は、1以上のV遺伝子セグメント、25の機能性ヒトD遺伝子セグメントおよび6つのヒトJ遺伝子セグメントを含む。
【0062】
重鎖定常領域をコードする各遺伝子セグメントは、重鎖定常領域遺伝子セグメントがCH1ドメインを発現しないという条件で、Cδ、Cγ1−4、Cμ、CεまたはCα1−2クラスの1個以上の重鎖定常領域エキソンを含み得る。重鎖定常領域遺伝子セグメントは、好ましいクラスまたは必要な抗体クラスの混合物に応じて選択される。場合により、異種重鎖遺伝子座はCμおよびCδを欠損している。
【0063】
従って、各VH重鎖遺伝子座は、インビボでエフェクター機能をもたらす少なくとも1つの重鎖定常領域をコードする遺伝子セグメントを含んでよく(例えばIgG、IgM、IgA、IgEもしくはIgDまたはそのアイソタイプ)、各定常領域はCH1ドメインを含まない。各遺伝子座は、1つの特定の定常領域をコードする遺伝子セグメントをただ1つ含有してよい。これは、エフェクター機能が必要とされる場合に有利である。あるいは、各遺伝子座は、それぞれが異なる定常領域をコードする1個よりも多くの遺伝子セグメントを含んでよい。これは、様々なクラスの重鎖のみ抗体が産生されることを可能とするような、特定のエフェクター機能についての要件がない場合に有利である。
【0064】
各遺伝子座は同じ定常領域コード遺伝子セグメントを含有してもよく、または各遺伝子座は異なる定常領域コード遺伝子セグメントまたはその異なる組合せを有してもよい。
【0065】
操作上、重鎖定常領域は、B細胞においてV遺伝子セグメント、D遺伝子セグメントおよびJ遺伝子セグメントと組換え可能な天然のまたは操作された遺伝子セグメントによってコードされる。
【0066】
各重鎖のみ抗体において、定常領域は、重鎖のみ抗体の生成が起こり得るように、CH1ドメインを含まずに発現される。CH1エキソンは、上記のVH重鎖のみ抗体の定常領域が機能性CH1ドメインを含有しないように除去され得る。
【0067】
多価の結合複合体を操作する際にクラス特異的な重鎖定常領域を含むことにより、インビボでエフェクター機能の治療上の利益が必要される機能性に応じてもたらされる。個々のエフェクター領域を操作することも、機能性の付加または欠失をもたらし得る[23]。従って、IgA定常領域の機能性を含むことにより、病原体に対して改良された粘膜機能をもたらされるが[24]、一方で、IgG1定常領域の機能性が存在することにより、インビボで強化された血清安定性がもたらされる。重鎖CH2およびCH3定常ドメインが存在することにより、自然抗体に見られるような安定的な二量体形成の基盤がもたらされ、翻訳後グリコシル化の認識部位がもたらされる。CH2およびCH3の存在はまた、二重特異性および多価の複合体を試薬および診断薬として用いる際に、二次抗体の認識も可能にする。
【0068】
例えば、IgM抗体は、マクロファージの活性化および補体経路の活性化に重要な役割を果たすことが公知である。その結合部位が近接していることに起因して、IgMは、ウイルスを含む病原体に高い親和力を有する。しかし、IgMは、迅速なイムノアッセイ技法での使用が困難であることも公知であり、その一方でIgG抗体はこれらの技法で容易に使用することができる。用途に応じて、好ましい抗体クラス、すなわちIgGまたはIgMを選択することが有用である。
【0069】
CH1を欠く異種の重鎖Cγ遺伝子座の全部または一部の発現は、異種のIgG遺伝子座に存在するIgG1、IgG2、IgG3およびIgG4アイソタイプに応じて、場合により一部または全てのIgGアイソタイプを生成する。あるいは、重鎖はCε遺伝子を含んでよい。得られるIgE分子も治療に使用してよい。
【0070】
あるいは、選択された抗体の混合物を得ることもできる。例えば、IgAおよびIgMは、重鎖定常領域がCαおよびCμ遺伝子を含む場合に得ることができる。
【0071】
好ましくは、重鎖定常領域は、特に重鎖のみ抗体がヒトにおける治療用途に使用される場合に、ヒト起源である。重鎖のみ抗体が獣医学目的に使用される場合は、重鎖定常領域は、獣医学治療が行われる標的生物、脊椎動物または哺乳類に由来することが好ましい。
【0072】
発現した場合、各重鎖定常領域はCH1ドメインを欠く。好ましくは、CH1エキソンが欠失している。場合により、CμおよびCδ定常領域は変異しているか、欠失しているか、または置換されていてよい。例えば、機能性CH1ドメインを含むIgMの存在は、B細胞の成熟を阻害し、その結果として同じ遺伝子座の中で重鎖のみのIgG(CH1を欠く)の生産的発現を制限する。
【0073】
本明細書に定義される「重鎖定常領域エキソン」(「CHエキソン」)には、天然の脊椎動物の配列が含まれるが、特に哺乳類のCHエキソンである。これはクラス特異的な方法で変動する。例えば、IgGおよびIgAは天然にCH4ドメインを欠く。用語「CHエキソン」は、CHエキソンが、それが重鎖定常領域の成分である場合に、本明細書に定義される機能性重鎖のみ抗体を形成することができる限り、その範囲内に誘導体、相同体およびその断片も含む。
【0074】
CH4またはその他の機能性ドメインは、存在する場合、細胞内分泌プロセス、B細胞の成熟または得られる抗体の結合活性を阻害しないならば、任意選択的に導入遺伝子内で操作されるかまたは除去されてよい。
【0075】
細胞表面の集合を妨げる分泌機構とその結果起こるB細胞成熟に操作が影響を及ぼさないならば、重鎖エフェクター分子は操作されて、機能性ドメイン、例えばカルボキシ末端CH4ドメインが取り除かれてよい。例えばグリコシル化を改良するかまたは機能を付加するために、さらなる特徴を遺伝子座に組み換えてもよい。
【0076】
従って、異種の重鎖遺伝子座は、基本的に正常にB細胞成熟した状態で、必要とされる抗体クラスに応じて、重鎖のみ抗体の好ましいクラスまたは混合物を生成するように設計される。ラクダ科動物V、DおよびJ遺伝子セグメントおよびラクダ科動物エフェクター領域を利用すると、ラクダ科動物抗体に特有の特長、例えば伸張したCDR3ループなどを有するラクダ科動物抗体が産生される。ヒトV、DおよびJ遺伝子セグメントを使用すると、伸張したCDR3ループを欠くヒト重鎖のみ抗体が産生される。
【0077】
本発明の状況において「VHドメイン」は、上記に定義されるように、D遺伝子セグメントおよびJ遺伝子セグメントと組み換えられる場合、V遺伝子セグメントの発現産物を指す。好ましくは、VHドメインは、VDJ組換えおよび体細胞突然変異の結果として抗原と結合する改良された能力を有する。ラクダ科動物種に特有の伸張したCDR3ループの存在または不在に対する依存性はない。VHドメインはモノマーとして抗原と結合することが可能である。可溶性重鎖のみ抗体複合体の一部として発現された場合に、VLドメインとの組換えの可能性は、CH1エキソンの除去により排除されている([25]参照)。
【0078】
VHドメインは、単独で、多様なタンパク質ドメインと遺伝子操作されて、例えば毒素、酵素および造影剤を含む、標的とされる治療および診断目的のための融合タンパク質を生成することもできる。
【0079】
VHドメインコード配列は、天然の供給源から誘導されてもよいし、それらは当業者の熟知の方法を用いて合成されてもよい。
【0080】
VHドメインの性質は、必要とされる特徴を有する配列をコードするV、Dおよび/またはJ遺伝子セグメントを選択するかまたは操作することにより変更または改良することができる。上記に示されるように、39個の機能性ヒトVH領域の一部は重鎖のみ抗体の産生に適していない可能性がある。様々な抗体特性を改良しようと試みて多数の研究が先行技術において行われた[26,29]。特定のVH領域特性に関して、Dolk et al.[30]は、ファージディスプレイ技術を用いて、フケ予防シャンプーに伴う厳しい条件下において改良された安定性を示す、重鎖のみ抗体を生成した。
【0081】
ヒトを除くトランスジェニック哺乳類は、好ましくは、げっ歯類、例えば、ウサギ、モルモット、ラットまたはマウスである。マウスが特に好ましい。代わりとなる哺乳類、例えばヤギ、ヒツジ、ネコ、イヌまたはその他の動物を用いてもよい。好ましくは、哺乳類はマウスである。
【0082】
好ましくは、トランスジェニック非ヒト動物は、確立された卵母細胞注射技術のみを用いて作製される。確立されていれば、ES細胞技術またはクローニングを用いてもよい。
【0083】
有利には、哺乳類に内在する抗体の重鎖遺伝子座および場合により軽鎖遺伝子座は、重鎖のみ抗体が本発明の方法に従って発現される場合に、削除されるか抑制される(silenced)。
【0084】
抗体の供給源とは異なる起源の脊椎動物の種への抗体の投与は、しばしば投与された抗体に対する免疫応答の発現をもたらすので、本発明の上記の態様に記載される重鎖のみ抗体を生成する方法は、ヒトの治療用途のための抗体の生成において特に有用であり得る。本発明に従って生成される抗体は、それらが実質的に単一または既知のクラスの抗体であり、好ましくは、ヒト起源であるという点で、先行技術のそれに優る利点を有する。抗体は、インビボでのVDJ組換えと親和性成熟の組合せにより高親和性である。
【0085】
従って、本発明のさらなる態様は、上記により定義された複数の異種のVH重鎖遺伝子座を含むヒトを除くトランスジェニック哺乳類を提供する。好ましくは、ヒトを除くトランスジェニック哺乳類は、軽鎖を含む抗体を産生する能力が低下するように遺伝子操作されてよい。
【0086】
抗体産生細胞は、本明細書に定義されるヒトを除くトランスジェニック哺乳類から誘導され、例えば、本明細書に定義される重鎖のみ抗体の産生のためのハイブリドーマの調製に使用されてよい。それに加えて、またあるいは、核酸配列をこれらのヒトを除くトランスジェニック哺乳類から単離し、当業者の熟知な組換えDNA技法を用いてVHドメイン重鎖のみ抗体またはその二重特異性/二機能性複合体を生成するために用いてよい。
【0087】
あるいは、またはそれに加えて、抗原特異的な重鎖のみ抗体は、本明細書に定義されるヒトを除くトランスジェニック哺乳類の免疫化によって生成されてよい。
【0088】
従って、本発明はまた、上に定義されるヒトを除くトランスジェニック哺乳類を抗原で免疫することによる、重鎖のみ抗体の産生のための方法を提供する。抗体およびその断片は、当業者に公知の確立された方法を用いて単離され、特性決定され、製造されてよい。これらの抗体は、PCT/GB2005/00292号に記載される方法に特に役立つ。
【0089】
本発明を、これから一例として、以下の図を参照する以下の詳細な説明において説明する。
〔実施例〕
【0090】
実施例1−重鎖のみ抗体遺伝子座はマウスにおいて完全に機能性であり、対立遺伝子排除に対して感受性である。
【0091】
上記に考察されるように、Janssens et al.[15]は、トランスジェニックマウスにおける重鎖のみ抗体の誘導のための方法を開発した。本方法および本明細書に記載される実験のさらなる詳細に関して、当業者は、参照により本明細書に組み込まれるJanssens et al.[15]を参照されたい。1
<方法>
・構築物
ゲノムコスミドライブラリーを、標準法を用いてラマ(Lama Glama)の末梢血細胞から作成した。2つの異なる生殖細胞系列VHHを、それらの配列、停止コドンを含まないオープンリーディングフレームおよびルフランの番号付け[32]に従う42、50および52位の親水性のアミノ酸コドンの存在、ならびに49位に親水性のアミノ酸を含むものと含まないものに基づいて選択した。一方はIGHV1S1(受託番号AF305944)と同一であり、もう一方はIGHV1S3(受託番号AF305946)と94%の同一性を有する。2つのクローン(ヒト重鎖DおよびJ領域、CμおよびCδを含有するクローン1065N8ならびにCγ3遺伝子を含有するクローン1115 N15)をヒトゲノムPacライブラリーRPCI−11(BACPAC Recource Center,USA)から選択した。異なるヒトゲノムライブラリー(Incyte Genomics,CA,USA)からのBacクローン11771をCγ2遺伝子およびIg重鎖LCRの供給源として用いた[33]。標準技法を用いて、Cγ3およびCγ2遺伝子を別々にpFastBac(Invitrogen)にサブクローニングした。単一の点突然変異(G〜A)[34]またはCH1エキソンの完全な欠失を相同組換えにより達成した[35]。同様にfrtおよびloxP部位をCμスイッチ領域の前に導入し、2番目のloxP部位をCγ2スイッチ領域の前に設置し、MGSまたはMGΔを得た。
【0092】
GSまたはGΔ構築物を得るために、MGSまたはMGΔベクター(図1)、2つのラマVHH遺伝子、その後にヒトDおよびJ重鎖領域、Cμ、Cδならびに修飾されたヒトCγ2およびCγ3遺伝子および3’LCRを含有し、16個の294Cre大腸菌株44に形質転換し、creに媒介される組換えによってGSまたはGΔ遺伝子座を得た(図1)。相同組換えによるCμCH1領域の欠失によりMGΔからMΔGΔを得た。
・トランスジェニックマウスの生成、育種および遺伝子型同定
220KbのMGSまたはMGΔまたはMΔGΔ断片、150KbのGSまたはGΔ断片(図1)をベクター配列から精製し、受精したFVB X B16/μMT−/−卵の前核に2ng/μlの濃度で注入した。5’および3’末端プローブを用いる尾由来DNAのサザンブロット分析により、トランスジェニック遺伝子座を、完全性およびコピーの数についてチェックした。トランスジェニックμMT+/−樹立系統をμMT−/−バックグラウンドの系統として育種した。以下のプライマー:Asp5IgMfw:5’−GCGGGTACCGAATGGTGGCAGGGATGGCTC−3’(配列番号1)と、HLL−MDには、Asp3’IgG2rv:5’−CGCGGTACCCTGCGGTGTGGGACAGAGCTG−3’(配列番号2)の組合せ、または、MGSには、Asp3’IgMrv:5’−CGCGGTACCACGGCCACGGCCACGCTGCTCGATTC−3’(配列番号3)の組合せ、およびμMT遺伝子型には、MIGMEMBINTRON1:fw:5’−CCAGTCAATACTACTCGCTAAGATTC−3’(配列番号4)とMIGMEMBEXON1rv:5’−CAGTGGTCCACAGTTTCTCAAAGC−3’(配列番号5)の組合せを用いるPCR(94℃にて45秒間の変性、60℃にて30秒間のアニーリングおよび72℃にて1分40秒間の伸張で30サイクル)により遺伝子型同定を行った。
・RT−PCR
Ultraspec RNA単離系統(Biotecx Laboratories,Houston)を用いて全RNAを単離した。逆転写酵素(Superscript II、Life technology)およびオリゴ(dT)プライマーを用いてcDNAを合成した。以下のプライマー:LVHHfw:5’−AGACTCTCCTGTGCAGCCTCTGG−3’(配列番号6)と、HINGEIgG2rv:5’CACTCGACACAACATTTGCGCTC−3’(配列番号7)またはhIgMCH2rv:CACTTTGGGAGGCAGCTCAGC−3’(配列番号8)の組み合わせを用いてPCRを行った。増幅は、94℃にて30秒間の変性、60℃にて30秒間のアニーリングおよび72℃にて1分間の伸張で30サイクル間であった。PCR産物をpGEM Tイージーベクター(Promega)にクローニングし、T7またはSP6プライマーを用いて配列決定した。
・フローサイトメトリー分析:
単細胞懸濁液を、既に記載したようにPBS中のリンパ器官から調製した45。約1×106個の細胞を、96ウェルプレート中のPBS/0.5%ウシ血清アルブミン(BSA)中の抗体を用いて4℃にて30分間インキュベートした。細胞をPBS/0.5% BSAで2回洗浄した。それぞれのサンプルについて、FACScanアナライザー(Becton Dickinson,Sunnyvale,CA)を用いて3×104イベントが記録された。CellQuestバージョン1.0コンピュータソフトウェアを用いてFACSデータを分析した。Becton Dickinson FACS Caliburで四色分析を行った。使用した大部分の抗体は記載されている[36];FITCコンジュゲート抗ヒトIgGおよび抗ヒトIgMはSigma(Zwijndrecht,NL)より購入した。
・Ig遺伝子再構成分析
wtマウスMΔGΔおよびGΔトランスジェニックマウスの脾臓および肝臓から単細胞懸濁液を作成した。MACS CD45(B220)MicroBeads(Miltenyi Biotec,Germany)をAutomacsセパレーターで製造業者の説明書に従って用いて 約90%の純度までB細胞を確実に選択した[37]。ゲノムDNAをHind III消化させ、Hybondナイロンフィルター上でブロットした。32P放射標識されたJκプローブ(Jκ1およびJκ5領域のゲノムDNAからのPCR増幅より入手)および32P放射標識された炭酸脱水酵素II(CAII)プローブを用いてフィルターをハイブリダイズした。肝臓DNAを流してκ生殖細胞系列の構成シグナルを示した(2.8Kbバンド)。4kbバンドとハイブリダイズするCAIIプローブを負荷対照として用いた。Tyfoon9200(Amerscham Biosciences)でフィルターを走査した。生殖細胞系Jκバンドの強度を、Image Quant5.2ソフトウェアを用いて定量し、負荷対照のそれに正規化し、対照肝臓DNA(100%とする)の割合で表した。異なる再構成イベントの増幅のためのハイブリドーマ由来のゲノムDNAで使用されるPCRプライマーは次の通りであった、
IGHJlR:5’−CCAGTGCTGGAAGTATTCAGC−3’(配列番号9)、
IGHJ2R:5’−CAGAGATCGAAGTACCAGTAG−3’(配列番号10)、
IGHJ3R:5’−GGCCCCAGAYATCAAAAGCAT−3’(配列番号11)、
IGHJ4R:5’−GGCCCCAGTAGTCAAAGTAGT−3’(配列番号12)、
IGHJ5R:5’−CCCAGGRGTCGAACCAGTTGT−3’(配列番号13)、
IGHJ6R:5’−CCAGAACGTCCATRYMGTAGTA−3’(配列番号14)。
・DNA FISH分析
標的DNAの調製:モノクローナルハイブリドーマ細胞をDMEM/10% FCS中で培養し、Heiskanen[38]に記載されるようにアガロースに包埋した。GΔトランスジェニック系統1のマウス由来の肺を収集し、単細胞懸濁液を作った。包埋した細胞をプロテイナーゼKおよびリボヌクレアーゼHで処理した。機械的に伸張されたDNAを、ポリ−L−リシンスライド(Sigma)上で電子レンジと別のスライドの端を用いて調製した。
【0093】
プローブ:GΔ導入遺伝子の再構成されたコピーおよび再構成されなかったコピーを検出するため、DNA断片を精製した。VHHとDの間の領域のハイブリダイズ用に2.3kB SpeIおよび3.6kB SpeI−BssHII断片、およびLCR検出用に5.9kB BamHI−SpeI断片またはIgH3’LCR(16kB)を含有する低コピー数のBluescriptプラスミド。ニックトランスレーションによりビオチン−11−dUTP(Roche)またはジゴキシゲニン−11−dUTP(Roche)でプローブを標識した。プローブをピペットでスライドの上に移す前に、90℃にて5分間、氷上で5分間、および37℃にて45分間でそれらを変性させた。インサイチューハイブリダイゼーション:ハイブリダイゼーション混合物は50%ホルムアミド、2×SSC、サケ精子DNA(200ng/μl)、5×デンハート液、1mM EDTA、および50mMリン酸ナトリウム、pH7.0を含有した。プローブのハイブリダイゼーションは、ピペットで25μlの混合物をスライド上に移し、24×32mmのカバーガラスで覆うことにより行われる。プローブおよび標的配列を変性させるため、スライドを80℃の加温板の上に2分間置いた。プローブは加湿チャンバー(加湿剤は50%ホルムアミド、2×SSCである)中で37℃にて一晩ハイブリダイズする。ハイブリダイゼーション後の洗浄は、記載される通り行った[39]。
【0094】
ハプテン標識されたプローブの免疫学的検出:ジゴキシゲニンプローブを、ヒツジ抗ジゴキシゲニン(1:500、Sigma)、フルオレセイン結合ウサギ抗ヒツジ(1:500、Sigma)、およびフルオレセイン結合ヤギ抗ウサギ(1:500、Sigma)を用いて検出した。ビオチンプローブは、テキサスレッド結合アビジン(1:500、Sigma)およびビオチン結合ヤギ抗アビジン(1:500、Boehringer)を用いて検出した。この段階を2回繰り返した。全てのインキュベーションおよび洗浄は記載される通り行った48。染色の後、スライドを段階的な一連のエタノール(70、90、および100%)中で段階ごとに5分間室温にて脱水した。細胞またはDNAを25μlの抗退色性包埋剤Vectashield(Vector Laboratories)に包埋した。100倍対物レンズを用いるLeica DMRBE 蛍光顕微鏡で視覚化を行った。
・免疫化およびハイブリドーマ作成
8週齢のマウスを、Specolアジュバント(IDDLO,Lelystadt,NL)または事前処方DKTPワクチンを含む5〜20μgの抗原で0、14、28、42日に皮下注射し、50日に腹腔内注射して免疫した。0、14および45日に採血した。56日にClonalCellTMHYキット(StemCell Technologies,Canada)を製造業者の説明書に従って用いて、脾臓細胞をSp2−O−Agl4骨髄腫細胞株(R.Haperenより寄贈)と融合させた。DKTPワクチンはNetherlands Vaccine Institute(Bilthoven,NL)より得られた。
・sdABライブラリーの構築およびスクリーニング
Ultraspec RNA単離系統(Biotecx Laboratories Inc,Houston,Texas,USA)を用いて、DKTP免疫単一コピーGΔおよびTNFα免疫MΔGΔマウスの脾臓から全RNAを単離した。cDNAはオリゴ(dT)プライマーを用いて作成した。VHHDJ断片をコードするDNA断片を、特異的プライマー:vh1 back Sfi Iプライマー[40〜42]を、hIgG2hingrevプライマー(5’−AATCTGGGCAGCGGCCGCCTCGACACAACATTTGCGCTC−3’(配列番号15))またはCH2huIgMrevプライマー(5’−TGGGACGAAGACGGCCGCTTTGGGAGGCAGCTCGGCAAT−3’(配列番号16))と組み合せたものを用いてPCRにより増幅させた。増幅したVHHDJ(約400bp)をSfi I/Not I消化させ、ゲル精製し、Sfi I/NotI消化ファージミドpHEN由来ベクターにクローニングした。TG1エレクトロコンピテント細胞(Stratagene La Jolla,USA)への形質転換により、ヒト単一のドメイン抗体ライブラリーが得られた。プラスチック(希釈されていないワクチンをコートしたイムノチューブ)に吸着させたDKTPワクチン抗原または精製されたヒトTNFα(Biosource International,USA)へのパニングを用いて、2ラウンドの選択を行った。
・免疫細胞化学およびウエスタンブロット
rtTAの存在に応答するマーカープラスミドでトランスフェクトされたtet−on細胞系統の細胞をスライド上で増殖させた。ドキシサイクリン誘導の24時間後、細胞を4%パラホルムアルデヒド/PBSに固定し、0.5% Triton−X−100/PBSで透過処理した。rtTAに対するHCAbを1:50希釈して使用し、その後ヤギ抗ヒトIgG FITC染色(Sigma,1:500希釈)に使用した。マーカータンパク質を前記に記載したように検出した51。ウエスタンブロットは、ヤギ抗ヒトIgG−HRP(Sigma,1/2500希釈)、ヒトIgM−HRP(Sigma,1/2500希釈)を用いる標準法であった。
・Superose6ゲル濾過
200mM KCl/20mM HEPES−KOH pH 7.9/1mM MgCl2/0.5mM EGTA/20%グリセロールで平衡したSuperose6 10/30カラムを備えるAKTA FPLC装置(Amersham Biosciences,Piscataway NJ)を用いて、MΔGΔマウス血清およびヒト対照血清のサイズ分画を行った。100μl/分の流量を用いて、500μlの画分を回収し、100%トリクロロ酢酸でで沈殿させ、SDS−PAGE(還元条件下)に続いてウエスタンブロット免疫ブロット法により解析した。
・BIAcore測定
実験は、BIAcore3000表面プラズモン共鳴バイオセンサー(Biacore AB,Uppsala,Sweden)にて行った。製造業者の提供する標準的なNHS−EDCキットを用いて3000共鳴単位(RU、任意の結合応答単位)のレベルまでCM5センサーチップ上に精製タンパク質を固定化した。10mM Hepes、pH7.4、150mM NaCl、2mM EDTA、0.005% Tween20中、40マイクログラム/mlの一定の流速で抗体を固定化抗原に通過させた。製造業者のBIA評価ソフトウェアを用いて平衡時の応答を記録し、カーブフィッティングを使用して平衡解離定数を得た。
<結果>
・ラクダ科動物様CH1スプライス変異は、ヒト重鎖遺伝子座におけるエキソンスキッピングには不十分である。
【0095】
ラクダ科動物におけるHCAb(IgG2およびIgG3)の生成にIgM+中間段階が必要かどうかは不明である。そのため、本発明者らは、まず、μMTバックグラウンドにおいて2つのハイブリッドヒト遺伝子座、1つはヒトCμ、Cδ、Cγ2およびCγ3定常領域を有する遺伝子座(MGS)、およびもう1つはヒトCγ2およびCγ3しか有さない1つの遺伝子座(図1)を生成した[43]。そのCγ領域に、まず、ラクダ科動物CH1スプライス変異を含有するように突然変異を起こさせた。μ鎖遺伝子の膜貫通ドメイン欠失によりB細胞がほぼ完全に欠けているμMT動物[43]がやはりB細胞小集団を有し、膜IgMの不在下で機能性IgG、IgAおよびIgEを産生することが証明され[44〜46]、(一部の)B細胞はIgMの表面発現なしに成長することが示唆されていることから、MGSを作製した。VH溶解度を向上させるためにヒトVHドメインを突然変異させる[47、48]代わりに、ラマ起源の2つのVHHを導入した。ラクダ科動物VHH領域は、42位、49位、50位および52位に特徴的なアミノ酸を含有する[49]。第1の、VHH1は、これらのVHHに特徴的なアミノ酸の全てを含有したが、基礎実験のこの証明における溶解度の重要性を検証するために、もう1つの、VHH2では、49位のGlu(E)の代わりに、これらの重大な「溶解」アミノ酸の1つであるGln(Q)を欠失させた。本発明者らは、さらに軽鎖可変部(VL)対合21に重要であると考えられることから、50位(Arg、R)ではなく49位を選択した。前記遺伝子座は、5つのヒト重鎖DおよびJ領域の全ておよびその遺伝子座の3’末端に遺伝子座制御領域(LCR)も含有した(図8)。驚くべきことに、スプライス変異によりトランスジェニックマウスに正しいCH1エキソンスキッピングは起こらず、ヒトIg発現の欠如につながらなかった(図8)。
・CH1領域を欠くヒト遺伝子座を含有するトランスジェニックマウスの解析
CH1のスプライシング問題を克服するために、本発明者らは3種類の新規構築物を生成した(図1)。全てがCH1を欠失させたCγ2およびCγ3を含有し、1つはCμおよびCδを含むもの(MGΔ)であり、1つはCμおよびCδを含まないもの(GΔ)であり、1つはCH1を欠失させたCμセグメントを有するもの(MΔGΔ)であった。μMTバックグラウンドにおいて、異なるコピー数(1〜5コピー)の3つのMGΔ、6つのGΔおよび4つのMΔGΔトランスジェニックマウス系統を得た。骨髄(BM)および脾臓においてB細胞の成長を解析した。異なるコピー数のマウスで同じ結果を得た。
・GΔおよびMΔGΔの発現はμMTマウスにおけるB細胞の成長を救済する
MGΔマウスはμMTバックグラウンドにおいてB細胞の成長を救済することができなかったが、一方、GΔおよびMΔGΔ構築物はB細胞の成長を効果的に救済した。B220/CD19陽性細胞の救済は、異なるリンパ系コンパートメントにおいて、コピー数に関係なく、30〜100%間であった(図2A)。これは、B220対ヒトIgMまたはヒトIgG染色を用いたBMのフローサイトメトリーによって確認される(図2B)。MΔGΔマウスは、野生型またはμMTマウスに存在しないヒトIgM産生細胞をBMにおいて含有する。従って、これらの細胞は、これらの細胞がヒトIgGを含有しないように、クラススイッチを受けなかった。GΔマウスはヒトIgG発現B細胞だけを含む。MGΔマウスは、細胞表面でヒトIgを発現するB細胞を含有するとしても非常に少なく、興味深いことに、B220細胞の一部は細胞内IgMを発現するが、IgGは発現しない(図2B)。MΔGΔおよびGΔマウス(下記参照)とは対照的に、MGΔマウスはマウスIg軽鎖を発現する(図6G)。これらの結果は、異なる構築物中でCμおよびCγ遺伝子が発現されることを示し、CH1が存在しないことがVHHに基づく抗体の細胞表面発現にとって重大であることが強く示唆される。
・ヒトHCAb IgGおよびIgMはBMでのB細胞の成長中にマウス(プレ)BCRに機能的に置き換わる
小型静止プレB細胞への大循環の発展進行中、特異的細胞表面マーカーの発現はプレBCR依存的にダウンレギュレートされる[36]。プレBCRに機能的に置き換わるヒトHCAbの能力を調査するために、様々なマーカーの発現をFACSにより解析した。プロB細胞は、高レベルの細胞質SLC、IL−7RおよびCD43を発現するが、これらはプレBCR発現時にダウンレギュレートされ、成熟B細胞では存在しない(図2C、μMTマウスからのプロB細胞、wtマウスの表面IgM−プロB/プレB細胞画分および表面IgM+B細胞画分と比較)。
【0096】
MΔGΔ/μMTまたはGΔ/μMTマウスのヒトIg+B細胞はSLCおよびIL−7Rレベルが低く、これにより、ヒト単鎖IgGおよびIgM受容体は、SLCおよびIL−7RのダウンレギュレーションにおいてマウスプレBCRに機能的に置き換わることが示される。CD43では、これはGΔマウスにおける場合のみのようであるが、MΔGΔマウスにおけるCD43発現の持続はこれらのマウスにおけるB−1 B細胞分化の増加という知見に関連があるかもしれない。同様に、正常なプレBCRシグナル伝達のように、CD2およびMHCクラスII発現が誘導される。プレB細胞段階で一過性に誘導されたIL−2R/CD25のレベルは、成熟MΔGΔまたはGΔ/μMT B細胞では非常に低く、成熟wt B細胞のレベルと同程度である(図2C)。さらに、ic Igκ発現は成熟MΔGΔまたはGΔ/μMT B細胞では検出できず(図2C)、IL−7+培養物から5日後のIL−7除去時にはインビトロBM培養物においても誘導されていなかった(図なし)。最後に、MΔGΔまたはGΔトランスジェニックマウスにおけるヒトHCAb発現B細胞集団は、BMで生成された細胞(HSAhighおよびAA4.1/CD93high)から部分的になり、末梢で成熟し、再循環している細胞(HSAlowおよびCD93low)から部分的になり、正常マウスでの結果と同様であった。
【0097】
まとめると、これらの結果は、ヒトHCAbのIgGおよびIgMが、発達上調節されるマーカーの発現に関して、B細胞の成長中にマウス(プレ)BCRに機能的に置き換わることを示す。IgのL鎖は誘導されない(下記参照)。BM RNAのcDNA解析は、VDJ組換えのための両方のVHHセグメントの使用、CH1の不在、そして重要なことにCDR3領域の大きな多様性を示している(図2D)。
・複数の再配列および対立遺伝子排除
免疫後、MΔGΔおよびGΔ系統から、特に5コピーGΔ系統1で多数のハイブリドーマを作製した(下記参照)。配列解析により、多コピーGΔ遺伝子座において1つ以上の再配列が起こり得ることが示された。解析した5つの異なる5コピーハイブリドーマのうち、1つのハイブリドーマ(G20)はフレーム内にある1つのコピーを再配列し;2つのハイブリドーマ(T7およびT12)は各々2つの再配列を有し、そのうちの1つの再配列は両方のハイブリドーマにおいてフレーム外であり;1つのハイブリドーマ(T3)は2つのフレーム内再配列(J2およびJ4)を有し;一方、1つのハイブリドーマ(T1)はフレーム内の2つの再配列(J2およびJ4)を含む4つの再配列を有した。
【0098】
T1およびT3は2つの生産的mRNAを発現し、それらは分泌された抗体の質量分析によりcDNAと正確にマッチさせて確認された(図なし)。本発明者らは、2つの異なるハイブリドーマ:G20(1つの再配列)およびT1(4つの再配列)において、それぞれのコピーを検出するLCRプローブと、再配列されていないコピーだけを検出する、VHHおよびDセグメントの間に位置するプローブとを用いて、DNA伸張線維FISHおよび標準的なDNA−FISHも行った(図3A〜E)。対照肺細胞は、サザンブロットマッピング(図なし)に一致して、一方の末端に5つの完全なコピーと半分の導入遺伝子を示したが(図3A)、一方、ハイブリドーマではG20およびT1それぞれにおいて1つおよび4つの再配列されたコピーを確かに示している(図3B、C〜E)。従って、複数のコピーは同じ対立遺伝子上でうまく再配列することができる。次に、本発明者らは、HCAb遺伝子座に、正常なマウス遺伝子座に対する(不)利点があるかどうか、そして遺伝子座の対立遺伝子排除が起こっているかどうかを求めた。wtバックグラウンドにおけるGΔ系統1トランスジェニックマウスのB220/CD19陽性BM細胞をヒトIgGおよびマウスIgMの発現について解析した。明らかに、GΔB細胞はマウスIgまたはヒトIgのいずれかを発現し(図3F)、対立遺伝子排除を示している。
・GΔ、MΔGΔおよびMGΔトランスジェニックマウスにおける脾臓B細胞
導入遺伝子がμMTマウスにおけるB細胞分化に与える影響を評価するために、本発明者らはCD21/CD23プロフィールを用いてフローサイトメトリーにより脾臓B細胞亜集団サイズを調べた(図4A)。GΔマウスでは、CD21lowCD23low未成熟B細胞の割合は正常な範囲にあり、ヒト単鎖IgG発現細胞は濾胞(FO;CD21+CD23+)B細胞へも辺縁帯(MZ;CD21highCD23low)B細胞へも分化し得た。MΔGΔマウスでは、未成熟B細胞画分が増加し、HCAb−IgM発現細胞のFOおよびMZ−8 B細胞への分化がいくらか損なうことを示している。これらの脾臓では、CD23発現の減少は、B−1 B細胞系統への分化を示す、CD43およびCD5の表面発現レベルの増加を伴っていた。MGΔトランスジェニックマウスに存在する少数のヒトIgM発現B細胞(マウス軽鎖も発現する、図6参照)は、MΔGΔトランスジェニックマウスにおけるものと同様のFO/MZ分布を現した。MGΔマウスは違うが、MΔGΔおよびGΔマウスは、正常な脾臓構造を有し、白脾髄の外側境界に存在する濾胞および辺縁帯を含有するB細胞リッチな領域で囲まれた動脈周囲リンパ球鞘(PALS)に集中しているT細胞の分離を示す(図4B)。
【0099】
それらはまた、T細胞依存性抗体応答中のwtに匹敵する、二次リンパ組織のB細胞濾胞胚中心を形成する(図なし)。これらは、記憶形成と、体細胞超変異に起因する親和性に基づく選択の部位である。GΔマウスでは、これらの胚中心においてヒトIgG陽性細胞が検出される。本発明者らは、ヒトIgG遺伝子の超突然変異を、パイエル板(Payer′s patches)に存在するB細胞のcDNA解析により確認した(図4C)。VHH1およびVHH2の両方を使用する。総合すれば、これらの発見により、GΔおよびMΔGΔトランスジェニックマウスにおいて、BMから移動する未成熟B細胞は、脾臓においてFO−B細胞およびMZ−B細胞の両方へと分化し、抗原曝露後に体細胞超変異を受ける能力を有することが実証される。
・単一コピー遺伝子座は効果的に救済し、CH1の不在が必須である
GΔ系統1マウス(図2A)は5コピーのGΔ遺伝子座を含有するため、従って効果的な救済が遺伝子座のコピー数と関係がある可能性があった。しかし、5コピーGΔ系統1からFlpeR系統23との交配によるFlp組換えにより得られた単一コピートランスジェニック系統は、B細胞の成長を同程度まで救済した(図4D、図2A)。単一コピーの遺伝子座は救済に十分であり、そしてCH1領域を含むCμ定常領域の存在が阻害性であることを、非救済単一コピー系統MGΔ(系統3)から、単一コピーGΔ系統をもたらすCre組換え(cre発現系統への品種改良)によりCμおよびCδ領域を欠失させることによって確認した(図4D)。それまで非救済であった遺伝子座はその時点で他のGΔ系統と同じB細胞の成長9を与える。従って、コピー数は救済に影響を及ぼさず、Cμ領域にCH1が存在することによりB細胞救済が阻害される(下記も参照)。
・MΔGΔおよびGΔトランスジェニックマウスにおいてマウスIg軽鎖遺伝子座は再配列しない
マウスIg軽鎖タンパク質はMΔGΔおよびGΔマウスでは血清のウエスタンブロット(図なし、図2Cおよび6A参照)によってまたはFACSによって検出されず、マウス軽鎖遺伝子が再配列しないことが示唆された。これは、サザンブロット解析により選別された脾臓B220+細胞画分および肝臓細胞のDNAにおけるIgκ遺伝子座生殖細胞系列シグナルの密度を比較することによって確認され(図5AおよびB)、これによりマウス軽鎖は再配列せず、生殖細胞系列構成を保つことが示される。対照的に、MGΔ/μMTマウスの少数のヒトIg発現細胞において軽鎖が検出される(図6G参照)。
【0100】
従って、BMでの初期B細胞の成長におけるヒトHCAbの発現は軽鎖再配列を導くシグナルを与えることができない。これに関連して、HCAbはプレBCRよりもむしろBCRによく似ており、このことはHCAbがCH1の不在下で擬似軽鎖と結合できなかったことと関係があると考えられる[61]。
・GΔ/μMTおよびMΔGΔ/μMTマウスの血清分析
ヒトIgMはMΔGΔ血清に存在し、ヒトIgGはMΔGΔマウス血清およびGΔマウス血清の両方に存在した。免疫されていない成体動物では、ヒトIgM(<50μg/ml)およびIgG(200〜1000μg/ml)は、正常ヒトIgH遺伝子座を有する正常マウスまたはトランスジェニックマウスに見られるものと同様のレベルで存在する[65]。6種全てのGΔマウスの血清のゲル電気泳動では、非還元性条件下で約70kD、還元性条件下で約35kDのMWを有するHCAbのIgGが現われ、軽鎖を欠く重鎖二量体とCH1エキソンを欠く各重鎖と一致した(図6A、B)。
【0101】
MΔGΔマウスの血清は、多量体重鎖のみのヒトIgMを含有した。還元性条件下(図6C)では、4種全ての系統が、CH1のMWを減じた後にヒト対照IgMとしてのMWを有するIgM鎖を含有した。また、血清を非還元性条件下でも分画した(図6D、横方向の画分)後、それぞれの画分10を還元性条件下でゲル電気泳動により解析した(図6D、縦方向のレーン)。900kDのヒト血清五量体IgM対照と比較すると、トランスジェニックIgMは600kDで分別され、それが多量体であること、ならびに軽鎖およびCH1を欠くことに一致する。従って、MΔGΔマウスはHCAb多量体IgMおよび二量体IgGを産生するが、一方、GΔは血清中で二量体IgGを産生する。
・MGΔマウスの血清分析
MGΔ/μMT系統の末梢および脾臓では、B220陽性細胞はほとんど見られず(野生型の1%未満)、極まれに小さなB細胞集団が脾臓において見られる(図4B)。精製後にのみ血清中に少量のヒトIgMおよびIgGが検出された(図6E、F)。これらのマウスにおけるヒトIgMは、還元性条件下では通常のサイズであったが、一方、循環しているヒトIgGはより短く(CH1の欠失に一致する、見かけのMWは35kD)。興味深いことに、ヒトIgMと結合していると思われるマウスκ軽鎖も検出された(図6G)。
【0102】
ヒトIgMおよびIgGは定量ELISAアッセイでの検出レベルを下回っていたが、本発明者らは、それにもかかわらず、MGΔ/μMTマウスが免疫化に応答し得るかどうかを検証した。マウスをヒト腫瘍壊死因子α(TNF−α)で免疫し、wtマウスは強いTNF−α特異的抗体応答を発現したが、一方、用いた2種のMGΔ系統の3匹のマウスにおいては、ELISAまたはウエスタンブロット解析によって抗原特異的ヒトIgGを検出することができなかった(図なし)。
・MΔGΔおよびGΔマウスの免疫化は抗原特異的Ab産生をもたらす
GΔ/μMTマウスを大腸菌hsp70、DKTP(ジフテリア毒素、百日咳菌の全細胞溶解物、破傷風毒素ならびに不活化ポリオウイルス1型、2型および3型)およびrtTAで免疫し[50]、一方、MΔGΔマウスをヒトTNFαで免疫した。ELISAで陽性であった血清を有するマウスから、ファージディスプレイライブラリーによりハイブリドーマまたは単一ドメインAb(sdAb)を用いて個々の完全抗体を単離した。
【0103】
抗体RNAのRTPCR後にαhsp70特異的、破傷風毒素特異的およびrtTA特異的モノクローナルを配列決定した(図7A)。これにより、IgG2(8つのうちの7つ)およびIgG3(8つのうちの1つ)抗体の両方が産生されることが示された(sdAbはIgG2ライブラリーから単離した)。異なるDおよびJ領域を使用した。高頻度ではないが、11のHCAbのVHHは超突然変異を受けていた。3つのhTNFα特異的抗体(1つは陽性IgMハイブリドーマ、図6α−hTNFα#1および2つはsdAbα−hTNFα#2&3)は全て、CDR2領域に異なる超突然変異を有した。14全ての抗体の配列を比較すると、全てのJ領域が用いられているものの、ヒトと同じようにJH4が最もよく用いられていることが明らかであった。驚くべきことに、全ての抗体はラマVHH2(49位に典型なグルタミン酸ではなくグルタミン(Q)を有する[51])を含有した。最後に、CDR3領域が多様性の大部分をもたらしていることは明らかである27。それは、ラマおよびヒトにおいて通常見られるものと非常に類似している、10〜20アミノ酸長の間で変化する(平均は13.6aa)[52、53]。本発明者らは、次に、これらのHCAbが常用アッセイにおいてハイブリドーマ上清および細菌原形質周辺画分のsdAbとして機能するかどうかを検証した(図7)。ELISAアッセイでは抗体の全てが陽性であり、ウエスタンブロットでの抗原検出でも全てが陽性であった(例えばα−DKTPおよびα−rtTA血清とハイブリドーマ培地とα百日咳菌sdAb、図7B)。また、本発明者らは、rtTA発現プラスミドでトランスフェクトした細胞系統を用いて免疫細胞化学によりα−rtTA IgGも検証した(図7C、D)。rtTA HC−IgGは極めてはっきりとした特異的な核染色を示した。多数の抗体の親和性は高かったが、一部のもの(特にsdAb)ではかなり低かった。例えば、免疫細胞化学に用いたα−rtTA抗体(図7C、D)は、BiaCoreで決定されるように約3nMであった(図7E)。
<結論>
本明細書において、本発明者らは、μMTマウスにおけるB細胞の成長を救済し、結果的に機能性HCAb産生をもたらす修飾HCAb遺伝子座の発現を報告した。これらのマウスを免疫して、抗原特異的HCAbを産生させることができる。ラクダ科動物の場合、HCAbの分泌にはCH1の除去が不可欠であるが、CH1の排除には3’CH1/イントロン境界における単一ラクダ科動物(総説30)スプライス変異では不十分であり、従って少なくともヒト遺伝子座において、単にこの単一点突然変異以上のものが必要である。CH1をVHHとともに有するIgMはB細胞の成長を妨げるが、これは恐らく、プレBCRとの関連でIgM表面分子の効果のない会合に起因する。対照的に、HCD様ヒトμタンパク質を発現するマウスは、λ5とは無関係にSCIDバックグラウンドにおいて正常CD43プレB細胞を生じる[54]。末端切断型Cμタンパク質は、関連L鎖なしにB細胞表面で発現され、自己会合を介してシグナル伝達するプレBCRによく似ていると考えられる[55]。
【0104】
通常、BiPシャペロンは、後に鎖間接触に関与するIg鎖の疎水性表面と結合することによって抗体分子の折り畳みおよび会合を監視している31。VHHのFR2に親水性アミノ酸が存在することによって、VHHとのBiPの結合がほぼ確実に阻止され、この場合には可溶性にする(代理)軽鎖は必要でない。同時に、CH1は、会合(軽鎖によるBiPの置換)が完了するまでERに重鎖を保持するとされているBiPとの相互作用をもたらす。本発明者らの結果は、プレB細胞受容体レベルでは、λ5タンパク質との非共有結合における代理軽鎖(SLC)の一部としてのVpreタンパク質とのトランスジェニックμ重鎖対合は、CH1ドメインを含有するμ重鎖がVHHと結合されている場合には起こるのができないことを示唆している。そのため、MGSおよびMGΔトランスジェニックマウスにおけるヒトIgMは、この点で、増殖展開および発展進行をシグナル伝達するために不十分であることが公知である不完全プレBCR様複合体に似ている[56、57]。これにより、BMではB220陽性細胞の30%しか細胞内IgMを有さない理由を説明することができる(図2B)。これらのマウスの脾臓に少数の成熟B細胞が存在することは、μ重鎖を含有するが任意のSLCまたは通常の軽鎖が欠失している最近記載された新規な受容体複合体によって説明する[58]。
【0105】
CH1がCγ2および3に存在せず、IgMがその遺伝子座から除去されている場合には、B細胞の成長の救済が起こり、IgGがIgMに機能的に置き換わることが示される。Rag2−/−マウスにおける成熟CD21+B細胞の成長の支援において、プロB細胞段階以降に発現されるIgG1受容体をIgMの代わりにすることができる[59]。最近、事前に再配列したラクダ科動物IgG2aがμMT(およびCΔ−/−)バックグラウンドの2匹のトランスジェニックマウスのうちの1匹においてB細胞の成長を部分的に救済することができるということも示された13。本発明者らの場合では、CH1を欠くIgMまたはIgGは10種の独立したトランスジェニックマウス系統のうちの10種においてB細胞の成長を救済する。さらに、本発明者らは軽鎖再配列を観察しておらず、さらなるB細胞分化に軽鎖発現は必要でないと結論づけられる。本明細書で得られた結果とZou et al.の結果[60]における相違は、LCRが本発明者らの構築物に含まれていることによる重鎖遺伝子座の発現レベル(すなわちシグナル伝達)によって説明することができる。本発明者らの結果は、CH1[61]またはVHおよびCH1[62]を欠く末端切断型μ重鎖タンパク質が、SLCと結合することができず、κ遺伝子再配列を活性化することができないことを裏付ける。
【0106】
5コピーGΔ系統1(および他の多コピー系統)は、ゲノムの同じ位置に組み込まれた単一コピー系統と同じ程度までB細胞の成長を救済する。興味深いことに、多コピートランスジェニック遺伝子座では1以上の再配列が起こる(図3)。2種の単一脾細胞由来のハイブリドーマの2つは、2種の生産的HCAb転写物およびタンパク質をもたらした。この結果は、同じB細胞における2種の抗体の発現は無害であることを裏付けている[63]。しかし、二重抗体産生B細胞は抗原曝露下で単一抗体産生細胞との競争に敗れるという予測37は、抗原曝露後に得られた5つのハイブリドーマから2種の二重抗体発現細胞を見出した本発明者らの結果からは実証されない。
【0107】
BM細胞は細胞表面でマウスまたはヒトIgを発現するため、野生型マウスにおいて(多コピーの)遺伝子座は対立遺伝子排除を受けやすい。細胞表面で両方を発現する細胞の有意な集団はない(図3F、上のパネル)。恐らく最も興味深いのは、マウスIgおよびヒトIgを発現している細胞の数の対比である。wt/5コピーGΔマウスでは、再配列に利用できる3つの可能性ある対立遺伝子、1つのIg遺伝子座を有する2つのマウス対立遺伝子および5つのヒトHCAb遺伝子座を有する1つの対立遺伝子が存在する。確率的に選択されるならば、ヒト対立遺伝子は3回のうち1回しか選択されない。
【0108】
遺伝子座の数を計数すれば、ヒト遺伝子座はまず7細胞のうち5細胞で再配列する。しかし、内因性マウスおよびトランスジェニックヒトHCAbはほぼ同等に発現される(44/38、図3F 下のパネル)。これらの数はV領域のランダム使用によって起こり得るずれやトランスジェニック遺伝子座における位置効果の可能性を無視しているが、最初の選択が対立遺伝子の確率的選択であり、続いて1対立遺伝子につき複数の遺伝子の再配列が行われ得ることを強く示唆している。
【0109】
これは、本発明者らがGΔ遺伝子座の複数の再配列を頻繁に観察しているという事実と一致する。通常、生産的再配列は組換えをダウンレギュレートして、他の対立遺伝子の再配列を妨げる。しかし、トランスジェニック対立遺伝子における複数のコピーは同じオープンな遺伝子座に存在し、RAGダウンレギュレーション前に組み換えられる。これは、複数の再配列が同時に起こるためであるか、または空間要素(「コンパートメント」)を含むならば、RAGがダウンレギュレートされる前に接近していれば遺伝子座の別の遺伝子を再配列する十分な時間があり得る。
【0110】
wtマウスにおいて再配列が生産的ではない(シグナル伝達はなく、RAGはとどまる)場合に限り、第2の遺伝子座が活性化され、第1の遺伝子座に置き換わり、再配列されるための十分な時間がある。この論証を支持して、同じ染色体上に複数の遺伝子座を有する他の種には、2つのAbを発現する、より多くの細胞が存在するという見解が生じる[64]。
【0111】
重要なことには、これらの試験によってHCAb遺伝子座はマウスにおける発現に成功し得ることが分かる。抗原曝露は、遺伝子座の構成によって異なるクラスの高親和性抗原特異的ヒトHCAbの産生をもたらす。これらの抗体は、正常マウスまたは他の「正常」なヒトIgHトランスジェニックマウスでのレベルと同様のレベルで発現される[65]。本発明者らの実験では2つの可変領域だけを使用したが、本発明者らが検証した無関係なタンパク質のほぼ全てに対して様々な特異性を有する高親和性抗体の単離に成功し、CDR3によってもたらされる多様性の効率および有効性を立証した[66]。従って、V(D)J組換えおよびインビボ選択を行うことは、ファージディスプレイを用いたファージミドライブラリーからのヒト単鎖抗体断片の作製に対して非常に重要な利点をもたらす39。ハイブリドーマは容易に生成することができ、このことによって、重要なことに、ファージディスプレイやさらなるスクリーニングの必要なく、完全ヒトHCAbまたはsdAb断片の直接クローニングおよび発現を可能にする。
【0112】
従って、これらのマウスは、特にHCAbが通常の抗体、例えば酵素の活性部位に対してほとんど抗原性がないエピトープを認識し得るという証拠4に照らして、臨床目的またはその他の目的でのヒトHCAbの産生に関して全く新しい可能性を広げる。抗原の全てが認識されるわけではないことの理由は、限られた数の可変領域によって説明することができる(ポリオおよびジフテリアタンパク質はGΔマウスでは応答をもたらさなかったが、一方、wt対照マウスでは応答がもたらされた)。驚くべきことに、抗体の全てがラマVHH2領域を有した。これには、49位に保存アミノ酸[67]が含まれず、それを有し、溶解性がより高いはずであるVHH1とは対照的である。
【0113】
それでもなお、本発明者らは、遺伝子座により多くの可変領域を追加することによっていっそう広範囲にわたるレパートリーをもたらすものと予測する。単一対立遺伝子上では複数のコピーの遺伝子座を避けることが好ましいが、多様性を高めるために異なるVH領域の単一コピーをそれぞれが含む複対立遺伝子を含むマウスを生成することが有利。そのような新しい遺伝子座では、通常発生する(ヒト)VH領域または溶解度18および軽鎖対合を高めるために操作されたVH領域のいずれかを使用することができる。
【0114】
結論として、本発明者らは、潜在的に任意のクラスの抗原特異的高親和性HCAbがマウスで産生され得ることを立証した。この技術は、人における治療薬または診断薬として使用するための、抗原曝露に応答する任意のクラスの完全ヒトHCAbまたはそれらの断片の生成を可能にする。また、異なる脊椎動物遺伝子座を用いることによって、本発明者らの技術は、試薬、診断法として使用するための、または動物の治療のための任意の脊椎動物由来の高親和性成熟抗体の産生を可能にする。
【実施例1】
【0115】
Janssens et al.(2006)は、トランスジェニックVH遺伝子座が遺伝子導入によってコードされた重鎖のみ抗体を産生するように正確に再結合していること、そしてそのような遺伝子座は内因性(マウス)重鎖免疫グロブリン遺伝子座の存在下で対立遺伝子排除に対して感受性があることを示した。対立遺伝子排除プロセスを用いることによって、重鎖のみ抗体産生に使用されるVHトランスジェニック遺伝子座の数を増加させることができるということを示すために、異なる重鎖のみの遺伝子座を含む2匹のトランスジェニックマウスを交配させ、結果として両方の遺伝子座を含む子孫を得た。一方のマウスはMGΔ遺伝子座を含有した(IgMおよびIgG遺伝子座、Janssens et al., 2006)が、もう一方のマウスはGΔ遺伝子座(IgG遺伝子座単独、Janssens et al., 2006)を含有し、両方のマウスはμMTバックグラウンドを有するものであった。ダブルトランスジェニック子孫のB細胞からハイブリドーマを誘導し、標準的な方法によって培養下で増殖させた。多数の得られた個々のモノクローナル細胞系統をPCRおよびサザンブロットによって解析し、それによって生産的に再配列されたMGΔ遺伝子座を含有する系統が、再配列されていないGΔまたは生産的に再配列されていないGΔ遺伝子座を含むことが示された。逆に、生産的に再配列されたGΔ遺伝子座を含有する細胞系統は、再配列されていないまたは生産的に再配列されていないMGΔ遺伝子座を含有した。このように、利用できるVH領域の合計が組換えプロセスに使用される。
<実施例3および4>
本発明の第1の態様の好ましい実施形態では、前の実施例に記載される、当技術分野で公知の方法(Janssens et al 2006)を用いて、機能性ヒトVH領域全数を、ヒトVH領域(またはそれらの変異体)を、全DH領域、全JH領域ならびにCμ、Cγ2、Cγ3およびCα領域と3’LCRの組合せを含む複数の修飾ヒト遺伝子座にクローニングすることによって増加させる。この手順は、独立した遺伝子座の複数の同一VH領域または独立した遺伝子座の異なるVH領域を用いて行うことができる。異なる遺伝子座は同一重鎖領域または異なる重鎖領域を含有し得る。実施例3は、重鎖領域の異なる組合せを有する遺伝子座の同一VH領域を有する遺伝子座に関し、実施例4は同一重鎖定常領域と異なるVH領域を有する2つの遺伝子座に関する。両実施例において、さらに遺伝子座を追加してよいことは明らかである。
【実施例2】
【0116】
ヒトVH領域は、可能性ある39個の機能性ヒトVH領域(あるいはヒトVL領域またはTCRのV領域または突然変異誘発によって得られるこれらの全ての変異体を使用または追加してよい)の中から選択されたそれぞれのVHに特異的なプライマーを用いたヒトゲノムDNAのPCR増幅によって単離する。ヒトVH領域は、前記実施例に記載した、すなわちヒトDHおよびJH(または他のDおよびJ領域)とそれぞれCH1および3’LCRが欠失しているCμ、Cγ2、Cγ3を含む遺伝子座にセットでクローニングされる(図9)。CH1およびスイッチ領域が欠失しているCα領域はGΔ遺伝子座に別個にクローニングされる(図10)。このGΔ遺伝子座は、lox部位を含有せず、ラマVHH領域が標準的な相同組換えによって除去されて、特異PI−PspI部位を残している(図10)という点から原型遺伝子座の変異体である。
【0117】
機能性VH領域は、各遺伝子座の任意の複数に関して同時にクローニングすることができる。最初に、17個の機能性ヒトVH領域を、クローニングされた2個の遺伝子を1セットで同時にクローニングする。これらの最初の構築物のそれぞれに、第2のセットを従来の方法(例えばXhoI−SalI制限消化/連結を用いて、XhoIおよびSalI適合部位の連結により両方を破壊する)により追加する。4、4、4、3および2個の遺伝子を含有するセットがBACベクター中の17個の遺伝子の1セットに連結される。この手法は、他の数の遺伝子を含有するセットを組み合わせることによって行ってよいことは明らかである。前記方法は、所望の数のVH領域を得る任意の時点で終了してよく、またはより多数のVH領域を得るために延長してよい。
【0118】
1セット全て(例えば17個の遺伝子)を特異PI−PspI部位の修飾GΔ遺伝子座にクローニングする。Cα領域はこの遺伝子座のI−CeuI部位にクローニングされ、結果としてIgαおよびIgGを産生可能なAΔGΔ遺伝子座が生じる(図10)。また、他の重鎖領域もクローニングすることができた。
【0119】
これらの最終的な遺伝子座は、その後、前記実施例に記載されている独立したトランスジェニックマウス(好ましくは、μMTなどのIgH遺伝子座欠損マウス)に導入される。これらの独立した遺伝子座は、独立したマウス系統、IgGだけを作製する系統ならびにIgAおよび/またはIgGを作製する系統を生成するために用いられる。これらのマウスは、続いて、2つの異なる遺伝子座から34個のVH領域を提供できるようにVH領域全体を有するように交配される。これらのマウスを両方の遺伝子座に関してホモ接合性になるように交配することによって、組換えに利用できる68個のVH領域が作製されることになる。組み込まれた遺伝子座の複数のコピーを有することによって、この数はいっそうさらに増加する。これらのマウスのB細胞から作製されたハイブリドーマの解析は、生産的に再配列された遺伝子座が存在する場合に、作り出された他の遺伝子座が、標準的な手法による対立遺伝子排除の結果、再配列されていないかまたは生産的に再配列されていないことを示すために利用される。
【実施例3】
【0120】
前記領域(または突然変異誘発によって得られたそれらの変異体)に対して同様に単離された1以上のVH領域の異なるセットを、前記と同じ方法により2つの独立した遺伝子座にクローニングする。これは結果として2つの異なるGΔ遺伝子座(またはそれらの変異体、例えばCαが追加されたもの)をもたらすことになる。これらの遺伝子座は、独立したマウスに導入されて、独立したトランスジェニック系統(例えばそれぞれ10個のVHドメインを有する2つの遺伝子座、図11)をもたらす。これらのマウスは、その後、組換えプロセスに利用できる全ての使用VH領域を有するダブルトランスジェニックマウスを得るために交配されることになる。これらのマウスを両方の遺伝子座に関してホモ接合性なるように交配することによって、組換えに利用できるVH領域の数が倍増することになる(図12、染色体1に組み込まれた1つの遺伝子座と染色体8上の1つの遺伝子座に関するカリオグラム)。組み込まれた遺伝子座の複数のコピーを有することによって、この数はいっそうさらに増加する。これらのマウスのB細胞から作製されたハイブリドーマの解析は、生産的に再配列された遺伝子座が存在する場合に、作り出された他の遺伝子座が、対立遺伝子排除の結果、再配列されていないかまたは生産的に再配列されていないということを示すために利用される。
【0121】
前記プロセスは、所望の数のVH領域を得る任意の時点で終了してよい。これらのVH領域にD、JHおよび定常領域が追加される。これらの最終遺伝子座は、その後、前記実施例に記載されている独立したトランスジェニックマウス(好ましくは、IgH遺伝子座欠損マウスを含む)に導入され得る。あるいは、lox部位の位置により、Cμ(IgM)単独、またはCγ2およびCγ3(IgG2およびIgG3)単独、またはCα単独(IgA)またはそれらの組合せを含有する独立した遺伝子座を生成するための個々の定常領域の排除が可能となる。これらの独立した遺伝子座は、ヒトIgM単独、またはIgG単独またはIgA単独またはそれらの組合せのいずれかを作製する独立したマウス系統を生成するために用いられる。
[参考文献]
【0122】
【表2A】
【0123】
【表2B】
【0124】
【表2C】
【0125】
【表2D】
【0126】
【表2E】
【図面の簡単な説明】
【0127】
【図1】トランスジェニックマウスを生成するために使用されたDNA断片の模式図である。2つのラマVHHエキソンは、ヒト重鎖多様性(D)および結合(J)遺伝子セグメントと連結され、その後にCμ、Cδ、Cγ2およびCγ3ヒト定常領域遺伝子およびヒト重鎖Ig3’LCRが続く。ヒトCγ2およびCγ3遺伝子は、構築物MGΔおよびGΔ中のCγ2およびCγ3遺伝子からの、または同様に構築物MΔGΔ中のCμからのCH1エキソンの完全な欠失により修飾されていた。2つのLoxP位(赤色)が同じ方向に存在することにより、Creに媒介される組換えの際、CμおよびCδ遺伝子の除去が可能になる。Frt位(緑色)が存在することにより、Flpに媒介される組換えによる、複数コピー導入遺伝子アレイからの単一コピートランスジェニックマウスの生成が可能になる。
【図2】パネルA:異なる器官における総細胞のうちのB220/CD19陽性細胞の百分率として表されたBリンパ球のフローサイトメトリー分析の表である。パネルB:BM中のwt、μMT、MGΔ/μMT、MΔGΔ/μMTおよびGΔ/μMTマウスのB細胞集団のフローサイトメトリー分析。B220の前方および側方散乱ならびに表面発現に基づいてリンパ球系統細胞をゲートし、ヒトIgMまたはIgGをプロットした。データはドットプロットとして表示される。MGΔμMTマウスについて、B220+画分をゲートして細胞内(ic)ヒトIgμおよびγH鎖の発現について分析し、ヒストグラムオーバーレイ(赤色線)として、一緒にμMTマウス由来のB220+細胞のバックグラウンド染色(黒色線)を対照として表示した。陽性細胞の百分率が示されている。パネルC:MΔGΔまたはGΔ導入遺伝子の発現は、プレBCRおよびBCRの機能を救済する。μMTマウス由来の総CD19+画分(プロB細胞)、野生型マウス由来のCD19+表面IgM+画分(プロB/プレB細胞)およびCD19+表面IgM+画分(B細胞)、MΔ−GΔμMTマウス由来のCD19+表面ヒトIgM+画分(B細胞)およびGΔμMTマウス由来のCD19+表面ヒトIgG+画分(B細胞)において示されたマーカーの発現プロフィール。ic−Igκ=細胞内IgκL鎖。フローサイトメトリーデータはヒストグラムとして表示される。示されるデータは、各群内で試験された3〜8匹の動物の代表である。パネルD:VHH1およびVHH2特異的プライマーをヒトCγ2プライマーと組合せて用いてBM cDNAから得た、PCR産物の配列アラインメント、VDJ組換えを示す。配列はGΔに由来する。緑色は配列同一性を示す。
【図3】パネルA〜E:5つのコピーのヒトGΔ遺伝子座のDNA−FISH。パネルA:その半分をLCR(赤色)を含む遺伝子座に、その半分をVHHからJ領域(緑色)を有する遺伝子座に隣接されている5つのインタクトなGΔ遺伝子座のコピー(1〜5)を有するGΔ系統1トランスジェニックマウスの肺細胞由来の伸張されたクロマチン線維。パネルB:1つのコピーが再構成されている(白色矢印)GΔ系統1のB細胞に由来するハイブリドーマ(G20)の伸張されたクロマチン線維FISH。パネルC:LCRプローブを用いるハイブリドーマT1の伸張されていないDNA−FISH(赤色)。パネルD:VHH〜D間のプローブを用い、Cと同内容(緑色)。パネルE:パネルCとEのオーバーレイ。赤色シグナルの喪失がないのに対し、4つの緑色シグナルの喪失が見られることから、T1が4つの再構成を有することに留意されたい。パネルF:GΔトランスジェニックマウスにおいて対立遺伝子が排除されている。マウス表面または細胞内(ic)μH鎖および示されるマウス由来の総BM CD19+細胞画分上のトランスジェニックヒトIgGのフローサイトメトリー分析。データはドットプロットとして表示され、示される象限内の細胞の百分率が示される。示されるデータは、各群内で試験された4匹のマウスの代表的なデータである。
【図4】wt、μMT、GΔ、MΔGΔおよびMGΔマウスの脾臓のB細胞集団の分析を示す図である。示されるデータは、各群内で試験された4〜8匹のマウスの代表的なデータである。パネルA:上側、B220に対してマウスIgM、ヒトIgG、ヒトIgMについて染色された脾臓細胞のFACSデータ。下側、脾臓のB細胞集団のフローサイトメトリー分析。B220の前方および側方散乱ならびに表面発現に基づいてリンパ球系統細胞をゲートし、示されたIg(上部)またはCD21/CD23プロフィールはドットプロットとして示され、示されたゲート内の細胞の百分率が示される。CD21lowCD23low:未熟B細胞;CD21+CD23+:濾胞性B細胞;CD21highCD23low:辺縁帯B細胞。パネルB:wt、μMT、GΔ/μMT、MΔGΔ/μMTおよびMGΔ/μMTマウスの脾臓の組織像。免疫組織化学分析;5μmの凍結切片を、B細胞には抗B220(青色)で、樹状細胞には抗CD11c/N418(茶色)で染色した。矢印は、MGΔ脾臓におけるB細胞の小さなクラスターの位置を示す。パネルC:VHH1およびVHH2特異的プライマーをヒトCγ2プライマーと組合せて用いてパイエル板cDNAから得たPCR産物の配列アラインメント、トランスジェニック遺伝子座がCDR1および2領域で超突然変異を受けることを示す。配列は、CH1の欠失しているトランスジェニック遺伝子座GΔに由来する。パネルD:上側;脾臓細胞のFACSデータ、抗CD19および抗B220で染色。下側左:FlpeRトランスジェニック系統に交配することによるインビボFlp組換えの模式図、および5つのコピーGΔ系統1に由来する単一コピー組換え型の脾臓細胞に関するFACSデータ。下側右:MGΔ系統をCAGCreトランスジェニック系統に交配することによるインビボCre組換えの模式図、組換え型の脾臓細胞に関するFACSスキャンデータを支持し、直接生成した元のGΔ系統に見られるようなB細胞の救済を示す。
【図5】GΔ/μMT(パネルA)およびMΔGΔ/μMT(パネルB)トランスジェニック系統においてκ軽鎖再構成が行われないことを示すサザンブロットを示す図である。1匹のwtマウスおよび2匹のGΔまたは4匹のMΔGΔトランスジェニックマウス由来の肝臓DNA(L)およびB細胞DNA(B)をHind IIIで消化し、32P放射標識されたJκプローブおよび炭酸脱水酵素II(CAII)プローブを用いて分析した。4kbバンドにハイブリダイズするCAIIプローブを泳動対照として用いた。肝臓DNAを流してκ生殖細胞系列の配置を示した(2.8Kbバンド)。wtB細胞だけが、2.8kb断片の強度の低下で測定されるκ遺伝子座の再構成を示す(肝臓と比較するとシグナルの30%が残る)。
【図6】μMTバックグラウンドにおいて非還元条件下(A)および還元条件下(B〜G)、6種類の異なるGΔ系統(A、B)、4種類の異なるMΔGΔ系統(C)および2種類の異なるMGΔ系統(E〜G)の、Prot Gまたはコンカバリン(concavalin)精製された血清サンプルを泳動した。トランスジェニックヒトIgG(パネルB、F)およびIgM(パネルC、D)のサイズはCH1欠失と一致し、軽鎖が存在しない。マウスκ軽鎖は正常なサイズであった(G)。ヒト血清を陽性対照として用いた。パネルD:非還元条件下でヒトIgM対照に混合後のMΔGΔ血清のSuperose6サイズ分画。それぞれの画分を還元条件下でゲル電気泳動により分析した。Superose6カラムから回収された画分は左(高MW)から右(低MW)である。対照は、ヒト血清単独(第1レーン左)およびヒトIgM対照血清(レーンMΔGΔ血清)に混合する前のマウス血清である。サイズマーカーが示される。
【図7】パネルA:破傷風毒素;HSP70、rtTAおよびヒトTNFαに特異的なモノクローナル抗体cDNAの配列。上の配列は生殖細胞系列VHH2配列である。CDR1、2および3ならびにヒンジ領域が配列の上に示されている。異なるアイソタイプおよびクラスは右側に異なる色で示されている。用いられるJ領域は右側に示されている。パネルB:異なる重鎖のみ抗体(ハイブリドーマ、血清およびsdAb)を用いるウエスタンブロットの例。左側のパネル、それぞれ1/100および1/250希釈された、抗rtTA血清およびハイブリドーマ培地。中央のパネル、それぞれ1/200および1/100希釈された、wtおよびGΔマウス由来の抗DKTP血清。右側のパネル、百日咳菌抗原(DKTP)を含むかまたはそれを欠く(DTP、本発明者らは精製された百日咳菌抗原を購入することができなかったため)ワクチンに対する抗百日咳菌sdAb。パネルCおよびD:細胞質においてマーカータンパク質を発現することによりrtTAの存在に応答するマーカープラスミドを追加的にトランスフェクトされたTet−on細胞系統の1つの免疫染色51。パネルCは、rtTAを発現している核を示す(緑色)。パネルDは、rtTAに応答した細胞質中のマーカータンパク質のドキシサイクリン誘導発現(赤色)、および細胞のDAPIでの核染色(青色)を示す。パネルE:抗rtTA抗体のBiaCore分析の例。親和性が示されている。
【図8】パネルA:ラクダ科動物様スプライス変異を含むIg遺伝子座の概略図である。2つのヒトIgG定常領域(Cγ2およびCγ3)を、まず、Gγ2−SおよびGγ3−Sで表されるラクダ科動物IgG HCAbs5においてCH1エキソンのスキッピングをもたらすと考えられるG+1〜A+1のスプライスを変更することにより変異させた。遺伝子座は、2つのラマVHH領域、全てのヒトDおよびJH領域ならびにヒトCμ、CδおよびCγ2およびCγ3およびLCRを含有した(明細書本文も参照のこと)。これらの遺伝子座をμMTトランスジェニックマウスに導入し、ヒト遺伝子座の発現について分析した。パネルB:GSマウス由来の骨髄(BM)ヒトIgGcDNAの配列決定から、両方のVHHが異なるヒトDおよびJセグメントと組み換えられ、Cγ2定常領域で転写されることが示された。しかし、CH1エキソンは、まだ最後の16bpを残して存在し、それを切り出した。進行中であるが、イントロン1の4位におけるA〜Gの移行に起因する、CH1エキソンの同じ潜在スプライス部位が白血病患者において報告された[31]。MGSまたはGS系統のいずれもμMTマウスにおいてB細胞の成長を救済しなかった(図なし)。マウスのB細胞転写/翻訳機構は再構成されたヒトコブラクダVHH−γ2aをプロセシングすることができるが[34、60]、本発明者らのデータは、G〜A突然変異に加えて、その他の特徴がCH1エキソンスキッピングに重要であることを示す。
【図9】実施例3および4に説明される様々な遺伝子座でのヒトVH領域のクローニングの模式図である。
【図10】複数のVH遺伝子セグメント、全D領域、全JH領域、Cγ2、Cγ3およびCx領域および3’LCRを含む重鎖遺伝子座の例を示す図である。
【図11】複数のVH遺伝子セグメント、全D領域、全JH領域、Cγ2、Cγ3およびCx領域および3’LCRを含む重鎖遺伝子座の例を示す図である。
【図12】染色体1に組み込まれた1つの遺伝子座と染色体8に組み込まれた1つの遺伝子座を含むカリオグラムを示す図である。
【技術分野】
【0001】
本発明は、ヒトを除くトランスジェニック哺乳類における、抗原曝露に応答する親和性成熟した機能性重鎖のみ抗体の多様なレパートリーの改良された製造方法およびその使用に関する。本発明はまた、複数の遺伝子座からのクラス特異的重鎖のみ抗体の多様なレパートリーの製造に関する。
【0002】
特に、本発明は、任意のクラスまたは複数のクラスの混合物のヒト抗原特異的な高親和性の重鎖のみ抗体の生成、ならびに十分に機能性のVH抗原結合ドメインの単離および発現のための方法に関する。
【0003】
本発明の方法を用いて生成された重鎖のみ抗体も記載される。
【0004】
以下の説明において、全てのアミノ酸残基位置番号は、Kabat et al.により考案された番号付け方式に従って示される[1]。
【背景技術】
【0005】
<抗体>
抗体の構造は当技術分野で周知である。大部分の天然抗体は四量体であり、2本の重鎖と2本の軽鎖を含む。重鎖はそれぞれの重鎖に沿ってほぼ中間に位置するヒンジドメイン間のジスルフィド結合を介して相互に連結されている。軽鎖はそれぞれの重鎖とヒンジドメインのN末端側で会合している。それぞれの軽鎖は、通常、各重鎖とヒンジドメインに近いジスルフィド結合により結合している。
【0006】
抗体分子が正常に折り畳まれると、それぞれの鎖は、より線状のポリペプチド配列により連結された多数の別個の球状のドメインに折り畳まれる。例えば、軽鎖は、可変(VL)および定常(CL)ドメインに折り畳まれる。重鎖は、単一の可変ドメインVH、第1の定常ドメイン(CH1)、ヒンジドメインおよび2または3個のさらなる定常ドメインを有する。重鎖定常ドメインおよびヒンジドメインは一体となって抗体重鎖の定常領域として一般に公知な構造を形成する。重(VH)および軽(VL)鎖可変ドメインの相互作用は、抗原結合領域(Fv)の形成をもたらす。重鎖および軽鎖の相互作用は、重鎖のCH1ドメインと軽鎖のCκまたはCλドメインにより促進される。一般に、VHおよびVLの両方は抗原結合に必要とされるが、重鎖二量体およびアミノ末端断片は軽鎖の不在下で活性を保持することが示されている[2]。
【0007】
重(VH)および軽(VL)鎖の両方の可変ドメインの中で、一部の短いポリペプチドセグメントは例外的な可変性を示す。これらのセグメントは超可変領域または相補性決定領域(CDR)と称される。間にあるセグメントはフレームワーク領域(FR)と呼ばれる。VHおよびVLドメインのそれぞれの中に、3つのCDR(CDR1〜CDR3)がある。
【0008】
哺乳類において、5つのクラスの抗体(IgA、IgD、IgE、IgGおよびIgM)が存在し、ヒトには4つのIgGサブタイプと2つのIgAサブタイプが存在する。
【0009】
【表1】
抗体クラスはその生理学的機能において異なる。例えば、IgGは、成熟した免疫応答において主要な役割を果たす。IgMは、補体結合および凝集に関与する。IgAは、分泌物(涙、唾液、初乳、粘液)中の抗体の主要なクラスであり、従って局所免疫に役割を果たす。天然抗体のエフェクター機能は、重鎖定常領域によってもたらされる。
【0010】
IgAは、粘液を含有する部位(例えば、腸内、気道内または泌尿生殖器路内)に見出すことができ、病原体による粘膜部位のコロニー形成を防ぐ。IgDは主にB細胞上の抗原受容体として機能する。IgEはアレルゲンと結合して肥満細胞からのヒスタミン放出(アレルギーの基礎をなす機構)を誘発し、また、蠕虫(虫)に対する保護をもたらす。IgGは(その4つのアイソタイプにおいて)、侵入してくる病原体に対して抗体に基づく免疫性の大部分をもたらす。IgMは、B細胞の表面に発現され、B細胞の媒介する免疫性初期段階において(すなわち、病原体を排除するために十分なIgGが存在する前に)病原体を排除するための非常に高い親和性を有する分泌型としても存在する。
【0011】
正常なヒトB細胞は、単一の重鎖遺伝子座を14番染色体上に含有し、その遺伝子座から重鎖をコードする遺伝子が再構成により形成される。マウスにおいて、重鎖遺伝子座は12番染色体に位置する。正常な重鎖遺伝子座は、複数のV遺伝子セグメント、多数のD遺伝子セグメントおよび多数のJ遺伝子セグメントを含む。VHドメインの大部分はV遺伝子セグメントによってコードされるが、それぞれのVHドメインのC末端はD遺伝子セグメントとJ遺伝子セグメントによってコードされる。B細胞におけるVDJの再構成と、その後の親和性成熟は、それぞれのVHドメインにその抗原結合特異性をもたらす。正常なH2L2四量体の配列解析は、多様性が主にVDJ再構成と体細胞超変異の組合せに起因することを実証する[3]。ヒトゲノムには50個を超えるヒトV遺伝子セグメントが存在し、そのうちの39個のみが機能性である。
【0012】
完全なヒト抗体(H2L2)は、現在、トランスジェニックマウスから抗原曝露に応答して導くことができる。そのようなトランスジェニックマウスは単一のヒト重鎖遺伝子座および別個の軽鎖遺伝子座を含む。対応するマウス重鎖および軽鎖遺伝子座は、マウス抗体の不在下でヒト抗体のみが産生されるように欠失または抑制される([4〜10])。
【0013】
新しい分子生物学技法の出現で、重鎖のみ抗体(軽鎖を欠く)の存在が、人におけるB細胞増殖性障害において同定され(重鎖疾患)、さらにマウスモデル系においても同定された。分子レベルでの重鎖疾患の分析により、ゲノムレベルでの突然変異および欠失が重鎖CH1ドメインの不適切な発現をもたらし、軽鎖と結合する能力を欠く重鎖のみ抗体の発現を引き起こしたことが示された[11、12]。
【0014】
ラクダ科動物が、遺伝子の自然突然変異の結果として、軽鎖との結合を媒介するCH1ドメインが存在しないために軽鎖と結合することのできない機能性IgG2およびIgG3重鎖のみ二量体を生成することが示された[13]。ラクダ科動物重鎖のみ抗体の特性を特徴付けるのは、ヒトおよび正常なラクダ科動物VHドメインに対して改良された溶解度をもたらすラクダ科動物VHドメインの特定のサブセットである。この特定のサブセット中のラクダ科動物VHドメインは通常VHHドメインと称される。
【0015】
サメなどの種が、おそらく哺乳類T細胞受容体または抗体軽鎖に類縁であり、重鎖のみ様結合タンパク質ファミリーを生成することも示されている[14]。
【0016】
ラクダ科動物重鎖のみ抗体の産生のために、ラクダ科動物生殖細胞系列の重鎖遺伝子座は、可能性のある重鎖定常領域の一部または全部をコードする遺伝子セグメントを含む。成熟の間、再構成されたVHHDJ結合ドメインは、ヒンジドメインをコードする遺伝子セグメントの5’末端上にスプライスされて、CH1ドメインを欠き、従って軽鎖と会合することのできない重鎖をコードする、再構成された遺伝子がもたらされる。
【0017】
ラクダ科動物VHHドメインは、特徴的な多数のアミノ酸を37、44、45および47の位置に含有する[49]。これらの保存されたアミノ酸は、重鎖のみ抗体に溶解性を付与するために重要であると考えられる。特定のラクダ科動物VHドメインだけが、改良された溶解度特性を有するVHHドメインである。それらはVHサブファミリーに限定され、そのため限定された範囲の抗原だけに増殖的に応答する。
【0018】
重鎖のみモノクローナル抗体は、標準的なクローニング技術によりラクダ科動物脾臓のB細胞から、またはファージもしくはその他のディスプレイ技術によりB細胞mRNAから回復することができる[18]。ラクダ科動物に由来する重鎖のみ抗体は高親和性である。重鎖のみ抗体をコードするmRNAの配列解析は、正常な四量体抗体の産生においても観察されるように、多様性が主にVHHDJ再構成と体細胞超変異の組合せに起因することを実証する[49]。
【0019】
天然のラクダ科動物およびヒトVHドメインの重要かつ共通する特徴は、それぞれのドメインが、最適な溶解度および結合親和性のためにVLドメインでの二量化に依存することなく、モノマーとして結合することである。
【0020】
近年、ヒトを除くトランスジェニック哺乳類における重鎖のみ抗体の産生のための方法が開発された(WO02/085945号およびWO02/085944号参照)。潜在的に任意のクラス(IgM、IgG、IgD、IgAまたはIgE)の、任意の哺乳類に由来する機能性重鎖のみ抗体は、抗原曝露の結果としてヒトを除くトランスジェニック哺乳類(好ましくはマウス)から生成することができる。これらの初期研究は2つのラマV遺伝子セグメントの使用に頼り、抗体応答の限定されたレパートリーに悩まされた。
【0021】
Janssens et al.[15]は、トランスジェニックマウスにおける重鎖のみ抗体の誘導のための方法を開発した。B細胞での抗体成熟の結果として高い結合親和性を有するこれらの重鎖のみ抗体は、抗原曝露の結果として誘導され、確立されたハイブリドーマ技術を用いて選択され、軽鎖(例えばIgG、IgA、IgM)の不在下で任意のクラスの抗体として、またはVH結合ドメイン単独として生成され得る。これらの重鎖のみ抗体は、全てのヒトDおよびJ遺伝子セグメントと結合した2つのラマVHH(クラス3)遺伝子セグメントならびに全てのヒト定常領域をコードする遺伝子セグメントを含有する生殖細胞系列(すなわち非再構成)配置中の抗体重鎖遺伝子座から誘導された。それぞれの定常領域をコードする遺伝子セグメントは、CH1ドメインの欠失を有し、軽鎖の結合を妨げた。さらに、この遺伝子座は、3’末端に抗体LCRを含有し、B系列の細胞における高いレベルの発現を確保する。この遺伝子座を受精卵のマイクロインジェクションによりマウスに導入した。
<抗体を原料とした産物の製造>
抗体を原料とした産物の遺伝子工学による製造、特にヒトまたはヒト化抗体を原料とした産物の製造は、新規なクラスの医薬、診断法および試薬の生成をもたらすと同時に、新規な産業、雇用および富の創造の機会の創出をもたらした(www.drugresearcher.com、www.leaddiscovery.co.uk参照)。抗体を原料とした産物は通常、天然の四量体抗体から導かれる。抗体を原料とした産物の製造に関する特許および特許出願は多くある。これらの特許および特許出願は、(例えばトランスジェニックマウスからの)誘導の経路、製造の経路および産物に特異的な物質の事柄に関する。そのような抗体を原料とした産物は、抗体断片を介して完全な四量体抗体から一本鎖Fv(scFv)分子までさまざまである。
【0022】
抗体を原料とした産物は、21世紀に着手される新規な医薬品の高い割合を占める。モノクローナル抗体療法は、関節リウマチおよびクローン病の治療のための好ましい経路として既に容認され、癌の治療において見事な進展がある。また、抗体を原料とした産物は、心血管疾患および感染症の治療のために開発中である。大部分の販売されている抗体を原料とした産物は、標的リガンド(例えばTNFα)上の単一の明確なエピトープを認識し、結合する。
【0023】
近年、高親和性VHドメインは、ディスプレイライブラリー中の無作為化されたヒトVHドメインから選択されるか、またはラクダ科動物の抗原曝露によって自然に産生された重鎖のみ抗体から誘導されるか、またはラクダ科動物で作られたVHドメインライブラリーから誘導されている。これらの高親和性VHドメインは抗体を原料とした産物に組み込まれている。これらのVHドメインは、VHHドメインとも呼ばれ、多数の古典的なVHドメインとは違い、特に、軽鎖の不在下で重鎖の改良された溶解度および安定性を確保する多数の突然変異を提示する。これらの変化の中で最も顕著なものは、荷電アミノ酸が45位に存在することである[16]。
【0024】
多数のグループが、天然の四量体抗体から誘導される重鎖のみ抗体の作製に取り組んできた。Jaton et al.は(同文中に2およびその他の参照文献が引用される)、親和性精製された、特性が十分明らかであるウサギ抗体の縮小された重鎖成分の分離、それに続いてその後の個々の重鎖の再生を記載している。再生された重鎖の免疫学的特性解析により、軽鎖を含まない重鎖ホモ二量体が、単独で、能力のある結合抗原であることが実証された。
【0025】
その後、Ward et al.[18]は、クローン化されたマウスVH領域が、大腸菌発現系で可溶性タンパク質モノマーとして発現されると、高い親和性で抗原と結合する能力を保持することを明白に実証した。Ward et al.[18]は、VHドメインの単離および特性解析を記載し、古典的なモノクローナル抗体産生(最後の段落参照)と比較した場合のこのアプローチの潜在的な商業上の利益を示した。彼らはまた、通常は軽鎖と会合する重鎖から単離されたVHドメインが天然の四量体抗体の溶解性に欠けていることを認めている。従って、Ward et al.[18]は、これらの分子を表現するために「粘着性のある」という用語を用い、この「粘着性」は改良された溶解度特性を有するVHドメインの設計によって対処することができると提案した。
【0026】
VH溶解度の改良は、その後、ファージディスプレイを用いる無作為化された部位特異的なアプローチの組合せを用いて対処されてきた。例えば、Davies and Riechmann[17]は(WO92/01047号参照)、溶解度を改良すると同時に結合特異性を維持するためにラクダ科動物重鎖のみ抗体のVHドメインの特徴の一部をファージディスプレイと組み合わせて組み込んだ。
【0027】
ファージライブラリーに由来する単離されたヒトVHドメインに関する複数の独立した研究により、VHドメインの抗原特異的な結合が実証されたが、これらのVHドメインは溶解度が低いことが分かった。さらに、ファージアレイに提示される特異的結合特性を有するヒトVHドメインの選択が操作された抗体のためのビルディングブロックを形成することが示唆された[18]。
【0028】
ヒトVHドメインは改良された溶解度特性のために操作されてよく[19、20]、または溶解度はインビボで自然選択により獲得されてもよい[21]。しかし、VH結合ドメインがファージライブラリーから誘導されている場合、例えば親和性ホットスポットのランダム化を含む親和性改善策の適用にもかかわらず、抗原に対する固有の親和性は低マイクロモルから高ナノモルの範囲のままである[22]。
【0029】
ファージまたは代替ディスプレイ技術により生成されたヒトVHまたはラクダ科動物VHドメインは、体細胞突然変異の結果としての改良された特性の利点および正常な抗体結合部位のCDR3領域におけるDおよびJ遺伝子セグメントの組換えにより提供されるさらなる多様性を欠く。一部のラクダ科動物VH(VHH)ドメインは、ヒトVHに対して溶解度の点で利点を示すものの、ヒトにおいて抗原性であることが判明し、さらに、ラクダ科動物VHがラクダ科動物の免疫化またはファージディスプレイ技術により生成されねばならないという不利点を被る可能性がある。
【0030】
ファージ由来ヒトVH領域は、高い親和性結合特性を達成するためにそれらが多くの回のパニングおよびその後の変異誘発を必要とするので、使用することが困難である。ラクダ科動物VHHドメインは、ファージもしくは類似のディスプレイライブラリーから単離された場合に同じ困難な手順を必要とするか、または、古典的なハイブリドーマ技術に従わない大型動物(同様に古典的な抗体を作成するラマまたはラクダ)の免疫化を必要とする。さらに、ラクダ科動物の結合ドメインは抗原性であることが分かり、ヒト化を必要とし得る。
【0031】
抗原曝露の結果としてのヒトを除くトランスジェニック哺乳類における重鎖のみ抗体の産生(WO02/085945号およびWO02/085944号参照)は、これらの多くの問題を克服する。
【0032】
しかし、当技術分野において、重鎖のみ抗体の多様性およびインビボでB細胞の応答を最大化する、特に、クラス特異的ヒト重鎖のみ抗体の機能性レパートリーおよび多様な臨床、工業および研究用途で使用するための最大の抗原結合可能性を保持する機能性VH重鎖のみ結合ドメインを生成する必要性が残っている。
【発明の開示】
【0033】
本発明者らは、驚くべきことに、先行技術の制限を克服し、クラス特異的な重鎖のみ抗体を生成するために用いられるヒトを除くトランスジェニック哺乳類に存在する重鎖のみの遺伝子座の数を増加させることにより、抗体応答のレパートリーを大いに増大させることができることを示した。
【0034】
本発明は、ヒトを除くトランスジェニック哺乳類が複数の重鎖のみの遺伝子座を有する場合、それらの遺伝子座は対立遺伝子排除の対象となるという発見に依存する。その結果、ただ1つの遺伝子座が確率的に選択され、首尾よく組換えられ、結果として重鎖のみ抗体の産生がもたらされる。従って、複数のVH重鎖遺伝子座を同じヒトを除くトランスジェニック哺乳類に用いて、哺乳類から入手可能な抗体レパートリーおよび多様性を最大化することができる。抗原に曝露された場合、ヒトを除くトランスジェニック哺乳類は、残りの遺伝子座を除いて特異的抗原曝露に応答するために最も適しているV遺伝子セグメントを含む遺伝子座を「選択する」。
【0035】
本発明の方法により生成され得る重鎖のみ抗体は、ヒトを除くトランスジェニック哺乳類が一連の遺伝子座から「選択する」ことが可能であることの結果として高い結合親和性を示し、それから一般に伸張されたCDR3ループの不在下でV、DおよびJ遺伝子セグメントの再構成ならびに体細胞突然変異が起こり得る。本質的に正常なB細胞の成熟は、高いレベルの重鎖のみ抗体が(CH1ドメインが組換え遺伝子座に存在する全ての抗体クラスから排除されているという条件で)単離された血漿中に存在する状態で観察される。B細胞の成熟および構築された二量体(例えばIgG)または多量体(例えばIgM)の分泌は、軽鎖遺伝子の存在または発現に依存しない。
【0036】
従って、本発明の第1の態様では、その哺乳類に複数の異種のVH重鎖遺伝子座を供与する段階を含む、ヒトを除くトランスジェニック哺乳類におけるVH重鎖のみ抗体の産生のための方法が提供され、各VH重鎖遺伝子座は、1個以上のV遺伝子セグメント、1個以上のD遺伝子セグメント、1個以上のJ遺伝子セグメント、および発現した場合、CH1ドメインを含まず、前記遺伝子座の少なくとも1つからVH重鎖のみ抗体を発現する重鎖定常領域をコードする遺伝子セグメントを含む。
【0037】
好ましくは、それぞれのVH重鎖遺伝子座は、1つ以上のV遺伝子セグメント、例えば1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、45、50または60個のV遺伝子セグメントを含み、それは任意の脊椎動物種に由来してもよい。
【0038】
一実施形態では、各遺伝子座は、ただ1つのV遺伝子セグメントを含んでよい。本実施形態の一つの代替形態では、各V遺伝子セグメントは他の全てのV遺伝子セグメントとは異なっている。第2の代替形態では、各V遺伝子セグメントは、他の全てのV遺伝子セグメントと同一である。この第2の代替形態では、各遺伝子座中の残りの遺伝子セグメントは、他の全ての遺伝子座中の遺伝子セグメントと同じであってもよいし、異なっていてもよい。
【0039】
従って、非ヒト哺乳類は単一のVH重鎖遺伝子座の複数のコピーを含有し得ることが想定される。これは、B細胞において生産的な再構成が起こり、従って有用な重鎖のみ抗体の産生を可能にする機会を最適化するという利点を有する。
【0040】
非ヒト哺乳類が、多数の異なるVH重鎖遺伝子座を含有するならば、これは、所望の特異性を有する重鎖のみ抗体を得る機会をさらに最適化する。
【0041】
もう1つの実施形態では、それぞれの遺伝子座は複数のV遺伝子セグメントを含む。本実施形態では、任意の1つの遺伝子座中のV遺伝子セグメントは、全て同じ種の生物に由来してよい、例えば、全てのV遺伝子セグメントがヒト起源であってよい。あるいは、任意の1つの遺伝子座中のV遺伝子セグメントは異なる種の生物に由来してよい。例えば、一部のV遺伝子セグメントがヒトに由来し、他のものはラクダ科動物またはサメに由来してよい。
【0042】
好ましくは、V遺伝子セグメントはヒト起源である。
【0043】
用語「V遺伝子セグメント」は、ラクダ科動物およびヒトを含む脊椎動物に由来する任意の天然のV遺伝子セグメントを包含する。V遺伝子セグメントは、D遺伝子セグメント、J遺伝子セグメント、および重鎖定常(エフェクター)領域(CH1エキソン以外の数個のエキソンを含み得る)をコードする遺伝子セグメントと組み換えて、核酸が発現されるとVH重鎖のみ抗体を生成できなければならない。
【0044】
V遺伝子セグメントはまた、D遺伝子セグメント、J遺伝子セグメント、および重鎖定常領域(1個以上のエキソンを含むがCH1エキソンは含まない)をコードする遺伝子セグメントと組み換えて、本明細書に定義される重鎖のみ抗体を生成できる相同体、誘導体またはタンパク質断片をコードする任意の遺伝子配列をその範囲内に含む。V遺伝子セグメントは、例えばT細胞受容体遺伝子座または免疫グロブリン軽鎖遺伝子座に由来してよい。
【0045】
好ましくは、本発明の複数の重鎖遺伝子座は、複数の遺伝子座に分散された、改良された溶解度特性を有する39個の機能性ヒトV遺伝子セグメントおよび操作された前記ヒトV遺伝子セグメントの変異体のうちの、任意の数または任意の組合せを含む。これらは、任意の数の遺伝子座、例えば8つのV遺伝子セグメントを含む4つの遺伝子座に加えて7つのV遺伝子セグメントを含む1つの遺伝子座;4つのV遺伝子セグメントを含む7つの遺伝子座に加えて3つのV遺伝子セグメントを含む1つの遺伝子座;またはそれぞれ1つのV遺伝子セグメントを含む39の遺伝子座上であってよい。
【0046】
ヒトV遺伝子は7つのファミリー、VH1〜VH7に分類され、各ファミリー内の個々の遺伝子に番号が付けられている。それぞれの遺伝子が使用される頻度は、特定の免疫応答の変動する要件に依存する。例えば、ファミリーVH3の遺伝子は、細菌性抗原に応答する際にファミリーVH5の遺伝子と比較して選択的に使用され得る。従って、本発明のさらに好ましい実施形態では、特定の抗原に対する抗体応答を生成するために有用であることが示されたV遺伝子セグメントのグループを別々のヒトを除くトランスジェニック哺乳類の系へ分類する。V遺伝子セグメントはファミリーに従って分類されるか、またはそれらは個々の機能に従って分類される。例えば、ファミリーVH3のV遺伝子が細菌性抗原に対する抗体応答を生成するために有用であることが示されるならば、細菌性抗原に対する重鎖のみ抗体を生成するために特に有用であるヒトを除くトランスジェニック哺乳類を生成するためにファミリーVH3のV遺伝子を用いることができる。あるいは、ファミリーVH3およびVH5の数個の個別の遺伝子が細菌性抗原に対する免疫応答を生成するために有用であることが示されるならば、それらは一緒に分類されて、細菌性抗原に対する重鎖のみ抗体を生成するために特に有用であるヒトを除くトランスジェニック哺乳類を生成するために用いることができる。
【0047】
本発明の状況において、用語「異種の」とは、本明細書に記載される核酸配列または遺伝子座が、それが位置する哺乳類に対して内在性でないことを意味する。
【0048】
「VH重鎖遺伝子座」は、本発明の状況において、重鎖エフェクター領域をコードする1つ以上の遺伝子セグメント(それぞれCH1ドメインを欠く)と操作によって連結されている、1つ以上のV遺伝子セグメント、1つ以上のD遺伝子セグメント、および1つ以上のJ遺伝子セグメントを含む、VHドメインをコードする最小のマイクロ遺伝子座に関する。好ましくは、抗体レパートリーの可変性の主な原因は、V、DおよびJ遺伝子セグメントの選択により、さらにV−DおよびD−J接合により形成されるCDR3領域である。
【0049】
本発明の利点は、再構成されたVH遺伝子配列において得られた抗体レパートリーおよび多様性が、同じヒトを除くトランスジェニック哺乳類における複数のVH重鎖遺伝子座の使用によって最大化することができることである。Janssens et al.、2006[15]は、上に記載されるトランスジェニック遺伝子座が、再構成および対立遺伝子排除に関して正常な免疫グロブリン遺伝子座のような挙動をすることを示した。このことは、同じ動物において(異なる染色体上に)複数の遺伝子座を有して、対立遺伝子排除を利用することにより起こり得るVH組換えの数を最大化する可能性を開いた。各トランスジェニック遺伝子座は、1から40を超えるVH領域を含有する。遺伝子座の1つを無作為に選択して組換えを開始し、最初の組換えが非生産的などであった場合に生産的な組換えが遺伝子座の1つから産出されるまで次の遺伝子座がその後に続く、対立遺伝子排除のプロセスは、組み合わせた遺伝子座中に存在する全てのVH領域が実際に全体的な組換えプロセスの一部となることを確保するものである。
【0050】
Janssens et al.、[15]はまた、驚くべきことに、同じヒトを除くトランスジェニック哺乳類において複数の重鎖遺伝子座が同じ染色体上で直列になっている場合、その結果、生産的な再構成および抗体の発現が複数の重鎖遺伝子座から起こり得ることを見出した。そのような場合、その後のハイブリドーマクローンはポリクローナルである。当業者であれば、これが抗体の多様性を増加させるためのもう一つの機構であることを理解する。
【0051】
従って、本発明の別の態様では、本明細書に記載されるように、ヒトを除くトランスジェニック哺乳類において少なくとも2つ以上のVH重鎖遺伝子座が同じ染色体上で直列になって存在し、その結果、生産的な再構成および抗体の発現が2つ以上の遺伝子座から同時に起こり得る方法が提供される。
【0052】
好ましくは、多数の重鎖遺伝子座は最初に別々にヒトを除くトランスジェニック哺乳類に導入され、単一の重鎖遺伝子座を有するヒトを除くトランスジェニック哺乳類を作り出す。次に、VH領域の数を最大化するためにこれらの動物を交配させて複数の重鎖遺伝子座を有する後代を生成し、最大の多様性をもたらす。遺伝子座はまた、新しい1ラウンドの遺伝子組換えによって加えてもよい。新しい遺伝子座は、既に1以上の異種のVH重鎖遺伝子座を含むヒトを除く哺乳類由来の卵に注入される。異種のVH重鎖遺伝子座の安定な融合により、利用可能なVH領域の増加および、従って多様性がもたらされる。異種のVH重鎖遺伝子座を、さらなる異種のVH重鎖遺伝子座とともに含む、ヒトを除くトランスジェニック哺乳類に由来するES細胞の安定なトランスフェクションは、ES細胞技術を胚融合および胚盤胞注入による遺伝子組換えに使用することのできるヒトを除く哺乳類(例えばマウス)において多様性を増加させる代替経路を提供する。
【0053】
好ましくは、それぞれの異なる重鎖遺伝子座は、ヒトを除くトランスジェニック哺乳類のゲノム中に単一コピーとして存在する。
【0054】
さらに、複数のV、DおよびJ遺伝子セグメントを使用することにより、得られる抗体レパートリーおよび多様性のさらなる増加がもたらされる。その後の体細胞突然変異は、VLおよびLc(軽鎖)抗体遺伝子座の必要なく最小の遺伝子座(マイクロ遺伝子座)を使用しながら達成される。
【0055】
本発明の状況において、用語「D遺伝子セグメント」および「J遺伝子セグメント」には、DおよびJ遺伝子セグメントの天然の配列が含まれる。好ましくは、DおよびJ遺伝子セグメントは、V遺伝子セグメントと同じ脊椎動物に由来する。例えば、V遺伝子セグメントがヒトに由来する場合、必要に応じて可溶化されたまたは操作されたDおよびJ遺伝子セグメントも、ヒトに由来することが好ましい。あるいは、V遺伝子セグメントは、例えばラクダ科動物に由来し、DおよびJ遺伝子セグメントはヒトに由来してもよく、逆も同様である。
【0056】
D遺伝子セグメントおよびJ遺伝子セグメントという用語はまた、得られるセグメントが本明細書に記載される重鎖抗体遺伝子座の残りの成分と組み換えられて本明細書に記載の重鎖のみ抗体を生成することができる限り、誘導体、相同体およびその断片をその範囲内に含む。DおよびJ遺伝子セグメントは天然の供給源から誘導されてもよいし、それらは当業者が熟知ある、本明細書に記載される方法を用いて合成されてもよい。V、DおよびJ遺伝子セグメントは組換えが可能であり、好ましくは、体細胞突然変異を受ける。
【0057】
DおよびJ遺伝子セグメントは、単一の脊椎動物種に由来することが好ましい。これはいずれの脊椎動物種であってもよいが、好ましくはヒトである。
【0058】
好ましくは、各VH重鎖遺伝子座は、1〜40(2、3、4、5、6、7、8、9、10、12、14、16、18、20、30または40)以上のD遺伝子セグメントを含む。D遺伝子セグメントはいずれの脊椎動物種に由来してもよいが、最も好ましくは、D遺伝子セグメントはヒトD遺伝子セグメント(通常25個の機能性D遺伝子セグメント)である。
【0059】
好ましくは、それぞれのVH重鎖遺伝子座は、1〜20(2、3、4、5、6、7、8、9、10、12、14、16、18または20)以上のJ遺伝子セグメントを含む。J遺伝子セグメントはいずれの脊椎動物種に由来してもよいが、最も好ましくは、J遺伝子セグメントはヒトJ遺伝子セグメント(通常6つのJ遺伝子セグメント)である。
【0060】
各VH重鎖遺伝子座は、同一のDおよびJ遺伝子セグメントを含有してよい。あるいは、各VH重鎖遺伝子座は、DおよびJ遺伝子セグメントの異なる組合せを含有してよい。例えば、各VH重鎖遺伝子座がただ1つのV遺伝子セグメントを含有し、このセグメントが各遺伝子座において同一である場合、生産的な再構成を得るための機会をさらに最適化するために、各遺伝子座においてDおよびJ遺伝子セグメントの異なる組合せを使用することが有利である。しかし、各VH重鎖遺伝子座が1つ以上の異なるV遺伝子セグメントを含有する場合、各遺伝子座においてDおよびJ遺伝子セグメントの同一の組合せを使用することが有利であろう。
【0061】
好ましくは、VH重鎖遺伝子座は、1以上のV遺伝子セグメント、25の機能性ヒトD遺伝子セグメントおよび6つのヒトJ遺伝子セグメントを含む。
【0062】
重鎖定常領域をコードする各遺伝子セグメントは、重鎖定常領域遺伝子セグメントがCH1ドメインを発現しないという条件で、Cδ、Cγ1−4、Cμ、CεまたはCα1−2クラスの1個以上の重鎖定常領域エキソンを含み得る。重鎖定常領域遺伝子セグメントは、好ましいクラスまたは必要な抗体クラスの混合物に応じて選択される。場合により、異種重鎖遺伝子座はCμおよびCδを欠損している。
【0063】
従って、各VH重鎖遺伝子座は、インビボでエフェクター機能をもたらす少なくとも1つの重鎖定常領域をコードする遺伝子セグメントを含んでよく(例えばIgG、IgM、IgA、IgEもしくはIgDまたはそのアイソタイプ)、各定常領域はCH1ドメインを含まない。各遺伝子座は、1つの特定の定常領域をコードする遺伝子セグメントをただ1つ含有してよい。これは、エフェクター機能が必要とされる場合に有利である。あるいは、各遺伝子座は、それぞれが異なる定常領域をコードする1個よりも多くの遺伝子セグメントを含んでよい。これは、様々なクラスの重鎖のみ抗体が産生されることを可能とするような、特定のエフェクター機能についての要件がない場合に有利である。
【0064】
各遺伝子座は同じ定常領域コード遺伝子セグメントを含有してもよく、または各遺伝子座は異なる定常領域コード遺伝子セグメントまたはその異なる組合せを有してもよい。
【0065】
操作上、重鎖定常領域は、B細胞においてV遺伝子セグメント、D遺伝子セグメントおよびJ遺伝子セグメントと組換え可能な天然のまたは操作された遺伝子セグメントによってコードされる。
【0066】
各重鎖のみ抗体において、定常領域は、重鎖のみ抗体の生成が起こり得るように、CH1ドメインを含まずに発現される。CH1エキソンは、上記のVH重鎖のみ抗体の定常領域が機能性CH1ドメインを含有しないように除去され得る。
【0067】
多価の結合複合体を操作する際にクラス特異的な重鎖定常領域を含むことにより、インビボでエフェクター機能の治療上の利益が必要される機能性に応じてもたらされる。個々のエフェクター領域を操作することも、機能性の付加または欠失をもたらし得る[23]。従って、IgA定常領域の機能性を含むことにより、病原体に対して改良された粘膜機能をもたらされるが[24]、一方で、IgG1定常領域の機能性が存在することにより、インビボで強化された血清安定性がもたらされる。重鎖CH2およびCH3定常ドメインが存在することにより、自然抗体に見られるような安定的な二量体形成の基盤がもたらされ、翻訳後グリコシル化の認識部位がもたらされる。CH2およびCH3の存在はまた、二重特異性および多価の複合体を試薬および診断薬として用いる際に、二次抗体の認識も可能にする。
【0068】
例えば、IgM抗体は、マクロファージの活性化および補体経路の活性化に重要な役割を果たすことが公知である。その結合部位が近接していることに起因して、IgMは、ウイルスを含む病原体に高い親和力を有する。しかし、IgMは、迅速なイムノアッセイ技法での使用が困難であることも公知であり、その一方でIgG抗体はこれらの技法で容易に使用することができる。用途に応じて、好ましい抗体クラス、すなわちIgGまたはIgMを選択することが有用である。
【0069】
CH1を欠く異種の重鎖Cγ遺伝子座の全部または一部の発現は、異種のIgG遺伝子座に存在するIgG1、IgG2、IgG3およびIgG4アイソタイプに応じて、場合により一部または全てのIgGアイソタイプを生成する。あるいは、重鎖はCε遺伝子を含んでよい。得られるIgE分子も治療に使用してよい。
【0070】
あるいは、選択された抗体の混合物を得ることもできる。例えば、IgAおよびIgMは、重鎖定常領域がCαおよびCμ遺伝子を含む場合に得ることができる。
【0071】
好ましくは、重鎖定常領域は、特に重鎖のみ抗体がヒトにおける治療用途に使用される場合に、ヒト起源である。重鎖のみ抗体が獣医学目的に使用される場合は、重鎖定常領域は、獣医学治療が行われる標的生物、脊椎動物または哺乳類に由来することが好ましい。
【0072】
発現した場合、各重鎖定常領域はCH1ドメインを欠く。好ましくは、CH1エキソンが欠失している。場合により、CμおよびCδ定常領域は変異しているか、欠失しているか、または置換されていてよい。例えば、機能性CH1ドメインを含むIgMの存在は、B細胞の成熟を阻害し、その結果として同じ遺伝子座の中で重鎖のみのIgG(CH1を欠く)の生産的発現を制限する。
【0073】
本明細書に定義される「重鎖定常領域エキソン」(「CHエキソン」)には、天然の脊椎動物の配列が含まれるが、特に哺乳類のCHエキソンである。これはクラス特異的な方法で変動する。例えば、IgGおよびIgAは天然にCH4ドメインを欠く。用語「CHエキソン」は、CHエキソンが、それが重鎖定常領域の成分である場合に、本明細書に定義される機能性重鎖のみ抗体を形成することができる限り、その範囲内に誘導体、相同体およびその断片も含む。
【0074】
CH4またはその他の機能性ドメインは、存在する場合、細胞内分泌プロセス、B細胞の成熟または得られる抗体の結合活性を阻害しないならば、任意選択的に導入遺伝子内で操作されるかまたは除去されてよい。
【0075】
細胞表面の集合を妨げる分泌機構とその結果起こるB細胞成熟に操作が影響を及ぼさないならば、重鎖エフェクター分子は操作されて、機能性ドメイン、例えばカルボキシ末端CH4ドメインが取り除かれてよい。例えばグリコシル化を改良するかまたは機能を付加するために、さらなる特徴を遺伝子座に組み換えてもよい。
【0076】
従って、異種の重鎖遺伝子座は、基本的に正常にB細胞成熟した状態で、必要とされる抗体クラスに応じて、重鎖のみ抗体の好ましいクラスまたは混合物を生成するように設計される。ラクダ科動物V、DおよびJ遺伝子セグメントおよびラクダ科動物エフェクター領域を利用すると、ラクダ科動物抗体に特有の特長、例えば伸張したCDR3ループなどを有するラクダ科動物抗体が産生される。ヒトV、DおよびJ遺伝子セグメントを使用すると、伸張したCDR3ループを欠くヒト重鎖のみ抗体が産生される。
【0077】
本発明の状況において「VHドメイン」は、上記に定義されるように、D遺伝子セグメントおよびJ遺伝子セグメントと組み換えられる場合、V遺伝子セグメントの発現産物を指す。好ましくは、VHドメインは、VDJ組換えおよび体細胞突然変異の結果として抗原と結合する改良された能力を有する。ラクダ科動物種に特有の伸張したCDR3ループの存在または不在に対する依存性はない。VHドメインはモノマーとして抗原と結合することが可能である。可溶性重鎖のみ抗体複合体の一部として発現された場合に、VLドメインとの組換えの可能性は、CH1エキソンの除去により排除されている([25]参照)。
【0078】
VHドメインは、単独で、多様なタンパク質ドメインと遺伝子操作されて、例えば毒素、酵素および造影剤を含む、標的とされる治療および診断目的のための融合タンパク質を生成することもできる。
【0079】
VHドメインコード配列は、天然の供給源から誘導されてもよいし、それらは当業者の熟知の方法を用いて合成されてもよい。
【0080】
VHドメインの性質は、必要とされる特徴を有する配列をコードするV、Dおよび/またはJ遺伝子セグメントを選択するかまたは操作することにより変更または改良することができる。上記に示されるように、39個の機能性ヒトVH領域の一部は重鎖のみ抗体の産生に適していない可能性がある。様々な抗体特性を改良しようと試みて多数の研究が先行技術において行われた[26,29]。特定のVH領域特性に関して、Dolk et al.[30]は、ファージディスプレイ技術を用いて、フケ予防シャンプーに伴う厳しい条件下において改良された安定性を示す、重鎖のみ抗体を生成した。
【0081】
ヒトを除くトランスジェニック哺乳類は、好ましくは、げっ歯類、例えば、ウサギ、モルモット、ラットまたはマウスである。マウスが特に好ましい。代わりとなる哺乳類、例えばヤギ、ヒツジ、ネコ、イヌまたはその他の動物を用いてもよい。好ましくは、哺乳類はマウスである。
【0082】
好ましくは、トランスジェニック非ヒト動物は、確立された卵母細胞注射技術のみを用いて作製される。確立されていれば、ES細胞技術またはクローニングを用いてもよい。
【0083】
有利には、哺乳類に内在する抗体の重鎖遺伝子座および場合により軽鎖遺伝子座は、重鎖のみ抗体が本発明の方法に従って発現される場合に、削除されるか抑制される(silenced)。
【0084】
抗体の供給源とは異なる起源の脊椎動物の種への抗体の投与は、しばしば投与された抗体に対する免疫応答の発現をもたらすので、本発明の上記の態様に記載される重鎖のみ抗体を生成する方法は、ヒトの治療用途のための抗体の生成において特に有用であり得る。本発明に従って生成される抗体は、それらが実質的に単一または既知のクラスの抗体であり、好ましくは、ヒト起源であるという点で、先行技術のそれに優る利点を有する。抗体は、インビボでのVDJ組換えと親和性成熟の組合せにより高親和性である。
【0085】
従って、本発明のさらなる態様は、上記により定義された複数の異種のVH重鎖遺伝子座を含むヒトを除くトランスジェニック哺乳類を提供する。好ましくは、ヒトを除くトランスジェニック哺乳類は、軽鎖を含む抗体を産生する能力が低下するように遺伝子操作されてよい。
【0086】
抗体産生細胞は、本明細書に定義されるヒトを除くトランスジェニック哺乳類から誘導され、例えば、本明細書に定義される重鎖のみ抗体の産生のためのハイブリドーマの調製に使用されてよい。それに加えて、またあるいは、核酸配列をこれらのヒトを除くトランスジェニック哺乳類から単離し、当業者の熟知な組換えDNA技法を用いてVHドメイン重鎖のみ抗体またはその二重特異性/二機能性複合体を生成するために用いてよい。
【0087】
あるいは、またはそれに加えて、抗原特異的な重鎖のみ抗体は、本明細書に定義されるヒトを除くトランスジェニック哺乳類の免疫化によって生成されてよい。
【0088】
従って、本発明はまた、上に定義されるヒトを除くトランスジェニック哺乳類を抗原で免疫することによる、重鎖のみ抗体の産生のための方法を提供する。抗体およびその断片は、当業者に公知の確立された方法を用いて単離され、特性決定され、製造されてよい。これらの抗体は、PCT/GB2005/00292号に記載される方法に特に役立つ。
【0089】
本発明を、これから一例として、以下の図を参照する以下の詳細な説明において説明する。
〔実施例〕
【0090】
実施例1−重鎖のみ抗体遺伝子座はマウスにおいて完全に機能性であり、対立遺伝子排除に対して感受性である。
【0091】
上記に考察されるように、Janssens et al.[15]は、トランスジェニックマウスにおける重鎖のみ抗体の誘導のための方法を開発した。本方法および本明細書に記載される実験のさらなる詳細に関して、当業者は、参照により本明細書に組み込まれるJanssens et al.[15]を参照されたい。1
<方法>
・構築物
ゲノムコスミドライブラリーを、標準法を用いてラマ(Lama Glama)の末梢血細胞から作成した。2つの異なる生殖細胞系列VHHを、それらの配列、停止コドンを含まないオープンリーディングフレームおよびルフランの番号付け[32]に従う42、50および52位の親水性のアミノ酸コドンの存在、ならびに49位に親水性のアミノ酸を含むものと含まないものに基づいて選択した。一方はIGHV1S1(受託番号AF305944)と同一であり、もう一方はIGHV1S3(受託番号AF305946)と94%の同一性を有する。2つのクローン(ヒト重鎖DおよびJ領域、CμおよびCδを含有するクローン1065N8ならびにCγ3遺伝子を含有するクローン1115 N15)をヒトゲノムPacライブラリーRPCI−11(BACPAC Recource Center,USA)から選択した。異なるヒトゲノムライブラリー(Incyte Genomics,CA,USA)からのBacクローン11771をCγ2遺伝子およびIg重鎖LCRの供給源として用いた[33]。標準技法を用いて、Cγ3およびCγ2遺伝子を別々にpFastBac(Invitrogen)にサブクローニングした。単一の点突然変異(G〜A)[34]またはCH1エキソンの完全な欠失を相同組換えにより達成した[35]。同様にfrtおよびloxP部位をCμスイッチ領域の前に導入し、2番目のloxP部位をCγ2スイッチ領域の前に設置し、MGSまたはMGΔを得た。
【0092】
GSまたはGΔ構築物を得るために、MGSまたはMGΔベクター(図1)、2つのラマVHH遺伝子、その後にヒトDおよびJ重鎖領域、Cμ、Cδならびに修飾されたヒトCγ2およびCγ3遺伝子および3’LCRを含有し、16個の294Cre大腸菌株44に形質転換し、creに媒介される組換えによってGSまたはGΔ遺伝子座を得た(図1)。相同組換えによるCμCH1領域の欠失によりMGΔからMΔGΔを得た。
・トランスジェニックマウスの生成、育種および遺伝子型同定
220KbのMGSまたはMGΔまたはMΔGΔ断片、150KbのGSまたはGΔ断片(図1)をベクター配列から精製し、受精したFVB X B16/μMT−/−卵の前核に2ng/μlの濃度で注入した。5’および3’末端プローブを用いる尾由来DNAのサザンブロット分析により、トランスジェニック遺伝子座を、完全性およびコピーの数についてチェックした。トランスジェニックμMT+/−樹立系統をμMT−/−バックグラウンドの系統として育種した。以下のプライマー:Asp5IgMfw:5’−GCGGGTACCGAATGGTGGCAGGGATGGCTC−3’(配列番号1)と、HLL−MDには、Asp3’IgG2rv:5’−CGCGGTACCCTGCGGTGTGGGACAGAGCTG−3’(配列番号2)の組合せ、または、MGSには、Asp3’IgMrv:5’−CGCGGTACCACGGCCACGGCCACGCTGCTCGATTC−3’(配列番号3)の組合せ、およびμMT遺伝子型には、MIGMEMBINTRON1:fw:5’−CCAGTCAATACTACTCGCTAAGATTC−3’(配列番号4)とMIGMEMBEXON1rv:5’−CAGTGGTCCACAGTTTCTCAAAGC−3’(配列番号5)の組合せを用いるPCR(94℃にて45秒間の変性、60℃にて30秒間のアニーリングおよび72℃にて1分40秒間の伸張で30サイクル)により遺伝子型同定を行った。
・RT−PCR
Ultraspec RNA単離系統(Biotecx Laboratories,Houston)を用いて全RNAを単離した。逆転写酵素(Superscript II、Life technology)およびオリゴ(dT)プライマーを用いてcDNAを合成した。以下のプライマー:LVHHfw:5’−AGACTCTCCTGTGCAGCCTCTGG−3’(配列番号6)と、HINGEIgG2rv:5’CACTCGACACAACATTTGCGCTC−3’(配列番号7)またはhIgMCH2rv:CACTTTGGGAGGCAGCTCAGC−3’(配列番号8)の組み合わせを用いてPCRを行った。増幅は、94℃にて30秒間の変性、60℃にて30秒間のアニーリングおよび72℃にて1分間の伸張で30サイクル間であった。PCR産物をpGEM Tイージーベクター(Promega)にクローニングし、T7またはSP6プライマーを用いて配列決定した。
・フローサイトメトリー分析:
単細胞懸濁液を、既に記載したようにPBS中のリンパ器官から調製した45。約1×106個の細胞を、96ウェルプレート中のPBS/0.5%ウシ血清アルブミン(BSA)中の抗体を用いて4℃にて30分間インキュベートした。細胞をPBS/0.5% BSAで2回洗浄した。それぞれのサンプルについて、FACScanアナライザー(Becton Dickinson,Sunnyvale,CA)を用いて3×104イベントが記録された。CellQuestバージョン1.0コンピュータソフトウェアを用いてFACSデータを分析した。Becton Dickinson FACS Caliburで四色分析を行った。使用した大部分の抗体は記載されている[36];FITCコンジュゲート抗ヒトIgGおよび抗ヒトIgMはSigma(Zwijndrecht,NL)より購入した。
・Ig遺伝子再構成分析
wtマウスMΔGΔおよびGΔトランスジェニックマウスの脾臓および肝臓から単細胞懸濁液を作成した。MACS CD45(B220)MicroBeads(Miltenyi Biotec,Germany)をAutomacsセパレーターで製造業者の説明書に従って用いて 約90%の純度までB細胞を確実に選択した[37]。ゲノムDNAをHind III消化させ、Hybondナイロンフィルター上でブロットした。32P放射標識されたJκプローブ(Jκ1およびJκ5領域のゲノムDNAからのPCR増幅より入手)および32P放射標識された炭酸脱水酵素II(CAII)プローブを用いてフィルターをハイブリダイズした。肝臓DNAを流してκ生殖細胞系列の構成シグナルを示した(2.8Kbバンド)。4kbバンドとハイブリダイズするCAIIプローブを負荷対照として用いた。Tyfoon9200(Amerscham Biosciences)でフィルターを走査した。生殖細胞系Jκバンドの強度を、Image Quant5.2ソフトウェアを用いて定量し、負荷対照のそれに正規化し、対照肝臓DNA(100%とする)の割合で表した。異なる再構成イベントの増幅のためのハイブリドーマ由来のゲノムDNAで使用されるPCRプライマーは次の通りであった、
IGHJlR:5’−CCAGTGCTGGAAGTATTCAGC−3’(配列番号9)、
IGHJ2R:5’−CAGAGATCGAAGTACCAGTAG−3’(配列番号10)、
IGHJ3R:5’−GGCCCCAGAYATCAAAAGCAT−3’(配列番号11)、
IGHJ4R:5’−GGCCCCAGTAGTCAAAGTAGT−3’(配列番号12)、
IGHJ5R:5’−CCCAGGRGTCGAACCAGTTGT−3’(配列番号13)、
IGHJ6R:5’−CCAGAACGTCCATRYMGTAGTA−3’(配列番号14)。
・DNA FISH分析
標的DNAの調製:モノクローナルハイブリドーマ細胞をDMEM/10% FCS中で培養し、Heiskanen[38]に記載されるようにアガロースに包埋した。GΔトランスジェニック系統1のマウス由来の肺を収集し、単細胞懸濁液を作った。包埋した細胞をプロテイナーゼKおよびリボヌクレアーゼHで処理した。機械的に伸張されたDNAを、ポリ−L−リシンスライド(Sigma)上で電子レンジと別のスライドの端を用いて調製した。
【0093】
プローブ:GΔ導入遺伝子の再構成されたコピーおよび再構成されなかったコピーを検出するため、DNA断片を精製した。VHHとDの間の領域のハイブリダイズ用に2.3kB SpeIおよび3.6kB SpeI−BssHII断片、およびLCR検出用に5.9kB BamHI−SpeI断片またはIgH3’LCR(16kB)を含有する低コピー数のBluescriptプラスミド。ニックトランスレーションによりビオチン−11−dUTP(Roche)またはジゴキシゲニン−11−dUTP(Roche)でプローブを標識した。プローブをピペットでスライドの上に移す前に、90℃にて5分間、氷上で5分間、および37℃にて45分間でそれらを変性させた。インサイチューハイブリダイゼーション:ハイブリダイゼーション混合物は50%ホルムアミド、2×SSC、サケ精子DNA(200ng/μl)、5×デンハート液、1mM EDTA、および50mMリン酸ナトリウム、pH7.0を含有した。プローブのハイブリダイゼーションは、ピペットで25μlの混合物をスライド上に移し、24×32mmのカバーガラスで覆うことにより行われる。プローブおよび標的配列を変性させるため、スライドを80℃の加温板の上に2分間置いた。プローブは加湿チャンバー(加湿剤は50%ホルムアミド、2×SSCである)中で37℃にて一晩ハイブリダイズする。ハイブリダイゼーション後の洗浄は、記載される通り行った[39]。
【0094】
ハプテン標識されたプローブの免疫学的検出:ジゴキシゲニンプローブを、ヒツジ抗ジゴキシゲニン(1:500、Sigma)、フルオレセイン結合ウサギ抗ヒツジ(1:500、Sigma)、およびフルオレセイン結合ヤギ抗ウサギ(1:500、Sigma)を用いて検出した。ビオチンプローブは、テキサスレッド結合アビジン(1:500、Sigma)およびビオチン結合ヤギ抗アビジン(1:500、Boehringer)を用いて検出した。この段階を2回繰り返した。全てのインキュベーションおよび洗浄は記載される通り行った48。染色の後、スライドを段階的な一連のエタノール(70、90、および100%)中で段階ごとに5分間室温にて脱水した。細胞またはDNAを25μlの抗退色性包埋剤Vectashield(Vector Laboratories)に包埋した。100倍対物レンズを用いるLeica DMRBE 蛍光顕微鏡で視覚化を行った。
・免疫化およびハイブリドーマ作成
8週齢のマウスを、Specolアジュバント(IDDLO,Lelystadt,NL)または事前処方DKTPワクチンを含む5〜20μgの抗原で0、14、28、42日に皮下注射し、50日に腹腔内注射して免疫した。0、14および45日に採血した。56日にClonalCellTMHYキット(StemCell Technologies,Canada)を製造業者の説明書に従って用いて、脾臓細胞をSp2−O−Agl4骨髄腫細胞株(R.Haperenより寄贈)と融合させた。DKTPワクチンはNetherlands Vaccine Institute(Bilthoven,NL)より得られた。
・sdABライブラリーの構築およびスクリーニング
Ultraspec RNA単離系統(Biotecx Laboratories Inc,Houston,Texas,USA)を用いて、DKTP免疫単一コピーGΔおよびTNFα免疫MΔGΔマウスの脾臓から全RNAを単離した。cDNAはオリゴ(dT)プライマーを用いて作成した。VHHDJ断片をコードするDNA断片を、特異的プライマー:vh1 back Sfi Iプライマー[40〜42]を、hIgG2hingrevプライマー(5’−AATCTGGGCAGCGGCCGCCTCGACACAACATTTGCGCTC−3’(配列番号15))またはCH2huIgMrevプライマー(5’−TGGGACGAAGACGGCCGCTTTGGGAGGCAGCTCGGCAAT−3’(配列番号16))と組み合せたものを用いてPCRにより増幅させた。増幅したVHHDJ(約400bp)をSfi I/Not I消化させ、ゲル精製し、Sfi I/NotI消化ファージミドpHEN由来ベクターにクローニングした。TG1エレクトロコンピテント細胞(Stratagene La Jolla,USA)への形質転換により、ヒト単一のドメイン抗体ライブラリーが得られた。プラスチック(希釈されていないワクチンをコートしたイムノチューブ)に吸着させたDKTPワクチン抗原または精製されたヒトTNFα(Biosource International,USA)へのパニングを用いて、2ラウンドの選択を行った。
・免疫細胞化学およびウエスタンブロット
rtTAの存在に応答するマーカープラスミドでトランスフェクトされたtet−on細胞系統の細胞をスライド上で増殖させた。ドキシサイクリン誘導の24時間後、細胞を4%パラホルムアルデヒド/PBSに固定し、0.5% Triton−X−100/PBSで透過処理した。rtTAに対するHCAbを1:50希釈して使用し、その後ヤギ抗ヒトIgG FITC染色(Sigma,1:500希釈)に使用した。マーカータンパク質を前記に記載したように検出した51。ウエスタンブロットは、ヤギ抗ヒトIgG−HRP(Sigma,1/2500希釈)、ヒトIgM−HRP(Sigma,1/2500希釈)を用いる標準法であった。
・Superose6ゲル濾過
200mM KCl/20mM HEPES−KOH pH 7.9/1mM MgCl2/0.5mM EGTA/20%グリセロールで平衡したSuperose6 10/30カラムを備えるAKTA FPLC装置(Amersham Biosciences,Piscataway NJ)を用いて、MΔGΔマウス血清およびヒト対照血清のサイズ分画を行った。100μl/分の流量を用いて、500μlの画分を回収し、100%トリクロロ酢酸でで沈殿させ、SDS−PAGE(還元条件下)に続いてウエスタンブロット免疫ブロット法により解析した。
・BIAcore測定
実験は、BIAcore3000表面プラズモン共鳴バイオセンサー(Biacore AB,Uppsala,Sweden)にて行った。製造業者の提供する標準的なNHS−EDCキットを用いて3000共鳴単位(RU、任意の結合応答単位)のレベルまでCM5センサーチップ上に精製タンパク質を固定化した。10mM Hepes、pH7.4、150mM NaCl、2mM EDTA、0.005% Tween20中、40マイクログラム/mlの一定の流速で抗体を固定化抗原に通過させた。製造業者のBIA評価ソフトウェアを用いて平衡時の応答を記録し、カーブフィッティングを使用して平衡解離定数を得た。
<結果>
・ラクダ科動物様CH1スプライス変異は、ヒト重鎖遺伝子座におけるエキソンスキッピングには不十分である。
【0095】
ラクダ科動物におけるHCAb(IgG2およびIgG3)の生成にIgM+中間段階が必要かどうかは不明である。そのため、本発明者らは、まず、μMTバックグラウンドにおいて2つのハイブリッドヒト遺伝子座、1つはヒトCμ、Cδ、Cγ2およびCγ3定常領域を有する遺伝子座(MGS)、およびもう1つはヒトCγ2およびCγ3しか有さない1つの遺伝子座(図1)を生成した[43]。そのCγ領域に、まず、ラクダ科動物CH1スプライス変異を含有するように突然変異を起こさせた。μ鎖遺伝子の膜貫通ドメイン欠失によりB細胞がほぼ完全に欠けているμMT動物[43]がやはりB細胞小集団を有し、膜IgMの不在下で機能性IgG、IgAおよびIgEを産生することが証明され[44〜46]、(一部の)B細胞はIgMの表面発現なしに成長することが示唆されていることから、MGSを作製した。VH溶解度を向上させるためにヒトVHドメインを突然変異させる[47、48]代わりに、ラマ起源の2つのVHHを導入した。ラクダ科動物VHH領域は、42位、49位、50位および52位に特徴的なアミノ酸を含有する[49]。第1の、VHH1は、これらのVHHに特徴的なアミノ酸の全てを含有したが、基礎実験のこの証明における溶解度の重要性を検証するために、もう1つの、VHH2では、49位のGlu(E)の代わりに、これらの重大な「溶解」アミノ酸の1つであるGln(Q)を欠失させた。本発明者らは、さらに軽鎖可変部(VL)対合21に重要であると考えられることから、50位(Arg、R)ではなく49位を選択した。前記遺伝子座は、5つのヒト重鎖DおよびJ領域の全ておよびその遺伝子座の3’末端に遺伝子座制御領域(LCR)も含有した(図8)。驚くべきことに、スプライス変異によりトランスジェニックマウスに正しいCH1エキソンスキッピングは起こらず、ヒトIg発現の欠如につながらなかった(図8)。
・CH1領域を欠くヒト遺伝子座を含有するトランスジェニックマウスの解析
CH1のスプライシング問題を克服するために、本発明者らは3種類の新規構築物を生成した(図1)。全てがCH1を欠失させたCγ2およびCγ3を含有し、1つはCμおよびCδを含むもの(MGΔ)であり、1つはCμおよびCδを含まないもの(GΔ)であり、1つはCH1を欠失させたCμセグメントを有するもの(MΔGΔ)であった。μMTバックグラウンドにおいて、異なるコピー数(1〜5コピー)の3つのMGΔ、6つのGΔおよび4つのMΔGΔトランスジェニックマウス系統を得た。骨髄(BM)および脾臓においてB細胞の成長を解析した。異なるコピー数のマウスで同じ結果を得た。
・GΔおよびMΔGΔの発現はμMTマウスにおけるB細胞の成長を救済する
MGΔマウスはμMTバックグラウンドにおいてB細胞の成長を救済することができなかったが、一方、GΔおよびMΔGΔ構築物はB細胞の成長を効果的に救済した。B220/CD19陽性細胞の救済は、異なるリンパ系コンパートメントにおいて、コピー数に関係なく、30〜100%間であった(図2A)。これは、B220対ヒトIgMまたはヒトIgG染色を用いたBMのフローサイトメトリーによって確認される(図2B)。MΔGΔマウスは、野生型またはμMTマウスに存在しないヒトIgM産生細胞をBMにおいて含有する。従って、これらの細胞は、これらの細胞がヒトIgGを含有しないように、クラススイッチを受けなかった。GΔマウスはヒトIgG発現B細胞だけを含む。MGΔマウスは、細胞表面でヒトIgを発現するB細胞を含有するとしても非常に少なく、興味深いことに、B220細胞の一部は細胞内IgMを発現するが、IgGは発現しない(図2B)。MΔGΔおよびGΔマウス(下記参照)とは対照的に、MGΔマウスはマウスIg軽鎖を発現する(図6G)。これらの結果は、異なる構築物中でCμおよびCγ遺伝子が発現されることを示し、CH1が存在しないことがVHHに基づく抗体の細胞表面発現にとって重大であることが強く示唆される。
・ヒトHCAb IgGおよびIgMはBMでのB細胞の成長中にマウス(プレ)BCRに機能的に置き換わる
小型静止プレB細胞への大循環の発展進行中、特異的細胞表面マーカーの発現はプレBCR依存的にダウンレギュレートされる[36]。プレBCRに機能的に置き換わるヒトHCAbの能力を調査するために、様々なマーカーの発現をFACSにより解析した。プロB細胞は、高レベルの細胞質SLC、IL−7RおよびCD43を発現するが、これらはプレBCR発現時にダウンレギュレートされ、成熟B細胞では存在しない(図2C、μMTマウスからのプロB細胞、wtマウスの表面IgM−プロB/プレB細胞画分および表面IgM+B細胞画分と比較)。
【0096】
MΔGΔ/μMTまたはGΔ/μMTマウスのヒトIg+B細胞はSLCおよびIL−7Rレベルが低く、これにより、ヒト単鎖IgGおよびIgM受容体は、SLCおよびIL−7RのダウンレギュレーションにおいてマウスプレBCRに機能的に置き換わることが示される。CD43では、これはGΔマウスにおける場合のみのようであるが、MΔGΔマウスにおけるCD43発現の持続はこれらのマウスにおけるB−1 B細胞分化の増加という知見に関連があるかもしれない。同様に、正常なプレBCRシグナル伝達のように、CD2およびMHCクラスII発現が誘導される。プレB細胞段階で一過性に誘導されたIL−2R/CD25のレベルは、成熟MΔGΔまたはGΔ/μMT B細胞では非常に低く、成熟wt B細胞のレベルと同程度である(図2C)。さらに、ic Igκ発現は成熟MΔGΔまたはGΔ/μMT B細胞では検出できず(図2C)、IL−7+培養物から5日後のIL−7除去時にはインビトロBM培養物においても誘導されていなかった(図なし)。最後に、MΔGΔまたはGΔトランスジェニックマウスにおけるヒトHCAb発現B細胞集団は、BMで生成された細胞(HSAhighおよびAA4.1/CD93high)から部分的になり、末梢で成熟し、再循環している細胞(HSAlowおよびCD93low)から部分的になり、正常マウスでの結果と同様であった。
【0097】
まとめると、これらの結果は、ヒトHCAbのIgGおよびIgMが、発達上調節されるマーカーの発現に関して、B細胞の成長中にマウス(プレ)BCRに機能的に置き換わることを示す。IgのL鎖は誘導されない(下記参照)。BM RNAのcDNA解析は、VDJ組換えのための両方のVHHセグメントの使用、CH1の不在、そして重要なことにCDR3領域の大きな多様性を示している(図2D)。
・複数の再配列および対立遺伝子排除
免疫後、MΔGΔおよびGΔ系統から、特に5コピーGΔ系統1で多数のハイブリドーマを作製した(下記参照)。配列解析により、多コピーGΔ遺伝子座において1つ以上の再配列が起こり得ることが示された。解析した5つの異なる5コピーハイブリドーマのうち、1つのハイブリドーマ(G20)はフレーム内にある1つのコピーを再配列し;2つのハイブリドーマ(T7およびT12)は各々2つの再配列を有し、そのうちの1つの再配列は両方のハイブリドーマにおいてフレーム外であり;1つのハイブリドーマ(T3)は2つのフレーム内再配列(J2およびJ4)を有し;一方、1つのハイブリドーマ(T1)はフレーム内の2つの再配列(J2およびJ4)を含む4つの再配列を有した。
【0098】
T1およびT3は2つの生産的mRNAを発現し、それらは分泌された抗体の質量分析によりcDNAと正確にマッチさせて確認された(図なし)。本発明者らは、2つの異なるハイブリドーマ:G20(1つの再配列)およびT1(4つの再配列)において、それぞれのコピーを検出するLCRプローブと、再配列されていないコピーだけを検出する、VHHおよびDセグメントの間に位置するプローブとを用いて、DNA伸張線維FISHおよび標準的なDNA−FISHも行った(図3A〜E)。対照肺細胞は、サザンブロットマッピング(図なし)に一致して、一方の末端に5つの完全なコピーと半分の導入遺伝子を示したが(図3A)、一方、ハイブリドーマではG20およびT1それぞれにおいて1つおよび4つの再配列されたコピーを確かに示している(図3B、C〜E)。従って、複数のコピーは同じ対立遺伝子上でうまく再配列することができる。次に、本発明者らは、HCAb遺伝子座に、正常なマウス遺伝子座に対する(不)利点があるかどうか、そして遺伝子座の対立遺伝子排除が起こっているかどうかを求めた。wtバックグラウンドにおけるGΔ系統1トランスジェニックマウスのB220/CD19陽性BM細胞をヒトIgGおよびマウスIgMの発現について解析した。明らかに、GΔB細胞はマウスIgまたはヒトIgのいずれかを発現し(図3F)、対立遺伝子排除を示している。
・GΔ、MΔGΔおよびMGΔトランスジェニックマウスにおける脾臓B細胞
導入遺伝子がμMTマウスにおけるB細胞分化に与える影響を評価するために、本発明者らはCD21/CD23プロフィールを用いてフローサイトメトリーにより脾臓B細胞亜集団サイズを調べた(図4A)。GΔマウスでは、CD21lowCD23low未成熟B細胞の割合は正常な範囲にあり、ヒト単鎖IgG発現細胞は濾胞(FO;CD21+CD23+)B細胞へも辺縁帯(MZ;CD21highCD23low)B細胞へも分化し得た。MΔGΔマウスでは、未成熟B細胞画分が増加し、HCAb−IgM発現細胞のFOおよびMZ−8 B細胞への分化がいくらか損なうことを示している。これらの脾臓では、CD23発現の減少は、B−1 B細胞系統への分化を示す、CD43およびCD5の表面発現レベルの増加を伴っていた。MGΔトランスジェニックマウスに存在する少数のヒトIgM発現B細胞(マウス軽鎖も発現する、図6参照)は、MΔGΔトランスジェニックマウスにおけるものと同様のFO/MZ分布を現した。MGΔマウスは違うが、MΔGΔおよびGΔマウスは、正常な脾臓構造を有し、白脾髄の外側境界に存在する濾胞および辺縁帯を含有するB細胞リッチな領域で囲まれた動脈周囲リンパ球鞘(PALS)に集中しているT細胞の分離を示す(図4B)。
【0099】
それらはまた、T細胞依存性抗体応答中のwtに匹敵する、二次リンパ組織のB細胞濾胞胚中心を形成する(図なし)。これらは、記憶形成と、体細胞超変異に起因する親和性に基づく選択の部位である。GΔマウスでは、これらの胚中心においてヒトIgG陽性細胞が検出される。本発明者らは、ヒトIgG遺伝子の超突然変異を、パイエル板(Payer′s patches)に存在するB細胞のcDNA解析により確認した(図4C)。VHH1およびVHH2の両方を使用する。総合すれば、これらの発見により、GΔおよびMΔGΔトランスジェニックマウスにおいて、BMから移動する未成熟B細胞は、脾臓においてFO−B細胞およびMZ−B細胞の両方へと分化し、抗原曝露後に体細胞超変異を受ける能力を有することが実証される。
・単一コピー遺伝子座は効果的に救済し、CH1の不在が必須である
GΔ系統1マウス(図2A)は5コピーのGΔ遺伝子座を含有するため、従って効果的な救済が遺伝子座のコピー数と関係がある可能性があった。しかし、5コピーGΔ系統1からFlpeR系統23との交配によるFlp組換えにより得られた単一コピートランスジェニック系統は、B細胞の成長を同程度まで救済した(図4D、図2A)。単一コピーの遺伝子座は救済に十分であり、そしてCH1領域を含むCμ定常領域の存在が阻害性であることを、非救済単一コピー系統MGΔ(系統3)から、単一コピーGΔ系統をもたらすCre組換え(cre発現系統への品種改良)によりCμおよびCδ領域を欠失させることによって確認した(図4D)。それまで非救済であった遺伝子座はその時点で他のGΔ系統と同じB細胞の成長9を与える。従って、コピー数は救済に影響を及ぼさず、Cμ領域にCH1が存在することによりB細胞救済が阻害される(下記も参照)。
・MΔGΔおよびGΔトランスジェニックマウスにおいてマウスIg軽鎖遺伝子座は再配列しない
マウスIg軽鎖タンパク質はMΔGΔおよびGΔマウスでは血清のウエスタンブロット(図なし、図2Cおよび6A参照)によってまたはFACSによって検出されず、マウス軽鎖遺伝子が再配列しないことが示唆された。これは、サザンブロット解析により選別された脾臓B220+細胞画分および肝臓細胞のDNAにおけるIgκ遺伝子座生殖細胞系列シグナルの密度を比較することによって確認され(図5AおよびB)、これによりマウス軽鎖は再配列せず、生殖細胞系列構成を保つことが示される。対照的に、MGΔ/μMTマウスの少数のヒトIg発現細胞において軽鎖が検出される(図6G参照)。
【0100】
従って、BMでの初期B細胞の成長におけるヒトHCAbの発現は軽鎖再配列を導くシグナルを与えることができない。これに関連して、HCAbはプレBCRよりもむしろBCRによく似ており、このことはHCAbがCH1の不在下で擬似軽鎖と結合できなかったことと関係があると考えられる[61]。
・GΔ/μMTおよびMΔGΔ/μMTマウスの血清分析
ヒトIgMはMΔGΔ血清に存在し、ヒトIgGはMΔGΔマウス血清およびGΔマウス血清の両方に存在した。免疫されていない成体動物では、ヒトIgM(<50μg/ml)およびIgG(200〜1000μg/ml)は、正常ヒトIgH遺伝子座を有する正常マウスまたはトランスジェニックマウスに見られるものと同様のレベルで存在する[65]。6種全てのGΔマウスの血清のゲル電気泳動では、非還元性条件下で約70kD、還元性条件下で約35kDのMWを有するHCAbのIgGが現われ、軽鎖を欠く重鎖二量体とCH1エキソンを欠く各重鎖と一致した(図6A、B)。
【0101】
MΔGΔマウスの血清は、多量体重鎖のみのヒトIgMを含有した。還元性条件下(図6C)では、4種全ての系統が、CH1のMWを減じた後にヒト対照IgMとしてのMWを有するIgM鎖を含有した。また、血清を非還元性条件下でも分画した(図6D、横方向の画分)後、それぞれの画分10を還元性条件下でゲル電気泳動により解析した(図6D、縦方向のレーン)。900kDのヒト血清五量体IgM対照と比較すると、トランスジェニックIgMは600kDで分別され、それが多量体であること、ならびに軽鎖およびCH1を欠くことに一致する。従って、MΔGΔマウスはHCAb多量体IgMおよび二量体IgGを産生するが、一方、GΔは血清中で二量体IgGを産生する。
・MGΔマウスの血清分析
MGΔ/μMT系統の末梢および脾臓では、B220陽性細胞はほとんど見られず(野生型の1%未満)、極まれに小さなB細胞集団が脾臓において見られる(図4B)。精製後にのみ血清中に少量のヒトIgMおよびIgGが検出された(図6E、F)。これらのマウスにおけるヒトIgMは、還元性条件下では通常のサイズであったが、一方、循環しているヒトIgGはより短く(CH1の欠失に一致する、見かけのMWは35kD)。興味深いことに、ヒトIgMと結合していると思われるマウスκ軽鎖も検出された(図6G)。
【0102】
ヒトIgMおよびIgGは定量ELISAアッセイでの検出レベルを下回っていたが、本発明者らは、それにもかかわらず、MGΔ/μMTマウスが免疫化に応答し得るかどうかを検証した。マウスをヒト腫瘍壊死因子α(TNF−α)で免疫し、wtマウスは強いTNF−α特異的抗体応答を発現したが、一方、用いた2種のMGΔ系統の3匹のマウスにおいては、ELISAまたはウエスタンブロット解析によって抗原特異的ヒトIgGを検出することができなかった(図なし)。
・MΔGΔおよびGΔマウスの免疫化は抗原特異的Ab産生をもたらす
GΔ/μMTマウスを大腸菌hsp70、DKTP(ジフテリア毒素、百日咳菌の全細胞溶解物、破傷風毒素ならびに不活化ポリオウイルス1型、2型および3型)およびrtTAで免疫し[50]、一方、MΔGΔマウスをヒトTNFαで免疫した。ELISAで陽性であった血清を有するマウスから、ファージディスプレイライブラリーによりハイブリドーマまたは単一ドメインAb(sdAb)を用いて個々の完全抗体を単離した。
【0103】
抗体RNAのRTPCR後にαhsp70特異的、破傷風毒素特異的およびrtTA特異的モノクローナルを配列決定した(図7A)。これにより、IgG2(8つのうちの7つ)およびIgG3(8つのうちの1つ)抗体の両方が産生されることが示された(sdAbはIgG2ライブラリーから単離した)。異なるDおよびJ領域を使用した。高頻度ではないが、11のHCAbのVHHは超突然変異を受けていた。3つのhTNFα特異的抗体(1つは陽性IgMハイブリドーマ、図6α−hTNFα#1および2つはsdAbα−hTNFα#2&3)は全て、CDR2領域に異なる超突然変異を有した。14全ての抗体の配列を比較すると、全てのJ領域が用いられているものの、ヒトと同じようにJH4が最もよく用いられていることが明らかであった。驚くべきことに、全ての抗体はラマVHH2(49位に典型なグルタミン酸ではなくグルタミン(Q)を有する[51])を含有した。最後に、CDR3領域が多様性の大部分をもたらしていることは明らかである27。それは、ラマおよびヒトにおいて通常見られるものと非常に類似している、10〜20アミノ酸長の間で変化する(平均は13.6aa)[52、53]。本発明者らは、次に、これらのHCAbが常用アッセイにおいてハイブリドーマ上清および細菌原形質周辺画分のsdAbとして機能するかどうかを検証した(図7)。ELISAアッセイでは抗体の全てが陽性であり、ウエスタンブロットでの抗原検出でも全てが陽性であった(例えばα−DKTPおよびα−rtTA血清とハイブリドーマ培地とα百日咳菌sdAb、図7B)。また、本発明者らは、rtTA発現プラスミドでトランスフェクトした細胞系統を用いて免疫細胞化学によりα−rtTA IgGも検証した(図7C、D)。rtTA HC−IgGは極めてはっきりとした特異的な核染色を示した。多数の抗体の親和性は高かったが、一部のもの(特にsdAb)ではかなり低かった。例えば、免疫細胞化学に用いたα−rtTA抗体(図7C、D)は、BiaCoreで決定されるように約3nMであった(図7E)。
<結論>
本明細書において、本発明者らは、μMTマウスにおけるB細胞の成長を救済し、結果的に機能性HCAb産生をもたらす修飾HCAb遺伝子座の発現を報告した。これらのマウスを免疫して、抗原特異的HCAbを産生させることができる。ラクダ科動物の場合、HCAbの分泌にはCH1の除去が不可欠であるが、CH1の排除には3’CH1/イントロン境界における単一ラクダ科動物(総説30)スプライス変異では不十分であり、従って少なくともヒト遺伝子座において、単にこの単一点突然変異以上のものが必要である。CH1をVHHとともに有するIgMはB細胞の成長を妨げるが、これは恐らく、プレBCRとの関連でIgM表面分子の効果のない会合に起因する。対照的に、HCD様ヒトμタンパク質を発現するマウスは、λ5とは無関係にSCIDバックグラウンドにおいて正常CD43プレB細胞を生じる[54]。末端切断型Cμタンパク質は、関連L鎖なしにB細胞表面で発現され、自己会合を介してシグナル伝達するプレBCRによく似ていると考えられる[55]。
【0104】
通常、BiPシャペロンは、後に鎖間接触に関与するIg鎖の疎水性表面と結合することによって抗体分子の折り畳みおよび会合を監視している31。VHHのFR2に親水性アミノ酸が存在することによって、VHHとのBiPの結合がほぼ確実に阻止され、この場合には可溶性にする(代理)軽鎖は必要でない。同時に、CH1は、会合(軽鎖によるBiPの置換)が完了するまでERに重鎖を保持するとされているBiPとの相互作用をもたらす。本発明者らの結果は、プレB細胞受容体レベルでは、λ5タンパク質との非共有結合における代理軽鎖(SLC)の一部としてのVpreタンパク質とのトランスジェニックμ重鎖対合は、CH1ドメインを含有するμ重鎖がVHHと結合されている場合には起こるのができないことを示唆している。そのため、MGSおよびMGΔトランスジェニックマウスにおけるヒトIgMは、この点で、増殖展開および発展進行をシグナル伝達するために不十分であることが公知である不完全プレBCR様複合体に似ている[56、57]。これにより、BMではB220陽性細胞の30%しか細胞内IgMを有さない理由を説明することができる(図2B)。これらのマウスの脾臓に少数の成熟B細胞が存在することは、μ重鎖を含有するが任意のSLCまたは通常の軽鎖が欠失している最近記載された新規な受容体複合体によって説明する[58]。
【0105】
CH1がCγ2および3に存在せず、IgMがその遺伝子座から除去されている場合には、B細胞の成長の救済が起こり、IgGがIgMに機能的に置き換わることが示される。Rag2−/−マウスにおける成熟CD21+B細胞の成長の支援において、プロB細胞段階以降に発現されるIgG1受容体をIgMの代わりにすることができる[59]。最近、事前に再配列したラクダ科動物IgG2aがμMT(およびCΔ−/−)バックグラウンドの2匹のトランスジェニックマウスのうちの1匹においてB細胞の成長を部分的に救済することができるということも示された13。本発明者らの場合では、CH1を欠くIgMまたはIgGは10種の独立したトランスジェニックマウス系統のうちの10種においてB細胞の成長を救済する。さらに、本発明者らは軽鎖再配列を観察しておらず、さらなるB細胞分化に軽鎖発現は必要でないと結論づけられる。本明細書で得られた結果とZou et al.の結果[60]における相違は、LCRが本発明者らの構築物に含まれていることによる重鎖遺伝子座の発現レベル(すなわちシグナル伝達)によって説明することができる。本発明者らの結果は、CH1[61]またはVHおよびCH1[62]を欠く末端切断型μ重鎖タンパク質が、SLCと結合することができず、κ遺伝子再配列を活性化することができないことを裏付ける。
【0106】
5コピーGΔ系統1(および他の多コピー系統)は、ゲノムの同じ位置に組み込まれた単一コピー系統と同じ程度までB細胞の成長を救済する。興味深いことに、多コピートランスジェニック遺伝子座では1以上の再配列が起こる(図3)。2種の単一脾細胞由来のハイブリドーマの2つは、2種の生産的HCAb転写物およびタンパク質をもたらした。この結果は、同じB細胞における2種の抗体の発現は無害であることを裏付けている[63]。しかし、二重抗体産生B細胞は抗原曝露下で単一抗体産生細胞との競争に敗れるという予測37は、抗原曝露後に得られた5つのハイブリドーマから2種の二重抗体発現細胞を見出した本発明者らの結果からは実証されない。
【0107】
BM細胞は細胞表面でマウスまたはヒトIgを発現するため、野生型マウスにおいて(多コピーの)遺伝子座は対立遺伝子排除を受けやすい。細胞表面で両方を発現する細胞の有意な集団はない(図3F、上のパネル)。恐らく最も興味深いのは、マウスIgおよびヒトIgを発現している細胞の数の対比である。wt/5コピーGΔマウスでは、再配列に利用できる3つの可能性ある対立遺伝子、1つのIg遺伝子座を有する2つのマウス対立遺伝子および5つのヒトHCAb遺伝子座を有する1つの対立遺伝子が存在する。確率的に選択されるならば、ヒト対立遺伝子は3回のうち1回しか選択されない。
【0108】
遺伝子座の数を計数すれば、ヒト遺伝子座はまず7細胞のうち5細胞で再配列する。しかし、内因性マウスおよびトランスジェニックヒトHCAbはほぼ同等に発現される(44/38、図3F 下のパネル)。これらの数はV領域のランダム使用によって起こり得るずれやトランスジェニック遺伝子座における位置効果の可能性を無視しているが、最初の選択が対立遺伝子の確率的選択であり、続いて1対立遺伝子につき複数の遺伝子の再配列が行われ得ることを強く示唆している。
【0109】
これは、本発明者らがGΔ遺伝子座の複数の再配列を頻繁に観察しているという事実と一致する。通常、生産的再配列は組換えをダウンレギュレートして、他の対立遺伝子の再配列を妨げる。しかし、トランスジェニック対立遺伝子における複数のコピーは同じオープンな遺伝子座に存在し、RAGダウンレギュレーション前に組み換えられる。これは、複数の再配列が同時に起こるためであるか、または空間要素(「コンパートメント」)を含むならば、RAGがダウンレギュレートされる前に接近していれば遺伝子座の別の遺伝子を再配列する十分な時間があり得る。
【0110】
wtマウスにおいて再配列が生産的ではない(シグナル伝達はなく、RAGはとどまる)場合に限り、第2の遺伝子座が活性化され、第1の遺伝子座に置き換わり、再配列されるための十分な時間がある。この論証を支持して、同じ染色体上に複数の遺伝子座を有する他の種には、2つのAbを発現する、より多くの細胞が存在するという見解が生じる[64]。
【0111】
重要なことには、これらの試験によってHCAb遺伝子座はマウスにおける発現に成功し得ることが分かる。抗原曝露は、遺伝子座の構成によって異なるクラスの高親和性抗原特異的ヒトHCAbの産生をもたらす。これらの抗体は、正常マウスまたは他の「正常」なヒトIgHトランスジェニックマウスでのレベルと同様のレベルで発現される[65]。本発明者らの実験では2つの可変領域だけを使用したが、本発明者らが検証した無関係なタンパク質のほぼ全てに対して様々な特異性を有する高親和性抗体の単離に成功し、CDR3によってもたらされる多様性の効率および有効性を立証した[66]。従って、V(D)J組換えおよびインビボ選択を行うことは、ファージディスプレイを用いたファージミドライブラリーからのヒト単鎖抗体断片の作製に対して非常に重要な利点をもたらす39。ハイブリドーマは容易に生成することができ、このことによって、重要なことに、ファージディスプレイやさらなるスクリーニングの必要なく、完全ヒトHCAbまたはsdAb断片の直接クローニングおよび発現を可能にする。
【0112】
従って、これらのマウスは、特にHCAbが通常の抗体、例えば酵素の活性部位に対してほとんど抗原性がないエピトープを認識し得るという証拠4に照らして、臨床目的またはその他の目的でのヒトHCAbの産生に関して全く新しい可能性を広げる。抗原の全てが認識されるわけではないことの理由は、限られた数の可変領域によって説明することができる(ポリオおよびジフテリアタンパク質はGΔマウスでは応答をもたらさなかったが、一方、wt対照マウスでは応答がもたらされた)。驚くべきことに、抗体の全てがラマVHH2領域を有した。これには、49位に保存アミノ酸[67]が含まれず、それを有し、溶解性がより高いはずであるVHH1とは対照的である。
【0113】
それでもなお、本発明者らは、遺伝子座により多くの可変領域を追加することによっていっそう広範囲にわたるレパートリーをもたらすものと予測する。単一対立遺伝子上では複数のコピーの遺伝子座を避けることが好ましいが、多様性を高めるために異なるVH領域の単一コピーをそれぞれが含む複対立遺伝子を含むマウスを生成することが有利。そのような新しい遺伝子座では、通常発生する(ヒト)VH領域または溶解度18および軽鎖対合を高めるために操作されたVH領域のいずれかを使用することができる。
【0114】
結論として、本発明者らは、潜在的に任意のクラスの抗原特異的高親和性HCAbがマウスで産生され得ることを立証した。この技術は、人における治療薬または診断薬として使用するための、抗原曝露に応答する任意のクラスの完全ヒトHCAbまたはそれらの断片の生成を可能にする。また、異なる脊椎動物遺伝子座を用いることによって、本発明者らの技術は、試薬、診断法として使用するための、または動物の治療のための任意の脊椎動物由来の高親和性成熟抗体の産生を可能にする。
【実施例1】
【0115】
Janssens et al.(2006)は、トランスジェニックVH遺伝子座が遺伝子導入によってコードされた重鎖のみ抗体を産生するように正確に再結合していること、そしてそのような遺伝子座は内因性(マウス)重鎖免疫グロブリン遺伝子座の存在下で対立遺伝子排除に対して感受性があることを示した。対立遺伝子排除プロセスを用いることによって、重鎖のみ抗体産生に使用されるVHトランスジェニック遺伝子座の数を増加させることができるということを示すために、異なる重鎖のみの遺伝子座を含む2匹のトランスジェニックマウスを交配させ、結果として両方の遺伝子座を含む子孫を得た。一方のマウスはMGΔ遺伝子座を含有した(IgMおよびIgG遺伝子座、Janssens et al., 2006)が、もう一方のマウスはGΔ遺伝子座(IgG遺伝子座単独、Janssens et al., 2006)を含有し、両方のマウスはμMTバックグラウンドを有するものであった。ダブルトランスジェニック子孫のB細胞からハイブリドーマを誘導し、標準的な方法によって培養下で増殖させた。多数の得られた個々のモノクローナル細胞系統をPCRおよびサザンブロットによって解析し、それによって生産的に再配列されたMGΔ遺伝子座を含有する系統が、再配列されていないGΔまたは生産的に再配列されていないGΔ遺伝子座を含むことが示された。逆に、生産的に再配列されたGΔ遺伝子座を含有する細胞系統は、再配列されていないまたは生産的に再配列されていないMGΔ遺伝子座を含有した。このように、利用できるVH領域の合計が組換えプロセスに使用される。
<実施例3および4>
本発明の第1の態様の好ましい実施形態では、前の実施例に記載される、当技術分野で公知の方法(Janssens et al 2006)を用いて、機能性ヒトVH領域全数を、ヒトVH領域(またはそれらの変異体)を、全DH領域、全JH領域ならびにCμ、Cγ2、Cγ3およびCα領域と3’LCRの組合せを含む複数の修飾ヒト遺伝子座にクローニングすることによって増加させる。この手順は、独立した遺伝子座の複数の同一VH領域または独立した遺伝子座の異なるVH領域を用いて行うことができる。異なる遺伝子座は同一重鎖領域または異なる重鎖領域を含有し得る。実施例3は、重鎖領域の異なる組合せを有する遺伝子座の同一VH領域を有する遺伝子座に関し、実施例4は同一重鎖定常領域と異なるVH領域を有する2つの遺伝子座に関する。両実施例において、さらに遺伝子座を追加してよいことは明らかである。
【実施例2】
【0116】
ヒトVH領域は、可能性ある39個の機能性ヒトVH領域(あるいはヒトVL領域またはTCRのV領域または突然変異誘発によって得られるこれらの全ての変異体を使用または追加してよい)の中から選択されたそれぞれのVHに特異的なプライマーを用いたヒトゲノムDNAのPCR増幅によって単離する。ヒトVH領域は、前記実施例に記載した、すなわちヒトDHおよびJH(または他のDおよびJ領域)とそれぞれCH1および3’LCRが欠失しているCμ、Cγ2、Cγ3を含む遺伝子座にセットでクローニングされる(図9)。CH1およびスイッチ領域が欠失しているCα領域はGΔ遺伝子座に別個にクローニングされる(図10)。このGΔ遺伝子座は、lox部位を含有せず、ラマVHH領域が標準的な相同組換えによって除去されて、特異PI−PspI部位を残している(図10)という点から原型遺伝子座の変異体である。
【0117】
機能性VH領域は、各遺伝子座の任意の複数に関して同時にクローニングすることができる。最初に、17個の機能性ヒトVH領域を、クローニングされた2個の遺伝子を1セットで同時にクローニングする。これらの最初の構築物のそれぞれに、第2のセットを従来の方法(例えばXhoI−SalI制限消化/連結を用いて、XhoIおよびSalI適合部位の連結により両方を破壊する)により追加する。4、4、4、3および2個の遺伝子を含有するセットがBACベクター中の17個の遺伝子の1セットに連結される。この手法は、他の数の遺伝子を含有するセットを組み合わせることによって行ってよいことは明らかである。前記方法は、所望の数のVH領域を得る任意の時点で終了してよく、またはより多数のVH領域を得るために延長してよい。
【0118】
1セット全て(例えば17個の遺伝子)を特異PI−PspI部位の修飾GΔ遺伝子座にクローニングする。Cα領域はこの遺伝子座のI−CeuI部位にクローニングされ、結果としてIgαおよびIgGを産生可能なAΔGΔ遺伝子座が生じる(図10)。また、他の重鎖領域もクローニングすることができた。
【0119】
これらの最終的な遺伝子座は、その後、前記実施例に記載されている独立したトランスジェニックマウス(好ましくは、μMTなどのIgH遺伝子座欠損マウス)に導入される。これらの独立した遺伝子座は、独立したマウス系統、IgGだけを作製する系統ならびにIgAおよび/またはIgGを作製する系統を生成するために用いられる。これらのマウスは、続いて、2つの異なる遺伝子座から34個のVH領域を提供できるようにVH領域全体を有するように交配される。これらのマウスを両方の遺伝子座に関してホモ接合性になるように交配することによって、組換えに利用できる68個のVH領域が作製されることになる。組み込まれた遺伝子座の複数のコピーを有することによって、この数はいっそうさらに増加する。これらのマウスのB細胞から作製されたハイブリドーマの解析は、生産的に再配列された遺伝子座が存在する場合に、作り出された他の遺伝子座が、標準的な手法による対立遺伝子排除の結果、再配列されていないかまたは生産的に再配列されていないことを示すために利用される。
【実施例3】
【0120】
前記領域(または突然変異誘発によって得られたそれらの変異体)に対して同様に単離された1以上のVH領域の異なるセットを、前記と同じ方法により2つの独立した遺伝子座にクローニングする。これは結果として2つの異なるGΔ遺伝子座(またはそれらの変異体、例えばCαが追加されたもの)をもたらすことになる。これらの遺伝子座は、独立したマウスに導入されて、独立したトランスジェニック系統(例えばそれぞれ10個のVHドメインを有する2つの遺伝子座、図11)をもたらす。これらのマウスは、その後、組換えプロセスに利用できる全ての使用VH領域を有するダブルトランスジェニックマウスを得るために交配されることになる。これらのマウスを両方の遺伝子座に関してホモ接合性なるように交配することによって、組換えに利用できるVH領域の数が倍増することになる(図12、染色体1に組み込まれた1つの遺伝子座と染色体8上の1つの遺伝子座に関するカリオグラム)。組み込まれた遺伝子座の複数のコピーを有することによって、この数はいっそうさらに増加する。これらのマウスのB細胞から作製されたハイブリドーマの解析は、生産的に再配列された遺伝子座が存在する場合に、作り出された他の遺伝子座が、対立遺伝子排除の結果、再配列されていないかまたは生産的に再配列されていないということを示すために利用される。
【0121】
前記プロセスは、所望の数のVH領域を得る任意の時点で終了してよい。これらのVH領域にD、JHおよび定常領域が追加される。これらの最終遺伝子座は、その後、前記実施例に記載されている独立したトランスジェニックマウス(好ましくは、IgH遺伝子座欠損マウスを含む)に導入され得る。あるいは、lox部位の位置により、Cμ(IgM)単独、またはCγ2およびCγ3(IgG2およびIgG3)単独、またはCα単独(IgA)またはそれらの組合せを含有する独立した遺伝子座を生成するための個々の定常領域の排除が可能となる。これらの独立した遺伝子座は、ヒトIgM単独、またはIgG単独またはIgA単独またはそれらの組合せのいずれかを作製する独立したマウス系統を生成するために用いられる。
[参考文献]
【0122】
【表2A】
【0123】
【表2B】
【0124】
【表2C】
【0125】
【表2D】
【0126】
【表2E】
【図面の簡単な説明】
【0127】
【図1】トランスジェニックマウスを生成するために使用されたDNA断片の模式図である。2つのラマVHHエキソンは、ヒト重鎖多様性(D)および結合(J)遺伝子セグメントと連結され、その後にCμ、Cδ、Cγ2およびCγ3ヒト定常領域遺伝子およびヒト重鎖Ig3’LCRが続く。ヒトCγ2およびCγ3遺伝子は、構築物MGΔおよびGΔ中のCγ2およびCγ3遺伝子からの、または同様に構築物MΔGΔ中のCμからのCH1エキソンの完全な欠失により修飾されていた。2つのLoxP位(赤色)が同じ方向に存在することにより、Creに媒介される組換えの際、CμおよびCδ遺伝子の除去が可能になる。Frt位(緑色)が存在することにより、Flpに媒介される組換えによる、複数コピー導入遺伝子アレイからの単一コピートランスジェニックマウスの生成が可能になる。
【図2】パネルA:異なる器官における総細胞のうちのB220/CD19陽性細胞の百分率として表されたBリンパ球のフローサイトメトリー分析の表である。パネルB:BM中のwt、μMT、MGΔ/μMT、MΔGΔ/μMTおよびGΔ/μMTマウスのB細胞集団のフローサイトメトリー分析。B220の前方および側方散乱ならびに表面発現に基づいてリンパ球系統細胞をゲートし、ヒトIgMまたはIgGをプロットした。データはドットプロットとして表示される。MGΔμMTマウスについて、B220+画分をゲートして細胞内(ic)ヒトIgμおよびγH鎖の発現について分析し、ヒストグラムオーバーレイ(赤色線)として、一緒にμMTマウス由来のB220+細胞のバックグラウンド染色(黒色線)を対照として表示した。陽性細胞の百分率が示されている。パネルC:MΔGΔまたはGΔ導入遺伝子の発現は、プレBCRおよびBCRの機能を救済する。μMTマウス由来の総CD19+画分(プロB細胞)、野生型マウス由来のCD19+表面IgM+画分(プロB/プレB細胞)およびCD19+表面IgM+画分(B細胞)、MΔ−GΔμMTマウス由来のCD19+表面ヒトIgM+画分(B細胞)およびGΔμMTマウス由来のCD19+表面ヒトIgG+画分(B細胞)において示されたマーカーの発現プロフィール。ic−Igκ=細胞内IgκL鎖。フローサイトメトリーデータはヒストグラムとして表示される。示されるデータは、各群内で試験された3〜8匹の動物の代表である。パネルD:VHH1およびVHH2特異的プライマーをヒトCγ2プライマーと組合せて用いてBM cDNAから得た、PCR産物の配列アラインメント、VDJ組換えを示す。配列はGΔに由来する。緑色は配列同一性を示す。
【図3】パネルA〜E:5つのコピーのヒトGΔ遺伝子座のDNA−FISH。パネルA:その半分をLCR(赤色)を含む遺伝子座に、その半分をVHHからJ領域(緑色)を有する遺伝子座に隣接されている5つのインタクトなGΔ遺伝子座のコピー(1〜5)を有するGΔ系統1トランスジェニックマウスの肺細胞由来の伸張されたクロマチン線維。パネルB:1つのコピーが再構成されている(白色矢印)GΔ系統1のB細胞に由来するハイブリドーマ(G20)の伸張されたクロマチン線維FISH。パネルC:LCRプローブを用いるハイブリドーマT1の伸張されていないDNA−FISH(赤色)。パネルD:VHH〜D間のプローブを用い、Cと同内容(緑色)。パネルE:パネルCとEのオーバーレイ。赤色シグナルの喪失がないのに対し、4つの緑色シグナルの喪失が見られることから、T1が4つの再構成を有することに留意されたい。パネルF:GΔトランスジェニックマウスにおいて対立遺伝子が排除されている。マウス表面または細胞内(ic)μH鎖および示されるマウス由来の総BM CD19+細胞画分上のトランスジェニックヒトIgGのフローサイトメトリー分析。データはドットプロットとして表示され、示される象限内の細胞の百分率が示される。示されるデータは、各群内で試験された4匹のマウスの代表的なデータである。
【図4】wt、μMT、GΔ、MΔGΔおよびMGΔマウスの脾臓のB細胞集団の分析を示す図である。示されるデータは、各群内で試験された4〜8匹のマウスの代表的なデータである。パネルA:上側、B220に対してマウスIgM、ヒトIgG、ヒトIgMについて染色された脾臓細胞のFACSデータ。下側、脾臓のB細胞集団のフローサイトメトリー分析。B220の前方および側方散乱ならびに表面発現に基づいてリンパ球系統細胞をゲートし、示されたIg(上部)またはCD21/CD23プロフィールはドットプロットとして示され、示されたゲート内の細胞の百分率が示される。CD21lowCD23low:未熟B細胞;CD21+CD23+:濾胞性B細胞;CD21highCD23low:辺縁帯B細胞。パネルB:wt、μMT、GΔ/μMT、MΔGΔ/μMTおよびMGΔ/μMTマウスの脾臓の組織像。免疫組織化学分析;5μmの凍結切片を、B細胞には抗B220(青色)で、樹状細胞には抗CD11c/N418(茶色)で染色した。矢印は、MGΔ脾臓におけるB細胞の小さなクラスターの位置を示す。パネルC:VHH1およびVHH2特異的プライマーをヒトCγ2プライマーと組合せて用いてパイエル板cDNAから得たPCR産物の配列アラインメント、トランスジェニック遺伝子座がCDR1および2領域で超突然変異を受けることを示す。配列は、CH1の欠失しているトランスジェニック遺伝子座GΔに由来する。パネルD:上側;脾臓細胞のFACSデータ、抗CD19および抗B220で染色。下側左:FlpeRトランスジェニック系統に交配することによるインビボFlp組換えの模式図、および5つのコピーGΔ系統1に由来する単一コピー組換え型の脾臓細胞に関するFACSデータ。下側右:MGΔ系統をCAGCreトランスジェニック系統に交配することによるインビボCre組換えの模式図、組換え型の脾臓細胞に関するFACSスキャンデータを支持し、直接生成した元のGΔ系統に見られるようなB細胞の救済を示す。
【図5】GΔ/μMT(パネルA)およびMΔGΔ/μMT(パネルB)トランスジェニック系統においてκ軽鎖再構成が行われないことを示すサザンブロットを示す図である。1匹のwtマウスおよび2匹のGΔまたは4匹のMΔGΔトランスジェニックマウス由来の肝臓DNA(L)およびB細胞DNA(B)をHind IIIで消化し、32P放射標識されたJκプローブおよび炭酸脱水酵素II(CAII)プローブを用いて分析した。4kbバンドにハイブリダイズするCAIIプローブを泳動対照として用いた。肝臓DNAを流してκ生殖細胞系列の配置を示した(2.8Kbバンド)。wtB細胞だけが、2.8kb断片の強度の低下で測定されるκ遺伝子座の再構成を示す(肝臓と比較するとシグナルの30%が残る)。
【図6】μMTバックグラウンドにおいて非還元条件下(A)および還元条件下(B〜G)、6種類の異なるGΔ系統(A、B)、4種類の異なるMΔGΔ系統(C)および2種類の異なるMGΔ系統(E〜G)の、Prot Gまたはコンカバリン(concavalin)精製された血清サンプルを泳動した。トランスジェニックヒトIgG(パネルB、F)およびIgM(パネルC、D)のサイズはCH1欠失と一致し、軽鎖が存在しない。マウスκ軽鎖は正常なサイズであった(G)。ヒト血清を陽性対照として用いた。パネルD:非還元条件下でヒトIgM対照に混合後のMΔGΔ血清のSuperose6サイズ分画。それぞれの画分を還元条件下でゲル電気泳動により分析した。Superose6カラムから回収された画分は左(高MW)から右(低MW)である。対照は、ヒト血清単独(第1レーン左)およびヒトIgM対照血清(レーンMΔGΔ血清)に混合する前のマウス血清である。サイズマーカーが示される。
【図7】パネルA:破傷風毒素;HSP70、rtTAおよびヒトTNFαに特異的なモノクローナル抗体cDNAの配列。上の配列は生殖細胞系列VHH2配列である。CDR1、2および3ならびにヒンジ領域が配列の上に示されている。異なるアイソタイプおよびクラスは右側に異なる色で示されている。用いられるJ領域は右側に示されている。パネルB:異なる重鎖のみ抗体(ハイブリドーマ、血清およびsdAb)を用いるウエスタンブロットの例。左側のパネル、それぞれ1/100および1/250希釈された、抗rtTA血清およびハイブリドーマ培地。中央のパネル、それぞれ1/200および1/100希釈された、wtおよびGΔマウス由来の抗DKTP血清。右側のパネル、百日咳菌抗原(DKTP)を含むかまたはそれを欠く(DTP、本発明者らは精製された百日咳菌抗原を購入することができなかったため)ワクチンに対する抗百日咳菌sdAb。パネルCおよびD:細胞質においてマーカータンパク質を発現することによりrtTAの存在に応答するマーカープラスミドを追加的にトランスフェクトされたTet−on細胞系統の1つの免疫染色51。パネルCは、rtTAを発現している核を示す(緑色)。パネルDは、rtTAに応答した細胞質中のマーカータンパク質のドキシサイクリン誘導発現(赤色)、および細胞のDAPIでの核染色(青色)を示す。パネルE:抗rtTA抗体のBiaCore分析の例。親和性が示されている。
【図8】パネルA:ラクダ科動物様スプライス変異を含むIg遺伝子座の概略図である。2つのヒトIgG定常領域(Cγ2およびCγ3)を、まず、Gγ2−SおよびGγ3−Sで表されるラクダ科動物IgG HCAbs5においてCH1エキソンのスキッピングをもたらすと考えられるG+1〜A+1のスプライスを変更することにより変異させた。遺伝子座は、2つのラマVHH領域、全てのヒトDおよびJH領域ならびにヒトCμ、CδおよびCγ2およびCγ3およびLCRを含有した(明細書本文も参照のこと)。これらの遺伝子座をμMTトランスジェニックマウスに導入し、ヒト遺伝子座の発現について分析した。パネルB:GSマウス由来の骨髄(BM)ヒトIgGcDNAの配列決定から、両方のVHHが異なるヒトDおよびJセグメントと組み換えられ、Cγ2定常領域で転写されることが示された。しかし、CH1エキソンは、まだ最後の16bpを残して存在し、それを切り出した。進行中であるが、イントロン1の4位におけるA〜Gの移行に起因する、CH1エキソンの同じ潜在スプライス部位が白血病患者において報告された[31]。MGSまたはGS系統のいずれもμMTマウスにおいてB細胞の成長を救済しなかった(図なし)。マウスのB細胞転写/翻訳機構は再構成されたヒトコブラクダVHH−γ2aをプロセシングすることができるが[34、60]、本発明者らのデータは、G〜A突然変異に加えて、その他の特徴がCH1エキソンスキッピングに重要であることを示す。
【図9】実施例3および4に説明される様々な遺伝子座でのヒトVH領域のクローニングの模式図である。
【図10】複数のVH遺伝子セグメント、全D領域、全JH領域、Cγ2、Cγ3およびCx領域および3’LCRを含む重鎖遺伝子座の例を示す図である。
【図11】複数のVH遺伝子セグメント、全D領域、全JH領域、Cγ2、Cγ3およびCx領域および3’LCRを含む重鎖遺伝子座の例を示す図である。
【図12】染色体1に組み込まれた1つの遺伝子座と染色体8に組み込まれた1つの遺伝子座を含むカリオグラムを示す図である。
【特許請求の範囲】
【請求項1】
複数の異種VH重鎖遺伝子座を哺乳類に供与するステップを含み、各VH重鎖遺伝子座が、1つ以上のV遺伝子セグメントと、1つ以上のD遺伝子セグメントと、1つ以上のJ遺伝子セグメントと、発現されたときにCH1ドメインを含まずに前記遺伝子座の少なくとも1つからVH重鎖のみ抗体を発現する重鎖定常領域をコードする遺伝子セグメントとを含む、ヒトを除くトランスジェニック哺乳類におけるVH重鎖のみ抗体の産生方法。
【請求項2】
各VH重鎖遺伝子座が1つ以上のV遺伝子セグメントを含む、請求項1に記載の方法。
【請求項3】
各遺伝子座がV遺伝子セグメントのみを含む、請求項1または請求項2に記載の方法。
【請求項4】
各V遺伝子セグメントが、他の全てのV遺伝子セグメントと異なっている、請求項3に記載の方法。
【請求項5】
各V遺伝子セグメントが他の全てのV遺伝子セグメントと同一である、請求項3に記載の方法。
【請求項6】
各遺伝子座中の残りの遺伝子セグメントが他の全ての遺伝子座中の遺伝子セグメントと同じである、請求項5に記載の方法。
【請求項7】
各遺伝子座中の残りの遺伝子セグメントが他の全ての遺伝子座中の遺伝子セグメントと異なる、請求項5に記載の方法。
【請求項8】
各遺伝子座が複数のV遺伝子セグメントを含む、請求項1または請求項2に記載の方法。
【請求項9】
任意の1つの遺伝子座中のV遺伝子セグメントが全て同じ種の生物に由来する、請求項8に記載の方法。
【請求項10】
任意の1つの遺伝子座中のV遺伝子セグメントが異なる種の生物に由来する、請求項8に記載の方法。
【請求項11】
V遺伝子セグメントがヒト起源のものである、請求項9に記載の方法。
【請求項12】
複数の重鎖遺伝子座が、複数の遺伝子座に分散された、改良された溶解度特性を有する39個の機能性ヒトV遺伝子セグメントおよび操作された前記ヒトV遺伝子セグメントの変異体のうちの、任意の数または任意の組合せを含む、請求項1〜4および8〜11のいずれか一項に記載の方法。
【請求項13】
各異なる重鎖遺伝子座がヒトを除くトランスジェニック哺乳類のゲノム中の単一コピーとして存在する、請求項1〜12のいずれか一項に記載の方法。
【請求項14】
各VH重鎖遺伝子座が1〜40個のD遺伝子セグメントを含む、請求項1〜13のいずれか一項に記載の方法。
【請求項15】
D遺伝子セグメントがヒトD遺伝子セグメントである、請求項1〜14のいずれか一項に記載の方法。
【請求項16】
それぞれのVH重鎖遺伝子座が1〜20個のJ遺伝子セグメントを含む、請求項1〜15のいずれか一項に記載の方法。
【請求項17】
J遺伝子セグメントがヒトJ遺伝子セグメントである、請求項16に記載の方法。
【請求項18】
各VH重鎖遺伝子座が同一のDおよびJ遺伝子セグメントを含有する、請求項1〜17のいずれか一項に記載の方法。
【請求項19】
各VH重鎖遺伝子座が、DおよびJ遺伝子セグメントの異なる組合せを含有する、請求項1〜17のいずれか一項に記載の方法。
【請求項20】
各VH重鎖遺伝子座が1個以上のV遺伝子セグメント、25個の機能性ヒトD遺伝子セグメントおよび6個のヒトJ遺伝子セグメントを含む、請求項1〜19のいずれか一項に記載の方法。
【請求項21】
各VH重鎖遺伝子座が、インビボでエフェクター機能をもたらす少なくとも1つの重鎖定常領域をコードする遺伝子セグメントを含み、定常領域は、発現されたときに、抗体中にCH1ドメインを含まない、請求項1〜20のいずれか一項に記載の方法。
【請求項22】
各遺伝子座が、1つの特定の定常領域をコードする遺伝子セグメントを1つのみ含む、請求項21に記載の方法。
【請求項23】
各遺伝子座が、それぞれ異なる定常領域をコードする、1個よりも多くの遺伝子セグメントを含む、請求項21に記載の方法。
【請求項24】
各遺伝子座が、同一の定常領域をコードする遺伝子セグメントを含有する、請求項21〜23のいずれか一項に記載の方法。
【請求項25】
各遺伝子座が、異なる定常領域コード遺伝子セグメントまたは定常領域コード遺伝子セグメントの異なる組合せを有する、請求項21〜23のいずれか一項に記載の方法。
【請求項26】
重鎖定常領域をコードする各遺伝子セグメントが、Cδ、Cγ1−4、Cμ、CεまたはCα1−2クラスの1個以上の重鎖定常領域エキソンを含むが、但し重鎖定常領域遺伝子セグメントがCH1ドメインを発現しない、請求項21〜25のいずれか一項に記載の方法。
【請求項27】
重鎖定常領域がヒト起源のものである、請求項21〜26のいずれか一項に記載の方法。
【請求項28】
ヒトを除くトランスジェニック哺乳類における2以上の異種のVH重鎖遺伝子座が同一の染色体上に直列に存在する、請求項1〜27のいずれか一項に記載の方法。
【請求項29】
ヒトを除くトランスジェニック哺乳類における1つ以上の異種のVH重鎖遺伝子座が異なる染色体上に存在する、請求項1〜27のいずれか一項に記載の方法。
【請求項30】
ヒトを除くトランスジェニック哺乳類がげっ歯類である、請求項1〜29のいずれか一項に記載の方法。
【請求項31】
げっ歯類がマウスである、請求項30に記載の方法。
【請求項32】
請求項1〜31のいずれか一項に記載の複数の異種のVH重鎖遺伝子座を含む、ヒトを除くトランスジェニック哺乳類。
【請求項33】
請求項32に記載のヒトを除くトランスジェニック哺乳類を抗原で免疫することによる、重鎖のみ抗体の産生方法。
【請求項34】
請求項32に記載のヒトを除くトランスジェニック哺乳類を抗原で免疫するステップと、
B細胞ハイブリドーマを生成するステップと、
抗原特異的な重鎖のみ抗体を発現している細胞を選択するステップと、
抗原特異的な組換え親和性成熟VH重鎖のみ抗体を単離するステップと
を含む、高親和性の抗原特異的VH重鎖のみ抗体の産生方法。
【請求項1】
複数の異種VH重鎖遺伝子座を哺乳類に供与するステップを含み、各VH重鎖遺伝子座が、1つ以上のV遺伝子セグメントと、1つ以上のD遺伝子セグメントと、1つ以上のJ遺伝子セグメントと、発現されたときにCH1ドメインを含まずに前記遺伝子座の少なくとも1つからVH重鎖のみ抗体を発現する重鎖定常領域をコードする遺伝子セグメントとを含む、ヒトを除くトランスジェニック哺乳類におけるVH重鎖のみ抗体の産生方法。
【請求項2】
各VH重鎖遺伝子座が1つ以上のV遺伝子セグメントを含む、請求項1に記載の方法。
【請求項3】
各遺伝子座がV遺伝子セグメントのみを含む、請求項1または請求項2に記載の方法。
【請求項4】
各V遺伝子セグメントが、他の全てのV遺伝子セグメントと異なっている、請求項3に記載の方法。
【請求項5】
各V遺伝子セグメントが他の全てのV遺伝子セグメントと同一である、請求項3に記載の方法。
【請求項6】
各遺伝子座中の残りの遺伝子セグメントが他の全ての遺伝子座中の遺伝子セグメントと同じである、請求項5に記載の方法。
【請求項7】
各遺伝子座中の残りの遺伝子セグメントが他の全ての遺伝子座中の遺伝子セグメントと異なる、請求項5に記載の方法。
【請求項8】
各遺伝子座が複数のV遺伝子セグメントを含む、請求項1または請求項2に記載の方法。
【請求項9】
任意の1つの遺伝子座中のV遺伝子セグメントが全て同じ種の生物に由来する、請求項8に記載の方法。
【請求項10】
任意の1つの遺伝子座中のV遺伝子セグメントが異なる種の生物に由来する、請求項8に記載の方法。
【請求項11】
V遺伝子セグメントがヒト起源のものである、請求項9に記載の方法。
【請求項12】
複数の重鎖遺伝子座が、複数の遺伝子座に分散された、改良された溶解度特性を有する39個の機能性ヒトV遺伝子セグメントおよび操作された前記ヒトV遺伝子セグメントの変異体のうちの、任意の数または任意の組合せを含む、請求項1〜4および8〜11のいずれか一項に記載の方法。
【請求項13】
各異なる重鎖遺伝子座がヒトを除くトランスジェニック哺乳類のゲノム中の単一コピーとして存在する、請求項1〜12のいずれか一項に記載の方法。
【請求項14】
各VH重鎖遺伝子座が1〜40個のD遺伝子セグメントを含む、請求項1〜13のいずれか一項に記載の方法。
【請求項15】
D遺伝子セグメントがヒトD遺伝子セグメントである、請求項1〜14のいずれか一項に記載の方法。
【請求項16】
それぞれのVH重鎖遺伝子座が1〜20個のJ遺伝子セグメントを含む、請求項1〜15のいずれか一項に記載の方法。
【請求項17】
J遺伝子セグメントがヒトJ遺伝子セグメントである、請求項16に記載の方法。
【請求項18】
各VH重鎖遺伝子座が同一のDおよびJ遺伝子セグメントを含有する、請求項1〜17のいずれか一項に記載の方法。
【請求項19】
各VH重鎖遺伝子座が、DおよびJ遺伝子セグメントの異なる組合せを含有する、請求項1〜17のいずれか一項に記載の方法。
【請求項20】
各VH重鎖遺伝子座が1個以上のV遺伝子セグメント、25個の機能性ヒトD遺伝子セグメントおよび6個のヒトJ遺伝子セグメントを含む、請求項1〜19のいずれか一項に記載の方法。
【請求項21】
各VH重鎖遺伝子座が、インビボでエフェクター機能をもたらす少なくとも1つの重鎖定常領域をコードする遺伝子セグメントを含み、定常領域は、発現されたときに、抗体中にCH1ドメインを含まない、請求項1〜20のいずれか一項に記載の方法。
【請求項22】
各遺伝子座が、1つの特定の定常領域をコードする遺伝子セグメントを1つのみ含む、請求項21に記載の方法。
【請求項23】
各遺伝子座が、それぞれ異なる定常領域をコードする、1個よりも多くの遺伝子セグメントを含む、請求項21に記載の方法。
【請求項24】
各遺伝子座が、同一の定常領域をコードする遺伝子セグメントを含有する、請求項21〜23のいずれか一項に記載の方法。
【請求項25】
各遺伝子座が、異なる定常領域コード遺伝子セグメントまたは定常領域コード遺伝子セグメントの異なる組合せを有する、請求項21〜23のいずれか一項に記載の方法。
【請求項26】
重鎖定常領域をコードする各遺伝子セグメントが、Cδ、Cγ1−4、Cμ、CεまたはCα1−2クラスの1個以上の重鎖定常領域エキソンを含むが、但し重鎖定常領域遺伝子セグメントがCH1ドメインを発現しない、請求項21〜25のいずれか一項に記載の方法。
【請求項27】
重鎖定常領域がヒト起源のものである、請求項21〜26のいずれか一項に記載の方法。
【請求項28】
ヒトを除くトランスジェニック哺乳類における2以上の異種のVH重鎖遺伝子座が同一の染色体上に直列に存在する、請求項1〜27のいずれか一項に記載の方法。
【請求項29】
ヒトを除くトランスジェニック哺乳類における1つ以上の異種のVH重鎖遺伝子座が異なる染色体上に存在する、請求項1〜27のいずれか一項に記載の方法。
【請求項30】
ヒトを除くトランスジェニック哺乳類がげっ歯類である、請求項1〜29のいずれか一項に記載の方法。
【請求項31】
げっ歯類がマウスである、請求項30に記載の方法。
【請求項32】
請求項1〜31のいずれか一項に記載の複数の異種のVH重鎖遺伝子座を含む、ヒトを除くトランスジェニック哺乳類。
【請求項33】
請求項32に記載のヒトを除くトランスジェニック哺乳類を抗原で免疫することによる、重鎖のみ抗体の産生方法。
【請求項34】
請求項32に記載のヒトを除くトランスジェニック哺乳類を抗原で免疫するステップと、
B細胞ハイブリドーマを生成するステップと、
抗原特異的な重鎖のみ抗体を発現している細胞を選択するステップと、
抗原特異的な組換え親和性成熟VH重鎖のみ抗体を単離するステップと
を含む、高親和性の抗原特異的VH重鎖のみ抗体の産生方法。
【図1】

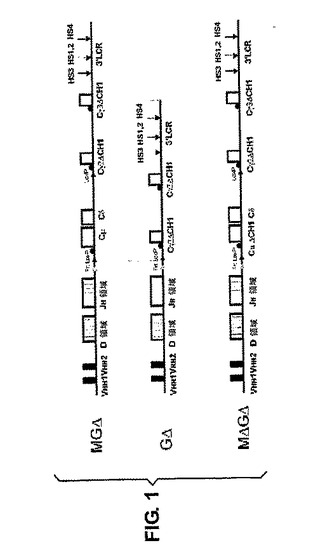
【図2A.2B】
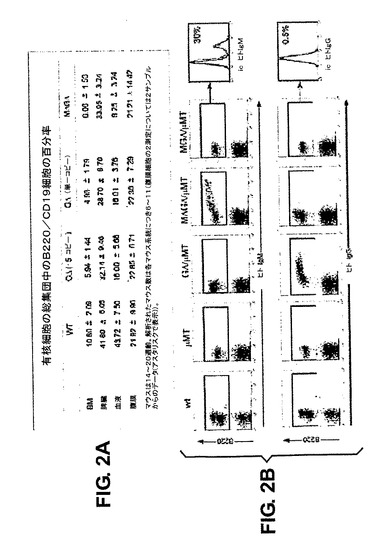
【図2C.2D】
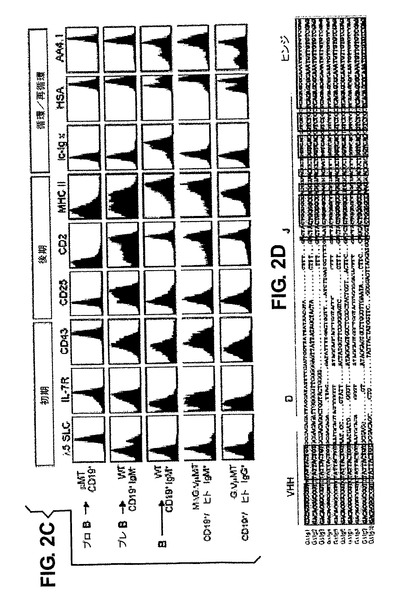
【図3A−3E】
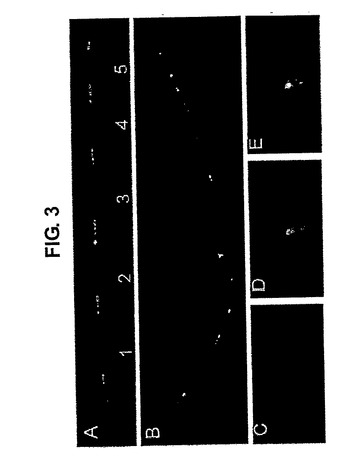
【図3F】
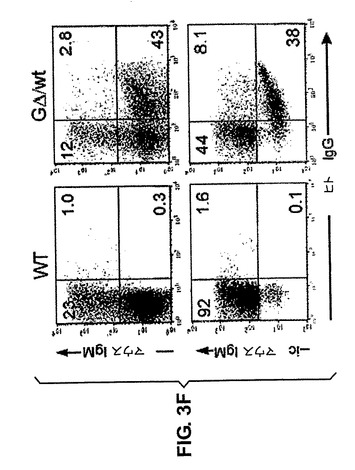
【図4A.4B】

【図4C】
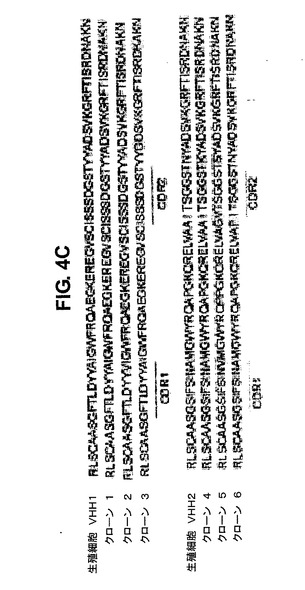
【図4D】
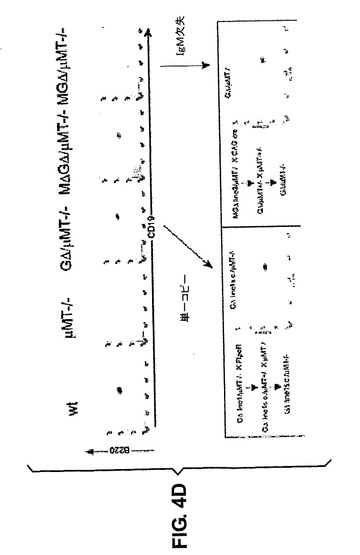
【図5】
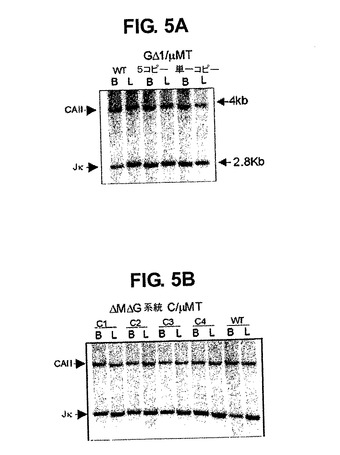
【図6】
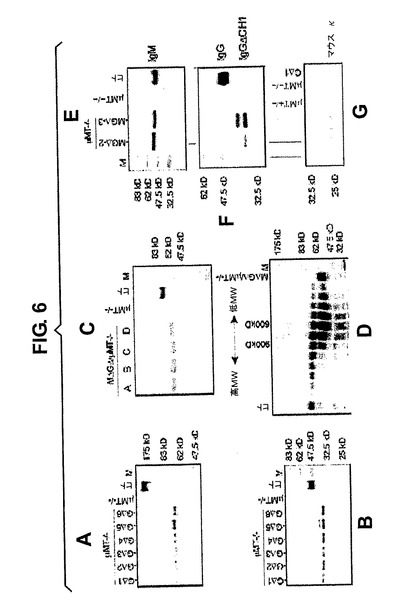
【図7A】
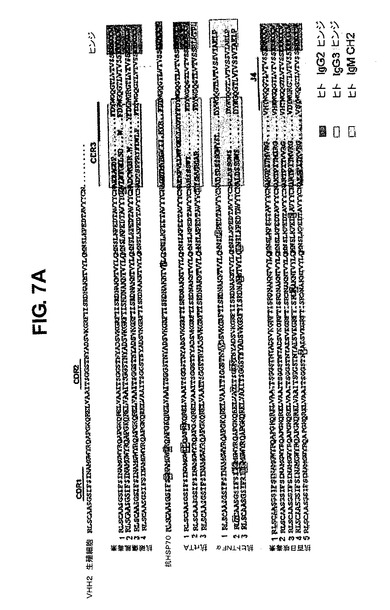
【図7B−7E】
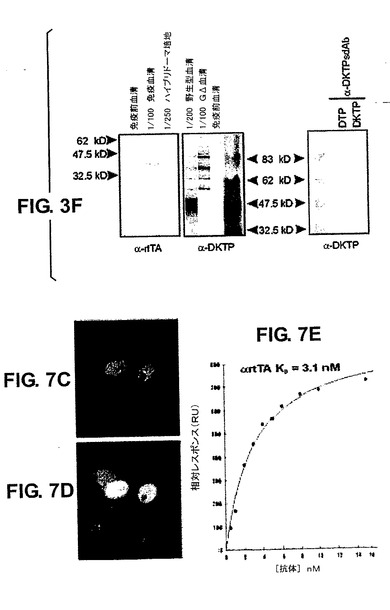
【図8】
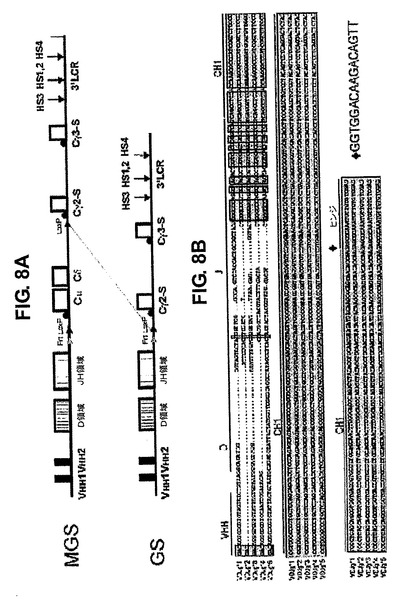
【図9】
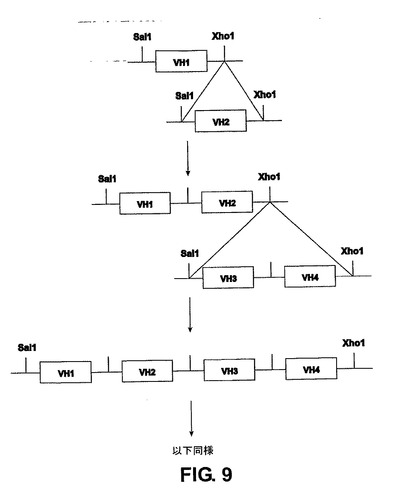
【図10】
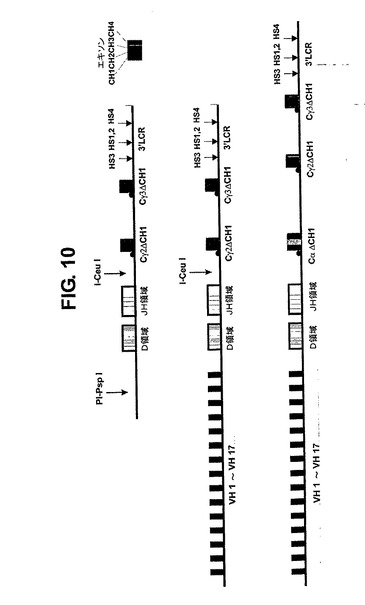
【図11】
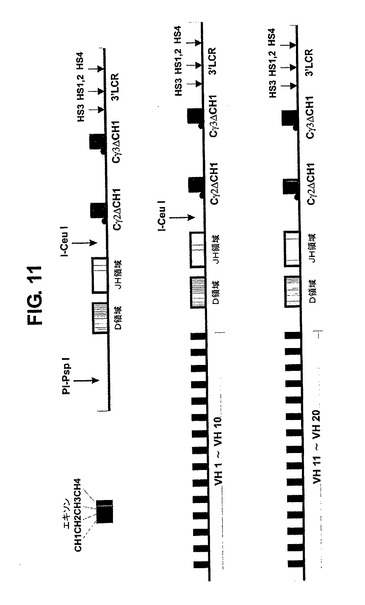
【図12】

【図2A.2B】
【図2C.2D】
【図3A−3E】
【図3F】
【図4A.4B】
【図4C】
【図4D】
【図5】
【図6】
【図7A】
【図7B−7E】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2009−524641(P2009−524641A)
【公表日】平成21年7月2日(2009.7.2)
【国際特許分類】
【出願番号】特願2008−551908(P2008−551908)
【出願日】平成19年1月25日(2007.1.25)
【国際出願番号】PCT/IB2007/001491
【国際公開番号】WO2007/096779
【国際公開日】平成19年8月30日(2007.8.30)
【出願人】(508226300)エラスムス・ユニヴァーシティ・メディカル・センター・ロッテルダム (2)
【Fターム(参考)】
【公表日】平成21年7月2日(2009.7.2)
【国際特許分類】
【出願日】平成19年1月25日(2007.1.25)
【国際出願番号】PCT/IB2007/001491
【国際公開番号】WO2007/096779
【国際公開日】平成19年8月30日(2007.8.30)
【出願人】(508226300)エラスムス・ユニヴァーシティ・メディカル・センター・ロッテルダム (2)
【Fターム(参考)】
[ Back to top ]