
微生物のグリセロール代謝促進方法

【課題】グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物について、グリセロール代謝を促進する。
【解決手段】グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液に投入し、培養液に接触させた電極に還元電位を印加しながら培養して、パエニバチルス属の微生物のグリセロール代謝を促進するようにした。
【解決手段】グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液に投入し、培養液に接触させた電極に還元電位を印加しながら培養して、パエニバチルス属の微生物のグリセロール代謝を促進するようにした。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、微生物のグリセロール代謝促進方法に関する。さらに詳述すると、本発明は、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物のグリセロール代謝を促進する方法、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を利用したエタノール生産方法、及びグリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を雑多な微生物群集から優占的に増殖させる培養方法に関する。
【背景技術】
【0002】
近年、温室効果ガスの排出量削減や化石燃料依存からの脱却に向けた動きの一つとして、バイオマスを利用したバイオディーゼル燃料の生産が進められつつある。バイオディーゼル燃料とは、バイオマス由来の油脂類の主成分であるトリグリセライドをメタノールとエステル交換反応させることによって得られる液体燃料であり、脂肪酸メチルエステルを主体とするものである。
【0003】
ここで、図19に示すように、バイオディーゼル燃料の生成過程においては、グリセロールが副産物として生じる。アルカリ触媒を用いるバイオディーゼル燃料の生産方法においては、バイオディーゼル燃料に対して約10%のグリセロールが生成されるとされており(非特許文献1を参照)、その処理が懸案事項となっている。
【0004】
ところで、最近では、グリセロールからエタノールやポリエステルの原料となる1,3−プロパンジオール等の有用物質を産生するグリセロール代謝能を有する微生物の存在に関する報告がいくつかなされている(例えば、特許文献1及び非特許得文献2を参照)。このような微生物を利用してグリセロールを有用物質に変換することで、グリセロールを有効利用してその処理を促進することができる。したがって、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物は、上記懸案事項を解決に導き得る可能性を秘めていると言える。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2010−226959号公報
【非特許文献】
【0006】
【非特許文献1】横山伸也他、「バイオマスエネルギー」、森北出版株式会社(2009)
【非特許文献2】伊藤武志他「通性嫌気性細菌を利用した廃棄物処理と水素生産の検討」、弓削商船高等専門学校紀要、29(2007)
【発明の概要】
【発明が解決しようとする課題】
【0007】
グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物を利用して実際にグリセロールを処理する際には、グリセロール代謝能をできる限り高めて、処理効率を向上させることが望ましいと言える。しかしながら、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物に関する知見の蓄積は未だ乏しく、グリセロール代謝能を高めて、処理効率を向上させるような検討は殆ど行われていない状況である。
【0008】
また、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物が、雑多な微生物群集の中に埋もれていることがある。このような状況において、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物を優占的に増殖させる技術を確立できれば、非常に有用であると考えられる。
【0009】
そこで、本発明は、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物について、グリセロール代謝を促進する方法を提供することを目的とする。
【0010】
また、本発明は、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物を利用して、グリセロールを効率よく処理すると共に、有用物質を効率よく生産する方法を提供することを目的とする。
【0011】
さらに、本発明は、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物を、雑多な微生物群集の中から優占的に増殖させることのできる方法を提供することを目的とする。
【課題を解決するための手段】
【0012】
かかる課題を解決するため、本願発明者等が鋭意検討を行った結果、雑多な微生物群集からなる微生物試料を、グリセロールとセルロースを含む培養液に投入し、この培養液に接触させた電極に銀・塩化銀電極電位基準で−0.6Vの電位を印加して培養試験を行ったところ、ある種の微生物が優占的に増殖すると共に、グリセロールがエタノールに変換されていることを知見した。
【0013】
そこで、優占的に増殖した微生物について解析を行った結果、驚くべきことに、この微生物が、微生物試料に僅か0.01%の割合でした含まれていなかったパエニバチルス(Paenibacillus)属の微生物であることが判明した。
【0014】
そして、このパエニバチルス(Paenibacillus)属の微生物を単離し、資化性について検討したところ、グリセロール資化性を有していることが判明した。
【0015】
ところが、上記と同様の微生物試料を、グリセロールを含みセルロースを含まない培養液に投入して電極に通電せずに培養試験を行った場合には、パエニバチルス(Paenibacillus)属の微生物の優占的な増殖は見られなかった。このことは、グリセロールとセルロースを含む培養液に投入して電極に通電を行わずに培養試験を行った場合、グリセロールを含みセルロースを含まない培養液に投入して電極に銀・塩化銀電極電位基準で−0.6Vの電位を印加して培養試験を行った場合においても同様であった。
【0016】
このことから、培養液中のセルロースの存在及び電極への還元電位の印加が、パエニバチルス(Paenibacillus)属の微生物のグリセロール代謝の促進に寄与し、その結果としてパエニバチルス(Paenibacillus)属の微生物の優占的な増殖が起こっているものと考えられた。
【0017】
また、上記培養試験で用いたセルロースをセロビオースに替えて培養試験を実施したところ、セルロースを用いた培養試験結果とほぼ同じ結果が得られた。このことから、培養液中のグルコース多量体の存在、及び電極への還元電位の印加が、パエニバチルス(Paenibacillus)属の微生物のグリセロール代謝の促進に寄与し、その結果としてパエニバチルス(Paenibacillus)属の微生物の優占的な増殖が起こっているものと考えられた。
【0018】
以上のことから、本願発明者等は、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物について、グリセロールとグルコース多量体及び/又はグルコースとの存在下で、培養液に接触させた電位に還元電位を印加することで、グリセロール代謝を促進できる可能性が導かれることを知見するに至り、この知見に基づきさらに種々検討を重ねて、本発明を完成するに至った。
【0019】
即ち、本発明のグリセロール代謝促進方法は、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液に投入し、培養液に接触させた電極に還元電位を印加しながら培養して、パエニバチルス属の微生物のグリセロール代謝を促進するようにしている。
【0020】
ここで、本発明のグリセロール代謝促進方法において、パエニバチルス属の微生物は、16S rRNA遺伝子の塩基配列が配列番号1に示される塩基配列からなる微生物であることが好ましい。
【0021】
また、本発明のグリセロール代謝促進方法において、パエニバチルス属の微生物は、パエニバチルス マセランス(Paenibacillus macerans)NS−1(寄託番号FERM P−22124)であることが好ましい。
【0022】
次に、本発明の有用物質生産方法は、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液に投入し、培養液に接触させた電極に還元電位を印加しながら培養する工程と、有用物質を回収する工程とを含むようにしている。
【0023】
ここで、本発明の有用物質生産方法において、パエニバチルス属の微生物は、16S rRNA遺伝子の塩基配列が配列番号1に示される塩基配列からなる微生物であり、有用物質は、エタノールであることが好ましい。
【0024】
また、本発明の有用物質生産方法において、パエニバチルス属の微生物は、パエニバチルス マセランス(Paenibacillus macerans)NS−1(寄託番号FERM P−22124)であり、有用物質はエタノールであることが好ましい。
【0025】
次に、本発明の培養方法は、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を含む微生物群集を、グリセロールとグルコース多量体を含む培養液に投入し、グルコース多量体として二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物を用い、培養液に接触させた電極に還元電位を印加して、パエニバチルス属の微生物を優占的に増殖させるようにしている。
【発明の効果】
【0026】
本発明のグリセロール代謝促進方法によれば、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物について、グリセロール代謝を促進させることができる。したがって、グリセロールの処理速度を向上させて、グリセロール処理効率の向上を図ることができる。
【0027】
また、本発明の有用物質生産方法によれば、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を利用して、グリセロール代謝を促進させ、エタノール等の有用物質の生産速度を向上させることができる。したがって、グリセロールの処理効率を向上させると共にエタノール等の有用物質の生産効率を向上させることができる。
【0028】
さらに、本発明の培養方法によれば、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、雑多な微生物群集の中から優占的に増殖させることができる。したがって、他の微生物によるグリセロール代謝の阻害を受けることなく、グリセロール代謝能を最大限に発揮できる微生物環境を構築することが可能となる。
【図面の簡単な説明】
【0029】
【図1】本発明を実施するための電気培養装置の一例を示す図ある。
【図2】実施例において使用した電気培養槽の模式図である。
【図3】電気培養試験において使用した嫌気チャンバーを示す図である。
【図4】実施例1の培養試験における菌体密度変化を示す図である。
【図5】実施例1の培養試験における培養液中のグリセロール濃度の経時変化を示す図である。
【図6】実施例1の培養試験における培養液中のエタノール濃度の経時変化を示す図である。
【図7】菌叢解析結果を示す電気泳動写真である。
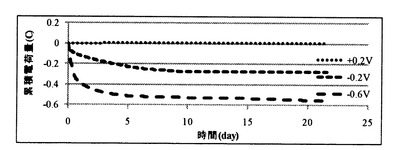
【図8A】実施例1の培養試験において、設定電位を+0.6Vとしたときの累積電荷量を示す図である。
【図8B】実施例1の培養試験において、設定電位を+0.2V、−0.2V、−0.6Vとした場合の累積電荷量を示す図である。
【図9】実施例2の培養試験におけるグリセロール濃度の経時変化を示す図である。
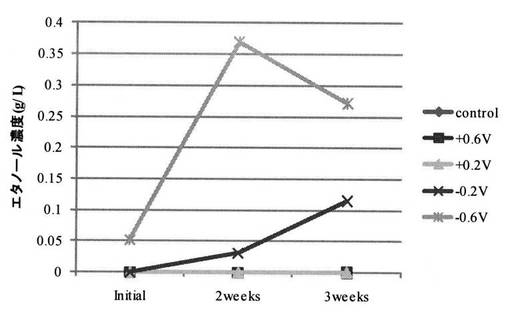
【図10】実施例2の培養試験におけるエタノール濃度の経時変化を示す図である。
【図11】実施例3の培養試験結果(グリセロール濃度変化とエタノール濃度変化)を示す図である。
【図12】実施例4の培養試験における菌体密度変化を示す図である。
【図13】実施例4の培養試験におけるグリセロール濃度の経時変化を示す図である。
【図14】実施例4の培養試験におけるセルロース濃度の経時変化を示す図である。
【図15】実施例5の培養試験における菌体密度変化を示す図である。
【図16】実施例5の培養試験(セロビオース使用)の各種測定結果を示す図である。
【図17】実施例5の培養試験(セルロース使用)の各種試験結果を示す図である。
【図18】微生物試料に含まれる微生物を網羅的に解析した結果を示す図である(実施例1)。
【図19】油脂からバイオディーゼルを生成する過程を示す化学反応式である。
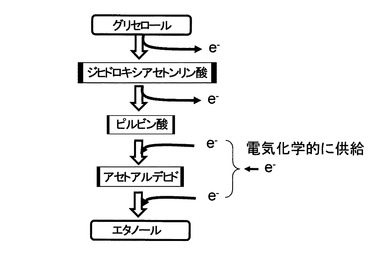
【図20】グリセロールからのエタノールの生成促進メカニズムを示す図である。
【発明を実施するための形態】
【0030】
以下、本発明を実施するための形態について、図面に基づいて詳細に説明する。
【0031】
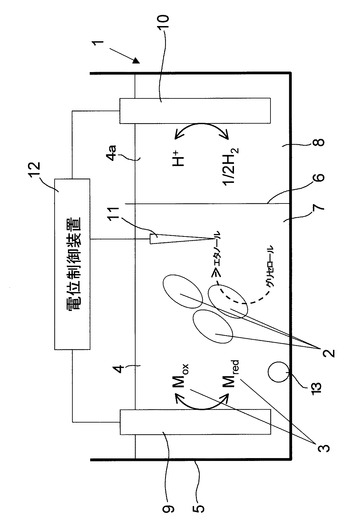
本発明のグリセロール代謝促進方法、有用物質生産方法及び培養方法は、例えば図1に示す電気培養装置1により実施することができる。図1に示す電気培養装置1は、大まかには、培養容器5、培養容器5内を二槽に区画するイオン交換膜6、培養容器5の一方の槽(培養槽7)に収容された培養液4、培養容器5の他方の槽(電解質槽8)に収容された電解液4a、培養液4に浸された作用極9及び参照電極11、電解液4aに浸された対極10、作用極9と対極10と参照電極10を結線して三電極方式で作用極9の電位を制御する定電位設定装置(ポテンシオスタット)12、培養液4を撹拌するための気泡発生部13により構成されている。
【0032】
三電極方式で作用極9の電位を制御することで、作用極9の電位を厳密に設定電位に制御することができる。詳細には、定電位設定装置12により、作用極9と対極10との間に電流を流し、基準となる参照電極11には一切電流が流れないようにしている。尚、三電極方式による電位制御については、例えば、電気化学測定法(上)、技報動出版株式会社、第1版15刷、2004年6月発行の6〜9ページにその詳細が記載されている。但し、作用極9と対極10の極間電圧のみで作用極9の電位を制御できる場合には、三電極方式とせずともよい。
【0033】
培養液4としては、パエニバチルス属の微生物2の培養に適した培地成分を含む培養液を適宜使用することができる。例えば、NH4Cl、K2HPO4、MgCl2・6H2O、CaCl2・2H2O、KCl等の無機塩類を主体とする培地成分を含む培養液等を適宜用いることができる。また、土壌成分を抽出した土壌抽出液を培養液に添加してもよい。これにより、パエニバチルス属の微生物の生育が促進され得る。
【0034】
電解液4aとしては、例えばNaClやKCl等の電解質を含む溶液を適宜用いることができる。したがって、培養液4を電解液4aとして用いることも可能である。
【0035】
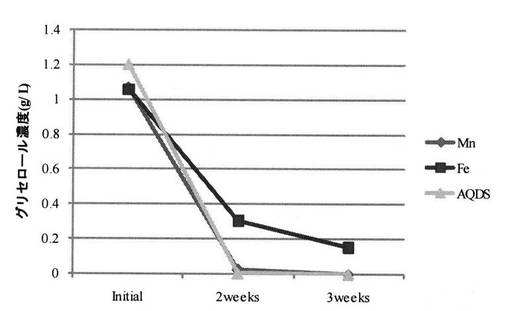
ここで、培養液4には、酸化還元物質3を添加することが好ましい。酸化還元物質3を培養液4に添加することで、培養液4の電位制御性を良好なものとすることができる。本発明に使用することができる酸化還元物質としては、パエニバチルス属の微生物2のグリセロール代謝能を阻害することのないものを適宜用いることができる。具体的には、鉄イオン、マンガンイオン等の金属イオン、アントラキノン−2,6−ジスルホン酸二ナトリウム等のキノン化合物が挙げられるが、これらに限定されるものではない。尚、金属イオンを用いる場合には、金属イオンをキレート剤に配位させて、金属イオンを培養液4中で安定に存在させるようにすることが好ましい。キレート剤としては、例えばジエチレントリアミンペンタ酢酸(DTPA)、エチレンジアミンテトラ酢酸(EDTA)、テトラエチレントリアミン(TET)、エチレンジアミン(EDA)、ジエチレントリアミン(DETA)、クエン酸、シュウ酸、クラウンエーテル、ニトリロテトラ酢酸、エデト酸二ナトリウム、エデト酸ナトリウム、エデト酸三ナトリウム、ペニシラミン、ペンテテートカルシウム三ナトリウム、ペンテト酸、スクシメルおよびエデト酸トリエンチン等、金属イオンを配位し得る任意のキレート剤を挙げることができる。
【0036】
酸化還元物質3の添加量は特に限定されるものではないが、0.1〜10mMとすることが好適であり、0.1〜5mMとすることがより好適であり、1〜3mMとすることがさらに好適である。
【0037】
イオン交換膜6は、培養液4に投入される微生物2、グリセロール、グリセロール多量体、酸化還元物質3を、電解質槽8に透過させることなく、培養液4中に留まらせ、且つ培養液4に含まれるイオンまたは電解液4aに含まれるイオンを透過させてイオン電流を生じさせ、作用極9において生じる還元反応を補完する酸化反応を対極10にて良好に生じさせるものである。また、酸化還元物質が培養液4に留まることによって、培養液4の電位制御性を長期に亘って良好なものとすることができる。
【0038】
作用極9は、酸化還元物質の酸化体を還元体に還元することが可能な材質の電極、例えば炭素板やグラッシーカーボン等の炭素電極、白金電極等により構成される。但し、作用極9の材質は、これらに限定されるものではない。
【0039】
対極10は、作用極9における還元反応を補完する酸化反応を進行させ得る材質の電極、例えば作用極9と同様、炭素板やグラッシーカーボン等の炭素電極、白金電極等により構成される。但し、対極10の材質は、これらに限定されるものではない。
【0040】
気泡発生部13は、培養液4中に気泡を発生させて、培養液4を撹拌するものである。本実施形態では、水素ガスを含む不活性ガス(例えばN2ガス)を気泡発生部13に導入し、気泡発生部13の孔隙を介して水素ガスを含む不活性ガスの気泡を培養液4に供給し、この気泡により培養液4を撹拌するようにしている。但し、培養液4の撹拌は、本実施形態のように気泡発生部13を用いる形態には限定されず、例えば撹拌子を用いた撹拌としてもよい。また、培養液4の撹拌を行わずに本発明を実施するようにしてもよい。
【0041】
ここで、本発明のグリセロール代謝促進方法、エタノール生産方法及び培養方法は、パエニバチルス属の微生物が嫌気性微生物であることから、嫌気条件下にて実施される。嫌気環境は、例えば電気培養装置1を密閉構造のボックス(不図示)内に入れ、当該ボックス内に水素ガス、不活性ガス(窒素、希ガス)、またはそれらの混合ガスを充填することにより形成することができるが、嫌気環境の形成方法は、このような方法に限定されるものではない。例えば、図1に示す電気培養装置1の培養容器5を密閉構造とし、培養容器5内に水素ガス、不活性ガス、またはそれらの混合ガスを充填して嫌気環境を形成するようにしても勿論よい。
【0042】
以下、図1に示す電気培養装置1を利用した本発明の実施形態の一例を説明する。
【0043】
<グリセロール代謝促進方法>
本発明のグリセロール代謝促進方法は、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物2を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液4に投入して実施される。
【0044】
グリセロールは、不純物を含まない単体として培養液4に投入してもよいし、不純物が混入した実廃棄物をそのまま培養液4に投入してもよい。培養液4中のグリセロール濃度は、10g/L程度とすることが好適であるが、15g/L、さらには20g/L程度まで濃度を上昇させても十分に処理可能と考えられる。
【0045】
グルコース多量体は、グルコースが二分子以上結合した多量体であれば特に限定されるものではない。具体的には、二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物であるセロビオースやセルロース、二分子以上のグルコースが1,4’−α−グリコシド結合した炭化水素化合物であるマルトースやアミロース等が挙げられる。尚、グルコース多量体を構成するグルコース分子は、置換基等が付加された誘導体を構成分子としたものを適宜使用してもよい。例えば、セルロースに関して言えば、水溶性セルロースであるカルボキシメチルセルロースやヒドロキシエチルセルロースを用いてもよいし、不溶性セルロースであるアビセル(結晶性セルロース)やろ紙を用いてもよい。
【0046】
また、グルコース多量体は、不純物を含まない単体として培養液4に投入してもよいし、不純物が混入した実廃棄物をそのまま培養液4に投入してもよい。例えば、セルロースを含む実廃棄物等をそのまま培養液4に投入してもよい。
【0047】
培養液4中のグルコース多量体の濃度は、1g/L程度とすれば十分にグリセロール代謝を促進することができる。但し、培養液中のグルコース多量体の濃度は、グリセロール代謝を促進し得る濃度であれば特に限定されるものではなく、1g/Lの濃度に限定されるものではない。
【0048】
グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス属の微生物2としては、例えば16S rRNA遺伝子の塩基配列が配列番号1に示す塩基配列からなるパエニバチルス属の微生物を挙げることができる。このパエニバチルス属の微生物は、グリセロールからエタノールを産生するグリセロール代謝能を有している。
【0049】
16S rRNA遺伝子の塩基配列が配列番号1に示す塩基配列からなるパエニバチルス属の微生物としては、例えば、Paenibacillus macerans NS-1が挙げられる。この微生物は、独立行政法人 産業技術研究所 特許生物寄託センターに2011年5月19日付けで寄託されている(寄託番号FERM P−22124)。
【0050】
但し、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス属の微生物は、上記の微生物に限定されるものではなく、他のパエニバチルス属の微生物を用いることもできる。また、パエニバチルス属の微生物は、自然界に存在するものに限られず、遺伝子組み換えしてグリセロール代謝能を付与したものを用いてもよい。さらに、パエニバチルス属の微生物は、エタノール以外の代謝産物、例えばD−乳酸やアセトアルデヒド等を高効率に産生するものとしてもよい。
【0051】
また、パエニバチルス属の微生物は、1種または2種以上を培養液4に投入してもよいし、1種または2種以上のパエニバチルス属の微生物を他の微生物または微生物群集に混入している状態(例えば環境中から採取した土壌試料等の形態)で培養液4に投入してもよい。後者の場合においても、パエニバチルス属の微生物のグリセロール代謝の促進を図ることができる。
【0052】
ここで、パエニバチルス属の微生物を他の微生物または微生物群集に混入している状態で培養液4に投入する場合には、グルコース多量体として、二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物であるセロビオースやセルロース等を用いることが好ましい。二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物を炭素源として使用できる微生物は、パエニバチルス属の微生物以外では稀である。したがって、本発明の方法により、パエニバチルス属の微生物のグリセロール代謝を促進させれば、パエニバチルス以外の微生物に使用可能な炭素源を遮断することができ、パエニバチルス属の微生物を優占的に増殖させ易くして、パエニバチルス属の微生物のグリセロール代謝の促進を図り易くすることができる。
【0053】
尚、パエニバチルス属の微生物を他の微生物または微生物群集に混入している状態で培養液4に投入する場合とは、パエニバチルス属の微生物が他の微生物または微生物群集に混入している微生物試料を意図的に用いることを意味しているが、本発明においては、パエニバチルス属の微生物以外の他の微生物が非意図的に混入する場合にも適用できる。即ち、パエニバチルス属の微生物を培養する培養槽の滅菌が不十分で他の微生物が混在してしまった場合や、培養期間中に外部から培養槽に微生物が侵入して混在してしまうような場合(例えば非密閉系での培養)にも、パエニバチルス属の微生物を優占的に増殖させて、パエニバチルス属の微生物のグリセロール代謝の促進を図り易いものとできる。
【0054】
逆に、培養液4に実質的にパエニバチルス属の微生物のみしか投入しないような場合には、グルコース多量体として、二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物に限らず、二分子以上のグルコースが1,4’−α−グリコシド結合した炭化水素化合物を用いてもよいし、さらには、グルコース多量体の単位ユニットを構成するグルコースを用いてもグルコース多量体を用いた場合と同様の効果を奏し得る。
【0055】
次に、培養液4に接触させた電極(作用極9)に還元電位を印加する。還元電位とは、非通電時の培養液の酸化還元電位よりも低い電位を意味している。例えば、非通電時の培養液の酸化還元電位が+0.2Vであれば、+0.2Vよりも低い電位(X<+0.2V)が還元電位となる。還元電位を印加することで、電極から培養液に電子が供給され、これがパエニバチルス属の微生物に供給されて、グリセロール代謝の促進に何らかの形で寄与しているものと考えられる。
【0056】
還元電位の好適な範囲としては、銀・塩化銀電極電位基準で、−0.2V以下、より好適には−0.4V以下、さらに好適には−0.6V以下である。但し、電位値が低すぎると、水の電気分解等の目的外の反応が進行して、投入した電気エネルギーの一部がグリセロール代謝に寄与しなくなり、無駄となり得るので、−1.2V以上とすることが好適であり、−1.0V以上とすることがより好適であり、−0.8V以上とすることがさらに好適である。
【0057】
電極(作用極9)に還元電位を印加しながら培養を行うと、パエニバチルス属の微生物のグリセロール代謝が促進される。したがって、グリセロールの消費速度が向上する。これにより、未利用バイオマスとしてその処理に苦慮していたバイオディーゼル燃料生成時の副産物であるグリセロールを、微生物を利用して効率よく処理することができる。
【0058】
<有用物質生産方法>
本発明の有用物質生産方法は、上記のグリセロール代謝促進方法によりパエニバチルス属の微生物のグリセロール代謝を促進し、この際に産生される有用物質を回収するものである。
【0059】
例えば、パエニバチルス属の微生物としてPaenibacillus macerans NS-1を用いた場合には、グリセロール代謝によりエタノールが産生される。このエタノールを培養液4から例えば蒸留等により回収する。これにより、グリセロールをエタノール等の有用物質に変換して回収することができる。
【0060】
本発明の有用物質生産方法によれば、パエニバチルス属の微生物のグリセロール代謝を促進させて、グリセロールの消費速度を向上させ、且つエタノール等の有用物質の生産速度を向上させることができる。これにより、未利用バイオマスとしてその処理に苦慮していたバイオディーゼル燃料生成時の副産物であるグリセロールを、微生物を利用して効率よく処理しながらも、グリセロールを有用物質に効率よく変換して回収することができる。つまり、未利用バイオマスであるグリセロールにエタノール等の有用物質の原料としての付加価値を与えて、その利用の拡大・促進を図ることができる。
【0061】
<培養方法>
本発明の培養方法は、パエニバチルス属の微生物を他の微生物群集に混入した状態で培養液に投入することにより投入し、上記のグリセロール代謝促進方法と同様の操作を行うことで実施される。
【0062】
但し、本発明の培養方法においては、グルコース多量体として、二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物であるセロビオースやセルロース等を用いることが好ましい。二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物を炭素源として使用できる微生物は、パエニバチルス属の微生物以外では稀である。したがって、本発明の方法により、パエニバチルス属の微生物のグリセロール代謝を促進させれば、パエニバチルス以外の微生物に使用可能な炭素源を遮断することができ、パエニバチルス属の微生物を優占的に増殖させることができる。
【0063】
上述の形態は本発明の好適な形態の一例ではあるがこれに限定されるものではなく本発明の要旨を逸脱しない範囲において種々変形実施可能である。
【0064】
例えば、上述の実施形態では、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体を含む培養液に投入し、培養液に接触させた電極に還元電位を印加しながら培養して、パエニバチルス属の微生物のグリセロール代謝を促進するようにしていたが、電極に還元電位を印加することなく(つまり、通電することなく)、培養を行うようにしてもよい。この場合にも、電極に還元電位を印加したとき程ではないが、パエニバチルス属の微生物のグリセロール代謝を促進し得る。
【実施例】
【0065】
以下に本発明の実施例を説明するが、本発明はこれら実施例に限られるものではない。
【0066】
<実施例1>
1.実験方法
(1)微生物試料の調製
千葉県我孫子市の手賀沼の底泥に生息する環境微生物群を使用した。採取した汚泥10gを、純水100g中に懸濁させ、撹拌子により約30分間撹拌した。その後、ろ紙(No.1、ADVANTEC)により土粒子成分をろ過分離し、得られたろ液を微生物試料とした。
【0067】
尚、微生物試料に含まれる微生物を網羅的に解析した結果を図18に示す。パエニバチルス(Paenibacillus)属の微生物は、微生物試料中に存在する微生物の全量のうち、僅か0.01%しか存在していなかった(94位)。
【0068】
(2)培養液の調製
本実施例において使用した培養液の基本組成を以下に示す。
[培養液の基本組成:g/L]
・NH4Cl :0.5
・K2HPO4 :0.4
・MgCl2・6H2O :0.49
・CaCl2・2H2O :0.05
・KCl :0.052
【0069】
本実施例では、この基本組成の培養液に、酸化還元物質、セルロース、グリセロール、土壌抽出液を添加し、リン酸緩衝液にてpH7.2に調製したものを培養液として使用した。
【0070】
酸化還元物質は、Mn(II)−EDTAとした。添加量は2mMとした。
【0071】
セルロースは、水溶性セルロースであるヒドロキシエチルセルロース(和光純薬製、LotCEH0259)とした。添加量は1g/Lとした。
【0072】
グリセロールの添加量は1g/Lとした。
【0073】
土壌抽出液は、上記1の(1)欄で説明した底泥10gを純水100gに懸濁し、加熱型の撹拌器を用いて約1時間煮沸撹拌することで、土中に含まれる成分を抽出し、これを常温まで自然冷却させた後、ろ紙(No.1、ADVANTEC)により土粒子成分をろ過分離し、得られたろ液を使用した。添加量は26mL/Lとした。
【0074】
尚、電気培養の際に用いる電解液は、上記基本組成の培養液とした。
【0075】
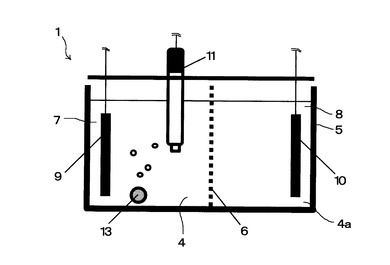
(3)電気培養槽
本実施例において使用した電気培養槽の模式図を図2に示す。ポリカーボネート製の容器(Kartell 351, Italy、幅70mm、奥行85mm、深さ45mm)5の内部を、イオン交換膜(Nafion(登録商標)N−117、デュポン社製)6で二層に仕切り、一方を培養槽7(容積65mL)、他方を電解質槽8(容積55mL)とした。
【0076】
各槽には、電極(作用極9、対極10)として炭素板(長さ75mm、幅35mm、厚さ2mm)を設置した。培養槽7には参照電極11(RE-1B, BAS株式会社)を設置した。
【0077】
さらに、培養液撹拌用の気泡器13を設置した。
【0078】
以上の各部品は、プラズマ滅菌装置(STERAD(登録商標)50、ジョンソン・エンド・ジョンソン社製)を用いて低温ガス滅菌をおこなった。
【0079】
培養の際には、別途殺菌処理を施した電位制御用の参照電極(銀・塩化銀電極 RE−1B、BAS社製)を培養槽に設置した。
【0080】
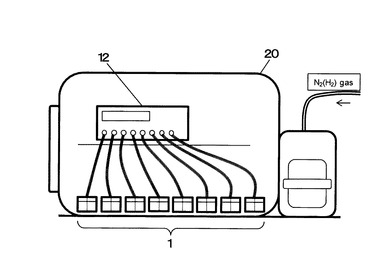
(4)電気培養試験
電気培養試験は、嫌気性ガス(4%H2含有N2ガス)を充填した嫌気チャンバー(AIRLOCK、COY社製)20内にて実施した(図3)。
【0081】
嫌気チャンバー内の温度は30℃に保持した。
【0082】
上記1の(3)欄にて説明した電気培養槽1を最大で8組、嫌気チャンバー20内に配置し、それぞれの電気培養槽1の培養槽7に培養液4を63mLを充填し、電解質槽8に電解液4aを55mLを充填した。
【0083】
続いて、多チャンネル型の電位制御装置12(8ch POTENTIOSTAT/GALVANOSTAT Model PS-08、東方技研社製)に各電極を結線するとともに、嫌気チャンバー内の嫌気ガスをポンプにより吸引し、各培養槽底部に設置した気泡器13から供給した。
【0084】
電位制御装置12によって、作用極9の電位を所定の電位(銀・塩化銀電極基準で+0.6V、+0.2V、−0.2V、−0.6V、通電無し(コントロール))に設定した後、培養液4全体の電位が安定するまで、約8時間放置した。その後、各培養槽7に微生物試料1.8mLを植菌し、3週間の電気培養を実施した。培養中、培養液の揮発を補うため、適宜、滅菌した純水を培養槽7に補充した。
【0085】
尚、以降の説明における電位の値は、全て銀・塩化銀電極を基準とする値である。
【0086】
(5)菌数計測
培養液中に生息する微生物の計測は、微生物計算盤(A−161、サンリード硝子社製)を使用し、顕微鏡観察により行った。
【0087】
(6)成分分析
培養液に含まれる有機物の成分分析は、液体クロマトグラフィー(Lachrome Elite、日立製)で測定した。カラムは、Gelpack(登録商標)(GL−C610H−S、7.8φ×300mm)を使用し、0.1%リン酸をバッファーとして用い、RI検出器により信号を得た。
【0088】
尚、計測時のオーブン温度は40℃とし、カラム内の流速は0.5mL/分とした。
【0089】
測定には、フィルターを通過し、微生物及び含有固形分を除去した培養液10μLを使用した。
【0090】
(7)菌叢解析
培養液に含まれる微生物の菌叢解析を、変性剤濃度勾配ゲル電気泳動法(PCR−DGGE:Polymerase Chain Reaction Denaturing Gradient Gel Electrophoresis)により、以下の手順で実施した。
【0091】
培養試験終了後の培養液(微生物懸濁液)0.5mLについて、12,000rpmで3分間遠心分離を行い、上澄みを取り除いて、沈殿物を得た。この沈殿物に細胞壁分解酵素を含むDNA抽出用バッファー(500U/mL アクロモペプターゼ、10mM Tris−HCl(pH8.0)、1mM EDTA、10mM NaCl)を100μL加えて微生物の細胞壁を破壊した。
【0092】
次に、InstaGene Matrix (Bio-Rad社) を100μL加え、精製したDNAを得た(参考文献1:Nagashima,K., et al., Food Sci. Technol. Res., 6, 115-118 (2000))。
【0093】
このDNAを鋳型とし、PCR法により16S rRNAをコードする約200bpのDNAを増殖した。
【0094】
DNAポリメラーゼにはAmpliTaqGold(Applied Biosystem社)を、サーマルサイクラーにはABI9700(Applied Biosystem社)を使用した。
【0095】
プライマーにはGC−341f及び534rを用いた(参考文献2:電力中央研究所報告、V08038(2009))。
【0096】
PCR条件は、94℃, 7 min, (94℃ 1 min, 65℃→55℃ 1 min (-1℃/2 cycle), 72℃ 2 min) × 20 cycle, (94℃ 1 min, 55℃ 1 min, 72℃ 2min) × 15 cycle, 72℃ 10 min とした。
【0097】
得られたPCR産物を、Dcode DGGEコンプリートシステム(Bio-Rad社)を用いてDGGE解析した。変性剤の濃度勾配は電気泳動方向に25%から65%とし、ポリアクリルアミドの濃度は8%とした。泳動条件は、電圧100V、12時間とした。
【0098】
PCR−DGGEにより得られた特徴的なバンドについて、ゲルより切り出し、以下の方法で微生物の同定をおこなった。
【0099】
切り出した切片より抽出したDNAを、BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystem社)のプロトコルに従い精製した。続いて、ABI PRISM 3100 Genetic Analyzer (Applied Biosystem社)を使用し、dideoxy nucleotide chain terimination法(参考資料3:Sanger, F., et al., Proc. Natl. Acad. Sci. USA., 74, 5463-5467 (1977))により塩基配列を決定した。得られた配列を、微生物同定用DNAデータベース系統解析システム「アポロン」(テクノスルガ・ラボ社)に供することで、微生物種を推定した。
【0100】
2.実験結果
(1)電気培養試験結果
電気培養試験期間における微生物密度変化を図4に示す。作用極への印加電位を+0.6V、+0.2V、通電無し(コントロール)とした場合には、微生物の増殖は見られなかった。これに対し、作用極への印加電位を−0.2Vとした場合には、微生物の若干の増殖が見られ、作用極への印加電位を−0.6Vとした場合には、顕著な微生物の増殖が見られ、3週間で約20倍に増殖した。
【0101】
次に、電気培養試験期間における培養液中のグリセロール濃度の経時変化を図5に示す。作用極への印加電位を+0.6V、+0.2V、通電無し(コントロール)とした場合には、グリセロール濃度の減少がほとんど見られなかった。これに対し、作用極への印加電位を−0.2Vとした場合には、グリセロール濃度が減少する傾向が見られた。そして、作用極への印加電位を−0.6Vとすることで、グリセロール濃度がさらに顕著に減少し、培養開始後2週間目にはグリセロールがほぼ消失した。
【0102】
次に、電気培養試験期間中における培養液中のエタノール濃度の経時変化を図6に示す。作用極への印加電位を+0.6V、+0.2V、通電無し(コントロール)とした場合には、エタノール濃度の増加が見られなかった。これに対し、作用極への印加電位を−0.2Vとした場合には、エタノール濃度が増加する傾向が見られた。そして、作用極への印加電位を−0.6Vとすることで、エタノール濃度がさらに顕著に増加した。
【0103】
ここで、電気培養試験期間中において、作用極への印加電位を−0.2V、−0.6Vとした場合には、セルロース濃度が若干減少する傾向が見られたものの、消費された有機物のほとんどがグリセロールであった。また、エタノール濃度の増加は、グリセロール濃度の減少に対して正の相関を有していた。これらのことからすれば、本実施例において増殖した微生物は、グリセロールをエタノールに変換する機能を有する微生物であることが明らかとなった。
【0104】
(2)菌叢解析結果
グリセロールからエタノールへの変換が顕著に見られた−0.6V条件での電気培養試験における3週間の培養後の微生物について、菌叢解析を実施した。結果を図7に示す。図7において、左はマーカーであり、中央は通電無し条件での電気培養試験における3週間の培養後の微生物について菌叢解析を実施した結果であり、右は−0.6V条件での電気培養試験における3週間の培養後の微生物について菌叢解析を実施した結果である。
【0105】
通電無し条件(中央)と−0.6V条件(右)を比較すると、−0.6V条件(右)では、通電無し条件(中央)では見られない複数のバンドが生じることが明らかとなった。
【0106】
そして、相同性検索を行った結果、−0.6V条件(右)に特有の3つのバンドは、全てパエニバチルス(Paenibacillus)属の近縁の微生物に帰属された。
【0107】
さらに解析を進めた結果、上2つのバンドは、Uncultured Paenibacillus(単離培養できておらず、遺伝子情報のみが登録されている)と97.4%の相同性を有していた。
【0108】
また、一番下のバンドは、Paenibacillus sp. FZ-1と100%の相同性を有していた。
【0109】
さらに、−0.6V条件での電気培養試験における3週間の培養後の微生物について、本願発明者等が、グリセロール変換活性を有する微生物の単離を試みたところ、一つの菌株を純粋分離することに成功した。この菌株は、Paenibacillus macerans NS-1と名付け、独立行政法人 産業技術研究所 特許生物寄託センターに2011年5月19日付けで寄託した(寄託番号FERM P−22124)。以下、この微生物をNS−1株と呼ぶこととする。表1〜3にNS−1株の菌学的性質を示す。
【0110】
【表1】

【0111】
【表2】
【0112】
【表3】
【0113】
尚、NS−1株とPaenibacillus sp. FZ-1との相同性について、配列番号1に示す1519bpの16S rRNA遺伝子の塩基配列に基づき、より高精度に比較検討を行った結果、相同性は98%であり、NS−1株とPaenibacillus sp. FZ-1が異なる菌株であることが明らかとなった。
【0114】
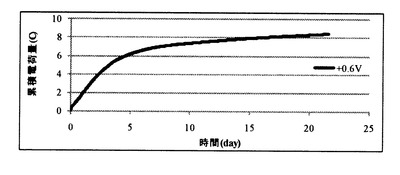
(3)作用極への印加電位と累積電荷量との関係
図8A及び図8Bに、種々の電位条件での電気培養試験において、培養液と電極(作用極)との間で移動した累積電荷量を示す。設定電位を+0.6Vとした場合には、8.4クーロンの電荷(電子)が培養液から引き抜かれていた。これに対し、設定電位を+0.2Vとした場合には、培養液からの電荷(電子)の引き抜きも培養液への電荷(電子)の供給も起こっておらず、設定電位を−0.2V、−0.6Vとした場合には、それぞれ約0.3、0.6クーロンの電荷(電子)が培養液に供給されていた。
【0115】
ここで、通電無し(コントロール)条件においては、培養期間中の酸化還元電位がほぼ+0.2Vで一定であったことから、設定電位+0.2Vでは、電荷の移動が生じなかったものと考えられる。
【0116】
また、パエニバチルス(Paenibacillus)属の微生物の増殖、グリセロールの減少及びエタノールの増加が見られた、設定電位−0.2V、−0.6V条件では、培養液に電荷(電子)が供給される条件であったことから、増殖したパエニバチルス(Paenibacillus)属の微生物において、グリセロール代謝が、還元電位を与えることによる電子の供給によって促進されたことが示唆された。
【0117】
本発明におけるグリセロール代謝促進メカニズムについて、図20に示すグリセロールからエタノールへの代謝経路に基づいて説明する。グリセロールからジヒドロキシアセトンリン酸への経路及びジヒドロキシアセトンリン酸からピルビン酸への経路は電子を放出する経路である。これに対し、ピルビン酸からアセトアルデヒドへの経路及びアセトアルデヒドからエタノールへの経路は電子が与えられる経路である。本発明では、還元電位を与えることによる電子の供給によって、ピルビン酸からアセトアルデヒドへの経路及びアセトアルデヒドからエタノールへの経路に電子が供給されてグリセロール代謝が促進されたものと推測された。
【0118】
<実施例2>
酸化還元物質がグリセロール変換活性に与える影響について検討した。
【0119】
実験方法は、実施例1と同様とし、酸化還元物質をFe(III)−EDTA、AQDS(アントラキノン−2,6−ジスルホン酸二ナトリウム)に変更した場合について、設定電位を−0.6Vとして検討した。
【0120】
グリセロール濃度の経時変化を図9に示し、エタノール濃度の経時変化を図10に示す。図中、◆は酸化還元物質をMn(II)−EDTA(以下、単にMnと呼ぶこともある)とした場合の結果であり、■は酸化還元物質をFe(III)−EDTA(以下、単にFeと呼ぶこともある)とした場合の結果であり、▲は酸化還元物質をAQDSとした場合の結果である。
【0121】
MnとAQDSではほぼ同じ傾向が見られ、培養開始2週間で溶存グリセロールはほぼ消失した。Feを用いた場合には、Mn、AQDSを用いた場合よりもグリセロール変換速度が低下する傾向が見られた。
【0122】
ここで、サイクリックボルタンメトリー測定におけるFe(III)−EDTAの還元電流ピークは−0.25V付近に存在する。一方、AQDSとMn(II)−EDTAの還元電流ピークは−0.6V付近に存在する(参考文献2:電力中央研究所報告、V08038(2009))。−0.6Vにおけるグリセロール変換速度の酸化還元物質による相違は、電極(作用極)から微生物への電子授受に起因している可能性が示唆された。即ち、AQDSとMn(II)−EDTAでは、−0.6V付近に還元電流ピークが存在することから、電極(作用極)から微生物への電子授受がスムーズに進行し、その結果として、Fe(III)−EDTAを用いた場合よりもグリセロール変換速度が加速された可能性が示唆された。
【0123】
但し、いずれの酸化還元物質を使用した際にも、エタノールの生成量とグリセロールの減少量との間に相関が見られたことから、酸化還元物質の違いによって代謝経路に変化が生じることはなかったと考えられる。
【0124】
<実施例3>
初期グリセロール濃度とエタノール生成量との関係について検討した。
【0125】
実験方法は、グリセロール濃度を以下の濃度とした以外には、同様とした。設定電位は−0.6Vとした。
[グリセロール濃度]
・A:1g/L(実施例1と同じ条件)
・B:2g/L
・C:4g/L
・D:9g/L
【0126】
培養液中のグリセロールとエタノールの初期濃度、及び培養3週間後の培養液中のグリセロールとエタノールの濃度を測定した結果を図11に示す。
【0127】
全ての試験区において、グリセロールはほぼ完全に消費されていた。そして、初期グリセロール濃度に比例したエタノールの生成が見られることが明らかとなった。この結果から、グリセロールの初期濃度が9g/Lであっても、グリセロールによる基質阻害や、エタノールによる生成物阻害を受けることなく、グリセロールの変換を生じさせ得ることが明らかとなった。
【0128】
<実施例4>
セルロースの有無の通電の有無による微生物のグリセロール代謝への影響について検討した。
【0129】
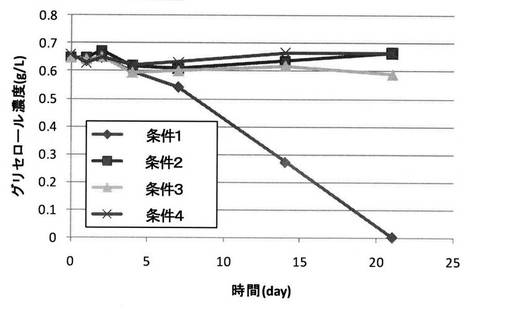
実験方法は、以下の条件以外は、実施例1と同様とした。
・条件1:セルロース有り(1g/L)、設定電位−0.6V
・条件2:セルロース有り(1g/L)、通電無し
・条件3:セルロース無し、設定電位−0.6V
・条件4:セルロール無し、通電無し
【0130】
菌体密度変化を図12に示し、グリセロール濃度の経時変化を図13に示し、セルロース濃度の経時変化を図14に示す。
【0131】
図12に示される結果から、条件1(セルロース有り(1g/L)、設定電位−0.6V)とすることで菌数の増加が確認できたものの、それ以外の条件では菌数の増加が確認できなかった。
【0132】
また、図13に示される結果から、条件1(セルロース有り(1g/L)、設定電位−0.6V)とすることでグルコースの減少(消費)が確認できたものの、それ以外の条件ではグルコースの減少(消費)が確認できなかった。
【0133】
これらの結果から、菌数を増加させてグリセロール変換を行うためには、培養液中のセルロースの存在と電極への還元電位の印加が必須であることが明らかとなった。
【0134】
尚、図14に示される結果から、通電の有無によらず、セルロースの減少(消費)に関してはほとんど相違が無いことが確認できたが、セルロースは数千のグルコースが結合した多量体であることから、セルロースを構成するグルコース単位で見れば、消費量に差がある可能性があるものと考えられた。
【0135】
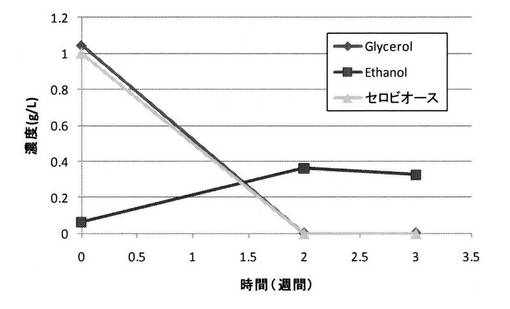
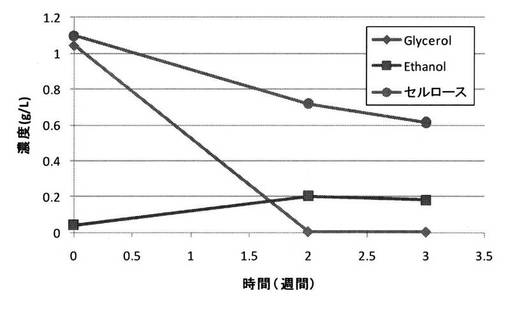
<実施例5>
セルロースに替えて、グルコースの二量体(より詳細には、二分子のグルコースが1,4’−β−グリコシド結合した炭化水素化合物)であるセロビオースを用いた場合にもグリセロールの変換が生じるかどうか検討した。
【0136】
実験方法は、以下の条件以外は、実施例1と同様とした。
・条件X:セルロースに替えてセロビオースを1g/L使用
・条件Y:セルロースを1g/L使用(実施例1と同じ条件)
※設定電位は共に−0.6V
【0137】
菌体密度変化の比較図を図15に示し、セロビオースを用いた場合(条件X)のセロビオース濃度、グリセロール濃度及びエタノール濃度の経時変化を図16に示し、セルロースを用いた場合(条件Y)のセルロース濃度、グリセロール濃度及びエタノール濃度の経時変化を図17に示す。
【0138】
図15に示される結果から、セロビオースを用いた場合にも菌数を増加させることができ、しかもその効果はセルロースを用いた場合よりも大きいことが明らかとなった。
【0139】
また、図16に示される結果から、セロビオースを用いた場合にも、セルロースを用いた場合(図17)と同様に、グリセロール変換を行うことが可能であることが明らかとなった。
【0140】
以上の結果から、セルロースに限らず、二分子のグルコースが1,4’−β−グリコシド結合した炭化水素化合物等といったグルコース多量体を用いることで、菌数を増加させてグリセロール変換を行うことが可能であることが明らかとなった。
【符号の説明】
【0141】
4 培養液
9 電極(作用極)
【技術分野】
【0001】
本発明は、微生物のグリセロール代謝促進方法に関する。さらに詳述すると、本発明は、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物のグリセロール代謝を促進する方法、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を利用したエタノール生産方法、及びグリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を雑多な微生物群集から優占的に増殖させる培養方法に関する。
【背景技術】
【0002】
近年、温室効果ガスの排出量削減や化石燃料依存からの脱却に向けた動きの一つとして、バイオマスを利用したバイオディーゼル燃料の生産が進められつつある。バイオディーゼル燃料とは、バイオマス由来の油脂類の主成分であるトリグリセライドをメタノールとエステル交換反応させることによって得られる液体燃料であり、脂肪酸メチルエステルを主体とするものである。
【0003】
ここで、図19に示すように、バイオディーゼル燃料の生成過程においては、グリセロールが副産物として生じる。アルカリ触媒を用いるバイオディーゼル燃料の生産方法においては、バイオディーゼル燃料に対して約10%のグリセロールが生成されるとされており(非特許文献1を参照)、その処理が懸案事項となっている。
【0004】
ところで、最近では、グリセロールからエタノールやポリエステルの原料となる1,3−プロパンジオール等の有用物質を産生するグリセロール代謝能を有する微生物の存在に関する報告がいくつかなされている(例えば、特許文献1及び非特許得文献2を参照)。このような微生物を利用してグリセロールを有用物質に変換することで、グリセロールを有効利用してその処理を促進することができる。したがって、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物は、上記懸案事項を解決に導き得る可能性を秘めていると言える。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2010−226959号公報
【非特許文献】
【0006】
【非特許文献1】横山伸也他、「バイオマスエネルギー」、森北出版株式会社(2009)
【非特許文献2】伊藤武志他「通性嫌気性細菌を利用した廃棄物処理と水素生産の検討」、弓削商船高等専門学校紀要、29(2007)
【発明の概要】
【発明が解決しようとする課題】
【0007】
グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物を利用して実際にグリセロールを処理する際には、グリセロール代謝能をできる限り高めて、処理効率を向上させることが望ましいと言える。しかしながら、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物に関する知見の蓄積は未だ乏しく、グリセロール代謝能を高めて、処理効率を向上させるような検討は殆ど行われていない状況である。
【0008】
また、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物が、雑多な微生物群集の中に埋もれていることがある。このような状況において、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物を優占的に増殖させる技術を確立できれば、非常に有用であると考えられる。
【0009】
そこで、本発明は、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物について、グリセロール代謝を促進する方法を提供することを目的とする。
【0010】
また、本発明は、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物を利用して、グリセロールを効率よく処理すると共に、有用物質を効率よく生産する方法を提供することを目的とする。
【0011】
さらに、本発明は、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物を、雑多な微生物群集の中から優占的に増殖させることのできる方法を提供することを目的とする。
【課題を解決するための手段】
【0012】
かかる課題を解決するため、本願発明者等が鋭意検討を行った結果、雑多な微生物群集からなる微生物試料を、グリセロールとセルロースを含む培養液に投入し、この培養液に接触させた電極に銀・塩化銀電極電位基準で−0.6Vの電位を印加して培養試験を行ったところ、ある種の微生物が優占的に増殖すると共に、グリセロールがエタノールに変換されていることを知見した。
【0013】
そこで、優占的に増殖した微生物について解析を行った結果、驚くべきことに、この微生物が、微生物試料に僅か0.01%の割合でした含まれていなかったパエニバチルス(Paenibacillus)属の微生物であることが判明した。
【0014】
そして、このパエニバチルス(Paenibacillus)属の微生物を単離し、資化性について検討したところ、グリセロール資化性を有していることが判明した。
【0015】
ところが、上記と同様の微生物試料を、グリセロールを含みセルロースを含まない培養液に投入して電極に通電せずに培養試験を行った場合には、パエニバチルス(Paenibacillus)属の微生物の優占的な増殖は見られなかった。このことは、グリセロールとセルロースを含む培養液に投入して電極に通電を行わずに培養試験を行った場合、グリセロールを含みセルロースを含まない培養液に投入して電極に銀・塩化銀電極電位基準で−0.6Vの電位を印加して培養試験を行った場合においても同様であった。
【0016】
このことから、培養液中のセルロースの存在及び電極への還元電位の印加が、パエニバチルス(Paenibacillus)属の微生物のグリセロール代謝の促進に寄与し、その結果としてパエニバチルス(Paenibacillus)属の微生物の優占的な増殖が起こっているものと考えられた。
【0017】
また、上記培養試験で用いたセルロースをセロビオースに替えて培養試験を実施したところ、セルロースを用いた培養試験結果とほぼ同じ結果が得られた。このことから、培養液中のグルコース多量体の存在、及び電極への還元電位の印加が、パエニバチルス(Paenibacillus)属の微生物のグリセロール代謝の促進に寄与し、その結果としてパエニバチルス(Paenibacillus)属の微生物の優占的な増殖が起こっているものと考えられた。
【0018】
以上のことから、本願発明者等は、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有する微生物について、グリセロールとグルコース多量体及び/又はグルコースとの存在下で、培養液に接触させた電位に還元電位を印加することで、グリセロール代謝を促進できる可能性が導かれることを知見するに至り、この知見に基づきさらに種々検討を重ねて、本発明を完成するに至った。
【0019】
即ち、本発明のグリセロール代謝促進方法は、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液に投入し、培養液に接触させた電極に還元電位を印加しながら培養して、パエニバチルス属の微生物のグリセロール代謝を促進するようにしている。
【0020】
ここで、本発明のグリセロール代謝促進方法において、パエニバチルス属の微生物は、16S rRNA遺伝子の塩基配列が配列番号1に示される塩基配列からなる微生物であることが好ましい。
【0021】
また、本発明のグリセロール代謝促進方法において、パエニバチルス属の微生物は、パエニバチルス マセランス(Paenibacillus macerans)NS−1(寄託番号FERM P−22124)であることが好ましい。
【0022】
次に、本発明の有用物質生産方法は、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液に投入し、培養液に接触させた電極に還元電位を印加しながら培養する工程と、有用物質を回収する工程とを含むようにしている。
【0023】
ここで、本発明の有用物質生産方法において、パエニバチルス属の微生物は、16S rRNA遺伝子の塩基配列が配列番号1に示される塩基配列からなる微生物であり、有用物質は、エタノールであることが好ましい。
【0024】
また、本発明の有用物質生産方法において、パエニバチルス属の微生物は、パエニバチルス マセランス(Paenibacillus macerans)NS−1(寄託番号FERM P−22124)であり、有用物質はエタノールであることが好ましい。
【0025】
次に、本発明の培養方法は、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を含む微生物群集を、グリセロールとグルコース多量体を含む培養液に投入し、グルコース多量体として二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物を用い、培養液に接触させた電極に還元電位を印加して、パエニバチルス属の微生物を優占的に増殖させるようにしている。
【発明の効果】
【0026】
本発明のグリセロール代謝促進方法によれば、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物について、グリセロール代謝を促進させることができる。したがって、グリセロールの処理速度を向上させて、グリセロール処理効率の向上を図ることができる。
【0027】
また、本発明の有用物質生産方法によれば、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を利用して、グリセロール代謝を促進させ、エタノール等の有用物質の生産速度を向上させることができる。したがって、グリセロールの処理効率を向上させると共にエタノール等の有用物質の生産効率を向上させることができる。
【0028】
さらに、本発明の培養方法によれば、グリセロールからエタノール等の有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、雑多な微生物群集の中から優占的に増殖させることができる。したがって、他の微生物によるグリセロール代謝の阻害を受けることなく、グリセロール代謝能を最大限に発揮できる微生物環境を構築することが可能となる。
【図面の簡単な説明】
【0029】
【図1】本発明を実施するための電気培養装置の一例を示す図ある。
【図2】実施例において使用した電気培養槽の模式図である。
【図3】電気培養試験において使用した嫌気チャンバーを示す図である。
【図4】実施例1の培養試験における菌体密度変化を示す図である。
【図5】実施例1の培養試験における培養液中のグリセロール濃度の経時変化を示す図である。
【図6】実施例1の培養試験における培養液中のエタノール濃度の経時変化を示す図である。
【図7】菌叢解析結果を示す電気泳動写真である。
【図8A】実施例1の培養試験において、設定電位を+0.6Vとしたときの累積電荷量を示す図である。
【図8B】実施例1の培養試験において、設定電位を+0.2V、−0.2V、−0.6Vとした場合の累積電荷量を示す図である。
【図9】実施例2の培養試験におけるグリセロール濃度の経時変化を示す図である。
【図10】実施例2の培養試験におけるエタノール濃度の経時変化を示す図である。
【図11】実施例3の培養試験結果(グリセロール濃度変化とエタノール濃度変化)を示す図である。
【図12】実施例4の培養試験における菌体密度変化を示す図である。
【図13】実施例4の培養試験におけるグリセロール濃度の経時変化を示す図である。
【図14】実施例4の培養試験におけるセルロース濃度の経時変化を示す図である。
【図15】実施例5の培養試験における菌体密度変化を示す図である。
【図16】実施例5の培養試験(セロビオース使用)の各種測定結果を示す図である。
【図17】実施例5の培養試験(セルロース使用)の各種試験結果を示す図である。
【図18】微生物試料に含まれる微生物を網羅的に解析した結果を示す図である(実施例1)。
【図19】油脂からバイオディーゼルを生成する過程を示す化学反応式である。
【図20】グリセロールからのエタノールの生成促進メカニズムを示す図である。
【発明を実施するための形態】
【0030】
以下、本発明を実施するための形態について、図面に基づいて詳細に説明する。
【0031】
本発明のグリセロール代謝促進方法、有用物質生産方法及び培養方法は、例えば図1に示す電気培養装置1により実施することができる。図1に示す電気培養装置1は、大まかには、培養容器5、培養容器5内を二槽に区画するイオン交換膜6、培養容器5の一方の槽(培養槽7)に収容された培養液4、培養容器5の他方の槽(電解質槽8)に収容された電解液4a、培養液4に浸された作用極9及び参照電極11、電解液4aに浸された対極10、作用極9と対極10と参照電極10を結線して三電極方式で作用極9の電位を制御する定電位設定装置(ポテンシオスタット)12、培養液4を撹拌するための気泡発生部13により構成されている。
【0032】
三電極方式で作用極9の電位を制御することで、作用極9の電位を厳密に設定電位に制御することができる。詳細には、定電位設定装置12により、作用極9と対極10との間に電流を流し、基準となる参照電極11には一切電流が流れないようにしている。尚、三電極方式による電位制御については、例えば、電気化学測定法(上)、技報動出版株式会社、第1版15刷、2004年6月発行の6〜9ページにその詳細が記載されている。但し、作用極9と対極10の極間電圧のみで作用極9の電位を制御できる場合には、三電極方式とせずともよい。
【0033】
培養液4としては、パエニバチルス属の微生物2の培養に適した培地成分を含む培養液を適宜使用することができる。例えば、NH4Cl、K2HPO4、MgCl2・6H2O、CaCl2・2H2O、KCl等の無機塩類を主体とする培地成分を含む培養液等を適宜用いることができる。また、土壌成分を抽出した土壌抽出液を培養液に添加してもよい。これにより、パエニバチルス属の微生物の生育が促進され得る。
【0034】
電解液4aとしては、例えばNaClやKCl等の電解質を含む溶液を適宜用いることができる。したがって、培養液4を電解液4aとして用いることも可能である。
【0035】
ここで、培養液4には、酸化還元物質3を添加することが好ましい。酸化還元物質3を培養液4に添加することで、培養液4の電位制御性を良好なものとすることができる。本発明に使用することができる酸化還元物質としては、パエニバチルス属の微生物2のグリセロール代謝能を阻害することのないものを適宜用いることができる。具体的には、鉄イオン、マンガンイオン等の金属イオン、アントラキノン−2,6−ジスルホン酸二ナトリウム等のキノン化合物が挙げられるが、これらに限定されるものではない。尚、金属イオンを用いる場合には、金属イオンをキレート剤に配位させて、金属イオンを培養液4中で安定に存在させるようにすることが好ましい。キレート剤としては、例えばジエチレントリアミンペンタ酢酸(DTPA)、エチレンジアミンテトラ酢酸(EDTA)、テトラエチレントリアミン(TET)、エチレンジアミン(EDA)、ジエチレントリアミン(DETA)、クエン酸、シュウ酸、クラウンエーテル、ニトリロテトラ酢酸、エデト酸二ナトリウム、エデト酸ナトリウム、エデト酸三ナトリウム、ペニシラミン、ペンテテートカルシウム三ナトリウム、ペンテト酸、スクシメルおよびエデト酸トリエンチン等、金属イオンを配位し得る任意のキレート剤を挙げることができる。
【0036】
酸化還元物質3の添加量は特に限定されるものではないが、0.1〜10mMとすることが好適であり、0.1〜5mMとすることがより好適であり、1〜3mMとすることがさらに好適である。
【0037】
イオン交換膜6は、培養液4に投入される微生物2、グリセロール、グリセロール多量体、酸化還元物質3を、電解質槽8に透過させることなく、培養液4中に留まらせ、且つ培養液4に含まれるイオンまたは電解液4aに含まれるイオンを透過させてイオン電流を生じさせ、作用極9において生じる還元反応を補完する酸化反応を対極10にて良好に生じさせるものである。また、酸化還元物質が培養液4に留まることによって、培養液4の電位制御性を長期に亘って良好なものとすることができる。
【0038】
作用極9は、酸化還元物質の酸化体を還元体に還元することが可能な材質の電極、例えば炭素板やグラッシーカーボン等の炭素電極、白金電極等により構成される。但し、作用極9の材質は、これらに限定されるものではない。
【0039】
対極10は、作用極9における還元反応を補完する酸化反応を進行させ得る材質の電極、例えば作用極9と同様、炭素板やグラッシーカーボン等の炭素電極、白金電極等により構成される。但し、対極10の材質は、これらに限定されるものではない。
【0040】
気泡発生部13は、培養液4中に気泡を発生させて、培養液4を撹拌するものである。本実施形態では、水素ガスを含む不活性ガス(例えばN2ガス)を気泡発生部13に導入し、気泡発生部13の孔隙を介して水素ガスを含む不活性ガスの気泡を培養液4に供給し、この気泡により培養液4を撹拌するようにしている。但し、培養液4の撹拌は、本実施形態のように気泡発生部13を用いる形態には限定されず、例えば撹拌子を用いた撹拌としてもよい。また、培養液4の撹拌を行わずに本発明を実施するようにしてもよい。
【0041】
ここで、本発明のグリセロール代謝促進方法、エタノール生産方法及び培養方法は、パエニバチルス属の微生物が嫌気性微生物であることから、嫌気条件下にて実施される。嫌気環境は、例えば電気培養装置1を密閉構造のボックス(不図示)内に入れ、当該ボックス内に水素ガス、不活性ガス(窒素、希ガス)、またはそれらの混合ガスを充填することにより形成することができるが、嫌気環境の形成方法は、このような方法に限定されるものではない。例えば、図1に示す電気培養装置1の培養容器5を密閉構造とし、培養容器5内に水素ガス、不活性ガス、またはそれらの混合ガスを充填して嫌気環境を形成するようにしても勿論よい。
【0042】
以下、図1に示す電気培養装置1を利用した本発明の実施形態の一例を説明する。
【0043】
<グリセロール代謝促進方法>
本発明のグリセロール代謝促進方法は、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物2を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液4に投入して実施される。
【0044】
グリセロールは、不純物を含まない単体として培養液4に投入してもよいし、不純物が混入した実廃棄物をそのまま培養液4に投入してもよい。培養液4中のグリセロール濃度は、10g/L程度とすることが好適であるが、15g/L、さらには20g/L程度まで濃度を上昇させても十分に処理可能と考えられる。
【0045】
グルコース多量体は、グルコースが二分子以上結合した多量体であれば特に限定されるものではない。具体的には、二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物であるセロビオースやセルロース、二分子以上のグルコースが1,4’−α−グリコシド結合した炭化水素化合物であるマルトースやアミロース等が挙げられる。尚、グルコース多量体を構成するグルコース分子は、置換基等が付加された誘導体を構成分子としたものを適宜使用してもよい。例えば、セルロースに関して言えば、水溶性セルロースであるカルボキシメチルセルロースやヒドロキシエチルセルロースを用いてもよいし、不溶性セルロースであるアビセル(結晶性セルロース)やろ紙を用いてもよい。
【0046】
また、グルコース多量体は、不純物を含まない単体として培養液4に投入してもよいし、不純物が混入した実廃棄物をそのまま培養液4に投入してもよい。例えば、セルロースを含む実廃棄物等をそのまま培養液4に投入してもよい。
【0047】
培養液4中のグルコース多量体の濃度は、1g/L程度とすれば十分にグリセロール代謝を促進することができる。但し、培養液中のグルコース多量体の濃度は、グリセロール代謝を促進し得る濃度であれば特に限定されるものではなく、1g/Lの濃度に限定されるものではない。
【0048】
グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス属の微生物2としては、例えば16S rRNA遺伝子の塩基配列が配列番号1に示す塩基配列からなるパエニバチルス属の微生物を挙げることができる。このパエニバチルス属の微生物は、グリセロールからエタノールを産生するグリセロール代謝能を有している。
【0049】
16S rRNA遺伝子の塩基配列が配列番号1に示す塩基配列からなるパエニバチルス属の微生物としては、例えば、Paenibacillus macerans NS-1が挙げられる。この微生物は、独立行政法人 産業技術研究所 特許生物寄託センターに2011年5月19日付けで寄託されている(寄託番号FERM P−22124)。
【0050】
但し、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス属の微生物は、上記の微生物に限定されるものではなく、他のパエニバチルス属の微生物を用いることもできる。また、パエニバチルス属の微生物は、自然界に存在するものに限られず、遺伝子組み換えしてグリセロール代謝能を付与したものを用いてもよい。さらに、パエニバチルス属の微生物は、エタノール以外の代謝産物、例えばD−乳酸やアセトアルデヒド等を高効率に産生するものとしてもよい。
【0051】
また、パエニバチルス属の微生物は、1種または2種以上を培養液4に投入してもよいし、1種または2種以上のパエニバチルス属の微生物を他の微生物または微生物群集に混入している状態(例えば環境中から採取した土壌試料等の形態)で培養液4に投入してもよい。後者の場合においても、パエニバチルス属の微生物のグリセロール代謝の促進を図ることができる。
【0052】
ここで、パエニバチルス属の微生物を他の微生物または微生物群集に混入している状態で培養液4に投入する場合には、グルコース多量体として、二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物であるセロビオースやセルロース等を用いることが好ましい。二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物を炭素源として使用できる微生物は、パエニバチルス属の微生物以外では稀である。したがって、本発明の方法により、パエニバチルス属の微生物のグリセロール代謝を促進させれば、パエニバチルス以外の微生物に使用可能な炭素源を遮断することができ、パエニバチルス属の微生物を優占的に増殖させ易くして、パエニバチルス属の微生物のグリセロール代謝の促進を図り易くすることができる。
【0053】
尚、パエニバチルス属の微生物を他の微生物または微生物群集に混入している状態で培養液4に投入する場合とは、パエニバチルス属の微生物が他の微生物または微生物群集に混入している微生物試料を意図的に用いることを意味しているが、本発明においては、パエニバチルス属の微生物以外の他の微生物が非意図的に混入する場合にも適用できる。即ち、パエニバチルス属の微生物を培養する培養槽の滅菌が不十分で他の微生物が混在してしまった場合や、培養期間中に外部から培養槽に微生物が侵入して混在してしまうような場合(例えば非密閉系での培養)にも、パエニバチルス属の微生物を優占的に増殖させて、パエニバチルス属の微生物のグリセロール代謝の促進を図り易いものとできる。
【0054】
逆に、培養液4に実質的にパエニバチルス属の微生物のみしか投入しないような場合には、グルコース多量体として、二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物に限らず、二分子以上のグルコースが1,4’−α−グリコシド結合した炭化水素化合物を用いてもよいし、さらには、グルコース多量体の単位ユニットを構成するグルコースを用いてもグルコース多量体を用いた場合と同様の効果を奏し得る。
【0055】
次に、培養液4に接触させた電極(作用極9)に還元電位を印加する。還元電位とは、非通電時の培養液の酸化還元電位よりも低い電位を意味している。例えば、非通電時の培養液の酸化還元電位が+0.2Vであれば、+0.2Vよりも低い電位(X<+0.2V)が還元電位となる。還元電位を印加することで、電極から培養液に電子が供給され、これがパエニバチルス属の微生物に供給されて、グリセロール代謝の促進に何らかの形で寄与しているものと考えられる。
【0056】
還元電位の好適な範囲としては、銀・塩化銀電極電位基準で、−0.2V以下、より好適には−0.4V以下、さらに好適には−0.6V以下である。但し、電位値が低すぎると、水の電気分解等の目的外の反応が進行して、投入した電気エネルギーの一部がグリセロール代謝に寄与しなくなり、無駄となり得るので、−1.2V以上とすることが好適であり、−1.0V以上とすることがより好適であり、−0.8V以上とすることがさらに好適である。
【0057】
電極(作用極9)に還元電位を印加しながら培養を行うと、パエニバチルス属の微生物のグリセロール代謝が促進される。したがって、グリセロールの消費速度が向上する。これにより、未利用バイオマスとしてその処理に苦慮していたバイオディーゼル燃料生成時の副産物であるグリセロールを、微生物を利用して効率よく処理することができる。
【0058】
<有用物質生産方法>
本発明の有用物質生産方法は、上記のグリセロール代謝促進方法によりパエニバチルス属の微生物のグリセロール代謝を促進し、この際に産生される有用物質を回収するものである。
【0059】
例えば、パエニバチルス属の微生物としてPaenibacillus macerans NS-1を用いた場合には、グリセロール代謝によりエタノールが産生される。このエタノールを培養液4から例えば蒸留等により回収する。これにより、グリセロールをエタノール等の有用物質に変換して回収することができる。
【0060】
本発明の有用物質生産方法によれば、パエニバチルス属の微生物のグリセロール代謝を促進させて、グリセロールの消費速度を向上させ、且つエタノール等の有用物質の生産速度を向上させることができる。これにより、未利用バイオマスとしてその処理に苦慮していたバイオディーゼル燃料生成時の副産物であるグリセロールを、微生物を利用して効率よく処理しながらも、グリセロールを有用物質に効率よく変換して回収することができる。つまり、未利用バイオマスであるグリセロールにエタノール等の有用物質の原料としての付加価値を与えて、その利用の拡大・促進を図ることができる。
【0061】
<培養方法>
本発明の培養方法は、パエニバチルス属の微生物を他の微生物群集に混入した状態で培養液に投入することにより投入し、上記のグリセロール代謝促進方法と同様の操作を行うことで実施される。
【0062】
但し、本発明の培養方法においては、グルコース多量体として、二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物であるセロビオースやセルロース等を用いることが好ましい。二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物を炭素源として使用できる微生物は、パエニバチルス属の微生物以外では稀である。したがって、本発明の方法により、パエニバチルス属の微生物のグリセロール代謝を促進させれば、パエニバチルス以外の微生物に使用可能な炭素源を遮断することができ、パエニバチルス属の微生物を優占的に増殖させることができる。
【0063】
上述の形態は本発明の好適な形態の一例ではあるがこれに限定されるものではなく本発明の要旨を逸脱しない範囲において種々変形実施可能である。
【0064】
例えば、上述の実施形態では、グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体を含む培養液に投入し、培養液に接触させた電極に還元電位を印加しながら培養して、パエニバチルス属の微生物のグリセロール代謝を促進するようにしていたが、電極に還元電位を印加することなく(つまり、通電することなく)、培養を行うようにしてもよい。この場合にも、電極に還元電位を印加したとき程ではないが、パエニバチルス属の微生物のグリセロール代謝を促進し得る。
【実施例】
【0065】
以下に本発明の実施例を説明するが、本発明はこれら実施例に限られるものではない。
【0066】
<実施例1>
1.実験方法
(1)微生物試料の調製
千葉県我孫子市の手賀沼の底泥に生息する環境微生物群を使用した。採取した汚泥10gを、純水100g中に懸濁させ、撹拌子により約30分間撹拌した。その後、ろ紙(No.1、ADVANTEC)により土粒子成分をろ過分離し、得られたろ液を微生物試料とした。
【0067】
尚、微生物試料に含まれる微生物を網羅的に解析した結果を図18に示す。パエニバチルス(Paenibacillus)属の微生物は、微生物試料中に存在する微生物の全量のうち、僅か0.01%しか存在していなかった(94位)。
【0068】
(2)培養液の調製
本実施例において使用した培養液の基本組成を以下に示す。
[培養液の基本組成:g/L]
・NH4Cl :0.5
・K2HPO4 :0.4
・MgCl2・6H2O :0.49
・CaCl2・2H2O :0.05
・KCl :0.052
【0069】
本実施例では、この基本組成の培養液に、酸化還元物質、セルロース、グリセロール、土壌抽出液を添加し、リン酸緩衝液にてpH7.2に調製したものを培養液として使用した。
【0070】
酸化還元物質は、Mn(II)−EDTAとした。添加量は2mMとした。
【0071】
セルロースは、水溶性セルロースであるヒドロキシエチルセルロース(和光純薬製、LotCEH0259)とした。添加量は1g/Lとした。
【0072】
グリセロールの添加量は1g/Lとした。
【0073】
土壌抽出液は、上記1の(1)欄で説明した底泥10gを純水100gに懸濁し、加熱型の撹拌器を用いて約1時間煮沸撹拌することで、土中に含まれる成分を抽出し、これを常温まで自然冷却させた後、ろ紙(No.1、ADVANTEC)により土粒子成分をろ過分離し、得られたろ液を使用した。添加量は26mL/Lとした。
【0074】
尚、電気培養の際に用いる電解液は、上記基本組成の培養液とした。
【0075】
(3)電気培養槽
本実施例において使用した電気培養槽の模式図を図2に示す。ポリカーボネート製の容器(Kartell 351, Italy、幅70mm、奥行85mm、深さ45mm)5の内部を、イオン交換膜(Nafion(登録商標)N−117、デュポン社製)6で二層に仕切り、一方を培養槽7(容積65mL)、他方を電解質槽8(容積55mL)とした。
【0076】
各槽には、電極(作用極9、対極10)として炭素板(長さ75mm、幅35mm、厚さ2mm)を設置した。培養槽7には参照電極11(RE-1B, BAS株式会社)を設置した。
【0077】
さらに、培養液撹拌用の気泡器13を設置した。
【0078】
以上の各部品は、プラズマ滅菌装置(STERAD(登録商標)50、ジョンソン・エンド・ジョンソン社製)を用いて低温ガス滅菌をおこなった。
【0079】
培養の際には、別途殺菌処理を施した電位制御用の参照電極(銀・塩化銀電極 RE−1B、BAS社製)を培養槽に設置した。
【0080】
(4)電気培養試験
電気培養試験は、嫌気性ガス(4%H2含有N2ガス)を充填した嫌気チャンバー(AIRLOCK、COY社製)20内にて実施した(図3)。
【0081】
嫌気チャンバー内の温度は30℃に保持した。
【0082】
上記1の(3)欄にて説明した電気培養槽1を最大で8組、嫌気チャンバー20内に配置し、それぞれの電気培養槽1の培養槽7に培養液4を63mLを充填し、電解質槽8に電解液4aを55mLを充填した。
【0083】
続いて、多チャンネル型の電位制御装置12(8ch POTENTIOSTAT/GALVANOSTAT Model PS-08、東方技研社製)に各電極を結線するとともに、嫌気チャンバー内の嫌気ガスをポンプにより吸引し、各培養槽底部に設置した気泡器13から供給した。
【0084】
電位制御装置12によって、作用極9の電位を所定の電位(銀・塩化銀電極基準で+0.6V、+0.2V、−0.2V、−0.6V、通電無し(コントロール))に設定した後、培養液4全体の電位が安定するまで、約8時間放置した。その後、各培養槽7に微生物試料1.8mLを植菌し、3週間の電気培養を実施した。培養中、培養液の揮発を補うため、適宜、滅菌した純水を培養槽7に補充した。
【0085】
尚、以降の説明における電位の値は、全て銀・塩化銀電極を基準とする値である。
【0086】
(5)菌数計測
培養液中に生息する微生物の計測は、微生物計算盤(A−161、サンリード硝子社製)を使用し、顕微鏡観察により行った。
【0087】
(6)成分分析
培養液に含まれる有機物の成分分析は、液体クロマトグラフィー(Lachrome Elite、日立製)で測定した。カラムは、Gelpack(登録商標)(GL−C610H−S、7.8φ×300mm)を使用し、0.1%リン酸をバッファーとして用い、RI検出器により信号を得た。
【0088】
尚、計測時のオーブン温度は40℃とし、カラム内の流速は0.5mL/分とした。
【0089】
測定には、フィルターを通過し、微生物及び含有固形分を除去した培養液10μLを使用した。
【0090】
(7)菌叢解析
培養液に含まれる微生物の菌叢解析を、変性剤濃度勾配ゲル電気泳動法(PCR−DGGE:Polymerase Chain Reaction Denaturing Gradient Gel Electrophoresis)により、以下の手順で実施した。
【0091】
培養試験終了後の培養液(微生物懸濁液)0.5mLについて、12,000rpmで3分間遠心分離を行い、上澄みを取り除いて、沈殿物を得た。この沈殿物に細胞壁分解酵素を含むDNA抽出用バッファー(500U/mL アクロモペプターゼ、10mM Tris−HCl(pH8.0)、1mM EDTA、10mM NaCl)を100μL加えて微生物の細胞壁を破壊した。
【0092】
次に、InstaGene Matrix (Bio-Rad社) を100μL加え、精製したDNAを得た(参考文献1:Nagashima,K., et al., Food Sci. Technol. Res., 6, 115-118 (2000))。
【0093】
このDNAを鋳型とし、PCR法により16S rRNAをコードする約200bpのDNAを増殖した。
【0094】
DNAポリメラーゼにはAmpliTaqGold(Applied Biosystem社)を、サーマルサイクラーにはABI9700(Applied Biosystem社)を使用した。
【0095】
プライマーにはGC−341f及び534rを用いた(参考文献2:電力中央研究所報告、V08038(2009))。
【0096】
PCR条件は、94℃, 7 min, (94℃ 1 min, 65℃→55℃ 1 min (-1℃/2 cycle), 72℃ 2 min) × 20 cycle, (94℃ 1 min, 55℃ 1 min, 72℃ 2min) × 15 cycle, 72℃ 10 min とした。
【0097】
得られたPCR産物を、Dcode DGGEコンプリートシステム(Bio-Rad社)を用いてDGGE解析した。変性剤の濃度勾配は電気泳動方向に25%から65%とし、ポリアクリルアミドの濃度は8%とした。泳動条件は、電圧100V、12時間とした。
【0098】
PCR−DGGEにより得られた特徴的なバンドについて、ゲルより切り出し、以下の方法で微生物の同定をおこなった。
【0099】
切り出した切片より抽出したDNAを、BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystem社)のプロトコルに従い精製した。続いて、ABI PRISM 3100 Genetic Analyzer (Applied Biosystem社)を使用し、dideoxy nucleotide chain terimination法(参考資料3:Sanger, F., et al., Proc. Natl. Acad. Sci. USA., 74, 5463-5467 (1977))により塩基配列を決定した。得られた配列を、微生物同定用DNAデータベース系統解析システム「アポロン」(テクノスルガ・ラボ社)に供することで、微生物種を推定した。
【0100】
2.実験結果
(1)電気培養試験結果
電気培養試験期間における微生物密度変化を図4に示す。作用極への印加電位を+0.6V、+0.2V、通電無し(コントロール)とした場合には、微生物の増殖は見られなかった。これに対し、作用極への印加電位を−0.2Vとした場合には、微生物の若干の増殖が見られ、作用極への印加電位を−0.6Vとした場合には、顕著な微生物の増殖が見られ、3週間で約20倍に増殖した。
【0101】
次に、電気培養試験期間における培養液中のグリセロール濃度の経時変化を図5に示す。作用極への印加電位を+0.6V、+0.2V、通電無し(コントロール)とした場合には、グリセロール濃度の減少がほとんど見られなかった。これに対し、作用極への印加電位を−0.2Vとした場合には、グリセロール濃度が減少する傾向が見られた。そして、作用極への印加電位を−0.6Vとすることで、グリセロール濃度がさらに顕著に減少し、培養開始後2週間目にはグリセロールがほぼ消失した。
【0102】
次に、電気培養試験期間中における培養液中のエタノール濃度の経時変化を図6に示す。作用極への印加電位を+0.6V、+0.2V、通電無し(コントロール)とした場合には、エタノール濃度の増加が見られなかった。これに対し、作用極への印加電位を−0.2Vとした場合には、エタノール濃度が増加する傾向が見られた。そして、作用極への印加電位を−0.6Vとすることで、エタノール濃度がさらに顕著に増加した。
【0103】
ここで、電気培養試験期間中において、作用極への印加電位を−0.2V、−0.6Vとした場合には、セルロース濃度が若干減少する傾向が見られたものの、消費された有機物のほとんどがグリセロールであった。また、エタノール濃度の増加は、グリセロール濃度の減少に対して正の相関を有していた。これらのことからすれば、本実施例において増殖した微生物は、グリセロールをエタノールに変換する機能を有する微生物であることが明らかとなった。
【0104】
(2)菌叢解析結果
グリセロールからエタノールへの変換が顕著に見られた−0.6V条件での電気培養試験における3週間の培養後の微生物について、菌叢解析を実施した。結果を図7に示す。図7において、左はマーカーであり、中央は通電無し条件での電気培養試験における3週間の培養後の微生物について菌叢解析を実施した結果であり、右は−0.6V条件での電気培養試験における3週間の培養後の微生物について菌叢解析を実施した結果である。
【0105】
通電無し条件(中央)と−0.6V条件(右)を比較すると、−0.6V条件(右)では、通電無し条件(中央)では見られない複数のバンドが生じることが明らかとなった。
【0106】
そして、相同性検索を行った結果、−0.6V条件(右)に特有の3つのバンドは、全てパエニバチルス(Paenibacillus)属の近縁の微生物に帰属された。
【0107】
さらに解析を進めた結果、上2つのバンドは、Uncultured Paenibacillus(単離培養できておらず、遺伝子情報のみが登録されている)と97.4%の相同性を有していた。
【0108】
また、一番下のバンドは、Paenibacillus sp. FZ-1と100%の相同性を有していた。
【0109】
さらに、−0.6V条件での電気培養試験における3週間の培養後の微生物について、本願発明者等が、グリセロール変換活性を有する微生物の単離を試みたところ、一つの菌株を純粋分離することに成功した。この菌株は、Paenibacillus macerans NS-1と名付け、独立行政法人 産業技術研究所 特許生物寄託センターに2011年5月19日付けで寄託した(寄託番号FERM P−22124)。以下、この微生物をNS−1株と呼ぶこととする。表1〜3にNS−1株の菌学的性質を示す。
【0110】
【表1】
【0111】
【表2】
【0112】
【表3】
【0113】
尚、NS−1株とPaenibacillus sp. FZ-1との相同性について、配列番号1に示す1519bpの16S rRNA遺伝子の塩基配列に基づき、より高精度に比較検討を行った結果、相同性は98%であり、NS−1株とPaenibacillus sp. FZ-1が異なる菌株であることが明らかとなった。
【0114】
(3)作用極への印加電位と累積電荷量との関係
図8A及び図8Bに、種々の電位条件での電気培養試験において、培養液と電極(作用極)との間で移動した累積電荷量を示す。設定電位を+0.6Vとした場合には、8.4クーロンの電荷(電子)が培養液から引き抜かれていた。これに対し、設定電位を+0.2Vとした場合には、培養液からの電荷(電子)の引き抜きも培養液への電荷(電子)の供給も起こっておらず、設定電位を−0.2V、−0.6Vとした場合には、それぞれ約0.3、0.6クーロンの電荷(電子)が培養液に供給されていた。
【0115】
ここで、通電無し(コントロール)条件においては、培養期間中の酸化還元電位がほぼ+0.2Vで一定であったことから、設定電位+0.2Vでは、電荷の移動が生じなかったものと考えられる。
【0116】
また、パエニバチルス(Paenibacillus)属の微生物の増殖、グリセロールの減少及びエタノールの増加が見られた、設定電位−0.2V、−0.6V条件では、培養液に電荷(電子)が供給される条件であったことから、増殖したパエニバチルス(Paenibacillus)属の微生物において、グリセロール代謝が、還元電位を与えることによる電子の供給によって促進されたことが示唆された。
【0117】
本発明におけるグリセロール代謝促進メカニズムについて、図20に示すグリセロールからエタノールへの代謝経路に基づいて説明する。グリセロールからジヒドロキシアセトンリン酸への経路及びジヒドロキシアセトンリン酸からピルビン酸への経路は電子を放出する経路である。これに対し、ピルビン酸からアセトアルデヒドへの経路及びアセトアルデヒドからエタノールへの経路は電子が与えられる経路である。本発明では、還元電位を与えることによる電子の供給によって、ピルビン酸からアセトアルデヒドへの経路及びアセトアルデヒドからエタノールへの経路に電子が供給されてグリセロール代謝が促進されたものと推測された。
【0118】
<実施例2>
酸化還元物質がグリセロール変換活性に与える影響について検討した。
【0119】
実験方法は、実施例1と同様とし、酸化還元物質をFe(III)−EDTA、AQDS(アントラキノン−2,6−ジスルホン酸二ナトリウム)に変更した場合について、設定電位を−0.6Vとして検討した。
【0120】
グリセロール濃度の経時変化を図9に示し、エタノール濃度の経時変化を図10に示す。図中、◆は酸化還元物質をMn(II)−EDTA(以下、単にMnと呼ぶこともある)とした場合の結果であり、■は酸化還元物質をFe(III)−EDTA(以下、単にFeと呼ぶこともある)とした場合の結果であり、▲は酸化還元物質をAQDSとした場合の結果である。
【0121】
MnとAQDSではほぼ同じ傾向が見られ、培養開始2週間で溶存グリセロールはほぼ消失した。Feを用いた場合には、Mn、AQDSを用いた場合よりもグリセロール変換速度が低下する傾向が見られた。
【0122】
ここで、サイクリックボルタンメトリー測定におけるFe(III)−EDTAの還元電流ピークは−0.25V付近に存在する。一方、AQDSとMn(II)−EDTAの還元電流ピークは−0.6V付近に存在する(参考文献2:電力中央研究所報告、V08038(2009))。−0.6Vにおけるグリセロール変換速度の酸化還元物質による相違は、電極(作用極)から微生物への電子授受に起因している可能性が示唆された。即ち、AQDSとMn(II)−EDTAでは、−0.6V付近に還元電流ピークが存在することから、電極(作用極)から微生物への電子授受がスムーズに進行し、その結果として、Fe(III)−EDTAを用いた場合よりもグリセロール変換速度が加速された可能性が示唆された。
【0123】
但し、いずれの酸化還元物質を使用した際にも、エタノールの生成量とグリセロールの減少量との間に相関が見られたことから、酸化還元物質の違いによって代謝経路に変化が生じることはなかったと考えられる。
【0124】
<実施例3>
初期グリセロール濃度とエタノール生成量との関係について検討した。
【0125】
実験方法は、グリセロール濃度を以下の濃度とした以外には、同様とした。設定電位は−0.6Vとした。
[グリセロール濃度]
・A:1g/L(実施例1と同じ条件)
・B:2g/L
・C:4g/L
・D:9g/L
【0126】
培養液中のグリセロールとエタノールの初期濃度、及び培養3週間後の培養液中のグリセロールとエタノールの濃度を測定した結果を図11に示す。
【0127】
全ての試験区において、グリセロールはほぼ完全に消費されていた。そして、初期グリセロール濃度に比例したエタノールの生成が見られることが明らかとなった。この結果から、グリセロールの初期濃度が9g/Lであっても、グリセロールによる基質阻害や、エタノールによる生成物阻害を受けることなく、グリセロールの変換を生じさせ得ることが明らかとなった。
【0128】
<実施例4>
セルロースの有無の通電の有無による微生物のグリセロール代謝への影響について検討した。
【0129】
実験方法は、以下の条件以外は、実施例1と同様とした。
・条件1:セルロース有り(1g/L)、設定電位−0.6V
・条件2:セルロース有り(1g/L)、通電無し
・条件3:セルロース無し、設定電位−0.6V
・条件4:セルロール無し、通電無し
【0130】
菌体密度変化を図12に示し、グリセロール濃度の経時変化を図13に示し、セルロース濃度の経時変化を図14に示す。
【0131】
図12に示される結果から、条件1(セルロース有り(1g/L)、設定電位−0.6V)とすることで菌数の増加が確認できたものの、それ以外の条件では菌数の増加が確認できなかった。
【0132】
また、図13に示される結果から、条件1(セルロース有り(1g/L)、設定電位−0.6V)とすることでグルコースの減少(消費)が確認できたものの、それ以外の条件ではグルコースの減少(消費)が確認できなかった。
【0133】
これらの結果から、菌数を増加させてグリセロール変換を行うためには、培養液中のセルロースの存在と電極への還元電位の印加が必須であることが明らかとなった。
【0134】
尚、図14に示される結果から、通電の有無によらず、セルロースの減少(消費)に関してはほとんど相違が無いことが確認できたが、セルロースは数千のグルコースが結合した多量体であることから、セルロースを構成するグルコース単位で見れば、消費量に差がある可能性があるものと考えられた。
【0135】
<実施例5>
セルロースに替えて、グルコースの二量体(より詳細には、二分子のグルコースが1,4’−β−グリコシド結合した炭化水素化合物)であるセロビオースを用いた場合にもグリセロールの変換が生じるかどうか検討した。
【0136】
実験方法は、以下の条件以外は、実施例1と同様とした。
・条件X:セルロースに替えてセロビオースを1g/L使用
・条件Y:セルロースを1g/L使用(実施例1と同じ条件)
※設定電位は共に−0.6V
【0137】
菌体密度変化の比較図を図15に示し、セロビオースを用いた場合(条件X)のセロビオース濃度、グリセロール濃度及びエタノール濃度の経時変化を図16に示し、セルロースを用いた場合(条件Y)のセルロース濃度、グリセロール濃度及びエタノール濃度の経時変化を図17に示す。
【0138】
図15に示される結果から、セロビオースを用いた場合にも菌数を増加させることができ、しかもその効果はセルロースを用いた場合よりも大きいことが明らかとなった。
【0139】
また、図16に示される結果から、セロビオースを用いた場合にも、セルロースを用いた場合(図17)と同様に、グリセロール変換を行うことが可能であることが明らかとなった。
【0140】
以上の結果から、セルロースに限らず、二分子のグルコースが1,4’−β−グリコシド結合した炭化水素化合物等といったグルコース多量体を用いることで、菌数を増加させてグリセロール変換を行うことが可能であることが明らかとなった。
【符号の説明】
【0141】
4 培養液
9 電極(作用極)
【特許請求の範囲】
【請求項1】
グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液に投入し、前記培養液に接触させた電極に還元電位を印加しながら培養して、前記パエニバチルス属の微生物のグリセロール代謝を促進することを特徴とするグリセロール代謝促進方法。
【請求項2】
前記パエニバチルス属の微生物は、16S rRNA遺伝子の塩基配列が配列番号1に示される塩基配列からなる微生物である請求項1に記載のグリセロール代謝促進方法。
【請求項3】
前記パエニバチルス属の微生物は、パエニバチルス マセランス(Paenibacillus macerans)NS−1(寄託番号FERM P−22124)である請求項1に記載のグリセロール代謝促進方法。
【請求項4】
グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液に投入し、前記培養液に接触させた電極に還元電位を印加しながら培養する工程と、
前記有用物質を回収する工程と
を含むことを特徴とする有用物質生産方法。
【請求項5】
前記パエニバチルス属の微生物は、16S rRNA遺伝子の塩基配列が配列番号1に示される塩基配列からなる微生物であり、
前記有用物質はエタノールである
請求項4に記載の有用物質生産方法。
【請求項6】
前記パエニバチルス属の微生物は、パエニバチルス マセランス(Paenibacillus macerans)NS−1(寄託番号FERM P−22124)であり、
前記有用物質はエタノールである
請求項4に記載の有用物質生産方法。
【請求項7】
グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を含む微生物群集を、グリセロールとグルコース多量体を含む培養液に投入し、前記グルコース多量体として二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物を用い、前記培養液に接触させた電極に還元電位を印加して、前記パエニバチルス属の微生物を優占的に増殖させることを特徴とする培養方法。
【請求項1】
グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液に投入し、前記培養液に接触させた電極に還元電位を印加しながら培養して、前記パエニバチルス属の微生物のグリセロール代謝を促進することを特徴とするグリセロール代謝促進方法。
【請求項2】
前記パエニバチルス属の微生物は、16S rRNA遺伝子の塩基配列が配列番号1に示される塩基配列からなる微生物である請求項1に記載のグリセロール代謝促進方法。
【請求項3】
前記パエニバチルス属の微生物は、パエニバチルス マセランス(Paenibacillus macerans)NS−1(寄託番号FERM P−22124)である請求項1に記載のグリセロール代謝促進方法。
【請求項4】
グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を、グリセロールとグルコース多量体及び/又はグルコースとを含む培養液に投入し、前記培養液に接触させた電極に還元電位を印加しながら培養する工程と、
前記有用物質を回収する工程と
を含むことを特徴とする有用物質生産方法。
【請求項5】
前記パエニバチルス属の微生物は、16S rRNA遺伝子の塩基配列が配列番号1に示される塩基配列からなる微生物であり、
前記有用物質はエタノールである
請求項4に記載の有用物質生産方法。
【請求項6】
前記パエニバチルス属の微生物は、パエニバチルス マセランス(Paenibacillus macerans)NS−1(寄託番号FERM P−22124)であり、
前記有用物質はエタノールである
請求項4に記載の有用物質生産方法。
【請求項7】
グリセロールから有用物質を産生するグリセロール代謝能を有するパエニバチルス(Paenibacillus)属の微生物を含む微生物群集を、グリセロールとグルコース多量体を含む培養液に投入し、前記グルコース多量体として二分子以上のグルコースが1,4’−β−グリコシド結合した炭化水素化合物を用い、前記培養液に接触させた電極に還元電位を印加して、前記パエニバチルス属の微生物を優占的に増殖させることを特徴とする培養方法。
【図1】

【図2】
【図3】
【図8A】
【図8B】
【図19】

【図20】
【図4】

【図5】

【図6】
【図7】

【図9】
【図10】

【図11】

【図12】

【図13】
【図14】

【図15】

【図16】
【図17】
【図18】

【図2】
【図3】
【図8A】
【図8B】
【図19】
【図20】
【図4】
【図5】
【図6】
【図7】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公開番号】特開2013−16(P2013−16A)
【公開日】平成25年1月7日(2013.1.7)
【国際特許分類】
【出願番号】特願2011−131899(P2011−131899)
【出願日】平成23年6月14日(2011.6.14)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成23年3月5日 社団法人日本農芸化学会発行の「日本農芸化学会 2011年度大会 講演要旨集」に発表
【出願人】(000173809)一般財団法人電力中央研究所 (1,040)
【Fターム(参考)】
【公開日】平成25年1月7日(2013.1.7)
【国際特許分類】
【出願日】平成23年6月14日(2011.6.14)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 平成23年3月5日 社団法人日本農芸化学会発行の「日本農芸化学会 2011年度大会 講演要旨集」に発表
【出願人】(000173809)一般財団法人電力中央研究所 (1,040)
【Fターム(参考)】
[ Back to top ]