
微生物由来の細胞外プロテアーゼの産生ならびにその用途
【課題】 目的とするタンパク質、特に中性プロテアーゼの宿主細胞外への分泌産生を可能とするシグナル配列及びプロ配列を提供すること。
【解決手段】 クロストリディウム・ヒストリティカムのゲノム遺伝子を精査し、中性プロテアーゼのシグナル配列及びプロ配列を同定した。また、これら配列を連結させた中性プロテアーゼの成熟配列を宿主細胞内にて発現させたところ、中性プロテアーゼが活性を有した成熟型として、宿主細胞外に分泌産生されることが実証された。
【解決手段】 クロストリディウム・ヒストリティカムのゲノム遺伝子を精査し、中性プロテアーゼのシグナル配列及びプロ配列を同定した。また、これら配列を連結させた中性プロテアーゼの成熟配列を宿主細胞内にて発現させたところ、中性プロテアーゼが活性を有した成熟型として、宿主細胞外に分泌産生されることが実証された。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、シグナル配列及びプロ配列に関し、より詳しくは、シグナル配列及びプロ配列をコードするポリヌクレオチド、シグナル配列と、プロ配列と、中性プロテアーゼの成熟配列とが連結しているタンパク質をコードするポリヌクレオチド、該ポリヌクレオチドを含有する発現ベクター、該発現ベクターで形質転換された宿主細胞、並びに、該宿主細胞から分泌された中性プロテアーゼを含む培養物に関する。また、本発明は、前記形質転換された宿主細胞を用いた中性プロテアーゼの製造方法、並びに、該製造方法によって製造された中性プロテアーゼを含むプロテアーゼ調製物に関する。
【背景技術】
【0002】
微生物から菌体外に分泌されるプロテアーゼ(菌体外プロテアーゼ)は、グラム陰性菌に関しては、最初に菌体内でシグナル配列、プロ配列及び成熟配列が連結した不活性のプレプロ型として合成される。その後、シグナル配列により細胞内膜を通過してペリプラズムに移行し、シグナル配列は切断されプロ型に変換される。プロ配列は主に成熟配列部分をフォールディングする分子内シャペロンとして機能している。このプロ配列が切断されることにより、活性を有した成熟型プロテアーゼとして菌体外へ分泌される。また、グラム陽性菌に関しては、最初に菌体内で不活性のプレプロ型として合成された後、細胞壁付近でシグナル配列が細菌内酵素により切断され、菌体外に分泌される。そして、菌体外に分泌されると直ぐに自己触媒的切断(autocatalytic cleavage)により速やかにプロ配列が切断され、活性を有した成熟配列となることが想定される。
【0003】
一方、糖尿病の根治療法として、膵臓から単離される膵島(インスリン産生細胞)を、糖尿病患者の門脈へ点滴にて移植する膵島移植が知られている。本移植法は、移植の際に患者の開腹手術が必要ないため、安全かつ簡便な糖尿病の根治療法として近年注目されている。通常、膵臓からの膵島の分離は、膵臓をコラゲナーゼ及び中性プロテアーゼにて処理することにより実施されるが、本目的に使用される中性プロテアーゼの一つとしてクロストリディウム・ヒストリティカム(Clostridium histolyticum)由来の中性プロテアーゼ(亜鉛金属プロテアーゼ)がある。
【0004】
これまでクロストリディウム・ヒストリティカム由来の中性プロテアーゼは、前記菌体外プロテアーゼの1種であるため、クロストリディウム・ヒストリティカムの培養上清より製造されているが、クロストリディウム・ヒストリティカムの培養液には非常に多くのプロテアーゼ活性を有するタンパク質が含まれており、調製物にその他のプロテアーゼ様活性が混入する懸念があった。
【0005】
そこで、他の中性プロテアーゼ様活性を混入させる恐れがなく、宿主細胞外に活性を有したまま多量に発現させるクロストリディウム・ヒストリティカム由来中性プロテアーゼの製造法が所望されていた。さらには、クロストリディウム・ヒストリティカム由来中性プロテアーゼの分泌機構が解明されることにより、中性プロテアーゼに限らず、目的とするタンパク質を中性プロテアーゼ由来のシグナル配列やプロ配列と融合させた形でクロストリディウム・ヒストリティカム等の宿主細胞内にて発現させ、その活性を維持しつつ、宿主細胞外に分泌された該タンパク質を効率良く調製することができるようになるため、クロストリディウム・ヒストリティカム由来中性プロテアーゼの分泌機構、特にシグナル配列及びプロ配列の同定が所望されていた。
【0006】
そして、このような背景の中、特許文献1には、クロストリディウム・ヒストリティカム由来中性プロテアーゼをコードする核酸の塩基配列、並びに該配列を用いることによって、そのプロテアーゼの活性を有するタンパク質を製造できることも開示されている。
【0007】
しかしながら、特許文献1において、クロストリディウム・ヒストリティカム由来中性プロテアーゼの遺伝子組換え体を用いた製造法に関する記載はあるものの、実施例中には遺伝子組換え体による中性プロテアーゼの発現の結果ならびにその活性についての記載はない。また、特許文献1において開示されている配列は、成熟型中性プロテアーゼ部分のアミノ酸配列だけであり、シグナル配列及びプロ配列は開示されていない。そのため、特許文献1に記載のアミノ酸配列をコードするポリヌクレオチドを用いても、宿主細胞外に活性を有した成熟型中性プロテアーゼを分泌できないことが想定され、実際、後述の比較例1において示す通り、特許文献1に記載の方法ではクロストリディウム・ヒストリティカム由来中性プロテアーゼを製造することはできなかった。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特表平10−500581号公報
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、前記従来技術の有する課題に鑑みてなされたものであり、目的とするタンパク質、特に中性プロテアーゼの宿主細胞外への分泌産生を可能とするシグナル配列及びプロ配列を提供することを目的とする。
【課題を解決するための手段】
【0010】
本発明者らは、前記目的を達成すべく鋭意研究を重ねた結果、クロストリディウム・ヒストリティカムのゲノム遺伝子を精査し、中性プロテアーゼのシグナル配列の開始コドンとなりうる部分を複数見出した。これらの配列について順に遺伝子組み換え体を造出し、発現を確認したところ、成熟配列のN末端からもっとも遠い位置に存在する開始コドンのみで中性プロテアーゼの発現が見られた。よって、配列番号1に示すシグナル配列及び配列番号3に示すプロ配列を含むポリヌクレオチドを用いることにより、クロストリディウム・ヒストリティカム以外の宿主においてもクロストリディウム・ヒストリティカム由来中性プロテアーゼ等のタンパク質を、宿主細胞外に分泌産生できることを見出した。なお、配列番号1に示すシグナル配列及び配列番号3に示すプロ配列は、他の遺伝子との相同性をほとんど持たない新規な配列であった。さらに、本発明者らは、当該シグナル配列と、当該プロ配列と、中性プロテアーゼの成熟配列とが連結しているタンパク質をコードするポリヌクレオチドを用いて形質転換された宿主細胞を用いることにより、中性プロテアーゼを活性を有した成熟型として宿主細胞外に分泌産生され、取得することができることを見出した。
【0011】
本発明は、より詳しくは以下を提供するものである。
<1> シグナル配列をコードする、下記(i)から(v)のいずれかに記載のポリヌクレオチド。
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:1に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:2に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:2に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:1に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
<2> プロ配列をコードする、下記(i)から(v)のいずれかに記載のポリヌクレオチド。
(i)配列番号:4に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:3に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:4に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:4に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:3に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
<3> クロストリディウム・ヒストリティカム(Clostridium histolyticum)由来である、<1>又は<2>に記載のポリヌクレオチド。
<4> <1>に記載のポリヌクレオチドと、<2>に記載のポリヌクレオチドと、中性プロテアーゼの成熟配列をコードする下記(i)から(v)のいずれかに記載のポリヌクレオチドとが連結しているポリヌクレオチド。
(i)配列番号:6に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:5に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:6に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:6に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:5に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
<5> クロストリディウム・ヒストリティカム(Clostridium histolyticum)由来である、<4>に記載のポリヌクレオチド。
<6> <4>又は<5>に記載のポリヌクレオチドを含有する発現ベクター。
<7> <6>に記載の発現ベクターで形質転換された宿主細胞。
<8> <7>に記載の形質転換された宿主細胞から分泌された中性プロテアーゼを含む培養物。
<9> <7>に記載の形質転換された宿主細胞を培養し、該宿主細胞および/またはその培養物から発現したタンパク質を採取する工程を含む、中性プロテアーゼの製造方法。
<10> <9>に記載の方法によって製造された中性プロテアーゼを含む、プロテアーゼ調製物。
【発明の効果】
【0012】
本発明によれば、目的とするタンパク質、特に中性プロテアーゼの宿主細胞外への分泌産生が可能となる。
【図面の簡単な説明】
【0013】
【図1】プラスミドpNCMO2−P2−NP(F1)を導入したBrevibacillus choshinensis SP−3株(pNCMO2−P2−NP(F1)形質転換体)の培養上清を電気泳動分離によって分析した結果を示すゲルの写真である。図中、1のレーンはサイズマーカーを泳動した結果を示し、2のレーンはpNCMO2−P2−NP(F1)形質転換体の培養上清を泳動した結果を示す。
【図2】pNCMO2−P2−NP(F1)形質転換体と、プラスミドpNCMO−P2−NPを導入したBrevibacillus choshinensis SP−3株(pNCMO2−P2−NP形質転換体)とをスキムミルクプレートに各々3点ずつ接種して培養し、スキムミルクの分解の程度(ハロー)を調べた結果を示す写真である。図中、AはpNCMO2−P2−NP(F1)形質転換体を培養した結果を示し、BはpNCMO2−P2−NP形質転換体を培養した結果を示す。
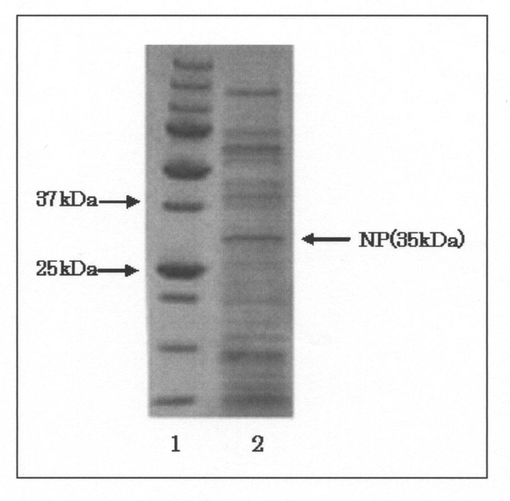
【図3】pNCMO2−P2−NP形質転換体の培養上清を電気泳動分離によって分析した結果を示すゲルの写真である。図中、1のレーンはサイズマーカーを泳動した結果を示し、2のレーンはpNCMO2−P2−NP形質転換体の培養上清を泳動した結果を示す。
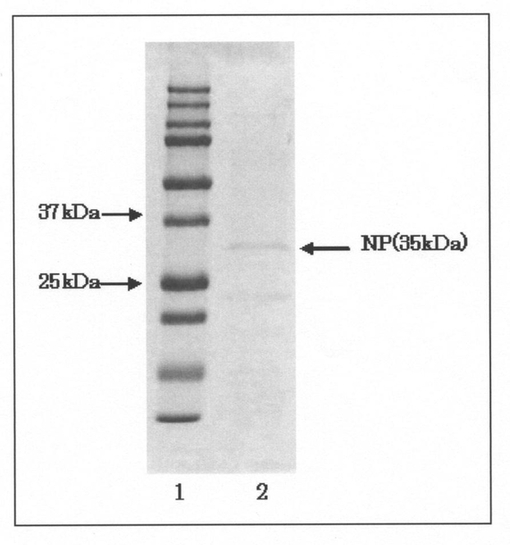
【図4】プラスミドpHT01−NPを導入したBacillus subtilis KN−2株(pHT01−NP形質転換体)の培養上清を電気泳動分離によって分析した結果を示すゲルの写真である。図中、1のレーンはサイズマーカーを泳動した結果を示し、2のレーンはpHT01−NP形質転換体の培養上清を泳動した結果を示す。
【発明を実施するための形態】
【0014】
<ポリヌクレオチド>
本発明のシグナル配列をコードするポリヌクレオチドは、下記(i)から(v)のいずれかに記載のポリヌクレオチドである。
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:1に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:2に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:2に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:1に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド。
【0015】
また、本発明のプロ配列をコードするポリヌクレオチドは、下記(i)から(v)のいずれかに記載のポリヌクレオチドである。
(i)配列番号:4に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:3に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:4に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:4に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:3に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド。
【0016】
本発明において、「シグナル配列」とは、その配列のC末端側に連結させる形で任意のタンパク質を宿主内において発現させた場合、該タンパク質を該宿主の外に分泌させる活性を有するタンパク質のことである。
【0017】
また、本発明において、「プロ配列」とは、その配列のC末端側に連結させる形で任意のタンパク質の成熟配列部分を宿主内において発現させた場合、主に、該成熟配列部分をフォールディングする分子内シャペロンとして機能するタンパク質のことである。
【0018】
本発明は、本発明のシグナル配列をコードするポリヌクレオチドと、本発明のプロ配列をコードするポリヌクレオチドと、中性プロテアーゼの成熟配列をコードする下記(i)から(v)のいずれかに記載のポリヌクレオチドとが連結しているポリヌクレオチドも提供するものである。
(i)配列番号:6に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:5に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:6に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:6に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:5に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド。
【0019】
本発明において、「中性プロテアーゼ」とは、その最適活性が中性pH範囲内にあり、カゼイン及び変性コラーゲンを解裂させる活性を有するタンパク質のことである。
【0020】
後述の実施例において示す通り、N末端側から順に、配列番号:2に記載のアミノ酸配列を含むタンパク質と、配列番号:4に記載のアミノ酸配列を含むタンパク質と、配列番号:6に記載のアミノ酸配列を含むタンパク質(クロストリディウム・ヒストリティカム由来中性プロテアーゼの成熟配列部分)とが連結しているタンパク質を宿主内において発現させた場合、活性を有した成熟型の中性プロテアーゼが該宿主の外に分泌された。従って、本発明のシグナル配列としては、配列番号:2に記載のアミノ酸配列を含むタンパク質(配列番号:3に記載の塩基配列がコードするタンパク質)が挙げられ、本発明のプロ配列としては、配列番号:4に記載のアミノ酸配列を含むタンパク質(配列番号:3に記載の塩基配列がコードするタンパク質)が挙げられ、本発明の中性プロテアーゼの成熟配列としては、配列番号:6に記載のアミノ酸配列を含むタンパク質(配列番号:5に記載の塩基配列がコードするタンパク質)が挙げられる。
【0021】
また、現在の技術水準においては、当業者であれば、かかるタンパク質をコードする塩基配列の情報が得られた場合、その塩基配列に対して種々の改変を行い、各活性や機能を有する、シグナル配列やプロ配列や中性プロテアーゼの製造を行うことが可能である。また、自然界においても、塩基配列が変異することは起こり得ることである。従って、本発明のシグナル配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:2に記載のアミノ酸配列において1もしくは複数個のアミノ酸が置換、欠失、挿入、および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチドも含まれ、本発明のプロ配列をコードするポリヌクレオチドには、前記機能を有するタンパク質をコードする限り、配列番号:4に記載のアミノ酸配列において1もしくは複数個のアミノ酸が置換、欠失、挿入、および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチドも含まれ、本発明の中性プロテアーゼの成熟配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:6に記載のアミノ酸配列において1もしくは複数個のアミノ酸が置換、欠失、挿入、および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチドも含まれる。
【0022】
「1もしくは複数個のアミノ酸が欠失、置換、挿入もしくは付加されたアミノ酸配列」とは、部位特異的突然変異誘発法等の周知の方法により、または天然に生じ得る程度の複数個の数のアミノ酸の置換等により改変がなされたことを意味する。アミノ酸の改変の個数は、好ましくは1〜10個、より好ましくは1〜5個、最も好ましくは1〜2個である。
【0023】
改変アミノ酸配列の例は、好ましくは、そのアミノ酸が、1または複数個(好ましくは、1ないし数個あるいは1、2、3、または4個)の保存的置換を有するアミノ酸配列であることができる。ここで、「保存的置換」とは、1または複数個のアミノ酸残基を、別の化学的に類似したアミノ酸残基で置き換えることを意味する。例えば、ある疎水性残基を別の疎水性残基によって置換する場合、ある極性残基を同じ電荷を有する別の極性残基によって置換する場合などが挙げられる。このような置換を行うことができる機能的に類似のアミノ酸は、アミノ酸毎に当該技術分野において公知である。具体例を挙げると、非極性(疎水性)アミノ酸としては、アラニン、バリン、イソロイシン、ロイシン、プロリン、トリプトファン、フェニルアラニン、メチオニンなどが挙げられる。極性(中性)アミノ酸としては、グリシン、セリン、スレオニン、チロシン、グルタミン、アスパラギン、システインなどが挙げられる。陽電荷をもつ(塩基性)アミノ酸としては、アルギニン、ヒスチジン、リジンなどが挙げられる。また、負電荷をもつ(酸性)アミノ酸としては、アスパラギン酸、グルタミン酸などが挙げられる。
【0024】
さらに、現在の技術水準においては、当業者であれば、かかるタンパク質をコードする特定の塩基配列の情報が得られた場合、それらの塩基配列情報を利用して、同種若しくは他の菌種等から、各活性や機能を有するタンパク質をコードする相同遺伝子を単離することが可能である。このような相同遺伝子を取得するための菌種等としては枯草菌(例えばBacillus subtilisやBacillus thermoproteolyticus rokko)や麹菌(例えばAspergillus oryzae)などが好ましい。
【0025】
相同遺伝子を取得するための方法としては、例えば、ハイブリダイゼーション技術(Southern,E.M.,J.Mol.Biol.,98:503,1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki,R.K.,et al.Science,230:1350−1354,1985、Saiki,R.K.et al.Science,239:487−491,1988)が挙げられる。相同遺伝子を単離するためには、通常、ストリンジェントな条件下でハイブリダイゼーション反応を行なう。
【0026】
ここで、「ストリンジェントな条件」とは、ハイブリダイゼーション後のメンブレンの洗浄操作を、高温度低塩濃度溶液中で行うことを意味し、例えば、2×SSC濃度(1×SSC:15mmol/Lクエン酸3ナトリウム、150mmol/L塩化ナトリウム)、0.5%SDS溶液中で、60℃、20分間の洗浄条件を意味する。
【0027】
従って、本発明のシグナル配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:1に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチドも含まれ、本発明のプロ配列をコードするポリヌクレオチドには、前記機能を有するタンパク質をコードする限り、配列番号:3に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチドも含まれ、本発明の中性プロテアーゼの成熟配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:5に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチドも含まれる。
【0028】
また、取得された相同遺伝子がコードするタンパク質は、通常、本発明のシグナル配列やプロ配列や中性プロテアーゼの成熟配列のアミノ酸配列と高い同一性を有するアミノ酸配列を含むタンパク質である。本発明において、高い同一性とは、好ましくは、95%以上、より好ましくは96%以上、特に好ましくは98%以上、そして最も好ましくは99%以上の同一性を有する。
【0029】
従って、本発明のシグナル配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:2に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチドも含まれ、本発明のプロ配列をコードするポリヌクレオチドには、前記機能を有するタンパク質をコードする限り、配列番号:4に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチドも含まれ、本発明の中性プロテアーゼの成熟配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:6に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチドも含まれる。
【0030】
塩基配列またはアミノ酸配列についての「同一性」は、比較される配列間において、各々の配列を構成する塩基またはアミノ酸残基の一致の程度の意味で用いられる。本明細書において示した「同一性」の数値はいずれも、当業者に公知の相同性検索プログラムを用いて算出される数値であればよく、例えばFASTA、BLAST等においてデフォルト(初期設定)のパラメータを用いることにより、容易に算出することができる。より具体的には、配列の同一性は、BLASTX(アミノ酸レベル)のプログラム(Altschul et al.J.Mol.Biol.,215:403−410,1990)を利用して決定することができる。該プログラムは、KarlinおよびAltschulによるアルゴリズムBLAST(Proc.Natl.Acad.Sci.USA,87:2264−2268,1990,Proc.Natl.Acad.Sci.USA,90:5873−5877,1993)に基づいている。BLASTXによってアミノ酸配列を解析する場合には、パラメーターは、例えばscore=50、wordlength=3とする。また、Gapped BLASTプログラムを用いて、アミノ酸配列を解析する場合は、Altschulら(Nucleic Acids Res.25:3389−3402,1997)に記載されているように行うことができる。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。
【0031】
なお、特定のポリヌクレオチドがコードするタンパク質が、前記活性や機能を有するタンパク質をコードするか否かは、例えば、後述の実施例に記載するように、特定のポリヌクレオチドがコードするタンパク質が発現するような形で、プラスミドベクターにクローニングし、該発現ベクターを宿主細胞(例えば、Brevibacillus choshinensis SP−3株、Bacillus subtilis KN−2株)に導入することにより得られた培養上清をSDS−PAGEにて分析し、予想される分子量を有するタンパク質のバンド(本発明にかかる中性プロテアーゼの成熟配列部分であれば、分子量 約35kDaのタンパク質が示すバンド)を検出できるか否かを検定することにより判定することができる(実施例2 参照)。また、前記発現ベクターを導入した宿主細胞(形質転換体)や該形質転換体の培養上清を用いて、スキムミルク(カゼイン)の分解の程度(例えば、ハローの有無やその大きさ)を培地(スキムミルクプレート)上で検出できるか否かを検定することにより判定することができる(実施例2 参照)。さらに、前記形質転換体や該形質転換体の培養上清を用い、アゾカゼインを基質とし、基質の分解によって生じる遊離チロシン量を335nmの吸光度で測定することによって、アゾカゼイン分解活性を検出できるか否かを検定することにより判定することができる(実施例3 参照)。
【0032】
また、本発明のポリヌクレオチドとしては、その形態に特に制限はなく、cDNAの他、ゲノムDNA、および化学合成DNAが含まれる。これらDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。ゲノムDNAは、例えば、クロストリディウム・ヒストリティカムからゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、BAC、PACなどが利用できる)を作製し、これを展開して、クロストリディウム・ヒストリティカム由来中性プロテアーゼのシグナル配列、プロ配列、成熟配列をコードする塩基配列(例えば、配列番号:1、3、5に記載のDNA)を基に調製したプローブを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより調製することが可能である。また、クロストリディウム・ヒストリティカム由来中性プロテアーゼのシグナル配列、プロ配列、成熟配列をコードする塩基配列に特異的なプライマーを作製し、これを利用したPCRを行うことによって調製することも可能である。また、cDNAは、例えば、クロストリディウム・ヒストリティカムから抽出したmRNAを基にcDNAを合成し、これをλZAP等のベクターに挿入してcDNAライブラリーを作成し、これを展開して、上記と同様にコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより、また、PCRを行うことにより調製することが可能である。さらに、市販のDNA合成機を用いれば、目的のDNAを合成により調製することも可能である。また、前記配列番号:1等に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチドや、前記配列番号:2等に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチドは、前記ハイブリダイゼーション技術やPCR技術の他に、特定の塩基配列やアミノ酸配列に基づき、当業者に公知の部位特異的変異誘発(site−directed mutagenesis)法(Kramer,W.&Fritz,HJ.,Methods Enzymol,1987,154,350.)等の遺伝子増幅技術や組換え技術を利用して調製することも可能である。
【0033】
また、本発明のポリヌクレオチドとしては、クロストリディウム・ヒストリティカム(Clostridium histolyticum)由来のポリヌクレオチド、すなわち、シグナル配列をコードするポリヌクレオチドとしては、配列番号:1に記載の塩基配列のコード領域を含むポリヌクレオチドであり、プロ配列をコードするポリヌクレオチドとしては、配列番号:3に記載の塩基配列のコード領域を含むポリヌクレオチドであり、中性プロテアーゼの成熟配列をコードするポリヌクレオチドとしては、配列番号:4に記載の塩基配列のコード領域を含むポリヌクレオチドであることが好ましい。
【0034】
以上、本発明のポリヌクレオチドの好適な実施形態について説明したが、本発明のポリヌクレオチドは上記実施形態に限定されるものではない。例えば、本発明のポリヌクレオチドとしては、前記本発明のシグナル配列をコードするポリヌクレオチドの代わりに、公知のシグナル配列をコードするポリヌクレオチドと、前記本発明のプロ配列をコードするポリヌクレオチドと、前記本発明にかかる中性プロテアーゼの成熟配列をコードするポリヌクレオチドとが連結しているポリヌクレオチドであってもよい。かかる公知のシグナル配列としては、その配列に連結させる形で任意のタンパク質を宿主内において発現させた場合、該タンパク質を該宿主の外に分泌させる活性を有するタンパク質であればよく、例えば、大腸菌ではpelB、OmpA、PhoA、バチルス属ではα−アミラーゼまたはβ−ラクタマーゼのシグナル配列、ブレビバチルス属ではHWPのシグナル配列、麹菌ではAmyBのシグナル配列が挙げられる。
【0035】
また、本発明のポリヌクレオチドとして、前記本発明のシグナル配列をコードするポリヌクレオチドと、前記本発明にかかる中性プロテアーゼの成熟配列やその他の任意のタンパク質をコードするポリヌクレオチドとが連結しているポリヌクレオチドであってもよい。
【0036】
さらに、本発明のポリヌクレオチドとして、任意のシグナル配列をコードするポリヌクレオチド及び本発明にかかる中性プロテアーゼの成熟配列をコードするポリヌクレオチドが連結しているポリヌクレオチドと、任意のシグナル配列をコードするポリヌクレオチド及び本発明のプロ配列が連結しているポリヌクレオチドとの組み合わせであってもよい。かかる場合、「任意のシグナル配列」は本発明のシグナル配列であってもよく、前記公知のシグナル配列であってもよい。そして、これらのポリヌクレオチドの組み合わせを同一の宿主細胞内にて別々のタンパク質として発現させても、「Yasukawa K.ら、Protein Engineering,Design&Selection、2007年、20巻、8号、375〜383ページ」に記載されているように、活性を有した成熟型の中性プロテアーゼとして宿主細胞外に分泌産生させることができる。
【0037】
また、本発明のポリヌクレオチドがコードするタンパク質としては、本発明にかかる中性プロテアーゼの成熟配列やその他の任意のタンパク質のN末端及び/またはC末端に、その活性(本発明にかかる中性プロテアーゼの成熟配列であれば、中性プロテアーゼ活性)に影響を与えない範囲で、任意のポリペプチド配列を付与することができる。このようなポリペプチド配列としては、例えば、検出用マーカー(例えば、FLAGタグ)、精製用ポリペプチド(例えば、グルタチオンS−トランスフェラーゼ)を挙げることができる。
【0038】
また、本発明のポリヌクレオチドがコードするタンパク質における、各配列間の「連結」は、各配列どうしを直接結合するものであってもよく、リンカーペプチドを介して結合するものであってもよい。かかるリンカーペプチドとしては、その両端に連結されたプロ配列やシグナル配列や中性プロテアーゼの成熟配列等の発現や、これらの機能や活性を抑制するものでなければ特に制限されない。リンカーペプチドの長さとしては、通常、1〜100アミノ酸、好ましくは1〜50アミノ酸、より好ましくは1〜30アミノ酸、特に好ましくは12〜18アミノ酸(例えば15アミノ酸)である。
【0039】
<発現ベクター、並びに該発現ベクターにより形質転換された宿主細胞>
本発明は、本発明のシグナル配列をコードするポリヌクレオチドと、本発明のプロ配列をコードするポリヌクレオチドと、本発明の中性プロテアーゼの成熟配列をコードするポリヌクレオチドとが連結しているポリヌクレオチドを含有する発現ベクターも提供するものである。
【0040】
すなわち、本発明においては、前記シグナル配列、プロ配列及び中性プロテアーゼの成熟配列(以下、「中性プロテアーゼのプレプロ配列」とも称する)をコードするポリヌクレオチドを、宿主細胞内において複製することができ、かつ、該ポリヌクレオチドがコードするタンパク質を発現可能な状態で含んでなる発現ベクターが提供される。
【0041】
本発明において、「発現ベクター」は、自己複製ベクター、すなわち、染色体外の独立体として存在し、その複製が染色体の複製に依存しない、例えば、プラスミドを基本に構築することができる。また、本発現ベクターは、宿主細胞に導入されたとき、その宿主細胞のゲノム中に組み込まれ、それが組み込まれた染色体と一緒に複製されるものであってもよい。本発明によるベクター構築の手順及び方法は、遺伝子工学の分野で慣用されているものを用いることができる。
【0042】
本発明の発現ベクターは、これを宿主細胞に導入して中性プロテアーゼを発現させるために、前記の中性プロテアーゼのプレプロ配列をコードするポリヌクレオチドの他に、その発現を制御するDNA配列や形質転換された宿主細胞を選択するための遺伝子マーカー等を含んでいるのが望ましい。発現を制御するDNA配列としては、プロモーター、エンハンサー及びターミネーター等がこれに含まれる。プロモーターは宿主細胞において転写活性を示すものであれば特に限定されず、宿主細胞と同種若しくは異種のいずれかのタンパク質をコードする遺伝子の発現を制御するDNA配列として得ることができる。また、前記発現を制御するDNA配列以外に発現を誘導するDNA配列を含んでいても良い。かかる発現を誘導するDNA配列としては、宿主細胞が細菌である場合には、イソプロピル−β−D−チオガラクトピラノシド(IPTG)の添加により、下流に配置された遺伝子の発現を誘導することのできるラクトースオペロンが挙げられる。本発明における遺伝子マーカーは、形質転換された宿主細胞の選択の方法に応じて適宜選択されてよいが、例えば薬剤耐性をコードする遺伝子、栄養要求性を相補する遺伝子を利用することができる。
【0043】
更に本発明によれば、この発現ベクターによって形質転換された宿主細胞(形質転換体)が提供される。この宿主細胞は特に限定されず、例えば、大腸菌、放線菌、酵母、糸状菌、植物細胞、昆虫細胞、動物細胞が挙げられるが、特に発現させた中性プロテアーゼによる溶菌の問題を回避するという観点から、グラム陽性菌を宿主細胞として利用するのが好ましく、ブレビバチルス・コシネンシス(Brevibacillus choshinensis)、バチルス・ズブチリス(Bacillus subtilis)を利用するのがより好ましい。
【0044】
また、これらの発現ベクターによる宿主細胞の形質転換も、この分野で慣用されている方法に従い実施することができる。例えば、大腸菌、放線菌、酵母、糸状菌、グラム陽性菌等の細菌の形質転換の方法としては、ヒートショック法、エレクトロポレーション法、スフェロプラスト法、酢酸リチウム法が挙げられ、植物細胞の形質転換の方法としては、アグロバクテリウムを用いる方法やパーティクルガン法が挙げられ、昆虫細胞の形質転換の方法としては、バキュロウィルスを用いる方法やエレクトロポレーション法が挙げられ、動物細胞の形質転換の方法としては、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法が挙げられる。
【0045】
<形質転換体の培養物、中性プロテアーゼの製造方法、並びにプロテアーゼ調製物>
前述の通り、また後述の実施例に示す通り、本発明の発現ベクターを宿主細胞に導入し、該宿主細胞内で発現させ、該宿主細胞を培養することにより、活性を有した成熟型の中性プロテアーゼが該宿主細胞の外へ分泌されることになる。従って、本発明は、本発明の形質転換体から分泌された中性プロテアーゼを含む培養物をも提供することができる。
【0046】
本発明において「培養物」とは、前記本発明の形質転換体を、その宿主細胞に対して適当な培地で培養することによって得られる、増殖した形質転換体、該形質転換体の分泌産物及び該形質転換体の代謝産物等を含有する培地のことであり、それらの希釈物、濃縮物を含む。
【0047】
かかる培地としては、宿主細胞が資化し得るものが含有されていればよく、炭素源、窒素源、硫黄源、無機塩類、金属、ペプトン、酵母エキス、肉エキス、カゼイン加水分解物、血清等が含有物として挙げられる。また、かかる培地には、例えば、中性プロテアーゼのプレプロ配列をコードするポリヌクレオチドの発現を誘導するためのIPTGや、本発明の発現ベクターがコードし得る薬剤耐性遺伝子に対応する抗生物質(例えば、ネオマイシン、クロラムフェニコール)や、本発明の発現ベクターがコードし得る栄養要求性を相補する遺伝子に対応する栄養物(例えば、アルギニン、ヒスチジン)を添加してもよい。
【0048】
本発明にかかる培養の条件としては、本発明の形質転換体が前記培地中に活性を有した成熟型の中性プロテアーゼを分泌産生できる条件であればよく、当業者であれば、宿主細胞の種類、用いる培地等に合わせて、温度、空気の添加の有無、酸素の濃度、二酸化炭素の濃度、培地のpH、培養温度、培養時間、湿度等を適宜調整し、設定することができる。
【0049】
前述の通り、また後述の実施例に示す通り、本発明の形質転換体を培養することにより、活性を有した成熟型の中性プロテアーゼが該宿主細胞の外へ分泌されることになる。従って、本発明は、本発明の形質転換体を培養し、該形質転換体および/またはその培養物から発現したタンパク質を採取する工程を含む、中性プロテアーゼの製造方法をも提供することができる。
【0050】
「本発明の形質転換体を培養」とは前記の通りである。また、「本発明の形質転換体および/またはその培養物から発現したタンパク質を採取する」方法としては、フィルター(例えば、孔径0.2μm以下のフィルター)によって宿主細胞を除去する方法や、脱塩用フィルターを用いた脱塩濃縮により、培養物中のタンパク質の濃度を高める方法や、カラム(例えば、陰イオン交換カラムや疎水性カラム)を用いたタンパク質の濃縮方法が挙げられる。
【0051】
また、本発明は前記製造方法によって製造された中性プロテアーゼを含む、プロテアーゼ調製物をも提供する。本発明のプロテアーゼ調製物としては、前記製造方法によって製造された中性プロテアーゼを含んでいればよいが、本発明の中性プロテアーゼの他、プロテアーゼ調製物として許容される他の成分を含むことができる。このような他の成分としては、例えば、担体、賦形剤、崩壊剤、緩衝剤、乳化剤、懸濁剤、安定剤、保存剤、防腐剤、生理食塩が挙げられる。賦形剤としては乳糖、デンプン、ソルビトール、D−マンニトール、白糖等を用いることができる。崩壊剤としてはデンプン、カルボキシメチルセルロース、炭酸カルシウム等を用いることができる。緩衝剤としてはリン酸塩、クエン酸塩、酢酸塩等を用いることができる。乳化剤としてはアラビアゴム、アルギン酸ナトリウム、トラガント等を用いることができる。懸濁剤としてはモノステアリン酸グリセリン、モノステアリン酸アルミニウム、メチルセルロース、カルボキシメチルセルロース、ヒドロキシメチルセルロース、ラウリル硫酸ナトリウム等を用いることができる。安定剤としてはプロピレングリコール、ジエチリン亜硫酸塩、アスコルビン酸等を用いることができる。保存剤としてはフェノール、塩化ベンザルコニウム、ベンジルアルコール、クロロブタノール、メチルパラベン等を用いることができる。防腐剤としてはアジ化ナトリウム、塩化ベンザルコニウム、パラオキシ安息香酸、クロロブタノール等を用いることができる。
【0052】
さらに、本発明のプロテアーゼ調製物を、糖尿病等の治療を目的とし、膵臓等の臓器から膵島等の細胞群やβ細胞等の細胞を単離するための薬剤として利用する場合には、例えばコラゲナーゼ、エラステラーゼ、トリプシン、キモトリプシン等の他の酵素を含有していてもよい。また、本発明のプロテアーゼ調製物を、中性洗剤として利用する場合には、例えば、プロテアーゼ、アミラーゼ、リパーゼ、マンナーゼ、ペクチナーゼ、クチナーゼ、オキシドレダクターゼ、ヘミセルラーゼ、セルラーゼ等の他の酵素を含有していてもよい。
【実施例】
【0053】
以下、実施例及び比較例に基づいて本発明をより具体的に説明するが、本発明は以下の実施例に限定されるものではない。また、各実施例及び比較例で用いたプライマーの配列を表1に示す。
【0054】
【表1】

【0055】
(比較例1)
<特許文献1に記載の遺伝子配列を用いた中性プロテアーゼの発現>
(1−1) 特許文献1に記載の塩基配列を用いた中性プロテアーゼ発現用プラスミドの構築
Brevibacillusにおいて強力に作用するP2プロモータの配列についてはpNCMO2ベクター(タカラバイオ株式会社製)を鋳型としてP2−F及びP2−Rプライマーを用いて、中性プロテアーゼ遺伝子についてはClostridium histolyticumのゲノムDNAを鋳型としてNP−F1及びNP−Rプライマーを用いてPCRにより増幅した。PCRはPrimeSTAR Max DNA Polymerase(タカラバイオ社製)を用いて、98℃で10秒、55℃で5秒、72℃で30秒を40サイクル実施した。増幅された約700bpのDNA断片及び1kbpのDNA断片(配列番号:7に記載の601〜1599番目の塩基配列を含むDNA断片)をQIAquick PCR Purification Kitにより回収した。回収したDNA断片をそれぞれ1μLずつ鋳型としてP2−F及びNP−Rプライマーを用いてPCRを行った。なお、NP−F1プライマーの1〜20位はP2−Rプライマーと相補的な配列となっており、該相補的な配列を介して回収したDNA断片どうしはアニーリングすることができる。そのため、回収したDNA断片を鋳型としてP2−F及びNP−Rプライマーを用いてPCRを行うことにより、P2プロモータ−の下流に中性プロテアーゼをコードする塩基配列が連結している配列を増幅することができる。そして、このようにして増幅して得られた約1.7kbpのDNA断片、及びBrevibacillus用発現ベクターであるpNCMO2をBalI及びBamHIで制限酵素処理した後に、ライゲーション反応を行い、プラスミドpNCMO2−P2−NP(F1)を得た。
【0056】
(1−2) 特許文献1に記載の遺伝子配列を用いた中性プロテアーゼの発現
プラスミドpNCMO2−P2−NP(F1)をエレクトロポレーションによりBrevibacillus choshinensis SP−3株に導入し、ネオマイシン耐性株(pNCMO2−P2−NP(F1)形質転換体)を得た。次いで、TMNM液体培地(グルコース10.0g/L、ポリペプトン10.0g/L、肉エキス5.0g/L、酵母エキス2.0g/L、FeSO4・7H2O 10mg/L、MnSO4・4H2O 10mg/L、ZnSO4・7H2O 1mg/L、ネオマイシン 10μg/mL)にて、pNCMO2−P2−NP(F1)形質転換体の培養を行った。そして、得られた培養液を遠心し、菌体と培養上清に分離した後、該培養上清をフィルター(孔径:0.2μm)に通してろ過滅菌することにより、電気泳動用のサンプルとして調製した。次いで、このサンプルをXV PANTERA Gel 15%(DRC社製)に供し、電気泳動分離を行った後、クマシーブリリアントブルー(CBB)染色を行った。得られた結果を図1に示す。
【0057】
また、pNCMO2−P2−NP(F1)形質転換体を、1%スキムミルクを含むLB培地(スキムミルクプレート)に接種して37℃で一夜培養し、スキムミルクの分解の程度を調べた。得られた結果を図2のAに示す。
【0058】
図1に示した結果から明らかなように、pNCMO2−P2−NP(F1)形質転換体の培養上清中に約35kDaの中性プロテアーゼとおもわれるタンパク質は検出されなかった。また、図2のAに示した結果から明らかなように、当該形質転換体をスキムミルクプレートで培養しても、スキムミルクの分解に起因するハローは検出されなかった。
【0059】
(実施例1)
<中性プロテアーゼのシグナル配列ならびにプロ配列のクローニング>
(2−1) 中性プロテアーゼのシグナル配列ならびにプロ配列の取得
中性プロテアーゼ成熟配列の上流部分を同定するために、Clostridium histolyticumのゲノムDNAを鋳型としてNP5−1及びNP3プライマーを用いてPCRにより増幅した。PCRはPrimeSTAR Max DNA Polymerase(タカラバイオ社製)を用いて、98℃で10秒、55℃で5秒、72℃で30秒を30サイクル実施した。増幅された約1.1kbpのDNA断片をQIAquick PCR Purification Kitにより回収した。PCRにより取得したDNA断片のシークエンスは、BigDye Terminator v3.1 Cycle Sequebcing Kit(アプライドバイオシステムズ社製)とABI PRISMジェネティックアナライザー(アプライドバイオシステムズ社製)を用い、添付のプロトコールに従って決定した。そして、得られた塩基配列をアミノ酸配列に翻訳し、そのアミノ酸配列から4箇所のメチオニン(開始コドン:配列番号7に記載の1〜3番目、61〜63番目、295〜297番目及び601〜603番目のatgがコードするメチオニン)を見出した。
【0060】
(実施例2)
<シグナル配列及びプロ配列を含む中性プロテアーゼ遺伝子を用いたBrevibacillus choshinensis SP−3株での発現>
(3−1) シグナル配列及びプロ配列を含む中性プロテアーゼ遺伝子発現用プラスミドの構築
シグナル配列及びプロ配列を含む中性プロテアーゼ遺伝子をClostridium histolyticumのゲノムDNAを鋳型としてNP−F2及びNP−Rプライマーを用いてPCRにより増幅した。PCRはPrimeSTAR Max DNA Polymerase(タカラバイオ社製)を用いて、98℃で10秒、55℃で5秒、72℃で30秒を40サイクル実施した。増幅された1.6kbpのDNA断片(配列番号:7に記載の塩基配列を含むDNA断片)をQIAquick PCR Purification Kitにより回収した。回収したDNA断片と比較例1(1−1)で作製したP2プロモータ配列の断片をそれぞれ1μLずつ鋳型としてP2−F及びNP−Rプライマーを用いてPCRを行い、P2プロモータ−の下流にシグナル配列ならびにプロ配列を含む中性プロテアーゼをコードする塩基配列が連結している配列を増幅した。増幅された約2.3kbpのDNA断片を、TOPO TAクローニングキット(インビトロジェン社製)により、添付のプロトコールに従ってpCR2.1 TOPOプラスミドベクターに挿入しプラスミドpCR−P2−NPを得た。そして、pCR−P2−NP及びpNCMO2を、BalI及びBamHIで制限酵素処理した後に、pCR−P2−NPのインサート部分(約2.3kbpのDNA断片)とpNCMOとのライゲーション反応を行い、プラスミドpNCMO2−P2−NPを得た。
【0061】
(3−2) 中性プロテアーゼの発現
プラスミドpNCMO−P2−NPをエレクトロポレーション法によりBrevibacillus choshinensis SP−3株に導入し、ネオマイシン耐性株(pNCMO2−P2−NP形質転換体)を得た。そして、pNCMO2−P2−NP形質転換体の培養をTMNM液体培地にて行い、得られた培養液を遠心し、菌体と培養上清に分離した後、該培養上清をフィルター(孔径:0.2μm)に通してろ過滅菌することにより、電気泳動用のサンプルとして調製した。次いで、このサンプルをXV PANTERA Gel 15%(DRC社製)に供し、電気泳動分離を行った後、クマシーブリリアントブルー(CBB)染色を行った。得られた結果を図3に示す。
【0062】
また、pNCMO2−P2−NP形質転換体を、1%スキムミルクを含むLB培地(スキムミルクプレート)に接種して37℃で一夜培養し、スキムミルクの分解の程度を調べた。得られた結果を図2のBに示す。
【0063】
図3に示した結果から明らかなように、pNCMO2−P2−NP形質転換体の培養上清中に約35kDaの中性プロテアーゼとおもわれるタンパク質が検出された。また、図2のBに示した結果から明らかなように、当該形質転換体をスキムミルクプレートで培養したところ、スキムミルクの分解に起因するハローが検出された。
【0064】
また、図には示さないが、配列番号7に記載の61〜63番目のatgを開始コドンとする中性プロテアーゼをコードする塩基配列及び配列番号7に記載の295〜297番目のatgを開始コドンとする中性プロテアーゼをコードする塩基配列についても前記同様に各々発現ベクターを作製し、これらの発現ベクターを導入した宿主細胞を用いて、中性プロテアーゼの活性を評価した。しかし、比較例1に示した配列番号7に記載の601〜603番目のatgを開始コドンとする中性プロテアーゼをコードする塩基配列を用いた場合と同じく、いずれも中性プロテアーゼの活性は検出されなかった。
【0065】
(実施例3)
<シグナル配列及びプロ配列を含む中性プロテアーゼ遺伝子を用いたBacillus subtilis KN−2株での発現>
(4−1) シグナル配列及びプロ配列を含む中性プロテアーゼ遺伝子発現用プラスミドの構築
両端にBamHI及びSmaIの制限酵素サイトを導入するため、プラスミドpCR−P2−NPを鋳型としてNP−BamHI−N0及びNP−SmaI−C0プライマーを用いて、PCRにより増幅した。PCRはPrimeSTAR Max DNA Polymerase(タカラバイオ社製)を用いて、98℃で10秒、55℃で5秒、72℃で30秒を35サイクル実施した。増幅された1.6kbpのDNA断片(配列番号:7に記載の塩基配列を含むDNA断片)をQIAquick PCR Purification Kitにより回収した。回収したDNA断片及びBacillus subtilis発現用ベクターpHT01(Mo Bi Tec社)をBamHI及びSmaIで制限酵素処理した後に、ライゲーション反応を行い、プラスミドpHT01−NPを得た。
【0066】
(4−2) Bacillus subtilis KN−2株での中性プロテアーゼ発現
プラスミドpHT01−NPをBacillus subtilis KN−2株のコンピテントセルに導入し、クロラムフェニコール耐性株(pHT01−NP形質転換体)を得た。クロラムフェニコールを含むLB液体培地(バクトトリプトン 10.0g/L、イースト 5.0g/L、NaCl 10.0g/L、クロラムフェニコール 25μg/mL)にて37℃で24時間、pHT01−NP形質転換体を培養した後、IPTGにてタンパク質の発現誘導を37℃で4時間行なった。
【0067】
そして、得られた培養液を遠心し、菌体と培養上清に分離した後、該培養上清をフィルター(孔径:0.2μm)に通してろ過滅菌することにより、電気泳動用のサンプルとして調製した。次いで、このサンプルを12% Gel SDS−PAGE mini(テフコ社製)に供し、電気泳動分離を行った後、クマシーブリリアントブルー(CBB)染色を行った。得られた結果を図4に示す。
【0068】
図4に示した結果から明らかなように、宿主をBrevibacillus choshinensis SP−3株からBacillus subtilis KN−2株に変えても、実施例2の結果と同様に、pHT01−NP形質転換体の培養上清においても約35kDaの中性プロテアーゼと思われるタンパク質が検出された。
【0069】
(4−3) 発現した中性プロテアーゼのN末端アミノ酸配列の同定
実施例3の(4−2)で発現させたタンパク質がクロストリディウム・ヒストリティカム由来中性プロテアーゼであることを確認するために、N末端アミノ酸配列を決定した。まず、培養上清中のタンパク質について12% Gel SDS−PAGE mini(テフコ社製)を用いて電気泳動による分離を行い、PVDF膜(ミリポア社製)上にブロットした。得られた約35kDaのバンドを切り出し、プロテインシークエンサーModel 492(アプライドバイオシステムズ社製)を用いて、N末端アミノ酸配列を決定した。その結果、クロストリディウム・ヒストリティカム由来中性プロテアーゼのN末端アミノ酸配列(配列番号:6に記載の1〜5番目のアミノ酸配列)と一致した。
【0070】
(4−4) 形質転換体における中性プロテアーゼ活性の測定
実施例3の(4−2)で得られたpHT01−NP形質転換体の培養上清を用いてアゾカゼイン分解活性を測定した。測定は、アゾカゼインを基質とし、基質の分解によって生じる遊離チロシン量を335nmの吸光度で測定する方法を用い、文献(Nagase H, Harris ED Jr;J Biol Chem. 1983 Jun 25;258(12):7490−8)に記載の方法に準じて行った。
【0071】
なお、アゾカゼイン分解活性は、反応条件においてO.D.(335nm)=1.0になるのに必要な酵素量を1 unitと定義し、培養上清1mL当りの活性(U/mL)として表した。得られた結果を表2に示す。表2に示した結果から明らかなように、pHT01−NP形質転換体は親株(遺伝子を導入していない株)の約19倍の活性を示した。
【0072】
【表2】
【0073】
以上の結果から、配列番号7:に記載の1〜657番目の塩基配列がコードするポリペプチド(配列番号8:に記載の1〜219番目のアミノ酸配列からなるポリペプチド)が、中性プロテアーゼの宿主細胞外への分泌産生を可能としていることが明らかになった。また、配列番号8:に記載のアミノ酸配列をシグナル配列予測ソフトウェア(SignalP 3.0、http://www.cbs.dtu.dk/services/SignalP/)を用いて解析したところ、配列番号:2に記載のアミノ酸配列(配列番号:1に記載の塩基配列がコ―ドするアミノ酸配列)がシグナル配列と推定され、結果、配列番号:4に記載のアミノ酸配列(配列番号:3に記載の塩基配列がコ―ドするアミノ酸配列)がプロ配列として機能していることが推定された。
【産業上の利用可能性】
【0074】
以上説明したように、本発明によれば、目的とするタンパク質の宿主細胞外への分泌産生が可能となる。特に、本発明のシグナル配列やプロ配列をコードするポリヌクレオチドを利用して得られる中性プロテアーゼは、クロストリディウム・ヒストリティカム以外の宿主細胞から、活性を有した成熟型として分泌産生することができる。
【0075】
したがって、クロストリディウム・ヒストリティカムの培養液に含まれる非常に多くのプロテアーゼ活性を有するタンパク質を混入させることなく、中性プロテアーゼを調製することができるため、本発明のシグナル配列やプロ配列を利用して得られる中性プロテアーゼは、糖尿病等の治療を目的とし、膵臓等の臓器から膵島等の細胞群やβ細胞等の細胞を単離するための薬剤や、中性洗剤等として有用である。
【配列表フリーテキスト】
【0076】
配列番号9〜17
<223> 人工的に合成されたプライマーの配列
【技術分野】
【0001】
本発明は、シグナル配列及びプロ配列に関し、より詳しくは、シグナル配列及びプロ配列をコードするポリヌクレオチド、シグナル配列と、プロ配列と、中性プロテアーゼの成熟配列とが連結しているタンパク質をコードするポリヌクレオチド、該ポリヌクレオチドを含有する発現ベクター、該発現ベクターで形質転換された宿主細胞、並びに、該宿主細胞から分泌された中性プロテアーゼを含む培養物に関する。また、本発明は、前記形質転換された宿主細胞を用いた中性プロテアーゼの製造方法、並びに、該製造方法によって製造された中性プロテアーゼを含むプロテアーゼ調製物に関する。
【背景技術】
【0002】
微生物から菌体外に分泌されるプロテアーゼ(菌体外プロテアーゼ)は、グラム陰性菌に関しては、最初に菌体内でシグナル配列、プロ配列及び成熟配列が連結した不活性のプレプロ型として合成される。その後、シグナル配列により細胞内膜を通過してペリプラズムに移行し、シグナル配列は切断されプロ型に変換される。プロ配列は主に成熟配列部分をフォールディングする分子内シャペロンとして機能している。このプロ配列が切断されることにより、活性を有した成熟型プロテアーゼとして菌体外へ分泌される。また、グラム陽性菌に関しては、最初に菌体内で不活性のプレプロ型として合成された後、細胞壁付近でシグナル配列が細菌内酵素により切断され、菌体外に分泌される。そして、菌体外に分泌されると直ぐに自己触媒的切断(autocatalytic cleavage)により速やかにプロ配列が切断され、活性を有した成熟配列となることが想定される。
【0003】
一方、糖尿病の根治療法として、膵臓から単離される膵島(インスリン産生細胞)を、糖尿病患者の門脈へ点滴にて移植する膵島移植が知られている。本移植法は、移植の際に患者の開腹手術が必要ないため、安全かつ簡便な糖尿病の根治療法として近年注目されている。通常、膵臓からの膵島の分離は、膵臓をコラゲナーゼ及び中性プロテアーゼにて処理することにより実施されるが、本目的に使用される中性プロテアーゼの一つとしてクロストリディウム・ヒストリティカム(Clostridium histolyticum)由来の中性プロテアーゼ(亜鉛金属プロテアーゼ)がある。
【0004】
これまでクロストリディウム・ヒストリティカム由来の中性プロテアーゼは、前記菌体外プロテアーゼの1種であるため、クロストリディウム・ヒストリティカムの培養上清より製造されているが、クロストリディウム・ヒストリティカムの培養液には非常に多くのプロテアーゼ活性を有するタンパク質が含まれており、調製物にその他のプロテアーゼ様活性が混入する懸念があった。
【0005】
そこで、他の中性プロテアーゼ様活性を混入させる恐れがなく、宿主細胞外に活性を有したまま多量に発現させるクロストリディウム・ヒストリティカム由来中性プロテアーゼの製造法が所望されていた。さらには、クロストリディウム・ヒストリティカム由来中性プロテアーゼの分泌機構が解明されることにより、中性プロテアーゼに限らず、目的とするタンパク質を中性プロテアーゼ由来のシグナル配列やプロ配列と融合させた形でクロストリディウム・ヒストリティカム等の宿主細胞内にて発現させ、その活性を維持しつつ、宿主細胞外に分泌された該タンパク質を効率良く調製することができるようになるため、クロストリディウム・ヒストリティカム由来中性プロテアーゼの分泌機構、特にシグナル配列及びプロ配列の同定が所望されていた。
【0006】
そして、このような背景の中、特許文献1には、クロストリディウム・ヒストリティカム由来中性プロテアーゼをコードする核酸の塩基配列、並びに該配列を用いることによって、そのプロテアーゼの活性を有するタンパク質を製造できることも開示されている。
【0007】
しかしながら、特許文献1において、クロストリディウム・ヒストリティカム由来中性プロテアーゼの遺伝子組換え体を用いた製造法に関する記載はあるものの、実施例中には遺伝子組換え体による中性プロテアーゼの発現の結果ならびにその活性についての記載はない。また、特許文献1において開示されている配列は、成熟型中性プロテアーゼ部分のアミノ酸配列だけであり、シグナル配列及びプロ配列は開示されていない。そのため、特許文献1に記載のアミノ酸配列をコードするポリヌクレオチドを用いても、宿主細胞外に活性を有した成熟型中性プロテアーゼを分泌できないことが想定され、実際、後述の比較例1において示す通り、特許文献1に記載の方法ではクロストリディウム・ヒストリティカム由来中性プロテアーゼを製造することはできなかった。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】特表平10−500581号公報
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、前記従来技術の有する課題に鑑みてなされたものであり、目的とするタンパク質、特に中性プロテアーゼの宿主細胞外への分泌産生を可能とするシグナル配列及びプロ配列を提供することを目的とする。
【課題を解決するための手段】
【0010】
本発明者らは、前記目的を達成すべく鋭意研究を重ねた結果、クロストリディウム・ヒストリティカムのゲノム遺伝子を精査し、中性プロテアーゼのシグナル配列の開始コドンとなりうる部分を複数見出した。これらの配列について順に遺伝子組み換え体を造出し、発現を確認したところ、成熟配列のN末端からもっとも遠い位置に存在する開始コドンのみで中性プロテアーゼの発現が見られた。よって、配列番号1に示すシグナル配列及び配列番号3に示すプロ配列を含むポリヌクレオチドを用いることにより、クロストリディウム・ヒストリティカム以外の宿主においてもクロストリディウム・ヒストリティカム由来中性プロテアーゼ等のタンパク質を、宿主細胞外に分泌産生できることを見出した。なお、配列番号1に示すシグナル配列及び配列番号3に示すプロ配列は、他の遺伝子との相同性をほとんど持たない新規な配列であった。さらに、本発明者らは、当該シグナル配列と、当該プロ配列と、中性プロテアーゼの成熟配列とが連結しているタンパク質をコードするポリヌクレオチドを用いて形質転換された宿主細胞を用いることにより、中性プロテアーゼを活性を有した成熟型として宿主細胞外に分泌産生され、取得することができることを見出した。
【0011】
本発明は、より詳しくは以下を提供するものである。
<1> シグナル配列をコードする、下記(i)から(v)のいずれかに記載のポリヌクレオチド。
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:1に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:2に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:2に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:1に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
<2> プロ配列をコードする、下記(i)から(v)のいずれかに記載のポリヌクレオチド。
(i)配列番号:4に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:3に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:4に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:4に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:3に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
<3> クロストリディウム・ヒストリティカム(Clostridium histolyticum)由来である、<1>又は<2>に記載のポリヌクレオチド。
<4> <1>に記載のポリヌクレオチドと、<2>に記載のポリヌクレオチドと、中性プロテアーゼの成熟配列をコードする下記(i)から(v)のいずれかに記載のポリヌクレオチドとが連結しているポリヌクレオチド。
(i)配列番号:6に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:5に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:6に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:6に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:5に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
<5> クロストリディウム・ヒストリティカム(Clostridium histolyticum)由来である、<4>に記載のポリヌクレオチド。
<6> <4>又は<5>に記載のポリヌクレオチドを含有する発現ベクター。
<7> <6>に記載の発現ベクターで形質転換された宿主細胞。
<8> <7>に記載の形質転換された宿主細胞から分泌された中性プロテアーゼを含む培養物。
<9> <7>に記載の形質転換された宿主細胞を培養し、該宿主細胞および/またはその培養物から発現したタンパク質を採取する工程を含む、中性プロテアーゼの製造方法。
<10> <9>に記載の方法によって製造された中性プロテアーゼを含む、プロテアーゼ調製物。
【発明の効果】
【0012】
本発明によれば、目的とするタンパク質、特に中性プロテアーゼの宿主細胞外への分泌産生が可能となる。
【図面の簡単な説明】
【0013】
【図1】プラスミドpNCMO2−P2−NP(F1)を導入したBrevibacillus choshinensis SP−3株(pNCMO2−P2−NP(F1)形質転換体)の培養上清を電気泳動分離によって分析した結果を示すゲルの写真である。図中、1のレーンはサイズマーカーを泳動した結果を示し、2のレーンはpNCMO2−P2−NP(F1)形質転換体の培養上清を泳動した結果を示す。
【図2】pNCMO2−P2−NP(F1)形質転換体と、プラスミドpNCMO−P2−NPを導入したBrevibacillus choshinensis SP−3株(pNCMO2−P2−NP形質転換体)とをスキムミルクプレートに各々3点ずつ接種して培養し、スキムミルクの分解の程度(ハロー)を調べた結果を示す写真である。図中、AはpNCMO2−P2−NP(F1)形質転換体を培養した結果を示し、BはpNCMO2−P2−NP形質転換体を培養した結果を示す。
【図3】pNCMO2−P2−NP形質転換体の培養上清を電気泳動分離によって分析した結果を示すゲルの写真である。図中、1のレーンはサイズマーカーを泳動した結果を示し、2のレーンはpNCMO2−P2−NP形質転換体の培養上清を泳動した結果を示す。
【図4】プラスミドpHT01−NPを導入したBacillus subtilis KN−2株(pHT01−NP形質転換体)の培養上清を電気泳動分離によって分析した結果を示すゲルの写真である。図中、1のレーンはサイズマーカーを泳動した結果を示し、2のレーンはpHT01−NP形質転換体の培養上清を泳動した結果を示す。
【発明を実施するための形態】
【0014】
<ポリヌクレオチド>
本発明のシグナル配列をコードするポリヌクレオチドは、下記(i)から(v)のいずれかに記載のポリヌクレオチドである。
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:1に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:2に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:2に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:1に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド。
【0015】
また、本発明のプロ配列をコードするポリヌクレオチドは、下記(i)から(v)のいずれかに記載のポリヌクレオチドである。
(i)配列番号:4に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:3に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:4に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:4に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:3に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド。
【0016】
本発明において、「シグナル配列」とは、その配列のC末端側に連結させる形で任意のタンパク質を宿主内において発現させた場合、該タンパク質を該宿主の外に分泌させる活性を有するタンパク質のことである。
【0017】
また、本発明において、「プロ配列」とは、その配列のC末端側に連結させる形で任意のタンパク質の成熟配列部分を宿主内において発現させた場合、主に、該成熟配列部分をフォールディングする分子内シャペロンとして機能するタンパク質のことである。
【0018】
本発明は、本発明のシグナル配列をコードするポリヌクレオチドと、本発明のプロ配列をコードするポリヌクレオチドと、中性プロテアーゼの成熟配列をコードする下記(i)から(v)のいずれかに記載のポリヌクレオチドとが連結しているポリヌクレオチドも提供するものである。
(i)配列番号:6に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:5に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:6に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:6に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:5に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド。
【0019】
本発明において、「中性プロテアーゼ」とは、その最適活性が中性pH範囲内にあり、カゼイン及び変性コラーゲンを解裂させる活性を有するタンパク質のことである。
【0020】
後述の実施例において示す通り、N末端側から順に、配列番号:2に記載のアミノ酸配列を含むタンパク質と、配列番号:4に記載のアミノ酸配列を含むタンパク質と、配列番号:6に記載のアミノ酸配列を含むタンパク質(クロストリディウム・ヒストリティカム由来中性プロテアーゼの成熟配列部分)とが連結しているタンパク質を宿主内において発現させた場合、活性を有した成熟型の中性プロテアーゼが該宿主の外に分泌された。従って、本発明のシグナル配列としては、配列番号:2に記載のアミノ酸配列を含むタンパク質(配列番号:3に記載の塩基配列がコードするタンパク質)が挙げられ、本発明のプロ配列としては、配列番号:4に記載のアミノ酸配列を含むタンパク質(配列番号:3に記載の塩基配列がコードするタンパク質)が挙げられ、本発明の中性プロテアーゼの成熟配列としては、配列番号:6に記載のアミノ酸配列を含むタンパク質(配列番号:5に記載の塩基配列がコードするタンパク質)が挙げられる。
【0021】
また、現在の技術水準においては、当業者であれば、かかるタンパク質をコードする塩基配列の情報が得られた場合、その塩基配列に対して種々の改変を行い、各活性や機能を有する、シグナル配列やプロ配列や中性プロテアーゼの製造を行うことが可能である。また、自然界においても、塩基配列が変異することは起こり得ることである。従って、本発明のシグナル配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:2に記載のアミノ酸配列において1もしくは複数個のアミノ酸が置換、欠失、挿入、および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチドも含まれ、本発明のプロ配列をコードするポリヌクレオチドには、前記機能を有するタンパク質をコードする限り、配列番号:4に記載のアミノ酸配列において1もしくは複数個のアミノ酸が置換、欠失、挿入、および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチドも含まれ、本発明の中性プロテアーゼの成熟配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:6に記載のアミノ酸配列において1もしくは複数個のアミノ酸が置換、欠失、挿入、および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチドも含まれる。
【0022】
「1もしくは複数個のアミノ酸が欠失、置換、挿入もしくは付加されたアミノ酸配列」とは、部位特異的突然変異誘発法等の周知の方法により、または天然に生じ得る程度の複数個の数のアミノ酸の置換等により改変がなされたことを意味する。アミノ酸の改変の個数は、好ましくは1〜10個、より好ましくは1〜5個、最も好ましくは1〜2個である。
【0023】
改変アミノ酸配列の例は、好ましくは、そのアミノ酸が、1または複数個(好ましくは、1ないし数個あるいは1、2、3、または4個)の保存的置換を有するアミノ酸配列であることができる。ここで、「保存的置換」とは、1または複数個のアミノ酸残基を、別の化学的に類似したアミノ酸残基で置き換えることを意味する。例えば、ある疎水性残基を別の疎水性残基によって置換する場合、ある極性残基を同じ電荷を有する別の極性残基によって置換する場合などが挙げられる。このような置換を行うことができる機能的に類似のアミノ酸は、アミノ酸毎に当該技術分野において公知である。具体例を挙げると、非極性(疎水性)アミノ酸としては、アラニン、バリン、イソロイシン、ロイシン、プロリン、トリプトファン、フェニルアラニン、メチオニンなどが挙げられる。極性(中性)アミノ酸としては、グリシン、セリン、スレオニン、チロシン、グルタミン、アスパラギン、システインなどが挙げられる。陽電荷をもつ(塩基性)アミノ酸としては、アルギニン、ヒスチジン、リジンなどが挙げられる。また、負電荷をもつ(酸性)アミノ酸としては、アスパラギン酸、グルタミン酸などが挙げられる。
【0024】
さらに、現在の技術水準においては、当業者であれば、かかるタンパク質をコードする特定の塩基配列の情報が得られた場合、それらの塩基配列情報を利用して、同種若しくは他の菌種等から、各活性や機能を有するタンパク質をコードする相同遺伝子を単離することが可能である。このような相同遺伝子を取得するための菌種等としては枯草菌(例えばBacillus subtilisやBacillus thermoproteolyticus rokko)や麹菌(例えばAspergillus oryzae)などが好ましい。
【0025】
相同遺伝子を取得するための方法としては、例えば、ハイブリダイゼーション技術(Southern,E.M.,J.Mol.Biol.,98:503,1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki,R.K.,et al.Science,230:1350−1354,1985、Saiki,R.K.et al.Science,239:487−491,1988)が挙げられる。相同遺伝子を単離するためには、通常、ストリンジェントな条件下でハイブリダイゼーション反応を行なう。
【0026】
ここで、「ストリンジェントな条件」とは、ハイブリダイゼーション後のメンブレンの洗浄操作を、高温度低塩濃度溶液中で行うことを意味し、例えば、2×SSC濃度(1×SSC:15mmol/Lクエン酸3ナトリウム、150mmol/L塩化ナトリウム)、0.5%SDS溶液中で、60℃、20分間の洗浄条件を意味する。
【0027】
従って、本発明のシグナル配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:1に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチドも含まれ、本発明のプロ配列をコードするポリヌクレオチドには、前記機能を有するタンパク質をコードする限り、配列番号:3に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチドも含まれ、本発明の中性プロテアーゼの成熟配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:5に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチドも含まれる。
【0028】
また、取得された相同遺伝子がコードするタンパク質は、通常、本発明のシグナル配列やプロ配列や中性プロテアーゼの成熟配列のアミノ酸配列と高い同一性を有するアミノ酸配列を含むタンパク質である。本発明において、高い同一性とは、好ましくは、95%以上、より好ましくは96%以上、特に好ましくは98%以上、そして最も好ましくは99%以上の同一性を有する。
【0029】
従って、本発明のシグナル配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:2に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチドも含まれ、本発明のプロ配列をコードするポリヌクレオチドには、前記機能を有するタンパク質をコードする限り、配列番号:4に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチドも含まれ、本発明の中性プロテアーゼの成熟配列をコードするポリヌクレオチドには、前記活性を有するタンパク質をコードする限り、配列番号:6に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチドも含まれる。
【0030】
塩基配列またはアミノ酸配列についての「同一性」は、比較される配列間において、各々の配列を構成する塩基またはアミノ酸残基の一致の程度の意味で用いられる。本明細書において示した「同一性」の数値はいずれも、当業者に公知の相同性検索プログラムを用いて算出される数値であればよく、例えばFASTA、BLAST等においてデフォルト(初期設定)のパラメータを用いることにより、容易に算出することができる。より具体的には、配列の同一性は、BLASTX(アミノ酸レベル)のプログラム(Altschul et al.J.Mol.Biol.,215:403−410,1990)を利用して決定することができる。該プログラムは、KarlinおよびAltschulによるアルゴリズムBLAST(Proc.Natl.Acad.Sci.USA,87:2264−2268,1990,Proc.Natl.Acad.Sci.USA,90:5873−5877,1993)に基づいている。BLASTXによってアミノ酸配列を解析する場合には、パラメーターは、例えばscore=50、wordlength=3とする。また、Gapped BLASTプログラムを用いて、アミノ酸配列を解析する場合は、Altschulら(Nucleic Acids Res.25:3389−3402,1997)に記載されているように行うことができる。BLASTとGapped BLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。
【0031】
なお、特定のポリヌクレオチドがコードするタンパク質が、前記活性や機能を有するタンパク質をコードするか否かは、例えば、後述の実施例に記載するように、特定のポリヌクレオチドがコードするタンパク質が発現するような形で、プラスミドベクターにクローニングし、該発現ベクターを宿主細胞(例えば、Brevibacillus choshinensis SP−3株、Bacillus subtilis KN−2株)に導入することにより得られた培養上清をSDS−PAGEにて分析し、予想される分子量を有するタンパク質のバンド(本発明にかかる中性プロテアーゼの成熟配列部分であれば、分子量 約35kDaのタンパク質が示すバンド)を検出できるか否かを検定することにより判定することができる(実施例2 参照)。また、前記発現ベクターを導入した宿主細胞(形質転換体)や該形質転換体の培養上清を用いて、スキムミルク(カゼイン)の分解の程度(例えば、ハローの有無やその大きさ)を培地(スキムミルクプレート)上で検出できるか否かを検定することにより判定することができる(実施例2 参照)。さらに、前記形質転換体や該形質転換体の培養上清を用い、アゾカゼインを基質とし、基質の分解によって生じる遊離チロシン量を335nmの吸光度で測定することによって、アゾカゼイン分解活性を検出できるか否かを検定することにより判定することができる(実施例3 参照)。
【0032】
また、本発明のポリヌクレオチドとしては、その形態に特に制限はなく、cDNAの他、ゲノムDNA、および化学合成DNAが含まれる。これらDNAの調製は、当業者にとって常套手段を利用して行うことが可能である。ゲノムDNAは、例えば、クロストリディウム・ヒストリティカムからゲノムDNAを抽出し、ゲノミックライブラリー(ベクターとしては、プラスミド、ファージ、コスミド、BAC、PACなどが利用できる)を作製し、これを展開して、クロストリディウム・ヒストリティカム由来中性プロテアーゼのシグナル配列、プロ配列、成熟配列をコードする塩基配列(例えば、配列番号:1、3、5に記載のDNA)を基に調製したプローブを用いてコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより調製することが可能である。また、クロストリディウム・ヒストリティカム由来中性プロテアーゼのシグナル配列、プロ配列、成熟配列をコードする塩基配列に特異的なプライマーを作製し、これを利用したPCRを行うことによって調製することも可能である。また、cDNAは、例えば、クロストリディウム・ヒストリティカムから抽出したmRNAを基にcDNAを合成し、これをλZAP等のベクターに挿入してcDNAライブラリーを作成し、これを展開して、上記と同様にコロニーハイブリダイゼーションあるいはプラークハイブリダイゼーションを行うことにより、また、PCRを行うことにより調製することが可能である。さらに、市販のDNA合成機を用いれば、目的のDNAを合成により調製することも可能である。また、前記配列番号:1等に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチドや、前記配列番号:2等に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチドは、前記ハイブリダイゼーション技術やPCR技術の他に、特定の塩基配列やアミノ酸配列に基づき、当業者に公知の部位特異的変異誘発(site−directed mutagenesis)法(Kramer,W.&Fritz,HJ.,Methods Enzymol,1987,154,350.)等の遺伝子増幅技術や組換え技術を利用して調製することも可能である。
【0033】
また、本発明のポリヌクレオチドとしては、クロストリディウム・ヒストリティカム(Clostridium histolyticum)由来のポリヌクレオチド、すなわち、シグナル配列をコードするポリヌクレオチドとしては、配列番号:1に記載の塩基配列のコード領域を含むポリヌクレオチドであり、プロ配列をコードするポリヌクレオチドとしては、配列番号:3に記載の塩基配列のコード領域を含むポリヌクレオチドであり、中性プロテアーゼの成熟配列をコードするポリヌクレオチドとしては、配列番号:4に記載の塩基配列のコード領域を含むポリヌクレオチドであることが好ましい。
【0034】
以上、本発明のポリヌクレオチドの好適な実施形態について説明したが、本発明のポリヌクレオチドは上記実施形態に限定されるものではない。例えば、本発明のポリヌクレオチドとしては、前記本発明のシグナル配列をコードするポリヌクレオチドの代わりに、公知のシグナル配列をコードするポリヌクレオチドと、前記本発明のプロ配列をコードするポリヌクレオチドと、前記本発明にかかる中性プロテアーゼの成熟配列をコードするポリヌクレオチドとが連結しているポリヌクレオチドであってもよい。かかる公知のシグナル配列としては、その配列に連結させる形で任意のタンパク質を宿主内において発現させた場合、該タンパク質を該宿主の外に分泌させる活性を有するタンパク質であればよく、例えば、大腸菌ではpelB、OmpA、PhoA、バチルス属ではα−アミラーゼまたはβ−ラクタマーゼのシグナル配列、ブレビバチルス属ではHWPのシグナル配列、麹菌ではAmyBのシグナル配列が挙げられる。
【0035】
また、本発明のポリヌクレオチドとして、前記本発明のシグナル配列をコードするポリヌクレオチドと、前記本発明にかかる中性プロテアーゼの成熟配列やその他の任意のタンパク質をコードするポリヌクレオチドとが連結しているポリヌクレオチドであってもよい。
【0036】
さらに、本発明のポリヌクレオチドとして、任意のシグナル配列をコードするポリヌクレオチド及び本発明にかかる中性プロテアーゼの成熟配列をコードするポリヌクレオチドが連結しているポリヌクレオチドと、任意のシグナル配列をコードするポリヌクレオチド及び本発明のプロ配列が連結しているポリヌクレオチドとの組み合わせであってもよい。かかる場合、「任意のシグナル配列」は本発明のシグナル配列であってもよく、前記公知のシグナル配列であってもよい。そして、これらのポリヌクレオチドの組み合わせを同一の宿主細胞内にて別々のタンパク質として発現させても、「Yasukawa K.ら、Protein Engineering,Design&Selection、2007年、20巻、8号、375〜383ページ」に記載されているように、活性を有した成熟型の中性プロテアーゼとして宿主細胞外に分泌産生させることができる。
【0037】
また、本発明のポリヌクレオチドがコードするタンパク質としては、本発明にかかる中性プロテアーゼの成熟配列やその他の任意のタンパク質のN末端及び/またはC末端に、その活性(本発明にかかる中性プロテアーゼの成熟配列であれば、中性プロテアーゼ活性)に影響を与えない範囲で、任意のポリペプチド配列を付与することができる。このようなポリペプチド配列としては、例えば、検出用マーカー(例えば、FLAGタグ)、精製用ポリペプチド(例えば、グルタチオンS−トランスフェラーゼ)を挙げることができる。
【0038】
また、本発明のポリヌクレオチドがコードするタンパク質における、各配列間の「連結」は、各配列どうしを直接結合するものであってもよく、リンカーペプチドを介して結合するものであってもよい。かかるリンカーペプチドとしては、その両端に連結されたプロ配列やシグナル配列や中性プロテアーゼの成熟配列等の発現や、これらの機能や活性を抑制するものでなければ特に制限されない。リンカーペプチドの長さとしては、通常、1〜100アミノ酸、好ましくは1〜50アミノ酸、より好ましくは1〜30アミノ酸、特に好ましくは12〜18アミノ酸(例えば15アミノ酸)である。
【0039】
<発現ベクター、並びに該発現ベクターにより形質転換された宿主細胞>
本発明は、本発明のシグナル配列をコードするポリヌクレオチドと、本発明のプロ配列をコードするポリヌクレオチドと、本発明の中性プロテアーゼの成熟配列をコードするポリヌクレオチドとが連結しているポリヌクレオチドを含有する発現ベクターも提供するものである。
【0040】
すなわち、本発明においては、前記シグナル配列、プロ配列及び中性プロテアーゼの成熟配列(以下、「中性プロテアーゼのプレプロ配列」とも称する)をコードするポリヌクレオチドを、宿主細胞内において複製することができ、かつ、該ポリヌクレオチドがコードするタンパク質を発現可能な状態で含んでなる発現ベクターが提供される。
【0041】
本発明において、「発現ベクター」は、自己複製ベクター、すなわち、染色体外の独立体として存在し、その複製が染色体の複製に依存しない、例えば、プラスミドを基本に構築することができる。また、本発現ベクターは、宿主細胞に導入されたとき、その宿主細胞のゲノム中に組み込まれ、それが組み込まれた染色体と一緒に複製されるものであってもよい。本発明によるベクター構築の手順及び方法は、遺伝子工学の分野で慣用されているものを用いることができる。
【0042】
本発明の発現ベクターは、これを宿主細胞に導入して中性プロテアーゼを発現させるために、前記の中性プロテアーゼのプレプロ配列をコードするポリヌクレオチドの他に、その発現を制御するDNA配列や形質転換された宿主細胞を選択するための遺伝子マーカー等を含んでいるのが望ましい。発現を制御するDNA配列としては、プロモーター、エンハンサー及びターミネーター等がこれに含まれる。プロモーターは宿主細胞において転写活性を示すものであれば特に限定されず、宿主細胞と同種若しくは異種のいずれかのタンパク質をコードする遺伝子の発現を制御するDNA配列として得ることができる。また、前記発現を制御するDNA配列以外に発現を誘導するDNA配列を含んでいても良い。かかる発現を誘導するDNA配列としては、宿主細胞が細菌である場合には、イソプロピル−β−D−チオガラクトピラノシド(IPTG)の添加により、下流に配置された遺伝子の発現を誘導することのできるラクトースオペロンが挙げられる。本発明における遺伝子マーカーは、形質転換された宿主細胞の選択の方法に応じて適宜選択されてよいが、例えば薬剤耐性をコードする遺伝子、栄養要求性を相補する遺伝子を利用することができる。
【0043】
更に本発明によれば、この発現ベクターによって形質転換された宿主細胞(形質転換体)が提供される。この宿主細胞は特に限定されず、例えば、大腸菌、放線菌、酵母、糸状菌、植物細胞、昆虫細胞、動物細胞が挙げられるが、特に発現させた中性プロテアーゼによる溶菌の問題を回避するという観点から、グラム陽性菌を宿主細胞として利用するのが好ましく、ブレビバチルス・コシネンシス(Brevibacillus choshinensis)、バチルス・ズブチリス(Bacillus subtilis)を利用するのがより好ましい。
【0044】
また、これらの発現ベクターによる宿主細胞の形質転換も、この分野で慣用されている方法に従い実施することができる。例えば、大腸菌、放線菌、酵母、糸状菌、グラム陽性菌等の細菌の形質転換の方法としては、ヒートショック法、エレクトロポレーション法、スフェロプラスト法、酢酸リチウム法が挙げられ、植物細胞の形質転換の方法としては、アグロバクテリウムを用いる方法やパーティクルガン法が挙げられ、昆虫細胞の形質転換の方法としては、バキュロウィルスを用いる方法やエレクトロポレーション法が挙げられ、動物細胞の形質転換の方法としては、リン酸カルシウム法、リポフェクション法、エレクトロポレーション法が挙げられる。
【0045】
<形質転換体の培養物、中性プロテアーゼの製造方法、並びにプロテアーゼ調製物>
前述の通り、また後述の実施例に示す通り、本発明の発現ベクターを宿主細胞に導入し、該宿主細胞内で発現させ、該宿主細胞を培養することにより、活性を有した成熟型の中性プロテアーゼが該宿主細胞の外へ分泌されることになる。従って、本発明は、本発明の形質転換体から分泌された中性プロテアーゼを含む培養物をも提供することができる。
【0046】
本発明において「培養物」とは、前記本発明の形質転換体を、その宿主細胞に対して適当な培地で培養することによって得られる、増殖した形質転換体、該形質転換体の分泌産物及び該形質転換体の代謝産物等を含有する培地のことであり、それらの希釈物、濃縮物を含む。
【0047】
かかる培地としては、宿主細胞が資化し得るものが含有されていればよく、炭素源、窒素源、硫黄源、無機塩類、金属、ペプトン、酵母エキス、肉エキス、カゼイン加水分解物、血清等が含有物として挙げられる。また、かかる培地には、例えば、中性プロテアーゼのプレプロ配列をコードするポリヌクレオチドの発現を誘導するためのIPTGや、本発明の発現ベクターがコードし得る薬剤耐性遺伝子に対応する抗生物質(例えば、ネオマイシン、クロラムフェニコール)や、本発明の発現ベクターがコードし得る栄養要求性を相補する遺伝子に対応する栄養物(例えば、アルギニン、ヒスチジン)を添加してもよい。
【0048】
本発明にかかる培養の条件としては、本発明の形質転換体が前記培地中に活性を有した成熟型の中性プロテアーゼを分泌産生できる条件であればよく、当業者であれば、宿主細胞の種類、用いる培地等に合わせて、温度、空気の添加の有無、酸素の濃度、二酸化炭素の濃度、培地のpH、培養温度、培養時間、湿度等を適宜調整し、設定することができる。
【0049】
前述の通り、また後述の実施例に示す通り、本発明の形質転換体を培養することにより、活性を有した成熟型の中性プロテアーゼが該宿主細胞の外へ分泌されることになる。従って、本発明は、本発明の形質転換体を培養し、該形質転換体および/またはその培養物から発現したタンパク質を採取する工程を含む、中性プロテアーゼの製造方法をも提供することができる。
【0050】
「本発明の形質転換体を培養」とは前記の通りである。また、「本発明の形質転換体および/またはその培養物から発現したタンパク質を採取する」方法としては、フィルター(例えば、孔径0.2μm以下のフィルター)によって宿主細胞を除去する方法や、脱塩用フィルターを用いた脱塩濃縮により、培養物中のタンパク質の濃度を高める方法や、カラム(例えば、陰イオン交換カラムや疎水性カラム)を用いたタンパク質の濃縮方法が挙げられる。
【0051】
また、本発明は前記製造方法によって製造された中性プロテアーゼを含む、プロテアーゼ調製物をも提供する。本発明のプロテアーゼ調製物としては、前記製造方法によって製造された中性プロテアーゼを含んでいればよいが、本発明の中性プロテアーゼの他、プロテアーゼ調製物として許容される他の成分を含むことができる。このような他の成分としては、例えば、担体、賦形剤、崩壊剤、緩衝剤、乳化剤、懸濁剤、安定剤、保存剤、防腐剤、生理食塩が挙げられる。賦形剤としては乳糖、デンプン、ソルビトール、D−マンニトール、白糖等を用いることができる。崩壊剤としてはデンプン、カルボキシメチルセルロース、炭酸カルシウム等を用いることができる。緩衝剤としてはリン酸塩、クエン酸塩、酢酸塩等を用いることができる。乳化剤としてはアラビアゴム、アルギン酸ナトリウム、トラガント等を用いることができる。懸濁剤としてはモノステアリン酸グリセリン、モノステアリン酸アルミニウム、メチルセルロース、カルボキシメチルセルロース、ヒドロキシメチルセルロース、ラウリル硫酸ナトリウム等を用いることができる。安定剤としてはプロピレングリコール、ジエチリン亜硫酸塩、アスコルビン酸等を用いることができる。保存剤としてはフェノール、塩化ベンザルコニウム、ベンジルアルコール、クロロブタノール、メチルパラベン等を用いることができる。防腐剤としてはアジ化ナトリウム、塩化ベンザルコニウム、パラオキシ安息香酸、クロロブタノール等を用いることができる。
【0052】
さらに、本発明のプロテアーゼ調製物を、糖尿病等の治療を目的とし、膵臓等の臓器から膵島等の細胞群やβ細胞等の細胞を単離するための薬剤として利用する場合には、例えばコラゲナーゼ、エラステラーゼ、トリプシン、キモトリプシン等の他の酵素を含有していてもよい。また、本発明のプロテアーゼ調製物を、中性洗剤として利用する場合には、例えば、プロテアーゼ、アミラーゼ、リパーゼ、マンナーゼ、ペクチナーゼ、クチナーゼ、オキシドレダクターゼ、ヘミセルラーゼ、セルラーゼ等の他の酵素を含有していてもよい。
【実施例】
【0053】
以下、実施例及び比較例に基づいて本発明をより具体的に説明するが、本発明は以下の実施例に限定されるものではない。また、各実施例及び比較例で用いたプライマーの配列を表1に示す。
【0054】
【表1】
【0055】
(比較例1)
<特許文献1に記載の遺伝子配列を用いた中性プロテアーゼの発現>
(1−1) 特許文献1に記載の塩基配列を用いた中性プロテアーゼ発現用プラスミドの構築
Brevibacillusにおいて強力に作用するP2プロモータの配列についてはpNCMO2ベクター(タカラバイオ株式会社製)を鋳型としてP2−F及びP2−Rプライマーを用いて、中性プロテアーゼ遺伝子についてはClostridium histolyticumのゲノムDNAを鋳型としてNP−F1及びNP−Rプライマーを用いてPCRにより増幅した。PCRはPrimeSTAR Max DNA Polymerase(タカラバイオ社製)を用いて、98℃で10秒、55℃で5秒、72℃で30秒を40サイクル実施した。増幅された約700bpのDNA断片及び1kbpのDNA断片(配列番号:7に記載の601〜1599番目の塩基配列を含むDNA断片)をQIAquick PCR Purification Kitにより回収した。回収したDNA断片をそれぞれ1μLずつ鋳型としてP2−F及びNP−Rプライマーを用いてPCRを行った。なお、NP−F1プライマーの1〜20位はP2−Rプライマーと相補的な配列となっており、該相補的な配列を介して回収したDNA断片どうしはアニーリングすることができる。そのため、回収したDNA断片を鋳型としてP2−F及びNP−Rプライマーを用いてPCRを行うことにより、P2プロモータ−の下流に中性プロテアーゼをコードする塩基配列が連結している配列を増幅することができる。そして、このようにして増幅して得られた約1.7kbpのDNA断片、及びBrevibacillus用発現ベクターであるpNCMO2をBalI及びBamHIで制限酵素処理した後に、ライゲーション反応を行い、プラスミドpNCMO2−P2−NP(F1)を得た。
【0056】
(1−2) 特許文献1に記載の遺伝子配列を用いた中性プロテアーゼの発現
プラスミドpNCMO2−P2−NP(F1)をエレクトロポレーションによりBrevibacillus choshinensis SP−3株に導入し、ネオマイシン耐性株(pNCMO2−P2−NP(F1)形質転換体)を得た。次いで、TMNM液体培地(グルコース10.0g/L、ポリペプトン10.0g/L、肉エキス5.0g/L、酵母エキス2.0g/L、FeSO4・7H2O 10mg/L、MnSO4・4H2O 10mg/L、ZnSO4・7H2O 1mg/L、ネオマイシン 10μg/mL)にて、pNCMO2−P2−NP(F1)形質転換体の培養を行った。そして、得られた培養液を遠心し、菌体と培養上清に分離した後、該培養上清をフィルター(孔径:0.2μm)に通してろ過滅菌することにより、電気泳動用のサンプルとして調製した。次いで、このサンプルをXV PANTERA Gel 15%(DRC社製)に供し、電気泳動分離を行った後、クマシーブリリアントブルー(CBB)染色を行った。得られた結果を図1に示す。
【0057】
また、pNCMO2−P2−NP(F1)形質転換体を、1%スキムミルクを含むLB培地(スキムミルクプレート)に接種して37℃で一夜培養し、スキムミルクの分解の程度を調べた。得られた結果を図2のAに示す。
【0058】
図1に示した結果から明らかなように、pNCMO2−P2−NP(F1)形質転換体の培養上清中に約35kDaの中性プロテアーゼとおもわれるタンパク質は検出されなかった。また、図2のAに示した結果から明らかなように、当該形質転換体をスキムミルクプレートで培養しても、スキムミルクの分解に起因するハローは検出されなかった。
【0059】
(実施例1)
<中性プロテアーゼのシグナル配列ならびにプロ配列のクローニング>
(2−1) 中性プロテアーゼのシグナル配列ならびにプロ配列の取得
中性プロテアーゼ成熟配列の上流部分を同定するために、Clostridium histolyticumのゲノムDNAを鋳型としてNP5−1及びNP3プライマーを用いてPCRにより増幅した。PCRはPrimeSTAR Max DNA Polymerase(タカラバイオ社製)を用いて、98℃で10秒、55℃で5秒、72℃で30秒を30サイクル実施した。増幅された約1.1kbpのDNA断片をQIAquick PCR Purification Kitにより回収した。PCRにより取得したDNA断片のシークエンスは、BigDye Terminator v3.1 Cycle Sequebcing Kit(アプライドバイオシステムズ社製)とABI PRISMジェネティックアナライザー(アプライドバイオシステムズ社製)を用い、添付のプロトコールに従って決定した。そして、得られた塩基配列をアミノ酸配列に翻訳し、そのアミノ酸配列から4箇所のメチオニン(開始コドン:配列番号7に記載の1〜3番目、61〜63番目、295〜297番目及び601〜603番目のatgがコードするメチオニン)を見出した。
【0060】
(実施例2)
<シグナル配列及びプロ配列を含む中性プロテアーゼ遺伝子を用いたBrevibacillus choshinensis SP−3株での発現>
(3−1) シグナル配列及びプロ配列を含む中性プロテアーゼ遺伝子発現用プラスミドの構築
シグナル配列及びプロ配列を含む中性プロテアーゼ遺伝子をClostridium histolyticumのゲノムDNAを鋳型としてNP−F2及びNP−Rプライマーを用いてPCRにより増幅した。PCRはPrimeSTAR Max DNA Polymerase(タカラバイオ社製)を用いて、98℃で10秒、55℃で5秒、72℃で30秒を40サイクル実施した。増幅された1.6kbpのDNA断片(配列番号:7に記載の塩基配列を含むDNA断片)をQIAquick PCR Purification Kitにより回収した。回収したDNA断片と比較例1(1−1)で作製したP2プロモータ配列の断片をそれぞれ1μLずつ鋳型としてP2−F及びNP−Rプライマーを用いてPCRを行い、P2プロモータ−の下流にシグナル配列ならびにプロ配列を含む中性プロテアーゼをコードする塩基配列が連結している配列を増幅した。増幅された約2.3kbpのDNA断片を、TOPO TAクローニングキット(インビトロジェン社製)により、添付のプロトコールに従ってpCR2.1 TOPOプラスミドベクターに挿入しプラスミドpCR−P2−NPを得た。そして、pCR−P2−NP及びpNCMO2を、BalI及びBamHIで制限酵素処理した後に、pCR−P2−NPのインサート部分(約2.3kbpのDNA断片)とpNCMOとのライゲーション反応を行い、プラスミドpNCMO2−P2−NPを得た。
【0061】
(3−2) 中性プロテアーゼの発現
プラスミドpNCMO−P2−NPをエレクトロポレーション法によりBrevibacillus choshinensis SP−3株に導入し、ネオマイシン耐性株(pNCMO2−P2−NP形質転換体)を得た。そして、pNCMO2−P2−NP形質転換体の培養をTMNM液体培地にて行い、得られた培養液を遠心し、菌体と培養上清に分離した後、該培養上清をフィルター(孔径:0.2μm)に通してろ過滅菌することにより、電気泳動用のサンプルとして調製した。次いで、このサンプルをXV PANTERA Gel 15%(DRC社製)に供し、電気泳動分離を行った後、クマシーブリリアントブルー(CBB)染色を行った。得られた結果を図3に示す。
【0062】
また、pNCMO2−P2−NP形質転換体を、1%スキムミルクを含むLB培地(スキムミルクプレート)に接種して37℃で一夜培養し、スキムミルクの分解の程度を調べた。得られた結果を図2のBに示す。
【0063】
図3に示した結果から明らかなように、pNCMO2−P2−NP形質転換体の培養上清中に約35kDaの中性プロテアーゼとおもわれるタンパク質が検出された。また、図2のBに示した結果から明らかなように、当該形質転換体をスキムミルクプレートで培養したところ、スキムミルクの分解に起因するハローが検出された。
【0064】
また、図には示さないが、配列番号7に記載の61〜63番目のatgを開始コドンとする中性プロテアーゼをコードする塩基配列及び配列番号7に記載の295〜297番目のatgを開始コドンとする中性プロテアーゼをコードする塩基配列についても前記同様に各々発現ベクターを作製し、これらの発現ベクターを導入した宿主細胞を用いて、中性プロテアーゼの活性を評価した。しかし、比較例1に示した配列番号7に記載の601〜603番目のatgを開始コドンとする中性プロテアーゼをコードする塩基配列を用いた場合と同じく、いずれも中性プロテアーゼの活性は検出されなかった。
【0065】
(実施例3)
<シグナル配列及びプロ配列を含む中性プロテアーゼ遺伝子を用いたBacillus subtilis KN−2株での発現>
(4−1) シグナル配列及びプロ配列を含む中性プロテアーゼ遺伝子発現用プラスミドの構築
両端にBamHI及びSmaIの制限酵素サイトを導入するため、プラスミドpCR−P2−NPを鋳型としてNP−BamHI−N0及びNP−SmaI−C0プライマーを用いて、PCRにより増幅した。PCRはPrimeSTAR Max DNA Polymerase(タカラバイオ社製)を用いて、98℃で10秒、55℃で5秒、72℃で30秒を35サイクル実施した。増幅された1.6kbpのDNA断片(配列番号:7に記載の塩基配列を含むDNA断片)をQIAquick PCR Purification Kitにより回収した。回収したDNA断片及びBacillus subtilis発現用ベクターpHT01(Mo Bi Tec社)をBamHI及びSmaIで制限酵素処理した後に、ライゲーション反応を行い、プラスミドpHT01−NPを得た。
【0066】
(4−2) Bacillus subtilis KN−2株での中性プロテアーゼ発現
プラスミドpHT01−NPをBacillus subtilis KN−2株のコンピテントセルに導入し、クロラムフェニコール耐性株(pHT01−NP形質転換体)を得た。クロラムフェニコールを含むLB液体培地(バクトトリプトン 10.0g/L、イースト 5.0g/L、NaCl 10.0g/L、クロラムフェニコール 25μg/mL)にて37℃で24時間、pHT01−NP形質転換体を培養した後、IPTGにてタンパク質の発現誘導を37℃で4時間行なった。
【0067】
そして、得られた培養液を遠心し、菌体と培養上清に分離した後、該培養上清をフィルター(孔径:0.2μm)に通してろ過滅菌することにより、電気泳動用のサンプルとして調製した。次いで、このサンプルを12% Gel SDS−PAGE mini(テフコ社製)に供し、電気泳動分離を行った後、クマシーブリリアントブルー(CBB)染色を行った。得られた結果を図4に示す。
【0068】
図4に示した結果から明らかなように、宿主をBrevibacillus choshinensis SP−3株からBacillus subtilis KN−2株に変えても、実施例2の結果と同様に、pHT01−NP形質転換体の培養上清においても約35kDaの中性プロテアーゼと思われるタンパク質が検出された。
【0069】
(4−3) 発現した中性プロテアーゼのN末端アミノ酸配列の同定
実施例3の(4−2)で発現させたタンパク質がクロストリディウム・ヒストリティカム由来中性プロテアーゼであることを確認するために、N末端アミノ酸配列を決定した。まず、培養上清中のタンパク質について12% Gel SDS−PAGE mini(テフコ社製)を用いて電気泳動による分離を行い、PVDF膜(ミリポア社製)上にブロットした。得られた約35kDaのバンドを切り出し、プロテインシークエンサーModel 492(アプライドバイオシステムズ社製)を用いて、N末端アミノ酸配列を決定した。その結果、クロストリディウム・ヒストリティカム由来中性プロテアーゼのN末端アミノ酸配列(配列番号:6に記載の1〜5番目のアミノ酸配列)と一致した。
【0070】
(4−4) 形質転換体における中性プロテアーゼ活性の測定
実施例3の(4−2)で得られたpHT01−NP形質転換体の培養上清を用いてアゾカゼイン分解活性を測定した。測定は、アゾカゼインを基質とし、基質の分解によって生じる遊離チロシン量を335nmの吸光度で測定する方法を用い、文献(Nagase H, Harris ED Jr;J Biol Chem. 1983 Jun 25;258(12):7490−8)に記載の方法に準じて行った。
【0071】
なお、アゾカゼイン分解活性は、反応条件においてO.D.(335nm)=1.0になるのに必要な酵素量を1 unitと定義し、培養上清1mL当りの活性(U/mL)として表した。得られた結果を表2に示す。表2に示した結果から明らかなように、pHT01−NP形質転換体は親株(遺伝子を導入していない株)の約19倍の活性を示した。
【0072】
【表2】
【0073】
以上の結果から、配列番号7:に記載の1〜657番目の塩基配列がコードするポリペプチド(配列番号8:に記載の1〜219番目のアミノ酸配列からなるポリペプチド)が、中性プロテアーゼの宿主細胞外への分泌産生を可能としていることが明らかになった。また、配列番号8:に記載のアミノ酸配列をシグナル配列予測ソフトウェア(SignalP 3.0、http://www.cbs.dtu.dk/services/SignalP/)を用いて解析したところ、配列番号:2に記載のアミノ酸配列(配列番号:1に記載の塩基配列がコ―ドするアミノ酸配列)がシグナル配列と推定され、結果、配列番号:4に記載のアミノ酸配列(配列番号:3に記載の塩基配列がコ―ドするアミノ酸配列)がプロ配列として機能していることが推定された。
【産業上の利用可能性】
【0074】
以上説明したように、本発明によれば、目的とするタンパク質の宿主細胞外への分泌産生が可能となる。特に、本発明のシグナル配列やプロ配列をコードするポリヌクレオチドを利用して得られる中性プロテアーゼは、クロストリディウム・ヒストリティカム以外の宿主細胞から、活性を有した成熟型として分泌産生することができる。
【0075】
したがって、クロストリディウム・ヒストリティカムの培養液に含まれる非常に多くのプロテアーゼ活性を有するタンパク質を混入させることなく、中性プロテアーゼを調製することができるため、本発明のシグナル配列やプロ配列を利用して得られる中性プロテアーゼは、糖尿病等の治療を目的とし、膵臓等の臓器から膵島等の細胞群やβ細胞等の細胞を単離するための薬剤や、中性洗剤等として有用である。
【配列表フリーテキスト】
【0076】
配列番号9〜17
<223> 人工的に合成されたプライマーの配列
【特許請求の範囲】
【請求項1】
シグナル配列をコードする、下記(i)から(v)のいずれかに記載のポリヌクレオチド。
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:1に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:2に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:2に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:1に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
【請求項2】
プロ配列をコードする、下記(i)から(v)のいずれかに記載のポリヌクレオチド。
(i)配列番号:4に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:3に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:4に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:4に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:3に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
【請求項3】
クロストリディウム・ヒストリティカム(Clostridium histolyticum)由来である、請求項1又は2に記載のポリヌクレオチド。
【請求項4】
請求項1に記載のポリヌクレオチドと、請求項2に記載のポリヌクレオチドと、中性プロテアーゼの成熟配列をコードする下記(i)から(v)のいずれかに記載のポリヌクレオチドとが連結しているポリヌクレオチド。
(i)配列番号:6に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:5に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:6に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:6に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:5に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
【請求項5】
クロストリディウム・ヒストリティカム(Clostridium histolyticum)由来である、請求項4に記載のポリヌクレオチド。
【請求項6】
請求項4又は5に記載のポリヌクレオチドを含有する発現ベクター。
【請求項7】
請求項6に記載の発現ベクターで形質転換された宿主細胞。
【請求項8】
請求項7に記載の形質転換された宿主細胞から分泌された中性プロテアーゼを含む培養物。
【請求項9】
請求項7に記載の形質転換された宿主細胞を培養し、該宿主細胞および/またはその培養物から発現したタンパク質を採取する工程を含む、中性プロテアーゼの製造方法。
【請求項10】
請求項9に記載の方法によって製造された中性プロテアーゼを含む、プロテアーゼ調製物。
【請求項1】
シグナル配列をコードする、下記(i)から(v)のいずれかに記載のポリヌクレオチド。
(i)配列番号:2に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:1に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:2に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:2に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:1に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
【請求項2】
プロ配列をコードする、下記(i)から(v)のいずれかに記載のポリヌクレオチド。
(i)配列番号:4に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:3に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:4に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:4に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:3に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
【請求項3】
クロストリディウム・ヒストリティカム(Clostridium histolyticum)由来である、請求項1又は2に記載のポリヌクレオチド。
【請求項4】
請求項1に記載のポリヌクレオチドと、請求項2に記載のポリヌクレオチドと、中性プロテアーゼの成熟配列をコードする下記(i)から(v)のいずれかに記載のポリヌクレオチドとが連結しているポリヌクレオチド。
(i)配列番号:6に記載のアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(ii)配列番号:5に記載の塩基配列のコード領域を含むポリヌクレオチド
(iii)配列番号:6に記載のアミノ酸配列において、1もしくは複数個のアミノ酸が置換、欠失、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするポリヌクレオチド
(iv)配列番号:6に記載のアミノ酸配列と95%以上の同一性を有するアミノ酸配列を含むタンパク質をコードするポリヌクレオチド
(v)配列番号:5に記載の塩基配列からなるポリヌクレオチドとストリンジェントな条件でハイブリダイズするポリヌクレオチド
【請求項5】
クロストリディウム・ヒストリティカム(Clostridium histolyticum)由来である、請求項4に記載のポリヌクレオチド。
【請求項6】
請求項4又は5に記載のポリヌクレオチドを含有する発現ベクター。
【請求項7】
請求項6に記載の発現ベクターで形質転換された宿主細胞。
【請求項8】
請求項7に記載の形質転換された宿主細胞から分泌された中性プロテアーゼを含む培養物。
【請求項9】
請求項7に記載の形質転換された宿主細胞を培養し、該宿主細胞および/またはその培養物から発現したタンパク質を採取する工程を含む、中性プロテアーゼの製造方法。
【請求項10】
請求項9に記載の方法によって製造された中性プロテアーゼを含む、プロテアーゼ調製物。
【図1】


【図2】
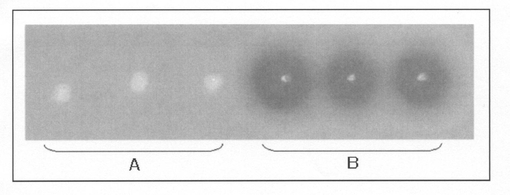
【図3】
【図4】
【図2】
【図3】
【図4】
【公開番号】特開2013−5795(P2013−5795A)
【公開日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願番号】特願2012−117169(P2012−117169)
【出願日】平成24年5月23日(2012.5.23)
【出願人】(000006091)Meiji Seikaファルマ株式会社 (180)
【出願人】(504157024)国立大学法人東北大学 (2,297)
【Fターム(参考)】
【公開日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願日】平成24年5月23日(2012.5.23)
【出願人】(000006091)Meiji Seikaファルマ株式会社 (180)
【出願人】(504157024)国立大学法人東北大学 (2,297)
【Fターム(参考)】
[ Back to top ]