
形質転換体、プラスミドベクター及びイタコン酸の製造方法
【課題】特定のDNAからなるシス−アコニット酸デカルボキシラーゼ(CAD)をコードするDNAを含有するプラスミドベクターが導入された形質転換体、及び、宿主にイタコン酸高生産能を与えるプラスミドベクター、並びに、イタコン酸を効率的に製造することができるイタコン酸の製造方法を提供する。
【解決手段】本形質転換体は、(a)〜(d)のいずれかのDNAからなるCADをコードするDNAを含有するプラスミドが導入されたものであり、宿主は糸状菌である。(a)特定な配列のアミノ酸配列からなるタンパク質をコードするDNA、(b)特定な配列の塩基配列からなるDNA、(c)(b)とは異なる、特定な配列の塩基配列からなるDNA、(d)(b)又は(c)の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA。
【解決手段】本形質転換体は、(a)〜(d)のいずれかのDNAからなるCADをコードするDNAを含有するプラスミドが導入されたものであり、宿主は糸状菌である。(a)特定な配列のアミノ酸配列からなるタンパク質をコードするDNA、(b)特定な配列の塩基配列からなるDNA、(c)(b)とは異なる、特定な配列の塩基配列からなるDNA、(d)(b)又は(c)の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、形質転換体、プラスミドベクター及びイタコン酸の製造方法に関する。更に詳しくは、本発明は、特定のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターが導入された形質転換体、及び、宿主にイタコン酸高生産能を与えるプラスミドベクター、並びに、イタコン酸を効率的に製造することができるイタコン酸の製造方法に関する。
【背景技術】
【0002】
イタコン酸は不飽和基を有する二価の水溶性有機酸であり、その構造から重合や共重合によって線状高分子を形成できる特性がある。また、その安全性も高いことから、合成樹脂や合成繊維、接着剤や印刷用インキ等、工業製品の原料や添加剤として幅広く利用されている。イタコン酸は、Aspergillus terreus、Aspergillus itaconicusのようなアスペルギルス属等の糸状菌や、Candida tsukubaensisのような酵母において生産できることが知られている。イタコン酸の工業生産においてはもっぱら生育速度の観点からA.terreusが使用されている。より効率的なイタコン酸製造のため、イタコン酸高性生産菌の開発が求められているが、イタコン酸高生産菌の育種は、変異原性物質による処理や紫外線照射等の古典的なランダム変異導入法に頼っているのが現状である(特許文献1参照)。
【0003】
Bentleyらは、A.terreusにおいて、クエン酸回路で生成するシス−アコニット酸からシス−アコニット酸デカルボキシラーゼによりイタコン酸が生合成されるという説を提唱した(非特許文献1参照)。また、その後、岡部らにより、A.terreus由来のシス−アコニット酸デカルボキシラーゼタンパク質が単離精製されている(非特許文献2参照)。更に、その後、本願発明者らにより、シス−アコニット酸デカルボキシラーゼをコードするDNAについて単離同定がなされており、このDNAを含むプラスミドを酵母細胞に導入した微生物を用いたイタコン酸の製造方法が提案されている(特許文献2参照)。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特開平6−38774号公報
【特許文献2】特開2009−27999号公報
【非特許文献】
【0005】
【非特許文献1】Bentley,R.and Thiessen C.P.,J.Biol.Chem.226:703−720(1957)
【非特許文献2】Dwiarti L.,Yamane K.,Yamatani H.,Kahar P.,Okabe M.,J.Biosci.Bioeng.94:29−33(2002)
【発明の概要】
【発明が解決しようとする課題】
【0006】
しかしながら、従来の微生物を用いたイタコン酸の製造方法では、イタコン酸の生産効率が未だ十分とはいえず、更なる効率の向上が求められているのが現状である。
本発明は、上記現状に鑑みてなされたものであり、特定のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターが導入された形質転換体、及び、宿主にイタコン酸高生産能を与えるプラスミドベクター、並びに、イタコン酸を効率的に製造することができるイタコン酸の製造方法を提供することを目的とする。
【課題を解決するための手段】
【0007】
本発明は以下の通りである。
[1]宿主に、下記(a)〜(d)のいずれかに記載のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターが導入された形質転換体であって、
前記宿主は糸状菌であることを特徴とする形質転換体。
(a)配列番号1に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号2に記載の塩基配列からなるDNA
(c)配列番号3に記載の塩基配列からなるDNA
(d)配列番号2又は配列番号3に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA
[2]前記宿主がアスペルギルス(Aspergillus)属の糸状菌である上記[1]に記載の形質転換体。
[3]前記宿主がアスペルギルス・テレウス(Aspergillus terreus)である上記[1]に記載の形質転換体。
[4]宿主にイタコン酸高生産能を与えるプラスミドベクターであって、
下記(a)〜(d)のいずれかに記載のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAと、
配列番号4に記載の塩基配列からなるプロモーターDNAと、を含有することを特徴とするプラスミドベクター。
(a)配列番号1に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号2に記載の塩基配列からなるDNA
(c)配列番号3に記載の塩基配列からなるDNA
(d)配列番号2又は配列番号3に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA
[5]更に、配列番号5に記載の塩基配列からなるターミネーターDNAを含有する上記[4]に記載のプラスミドベクター。
[6]上記[1]乃至[3]のいずれかに記載の形質転換体を培養する工程と、
前記形質転換体が生成したイタコン酸を回収する工程と、を備えることを特徴とするイタコン酸の製造方法。
【発明の効果】
【0008】
本発明の形質転換体は、宿主である糸状菌に、特定のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターが導入されているため、優れたイタコン酸高生産能を備えている。そのため、微生物を用いたイタコン酸の製造方法に好適に用いることができる。
本発明のプラスミドベクターは、特定のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターと、配列番号4に記載の特定のプロモーターDNAとを含有しているため、宿主にイタコン酸高生産能を付与することができる。
また、プラスミドベクターが、配列番号5に記載の特定の塩基配列からなるターミネーターDNAを含有する場合には、宿主にイタコン酸高生産能を確実に付与することができる。
本発明のイタコン酸の製造方法によれば、イタコン酸を従来よりも効率的に生産することができる。
【図面の簡単な説明】
【0009】
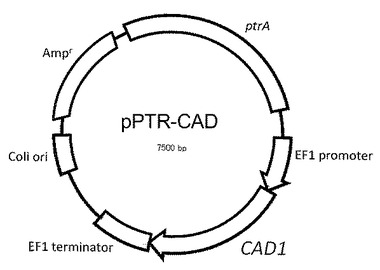
【図1】実施例1において作製したプラスミドベクターを示す。
【図2】イタコン酸生産量を示すグラフである。
【発明を実施するための形態】
【0010】
以下、本発明を詳細に説明する。
[1]形質転換体
本発明の形質転換体は、宿主に、シス−アコニット酸デカルボキシラーゼ(以下、「CAD」ともいう。)をコードするDNAを含有するプラスミドベクターが導入されたものであって、上記宿主が糸状菌であることを特徴とする。
【0011】
本発明の形質転換体における宿主細胞である上記「糸状菌」としては、例えば、アスペルギルス属、ウスチラゴ属等のイタコン酸生産菌が挙げられる。これらのなかでも、アスペルギルス属の糸状菌が好ましい。
上記アスペルギルス属の糸状菌としては、例えば、アスペルギルス・テレウス(Aspergillus tereus)、アスペルギルス・イタコニカス(Aspergillus itaconicus)等が挙げられる。これらのなかでも、アスペルギルス・テレウスが好ましく、アスペルギルス・テレウスIFO6365株が特に好ましい。
【0012】
また、上記「プラスミドベクター」が含有しているCADをコードするDNAは下記(a)〜(d)のいずれかに記載のDNAからなる。
(a)配列番号1(表1参照)に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号2(表2参照)に記載の塩基配列からなるDNA
(c)配列番号3(表3参照)に記載の塩基配列からなるDNA
(d)配列番号2(表2参照)又は配列番号3(表3参照)に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA
【0013】
【表1】

【0014】
【表2】
【0015】
【表3】
【0016】
本発明において、上記(a)に記載のDNAには、配列番号1に記載のアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、CAD活性を有するタンパク質をコードするDNAも含まれる。
【0017】
配列番号1に記載のアミノ酸配列の1若しくは数個のアミノ酸が置換されたアミノ酸配列におけるアミノ酸置換の具体例としては、例えば、配列番号1に記載のアミノ酸配列のうち、N末端から16番目のSerの他のアミノ酸(例えばAla)への置換、236番目のHisの他のアミノ酸(例えばTyr)への置換、289番目のLeuの他のアミノ酸(例えばTyr)への置換、290番目のTyrの他のアミノ酸(例えばLeu)への置換、305番目のAsnの他のアミノ酸(例えばLys)への置換、308番目のGlyの他のアミノ酸(例えばArg)への置換、326番目のHisの他のアミノ酸(例えばTyr)への置換、371番目のSerの他のアミノ酸(例えばAla)への置換、394番目のSerの他のアミノ酸(例えばPro)への置換、396番目のGlnの他のアミノ酸(例えばHis)への置換、422番目のIleの他のアミノ酸(例えばVal)への置換、425番目のSerの他のアミノ酸(例えばThr)への置換、467番目のGlyの他のアミノ酸(例えばSer)への置換、475番目のSerの他のアミノ酸(例えばThr)への置換等を挙げることができる。
【0018】
配列番号1に記載のアミノ酸配列からなるタンパク質をコードするDNAは、既知のアミノ酸配列からそれをコードするDNAを得る従来の方法をそのまま用いて得ることができる。例えば、上記アミノ酸配列からDNAの塩基配列を決定して得ることができる。また、上記アミノ酸配列に対応する部分塩基配列に基づき、PCRにより全塩基配列を得ることができる。
【0019】
配列番号2に記載の塩基配列からなるDNAは、CADをコードするゲノム配列であることから、アミノ酸配列に翻訳されないイントロン領域を含んでいる。
配列番号3に記載の塩基配列は、配列番号2に記載の塩基配列からイントロン領域を除外した塩基配列である。配列番号3に記載の塩基配列は、例えば、配列番号1に記載のアミノ酸配列からなるタンパク質のcDNA塩基配列である。
【0020】
また、上記(d)に記載のDNAにおける、ストリンジェントな条件としては、例えば、65℃で4×SSCにおけるハイブリダイゼーション、次いで65℃で1時間0.1×SSCでの洗浄という条件を挙げることができる。
【0021】
配列番号2又は配列番号3に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNAは、例えば、上記の配列番号2又は配列番号3に記載の塩基配列をもとにPCRプライマーを設計し、種々のイタコン酸生産菌に由来する染色体DNA又はcDNAライブラリーを鋳型としてPCRを行うことで得ることができる。
適用可能なイタコン酸生産菌については特に制限はなく、種々のアスペルギルス属糸状菌や酵母を用いることができる。具体的には種々のAspergillus tereus、Aspergillus itaconicus、及び、Candida tsukubaensis等のイタコン酸生産菌を挙げることができる。
【0022】
上記プラスミドベクターは、CADをコードするDNAを公知の方法で糸状菌用ベクター等に挿入することで得ることができる。
また、このプラスミドベクターは、後段で説明する特定のプロモーターDNAとCADをコードするDNAとを含有するプラスミドベクターであることが好ましい。
【0023】
本発明の形質転換体は、宿主(糸状菌)に適した方法で、糸状菌細胞にプラスミドベクターを導入することで得ることができる。例えば、プロトプラスト−ポリエチレングリコール法、電気穿孔法、アグロバクテリウム法、パーティクル・ガン法等の導入方法を用いることができる。
【0024】
[2]プラスミドベクター
本発明のプラスミドベクターは、宿主にイタコン酸高生産能を与えるものであって、CADをコードするDNAと、配列番号4に記載の塩基配列からなるプロモーターDNAと、を含有することを特徴とする。
尚、「シス−アコニット酸デカルボキシラーゼ(CAD)をコードするDNA」については、上述の形質転換体における説明をそのまま適用することができる。
【0025】
上記「プロモーターDNA」は、配列番号4(後述の表5参照)に記載の塩基配列からなる。
【0026】
また、本発明におけるプラスミドベクターは、上記CADをコードするDNA及び上記プロモーターDNA以外に、配列番号5(後述の表7参照)に記載の塩基配列からなるターミネーターDNAを更に含有するものとすることができる。
【0027】
更に、本発明におけるプラスミドベクターは、上記CADをコードするDNA及び上記プロモーターDNA以外に、ピリチアミン耐性マーカー遺伝子を更に含有するものとすることができる。
【0028】
上記プラスミドベクターは、CADをコードするDNA、プロモーターDNA等のDNAを公知の方法で発現ベクター等に挿入することで得ることができる。尚、用いられる発現ベクターは、シャトルベクターであってもよい。
上記発現ベクターとしては、糸状菌用ベクター等が挙げられる。具体的には、例えば、「pPTR I」(タカラバイオ株式会社製)、「pPTR II」(タカラバイオ株式会社製)、「pAUR123」(タカラバイオ株式会社製)等を挙げることができる。
【0029】
[3]イタコン酸の製造方法
本発明のイタコン酸の製造方法は、上述の形質転換体を培養する工程と、形質転換体が生成したイタコン酸を回収する工程と、を備えることを特徴とする。
【0030】
形質転換体を培養する工程では、宿主である糸状菌に適した方法で宿主細胞を培養すればよい。
特に、この培養工程では、培地のpHを4以下(特に3以下、更には2以下)とすることができる。このように培地のpHが低い場合、炭素源として、従来用いられているブドウ糖よりも安価なデンプンの酸分解物をそのまま用いることができる。また、植物から抽出されたデンプンを硝酸によって酸分解すれば、糸状菌は硝酸を炭素源として利用できるため、その他の栄養となる物質を一切添加することなく培養が可能である。更には、培地が酸性であるため、培養の際に細菌や酵母が混入しても、その影響を最小限に留めることができる。
また、この培養工程では、培養温度を15〜38℃(特に25〜34℃、更には28〜32℃)とすることができる。培養温度が上記範囲である場合には、宿主細胞を十分に培養することができる。
【0031】
また、形質転換体が生成したイタコン酸の回収工程では、微生物の培養によって生成する有機酸類をその培養物から分離するのに用いられる方法を特に制限なく用いることができる。例えば、遠心分離や濾過等による菌体の分離、上清の濃縮、各種のカラムクロマトグラフィー、活性炭処理、濾過、及び、再結晶等の手段を適宜組合せて用いることにより、分離、精製することができる。
【実施例】
【0032】
以下、実施例を挙げて、本発明を更に具体的に説明する。但し、本発明は、これらの実施例に何ら制約されるものではない。ここで、「部」及び「%」は、特記しない限り質量基準である。
【0033】
<実施例1>
[1]プラスミドベクターの構築
(1−1)糸状菌用ベクターへのプロモーターDNAの挿入
糸状菌を宿主としてCAD1遺伝子を発現させるために、アスペルギルス・ニドランス由来伸長因子遺伝子プロモーターDNAを用いた。
このプロモーターDNAを得るために、下記表4に示すプライマーを用い、アスペルギルス・ニドランスの染色体DNAを鋳型としてPCRを行った。これにより、配列番号4(表5参照)に記載の塩基配列からなるDNAが得られた。
尚、PCRの条件は、94℃、2分の後、94℃ 30秒、57℃ 30秒、72℃ 2分を30サイクル行った。
【0034】
【表4】
【0035】
【表5】
【0036】
表4における配列番号6には制限酵素KpnI、配列番号7にはHindIII及びSpeIの認識配列を付加しており、増幅したプロモーターDNAを制限酵素KpnI及びHindIIIにて消化した。
次いで、同じく制限酵素KpnI及びHindIIIにて消化した糸状菌用ベクター(タカラバイオ株式会社製、「pPTR I」)と結合した。DNAの結合にはライゲーション試薬(東洋紡績株式会社製、「Ligation high」)を用いた。得られたプラスミドDNAをpPTR−Pとする。
【0037】
(1−2)ターミネーターDNAの挿入
アスペルギルス・ニドランス由来伸長因子遺伝子ターミネーターDNAを得るために、下記表6に示すプライマーを用い、アスペルギルス・ニドランスの染色体DNAを鋳型としてPCRを行った。これにより、配列番号5(表7参照)に記載の塩基配列からなるDNAが得られた。
尚、PCRの条件は、94℃、2分の後、94℃ 30秒、57℃ 30秒、72℃ 2分を30サイクル行った。
【0038】
【表6】
【0039】
【表7】
【0040】
表6のプライマーには、In−fusion Advantage PCR Cloning kit(タカラバイオ株式会社製)の取扱説明書に従い、次に結合するベクター配列に同一性のあるDNAを付加しており、取扱説明書に従いこのキットを用いて配列番号5のDNAを制限酵素HindIIIで消化したpPTR−Pと結合した。得られたプラスミドDNAをpPTR−PTとする。
【0041】
(1−3)CAD1遺伝子の挿入
アスペルギルス・テレウス由来CAD1遺伝子発現用DNAを得るために、下記表8に示すプライマーを用い、アスペルギルス・テレウス IFO6365株の染色体DNAを鋳型としてPCRを行った。これにより、配列番号12(表9参照)に記載の塩基配列からなるDNAが得られた。このCAD1遺伝子配列は特開2009−27999や、Appl.Microbiol.Biotechnol.(2008)80,223−229に記述され、DDBJにAccession Number AB326105として登録されたものである。
尚、PCRの条件は、98℃ 10秒、60℃ 15秒、68℃ 2分を30サイクル行った。
【0042】
【表8】
【0043】
【表9】
【0044】
表8のプライマーには、In−fusion Advantage PCR Cloning kit(タカラバイオ株式会社製)の取扱説明書に従い、次に結合するベクター配列に同一性のあるDNAを付加しており、取扱説明書に従いこのキットを用いて配列番号12のDNAを制限酵素SpeIで消化したpPTR−PTと結合した。得られたプラスミドベクター(プラスミドDNA)をpPTR−CADとする(図1参照)。
【0045】
[2]宿主(アスペルギルス・テレウス)へのプラスミドベクターの導入(形質転換体の製造)
上記[1]で得られたプラスミドベクター(pPTR−CAD)を、糸状菌細胞(アスペルギルス・テレウス IFO6365株)に、プロトプラスト−ポリエチレングリコール法により導入した。アスペルギルス・テレウスの形質転換法についてはBiosci.Biotechnol.Biochem.(2002)66,404−406に報告があるが、これに記載の方法をアスペルギルス・テレウス IFO6365株に適用すると、形質転換体の判別が困難であり、また菌の生育も不十分であった。そこで、形質転換体を分離するために培地に加えるピリチアミン臭化水素酸塩の濃度を0.2μg/mlにしたところ、形質転換体を明確に判別が可能になった。菌の生育は、培地1リットルあたりにビオチン2.5mg、ニコチン酸2.5mg、パラアミノ安息香酸0.8mg、ピリドキシン塩酸塩1.0mg、パントテン酸2.0mg、リボフラビン2.5mg、塩化コリン20mgを添加することにより改善した。常法であるプロトプラスト−ポリエチレングリコール法にこれらの工夫を加えることによって、pPTR−CADが導入された形質転換体(アスペルギルス・テレウス AtCAD株)を得た。
【0046】
[3]形質転換体のイタコン酸生産性評価
上記[2]で得られた形質転換体(アスペルギルス・テレウス AtCAD株)を糸状菌用最少培地にて13日間振とう培養した。糸状菌最少培地の組成は1リットルあたり、グルコース10g、硝酸ナトリウム3g、塩化カリウム1.3g、硫酸マグネシウム七水和物1.3g、リン酸二水素カリウム3.8g、七モリブデン酸六アンモニウム四水和物1mg、ホウ酸10mg、塩化コバルト六水和物2mg、硫酸銅五水和物2mg、硫酸鉄七水和物5mg、塩化マンガン四水和物5mg、硫酸亜鉛七水和物20mg、エチレンジアミン四酢酸四ナトリウム50mg、ピリチアミン臭化水素酸塩0.2mgを基本とし、今回生育速度を改善させるためにビオチン2.5mg、ニコチン酸2.5mg、パラアミノ安息香酸0.8mg、ピリドキシン塩酸塩1mg、パントテン酸2mg、リボフラビン2.5mg、塩化コリン20mgを加えた。尚、培地のpHは2.0、培養温度は30℃とした。
そして、培養開始3日目から2日毎に培養液の一部を採取し、0.20μmの孔径を有するフィルターにてろ過することにより試料を調製した。試料に含まれるイタコン酸は高速液体クロマトグラフィ(HPLC)(カラム:カプセルパックC18 MG(5.0×150mm、株式会社資生堂製)、溶媒:0.1%リン酸溶液、温度:60℃、流速:1.5ml/分、検出波長:210nm)で定量した。
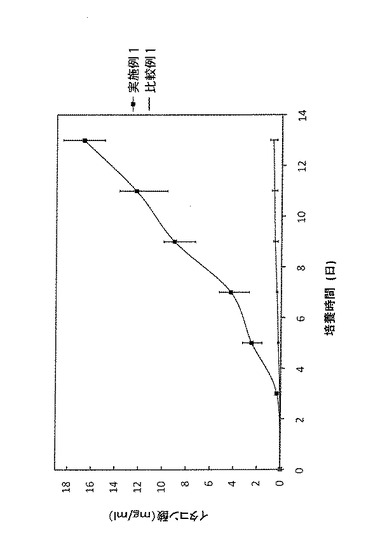
尚、コントロール(比較例1)には、CADをコードするDNAを挿入していないpPTR−PTが導入された形質転換体(アスペルギルス・テレウス AtPT株)を用いた。そして、培養から13日間の結果を表10及び図2に示した。
【0047】
【表10】
【0048】
表10及び図2によれば、実施例1の形質転換体(アスペルギルス・テレウス AtCAD株)から得られた培養液においては、CADをコードするDNAを挿入していない比較例1のコントロール(アスペルギルス・テレウス AtPT株)と比較すると、培養13日でイタコン酸濃度が約26倍に上昇していることが確認できた。
以上より、シス−アコニット酸デカルボキシラーゼ(CAD)をコードするDNAを糸状菌(アスペルギルス・テレウス)にて高発現させることにより、イタコン酸をより効率的に製造することができることが分かった。
【技術分野】
【0001】
本発明は、形質転換体、プラスミドベクター及びイタコン酸の製造方法に関する。更に詳しくは、本発明は、特定のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターが導入された形質転換体、及び、宿主にイタコン酸高生産能を与えるプラスミドベクター、並びに、イタコン酸を効率的に製造することができるイタコン酸の製造方法に関する。
【背景技術】
【0002】
イタコン酸は不飽和基を有する二価の水溶性有機酸であり、その構造から重合や共重合によって線状高分子を形成できる特性がある。また、その安全性も高いことから、合成樹脂や合成繊維、接着剤や印刷用インキ等、工業製品の原料や添加剤として幅広く利用されている。イタコン酸は、Aspergillus terreus、Aspergillus itaconicusのようなアスペルギルス属等の糸状菌や、Candida tsukubaensisのような酵母において生産できることが知られている。イタコン酸の工業生産においてはもっぱら生育速度の観点からA.terreusが使用されている。より効率的なイタコン酸製造のため、イタコン酸高性生産菌の開発が求められているが、イタコン酸高生産菌の育種は、変異原性物質による処理や紫外線照射等の古典的なランダム変異導入法に頼っているのが現状である(特許文献1参照)。
【0003】
Bentleyらは、A.terreusにおいて、クエン酸回路で生成するシス−アコニット酸からシス−アコニット酸デカルボキシラーゼによりイタコン酸が生合成されるという説を提唱した(非特許文献1参照)。また、その後、岡部らにより、A.terreus由来のシス−アコニット酸デカルボキシラーゼタンパク質が単離精製されている(非特許文献2参照)。更に、その後、本願発明者らにより、シス−アコニット酸デカルボキシラーゼをコードするDNAについて単離同定がなされており、このDNAを含むプラスミドを酵母細胞に導入した微生物を用いたイタコン酸の製造方法が提案されている(特許文献2参照)。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特開平6−38774号公報
【特許文献2】特開2009−27999号公報
【非特許文献】
【0005】
【非特許文献1】Bentley,R.and Thiessen C.P.,J.Biol.Chem.226:703−720(1957)
【非特許文献2】Dwiarti L.,Yamane K.,Yamatani H.,Kahar P.,Okabe M.,J.Biosci.Bioeng.94:29−33(2002)
【発明の概要】
【発明が解決しようとする課題】
【0006】
しかしながら、従来の微生物を用いたイタコン酸の製造方法では、イタコン酸の生産効率が未だ十分とはいえず、更なる効率の向上が求められているのが現状である。
本発明は、上記現状に鑑みてなされたものであり、特定のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターが導入された形質転換体、及び、宿主にイタコン酸高生産能を与えるプラスミドベクター、並びに、イタコン酸を効率的に製造することができるイタコン酸の製造方法を提供することを目的とする。
【課題を解決するための手段】
【0007】
本発明は以下の通りである。
[1]宿主に、下記(a)〜(d)のいずれかに記載のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターが導入された形質転換体であって、
前記宿主は糸状菌であることを特徴とする形質転換体。
(a)配列番号1に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号2に記載の塩基配列からなるDNA
(c)配列番号3に記載の塩基配列からなるDNA
(d)配列番号2又は配列番号3に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA
[2]前記宿主がアスペルギルス(Aspergillus)属の糸状菌である上記[1]に記載の形質転換体。
[3]前記宿主がアスペルギルス・テレウス(Aspergillus terreus)である上記[1]に記載の形質転換体。
[4]宿主にイタコン酸高生産能を与えるプラスミドベクターであって、
下記(a)〜(d)のいずれかに記載のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAと、
配列番号4に記載の塩基配列からなるプロモーターDNAと、を含有することを特徴とするプラスミドベクター。
(a)配列番号1に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号2に記載の塩基配列からなるDNA
(c)配列番号3に記載の塩基配列からなるDNA
(d)配列番号2又は配列番号3に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA
[5]更に、配列番号5に記載の塩基配列からなるターミネーターDNAを含有する上記[4]に記載のプラスミドベクター。
[6]上記[1]乃至[3]のいずれかに記載の形質転換体を培養する工程と、
前記形質転換体が生成したイタコン酸を回収する工程と、を備えることを特徴とするイタコン酸の製造方法。
【発明の効果】
【0008】
本発明の形質転換体は、宿主である糸状菌に、特定のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターが導入されているため、優れたイタコン酸高生産能を備えている。そのため、微生物を用いたイタコン酸の製造方法に好適に用いることができる。
本発明のプラスミドベクターは、特定のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターと、配列番号4に記載の特定のプロモーターDNAとを含有しているため、宿主にイタコン酸高生産能を付与することができる。
また、プラスミドベクターが、配列番号5に記載の特定の塩基配列からなるターミネーターDNAを含有する場合には、宿主にイタコン酸高生産能を確実に付与することができる。
本発明のイタコン酸の製造方法によれば、イタコン酸を従来よりも効率的に生産することができる。
【図面の簡単な説明】
【0009】
【図1】実施例1において作製したプラスミドベクターを示す。
【図2】イタコン酸生産量を示すグラフである。
【発明を実施するための形態】
【0010】
以下、本発明を詳細に説明する。
[1]形質転換体
本発明の形質転換体は、宿主に、シス−アコニット酸デカルボキシラーゼ(以下、「CAD」ともいう。)をコードするDNAを含有するプラスミドベクターが導入されたものであって、上記宿主が糸状菌であることを特徴とする。
【0011】
本発明の形質転換体における宿主細胞である上記「糸状菌」としては、例えば、アスペルギルス属、ウスチラゴ属等のイタコン酸生産菌が挙げられる。これらのなかでも、アスペルギルス属の糸状菌が好ましい。
上記アスペルギルス属の糸状菌としては、例えば、アスペルギルス・テレウス(Aspergillus tereus)、アスペルギルス・イタコニカス(Aspergillus itaconicus)等が挙げられる。これらのなかでも、アスペルギルス・テレウスが好ましく、アスペルギルス・テレウスIFO6365株が特に好ましい。
【0012】
また、上記「プラスミドベクター」が含有しているCADをコードするDNAは下記(a)〜(d)のいずれかに記載のDNAからなる。
(a)配列番号1(表1参照)に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号2(表2参照)に記載の塩基配列からなるDNA
(c)配列番号3(表3参照)に記載の塩基配列からなるDNA
(d)配列番号2(表2参照)又は配列番号3(表3参照)に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA
【0013】
【表1】
【0014】
【表2】
【0015】
【表3】
【0016】
本発明において、上記(a)に記載のDNAには、配列番号1に記載のアミノ酸配列の1個又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列からなり、CAD活性を有するタンパク質をコードするDNAも含まれる。
【0017】
配列番号1に記載のアミノ酸配列の1若しくは数個のアミノ酸が置換されたアミノ酸配列におけるアミノ酸置換の具体例としては、例えば、配列番号1に記載のアミノ酸配列のうち、N末端から16番目のSerの他のアミノ酸(例えばAla)への置換、236番目のHisの他のアミノ酸(例えばTyr)への置換、289番目のLeuの他のアミノ酸(例えばTyr)への置換、290番目のTyrの他のアミノ酸(例えばLeu)への置換、305番目のAsnの他のアミノ酸(例えばLys)への置換、308番目のGlyの他のアミノ酸(例えばArg)への置換、326番目のHisの他のアミノ酸(例えばTyr)への置換、371番目のSerの他のアミノ酸(例えばAla)への置換、394番目のSerの他のアミノ酸(例えばPro)への置換、396番目のGlnの他のアミノ酸(例えばHis)への置換、422番目のIleの他のアミノ酸(例えばVal)への置換、425番目のSerの他のアミノ酸(例えばThr)への置換、467番目のGlyの他のアミノ酸(例えばSer)への置換、475番目のSerの他のアミノ酸(例えばThr)への置換等を挙げることができる。
【0018】
配列番号1に記載のアミノ酸配列からなるタンパク質をコードするDNAは、既知のアミノ酸配列からそれをコードするDNAを得る従来の方法をそのまま用いて得ることができる。例えば、上記アミノ酸配列からDNAの塩基配列を決定して得ることができる。また、上記アミノ酸配列に対応する部分塩基配列に基づき、PCRにより全塩基配列を得ることができる。
【0019】
配列番号2に記載の塩基配列からなるDNAは、CADをコードするゲノム配列であることから、アミノ酸配列に翻訳されないイントロン領域を含んでいる。
配列番号3に記載の塩基配列は、配列番号2に記載の塩基配列からイントロン領域を除外した塩基配列である。配列番号3に記載の塩基配列は、例えば、配列番号1に記載のアミノ酸配列からなるタンパク質のcDNA塩基配列である。
【0020】
また、上記(d)に記載のDNAにおける、ストリンジェントな条件としては、例えば、65℃で4×SSCにおけるハイブリダイゼーション、次いで65℃で1時間0.1×SSCでの洗浄という条件を挙げることができる。
【0021】
配列番号2又は配列番号3に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNAは、例えば、上記の配列番号2又は配列番号3に記載の塩基配列をもとにPCRプライマーを設計し、種々のイタコン酸生産菌に由来する染色体DNA又はcDNAライブラリーを鋳型としてPCRを行うことで得ることができる。
適用可能なイタコン酸生産菌については特に制限はなく、種々のアスペルギルス属糸状菌や酵母を用いることができる。具体的には種々のAspergillus tereus、Aspergillus itaconicus、及び、Candida tsukubaensis等のイタコン酸生産菌を挙げることができる。
【0022】
上記プラスミドベクターは、CADをコードするDNAを公知の方法で糸状菌用ベクター等に挿入することで得ることができる。
また、このプラスミドベクターは、後段で説明する特定のプロモーターDNAとCADをコードするDNAとを含有するプラスミドベクターであることが好ましい。
【0023】
本発明の形質転換体は、宿主(糸状菌)に適した方法で、糸状菌細胞にプラスミドベクターを導入することで得ることができる。例えば、プロトプラスト−ポリエチレングリコール法、電気穿孔法、アグロバクテリウム法、パーティクル・ガン法等の導入方法を用いることができる。
【0024】
[2]プラスミドベクター
本発明のプラスミドベクターは、宿主にイタコン酸高生産能を与えるものであって、CADをコードするDNAと、配列番号4に記載の塩基配列からなるプロモーターDNAと、を含有することを特徴とする。
尚、「シス−アコニット酸デカルボキシラーゼ(CAD)をコードするDNA」については、上述の形質転換体における説明をそのまま適用することができる。
【0025】
上記「プロモーターDNA」は、配列番号4(後述の表5参照)に記載の塩基配列からなる。
【0026】
また、本発明におけるプラスミドベクターは、上記CADをコードするDNA及び上記プロモーターDNA以外に、配列番号5(後述の表7参照)に記載の塩基配列からなるターミネーターDNAを更に含有するものとすることができる。
【0027】
更に、本発明におけるプラスミドベクターは、上記CADをコードするDNA及び上記プロモーターDNA以外に、ピリチアミン耐性マーカー遺伝子を更に含有するものとすることができる。
【0028】
上記プラスミドベクターは、CADをコードするDNA、プロモーターDNA等のDNAを公知の方法で発現ベクター等に挿入することで得ることができる。尚、用いられる発現ベクターは、シャトルベクターであってもよい。
上記発現ベクターとしては、糸状菌用ベクター等が挙げられる。具体的には、例えば、「pPTR I」(タカラバイオ株式会社製)、「pPTR II」(タカラバイオ株式会社製)、「pAUR123」(タカラバイオ株式会社製)等を挙げることができる。
【0029】
[3]イタコン酸の製造方法
本発明のイタコン酸の製造方法は、上述の形質転換体を培養する工程と、形質転換体が生成したイタコン酸を回収する工程と、を備えることを特徴とする。
【0030】
形質転換体を培養する工程では、宿主である糸状菌に適した方法で宿主細胞を培養すればよい。
特に、この培養工程では、培地のpHを4以下(特に3以下、更には2以下)とすることができる。このように培地のpHが低い場合、炭素源として、従来用いられているブドウ糖よりも安価なデンプンの酸分解物をそのまま用いることができる。また、植物から抽出されたデンプンを硝酸によって酸分解すれば、糸状菌は硝酸を炭素源として利用できるため、その他の栄養となる物質を一切添加することなく培養が可能である。更には、培地が酸性であるため、培養の際に細菌や酵母が混入しても、その影響を最小限に留めることができる。
また、この培養工程では、培養温度を15〜38℃(特に25〜34℃、更には28〜32℃)とすることができる。培養温度が上記範囲である場合には、宿主細胞を十分に培養することができる。
【0031】
また、形質転換体が生成したイタコン酸の回収工程では、微生物の培養によって生成する有機酸類をその培養物から分離するのに用いられる方法を特に制限なく用いることができる。例えば、遠心分離や濾過等による菌体の分離、上清の濃縮、各種のカラムクロマトグラフィー、活性炭処理、濾過、及び、再結晶等の手段を適宜組合せて用いることにより、分離、精製することができる。
【実施例】
【0032】
以下、実施例を挙げて、本発明を更に具体的に説明する。但し、本発明は、これらの実施例に何ら制約されるものではない。ここで、「部」及び「%」は、特記しない限り質量基準である。
【0033】
<実施例1>
[1]プラスミドベクターの構築
(1−1)糸状菌用ベクターへのプロモーターDNAの挿入
糸状菌を宿主としてCAD1遺伝子を発現させるために、アスペルギルス・ニドランス由来伸長因子遺伝子プロモーターDNAを用いた。
このプロモーターDNAを得るために、下記表4に示すプライマーを用い、アスペルギルス・ニドランスの染色体DNAを鋳型としてPCRを行った。これにより、配列番号4(表5参照)に記載の塩基配列からなるDNAが得られた。
尚、PCRの条件は、94℃、2分の後、94℃ 30秒、57℃ 30秒、72℃ 2分を30サイクル行った。
【0034】
【表4】
【0035】
【表5】
【0036】
表4における配列番号6には制限酵素KpnI、配列番号7にはHindIII及びSpeIの認識配列を付加しており、増幅したプロモーターDNAを制限酵素KpnI及びHindIIIにて消化した。
次いで、同じく制限酵素KpnI及びHindIIIにて消化した糸状菌用ベクター(タカラバイオ株式会社製、「pPTR I」)と結合した。DNAの結合にはライゲーション試薬(東洋紡績株式会社製、「Ligation high」)を用いた。得られたプラスミドDNAをpPTR−Pとする。
【0037】
(1−2)ターミネーターDNAの挿入
アスペルギルス・ニドランス由来伸長因子遺伝子ターミネーターDNAを得るために、下記表6に示すプライマーを用い、アスペルギルス・ニドランスの染色体DNAを鋳型としてPCRを行った。これにより、配列番号5(表7参照)に記載の塩基配列からなるDNAが得られた。
尚、PCRの条件は、94℃、2分の後、94℃ 30秒、57℃ 30秒、72℃ 2分を30サイクル行った。
【0038】
【表6】
【0039】
【表7】
【0040】
表6のプライマーには、In−fusion Advantage PCR Cloning kit(タカラバイオ株式会社製)の取扱説明書に従い、次に結合するベクター配列に同一性のあるDNAを付加しており、取扱説明書に従いこのキットを用いて配列番号5のDNAを制限酵素HindIIIで消化したpPTR−Pと結合した。得られたプラスミドDNAをpPTR−PTとする。
【0041】
(1−3)CAD1遺伝子の挿入
アスペルギルス・テレウス由来CAD1遺伝子発現用DNAを得るために、下記表8に示すプライマーを用い、アスペルギルス・テレウス IFO6365株の染色体DNAを鋳型としてPCRを行った。これにより、配列番号12(表9参照)に記載の塩基配列からなるDNAが得られた。このCAD1遺伝子配列は特開2009−27999や、Appl.Microbiol.Biotechnol.(2008)80,223−229に記述され、DDBJにAccession Number AB326105として登録されたものである。
尚、PCRの条件は、98℃ 10秒、60℃ 15秒、68℃ 2分を30サイクル行った。
【0042】
【表8】
【0043】
【表9】
【0044】
表8のプライマーには、In−fusion Advantage PCR Cloning kit(タカラバイオ株式会社製)の取扱説明書に従い、次に結合するベクター配列に同一性のあるDNAを付加しており、取扱説明書に従いこのキットを用いて配列番号12のDNAを制限酵素SpeIで消化したpPTR−PTと結合した。得られたプラスミドベクター(プラスミドDNA)をpPTR−CADとする(図1参照)。
【0045】
[2]宿主(アスペルギルス・テレウス)へのプラスミドベクターの導入(形質転換体の製造)
上記[1]で得られたプラスミドベクター(pPTR−CAD)を、糸状菌細胞(アスペルギルス・テレウス IFO6365株)に、プロトプラスト−ポリエチレングリコール法により導入した。アスペルギルス・テレウスの形質転換法についてはBiosci.Biotechnol.Biochem.(2002)66,404−406に報告があるが、これに記載の方法をアスペルギルス・テレウス IFO6365株に適用すると、形質転換体の判別が困難であり、また菌の生育も不十分であった。そこで、形質転換体を分離するために培地に加えるピリチアミン臭化水素酸塩の濃度を0.2μg/mlにしたところ、形質転換体を明確に判別が可能になった。菌の生育は、培地1リットルあたりにビオチン2.5mg、ニコチン酸2.5mg、パラアミノ安息香酸0.8mg、ピリドキシン塩酸塩1.0mg、パントテン酸2.0mg、リボフラビン2.5mg、塩化コリン20mgを添加することにより改善した。常法であるプロトプラスト−ポリエチレングリコール法にこれらの工夫を加えることによって、pPTR−CADが導入された形質転換体(アスペルギルス・テレウス AtCAD株)を得た。
【0046】
[3]形質転換体のイタコン酸生産性評価
上記[2]で得られた形質転換体(アスペルギルス・テレウス AtCAD株)を糸状菌用最少培地にて13日間振とう培養した。糸状菌最少培地の組成は1リットルあたり、グルコース10g、硝酸ナトリウム3g、塩化カリウム1.3g、硫酸マグネシウム七水和物1.3g、リン酸二水素カリウム3.8g、七モリブデン酸六アンモニウム四水和物1mg、ホウ酸10mg、塩化コバルト六水和物2mg、硫酸銅五水和物2mg、硫酸鉄七水和物5mg、塩化マンガン四水和物5mg、硫酸亜鉛七水和物20mg、エチレンジアミン四酢酸四ナトリウム50mg、ピリチアミン臭化水素酸塩0.2mgを基本とし、今回生育速度を改善させるためにビオチン2.5mg、ニコチン酸2.5mg、パラアミノ安息香酸0.8mg、ピリドキシン塩酸塩1mg、パントテン酸2mg、リボフラビン2.5mg、塩化コリン20mgを加えた。尚、培地のpHは2.0、培養温度は30℃とした。
そして、培養開始3日目から2日毎に培養液の一部を採取し、0.20μmの孔径を有するフィルターにてろ過することにより試料を調製した。試料に含まれるイタコン酸は高速液体クロマトグラフィ(HPLC)(カラム:カプセルパックC18 MG(5.0×150mm、株式会社資生堂製)、溶媒:0.1%リン酸溶液、温度:60℃、流速:1.5ml/分、検出波長:210nm)で定量した。
尚、コントロール(比較例1)には、CADをコードするDNAを挿入していないpPTR−PTが導入された形質転換体(アスペルギルス・テレウス AtPT株)を用いた。そして、培養から13日間の結果を表10及び図2に示した。
【0047】
【表10】
【0048】
表10及び図2によれば、実施例1の形質転換体(アスペルギルス・テレウス AtCAD株)から得られた培養液においては、CADをコードするDNAを挿入していない比較例1のコントロール(アスペルギルス・テレウス AtPT株)と比較すると、培養13日でイタコン酸濃度が約26倍に上昇していることが確認できた。
以上より、シス−アコニット酸デカルボキシラーゼ(CAD)をコードするDNAを糸状菌(アスペルギルス・テレウス)にて高発現させることにより、イタコン酸をより効率的に製造することができることが分かった。
【特許請求の範囲】
【請求項1】
宿主に、下記(a)〜(d)のいずれかに記載のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターが導入された形質転換体であって、
前記宿主は糸状菌であることを特徴とする形質転換体。
(a)配列番号1に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号2に記載の塩基配列からなるDNA
(c)配列番号3に記載の塩基配列からなるDNA
(d)配列番号2又は配列番号3に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA
【請求項2】
前記宿主がアスペルギルス(Aspergillus)属の糸状菌である請求項1に記載の形質転換体。
【請求項3】
前記宿主がアスペルギルス・テレウス(Aspergillus terreus)である請求項1に記載の形質転換体。
【請求項4】
宿主にイタコン酸高生産能を与えるプラスミドベクターであって、
下記(a)〜(d)のいずれかに記載のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAと、
配列番号4に記載の塩基配列からなるプロモーターDNAと、を含有することを特徴とするプラスミドベクター。
(a)配列番号1に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号2に記載の塩基配列からなるDNA
(c)配列番号3に記載の塩基配列からなるDNA
(d)配列番号2又は配列番号3に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA
【請求項5】
更に、配列番号5に記載の塩基配列からなるターミネーターDNAを含有する請求項4に記載のプラスミドベクター。
【請求項6】
請求項1乃至3のいずれかに記載の形質転換体を培養する工程と、
前記形質転換体が生成したイタコン酸を回収する工程と、を備えることを特徴とするイタコン酸の製造方法。
【請求項1】
宿主に、下記(a)〜(d)のいずれかに記載のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAを含有するプラスミドベクターが導入された形質転換体であって、
前記宿主は糸状菌であることを特徴とする形質転換体。
(a)配列番号1に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号2に記載の塩基配列からなるDNA
(c)配列番号3に記載の塩基配列からなるDNA
(d)配列番号2又は配列番号3に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA
【請求項2】
前記宿主がアスペルギルス(Aspergillus)属の糸状菌である請求項1に記載の形質転換体。
【請求項3】
前記宿主がアスペルギルス・テレウス(Aspergillus terreus)である請求項1に記載の形質転換体。
【請求項4】
宿主にイタコン酸高生産能を与えるプラスミドベクターであって、
下記(a)〜(d)のいずれかに記載のDNAからなるシス−アコニット酸デカルボキシラーゼをコードするDNAと、
配列番号4に記載の塩基配列からなるプロモーターDNAと、を含有することを特徴とするプラスミドベクター。
(a)配列番号1に記載のアミノ酸配列からなるタンパク質をコードするDNA
(b)配列番号2に記載の塩基配列からなるDNA
(c)配列番号3に記載の塩基配列からなるDNA
(d)配列番号2又は配列番号3に記載の塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、シス−アコニット酸デカルボキシラーゼ活性を有するタンパク質をコードするDNA
【請求項5】
更に、配列番号5に記載の塩基配列からなるターミネーターDNAを含有する請求項4に記載のプラスミドベクター。
【請求項6】
請求項1乃至3のいずれかに記載の形質転換体を培養する工程と、
前記形質転換体が生成したイタコン酸を回収する工程と、を備えることを特徴とするイタコン酸の製造方法。
【図1】

【図2】
【図2】
【公開番号】特開2013−51900(P2013−51900A)
【公開日】平成25年3月21日(2013.3.21)
【国際特許分類】
【出願番号】特願2011−191055(P2011−191055)
【出願日】平成23年9月1日(2011.9.1)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 刊行物に発表 発行者名:社団法人日本農芸化学会 刊行物名:日本農芸化学会2011年度大会講演要旨集 発行年月日:平成23年3月5日
【出願人】(500433225)学校法人中部大学 (105)
【Fターム(参考)】
【公開日】平成25年3月21日(2013.3.21)
【国際特許分類】
【出願日】平成23年9月1日(2011.9.1)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 刊行物に発表 発行者名:社団法人日本農芸化学会 刊行物名:日本農芸化学会2011年度大会講演要旨集 発行年月日:平成23年3月5日
【出願人】(500433225)学校法人中部大学 (105)
【Fターム(参考)】
[ Back to top ]