
幹細胞/前駆細胞の多分化能の維持方法
本発明は、幹細胞/前駆細胞の多分化能または自己再生特性のうち少なくともいずれか一方を維持する方法に関する。本発明は、細胞中における遺伝子発現を調節する方法にも関する。本方法は、少なくとも2つの転写因子またはその機能的断片を、nanog遺伝子のプロモータ領域と接触させることを含む。少なくとも2つの転写因子の一方はPOUおよびホメオドメイン含有転写因子から選択される。少なくとも2つの転写因子の他方はHMGドメイン含有転写因子から選択される。本方法は、さらに、少なくとも2つの転写因子がnanogプロモータ内の特異的結合要素と複合体を形成できるようにすることを含む。このようにして形成された複合体は転写活性化を介してnanog遺伝子発現を調節する。
【発明の詳細な説明】
【技術分野】
【0001】
(0001)本発明は、幹細胞/前駆細胞の多分化能または自己再生特性のうち少なくともいずれか一方を維持する方法に関する。本発明は、細胞内の遺伝子発現を調節するための方法にも関する。本方法は、少なくとも2つの転写因子またはその機能的断片を、nanog遺伝子のプロモータ領域と接触させることを含む。少なくとも2つの転写因子のうちの一方は、POUドメインおよびホメオドメイン含有転写因子から選択される。少なくとも2つの転写因子のうちの他方は、HMGドメイン含有転写因子から選択される。本方法は、少なくとも2つの転写因子が、nanogのプロモータ内の特異的結合要素と複合体を形成できるようにすることをさらに含む。このようにして形成される複合体は、転写活性化を介してnanog遺伝子の発現を調節する。
【背景技術】
【0002】
(0002)幹細胞は、再生医療に対する鍵を握っていることがわかってきている。というのも幹細胞は、疾患、感染または先天性異常によって損傷した対応組織の代わりとなる細胞源を供給できるためである。これは、幹細胞が、無制限に増殖できる能力を持つとともに、心臓、肝臓、脳細胞などの機能的な成熟細胞に分化しうる未分化細胞であるためである。ギュンター病、ハンター症候群、およびハーラー症候群は、たとえば、幹細胞を用いて治療されてきた。白血病やリンパ腫などの病状を有するがん患者の治療は、骨髄からの成人幹細胞を用いて行われてきた。化学療法投薬量の段階的増大による血球減少に苦しむがん患者の造血系は、幹細胞の静脈内注射によって回復させることができた。韓国の研究者は、臍帯血からの成人幹細胞をうまく用いて、脊髄損傷を煩う麻痺をもつ女性が、歩行器の助けを借りて歩行できるようになったことを報告している。パーキンソン病、心筋梗塞、若年発症糖尿病をはじめとするさらに多くの疾病が、現在のところ、幹細胞または幹細胞由来の細胞の治療的移植によって治療可能であると考えられている。
【0003】
(0003)この点に関する特に実現可能なツールは、哺乳類の多分化性幹細胞であるが、これは当該細胞が、任意の器官、細胞タイプまたは組織タイプへ分化可能であり、少なくとも潜在的には完全な生命体へと分化することができるためである。このような多分化性幹細胞は、着床前胚および多くの成人組織のいずれにおいても見られる。特に優れているのは、哺乳類の胚盤胞の段階にある胚の内細胞塊(ICM)に由来する胚幹細胞(ESC)である。ESCは、特定の細胞培養条件下で、より長い時間、自己再生的に細胞分裂することにより、その多分化能を維持する(たとえば、非特許文献1または非特許文献2を参照のこと)。
【0004】
(0004)ESCは、たとえば、神経成長因子とレチノイン酸の共存下に、ニューロンへと制御的に分化させることができるが(非特許文献3)、そのように容易に分化できることが、実践上の大きな難問をつきつけるのである。ESCを多分化性状態に維持するためには、取扱時および培養増殖時の分化を防止しなければならない。こうした理由から、ESCは、伝統的にはフィーダー細胞層上でウシ胎仔血清の共存下で(たとえば、特許文献1および特許文献2を参照のこと)、または線維芽細胞調整培地(CM)中で培養されてきた。にもかかわらず、念入りに制御された条件下においてさえ、ESCは、in vitroでの増殖のあいだに自発的に分化することがある。マウスESCにおける自己再生を媒介する因子である白血病抑制因子(LIF)は、マウスESCの分化を阻害することがわかっているが、ヒトESCの分化を阻害する際のフィーダー細胞の役割に変わるものではない。したがって、ECSの多分化能または自己再生特性のうち少なくともいずれか一方を維持する手段は、幹細胞治療の完全な商業的潜在性の実現に向けての多大な功績となるであろう。
【0005】
(0005)成人幹細胞は、ESCのような多分化性ではないものの、自己再生可能であり、その発生能を、より未成熟な多分化性ESCの発生能と同等にする可塑性を持つことがわかっている。一例として、成人幹細胞は、その元となる組織とは異なる細胞系統に分化することができる。
【0006】
(0006)幹細胞は、2以上の発生学上の層が混ざり合った組織を生み出す、様々な組織(精巣および卵巣のものであることが多い)の奇形腫と呼ばれる癌においても見つかっている。このような癌の悪性型は、奇形癌とも呼ばれている。マウス奇形癌における幹細胞の発達は、正常な胚における事象と平行したものである。これらの存在は、化学治療で大量の腫瘍塊を除去しても、腫瘍の再発を防止できない場合が多いことの説明となる(非特許文献4)。したがって、このような腫瘍において幹細胞の多分化能または自己再生特性のうち少なくともいずれか一方を無効にする手段は、典型的にはこのような癌の永久的除去のための前提条件となるであろう。
【0007】
(0007)いまのところ数種のタンパク質が、多分化性細胞の正常な発達のために、かつ/または多分化性の細胞状態を維持するために重要であることが確認されている。そうしたタンパク質としては、Oct4(非特許文献5)、Sox2(非特許文献6)、Nanog(非特許文献7、非特許文献8)、およびFoxD3(非特許文献9)が挙げられる。他のタンパク質、たとえば、LIF(上記参照)またはRexl(Zfp42)も、このような機能に関与することが示唆されている。しかしながら、それらのタンパク質の作用機構、ならびに多分化性細胞の発生または多分化性細胞状態の維持における、当該タンパク質の特異的役割については今のところ知られていない。
【特許文献1】米国特許第5,843,780号明細書
【特許文献2】米国特許第6,090,622号明細書
【非特許文献1】ロエベル、ディー.エイ.(Loebel,D.A.)ら、2003年、Dev.Biol.第264巻、p.1〜14
【非特許文献2】スミス、エイ.ジー.(Smith,A.G.)、2001年、Annu.Rev.Cell Dev.Biol.第17巻、p.435〜462
【非特許文献3】シュルディナー(Schuldiner)ら、2001年、Br.Res.第913巻、p.201〜205
【非特許文献4】チャンバース、アイ.、スミス、エイ.(Chambers,I.Smith,A)、2004年、Oncogene 第23巻、p.7150〜7160
【非特許文献5】ニコルス、ジェイ.(Nichols,J.)ら、1998年、Cell 第95巻、p.379〜391
【非特許文献6】アビリオン、エイ.エイ.(Avilion,A.A.)ら、2003年、Genes Dev.第17巻、p.126〜140
【非特許文献7】ミツイ(Mitsui)ら、2003年、Cell 第113巻、p.631〜642
【非特許文献8】チャンバース(Chambers)ら、2003年、Cell 第113巻、p.643〜655
【非特許文献9】ハンナ(Hanna)ら、2002年、Genes Dev.第16巻、p.2650〜2651
【発明の開示】
【発明が解決しようとする課題】
【0008】
(0008)したがって、本発明の目的は、幹細胞/前駆細胞の多分化能または自己再生特性のうち少なくともいずれか一方を制御的に維持する方法、ならびに、たとえば、奇形腫中の幹細胞の多分化能または自己再生特性のうち少なくともいずれか一方を無効にするために、細胞内の遺伝子発現を調節する方法を提供することにある。
【課題を解決するための手段】
【0009】
(0009)一つの態様において、本発明は、幹細胞/前駆細胞の多分化能または自己再生特性のうち少なくともいずれか一方を維持する方法を提供する。本方法は、少なくとも2つの転写因子またはその機能的断片を、nanog遺伝子のプロモータ領域と接触させることを含む。少なくとも2つの転写因子のうちの一方は、POUドメインおよびホメオドメイン含有転写因子から選択される。少なくとも2つの転写因子のうちの他方は、HMGドメイン含有転写因子から選択される。本方法は、少なくとも2つの転写因子が、nanogプロモータ内の特異的結合要素と複合体を形成できるようにすることをさらに含む。このように形成された複合体は、転写活性化を介してnanog遺伝子発現を調節する。
【0010】
(0010)別の態様において、本発明は、細胞内の遺伝子発現を調節する方法を提供する。本方法は、少なくとも2つの転写因子またはその機能的断片をnanog遺伝子のプロモータ領域と接触させることを含む。少なくとも2つの転写因子のうちの一方は、POUドメインおよびホメオドメイン含有転写因子から選択される。少なくとも2つの転写因子のうちの他方は、HMGドメイン含有転写因子から選択される。本方法は、少なくとも2つの転写因子が、nanogプロモータ内の特異的結合要素と複合体を形成できるようにすることをさらに含む。このように形成された複合体は、転写活性化を介してnanog遺伝子発現を調節する。
【発明を実施するための最良の形態】
【0011】
(0011)本発明は、非限定的な実施例および図面と組み合わせて、以下の詳細な説明を参照してより良く理解されるであろう。
(0019)本発明は、POU特異的ドメインおよびホメオドメインを含む転写因子、ならびにHMGドメインを含む転写因子が、in vitroおよびin vivoのいずれにおいてもnanogのプロモータと複合体を形成し、さらに、この複合体の形成がタンパク質Nanogの発現を調節するという驚くべき発見に基づいている。さらに驚くべきことに、このような結合領域の少なくとも1つが累積して2億五千万年以上もの進化の間、不変のままであることが判った。
【0012】
(0020)本願発明者の知見は、特に実用的に関係がある。というのも、POU特異的ドメインおよびホメオドメインを含む転写因子がタンパク質Oct4であり、HMGドメインを含む転写因子がタンパク質Sox2(参考:上記および下記)であるためである。Oct3としても知られるOct4は、細胞の未分化状態に関するマーカーとしてよく使用される。Oct4は、ヒトおよびマウスESCにおいて高度に発現され、その発現は、これらの細胞が分化して多分化能を失うときに減衰する(パルミエリ(Palmieri)ら、1994年、Dev.Biol.第166巻、p.259〜267)。また、Oct4は成人幹細胞、腫瘍細胞、および不死化非腫瘍原性細胞においても発現されるが、分化した組織の細胞においては発現されない(たとえば、モンク、エム.(Monk,M.)、ホールディング、シー.(Holding,C.)、2001年、Oncogene 第20巻、p.8085〜8091を参照のこと)。
【0013】
(0021)Sox2は転写因子であり、Oct4と同様に、分化中のESCと比べて、未分化ヒトESC内で強くアップレギュレートされる(ブランデンバーガー、アール(Brandenberger,R)ら、2004年、Nature Biotechnology 第22巻、p.707〜716)。Sox2は、たとえば、胚発生の初期段階における全CNSの神経上皮の未熟な未分化細胞においても発現される。分化が起こると、Sox2がダウンレギュレートされる(ステバノヴィク、エム(Stevanovic,M)、2003年、Molecular Biology Reports 第30巻
第2号、p.127〜132)。Sox2とOct4のいずれも、他の細胞タイプを決定する際に独立した役割を果たし(ニワ、エイチ.(Niwa,H.)ら、2000年、Nat Genet 第24巻、p.372〜37;非特許文献6)、多分化性細胞におけるそれらの機能の少なくとも一部は、標的遺伝子の転写を駆動するために2者の間の相乗作用を介する。
【0014】
(0022)転写因子という用語は、DNAからのRNAの合成(転写)のレベルを変えるタンパク質の能力のことをいう。典型的には、このような因子は、エンハンサ要素またはプロモータ要素などのような、各DNA上に見られる遺伝子の所定の領域に結合して、該DNAと複合体を形成する、細胞質タンパク質または核タンパク質である。
【0015】
(0023)Oct4およびSox2の相乗作用の標的としては現在、fgf4,utfl,およびfbx15の3種が知られており(ユアン、エイチ.(Yuan,H.)ら、1995年、Genes Dev.第9巻、p.2635〜2645;ニシモト、エム.(Nishimoto,M.)ら、1999年、Mol.Cell.Biol.第19巻、p.5453〜5465;トクザワ、ワイ.(Tokuzawa,Y.)ら、2003年、Mol.Cell.Biol.第23巻、p.2699〜2708)、4つ目は、sox2自身であると思われる(トミオカ、エム.(Tomioka,M.)ら、2002年、Nucleic Acids Res.第30巻、p.3202〜3213)が、対応するシス要素は多分化性細胞発現に対しては省略可能であると思われる(ハヤシ、エス.(Hayashi,S.)ら、2002年、Mech.Dev.第119巻、追補1,S97〜S101)。これらの標的遺伝子のそれぞれは、8量体およびsox結合部位を含む複合要素を有する。
【0016】
(0024)本発明の方法において、上述のような転写因子(以下も参照)は、新しく同定されNanogと命名されたタンパク質をコードする遺伝子のプロモータとともに複合体を形成する。これは、細胞の多分化性状態の維持、ならびに初期胚盤葉上層/内細胞塊(ICM)の多分化性細胞の正常な発達、およびこの細胞集団からのES細胞の派生にも必要なタンパク質である(ミツイ(Mitsui)ら、前掲)。またこのタンパク質は、たとえば分化中のESC中よりも未分化のESC中にはるかに多く存在することから、未分化状態の細胞、たとえば、幹細胞または胚性生殖細胞に対するマーカーとして使われることも多い(ブランデンバーガー、アール(Brandenberger,R)ら、前掲)。Nanogは、成人骨髄中でも発現されており、腎細胞やある種のがんなどの様々な成人組織において、低レベルで発現されている。Oct−4と同様に、Nanogは、成人の成熟細胞にいくらかの胚様の可塑性を回復させうることで知られている(たとえば、シース、エヌエックス.(Theise,NX.)、ウィルマット、アイ.(Wilmut,I)、2003年、Nature 第425巻、p.21を参照)。
【0017】
(0025)Nanogはホメオボックスを含み、転写因子として作用する場合もある。nanogの過剰発現により、通常は分化を誘発する培養条件において、ESCの多分化能および自己再生特性を維持することができる(チャンバース、アイ.(Chambers,I.)ら、前掲)。標準増殖条件下でnanog発現をダウンレギュレートすると、ESCにおける分化が誘導される(図1を参照)。多分化性細胞の維持においてこの必須機能に付随するのが、nanogの限局的な発現パターンである。nanog転写物は、胚盤胞形成前に、まず桑実胚の内部細胞内に現れ(チャンバース、アイ.(Chambers,I.)ら、前掲;ミツイ、ケイ.(Mitsui,K.)ら、前掲)、胚盤胞内では発現は内細胞塊(ICM)に限られ(ワン、エス.エイチ.(Wang,S.H.)ら、2003年、Gene Expr.Patterns 第3巻、p.99〜103)、着床時にはすでに検出不能である。
【0018】
(0026)本願発明者による知見を考慮すると、Oct4およびSox2がマウスの着床前発生において類似の発現パターンを有するという最近発表された研究結果(アビロン、エイ.エイ.(Avillon,A.A.)ら、2003年、Genes Dev.第17巻、p.126〜140)が容易に理解できるようになる。Nanogは、幹細胞の多分化性細胞状態および自己再生特性の維持のための鍵となる要素であるという知識に基づいて、本発明者らは、幹細胞および前駆細胞の多分化能または自己再生特性のうち少なくともいずれか一方を維持する方法を確立することができた。本発明者らは、さらに、それらの方法が細胞内の遺伝子発現を調節するのに適していることを発見した。
【0019】
(0027)nanog遺伝子は、凝縮後に現れ桑実胚の内部細胞に特異的な第1の既知の転写因子をコードし、Oct4およびSox2のいずれも、すべての割球において凝縮の前に発現される。本明細書中で用いる「nanog遺伝子」という用語は、あらゆる哺乳類のnanog遺伝子、たとえば、ハート、エイ.エイチ.(Hart,A.H.)ら(2004年、Dev.Dynamics 第230巻、p.187〜198)に記載されるようなマウスおよびヒトのnanog遺伝子、ウシの遺伝子(NCBI遺伝子ID:538951)、ブタの遺伝子(EMBLアクセス番号Q5GMQ0およびQ64HX3)、ディンゴの遺伝子(NCBI遺伝子ID:486701)、チンパンジーの遺伝子(NCBI遺伝子ID:452438)、マカクの遺伝子(EMBLアクセス番号Q5TM84)、ラットの遺伝子(NCBI遺伝子ID:414065(Locusタグ:RGD:1303178))、NCBIアクセス番号AY786437のRNAを転写し、NCBIアクセス番号AAW50709の転写因子をコードするヤギ遺伝子、ならびにこれらのオーソログを含む。また、まだ同定されてないnanog遺伝子も含まれる。また、たとえばアンネ(Anne)ら(2004年、Genomics 第84巻、p.229〜238)によって同定され、当技術分野で周知の標準的な方法(たとえば、部位特異的変異法)によって機能するように改変されたもののようなnanogの偽遺伝子も含まれる。さらに、既知のnanog配列と比較すると核酸配列に逸脱が見られる形態の遺伝子も含まれる。その差異は、たとえば、多型、1つのヌクレオチドの変化または修飾、(連続する一続きの)置換、欠失または挿入、および対応する天然の核酸配列に導入されたN末端および/またはC末端への付加によるものであってよい。本発明の方法において用いるnanog遺伝子のプロモータ領域は、任意の生物種のもの、または任意の生物種に由来するものであってよい。2つの例示的実施例は、ヒトおよびマウスのnanog遺伝子のプロモータ領域である。
【0020】
(0028)Nanogは、マウスおよびヒトの多分化性細胞のいずれにおいても発現されるので、この遺伝子の多分化性転写は、シス調節要素の機能保存を通して維持されており、同時に、これらの要素の位置および配列は、浄化選択を経ても保存されると思われる。したがって、マウスおよびヒトのnanogゲノム配列を、配列比較のために公開データベースから得た。驚いたことに、マウスnanogおよびslc2a3のタンデム重複の結果として、2コピーのヒトnanog遺伝子が同定された(ブース、エイチ.エイ.(Booth,H.A.)およびホランド、ピー.ダブリュ.(Holland,P.W.)、2004年、Genomics 第84巻、p.229〜238;ハート、エイ.エイチ.(Hart,A.H.)ら、前掲)。ヒトESTデータ(Hs.329296)に基づけば、NANOG/NANOGlは活発に転写されるが、NANOGP1/NANOG2は検出不能レベルを維持している。以下の実施例においては、活発に転写されるNANOGを用いて、マウスのホモログnanogと比較した。
【0021】
(0029)本発明の方法は、少なくとも2つの転写因子、またはその機能的断片を、nanog遺伝子のプロモータ領域、すなわち、転写開始時にRNAポリメラーゼによって認識され結合されるnanog遺伝子の核酸配列と接触させることを含む。これらの転写因子のうちの一方は、Pit,Oct,Unc(POU)特異的ドメインおよびホメオ
ドメインを含む転写因子から選択される。多くの実施形態において、ホメオドメインはPOU−ホメオドメインである。POU特異的ドメインは一般には75〜82アミノ酸長であり、POU−ホメオドメインは約60アミノ酸長である。天然の転写因子において、これらの2つのドメインは典型的には一緒になっていわゆるPOUドメインを構成し、この場合、POU特異的ドメインはN末端領域を形成し、POU−ホメオドメインはC末端領域を形成する。POU特異的ドメインおよびPOUホメオドメインのいずれも本発明において必要ではあるが、nanog遺伝子のプロモータとの複合体形成が起こりさえすれば、該ドメインを任意の配置構成で選択することができる。多くの実施形態において、該ドメインは適切な長さのリンカー領域によって連結される。当業者であれば、POUドメイン外の非保存配列が標的配列のトランス活性化に参加するとの報告を認識しているであろう。したがって、人工的に作成した転写因子、またはその機能的断片の、本発明の方法への適合性は、該転写因子または断片のnanogへの結合に関して調べる必要がある(下記およびたとえば、実施例2または3、図2または図3を参照)。適切な転写因子の例としては、限定はされないが、Pit−1,Oct−1,Oct−2、Oct−4、Oct−6、Oct−11、Brn−3、Brn−3A、BおよびC、肝細胞核因子lαおよびβ、網膜由来POUドメイン因子−1、精子1POU−ドメイン転写因子、ラットタンパク質Rov−1、アフリカツメガエル(Xenopus laevis)の転写因子POUl、POU2、GBX−2およびOct−1.5、キイロショウジョウバエ(Drosophila melanogaster)の転写因子Cfla、カタユウレイボヤ(Ciona intestinalis)の因子(NCBIアクセス番号BAE06650)、線虫(C.elegans)遺伝子unc−86によってコードされるタンパク質、線虫の因子ceh−6、ショウジョウバエ(Drosophila)タンパク質dPOU−19/pdm−1およびdPOU−19/pdm−2、およびゼブラフィッシュの因子pou5fl/pou2が挙げられる。説明的な例としては、転写因子Oct−1、Oct−4およびOct−6が、nanogプロモータ上の領域に結合することが示されている(ウー、ディー.(Wu,D)、ヤオ、ズィー.(Yao,Z)、2005年、Cell Research 第15巻、第5号、p.317〜324)。
【0022】
(0030)上記少なくとも2つの転写因子またはその機能的断片の他方は、HMG(高移動度群)ドメイン含有転写因子から選択される。HMG DNA結合ドメインは、4方向接合(4‐way junctions)など湾曲DNAに対する著しい優先性を与える。このような転写因子の例としては、限定はされないが、Sry HMGボックス(Sox)転写因子、HrSoxB1、リンパ球エンハンサ結合因子1(LEF−I,T細胞特異的転写因子1−α)、HMGB1aタンパク質、HMGB1bタンパク質、上流結合因子(UBF)、CCAAT結合転写因子2(CTF2)、活性化転写因子4(ATF4)、YABBYファミリーのタンパク質、ミトコンドリア転写因子A(mtTFA)、顆粒膜細胞高移動度群ボックスタンパク質1(GCX−1)、T細胞特異的転写因子7、黒穂菌Ustilago maydisのPrfl、分裂酵母(Schizosaccharomyces pombe)のStellp因子および未命名の因子(NCBIアクセス番号NP595672)、線虫(C.elegans)のTCF/LEF−1、アフリカツメガエル(Xenopus)のXTcf3、睾丸決定因子SRY、XSOX3、ウニ遺伝子Unichromの転写産物、クルイベロマイセス・ラクティス(Kluyveromyces lactis)由来の交配型タンパク質A2、キイロショウジョウバエ(Drosophila melanogaster)のタンパク質CG17964−PHアイソフォームH(NCBIアクセス番号NP001014685)、およびNCBIアクセス番号T12113を有するソラマメの転写因子が挙げられる。
【0023】
(0031)いくつかの実施形態において、第2の転写因子は、SOX(SRY関連HMGボックス)タンパク質であり、例として、限定はされないが、Sox−1,Sox−2,Sox−3,Sox−4,Sox6,Sox7,Sox8,Sox9,Sox10,
Sox11,Sox−13,Sox14,Sox15,Sox18,Sox20,Sox21,Sox30,Sox32またはアフリカツメガエル(Xenopus laevis)の因子Sox−11−D、またはこれらの機能的断片が挙げられる。
【0024】
(0032)各転写因子のうち一方の機能的断片は、任意の長さのものであってよく、2つの基準によって定義することができる。第1に、機能的断片は、nanogプロモータの結合領域と結合して複合体を形成することができ、前記複合体は、たとえばnanog遺伝子などの当該プロモータによって駆動されるそれぞれの遺伝子の活性に影響するのに十分な安定性を有する。第2に、このような機能的断片は、POUおよびホメオドメイン(たとえばPOU−ホメオドメイン)またはHMGドメインをそれぞれ含む天然に存在する転写因子の対応するアミノ酸配列と少なくとも60%の配列同一性を有しうる。いくつかの実施形態において、各断片は、既知の各転写因子の対応するアミノ酸配列と少なくとも80%の配列同一性を、一部の実施形態においては少なくとも95%の配列同一性を有する。「配列同一性」という用語は、既知のPOUおよびホメオドメイン含有転写因子またはHMGドメイン含有転写因子それぞれのアミノ酸配列と、問題とするアミノ酸配列との相同性アラインメント後に得られる、ペアワイズな同一残基の割合(%)を意味し、%の数字は、2つの配列のうち長い方の残基数に対するものである。
【0025】
(0033)本発明の方法において用いる転写因子の機能的断片には、たとえば融合タンパク質の形で、追加の配列がさらに結合されてもよい。また、たとえば、いわゆるアフィニティタグまたはラベルなどの天然または人工の化学修飾を含んでいてもよい。アフィニティタグの例としては、限定はされないが、ビオチン、ジニトロフェノール、ジゴキシゲニン、オリゴヒスチジン、ポリヒスチジン、免疫グロブリンドメイン、マルトース結合タンパク質、グルタチオン−S−トランスフェラーゼ(GST)、カルモジュリン結合ペプチド(CBP)、FLAGのペプチド、T7エピトープ(Ala−Ser−Met−Thr−Gly−Gly−Gln−Gln−Met−Gly)、マルトース結合タンパク質(MBP)、単純ヘルペスウィルス糖タンパク質Dの配列Gln−Pro−Glu−Leu−Ala−Pro−Glu−Asp−Pro−Glu−AspからなるHSVエピトープ、配列Tyr−Pro−Tyr−Asp−Val−Pro−Asp−Tyr−Alaからなるヘマグルチニン(HA)エピトープおよび転写因子c−mycの配列Glu−Gln−Lys−Leu−Ile−Ser−Glu−Glu−Asp−Leuからなる“myc”エピトープが挙げられる。ラベルはさらに、たとえば本発明の方法において望ましい場合には、検出を助ける部分であってもよい。一例としては、限定はされないが、放射活性アミノ酸、フルオレセインイソチオシアネート、5,6−カルボキシメチルフルオレセイン、Texas Red(登録商標)、ニトロベンズ−2−オキサ−1,3−ジアゾール−4−イル、クマリン、ダンシルクロライド、ローダミン、アミノ−メチルクマリン、エオシン、エリスロシン、BODIPY(登録商標)、Cascade Blue(登録商標),Oregon Green(登録商標)、ピレン、リサミン、キサンテン、アクリジン、オキサジン類、フィコエリスリン、Cy3、Cy3.5、Cy5、Cy5.5、およびCy7酵素が挙げられる。さらに適した酵素としては、限定はされないが、アルカリホスファターゼ、大豆ペルオキシダーゼ、またはサイヨウワサビペルオキシダーゼが挙げられる。
【0026】
(0034)各アフィニティタグまたはラベルは、選択された転写因子の任意の部分内に配置されてもよいし、任意の部分に付加されてもよい。説明的な例として、任意のPOUドメインおよびホメオドメイン含有転写因子またはHMGドメイン含有転写因子のアミノ末端またはカルボキシ末端に機能可能なように融合させるとよい。
【0027】
(0035)本発明の方法は、少なくとも2つの転写因子(上記参照)またはその機能的断片が、nanogプロモータ内の特異的結合要素との複合体を形成できるようにする
ことをさらに含む。
【0028】
(0036)本発明の方法のいくつかの実施形態において、少なくとも2つの転写因子またはその機能的断片は、nanogプロモータ上にヘテロ三量体複合体を形成する。それぞれの複合体の形成は、当業者周知の様々な分析方法によって検出することができ、そのような方法としては、表面プラズモン共鳴(たとえば、Biacore(登録商標)技術)、核磁気共鳴または結晶化とそれに続くX線解析が挙げられる。本発明の方法のいくつかの実施形態において、上記で詳述したような転写因子またはその機能的断片は、ヘテロ二量体を形成する。このヘテロ二量体は、その後、nanogプロモータの結合領域を含む核酸とのヘテロ三量体複合体を形成する。ヘテロ三量体複合体を形成するそれぞれの二量体の例は、Oct4およびSox2のnanogへの結合である。結晶学的データは、これらの2つの関係する転写因子がDNA上の結合要素と三重複合体を形成することを示している(レメニ(Remenyi)ら、2003年、Genes&Development 第17巻、p.2048〜2059)。Octl構造の上にOct4を用いて作成した相同性モデルは、Oct4とSox2の間で形成された複合体が、Oct1とSox2の間の複合体と類似していることを示す。Sox2は、Oct4との相互作用のための2つの表面パッチを有する。そのうちの一方は、DNAとも相互作用できることがわかっているSox2のHMGドメインのC末端である。様々なSOXおよびPOU転写因子の間での複合体形成のキャラクタリゼーションが進んでいることに鑑みて(たとえばレメニ(Remenyi)ら、前掲を参照)、様々なPOUドメインおよびホメオドメインを含む転写因子、ならびにHMGドメインを含む転写因子が、核酸、たとえばnanog遺伝子上の結合領域とヘテロ三量体複合体を形成することが予想される。
【0029】
(0037)各結合要素は、当業者によって知られる様々な手段によって同定することができる。実施例4は、配列比較を用いたNANOG上の結合要素の同定を示したものである。このような手法は典型的には、それぞれの転写因子に対する結合要素が存在するかどうかについての情報が全く得られない場合に行われる。本発明者らは少なくとも1つのそのような結合要素をすでに同定していたので、総じてこの手順を用いることは必要でないことになる。代替の手段は、たとえば米国特許第6,735,530号明細書に開示されるようなコンピュータによって実施する方法である。さらに別の結合要素同定手段としては、たとえば実施例3に記載するようなクロマチン免疫沈降アッセイがある。この方法はさらに、活性転写領域を判定したり、ヒストン結合によるゲノムの修飾を評価したりするために使用することもできる。さらに別の手段は、実施例2および図2に記載するような電気泳動移動度シフトアッセイ(EMSA)である。nanogプロモータ内の結合要素を同定するさらなる方法は、たとえば蛍光タンパク質などのマーカーの発現のために該プロモータを使用することである。この手法がどのように実施されるかの例を図4に示す。強化緑色蛍光タンパク質(EGFP)の発現を駆動するnanog遺伝子の−289から+117(転写開始部位(TSS)に対して)のマウス配列を含むプラスミドベクターで、マウスESC株をトランスフェクトした。レチノイン酸による分化誘導によってnanog遺伝子をダウンレギュレートし、これにより、処理3日後のEGFP発現が劇的に減少した(図4E、Fを参照)。リアルタイムPCRによる、NanogEGFPと、内在性Nanogとの比較(図4G)から、該構築物は、内在性Nanogの発現を再現することができたことがわかる。当業者であれば、これらの方法は、さらに他の方法、たとえばin vitro突然変異誘発と組み合わせて、特定の領域についての結合要素の一部としてのさらなる検証に用いることができることに気づくであろう。
【0030】
(0038)本発明のいくつかの実施形態において、少なくとも2つの転写因子は、nonogプロモータ内の複合要素に結合する。このように、これらの実施形態において、POUドメインおよびホメオドメイン含有転写因子は、複合要素の一方部分に結合し、HMGドメイン含有転写因子は、複合要素の他方部分に結合する。各結合要素は、それぞれ
の転写因子について任意の数の結合領域を有していてもよい。たとえば、2つの部分からなる結合部位であってもよい。そのような結合要素は、たとえば、2つの転写因子Sox2およびOct4に対して存在することが知られている。8量体とsox結合部位を含み、Sox2およびOct4の標的となる複合要素を含む遺伝子は、Fgf4,Utfl,Fbx15およびSox2、ならびにPou5fl(Oct4をコードする遺伝子)自体である(ユアン、エイチ.(Yuan,H.)ら、前掲;ニシモト、エム.(Nishimoto,M.)ら、前掲;トクザワ、ワイ.(Tokuzawa,Y.)ら、前掲;トミオカ、エム.(Tomioka,M.)ら、前掲;チュー、ジェイ.エル.(Chew,J.−L.)ら、2005年、Mol.Cell.Biol.第25巻、第14号、p.6031〜6046;オクムラ‐ナカニシ、エス.(Okumura−Nakanishi,S.)ら、2005年、J.Biol.Chem.第280巻、p.5307〜5317)である。本願発明者は、nanogプロモータ内の各複合要素を同定した(たとえば、図5Aを参照)。いくつかの実施形態において、複合結合要素はoct4/Sox2結合部位である。oct4/Sox2結合部位内では、oct4およびSox2に対する結合領域は、Sox2およびoct4の両者が同時に該複合結合部位に結合できさえすれば、任意の配向、かつ任意の相対位置に配置されていてよい。2つの領域は、たとえば、互いに直接隣接していてもよい(図5Aを参照)。
【0031】
(0039)本発明のいくつかの実施形態において、nanog遺伝子のプロモータ領域内の結合要素と、少なくとも2つの転写因子またはその機能的断片との間での複合体の形成も検出される。この目的のために、核酸へのタンパク質の結合を検出するのに十分な感度をもつ任意の方法を用いることができる。このような方法は、たとえば、分光学的、光化学的、光分析的、蛍光分析的、放射線学的、酵素的もしくは熱力学的手段、または細胞効果を利用するものである。分光学的検出方法の例としては、蛍光相関分光法(トンプソン、エヌ.エル.(Thompson,N.L.)ら、2002年、Curr.Opin.Struct.Biol.第12巻、第5号、p.634〜641)が挙げられる。光化学的方法は、たとえば、光化学的架橋である(スティーン、エイチ.(Steen,H.)、ジェンセン、オー.エヌ.(Jensen,O.N.)、2002年、Mass.Spectrom.Rev.第21巻、第3号、p.163〜182)。光活性、蛍光、放射活性、または酵素的ラベルの使用(総覧として、リッペ、アール.エイ.(Rippe R.A.)ら、2001年、Methods Mol Biol.第160巻、p.459〜479を参照のこと)は、それぞれ光分析、蛍光分析、放射線学および酵素による検出法に対する例である。熱力学的検出方法の例は、等温滴定熱量分析(ITC、総覧として、ベラツケツ‐カンポイ、エイ.(Velazquez−Campoy,A.)ら、2004年、Methods Mol Biol.第261巻、p.35〜54を参照のこと)である。任意の細胞効果、たとえば、表現型の変化などは、nanog遺伝子プロモータ領域制御下の組換え因子の発現によって引き起こされうる。細胞効果を用いた方法の例には、マーカータンパク質のパターンの判定を含む、細胞分化状態の測定が含まれていてもよい(たとえば、ノークソン、ケイ.(Noaksson,K.)ら、2005年、Stem Cell Express doi:10.1634/stemcells.2005−0093を参照のこと)。これらの方法のいくつかは、電気泳動またはHPLCなどの追加の分離技術を含んでいてもよい。詳細には、ラベルの使用についての例としては、限定はされないが、プローブとしての化合物、または触媒する反応が検出可能なシグナルをもたらすような酵素を付加した免役グロブリンが挙げられる。放射活性ラベルおよび電気泳動による分離を用いた方法の例としては、電気泳動移動度シフトアッセイが挙げられる。
【0032】
(0040)したがって、nanog遺伝子の結合要素と、少なくとも2つの転写因子またはその機能的断片との間の複合体の形成を検出する例としては、限定はされないが、免役沈降(もしくはウェスタンブロットハイブリダイゼーション)またはクロマチン免疫
沈降アッセイ、電気泳動移動度シフトアッセイ、あるいは表面プラズモン共鳴などの結合要素の同定に適した例も挙げられる。
【0033】
(0041)いくつかの実施形態において、少なくとも2つの転写因子が結合するそれぞれのnanog遺伝子の領域は、ヒトnanog遺伝子配列の位置−289〜+117を含むヒトnanog遺伝子領域に相当する領域内に位置する。一実施形態において、この領域は、ヒトnanog遺伝子配列の位置−212〜−119を含むヒトnanog遺伝子領域に相当する。
【0034】
(0042)上記詳述したように、本発明の方法において、複合体は、少なくとも2つの転写因子またはその機能的断片と、nanog遺伝子のプロモータ領域内の結合要素との間で形成される。この複合体の形成は、さらに、nanog遺伝子の発現を調節する。この効果は、転写活性化を調節することによって達成される。このように、少なくとも2つの転写因子の結合が、nanog遺伝子の転写を増大させる。nanog遺伝子の転写の増大によって、ESCの多分化性細胞状態が維持されることがわかっている(前掲)。nanog遺伝子の転写の抑制により、ヒトECおよびヒト胚性癌細胞における分化が誘導されることがわかっている(図1Eおよびヒスロップ、エル.(Hyslop,L.)ら、2005年、Stem Cell Express doi:10.1634/stemcells.2005−0080を参照のこと)。
【0035】
(0043)本発明のいくつかの実施形態において、nanog遺伝子の転写の増大は、nanog遺伝子発現に換算して測定される。これは、たとえば、nanogプロモータの制御下にある遺伝子から転写されたRNA分子の数を測定することによって行える。当分野で一般的に用いられる方法は、RNA分子転写後の逆転写酵素を用いたRNAからcDNAへのコピーと、該cDNA分子の蛍光染料とのカップリングである。分析は典型的には、DNAマイクロアレイの形で行われる。アフィメトリクス(Affymetrix)製のGeneChip(登録商標)発現アレイといった数々のサービスやキットが市販されている。nanog遺伝子の発現を判定するための他の手段としては、限定はされないが、オリゴヌクレオチドアレイ、および定量的リアルタイムポリメラーゼ連鎖反応(RT−PCR)が挙げられる。
【0036】
(0044)実施例5では、nanog遺伝子発現のさらなる判定手段であるルシフェラーゼレポーターベクターの使用について説明する。この方法において、発現レベルは、nanogプロモータを含むベクターを発現している細胞のルシフェラーゼ活性に反映される。ルシフェラーゼ活性は、市販のキットを用いて照度計で検出することができる(たとえば、実施例5を参照)。
【0037】
(0045)いくつかの実施形態において、nanog遺伝子発現データを較正するか、ランク付けすることが有益または望ましいかもしれない。このように、いくつかの実施形態において、本発明の方法は、得られた結果を、1つまたはそれ以上の対照測定の結果と比較することをさらに含む。
【0038】
(0046)このような対照測定は、主要な測定自体とは異なる任意の条件を含んでいてもよい。たとえば、nanogプロモータの制御下での全く発現が起こらない条件下、または転写因子とnanogプロモータとの間の複合体形成が生じ得ないか、複合体形成の調節ができないような条件下での方法の条件が含まれる。いくつかの実施形態において、nanogプロモータの活性を所定のレベルに調節する化合物を使用することを含んでいてもよい。他の実施形態において、各化合物は、nanogプロモータと2つの転写因子のうちの一方との間での複合体形成を防止するものであってもよい。
【0039】
(0047)対照測定のさらなる手段は、上述の転写因子の一方または両方に結合することができないか、または周知レベルよりも低い親和性または高い親和性で結合する、変異nanogプロモータを使用することである。図5Cは、変異に用いることのできる各部位の同定例である。3つの部位が、変異導入された場合に、多分化性プロモータの機能にルシフェラーゼ活性を野生型構築物の20%以下に低下させるという劇的な影響を及ぼした(図5Cを参照)。これには、oct部位およびsox部位に影響する変異が含まれていた。
【0040】
(0048)本発明の方法のいくつかの実施形態において、2つの転写因子は相乗的に作用して、nanog遺伝子の転写を活性化する。2つの転写因子のそれぞれの相乗作用は、nanog遺伝子の転写に対する個々の転写因子の作用によって得られるデータを、転写に対する2つ(またはそれ以上)の転写因子の作用の組み合わせと比較することによって評定できる。たとえば上述したような、遺伝子発現を判定するのに適した任意の方法をこの目的のために使用することができる。説明的な例として、図5Cは、ルシフェラーゼ活性アッセイ(上記参照)によるnanog遺伝子発現の検出を示している。同一の構築物中でoct部位およびsox部位の両方が変異していると、活性は、(野生型構築物の)約20%の活性に対して、野生型の6%にまで低下する。
【0041】
(0049)多分化性または自己再生特性のうち少なくともいずれか一方を維持するための本発明の方法は、nanogプロモータの結合要素と複合体を形成することのできる、POUドメインおよびPOUホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子を発現できさえすれば、任意の幹細胞、前駆細胞、奇形腫細胞またはそれらに由来する任意の細胞に適している。説明的な例として、任意の多分化性ヒトESCまたはそれぞれの細胞株をそれぞれの方法において用いることができる。そのような細胞の集団を得る手段は、当該技術分野において十分に確立されている(たとえば、トンプソン、ジェイ.エイ.(Thomson,J.A.)ら、1998年、Science 第282巻、p.1145〜1147またはカウアン、シー.エイ.(Cowan,C.A.)ら、2004年、N.Engl.J.Med.第350巻、p.1353〜1356を参照のこと)。さらに、たとえば71の独立したヒトESC株が存在することが知られており、そのうちGE01、GE09、BG01、BG02、TE06またはWA09などの11の細胞株が研究目的で入手可能である(たとえば、NIHヒト胚幹細胞レジストリー(http://stemcells.nih.gov/research/registry/eligibilityCriteria.asp)を参照のこと)。成人幹細胞は、たとえば、分娩後に残った胎盤および臍帯由来の血液から、または成人幹細胞が「衛星細胞」として会合している筋原繊維から、単離することができる(コリンズ、シー.エイ.(Collins,C.A.)ら、2005年、Cell 第122巻、p.289〜301、また、ランド、ティー.エイ.(Rando,T.A.)、2005年、Nature Medicine 第11巻、第8号、p.829〜831も参照のこと)。
【0042】
(0050)本方法が前駆細胞、すなわち成熟体細胞を生じる細胞に対して使用されることが意図される場合、本発明の本方法においては任意の前駆細胞を用いることができる。適切な前駆細胞の例としては、限定はされないが、神経前駆細胞、内皮前駆細胞、赤血球前駆細胞、心臓前駆細胞、オリゴデンドロサイト前駆細胞、網膜前駆細胞、または造血前駆細胞が挙げられる。前駆細胞を得る方法は当該技術分野において周知である。2つの説明的な例として、巨核球前駆細胞を得る方法が米国特許出願第2005/0176142号明細書に開示されており、マウス肝前駆細胞株を得る方法が、リ(Li)ら(2005年、Stem Cell Express,doi:10.1634/stemcells.2005−0108)によって記載されている。いまのところ限られたデータしか入手できないが、前駆細胞は、幹細胞と同様に、POUドメインおよびホメオドメイン含有転写因子ならびにHMGドメイン含有転写因子を発現しているようである。たとえば末
梢血から得られる内皮前駆細胞などの前駆細胞は、NanogやOct−4の高い発現レベルを示すことが発見されている(ロマグナニ、ピー.(Romagnani,P.)ら、2005年、Circ.Res.第97巻、p.314)。網膜前駆細胞などのCNS前駆細胞は、Sox2を高レベルで発現することが報告されている(グラハム、ブイ.(Graham,V.)ら、2003年、Neuron 第39巻、第5号、p.749〜765;クラッセン、エイチ.(Klassen,H.)ら、2004年、J.Neurosci.Res.第77巻、第3号、p.334〜343)。
【0043】
(0051)上に示したように、本発明の1つの方法は、細胞内の遺伝子発現を調節するための方法である。上述の少なくとも2つの転写因子を発現し、かつ機能的nanog遺伝子を含む任意の細胞を用いることができる。いくつかの実施形態において、内在性nanog遺伝子は機能的に活性である。これらの実施形態の一部において、それぞれの細胞は幹細胞または前駆細胞である。本発明の方法において用いることのできる幹細胞の例としては、限定はされないが、胚幹細胞、トロホブラスト幹細胞および胚外幹細胞が挙げられる。本発明の方法のいくつかの実施形態において、ヒト由来のESCなどのESC(胚幹細胞)、すなわちヒトESCを用いることができる。他の実施形態においては、細胞は前駆細胞である(上記参照)。さらに他の実施形態において、細胞はがん細胞である。がん細胞の例示的例としては、奇形腫がん細胞、たとえば、F9、NTERA2、C3H、TES−1、1246(1246−3Aなど)、SuSa(SuSa/DXR10およびSKOV−3/DXR10など)、AT805(ATDC5など)、HTST、HGRT、PC(たとえば、PCC3/A/1)またはGCT27などが挙げられる。がん細胞のさらなる2つの例示的な例としては、HeLa細胞およびMCF−7細胞が挙げられる。いくつかの実施形態において、細胞は幹細胞と体細胞のハイブリッド細胞である。nanog遺伝子は、このようなハイブリッド細胞内で機能的に活性であることが示されている(たとえば、ハタノ(Hatano)ら、2005年、Mech.Dev.第122巻、p.67〜79を参照)。他の実施形態において、内在性nanog遺伝子は機能的に不活性である。これらの実施形態のいくつかにおいて、確立された真核細胞株の任意の細胞、たとえばHEK、COS、CHO、CRE、MT4、DE(アヒル胚)、QF(ウズラ線維肉腫)、NS0、BHK、Sf9、PC12、またはHigh5が選択される。説明的な例は、HEK293T細胞である。さらなる実施形態において、外来性nanog遺伝子は、組換え技術によって、たとえばnanog遺伝子を担持するベクターを用いて導入する(以下も参照されたい)。
【0044】
(0052)いくつかの実施形態において、選択された転写因子またはその機能的断片は、本発明の本方法を行うのに十分な量で内在的に発現される。他の実施形態において、選択された転写因子は、細胞のプロテオーム由来のものはほとんどまたは全く存在しない。いずれの場合も、各転写因子またはその機能的断片を、所望の転写因子をコードする遺伝子を含む1つ以上の組換えベクターを用いて細胞中に導入することができる。細胞が当該転写因子をすでに内在的に発現している場合には、いくつかの実施形態においては、例えばスクリーニングアッセイにおけるシグナル/ノイズ比を改善する目的で、細胞内でのそれぞれの転写ベクターの量を増加させることが望ましいかもしれない。例示的な例として、Oct4および他の転写因子(nanogを含む)が、アデノウィルスベクターを用いてESC中で発現されている(カワタバ、ケイ.(Kawataba,K.)ら、2005年、Mol.Therapy 第12巻、第3号、p.547〜553)。必然ではないが、典型的には、各遺伝子は活性プロモータまたは外部刺激によって都合良く活性化可能なプロモータの制御下におかれる。
【0045】
(0053)目的の細胞内に、各細胞の内在性遺伝子に加えて、nanogプロモータまたは完全なnanog遺伝子のさらなるコピーを含めることが望まれる場合には、これも同様に組換えベクターによって達成することができる(たとえば、カワタバ、ケイ.(
Kawataba,K)ら、前掲)。nanogプロモータ(内在性または外来性のいずれの起源であっても)の活性化を容易にするために、nanogプロモータを含むベクターを細胞内に導入することが有益であるかもしれない。さらに、たとえばタンパク質を得るために、nanogプロモータを用いて外来性遺伝子を発現することが望ましい場合がある。この場合、典型的には、nanogプロモータ制御下に所望の外来性遺伝子を含むベクターが選ばれる。この目的のために、任意の所望の遺伝子の配列を各ベクターに含めることができる。当業者であれば、それぞれ転写されたタンパク質が本発明の方法において用いられる細胞内で翻訳後修飾を受けることが望ましい場合には、さらに酵素を同時発現することが必要なこともあることに気づくであろう。
【0046】
(0054)本方法においてnanogプロモータの制御下で用いることのできる外来性遺伝子の例としては、限定はされないが、レポータ遺伝子、薬剤耐性遺伝子、アポトーシス遺伝子(いわゆる「細胞死」遺伝子)または各細胞内で所望の発現をする任意の他の遺伝子が挙げられる。いくつかの実施形態において、各遺伝子は、目的のタンパク質をコードするものであってもよい。そのような場合、本発明の方法は、各タンパク質を発現して得るために用いてもよい。nanogプロモータの制御下にある遺伝子がアポトーシス遺伝子の場合、本方法は、たとえば、組織から多分化性細胞を除外するために用いることができる。
【0047】
(0055)例示的な例としては、nanogプロモータの制御下にあるアポトーシス遺伝子を含むベクターを組織の細胞内に導入してもよい。組織は、幹細胞/前駆細胞などの多分化性細胞の分化によって得られたものであってもよい。このように本発明の方法は、各組織から、残っている全ての未分化の細胞、たとえば多分化性細胞を除外するために用いることもできる。本実施形態において、本方法は、このような未分化の細胞が生物体に移植されるのを防止するために用いることもできる。したがって、本方法の本実施形態は、たとえば、移植または埋め込み後の奇形腫の発生を防ぐために用いることもできる。さらなる説明的な例として、nanogプロモータによって駆動される薬剤耐性遺伝子(たとえば、抗生物質耐性遺伝子)を含むベクターを、幹細胞/前駆細胞の選択のために用いてもよい。
【0048】
(0056)他の実施形態において、本方法は、上述の少なくとも2つの転写因子またはその機能的断片の、nanogプロモータの結合要素との複合体形成を調節する化合物を各細胞内に導入することをさらに含む。これらの実施形態のいくつかにおいて、本方法は、転写因子、そのヘテロ二量体複合体、またはnanogプロモータの結合要素を、上記のような化合物(すなわち、少なくとも2つの転写因子の、nanogプロモータの結合要素との複合体形成を調節する化合物)と接触させることをさらに含む。
【0049】
(0057)これらの実施形態のうちのいくつかにおいて、本方法は、少なくとも2つの転写因子またはその機能的断片と、nanogプロモータの結合要素との間の上述の複合体の形成を調節する適切な化合物を同定するためのin−vitro方法である。一実施形態において、本発明の方法は、そのような化合物を同定または選択することを目的としたスクリーニング方法として用いられる。そうしたスクリーニング方法は、自動作業ステーションを用いたマルチウェルミクロプレート(たとえば、従来の48ウェル、96ウェル、384ウェルまたは1536ウェルのプレート)上での化合物ライブラリの同時スクリーニングを含んでいてもよい。説明的な例として、上述のようなnanogプロモータ内の結合領域と少なくとも2つの転写因子との間の複合体形成を阻害する化合物を同定することが望ましい場合がある。そのような化合物は、たとえば胚細胞腫瘍を標的とするか、ある種の組織の細胞の均一集団を生じさせるために、たとえば細胞分化を開始または補助することが望まれるかもしれない。このような化合物を、細胞を除去または分化させるために用いることが望ましい場合もある。この特性は、たとえば、上述の少なくとも2
つの転写因子またはその機能的断片と、nanogプロモータの各結合要素との複合体形成の阻害によるものかもしれない(上記参照)。
【0050】
(0058)他の実施形態において、本方法はin−vivo方法であり、本方法において用いられる細胞は、哺乳類または無脊椎動物種、または微生物の一部であるか、これらに含まれる。使用しうる哺乳類の例は、限定はされないが、ラット、マウス、ディンゴ、ウシ、ブタ、ヤギ、チンパンジー、マカウ、およびヒトである。各in−vivo法は、少なくとも2つの転写因子とnanog遺伝子プロモータ内の結合領域との間の上述の複合体の形成を調節する化合物を投与することを含みうる。
【0051】
(0059)さらなる実施形態において、本発明の方法は、哺乳類(たとえば上記)または無脊椎動物において、nanogプロモータ内の結合要素と上述の転写因子またはその機能的断片との間の複合体形成を調節することが判っている化合物を使用することを含む。典型的な実施形態において、上記のような使用には、たとえばヒトなどの哺乳類に投与可能な医薬品または医薬組成物の製造が含まれる。
【0052】
(0060)上述の複合体形成を調節する化合物は、任意の適切な手段によって投与することができる。細胞が哺乳類に含まれるか、その一部である場合には、化合物は非経口投与でも経口(経腸)投与でもよい。哺乳類への投与に対する典型的な実施形態において、投与は、例えば化合物の調製物を経口投与、静脈内投与、または吸入によって、血液および肝臓へ確実に送達できるものとする。経口投与のための調製物の例としては、錠剤、丸剤または飲用液が挙げられ、静脈内投与に対する調製物の例としては、注射液または注入液が挙げられ、吸入による投与に対する調製物の例としては、エアロゾル混合物または噴霧剤が挙げられる。細胞が、微生物に含まれるかその一部である場合、または個々の細胞(たとえば、培養中の組換え細胞)として用いられる場合、投与の例は、化合物を該細胞の環境中に注入または添加することである。個々の細胞を用いる場合には、後者の投与形態が、場合によっては微生物を改変する技術と組み合わせて行われることもある。そのような技術としては、エレクトロポレーションまたは細胞膜の透過化が挙げられる。
【0053】
(0061)本発明のin−vivo方法、またはnanogプロモータ内の結合要素と上述の転写因子との間の複合体形成を調節するための化合物の使用は、様々な目的のために用いることができる。そのような目的の例としては、治療、診断または検査の目的がある。検査目的においては、いくつかの方法は、上述の少なくとも2つの転写因子とnanog遺伝子プロモータ内の結合領域との間の複合体形成を調節することができるとしてすでに同定されている化合物を投与することを含んでいてもよく、他の方法は、そのような化合物の同定を目的としたものであってもよい。治療目的の説明的な例は、奇形腫の治療である。
【0054】
(0062)すでに上に示したように、本発明の本方法は、in vitroおよびin vivoのいずれにおいても、たとえば幹細胞/前駆細胞の分化を開始または補助するためにさらに用いることができる。
【0055】
(0063)上述の複合体形成を調節するために用いられる化合物は、任意の性質を有するものであってよい。たとえば、生物供給源もしくは非生物供給源から単離されてもよいし、または化学的もしくは生物工学的に生産されてもよい。そのような化合物の例としては、限定はされないが、有機小分子またはポリペプチドなどの生物活性ポリマー、たとえば、免役グロブリンまたは免疫グロブリン様の機能を有した結合タンパク質、またはオリゴヌクレオチドが挙げられる。
【0056】
(0064)各化合物の例示的実施形態は、核酸のメチル化状態を変化させる分子、ま
たは、POUドメインおよびホメオドメインを含む転写因子および/またはHMGドメインを含む転写因子のプロモータのメチル化状態を調節する化合物である。説明的な例として、脱メチル化分子5−アザシチジンが、Sox2およびOct4の発現を増加させることが発見されている(ツジ‐タカヤマ(Tsuji−Takayama)ら、2004年、Biochem.Biophys.Res.Commun.第323巻、p.86〜90)。
【0057】
(0065)上述の複合体形成を調節するために用いられる化合物の別の実施形態は、複合体に参加している転写因子の少なくとも一方の翻訳後修飾のパターンを変化させる分子である。転写因子の活性は、リン酸化、アセチル化またはユビキチン化を含むいくつかの翻訳後修飾によって調節されていることが知られている。
【0058】
(0066)このような化合物のさらなる実施形態は、核酸分子である。本明細書中で用いる「核酸分子」という用語は、一本鎖、二本鎖またはその組み合わせなど、任意の可能な立体構造にある任意の核酸のことをさす。核酸には、たとえば、DNA分子(たとえばcDNAまたはゲノムDNA)、RNA分子(たとえばmRNA)、ヌクレオチドアナログを用いて、または核酸化学を用いて作製されたDNAまたはRNAのアナログ、およびPNA(タンパク質核酸)が含まれる。DNAまたはRNAは、ゲノム由来でも合成されたものでもよいし、一本鎖または二本鎖のいずれであってもよい。本発明の本方法において、必然ではないが、典型的にはRNA分子またはDNA分子が用いられる。このような核酸は、たとえば、mRNA、cRNA、合成RNA、ゲノムDNA、cDNA、合成DNA、DNAとRNAのコポリマー、オリゴヌクレオチドなどであってよい。各核酸は、非天然のヌクレオチドアナログをさらに含んでいてもよいし、かつ/またはアフィニティタグもしくはラベルが結合されていてもよい(上記参照)。
【0059】
(0067)多くのヌクレオチドアナログが知られており、本発明の本方法において用いられる核酸およびオリゴヌクオチドにおいて使用することができる。ヌクレオチドアナログは、たとえば塩基、糖、またはリン酸部分に修飾を含むヌクレオチドである。塩基部分の修飾には、A、C、G、およびT/Uの天然または合成の修飾体、別のプリンまたはピリミジン塩基(たとえばウラシル−5−イル、ヒポキサチン−9−イル、および2−アミノアデニン−9−イル)、ならびに非プリンまたは非ピリミジンヌクレオチド塩基が含まれる。他のヌクレオチドアナログは、ユニバーサル塩基として機能する。ユニバーサル塩基には、3−ニトロピロールおよび5−ニトロインドールが挙げられる。ユニバーサル塩基は任意の他の塩基と塩基対を形成することができる。塩基修飾は、二本鎖の安定性を高めるなどの固有の特性を達成するために、たとえば糖修飾(2’−O−メトキシエチルなど)と組み合わせることができる場合が多い。
【0060】
(0068)本発明の本方法のいくつかの実施形態において、核酸分子は、アプタマー、Spiegelmer(登録商標)(国際公開公報第01/92655号パンフレットに記載)、ミクロRNA(miRNA)分子、短鎖ヘアピンRNA(shRNA)分子、または短鎖干渉核酸(siNA)分子、たとえば、短鎖干渉RNA(si−RNA)分子もしくは反復関連短鎖干渉RNA(rasiRNA)分子である。
【0061】
(0069)短鎖干渉RNAの使用は、特定の遺伝子を「ノックダウン」するためのツールとなっている。これは、転写後レベルで起こり、mRNAの分解を伴うRNA干渉(RNAi)を介した、遺伝子のサイレンシングまたは遺伝子抑圧を利用する。RNA干渉は、ゲノムを保護する細胞機構に相当する。siRNA分子は、該siRNAが多酵素複合体と会合していわゆるRNA誘導サイレンシング複合体(RISC)を形成することにより、該siRNAに相補的なRNAの分解を媒介する。siRNAは、RISCの一部となって相補的RNA種を標的とし、次いで相補的RNAが開裂される。これが、各遺伝
子の発現の損失をもたらす(簡単な概説として、ザモア、ピーディー(Zamore,PD)、ハーレイ、ビー(Haley,B)、2005年、Science 第309巻、p.1519〜1524を参照されたい)。この技術は、たとえば、米国特許出願第2005/0191618号明細書に開示されるように、寄生DNA配列のサイレンシング、たとえばHIV RNAの開裂に応用されてきた。
【0062】
(0070)本発明に対するこのようなsiRNAの典型的な実施形態は、10〜35ヌクレオチド、一部の実施形態においては15〜25ヌクレオチドのin vitroまたはin vivoで合成された分子を含む。転写によって(たとえば、生きた細胞の転写機構を用いて)si−RNA分子を生産するために用いることのできる核酸配列の説明的な2例が、配列番号31および配列番号32である(実施例6を参照)。各si−RNA分子を対象とする細胞(微生物および動物の一部である細胞を含む)内で直接合成してもよい。また、各細胞に導入してもよいし、かつ/または該細胞に送達してもよい。siRNA分子をin vivoで選択された細胞に送達する説明的例は、重鎖抗体断片(Fab)と核酸結合タンパク質プロタミンとの融合タンパク質にsiRNA分子を非共有結合により結合させることである。(ソング、イー.(Song,E.)ら、2005年、Nature Biotech.第23巻、第6号、p.709〜717)。本発明の一実施形態では、siRNA分子を用いて、1つまたはそれ以上のPOUドメインおよびホメオドメイン含有転写因子および/または1つまたはそれ以上のHMGドメイン含有転写因子をコードするmRNA分子の分解を誘導する(たとえば、実施例6参照)。
【0063】
(0071)本発明の方法は、nanog遺伝子上の領域に特異的に結合することのできるさらなる転写因子を使用することを含んでいてもよい。したがって、いくつかの実施形態において、該方法は、少なくとも第3の転写因子をnanog遺伝子と接触させることを含む。このような転写因子の例としては、限定はされないが、SMADタンパク質ファミリーのメンバー(たとえば、SMAD1、SMAD2、SMAD3、SMAD4、SMAD5、SMAD7、およびSMAD9、またはキツネの条虫である多包条虫(Echinococcus multilocularis)由来のSmadAおよびSmadB)、AP1(アクチベータタンパク質1)ファミリーのメンバー、hand1(心臓および神経冠誘導体発現タンパク質1)転写因子またはhand1関連転写因子が含まれる。
【0064】
(0072)本発明を、以下の非限定的な実施例によってさらに説明する。
【実施例】
【0065】
配列解析およびプロモータ構築物
(0073)すべての配列は、www.ncbi.nlm.nih.govおよび/またはwww.ensembl.org.にある公的に利用可能なデータベースから得た。ウシ(Bos taurus)およびアフィリカゾウ(Loxodonta africana)のNanogプロモータ領域は、NCBIにおいて入手可能なトレースファイル配列から構築した。ウェブ上(www-gsd.lbl.gov/vista )のVistaゲノム配列アラインメントツール(メイヤー(Mayor)ら、2000年)を用いて、マウスとヒトのNanogゲノム配列を比較した。マウスnanogおよびヒトNanogおよびNanogGP1に対するプロモータ領域を適当な生物種のゲノムDNAから増幅した。増幅のためのプライマーは、クローニング用の制限酵素切断部位(小文字で示す)を備えた下記の
nanog順方向 5’−CGCgtcgacTAAAGTGAAATGAGGTAAAGCC−3’(配列番号1)、
nanog逆方向 5’−CGCggatccGGAAAGATCATAGAAAGAAGAG−3’(配列番号2)、
Nanog順方向 5’−CGGctcgagTTGCTCGGTTTTCTAGTTC
C−3’(配列番号3)、
Nanog逆方向 5’−CGGctcgagCAAGAAATTGGGATAAAGTGAG−3’(配列番号4)、
NanogGP1順方向 5’−CGGctcgagTTGCTCGGTTTTCTAGTTCC−3’(配列番号5)、
NanogGP1逆方向 5’−CGGctcgagCAAGAAGTTGTGATGAAGTGAG−3’(配列番号6)
とした。マウスnanogプロモータ断片を、pGL3−Basicベクター(プロメガ(Promega))およびpEGFP1ベクター(クロンテック(Clontech))の両方にクローニングし、ヒトプロモータはpGL3−Basicだけにクローニングした。すべての構築物について配列確認を行った。
【0066】
胚幹細胞培養およびレポータの系
(0074)E14マウスESCを、ダルベッコ改変イーグル培地、20%ウシ胎仔血清、1×非必須アミノ酸、0.1mMの2−メルカプトエタノール、および組換えLIF調整培地のアリコート中で培養した。標準的なプロトコールを用いてNanogEGFP構築物でESCを安定的にトランスフェクトし、ネオマイシン耐性マウス胚線維芽細胞上で培養した細胞を用いて300μg/mLのG418で10日間選択した後に、個々のコロニーを採取した。ESCを、LIF調整培地の回収、胚様体への自発的分化、またはレチノイン酸の添加により分化させた。蛍光活性化細胞選別(FACS)は、FACSCalibur(商標)(ビーディーバイオサイエンシズ(BD Biosciences))で行った。内在性Nanogおよび強化緑色蛍光タンパク質(EGFP)レポータの発現の定量は、ABI PRISM(登録商標)7900配列検出システム(アプライドバイオシステムズ(Applied Biosystems))を用いてリアルタイムの逆転写PCR分析によって行った。この分析のためにProligo(登録商標)により合成されたプライマーとプローブのセットは以下の
Nanog順方向、5’−GGTTGAAGACTAGCAATGGTCTGA−3’(配列番号7)
Nanog逆方向、5’−TGCAATGGATGCTGGGATACTC−3’(配列番号8)
Nanogプローブ、5’−TTCAGAAGGGCTCAGCACCA−3’(配列番号9)
EGFP順方向、5’−CGACAACCACTACCTGAGCAC−3’(配列番号10)
EGFP逆方向、5’−TCGTCCATGCCGAGAGTGAT−3’(配列番号11)
EGFPプローブ、5’−CGGCGGCGGTCACGAACTCCAGC−3’(配列番号12)
である。発現はa−アクチン対照(アプライドバイオシステムズ(Applied Biosystems))に対して正規化した。ヒトESC株HUES−6(ハーバード大学のダグ・メルトン氏(Doug Melton)より入手)は、カウアン(Cowan)ら(2004年、N.Engl.J.Med.第350巻、p.1353〜1356)に従って継代した。
【0067】
実施例1:Nanogのダウンレギュレーション効果の分析
(a)RNAi分析
(0075)本実施例では、nanog遺伝子活性のダウンレギュレートによる細胞効果をどのようにして分析するかについて説明する。実施例6(下記参照)に示すのと同様の方法から、NanogのダウンレギュレーションがESCの分化に繋がることが判った。
【0068】
(0076)RNAiのための遺伝子特異的オリゴヌクレオチドを、レイノルド(Reynolds)らの文献(2004年)およびウイ‐タイ(Ui−Tei)らの文献(2004年)にしたがって設計した。9ヌクレオチドのループを有する19ヌクレオチドのヘアピン型shRNAsを、pSUPER.puro(BglIIおよびHindIII部位、オリゴエンジン(Oligoengine))中にクローニングした。使用したオリゴヌクレオチドは以下の通り、すなわち
GFP RNAi(対照)用として、
5’−GATCCCCGAACGGCATCAAGGTGAACTTCAAGAGAGTTCACCTTGATGCCGTTCTTTTTA−3’(配列番号13)および
5’−AGCTTAAAAAGAACGGCATCAAGGTGAACTCTCTTGAAGTTCACCTTGATGCCGTTCGGG−3’(配列番号14)
Nanog RNAi用として、
5’−GATCCCCGAACTATTCTTGCTTACAATTCAAGAGATTGTAAGCAGAATAGTTCTTTTTA−3’(配列番号15)および
5’−AGCTTAAAAAGAACTATTCTTGCTTACAATCTCTTGAATTGTAAGCAAGAATAGTTCGGG−3’(配列番号16)
とした。E14ES細胞を、50%コンフルエントの状態でshRNAプラスミドおよびGFPプラスミドで同時にトランスフェクションした。
【0069】
(b)ウェスタンブロット分析
(0077)NanogRNAiが、Sox2またはOct4などの他の転写因子に影響を与えることなくNanog発現を特異的にダウンレギュレートするかどうかを調べるために、SDS−PAGEおよび続くウェスタンブロットによってタンパク質レベルを分析した。この2つの方法は当業者に周知である。Nanog発現ベクターを、対照、Nanog、Oct4またはSox2のsiRNAを発現する構築物とともに、293T細胞に同時トランスフェクトした。細胞溶解物のウェスタンブロット分析は、抗Nanog抗体または抗βアクチン抗体を用いて行い、βアクチンを装荷量の対照とした。
【0070】
(0078)タンパク質を12%SDS−PAGEゲルで分離し、当該技術分野においてよく確立されている標準的な方法によってImmobilon(商標)Pブロッティング膜(ミリポア(Millipore))に移した。膜をPBS+0.1%tween20および5%ミルクでブロッキングし、特異的抗体をプローブとして調べた。その後、膜をセイヨウワサビペルオキシダーゼ共役二次抗体とともにインキュベートした。このブロットをECL Advance(商標)ウェスタンブロッティング検出キット(Amersham)によって現像した。
【0071】
(0079)図1Aに示すように、Nanog siRNAだけが、Nanogタンパク質レベルを低減することができた。対照RNAi、Oct4 RNAiおよびSox2
RNAi(実施例6参照)は、Nanogの発現に影響を及ぼさなかった。このNanog siRNAは、同時発現させたSox2またはOct4のレベルに影響を及ぼさなかった(図1B、Cを参照)。これらのデータから、Nanog siRNAがNanogに対して特異的であり、POUドメインおよびホメオドメイン含有転写因子ならびにHMGドメイン含有転写因子に対するモデルタンパク質として選ばれた他の2つの転写因子には影響を及ぼさないことがわかった。
【0072】
(0080)ES細胞を、NanogまたはGFPのsiRNAを発現する構築物でトランスフェクトした(下記も参照のこと)。トランスフェクトされた細胞をピューロマイシンで選択し、ウェスタンブロットから、Nanogのレベルが大きく減少、すなわち、検出限界のタンパク質レベルまで減少したことがわかった(図1D)。
【0073】
(0081)さらに、未分化ES細胞の典型的な形態が、Nanogノックダウン後に失われていた(図1E)。各写真を撮るために従来の顕微鏡を用いた。アルカリホスファターゼ染色もNanogノックダウン細胞において劇的に減少していた。
【0074】
実施例2:NANOGと、(POU)特異的ドメインおよびホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子との間の、in vitroでの複合体形成についての検証
(0082)当該技術分野において電気泳動移動度シフトアッセイ(EMSA)として知られるアッセイを用いて、Oct4およびSox2が、nanogプロモータ内の要素と複合体を形成することができるかを判定した。
【0075】
(0083)この目的のために、核抽出物を、ディグナム(Dignam)ら(1983年、Nucleic Acids Res.第11巻、p.1475〜1489)の方法を次のように改変したものを用いて、マウス胚線維芽細胞フィーダー上で成長させたE14マウス胚幹細胞から調製した。すなわち、細胞を洗浄し、PBS中で掻き取ることにより回収し、ペレットの5倍容のバッファA(10mM Hepes(pH7.9)、1.5mM MgCl2、10mM KCl、0.5mM DTT、1%プロテアーゼインヒビターカクテル(シグマ(Sigma)))中に再懸濁し、氷上で10分間インキュベートした。細胞を遠心分離し、ペレットの2倍容のバッファAに再懸濁し、Dounceホモジナイザー(タイプBペッスル)を20ストロークとして細胞膜を溶解した。遠心分離によって核を回収し、核ペレットの0.6倍容のバッファC(20mM Hepes(pH7.9)、25%グリセロール、420mM NaCl、1.5mM MgCl2、0.2mM EDTA、0.5mM DTT、1%プロテアーゼインヒビターカクテル)に再懸濁してから、回転させながら4℃で30分間インキュベートした。次に核を13000rpmで5分間遠心分離し、上清を回収して透析バッファ(20mM Hepes(pH7.9)、20%グリセロール、100mM KCl、0.83mM EDTA、1.66mM DTT、1%プロテアーゼインヒビターカクテル)に対して4℃で2時間透析した。抽出物は−80℃で保存した。
【0076】
(0084)EMSAは、DNAプローブとして、NOSと呼ばれる推定上の要素と重複するnanogプロモータ由来の配列を含む37塩基対の二本鎖オリゴヌクレオチドを用いて行った。(POU)特異的ドメインおよびホメオドメインを含む転写因子としてSox2を選び、HMGドメインを含む転写因子としてOct4を選んだ。陽性対照として、FGF4エンハンサ由来の既知のOct4/Sox2複合結合部位(FOS)を用いた。これはnanog要素とは違って、8量体とHMGモチーフとの間に3塩基対の間隔があいている。
【0077】
(0085)両鎖の5’末端がcy5によって標識された二本鎖DNAオリゴヌクレオチドを用いた。一本鎖の5’標識オリゴヌクレオチドをプロリゴ(Proligo)より購入し、95℃から室温に徐々に冷却することにより、相補的な5’標識オリゴヌクレオチドにアニールさせた。使用したオリゴヌクレオチドの配列は以下のとおり、すなわち
FOS、5’−TTTAAGTATCCCATTAGCATCCAAACAAAGAGTTTTC−3’(配列番号17);
NOS、5’−CTTACAGCTTCTTTTGCATTACAATGTCCATGGTGGA−3’(配列番号18);
NmOS、5’−CTTACAGCTTCTTTCAAATTACAATGTCCATGGTGGA−3’(配列番号19);
NOmS、5’−CTTACAGCTTCTTTTGCATTAACCTGTCCATGGTGGA−3’(配列番号20);
NmOmS、5’−CTTACAGCTTCTTTCAAATTAACCTGTCCATGGTGGA−3’(配列番号21);
NNS、5’−CTGCAGGTGGGATTAACTGTGAATTCA−3’(配列番号22)
である(図4Aを参照)。DNA結合反応のためには、2μl(約16μg)の核抽出物を、50nM二本鎖標識オリゴヌクレオチドと5μgのポリdGdC(アマシャム(Amersham))とを含む反応物に添加して反応体積10μl(最終)とした。最終的な結合バッファ組成は、60%透析バッファとした。核抽出物を加える前に、所定の1μM非標識オリゴヌクレオチド競合物も含めた。結合反応は、室温(RT)で約20分間行った。所定の2μl(抗Oct4および抗JunB)または8μl(抗Sox2および抗Sox4)の抗体を最初の結合反応に続いて付加し、さらに20分間インキュベートした。サンタクルーズ(Santa Cruz)より購入した抗体は以下の通りである。ウサギ抗Oct4ポリクローナル抗体(H−134)sc−9081x;ヤギ抗Sox2ポリクローナル抗体(Y−17)sc−17320;ウサギ抗JunBポリクローナル抗体(N−17)sc−46x;ヤギ抗Sox4ポリクローナル抗体(C−20)sc−17326。結合反応物を、プレラン済みの6%非変性ポリアクリルアミドゲルで、0.5×TBE中、バイオラッド(Bio−Rad)protean(登録商標)II xi装置で300Vにて2時間かけて分離した。ゲルは、モレキュラーダイナミクス(Molecular Dynamics)のTyphoon(商標)9140型画像解析装置を用いて、赤色レーザ(633nm)ならびに、設定を600V、通常感度および焦点面+3mmとしたPMTとともにcy5エミッションフィルタを用いて、ガラス板中で直接撮影した。マウス胚線維芽細胞フィーダーのみについて行ったEMSAでは、有意な移動度シフトは生じなかったことから、それらが観察されるタンパク質DNA複合体に寄与しないことが示された。
【0078】
(0086)最初に、E14マウス胚幹細胞の核抽出物中のDNA結合タンパク質が、NOS要素を認識することができるかどうかを調べた。図2に示すように、EMSAにおいて、NOS要素に結合する4つの主要タンパク質複合体が観察された。FOS要素に結合する類似の移動度を示す4つの複合体が観察されたが、この配列に結合するもう1つの複合体も観察された(レーン12)。さらに、複合体Cは、FOS要素上よりもNOS要素上において僅かに速い移動度を有するようにみえた。
【0079】
(0087)これらのバンドのいずれがOct4またはSox2に相当するかを判定するために、これらのタンパク質に対する抗体を結合反応物に加えることによってスーパーシフトが起こるかどうかを調べた。実際には、抗Oct4抗体を添加した後、複合体のうちの2つ、AおよびCが消失し、標識されたプローブがウェル中に蓄積した(標識Oct4のスーパーシフト)。これは、NOSおよびFOSプローブの両方で観察され(レーン2および13)、いずれの配列上の複合体AおよびCもOct4を含んでいたことがわかった。同様に、抗Sox2抗体の添加により、NOSプローブとの複合体C、およびFOSプローブとの複合体CおよびEが消失した(レーン3および14)。いずれの場合も、より移動の遅い複合体(標識Sox2のスーパーシフト)が観察された。このことは、複合体CおよびEがSox2を含んでいたことを示す。重要なのは、JunBまたはSox4のいずれかに対する抗体を加えてもバンドの移動度に全く影響がなく、Oct4およびSox2抗体の影響が特異的であることが示されたことである。したがって、複合体CがOct4抗体およびSox2抗体の両方によってスーパーシフトしたことから、このバンドは複合要素に結合したこれらのタンパク質のヘテロ二量体を示す。複合体Aは該ヘテロ二量体よりもより迅速に移動し、Oct4抗体によってのみ認識されたことから、この複合体はOct4単量体であることが強く示唆される。同様に、FOS配列上の複合体Eは、ヘテロ二量体よりも速く移動し、Sox2抗体によってのみ認識されたことから、これはSox2単量体に相当することが示された。これらのバンドの相対移動度は、以前に公
表されている、FGF4要素に結合するF9テラトカルシノーマ細胞抽出物中のOct4およびSox2を同定した結果と矛盾するものではない。
【0080】
(0088)現在、複合体BおよびD中の結合因子は同定されていない。複合体Dの有力候補は転写因子Oct1であり、該転写因子Oct1は、FGF4要素内の8量体モチーフを認識し、移動度が同様に遅いことが示されている。さらに、Oct1は、FGF4複合要素上でSox2とヘテロ二量体を形成することが示されている。しかしながら、さらに遅く移動する複合体が無いことから、E14核抽出物においては、少なくともEMSAによって検出されるレベルではヘテロ二量体の形成が起こっていないことが示唆される。
【0081】
(0089)NOSに結合したOct4/Sox2ヘテロ二量体が、FOSに結合したものよりも移動がわずかに速い(レーン1と12の複合体Cを比較)ことは、FOS配列の場合のように8量体モチーフをHMFモチーフから隔てるヌクレオチドが存在するわけではないために、NOS配列上でより緊密なコンフォメーションが生じることもありうるという考えと矛盾しない。このようなコンフォメーションの違いが転写活性の差異や共調節因子の動員の差異をもたらすと考えると面白い。
【0082】
(0090)これらの因子のDNA結合特異性を確立するために、オリゴヌクレオチド競合を行った。いずれの因子によるNOSへの結合も、20倍過剰の同一の非標識NOSを加えることによって強く競合したが(レーン7)、nanogプロモータのすぐ下流の配列を含む非特異的非標識オリゴヌクレオチド(NNS)は、結合に対して有効な競合を示さなかった(レーン11)。これらの結果は、これらの因子のNOSに対する結合が特異的であることを示す。同様に、非標識FOSが標識FOSに対するすべての結合と競合可能であり(レーン17)、NSSでは競合できなかった(レーン22)ことから、FOS配列に対する結合もまた特異的である。さらに、非標識のFOSおよびNOSのいずれも相互に競合可能である(レーン6および18)ことから、0または3塩基対の間隔を有する競合物のいずれが20倍過剰に加えられた場合にも、同等に有効であることがわかる。NOSおよびFOSのいずれの場合も、8量体モチーフ内に3塩基対の置換を有する競合物の存在下で結合が行われる場合、Oct4単量体およびOct4/Sox2ヘテロ二量体による結合に対する競合は起こらなくなるが(レーン8および19)、Sox2単量体に対する競合は依然として起こる(レーン19)。このことは、Oct4単量体およびOct4/Sox2ヘテロ二量体のいずれの結合に対しても完全な8量体モチーフが必要であることを示している。同様に、競合物のHMGモチーフ内を3塩基対置換することにより、NOSおよびFOSに関するOct4/Sox2ヘテロ二量体との競合(レーン9および20)およびFOSに関するSox2単量体との競合(レーン20)が消失する。いずれの場合も、Oct4単量体は依然として競合を受けている。このことは、Sox2単量体およびSox2/Oct4ヘテロ二量体の結合には完全なHMGドメインが必要であることを示している。最後に、同じ競合物中の8量体およびHMGモチーフの両方において2塩基対を置換することにより、Oct4単量体およびOct4/Sox2ヘテロ二量体のNOSおよびFOS両者への結合に対する競合が消失し(レーン10および21)、かつSox2単量体のFOSへの結合に対する競合が消失した(レーン21)。図2からわかるように、EMSAは転写因子とNanogとの間の形成を検証する便利な手段である。しかしながら、例えばGSTプルダウンアッセイまたはBiacoreなどの他の手段を利用してもよい。
【0083】
実施例3:NANOGと(POU)特異的ドメインおよびホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子との間のin vivo複合体形成の検証
(0091)in vivoにおけるNanogと転写因子との間の相互作用を分析する方法は、当業者には「クロマチン免役沈降アッセイ」または「ChIPアッセイ」とし
て知られている。
【0084】
(0092)本実施例では、Oct4抗体およびSox2抗体ならびにマウスおよびヒトのESCからの核抽出物とともに、またはレチノイン酸で分化させたマウスESCにおいて、上記方法を使用することについて説明する。
【0085】
(0093)E14マウスESCを用いたChIPアッセイは、既述(ウェルズ、ジェイ.(Wells,J.)およびファーナム、ピー.ジェイ.(Farnham,P.J.)、2002年、Methods(Orlando)第26巻、p.48〜56)のようにして行った。簡潔に述べると、細胞を1%ホルムアルデヒドで10分間室温にて架橋し、125mMグリシンを添加することによってホルムアルデヒドを不活性化した。平均500塩基対の大きさのDNA断片を含んだクロマチン抽出物を、Oct4ポリクローナル抗体(N19)またはSox2ポリクローナル抗体(Y17)(サンタクルーズ・バイオテクノロジー(Santa Cruz Biotechnology))またはSox2ポリクローナル抗体(AB5603)(ケミコン(Chemicon))を用いて免役沈降させた。すべてのChIP実験に対して、定量的PCR分析を、ABI PRISM(登録商標)7900配列検出システム(アプライドバイオシステムズ(Applied
Biosystems))およびSYBR Greenマスターミックス(アプライドバイオシステムズ)を用いて、既述(ン、エイチ.エイチ.(Ng,H.H.)ら、2003年、Mol Cell 第11巻、p.709〜719)のようにリアルタイムで行った。相対占有率は、見かけ上の免役沈降効率(入れた試料の量に対する免役沈降したDNA量の比)を測定し、対照の領域について観察されたレベルを1.0として正規化することによって算出した。プライマー対は以下のとおり、すなわち
1、5’−GGCAAACTTTGAACTTGGGATGTGGAAATA−3’(配列番号23)、
5’−CTCAGCCGTCTAAGCAATGGAAGAAGAAAT−3’(配列番号24);
2、5’−GAGGATGCCCCCTAAGCTTTCCCTCCC−3’(配列番号25)、
5’−CCTCCTACCCTACCCACCCCCTATTCTCCC−3’(配列番号26);
3、5’−GGGTCACCTTACAGCTTCTTTTGCATTA−3’(配列番号27)、
5’−GGCTCAAGGCGATAGATTTAAAGGGTAG−3’(配列番号28);
4、5’−GGTGATACGTTGGCCTTCTAGTCTGAA−3’(配列番号29)、
5’−GGGCAAATTGCAAACTAACTGTATAACCTC−3’(配列番号30)
とした。
【0086】
(0094)フィーダー無しの条件で増殖させた未分化のマウスESCからは、複合oct−sox要素を含むDNA断片が、それぞれOct4抗体およびSox2抗体による免役沈降で43倍および37倍にまで富化された(図3A、アンプリコン2)。oct−sox複合要素を含まない2つの隣接領域には有意な富化はみられなかった(図3A、アンプリコン1および3)。さらに、マウスESCをレチノイン酸によって分化誘導すると、このoct−sox含有断片の富化は、その分化の程度に応じて低減された。分化の3日後、Oct4抗体およびSox2抗体を両方用いた場合にのみ富化は最大でバックグラウンドの10倍にまで達したが、分化の6日後には、有意な富化は検出できなかった(図3A)。Mllに対する抗体を陰性対照として用いた場合、分析した3つのESC状態の
いずれからもアンプリコンのいずれについても有意な富化は見られなかった(データ示さず)。ここで同定された富化のレベルは、EGFPレポータ系を用いた上述の類似の分化プロトコールにおけるNanogプロモータの活性の場合とよく対応している。
【0087】
(0095)OCT4およびSOX2がヒトESCのNANOGプロモータとも相互作用することを検証するために、類似のChIP分析を行ってもよい。本実施例においては、不活性化マウス胚線維芽細胞上で増殖させたヒトESC株HUES−6を用いた。OCT4抗体およびSOX2抗体でクロマチン免役沈降したDNAの富化を検出するために、いずれもNANOGのエキソン1に近接する6つの異なるアンプリコンを用いた(図3B)。複合sox−oct要素と近いほうの3つのアンプリコンだけが、両抗体による有意な富化を示した(図3B)。第2のSox2抗体(Y17)から、類似の結果が得られた(データ示さず)。マウスESCにおいて見られるものと比較して3つすべての抗体について富化率が低い(6倍)のは、未分化なヒトESCを培養中に維持するのがより難しい傾向にあるので、ヒト培養物に分化した細胞がより多く存在することによるのかもしれない。グルタチオンS−トランスフェラーゼ抗体を陰性対照として用いたところ、これらのヒトESC中のアンプリコンのいずれに対しても有意な富化は見られなかった(データ示さず)。まとめると、これらのChIPデータのすべてが、in vivoにおいて、未分化なマウスおよびヒトESC中のNanog/NANOGプロモータ上をOct4/OCT4およびSox2/SOX2が占有することを明確に示している。
【0088】
実施例4:NANOG上の結合要素の同定
(0096)本実施例では、Nanog遺伝子上の結合領域を、その配列保存性に基づいてどのようにして同定できるかを説明する。
【0089】
(a)シス調節情報を含む非コーディング領域の配列比較
(0097)本発明者らは、nanogの高度に保存された結合領域、sox−oct複合要素を同定済みである。マウスおよびヒト遺伝子配列のペアワイズなアラインメントを、エルビーエル・バークレー(LBL Berkley)のVistaオンラインツール(メイヤー、シー.(Mayor,C.)ら、2000年、Bioinformatics 第16巻、p.1046〜1047)を用いて作成し、作成にあたっては、最適なアラインメントの作成時に種特異的な反復要素を考慮した。このアラインメントに対しては、マウスnanogESTの最も5’側の10kb上流から、3’マウスnanogESTの最も3’側の10kb下流までのゲノム領域を選択した。配列類似性のピークがnanogの4つのエキソンに対応して同定されたが、ホメオドメインをコードするエキソン2および3で最も顕著であり、エキソン1で最も顕著でなかった(データ示さず)。非コーディング領域のなかでは、ESTの最も5’側のすぐ上流に、近位プロモータと定義される、有意な配列保存性を有する唯一の領域がみつかった。
【0090】
(0098)非コーディング配列に有意な保存性が認められないことは、多分化性発現を指示するシス調節要素が、すべてこの近位プロモータ領域内にあることを示唆することに他ならない。このことを調べるために、レポータEGFPの発現を駆動するマウス近位プロモータ領域を含むプラスミドベクターで、安定にトランスフェクトされたマウスES細胞株を作成した。406塩基対のプロモータ断片は、優勢な転写開始部位として定義されてきたことから、最も遠い5’共通(public)ESTの位置として同定されている位置に対して−289から+117の配列で構成した(ハート、エイ.エイチ.(Hart,A.H.)ら、前掲)。選択後、これらのneo耐性コロニーの大半が、緑色の蛍光を発し、nanogプロモータのこの領域が多分化性細胞において実際に活性を有することを示した(図4Bを参照)。これらのnanogEGΕVコロニーのいくつかをさらなる分析のために選択した。
【0091】
(0099)このプロモータ断片内の明らかに多分化性ポジティブなシス調節要素に加えて、発明者らは、内在性NanogのmRNAレベルでは起こるような、ES細胞分化時の転写のダウンレギュレーションに十分なシス調節情報を該断片が含んでいるかを判定することに関心を持った。ES細胞分化の強力な誘発剤であるレチノイン酸を0.1μMで使用して分化を誘発させた。この処理により、3日後にEGFP発現が劇的に減少し(図4E−F参照)、分化から4日後までにはFACS分析による測定でEGFPポジティブのままの細胞はわずか9%にすぎなかった(図4Gを参照)。これは同じ分化プロトコールにおける内在性nanogのmRNAレベルと非常に似ている(チャンバース、アイ.(Chambers,I.)ら、前掲)。
【0092】
(0100)より明敏な分化プロトコールにおいても、このプロモータ構築物の活性はやはり内在性Nanog mRNAレベルと類似していた。nanogEGFP細胞をLIFの非共存下で培養したところ、EGFP発現が培養の3日後に顕著に低下し(図4C〜Dを参照)、4日目までには、EGFPポジティブであったのは、フィーダーを含まない未分化nanogEGFP細胞では74%であるのに対して、48%の細胞にすぎなかった(図4Gを参照)。胚様体形成もまた、FACSCalibur(商標)(ビーディーバイオサイエンシズ(BD Biosciences))上でのFACS分析によって測定されるように、EGFP発現のダウンレギュレーションを誘発した。内在性NanogおよびEGFPレポータの発現の定量化は、上述のようなRT−PCR分析によって行った(「胚幹細胞培養およびレポータの系」を参照)。分化の最初の2日の後、EGFP発現は次第に減少し、8日目までには、バックグラウンドレベルの発現しか認められず(図4Gを参照)、これは、内在性Nanog mRNAの発現と類似している(ハート(Hart)ら、2004年)。よってこのnanogの406塩基対断片は、少なくともES細胞を用いるin vitroの系において試験した場合には、内在性nanog発現を再現するのに十分なシス調節情報を含んでいる。
【0093】
(b)系統発生学的フットプリンティングによる結合要素の同定
(0101)広範囲の哺乳類から近位プロモータ領域のアラインメントを作成することにより、同定を強力な系統発生学的フットプリントにすることができる。このアラインメントの構築にあたって使用した5つの種の間の分岐年代は、控えめに見積もっても、マウスとラットの分岐については千2百万年前であり、マウス、ラット、ヒト、ウシおよびゾウの間では六千万年前であるが、この見積もりから、フットプリントを同定するための累積2億5千2百万年の浄化選択が提供される。最も大きい配列保存の長さは、nanog転写開始部位との相対位置−212から−119までに位置する94塩基対領域にわたり、このうち64位置が不変である(図5Aを参照)。
【0094】
(0102)最も長い連続不変配列は16塩基対長であり、面白いことに、この配列の中に、fgf4,utf1,sox2,およびfbx15の多分化性発現において同定された要素もしくは前記発現に対して機能的であることが知られている要素と類似した、15塩基対のoct−sox複合要素(図5Aを参照)がある(ユアン、エイチ.(Yuan,H.)ら、前掲;ニシモト、エム.(Nishimoto,M.)ら、前掲;トミオカ、エム.(Tomioka,M.)ら、前掲;トクザワ、ワイ.(Tokuzawa,Y.)ら、前掲)。sox要素とoct要素の相対位置は、上記5つの遺伝子全部において同じであり、このことは、多分化性細胞においてこれらの要素と結合することが知られている対応する転写因子(Sox2およびOct4)の間の特異的なタンパク質−タンパク質接触を示すものである(レメニ、エイ.(Remenyi,A.)ら、2003年、Genes Dev.第17巻、p.2048〜2059)。utfl,sox2,およびfbx15と同様に、nanog内のsoxシス要素およびoctシス要素は互いに直接隣接しており、fgf4においては該2つの要素は3つの塩基で隔てられている。
【0095】
(0103)nanogプロモータ内の多数の部位におけるこのような強力な浄化選択は、転写の調節におけるこれらの部位の機能的役割を示唆するものである。これらの部位のいくつか、たとえば、oct−sox複合要素などは、多分化性発現において重要でありそうだし、他のものは、nanogの移植後発現において機能するシス調節要素に相当するかもしれない。さらに、マウスESCにおける超並列シグニチャシーケンシング(ウェイ、シー.エル.(Wei,C.L.)ら、2005年、Stem Cells 第23巻、p.166〜185)によって測定されたように、sox−oct複合要素の位置および向きと、対応する遺伝子の発現レベルとの間には、明確な関係はない(表1)。
【0096】
実施例5:NANOGと、(POU)特異的ドメインおよびホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子との複合体によるnanog遺伝子発現の調節
(0104)本実施例では、NANOGと、(POU)特異的ドメインおよびホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子との複合体形成の、多分化性細胞の転写活性化に対する影響の分析について説明する。
【0097】
(0105)上記保存プロモータ内の特定の配列要素の役割を調べるために、一連のルシフェラーゼレポータ構築物を構築し、F9テラトカルシノーマ細胞内にトランスフェクトした。
【0098】
(0106)DNA断片(−289〜+117)は、プライマーpNanogFおよびpNanogRを用いてPCRによりマウスゲノムDNA(2B gDNA)から増幅した(プロメガ(Promega)Pfuポリメラーゼ)。そのSalI/BamHI消化断片をpEGFP−1(ビーディーバイオサイエンシズ・クロンテック(BD Biosciences Clontech))のSalI/BamHI部位にクローニングした。この完全な構築物に続いて、pEGFP−1マルチクローニングサイト由来の22塩基対を含むSacI/BamHI断片をpGL3 Basic(プロメガ)のSacI/BglII部位にクローニングした。得られたプラスミドをシーケンシングにより確認し、pGL3Basic pNanogと名付け、次の突然変異誘発実験に用いた。レポータプラスミドは、Transformer(商標)部位特異的突然変異誘発キット(クロンテック(Clontech))を用いて3塩基対の変異が組み込まれるように改変した。修飾プラスミドはシーケンシングにより確認した。F9細胞(ATCC)を、15%標準ウシ胎仔血清(ハイクローン(Hyclone))および1%pen/strep(ペニシリン‐ストレプトマイシン)を含む高グルコースのダルベッコ改変イーグル培地(Gibco)を用いて培養した。細胞は、5%CO2で37℃に維持した。DNAは、Lipofectamine(商標)2000(インビトロジェン(Invitrogen))を使用し、24ウェル形式の細胞培養に対する同社のプロトコールを用いてF9細胞中にトランスフェクトした。ウミシイタケルシフェラーゼプラスミド(プロメガより入手したpRL−TK)を内部対照として同時トランスフェクトした。24時間後、細胞を溶解し、溶解物のルシフェラーゼ活性を、Dual−Luciferase(登録商標)レポータアッセイシステム(プロメガ)により、Centro LB960型96ウェル照度計(ベルトールドテクノロジーズ(Berthold Technologies))を用いて測定した。少なくとも、トランスフェクションは2回ずつ、2回の独立した状況で行った。レポータプラスミドは、Transformer(商標)部位特異的変異誘発キット(クロンテック)を用いて、3塩基対の変異が組み込まれるように改変し、変異はシーケンシングによって確認した。
【0099】
(0107)nanogEGF ES細胞を作製するために用いた野生型406塩基対プロモータ断片は、予想どおり顕著な転写活性を有し、すべての比較において該活性を100%と設定しポジティブコントロールとして使用した(図5B、2Cを参照)。図5A
中の保存配列のすぐ5’側から始まるこの野生型プロモータのより短いもの(−230〜+63)は、元のプロモータ活性の約130%を維持していた。このことは、この短いプロモータから除外された配列内に潜在的な負の調節要素が含まれていることを示唆するだけでなく、多分化能エンハンサ要素が−230〜+63の領域内に含まれることを示している。
【0100】
(0108)この保存されたモジュールの方向依存性を調べるために、この領域(−289〜−94)が直近位プロモータ(−93〜+117)に対して順方向または逆方向のいずれかの方向の相同な構築物を構築した。野生型活性の79%が順方向構築物において維持され、48%が逆方向構築物において維持されていた。有意な転写活性は、この保存領域を逆向きにした場合にも該領域に維持されていたことから、わずかに低い活性が向きの微妙な影響を示すかもしれないものの、エンハンサ活性は向きに依存しないことが示唆された。
【0101】
(0109)保存領域の機能的重要性の別の尺度として、第2の種からの、すなわち、ヒトnanogを用いて、相同領域を調べた。マウスF9細胞にトランスフェクトされたこの構築物は、マウス転写活性の60%を維持していた(Hs.1、図5Bを参照)ことから、実質的な機能保存が示された。本発明者らは、NanogGPlの、ヒトnanogのタンデム重複コピーからの相同領域についても調べた。おもしろいことに、ヒト多分化性細胞においてこの転写物のレベルは非常に低いにもかからわず、該領域は有意な転写活性を維持しており(Hs.2、図5Bを参照)、このことは、NanogGPlの不活性化変異がシス調節要素中ではなく、コーディング配列中にストップコドン8アミノ酸を生じることが判っているものでありそうなことを示唆している(ブース、エイチ.エイ.(Booth,H.A.)およびホランド、ピー.ダブリュ.(Holland,P.W)、前掲)。
【0102】
実施例6:Nanog活性RNAi分析の調節の分析
(0110)本実施例では、NANOGと、(POU)特異的ドメインおよびホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子とのあいだの複合体形成による、Nanogプロモータの活性調節の分析について示す。本実施例においては、Oct4およびSox2がNanogプロモータの多分化性活性に与える影響を分析した。
【0103】
(0111)この目的のために、RNAi実験を実施して、Oct4またはSox2のレベルを低下させた。この実験で用いたPou5flおよびSox2 RNAi構築物のいずれも、それぞれのmRNAを特異的にノックダウンすることがすでに知られているものである。Nanog−ルシフェラーゼレポータ構築物を、マウスESCにPou5fl、Sox2、GFP、または空のRNAi構築物と同時トランスフェクトし(図6A)、その後ルシフェラーゼ活性のアッセイを行った。
【0104】
(0112)オリゴヌクレオチドを、既述(ブルメルカンプ(Brummelkamp)ら、2002年、Science 第296巻、p.550〜553)のようにして、9ヌクレオチドループを有する19ヌクレオチドのヘアピン型短鎖ヘアピンRNA(shRNA)を発現するpSUPER.puro(BglIIおよびHindIII部位;オリゴエンジン(Oligoengine))にクローニングした。すべての配列について、BLAST検索で解析し、他の遺伝子と有意な配列類似性を有していないことを確認した。E14マウスESCは、トランスフェクションの1日前に、96ウェルプレートに細胞が20,000〜30,000個/ウェルの密度で(1000個/ウェルの初代胚線維芽細胞とともに)播種した。トランスフェクションは、Lipofectamine2000を用いて行った。50ngのNanog−pGL3(ホタルルシフェラーゼ構築物),150ngのpSuperまたはpSuper_RNAi,およびphRLSV40(
ウミシイタケルシフェラーゼを発現する正規化対照)をトランスフェクトした。
【0105】
(0113)トランスフェクションから24時間後に、ピューロマイシン耐性でさらに2日間細胞を選択してから、ルシフェラーゼ活性を測定した。RNAiに用いた配列は以下のとおり、すなわち
Sox2、5’−GAAGGAGCACCCGGATTA−3’(配列番号31);
Oct4、5’−GAAGGATGTGGTTCGAGT−3’(配列番号32)
である。
【0106】
(0114)図6Bに見られるように、ルシフェラーゼ活性は、Pou5flおよびSox2のRNAiによってほとんどバックグラウンドレベルにまで低減されたが、GFPおよび空のRNAi対照では全く影響がなかった。
【0107】
(0115)本明細書で例示的に記載した発明は、本明細書に具体的に開示されていないような任意の要素または限定なしで適切に実施してもよい。したがって、たとえば、用語「〜を含む(“comprising”,“including”,“containing”,など)」という用語は、限定的ではなく開放的に読まれることになる。さらに、本明細書で用いる用語および表現は、限定ではなく説明の用語として用いられており、開示および説明した特徴およびその一部の任意の均等物を除外するような用語および表現を使用する意図は全くなく、特許請求の範囲に記載の本発明の範囲内で様々な修正が可能であるものと理解される。したがって、本発明を説明的実施形態および任意選択の特徴によって具体的に開示してきたものの、本明細書に開示する具現された発明の修正および変更も、当業者によって行使されることができ、そのような修正や変更も本発明の範囲に含まれるとみなされると理解すべきである。
【0108】
(0116)本発明について、広範かつ一般的に本明細書において説明してきた。包括的開示の範囲内に入るより狭い種類のものおよび準包括的な群もまた、本発明の一部を構成する。これには、除外されたものが本明細書に具体的に列挙されているかどうかに関係なく、その分類から任意の特徴を除外する条件または負の限定を有する本発明の包括的説明も含まれる。
【0109】
(0117)他の実施形態は、添付の特許請求の範囲の中にある。さらに、本発明の特徴または態様がマーカッシュ形式で記載されている場合、当業者であれば、本発明が、そのマーカッシュ群の任意の個々のメンバーまたはメンバーのサブグループについても記載されていることを理解できるであろう。
【0110】
【表1】

上記表中、
a参照文献:Fgf4:ハート(Hart)ら、前掲;
Utfl:ニシモト(Nishimoto)ら、前掲;
Sox2:トミオカ(Tomioka)ら、前掲;
Fbxl5:トクザワ(Tokuzawa)ら、前掲;
Nanog:本願発明者;
Pou5fl:本願発明者,オクムラ‐ナカニシ(Okumura−Nakanishi)ら、前掲
b転写開始部位から
cMPSS:超並列シグニチャシーケンシング(ウェイ(Wei)ら、前掲)
dtpm,マウスESCにおける100万あたりのタグ数
【図面の簡単な説明】
【0111】
【図1】NanogRNAiを用いた胚幹細胞(ESC)におけるNanogダウンレギュレーションの効果を示す図。(A)使用したNanogRNAiの特異性について、Nanog発現ベクターを、対照、Nanog,Oct4またはSox2のsiRNAを発現する構築物とともに293T細胞中に同時トランスフェクトすることによって調べた。細胞溶解物を、抗Nanog抗体または抗βアクチン抗体を用いたウェスタンブロットによって分析した。βアクチンは装荷量の対照とした。(B)nanog siRNAは、Sox2の発現に影響を及ぼさなかった。nanogまたは対照のsiRNA構築物を、Sox2発現ベクターとともに293T細胞中に同時トランスフェクトした。溶解物を、抗Sox2抗体または抗βアクチン抗体を用いたウェスタンブロットによって分析した。(C)nanog siRNAは、Oct4の発現に影響を及ぼさなかった。nanogまたは対照のsiRNA構築物を、Oct4発現ベクターとともに293T細胞中に同時トランスフェクトした。溶解物を、抗Oct4抗体または抗βアクチン抗体を用いたウェスタンブロットによって分析した。(D)nanogのノックダウンにより、ES細胞中のNanogが減少した。nanogまたは対照のsiRNA構築物を、ES細胞中にトランスフェクトした。溶解物を、抗Nanog抗体または抗βアクチン抗体を用いて調べた。(E)nanogのノックダウンにより、ES細胞の分化が誘発された。nanogおよび対照のノックダウン細胞をアルカリホスファターゼについて染色した。写真は、対照siRNAでトランスフェクトしたESC(I)およびnanog siRNAでトランスフェクトしたESC(II)を同時に撮影したものである。ノックダウンした細胞の扁平な上皮様細胞の存在は、ベクター対照のES細胞においては全く認められないことに注目されたい。
【図2】POUおよびホメオドメイン含有転写因子ならびにHMGドメイン含有転写因子の、nanogプロモータ内の特異的結合要素との複合体形成を分析するための例示的方法として、電気泳動移動度シフトアッセイ(EMSA)を示す図。本実施例はOct4およびSox2の、Nanog8量体/HMG複合要素への結合を説明する。EMSAは、nanogプロモータ由来の標識した推定Oct4/Sox2要素、または陽性対照としてのFGF4エンハンサ由来の既知のOct4/Sox2結合部位のいずれかを用いて実施した。因子の結合は、E14胚幹細胞の粗製核抽出物中で調べた。Oct4およびSox2を含むタンパク質−DNA複合体は、示したバンドをスーパーシフトさせる抗体の添加によって同定した。結合特異性を、オリゴ競合物(PAGEゲルの一番上に表示)を用いて調べたところ、Oct4の結合には完全な8量体モチーフを必要とし、Sox2の結合には完全なHMG要素を必要とすることが示された。ヘテロ二量化には、両方が完全であることが必要であった。競合物についての記号は次のとおりである。N=Nanog,F=FGF4,O=Oct4結合部位、S=Sox2結合部位、mO=変異型Oct4結合部位、mS=変異型Sox2結合部位、Gbx=非特異的競合物。
【図3】Oct4/OCT4およびSox2/SOX2の、Nanog/NANOGプロモータとのin vivo相互作用を示す図。(A)クロマチン免役沈降(ChIP)は、未分化のマウスESC(第0日、レチノイン酸、RA)およびレチノイン酸(RA)の共存下で3日間および6日間培養したESCについて行った。Oct4(N19)およびSox2(Y17)に対する抗体による免疫沈降を示す。リアルタイムPCRによって測定した富化の倍率を3つの異なるアンプリコンで比較した。このうち1つ(アンプリコン2)は、複合oct−sox要素(概略図では白抜きのボックスで示す)を含み、残りの2つ(アンプリコン1および3)は含まなかった。各アンプリコンの転写開始部位に対する位置が概略図に示されており、Nanogのエキソン1が黒塗りのボックスで示されている。(B)ヒトESCにおけるNANOGのOCT4およびSOX2 ChIP。未分化細胞からの特定のアンプリコンの富化の倍率が示されている。このデータから、複合sox−oct要素に最も近い3つのアンプリコンだけが、使用した両抗体によって富化を示したことがわかる。すべて標準偏差が示されている。
【図4】nanog近位プロモータが、未分化ES細胞においてはEGFP(強化緑色蛍光タンパク質)発現を駆動するが、同細胞の分化誘導体においては駆動しないことを示す図。EGFP発現を駆動するnanogプロモータ(転写開始部位に対して−289〜+117)を含むベクター構築物で安定的にトランスフェクトされたマウスESC株(NanogEGFP)を、異なる培養条件において比較した。光学顕微鏡写真(A,C,E)および蛍光顕微鏡写真(B,D,F)は、未分化のESC(A,B)、LIFなしで3日間培養したESC(C,D)、および0.1μMレチノイン酸(RA)で3日間処理したESC(E,F)のものである。LIFを回収すると、RAへの曝露よりも分化誘発の積極性は小さいが、EGFP発現は大きく減少した。ESC分化の強力な誘発剤であるRAは、EGFP発現を劇的に減少させた。(G)リアルタイムPCRでの検出により、内在性Nanog転写物のレベルを、未分化および分化条件下においてNanogプロモータによって誘導されるEGFPのレベルと比較して示すグラフ。グラフ中の数値は、FACS分析によって測定したEGFP陽性細胞の割合(%)を示す。EBは胚様体であり、RAはレチノイン酸である。
【図5】(A)系統発生学的フットプリンティングによる結合要素の同定の例を示す図。nanog配列は、マウス、ラット、ヒト、ウシおよびゾウのゲノムシーケンシングプロジェクトから得た。2億5千万年以上の累積進化あいだ不変のままである、転写開始位置から−212〜−119の領域に影をつけてある。oct−sox複合要素は、3塩基対の置換変異体として示してあり、該変異体の対応名は下に表示してある。 (B、C)Nanogと、(POU)特異的ドメインおよびホメオドメインを含む転写遺伝子ならびにHMGドメインを含む転写因子との間の複合体形成の影響の分析を示す図。F9テラトカルシノーマ細胞に一時的にトランスフェクトされた様々なnanogプロモータ構築物のプロモータ−ルシフェラーゼレポータアッセイを示している。−289〜+117のマウスnanogプロモータ断片(wt)のルシフェラーゼ活性を任意で100%に設定する。(B)変更を加えたマウスプロモータ構築物の活性を、野生型(wt)、短縮型(short)(短い野生型プロモータ、−230〜+63の領域)と比較したもの。ForおよびRevは、互いに同じであるが、保存領域(−93〜+117)の転写開始位置に対する向きが逆である。Hs.1およびHs.2は、それぞれヒトNanogおよびNanogGPl由来の、マウスwt構築物に相同な領域である。(C)置換変異のマウスプロモータ活性への影響。変異は、oct/sox要素がいずれも変異を有するoct/sox構築物に関して、(A)に示したものに相当する。
【図6】Nanogプロモータ活性に対する、Oct4またはSox2のRNAiノックダウンの結果を示す図。図2Aは、Sox2、Oct4、およびEGFP(対照)を標的とするRNAi構築物の、Nanogプロモータ−ルシフェラーゼレポータ構築物とともにマウスESC中への同時トランスフェクションの概略図である。図2Aは、トランスフェクションから3日後に測定されたルシフェラーゼ活性を示す。(B)ルシフェラーゼ活性を、ウミシイタケルシフェラーゼ内部対照と比較して測定した。阻害性RNAを含まない構築物を陰性対照(−)として用いた。示したとおり、Nanogプロモータ活性は、Sox2およびOct4の両者を標的としたRNAi構築物によって低減される。標準偏差も示してある。
【技術分野】
【0001】
(0001)本発明は、幹細胞/前駆細胞の多分化能または自己再生特性のうち少なくともいずれか一方を維持する方法に関する。本発明は、細胞内の遺伝子発現を調節するための方法にも関する。本方法は、少なくとも2つの転写因子またはその機能的断片を、nanog遺伝子のプロモータ領域と接触させることを含む。少なくとも2つの転写因子のうちの一方は、POUドメインおよびホメオドメイン含有転写因子から選択される。少なくとも2つの転写因子のうちの他方は、HMGドメイン含有転写因子から選択される。本方法は、少なくとも2つの転写因子が、nanogのプロモータ内の特異的結合要素と複合体を形成できるようにすることをさらに含む。このようにして形成される複合体は、転写活性化を介してnanog遺伝子の発現を調節する。
【背景技術】
【0002】
(0002)幹細胞は、再生医療に対する鍵を握っていることがわかってきている。というのも幹細胞は、疾患、感染または先天性異常によって損傷した対応組織の代わりとなる細胞源を供給できるためである。これは、幹細胞が、無制限に増殖できる能力を持つとともに、心臓、肝臓、脳細胞などの機能的な成熟細胞に分化しうる未分化細胞であるためである。ギュンター病、ハンター症候群、およびハーラー症候群は、たとえば、幹細胞を用いて治療されてきた。白血病やリンパ腫などの病状を有するがん患者の治療は、骨髄からの成人幹細胞を用いて行われてきた。化学療法投薬量の段階的増大による血球減少に苦しむがん患者の造血系は、幹細胞の静脈内注射によって回復させることができた。韓国の研究者は、臍帯血からの成人幹細胞をうまく用いて、脊髄損傷を煩う麻痺をもつ女性が、歩行器の助けを借りて歩行できるようになったことを報告している。パーキンソン病、心筋梗塞、若年発症糖尿病をはじめとするさらに多くの疾病が、現在のところ、幹細胞または幹細胞由来の細胞の治療的移植によって治療可能であると考えられている。
【0003】
(0003)この点に関する特に実現可能なツールは、哺乳類の多分化性幹細胞であるが、これは当該細胞が、任意の器官、細胞タイプまたは組織タイプへ分化可能であり、少なくとも潜在的には完全な生命体へと分化することができるためである。このような多分化性幹細胞は、着床前胚および多くの成人組織のいずれにおいても見られる。特に優れているのは、哺乳類の胚盤胞の段階にある胚の内細胞塊(ICM)に由来する胚幹細胞(ESC)である。ESCは、特定の細胞培養条件下で、より長い時間、自己再生的に細胞分裂することにより、その多分化能を維持する(たとえば、非特許文献1または非特許文献2を参照のこと)。
【0004】
(0004)ESCは、たとえば、神経成長因子とレチノイン酸の共存下に、ニューロンへと制御的に分化させることができるが(非特許文献3)、そのように容易に分化できることが、実践上の大きな難問をつきつけるのである。ESCを多分化性状態に維持するためには、取扱時および培養増殖時の分化を防止しなければならない。こうした理由から、ESCは、伝統的にはフィーダー細胞層上でウシ胎仔血清の共存下で(たとえば、特許文献1および特許文献2を参照のこと)、または線維芽細胞調整培地(CM)中で培養されてきた。にもかかわらず、念入りに制御された条件下においてさえ、ESCは、in vitroでの増殖のあいだに自発的に分化することがある。マウスESCにおける自己再生を媒介する因子である白血病抑制因子(LIF)は、マウスESCの分化を阻害することがわかっているが、ヒトESCの分化を阻害する際のフィーダー細胞の役割に変わるものではない。したがって、ECSの多分化能または自己再生特性のうち少なくともいずれか一方を維持する手段は、幹細胞治療の完全な商業的潜在性の実現に向けての多大な功績となるであろう。
【0005】
(0005)成人幹細胞は、ESCのような多分化性ではないものの、自己再生可能であり、その発生能を、より未成熟な多分化性ESCの発生能と同等にする可塑性を持つことがわかっている。一例として、成人幹細胞は、その元となる組織とは異なる細胞系統に分化することができる。
【0006】
(0006)幹細胞は、2以上の発生学上の層が混ざり合った組織を生み出す、様々な組織(精巣および卵巣のものであることが多い)の奇形腫と呼ばれる癌においても見つかっている。このような癌の悪性型は、奇形癌とも呼ばれている。マウス奇形癌における幹細胞の発達は、正常な胚における事象と平行したものである。これらの存在は、化学治療で大量の腫瘍塊を除去しても、腫瘍の再発を防止できない場合が多いことの説明となる(非特許文献4)。したがって、このような腫瘍において幹細胞の多分化能または自己再生特性のうち少なくともいずれか一方を無効にする手段は、典型的にはこのような癌の永久的除去のための前提条件となるであろう。
【0007】
(0007)いまのところ数種のタンパク質が、多分化性細胞の正常な発達のために、かつ/または多分化性の細胞状態を維持するために重要であることが確認されている。そうしたタンパク質としては、Oct4(非特許文献5)、Sox2(非特許文献6)、Nanog(非特許文献7、非特許文献8)、およびFoxD3(非特許文献9)が挙げられる。他のタンパク質、たとえば、LIF(上記参照)またはRexl(Zfp42)も、このような機能に関与することが示唆されている。しかしながら、それらのタンパク質の作用機構、ならびに多分化性細胞の発生または多分化性細胞状態の維持における、当該タンパク質の特異的役割については今のところ知られていない。
【特許文献1】米国特許第5,843,780号明細書
【特許文献2】米国特許第6,090,622号明細書
【非特許文献1】ロエベル、ディー.エイ.(Loebel,D.A.)ら、2003年、Dev.Biol.第264巻、p.1〜14
【非特許文献2】スミス、エイ.ジー.(Smith,A.G.)、2001年、Annu.Rev.Cell Dev.Biol.第17巻、p.435〜462
【非特許文献3】シュルディナー(Schuldiner)ら、2001年、Br.Res.第913巻、p.201〜205
【非特許文献4】チャンバース、アイ.、スミス、エイ.(Chambers,I.Smith,A)、2004年、Oncogene 第23巻、p.7150〜7160
【非特許文献5】ニコルス、ジェイ.(Nichols,J.)ら、1998年、Cell 第95巻、p.379〜391
【非特許文献6】アビリオン、エイ.エイ.(Avilion,A.A.)ら、2003年、Genes Dev.第17巻、p.126〜140
【非特許文献7】ミツイ(Mitsui)ら、2003年、Cell 第113巻、p.631〜642
【非特許文献8】チャンバース(Chambers)ら、2003年、Cell 第113巻、p.643〜655
【非特許文献9】ハンナ(Hanna)ら、2002年、Genes Dev.第16巻、p.2650〜2651
【発明の開示】
【発明が解決しようとする課題】
【0008】
(0008)したがって、本発明の目的は、幹細胞/前駆細胞の多分化能または自己再生特性のうち少なくともいずれか一方を制御的に維持する方法、ならびに、たとえば、奇形腫中の幹細胞の多分化能または自己再生特性のうち少なくともいずれか一方を無効にするために、細胞内の遺伝子発現を調節する方法を提供することにある。
【課題を解決するための手段】
【0009】
(0009)一つの態様において、本発明は、幹細胞/前駆細胞の多分化能または自己再生特性のうち少なくともいずれか一方を維持する方法を提供する。本方法は、少なくとも2つの転写因子またはその機能的断片を、nanog遺伝子のプロモータ領域と接触させることを含む。少なくとも2つの転写因子のうちの一方は、POUドメインおよびホメオドメイン含有転写因子から選択される。少なくとも2つの転写因子のうちの他方は、HMGドメイン含有転写因子から選択される。本方法は、少なくとも2つの転写因子が、nanogプロモータ内の特異的結合要素と複合体を形成できるようにすることをさらに含む。このように形成された複合体は、転写活性化を介してnanog遺伝子発現を調節する。
【0010】
(0010)別の態様において、本発明は、細胞内の遺伝子発現を調節する方法を提供する。本方法は、少なくとも2つの転写因子またはその機能的断片をnanog遺伝子のプロモータ領域と接触させることを含む。少なくとも2つの転写因子のうちの一方は、POUドメインおよびホメオドメイン含有転写因子から選択される。少なくとも2つの転写因子のうちの他方は、HMGドメイン含有転写因子から選択される。本方法は、少なくとも2つの転写因子が、nanogプロモータ内の特異的結合要素と複合体を形成できるようにすることをさらに含む。このように形成された複合体は、転写活性化を介してnanog遺伝子発現を調節する。
【発明を実施するための最良の形態】
【0011】
(0011)本発明は、非限定的な実施例および図面と組み合わせて、以下の詳細な説明を参照してより良く理解されるであろう。
(0019)本発明は、POU特異的ドメインおよびホメオドメインを含む転写因子、ならびにHMGドメインを含む転写因子が、in vitroおよびin vivoのいずれにおいてもnanogのプロモータと複合体を形成し、さらに、この複合体の形成がタンパク質Nanogの発現を調節するという驚くべき発見に基づいている。さらに驚くべきことに、このような結合領域の少なくとも1つが累積して2億五千万年以上もの進化の間、不変のままであることが判った。
【0012】
(0020)本願発明者の知見は、特に実用的に関係がある。というのも、POU特異的ドメインおよびホメオドメインを含む転写因子がタンパク質Oct4であり、HMGドメインを含む転写因子がタンパク質Sox2(参考:上記および下記)であるためである。Oct3としても知られるOct4は、細胞の未分化状態に関するマーカーとしてよく使用される。Oct4は、ヒトおよびマウスESCにおいて高度に発現され、その発現は、これらの細胞が分化して多分化能を失うときに減衰する(パルミエリ(Palmieri)ら、1994年、Dev.Biol.第166巻、p.259〜267)。また、Oct4は成人幹細胞、腫瘍細胞、および不死化非腫瘍原性細胞においても発現されるが、分化した組織の細胞においては発現されない(たとえば、モンク、エム.(Monk,M.)、ホールディング、シー.(Holding,C.)、2001年、Oncogene 第20巻、p.8085〜8091を参照のこと)。
【0013】
(0021)Sox2は転写因子であり、Oct4と同様に、分化中のESCと比べて、未分化ヒトESC内で強くアップレギュレートされる(ブランデンバーガー、アール(Brandenberger,R)ら、2004年、Nature Biotechnology 第22巻、p.707〜716)。Sox2は、たとえば、胚発生の初期段階における全CNSの神経上皮の未熟な未分化細胞においても発現される。分化が起こると、Sox2がダウンレギュレートされる(ステバノヴィク、エム(Stevanovic,M)、2003年、Molecular Biology Reports 第30巻
第2号、p.127〜132)。Sox2とOct4のいずれも、他の細胞タイプを決定する際に独立した役割を果たし(ニワ、エイチ.(Niwa,H.)ら、2000年、Nat Genet 第24巻、p.372〜37;非特許文献6)、多分化性細胞におけるそれらの機能の少なくとも一部は、標的遺伝子の転写を駆動するために2者の間の相乗作用を介する。
【0014】
(0022)転写因子という用語は、DNAからのRNAの合成(転写)のレベルを変えるタンパク質の能力のことをいう。典型的には、このような因子は、エンハンサ要素またはプロモータ要素などのような、各DNA上に見られる遺伝子の所定の領域に結合して、該DNAと複合体を形成する、細胞質タンパク質または核タンパク質である。
【0015】
(0023)Oct4およびSox2の相乗作用の標的としては現在、fgf4,utfl,およびfbx15の3種が知られており(ユアン、エイチ.(Yuan,H.)ら、1995年、Genes Dev.第9巻、p.2635〜2645;ニシモト、エム.(Nishimoto,M.)ら、1999年、Mol.Cell.Biol.第19巻、p.5453〜5465;トクザワ、ワイ.(Tokuzawa,Y.)ら、2003年、Mol.Cell.Biol.第23巻、p.2699〜2708)、4つ目は、sox2自身であると思われる(トミオカ、エム.(Tomioka,M.)ら、2002年、Nucleic Acids Res.第30巻、p.3202〜3213)が、対応するシス要素は多分化性細胞発現に対しては省略可能であると思われる(ハヤシ、エス.(Hayashi,S.)ら、2002年、Mech.Dev.第119巻、追補1,S97〜S101)。これらの標的遺伝子のそれぞれは、8量体およびsox結合部位を含む複合要素を有する。
【0016】
(0024)本発明の方法において、上述のような転写因子(以下も参照)は、新しく同定されNanogと命名されたタンパク質をコードする遺伝子のプロモータとともに複合体を形成する。これは、細胞の多分化性状態の維持、ならびに初期胚盤葉上層/内細胞塊(ICM)の多分化性細胞の正常な発達、およびこの細胞集団からのES細胞の派生にも必要なタンパク質である(ミツイ(Mitsui)ら、前掲)。またこのタンパク質は、たとえば分化中のESC中よりも未分化のESC中にはるかに多く存在することから、未分化状態の細胞、たとえば、幹細胞または胚性生殖細胞に対するマーカーとして使われることも多い(ブランデンバーガー、アール(Brandenberger,R)ら、前掲)。Nanogは、成人骨髄中でも発現されており、腎細胞やある種のがんなどの様々な成人組織において、低レベルで発現されている。Oct−4と同様に、Nanogは、成人の成熟細胞にいくらかの胚様の可塑性を回復させうることで知られている(たとえば、シース、エヌエックス.(Theise,NX.)、ウィルマット、アイ.(Wilmut,I)、2003年、Nature 第425巻、p.21を参照)。
【0017】
(0025)Nanogはホメオボックスを含み、転写因子として作用する場合もある。nanogの過剰発現により、通常は分化を誘発する培養条件において、ESCの多分化能および自己再生特性を維持することができる(チャンバース、アイ.(Chambers,I.)ら、前掲)。標準増殖条件下でnanog発現をダウンレギュレートすると、ESCにおける分化が誘導される(図1を参照)。多分化性細胞の維持においてこの必須機能に付随するのが、nanogの限局的な発現パターンである。nanog転写物は、胚盤胞形成前に、まず桑実胚の内部細胞内に現れ(チャンバース、アイ.(Chambers,I.)ら、前掲;ミツイ、ケイ.(Mitsui,K.)ら、前掲)、胚盤胞内では発現は内細胞塊(ICM)に限られ(ワン、エス.エイチ.(Wang,S.H.)ら、2003年、Gene Expr.Patterns 第3巻、p.99〜103)、着床時にはすでに検出不能である。
【0018】
(0026)本願発明者による知見を考慮すると、Oct4およびSox2がマウスの着床前発生において類似の発現パターンを有するという最近発表された研究結果(アビロン、エイ.エイ.(Avillon,A.A.)ら、2003年、Genes Dev.第17巻、p.126〜140)が容易に理解できるようになる。Nanogは、幹細胞の多分化性細胞状態および自己再生特性の維持のための鍵となる要素であるという知識に基づいて、本発明者らは、幹細胞および前駆細胞の多分化能または自己再生特性のうち少なくともいずれか一方を維持する方法を確立することができた。本発明者らは、さらに、それらの方法が細胞内の遺伝子発現を調節するのに適していることを発見した。
【0019】
(0027)nanog遺伝子は、凝縮後に現れ桑実胚の内部細胞に特異的な第1の既知の転写因子をコードし、Oct4およびSox2のいずれも、すべての割球において凝縮の前に発現される。本明細書中で用いる「nanog遺伝子」という用語は、あらゆる哺乳類のnanog遺伝子、たとえば、ハート、エイ.エイチ.(Hart,A.H.)ら(2004年、Dev.Dynamics 第230巻、p.187〜198)に記載されるようなマウスおよびヒトのnanog遺伝子、ウシの遺伝子(NCBI遺伝子ID:538951)、ブタの遺伝子(EMBLアクセス番号Q5GMQ0およびQ64HX3)、ディンゴの遺伝子(NCBI遺伝子ID:486701)、チンパンジーの遺伝子(NCBI遺伝子ID:452438)、マカクの遺伝子(EMBLアクセス番号Q5TM84)、ラットの遺伝子(NCBI遺伝子ID:414065(Locusタグ:RGD:1303178))、NCBIアクセス番号AY786437のRNAを転写し、NCBIアクセス番号AAW50709の転写因子をコードするヤギ遺伝子、ならびにこれらのオーソログを含む。また、まだ同定されてないnanog遺伝子も含まれる。また、たとえばアンネ(Anne)ら(2004年、Genomics 第84巻、p.229〜238)によって同定され、当技術分野で周知の標準的な方法(たとえば、部位特異的変異法)によって機能するように改変されたもののようなnanogの偽遺伝子も含まれる。さらに、既知のnanog配列と比較すると核酸配列に逸脱が見られる形態の遺伝子も含まれる。その差異は、たとえば、多型、1つのヌクレオチドの変化または修飾、(連続する一続きの)置換、欠失または挿入、および対応する天然の核酸配列に導入されたN末端および/またはC末端への付加によるものであってよい。本発明の方法において用いるnanog遺伝子のプロモータ領域は、任意の生物種のもの、または任意の生物種に由来するものであってよい。2つの例示的実施例は、ヒトおよびマウスのnanog遺伝子のプロモータ領域である。
【0020】
(0028)Nanogは、マウスおよびヒトの多分化性細胞のいずれにおいても発現されるので、この遺伝子の多分化性転写は、シス調節要素の機能保存を通して維持されており、同時に、これらの要素の位置および配列は、浄化選択を経ても保存されると思われる。したがって、マウスおよびヒトのnanogゲノム配列を、配列比較のために公開データベースから得た。驚いたことに、マウスnanogおよびslc2a3のタンデム重複の結果として、2コピーのヒトnanog遺伝子が同定された(ブース、エイチ.エイ.(Booth,H.A.)およびホランド、ピー.ダブリュ.(Holland,P.W.)、2004年、Genomics 第84巻、p.229〜238;ハート、エイ.エイチ.(Hart,A.H.)ら、前掲)。ヒトESTデータ(Hs.329296)に基づけば、NANOG/NANOGlは活発に転写されるが、NANOGP1/NANOG2は検出不能レベルを維持している。以下の実施例においては、活発に転写されるNANOGを用いて、マウスのホモログnanogと比較した。
【0021】
(0029)本発明の方法は、少なくとも2つの転写因子、またはその機能的断片を、nanog遺伝子のプロモータ領域、すなわち、転写開始時にRNAポリメラーゼによって認識され結合されるnanog遺伝子の核酸配列と接触させることを含む。これらの転写因子のうちの一方は、Pit,Oct,Unc(POU)特異的ドメインおよびホメオ
ドメインを含む転写因子から選択される。多くの実施形態において、ホメオドメインはPOU−ホメオドメインである。POU特異的ドメインは一般には75〜82アミノ酸長であり、POU−ホメオドメインは約60アミノ酸長である。天然の転写因子において、これらの2つのドメインは典型的には一緒になっていわゆるPOUドメインを構成し、この場合、POU特異的ドメインはN末端領域を形成し、POU−ホメオドメインはC末端領域を形成する。POU特異的ドメインおよびPOUホメオドメインのいずれも本発明において必要ではあるが、nanog遺伝子のプロモータとの複合体形成が起こりさえすれば、該ドメインを任意の配置構成で選択することができる。多くの実施形態において、該ドメインは適切な長さのリンカー領域によって連結される。当業者であれば、POUドメイン外の非保存配列が標的配列のトランス活性化に参加するとの報告を認識しているであろう。したがって、人工的に作成した転写因子、またはその機能的断片の、本発明の方法への適合性は、該転写因子または断片のnanogへの結合に関して調べる必要がある(下記およびたとえば、実施例2または3、図2または図3を参照)。適切な転写因子の例としては、限定はされないが、Pit−1,Oct−1,Oct−2、Oct−4、Oct−6、Oct−11、Brn−3、Brn−3A、BおよびC、肝細胞核因子lαおよびβ、網膜由来POUドメイン因子−1、精子1POU−ドメイン転写因子、ラットタンパク質Rov−1、アフリカツメガエル(Xenopus laevis)の転写因子POUl、POU2、GBX−2およびOct−1.5、キイロショウジョウバエ(Drosophila melanogaster)の転写因子Cfla、カタユウレイボヤ(Ciona intestinalis)の因子(NCBIアクセス番号BAE06650)、線虫(C.elegans)遺伝子unc−86によってコードされるタンパク質、線虫の因子ceh−6、ショウジョウバエ(Drosophila)タンパク質dPOU−19/pdm−1およびdPOU−19/pdm−2、およびゼブラフィッシュの因子pou5fl/pou2が挙げられる。説明的な例としては、転写因子Oct−1、Oct−4およびOct−6が、nanogプロモータ上の領域に結合することが示されている(ウー、ディー.(Wu,D)、ヤオ、ズィー.(Yao,Z)、2005年、Cell Research 第15巻、第5号、p.317〜324)。
【0022】
(0030)上記少なくとも2つの転写因子またはその機能的断片の他方は、HMG(高移動度群)ドメイン含有転写因子から選択される。HMG DNA結合ドメインは、4方向接合(4‐way junctions)など湾曲DNAに対する著しい優先性を与える。このような転写因子の例としては、限定はされないが、Sry HMGボックス(Sox)転写因子、HrSoxB1、リンパ球エンハンサ結合因子1(LEF−I,T細胞特異的転写因子1−α)、HMGB1aタンパク質、HMGB1bタンパク質、上流結合因子(UBF)、CCAAT結合転写因子2(CTF2)、活性化転写因子4(ATF4)、YABBYファミリーのタンパク質、ミトコンドリア転写因子A(mtTFA)、顆粒膜細胞高移動度群ボックスタンパク質1(GCX−1)、T細胞特異的転写因子7、黒穂菌Ustilago maydisのPrfl、分裂酵母(Schizosaccharomyces pombe)のStellp因子および未命名の因子(NCBIアクセス番号NP595672)、線虫(C.elegans)のTCF/LEF−1、アフリカツメガエル(Xenopus)のXTcf3、睾丸決定因子SRY、XSOX3、ウニ遺伝子Unichromの転写産物、クルイベロマイセス・ラクティス(Kluyveromyces lactis)由来の交配型タンパク質A2、キイロショウジョウバエ(Drosophila melanogaster)のタンパク質CG17964−PHアイソフォームH(NCBIアクセス番号NP001014685)、およびNCBIアクセス番号T12113を有するソラマメの転写因子が挙げられる。
【0023】
(0031)いくつかの実施形態において、第2の転写因子は、SOX(SRY関連HMGボックス)タンパク質であり、例として、限定はされないが、Sox−1,Sox−2,Sox−3,Sox−4,Sox6,Sox7,Sox8,Sox9,Sox10,
Sox11,Sox−13,Sox14,Sox15,Sox18,Sox20,Sox21,Sox30,Sox32またはアフリカツメガエル(Xenopus laevis)の因子Sox−11−D、またはこれらの機能的断片が挙げられる。
【0024】
(0032)各転写因子のうち一方の機能的断片は、任意の長さのものであってよく、2つの基準によって定義することができる。第1に、機能的断片は、nanogプロモータの結合領域と結合して複合体を形成することができ、前記複合体は、たとえばnanog遺伝子などの当該プロモータによって駆動されるそれぞれの遺伝子の活性に影響するのに十分な安定性を有する。第2に、このような機能的断片は、POUおよびホメオドメイン(たとえばPOU−ホメオドメイン)またはHMGドメインをそれぞれ含む天然に存在する転写因子の対応するアミノ酸配列と少なくとも60%の配列同一性を有しうる。いくつかの実施形態において、各断片は、既知の各転写因子の対応するアミノ酸配列と少なくとも80%の配列同一性を、一部の実施形態においては少なくとも95%の配列同一性を有する。「配列同一性」という用語は、既知のPOUおよびホメオドメイン含有転写因子またはHMGドメイン含有転写因子それぞれのアミノ酸配列と、問題とするアミノ酸配列との相同性アラインメント後に得られる、ペアワイズな同一残基の割合(%)を意味し、%の数字は、2つの配列のうち長い方の残基数に対するものである。
【0025】
(0033)本発明の方法において用いる転写因子の機能的断片には、たとえば融合タンパク質の形で、追加の配列がさらに結合されてもよい。また、たとえば、いわゆるアフィニティタグまたはラベルなどの天然または人工の化学修飾を含んでいてもよい。アフィニティタグの例としては、限定はされないが、ビオチン、ジニトロフェノール、ジゴキシゲニン、オリゴヒスチジン、ポリヒスチジン、免疫グロブリンドメイン、マルトース結合タンパク質、グルタチオン−S−トランスフェラーゼ(GST)、カルモジュリン結合ペプチド(CBP)、FLAGのペプチド、T7エピトープ(Ala−Ser−Met−Thr−Gly−Gly−Gln−Gln−Met−Gly)、マルトース結合タンパク質(MBP)、単純ヘルペスウィルス糖タンパク質Dの配列Gln−Pro−Glu−Leu−Ala−Pro−Glu−Asp−Pro−Glu−AspからなるHSVエピトープ、配列Tyr−Pro−Tyr−Asp−Val−Pro−Asp−Tyr−Alaからなるヘマグルチニン(HA)エピトープおよび転写因子c−mycの配列Glu−Gln−Lys−Leu−Ile−Ser−Glu−Glu−Asp−Leuからなる“myc”エピトープが挙げられる。ラベルはさらに、たとえば本発明の方法において望ましい場合には、検出を助ける部分であってもよい。一例としては、限定はされないが、放射活性アミノ酸、フルオレセインイソチオシアネート、5,6−カルボキシメチルフルオレセイン、Texas Red(登録商標)、ニトロベンズ−2−オキサ−1,3−ジアゾール−4−イル、クマリン、ダンシルクロライド、ローダミン、アミノ−メチルクマリン、エオシン、エリスロシン、BODIPY(登録商標)、Cascade Blue(登録商標),Oregon Green(登録商標)、ピレン、リサミン、キサンテン、アクリジン、オキサジン類、フィコエリスリン、Cy3、Cy3.5、Cy5、Cy5.5、およびCy7酵素が挙げられる。さらに適した酵素としては、限定はされないが、アルカリホスファターゼ、大豆ペルオキシダーゼ、またはサイヨウワサビペルオキシダーゼが挙げられる。
【0026】
(0034)各アフィニティタグまたはラベルは、選択された転写因子の任意の部分内に配置されてもよいし、任意の部分に付加されてもよい。説明的な例として、任意のPOUドメインおよびホメオドメイン含有転写因子またはHMGドメイン含有転写因子のアミノ末端またはカルボキシ末端に機能可能なように融合させるとよい。
【0027】
(0035)本発明の方法は、少なくとも2つの転写因子(上記参照)またはその機能的断片が、nanogプロモータ内の特異的結合要素との複合体を形成できるようにする
ことをさらに含む。
【0028】
(0036)本発明の方法のいくつかの実施形態において、少なくとも2つの転写因子またはその機能的断片は、nanogプロモータ上にヘテロ三量体複合体を形成する。それぞれの複合体の形成は、当業者周知の様々な分析方法によって検出することができ、そのような方法としては、表面プラズモン共鳴(たとえば、Biacore(登録商標)技術)、核磁気共鳴または結晶化とそれに続くX線解析が挙げられる。本発明の方法のいくつかの実施形態において、上記で詳述したような転写因子またはその機能的断片は、ヘテロ二量体を形成する。このヘテロ二量体は、その後、nanogプロモータの結合領域を含む核酸とのヘテロ三量体複合体を形成する。ヘテロ三量体複合体を形成するそれぞれの二量体の例は、Oct4およびSox2のnanogへの結合である。結晶学的データは、これらの2つの関係する転写因子がDNA上の結合要素と三重複合体を形成することを示している(レメニ(Remenyi)ら、2003年、Genes&Development 第17巻、p.2048〜2059)。Octl構造の上にOct4を用いて作成した相同性モデルは、Oct4とSox2の間で形成された複合体が、Oct1とSox2の間の複合体と類似していることを示す。Sox2は、Oct4との相互作用のための2つの表面パッチを有する。そのうちの一方は、DNAとも相互作用できることがわかっているSox2のHMGドメインのC末端である。様々なSOXおよびPOU転写因子の間での複合体形成のキャラクタリゼーションが進んでいることに鑑みて(たとえばレメニ(Remenyi)ら、前掲を参照)、様々なPOUドメインおよびホメオドメインを含む転写因子、ならびにHMGドメインを含む転写因子が、核酸、たとえばnanog遺伝子上の結合領域とヘテロ三量体複合体を形成することが予想される。
【0029】
(0037)各結合要素は、当業者によって知られる様々な手段によって同定することができる。実施例4は、配列比較を用いたNANOG上の結合要素の同定を示したものである。このような手法は典型的には、それぞれの転写因子に対する結合要素が存在するかどうかについての情報が全く得られない場合に行われる。本発明者らは少なくとも1つのそのような結合要素をすでに同定していたので、総じてこの手順を用いることは必要でないことになる。代替の手段は、たとえば米国特許第6,735,530号明細書に開示されるようなコンピュータによって実施する方法である。さらに別の結合要素同定手段としては、たとえば実施例3に記載するようなクロマチン免疫沈降アッセイがある。この方法はさらに、活性転写領域を判定したり、ヒストン結合によるゲノムの修飾を評価したりするために使用することもできる。さらに別の手段は、実施例2および図2に記載するような電気泳動移動度シフトアッセイ(EMSA)である。nanogプロモータ内の結合要素を同定するさらなる方法は、たとえば蛍光タンパク質などのマーカーの発現のために該プロモータを使用することである。この手法がどのように実施されるかの例を図4に示す。強化緑色蛍光タンパク質(EGFP)の発現を駆動するnanog遺伝子の−289から+117(転写開始部位(TSS)に対して)のマウス配列を含むプラスミドベクターで、マウスESC株をトランスフェクトした。レチノイン酸による分化誘導によってnanog遺伝子をダウンレギュレートし、これにより、処理3日後のEGFP発現が劇的に減少した(図4E、Fを参照)。リアルタイムPCRによる、NanogEGFPと、内在性Nanogとの比較(図4G)から、該構築物は、内在性Nanogの発現を再現することができたことがわかる。当業者であれば、これらの方法は、さらに他の方法、たとえばin vitro突然変異誘発と組み合わせて、特定の領域についての結合要素の一部としてのさらなる検証に用いることができることに気づくであろう。
【0030】
(0038)本発明のいくつかの実施形態において、少なくとも2つの転写因子は、nonogプロモータ内の複合要素に結合する。このように、これらの実施形態において、POUドメインおよびホメオドメイン含有転写因子は、複合要素の一方部分に結合し、HMGドメイン含有転写因子は、複合要素の他方部分に結合する。各結合要素は、それぞれ
の転写因子について任意の数の結合領域を有していてもよい。たとえば、2つの部分からなる結合部位であってもよい。そのような結合要素は、たとえば、2つの転写因子Sox2およびOct4に対して存在することが知られている。8量体とsox結合部位を含み、Sox2およびOct4の標的となる複合要素を含む遺伝子は、Fgf4,Utfl,Fbx15およびSox2、ならびにPou5fl(Oct4をコードする遺伝子)自体である(ユアン、エイチ.(Yuan,H.)ら、前掲;ニシモト、エム.(Nishimoto,M.)ら、前掲;トクザワ、ワイ.(Tokuzawa,Y.)ら、前掲;トミオカ、エム.(Tomioka,M.)ら、前掲;チュー、ジェイ.エル.(Chew,J.−L.)ら、2005年、Mol.Cell.Biol.第25巻、第14号、p.6031〜6046;オクムラ‐ナカニシ、エス.(Okumura−Nakanishi,S.)ら、2005年、J.Biol.Chem.第280巻、p.5307〜5317)である。本願発明者は、nanogプロモータ内の各複合要素を同定した(たとえば、図5Aを参照)。いくつかの実施形態において、複合結合要素はoct4/Sox2結合部位である。oct4/Sox2結合部位内では、oct4およびSox2に対する結合領域は、Sox2およびoct4の両者が同時に該複合結合部位に結合できさえすれば、任意の配向、かつ任意の相対位置に配置されていてよい。2つの領域は、たとえば、互いに直接隣接していてもよい(図5Aを参照)。
【0031】
(0039)本発明のいくつかの実施形態において、nanog遺伝子のプロモータ領域内の結合要素と、少なくとも2つの転写因子またはその機能的断片との間での複合体の形成も検出される。この目的のために、核酸へのタンパク質の結合を検出するのに十分な感度をもつ任意の方法を用いることができる。このような方法は、たとえば、分光学的、光化学的、光分析的、蛍光分析的、放射線学的、酵素的もしくは熱力学的手段、または細胞効果を利用するものである。分光学的検出方法の例としては、蛍光相関分光法(トンプソン、エヌ.エル.(Thompson,N.L.)ら、2002年、Curr.Opin.Struct.Biol.第12巻、第5号、p.634〜641)が挙げられる。光化学的方法は、たとえば、光化学的架橋である(スティーン、エイチ.(Steen,H.)、ジェンセン、オー.エヌ.(Jensen,O.N.)、2002年、Mass.Spectrom.Rev.第21巻、第3号、p.163〜182)。光活性、蛍光、放射活性、または酵素的ラベルの使用(総覧として、リッペ、アール.エイ.(Rippe R.A.)ら、2001年、Methods Mol Biol.第160巻、p.459〜479を参照のこと)は、それぞれ光分析、蛍光分析、放射線学および酵素による検出法に対する例である。熱力学的検出方法の例は、等温滴定熱量分析(ITC、総覧として、ベラツケツ‐カンポイ、エイ.(Velazquez−Campoy,A.)ら、2004年、Methods Mol Biol.第261巻、p.35〜54を参照のこと)である。任意の細胞効果、たとえば、表現型の変化などは、nanog遺伝子プロモータ領域制御下の組換え因子の発現によって引き起こされうる。細胞効果を用いた方法の例には、マーカータンパク質のパターンの判定を含む、細胞分化状態の測定が含まれていてもよい(たとえば、ノークソン、ケイ.(Noaksson,K.)ら、2005年、Stem Cell Express doi:10.1634/stemcells.2005−0093を参照のこと)。これらの方法のいくつかは、電気泳動またはHPLCなどの追加の分離技術を含んでいてもよい。詳細には、ラベルの使用についての例としては、限定はされないが、プローブとしての化合物、または触媒する反応が検出可能なシグナルをもたらすような酵素を付加した免役グロブリンが挙げられる。放射活性ラベルおよび電気泳動による分離を用いた方法の例としては、電気泳動移動度シフトアッセイが挙げられる。
【0032】
(0040)したがって、nanog遺伝子の結合要素と、少なくとも2つの転写因子またはその機能的断片との間の複合体の形成を検出する例としては、限定はされないが、免役沈降(もしくはウェスタンブロットハイブリダイゼーション)またはクロマチン免疫
沈降アッセイ、電気泳動移動度シフトアッセイ、あるいは表面プラズモン共鳴などの結合要素の同定に適した例も挙げられる。
【0033】
(0041)いくつかの実施形態において、少なくとも2つの転写因子が結合するそれぞれのnanog遺伝子の領域は、ヒトnanog遺伝子配列の位置−289〜+117を含むヒトnanog遺伝子領域に相当する領域内に位置する。一実施形態において、この領域は、ヒトnanog遺伝子配列の位置−212〜−119を含むヒトnanog遺伝子領域に相当する。
【0034】
(0042)上記詳述したように、本発明の方法において、複合体は、少なくとも2つの転写因子またはその機能的断片と、nanog遺伝子のプロモータ領域内の結合要素との間で形成される。この複合体の形成は、さらに、nanog遺伝子の発現を調節する。この効果は、転写活性化を調節することによって達成される。このように、少なくとも2つの転写因子の結合が、nanog遺伝子の転写を増大させる。nanog遺伝子の転写の増大によって、ESCの多分化性細胞状態が維持されることがわかっている(前掲)。nanog遺伝子の転写の抑制により、ヒトECおよびヒト胚性癌細胞における分化が誘導されることがわかっている(図1Eおよびヒスロップ、エル.(Hyslop,L.)ら、2005年、Stem Cell Express doi:10.1634/stemcells.2005−0080を参照のこと)。
【0035】
(0043)本発明のいくつかの実施形態において、nanog遺伝子の転写の増大は、nanog遺伝子発現に換算して測定される。これは、たとえば、nanogプロモータの制御下にある遺伝子から転写されたRNA分子の数を測定することによって行える。当分野で一般的に用いられる方法は、RNA分子転写後の逆転写酵素を用いたRNAからcDNAへのコピーと、該cDNA分子の蛍光染料とのカップリングである。分析は典型的には、DNAマイクロアレイの形で行われる。アフィメトリクス(Affymetrix)製のGeneChip(登録商標)発現アレイといった数々のサービスやキットが市販されている。nanog遺伝子の発現を判定するための他の手段としては、限定はされないが、オリゴヌクレオチドアレイ、および定量的リアルタイムポリメラーゼ連鎖反応(RT−PCR)が挙げられる。
【0036】
(0044)実施例5では、nanog遺伝子発現のさらなる判定手段であるルシフェラーゼレポーターベクターの使用について説明する。この方法において、発現レベルは、nanogプロモータを含むベクターを発現している細胞のルシフェラーゼ活性に反映される。ルシフェラーゼ活性は、市販のキットを用いて照度計で検出することができる(たとえば、実施例5を参照)。
【0037】
(0045)いくつかの実施形態において、nanog遺伝子発現データを較正するか、ランク付けすることが有益または望ましいかもしれない。このように、いくつかの実施形態において、本発明の方法は、得られた結果を、1つまたはそれ以上の対照測定の結果と比較することをさらに含む。
【0038】
(0046)このような対照測定は、主要な測定自体とは異なる任意の条件を含んでいてもよい。たとえば、nanogプロモータの制御下での全く発現が起こらない条件下、または転写因子とnanogプロモータとの間の複合体形成が生じ得ないか、複合体形成の調節ができないような条件下での方法の条件が含まれる。いくつかの実施形態において、nanogプロモータの活性を所定のレベルに調節する化合物を使用することを含んでいてもよい。他の実施形態において、各化合物は、nanogプロモータと2つの転写因子のうちの一方との間での複合体形成を防止するものであってもよい。
【0039】
(0047)対照測定のさらなる手段は、上述の転写因子の一方または両方に結合することができないか、または周知レベルよりも低い親和性または高い親和性で結合する、変異nanogプロモータを使用することである。図5Cは、変異に用いることのできる各部位の同定例である。3つの部位が、変異導入された場合に、多分化性プロモータの機能にルシフェラーゼ活性を野生型構築物の20%以下に低下させるという劇的な影響を及ぼした(図5Cを参照)。これには、oct部位およびsox部位に影響する変異が含まれていた。
【0040】
(0048)本発明の方法のいくつかの実施形態において、2つの転写因子は相乗的に作用して、nanog遺伝子の転写を活性化する。2つの転写因子のそれぞれの相乗作用は、nanog遺伝子の転写に対する個々の転写因子の作用によって得られるデータを、転写に対する2つ(またはそれ以上)の転写因子の作用の組み合わせと比較することによって評定できる。たとえば上述したような、遺伝子発現を判定するのに適した任意の方法をこの目的のために使用することができる。説明的な例として、図5Cは、ルシフェラーゼ活性アッセイ(上記参照)によるnanog遺伝子発現の検出を示している。同一の構築物中でoct部位およびsox部位の両方が変異していると、活性は、(野生型構築物の)約20%の活性に対して、野生型の6%にまで低下する。
【0041】
(0049)多分化性または自己再生特性のうち少なくともいずれか一方を維持するための本発明の方法は、nanogプロモータの結合要素と複合体を形成することのできる、POUドメインおよびPOUホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子を発現できさえすれば、任意の幹細胞、前駆細胞、奇形腫細胞またはそれらに由来する任意の細胞に適している。説明的な例として、任意の多分化性ヒトESCまたはそれぞれの細胞株をそれぞれの方法において用いることができる。そのような細胞の集団を得る手段は、当該技術分野において十分に確立されている(たとえば、トンプソン、ジェイ.エイ.(Thomson,J.A.)ら、1998年、Science 第282巻、p.1145〜1147またはカウアン、シー.エイ.(Cowan,C.A.)ら、2004年、N.Engl.J.Med.第350巻、p.1353〜1356を参照のこと)。さらに、たとえば71の独立したヒトESC株が存在することが知られており、そのうちGE01、GE09、BG01、BG02、TE06またはWA09などの11の細胞株が研究目的で入手可能である(たとえば、NIHヒト胚幹細胞レジストリー(http://stemcells.nih.gov/research/registry/eligibilityCriteria.asp)を参照のこと)。成人幹細胞は、たとえば、分娩後に残った胎盤および臍帯由来の血液から、または成人幹細胞が「衛星細胞」として会合している筋原繊維から、単離することができる(コリンズ、シー.エイ.(Collins,C.A.)ら、2005年、Cell 第122巻、p.289〜301、また、ランド、ティー.エイ.(Rando,T.A.)、2005年、Nature Medicine 第11巻、第8号、p.829〜831も参照のこと)。
【0042】
(0050)本方法が前駆細胞、すなわち成熟体細胞を生じる細胞に対して使用されることが意図される場合、本発明の本方法においては任意の前駆細胞を用いることができる。適切な前駆細胞の例としては、限定はされないが、神経前駆細胞、内皮前駆細胞、赤血球前駆細胞、心臓前駆細胞、オリゴデンドロサイト前駆細胞、網膜前駆細胞、または造血前駆細胞が挙げられる。前駆細胞を得る方法は当該技術分野において周知である。2つの説明的な例として、巨核球前駆細胞を得る方法が米国特許出願第2005/0176142号明細書に開示されており、マウス肝前駆細胞株を得る方法が、リ(Li)ら(2005年、Stem Cell Express,doi:10.1634/stemcells.2005−0108)によって記載されている。いまのところ限られたデータしか入手できないが、前駆細胞は、幹細胞と同様に、POUドメインおよびホメオドメイン含有転写因子ならびにHMGドメイン含有転写因子を発現しているようである。たとえば末
梢血から得られる内皮前駆細胞などの前駆細胞は、NanogやOct−4の高い発現レベルを示すことが発見されている(ロマグナニ、ピー.(Romagnani,P.)ら、2005年、Circ.Res.第97巻、p.314)。網膜前駆細胞などのCNS前駆細胞は、Sox2を高レベルで発現することが報告されている(グラハム、ブイ.(Graham,V.)ら、2003年、Neuron 第39巻、第5号、p.749〜765;クラッセン、エイチ.(Klassen,H.)ら、2004年、J.Neurosci.Res.第77巻、第3号、p.334〜343)。
【0043】
(0051)上に示したように、本発明の1つの方法は、細胞内の遺伝子発現を調節するための方法である。上述の少なくとも2つの転写因子を発現し、かつ機能的nanog遺伝子を含む任意の細胞を用いることができる。いくつかの実施形態において、内在性nanog遺伝子は機能的に活性である。これらの実施形態の一部において、それぞれの細胞は幹細胞または前駆細胞である。本発明の方法において用いることのできる幹細胞の例としては、限定はされないが、胚幹細胞、トロホブラスト幹細胞および胚外幹細胞が挙げられる。本発明の方法のいくつかの実施形態において、ヒト由来のESCなどのESC(胚幹細胞)、すなわちヒトESCを用いることができる。他の実施形態においては、細胞は前駆細胞である(上記参照)。さらに他の実施形態において、細胞はがん細胞である。がん細胞の例示的例としては、奇形腫がん細胞、たとえば、F9、NTERA2、C3H、TES−1、1246(1246−3Aなど)、SuSa(SuSa/DXR10およびSKOV−3/DXR10など)、AT805(ATDC5など)、HTST、HGRT、PC(たとえば、PCC3/A/1)またはGCT27などが挙げられる。がん細胞のさらなる2つの例示的な例としては、HeLa細胞およびMCF−7細胞が挙げられる。いくつかの実施形態において、細胞は幹細胞と体細胞のハイブリッド細胞である。nanog遺伝子は、このようなハイブリッド細胞内で機能的に活性であることが示されている(たとえば、ハタノ(Hatano)ら、2005年、Mech.Dev.第122巻、p.67〜79を参照)。他の実施形態において、内在性nanog遺伝子は機能的に不活性である。これらの実施形態のいくつかにおいて、確立された真核細胞株の任意の細胞、たとえばHEK、COS、CHO、CRE、MT4、DE(アヒル胚)、QF(ウズラ線維肉腫)、NS0、BHK、Sf9、PC12、またはHigh5が選択される。説明的な例は、HEK293T細胞である。さらなる実施形態において、外来性nanog遺伝子は、組換え技術によって、たとえばnanog遺伝子を担持するベクターを用いて導入する(以下も参照されたい)。
【0044】
(0052)いくつかの実施形態において、選択された転写因子またはその機能的断片は、本発明の本方法を行うのに十分な量で内在的に発現される。他の実施形態において、選択された転写因子は、細胞のプロテオーム由来のものはほとんどまたは全く存在しない。いずれの場合も、各転写因子またはその機能的断片を、所望の転写因子をコードする遺伝子を含む1つ以上の組換えベクターを用いて細胞中に導入することができる。細胞が当該転写因子をすでに内在的に発現している場合には、いくつかの実施形態においては、例えばスクリーニングアッセイにおけるシグナル/ノイズ比を改善する目的で、細胞内でのそれぞれの転写ベクターの量を増加させることが望ましいかもしれない。例示的な例として、Oct4および他の転写因子(nanogを含む)が、アデノウィルスベクターを用いてESC中で発現されている(カワタバ、ケイ.(Kawataba,K.)ら、2005年、Mol.Therapy 第12巻、第3号、p.547〜553)。必然ではないが、典型的には、各遺伝子は活性プロモータまたは外部刺激によって都合良く活性化可能なプロモータの制御下におかれる。
【0045】
(0053)目的の細胞内に、各細胞の内在性遺伝子に加えて、nanogプロモータまたは完全なnanog遺伝子のさらなるコピーを含めることが望まれる場合には、これも同様に組換えベクターによって達成することができる(たとえば、カワタバ、ケイ.(
Kawataba,K)ら、前掲)。nanogプロモータ(内在性または外来性のいずれの起源であっても)の活性化を容易にするために、nanogプロモータを含むベクターを細胞内に導入することが有益であるかもしれない。さらに、たとえばタンパク質を得るために、nanogプロモータを用いて外来性遺伝子を発現することが望ましい場合がある。この場合、典型的には、nanogプロモータ制御下に所望の外来性遺伝子を含むベクターが選ばれる。この目的のために、任意の所望の遺伝子の配列を各ベクターに含めることができる。当業者であれば、それぞれ転写されたタンパク質が本発明の方法において用いられる細胞内で翻訳後修飾を受けることが望ましい場合には、さらに酵素を同時発現することが必要なこともあることに気づくであろう。
【0046】
(0054)本方法においてnanogプロモータの制御下で用いることのできる外来性遺伝子の例としては、限定はされないが、レポータ遺伝子、薬剤耐性遺伝子、アポトーシス遺伝子(いわゆる「細胞死」遺伝子)または各細胞内で所望の発現をする任意の他の遺伝子が挙げられる。いくつかの実施形態において、各遺伝子は、目的のタンパク質をコードするものであってもよい。そのような場合、本発明の方法は、各タンパク質を発現して得るために用いてもよい。nanogプロモータの制御下にある遺伝子がアポトーシス遺伝子の場合、本方法は、たとえば、組織から多分化性細胞を除外するために用いることができる。
【0047】
(0055)例示的な例としては、nanogプロモータの制御下にあるアポトーシス遺伝子を含むベクターを組織の細胞内に導入してもよい。組織は、幹細胞/前駆細胞などの多分化性細胞の分化によって得られたものであってもよい。このように本発明の方法は、各組織から、残っている全ての未分化の細胞、たとえば多分化性細胞を除外するために用いることもできる。本実施形態において、本方法は、このような未分化の細胞が生物体に移植されるのを防止するために用いることもできる。したがって、本方法の本実施形態は、たとえば、移植または埋め込み後の奇形腫の発生を防ぐために用いることもできる。さらなる説明的な例として、nanogプロモータによって駆動される薬剤耐性遺伝子(たとえば、抗生物質耐性遺伝子)を含むベクターを、幹細胞/前駆細胞の選択のために用いてもよい。
【0048】
(0056)他の実施形態において、本方法は、上述の少なくとも2つの転写因子またはその機能的断片の、nanogプロモータの結合要素との複合体形成を調節する化合物を各細胞内に導入することをさらに含む。これらの実施形態のいくつかにおいて、本方法は、転写因子、そのヘテロ二量体複合体、またはnanogプロモータの結合要素を、上記のような化合物(すなわち、少なくとも2つの転写因子の、nanogプロモータの結合要素との複合体形成を調節する化合物)と接触させることをさらに含む。
【0049】
(0057)これらの実施形態のうちのいくつかにおいて、本方法は、少なくとも2つの転写因子またはその機能的断片と、nanogプロモータの結合要素との間の上述の複合体の形成を調節する適切な化合物を同定するためのin−vitro方法である。一実施形態において、本発明の方法は、そのような化合物を同定または選択することを目的としたスクリーニング方法として用いられる。そうしたスクリーニング方法は、自動作業ステーションを用いたマルチウェルミクロプレート(たとえば、従来の48ウェル、96ウェル、384ウェルまたは1536ウェルのプレート)上での化合物ライブラリの同時スクリーニングを含んでいてもよい。説明的な例として、上述のようなnanogプロモータ内の結合領域と少なくとも2つの転写因子との間の複合体形成を阻害する化合物を同定することが望ましい場合がある。そのような化合物は、たとえば胚細胞腫瘍を標的とするか、ある種の組織の細胞の均一集団を生じさせるために、たとえば細胞分化を開始または補助することが望まれるかもしれない。このような化合物を、細胞を除去または分化させるために用いることが望ましい場合もある。この特性は、たとえば、上述の少なくとも2
つの転写因子またはその機能的断片と、nanogプロモータの各結合要素との複合体形成の阻害によるものかもしれない(上記参照)。
【0050】
(0058)他の実施形態において、本方法はin−vivo方法であり、本方法において用いられる細胞は、哺乳類または無脊椎動物種、または微生物の一部であるか、これらに含まれる。使用しうる哺乳類の例は、限定はされないが、ラット、マウス、ディンゴ、ウシ、ブタ、ヤギ、チンパンジー、マカウ、およびヒトである。各in−vivo法は、少なくとも2つの転写因子とnanog遺伝子プロモータ内の結合領域との間の上述の複合体の形成を調節する化合物を投与することを含みうる。
【0051】
(0059)さらなる実施形態において、本発明の方法は、哺乳類(たとえば上記)または無脊椎動物において、nanogプロモータ内の結合要素と上述の転写因子またはその機能的断片との間の複合体形成を調節することが判っている化合物を使用することを含む。典型的な実施形態において、上記のような使用には、たとえばヒトなどの哺乳類に投与可能な医薬品または医薬組成物の製造が含まれる。
【0052】
(0060)上述の複合体形成を調節する化合物は、任意の適切な手段によって投与することができる。細胞が哺乳類に含まれるか、その一部である場合には、化合物は非経口投与でも経口(経腸)投与でもよい。哺乳類への投与に対する典型的な実施形態において、投与は、例えば化合物の調製物を経口投与、静脈内投与、または吸入によって、血液および肝臓へ確実に送達できるものとする。経口投与のための調製物の例としては、錠剤、丸剤または飲用液が挙げられ、静脈内投与に対する調製物の例としては、注射液または注入液が挙げられ、吸入による投与に対する調製物の例としては、エアロゾル混合物または噴霧剤が挙げられる。細胞が、微生物に含まれるかその一部である場合、または個々の細胞(たとえば、培養中の組換え細胞)として用いられる場合、投与の例は、化合物を該細胞の環境中に注入または添加することである。個々の細胞を用いる場合には、後者の投与形態が、場合によっては微生物を改変する技術と組み合わせて行われることもある。そのような技術としては、エレクトロポレーションまたは細胞膜の透過化が挙げられる。
【0053】
(0061)本発明のin−vivo方法、またはnanogプロモータ内の結合要素と上述の転写因子との間の複合体形成を調節するための化合物の使用は、様々な目的のために用いることができる。そのような目的の例としては、治療、診断または検査の目的がある。検査目的においては、いくつかの方法は、上述の少なくとも2つの転写因子とnanog遺伝子プロモータ内の結合領域との間の複合体形成を調節することができるとしてすでに同定されている化合物を投与することを含んでいてもよく、他の方法は、そのような化合物の同定を目的としたものであってもよい。治療目的の説明的な例は、奇形腫の治療である。
【0054】
(0062)すでに上に示したように、本発明の本方法は、in vitroおよびin vivoのいずれにおいても、たとえば幹細胞/前駆細胞の分化を開始または補助するためにさらに用いることができる。
【0055】
(0063)上述の複合体形成を調節するために用いられる化合物は、任意の性質を有するものであってよい。たとえば、生物供給源もしくは非生物供給源から単離されてもよいし、または化学的もしくは生物工学的に生産されてもよい。そのような化合物の例としては、限定はされないが、有機小分子またはポリペプチドなどの生物活性ポリマー、たとえば、免役グロブリンまたは免疫グロブリン様の機能を有した結合タンパク質、またはオリゴヌクレオチドが挙げられる。
【0056】
(0064)各化合物の例示的実施形態は、核酸のメチル化状態を変化させる分子、ま
たは、POUドメインおよびホメオドメインを含む転写因子および/またはHMGドメインを含む転写因子のプロモータのメチル化状態を調節する化合物である。説明的な例として、脱メチル化分子5−アザシチジンが、Sox2およびOct4の発現を増加させることが発見されている(ツジ‐タカヤマ(Tsuji−Takayama)ら、2004年、Biochem.Biophys.Res.Commun.第323巻、p.86〜90)。
【0057】
(0065)上述の複合体形成を調節するために用いられる化合物の別の実施形態は、複合体に参加している転写因子の少なくとも一方の翻訳後修飾のパターンを変化させる分子である。転写因子の活性は、リン酸化、アセチル化またはユビキチン化を含むいくつかの翻訳後修飾によって調節されていることが知られている。
【0058】
(0066)このような化合物のさらなる実施形態は、核酸分子である。本明細書中で用いる「核酸分子」という用語は、一本鎖、二本鎖またはその組み合わせなど、任意の可能な立体構造にある任意の核酸のことをさす。核酸には、たとえば、DNA分子(たとえばcDNAまたはゲノムDNA)、RNA分子(たとえばmRNA)、ヌクレオチドアナログを用いて、または核酸化学を用いて作製されたDNAまたはRNAのアナログ、およびPNA(タンパク質核酸)が含まれる。DNAまたはRNAは、ゲノム由来でも合成されたものでもよいし、一本鎖または二本鎖のいずれであってもよい。本発明の本方法において、必然ではないが、典型的にはRNA分子またはDNA分子が用いられる。このような核酸は、たとえば、mRNA、cRNA、合成RNA、ゲノムDNA、cDNA、合成DNA、DNAとRNAのコポリマー、オリゴヌクレオチドなどであってよい。各核酸は、非天然のヌクレオチドアナログをさらに含んでいてもよいし、かつ/またはアフィニティタグもしくはラベルが結合されていてもよい(上記参照)。
【0059】
(0067)多くのヌクレオチドアナログが知られており、本発明の本方法において用いられる核酸およびオリゴヌクオチドにおいて使用することができる。ヌクレオチドアナログは、たとえば塩基、糖、またはリン酸部分に修飾を含むヌクレオチドである。塩基部分の修飾には、A、C、G、およびT/Uの天然または合成の修飾体、別のプリンまたはピリミジン塩基(たとえばウラシル−5−イル、ヒポキサチン−9−イル、および2−アミノアデニン−9−イル)、ならびに非プリンまたは非ピリミジンヌクレオチド塩基が含まれる。他のヌクレオチドアナログは、ユニバーサル塩基として機能する。ユニバーサル塩基には、3−ニトロピロールおよび5−ニトロインドールが挙げられる。ユニバーサル塩基は任意の他の塩基と塩基対を形成することができる。塩基修飾は、二本鎖の安定性を高めるなどの固有の特性を達成するために、たとえば糖修飾(2’−O−メトキシエチルなど)と組み合わせることができる場合が多い。
【0060】
(0068)本発明の本方法のいくつかの実施形態において、核酸分子は、アプタマー、Spiegelmer(登録商標)(国際公開公報第01/92655号パンフレットに記載)、ミクロRNA(miRNA)分子、短鎖ヘアピンRNA(shRNA)分子、または短鎖干渉核酸(siNA)分子、たとえば、短鎖干渉RNA(si−RNA)分子もしくは反復関連短鎖干渉RNA(rasiRNA)分子である。
【0061】
(0069)短鎖干渉RNAの使用は、特定の遺伝子を「ノックダウン」するためのツールとなっている。これは、転写後レベルで起こり、mRNAの分解を伴うRNA干渉(RNAi)を介した、遺伝子のサイレンシングまたは遺伝子抑圧を利用する。RNA干渉は、ゲノムを保護する細胞機構に相当する。siRNA分子は、該siRNAが多酵素複合体と会合していわゆるRNA誘導サイレンシング複合体(RISC)を形成することにより、該siRNAに相補的なRNAの分解を媒介する。siRNAは、RISCの一部となって相補的RNA種を標的とし、次いで相補的RNAが開裂される。これが、各遺伝
子の発現の損失をもたらす(簡単な概説として、ザモア、ピーディー(Zamore,PD)、ハーレイ、ビー(Haley,B)、2005年、Science 第309巻、p.1519〜1524を参照されたい)。この技術は、たとえば、米国特許出願第2005/0191618号明細書に開示されるように、寄生DNA配列のサイレンシング、たとえばHIV RNAの開裂に応用されてきた。
【0062】
(0070)本発明に対するこのようなsiRNAの典型的な実施形態は、10〜35ヌクレオチド、一部の実施形態においては15〜25ヌクレオチドのin vitroまたはin vivoで合成された分子を含む。転写によって(たとえば、生きた細胞の転写機構を用いて)si−RNA分子を生産するために用いることのできる核酸配列の説明的な2例が、配列番号31および配列番号32である(実施例6を参照)。各si−RNA分子を対象とする細胞(微生物および動物の一部である細胞を含む)内で直接合成してもよい。また、各細胞に導入してもよいし、かつ/または該細胞に送達してもよい。siRNA分子をin vivoで選択された細胞に送達する説明的例は、重鎖抗体断片(Fab)と核酸結合タンパク質プロタミンとの融合タンパク質にsiRNA分子を非共有結合により結合させることである。(ソング、イー.(Song,E.)ら、2005年、Nature Biotech.第23巻、第6号、p.709〜717)。本発明の一実施形態では、siRNA分子を用いて、1つまたはそれ以上のPOUドメインおよびホメオドメイン含有転写因子および/または1つまたはそれ以上のHMGドメイン含有転写因子をコードするmRNA分子の分解を誘導する(たとえば、実施例6参照)。
【0063】
(0071)本発明の方法は、nanog遺伝子上の領域に特異的に結合することのできるさらなる転写因子を使用することを含んでいてもよい。したがって、いくつかの実施形態において、該方法は、少なくとも第3の転写因子をnanog遺伝子と接触させることを含む。このような転写因子の例としては、限定はされないが、SMADタンパク質ファミリーのメンバー(たとえば、SMAD1、SMAD2、SMAD3、SMAD4、SMAD5、SMAD7、およびSMAD9、またはキツネの条虫である多包条虫(Echinococcus multilocularis)由来のSmadAおよびSmadB)、AP1(アクチベータタンパク質1)ファミリーのメンバー、hand1(心臓および神経冠誘導体発現タンパク質1)転写因子またはhand1関連転写因子が含まれる。
【0064】
(0072)本発明を、以下の非限定的な実施例によってさらに説明する。
【実施例】
【0065】
配列解析およびプロモータ構築物
(0073)すべての配列は、www.ncbi.nlm.nih.govおよび/またはwww.ensembl.org.にある公的に利用可能なデータベースから得た。ウシ(Bos taurus)およびアフィリカゾウ(Loxodonta africana)のNanogプロモータ領域は、NCBIにおいて入手可能なトレースファイル配列から構築した。ウェブ上(www-gsd.lbl.gov/vista )のVistaゲノム配列アラインメントツール(メイヤー(Mayor)ら、2000年)を用いて、マウスとヒトのNanogゲノム配列を比較した。マウスnanogおよびヒトNanogおよびNanogGP1に対するプロモータ領域を適当な生物種のゲノムDNAから増幅した。増幅のためのプライマーは、クローニング用の制限酵素切断部位(小文字で示す)を備えた下記の
nanog順方向 5’−CGCgtcgacTAAAGTGAAATGAGGTAAAGCC−3’(配列番号1)、
nanog逆方向 5’−CGCggatccGGAAAGATCATAGAAAGAAGAG−3’(配列番号2)、
Nanog順方向 5’−CGGctcgagTTGCTCGGTTTTCTAGTTC
C−3’(配列番号3)、
Nanog逆方向 5’−CGGctcgagCAAGAAATTGGGATAAAGTGAG−3’(配列番号4)、
NanogGP1順方向 5’−CGGctcgagTTGCTCGGTTTTCTAGTTCC−3’(配列番号5)、
NanogGP1逆方向 5’−CGGctcgagCAAGAAGTTGTGATGAAGTGAG−3’(配列番号6)
とした。マウスnanogプロモータ断片を、pGL3−Basicベクター(プロメガ(Promega))およびpEGFP1ベクター(クロンテック(Clontech))の両方にクローニングし、ヒトプロモータはpGL3−Basicだけにクローニングした。すべての構築物について配列確認を行った。
【0066】
胚幹細胞培養およびレポータの系
(0074)E14マウスESCを、ダルベッコ改変イーグル培地、20%ウシ胎仔血清、1×非必須アミノ酸、0.1mMの2−メルカプトエタノール、および組換えLIF調整培地のアリコート中で培養した。標準的なプロトコールを用いてNanogEGFP構築物でESCを安定的にトランスフェクトし、ネオマイシン耐性マウス胚線維芽細胞上で培養した細胞を用いて300μg/mLのG418で10日間選択した後に、個々のコロニーを採取した。ESCを、LIF調整培地の回収、胚様体への自発的分化、またはレチノイン酸の添加により分化させた。蛍光活性化細胞選別(FACS)は、FACSCalibur(商標)(ビーディーバイオサイエンシズ(BD Biosciences))で行った。内在性Nanogおよび強化緑色蛍光タンパク質(EGFP)レポータの発現の定量は、ABI PRISM(登録商標)7900配列検出システム(アプライドバイオシステムズ(Applied Biosystems))を用いてリアルタイムの逆転写PCR分析によって行った。この分析のためにProligo(登録商標)により合成されたプライマーとプローブのセットは以下の
Nanog順方向、5’−GGTTGAAGACTAGCAATGGTCTGA−3’(配列番号7)
Nanog逆方向、5’−TGCAATGGATGCTGGGATACTC−3’(配列番号8)
Nanogプローブ、5’−TTCAGAAGGGCTCAGCACCA−3’(配列番号9)
EGFP順方向、5’−CGACAACCACTACCTGAGCAC−3’(配列番号10)
EGFP逆方向、5’−TCGTCCATGCCGAGAGTGAT−3’(配列番号11)
EGFPプローブ、5’−CGGCGGCGGTCACGAACTCCAGC−3’(配列番号12)
である。発現はa−アクチン対照(アプライドバイオシステムズ(Applied Biosystems))に対して正規化した。ヒトESC株HUES−6(ハーバード大学のダグ・メルトン氏(Doug Melton)より入手)は、カウアン(Cowan)ら(2004年、N.Engl.J.Med.第350巻、p.1353〜1356)に従って継代した。
【0067】
実施例1:Nanogのダウンレギュレーション効果の分析
(a)RNAi分析
(0075)本実施例では、nanog遺伝子活性のダウンレギュレートによる細胞効果をどのようにして分析するかについて説明する。実施例6(下記参照)に示すのと同様の方法から、NanogのダウンレギュレーションがESCの分化に繋がることが判った。
【0068】
(0076)RNAiのための遺伝子特異的オリゴヌクレオチドを、レイノルド(Reynolds)らの文献(2004年)およびウイ‐タイ(Ui−Tei)らの文献(2004年)にしたがって設計した。9ヌクレオチドのループを有する19ヌクレオチドのヘアピン型shRNAsを、pSUPER.puro(BglIIおよびHindIII部位、オリゴエンジン(Oligoengine))中にクローニングした。使用したオリゴヌクレオチドは以下の通り、すなわち
GFP RNAi(対照)用として、
5’−GATCCCCGAACGGCATCAAGGTGAACTTCAAGAGAGTTCACCTTGATGCCGTTCTTTTTA−3’(配列番号13)および
5’−AGCTTAAAAAGAACGGCATCAAGGTGAACTCTCTTGAAGTTCACCTTGATGCCGTTCGGG−3’(配列番号14)
Nanog RNAi用として、
5’−GATCCCCGAACTATTCTTGCTTACAATTCAAGAGATTGTAAGCAGAATAGTTCTTTTTA−3’(配列番号15)および
5’−AGCTTAAAAAGAACTATTCTTGCTTACAATCTCTTGAATTGTAAGCAAGAATAGTTCGGG−3’(配列番号16)
とした。E14ES細胞を、50%コンフルエントの状態でshRNAプラスミドおよびGFPプラスミドで同時にトランスフェクションした。
【0069】
(b)ウェスタンブロット分析
(0077)NanogRNAiが、Sox2またはOct4などの他の転写因子に影響を与えることなくNanog発現を特異的にダウンレギュレートするかどうかを調べるために、SDS−PAGEおよび続くウェスタンブロットによってタンパク質レベルを分析した。この2つの方法は当業者に周知である。Nanog発現ベクターを、対照、Nanog、Oct4またはSox2のsiRNAを発現する構築物とともに、293T細胞に同時トランスフェクトした。細胞溶解物のウェスタンブロット分析は、抗Nanog抗体または抗βアクチン抗体を用いて行い、βアクチンを装荷量の対照とした。
【0070】
(0078)タンパク質を12%SDS−PAGEゲルで分離し、当該技術分野においてよく確立されている標準的な方法によってImmobilon(商標)Pブロッティング膜(ミリポア(Millipore))に移した。膜をPBS+0.1%tween20および5%ミルクでブロッキングし、特異的抗体をプローブとして調べた。その後、膜をセイヨウワサビペルオキシダーゼ共役二次抗体とともにインキュベートした。このブロットをECL Advance(商標)ウェスタンブロッティング検出キット(Amersham)によって現像した。
【0071】
(0079)図1Aに示すように、Nanog siRNAだけが、Nanogタンパク質レベルを低減することができた。対照RNAi、Oct4 RNAiおよびSox2
RNAi(実施例6参照)は、Nanogの発現に影響を及ぼさなかった。このNanog siRNAは、同時発現させたSox2またはOct4のレベルに影響を及ぼさなかった(図1B、Cを参照)。これらのデータから、Nanog siRNAがNanogに対して特異的であり、POUドメインおよびホメオドメイン含有転写因子ならびにHMGドメイン含有転写因子に対するモデルタンパク質として選ばれた他の2つの転写因子には影響を及ぼさないことがわかった。
【0072】
(0080)ES細胞を、NanogまたはGFPのsiRNAを発現する構築物でトランスフェクトした(下記も参照のこと)。トランスフェクトされた細胞をピューロマイシンで選択し、ウェスタンブロットから、Nanogのレベルが大きく減少、すなわち、検出限界のタンパク質レベルまで減少したことがわかった(図1D)。
【0073】
(0081)さらに、未分化ES細胞の典型的な形態が、Nanogノックダウン後に失われていた(図1E)。各写真を撮るために従来の顕微鏡を用いた。アルカリホスファターゼ染色もNanogノックダウン細胞において劇的に減少していた。
【0074】
実施例2:NANOGと、(POU)特異的ドメインおよびホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子との間の、in vitroでの複合体形成についての検証
(0082)当該技術分野において電気泳動移動度シフトアッセイ(EMSA)として知られるアッセイを用いて、Oct4およびSox2が、nanogプロモータ内の要素と複合体を形成することができるかを判定した。
【0075】
(0083)この目的のために、核抽出物を、ディグナム(Dignam)ら(1983年、Nucleic Acids Res.第11巻、p.1475〜1489)の方法を次のように改変したものを用いて、マウス胚線維芽細胞フィーダー上で成長させたE14マウス胚幹細胞から調製した。すなわち、細胞を洗浄し、PBS中で掻き取ることにより回収し、ペレットの5倍容のバッファA(10mM Hepes(pH7.9)、1.5mM MgCl2、10mM KCl、0.5mM DTT、1%プロテアーゼインヒビターカクテル(シグマ(Sigma)))中に再懸濁し、氷上で10分間インキュベートした。細胞を遠心分離し、ペレットの2倍容のバッファAに再懸濁し、Dounceホモジナイザー(タイプBペッスル)を20ストロークとして細胞膜を溶解した。遠心分離によって核を回収し、核ペレットの0.6倍容のバッファC(20mM Hepes(pH7.9)、25%グリセロール、420mM NaCl、1.5mM MgCl2、0.2mM EDTA、0.5mM DTT、1%プロテアーゼインヒビターカクテル)に再懸濁してから、回転させながら4℃で30分間インキュベートした。次に核を13000rpmで5分間遠心分離し、上清を回収して透析バッファ(20mM Hepes(pH7.9)、20%グリセロール、100mM KCl、0.83mM EDTA、1.66mM DTT、1%プロテアーゼインヒビターカクテル)に対して4℃で2時間透析した。抽出物は−80℃で保存した。
【0076】
(0084)EMSAは、DNAプローブとして、NOSと呼ばれる推定上の要素と重複するnanogプロモータ由来の配列を含む37塩基対の二本鎖オリゴヌクレオチドを用いて行った。(POU)特異的ドメインおよびホメオドメインを含む転写因子としてSox2を選び、HMGドメインを含む転写因子としてOct4を選んだ。陽性対照として、FGF4エンハンサ由来の既知のOct4/Sox2複合結合部位(FOS)を用いた。これはnanog要素とは違って、8量体とHMGモチーフとの間に3塩基対の間隔があいている。
【0077】
(0085)両鎖の5’末端がcy5によって標識された二本鎖DNAオリゴヌクレオチドを用いた。一本鎖の5’標識オリゴヌクレオチドをプロリゴ(Proligo)より購入し、95℃から室温に徐々に冷却することにより、相補的な5’標識オリゴヌクレオチドにアニールさせた。使用したオリゴヌクレオチドの配列は以下のとおり、すなわち
FOS、5’−TTTAAGTATCCCATTAGCATCCAAACAAAGAGTTTTC−3’(配列番号17);
NOS、5’−CTTACAGCTTCTTTTGCATTACAATGTCCATGGTGGA−3’(配列番号18);
NmOS、5’−CTTACAGCTTCTTTCAAATTACAATGTCCATGGTGGA−3’(配列番号19);
NOmS、5’−CTTACAGCTTCTTTTGCATTAACCTGTCCATGGTGGA−3’(配列番号20);
NmOmS、5’−CTTACAGCTTCTTTCAAATTAACCTGTCCATGGTGGA−3’(配列番号21);
NNS、5’−CTGCAGGTGGGATTAACTGTGAATTCA−3’(配列番号22)
である(図4Aを参照)。DNA結合反応のためには、2μl(約16μg)の核抽出物を、50nM二本鎖標識オリゴヌクレオチドと5μgのポリdGdC(アマシャム(Amersham))とを含む反応物に添加して反応体積10μl(最終)とした。最終的な結合バッファ組成は、60%透析バッファとした。核抽出物を加える前に、所定の1μM非標識オリゴヌクレオチド競合物も含めた。結合反応は、室温(RT)で約20分間行った。所定の2μl(抗Oct4および抗JunB)または8μl(抗Sox2および抗Sox4)の抗体を最初の結合反応に続いて付加し、さらに20分間インキュベートした。サンタクルーズ(Santa Cruz)より購入した抗体は以下の通りである。ウサギ抗Oct4ポリクローナル抗体(H−134)sc−9081x;ヤギ抗Sox2ポリクローナル抗体(Y−17)sc−17320;ウサギ抗JunBポリクローナル抗体(N−17)sc−46x;ヤギ抗Sox4ポリクローナル抗体(C−20)sc−17326。結合反応物を、プレラン済みの6%非変性ポリアクリルアミドゲルで、0.5×TBE中、バイオラッド(Bio−Rad)protean(登録商標)II xi装置で300Vにて2時間かけて分離した。ゲルは、モレキュラーダイナミクス(Molecular Dynamics)のTyphoon(商標)9140型画像解析装置を用いて、赤色レーザ(633nm)ならびに、設定を600V、通常感度および焦点面+3mmとしたPMTとともにcy5エミッションフィルタを用いて、ガラス板中で直接撮影した。マウス胚線維芽細胞フィーダーのみについて行ったEMSAでは、有意な移動度シフトは生じなかったことから、それらが観察されるタンパク質DNA複合体に寄与しないことが示された。
【0078】
(0086)最初に、E14マウス胚幹細胞の核抽出物中のDNA結合タンパク質が、NOS要素を認識することができるかどうかを調べた。図2に示すように、EMSAにおいて、NOS要素に結合する4つの主要タンパク質複合体が観察された。FOS要素に結合する類似の移動度を示す4つの複合体が観察されたが、この配列に結合するもう1つの複合体も観察された(レーン12)。さらに、複合体Cは、FOS要素上よりもNOS要素上において僅かに速い移動度を有するようにみえた。
【0079】
(0087)これらのバンドのいずれがOct4またはSox2に相当するかを判定するために、これらのタンパク質に対する抗体を結合反応物に加えることによってスーパーシフトが起こるかどうかを調べた。実際には、抗Oct4抗体を添加した後、複合体のうちの2つ、AおよびCが消失し、標識されたプローブがウェル中に蓄積した(標識Oct4のスーパーシフト)。これは、NOSおよびFOSプローブの両方で観察され(レーン2および13)、いずれの配列上の複合体AおよびCもOct4を含んでいたことがわかった。同様に、抗Sox2抗体の添加により、NOSプローブとの複合体C、およびFOSプローブとの複合体CおよびEが消失した(レーン3および14)。いずれの場合も、より移動の遅い複合体(標識Sox2のスーパーシフト)が観察された。このことは、複合体CおよびEがSox2を含んでいたことを示す。重要なのは、JunBまたはSox4のいずれかに対する抗体を加えてもバンドの移動度に全く影響がなく、Oct4およびSox2抗体の影響が特異的であることが示されたことである。したがって、複合体CがOct4抗体およびSox2抗体の両方によってスーパーシフトしたことから、このバンドは複合要素に結合したこれらのタンパク質のヘテロ二量体を示す。複合体Aは該ヘテロ二量体よりもより迅速に移動し、Oct4抗体によってのみ認識されたことから、この複合体はOct4単量体であることが強く示唆される。同様に、FOS配列上の複合体Eは、ヘテロ二量体よりも速く移動し、Sox2抗体によってのみ認識されたことから、これはSox2単量体に相当することが示された。これらのバンドの相対移動度は、以前に公
表されている、FGF4要素に結合するF9テラトカルシノーマ細胞抽出物中のOct4およびSox2を同定した結果と矛盾するものではない。
【0080】
(0088)現在、複合体BおよびD中の結合因子は同定されていない。複合体Dの有力候補は転写因子Oct1であり、該転写因子Oct1は、FGF4要素内の8量体モチーフを認識し、移動度が同様に遅いことが示されている。さらに、Oct1は、FGF4複合要素上でSox2とヘテロ二量体を形成することが示されている。しかしながら、さらに遅く移動する複合体が無いことから、E14核抽出物においては、少なくともEMSAによって検出されるレベルではヘテロ二量体の形成が起こっていないことが示唆される。
【0081】
(0089)NOSに結合したOct4/Sox2ヘテロ二量体が、FOSに結合したものよりも移動がわずかに速い(レーン1と12の複合体Cを比較)ことは、FOS配列の場合のように8量体モチーフをHMFモチーフから隔てるヌクレオチドが存在するわけではないために、NOS配列上でより緊密なコンフォメーションが生じることもありうるという考えと矛盾しない。このようなコンフォメーションの違いが転写活性の差異や共調節因子の動員の差異をもたらすと考えると面白い。
【0082】
(0090)これらの因子のDNA結合特異性を確立するために、オリゴヌクレオチド競合を行った。いずれの因子によるNOSへの結合も、20倍過剰の同一の非標識NOSを加えることによって強く競合したが(レーン7)、nanogプロモータのすぐ下流の配列を含む非特異的非標識オリゴヌクレオチド(NNS)は、結合に対して有効な競合を示さなかった(レーン11)。これらの結果は、これらの因子のNOSに対する結合が特異的であることを示す。同様に、非標識FOSが標識FOSに対するすべての結合と競合可能であり(レーン17)、NSSでは競合できなかった(レーン22)ことから、FOS配列に対する結合もまた特異的である。さらに、非標識のFOSおよびNOSのいずれも相互に競合可能である(レーン6および18)ことから、0または3塩基対の間隔を有する競合物のいずれが20倍過剰に加えられた場合にも、同等に有効であることがわかる。NOSおよびFOSのいずれの場合も、8量体モチーフ内に3塩基対の置換を有する競合物の存在下で結合が行われる場合、Oct4単量体およびOct4/Sox2ヘテロ二量体による結合に対する競合は起こらなくなるが(レーン8および19)、Sox2単量体に対する競合は依然として起こる(レーン19)。このことは、Oct4単量体およびOct4/Sox2ヘテロ二量体のいずれの結合に対しても完全な8量体モチーフが必要であることを示している。同様に、競合物のHMGモチーフ内を3塩基対置換することにより、NOSおよびFOSに関するOct4/Sox2ヘテロ二量体との競合(レーン9および20)およびFOSに関するSox2単量体との競合(レーン20)が消失する。いずれの場合も、Oct4単量体は依然として競合を受けている。このことは、Sox2単量体およびSox2/Oct4ヘテロ二量体の結合には完全なHMGドメインが必要であることを示している。最後に、同じ競合物中の8量体およびHMGモチーフの両方において2塩基対を置換することにより、Oct4単量体およびOct4/Sox2ヘテロ二量体のNOSおよびFOS両者への結合に対する競合が消失し(レーン10および21)、かつSox2単量体のFOSへの結合に対する競合が消失した(レーン21)。図2からわかるように、EMSAは転写因子とNanogとの間の形成を検証する便利な手段である。しかしながら、例えばGSTプルダウンアッセイまたはBiacoreなどの他の手段を利用してもよい。
【0083】
実施例3:NANOGと(POU)特異的ドメインおよびホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子との間のin vivo複合体形成の検証
(0091)in vivoにおけるNanogと転写因子との間の相互作用を分析する方法は、当業者には「クロマチン免役沈降アッセイ」または「ChIPアッセイ」とし
て知られている。
【0084】
(0092)本実施例では、Oct4抗体およびSox2抗体ならびにマウスおよびヒトのESCからの核抽出物とともに、またはレチノイン酸で分化させたマウスESCにおいて、上記方法を使用することについて説明する。
【0085】
(0093)E14マウスESCを用いたChIPアッセイは、既述(ウェルズ、ジェイ.(Wells,J.)およびファーナム、ピー.ジェイ.(Farnham,P.J.)、2002年、Methods(Orlando)第26巻、p.48〜56)のようにして行った。簡潔に述べると、細胞を1%ホルムアルデヒドで10分間室温にて架橋し、125mMグリシンを添加することによってホルムアルデヒドを不活性化した。平均500塩基対の大きさのDNA断片を含んだクロマチン抽出物を、Oct4ポリクローナル抗体(N19)またはSox2ポリクローナル抗体(Y17)(サンタクルーズ・バイオテクノロジー(Santa Cruz Biotechnology))またはSox2ポリクローナル抗体(AB5603)(ケミコン(Chemicon))を用いて免役沈降させた。すべてのChIP実験に対して、定量的PCR分析を、ABI PRISM(登録商標)7900配列検出システム(アプライドバイオシステムズ(Applied
Biosystems))およびSYBR Greenマスターミックス(アプライドバイオシステムズ)を用いて、既述(ン、エイチ.エイチ.(Ng,H.H.)ら、2003年、Mol Cell 第11巻、p.709〜719)のようにリアルタイムで行った。相対占有率は、見かけ上の免役沈降効率(入れた試料の量に対する免役沈降したDNA量の比)を測定し、対照の領域について観察されたレベルを1.0として正規化することによって算出した。プライマー対は以下のとおり、すなわち
1、5’−GGCAAACTTTGAACTTGGGATGTGGAAATA−3’(配列番号23)、
5’−CTCAGCCGTCTAAGCAATGGAAGAAGAAAT−3’(配列番号24);
2、5’−GAGGATGCCCCCTAAGCTTTCCCTCCC−3’(配列番号25)、
5’−CCTCCTACCCTACCCACCCCCTATTCTCCC−3’(配列番号26);
3、5’−GGGTCACCTTACAGCTTCTTTTGCATTA−3’(配列番号27)、
5’−GGCTCAAGGCGATAGATTTAAAGGGTAG−3’(配列番号28);
4、5’−GGTGATACGTTGGCCTTCTAGTCTGAA−3’(配列番号29)、
5’−GGGCAAATTGCAAACTAACTGTATAACCTC−3’(配列番号30)
とした。
【0086】
(0094)フィーダー無しの条件で増殖させた未分化のマウスESCからは、複合oct−sox要素を含むDNA断片が、それぞれOct4抗体およびSox2抗体による免役沈降で43倍および37倍にまで富化された(図3A、アンプリコン2)。oct−sox複合要素を含まない2つの隣接領域には有意な富化はみられなかった(図3A、アンプリコン1および3)。さらに、マウスESCをレチノイン酸によって分化誘導すると、このoct−sox含有断片の富化は、その分化の程度に応じて低減された。分化の3日後、Oct4抗体およびSox2抗体を両方用いた場合にのみ富化は最大でバックグラウンドの10倍にまで達したが、分化の6日後には、有意な富化は検出できなかった(図3A)。Mllに対する抗体を陰性対照として用いた場合、分析した3つのESC状態の
いずれからもアンプリコンのいずれについても有意な富化は見られなかった(データ示さず)。ここで同定された富化のレベルは、EGFPレポータ系を用いた上述の類似の分化プロトコールにおけるNanogプロモータの活性の場合とよく対応している。
【0087】
(0095)OCT4およびSOX2がヒトESCのNANOGプロモータとも相互作用することを検証するために、類似のChIP分析を行ってもよい。本実施例においては、不活性化マウス胚線維芽細胞上で増殖させたヒトESC株HUES−6を用いた。OCT4抗体およびSOX2抗体でクロマチン免役沈降したDNAの富化を検出するために、いずれもNANOGのエキソン1に近接する6つの異なるアンプリコンを用いた(図3B)。複合sox−oct要素と近いほうの3つのアンプリコンだけが、両抗体による有意な富化を示した(図3B)。第2のSox2抗体(Y17)から、類似の結果が得られた(データ示さず)。マウスESCにおいて見られるものと比較して3つすべての抗体について富化率が低い(6倍)のは、未分化なヒトESCを培養中に維持するのがより難しい傾向にあるので、ヒト培養物に分化した細胞がより多く存在することによるのかもしれない。グルタチオンS−トランスフェラーゼ抗体を陰性対照として用いたところ、これらのヒトESC中のアンプリコンのいずれに対しても有意な富化は見られなかった(データ示さず)。まとめると、これらのChIPデータのすべてが、in vivoにおいて、未分化なマウスおよびヒトESC中のNanog/NANOGプロモータ上をOct4/OCT4およびSox2/SOX2が占有することを明確に示している。
【0088】
実施例4:NANOG上の結合要素の同定
(0096)本実施例では、Nanog遺伝子上の結合領域を、その配列保存性に基づいてどのようにして同定できるかを説明する。
【0089】
(a)シス調節情報を含む非コーディング領域の配列比較
(0097)本発明者らは、nanogの高度に保存された結合領域、sox−oct複合要素を同定済みである。マウスおよびヒト遺伝子配列のペアワイズなアラインメントを、エルビーエル・バークレー(LBL Berkley)のVistaオンラインツール(メイヤー、シー.(Mayor,C.)ら、2000年、Bioinformatics 第16巻、p.1046〜1047)を用いて作成し、作成にあたっては、最適なアラインメントの作成時に種特異的な反復要素を考慮した。このアラインメントに対しては、マウスnanogESTの最も5’側の10kb上流から、3’マウスnanogESTの最も3’側の10kb下流までのゲノム領域を選択した。配列類似性のピークがnanogの4つのエキソンに対応して同定されたが、ホメオドメインをコードするエキソン2および3で最も顕著であり、エキソン1で最も顕著でなかった(データ示さず)。非コーディング領域のなかでは、ESTの最も5’側のすぐ上流に、近位プロモータと定義される、有意な配列保存性を有する唯一の領域がみつかった。
【0090】
(0098)非コーディング配列に有意な保存性が認められないことは、多分化性発現を指示するシス調節要素が、すべてこの近位プロモータ領域内にあることを示唆することに他ならない。このことを調べるために、レポータEGFPの発現を駆動するマウス近位プロモータ領域を含むプラスミドベクターで、安定にトランスフェクトされたマウスES細胞株を作成した。406塩基対のプロモータ断片は、優勢な転写開始部位として定義されてきたことから、最も遠い5’共通(public)ESTの位置として同定されている位置に対して−289から+117の配列で構成した(ハート、エイ.エイチ.(Hart,A.H.)ら、前掲)。選択後、これらのneo耐性コロニーの大半が、緑色の蛍光を発し、nanogプロモータのこの領域が多分化性細胞において実際に活性を有することを示した(図4Bを参照)。これらのnanogEGΕVコロニーのいくつかをさらなる分析のために選択した。
【0091】
(0099)このプロモータ断片内の明らかに多分化性ポジティブなシス調節要素に加えて、発明者らは、内在性NanogのmRNAレベルでは起こるような、ES細胞分化時の転写のダウンレギュレーションに十分なシス調節情報を該断片が含んでいるかを判定することに関心を持った。ES細胞分化の強力な誘発剤であるレチノイン酸を0.1μMで使用して分化を誘発させた。この処理により、3日後にEGFP発現が劇的に減少し(図4E−F参照)、分化から4日後までにはFACS分析による測定でEGFPポジティブのままの細胞はわずか9%にすぎなかった(図4Gを参照)。これは同じ分化プロトコールにおける内在性nanogのmRNAレベルと非常に似ている(チャンバース、アイ.(Chambers,I.)ら、前掲)。
【0092】
(0100)より明敏な分化プロトコールにおいても、このプロモータ構築物の活性はやはり内在性Nanog mRNAレベルと類似していた。nanogEGFP細胞をLIFの非共存下で培養したところ、EGFP発現が培養の3日後に顕著に低下し(図4C〜Dを参照)、4日目までには、EGFPポジティブであったのは、フィーダーを含まない未分化nanogEGFP細胞では74%であるのに対して、48%の細胞にすぎなかった(図4Gを参照)。胚様体形成もまた、FACSCalibur(商標)(ビーディーバイオサイエンシズ(BD Biosciences))上でのFACS分析によって測定されるように、EGFP発現のダウンレギュレーションを誘発した。内在性NanogおよびEGFPレポータの発現の定量化は、上述のようなRT−PCR分析によって行った(「胚幹細胞培養およびレポータの系」を参照)。分化の最初の2日の後、EGFP発現は次第に減少し、8日目までには、バックグラウンドレベルの発現しか認められず(図4Gを参照)、これは、内在性Nanog mRNAの発現と類似している(ハート(Hart)ら、2004年)。よってこのnanogの406塩基対断片は、少なくともES細胞を用いるin vitroの系において試験した場合には、内在性nanog発現を再現するのに十分なシス調節情報を含んでいる。
【0093】
(b)系統発生学的フットプリンティングによる結合要素の同定
(0101)広範囲の哺乳類から近位プロモータ領域のアラインメントを作成することにより、同定を強力な系統発生学的フットプリントにすることができる。このアラインメントの構築にあたって使用した5つの種の間の分岐年代は、控えめに見積もっても、マウスとラットの分岐については千2百万年前であり、マウス、ラット、ヒト、ウシおよびゾウの間では六千万年前であるが、この見積もりから、フットプリントを同定するための累積2億5千2百万年の浄化選択が提供される。最も大きい配列保存の長さは、nanog転写開始部位との相対位置−212から−119までに位置する94塩基対領域にわたり、このうち64位置が不変である(図5Aを参照)。
【0094】
(0102)最も長い連続不変配列は16塩基対長であり、面白いことに、この配列の中に、fgf4,utf1,sox2,およびfbx15の多分化性発現において同定された要素もしくは前記発現に対して機能的であることが知られている要素と類似した、15塩基対のoct−sox複合要素(図5Aを参照)がある(ユアン、エイチ.(Yuan,H.)ら、前掲;ニシモト、エム.(Nishimoto,M.)ら、前掲;トミオカ、エム.(Tomioka,M.)ら、前掲;トクザワ、ワイ.(Tokuzawa,Y.)ら、前掲)。sox要素とoct要素の相対位置は、上記5つの遺伝子全部において同じであり、このことは、多分化性細胞においてこれらの要素と結合することが知られている対応する転写因子(Sox2およびOct4)の間の特異的なタンパク質−タンパク質接触を示すものである(レメニ、エイ.(Remenyi,A.)ら、2003年、Genes Dev.第17巻、p.2048〜2059)。utfl,sox2,およびfbx15と同様に、nanog内のsoxシス要素およびoctシス要素は互いに直接隣接しており、fgf4においては該2つの要素は3つの塩基で隔てられている。
【0095】
(0103)nanogプロモータ内の多数の部位におけるこのような強力な浄化選択は、転写の調節におけるこれらの部位の機能的役割を示唆するものである。これらの部位のいくつか、たとえば、oct−sox複合要素などは、多分化性発現において重要でありそうだし、他のものは、nanogの移植後発現において機能するシス調節要素に相当するかもしれない。さらに、マウスESCにおける超並列シグニチャシーケンシング(ウェイ、シー.エル.(Wei,C.L.)ら、2005年、Stem Cells 第23巻、p.166〜185)によって測定されたように、sox−oct複合要素の位置および向きと、対応する遺伝子の発現レベルとの間には、明確な関係はない(表1)。
【0096】
実施例5:NANOGと、(POU)特異的ドメインおよびホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子との複合体によるnanog遺伝子発現の調節
(0104)本実施例では、NANOGと、(POU)特異的ドメインおよびホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子との複合体形成の、多分化性細胞の転写活性化に対する影響の分析について説明する。
【0097】
(0105)上記保存プロモータ内の特定の配列要素の役割を調べるために、一連のルシフェラーゼレポータ構築物を構築し、F9テラトカルシノーマ細胞内にトランスフェクトした。
【0098】
(0106)DNA断片(−289〜+117)は、プライマーpNanogFおよびpNanogRを用いてPCRによりマウスゲノムDNA(2B gDNA)から増幅した(プロメガ(Promega)Pfuポリメラーゼ)。そのSalI/BamHI消化断片をpEGFP−1(ビーディーバイオサイエンシズ・クロンテック(BD Biosciences Clontech))のSalI/BamHI部位にクローニングした。この完全な構築物に続いて、pEGFP−1マルチクローニングサイト由来の22塩基対を含むSacI/BamHI断片をpGL3 Basic(プロメガ)のSacI/BglII部位にクローニングした。得られたプラスミドをシーケンシングにより確認し、pGL3Basic pNanogと名付け、次の突然変異誘発実験に用いた。レポータプラスミドは、Transformer(商標)部位特異的突然変異誘発キット(クロンテック(Clontech))を用いて3塩基対の変異が組み込まれるように改変した。修飾プラスミドはシーケンシングにより確認した。F9細胞(ATCC)を、15%標準ウシ胎仔血清(ハイクローン(Hyclone))および1%pen/strep(ペニシリン‐ストレプトマイシン)を含む高グルコースのダルベッコ改変イーグル培地(Gibco)を用いて培養した。細胞は、5%CO2で37℃に維持した。DNAは、Lipofectamine(商標)2000(インビトロジェン(Invitrogen))を使用し、24ウェル形式の細胞培養に対する同社のプロトコールを用いてF9細胞中にトランスフェクトした。ウミシイタケルシフェラーゼプラスミド(プロメガより入手したpRL−TK)を内部対照として同時トランスフェクトした。24時間後、細胞を溶解し、溶解物のルシフェラーゼ活性を、Dual−Luciferase(登録商標)レポータアッセイシステム(プロメガ)により、Centro LB960型96ウェル照度計(ベルトールドテクノロジーズ(Berthold Technologies))を用いて測定した。少なくとも、トランスフェクションは2回ずつ、2回の独立した状況で行った。レポータプラスミドは、Transformer(商標)部位特異的変異誘発キット(クロンテック)を用いて、3塩基対の変異が組み込まれるように改変し、変異はシーケンシングによって確認した。
【0099】
(0107)nanogEGF ES細胞を作製するために用いた野生型406塩基対プロモータ断片は、予想どおり顕著な転写活性を有し、すべての比較において該活性を100%と設定しポジティブコントロールとして使用した(図5B、2Cを参照)。図5A
中の保存配列のすぐ5’側から始まるこの野生型プロモータのより短いもの(−230〜+63)は、元のプロモータ活性の約130%を維持していた。このことは、この短いプロモータから除外された配列内に潜在的な負の調節要素が含まれていることを示唆するだけでなく、多分化能エンハンサ要素が−230〜+63の領域内に含まれることを示している。
【0100】
(0108)この保存されたモジュールの方向依存性を調べるために、この領域(−289〜−94)が直近位プロモータ(−93〜+117)に対して順方向または逆方向のいずれかの方向の相同な構築物を構築した。野生型活性の79%が順方向構築物において維持され、48%が逆方向構築物において維持されていた。有意な転写活性は、この保存領域を逆向きにした場合にも該領域に維持されていたことから、わずかに低い活性が向きの微妙な影響を示すかもしれないものの、エンハンサ活性は向きに依存しないことが示唆された。
【0101】
(0109)保存領域の機能的重要性の別の尺度として、第2の種からの、すなわち、ヒトnanogを用いて、相同領域を調べた。マウスF9細胞にトランスフェクトされたこの構築物は、マウス転写活性の60%を維持していた(Hs.1、図5Bを参照)ことから、実質的な機能保存が示された。本発明者らは、NanogGPlの、ヒトnanogのタンデム重複コピーからの相同領域についても調べた。おもしろいことに、ヒト多分化性細胞においてこの転写物のレベルは非常に低いにもかからわず、該領域は有意な転写活性を維持しており(Hs.2、図5Bを参照)、このことは、NanogGPlの不活性化変異がシス調節要素中ではなく、コーディング配列中にストップコドン8アミノ酸を生じることが判っているものでありそうなことを示唆している(ブース、エイチ.エイ.(Booth,H.A.)およびホランド、ピー.ダブリュ.(Holland,P.W)、前掲)。
【0102】
実施例6:Nanog活性RNAi分析の調節の分析
(0110)本実施例では、NANOGと、(POU)特異的ドメインおよびホメオドメインを含む転写因子ならびにHMGドメインを含む転写因子とのあいだの複合体形成による、Nanogプロモータの活性調節の分析について示す。本実施例においては、Oct4およびSox2がNanogプロモータの多分化性活性に与える影響を分析した。
【0103】
(0111)この目的のために、RNAi実験を実施して、Oct4またはSox2のレベルを低下させた。この実験で用いたPou5flおよびSox2 RNAi構築物のいずれも、それぞれのmRNAを特異的にノックダウンすることがすでに知られているものである。Nanog−ルシフェラーゼレポータ構築物を、マウスESCにPou5fl、Sox2、GFP、または空のRNAi構築物と同時トランスフェクトし(図6A)、その後ルシフェラーゼ活性のアッセイを行った。
【0104】
(0112)オリゴヌクレオチドを、既述(ブルメルカンプ(Brummelkamp)ら、2002年、Science 第296巻、p.550〜553)のようにして、9ヌクレオチドループを有する19ヌクレオチドのヘアピン型短鎖ヘアピンRNA(shRNA)を発現するpSUPER.puro(BglIIおよびHindIII部位;オリゴエンジン(Oligoengine))にクローニングした。すべての配列について、BLAST検索で解析し、他の遺伝子と有意な配列類似性を有していないことを確認した。E14マウスESCは、トランスフェクションの1日前に、96ウェルプレートに細胞が20,000〜30,000個/ウェルの密度で(1000個/ウェルの初代胚線維芽細胞とともに)播種した。トランスフェクションは、Lipofectamine2000を用いて行った。50ngのNanog−pGL3(ホタルルシフェラーゼ構築物),150ngのpSuperまたはpSuper_RNAi,およびphRLSV40(
ウミシイタケルシフェラーゼを発現する正規化対照)をトランスフェクトした。
【0105】
(0113)トランスフェクションから24時間後に、ピューロマイシン耐性でさらに2日間細胞を選択してから、ルシフェラーゼ活性を測定した。RNAiに用いた配列は以下のとおり、すなわち
Sox2、5’−GAAGGAGCACCCGGATTA−3’(配列番号31);
Oct4、5’−GAAGGATGTGGTTCGAGT−3’(配列番号32)
である。
【0106】
(0114)図6Bに見られるように、ルシフェラーゼ活性は、Pou5flおよびSox2のRNAiによってほとんどバックグラウンドレベルにまで低減されたが、GFPおよび空のRNAi対照では全く影響がなかった。
【0107】
(0115)本明細書で例示的に記載した発明は、本明細書に具体的に開示されていないような任意の要素または限定なしで適切に実施してもよい。したがって、たとえば、用語「〜を含む(“comprising”,“including”,“containing”,など)」という用語は、限定的ではなく開放的に読まれることになる。さらに、本明細書で用いる用語および表現は、限定ではなく説明の用語として用いられており、開示および説明した特徴およびその一部の任意の均等物を除外するような用語および表現を使用する意図は全くなく、特許請求の範囲に記載の本発明の範囲内で様々な修正が可能であるものと理解される。したがって、本発明を説明的実施形態および任意選択の特徴によって具体的に開示してきたものの、本明細書に開示する具現された発明の修正および変更も、当業者によって行使されることができ、そのような修正や変更も本発明の範囲に含まれるとみなされると理解すべきである。
【0108】
(0116)本発明について、広範かつ一般的に本明細書において説明してきた。包括的開示の範囲内に入るより狭い種類のものおよび準包括的な群もまた、本発明の一部を構成する。これには、除外されたものが本明細書に具体的に列挙されているかどうかに関係なく、その分類から任意の特徴を除外する条件または負の限定を有する本発明の包括的説明も含まれる。
【0109】
(0117)他の実施形態は、添付の特許請求の範囲の中にある。さらに、本発明の特徴または態様がマーカッシュ形式で記載されている場合、当業者であれば、本発明が、そのマーカッシュ群の任意の個々のメンバーまたはメンバーのサブグループについても記載されていることを理解できるであろう。
【0110】
【表1】
上記表中、
a参照文献:Fgf4:ハート(Hart)ら、前掲;
Utfl:ニシモト(Nishimoto)ら、前掲;
Sox2:トミオカ(Tomioka)ら、前掲;
Fbxl5:トクザワ(Tokuzawa)ら、前掲;
Nanog:本願発明者;
Pou5fl:本願発明者,オクムラ‐ナカニシ(Okumura−Nakanishi)ら、前掲
b転写開始部位から
cMPSS:超並列シグニチャシーケンシング(ウェイ(Wei)ら、前掲)
dtpm,マウスESCにおける100万あたりのタグ数
【図面の簡単な説明】
【0111】
【図1】NanogRNAiを用いた胚幹細胞(ESC)におけるNanogダウンレギュレーションの効果を示す図。(A)使用したNanogRNAiの特異性について、Nanog発現ベクターを、対照、Nanog,Oct4またはSox2のsiRNAを発現する構築物とともに293T細胞中に同時トランスフェクトすることによって調べた。細胞溶解物を、抗Nanog抗体または抗βアクチン抗体を用いたウェスタンブロットによって分析した。βアクチンは装荷量の対照とした。(B)nanog siRNAは、Sox2の発現に影響を及ぼさなかった。nanogまたは対照のsiRNA構築物を、Sox2発現ベクターとともに293T細胞中に同時トランスフェクトした。溶解物を、抗Sox2抗体または抗βアクチン抗体を用いたウェスタンブロットによって分析した。(C)nanog siRNAは、Oct4の発現に影響を及ぼさなかった。nanogまたは対照のsiRNA構築物を、Oct4発現ベクターとともに293T細胞中に同時トランスフェクトした。溶解物を、抗Oct4抗体または抗βアクチン抗体を用いたウェスタンブロットによって分析した。(D)nanogのノックダウンにより、ES細胞中のNanogが減少した。nanogまたは対照のsiRNA構築物を、ES細胞中にトランスフェクトした。溶解物を、抗Nanog抗体または抗βアクチン抗体を用いて調べた。(E)nanogのノックダウンにより、ES細胞の分化が誘発された。nanogおよび対照のノックダウン細胞をアルカリホスファターゼについて染色した。写真は、対照siRNAでトランスフェクトしたESC(I)およびnanog siRNAでトランスフェクトしたESC(II)を同時に撮影したものである。ノックダウンした細胞の扁平な上皮様細胞の存在は、ベクター対照のES細胞においては全く認められないことに注目されたい。
【図2】POUおよびホメオドメイン含有転写因子ならびにHMGドメイン含有転写因子の、nanogプロモータ内の特異的結合要素との複合体形成を分析するための例示的方法として、電気泳動移動度シフトアッセイ(EMSA)を示す図。本実施例はOct4およびSox2の、Nanog8量体/HMG複合要素への結合を説明する。EMSAは、nanogプロモータ由来の標識した推定Oct4/Sox2要素、または陽性対照としてのFGF4エンハンサ由来の既知のOct4/Sox2結合部位のいずれかを用いて実施した。因子の結合は、E14胚幹細胞の粗製核抽出物中で調べた。Oct4およびSox2を含むタンパク質−DNA複合体は、示したバンドをスーパーシフトさせる抗体の添加によって同定した。結合特異性を、オリゴ競合物(PAGEゲルの一番上に表示)を用いて調べたところ、Oct4の結合には完全な8量体モチーフを必要とし、Sox2の結合には完全なHMG要素を必要とすることが示された。ヘテロ二量化には、両方が完全であることが必要であった。競合物についての記号は次のとおりである。N=Nanog,F=FGF4,O=Oct4結合部位、S=Sox2結合部位、mO=変異型Oct4結合部位、mS=変異型Sox2結合部位、Gbx=非特異的競合物。
【図3】Oct4/OCT4およびSox2/SOX2の、Nanog/NANOGプロモータとのin vivo相互作用を示す図。(A)クロマチン免役沈降(ChIP)は、未分化のマウスESC(第0日、レチノイン酸、RA)およびレチノイン酸(RA)の共存下で3日間および6日間培養したESCについて行った。Oct4(N19)およびSox2(Y17)に対する抗体による免疫沈降を示す。リアルタイムPCRによって測定した富化の倍率を3つの異なるアンプリコンで比較した。このうち1つ(アンプリコン2)は、複合oct−sox要素(概略図では白抜きのボックスで示す)を含み、残りの2つ(アンプリコン1および3)は含まなかった。各アンプリコンの転写開始部位に対する位置が概略図に示されており、Nanogのエキソン1が黒塗りのボックスで示されている。(B)ヒトESCにおけるNANOGのOCT4およびSOX2 ChIP。未分化細胞からの特定のアンプリコンの富化の倍率が示されている。このデータから、複合sox−oct要素に最も近い3つのアンプリコンだけが、使用した両抗体によって富化を示したことがわかる。すべて標準偏差が示されている。
【図4】nanog近位プロモータが、未分化ES細胞においてはEGFP(強化緑色蛍光タンパク質)発現を駆動するが、同細胞の分化誘導体においては駆動しないことを示す図。EGFP発現を駆動するnanogプロモータ(転写開始部位に対して−289〜+117)を含むベクター構築物で安定的にトランスフェクトされたマウスESC株(NanogEGFP)を、異なる培養条件において比較した。光学顕微鏡写真(A,C,E)および蛍光顕微鏡写真(B,D,F)は、未分化のESC(A,B)、LIFなしで3日間培養したESC(C,D)、および0.1μMレチノイン酸(RA)で3日間処理したESC(E,F)のものである。LIFを回収すると、RAへの曝露よりも分化誘発の積極性は小さいが、EGFP発現は大きく減少した。ESC分化の強力な誘発剤であるRAは、EGFP発現を劇的に減少させた。(G)リアルタイムPCRでの検出により、内在性Nanog転写物のレベルを、未分化および分化条件下においてNanogプロモータによって誘導されるEGFPのレベルと比較して示すグラフ。グラフ中の数値は、FACS分析によって測定したEGFP陽性細胞の割合(%)を示す。EBは胚様体であり、RAはレチノイン酸である。
【図5】(A)系統発生学的フットプリンティングによる結合要素の同定の例を示す図。nanog配列は、マウス、ラット、ヒト、ウシおよびゾウのゲノムシーケンシングプロジェクトから得た。2億5千万年以上の累積進化あいだ不変のままである、転写開始位置から−212〜−119の領域に影をつけてある。oct−sox複合要素は、3塩基対の置換変異体として示してあり、該変異体の対応名は下に表示してある。 (B、C)Nanogと、(POU)特異的ドメインおよびホメオドメインを含む転写遺伝子ならびにHMGドメインを含む転写因子との間の複合体形成の影響の分析を示す図。F9テラトカルシノーマ細胞に一時的にトランスフェクトされた様々なnanogプロモータ構築物のプロモータ−ルシフェラーゼレポータアッセイを示している。−289〜+117のマウスnanogプロモータ断片(wt)のルシフェラーゼ活性を任意で100%に設定する。(B)変更を加えたマウスプロモータ構築物の活性を、野生型(wt)、短縮型(short)(短い野生型プロモータ、−230〜+63の領域)と比較したもの。ForおよびRevは、互いに同じであるが、保存領域(−93〜+117)の転写開始位置に対する向きが逆である。Hs.1およびHs.2は、それぞれヒトNanogおよびNanogGPl由来の、マウスwt構築物に相同な領域である。(C)置換変異のマウスプロモータ活性への影響。変異は、oct/sox要素がいずれも変異を有するoct/sox構築物に関して、(A)に示したものに相当する。
【図6】Nanogプロモータ活性に対する、Oct4またはSox2のRNAiノックダウンの結果を示す図。図2Aは、Sox2、Oct4、およびEGFP(対照)を標的とするRNAi構築物の、Nanogプロモータ−ルシフェラーゼレポータ構築物とともにマウスESC中への同時トランスフェクションの概略図である。図2Aは、トランスフェクションから3日後に測定されたルシフェラーゼ活性を示す。(B)ルシフェラーゼ活性を、ウミシイタケルシフェラーゼ内部対照と比較して測定した。阻害性RNAを含まない構築物を陰性対照(−)として用いた。示したとおり、Nanogプロモータ活性は、Sox2およびOct4の両者を標的としたRNAi構築物によって低減される。標準偏差も示してある。
【特許請求の範囲】
【請求項1】
幹細胞または前駆細胞のうち少なくともいずれか一方の、多分化能または自己再生特性のうち少なくともいずれか一方を維持する方法であって、
(a)少なくとも2つの転写因子またはその機能的断片を、nanog遺伝子のプロモータ領域と接触させる工程であり、前記少なくとも2つの転写因子の一方はPOUドメインおよびホメオドメイン含有転写因子から選択され、前記少なくとも2つの転写因子の他方はHMGドメイン含有転写因子から選択される接触工程と、
(b)前記少なくとも2つの転写因子がnanogプロモータ内の特異的結合要素と複合体を形成できるようにする工程であり、前記複合体は転写活性化を介してnanog遺伝子発現を調節することを特徴とする工程と
を含む方法。
【請求項2】
前記POUドメインおよびホメオドメイン含有転写因子のホメオドメインは、POU−ホメオドメインであることを特徴とする請求項1に記載の方法。
【請求項3】
前記複合体形成を検出する工程をさらに含む請求項1または2に記載の方法。
【請求項4】
(c)幹細胞または前駆細胞のうち少なくともいずれか一方におけるnanog遺伝子発現を測定する工程をさらに含む、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
(d)工程(c)において得られた測定結果を、対照測定の結果と比較する工程をさらに含む、請求項4に記載の方法。
【請求項6】
少なくとも2つの転写因子は、nanogプロモータ上でヘテロ複合体を形成することを特徴とする請求項1〜5のいずれか1項に記載の方法。
【請求項7】
ヘテロ複合体は、2つの転写因子からなるヘテロ二量体であることを特徴とする請求項6に記載の方法。
【請求項8】
少なくとも2つの転写因子は相乗的に作用して、nanog遺伝子の転写を活性化することを特徴とする請求項1〜7のいずれか1項に記載の方法。
【請求項9】
POUドメインおよびホメオドメイン含有転写因子はOct4であることを特徴とする請求項1〜8のいずれか1項に記載の方法。
【請求項10】
HMGドメイン含有転写因子はSox2であることを特徴とする請求項1〜9のいずれか1項に記載の方法。
【請求項11】
少なくとも2つの転写因子は、nanogプロモータ内の複合結合要素に結合し、前記POUドメインおよびホメオドメイン含有転写因子は複合結合要素の一方部分に結合し、前記HMGドメイン含有転写因子は複合結合要素の他方部分に結合することを特徴とする請求項1〜10のいずれか1項に記載の方法。
【請求項12】
複合結合要素は2つの部分からなる結合部位であることを特徴とする請求項11に記載の方法。
【請求項13】
複合結合要素は、Oct4/Sox2結合部位であることを特徴とする請求項12に記載の方法。
【請求項14】
Oct4結合部位およびSox2結合部位は互いに直接隣接していることを特徴とする請求項13に記載の方法。
【請求項15】
幹細胞/前駆細胞は、胚幹細胞であることを特徴とする請求項1〜14のいずれか1項に記載の方法。
【請求項16】
胚幹細胞はヒト由来であることを特徴とする請求項15に記載の方法。
【請求項17】
少なくとも第3の転写因子をnanog遺伝子と接触させる工程をさらに含み、前記第3の転写因子は、SMADタンパク質ファミリーのメンバー、AP1ファミリーのメンバー、hand1転写因子およびhand1関連転写因子からなる群より選択されることを特徴とする請求項1〜16のいずれか1項に記載の方法。
【請求項18】
少なくとも2つの転写因子のそれぞれが、ヒトnanog遺伝子配列の位置−289〜+117からなるヒトnanog遺伝子の領域に対応するそれぞれのnanog遺伝子の領域内に位置する結合部位に結合することを特徴とする請求項1〜17のいずれか1項に記載の方法。
【請求項19】
3つの転写因子のそれぞれに対する結合部位が、ヒトnanog遺伝子配列の位置−212〜−119からなるヒトnanog遺伝子の領域に対応するnano遺伝子の領域内に位置することを特徴とする請求項18に記載の方法。
【請求項20】
細胞内の遺伝子発現を調節する方法であって、
(a)少なくとも2つの転写因子またはその機能的断片を、nanog遺伝子のプロモータ領域と接触させる工程であり、前記少なくとも2つの転写因子の一方はPOUドメインおよびホメオドメイン含有転写因子から選択され、前記少なくとも2つの転写因子の他方はHMGドメイン含有転写因子から選択される接触工程と、
(b)前記少なくとも2つの転写因子がnanogプロモータ内の特異的結合要素と複合体を形成できるようにする工程であり、前記複合体は転写活性化を介してnanog遺伝子発現を調節することを特徴とする工程と
を含む方法。
【請求項21】
前記POUドメインおよびホメオドメイン含有転写因子のホメオドメインは、POU−ホメオドメインであることを特徴とする請求項20に記載の方法。
【請求項22】
細胞は、幹細胞または前駆細胞のうち少なくともいずれか一方であることを特徴とする請求項20または21に記載の方法。
【請求項23】
細胞は、胚幹細胞であることを特徴とする請求項20〜22のいずれか1項に記載の方法。
【請求項24】
細胞は、がん細胞であることを特徴とする請求項20または21に記載の方法。
【請求項25】
がんは、奇形腫であることを特徴とする請求項24に記載の方法。
【請求項26】
細胞は、nanogプロモータを含み少なくとも2つの転写因子を発現する組換え細胞であり、少なくとも2つの転写因子の一方はPOUドメインおよびホメオドメイン含有転写因子またはその機能的断片から選択され、少なくとも2つの転写因子の他方はHMGドメイン含有転写因子またはその機能的断片から選択されることを特徴とする請求項20〜25のいずれか1項に記載の方法。
【請求項27】
発現が調節される遺伝子は、レポータ遺伝子、薬剤耐性遺伝子、アポトーシス遺伝子(いわゆる「細胞死」遺伝子)、または各細胞における所望の発現を有する任意の他の遺伝子からなる群より選択されることを特徴とする請求項20〜26のいずれか1項に記載の方法。
【請求項28】
前記転写因子を、前記少なくとも2つの転写因子とnanogプロモータの前記要素との複合体形成を調節する化合物と接触させる工程をさらに含む請求項20〜27のいずれか1項に記載の方法。
【請求項29】
前記少なくとも2つの転写因子またはその機能的断片と、nanogプロモータの前記要素との複合体形成が、核酸分子によって低減されることを特徴とする請求項28に記載の方法。
【請求項30】
核酸分子はRNAまたはDNAであることを特徴とする請求項29に記載の方法。
【請求項31】
核酸分子はアプタマー、ミクロRNA(miRNA)分子、短鎖干渉RNA(si−RNA)分子、または反復関連短鎖干渉RNA(rasiRNA)分子であることを特徴とする請求項30に記載の方法。
【請求項32】
核酸分子は、配列番号31の配列から転写されたsi−RNA分子、または配列番号32の配列から転写されたsi−RNA分子のうち少なくともいずれか一方であることを特徴とする請求項31に記載の方法。
【請求項33】
方法は、前記複合体形成を調節する適切な化合物を同定するためのin vitro方法であることを特徴とする請求項27〜32のいずれか1項に記載の方法。
【請求項34】
前記少なくとも2つの転写因子またはその機能的断片と、nanogプロモータの前記要素との間の複合体形成の阻害により細胞を除去するかまたは分化させるのに有用である潜在的化合物を、in−vitroでスクリーニングするための請求項33に記載の方法であり、自動化作業ステーションを用いたマルチウェルマイクロプレート上での化合物ライブラリの同時スクリーニングを含む方法。
【請求項35】
前記細胞は哺乳類に含まれており、方法はin−vivo方法であることを特徴とする請求項20〜26のいずれか1項に記載の方法。
【請求項36】
哺乳類は、ラット、マウス、ディンゴ、ウシ、ヤギ、ブタ、チンパンジー、マカウおよびヒトからなる群より選択されることを特徴とする請求項35に記載の方法。
【請求項37】
少なくとも2つの転写因子と、nanogプロモータの結合領域との間の前記複合体の形成を調節する化合物を投与することを含む、請求項35または36のいずれか1項に記載の方法。
【請求項38】
奇形腫の治療のための方法であるか、または奇形腫の治療の一部である請求項35〜37のいずれか1項に記載の方法。
【請求項39】
複合体形成を検出することをさらに含む請求項20〜38のいずれか1項に記載の方法。
【請求項40】
(c)細胞内でのnanog遺伝子の発現を測定する工程をさらに含む請求項20〜3
9のいずれか1項に記載の方法。
【請求項41】
(d)工程(c)において得られる測定結果を、対照測定の結果と比較することをさらに含む請求項40に記載の方法。
【請求項42】
検出は、適切な分光学的、光化学的、光分析的、蛍光分析的、放射線学的、酵素的または熱力学的方法によって行うか、または細胞効果に基づいて行うことを特徴とする請求項3または39に記載の方法。
【請求項43】
少なくとも2つの転写因子が相乗的に作用して、nanog遺伝子の転写を活性化することを特徴とする請求項20〜42のいずれか1項に記載の方法。
【請求項44】
POUドメインおよびホメオドメイン含有転写因子はOct4であることを特徴とする請求項20〜43のいずれか1項に記載の方法。
【請求項1】
幹細胞または前駆細胞のうち少なくともいずれか一方の、多分化能または自己再生特性のうち少なくともいずれか一方を維持する方法であって、
(a)少なくとも2つの転写因子またはその機能的断片を、nanog遺伝子のプロモータ領域と接触させる工程であり、前記少なくとも2つの転写因子の一方はPOUドメインおよびホメオドメイン含有転写因子から選択され、前記少なくとも2つの転写因子の他方はHMGドメイン含有転写因子から選択される接触工程と、
(b)前記少なくとも2つの転写因子がnanogプロモータ内の特異的結合要素と複合体を形成できるようにする工程であり、前記複合体は転写活性化を介してnanog遺伝子発現を調節することを特徴とする工程と
を含む方法。
【請求項2】
前記POUドメインおよびホメオドメイン含有転写因子のホメオドメインは、POU−ホメオドメインであることを特徴とする請求項1に記載の方法。
【請求項3】
前記複合体形成を検出する工程をさらに含む請求項1または2に記載の方法。
【請求項4】
(c)幹細胞または前駆細胞のうち少なくともいずれか一方におけるnanog遺伝子発現を測定する工程をさらに含む、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
(d)工程(c)において得られた測定結果を、対照測定の結果と比較する工程をさらに含む、請求項4に記載の方法。
【請求項6】
少なくとも2つの転写因子は、nanogプロモータ上でヘテロ複合体を形成することを特徴とする請求項1〜5のいずれか1項に記載の方法。
【請求項7】
ヘテロ複合体は、2つの転写因子からなるヘテロ二量体であることを特徴とする請求項6に記載の方法。
【請求項8】
少なくとも2つの転写因子は相乗的に作用して、nanog遺伝子の転写を活性化することを特徴とする請求項1〜7のいずれか1項に記載の方法。
【請求項9】
POUドメインおよびホメオドメイン含有転写因子はOct4であることを特徴とする請求項1〜8のいずれか1項に記載の方法。
【請求項10】
HMGドメイン含有転写因子はSox2であることを特徴とする請求項1〜9のいずれか1項に記載の方法。
【請求項11】
少なくとも2つの転写因子は、nanogプロモータ内の複合結合要素に結合し、前記POUドメインおよびホメオドメイン含有転写因子は複合結合要素の一方部分に結合し、前記HMGドメイン含有転写因子は複合結合要素の他方部分に結合することを特徴とする請求項1〜10のいずれか1項に記載の方法。
【請求項12】
複合結合要素は2つの部分からなる結合部位であることを特徴とする請求項11に記載の方法。
【請求項13】
複合結合要素は、Oct4/Sox2結合部位であることを特徴とする請求項12に記載の方法。
【請求項14】
Oct4結合部位およびSox2結合部位は互いに直接隣接していることを特徴とする請求項13に記載の方法。
【請求項15】
幹細胞/前駆細胞は、胚幹細胞であることを特徴とする請求項1〜14のいずれか1項に記載の方法。
【請求項16】
胚幹細胞はヒト由来であることを特徴とする請求項15に記載の方法。
【請求項17】
少なくとも第3の転写因子をnanog遺伝子と接触させる工程をさらに含み、前記第3の転写因子は、SMADタンパク質ファミリーのメンバー、AP1ファミリーのメンバー、hand1転写因子およびhand1関連転写因子からなる群より選択されることを特徴とする請求項1〜16のいずれか1項に記載の方法。
【請求項18】
少なくとも2つの転写因子のそれぞれが、ヒトnanog遺伝子配列の位置−289〜+117からなるヒトnanog遺伝子の領域に対応するそれぞれのnanog遺伝子の領域内に位置する結合部位に結合することを特徴とする請求項1〜17のいずれか1項に記載の方法。
【請求項19】
3つの転写因子のそれぞれに対する結合部位が、ヒトnanog遺伝子配列の位置−212〜−119からなるヒトnanog遺伝子の領域に対応するnano遺伝子の領域内に位置することを特徴とする請求項18に記載の方法。
【請求項20】
細胞内の遺伝子発現を調節する方法であって、
(a)少なくとも2つの転写因子またはその機能的断片を、nanog遺伝子のプロモータ領域と接触させる工程であり、前記少なくとも2つの転写因子の一方はPOUドメインおよびホメオドメイン含有転写因子から選択され、前記少なくとも2つの転写因子の他方はHMGドメイン含有転写因子から選択される接触工程と、
(b)前記少なくとも2つの転写因子がnanogプロモータ内の特異的結合要素と複合体を形成できるようにする工程であり、前記複合体は転写活性化を介してnanog遺伝子発現を調節することを特徴とする工程と
を含む方法。
【請求項21】
前記POUドメインおよびホメオドメイン含有転写因子のホメオドメインは、POU−ホメオドメインであることを特徴とする請求項20に記載の方法。
【請求項22】
細胞は、幹細胞または前駆細胞のうち少なくともいずれか一方であることを特徴とする請求項20または21に記載の方法。
【請求項23】
細胞は、胚幹細胞であることを特徴とする請求項20〜22のいずれか1項に記載の方法。
【請求項24】
細胞は、がん細胞であることを特徴とする請求項20または21に記載の方法。
【請求項25】
がんは、奇形腫であることを特徴とする請求項24に記載の方法。
【請求項26】
細胞は、nanogプロモータを含み少なくとも2つの転写因子を発現する組換え細胞であり、少なくとも2つの転写因子の一方はPOUドメインおよびホメオドメイン含有転写因子またはその機能的断片から選択され、少なくとも2つの転写因子の他方はHMGドメイン含有転写因子またはその機能的断片から選択されることを特徴とする請求項20〜25のいずれか1項に記載の方法。
【請求項27】
発現が調節される遺伝子は、レポータ遺伝子、薬剤耐性遺伝子、アポトーシス遺伝子(いわゆる「細胞死」遺伝子)、または各細胞における所望の発現を有する任意の他の遺伝子からなる群より選択されることを特徴とする請求項20〜26のいずれか1項に記載の方法。
【請求項28】
前記転写因子を、前記少なくとも2つの転写因子とnanogプロモータの前記要素との複合体形成を調節する化合物と接触させる工程をさらに含む請求項20〜27のいずれか1項に記載の方法。
【請求項29】
前記少なくとも2つの転写因子またはその機能的断片と、nanogプロモータの前記要素との複合体形成が、核酸分子によって低減されることを特徴とする請求項28に記載の方法。
【請求項30】
核酸分子はRNAまたはDNAであることを特徴とする請求項29に記載の方法。
【請求項31】
核酸分子はアプタマー、ミクロRNA(miRNA)分子、短鎖干渉RNA(si−RNA)分子、または反復関連短鎖干渉RNA(rasiRNA)分子であることを特徴とする請求項30に記載の方法。
【請求項32】
核酸分子は、配列番号31の配列から転写されたsi−RNA分子、または配列番号32の配列から転写されたsi−RNA分子のうち少なくともいずれか一方であることを特徴とする請求項31に記載の方法。
【請求項33】
方法は、前記複合体形成を調節する適切な化合物を同定するためのin vitro方法であることを特徴とする請求項27〜32のいずれか1項に記載の方法。
【請求項34】
前記少なくとも2つの転写因子またはその機能的断片と、nanogプロモータの前記要素との間の複合体形成の阻害により細胞を除去するかまたは分化させるのに有用である潜在的化合物を、in−vitroでスクリーニングするための請求項33に記載の方法であり、自動化作業ステーションを用いたマルチウェルマイクロプレート上での化合物ライブラリの同時スクリーニングを含む方法。
【請求項35】
前記細胞は哺乳類に含まれており、方法はin−vivo方法であることを特徴とする請求項20〜26のいずれか1項に記載の方法。
【請求項36】
哺乳類は、ラット、マウス、ディンゴ、ウシ、ヤギ、ブタ、チンパンジー、マカウおよびヒトからなる群より選択されることを特徴とする請求項35に記載の方法。
【請求項37】
少なくとも2つの転写因子と、nanogプロモータの結合領域との間の前記複合体の形成を調節する化合物を投与することを含む、請求項35または36のいずれか1項に記載の方法。
【請求項38】
奇形腫の治療のための方法であるか、または奇形腫の治療の一部である請求項35〜37のいずれか1項に記載の方法。
【請求項39】
複合体形成を検出することをさらに含む請求項20〜38のいずれか1項に記載の方法。
【請求項40】
(c)細胞内でのnanog遺伝子の発現を測定する工程をさらに含む請求項20〜3
9のいずれか1項に記載の方法。
【請求項41】
(d)工程(c)において得られる測定結果を、対照測定の結果と比較することをさらに含む請求項40に記載の方法。
【請求項42】
検出は、適切な分光学的、光化学的、光分析的、蛍光分析的、放射線学的、酵素的または熱力学的方法によって行うか、または細胞効果に基づいて行うことを特徴とする請求項3または39に記載の方法。
【請求項43】
少なくとも2つの転写因子が相乗的に作用して、nanog遺伝子の転写を活性化することを特徴とする請求項20〜42のいずれか1項に記載の方法。
【請求項44】
POUドメインおよびホメオドメイン含有転写因子はOct4であることを特徴とする請求項20〜43のいずれか1項に記載の方法。
【図1】

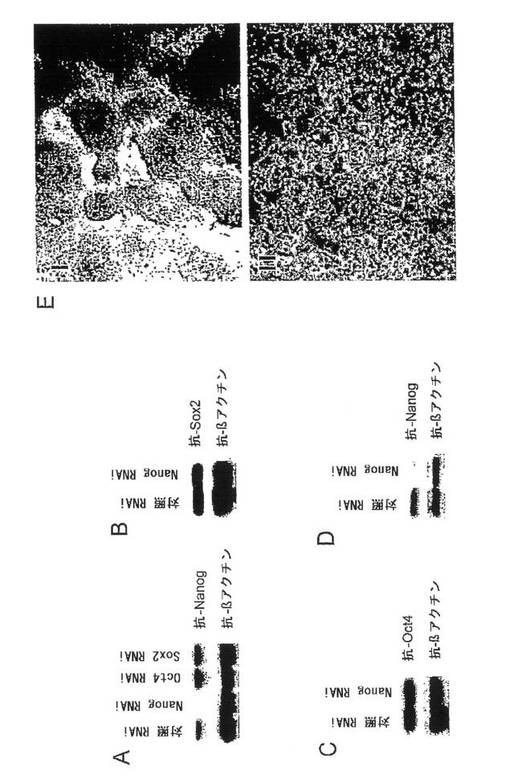
【図2】
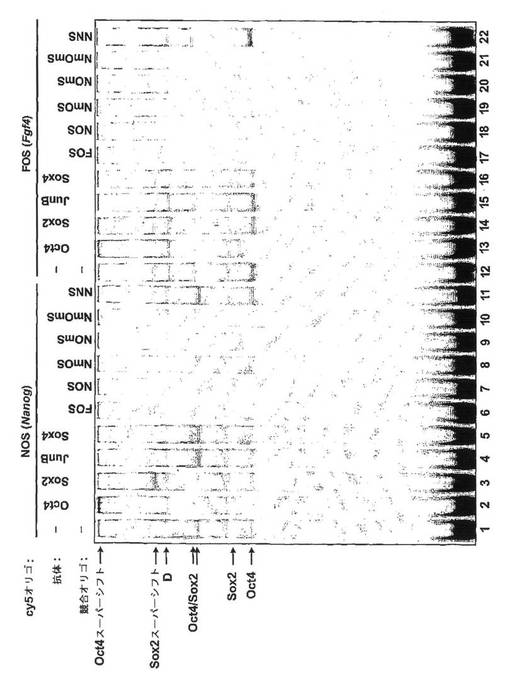
【図3】
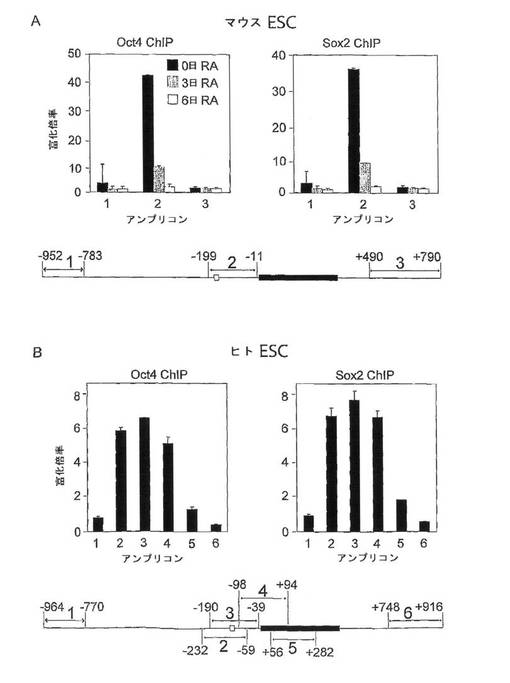
【図4】
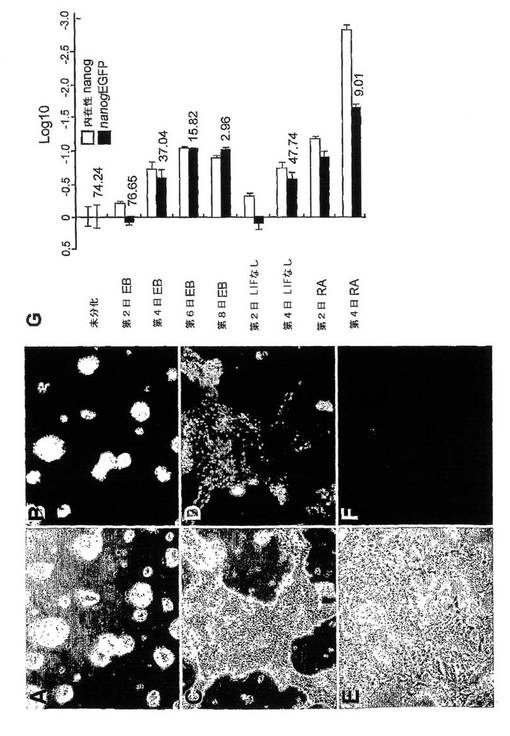
【図5】

【図6】
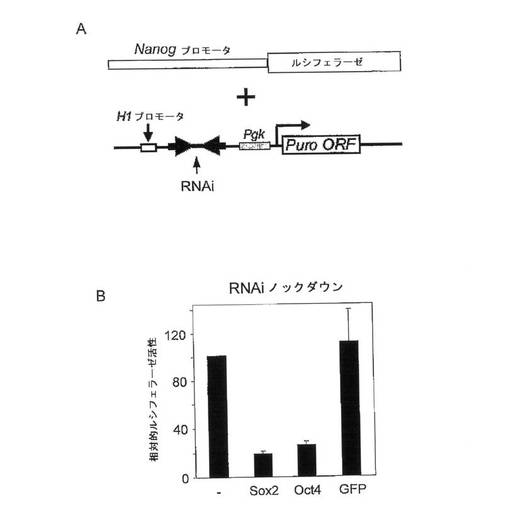
【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2008−511321(P2008−511321A)
【公表日】平成20年4月17日(2008.4.17)
【国際特許分類】
【出願番号】特願2007−529787(P2007−529787)
【出願日】平成17年9月2日(2005.9.2)
【国際出願番号】PCT/SG2005/000304
【国際公開番号】WO2006/025802
【国際公開日】平成18年3月9日(2006.3.9)
【出願人】(506205103)エージェンシー フォー サイエンス,テクノロジー アンド リサーチ (26)
【氏名又は名称原語表記】AGENCY FOR SCIENCE,TECHNOLOGY AND RESEARCH
【Fターム(参考)】
【公表日】平成20年4月17日(2008.4.17)
【国際特許分類】
【出願日】平成17年9月2日(2005.9.2)
【国際出願番号】PCT/SG2005/000304
【国際公開番号】WO2006/025802
【国際公開日】平成18年3月9日(2006.3.9)
【出願人】(506205103)エージェンシー フォー サイエンス,テクノロジー アンド リサーチ (26)
【氏名又は名称原語表記】AGENCY FOR SCIENCE,TECHNOLOGY AND RESEARCH
【Fターム(参考)】
[ Back to top ]