
対立遺伝子特異的RNA干渉
優性の機能獲得型遺伝子によって引き起こされるヒト疾患は、遺伝子の1つの突然変異体と1つの野生種コピーを有するヘテロ接合体に生じる。前記野生種遺伝子は重要な機能を果たすことが多い一方で、突然変異遺伝子は有毒であることが多いため、いずれの治療戦略も野生種遺伝子の発現を維持しながら突然変異体を選択的に阻害しなければならない。本願発明には、RNAiを用いた同時発現した野生種対立遺伝子の発現を保護しながら突然変異対立遺伝子の発現を特異的に阻害する方法であって、優性の機能獲得遺伝子突然変異に関連する遺伝子障害を治療する治療戦略が含まれる。本願発明にはまた、遺伝性筋萎縮性側策硬化症(ALS)を引き起こす、銅亜鉛スーパーオキシドジスムターゼ(SOD1)の、野生種ではなく突然変異種の発現を選択的に抑制する、低分子干渉RNA(siRNA)および低分子ヘアピンRNA(shRNA)が含まれる。
【発明の詳細な説明】
【背景技術】
【0001】
関連出願
本願明細書は、2002年11月4日に提出された米国特許仮出願第60/423,507号「Allele-Specific RNA Interference(対立特異的RNA干渉)」、および2003年7月18日に提出された米国特許仮出願60/488,283号「Allele-Specific RNA
Interference(対立特異的RNA干渉)」の利益を主張する。参照された特許仮出願の内容全体は、引用をもって本願明細書に援用するものとする。
【0002】
連邦政府の資金による研究に関する記載
米国政府は、国立衛生研究所(NIH)により授与された認可番号GM62862およびGM53874、ならびに国立神経疾患・卒中研究所(NINDS)により授与された認可番号NS35750にしたがった本願発明の一定の権利を有する可能性がある。
【0003】
背景
優性の機能獲得型遺伝子によって引き起こされる疾患は、1つの突然変異体と1つのその遺伝子の野生種コピーを有するヘテロ接合体に生じる。この種の疾患で最も知られるものの一部は、共通の神経変性疾患で、アルツハイマー病、ハンチントン病、パーキンソン病、および筋萎縮性側策硬化症(ALS; 「ルー・ゲーリック病」)が含まれる(Taylor
et al., 2002)。これらの疾患では、突然変異タンパク質が細胞変性を引き起こす正確な経路が明らかになってはいないが、突然変異タンパク質となる細胞毒性の起源は知られている。
【0004】
SOD1における突然変異は運動ニューロンの変性を生じて、ALSを引き起こす。それは、前記突然変異タンパク質がいくらかの毒性を獲得したためである(Cleveland et al., 2001)。この毒性の性質も、最終的に運動ニューロンの変性に至る下流経路も解明されていない。マウスでは、突然変異体SOD1の発現だけでなく、遺伝子ノックアウトによるSOD1の削除がALSを引き起こす。しかし、遺伝子ノックアウトマウスは、繁殖性の減退(Matzuk et al., 1990)、運動軸索障害(Shefner et al., 1999)、蝸牛有毛細胞の加齢に関連する喪失(McFadden et al., 2001)、および神経筋接合部シナプス(Flood et al.,
1999)を含む様々な異常、ならびに、CNSおよびその他の系への、興奮毒性、虚血、神経毒、および放射線照射など各種の有害な攻撃への高い感受性を生じる。突然変異種の毒性、および野生種タンパク質の機能的重要性を考えると、この疾患のための理想的な治療法は、前記突然変異タンパク質の発現を選択的に阻害する一方で野生種の発現を維持することである。
【0005】
要約
本願発明は、優性の機能獲得型疾患を治療する新規の方法に関連する。特に、本願発明は、機能獲得型遺伝子から転写された突然変異mRNAを選択的に破壊し、それによりそのような遺伝子によってコードされる突然変異タンパク質の産生を抑制する方法を提供する。本願発明は、低分子干渉RNA(siRNA)および低分子ヘアピンRNA(shRNA)の両方のがたとえばG85R SOD1またはG93A SOD1などの突然変異対立遺伝子の発現を選択的に阻害する一方で、1ヌクレオチド特異性で前記野生種タンパク質の発現を阻害するようにデザインすることができるという発見に、部分的に基づいている。
【0006】
本願発明の方法は、たとえば、筋萎縮性側索硬化症(ALS)に関連する銅亜鉛スーパーオキシドジスムターゼ(SOD1)遺伝子にある点突然変位など、突然変異遺伝子における単一の対立遺伝子に生じる選択的な点変異に対して、RNA干渉技術(RNAi)を用いる。RNAiは多様な真核生物の遺伝子発現を、標的遺伝子への配列類似性を有する短い2本鎖RNA(低分子干渉RNAまたはsiRNAとよばれる)を導入することによって配列選択的に抑制するのを媒介する(Caplen et al.,
2001; Elbashir et al.2001c)。本願発明の2本鎖siRNAまたはshRNA発現ベクタを用いて、たとえばSOD1突然変異タンパク質などの毒性突然変異遺伝子の発現を選択的にサイレンシングし、それにより野生種SOD1対立遺伝子を継続的に機能させることができる。
【0007】
本願発明はまた、たとえばin vivoなどにおいて細胞で発現する場合、細胞自体のRNAi経路を用いて標的遺伝子の突然変異対立遺伝子を選択的にサイレンシングする標的siRNAを生じるように細胞によって処理された、新規の人為的操作RNA前駆体の発見に基づく。これらの操作したRNA前駆体をコードする核酸分子を、in vivoにおいて適切な制御配列(たとえば、プラスミドなどのベクタにある導入遺伝子)を用いて細胞に導入することによって、操作したRNA前駆体の発現は、時間的および空間的に、すなわち、特定の時間および/または特定の組織、器官、または細胞において選択的に制御することができる。
【0008】
ある局面では、本願発明は、遺伝子の少なくとも2つの異なる対立遺伝子を含む、細胞中の標的対立遺伝子の発現を、前記標的対立遺伝子に特異的なsiRNAを前記細胞に投与することによって阻害する方法を特徴とする。ある実施態様では、前記標的対立遺伝子は、優性の機能獲得型突然変異に関連する障害と相関する。別の実施態様では、前記障害は筋萎縮性軸索硬化症、ハンチントン病、アルツハイマー病、またはパーキンソン病である。
【0009】
別の局面では、本願発明は、優性の機能獲得型突然変異対立遺伝子の存在と相関する疾患を有する対象の治療方法であって、当該方法が前記突然変異対立遺伝子に特異的なsiRNAを治療上有効な量、前記対象に投与するステップを含む方法を特徴とする。ある実施態様では、前記siRNAは機能獲得型突然変異を標的とする。別の実施態様では、前記障害は筋萎縮性軸索硬化症、ハンチントン病、アルツハイマー病、またはパーキンソン病である。
【0010】
ある実施態様では、前記疾患は筋萎縮性軸索硬化症である。さらなる実施態様では、前記対立遺伝子はSOD1突然変異対立遺伝子である。
【0011】
ある実施態様では、前記siRNAは突然変異SOD1対立遺伝子(配列番号8)を標的とし、好ましくはP10(配列番号4)、次に好ましくはP9(配列番号2)、次に好ましくはP11(配列番号6)を有する図1Aに記載の突然変異siRNA配列を含む、またはそれから成る。
【0012】
別の実施態様では、前記siRNA(たとえば制御siRNA)は野生種SOD1対立遺伝子を標的とし、好ましくはP9(配列番号14)またはP10(配列番号12)、次に好ましいのはP11(配列番号10)を有する図1Aに記載の野生種siRNA配列を含む、またはそれから成る。
【0013】
別の局面では、本願発明は図1Aに記載の配列を含むsiRNAを提供する。
【0014】
別の局面では、本願発明は、図1Aに記載の配列(配列番号4)を含むp10突然変異siRNAを提供する。
【0015】
別の局面では、本願発明は、図1Aに記載の配列(配列番号2)を含むp9突然変異siRNAを提供する。
【0016】
別の局面では、本願発明は、図3Aに記載の配列(配列番号16)を含むG93A SOD1 shRNA、および本願発明のshRNAを含む発現作製物を提供する。
別の局面では、本願発明は本願発明のsiRNAおよび/またはshRNAを含む治療用組成物、および薬学的に許容な担体を提供する。
【0017】
本願発明のその他の特徴および利点は、以下の発明の詳細な説明および特許請求の範囲から明らかになるであろう。
【0018】
発明の詳細な説明
銅亜鉛スーパーオキシドジスムターゼ(SOD1)遺伝子の突然変異は、筋萎縮性側策硬化症のサブセット、運動ニューロンの変性を来す神経変性疾患、麻痺および死を引き起こす(Brown and Robberecht, 2001;Siddique and Lalani, 2002)。突然変異SOD1が毒性を獲得することによって運動ニューロン変性を引き起こすことは、既知である。しかし、この毒性の分子機序も、運動ニューロンの死滅を来す機序も、いずれも解明されていない。疾患の機序の理解が不十分なことから、合理的なデザインの治療法が着実で有効な転帰を生じたことがない。その一方で、運動ニューロンを死滅させる毒性が突然変異したタンパク質に起因することから(Cleveland and Rothstein, 2001)、突然変異タンパク質の減少によって、その疾患を緩和または予防できるはずである。RNA干渉(RNAi)技術を用いてこの目的を達成することができる。
【0019】
本願発明は、siRNAおよびshRNAが突然変異対立遺伝子の発現を選択的に阻害できるという発見であって、たとえばALSと相関するSOD1の突然変異など、特定の突然変異についてのケースに見られるとおり、突然変異mRNAと野生種と違いが1ヌクレオチドだけの場合であっても阻害できるという発見に基づく。これらの方法は、ALSに限定しないがそれを含む、優性で機能獲得型の遺伝子突然変異によって生じる疾患の治療に適用することができる。本願発明のsiRNAは、1ヌクレオチドの識別、およびその標的遺伝子の発現の選択的下方制御を可能にする。
【0020】
本願発明の方法は、たとえば、筋萎縮性側索硬化症(ALS)に関連する銅亜鉛スーパーオキシドジスムターゼ(SOD1)遺伝子の中ある点突然変位など、突然変異遺伝子における単一の対立遺伝子に生じる選択的な点変異に対して、RNA干渉技術(RNAi)を用いる。RNAiは多様な真核生物の遺伝子発現を、標的遺伝子への配列相同性を有する短い2本鎖RNA(低分子干渉RNAまたはsiRNAとよばれる)を導入することによって配列選択的に抑制するのを媒介する(Caplen et al.,
2001; Elbashir et al.2001c)。本願発明の2本鎖siRNAまたはshRNA発現ベクタを用いて、たとえばSOD1突然変異タンパク質などの毒性突然変異遺伝子の発現を選択的にサイレンシングし、それにより野生種SOD1対立遺伝子を継続的に機能させることができる。
【0021】
遺伝子発現の配列選択的、転写後不活性化は、RNAiと呼ばれる現象である、標的遺伝子に相当する2本鎖RNAの導入によって、多様な原核生物に実現させることが可能である(Hutvagner and Zamore, 2002; Hannon, G. J., 2002; McManus and Sharp,
2002)。RNAiの方法論は、培養哺乳類細胞にも拡張されている(Caplen et al, 2001; Elbashir
et al., 2001)。このアプローチは、RNAi経路の中間体であるsiRNAが、当該siRNA配列に相当するmRNAの分解を引き起こすことがあるという発見を活用している。さらに、in vivoにおいて転写されたshRNAは、shRNAステムの配列に相補的な標的RNAの分解を引き起こすことがある。それは、細胞内でshRNAがsiRNAに加工されるためである(Paul et al., 2002; Lee et al., 2002;
Paddison et al., 2002; Sui et al., 2002; Yu et al., 2002; McManus et al., 2002;
Zeng et al., 2002; Brummelkamp et al., 2002; Miyagishi et al., 2002; Jacque et al.,
2002)。本願出願人らは、siRNA 2本鎖またはshRNAを発現するウイルスを用いて、突然変異体率遺伝子の発現を選択的に阻害しながら、同時に発現する野生種対立遺伝子の発現を保護することができることを実証する。
【0022】
ALS関連SOD1突然変異の大半は、1アミノ酸の変化を生じる1ヌクレオチド点変異である(ALS遺伝子(SOD1, ALS など)突然変異のALSオンラインデータベース)。したがって、突然変異体の発現だけを選択的にサイレンシングを生じさせ、野生種にはサイレンシングを生じさせないために、1ヌクレオチド特異性が必要である。本願出願人らは、哺乳類細胞において1ヌクレオチドの認識が実現可能であることを証明している。
【0023】
本願発明をより容易に理解するため、まず特定の用語を定義する。
【0024】
「単離された核酸分子または配列」は、由来する生物の天然のゲノムにおいて、(片方は5’末端でもう片方が3’末端に)ただちに連続するコード配列の両側に、ただちに連続しない核酸分子または配列である。したがって、当該用語には、たとえば、ベクター、自己複製プラスミドもしくはウイルス、または原核生物もしくは真核生物のゲノムDNAに組み込まれた、または分離可能な分子として存在する組換えDNAまたはRNAが含まれる。また、さらなるポリペプチド配列をコードするハイブリッド遺伝子の一部分である組換えDNAも含まれる。
【0025】
「ヌクレオシド」という用語は、リボースまたはデオキシリボース糖に共有結合したプリンまたはピリミジンを有する分子を意味する。例示的なヌクレオチドには、アデノシン、グアシン、シチジン、ウリジン、およびチミジンが含まれる。さらなる例示的なヌクレオシドには、イノシン、1-メチルイノシン、シュードウリジン、5,6-ジヒドロウリジン、リボチミジン、2N-メチルグアノシン、および2,2N,N-ジメチルグアノシン(「稀少」ヌクレオチドともよばれる)が含まれる。「ヌクレオチド」という用語は、糖部分にエステル結合したリン酸基を1つ以上有するヌクレオシドを意味する。例示的なヌクレオシドには、1リン酸ヌクレオシド、2リン酸ヌクレオシド、および3リン酸ヌクレオシドが含まれる。「ポリヌクレオチド」および「核酸分子」という用語は、本願明細書では交換可能に用いられ、5’および3’炭素原子の間のホスホジエステル結合によって結合したヌクレオシドのポリマーを意味する。
【0026】
「RNA」または「RNA分子」または「リボ核酸分子」という用語は、リボヌクレオチドのポリマーを意味する。「DNA」または「DNA分子」または「デオキシリボ核酸分子」という用語は、デオキシリボヌクレオチドのポリマーを意味する。DNAおよびRNAは自然に合成することができる(たとえば、それぞれDNA複製またはDNAの転写によって)。RNAは転写後に修飾されることがある。DNAおよびRNAは化学的にも合成することができる。DNAおよびRNAは1本鎖であっても(すなわちそれぞれssRNAおよびssDNA)、または多重鎖(たとえば2本鎖、すなわちそれぞれdsRNAおよびdsDNA)であってもよい。「mRNA」または「メッセンジャRNA」は、1つ以上のポリペプチド鎖のアミノ酸配列を特定する1本鎖RNAである。この情報は、リボソームがmRNAに結合する際にタンパク質合成中に翻訳される。
【0027】
組換えRNA前駆体、または組換え核酸分子の中などに使われる「組換え」という用語は、その前駆体または分子が天然には認められず、その前駆体または分子の核酸配列全体または一部分が人工的に作成された、または選択されたものであることを示す。ひとたび作製または選択されると、その配列は、細胞内のメカニズムによって複製、翻訳、または転写などの処理を受けることができる。したがって、組換え核酸分子を含む導入遺伝子から細胞内で産生されたRNA前駆体は組換えRNA前駆体である。
【0028】
本願明細書に記載の「低分子干渉RNA」(「siRNA」)という用語(「短分子干渉RNA」ともいう)は、RNA干渉を方向付けるまたは媒介する能力のある約10乃至50ヌクレオチドを含むRNA(またはRNA類似体)を意味する。好ましくは、siRNAは、約15乃至30ヌクレオチドまたはヌクレオチド類似体、さらに好ましくは約16乃至25ヌクレオチド(またはヌクレオチド類似体)、よりさらに好ましくは約18乃至23ヌクレオチド(またはヌクレオチド類似体)および、よりさらに好ましくは約19乃至22ヌクレオチド(またはヌクレオチド類似体)(たとえば、19、20、21、または22ヌクレオチドまたはヌクレオチド類似体)を含む。「短分子」siRNAとは、たとえば19、20、21、または22ヌクレオチドなど、約21ヌクレオチド(またはヌクレオチド類似体)を含む、siRNAを意味する。「長分子」siRNAとは、たとえば23、24、24、または26ヌクレオチドなど、約24乃至25ヌクレオチド(またはヌクレオチド類似体)を含む、siRNAを意味する。短いsiRNAほどRNAiを媒介する能力を維持しているとすると、短分子siRNAには、たとえば16、17、または18ヌクレオチドなど、19ヌクレオチド未満が含まれてよい。同様に、長分子siRNAは、短分子siRNAに対して、たとえば酵素的処理など、RNAiが不在のさらなる処理を媒介する能力を維持しているとすると、ある場合には、長分子siRNAには26ヌクレオチド超が含まれてもよい。
【0029】
「ヌクレオチド類似体」または「改変ヌクレオチド」または「修飾ヌクレオチド」という用語は、天然に存在しないリボヌクレオチドまたはデオキシリボヌクレオチドを含む、標準的ではないヌクレオチドを意味する。好ましいヌクレオチド類似体は、任意の位置を修飾し、ヌクレオチドの特定の化学的特性を変えつつ、核酸類似体の目的の機能を実行する能力を維持できるようにする。誘導可能なヌクレオチドの位置の例には、たとえば5-(2-アミノ)プロピルウリジン、5-ブロモウリジン、5-プロピンウリジン、5-プロペニルウリジンなどの5位、たとえば6-(2-アミノ)プロピルウリジンなどの6位、たとえば8-ブロモグアノシン、8-クロログアノシン、8-フルオログアノシンなどのアデノシンおよび/またはグアノシンの8位が含まれる。ヌクレオチド類似体も、たとえば7-デアザアデノシンなどのデアザヌクレオチド、O-およびN修飾(たとえばアルカリ化、たとえばN6-メチルアデノシン、またはその他の当業に知られる)ヌクレオチド、およびたとえばHerdewijn, Antisense Nucleic Acid Drug
Dev., 2000 Aug. 10(4):297-310に記載されているなどのその他のヘテロ周期性修飾ヌクレオチド類似体も含まれる。
【0030】
ヌクレオチド類似体は、ヌクレオチドの糖部分への修飾も含んでもよい。たとえば、2’OH基はH、OR、R、F、Cl、Br、I、SH、SR、NH2、NHR、NR2、COOR、またはORであって、当該化学基においてRは置換または非置換C1乃至C6
アルキル、アルケニル、アルキニル、アリールなどから選択される化学基と置換してもよい。その他の可能な修飾には、米国特許第5,858,988号および6,291,438号に記載されるものが含まれる。
【0031】
ヌクレオチドのリン酸基は、たとえばリン酸基の1つ以上の酸素を硫黄で置換する(たとえばホスホロチオ酸)ことにより、またはヌクレオチドがたとえばEckstein, Antisense Nucleic Acid Drug Dev. 2000
Apr. 10(2):117-21、
Rusckowski et al. Antisense Nucleic Acid Drug Dev. 2000
Oct. 10(5):333-45、 Stein, Antisense
Nucleic Acid Drug Dev. 2001 Oct. 11(5): 317-25、 Vorobjev et al. Antisense
Nucleic Acid Drug Dev. 2001 Apr. 11(2):77-85、 および米国特許第5,684,143号などに記載される目的の機能を実施できるようなその他の置換を行うことによって修飾してもよい。上に参照した修飾の一部は、好ましくは、たとえばin vivo またはin vitroにおいて前記類似体を含むポリヌクレオチドの加水分解速度を低減させる。
【0032】
本願明細書に記載の、たとえばsiRNA2本鎖またはsiRNA配列のアンチセンス鎖など、siRNAまたはRNAi剤の「アンチセンス鎖」という用語は、たとえばサイレンシングのターゲットである遺伝子のmRNAの約15乃至30、16乃至25、18乃至23または19乃至22ヌクレオチドなど、 約10乃至50ヌクレオチドの部分に実質的に相補的な鎖を意味する。第1の鎖のアンチセンス鎖は、たとえばRNAi機構またはプロセスによる望ましい標的mRNAの破壊を引き起こすのに十分な相補性など、標的特異的RNA干渉(RNAi)を方向付ける望ましい標的mRNA配列に十分相補的な配列を有する。たとえばsiRNA2本鎖またはsiRNA配列のアンチセンス鎖など、siRNAまたはRNAi剤の「センス鎖」または「第2の鎖」という用語は、アンチセンス鎖または第1の鎖に相補的な鎖を意味する。アンチセンスまたはセンス鎖は、第1鎖または第2鎖とも呼ばれ、標的配列に相補性を有する第1鎖または第2鎖、および前記第1鎖または第2鎖に相補性を有する第2鎖または第1鎖である。
【0033】
本願明細書に記載の「ガイド鎖」という用語は、たとえばsiRNA2本鎖またはsiRNA配列のアンチセンス鎖など、RNAi剤の鎖であって、RISC複合体に進入し、標的mRNAの切断を方向付ける、鎖を意味する。
【0034】
本願明細書に記載のアンチセンス鎖の5’末端としての「5’末端」は、たとえばアンチセンス鎖の5’末端における1乃至約5ヌクレオチドなどの5’末端ヌクレオチドを意味する。本願明細書に記載のセンス鎖の3’末端としての「3’末端」は、たとえば相補的アンチセンス鎖の5’末端のヌクレオチドに相補的な1乃至約5ヌクレオチドの領域などの、領域を意味する。
【0035】
「オリゴヌクレオチド」という用語は、ヌクレオチドおよび/またはヌクレオチド類似体の短いポリマーを意味する。「RNA類似体」という用語は、相当する未改変または未修飾RNAと比較して少なくとも1つの改変またな修飾ヌクレオチドを有するが、前記の相当する未改変または未修飾RNAと同一のまたは類似する性質または機能を保持する、ポリヌクレオチド(たとえば化学合成されたポリヌクレオチドなど)を意味する。上述の通り、前記オリゴヌクレオチドは、ホスホジエステル結合を有するRNA分子と比較してRNA類似体の加水分解が低速度になる結合で結合していてよい。たとえば、前記類似体のヌクレオチドはメチレンジオール、エチレンジオール、オキシメチルチオ、オキシエチルチオ、オキシカルボニルオキシ、ホスホロジアミド酸、ホホロアミド酸、および/またはホスホロチオ酸結合を含んでよい。好ましいRNA類似体には、糖および/または骨格修飾リボヌクレオチドおよび/またはデオキシリボヌクレオチドが含まれる。そのような改変または修飾にはさらに、RNA末端または内部などへの非ヌクレオチド物質の付加(RNAの1つ以上のヌクレオチド)が含まれる。RNA類似体は、RNA干渉を媒介する能力を有する天然のRNAに十分類似しているだけでよい。
【0036】
本願明細書に記載の通り、「RNA干渉」(「RNAi」)という用語は、RNAの選択的な細胞内分解を意味する。RNAi は、細胞内で外来のRNA(たとえばウイルス性RNA)を除去するため自然にに生じる。天然のRNAiは、分解メカニズムをその他の同様のRNA配列に方向付ける遊離dsRNAから切断された断片によって進行する。代替的には、RNAiは、たとえば標的遺伝子の発現をサイレンシングさせるなど、人工的に開始させることもできる。
【0037】
「標的特異的RNA干渉(RNAi)を方向付ける望ましい標的mRNA配列に十分相補的な配列」である鎖を有するRNAi作用剤は、その鎖がRNAi機構またはプロセスによる、標的mRNAの破壊を惹起するのに十分な配列を有することを意味する。
本願明細書に記載の「単離されたRNA」(たとえば「単離されたsiRNA」または「単離されたsiRNA前駆体」)という用語は、組換え技術によって生成した場合には他の細胞物質または培養培地を実質的に含まない、または化学的に合成された場合には、化学前駆体またはその他の化学物質を実質的に含まない、RNA分子を意味する。
【0038】
「標的遺伝子」は、発現が選択的に阻害または「サイレンシング」されている遺伝子である。このサイレンシングは、細胞のRNAiシステムによって人工的に作製したRNAからつくり出されたsiRNAによる、標的遺伝子のmRNAの切断によって実現する。RNA前駆体の2本鎖ステムの一部分またはセグメントは、標的遺伝子のmRNAの約18乃至約40ヌクレオチド、またはそれ以上のヌクレオチドの部分に、たとえば完全に相補的など、相補的なアンチセンス鎖である。
【0039】
本願明細書に記載の「導入遺伝子」という用語は、細胞内に人為的に挿入した任意の核酸分子を意味する。そのような導入遺伝子には、トランスジェニック生物に部分的または完全に非相同的(すなわち外来性)である遺伝子を含むこともある。「導入遺伝子」という用語はまた、たとえばDNAなどの選択された1つ以上の核酸配列であって、その核酸配列は、たとえば動物などのトランスジェニック生物に発現させるために1つ以上の作製されたRNA前駆体をコードする核酸配列であって、その生物は部分的または完全にトランスジェニック動物に非相同的または外来性であるか、またはトランスジェニック動物の内因性遺伝子に相同的であるが、天然遺伝子とは異なる部分にある前記動物のゲノムに挿入するようにデザインされたトランスジェニック生物である、核酸配列を意味する。導入遺伝子には、1つ以上のプロモータ、および特定の核酸配列の発現に必要なイントロンなどのその他の任意のDNAであって、すべて特定の配列に機能的に連結されているプロモータまたはその他の任意のDNAが含まれ、およびエンハンサ配列が含まれてもよい。
【0040】
疾患または障害に「関与する」遺伝子には、前記疾患または障害もしくは前記疾患または障害の少なくとも1つの症状に作用するまたはそれを生じる、正常なまたは異常な発現または機能の遺伝子が含まれる。
【0041】
「発現の対立遺伝子特異的阻害」とは、たとえば両方の対立遺伝子が同一の細胞に存在する場合など、1つの対立遺伝子の発現をもう1つの対立遺伝子の発現よりも著しく阻害する能力を意味する。たとえば、その対立遺伝子は1、2、3またはそれ以上のヌクレオチドが異なっていてもよい。ある場合には、1つの対立遺伝子は、たとえば、優性の機能獲得型突然変異に相関する疾患など、疾患の原因に関連する。
【0042】
本願明細書に記載の「機能獲得型突然変異」という用語は、遺伝子中の任意の突然変異であって、当該突然変異において、前記遺伝子によってコードされたタンパク質(つまり突然変異タンパク質)は、通常タンパク質(つまり野生種タンパク質)に関連していない機能を獲得するか、または疾患もしくは障害を引き起こすまたはそれに寄与する機能を獲得する、突然変異を意味する。機能獲得突然変異は、コードされたタンパク質の機能に変化を生じる、遺伝子中の1つ以上のヌクレオチドの欠失、付加、または置換であってよい。ある実施態様では、機能獲得突然変異は前記突然変異の機能を変更するか、またはその他のタンパク質と相互作用を引き起こす。別の実施態様では、機能獲得突然変異は、たとえば改変した突然変異タンパク質と前記正常野生種タンパク質との相互作用によって、正常野生種タンパク質の減少、または除去を引き起こす。
【0043】
「細胞または組織における遺伝子機能の検証」という文言は、そこから生じる発現、活性、機能、または表現型の検証または研究を意味する。
【0044】
本願発明の様々な方法論には、本願明細書では「適切な対照」とも交換可能な意味である「好適な対照」と、価値、レベル、特色、特徴、性質などとの比較に関与するステップを含む。「好適な対照」または「適切な対照」は、比較の目的で有用な当業者が精通している任意の対照または標準である。ある実施態様では、「好適な対照」または「適切な対照」は、本願明細書に記載の通り、RNAi方法論を実行する前に決定する、価値、レベル、特色、特徴、性質などである。たとえば、転写速度、mRNAレベル、翻訳速度、タンパク質レベル、生物学的活性、細胞の特徴または性質、ゲノタイプ、表現型などは、細胞または生物に本願発明のRNAiを導入する前に決定することができる。別の実施態様では、「好適な対照」または「適切な対照」は、細胞、またはたとえば正常な形質を示す対象、または正常細胞もしくは生物などの生物において決定される価値、レベル、特色、特徴、性質などである。さらに別の実施態様では、「好適な対照」または「適切な対照」は、予め決定された価値、レベル、特色、特徴、性質などである。
【0045】
他に定義のない限り、本願明細書に用いられたすべての技術および科学用語は、本願発明が属する当業者によって普通に理解されるものと同じ意味を有する。本願明細書に記載のものに類似するまたはそれに相当する方法および材料は、本願発明の実施またはテストに用いることができ、好適な方法および材料を以下に説明する。 本願明細書に引用されるすべての出版物、特許出願、特許、およびその他の参考文献は、その全体を引用によりここに援用する。紛争の場合、定義を含む本願明細書が支配するだろう。さらに、当該材料、方法、および例は、実例のみで、制限を意図しない
【0046】
本願発明のさまざまな局面は、以下のサブセクションでさらに詳細に説明する。
【0047】
I. 筋萎縮性側策硬化症(ALS)
筋萎縮性側策硬化症(ALS)は、ルー・ゲーリック病としても知られ、大脳皮質、脳幹、および脊髄の運動ニューロンが関与する、進行性、致死性の神経変性障害である(Hirano, A., 1996, Neurology 47 (Suppl. 2), S63-S66)。この疾患は、たとえばSOD1など、前記遺伝子の1つの突然変異種と1つの野生種コピーを有する人間において生じる、優性の機能獲得型遺伝子によって引き起こされる。SOD1突然変異を引き起こすALSは、前記タンパク質中の1つのアミノ酸を変化させる1ヌクレオチド点突然変異である。この疾患はさらに、進行性の運動ニューロン変性によって特徴づけられ、麻痺、運動神経および呼吸機能の完全喪失、および最終的には最初の臨床徴候の出現から2乃至8年(発症後の平均期間は3年)に死に至るALSは、患者の10%が遺伝子に由来しており、症例の90%が特発性である。染色体21 q22-1に位置する銅亜鉛スーパーオキシドジスムターゼ(SOD1)をコードする遺伝子における点突然変異は、家族性の症例の20%の病変に関与している(Rosen et al., Mutations in Cu/Zn
superoxide dismutase gene are associated with familial amyotrophic lateral sclerosis,
Nature, 362, 59-62, 1993, review in Rowland, Amyotrophic lateral sclerosis:
Human challenge for neuroscience, Proc. Natl. Acad. Sci. USA, 92, 1251-1253,
1995)。したがって、欠陥のあるSOD1は、運動ニューロン死に関連しており、家族性筋萎縮性側策硬化症の理解および可能な治療についての示唆を有する。
【0048】
II. SOD-1遺伝子
SOD1は銅原子1個、亜鉛原子1個を含む金属酵素で、ホモ二量体として細胞質の中に存在している。銅は酵素活性に必要であり、その一方で亜鉛はタンパク質の構造を安定化させる(Hirano, A. 1996, Neurology 47 (Suppl. 2). S63-S66)。SOD1はすべての真核細胞で発現し、マンガン依存性ミトコンドリアSOD(SOD2)および銅/亜鉛細胞外(SOD3)を含む、3種類のSOD酵素のファミリの1種類である(I Fridovich, 1986,
"Superoxide dismutases," Advances in Enzymology 58: 61-97)。SOD1の主な天然の機能は、スーパーオキシドジスムターゼで、スーパーオキシド(O2-)が過酸化水素(H2O2)および酸素に転換する。下流酵素のカタラーゼおよびグルタチオンペルオキシダーゼ(H2O2 を水と酸素に転換する)とともに、SOD1は細胞のフリーラジカルを解毒する。この機能の重要性は、生殖性の減退(Matzuk et al., 1998)、運動軸索障害(Shefner et al., 1999)、蝸牛有毛細胞の加齢に関連する喪失(McFadden et al., 2001)、および神経筋接合部シナプス(Flood et al., 1999)を含む、SOD1遺伝子が欠損するマウスの様々な異常、ならびに、軸索損傷(Reaume et al., 1996)、虚血(Kondo
et al., 1997; Kawase et al., 1999)、溶血血液の曝露(Matz et al., 2000)、および放射線照射(Behndig et al., 2001)など各種の有害な攻撃への高い感受性によって理解される。突然変異タンパク質の毒性、および野生種の機能的重要性を考えると、ALSの理想的な治療法は、前記突然変異SOD1タンパク質の発現を選択的に阻害する一方で野生種SOD1タンパク質の発現を維持することである。
【0049】
本願発明は、RNAiを用いた突然変異SOD1を標的とする。RNAiに用いる方法は、突然変異SOD1 mRNA内に点突然変異を含有する領域を補完する2本鎖RNA(siRNA)の1本を含む。siRNAをニューロンに導入後、siRNAは部分的に巻き戻され、部位特異的にSOD1 mRNA内に点突然変異を含む領域に結合し、mRNAヌクレアーゼを活性化する。このヌクレアーゼはSOD1 mRNAを切断し、突然変異SOD1の翻訳を中断させる。細胞は、部分的に切断されたmRNAを切り離し、その結果、翻訳を妨げるか、または細胞は部分的に翻訳されたタンパク質を切断する。ニューロンは野生種SOD1(正常対立遺伝子の)上で生存するため、このアプローチは、突然変異SOD1の産生物を除外することによって、突然変異SOD1の破壊を防ぐ。
【0050】
ヒト野生種SOD1タンパク質のアミノ酸配列は、図1(配列番号18)に記載されている。ヒト野生種SOD1遺伝子のコンセンサスヌクレオチド配列は、図2(配列番号17)に記載されている。
【0051】
III SOD-1突然変異遺伝子
100を超えるSOD1突然変異が特定されている。これらの突然変異の大半が、アミノ酸のスーパーオキシドジスムターゼの酵素鎖において1アミノ酸の置換を生じる。最も頻発する置換は、アミノ酸鎖の4位で生じるバリンによるアルギニンの置換(Arg4Valとも書く)であって、1型筋萎縮性側策硬化症を罹患するアメリカ人患者の50%に生じる。
【0052】
SOD1突然変異は、1型筋萎縮性側策硬化症の症状が始まる年齢、および疾患の進行の速さに影響を与える。たとえば、Arg4Val突然変異は、生存期間が疾患の発症から2年間未満という、侵攻性の障害を生じる。37位におけるグリシンのアルギニンへの置換(Gly37Arg)は、疾患の発症は早期だが生存期間は長い。さらに、その他の因子がSOD1突然変異と組み合わさると、1型筋萎縮性側策硬化症の経過が変化する。たとえば、SOD1遺伝子とCNTFとして知られる遺伝子の両方に生じる突然変異は、前記疾患の発症を早めるようである。CNTF突然変異だけでは病的影響はないが、SOD1突然変異と組み合わさると、疾患の症状は他の影響を受けたファミリメンバと比較すると何十年も早まるようである。
【0053】
SOD1突然変異が、筋肉の運動を制御する働きに特化した脳および脊髄の神経細胞である運動ニューロンに、選択的死滅を引き起こす機序は明らかではない。スーパーオキシドジスムターゼ酵素が、SOD1遺伝子において生じた変化が原因で、新しい(だがまだ未同定の)毒性機能を獲得すると考えられている。機能が異常な酵素は、有害なスーパーオキシドラジカルの蓄積、他の種類の有毒ラジカルの異常産生、細胞死(アポトーシス)の促進、前記酵素とその他の細胞タンパク質との凝集、または消耗と死滅を引き起こすような運動ニューロンへの連続的な刺激によって運動ニューロンの死滅を引き起こすこともある。
【0054】
【表1】

【0055】
IV RNA干渉
RNAiは2本鎖RNA(dsRNA)が動物および植物細胞において相同的なmRNAの配列特異的分解を誘発する著しく有効なプロセスである(Hutvagner and Zamore
(2002), Curr. Opin. Genet. Dev., 12, 225-232; Sharp (2001), Genes Dev., 15,
485-490)。哺乳類細胞では、RNAiは低分子干渉RNA(siRNA)の21ヌクレオチド(nt)2本鎖 (Chiu et al. (2002), Mol.
Cell., 10, 549-561; Elbashir et al. (2001),
Nature, 411, 494-498)、またはDNAテンプレートを用いてRNAポリメラーゼIIIプロモータとともにin vivoで発現するその他のdsRNA(Zeng et al. (2002), Mol. Cell, 9, 1327-1333; Paddison et al. (2002), Genes Dev.,
16, 948-958; Lee et al. (2002), Nature Biotechnol., 20, 500-505; Paul et al.
(2002), Nature Biotechnol., 20, 505-508; Tuschl, T. (2002), Nature Biotechnol.,
20, 440-448; Yu et al. (2002), Proc. Natl. Acad. Sci. USA, 99(9), 6047-6052;
McManus et al. (2002), RNA, 8, 842-850; Sui et al. (2002), Proc. Natl. Acad.
Sci. USA, 99(6), 5515-5520.)によって引き起こすことができる
【0056】
V. RNA分子および薬剤
本願発明は「低分子干渉RNA分子」(「siRNA分子」または「siRNA」)、前記siRNA分子を作製する方法、および前記siRNA分子を使用する方法(たとえば研究および/または治療方法)を特徴とする。本願発明のsiRNA分子はセンス鎖および相補的なアンチセンス鎖からなる2本鎖であって、アンチセンス鎖はRNAiを媒介するのために十分な標的mRNAへの相補性を有している。好ましくは、前記鎖は、鎖がアニーリングされる時に2本鎖末端の片方または両方に1、2、または3残基のオーバーハングが生じるように、鎖末端に、アラインメントされない(つまり相補的な塩基が反対側の鎖に生じない)少なくとも1、2、または3塩基があるようにアラインメントされる。好ましくは、前記siRNA分子は約10乃至50またはそれを超えるヌクレオチドの長さを有し、つまり、各鎖が10乃至50ヌクレオチド(またはヌクレオチド類似体)を含むより好ましくは、前記siRNA分子は、各鎖にたとえば16、17、18、19、20、21、22、23、24、25、26、27、28、29、または30 ヌクレオチドなど約16乃至30の長さを有し、当該分子において片方の鎖が標的領域に実質的にたとえば少なくとも80%(またはそれを超える、たとえば85%、90%、95%、または100%)相補的であって、たとえば3、2、1または0のミスマッチヌクレオチドを有し、当該標的領域が野生種と突然変異の対立遺伝子の差が少なくとも1塩基対はある、たとえば機能獲得型突然変異を含む標的領域などの標的領域であって、もう片方の鎖は第1の鎖と実質的に同一である。
【0057】
一般的に、siRNAは当業に知られる任意の方法、たとえば以下のプロトコルを用いることによってデザインすることができる。
【0058】
1. AUG開始コドンから始めてAAジヌクレオチド配列を探す。各AAおよび3’側に16ヌクレオチド以上離れたところが、siRNA標的の候補である。siRNAは機能獲得型突然変異を含む標的領域など、野生種と突然変異の対立遺伝子の差が少なくとも1塩基対の標的領域に特異的でなければならない。第1の鎖は、この配列に相補的でなければならず、その他の鎖は第1の鎖に同一であるか実質的に同一である。ある実施態様では、前記核酸分子は、例えばSOD1の配列など、開始コドンの少なくとも50乃至100nt下流から始まる標的対立遺伝子配列の領域から選択される。さらに、G/C含有量が少ないsiRNA(35乃至55%)は、G/C含有量が55%を超えるものよりも活性であることもある。したがってある実施態様では、前記発明には35乃至55%のG/C含有量を有する核酸分子が含まれる。さらに、siRNAの鎖は、たとえば2ヌクレオチドなど、1乃至4ヌクレオチドの3’オーバーハングを有するように対を形成させることができる。したがって別の実施態様では、前記核酸分子はTTなどの2ヌクレオチドの3’オーバーハングを有してもよい。前記のオーバーハングしているヌクレオチドは、RNAまたはDNAのいずれかである。上述のように、突然変異:野生種ミスマッチがプリン:プリンミスマッチである標的領域を選択することが望ましい。
【0059】
2.
当業に知られる任意の方法を用いて、標的候補と適切なゲノムデータベース(ヒト、マウス、ラットなど)を比較し、他のコード配列に顕著に相同性を有する任意の標的配列を考慮から除外する。そのような配列相同性の探索に使われる1つの方法はBLASTとして知られ、国立バイオテクノロジー情報センターのウェブサイトで入手することができる。
【0060】
3.
評価基準に合う1つ以上の配列を選択する。
さらに、siRNAのデザインおよび使用についての一般的な情報は、マックスプランク生物物理化学研究所のウエブサイトで入手できる「The siRNA User Guide(siRNAユーザーガイド)」の中に見つけることもできる。
【0061】
陰性対照siRNAは、選択されたsiRNAと同一のヌクレオチド組成を有するべきだが、適切なゲノムに相補的な重要な配列は有さない。そのような陰性対照は、選択されたsiRNAのヌクレオチド配列をランダムにスクランブルしてデザインしてもよく、相同性探索は、前記陰性対照が適切なゲノムにおける任意のその他の遺伝子に対する相同性を欠損していることを確認するために行うことができる。さらに、陰性対照siRNAは、前記配列に1つ以上の塩基ミスマッチを導入することによってデザインすることができる。
【0062】
1ヌクレオチド特異性を有するsiRNAは、以下の通りデザインすることができる。
【0063】
標的mRNAは、参照mRNA配列(たとえば野生種対立遺伝子またはmRNA配列)と比較したミスマッチ(たとえば点突然変異などの1ヌクレオチドミスマッチなど)を有するものが選択される(たとえば突然変異対立遺伝子またはmRNA)。siRNAは、1ヌクレオチドのところに、siRNAと標的mRNA(たとえば突然変異mRNA)の間に完全な相補性が存在し、したがって、siRNA(たとえばアライニングされた)を参照配列(たとえば野生種対立遺伝子またはmRNA配列)と比較する場合、ミスマッチが存在するようにデザインされる。好ましくは、前記siRNAは、1ヌクレオチド(たとえば点突然変異)が意図される切断部位、またはその近傍に存在するようにデザインされる。好ましくは、前記siRNAが、標的とされる1ヌクレオチド(たとえば点突然変異)が完全にまたは正確にsiRNA(たとえばsiRNAのアンチセンス鎖)の中央に配置されるようにデザインされる。完全に中央に配置するという文言は、前記1ヌクレオチド(たとえば点突然変異)の両側に並んでいるヌクレオチドの数が同一であるが(つまり8、9、10、11、または12)、たとえばdTdT末端など任意のオーバーハングは含まないことを意味する。たとえば、2ヌクレオチドの3’オーバーハング(たとえばアンチセンス鎖の3’末端にあるオーバーハング)を有する21ヌクレオチドsiRNAが選択される場合、前記1ヌクレオチド(たとえば点突然変異)の両側には9ヌクレオチドが存在する。たとえば、2ヌクレオチドの3’オーバーハング(たとえばアンチセンス鎖の3’末端にあるオーバーハング)を有する22ヌクレオチドsiRNAの場合、前記1ヌクレオチド(たとえば点突然変異)の両側には9および10ヌクレオチドが存在する。23ヌクレオチドsiRNAの場合、前記1ヌクレオチド(例えば点突然変異)の両側に10ヌクレオチドが存在する。24ヌクレオチドsiRNAの場合、前記1ヌクレオチド(例えば点突然変異)の両側に10および11ヌクレオチドが存在する。例示した数は、2ヌクレオチド3’オーバーハングを有するsiRNAの場合であるが、より長いまたは短いオーバーハングを有するか、またはオーバーハングを有さないsiRNAにも容易に当てはめることができる。前記1ヌクレオチド(たとえば点突然変異)がsiRNAの中央に位置しないようなsiRNAをデザインする場合、siRNAによる切断の有効性が減少する場合もある。
【0064】
1ヌクレオチド特異性を有するsiRNAは、好ましくは、相当する参照(たとえば野生種)配列における塩基対の形成が生じにくいようにデザインされる。たとえば、siRNAをプリン:プリン対形成が前記1ヌクレオチドの位置でsiRNAと野生種mRNAの間に存在するようにデザインすると、1ヌクレオチド特異性が促進される。プリン:プリン対形成は、たとえばG:G、A:G、G:AおよびA:A対形成のグループから選択される。 さらに、前記1ヌクレオチドの位置でのsiRNAと突然変異mRNAの間のプリン:ピリミジン対形成は、1ヌクレオチド特異性を促進する。プリン:ピリミジン対形成は、たとえばG:C、C:G、A:U、U:A、C:A、A:C、U:AおよびA:U
のグループから選択される。
【0065】
本願発明の核酸組成物には、本願明細書に記載のsiRNAおよびsiRNA誘導体の両方が含まれる。たとえば、たとえば体内の半減期を延長するためなど、前記組成物の薬物動態学を変化させるために、架橋を用いることができる。したがって、本願発明には、2本の鎖が架橋されるように、核酸の相補的な2本の鎖を有するsiRNA誘導体を含む、siRNA誘導体が含まれる。本願発明にはまた、3’末端(たとえばペプチド)、または有機化合物(たとえば色素)などに結合する非核酸部分を有するsiRNA誘導体も含まれる。このようなsiRNA誘導体の修飾は、細胞取り込みが改善するか、もしくは相当するsiRNAと比較して、得られたsiRNA誘導体の細胞の標的活性を促進するか、前記細胞のsiRNA誘導体を追跡するのに有用であるか、または相当するsiRNAと比較したsiRNA誘導体の安定性を向上することもある。
【0066】
本願発明のsiRNA分子は、化学的に合成することができ、またはin vitroではDNAテンプレートから、もしくはin vivoではたとえばshRNAから、もしくはin vitro転写dsRNAテンプレートをRNAiを切断して媒介する20、21、もしくは23bp 2本鎖RNAのプールをつくるためにヒトDICER酵素を用いて、転写することができる。前記siRNA分子は、当業に知られる任意の方法を用いてデザインすることができる。
【0067】
ある局面では、RNAi剤を本願明細書に記載のたとえばsiRNAまたはshRNAなどの干渉リボ核酸ではなく、前記RNAi剤は本願明細書に記載のたとえばshRNAなどの干渉ヌクレアーゼをコードしてもよい。つまり、前記RNAi剤は前記干渉リボ核酸の転写テンプレートであってもよい。したがって、本願発明のRNAi剤には、低分子ヘアピンRNA(shRNA)、およびshRNAを発現するためにつくられた発現作製物が含まれてよい。shRNAの転写は、ポリメラーゼIII(pol III)プロモータから開始し、4-5チミン転写終了部位の2位で終了すると考えられている。発現後、shRNAは3’UUオーバーハングを有するステム-ループ構造に折り畳まれると考えられ、その後、これらのshRNAの末端が処理されて、shRNAが約21乃至23ヌクレオチドのsiRNA様分子に転換される。 Brummelkamp et al.,
Science 296:550-553 (2002); Lee et al, (2002). supra; Miyagishi and
Taira, Nature Biotechnol. 20:497-500 (2002); Paddison et al. (2002), supra;
Paul (2002), supra; Sui (2002) supra; Yu et al. (2002), supra。
【0068】
本願発明の発現作製物には、適切な発現システムに使用するのに好適な任意の作製物が含まれ、当業に知られるレトロウイルスベクタ、直線状発現カセット、プラスミドおよびウイルスまたはウイルス由来ベクタがそれらに限定されずに含まれる。そのような発現作製物には、1つ以上の誘導プロモータ、U6 snRNAプロモータもしくはH1 RNAポリメラーゼIIIプロモータなどのRNA Pol IIIプロモータシステム、または当業に知られるその他のプロモータが含まれてよい。両方の鎖を発現する発現作製物には、両方の鎖を連結するループ構造が含まれてもよく、または各鎖は同一の作製物内の個別のプロモータから別々に転写されてもよい。各鎖は個別の発現作製物から転写されてもよい。 (Tuschl (2002), supra)。
【0069】
合成siRNAは、カチオン性リポソームトランスフェクションおよび電気穿孔を含む、当業に知られる方法によって細胞に送達される。しかし、これらの外来性siRNAは、一般に、サイレンシング効果が短寿命で(培養細胞中で4乃至5日)、特定の実施態様でしか有益でない場合もある。前記標的遺伝子(つまり突然変異遺伝子)を長期間抑制し、特定の環境下での送達を容易にするために、細胞内で1つ以上のsiRNAを組換えDNA作製物から発現させることができる。細胞内における長期間の標的遺伝子の抑制を可能にするために、細胞内で組換えDNA作製物からsiRNA
2本差を発現させる方法は当業に知られており、機能的な2本鎖siRNAを発現することが可能な哺乳類Pol III プロモータシステム(たとえばH1またはU6/snRNAプロモータシステム(Tuschl (2002), supra)
)が含まれる(Bagella et
al., J. Cell. Physiol. 177:206−213 (1998); Lee et al. (2002), supra;
Miyagishi et al. (2002), supra; Paul et al. (2002), supra;
Yu et al. (2002), supra; Sui et al. (2002), supra)。RNA Pol IIIによる転写終了は、一連のDNAテンプレートの連続する4つのT残基で生じ、特定の配列においてsiRNA転写物を終了させるメカニズムを提供する。前記siRNAは5’-3’および3’-5’の方向に向いた前記標的遺伝子の配列に相補的であって、siRNAの2本鎖は同一の作製物または別々の作製物中に発現することができる。H1またはU6 snRNAプロモータによって駆動され、細胞内で発現するヘアピンsiRNAは、標的遺伝子の発現を阻害することができる(Bagella et al. (1998),
supra; Lee et al.
(2002), supra; Miyagishi et al. (2002), supra; Paul et
al. (2002), supra; Yu et al. (2002), supra; Sui et
al. (2002) supra)。T7プロモータの制御下にあるsiRNA配列を含有する作製物もまた、T7 RNA ポリメラーゼを発現するベクタを有する細胞に同時トランスフェクトされると、機能性siRNAをつくる。1つの作製物は、同一の遺伝子または複数の遺伝子に方向付けられている突然変異SOD1をコードする遺伝子の複数の領域など、siRNAをコードする複数の配列を含有することがあり、たとえば、別々のPolIIIプロモータ部位によって駆動することができる。
【0070】
動物細胞はミクロRNA(miRNA)とよばれる約22ヌクレオチドの非コードRNAの領域を発現する。ミクロRNAは動物の成長中に転写後または転写レベルで遺伝子発現を制御することができる。miRNAのある共通の性質は、おそらくダイサー、RNase III型酵素、またはその相同体によって、約70ヌクレオチドの前駆体RNAステム-ループからすべて切除されることである。標的mRNAに相補的な配列を有するmiRNA前駆体のステム配列を置換することによって、組換えされた前駆体を発現するベクター作製物を用いて、哺乳類細胞の中で特異的mRNA標的に対してRNAiを開始するためにsiRNAを産生することができる(Zeng (2002), supra)。ポリメラーゼIIIプロモータを含有するDNAベクタによって発現する場合、ミクロRNAでデザインされたヘアピンは遺伝子発現をサイレンシングすることができる(McManus (2002), supra)。ミクロRNA標的多型が、siRNA媒介遺伝子サイレンシングの欠如下において突然変異タンパク質の翻訳を阻害するために有用な場合もある。そのような適用は、たとえばデザインされたsiRNAが野生種タンパク質の標的外サイレンシングを生じるような状況で有用である場合がある。
【0071】
ウイルス媒介送達メカニズムを用いて、たとえばRNA Pol IIプロモータ転写制御下におけるsiRNAを有する組換えアデノウイルスの作製などによるsiRNAの発現を通して、標的遺伝子の特異的サイレンシングを誘導することができる(Xia et al. (2002), supra)。 これらの組換えアデノウイルスによるHeLa細胞の感染によって、内因性標的遺伝子発現を低減することができる。siRNAの標的遺伝子を発現するトランスジェニックマウスに組換えアデノウイルスベクタを注射すると、in vivoにおいて標的遺伝子の発現が減少する。同文献。動物モデルにおいて、全胚電気穿孔は、合成siRNAを効率的に移植後マウス胚合成siRNAに送達させることができる。成体マウスにおいて、siRNAの効率的な送達は、「高圧」送達技術によって実現することができる。これは溶液を含有する大量のsiRNAを、尾部静脈を介して動物体内へ迅速に(5秒以内に)注入する技術である。ナノ粒子およびリポソームを用いて、siRNAを動物体内へ送達することもできる。
【0072】
本願発明の核酸組成物には、非修飾siRNA、および架橋siRNA誘導体、または3’もしくは5’末端などに結合させた非ヌクレオチド部分を有する誘導体など、当業に知られる修飾RNAの両方が含まれる。このようなsiRNA誘導体の修飾は、細胞取り込みが改善するか、もしくは相当するsiRNAと比較して、得られたsiRNA誘導体の細胞の標的活性を促進するか、前記細胞のsiRNA誘導体を追跡するのに有用であるか、または相当するsiRNAと比較したsiRNA誘導体の安定性を向上することもある。
【0073】
本願明細書に記載の細胞または組織全体に導入された組換えRNA前駆体によって、望ましいsiRNA分子の産生が引き起こされるだろう。そこで、そのようなsiRNA分子は、切断および破壊のための特異的なmRNA配列に結合し標的とするRNAi経路の内因性タンパク質要素に関連するだろう。このようにすれば、組換えRNA前駆体から作製されたsiRNAが標的とするmRNAは、前記細胞および組織から大幅に減少し、前記細胞または組織中のmRNAによってコードされるタンパク質の濃度が減少するだろう。RNA前駆体は、典型的にはdsRNAの1本鎖を個別にコードするか、またはRNAヘアピンループ構造のヌクレオチド配列全体をコードする核酸分子である。
【0074】
本願発明の核酸組成物は、たとえば吸収、有効性、バイオアベイラビリティ、および/または半減期などの薬物動態学的パラメタなどの前記組成物の性質を向上させるナノ粒子などの別の部分と結合させることもでき、または結合させないでもよい。その結合は、たとえば、Lambert et al., Drug Deliv. Rev.:47(1), 99-112 (2001) (ポリアルキルシアノアクリル酸(PACA)ナノ粒子に装着された核酸について記載)、Fattal et al., J. Control
Release 53(1-3):137-43 (1998) (ナノ粒子に結合させた核酸について記載)、 Schwab et al., Ann. Oncol.
5 Suppl. 4:55-8 (1994) (挿入剤、疎水基、ポリカチオンPACAナノ粒子に結合させた核酸について記載)、およびGodard et al., Eur. J. Biochem. 232(2):404-10 (1995) (ナノ粒子に結合させた核酸について記載)の方法を用いるなど、当業に知られる方法によって実現することができる。
【0075】
本願発明の核酸分子は、当業に知られる任意の方法を用いてラベルすることもでき、たとえば前記核酸分子組成物がたとえばCy3、フルオレセイン、ローダミンなどのフルオロフォアで標識することができる。その標識は、たとえばSILENCERTM siRNA ラベリングキット(アンビオン)など、キットを用いて行うことができる。さらに、前記siRNAは、たとえば3H、32Pまたはその他の適切な同位体などを用いて放射標識することができる。
【0076】
さらに、RNAiは、少なくとも1つの1本鎖RNA中間体を経る過程であると考えられるため、当業者は、ss-siRNA(たとえばds-siRNAのアンチセンス鎖など、本願明細書に記載のとおりデザイン(たとえば化学合成用に)、作製(たとえば酵素的に作製)または発現(たとえばベクターまたはプラスミドから)することもでき、本願特許請求の方法論にしたがって利用することができるということは理解されよう。さらに、無脊椎動物では、RNAiは長いdsRNA(たとえば長さが約100乃至1000ヌクレオチド、好ましくは約200乃至500であって、たとえば、約250、300、350、400、または450ヌクレオチドのdsRNA)であって、RNAiのエフェクタT細胞として作用するRNAiは長いdsRNAによって効果的に引き起こすことができる。
【0077】
本願発明のsiRNA分子は、突然変異siRNA P11 (配列番号5, センス; 配列番号6, アンチセンスまたはガイド), 突然変異 siRNA P10 (配列番号3, センス;
配列番号4, アンチセンス、またはガイド), 突然変異 siRNA P9 (配列番号1, センス; 配列番号2 アンチセンス、またはガイド), SOD1 野生種標的(配列番号7), SOD1 突然変異 標的(配列番号8), 野生種siRNA
P11 (配列番号9 センス; 配列番号10, アンチセンス、またはガイド), 野生種siRNA P10 (配列番号11, センス; 配列番号12, アンチセンス、またはガイド), 野生種siRNA P9 (配列番号13, センス;
配列番号14, アンチセンス、またはガイド)が含まれる図1に記載の配列、G93A SOD1 siRNA (SEQ ID
NO:16)が含まれる図3に記載の配列、およびその対立遺伝子変異種などを含む、またはそれらからなっていてもよい。
【0078】
VI. RNAiを誘導するための組換えRNA前駆体の使用
本願明細書に記載の細胞または組織全体に導入された組換えRNA前駆体によって、望ましいsiRNA分子の産生が引き起こされるだろう。そこで、そのようなsiRNA分子は、切断および破壊のための特異的なmRNA配列に結合し、標的とするRNAi経路の内因性タンパク質要素に関連するだろう。このようにすれば、組換えRNA前駆体から作製されたsiRNAが標的とするmRNAは、前記細胞および組織から大幅に減少し、前記細胞または組織中のmRNAによってコードされるタンパク質の濃度が減少するだろう。
【0079】
VII. 薬学的組成物および投与方法
本願発明のsiRNA分子は薬学的組成物に組み込むことができる。そのような組成物には、典型的には、核酸分子および薬学的に許容な担体が含まれる。本願明細書に記載の「薬学的に許容な担体」という用語は、薬学的投与に適合する、食塩水、溶媒、分散媒、コーティング、抗細菌および抗真菌剤、ならびに等張および吸収遅延剤などを意味する。補助的な活性化合物も、当該組成物に組み込むことができる。
【0080】
薬学的組成物は、意図する投与経路と適合するように調剤される。投与経路の例には、たとえば静脈内、皮内、皮下、口腔内(たとえば吸入)、経皮(局所)、経粘膜、および直腸投与などの非経口投与が含まれる。非経口、皮内、または皮下投与に使用する溶液または懸濁液には、以下の成分、注射用の水、食塩水、固定油、ポリエチレングリコール、グリセリン、プロピレングリコール、またはその他の合成溶媒などの滅菌希釈剤、ベンジルアルコールまたはメチルパラベンなどの抗菌剤、アスコルビン酸または亜硫酸ナトリウムなどの抗酸化剤、エチレンジアミン四酢酸などおキレート剤、酢酸塩、クエン酸塩、またはリン酸塩などのバッファ、および塩化ナトリウムまたはデキストロースなどの張力調整剤などが含まれていてよい。非経口製剤は、アンプル、ガラスまたはプラスチック製の使い捨て用シリンジ、または複数の投与用バイアルの中に封入されていてよい。
【0081】
注射に使用するために好適な薬学的組成物には、滅菌水溶液(水溶性)、または分散液、および滅菌注射用溶液または分散液の即時調整用滅菌粉末が含まれる。静脈内投与に好適な担体には、生理食塩水、静菌水、クレモホールELTM(BASF、ニュージャージー州パシッパニー)、またはリン酸緩衝食塩水(PBS)が含まれる。すべての場合において、前記組成物は滅菌状態であって、注射容器への充填が容易な程度、流動的でなければならない。前記組成物は、製造および保存の条件下において安定でなければならず、細菌および真菌などの微生物の汚染作用から保護されなければならない。前記担体は、たとえば水、エタノール、ポリオール(たとえばグリセロール、ポリエチレングリコール、および液体状ポリエチレングリコールなど)、それらの好適な混合物、ならびに植物油などを含む溶媒又は分散媒であってよい。適切な流動性は、たとえばレシチンなどのコーティングの使用、分散液の場合は所要の粒子サイズの維持、および表面活性剤の使用などによって維持することができる。微生物の作用は、たとえばパラベン、クロロブタノール、フェノール、アスコルビン酸、チメロサールなどの各種抗細菌剤および抗真菌剤によって防ぐことができる。多くの場合、たとえば糖、マニトールなどのポリアルコール、ソルビトール、塩化ナトリウムなどの等張剤を前記化合物に含めることが望ましいだろう。注射可能な組成物の吸収を持続させるために、たとえばモノステアリン酸アルミニウムおよびゼラチンなどの吸収遅延剤を組成物に含ませてもよい。
【0082】
滅菌された注射可能な溶液は、適切な溶媒中に入れた必要量の前記活性化合物を、必要に応じて上述の成分1つまたはそれらの組み合わせに組み入れてからろ過滅菌することによって調製することができる。一般に、分散液は、塩基性分散媒および上述の物質のうちの必要なその他の成分を含む滅菌担体に前記活性化合物を組み入れて調製する。滅菌注射用溶液の調製のための滅菌粉末の場合、好ましい調製方法は、前記活性成分の粉末に加えて予め滅菌ろ過されたその溶液から得られた任意の追加の望ましい成分を得られる、真空乾燥および凍結乾燥である。
【0083】
経口組成物には一般に、不活性な希釈剤または食用担体が含まれる。治療用の経口投与の場合、前記活性化合物を賦形剤に組み込み、錠剤、トローチ、またはたとえばゼラチンカプセルなどのカプセルなどの形状で用いることができる。経口組成物はまた、洗口剤として用いられる液体担体を用いて調製することもできる。薬学的に適合な結合剤、および/またはアジュバント物質は、前記組成物の一部分として含めることもできる。前記錠剤、丸剤、カプセル、およびトローチなどは、以下の成分、もしくは同様の性質の化合物のいずれかを含むことができる。それは、微結晶性セルロース、ガムトラガカント、もしくはゼラチンなどの結合剤、デンプンもしくはラクトースなどの賦形剤、アルギン酸、プリモゲル、もしくはトウモロコシデンプンなどの崩壊剤、ステアリン酸マグネシウムもしくはステロート(Sterote)などの潤滑剤、コロイド状二酸化ケイ素などの流動促進剤、ショ糖もしくはサッカリンなどの甘味剤、またはペパーミント、サリチル酸メチル、もしくはオレンジ香味料などの香味剤である。
【0084】
吸入による投与の場合、前記化合物は、たとえば二酸化炭素などの好適な高圧ガスを含む加圧容器もしくはディスペンサ、またはネビュライザからのエアロゾルスプレーの形状で送達する。そのような方法には、米国特許第6,468,798号に記載されるものが含まれる。
【0085】
全身投与はまた、経粘膜または経皮手段でもよい。経粘膜または経皮投与の場合、透過させようとする関門に適切な浸透剤を、前記製剤に用いる。そのような浸透剤は一般に当業に知られており、たとえば経粘膜投与の場合、界面活性剤、胆汁塩、およびフシジン酸誘導体が含まれる。経粘膜投与は、経鼻スプレーまたは坐剤の使用によって実施することができる。経皮投与の場合、前記活性化合物は、一般に当業に知られる軟膏(ointment)、軟膏(salve)、ゲル、またはクリームに調製する。
【0086】
前記化合物はまた、坐剤(たとえばカカオバターおよびその他のグリセリドなどの従来の坐剤ベース)または直腸送達用の停留浣腸の形状に調製してもよい。
【0087】
前記化合物はまた、McCaffrey et al.
(2002), Nature, 418(6893), 38-9 (流体力学トランスフェクション); Xia et al. (2002), Nature Biotechnol., 20(10), 1006-10 (ウイルス媒介送達); またはPutnam
(1996), Am. J. Health Syst. Pharm.53(2), 151-160, 誤字 Am. J. Health Syst. Pharm. 53(3), 325 (1996)に記載の方法が限定されずに含まれる、当業に知られる方法を用いてトランスフェクションまたはインフェクションによって投与することができる。
【0088】
前記化合物はまた、DNAワクチンなど、核酸剤の投与に好適な任意の方法によって投与することもできる。前記方法には、遺伝子銃、バイオ注射器、および皮膚用添付剤、ならびに米国特許第6,194,389号に開示されている微粒子DNAワクチン技術などの無針法、米国特許第6,168,587号に開示されるパウダー状ワクチンを有する哺乳類経皮無心ワクチン投与が含まれる。さらに、とりわけHamajima et al. (1998), Clin. Immunol. Immunopathol., 88(2),
205-10に記載されているものなど、経鼻送達も可能である。リポソーム(たとえば米国特許第6,472,375号に記載されているなど)およびマイクロカプセル封入も使用することができる。生分解性標的化可能微粒子送達システムも使用することができる(たとえば米国特許第6,471,996号に記載されているなど)。
【0089】
ある実施態様では、前記活性化合物は、インプラントおよびマイクロカプセル封入送達システムを含む、制御放出製剤などの、体外への急速な排出から前記化合物を保護する担体とともに調製する。エチレン酢酸ビニル、ポリ無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、およびポリ乳酸などの生分解性、生体適合性ポリマーを用いることができる。そのような製剤の調製は、標準技術を用いて行うことができる。前記材料はアルザ社およびノバファーマシューティカルズ社から購入することもできる。リポソーム懸濁液(ウイルス抗原に対するモノクローナル抗体で感染細胞を狙ったリポソームを含む)を薬学的に許容な担体として用いることもできる。これらは、米国特許第4,522,811号などに記載の、当業に知られる方法に従って調製することができる。
【0090】
当該化合物の毒性および治療上の有効性は、たとえばLD50(個体群の50%に致死的な投与量)およびED50(個体群の50%に治療上有効な投与量)など、細胞培養物または実験動物における標準的な薬学的方法によって決定することができる。毒性作用および治療上有効な作用の投与量の比は治療指数であって、LD50/ED50の比で表すことができる。高い治療指数を示す化合物が好ましい。有毒な副作用を示す化合物を用いる場合、感染していない細胞に与える可能性のある損傷を最小にして副作用を減少するよう、そのような化合物を罹患組織の部位に狙わせる送達システムをデザインするように、注意が払われなければならない。
【0091】
細胞培養アッセイおよび動物研究から得られるデータは、ヒトにおける使用のための用量の範囲を決定する際に用いることができる。そのような化合物の用量は、好ましくは毒性が小さいかまたは皆無のED50を含む循環濃度の範囲内にある。前記用量は、用いられた投与剤形および使用された投与経路によって、この範囲内で変化させてもよい。本願発明の方法に用いられるいずれの化合物の場合も、治療上有効な投与量は、最初は細胞培養アッセイで推定することができる。1回投与量は、動物モデルにおいて、細胞培養物で決定したIC50(すなわち症状の最大半減抑制を達成する前記テスト化合物の濃度)を含む循環血漿濃度の範囲になるように調製してもよい。このような知見は、ヒトにおける有用な投与量をより正確に決定するために用いることができる。血漿濃度は、たとえば高性能液体クロマトグラフィによって測定することもできる。
【0092】
本願明細書に記載の、核酸分子の治療上有効な量(すなわち有効投与量)は、選択された核酸による。たとえば、shRNAをコードするプラスミドを選択する場合、単回投与は約1μg乃至100mgの範囲内で投与してもよく、ある実施態様では10、30、100または1000 μgを投与することがある。 ある実施態様では、1乃至5gの前記組成物を投与することができる。前記組成物は、2日に1回など、1日1回乃至1回以上、または1週間に1回以上投与することができる。当業者は、特定の要因が、疾患またな障害の重篤度、治療歴、対象の全般的な健康状態、およびその他に罹患する疾患をそれらに限定せずに含む、前記対象を有効に治療するために必要な用量および時間に影響することもあるということを理解するであろう。さらに、対象の治療上有効な量のタンパク質、ポリペプチド、または抗体での治療は、単回の治療を含んでもよく、または好ましくは、一連の治療を含んでもよい。
【0093】
本願発明の核酸分子は、たとえば上述のXia et al.,
(2002)に記載の方法が限定されずに含まれる、当業に知られる方法を用いて、たとえばウイルスベクタ、レトロウイルスベクタ、発現カセット、またはプラスミドウイルスベクタなどの発現作製物に挿入することができる。発現作製物は、たとえば吸入、経口、静脈内投与、局所投与(米国特許第5,328,470号参照)、または定位固定注射(たとえばChen et al. (1994), Proc.
Natl. Acad. Sci. USA, 91, 3054-3057参照)によって対照に送達することができる。前記輸送ベクタの薬学的調製には許容な希釈剤に入れたベクタが含まれてもよく、または輸送用賦形剤が埋め込まれた徐放性基質を含んでもよい。代替的には、たとえばレトロウイルスベクタなど、完全な輸送ベクタを組換え細胞から無傷で作製できる場合、前記薬学的調製物は遺伝子送達システムをつくる1つ以上の細胞を含むことができる。
【0094】
本願発明の核酸分子には、低分子ヘアピンRNA(shRNA)、およびshRNAを発現するためにつくられた発現作製物が含まれてよい。shRNAの転写は、ポリメラーゼIII(pol III)プロモータから開始し、4-5チミン転写終了部位の2位で終了すると考えられている。発現後、shRNAは3’UUオーバーハングを有するステム-ループ構造に折り畳まれると考えられ、その後、これらのshRNAの末端が処理されて、shRNAが約21ヌクレオチドのsiRNA様分子に転換される。
Brummelkamp et al. (2002), Science, 296, 550-553; Lee et al, (2002). supra;
Miyagishi and Taira (2002), Nature Biotechnol., 20, 497-500; Paddison et al.
(2002), supra; Paul (2002), supra; Sui (2002) supra; Yu et
al. (2002), supra。
【0095】
前記発現作製物は、適切な発現システムに使用するのに好適な任意の作製物であってよく、当業に知られるレトロウイルスベクタ、直線状発現カセット、プラスミドおよびウイルスまたはウイルス由来ベクタがそれらに限定されずに含まれる。そのような発現作製物には、1つ以上の誘導プロモータ、U6 snRNAプロモータもしくはH1 RNAポリメラーゼIIIプロモータなどのRNA Pol IIIプロモータシステム、または当業に知られるその他のプロモータが含まれてよい。両方の鎖を発現する発現作製物には、両方の鎖を連結するループ構造が含まれてもよく、または各鎖は同一の作製物内の個別のプロモータから別々に転写されてもよい。各鎖は個別の発現作製物から転写されてもよい。上述のTuschl (2002)。
【0096】
薬学的組成物は、投与法の説明書とともに、容器、パック、またはディスペンサに包含されていてよい。
【0097】
VIII. 治療方法
本願発明は、機能獲得型突然変異タンパク質によって、全体的にまたは部分的に引き起こされた疾患または障害のリスクがある(または感受性がある)を治療するための予防および治療方法の両方を提供する。ある実施態様では、前記疾患または障害は優性の機能獲得型疾患である。好ましい実施態様では、前記疾患または障害はSOD1遺伝子の改変、特にSOD1突然変異対立遺伝子における点突然変異に関連する疾患であって、臨床徴候にはALS患者に認められるものが含まれるような、SOD1遺伝子(構造または機能)またはSOD1タンパク質(構造または機能または発現)に異常を引き起こす疾患である。
【0098】
本願明細書に記載の「治療」または「治療する」は、疾患もしくは障害、疾患または障害の症状、または疾患もしくは障害の素因を有する患者への治療用の物質(たとえばRNA剤またはベクターまたはそれをコードする導入遺伝子)の適用もしくは投与、または前記患者から単離された組織または細胞株への治療用物質の適用もしくは投与であって、当該適用もしくは投与の目的が疾患もしくは障害、疾患もしくは障害の症状、または疾患の素因の治療、治癒、緩和、軽減、改変、改善、改良、向上、または作用である、適用もしくは投与として定義される。
【0099】
ある局面では、本願発明は、対象において、前記対照に治療剤を投与することによって、上述の疾患または障害を予防する方法を提供する。前記疾患のリスクがある対照は、たとえば本願明細書に記載の任意のまたは組み合わせた診断アッセイまたは予後アッセイによって同定することができる。予防剤は、前記疾患または障害を予防、または代替的にはその進行を遅らせるように、前記疾患または障害に特徴的な症状の徴候の前に投与することができる。
【0100】
本願発明の別の局面は、対照を治療的に処理する、つまり前記疾患または障害の症状の発症を改変させる方法に関連する。ある例示的な実施態様では、本願発明の調節方法は、機能獲得型突然変異を発現する細胞と、前記遺伝子の配列特異的な干渉を実現するような、前記遺伝子内の突然変異に特異的な治療剤(たとえばRNAi剤またはベクターまたはそれをコードする導入遺伝子)と接触させるステップに関与する。これらの方法は、in vitro(たとえば前記細胞を前記作用剤で培養するなど)または代替的にはin vivoにおいて(たとえばその物質を対象に「投与する」ステップによって)実行することができる。
【0101】
処置の予防的および治療的な方法の両方に関して、そのような処置は薬理ゲノム学分野から得られた知識を本に、特別にあつらえるか、または修正してもよい。本願明細書に記載の「薬理ゲノム学」は、臨床開発用および市販用薬物の遺伝子配列解析、統計学的遺伝子学、および遺伝子発現解析などのゲノミクス技術の適用を意味する。より具体的には、この用語は、患者の遺伝子が薬物に対する応答(たとえば患者の「薬物応答表現型」または「薬物応答遺伝子型」など)をどのように決定するかを研究することを意味する。したがって、本願発明の別の局面は、本願発明の標的遺伝子分子、または個人の薬物応答遺伝子型にしたがった標的遺伝子モジュレータのいずれかによる、個人の予防的または治療的処置をあつらえる方法を提供する。薬理ゲノム学は、臨床医または医師が、予防的または治療的処置から最も利益を得るであろう患者のみを前記処置の対象にし、有毒な薬物関連の副作用を経験するであろう患者の治療を避けることを可能にする。
【0102】
治療剤は適切な動物モデルにおいてテストすることができる。たとえば、本願明細書に記載のRNAi剤(または発現ベクタもしくはそれをコードする導入遺伝子)を動物モデルに用いて、前記薬剤での処置の有効性、毒性、または副作用を決定することができる。 代替的には、治療剤を動物モデルに用い、そのような作用薬の作用機序を決定することができる。たとえば、薬剤を動物モデルに用いて、前記薬剤での処置の有効性、毒性、または副作用を決定することができる。 代替的には、薬剤を動物モデルに用い、そのような作用薬の作用機序を決定することができる。
【実施例】
【0103】
以下の材料、方法、および例は、具体例を示すのみで、制限を意図しない
材料および方法
RNAおよびDNA作製物
21本のヌクレオチド1本鎖RNA(図1)は、ダーマコン・リサーチ社から購入し、製造者の指示に従って脱保護して記載の通りアニーリングした(Nykanen et al., 2001)。野生種および突然変異SOD1-GFP融合タンパク質を作製するために、SOD1 wt(Genbank Accession No. ジェンバンク受入番号NP_000445; 図7; 配列番号18)、SOD1G85R、およびSOD1 G93A(配列番号16)cDNAを、pCMV/myc/mito/GFP(インビトロゲン社)のPmlI およびPstI部位の間でPCRクローニングした。このクローニングステップによって、ミトコンドリア標的配列が削除された。mycタグ付き野生種SOD1を作製するために、SOD1wt cDNA (配列番号17)をpCMV/myc/mito/GFPのPstIおよびXhoI部位の間でPCRクローニングした。ミトコンドリア標的配列は、BssHIIおよびPmlIで切断し、平滑末端化ライゲーションすることによって削除した。すべての作製物を、シーケンシングによって確認した。DsRed (pDsRed2-C1) は、クロンテック社(カリフォルニア州パロアルト)から購入した。U6-G93A は上述の通り作製した(Sui et al., 2002)。3’ブロックsiRNAは標準技術によって合成された。
【0104】
In-vitro RNAiアッセイ
野生種または突然変異SOD1のいずれかをコードする配列を有する560ヌクレオチドヒトSOD1標的RNAを、既報の通り調製する(Zamore et al., 2000)。標的切断は、濃度約5nMのショウジョウバエ胚ライセートを含有する標準in vitro RNAi反応において25乃至100nM siRNAを有する5’32P-キャップ放射標識標的をインキュベートすることによって決定した。
【0105】
細胞培養およびトランスフェクション
Hela細胞を、10%ウシ胎仔血清(FBS)、100ユニット/mlペニシリン、および100μg/mlストレプトマイシンをDMEMならびにDMEDおよびOpti-MEM(1:1)に入れたN2A 細胞の両方ともに添加し、その中で培養した。
トランスフェクションの24時間前、細胞(集密度70乃至90% )を倍散で脱離させ、6穴プレートに移して、抗生物質を含まない新鮮培地で培養した。トランスフェクションは、リポフェクタミン2000(インビトロゲン社)を、製造者の指示に従って用いて行った。トランスフェクションに用いた作製物の量は、突然変異または野生種SOD1-GFPおよびDsRedプラスミドそれぞれ4μg 、siRNAを4x10-11または4x10-12モル、ならびにU6-G93Aを20または8μgである。
【0106】
In vivoトランスフェクション
生後6乃至8週間のマウス24匹を3つのグループに分けた。第1のグループはshRNAベクタの投与を受けておらず、第2のグループは20μgの空ベクタの投与を受け、第3のグループはSOD1 G93A (配列番号16)に対するU6-hpRNA ベクタ20μgを投与された。すべてのグループは、mycでタグしたヒト野生種SOD1 (SEQ ID
NO: 7) 20μg、およびGFPでタグしたSOD1 20μgの両方が投与された。前記ベクタを合計体積がマウス1匹あたり2.5mlになるように、リンガ液で希釈した。マウスをアバーチン(240mg/kg)で麻酔し、前記ベクタを26ゲージ針を用いて10病未満で尾部静脈から注入する。注入から48時間後、肝臓から血液を除去するために、動物を5ml PBSで灌流した。肝臓を切り取り、素速くドライアイス上で冷凍させた。サンプルを、1% SDS、1mM DTT、1mMフェニルメチルスルホニルフルオリド(PMSF)、およびプロテアーゼ阻害物質カクテル(シグマ社1:100希釈)を含有する25mM PBSバッファ(pH 7.2)中に入れ、携帯用ポリトロン(プロサイエンティフィック社)を用いてホモジナイズした。
【0107】
ウエスタンブロット分析法
タンパク質濃度は、BCAタンパク質アッセイキット(ピアース社、イリノイ州ロックビル)を用いて決定した。Hela細胞タンパク質25μg、または100μg肝タンパク質を15% SDS-PAGE 上で分離し、ジーンスクリーン・プラス(パーキンエルマー社)上に移動させた。ウサギ抗SOD1(バイオデザイン社)またはヒツジ抗SOD1は1次抗体、およびHRP標識ヤギ抗ウサギIgG(アマシャム社)またはロバ抗ヒツジIgGは2次抗体だった。タンパク質のバンドの可視化には、スーパーシグナル・キット(ピアース社)およびコダック・デジタルイメージステーション440CFを用いた。そのバンドの強度の定量化には、コダック1Dソフトウェアを用いた。
【0108】
例I.
例I乃至VIIIは、siRNAが、まずsiRNA活性をショウジョウバエ胚ライセートを含む無細胞RNAI反応でテストし、次に培養哺乳類細胞中の活性1ヌクレオチド選択性siRNAを分析することによって、1ヌクレオチド選択性を有するようにデザインされたことを明らかにする。結果は、2つのmRNAの差が1ヌクレオチドしかなく同一の細胞に存在するとしても、21ヌクレオチドsiRNAおよびshRNAの両方とも、選択的に突然変異SOD1(配列番号8)の発現を阻害するが、野生種SOD1(配列番号7)ではそのようなことがないようにデザインすることができることを示した。したがって、RNAiはとりわけ、優性機能獲得型突然経によって生じる疾患の治療法として有用である。
【0109】
例I. siRNA 2本鎖は突然変異SOD1を区別することができる
それぞれ野生種または突然変異mRNAのいずれかを狙う3つのsiRNA
2セット(図1A、配列番号8)を、標的の切断部位またはその近傍のミスマッチが要求されるA型らせん形を乱しているかどうかをテストするように方向付けた。グアニン256(たとえばジェンバンク受入番号K00056などの、翻訳開始に関連するG256)SOD1の対立遺伝子がシトシンに突然変異し、グリシン-アルギニン突然変異(G85R) を生じているSOD1の対立遺伝子を選択した。突然変異したヌクレオチドは、SOD1 mRNA切断の予測された部位の近傍、すなわち、siRNAのアンチセンス鎖の5’末端に対して9位(P9)、10位(P10)、11位(P11)(図1A)に位置した。SOD1 mRNA切断のこの予測された部位は、前記siRNAとRNAiエンドヌクレアーゼの活性部位の中または近傍の非同族標的RNAとの間にミスマッチを配置する。これらのsiRNAのテストは、in vitroにおいてRNAを再利用する、確立されたショウジョウバエ胚性ライセートで行った(Zamore et al., 2000; Tuschl et al., 1999)。予想通り、6つのsiRNAのそれぞれが相当する標的RNAを切断したが、その有効性の差は劇的だった。たとえば、P11突然変異種および野生種siRNA(配列番号6, 10)は、それぞれの標的RNAを効率的に切断できなかった。反対に、P10突然変異種siRNA(配列番号4)は前記突然変異種標的RNAを効率的に切断した。完全長突然変異種SOD1標的mRNAの切断は、相当する約288ヌクレオチドの5’切断産生物の蓄積によって行われ、結果は、前記標的mRNAの非特異的分解ではなくRNAiを示唆している。siRNAの欠如下、またはルシフェラーゼに対するsiRNAの存在下において、前記突然変異標的RNA(配列番号8)はショウジョウバエ胚性ライセート中で安定だった(データは非表示)。標的RNAの切断および5’切断産生物の蓄積のデータは両方とも、1つの指数方程式に良くフィットし、この反応は擬1次動力学にしたがっていることを示している(図1B)。
【0110】
例I. siRNA 2本鎖は野生種SOD1を区別することができる
6つのsiRNAの特異性を決定するために、突然変異SOD1配列(配列番号8)に相当する各siRNAを、野生種SOD1 RNA (配列番号7)を切断する能力についてテストし、各野生種siRNAについて突然変異RNAを切断する能力をテストした。すべてではないが一部のsiRNA 2本鎖が、効率的に、完全にマッチする標的と1ヌクレオチドのミスマッチを有する標的を区別した。たとえば、突然変異と野生種のsiRNA(配列番号6、10)の両方のP11は、完全にマッチした標的RNAもミスマッチの標的RNAのいずれにも効率的な切断を惹起しなかった(図1A)。したがって、これらのsiRNA配列は本質的にRNAiを惹起する能力が乏しい。反対に、P9 (配列番号 14)およびP10野生種(配列番号12) siRNAは、相当する野生種標的の迅速な切断を惹起しただけでなく、突然変異RNAの著しい切断も生じた(図1A)。これらのsiRNAはRNAiを惹起する優れた能力があるが、乏しい選択性を示す。P10突然変異種siRNA(配列番号4)が、突然変異種および野生種SOD1 RNA (配列番号7、8)を効率的および堅固に識別し、突然変異RNAを野生種よりもはるかに効率的に切断することが示された。最も重要なのは、P10突然変異siRNA(配列番号4)は、突然変異および野生種SOD1 mRNA 標的を実質的に完全に識別することが示された(Fig. 1A)。このP10突然変異siRNAは、突然変異SOD1標的の切断を効率的に媒介したが、野生種SOD1 mRNAの切断はほとんど媒介せず(Fig. 1B)、このsiRNAは治療用途に理想的であることが示唆された。
【0111】
例III. Hela細胞における突然変異SOD1 G85R発現の選択的阻害
哺乳類細胞において、無細胞反応が正確、効率的、および選択的にsiRNAを予想できるかどうかをテストするために、カルボキシ末端を融合させたGFPを有する野生種または突然変異種SOD1 G85Rを発現したプラスミド作製物を作製した。各作製物を、dsRed発現ベクターを用いてHela細胞にトランスフェクトし、トランスフェクション対照とした。突然変異または野生種のSOD1 (配列番号7、8)のいずれかの発現を、蛍光標示式細胞分取法(FACS)で緑色および赤色細胞を定量することによって監視した。P9、P10およびP11 siRNAを相当する突然変異種または野生種標的でトランスフェクションしたところ、遺伝子発現を抑制したが、効率および選択性には差があった(図2)。反対に、ホタルルシフェラーゼに相補的なsiRNAによるコトランスフェクションでは、突然変異または野生種SOD1の発現のいずれも抑制しなかった(図2)。野生種に対するsiRNA p10以外のすべてのsiRNAは1ヌクレオチドミスマッチを有するmRNAを抑制しなかったが、野生種に対するsiRNA
p10は野生種および突然変異SOD1の発現の両方を効率的に抑制した(図2)。この結果は全般的にin vitroデータ(図1)と一致しており、すべてではないが一部のsiRNAは1ヌクレオチドの差を有するmRNA標的を効率的に区別することができることを示唆した。
【0112】
例IV. Hela細胞におけるU6-G93A ベクタによる突然変異SOD1 G93A発現の選択的阻害
最近、shRNAがin vivoにおいてRNAiを惹起することができることが示された。突然変異種SOD1に対するshRNAが、選択的に野生種SOD1発現ではなく突然変異種SOD1の発現を阻害することができるかどうかをテストするために、別の疾患の原因となる突然変異種SOD1
G93A に相同なshRNA(ヌクレオチド281位のGをCに交換し、shRNAと野生種SOD1G:Gミスマッチを配置するヌクレオチド交換;配列番号16)を、RNAポリメラーゼIII(U6)プロモータの制御下において合成するプラスミドを作製した(Sui et al., 2002)。結果から、野生種または突然変異SOD1-GFPプラスミドのいずれかとコトランスフェクトした場合、この作製物を用いて、培養細胞中の突然変異種SOD1の1ヌクレオチド選択的RNAiを惹起することができるということが示された(図3)。
【0113】
例V. in vivoにおけるsiRNAおよびU6-G93A ベクタによる突然変異SOD1発現の選択的阻害
突然変異種の選択的阻害が神経細胞で実現するかどうかをテストするために、野生種および突然変異SOD1-GFP作製物を、SOD1 G85R に対するsiRNA P10、またはSOD G93A (配列番号16)に対するshRNA合成ベクタのいずれかを用いて、神経芽細胞腫細胞株N2aにコトランスフェクトした。Hela細胞と同様に、合成siRNAおよびshRNA作製物が、N2a細胞における突然変異SOD1発現の選択的阻害を方向付けた(図4A、B)。
【0114】
例VI. in vivoにおける突然変異SOD1 G85Rの選択的阻害
同一の細胞に突然変異および野生種SOD1 mRNA1が両方とも存在する場合、ヌクレオチド選択的siRNAがその突然変異および野生種SOD1を識別できるかどうかを明らかにするために、Hela細胞にP10 siRNAおよび突然変異SOD1 G85R-GFPでトランスフェクトした。抗SOD1抗体で免疫ブロットすることによって、トランスフェクト融合SOD1-GFPおよび内因性野生種ヒトSOD1の両方の検出が可能になった。内因性野生種SOD1発現の約50%阻害は、約50%のトランスフェクション効率を反映した。P10野生種siRNAに対して、突然変異種に対するP10 siRNAは2種類の異なる用量において突然変異種の発現を阻害したが、内因性野生種SOD1の発現には効果がなかった。
【0115】
例VII. in vivoにおけるU6-G93A ベクタによる突然変異種SOD1発現の選択的阻害
選択的阻害をin vivoで生じさせることができるかどうかをテストするために、SOD1レポータおよびshRNAプラスミドを、水力学的トランスフェクションプロトコルを用いてマウスにトランスフェクトした。突然変異SOD1 G93A-GFP プラスミドおよびmycでタグした野生種ヒトSOD1(ゲル上においてトランスフェクトしたヒトSOD1を内因性マウスSOD1からよりよく分離することができる)をU6空ベクトルまたはU6-G93Aベクタでコトランスフェクトした。SOD1
G93A-GFP およびSOD1mycの肝臓における発現を、ウエスタンブロットによって検証した。結果から、U6-G93Aで選択的にコトランスフェクトする場合のみ、G93A発現が減少することが示された(図6)。
【0116】
例VIII. トランスジェニックマウスを用いたin vivoにおける突然変異SOD1のshRNA抑制
突然変異SOD1に対するshRNAがin
vivoにおいて突然変異SOD1の発現を抑制することができるかどうかを明らかにするために、RNAポリメラーゼIII(Pol
III)プロモータU6の制御下におけるSOD1G93A に対するshRNAを発現するトランスジェニックマウス(U6-G93A マウス)を、C57BL/6JおよびSJLのハイブリッド背景において作製した。
【0117】
マウスU6プロモータの制御下におけるshRNA相同的突然変異SOD1G93A (shG93A) を合成するプラスミドは、公表されているプロトコルにしたがって作製した(Sui et al., 2002 Proc Natl Acad Sci USA 99:5515-5520)。前記マウスを作製するために、Kpn IおよびSac Iを用いた消化によって導入遺伝子を直線化し、精製して、マサチューセッツ大学医学部(UMMS)トランスジェニック・コアでマウスの有精卵に注入した。
【0118】
U6-G93A トランスジェニックマウスをスクリーニングするために、前記導入遺伝子の配列を選択的に増幅するPCRプライマをデザインし、前記トランスジェニックマウスの同定に用いた。合計7匹の第0世代(F0)が同定された。これらの第0世代を、FVB背景において、突然変異種SOD1G93A
導入マウスと交雑させた。
【0119】
F1マウスを、既報のとおりのサザンブロット法を用いて導入遺伝子コピー数について解析した(Xu et al., 1993 Cell 73:23-33)。末端DNAをBam H1で消化し、388ヌクレオチドの導入遺伝子断片を作製した。内因性マウスU6プロモータにはBamHI部位が1つしかないため、BamHI消化によって、内因性マウスU6遺伝子からより大きな断片を作製できた。前記U6プロモータ領域に相補的な32P標識RNAオリゴヌクレオチドプローブを、ハイブリダイゼーションに用いた。前記U6領域を標的として用いた。これは、前記内因性マウスU6バンドは同一ブロット上で前記導入遺伝子とともに検出でき、導入遺伝子のコピー数を定量化するための参照として前記内因性バンドを用いることができるためである。
【0120】
ノーザンブロット法を用いて測定したとおり、前記U6-G93A
shRNA作製物を、2種類の遺伝子を導入したトランスジェニックマウスから採取した細胞に発現させた。ウエスタンブロット法を用いて測定した結果、前記U6-G93A
shRNA 作製物は、2種類の遺伝子を導入したトランスジェニックマウスにおける突然変異種SOD1G93A
の発現(U6-G93A shRNA 作製物および突然変異種SOD1G93Aを発現)をサイレンシングすることが明らかになった。
【0121】
例I乃至VIIIの結果の考察
RNAiを用いた優性突然変異種ALS遺伝子を選択的にサイレンシングする可能性を調べた。野生種または突然変異種のSOD1のいずれかにマッチする複数のsiRNAを用いた結果、in vitroにおいて、突然変異種SOD1 G85R に対するsiRNAは前記突然変異種を切断するが、野生種SOD1 RNAの切断は効率的ではないことが示された(図1)。さらに、同一の細胞に突然変異および野生種SOD1タンパク質の両方が存在する場合でも(図4)、これらのsiRNAは、哺乳類の細胞において突然変異SOD1タンパク質発現を選択的に阻害するが野生種については阻害しなかった(図2)。in vivoにおいてsiRNAに処理されたヘアピンを発現するベクタも、マウス肝において突然変異SOD1の発現を選択的に阻害するが、野生種SOD1の発現は阻害しなかった。これらの結果から、優性突然変異対立遺伝子の選択的阻害はRNAiを用いて実現することができ、in vitroまたはin vivoにおいて、前臨床スクリーニングによって最適化したsiRNAおよびshRNA配列を同定することができることが明らかになった。
【0122】
哺乳類細胞においてSOD1 1ヌクレオチドの識別を実現することはできるが、この識別は確実なものではない。一部のsiRNAは1ヌクレオチドの差を有する対立遺伝子間の識別が可能であるが、その他のsiRNAでは識別することができない。結果は、突然変異病因対立遺伝子に方向付けられるようにデザインされたsiRNAに2種類の異なる欠陥があることを示している。第1に、標的に完全にマッチするsiRNAは、その標的を切断し、前記標的遺伝子からのタンパク質発現を阻害することができるが、すべてのsiRNAが同一の効率でサイレンシングを生じるわけではない。たとえば、前記野生種に対するsiRNAの中で、in vitroにおいて、P9およびP10はP11よりも効率的に標的を切断した。P10はまた、哺乳類細胞において、標的遺伝子発現を最も効率的に阻害した(図2)。同様に、前記突然変異SOD1 G85Rに対するsiRNAの中で、P9およびP10はP11よりも効率的に突然変異RNAを切断した。P10はまた、哺乳類細胞において、突然変異SOD1発現の阻害について最も効率的だった(図2)。前記標的に対するsiRNA配列の1ヌクレオチドのシフトの結果、サイレンシングの効率においてこのような顕著な変化を生じる。
第2に、完全にマッチした標的RNAと1ヌクレオチドのミスマッチを有するRNAの間の選択性の差が、使用された6つのsiRNAの中で認められた。たとえば、野生種P10 siRNAは乏しい選択性を付与した。野生種P10は、無細胞アッセイにおいて野生種および突然変異SOD1 RNA の両方を切断し、哺乳類動物の両方の対立遺伝子の発現を高い効率で阻害した。反対に、突然変異SOD1に対して方向付けられたP10 siRNAは最高の選択性を付与した。それは突然変異SOD1 RNA を切断し、無細胞アッセイにおいて突然変異SOD1の発現を阻害し、哺乳類細胞における突然変異SOD1の発現は阻害したが野生種SOD1は阻害しなかった(図1乃至6)。
【0123】
突然変異と野生種のP10 siRNAの選択性の差についての説明は、以下の通りである。突然変異種P10 siRNAと野生種SOD1 mRNA の間のミスマッチがG:G衝突をつくり出したが、野生種P10 siRNA と突然変異種G85R mRNA の間のミスマッチはC:C衝突を生じた(図1A参照)。したがって、特定の配列の1つの対立遺伝子に選択的に作用するsiRNAをデザインする場合、以下が考慮された。理論によって制限したいとは考えないが、プリン:プリンミスマッチは、siRNAのアンチセンス鎖とそのmRNA標的の間に要求されるA型らせんを妨げる(Chiu et al., 2002)。反対に、ピリミジン:ピリミジンミスマッチはA型らせんの中に容易に収容することができる。したがって、siRNAと野生種標的RNAの間のG:G衝突は前記野生種標的を排除し、前記突然変異標的の選択性を高める。顕著なことに、突然変異種SOD1G93Aについて優れた選択性を示す、G93Aに対するsiRNAヘアピンベクタも、野生種SOD1 mRNAを有するG:G衝突をつくり出した。これらの結果は、プリン:プリンミスマッチはピリジン:ピリジンミスマッチよりも大きな選択性を付与することを示唆した。ピリジン:ピリジンミスマッチを含有する本願発明の方法に用いたsiRNAのデザインに加えて、前記siRNAは当業に知られる方法を用いてデザインされる。
【0124】
参考文献
1. Amarzguioui, M., Holen, T., Babaie, E., and
Prydz, H. (2003). Tolerance for mutations and chemical modifications in a
siRNA. Nucleic Acids Res 31, 589-595.
2. Behndig,
A., Karlsson, K., Reaume, A. G., Sentman, M. L. & Marklund, S. L. In
vitro photochemical cataract in mice lacking copper-zinc superoxide dismutase.
Free Radic Biol Med 31, 738-44. (2001).
3. Boutla,
A., Delidakis, C., Livadaras, I., Tsagris, M. & Tabler, M. Short
5'-phosphorylated double-stranded RNAs induce RNA interference in Drosophila.
Curr Biol 11, 1776-80. (2001).
4. Brummelkamp,
T. R., Bernards, R. & Agami, R. A system for stable expression of short
interfering RNAs in mammalian cells. Science 296, 550-3. (2002).
5. Brummelkamp,
T., Bernards, R. & Agami, R. Stable suppression of tumorigenicity by
virus-mediated RNA interference. Cancer Cell 2, 243. (2002).
6. Caplen,
N. J., Parrish, S., Imani, F., Fire, A. & Morgan, R. A. Specific inhibition
of gene expression by small double-stranded RNAs in invertebrate and vertebrate
systems. Proc Natl Acad Sci U S A 98, 9742-7. (2001).
7. Caplen,
N. J. et al. Rescue of polyglutamine-mediated cytotoxicity by double-stranded
RNA-mediated RNA interference. Hum Mol Genet 11, 175-84. (2002).
8. Chiu,
Y.-L. & Rana, T. M. RNAi in human cells: basic structural and functional
features of small interfering RNA. Molecular Cell 10, 549-561 (2002).
9. Cleveland,
D. W. & Rothstein, J. D. From Charcot to Lou Gehrig: deciphering selective
motor neuron death in ALS. Nat Rev Neurosci 2, 806-19. (2001).
10. Devroe,
E. & Silver, P. A. Retrovirus-delivered siRNA. BMC Biotechnol 2, 15.
(2002).
11. Elbashir,
S. M., Lendeckel, W. & Tuschl, T. RNA interference is mediated by 21- and
22-nucleotide RNAs. Genes Dev 15, 188-200. (2001).
12. Elbashir,
S. M. et al. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured
mammalian cells. Nature 411, 494-8. (2001).
13. Elbashir,
S. M., Martinez, J., Patkaniowska, A., Lendeckel, W. & Tuschl, T.
Functional anatomy of siRNAs for mediating efficient RNAi in Drosophila
melanogaster embryo lysate. Embo J 20, 6877-88. (2001).
14. Flood,
D. G. et al. Hindlimb motor neurons require Cu/Zn superoxide dismutase for
maintenance of neuromuscular junctions. Am J Pathol 155, 663-72. (1999).
15. Hamilton,
A. J. & Baulcombe, D. C. A species of small antisense RNA in
posttranscriptional gene silencing in plants. Science 286, 950-2. (1999).
16. Hammond,
S. M., Bernstein, E., Beach, D. & Hannon, G. J. An RNA-directed nuclease
mediates post-transcriptional gene silencing in Drosophila cells. Nature 404,
293-6. (2000).
17. Hannon,
G. J. RNA interference. Nature 418, 244-51. (2002).
18. Hutvagner,
G. & Zamore, P. D. RNAi: nature abhors a double-strand. Curr Opin Genet Dev
12, 225-32. (2002).
19. Jacque,
J. M., Triques, K. & Stevenson, M. Modulation of HIV-1 replication by RNA
interference. Nature 418, 435-8. (2002).
20. Kawase,
M. et al. Exacerbation of delayed cell injury after transient global ischemia
in mutant mice with CuZn superoxide dismutase deficiency. Stroke 30, 1962-8.
(1999).
21. Kondo,
T. et al. Reduction of CuZn-Superoxide Dismutase Activity Exacerbates Neuronal
Cell Injury and Edema Formation after Transient Focal Cerebral Ischemia.
Journal of Neuroscience 17, 4180-9 (1997).
22. Lee,
N. S. et al. Expression of small interfering RNAs targeted against HIV-1 rev
transcripts in human cells. Nat Biotechnol 20, 500-5. (2002).
23. Lipardi,
C., Wei, Q. & Paterson, B. M. RNAi as random degradative PCR: siRNA primers
convert mRNA into dsRNAs that are degraded to generate new siRNAs. Cell 107,
297-307. (2001).
24. Matz,
P. G., Copin, J. C. & Chan, P. H. Cell death after exposure to subarachnoid
hemolysate correlates inversely with expression of CuZn-superoxide dismutase.
Stroke 31, 2450-9. (2000).
25. Matzuk,
M. M., Dionne, L., Guo, Q., Kumar, T. R. & Lebovitz, R. M. Ovarian function
in superoxide dismutase 1 and 2 knockout mice. Endocrinology 139, 4008-11.
(1998).
26. McFadden,
S. L., Ding, D. & Salvi, R. Anatomical, metabolic and genetic aspects of
age-related hearing loss in mice. Audiology 40, 313-21. (2001).
27. McManus,
M. T. & Sharp, P. A. Gene silencing in mammals by small interfering RNAs.
Nat Rev Genet 3, 737-47. (2002).
28. McManus,
M. T., Petersen, C. P., Haines, B. B., Chen, J. & Sharp, P. A. Gene
silencing using micro-RNA designed hairpins. Rna 8, 842-50. (2002).
29. Miyagishi,
M. & Taira, K. U6 promoter-driven siRNAs with four uridine 3' overhangs
efficiently suppress targeted gene expression in mammalian cells. Nat
Biotechnol 20, 497-500. (2002).
30. Nykanen,
A., Haley, B. & Zamore, P. D. ATP requirements and small interfering RNA
structure in the RNA interference pathway. Cell 107, 309-21. (2001).
31. Paddison,
P. J., Caudy, A. A., Bernstein, E., Hannon, G. J. & Conklin, D. S. Short
hairpin RNAs (shRNAs) induce sequence-specific silencing in mammalian cells. Genes
Dev 16, 948-58. (2002).
32. Paul,
C. P., Good, P. D., Winer, I. & Engelke, D. R. Effective expression of
small interfering RNA in human cells. Nat Biotechnol 20, 505-8. (2002).
33. Shefner,
J. M. et al. Mice lacking cytosolic copper/zinc superoxide dismutase display a
distinctive motor axonopathy. Neurology 53, 1239-46. (1999).
34. Schwarz,
D. S., Hutvagner, G., Haley, B. & Zamore, P. D. Evidence that siRNAs
function as guides, not primers, in the Drosophila and human RNAi pathways.
Molecular Cell 10, 537-548 (2002).
35. Sui,
G. et al. A DNA vector-based RNAi technology to suppress gene expression in
mammalian cells. Proc Natl Acad Sci U S A 99, 5515-20. (2002).
36. Taylor,
J. P., Hardy, J. & Fischbeck, K. H. Toxic proteins in neurodegenerative disease.
Science 296, 1991-5. (2002).
37.
Tuschl, T., Zamore, P. D., Lehmann, R., Bartel, D. P. & Sharp, P. A.
Targeted mRNA degradation by double-stranded RNA in vitro. Genes Dev 13,
3191-7. (1999).
38. Xia,
H., Mao, Q., Paulson, H. L. & Davidson, B. L. siRNA-mediated gene silencing
in vitro and in vivo. Nat Biotechnol 20, 1006-10. (2002).
39. Xu, P., Vernooy, S. Y., Guo, M., and Hay, B. A.
(2003). The Drosophila MicroRNA Mir-14 Suppresses Cell Death and Is Required
for Normal Fat Metabolism. Curr Biol 13, 790-795.
40. Yu, J. Y., DeRuiter, S. L. & Turner, D. L. RNA interference by
expression of short-interfering RNAs and hairpin RNAs in mammalian cells. Proc
Natl Acad Sci U S A 99, 6047-52. (2002).
41. Zamore,
P. D., Tuschl, T., Sharp, P. A. & Bartel, D. P. RNAi: double-stranded RNA
directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals.
Cell 101, 25-33. (2000).
42. Zeng,
Y., Wagner, E. J. & Cullen, B. R. Both natural and designed micro RNAs can
inhibit the expression of cognate mRNAs when expressed in human cells. Mol Cell
9, 1327-33. (2002).
【0125】
その他の実施態様
本願発明は、発明の詳細な説明に関連して説明されているが、上述の説明は具体例を示すことを意図しており本願発明の範囲を制限することは意図しておらず、それは添付の特許請求の範囲によって定義されると理解される。その他の局面、利点、および改善点は、添付の特許請求の範囲内である。
【図面の簡単な説明】
【0126】
【図1】siRNA 2本鎖は、in vitroにおいて突然変異および野生種のSOD1を区別することができる(A) 使用したsiRNA 2本鎖:突然変異siRNA P11 (配列番号 5、センス; 配列番号 6、アンチセンスまたはガイド)、突然変異 siRNA P10 (SEQ ID NO: 3、センス; SEQ ID NO: 4、アンチセンスまたはガイド)、突然変異 siRNA P9 (SEQ ID NO: 1、センス; SEQ ID NO: 2 アンチセンスまたはガイド)、SOD1 野生種標的 (SEQ ID NO: 7)、SOD1 突然変異標的 (SEQ ID NO: 8)、野生種 siRNA P11 (SEQ ID NO: 9 センス; SEQ ID NO: 10、アンチセンスまたはガイド)、野生種 siRNA P10 (SEQ ID NO: 11、センス; SEQ ID NO: 12、アンチセンスまたはガイド)、野生種 siRNA P9 (SEQ ID NO: 13、センス; SEQ ID NO: 14、アンチセンスまたはガイド) (B) 突然変異siRNA p10は、野生種SOD1 mRNA ではなく突然変異SOD1 mRNA をRNAi経路による破壊に方向付ける。
【図2】Hela細胞におけるsiRNAによる突然変異SOD1 G85R発現の選択的阻害。SOD1wtGFPまたはG85R-GFPを各種のsiRNAとともにコトランスフェクトした。dsRedをトランスフェクション対照としてコトランスフェクトした。緑色および赤色蛍光細胞をFACSを用いて定量した。(A) トランスフェクションにおける緑色(黒線)および赤色細胞(白線)の相対数(n=3)。エラーバーは標準偏差を示す。
【図3】Hela細胞におけるU6-G93A ベクタによる突然変異SOD1 G93A発現の選択的阻害。(A) G93A shRNA のデザイン(配列番号16)、(B) SOD1wtGFPまたはSOD1G93A-GFPをU6空またはU6-G93Aとともにコトランスフェクトした。dsRedをトランスフェクション対照としてコトランスフェクトした。緑色および赤色傾向細胞をFACSを用いて定量した。4回の実験の結果を平均した。エラーバーは標準偏差を示す。
【図4】神経細胞芽腫N2a細胞におけるsiRNAおよびU6-G93Aベクタによる突然変異SOD1発現の選択的阻害。(A) G85Rに対するsiRNA (n=4)、(B) U6-G93Aベクタ(n=3)。エラーバーは標準偏差を示す。
【図5】同一の細胞におけるsiRNAによる、野生種SOD1ではなく、突然変異SOD1 G85R発現の選択的阻害。(A) 突然変異SOD1 G85R-GFP を検出するトランスフェクトHela細胞のタンパク質ブロットから測定した、SOD1の相対的レベル(4回のトランスフェクションの平均)。エラーバーは標準誤差を示す。
【図6】in vivoにおけるU6-G93A ベクタによる突然変異SOD1発現の選択的阻害(A) 水力学的トランスフェクション法を用い、マウスにおいてSOD1 G93A-GFP をC末端mycタグ付き野生種ヒトSOD1とともにコトランスフェクトした。SDA-PAGE上の相対バンド強度を定量した。野生種SOD1mycに対するSOD1 G93A-GFPの割合を示す。各グループにおいて8匹の動物を用いた。U6-G93A グループは、他の2つのグループとは有意に異なる(p < 0.05)。
【図7】ヒトSOD-1タンパク質のジェンバンク登録。受入番号NP_000445。野生種SOD-1 (配列番号18)の推定アミノ酸配列を示す。
【図8】ヒトSOD-1 mRNAのジェンバンク登録。受入番号NM_000454。野生種SOD-1 (配列番号17)のヌクレオチド配列を示す。
【図9】図9はSOD1ゲノム配列(配列番号19)である。
【配列表】
【背景技術】
【0001】
関連出願
本願明細書は、2002年11月4日に提出された米国特許仮出願第60/423,507号「Allele-Specific RNA Interference(対立特異的RNA干渉)」、および2003年7月18日に提出された米国特許仮出願60/488,283号「Allele-Specific RNA
Interference(対立特異的RNA干渉)」の利益を主張する。参照された特許仮出願の内容全体は、引用をもって本願明細書に援用するものとする。
【0002】
連邦政府の資金による研究に関する記載
米国政府は、国立衛生研究所(NIH)により授与された認可番号GM62862およびGM53874、ならびに国立神経疾患・卒中研究所(NINDS)により授与された認可番号NS35750にしたがった本願発明の一定の権利を有する可能性がある。
【0003】
背景
優性の機能獲得型遺伝子によって引き起こされる疾患は、1つの突然変異体と1つのその遺伝子の野生種コピーを有するヘテロ接合体に生じる。この種の疾患で最も知られるものの一部は、共通の神経変性疾患で、アルツハイマー病、ハンチントン病、パーキンソン病、および筋萎縮性側策硬化症(ALS; 「ルー・ゲーリック病」)が含まれる(Taylor
et al., 2002)。これらの疾患では、突然変異タンパク質が細胞変性を引き起こす正確な経路が明らかになってはいないが、突然変異タンパク質となる細胞毒性の起源は知られている。
【0004】
SOD1における突然変異は運動ニューロンの変性を生じて、ALSを引き起こす。それは、前記突然変異タンパク質がいくらかの毒性を獲得したためである(Cleveland et al., 2001)。この毒性の性質も、最終的に運動ニューロンの変性に至る下流経路も解明されていない。マウスでは、突然変異体SOD1の発現だけでなく、遺伝子ノックアウトによるSOD1の削除がALSを引き起こす。しかし、遺伝子ノックアウトマウスは、繁殖性の減退(Matzuk et al., 1990)、運動軸索障害(Shefner et al., 1999)、蝸牛有毛細胞の加齢に関連する喪失(McFadden et al., 2001)、および神経筋接合部シナプス(Flood et al.,
1999)を含む様々な異常、ならびに、CNSおよびその他の系への、興奮毒性、虚血、神経毒、および放射線照射など各種の有害な攻撃への高い感受性を生じる。突然変異種の毒性、および野生種タンパク質の機能的重要性を考えると、この疾患のための理想的な治療法は、前記突然変異タンパク質の発現を選択的に阻害する一方で野生種の発現を維持することである。
【0005】
要約
本願発明は、優性の機能獲得型疾患を治療する新規の方法に関連する。特に、本願発明は、機能獲得型遺伝子から転写された突然変異mRNAを選択的に破壊し、それによりそのような遺伝子によってコードされる突然変異タンパク質の産生を抑制する方法を提供する。本願発明は、低分子干渉RNA(siRNA)および低分子ヘアピンRNA(shRNA)の両方のがたとえばG85R SOD1またはG93A SOD1などの突然変異対立遺伝子の発現を選択的に阻害する一方で、1ヌクレオチド特異性で前記野生種タンパク質の発現を阻害するようにデザインすることができるという発見に、部分的に基づいている。
【0006】
本願発明の方法は、たとえば、筋萎縮性側索硬化症(ALS)に関連する銅亜鉛スーパーオキシドジスムターゼ(SOD1)遺伝子にある点突然変位など、突然変異遺伝子における単一の対立遺伝子に生じる選択的な点変異に対して、RNA干渉技術(RNAi)を用いる。RNAiは多様な真核生物の遺伝子発現を、標的遺伝子への配列類似性を有する短い2本鎖RNA(低分子干渉RNAまたはsiRNAとよばれる)を導入することによって配列選択的に抑制するのを媒介する(Caplen et al.,
2001; Elbashir et al.2001c)。本願発明の2本鎖siRNAまたはshRNA発現ベクタを用いて、たとえばSOD1突然変異タンパク質などの毒性突然変異遺伝子の発現を選択的にサイレンシングし、それにより野生種SOD1対立遺伝子を継続的に機能させることができる。
【0007】
本願発明はまた、たとえばin vivoなどにおいて細胞で発現する場合、細胞自体のRNAi経路を用いて標的遺伝子の突然変異対立遺伝子を選択的にサイレンシングする標的siRNAを生じるように細胞によって処理された、新規の人為的操作RNA前駆体の発見に基づく。これらの操作したRNA前駆体をコードする核酸分子を、in vivoにおいて適切な制御配列(たとえば、プラスミドなどのベクタにある導入遺伝子)を用いて細胞に導入することによって、操作したRNA前駆体の発現は、時間的および空間的に、すなわち、特定の時間および/または特定の組織、器官、または細胞において選択的に制御することができる。
【0008】
ある局面では、本願発明は、遺伝子の少なくとも2つの異なる対立遺伝子を含む、細胞中の標的対立遺伝子の発現を、前記標的対立遺伝子に特異的なsiRNAを前記細胞に投与することによって阻害する方法を特徴とする。ある実施態様では、前記標的対立遺伝子は、優性の機能獲得型突然変異に関連する障害と相関する。別の実施態様では、前記障害は筋萎縮性軸索硬化症、ハンチントン病、アルツハイマー病、またはパーキンソン病である。
【0009】
別の局面では、本願発明は、優性の機能獲得型突然変異対立遺伝子の存在と相関する疾患を有する対象の治療方法であって、当該方法が前記突然変異対立遺伝子に特異的なsiRNAを治療上有効な量、前記対象に投与するステップを含む方法を特徴とする。ある実施態様では、前記siRNAは機能獲得型突然変異を標的とする。別の実施態様では、前記障害は筋萎縮性軸索硬化症、ハンチントン病、アルツハイマー病、またはパーキンソン病である。
【0010】
ある実施態様では、前記疾患は筋萎縮性軸索硬化症である。さらなる実施態様では、前記対立遺伝子はSOD1突然変異対立遺伝子である。
【0011】
ある実施態様では、前記siRNAは突然変異SOD1対立遺伝子(配列番号8)を標的とし、好ましくはP10(配列番号4)、次に好ましくはP9(配列番号2)、次に好ましくはP11(配列番号6)を有する図1Aに記載の突然変異siRNA配列を含む、またはそれから成る。
【0012】
別の実施態様では、前記siRNA(たとえば制御siRNA)は野生種SOD1対立遺伝子を標的とし、好ましくはP9(配列番号14)またはP10(配列番号12)、次に好ましいのはP11(配列番号10)を有する図1Aに記載の野生種siRNA配列を含む、またはそれから成る。
【0013】
別の局面では、本願発明は図1Aに記載の配列を含むsiRNAを提供する。
【0014】
別の局面では、本願発明は、図1Aに記載の配列(配列番号4)を含むp10突然変異siRNAを提供する。
【0015】
別の局面では、本願発明は、図1Aに記載の配列(配列番号2)を含むp9突然変異siRNAを提供する。
【0016】
別の局面では、本願発明は、図3Aに記載の配列(配列番号16)を含むG93A SOD1 shRNA、および本願発明のshRNAを含む発現作製物を提供する。
別の局面では、本願発明は本願発明のsiRNAおよび/またはshRNAを含む治療用組成物、および薬学的に許容な担体を提供する。
【0017】
本願発明のその他の特徴および利点は、以下の発明の詳細な説明および特許請求の範囲から明らかになるであろう。
【0018】
発明の詳細な説明
銅亜鉛スーパーオキシドジスムターゼ(SOD1)遺伝子の突然変異は、筋萎縮性側策硬化症のサブセット、運動ニューロンの変性を来す神経変性疾患、麻痺および死を引き起こす(Brown and Robberecht, 2001;Siddique and Lalani, 2002)。突然変異SOD1が毒性を獲得することによって運動ニューロン変性を引き起こすことは、既知である。しかし、この毒性の分子機序も、運動ニューロンの死滅を来す機序も、いずれも解明されていない。疾患の機序の理解が不十分なことから、合理的なデザインの治療法が着実で有効な転帰を生じたことがない。その一方で、運動ニューロンを死滅させる毒性が突然変異したタンパク質に起因することから(Cleveland and Rothstein, 2001)、突然変異タンパク質の減少によって、その疾患を緩和または予防できるはずである。RNA干渉(RNAi)技術を用いてこの目的を達成することができる。
【0019】
本願発明は、siRNAおよびshRNAが突然変異対立遺伝子の発現を選択的に阻害できるという発見であって、たとえばALSと相関するSOD1の突然変異など、特定の突然変異についてのケースに見られるとおり、突然変異mRNAと野生種と違いが1ヌクレオチドだけの場合であっても阻害できるという発見に基づく。これらの方法は、ALSに限定しないがそれを含む、優性で機能獲得型の遺伝子突然変異によって生じる疾患の治療に適用することができる。本願発明のsiRNAは、1ヌクレオチドの識別、およびその標的遺伝子の発現の選択的下方制御を可能にする。
【0020】
本願発明の方法は、たとえば、筋萎縮性側索硬化症(ALS)に関連する銅亜鉛スーパーオキシドジスムターゼ(SOD1)遺伝子の中ある点突然変位など、突然変異遺伝子における単一の対立遺伝子に生じる選択的な点変異に対して、RNA干渉技術(RNAi)を用いる。RNAiは多様な真核生物の遺伝子発現を、標的遺伝子への配列相同性を有する短い2本鎖RNA(低分子干渉RNAまたはsiRNAとよばれる)を導入することによって配列選択的に抑制するのを媒介する(Caplen et al.,
2001; Elbashir et al.2001c)。本願発明の2本鎖siRNAまたはshRNA発現ベクタを用いて、たとえばSOD1突然変異タンパク質などの毒性突然変異遺伝子の発現を選択的にサイレンシングし、それにより野生種SOD1対立遺伝子を継続的に機能させることができる。
【0021】
遺伝子発現の配列選択的、転写後不活性化は、RNAiと呼ばれる現象である、標的遺伝子に相当する2本鎖RNAの導入によって、多様な原核生物に実現させることが可能である(Hutvagner and Zamore, 2002; Hannon, G. J., 2002; McManus and Sharp,
2002)。RNAiの方法論は、培養哺乳類細胞にも拡張されている(Caplen et al, 2001; Elbashir
et al., 2001)。このアプローチは、RNAi経路の中間体であるsiRNAが、当該siRNA配列に相当するmRNAの分解を引き起こすことがあるという発見を活用している。さらに、in vivoにおいて転写されたshRNAは、shRNAステムの配列に相補的な標的RNAの分解を引き起こすことがある。それは、細胞内でshRNAがsiRNAに加工されるためである(Paul et al., 2002; Lee et al., 2002;
Paddison et al., 2002; Sui et al., 2002; Yu et al., 2002; McManus et al., 2002;
Zeng et al., 2002; Brummelkamp et al., 2002; Miyagishi et al., 2002; Jacque et al.,
2002)。本願出願人らは、siRNA 2本鎖またはshRNAを発現するウイルスを用いて、突然変異体率遺伝子の発現を選択的に阻害しながら、同時に発現する野生種対立遺伝子の発現を保護することができることを実証する。
【0022】
ALS関連SOD1突然変異の大半は、1アミノ酸の変化を生じる1ヌクレオチド点変異である(ALS遺伝子(SOD1, ALS など)突然変異のALSオンラインデータベース)。したがって、突然変異体の発現だけを選択的にサイレンシングを生じさせ、野生種にはサイレンシングを生じさせないために、1ヌクレオチド特異性が必要である。本願出願人らは、哺乳類細胞において1ヌクレオチドの認識が実現可能であることを証明している。
【0023】
本願発明をより容易に理解するため、まず特定の用語を定義する。
【0024】
「単離された核酸分子または配列」は、由来する生物の天然のゲノムにおいて、(片方は5’末端でもう片方が3’末端に)ただちに連続するコード配列の両側に、ただちに連続しない核酸分子または配列である。したがって、当該用語には、たとえば、ベクター、自己複製プラスミドもしくはウイルス、または原核生物もしくは真核生物のゲノムDNAに組み込まれた、または分離可能な分子として存在する組換えDNAまたはRNAが含まれる。また、さらなるポリペプチド配列をコードするハイブリッド遺伝子の一部分である組換えDNAも含まれる。
【0025】
「ヌクレオシド」という用語は、リボースまたはデオキシリボース糖に共有結合したプリンまたはピリミジンを有する分子を意味する。例示的なヌクレオチドには、アデノシン、グアシン、シチジン、ウリジン、およびチミジンが含まれる。さらなる例示的なヌクレオシドには、イノシン、1-メチルイノシン、シュードウリジン、5,6-ジヒドロウリジン、リボチミジン、2N-メチルグアノシン、および2,2N,N-ジメチルグアノシン(「稀少」ヌクレオチドともよばれる)が含まれる。「ヌクレオチド」という用語は、糖部分にエステル結合したリン酸基を1つ以上有するヌクレオシドを意味する。例示的なヌクレオシドには、1リン酸ヌクレオシド、2リン酸ヌクレオシド、および3リン酸ヌクレオシドが含まれる。「ポリヌクレオチド」および「核酸分子」という用語は、本願明細書では交換可能に用いられ、5’および3’炭素原子の間のホスホジエステル結合によって結合したヌクレオシドのポリマーを意味する。
【0026】
「RNA」または「RNA分子」または「リボ核酸分子」という用語は、リボヌクレオチドのポリマーを意味する。「DNA」または「DNA分子」または「デオキシリボ核酸分子」という用語は、デオキシリボヌクレオチドのポリマーを意味する。DNAおよびRNAは自然に合成することができる(たとえば、それぞれDNA複製またはDNAの転写によって)。RNAは転写後に修飾されることがある。DNAおよびRNAは化学的にも合成することができる。DNAおよびRNAは1本鎖であっても(すなわちそれぞれssRNAおよびssDNA)、または多重鎖(たとえば2本鎖、すなわちそれぞれdsRNAおよびdsDNA)であってもよい。「mRNA」または「メッセンジャRNA」は、1つ以上のポリペプチド鎖のアミノ酸配列を特定する1本鎖RNAである。この情報は、リボソームがmRNAに結合する際にタンパク質合成中に翻訳される。
【0027】
組換えRNA前駆体、または組換え核酸分子の中などに使われる「組換え」という用語は、その前駆体または分子が天然には認められず、その前駆体または分子の核酸配列全体または一部分が人工的に作成された、または選択されたものであることを示す。ひとたび作製または選択されると、その配列は、細胞内のメカニズムによって複製、翻訳、または転写などの処理を受けることができる。したがって、組換え核酸分子を含む導入遺伝子から細胞内で産生されたRNA前駆体は組換えRNA前駆体である。
【0028】
本願明細書に記載の「低分子干渉RNA」(「siRNA」)という用語(「短分子干渉RNA」ともいう)は、RNA干渉を方向付けるまたは媒介する能力のある約10乃至50ヌクレオチドを含むRNA(またはRNA類似体)を意味する。好ましくは、siRNAは、約15乃至30ヌクレオチドまたはヌクレオチド類似体、さらに好ましくは約16乃至25ヌクレオチド(またはヌクレオチド類似体)、よりさらに好ましくは約18乃至23ヌクレオチド(またはヌクレオチド類似体)および、よりさらに好ましくは約19乃至22ヌクレオチド(またはヌクレオチド類似体)(たとえば、19、20、21、または22ヌクレオチドまたはヌクレオチド類似体)を含む。「短分子」siRNAとは、たとえば19、20、21、または22ヌクレオチドなど、約21ヌクレオチド(またはヌクレオチド類似体)を含む、siRNAを意味する。「長分子」siRNAとは、たとえば23、24、24、または26ヌクレオチドなど、約24乃至25ヌクレオチド(またはヌクレオチド類似体)を含む、siRNAを意味する。短いsiRNAほどRNAiを媒介する能力を維持しているとすると、短分子siRNAには、たとえば16、17、または18ヌクレオチドなど、19ヌクレオチド未満が含まれてよい。同様に、長分子siRNAは、短分子siRNAに対して、たとえば酵素的処理など、RNAiが不在のさらなる処理を媒介する能力を維持しているとすると、ある場合には、長分子siRNAには26ヌクレオチド超が含まれてもよい。
【0029】
「ヌクレオチド類似体」または「改変ヌクレオチド」または「修飾ヌクレオチド」という用語は、天然に存在しないリボヌクレオチドまたはデオキシリボヌクレオチドを含む、標準的ではないヌクレオチドを意味する。好ましいヌクレオチド類似体は、任意の位置を修飾し、ヌクレオチドの特定の化学的特性を変えつつ、核酸類似体の目的の機能を実行する能力を維持できるようにする。誘導可能なヌクレオチドの位置の例には、たとえば5-(2-アミノ)プロピルウリジン、5-ブロモウリジン、5-プロピンウリジン、5-プロペニルウリジンなどの5位、たとえば6-(2-アミノ)プロピルウリジンなどの6位、たとえば8-ブロモグアノシン、8-クロログアノシン、8-フルオログアノシンなどのアデノシンおよび/またはグアノシンの8位が含まれる。ヌクレオチド類似体も、たとえば7-デアザアデノシンなどのデアザヌクレオチド、O-およびN修飾(たとえばアルカリ化、たとえばN6-メチルアデノシン、またはその他の当業に知られる)ヌクレオチド、およびたとえばHerdewijn, Antisense Nucleic Acid Drug
Dev., 2000 Aug. 10(4):297-310に記載されているなどのその他のヘテロ周期性修飾ヌクレオチド類似体も含まれる。
【0030】
ヌクレオチド類似体は、ヌクレオチドの糖部分への修飾も含んでもよい。たとえば、2’OH基はH、OR、R、F、Cl、Br、I、SH、SR、NH2、NHR、NR2、COOR、またはORであって、当該化学基においてRは置換または非置換C1乃至C6
アルキル、アルケニル、アルキニル、アリールなどから選択される化学基と置換してもよい。その他の可能な修飾には、米国特許第5,858,988号および6,291,438号に記載されるものが含まれる。
【0031】
ヌクレオチドのリン酸基は、たとえばリン酸基の1つ以上の酸素を硫黄で置換する(たとえばホスホロチオ酸)ことにより、またはヌクレオチドがたとえばEckstein, Antisense Nucleic Acid Drug Dev. 2000
Apr. 10(2):117-21、
Rusckowski et al. Antisense Nucleic Acid Drug Dev. 2000
Oct. 10(5):333-45、 Stein, Antisense
Nucleic Acid Drug Dev. 2001 Oct. 11(5): 317-25、 Vorobjev et al. Antisense
Nucleic Acid Drug Dev. 2001 Apr. 11(2):77-85、 および米国特許第5,684,143号などに記載される目的の機能を実施できるようなその他の置換を行うことによって修飾してもよい。上に参照した修飾の一部は、好ましくは、たとえばin vivo またはin vitroにおいて前記類似体を含むポリヌクレオチドの加水分解速度を低減させる。
【0032】
本願明細書に記載の、たとえばsiRNA2本鎖またはsiRNA配列のアンチセンス鎖など、siRNAまたはRNAi剤の「アンチセンス鎖」という用語は、たとえばサイレンシングのターゲットである遺伝子のmRNAの約15乃至30、16乃至25、18乃至23または19乃至22ヌクレオチドなど、 約10乃至50ヌクレオチドの部分に実質的に相補的な鎖を意味する。第1の鎖のアンチセンス鎖は、たとえばRNAi機構またはプロセスによる望ましい標的mRNAの破壊を引き起こすのに十分な相補性など、標的特異的RNA干渉(RNAi)を方向付ける望ましい標的mRNA配列に十分相補的な配列を有する。たとえばsiRNA2本鎖またはsiRNA配列のアンチセンス鎖など、siRNAまたはRNAi剤の「センス鎖」または「第2の鎖」という用語は、アンチセンス鎖または第1の鎖に相補的な鎖を意味する。アンチセンスまたはセンス鎖は、第1鎖または第2鎖とも呼ばれ、標的配列に相補性を有する第1鎖または第2鎖、および前記第1鎖または第2鎖に相補性を有する第2鎖または第1鎖である。
【0033】
本願明細書に記載の「ガイド鎖」という用語は、たとえばsiRNA2本鎖またはsiRNA配列のアンチセンス鎖など、RNAi剤の鎖であって、RISC複合体に進入し、標的mRNAの切断を方向付ける、鎖を意味する。
【0034】
本願明細書に記載のアンチセンス鎖の5’末端としての「5’末端」は、たとえばアンチセンス鎖の5’末端における1乃至約5ヌクレオチドなどの5’末端ヌクレオチドを意味する。本願明細書に記載のセンス鎖の3’末端としての「3’末端」は、たとえば相補的アンチセンス鎖の5’末端のヌクレオチドに相補的な1乃至約5ヌクレオチドの領域などの、領域を意味する。
【0035】
「オリゴヌクレオチド」という用語は、ヌクレオチドおよび/またはヌクレオチド類似体の短いポリマーを意味する。「RNA類似体」という用語は、相当する未改変または未修飾RNAと比較して少なくとも1つの改変またな修飾ヌクレオチドを有するが、前記の相当する未改変または未修飾RNAと同一のまたは類似する性質または機能を保持する、ポリヌクレオチド(たとえば化学合成されたポリヌクレオチドなど)を意味する。上述の通り、前記オリゴヌクレオチドは、ホスホジエステル結合を有するRNA分子と比較してRNA類似体の加水分解が低速度になる結合で結合していてよい。たとえば、前記類似体のヌクレオチドはメチレンジオール、エチレンジオール、オキシメチルチオ、オキシエチルチオ、オキシカルボニルオキシ、ホスホロジアミド酸、ホホロアミド酸、および/またはホスホロチオ酸結合を含んでよい。好ましいRNA類似体には、糖および/または骨格修飾リボヌクレオチドおよび/またはデオキシリボヌクレオチドが含まれる。そのような改変または修飾にはさらに、RNA末端または内部などへの非ヌクレオチド物質の付加(RNAの1つ以上のヌクレオチド)が含まれる。RNA類似体は、RNA干渉を媒介する能力を有する天然のRNAに十分類似しているだけでよい。
【0036】
本願明細書に記載の通り、「RNA干渉」(「RNAi」)という用語は、RNAの選択的な細胞内分解を意味する。RNAi は、細胞内で外来のRNA(たとえばウイルス性RNA)を除去するため自然にに生じる。天然のRNAiは、分解メカニズムをその他の同様のRNA配列に方向付ける遊離dsRNAから切断された断片によって進行する。代替的には、RNAiは、たとえば標的遺伝子の発現をサイレンシングさせるなど、人工的に開始させることもできる。
【0037】
「標的特異的RNA干渉(RNAi)を方向付ける望ましい標的mRNA配列に十分相補的な配列」である鎖を有するRNAi作用剤は、その鎖がRNAi機構またはプロセスによる、標的mRNAの破壊を惹起するのに十分な配列を有することを意味する。
本願明細書に記載の「単離されたRNA」(たとえば「単離されたsiRNA」または「単離されたsiRNA前駆体」)という用語は、組換え技術によって生成した場合には他の細胞物質または培養培地を実質的に含まない、または化学的に合成された場合には、化学前駆体またはその他の化学物質を実質的に含まない、RNA分子を意味する。
【0038】
「標的遺伝子」は、発現が選択的に阻害または「サイレンシング」されている遺伝子である。このサイレンシングは、細胞のRNAiシステムによって人工的に作製したRNAからつくり出されたsiRNAによる、標的遺伝子のmRNAの切断によって実現する。RNA前駆体の2本鎖ステムの一部分またはセグメントは、標的遺伝子のmRNAの約18乃至約40ヌクレオチド、またはそれ以上のヌクレオチドの部分に、たとえば完全に相補的など、相補的なアンチセンス鎖である。
【0039】
本願明細書に記載の「導入遺伝子」という用語は、細胞内に人為的に挿入した任意の核酸分子を意味する。そのような導入遺伝子には、トランスジェニック生物に部分的または完全に非相同的(すなわち外来性)である遺伝子を含むこともある。「導入遺伝子」という用語はまた、たとえばDNAなどの選択された1つ以上の核酸配列であって、その核酸配列は、たとえば動物などのトランスジェニック生物に発現させるために1つ以上の作製されたRNA前駆体をコードする核酸配列であって、その生物は部分的または完全にトランスジェニック動物に非相同的または外来性であるか、またはトランスジェニック動物の内因性遺伝子に相同的であるが、天然遺伝子とは異なる部分にある前記動物のゲノムに挿入するようにデザインされたトランスジェニック生物である、核酸配列を意味する。導入遺伝子には、1つ以上のプロモータ、および特定の核酸配列の発現に必要なイントロンなどのその他の任意のDNAであって、すべて特定の配列に機能的に連結されているプロモータまたはその他の任意のDNAが含まれ、およびエンハンサ配列が含まれてもよい。
【0040】
疾患または障害に「関与する」遺伝子には、前記疾患または障害もしくは前記疾患または障害の少なくとも1つの症状に作用するまたはそれを生じる、正常なまたは異常な発現または機能の遺伝子が含まれる。
【0041】
「発現の対立遺伝子特異的阻害」とは、たとえば両方の対立遺伝子が同一の細胞に存在する場合など、1つの対立遺伝子の発現をもう1つの対立遺伝子の発現よりも著しく阻害する能力を意味する。たとえば、その対立遺伝子は1、2、3またはそれ以上のヌクレオチドが異なっていてもよい。ある場合には、1つの対立遺伝子は、たとえば、優性の機能獲得型突然変異に相関する疾患など、疾患の原因に関連する。
【0042】
本願明細書に記載の「機能獲得型突然変異」という用語は、遺伝子中の任意の突然変異であって、当該突然変異において、前記遺伝子によってコードされたタンパク質(つまり突然変異タンパク質)は、通常タンパク質(つまり野生種タンパク質)に関連していない機能を獲得するか、または疾患もしくは障害を引き起こすまたはそれに寄与する機能を獲得する、突然変異を意味する。機能獲得突然変異は、コードされたタンパク質の機能に変化を生じる、遺伝子中の1つ以上のヌクレオチドの欠失、付加、または置換であってよい。ある実施態様では、機能獲得突然変異は前記突然変異の機能を変更するか、またはその他のタンパク質と相互作用を引き起こす。別の実施態様では、機能獲得突然変異は、たとえば改変した突然変異タンパク質と前記正常野生種タンパク質との相互作用によって、正常野生種タンパク質の減少、または除去を引き起こす。
【0043】
「細胞または組織における遺伝子機能の検証」という文言は、そこから生じる発現、活性、機能、または表現型の検証または研究を意味する。
【0044】
本願発明の様々な方法論には、本願明細書では「適切な対照」とも交換可能な意味である「好適な対照」と、価値、レベル、特色、特徴、性質などとの比較に関与するステップを含む。「好適な対照」または「適切な対照」は、比較の目的で有用な当業者が精通している任意の対照または標準である。ある実施態様では、「好適な対照」または「適切な対照」は、本願明細書に記載の通り、RNAi方法論を実行する前に決定する、価値、レベル、特色、特徴、性質などである。たとえば、転写速度、mRNAレベル、翻訳速度、タンパク質レベル、生物学的活性、細胞の特徴または性質、ゲノタイプ、表現型などは、細胞または生物に本願発明のRNAiを導入する前に決定することができる。別の実施態様では、「好適な対照」または「適切な対照」は、細胞、またはたとえば正常な形質を示す対象、または正常細胞もしくは生物などの生物において決定される価値、レベル、特色、特徴、性質などである。さらに別の実施態様では、「好適な対照」または「適切な対照」は、予め決定された価値、レベル、特色、特徴、性質などである。
【0045】
他に定義のない限り、本願明細書に用いられたすべての技術および科学用語は、本願発明が属する当業者によって普通に理解されるものと同じ意味を有する。本願明細書に記載のものに類似するまたはそれに相当する方法および材料は、本願発明の実施またはテストに用いることができ、好適な方法および材料を以下に説明する。 本願明細書に引用されるすべての出版物、特許出願、特許、およびその他の参考文献は、その全体を引用によりここに援用する。紛争の場合、定義を含む本願明細書が支配するだろう。さらに、当該材料、方法、および例は、実例のみで、制限を意図しない
【0046】
本願発明のさまざまな局面は、以下のサブセクションでさらに詳細に説明する。
【0047】
I. 筋萎縮性側策硬化症(ALS)
筋萎縮性側策硬化症(ALS)は、ルー・ゲーリック病としても知られ、大脳皮質、脳幹、および脊髄の運動ニューロンが関与する、進行性、致死性の神経変性障害である(Hirano, A., 1996, Neurology 47 (Suppl. 2), S63-S66)。この疾患は、たとえばSOD1など、前記遺伝子の1つの突然変異種と1つの野生種コピーを有する人間において生じる、優性の機能獲得型遺伝子によって引き起こされる。SOD1突然変異を引き起こすALSは、前記タンパク質中の1つのアミノ酸を変化させる1ヌクレオチド点突然変異である。この疾患はさらに、進行性の運動ニューロン変性によって特徴づけられ、麻痺、運動神経および呼吸機能の完全喪失、および最終的には最初の臨床徴候の出現から2乃至8年(発症後の平均期間は3年)に死に至るALSは、患者の10%が遺伝子に由来しており、症例の90%が特発性である。染色体21 q22-1に位置する銅亜鉛スーパーオキシドジスムターゼ(SOD1)をコードする遺伝子における点突然変異は、家族性の症例の20%の病変に関与している(Rosen et al., Mutations in Cu/Zn
superoxide dismutase gene are associated with familial amyotrophic lateral sclerosis,
Nature, 362, 59-62, 1993, review in Rowland, Amyotrophic lateral sclerosis:
Human challenge for neuroscience, Proc. Natl. Acad. Sci. USA, 92, 1251-1253,
1995)。したがって、欠陥のあるSOD1は、運動ニューロン死に関連しており、家族性筋萎縮性側策硬化症の理解および可能な治療についての示唆を有する。
【0048】
II. SOD-1遺伝子
SOD1は銅原子1個、亜鉛原子1個を含む金属酵素で、ホモ二量体として細胞質の中に存在している。銅は酵素活性に必要であり、その一方で亜鉛はタンパク質の構造を安定化させる(Hirano, A. 1996, Neurology 47 (Suppl. 2). S63-S66)。SOD1はすべての真核細胞で発現し、マンガン依存性ミトコンドリアSOD(SOD2)および銅/亜鉛細胞外(SOD3)を含む、3種類のSOD酵素のファミリの1種類である(I Fridovich, 1986,
"Superoxide dismutases," Advances in Enzymology 58: 61-97)。SOD1の主な天然の機能は、スーパーオキシドジスムターゼで、スーパーオキシド(O2-)が過酸化水素(H2O2)および酸素に転換する。下流酵素のカタラーゼおよびグルタチオンペルオキシダーゼ(H2O2 を水と酸素に転換する)とともに、SOD1は細胞のフリーラジカルを解毒する。この機能の重要性は、生殖性の減退(Matzuk et al., 1998)、運動軸索障害(Shefner et al., 1999)、蝸牛有毛細胞の加齢に関連する喪失(McFadden et al., 2001)、および神経筋接合部シナプス(Flood et al., 1999)を含む、SOD1遺伝子が欠損するマウスの様々な異常、ならびに、軸索損傷(Reaume et al., 1996)、虚血(Kondo
et al., 1997; Kawase et al., 1999)、溶血血液の曝露(Matz et al., 2000)、および放射線照射(Behndig et al., 2001)など各種の有害な攻撃への高い感受性によって理解される。突然変異タンパク質の毒性、および野生種の機能的重要性を考えると、ALSの理想的な治療法は、前記突然変異SOD1タンパク質の発現を選択的に阻害する一方で野生種SOD1タンパク質の発現を維持することである。
【0049】
本願発明は、RNAiを用いた突然変異SOD1を標的とする。RNAiに用いる方法は、突然変異SOD1 mRNA内に点突然変異を含有する領域を補完する2本鎖RNA(siRNA)の1本を含む。siRNAをニューロンに導入後、siRNAは部分的に巻き戻され、部位特異的にSOD1 mRNA内に点突然変異を含む領域に結合し、mRNAヌクレアーゼを活性化する。このヌクレアーゼはSOD1 mRNAを切断し、突然変異SOD1の翻訳を中断させる。細胞は、部分的に切断されたmRNAを切り離し、その結果、翻訳を妨げるか、または細胞は部分的に翻訳されたタンパク質を切断する。ニューロンは野生種SOD1(正常対立遺伝子の)上で生存するため、このアプローチは、突然変異SOD1の産生物を除外することによって、突然変異SOD1の破壊を防ぐ。
【0050】
ヒト野生種SOD1タンパク質のアミノ酸配列は、図1(配列番号18)に記載されている。ヒト野生種SOD1遺伝子のコンセンサスヌクレオチド配列は、図2(配列番号17)に記載されている。
【0051】
III SOD-1突然変異遺伝子
100を超えるSOD1突然変異が特定されている。これらの突然変異の大半が、アミノ酸のスーパーオキシドジスムターゼの酵素鎖において1アミノ酸の置換を生じる。最も頻発する置換は、アミノ酸鎖の4位で生じるバリンによるアルギニンの置換(Arg4Valとも書く)であって、1型筋萎縮性側策硬化症を罹患するアメリカ人患者の50%に生じる。
【0052】
SOD1突然変異は、1型筋萎縮性側策硬化症の症状が始まる年齢、および疾患の進行の速さに影響を与える。たとえば、Arg4Val突然変異は、生存期間が疾患の発症から2年間未満という、侵攻性の障害を生じる。37位におけるグリシンのアルギニンへの置換(Gly37Arg)は、疾患の発症は早期だが生存期間は長い。さらに、その他の因子がSOD1突然変異と組み合わさると、1型筋萎縮性側策硬化症の経過が変化する。たとえば、SOD1遺伝子とCNTFとして知られる遺伝子の両方に生じる突然変異は、前記疾患の発症を早めるようである。CNTF突然変異だけでは病的影響はないが、SOD1突然変異と組み合わさると、疾患の症状は他の影響を受けたファミリメンバと比較すると何十年も早まるようである。
【0053】
SOD1突然変異が、筋肉の運動を制御する働きに特化した脳および脊髄の神経細胞である運動ニューロンに、選択的死滅を引き起こす機序は明らかではない。スーパーオキシドジスムターゼ酵素が、SOD1遺伝子において生じた変化が原因で、新しい(だがまだ未同定の)毒性機能を獲得すると考えられている。機能が異常な酵素は、有害なスーパーオキシドラジカルの蓄積、他の種類の有毒ラジカルの異常産生、細胞死(アポトーシス)の促進、前記酵素とその他の細胞タンパク質との凝集、または消耗と死滅を引き起こすような運動ニューロンへの連続的な刺激によって運動ニューロンの死滅を引き起こすこともある。
【0054】
【表1】
【0055】
IV RNA干渉
RNAiは2本鎖RNA(dsRNA)が動物および植物細胞において相同的なmRNAの配列特異的分解を誘発する著しく有効なプロセスである(Hutvagner and Zamore
(2002), Curr. Opin. Genet. Dev., 12, 225-232; Sharp (2001), Genes Dev., 15,
485-490)。哺乳類細胞では、RNAiは低分子干渉RNA(siRNA)の21ヌクレオチド(nt)2本鎖 (Chiu et al. (2002), Mol.
Cell., 10, 549-561; Elbashir et al. (2001),
Nature, 411, 494-498)、またはDNAテンプレートを用いてRNAポリメラーゼIIIプロモータとともにin vivoで発現するその他のdsRNA(Zeng et al. (2002), Mol. Cell, 9, 1327-1333; Paddison et al. (2002), Genes Dev.,
16, 948-958; Lee et al. (2002), Nature Biotechnol., 20, 500-505; Paul et al.
(2002), Nature Biotechnol., 20, 505-508; Tuschl, T. (2002), Nature Biotechnol.,
20, 440-448; Yu et al. (2002), Proc. Natl. Acad. Sci. USA, 99(9), 6047-6052;
McManus et al. (2002), RNA, 8, 842-850; Sui et al. (2002), Proc. Natl. Acad.
Sci. USA, 99(6), 5515-5520.)によって引き起こすことができる
【0056】
V. RNA分子および薬剤
本願発明は「低分子干渉RNA分子」(「siRNA分子」または「siRNA」)、前記siRNA分子を作製する方法、および前記siRNA分子を使用する方法(たとえば研究および/または治療方法)を特徴とする。本願発明のsiRNA分子はセンス鎖および相補的なアンチセンス鎖からなる2本鎖であって、アンチセンス鎖はRNAiを媒介するのために十分な標的mRNAへの相補性を有している。好ましくは、前記鎖は、鎖がアニーリングされる時に2本鎖末端の片方または両方に1、2、または3残基のオーバーハングが生じるように、鎖末端に、アラインメントされない(つまり相補的な塩基が反対側の鎖に生じない)少なくとも1、2、または3塩基があるようにアラインメントされる。好ましくは、前記siRNA分子は約10乃至50またはそれを超えるヌクレオチドの長さを有し、つまり、各鎖が10乃至50ヌクレオチド(またはヌクレオチド類似体)を含むより好ましくは、前記siRNA分子は、各鎖にたとえば16、17、18、19、20、21、22、23、24、25、26、27、28、29、または30 ヌクレオチドなど約16乃至30の長さを有し、当該分子において片方の鎖が標的領域に実質的にたとえば少なくとも80%(またはそれを超える、たとえば85%、90%、95%、または100%)相補的であって、たとえば3、2、1または0のミスマッチヌクレオチドを有し、当該標的領域が野生種と突然変異の対立遺伝子の差が少なくとも1塩基対はある、たとえば機能獲得型突然変異を含む標的領域などの標的領域であって、もう片方の鎖は第1の鎖と実質的に同一である。
【0057】
一般的に、siRNAは当業に知られる任意の方法、たとえば以下のプロトコルを用いることによってデザインすることができる。
【0058】
1. AUG開始コドンから始めてAAジヌクレオチド配列を探す。各AAおよび3’側に16ヌクレオチド以上離れたところが、siRNA標的の候補である。siRNAは機能獲得型突然変異を含む標的領域など、野生種と突然変異の対立遺伝子の差が少なくとも1塩基対の標的領域に特異的でなければならない。第1の鎖は、この配列に相補的でなければならず、その他の鎖は第1の鎖に同一であるか実質的に同一である。ある実施態様では、前記核酸分子は、例えばSOD1の配列など、開始コドンの少なくとも50乃至100nt下流から始まる標的対立遺伝子配列の領域から選択される。さらに、G/C含有量が少ないsiRNA(35乃至55%)は、G/C含有量が55%を超えるものよりも活性であることもある。したがってある実施態様では、前記発明には35乃至55%のG/C含有量を有する核酸分子が含まれる。さらに、siRNAの鎖は、たとえば2ヌクレオチドなど、1乃至4ヌクレオチドの3’オーバーハングを有するように対を形成させることができる。したがって別の実施態様では、前記核酸分子はTTなどの2ヌクレオチドの3’オーバーハングを有してもよい。前記のオーバーハングしているヌクレオチドは、RNAまたはDNAのいずれかである。上述のように、突然変異:野生種ミスマッチがプリン:プリンミスマッチである標的領域を選択することが望ましい。
【0059】
2.
当業に知られる任意の方法を用いて、標的候補と適切なゲノムデータベース(ヒト、マウス、ラットなど)を比較し、他のコード配列に顕著に相同性を有する任意の標的配列を考慮から除外する。そのような配列相同性の探索に使われる1つの方法はBLASTとして知られ、国立バイオテクノロジー情報センターのウェブサイトで入手することができる。
【0060】
3.
評価基準に合う1つ以上の配列を選択する。
さらに、siRNAのデザインおよび使用についての一般的な情報は、マックスプランク生物物理化学研究所のウエブサイトで入手できる「The siRNA User Guide(siRNAユーザーガイド)」の中に見つけることもできる。
【0061】
陰性対照siRNAは、選択されたsiRNAと同一のヌクレオチド組成を有するべきだが、適切なゲノムに相補的な重要な配列は有さない。そのような陰性対照は、選択されたsiRNAのヌクレオチド配列をランダムにスクランブルしてデザインしてもよく、相同性探索は、前記陰性対照が適切なゲノムにおける任意のその他の遺伝子に対する相同性を欠損していることを確認するために行うことができる。さらに、陰性対照siRNAは、前記配列に1つ以上の塩基ミスマッチを導入することによってデザインすることができる。
【0062】
1ヌクレオチド特異性を有するsiRNAは、以下の通りデザインすることができる。
【0063】
標的mRNAは、参照mRNA配列(たとえば野生種対立遺伝子またはmRNA配列)と比較したミスマッチ(たとえば点突然変異などの1ヌクレオチドミスマッチなど)を有するものが選択される(たとえば突然変異対立遺伝子またはmRNA)。siRNAは、1ヌクレオチドのところに、siRNAと標的mRNA(たとえば突然変異mRNA)の間に完全な相補性が存在し、したがって、siRNA(たとえばアライニングされた)を参照配列(たとえば野生種対立遺伝子またはmRNA配列)と比較する場合、ミスマッチが存在するようにデザインされる。好ましくは、前記siRNAは、1ヌクレオチド(たとえば点突然変異)が意図される切断部位、またはその近傍に存在するようにデザインされる。好ましくは、前記siRNAが、標的とされる1ヌクレオチド(たとえば点突然変異)が完全にまたは正確にsiRNA(たとえばsiRNAのアンチセンス鎖)の中央に配置されるようにデザインされる。完全に中央に配置するという文言は、前記1ヌクレオチド(たとえば点突然変異)の両側に並んでいるヌクレオチドの数が同一であるが(つまり8、9、10、11、または12)、たとえばdTdT末端など任意のオーバーハングは含まないことを意味する。たとえば、2ヌクレオチドの3’オーバーハング(たとえばアンチセンス鎖の3’末端にあるオーバーハング)を有する21ヌクレオチドsiRNAが選択される場合、前記1ヌクレオチド(たとえば点突然変異)の両側には9ヌクレオチドが存在する。たとえば、2ヌクレオチドの3’オーバーハング(たとえばアンチセンス鎖の3’末端にあるオーバーハング)を有する22ヌクレオチドsiRNAの場合、前記1ヌクレオチド(たとえば点突然変異)の両側には9および10ヌクレオチドが存在する。23ヌクレオチドsiRNAの場合、前記1ヌクレオチド(例えば点突然変異)の両側に10ヌクレオチドが存在する。24ヌクレオチドsiRNAの場合、前記1ヌクレオチド(例えば点突然変異)の両側に10および11ヌクレオチドが存在する。例示した数は、2ヌクレオチド3’オーバーハングを有するsiRNAの場合であるが、より長いまたは短いオーバーハングを有するか、またはオーバーハングを有さないsiRNAにも容易に当てはめることができる。前記1ヌクレオチド(たとえば点突然変異)がsiRNAの中央に位置しないようなsiRNAをデザインする場合、siRNAによる切断の有効性が減少する場合もある。
【0064】
1ヌクレオチド特異性を有するsiRNAは、好ましくは、相当する参照(たとえば野生種)配列における塩基対の形成が生じにくいようにデザインされる。たとえば、siRNAをプリン:プリン対形成が前記1ヌクレオチドの位置でsiRNAと野生種mRNAの間に存在するようにデザインすると、1ヌクレオチド特異性が促進される。プリン:プリン対形成は、たとえばG:G、A:G、G:AおよびA:A対形成のグループから選択される。 さらに、前記1ヌクレオチドの位置でのsiRNAと突然変異mRNAの間のプリン:ピリミジン対形成は、1ヌクレオチド特異性を促進する。プリン:ピリミジン対形成は、たとえばG:C、C:G、A:U、U:A、C:A、A:C、U:AおよびA:U
のグループから選択される。
【0065】
本願発明の核酸組成物には、本願明細書に記載のsiRNAおよびsiRNA誘導体の両方が含まれる。たとえば、たとえば体内の半減期を延長するためなど、前記組成物の薬物動態学を変化させるために、架橋を用いることができる。したがって、本願発明には、2本の鎖が架橋されるように、核酸の相補的な2本の鎖を有するsiRNA誘導体を含む、siRNA誘導体が含まれる。本願発明にはまた、3’末端(たとえばペプチド)、または有機化合物(たとえば色素)などに結合する非核酸部分を有するsiRNA誘導体も含まれる。このようなsiRNA誘導体の修飾は、細胞取り込みが改善するか、もしくは相当するsiRNAと比較して、得られたsiRNA誘導体の細胞の標的活性を促進するか、前記細胞のsiRNA誘導体を追跡するのに有用であるか、または相当するsiRNAと比較したsiRNA誘導体の安定性を向上することもある。
【0066】
本願発明のsiRNA分子は、化学的に合成することができ、またはin vitroではDNAテンプレートから、もしくはin vivoではたとえばshRNAから、もしくはin vitro転写dsRNAテンプレートをRNAiを切断して媒介する20、21、もしくは23bp 2本鎖RNAのプールをつくるためにヒトDICER酵素を用いて、転写することができる。前記siRNA分子は、当業に知られる任意の方法を用いてデザインすることができる。
【0067】
ある局面では、RNAi剤を本願明細書に記載のたとえばsiRNAまたはshRNAなどの干渉リボ核酸ではなく、前記RNAi剤は本願明細書に記載のたとえばshRNAなどの干渉ヌクレアーゼをコードしてもよい。つまり、前記RNAi剤は前記干渉リボ核酸の転写テンプレートであってもよい。したがって、本願発明のRNAi剤には、低分子ヘアピンRNA(shRNA)、およびshRNAを発現するためにつくられた発現作製物が含まれてよい。shRNAの転写は、ポリメラーゼIII(pol III)プロモータから開始し、4-5チミン転写終了部位の2位で終了すると考えられている。発現後、shRNAは3’UUオーバーハングを有するステム-ループ構造に折り畳まれると考えられ、その後、これらのshRNAの末端が処理されて、shRNAが約21乃至23ヌクレオチドのsiRNA様分子に転換される。 Brummelkamp et al.,
Science 296:550-553 (2002); Lee et al, (2002). supra; Miyagishi and
Taira, Nature Biotechnol. 20:497-500 (2002); Paddison et al. (2002), supra;
Paul (2002), supra; Sui (2002) supra; Yu et al. (2002), supra。
【0068】
本願発明の発現作製物には、適切な発現システムに使用するのに好適な任意の作製物が含まれ、当業に知られるレトロウイルスベクタ、直線状発現カセット、プラスミドおよびウイルスまたはウイルス由来ベクタがそれらに限定されずに含まれる。そのような発現作製物には、1つ以上の誘導プロモータ、U6 snRNAプロモータもしくはH1 RNAポリメラーゼIIIプロモータなどのRNA Pol IIIプロモータシステム、または当業に知られるその他のプロモータが含まれてよい。両方の鎖を発現する発現作製物には、両方の鎖を連結するループ構造が含まれてもよく、または各鎖は同一の作製物内の個別のプロモータから別々に転写されてもよい。各鎖は個別の発現作製物から転写されてもよい。 (Tuschl (2002), supra)。
【0069】
合成siRNAは、カチオン性リポソームトランスフェクションおよび電気穿孔を含む、当業に知られる方法によって細胞に送達される。しかし、これらの外来性siRNAは、一般に、サイレンシング効果が短寿命で(培養細胞中で4乃至5日)、特定の実施態様でしか有益でない場合もある。前記標的遺伝子(つまり突然変異遺伝子)を長期間抑制し、特定の環境下での送達を容易にするために、細胞内で1つ以上のsiRNAを組換えDNA作製物から発現させることができる。細胞内における長期間の標的遺伝子の抑制を可能にするために、細胞内で組換えDNA作製物からsiRNA
2本差を発現させる方法は当業に知られており、機能的な2本鎖siRNAを発現することが可能な哺乳類Pol III プロモータシステム(たとえばH1またはU6/snRNAプロモータシステム(Tuschl (2002), supra)
)が含まれる(Bagella et
al., J. Cell. Physiol. 177:206−213 (1998); Lee et al. (2002), supra;
Miyagishi et al. (2002), supra; Paul et al. (2002), supra;
Yu et al. (2002), supra; Sui et al. (2002), supra)。RNA Pol IIIによる転写終了は、一連のDNAテンプレートの連続する4つのT残基で生じ、特定の配列においてsiRNA転写物を終了させるメカニズムを提供する。前記siRNAは5’-3’および3’-5’の方向に向いた前記標的遺伝子の配列に相補的であって、siRNAの2本鎖は同一の作製物または別々の作製物中に発現することができる。H1またはU6 snRNAプロモータによって駆動され、細胞内で発現するヘアピンsiRNAは、標的遺伝子の発現を阻害することができる(Bagella et al. (1998),
supra; Lee et al.
(2002), supra; Miyagishi et al. (2002), supra; Paul et
al. (2002), supra; Yu et al. (2002), supra; Sui et
al. (2002) supra)。T7プロモータの制御下にあるsiRNA配列を含有する作製物もまた、T7 RNA ポリメラーゼを発現するベクタを有する細胞に同時トランスフェクトされると、機能性siRNAをつくる。1つの作製物は、同一の遺伝子または複数の遺伝子に方向付けられている突然変異SOD1をコードする遺伝子の複数の領域など、siRNAをコードする複数の配列を含有することがあり、たとえば、別々のPolIIIプロモータ部位によって駆動することができる。
【0070】
動物細胞はミクロRNA(miRNA)とよばれる約22ヌクレオチドの非コードRNAの領域を発現する。ミクロRNAは動物の成長中に転写後または転写レベルで遺伝子発現を制御することができる。miRNAのある共通の性質は、おそらくダイサー、RNase III型酵素、またはその相同体によって、約70ヌクレオチドの前駆体RNAステム-ループからすべて切除されることである。標的mRNAに相補的な配列を有するmiRNA前駆体のステム配列を置換することによって、組換えされた前駆体を発現するベクター作製物を用いて、哺乳類細胞の中で特異的mRNA標的に対してRNAiを開始するためにsiRNAを産生することができる(Zeng (2002), supra)。ポリメラーゼIIIプロモータを含有するDNAベクタによって発現する場合、ミクロRNAでデザインされたヘアピンは遺伝子発現をサイレンシングすることができる(McManus (2002), supra)。ミクロRNA標的多型が、siRNA媒介遺伝子サイレンシングの欠如下において突然変異タンパク質の翻訳を阻害するために有用な場合もある。そのような適用は、たとえばデザインされたsiRNAが野生種タンパク質の標的外サイレンシングを生じるような状況で有用である場合がある。
【0071】
ウイルス媒介送達メカニズムを用いて、たとえばRNA Pol IIプロモータ転写制御下におけるsiRNAを有する組換えアデノウイルスの作製などによるsiRNAの発現を通して、標的遺伝子の特異的サイレンシングを誘導することができる(Xia et al. (2002), supra)。 これらの組換えアデノウイルスによるHeLa細胞の感染によって、内因性標的遺伝子発現を低減することができる。siRNAの標的遺伝子を発現するトランスジェニックマウスに組換えアデノウイルスベクタを注射すると、in vivoにおいて標的遺伝子の発現が減少する。同文献。動物モデルにおいて、全胚電気穿孔は、合成siRNAを効率的に移植後マウス胚合成siRNAに送達させることができる。成体マウスにおいて、siRNAの効率的な送達は、「高圧」送達技術によって実現することができる。これは溶液を含有する大量のsiRNAを、尾部静脈を介して動物体内へ迅速に(5秒以内に)注入する技術である。ナノ粒子およびリポソームを用いて、siRNAを動物体内へ送達することもできる。
【0072】
本願発明の核酸組成物には、非修飾siRNA、および架橋siRNA誘導体、または3’もしくは5’末端などに結合させた非ヌクレオチド部分を有する誘導体など、当業に知られる修飾RNAの両方が含まれる。このようなsiRNA誘導体の修飾は、細胞取り込みが改善するか、もしくは相当するsiRNAと比較して、得られたsiRNA誘導体の細胞の標的活性を促進するか、前記細胞のsiRNA誘導体を追跡するのに有用であるか、または相当するsiRNAと比較したsiRNA誘導体の安定性を向上することもある。
【0073】
本願明細書に記載の細胞または組織全体に導入された組換えRNA前駆体によって、望ましいsiRNA分子の産生が引き起こされるだろう。そこで、そのようなsiRNA分子は、切断および破壊のための特異的なmRNA配列に結合し標的とするRNAi経路の内因性タンパク質要素に関連するだろう。このようにすれば、組換えRNA前駆体から作製されたsiRNAが標的とするmRNAは、前記細胞および組織から大幅に減少し、前記細胞または組織中のmRNAによってコードされるタンパク質の濃度が減少するだろう。RNA前駆体は、典型的にはdsRNAの1本鎖を個別にコードするか、またはRNAヘアピンループ構造のヌクレオチド配列全体をコードする核酸分子である。
【0074】
本願発明の核酸組成物は、たとえば吸収、有効性、バイオアベイラビリティ、および/または半減期などの薬物動態学的パラメタなどの前記組成物の性質を向上させるナノ粒子などの別の部分と結合させることもでき、または結合させないでもよい。その結合は、たとえば、Lambert et al., Drug Deliv. Rev.:47(1), 99-112 (2001) (ポリアルキルシアノアクリル酸(PACA)ナノ粒子に装着された核酸について記載)、Fattal et al., J. Control
Release 53(1-3):137-43 (1998) (ナノ粒子に結合させた核酸について記載)、 Schwab et al., Ann. Oncol.
5 Suppl. 4:55-8 (1994) (挿入剤、疎水基、ポリカチオンPACAナノ粒子に結合させた核酸について記載)、およびGodard et al., Eur. J. Biochem. 232(2):404-10 (1995) (ナノ粒子に結合させた核酸について記載)の方法を用いるなど、当業に知られる方法によって実現することができる。
【0075】
本願発明の核酸分子は、当業に知られる任意の方法を用いてラベルすることもでき、たとえば前記核酸分子組成物がたとえばCy3、フルオレセイン、ローダミンなどのフルオロフォアで標識することができる。その標識は、たとえばSILENCERTM siRNA ラベリングキット(アンビオン)など、キットを用いて行うことができる。さらに、前記siRNAは、たとえば3H、32Pまたはその他の適切な同位体などを用いて放射標識することができる。
【0076】
さらに、RNAiは、少なくとも1つの1本鎖RNA中間体を経る過程であると考えられるため、当業者は、ss-siRNA(たとえばds-siRNAのアンチセンス鎖など、本願明細書に記載のとおりデザイン(たとえば化学合成用に)、作製(たとえば酵素的に作製)または発現(たとえばベクターまたはプラスミドから)することもでき、本願特許請求の方法論にしたがって利用することができるということは理解されよう。さらに、無脊椎動物では、RNAiは長いdsRNA(たとえば長さが約100乃至1000ヌクレオチド、好ましくは約200乃至500であって、たとえば、約250、300、350、400、または450ヌクレオチドのdsRNA)であって、RNAiのエフェクタT細胞として作用するRNAiは長いdsRNAによって効果的に引き起こすことができる。
【0077】
本願発明のsiRNA分子は、突然変異siRNA P11 (配列番号5, センス; 配列番号6, アンチセンスまたはガイド), 突然変異 siRNA P10 (配列番号3, センス;
配列番号4, アンチセンス、またはガイド), 突然変異 siRNA P9 (配列番号1, センス; 配列番号2 アンチセンス、またはガイド), SOD1 野生種標的(配列番号7), SOD1 突然変異 標的(配列番号8), 野生種siRNA
P11 (配列番号9 センス; 配列番号10, アンチセンス、またはガイド), 野生種siRNA P10 (配列番号11, センス; 配列番号12, アンチセンス、またはガイド), 野生種siRNA P9 (配列番号13, センス;
配列番号14, アンチセンス、またはガイド)が含まれる図1に記載の配列、G93A SOD1 siRNA (SEQ ID
NO:16)が含まれる図3に記載の配列、およびその対立遺伝子変異種などを含む、またはそれらからなっていてもよい。
【0078】
VI. RNAiを誘導するための組換えRNA前駆体の使用
本願明細書に記載の細胞または組織全体に導入された組換えRNA前駆体によって、望ましいsiRNA分子の産生が引き起こされるだろう。そこで、そのようなsiRNA分子は、切断および破壊のための特異的なmRNA配列に結合し、標的とするRNAi経路の内因性タンパク質要素に関連するだろう。このようにすれば、組換えRNA前駆体から作製されたsiRNAが標的とするmRNAは、前記細胞および組織から大幅に減少し、前記細胞または組織中のmRNAによってコードされるタンパク質の濃度が減少するだろう。
【0079】
VII. 薬学的組成物および投与方法
本願発明のsiRNA分子は薬学的組成物に組み込むことができる。そのような組成物には、典型的には、核酸分子および薬学的に許容な担体が含まれる。本願明細書に記載の「薬学的に許容な担体」という用語は、薬学的投与に適合する、食塩水、溶媒、分散媒、コーティング、抗細菌および抗真菌剤、ならびに等張および吸収遅延剤などを意味する。補助的な活性化合物も、当該組成物に組み込むことができる。
【0080】
薬学的組成物は、意図する投与経路と適合するように調剤される。投与経路の例には、たとえば静脈内、皮内、皮下、口腔内(たとえば吸入)、経皮(局所)、経粘膜、および直腸投与などの非経口投与が含まれる。非経口、皮内、または皮下投与に使用する溶液または懸濁液には、以下の成分、注射用の水、食塩水、固定油、ポリエチレングリコール、グリセリン、プロピレングリコール、またはその他の合成溶媒などの滅菌希釈剤、ベンジルアルコールまたはメチルパラベンなどの抗菌剤、アスコルビン酸または亜硫酸ナトリウムなどの抗酸化剤、エチレンジアミン四酢酸などおキレート剤、酢酸塩、クエン酸塩、またはリン酸塩などのバッファ、および塩化ナトリウムまたはデキストロースなどの張力調整剤などが含まれていてよい。非経口製剤は、アンプル、ガラスまたはプラスチック製の使い捨て用シリンジ、または複数の投与用バイアルの中に封入されていてよい。
【0081】
注射に使用するために好適な薬学的組成物には、滅菌水溶液(水溶性)、または分散液、および滅菌注射用溶液または分散液の即時調整用滅菌粉末が含まれる。静脈内投与に好適な担体には、生理食塩水、静菌水、クレモホールELTM(BASF、ニュージャージー州パシッパニー)、またはリン酸緩衝食塩水(PBS)が含まれる。すべての場合において、前記組成物は滅菌状態であって、注射容器への充填が容易な程度、流動的でなければならない。前記組成物は、製造および保存の条件下において安定でなければならず、細菌および真菌などの微生物の汚染作用から保護されなければならない。前記担体は、たとえば水、エタノール、ポリオール(たとえばグリセロール、ポリエチレングリコール、および液体状ポリエチレングリコールなど)、それらの好適な混合物、ならびに植物油などを含む溶媒又は分散媒であってよい。適切な流動性は、たとえばレシチンなどのコーティングの使用、分散液の場合は所要の粒子サイズの維持、および表面活性剤の使用などによって維持することができる。微生物の作用は、たとえばパラベン、クロロブタノール、フェノール、アスコルビン酸、チメロサールなどの各種抗細菌剤および抗真菌剤によって防ぐことができる。多くの場合、たとえば糖、マニトールなどのポリアルコール、ソルビトール、塩化ナトリウムなどの等張剤を前記化合物に含めることが望ましいだろう。注射可能な組成物の吸収を持続させるために、たとえばモノステアリン酸アルミニウムおよびゼラチンなどの吸収遅延剤を組成物に含ませてもよい。
【0082】
滅菌された注射可能な溶液は、適切な溶媒中に入れた必要量の前記活性化合物を、必要に応じて上述の成分1つまたはそれらの組み合わせに組み入れてからろ過滅菌することによって調製することができる。一般に、分散液は、塩基性分散媒および上述の物質のうちの必要なその他の成分を含む滅菌担体に前記活性化合物を組み入れて調製する。滅菌注射用溶液の調製のための滅菌粉末の場合、好ましい調製方法は、前記活性成分の粉末に加えて予め滅菌ろ過されたその溶液から得られた任意の追加の望ましい成分を得られる、真空乾燥および凍結乾燥である。
【0083】
経口組成物には一般に、不活性な希釈剤または食用担体が含まれる。治療用の経口投与の場合、前記活性化合物を賦形剤に組み込み、錠剤、トローチ、またはたとえばゼラチンカプセルなどのカプセルなどの形状で用いることができる。経口組成物はまた、洗口剤として用いられる液体担体を用いて調製することもできる。薬学的に適合な結合剤、および/またはアジュバント物質は、前記組成物の一部分として含めることもできる。前記錠剤、丸剤、カプセル、およびトローチなどは、以下の成分、もしくは同様の性質の化合物のいずれかを含むことができる。それは、微結晶性セルロース、ガムトラガカント、もしくはゼラチンなどの結合剤、デンプンもしくはラクトースなどの賦形剤、アルギン酸、プリモゲル、もしくはトウモロコシデンプンなどの崩壊剤、ステアリン酸マグネシウムもしくはステロート(Sterote)などの潤滑剤、コロイド状二酸化ケイ素などの流動促進剤、ショ糖もしくはサッカリンなどの甘味剤、またはペパーミント、サリチル酸メチル、もしくはオレンジ香味料などの香味剤である。
【0084】
吸入による投与の場合、前記化合物は、たとえば二酸化炭素などの好適な高圧ガスを含む加圧容器もしくはディスペンサ、またはネビュライザからのエアロゾルスプレーの形状で送達する。そのような方法には、米国特許第6,468,798号に記載されるものが含まれる。
【0085】
全身投与はまた、経粘膜または経皮手段でもよい。経粘膜または経皮投与の場合、透過させようとする関門に適切な浸透剤を、前記製剤に用いる。そのような浸透剤は一般に当業に知られており、たとえば経粘膜投与の場合、界面活性剤、胆汁塩、およびフシジン酸誘導体が含まれる。経粘膜投与は、経鼻スプレーまたは坐剤の使用によって実施することができる。経皮投与の場合、前記活性化合物は、一般に当業に知られる軟膏(ointment)、軟膏(salve)、ゲル、またはクリームに調製する。
【0086】
前記化合物はまた、坐剤(たとえばカカオバターおよびその他のグリセリドなどの従来の坐剤ベース)または直腸送達用の停留浣腸の形状に調製してもよい。
【0087】
前記化合物はまた、McCaffrey et al.
(2002), Nature, 418(6893), 38-9 (流体力学トランスフェクション); Xia et al. (2002), Nature Biotechnol., 20(10), 1006-10 (ウイルス媒介送達); またはPutnam
(1996), Am. J. Health Syst. Pharm.53(2), 151-160, 誤字 Am. J. Health Syst. Pharm. 53(3), 325 (1996)に記載の方法が限定されずに含まれる、当業に知られる方法を用いてトランスフェクションまたはインフェクションによって投与することができる。
【0088】
前記化合物はまた、DNAワクチンなど、核酸剤の投与に好適な任意の方法によって投与することもできる。前記方法には、遺伝子銃、バイオ注射器、および皮膚用添付剤、ならびに米国特許第6,194,389号に開示されている微粒子DNAワクチン技術などの無針法、米国特許第6,168,587号に開示されるパウダー状ワクチンを有する哺乳類経皮無心ワクチン投与が含まれる。さらに、とりわけHamajima et al. (1998), Clin. Immunol. Immunopathol., 88(2),
205-10に記載されているものなど、経鼻送達も可能である。リポソーム(たとえば米国特許第6,472,375号に記載されているなど)およびマイクロカプセル封入も使用することができる。生分解性標的化可能微粒子送達システムも使用することができる(たとえば米国特許第6,471,996号に記載されているなど)。
【0089】
ある実施態様では、前記活性化合物は、インプラントおよびマイクロカプセル封入送達システムを含む、制御放出製剤などの、体外への急速な排出から前記化合物を保護する担体とともに調製する。エチレン酢酸ビニル、ポリ無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、およびポリ乳酸などの生分解性、生体適合性ポリマーを用いることができる。そのような製剤の調製は、標準技術を用いて行うことができる。前記材料はアルザ社およびノバファーマシューティカルズ社から購入することもできる。リポソーム懸濁液(ウイルス抗原に対するモノクローナル抗体で感染細胞を狙ったリポソームを含む)を薬学的に許容な担体として用いることもできる。これらは、米国特許第4,522,811号などに記載の、当業に知られる方法に従って調製することができる。
【0090】
当該化合物の毒性および治療上の有効性は、たとえばLD50(個体群の50%に致死的な投与量)およびED50(個体群の50%に治療上有効な投与量)など、細胞培養物または実験動物における標準的な薬学的方法によって決定することができる。毒性作用および治療上有効な作用の投与量の比は治療指数であって、LD50/ED50の比で表すことができる。高い治療指数を示す化合物が好ましい。有毒な副作用を示す化合物を用いる場合、感染していない細胞に与える可能性のある損傷を最小にして副作用を減少するよう、そのような化合物を罹患組織の部位に狙わせる送達システムをデザインするように、注意が払われなければならない。
【0091】
細胞培養アッセイおよび動物研究から得られるデータは、ヒトにおける使用のための用量の範囲を決定する際に用いることができる。そのような化合物の用量は、好ましくは毒性が小さいかまたは皆無のED50を含む循環濃度の範囲内にある。前記用量は、用いられた投与剤形および使用された投与経路によって、この範囲内で変化させてもよい。本願発明の方法に用いられるいずれの化合物の場合も、治療上有効な投与量は、最初は細胞培養アッセイで推定することができる。1回投与量は、動物モデルにおいて、細胞培養物で決定したIC50(すなわち症状の最大半減抑制を達成する前記テスト化合物の濃度)を含む循環血漿濃度の範囲になるように調製してもよい。このような知見は、ヒトにおける有用な投与量をより正確に決定するために用いることができる。血漿濃度は、たとえば高性能液体クロマトグラフィによって測定することもできる。
【0092】
本願明細書に記載の、核酸分子の治療上有効な量(すなわち有効投与量)は、選択された核酸による。たとえば、shRNAをコードするプラスミドを選択する場合、単回投与は約1μg乃至100mgの範囲内で投与してもよく、ある実施態様では10、30、100または1000 μgを投与することがある。 ある実施態様では、1乃至5gの前記組成物を投与することができる。前記組成物は、2日に1回など、1日1回乃至1回以上、または1週間に1回以上投与することができる。当業者は、特定の要因が、疾患またな障害の重篤度、治療歴、対象の全般的な健康状態、およびその他に罹患する疾患をそれらに限定せずに含む、前記対象を有効に治療するために必要な用量および時間に影響することもあるということを理解するであろう。さらに、対象の治療上有効な量のタンパク質、ポリペプチド、または抗体での治療は、単回の治療を含んでもよく、または好ましくは、一連の治療を含んでもよい。
【0093】
本願発明の核酸分子は、たとえば上述のXia et al.,
(2002)に記載の方法が限定されずに含まれる、当業に知られる方法を用いて、たとえばウイルスベクタ、レトロウイルスベクタ、発現カセット、またはプラスミドウイルスベクタなどの発現作製物に挿入することができる。発現作製物は、たとえば吸入、経口、静脈内投与、局所投与(米国特許第5,328,470号参照)、または定位固定注射(たとえばChen et al. (1994), Proc.
Natl. Acad. Sci. USA, 91, 3054-3057参照)によって対照に送達することができる。前記輸送ベクタの薬学的調製には許容な希釈剤に入れたベクタが含まれてもよく、または輸送用賦形剤が埋め込まれた徐放性基質を含んでもよい。代替的には、たとえばレトロウイルスベクタなど、完全な輸送ベクタを組換え細胞から無傷で作製できる場合、前記薬学的調製物は遺伝子送達システムをつくる1つ以上の細胞を含むことができる。
【0094】
本願発明の核酸分子には、低分子ヘアピンRNA(shRNA)、およびshRNAを発現するためにつくられた発現作製物が含まれてよい。shRNAの転写は、ポリメラーゼIII(pol III)プロモータから開始し、4-5チミン転写終了部位の2位で終了すると考えられている。発現後、shRNAは3’UUオーバーハングを有するステム-ループ構造に折り畳まれると考えられ、その後、これらのshRNAの末端が処理されて、shRNAが約21ヌクレオチドのsiRNA様分子に転換される。
Brummelkamp et al. (2002), Science, 296, 550-553; Lee et al, (2002). supra;
Miyagishi and Taira (2002), Nature Biotechnol., 20, 497-500; Paddison et al.
(2002), supra; Paul (2002), supra; Sui (2002) supra; Yu et
al. (2002), supra。
【0095】
前記発現作製物は、適切な発現システムに使用するのに好適な任意の作製物であってよく、当業に知られるレトロウイルスベクタ、直線状発現カセット、プラスミドおよびウイルスまたはウイルス由来ベクタがそれらに限定されずに含まれる。そのような発現作製物には、1つ以上の誘導プロモータ、U6 snRNAプロモータもしくはH1 RNAポリメラーゼIIIプロモータなどのRNA Pol IIIプロモータシステム、または当業に知られるその他のプロモータが含まれてよい。両方の鎖を発現する発現作製物には、両方の鎖を連結するループ構造が含まれてもよく、または各鎖は同一の作製物内の個別のプロモータから別々に転写されてもよい。各鎖は個別の発現作製物から転写されてもよい。上述のTuschl (2002)。
【0096】
薬学的組成物は、投与法の説明書とともに、容器、パック、またはディスペンサに包含されていてよい。
【0097】
VIII. 治療方法
本願発明は、機能獲得型突然変異タンパク質によって、全体的にまたは部分的に引き起こされた疾患または障害のリスクがある(または感受性がある)を治療するための予防および治療方法の両方を提供する。ある実施態様では、前記疾患または障害は優性の機能獲得型疾患である。好ましい実施態様では、前記疾患または障害はSOD1遺伝子の改変、特にSOD1突然変異対立遺伝子における点突然変異に関連する疾患であって、臨床徴候にはALS患者に認められるものが含まれるような、SOD1遺伝子(構造または機能)またはSOD1タンパク質(構造または機能または発現)に異常を引き起こす疾患である。
【0098】
本願明細書に記載の「治療」または「治療する」は、疾患もしくは障害、疾患または障害の症状、または疾患もしくは障害の素因を有する患者への治療用の物質(たとえばRNA剤またはベクターまたはそれをコードする導入遺伝子)の適用もしくは投与、または前記患者から単離された組織または細胞株への治療用物質の適用もしくは投与であって、当該適用もしくは投与の目的が疾患もしくは障害、疾患もしくは障害の症状、または疾患の素因の治療、治癒、緩和、軽減、改変、改善、改良、向上、または作用である、適用もしくは投与として定義される。
【0099】
ある局面では、本願発明は、対象において、前記対照に治療剤を投与することによって、上述の疾患または障害を予防する方法を提供する。前記疾患のリスクがある対照は、たとえば本願明細書に記載の任意のまたは組み合わせた診断アッセイまたは予後アッセイによって同定することができる。予防剤は、前記疾患または障害を予防、または代替的にはその進行を遅らせるように、前記疾患または障害に特徴的な症状の徴候の前に投与することができる。
【0100】
本願発明の別の局面は、対照を治療的に処理する、つまり前記疾患または障害の症状の発症を改変させる方法に関連する。ある例示的な実施態様では、本願発明の調節方法は、機能獲得型突然変異を発現する細胞と、前記遺伝子の配列特異的な干渉を実現するような、前記遺伝子内の突然変異に特異的な治療剤(たとえばRNAi剤またはベクターまたはそれをコードする導入遺伝子)と接触させるステップに関与する。これらの方法は、in vitro(たとえば前記細胞を前記作用剤で培養するなど)または代替的にはin vivoにおいて(たとえばその物質を対象に「投与する」ステップによって)実行することができる。
【0101】
処置の予防的および治療的な方法の両方に関して、そのような処置は薬理ゲノム学分野から得られた知識を本に、特別にあつらえるか、または修正してもよい。本願明細書に記載の「薬理ゲノム学」は、臨床開発用および市販用薬物の遺伝子配列解析、統計学的遺伝子学、および遺伝子発現解析などのゲノミクス技術の適用を意味する。より具体的には、この用語は、患者の遺伝子が薬物に対する応答(たとえば患者の「薬物応答表現型」または「薬物応答遺伝子型」など)をどのように決定するかを研究することを意味する。したがって、本願発明の別の局面は、本願発明の標的遺伝子分子、または個人の薬物応答遺伝子型にしたがった標的遺伝子モジュレータのいずれかによる、個人の予防的または治療的処置をあつらえる方法を提供する。薬理ゲノム学は、臨床医または医師が、予防的または治療的処置から最も利益を得るであろう患者のみを前記処置の対象にし、有毒な薬物関連の副作用を経験するであろう患者の治療を避けることを可能にする。
【0102】
治療剤は適切な動物モデルにおいてテストすることができる。たとえば、本願明細書に記載のRNAi剤(または発現ベクタもしくはそれをコードする導入遺伝子)を動物モデルに用いて、前記薬剤での処置の有効性、毒性、または副作用を決定することができる。 代替的には、治療剤を動物モデルに用い、そのような作用薬の作用機序を決定することができる。たとえば、薬剤を動物モデルに用いて、前記薬剤での処置の有効性、毒性、または副作用を決定することができる。 代替的には、薬剤を動物モデルに用い、そのような作用薬の作用機序を決定することができる。
【実施例】
【0103】
以下の材料、方法、および例は、具体例を示すのみで、制限を意図しない
材料および方法
RNAおよびDNA作製物
21本のヌクレオチド1本鎖RNA(図1)は、ダーマコン・リサーチ社から購入し、製造者の指示に従って脱保護して記載の通りアニーリングした(Nykanen et al., 2001)。野生種および突然変異SOD1-GFP融合タンパク質を作製するために、SOD1 wt(Genbank Accession No. ジェンバンク受入番号NP_000445; 図7; 配列番号18)、SOD1G85R、およびSOD1 G93A(配列番号16)cDNAを、pCMV/myc/mito/GFP(インビトロゲン社)のPmlI およびPstI部位の間でPCRクローニングした。このクローニングステップによって、ミトコンドリア標的配列が削除された。mycタグ付き野生種SOD1を作製するために、SOD1wt cDNA (配列番号17)をpCMV/myc/mito/GFPのPstIおよびXhoI部位の間でPCRクローニングした。ミトコンドリア標的配列は、BssHIIおよびPmlIで切断し、平滑末端化ライゲーションすることによって削除した。すべての作製物を、シーケンシングによって確認した。DsRed (pDsRed2-C1) は、クロンテック社(カリフォルニア州パロアルト)から購入した。U6-G93A は上述の通り作製した(Sui et al., 2002)。3’ブロックsiRNAは標準技術によって合成された。
【0104】
In-vitro RNAiアッセイ
野生種または突然変異SOD1のいずれかをコードする配列を有する560ヌクレオチドヒトSOD1標的RNAを、既報の通り調製する(Zamore et al., 2000)。標的切断は、濃度約5nMのショウジョウバエ胚ライセートを含有する標準in vitro RNAi反応において25乃至100nM siRNAを有する5’32P-キャップ放射標識標的をインキュベートすることによって決定した。
【0105】
細胞培養およびトランスフェクション
Hela細胞を、10%ウシ胎仔血清(FBS)、100ユニット/mlペニシリン、および100μg/mlストレプトマイシンをDMEMならびにDMEDおよびOpti-MEM(1:1)に入れたN2A 細胞の両方ともに添加し、その中で培養した。
トランスフェクションの24時間前、細胞(集密度70乃至90% )を倍散で脱離させ、6穴プレートに移して、抗生物質を含まない新鮮培地で培養した。トランスフェクションは、リポフェクタミン2000(インビトロゲン社)を、製造者の指示に従って用いて行った。トランスフェクションに用いた作製物の量は、突然変異または野生種SOD1-GFPおよびDsRedプラスミドそれぞれ4μg 、siRNAを4x10-11または4x10-12モル、ならびにU6-G93Aを20または8μgである。
【0106】
In vivoトランスフェクション
生後6乃至8週間のマウス24匹を3つのグループに分けた。第1のグループはshRNAベクタの投与を受けておらず、第2のグループは20μgの空ベクタの投与を受け、第3のグループはSOD1 G93A (配列番号16)に対するU6-hpRNA ベクタ20μgを投与された。すべてのグループは、mycでタグしたヒト野生種SOD1 (SEQ ID
NO: 7) 20μg、およびGFPでタグしたSOD1 20μgの両方が投与された。前記ベクタを合計体積がマウス1匹あたり2.5mlになるように、リンガ液で希釈した。マウスをアバーチン(240mg/kg)で麻酔し、前記ベクタを26ゲージ針を用いて10病未満で尾部静脈から注入する。注入から48時間後、肝臓から血液を除去するために、動物を5ml PBSで灌流した。肝臓を切り取り、素速くドライアイス上で冷凍させた。サンプルを、1% SDS、1mM DTT、1mMフェニルメチルスルホニルフルオリド(PMSF)、およびプロテアーゼ阻害物質カクテル(シグマ社1:100希釈)を含有する25mM PBSバッファ(pH 7.2)中に入れ、携帯用ポリトロン(プロサイエンティフィック社)を用いてホモジナイズした。
【0107】
ウエスタンブロット分析法
タンパク質濃度は、BCAタンパク質アッセイキット(ピアース社、イリノイ州ロックビル)を用いて決定した。Hela細胞タンパク質25μg、または100μg肝タンパク質を15% SDS-PAGE 上で分離し、ジーンスクリーン・プラス(パーキンエルマー社)上に移動させた。ウサギ抗SOD1(バイオデザイン社)またはヒツジ抗SOD1は1次抗体、およびHRP標識ヤギ抗ウサギIgG(アマシャム社)またはロバ抗ヒツジIgGは2次抗体だった。タンパク質のバンドの可視化には、スーパーシグナル・キット(ピアース社)およびコダック・デジタルイメージステーション440CFを用いた。そのバンドの強度の定量化には、コダック1Dソフトウェアを用いた。
【0108】
例I.
例I乃至VIIIは、siRNAが、まずsiRNA活性をショウジョウバエ胚ライセートを含む無細胞RNAI反応でテストし、次に培養哺乳類細胞中の活性1ヌクレオチド選択性siRNAを分析することによって、1ヌクレオチド選択性を有するようにデザインされたことを明らかにする。結果は、2つのmRNAの差が1ヌクレオチドしかなく同一の細胞に存在するとしても、21ヌクレオチドsiRNAおよびshRNAの両方とも、選択的に突然変異SOD1(配列番号8)の発現を阻害するが、野生種SOD1(配列番号7)ではそのようなことがないようにデザインすることができることを示した。したがって、RNAiはとりわけ、優性機能獲得型突然経によって生じる疾患の治療法として有用である。
【0109】
例I. siRNA 2本鎖は突然変異SOD1を区別することができる
それぞれ野生種または突然変異mRNAのいずれかを狙う3つのsiRNA
2セット(図1A、配列番号8)を、標的の切断部位またはその近傍のミスマッチが要求されるA型らせん形を乱しているかどうかをテストするように方向付けた。グアニン256(たとえばジェンバンク受入番号K00056などの、翻訳開始に関連するG256)SOD1の対立遺伝子がシトシンに突然変異し、グリシン-アルギニン突然変異(G85R) を生じているSOD1の対立遺伝子を選択した。突然変異したヌクレオチドは、SOD1 mRNA切断の予測された部位の近傍、すなわち、siRNAのアンチセンス鎖の5’末端に対して9位(P9)、10位(P10)、11位(P11)(図1A)に位置した。SOD1 mRNA切断のこの予測された部位は、前記siRNAとRNAiエンドヌクレアーゼの活性部位の中または近傍の非同族標的RNAとの間にミスマッチを配置する。これらのsiRNAのテストは、in vitroにおいてRNAを再利用する、確立されたショウジョウバエ胚性ライセートで行った(Zamore et al., 2000; Tuschl et al., 1999)。予想通り、6つのsiRNAのそれぞれが相当する標的RNAを切断したが、その有効性の差は劇的だった。たとえば、P11突然変異種および野生種siRNA(配列番号6, 10)は、それぞれの標的RNAを効率的に切断できなかった。反対に、P10突然変異種siRNA(配列番号4)は前記突然変異種標的RNAを効率的に切断した。完全長突然変異種SOD1標的mRNAの切断は、相当する約288ヌクレオチドの5’切断産生物の蓄積によって行われ、結果は、前記標的mRNAの非特異的分解ではなくRNAiを示唆している。siRNAの欠如下、またはルシフェラーゼに対するsiRNAの存在下において、前記突然変異標的RNA(配列番号8)はショウジョウバエ胚性ライセート中で安定だった(データは非表示)。標的RNAの切断および5’切断産生物の蓄積のデータは両方とも、1つの指数方程式に良くフィットし、この反応は擬1次動力学にしたがっていることを示している(図1B)。
【0110】
例I. siRNA 2本鎖は野生種SOD1を区別することができる
6つのsiRNAの特異性を決定するために、突然変異SOD1配列(配列番号8)に相当する各siRNAを、野生種SOD1 RNA (配列番号7)を切断する能力についてテストし、各野生種siRNAについて突然変異RNAを切断する能力をテストした。すべてではないが一部のsiRNA 2本鎖が、効率的に、完全にマッチする標的と1ヌクレオチドのミスマッチを有する標的を区別した。たとえば、突然変異と野生種のsiRNA(配列番号6、10)の両方のP11は、完全にマッチした標的RNAもミスマッチの標的RNAのいずれにも効率的な切断を惹起しなかった(図1A)。したがって、これらのsiRNA配列は本質的にRNAiを惹起する能力が乏しい。反対に、P9 (配列番号 14)およびP10野生種(配列番号12) siRNAは、相当する野生種標的の迅速な切断を惹起しただけでなく、突然変異RNAの著しい切断も生じた(図1A)。これらのsiRNAはRNAiを惹起する優れた能力があるが、乏しい選択性を示す。P10突然変異種siRNA(配列番号4)が、突然変異種および野生種SOD1 RNA (配列番号7、8)を効率的および堅固に識別し、突然変異RNAを野生種よりもはるかに効率的に切断することが示された。最も重要なのは、P10突然変異siRNA(配列番号4)は、突然変異および野生種SOD1 mRNA 標的を実質的に完全に識別することが示された(Fig. 1A)。このP10突然変異siRNAは、突然変異SOD1標的の切断を効率的に媒介したが、野生種SOD1 mRNAの切断はほとんど媒介せず(Fig. 1B)、このsiRNAは治療用途に理想的であることが示唆された。
【0111】
例III. Hela細胞における突然変異SOD1 G85R発現の選択的阻害
哺乳類細胞において、無細胞反応が正確、効率的、および選択的にsiRNAを予想できるかどうかをテストするために、カルボキシ末端を融合させたGFPを有する野生種または突然変異種SOD1 G85Rを発現したプラスミド作製物を作製した。各作製物を、dsRed発現ベクターを用いてHela細胞にトランスフェクトし、トランスフェクション対照とした。突然変異または野生種のSOD1 (配列番号7、8)のいずれかの発現を、蛍光標示式細胞分取法(FACS)で緑色および赤色細胞を定量することによって監視した。P9、P10およびP11 siRNAを相当する突然変異種または野生種標的でトランスフェクションしたところ、遺伝子発現を抑制したが、効率および選択性には差があった(図2)。反対に、ホタルルシフェラーゼに相補的なsiRNAによるコトランスフェクションでは、突然変異または野生種SOD1の発現のいずれも抑制しなかった(図2)。野生種に対するsiRNA p10以外のすべてのsiRNAは1ヌクレオチドミスマッチを有するmRNAを抑制しなかったが、野生種に対するsiRNA
p10は野生種および突然変異SOD1の発現の両方を効率的に抑制した(図2)。この結果は全般的にin vitroデータ(図1)と一致しており、すべてではないが一部のsiRNAは1ヌクレオチドの差を有するmRNA標的を効率的に区別することができることを示唆した。
【0112】
例IV. Hela細胞におけるU6-G93A ベクタによる突然変異SOD1 G93A発現の選択的阻害
最近、shRNAがin vivoにおいてRNAiを惹起することができることが示された。突然変異種SOD1に対するshRNAが、選択的に野生種SOD1発現ではなく突然変異種SOD1の発現を阻害することができるかどうかをテストするために、別の疾患の原因となる突然変異種SOD1
G93A に相同なshRNA(ヌクレオチド281位のGをCに交換し、shRNAと野生種SOD1G:Gミスマッチを配置するヌクレオチド交換;配列番号16)を、RNAポリメラーゼIII(U6)プロモータの制御下において合成するプラスミドを作製した(Sui et al., 2002)。結果から、野生種または突然変異SOD1-GFPプラスミドのいずれかとコトランスフェクトした場合、この作製物を用いて、培養細胞中の突然変異種SOD1の1ヌクレオチド選択的RNAiを惹起することができるということが示された(図3)。
【0113】
例V. in vivoにおけるsiRNAおよびU6-G93A ベクタによる突然変異SOD1発現の選択的阻害
突然変異種の選択的阻害が神経細胞で実現するかどうかをテストするために、野生種および突然変異SOD1-GFP作製物を、SOD1 G85R に対するsiRNA P10、またはSOD G93A (配列番号16)に対するshRNA合成ベクタのいずれかを用いて、神経芽細胞腫細胞株N2aにコトランスフェクトした。Hela細胞と同様に、合成siRNAおよびshRNA作製物が、N2a細胞における突然変異SOD1発現の選択的阻害を方向付けた(図4A、B)。
【0114】
例VI. in vivoにおける突然変異SOD1 G85Rの選択的阻害
同一の細胞に突然変異および野生種SOD1 mRNA1が両方とも存在する場合、ヌクレオチド選択的siRNAがその突然変異および野生種SOD1を識別できるかどうかを明らかにするために、Hela細胞にP10 siRNAおよび突然変異SOD1 G85R-GFPでトランスフェクトした。抗SOD1抗体で免疫ブロットすることによって、トランスフェクト融合SOD1-GFPおよび内因性野生種ヒトSOD1の両方の検出が可能になった。内因性野生種SOD1発現の約50%阻害は、約50%のトランスフェクション効率を反映した。P10野生種siRNAに対して、突然変異種に対するP10 siRNAは2種類の異なる用量において突然変異種の発現を阻害したが、内因性野生種SOD1の発現には効果がなかった。
【0115】
例VII. in vivoにおけるU6-G93A ベクタによる突然変異種SOD1発現の選択的阻害
選択的阻害をin vivoで生じさせることができるかどうかをテストするために、SOD1レポータおよびshRNAプラスミドを、水力学的トランスフェクションプロトコルを用いてマウスにトランスフェクトした。突然変異SOD1 G93A-GFP プラスミドおよびmycでタグした野生種ヒトSOD1(ゲル上においてトランスフェクトしたヒトSOD1を内因性マウスSOD1からよりよく分離することができる)をU6空ベクトルまたはU6-G93Aベクタでコトランスフェクトした。SOD1
G93A-GFP およびSOD1mycの肝臓における発現を、ウエスタンブロットによって検証した。結果から、U6-G93Aで選択的にコトランスフェクトする場合のみ、G93A発現が減少することが示された(図6)。
【0116】
例VIII. トランスジェニックマウスを用いたin vivoにおける突然変異SOD1のshRNA抑制
突然変異SOD1に対するshRNAがin
vivoにおいて突然変異SOD1の発現を抑制することができるかどうかを明らかにするために、RNAポリメラーゼIII(Pol
III)プロモータU6の制御下におけるSOD1G93A に対するshRNAを発現するトランスジェニックマウス(U6-G93A マウス)を、C57BL/6JおよびSJLのハイブリッド背景において作製した。
【0117】
マウスU6プロモータの制御下におけるshRNA相同的突然変異SOD1G93A (shG93A) を合成するプラスミドは、公表されているプロトコルにしたがって作製した(Sui et al., 2002 Proc Natl Acad Sci USA 99:5515-5520)。前記マウスを作製するために、Kpn IおよびSac Iを用いた消化によって導入遺伝子を直線化し、精製して、マサチューセッツ大学医学部(UMMS)トランスジェニック・コアでマウスの有精卵に注入した。
【0118】
U6-G93A トランスジェニックマウスをスクリーニングするために、前記導入遺伝子の配列を選択的に増幅するPCRプライマをデザインし、前記トランスジェニックマウスの同定に用いた。合計7匹の第0世代(F0)が同定された。これらの第0世代を、FVB背景において、突然変異種SOD1G93A
導入マウスと交雑させた。
【0119】
F1マウスを、既報のとおりのサザンブロット法を用いて導入遺伝子コピー数について解析した(Xu et al., 1993 Cell 73:23-33)。末端DNAをBam H1で消化し、388ヌクレオチドの導入遺伝子断片を作製した。内因性マウスU6プロモータにはBamHI部位が1つしかないため、BamHI消化によって、内因性マウスU6遺伝子からより大きな断片を作製できた。前記U6プロモータ領域に相補的な32P標識RNAオリゴヌクレオチドプローブを、ハイブリダイゼーションに用いた。前記U6領域を標的として用いた。これは、前記内因性マウスU6バンドは同一ブロット上で前記導入遺伝子とともに検出でき、導入遺伝子のコピー数を定量化するための参照として前記内因性バンドを用いることができるためである。
【0120】
ノーザンブロット法を用いて測定したとおり、前記U6-G93A
shRNA作製物を、2種類の遺伝子を導入したトランスジェニックマウスから採取した細胞に発現させた。ウエスタンブロット法を用いて測定した結果、前記U6-G93A
shRNA 作製物は、2種類の遺伝子を導入したトランスジェニックマウスにおける突然変異種SOD1G93A
の発現(U6-G93A shRNA 作製物および突然変異種SOD1G93Aを発現)をサイレンシングすることが明らかになった。
【0121】
例I乃至VIIIの結果の考察
RNAiを用いた優性突然変異種ALS遺伝子を選択的にサイレンシングする可能性を調べた。野生種または突然変異種のSOD1のいずれかにマッチする複数のsiRNAを用いた結果、in vitroにおいて、突然変異種SOD1 G85R に対するsiRNAは前記突然変異種を切断するが、野生種SOD1 RNAの切断は効率的ではないことが示された(図1)。さらに、同一の細胞に突然変異および野生種SOD1タンパク質の両方が存在する場合でも(図4)、これらのsiRNAは、哺乳類の細胞において突然変異SOD1タンパク質発現を選択的に阻害するが野生種については阻害しなかった(図2)。in vivoにおいてsiRNAに処理されたヘアピンを発現するベクタも、マウス肝において突然変異SOD1の発現を選択的に阻害するが、野生種SOD1の発現は阻害しなかった。これらの結果から、優性突然変異対立遺伝子の選択的阻害はRNAiを用いて実現することができ、in vitroまたはin vivoにおいて、前臨床スクリーニングによって最適化したsiRNAおよびshRNA配列を同定することができることが明らかになった。
【0122】
哺乳類細胞においてSOD1 1ヌクレオチドの識別を実現することはできるが、この識別は確実なものではない。一部のsiRNAは1ヌクレオチドの差を有する対立遺伝子間の識別が可能であるが、その他のsiRNAでは識別することができない。結果は、突然変異病因対立遺伝子に方向付けられるようにデザインされたsiRNAに2種類の異なる欠陥があることを示している。第1に、標的に完全にマッチするsiRNAは、その標的を切断し、前記標的遺伝子からのタンパク質発現を阻害することができるが、すべてのsiRNAが同一の効率でサイレンシングを生じるわけではない。たとえば、前記野生種に対するsiRNAの中で、in vitroにおいて、P9およびP10はP11よりも効率的に標的を切断した。P10はまた、哺乳類細胞において、標的遺伝子発現を最も効率的に阻害した(図2)。同様に、前記突然変異SOD1 G85Rに対するsiRNAの中で、P9およびP10はP11よりも効率的に突然変異RNAを切断した。P10はまた、哺乳類細胞において、突然変異SOD1発現の阻害について最も効率的だった(図2)。前記標的に対するsiRNA配列の1ヌクレオチドのシフトの結果、サイレンシングの効率においてこのような顕著な変化を生じる。
第2に、完全にマッチした標的RNAと1ヌクレオチドのミスマッチを有するRNAの間の選択性の差が、使用された6つのsiRNAの中で認められた。たとえば、野生種P10 siRNAは乏しい選択性を付与した。野生種P10は、無細胞アッセイにおいて野生種および突然変異SOD1 RNA の両方を切断し、哺乳類動物の両方の対立遺伝子の発現を高い効率で阻害した。反対に、突然変異SOD1に対して方向付けられたP10 siRNAは最高の選択性を付与した。それは突然変異SOD1 RNA を切断し、無細胞アッセイにおいて突然変異SOD1の発現を阻害し、哺乳類細胞における突然変異SOD1の発現は阻害したが野生種SOD1は阻害しなかった(図1乃至6)。
【0123】
突然変異と野生種のP10 siRNAの選択性の差についての説明は、以下の通りである。突然変異種P10 siRNAと野生種SOD1 mRNA の間のミスマッチがG:G衝突をつくり出したが、野生種P10 siRNA と突然変異種G85R mRNA の間のミスマッチはC:C衝突を生じた(図1A参照)。したがって、特定の配列の1つの対立遺伝子に選択的に作用するsiRNAをデザインする場合、以下が考慮された。理論によって制限したいとは考えないが、プリン:プリンミスマッチは、siRNAのアンチセンス鎖とそのmRNA標的の間に要求されるA型らせんを妨げる(Chiu et al., 2002)。反対に、ピリミジン:ピリミジンミスマッチはA型らせんの中に容易に収容することができる。したがって、siRNAと野生種標的RNAの間のG:G衝突は前記野生種標的を排除し、前記突然変異標的の選択性を高める。顕著なことに、突然変異種SOD1G93Aについて優れた選択性を示す、G93Aに対するsiRNAヘアピンベクタも、野生種SOD1 mRNAを有するG:G衝突をつくり出した。これらの結果は、プリン:プリンミスマッチはピリジン:ピリジンミスマッチよりも大きな選択性を付与することを示唆した。ピリジン:ピリジンミスマッチを含有する本願発明の方法に用いたsiRNAのデザインに加えて、前記siRNAは当業に知られる方法を用いてデザインされる。
【0124】
参考文献
1. Amarzguioui, M., Holen, T., Babaie, E., and
Prydz, H. (2003). Tolerance for mutations and chemical modifications in a
siRNA. Nucleic Acids Res 31, 589-595.
2. Behndig,
A., Karlsson, K., Reaume, A. G., Sentman, M. L. & Marklund, S. L. In
vitro photochemical cataract in mice lacking copper-zinc superoxide dismutase.
Free Radic Biol Med 31, 738-44. (2001).
3. Boutla,
A., Delidakis, C., Livadaras, I., Tsagris, M. & Tabler, M. Short
5'-phosphorylated double-stranded RNAs induce RNA interference in Drosophila.
Curr Biol 11, 1776-80. (2001).
4. Brummelkamp,
T. R., Bernards, R. & Agami, R. A system for stable expression of short
interfering RNAs in mammalian cells. Science 296, 550-3. (2002).
5. Brummelkamp,
T., Bernards, R. & Agami, R. Stable suppression of tumorigenicity by
virus-mediated RNA interference. Cancer Cell 2, 243. (2002).
6. Caplen,
N. J., Parrish, S., Imani, F., Fire, A. & Morgan, R. A. Specific inhibition
of gene expression by small double-stranded RNAs in invertebrate and vertebrate
systems. Proc Natl Acad Sci U S A 98, 9742-7. (2001).
7. Caplen,
N. J. et al. Rescue of polyglutamine-mediated cytotoxicity by double-stranded
RNA-mediated RNA interference. Hum Mol Genet 11, 175-84. (2002).
8. Chiu,
Y.-L. & Rana, T. M. RNAi in human cells: basic structural and functional
features of small interfering RNA. Molecular Cell 10, 549-561 (2002).
9. Cleveland,
D. W. & Rothstein, J. D. From Charcot to Lou Gehrig: deciphering selective
motor neuron death in ALS. Nat Rev Neurosci 2, 806-19. (2001).
10. Devroe,
E. & Silver, P. A. Retrovirus-delivered siRNA. BMC Biotechnol 2, 15.
(2002).
11. Elbashir,
S. M., Lendeckel, W. & Tuschl, T. RNA interference is mediated by 21- and
22-nucleotide RNAs. Genes Dev 15, 188-200. (2001).
12. Elbashir,
S. M. et al. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured
mammalian cells. Nature 411, 494-8. (2001).
13. Elbashir,
S. M., Martinez, J., Patkaniowska, A., Lendeckel, W. & Tuschl, T.
Functional anatomy of siRNAs for mediating efficient RNAi in Drosophila
melanogaster embryo lysate. Embo J 20, 6877-88. (2001).
14. Flood,
D. G. et al. Hindlimb motor neurons require Cu/Zn superoxide dismutase for
maintenance of neuromuscular junctions. Am J Pathol 155, 663-72. (1999).
15. Hamilton,
A. J. & Baulcombe, D. C. A species of small antisense RNA in
posttranscriptional gene silencing in plants. Science 286, 950-2. (1999).
16. Hammond,
S. M., Bernstein, E., Beach, D. & Hannon, G. J. An RNA-directed nuclease
mediates post-transcriptional gene silencing in Drosophila cells. Nature 404,
293-6. (2000).
17. Hannon,
G. J. RNA interference. Nature 418, 244-51. (2002).
18. Hutvagner,
G. & Zamore, P. D. RNAi: nature abhors a double-strand. Curr Opin Genet Dev
12, 225-32. (2002).
19. Jacque,
J. M., Triques, K. & Stevenson, M. Modulation of HIV-1 replication by RNA
interference. Nature 418, 435-8. (2002).
20. Kawase,
M. et al. Exacerbation of delayed cell injury after transient global ischemia
in mutant mice with CuZn superoxide dismutase deficiency. Stroke 30, 1962-8.
(1999).
21. Kondo,
T. et al. Reduction of CuZn-Superoxide Dismutase Activity Exacerbates Neuronal
Cell Injury and Edema Formation after Transient Focal Cerebral Ischemia.
Journal of Neuroscience 17, 4180-9 (1997).
22. Lee,
N. S. et al. Expression of small interfering RNAs targeted against HIV-1 rev
transcripts in human cells. Nat Biotechnol 20, 500-5. (2002).
23. Lipardi,
C., Wei, Q. & Paterson, B. M. RNAi as random degradative PCR: siRNA primers
convert mRNA into dsRNAs that are degraded to generate new siRNAs. Cell 107,
297-307. (2001).
24. Matz,
P. G., Copin, J. C. & Chan, P. H. Cell death after exposure to subarachnoid
hemolysate correlates inversely with expression of CuZn-superoxide dismutase.
Stroke 31, 2450-9. (2000).
25. Matzuk,
M. M., Dionne, L., Guo, Q., Kumar, T. R. & Lebovitz, R. M. Ovarian function
in superoxide dismutase 1 and 2 knockout mice. Endocrinology 139, 4008-11.
(1998).
26. McFadden,
S. L., Ding, D. & Salvi, R. Anatomical, metabolic and genetic aspects of
age-related hearing loss in mice. Audiology 40, 313-21. (2001).
27. McManus,
M. T. & Sharp, P. A. Gene silencing in mammals by small interfering RNAs.
Nat Rev Genet 3, 737-47. (2002).
28. McManus,
M. T., Petersen, C. P., Haines, B. B., Chen, J. & Sharp, P. A. Gene
silencing using micro-RNA designed hairpins. Rna 8, 842-50. (2002).
29. Miyagishi,
M. & Taira, K. U6 promoter-driven siRNAs with four uridine 3' overhangs
efficiently suppress targeted gene expression in mammalian cells. Nat
Biotechnol 20, 497-500. (2002).
30. Nykanen,
A., Haley, B. & Zamore, P. D. ATP requirements and small interfering RNA
structure in the RNA interference pathway. Cell 107, 309-21. (2001).
31. Paddison,
P. J., Caudy, A. A., Bernstein, E., Hannon, G. J. & Conklin, D. S. Short
hairpin RNAs (shRNAs) induce sequence-specific silencing in mammalian cells. Genes
Dev 16, 948-58. (2002).
32. Paul,
C. P., Good, P. D., Winer, I. & Engelke, D. R. Effective expression of
small interfering RNA in human cells. Nat Biotechnol 20, 505-8. (2002).
33. Shefner,
J. M. et al. Mice lacking cytosolic copper/zinc superoxide dismutase display a
distinctive motor axonopathy. Neurology 53, 1239-46. (1999).
34. Schwarz,
D. S., Hutvagner, G., Haley, B. & Zamore, P. D. Evidence that siRNAs
function as guides, not primers, in the Drosophila and human RNAi pathways.
Molecular Cell 10, 537-548 (2002).
35. Sui,
G. et al. A DNA vector-based RNAi technology to suppress gene expression in
mammalian cells. Proc Natl Acad Sci U S A 99, 5515-20. (2002).
36. Taylor,
J. P., Hardy, J. & Fischbeck, K. H. Toxic proteins in neurodegenerative disease.
Science 296, 1991-5. (2002).
37.
Tuschl, T., Zamore, P. D., Lehmann, R., Bartel, D. P. & Sharp, P. A.
Targeted mRNA degradation by double-stranded RNA in vitro. Genes Dev 13,
3191-7. (1999).
38. Xia,
H., Mao, Q., Paulson, H. L. & Davidson, B. L. siRNA-mediated gene silencing
in vitro and in vivo. Nat Biotechnol 20, 1006-10. (2002).
39. Xu, P., Vernooy, S. Y., Guo, M., and Hay, B. A.
(2003). The Drosophila MicroRNA Mir-14 Suppresses Cell Death and Is Required
for Normal Fat Metabolism. Curr Biol 13, 790-795.
40. Yu, J. Y., DeRuiter, S. L. & Turner, D. L. RNA interference by
expression of short-interfering RNAs and hairpin RNAs in mammalian cells. Proc
Natl Acad Sci U S A 99, 6047-52. (2002).
41. Zamore,
P. D., Tuschl, T., Sharp, P. A. & Bartel, D. P. RNAi: double-stranded RNA
directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals.
Cell 101, 25-33. (2000).
42. Zeng,
Y., Wagner, E. J. & Cullen, B. R. Both natural and designed micro RNAs can
inhibit the expression of cognate mRNAs when expressed in human cells. Mol Cell
9, 1327-33. (2002).
【0125】
その他の実施態様
本願発明は、発明の詳細な説明に関連して説明されているが、上述の説明は具体例を示すことを意図しており本願発明の範囲を制限することは意図しておらず、それは添付の特許請求の範囲によって定義されると理解される。その他の局面、利点、および改善点は、添付の特許請求の範囲内である。
【図面の簡単な説明】
【0126】
【図1】siRNA 2本鎖は、in vitroにおいて突然変異および野生種のSOD1を区別することができる(A) 使用したsiRNA 2本鎖:突然変異siRNA P11 (配列番号 5、センス; 配列番号 6、アンチセンスまたはガイド)、突然変異 siRNA P10 (SEQ ID NO: 3、センス; SEQ ID NO: 4、アンチセンスまたはガイド)、突然変異 siRNA P9 (SEQ ID NO: 1、センス; SEQ ID NO: 2 アンチセンスまたはガイド)、SOD1 野生種標的 (SEQ ID NO: 7)、SOD1 突然変異標的 (SEQ ID NO: 8)、野生種 siRNA P11 (SEQ ID NO: 9 センス; SEQ ID NO: 10、アンチセンスまたはガイド)、野生種 siRNA P10 (SEQ ID NO: 11、センス; SEQ ID NO: 12、アンチセンスまたはガイド)、野生種 siRNA P9 (SEQ ID NO: 13、センス; SEQ ID NO: 14、アンチセンスまたはガイド) (B) 突然変異siRNA p10は、野生種SOD1 mRNA ではなく突然変異SOD1 mRNA をRNAi経路による破壊に方向付ける。
【図2】Hela細胞におけるsiRNAによる突然変異SOD1 G85R発現の選択的阻害。SOD1wtGFPまたはG85R-GFPを各種のsiRNAとともにコトランスフェクトした。dsRedをトランスフェクション対照としてコトランスフェクトした。緑色および赤色蛍光細胞をFACSを用いて定量した。(A) トランスフェクションにおける緑色(黒線)および赤色細胞(白線)の相対数(n=3)。エラーバーは標準偏差を示す。
【図3】Hela細胞におけるU6-G93A ベクタによる突然変異SOD1 G93A発現の選択的阻害。(A) G93A shRNA のデザイン(配列番号16)、(B) SOD1wtGFPまたはSOD1G93A-GFPをU6空またはU6-G93Aとともにコトランスフェクトした。dsRedをトランスフェクション対照としてコトランスフェクトした。緑色および赤色傾向細胞をFACSを用いて定量した。4回の実験の結果を平均した。エラーバーは標準偏差を示す。
【図4】神経細胞芽腫N2a細胞におけるsiRNAおよびU6-G93Aベクタによる突然変異SOD1発現の選択的阻害。(A) G85Rに対するsiRNA (n=4)、(B) U6-G93Aベクタ(n=3)。エラーバーは標準偏差を示す。
【図5】同一の細胞におけるsiRNAによる、野生種SOD1ではなく、突然変異SOD1 G85R発現の選択的阻害。(A) 突然変異SOD1 G85R-GFP を検出するトランスフェクトHela細胞のタンパク質ブロットから測定した、SOD1の相対的レベル(4回のトランスフェクションの平均)。エラーバーは標準誤差を示す。
【図6】in vivoにおけるU6-G93A ベクタによる突然変異SOD1発現の選択的阻害(A) 水力学的トランスフェクション法を用い、マウスにおいてSOD1 G93A-GFP をC末端mycタグ付き野生種ヒトSOD1とともにコトランスフェクトした。SDA-PAGE上の相対バンド強度を定量した。野生種SOD1mycに対するSOD1 G93A-GFPの割合を示す。各グループにおいて8匹の動物を用いた。U6-G93A グループは、他の2つのグループとは有意に異なる(p < 0.05)。
【図7】ヒトSOD-1タンパク質のジェンバンク登録。受入番号NP_000445。野生種SOD-1 (配列番号18)の推定アミノ酸配列を示す。
【図8】ヒトSOD-1 mRNAのジェンバンク登録。受入番号NM_000454。野生種SOD-1 (配列番号17)のヌクレオチド配列を示す。
【図9】図9はSOD1ゲノム配列(配列番号19)である。
【配列表】
【特許請求の範囲】
【請求項1】
遺伝子の少なくとも2つの異なる対立遺伝子を含む細胞中の標的対立遺伝子の発現を阻害する方法であって、当該方法が前記標的対立遺伝子に特異的なsiRNAを前記細胞に投与するステップを含む、方法。
【請求項2】
請求項1に記載の方法であって、当該方法において、前記標的対立遺伝子が、優性の機能獲得型突然変異に関連する障害と相関する、方法。
【請求項3】
請求項2に記載の方法であって、当該方法において、前記障害が筋萎縮性軸索硬化症、ハンチントン病、アルツハイマー病、およびパーキンソン病からなるグループから選択される、方法。
【請求項4】
優性の機能獲得型突然変異対立遺伝子の存在と相関する疾患を有する対象の治療方法であって、当該方法が前記突然変異対立遺伝子に特異的なsiRNAを治療上有効な量、前記対象に投与するステップを含む、方法。
【請求項5】
請求項4に記載の方法であって、当該方法において、前記siRNAが機能獲得型突然変異を標的とする、方法。
【請求項6】
請求項4に記載の方法であって、当該方法において、前記障害が筋萎縮性軸索硬化症、ハンチントン病、アルツハイマー病、およびパーキンソン病からなるグループから選択される、方法。
【請求項7】
請求項4に記載の方法であって、当該方法において前記疾患は筋萎縮性側策硬化症である、方法。
【請求項8】
請求項7に記載の方法であって、当該方法において前記対立遺伝子はSOD1である、方法。
【請求項9】
請求項8に記載の方法であって、当該方法において、前記突然変異対立遺伝子は点変異を含む、方法。
【請求項10】
請求項8に記載の方法であって、当該方法において、前記点突然変異はグアニン:シトシン突然変異である、方法。
【請求項11】
請求項8に記載の方法であって、当該方法において、前記突然変異はG256Cである、方法。
【請求項12】
請求項8に記載の方法であって、当該方法において、前記突然変異はG281Cである、方法。
【請求項13】
請求項8に記載の方法であって、当該方法において、前記siRNAは図1Aに記載の配列を含む、方法。
【請求項14】
図1Aに記載の配列を含むsiRNA。
【請求項15】
図1Aに記載の前記配列を含むp10突然変異siRNA。
【請求項16】
図1Aに記載の前記配列を含むp9突然変異siRNA。
【請求項17】
図3Aに記載の前記配列を含むG93A SOD1 shRNA 。
【請求項18】
請求項13に記載のshRNAを含む発現作製物。
【請求項19】
請求項10乃至12に記載の前記siRNAを含む治療用組成物、および薬学的に許容の担体。
【請求項20】
請求項13に記載の前記shRNAを含む治療用組成物、および薬学的に許容の担体。
【請求項1】
遺伝子の少なくとも2つの異なる対立遺伝子を含む細胞中の標的対立遺伝子の発現を阻害する方法であって、当該方法が前記標的対立遺伝子に特異的なsiRNAを前記細胞に投与するステップを含む、方法。
【請求項2】
請求項1に記載の方法であって、当該方法において、前記標的対立遺伝子が、優性の機能獲得型突然変異に関連する障害と相関する、方法。
【請求項3】
請求項2に記載の方法であって、当該方法において、前記障害が筋萎縮性軸索硬化症、ハンチントン病、アルツハイマー病、およびパーキンソン病からなるグループから選択される、方法。
【請求項4】
優性の機能獲得型突然変異対立遺伝子の存在と相関する疾患を有する対象の治療方法であって、当該方法が前記突然変異対立遺伝子に特異的なsiRNAを治療上有効な量、前記対象に投与するステップを含む、方法。
【請求項5】
請求項4に記載の方法であって、当該方法において、前記siRNAが機能獲得型突然変異を標的とする、方法。
【請求項6】
請求項4に記載の方法であって、当該方法において、前記障害が筋萎縮性軸索硬化症、ハンチントン病、アルツハイマー病、およびパーキンソン病からなるグループから選択される、方法。
【請求項7】
請求項4に記載の方法であって、当該方法において前記疾患は筋萎縮性側策硬化症である、方法。
【請求項8】
請求項7に記載の方法であって、当該方法において前記対立遺伝子はSOD1である、方法。
【請求項9】
請求項8に記載の方法であって、当該方法において、前記突然変異対立遺伝子は点変異を含む、方法。
【請求項10】
請求項8に記載の方法であって、当該方法において、前記点突然変異はグアニン:シトシン突然変異である、方法。
【請求項11】
請求項8に記載の方法であって、当該方法において、前記突然変異はG256Cである、方法。
【請求項12】
請求項8に記載の方法であって、当該方法において、前記突然変異はG281Cである、方法。
【請求項13】
請求項8に記載の方法であって、当該方法において、前記siRNAは図1Aに記載の配列を含む、方法。
【請求項14】
図1Aに記載の配列を含むsiRNA。
【請求項15】
図1Aに記載の前記配列を含むp10突然変異siRNA。
【請求項16】
図1Aに記載の前記配列を含むp9突然変異siRNA。
【請求項17】
図3Aに記載の前記配列を含むG93A SOD1 shRNA 。
【請求項18】
請求項13に記載のshRNAを含む発現作製物。
【請求項19】
請求項10乃至12に記載の前記siRNAを含む治療用組成物、および薬学的に許容の担体。
【請求項20】
請求項13に記載の前記shRNAを含む治療用組成物、および薬学的に許容の担体。
【図1】

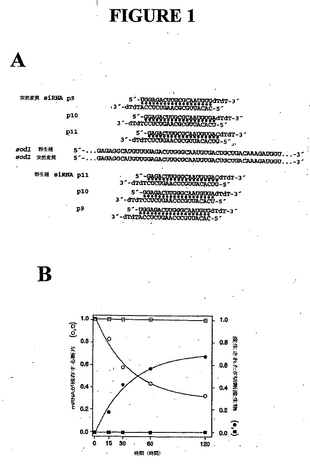
【図2】
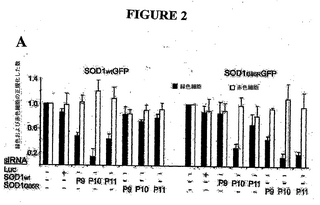
【図3】
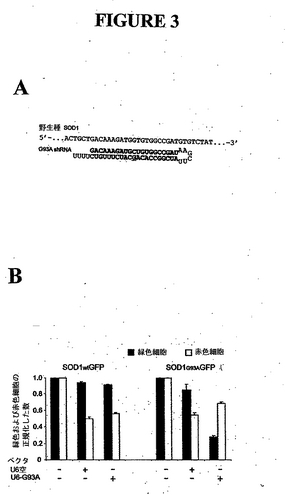
【図4】
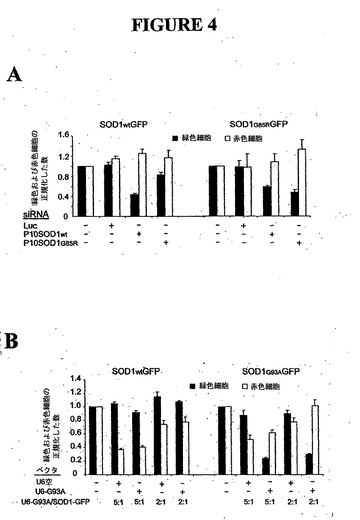
【図5】
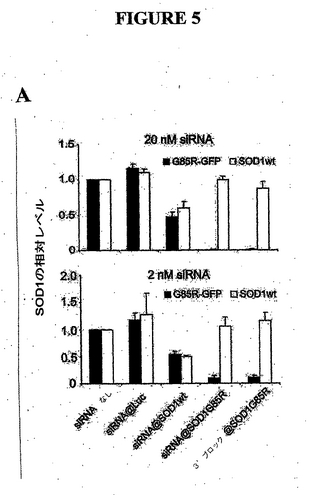
【図6】
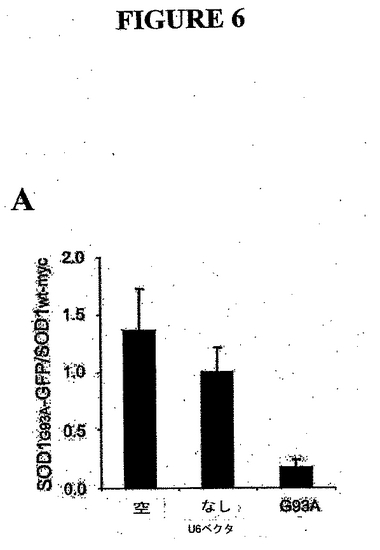
【図7】

【図8】

【図9】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2006−505288(P2006−505288A)
【公表日】平成18年2月16日(2006.2.16)
【国際特許分類】
【出願番号】特願2005−502260(P2005−502260)
【出願日】平成15年11月4日(2003.11.4)
【国際出願番号】PCT/US2003/035009
【国際公開番号】WO2004/042027
【国際公開日】平成16年5月21日(2004.5.21)
【出願人】(500143324)ユニバーシティー オブ マサチューセッツ (4)
【Fターム(参考)】
【公表日】平成18年2月16日(2006.2.16)
【国際特許分類】
【出願日】平成15年11月4日(2003.11.4)
【国際出願番号】PCT/US2003/035009
【国際公開番号】WO2004/042027
【国際公開日】平成16年5月21日(2004.5.21)
【出願人】(500143324)ユニバーシティー オブ マサチューセッツ (4)
【Fターム(参考)】
[ Back to top ]