
幹細胞増殖の阻害剤および刺激剤ならびにその使用
【課題】レトロウイルスベクターまたは遺伝子導入の物理的技術のいずれかを用いたヒト造血細胞への遺伝子導入に伴ういくつかの問題を改善すること。
【解決手段】本発明は、例えば、C末端疎水性ドメインが置換または欠失されているヘモグロビンα鎖を含むポリペプチド、C末端ハプトグロビン結合ドメインが置換または欠失されているヘモグロビンα鎖を含むポリペプチド、またはヒトαヘモグロビン鎖のアミノ酸1-97を含むポリペプチドなどを提供する。
【解決手段】本発明は、例えば、C末端疎水性ドメインが置換または欠失されているヘモグロビンα鎖を含むポリペプチド、C末端ハプトグロビン結合ドメインが置換または欠失されているヘモグロビンα鎖を含むポリペプチド、またはヒトαヘモグロビン鎖のアミノ酸1-97を含むポリペプチドなどを提供する。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、自己免疫疾患、加齢、ガン、脊髄形成異常、前白血病、白血病、乾癬、後天性免疫不全症候群(AIDS)、脊髄形成異常症候群、再生不良性貧血、または過剰または過少増殖状態を含む他の疾患を有するヒトまたは動物の処置における幹細胞の細胞周期を調節するための幹細胞増殖モジュレーターの使用、ならびに鎮痛のためのこのような化合物の使用に関する。本発明はまた、化学療法剤または細胞周期内にある幹細胞に損傷を与える他の薬剤への曝露、あるいは放射線の照射を受けることが予想されるかまたは受けているヒトまたは動物の処置、ならびにエクスビボでの処置の間のそのような薬剤に対する防御の方法に関する。最後に、本発明は、自家移植および同種移植の手法あるいは遺伝子導入のための幹細胞の維持または拡大培養の改良、ならびにそのような手法の改良のためのインビボでの処置に関する。
【背景技術】
【0002】
発明の背景
再生系のほとんどの終期細胞は、短命であり、そして一生を通じて連続的に置き換えられなければならない。例えば、血液細胞は、多能性造血幹細胞(HSC)の自己再生集団から生じる。造血幹細胞は造血細胞の亜集団である。造血細胞は、例えば、骨髄、臍帯血、または末梢血から得られ得る(造血細胞は、移動しないか、G-CSFのような因子によって移動される)。造血細胞は、幹細胞集団、前駆細胞、分化細胞、補助細胞、間質細胞、および成熟血液細胞の産生に必要な環境を寄与する他の細胞を含む。造血前駆細胞は、それらの発生効力がさらに制限される幹細胞の部分集合である。前駆細胞は、1または2系統にだけ分化し得る(例えば、赤血球のみを生じるBFU-EおよびCFU-E、または顆粒球およびマクロファージを生じるCFU-GM)が、幹細胞(例えば、CFU-MIXまたはCFU-GEMM)は、複数の系統および/または他の幹細胞を生成し得る。造血幹細胞は造血系および免疫系のすべての成熟細胞の発生に必要であるので、化学療法剤または他の薬剤で処置される被験体において十分に機能的な宿主防御系を再確立するために、造血幹細胞の生存は必須である。
【0003】
造血細胞の産生は、造血細胞の増殖および分化を刺激する一連の因子によって調節され、それらのいくつか(例えば、エリスロポエチン、GM-CSF、およびG-CSF)は、現在では臨床的な実用に用いられている。しかし、広範に特徴付けられていない制御ネットワークの一部分は、幹細胞の細胞周期の状態を制御する生理学的機構である(非特許文献1;非特許文献2)。
【0004】
Lordおよびその共同研究者による初期の研究は、正常および再生骨髄の抽出物中の、幹細胞の増殖を阻害または刺激のいずれかをし得る可溶性タンパク質因子の存在を示した(非特許文献3;WrightおよびLorimore,Cell Tissue Kinet.20:191-203,1987;MarshallおよびLord,Int Rev.Cyt.167:185-261,1996に概説される)。これらの活性は、それぞれ幹細胞阻害剤(SCI)および幹細胞刺激剤(SCS)と称された。
【0005】
現在まで、Lordら(上記を参照)に記載されるように、調製された骨髄抽出物からは、SCSの候補となる分子は精製されていない。一次供給源からのSCSまたはSCIのいずれかの精製は、放射線照射された多数のマウスを必要とするインビボアッセイに固有の困難さにより達成されなかった。これらの問題を克服するための試みにおいて、Pragnellおよびその共同研究者は、原始造血細胞(CFU-A)のインビトロアッセイを開発し、そして阻害活性源としての細胞株をスクリーニングした(Grahamら、Nature 344:442-444,1990を参照)。初期の研究はマクロファージをSCIの可能な供給源として同定していたので(非特許文献3)、マウスマクロファージ細胞株J774.2が選択された(Grahamら、Nature 344:442-444、1990)。この細胞株由来の馴化培地が精製のために、Grahamらによって用いられた。阻害ペプチドが単離され、それは、前述のサイトカインマクロファージ炎症性タンパク質1-α(MIP-1α)と同一であることが証明された。他のケモカインレセプターのように、MIP-1αのレセプターがクローニングされ、これらのMIP-1αは、G阻害性サブクラス(「Gi」)のグアニンヌクレオチド(GTP)結合タンパク質に結合する7回膜貫通ドメイン(または「G-関連」)レセプターである(Murphy,Cytokine & Growth Factor Rev.7:47-64,1996に概説される)。Giサブクラスに対する「inhibitory」の名称は、そのアデニル酸シクラーゼの阻害活性をいう。
【0006】
MIP-1αは、細胞株から単離されたのであって、一次材料から単離されたのではない。Grahamらは、MIP-1αに対する抗体が骨髄の粗抽出物の活性を失わせることを観察したが、他の研究者は、他の阻害活性が重要であることを示した。例えば、Grahamら(J.Exp.Med.178:925-32、1993)は、MIP-1αではなくTGFβが造血幹細胞の主要な阻害剤であることを示唆した。さらにEavesら(PNAS 90:12015-19、1993)は、MIP-1αおよびTGFβの両方がやや最適のレベルで正常骨髄中に存在し、そして阻害にはこれら2つの因子間の相乗作用が必要であることを示唆した。
【0007】
最近、相同組換えによってMIP-1α遺伝子が欠失したマウスが作製された。(Cookら,Science 269:1583-5,1995)。そのようなマウスは、その造血系に明白な混乱は見られず、普通の恒常性条件下における幹細胞の細胞周期の生理学的調節因子としてのMIP-1αの役割に異議を唱えられている。同様に、トランスフォーミング成長因子β(TGFβ)もまた幹細胞阻害効果を有するが、長い間、幹細胞がこのサイトカインに反応する思われており、さらに、これが骨髄抽出物内に存在する内因性因子ではないこと、さらに、TGFβに対する中和抗体が、骨髄上清のSCI活性を無効にできないことを示唆する(Hampsonら、Exp.Hemat.19:245-249,1991)。
【0008】
他の研究者は、さらなる幹細胞阻害剤を記載した。Frindelおよびその共同研究者は、ウシ胎児骨髄および肝臓抽出物から幹細胞阻害活性を有するテトラペプチドを単離した(Lenfantら、PNAS 86:779-782、1989)。Paukovitsら(Cancer Res.50:328-332、1990)は、ペンタペプチドを特徴付け、それはモノマー形体では阻害剤であり、そしてダイマー形体では幹細胞の細胞周期の刺激剤であるとしている。他の因子もまた、種々のインビトロ系において阻害性であることが主張されている(WrightおよびPragnell、Bailliere’s Clinical Haematology 第5巻、723〜39頁、1992(Bailliere Tinadall、Paris):MarshallおよびLord,Int Rev.Cyt.167:185-261,1996を参照)。
【0009】
Tsyrlovaら、SU 1561261 A1は、幹細胞増殖阻害剤のための精製プロセスを開示した。
【0010】
共有に係る出願WO 94/22915およびWO96/10634は、幹細胞増殖の阻害剤を開示し、またそこにその全体が本明細書中に参考として援用される。
【0011】
現在まで、これらの因子のいずれもが臨床的使用に対して認可されていない。しかし、効果的な幹細胞阻害剤の必要性は存在している。化学療法または放射線処置に関連する主要な毒性は正常増殖細胞の破壊であり、これにより骨髄の抑制または胃腸に対する毒性を生じ得る。効果的な幹細胞阻害剤は、これらの細胞を保護し、そしてこれらの治療レジメを最適化し得る。臨床的な症状に応じて種々の刺激性サイトカイン(すなわち、IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-11、IL-13、IL-14、IL-15、G-CSF、GM-CSF、エリスロポエチン、トロンボポエチン、幹細胞因子、flk2/flt3リガンドなどのようなサイトカイン、これらは造血細胞の細胞周期を刺激する)が必要とされることが実証され、同様に異なる臨床的な必要性に関して種々の阻害剤が必要とされるようである。
【0012】
さらに、そのような阻害剤の活性を迅速に無効にする必要がある。Lordらの最初の研究(上記を参照)は、阻害活性が刺激活性の付加により逆転され得ることを実証した。種々の幹細胞刺激性サイトカインが同定されている(上記を参照)一方、Lordおよび共同研究者らによって記載された、骨髄抽出物に存在し、そして阻害剤の活性を逆転させ得る活性を示すものがないことが示されている。
【0013】
造血前駆細胞および幹細胞は健常な成人の骨髄中に主に存在する。例えば、化学療法またはG-CSFのようなサイトカインでの処置のような特定の条件下で、大量の前駆細胞および幹細胞が末梢血中に出ていくプロセスは、「移動(mobilization)」と呼ばれる。(Simmonsら,Stem Cells 12(補遺1):187-202,1994;Schedingら,Stem Cells 12(補遺1):203-11,1994;Mangan,Sem.Oncology 22:202-9,1995;Moolten,Sem.Oncology 22:271-90,1995)。最近公表されたデータは、大部分の移動した前駆細胞が細胞周期中は活性でないことを示唆する(RobertsおよびMetcalf,Blood 86:1600-,1995;Donahueら,Blood 87:1644-,1996;SiegertおよびSerke,Bone Marrow Trans.17:467-1996;UchidaらBlood 89:465-72,1997)。
【0014】
ヘモグロビンは、高度に保存された四量体タンパク質であり、その分子量は約64,000ダルトンである。これは、2つのα鎖および2つのβ鎖からなる。各鎖は、ヘムの単一分子(フェロプロトポルフィリンIX)(鉄含有補欠分子族)に結合する。脊椎動物α鎖およびβ鎖は、おそらく、複製し、次いで分岐した1つの単一祖先遺伝子に由来した。この2つの鎖は、それら鎖間および種々の脊椎動物間で高度の配列同一性を保持している(図16Aを参照)。ヒトにおいて、第16染色体上のα鎖クラスターは、同一のポリペプチドをコードする2つのα遺伝子(α1およびα2)、ならびに他のα様鎖をコードする遺伝子(ζ、θ)およびいくつかの非転写偽遺伝子を含有する(ヒトα鎖のcDNA配列およびアミノ酸配列については図16Bを参照)。第11染色体上のβ鎖クラスターは、1つのβ鎖遺伝子およびいくつかのβ様鎖遺伝子(δ、ε、Gγ、およびAγ)、ならびに少なくとも2つの非発現偽遺伝子からなる(ヒトβ鎖のcDNA配列およびアミノ酸配列については図16Cを参照)。
【0015】
これらの遺伝子の発現は、発育の間で変化する。ヒトの造血(これは十分に特徴付けられている)において、胚赤芽球は、2つのζ鎖および2つのε鎖(Gower I)、2つのα鎖および2つのε鎖(Gower II)、または2つのζ鎖および2つのγ鎖(Hb Portland)からなる四量体を連続的に合成する。胚形成が進行するにつれて、優勢型は、2つのα鎖および2つのγ鎖で構成される胎児ヘモグロビン(Hb F)からなる。成人ヘモグロビン(2つのα鎖および2つのβ鎖)は、胎児期の間に合成が開始される。誕生時には、約50%のヘモグロビンが成人型であり、そしてその移行は約6カ月齢までに完了する。成人におけるヘモグロビンの大部分(約97%)は、2つのα鎖および2つのβ鎖からなる種(Hb A)であり、少量のHb Fまたはδ鎖(Hb A2)もまた検出され得る。
【0016】
いくつかの方法が、E.coli内および酵母内で組換え体ヘモグロビン鎖を発現するために使用された(例えば、Jessenら,Methods Enz.231:347-364,1994;Lookerら,Methods Enz.231:364-374,1994;Ogdenら,Methods Enz.231:374-390,1994;Martin de Llanoら,Methods Enz.231:364-374,1994)。現在まで、組換えの方法では、単離されたヒトα鎖を高収率で発現することは可能ではない(例えば、Hoffmanら,PNAS 87:8521-25,1990;Hernanら,Biochem.31:8619-28,1992)。明らかに、単離されたアルファ鎖は安定した高次構造であるとは考えられず、E.coli内では速やかに分解される。β鎖とα鎖との同時発現は両方の発現が増加を生じた(Hoffmanら、およびHernanら、前出)。α鎖はβ鎖部分とXa因子の認識部位(NagaiおよびThorgersen,Methods Enz.231:347-364,1994)との融合タンパク質として発現されているが、これらの状態下では、不溶性封入体として発現される。
【0017】
β鎖およびα鎖の両方がハプトグロビンに対する結合部位を含む。ハプトグロビンはヘモグロビンに対して極端に高い親和性を有する血清タンパク質である(例えばPutnam in The Plasma Proteins - Structure,Function and Genetic Control(F.W.Putnam,Ed.)Vol.2,1-49頁(Academic Press,NY);HwangおよびGreer,JBC 255:3038-3041,1980)。肝臓のハプトグロビン輸送は、循環しているヘモグロビンの主要な代謝経路である。これらはα鎖に、ハプトグロビンへの1つの結合部位(アミノ酸121-127)があり、そしてβ鎖には2つの結合部位(アミノ酸領域11-25および131-146)がある(KazimおよびAtassi,Biochem J.197:507-510,1981;McCormickおよびAtassi,J.Prot.Chem.9:735-742,1990)。
【0018】
生物学的に活性なオピエート活性を有するペプチドは、ヘモグロビンのタンパク質分解性の分解によって得られている(Karelinら,Peptides 16:693-697,1995に概説される)。ヘモグロビンα鎖はアミノ酸94-95間に酸に不安定(acid-labile)な切断部位を有する(Shaeffer,J.Biol.Chem.269-29530-29536,1994)。
【0019】
Kreglerら(Exp.Hemat.9:11-21,1981)は、ヘモグロビンがマウス骨髄のCFU-C前駆細胞コロニーに対して増強活性を有することを開示した。そのようなアッセイは、CFU-MIXのような幹細胞に対して、CFU-GMおよびCFU-M前駆細胞集団に対する効果を実証する。著者らは、ヘモグロビンの単離されたα鎖およびβ鎖の両方における活性を観察した。この活性は、N-エチルマレイミドでの処理により消滅し、このことはKreglerらに、スルフヒドリル基が必要であることを示唆した。この観察は、刺激活性がトリプシン消化に対する耐性であるという事実と合わせて、KreglerらにC-末端疎水性ドメインまたは「核」領域が活性を担っていることを示唆した。Moqattashら(Acta.Haematol.92:182-186,1994)は、組換えヘモグロビンがCFU-E、BFU-EおよびCFU-GM前駆細胞の数に対して刺激効果を有することを開示し、これは、ヘミンで観察されたのと同様である。Muellerら(Blood 86:1974,1955)は、精製された成人のヘモグロビンがヘミンと同様の様式で赤血球系の前駆細胞を刺激することを開示した。
【0020】
Petrovら(Bioscience Reports 15:1-14,1995)は、「非同定ミエロペプチド混合物」のWv/Wvマウスでの先天性貧血症の処置での使用を開示した。この混合物は、脾臓コロニー、特にその赤血球型の数を増加させた。
【0021】
ヘムおよびヘミンは、造血に対するその影響に関して十分に試験されてきた(概説についてS.Sassa、Seminars Hemat.25:312-20,1988およびN.Abrahamら、Int.J.Cell Cloning 9:185-210,1991を参照のこと)。ヘムは、赤芽球の成熟に必要とされる。インビトロにおいて、ヘミン(クロロフェロプロトポルフィリンIX−すなわち、付加塩化物イオンを有するヘム)は、CFU-GEMM、BFU-E、およびCFU-Eの増殖を増大させる。同様に、ヘミンは、長期骨髄培養物において細胞充実性を増大させる。
【0022】
「オピエート」は、アヘンの主要な作用物質であるモルヒネと類似する鎮痛性の性質を持つ物質である。オピエートは、モルヒネおよび他のアルカロイドもしくは合成化合物のような有機小分子、またはエンケファリン、エンドルフィン、ダイノルフィンおよびその合成誘導体のような内因性のペプチドであり得る。内因性のオピエートペプチドは、より大きな前駆体(Met-またはLeu-エンケファリンに対するプレプロエンケファリンA、α、βおよびγエンドルフィンに対するプレプロオピオメラノコルチン、ダイノルフィンAおよびBに対するプレプロダイノルフィン、βネオエンドルフィンに対するαネオエンドルフィン)よりインビボで生成される。さらに、オピエート活性を有するペプチドは、αまたはβカゼイン、小麦グルテン、ラクトアルブミン、チトクローム、またはヘモグロビンのようなタンパク質のタンパク質分解または加水分解のような非古典的な供給源、またはカエルの皮膚(デルモルフィン)またはウシ副腎髄質のような他の種から得られ得る。このようなペプチドは、古典的なエンドルフィンに対して「エキソルフィン(exorphin)」と呼ばれ、それらはまた異型オピエートペプチドとも呼ばれる(非特許文献4;非特許文献5;非特許文献6;Brantl,Eur.J.Pharm.106:213-14,1984;Brantlら,Eur.J.Pharm.111:293-4,1985;Brantlら,Eur.J.Pharm.125:309-10,1986;BlantlおよびNeubert,TIPS 7:6-7,1986;Glamstaら,BBRC 184:1060-6,1992;Teschemacher,Handbook Exp.Pharm.104:499-28,1993;Petrovら,Bioscience Reports 15:1-14,1995;Karelinら,Peptides 16:693-7,1995)。Tyr-MIF-1ファミリーのような他の内因性ペプチドはまた、オピエート活性を有することが示されている(Reedら,Neurosci.and Biobehav.Rev.18:519-25,1994)。
【0023】
オピエートは、内因性オピエートレセプターの3つの主要な薬理学的クラス(μ、δ、およびκ)に結合することによって効果を発揮する。各薬理学的クラスを代表するレセプターは、クローン化され、そしてGiに結合したG関連レセプターであることが示されている。(ReisineおよびBell,TINS 16:506-510,1993;Uhlら,TINS 17:89-93,1994;Knappら,FASEB J.9:516-525,1995;SatohおよびMinami,Pharm.Ther.68:343-64,1995;Kieffer,Cell.Mol.Neurobiol.15:615-635,1995;Reisine,Neuropharm.34:463-472,1995;Zakiら,Ann.Rev.Pharm.Toxicol.,36:379-401,1996に概説されている)。
【0024】
特定のアゴニストおよびアンタゴニストが、各レセプタータイプについて入手可能である。レセプタータイプには、例えば、μレセプター(これは、DAMGOおよびDALDAによって選択的に活性化され、CTOPおよびナロキソナジンによって選択的に拮抗される)、κレセプター(これは、GR 89696フマル酸塩またはU-69593で選択的に活性化され、そしてノルビナルトルフィミンハイドロクロリドで選択的に拮抗される)、およびδレセプター(これは、DADLEおよびDPDPEで選択的に活性化され、そしてナトリンドール(natrindole)で選択的に拮抗される)がある。さらに、3つのレセプターサブタイプの全てに作用する広域スペクトルのナロキソンのようなアンタゴニストおよびエトルフィンのようなアゴニストがある。
【0025】
古典的かつ異型のオピエートペプチドは、化学的に変化または誘導体化され、その特異的オピエートレセプター結合特性を変化され得る(HrubyおよびGehrig,Med.Res.Rev.9:343-401,1989;Schiller,Prog.Med.Chem.28:301-40,1991;Teschemacher,Handbook Exp.Pharm.104:499-28,1993;Handbook of Experimental Pharmacology,A.Hertz(編)第104/Iおよび104/II刊、1993,Springer Verlag,Berlin;Karelinら,Peptides 16:693-7,1995)。例には、デルモルフィン(例えば、DALDA)およびエンケファリン(例えば、DADLE,DAMGO、またはDAMME)の誘導体が挙げられる。ソマトスタチンのようなオピエートレセプターに通常結合しないペプチドはまた、特異的オピエートレセプター結合を示すように誘導体化され得る(例えば、CTOP(Hawkinsら,J.Pharm.Exp.Ther.248:73,1989))。アナログもまた、改変したレセプター結合特性を有するモルヒネのようなアルカロイドから誘導され得る(例えば、ヘロイン、コデイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノールおよびナルブフィン);さらに、モルヒネに構造的に関連のない小分子もまた、オピエートレセプターに作用し得る(例えば、メペリジンおよびその同族種アルファプロジン、ジフェノキシレート、およびフェンタニール、)(Handbook of Experimental Pharmacology,前掲書、GoodmanおよびGilmanのThe Pharmacological Basis of Therapeutics,第7版、A.G.Gilman,L.S.Goodman,T.W.Rall and F.Murad(編)1985 Macmillan Publishing Co.NYを参照のこと)。
【0026】
内因性オピエートペプチド(エンケファリン、エンドルフィン、およびダイノルフィン)は、保存されたN末端テトラペプチド Tyr-Gly-Gly-Phe、続いてLeuまたはMetおよびいくらかの残りのC末端配列を有する。N末端Tyrのヒドロキシル基を除去することにより(N末端Pheを生じる)、Metエンケファリンに対する活性を劇的に損失する。これらの構造的データは、「メッセージ−アドレス(message-address)」仮説を導き、それによりN末端「メッセージ」は、生物学的活性を与え、一方C末端「アドレス」は特異性および増強された効力を与える。(ChavkinおよびGoldstein,PNAS 78:6543-7,1981)。エキソルフィンは、一般に、古典的なオピエートペプチドのN末端Tyr-Glyを置き換えたTyr-Proを有する。このプロリン残基は、アミノペプチダーゼ分解に対するより高度の安定性を与えると考えられている(Shippら,PNAS 86:287-,1989;Glamstaら,BBRC 184:1060-6,1992)。
【0027】
最近、オーファンレセプター(「ORL1」)が、μ、δ、およびκオピエートレセプターに対する配列関連性によってクローン化された(Mollereauら,FEBS 341:33-38,1994;Fukudaら,FEBS 343:42-46,1994;Bunzowら,FEBS 347:284-8,1994;Chenら,FEBS 347:279-83,1994;Wangら,FEBS 348:75-79,1994;Keithら,Reg.Peptides 54 143-4,1994;Wickら,Mol.Brain Res.27:37-44,1994;Halfordら,J.Neuroimmun.59:91-101,1995)。ノシセプチン(nociceptin)またはオーファニン(orphanin)FQ(本明細書中以下「ノシセプチン」という)と様々に呼ばれるこのレセプターに対するリガンドは、クローン化され、そしてより大きな前駆体由来のヘプタデカペプチドであることが示された(Meunierら,Nature 377:532-535,1995;Reinscheidら,Science 270:792-794,1995)。これは、鎮痛特性を有する古典的なオピエートに反して、インビボで前侵害性の痛覚過敏機能を有することが示された。ノシセプチンは、先に議論した古典的なオピエートペプチドのTyr-Gly-Gly-Phe...というN末端モチーフと対照的に、Phe-Gly-Gly-Phe...というN末端モチーフを有する。古典的なオピエートペプチドのオピエート活性についてのN末端Tyrの要件と一致して、ノシセプチンはμ、κまたはδオピエートレセプターに対してほとんどまたは全く親和性を示さない。同様に、広域スペクトルオピエートアンタゴニストのナロキソンは、ORL1に対して感知できるほどの親和性を有さない。
【0028】
エンケファリンは、固定化ストレスの状況下で、インビボでマウス造血に効果を有することが観察された(Goldbergら,Folia Biol.(Praha)36:319-331,1990)。骨髄の造血を、Leuエンケファリンは阻害し、Metエンケファリンは刺激した。これらの効果は、間接的に、グルココルチコイドレベルおよびTリンパ球遊走に対する効果によると、Goldbergらは考えた。Krizanac-Bengezら(Biomed.& Pharmacother.46:367-43,1992;Biomed.& Pharmacother.49:27-31,1995;Biomed.& Phrmacother.50:85-91,1996)は、インビトロでのこれらの化合物の効果を観察した。マウス骨髄のMet-またはLeu-エンケファリンまたはナロキソンでの前処理は、コロニーアッセイで観察されたGM前駆細胞の数に影響を及ぼした。この効果は、高度に可変性であり、そして抑制、刺激、または効果なしという結果を生じた。さらに、明らかな用量応答は存在しなかった。この可変性は、Krizanac-Bengezらは、概日性リズムおよび補助細胞のためであるとしている。
【0029】
最近、μオピエートレセプターが相同性組換えにより欠失したマウスは、大腿あたりのCFU-GM、BFU-EおよびCFU-GEMMの数が上昇することが実証された。骨髄および脾臓前駆細胞は、正常マウスと比較してこれらのμレセプターノックアウトマウスではより急速に循環していた。これらの効果が、これらの動物におけるμレセプターの不在から生じる骨髄幹細胞に対する直接的または間接的効果に起因するかどうかは決定されなかった(Broxmeyerら,Blood 88:338a,1997)。
【0030】
I.ガンの化学療法および放射線療法
刺激性増殖因子に関する生産的な研究は、多くのこれらの因子(エリスロポエチン、G-CSF、GM-CSFなど)の臨床的な使用をもたらした。これらの因子は、化学療法および放射線処置に関連した死亡率および罹患率を低下させた。化学療法または放射線照射を受けている患者に対するさらなる臨床的利点が、幹細胞が細胞周期に入ることをブロックし、それによって幹細胞を有毒な副作用から保護する代替のストラテジーにより実現され得る。この保護の逆は、化学または放射線処置後の、骨髄機能の迅速な回復を可能にする。
【0031】
II.骨髄および幹細胞移植、エキソビボ幹細胞拡大、ならびに腫瘍パージング
骨髄移植(BMT)は、種々の血液疾患、自己免疫疾患および悪性疾患にとって有用な処置である。現在の治療法は、臍帯血、胎児肝臓から、または末梢血(移動化していないか、またはG-CSFもしくはシクロホスファミドのような因子によって移動化される)から、ならびに骨髄から得られた造血細胞を含む;幹細胞は未精製、部分的に精製(例えば、CD34+集団のアフィニティー精製)、または高度に精製(例えば、CD34,CD38、またはローダミンのようなマーカーを使用する、蛍光活性化細胞選別により)され得る。造血細胞のエクスビボ操作は、現在では、原始幹細胞を移植に適した集団に拡大するために用いられている。この手順を最適化するには以下のことが必要である:(1)造血の長期の再構築を維持し得るのに十分な数の幹細胞;(2)移植片対宿主誘導Tリンパ球の涸渇、および(3)悪性細胞の残留がないこと。この手法は、幹細胞阻害剤および/または幹細胞刺激剤を含むことにより最適化され得る。
【0032】
残留の悪性細胞を排除するために細胞傷害性薬物を用いて造血細胞をパージ(purge)することの効果は、正常な造血細胞および特に幹細胞に対するこれらの化合物の毒性により制限される。パージ中は、正常細胞を効果的に保護する必要性がある。保護は、効果的な阻害剤を用いて、幹細胞をその細胞周期からはずすことにより与えられ得る。
【0033】
III.末梢幹細胞の採取
末梢血幹細胞(PBSC)は、自家移植に対して骨髄よりも多くの潜在的な利点を提供する。腫瘍の保持、または以前の放射線処置により適切な骨髄採取部位を持たない患者は、さらにPBSC採集を受け得る。血液幹細胞の使用は、一般的な麻酔法および外科的手段に十分に耐えられない患者におけるそれらの必要性を排除する。血液細胞を採集するのに必要な血液成分分離技術は、効率的であり、そしてほとんどの主要な医療センターで広く利用可能である。この方法の主な制限事項は、末梢血中の幹細胞が正常な定常状態にある頻度が低いこと、および薬物または増殖因子(例えば、シクロホスファミド、G-CSF、幹細胞因子)を用いた移動手順の後では細胞周期の進行が速いことの両方である。効果的な幹細胞阻害剤は、そのような細胞を休止状態に戻すために有用であり、それによって分化の間にそれら細胞が消失することを防ぐ。
【0034】
IV.過剰増殖性疾患の処置
多くの疾患は、調節異常の幹細胞が、末期細胞の過剰生産を引き起こす過剰増殖状態によって特徴付けられる。このような病態は、上皮細胞の過剰生産である乾癬、および腸ポリープ出現を特徴とする胃腸管における前悪性状態,および初期幹細胞がHIVによって感染されていないが迅速に循環し、幹細胞の消耗を生じる後天性免疫不全症候群(AIDS)を含むが、それらに限定されない。幹細胞阻害剤はこのような状態の処置に有用である。
【0035】
V.過剰増殖障害の処置
多くの疾患は、調節異常の幹細胞が末期細胞の生産不足を引き起こす過剰増殖状態によって特徴づけられる。このような病態は、血液細胞の生産不足である脊髄形成異常症候群または再生不良性貧血、および細胞の再生および置換の欠乏が存在する加齢と関連する状態を含む。幹細胞刺激剤は、このような状態の処置に有用である。
【0036】
VI.遺伝子導入
遺伝情報を造血細胞に導入する能力は、現在では、臨床的設定に利用されている。利用の容易さ、エクスビボでこの組織を操作し処理する幅広い経験のため、および血液細胞が組織に浸透する能力のため、造血細胞は遺伝子療法の有用な標的である。さらに、特定のヒトの遺伝子欠損の修正は、機能的遺伝子をヒト造血系の原始幹細胞に挿入することにより可能であり得る。
【非特許文献1】Eavesら、Blood 78:110-117、1991
【非特許文献2】Lord,Stem Cells(C.S.Potten,Ed.)pp 401-22,1997(Academic Press,NY)
【非特許文献3】LordおよびWright,Blood Cells 6:581-593,1980
【非特許文献4】Zioudrouら,JBC 254:2446-9,1979
【非特許文献5】QuirionおよびWeiss,Peptides 4:445,1983
【非特許文献6】Loukasら,Biochem.22:4567,1983
【発明の開示】
【発明が解決しようとする課題】
【0037】
レトロウイルスベクターまたは遺伝子導入の物理的技術のいずれかを用いて、遺伝子をヒト造血細胞に導入することにはいくつかの制限がある:(1)造血組織中の幹細胞の頻度の低さは、効率の高い遺伝子導入技術の開発を必要とする;および(2)より迅速に細胞周期の進行している幹細胞はベクターの感染をより受けやすくなることが証明されたが、成長因子による幹細胞増殖の刺激によって感染頻度が増加することは、導入遺伝子を含む細胞が不可逆的に分化させられ、そしてそれらの自己再生ができなくなるので、長期の遺伝子発現にネガティブな影響を与える。これらの問題は、分化および自己再生の損失を妨げるための幹細胞阻害剤、および幹細胞の細胞周期への進入を調節するための幹細胞刺激剤の使用によって改善され、それによってレトロウィルス媒介遺伝子導入を促進し得る。
【課題を解決するための手段】
【0038】
発明の要旨
本発明は、幹細胞増殖阻害剤および/または刺激剤(INPROLおよびオピエート化合物)であるペプチドおよびポリペプチドを含む化合物、およびそれらの使用に関する。
【0039】
本発明は、以下の性質により特徴付けられる幹細胞増殖阻害剤を包含する:
(a)マウスコロニー形成脾臓(CFU-S)アッセイにおいて比活性(IC50)が20ng/ml以下である(実施例4を参照のこと)、
(b)(限外濾過により)分子量が10,000ダルトンより大きく、かつ100,000ダルトンより小さい、
(c)活性がトリプシンによる分解に対して感受性である、
(d)MIP-1αまたはTGFβよりも疎水性であり、そして逆相クロマトグラフィーにより両方から分離可能である(実施例12を参照のこと)、
(e)水溶液中で37℃、55℃または75℃で1時間加熱した後でも生物学的活性が保持されている、および
(f)1%塩酸のアセトン溶液を用いて沈澱させた後でも生物学的活性が保持されている。
【0040】
本発明はさらに、短時間のプレインキュベーションの後のインビトロアッセイにおける阻害の達成能力によって特徴付けられ、そして他の幹細胞阻害剤候補(例えば、MIP-1α、TGFβ、および種々のオリゴペプチド)と区別される。
【0041】
本発明はまた、種々の疾患の処置のためのINPROLを含有する薬学的組成物を包含する。
【0042】
本発明は、幹細胞を死滅させるかまたは損傷し得る薬剤への曝露が予想される被験体に有効量の幹細胞阻害性組成物を投与することにより、その被験体を処置する方法を提供する。この方法によって保護された幹細胞は、通常は骨髄、臍帯血、胎児肝に存在し、分裂するか、または末梢血循環へ移動した造血幹細胞であり得る。蛍光活性化セルソーター(FACS)分析により、大部分の移動した幹細胞は静止状態であり、多分化能の幹細胞は細胞周期中であり、幹細胞阻害量のINPROLにより阻害され得ることが実証された。あるいは、幹細胞は上皮性(例えば、腸管または頭皮または身体の他の領域に位置する)であり得るか、あるいは生殖器官内に位置する生殖細胞であり得る。本発明の方法は、望ましくは、ヒトに用いられ得るが、動物の処置もまた、本方法に包含される。本明細書で用いられる用語「被験体」または「患者」は、動物(例えば哺乳動物(ヒトを含む))を指す。
【0043】
本発明はまた、低増殖性の幹細胞を有する被験体を、有効量の幹細胞刺激性組成物をその被験体に投与することにより処置する方法を提供する。この方法により刺激された幹細胞は、通常は骨髄、臍帯血、胎児肝に存在するか、または末梢血循環へ移動した造血幹細胞であり得る;このような幹細胞は、以前に幹細胞阻害量のINPROLの使用により、静止状態へ置かれ得る。幹細胞刺激量のINPROLは、所望の場合、例えばエクスビボ拡大の間の使用のために幹細胞を収集後、またはその後のインビボでの幹細胞の移植(transplantation)および移植(engraftment)後に、幹細胞の細胞周期の刺激を可能にする。あるいは、幹細胞は;上皮性(例えば小腸、もしくは頭皮または身体の他の領域)であり得るか、あるいは生殖器官内に位置する生殖細胞であり得る。
【0044】
他の局面では、本発明は、化学療法を受けている患者の造血幹細胞系、免疫幹細胞系または他の幹細胞系を保護し、かつ回復させる方法を提供する。この方法は患者に有効な幹細胞阻害量のINPROLを投与する工程、および/または、有効な幹細胞刺激量のINPROLを投与することによって、化学療法または放射線照射後の回復を刺激する工程を含む。
【0045】
なおさらなる局面では、本発明は、ガンを有する患者に有効な幹細胞阻害量のINPROLを投与して、骨髄、胃腸管、または他の器官の幹細胞を、化学療法または放射線療法の有害な作用から保護することにより、および/または幹細胞刺激量のINPROLを投与することによって化学療法または放射線照射後の回復を刺激することにより、固形腫瘍(例えば、乳、大腸、肺、精巣、卵巣、肝臓、腎臓、膵臓、脳、肉腫)によって特徴付けられるガンを含む、任意のガンを補助的に処置する方法を包含する。
【0046】
本発明のさらに別の局面は、正常幹細胞の増殖を阻害するために、増殖性白血病細胞を内部に有する造血細胞を有効量のINPROLで処理する工程、および白血病細胞を破壊するために、細胞傷害性薬剤で骨髄を処理する工程を含む、白血病(例えば、慢性骨髄性白血病、急性骨髄性白血病、慢性リンパ性白血病、急性リンパ性白血病、骨髄腫、ホジキン病)の処置を包含する。この方法は、骨髄の増殖を刺激する他の薬剤(例えば、コロニー刺激剤、および/または幹細胞刺激量でのINPROL)で骨髄を継続処置することによって増強され得る。1つの実施態様において、この方法はインビボで行われる。あるいは、この方法はまた、移植のためにエクスビボで造血細胞をパージし拡大するのに有用である。
【0047】
なおさらなる別の局面では、この方法は、増殖性幹細胞によって引き起こされる任意の疾患を有する被験体を処置する工程を含む。乾癬、脊髄形成異常、いくつかのの自己免疫疾患、加齢における免疫低下、脊髄形成異常症候群、再生不良性貧血またはAIDSによる幹細胞の消耗のような疾患は、被験体に有効量のINPROLを投与して問題の幹細胞の増殖を阻害または刺激することにより処置される。
【0048】
本発明は、幹細胞を死滅させるかまたは損傷し得る細胞傷害性薬剤由来の損傷から幹細胞を可逆的に保護する方法を提供する。この方法は、そのような薬剤への暴露が予想される被験体に有効な幹細胞阻害量のINPROLを投与する工程を含む。
【0049】
本発明はまた、化学療法または放射線照射後の回復期の間に幹細胞の増殖を可逆的に刺激する方法を提供する。この方法は、このような薬剤への暴露が予想される被験体に、有効な幹細胞刺激量のINPROLを投与する工程を包含する。
【0050】
本発明はまた以下を提供する:
以下の手順によってブタまたは他の骨髄から単離された幹細胞増殖阻害剤(実施例12を参照のこと):
(a)骨髄を抽出し、そして濾過によって粒子状物質を除去する;
(b)56℃で40分間熱処理した後、氷浴で冷却する;
(c)4℃で30分間10,000gで遠心分離することによって沈澱物を除去する;
(d)10容量の撹拌された氷冷アセトン(1容量%の濃塩酸を含む)に上清を添加し、そして4℃で16時間インキュベーションすることにより酸沈澱させる;
(e)4℃で30分間20,000gで遠心分離することにより沈澱物を単離し、そして冷アセトンで洗浄した後、乾燥させる;
(f)逆相クロマトグラフィーによって単離し、そして5-フルオロウラシルで前処理し、マウスIL-3存在下でインキュベートした骨髄細胞によるコロニー形成の阻害、ならびに280nmにおける吸光度およびSDS-PAGEにより活性をモニターする。
【0051】
本発明はまた以下を提供する:
実質的に他のタンパク性物質を含まない幹細胞増殖阻害剤を精製する方法であって、前記の工程を包含し、さらに以下により詳細に記載される方法。
【0052】
本発明はまた以下を提供する:
幹細胞増殖阻害剤が、幹細胞の過剰増殖によって引き起こされる免疫抑制を改善するように作用する、ヒトまたは動物を処置する方法。
【0053】
本発明はまた以下を提供する:
幹細胞刺激量のINPROLが、幹細胞の低増殖によって引き起こされる骨髄抑制を改善する、ヒトまたは動物を処置する方法。
【0054】
本発明はまた以下を提供する:
前記幹細胞増殖阻害剤が、細胞傷害性薬物または放射線照射手順に曝されることによって幹細胞が増殖するように誘導された後、投与される、ヒトまたは動物を処置する方法。幹細胞は通常は休止しているが、化学療法後、細胞周期に入るように刺激される。これによって幹細胞は2回目の化学療法剤の投与に対してより感受性となる;本方法はこの処置から幹細胞を保護する。
【0055】
本発明はまた以下を提供する:
幹細胞増殖の刺激剤(例えば、幹細胞刺激量でのINPROL)が、骨髄の再生を促進するために、幹細胞阻害量のINPROL前後に投与される、ヒトまたは動物を処置する方法。幹細胞阻害量のINPROLは、幹細胞の細胞周期への移行速度を遅らせ、そして化学療法や放射線照射から保護する;幹細胞刺激量のINPROLはこの阻害を逆転し、そして骨髄の回復を促進する。逆に、幹細胞刺激量のINPROLは骨髄の回復に使用され得るが、幹細胞阻害量のINPROLは一旦骨髄の回復が達成すると、次に幹細胞を静止状態に戻すために使用される。
【0056】
本発明はまた以下を提供する:
前記幹細胞増殖阻害剤が、免疫応答を増大させるためにワクチン接種の前または同時にアジュバントとして投与される、ヒトまたは動物を処置する方法。
【0057】
本発明はまた以下を提供する:
前記哺乳動物に免疫刺激的量のINPROLを投与する工程を含む、哺乳動物の免疫不全を処置する方法。
【0058】
本発明はまた以下を提供する:
前記哺乳動物に鎮痛誘導量のINPROLを投与する工程を含む、哺乳動物の疼痛を処置する方法。
【0059】
本発明はまた以下を提供する:
幹細胞を損傷から保護するために有効量の幹細胞増殖阻害剤を投与する工程を含む、細胞傷害性薬物または放射線処置を受けたヒトまたは動物を処置する方法。
【0060】
本発明はまた以下を提供する:
処置後の回復を増強するために、有効な幹細胞刺激量のINPROLを投与する工程を含む、細胞傷害性薬物または放射線処置を受けたヒトまたは動物を処置する方法。
【0061】
本発明はまた、ヘモグロビンおよび薬学的に受容可能なキャリアを含む薬学的組成物を含む。
【0062】
本発明はまた、(a)ヘモグロビンのα鎖、ヘモグロビンのβ鎖、ヘモグロビンのγ鎖、ヘモグロビンのδ鎖、ヘモグロビンのε鎖、およびヘモグロビンのζ鎖からなる群から選択されるポリペプチドであって、ポリペプチドはヒトαヘモグロビン鎖のアミノ酸1-97(「ペプチド1-97」)を含み、そしてポリペプチドはヒトαヘモグロビン鎖のアミノ酸1-94(「ペプチド1-94」)を含む、ならびに(b)薬学的に受容可能なキャリアを含む薬学的組成物を包含する。このような薬学的組成物は、ヘムと共にまたはヘムを有さずに、上記群から選択される単一ポリペプチド、上記群から選択されるポリペプチドの混合物、または上記群から選択される二量体または多量体の形態のポリペプチドで構成され得る。
【0063】
本発明はまた、以下の配列を有するペプチドを包含する:
【0064】
【化1】

ここで、2つのCys残基はジスルフィド結合を形成する(「環状ペプチド43-55」)
【0065】
【化2】
ここで、2つのCys残基は炭素架橋により結合されている、
【0066】
【化3】
および
図16Aに示すように、ヒトαヘモグロビンの最初の97 N末端アミノ酸を含むペプチド。
【0067】
本発明はまた、幹細胞刺激および/または刺激特性を保持した、ヒトα鎖の修飾型からなるタンパク質およびペプチド配列を含む。そのような修飾は、C末端疎水性ドメインの除去または修飾(改善された溶解特性を生じる)、および/またはハプトグロビン結合ドメイン除去または修飾(改善された薬物動態学的特性を生じる)を含むが、これらに限定されない。ヒトαヘモグロビンのC末端疎水性ドメインは、アミノ酸98(フェニルアラニン)から141(アルギニン)までの領域を含み、総数44のうち23の疎水性アミノ酸を含む。ドメイン全体または1つ以上のこれらの疎水性アミノ酸(6つのアラニン、4つのバリン、8つのロイシン、2つのプロリンおよび3つのフェニルアラニン)は、欠失(deletion)(C末端疎水性ドメインの「欠失」)によって除去され得る。あるいは、1つ以上のこれらのアミノ酸は、非極性アミノ酸(例えば、グリシン、セリン、トレオニン、システイン、チロシン、アスパラギンまたはグルタミン)で置換され得る(「置換」C末端疎水性ドメイン)。
【0068】
別の実施態様では、カルボキシメチル化のような、この領域の疎水的特性を減じ、溶解性を増加する化学的修飾が使用される。
【0069】
別の実施態様では、疎水性残基はヒトβヘモグロビン配列の対応する親水性の領域と置換される。例えば、ヒトβヘモグロビン配列では、アミノ酸107(グリシン)から117(ヒスチジン)間の領域またはアミノ酸130(チロシン)から139(ヒスチジン)間の領域はそれぞれ比較的親水性で、それぞれまたは共にヒトαヘモグロビン内の等価な疎水性の領域について置換され得る。
【0070】
ハプトグロビン結合ドメインはC末端疎水性領域に含まれ、アミノ酸121-127からなる。この領域は全体的に欠失により除去され得るか、またはこの領域の1つ以上のアミノ酸が欠失され得る(「欠失」C末端ハプトグロビン結合ドメイン)。この領域またはこの領域の1つ以上のアミノ酸は、例えばポリアラニンもしくはポリグリシン、またはポリペプチドのハプトグロビンへの結合を無効にする効果を有するが、幹細胞阻害活性は維持するような他のアミノ酸に置換され得る(「置換」C末端ハプトグロビン結合ドメイン)。
【0071】
本発明の他の実施態様は、βヘモグロビン鎖(C末端疎水性領域内および/または1つまたは両ハプトグロビン結合ドメイン(アミノ酸11-25および136-146))に対する相当する修飾、ならびにδ、γ、ε、および/またはζヘモグロビン鎖に対する相当する修飾を包含する。
【0072】
上記の同定されたペプチドをコードするDNA配列、そのDNA配列を含むベクター、およびそのベクターを含む宿主細胞もまた、本発明に包含される。これらのペプチドは、標準的な化学技術(例えば、固相合成)を用いるか、または組換え技術(融合システム(例えば、グルタチオン-S-トランスフェラーゼ(D.B.SmithおよびK.S.Johnson、Gene 67:31-40,1988)、チオレドキシン(La Vallieら、Biotechnology 11:187-193、1993)、またはユビキチン(Buttら、PNAS 86:2540-4、1989;Cherneyら,Biochem.30:10420-7,1991;Bakerら,JBC 269:25381-6,1994;米国特許第5,132,213号;第5,196,321号および第5,391,490号およびPCT WO 91/17245))を用いる融合システム)を含む)を用いることにより合成され得る。これらの論文、適用、および特許のそれぞれは、本明細書中に参考として援用される。
【0073】
さらに、本発明は、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、オピエートレセプター、好都合には、オピエートレセプターのμサブクラスに結合し得る化合物とを接触させる工程を含む方法を包含する。さらに、本発明は、幹細胞増殖を阻害または刺激する方法であって、造血細胞とノシセプチン(nociceptin)レセプター(例えば、ORL1)に結合し得る化合物とを接触させる工程を含む方法を包含する。さらに、本発明は、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、「オピエート様」レセプター結合し得る化合物とを接触させる工程を含む方法を包含する。
【0074】
オピエート活性を示すペプチド(「ヘモルフィン(hemorphin)」と呼ばれる)が、ヘモグロビンから単離されている(例えば、Brantlら、Eur.J.Pharm.、125:309-10、1986;Davisら、Peptides 10:747-51、1989;Hoffmanら,PNAS 87:8721-25,1990;Hernanら,Biochem.31:8619-28,1992;Karelinら、Bioch.Biophys.Res.Comm、202:410-5、1994;Zhaoら、Ann.N.Y.Acad.Science 750:452-8、1995;Petrovら,Bioscience Reports,15:1-14,1995;Karelinら,16:693-697,1995)。これらの論文のそれぞれは、本明細書中に参考として援用される。他の異型のオピエートペプチドおよび小分子もまた存在する(Zioudrouら,JBC 254:2446-9,1979;QuirionおよびWeiss,Peptides 4:445,1983;Loukasら,Biochem.22:4657,1983;Brantl,Eur.J.Pharm.106:213-14,1984;Brantlら,Eur.J.Pharm.111:293-4,1985;BrantlおよびNeubert,TIPS 7:6-7,1986;HurbyおよびGehrig,Med.Res.Rev.9:343-401,1989;Schiller,Prog.Med.Chem.28:301-40,1991;Glamstaら,BBRC 184:1060-6,1992;Teschemacher,Handbook Exp.Pharm.104:499-28,1993;Handbook of Experimental Pharmacology,A.Hertz(編)volumes 104/Iおよび104/II,1993,Springer Verlag,Berlin;Reedら,Neurosci.and Biobehav.Rev.18:519-25,1994;Karelinら,Peptides 16:693-7,1995)。これらの論文のそれぞれは、本明細書中に参考として援用される。本明細書中で使用されたように、「オピエート様レセプター」は、オピエート、INPROL、ヘモルフィン(hemorphins)、エキソルフィン(exorphins)、ノイセプチン(noiceptin)、Tyr-MIF-1ファミリーメンバー、アルカロイド、および/または適当量のナロキソン(naloxone)を含ませることにより拮抗されるように幹細胞増殖を阻害または刺激する他の化合物に結合するそれらの能力によって定義される(実施例29および38を参照のこと)。
【0075】
さらに、本発明は、レセプター(単数または複数)およびリガンドを同定する方法であって、結合アッセイにおいてINPROL(好都合には、ペプチド1-94、1-97、43-55または64-82のようなペプチド形態)を使用する工程を含む方法を包含する。さらに、本発明は、レセプター(単数または複数)およびリガンドを同定する方法であって、アデニル酸シクラーゼアッセイにおいてINPROLを使用する工程を含む方法を包含する。
【0076】
さらに、本発明は、幹細胞増殖を阻害または刺激する方法であって、造血細胞とGTP結合タンパク質(好都合には、これらのG阻害性サブタイプのタンパク質)を活性化し得る化合物(例えば、マストパラン(mastoparan))とを接触させる工程を含む方法を包含する。
【0077】
本発明はまた、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、以下の配列を有するヘモルフィンペプチドの群から選択されるペプチドとを接触させる工程を含む方法を包含する:
【0078】
【化4】
。
【0079】
上記ペプチドは、他の異型オピエートペプチド(例えば、Tyr-MIF-1ファミリー(概説についてはReedら、Neurosci.Biobehav.Rev.18:519-25、1994を参照のこと)、カゼイン由来カソモルフィン(casomorphin)(Brantlら、Hoppe-Seyler’s Z.Physiol.Chem.360:1211-16、1979;Loukasら、Biochem.22:4567-4573、1983;FiatおよびJolles、Mol.Cell.Biochem.87:5-30、1989)、シトクロームb由来のペプチド(シトクロフィン(cytochrophin)と称される)(Brantlら、Eur.J.Pharm.111:293-4、1985)、ヒトおよびヒト以外の種由来の種々なエキソルフィンおよびオピエートペプチド(Zioudrouら,JBC 254:2446-9,1979;Brantl,Eur.J.Pharm.106:213-14,1984;Brantlら,Eur.J.Pharm.125:309-10,1986;BrantlおよびNeubert,TIPS 7:6-7,1986;Glamstaら,BBRC 184:1060-6,1992;Teschemacher,Handbook Exp.Pharm.104:499-28,1993;Karelinら,Peptides 16:693-7,1995)、ならびにオピエートレセプターへの結合についてスクリーニングされたコンビナトリアルライブラリー由来ペプチド(概説についてはDooleyら、Peptide Research 8:124-137、1995を参照のこと))に類似の配列類似性および/または生物学的活性を有する。これらの論文のそれぞれは、本明細書中に参考として援用される。
【0080】
本発明はまた、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、Tyr-MIF-1関連ペプチド、カソモルフィン、シトクロフィンおよびエキソルフィンからなる群から選択されるペプチドとを接触させる工程を含む方法を包含する。以下の配列を有するTyr-MIF-1ペプチドが特に含まれる:
【0081】
【化5】
。
【0082】
本発明はまた、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、以下の群から選択されるオピエートペプチドとを接触させる工程を含む方法を包含する:
【0083】
【化6】
。
【0084】
本発明はまた、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、モルヒネ、コデイン、メタドン、ヘロイン、メペリジン(meperidine)、アルファプロジン、ジフェノキシラート、フェンタニール、スフェンタニル、アルフェンタニル、レボルファノール、ハイドロコドン、ジヒドロコデイン、オキシコドン、ヒドロモルフォン、プロポキシフェン、ブプレノルフィン、エトルフィン、オキシモルフォン デキストプロポキシフェン(oxymorphone dextopropoxyphene)およびメプタジノールからなる群から選択されるオピエートアゴニスト化合物とを接触させる工程を含む方法を包含する。特に、10-7モル未満の阻害量のモルヒネが含まれる。
【0085】
本発明はまた、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、ナロキソン、ナルトレキソン、ナロルフィン、ペンタゾシン、ナルブフィン、ブトルファノールからなる群から選択されるオピエートアンタゴニストまたは混合アゴニスト/アンタゴニストとを接触させる工程を含む方法を包含する。特に、10-8モル未満の阻害量のナロキソンが含まれる。
【0086】
本発明はまた、幹細胞増殖を刺激する方法であって、造血細胞と、INPROL、ミオグロビン、DAMGO、およびDALDAからなる群から選択されるタンパク質またはペプチドの幹細胞刺激量とを接触させる工程を含む方法を包含する。
【0087】
本発明はまた、哺乳動物において遺伝子療法を実施する方法を包含し、この方法は以下の工程を含む:
a)上記哺乳動物から造血細胞を取り出す工程、
b)エクスビボで上記造血細胞を幹細胞刺激量のINPROLおよび/またはオピエート化合物で処理する工程、
c)予め決定された遺伝子で上記造血細胞をトランスフェクトまたは感染する工程、
d)エクスビボで上記トランスフェクト造血細胞と幹細胞阻害量のINPROLおよび/またはオピエート化合物とを接触させる工程、
e)上記哺乳動物に工程dの造血細胞を移植する工程、
f)必要に応じて、インビボで上記哺乳動物を幹細胞を阻害または刺激する量のINPROLおよび/またはオピエート化合物で処理する工程。
【0088】
本発明はまた、エクスビボで幹細胞拡大を実施する方法であって、この造血細胞を幹細胞阻害量のINPROLおよび少なくとも1つの刺激性サイトカインで処理する工程を含む方法を包含する。INPROLは、刺激性サイトカインと接触させる前、間、および/または後で、造血細胞と接触させられる。エクスビボでの幹細胞の拡大は、臍帯血、胎児肝、化学療法などの後または精製後(例えば、CD34、CD38またはローダミンのようなマーカーを使用する蛍光活性化細胞選別による)の自家骨髄のような限られた供給源から十分量の幹細胞の産生が可能となる。特定の造血系列を選択的に増殖する能力はまた、臨床家が個々の患者の必要性に従って幹細胞移植を特別に設計することを可能とする。
【0089】
本発明はまた、エクスビボでの幹細胞の拡大を実施する方法であって、少なくとも1つさらなる刺激性サイトカインと共にまたはこれを伴わずに、造血細胞を幹細胞刺激量のINPROLで処理する工程を含む方法を包含する。INPROLは、刺激性サイトカイン(単数または複数)と接触させる前、間および/または後で、造血細胞と接触させられる。エクスビボで、幹細胞刺激剤は幹細胞および/または前駆細胞の拡大を可能にし、一方、幹細胞阻害剤は幹細胞の未分化状態に幹細胞を維持する。手順はまた、インビボで幹細胞阻害量でのINPROLの使用により、幹細胞を移植されるまで静止状態に維持することを最適化し得、後に幹細胞刺激量でのINPROLが、骨髄再生を刺激するために使用され得る。必要に応じて、造血細胞は2つの調製物に分割し得、そして幹細胞刺激量のINPROLで処置されたものは、幹細胞および/または前駆細胞の拡大を促進し、一方幹細胞阻害量のINPROLで処理された別のものは幹細胞をその未分化状態に維持する。次に、2つの調製物は組み合わされ得、そして患者に注入され得る。
【0090】
本発明はまた、(a)INPROLおよび(b)少なくとも1つの阻害性化合物(MIP-1α、TGFβ、TNFα、INFα、INFβ、INFγ、ペンタペプチドであるピロGlu-Glu-Asp-Cys-Lys、テトラペプチドであるN-アセチル-Ser-Asp-Lys-Pro、およびトリペプチドであるグルタチオン(Gly-Cys-γGlu)からなる群から選択される)を含む薬学的組成物を包含する。
【0091】
本発明はまた、(a)INPROLおよび(b)少なくとも1つの刺激性サイトカイン(IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-11、IL-13、IL-14、IL-15、G-CSF、GM-CSF、M-CSF、エリスロポエチン、トロンボポエチン、幹細胞因子、δ様タンパクおよびflk2/flt3リガンドからなる群から選択される)を含む薬学的組成物を包含する。
【0092】
本発明は、幹細胞阻害剤(INPROL)を記載する。この阻害剤は、MIP-1α、TGFβ、Frindelおよびその共同研究者のテトラペプチド、またはPaukovitsおよびその共同研究者のペンタペプチド(WrightおよびPragnell、1992(前出)参照)のような当該分野で公知の阻害剤とは異なる。天然に存在するネイティブINPROLは、INPROLとテトラペプチドおよびペンタペプチドとを区別する限外濾過によれば、10,000ダルトンを超える分子量を有する。ネイティブINPROLは、逆相クロマトグラフィーシステムにおいてはMIP-1αまたはTGFβよりも疎水性であり、これをそれらのサイトカインから区別する。さらに、その作用様式は、プレインキュベーションの期間でのみ用いられる場合に、インビトロアッセイで活性である点において、前記のいずれの阻害剤の作用様式とも異なる。例えば、MIP-1αは、プレインキュベーションの期間でのみ用いられる場合、効果的ではない(実施例5)。さらに、天然に存在するINPROLは、「高増殖性潜在細胞(high proliferative potential cells)」(HPP-PFC)を測定するアッセイにおいて活性であるが、MIP-1αは活性ではない(実施例6)。INPROLは、IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-10、IL-11、IL-12、IL-13、IL-14、IL-15、IL-16、G-CSF、GM-CSF、M-CSF、エリスロポエチン、トロンボポエチン、幹細胞因子、およびflk2/flt3リガンドのような当該分野では公知のこれらの刺激剤とは異なる。天然に存在するINPROLは、これらのサイトカインとはほとんどまたは全く配列類似性を示さない。
【0093】
本発明はまた、以下の項目を提供する:
項目1.C末端疎水性ドメインが置換または欠失されているヘモグロビンα鎖を含むポリペプチド。
項目2.C末端ハプトグロビン結合ドメインが置換または欠失されているヘモグロビンα鎖を含むポリペプチド。
項目3.ヒトαヘモグロビン鎖のアミノ酸1-97を含むポリペプチド。
項目4.(a)請求項1または2に記載のポリペプチドおよび(b)薬学的に受容可能なキャリアを含む薬学的組成物。
項目5.ヒトαヘモグロビン鎖のアミノ酸1-97からなるポリペプチドおよび薬学的に受容可能なキャリアを含む薬学的組成物。
項目6.ヒトαヘモグロビン鎖のアミノ酸1-94からなるポリペプチドおよび薬学的に受容可能なキャリアを含む薬学的組成物。
項目7.単位投与量形態の請求項4から6に記載の薬学的組成物。
項目8.0.1mgから6gの、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチドおよびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される1つまたは2つの化合物を含む、請求項7に記載の薬学的組成物。
項目9.幹細胞増殖を阻害する方法であって、造血細胞と幹細胞増殖阻害量の請求項1または2に記載のポリペプチドとを接触させる工程を包含する、方法。
項目10.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、および配列Phe-Leu-Gly-Phe-Pro-Thrを有するペプチドからなる群より選択される、請求項9に記載の方法。
項目11.B細胞の増殖を刺激する方法であって、造血細胞と増殖刺激量の請求項1または2に記載のポリペプチドとを接触させる工程を包含する、方法。
項目12.ガンに羅患している哺乳動物においてガンを処置する方法であって、該方法は以下の工程:
(a)放射線療法または化学療法を投与する工程、および
(b)幹細胞増殖阻害量の請求項1または2に記載のポリペプチドを投与する工程、
を包含する、方法。
項目13.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチドおよびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項12に記載の方法。
項目14.前記工程aおよびbが1回以上繰り返される、請求項12に記載の方法。
項目15.前記工程aが前記工程bの後に行われる、請求項12に記載の方法。
項目16.前記工程bが前記工程aの前または後の24時間以内に行われる、請求項12に記載の方法。
項目17.哺乳動物においてガンを処置する方法であって、該方法は以下の工程:
a)該哺乳動物から造血細胞を取り出す工程、
b)該造血細胞を、請求項1または2に記載のポリペプチドでエクスビボで処理する工程、
c)該工程bの造血細胞を、化学療法または放射線で処理する工程、
d)該哺乳動物において骨髄切除処置を行う工程、および
e)該工程cの造血細胞を該哺乳動物に移植する工程、
を包含する、方法。
項目18.前記工程(b)におけるポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチドおよびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項17に記載の方法。
項目19.幹細胞に損傷を与えるか、または破壊する薬剤に暴露された哺乳動物において幹細胞分裂を阻害する方法であって、幹細胞増殖阻害量の請求項1または2に記載のポリペプチドを投与する工程を包含する、方法。
項目20.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、および配列Phe-Leu-Gly-Phe-Pro-Thrを有するペプチドからなる群より選択される、請求項19に記載の方法。
項目21.前記薬剤が抗ウイルス剤である、請求項19に記載の方法。
項目22.哺乳動物造血幹細胞をエクスビボで維持する方法であって、造血細胞と幹細胞増殖阻害量の請求項1または2に記載のポリペプチドとを接触させる工程を包含する、方法。
項目23.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、および配列Phe-Leu-Gly-Phe-Pro-Thrを有するペプチドからなる群より選択される、請求項22に記載の方法。
項目24.前記造血細胞が、骨髄細胞、末梢血細胞、移動末梢血細胞、胎児肝臓、および臍帯血細胞からなる群より選択される、請求項22に記載の方法。
項目25.骨髄増殖性疾患もしくは自己免疫疾患または上皮幹細胞過剰増殖を、それらに羅患している哺乳動物において処置する方法であって、過剰増殖減少量の請求項1または2に記載のポリペプチドを投与する工程を包含する、方法。
項目26.前記骨髄増殖性疾患が脊髄形成異常症候群である、請求項25に記載の方法。
項目27.化学療法または放射線から哺乳動物における正常な幹細胞を区別して保護し、そしてガン細胞を保護しない方法であって、幹細胞保護量の請求項1または2に記載のポリペプチドを投与する工程を包含する、方法。
項目28.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、および配列Phe-Leu-Gly-Phe-Pro-Thrを有するペプチドからなる群より選択される、請求項27に記載の方法。
項目29.前記正常な幹細胞が細胞傷害性薬物または放射線への暴露により増殖するように誘導された後に、前記ポリペプチドが投与される、請求項27に記載の方法。
項目30.哺乳動物をワクチン接種する方法であって、ワクチン投与の前、間、または後に請求項1または2に記載のポリペプチドをアジュバントとして投与する工程を包含する、方法。
項目31.幹細胞過剰増殖により引き起こされる免疫抑制を有する哺乳動物を処置する方法であって、該哺乳動物に過剰増殖後退量の請求項1または2に記載のポリペプチドを投与する工程を包含する、方法。
項目32.哺乳動物において遺伝子療法を行う方法であって、該方法は以下の工程:
a)造血細胞を該哺乳動物から取り出す工程、
b)該造血細胞を予め決定された遺伝子でトランスフェクトする工程、
c)該トランスフェクトされた造血細胞と請求項1または2に記載のポリペプチドとをエクスビボで接触させる工程、および
d)該工程cの造血細胞を該哺乳動物に移植する工程、
を包含する、方法。
項目33.前記工程cにおけるポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチドおよびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項32に記載の方法。
項目34.前記工程(a)の後に、幹細胞増殖を誘導するために、前記造血細胞を少なくとも1つの刺激性サイトカインで処理する工程をさらに包含する、請求項32に記載の方法。
項目35.前記工程(d)の後に、前記哺乳動物を前記ポリペプチドでインビボで処置する工程をさらに包含する、請求項32に記載の方法。
項目36.幹細胞拡大をエクスビボで行う方法であって、造血細胞と請求項1または2に記載のポリペプチドおよび少なくとも1つの刺激性サイトカインとを接触させる工程を包含する、方法。
項目37.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、およびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項36に記載の方法。
項目38.前記造血細胞が、骨髄細胞、末梢血細胞、移動末梢血細胞、胎児肝臓、および臍帯血細胞からなる群より選択される細胞である、請求項36に記載の方法。
項目39.(a)請求項1または2に記載のポリペプチド、ならびに(b)MIP-1α、TGFβ、TNFα、INFα、INFβ、INFγ、ペンタペプチドピロGlu-Glu-Asp-Cys-Lys、テトラペプチドN-アセチル-Ser-Asp-Lys-Pro、およびトリペプチドグルタチオン(Gly-Cys-γGlu)からなる群より選択される少なくとも1つの阻害性化合物を含む、薬学的組成物。
項目40.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、およびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項39に記載の方法。
項目41.(a)請求項1または2に記載のポリペプチド、ならびに(b)IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-11、IL-13、IL-14、IL-15、G-CSF、GM-CSF、M-CSF、エリスロポエチン、トロンボポエチン、幹細胞因子、およびflk2/flt3リガンドからなる群より選択される少なくとも1つの刺激性化合物を含む、薬学的組成物。
項目42.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、およびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項41に記載の方法。
項目43.αヘモグロビンまたはその置換もしくは欠失アナログを発現させる方法であって、該αヘモグロビンまたは置換もしくは欠失アナログを、ユビキチン融合体として発現させる工程を包含する、方法。
項目44.前記発現させる工程がE.coliにおいて行われる、請求項43に記載の方法。
項目45.前記発現させる工程が、ユビキチン切断酵素を発現させる工程を包含する、請求項43に記載の方法。
項目46.ビオチン-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、(ヨード)Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、および(ヨード)Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Valからなる群より選択される配列を有するペプチド。
項目47.幹細胞増殖を刺激する方法であって、造血細胞と幹細胞増殖刺激量のINPROLおよび/またはオピエート化合物とを接触させる工程を包含する、方法。
項目48.前記INPROLが、ヘモグロビンα鎖、ヘモグロビンβ鎖、ヘモグロビンγ鎖、ヘモグロビンδ鎖、ヘモグロビンε鎖、ヘモグロビンζ鎖、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、およびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項47に記載の方法。
項目49.前記INPROLが、以下の配列:
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Cys-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val-Cys
(ここで、2つのCys残基はジスルフィド結合を形成する)、
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala、
Phe-Leu-Gly-Phe-Pro-Thr、
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln-Arg-Phe、
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln-Arg、
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln、
Leu-Val-Val-Tyr-Pro-Trp-Thr、
Leu-Val-Val-Tyr-Pro-Trp、
Leu-Val-Val-Tyr-Pro、
Val-Val-Tyr-Pro-Trp-Thr-Gln、
Tyr-Pro-Trp-Thr-Gln-Arg-Phe、
Tyr-Pro-Trp-Thr-Gln-Arg、
Tyr-Pro-Trp-Thr-Gln、および
Tyr-Pro-Trp-Thr、
からなる群より選択される、請求項47に記載の方法。
項目50.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項47に記載の方法。
項目51.幹細胞増殖を刺激する方法であって、造血細胞とオピエートレセプターに結合し得る化合物とを接触させる工程を包含する、方法。
項目52.前記化合物がμサブクラスのオピエートレセプターに対する選択性を有する、請求項51に記載の方法。
項目53.幹細胞増殖を刺激または阻害する方法であって、造血細胞とノシセプチンレセプターに結合し得る化合物とを接触させる工程を包含する、方法。
項目54.幹細胞増殖を刺激または阻害する方法であって、造血細胞とG阻害性サブクラスのGTP結合タンパク質を活性化し得る化合物とを接触させる工程を包含する、方法。
項目55.幹細胞増殖を刺激または阻害する方法であって、造血細胞と古典的μ、κ、もしくはδオピエートレセプターまたはORL1を含まないオピエート様レセプターに結合し得る化合物とを接触させる工程を包含し、ここで該レセプターは(a)幹細胞刺激および/または阻害特性を有し、そして(b)ナロキソンにより拮抗可能な該幹細胞刺激および/または阻害能力を有する、方法。
項目56.前記オピエート様レセプターが、1μM未満またはそれに等しい解離定数(Kd)で、ペプチドPhe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Valに結合する能力を有する、請求項55に記載の方法。
項目57.前記解離定数が10nM未満またはそれに等しい、請求項55に記載の方法。
項目58.INPROLに対するレセプターを同定する方法であって、レセプター結合アッセイにおいて該レセプターを含む物質とINPROLとを接触させる工程を包含する、方法。
項目59.前記INPROLが、ヘモグロビンα鎖、ヘモグロビンβ鎖、ヘモグロビンγ鎖、ヘモグロビンδ鎖、ヘモグロビンε鎖、ヘモグロビンζ鎖、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
ビオチン-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
(ヨード)Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
(ヨード)Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Cys-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val-Cys、および
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala、
からなる群より選択される、請求項58に記載の方法。
項目60.INPROLに対するレセプターを同定する方法であって、アデニル酸シクラーゼアッセイにおいて該レセプターを含む物質とINPROLとを接触させる工程を包含する、方法。
項目61.前記INPROLが、ヘモグロビンα鎖、ヘモグロビンβ鎖、ヘモグロビンγ鎖、ヘモグロビンδ鎖、ヘモグロビンε鎖、ヘモグロビンζ鎖、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
ビオチン-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
(ヨード)Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
(ヨード)Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Cys-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val-Cys、および
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala、
からなる群より選択される、請求項60に記載の方法。
項目62.ガンを羅患している哺乳動物においてガンを処置する方法であって、該方法は以下の工程:
(a)放射線療法および/または化学療法を投与する工程、および
(b)幹細胞増殖刺激量のINPROLおよび/またはオピエート化合物を投与する工程、
を包含する、方法。
項目63.前記工程aおよびbが1回以上繰り返される、請求項62に記載の方法。
項目64.前記工程aが前記工程bの前に行われる、請求項62に記載の方法。
項目65.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項62に記載の化合物。
項目66.幹細胞に損傷を与えるかまたは破壊する薬剤に暴露された哺乳動物において幹細胞分裂を刺激する方法であって、幹細胞増殖刺激量のINPROLおよび/またはオピエート化合物を投与する工程を包含する、方法。
項目67.前記薬剤が抗ウイルス剤または抗腫瘍剤である、請求項66に記載の方法。
項目68.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項66に記載の方法。
項目69.哺乳動物造血幹細胞をエクスビボで維持する方法であって、造血細胞と幹細胞増殖刺激量のINPROLおよび/またはオピエート化合物とを接触させる工程を包含する、方法。
項目70.前記造血細胞が、骨髄細胞、末梢血細胞、移動末梢血細胞、胎児肝臓、および臍帯血細胞からなる群より選択される、請求項69に記載の方法。
項目71.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項69に記載の方法。
項目72.骨髄増殖性疾患、造血幹細胞過剰増殖または上皮幹細胞過剰増殖を、それらを羅患している哺乳動物において処置する方法であって、刺激量のINPROLおよび/またはオピエート化合物を投与する工程を包含する、方法。
項目73.前記骨髄増殖性疾患が脊髄形成異常症候群または再生不良性貧血である、請求項72に記載の方法。
項目74.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項72に記載の方法。
項目75.幹細胞消耗を処置または予防する方法であって、幹細胞増殖阻害量のINPROLおよび/またはオピエート化合物を投与する工程を包含する、方法。
項目76.前記幹細胞消耗が後天性免疫不全症候群によるものである、請求項75に記載の方法。
項目77.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項75に記載の方法。
項目78.化学療法または放射線から哺乳動物における正常な幹細胞を区別して保護する方法であって、幹細胞保護量のオピエート化合物を投与する工程を包含する、方法。
項目79.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項78に記載の方法。
項目80.哺乳動物において遺伝子療法を行う方法であって、該方法は以下の工程:
a)該哺乳動物から造血細胞を取り出す工程、
b)該造血細胞を、幹細胞刺激量のINPROLおよび/またはオピエート化合物でエクスビボで処理する工程、
c)該造血細胞を予め決定された遺伝子でトランスフェクトまたは感染する工程、
d)該トランスフェクトされた造血細胞と、幹細胞阻害量のINPROLおよび/またはオピエート化合物とをエクスビボで接触させる工程、
e)該工程dの造血細胞を該哺乳動物に移植する工程、
f)必要に応じて、該哺乳動物を幹細胞阻害量または刺激量のINPROLおよび/またはオピエート化合物でインビボで処置する工程、
を包含する、方法。
項目81.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項80に記載の方法。
項目82.幹細胞拡大をエクスビボで行う方法であって、造血細胞と幹細胞刺激量のINPROLおよび/またはオピエート化合物とを接触させる工程を包含する、方法。
項目83.前記造血細胞が、骨髄細胞、末梢血細胞、移動末梢血細胞、胎児肝臓、および臍帯血細胞からなる群より選択される細胞である、請求項80に記載の方法。
項目84.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項80に記載の方法。
項目85.(a)オピエート化合物、ならびに(b)MIP-1α、TGFβ、TNFα、INFα、INFβ、INFγ、ペンタペプチドピロGlu-Glu-Asp-Cys-Lys、テトラペプチドN-アセチル-Ser-Asp-Lys-Pro、およびトリペプチドグルタチオン(Gly-Cys-γGlu)からなる群より選択される少なくとも1つの阻害性化合物を含む、薬学的組成物。
項目86.a)オピエート化合物、ならびに(b)IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-11、IL-13、IL-14、IL-15、G-CSF、GM-CSF、M-CSF、エリスロポエチン、トロンボポエチン、幹細胞因子、およびflk2/flt3リガンドからなる群より選択される少なくとも1つの刺激性化合物を含む、薬学的組成物。
項目87.哺乳動物において疼痛を処置する方法であって、鎮痛誘導量のINPROLを該哺乳動物に投与する工程を包含する、方法。
項目88.前記INPROLが、ヘモグロビンα鎖、ヘモグロビンβ鎖、ヘモグロビンγ鎖、ヘモグロビンδ鎖、ヘモグロビンε鎖、ヘモグロビンζ鎖、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Cys-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ara-Gln-Val-Cys、および
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala、
からなる群より選択される、請求項87に記載の方法。
項目89.哺乳動物における免疫不全を処置する方法であって、免疫刺激量のINPROLを該哺乳動物に投与する工程を包含する、方法。
項目90.前記INPROLが、ヘモグロビンα鎖、ヘモグロビンβ鎖、ヘモグロビンγ鎖、ヘモグロビンδ鎖、ヘモグロビンε鎖、ヘモグロビンζ鎖、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Cys-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ara-Gln-Val-Cys、および
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala、
からなる群より選択される、請求項89に記載の方法。
【0094】
本明細書中に記載される本発明がより十分に理解され得るために、以下の詳細な説明を記載する。この説明は、本発明の例示であるが、本発明を特に限定すると解釈されるべきではなく、そして当業者の理解範囲であるような変化は本発明の範囲に入ると考えられるべきである。
【発明を実施するための最良の形態】
【0095】
好適な実施態様の詳細な説明
INPROLは、幹細胞の分裂を可逆的に阻害または刺激する。特定の理論に縛られることを望まないが、幹細胞阻害剤および刺激剤は、幹細胞が細胞周期を通って移行する割合に影響を与えることによってそれらの効果を発揮すると考えられる。特に、INPROLは、使用量に依存して造血幹細胞の細胞分裂を一時的に阻害または刺激することに効果的である。幹細胞増殖を阻害または刺激し得る化合物を臨床的に使用する能力は、例えば化学療法、幹細胞移植または遺伝子療法プロトコルの間の、造血幹細胞の細胞周期進行の精巧な制御を可能にする。従って、本発明の方法は、ガン細胞もしくはウイルスに感染した細胞を破壊するために用いられる化学療法剤もしくは放射線によって引き起こされる損傷から幹細胞を保護することによって、またはそのような損傷後の回復を刺激することによって、患者の造血系、骨髄系および免疫系に対する化学療法の望ましくない副作用を軽減することに用いられ得る。本発明の一つの実施態様では、INPROLは、化学療法剤が疾患細胞に作用している間の幹細胞分裂を阻害するのに十分な用量で患者に投与される。化学療法剤がその機能を発揮した後、INPROLによって阻害された幹細胞は、さらなる処置を受けずに分裂細胞に戻る。造血再生を高めることが所望されるならば、刺激性成長因子、サイトカインまたは幹細胞刺激量のINPROLがさらに用いられ得る。
【0096】
本明細書中で用いられる用語「INPROL」は、実施例におけるように精製された哺乳動物および非哺乳動物タンパク質、ヘモグロビン、ヘモグロビンのα鎖(ヘム基を有するかまたは有しない)、ヘモグロビンのβ鎖(ヘム基を有するかまたは有しない)、α鎖およびβ鎖の混合物(ヘム基を有するかまたは有しない)、およびこれらのタンパク質のフラグメントまたはアナログ(例えば、α、β、γ、δ、ε、またはζ鎖、単独で、あるいは混合物、二量体、または多量体のいずれかとして、ヘム基を有するかまたは有しない)を包含する。このようなフラグメントまたはアナログは、胚型、胎児型、または成体型を包含し、幹細胞増殖の阻害能および/または刺激能を有する。用語「INPROL」は、これらのタンパク質の天然に存在する形態ならびに天然に存在しない(例えば、組換え的および/または合成的に生成された)形態を包含する。用語「INPROLポリペプチド」は、40以上のアミノ酸からなるINPROLをいう。
【0097】
本明細書中で用いられる用語「オピエート化合物」は、オピエートレセプター(または、オピエートレセプターに配列関連性のあるレセプター、例えばORL1)に結合し、そしてアゴニスト、アンタゴニスト、または混合アゴニスト/アンタゴニストのいずれかの活性を発揮する、INPROLではないオピエートを含む化合物である。例えば、特異的アゴニストおよびアンタゴニストが、μレセプター(これは、DAMGOおよびDALDAによって選択的に活性化され、そしてCTOPおよびナロキソナジン(naloxonazine)によって選択的に拮抗される)、κレセプター(これは、GR 89696フマル酸塩またはU-69593によって選択的に活性化され、そしてnor-ビナルトルフィミン(binaltorphimine)塩酸塩によって選択的に拮抗される)、δレセプター(これは、DADLEおよびDPDPEによって選択的に活性化され、そしてナトリンドール(natrindole)、によって選択的に拮抗される)について存在する。さらに、3つのレセプターサブタイプの全てに作用する、広域スペクトルアンタゴニスト(例えば、ナロキソン)およびアゴニスト(例えば、エトルフィン)が存在する。ノシセプチン(nociceptin)はORL1レセプターを特異的にアゴナイズする。幹細胞の刺激および/または阻害活性を有するオピエート化合物は、INPROLについて本明細書中に記載された適用のそれぞれに使用され得る。
【0098】
本明細書中で用いられる「幹細胞刺激量」は、幹細胞の増殖を誘導する量である。本明細書中で用いられる「幹細胞阻害量」は、幹細胞の増殖を阻害する量である。全ての場合において、インビボおよびエクスビボの両方で、選択される量は、選択された特定のINPROLまたはオピエート化合物および特定の条件または適用に依存する。とりわけ、INPROLのポリペプチドまたはフラグメントの等モル用量は、オピエートペプチドまたは小分子と等モルとして活性である。
【0099】
典型的な臨床状況においては、幹細胞の阻害を所望するのであれば、INPROLは、例えば、0.01〜100mg/kg、好都合には0.1〜1.0mg/kgのINPROLを、例えば幹細胞の細胞周期進行を阻害することが望ましい場合は、標準的化学療法または放射線照射処置の4〜60時間前に投与することを用いて、投与量単位形態で静脈注射または点滴により日毎のレジメで患者に投与される。
【0100】
化学療法または放射線照射後の回復を促進するために、幹細胞の細胞周期進行の刺激が所望される状況では、幹細胞刺激量でINPROLが使用される。そのような用量は、典型的には1-500mg/kg、好都合には10mg〜100mg/kgである。
【0101】
幹細胞の細胞周期進行を阻害または刺激するためにオピエート化合物(単数または複数)を使用することが所望される場合では、オピエート化合物(単数または複数)は、INPROLについて記載した濃度の等モルで使用される。
【0102】
本発明の別の実施態様では、幹細胞阻害量のINPROLによる前処置により、患者が通常耐えられる用量を上回る量への化学療法剤用量または放射線量の増大が可能になる。同様に、幹細胞刺激量でのINPROLを用いた化学療法後または放射線後の処置はまた、化学療法または放射線照射の通常耐えられる用量の増大を可能にする。
【0103】
大部分の造血幹細胞は通常静止(ゆっくり細胞周期を進行しているか、全く進行していない)している。しかし、化学療法誘導性造血損傷に対する代償応答として、多くの割合の幹細胞が化学療法後に細胞周期進行に入る。それにより、細胞は、特に、次の用量の細胞傷害性化学療法または治療的放射線照射に対して抵抗力のないものとなる。このような幹細胞の細胞周期進行を阻害することにより、INPROL処置によって、次の用量の細胞傷害性化学療法をより早くまたはより頻度を高くして投与すること(従来用量または高用量のいずれかで)が可能になる。
【0104】
いくつかの正常な個体は、自然にすばやく細胞周期に入る幹細胞を有する。幹細胞阻害量のINPROLは、たとえ初回の用量の放射線照射または化学療法の前に与えるとしてもそのような個体には有用である。
【0105】
本発明の1つの実施態様では、INPROL(0.1mg〜6g/kg体重−好都合には1.0〜60mg/kg)は、化学療法の初回用量の投与から約24時間〜10日後に投与される。さらに4〜60時間後、好都合には24〜48時間後、次回の用量の化学療法が投与される。この化学療法とINPROLとを交互に行うサイクルは、治療上の利点に従って続けられる。化学療法剤および投与プロトコルは、標準的な医療実施における特定の腫瘍タイプに対する適合性に従って選択される。必要に応じて、G-CSF、幹細胞因子、または幹細胞刺激量のINPROLのような刺激性成長因子が、化学療法または放射線照射処置の後に用いられて、さらに造血再構築を改善する。
【0106】
エクスビボ適用のためには、幹細胞増殖の阻害が所望される場合では、106細胞/ml当たり0.1ng〜100ng、好都合には106細胞/ml当たり2〜50ngのINPROLが用いられる。幹細胞の刺激が所望される場合では、106細胞/ml当たり10ng〜100μg、好都合には106細胞/ml当たり1〜100μgのINPROLが用いられる。
【0107】
オピエート化合物を幹細胞の細胞周期進行の阻害または刺激のために使用することが所望される場合は、オピエート化合物(単数または複数)はINPROLについて記載された濃度と等モル濃度で用いられる。
【0108】
本発明の別の実施態様では、INPROLは、移植のための自家造血細胞を調製するための方法において用いられる。造血細胞は、有効量のINPROLでエクスビボで処理されて、幹細胞分裂を阻害し、次いで骨髄培養物に有効量の化学療法剤または放射線照射を投与することによってガン性細胞をパージする。細胞周期が進行している細胞に特異性を有する化学療法剤が好ましい。このように処理された骨髄は、自家ドナー内に再注射される。必要に応じて、患者は、幹細胞刺激量のINPROLおよび/または造血を刺激することが知られている別の薬剤で処置されて、患者の造血再構築を改善する。このような技術によって自家骨髄移植片の間の腫瘍細胞を効果的にパージするが、一方造血幹細胞を保護することが可能になる。このような保護は、エクスビボまたはインビボのいずれかのパージングのプロトコルを与え得る。一旦首尾良く移植されたら、幹細胞を迅速に増殖させて、正常な骨髄機能を再生させる必要がある。これは、幹細胞の細胞周期進行を刺激しそして骨髄機能の回復を増強する幹細胞刺激量でのINPROLの使用によって与えられ得る。
【0109】
本発明の別の実施態様では、INPROLは、遺伝子療法のための造血細胞を調製するための方法において用いられる。造血細胞はエクスビボで幹細胞刺激量のINPROLおよび/または他の刺激性サイトカイン(単数または複数)で処理されて幹細胞の分裂を刺激し、次に目的の遺伝子(単数または複数)でトランスフェクトする(好都合には、例えばレトロウイルスベクターを用いて感染させる)。トランスフェクションが達成された後、細胞を洗浄し、そして幹細胞阻害量のINPROLで処理して幹細胞を静止状態に戻す。このように処理された骨髄は、ドナーに再注入される。必要に応じて、患者は幹細胞阻害量のINPROLでインビボで処理されて、幹細胞がそれらの静止形態に維持され、そしてそれらの骨髄再生能が増大する。
【0110】
本発明の別の実施態様では、INPROLは、白血病処置の補助的な処置として用いられる。例えば、白血病細胞がINPROLに応答しない病態において、造血細胞はエクスビボで幹細胞阻害量のINPROLにより処理される。正常幹細胞の増殖はINPROLの投与により妨げられる。従って、増殖する白血病細胞が細胞周期に特異的な細胞傷害性薬剤により処理されている間は、正常幹細胞の集団は損傷から保護される。さらに、IL-3、GM-CSFのような刺激性サイトカインは、薬物処置または放射線照射処置の間に必要に応じて投与されて白血病細胞における細胞周期の進行を誘導するが、正常幹細胞はINPROLにより保護される。患者は、白血病細胞を破壊するために化学療法剤または放射線により処置され、次いでパージされた骨髄は、造血再構築を確立するために患者内に移植により戻される。
【0111】
同様に、血液細胞またはリンパ球に関与する重篤なウイルス感染(例えば、HIV感染)にかかった患者を処置するための本発明の別の実施態様においては、造血細胞はエクスビボまたはインビボでINPROLにより処理され、その後抗ウイルス剤、感染細胞を破壊する薬物、または感染細胞を排除するための抗体に基づく系により処理される。患者からウイルス性宿主細胞を根絶するための骨髄切除(myeloablative)抗ウイルスまたは骨髄切除化学療法の後、INPROL処理した骨髄は患者に戻される。
【0112】
本発明の別の実施態様では、INPROLは過剰増殖性幹細胞に関連した疾患を処置するために用いられる。例えば、乾癬は、皮膚の過剰増殖する上皮細胞によって引き起こされる障害であり、時には細胞傷害性薬物により処置される。幹細胞増殖を含む他の前新生物性病変はまた、幹細胞の増殖を阻害するために用いられる有効量のINPROLに影響されやすい。後天性免疫不全症候群の患者は、幹細胞の枯渇という結果となる異常に速い速度の幹細胞の細胞周期進行を有する。これらの患者はまた、幹細胞の細胞周期進行を阻害するのに有効な量のINPROLで処置することから恩恵を受ける。これらの使用のため、INPROLを含む局所的または経皮的な送達組成物(例えば、軟膏、ローション剤、ゲル剤、またはパッチ剤)は、非経口投与の代わりとして、適切な箇所で用いられる。
【0113】
ほとんどの白血病の場合、白血病前駆細胞は、INPROLにより影響を受けない分化細胞集団であり、従って上記のようなINPROLを用いる方法によって処理される分化細胞集団である。白血病前駆細胞が非常に原始期のものであり、INPROLによる阻害に対して直接的に感受性である場合には、白血病細胞の増殖は有効量のINPROLの投与によって弱められる。
【0114】
本発明の別の実施態様では、INPROLは、過剰増殖性幹細胞に関する障害を処置するために用いられる。例えば、骨髄形成異常症候群および再生不良性貧血は、骨髄の幹細胞の過剰増殖により引き起こされる障害である。幹細胞の過剰増殖に関与する他の症候群は、幹細胞刺激量のINPROLで処置可能である。
【0115】
モノクローナルまたはポリクローナルである抗体は、INPROLペプチドもしくはポリペプチドに対して標準的な技術によって生成される。これらの抗体またはINPROLペプチドもしくはポリペプチドは、当該分野で公知の多くのタイプの検出可能な標識により標識される。次いで、標識されたINPROLまたは抗INPROL抗体は、幹細胞マーカーとして用いられ、診断を目的として患者に直接それらを投与することによって幹細胞を同定および単離する。あるいは、これらの標識されたペプチド、ポリペプチドまたは抗体はエクスビボで用いられ、骨髄中の新生物形成細胞を除去する前にそれらを除き得るように造血細胞調製物中の幹細胞を同定する。同様の方法で、このような標識されたペプチド、ポリペプチドまたは抗体は、上皮性または他の幹細胞を単離および同定するために用いられる。さらに、標識されるかまたは標識されていないこのような抗体は、INPROL活性の中和によって治療的に、または循環しているINPROLレベルを検出することによって診断的に用いられる。
【0116】
INPROLは、標準的な技術を用いて、ヒト遺伝子または組換えヒトINPROLの発現用のcDNAライブラリーからクローニングされ得る。例えば、精製タンパク質から得られる配列情報を用いて、例えば32-リンで標識され得るオリゴヌクレオチドプローブが構築され、そして適切なcDNAライブラリー(例えば、骨髄由来)をスクリーニングするために用いられる。あるいは、適切な供給源(例えば、骨髄)由来の発現ライブラリーは、抗体を用いて、または適切な機能的アッセイ(例えば、実施例2に記載されるアッセイ)を用いて、INPROLをコードするcDNAについてスクリーニングされる。ヘモグロビン自体、ならびに個々のα鎖およびβ鎖は、当該分野において公知の方法を用いてクローニングおよび発現されている(Pagnierら、Rev.Fr.Transfus.Hemobiol.35:407-15,1992;Lookerら、Nature 356:258-60,1992;Methods in Enzymology 231巻,1994)。
【0117】
本発明は、以下を含むDNA配列を包含する:選択された非哺乳動物宿主による発現に「好ましい」コドンの取り込み;制限エンドヌクレアーゼ酵素による切断部位の供給;および付加的な開始、末端、または中間のDNA配列の供給、これらの配列は、容易に発現されるベクターの構築、あるいはヘモグロビンのα鎖、β鎖、γ鎖、δ鎖、ε鎖、および/またはζ鎖の生成または精製を容易にする。
【0118】
本発明はまた、ヘモグロビンα鎖、β鎖、γ鎖、δ鎖、ε鎖、および/またはζ鎖のポリペプチドアナログまたは誘導体をコードするDNA配列を提供する。これらアナログまたは誘導体は、1つ以上のアミノ酸残基の同一性または位置に関して天然に存在する形態と異なり(すなわち、欠失アナログ、記載の全残基より少ない残基を含有する;置換アナログ、記載の1つ以上の残基が他の残基により置換される;および付加アナログ、1つ以上のアミノ酸残基がポリペプチドの末端または中間部分に付加される)、かつ天然に存在する形態の一部または全ての特性を共有する。
【0119】
好都合な実施態様では、INPROLは、ゲノムクローニングもしくはcDNAクローニング、または遺伝子合成により得られた外因性DNA配列の原核生物または真核生物宿主発現(例えば、細菌、酵母、高等植物、昆虫、および哺乳動物培養細胞による)の産物である。すなわち、好都合な実施態様では、INPROLは「組換えINPROL」である。代表的な酵母(例えば、Saccharomyces cerevisiae)または原核生物(例えば、E.coli)宿主細胞における発現産物は、いずれの哺乳動物タンパク質とも関連しない。脊椎動物(例えば、ヒトではない哺乳動物(例えば、COSまたはCHO)および鳥類)細胞における発現産物は、いずれのヒトタンパク質とも関連しない。用いられる宿主に依存して、本発明のポリペプチドは、グリコシル化され得るか、またはグリコシル化され得ない。本発明のポリペプチドは、必要に応じて開始メチオニンアミノ酸残基(−1位)もまた含む。
【0120】
本発明は、ヘモグロビンのα鎖、β鎖、γ鎖、δ鎖、ε鎖、および/またはζ鎖のポリペプチドアナログのような他の産物もまた包含する。このようなアナログは、ヘモグロビンのα鎖、β鎖、γ鎖、δ鎖、ε鎖、および/またはζ鎖のフラグメントを包含する。周知の手順に従って、当業者は、本明細書中に記載の1つ以上の残基の同一性または位置に関して天然に存在する形態と異なる(例えば、置換、末端および中間における付加、および欠失)一次配列を有する微生物発現ポリペプチドをコードする遺伝子を容易に設計および製造し得る。あるいは、cDNA遺伝子およびゲノム遺伝子の改変は、周知の部位特異的変異誘発技術により容易に達成され得、そしてヘモグロビンのα鎖、β鎖、γ鎖、δ鎖、ε鎖、またはζ鎖のアナログおよび誘導体を生成するために用いられ得る。このような産物は、少なくとも1つのINPROLの生物学的特性を共有するが、他は異なり得る。例として、本発明の産物は、以下のようなα鎖、β鎖、γ鎖、δ鎖、ε鎖、またはζ鎖を包含する:例えば、欠失により短縮される;または加水分解に対してより安定である(従って、天然に存在するよりもより顕著なまたはより長期間持続する効果を有し得る);あるいはO-グリコシル化および/またはN-グリコシル化のための1つ以上の潜在的な部位を欠失または付加するように改変されたか、あるいは欠失または置換(例えば、アラニンまたはセリン残基により)された1つ以上のシステイン残基を有しており、そして微生物系から活性形態でより容易に単離される;あるいはフェニルアラニンにより置換される1つ以上のチロシン残基を有し、そして標的タンパク質または標的細胞上のレセプターにある程度容易に結合する。α鎖、β鎖、γ鎖、δ鎖、ε鎖、またはζ鎖内の連続アミノ酸配列または二次コンフォメーションの一部のみを複製するペプチドフラグメントもしくはポリペプチドフラグメントもまた包含される。これらのフラグメントは、INPROLの1つの特性(例えば、レセプター結合性)を有し、そして他(例えば、幹細胞阻害活性)は有さない。本発明の産物のいずれか1つ以上が治療有用性(Weilandら、Blut 44:173-5,1982を参照のこと)または他の情況(例えば、阻害剤拮抗作用のアッセイ)における有用性を有するために、活性は必要でないことは注目すべきことである。競合的アンタゴニストは、幹細胞阻害剤またはそのレセプターの過剰生産の場合において有用である。
【0121】
さらに、生物学的活性を保持するタンパク質配列に由来するペプチドは、標準的な方法に従って化学的に合成され得る。本発明はまた、ヘモグロビンα鎖、β鎖、γ鎖、δ鎖、ε鎖、および/またはζ鎖のペプチドアナログまたは誘導体をコードする配列を提供する。これらアナログまたは誘導体は、1つ以上のアミノ酸残基の同一性または位置に関して天然に存在する形態と異なり(例えば、欠失アナログ、記載の全残基より少ない残基を含有する;置換アナログ、記載の1つ以上の残基が他の残基、天然に存在するかまたは当該分野で公知の他のアナログ(例えば、D-アミノ酸)のいずれかにより置換される;および付加アナログ、1つ以上のアミノ酸残基が、安定性、溶解度、および/またはタンパク質分解に対する耐性を増大させるように化学修飾される)、かつ天然に存在する形態の特性の一部または全ての特性を共有する。
【0122】
上記のようなペプチド配列は、種々の方法により同定され得る。アッセイにおいて活性な天然ヘモグロビン鎖(例えば、α鎖)の三次元構造と不活性な構造的に関連したタンパク質(例えば、ミオグロビン)との比較は、三次元空間において異なるコンフォメーションを有し、従って活性ペプチドの候補領域である領域を同定し得る。別のアプローチは選択的タンパク質分解を用いる。このタンパク質分解では、タンパク質分解酵素は、ヘモグロビン鎖の限定消化において用いられる。この消化は、例えば逆相HPLCにより分離され、次いで幹細胞阻害についてアッセイされ得るペプチドを生じる。ペプチドはまた、化学合成(例えば、固相合成)によっても生成され得る;目的のヘモグロビン鎖(例えば、α鎖)の配列を含む一連の重複ペプチド(例えば、15マー)は、容易に生成され得、そして幹細胞アッセイにおいて試験され得る。コンビナトリアルライブラリーが生成され得、このライブラリーでは、複数の化学合成が実施され、かつ選択されたアミノ酸位置を変えることが可能であり、その結果、多くのスクリーニング用ペプチドアナログを生じる(例えば、Dooleyら、Peptide Research 8:124-137,1995)。あるいは、組換え法が用いられ得る。部位特異的変異誘発は、特定のヘモグロビン鎖の活性に必要な必須残基を同定するために用いられ得る。幹細胞阻害剤として活性であると知られる鎖(例えば、α鎖)の領域は、関連するが不活性なタンパク質(例えば、ミオグロビン)の領域と置換され得、そして幹細胞アッセイにおいて試験され得、これにより活性に必要な領域の同定が可能になる。このような同定された領域は、ペプチドとして発現され得、そして幹細胞周期アッセイにおいて活性について試験され得る。
【0123】
他の種に由来するINPROLの同種または類似のバージョンは、上記の本発明の治療的実施態様と同様に、種々の獣医学的使用に用いられる。
【0124】
幹細胞刺激量のINPROLは、細胞周期の進行している幹細胞を分裂しないかまたは緩徐にに分裂する「休止」期に可逆的に移行させることによって、それらに作用する。休止している幹細胞を刺激して分裂させることが望ましい場合(例えば、ガンの化学療法剤または放射線により患者を処置した後)、幹細胞刺激量のINPROLが用いられ得る。あるいは、またはさらに、コロニー刺激剤および他の造血刺激剤が被験体に投与される。このような因子の例としては以下のものが挙げられるが、それらに限定されない:M-CSF(CSF-1)、GM-CSF、G-CSF、巨核球-CSF、トロンボポエチン、幹細胞因子、または他のサイトカイン(例えば、IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-11、IL-12、IL-13、IL-14、あるいはエリスロポエチン)。
【0125】
INPROLポリペプチドまたは幹細胞阻害活性を有する活性フラグメントは、下記のプロトコルで例示されるように、精製されるか、または幹細胞阻害活性用の適切なバイオアッセイを組み合わせた従来の化学的プロセスによって合成される。
【0126】
本発明の1つの実施態様では、治療的に有効な量のINPROLタンパク質または治療的に有効なそれらのフラグメントは、薬学的に受容可能なキャリアと混合して用いられる。このINPROL組成物は、一般的に、非経口的注射または点滴によって投与される。皮下注射、静脈注射、または筋肉内注射の経路が、達成される治療的効果によって選択される。
【0127】
全身投与される場合、本発明における使用のための治療的組成物は、発熱物質非含有の非経口的に受容可能な水溶液の形態である。薬学的に受容可能な滅菌タンパク質溶液は、pH、等張性、安定性、キャリアタンパク質などに関する要件を有し、当該分野の技術に属する。
【0128】
INPROL療法において有用な適切な希釈剤、保存剤、安定化剤、乳化剤、アジュバント、および/またはキャリアと共に、治療的に有効な量の本発明のポリペプチド産物を含む薬学的組成物もまた、本発明により包含される。本明細書中で用いられる「治療的に有効な量」は、所定の条件および投与レジメで治療効果を提供する量を指す。このような組成物は、液剤、ゲル剤、軟膏、または凍結乾燥処方物もしくは他の乾燥処方物であり、そして種々の緩衝液内容物(例えば、Tris-HCl、酢酸、リン酸)の希釈物、pHおよびイオン強度、表面吸着防止のための添加剤(例えば、アルブミンまたはゼラチン)、界面活性剤(例えば、Tween 20、Tween 80、Pluronic F68、胆汁酸塩)、可溶化剤(例えば、グリセロール、ポリエチレングリコール)、抗酸化剤(例えば、アスコルビン酸、メタ重亜硫酸ナトリウム)、保存剤(例えば、チメロサール、ベンジルアルコール、パラベン)、増量物または張度改変剤(例えば、ラクトース、マンニトール)を含み、ポリマー(例えば、ポリエチレングリコール)のタンパク質への共有結合、金属イオンとの複合体化、あるいは物質のポリマー性化合物(例えば、ポリ乳酸、ポリグリコール酸、水性ゲルなど)の粒状調製物中または上への取り込み、またはリポソーム、ニオソーム(niosome)、マイクロエマルジョン、ミセル、単ラメラ小胞または多重ラメラ小胞、生分解性注入マイクロカプセルまたはマイクロスフェア、またはタンパク質マトリックス、赤血球影、スフェロプラスト、皮膚パッチ、または薬剤を放出またはパッケージングする他の公知の方法中への取り込みを含む。このような組成物は、INPROLの物理的状態、溶解度、安定度、インビボ放出速度、およびインビボ消失速度に影響を及ぼす。徐放性組成物または持続放出性組成物は、親油性デポ剤(例えば、脂肪酸、ワックス、オイル)中の処方物を包含する。ポリマー(例えば、ポロキサマーまたはポロキサミン)で被覆された粒状組成物および組織特異的レセプター、リガンド、または抗原に対する抗体に結合されたINPROL、あるいは組織特異的レセプターのリガンドに結合されたINPROLもまた、本発明により包含される。本発明の組成物の他の実施態様は、種々の投与経路のために、粒状形態の保護コーティング、プロテアーゼ阻害性因子、または浸透増強剤を取り込む。この投与経路としては、非経口投与、経肺投与、鼻内投与、局所(皮膚または粘膜)投与、および経口投与が挙げられる。別の実施態様では、INPROLを含有する組成物は、局所的にまたは経皮パッチを介して投与される。
【0129】
1つの実施態様では、本発明の組成物は、単位投与量形態で滅菌バイアルまたはアンプル中にパッケージされる。
【0130】
本発明は、1つ以上の付加因子(例えば、化学療法剤(例えば、5-フルオロウラシル(5FU)、シトシンアラビノシド、シクロホスファミド、シスプラチン、カルボプラチン、ドキソルビシン(doxyrubicin)、エトポシド、タキソール、アルキル化剤)、抗ウイルス剤(例えば、AZT、アシクロビル)、TNF、サイトカイン(例えば、インターロイキン)、抗増殖薬、代謝拮抗物質、およびDNA代謝に干渉する薬物)を含む組成物もまた包含する。
【0131】
そのような細胞傷害性薬剤に曝されることが予想される被験体を処置する方法または過剰増殖性幹細胞を処理する方法に含まれる投与量レジメは、担当の医師により、薬物の作用を改変する種々の要因(例えば、症状、体重、性別、患者の規定食、感染の重篤度、投与時間、および他の臨床的要因)を考慮して決定される。
【0132】
被験者が細胞傷害性薬剤または放射線に曝された後、本発明の治療方法は、1種以上のリンホカイン、コロニー刺激剤、または他のサイトカイン、造血素、インターロイキン、またはINPROLによる前処理により阻害される幹細胞(およびそれらの派生物)の成長および分裂を一般に刺激する成長因子を必要に応じて含む、幹細胞刺激量のINPROLを被験体に投与することを用いる。造血を促進するこのような治療剤としては、IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、Meg-CSF、M-CSF(CSF-1)、GM-CSF、G-CSF、またはエリスロポエチンが挙げられる。これらの薬剤の投与量は、化学療法または造血幹細胞移植後の造血再構築の促進における効能に対する臨床試験にそれらを用いたときに得られる知見によって選択される。これらの投与量は、患者の身体状態の変化および被験体が曝された化学療法剤または放射線の量およびタイプを補償するように調整される。処置を受けた患者へのINPROLの投与によって引き起こされる幹細胞の阻害の回復の進行は、従来の方法によってモニターされる。
【0133】
白血病の処置では、正常な幹細胞の細胞周期の進行を阻害するINPROLおよびIL-3またはGM-CSFのような白血病細胞の成長の刺激剤の両方を、細胞傷害性薬物処置または放射線照射の間に同時に投与することが有利である。このプロトコルにより、正常細胞および白血病細胞の細胞周期の進行状態と薬物感受性との間の最大の差違を達成し得る。
【実施例】
【0134】
実施例1:インビボ幹細胞増殖阻害アッセイ
幹細胞増殖の検出のために、細胞周期のS期にあるCFU-Sの数を3H-チミジン「自殺(suicide)」法(Beckerら、Blood 26:296-308、1965)により測定した。
【0135】
未成熟の造血前駆細胞--脾コロニー形成単位(CFU-S)--は、致死量の放射線を照射したマウスの脾臓において肉眼で見えるコロニーを形成させることにより、インビボで、造血細胞の静脈注射の8〜12日後に検出され得る(Till & McCulloch、1961)。
【0136】
標準的CFU-S増殖アッセイに関しては、3H-チミジン「自殺」の方法が通常適用される(Beckerら、1965)。この方法は、DNAの前駆体である放射標識したチミジン(3H-チミジン)をDNA合成中の細胞内に取り込むことに基づいている。試験の時点で細胞周期のS期にあるこのCFU-Sは、高い放射活性により死滅しており、そのため脾臓ではコロニーを形成できない。従って、3H-チミジンなしでインキュベーションされた細胞サンプルと3H-チミジンとともにインキュベーションされた同じ細胞との間の、注射によって形成されるCFU-Sの数の違いは、元のサンプルにあった増殖性CFU-Sのパーセントを示す。
【0137】
阻害剤試験は、阻害剤が細胞周期が進行しているCFU-S(正常マウスの骨髄では全CFU-S集団の7〜10%程の低さ)にのみ影響する限り、刺激を受けていない動物由来の骨髄幹細胞集団を用いて行うことはできない。
【0138】
CFU-Sの増殖を刺激するために、フェニルヒドラジン(PHZ)、または致死量以下(sublethal)の放射線照射を用いた(Lord、1976)。
【0139】
本発明者らは、テストステロンプロピオネート(TSP)がCFU-Sの細胞周期の進行に刺激効果を及ぼすこと(Byronら、Nature228:1204、1970)に基づいて、TSPを用いる方法を開発し、それによって試験は簡素化され、いかなる副作用も引き起こされなかった。TSPは、注射後20〜24時間以内にCFU-S増殖の刺激を誘導し、そしてその効果は少なくとも7日間見られ得た。
【0140】
阻害剤の精製の間に画分のスクリーニングに用いられた手順は以下の通りであった:
マウス:全ての試験を通じてBDF1またはCBF1マウス種を用いた。
【0141】
30〜35%のCFU-SをS期に誘導するために、ドナーマウスを、0.2ml/マウスの腹腔内注射によって、10mg/100gの用量のTSPで処理した。
【0142】
24時間後、細胞懸濁調製のため骨髄を大腿骨から取り出す。次いで、1ml当たり5×106から10×106個の細胞を異なるコントロールおよび試験画分と、37℃で3.5時間水浴中でインキュベーションする(各グループにつき試験管2本ずつ、1つはホット(標識したもの)用およびもう1つはコールド(標識していないもの)用)。
【0143】
3.5時間後、3H-チミジン(1mCi/ml、比活性18〜25Ci/mmole)を、ホット用の各試験管に細胞懸濁液1ml当たり200μlずつの容量で添加し、コールド用の試験管には何も添加しない。37℃でさらに30分間インキュベーションし続ける。
【0144】
30分間のインキュベーションの後、400μg/mlの放射能活性のないチミジンを含む10mlの冷培地(4℃)を添加することにより、死滅反応を停止させる。細胞を十分に洗浄する(3回)。
【0145】
細胞を再懸濁し、そして注射に望ましい濃度、通常マウス1匹当たり0.3〜0.5ml中2〜4×104個細胞、まで希釈する。
【0146】
1グループ当たり8〜10匹のレシピエントマウスを注射の6時間以上前に放射線照射する。
【0147】
9〜12日目にレシピエントの脾臓を集め、そしてTellesnitsky溶液中で固定して、コロニー数を目で数える。下式を用いてS期にある細胞のパーセントを計算する。
【0148】
【化7】
ここで、a--3H-チミジンを有さないCFU-Sの数
ここで、b--3H-チミジンを有するCFU-Sの数
表1に示されるINPROLの試験データは、INPROLでの処理の後に細胞周期の進行している幹細胞が3H-チミジンの作用に対して耐性になることを示している。実施例1および以下の実施例の全てについて、用語「pINPROL」は、ブタ骨髄由来の精製タンパク質をいう。同じ保護が、S期特異的細胞障害性薬物であるシトシンアラビノシドおよびヒドロキシ尿素にも見られる(データは示していない)。次いで、処理された幹細胞を放射能活性のないチミジンを含む冷培地で洗浄した場合、生き残った幹細胞はマウス脾臓内で増殖して、正常にコロニーを形成する。
【0149】
表1
骨髄細胞との4時間のインキュベーションの間に
pINPROLがCFU-S増殖に及ぼす阻害活性
【0150】
【表1】
*2×104細胞当たりのCFU-S。
【0151】
実施例2:インビトロ幹細胞増殖阻害アッセイ
以下の試験系(Lordら、The Inhibitors of Hematopoiesis 227〜239頁、1987)を用いて、INPROLの直接的効果を示した。多系統(multilineage)因子(IL-3)依存性幹細胞株であるFDCP mix A4(A4)を、コロニー刺激性IL-3の供給源として、20%ウマ血清および10%WEHI-3馴化培地を補充したIMDM培地中で維持した。
【0152】
トリチウム化チミジン取り込みアッセイを使用して増殖を測定した:A4細胞(20%ウマ血清および50%WEHI-3 CMを有する100μlの培地中の5×104個)を、5%CO2中で37℃、16時間インキュベーションした。
【0153】
pINPROLまたは粗製のBME(画分IV)を開始時点で添加した。次いで、トリチウム化チミジン((3H-Tdr)50μl中の3.7KBq、740 GBq/mmole)を各グループに添加し、さらに3時間インキュベーションした。増殖レベルを集めた細胞によって測定し、そして阻害%を下記の式を用いて計算した。
【0154】
【化8】
。
【0155】
正常骨髄抽出物またはpINPROLを段階的な用量で存在させて増殖させたFDCPmix-A4細胞によるトリチウム化チミジン(3H-Tdr)取り込みを図6に示す。pINPROLの精製組成物は出発物質より少なくとも1000倍活性が高いことがわかった。曝露時間(16時間)は、効果的な阻害についての重要な因子であり、そしてA4細胞株の幹細胞に、pINPROLの直接的な効果を及ぼしている証拠を示している。
【0156】
実施例3:インビボのINPROL注射によるCFU-S増殖の阻害:用量および効果の持続期間
インビボで注射されたINPROLの効果の研究は、INPROLがCFU-Sの細胞周期への加入を効果的にブロックし得、これによりそれらの細胞をさらなる処理の細胞障害効果から保護し、その臨床使用への可能性を示すことを示した。
【0157】
実験プロトコルには2つの目的がある:インビボで注射された場合にCFU-Sに対するINPROLの効果をチェックすること、および、細胞周期の進行している幹細胞に関してINPROL活性の有効持続期間を明確にすること。
【0158】
CFU-Sの増殖を刺激するために、上記の実施例1で述べた効果に基づいて、テストステロン-プロピオネートの注射を用いた。
【0159】
マウスBDF1をTSP(10mg/100g)と共に第0日に注射した;24時間後に、各実験グループのマウス(各グループあたり4匹)に0μg、5μg、10μg、および15μg/マウスの用量で腹腔内にpINPROL注射を1回行った。
【0160】
pINPROL注射の24時間後、マウスを屠殺し、そして細胞周期の進行しているCFU-Sのパーセントを実施例1に記載されたアッセイにより測定した。TSP注射は、約50%のCFU-Sを細胞周期に誘導したが、未処理のマウスでは7%であった。2μg/マウス程度に低い用量のpINPROLは、TSPで誘導される増殖を阻害して正常レベルまで下げ得た。
【0161】
効果評価の持続期間の間、一方のグループのマウス(1グループ当たりマウス21匹)にTSPのみを注射し、そして他方のグループにTSPとpINPROL(TSPの24時間後)の両方を注射した。CFU-Sの細胞周期の進行は、各グループから3匹ずつドナーを取り出して、記載した方法(実施例1参照)で骨髄のCFU-Sの細胞周期の状態を測定することにより、1週間、24時間毎に測定された。図7に示すデータは、TSPの有効持続期間は少なくとも7日間であるが、1回のINPROLの注射によりCFU-Sは休止期に入り、わずか48〜72時間は細胞周期に入らないようにすることができることを示している。ガンおよび白血病の化学療法に用いられる化学療法剤の大多数はインビボでの半減期は比較的短い(通常24時間より短い)ので、得られたデータによれば、INPROLの効果はシトシンアラビノシドまたはヒドロキシ尿素のような化学療法剤がインビボで活性である間は有効期間よりも長く維持される。さらに重要なことは、第1回目の処理(幹細胞に損傷を与えない)と第2回目の処理(CFU-Sに損傷を与える)との間により長い間隔(24時間より長く96時間より短い)を有する化学療法および放射線療法に対しては、化学療法剤または放射線照射の2回の適用の間の間隔にINPROLを1回注射すれば十分である。繰り返し行う数回の細胞傷害性治療または放射線照射に関しても、INPROL効果の持続期間に基づいて同じストラテジーを適用し得た。
【0162】
実施例4:5-FUによる処理の後、迅速に周期に入るように刺激された最原始造血幹細胞は、INPROLにより第2回目の5-FU被曝から保護される
薬剤5-フルオロウラシル(5-FU)は、骨髄およびリンパ球の区画(compartment)中の細胞数を劇的に減少させる。それは通常、細胞周期特異的であり、細胞周期のS期の間または前にヌクレオチドアナログをDNA内に取り込むことは細胞死となるので、増殖細胞を迅速に標的とすると考えられている。マウスの骨髄の長期生存および免疫造血再構築は、5-FUの1回の用量では影響を受けない;しかし、多能性造血幹細胞(PHSC)は最初の投薬後約3〜5日間という短い期間に第2回目の5-FUの投薬に対して攻撃されやすくなるということが示された(Harrisonら、Blood 78:1237〜1240、1991)。PHSCは、通常、5-FUの1回の投薬に対して細胞周期が遅すぎて効果的ではなく、そして第1回目の5-FU処理の結果起こる刺激により刺激されて迅速に細胞周期に入ることが説明され得る。本発明者らは、PHSCは、INPROLにより遅い細胞周期状態に戻り得、これにより2回目の5-FU処理から保護され得るということを提案した。
【0163】
これらの実験に用いられたマウスは、BDF1雄マウスであった。5-FU(Sigma)の貯蔵液を生理食塩水中で10μg/mlの濃度に調製した。各処理マウスは、実験0日目に尾部静脈を介して体重10g当たり2mgの5-FUを受けた;24時間後、マウスにpINPROL(10μg/体重100g)を腹腔内注射し、そして3日目に第2回目の5-FUの用量を注射した。生存研究は、各々30匹のマウスからなる実験グループ(pINPROL処理)およびコントロールグループにおいて、マウスの死亡をモニターすることにより行った。生存曲線を図8に示す。
【0164】
実施例5:骨髄細胞におけるINPROLでプレインキュベーションした場合とMIP1αでプレインキュベーションした場合の効果
この実験の目的は、インビトロでのマウス骨髄細胞に対するpINPROLおよびMIP-1αでプレインキュベーションした場合の阻害効果を比較することであった。
【0165】
以下の手順を用いた:
インビボ:大腿骨から骨髄を採取する48時間前に、6〜15週齢のBDF1マウスに200mg/kgの5-FUを腹腔内に注射する。
【0166】
インビトロ:単一の細胞をプールした懸濁液を計数し、そして5×106細胞を、pINPROLまたはMIP-1αとともにまたはなしで、5%ウマ血清、L-グルタミン添加IMDM培地とともに総容量2mlとして、37℃および5%CO2で4時間インキュベーションする。次いで、細胞を2回洗浄し、そして再計数した。それらを以下の最終条件下でメチルセルロースにプレートする:
0.8% メチルセルロース
25% ウマ血清
20ng/ml 組換え型マウスIL3
L-グルタミン添加
5×105細胞/ml
IMDM培地
プレートを11日間、37℃および湿度100%中5%CO2で、インキュベーションした。50細胞より多いコロニーを計数した。
【0167】
【表2】
。
【0168】
実施例6:INPROLはHPP-CFC増殖を阻害する
マウス再構築幹細胞および初期前駆体を評価するインビトロアッセイは、高増殖性潜在コロニー(HPP-PFC)アッセイである;他の関連アッセイ、例えばCFU-A、CFU-GM、CFU-EおよびCFU-GEMMは、次第に制限された前駆細胞の集団を検出する(M.Moore、Blood 177:2122-2128、1991)。この例は、pINPROLでの細胞の前処理が、それらの増殖を阻害するのに対し、MIP-1αは、これらの実験条件下では阻害しないということを示す。
【0169】
BDF1マウスを、それらの骨髄をHPP-CFC数に関してアッセイする前に、5-フルオロウラシル(200mg/kg腹腔内)で処置した。細胞を遠心分離により洗浄し、そして添加剤のない培地(コントロール)、pINPROL(25ng/ml)またはMIP-1α(200ng/ml)を含む培地のいずれかにおいて、106〜5×106/mlの密度で4時間インキュベーションした。インキュベーション後、細胞を洗浄し、そして30%FCSならびに5637細胞株およびWEHI-3B細胞株由来の組合せ馴化培地(7.5%の各馴化培地、Sharpら、1991の推奨による)を有する寒天(0.3%)にプレートした。プレート濃度は、60mmの皿に5×104細胞/mlであった。14日目にコロニー数を計数し、そして結果を以下に示す。
【0170】
【表3】
。
【0171】
これらの結果により、MIP-1αは、プレインキュベーションの期間中にのみ存在した場合には、ほとんどの未成熟な前駆体の増殖を阻害しなかった。pINPROLは、これらの条件下で増殖を効果的に阻害し、生物学的活性という点から見るとpINPROLとMIP-1αとの間には基本的な違いがあることを示している。
【0172】
実施例7:放射線誘導性骨髄形成不全からの回復に対するINPROL処理の効果
骨髄形成不全は、放射線ガン療法の一次的制限毒性(primary limiting toxicity)である。いくつかの増殖因子(例えば、G-CSF、GM-CSF、エリスロポエチン)は、放射線誘導性骨髄形成不全からの回復を促進し得るということが示されてきた。幹細胞増殖の阻害剤の使用による保護という概念は、血液学的な損傷に上手く対処することにおける異なった補足的なアプローチである。初期に開発された処置手順(実施例3、4)に従い、致死量の放射線を照射されたマウスのモデルを確立した。9Gyのコバルト60を受けたマウスは10〜14日後に死亡し始め;30日目までには死亡率は約50%となる、ということは当該分野では公知である。この致死用量を、それを各4.5Gyずつの連続する2回の適用に分けて、投薬の間は3日間の間隔を置いて、本発明者らのモデルに用いた。予備データは、そのモデルの生存曲線が、9Gyによる1回の放射線照射で知られた生存曲線と非常に近いことを示した;さらに、CFU-S増殖に関する試験は、第1回目の放射線照射の24時間後に35〜50%のCFU-Sが増殖するように誘導されることを示した。このような細胞は、第2回目の投薬の前に送達される幹細胞阻害剤により保護され得る。
【0173】
この可能性を試験するために、マウス(50匹/グループ)に0日目に4.5Gyを照射し、その24時間後に、1つのグループにはpINPROL(2μg/マウス、腹腔内)を投与し、別のコントロールグループには生理食塩水を注射した。そして第2回目の放射線の照射用量(4.5Gy)を3日目に与えた。
【0174】
図9は、pINPROLの1回の用量を投与した後の増大した生存を示す。モデルの状態は、固形腫瘍により特徴付けられるガンを含むいかなるガンの処置にも臨床的に有意義であり、このような処置は、放射線照射の2回続けた投与の間に有効用量のINPROLを送達することによりガンを有する患者に投与し、それによってガンの処置に用いられる放射線の照射線量をより多くすることができる。この様式は化学療法剤にも広げることが可能なはずである。
【0175】
実施例8:自己骨髄移植へのINPROLの使用
骨髄移植は、いくつかの白血病(CML、AML、およびその他)にとっては唯一知られた治療的な療法である。輸注のための自己BMTのエクスビボ調整により、白血病細胞の混入のない正常幹細胞の潜在的な自己供給源が提供され、そしてレシピエントの造血系を再生させて(repopulate)、積極的で効果的な療法を可能にする。
【0176】
1.INPROL効果の研究用の長期骨髄培養物L1210白血病モデルはAraCによるパージの間、正常な造血を保持する
長期骨髄培養物(LTBMC)をToksozら(Blood 55;931-936、1980)に従って確立し、そして白血病細胞株L1210を2週間LTBMCと共培養した。正常前駆細胞および白血病前駆細胞の同時増殖は、これらのLTBMC/L1210の混合培養物内で起こり、それは白血病患者の骨髄内と同様の状況である。正常なコロニー形成単位CFUと白血病CFUとの間の区別は、WEHI-3(マウスIL-3産生細胞株)由来の馴化培地の存在下または非存在下での寒天コロニーとしてそれらを増殖させることにより可能であった。正常細胞は、IL-3非存在下で、アポトーシスを受けるのに対し、白血病細胞はIL-3非存在下でコロニーを形成し得る。LTBMC-L1210組成物由来の懸濁細胞は、プレートされた50,000細胞当たり、IL-3存在下で約150個のコロニー(正常造血クローン)を生じ、IL-3なしで増殖した場合は70コロニー(白血病クローン)を生じた。
【0177】
パージ手順は、以下の通りであった:0日目に、全ての懸濁細胞および培地(10ml/フラスコ)をLTBMC-L1210の入っているフラスコから取り出し、そして200μgのシトシンアラビノシド(AraC)を含む2mlの培地で置き換えた(Tsyrlovaら、Leukemia:Advances in Biology and Therapy、第35巻、1988);20時間のインキュベーション後、フラスコを洗浄し、そして2mlの新鮮な培地のみ(コントロールグループ)または25ng/mlのpINPROLを含む培地で4時間置き換えた。このプレインキュベーションの後、細胞を100μg/フラスコのAraCとともに、37℃で3時間再びインキュベーションした。各グループは4本のフラスコを含んだ。LTBMC-L1210培養物を3回洗浄し、そして新鮮なLTBC培地で置き換えた;それらを再生研究のため前述のように3〜4週間維持した。
【0178】
データを図10に示す。AraCのみで処理されたコントロール培養物には細胞の増殖が見られなかったが、INPROLで保護されたフラスコ内では、接着層由来の前駆細胞の増殖のため、造血再生がさらにより急速に起こった。さらに、実験グループ由来の細胞は、寒天にプレートされた場合、IL-3存在下では50,000細胞当たりわずか約100 CFUしか増殖しなかった;少なくとも4週間の間には白血病細胞の増殖は観察されなかった。従って、エクスビボでINPROLと組み合わせて、有効量のAraCにより処理された骨髄は、ガン細胞はパージされ得るが、幹細胞は保護される。この様式は他の形態の化学療法または放射線処置に拡げることが可能なはずである。
【0179】
2.骨髄再生能(MRA)および30日間放射線防御は、インビトロのINPROL処理により増大する
MRAは、致死量の放射線を照射されたマウスの骨髄を再生する細胞の能力であり、30日間の放射線防御を付与する能力を伴っていて、骨髄抑制動物を救う潜在能力の直接的なインビボ測定である(Visserら、Blood Cells 14:369〜384、1988)。
【0180】
放射線防御研究に関して、BDF1マウスは9.5Gyで放射線照射され、テストステロンで刺激されたドナー由来の骨髄の移植により回復した。レシピエントの1グループは培地で(コントロール−グループA)、そして別のグループ(グループB)は25ng/mlのpINPROLで、4時間プレインキュベーションされた骨髄細胞により回復した。両グループの細胞を洗浄し、そしてマウス1匹当たり30,000個の細胞を放射線照射された動物に移植した。生存データを示す(図11)。3つの実験の合計を、コントロールを100%に正規化して示す。30日目までに、pINPROLによるインキュベーションにより、マウスの生存をコントロールグループの36.5%から61.8%まで増加させた。
【0181】
INPROLによるプレインキュベーションによって誘導されるMRAの増加は、放射線防御を改善するメカニズムの1つであり得る。この仮説を試験するため、Visserら(前出)に従ってMRAを測定した。簡潔に記載すると、ドナーBDF1マウスをテストステロンで前処理し、それらの骨髄を培地またはpINPROLで4時間プレインキュベーションし、そして放射線照射された動物に注射した。13日目に、レシピエントの大腿骨由来の骨髄細胞を、20%ウマ血清、および10%WEHI-CMの存在下で、3つの異なる濃度(0.01、0.05、0.1当量の大腿骨)で寒天に接種した。7日目のコロニー数は、その時点のレシピエントの骨髄中のコロニー形成細胞がドナーの未成熟幹細胞の前駆細胞である限り、MRAを表す。
【0182】
図12に見られ得るように、INPROLでプレインキュベーションされた細胞集団のMRAは、コントロールグループ(B)の場合より多い。
【0183】
実施例9:INPROLの幹細胞への抗過剰増殖効果は、それらの分化の異常を変化させ得る
CFU-Sの過剰増殖は、細胞障害性薬物または放射線照射からの回復中に見られるだけでなく、正常な加齢の結果としても見られ、そして骨髄異形成症候群(MDS)の主要な特徴であると考えられている。それは、顆粒球経路に沿った分化が減少する一方で、赤血球分化が広まるような分化障害を伴う。
【0184】
骨髄細胞を25ng/mlのpINPROLまたは培地(コントロール)とともに37℃で4時間インキュベーションし、洗浄し、次いで20%ウマ血清、2U/mlエリスロポエチン、および10%WEHI-CMを有する寒天にプレートした。BFU-EおよびGM-CFUコロニーの数を7日目に計数した。表4に示すデータは、3つの実験をまとめたものであり、各グループから各時点で4匹を取り出し;4枚の皿にプレートした。
【0185】
表4から明らかなように、無傷の幼動物(8〜12週齢のBDF1)由来の正常骨髄(NBM)をINPROLとともにインキュベーションすることにより、異なるタイプのコロニーの数または比率は変化しなかった。テストステロンプロピオネート(TSP)で前処理されたBDF1ドナーは、以前に観察したように(実施例1、3、4)、CFU-S増殖における同じ増加、赤血球前駆細胞数(BFU-Eコロニー)におけるわずかな増加、およびGM-CFUにおける減少を示し、それはINPROLとのインキュベーションにより完全に妨げられた。さらに、異常に高いレベルのCFU-S増殖は、細胞周期のS期に10%のCFU-Sに戻った。CFU-Sの過剰増殖は、例えばBalb/cマウス(表4)のようにウイルス性白血病誘発を受けやすいある種のマウス系統の特徴であることが知られており、そしてより高齢の動物においても観察され得る(表4)。TSP処理したBDF1マウスに見られた分化方向の決定付けられた(committed)前駆細胞と同じ再分布が、Balb/cおよびより高齢(23〜25ヶ月齢)のBDF1において観察され、それは一般に異常に高いレベルのCFU-Sの増殖を有する。CFU-Sの増殖および分化の両方の修正は、INPROLとのインキュベーションによって誘導された。臨床的により関連したことに、研究によって、INPROLのインビボ注射(2μg/マウス)がCFU-Sの増殖ならびに赤血球(BFU-E)とGM-コロニーとの比率の両方に影響することが示された(表4)。
【0186】
表4
分化方向の決定付けられた前駆細胞BFU-EおよびCFU-GMへの
CFU-Sの分化に対するINPROLの効果
【0187】
【表4】
。
【0188】
実施例10:INPROLの免疫刺激活性
増殖CFU-Sを高い比率で含む骨髄細胞とINPROLとのインキュベーションは、CFU-Sの周期を変えるだけでなく、それらの分化を、優勢な赤血球の分化から顆粒球およびリンパ球の前駆細胞へと切り換える。INPROLのこの特性は、細胞障害性の化学療法または放射線療法の免疫抑制的な副作用および過剰増殖幹細胞障害および加齢に伴う免疫抑制のために重要である。
【0189】
本実施例は、Wittlock & Witte(Ann.Rev.Immun.3:213〜35、1985)によって確立されたリンパ球長期培養(LLTC)由来の未成熟前駆体の前B前駆細胞への分化に対するINPROLの直接的な効果を示し、これはIL-7を含むメチルセルロース中のコロニー形成によって測定される。
【0190】
LLTCを、記載されたように確立し、そして新鮮なLLTC培地(Terry Fox Labs.,Vancouver,Canada)を1週間に2回与えた。非接着細胞を1週間に1度採取し、因子を洗い流し、そして25ng/mlのpINPROL、またはコントロールとして培地のみと4時間インキュベーションした。インキュベーションの後、細胞を洗浄し、そして105細胞/mlの濃度で、30%FCSおよび10ng/mlのIL-7を含むメチルセルロース中にプレートした。3週間のデータを図13に示す。大きなpre-Bコロニーの数はコントロール中で変化し、時間とともに増加したが、INPROLとのプレインキュベーションは、常にコロニーの増殖をコントロールレベルの4〜8倍に刺激した。このことは、免疫不全状態を修正し、そして例えばワクチン投与に対して所望の免疫応答を増加させるために有用であるINPROLの免疫刺激特性を示す。
【0191】
実施例11:INPROLは幹細胞の再生能を改善する--CMLの患者由来の長期培養開始細胞
慢性骨髄性白血病(CML)は、造血幹細胞の致死的な悪性疾患である。慢性期にあるCMLを単一薬剤による化学療法、組み合わせ化学療法、脾摘出、または脾臓への放射線照射を用いて処置することは、臨床的徴候および症状を制御し得るが、生存を有意に伸ばすことはない。CMLが慢性状態から促進状態に進むと、標準的な療法は効果的でない。現在では、骨髄移植(BMT)がCMLに対する唯一の既知の治療的療法である。血縁でないドナーのBMTを用いる治療は、組織不適合性の問題により困難である。適格なCML患者の40%未満にしか適切に適合する血縁のドナーがいない;従って自己移植が好ましい。長期培養(LTC)において増殖するPh陽性患者由来の非白血病(Ph-陰性)骨髄前駆細胞を選択する能力とともに、輸注のための自己BMTのエクスビボ調整は、正常幹細胞の自己供給源がCMLの積極的で効果的な治療を行い得る可能性を示唆する。
【0192】
BMTの状況においては、造血幹細胞は、長期間にわたって成熟した血液細胞を生成する能力を有する細胞として定義され得る。本発明者らは、C.Eaves & A.Eavesによって開発されたヒトLTC系を、幹細胞数の定量用およびそれらの治療的使用のための操作手段の両方で用いた。これは、予め確立され、放射線照射されたヒト骨髄接着層上に細胞を接種することを包含し;次いでこれらの培養物を5週間維持する。エンドポイント(end point)は、この期間の終わりに採取された培養物の総クローン原性細胞内容物(接着および非接着)である。このような条件下で生じたクローン原性細胞は、最初に添加された前駆細胞(長期培養開始細胞(LTC-IC))の数と直線関係にある;個々のヒトLTC-ICから生じた平均は、1個のLTC-ICにつき4クローン原性前駆細胞である。CMLの患者由来の骨髄が同様の条件下に置かれた場合、白血病性(Ph陽性)クローン原性細胞は迅速に衰退することが以前に示されていた。CML患者の残った正常LTC-ICの定量を用いることにより、培養自己移植片の移植により支持される集中治療から利益を得やすい細胞を選択することが可能である(Phillipsら、Bone Marrow Transplantation 8:477〜487、1991)。
【0193】
以下の手順を用いて、CMLの患者の末梢血から確立された骨髄移植細胞中のクローン原性細胞(LTC-IC)の数に対するINPROLの効果を試験した。
【0194】
予め放射線照射した間質上の長期培養と同様に培養を開始した。健常ドナーの末梢血をコントロールとして用いた。CML患者由来の末梢血細胞を、pINPROL(25ng/ml)とともに、またはなしで4時間プレインキュベーションし、洗浄し、そしてLTC-IC系に5週間置き、LTC-ICのコントロール数を測定した。実験のために、別の平行培養を10日間確立した。10日間増殖させた培養物からの接着細胞および非接着細胞の混合物を、pINPROLとともに、またはなしでプレインキュベーションし、そしてさらに8週間予め確立したフィーダー上に置いた。各実験培養物由来のLTC-ICの数は、接着細胞および非接着細胞の両方を適当な増殖因子(Terry Fox Laboratories、Vancouver、Canada)を有するメチルセルロース中にプレートし、そして得られた全コロニー形成細胞の数を計数することにより概算した。この手順を用いて得られたLTC-IC値は、下記の式を用いた全クローン原性細胞(CFC)内容物の評価に由来した:
#LTC-IC=#CFC/4。
【0195】
図14に示したデータは、健常ドナーの骨髄から始めた最初の10日間の培養中ではLTC-ICに損失がなく、そして投入したLTC-IC数の約30%が5週間たってもなお培養中に存在していたことを示す。CML患者のLTC-IC数は、10日間の間に約8%にまで劇的に低減し、そしてさらなるインキュベーション中、元には戻らなかったが、INPROLとともに細胞をプレインキュベーションすると、最初の数の30%までLTC-ICレベルが増加し、そしてそれは8週間維持された。
【0196】
これらの予備データによって予想されるINPROLの臨床関連の適用は、新鮮なまたは培養されたの骨髄移植物の正常幹細胞内容物を選択的に改善するためのストラテジー、インビボの残存正常幹細胞の補充を高めるためのストラテジー、さらに患者へのさらなる移植のために新しい遺伝物質をヒト骨髄幹細胞に転移させるためのプロトコルにおけるそれらの使用を包含する。
【0197】
実施例12A:骨髄調製物由来の幹細胞増殖の免疫活性阻害剤の単離方法
骨髄はブタの肋骨から単離した。ブタの胴体から肋骨を分離して筋繊維および脂肪を除き、細片に刻んで骨髄をBiophyzpribor社製の水圧プレスによって抽出した。骨髄細胞を、遠心機K-70中で2,000rpmで20分間遠心分離することにより分離する。次いで、引き続き抽出上清をAmicon USAメンブレンXM-100、PM30、PM-50により連続的に限外濾過にかける。電気泳動による分析によると、この生成物の主要な成分はアルブミンである(図1参照)。
【0198】
生物化学的精製
画分の骨髄抽出物およびタンパク質成分を、精製の各段階で、0.1%ドデシル硫酸ナトリウムを含む10%ポリアクリルアミドゲル電気泳動により分析した。7%までのドデシル硫酸ナトリウムおよび0.5〜1%までのメルカプトエタノールをサンプルに添加し、そしてそのサンプルをゲルにロードする前に70℃で5分間インキュベーションした。
【0199】
電気泳動は20Ycmのゲルで5時間行った。次いで、ゲルを、エタノール:水:酢酸が5:5:1の混合物中の0.25%クーマシーCBBC250で20℃で1時間染色し、そして7%酢酸を数回交換して洗浄した。生成物の活性を造血幹細胞(CFU-S)増殖の阻害方法により評価した。この方法を以下に詳細に述べる。
【0200】
ステージ1.硫安沈澱による精製
活性を、表5の結果に基づいて選択された飽和度40〜80%で25%の硫安を用いて沈澱させることにより精製した。
【0201】
【表5】
。
【0202】
精製の各工程の後の試験に用いられる調製物の量は、精製レベルによって決定し、始めの生成物の2×10-2mgの用量と同じであった。活性を下式により決定した:
変化%=Sa%−Sb%
ここで、Sa%は、コントロールのS%であり、
Sb%は、試験画分とのインキュベーション後のS%である。
【0203】
画分は、各活性試験の前、および次の各精製工程の前に硫安濃度を20倍低くするために脱塩した。
【0204】
ステージ2.ステージ1由来の不純物のある阻害剤を、脱塩後にイオン交換クロマトグラフィー(この場合DEAE23セルロース)を利用して分画し、次いで酢酸ナトリウム緩衝液(pH6.0)の勾配を用いて溶出する。
【0205】
阻害剤の活性画分は3〜5mMの間に溶出する。
【0206】
カラム容積は1mlであり、溶出速度は4ml/時間であった。検出はクロマトグラフMillicromeにより230および280nmで行った。最も高い活性を示した画分1(図2参照)を単離し、5mM酢酸ナトリウム緩衝液中に溶出した(表6参照)。
【0207】
【表6】
。
【0208】
電気泳動のデータは、主なタンパク質混入物であるアルブミン(図3参照)がこの画分から除かれることを示し、さらに4回の精製に続く。
【0209】
ステージ3.ステージ2由来の部分精製された阻害剤を直接G-75Sephadexカラムにかける。
【0210】
カラム体積は20ml(20×1)であり、溶出速度は2ml/時間である。溶出緩衝液は50mM NaCl、10mM Tris-HCl、pH7.5である。検出はクロマトグラフMillichromeにより230および280nmで行った。最も高い活性を示した画分5を単離した。
【0211】
【表7】
。
【0212】
ステージ4.Pro-RECカラムを利用した逆相クロマトグラフィー(Pharmacia FPLC System)をUltrasfera matrix 上で行った。タンパク質は0.1%トリフルオロ酢酸のアセトニトリル勾配を用いて溶出した。
【0213】
分子量16〜17kDの生成物の均質度は、アクリルアミド/ドデシル硫酸ナトリウムゲルの分析において示されるように、90%に等しい(図6参照)。この結果を図4に示す。画分5についての活性を測定する。生成物の最終回収率は5%である。結果として、精製後の分子量16kDのタンパク質の総量は始めの生成物の650ng/mlである。精製プロセスの間、生成物は70℃の加熱インキュベーションを数分受けたが、生物学的活性の損失は検出されなかった。
【0214】
実施例12B:より大量のINPROLを単離するための代替方法
最初の単離
新鮮なブタの胴体から採取した肋骨から筋繊維および脂肪を除き、次いでこれを刻んで細かくし、リン酸緩化衝生理食塩水に1:1(重量/容量)の比率で浸漬する。得られた混合物を水圧プレスで粉砕し、固体骨物質から骨髄を分離する。
【0215】
骨髄細胞懸濁液を回収し、そして4層のチーズ・クロス(cheese-cloth)を通して固形粒子を濾過する。濾液を56℃で40分間インキュベーションし、次いで氷浴で4℃まで冷却する。得られた沈澱物を10,000gで30分間4℃で遠心分離することにより除去し、廃棄する。
【0216】
清澄化した上清を、1容量%の濃塩酸を含む10容量の撹拌された氷冷アセトンに30分間滴下して添加する。得られた混合物を4℃に16時間保存し、完全な沈澱物を形成させる。次いで、沈澱物を20,000gで30分間4℃で遠心分離することによりペレット化する。このペレットを冷アセトンにより洗浄し、そして乾燥する。
【0217】
HPLC精製
ペレットを、5%アセトニトリル(MeCN)および0.1%トリフルオロ酢酸(TFA)を含むHPLC溶出緩衝液Aに溶かし、最終タンパク質濃度を8〜10mg/mlとする。この溶液(0.5〜0.6ml)を、PolisilODS-300(10mcm)が充填され、そして同じ緩衝液Aにより平衡化された250×4.6mmのHPLCカラムにロードする。
【0218】
溶出は緩衝液A中の緩衝液B(90%MeCN、0.1%TFA)の勾配により、流速1ml/分で、以下のプログラムに従って行う。
【0219】
時間、分 Bの%
0 0
4 0
5 25
25 90
。
【0220】
100%の緩衝液Bを5分間流すさらなる工程を用いて再平衡化の前にカラムを洗浄する。次いで、カラムを開始時の状態に戻すため再び平衡化し、そして次の部分のタンパク質溶液をカラムにロードし得る。代表的なクロマトグラムを表5に示す。
【0221】
分離の間、カラム溶出物をタンパク質ピークの検出のため280nmでモニターする。タンパク質物質を含む画分を回収し、分離したピークをプールして、30℃でロータリーエバポレーターにかけて乾燥させる。得られた残渣を蒸留水に溶解し、生物活性試験およびSDS-PAGE(14%ゲル、還元条件)によりアッセイする。活性物質を含むピークは、緩衝液Bが70%と80%との間に溶出され、SDS-PAGEによってアッセイされたように、16kDの主要なタンパク質バンドおよび速く移動するタンパク質の痕跡を含む。
【0222】
第2の主要なHPLCピークのみを回収することにより得られた物質の分析を、図15(AおよびC)に示す。両方のピークを含む物質(例えば、表5)を、本明細書中ではpINPROL調製物1といい、そして第2のピークのみからなる物質をpINPROL調製物2という。500μgのこの活性な、精製pINPROL調製物2をC4逆相カラム(Vydac)にロードし、そして0.1%トリフルオロ酢酸中の5〜95%アセトニトリルの直線勾配を用いて溶出した。その物質は53%アセトニトリルで単一ピークとして溶出した(図15A)。しかし、250μgのMIP-1α(R & D Systems)を同じ条件で流した場合、それは43.9%アセトニトリルで溶出した(図15B−14%アセトニトリルより前に溶出する初期のピークは人工的で検出器内の気泡によることを銘記されたい)。従って、天然に存在するINPROLは、実質的には、これらのような条件下ではMIP-1αより疎水性である。TGF-βはこれらの条件下ではpINPROLで観察される濃度より低い濃度で溶出することが知られている(Miyazonoら、J.Biol.Chem.263:6407〜15、1988)。
【0223】
溶出したpINPROL物質のゲルを図15Cに示す。レーン1は粗物質、レーン2は、分子量マーカー、そしてレーン3は精製物質である。2本の主要なバンドがあり、1本は約14kD、そしてもう1本は約35kDである。この35kDのバンドは14kDのバンドのマルチマー形態であると考えられる。
【0224】
実施例13A:活性INPROL調製物は、ヘモグロビンβ鎖を含む
pINPROLを、図5に示すように調製した(すなわち、pINPROL調製物1(実施例12Bを参照のこと))。物質をSDS-PAGEにかけ、そして標準的な技法を用いてニトロセルロースに移した。物質を、標準的な技術を用いて120AオンラインPTH-AA分析器を有するABI 477Aタンパク質シーケンサーを使用して、N末端配列分析に供した。以下のN末端配列が得られた:
VHLSAEEKEAVLGLWGKVNVDEV....
タンパク質データベースのコンピューターサーチは、この配列がブタヘモグロビンのβ鎖のN末端配列と同一性を有することを明らかにする(図16Cを参照のこと)。
【0225】
実施例13B:活性INPROL調製物はヘモグロビンα鎖を含む
図15Cにおいて示されるように、図5に示される第2の主要なピークを回収することにより精製したタンパク質(すなわち、pINPROL調製物2)は、分子量約15Kおよび30Kに対応する2つの主要なバンド、ならびに幾つかのマイナーバンドを生じた。SDS-PAGEゲルを、標準的な技術を用いてニトロセルロースに移し、そして個々のバンドを切り出して、実施例13Aに記載のように、N末端配列決定分析に供した。15kDのバンドについては以下のN末端配列が得られた:
VLSAADKANVKAAWGKVGGQ......
30kDのバンドを、制限タンパク質分解消化に供し、そして以下の内部の配列を得た:
**FPHFNLSHGSDQVK....
。
【0226】
第1の配列は、ブタヘモグロビンα鎖のN末端配列と同一性を示し、一方第2の配列はブタヘモグロビンα鎖の残基43〜56と同一性を示した(ヒト、マウス、およびブタのαヘモグロビン鎖およびβヘモグロビン鎖の配列比較については図16Cを参照のこと)。これらのバンドおよびいくつかのマイナーバンドの反復配列決定により、一貫して、αグロビン配列の部分を得た。従って、図15Cにおいて観察された種々のバンドは、ブタヘモグロビンα鎖のフラグメントまたは凝集体のいずれかを示す。
【0227】
実施例13C:ブタINPROLのさらなる特徴付け
pINPROLとブタヘモグロビンとをさらに比較するために、2回結晶化したブタヘモグロビンをSigma Chemical Companyから入手し、そして実施例12Bにおいて図15について記載したように、逆相HPLCに供した。図17において示され得るように、完全なヘモグロビンのHPLCクロマトグラムは、pINPROL調製物1について示されるクロマトグラムに類似する。さらに、直接比較において、図17Aにおいて示されるpINPROL調製物2(すなわち、図5の第2のピークに由来する)は、ブタヘモグロビンの第2の2つのピークの調製物(図17B)に重複することが見られ、主要ピークについての保持時間はそれぞれ52.51分および52.25分であった。ヘムが、この場合、49.153分で、ヘモグロビンにおける第1の主要ピークと共移動(co-migrate)することに注意すべきである;それゆえ、ヘムはpINPROL調製物1の成分であるが、調製物2の成分ではない。このことは、ヘムの存在について診断する波長575nmでのこのpINPROL調製物の吸光の欠失により確認される。
【0228】
ブタヘモグロビンα鎖およびβ鎖の推定分子量は、それぞれ、15038ダルトンおよび16034ダルトンである。図18のSDS-PAGEクロマトグラムにおいて示され得るように、第1の2つのピークはより高い分子量鎖から構成され、および第2の2つのピークはより低い分子量鎖からなる。したがって、第1の2つのピークはヘモグロビンβ鎖を示すようであり、および第2の2つのピークはヘモグロビンα鎖を示すようであった。
【0229】
ブタヘモグロビンのさらなる分離を、ゆるやかな溶出勾配を用いて行った(図21)。これらのピークのN末端分析は、第1のピークがブタα鎖であり、および第2のピークがブタβ鎖であることを示した。バイオアッセイ結果により、単離されたヘモグロビン鎖の両方が生物学的に活性であることが確認される(例えば、実施例14および15を参照のこと)。
【0230】
pINPROL調製物2およびヘモグロビンβ鎖のさらなる比較のために、2次元電気泳動を行った(図19)。1次元について、等電点を、9600ボルト時間について2%pH4〜8アンホラインを用いて、ガラスチューブ中で行った。トロポミオシン(分子量33kD、pI5.2)を、内部標準として用いた;その位置を最終的な2Dゲル上で矢印で記した。チューブゲルを緩衝液中で平衡化させ、そして12.5%アクリルアミドスラブゲルの上部のスタッキングゲルの上部にシールした。SDSスラブゲル電気泳動を4時間、12.5mA/ゲルで行った。ゲルを銀染色し、そして乾燥させた。
【0231】
2D電気泳動のパターンの比較により、HPLC精製ヘモグロビンβ鎖とpINPROL調製物2との間で異なるスポットは、1つまたは2つの少数のスポットのみであることを示した。抗ブタヘモグロビン抗体、および1Dまたは2D電気泳動のいずれかを用いたウエスタン分析により、調製物中にβヘモグロビンが存在することを確認する。したがって、活性pINPROL調製物2(実施例12Bにしたがって調製される)は、実質的にブタヘモグロビンβ鎖である。
【0232】
実施例14:ヘモグロビンα鎖、ヘモグロビンβ鎖、または完全なヘモグロビンは、幹細胞阻害活性を示す
ヘモグロビンβ鎖がINPROL活性を有することを確認するために、実施例1に記載したテストステロン処理したマウス由来の骨髄を用いた自殺アッセイを、実施例12Bにおけるように精製した物質を使用して実施例1に記載の方法論を用いて行った。表8に示すように、正常なマウスの骨髄細胞の15%が、テストステロン処理動物の36%とは対照的に死滅された。予想されたように、このことはテストロン処理が、細胞周期における細胞のパーセントを増加させる(したがって、細胞がより死滅されやすくする、例えば、実施例1)ことを示している。より際だった対照において、40ng/mlでpINPROLまたは精製ヘモグロビンβ鎖のいずれかとともにインキュベーションしたテストステロン処理した動物由来の細胞は、細胞周期における細胞のパーセントが、36%から0%および7%に、それぞれ、劇的に減少することを示した。200ngのより高い用量は、両方のタンパク質についてほとんど効果がなかった。ポジティブコントロールとして、以前に特徴付けられた幹細胞阻害剤MIP-1αは、13%に細胞周期の進行を減少させた。
【0233】
同様のアッセイを、CFU-Sの代わりにCFU-MIXの細胞周期の進行状態を用いて、インビトロで行い得る。高用量のトリチウム化されたチミジン(B.I.Lord、Haemopoiesis-A Practical Approach,N.G.TestaおよびG.Molineux(編)、IRL出版、1993;Pragnellら、Culture of Hematopoietic Cells,R.I.Freshney,I.B.PragnellおよびM.G.Freshney(編)、Wiley Liss 1994)の代わりに、シトシンアラビノシド(Ara C,30mg/ml)を細胞周期特異的毒性薬剤として用いた以外は、CFU-Sアッセイについて上述したようにアッセイを行い、そして高い内在性細胞周期速度を有するマウス系統(Balb/c)をテストステロンで処理したBDF1マウスの代わりに用いる。表9において示すように、高度に精製されたブタβ鎖、または高度に精製されたブタα鎖は、このアッセイにおいて両方とも活性であった。このアッセイにおいて注目すべき点は、pINPROLで処理した細胞についての細胞周期の進行レベルが、場合によってはネガティブな数を有し、このことは、非処理のプールよりもAra Cで処理したプールにおいてより多くのコロニーが存在することを示している。
【0234】
実施例2に記載したように、pINPROLは、トリチウム化したチミジン取り込みアッセイにおいて、マウス幹細胞FDCP-MIX株の増殖を阻害する。図20は精製ヘモグロビンα鎖およびβ鎖が、2ng/ml未満で示される阻害を伴いつつ、このアッセイにおいて両方とも活性であることを示している。
【0235】
前述は、ブタヘモグロビンのβ鎖がINPROL活性を示すという証拠を提供する。他のデータ(例えば、表9、図20)は、単離されたα鎖、ならびに完全なヘモグロビンがまた、幹細胞阻害剤として活性であることを示す。活性な調製物にはまた、α鎖およびβ鎖の混合物が含まれる(例えば、図5)。
【0236】
単離されたαグロビン鎖および/または単離されたβグロビン鎖が活性であるという観察は、本実施例で記載した活性が、完全な3次元ヘモグロビン構造を必要としないことを示している。α鎖およびβ鎖のフラグメントはまた、幹細胞阻害剤として活性である。
【0237】
【表8】
1NBM=正常な骨髄
2TPBM=テストステロン-処理マウス由来の骨髄
3Hbg=C4逆相精製ブタヘモグロビンβ鎖(2回結晶化したブタヘモグロビン由来)
【0238】
【表9】
1コントロール-Balb/cマウス由来の骨髄
2100ng/ml(図21のように精製)。
【0239】
実施例15:精製INPROL、精製ブタαヘモグロビン、または精製ブタβヘモグロビンは、インビボで活性である
精製したブタヘモグロビン鎖のインビボでの作用能力を試験するために、BDF1マウスに実施例1に記載のようにテストステロンプロピオネートを用いて注射した。24時間後、マウスに、pINPROL、ブタヘモグロビンα鎖(図21のように末梢血赤血球から精製した)、ブタヘモグロビンβ鎖(図21のように末梢赤血球から精製した)のいずれか500ng、あるいは等容量のキャリアを静脈内投与した。48時間後、各マウス由来の骨髄を採取し、そしてCFU-MIXアッセイを実施例14に記載のように行った。表10において示されるように、pINPROL、ブタα鎖およびブタβ鎖は、細胞周期におけるCFU-MIXのパーセントを基底レベルまで減少させ、全てインビボで活性であった。
【0240】
【表10】
1コントロール-テストステロン処理BDF1マウス由来の骨髄
2100ng/ml
3基底-未処理のBDF1マウス由来の骨髄。
【0241】
実施例16:精製ヒトヘモグロビンα鎖、ビオチン化ヒトヘモグロビンα鎖、ビオチン化ヒトヘモグロビンβ鎖、ヒトヘモグロビンγ鎖、およびヒトヘモグロビンδ鎖は、全てインビトロで幹細胞阻害活性を示す
ヒトヘモグロビンを、Sigma Chemical Corporationから入手するか、あるいは成人ヒト末梢血または臍帯血から標準的な手段により単離した。個々の鎖を、ブタα鎖およびβ鎖について上述したのと同様の様式で逆相HPLCにより単離した(B.MasalaおよびL.Manca,Methods in Enzymology、第231巻、21〜44頁、1994を参照のこと)。精製α、β、γ、およびδ鎖を得た。ビオチン化したα鎖およびβ鎖について、1mgの成人ヒトヘモグロビンを、37μgのNHS LC ビオチン(Pierce)を用いて処理し、そして上記のように逆相クロマトグラフィーにより鎖を分離した。
【0242】
表11、12、および13において示すように、精製ヒトαヘモグロビン鎖、ビオチン化ヒトαヘモグロビン鎖、ビオチン化ヒトβヘモグロビン鎖、ヒトγ鎖ヘモグロビン鎖、およびヒトδヘモグロビン鎖は、全て、CFU-MIX細胞周期の進行アッセイにおいて活性である。
【0243】
【表11】
1コントロール-Balb/cマウス由来の骨髄
2100ng/ml
【0244】
【表12】
1コントロール-Balb/cマウス由来の骨髄
2100ng/ml
【0245】
【表13】
1コントロール-Balb/cマウス由来の骨髄
2100ng/ml。
【0246】
実施例17:精製ヒトα鎖、α-βダイマー、またはヘモグロビンは、インビボで活性である
精製ヒトα鎖、α-βダイマー、またはヘモグロビンを実施例15に記載のようにインビボアッセイで試験した。表14に示したように、これらのそれぞれは濃度500ng/マウスで活性であった。
【0247】
【表14】
1コントロール-テストステロン処理BDF1マウス由来骨髄。
【0248】
実施例18:ブタINPROLは、インビトロでヒト単核細胞またはCD34+臍帯血細胞において活性である
ブタ骨髄由来の精製INPROLが、ヒト前駆細胞における細胞周期の進行に影響する能力を調べるために、臍帯血細胞を得た。Ficoll上で分離した後に得られた総単核細胞画分または抗CD34親和性カラム(CellPro Inc.)上での分画後に得られたCD34+画分のいずれかを使用した。初期の幹細胞が細胞周期にあることを確認するために細胞を、インターロイキン3(IL-3)および幹細胞因子(SCF)(各100ng/ml)の存在下でインビトロにおいて48時間インキュベーションした。このプレインキュベーション後、細胞周期の進行アッセイを、CFU-GEMM(CFU-MIXの代わりに)をプレート後18日目に計数した以外は、実施例14に記載のようにマウスについて行った。表15において示すように、ブタINPROLは、大多数の単核細胞またはCD34+画分のいずれかにおいて、CFU-GEMMの細胞周期の進行を阻害した。
【0249】
【表15】
。
【0250】
実施例19:精製ヒトαヘモグロビンは、ヒトCFU-GEMMにおいて活性である
ヒト臍帯血単核細胞を得、そしてIL-3およびSCF中でインキュベーションし、そして実施例18に記載のように細胞周期の進行アッセイに使用した。表16に示すように、骨髄から精製したブタINPROLおよび末梢血から精製したヒトαヘモグロビンは、このアッセイにおいて活性であった。
【0251】
【表16】
。
【0252】
実施例20:ヒトαヘモグロビンおよびヒトβヘモグロビン配列から得られたペプチドは活性である
活性なペプチド配列を同定するために、ミオグロビンの3次元構造(本アッセイにおいて不活性である)を、コンピューターモデリングプログラムを用いて成人ヒトヘモグロビンに存在するα鎖のネイティブな3次元構造に重ね合わせた。2つのペプチド(アミノ酸43〜55および64〜82で表される、これらは3次元空間においてミオグロビンと構造的に異なる領域である)を、CFU-MIX細胞周期の進行アッセイにおいて活性を有するとして同定した。ネイティブなα鎖において見出されるループをより近接に近づけるために、ペプチド43〜55の環状誘導体(c43〜55)(ジスルフィド結合を利用)をまた合成し、そして活性であることが見出された。
【0253】
これらのペプチドの配列は以下のようである:
【0254】
【化9】
2つのヘモルフィン配列、ヘモルフィン10(β鎖配列のアミノ酸32〜41)およびヘモルフィン7(アミノ酸33〜40)を試験し、そして活性であることを見出した。
【0255】
配列は以下のようである:
【0256】
【化10】
。
【0257】
これらの配列の活性を試験するために、CFU-MIX細胞周期の進行アッセイを実施例14に記載のように行った。表17〜19に示すように、これらのペプチドは全てこのアッセイにおいて活性である。
【0258】
【表17】
【0259】
【表18】
1全てのペプチドは100ng/mlで試験された。
【0260】
【表19】
1100ng/mlで試験された。
【0261】
実施例21:蟻酸切断によりヒトαヘモグロビンから得られたペプチドフラグメントは活性である
ヒトαヘモグロビン鎖は、アミノ酸94位と95位(Asp-Pro)との間に蟻酸切断部位を有する。切断を、精製ヒトα鎖(実施例16と同様)を70%蟻酸中1mg/mlの濃度で37℃で72時間インキュベートすることにより得た。1-94フラグメントを未切断α鎖から精製し、そして95-141フラグメントを実施例16のような逆相HPLCにより精製した;その後、分画をSDS-PAGEを用いて行った(実施例22と同様)。精製1-94タンパク質フラグメントの同一性を、エレクトロスプレーイオン化質量分析により確認した。
【0262】
このフラグメントの幹細胞阻害活性を評価するために、CFU-MIX細胞周期進行アッセイを実施例14と同様に使用する:
【0263】
【表20】
1Balb/c骨髄
2精製非組換えヒトαヘモグロビン(実施例16と同様)(100ng/ml)
3精製蟻酸切断タンパク質(本実施例と同様)(100ng/ml)。
【0264】
実施例22:ユビキチン融合体としてのE.coliにおけるヘモグロビンα鎖、ポリペプチド1-141、ポリペプチド1-97、ペプチド43-55、およびペプチドc(43-55)の発現
ペプチド43-55(「p13」)およびc43-55(「p15」)(実施例20と同様)の遺伝子を、最適E.coliコドン使用頻度(AnderssenおよびKurland,Micro.Reviews 54:198-210,1990)に従って対応するオリゴヌクレオチドをアニールすることにより合成した。インタクトなヒトαヘモグロビン鎖(「p141」)の遺伝子を、1セットのオリゴを設計して、ヒト骨髄cDNAプール(Clontech,Palo Alto,CA)からPCR増幅することにより得た。1-97フラグメント(「p1-97」)の遺伝子を、適切なサブクローニング後のp141遺伝子を含むプラスミドのPCR増幅により得た。
【0265】
上記の遺伝子を、ユビキチン融合タンパク質として発現した(米国特許第5,132,213号;同第5,196,321号;および同第5,391,490号、ならびにPCT WO 91/17245を参照のこと)。宿主株E.coli DH5αFIQ(Life Technologies,Inc.,Gaithersburg,MD)を、適切に合成した遺伝子(上記)を含むユビキチン発現ベクターpDSUbで形質転換した。pDSUbは、ヒトユビキチンを発現するpDS78/RBSIIの誘導体である(図22A)(Hochuliら、Biotechnology 6:1321-5,1988)。Loetscherら(JBC 266:11213-11220,1991)は、クロラムフェニコールアセチルトランスフェラーゼ(CAT)配列をHochuliプラスミドから切り出し、そしてプラスミドを再連結することによりpDS78/RBSIIを改変した(図22B)。合成ユビキチン遺伝子を、細菌発現に最適なコドン使用頻度を有するヒトユビキチンをコードするリン酸化合成オリゴヌクレオチドの対様アニーリングにより構築した。次いで、pDSUbを、組み立てたオリゴヌクレオチドからなる合成ユビキチン遺伝子を、誘導体化pDS78/RBSII のKlenow平滑化BamHI-BglI消化物に挿入することにより構築した。得られたプラスミドpDSUb(図22C)は、E.coliにおいて高レベルでユビキチンを発現することが示された。
【0266】
p97を含むプラスミドおよびp141を含むプラスミドを、指向性クローニングにおいて、p97またはp141タンパク質および融合接合部をコードするAflII-PstI消化PCR産物を、AflIIおよびPstIで消化したpDSUbに挿入することにより構築した。同様に、p13を含むプラスミドおよびp15を含むプラスミドを、適切な付着末端を有し、そしてペプチドおよび融合接合部をコードする、リン酸化およびアニールされたオリゴヌクレオチドを、AflII-PstI消化pDSUbに挿入することにより構築した。
【0267】
形質転換体を、100μg/mlアンピシリン、5μg/mlネオマイシンで選択した。コロニーは30℃で2日後に現れる。形質転換体を、挿入部位を横切るPCRによりスクリーニングした。次いで、正確なサイズの挿入物を含むコロニーを、SDS-PAGEにより適切なサイズの融合タンパク質の発現についてスクリーニングした(以下を参照のこと)。ユビキチン融合体を、lacリプレッサーを滴定し(titrate)、それをpDSUbのプロモーターから除去するIPTGの添加により過剰発現させた(DH5αF’IQは、10μg/mlネオマイシンで選択されるF’因子上にアップレギュレートされたlaqIq遺伝子を含む)。
【0268】
過剰発現された誘導ユビキチン融合タンパク質を示したクローン由来のプラスミドDNAを調製し、そしてジデオキシ法によりSequenase Version 2.0キット(United States Biochemical.)を用いて配列決定した。次いで、ポジティブクローンを凍結し、そして-80℃でグリセロール中に保存した。ポジティブクローンを、アンピシリン(100μg/ml)、ネオマイシン(10μg/ml)、および1%グルコースを含むLBプレート上で30℃で維持した。それらを10継代まで一週間毎に画線培養し(streak)、その後、新たな画線培養物を、連続培養のために凍結した種バイアルから採取して、株が本物であることを保証した。
【0269】
アッセイ用のタンパク質を得るために、250ml振盪フラスコ中の100ml接種菌培養物を、一晩のインキュベーション(16〜20時間)により、アンピシリン(100μg/ml)、ネオマイシン(10μg/ml)、および1%グルコースを有する2×YT培地中でシングルコロニーから増殖させた。振盪フラスコ培養物を、New Brunswick環境振盪インキュベーターにおいて30℃および250rpmで維持した。翌朝、培養物を、培地で1リットルに希釈した。細胞を、1mM(最終希釈)のIPTG添加によりOD600=0.5で誘導し、そしてOD600=0.8で遠心分離により採集した。採集細胞を低張溶解緩衝液(100μlの50mM Tris(pH10.0))に再懸濁した。細菌細胞を、その懸濁物を3サイクルの凍結-解凍(凍結のためのドライアイス-エタノール浴および解凍のための60℃)に供することにより溶解した。次いで、懸濁物を10分間超音波処理し、そして12,000gで10分間遠心分離した。得られた上清を「S1」と呼ぶ。細胞ペレットを、50mM Tris(pH10)および2×SDSトリシンローディング緩衝液(Novex,San Diego,CA)(1:1)に再懸濁した。次いで、混合物を95℃で15分間加熱し、そして12,000gで10分間遠心分離した。この方法で再可溶化され得る沈殿物の一部を「P1」と呼ぶ。残存するペレットから得られた沈殿物の一部を「P2」と呼ぶ。P2を、P1と同様にローディング緩衝液に再懸濁した。S1、P1、およびP2由来のサンプルを、SDS-PAGEにより分析した。
【0270】
SDS-PAGEゲルを、10〜20%トリシンゲル(Novex)と共に、ミニゲル装置における二緩衝液トリシン系を用いて泳動させた。陽極(下部)緩衝液は0.2M Tris(pH9.0)であった。陰極(上部)緩衝液は、0.1M Tris、0.1Mトリシン、0.1%SDS(pH8.25)であった。市販の分子量マーカー「Multi-Mark」(Novex)を使用した。標準として使用したウシユビキチンをSigmaから購入した。ゲルを、色素マーカーがゲルの下部に達するまで4mAの定電流で泳動させた。ゲルを、酢酸:メタノール(10%:40%)中の0.25%のCoomassie Blue R250(Sigma)で染色し、そして色素のない同じ溶液で脱染した。
【0271】
インタクトなp141ユビキチン融合タンパク質の大部分(>70%)を、細菌溶解物の遠心分離後の沈殿物(P1およびP2)において見出した。はっきりと対照的に、p97ユビキチン融合タンパク質の大部分(>70%)を、可溶性画分(S1)において見出した。これにより、C末端疎水性領域の除去は、改善された可溶性特性を有する産物を生じることが確認された。同様に、p13およびp15ペプチドもまた可溶性画分に含まれた。
【0272】
UCH-L3ユビキナーゼ(ubiquinase)酵素(Recksteiner,M.(編)Ubiquitin,Plenum Press(NY)1988;Wilkinsonら、Science 246:670-73,1989)を、宿主株BL21/DE3を形質転換するために使用されたpRSET(Invitrogen,San Diego,CA)において発現させた。UCH-L3は、ユビキチンC末端伸長を切断するユビキチン特異的プロテアーゼである。これを、35%(w/v)硫酸アンモニウム沈殿により細菌溶解物から部分的に精製した。使用した硫酸アンモニウムの正確な割合を、25.5kDバンドの存在についてSDS-PAGEによりモニターした。上清を50mM Tris(pH7.4)に対して透析し、そしてユビキチンペプチド融合基質に対してアッセイした。活性な上清を等分し、そして-20℃で凍結した。代表的な反応混合物は、3μlの溶解物、1μlの1M DTT、1μlのUCH-L3(上記)、および5μlの反応緩衝液(50mM Tris(pH7.4))を含む。反応を室温で20分間行った。大規模な消化については、300μlの溶解物を、100μlの1M DTT、20μl UCH-L3、および580μl反応緩衝液と混合した。
【0273】
可溶性(S1)画分に含まれるペプチドまたはタンパク質を、実施例16のように逆相HPLCによりさらに精製した;画分をSDS-PAGEによりモニターし、そして同一性を、エレクトロスプレーイオン化質量分析により確認した(以下を参照のこと)。精製したペプチドまたはタンパク質を、上記のUCH-L3によって酵素学的に消化し、非ユビキノン化最終生成物を得た。次いで、この切断物質を、逆相HPLCによって再精製した。精製を、SDS-PAGEによって行い、そして最終生成物の同一性をエレクトロスプレーイオン化質量分析によって確認した。
【0274】
上記のようなUCH-L3でのインビトロ切断に対する代替は、所望のユビキチン融合体と同じ細菌においてユビキチン切断酵素を同時発現させることである。この目的のために、ユビキナーゼUBP1(TobiasおよびVarshavsky,JBC 266:12021-12028,1991)を発現するベクター(pJT184)を使用した。P97ユビキチン融合体およびUBP1を同時発現する細菌は、インビボで融合タンパク質の完全な消化を示した;p141ユビキチン融合体およびUBP1を同時発現する細菌は、融合タンパク質の部分的な(約70%)消化を示した。インビボ消化したp97タンパク質を、硫酸アンモニウム沈殿、続いて上記のように逆相HPLCにより精製した。
【0275】
発現および精製されたポリペプチドの同一性を確認するために、エレクトロスプレーイオン化質量分析を、四重極分析器を有するVG Biotech BIO-Q器械を用いて行った。この器械の較正をするために、ミオグロビンを使用した。精製p97で得られた主な成分は、分子量10,339ダルトンの単一のピークであった;これは、計算された分子量10,347にほぼ匹敵し、組換えp97フラグメントの同一性を確認した。
【0276】
実施例23:組換えp1-97は、幹細胞阻害活性を維持する
組換えp1-97の生物活性を評価するために、GFU-GEMM細胞周期進行アッセイを実施例18と同様に使用した:
【0277】
【表21】
1ヒト骨髄単核細胞
2精製非組換えヒトαヘモグロビン(実施例16と同様)(100ng/ml)
3精製組換えp97(実施例22と同様)(100ng/ml)。
【0278】
実施例24:ヒトαヘモグロビンおよびペプチド43-55は、テストステロンまたは5-フルオロウラシル(5FU)処理マウスにおいて、インビボでCFU-MIX細胞周期進行を阻害する
ペプチド43-55のインビボでの幹細胞阻害活性を評価するために、B6D2 F1マウスを、実施例1に記載のテストステロンプロピオネートまたは5FUで前処理した。詳細には、テストステロンプロピオネート(100mg/kg体重)を、マウスに0日目にi.p.注射した。あるいは、5FU(200μg/kg体重)を、マウスに0日目に注射した。
【0279】
24時間後(1日目)、種々の用量のペプチド43-55またはビヒクルを、i.v.注射した。骨髄を2日目に採集し、そしてCFU-MIX細胞周期進行アッセイを、実施例14と同様に行った。詳細には、マウスを屠殺し、そして単一細胞骨髄懸濁物を大腿骨から調製した。細胞を1回洗浄し、そして濃度をFischer培地で5×106細胞/mlに調節した。各試験条件のために、1ミリリットルの細胞を、2つのポリプロピレンチューブのそれぞれに添加した。チューブを、適切な濃度の試験基質を共なわずに(「コントロール」)または共に(「実験」)で37℃で3時間インキュベートした。インキュベーションの終わりに、30μg/mlのシトシンアラビノシド(「Ara C」(Sigma))を、チューブの半分まで添加し、そして同じ容量のFischer培地をもう1つのチューブに添加した。チューブをさらに1時間37℃でインキュベートし、その後それらを氷上に置き、そして冷Fischer培地で2回洗浄した。
【0280】
細胞をFischer培地で5×104〜106/mlに再調節し、そして0.5mlの細胞懸濁物を、5mlのMethocult M3430メチルセルロース培地(Stem Cell Technologies,Vancouver,British Columbia)に添加した。混合物を、ボルテックスしながら力強く混合し、そして1mlを5つの35mmディッシュのそれぞれに分配した。35mmディッシュを、順に、滅菌水を含むふたのない35mmディッシュと共に、ふた付きの150mmディッシュに入れた。CFU-MIXコロニーを、37℃でのインキュベーションの7日後に倒立顕微鏡を用いて数えた。
【0281】
培地処理チューブとAra C処理チューブとの間のコロニー数の差異は、以下の式に従った条件下でのサイクルにおける細胞の割合を示す:
【0282】
【化11】
ここで、a=培地単独でインキュベートしたチューブ由来のCFU-MIXの数
b=Ara Cでインキュベートしたチューブ由来のCFU-MIXの数
【0283】
【表22】
1テストステロン前処理動物
220グラムマウスあたりのi.v.注射した量
【0284】
【表23】
15FU前処理動物
220グラムマウスあたりのi.v.注射した量。
【0285】
実施例25:ペプチド43-55は、N末端Phe(Phe43)でビオチン化またはPhe43もしくはPhe46でヨウ素化した場合、幹細胞阻害剤として活性である
ペプチド43-55を、固相ペプチド合成技術(American Peptide Co.,Sunnyvale,CA)により合成した。ペプチドアナログを、Phe43またはPhe46のパラ位でヨウ素を用いて合成した。ビオチン化ペプチド43-55を、C4炭素リンカーを有するビオチンのCOOHを、Phe43のN末端NH2に連結することにより合成した。
【0286】
【表24】
。
【0287】
実施例26:モルヒネは、マウスCFU-MIXの細胞周期進行をインビトロで阻害する
モルヒネを、実施例24と同様にBalb/cマウス由来の骨髄を用いてCFU-MIX細胞周期進行アッセイにおいて試験した。
【0288】
【表25】
。
【0289】
実施例27:オピエートペプチドDAMGOおよびDALDAは、マウスCFU-MIXの細胞周期進行をインビトロで阻害する
DAMGOおよびDALDAを、実施例24と同様にBalb/cマウス由来の骨髄を用いてCFU-MIX細胞周期進行アッセイにおいて試験した。
【0290】
【表26】
。
【0291】
実施例28:ノシセプチンは、マウスCFU-MIXの細胞周期進行をインビトロで阻害する
ノシセプチンを、実施例24と同様にBalb/cマウス由来の骨髄を用いてCFU-MIX細胞周期進行アッセイにおいて試験した。
【0292】
【表27】
。
【0293】
実施例29:ナロキソンは、ヒトαおよびδヘモグロビン鎖、へモルフィン10、ならびにペプチド43-55の阻害活性と拮抗する
CFU-MIX細胞周期進行アッセイを実施例24と同様に行う。試験基質を、それ自身でまたはナロキソン(10-5、10-7M)の存在下でアッセイした。ナロキソン自身は、これらの濃度でアッセイに効果を生じなかった。
【0294】
【表28】
110-5M最終濃度で使用した。
【0295】
【表29】
110-7M最終濃度で使用した。
【0296】
実施例30:低濃度のナロキソンは、マウスCFU-MIXの細胞周期進行を阻害する
CFU-MIXアッセイを実施例24と同様に行った。
【0297】
【表30】
。
【0298】
実施例31:μオピエートレセプターアンタゴニストCTOPは、ヒトヘモグロビンα鎖、ペプチド43-55、およびペプチド64-82の阻害活性と拮抗する
CTOP(H-D-Phe-Cys-Tyr-D-Trp-Orn-Thr-Pen-Thr-NH2、Cys2とPen7との間にジスルフィドを有する)は、μオピエートレセプター特異的アンタゴニストであるソマトスタチンのアナログである。試験基質を、それ自身またはCTOP(10-7M)の存在下でアッセイした。CTOP自身は、この濃度でアッセイでの影響を有さないが、αヘモグロビンまたはペプチド43-55により引き起こされる細胞周期阻害と拮抗する。
【0299】
【表31】
110-7Mの最終濃度で使用した。
【0300】
実施例32:ヒトα鎖での前処理は、後期形成コブルストーン(cobblestone)形成細胞の数を増加させる
コブルストーンアッセイを、Ploemacherおよびその同僚(Ploemacherら、Blood 74:2755-63,1989;van der Sluijsら、Exp.Hematol.18:893-6,1990;Ploemacherら、Blood 78:2527-33,1991;Ploemacherら、J.Tiss.Cult.Meth.13:63-68,1991;DownおよびPloemacher,Exp.Hematol.21:213-21,1993)により記載されたように行った。コブルストーンアッセイは、ストロマ細胞の単層内に細胞群(または「コブルストーン」)の出現を測定する。非常に初期の幹細胞は、軟寒天においてコロニーを形成しないが、ストロマ単層の存在下でコブルストーンを形成する。コブルストーンを形成する細胞は、「コブルストーン領域形成細胞」(CAFC)と呼ばれる。より分化した(例えば、GM-CFC)前駆体は、培養の最初の数週間以内に出現し、そして消滅する一過的なコブルストーンを形成するが、より初期の幹細胞(例えば、長期再増殖細胞)は、4〜5週間の培養後のみに出現するコブルストーンを形成する。従って、培養の7〜14日目に形成するCAFCはCFU-GMに富み、28〜35日目に形成するCAFCはCFU-MIXに富み、そして28〜35日目に形成するCAFCは長期再増殖細胞に富む。
【0301】
B6D2F1マウスを、実施例24と同様にテストステロンプロピオネートで処理した。その翌日、骨髄を取り出し、そしてそれらをブレストーンアッセイでプレーティングした後に、ヒトαヘモグロビン鎖(100ng/106細胞)と共にまたは共なわずに4時間インキュベートした。使用したアッセイは、96ウェルプレートにおける限界希釈長期骨髄培養(LTBMC)からなる。培養物を、FBMD-1マウスストロマ細胞株(Breemsら、Leukemia 11:142-50,1997)をコンフルエントになるまで増殖させることにより調製した;コンフルエントな単層を有する96ウェルプレートを、アッセイまで33℃で保存した。マウス骨髄細胞を単細胞懸濁物として調製し、そして以下の細胞希釈物:27,000;9000;3000;1000;333を、0.2mlのLTBMC培地(Stem Cell Technologies,Vancouver)にウェルあたりプレーティングした。20ウェルを、条件あたり各希釈についてプレーティングし、そして2プレート以上に分配した。
【0302】
コブルストーン領域形成細胞(CAFC)の頻度を、以前に記載されたように(Ploemacherら、Blood 78:2527-33,1991;Ploemacherら、J.Tiss.Cult.Meth.13:663-68,1991;Breemsら、Leukemia 8:1095-104,1994)計算した。結果を実施例23に示す。細胞周期進行幹細胞のヒトαヘモグロビン鎖とのプレインキュベーションは、約5倍まで後期形成CAFCの比を増大させた。非細胞周期進行細胞のα鎖での処理は効果がなかった。
【0303】
実施例33:ヒトαヘモグロビン、ならびにペプチド43-55、環状43-55および64-82は、ヒト臍帯血CFU-GEMMの細胞周期進行を阻害する
ヒト臍帯血CFU-GEMM細胞周期進行アッセイを、実施例19と同様に行った。詳細には、単核細胞を、ヒト臍帯血細胞から単離し、そして10%FBS、100ng/mlキットリガンド、および100ng/mlヒトIL-3を補充したIMDM組織培養培地で2〜4×104細胞/mlに調節した。細胞を37℃で48時間インキュベートした。
【0304】
インキュベーション後、細胞を洗浄し、そして106細胞/mlの濃度で無血清IMDMに再懸濁した。1mlの細胞を、条件あたり2つのポリプロピレンチューブのそれぞれに添加し、そして細胞周期進行アッセイを、マウス骨髄について実施例24と同様に行った。Ara Cインキュベーション後、細胞を冷IMDMで洗浄し、そして0.5ml IMDMあたり10,000〜20,000細胞に調節し、そして5mlのMethocult H4433(Stem Cell Technologies)と混合した。あるいは、細胞濃度を0.5ml IMDMあたり2500〜5000細胞に調節する場合に、Methocult H4435メチルセルロース培地(Stem Cell Technologies)を使用した。細胞を実施例24と同様にプレーティングし、そしてCFU-GEMMコロニーを14〜18日目にスコア付けした。
【0305】
【表32】
。
【0306】
実施例34:ヒトαヘモグロビンおよびペプチド43-55は、成人骨髄CFU-GEMMの細胞周期進行を阻害する
CD34+幹細胞を、CellProカラムの手段によるヒト骨髄からの精製の後、Poietic Technologies(Gaithersburg,MD)から得た。細胞を、キットリガンドおよびIL-3と48時間インキュベートし、そして実施例26におけるCFU-GEMM細胞周期進行アッセイにおいて使用した。
【0307】
【表33】
。
【0308】
実施例35:移動したヒト末梢血におけるCFU-GEMMは活発にサイクルし、そしてヒトαヘモグロビン、ペプチド43-55、DAMGO、またはモルヒネにより阻害可能である
末梢血を、標準的プロトコルに従ったシクロフォスファミドおよびG-CSFでの末梢幹細胞移動を受けている乳ガン患者から得た。赤血球をFicoll Hypaqueで除去した(細胞をIMDMで1:1に希釈し、そして20mlを16mlのFicollの上部に重層し、そして800gで30分間遠心分離した;単核細胞を中間層(interphase)から取り出し、そしてIMDM中で2回洗浄した)。1つの場合、単核細胞を、アッセイ前に液体窒素中で凍結保存した。単核細胞を、2.5〜5×105細胞をディッシュあたりにプレーティングした以外は、実施例26と同様にCFU-GEMM細胞周期進行アッセイのためにプレーティングした。
【0309】
【表34】
。
【0310】
【表35】
。
【0311】
【表36】
1アッセイ前に凍結保存した細胞に由来する。
【0312】
実施例36:高用量のヒトαまたはβヘモグロビン、ミオグロビン、ペプチド1-97、ペプチド43-55、ペプチド64-82、ノシセプチン、あるいはDALDAは、静止マウス幹細胞の細胞周期進行を刺激する
1μg/ml用量のヘモグロビン鎖、ミオグロビン、およびペプチドを、静止幹細胞の刺激についてアッセイした。骨髄を、実施例24と同様のCFU-MIX細胞周期進行アッセイにおいて試験した未処理B6D2F1から得た。未処理B6D2F1マウスから単離した幹細胞は、(例えば、テストステロンプロピオネート(実施例1を参照のこと)または5FUのような化学療法(実施例4を参照のこと)により)刺激されるまで、通常ゆっくりと細胞周期進行して、周期に入る。
【0313】
【表37】
。
【0314】
【表38】
。
【0315】
【表39】
。
【0316】
【表40】
。
【0317】
【表41】
。
【0318】
【表42】
。
【0319】
実施例37:高用量のヒトαヘモグロビンの静脈内投与は、静止マウス幹細胞の細胞周期進行を刺激する
ヒトαヘモグロビンを、未処理B6D2F1マウスにi.v.注射した。24時間後、マウスを屠殺し、骨髄を大腿骨から回収し、そしてCFU-MIXアッセイを実施例24と同様に行った。
【0320】
【表43】
。
【0321】
実施例38:ナロキソンは、高用量ヒトαヘモグロビン、ペプチド43-55の幹細胞刺激活性と拮抗する
【0322】
【表44】
110-7M最終濃度で使用した。
【0323】
***
本発明は好ましい実施態様によって記載されたが、変化および改変は当業者に考えられることが理解される。従って、添付の請求の範囲は、請求される本発明の範囲内にある全てのこのような等価な変化を含むことが意図される。
【図面の簡単な説明】
【0324】
【図1】図1〜図4は、各精製段階後の産物のSDSポリアクリルアミドゲル泳動を示す。図1−レーン1はキモトリプシノーゲンであり、レーン2はオボアルブミンであり、レーン3はBSAであり、レーン4は30kD未満の画分であり、レーン5は30〜50kDの画分であり、そしてレーン6は50〜100kDの画分である。
【図2】図1〜図4は、各精製段階後の産物のSDSポリアクリルアミドゲル泳動を示す。図2−レーン1は硫酸アンモニウム沈澱後(40〜80%)であり、そしてレーン2〜5はDEAE画分である(レーン番号2は活性画分を表す)。
【図3】図1〜図4は、各精製段階後の産物のSDSポリアクリルアミドゲル泳動を示す。図3−レーン1は硫酸アンモニウム沈澱後の上清であり、そしてレーン2は活性DEAE画分であり、レーン3〜5はゲル濾過画分を表す(レーン番号5は活性画分を表す)。
【図4】図1〜図4は、各精製段階後の産物のSDSポリアクリルアミドゲル泳動を示す。図4−レーン2は最終産物を表す。
【図5】図5は、最終精製物の逆相HPLCクロマトグラムを示す。
【図6】図6は、ブタ骨髄から精製されたINPROL(pINPROL)を含まない場合(コントロール=0%阻害)および、種々の量で含む場合のFDCP-mix細胞株内へのトリチウム化チミジンの取り込み(cpm)を示す。データは、コントロールの値に対して正規化されている。
【図7】図7は、テストステロンプロピオネート(TSP)、TSP+pINPROL、またはビヒクル(コントロール)を用いてマウスを処理した後の細胞周期のS期にある細胞の%を示す。各グループは25匹の動物を含んでいた(時点毎に3〜4匹)。
【図8】図8は、2用量の5-FUで処置され、pINPROL処理を受けたかまたは受けていないマウスの生存を示す。各グループは30匹の動物を含んでいた。
【図9】図9は、放射線照射され、pINPROL処置を受けた場合および受けていない場合のマウスの生存を示す。各グループは50匹の動物を含んでいた。
【図10A】図10Aおよび図10Bは、Ara-CまたはAra-C+pINPROLで処理した後、1週間目(図10A)および3週間目(図10B)の正常骨髄の長期培養細胞の再生を示す。
【図10B】図10Aおよび図10Bは、Ara-CまたはAra-C+pINPROLで処理した後、1週間目(図10A)および3週間目(図10B)の正常骨髄の長期培養細胞の再生を示す。
【図11】図11は、致死量の放射線照射を行い、培地(コントロール)またはpINPROL(25ng/ml)と4時間プレインキュベーションした後の3×104個の骨髄細胞を移植した後のマウス(グループ当たり75匹)の生存を示す。生存は30日間モニターされた。
【図12】図12は、致死量の放射線照射を行い、pINPROLまたは培地と4時間プレインキュベートされたドナーの骨髄細胞により回復した後のマウスの骨髄細胞により14日後に培養物中で形成されたCFU-GMの数を示す。
【図13】図13は、毎週取り出し、洗浄し、そして培地またはpINPROLと4時間プレインキュベートした後にIL-7(10ng/ml)と共にプレートしたリンパ球の長期培養物由来の懸濁細胞を示す。
【図14】図14は、pINPROLで処理された白血病の末梢血細胞の改善された再生能を示す。長期培養開始細胞(LTC-IC)は、粘着性および非粘着性LTC細胞をpINPROLと共に、およびpINPROLなしでプレートし、そして7日目にCFU-GMを数えることにより測定された。データはコントロールの値に対して正規化されている。
【図15A】図15Aは、53%アセトニトリルで溶出した精製pINPROLのC4逆相クロマトグラムを示す。レーン1は粗物質であり、レーン2は分子量マーカーであり、そしてレーン3は精製物質である。図15Bは、43.9%アセトニトリルで溶出したMIP-1αのC4逆相クロマトグラムを示す。図15Cは、pINPROLの粗調製物および逆相後の精製された調製物のSDS-PAGEクロマトグラムを示す。
【図15B】図15Aは、53%アセトニトリルで溶出した精製pINPROLのC4逆相クロマトグラムを示す。レーン1は粗物質であり、レーン2は分子量マーカーであり、そしてレーン3は精製物質である。図15Bは、43.9%アセトニトリルで溶出したMIP-1αのC4逆相クロマトグラムを示す。図15Cは、pINPROLの粗調製物および逆相後の精製された調製物のSDS-PAGEクロマトグラムを示す。
【図15C】図15Aは、53%アセトニトリルで溶出した精製pINPROLのC4逆相クロマトグラムを示す。レーン1は粗物質であり、レーン2は分子量マーカーであり、そしてレーン3は精製物質である。図15Bは、43.9%アセトニトリルで溶出したMIP-1αのC4逆相クロマトグラムを示す。図15Cは、pINPROLの粗調製物および逆相後の精製された調製物のSDS-PAGEクロマトグラムを示す。
【図16A】図16は、ヘモグロビン配列を示す:図16AはヒトαヘモグロビンのcDNA配列およびアミノ酸配列を示し、そして図16BはヒトβヘモグロビンのcDNA配列およびアミノ酸配列を示す。番号付けはアミノ酸に従う。図16Cは、ヒト、マウス、およびブタヘモグロビンのα鎖およびβ鎖のアミノ酸配列比較を示す。
【図16B】図16は、ヘモグロビン配列を示す:図16AはヒトαヘモグロビンのcDNA配列およびアミノ酸配列を示し、そして図16BはヒトβヘモグロビンのcDNA配列およびアミノ酸配列を示す。番号付けはアミノ酸に従う。図16Cは、ヒト、マウス、およびブタヘモグロビンのα鎖およびβ鎖のアミノ酸配列比較を示す。
【図16C】図16は、ヘモグロビン配列を示す:図16AはヒトαヘモグロビンのcDNA配列およびアミノ酸配列を示し、そして図16BはヒトβヘモグロビンのcDNA配列およびアミノ酸配列を示す。番号付けはアミノ酸に従う。図16Cは、ヒト、マウス、およびブタヘモグロビンのα鎖およびβ鎖のアミノ酸配列比較を示す。
【図17】図17は、pINPROL(図17A)および結晶化ブタヘモグロビン(図17B)のC4逆相HPLCトレースの比較を示す。
【図18】図18は、結晶化ブタヘモグロビンのC4逆相HPLC分離からの画分のSDS-PAGEゲルを示す。レーン1は分子量マーカーを示し、レーン2は第1ピーク(47.11分)由来の画分48〜49を示し、レーン3は第2ピーク(49.153分)由来の画分50〜51を示し、レーン4は第3ピーク(52.25分)由来の画分54〜55を示し、そしてレーン5は第4ピーク(53.613分)由来の画分56〜57を示す。
【図19】図19は、pINPROL(図19A)および精製ブタβヘモグロビン(図19B)の2次元ゲル電気泳動の比較を示す。
【図20】図20は、FDCP-MIXアッセイにおける精製ブタαヘモグロビン、βヘモグロビン、またはpINPROLの効果の比較を示す。
【図21】図21は、ゆるやかな溶出勾配を用いるブタヘモグロビンの逆相分離を示す。
【図22】図22Aは、Hochuliら(1988)からのプラスミドを示す;図22Bは、Loetscherら(1991)のプラスミドを示す;図22CはpDSUbプラスミドを示す。
【図23】図23は、cobblestoneアッセイでの、INPROLでの処置の結果を示す。
【技術分野】
【0001】
発明の分野
本発明は、自己免疫疾患、加齢、ガン、脊髄形成異常、前白血病、白血病、乾癬、後天性免疫不全症候群(AIDS)、脊髄形成異常症候群、再生不良性貧血、または過剰または過少増殖状態を含む他の疾患を有するヒトまたは動物の処置における幹細胞の細胞周期を調節するための幹細胞増殖モジュレーターの使用、ならびに鎮痛のためのこのような化合物の使用に関する。本発明はまた、化学療法剤または細胞周期内にある幹細胞に損傷を与える他の薬剤への曝露、あるいは放射線の照射を受けることが予想されるかまたは受けているヒトまたは動物の処置、ならびにエクスビボでの処置の間のそのような薬剤に対する防御の方法に関する。最後に、本発明は、自家移植および同種移植の手法あるいは遺伝子導入のための幹細胞の維持または拡大培養の改良、ならびにそのような手法の改良のためのインビボでの処置に関する。
【背景技術】
【0002】
発明の背景
再生系のほとんどの終期細胞は、短命であり、そして一生を通じて連続的に置き換えられなければならない。例えば、血液細胞は、多能性造血幹細胞(HSC)の自己再生集団から生じる。造血幹細胞は造血細胞の亜集団である。造血細胞は、例えば、骨髄、臍帯血、または末梢血から得られ得る(造血細胞は、移動しないか、G-CSFのような因子によって移動される)。造血細胞は、幹細胞集団、前駆細胞、分化細胞、補助細胞、間質細胞、および成熟血液細胞の産生に必要な環境を寄与する他の細胞を含む。造血前駆細胞は、それらの発生効力がさらに制限される幹細胞の部分集合である。前駆細胞は、1または2系統にだけ分化し得る(例えば、赤血球のみを生じるBFU-EおよびCFU-E、または顆粒球およびマクロファージを生じるCFU-GM)が、幹細胞(例えば、CFU-MIXまたはCFU-GEMM)は、複数の系統および/または他の幹細胞を生成し得る。造血幹細胞は造血系および免疫系のすべての成熟細胞の発生に必要であるので、化学療法剤または他の薬剤で処置される被験体において十分に機能的な宿主防御系を再確立するために、造血幹細胞の生存は必須である。
【0003】
造血細胞の産生は、造血細胞の増殖および分化を刺激する一連の因子によって調節され、それらのいくつか(例えば、エリスロポエチン、GM-CSF、およびG-CSF)は、現在では臨床的な実用に用いられている。しかし、広範に特徴付けられていない制御ネットワークの一部分は、幹細胞の細胞周期の状態を制御する生理学的機構である(非特許文献1;非特許文献2)。
【0004】
Lordおよびその共同研究者による初期の研究は、正常および再生骨髄の抽出物中の、幹細胞の増殖を阻害または刺激のいずれかをし得る可溶性タンパク質因子の存在を示した(非特許文献3;WrightおよびLorimore,Cell Tissue Kinet.20:191-203,1987;MarshallおよびLord,Int Rev.Cyt.167:185-261,1996に概説される)。これらの活性は、それぞれ幹細胞阻害剤(SCI)および幹細胞刺激剤(SCS)と称された。
【0005】
現在まで、Lordら(上記を参照)に記載されるように、調製された骨髄抽出物からは、SCSの候補となる分子は精製されていない。一次供給源からのSCSまたはSCIのいずれかの精製は、放射線照射された多数のマウスを必要とするインビボアッセイに固有の困難さにより達成されなかった。これらの問題を克服するための試みにおいて、Pragnellおよびその共同研究者は、原始造血細胞(CFU-A)のインビトロアッセイを開発し、そして阻害活性源としての細胞株をスクリーニングした(Grahamら、Nature 344:442-444,1990を参照)。初期の研究はマクロファージをSCIの可能な供給源として同定していたので(非特許文献3)、マウスマクロファージ細胞株J774.2が選択された(Grahamら、Nature 344:442-444、1990)。この細胞株由来の馴化培地が精製のために、Grahamらによって用いられた。阻害ペプチドが単離され、それは、前述のサイトカインマクロファージ炎症性タンパク質1-α(MIP-1α)と同一であることが証明された。他のケモカインレセプターのように、MIP-1αのレセプターがクローニングされ、これらのMIP-1αは、G阻害性サブクラス(「Gi」)のグアニンヌクレオチド(GTP)結合タンパク質に結合する7回膜貫通ドメイン(または「G-関連」)レセプターである(Murphy,Cytokine & Growth Factor Rev.7:47-64,1996に概説される)。Giサブクラスに対する「inhibitory」の名称は、そのアデニル酸シクラーゼの阻害活性をいう。
【0006】
MIP-1αは、細胞株から単離されたのであって、一次材料から単離されたのではない。Grahamらは、MIP-1αに対する抗体が骨髄の粗抽出物の活性を失わせることを観察したが、他の研究者は、他の阻害活性が重要であることを示した。例えば、Grahamら(J.Exp.Med.178:925-32、1993)は、MIP-1αではなくTGFβが造血幹細胞の主要な阻害剤であることを示唆した。さらにEavesら(PNAS 90:12015-19、1993)は、MIP-1αおよびTGFβの両方がやや最適のレベルで正常骨髄中に存在し、そして阻害にはこれら2つの因子間の相乗作用が必要であることを示唆した。
【0007】
最近、相同組換えによってMIP-1α遺伝子が欠失したマウスが作製された。(Cookら,Science 269:1583-5,1995)。そのようなマウスは、その造血系に明白な混乱は見られず、普通の恒常性条件下における幹細胞の細胞周期の生理学的調節因子としてのMIP-1αの役割に異議を唱えられている。同様に、トランスフォーミング成長因子β(TGFβ)もまた幹細胞阻害効果を有するが、長い間、幹細胞がこのサイトカインに反応する思われており、さらに、これが骨髄抽出物内に存在する内因性因子ではないこと、さらに、TGFβに対する中和抗体が、骨髄上清のSCI活性を無効にできないことを示唆する(Hampsonら、Exp.Hemat.19:245-249,1991)。
【0008】
他の研究者は、さらなる幹細胞阻害剤を記載した。Frindelおよびその共同研究者は、ウシ胎児骨髄および肝臓抽出物から幹細胞阻害活性を有するテトラペプチドを単離した(Lenfantら、PNAS 86:779-782、1989)。Paukovitsら(Cancer Res.50:328-332、1990)は、ペンタペプチドを特徴付け、それはモノマー形体では阻害剤であり、そしてダイマー形体では幹細胞の細胞周期の刺激剤であるとしている。他の因子もまた、種々のインビトロ系において阻害性であることが主張されている(WrightおよびPragnell、Bailliere’s Clinical Haematology 第5巻、723〜39頁、1992(Bailliere Tinadall、Paris):MarshallおよびLord,Int Rev.Cyt.167:185-261,1996を参照)。
【0009】
Tsyrlovaら、SU 1561261 A1は、幹細胞増殖阻害剤のための精製プロセスを開示した。
【0010】
共有に係る出願WO 94/22915およびWO96/10634は、幹細胞増殖の阻害剤を開示し、またそこにその全体が本明細書中に参考として援用される。
【0011】
現在まで、これらの因子のいずれもが臨床的使用に対して認可されていない。しかし、効果的な幹細胞阻害剤の必要性は存在している。化学療法または放射線処置に関連する主要な毒性は正常増殖細胞の破壊であり、これにより骨髄の抑制または胃腸に対する毒性を生じ得る。効果的な幹細胞阻害剤は、これらの細胞を保護し、そしてこれらの治療レジメを最適化し得る。臨床的な症状に応じて種々の刺激性サイトカイン(すなわち、IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-11、IL-13、IL-14、IL-15、G-CSF、GM-CSF、エリスロポエチン、トロンボポエチン、幹細胞因子、flk2/flt3リガンドなどのようなサイトカイン、これらは造血細胞の細胞周期を刺激する)が必要とされることが実証され、同様に異なる臨床的な必要性に関して種々の阻害剤が必要とされるようである。
【0012】
さらに、そのような阻害剤の活性を迅速に無効にする必要がある。Lordらの最初の研究(上記を参照)は、阻害活性が刺激活性の付加により逆転され得ることを実証した。種々の幹細胞刺激性サイトカインが同定されている(上記を参照)一方、Lordおよび共同研究者らによって記載された、骨髄抽出物に存在し、そして阻害剤の活性を逆転させ得る活性を示すものがないことが示されている。
【0013】
造血前駆細胞および幹細胞は健常な成人の骨髄中に主に存在する。例えば、化学療法またはG-CSFのようなサイトカインでの処置のような特定の条件下で、大量の前駆細胞および幹細胞が末梢血中に出ていくプロセスは、「移動(mobilization)」と呼ばれる。(Simmonsら,Stem Cells 12(補遺1):187-202,1994;Schedingら,Stem Cells 12(補遺1):203-11,1994;Mangan,Sem.Oncology 22:202-9,1995;Moolten,Sem.Oncology 22:271-90,1995)。最近公表されたデータは、大部分の移動した前駆細胞が細胞周期中は活性でないことを示唆する(RobertsおよびMetcalf,Blood 86:1600-,1995;Donahueら,Blood 87:1644-,1996;SiegertおよびSerke,Bone Marrow Trans.17:467-1996;UchidaらBlood 89:465-72,1997)。
【0014】
ヘモグロビンは、高度に保存された四量体タンパク質であり、その分子量は約64,000ダルトンである。これは、2つのα鎖および2つのβ鎖からなる。各鎖は、ヘムの単一分子(フェロプロトポルフィリンIX)(鉄含有補欠分子族)に結合する。脊椎動物α鎖およびβ鎖は、おそらく、複製し、次いで分岐した1つの単一祖先遺伝子に由来した。この2つの鎖は、それら鎖間および種々の脊椎動物間で高度の配列同一性を保持している(図16Aを参照)。ヒトにおいて、第16染色体上のα鎖クラスターは、同一のポリペプチドをコードする2つのα遺伝子(α1およびα2)、ならびに他のα様鎖をコードする遺伝子(ζ、θ)およびいくつかの非転写偽遺伝子を含有する(ヒトα鎖のcDNA配列およびアミノ酸配列については図16Bを参照)。第11染色体上のβ鎖クラスターは、1つのβ鎖遺伝子およびいくつかのβ様鎖遺伝子(δ、ε、Gγ、およびAγ)、ならびに少なくとも2つの非発現偽遺伝子からなる(ヒトβ鎖のcDNA配列およびアミノ酸配列については図16Cを参照)。
【0015】
これらの遺伝子の発現は、発育の間で変化する。ヒトの造血(これは十分に特徴付けられている)において、胚赤芽球は、2つのζ鎖および2つのε鎖(Gower I)、2つのα鎖および2つのε鎖(Gower II)、または2つのζ鎖および2つのγ鎖(Hb Portland)からなる四量体を連続的に合成する。胚形成が進行するにつれて、優勢型は、2つのα鎖および2つのγ鎖で構成される胎児ヘモグロビン(Hb F)からなる。成人ヘモグロビン(2つのα鎖および2つのβ鎖)は、胎児期の間に合成が開始される。誕生時には、約50%のヘモグロビンが成人型であり、そしてその移行は約6カ月齢までに完了する。成人におけるヘモグロビンの大部分(約97%)は、2つのα鎖および2つのβ鎖からなる種(Hb A)であり、少量のHb Fまたはδ鎖(Hb A2)もまた検出され得る。
【0016】
いくつかの方法が、E.coli内および酵母内で組換え体ヘモグロビン鎖を発現するために使用された(例えば、Jessenら,Methods Enz.231:347-364,1994;Lookerら,Methods Enz.231:364-374,1994;Ogdenら,Methods Enz.231:374-390,1994;Martin de Llanoら,Methods Enz.231:364-374,1994)。現在まで、組換えの方法では、単離されたヒトα鎖を高収率で発現することは可能ではない(例えば、Hoffmanら,PNAS 87:8521-25,1990;Hernanら,Biochem.31:8619-28,1992)。明らかに、単離されたアルファ鎖は安定した高次構造であるとは考えられず、E.coli内では速やかに分解される。β鎖とα鎖との同時発現は両方の発現が増加を生じた(Hoffmanら、およびHernanら、前出)。α鎖はβ鎖部分とXa因子の認識部位(NagaiおよびThorgersen,Methods Enz.231:347-364,1994)との融合タンパク質として発現されているが、これらの状態下では、不溶性封入体として発現される。
【0017】
β鎖およびα鎖の両方がハプトグロビンに対する結合部位を含む。ハプトグロビンはヘモグロビンに対して極端に高い親和性を有する血清タンパク質である(例えばPutnam in The Plasma Proteins - Structure,Function and Genetic Control(F.W.Putnam,Ed.)Vol.2,1-49頁(Academic Press,NY);HwangおよびGreer,JBC 255:3038-3041,1980)。肝臓のハプトグロビン輸送は、循環しているヘモグロビンの主要な代謝経路である。これらはα鎖に、ハプトグロビンへの1つの結合部位(アミノ酸121-127)があり、そしてβ鎖には2つの結合部位(アミノ酸領域11-25および131-146)がある(KazimおよびAtassi,Biochem J.197:507-510,1981;McCormickおよびAtassi,J.Prot.Chem.9:735-742,1990)。
【0018】
生物学的に活性なオピエート活性を有するペプチドは、ヘモグロビンのタンパク質分解性の分解によって得られている(Karelinら,Peptides 16:693-697,1995に概説される)。ヘモグロビンα鎖はアミノ酸94-95間に酸に不安定(acid-labile)な切断部位を有する(Shaeffer,J.Biol.Chem.269-29530-29536,1994)。
【0019】
Kreglerら(Exp.Hemat.9:11-21,1981)は、ヘモグロビンがマウス骨髄のCFU-C前駆細胞コロニーに対して増強活性を有することを開示した。そのようなアッセイは、CFU-MIXのような幹細胞に対して、CFU-GMおよびCFU-M前駆細胞集団に対する効果を実証する。著者らは、ヘモグロビンの単離されたα鎖およびβ鎖の両方における活性を観察した。この活性は、N-エチルマレイミドでの処理により消滅し、このことはKreglerらに、スルフヒドリル基が必要であることを示唆した。この観察は、刺激活性がトリプシン消化に対する耐性であるという事実と合わせて、KreglerらにC-末端疎水性ドメインまたは「核」領域が活性を担っていることを示唆した。Moqattashら(Acta.Haematol.92:182-186,1994)は、組換えヘモグロビンがCFU-E、BFU-EおよびCFU-GM前駆細胞の数に対して刺激効果を有することを開示し、これは、ヘミンで観察されたのと同様である。Muellerら(Blood 86:1974,1955)は、精製された成人のヘモグロビンがヘミンと同様の様式で赤血球系の前駆細胞を刺激することを開示した。
【0020】
Petrovら(Bioscience Reports 15:1-14,1995)は、「非同定ミエロペプチド混合物」のWv/Wvマウスでの先天性貧血症の処置での使用を開示した。この混合物は、脾臓コロニー、特にその赤血球型の数を増加させた。
【0021】
ヘムおよびヘミンは、造血に対するその影響に関して十分に試験されてきた(概説についてS.Sassa、Seminars Hemat.25:312-20,1988およびN.Abrahamら、Int.J.Cell Cloning 9:185-210,1991を参照のこと)。ヘムは、赤芽球の成熟に必要とされる。インビトロにおいて、ヘミン(クロロフェロプロトポルフィリンIX−すなわち、付加塩化物イオンを有するヘム)は、CFU-GEMM、BFU-E、およびCFU-Eの増殖を増大させる。同様に、ヘミンは、長期骨髄培養物において細胞充実性を増大させる。
【0022】
「オピエート」は、アヘンの主要な作用物質であるモルヒネと類似する鎮痛性の性質を持つ物質である。オピエートは、モルヒネおよび他のアルカロイドもしくは合成化合物のような有機小分子、またはエンケファリン、エンドルフィン、ダイノルフィンおよびその合成誘導体のような内因性のペプチドであり得る。内因性のオピエートペプチドは、より大きな前駆体(Met-またはLeu-エンケファリンに対するプレプロエンケファリンA、α、βおよびγエンドルフィンに対するプレプロオピオメラノコルチン、ダイノルフィンAおよびBに対するプレプロダイノルフィン、βネオエンドルフィンに対するαネオエンドルフィン)よりインビボで生成される。さらに、オピエート活性を有するペプチドは、αまたはβカゼイン、小麦グルテン、ラクトアルブミン、チトクローム、またはヘモグロビンのようなタンパク質のタンパク質分解または加水分解のような非古典的な供給源、またはカエルの皮膚(デルモルフィン)またはウシ副腎髄質のような他の種から得られ得る。このようなペプチドは、古典的なエンドルフィンに対して「エキソルフィン(exorphin)」と呼ばれ、それらはまた異型オピエートペプチドとも呼ばれる(非特許文献4;非特許文献5;非特許文献6;Brantl,Eur.J.Pharm.106:213-14,1984;Brantlら,Eur.J.Pharm.111:293-4,1985;Brantlら,Eur.J.Pharm.125:309-10,1986;BlantlおよびNeubert,TIPS 7:6-7,1986;Glamstaら,BBRC 184:1060-6,1992;Teschemacher,Handbook Exp.Pharm.104:499-28,1993;Petrovら,Bioscience Reports 15:1-14,1995;Karelinら,Peptides 16:693-7,1995)。Tyr-MIF-1ファミリーのような他の内因性ペプチドはまた、オピエート活性を有することが示されている(Reedら,Neurosci.and Biobehav.Rev.18:519-25,1994)。
【0023】
オピエートは、内因性オピエートレセプターの3つの主要な薬理学的クラス(μ、δ、およびκ)に結合することによって効果を発揮する。各薬理学的クラスを代表するレセプターは、クローン化され、そしてGiに結合したG関連レセプターであることが示されている。(ReisineおよびBell,TINS 16:506-510,1993;Uhlら,TINS 17:89-93,1994;Knappら,FASEB J.9:516-525,1995;SatohおよびMinami,Pharm.Ther.68:343-64,1995;Kieffer,Cell.Mol.Neurobiol.15:615-635,1995;Reisine,Neuropharm.34:463-472,1995;Zakiら,Ann.Rev.Pharm.Toxicol.,36:379-401,1996に概説されている)。
【0024】
特定のアゴニストおよびアンタゴニストが、各レセプタータイプについて入手可能である。レセプタータイプには、例えば、μレセプター(これは、DAMGOおよびDALDAによって選択的に活性化され、CTOPおよびナロキソナジンによって選択的に拮抗される)、κレセプター(これは、GR 89696フマル酸塩またはU-69593で選択的に活性化され、そしてノルビナルトルフィミンハイドロクロリドで選択的に拮抗される)、およびδレセプター(これは、DADLEおよびDPDPEで選択的に活性化され、そしてナトリンドール(natrindole)で選択的に拮抗される)がある。さらに、3つのレセプターサブタイプの全てに作用する広域スペクトルのナロキソンのようなアンタゴニストおよびエトルフィンのようなアゴニストがある。
【0025】
古典的かつ異型のオピエートペプチドは、化学的に変化または誘導体化され、その特異的オピエートレセプター結合特性を変化され得る(HrubyおよびGehrig,Med.Res.Rev.9:343-401,1989;Schiller,Prog.Med.Chem.28:301-40,1991;Teschemacher,Handbook Exp.Pharm.104:499-28,1993;Handbook of Experimental Pharmacology,A.Hertz(編)第104/Iおよび104/II刊、1993,Springer Verlag,Berlin;Karelinら,Peptides 16:693-7,1995)。例には、デルモルフィン(例えば、DALDA)およびエンケファリン(例えば、DADLE,DAMGO、またはDAMME)の誘導体が挙げられる。ソマトスタチンのようなオピエートレセプターに通常結合しないペプチドはまた、特異的オピエートレセプター結合を示すように誘導体化され得る(例えば、CTOP(Hawkinsら,J.Pharm.Exp.Ther.248:73,1989))。アナログもまた、改変したレセプター結合特性を有するモルヒネのようなアルカロイドから誘導され得る(例えば、ヘロイン、コデイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノールおよびナルブフィン);さらに、モルヒネに構造的に関連のない小分子もまた、オピエートレセプターに作用し得る(例えば、メペリジンおよびその同族種アルファプロジン、ジフェノキシレート、およびフェンタニール、)(Handbook of Experimental Pharmacology,前掲書、GoodmanおよびGilmanのThe Pharmacological Basis of Therapeutics,第7版、A.G.Gilman,L.S.Goodman,T.W.Rall and F.Murad(編)1985 Macmillan Publishing Co.NYを参照のこと)。
【0026】
内因性オピエートペプチド(エンケファリン、エンドルフィン、およびダイノルフィン)は、保存されたN末端テトラペプチド Tyr-Gly-Gly-Phe、続いてLeuまたはMetおよびいくらかの残りのC末端配列を有する。N末端Tyrのヒドロキシル基を除去することにより(N末端Pheを生じる)、Metエンケファリンに対する活性を劇的に損失する。これらの構造的データは、「メッセージ−アドレス(message-address)」仮説を導き、それによりN末端「メッセージ」は、生物学的活性を与え、一方C末端「アドレス」は特異性および増強された効力を与える。(ChavkinおよびGoldstein,PNAS 78:6543-7,1981)。エキソルフィンは、一般に、古典的なオピエートペプチドのN末端Tyr-Glyを置き換えたTyr-Proを有する。このプロリン残基は、アミノペプチダーゼ分解に対するより高度の安定性を与えると考えられている(Shippら,PNAS 86:287-,1989;Glamstaら,BBRC 184:1060-6,1992)。
【0027】
最近、オーファンレセプター(「ORL1」)が、μ、δ、およびκオピエートレセプターに対する配列関連性によってクローン化された(Mollereauら,FEBS 341:33-38,1994;Fukudaら,FEBS 343:42-46,1994;Bunzowら,FEBS 347:284-8,1994;Chenら,FEBS 347:279-83,1994;Wangら,FEBS 348:75-79,1994;Keithら,Reg.Peptides 54 143-4,1994;Wickら,Mol.Brain Res.27:37-44,1994;Halfordら,J.Neuroimmun.59:91-101,1995)。ノシセプチン(nociceptin)またはオーファニン(orphanin)FQ(本明細書中以下「ノシセプチン」という)と様々に呼ばれるこのレセプターに対するリガンドは、クローン化され、そしてより大きな前駆体由来のヘプタデカペプチドであることが示された(Meunierら,Nature 377:532-535,1995;Reinscheidら,Science 270:792-794,1995)。これは、鎮痛特性を有する古典的なオピエートに反して、インビボで前侵害性の痛覚過敏機能を有することが示された。ノシセプチンは、先に議論した古典的なオピエートペプチドのTyr-Gly-Gly-Phe...というN末端モチーフと対照的に、Phe-Gly-Gly-Phe...というN末端モチーフを有する。古典的なオピエートペプチドのオピエート活性についてのN末端Tyrの要件と一致して、ノシセプチンはμ、κまたはδオピエートレセプターに対してほとんどまたは全く親和性を示さない。同様に、広域スペクトルオピエートアンタゴニストのナロキソンは、ORL1に対して感知できるほどの親和性を有さない。
【0028】
エンケファリンは、固定化ストレスの状況下で、インビボでマウス造血に効果を有することが観察された(Goldbergら,Folia Biol.(Praha)36:319-331,1990)。骨髄の造血を、Leuエンケファリンは阻害し、Metエンケファリンは刺激した。これらの効果は、間接的に、グルココルチコイドレベルおよびTリンパ球遊走に対する効果によると、Goldbergらは考えた。Krizanac-Bengezら(Biomed.& Pharmacother.46:367-43,1992;Biomed.& Pharmacother.49:27-31,1995;Biomed.& Phrmacother.50:85-91,1996)は、インビトロでのこれらの化合物の効果を観察した。マウス骨髄のMet-またはLeu-エンケファリンまたはナロキソンでの前処理は、コロニーアッセイで観察されたGM前駆細胞の数に影響を及ぼした。この効果は、高度に可変性であり、そして抑制、刺激、または効果なしという結果を生じた。さらに、明らかな用量応答は存在しなかった。この可変性は、Krizanac-Bengezらは、概日性リズムおよび補助細胞のためであるとしている。
【0029】
最近、μオピエートレセプターが相同性組換えにより欠失したマウスは、大腿あたりのCFU-GM、BFU-EおよびCFU-GEMMの数が上昇することが実証された。骨髄および脾臓前駆細胞は、正常マウスと比較してこれらのμレセプターノックアウトマウスではより急速に循環していた。これらの効果が、これらの動物におけるμレセプターの不在から生じる骨髄幹細胞に対する直接的または間接的効果に起因するかどうかは決定されなかった(Broxmeyerら,Blood 88:338a,1997)。
【0030】
I.ガンの化学療法および放射線療法
刺激性増殖因子に関する生産的な研究は、多くのこれらの因子(エリスロポエチン、G-CSF、GM-CSFなど)の臨床的な使用をもたらした。これらの因子は、化学療法および放射線処置に関連した死亡率および罹患率を低下させた。化学療法または放射線照射を受けている患者に対するさらなる臨床的利点が、幹細胞が細胞周期に入ることをブロックし、それによって幹細胞を有毒な副作用から保護する代替のストラテジーにより実現され得る。この保護の逆は、化学または放射線処置後の、骨髄機能の迅速な回復を可能にする。
【0031】
II.骨髄および幹細胞移植、エキソビボ幹細胞拡大、ならびに腫瘍パージング
骨髄移植(BMT)は、種々の血液疾患、自己免疫疾患および悪性疾患にとって有用な処置である。現在の治療法は、臍帯血、胎児肝臓から、または末梢血(移動化していないか、またはG-CSFもしくはシクロホスファミドのような因子によって移動化される)から、ならびに骨髄から得られた造血細胞を含む;幹細胞は未精製、部分的に精製(例えば、CD34+集団のアフィニティー精製)、または高度に精製(例えば、CD34,CD38、またはローダミンのようなマーカーを使用する、蛍光活性化細胞選別により)され得る。造血細胞のエクスビボ操作は、現在では、原始幹細胞を移植に適した集団に拡大するために用いられている。この手順を最適化するには以下のことが必要である:(1)造血の長期の再構築を維持し得るのに十分な数の幹細胞;(2)移植片対宿主誘導Tリンパ球の涸渇、および(3)悪性細胞の残留がないこと。この手法は、幹細胞阻害剤および/または幹細胞刺激剤を含むことにより最適化され得る。
【0032】
残留の悪性細胞を排除するために細胞傷害性薬物を用いて造血細胞をパージ(purge)することの効果は、正常な造血細胞および特に幹細胞に対するこれらの化合物の毒性により制限される。パージ中は、正常細胞を効果的に保護する必要性がある。保護は、効果的な阻害剤を用いて、幹細胞をその細胞周期からはずすことにより与えられ得る。
【0033】
III.末梢幹細胞の採取
末梢血幹細胞(PBSC)は、自家移植に対して骨髄よりも多くの潜在的な利点を提供する。腫瘍の保持、または以前の放射線処置により適切な骨髄採取部位を持たない患者は、さらにPBSC採集を受け得る。血液幹細胞の使用は、一般的な麻酔法および外科的手段に十分に耐えられない患者におけるそれらの必要性を排除する。血液細胞を採集するのに必要な血液成分分離技術は、効率的であり、そしてほとんどの主要な医療センターで広く利用可能である。この方法の主な制限事項は、末梢血中の幹細胞が正常な定常状態にある頻度が低いこと、および薬物または増殖因子(例えば、シクロホスファミド、G-CSF、幹細胞因子)を用いた移動手順の後では細胞周期の進行が速いことの両方である。効果的な幹細胞阻害剤は、そのような細胞を休止状態に戻すために有用であり、それによって分化の間にそれら細胞が消失することを防ぐ。
【0034】
IV.過剰増殖性疾患の処置
多くの疾患は、調節異常の幹細胞が、末期細胞の過剰生産を引き起こす過剰増殖状態によって特徴付けられる。このような病態は、上皮細胞の過剰生産である乾癬、および腸ポリープ出現を特徴とする胃腸管における前悪性状態,および初期幹細胞がHIVによって感染されていないが迅速に循環し、幹細胞の消耗を生じる後天性免疫不全症候群(AIDS)を含むが、それらに限定されない。幹細胞阻害剤はこのような状態の処置に有用である。
【0035】
V.過剰増殖障害の処置
多くの疾患は、調節異常の幹細胞が末期細胞の生産不足を引き起こす過剰増殖状態によって特徴づけられる。このような病態は、血液細胞の生産不足である脊髄形成異常症候群または再生不良性貧血、および細胞の再生および置換の欠乏が存在する加齢と関連する状態を含む。幹細胞刺激剤は、このような状態の処置に有用である。
【0036】
VI.遺伝子導入
遺伝情報を造血細胞に導入する能力は、現在では、臨床的設定に利用されている。利用の容易さ、エクスビボでこの組織を操作し処理する幅広い経験のため、および血液細胞が組織に浸透する能力のため、造血細胞は遺伝子療法の有用な標的である。さらに、特定のヒトの遺伝子欠損の修正は、機能的遺伝子をヒト造血系の原始幹細胞に挿入することにより可能であり得る。
【非特許文献1】Eavesら、Blood 78:110-117、1991
【非特許文献2】Lord,Stem Cells(C.S.Potten,Ed.)pp 401-22,1997(Academic Press,NY)
【非特許文献3】LordおよびWright,Blood Cells 6:581-593,1980
【非特許文献4】Zioudrouら,JBC 254:2446-9,1979
【非特許文献5】QuirionおよびWeiss,Peptides 4:445,1983
【非特許文献6】Loukasら,Biochem.22:4567,1983
【発明の開示】
【発明が解決しようとする課題】
【0037】
レトロウイルスベクターまたは遺伝子導入の物理的技術のいずれかを用いて、遺伝子をヒト造血細胞に導入することにはいくつかの制限がある:(1)造血組織中の幹細胞の頻度の低さは、効率の高い遺伝子導入技術の開発を必要とする;および(2)より迅速に細胞周期の進行している幹細胞はベクターの感染をより受けやすくなることが証明されたが、成長因子による幹細胞増殖の刺激によって感染頻度が増加することは、導入遺伝子を含む細胞が不可逆的に分化させられ、そしてそれらの自己再生ができなくなるので、長期の遺伝子発現にネガティブな影響を与える。これらの問題は、分化および自己再生の損失を妨げるための幹細胞阻害剤、および幹細胞の細胞周期への進入を調節するための幹細胞刺激剤の使用によって改善され、それによってレトロウィルス媒介遺伝子導入を促進し得る。
【課題を解決するための手段】
【0038】
発明の要旨
本発明は、幹細胞増殖阻害剤および/または刺激剤(INPROLおよびオピエート化合物)であるペプチドおよびポリペプチドを含む化合物、およびそれらの使用に関する。
【0039】
本発明は、以下の性質により特徴付けられる幹細胞増殖阻害剤を包含する:
(a)マウスコロニー形成脾臓(CFU-S)アッセイにおいて比活性(IC50)が20ng/ml以下である(実施例4を参照のこと)、
(b)(限外濾過により)分子量が10,000ダルトンより大きく、かつ100,000ダルトンより小さい、
(c)活性がトリプシンによる分解に対して感受性である、
(d)MIP-1αまたはTGFβよりも疎水性であり、そして逆相クロマトグラフィーにより両方から分離可能である(実施例12を参照のこと)、
(e)水溶液中で37℃、55℃または75℃で1時間加熱した後でも生物学的活性が保持されている、および
(f)1%塩酸のアセトン溶液を用いて沈澱させた後でも生物学的活性が保持されている。
【0040】
本発明はさらに、短時間のプレインキュベーションの後のインビトロアッセイにおける阻害の達成能力によって特徴付けられ、そして他の幹細胞阻害剤候補(例えば、MIP-1α、TGFβ、および種々のオリゴペプチド)と区別される。
【0041】
本発明はまた、種々の疾患の処置のためのINPROLを含有する薬学的組成物を包含する。
【0042】
本発明は、幹細胞を死滅させるかまたは損傷し得る薬剤への曝露が予想される被験体に有効量の幹細胞阻害性組成物を投与することにより、その被験体を処置する方法を提供する。この方法によって保護された幹細胞は、通常は骨髄、臍帯血、胎児肝に存在し、分裂するか、または末梢血循環へ移動した造血幹細胞であり得る。蛍光活性化セルソーター(FACS)分析により、大部分の移動した幹細胞は静止状態であり、多分化能の幹細胞は細胞周期中であり、幹細胞阻害量のINPROLにより阻害され得ることが実証された。あるいは、幹細胞は上皮性(例えば、腸管または頭皮または身体の他の領域に位置する)であり得るか、あるいは生殖器官内に位置する生殖細胞であり得る。本発明の方法は、望ましくは、ヒトに用いられ得るが、動物の処置もまた、本方法に包含される。本明細書で用いられる用語「被験体」または「患者」は、動物(例えば哺乳動物(ヒトを含む))を指す。
【0043】
本発明はまた、低増殖性の幹細胞を有する被験体を、有効量の幹細胞刺激性組成物をその被験体に投与することにより処置する方法を提供する。この方法により刺激された幹細胞は、通常は骨髄、臍帯血、胎児肝に存在するか、または末梢血循環へ移動した造血幹細胞であり得る;このような幹細胞は、以前に幹細胞阻害量のINPROLの使用により、静止状態へ置かれ得る。幹細胞刺激量のINPROLは、所望の場合、例えばエクスビボ拡大の間の使用のために幹細胞を収集後、またはその後のインビボでの幹細胞の移植(transplantation)および移植(engraftment)後に、幹細胞の細胞周期の刺激を可能にする。あるいは、幹細胞は;上皮性(例えば小腸、もしくは頭皮または身体の他の領域)であり得るか、あるいは生殖器官内に位置する生殖細胞であり得る。
【0044】
他の局面では、本発明は、化学療法を受けている患者の造血幹細胞系、免疫幹細胞系または他の幹細胞系を保護し、かつ回復させる方法を提供する。この方法は患者に有効な幹細胞阻害量のINPROLを投与する工程、および/または、有効な幹細胞刺激量のINPROLを投与することによって、化学療法または放射線照射後の回復を刺激する工程を含む。
【0045】
なおさらなる局面では、本発明は、ガンを有する患者に有効な幹細胞阻害量のINPROLを投与して、骨髄、胃腸管、または他の器官の幹細胞を、化学療法または放射線療法の有害な作用から保護することにより、および/または幹細胞刺激量のINPROLを投与することによって化学療法または放射線照射後の回復を刺激することにより、固形腫瘍(例えば、乳、大腸、肺、精巣、卵巣、肝臓、腎臓、膵臓、脳、肉腫)によって特徴付けられるガンを含む、任意のガンを補助的に処置する方法を包含する。
【0046】
本発明のさらに別の局面は、正常幹細胞の増殖を阻害するために、増殖性白血病細胞を内部に有する造血細胞を有効量のINPROLで処理する工程、および白血病細胞を破壊するために、細胞傷害性薬剤で骨髄を処理する工程を含む、白血病(例えば、慢性骨髄性白血病、急性骨髄性白血病、慢性リンパ性白血病、急性リンパ性白血病、骨髄腫、ホジキン病)の処置を包含する。この方法は、骨髄の増殖を刺激する他の薬剤(例えば、コロニー刺激剤、および/または幹細胞刺激量でのINPROL)で骨髄を継続処置することによって増強され得る。1つの実施態様において、この方法はインビボで行われる。あるいは、この方法はまた、移植のためにエクスビボで造血細胞をパージし拡大するのに有用である。
【0047】
なおさらなる別の局面では、この方法は、増殖性幹細胞によって引き起こされる任意の疾患を有する被験体を処置する工程を含む。乾癬、脊髄形成異常、いくつかのの自己免疫疾患、加齢における免疫低下、脊髄形成異常症候群、再生不良性貧血またはAIDSによる幹細胞の消耗のような疾患は、被験体に有効量のINPROLを投与して問題の幹細胞の増殖を阻害または刺激することにより処置される。
【0048】
本発明は、幹細胞を死滅させるかまたは損傷し得る細胞傷害性薬剤由来の損傷から幹細胞を可逆的に保護する方法を提供する。この方法は、そのような薬剤への暴露が予想される被験体に有効な幹細胞阻害量のINPROLを投与する工程を含む。
【0049】
本発明はまた、化学療法または放射線照射後の回復期の間に幹細胞の増殖を可逆的に刺激する方法を提供する。この方法は、このような薬剤への暴露が予想される被験体に、有効な幹細胞刺激量のINPROLを投与する工程を包含する。
【0050】
本発明はまた以下を提供する:
以下の手順によってブタまたは他の骨髄から単離された幹細胞増殖阻害剤(実施例12を参照のこと):
(a)骨髄を抽出し、そして濾過によって粒子状物質を除去する;
(b)56℃で40分間熱処理した後、氷浴で冷却する;
(c)4℃で30分間10,000gで遠心分離することによって沈澱物を除去する;
(d)10容量の撹拌された氷冷アセトン(1容量%の濃塩酸を含む)に上清を添加し、そして4℃で16時間インキュベーションすることにより酸沈澱させる;
(e)4℃で30分間20,000gで遠心分離することにより沈澱物を単離し、そして冷アセトンで洗浄した後、乾燥させる;
(f)逆相クロマトグラフィーによって単離し、そして5-フルオロウラシルで前処理し、マウスIL-3存在下でインキュベートした骨髄細胞によるコロニー形成の阻害、ならびに280nmにおける吸光度およびSDS-PAGEにより活性をモニターする。
【0051】
本発明はまた以下を提供する:
実質的に他のタンパク性物質を含まない幹細胞増殖阻害剤を精製する方法であって、前記の工程を包含し、さらに以下により詳細に記載される方法。
【0052】
本発明はまた以下を提供する:
幹細胞増殖阻害剤が、幹細胞の過剰増殖によって引き起こされる免疫抑制を改善するように作用する、ヒトまたは動物を処置する方法。
【0053】
本発明はまた以下を提供する:
幹細胞刺激量のINPROLが、幹細胞の低増殖によって引き起こされる骨髄抑制を改善する、ヒトまたは動物を処置する方法。
【0054】
本発明はまた以下を提供する:
前記幹細胞増殖阻害剤が、細胞傷害性薬物または放射線照射手順に曝されることによって幹細胞が増殖するように誘導された後、投与される、ヒトまたは動物を処置する方法。幹細胞は通常は休止しているが、化学療法後、細胞周期に入るように刺激される。これによって幹細胞は2回目の化学療法剤の投与に対してより感受性となる;本方法はこの処置から幹細胞を保護する。
【0055】
本発明はまた以下を提供する:
幹細胞増殖の刺激剤(例えば、幹細胞刺激量でのINPROL)が、骨髄の再生を促進するために、幹細胞阻害量のINPROL前後に投与される、ヒトまたは動物を処置する方法。幹細胞阻害量のINPROLは、幹細胞の細胞周期への移行速度を遅らせ、そして化学療法や放射線照射から保護する;幹細胞刺激量のINPROLはこの阻害を逆転し、そして骨髄の回復を促進する。逆に、幹細胞刺激量のINPROLは骨髄の回復に使用され得るが、幹細胞阻害量のINPROLは一旦骨髄の回復が達成すると、次に幹細胞を静止状態に戻すために使用される。
【0056】
本発明はまた以下を提供する:
前記幹細胞増殖阻害剤が、免疫応答を増大させるためにワクチン接種の前または同時にアジュバントとして投与される、ヒトまたは動物を処置する方法。
【0057】
本発明はまた以下を提供する:
前記哺乳動物に免疫刺激的量のINPROLを投与する工程を含む、哺乳動物の免疫不全を処置する方法。
【0058】
本発明はまた以下を提供する:
前記哺乳動物に鎮痛誘導量のINPROLを投与する工程を含む、哺乳動物の疼痛を処置する方法。
【0059】
本発明はまた以下を提供する:
幹細胞を損傷から保護するために有効量の幹細胞増殖阻害剤を投与する工程を含む、細胞傷害性薬物または放射線処置を受けたヒトまたは動物を処置する方法。
【0060】
本発明はまた以下を提供する:
処置後の回復を増強するために、有効な幹細胞刺激量のINPROLを投与する工程を含む、細胞傷害性薬物または放射線処置を受けたヒトまたは動物を処置する方法。
【0061】
本発明はまた、ヘモグロビンおよび薬学的に受容可能なキャリアを含む薬学的組成物を含む。
【0062】
本発明はまた、(a)ヘモグロビンのα鎖、ヘモグロビンのβ鎖、ヘモグロビンのγ鎖、ヘモグロビンのδ鎖、ヘモグロビンのε鎖、およびヘモグロビンのζ鎖からなる群から選択されるポリペプチドであって、ポリペプチドはヒトαヘモグロビン鎖のアミノ酸1-97(「ペプチド1-97」)を含み、そしてポリペプチドはヒトαヘモグロビン鎖のアミノ酸1-94(「ペプチド1-94」)を含む、ならびに(b)薬学的に受容可能なキャリアを含む薬学的組成物を包含する。このような薬学的組成物は、ヘムと共にまたはヘムを有さずに、上記群から選択される単一ポリペプチド、上記群から選択されるポリペプチドの混合物、または上記群から選択される二量体または多量体の形態のポリペプチドで構成され得る。
【0063】
本発明はまた、以下の配列を有するペプチドを包含する:
【0064】
【化1】
ここで、2つのCys残基はジスルフィド結合を形成する(「環状ペプチド43-55」)
【0065】
【化2】
ここで、2つのCys残基は炭素架橋により結合されている、
【0066】
【化3】
および
図16Aに示すように、ヒトαヘモグロビンの最初の97 N末端アミノ酸を含むペプチド。
【0067】
本発明はまた、幹細胞刺激および/または刺激特性を保持した、ヒトα鎖の修飾型からなるタンパク質およびペプチド配列を含む。そのような修飾は、C末端疎水性ドメインの除去または修飾(改善された溶解特性を生じる)、および/またはハプトグロビン結合ドメイン除去または修飾(改善された薬物動態学的特性を生じる)を含むが、これらに限定されない。ヒトαヘモグロビンのC末端疎水性ドメインは、アミノ酸98(フェニルアラニン)から141(アルギニン)までの領域を含み、総数44のうち23の疎水性アミノ酸を含む。ドメイン全体または1つ以上のこれらの疎水性アミノ酸(6つのアラニン、4つのバリン、8つのロイシン、2つのプロリンおよび3つのフェニルアラニン)は、欠失(deletion)(C末端疎水性ドメインの「欠失」)によって除去され得る。あるいは、1つ以上のこれらのアミノ酸は、非極性アミノ酸(例えば、グリシン、セリン、トレオニン、システイン、チロシン、アスパラギンまたはグルタミン)で置換され得る(「置換」C末端疎水性ドメイン)。
【0068】
別の実施態様では、カルボキシメチル化のような、この領域の疎水的特性を減じ、溶解性を増加する化学的修飾が使用される。
【0069】
別の実施態様では、疎水性残基はヒトβヘモグロビン配列の対応する親水性の領域と置換される。例えば、ヒトβヘモグロビン配列では、アミノ酸107(グリシン)から117(ヒスチジン)間の領域またはアミノ酸130(チロシン)から139(ヒスチジン)間の領域はそれぞれ比較的親水性で、それぞれまたは共にヒトαヘモグロビン内の等価な疎水性の領域について置換され得る。
【0070】
ハプトグロビン結合ドメインはC末端疎水性領域に含まれ、アミノ酸121-127からなる。この領域は全体的に欠失により除去され得るか、またはこの領域の1つ以上のアミノ酸が欠失され得る(「欠失」C末端ハプトグロビン結合ドメイン)。この領域またはこの領域の1つ以上のアミノ酸は、例えばポリアラニンもしくはポリグリシン、またはポリペプチドのハプトグロビンへの結合を無効にする効果を有するが、幹細胞阻害活性は維持するような他のアミノ酸に置換され得る(「置換」C末端ハプトグロビン結合ドメイン)。
【0071】
本発明の他の実施態様は、βヘモグロビン鎖(C末端疎水性領域内および/または1つまたは両ハプトグロビン結合ドメイン(アミノ酸11-25および136-146))に対する相当する修飾、ならびにδ、γ、ε、および/またはζヘモグロビン鎖に対する相当する修飾を包含する。
【0072】
上記の同定されたペプチドをコードするDNA配列、そのDNA配列を含むベクター、およびそのベクターを含む宿主細胞もまた、本発明に包含される。これらのペプチドは、標準的な化学技術(例えば、固相合成)を用いるか、または組換え技術(融合システム(例えば、グルタチオン-S-トランスフェラーゼ(D.B.SmithおよびK.S.Johnson、Gene 67:31-40,1988)、チオレドキシン(La Vallieら、Biotechnology 11:187-193、1993)、またはユビキチン(Buttら、PNAS 86:2540-4、1989;Cherneyら,Biochem.30:10420-7,1991;Bakerら,JBC 269:25381-6,1994;米国特許第5,132,213号;第5,196,321号および第5,391,490号およびPCT WO 91/17245))を用いる融合システム)を含む)を用いることにより合成され得る。これらの論文、適用、および特許のそれぞれは、本明細書中に参考として援用される。
【0073】
さらに、本発明は、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、オピエートレセプター、好都合には、オピエートレセプターのμサブクラスに結合し得る化合物とを接触させる工程を含む方法を包含する。さらに、本発明は、幹細胞増殖を阻害または刺激する方法であって、造血細胞とノシセプチン(nociceptin)レセプター(例えば、ORL1)に結合し得る化合物とを接触させる工程を含む方法を包含する。さらに、本発明は、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、「オピエート様」レセプター結合し得る化合物とを接触させる工程を含む方法を包含する。
【0074】
オピエート活性を示すペプチド(「ヘモルフィン(hemorphin)」と呼ばれる)が、ヘモグロビンから単離されている(例えば、Brantlら、Eur.J.Pharm.、125:309-10、1986;Davisら、Peptides 10:747-51、1989;Hoffmanら,PNAS 87:8721-25,1990;Hernanら,Biochem.31:8619-28,1992;Karelinら、Bioch.Biophys.Res.Comm、202:410-5、1994;Zhaoら、Ann.N.Y.Acad.Science 750:452-8、1995;Petrovら,Bioscience Reports,15:1-14,1995;Karelinら,16:693-697,1995)。これらの論文のそれぞれは、本明細書中に参考として援用される。他の異型のオピエートペプチドおよび小分子もまた存在する(Zioudrouら,JBC 254:2446-9,1979;QuirionおよびWeiss,Peptides 4:445,1983;Loukasら,Biochem.22:4657,1983;Brantl,Eur.J.Pharm.106:213-14,1984;Brantlら,Eur.J.Pharm.111:293-4,1985;BrantlおよびNeubert,TIPS 7:6-7,1986;HurbyおよびGehrig,Med.Res.Rev.9:343-401,1989;Schiller,Prog.Med.Chem.28:301-40,1991;Glamstaら,BBRC 184:1060-6,1992;Teschemacher,Handbook Exp.Pharm.104:499-28,1993;Handbook of Experimental Pharmacology,A.Hertz(編)volumes 104/Iおよび104/II,1993,Springer Verlag,Berlin;Reedら,Neurosci.and Biobehav.Rev.18:519-25,1994;Karelinら,Peptides 16:693-7,1995)。これらの論文のそれぞれは、本明細書中に参考として援用される。本明細書中で使用されたように、「オピエート様レセプター」は、オピエート、INPROL、ヘモルフィン(hemorphins)、エキソルフィン(exorphins)、ノイセプチン(noiceptin)、Tyr-MIF-1ファミリーメンバー、アルカロイド、および/または適当量のナロキソン(naloxone)を含ませることにより拮抗されるように幹細胞増殖を阻害または刺激する他の化合物に結合するそれらの能力によって定義される(実施例29および38を参照のこと)。
【0075】
さらに、本発明は、レセプター(単数または複数)およびリガンドを同定する方法であって、結合アッセイにおいてINPROL(好都合には、ペプチド1-94、1-97、43-55または64-82のようなペプチド形態)を使用する工程を含む方法を包含する。さらに、本発明は、レセプター(単数または複数)およびリガンドを同定する方法であって、アデニル酸シクラーゼアッセイにおいてINPROLを使用する工程を含む方法を包含する。
【0076】
さらに、本発明は、幹細胞増殖を阻害または刺激する方法であって、造血細胞とGTP結合タンパク質(好都合には、これらのG阻害性サブタイプのタンパク質)を活性化し得る化合物(例えば、マストパラン(mastoparan))とを接触させる工程を含む方法を包含する。
【0077】
本発明はまた、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、以下の配列を有するヘモルフィンペプチドの群から選択されるペプチドとを接触させる工程を含む方法を包含する:
【0078】
【化4】
。
【0079】
上記ペプチドは、他の異型オピエートペプチド(例えば、Tyr-MIF-1ファミリー(概説についてはReedら、Neurosci.Biobehav.Rev.18:519-25、1994を参照のこと)、カゼイン由来カソモルフィン(casomorphin)(Brantlら、Hoppe-Seyler’s Z.Physiol.Chem.360:1211-16、1979;Loukasら、Biochem.22:4567-4573、1983;FiatおよびJolles、Mol.Cell.Biochem.87:5-30、1989)、シトクロームb由来のペプチド(シトクロフィン(cytochrophin)と称される)(Brantlら、Eur.J.Pharm.111:293-4、1985)、ヒトおよびヒト以外の種由来の種々なエキソルフィンおよびオピエートペプチド(Zioudrouら,JBC 254:2446-9,1979;Brantl,Eur.J.Pharm.106:213-14,1984;Brantlら,Eur.J.Pharm.125:309-10,1986;BrantlおよびNeubert,TIPS 7:6-7,1986;Glamstaら,BBRC 184:1060-6,1992;Teschemacher,Handbook Exp.Pharm.104:499-28,1993;Karelinら,Peptides 16:693-7,1995)、ならびにオピエートレセプターへの結合についてスクリーニングされたコンビナトリアルライブラリー由来ペプチド(概説についてはDooleyら、Peptide Research 8:124-137、1995を参照のこと))に類似の配列類似性および/または生物学的活性を有する。これらの論文のそれぞれは、本明細書中に参考として援用される。
【0080】
本発明はまた、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、Tyr-MIF-1関連ペプチド、カソモルフィン、シトクロフィンおよびエキソルフィンからなる群から選択されるペプチドとを接触させる工程を含む方法を包含する。以下の配列を有するTyr-MIF-1ペプチドが特に含まれる:
【0081】
【化5】
。
【0082】
本発明はまた、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、以下の群から選択されるオピエートペプチドとを接触させる工程を含む方法を包含する:
【0083】
【化6】
。
【0084】
本発明はまた、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、モルヒネ、コデイン、メタドン、ヘロイン、メペリジン(meperidine)、アルファプロジン、ジフェノキシラート、フェンタニール、スフェンタニル、アルフェンタニル、レボルファノール、ハイドロコドン、ジヒドロコデイン、オキシコドン、ヒドロモルフォン、プロポキシフェン、ブプレノルフィン、エトルフィン、オキシモルフォン デキストプロポキシフェン(oxymorphone dextopropoxyphene)およびメプタジノールからなる群から選択されるオピエートアゴニスト化合物とを接触させる工程を含む方法を包含する。特に、10-7モル未満の阻害量のモルヒネが含まれる。
【0085】
本発明はまた、幹細胞増殖を阻害または刺激する方法であって、造血細胞と、ナロキソン、ナルトレキソン、ナロルフィン、ペンタゾシン、ナルブフィン、ブトルファノールからなる群から選択されるオピエートアンタゴニストまたは混合アゴニスト/アンタゴニストとを接触させる工程を含む方法を包含する。特に、10-8モル未満の阻害量のナロキソンが含まれる。
【0086】
本発明はまた、幹細胞増殖を刺激する方法であって、造血細胞と、INPROL、ミオグロビン、DAMGO、およびDALDAからなる群から選択されるタンパク質またはペプチドの幹細胞刺激量とを接触させる工程を含む方法を包含する。
【0087】
本発明はまた、哺乳動物において遺伝子療法を実施する方法を包含し、この方法は以下の工程を含む:
a)上記哺乳動物から造血細胞を取り出す工程、
b)エクスビボで上記造血細胞を幹細胞刺激量のINPROLおよび/またはオピエート化合物で処理する工程、
c)予め決定された遺伝子で上記造血細胞をトランスフェクトまたは感染する工程、
d)エクスビボで上記トランスフェクト造血細胞と幹細胞阻害量のINPROLおよび/またはオピエート化合物とを接触させる工程、
e)上記哺乳動物に工程dの造血細胞を移植する工程、
f)必要に応じて、インビボで上記哺乳動物を幹細胞を阻害または刺激する量のINPROLおよび/またはオピエート化合物で処理する工程。
【0088】
本発明はまた、エクスビボで幹細胞拡大を実施する方法であって、この造血細胞を幹細胞阻害量のINPROLおよび少なくとも1つの刺激性サイトカインで処理する工程を含む方法を包含する。INPROLは、刺激性サイトカインと接触させる前、間、および/または後で、造血細胞と接触させられる。エクスビボでの幹細胞の拡大は、臍帯血、胎児肝、化学療法などの後または精製後(例えば、CD34、CD38またはローダミンのようなマーカーを使用する蛍光活性化細胞選別による)の自家骨髄のような限られた供給源から十分量の幹細胞の産生が可能となる。特定の造血系列を選択的に増殖する能力はまた、臨床家が個々の患者の必要性に従って幹細胞移植を特別に設計することを可能とする。
【0089】
本発明はまた、エクスビボでの幹細胞の拡大を実施する方法であって、少なくとも1つさらなる刺激性サイトカインと共にまたはこれを伴わずに、造血細胞を幹細胞刺激量のINPROLで処理する工程を含む方法を包含する。INPROLは、刺激性サイトカイン(単数または複数)と接触させる前、間および/または後で、造血細胞と接触させられる。エクスビボで、幹細胞刺激剤は幹細胞および/または前駆細胞の拡大を可能にし、一方、幹細胞阻害剤は幹細胞の未分化状態に幹細胞を維持する。手順はまた、インビボで幹細胞阻害量でのINPROLの使用により、幹細胞を移植されるまで静止状態に維持することを最適化し得、後に幹細胞刺激量でのINPROLが、骨髄再生を刺激するために使用され得る。必要に応じて、造血細胞は2つの調製物に分割し得、そして幹細胞刺激量のINPROLで処置されたものは、幹細胞および/または前駆細胞の拡大を促進し、一方幹細胞阻害量のINPROLで処理された別のものは幹細胞をその未分化状態に維持する。次に、2つの調製物は組み合わされ得、そして患者に注入され得る。
【0090】
本発明はまた、(a)INPROLおよび(b)少なくとも1つの阻害性化合物(MIP-1α、TGFβ、TNFα、INFα、INFβ、INFγ、ペンタペプチドであるピロGlu-Glu-Asp-Cys-Lys、テトラペプチドであるN-アセチル-Ser-Asp-Lys-Pro、およびトリペプチドであるグルタチオン(Gly-Cys-γGlu)からなる群から選択される)を含む薬学的組成物を包含する。
【0091】
本発明はまた、(a)INPROLおよび(b)少なくとも1つの刺激性サイトカイン(IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-11、IL-13、IL-14、IL-15、G-CSF、GM-CSF、M-CSF、エリスロポエチン、トロンボポエチン、幹細胞因子、δ様タンパクおよびflk2/flt3リガンドからなる群から選択される)を含む薬学的組成物を包含する。
【0092】
本発明は、幹細胞阻害剤(INPROL)を記載する。この阻害剤は、MIP-1α、TGFβ、Frindelおよびその共同研究者のテトラペプチド、またはPaukovitsおよびその共同研究者のペンタペプチド(WrightおよびPragnell、1992(前出)参照)のような当該分野で公知の阻害剤とは異なる。天然に存在するネイティブINPROLは、INPROLとテトラペプチドおよびペンタペプチドとを区別する限外濾過によれば、10,000ダルトンを超える分子量を有する。ネイティブINPROLは、逆相クロマトグラフィーシステムにおいてはMIP-1αまたはTGFβよりも疎水性であり、これをそれらのサイトカインから区別する。さらに、その作用様式は、プレインキュベーションの期間でのみ用いられる場合に、インビトロアッセイで活性である点において、前記のいずれの阻害剤の作用様式とも異なる。例えば、MIP-1αは、プレインキュベーションの期間でのみ用いられる場合、効果的ではない(実施例5)。さらに、天然に存在するINPROLは、「高増殖性潜在細胞(high proliferative potential cells)」(HPP-PFC)を測定するアッセイにおいて活性であるが、MIP-1αは活性ではない(実施例6)。INPROLは、IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-10、IL-11、IL-12、IL-13、IL-14、IL-15、IL-16、G-CSF、GM-CSF、M-CSF、エリスロポエチン、トロンボポエチン、幹細胞因子、およびflk2/flt3リガンドのような当該分野では公知のこれらの刺激剤とは異なる。天然に存在するINPROLは、これらのサイトカインとはほとんどまたは全く配列類似性を示さない。
【0093】
本発明はまた、以下の項目を提供する:
項目1.C末端疎水性ドメインが置換または欠失されているヘモグロビンα鎖を含むポリペプチド。
項目2.C末端ハプトグロビン結合ドメインが置換または欠失されているヘモグロビンα鎖を含むポリペプチド。
項目3.ヒトαヘモグロビン鎖のアミノ酸1-97を含むポリペプチド。
項目4.(a)請求項1または2に記載のポリペプチドおよび(b)薬学的に受容可能なキャリアを含む薬学的組成物。
項目5.ヒトαヘモグロビン鎖のアミノ酸1-97からなるポリペプチドおよび薬学的に受容可能なキャリアを含む薬学的組成物。
項目6.ヒトαヘモグロビン鎖のアミノ酸1-94からなるポリペプチドおよび薬学的に受容可能なキャリアを含む薬学的組成物。
項目7.単位投与量形態の請求項4から6に記載の薬学的組成物。
項目8.0.1mgから6gの、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチドおよびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される1つまたは2つの化合物を含む、請求項7に記載の薬学的組成物。
項目9.幹細胞増殖を阻害する方法であって、造血細胞と幹細胞増殖阻害量の請求項1または2に記載のポリペプチドとを接触させる工程を包含する、方法。
項目10.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、および配列Phe-Leu-Gly-Phe-Pro-Thrを有するペプチドからなる群より選択される、請求項9に記載の方法。
項目11.B細胞の増殖を刺激する方法であって、造血細胞と増殖刺激量の請求項1または2に記載のポリペプチドとを接触させる工程を包含する、方法。
項目12.ガンに羅患している哺乳動物においてガンを処置する方法であって、該方法は以下の工程:
(a)放射線療法または化学療法を投与する工程、および
(b)幹細胞増殖阻害量の請求項1または2に記載のポリペプチドを投与する工程、
を包含する、方法。
項目13.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチドおよびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項12に記載の方法。
項目14.前記工程aおよびbが1回以上繰り返される、請求項12に記載の方法。
項目15.前記工程aが前記工程bの後に行われる、請求項12に記載の方法。
項目16.前記工程bが前記工程aの前または後の24時間以内に行われる、請求項12に記載の方法。
項目17.哺乳動物においてガンを処置する方法であって、該方法は以下の工程:
a)該哺乳動物から造血細胞を取り出す工程、
b)該造血細胞を、請求項1または2に記載のポリペプチドでエクスビボで処理する工程、
c)該工程bの造血細胞を、化学療法または放射線で処理する工程、
d)該哺乳動物において骨髄切除処置を行う工程、および
e)該工程cの造血細胞を該哺乳動物に移植する工程、
を包含する、方法。
項目18.前記工程(b)におけるポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチドおよびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項17に記載の方法。
項目19.幹細胞に損傷を与えるか、または破壊する薬剤に暴露された哺乳動物において幹細胞分裂を阻害する方法であって、幹細胞増殖阻害量の請求項1または2に記載のポリペプチドを投与する工程を包含する、方法。
項目20.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、および配列Phe-Leu-Gly-Phe-Pro-Thrを有するペプチドからなる群より選択される、請求項19に記載の方法。
項目21.前記薬剤が抗ウイルス剤である、請求項19に記載の方法。
項目22.哺乳動物造血幹細胞をエクスビボで維持する方法であって、造血細胞と幹細胞増殖阻害量の請求項1または2に記載のポリペプチドとを接触させる工程を包含する、方法。
項目23.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、および配列Phe-Leu-Gly-Phe-Pro-Thrを有するペプチドからなる群より選択される、請求項22に記載の方法。
項目24.前記造血細胞が、骨髄細胞、末梢血細胞、移動末梢血細胞、胎児肝臓、および臍帯血細胞からなる群より選択される、請求項22に記載の方法。
項目25.骨髄増殖性疾患もしくは自己免疫疾患または上皮幹細胞過剰増殖を、それらに羅患している哺乳動物において処置する方法であって、過剰増殖減少量の請求項1または2に記載のポリペプチドを投与する工程を包含する、方法。
項目26.前記骨髄増殖性疾患が脊髄形成異常症候群である、請求項25に記載の方法。
項目27.化学療法または放射線から哺乳動物における正常な幹細胞を区別して保護し、そしてガン細胞を保護しない方法であって、幹細胞保護量の請求項1または2に記載のポリペプチドを投与する工程を包含する、方法。
項目28.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、および配列Phe-Leu-Gly-Phe-Pro-Thrを有するペプチドからなる群より選択される、請求項27に記載の方法。
項目29.前記正常な幹細胞が細胞傷害性薬物または放射線への暴露により増殖するように誘導された後に、前記ポリペプチドが投与される、請求項27に記載の方法。
項目30.哺乳動物をワクチン接種する方法であって、ワクチン投与の前、間、または後に請求項1または2に記載のポリペプチドをアジュバントとして投与する工程を包含する、方法。
項目31.幹細胞過剰増殖により引き起こされる免疫抑制を有する哺乳動物を処置する方法であって、該哺乳動物に過剰増殖後退量の請求項1または2に記載のポリペプチドを投与する工程を包含する、方法。
項目32.哺乳動物において遺伝子療法を行う方法であって、該方法は以下の工程:
a)造血細胞を該哺乳動物から取り出す工程、
b)該造血細胞を予め決定された遺伝子でトランスフェクトする工程、
c)該トランスフェクトされた造血細胞と請求項1または2に記載のポリペプチドとをエクスビボで接触させる工程、および
d)該工程cの造血細胞を該哺乳動物に移植する工程、
を包含する、方法。
項目33.前記工程cにおけるポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチドおよびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項32に記載の方法。
項目34.前記工程(a)の後に、幹細胞増殖を誘導するために、前記造血細胞を少なくとも1つの刺激性サイトカインで処理する工程をさらに包含する、請求項32に記載の方法。
項目35.前記工程(d)の後に、前記哺乳動物を前記ポリペプチドでインビボで処置する工程をさらに包含する、請求項32に記載の方法。
項目36.幹細胞拡大をエクスビボで行う方法であって、造血細胞と請求項1または2に記載のポリペプチドおよび少なくとも1つの刺激性サイトカインとを接触させる工程を包含する、方法。
項目37.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、およびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項36に記載の方法。
項目38.前記造血細胞が、骨髄細胞、末梢血細胞、移動末梢血細胞、胎児肝臓、および臍帯血細胞からなる群より選択される細胞である、請求項36に記載の方法。
項目39.(a)請求項1または2に記載のポリペプチド、ならびに(b)MIP-1α、TGFβ、TNFα、INFα、INFβ、INFγ、ペンタペプチドピロGlu-Glu-Asp-Cys-Lys、テトラペプチドN-アセチル-Ser-Asp-Lys-Pro、およびトリペプチドグルタチオン(Gly-Cys-γGlu)からなる群より選択される少なくとも1つの阻害性化合物を含む、薬学的組成物。
項目40.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、およびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項39に記載の方法。
項目41.(a)請求項1または2に記載のポリペプチド、ならびに(b)IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-11、IL-13、IL-14、IL-15、G-CSF、GM-CSF、M-CSF、エリスロポエチン、トロンボポエチン、幹細胞因子、およびflk2/flt3リガンドからなる群より選択される少なくとも1つの刺激性化合物を含む、薬学的組成物。
項目42.前記ポリペプチドが、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、およびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項41に記載の方法。
項目43.αヘモグロビンまたはその置換もしくは欠失アナログを発現させる方法であって、該αヘモグロビンまたは置換もしくは欠失アナログを、ユビキチン融合体として発現させる工程を包含する、方法。
項目44.前記発現させる工程がE.coliにおいて行われる、請求項43に記載の方法。
項目45.前記発現させる工程が、ユビキチン切断酵素を発現させる工程を包含する、請求項43に記載の方法。
項目46.ビオチン-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、(ヨード)Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、および(ヨード)Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Valからなる群より選択される配列を有するペプチド。
項目47.幹細胞増殖を刺激する方法であって、造血細胞と幹細胞増殖刺激量のINPROLおよび/またはオピエート化合物とを接触させる工程を包含する、方法。
項目48.前記INPROLが、ヘモグロビンα鎖、ヘモグロビンβ鎖、ヘモグロビンγ鎖、ヘモグロビンδ鎖、ヘモグロビンε鎖、ヘモグロビンζ鎖、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、およびヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチドからなる群より選択される、請求項47に記載の方法。
項目49.前記INPROLが、以下の配列:
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Cys-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val-Cys
(ここで、2つのCys残基はジスルフィド結合を形成する)、
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala、
Phe-Leu-Gly-Phe-Pro-Thr、
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln-Arg-Phe、
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln-Arg、
Leu-Val-Val-Tyr-Pro-Trp-Thr-Gln、
Leu-Val-Val-Tyr-Pro-Trp-Thr、
Leu-Val-Val-Tyr-Pro-Trp、
Leu-Val-Val-Tyr-Pro、
Val-Val-Tyr-Pro-Trp-Thr-Gln、
Tyr-Pro-Trp-Thr-Gln-Arg-Phe、
Tyr-Pro-Trp-Thr-Gln-Arg、
Tyr-Pro-Trp-Thr-Gln、および
Tyr-Pro-Trp-Thr、
からなる群より選択される、請求項47に記載の方法。
項目50.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項47に記載の方法。
項目51.幹細胞増殖を刺激する方法であって、造血細胞とオピエートレセプターに結合し得る化合物とを接触させる工程を包含する、方法。
項目52.前記化合物がμサブクラスのオピエートレセプターに対する選択性を有する、請求項51に記載の方法。
項目53.幹細胞増殖を刺激または阻害する方法であって、造血細胞とノシセプチンレセプターに結合し得る化合物とを接触させる工程を包含する、方法。
項目54.幹細胞増殖を刺激または阻害する方法であって、造血細胞とG阻害性サブクラスのGTP結合タンパク質を活性化し得る化合物とを接触させる工程を包含する、方法。
項目55.幹細胞増殖を刺激または阻害する方法であって、造血細胞と古典的μ、κ、もしくはδオピエートレセプターまたはORL1を含まないオピエート様レセプターに結合し得る化合物とを接触させる工程を包含し、ここで該レセプターは(a)幹細胞刺激および/または阻害特性を有し、そして(b)ナロキソンにより拮抗可能な該幹細胞刺激および/または阻害能力を有する、方法。
項目56.前記オピエート様レセプターが、1μM未満またはそれに等しい解離定数(Kd)で、ペプチドPhe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Valに結合する能力を有する、請求項55に記載の方法。
項目57.前記解離定数が10nM未満またはそれに等しい、請求項55に記載の方法。
項目58.INPROLに対するレセプターを同定する方法であって、レセプター結合アッセイにおいて該レセプターを含む物質とINPROLとを接触させる工程を包含する、方法。
項目59.前記INPROLが、ヘモグロビンα鎖、ヘモグロビンβ鎖、ヘモグロビンγ鎖、ヘモグロビンδ鎖、ヘモグロビンε鎖、ヘモグロビンζ鎖、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
ビオチン-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
(ヨード)Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
(ヨード)Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Cys-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val-Cys、および
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala、
からなる群より選択される、請求項58に記載の方法。
項目60.INPROLに対するレセプターを同定する方法であって、アデニル酸シクラーゼアッセイにおいて該レセプターを含む物質とINPROLとを接触させる工程を包含する、方法。
項目61.前記INPROLが、ヘモグロビンα鎖、ヘモグロビンβ鎖、ヘモグロビンγ鎖、ヘモグロビンδ鎖、ヘモグロビンε鎖、ヘモグロビンζ鎖、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
ビオチン-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
(ヨード)Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
(ヨード)Phe-Pro-His-(ヨード)Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Cys-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val-Cys、および
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala、
からなる群より選択される、請求項60に記載の方法。
項目62.ガンを羅患している哺乳動物においてガンを処置する方法であって、該方法は以下の工程:
(a)放射線療法および/または化学療法を投与する工程、および
(b)幹細胞増殖刺激量のINPROLおよび/またはオピエート化合物を投与する工程、
を包含する、方法。
項目63.前記工程aおよびbが1回以上繰り返される、請求項62に記載の方法。
項目64.前記工程aが前記工程bの前に行われる、請求項62に記載の方法。
項目65.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項62に記載の化合物。
項目66.幹細胞に損傷を与えるかまたは破壊する薬剤に暴露された哺乳動物において幹細胞分裂を刺激する方法であって、幹細胞増殖刺激量のINPROLおよび/またはオピエート化合物を投与する工程を包含する、方法。
項目67.前記薬剤が抗ウイルス剤または抗腫瘍剤である、請求項66に記載の方法。
項目68.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項66に記載の方法。
項目69.哺乳動物造血幹細胞をエクスビボで維持する方法であって、造血細胞と幹細胞増殖刺激量のINPROLおよび/またはオピエート化合物とを接触させる工程を包含する、方法。
項目70.前記造血細胞が、骨髄細胞、末梢血細胞、移動末梢血細胞、胎児肝臓、および臍帯血細胞からなる群より選択される、請求項69に記載の方法。
項目71.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項69に記載の方法。
項目72.骨髄増殖性疾患、造血幹細胞過剰増殖または上皮幹細胞過剰増殖を、それらを羅患している哺乳動物において処置する方法であって、刺激量のINPROLおよび/またはオピエート化合物を投与する工程を包含する、方法。
項目73.前記骨髄増殖性疾患が脊髄形成異常症候群または再生不良性貧血である、請求項72に記載の方法。
項目74.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項72に記載の方法。
項目75.幹細胞消耗を処置または予防する方法であって、幹細胞増殖阻害量のINPROLおよび/またはオピエート化合物を投与する工程を包含する、方法。
項目76.前記幹細胞消耗が後天性免疫不全症候群によるものである、請求項75に記載の方法。
項目77.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項75に記載の方法。
項目78.化学療法または放射線から哺乳動物における正常な幹細胞を区別して保護する方法であって、幹細胞保護量のオピエート化合物を投与する工程を包含する、方法。
項目79.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項78に記載の方法。
項目80.哺乳動物において遺伝子療法を行う方法であって、該方法は以下の工程:
a)該哺乳動物から造血細胞を取り出す工程、
b)該造血細胞を、幹細胞刺激量のINPROLおよび/またはオピエート化合物でエクスビボで処理する工程、
c)該造血細胞を予め決定された遺伝子でトランスフェクトまたは感染する工程、
d)該トランスフェクトされた造血細胞と、幹細胞阻害量のINPROLおよび/またはオピエート化合物とをエクスビボで接触させる工程、
e)該工程dの造血細胞を該哺乳動物に移植する工程、
f)必要に応じて、該哺乳動物を幹細胞阻害量または刺激量のINPROLおよび/またはオピエート化合物でインビボで処置する工程、
を包含する、方法。
項目81.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項80に記載の方法。
項目82.幹細胞拡大をエクスビボで行う方法であって、造血細胞と幹細胞刺激量のINPROLおよび/またはオピエート化合物とを接触させる工程を包含する、方法。
項目83.前記造血細胞が、骨髄細胞、末梢血細胞、移動末梢血細胞、胎児肝臓、および臍帯血細胞からなる群より選択される細胞である、請求項80に記載の方法。
項目84.前記オピエート化合物が、モルヒネ、エトルフィン、コデイン、ヘロイン、ヒドロモルフォン、オキシモルフォン、レボルファノール、レバロルファン、コデイン、ハイドロコドン、オキシコドン、ナロルフィン、ナロキソン、ナルトレキソン、ブプレノルフィン、ブタノルファノール、ナルブフィン、メペリジン、アルファプロジン、ジフェノキシレート、フェンタニル、DAMGO、DALDA、およびノシセプチンからなる群より選択される、請求項80に記載の方法。
項目85.(a)オピエート化合物、ならびに(b)MIP-1α、TGFβ、TNFα、INFα、INFβ、INFγ、ペンタペプチドピロGlu-Glu-Asp-Cys-Lys、テトラペプチドN-アセチル-Ser-Asp-Lys-Pro、およびトリペプチドグルタチオン(Gly-Cys-γGlu)からなる群より選択される少なくとも1つの阻害性化合物を含む、薬学的組成物。
項目86.a)オピエート化合物、ならびに(b)IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-11、IL-13、IL-14、IL-15、G-CSF、GM-CSF、M-CSF、エリスロポエチン、トロンボポエチン、幹細胞因子、およびflk2/flt3リガンドからなる群より選択される少なくとも1つの刺激性化合物を含む、薬学的組成物。
項目87.哺乳動物において疼痛を処置する方法であって、鎮痛誘導量のINPROLを該哺乳動物に投与する工程を包含する、方法。
項目88.前記INPROLが、ヘモグロビンα鎖、ヘモグロビンβ鎖、ヘモグロビンγ鎖、ヘモグロビンδ鎖、ヘモグロビンε鎖、ヘモグロビンζ鎖、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Cys-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ara-Gln-Val-Cys、および
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala、
からなる群より選択される、請求項87に記載の方法。
項目89.哺乳動物における免疫不全を処置する方法であって、免疫刺激量のINPROLを該哺乳動物に投与する工程を包含する、方法。
項目90.前記INPROLが、ヘモグロビンα鎖、ヘモグロビンβ鎖、ヘモグロビンγ鎖、ヘモグロビンδ鎖、ヘモグロビンε鎖、ヘモグロビンζ鎖、ヒトαヘモグロビン鎖のアミノ酸1-97の配列を有するポリペプチド、ヒトαヘモグロビン鎖のアミノ酸1-94の配列を有するポリペプチド、
Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ala-Gln-Val、
Cys-Phe-Pro-His-Phe-Asp-Leu-Ser-His-Gly-Ser-Ara-Gln-Val-Cys、および
Asp-Ala-Leu-Thr-Asn-Ala-Val-Ala-His-Val-Asp-Asp-Met-Pro-Asn-Ala-Leu-Ser-Ala、
からなる群より選択される、請求項89に記載の方法。
【0094】
本明細書中に記載される本発明がより十分に理解され得るために、以下の詳細な説明を記載する。この説明は、本発明の例示であるが、本発明を特に限定すると解釈されるべきではなく、そして当業者の理解範囲であるような変化は本発明の範囲に入ると考えられるべきである。
【発明を実施するための最良の形態】
【0095】
好適な実施態様の詳細な説明
INPROLは、幹細胞の分裂を可逆的に阻害または刺激する。特定の理論に縛られることを望まないが、幹細胞阻害剤および刺激剤は、幹細胞が細胞周期を通って移行する割合に影響を与えることによってそれらの効果を発揮すると考えられる。特に、INPROLは、使用量に依存して造血幹細胞の細胞分裂を一時的に阻害または刺激することに効果的である。幹細胞増殖を阻害または刺激し得る化合物を臨床的に使用する能力は、例えば化学療法、幹細胞移植または遺伝子療法プロトコルの間の、造血幹細胞の細胞周期進行の精巧な制御を可能にする。従って、本発明の方法は、ガン細胞もしくはウイルスに感染した細胞を破壊するために用いられる化学療法剤もしくは放射線によって引き起こされる損傷から幹細胞を保護することによって、またはそのような損傷後の回復を刺激することによって、患者の造血系、骨髄系および免疫系に対する化学療法の望ましくない副作用を軽減することに用いられ得る。本発明の一つの実施態様では、INPROLは、化学療法剤が疾患細胞に作用している間の幹細胞分裂を阻害するのに十分な用量で患者に投与される。化学療法剤がその機能を発揮した後、INPROLによって阻害された幹細胞は、さらなる処置を受けずに分裂細胞に戻る。造血再生を高めることが所望されるならば、刺激性成長因子、サイトカインまたは幹細胞刺激量のINPROLがさらに用いられ得る。
【0096】
本明細書中で用いられる用語「INPROL」は、実施例におけるように精製された哺乳動物および非哺乳動物タンパク質、ヘモグロビン、ヘモグロビンのα鎖(ヘム基を有するかまたは有しない)、ヘモグロビンのβ鎖(ヘム基を有するかまたは有しない)、α鎖およびβ鎖の混合物(ヘム基を有するかまたは有しない)、およびこれらのタンパク質のフラグメントまたはアナログ(例えば、α、β、γ、δ、ε、またはζ鎖、単独で、あるいは混合物、二量体、または多量体のいずれかとして、ヘム基を有するかまたは有しない)を包含する。このようなフラグメントまたはアナログは、胚型、胎児型、または成体型を包含し、幹細胞増殖の阻害能および/または刺激能を有する。用語「INPROL」は、これらのタンパク質の天然に存在する形態ならびに天然に存在しない(例えば、組換え的および/または合成的に生成された)形態を包含する。用語「INPROLポリペプチド」は、40以上のアミノ酸からなるINPROLをいう。
【0097】
本明細書中で用いられる用語「オピエート化合物」は、オピエートレセプター(または、オピエートレセプターに配列関連性のあるレセプター、例えばORL1)に結合し、そしてアゴニスト、アンタゴニスト、または混合アゴニスト/アンタゴニストのいずれかの活性を発揮する、INPROLではないオピエートを含む化合物である。例えば、特異的アゴニストおよびアンタゴニストが、μレセプター(これは、DAMGOおよびDALDAによって選択的に活性化され、そしてCTOPおよびナロキソナジン(naloxonazine)によって選択的に拮抗される)、κレセプター(これは、GR 89696フマル酸塩またはU-69593によって選択的に活性化され、そしてnor-ビナルトルフィミン(binaltorphimine)塩酸塩によって選択的に拮抗される)、δレセプター(これは、DADLEおよびDPDPEによって選択的に活性化され、そしてナトリンドール(natrindole)、によって選択的に拮抗される)について存在する。さらに、3つのレセプターサブタイプの全てに作用する、広域スペクトルアンタゴニスト(例えば、ナロキソン)およびアゴニスト(例えば、エトルフィン)が存在する。ノシセプチン(nociceptin)はORL1レセプターを特異的にアゴナイズする。幹細胞の刺激および/または阻害活性を有するオピエート化合物は、INPROLについて本明細書中に記載された適用のそれぞれに使用され得る。
【0098】
本明細書中で用いられる「幹細胞刺激量」は、幹細胞の増殖を誘導する量である。本明細書中で用いられる「幹細胞阻害量」は、幹細胞の増殖を阻害する量である。全ての場合において、インビボおよびエクスビボの両方で、選択される量は、選択された特定のINPROLまたはオピエート化合物および特定の条件または適用に依存する。とりわけ、INPROLのポリペプチドまたはフラグメントの等モル用量は、オピエートペプチドまたは小分子と等モルとして活性である。
【0099】
典型的な臨床状況においては、幹細胞の阻害を所望するのであれば、INPROLは、例えば、0.01〜100mg/kg、好都合には0.1〜1.0mg/kgのINPROLを、例えば幹細胞の細胞周期進行を阻害することが望ましい場合は、標準的化学療法または放射線照射処置の4〜60時間前に投与することを用いて、投与量単位形態で静脈注射または点滴により日毎のレジメで患者に投与される。
【0100】
化学療法または放射線照射後の回復を促進するために、幹細胞の細胞周期進行の刺激が所望される状況では、幹細胞刺激量でINPROLが使用される。そのような用量は、典型的には1-500mg/kg、好都合には10mg〜100mg/kgである。
【0101】
幹細胞の細胞周期進行を阻害または刺激するためにオピエート化合物(単数または複数)を使用することが所望される場合では、オピエート化合物(単数または複数)は、INPROLについて記載した濃度の等モルで使用される。
【0102】
本発明の別の実施態様では、幹細胞阻害量のINPROLによる前処置により、患者が通常耐えられる用量を上回る量への化学療法剤用量または放射線量の増大が可能になる。同様に、幹細胞刺激量でのINPROLを用いた化学療法後または放射線後の処置はまた、化学療法または放射線照射の通常耐えられる用量の増大を可能にする。
【0103】
大部分の造血幹細胞は通常静止(ゆっくり細胞周期を進行しているか、全く進行していない)している。しかし、化学療法誘導性造血損傷に対する代償応答として、多くの割合の幹細胞が化学療法後に細胞周期進行に入る。それにより、細胞は、特に、次の用量の細胞傷害性化学療法または治療的放射線照射に対して抵抗力のないものとなる。このような幹細胞の細胞周期進行を阻害することにより、INPROL処置によって、次の用量の細胞傷害性化学療法をより早くまたはより頻度を高くして投与すること(従来用量または高用量のいずれかで)が可能になる。
【0104】
いくつかの正常な個体は、自然にすばやく細胞周期に入る幹細胞を有する。幹細胞阻害量のINPROLは、たとえ初回の用量の放射線照射または化学療法の前に与えるとしてもそのような個体には有用である。
【0105】
本発明の1つの実施態様では、INPROL(0.1mg〜6g/kg体重−好都合には1.0〜60mg/kg)は、化学療法の初回用量の投与から約24時間〜10日後に投与される。さらに4〜60時間後、好都合には24〜48時間後、次回の用量の化学療法が投与される。この化学療法とINPROLとを交互に行うサイクルは、治療上の利点に従って続けられる。化学療法剤および投与プロトコルは、標準的な医療実施における特定の腫瘍タイプに対する適合性に従って選択される。必要に応じて、G-CSF、幹細胞因子、または幹細胞刺激量のINPROLのような刺激性成長因子が、化学療法または放射線照射処置の後に用いられて、さらに造血再構築を改善する。
【0106】
エクスビボ適用のためには、幹細胞増殖の阻害が所望される場合では、106細胞/ml当たり0.1ng〜100ng、好都合には106細胞/ml当たり2〜50ngのINPROLが用いられる。幹細胞の刺激が所望される場合では、106細胞/ml当たり10ng〜100μg、好都合には106細胞/ml当たり1〜100μgのINPROLが用いられる。
【0107】
オピエート化合物を幹細胞の細胞周期進行の阻害または刺激のために使用することが所望される場合は、オピエート化合物(単数または複数)はINPROLについて記載された濃度と等モル濃度で用いられる。
【0108】
本発明の別の実施態様では、INPROLは、移植のための自家造血細胞を調製するための方法において用いられる。造血細胞は、有効量のINPROLでエクスビボで処理されて、幹細胞分裂を阻害し、次いで骨髄培養物に有効量の化学療法剤または放射線照射を投与することによってガン性細胞をパージする。細胞周期が進行している細胞に特異性を有する化学療法剤が好ましい。このように処理された骨髄は、自家ドナー内に再注射される。必要に応じて、患者は、幹細胞刺激量のINPROLおよび/または造血を刺激することが知られている別の薬剤で処置されて、患者の造血再構築を改善する。このような技術によって自家骨髄移植片の間の腫瘍細胞を効果的にパージするが、一方造血幹細胞を保護することが可能になる。このような保護は、エクスビボまたはインビボのいずれかのパージングのプロトコルを与え得る。一旦首尾良く移植されたら、幹細胞を迅速に増殖させて、正常な骨髄機能を再生させる必要がある。これは、幹細胞の細胞周期進行を刺激しそして骨髄機能の回復を増強する幹細胞刺激量でのINPROLの使用によって与えられ得る。
【0109】
本発明の別の実施態様では、INPROLは、遺伝子療法のための造血細胞を調製するための方法において用いられる。造血細胞はエクスビボで幹細胞刺激量のINPROLおよび/または他の刺激性サイトカイン(単数または複数)で処理されて幹細胞の分裂を刺激し、次に目的の遺伝子(単数または複数)でトランスフェクトする(好都合には、例えばレトロウイルスベクターを用いて感染させる)。トランスフェクションが達成された後、細胞を洗浄し、そして幹細胞阻害量のINPROLで処理して幹細胞を静止状態に戻す。このように処理された骨髄は、ドナーに再注入される。必要に応じて、患者は幹細胞阻害量のINPROLでインビボで処理されて、幹細胞がそれらの静止形態に維持され、そしてそれらの骨髄再生能が増大する。
【0110】
本発明の別の実施態様では、INPROLは、白血病処置の補助的な処置として用いられる。例えば、白血病細胞がINPROLに応答しない病態において、造血細胞はエクスビボで幹細胞阻害量のINPROLにより処理される。正常幹細胞の増殖はINPROLの投与により妨げられる。従って、増殖する白血病細胞が細胞周期に特異的な細胞傷害性薬剤により処理されている間は、正常幹細胞の集団は損傷から保護される。さらに、IL-3、GM-CSFのような刺激性サイトカインは、薬物処置または放射線照射処置の間に必要に応じて投与されて白血病細胞における細胞周期の進行を誘導するが、正常幹細胞はINPROLにより保護される。患者は、白血病細胞を破壊するために化学療法剤または放射線により処置され、次いでパージされた骨髄は、造血再構築を確立するために患者内に移植により戻される。
【0111】
同様に、血液細胞またはリンパ球に関与する重篤なウイルス感染(例えば、HIV感染)にかかった患者を処置するための本発明の別の実施態様においては、造血細胞はエクスビボまたはインビボでINPROLにより処理され、その後抗ウイルス剤、感染細胞を破壊する薬物、または感染細胞を排除するための抗体に基づく系により処理される。患者からウイルス性宿主細胞を根絶するための骨髄切除(myeloablative)抗ウイルスまたは骨髄切除化学療法の後、INPROL処理した骨髄は患者に戻される。
【0112】
本発明の別の実施態様では、INPROLは過剰増殖性幹細胞に関連した疾患を処置するために用いられる。例えば、乾癬は、皮膚の過剰増殖する上皮細胞によって引き起こされる障害であり、時には細胞傷害性薬物により処置される。幹細胞増殖を含む他の前新生物性病変はまた、幹細胞の増殖を阻害するために用いられる有効量のINPROLに影響されやすい。後天性免疫不全症候群の患者は、幹細胞の枯渇という結果となる異常に速い速度の幹細胞の細胞周期進行を有する。これらの患者はまた、幹細胞の細胞周期進行を阻害するのに有効な量のINPROLで処置することから恩恵を受ける。これらの使用のため、INPROLを含む局所的または経皮的な送達組成物(例えば、軟膏、ローション剤、ゲル剤、またはパッチ剤)は、非経口投与の代わりとして、適切な箇所で用いられる。
【0113】
ほとんどの白血病の場合、白血病前駆細胞は、INPROLにより影響を受けない分化細胞集団であり、従って上記のようなINPROLを用いる方法によって処理される分化細胞集団である。白血病前駆細胞が非常に原始期のものであり、INPROLによる阻害に対して直接的に感受性である場合には、白血病細胞の増殖は有効量のINPROLの投与によって弱められる。
【0114】
本発明の別の実施態様では、INPROLは、過剰増殖性幹細胞に関する障害を処置するために用いられる。例えば、骨髄形成異常症候群および再生不良性貧血は、骨髄の幹細胞の過剰増殖により引き起こされる障害である。幹細胞の過剰増殖に関与する他の症候群は、幹細胞刺激量のINPROLで処置可能である。
【0115】
モノクローナルまたはポリクローナルである抗体は、INPROLペプチドもしくはポリペプチドに対して標準的な技術によって生成される。これらの抗体またはINPROLペプチドもしくはポリペプチドは、当該分野で公知の多くのタイプの検出可能な標識により標識される。次いで、標識されたINPROLまたは抗INPROL抗体は、幹細胞マーカーとして用いられ、診断を目的として患者に直接それらを投与することによって幹細胞を同定および単離する。あるいは、これらの標識されたペプチド、ポリペプチドまたは抗体はエクスビボで用いられ、骨髄中の新生物形成細胞を除去する前にそれらを除き得るように造血細胞調製物中の幹細胞を同定する。同様の方法で、このような標識されたペプチド、ポリペプチドまたは抗体は、上皮性または他の幹細胞を単離および同定するために用いられる。さらに、標識されるかまたは標識されていないこのような抗体は、INPROL活性の中和によって治療的に、または循環しているINPROLレベルを検出することによって診断的に用いられる。
【0116】
INPROLは、標準的な技術を用いて、ヒト遺伝子または組換えヒトINPROLの発現用のcDNAライブラリーからクローニングされ得る。例えば、精製タンパク質から得られる配列情報を用いて、例えば32-リンで標識され得るオリゴヌクレオチドプローブが構築され、そして適切なcDNAライブラリー(例えば、骨髄由来)をスクリーニングするために用いられる。あるいは、適切な供給源(例えば、骨髄)由来の発現ライブラリーは、抗体を用いて、または適切な機能的アッセイ(例えば、実施例2に記載されるアッセイ)を用いて、INPROLをコードするcDNAについてスクリーニングされる。ヘモグロビン自体、ならびに個々のα鎖およびβ鎖は、当該分野において公知の方法を用いてクローニングおよび発現されている(Pagnierら、Rev.Fr.Transfus.Hemobiol.35:407-15,1992;Lookerら、Nature 356:258-60,1992;Methods in Enzymology 231巻,1994)。
【0117】
本発明は、以下を含むDNA配列を包含する:選択された非哺乳動物宿主による発現に「好ましい」コドンの取り込み;制限エンドヌクレアーゼ酵素による切断部位の供給;および付加的な開始、末端、または中間のDNA配列の供給、これらの配列は、容易に発現されるベクターの構築、あるいはヘモグロビンのα鎖、β鎖、γ鎖、δ鎖、ε鎖、および/またはζ鎖の生成または精製を容易にする。
【0118】
本発明はまた、ヘモグロビンα鎖、β鎖、γ鎖、δ鎖、ε鎖、および/またはζ鎖のポリペプチドアナログまたは誘導体をコードするDNA配列を提供する。これらアナログまたは誘導体は、1つ以上のアミノ酸残基の同一性または位置に関して天然に存在する形態と異なり(すなわち、欠失アナログ、記載の全残基より少ない残基を含有する;置換アナログ、記載の1つ以上の残基が他の残基により置換される;および付加アナログ、1つ以上のアミノ酸残基がポリペプチドの末端または中間部分に付加される)、かつ天然に存在する形態の一部または全ての特性を共有する。
【0119】
好都合な実施態様では、INPROLは、ゲノムクローニングもしくはcDNAクローニング、または遺伝子合成により得られた外因性DNA配列の原核生物または真核生物宿主発現(例えば、細菌、酵母、高等植物、昆虫、および哺乳動物培養細胞による)の産物である。すなわち、好都合な実施態様では、INPROLは「組換えINPROL」である。代表的な酵母(例えば、Saccharomyces cerevisiae)または原核生物(例えば、E.coli)宿主細胞における発現産物は、いずれの哺乳動物タンパク質とも関連しない。脊椎動物(例えば、ヒトではない哺乳動物(例えば、COSまたはCHO)および鳥類)細胞における発現産物は、いずれのヒトタンパク質とも関連しない。用いられる宿主に依存して、本発明のポリペプチドは、グリコシル化され得るか、またはグリコシル化され得ない。本発明のポリペプチドは、必要に応じて開始メチオニンアミノ酸残基(−1位)もまた含む。
【0120】
本発明は、ヘモグロビンのα鎖、β鎖、γ鎖、δ鎖、ε鎖、および/またはζ鎖のポリペプチドアナログのような他の産物もまた包含する。このようなアナログは、ヘモグロビンのα鎖、β鎖、γ鎖、δ鎖、ε鎖、および/またはζ鎖のフラグメントを包含する。周知の手順に従って、当業者は、本明細書中に記載の1つ以上の残基の同一性または位置に関して天然に存在する形態と異なる(例えば、置換、末端および中間における付加、および欠失)一次配列を有する微生物発現ポリペプチドをコードする遺伝子を容易に設計および製造し得る。あるいは、cDNA遺伝子およびゲノム遺伝子の改変は、周知の部位特異的変異誘発技術により容易に達成され得、そしてヘモグロビンのα鎖、β鎖、γ鎖、δ鎖、ε鎖、またはζ鎖のアナログおよび誘導体を生成するために用いられ得る。このような産物は、少なくとも1つのINPROLの生物学的特性を共有するが、他は異なり得る。例として、本発明の産物は、以下のようなα鎖、β鎖、γ鎖、δ鎖、ε鎖、またはζ鎖を包含する:例えば、欠失により短縮される;または加水分解に対してより安定である(従って、天然に存在するよりもより顕著なまたはより長期間持続する効果を有し得る);あるいはO-グリコシル化および/またはN-グリコシル化のための1つ以上の潜在的な部位を欠失または付加するように改変されたか、あるいは欠失または置換(例えば、アラニンまたはセリン残基により)された1つ以上のシステイン残基を有しており、そして微生物系から活性形態でより容易に単離される;あるいはフェニルアラニンにより置換される1つ以上のチロシン残基を有し、そして標的タンパク質または標的細胞上のレセプターにある程度容易に結合する。α鎖、β鎖、γ鎖、δ鎖、ε鎖、またはζ鎖内の連続アミノ酸配列または二次コンフォメーションの一部のみを複製するペプチドフラグメントもしくはポリペプチドフラグメントもまた包含される。これらのフラグメントは、INPROLの1つの特性(例えば、レセプター結合性)を有し、そして他(例えば、幹細胞阻害活性)は有さない。本発明の産物のいずれか1つ以上が治療有用性(Weilandら、Blut 44:173-5,1982を参照のこと)または他の情況(例えば、阻害剤拮抗作用のアッセイ)における有用性を有するために、活性は必要でないことは注目すべきことである。競合的アンタゴニストは、幹細胞阻害剤またはそのレセプターの過剰生産の場合において有用である。
【0121】
さらに、生物学的活性を保持するタンパク質配列に由来するペプチドは、標準的な方法に従って化学的に合成され得る。本発明はまた、ヘモグロビンα鎖、β鎖、γ鎖、δ鎖、ε鎖、および/またはζ鎖のペプチドアナログまたは誘導体をコードする配列を提供する。これらアナログまたは誘導体は、1つ以上のアミノ酸残基の同一性または位置に関して天然に存在する形態と異なり(例えば、欠失アナログ、記載の全残基より少ない残基を含有する;置換アナログ、記載の1つ以上の残基が他の残基、天然に存在するかまたは当該分野で公知の他のアナログ(例えば、D-アミノ酸)のいずれかにより置換される;および付加アナログ、1つ以上のアミノ酸残基が、安定性、溶解度、および/またはタンパク質分解に対する耐性を増大させるように化学修飾される)、かつ天然に存在する形態の特性の一部または全ての特性を共有する。
【0122】
上記のようなペプチド配列は、種々の方法により同定され得る。アッセイにおいて活性な天然ヘモグロビン鎖(例えば、α鎖)の三次元構造と不活性な構造的に関連したタンパク質(例えば、ミオグロビン)との比較は、三次元空間において異なるコンフォメーションを有し、従って活性ペプチドの候補領域である領域を同定し得る。別のアプローチは選択的タンパク質分解を用いる。このタンパク質分解では、タンパク質分解酵素は、ヘモグロビン鎖の限定消化において用いられる。この消化は、例えば逆相HPLCにより分離され、次いで幹細胞阻害についてアッセイされ得るペプチドを生じる。ペプチドはまた、化学合成(例えば、固相合成)によっても生成され得る;目的のヘモグロビン鎖(例えば、α鎖)の配列を含む一連の重複ペプチド(例えば、15マー)は、容易に生成され得、そして幹細胞アッセイにおいて試験され得る。コンビナトリアルライブラリーが生成され得、このライブラリーでは、複数の化学合成が実施され、かつ選択されたアミノ酸位置を変えることが可能であり、その結果、多くのスクリーニング用ペプチドアナログを生じる(例えば、Dooleyら、Peptide Research 8:124-137,1995)。あるいは、組換え法が用いられ得る。部位特異的変異誘発は、特定のヘモグロビン鎖の活性に必要な必須残基を同定するために用いられ得る。幹細胞阻害剤として活性であると知られる鎖(例えば、α鎖)の領域は、関連するが不活性なタンパク質(例えば、ミオグロビン)の領域と置換され得、そして幹細胞アッセイにおいて試験され得、これにより活性に必要な領域の同定が可能になる。このような同定された領域は、ペプチドとして発現され得、そして幹細胞周期アッセイにおいて活性について試験され得る。
【0123】
他の種に由来するINPROLの同種または類似のバージョンは、上記の本発明の治療的実施態様と同様に、種々の獣医学的使用に用いられる。
【0124】
幹細胞刺激量のINPROLは、細胞周期の進行している幹細胞を分裂しないかまたは緩徐にに分裂する「休止」期に可逆的に移行させることによって、それらに作用する。休止している幹細胞を刺激して分裂させることが望ましい場合(例えば、ガンの化学療法剤または放射線により患者を処置した後)、幹細胞刺激量のINPROLが用いられ得る。あるいは、またはさらに、コロニー刺激剤および他の造血刺激剤が被験体に投与される。このような因子の例としては以下のものが挙げられるが、それらに限定されない:M-CSF(CSF-1)、GM-CSF、G-CSF、巨核球-CSF、トロンボポエチン、幹細胞因子、または他のサイトカイン(例えば、IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、IL-9、IL-11、IL-12、IL-13、IL-14、あるいはエリスロポエチン)。
【0125】
INPROLポリペプチドまたは幹細胞阻害活性を有する活性フラグメントは、下記のプロトコルで例示されるように、精製されるか、または幹細胞阻害活性用の適切なバイオアッセイを組み合わせた従来の化学的プロセスによって合成される。
【0126】
本発明の1つの実施態様では、治療的に有効な量のINPROLタンパク質または治療的に有効なそれらのフラグメントは、薬学的に受容可能なキャリアと混合して用いられる。このINPROL組成物は、一般的に、非経口的注射または点滴によって投与される。皮下注射、静脈注射、または筋肉内注射の経路が、達成される治療的効果によって選択される。
【0127】
全身投与される場合、本発明における使用のための治療的組成物は、発熱物質非含有の非経口的に受容可能な水溶液の形態である。薬学的に受容可能な滅菌タンパク質溶液は、pH、等張性、安定性、キャリアタンパク質などに関する要件を有し、当該分野の技術に属する。
【0128】
INPROL療法において有用な適切な希釈剤、保存剤、安定化剤、乳化剤、アジュバント、および/またはキャリアと共に、治療的に有効な量の本発明のポリペプチド産物を含む薬学的組成物もまた、本発明により包含される。本明細書中で用いられる「治療的に有効な量」は、所定の条件および投与レジメで治療効果を提供する量を指す。このような組成物は、液剤、ゲル剤、軟膏、または凍結乾燥処方物もしくは他の乾燥処方物であり、そして種々の緩衝液内容物(例えば、Tris-HCl、酢酸、リン酸)の希釈物、pHおよびイオン強度、表面吸着防止のための添加剤(例えば、アルブミンまたはゼラチン)、界面活性剤(例えば、Tween 20、Tween 80、Pluronic F68、胆汁酸塩)、可溶化剤(例えば、グリセロール、ポリエチレングリコール)、抗酸化剤(例えば、アスコルビン酸、メタ重亜硫酸ナトリウム)、保存剤(例えば、チメロサール、ベンジルアルコール、パラベン)、増量物または張度改変剤(例えば、ラクトース、マンニトール)を含み、ポリマー(例えば、ポリエチレングリコール)のタンパク質への共有結合、金属イオンとの複合体化、あるいは物質のポリマー性化合物(例えば、ポリ乳酸、ポリグリコール酸、水性ゲルなど)の粒状調製物中または上への取り込み、またはリポソーム、ニオソーム(niosome)、マイクロエマルジョン、ミセル、単ラメラ小胞または多重ラメラ小胞、生分解性注入マイクロカプセルまたはマイクロスフェア、またはタンパク質マトリックス、赤血球影、スフェロプラスト、皮膚パッチ、または薬剤を放出またはパッケージングする他の公知の方法中への取り込みを含む。このような組成物は、INPROLの物理的状態、溶解度、安定度、インビボ放出速度、およびインビボ消失速度に影響を及ぼす。徐放性組成物または持続放出性組成物は、親油性デポ剤(例えば、脂肪酸、ワックス、オイル)中の処方物を包含する。ポリマー(例えば、ポロキサマーまたはポロキサミン)で被覆された粒状組成物および組織特異的レセプター、リガンド、または抗原に対する抗体に結合されたINPROL、あるいは組織特異的レセプターのリガンドに結合されたINPROLもまた、本発明により包含される。本発明の組成物の他の実施態様は、種々の投与経路のために、粒状形態の保護コーティング、プロテアーゼ阻害性因子、または浸透増強剤を取り込む。この投与経路としては、非経口投与、経肺投与、鼻内投与、局所(皮膚または粘膜)投与、および経口投与が挙げられる。別の実施態様では、INPROLを含有する組成物は、局所的にまたは経皮パッチを介して投与される。
【0129】
1つの実施態様では、本発明の組成物は、単位投与量形態で滅菌バイアルまたはアンプル中にパッケージされる。
【0130】
本発明は、1つ以上の付加因子(例えば、化学療法剤(例えば、5-フルオロウラシル(5FU)、シトシンアラビノシド、シクロホスファミド、シスプラチン、カルボプラチン、ドキソルビシン(doxyrubicin)、エトポシド、タキソール、アルキル化剤)、抗ウイルス剤(例えば、AZT、アシクロビル)、TNF、サイトカイン(例えば、インターロイキン)、抗増殖薬、代謝拮抗物質、およびDNA代謝に干渉する薬物)を含む組成物もまた包含する。
【0131】
そのような細胞傷害性薬剤に曝されることが予想される被験体を処置する方法または過剰増殖性幹細胞を処理する方法に含まれる投与量レジメは、担当の医師により、薬物の作用を改変する種々の要因(例えば、症状、体重、性別、患者の規定食、感染の重篤度、投与時間、および他の臨床的要因)を考慮して決定される。
【0132】
被験者が細胞傷害性薬剤または放射線に曝された後、本発明の治療方法は、1種以上のリンホカイン、コロニー刺激剤、または他のサイトカイン、造血素、インターロイキン、またはINPROLによる前処理により阻害される幹細胞(およびそれらの派生物)の成長および分裂を一般に刺激する成長因子を必要に応じて含む、幹細胞刺激量のINPROLを被験体に投与することを用いる。造血を促進するこのような治療剤としては、IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-7、Meg-CSF、M-CSF(CSF-1)、GM-CSF、G-CSF、またはエリスロポエチンが挙げられる。これらの薬剤の投与量は、化学療法または造血幹細胞移植後の造血再構築の促進における効能に対する臨床試験にそれらを用いたときに得られる知見によって選択される。これらの投与量は、患者の身体状態の変化および被験体が曝された化学療法剤または放射線の量およびタイプを補償するように調整される。処置を受けた患者へのINPROLの投与によって引き起こされる幹細胞の阻害の回復の進行は、従来の方法によってモニターされる。
【0133】
白血病の処置では、正常な幹細胞の細胞周期の進行を阻害するINPROLおよびIL-3またはGM-CSFのような白血病細胞の成長の刺激剤の両方を、細胞傷害性薬物処置または放射線照射の間に同時に投与することが有利である。このプロトコルにより、正常細胞および白血病細胞の細胞周期の進行状態と薬物感受性との間の最大の差違を達成し得る。
【実施例】
【0134】
実施例1:インビボ幹細胞増殖阻害アッセイ
幹細胞増殖の検出のために、細胞周期のS期にあるCFU-Sの数を3H-チミジン「自殺(suicide)」法(Beckerら、Blood 26:296-308、1965)により測定した。
【0135】
未成熟の造血前駆細胞--脾コロニー形成単位(CFU-S)--は、致死量の放射線を照射したマウスの脾臓において肉眼で見えるコロニーを形成させることにより、インビボで、造血細胞の静脈注射の8〜12日後に検出され得る(Till & McCulloch、1961)。
【0136】
標準的CFU-S増殖アッセイに関しては、3H-チミジン「自殺」の方法が通常適用される(Beckerら、1965)。この方法は、DNAの前駆体である放射標識したチミジン(3H-チミジン)をDNA合成中の細胞内に取り込むことに基づいている。試験の時点で細胞周期のS期にあるこのCFU-Sは、高い放射活性により死滅しており、そのため脾臓ではコロニーを形成できない。従って、3H-チミジンなしでインキュベーションされた細胞サンプルと3H-チミジンとともにインキュベーションされた同じ細胞との間の、注射によって形成されるCFU-Sの数の違いは、元のサンプルにあった増殖性CFU-Sのパーセントを示す。
【0137】
阻害剤試験は、阻害剤が細胞周期が進行しているCFU-S(正常マウスの骨髄では全CFU-S集団の7〜10%程の低さ)にのみ影響する限り、刺激を受けていない動物由来の骨髄幹細胞集団を用いて行うことはできない。
【0138】
CFU-Sの増殖を刺激するために、フェニルヒドラジン(PHZ)、または致死量以下(sublethal)の放射線照射を用いた(Lord、1976)。
【0139】
本発明者らは、テストステロンプロピオネート(TSP)がCFU-Sの細胞周期の進行に刺激効果を及ぼすこと(Byronら、Nature228:1204、1970)に基づいて、TSPを用いる方法を開発し、それによって試験は簡素化され、いかなる副作用も引き起こされなかった。TSPは、注射後20〜24時間以内にCFU-S増殖の刺激を誘導し、そしてその効果は少なくとも7日間見られ得た。
【0140】
阻害剤の精製の間に画分のスクリーニングに用いられた手順は以下の通りであった:
マウス:全ての試験を通じてBDF1またはCBF1マウス種を用いた。
【0141】
30〜35%のCFU-SをS期に誘導するために、ドナーマウスを、0.2ml/マウスの腹腔内注射によって、10mg/100gの用量のTSPで処理した。
【0142】
24時間後、細胞懸濁調製のため骨髄を大腿骨から取り出す。次いで、1ml当たり5×106から10×106個の細胞を異なるコントロールおよび試験画分と、37℃で3.5時間水浴中でインキュベーションする(各グループにつき試験管2本ずつ、1つはホット(標識したもの)用およびもう1つはコールド(標識していないもの)用)。
【0143】
3.5時間後、3H-チミジン(1mCi/ml、比活性18〜25Ci/mmole)を、ホット用の各試験管に細胞懸濁液1ml当たり200μlずつの容量で添加し、コールド用の試験管には何も添加しない。37℃でさらに30分間インキュベーションし続ける。
【0144】
30分間のインキュベーションの後、400μg/mlの放射能活性のないチミジンを含む10mlの冷培地(4℃)を添加することにより、死滅反応を停止させる。細胞を十分に洗浄する(3回)。
【0145】
細胞を再懸濁し、そして注射に望ましい濃度、通常マウス1匹当たり0.3〜0.5ml中2〜4×104個細胞、まで希釈する。
【0146】
1グループ当たり8〜10匹のレシピエントマウスを注射の6時間以上前に放射線照射する。
【0147】
9〜12日目にレシピエントの脾臓を集め、そしてTellesnitsky溶液中で固定して、コロニー数を目で数える。下式を用いてS期にある細胞のパーセントを計算する。
【0148】
【化7】
ここで、a--3H-チミジンを有さないCFU-Sの数
ここで、b--3H-チミジンを有するCFU-Sの数
表1に示されるINPROLの試験データは、INPROLでの処理の後に細胞周期の進行している幹細胞が3H-チミジンの作用に対して耐性になることを示している。実施例1および以下の実施例の全てについて、用語「pINPROL」は、ブタ骨髄由来の精製タンパク質をいう。同じ保護が、S期特異的細胞障害性薬物であるシトシンアラビノシドおよびヒドロキシ尿素にも見られる(データは示していない)。次いで、処理された幹細胞を放射能活性のないチミジンを含む冷培地で洗浄した場合、生き残った幹細胞はマウス脾臓内で増殖して、正常にコロニーを形成する。
【0149】
表1
骨髄細胞との4時間のインキュベーションの間に
pINPROLがCFU-S増殖に及ぼす阻害活性
【0150】
【表1】
*2×104細胞当たりのCFU-S。
【0151】
実施例2:インビトロ幹細胞増殖阻害アッセイ
以下の試験系(Lordら、The Inhibitors of Hematopoiesis 227〜239頁、1987)を用いて、INPROLの直接的効果を示した。多系統(multilineage)因子(IL-3)依存性幹細胞株であるFDCP mix A4(A4)を、コロニー刺激性IL-3の供給源として、20%ウマ血清および10%WEHI-3馴化培地を補充したIMDM培地中で維持した。
【0152】
トリチウム化チミジン取り込みアッセイを使用して増殖を測定した:A4細胞(20%ウマ血清および50%WEHI-3 CMを有する100μlの培地中の5×104個)を、5%CO2中で37℃、16時間インキュベーションした。
【0153】
pINPROLまたは粗製のBME(画分IV)を開始時点で添加した。次いで、トリチウム化チミジン((3H-Tdr)50μl中の3.7KBq、740 GBq/mmole)を各グループに添加し、さらに3時間インキュベーションした。増殖レベルを集めた細胞によって測定し、そして阻害%を下記の式を用いて計算した。
【0154】
【化8】
。
【0155】
正常骨髄抽出物またはpINPROLを段階的な用量で存在させて増殖させたFDCPmix-A4細胞によるトリチウム化チミジン(3H-Tdr)取り込みを図6に示す。pINPROLの精製組成物は出発物質より少なくとも1000倍活性が高いことがわかった。曝露時間(16時間)は、効果的な阻害についての重要な因子であり、そしてA4細胞株の幹細胞に、pINPROLの直接的な効果を及ぼしている証拠を示している。
【0156】
実施例3:インビボのINPROL注射によるCFU-S増殖の阻害:用量および効果の持続期間
インビボで注射されたINPROLの効果の研究は、INPROLがCFU-Sの細胞周期への加入を効果的にブロックし得、これによりそれらの細胞をさらなる処理の細胞障害効果から保護し、その臨床使用への可能性を示すことを示した。
【0157】
実験プロトコルには2つの目的がある:インビボで注射された場合にCFU-Sに対するINPROLの効果をチェックすること、および、細胞周期の進行している幹細胞に関してINPROL活性の有効持続期間を明確にすること。
【0158】
CFU-Sの増殖を刺激するために、上記の実施例1で述べた効果に基づいて、テストステロン-プロピオネートの注射を用いた。
【0159】
マウスBDF1をTSP(10mg/100g)と共に第0日に注射した;24時間後に、各実験グループのマウス(各グループあたり4匹)に0μg、5μg、10μg、および15μg/マウスの用量で腹腔内にpINPROL注射を1回行った。
【0160】
pINPROL注射の24時間後、マウスを屠殺し、そして細胞周期の進行しているCFU-Sのパーセントを実施例1に記載されたアッセイにより測定した。TSP注射は、約50%のCFU-Sを細胞周期に誘導したが、未処理のマウスでは7%であった。2μg/マウス程度に低い用量のpINPROLは、TSPで誘導される増殖を阻害して正常レベルまで下げ得た。
【0161】
効果評価の持続期間の間、一方のグループのマウス(1グループ当たりマウス21匹)にTSPのみを注射し、そして他方のグループにTSPとpINPROL(TSPの24時間後)の両方を注射した。CFU-Sの細胞周期の進行は、各グループから3匹ずつドナーを取り出して、記載した方法(実施例1参照)で骨髄のCFU-Sの細胞周期の状態を測定することにより、1週間、24時間毎に測定された。図7に示すデータは、TSPの有効持続期間は少なくとも7日間であるが、1回のINPROLの注射によりCFU-Sは休止期に入り、わずか48〜72時間は細胞周期に入らないようにすることができることを示している。ガンおよび白血病の化学療法に用いられる化学療法剤の大多数はインビボでの半減期は比較的短い(通常24時間より短い)ので、得られたデータによれば、INPROLの効果はシトシンアラビノシドまたはヒドロキシ尿素のような化学療法剤がインビボで活性である間は有効期間よりも長く維持される。さらに重要なことは、第1回目の処理(幹細胞に損傷を与えない)と第2回目の処理(CFU-Sに損傷を与える)との間により長い間隔(24時間より長く96時間より短い)を有する化学療法および放射線療法に対しては、化学療法剤または放射線照射の2回の適用の間の間隔にINPROLを1回注射すれば十分である。繰り返し行う数回の細胞傷害性治療または放射線照射に関しても、INPROL効果の持続期間に基づいて同じストラテジーを適用し得た。
【0162】
実施例4:5-FUによる処理の後、迅速に周期に入るように刺激された最原始造血幹細胞は、INPROLにより第2回目の5-FU被曝から保護される
薬剤5-フルオロウラシル(5-FU)は、骨髄およびリンパ球の区画(compartment)中の細胞数を劇的に減少させる。それは通常、細胞周期特異的であり、細胞周期のS期の間または前にヌクレオチドアナログをDNA内に取り込むことは細胞死となるので、増殖細胞を迅速に標的とすると考えられている。マウスの骨髄の長期生存および免疫造血再構築は、5-FUの1回の用量では影響を受けない;しかし、多能性造血幹細胞(PHSC)は最初の投薬後約3〜5日間という短い期間に第2回目の5-FUの投薬に対して攻撃されやすくなるということが示された(Harrisonら、Blood 78:1237〜1240、1991)。PHSCは、通常、5-FUの1回の投薬に対して細胞周期が遅すぎて効果的ではなく、そして第1回目の5-FU処理の結果起こる刺激により刺激されて迅速に細胞周期に入ることが説明され得る。本発明者らは、PHSCは、INPROLにより遅い細胞周期状態に戻り得、これにより2回目の5-FU処理から保護され得るということを提案した。
【0163】
これらの実験に用いられたマウスは、BDF1雄マウスであった。5-FU(Sigma)の貯蔵液を生理食塩水中で10μg/mlの濃度に調製した。各処理マウスは、実験0日目に尾部静脈を介して体重10g当たり2mgの5-FUを受けた;24時間後、マウスにpINPROL(10μg/体重100g)を腹腔内注射し、そして3日目に第2回目の5-FUの用量を注射した。生存研究は、各々30匹のマウスからなる実験グループ(pINPROL処理)およびコントロールグループにおいて、マウスの死亡をモニターすることにより行った。生存曲線を図8に示す。
【0164】
実施例5:骨髄細胞におけるINPROLでプレインキュベーションした場合とMIP1αでプレインキュベーションした場合の効果
この実験の目的は、インビトロでのマウス骨髄細胞に対するpINPROLおよびMIP-1αでプレインキュベーションした場合の阻害効果を比較することであった。
【0165】
以下の手順を用いた:
インビボ:大腿骨から骨髄を採取する48時間前に、6〜15週齢のBDF1マウスに200mg/kgの5-FUを腹腔内に注射する。
【0166】
インビトロ:単一の細胞をプールした懸濁液を計数し、そして5×106細胞を、pINPROLまたはMIP-1αとともにまたはなしで、5%ウマ血清、L-グルタミン添加IMDM培地とともに総容量2mlとして、37℃および5%CO2で4時間インキュベーションする。次いで、細胞を2回洗浄し、そして再計数した。それらを以下の最終条件下でメチルセルロースにプレートする:
0.8% メチルセルロース
25% ウマ血清
20ng/ml 組換え型マウスIL3
L-グルタミン添加
5×105細胞/ml
IMDM培地
プレートを11日間、37℃および湿度100%中5%CO2で、インキュベーションした。50細胞より多いコロニーを計数した。
【0167】
【表2】
。
【0168】
実施例6:INPROLはHPP-CFC増殖を阻害する
マウス再構築幹細胞および初期前駆体を評価するインビトロアッセイは、高増殖性潜在コロニー(HPP-PFC)アッセイである;他の関連アッセイ、例えばCFU-A、CFU-GM、CFU-EおよびCFU-GEMMは、次第に制限された前駆細胞の集団を検出する(M.Moore、Blood 177:2122-2128、1991)。この例は、pINPROLでの細胞の前処理が、それらの増殖を阻害するのに対し、MIP-1αは、これらの実験条件下では阻害しないということを示す。
【0169】
BDF1マウスを、それらの骨髄をHPP-CFC数に関してアッセイする前に、5-フルオロウラシル(200mg/kg腹腔内)で処置した。細胞を遠心分離により洗浄し、そして添加剤のない培地(コントロール)、pINPROL(25ng/ml)またはMIP-1α(200ng/ml)を含む培地のいずれかにおいて、106〜5×106/mlの密度で4時間インキュベーションした。インキュベーション後、細胞を洗浄し、そして30%FCSならびに5637細胞株およびWEHI-3B細胞株由来の組合せ馴化培地(7.5%の各馴化培地、Sharpら、1991の推奨による)を有する寒天(0.3%)にプレートした。プレート濃度は、60mmの皿に5×104細胞/mlであった。14日目にコロニー数を計数し、そして結果を以下に示す。
【0170】
【表3】
。
【0171】
これらの結果により、MIP-1αは、プレインキュベーションの期間中にのみ存在した場合には、ほとんどの未成熟な前駆体の増殖を阻害しなかった。pINPROLは、これらの条件下で増殖を効果的に阻害し、生物学的活性という点から見るとpINPROLとMIP-1αとの間には基本的な違いがあることを示している。
【0172】
実施例7:放射線誘導性骨髄形成不全からの回復に対するINPROL処理の効果
骨髄形成不全は、放射線ガン療法の一次的制限毒性(primary limiting toxicity)である。いくつかの増殖因子(例えば、G-CSF、GM-CSF、エリスロポエチン)は、放射線誘導性骨髄形成不全からの回復を促進し得るということが示されてきた。幹細胞増殖の阻害剤の使用による保護という概念は、血液学的な損傷に上手く対処することにおける異なった補足的なアプローチである。初期に開発された処置手順(実施例3、4)に従い、致死量の放射線を照射されたマウスのモデルを確立した。9Gyのコバルト60を受けたマウスは10〜14日後に死亡し始め;30日目までには死亡率は約50%となる、ということは当該分野では公知である。この致死用量を、それを各4.5Gyずつの連続する2回の適用に分けて、投薬の間は3日間の間隔を置いて、本発明者らのモデルに用いた。予備データは、そのモデルの生存曲線が、9Gyによる1回の放射線照射で知られた生存曲線と非常に近いことを示した;さらに、CFU-S増殖に関する試験は、第1回目の放射線照射の24時間後に35〜50%のCFU-Sが増殖するように誘導されることを示した。このような細胞は、第2回目の投薬の前に送達される幹細胞阻害剤により保護され得る。
【0173】
この可能性を試験するために、マウス(50匹/グループ)に0日目に4.5Gyを照射し、その24時間後に、1つのグループにはpINPROL(2μg/マウス、腹腔内)を投与し、別のコントロールグループには生理食塩水を注射した。そして第2回目の放射線の照射用量(4.5Gy)を3日目に与えた。
【0174】
図9は、pINPROLの1回の用量を投与した後の増大した生存を示す。モデルの状態は、固形腫瘍により特徴付けられるガンを含むいかなるガンの処置にも臨床的に有意義であり、このような処置は、放射線照射の2回続けた投与の間に有効用量のINPROLを送達することによりガンを有する患者に投与し、それによってガンの処置に用いられる放射線の照射線量をより多くすることができる。この様式は化学療法剤にも広げることが可能なはずである。
【0175】
実施例8:自己骨髄移植へのINPROLの使用
骨髄移植は、いくつかの白血病(CML、AML、およびその他)にとっては唯一知られた治療的な療法である。輸注のための自己BMTのエクスビボ調整により、白血病細胞の混入のない正常幹細胞の潜在的な自己供給源が提供され、そしてレシピエントの造血系を再生させて(repopulate)、積極的で効果的な療法を可能にする。
【0176】
1.INPROL効果の研究用の長期骨髄培養物L1210白血病モデルはAraCによるパージの間、正常な造血を保持する
長期骨髄培養物(LTBMC)をToksozら(Blood 55;931-936、1980)に従って確立し、そして白血病細胞株L1210を2週間LTBMCと共培養した。正常前駆細胞および白血病前駆細胞の同時増殖は、これらのLTBMC/L1210の混合培養物内で起こり、それは白血病患者の骨髄内と同様の状況である。正常なコロニー形成単位CFUと白血病CFUとの間の区別は、WEHI-3(マウスIL-3産生細胞株)由来の馴化培地の存在下または非存在下での寒天コロニーとしてそれらを増殖させることにより可能であった。正常細胞は、IL-3非存在下で、アポトーシスを受けるのに対し、白血病細胞はIL-3非存在下でコロニーを形成し得る。LTBMC-L1210組成物由来の懸濁細胞は、プレートされた50,000細胞当たり、IL-3存在下で約150個のコロニー(正常造血クローン)を生じ、IL-3なしで増殖した場合は70コロニー(白血病クローン)を生じた。
【0177】
パージ手順は、以下の通りであった:0日目に、全ての懸濁細胞および培地(10ml/フラスコ)をLTBMC-L1210の入っているフラスコから取り出し、そして200μgのシトシンアラビノシド(AraC)を含む2mlの培地で置き換えた(Tsyrlovaら、Leukemia:Advances in Biology and Therapy、第35巻、1988);20時間のインキュベーション後、フラスコを洗浄し、そして2mlの新鮮な培地のみ(コントロールグループ)または25ng/mlのpINPROLを含む培地で4時間置き換えた。このプレインキュベーションの後、細胞を100μg/フラスコのAraCとともに、37℃で3時間再びインキュベーションした。各グループは4本のフラスコを含んだ。LTBMC-L1210培養物を3回洗浄し、そして新鮮なLTBC培地で置き換えた;それらを再生研究のため前述のように3〜4週間維持した。
【0178】
データを図10に示す。AraCのみで処理されたコントロール培養物には細胞の増殖が見られなかったが、INPROLで保護されたフラスコ内では、接着層由来の前駆細胞の増殖のため、造血再生がさらにより急速に起こった。さらに、実験グループ由来の細胞は、寒天にプレートされた場合、IL-3存在下では50,000細胞当たりわずか約100 CFUしか増殖しなかった;少なくとも4週間の間には白血病細胞の増殖は観察されなかった。従って、エクスビボでINPROLと組み合わせて、有効量のAraCにより処理された骨髄は、ガン細胞はパージされ得るが、幹細胞は保護される。この様式は他の形態の化学療法または放射線処置に拡げることが可能なはずである。
【0179】
2.骨髄再生能(MRA)および30日間放射線防御は、インビトロのINPROL処理により増大する
MRAは、致死量の放射線を照射されたマウスの骨髄を再生する細胞の能力であり、30日間の放射線防御を付与する能力を伴っていて、骨髄抑制動物を救う潜在能力の直接的なインビボ測定である(Visserら、Blood Cells 14:369〜384、1988)。
【0180】
放射線防御研究に関して、BDF1マウスは9.5Gyで放射線照射され、テストステロンで刺激されたドナー由来の骨髄の移植により回復した。レシピエントの1グループは培地で(コントロール−グループA)、そして別のグループ(グループB)は25ng/mlのpINPROLで、4時間プレインキュベーションされた骨髄細胞により回復した。両グループの細胞を洗浄し、そしてマウス1匹当たり30,000個の細胞を放射線照射された動物に移植した。生存データを示す(図11)。3つの実験の合計を、コントロールを100%に正規化して示す。30日目までに、pINPROLによるインキュベーションにより、マウスの生存をコントロールグループの36.5%から61.8%まで増加させた。
【0181】
INPROLによるプレインキュベーションによって誘導されるMRAの増加は、放射線防御を改善するメカニズムの1つであり得る。この仮説を試験するため、Visserら(前出)に従ってMRAを測定した。簡潔に記載すると、ドナーBDF1マウスをテストステロンで前処理し、それらの骨髄を培地またはpINPROLで4時間プレインキュベーションし、そして放射線照射された動物に注射した。13日目に、レシピエントの大腿骨由来の骨髄細胞を、20%ウマ血清、および10%WEHI-CMの存在下で、3つの異なる濃度(0.01、0.05、0.1当量の大腿骨)で寒天に接種した。7日目のコロニー数は、その時点のレシピエントの骨髄中のコロニー形成細胞がドナーの未成熟幹細胞の前駆細胞である限り、MRAを表す。
【0182】
図12に見られ得るように、INPROLでプレインキュベーションされた細胞集団のMRAは、コントロールグループ(B)の場合より多い。
【0183】
実施例9:INPROLの幹細胞への抗過剰増殖効果は、それらの分化の異常を変化させ得る
CFU-Sの過剰増殖は、細胞障害性薬物または放射線照射からの回復中に見られるだけでなく、正常な加齢の結果としても見られ、そして骨髄異形成症候群(MDS)の主要な特徴であると考えられている。それは、顆粒球経路に沿った分化が減少する一方で、赤血球分化が広まるような分化障害を伴う。
【0184】
骨髄細胞を25ng/mlのpINPROLまたは培地(コントロール)とともに37℃で4時間インキュベーションし、洗浄し、次いで20%ウマ血清、2U/mlエリスロポエチン、および10%WEHI-CMを有する寒天にプレートした。BFU-EおよびGM-CFUコロニーの数を7日目に計数した。表4に示すデータは、3つの実験をまとめたものであり、各グループから各時点で4匹を取り出し;4枚の皿にプレートした。
【0185】
表4から明らかなように、無傷の幼動物(8〜12週齢のBDF1)由来の正常骨髄(NBM)をINPROLとともにインキュベーションすることにより、異なるタイプのコロニーの数または比率は変化しなかった。テストステロンプロピオネート(TSP)で前処理されたBDF1ドナーは、以前に観察したように(実施例1、3、4)、CFU-S増殖における同じ増加、赤血球前駆細胞数(BFU-Eコロニー)におけるわずかな増加、およびGM-CFUにおける減少を示し、それはINPROLとのインキュベーションにより完全に妨げられた。さらに、異常に高いレベルのCFU-S増殖は、細胞周期のS期に10%のCFU-Sに戻った。CFU-Sの過剰増殖は、例えばBalb/cマウス(表4)のようにウイルス性白血病誘発を受けやすいある種のマウス系統の特徴であることが知られており、そしてより高齢の動物においても観察され得る(表4)。TSP処理したBDF1マウスに見られた分化方向の決定付けられた(committed)前駆細胞と同じ再分布が、Balb/cおよびより高齢(23〜25ヶ月齢)のBDF1において観察され、それは一般に異常に高いレベルのCFU-Sの増殖を有する。CFU-Sの増殖および分化の両方の修正は、INPROLとのインキュベーションによって誘導された。臨床的により関連したことに、研究によって、INPROLのインビボ注射(2μg/マウス)がCFU-Sの増殖ならびに赤血球(BFU-E)とGM-コロニーとの比率の両方に影響することが示された(表4)。
【0186】
表4
分化方向の決定付けられた前駆細胞BFU-EおよびCFU-GMへの
CFU-Sの分化に対するINPROLの効果
【0187】
【表4】
。
【0188】
実施例10:INPROLの免疫刺激活性
増殖CFU-Sを高い比率で含む骨髄細胞とINPROLとのインキュベーションは、CFU-Sの周期を変えるだけでなく、それらの分化を、優勢な赤血球の分化から顆粒球およびリンパ球の前駆細胞へと切り換える。INPROLのこの特性は、細胞障害性の化学療法または放射線療法の免疫抑制的な副作用および過剰増殖幹細胞障害および加齢に伴う免疫抑制のために重要である。
【0189】
本実施例は、Wittlock & Witte(Ann.Rev.Immun.3:213〜35、1985)によって確立されたリンパ球長期培養(LLTC)由来の未成熟前駆体の前B前駆細胞への分化に対するINPROLの直接的な効果を示し、これはIL-7を含むメチルセルロース中のコロニー形成によって測定される。
【0190】
LLTCを、記載されたように確立し、そして新鮮なLLTC培地(Terry Fox Labs.,Vancouver,Canada)を1週間に2回与えた。非接着細胞を1週間に1度採取し、因子を洗い流し、そして25ng/mlのpINPROL、またはコントロールとして培地のみと4時間インキュベーションした。インキュベーションの後、細胞を洗浄し、そして105細胞/mlの濃度で、30%FCSおよび10ng/mlのIL-7を含むメチルセルロース中にプレートした。3週間のデータを図13に示す。大きなpre-Bコロニーの数はコントロール中で変化し、時間とともに増加したが、INPROLとのプレインキュベーションは、常にコロニーの増殖をコントロールレベルの4〜8倍に刺激した。このことは、免疫不全状態を修正し、そして例えばワクチン投与に対して所望の免疫応答を増加させるために有用であるINPROLの免疫刺激特性を示す。
【0191】
実施例11:INPROLは幹細胞の再生能を改善する--CMLの患者由来の長期培養開始細胞
慢性骨髄性白血病(CML)は、造血幹細胞の致死的な悪性疾患である。慢性期にあるCMLを単一薬剤による化学療法、組み合わせ化学療法、脾摘出、または脾臓への放射線照射を用いて処置することは、臨床的徴候および症状を制御し得るが、生存を有意に伸ばすことはない。CMLが慢性状態から促進状態に進むと、標準的な療法は効果的でない。現在では、骨髄移植(BMT)がCMLに対する唯一の既知の治療的療法である。血縁でないドナーのBMTを用いる治療は、組織不適合性の問題により困難である。適格なCML患者の40%未満にしか適切に適合する血縁のドナーがいない;従って自己移植が好ましい。長期培養(LTC)において増殖するPh陽性患者由来の非白血病(Ph-陰性)骨髄前駆細胞を選択する能力とともに、輸注のための自己BMTのエクスビボ調整は、正常幹細胞の自己供給源がCMLの積極的で効果的な治療を行い得る可能性を示唆する。
【0192】
BMTの状況においては、造血幹細胞は、長期間にわたって成熟した血液細胞を生成する能力を有する細胞として定義され得る。本発明者らは、C.Eaves & A.Eavesによって開発されたヒトLTC系を、幹細胞数の定量用およびそれらの治療的使用のための操作手段の両方で用いた。これは、予め確立され、放射線照射されたヒト骨髄接着層上に細胞を接種することを包含し;次いでこれらの培養物を5週間維持する。エンドポイント(end point)は、この期間の終わりに採取された培養物の総クローン原性細胞内容物(接着および非接着)である。このような条件下で生じたクローン原性細胞は、最初に添加された前駆細胞(長期培養開始細胞(LTC-IC))の数と直線関係にある;個々のヒトLTC-ICから生じた平均は、1個のLTC-ICにつき4クローン原性前駆細胞である。CMLの患者由来の骨髄が同様の条件下に置かれた場合、白血病性(Ph陽性)クローン原性細胞は迅速に衰退することが以前に示されていた。CML患者の残った正常LTC-ICの定量を用いることにより、培養自己移植片の移植により支持される集中治療から利益を得やすい細胞を選択することが可能である(Phillipsら、Bone Marrow Transplantation 8:477〜487、1991)。
【0193】
以下の手順を用いて、CMLの患者の末梢血から確立された骨髄移植細胞中のクローン原性細胞(LTC-IC)の数に対するINPROLの効果を試験した。
【0194】
予め放射線照射した間質上の長期培養と同様に培養を開始した。健常ドナーの末梢血をコントロールとして用いた。CML患者由来の末梢血細胞を、pINPROL(25ng/ml)とともに、またはなしで4時間プレインキュベーションし、洗浄し、そしてLTC-IC系に5週間置き、LTC-ICのコントロール数を測定した。実験のために、別の平行培養を10日間確立した。10日間増殖させた培養物からの接着細胞および非接着細胞の混合物を、pINPROLとともに、またはなしでプレインキュベーションし、そしてさらに8週間予め確立したフィーダー上に置いた。各実験培養物由来のLTC-ICの数は、接着細胞および非接着細胞の両方を適当な増殖因子(Terry Fox Laboratories、Vancouver、Canada)を有するメチルセルロース中にプレートし、そして得られた全コロニー形成細胞の数を計数することにより概算した。この手順を用いて得られたLTC-IC値は、下記の式を用いた全クローン原性細胞(CFC)内容物の評価に由来した:
#LTC-IC=#CFC/4。
【0195】
図14に示したデータは、健常ドナーの骨髄から始めた最初の10日間の培養中ではLTC-ICに損失がなく、そして投入したLTC-IC数の約30%が5週間たってもなお培養中に存在していたことを示す。CML患者のLTC-IC数は、10日間の間に約8%にまで劇的に低減し、そしてさらなるインキュベーション中、元には戻らなかったが、INPROLとともに細胞をプレインキュベーションすると、最初の数の30%までLTC-ICレベルが増加し、そしてそれは8週間維持された。
【0196】
これらの予備データによって予想されるINPROLの臨床関連の適用は、新鮮なまたは培養されたの骨髄移植物の正常幹細胞内容物を選択的に改善するためのストラテジー、インビボの残存正常幹細胞の補充を高めるためのストラテジー、さらに患者へのさらなる移植のために新しい遺伝物質をヒト骨髄幹細胞に転移させるためのプロトコルにおけるそれらの使用を包含する。
【0197】
実施例12A:骨髄調製物由来の幹細胞増殖の免疫活性阻害剤の単離方法
骨髄はブタの肋骨から単離した。ブタの胴体から肋骨を分離して筋繊維および脂肪を除き、細片に刻んで骨髄をBiophyzpribor社製の水圧プレスによって抽出した。骨髄細胞を、遠心機K-70中で2,000rpmで20分間遠心分離することにより分離する。次いで、引き続き抽出上清をAmicon USAメンブレンXM-100、PM30、PM-50により連続的に限外濾過にかける。電気泳動による分析によると、この生成物の主要な成分はアルブミンである(図1参照)。
【0198】
生物化学的精製
画分の骨髄抽出物およびタンパク質成分を、精製の各段階で、0.1%ドデシル硫酸ナトリウムを含む10%ポリアクリルアミドゲル電気泳動により分析した。7%までのドデシル硫酸ナトリウムおよび0.5〜1%までのメルカプトエタノールをサンプルに添加し、そしてそのサンプルをゲルにロードする前に70℃で5分間インキュベーションした。
【0199】
電気泳動は20Ycmのゲルで5時間行った。次いで、ゲルを、エタノール:水:酢酸が5:5:1の混合物中の0.25%クーマシーCBBC250で20℃で1時間染色し、そして7%酢酸を数回交換して洗浄した。生成物の活性を造血幹細胞(CFU-S)増殖の阻害方法により評価した。この方法を以下に詳細に述べる。
【0200】
ステージ1.硫安沈澱による精製
活性を、表5の結果に基づいて選択された飽和度40〜80%で25%の硫安を用いて沈澱させることにより精製した。
【0201】
【表5】
。
【0202】
精製の各工程の後の試験に用いられる調製物の量は、精製レベルによって決定し、始めの生成物の2×10-2mgの用量と同じであった。活性を下式により決定した:
変化%=Sa%−Sb%
ここで、Sa%は、コントロールのS%であり、
Sb%は、試験画分とのインキュベーション後のS%である。
【0203】
画分は、各活性試験の前、および次の各精製工程の前に硫安濃度を20倍低くするために脱塩した。
【0204】
ステージ2.ステージ1由来の不純物のある阻害剤を、脱塩後にイオン交換クロマトグラフィー(この場合DEAE23セルロース)を利用して分画し、次いで酢酸ナトリウム緩衝液(pH6.0)の勾配を用いて溶出する。
【0205】
阻害剤の活性画分は3〜5mMの間に溶出する。
【0206】
カラム容積は1mlであり、溶出速度は4ml/時間であった。検出はクロマトグラフMillicromeにより230および280nmで行った。最も高い活性を示した画分1(図2参照)を単離し、5mM酢酸ナトリウム緩衝液中に溶出した(表6参照)。
【0207】
【表6】
。
【0208】
電気泳動のデータは、主なタンパク質混入物であるアルブミン(図3参照)がこの画分から除かれることを示し、さらに4回の精製に続く。
【0209】
ステージ3.ステージ2由来の部分精製された阻害剤を直接G-75Sephadexカラムにかける。
【0210】
カラム体積は20ml(20×1)であり、溶出速度は2ml/時間である。溶出緩衝液は50mM NaCl、10mM Tris-HCl、pH7.5である。検出はクロマトグラフMillichromeにより230および280nmで行った。最も高い活性を示した画分5を単離した。
【0211】
【表7】
。
【0212】
ステージ4.Pro-RECカラムを利用した逆相クロマトグラフィー(Pharmacia FPLC System)をUltrasfera matrix 上で行った。タンパク質は0.1%トリフルオロ酢酸のアセトニトリル勾配を用いて溶出した。
【0213】
分子量16〜17kDの生成物の均質度は、アクリルアミド/ドデシル硫酸ナトリウムゲルの分析において示されるように、90%に等しい(図6参照)。この結果を図4に示す。画分5についての活性を測定する。生成物の最終回収率は5%である。結果として、精製後の分子量16kDのタンパク質の総量は始めの生成物の650ng/mlである。精製プロセスの間、生成物は70℃の加熱インキュベーションを数分受けたが、生物学的活性の損失は検出されなかった。
【0214】
実施例12B:より大量のINPROLを単離するための代替方法
最初の単離
新鮮なブタの胴体から採取した肋骨から筋繊維および脂肪を除き、次いでこれを刻んで細かくし、リン酸緩化衝生理食塩水に1:1(重量/容量)の比率で浸漬する。得られた混合物を水圧プレスで粉砕し、固体骨物質から骨髄を分離する。
【0215】
骨髄細胞懸濁液を回収し、そして4層のチーズ・クロス(cheese-cloth)を通して固形粒子を濾過する。濾液を56℃で40分間インキュベーションし、次いで氷浴で4℃まで冷却する。得られた沈澱物を10,000gで30分間4℃で遠心分離することにより除去し、廃棄する。
【0216】
清澄化した上清を、1容量%の濃塩酸を含む10容量の撹拌された氷冷アセトンに30分間滴下して添加する。得られた混合物を4℃に16時間保存し、完全な沈澱物を形成させる。次いで、沈澱物を20,000gで30分間4℃で遠心分離することによりペレット化する。このペレットを冷アセトンにより洗浄し、そして乾燥する。
【0217】
HPLC精製
ペレットを、5%アセトニトリル(MeCN)および0.1%トリフルオロ酢酸(TFA)を含むHPLC溶出緩衝液Aに溶かし、最終タンパク質濃度を8〜10mg/mlとする。この溶液(0.5〜0.6ml)を、PolisilODS-300(10mcm)が充填され、そして同じ緩衝液Aにより平衡化された250×4.6mmのHPLCカラムにロードする。
【0218】
溶出は緩衝液A中の緩衝液B(90%MeCN、0.1%TFA)の勾配により、流速1ml/分で、以下のプログラムに従って行う。
【0219】
時間、分 Bの%
0 0
4 0
5 25
25 90
。
【0220】
100%の緩衝液Bを5分間流すさらなる工程を用いて再平衡化の前にカラムを洗浄する。次いで、カラムを開始時の状態に戻すため再び平衡化し、そして次の部分のタンパク質溶液をカラムにロードし得る。代表的なクロマトグラムを表5に示す。
【0221】
分離の間、カラム溶出物をタンパク質ピークの検出のため280nmでモニターする。タンパク質物質を含む画分を回収し、分離したピークをプールして、30℃でロータリーエバポレーターにかけて乾燥させる。得られた残渣を蒸留水に溶解し、生物活性試験およびSDS-PAGE(14%ゲル、還元条件)によりアッセイする。活性物質を含むピークは、緩衝液Bが70%と80%との間に溶出され、SDS-PAGEによってアッセイされたように、16kDの主要なタンパク質バンドおよび速く移動するタンパク質の痕跡を含む。
【0222】
第2の主要なHPLCピークのみを回収することにより得られた物質の分析を、図15(AおよびC)に示す。両方のピークを含む物質(例えば、表5)を、本明細書中ではpINPROL調製物1といい、そして第2のピークのみからなる物質をpINPROL調製物2という。500μgのこの活性な、精製pINPROL調製物2をC4逆相カラム(Vydac)にロードし、そして0.1%トリフルオロ酢酸中の5〜95%アセトニトリルの直線勾配を用いて溶出した。その物質は53%アセトニトリルで単一ピークとして溶出した(図15A)。しかし、250μgのMIP-1α(R & D Systems)を同じ条件で流した場合、それは43.9%アセトニトリルで溶出した(図15B−14%アセトニトリルより前に溶出する初期のピークは人工的で検出器内の気泡によることを銘記されたい)。従って、天然に存在するINPROLは、実質的には、これらのような条件下ではMIP-1αより疎水性である。TGF-βはこれらの条件下ではpINPROLで観察される濃度より低い濃度で溶出することが知られている(Miyazonoら、J.Biol.Chem.263:6407〜15、1988)。
【0223】
溶出したpINPROL物質のゲルを図15Cに示す。レーン1は粗物質、レーン2は、分子量マーカー、そしてレーン3は精製物質である。2本の主要なバンドがあり、1本は約14kD、そしてもう1本は約35kDである。この35kDのバンドは14kDのバンドのマルチマー形態であると考えられる。
【0224】
実施例13A:活性INPROL調製物は、ヘモグロビンβ鎖を含む
pINPROLを、図5に示すように調製した(すなわち、pINPROL調製物1(実施例12Bを参照のこと))。物質をSDS-PAGEにかけ、そして標準的な技法を用いてニトロセルロースに移した。物質を、標準的な技術を用いて120AオンラインPTH-AA分析器を有するABI 477Aタンパク質シーケンサーを使用して、N末端配列分析に供した。以下のN末端配列が得られた:
VHLSAEEKEAVLGLWGKVNVDEV....
タンパク質データベースのコンピューターサーチは、この配列がブタヘモグロビンのβ鎖のN末端配列と同一性を有することを明らかにする(図16Cを参照のこと)。
【0225】
実施例13B:活性INPROL調製物はヘモグロビンα鎖を含む
図15Cにおいて示されるように、図5に示される第2の主要なピークを回収することにより精製したタンパク質(すなわち、pINPROL調製物2)は、分子量約15Kおよび30Kに対応する2つの主要なバンド、ならびに幾つかのマイナーバンドを生じた。SDS-PAGEゲルを、標準的な技術を用いてニトロセルロースに移し、そして個々のバンドを切り出して、実施例13Aに記載のように、N末端配列決定分析に供した。15kDのバンドについては以下のN末端配列が得られた:
VLSAADKANVKAAWGKVGGQ......
30kDのバンドを、制限タンパク質分解消化に供し、そして以下の内部の配列を得た:
**FPHFNLSHGSDQVK....
。
【0226】
第1の配列は、ブタヘモグロビンα鎖のN末端配列と同一性を示し、一方第2の配列はブタヘモグロビンα鎖の残基43〜56と同一性を示した(ヒト、マウス、およびブタのαヘモグロビン鎖およびβヘモグロビン鎖の配列比較については図16Cを参照のこと)。これらのバンドおよびいくつかのマイナーバンドの反復配列決定により、一貫して、αグロビン配列の部分を得た。従って、図15Cにおいて観察された種々のバンドは、ブタヘモグロビンα鎖のフラグメントまたは凝集体のいずれかを示す。
【0227】
実施例13C:ブタINPROLのさらなる特徴付け
pINPROLとブタヘモグロビンとをさらに比較するために、2回結晶化したブタヘモグロビンをSigma Chemical Companyから入手し、そして実施例12Bにおいて図15について記載したように、逆相HPLCに供した。図17において示され得るように、完全なヘモグロビンのHPLCクロマトグラムは、pINPROL調製物1について示されるクロマトグラムに類似する。さらに、直接比較において、図17Aにおいて示されるpINPROL調製物2(すなわち、図5の第2のピークに由来する)は、ブタヘモグロビンの第2の2つのピークの調製物(図17B)に重複することが見られ、主要ピークについての保持時間はそれぞれ52.51分および52.25分であった。ヘムが、この場合、49.153分で、ヘモグロビンにおける第1の主要ピークと共移動(co-migrate)することに注意すべきである;それゆえ、ヘムはpINPROL調製物1の成分であるが、調製物2の成分ではない。このことは、ヘムの存在について診断する波長575nmでのこのpINPROL調製物の吸光の欠失により確認される。
【0228】
ブタヘモグロビンα鎖およびβ鎖の推定分子量は、それぞれ、15038ダルトンおよび16034ダルトンである。図18のSDS-PAGEクロマトグラムにおいて示され得るように、第1の2つのピークはより高い分子量鎖から構成され、および第2の2つのピークはより低い分子量鎖からなる。したがって、第1の2つのピークはヘモグロビンβ鎖を示すようであり、および第2の2つのピークはヘモグロビンα鎖を示すようであった。
【0229】
ブタヘモグロビンのさらなる分離を、ゆるやかな溶出勾配を用いて行った(図21)。これらのピークのN末端分析は、第1のピークがブタα鎖であり、および第2のピークがブタβ鎖であることを示した。バイオアッセイ結果により、単離されたヘモグロビン鎖の両方が生物学的に活性であることが確認される(例えば、実施例14および15を参照のこと)。
【0230】
pINPROL調製物2およびヘモグロビンβ鎖のさらなる比較のために、2次元電気泳動を行った(図19)。1次元について、等電点を、9600ボルト時間について2%pH4〜8アンホラインを用いて、ガラスチューブ中で行った。トロポミオシン(分子量33kD、pI5.2)を、内部標準として用いた;その位置を最終的な2Dゲル上で矢印で記した。チューブゲルを緩衝液中で平衡化させ、そして12.5%アクリルアミドスラブゲルの上部のスタッキングゲルの上部にシールした。SDSスラブゲル電気泳動を4時間、12.5mA/ゲルで行った。ゲルを銀染色し、そして乾燥させた。
【0231】
2D電気泳動のパターンの比較により、HPLC精製ヘモグロビンβ鎖とpINPROL調製物2との間で異なるスポットは、1つまたは2つの少数のスポットのみであることを示した。抗ブタヘモグロビン抗体、および1Dまたは2D電気泳動のいずれかを用いたウエスタン分析により、調製物中にβヘモグロビンが存在することを確認する。したがって、活性pINPROL調製物2(実施例12Bにしたがって調製される)は、実質的にブタヘモグロビンβ鎖である。
【0232】
実施例14:ヘモグロビンα鎖、ヘモグロビンβ鎖、または完全なヘモグロビンは、幹細胞阻害活性を示す
ヘモグロビンβ鎖がINPROL活性を有することを確認するために、実施例1に記載したテストステロン処理したマウス由来の骨髄を用いた自殺アッセイを、実施例12Bにおけるように精製した物質を使用して実施例1に記載の方法論を用いて行った。表8に示すように、正常なマウスの骨髄細胞の15%が、テストステロン処理動物の36%とは対照的に死滅された。予想されたように、このことはテストロン処理が、細胞周期における細胞のパーセントを増加させる(したがって、細胞がより死滅されやすくする、例えば、実施例1)ことを示している。より際だった対照において、40ng/mlでpINPROLまたは精製ヘモグロビンβ鎖のいずれかとともにインキュベーションしたテストステロン処理した動物由来の細胞は、細胞周期における細胞のパーセントが、36%から0%および7%に、それぞれ、劇的に減少することを示した。200ngのより高い用量は、両方のタンパク質についてほとんど効果がなかった。ポジティブコントロールとして、以前に特徴付けられた幹細胞阻害剤MIP-1αは、13%に細胞周期の進行を減少させた。
【0233】
同様のアッセイを、CFU-Sの代わりにCFU-MIXの細胞周期の進行状態を用いて、インビトロで行い得る。高用量のトリチウム化されたチミジン(B.I.Lord、Haemopoiesis-A Practical Approach,N.G.TestaおよびG.Molineux(編)、IRL出版、1993;Pragnellら、Culture of Hematopoietic Cells,R.I.Freshney,I.B.PragnellおよびM.G.Freshney(編)、Wiley Liss 1994)の代わりに、シトシンアラビノシド(Ara C,30mg/ml)を細胞周期特異的毒性薬剤として用いた以外は、CFU-Sアッセイについて上述したようにアッセイを行い、そして高い内在性細胞周期速度を有するマウス系統(Balb/c)をテストステロンで処理したBDF1マウスの代わりに用いる。表9において示すように、高度に精製されたブタβ鎖、または高度に精製されたブタα鎖は、このアッセイにおいて両方とも活性であった。このアッセイにおいて注目すべき点は、pINPROLで処理した細胞についての細胞周期の進行レベルが、場合によってはネガティブな数を有し、このことは、非処理のプールよりもAra Cで処理したプールにおいてより多くのコロニーが存在することを示している。
【0234】
実施例2に記載したように、pINPROLは、トリチウム化したチミジン取り込みアッセイにおいて、マウス幹細胞FDCP-MIX株の増殖を阻害する。図20は精製ヘモグロビンα鎖およびβ鎖が、2ng/ml未満で示される阻害を伴いつつ、このアッセイにおいて両方とも活性であることを示している。
【0235】
前述は、ブタヘモグロビンのβ鎖がINPROL活性を示すという証拠を提供する。他のデータ(例えば、表9、図20)は、単離されたα鎖、ならびに完全なヘモグロビンがまた、幹細胞阻害剤として活性であることを示す。活性な調製物にはまた、α鎖およびβ鎖の混合物が含まれる(例えば、図5)。
【0236】
単離されたαグロビン鎖および/または単離されたβグロビン鎖が活性であるという観察は、本実施例で記載した活性が、完全な3次元ヘモグロビン構造を必要としないことを示している。α鎖およびβ鎖のフラグメントはまた、幹細胞阻害剤として活性である。
【0237】
【表8】
1NBM=正常な骨髄
2TPBM=テストステロン-処理マウス由来の骨髄
3Hbg=C4逆相精製ブタヘモグロビンβ鎖(2回結晶化したブタヘモグロビン由来)
【0238】
【表9】
1コントロール-Balb/cマウス由来の骨髄
2100ng/ml(図21のように精製)。
【0239】
実施例15:精製INPROL、精製ブタαヘモグロビン、または精製ブタβヘモグロビンは、インビボで活性である
精製したブタヘモグロビン鎖のインビボでの作用能力を試験するために、BDF1マウスに実施例1に記載のようにテストステロンプロピオネートを用いて注射した。24時間後、マウスに、pINPROL、ブタヘモグロビンα鎖(図21のように末梢血赤血球から精製した)、ブタヘモグロビンβ鎖(図21のように末梢赤血球から精製した)のいずれか500ng、あるいは等容量のキャリアを静脈内投与した。48時間後、各マウス由来の骨髄を採取し、そしてCFU-MIXアッセイを実施例14に記載のように行った。表10において示されるように、pINPROL、ブタα鎖およびブタβ鎖は、細胞周期におけるCFU-MIXのパーセントを基底レベルまで減少させ、全てインビボで活性であった。
【0240】
【表10】
1コントロール-テストステロン処理BDF1マウス由来の骨髄
2100ng/ml
3基底-未処理のBDF1マウス由来の骨髄。
【0241】
実施例16:精製ヒトヘモグロビンα鎖、ビオチン化ヒトヘモグロビンα鎖、ビオチン化ヒトヘモグロビンβ鎖、ヒトヘモグロビンγ鎖、およびヒトヘモグロビンδ鎖は、全てインビトロで幹細胞阻害活性を示す
ヒトヘモグロビンを、Sigma Chemical Corporationから入手するか、あるいは成人ヒト末梢血または臍帯血から標準的な手段により単離した。個々の鎖を、ブタα鎖およびβ鎖について上述したのと同様の様式で逆相HPLCにより単離した(B.MasalaおよびL.Manca,Methods in Enzymology、第231巻、21〜44頁、1994を参照のこと)。精製α、β、γ、およびδ鎖を得た。ビオチン化したα鎖およびβ鎖について、1mgの成人ヒトヘモグロビンを、37μgのNHS LC ビオチン(Pierce)を用いて処理し、そして上記のように逆相クロマトグラフィーにより鎖を分離した。
【0242】
表11、12、および13において示すように、精製ヒトαヘモグロビン鎖、ビオチン化ヒトαヘモグロビン鎖、ビオチン化ヒトβヘモグロビン鎖、ヒトγ鎖ヘモグロビン鎖、およびヒトδヘモグロビン鎖は、全て、CFU-MIX細胞周期の進行アッセイにおいて活性である。
【0243】
【表11】
1コントロール-Balb/cマウス由来の骨髄
2100ng/ml
【0244】
【表12】
1コントロール-Balb/cマウス由来の骨髄
2100ng/ml
【0245】
【表13】
1コントロール-Balb/cマウス由来の骨髄
2100ng/ml。
【0246】
実施例17:精製ヒトα鎖、α-βダイマー、またはヘモグロビンは、インビボで活性である
精製ヒトα鎖、α-βダイマー、またはヘモグロビンを実施例15に記載のようにインビボアッセイで試験した。表14に示したように、これらのそれぞれは濃度500ng/マウスで活性であった。
【0247】
【表14】
1コントロール-テストステロン処理BDF1マウス由来骨髄。
【0248】
実施例18:ブタINPROLは、インビトロでヒト単核細胞またはCD34+臍帯血細胞において活性である
ブタ骨髄由来の精製INPROLが、ヒト前駆細胞における細胞周期の進行に影響する能力を調べるために、臍帯血細胞を得た。Ficoll上で分離した後に得られた総単核細胞画分または抗CD34親和性カラム(CellPro Inc.)上での分画後に得られたCD34+画分のいずれかを使用した。初期の幹細胞が細胞周期にあることを確認するために細胞を、インターロイキン3(IL-3)および幹細胞因子(SCF)(各100ng/ml)の存在下でインビトロにおいて48時間インキュベーションした。このプレインキュベーション後、細胞周期の進行アッセイを、CFU-GEMM(CFU-MIXの代わりに)をプレート後18日目に計数した以外は、実施例14に記載のようにマウスについて行った。表15において示すように、ブタINPROLは、大多数の単核細胞またはCD34+画分のいずれかにおいて、CFU-GEMMの細胞周期の進行を阻害した。
【0249】
【表15】
。
【0250】
実施例19:精製ヒトαヘモグロビンは、ヒトCFU-GEMMにおいて活性である
ヒト臍帯血単核細胞を得、そしてIL-3およびSCF中でインキュベーションし、そして実施例18に記載のように細胞周期の進行アッセイに使用した。表16に示すように、骨髄から精製したブタINPROLおよび末梢血から精製したヒトαヘモグロビンは、このアッセイにおいて活性であった。
【0251】
【表16】
。
【0252】
実施例20:ヒトαヘモグロビンおよびヒトβヘモグロビン配列から得られたペプチドは活性である
活性なペプチド配列を同定するために、ミオグロビンの3次元構造(本アッセイにおいて不活性である)を、コンピューターモデリングプログラムを用いて成人ヒトヘモグロビンに存在するα鎖のネイティブな3次元構造に重ね合わせた。2つのペプチド(アミノ酸43〜55および64〜82で表される、これらは3次元空間においてミオグロビンと構造的に異なる領域である)を、CFU-MIX細胞周期の進行アッセイにおいて活性を有するとして同定した。ネイティブなα鎖において見出されるループをより近接に近づけるために、ペプチド43〜55の環状誘導体(c43〜55)(ジスルフィド結合を利用)をまた合成し、そして活性であることが見出された。
【0253】
これらのペプチドの配列は以下のようである:
【0254】
【化9】
2つのヘモルフィン配列、ヘモルフィン10(β鎖配列のアミノ酸32〜41)およびヘモルフィン7(アミノ酸33〜40)を試験し、そして活性であることを見出した。
【0255】
配列は以下のようである:
【0256】
【化10】
。
【0257】
これらの配列の活性を試験するために、CFU-MIX細胞周期の進行アッセイを実施例14に記載のように行った。表17〜19に示すように、これらのペプチドは全てこのアッセイにおいて活性である。
【0258】
【表17】
【0259】
【表18】
1全てのペプチドは100ng/mlで試験された。
【0260】
【表19】
1100ng/mlで試験された。
【0261】
実施例21:蟻酸切断によりヒトαヘモグロビンから得られたペプチドフラグメントは活性である
ヒトαヘモグロビン鎖は、アミノ酸94位と95位(Asp-Pro)との間に蟻酸切断部位を有する。切断を、精製ヒトα鎖(実施例16と同様)を70%蟻酸中1mg/mlの濃度で37℃で72時間インキュベートすることにより得た。1-94フラグメントを未切断α鎖から精製し、そして95-141フラグメントを実施例16のような逆相HPLCにより精製した;その後、分画をSDS-PAGEを用いて行った(実施例22と同様)。精製1-94タンパク質フラグメントの同一性を、エレクトロスプレーイオン化質量分析により確認した。
【0262】
このフラグメントの幹細胞阻害活性を評価するために、CFU-MIX細胞周期進行アッセイを実施例14と同様に使用する:
【0263】
【表20】
1Balb/c骨髄
2精製非組換えヒトαヘモグロビン(実施例16と同様)(100ng/ml)
3精製蟻酸切断タンパク質(本実施例と同様)(100ng/ml)。
【0264】
実施例22:ユビキチン融合体としてのE.coliにおけるヘモグロビンα鎖、ポリペプチド1-141、ポリペプチド1-97、ペプチド43-55、およびペプチドc(43-55)の発現
ペプチド43-55(「p13」)およびc43-55(「p15」)(実施例20と同様)の遺伝子を、最適E.coliコドン使用頻度(AnderssenおよびKurland,Micro.Reviews 54:198-210,1990)に従って対応するオリゴヌクレオチドをアニールすることにより合成した。インタクトなヒトαヘモグロビン鎖(「p141」)の遺伝子を、1セットのオリゴを設計して、ヒト骨髄cDNAプール(Clontech,Palo Alto,CA)からPCR増幅することにより得た。1-97フラグメント(「p1-97」)の遺伝子を、適切なサブクローニング後のp141遺伝子を含むプラスミドのPCR増幅により得た。
【0265】
上記の遺伝子を、ユビキチン融合タンパク質として発現した(米国特許第5,132,213号;同第5,196,321号;および同第5,391,490号、ならびにPCT WO 91/17245を参照のこと)。宿主株E.coli DH5αFIQ(Life Technologies,Inc.,Gaithersburg,MD)を、適切に合成した遺伝子(上記)を含むユビキチン発現ベクターpDSUbで形質転換した。pDSUbは、ヒトユビキチンを発現するpDS78/RBSIIの誘導体である(図22A)(Hochuliら、Biotechnology 6:1321-5,1988)。Loetscherら(JBC 266:11213-11220,1991)は、クロラムフェニコールアセチルトランスフェラーゼ(CAT)配列をHochuliプラスミドから切り出し、そしてプラスミドを再連結することによりpDS78/RBSIIを改変した(図22B)。合成ユビキチン遺伝子を、細菌発現に最適なコドン使用頻度を有するヒトユビキチンをコードするリン酸化合成オリゴヌクレオチドの対様アニーリングにより構築した。次いで、pDSUbを、組み立てたオリゴヌクレオチドからなる合成ユビキチン遺伝子を、誘導体化pDS78/RBSII のKlenow平滑化BamHI-BglI消化物に挿入することにより構築した。得られたプラスミドpDSUb(図22C)は、E.coliにおいて高レベルでユビキチンを発現することが示された。
【0266】
p97を含むプラスミドおよびp141を含むプラスミドを、指向性クローニングにおいて、p97またはp141タンパク質および融合接合部をコードするAflII-PstI消化PCR産物を、AflIIおよびPstIで消化したpDSUbに挿入することにより構築した。同様に、p13を含むプラスミドおよびp15を含むプラスミドを、適切な付着末端を有し、そしてペプチドおよび融合接合部をコードする、リン酸化およびアニールされたオリゴヌクレオチドを、AflII-PstI消化pDSUbに挿入することにより構築した。
【0267】
形質転換体を、100μg/mlアンピシリン、5μg/mlネオマイシンで選択した。コロニーは30℃で2日後に現れる。形質転換体を、挿入部位を横切るPCRによりスクリーニングした。次いで、正確なサイズの挿入物を含むコロニーを、SDS-PAGEにより適切なサイズの融合タンパク質の発現についてスクリーニングした(以下を参照のこと)。ユビキチン融合体を、lacリプレッサーを滴定し(titrate)、それをpDSUbのプロモーターから除去するIPTGの添加により過剰発現させた(DH5αF’IQは、10μg/mlネオマイシンで選択されるF’因子上にアップレギュレートされたlaqIq遺伝子を含む)。
【0268】
過剰発現された誘導ユビキチン融合タンパク質を示したクローン由来のプラスミドDNAを調製し、そしてジデオキシ法によりSequenase Version 2.0キット(United States Biochemical.)を用いて配列決定した。次いで、ポジティブクローンを凍結し、そして-80℃でグリセロール中に保存した。ポジティブクローンを、アンピシリン(100μg/ml)、ネオマイシン(10μg/ml)、および1%グルコースを含むLBプレート上で30℃で維持した。それらを10継代まで一週間毎に画線培養し(streak)、その後、新たな画線培養物を、連続培養のために凍結した種バイアルから採取して、株が本物であることを保証した。
【0269】
アッセイ用のタンパク質を得るために、250ml振盪フラスコ中の100ml接種菌培養物を、一晩のインキュベーション(16〜20時間)により、アンピシリン(100μg/ml)、ネオマイシン(10μg/ml)、および1%グルコースを有する2×YT培地中でシングルコロニーから増殖させた。振盪フラスコ培養物を、New Brunswick環境振盪インキュベーターにおいて30℃および250rpmで維持した。翌朝、培養物を、培地で1リットルに希釈した。細胞を、1mM(最終希釈)のIPTG添加によりOD600=0.5で誘導し、そしてOD600=0.8で遠心分離により採集した。採集細胞を低張溶解緩衝液(100μlの50mM Tris(pH10.0))に再懸濁した。細菌細胞を、その懸濁物を3サイクルの凍結-解凍(凍結のためのドライアイス-エタノール浴および解凍のための60℃)に供することにより溶解した。次いで、懸濁物を10分間超音波処理し、そして12,000gで10分間遠心分離した。得られた上清を「S1」と呼ぶ。細胞ペレットを、50mM Tris(pH10)および2×SDSトリシンローディング緩衝液(Novex,San Diego,CA)(1:1)に再懸濁した。次いで、混合物を95℃で15分間加熱し、そして12,000gで10分間遠心分離した。この方法で再可溶化され得る沈殿物の一部を「P1」と呼ぶ。残存するペレットから得られた沈殿物の一部を「P2」と呼ぶ。P2を、P1と同様にローディング緩衝液に再懸濁した。S1、P1、およびP2由来のサンプルを、SDS-PAGEにより分析した。
【0270】
SDS-PAGEゲルを、10〜20%トリシンゲル(Novex)と共に、ミニゲル装置における二緩衝液トリシン系を用いて泳動させた。陽極(下部)緩衝液は0.2M Tris(pH9.0)であった。陰極(上部)緩衝液は、0.1M Tris、0.1Mトリシン、0.1%SDS(pH8.25)であった。市販の分子量マーカー「Multi-Mark」(Novex)を使用した。標準として使用したウシユビキチンをSigmaから購入した。ゲルを、色素マーカーがゲルの下部に達するまで4mAの定電流で泳動させた。ゲルを、酢酸:メタノール(10%:40%)中の0.25%のCoomassie Blue R250(Sigma)で染色し、そして色素のない同じ溶液で脱染した。
【0271】
インタクトなp141ユビキチン融合タンパク質の大部分(>70%)を、細菌溶解物の遠心分離後の沈殿物(P1およびP2)において見出した。はっきりと対照的に、p97ユビキチン融合タンパク質の大部分(>70%)を、可溶性画分(S1)において見出した。これにより、C末端疎水性領域の除去は、改善された可溶性特性を有する産物を生じることが確認された。同様に、p13およびp15ペプチドもまた可溶性画分に含まれた。
【0272】
UCH-L3ユビキナーゼ(ubiquinase)酵素(Recksteiner,M.(編)Ubiquitin,Plenum Press(NY)1988;Wilkinsonら、Science 246:670-73,1989)を、宿主株BL21/DE3を形質転換するために使用されたpRSET(Invitrogen,San Diego,CA)において発現させた。UCH-L3は、ユビキチンC末端伸長を切断するユビキチン特異的プロテアーゼである。これを、35%(w/v)硫酸アンモニウム沈殿により細菌溶解物から部分的に精製した。使用した硫酸アンモニウムの正確な割合を、25.5kDバンドの存在についてSDS-PAGEによりモニターした。上清を50mM Tris(pH7.4)に対して透析し、そしてユビキチンペプチド融合基質に対してアッセイした。活性な上清を等分し、そして-20℃で凍結した。代表的な反応混合物は、3μlの溶解物、1μlの1M DTT、1μlのUCH-L3(上記)、および5μlの反応緩衝液(50mM Tris(pH7.4))を含む。反応を室温で20分間行った。大規模な消化については、300μlの溶解物を、100μlの1M DTT、20μl UCH-L3、および580μl反応緩衝液と混合した。
【0273】
可溶性(S1)画分に含まれるペプチドまたはタンパク質を、実施例16のように逆相HPLCによりさらに精製した;画分をSDS-PAGEによりモニターし、そして同一性を、エレクトロスプレーイオン化質量分析により確認した(以下を参照のこと)。精製したペプチドまたはタンパク質を、上記のUCH-L3によって酵素学的に消化し、非ユビキノン化最終生成物を得た。次いで、この切断物質を、逆相HPLCによって再精製した。精製を、SDS-PAGEによって行い、そして最終生成物の同一性をエレクトロスプレーイオン化質量分析によって確認した。
【0274】
上記のようなUCH-L3でのインビトロ切断に対する代替は、所望のユビキチン融合体と同じ細菌においてユビキチン切断酵素を同時発現させることである。この目的のために、ユビキナーゼUBP1(TobiasおよびVarshavsky,JBC 266:12021-12028,1991)を発現するベクター(pJT184)を使用した。P97ユビキチン融合体およびUBP1を同時発現する細菌は、インビボで融合タンパク質の完全な消化を示した;p141ユビキチン融合体およびUBP1を同時発現する細菌は、融合タンパク質の部分的な(約70%)消化を示した。インビボ消化したp97タンパク質を、硫酸アンモニウム沈殿、続いて上記のように逆相HPLCにより精製した。
【0275】
発現および精製されたポリペプチドの同一性を確認するために、エレクトロスプレーイオン化質量分析を、四重極分析器を有するVG Biotech BIO-Q器械を用いて行った。この器械の較正をするために、ミオグロビンを使用した。精製p97で得られた主な成分は、分子量10,339ダルトンの単一のピークであった;これは、計算された分子量10,347にほぼ匹敵し、組換えp97フラグメントの同一性を確認した。
【0276】
実施例23:組換えp1-97は、幹細胞阻害活性を維持する
組換えp1-97の生物活性を評価するために、GFU-GEMM細胞周期進行アッセイを実施例18と同様に使用した:
【0277】
【表21】
1ヒト骨髄単核細胞
2精製非組換えヒトαヘモグロビン(実施例16と同様)(100ng/ml)
3精製組換えp97(実施例22と同様)(100ng/ml)。
【0278】
実施例24:ヒトαヘモグロビンおよびペプチド43-55は、テストステロンまたは5-フルオロウラシル(5FU)処理マウスにおいて、インビボでCFU-MIX細胞周期進行を阻害する
ペプチド43-55のインビボでの幹細胞阻害活性を評価するために、B6D2 F1マウスを、実施例1に記載のテストステロンプロピオネートまたは5FUで前処理した。詳細には、テストステロンプロピオネート(100mg/kg体重)を、マウスに0日目にi.p.注射した。あるいは、5FU(200μg/kg体重)を、マウスに0日目に注射した。
【0279】
24時間後(1日目)、種々の用量のペプチド43-55またはビヒクルを、i.v.注射した。骨髄を2日目に採集し、そしてCFU-MIX細胞周期進行アッセイを、実施例14と同様に行った。詳細には、マウスを屠殺し、そして単一細胞骨髄懸濁物を大腿骨から調製した。細胞を1回洗浄し、そして濃度をFischer培地で5×106細胞/mlに調節した。各試験条件のために、1ミリリットルの細胞を、2つのポリプロピレンチューブのそれぞれに添加した。チューブを、適切な濃度の試験基質を共なわずに(「コントロール」)または共に(「実験」)で37℃で3時間インキュベートした。インキュベーションの終わりに、30μg/mlのシトシンアラビノシド(「Ara C」(Sigma))を、チューブの半分まで添加し、そして同じ容量のFischer培地をもう1つのチューブに添加した。チューブをさらに1時間37℃でインキュベートし、その後それらを氷上に置き、そして冷Fischer培地で2回洗浄した。
【0280】
細胞をFischer培地で5×104〜106/mlに再調節し、そして0.5mlの細胞懸濁物を、5mlのMethocult M3430メチルセルロース培地(Stem Cell Technologies,Vancouver,British Columbia)に添加した。混合物を、ボルテックスしながら力強く混合し、そして1mlを5つの35mmディッシュのそれぞれに分配した。35mmディッシュを、順に、滅菌水を含むふたのない35mmディッシュと共に、ふた付きの150mmディッシュに入れた。CFU-MIXコロニーを、37℃でのインキュベーションの7日後に倒立顕微鏡を用いて数えた。
【0281】
培地処理チューブとAra C処理チューブとの間のコロニー数の差異は、以下の式に従った条件下でのサイクルにおける細胞の割合を示す:
【0282】
【化11】
ここで、a=培地単独でインキュベートしたチューブ由来のCFU-MIXの数
b=Ara Cでインキュベートしたチューブ由来のCFU-MIXの数
【0283】
【表22】
1テストステロン前処理動物
220グラムマウスあたりのi.v.注射した量
【0284】
【表23】
15FU前処理動物
220グラムマウスあたりのi.v.注射した量。
【0285】
実施例25:ペプチド43-55は、N末端Phe(Phe43)でビオチン化またはPhe43もしくはPhe46でヨウ素化した場合、幹細胞阻害剤として活性である
ペプチド43-55を、固相ペプチド合成技術(American Peptide Co.,Sunnyvale,CA)により合成した。ペプチドアナログを、Phe43またはPhe46のパラ位でヨウ素を用いて合成した。ビオチン化ペプチド43-55を、C4炭素リンカーを有するビオチンのCOOHを、Phe43のN末端NH2に連結することにより合成した。
【0286】
【表24】
。
【0287】
実施例26:モルヒネは、マウスCFU-MIXの細胞周期進行をインビトロで阻害する
モルヒネを、実施例24と同様にBalb/cマウス由来の骨髄を用いてCFU-MIX細胞周期進行アッセイにおいて試験した。
【0288】
【表25】
。
【0289】
実施例27:オピエートペプチドDAMGOおよびDALDAは、マウスCFU-MIXの細胞周期進行をインビトロで阻害する
DAMGOおよびDALDAを、実施例24と同様にBalb/cマウス由来の骨髄を用いてCFU-MIX細胞周期進行アッセイにおいて試験した。
【0290】
【表26】
。
【0291】
実施例28:ノシセプチンは、マウスCFU-MIXの細胞周期進行をインビトロで阻害する
ノシセプチンを、実施例24と同様にBalb/cマウス由来の骨髄を用いてCFU-MIX細胞周期進行アッセイにおいて試験した。
【0292】
【表27】
。
【0293】
実施例29:ナロキソンは、ヒトαおよびδヘモグロビン鎖、へモルフィン10、ならびにペプチド43-55の阻害活性と拮抗する
CFU-MIX細胞周期進行アッセイを実施例24と同様に行う。試験基質を、それ自身でまたはナロキソン(10-5、10-7M)の存在下でアッセイした。ナロキソン自身は、これらの濃度でアッセイに効果を生じなかった。
【0294】
【表28】
110-5M最終濃度で使用した。
【0295】
【表29】
110-7M最終濃度で使用した。
【0296】
実施例30:低濃度のナロキソンは、マウスCFU-MIXの細胞周期進行を阻害する
CFU-MIXアッセイを実施例24と同様に行った。
【0297】
【表30】
。
【0298】
実施例31:μオピエートレセプターアンタゴニストCTOPは、ヒトヘモグロビンα鎖、ペプチド43-55、およびペプチド64-82の阻害活性と拮抗する
CTOP(H-D-Phe-Cys-Tyr-D-Trp-Orn-Thr-Pen-Thr-NH2、Cys2とPen7との間にジスルフィドを有する)は、μオピエートレセプター特異的アンタゴニストであるソマトスタチンのアナログである。試験基質を、それ自身またはCTOP(10-7M)の存在下でアッセイした。CTOP自身は、この濃度でアッセイでの影響を有さないが、αヘモグロビンまたはペプチド43-55により引き起こされる細胞周期阻害と拮抗する。
【0299】
【表31】
110-7Mの最終濃度で使用した。
【0300】
実施例32:ヒトα鎖での前処理は、後期形成コブルストーン(cobblestone)形成細胞の数を増加させる
コブルストーンアッセイを、Ploemacherおよびその同僚(Ploemacherら、Blood 74:2755-63,1989;van der Sluijsら、Exp.Hematol.18:893-6,1990;Ploemacherら、Blood 78:2527-33,1991;Ploemacherら、J.Tiss.Cult.Meth.13:63-68,1991;DownおよびPloemacher,Exp.Hematol.21:213-21,1993)により記載されたように行った。コブルストーンアッセイは、ストロマ細胞の単層内に細胞群(または「コブルストーン」)の出現を測定する。非常に初期の幹細胞は、軟寒天においてコロニーを形成しないが、ストロマ単層の存在下でコブルストーンを形成する。コブルストーンを形成する細胞は、「コブルストーン領域形成細胞」(CAFC)と呼ばれる。より分化した(例えば、GM-CFC)前駆体は、培養の最初の数週間以内に出現し、そして消滅する一過的なコブルストーンを形成するが、より初期の幹細胞(例えば、長期再増殖細胞)は、4〜5週間の培養後のみに出現するコブルストーンを形成する。従って、培養の7〜14日目に形成するCAFCはCFU-GMに富み、28〜35日目に形成するCAFCはCFU-MIXに富み、そして28〜35日目に形成するCAFCは長期再増殖細胞に富む。
【0301】
B6D2F1マウスを、実施例24と同様にテストステロンプロピオネートで処理した。その翌日、骨髄を取り出し、そしてそれらをブレストーンアッセイでプレーティングした後に、ヒトαヘモグロビン鎖(100ng/106細胞)と共にまたは共なわずに4時間インキュベートした。使用したアッセイは、96ウェルプレートにおける限界希釈長期骨髄培養(LTBMC)からなる。培養物を、FBMD-1マウスストロマ細胞株(Breemsら、Leukemia 11:142-50,1997)をコンフルエントになるまで増殖させることにより調製した;コンフルエントな単層を有する96ウェルプレートを、アッセイまで33℃で保存した。マウス骨髄細胞を単細胞懸濁物として調製し、そして以下の細胞希釈物:27,000;9000;3000;1000;333を、0.2mlのLTBMC培地(Stem Cell Technologies,Vancouver)にウェルあたりプレーティングした。20ウェルを、条件あたり各希釈についてプレーティングし、そして2プレート以上に分配した。
【0302】
コブルストーン領域形成細胞(CAFC)の頻度を、以前に記載されたように(Ploemacherら、Blood 78:2527-33,1991;Ploemacherら、J.Tiss.Cult.Meth.13:663-68,1991;Breemsら、Leukemia 8:1095-104,1994)計算した。結果を実施例23に示す。細胞周期進行幹細胞のヒトαヘモグロビン鎖とのプレインキュベーションは、約5倍まで後期形成CAFCの比を増大させた。非細胞周期進行細胞のα鎖での処理は効果がなかった。
【0303】
実施例33:ヒトαヘモグロビン、ならびにペプチド43-55、環状43-55および64-82は、ヒト臍帯血CFU-GEMMの細胞周期進行を阻害する
ヒト臍帯血CFU-GEMM細胞周期進行アッセイを、実施例19と同様に行った。詳細には、単核細胞を、ヒト臍帯血細胞から単離し、そして10%FBS、100ng/mlキットリガンド、および100ng/mlヒトIL-3を補充したIMDM組織培養培地で2〜4×104細胞/mlに調節した。細胞を37℃で48時間インキュベートした。
【0304】
インキュベーション後、細胞を洗浄し、そして106細胞/mlの濃度で無血清IMDMに再懸濁した。1mlの細胞を、条件あたり2つのポリプロピレンチューブのそれぞれに添加し、そして細胞周期進行アッセイを、マウス骨髄について実施例24と同様に行った。Ara Cインキュベーション後、細胞を冷IMDMで洗浄し、そして0.5ml IMDMあたり10,000〜20,000細胞に調節し、そして5mlのMethocult H4433(Stem Cell Technologies)と混合した。あるいは、細胞濃度を0.5ml IMDMあたり2500〜5000細胞に調節する場合に、Methocult H4435メチルセルロース培地(Stem Cell Technologies)を使用した。細胞を実施例24と同様にプレーティングし、そしてCFU-GEMMコロニーを14〜18日目にスコア付けした。
【0305】
【表32】
。
【0306】
実施例34:ヒトαヘモグロビンおよびペプチド43-55は、成人骨髄CFU-GEMMの細胞周期進行を阻害する
CD34+幹細胞を、CellProカラムの手段によるヒト骨髄からの精製の後、Poietic Technologies(Gaithersburg,MD)から得た。細胞を、キットリガンドおよびIL-3と48時間インキュベートし、そして実施例26におけるCFU-GEMM細胞周期進行アッセイにおいて使用した。
【0307】
【表33】
。
【0308】
実施例35:移動したヒト末梢血におけるCFU-GEMMは活発にサイクルし、そしてヒトαヘモグロビン、ペプチド43-55、DAMGO、またはモルヒネにより阻害可能である
末梢血を、標準的プロトコルに従ったシクロフォスファミドおよびG-CSFでの末梢幹細胞移動を受けている乳ガン患者から得た。赤血球をFicoll Hypaqueで除去した(細胞をIMDMで1:1に希釈し、そして20mlを16mlのFicollの上部に重層し、そして800gで30分間遠心分離した;単核細胞を中間層(interphase)から取り出し、そしてIMDM中で2回洗浄した)。1つの場合、単核細胞を、アッセイ前に液体窒素中で凍結保存した。単核細胞を、2.5〜5×105細胞をディッシュあたりにプレーティングした以外は、実施例26と同様にCFU-GEMM細胞周期進行アッセイのためにプレーティングした。
【0309】
【表34】
。
【0310】
【表35】
。
【0311】
【表36】
1アッセイ前に凍結保存した細胞に由来する。
【0312】
実施例36:高用量のヒトαまたはβヘモグロビン、ミオグロビン、ペプチド1-97、ペプチド43-55、ペプチド64-82、ノシセプチン、あるいはDALDAは、静止マウス幹細胞の細胞周期進行を刺激する
1μg/ml用量のヘモグロビン鎖、ミオグロビン、およびペプチドを、静止幹細胞の刺激についてアッセイした。骨髄を、実施例24と同様のCFU-MIX細胞周期進行アッセイにおいて試験した未処理B6D2F1から得た。未処理B6D2F1マウスから単離した幹細胞は、(例えば、テストステロンプロピオネート(実施例1を参照のこと)または5FUのような化学療法(実施例4を参照のこと)により)刺激されるまで、通常ゆっくりと細胞周期進行して、周期に入る。
【0313】
【表37】
。
【0314】
【表38】
。
【0315】
【表39】
。
【0316】
【表40】
。
【0317】
【表41】
。
【0318】
【表42】
。
【0319】
実施例37:高用量のヒトαヘモグロビンの静脈内投与は、静止マウス幹細胞の細胞周期進行を刺激する
ヒトαヘモグロビンを、未処理B6D2F1マウスにi.v.注射した。24時間後、マウスを屠殺し、骨髄を大腿骨から回収し、そしてCFU-MIXアッセイを実施例24と同様に行った。
【0320】
【表43】
。
【0321】
実施例38:ナロキソンは、高用量ヒトαヘモグロビン、ペプチド43-55の幹細胞刺激活性と拮抗する
【0322】
【表44】
110-7M最終濃度で使用した。
【0323】
***
本発明は好ましい実施態様によって記載されたが、変化および改変は当業者に考えられることが理解される。従って、添付の請求の範囲は、請求される本発明の範囲内にある全てのこのような等価な変化を含むことが意図される。
【図面の簡単な説明】
【0324】
【図1】図1〜図4は、各精製段階後の産物のSDSポリアクリルアミドゲル泳動を示す。図1−レーン1はキモトリプシノーゲンであり、レーン2はオボアルブミンであり、レーン3はBSAであり、レーン4は30kD未満の画分であり、レーン5は30〜50kDの画分であり、そしてレーン6は50〜100kDの画分である。
【図2】図1〜図4は、各精製段階後の産物のSDSポリアクリルアミドゲル泳動を示す。図2−レーン1は硫酸アンモニウム沈澱後(40〜80%)であり、そしてレーン2〜5はDEAE画分である(レーン番号2は活性画分を表す)。
【図3】図1〜図4は、各精製段階後の産物のSDSポリアクリルアミドゲル泳動を示す。図3−レーン1は硫酸アンモニウム沈澱後の上清であり、そしてレーン2は活性DEAE画分であり、レーン3〜5はゲル濾過画分を表す(レーン番号5は活性画分を表す)。
【図4】図1〜図4は、各精製段階後の産物のSDSポリアクリルアミドゲル泳動を示す。図4−レーン2は最終産物を表す。
【図5】図5は、最終精製物の逆相HPLCクロマトグラムを示す。
【図6】図6は、ブタ骨髄から精製されたINPROL(pINPROL)を含まない場合(コントロール=0%阻害)および、種々の量で含む場合のFDCP-mix細胞株内へのトリチウム化チミジンの取り込み(cpm)を示す。データは、コントロールの値に対して正規化されている。
【図7】図7は、テストステロンプロピオネート(TSP)、TSP+pINPROL、またはビヒクル(コントロール)を用いてマウスを処理した後の細胞周期のS期にある細胞の%を示す。各グループは25匹の動物を含んでいた(時点毎に3〜4匹)。
【図8】図8は、2用量の5-FUで処置され、pINPROL処理を受けたかまたは受けていないマウスの生存を示す。各グループは30匹の動物を含んでいた。
【図9】図9は、放射線照射され、pINPROL処置を受けた場合および受けていない場合のマウスの生存を示す。各グループは50匹の動物を含んでいた。
【図10A】図10Aおよび図10Bは、Ara-CまたはAra-C+pINPROLで処理した後、1週間目(図10A)および3週間目(図10B)の正常骨髄の長期培養細胞の再生を示す。
【図10B】図10Aおよび図10Bは、Ara-CまたはAra-C+pINPROLで処理した後、1週間目(図10A)および3週間目(図10B)の正常骨髄の長期培養細胞の再生を示す。
【図11】図11は、致死量の放射線照射を行い、培地(コントロール)またはpINPROL(25ng/ml)と4時間プレインキュベーションした後の3×104個の骨髄細胞を移植した後のマウス(グループ当たり75匹)の生存を示す。生存は30日間モニターされた。
【図12】図12は、致死量の放射線照射を行い、pINPROLまたは培地と4時間プレインキュベートされたドナーの骨髄細胞により回復した後のマウスの骨髄細胞により14日後に培養物中で形成されたCFU-GMの数を示す。
【図13】図13は、毎週取り出し、洗浄し、そして培地またはpINPROLと4時間プレインキュベートした後にIL-7(10ng/ml)と共にプレートしたリンパ球の長期培養物由来の懸濁細胞を示す。
【図14】図14は、pINPROLで処理された白血病の末梢血細胞の改善された再生能を示す。長期培養開始細胞(LTC-IC)は、粘着性および非粘着性LTC細胞をpINPROLと共に、およびpINPROLなしでプレートし、そして7日目にCFU-GMを数えることにより測定された。データはコントロールの値に対して正規化されている。
【図15A】図15Aは、53%アセトニトリルで溶出した精製pINPROLのC4逆相クロマトグラムを示す。レーン1は粗物質であり、レーン2は分子量マーカーであり、そしてレーン3は精製物質である。図15Bは、43.9%アセトニトリルで溶出したMIP-1αのC4逆相クロマトグラムを示す。図15Cは、pINPROLの粗調製物および逆相後の精製された調製物のSDS-PAGEクロマトグラムを示す。
【図15B】図15Aは、53%アセトニトリルで溶出した精製pINPROLのC4逆相クロマトグラムを示す。レーン1は粗物質であり、レーン2は分子量マーカーであり、そしてレーン3は精製物質である。図15Bは、43.9%アセトニトリルで溶出したMIP-1αのC4逆相クロマトグラムを示す。図15Cは、pINPROLの粗調製物および逆相後の精製された調製物のSDS-PAGEクロマトグラムを示す。
【図15C】図15Aは、53%アセトニトリルで溶出した精製pINPROLのC4逆相クロマトグラムを示す。レーン1は粗物質であり、レーン2は分子量マーカーであり、そしてレーン3は精製物質である。図15Bは、43.9%アセトニトリルで溶出したMIP-1αのC4逆相クロマトグラムを示す。図15Cは、pINPROLの粗調製物および逆相後の精製された調製物のSDS-PAGEクロマトグラムを示す。
【図16A】図16は、ヘモグロビン配列を示す:図16AはヒトαヘモグロビンのcDNA配列およびアミノ酸配列を示し、そして図16BはヒトβヘモグロビンのcDNA配列およびアミノ酸配列を示す。番号付けはアミノ酸に従う。図16Cは、ヒト、マウス、およびブタヘモグロビンのα鎖およびβ鎖のアミノ酸配列比較を示す。
【図16B】図16は、ヘモグロビン配列を示す:図16AはヒトαヘモグロビンのcDNA配列およびアミノ酸配列を示し、そして図16BはヒトβヘモグロビンのcDNA配列およびアミノ酸配列を示す。番号付けはアミノ酸に従う。図16Cは、ヒト、マウス、およびブタヘモグロビンのα鎖およびβ鎖のアミノ酸配列比較を示す。
【図16C】図16は、ヘモグロビン配列を示す:図16AはヒトαヘモグロビンのcDNA配列およびアミノ酸配列を示し、そして図16BはヒトβヘモグロビンのcDNA配列およびアミノ酸配列を示す。番号付けはアミノ酸に従う。図16Cは、ヒト、マウス、およびブタヘモグロビンのα鎖およびβ鎖のアミノ酸配列比較を示す。
【図17】図17は、pINPROL(図17A)および結晶化ブタヘモグロビン(図17B)のC4逆相HPLCトレースの比較を示す。
【図18】図18は、結晶化ブタヘモグロビンのC4逆相HPLC分離からの画分のSDS-PAGEゲルを示す。レーン1は分子量マーカーを示し、レーン2は第1ピーク(47.11分)由来の画分48〜49を示し、レーン3は第2ピーク(49.153分)由来の画分50〜51を示し、レーン4は第3ピーク(52.25分)由来の画分54〜55を示し、そしてレーン5は第4ピーク(53.613分)由来の画分56〜57を示す。
【図19】図19は、pINPROL(図19A)および精製ブタβヘモグロビン(図19B)の2次元ゲル電気泳動の比較を示す。
【図20】図20は、FDCP-MIXアッセイにおける精製ブタαヘモグロビン、βヘモグロビン、またはpINPROLの効果の比較を示す。
【図21】図21は、ゆるやかな溶出勾配を用いるブタヘモグロビンの逆相分離を示す。
【図22】図22Aは、Hochuliら(1988)からのプラスミドを示す;図22Bは、Loetscherら(1991)のプラスミドを示す;図22CはpDSUbプラスミドを示す。
【図23】図23は、cobblestoneアッセイでの、INPROLでの処置の結果を示す。
【特許請求の範囲】
【請求項1】
本明細書中に記載のポリペプチド。
【請求項1】
本明細書中に記載のポリペプチド。
【図1】


【図2】
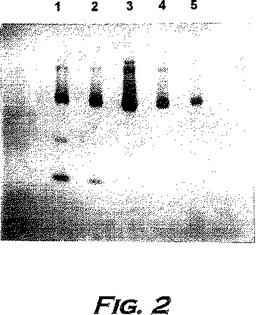
【図3】
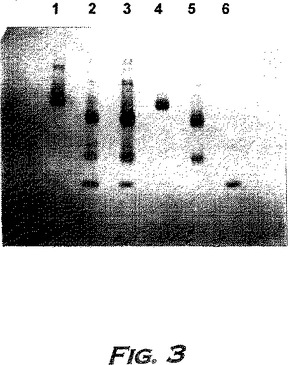
【図4】

【図5】
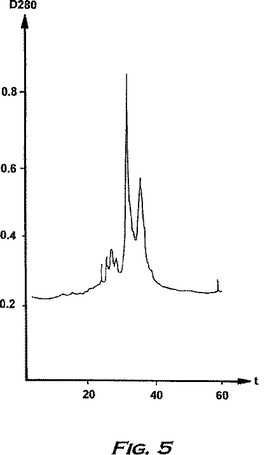
【図6】
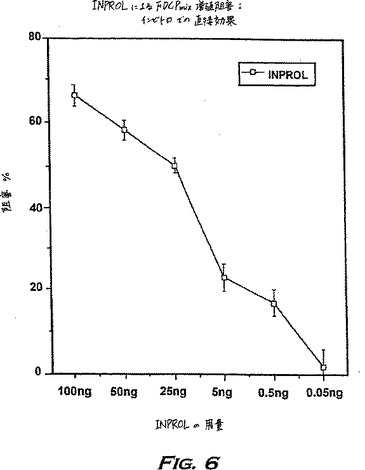
【図7】
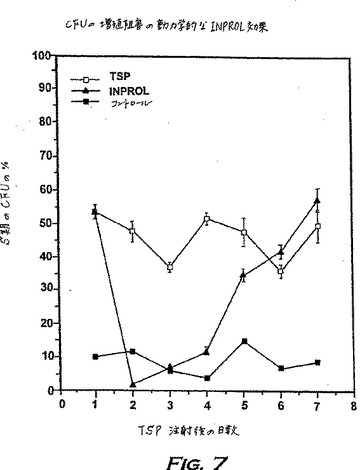
【図8】
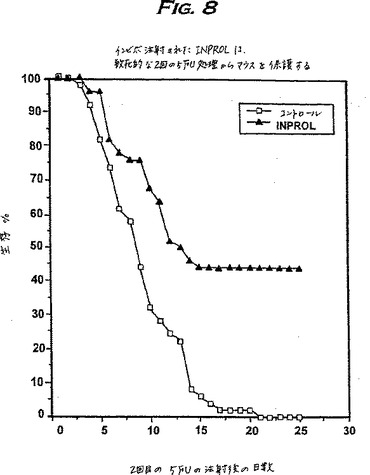
【図9】
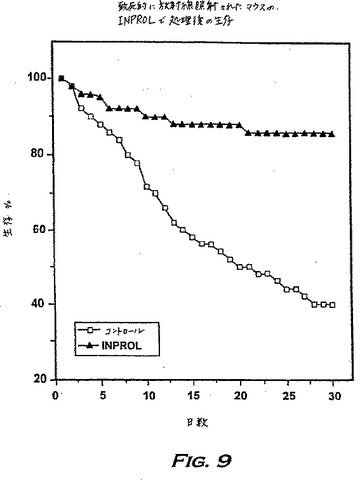
【図10A】
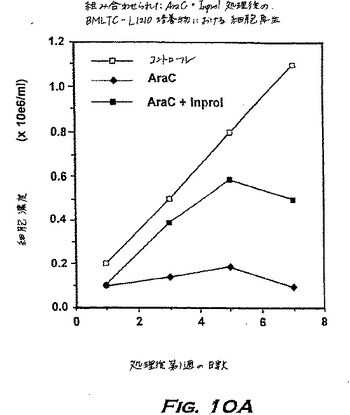
【図10B】
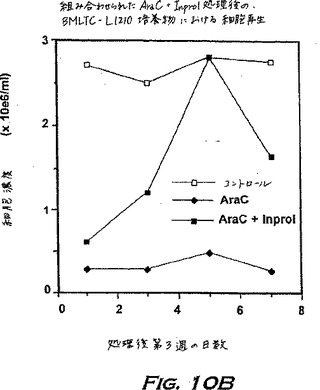
【図11】
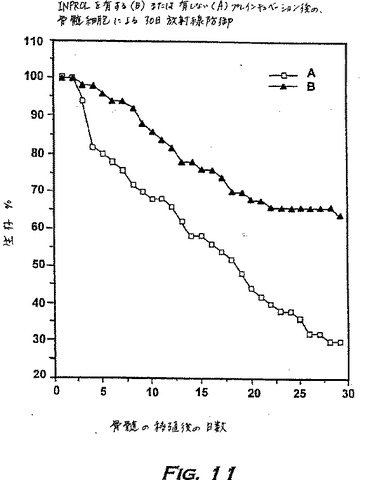
【図12】
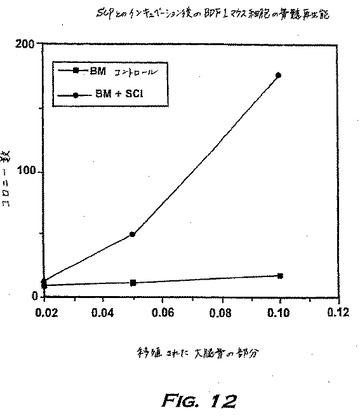
【図13】
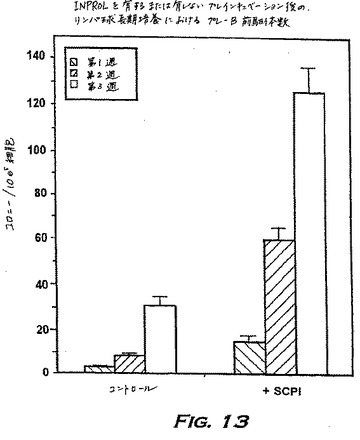
【図14】
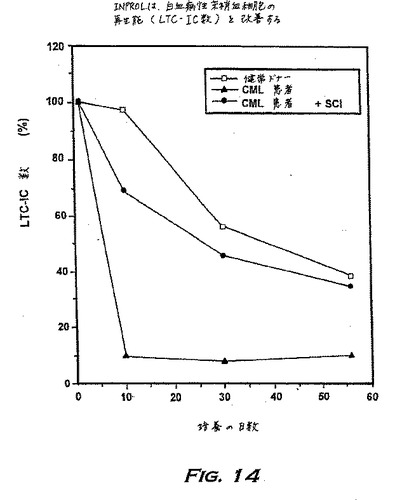
【図15A】
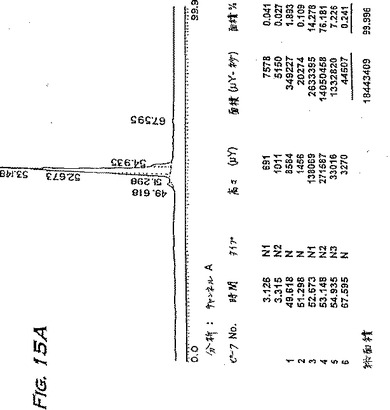
【図15B】
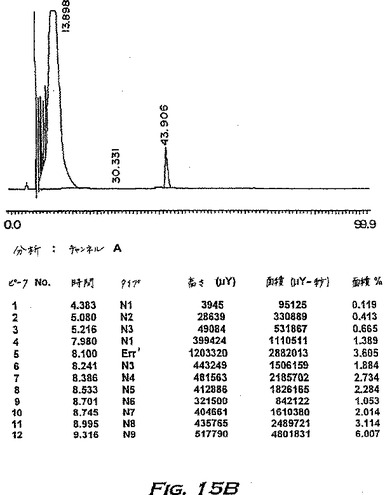
【図15C】

【図16A】

【図16B】

【図16C】

【図17】
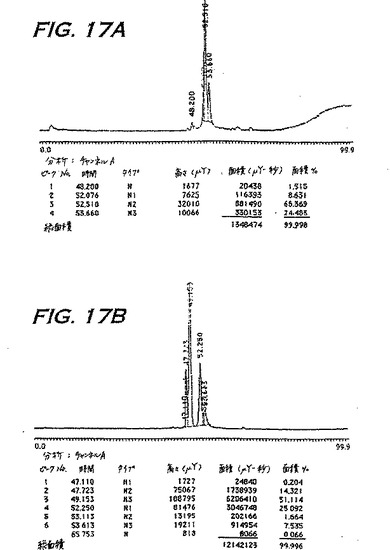
【図18】
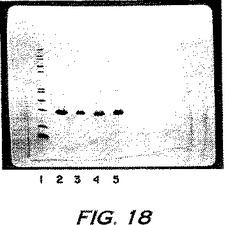
【図19】

【図20】
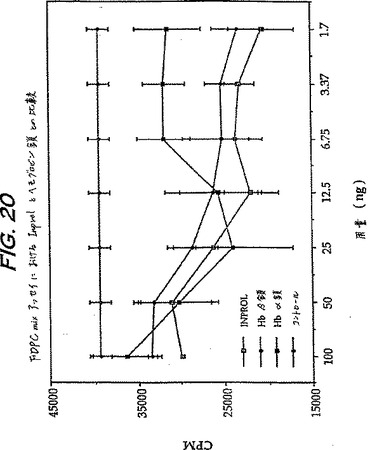
【図21】
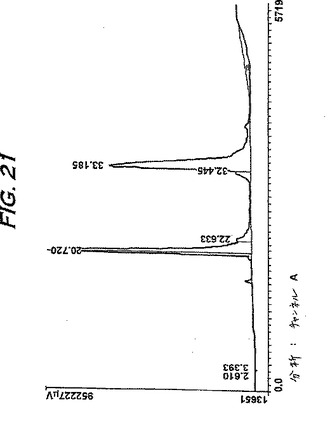
【図22】
【図23】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10A】
【図10B】
【図11】
【図12】
【図13】
【図14】
【図15A】
【図15B】
【図15C】
【図16A】
【図16B】
【図16C】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【公開番号】特開2009−13178(P2009−13178A)
【公開日】平成21年1月22日(2009.1.22)
【国際特許分類】
【出願番号】特願2008−201331(P2008−201331)
【出願日】平成20年8月4日(2008.8.4)
【分割の表示】特願平9−535611の分割
【原出願日】平成9年4月3日(1997.4.3)
【出願人】(594197621)プロ ー ニューロン, インコーポレーテッド (4)
【Fターム(参考)】
【公開日】平成21年1月22日(2009.1.22)
【国際特許分類】
【出願日】平成20年8月4日(2008.8.4)
【分割の表示】特願平9−535611の分割
【原出願日】平成9年4月3日(1997.4.3)
【出願人】(594197621)プロ ー ニューロン, インコーポレーテッド (4)
【Fターム(参考)】
[ Back to top ]