
微生物由来の新規コラゲナーゼ遺伝子

【課題】
活性の高いコラゲナーゼを得るために、微生物由来のコラゲナーゼをコードする新規な遺伝子配列を決定し、遺伝子工学的手法を用いた当該コラゲナーゼ生産のための技術を構築すること。
【解決手段】
グリモンティア・ホリセー由来のコラゲナーゼ遺伝子、該遺伝子が組み込まれた組換えベクター、該ベクターにより形質転換された宿主細胞、並びにこれらを利用してコラゲナーゼを製造する方法を提供する。
活性の高いコラゲナーゼを得るために、微生物由来のコラゲナーゼをコードする新規な遺伝子配列を決定し、遺伝子工学的手法を用いた当該コラゲナーゼ生産のための技術を構築すること。
【解決手段】
グリモンティア・ホリセー由来のコラゲナーゼ遺伝子、該遺伝子が組み込まれた組換えベクター、該ベクターにより形質転換された宿主細胞、並びにこれらを利用してコラゲナーゼを製造する方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は活性の高いコラゲナーゼを得るための、グリモンティア・ホリセー由来のコラゲナーゼ遺伝子、該遺伝子が組み込まれた組換えベクター、該ベクターにより形質転換された宿主細胞およびその利用に関する。
【背景技術】
【0002】
コラーゲンは細胞外マトリックスの主要タンパク質として各臓器に広く分布しており、哺乳動物では全タンパク質の約30%を占める。現在までに、遺伝子として29種類の型のコラーゲンが同定されており、分子構造や会合体の形成様式によりサブファミリーに分類されている。その中でもI型コラーゲンは皮膚や骨や腱を構成する線維性コラーゲンの主成分である。I型コラーゲンおよびその熱変性物であるゼラチンは工業的に生産され、食品、化粧品、医療等の分野で利用されている。I型コラーゲンは約10万の分子量をもつ3本のポリペプチド鎖が三重らせん構造をとった棒状の分子である。コラーゲン分子を構成する各ポリペプチドのアミノ酸配列はGly−X−Y(XおよびYは種々のアミノ酸残基を示す、Xはプロリン、Yはヒドロキシプロリンである頻度が高い)の繰返しであり、各ポリペプチドは分子内または分子間で架橋されている。I型コラーゲンは生体中で自己会合して線維を形成しており、組織に機械的特性と化学的安定性をもたらしている。コラーゲン特有の三重らせん構造は通常のプロテアーゼに対して抵抗性を示すが、コラゲナーゼには分解される。微生物由来のコラゲナーゼは、コラーゲンあるいはその変性物であるゼラチンに作用し、Gly−X−Yの単位にまで分解することが知られている。その特異的な性質を利用して生化学的研究や医療分野、さらに食品加工にも広く利用されている。
【0003】
従来、入手可能な微生物由来の主なコラゲナーゼ精製品は、クロストリジウム属由来、ビブリオ・アルギノリティカス・ケモバル・イオファガス(Vibrio alginolyticus chemovar.iophagus)由来、ストレプトミセス(Streptomyces)属由来のコラゲナーゼである。
【0004】
最も利用されているコラゲナーゼはクロストリジウム・ヒストリティカム(Clostridium histolyticum)由来のコラゲナーゼである。クロストリジウム・ヒストリティカムはガス壊疽菌として古くから知られている嫌気性の病原菌であり、毒素を産生する。従って該菌の大量培養によるコラゲナーゼの大量生産には適さないという問題があるが、近年は遺伝子組換え体が利用されている(特許第2634782、特表2001-510331)。クロストリジウム属のコラゲナーゼは種々の生化学的研究に研究試薬として使用されるだけでなく、たとえば組織の消化と個々の細胞または細胞群の単離(例えば膵臓から膵島(islet)の分離)に使用される(膵島:Suttonら、Transplantation 、42 (1986) 689-691;肝:Quibelら、Anal.Biochem.、154 (1986) 26-28;骨:Hefleyら、J.Bone Mineral Res.、2 (1987) 505-516;臍帯:Holzingerら、Immunol. Lett.、35 (1993) 109−118)。
【0005】
ビブリオ・アルギノリティカス・ケモバル・イオファガス(Vibrio alginolyticus chemovar.iophagus)由来のコラゲナーゼも種々の用途で使用されている。たとえば、コラーゲンに富む構造の無制限な基質が生じる火傷、潰瘍、痂皮、コラーゲンベースの白色硬痂、ケロイド、壊死とくに臥位または潰瘍による壊死の治療に使用される。虫歯の治療にも使用される。その他、肉質軟化剤としても使用できる。ビブリオ・アルギノリティカス由来のコラゲナーゼには安価な取得が困難であるという問題があるが、この酵素の遺伝子取得によるアミノ酸全配列が明らかにされたことで遺伝子工学的手法を用いることが可能となった(特許第3022984)。
【0006】
ストレプトミセス(Streptomyces)属由来コラゲナーゼは、公知の培養法により放線菌を好気的に培養した培養液より精製して得られる(特公平5-16832)。既存添加物名簿にストレプトミセス(Streptomyces)属のプロテアーゼが収載されており、同属由来のコラゲナーゼは他の2種とは異なって食品に使用することができる。
【0007】
既存のコラゲナーゼは安定性に難があるのに対して、安定性に優れさらに比活性が高い微生物由来コラゲナーゼである、ビブリオ・ホリセー(Vibrio hollisae)由来のコラゲナーゼが発見された(鈴木ら、皮革科学 44 (1998) 64-71、鈴木、皮革科学 45 (2000) 272-283、特開平8-70853)。しかし、この報告ではビブリオ・ホリセー(Vibrio hollisae)由来のコラゲナーゼをコードする遺伝子は特定されておらず、それが既存のコラゲナーゼとは異なる新規な酵素であることを示すには不十分であった。また、遺伝子工学的手法による同酵素の大量取得にも対応できていなかった。
【0008】
尚、ビブリオ・ホリセーは16S rRNAの相同性により、新たにグリモンティア・ホリセー(Grimontia hollisae)と分類し直された(Thompsonら、 Int. J. Syst. Evol. Microbiol.、 53 (2003) 1615-1617)。本明細書ではビブリオ・ホリセーをグリモンティア・ホリセーとして表記する。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】特許第2634782号公報
【特許文献2】特表2001-510331号公報
【特許文献3】特許第3022984号公報
【特許文献4】特公平5-16832号公報
【特許文献5】特開平8-70853号公報
【非特許文献】
【0010】
【非特許文献1】Suttonら、Transplantation、42 (1986) p.689-691
【非特許文献2】Quibelら、Anal. Biochem.、154 (1986) p.26-28
【非特許文献3】Hefleyら、J. Bone Mineral Res.、2 (1987) p.505-516
【非特許文献4】Holzingerら、Immunol. Lett.、35 (1993) p.109-118
【非特許文献5】鈴木ら、皮革科学 44 (1998) p.64-71
【非特許文献6】鈴木、皮革科学 45 (2000) p.272-283
【非特許文献7】Thompsonら、 Int. J. Syst. Evol. Microbiol.、53 (2003) p.1615-1617
【発明の概要】
【発明が解決しようとする課題】
【0011】
グリモンティア・ホリセー(Grimontia hollisae)由来コラゲナーゼは優れた比活性と安定性を有し、クロストリジウム・ヒストリティカム等由来の既存のコラゲナーゼに替わる新規コラゲナーゼとして研究用試薬や医療用途への利用が期待される。その一方で、安価な取得が困難であるという問題がある。当該コラゲナーゼはその産生菌であるグリモンティア・ホリセーを培養し、その培養液より回収・精製できるが、当該微生物のコラゲナーゼ産生量は1mg/Lと著しく低く、生産効率は実用的ではない。そのため、当該コラゲナーゼをコードする遺伝子配列を決定すること、そして決定した遺伝子配列をもとに遺伝子工学的手法を用いた当該コラゲナーゼ生産効率化のための技術を構築することが本発明の課題である。
【課題を解決するための手段】
【0012】
本発明者らはグリモンティア・ホリセーに属する微生物由来のコラゲナーゼ遺伝子(明細書中では、本発明のコラゲナーゼ遺伝子と呼ぶこともある)を取得し、そのアミノ酸配列を明らかにすることに成功し、本発明を完成した。これにより、遺伝子工学的手法を用いた当該コラゲナーゼの産生系を構築することが可能となった。すなわち、本発明の概略は以下の通りである。
【0013】
(1)少なくとも下記式(a)ないし(g)で示されるペプチド断片を有するグリモンティア・ホリセー(Grimontia hollisae)に属する微生物由来のコラゲナーゼをコードする遺伝子。
【0014】
式(a) AVEQCDL
式(b) GAVDAFV
式(c) IRSQDMSQAQHLAACDK
式(d) MAYEESFFHTTLE
式(e) WGYL
式(f) FSSNHMY
式(g) ASYAN
但し、式(a)ないし(g)中のアルファベットは以下のアミノ酸を示す。A:アラニン、C:システイン、D:アスパラギン酸、E:グルタミン酸 F:フェニルアラニン、G:グリシン、H:ヒスチジン、I:イソロイシン、K:リシン、L:ロイシン、M:メチオニン、N:アスパラギン、P:プロリン、Q:グルタミン、R:アルギニン、S:セリン、T:スレオニン、V:バリン、W:トリプトファン、Y:チロシン。
【0015】
(2)以下の(h)または(i)のタンパク質をコードする遺伝子。
(h)下記式(I)で表されるアミノ酸配列からなるグリモンティア・ホリセー(Grimontia hollisae)に属する微生物由来のコラゲナーゼタンパク質
式(I): X-
AVEQCDLSQFQTTSSNQLMAAIRQQGASCVNALFSADTGVQEAAFSSNHMYNVAQYTRTLAQQYAGGGSDELEALYLYLRAGYYAEFYNSNITFLSWVTPAVKGAVDAFVQNAHFYDNGDAHGKVLNEVIITMDSAGLQHAYLDVVTQWLTRWNAQYAEHWYMRNAVNGVFTLLFGGQWNNQYTSLIGEQTALVTALQAFALDRTKVNSPTEFMAANAARELGRLARYTDATIAPKVTEGLTAIFGQYPSYGDGDAIWLGAADTASYYADCSQFNICGFEDALRDAALNQTFICSDTIKIRSQDMSQAQHLAACDKMAYEESFFHTTLETGNQPVADDHNTQLQVNIFNSDTDYGKYAGPIFGIDTNNGGMYLEGNPANVGNIPNFIAYEASYANPDHFVWNLEHEYVHYLDGRFNMYGDFGTPTELVVWWSEGVAEYVSRVNDNPQAIATIQDGSTYTLAQVFDTTYDGFDVDRIYRWGYLAVRFMFERHPDEVQRMLSATRQGRWAEYKAIISGWANQYQSEFAQWTEALAKGDSGAGNGEGTGSGNEGGGESGGNTGLPENCAVLPKISDGRLALDEAACLADTASASDVLWFSIPAVSEYQTIAITAGNGTGDLTLEYSNLNWPDGTNVQASSANMGNSECIILEHQANYWGYLKVSGSFENAALLVEAGSNQCRQ
但し、式(I)中、Xは、水素原子または次式(II)で表されるポリペプチドを示す。
式(II):MELKALALTVSALLVGQSVYASEVFAHPGMPVQQKLQPNLLQQSTRLQPEQHIHGLERTDRQYRPTDATQQPEPPTLLKRQVSVQQD
また、式(I)、(II)中のアルファベットは、(1)と同義である。
(i)式(I)で表されるアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつコラゲナーゼ活性を有するタンパク質。
【0016】
(3)以下の(j)、(k)、(l)または(m)のDNAからなるコラゲナーゼ遺伝子:
(j)グリモンティア・ホリセーに属する微生物由来のコラゲナーゼをコードする、下記式(III)で表される塩基配列からなるDNA
式(III):Z-
GCGGTTGAACAGTGTGATCTCAGCCAGTTTCAGACGACCAGCAGTAACCAGTTGATGGCCGCGATTCGCCAGCAGGGCGCCAGTTGTGTCAATGCCTTGTTTAGTGCTGACACGGGAGTACAGGAAGCCGCGTTTTCGTCCAACCACATGTATAACGTCGCACAATATACCCGAACACTGGCGCAGCAATATGCGGGCGGGGGCAGTGATGAACTGGAAGCCTTATATCTGTATCTTCGGGCGGGGTATTACGCTGAGTTTTATAACAGCAATATCACGTTTCTGTCGTGGGTGACGCCGGCAGTGAAAGGTGCCGTGGATGCGTTTGTCCAGAATGCCCATTTTTATGACAACGGCGACGCTCATGGCAAGGTGCTGAACGAGGTGATCATCACGATGGACAGTGCTGGGCTACAGCATGCCTATCTCGATGTAGTGACGCAGTGGCTGACCCGTTGGAATGCGCAATATGCCGAGCATTGGTATATGCGAAACGCAGTCAATGGTGTCTTTACTCTTTTGTTTGGCGGGCAGTGGAACAATCAGTACACCAGCCTGATTGGCGAACAGACTGCGTTGGTGACGGCCTTACAGGCATTTGCGCTGGACCGTACGAAAGTGAACTCGCCAACGGAGTTTATGGCGGCCAATGCGGCCAGAGAACTGGGGCGGTTAGCTCGCTACACGGACGCGACCATTGCGCCTAAAGTCACGGAAGGATTAACCGCGATCTTCGGCCAGTATCCGTCCTATGGCGATGGAGATGCTATCTGGCTGGGGGCGGCGGATACGGCCTCTTATTATGCTGATTGCAGCCAGTTCAACATCTGTGGCTTTGAAGACGCGCTGCGTGATGCGGCGCTGAACCAGACTTTTATCTGTAGTGATACGATTAAAATTCGCTCACAGGATATGTCGCAGGCACAGCATCTCGCGGCTTGCGACAAAATGGCTTATGAAGAGTCATTTTTCCACACCACGCTTGAAACCGGTAATCAGCCGGTGGCTGATGATCATAATACGCAGCTGCAGGTGAATATTTTTAATTCCGATACCGATTACGGTAAATATGCCGGTCCGATATTTGGGATTGATACCAACAACGGCGGTATGTACCTCGAGGGGAATCCGGCCAATGTGGGCAATATTCCCAATTTCATCGCGTATGAAGCCAGCTATGCCAACCCGGACCATTTTGTCTGGAATCTTGAGCACGAGTACGTCCACTATTTGGATGGGCGGTTCAATATGTATGGCGATTTTGGTACGCCTACCGAGCTTGTGGTCTGGTGGAGCGAAGGGGTGGCCGAGTATGTGTCGCGGGTAAATGATAATCCTCAGGCGATTGCCACCATCCAGGATGGTAGCACGTACACTCTGGCGCAGGTGTTTGACACGACGTATGACGGTTTTGATGTGGATCGCATCTACCGATGGGGATATCTGGCGGTGCGATTCATGTTTGAACGTCATCCTGATGAAGTTCAACGTATGCTGAGTGCCACCCGACAGGGACGCTGGGCAGAATACAAGGCGATCATTAGCGGTTGGGCAAATCAGTATCAGTCAGAATTTGCCCAATGGACCGAGGCGCTGGCGAAGGGCGACAGTGGCGCTGGGAACGGTGAGGGGACAGGCTCCGGTAATGAAGGGGGCGGTGAATCTGGTGGTAATACCGGCTTGCCGGAAAACTGCGCAGTACTGCCAAAAATCAGTGATGGGCGTTTAGCACTGGATGAAGCGGCCTGTCTGGCCGACACGGCTTCAGCGTCTGACGTATTGTGGTTCAGTATTCCGGCTGTCAGTGAATATCAGACCATTGCCATTACGGCGGGCAACGGGACTGGCGACCTGACGCTGGAATACAGTAACCTGAATTGGCCAGATGGTACCAATGTGCAGGCATCATCGGCAAATATGGGTAACAGTGAATGCATTATTCTGGAACATCAGGCGAATTATTGGGGATATCTGAAAGTCTCGGGTTCCTTTGAAAATGCAGCGTTACTGGTGGAGGCTGGCAGTAACCAGTGTCGTCAG
但し、式(III)中、Zは無しか、または式(IV)で表される塩基配列を示す。
式(IV):
ATGGAACTTAAAGCACTTGCGTTAACGGTATCAGCCCTGCTTGTGGGTCAGTCGGTGTATGCATCTGAGGTGTTTGCTCACCCGGGCATGCCGGTTCAACAAAAGTTGCAGCCAAATTTGCTGCAGCAAAGTACCAGGCTTCAGCCAGAGCAACATATCCACGGGCTTGAACGCACGGATCGGCAATATCGTCCGACAGATGCTACTCAGCAGCCGGAACCGCCAACGTTACTGAAACGTCAGGTATCGGTTCAGCAGGAT
(k)遺伝子コドンの縮重の範囲内で(j)のDNAと同じアミノ酸をコードする塩基配列を有するDNA
(l)式(III)で表される塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつコラゲナーゼ活性を有するタンパク質をコードするDNA
(m)式(III)で表される塩基配列と生物学的に同等な塩基配列を有するDNA。
【0017】
(4)上記(1)〜(3)のいずれかに記載の遺伝子を含有する組換えベクター。
【0018】
(5)上記(4)に記載の組換えベクターにより形質転換された宿主細胞。
【0019】
(6)微生物由来のコラゲナーゼの製造方法であって、上記(5)記載の宿主細胞を培養し、培養物から産生されたコラゲナーゼを回収・精製することを特徴とする前記コラゲナーゼの製造方法。
【発明の効果】
【0020】
本発明によりグリモンティア・ホリセー由来コラゲナーゼの遺伝子およびアミノ酸の配列が明らかになり、物質としての新規性を証明・登録することができた。本発明により、当該コラゲナーゼを例えば適当な宿主を用いて大量に製造することや、遺伝子工学的手法により改良することなどの手段が講じられるようになった。
【0021】
最も利用されているクロストリジウム・ヒストリティカム由来のコラゲナーゼと比較して、当該コラゲナーゼは高い比活性と安定性を有し、細胞毒性も遜色なく低いことから、その遺伝子配列の同定は新規コラゲナーゼとしての研究用試薬や医療用途への利用等といった産業上の利用価値を持つ。
【図面の簡単な説明】
【0022】
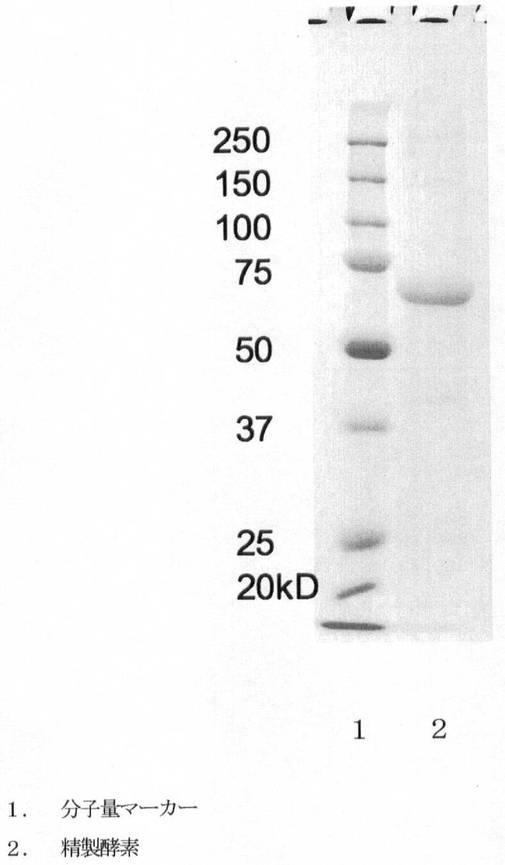
【図1】第1図は精製酵素のSDS-ポリアクリルアミドゲル電気泳動図である。レーン1が分子量マーカー、2が精製酵素である。
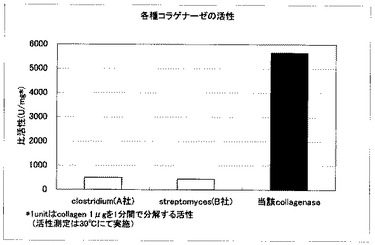
【図2】第2図は市販のクロストリジウム・ヒストリティカム(Clostridium histolyticum) 由来のコラゲナーゼおよびストレプトミセス(Streptomyces)由来のコラゲナーゼと、当該コラゲナーゼ(グリモンティア・ホリセー由来コラゲナーゼ)のFITC-コラーゲン分解酵素活性を比較したグラフである。
【図3】第3図はコラゲナーゼ遺伝子の全塩基配列と、推定されるコラゲナーゼのアミノ酸配列を示す。線で囲んだ部位は、精製コラゲナーゼの部分アミノ酸配列(実施例2参照)と一致する部分を示す。
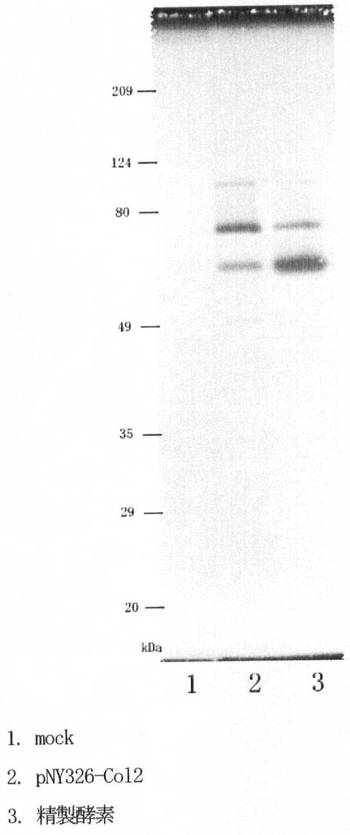
【図4】第4図はコラゲナーゼ活性を解析したゼラチンザイモグラムである。レーン1は空ベクター組換え体の培養上清、2は遺伝子組換え体の培養上清、3はグリモンティア・ホリセーの培養上清から精製した酵素である。
【発明を実施するための形態】
【0023】
本発明はグリモンティア・ホリセー由来コラゲナーゼに関するこれまでの知見(皮革科学 44 (1998) 64-71、皮革科学 45 (2000) 272-283、特開平8-70853)を基礎として完成された。
【0024】
本明細書中では、グリモンティア・ホリセー由来コラゲナーゼのことを「当該コラゲナーゼ」と称することがある。「当該コラゲナーゼ」の意味するところには、特開平8-70853等に記載の方法でグリモンティア・ホリセーに属する微生物を培養して得られるものと、本発明により遺伝子工学的手法で得られるものの両方が含まれる。
【0025】
グリモンティア・ホリセーに属する微生物の入手、微生物を培養し、その培養液からコラゲナーゼを単離する方法、および得られたコラゲナーゼの性状については特開平8-70853、Thompsonら、Int. J. Syst. Evol. Microbiol.、53 (2003) 1615-1617等に記載されている。
【0026】
本発明のコラゲナーゼ遺伝子は、グリモンティア・ホリセーに属する微生物由来のコラゲナーゼの活性(コラーゲン分解酵素活性)を有するタンパク質をコードする遺伝子である。そのようなコラゲナーゼは、好ましくは、特開平8-70853に記載された理化学的性質および酵素化学的性質を有するコラゲナーゼである。
【0027】
本発明のコラゲナーゼ遺伝子の一態様は、少なくとも下記式(a)ないし(g)で示されるペプチド断片を有するタンパク質をコードするものである。
【0028】
式(a) AVEQCDL (配列番号1)
式(b) GAVDAFV (配列番号2)
式(c) IRSQDMSQAQHLAACDK (配列番号3)
式(d) MAYEESFFHTTLE (配列番号4)
式(e) WGYL (配列番号5)
式(f) FSSNHMY (配列番号6)
式(g) ASYAN (配列番号7)
本明細書全体を通じて、アミノ酸の一文字表記は次の通りである。A:アラニン、C:システイン、D:アスパラギン酸、E:グルタミン酸 F:フェニルアラニン、G:グリシン、H:ヒスチジン、I:イソロイシン、K:リシン、L:ロイシン、M:メチオニン、N:アスパラギン、P:プロリン、Q:グルタミン、R:アルギニン、S:セリン、T:スレオニン、V:バリン、W:トリプトファン、Y:チロシン。
【0029】
本発明のコラゲナーゼ遺伝子の別の一態様は、グリモンティア・ホリセーに属する微生物のゲノムDNA由来の天然型の遺伝子をクローニングすることにより得られる。
【0030】
遺伝子のクローニング方法としては、例えば、グリモンティア・ホリセーに属する微生物の培養液から分離・精製されたコラゲナーゼの部分アミノ酸配列を決定した後、適当なDNAプローブを調製し、これを用いてグリモンティア・ホリセーの遺伝子ライブラリーをスクリーニングする方法が挙げられる。得られたDNAの塩基配列を部分アミノ酸配列の情報と比較することにより、グリモンティア・ホリセー由来の遺伝子が取得できたか否かを確認することができる。クローニングに用いる微生物DNAの単離、遺伝子ライブラリーの作製、スクリーニング、塩基配列の解析は当業者に周知の方法によって行うことができる。
【0031】
本発明者らが実際に行った手順は次の通りである。グリモンティア・ホリセーに属する微生物の培養液から分離・精製されたコラゲナーゼを加水分解し、その加水分解断片の部分アミノ酸配列の情報をもとにオリゴヌクレオチドプライマーを合成し、PCR反応を用いて当該コラゲナーゼをコードする遺伝子の部分配列を単離した。次いで、この部分配列を使ってDNAプローブを調製し、グリモンティア・ホリセーのゲノムライブラリーをスクリーニングし、コラゲナーゼを発現するクローンを選択し、目的とするコラゲナーゼタンパク質をコードする遺伝子配列を明らかにすることに成功した。
【0032】
当該コラゲナーゼに対応するオープンリーディングフレームは2304塩基対からなる767アミノ酸をコードする配列であり、分子量約70KDaのコラゲナーゼタンパク質を構成することが明らかとなった。(図3、配列番号14および15参照)。
【0033】
従って、本発明のコラゲナーゼ遺伝子の別の一態様は、下記式(I)で表されるアミノ酸配列からなるタンパク質をコードするものである。
式(I): X-
AVEQCDLSQFQTTSSNQLMAAIRQQGASCVNALFSADTGVQEAAFSSNHMYNVAQYTRTLAQQYAGGGSDELEALYLYLRAGYYAEFYNSNITFLSWVTPAVKGAVDAFVQNAHFYDNGDAHGKVLNEVIITMDSAGLQHAYLDVVTQWLTRWNAQYAEHWYMRNAVNGVFTLLFGGQWNNQYTSLIGEQTALVTALQAFALDRTKVNSPTEFMAANAARELGRLARYTDATIAPKVTEGLTAIFGQYPSYGDGDAIWLGAADTASYYADCSQFNICGFEDALRDAALNQTFICSDTIKIRSQDMSQAQHLAACDKMAYEESFFHTTLETGNQPVADDHNTQLQVNIFNSDTDYGKYAGPIFGIDTNNGGMYLEGNPANVGNIPNFIAYEASYANPDHFVWNLEHEYVHYLDGRFNMYGDFGTPTELVVWWSEGVAEYVSRVNDNPQAIATIQDGSTYTLAQVFDTTYDGFDVDRIYRWGYLAVRFMFERHPDEVQRMLSATRQGRWAEYKAIISGWANQYQSEFAQWTEALAKGDSGAGNGEGTGSGNEGGGESGGNTGLPENCAVLPKISDGRLALDEAACLADTASASDVLWFSIPAVSEYQTIAITAGNGTGDLTLEYSNLNWPDGTNVQASSANMGNSECIILEHQANYWGYLKVSGSFENAALLVEAGSNQCRQ (配列番号8)
但し、式(I)中、Xは、水素原子または次式(II)で表されるポリペプチドを示す
式(II):MELKALALTVSALLVGQSVYASEVFAHPGMPVQQKLQPNLLQQSTRLQPEQHIHGLERTDRQYRPTDATQQPEPPTLLKRQVSVQQD (配列番号9)
本発明のコラゲナーゼ遺伝子のさらに別の一態様は、下記式(III)で表される塩基配列を有するDNAである。
式(III):Z-
GCGGTTGAACAGTGTGATCTCAGCCAGTTTCAGACGACCAGCAGTAACCAGTTGATGGCCGCGATTCGCCAGCAGGGCGCCAGTTGTGTCAATGCCTTGTTTAGTGCTGACACGGGAGTACAGGAAGCCGCGTTTTCGTCCAACCACATGTATAACGTCGCACAATATACCCGAACACTGGCGCAGCAATATGCGGGCGGGGGCAGTGATGAACTGGAAGCCTTATATCTGTATCTTCGGGCGGGGTATTACGCTGAGTTTTATAACAGCAATATCACGTTTCTGTCGTGGGTGACGCCGGCAGTGAAAGGTGCCGTGGATGCGTTTGTCCAGAATGCCCATTTTTATGACAACGGCGACGCTCATGGCAAGGTGCTGAACGAGGTGATCATCACGATGGACAGTGCTGGGCTACAGCATGCCTATCTCGATGTAGTGACGCAGTGGCTGACCCGTTGGAATGCGCAATATGCCGAGCATTGGTATATGCGAAACGCAGTCAATGGTGTCTTTACTCTTTTGTTTGGCGGGCAGTGGAACAATCAGTACACCAGCCTGATTGGCGAACAGACTGCGTTGGTGACGGCCTTACAGGCATTTGCGCTGGACCGTACGAAAGTGAACTCGCCAACGGAGTTTATGGCGGCCAATGCGGCCAGAGAACTGGGGCGGTTAGCTCGCTACACGGACGCGACCATTGCGCCTAAAGTCACGGAAGGATTAACCGCGATCTTCGGCCAGTATCCGTCCTATGGCGATGGAGATGCTATCTGGCTGGGGGCGGCGGATACGGCCTCTTATTATGCTGATTGCAGCCAGTTCAACATCTGTGGCTTTGAAGACGCGCTGCGTGATGCGGCGCTGAACCAGACTTTTATCTGTAGTGATACGATTAAAATTCGCTCACAGGATATGTCGCAGGCACAGCATCTCGCGGCTTGCGACAAAATGGCTTATGAAGAGTCATTTTTCCACACCACGCTTGAAACCGGTAATCAGCCGGTGGCTGATGATCATAATACGCAGCTGCAGGTGAATATTTTTAATTCCGATACCGATTACGGTAAATATGCCGGTCCGATATTTGGGATTGATACCAACAACGGCGGTATGTACCTCGAGGGGAATCCGGCCAATGTGGGCAATATTCCCAATTTCATCGCGTATGAAGCCAGCTATGCCAACCCGGACCATTTTGTCTGGAATCTTGAGCACGAGTACGTCCACTATTTGGATGGGCGGTTCAATATGTATGGCGATTTTGGTACGCCTACCGAGCTTGTGGTCTGGTGGAGCGAAGGGGTGGCCGAGTATGTGTCGCGGGTAAATGATAATCCTCAGGCGATTGCCACCATCCAGGATGGTAGCACGTACACTCTGGCGCAGGTGTTTGACACGACGTATGACGGTTTTGATGTGGATCGCATCTACCGATGGGGATATCTGGCGGTGCGATTCATGTTTGAACGTCATCCTGATGAAGTTCAACGTATGCTGAGTGCCACCCGACAGGGACGCTGGGCAGAATACAAGGCGATCATTAGCGGTTGGGCAAATCAGTATCAGTCAGAATTTGCCCAATGGACCGAGGCGCTGGCGAAGGGCGACAGTGGCGCTGGGAACGGTGAGGGGACAGGCTCCGGTAATGAAGGGGGCGGTGAATCTGGTGGTAATACCGGCTTGCCGGAAAACTGCGCAGTACTGCCAAAAATCAGTGATGGGCGTTTAGCACTGGATGAAGCGGCCTGTCTGGCCGACACGGCTTCAGCGTCTGACGTATTGTGGTTCAGTATTCCGGCTGTCAGTGAATATCAGACCATTGCCATTACGGCGGGCAACGGGACTGGCGACCTGACGCTGGAATACAGTAACCTGAATTGGCCAGATGGTACCAATGTGCAGGCATCATCGGCAAATATGGGTAACAGTGAATGCATTATTCTGGAACATCAGGCGAATTATTGGGGATATCTGAAAGTCTCGGGTTCCTTTGAAAATGCAGCGTTACTGGTGGAGGCTGGCAGTAACCAGTGTCGTCAG (配列番号10)
但し、式(III)中、Zは無しか、または式(IV)で表される塩基配列を示す。
式(IV)
ATGGAACTTAAAGCACTTGCGTTAACGGTATCAGCCCTGCTTGTGGGTCAGTCGGTGTATGCATCTGAGGTGTTTGCTCACCCGGGCATGCCGGTTCAACAAAAGTTGCAGCCAAATTTGCTGCAGCAAAGTACCAGGCTTCAGCCAGAGCAACATATCCACGGGCTTGAACGCACGGATCGGCAATATCGTCCGACAGATGCTACTCAGCAGCCGGAACCGCCAACGTTACTGAAACGTCAGGTATCGGTTCAGCAGGAT (配列番号11)
本発明のコラゲナーゼ遺伝子のさらに別の一態様は、図3において示された核酸配列を有するDNAである。
【0034】
グリモンティア・ホリセーはビブリオ・アルギノリティカス(Vibrio alginolyticus)とは近縁種であるが、今回明らかにしたコラゲナーゼ遺伝子配列から両者のコラゲナーゼタンパク質のアミノ酸配列を比較した結果、その相同性は54%であった。ビブリオ・アルギノリティカス・コラゲナーゼと、同属のビブリオ・パラヘモリティカス(Vibrio parahaemolyticus)由来のコラゲナーゼの相同性が80%であることを考慮すると、ビブリオ・アルギノリティカスのコラゲナーゼとグリモンティア・ホリセーのコラゲナーゼとの相同性は低く、本発明のコラゲナーゼ遺伝子は新規のコラゲナーゼ遺伝子と考えられる。
【0035】
一般的に、ペプチドないしタンパク質は、そのアミノ酸配列中の1もしくは数個のアミノ酸が欠失、置換もしくは付加されていても同じ活性を有することが知られている。本発明において、「数個」のアミノ酸とは、コラゲナーゼタンパク質の構造におけるアミノ酸残基の位置または種類によっても異なるが、通常、例えば、2〜10個を意味する。また、一般的に、同じペプチドないしタンパク質をコードする遺伝子でもコドンの縮重により塩基配列が相違することが知られている。本明細書においては、これらの欠失、置換もしくは付加されたアミノ酸配列をコードする塩基配列や、コドンの縮重により相違する塩基配列も含めて「生物学的に同等」とする。
【0036】
さらに、「生物学的に同等」な塩基配列には、式(III)で表される塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつコラゲナーゼ活性を有するタンパク質をコードするDNAの配列も含まれる。ポリヌクレオチドのハイブリダイゼーション法は当業者に周知であり、例えば、Sambrookら,1989,Molecular Cloning: A Laboratory Manual,第2版,Nolan C.編,New York: Cold Spring Harbor Laboratory Pressに記載の方法により実施することができる。上記ストリンジェントな条件としては、例えば、6×SSC(1×SSCの組成:0.15M NaCl、0.015Mクエン酸ナトリウム、pH7.0)、0.5% SDS、5×デンハルト、100mg/mlニシン精子DNAを含む溶液中、プローブとともに65℃で一晩保温するという条件があげられる。
【0037】
このような生物学的に同等な塩基配列を有する遺伝子も、発現産物であるタンパク質がコラゲナーゼ活性を有する限り、グリモンティア・ホリセーに属する微生物由来のコラゲナーゼ遺伝子と等価なものとして本発明のコラゲナーゼ遺伝子の範囲内に包含される。
【0038】
本発明のコラゲナーゼ遺伝子を用いると、遺伝子組み換え技術によって当該コラゲナーゼを大量に製造することが可能である。
【0039】
すなわち、本発明のコラゲナーゼ遺伝子を適当なベクターに組み込むことにより、宿主細胞を形質転換することができる。これらのベクターに適当なプロモーターや形質発現にかかわる配列を導入することにより、それぞれの宿主細胞において本発明のコラゲナーゼ遺伝子を発現することが可能である。宿主細胞としては、エシェリヒア・コリ(Escherichia coli)やバチルス・ズブチリス(Bacillus subtilis)やバチルス・ブレビス(Bacillus brevis)を用いることができ、ベクターとしては、エシェリヒア・コリ内で複製できるpUC18、pUC19、pBR322、pGEM3、pGEM4など、バチルス・ズブチリス内で複製できるpUC110、pE194、pC194など、バチルス・ブレビス内で複製できるpNCMO2、pNY326などのプラスミドが使用できる。
【0040】
本発明のコラゲナーゼ遺伝子を含むベクターで形質転換された宿主細胞を培養し、培養物からコラゲナーゼを採取することにより、当該コラゲナーゼを製造することができる。宿主細胞の培養は、例えば、エイ・レクロイジーら、FEBSレターズ(A.Lecroiseyら、FEBS Lett.、59 (1975) 167−172)に記載される方法に従って、適当な炭素源、窒素源および微量の金属元素を含む培地中で液体培養することで行なえる。得られた培養物を回収し、例えば、培養上清や可溶化菌体上清を硫安沈澱(60%飽和)処理し、種々のカラムクロマトグラフィー(例えば、DEAEセルロースカラムクロマトグラフィーおよびセファデックスG−100カラムクロマトグラフィーあるいはキレートアフィニティクロマトグラフィーなど)を用いて精製することにより、所望のコラゲナーゼが得られる。
【0041】
以下に実施例を挙げて本発明をさらに詳しく説明するが、本発明はこれらに限定されるものではない。
【実施例1】
【0042】
[精製酵素の解析]
当該コラゲナーゼを特開平8-70853の方法で精製し、常法(例えば、細胞外マトリックス研究法、第2巻、77−85頁、コラーゲン技術研修会(1998))により蛍光標識したI型コラーゲンを用いてコラーゲン分解酵素活性を測定した。市販品のクロストリジウム・ヒストリティカム(Clostridium histolyticum) 由来のコラゲナーゼおよび、ストレプトミセス(Streptomyces) 由来のコラゲナーゼと比較した結果、グリモンティア・ホリセー由来のコラゲナーゼの活性が最も高かった。
【0043】
精製酵素をSDS-ポリアクリルアミド電気泳動により解析した。
【0044】
第1図に精製酵素のSDS-ポリアクリルアミド電気泳動像を、第2図にFITC-コラーゲンを分解する酵素活性を示す。
【実施例2】
【0045】
[アミノ酸配列の決定]
当該コラゲナーゼの部分アミノ酸配列は、以下のようにして決定した。精製した当該コラゲナーゼを常法(例えば、続生化学実験法、第2巻、260−270頁、日本生化学会編)によりトリプシン、プロテアーゼV8またはリジルエンドペプチダーゼで各々部分加水分解した。得られたペプチド断片を高速液体クロマトグラフィーおよびSDS-ポリアクリルアミド電気泳動法により精製した後、自動化エドマン分解法によりアミノ酸配列を決定した。その結果、本発明における当該コラゲナーゼは少なくとも以下の7種のペプチド断片のアミノ酸配列を有していた。
【0046】
1) AVEQCDL (配列番号1)
2) GAVDAFV (配列番号2)
3) IRSQDMSQAQHLAACDK (配列番号3)
4) MAYEESFFHTTLE (配列番号4)
5) WGYL (配列番号5)
6) FSSNHMY (配列番号6)
7) ASYAN (配列番号7)
【実施例3】
【0047】
[遺伝子ライブラリーの作製]
当該コラゲナーゼ産生菌グリモンティア・ホリセーから常法により全ゲノムDNAを単離した。微生物DNAの単離法としては、例えば、サイトウ・ミウラ法(H. Saito and K. Miura、Biochem. Biophys. Acta、72 (1963) 619-629)等が挙げられる。このDNAを制限酵素EcoRIで部分分解し、アガロースゲル電気泳動法により分画し、2kb以上のDNA断片を集めた。このDNA断片を、ベクターpCC1BAC(大腸菌バクミド(pBeloBAC11由来))のEcoRI挿入サイトにT4リガーゼで結合し、エシェリヒア・コリK12株(E.coli Trans for MaxTM EPI300TM)を形質転換して、クロラムフェニコール耐性形質転換体としてグリモンティア・ホリセーのゲノムライブラリーを作製した。エシェリヒア・コリの形質転換は常法(Dower, W. J.ら、Nucl. Acids Res.、16 (1988) 6127-6145)を用いた。
【実施例4】
【0048】
[遺伝子ライブラリーのスクリーニング]
精製した当該コラゲナーゼをプロテアーゼV8で消化して得た上記6) 7)のペプチド配列から誘導された当該コラゲナーゼ遺伝子の部分配列(長さ1.0kb)を、PCR反応を使って単離した。
【0049】
プライマー:
Fwd: GAGGCNATCTTTAGCTCCAATCATATGTAYAAY (配列番号12)
Rvs: ATCTAAGTAATGCACGTATTCATGYTCNAGRTT (配列番号13)
但し、配列番号12、13中のアルファベットは以下の塩基を示す。
N: A,C,G,T、 Y: C,T、 R: A,G
上記プライマーの組合わせがPCR産物(長さ1.0kb)を生じ、図3に示した全塩基配列の421〜1426番目の塩基に対応した。この部分塩基配列を使ってDIGシステム(Kruchen, B. and Rueger, B.、Biochemica、3 (2003)13-15)に記載されている方法によりDNAプローブを調製した。当該コラゲナーゼ遺伝子の全領域を単離する目的で、DNAプローブを用いたコロニーハイブリダイゼーション法により上記遺伝子ライブラリーの中から、コラゲナーゼを発現しているクローンとして、バクミドpCC1BAC-2、pCC1BAC-4を持つクローンを選択した。pCC1BAC-2は、50kb以上のグリモンティア・ホリセー由来のDNA挿入断片を持つ。
【0050】
なお、バクミドpCC1BAC-2は平成21(2009)年4月28日付けで独立行政法人製品評価技術基盤機構特許生物寄託センターに受託番号:NITE P-739として寄託した。
【実施例5】
【0051】
[DNA塩基配列の決定]
pCC1BAC-2の50kb以上の挿入断片の内、約2.4kbについてジデオキシ法(F.Sangerら、Proc.Nat,Acad,Sci,USA、74 (1977) 5963-5967)により塩基配列を決定した。決定されたオープンリーディングフレーム(図3、配列番号14、15)は、1番目のATGから始まり、2304番目のTAAで終わる2304塩基対からなり、当該コラゲナーゼの全領域を含んでいる。全塩基配列中において、決定された塩基配列から推測されるアミノ酸配列と、実施例2において決定された部分アミノ酸配列とを比較してみると、以下の7個のペプチド断片のアミノ酸配列を有していた。
【0052】
【化1】

【0053】
なお、(1)〜(7)の塩基配列(配列表において配列番号16〜22として示す)はそれぞれ配列番号1〜7のアミノ酸配列に対応する。
【実施例6】
【0054】
[コラゲナーゼ組換えタンパク質の発現および性状解析]
まず、コラゲナーゼ遺伝子をバチルス・ブレビス(Bacillus brevis)で大量に生産させるための組換えプラスミドを作製した。pCC1BAC-2を鋳型として挿入されているコラゲナーゼ遺伝子の成熟体領域2.0kbのDNA断片の5’側にNco Iサイトを、3’側にHind IIIサイトを付加し、Expand High Fidelity PCR System(Roche)により増幅した。増幅された断片をNco I-Hind IIIで処理して回収した当該DNA断片を、プラスミドベクターpNY326のサイトに挿入して、pNY326-Col2を作製した。さらに組換プラスミドpNY326-Col2をB.brevis S5株に形質転換して、コラゲナーゼを大量に生産するB.brevis組換え体を作製した。
【0055】
コラゲナーゼ遺伝子を含有するB.brevis組換え体を2SLN培地(4% ポリペプトンS、0.5% 酵母エキス、2% グルコース、0.001% FeSO4・7H2O、0.001% MnSO4・7H2O、0.0001% ZnSO4・7H2O、50μg/ml ネオマイシン)中で、30℃、48時間培養した。培養液を遠心し、上清を0.2μmのフィルターでろ過滅菌をした。このようにして得たB.brevis組換え体培養上清中に分泌されたグリモンティア・ホリセー由来コラゲナーゼ組換えタンパク質を、まずゼラチンザイモグラフィー法によって解析した。ゼラチンザイモグラフィー法は服部らの方法(Hattori, S., Anal. Biochem., 301,27-34,(2002))で行った。図4に示すように、組換え体培養上清にはグリモンティア・ホリセー由来の精製コラゲナーゼと同じ位置にバンドが認められた。対照となる非組換えプラスミド導入株(mock)培養上清にはバンドが認められなかった。
【0056】
次に組換えタンパク質のコラゲナーゼ活性を、蛍光標識したコラーゲン溶液を用いて測定した。測定法は常法(例えば細胞外マトリックス研究法、第2巻、77−85頁、コラーゲン技術研修会(1998))を用いた。第1表に示すように、B.brevis組換え体(pNY326-Col2)培養上清中には、高いコラゲナーゼ活性が認められた。一方、非組換えプラスミド導入株(mock)培養上清にはコラゲナーゼ活性は認められなかった。表中1ユニット(U)はコラーゲン1μgを、30℃、1分間で分解する活性を指す。
【0057】
【表1】
【受託番号】
【0058】
NITE P-739
【技術分野】
【0001】
本発明は活性の高いコラゲナーゼを得るための、グリモンティア・ホリセー由来のコラゲナーゼ遺伝子、該遺伝子が組み込まれた組換えベクター、該ベクターにより形質転換された宿主細胞およびその利用に関する。
【背景技術】
【0002】
コラーゲンは細胞外マトリックスの主要タンパク質として各臓器に広く分布しており、哺乳動物では全タンパク質の約30%を占める。現在までに、遺伝子として29種類の型のコラーゲンが同定されており、分子構造や会合体の形成様式によりサブファミリーに分類されている。その中でもI型コラーゲンは皮膚や骨や腱を構成する線維性コラーゲンの主成分である。I型コラーゲンおよびその熱変性物であるゼラチンは工業的に生産され、食品、化粧品、医療等の分野で利用されている。I型コラーゲンは約10万の分子量をもつ3本のポリペプチド鎖が三重らせん構造をとった棒状の分子である。コラーゲン分子を構成する各ポリペプチドのアミノ酸配列はGly−X−Y(XおよびYは種々のアミノ酸残基を示す、Xはプロリン、Yはヒドロキシプロリンである頻度が高い)の繰返しであり、各ポリペプチドは分子内または分子間で架橋されている。I型コラーゲンは生体中で自己会合して線維を形成しており、組織に機械的特性と化学的安定性をもたらしている。コラーゲン特有の三重らせん構造は通常のプロテアーゼに対して抵抗性を示すが、コラゲナーゼには分解される。微生物由来のコラゲナーゼは、コラーゲンあるいはその変性物であるゼラチンに作用し、Gly−X−Yの単位にまで分解することが知られている。その特異的な性質を利用して生化学的研究や医療分野、さらに食品加工にも広く利用されている。
【0003】
従来、入手可能な微生物由来の主なコラゲナーゼ精製品は、クロストリジウム属由来、ビブリオ・アルギノリティカス・ケモバル・イオファガス(Vibrio alginolyticus chemovar.iophagus)由来、ストレプトミセス(Streptomyces)属由来のコラゲナーゼである。
【0004】
最も利用されているコラゲナーゼはクロストリジウム・ヒストリティカム(Clostridium histolyticum)由来のコラゲナーゼである。クロストリジウム・ヒストリティカムはガス壊疽菌として古くから知られている嫌気性の病原菌であり、毒素を産生する。従って該菌の大量培養によるコラゲナーゼの大量生産には適さないという問題があるが、近年は遺伝子組換え体が利用されている(特許第2634782、特表2001-510331)。クロストリジウム属のコラゲナーゼは種々の生化学的研究に研究試薬として使用されるだけでなく、たとえば組織の消化と個々の細胞または細胞群の単離(例えば膵臓から膵島(islet)の分離)に使用される(膵島:Suttonら、Transplantation 、42 (1986) 689-691;肝:Quibelら、Anal.Biochem.、154 (1986) 26-28;骨:Hefleyら、J.Bone Mineral Res.、2 (1987) 505-516;臍帯:Holzingerら、Immunol. Lett.、35 (1993) 109−118)。
【0005】
ビブリオ・アルギノリティカス・ケモバル・イオファガス(Vibrio alginolyticus chemovar.iophagus)由来のコラゲナーゼも種々の用途で使用されている。たとえば、コラーゲンに富む構造の無制限な基質が生じる火傷、潰瘍、痂皮、コラーゲンベースの白色硬痂、ケロイド、壊死とくに臥位または潰瘍による壊死の治療に使用される。虫歯の治療にも使用される。その他、肉質軟化剤としても使用できる。ビブリオ・アルギノリティカス由来のコラゲナーゼには安価な取得が困難であるという問題があるが、この酵素の遺伝子取得によるアミノ酸全配列が明らかにされたことで遺伝子工学的手法を用いることが可能となった(特許第3022984)。
【0006】
ストレプトミセス(Streptomyces)属由来コラゲナーゼは、公知の培養法により放線菌を好気的に培養した培養液より精製して得られる(特公平5-16832)。既存添加物名簿にストレプトミセス(Streptomyces)属のプロテアーゼが収載されており、同属由来のコラゲナーゼは他の2種とは異なって食品に使用することができる。
【0007】
既存のコラゲナーゼは安定性に難があるのに対して、安定性に優れさらに比活性が高い微生物由来コラゲナーゼである、ビブリオ・ホリセー(Vibrio hollisae)由来のコラゲナーゼが発見された(鈴木ら、皮革科学 44 (1998) 64-71、鈴木、皮革科学 45 (2000) 272-283、特開平8-70853)。しかし、この報告ではビブリオ・ホリセー(Vibrio hollisae)由来のコラゲナーゼをコードする遺伝子は特定されておらず、それが既存のコラゲナーゼとは異なる新規な酵素であることを示すには不十分であった。また、遺伝子工学的手法による同酵素の大量取得にも対応できていなかった。
【0008】
尚、ビブリオ・ホリセーは16S rRNAの相同性により、新たにグリモンティア・ホリセー(Grimontia hollisae)と分類し直された(Thompsonら、 Int. J. Syst. Evol. Microbiol.、 53 (2003) 1615-1617)。本明細書ではビブリオ・ホリセーをグリモンティア・ホリセーとして表記する。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】特許第2634782号公報
【特許文献2】特表2001-510331号公報
【特許文献3】特許第3022984号公報
【特許文献4】特公平5-16832号公報
【特許文献5】特開平8-70853号公報
【非特許文献】
【0010】
【非特許文献1】Suttonら、Transplantation、42 (1986) p.689-691
【非特許文献2】Quibelら、Anal. Biochem.、154 (1986) p.26-28
【非特許文献3】Hefleyら、J. Bone Mineral Res.、2 (1987) p.505-516
【非特許文献4】Holzingerら、Immunol. Lett.、35 (1993) p.109-118
【非特許文献5】鈴木ら、皮革科学 44 (1998) p.64-71
【非特許文献6】鈴木、皮革科学 45 (2000) p.272-283
【非特許文献7】Thompsonら、 Int. J. Syst. Evol. Microbiol.、53 (2003) p.1615-1617
【発明の概要】
【発明が解決しようとする課題】
【0011】
グリモンティア・ホリセー(Grimontia hollisae)由来コラゲナーゼは優れた比活性と安定性を有し、クロストリジウム・ヒストリティカム等由来の既存のコラゲナーゼに替わる新規コラゲナーゼとして研究用試薬や医療用途への利用が期待される。その一方で、安価な取得が困難であるという問題がある。当該コラゲナーゼはその産生菌であるグリモンティア・ホリセーを培養し、その培養液より回収・精製できるが、当該微生物のコラゲナーゼ産生量は1mg/Lと著しく低く、生産効率は実用的ではない。そのため、当該コラゲナーゼをコードする遺伝子配列を決定すること、そして決定した遺伝子配列をもとに遺伝子工学的手法を用いた当該コラゲナーゼ生産効率化のための技術を構築することが本発明の課題である。
【課題を解決するための手段】
【0012】
本発明者らはグリモンティア・ホリセーに属する微生物由来のコラゲナーゼ遺伝子(明細書中では、本発明のコラゲナーゼ遺伝子と呼ぶこともある)を取得し、そのアミノ酸配列を明らかにすることに成功し、本発明を完成した。これにより、遺伝子工学的手法を用いた当該コラゲナーゼの産生系を構築することが可能となった。すなわち、本発明の概略は以下の通りである。
【0013】
(1)少なくとも下記式(a)ないし(g)で示されるペプチド断片を有するグリモンティア・ホリセー(Grimontia hollisae)に属する微生物由来のコラゲナーゼをコードする遺伝子。
【0014】
式(a) AVEQCDL
式(b) GAVDAFV
式(c) IRSQDMSQAQHLAACDK
式(d) MAYEESFFHTTLE
式(e) WGYL
式(f) FSSNHMY
式(g) ASYAN
但し、式(a)ないし(g)中のアルファベットは以下のアミノ酸を示す。A:アラニン、C:システイン、D:アスパラギン酸、E:グルタミン酸 F:フェニルアラニン、G:グリシン、H:ヒスチジン、I:イソロイシン、K:リシン、L:ロイシン、M:メチオニン、N:アスパラギン、P:プロリン、Q:グルタミン、R:アルギニン、S:セリン、T:スレオニン、V:バリン、W:トリプトファン、Y:チロシン。
【0015】
(2)以下の(h)または(i)のタンパク質をコードする遺伝子。
(h)下記式(I)で表されるアミノ酸配列からなるグリモンティア・ホリセー(Grimontia hollisae)に属する微生物由来のコラゲナーゼタンパク質
式(I): X-
AVEQCDLSQFQTTSSNQLMAAIRQQGASCVNALFSADTGVQEAAFSSNHMYNVAQYTRTLAQQYAGGGSDELEALYLYLRAGYYAEFYNSNITFLSWVTPAVKGAVDAFVQNAHFYDNGDAHGKVLNEVIITMDSAGLQHAYLDVVTQWLTRWNAQYAEHWYMRNAVNGVFTLLFGGQWNNQYTSLIGEQTALVTALQAFALDRTKVNSPTEFMAANAARELGRLARYTDATIAPKVTEGLTAIFGQYPSYGDGDAIWLGAADTASYYADCSQFNICGFEDALRDAALNQTFICSDTIKIRSQDMSQAQHLAACDKMAYEESFFHTTLETGNQPVADDHNTQLQVNIFNSDTDYGKYAGPIFGIDTNNGGMYLEGNPANVGNIPNFIAYEASYANPDHFVWNLEHEYVHYLDGRFNMYGDFGTPTELVVWWSEGVAEYVSRVNDNPQAIATIQDGSTYTLAQVFDTTYDGFDVDRIYRWGYLAVRFMFERHPDEVQRMLSATRQGRWAEYKAIISGWANQYQSEFAQWTEALAKGDSGAGNGEGTGSGNEGGGESGGNTGLPENCAVLPKISDGRLALDEAACLADTASASDVLWFSIPAVSEYQTIAITAGNGTGDLTLEYSNLNWPDGTNVQASSANMGNSECIILEHQANYWGYLKVSGSFENAALLVEAGSNQCRQ
但し、式(I)中、Xは、水素原子または次式(II)で表されるポリペプチドを示す。
式(II):MELKALALTVSALLVGQSVYASEVFAHPGMPVQQKLQPNLLQQSTRLQPEQHIHGLERTDRQYRPTDATQQPEPPTLLKRQVSVQQD
また、式(I)、(II)中のアルファベットは、(1)と同義である。
(i)式(I)で表されるアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつコラゲナーゼ活性を有するタンパク質。
【0016】
(3)以下の(j)、(k)、(l)または(m)のDNAからなるコラゲナーゼ遺伝子:
(j)グリモンティア・ホリセーに属する微生物由来のコラゲナーゼをコードする、下記式(III)で表される塩基配列からなるDNA
式(III):Z-
GCGGTTGAACAGTGTGATCTCAGCCAGTTTCAGACGACCAGCAGTAACCAGTTGATGGCCGCGATTCGCCAGCAGGGCGCCAGTTGTGTCAATGCCTTGTTTAGTGCTGACACGGGAGTACAGGAAGCCGCGTTTTCGTCCAACCACATGTATAACGTCGCACAATATACCCGAACACTGGCGCAGCAATATGCGGGCGGGGGCAGTGATGAACTGGAAGCCTTATATCTGTATCTTCGGGCGGGGTATTACGCTGAGTTTTATAACAGCAATATCACGTTTCTGTCGTGGGTGACGCCGGCAGTGAAAGGTGCCGTGGATGCGTTTGTCCAGAATGCCCATTTTTATGACAACGGCGACGCTCATGGCAAGGTGCTGAACGAGGTGATCATCACGATGGACAGTGCTGGGCTACAGCATGCCTATCTCGATGTAGTGACGCAGTGGCTGACCCGTTGGAATGCGCAATATGCCGAGCATTGGTATATGCGAAACGCAGTCAATGGTGTCTTTACTCTTTTGTTTGGCGGGCAGTGGAACAATCAGTACACCAGCCTGATTGGCGAACAGACTGCGTTGGTGACGGCCTTACAGGCATTTGCGCTGGACCGTACGAAAGTGAACTCGCCAACGGAGTTTATGGCGGCCAATGCGGCCAGAGAACTGGGGCGGTTAGCTCGCTACACGGACGCGACCATTGCGCCTAAAGTCACGGAAGGATTAACCGCGATCTTCGGCCAGTATCCGTCCTATGGCGATGGAGATGCTATCTGGCTGGGGGCGGCGGATACGGCCTCTTATTATGCTGATTGCAGCCAGTTCAACATCTGTGGCTTTGAAGACGCGCTGCGTGATGCGGCGCTGAACCAGACTTTTATCTGTAGTGATACGATTAAAATTCGCTCACAGGATATGTCGCAGGCACAGCATCTCGCGGCTTGCGACAAAATGGCTTATGAAGAGTCATTTTTCCACACCACGCTTGAAACCGGTAATCAGCCGGTGGCTGATGATCATAATACGCAGCTGCAGGTGAATATTTTTAATTCCGATACCGATTACGGTAAATATGCCGGTCCGATATTTGGGATTGATACCAACAACGGCGGTATGTACCTCGAGGGGAATCCGGCCAATGTGGGCAATATTCCCAATTTCATCGCGTATGAAGCCAGCTATGCCAACCCGGACCATTTTGTCTGGAATCTTGAGCACGAGTACGTCCACTATTTGGATGGGCGGTTCAATATGTATGGCGATTTTGGTACGCCTACCGAGCTTGTGGTCTGGTGGAGCGAAGGGGTGGCCGAGTATGTGTCGCGGGTAAATGATAATCCTCAGGCGATTGCCACCATCCAGGATGGTAGCACGTACACTCTGGCGCAGGTGTTTGACACGACGTATGACGGTTTTGATGTGGATCGCATCTACCGATGGGGATATCTGGCGGTGCGATTCATGTTTGAACGTCATCCTGATGAAGTTCAACGTATGCTGAGTGCCACCCGACAGGGACGCTGGGCAGAATACAAGGCGATCATTAGCGGTTGGGCAAATCAGTATCAGTCAGAATTTGCCCAATGGACCGAGGCGCTGGCGAAGGGCGACAGTGGCGCTGGGAACGGTGAGGGGACAGGCTCCGGTAATGAAGGGGGCGGTGAATCTGGTGGTAATACCGGCTTGCCGGAAAACTGCGCAGTACTGCCAAAAATCAGTGATGGGCGTTTAGCACTGGATGAAGCGGCCTGTCTGGCCGACACGGCTTCAGCGTCTGACGTATTGTGGTTCAGTATTCCGGCTGTCAGTGAATATCAGACCATTGCCATTACGGCGGGCAACGGGACTGGCGACCTGACGCTGGAATACAGTAACCTGAATTGGCCAGATGGTACCAATGTGCAGGCATCATCGGCAAATATGGGTAACAGTGAATGCATTATTCTGGAACATCAGGCGAATTATTGGGGATATCTGAAAGTCTCGGGTTCCTTTGAAAATGCAGCGTTACTGGTGGAGGCTGGCAGTAACCAGTGTCGTCAG
但し、式(III)中、Zは無しか、または式(IV)で表される塩基配列を示す。
式(IV):
ATGGAACTTAAAGCACTTGCGTTAACGGTATCAGCCCTGCTTGTGGGTCAGTCGGTGTATGCATCTGAGGTGTTTGCTCACCCGGGCATGCCGGTTCAACAAAAGTTGCAGCCAAATTTGCTGCAGCAAAGTACCAGGCTTCAGCCAGAGCAACATATCCACGGGCTTGAACGCACGGATCGGCAATATCGTCCGACAGATGCTACTCAGCAGCCGGAACCGCCAACGTTACTGAAACGTCAGGTATCGGTTCAGCAGGAT
(k)遺伝子コドンの縮重の範囲内で(j)のDNAと同じアミノ酸をコードする塩基配列を有するDNA
(l)式(III)で表される塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつコラゲナーゼ活性を有するタンパク質をコードするDNA
(m)式(III)で表される塩基配列と生物学的に同等な塩基配列を有するDNA。
【0017】
(4)上記(1)〜(3)のいずれかに記載の遺伝子を含有する組換えベクター。
【0018】
(5)上記(4)に記載の組換えベクターにより形質転換された宿主細胞。
【0019】
(6)微生物由来のコラゲナーゼの製造方法であって、上記(5)記載の宿主細胞を培養し、培養物から産生されたコラゲナーゼを回収・精製することを特徴とする前記コラゲナーゼの製造方法。
【発明の効果】
【0020】
本発明によりグリモンティア・ホリセー由来コラゲナーゼの遺伝子およびアミノ酸の配列が明らかになり、物質としての新規性を証明・登録することができた。本発明により、当該コラゲナーゼを例えば適当な宿主を用いて大量に製造することや、遺伝子工学的手法により改良することなどの手段が講じられるようになった。
【0021】
最も利用されているクロストリジウム・ヒストリティカム由来のコラゲナーゼと比較して、当該コラゲナーゼは高い比活性と安定性を有し、細胞毒性も遜色なく低いことから、その遺伝子配列の同定は新規コラゲナーゼとしての研究用試薬や医療用途への利用等といった産業上の利用価値を持つ。
【図面の簡単な説明】
【0022】
【図1】第1図は精製酵素のSDS-ポリアクリルアミドゲル電気泳動図である。レーン1が分子量マーカー、2が精製酵素である。
【図2】第2図は市販のクロストリジウム・ヒストリティカム(Clostridium histolyticum) 由来のコラゲナーゼおよびストレプトミセス(Streptomyces)由来のコラゲナーゼと、当該コラゲナーゼ(グリモンティア・ホリセー由来コラゲナーゼ)のFITC-コラーゲン分解酵素活性を比較したグラフである。
【図3】第3図はコラゲナーゼ遺伝子の全塩基配列と、推定されるコラゲナーゼのアミノ酸配列を示す。線で囲んだ部位は、精製コラゲナーゼの部分アミノ酸配列(実施例2参照)と一致する部分を示す。
【図4】第4図はコラゲナーゼ活性を解析したゼラチンザイモグラムである。レーン1は空ベクター組換え体の培養上清、2は遺伝子組換え体の培養上清、3はグリモンティア・ホリセーの培養上清から精製した酵素である。
【発明を実施するための形態】
【0023】
本発明はグリモンティア・ホリセー由来コラゲナーゼに関するこれまでの知見(皮革科学 44 (1998) 64-71、皮革科学 45 (2000) 272-283、特開平8-70853)を基礎として完成された。
【0024】
本明細書中では、グリモンティア・ホリセー由来コラゲナーゼのことを「当該コラゲナーゼ」と称することがある。「当該コラゲナーゼ」の意味するところには、特開平8-70853等に記載の方法でグリモンティア・ホリセーに属する微生物を培養して得られるものと、本発明により遺伝子工学的手法で得られるものの両方が含まれる。
【0025】
グリモンティア・ホリセーに属する微生物の入手、微生物を培養し、その培養液からコラゲナーゼを単離する方法、および得られたコラゲナーゼの性状については特開平8-70853、Thompsonら、Int. J. Syst. Evol. Microbiol.、53 (2003) 1615-1617等に記載されている。
【0026】
本発明のコラゲナーゼ遺伝子は、グリモンティア・ホリセーに属する微生物由来のコラゲナーゼの活性(コラーゲン分解酵素活性)を有するタンパク質をコードする遺伝子である。そのようなコラゲナーゼは、好ましくは、特開平8-70853に記載された理化学的性質および酵素化学的性質を有するコラゲナーゼである。
【0027】
本発明のコラゲナーゼ遺伝子の一態様は、少なくとも下記式(a)ないし(g)で示されるペプチド断片を有するタンパク質をコードするものである。
【0028】
式(a) AVEQCDL (配列番号1)
式(b) GAVDAFV (配列番号2)
式(c) IRSQDMSQAQHLAACDK (配列番号3)
式(d) MAYEESFFHTTLE (配列番号4)
式(e) WGYL (配列番号5)
式(f) FSSNHMY (配列番号6)
式(g) ASYAN (配列番号7)
本明細書全体を通じて、アミノ酸の一文字表記は次の通りである。A:アラニン、C:システイン、D:アスパラギン酸、E:グルタミン酸 F:フェニルアラニン、G:グリシン、H:ヒスチジン、I:イソロイシン、K:リシン、L:ロイシン、M:メチオニン、N:アスパラギン、P:プロリン、Q:グルタミン、R:アルギニン、S:セリン、T:スレオニン、V:バリン、W:トリプトファン、Y:チロシン。
【0029】
本発明のコラゲナーゼ遺伝子の別の一態様は、グリモンティア・ホリセーに属する微生物のゲノムDNA由来の天然型の遺伝子をクローニングすることにより得られる。
【0030】
遺伝子のクローニング方法としては、例えば、グリモンティア・ホリセーに属する微生物の培養液から分離・精製されたコラゲナーゼの部分アミノ酸配列を決定した後、適当なDNAプローブを調製し、これを用いてグリモンティア・ホリセーの遺伝子ライブラリーをスクリーニングする方法が挙げられる。得られたDNAの塩基配列を部分アミノ酸配列の情報と比較することにより、グリモンティア・ホリセー由来の遺伝子が取得できたか否かを確認することができる。クローニングに用いる微生物DNAの単離、遺伝子ライブラリーの作製、スクリーニング、塩基配列の解析は当業者に周知の方法によって行うことができる。
【0031】
本発明者らが実際に行った手順は次の通りである。グリモンティア・ホリセーに属する微生物の培養液から分離・精製されたコラゲナーゼを加水分解し、その加水分解断片の部分アミノ酸配列の情報をもとにオリゴヌクレオチドプライマーを合成し、PCR反応を用いて当該コラゲナーゼをコードする遺伝子の部分配列を単離した。次いで、この部分配列を使ってDNAプローブを調製し、グリモンティア・ホリセーのゲノムライブラリーをスクリーニングし、コラゲナーゼを発現するクローンを選択し、目的とするコラゲナーゼタンパク質をコードする遺伝子配列を明らかにすることに成功した。
【0032】
当該コラゲナーゼに対応するオープンリーディングフレームは2304塩基対からなる767アミノ酸をコードする配列であり、分子量約70KDaのコラゲナーゼタンパク質を構成することが明らかとなった。(図3、配列番号14および15参照)。
【0033】
従って、本発明のコラゲナーゼ遺伝子の別の一態様は、下記式(I)で表されるアミノ酸配列からなるタンパク質をコードするものである。
式(I): X-
AVEQCDLSQFQTTSSNQLMAAIRQQGASCVNALFSADTGVQEAAFSSNHMYNVAQYTRTLAQQYAGGGSDELEALYLYLRAGYYAEFYNSNITFLSWVTPAVKGAVDAFVQNAHFYDNGDAHGKVLNEVIITMDSAGLQHAYLDVVTQWLTRWNAQYAEHWYMRNAVNGVFTLLFGGQWNNQYTSLIGEQTALVTALQAFALDRTKVNSPTEFMAANAARELGRLARYTDATIAPKVTEGLTAIFGQYPSYGDGDAIWLGAADTASYYADCSQFNICGFEDALRDAALNQTFICSDTIKIRSQDMSQAQHLAACDKMAYEESFFHTTLETGNQPVADDHNTQLQVNIFNSDTDYGKYAGPIFGIDTNNGGMYLEGNPANVGNIPNFIAYEASYANPDHFVWNLEHEYVHYLDGRFNMYGDFGTPTELVVWWSEGVAEYVSRVNDNPQAIATIQDGSTYTLAQVFDTTYDGFDVDRIYRWGYLAVRFMFERHPDEVQRMLSATRQGRWAEYKAIISGWANQYQSEFAQWTEALAKGDSGAGNGEGTGSGNEGGGESGGNTGLPENCAVLPKISDGRLALDEAACLADTASASDVLWFSIPAVSEYQTIAITAGNGTGDLTLEYSNLNWPDGTNVQASSANMGNSECIILEHQANYWGYLKVSGSFENAALLVEAGSNQCRQ (配列番号8)
但し、式(I)中、Xは、水素原子または次式(II)で表されるポリペプチドを示す
式(II):MELKALALTVSALLVGQSVYASEVFAHPGMPVQQKLQPNLLQQSTRLQPEQHIHGLERTDRQYRPTDATQQPEPPTLLKRQVSVQQD (配列番号9)
本発明のコラゲナーゼ遺伝子のさらに別の一態様は、下記式(III)で表される塩基配列を有するDNAである。
式(III):Z-
GCGGTTGAACAGTGTGATCTCAGCCAGTTTCAGACGACCAGCAGTAACCAGTTGATGGCCGCGATTCGCCAGCAGGGCGCCAGTTGTGTCAATGCCTTGTTTAGTGCTGACACGGGAGTACAGGAAGCCGCGTTTTCGTCCAACCACATGTATAACGTCGCACAATATACCCGAACACTGGCGCAGCAATATGCGGGCGGGGGCAGTGATGAACTGGAAGCCTTATATCTGTATCTTCGGGCGGGGTATTACGCTGAGTTTTATAACAGCAATATCACGTTTCTGTCGTGGGTGACGCCGGCAGTGAAAGGTGCCGTGGATGCGTTTGTCCAGAATGCCCATTTTTATGACAACGGCGACGCTCATGGCAAGGTGCTGAACGAGGTGATCATCACGATGGACAGTGCTGGGCTACAGCATGCCTATCTCGATGTAGTGACGCAGTGGCTGACCCGTTGGAATGCGCAATATGCCGAGCATTGGTATATGCGAAACGCAGTCAATGGTGTCTTTACTCTTTTGTTTGGCGGGCAGTGGAACAATCAGTACACCAGCCTGATTGGCGAACAGACTGCGTTGGTGACGGCCTTACAGGCATTTGCGCTGGACCGTACGAAAGTGAACTCGCCAACGGAGTTTATGGCGGCCAATGCGGCCAGAGAACTGGGGCGGTTAGCTCGCTACACGGACGCGACCATTGCGCCTAAAGTCACGGAAGGATTAACCGCGATCTTCGGCCAGTATCCGTCCTATGGCGATGGAGATGCTATCTGGCTGGGGGCGGCGGATACGGCCTCTTATTATGCTGATTGCAGCCAGTTCAACATCTGTGGCTTTGAAGACGCGCTGCGTGATGCGGCGCTGAACCAGACTTTTATCTGTAGTGATACGATTAAAATTCGCTCACAGGATATGTCGCAGGCACAGCATCTCGCGGCTTGCGACAAAATGGCTTATGAAGAGTCATTTTTCCACACCACGCTTGAAACCGGTAATCAGCCGGTGGCTGATGATCATAATACGCAGCTGCAGGTGAATATTTTTAATTCCGATACCGATTACGGTAAATATGCCGGTCCGATATTTGGGATTGATACCAACAACGGCGGTATGTACCTCGAGGGGAATCCGGCCAATGTGGGCAATATTCCCAATTTCATCGCGTATGAAGCCAGCTATGCCAACCCGGACCATTTTGTCTGGAATCTTGAGCACGAGTACGTCCACTATTTGGATGGGCGGTTCAATATGTATGGCGATTTTGGTACGCCTACCGAGCTTGTGGTCTGGTGGAGCGAAGGGGTGGCCGAGTATGTGTCGCGGGTAAATGATAATCCTCAGGCGATTGCCACCATCCAGGATGGTAGCACGTACACTCTGGCGCAGGTGTTTGACACGACGTATGACGGTTTTGATGTGGATCGCATCTACCGATGGGGATATCTGGCGGTGCGATTCATGTTTGAACGTCATCCTGATGAAGTTCAACGTATGCTGAGTGCCACCCGACAGGGACGCTGGGCAGAATACAAGGCGATCATTAGCGGTTGGGCAAATCAGTATCAGTCAGAATTTGCCCAATGGACCGAGGCGCTGGCGAAGGGCGACAGTGGCGCTGGGAACGGTGAGGGGACAGGCTCCGGTAATGAAGGGGGCGGTGAATCTGGTGGTAATACCGGCTTGCCGGAAAACTGCGCAGTACTGCCAAAAATCAGTGATGGGCGTTTAGCACTGGATGAAGCGGCCTGTCTGGCCGACACGGCTTCAGCGTCTGACGTATTGTGGTTCAGTATTCCGGCTGTCAGTGAATATCAGACCATTGCCATTACGGCGGGCAACGGGACTGGCGACCTGACGCTGGAATACAGTAACCTGAATTGGCCAGATGGTACCAATGTGCAGGCATCATCGGCAAATATGGGTAACAGTGAATGCATTATTCTGGAACATCAGGCGAATTATTGGGGATATCTGAAAGTCTCGGGTTCCTTTGAAAATGCAGCGTTACTGGTGGAGGCTGGCAGTAACCAGTGTCGTCAG (配列番号10)
但し、式(III)中、Zは無しか、または式(IV)で表される塩基配列を示す。
式(IV)
ATGGAACTTAAAGCACTTGCGTTAACGGTATCAGCCCTGCTTGTGGGTCAGTCGGTGTATGCATCTGAGGTGTTTGCTCACCCGGGCATGCCGGTTCAACAAAAGTTGCAGCCAAATTTGCTGCAGCAAAGTACCAGGCTTCAGCCAGAGCAACATATCCACGGGCTTGAACGCACGGATCGGCAATATCGTCCGACAGATGCTACTCAGCAGCCGGAACCGCCAACGTTACTGAAACGTCAGGTATCGGTTCAGCAGGAT (配列番号11)
本発明のコラゲナーゼ遺伝子のさらに別の一態様は、図3において示された核酸配列を有するDNAである。
【0034】
グリモンティア・ホリセーはビブリオ・アルギノリティカス(Vibrio alginolyticus)とは近縁種であるが、今回明らかにしたコラゲナーゼ遺伝子配列から両者のコラゲナーゼタンパク質のアミノ酸配列を比較した結果、その相同性は54%であった。ビブリオ・アルギノリティカス・コラゲナーゼと、同属のビブリオ・パラヘモリティカス(Vibrio parahaemolyticus)由来のコラゲナーゼの相同性が80%であることを考慮すると、ビブリオ・アルギノリティカスのコラゲナーゼとグリモンティア・ホリセーのコラゲナーゼとの相同性は低く、本発明のコラゲナーゼ遺伝子は新規のコラゲナーゼ遺伝子と考えられる。
【0035】
一般的に、ペプチドないしタンパク質は、そのアミノ酸配列中の1もしくは数個のアミノ酸が欠失、置換もしくは付加されていても同じ活性を有することが知られている。本発明において、「数個」のアミノ酸とは、コラゲナーゼタンパク質の構造におけるアミノ酸残基の位置または種類によっても異なるが、通常、例えば、2〜10個を意味する。また、一般的に、同じペプチドないしタンパク質をコードする遺伝子でもコドンの縮重により塩基配列が相違することが知られている。本明細書においては、これらの欠失、置換もしくは付加されたアミノ酸配列をコードする塩基配列や、コドンの縮重により相違する塩基配列も含めて「生物学的に同等」とする。
【0036】
さらに、「生物学的に同等」な塩基配列には、式(III)で表される塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつコラゲナーゼ活性を有するタンパク質をコードするDNAの配列も含まれる。ポリヌクレオチドのハイブリダイゼーション法は当業者に周知であり、例えば、Sambrookら,1989,Molecular Cloning: A Laboratory Manual,第2版,Nolan C.編,New York: Cold Spring Harbor Laboratory Pressに記載の方法により実施することができる。上記ストリンジェントな条件としては、例えば、6×SSC(1×SSCの組成:0.15M NaCl、0.015Mクエン酸ナトリウム、pH7.0)、0.5% SDS、5×デンハルト、100mg/mlニシン精子DNAを含む溶液中、プローブとともに65℃で一晩保温するという条件があげられる。
【0037】
このような生物学的に同等な塩基配列を有する遺伝子も、発現産物であるタンパク質がコラゲナーゼ活性を有する限り、グリモンティア・ホリセーに属する微生物由来のコラゲナーゼ遺伝子と等価なものとして本発明のコラゲナーゼ遺伝子の範囲内に包含される。
【0038】
本発明のコラゲナーゼ遺伝子を用いると、遺伝子組み換え技術によって当該コラゲナーゼを大量に製造することが可能である。
【0039】
すなわち、本発明のコラゲナーゼ遺伝子を適当なベクターに組み込むことにより、宿主細胞を形質転換することができる。これらのベクターに適当なプロモーターや形質発現にかかわる配列を導入することにより、それぞれの宿主細胞において本発明のコラゲナーゼ遺伝子を発現することが可能である。宿主細胞としては、エシェリヒア・コリ(Escherichia coli)やバチルス・ズブチリス(Bacillus subtilis)やバチルス・ブレビス(Bacillus brevis)を用いることができ、ベクターとしては、エシェリヒア・コリ内で複製できるpUC18、pUC19、pBR322、pGEM3、pGEM4など、バチルス・ズブチリス内で複製できるpUC110、pE194、pC194など、バチルス・ブレビス内で複製できるpNCMO2、pNY326などのプラスミドが使用できる。
【0040】
本発明のコラゲナーゼ遺伝子を含むベクターで形質転換された宿主細胞を培養し、培養物からコラゲナーゼを採取することにより、当該コラゲナーゼを製造することができる。宿主細胞の培養は、例えば、エイ・レクロイジーら、FEBSレターズ(A.Lecroiseyら、FEBS Lett.、59 (1975) 167−172)に記載される方法に従って、適当な炭素源、窒素源および微量の金属元素を含む培地中で液体培養することで行なえる。得られた培養物を回収し、例えば、培養上清や可溶化菌体上清を硫安沈澱(60%飽和)処理し、種々のカラムクロマトグラフィー(例えば、DEAEセルロースカラムクロマトグラフィーおよびセファデックスG−100カラムクロマトグラフィーあるいはキレートアフィニティクロマトグラフィーなど)を用いて精製することにより、所望のコラゲナーゼが得られる。
【0041】
以下に実施例を挙げて本発明をさらに詳しく説明するが、本発明はこれらに限定されるものではない。
【実施例1】
【0042】
[精製酵素の解析]
当該コラゲナーゼを特開平8-70853の方法で精製し、常法(例えば、細胞外マトリックス研究法、第2巻、77−85頁、コラーゲン技術研修会(1998))により蛍光標識したI型コラーゲンを用いてコラーゲン分解酵素活性を測定した。市販品のクロストリジウム・ヒストリティカム(Clostridium histolyticum) 由来のコラゲナーゼおよび、ストレプトミセス(Streptomyces) 由来のコラゲナーゼと比較した結果、グリモンティア・ホリセー由来のコラゲナーゼの活性が最も高かった。
【0043】
精製酵素をSDS-ポリアクリルアミド電気泳動により解析した。
【0044】
第1図に精製酵素のSDS-ポリアクリルアミド電気泳動像を、第2図にFITC-コラーゲンを分解する酵素活性を示す。
【実施例2】
【0045】
[アミノ酸配列の決定]
当該コラゲナーゼの部分アミノ酸配列は、以下のようにして決定した。精製した当該コラゲナーゼを常法(例えば、続生化学実験法、第2巻、260−270頁、日本生化学会編)によりトリプシン、プロテアーゼV8またはリジルエンドペプチダーゼで各々部分加水分解した。得られたペプチド断片を高速液体クロマトグラフィーおよびSDS-ポリアクリルアミド電気泳動法により精製した後、自動化エドマン分解法によりアミノ酸配列を決定した。その結果、本発明における当該コラゲナーゼは少なくとも以下の7種のペプチド断片のアミノ酸配列を有していた。
【0046】
1) AVEQCDL (配列番号1)
2) GAVDAFV (配列番号2)
3) IRSQDMSQAQHLAACDK (配列番号3)
4) MAYEESFFHTTLE (配列番号4)
5) WGYL (配列番号5)
6) FSSNHMY (配列番号6)
7) ASYAN (配列番号7)
【実施例3】
【0047】
[遺伝子ライブラリーの作製]
当該コラゲナーゼ産生菌グリモンティア・ホリセーから常法により全ゲノムDNAを単離した。微生物DNAの単離法としては、例えば、サイトウ・ミウラ法(H. Saito and K. Miura、Biochem. Biophys. Acta、72 (1963) 619-629)等が挙げられる。このDNAを制限酵素EcoRIで部分分解し、アガロースゲル電気泳動法により分画し、2kb以上のDNA断片を集めた。このDNA断片を、ベクターpCC1BAC(大腸菌バクミド(pBeloBAC11由来))のEcoRI挿入サイトにT4リガーゼで結合し、エシェリヒア・コリK12株(E.coli Trans for MaxTM EPI300TM)を形質転換して、クロラムフェニコール耐性形質転換体としてグリモンティア・ホリセーのゲノムライブラリーを作製した。エシェリヒア・コリの形質転換は常法(Dower, W. J.ら、Nucl. Acids Res.、16 (1988) 6127-6145)を用いた。
【実施例4】
【0048】
[遺伝子ライブラリーのスクリーニング]
精製した当該コラゲナーゼをプロテアーゼV8で消化して得た上記6) 7)のペプチド配列から誘導された当該コラゲナーゼ遺伝子の部分配列(長さ1.0kb)を、PCR反応を使って単離した。
【0049】
プライマー:
Fwd: GAGGCNATCTTTAGCTCCAATCATATGTAYAAY (配列番号12)
Rvs: ATCTAAGTAATGCACGTATTCATGYTCNAGRTT (配列番号13)
但し、配列番号12、13中のアルファベットは以下の塩基を示す。
N: A,C,G,T、 Y: C,T、 R: A,G
上記プライマーの組合わせがPCR産物(長さ1.0kb)を生じ、図3に示した全塩基配列の421〜1426番目の塩基に対応した。この部分塩基配列を使ってDIGシステム(Kruchen, B. and Rueger, B.、Biochemica、3 (2003)13-15)に記載されている方法によりDNAプローブを調製した。当該コラゲナーゼ遺伝子の全領域を単離する目的で、DNAプローブを用いたコロニーハイブリダイゼーション法により上記遺伝子ライブラリーの中から、コラゲナーゼを発現しているクローンとして、バクミドpCC1BAC-2、pCC1BAC-4を持つクローンを選択した。pCC1BAC-2は、50kb以上のグリモンティア・ホリセー由来のDNA挿入断片を持つ。
【0050】
なお、バクミドpCC1BAC-2は平成21(2009)年4月28日付けで独立行政法人製品評価技術基盤機構特許生物寄託センターに受託番号:NITE P-739として寄託した。
【実施例5】
【0051】
[DNA塩基配列の決定]
pCC1BAC-2の50kb以上の挿入断片の内、約2.4kbについてジデオキシ法(F.Sangerら、Proc.Nat,Acad,Sci,USA、74 (1977) 5963-5967)により塩基配列を決定した。決定されたオープンリーディングフレーム(図3、配列番号14、15)は、1番目のATGから始まり、2304番目のTAAで終わる2304塩基対からなり、当該コラゲナーゼの全領域を含んでいる。全塩基配列中において、決定された塩基配列から推測されるアミノ酸配列と、実施例2において決定された部分アミノ酸配列とを比較してみると、以下の7個のペプチド断片のアミノ酸配列を有していた。
【0052】
【化1】
【0053】
なお、(1)〜(7)の塩基配列(配列表において配列番号16〜22として示す)はそれぞれ配列番号1〜7のアミノ酸配列に対応する。
【実施例6】
【0054】
[コラゲナーゼ組換えタンパク質の発現および性状解析]
まず、コラゲナーゼ遺伝子をバチルス・ブレビス(Bacillus brevis)で大量に生産させるための組換えプラスミドを作製した。pCC1BAC-2を鋳型として挿入されているコラゲナーゼ遺伝子の成熟体領域2.0kbのDNA断片の5’側にNco Iサイトを、3’側にHind IIIサイトを付加し、Expand High Fidelity PCR System(Roche)により増幅した。増幅された断片をNco I-Hind IIIで処理して回収した当該DNA断片を、プラスミドベクターpNY326のサイトに挿入して、pNY326-Col2を作製した。さらに組換プラスミドpNY326-Col2をB.brevis S5株に形質転換して、コラゲナーゼを大量に生産するB.brevis組換え体を作製した。
【0055】
コラゲナーゼ遺伝子を含有するB.brevis組換え体を2SLN培地(4% ポリペプトンS、0.5% 酵母エキス、2% グルコース、0.001% FeSO4・7H2O、0.001% MnSO4・7H2O、0.0001% ZnSO4・7H2O、50μg/ml ネオマイシン)中で、30℃、48時間培養した。培養液を遠心し、上清を0.2μmのフィルターでろ過滅菌をした。このようにして得たB.brevis組換え体培養上清中に分泌されたグリモンティア・ホリセー由来コラゲナーゼ組換えタンパク質を、まずゼラチンザイモグラフィー法によって解析した。ゼラチンザイモグラフィー法は服部らの方法(Hattori, S., Anal. Biochem., 301,27-34,(2002))で行った。図4に示すように、組換え体培養上清にはグリモンティア・ホリセー由来の精製コラゲナーゼと同じ位置にバンドが認められた。対照となる非組換えプラスミド導入株(mock)培養上清にはバンドが認められなかった。
【0056】
次に組換えタンパク質のコラゲナーゼ活性を、蛍光標識したコラーゲン溶液を用いて測定した。測定法は常法(例えば細胞外マトリックス研究法、第2巻、77−85頁、コラーゲン技術研修会(1998))を用いた。第1表に示すように、B.brevis組換え体(pNY326-Col2)培養上清中には、高いコラゲナーゼ活性が認められた。一方、非組換えプラスミド導入株(mock)培養上清にはコラゲナーゼ活性は認められなかった。表中1ユニット(U)はコラーゲン1μgを、30℃、1分間で分解する活性を指す。
【0057】
【表1】
【受託番号】
【0058】
NITE P-739
【特許請求の範囲】
【請求項1】
少なくとも下記式(a)ないし(g)で示されるペプチド断片を有するグリモンティア・ホリセー(Grimontia hollisae)に属する微生物由来のコラゲナーゼをコードする遺伝子。
式(a) AVEQCDL
式(b) GAVDAFV
式(c) IRSQDMSQAQHLAACDK
式(d) MAYEESFFHTTLE
式(e) WGYL
式(f) FSSNHMY
式(g) ASYAN
但し、式(a)ないし(g)中のアルファベットは以下のアミノ酸を示す。A:アラニン、C:システイン、D:アスパラギン酸、E:グルタミン酸 F:フェニルアラニン、G:グリシン、H:ヒスチジン、I:イソロイシン、K:リシン、L:ロイシン、M:メチオニン、N:アスパラギン、P:プロリン、Q:グルタミン、R:アルギニン、S:セリン、T:スレオニン、V:バリン、W:トリプトファン、Y:チロシン。
【請求項2】
以下の(h)または(i)のタンパク質をコードする遺伝子。
(h)下記式(I)で表されるアミノ酸配列からなるグリモンティア・ホリセー(Grimontia hollisae)に属する微生物由来のコラゲナーゼタンパク質
式(I): X-
AVEQCDLSQFQTTSSNQLMAAIRQQGASCVNALFSADTGVQEAAFSSNHMYNVAQYTRTLAQQYAGGGSDELEALYLYLRAGYYAEFYNSNITFLSWVTPAVKGAVDAFVQNAHFYDNGDAHGKVLNEVIITMDSAGLQHAYLDVVTQWLTRWNAQYAEHWYMRNAVNGVFTLLFGGQWNNQYTSLIGEQTALVTALQAFALDRTKVNSPTEFMAANAARELGRLARYTDATIAPKVTEGLTAIFGQYPSYGDGDAIWLGAADTASYYADCSQFNICGFEDALRDAALNQTFICSDTIKIRSQDMSQAQHLAACDKMAYEESFFHTTLETGNQPVADDHNTQLQVNIFNSDTDYGKYAGPIFGIDTNNGGMYLEGNPANVGNIPNFIAYEASYANPDHFVWNLEHEYVHYLDGRFNMYGDFGTPTELVVWWSEGVAEYVSRVNDNPQAIATIQDGSTYTLAQVFDTTYDGFDVDRIYRWGYLAVRFMFERHPDEVQRMLSATRQGRWAEYKAIISGWANQYQSEFAQWTEALAKGDSGAGNGEGTGSGNEGGGESGGNTGLPENCAVLPKISDGRLALDEAACLADTASASDVLWFSIPAVSEYQTIAITAGNGTGDLTLEYSNLNWPDGTNVQASSANMGNSECIILEHQANYWGYLKVSGSFENAALLVEAGSNQCRQ
但し、式(I)中、Xは、水素原子または次式(II)で表されるポリペプチドを示す。
式(II):MELKALALTVSALLVGQSVYASEVFAHPGMPVQQKLQPNLLQQSTRLQPEQHIHGLERTDRQYRPTDATQQPEPPTLLKRQVSVQQD
また、式(I)、(II)中のアルファベットは、請求項1と同義である。
(i)式(I)で表されるアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつコラゲナーゼ活性を有するタンパク質。
【請求項3】
以下の(j)、(k)、(l)または(m)のDNAからなるコラゲナーゼ遺伝子:
(j)グリモンティア・ホリセーに属する微生物由来のコラゲナーゼをコードする、下記式(III)で表される塩基配列からなるDNA
式(III):Z-
GCGGTTGAACAGTGTGATCTCAGCCAGTTTCAGACGACCAGCAGTAACCAGTTGATGGCCGCGATTCGCCAGCAGGGCGCCAGTTGTGTCAATGCCTTGTTTAGTGCTGACACGGGAGTACAGGAAGCCGCGTTTTCGTCCAACCACATGTATAACGTCGCACAATATACCCGAACACTGGCGCAGCAATATGCGGGCGGGGGCAGTGATGAACTGGAAGCCTTATATCTGTATCTTCGGGCGGGGTATTACGCTGAGTTTTATAACAGCAATATCACGTTTCTGTCGTGGGTGACGCCGGCAGTGAAAGGTGCCGTGGATGCGTTTGTCCAGAATGCCCATTTTTATGACAACGGCGACGCTCATGGCAAGGTGCTGAACGAGGTGATCATCACGATGGACAGTGCTGGGCTACAGCATGCCTATCTCGATGTAGTGACGCAGTGGCTGACCCGTTGGAATGCGCAATATGCCGAGCATTGGTATATGCGAAACGCAGTCAATGGTGTCTTTACTCTTTTGTTTGGCGGGCAGTGGAACAATCAGTACACCAGCCTGATTGGCGAACAGACTGCGTTGGTGACGGCCTTACAGGCATTTGCGCTGGACCGTACGAAAGTGAACTCGCCAACGGAGTTTATGGCGGCCAATGCGGCCAGAGAACTGGGGCGGTTAGCTCGCTACACGGACGCGACCATTGCGCCTAAAGTCACGGAAGGATTAACCGCGATCTTCGGCCAGTATCCGTCCTATGGCGATGGAGATGCTATCTGGCTGGGGGCGGCGGATACGGCCTCTTATTATGCTGATTGCAGCCAGTTCAACATCTGTGGCTTTGAAGACGCGCTGCGTGATGCGGCGCTGAACCAGACTTTTATCTGTAGTGATACGATTAAAATTCGCTCACAGGATATGTCGCAGGCACAGCATCTCGCGGCTTGCGACAAAATGGCTTATGAAGAGTCATTTTTCCACACCACGCTTGAAACCGGTAATCAGCCGGTGGCTGATGATCATAATACGCAGCTGCAGGTGAATATTTTTAATTCCGATACCGATTACGGTAAATATGCCGGTCCGATATTTGGGATTGATACCAACAACGGCGGTATGTACCTCGAGGGGAATCCGGCCAATGTGGGCAATATTCCCAATTTCATCGCGTATGAAGCCAGCTATGCCAACCCGGACCATTTTGTCTGGAATCTTGAGCACGAGTACGTCCACTATTTGGATGGGCGGTTCAATATGTATGGCGATTTTGGTACGCCTACCGAGCTTGTGGTCTGGTGGAGCGAAGGGGTGGCCGAGTATGTGTCGCGGGTAAATGATAATCCTCAGGCGATTGCCACCATCCAGGATGGTAGCACGTACACTCTGGCGCAGGTGTTTGACACGACGTATGACGGTTTTGATGTGGATCGCATCTACCGATGGGGATATCTGGCGGTGCGATTCATGTTTGAACGTCATCCTGATGAAGTTCAACGTATGCTGAGTGCCACCCGACAGGGACGCTGGGCAGAATACAAGGCGATCATTAGCGGTTGGGCAAATCAGTATCAGTCAGAATTTGCCCAATGGACCGAGGCGCTGGCGAAGGGCGACAGTGGCGCTGGGAACGGTGAGGGGACAGGCTCCGGTAATGAAGGGGGCGGTGAATCTGGTGGTAATACCGGCTTGCCGGAAAACTGCGCAGTACTGCCAAAAATCAGTGATGGGCGTTTAGCACTGGATGAAGCGGCCTGTCTGGCCGACACGGCTTCAGCGTCTGACGTATTGTGGTTCAGTATTCCGGCTGTCAGTGAATATCAGACCATTGCCATTACGGCGGGCAACGGGACTGGCGACCTGACGCTGGAATACAGTAACCTGAATTGGCCAGATGGTACCAATGTGCAGGCATCATCGGCAAATATGGGTAACAGTGAATGCATTATTCTGGAACATCAGGCGAATTATTGGGGATATCTGAAAGTCTCGGGTTCCTTTGAAAATGCAGCGTTACTGGTGGAGGCTGGCAGTAACCAGTGTCGTCAG
但し、式(III)中、Zは無しか、または式(IV)で表される塩基配列を示す。
式(IV):
ATGGAACTTAAAGCACTTGCGTTAACGGTATCAGCCCTGCTTGTGGGTCAGTCGGTGTATGCATCTGAGGTGTTTGCTCACCCGGGCATGCCGGTTCAACAAAAGTTGCAGCCAAATTTGCTGCAGCAAAGTACCAGGCTTCAGCCAGAGCAACATATCCACGGGCTTGAACGCACGGATCGGCAATATCGTCCGACAGATGCTACTCAGCAGCCGGAACCGCCAACGTTACTGAAACGTCAGGTATCGGTTCAGCAGGAT
(k)遺伝子コドンの縮重の範囲内で(j)のDNAと同じアミノ酸をコードする塩基配列を有するDNA
(l)式(III)で表される塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつコラゲナーゼ活性を有するタンパク質をコードするDNA
(m)式(III)で表される塩基配列と生物学的に同等な塩基配列を有するDNA。
【請求項4】
請求項1〜3のいずれか1項に記載の遺伝子を含有する組換えベクター。
【請求項5】
請求項4に記載の組換えベクターにより形質転換された宿主細胞。
【請求項6】
微生物由来のコラゲナーゼの製造方法であって、請求項5記載の宿主細胞を培養し、培養物から産生されたコラゲナーゼを回収・精製することを特徴とする前記コラゲナーゼの製造方法。
【請求項1】
少なくとも下記式(a)ないし(g)で示されるペプチド断片を有するグリモンティア・ホリセー(Grimontia hollisae)に属する微生物由来のコラゲナーゼをコードする遺伝子。
式(a) AVEQCDL
式(b) GAVDAFV
式(c) IRSQDMSQAQHLAACDK
式(d) MAYEESFFHTTLE
式(e) WGYL
式(f) FSSNHMY
式(g) ASYAN
但し、式(a)ないし(g)中のアルファベットは以下のアミノ酸を示す。A:アラニン、C:システイン、D:アスパラギン酸、E:グルタミン酸 F:フェニルアラニン、G:グリシン、H:ヒスチジン、I:イソロイシン、K:リシン、L:ロイシン、M:メチオニン、N:アスパラギン、P:プロリン、Q:グルタミン、R:アルギニン、S:セリン、T:スレオニン、V:バリン、W:トリプトファン、Y:チロシン。
【請求項2】
以下の(h)または(i)のタンパク質をコードする遺伝子。
(h)下記式(I)で表されるアミノ酸配列からなるグリモンティア・ホリセー(Grimontia hollisae)に属する微生物由来のコラゲナーゼタンパク質
式(I): X-
AVEQCDLSQFQTTSSNQLMAAIRQQGASCVNALFSADTGVQEAAFSSNHMYNVAQYTRTLAQQYAGGGSDELEALYLYLRAGYYAEFYNSNITFLSWVTPAVKGAVDAFVQNAHFYDNGDAHGKVLNEVIITMDSAGLQHAYLDVVTQWLTRWNAQYAEHWYMRNAVNGVFTLLFGGQWNNQYTSLIGEQTALVTALQAFALDRTKVNSPTEFMAANAARELGRLARYTDATIAPKVTEGLTAIFGQYPSYGDGDAIWLGAADTASYYADCSQFNICGFEDALRDAALNQTFICSDTIKIRSQDMSQAQHLAACDKMAYEESFFHTTLETGNQPVADDHNTQLQVNIFNSDTDYGKYAGPIFGIDTNNGGMYLEGNPANVGNIPNFIAYEASYANPDHFVWNLEHEYVHYLDGRFNMYGDFGTPTELVVWWSEGVAEYVSRVNDNPQAIATIQDGSTYTLAQVFDTTYDGFDVDRIYRWGYLAVRFMFERHPDEVQRMLSATRQGRWAEYKAIISGWANQYQSEFAQWTEALAKGDSGAGNGEGTGSGNEGGGESGGNTGLPENCAVLPKISDGRLALDEAACLADTASASDVLWFSIPAVSEYQTIAITAGNGTGDLTLEYSNLNWPDGTNVQASSANMGNSECIILEHQANYWGYLKVSGSFENAALLVEAGSNQCRQ
但し、式(I)中、Xは、水素原子または次式(II)で表されるポリペプチドを示す。
式(II):MELKALALTVSALLVGQSVYASEVFAHPGMPVQQKLQPNLLQQSTRLQPEQHIHGLERTDRQYRPTDATQQPEPPTLLKRQVSVQQD
また、式(I)、(II)中のアルファベットは、請求項1と同義である。
(i)式(I)で表されるアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつコラゲナーゼ活性を有するタンパク質。
【請求項3】
以下の(j)、(k)、(l)または(m)のDNAからなるコラゲナーゼ遺伝子:
(j)グリモンティア・ホリセーに属する微生物由来のコラゲナーゼをコードする、下記式(III)で表される塩基配列からなるDNA
式(III):Z-
GCGGTTGAACAGTGTGATCTCAGCCAGTTTCAGACGACCAGCAGTAACCAGTTGATGGCCGCGATTCGCCAGCAGGGCGCCAGTTGTGTCAATGCCTTGTTTAGTGCTGACACGGGAGTACAGGAAGCCGCGTTTTCGTCCAACCACATGTATAACGTCGCACAATATACCCGAACACTGGCGCAGCAATATGCGGGCGGGGGCAGTGATGAACTGGAAGCCTTATATCTGTATCTTCGGGCGGGGTATTACGCTGAGTTTTATAACAGCAATATCACGTTTCTGTCGTGGGTGACGCCGGCAGTGAAAGGTGCCGTGGATGCGTTTGTCCAGAATGCCCATTTTTATGACAACGGCGACGCTCATGGCAAGGTGCTGAACGAGGTGATCATCACGATGGACAGTGCTGGGCTACAGCATGCCTATCTCGATGTAGTGACGCAGTGGCTGACCCGTTGGAATGCGCAATATGCCGAGCATTGGTATATGCGAAACGCAGTCAATGGTGTCTTTACTCTTTTGTTTGGCGGGCAGTGGAACAATCAGTACACCAGCCTGATTGGCGAACAGACTGCGTTGGTGACGGCCTTACAGGCATTTGCGCTGGACCGTACGAAAGTGAACTCGCCAACGGAGTTTATGGCGGCCAATGCGGCCAGAGAACTGGGGCGGTTAGCTCGCTACACGGACGCGACCATTGCGCCTAAAGTCACGGAAGGATTAACCGCGATCTTCGGCCAGTATCCGTCCTATGGCGATGGAGATGCTATCTGGCTGGGGGCGGCGGATACGGCCTCTTATTATGCTGATTGCAGCCAGTTCAACATCTGTGGCTTTGAAGACGCGCTGCGTGATGCGGCGCTGAACCAGACTTTTATCTGTAGTGATACGATTAAAATTCGCTCACAGGATATGTCGCAGGCACAGCATCTCGCGGCTTGCGACAAAATGGCTTATGAAGAGTCATTTTTCCACACCACGCTTGAAACCGGTAATCAGCCGGTGGCTGATGATCATAATACGCAGCTGCAGGTGAATATTTTTAATTCCGATACCGATTACGGTAAATATGCCGGTCCGATATTTGGGATTGATACCAACAACGGCGGTATGTACCTCGAGGGGAATCCGGCCAATGTGGGCAATATTCCCAATTTCATCGCGTATGAAGCCAGCTATGCCAACCCGGACCATTTTGTCTGGAATCTTGAGCACGAGTACGTCCACTATTTGGATGGGCGGTTCAATATGTATGGCGATTTTGGTACGCCTACCGAGCTTGTGGTCTGGTGGAGCGAAGGGGTGGCCGAGTATGTGTCGCGGGTAAATGATAATCCTCAGGCGATTGCCACCATCCAGGATGGTAGCACGTACACTCTGGCGCAGGTGTTTGACACGACGTATGACGGTTTTGATGTGGATCGCATCTACCGATGGGGATATCTGGCGGTGCGATTCATGTTTGAACGTCATCCTGATGAAGTTCAACGTATGCTGAGTGCCACCCGACAGGGACGCTGGGCAGAATACAAGGCGATCATTAGCGGTTGGGCAAATCAGTATCAGTCAGAATTTGCCCAATGGACCGAGGCGCTGGCGAAGGGCGACAGTGGCGCTGGGAACGGTGAGGGGACAGGCTCCGGTAATGAAGGGGGCGGTGAATCTGGTGGTAATACCGGCTTGCCGGAAAACTGCGCAGTACTGCCAAAAATCAGTGATGGGCGTTTAGCACTGGATGAAGCGGCCTGTCTGGCCGACACGGCTTCAGCGTCTGACGTATTGTGGTTCAGTATTCCGGCTGTCAGTGAATATCAGACCATTGCCATTACGGCGGGCAACGGGACTGGCGACCTGACGCTGGAATACAGTAACCTGAATTGGCCAGATGGTACCAATGTGCAGGCATCATCGGCAAATATGGGTAACAGTGAATGCATTATTCTGGAACATCAGGCGAATTATTGGGGATATCTGAAAGTCTCGGGTTCCTTTGAAAATGCAGCGTTACTGGTGGAGGCTGGCAGTAACCAGTGTCGTCAG
但し、式(III)中、Zは無しか、または式(IV)で表される塩基配列を示す。
式(IV):
ATGGAACTTAAAGCACTTGCGTTAACGGTATCAGCCCTGCTTGTGGGTCAGTCGGTGTATGCATCTGAGGTGTTTGCTCACCCGGGCATGCCGGTTCAACAAAAGTTGCAGCCAAATTTGCTGCAGCAAAGTACCAGGCTTCAGCCAGAGCAACATATCCACGGGCTTGAACGCACGGATCGGCAATATCGTCCGACAGATGCTACTCAGCAGCCGGAACCGCCAACGTTACTGAAACGTCAGGTATCGGTTCAGCAGGAT
(k)遺伝子コドンの縮重の範囲内で(j)のDNAと同じアミノ酸をコードする塩基配列を有するDNA
(l)式(III)で表される塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつコラゲナーゼ活性を有するタンパク質をコードするDNA
(m)式(III)で表される塩基配列と生物学的に同等な塩基配列を有するDNA。
【請求項4】
請求項1〜3のいずれか1項に記載の遺伝子を含有する組換えベクター。
【請求項5】
請求項4に記載の組換えベクターにより形質転換された宿主細胞。
【請求項6】
微生物由来のコラゲナーゼの製造方法であって、請求項5記載の宿主細胞を培養し、培養物から産生されたコラゲナーゼを回収・精製することを特徴とする前記コラゲナーゼの製造方法。
【図2】

【図1】
【図3】

【図4】
【図1】
【図3】
【図4】
【公開番号】特開2010−263880(P2010−263880A)
【公開日】平成22年11月25日(2010.11.25)
【国際特許分類】
【出願番号】特願2009−203743(P2009−203743)
【出願日】平成21年9月3日(2009.9.3)
【出願人】(000135151)株式会社ニッピ (18)
【Fターム(参考)】
【公開日】平成22年11月25日(2010.11.25)
【国際特許分類】
【出願日】平成21年9月3日(2009.9.3)
【出願人】(000135151)株式会社ニッピ (18)
【Fターム(参考)】
[ Back to top ]