
幹細胞の分化誘導方法
【課題】ウイルスや遺伝子を用いることなくES細胞又はiPS細胞などの幹細胞から膵β細
胞に分化させる方法を提供すること。
【解決手段】哺乳動物由来の幹細胞の培養開始後5日目から7日目の間にPdx-1蛋白質を
1回以上導入し、9日目から11日目の間にNeuroD蛋白質を1回以上導入し、13日目か
ら15日目の間にMafA 蛋白質を1回以上導入することを含む、幹細胞から膵臓系細胞へ
と分化誘導する方法。
胞に分化させる方法を提供すること。
【解決手段】哺乳動物由来の幹細胞の培養開始後5日目から7日目の間にPdx-1蛋白質を
1回以上導入し、9日目から11日目の間にNeuroD蛋白質を1回以上導入し、13日目か
ら15日目の間にMafA 蛋白質を1回以上導入することを含む、幹細胞から膵臓系細胞へ
と分化誘導する方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ES細胞又はiPS細胞などの幹細胞の分化誘導方法に関する。より詳細には、
本発明は、3種類の転写調節因子の組換え蛋白質を幹細胞に導入することによって膵β細
胞に分化誘導させる方法に関する。
【背景技術】
【0002】
マウス膵臓へPdx-1遺伝子、Ngn3遺伝子及び MafA遺伝子をウイルスベクターで導入する
ことにより、膵β細胞に類似した細胞を誘導できることが報告されている(Nature 455,
627, 2008)。しかし、ウィルスベクターによる遺伝子導入は、染色体に組み込まれるこ
とによる腫瘍化などの問題点があり、臨床応用には適さない。他に、TATペプチドを付加
したNgn3蛋白質を胎生膵組織に導入することにより、膵内分泌細胞への分化を促進したと
の報告がある(Diabetes, 54, 720, 2005)。しかし、胎生膵組織を用いた膵β細胞分化
誘導技術は、臨床応用が困難である。
【0003】
胚性幹(ES)細胞は、胚盤胞の内細胞塊に由来する多能性細胞である。ES細胞は未分化
状態では無制限に培養でき、刺激されると多様な細胞型に分化でき、例えば、神経細胞、
造血性細胞、膵臓や肝臓などの内胚葉系の細胞への誘導が可能である。また、iPS細胞(
人工多能性幹細胞:induced pluripotent stem cell)は、体細胞を初期化することによ
って得られる多能性を有する細胞である。しかし、これまで、蛋白質を導入することによ
りES細胞から膵β細胞へ分化誘導を成功させたとの報告は無い。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Nature 455, 627, 2008
【非特許文献2】Diabetes, 54, 720, 2005
【発明の開示】
【発明が解決しようとする課題】
【0005】
本発明は、ウイルスや遺伝子を用いることなくES細胞又はiPS細胞などの幹細胞から膵
β細胞に分化させる方法を提供することを解決すべき課題とした。
【課題を解決するための手段】
【0006】
本発明者らは上記課題を解決するために鋭意検討した結果、3種類の転写調節因子の組
換え蛋白質(Pdx-1蛋白質、NeuroD蛋白質、及びポリアルギニンを付加したMafA蛋白質)
をES細胞に導入することによって膵β細胞に分化誘導させることに成功し、本発明を完成
するに至った。
【0007】
即ち、本発明によれば、哺乳動物由来の幹細胞の培養開始後5日目から7日目の間にP
dx-1蛋白質を1回以上導入し、9日目から11日目の間にNeuroD蛋白質を1回以上導入し
、13日目から15日目の間にMafA 蛋白質を1回以上導入することを含む、幹細胞から
膵臓系細胞へと分化誘導する方法が提供される。
好ましくは、哺乳動物由来の幹細胞はマウス、サル又はヒト由来の幹細胞である。
好ましくは、幹細胞はES細胞又はiPS細胞である。
好ましくは、膵臓系細胞はインスリン産生細胞である。
【0008】
好ましくは、MafA蛋白質は、ポリアルギニンが付加されているMafA蛋白質である。
好ましくは、ラット膀胱癌由来804G細胞由来の細胞外マトリックス上で培養している哺
乳動物由来の幹細胞に、Pdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質を導入する。
【0009】
本発明によれば、上記した本発明の方法により得られる、ES細胞から分化誘導された膵
臓系細胞が提供される。
本発明によればさらに、上記した本発明の方法により幹細胞から膵臓系細胞へと分化誘
導する工程、及び分化誘導された膵臓系細胞を分離する工程を含む、ES細胞から分化誘導
された膵臓系細胞を取得する方法が提供される。
【発明の効果】
【0010】
本発明によれば、ウイルスや遺伝子を用いることなくES細胞又はiPS細胞などの幹細胞
を膵β細胞に分化させることができる。本発明の方法は、糖尿病患者に対する膵β細胞移
植治療に利用する膵β細胞の作製方法として有用である。
【図面の簡単な説明】
【0011】
【図1】図1は、全長又は11個のアルギニンを付加した組み換え蛋白質の模式図を示す。
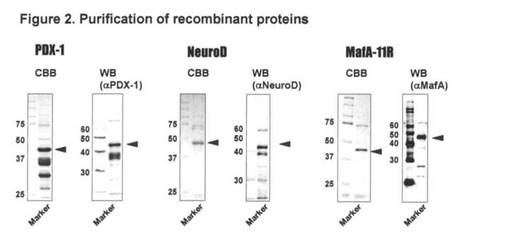
【図2】図2は、組み換え蛋白質の精製を示す。
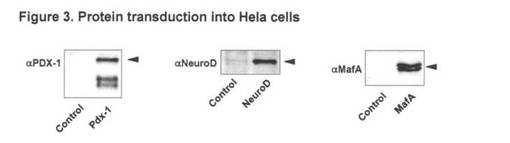
【図3】図3は、HeLa細胞への蛋白質の導入を示す。
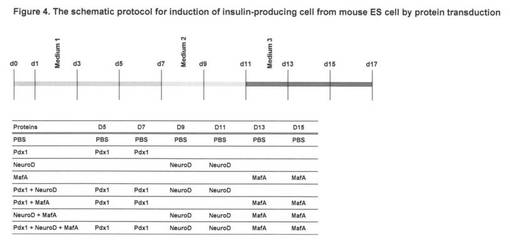
【図4】図4は、蛋白質の導入によるマウスES細胞からのインスリン産生細胞の誘導の手順を示す。
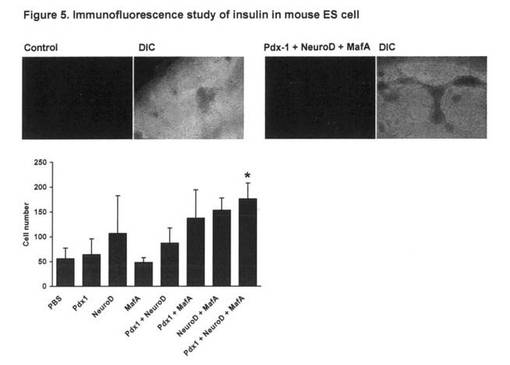
【図5】図5は、マウスES細胞におけるインスリンの免疫蛍光分析を示す。
【図6】図6は、インスリン1遺伝子発現のリアルタイムRT-PCR解析を示す。
【図7】図7は、9日目からはインスリン遺伝子発現は蛋白質導入により増加しないことを示す。
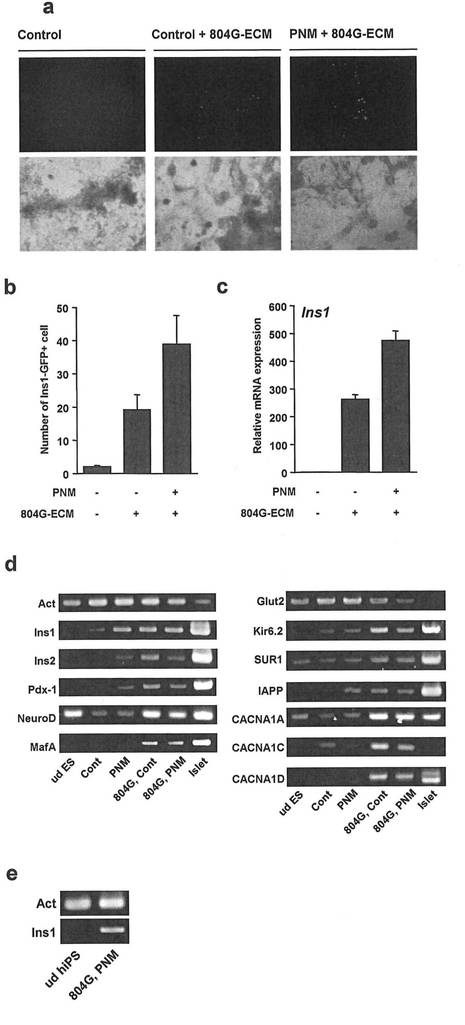
【図8】図8は、804G細胞由来の細胞外マトリックスを用いた、3因子の蛋白質導入によるマウスES細胞からインスリン産生細胞への分化を示す。
【発明を実施するための形態】
【0012】
以下、本発明の実施の形態についてさらに詳細に説明する。
本発明による幹細胞から膵臓系細胞へと分化誘導する方法は、哺乳動物由来の幹細胞に
Pdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質を導入することを特徴とする方法である。
【0013】
膵β細胞の分化の過程では、上述の3因子を含む転写因子が一過性に発現することが重
要である。本発明で用いる蛋白質導入法では、添加された蛋白質は細胞内に短時間で移行
する。また、導入後1日ないし2日で完全に分解され消失し、一過性の発現が期待できる
。さらに、発現プラスミドやウイルスベクターを用いて導入した場合のように発現した蛋
白質が長時間細胞に影響することもなく、外来遺伝子が染色体に組み込まれることもない
。本発明による方法は、臨床応用に不可欠な安全性の面をクリアしたものである。
【0014】
1型ならびに2型糖尿病に対して、インスリン産生細胞を移植する再生医療が期待され
ている。本治療法を臨床応用するためには、安全にES細胞またはiPS細胞から膵β細胞を
分化させる技術が必須である。本発明で用いた技術は、ウイルスベクターを使用せず安全
であり、移植に必要な膵β細胞の作製方法として利用できる。
【0015】
本発明で用いるES 細胞は、哺乳動物由来のES細胞であればよく、その種類などは特に
限定されず、例えば、マウス、サル又はヒト由来のES細胞などを使用することができる。
ES細胞としては、例えば、その分化の程度の確認を容易とするために、Pdx1遺伝子付近に
レポーター遺伝子を導入した細胞を用いることができる。例えば、Pdx1座位にlacZ遺伝子
を組み込んだ129/Sv由来ES細胞株R1、J1又は、Pdx1プロモーター制御下のGFPレポーター
トランスジーンをもつES細胞SK7株などを使用することができる。あるいは、Hnf3β内胚
葉特異的エンハンサー断片制御下のmRFP1レポータートランスジーン及びPdx1プロモータ
ー制御下のGFPレポータートランスジーンを有するES細胞PH3株を使用することもできる。
【0016】
哺乳動物由来のES細胞の培養方法は常法により行うことができ、例えば、所望によりフ
ィーダー細胞としてのマイトマイシンC処理マウス胚線維芽細胞(MEF)の存在下において
、白血病阻害因子(LIF,ESGRO 1000単位/ml、Chemicon製)を添加した分化培地(10%ウ
シ胎児血清(FBS)、0.1mM 2-メルカプトエタノール、100μM非必須アミノ酸、2mM L-グ
ルタミンを補充したダルベッコ改変イーグル培地(DMEM、シグマ製))で維持することが
できる。
【0017】
iPS細胞(人工多能性幹細胞:induced pluripotent stem cell)とは、体細胞を初期化
することによって得られる多能性を有する細胞である。人工多能性幹細胞の作製は、京都
大学の山中伸弥教授らのグループ、マサチューセッツ工科大学のルドルフ・ヤニッシュ(
Rudolf Jaenisch)らのグループ、ウイスコンシン大学のジェームス・トムソン(James T
homson)らのグループ、ハーバード大学のコンラッド・ホッケドリンガー(Konrad Hoch
edlinger)らのグループなどを含む複数のグループが成功している。人工多能性幹細胞は
、拒絶反応や倫理的問題のない理想的な多能性細胞として大きな期待を集めている。例え
ば、国際公開WO2007/069666号公報には、Octファミリー遺伝子、Klfファミ
リー遺伝子、及びMycファミリー遺伝子の遺伝子産物を含む体細胞の核初期化因子、並び
にOctファミリー遺伝子、Klfファミリー遺伝子、Soxファミリー遺伝子及びMycファミリー
遺伝子の遺伝子産物を含む体細胞の核初期化因子が記載されており、さらに体細胞に上記
核初期化因子を接触させる工程を含む、体細胞の核初期化により誘導多能性幹細胞を製造
する方法が記載されている。
【0018】
本発明で用いるiPS細胞は、体細胞を初期化することにより製造することができる。こ
こで用いる体細胞の種類は特に限定されず、任意の体細胞を用いることができる。即ち、
本発明で言う体細胞とは、生体を構成する細胞の内生殖細胞以外の全ての細胞を包含し、
分化した体細胞でもよいし、未分化の幹細胞でもよい。体細胞の由来は、哺乳動物、鳥類
、魚類、爬虫類、両生類の何れでもよく特に限定されないが、好ましくは哺乳動物(例え
ば、マウスなどのげっ歯類、またはヒトなどの霊長類)であり、特に好ましくはマウス又
はヒトである。また、ヒトの体細胞を用いる場合、胎児、新生児又は成人の何れの体細胞
を用いてもよい。
【0019】
本発明で言うiPS細胞は、所定の培養条件下(例えば、ES細胞を培養する条件下)に
おいて長期にわたって自己複製能を有し、また所定の分化誘導条件下において外胚葉、中
胚葉及び内胚葉への多分化能を有する幹細胞のことを言う。また、本発明における人工多
能性幹細胞はマウスなどの試験動物に移植した場合にテラトーマを形成する能力を有する
幹細胞でもよい。
【0020】
体細胞からiPS細胞を製造するためには、まず、少なくとも1種類以上の初期化遺伝子
を体細胞に導入する。初期化遺伝子とは、体細胞を初期化してiPS細胞とする作用を有す
る初期化因子をコードする遺伝子である。初期化遺伝子の組み合わせの具体例としては、
以下の組み合わせをあげることができるが、これらに限定されるものではない。
(i)Oct遺伝子、Klf遺伝子、Sox遺伝子、Myc遺伝子
(ii)Oct遺伝子、Sox遺伝子、NANOG遺伝子、LIN28遺伝子
(iii)Oct遺伝子、Klf遺伝子、Sox遺伝子、Myc遺伝子、hTERT遺伝子、SV40 large T遺伝
子
(iv)Oct遺伝子、Klf遺伝子、Sox遺伝子
【0021】
本発明では、哺乳動物由来の幹細胞にPdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質を
導入する。幹細胞に対して、上記3種類の蛋白質を導入する手順は特に限定されず、同時
に導入してもよいし、順番に逐次に導入してもよい。好ましくはPdx-1蛋白質、NeuroD蛋
白質、及びMafA蛋白質の順番で、各蛋白質を導入することができる。特に好ましくは、幹
細胞の培養開始後5日目から7日目の間にPdx-1蛋白質を1回以上導入し、9日目から1
1日目の間にNeuroD蛋白質を1回以上導入し、13日目から15日目の間にMafA 蛋白質
を1回以上導入することができる。
【0022】
本発明で用いるPdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質の由来は特に限定されず
、導入する幹細胞(ES細胞又はiPS細胞など)が由来する哺乳動物の種類と同種又は近い
種類の哺乳動物のPdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質を用いることができる。
例えば、マウス由来の幹細胞に導入する場合には、ラット又はマウスの組み換えPdx-1蛋
白質、NeuroD蛋白質及びMafA蛋白質を用いることができる。また、ヒト由来の幹細胞に導
入する場合には、ヒトの組み換えPdx-1蛋白質、NeuroD蛋白質及びMafA蛋白質を用いるこ
とができる。また、Pdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質としては、組み換え蛋
白質を用いることが好ましい。即ち、Pdx-1遺伝子、NeuroD遺伝子、及びMafA遺伝子をそ
れぞれコードする発現ベクターを構築し、これを適当な宿主(大腸菌など)に導入し、得
られた形質転換体を、導入された遺伝子を発現するような適当な条件下で培養することに
よって所望の組み換え蛋白質を製造することができる。また、本発明で用いるMafA蛋白質
としては、ポリアルギニンが付加されているMafA蛋白質を用いることが好ましい。
【0023】
本発明で用いることができるPdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質の各々の塩
基配列及びアミノ酸配列は公知であり、以下の通り配列表にも記載する。
【表1】

【0024】
哺乳動物由来の幹細胞(ES細胞又はiPS細胞など)を培養する方法は特に限定されず、
当業者に公知の定法により行うことができる(例えば、白木伸明,粂昭苑「ES細胞からの
内胚葉系細胞の分化誘導」実験医学別冊 培養細胞実験ハンドブック 297-301, 2008;
及びNature protocols, 2, 3081, 2007を参照)。
【0025】
本発明によれば、幹細胞から膵臓系細胞へと分化誘導することができる。分化誘導され
た膵臓系細胞は、好ましくはインスリン産生細胞である。
【0026】
さらに本発明によれば、ES細胞又はiPS細胞などの幹細胞から膵臓系細胞へと分化誘導
する際に、被験物質の存在下で幹細胞を培養し、被験物質の非存在下で幹細胞を培養した
場合における膵臓系細胞への分化誘導の程度と被験物質の存在下で幹細胞を培養した場合
における膵臓系細胞への分化誘導の程度とを比較することによって、幹細胞から膵臓系細
胞へと分化誘導を促進又は阻害する物質をスクリーニングする方法が提供される。被験物
質としては、成長因子又は低分子化合物などを使用することができる。この際、インスリ
ンの発現量を指標として、膵臓系細胞へと分化誘導の程度を測定することが可能である。
【0027】
以下の実施例により本発明をさらに具体的に説明するが、本発明は実施例によって限定
されるものではない。
【実施例】
【0028】
実施例1:ラット組換えPdx-1蛋白質、ラット組み換え NeuroD蛋白質、及びポリアルギニ
ンを付加したマウス組換えMafA蛋白質の大腸菌での発現及び精製(図1及び図2)
ラットPdx-1遺伝子、ラットNeuroD遺伝子およびマウスMafA遺伝子は米国ベイラー研究
所の野口洋文先生から提供されたものを用いた(Diabetes, 52, 1732, 2003; Diabetes,
54, 2859, 2005)。制限酵素サイトを付加したプライマーを用い、PCR法により、Pdx-1
、NeuroDの両遺伝子をpET21a(+)ベクターにそれぞれ組み込んだ。MafAは、あらかじめポ
リアルギニンを付加してあるpGEX-6p-1ベクターに組み込んだ。大腸菌BL21(DE3)株に、形
質転換により上記plasmidを導入し、37℃で培養する。対数増殖期にある菌体にIPTGを添
加し、24℃で12時間培養する。遠心により回収した菌体を、細胞溶解液により溶解し、P
dx-1およびNeuroDはTALON resinにより精製した。MafAはGlutathione Sepharose 4Bによ
り精製した後、PreScission ProteaseによりGSTタグを切断した。精製後の蛋白質はSDS-
PAGEの後、クマシー染色およびウエスタンブロット法により発現を確認した。
【0029】
結果を図2に示す。図2に示す通り、ラット組換えPdx-1蛋白質、ラット組み換え Neu
roD蛋白質、及びポリアルギニンを付加したマウス組換えMafA蛋白質の大腸菌での発現を
確認できた。
【0030】
実施例2:HeLa細胞への組み換え蛋白質の導入(図3)
実施例1で精製した組み換え蛋白質を細胞内に導入されることをHela細胞を用いて確認
した。Hela細胞にそれぞれの蛋白質を終濃度1 μMとなるよう培地に添加した。蛋白質添
加の6時間後にSDSサンプルバッファーで細胞を回収した。導入の確認はそれぞれの蛋白
質を認識する抗体を用いてウエスタンブロット法により行なった。
【0031】
結果を図3に示す。Pdx-1およびNeuroDはそれ自身PTD(Protein transduction domain
)を持ち、細胞内に導入されることはすでに報告されているが、MafAがポリアルギニンを
付加することにより、細胞内に導入されることは本実験では初めて実証された。
【0032】
実施例3:マウスES細胞から膵β細胞への分化誘導(図4〜7)
マウスES細胞から膵β細胞への分化誘導方法は粂らの方法に従った(図4の上)。粂ら
の方法では、ES細胞を3種類の培地条件で17日間培養するとインスリン産生細胞が増加
することを提唱している。具体的には、マウスES細胞からインスリン産生細胞への分化は
図4上に示すように17日間で行なった。Medium 1はActivin A, basic FGFを含むDMEM培
地、Medium 2はB27添加物, レチノイン酸, KAAD-Cyclopamine, FGF10を含むRPMI培地、M
edium 3はNicotinamide, GLP-1を含むDMEM培地であり、図に示すように段階的に培地を交
換し分化を誘導した。
【0033】
ES細胞への蛋白質の導入は、図4下表に示すようにDay5, 7, 9, 11, 13, 15に蛋白質の
終濃度1μMとなるようそれぞれ培地に直接添加した。Day17の時点で、細胞を固定し抗イ
ンスリン抗体で免疫染色を行なった(図5上)。インスリン陽性細胞の数はカウンターで
計数し、図4下表に示すグループ間で比較した(図5下)。さらに、Day17の分化細胞か
らRNAを抽出し、RT-PCR法によりcDNAを合成した。以下のプライマーを用いてインスリン
1遺伝子の発現を定量RT-PCR法により測定した(図6,7)。
センス:CAGCCCTTAGTGACCAGCTA (配列番号19)
アンチセンス:ATGCTGGTGCAGCACTGATC(配列番号20)
【0034】
本実施例では、図4の下に示すようにDay5からDay13の間で、導入する蛋白質の種類を
検討した。その結果、Day5および7にPdx-1をDay9および11にNeuroDをDay13および15にMa
fAを導入した細胞でInsulin陽性細胞の数が最も多かった(図5)。一方で、3因子それ
ぞれ単独での処理では対照群との間に差はみられなかった。
【0035】
さらに、3因子の導入はInsulin遺伝子の発現も著明に増加した(図6)。また、図7
に示すように、Day9からの蛋白質導入ではInsulin遺伝子の発現の増加が起こらないこと
から、3因子をそれぞれ特定の時期に導入することが分化誘導に重要であることが示唆さ
れた。
【0036】
実施例4:804G細胞由来の細胞外マトリックスを用いた、3因子の蛋白質導入によるマウ
スES細胞からインスリン産生細胞への分化
ラット膀胱癌由来の細胞株804Gはラミニン-5およびフィブロネクチンを含む細胞外マト
リックスを産生する。このマトリックスは膵β細胞の生育およびグルコース応答性のイン
スリン分泌を改善する(Diabetes, 49, 233, 2000; Diabetes, 53, 2034, 2004)。
【0037】
培養プレートを予め804G細胞の培養上清で一晩コーティングし、ES細胞を播種した。本
実験では、Ins-GFPトランスジェニックマウス由来のES細胞(マウスES細胞株ING112)を
用い、GFPの蛍光をインスリン産生細胞への分化の指標とした。分化誘導方法および蛋白
質導入方法は上記と同様に行ない、培養17日目でIns-GFP陽性細胞の数をカウンターで
計数し、804Gマトリックス処理群および3因子導入群を比較した(図8のa、b)。図8
のa及びbに示すように、マウスES細胞株ING112において、804G細胞由来の細胞外マトリ
ックスはIns-GFP陽性の細胞数を著明に増加し、さらに3因子を導入した群は、導入して
いない群に比べIns-GFP陽性の細胞数を増加した。
【0038】
Insulin 1遺伝子の発現を比較した結果を図8のcに示す。
培養17日目で分化細胞からRNAを抽出し、RT-PCR法によりcDNAを合成した。以下のプ
ライマーを用いてInsulin 1遺伝子の発現を定量RT-PCR法により測定し、対照群、804Gマ
トリックス処理群および3因子導入群を比較した。
センス:CAGCCCTTAGTGACCAGCTA (配列番号19)
アンチセンス:ATGCTGGTGCAGCACTGATC(配列番号20)
図8のcに示す通り、804G細胞外マトリックスおよび蛋白質導入によりInsulin 1遺伝子
の発現は増加した。
【0039】
膵臓特異的遺伝子の発現をRT-PCR法により検討した結果を図8のdに示す。
培養17日目で分化細胞からRNAを抽出し、RT-PCR法によりcDNAを合成し、以下のプラ
イマーを用いて膵臓特異的遺伝子の発現を解析した。
Act,
センス:CCTCATGAAGATCCTGACCGA (配列番号21)
アンチセンス:TTGCCAATAGTGATGACCTGG (配列番号22)
Ins1,
センス:CAGCCCTTAGTGACCAGCTA (配列番号19)
アンチセンス:ATGCTGGTGCAGCACTGATC(配列番号20)
Ins2,
センス:TCCGCAACAATCAAAAACCAT (配列番号23)
アンチセンス:GCTGGGTAGTGGTGGGTCTA (配列番号24)
Pdx-1,
センス:CCAAAACCGTCGCATGAAGTG (配列番号25)
アンチセンス:CTCTCGTGCCCTCAAGAATTTTC (配列番号26)
NeuroD,
センス:CTTGGCCAAGAACTACATCTGG (配列番号27)
アンチセンス:GGAGTAGGGATGCACCGGGAA (配列番号28)
MafA,
センス:ACACTGCCCACCATCATCACTC (配列番号29)
アンチセンス:CTTCTCGCTCTCCAGAATGTGC (配列番号30)
Glut2,
センス:ACAGAGCTACAATGCAACGTGG (配列番号31)
アンチセンス:CAACCAGAATGCCAATGACGAT (配列番号32)
Kir6.2,
センス:GGCTCCTAGTGACCTGCACCA (配列番号33)
アンチセンス:CCACAGCCACACTGCGCTTGCG (配列番号34)
SUR1,
センス:CCAGACCAAGGGAAGATTCA (配列番号35)
アンチセンス:GTCCTGTAGGATGATGGACA (配列番号36)
IAPP,
センス:TGATATTGCTGCCTCGGACC (配列番号37)
アンチセンス:GGAGGACTGGACCAAGGTTG (配列番号38)
CACNA1A,
センス:GGTCACACCTCACAAGTCCAC (配列番号39)
アンチセンス:CCAGTCTTCTGGAACATCTCTTG (配列番号40)
CACNA1C,
センス:ATGCAAGACGCTATGGGCTAT (配列番号41)
アンチセンス:CAGGTAGCCTTTGAGATCTTCTTC (配列番号42)
CACNA1D,
センス:ACATTCTGAACATGGTCTTCACAG (配列番号43)
アンチセンス:AGGACTTGATGAAGGTCCACAG (配列番号44)
図8のdに示す通り、804G細胞外マトリックスおよび蛋白質導入により、検討したすべて
の遺伝子の発現が認められた。
【0040】
ヒトiPS細胞から分化誘導した細胞のInsulin遺伝子の発現を調べた結果を図8のeに示
す。ヒトiPS細胞への蛋白質の導入は、Day15, 17, 19, 21, 23, 25に蛋白質の終濃度1μ
Mとなるようそれぞれ培地に直接添加した。
培養28日目で分化細胞からRNAを抽出し、RT-PCR法によりcDNAを合成し、以下のプラ
イマーを用いてInsulin遺伝子の発現を解析した。
センス:GAGGCCATCAAGCAGATCAC (配列番号45)
アンチセンス:GGCTGCGTCTAGTTGCAGTA (配列番号46)
図8のeに示す通り、ヒトiPS細胞においても、804G細胞外マトリックスおよび蛋白質
導入によりInsulin遺伝子の発現が認められた。
【技術分野】
【0001】
本発明は、ES細胞又はiPS細胞などの幹細胞の分化誘導方法に関する。より詳細には、
本発明は、3種類の転写調節因子の組換え蛋白質を幹細胞に導入することによって膵β細
胞に分化誘導させる方法に関する。
【背景技術】
【0002】
マウス膵臓へPdx-1遺伝子、Ngn3遺伝子及び MafA遺伝子をウイルスベクターで導入する
ことにより、膵β細胞に類似した細胞を誘導できることが報告されている(Nature 455,
627, 2008)。しかし、ウィルスベクターによる遺伝子導入は、染色体に組み込まれるこ
とによる腫瘍化などの問題点があり、臨床応用には適さない。他に、TATペプチドを付加
したNgn3蛋白質を胎生膵組織に導入することにより、膵内分泌細胞への分化を促進したと
の報告がある(Diabetes, 54, 720, 2005)。しかし、胎生膵組織を用いた膵β細胞分化
誘導技術は、臨床応用が困難である。
【0003】
胚性幹(ES)細胞は、胚盤胞の内細胞塊に由来する多能性細胞である。ES細胞は未分化
状態では無制限に培養でき、刺激されると多様な細胞型に分化でき、例えば、神経細胞、
造血性細胞、膵臓や肝臓などの内胚葉系の細胞への誘導が可能である。また、iPS細胞(
人工多能性幹細胞:induced pluripotent stem cell)は、体細胞を初期化することによ
って得られる多能性を有する細胞である。しかし、これまで、蛋白質を導入することによ
りES細胞から膵β細胞へ分化誘導を成功させたとの報告は無い。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Nature 455, 627, 2008
【非特許文献2】Diabetes, 54, 720, 2005
【発明の開示】
【発明が解決しようとする課題】
【0005】
本発明は、ウイルスや遺伝子を用いることなくES細胞又はiPS細胞などの幹細胞から膵
β細胞に分化させる方法を提供することを解決すべき課題とした。
【課題を解決するための手段】
【0006】
本発明者らは上記課題を解決するために鋭意検討した結果、3種類の転写調節因子の組
換え蛋白質(Pdx-1蛋白質、NeuroD蛋白質、及びポリアルギニンを付加したMafA蛋白質)
をES細胞に導入することによって膵β細胞に分化誘導させることに成功し、本発明を完成
するに至った。
【0007】
即ち、本発明によれば、哺乳動物由来の幹細胞の培養開始後5日目から7日目の間にP
dx-1蛋白質を1回以上導入し、9日目から11日目の間にNeuroD蛋白質を1回以上導入し
、13日目から15日目の間にMafA 蛋白質を1回以上導入することを含む、幹細胞から
膵臓系細胞へと分化誘導する方法が提供される。
好ましくは、哺乳動物由来の幹細胞はマウス、サル又はヒト由来の幹細胞である。
好ましくは、幹細胞はES細胞又はiPS細胞である。
好ましくは、膵臓系細胞はインスリン産生細胞である。
【0008】
好ましくは、MafA蛋白質は、ポリアルギニンが付加されているMafA蛋白質である。
好ましくは、ラット膀胱癌由来804G細胞由来の細胞外マトリックス上で培養している哺
乳動物由来の幹細胞に、Pdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質を導入する。
【0009】
本発明によれば、上記した本発明の方法により得られる、ES細胞から分化誘導された膵
臓系細胞が提供される。
本発明によればさらに、上記した本発明の方法により幹細胞から膵臓系細胞へと分化誘
導する工程、及び分化誘導された膵臓系細胞を分離する工程を含む、ES細胞から分化誘導
された膵臓系細胞を取得する方法が提供される。
【発明の効果】
【0010】
本発明によれば、ウイルスや遺伝子を用いることなくES細胞又はiPS細胞などの幹細胞
を膵β細胞に分化させることができる。本発明の方法は、糖尿病患者に対する膵β細胞移
植治療に利用する膵β細胞の作製方法として有用である。
【図面の簡単な説明】
【0011】
【図1】図1は、全長又は11個のアルギニンを付加した組み換え蛋白質の模式図を示す。
【図2】図2は、組み換え蛋白質の精製を示す。
【図3】図3は、HeLa細胞への蛋白質の導入を示す。
【図4】図4は、蛋白質の導入によるマウスES細胞からのインスリン産生細胞の誘導の手順を示す。
【図5】図5は、マウスES細胞におけるインスリンの免疫蛍光分析を示す。
【図6】図6は、インスリン1遺伝子発現のリアルタイムRT-PCR解析を示す。
【図7】図7は、9日目からはインスリン遺伝子発現は蛋白質導入により増加しないことを示す。
【図8】図8は、804G細胞由来の細胞外マトリックスを用いた、3因子の蛋白質導入によるマウスES細胞からインスリン産生細胞への分化を示す。
【発明を実施するための形態】
【0012】
以下、本発明の実施の形態についてさらに詳細に説明する。
本発明による幹細胞から膵臓系細胞へと分化誘導する方法は、哺乳動物由来の幹細胞に
Pdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質を導入することを特徴とする方法である。
【0013】
膵β細胞の分化の過程では、上述の3因子を含む転写因子が一過性に発現することが重
要である。本発明で用いる蛋白質導入法では、添加された蛋白質は細胞内に短時間で移行
する。また、導入後1日ないし2日で完全に分解され消失し、一過性の発現が期待できる
。さらに、発現プラスミドやウイルスベクターを用いて導入した場合のように発現した蛋
白質が長時間細胞に影響することもなく、外来遺伝子が染色体に組み込まれることもない
。本発明による方法は、臨床応用に不可欠な安全性の面をクリアしたものである。
【0014】
1型ならびに2型糖尿病に対して、インスリン産生細胞を移植する再生医療が期待され
ている。本治療法を臨床応用するためには、安全にES細胞またはiPS細胞から膵β細胞を
分化させる技術が必須である。本発明で用いた技術は、ウイルスベクターを使用せず安全
であり、移植に必要な膵β細胞の作製方法として利用できる。
【0015】
本発明で用いるES 細胞は、哺乳動物由来のES細胞であればよく、その種類などは特に
限定されず、例えば、マウス、サル又はヒト由来のES細胞などを使用することができる。
ES細胞としては、例えば、その分化の程度の確認を容易とするために、Pdx1遺伝子付近に
レポーター遺伝子を導入した細胞を用いることができる。例えば、Pdx1座位にlacZ遺伝子
を組み込んだ129/Sv由来ES細胞株R1、J1又は、Pdx1プロモーター制御下のGFPレポーター
トランスジーンをもつES細胞SK7株などを使用することができる。あるいは、Hnf3β内胚
葉特異的エンハンサー断片制御下のmRFP1レポータートランスジーン及びPdx1プロモータ
ー制御下のGFPレポータートランスジーンを有するES細胞PH3株を使用することもできる。
【0016】
哺乳動物由来のES細胞の培養方法は常法により行うことができ、例えば、所望によりフ
ィーダー細胞としてのマイトマイシンC処理マウス胚線維芽細胞(MEF)の存在下において
、白血病阻害因子(LIF,ESGRO 1000単位/ml、Chemicon製)を添加した分化培地(10%ウ
シ胎児血清(FBS)、0.1mM 2-メルカプトエタノール、100μM非必須アミノ酸、2mM L-グ
ルタミンを補充したダルベッコ改変イーグル培地(DMEM、シグマ製))で維持することが
できる。
【0017】
iPS細胞(人工多能性幹細胞:induced pluripotent stem cell)とは、体細胞を初期化
することによって得られる多能性を有する細胞である。人工多能性幹細胞の作製は、京都
大学の山中伸弥教授らのグループ、マサチューセッツ工科大学のルドルフ・ヤニッシュ(
Rudolf Jaenisch)らのグループ、ウイスコンシン大学のジェームス・トムソン(James T
homson)らのグループ、ハーバード大学のコンラッド・ホッケドリンガー(Konrad Hoch
edlinger)らのグループなどを含む複数のグループが成功している。人工多能性幹細胞は
、拒絶反応や倫理的問題のない理想的な多能性細胞として大きな期待を集めている。例え
ば、国際公開WO2007/069666号公報には、Octファミリー遺伝子、Klfファミ
リー遺伝子、及びMycファミリー遺伝子の遺伝子産物を含む体細胞の核初期化因子、並び
にOctファミリー遺伝子、Klfファミリー遺伝子、Soxファミリー遺伝子及びMycファミリー
遺伝子の遺伝子産物を含む体細胞の核初期化因子が記載されており、さらに体細胞に上記
核初期化因子を接触させる工程を含む、体細胞の核初期化により誘導多能性幹細胞を製造
する方法が記載されている。
【0018】
本発明で用いるiPS細胞は、体細胞を初期化することにより製造することができる。こ
こで用いる体細胞の種類は特に限定されず、任意の体細胞を用いることができる。即ち、
本発明で言う体細胞とは、生体を構成する細胞の内生殖細胞以外の全ての細胞を包含し、
分化した体細胞でもよいし、未分化の幹細胞でもよい。体細胞の由来は、哺乳動物、鳥類
、魚類、爬虫類、両生類の何れでもよく特に限定されないが、好ましくは哺乳動物(例え
ば、マウスなどのげっ歯類、またはヒトなどの霊長類)であり、特に好ましくはマウス又
はヒトである。また、ヒトの体細胞を用いる場合、胎児、新生児又は成人の何れの体細胞
を用いてもよい。
【0019】
本発明で言うiPS細胞は、所定の培養条件下(例えば、ES細胞を培養する条件下)に
おいて長期にわたって自己複製能を有し、また所定の分化誘導条件下において外胚葉、中
胚葉及び内胚葉への多分化能を有する幹細胞のことを言う。また、本発明における人工多
能性幹細胞はマウスなどの試験動物に移植した場合にテラトーマを形成する能力を有する
幹細胞でもよい。
【0020】
体細胞からiPS細胞を製造するためには、まず、少なくとも1種類以上の初期化遺伝子
を体細胞に導入する。初期化遺伝子とは、体細胞を初期化してiPS細胞とする作用を有す
る初期化因子をコードする遺伝子である。初期化遺伝子の組み合わせの具体例としては、
以下の組み合わせをあげることができるが、これらに限定されるものではない。
(i)Oct遺伝子、Klf遺伝子、Sox遺伝子、Myc遺伝子
(ii)Oct遺伝子、Sox遺伝子、NANOG遺伝子、LIN28遺伝子
(iii)Oct遺伝子、Klf遺伝子、Sox遺伝子、Myc遺伝子、hTERT遺伝子、SV40 large T遺伝
子
(iv)Oct遺伝子、Klf遺伝子、Sox遺伝子
【0021】
本発明では、哺乳動物由来の幹細胞にPdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質を
導入する。幹細胞に対して、上記3種類の蛋白質を導入する手順は特に限定されず、同時
に導入してもよいし、順番に逐次に導入してもよい。好ましくはPdx-1蛋白質、NeuroD蛋
白質、及びMafA蛋白質の順番で、各蛋白質を導入することができる。特に好ましくは、幹
細胞の培養開始後5日目から7日目の間にPdx-1蛋白質を1回以上導入し、9日目から1
1日目の間にNeuroD蛋白質を1回以上導入し、13日目から15日目の間にMafA 蛋白質
を1回以上導入することができる。
【0022】
本発明で用いるPdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質の由来は特に限定されず
、導入する幹細胞(ES細胞又はiPS細胞など)が由来する哺乳動物の種類と同種又は近い
種類の哺乳動物のPdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質を用いることができる。
例えば、マウス由来の幹細胞に導入する場合には、ラット又はマウスの組み換えPdx-1蛋
白質、NeuroD蛋白質及びMafA蛋白質を用いることができる。また、ヒト由来の幹細胞に導
入する場合には、ヒトの組み換えPdx-1蛋白質、NeuroD蛋白質及びMafA蛋白質を用いるこ
とができる。また、Pdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質としては、組み換え蛋
白質を用いることが好ましい。即ち、Pdx-1遺伝子、NeuroD遺伝子、及びMafA遺伝子をそ
れぞれコードする発現ベクターを構築し、これを適当な宿主(大腸菌など)に導入し、得
られた形質転換体を、導入された遺伝子を発現するような適当な条件下で培養することに
よって所望の組み換え蛋白質を製造することができる。また、本発明で用いるMafA蛋白質
としては、ポリアルギニンが付加されているMafA蛋白質を用いることが好ましい。
【0023】
本発明で用いることができるPdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質の各々の塩
基配列及びアミノ酸配列は公知であり、以下の通り配列表にも記載する。
【表1】
【0024】
哺乳動物由来の幹細胞(ES細胞又はiPS細胞など)を培養する方法は特に限定されず、
当業者に公知の定法により行うことができる(例えば、白木伸明,粂昭苑「ES細胞からの
内胚葉系細胞の分化誘導」実験医学別冊 培養細胞実験ハンドブック 297-301, 2008;
及びNature protocols, 2, 3081, 2007を参照)。
【0025】
本発明によれば、幹細胞から膵臓系細胞へと分化誘導することができる。分化誘導され
た膵臓系細胞は、好ましくはインスリン産生細胞である。
【0026】
さらに本発明によれば、ES細胞又はiPS細胞などの幹細胞から膵臓系細胞へと分化誘導
する際に、被験物質の存在下で幹細胞を培養し、被験物質の非存在下で幹細胞を培養した
場合における膵臓系細胞への分化誘導の程度と被験物質の存在下で幹細胞を培養した場合
における膵臓系細胞への分化誘導の程度とを比較することによって、幹細胞から膵臓系細
胞へと分化誘導を促進又は阻害する物質をスクリーニングする方法が提供される。被験物
質としては、成長因子又は低分子化合物などを使用することができる。この際、インスリ
ンの発現量を指標として、膵臓系細胞へと分化誘導の程度を測定することが可能である。
【0027】
以下の実施例により本発明をさらに具体的に説明するが、本発明は実施例によって限定
されるものではない。
【実施例】
【0028】
実施例1:ラット組換えPdx-1蛋白質、ラット組み換え NeuroD蛋白質、及びポリアルギニ
ンを付加したマウス組換えMafA蛋白質の大腸菌での発現及び精製(図1及び図2)
ラットPdx-1遺伝子、ラットNeuroD遺伝子およびマウスMafA遺伝子は米国ベイラー研究
所の野口洋文先生から提供されたものを用いた(Diabetes, 52, 1732, 2003; Diabetes,
54, 2859, 2005)。制限酵素サイトを付加したプライマーを用い、PCR法により、Pdx-1
、NeuroDの両遺伝子をpET21a(+)ベクターにそれぞれ組み込んだ。MafAは、あらかじめポ
リアルギニンを付加してあるpGEX-6p-1ベクターに組み込んだ。大腸菌BL21(DE3)株に、形
質転換により上記plasmidを導入し、37℃で培養する。対数増殖期にある菌体にIPTGを添
加し、24℃で12時間培養する。遠心により回収した菌体を、細胞溶解液により溶解し、P
dx-1およびNeuroDはTALON resinにより精製した。MafAはGlutathione Sepharose 4Bによ
り精製した後、PreScission ProteaseによりGSTタグを切断した。精製後の蛋白質はSDS-
PAGEの後、クマシー染色およびウエスタンブロット法により発現を確認した。
【0029】
結果を図2に示す。図2に示す通り、ラット組換えPdx-1蛋白質、ラット組み換え Neu
roD蛋白質、及びポリアルギニンを付加したマウス組換えMafA蛋白質の大腸菌での発現を
確認できた。
【0030】
実施例2:HeLa細胞への組み換え蛋白質の導入(図3)
実施例1で精製した組み換え蛋白質を細胞内に導入されることをHela細胞を用いて確認
した。Hela細胞にそれぞれの蛋白質を終濃度1 μMとなるよう培地に添加した。蛋白質添
加の6時間後にSDSサンプルバッファーで細胞を回収した。導入の確認はそれぞれの蛋白
質を認識する抗体を用いてウエスタンブロット法により行なった。
【0031】
結果を図3に示す。Pdx-1およびNeuroDはそれ自身PTD(Protein transduction domain
)を持ち、細胞内に導入されることはすでに報告されているが、MafAがポリアルギニンを
付加することにより、細胞内に導入されることは本実験では初めて実証された。
【0032】
実施例3:マウスES細胞から膵β細胞への分化誘導(図4〜7)
マウスES細胞から膵β細胞への分化誘導方法は粂らの方法に従った(図4の上)。粂ら
の方法では、ES細胞を3種類の培地条件で17日間培養するとインスリン産生細胞が増加
することを提唱している。具体的には、マウスES細胞からインスリン産生細胞への分化は
図4上に示すように17日間で行なった。Medium 1はActivin A, basic FGFを含むDMEM培
地、Medium 2はB27添加物, レチノイン酸, KAAD-Cyclopamine, FGF10を含むRPMI培地、M
edium 3はNicotinamide, GLP-1を含むDMEM培地であり、図に示すように段階的に培地を交
換し分化を誘導した。
【0033】
ES細胞への蛋白質の導入は、図4下表に示すようにDay5, 7, 9, 11, 13, 15に蛋白質の
終濃度1μMとなるようそれぞれ培地に直接添加した。Day17の時点で、細胞を固定し抗イ
ンスリン抗体で免疫染色を行なった(図5上)。インスリン陽性細胞の数はカウンターで
計数し、図4下表に示すグループ間で比較した(図5下)。さらに、Day17の分化細胞か
らRNAを抽出し、RT-PCR法によりcDNAを合成した。以下のプライマーを用いてインスリン
1遺伝子の発現を定量RT-PCR法により測定した(図6,7)。
センス:CAGCCCTTAGTGACCAGCTA (配列番号19)
アンチセンス:ATGCTGGTGCAGCACTGATC(配列番号20)
【0034】
本実施例では、図4の下に示すようにDay5からDay13の間で、導入する蛋白質の種類を
検討した。その結果、Day5および7にPdx-1をDay9および11にNeuroDをDay13および15にMa
fAを導入した細胞でInsulin陽性細胞の数が最も多かった(図5)。一方で、3因子それ
ぞれ単独での処理では対照群との間に差はみられなかった。
【0035】
さらに、3因子の導入はInsulin遺伝子の発現も著明に増加した(図6)。また、図7
に示すように、Day9からの蛋白質導入ではInsulin遺伝子の発現の増加が起こらないこと
から、3因子をそれぞれ特定の時期に導入することが分化誘導に重要であることが示唆さ
れた。
【0036】
実施例4:804G細胞由来の細胞外マトリックスを用いた、3因子の蛋白質導入によるマウ
スES細胞からインスリン産生細胞への分化
ラット膀胱癌由来の細胞株804Gはラミニン-5およびフィブロネクチンを含む細胞外マト
リックスを産生する。このマトリックスは膵β細胞の生育およびグルコース応答性のイン
スリン分泌を改善する(Diabetes, 49, 233, 2000; Diabetes, 53, 2034, 2004)。
【0037】
培養プレートを予め804G細胞の培養上清で一晩コーティングし、ES細胞を播種した。本
実験では、Ins-GFPトランスジェニックマウス由来のES細胞(マウスES細胞株ING112)を
用い、GFPの蛍光をインスリン産生細胞への分化の指標とした。分化誘導方法および蛋白
質導入方法は上記と同様に行ない、培養17日目でIns-GFP陽性細胞の数をカウンターで
計数し、804Gマトリックス処理群および3因子導入群を比較した(図8のa、b)。図8
のa及びbに示すように、マウスES細胞株ING112において、804G細胞由来の細胞外マトリ
ックスはIns-GFP陽性の細胞数を著明に増加し、さらに3因子を導入した群は、導入して
いない群に比べIns-GFP陽性の細胞数を増加した。
【0038】
Insulin 1遺伝子の発現を比較した結果を図8のcに示す。
培養17日目で分化細胞からRNAを抽出し、RT-PCR法によりcDNAを合成した。以下のプ
ライマーを用いてInsulin 1遺伝子の発現を定量RT-PCR法により測定し、対照群、804Gマ
トリックス処理群および3因子導入群を比較した。
センス:CAGCCCTTAGTGACCAGCTA (配列番号19)
アンチセンス:ATGCTGGTGCAGCACTGATC(配列番号20)
図8のcに示す通り、804G細胞外マトリックスおよび蛋白質導入によりInsulin 1遺伝子
の発現は増加した。
【0039】
膵臓特異的遺伝子の発現をRT-PCR法により検討した結果を図8のdに示す。
培養17日目で分化細胞からRNAを抽出し、RT-PCR法によりcDNAを合成し、以下のプラ
イマーを用いて膵臓特異的遺伝子の発現を解析した。
Act,
センス:CCTCATGAAGATCCTGACCGA (配列番号21)
アンチセンス:TTGCCAATAGTGATGACCTGG (配列番号22)
Ins1,
センス:CAGCCCTTAGTGACCAGCTA (配列番号19)
アンチセンス:ATGCTGGTGCAGCACTGATC(配列番号20)
Ins2,
センス:TCCGCAACAATCAAAAACCAT (配列番号23)
アンチセンス:GCTGGGTAGTGGTGGGTCTA (配列番号24)
Pdx-1,
センス:CCAAAACCGTCGCATGAAGTG (配列番号25)
アンチセンス:CTCTCGTGCCCTCAAGAATTTTC (配列番号26)
NeuroD,
センス:CTTGGCCAAGAACTACATCTGG (配列番号27)
アンチセンス:GGAGTAGGGATGCACCGGGAA (配列番号28)
MafA,
センス:ACACTGCCCACCATCATCACTC (配列番号29)
アンチセンス:CTTCTCGCTCTCCAGAATGTGC (配列番号30)
Glut2,
センス:ACAGAGCTACAATGCAACGTGG (配列番号31)
アンチセンス:CAACCAGAATGCCAATGACGAT (配列番号32)
Kir6.2,
センス:GGCTCCTAGTGACCTGCACCA (配列番号33)
アンチセンス:CCACAGCCACACTGCGCTTGCG (配列番号34)
SUR1,
センス:CCAGACCAAGGGAAGATTCA (配列番号35)
アンチセンス:GTCCTGTAGGATGATGGACA (配列番号36)
IAPP,
センス:TGATATTGCTGCCTCGGACC (配列番号37)
アンチセンス:GGAGGACTGGACCAAGGTTG (配列番号38)
CACNA1A,
センス:GGTCACACCTCACAAGTCCAC (配列番号39)
アンチセンス:CCAGTCTTCTGGAACATCTCTTG (配列番号40)
CACNA1C,
センス:ATGCAAGACGCTATGGGCTAT (配列番号41)
アンチセンス:CAGGTAGCCTTTGAGATCTTCTTC (配列番号42)
CACNA1D,
センス:ACATTCTGAACATGGTCTTCACAG (配列番号43)
アンチセンス:AGGACTTGATGAAGGTCCACAG (配列番号44)
図8のdに示す通り、804G細胞外マトリックスおよび蛋白質導入により、検討したすべて
の遺伝子の発現が認められた。
【0040】
ヒトiPS細胞から分化誘導した細胞のInsulin遺伝子の発現を調べた結果を図8のeに示
す。ヒトiPS細胞への蛋白質の導入は、Day15, 17, 19, 21, 23, 25に蛋白質の終濃度1μ
Mとなるようそれぞれ培地に直接添加した。
培養28日目で分化細胞からRNAを抽出し、RT-PCR法によりcDNAを合成し、以下のプラ
イマーを用いてInsulin遺伝子の発現を解析した。
センス:GAGGCCATCAAGCAGATCAC (配列番号45)
アンチセンス:GGCTGCGTCTAGTTGCAGTA (配列番号46)
図8のeに示す通り、ヒトiPS細胞においても、804G細胞外マトリックスおよび蛋白質
導入によりInsulin遺伝子の発現が認められた。
【特許請求の範囲】
【請求項1】
哺乳動物由来の幹細胞の培養開始後5日目から7日目の間にPdx-1蛋白質
を1回以上導入し、9日目から11日目の間にNeuroD蛋白質を1回以上導入し、13日目
から15日目の間にMafA 蛋白質を1回以上導入することを含む、幹細胞から膵臓系細胞
へと分化誘導する方法。
【請求項2】
哺乳動物由来の幹細胞がマウス、サル又はヒト由来の幹細胞である、請求
項1に記載の方法。
【請求項3】
幹細胞がES細胞又はiPS細胞である、請求項1又は2に記載の方法。
【請求項4】
膵臓系細胞がインスリン産生細胞である、請求項1から3の何れか1項に
記載の方法。
【請求項5】
MafA蛋白質が、ポリアルギニンが付加されているMafA蛋白質である、請求
項1から4の何れか1項に記載の方法。
【請求項6】
ラット膀胱癌由来804G細胞由来の細胞外マトリックス上で培養している哺
乳動物由来の幹細胞に、Pdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質を導入する、請求
項1から5の何れか1項に記載の方法。
【請求項7】
請求項1から6の何れか1項に記載の方法により得られる、ES細胞から分
化誘導された膵臓系細胞。
【請求項8】
請求項1から6の何れか1項に記載の方法により幹細胞から膵臓系細胞へ
と分化誘導する工程、及び分化誘導された膵臓系細胞を分離する工程を含む、ES細胞から
分化誘導された膵臓系細胞を取得する方法。
【請求項1】
哺乳動物由来の幹細胞の培養開始後5日目から7日目の間にPdx-1蛋白質
を1回以上導入し、9日目から11日目の間にNeuroD蛋白質を1回以上導入し、13日目
から15日目の間にMafA 蛋白質を1回以上導入することを含む、幹細胞から膵臓系細胞
へと分化誘導する方法。
【請求項2】
哺乳動物由来の幹細胞がマウス、サル又はヒト由来の幹細胞である、請求
項1に記載の方法。
【請求項3】
幹細胞がES細胞又はiPS細胞である、請求項1又は2に記載の方法。
【請求項4】
膵臓系細胞がインスリン産生細胞である、請求項1から3の何れか1項に
記載の方法。
【請求項5】
MafA蛋白質が、ポリアルギニンが付加されているMafA蛋白質である、請求
項1から4の何れか1項に記載の方法。
【請求項6】
ラット膀胱癌由来804G細胞由来の細胞外マトリックス上で培養している哺
乳動物由来の幹細胞に、Pdx-1蛋白質、NeuroD蛋白質、及びMafA蛋白質を導入する、請求
項1から5の何れか1項に記載の方法。
【請求項7】
請求項1から6の何れか1項に記載の方法により得られる、ES細胞から分
化誘導された膵臓系細胞。
【請求項8】
請求項1から6の何れか1項に記載の方法により幹細胞から膵臓系細胞へ
と分化誘導する工程、及び分化誘導された膵臓系細胞を分離する工程を含む、ES細胞から
分化誘導された膵臓系細胞を取得する方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】

【図7】

【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2011−223993(P2011−223993A)
【公開日】平成23年11月10日(2011.11.10)
【国際特許分類】
【出願番号】特願2011−71530(P2011−71530)
【出願日】平成23年3月29日(2011.3.29)
【出願人】(504159235)国立大学法人 熊本大学 (314)
【Fターム(参考)】
【公開日】平成23年11月10日(2011.11.10)
【国際特許分類】
【出願日】平成23年3月29日(2011.3.29)
【出願人】(504159235)国立大学法人 熊本大学 (314)
【Fターム(参考)】
[ Back to top ]