
悪性胸膜中皮腫の検出のための方法及びキット
【課題】悪性胸膜中皮腫を検出するための方法を提供する。
【解決手段】悪性胸膜中皮腫を検出するための方法あるいは悪性胸膜中皮腫に罹患した被験者の予後を測定する方法であって、被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化またはメチル化程度を検出することを含み、該遺伝子がメチル化されているとき悪性胸膜中皮腫であると判定する方法、あるいは、該遺伝子のメチル化程度が、予後不良被験者のメチル化程度と比べて低いとき、該疾患の予後が良好であると決定する方法、ならびに、これらの方法に使用するためのキット。
【解決手段】悪性胸膜中皮腫を検出するための方法あるいは悪性胸膜中皮腫に罹患した被験者の予後を測定する方法であって、被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化またはメチル化程度を検出することを含み、該遺伝子がメチル化されているとき悪性胸膜中皮腫であると判定する方法、あるいは、該遺伝子のメチル化程度が、予後不良被験者のメチル化程度と比べて低いとき、該疾患の予後が良好であると決定する方法、ならびに、これらの方法に使用するためのキット。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、DNAメチル化異常に基づいた悪性胸膜中皮腫の検出のための方法およびキットに関する。
【背景技術】
【0002】
悪性胸膜中皮腫(Malignant Pleural Mesothelioma (MPM))は、アスベスト曝露と関連のある侵襲性腫瘍である(非特許文献1, 2)。世界で毎年、約10,000〜15,000名の患者が、新たにMPMと診断され、今後20年以内に患者数は増加すると予測されている(非特許文献3〜5)。アスベストの吸入は、周知の危険因子であり、MPMの早期診断が望まれているが、早期のステージで臨床症状がないことと有用な診断マーカーがないことが、早期診断を困難にしている。さらにまた、MPMは、時に病理学的に肺腺癌(AdCa)との区別が難しく、抗体パネルによる追加の免疫組織化学的染色を必要とする場合がある(非特許文献6)。これらの困難性と有効な治療がいまだないために、治療にも拘らず、多くの患者が、診断から2年以内に死亡している(非特許文献4, 7, 8)。したがって、精確な早期診断に有効なマーカーを同定するために、MPMのさらなる分子分析が緊急に必要とされている。
【0003】
MPMが発症する中心的な機序は依然としてはっきりしていない。いくつかの遺伝子異常がMPMに関与していると考えられており、そのような異常には、例えばp16 遺伝子座の欠損やNF2 遺伝子の変異が挙げられる(非特許文献9〜11)。最近の全トランスクリプトーム配列決定法により、転写された約6個の遺伝子が個々のMPMで突然変異を受けていると推定されたが、このことは限られた数個の遺伝子変異がMPM発症に関与していることを示唆している(非特許文献12)。以上の報告から遺伝子変異はMPMにおいて低頻度であり、遺伝子変異以外の機序がMPMの腫瘍形成に関与すると考えられる。
【0004】
エピジェネティックな転写制御、特にプロモーターDNAメチル化の調節異常は、ヒト悪性腫瘍に高頻度に観察される(非特許文献13)。さらにDNAの異常メチル化と炎症との関係がこれまで多くの癌で記載されている(非特許文献14)。アスベストは、ヒト中皮培養細胞の悪性転換を直接誘導しない(非特許文献15, 16)。またアスベスト曝露により慢性炎症が引き起こされることがわかっている(非特許文献17)。すなわちMPM発症にDNAメチル化が関与している可能性が強く疑われる。しかし、これまでにMPMにおけるDNAメチル化異常についての情報は限られているのが現状である(非特許文献18〜20)。
【0005】
ちなみに、中皮腫の診断または予後予測に関係する文献には、特定のポリペプチドに対する抗体、特定の遺伝子の発現量の差を利用する、特許文献1、2および3などが知られている。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2007-071849
【特許文献2】特表2008-505862
【特許文献3】特表2003-524021
【非特許文献】
【0007】
【非特許文献1】Robinson BW, Musk AW, Lake RA. Malignant mesothelioma. Lancet 2005;366:397-408
【非特許文献2】Pass HI, Vogelzang N, Hahn S, Carbone M. Malignant pleural mesothelioma. Curr Probl Cancer 2004;28:93-174
【非特許文献3】Tsiouris A, Walesby RK. Malignant pleural mesothelioma: current concepts in treatment. Nat Clin Pract Oncol 2007;4:344-52
【非特許文献4】Robinson BW, Lake RA. Advances in malignant mesothelioma. N Engl J Med 2005;353:1591-603
【非特許文献5】Bianchi C, Bianchi T. Malignant mesothelioma: global incidence and relationship with asbestos. IndHealth 2007;45:379-87
【非特許文献6】Abutaily AS, Addis BJ, Roche WR. Immunohistochemistry in the distinction between malignant mesothelioma and pulmonary adenocarcinoma: a critical evaluation of new antibodies. J Clin Pathol 2002;55:662-8
【非特許文献7】Vogelzang NJ, Rusthoven JJ, Symanowski J, et al. Phase III study of pemetrexed in combination with cisplatin versus cisplatin alone in patients with malignant pleural mesothelioma. J Clin Oncol 2003;21:2636-44
【非特許文献8】van Meerbeeck JP, Gaafar R, Manegold C, et al. Randomized phase III study of cisplatin with or without raltitrexed in patients with malignant pleural mesothelioma: an intergroup study of the European Organisation for Research and Treatment of Cancer Lung Cancer Group and the National Cancer Institute of Canada. J Clin Oncol 2005;23:6881-9
【非特許文献9】Cheng JQ, Jhanwar SC, Klein WM, et al. p16 alterations and deletion mapping of 9p21-p22 in malignant mesothelioma. Cancer Res 1994;54:5547-51.10. Sekido Y, Pass HI, Bader S, et al. Neurofibromatosis type 2 (NF2) gene is somatically mutated in mesothelioma but not in lung cancer. Cancer Res 1995;55:1227-31
【非特許文献10】Sekido Y, Pass HI, Bader S, et al. Neurofibromatosis type 2 (NF2) gene is somatically mutated in mesothelioma but not in lung cancer. Cancer Res 1995;55:1227-31
【非特許文献11】Bianchi AB, Mitsunaga SI, Cheng JQ, et al. High frequency of inactivating mutations in the neurofibromatosis type 2 gene (NF2) in primary malignant mesotheliomas. Proc Natl Acad Sci U S A 1995;92:10854-8
【非特許文献12】Sugarbaker DJ, Richards WG, Gordon GJ, et al. Transcriptome sequencing of malignant pleural mesothelioma tumors. Proc Natl Acad Sci U S A 2008;105:3521-6
【非特許文献13】Jones PA, Baylin SB. The fundamental role of epigenetic events in cancer. Nat Rev Genet 2002;3:415-28
【非特許文献14】Perwez Hussain S, Harris CC. Inflammation and cancer: an ancient link with novel potentials. Int J Cancer 2007;121:2373-80
【非特許文献15】Xu L, Flynn BJ, Ungar S, et al. Asbestos induction of extended lifespan in normal human mesothelial cells: interindividual susceptibility and SV40 T antigen. Carcinogenesis 1999;20:773-83
【非特許文献16】Bocchetta M, Di Resta I, Powers A, et al. Human mesothelial cells are unusually susceptible to simian virus 40-mediated transformation and asbestos cocarcinogenicity. Proc Natl Acad Sci U S A 2000;97:10214-9
【非特許文献17】Yang H, Bocchetta M, Kroczynska B, et al. TNF-alpha inhibits asbestos-induced cytotoxicity via a NF-kappaB-dependent pathway, a possible mechanism for asbestos-induced oncogenesis. Proc Natl Acad Sci U S A 2006;103:10397-402
【非特許文献18】Toyooka S, Pass HI, Shivapurkar N, et al. Aberrant methylation and simian virus 40 tag sequences in malignant mesothelioma. Cancer Res 2001;61:5727-30
【非特許文献19】Tsou JA, Galler JS, Wali A, et al. DNA methylation profile of 28 potential marker loci in malignant mesothelioma. Lung Cancer 2007;58:220-30
【非特許文献20】Tsou JA, Shen LY, Siegmund KD, et al. Distinct DNA methylation profiles in malignant mesothelioma, lung adenocarcinoma, and non-tumor lung. Lung Cancer 2005;47:193-204
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の目的は、悪性胸膜中皮腫の検出のための方法を提供することである。
【課題を解決するための手段】
【0009】
MPMでのDNAメチル化についてより多くのことを理解するために、本発明者らは、メチル化CpGアイランド増幅マイクロアレイ(MCAM) ( Shen L, Kondo Y, Guo Y, et al. PLoS Genet 2007;3:2023-36)を用いてMPM中の異常にメチル化された遺伝子について広域に及ぶスクリーニングを行った。MCAMは、癌組織だけでなく正常組織においてもメチル化遺伝子を成功裏に検出可能にし、また、高有効率で再現性のある結果を提供する。さらに、本発明者らは、2種類の胸部癌におけるDNAメチル化異常の役割を比較するために、AdCaのメチル化を調べた。DNAのメチル化プロフィールの分析により、異常なDNAメチル化がAdCaと比べてMPMで頻度が少ないこと、ならびにAdCaは症例間で共通の遺伝子が高頻度にメチル化するが、MPMで症例間での共通メチル化遺伝子は少ないことが明示された。以上の結果を詳細に検討しMCAMで見出された高メチル化遺伝子のうち、MPMで特異的にメチル化されるがAdCaでメチル化されない、3つの遺伝子を同定した。これらの遺伝子は、MPMの早期診断マーカーとして役立つはずである。
【0010】
したがって、本発明は、要約すると、以下の特徴を含む。
本発明は、第1の態様において、悪性胸膜中皮腫を検出するための方法であって、被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を検出することを含み、該遺伝子がメチル化されているとき悪性胸膜中皮腫であると判定する、上記方法を提供する。
【0011】
本発明はまた、第2の態様において、悪性胸膜中皮腫に罹患した被験者の予後を判定する方法であって、該被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化の程度を測定することを含み、該遺伝子のメチル化程度が、予後不良被験者のメチル化程度と比べて低いとき、該疾患の予後が良好であると決定する、上記方法を提供する。
【0012】
上記2つの各発明の実施形態において、上記の方法が、上記TMEM30B遺伝子、KAZALD1遺伝子またはMAPK13遺伝子のプロモーターのCpG領域の塩基配列のメチル化を検出または測定する。
【0013】
別の実施形態において、上記プロモーターのCpG領域の塩基配列が、配列番号78、79または80に示される塩基配列、あるいは、その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含み、かつ、プロモーター活性を有する塩基配列からなる。
【0014】
別の実施形態において、上記メチル化が、メチル化特異的PCR法によって検出される。
別の実施形態において、上記PCR法で使用するためのプライマーを、配列番号78、79または80に示される塩基配列中の、あるいは、その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含み、かつ、プロモーター活性を有する塩基配列中の、連続する17〜35塩基からなる配列を、メチル化特異的プライマーおよび/または非メチル化特異的プライマーとして選択する。
【0015】
また、別の実施形態において、前記サンプルが血清、胸水、気管支洗浄液または患部組織である。
【0016】
本発明はさらに、第3の態様において、上記のいずれかの方法で使用するためのキットであって、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を測定することが可能である、メチル化特異的PCR用の下記のプライマーセットを含むキットを提供する。
TMEM30B遺伝子のメチル化特異的プライマー
TTGGGAGGGTGTGGGTATTGTTC(配列番号66)
ACGCGCGAACTAACCGAAT(配列番号67)
CTGGGAGGGTGTGGGCACTGCTCG(配列番号81)
CGAGTAGTTTTCGGGTTTGC(配列番号95)
ACCGACGCGCGAACTAAC(配列番号96)
TMEM30B遺伝子の非メチル化特異的プライマー
GGAATTGGGAGGGTGTGGGTATTGTTT(配列番号68)
ACCTCCCTCTCCACACCAC(配列番号69)
GGAACTGGGAGGGTGTGGGCACTGCT(配列番号82)
TGAGTAGTTTTTGGGTTTGT(配列番号97)
ACCATAACACTCCAAATCATAACA(配列番号98)
KAZALD1遺伝子のメチル化特異的プライマー
GCGGGTCGTAGTATTTGTCG(配列番号70)
AACAAAACCAACCACCTTCG(配列番号71)
CGGGTCGTAGTATTTGTC(配列番号85)
GGTAAGTTTGCGGTCGTTTTC(配列番号99)
AACAAAACCAACCACCTTCG(配列番号100)
KAZALD1遺伝子の非メチル化特異的プライマー
TTATTTGTGGGTTGTAGTATTTGTT(配列番号72)
AAACCAACCACCTTCATCTCC(配列番号73)
TTATTCGCGGGTCGTAGTATTTGTC(配列番号86)
GTGGGTAAGTTTGTGGTTG(配列番号101)
AACAAAACCAACCACCTTCATCT(配列番号102)
MAPK13遺伝子のメチル化特異的プライマー
CGTGTTTTCGACGTACGTCGGTAGC(配列番号74)
ACCGCTAAATAACCCCGAAA(配列番号75)
CGTGTCCCCGACGCACGTCGGCAGC(配列番号89)
MAPK13遺伝子の非メチル化特異的プライマー
GGTTTTGTTGGGTAGTTTT(配列番号76)
TCCCACCTACCAACTCACC(配列番号77)
GGTCCCGCTGGGCAGCCC(配列番号90)
【発明の効果】
【0017】
本発明により、悪性胸膜中皮腫を肺癌から区別可能とする悪性胸膜中皮腫検出方法が提供される。この方法は、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を測定することに基づいている。
【図面の簡単な説明】
【0018】
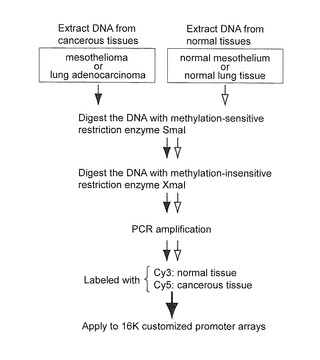
【図1A】この図は、悪性中皮腫細胞系におけるMCAM分析とその有効性を示す。図1AはMCAMの概説である。癌又は正常組織由来のDNAをSmaI及びXmaIで連続的に消化した。メチル化DNA断片を増幅したのち、MCA産物を、癌組織に対してはCy5(赤色)で、正常組織に対してはCy3(緑色)で それぞれ標識し、その後、プロモーターマイクロアレイに使用した。
【図1B】この図は、悪性中皮腫細胞系におけるMCAM分析とその有効性を示す。図1Bは、MCAMシグナル比とメチル化レベルの比較を示す。Y軸は、各遺伝子におけるMCAMのシグナル比を示す。pyrosequencing分析によって決定されたメチル化レベルを絶対的変数として表した。 水平線は、MCAMシグナル比2.0を示す。
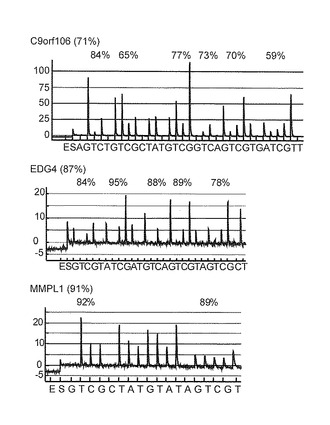
【図1C】この図は、悪性中皮腫細胞系におけるMCAM分析とその有効性を示す。図1Cは、3種の高メチル化遺伝子の代表的パイログラムを示す。灰色で示した領域は、各遺伝子で定量化されたCpG部位を示し、各部位の%メチル化は、ピークの上に示されている。CpG部位の平均メチル化の計算値をカッコのなかに示す。
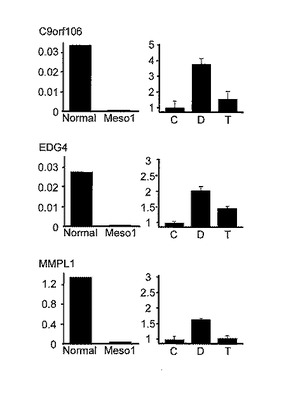
【図1D】この図は、悪性中皮腫細胞系におけるMCAM分析とその有効性を示す。図1Dは、DNAメチル化又はヒストン脱アセチル化の阻害による高メチル化遺伝子の再活性化を示す。PBS (コントロール, C)又は5-Aza-dC (D)、或いはトリコスタチン(Trichostatin) A (T)で処理したあとの、正常中皮組織(Normal)及びACC-MESO-1 (Meso1)における遺伝子発現を定量的-PCRによって測定した。GAPDHに対する各遺伝子のmRNA発現の相対値をY軸(左カラム) に示す。5-Aza-dC処理によって、サイレンシングされた遺伝子が有効に再活性化された。右カラムのY軸は、コントロールに対する処理細胞のRNA発現の相対値を示す。グラフ上の誤差バーは、3回の実験からの標準偏差を示す。
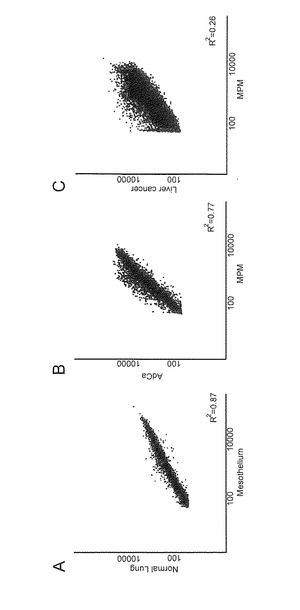
【図2】この図は、正常中皮及び正常肺組織間(A)、MPM及びAdCa 間(B)、並びにMPM及び肝臓癌間(C)、のシグナル強度(logスケール)のスキャッタープロット分析を示す。線形回帰モデルの重相関決定係数(R2) を各分析で示す。
【図3A】この図は、MPM及びAdCa患者でのMCAM分析結果を示す。図3Aの上段のカラムは、全6,161遺伝子(Y軸)を用いて行った、40サンプル(20サンプルのMPMを青色ボックスで示し、20サンプルのAdCaを白色ボックスで示す。)からのDNAメチル化データの階層的クラスター分析のデンドログラムとヒートマップの概観を示す。マトリックス中の各セルは、個々のサンプル中の遺伝子のDNAメチル化状態を表す。セル中の赤色と緑色はそれぞれ、高メチル化レベル、低メチル化レベルを反映している。ここで、メチル化レベルは、マトリックスの下にスケールバーとして示されている(log2変換スケール)。MPMでメチル化された遺伝子数が300未満の症例を黒丸で示す。AdCa症例のうち、950以上の遺伝子がメチル化されていた症例を白抜き菱形ボックスで示す。中段のカラムは、階層型クラスターを用いて各腫瘍におけるサブクラス分類を示す。全6,161遺伝子を用いて20サンプルのMPMの分析を行った。Y軸は、症例間の類似性を示す。黒丸は、上の図と同じMPM症例を示す。ボックスは、MPMの組織学的サブタイプを示し、細線のハッチング、太線のハッチング、黒色及び白色ボックスはそれぞれ、上皮、二相性(biphasic)、肉腫様、及び特殊型を表す。下段のカラムは、AdCaクラスター化を示し、ここで灰色及び白色ボックスはそれぞれ喫煙者及び非喫煙者を示す。Y軸は症例間の類似性を示す。白抜き菱形ボックスは、上の図と同じAdCa症例を示す。
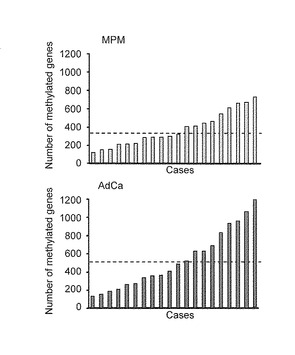
【図3B】この図は、MPM及びAdCa患者でのMCAM分析結果を示す。図3Bは、各症例におけるメチル化遺伝子数を示す。破線は、平均メチル化遺伝子数 (MPM及びAdCaでそれぞれ、387遺伝子及び544遺伝子)を表す。
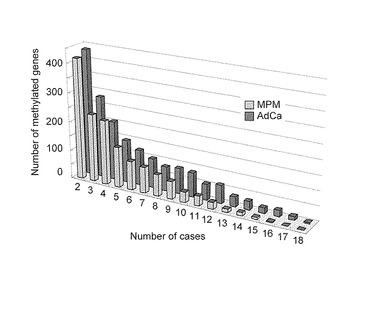
【図3C】この図は、MPM及びAdCa患者でのMCAM分析結果を示す。図3Cは、MPM (灰色バー)又はAdCa (黒色バー)において、症例数x(X軸)で共通にメチル化された遺伝子数(Y軸) を示す。
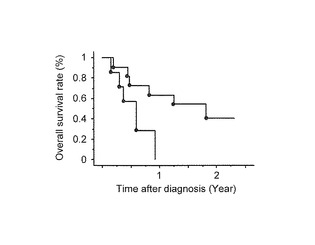
【図3D】この図は、MPM及びAdCa患者でのMCAM分析結果を示す。図3Dは、MPM患者の生存率のKaplan-Meier分析を示す。MPM患者は、メチル化遺伝子数によって二群に分けられた。二群間のlog順位統計による生存率の統計学的比較は、メチル化が高頻度である症例(破線, 387以上の遺伝子をもつ患者)が、メチル化が低頻度である症例(実線, 387未満の遺伝子をもつ患者; P=0.01)と比べて生存率が悪いことを示す。
【図4A】この図は、MPMに特異的なメチル化の同定を示す。図4Aは、各腫瘍での42個の特異的にメチル化された遺伝子を用いて行った階層型クラスター化(MPM及びAdCaでそれぞれ8遺伝子及び34遺伝子)を示す。
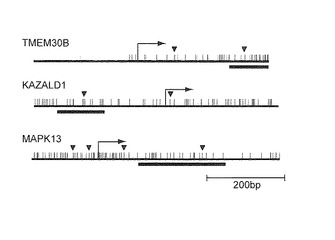
【図4B】この図は、MPMに特異的なメチル化の同定を示す。図4Bは、選択した3つの遺伝子のプロモーターの図を示す。各垂線は、単一のCpG部位を表す。転写開始部位(矢印)、エクソン1の位置(灰色ボックス)、制限酵素部位(矢頭)、並びに、MSPアッセイの位置(バー)を示す。
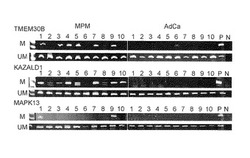
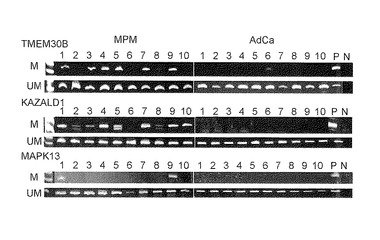
【図4C】この図は、MPMに特異的なメチル化の同定を示す。図4Cは、MCAM分析に使用された、MPMの10サンプル及びAdCaの10サンプルでの、TMEM30B、KAZALD1及びMAPK13のMSP分析を示す。DNAメチル化は、MPM症例で検出された。これに対して、わずか1個の不明瞭なメチル化バンドが、患者6のTMEM30B遺伝子で検出された。M, メチル化型; UM, 非メチル化型; P, メチル化型の陽性コントロール(M) がSssI処理DNAであり、非メチル化コントロール(UM)が正常リンパ球由来のDNAである; N, DNA鋳型を含まない。
【図4D】この図は、MPMに特異的なメチル化の同定を示す。図4Dは、別のコホートの患者のMSP分析(10 MPM及び30 AdCa)を示す。M, UM及びNは、上記4Cと同じ意味である;S, SssI処理DNA。
【発明を実施するための形態】
【0019】
(悪性胸膜中皮腫の検出)
本発明は、悪性胸膜中皮腫を検出するための方法であって、被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を検出することを含み、該遺伝子がメチル化されているとき悪性胸膜中皮腫であると判定する、上記方法を提供する。
【0020】
本明細書において、悪性胸膜中皮腫は、胸膜を構成する中皮細胞の腫瘍であり、アスベスト(石綿)への長期間の曝露に伴うアスベストの吸入が原因で発症すると考えられている。悪性胸膜中皮腫の一般的な症状として、胸水がたまり、胸痛、咳、呼吸困難などの症状が現れる。また、胸水中のヒアルロン酸含量が高値となる傾向がある。
【0021】
DNAメチル化は、MPM診断の強力なマーカーとして提案されている(Toyooka S, Pass HI, Shivapurkar N, et al., Cancer Res 2001;61:5727-30; Tsou JA, Shen LY, Siegmund KD, et al., Lung Cancer 2005;47:193-204.18, 20)。しかしながら、悪性胸膜中皮腫(MPM)と肺腺癌(AdCa)のメチル化状態のこれまでの検討により、その頻度に違いはあるが、候補マーカーの異常メチル化がある程度MPMとAdCaの両方で検出されることが示された。さらに、MPMの特定のメチル化マーカーがこれまで報告されたことはなかった。今回、本発明者らは、MPMとAdCaで全ゲノム領域におけるDNAメチル化プロフィールを同時に分析し、特定の遺伝子座の異常メチル化がMPMで頻繁に検出されること、および、3つの遺伝子(MEM30B、KAZALD1およびMAPK13遺伝子)がMPMで特異的にメチル化されていることを見出した。
【0022】
TMEM30Bは、機能が不明であり、methylation-sensitive representational difference分析法によりヒトメラノーマ細胞系で高メチル化された遺伝子として報告された (Furuta J, Nobeyama Y, Umebayashi Y, Otsuka F, Kikuchi K, Ushijima T., Cancer Res 2006;66:6080-637)。
【0023】
KAZALD1は、骨の形成と再生の間の骨芽細胞の増殖に関与する( Shibata Y, Tsukazaki T, Hirata K, Xin C, Yamaguchi A., Biochem Biophys Res Commun 2004;325:1194-20038)。
【0024】
MAPK13は、MAPキナーゼファミリーのメンバーであり、免疫欠陥、セントロメア領域の不安定化、および顔面異常症候群で調節異常が存在することが認められた(Ehrlich M, Sanchez C, Shao C, et al., Autoimmunity 2008;41: 253-71)。
【0025】
本発明は、特定の3つの遺伝子(TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子)のメチル化を検出することによって、これまで悪性胸膜中皮腫であるのか、それとも肺腺癌であるのか、容易には鑑別不能であったこれらの癌のうち悪性胸膜中皮腫を特異的に検出可能にした。すなわち、本発明は、悪性胸膜中皮腫を検出可能にする遺伝子マーカーを提供する。これらの遺伝子のうち特にKAZALD1およびMAPK13遺伝子のメチル化異常はヒト癌でこれまで報告されたことはなかったことに注目されるべきである。
【0026】
本明細書において、被験者は、悪性胸膜中皮腫に罹患したヒト患者、あるいは、該疾患の疑いのあるヒトを含む。
【0027】
本明細書において、生物学的サンプルは、被験者から生体外に採取された、必要により前処理された、例えば血清、胸水、気管支洗浄液または患部組織(特に、胸膜組織)などが挙げられる。
【0028】
本発明の上記3つの遺伝子の塩基配列は、例えばGenBank(米国NCBI)から下記の登録番号にアクセスすることによって入手可能である。
【0029】
ヒトTMEM30B遺伝子については、NW_925561、NT_025437など。
ヒトKAZALD1遺伝子については、NW_924884、NT_030059など。
ヒトMAPK13遺伝子については、NW_923073、NT_007592など。
【0030】
本発明の方法では、上記遺伝子のうちの少なくとも1つの遺伝子のメチル化を検出する。
【0031】
遺伝子上の好ましいメチル化部位は、上記TMEM30B遺伝子、KAZALD1遺伝子またはMAPK13遺伝子のプロモーターのCpG領域であり、この領域の塩基配列のメチル化が検出または測定される。これら遺伝子のプロモーターのCpG領域は、例えば、(1) 配列番号78、79または80に示される塩基配列を含むか、あるいは、(2) その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含む塩基配列、あるいは、その塩基配列と少なくとも95%、好ましくは少なくとも97%、より好ましくは少なくとも98%の同一性を有する塩基配列であって、かつ、プロモーター活性を有する塩基配列を含む。
【0032】
本明細書で使用する「プロモーターのCpG領域」は、CG配列に富む領域であり、例えば約20〜35%またはそれ以上のC+Gの塩基を含む領域である。また、「プロモーター活性」は、遺伝子の転写を制御する活性をいう。
【0033】
本明細書で使用する「数個」は、2〜10の範囲の整数をいう。
本明細書で使用する「同一性」は、2つの塩基配列を、一致度が最大になるように、ギャップを導入するかギャップを導入しないで、整列させたときに、ギャップの数を含めた総塩基数に対する同一(すなわち、一致する)塩基数のパーセンテージ(%)を意味する。
【0034】
本発明の方法におけるメチル化は、例えばメチル化特異的PCR法(J.G. Herman et al., Proc. Natl. Acad. Sci. USA 1996, 93:9821-9826; C.A. Eads et al., Nucleic Acids Res. 2000, 28:E32; K. Rand et al., Methods 2002, 27:114-120; D.T. Akey et al., Genomics 2002, 80:376-384)によって検出することができる。また、メチル化のレベルは、例えばpyrosequencing法(S. Colella et al., Biotechniques 2003, 35:146-150; J. Tost et al., Biotechniques 2003, 35:152-156)によって測定することができる。
【0035】
メチル化特異的PCR法(Methylation Specific PCR; MSP)は、重亜硫酸ナトリウム(sodium bisulfite)で処理したDNAを鋳型にし、PCRを行うことによってメチル化の有無を調べる方法である。重亜硫酸ナトリウムは、DNA上のシトシンをウラシルに変換するが、メチル化シトシン(CpGのシトシン残基の5'位のメチル化)ではこの変換反応が起こらない。
【0036】
PCR用プライマーとして、メチル化されている場合に増幅されるメチル化特異的プライマーセットと、メチル化されていない場合に増幅される非メチル化特異的プライマーセットの2種類が使用される。具体的には、これらのプライマーは、上記のとおり、転写を制御するプロモーターCpG領域の遺伝子配列を基準にして選択し、例えばDNA自動合成機を使用することによって作製することができる。
【0037】
上記PCR法で使用するためのプライマーについては、例えば、配列番号78、79または80に示される塩基配列、あるいは、その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含む塩基配列中の、あるいはその塩基配列と少なくとも95%、好ましくは少なくとも97%、より好ましくは少なくとも98%の同一性を有する塩基配列であって、かつ、プロモーター活性を有する塩基配列中の、連続する17〜35塩基、好ましくは20〜30塩基からなる配列を、メチル化特異的プライマーおよび/または非メチル化特異的プライマーとして選択することができる。
【0038】
PCRによる増幅後に、この2種類のプライマーセットからの増幅産物の比を比較することによってメチル化の有無を決定する。すなわち、メチル化特異的プライマーによる増幅産物が、試験検体で検出された時、問題の遺伝子がメチル化されていると判定する。このプライマーは、元の配列に近い配列を増幅し検出する際に有効である。一方、非メチル化特異的プライマーは、重亜硫酸ナトリウムの処理で変化した配列を検出する際に有効である。さらにまた、試験検体の重亜硫酸ナトリウムで処理後のDNAの質的評価としてメチル化の有無に関わらずPCR増幅が行われる例えばLINE1プライマー(例えば、LINE1−Fwd:TTTTAAAGTTGTTAGATAGGGATATTTAAGTTTGTA(配列番号91); LINE1−Rev: AAAAACCTACCTACCTCTATAAACTCCACC(配列番号92))などのプライマーを用いるときには、(メチル化特異的プライマーによる増幅産物)対(LINE1プライマーによる増幅産物)の比によりメチル化DNAの量的診断も可能となる。
【0039】
さらにまた、上記のPCRに加えて、定量的PCRを実施することも可能であり、正常のDNAが非特異的に増幅される場合に、それによる影響を排除するために利用しうる。定量的PCR用のプライマーの例は、TMEM30B遺伝子について、配列番号83(メチル化プライマー)、配列番号84(非メチル化プライマー)、KAZALD1遺伝子について、配列番号87(メチル化プライマー)、配列番号88(非メチル化プライマー)である。
【0040】
本発明で使用可能なプライマーセットは、ヒトTMEM30B遺伝子(NW_925561、NT_025437など)、ヒトKAZALD1遺伝子(NW_924884、NT_030059など)、またはヒトMAPK13遺伝子(NW_923073、NT_007592など)の塩基配列、好ましくはこれらの各遺伝子のプロモーターのCpG領域の塩基配列、さらに好ましくは配列番号78、79または80を含む塩基配列、において、その中の少なくとも1つのメチル化CpGを含む約17〜約35塩基、好ましくは約20〜30塩基の塩基配列からなる、任意に選択されたプライマーを含む。
【0041】
上記プライマーセットの非限定的な例は、以下のとおりである。
TMEM30B遺伝子のメチル化特異的プライマー:
TTGGGAGGGTGTGGGTATTGTTC(配列番号66)
ACGCGCGAACTAACCGAAT(配列番号67)
CTGGGAGGGTGTGGGCACTGCTCG(配列番号81)
CGAGTAGTTTTCGGGTTTGC(配列番号95)
ACCGACGCGCGAACTAAC(配列番号96)
TMEM30B遺伝子の非メチル化特異的プライマー
GGAATTGGGAGGGTGTGGGTATTGTTT(配列番号68)
ACCTCCCTCTCCACACCAC(配列番号69)
GGAACTGGGAGGGTGTGGGCACTGCT(配列番号82)
TGAGTAGTTTTTGGGTTTGT(配列番号97)
ACCATAACACTCCAAATCATAACA(配列番号98)
KAZALD1遺伝子のメチル化特異的プライマー
GCGGGTCGTAGTATTTGTCG(配列番号70)
AACAAAACCAACCACCTTCG(配列番号71)
CGGGTCGTAGTATTTGTC(配列番号85)
GGTAAGTTTGCGGTCGTTTTC(配列番号99)
AACAAAACCAACCACCTTCG(配列番号100)
KAZALD1遺伝子の非メチル化特異的プライマー
TTATTTGTGGGTTGTAGTATTTGTT(配列番号72)
AAACCAACCACCTTCATCTCC(配列番号73)
TTATTCGCGGGTCGTAGTATTTGTC(配列番号86)
GTGGGTAAGTTTGTGGTTG(配列番号101)
AACAAAACCAACCACCTTCATCT(配列番号102)
MAPK13遺伝子のメチル化特異的プライマー
CGTGTTTTCGACGTACGTCGGTAGC(配列番号74)
ACCGCTAAATAACCCCGAAA(配列番号75)
CGTGTCCCCGACGCACGTCGGCAGC(配列番号89)
MAPK13遺伝子の非メチル化特異的プライマー
GGTTTTGTTGGGTAGTTTT(配列番号76)
TCCCACCTACCAACTCACC(配列番号77)
GGTCCCGCTGGGCAGCCC(配列番号90)
【0042】
一方、pyrosequencing法は、遺伝子上の目的の領域をPCR増幅したのち、この領域内の特定のCpG部位の重亜硫酸塩変換配列を決定する手法である。この方法では、個々の部位のC対Tの比を、配列伸長の際のCおよびTの取り込み量に基づいて定量的に決定する。具体的には、この方法は、キアゲン社パイロシークエンサーによって行うことができる。
【0043】
PCR条件は、一般的手法を使用することができる。すなわち、PCRでは、変性、アニーリング及び伸長反応を1サイクルとして約20〜40サイクルが実施される。変性は、二本鎖の鋳型DNAを例えば94〜96℃、約5秒〜約10分加熱して一本鎖に分離させる工程である。アニーリングは、一本鎖の鋳型DNAのそれぞれの側面に、正方向プライマーまたは逆方向プライマーをアニールさせる工程であり、約50〜65℃、約5秒〜60秒の加熱によって行われる。伸長反応は、鋳型DNAに沿ってプライマーを伸長してDNAを合成する工程であり、通常、72〜77℃、約30秒〜10分の加熱によって行われる。PCR用溶媒は、Mg2+含有緩衝液であり、これに耐熱性DNAポリメラーゼ酵素(例えばTaqポリメラーゼなど)およびdNTPs(N=A,T,G,C)を添加する。PCRサイクルの初めに、例えば94〜96℃、約30秒〜10分の加熱を行ってもよいし、また、最終サイクルの終了時に72〜77℃、約2〜10分の加熱を行ってもよい。好適なPCR条件はプライマーにより異なり、例えばTMEM30Bメチル化プライマーのアニーリング温度は55℃、非メチル化プライマーは65℃である。PCRは、サーマルサイクラーのような増幅装置を使用すると便利である。
【0044】
PCR産物は、臭化エチジウム染色された、6%ポリアクリルアミドゲルまたは3%アガロースゲル上で視覚化することができる。
【0045】
本発明の方法によれば、上記遺伝子の少なくとも1つの遺伝子がメチル化されているとき、悪性胸膜中皮腫であると判定する。これらの遺伝子は、肺腺癌ではメチル化されていないため、これら遺伝子のメチル化を検出することによって悪性胸膜中皮腫を肺腺癌から容易に鑑別することができる。
【0046】
(悪性胸膜中皮腫に罹患した被験者の予後の測定)
本発明はさらに、悪性胸膜中皮腫に罹患した被験者の予後を判定する方法であって、該被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化の程度を測定することを含み、該遺伝子のメチル化程度が、予後不良被験者のメチル化程度と比べて低いとき、該疾患の予後が良好であると決定する、上記方法を提供する。
【0047】
本発明によれば、悪性胸膜中皮腫に罹患した被験者の予後を、上記3つの遺伝子のいずれかのメチル化の程度、すなわちメチル化のレベルを測定することによって判定することができる。これは、上記腫瘍に関する予後が不良の場合、該遺伝子が異常にメチル化されていることに基づく。逆に、予後が良好の場合、該遺伝子のメチル化のレベルが予後不良の場合と比べて相対的に低い。
【0048】
予後を判定する方法として、上記MEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つのヒト遺伝子のメチル化程度を測定するために、例えば、上記のメチル化特異的PCR法および/またはpyrosequencing法を使用できる。これらの方法およびプライマーの例については、上で説明したとおりである。
【0049】
また、生物学的サンプルについても、上で説明したように、具体的には、血清、胸水、気管支洗浄液または患部組織(特に、胸膜組織)である。
【0050】
(キット)
本発明はさらに、上記のいずれかの方法で使用するためのキットであって、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を測定することが可能である、メチル化特異的PCR用の下記のプライマーセットを含むキットを提供する。
プライマーセット:
TMEM30B遺伝子のメチル化特異的プライマー
TTGGGAGGGTGTGGGTATTGTTC(配列番号66)
ACGCGCGAACTAACCGAAT(配列番号67)
CTGGGAGGGTGTGGGCACTGCTCG(配列番号81)
CGAGTAGTTTTCGGGTTTGC(配列番号95)
ACCGACGCGCGAACTAAC(配列番号96)
TMEM30B遺伝子の非メチル化特異的プライマー
GGAATTGGGAGGGTGTGGGTATTGTTT(配列番号68)
ACCTCCCTCTCCACACCAC(配列番号69)
GGAACTGGGAGGGTGTGGGCACTGCT(配列番号82)
TGAGTAGTTTTTGGGTTTGT(配列番号97)
ACCATAACACTCCAAATCATAACA(配列番号98)
KAZALD1遺伝子のメチル化特異的プライマー
GCGGGTCGTAGTATTTGTCG(配列番号70)
AACAAAACCAACCACCTTCG(配列番号71)
CGGGTCGTAGTATTTGTC(配列番号85)
GGTAAGTTTGCGGTCGTTTTC(配列番号99)
AACAAAACCAACCACCTTCG(配列番号100)
KAZALD1遺伝子の非メチル化特異的プライマー
TTATTTGTGGGTTGTAGTATTTGTT(配列番号72)
AAACCAACCACCTTCATCTCC(配列番号73)
TTATTCGCGGGTCGTAGTATTTGTC(配列番号86)
GTGGGTAAGTTTGTGGTTG(配列番号101)
AACAAAACCAACCACCTTCATCT(配列番号102)
MAPK13遺伝子のメチル化特異的プライマー
CGTGTTTTCGACGTACGTCGGTAGC(配列番号74)
ACCGCTAAATAACCCCGAAA(配列番号75)
CGTGTCCCCGACGCACGTCGGCAGC(配列番号89)
MAPK13遺伝子の非メチル化特異的プライマー
GGTTTTGTTGGGTAGTTTT(配列番号76)
TCCCACCTACCAACTCACC(配列番号77)
GGTCCCGCTGGGCAGCCC(配列番号90)
【0051】
本発明のキットには、上記プライマーの他に、PCRの実施に必要な、例えば、緩衝液、耐熱性DNAポリメラーゼ酵素、dNTPs(N=A,T,C,G)、使用説明書などを含むことができる。プライマー類は、単独で、あるいは、組み合わせて包装することができる。
【0052】
(実施例)
本発明を以下の実施例によってされに具体的に説明するが、本発明の範囲は、これらの実施例によって制限されないものとする。
【実施例1】
【0053】
<材料及び方法>
細胞
MPM細胞 (ACC-MESO-1)(Usami N, Fukui T, Kondo M, et al. Cancer Sci 2006;97:387-94)及び非悪性の中皮細胞(MeT-5A)を解析のために使用した。MeT-5Aは、ATCC (American Type Culture Collection, Rockville, MD, USA)からATCC CRL-9444として入手可能である。ACC-MESO-1は、10%牛胎仔血清(Invitrogen, Carlsbad, CA, USA)及びantibiotic-antimycotic (Invitrogen)を添加したRPMI-1640培地(Sigma-Aldrich, St Louis, MO)中、37℃、5% CO2を含む湿気のあるインキュベーター中で培養された。MeT-5Aは、ATCCの説明書の条件にしたがって培養された。
【0054】
組織サンプル
31個のMPMサンプル及び51個のAdCaサンプル(いずれも日本人患者由来)を、愛知県がんセンター病院、名古屋大学病院、名古屋第一赤十字病院、名古屋第二赤十字病院、春日井市民病院、大垣市営病院、及び岡山労災病院から入手した。サンプル及び臨床データは、適正機関の検討部会の承認と、すべての患者からのインフォームドコンセントを書面で得た後、集められた。
【0055】
DNA調製
ゲノムDNAを、標準フェノール−クロロホルム法を用いて抽出した。ゲノムDNAをM. SssIメチラーゼ(New England Biolabs, Ipswich, MA)で処理することによって完全メチル化DNAを調製した。簡潔に説明すると、5μg のDNAを、20 UのM. SssIメチラーゼ、320μMのS-アデノシルメチオニン(SAM, New England Biolabs)及びNEBバッファー2 (New England Biolabs)を添加して計300 μlとし37℃でインキュベーションした。インキュベーションの間、同量のM. SssIメチラーゼ及びSAM をさらに1回添加し、DNAのメチル化反応を確実にした。非メチル化DNAは、製造元の指示書に従って、phi29 DNAポリメラーゼ(GenomiPhi DNA Amplification Kit; Amersham Biosciences, Uppsala, Sweden)を用いてゲノムDNAを処理することによって調製した。
【0056】
メチル化CpGアイランド増幅マイクロアレイ(MCAM)
本発明者らは、20サンプルの MPM (45〜78 歳、平均年齢59.1歳)及び20サンプルの AdCa(44〜76歳、平均年齢62.8歳)をMCAM分析にかけた。MCMの詳細なプロトコルは、これまでに記載された方法に従った(Shen L, Kondo Y, Guo Y, et al. PLoS Genet 2007;3:2023-3621; Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)。簡単に説明すると、2 μgのゲノムDNAを、100Uのメチル化感受性制限エンドヌクレアーゼSmaI (New England Biolabs)で消化した。この酵素は、非メチル化DNAを切断して平滑末端(CCC/GGG)を残す。つぎに、このDNAを、50 Uの メチル化非感受性制限エンドヌクレアーゼXmaIで消化した。この酵素は、粘着末端(C/CCGGG)を残す。この粘着末端にアダプターを結合した。アダプターは、オリゴヌクレオチド RMCA12 (5’-CCGGGCAGAAAG-3’(配列番号93))及びRMCA24(5’-CCACCGCCATCCGAGCCTTTCTGC-3’(配列番号94))(Toyota, M. et al., Cancer Res. 59:2307-2312, 1999)を65℃で2分間インキュベーションし、その後、60分間で室温まで冷却することによって作製した。消化されたDNAを、T4 DNAリガーゼ (Invitrogen, Carlsbad, CA)を用いてアダプターに結合し、RMCA24プライマーを用いるPCRによって増幅した。PCR条件は、72℃、5分の加熱により、アダプター結合DNA断片のオーバーハング末端の伸長を行い、さらに、95℃、3分の加熱を行ったのち、95℃、1分及び77℃、3分を25サイクル行い、最後に72℃、10分の伸長反応を行った。本発明者らは、6,161個のユニークな遺伝子に対応した15,134個のプローブを含むAgilent Technologies 社のカスタムプロモーターアレイ(G4497A, Agilent Technologies, Santa Clara, CA)を使用した。このアレイは、過去のMCAMを用いた研究で有効性について証明されている(Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)。マイクロアレイプロトコルは、これまで記載されたものを使用した(Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)。簡単に説明すると、MCA産物を、癌組織由来DNAに対してCy5(赤色)で標識し、 正常組織由来DNAに対してCy3(緑色)で標識した。正常対照として、MPMについては2サンプルの正常中皮組織由来のDNA、または、AdCaについては5サンプルの正常肺組織のDNAが使用された。色素で標識後、ヒトCot-1 DNA(Invitrogen)とともに、アレイに標識サンプルをハイブリダイズした。製造者のプロトコルに従ってアレイを洗浄し、ついで、Agilentスキャナーを用いアレイをスキャンし、Feature Extractionソフトウエアを用いて分析した。正規化は、製造者のプロトコル(Agilent Technologies)により、linear per-array アルゴリスムを用いて行った。
【0057】
階層型クラスタリング分析法
クラスタリング分析は、階層型クラスター化法(Eisen MB, Spellman PT, Brown PO, Botstein D., Proc Natl Acad Sci U S A 1998;95:14863-8)を用いて行った。検体のクラスタリングについては、Cluster 3.0ソフトウエア (http://rana.lbl.gov/EisenSoftware.htm)またはMinitab 15統計ソフトウエア (http://www.minitab.com)を用いて、検体間で、すべての遺伝子におけるDNAメチル化シグナル強度の測定値に基づき、類似性を計算した。デンドログラムおよびヒートマップは、TreeViewソフトウエア (http://rana.lbl.gov/EisenSoftware.htm)を用いて作成した。
【0058】
メチル化分析
本発明者らは、これまで報告された手法によって重亜硫酸塩(bisulfite)処理を行った(Kondo Y, Shen L, Suzuki S, et al. Hepatol Res 2007;37:974-83; Yang AS, Estecio MR, Doshi K, Kondo Y, Tajara EH, Issa JP. Nucleic Acids Res 2004;32:e38)。簡単に説明すると、2μgのゲノムDNAに、3M重亜硫酸ナトリウム(pH 5.0)を添加し50℃で16時間インキュベートした。Wizard Miniprep Column(Promega, Madison, WI)を用いてDNAを精製し、エタノールで沈殿させ、30μlの精製水に再溶解させた。DNAメチル化レベルは、Pyrosequencing法(Pyrosequencing AB, Uppsala, Sweden)(Colella S, Shen L, Baggerly KA, Issa JP, Krahe R., Biotechniques 2003;35:146-5027)を用いる定量的メチル化解析法によって解析された。各アッセイで、セットアップの段階で、陽性コントロール(SssI処理後のサンプル) と陰性コントロール(GenomiPhi V2を用いてゲノム全体を増幅したあとのサンプル)を混合した実験を行い、PCRバイアスが生じないアニーリングの至適温度を決定した。また実験を反復し、再現性を評価した。(Shen L, Guo Y, Chen X, Ahmed S, Issa JP. Biotechniques 2007;42:48-5828)。また、メチル化特異的PCR (MSP)を、 TMEM30B、KAZALD1およびMAPK13遺伝子において行った。PCR産物は、臭化エチジウム染色された、6%ポリアクリルアミドゲルまたは3%アガロースゲル上で視覚化した。pyrosequencingおよびMSPのプライマー配列およびPCR条件を、表1A〜表1Cに示した。
【0059】
【表1A】

【0060】
【表1B】
【0061】
【表1C】
【0062】
プライマーはすべて、転写開始部位から0.5kb以内のCpGのメチル化状態をアッセイ可能であるように設計した。
【0063】
細胞のトリコスタチンAおよび5Aza-dC 処理
細胞を12〜24時間培地で培養した後、細胞を、1) 5μM 5-アザ-2’-デオキシシチジン(5Aza-dC; Sigma-Aldrich)またはリン酸緩衝塩水で72時間処理する(このとき、5Aza-dCまたはリン酸緩衝塩水を含有する培地を24時間おきに交換した。)か、あるいは、2) 300nMのトリコスタチンA (TSA; MP Biomedicals, Solon, OH)または等容量のエタノールで24時間処理した。
【0064】
定量逆転写−ポリメラーゼ連鎖反応分析
全RNAをTRIzol (Invitrogen)を用いてRNAを抽出した。2μgのRNAをMPMLV (Promega)を用いて逆転写した。SYBR GREEN Quantitative逆転写-PCRを、標的遺伝子に対して実験を3回おこなった (Applied Biosystems, Foster City, CA)。Q-PCRのプライマー配列を表1Bに示した。
【0065】
統計分析
メチル化状態と臨床病理学的変化との間の関係を、StadView ソフトウエア (version 5.0; Abacus Concepts, Berkeley, CA)を用いて、Mann-Whitney U test、Fisher’s exact testまたはKruskal-Wallis testを用いて解析した。線形回帰モデルはStata version 8 (College Station, TX)を使用した。P値のすべてがtwo-sidedであり、P<0.05が統計的に有意であるとみなした。生存率曲線を、Kaplan-Meier法を用いて作成した。全生存率は、外科手術日から開始し死亡までの間で計算した。
【実施例2】
【0066】
<結果>
本発明者らは、はじめに、悪性中皮腫細胞系ACC-MESO-1のメチル化プロフィールを分析するためにMCAMを実施した。MCAによってメチル化断片を増幅したのち、MCA産物を、Cy5 (ACC-MESO-1)またはCy3 (正常中皮組織)で標識し、ともにマイクロアレイ(6,161遺伝子に対応する15,134プローブを含む)に共ハイブリダイズした(図1A)。これまで、本発明者らは、MCAMにおいて2.0以上をしめすCy5/Cy3シグナル比は、pyrosequencing分析でのメチル化陽性と一致した (Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)。ACC-MESO-1細胞でのMCAM解析の有効性を評価するために、 高、中、または低シグナル強度を示す19遺伝子をランダムに選択し、pyrosequencing法によってメチル化レベルを調べた。2.0以上のCy5/Cy3シグナル比を示す14遺伝子のうち11遺伝子がpyrosequencing分析でメチル化陽性(ACC-MESO-1で15%以上のメチル化レベル、および、正常中皮組織のメチル化レベルの少なくとも1.5倍以上のメチル化レベル)であったが、2.0以下の Cy5/Cy3シグナル比をもつ5遺伝子のうち4遺伝子がメチル化陰性(MPM細胞で15%未満のメチル化レベル、および/または正常中皮組織の1.5分の1以下のメチル化レベル)であった。MCAM分析とpyrosequencing分析との間の高い相関関係(特異性80%; 感度86%)は、これまでの研究のなかで示されているのと同様に(Shen L, Kondo Y, Guo Y, et al., PLoS Genet 2007;3:2023-36.: Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)、この細胞で確認された(図1Bおよび表2)。この規準を用いてACC-MESO-1でメチル化された971遺伝子を同定した。
【0067】
【表2】
【0068】
つぎに、MPMでのこれらの遺伝子のDNAメチル化と遺伝子発現との関係を調べるために、ACC-MESO-1をDNAメチルトランスフェラーゼ阻害剤、5-アザ-2’-デオキシシチジン(5-Aza-dC)もしくはヒストンデアセチラーゼ阻害剤、トリコスタチンA (TSA)で処理し、3つの同定された遺伝子C9orf106、EDG4およびMPMPL1 の発現を検討した(図1C および図1D)。ACC-MESO-1ではこれらの遺伝子発現は抑制されていたのに対して、正常中皮組織では高い発現レベルであった。本発明者らは、5-Aza-dCによる有意な遺伝子再発現を観察したがTSA処理では有効性が乏しかった。このことは、これらの遺伝子の発現抑制がDNAメチル化によることを示している。
【0069】
正常中皮DNA(2症例の混合)および正常肺組織DNA(4症例の混合)からのMCA産物の共ハイブリダイゼーションにおいて、本発明者らは、メチル化状態は高い一致(R2=0.87, 図2A)を示すことを確認した。このことは、組織特異的メチル化は、これら2つの正常組織ではまれであることを示している。つぎに、MPMおよびAdCaのメチル化プロフィールの違いを評価するために、MPM DNA (4症例の混合物)およびAdCa DNA (4症例の混合物)からのMCA産物を共ハイブリダイズし、遺伝子の一部はそれぞれの腫瘍で異なるメチル化プロファイルを示すことを見出した(R2=0.77, 図2B)。興味深いことに、本発明者らは、肝臓癌(4症例の混合)とMPMとのメチル化プロファイルを検討したところ、肺と肝臓はともに内胚葉から発生するのにもかかわらず、MPMとAdCaとのメチル化標的遺伝子の差に比べて、MPMと肝臓癌とではより多くの異なった遺伝子がメチル化していること (R2=0.26)を見出した(図2C)。
【0070】
MPMおよびAdCaの全ゲノム領域におけるDNAメチル化プロフィールを比較するために、本発明者らは、MCAMを使用して20症例のMPMと20症例のAdCaを分析した。6,161遺伝子を用いてのMPMおよびAdCaのすべてのヒートマップから、AdCaがMPMよりも高頻度でメチル化されているように見られること、ならびに、AdCaの一部の症例群が広範にメチル化されていることが示された(図3A)。MPMと AdCaは、DNAメチル化プロファイルによって異なるサブグループに分類された。MPMのなかには主にAdCaからなるサブグループに入る2つの例外症例が存在した。
【0071】
つぎに、DNAメチル化状態がMPMおよびAdCaにおいて臨床病理学的な特性に影響を与えるかどうかを調べるために、6,161遺伝子を用いる階層型クラスタリングによって個々にMPMおよびAdCaを分析した。DNAメチル化遺伝子数によって、MPMを2つのサブグループに分けた。すなわち、その1つのサブグループは、メチル化遺伝子数が300遺伝子以下と少ない4症例であり、そのすべての症例が上皮型であった。これらのデータは、DNAメチル化が腫瘍形成の際にMPMの表現型に影響することを示している。階層型クラスタリング分析によって、AdCaは、4つのサブグループに分けることができた。1つのサブグループは、6症例のAdCaからなり、それらは、他のAdCa症例と比べてメチル化遺伝子数を多く有する症例であった (911遺伝子vs 387遺伝子, P=0.001)。また、そのサブグループの症例のほとんどが喫煙者であった(喫煙者5/6症例;喫煙した平均パック年数, 68.6± 22.9)。
【0072】
本発明者らは、MPMおよびAdCaにおいてメチル化している遺伝子を数えた。MPMでメチル化されている遺伝子は700未満であり、これはマイクロアレイ上に存在する6,161 CpGの約11%を占める。また、MPM症例のメチル化遺伝子数の平均が387(範囲, 120 〜755; 図3B)であり、一方、AdCaでは544遺伝子 (範囲, 133〜1212)であった。
【0073】
AdCaとMPMのそれぞれで共通してメチル化されている遺伝子を比較した。10症例の MPMで共通してメチル化されている遺伝子は非常にまれ (40未満の遺伝子)であるが、10症例の AdCaでは、80以上の遺伝子が共通してメチル化されていた。このことは各MPM症例では共通してメチル化する遺伝子は少ない(P=0.001, 図3C)ことを示している。各腫瘍のメチル化遺伝子のリストを表3および表4に示した。
【0074】
【表3】
【0075】
【表4】
【0076】
MPMおよびAdCaにおけるメチル化遺伝子数と臨床病理学的パラメーターとの関係を表5に示した。
【0077】
【表5】
【0078】
AdCaでは、喫煙者は非喫煙者と比べて有意にメチル化遺伝子が多かった (728遺伝子vs 360遺伝子, P=0.02)。MPMでの喫煙者もまた、非喫煙者と比べてメチル化遺伝子が多い傾向であり(366遺伝子vs 261遺伝子, P=0.2)、このことは喫煙がMPMのDNAメチル化に影響していることを示している。これに対し、アスベスト曝露は、MPMではDNAメチル化に対する影響は少なかった(MPMでのアスベスト曝露あり対アスベスト曝露なしは、386遺伝子vs 320遺伝子であった。)。
【0079】
進行性大腸直腸癌患者でDNAメチル化が高頻度に存在する症例では予後不良であることが報告されている( Shen L, Catalano PJ, Benson AB, 3rd, O'Dwyer P, Hamilton SR, Issa JPClin Cancer Res 2007;13:6093-829)。本発明者らは、MPM患者を、2つの群、すなわち、メチル化遺伝子数が平均(387遺伝子)より多い群と少ない群に分けて、生存率とメチル化との関係を検討した。外科的合併症が原因で死亡した患者、または、手術合併症のため死亡した2人の患者は除かれた。平均メチル化遺伝子数よりメチル化遺伝子数の少ない群の患者(n=11)は、平均よりメチル化遺伝子数の多い群の患者(n=7)と比べて有意に生存率が長かった (平均生存時間, 22ヶ月vs 8ヶ月, P=0.01)(図3D)。MPM患者全集団の全体の平均生存期間は11.7ヶ月であった。
【0080】
DNAメチル化は、癌診断のための強力なマーカーとなりうるとされている(Belinsky SA, Nat Rev Cancer 2004;4:707-1730)。MPM診断のための特異的なDNAメチル化マーカーを同定するために、はじめに、本発明者らは、MCAM分析から8遺伝子を選択した。この分析では、少なくとも4症例のMPMでメチル化陽性(Cy5/Cy3>2.0)を示し、AdCaの全症例でメチル化陰性を示すものを選んだ(図4A)。興味深いことに、少なくとも4症例のAdCaでメチル化陽性、ならびに、MPMの全症例でメチル化陰性という同様の規準を用いて遺伝子を選ぶと、AdCaでより多くの遺伝子(34遺伝子)が存在し、このことは、メチル化した遺伝子がAdCaでより多く存在することを示している(図4A)。本発明者らは、MSPによって遺伝子のメチル化を確認し、3つの遺伝子、すなわち、膜貫通蛋白質30B (TMEM30B)、Kazal型セリンプロテアーゼ阻害剤ドメイン1 (KAZALD1)及び有糸分裂活性化プロテインキナーゼ13 (MAPK13)がMPMに特異的なメチル化マーカーであることを見出した(図4Bおよび4C)。MCAM解析で解析されたMPM症例では、DNAメチル化が、MSPによりDNAメチル化が、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子のそれぞれにおいて19症例のMPMのうち11 (58%)症例、8 (42%)症例および2 (11%)症例で検出された。しかしながら、AdCaでは、TMEM30B遺伝子でわずか1つの不明瞭なメチル化バンドが検出されたにすぎなかった。
【0081】
これらのメチル化マーカーがMPM患者の別の集団で鑑別可能であるかを確認するために、本発明者らは、別の機関からさらに10症例のMPMを入手し分析した(図4D)。DNAメチル化は、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子のそれぞれで、10症例のMPMのうち5 (50%)、7 (70%)および2 (20%)症例で検出された。これら3つの遺伝子のDNAメチル化を、合計31症例のMPMと51症例のAdCa(19症例のMPMサンプルおよび19症例のAdCaサンプルは、MCAM解析で用いた集団であった)で分析した。DNAメチル化は、MPMでは、TMEM30Bについて17症例(55%)で、KAZALD1について16症例(52%)で、MAPK13について4症例(13%)で検出されたが、AdCaでは、DNAメチル化は検出されなかった (図4Cおよび4D)。AdCaとMPMの鑑別診断として、3つの遺伝子のうち少なくとも1遺伝子がメチル化している場合にMPMと診断すると、その感度と特異性はそれぞれ77%および100%であった。
【0082】
さらにまた、上記3種の遺伝子の各配列のなかで、特にそれら遺伝子のプロモーターのCpGアイランド領域の配列(配列番号78、79又は80)のメチル化又はメチル化程度を検出又は測定することによって、MPMをより特異的に検出できることが分かった。また、そのための分析に使用されるPCR用プライマーとして有効なものは、TMEM30B遺伝子のメチル化特異的プライマーの場合、例えば配列番号66、配列番号67、配列番号81、配列番号95、及び/又は配列番号96のプライマーであり、TMEM30B遺伝子の非メチル化特異的プライマーの場合、例えば配列番号68、配列番号69、配列番号82、配列番号97、及び/又は配列番号98のプライマーであり、KAZALD1遺伝子のメチル化特異的プライマーの場合、例えば配列番号70、配列番号71、配列番号85、配列番号99、及び/又は配列番号100のプライマーであり、KAZALD1遺伝子の非メチル化特異的プライマーの場合、例えば配列番号72、配列番号73、配列番号86、配列番号101、及び/又は配列番号102のプライマーであり、MAPK13遺伝子のメチル化特異的プライマーの場合、例えば配列番号74、配列番号75、及び/又は配列番号89のプライマーであり、MAPK13遺伝子の非メチル化特異的プライマーの場合、例えば配列番号76、配列番号77、及び/又は配列番号90のプライマーであった。
【0083】
<考察>
この研究のなかで、本発明者らは、MCAMを使用してMPMおよびAdCaの全ゲノム領域におけるメチル化プロフィールを解析、比較し、ならびに、MPMにおいて特定の遺伝子のDNAメチル化の頻度を検討した。MCAMが高い特異性と感度をもって全ゲノム領域のメチル化の状態を評価することが可能であることは以前の報告で示されている( Shen L, Kondo Y, Guo Y, et al., PLoS Genet 2007;3:2023-36; Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10); Estecio MR, Yan PS, Ibrahim AE, et al., Nat Genet 2005;37:853-62)。しかしながら、MCAMは組織サンプルなかの異常にメチル化された遺伝子をより有意に検出する可能性があるので、本発明者らは、高メチル化遺伝子に照準を合わせた。実際、MPM細胞であるACC-MESO-1を使用したときの有効性は、MCAMからのシグナル比と、bisulfite-pyrosequencingによってアッセイされたメチル化レベルとの間で良好な相関性を示し、従来の研究結果と一致している( Shen L, Kondo Y, Guo Y, et al., PLoS Genet 2007;3:2023-36; Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)。ACC-MESO-1でMCAM分析によって検出された標的遺伝子は、 5-Aza-dCによって再活性化され、また、MPM患者で高メチル化されていることが見出された。このことは、MPMでのこれら遺伝子のDNAメチル化により遺伝子発現抑制されていることを示している。
【0084】
正常中皮と正常肺組織は異なる胚葉(それぞれ、中胚葉、内胚葉)から発生するにもかかわらず、それらのDNAメチル化プロフィールは非常に類似しており(R2=0.87)、このことは、これら2つの正常組織での組織特異的メチル化の差は少ないことを示している。これまでの種々の正常組織のゲノムワイドなメチル化分析では、組織特異的なメチル化が非常にまれであることを示しており、本発明者らの知見と一致する(Shen L, Kondo Y, Guo Y, et al., PLoS Genet 2007;3:2023-36; Estecio MR, Yan PS, Ibrahim AE, et al., Genome Res 2007;17:1529-36; Weber M, Davies JJ, Wittig D, et al., Nat Genet 2005;37:853-6221, 31, 32)。これに対して、MPMとAdCaとの間のDNAメチル化標的遺伝子の差は正常組織と比べてかなり多く、これは種々の組織での癌が異なるメチル化プロフィールを表すためと推測される(Esteller M, Corn PG, Baylin SB, Herman JG. Cancer Res 2001;61:3225-933)。興味深いことに、MPMのメチル化プロフィールは、肝臓癌と比べると、肺及び肝臓がともに内胚葉から発生するにも関わらず、AdCaの方により類似していた。MPMとAdCaとの間のDNAメチル化の類似性の理由は解っていない。喫煙などの腫瘍形成に寄与する特定の環境因子が、MPMとAdCaの両方のDNAメチル化異常に同一の影響を与えている可能性はある(Feinberg AP., Nature 2007;447:433-40)。
【0085】
いくつかの遺伝子座のDNAメチル化異常のこれまでの研究により、メチル化がAdCaと比較してMPMでは広範囲の遺伝子に起こらないことが示されてきた(Toyooka S, Pass HI, Shivapurkar N, et al., Cancer Res 2001;61:5727-30)。今回のゲノムワイドな(広域の)DNAメチル化解析により、DNAメチル化がAdCaと比べてMPMでは低頻度でありかつ共通性に乏しいことが示された。大多数のMPMではメチル化されていた遺伝子は700以下であった(平均メチル化遺伝子数, 387±196遺伝子)。これは、AdCa患者の一部の症例群(ほとんどが喫煙者による症例(喫煙した平均パック年数67.3± 14.2)であり、広範囲の遺伝子がメチル化されていた(>950遺伝子))と対比的であった。特定の肺腺癌は、メチル化されやすく、また、CpG island methylator phenotype (CIMP)と称される表現型になりやすいことが示唆されている (Toyota M, Ahuja N, Ohe-Toyota M, Herman JG, Baylin SB, Issa JP., Proc Natl Acad Sci U S A 1999;96:8681-6; Suzuki M, Shigematsu H, Iizasa T, et al., Cancer 2006;106:2200-7)。本発明者らの結果は、AdCaのサブセットがCIPMを示したのに対し、MPMではそのような症例群は存在しなかったことを示した。MPMで異常メチル化がおこる際に、アスベストによる継続的炎症は誘導因子であるように思われるが、AdCaでの喫煙ほどは強くない。これらのデータは、MPMとAdCaの形成の間の異常なDNAメチル化の獲得に対する異なる機序を反映しているかもしれない。
【0086】
DNAメチル化は、MPM診断の強力なマーカーとして提案されている(Toyooka S, Pass HI, Shivapurkar N, et al., Cancer Res 2001;61:5727-30; Tsou JA, Shen LY, Siegmund KD, et al., Lung Cancer 2005;47:193-204)。しかしながら、MPMとAdCaのメチル化状態のこれまでの検討により、その頻度に違いはあるが、候補マーカーの異常メチル化がある程度MPMとAdCaの両方で検出されることが示されている。さらに、MPMに特異的なメチル化マーカーがこれまで報告されたことはなかった。今回、本発明者らは、MPMとAdCaで全ゲノム領域のDNAメチル化プロフィールを解析し、特定の遺伝子座のDNAメチル化がMPMで頻繁に検出され、3つの遺伝子、すなわちTMEM30B、KAZALD1およびMAPK13遺伝子がMPMで特異的にメチル化されていたことを示した。特に、KAZALD1およびMAPK13遺伝子のDNAメチル化はヒト癌でこれまで報告されたことはなかったが、それらの異常なメチル化がMPMをAdCaから鑑別する有益なマーカーとして役立つはずである。
【0087】
まとめると、MPMとAdCaを全ゲノム領域のメチル化分析によって比較すると、これら2つの悪性腫瘍ではそれぞれ特徴的なDNAメチル化パターンを示し、そのパターンは、病態生理学的過程の違いから生じているようである。本発明者らは、今回、MPMの診断のための可能なマーカーを提案する。さらに、病理学的知見と関連するメチル化プロフィールによって分類されるMPMのサブセットを見出した。また、DNAメチル化が低頻度のMPM患者の予後が良好であることを見出した。これらの事実は、DNAメチル化異常がMPMの臨床上の特徴に影響を与えることを示している。したがって、プロモーターCpGアイランドのDNAメチル化およびそれと関連する遺伝子発現抑制がMPM発症に重要な役割を果たしている。
【産業上の利用可能性】
【0088】
本発明は、悪性胸膜中皮腫を肺癌から区別可能とする悪性胸膜中皮腫検出方法を提供するため、特にアスベストを原因とするような悪性胸膜中皮腫の確定診断のために医療上有用である。
【技術分野】
【0001】
本発明は、DNAメチル化異常に基づいた悪性胸膜中皮腫の検出のための方法およびキットに関する。
【背景技術】
【0002】
悪性胸膜中皮腫(Malignant Pleural Mesothelioma (MPM))は、アスベスト曝露と関連のある侵襲性腫瘍である(非特許文献1, 2)。世界で毎年、約10,000〜15,000名の患者が、新たにMPMと診断され、今後20年以内に患者数は増加すると予測されている(非特許文献3〜5)。アスベストの吸入は、周知の危険因子であり、MPMの早期診断が望まれているが、早期のステージで臨床症状がないことと有用な診断マーカーがないことが、早期診断を困難にしている。さらにまた、MPMは、時に病理学的に肺腺癌(AdCa)との区別が難しく、抗体パネルによる追加の免疫組織化学的染色を必要とする場合がある(非特許文献6)。これらの困難性と有効な治療がいまだないために、治療にも拘らず、多くの患者が、診断から2年以内に死亡している(非特許文献4, 7, 8)。したがって、精確な早期診断に有効なマーカーを同定するために、MPMのさらなる分子分析が緊急に必要とされている。
【0003】
MPMが発症する中心的な機序は依然としてはっきりしていない。いくつかの遺伝子異常がMPMに関与していると考えられており、そのような異常には、例えばp16 遺伝子座の欠損やNF2 遺伝子の変異が挙げられる(非特許文献9〜11)。最近の全トランスクリプトーム配列決定法により、転写された約6個の遺伝子が個々のMPMで突然変異を受けていると推定されたが、このことは限られた数個の遺伝子変異がMPM発症に関与していることを示唆している(非特許文献12)。以上の報告から遺伝子変異はMPMにおいて低頻度であり、遺伝子変異以外の機序がMPMの腫瘍形成に関与すると考えられる。
【0004】
エピジェネティックな転写制御、特にプロモーターDNAメチル化の調節異常は、ヒト悪性腫瘍に高頻度に観察される(非特許文献13)。さらにDNAの異常メチル化と炎症との関係がこれまで多くの癌で記載されている(非特許文献14)。アスベストは、ヒト中皮培養細胞の悪性転換を直接誘導しない(非特許文献15, 16)。またアスベスト曝露により慢性炎症が引き起こされることがわかっている(非特許文献17)。すなわちMPM発症にDNAメチル化が関与している可能性が強く疑われる。しかし、これまでにMPMにおけるDNAメチル化異常についての情報は限られているのが現状である(非特許文献18〜20)。
【0005】
ちなみに、中皮腫の診断または予後予測に関係する文献には、特定のポリペプチドに対する抗体、特定の遺伝子の発現量の差を利用する、特許文献1、2および3などが知られている。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2007-071849
【特許文献2】特表2008-505862
【特許文献3】特表2003-524021
【非特許文献】
【0007】
【非特許文献1】Robinson BW, Musk AW, Lake RA. Malignant mesothelioma. Lancet 2005;366:397-408
【非特許文献2】Pass HI, Vogelzang N, Hahn S, Carbone M. Malignant pleural mesothelioma. Curr Probl Cancer 2004;28:93-174
【非特許文献3】Tsiouris A, Walesby RK. Malignant pleural mesothelioma: current concepts in treatment. Nat Clin Pract Oncol 2007;4:344-52
【非特許文献4】Robinson BW, Lake RA. Advances in malignant mesothelioma. N Engl J Med 2005;353:1591-603
【非特許文献5】Bianchi C, Bianchi T. Malignant mesothelioma: global incidence and relationship with asbestos. IndHealth 2007;45:379-87
【非特許文献6】Abutaily AS, Addis BJ, Roche WR. Immunohistochemistry in the distinction between malignant mesothelioma and pulmonary adenocarcinoma: a critical evaluation of new antibodies. J Clin Pathol 2002;55:662-8
【非特許文献7】Vogelzang NJ, Rusthoven JJ, Symanowski J, et al. Phase III study of pemetrexed in combination with cisplatin versus cisplatin alone in patients with malignant pleural mesothelioma. J Clin Oncol 2003;21:2636-44
【非特許文献8】van Meerbeeck JP, Gaafar R, Manegold C, et al. Randomized phase III study of cisplatin with or without raltitrexed in patients with malignant pleural mesothelioma: an intergroup study of the European Organisation for Research and Treatment of Cancer Lung Cancer Group and the National Cancer Institute of Canada. J Clin Oncol 2005;23:6881-9
【非特許文献9】Cheng JQ, Jhanwar SC, Klein WM, et al. p16 alterations and deletion mapping of 9p21-p22 in malignant mesothelioma. Cancer Res 1994;54:5547-51.10. Sekido Y, Pass HI, Bader S, et al. Neurofibromatosis type 2 (NF2) gene is somatically mutated in mesothelioma but not in lung cancer. Cancer Res 1995;55:1227-31
【非特許文献10】Sekido Y, Pass HI, Bader S, et al. Neurofibromatosis type 2 (NF2) gene is somatically mutated in mesothelioma but not in lung cancer. Cancer Res 1995;55:1227-31
【非特許文献11】Bianchi AB, Mitsunaga SI, Cheng JQ, et al. High frequency of inactivating mutations in the neurofibromatosis type 2 gene (NF2) in primary malignant mesotheliomas. Proc Natl Acad Sci U S A 1995;92:10854-8
【非特許文献12】Sugarbaker DJ, Richards WG, Gordon GJ, et al. Transcriptome sequencing of malignant pleural mesothelioma tumors. Proc Natl Acad Sci U S A 2008;105:3521-6
【非特許文献13】Jones PA, Baylin SB. The fundamental role of epigenetic events in cancer. Nat Rev Genet 2002;3:415-28
【非特許文献14】Perwez Hussain S, Harris CC. Inflammation and cancer: an ancient link with novel potentials. Int J Cancer 2007;121:2373-80
【非特許文献15】Xu L, Flynn BJ, Ungar S, et al. Asbestos induction of extended lifespan in normal human mesothelial cells: interindividual susceptibility and SV40 T antigen. Carcinogenesis 1999;20:773-83
【非特許文献16】Bocchetta M, Di Resta I, Powers A, et al. Human mesothelial cells are unusually susceptible to simian virus 40-mediated transformation and asbestos cocarcinogenicity. Proc Natl Acad Sci U S A 2000;97:10214-9
【非特許文献17】Yang H, Bocchetta M, Kroczynska B, et al. TNF-alpha inhibits asbestos-induced cytotoxicity via a NF-kappaB-dependent pathway, a possible mechanism for asbestos-induced oncogenesis. Proc Natl Acad Sci U S A 2006;103:10397-402
【非特許文献18】Toyooka S, Pass HI, Shivapurkar N, et al. Aberrant methylation and simian virus 40 tag sequences in malignant mesothelioma. Cancer Res 2001;61:5727-30
【非特許文献19】Tsou JA, Galler JS, Wali A, et al. DNA methylation profile of 28 potential marker loci in malignant mesothelioma. Lung Cancer 2007;58:220-30
【非特許文献20】Tsou JA, Shen LY, Siegmund KD, et al. Distinct DNA methylation profiles in malignant mesothelioma, lung adenocarcinoma, and non-tumor lung. Lung Cancer 2005;47:193-204
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の目的は、悪性胸膜中皮腫の検出のための方法を提供することである。
【課題を解決するための手段】
【0009】
MPMでのDNAメチル化についてより多くのことを理解するために、本発明者らは、メチル化CpGアイランド増幅マイクロアレイ(MCAM) ( Shen L, Kondo Y, Guo Y, et al. PLoS Genet 2007;3:2023-36)を用いてMPM中の異常にメチル化された遺伝子について広域に及ぶスクリーニングを行った。MCAMは、癌組織だけでなく正常組織においてもメチル化遺伝子を成功裏に検出可能にし、また、高有効率で再現性のある結果を提供する。さらに、本発明者らは、2種類の胸部癌におけるDNAメチル化異常の役割を比較するために、AdCaのメチル化を調べた。DNAのメチル化プロフィールの分析により、異常なDNAメチル化がAdCaと比べてMPMで頻度が少ないこと、ならびにAdCaは症例間で共通の遺伝子が高頻度にメチル化するが、MPMで症例間での共通メチル化遺伝子は少ないことが明示された。以上の結果を詳細に検討しMCAMで見出された高メチル化遺伝子のうち、MPMで特異的にメチル化されるがAdCaでメチル化されない、3つの遺伝子を同定した。これらの遺伝子は、MPMの早期診断マーカーとして役立つはずである。
【0010】
したがって、本発明は、要約すると、以下の特徴を含む。
本発明は、第1の態様において、悪性胸膜中皮腫を検出するための方法であって、被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を検出することを含み、該遺伝子がメチル化されているとき悪性胸膜中皮腫であると判定する、上記方法を提供する。
【0011】
本発明はまた、第2の態様において、悪性胸膜中皮腫に罹患した被験者の予後を判定する方法であって、該被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化の程度を測定することを含み、該遺伝子のメチル化程度が、予後不良被験者のメチル化程度と比べて低いとき、該疾患の予後が良好であると決定する、上記方法を提供する。
【0012】
上記2つの各発明の実施形態において、上記の方法が、上記TMEM30B遺伝子、KAZALD1遺伝子またはMAPK13遺伝子のプロモーターのCpG領域の塩基配列のメチル化を検出または測定する。
【0013】
別の実施形態において、上記プロモーターのCpG領域の塩基配列が、配列番号78、79または80に示される塩基配列、あるいは、その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含み、かつ、プロモーター活性を有する塩基配列からなる。
【0014】
別の実施形態において、上記メチル化が、メチル化特異的PCR法によって検出される。
別の実施形態において、上記PCR法で使用するためのプライマーを、配列番号78、79または80に示される塩基配列中の、あるいは、その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含み、かつ、プロモーター活性を有する塩基配列中の、連続する17〜35塩基からなる配列を、メチル化特異的プライマーおよび/または非メチル化特異的プライマーとして選択する。
【0015】
また、別の実施形態において、前記サンプルが血清、胸水、気管支洗浄液または患部組織である。
【0016】
本発明はさらに、第3の態様において、上記のいずれかの方法で使用するためのキットであって、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を測定することが可能である、メチル化特異的PCR用の下記のプライマーセットを含むキットを提供する。
TMEM30B遺伝子のメチル化特異的プライマー
TTGGGAGGGTGTGGGTATTGTTC(配列番号66)
ACGCGCGAACTAACCGAAT(配列番号67)
CTGGGAGGGTGTGGGCACTGCTCG(配列番号81)
CGAGTAGTTTTCGGGTTTGC(配列番号95)
ACCGACGCGCGAACTAAC(配列番号96)
TMEM30B遺伝子の非メチル化特異的プライマー
GGAATTGGGAGGGTGTGGGTATTGTTT(配列番号68)
ACCTCCCTCTCCACACCAC(配列番号69)
GGAACTGGGAGGGTGTGGGCACTGCT(配列番号82)
TGAGTAGTTTTTGGGTTTGT(配列番号97)
ACCATAACACTCCAAATCATAACA(配列番号98)
KAZALD1遺伝子のメチル化特異的プライマー
GCGGGTCGTAGTATTTGTCG(配列番号70)
AACAAAACCAACCACCTTCG(配列番号71)
CGGGTCGTAGTATTTGTC(配列番号85)
GGTAAGTTTGCGGTCGTTTTC(配列番号99)
AACAAAACCAACCACCTTCG(配列番号100)
KAZALD1遺伝子の非メチル化特異的プライマー
TTATTTGTGGGTTGTAGTATTTGTT(配列番号72)
AAACCAACCACCTTCATCTCC(配列番号73)
TTATTCGCGGGTCGTAGTATTTGTC(配列番号86)
GTGGGTAAGTTTGTGGTTG(配列番号101)
AACAAAACCAACCACCTTCATCT(配列番号102)
MAPK13遺伝子のメチル化特異的プライマー
CGTGTTTTCGACGTACGTCGGTAGC(配列番号74)
ACCGCTAAATAACCCCGAAA(配列番号75)
CGTGTCCCCGACGCACGTCGGCAGC(配列番号89)
MAPK13遺伝子の非メチル化特異的プライマー
GGTTTTGTTGGGTAGTTTT(配列番号76)
TCCCACCTACCAACTCACC(配列番号77)
GGTCCCGCTGGGCAGCCC(配列番号90)
【発明の効果】
【0017】
本発明により、悪性胸膜中皮腫を肺癌から区別可能とする悪性胸膜中皮腫検出方法が提供される。この方法は、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を測定することに基づいている。
【図面の簡単な説明】
【0018】
【図1A】この図は、悪性中皮腫細胞系におけるMCAM分析とその有効性を示す。図1AはMCAMの概説である。癌又は正常組織由来のDNAをSmaI及びXmaIで連続的に消化した。メチル化DNA断片を増幅したのち、MCA産物を、癌組織に対してはCy5(赤色)で、正常組織に対してはCy3(緑色)で それぞれ標識し、その後、プロモーターマイクロアレイに使用した。
【図1B】この図は、悪性中皮腫細胞系におけるMCAM分析とその有効性を示す。図1Bは、MCAMシグナル比とメチル化レベルの比較を示す。Y軸は、各遺伝子におけるMCAMのシグナル比を示す。pyrosequencing分析によって決定されたメチル化レベルを絶対的変数として表した。 水平線は、MCAMシグナル比2.0を示す。
【図1C】この図は、悪性中皮腫細胞系におけるMCAM分析とその有効性を示す。図1Cは、3種の高メチル化遺伝子の代表的パイログラムを示す。灰色で示した領域は、各遺伝子で定量化されたCpG部位を示し、各部位の%メチル化は、ピークの上に示されている。CpG部位の平均メチル化の計算値をカッコのなかに示す。
【図1D】この図は、悪性中皮腫細胞系におけるMCAM分析とその有効性を示す。図1Dは、DNAメチル化又はヒストン脱アセチル化の阻害による高メチル化遺伝子の再活性化を示す。PBS (コントロール, C)又は5-Aza-dC (D)、或いはトリコスタチン(Trichostatin) A (T)で処理したあとの、正常中皮組織(Normal)及びACC-MESO-1 (Meso1)における遺伝子発現を定量的-PCRによって測定した。GAPDHに対する各遺伝子のmRNA発現の相対値をY軸(左カラム) に示す。5-Aza-dC処理によって、サイレンシングされた遺伝子が有効に再活性化された。右カラムのY軸は、コントロールに対する処理細胞のRNA発現の相対値を示す。グラフ上の誤差バーは、3回の実験からの標準偏差を示す。
【図2】この図は、正常中皮及び正常肺組織間(A)、MPM及びAdCa 間(B)、並びにMPM及び肝臓癌間(C)、のシグナル強度(logスケール)のスキャッタープロット分析を示す。線形回帰モデルの重相関決定係数(R2) を各分析で示す。
【図3A】この図は、MPM及びAdCa患者でのMCAM分析結果を示す。図3Aの上段のカラムは、全6,161遺伝子(Y軸)を用いて行った、40サンプル(20サンプルのMPMを青色ボックスで示し、20サンプルのAdCaを白色ボックスで示す。)からのDNAメチル化データの階層的クラスター分析のデンドログラムとヒートマップの概観を示す。マトリックス中の各セルは、個々のサンプル中の遺伝子のDNAメチル化状態を表す。セル中の赤色と緑色はそれぞれ、高メチル化レベル、低メチル化レベルを反映している。ここで、メチル化レベルは、マトリックスの下にスケールバーとして示されている(log2変換スケール)。MPMでメチル化された遺伝子数が300未満の症例を黒丸で示す。AdCa症例のうち、950以上の遺伝子がメチル化されていた症例を白抜き菱形ボックスで示す。中段のカラムは、階層型クラスターを用いて各腫瘍におけるサブクラス分類を示す。全6,161遺伝子を用いて20サンプルのMPMの分析を行った。Y軸は、症例間の類似性を示す。黒丸は、上の図と同じMPM症例を示す。ボックスは、MPMの組織学的サブタイプを示し、細線のハッチング、太線のハッチング、黒色及び白色ボックスはそれぞれ、上皮、二相性(biphasic)、肉腫様、及び特殊型を表す。下段のカラムは、AdCaクラスター化を示し、ここで灰色及び白色ボックスはそれぞれ喫煙者及び非喫煙者を示す。Y軸は症例間の類似性を示す。白抜き菱形ボックスは、上の図と同じAdCa症例を示す。
【図3B】この図は、MPM及びAdCa患者でのMCAM分析結果を示す。図3Bは、各症例におけるメチル化遺伝子数を示す。破線は、平均メチル化遺伝子数 (MPM及びAdCaでそれぞれ、387遺伝子及び544遺伝子)を表す。
【図3C】この図は、MPM及びAdCa患者でのMCAM分析結果を示す。図3Cは、MPM (灰色バー)又はAdCa (黒色バー)において、症例数x(X軸)で共通にメチル化された遺伝子数(Y軸) を示す。
【図3D】この図は、MPM及びAdCa患者でのMCAM分析結果を示す。図3Dは、MPM患者の生存率のKaplan-Meier分析を示す。MPM患者は、メチル化遺伝子数によって二群に分けられた。二群間のlog順位統計による生存率の統計学的比較は、メチル化が高頻度である症例(破線, 387以上の遺伝子をもつ患者)が、メチル化が低頻度である症例(実線, 387未満の遺伝子をもつ患者; P=0.01)と比べて生存率が悪いことを示す。
【図4A】この図は、MPMに特異的なメチル化の同定を示す。図4Aは、各腫瘍での42個の特異的にメチル化された遺伝子を用いて行った階層型クラスター化(MPM及びAdCaでそれぞれ8遺伝子及び34遺伝子)を示す。
【図4B】この図は、MPMに特異的なメチル化の同定を示す。図4Bは、選択した3つの遺伝子のプロモーターの図を示す。各垂線は、単一のCpG部位を表す。転写開始部位(矢印)、エクソン1の位置(灰色ボックス)、制限酵素部位(矢頭)、並びに、MSPアッセイの位置(バー)を示す。
【図4C】この図は、MPMに特異的なメチル化の同定を示す。図4Cは、MCAM分析に使用された、MPMの10サンプル及びAdCaの10サンプルでの、TMEM30B、KAZALD1及びMAPK13のMSP分析を示す。DNAメチル化は、MPM症例で検出された。これに対して、わずか1個の不明瞭なメチル化バンドが、患者6のTMEM30B遺伝子で検出された。M, メチル化型; UM, 非メチル化型; P, メチル化型の陽性コントロール(M) がSssI処理DNAであり、非メチル化コントロール(UM)が正常リンパ球由来のDNAである; N, DNA鋳型を含まない。
【図4D】この図は、MPMに特異的なメチル化の同定を示す。図4Dは、別のコホートの患者のMSP分析(10 MPM及び30 AdCa)を示す。M, UM及びNは、上記4Cと同じ意味である;S, SssI処理DNA。
【発明を実施するための形態】
【0019】
(悪性胸膜中皮腫の検出)
本発明は、悪性胸膜中皮腫を検出するための方法であって、被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を検出することを含み、該遺伝子がメチル化されているとき悪性胸膜中皮腫であると判定する、上記方法を提供する。
【0020】
本明細書において、悪性胸膜中皮腫は、胸膜を構成する中皮細胞の腫瘍であり、アスベスト(石綿)への長期間の曝露に伴うアスベストの吸入が原因で発症すると考えられている。悪性胸膜中皮腫の一般的な症状として、胸水がたまり、胸痛、咳、呼吸困難などの症状が現れる。また、胸水中のヒアルロン酸含量が高値となる傾向がある。
【0021】
DNAメチル化は、MPM診断の強力なマーカーとして提案されている(Toyooka S, Pass HI, Shivapurkar N, et al., Cancer Res 2001;61:5727-30; Tsou JA, Shen LY, Siegmund KD, et al., Lung Cancer 2005;47:193-204.18, 20)。しかしながら、悪性胸膜中皮腫(MPM)と肺腺癌(AdCa)のメチル化状態のこれまでの検討により、その頻度に違いはあるが、候補マーカーの異常メチル化がある程度MPMとAdCaの両方で検出されることが示された。さらに、MPMの特定のメチル化マーカーがこれまで報告されたことはなかった。今回、本発明者らは、MPMとAdCaで全ゲノム領域におけるDNAメチル化プロフィールを同時に分析し、特定の遺伝子座の異常メチル化がMPMで頻繁に検出されること、および、3つの遺伝子(MEM30B、KAZALD1およびMAPK13遺伝子)がMPMで特異的にメチル化されていることを見出した。
【0022】
TMEM30Bは、機能が不明であり、methylation-sensitive representational difference分析法によりヒトメラノーマ細胞系で高メチル化された遺伝子として報告された (Furuta J, Nobeyama Y, Umebayashi Y, Otsuka F, Kikuchi K, Ushijima T., Cancer Res 2006;66:6080-637)。
【0023】
KAZALD1は、骨の形成と再生の間の骨芽細胞の増殖に関与する( Shibata Y, Tsukazaki T, Hirata K, Xin C, Yamaguchi A., Biochem Biophys Res Commun 2004;325:1194-20038)。
【0024】
MAPK13は、MAPキナーゼファミリーのメンバーであり、免疫欠陥、セントロメア領域の不安定化、および顔面異常症候群で調節異常が存在することが認められた(Ehrlich M, Sanchez C, Shao C, et al., Autoimmunity 2008;41: 253-71)。
【0025】
本発明は、特定の3つの遺伝子(TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子)のメチル化を検出することによって、これまで悪性胸膜中皮腫であるのか、それとも肺腺癌であるのか、容易には鑑別不能であったこれらの癌のうち悪性胸膜中皮腫を特異的に検出可能にした。すなわち、本発明は、悪性胸膜中皮腫を検出可能にする遺伝子マーカーを提供する。これらの遺伝子のうち特にKAZALD1およびMAPK13遺伝子のメチル化異常はヒト癌でこれまで報告されたことはなかったことに注目されるべきである。
【0026】
本明細書において、被験者は、悪性胸膜中皮腫に罹患したヒト患者、あるいは、該疾患の疑いのあるヒトを含む。
【0027】
本明細書において、生物学的サンプルは、被験者から生体外に採取された、必要により前処理された、例えば血清、胸水、気管支洗浄液または患部組織(特に、胸膜組織)などが挙げられる。
【0028】
本発明の上記3つの遺伝子の塩基配列は、例えばGenBank(米国NCBI)から下記の登録番号にアクセスすることによって入手可能である。
【0029】
ヒトTMEM30B遺伝子については、NW_925561、NT_025437など。
ヒトKAZALD1遺伝子については、NW_924884、NT_030059など。
ヒトMAPK13遺伝子については、NW_923073、NT_007592など。
【0030】
本発明の方法では、上記遺伝子のうちの少なくとも1つの遺伝子のメチル化を検出する。
【0031】
遺伝子上の好ましいメチル化部位は、上記TMEM30B遺伝子、KAZALD1遺伝子またはMAPK13遺伝子のプロモーターのCpG領域であり、この領域の塩基配列のメチル化が検出または測定される。これら遺伝子のプロモーターのCpG領域は、例えば、(1) 配列番号78、79または80に示される塩基配列を含むか、あるいは、(2) その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含む塩基配列、あるいは、その塩基配列と少なくとも95%、好ましくは少なくとも97%、より好ましくは少なくとも98%の同一性を有する塩基配列であって、かつ、プロモーター活性を有する塩基配列を含む。
【0032】
本明細書で使用する「プロモーターのCpG領域」は、CG配列に富む領域であり、例えば約20〜35%またはそれ以上のC+Gの塩基を含む領域である。また、「プロモーター活性」は、遺伝子の転写を制御する活性をいう。
【0033】
本明細書で使用する「数個」は、2〜10の範囲の整数をいう。
本明細書で使用する「同一性」は、2つの塩基配列を、一致度が最大になるように、ギャップを導入するかギャップを導入しないで、整列させたときに、ギャップの数を含めた総塩基数に対する同一(すなわち、一致する)塩基数のパーセンテージ(%)を意味する。
【0034】
本発明の方法におけるメチル化は、例えばメチル化特異的PCR法(J.G. Herman et al., Proc. Natl. Acad. Sci. USA 1996, 93:9821-9826; C.A. Eads et al., Nucleic Acids Res. 2000, 28:E32; K. Rand et al., Methods 2002, 27:114-120; D.T. Akey et al., Genomics 2002, 80:376-384)によって検出することができる。また、メチル化のレベルは、例えばpyrosequencing法(S. Colella et al., Biotechniques 2003, 35:146-150; J. Tost et al., Biotechniques 2003, 35:152-156)によって測定することができる。
【0035】
メチル化特異的PCR法(Methylation Specific PCR; MSP)は、重亜硫酸ナトリウム(sodium bisulfite)で処理したDNAを鋳型にし、PCRを行うことによってメチル化の有無を調べる方法である。重亜硫酸ナトリウムは、DNA上のシトシンをウラシルに変換するが、メチル化シトシン(CpGのシトシン残基の5'位のメチル化)ではこの変換反応が起こらない。
【0036】
PCR用プライマーとして、メチル化されている場合に増幅されるメチル化特異的プライマーセットと、メチル化されていない場合に増幅される非メチル化特異的プライマーセットの2種類が使用される。具体的には、これらのプライマーは、上記のとおり、転写を制御するプロモーターCpG領域の遺伝子配列を基準にして選択し、例えばDNA自動合成機を使用することによって作製することができる。
【0037】
上記PCR法で使用するためのプライマーについては、例えば、配列番号78、79または80に示される塩基配列、あるいは、その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含む塩基配列中の、あるいはその塩基配列と少なくとも95%、好ましくは少なくとも97%、より好ましくは少なくとも98%の同一性を有する塩基配列であって、かつ、プロモーター活性を有する塩基配列中の、連続する17〜35塩基、好ましくは20〜30塩基からなる配列を、メチル化特異的プライマーおよび/または非メチル化特異的プライマーとして選択することができる。
【0038】
PCRによる増幅後に、この2種類のプライマーセットからの増幅産物の比を比較することによってメチル化の有無を決定する。すなわち、メチル化特異的プライマーによる増幅産物が、試験検体で検出された時、問題の遺伝子がメチル化されていると判定する。このプライマーは、元の配列に近い配列を増幅し検出する際に有効である。一方、非メチル化特異的プライマーは、重亜硫酸ナトリウムの処理で変化した配列を検出する際に有効である。さらにまた、試験検体の重亜硫酸ナトリウムで処理後のDNAの質的評価としてメチル化の有無に関わらずPCR増幅が行われる例えばLINE1プライマー(例えば、LINE1−Fwd:TTTTAAAGTTGTTAGATAGGGATATTTAAGTTTGTA(配列番号91); LINE1−Rev: AAAAACCTACCTACCTCTATAAACTCCACC(配列番号92))などのプライマーを用いるときには、(メチル化特異的プライマーによる増幅産物)対(LINE1プライマーによる増幅産物)の比によりメチル化DNAの量的診断も可能となる。
【0039】
さらにまた、上記のPCRに加えて、定量的PCRを実施することも可能であり、正常のDNAが非特異的に増幅される場合に、それによる影響を排除するために利用しうる。定量的PCR用のプライマーの例は、TMEM30B遺伝子について、配列番号83(メチル化プライマー)、配列番号84(非メチル化プライマー)、KAZALD1遺伝子について、配列番号87(メチル化プライマー)、配列番号88(非メチル化プライマー)である。
【0040】
本発明で使用可能なプライマーセットは、ヒトTMEM30B遺伝子(NW_925561、NT_025437など)、ヒトKAZALD1遺伝子(NW_924884、NT_030059など)、またはヒトMAPK13遺伝子(NW_923073、NT_007592など)の塩基配列、好ましくはこれらの各遺伝子のプロモーターのCpG領域の塩基配列、さらに好ましくは配列番号78、79または80を含む塩基配列、において、その中の少なくとも1つのメチル化CpGを含む約17〜約35塩基、好ましくは約20〜30塩基の塩基配列からなる、任意に選択されたプライマーを含む。
【0041】
上記プライマーセットの非限定的な例は、以下のとおりである。
TMEM30B遺伝子のメチル化特異的プライマー:
TTGGGAGGGTGTGGGTATTGTTC(配列番号66)
ACGCGCGAACTAACCGAAT(配列番号67)
CTGGGAGGGTGTGGGCACTGCTCG(配列番号81)
CGAGTAGTTTTCGGGTTTGC(配列番号95)
ACCGACGCGCGAACTAAC(配列番号96)
TMEM30B遺伝子の非メチル化特異的プライマー
GGAATTGGGAGGGTGTGGGTATTGTTT(配列番号68)
ACCTCCCTCTCCACACCAC(配列番号69)
GGAACTGGGAGGGTGTGGGCACTGCT(配列番号82)
TGAGTAGTTTTTGGGTTTGT(配列番号97)
ACCATAACACTCCAAATCATAACA(配列番号98)
KAZALD1遺伝子のメチル化特異的プライマー
GCGGGTCGTAGTATTTGTCG(配列番号70)
AACAAAACCAACCACCTTCG(配列番号71)
CGGGTCGTAGTATTTGTC(配列番号85)
GGTAAGTTTGCGGTCGTTTTC(配列番号99)
AACAAAACCAACCACCTTCG(配列番号100)
KAZALD1遺伝子の非メチル化特異的プライマー
TTATTTGTGGGTTGTAGTATTTGTT(配列番号72)
AAACCAACCACCTTCATCTCC(配列番号73)
TTATTCGCGGGTCGTAGTATTTGTC(配列番号86)
GTGGGTAAGTTTGTGGTTG(配列番号101)
AACAAAACCAACCACCTTCATCT(配列番号102)
MAPK13遺伝子のメチル化特異的プライマー
CGTGTTTTCGACGTACGTCGGTAGC(配列番号74)
ACCGCTAAATAACCCCGAAA(配列番号75)
CGTGTCCCCGACGCACGTCGGCAGC(配列番号89)
MAPK13遺伝子の非メチル化特異的プライマー
GGTTTTGTTGGGTAGTTTT(配列番号76)
TCCCACCTACCAACTCACC(配列番号77)
GGTCCCGCTGGGCAGCCC(配列番号90)
【0042】
一方、pyrosequencing法は、遺伝子上の目的の領域をPCR増幅したのち、この領域内の特定のCpG部位の重亜硫酸塩変換配列を決定する手法である。この方法では、個々の部位のC対Tの比を、配列伸長の際のCおよびTの取り込み量に基づいて定量的に決定する。具体的には、この方法は、キアゲン社パイロシークエンサーによって行うことができる。
【0043】
PCR条件は、一般的手法を使用することができる。すなわち、PCRでは、変性、アニーリング及び伸長反応を1サイクルとして約20〜40サイクルが実施される。変性は、二本鎖の鋳型DNAを例えば94〜96℃、約5秒〜約10分加熱して一本鎖に分離させる工程である。アニーリングは、一本鎖の鋳型DNAのそれぞれの側面に、正方向プライマーまたは逆方向プライマーをアニールさせる工程であり、約50〜65℃、約5秒〜60秒の加熱によって行われる。伸長反応は、鋳型DNAに沿ってプライマーを伸長してDNAを合成する工程であり、通常、72〜77℃、約30秒〜10分の加熱によって行われる。PCR用溶媒は、Mg2+含有緩衝液であり、これに耐熱性DNAポリメラーゼ酵素(例えばTaqポリメラーゼなど)およびdNTPs(N=A,T,G,C)を添加する。PCRサイクルの初めに、例えば94〜96℃、約30秒〜10分の加熱を行ってもよいし、また、最終サイクルの終了時に72〜77℃、約2〜10分の加熱を行ってもよい。好適なPCR条件はプライマーにより異なり、例えばTMEM30Bメチル化プライマーのアニーリング温度は55℃、非メチル化プライマーは65℃である。PCRは、サーマルサイクラーのような増幅装置を使用すると便利である。
【0044】
PCR産物は、臭化エチジウム染色された、6%ポリアクリルアミドゲルまたは3%アガロースゲル上で視覚化することができる。
【0045】
本発明の方法によれば、上記遺伝子の少なくとも1つの遺伝子がメチル化されているとき、悪性胸膜中皮腫であると判定する。これらの遺伝子は、肺腺癌ではメチル化されていないため、これら遺伝子のメチル化を検出することによって悪性胸膜中皮腫を肺腺癌から容易に鑑別することができる。
【0046】
(悪性胸膜中皮腫に罹患した被験者の予後の測定)
本発明はさらに、悪性胸膜中皮腫に罹患した被験者の予後を判定する方法であって、該被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化の程度を測定することを含み、該遺伝子のメチル化程度が、予後不良被験者のメチル化程度と比べて低いとき、該疾患の予後が良好であると決定する、上記方法を提供する。
【0047】
本発明によれば、悪性胸膜中皮腫に罹患した被験者の予後を、上記3つの遺伝子のいずれかのメチル化の程度、すなわちメチル化のレベルを測定することによって判定することができる。これは、上記腫瘍に関する予後が不良の場合、該遺伝子が異常にメチル化されていることに基づく。逆に、予後が良好の場合、該遺伝子のメチル化のレベルが予後不良の場合と比べて相対的に低い。
【0048】
予後を判定する方法として、上記MEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つのヒト遺伝子のメチル化程度を測定するために、例えば、上記のメチル化特異的PCR法および/またはpyrosequencing法を使用できる。これらの方法およびプライマーの例については、上で説明したとおりである。
【0049】
また、生物学的サンプルについても、上で説明したように、具体的には、血清、胸水、気管支洗浄液または患部組織(特に、胸膜組織)である。
【0050】
(キット)
本発明はさらに、上記のいずれかの方法で使用するためのキットであって、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を測定することが可能である、メチル化特異的PCR用の下記のプライマーセットを含むキットを提供する。
プライマーセット:
TMEM30B遺伝子のメチル化特異的プライマー
TTGGGAGGGTGTGGGTATTGTTC(配列番号66)
ACGCGCGAACTAACCGAAT(配列番号67)
CTGGGAGGGTGTGGGCACTGCTCG(配列番号81)
CGAGTAGTTTTCGGGTTTGC(配列番号95)
ACCGACGCGCGAACTAAC(配列番号96)
TMEM30B遺伝子の非メチル化特異的プライマー
GGAATTGGGAGGGTGTGGGTATTGTTT(配列番号68)
ACCTCCCTCTCCACACCAC(配列番号69)
GGAACTGGGAGGGTGTGGGCACTGCT(配列番号82)
TGAGTAGTTTTTGGGTTTGT(配列番号97)
ACCATAACACTCCAAATCATAACA(配列番号98)
KAZALD1遺伝子のメチル化特異的プライマー
GCGGGTCGTAGTATTTGTCG(配列番号70)
AACAAAACCAACCACCTTCG(配列番号71)
CGGGTCGTAGTATTTGTC(配列番号85)
GGTAAGTTTGCGGTCGTTTTC(配列番号99)
AACAAAACCAACCACCTTCG(配列番号100)
KAZALD1遺伝子の非メチル化特異的プライマー
TTATTTGTGGGTTGTAGTATTTGTT(配列番号72)
AAACCAACCACCTTCATCTCC(配列番号73)
TTATTCGCGGGTCGTAGTATTTGTC(配列番号86)
GTGGGTAAGTTTGTGGTTG(配列番号101)
AACAAAACCAACCACCTTCATCT(配列番号102)
MAPK13遺伝子のメチル化特異的プライマー
CGTGTTTTCGACGTACGTCGGTAGC(配列番号74)
ACCGCTAAATAACCCCGAAA(配列番号75)
CGTGTCCCCGACGCACGTCGGCAGC(配列番号89)
MAPK13遺伝子の非メチル化特異的プライマー
GGTTTTGTTGGGTAGTTTT(配列番号76)
TCCCACCTACCAACTCACC(配列番号77)
GGTCCCGCTGGGCAGCCC(配列番号90)
【0051】
本発明のキットには、上記プライマーの他に、PCRの実施に必要な、例えば、緩衝液、耐熱性DNAポリメラーゼ酵素、dNTPs(N=A,T,C,G)、使用説明書などを含むことができる。プライマー類は、単独で、あるいは、組み合わせて包装することができる。
【0052】
(実施例)
本発明を以下の実施例によってされに具体的に説明するが、本発明の範囲は、これらの実施例によって制限されないものとする。
【実施例1】
【0053】
<材料及び方法>
細胞
MPM細胞 (ACC-MESO-1)(Usami N, Fukui T, Kondo M, et al. Cancer Sci 2006;97:387-94)及び非悪性の中皮細胞(MeT-5A)を解析のために使用した。MeT-5Aは、ATCC (American Type Culture Collection, Rockville, MD, USA)からATCC CRL-9444として入手可能である。ACC-MESO-1は、10%牛胎仔血清(Invitrogen, Carlsbad, CA, USA)及びantibiotic-antimycotic (Invitrogen)を添加したRPMI-1640培地(Sigma-Aldrich, St Louis, MO)中、37℃、5% CO2を含む湿気のあるインキュベーター中で培養された。MeT-5Aは、ATCCの説明書の条件にしたがって培養された。
【0054】
組織サンプル
31個のMPMサンプル及び51個のAdCaサンプル(いずれも日本人患者由来)を、愛知県がんセンター病院、名古屋大学病院、名古屋第一赤十字病院、名古屋第二赤十字病院、春日井市民病院、大垣市営病院、及び岡山労災病院から入手した。サンプル及び臨床データは、適正機関の検討部会の承認と、すべての患者からのインフォームドコンセントを書面で得た後、集められた。
【0055】
DNA調製
ゲノムDNAを、標準フェノール−クロロホルム法を用いて抽出した。ゲノムDNAをM. SssIメチラーゼ(New England Biolabs, Ipswich, MA)で処理することによって完全メチル化DNAを調製した。簡潔に説明すると、5μg のDNAを、20 UのM. SssIメチラーゼ、320μMのS-アデノシルメチオニン(SAM, New England Biolabs)及びNEBバッファー2 (New England Biolabs)を添加して計300 μlとし37℃でインキュベーションした。インキュベーションの間、同量のM. SssIメチラーゼ及びSAM をさらに1回添加し、DNAのメチル化反応を確実にした。非メチル化DNAは、製造元の指示書に従って、phi29 DNAポリメラーゼ(GenomiPhi DNA Amplification Kit; Amersham Biosciences, Uppsala, Sweden)を用いてゲノムDNAを処理することによって調製した。
【0056】
メチル化CpGアイランド増幅マイクロアレイ(MCAM)
本発明者らは、20サンプルの MPM (45〜78 歳、平均年齢59.1歳)及び20サンプルの AdCa(44〜76歳、平均年齢62.8歳)をMCAM分析にかけた。MCMの詳細なプロトコルは、これまでに記載された方法に従った(Shen L, Kondo Y, Guo Y, et al. PLoS Genet 2007;3:2023-3621; Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)。簡単に説明すると、2 μgのゲノムDNAを、100Uのメチル化感受性制限エンドヌクレアーゼSmaI (New England Biolabs)で消化した。この酵素は、非メチル化DNAを切断して平滑末端(CCC/GGG)を残す。つぎに、このDNAを、50 Uの メチル化非感受性制限エンドヌクレアーゼXmaIで消化した。この酵素は、粘着末端(C/CCGGG)を残す。この粘着末端にアダプターを結合した。アダプターは、オリゴヌクレオチド RMCA12 (5’-CCGGGCAGAAAG-3’(配列番号93))及びRMCA24(5’-CCACCGCCATCCGAGCCTTTCTGC-3’(配列番号94))(Toyota, M. et al., Cancer Res. 59:2307-2312, 1999)を65℃で2分間インキュベーションし、その後、60分間で室温まで冷却することによって作製した。消化されたDNAを、T4 DNAリガーゼ (Invitrogen, Carlsbad, CA)を用いてアダプターに結合し、RMCA24プライマーを用いるPCRによって増幅した。PCR条件は、72℃、5分の加熱により、アダプター結合DNA断片のオーバーハング末端の伸長を行い、さらに、95℃、3分の加熱を行ったのち、95℃、1分及び77℃、3分を25サイクル行い、最後に72℃、10分の伸長反応を行った。本発明者らは、6,161個のユニークな遺伝子に対応した15,134個のプローブを含むAgilent Technologies 社のカスタムプロモーターアレイ(G4497A, Agilent Technologies, Santa Clara, CA)を使用した。このアレイは、過去のMCAMを用いた研究で有効性について証明されている(Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)。マイクロアレイプロトコルは、これまで記載されたものを使用した(Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)。簡単に説明すると、MCA産物を、癌組織由来DNAに対してCy5(赤色)で標識し、 正常組織由来DNAに対してCy3(緑色)で標識した。正常対照として、MPMについては2サンプルの正常中皮組織由来のDNA、または、AdCaについては5サンプルの正常肺組織のDNAが使用された。色素で標識後、ヒトCot-1 DNA(Invitrogen)とともに、アレイに標識サンプルをハイブリダイズした。製造者のプロトコルに従ってアレイを洗浄し、ついで、Agilentスキャナーを用いアレイをスキャンし、Feature Extractionソフトウエアを用いて分析した。正規化は、製造者のプロトコル(Agilent Technologies)により、linear per-array アルゴリスムを用いて行った。
【0057】
階層型クラスタリング分析法
クラスタリング分析は、階層型クラスター化法(Eisen MB, Spellman PT, Brown PO, Botstein D., Proc Natl Acad Sci U S A 1998;95:14863-8)を用いて行った。検体のクラスタリングについては、Cluster 3.0ソフトウエア (http://rana.lbl.gov/EisenSoftware.htm)またはMinitab 15統計ソフトウエア (http://www.minitab.com)を用いて、検体間で、すべての遺伝子におけるDNAメチル化シグナル強度の測定値に基づき、類似性を計算した。デンドログラムおよびヒートマップは、TreeViewソフトウエア (http://rana.lbl.gov/EisenSoftware.htm)を用いて作成した。
【0058】
メチル化分析
本発明者らは、これまで報告された手法によって重亜硫酸塩(bisulfite)処理を行った(Kondo Y, Shen L, Suzuki S, et al. Hepatol Res 2007;37:974-83; Yang AS, Estecio MR, Doshi K, Kondo Y, Tajara EH, Issa JP. Nucleic Acids Res 2004;32:e38)。簡単に説明すると、2μgのゲノムDNAに、3M重亜硫酸ナトリウム(pH 5.0)を添加し50℃で16時間インキュベートした。Wizard Miniprep Column(Promega, Madison, WI)を用いてDNAを精製し、エタノールで沈殿させ、30μlの精製水に再溶解させた。DNAメチル化レベルは、Pyrosequencing法(Pyrosequencing AB, Uppsala, Sweden)(Colella S, Shen L, Baggerly KA, Issa JP, Krahe R., Biotechniques 2003;35:146-5027)を用いる定量的メチル化解析法によって解析された。各アッセイで、セットアップの段階で、陽性コントロール(SssI処理後のサンプル) と陰性コントロール(GenomiPhi V2を用いてゲノム全体を増幅したあとのサンプル)を混合した実験を行い、PCRバイアスが生じないアニーリングの至適温度を決定した。また実験を反復し、再現性を評価した。(Shen L, Guo Y, Chen X, Ahmed S, Issa JP. Biotechniques 2007;42:48-5828)。また、メチル化特異的PCR (MSP)を、 TMEM30B、KAZALD1およびMAPK13遺伝子において行った。PCR産物は、臭化エチジウム染色された、6%ポリアクリルアミドゲルまたは3%アガロースゲル上で視覚化した。pyrosequencingおよびMSPのプライマー配列およびPCR条件を、表1A〜表1Cに示した。
【0059】
【表1A】
【0060】
【表1B】
【0061】
【表1C】
【0062】
プライマーはすべて、転写開始部位から0.5kb以内のCpGのメチル化状態をアッセイ可能であるように設計した。
【0063】
細胞のトリコスタチンAおよび5Aza-dC 処理
細胞を12〜24時間培地で培養した後、細胞を、1) 5μM 5-アザ-2’-デオキシシチジン(5Aza-dC; Sigma-Aldrich)またはリン酸緩衝塩水で72時間処理する(このとき、5Aza-dCまたはリン酸緩衝塩水を含有する培地を24時間おきに交換した。)か、あるいは、2) 300nMのトリコスタチンA (TSA; MP Biomedicals, Solon, OH)または等容量のエタノールで24時間処理した。
【0064】
定量逆転写−ポリメラーゼ連鎖反応分析
全RNAをTRIzol (Invitrogen)を用いてRNAを抽出した。2μgのRNAをMPMLV (Promega)を用いて逆転写した。SYBR GREEN Quantitative逆転写-PCRを、標的遺伝子に対して実験を3回おこなった (Applied Biosystems, Foster City, CA)。Q-PCRのプライマー配列を表1Bに示した。
【0065】
統計分析
メチル化状態と臨床病理学的変化との間の関係を、StadView ソフトウエア (version 5.0; Abacus Concepts, Berkeley, CA)を用いて、Mann-Whitney U test、Fisher’s exact testまたはKruskal-Wallis testを用いて解析した。線形回帰モデルはStata version 8 (College Station, TX)を使用した。P値のすべてがtwo-sidedであり、P<0.05が統計的に有意であるとみなした。生存率曲線を、Kaplan-Meier法を用いて作成した。全生存率は、外科手術日から開始し死亡までの間で計算した。
【実施例2】
【0066】
<結果>
本発明者らは、はじめに、悪性中皮腫細胞系ACC-MESO-1のメチル化プロフィールを分析するためにMCAMを実施した。MCAによってメチル化断片を増幅したのち、MCA産物を、Cy5 (ACC-MESO-1)またはCy3 (正常中皮組織)で標識し、ともにマイクロアレイ(6,161遺伝子に対応する15,134プローブを含む)に共ハイブリダイズした(図1A)。これまで、本発明者らは、MCAMにおいて2.0以上をしめすCy5/Cy3シグナル比は、pyrosequencing分析でのメチル化陽性と一致した (Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)。ACC-MESO-1細胞でのMCAM解析の有効性を評価するために、 高、中、または低シグナル強度を示す19遺伝子をランダムに選択し、pyrosequencing法によってメチル化レベルを調べた。2.0以上のCy5/Cy3シグナル比を示す14遺伝子のうち11遺伝子がpyrosequencing分析でメチル化陽性(ACC-MESO-1で15%以上のメチル化レベル、および、正常中皮組織のメチル化レベルの少なくとも1.5倍以上のメチル化レベル)であったが、2.0以下の Cy5/Cy3シグナル比をもつ5遺伝子のうち4遺伝子がメチル化陰性(MPM細胞で15%未満のメチル化レベル、および/または正常中皮組織の1.5分の1以下のメチル化レベル)であった。MCAM分析とpyrosequencing分析との間の高い相関関係(特異性80%; 感度86%)は、これまでの研究のなかで示されているのと同様に(Shen L, Kondo Y, Guo Y, et al., PLoS Genet 2007;3:2023-36.: Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)、この細胞で確認された(図1Bおよび表2)。この規準を用いてACC-MESO-1でメチル化された971遺伝子を同定した。
【0067】
【表2】
【0068】
つぎに、MPMでのこれらの遺伝子のDNAメチル化と遺伝子発現との関係を調べるために、ACC-MESO-1をDNAメチルトランスフェラーゼ阻害剤、5-アザ-2’-デオキシシチジン(5-Aza-dC)もしくはヒストンデアセチラーゼ阻害剤、トリコスタチンA (TSA)で処理し、3つの同定された遺伝子C9orf106、EDG4およびMPMPL1 の発現を検討した(図1C および図1D)。ACC-MESO-1ではこれらの遺伝子発現は抑制されていたのに対して、正常中皮組織では高い発現レベルであった。本発明者らは、5-Aza-dCによる有意な遺伝子再発現を観察したがTSA処理では有効性が乏しかった。このことは、これらの遺伝子の発現抑制がDNAメチル化によることを示している。
【0069】
正常中皮DNA(2症例の混合)および正常肺組織DNA(4症例の混合)からのMCA産物の共ハイブリダイゼーションにおいて、本発明者らは、メチル化状態は高い一致(R2=0.87, 図2A)を示すことを確認した。このことは、組織特異的メチル化は、これら2つの正常組織ではまれであることを示している。つぎに、MPMおよびAdCaのメチル化プロフィールの違いを評価するために、MPM DNA (4症例の混合物)およびAdCa DNA (4症例の混合物)からのMCA産物を共ハイブリダイズし、遺伝子の一部はそれぞれの腫瘍で異なるメチル化プロファイルを示すことを見出した(R2=0.77, 図2B)。興味深いことに、本発明者らは、肝臓癌(4症例の混合)とMPMとのメチル化プロファイルを検討したところ、肺と肝臓はともに内胚葉から発生するのにもかかわらず、MPMとAdCaとのメチル化標的遺伝子の差に比べて、MPMと肝臓癌とではより多くの異なった遺伝子がメチル化していること (R2=0.26)を見出した(図2C)。
【0070】
MPMおよびAdCaの全ゲノム領域におけるDNAメチル化プロフィールを比較するために、本発明者らは、MCAMを使用して20症例のMPMと20症例のAdCaを分析した。6,161遺伝子を用いてのMPMおよびAdCaのすべてのヒートマップから、AdCaがMPMよりも高頻度でメチル化されているように見られること、ならびに、AdCaの一部の症例群が広範にメチル化されていることが示された(図3A)。MPMと AdCaは、DNAメチル化プロファイルによって異なるサブグループに分類された。MPMのなかには主にAdCaからなるサブグループに入る2つの例外症例が存在した。
【0071】
つぎに、DNAメチル化状態がMPMおよびAdCaにおいて臨床病理学的な特性に影響を与えるかどうかを調べるために、6,161遺伝子を用いる階層型クラスタリングによって個々にMPMおよびAdCaを分析した。DNAメチル化遺伝子数によって、MPMを2つのサブグループに分けた。すなわち、その1つのサブグループは、メチル化遺伝子数が300遺伝子以下と少ない4症例であり、そのすべての症例が上皮型であった。これらのデータは、DNAメチル化が腫瘍形成の際にMPMの表現型に影響することを示している。階層型クラスタリング分析によって、AdCaは、4つのサブグループに分けることができた。1つのサブグループは、6症例のAdCaからなり、それらは、他のAdCa症例と比べてメチル化遺伝子数を多く有する症例であった (911遺伝子vs 387遺伝子, P=0.001)。また、そのサブグループの症例のほとんどが喫煙者であった(喫煙者5/6症例;喫煙した平均パック年数, 68.6± 22.9)。
【0072】
本発明者らは、MPMおよびAdCaにおいてメチル化している遺伝子を数えた。MPMでメチル化されている遺伝子は700未満であり、これはマイクロアレイ上に存在する6,161 CpGの約11%を占める。また、MPM症例のメチル化遺伝子数の平均が387(範囲, 120 〜755; 図3B)であり、一方、AdCaでは544遺伝子 (範囲, 133〜1212)であった。
【0073】
AdCaとMPMのそれぞれで共通してメチル化されている遺伝子を比較した。10症例の MPMで共通してメチル化されている遺伝子は非常にまれ (40未満の遺伝子)であるが、10症例の AdCaでは、80以上の遺伝子が共通してメチル化されていた。このことは各MPM症例では共通してメチル化する遺伝子は少ない(P=0.001, 図3C)ことを示している。各腫瘍のメチル化遺伝子のリストを表3および表4に示した。
【0074】
【表3】
【0075】
【表4】
【0076】
MPMおよびAdCaにおけるメチル化遺伝子数と臨床病理学的パラメーターとの関係を表5に示した。
【0077】
【表5】
【0078】
AdCaでは、喫煙者は非喫煙者と比べて有意にメチル化遺伝子が多かった (728遺伝子vs 360遺伝子, P=0.02)。MPMでの喫煙者もまた、非喫煙者と比べてメチル化遺伝子が多い傾向であり(366遺伝子vs 261遺伝子, P=0.2)、このことは喫煙がMPMのDNAメチル化に影響していることを示している。これに対し、アスベスト曝露は、MPMではDNAメチル化に対する影響は少なかった(MPMでのアスベスト曝露あり対アスベスト曝露なしは、386遺伝子vs 320遺伝子であった。)。
【0079】
進行性大腸直腸癌患者でDNAメチル化が高頻度に存在する症例では予後不良であることが報告されている( Shen L, Catalano PJ, Benson AB, 3rd, O'Dwyer P, Hamilton SR, Issa JPClin Cancer Res 2007;13:6093-829)。本発明者らは、MPM患者を、2つの群、すなわち、メチル化遺伝子数が平均(387遺伝子)より多い群と少ない群に分けて、生存率とメチル化との関係を検討した。外科的合併症が原因で死亡した患者、または、手術合併症のため死亡した2人の患者は除かれた。平均メチル化遺伝子数よりメチル化遺伝子数の少ない群の患者(n=11)は、平均よりメチル化遺伝子数の多い群の患者(n=7)と比べて有意に生存率が長かった (平均生存時間, 22ヶ月vs 8ヶ月, P=0.01)(図3D)。MPM患者全集団の全体の平均生存期間は11.7ヶ月であった。
【0080】
DNAメチル化は、癌診断のための強力なマーカーとなりうるとされている(Belinsky SA, Nat Rev Cancer 2004;4:707-1730)。MPM診断のための特異的なDNAメチル化マーカーを同定するために、はじめに、本発明者らは、MCAM分析から8遺伝子を選択した。この分析では、少なくとも4症例のMPMでメチル化陽性(Cy5/Cy3>2.0)を示し、AdCaの全症例でメチル化陰性を示すものを選んだ(図4A)。興味深いことに、少なくとも4症例のAdCaでメチル化陽性、ならびに、MPMの全症例でメチル化陰性という同様の規準を用いて遺伝子を選ぶと、AdCaでより多くの遺伝子(34遺伝子)が存在し、このことは、メチル化した遺伝子がAdCaでより多く存在することを示している(図4A)。本発明者らは、MSPによって遺伝子のメチル化を確認し、3つの遺伝子、すなわち、膜貫通蛋白質30B (TMEM30B)、Kazal型セリンプロテアーゼ阻害剤ドメイン1 (KAZALD1)及び有糸分裂活性化プロテインキナーゼ13 (MAPK13)がMPMに特異的なメチル化マーカーであることを見出した(図4Bおよび4C)。MCAM解析で解析されたMPM症例では、DNAメチル化が、MSPによりDNAメチル化が、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子のそれぞれにおいて19症例のMPMのうち11 (58%)症例、8 (42%)症例および2 (11%)症例で検出された。しかしながら、AdCaでは、TMEM30B遺伝子でわずか1つの不明瞭なメチル化バンドが検出されたにすぎなかった。
【0081】
これらのメチル化マーカーがMPM患者の別の集団で鑑別可能であるかを確認するために、本発明者らは、別の機関からさらに10症例のMPMを入手し分析した(図4D)。DNAメチル化は、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子のそれぞれで、10症例のMPMのうち5 (50%)、7 (70%)および2 (20%)症例で検出された。これら3つの遺伝子のDNAメチル化を、合計31症例のMPMと51症例のAdCa(19症例のMPMサンプルおよび19症例のAdCaサンプルは、MCAM解析で用いた集団であった)で分析した。DNAメチル化は、MPMでは、TMEM30Bについて17症例(55%)で、KAZALD1について16症例(52%)で、MAPK13について4症例(13%)で検出されたが、AdCaでは、DNAメチル化は検出されなかった (図4Cおよび4D)。AdCaとMPMの鑑別診断として、3つの遺伝子のうち少なくとも1遺伝子がメチル化している場合にMPMと診断すると、その感度と特異性はそれぞれ77%および100%であった。
【0082】
さらにまた、上記3種の遺伝子の各配列のなかで、特にそれら遺伝子のプロモーターのCpGアイランド領域の配列(配列番号78、79又は80)のメチル化又はメチル化程度を検出又は測定することによって、MPMをより特異的に検出できることが分かった。また、そのための分析に使用されるPCR用プライマーとして有効なものは、TMEM30B遺伝子のメチル化特異的プライマーの場合、例えば配列番号66、配列番号67、配列番号81、配列番号95、及び/又は配列番号96のプライマーであり、TMEM30B遺伝子の非メチル化特異的プライマーの場合、例えば配列番号68、配列番号69、配列番号82、配列番号97、及び/又は配列番号98のプライマーであり、KAZALD1遺伝子のメチル化特異的プライマーの場合、例えば配列番号70、配列番号71、配列番号85、配列番号99、及び/又は配列番号100のプライマーであり、KAZALD1遺伝子の非メチル化特異的プライマーの場合、例えば配列番号72、配列番号73、配列番号86、配列番号101、及び/又は配列番号102のプライマーであり、MAPK13遺伝子のメチル化特異的プライマーの場合、例えば配列番号74、配列番号75、及び/又は配列番号89のプライマーであり、MAPK13遺伝子の非メチル化特異的プライマーの場合、例えば配列番号76、配列番号77、及び/又は配列番号90のプライマーであった。
【0083】
<考察>
この研究のなかで、本発明者らは、MCAMを使用してMPMおよびAdCaの全ゲノム領域におけるメチル化プロフィールを解析、比較し、ならびに、MPMにおいて特定の遺伝子のDNAメチル化の頻度を検討した。MCAMが高い特異性と感度をもって全ゲノム領域のメチル化の状態を評価することが可能であることは以前の報告で示されている( Shen L, Kondo Y, Guo Y, et al., PLoS Genet 2007;3:2023-36; Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10); Estecio MR, Yan PS, Ibrahim AE, et al., Nat Genet 2005;37:853-62)。しかしながら、MCAMは組織サンプルなかの異常にメチル化された遺伝子をより有意に検出する可能性があるので、本発明者らは、高メチル化遺伝子に照準を合わせた。実際、MPM細胞であるACC-MESO-1を使用したときの有効性は、MCAMからのシグナル比と、bisulfite-pyrosequencingによってアッセイされたメチル化レベルとの間で良好な相関性を示し、従来の研究結果と一致している( Shen L, Kondo Y, Guo Y, et al., PLoS Genet 2007;3:2023-36; Gao W, Kondo Y, Shen L, et al., Carcinogenesis 2008;29:1901-10)。ACC-MESO-1でMCAM分析によって検出された標的遺伝子は、 5-Aza-dCによって再活性化され、また、MPM患者で高メチル化されていることが見出された。このことは、MPMでのこれら遺伝子のDNAメチル化により遺伝子発現抑制されていることを示している。
【0084】
正常中皮と正常肺組織は異なる胚葉(それぞれ、中胚葉、内胚葉)から発生するにもかかわらず、それらのDNAメチル化プロフィールは非常に類似しており(R2=0.87)、このことは、これら2つの正常組織での組織特異的メチル化の差は少ないことを示している。これまでの種々の正常組織のゲノムワイドなメチル化分析では、組織特異的なメチル化が非常にまれであることを示しており、本発明者らの知見と一致する(Shen L, Kondo Y, Guo Y, et al., PLoS Genet 2007;3:2023-36; Estecio MR, Yan PS, Ibrahim AE, et al., Genome Res 2007;17:1529-36; Weber M, Davies JJ, Wittig D, et al., Nat Genet 2005;37:853-6221, 31, 32)。これに対して、MPMとAdCaとの間のDNAメチル化標的遺伝子の差は正常組織と比べてかなり多く、これは種々の組織での癌が異なるメチル化プロフィールを表すためと推測される(Esteller M, Corn PG, Baylin SB, Herman JG. Cancer Res 2001;61:3225-933)。興味深いことに、MPMのメチル化プロフィールは、肝臓癌と比べると、肺及び肝臓がともに内胚葉から発生するにも関わらず、AdCaの方により類似していた。MPMとAdCaとの間のDNAメチル化の類似性の理由は解っていない。喫煙などの腫瘍形成に寄与する特定の環境因子が、MPMとAdCaの両方のDNAメチル化異常に同一の影響を与えている可能性はある(Feinberg AP., Nature 2007;447:433-40)。
【0085】
いくつかの遺伝子座のDNAメチル化異常のこれまでの研究により、メチル化がAdCaと比較してMPMでは広範囲の遺伝子に起こらないことが示されてきた(Toyooka S, Pass HI, Shivapurkar N, et al., Cancer Res 2001;61:5727-30)。今回のゲノムワイドな(広域の)DNAメチル化解析により、DNAメチル化がAdCaと比べてMPMでは低頻度でありかつ共通性に乏しいことが示された。大多数のMPMではメチル化されていた遺伝子は700以下であった(平均メチル化遺伝子数, 387±196遺伝子)。これは、AdCa患者の一部の症例群(ほとんどが喫煙者による症例(喫煙した平均パック年数67.3± 14.2)であり、広範囲の遺伝子がメチル化されていた(>950遺伝子))と対比的であった。特定の肺腺癌は、メチル化されやすく、また、CpG island methylator phenotype (CIMP)と称される表現型になりやすいことが示唆されている (Toyota M, Ahuja N, Ohe-Toyota M, Herman JG, Baylin SB, Issa JP., Proc Natl Acad Sci U S A 1999;96:8681-6; Suzuki M, Shigematsu H, Iizasa T, et al., Cancer 2006;106:2200-7)。本発明者らの結果は、AdCaのサブセットがCIPMを示したのに対し、MPMではそのような症例群は存在しなかったことを示した。MPMで異常メチル化がおこる際に、アスベストによる継続的炎症は誘導因子であるように思われるが、AdCaでの喫煙ほどは強くない。これらのデータは、MPMとAdCaの形成の間の異常なDNAメチル化の獲得に対する異なる機序を反映しているかもしれない。
【0086】
DNAメチル化は、MPM診断の強力なマーカーとして提案されている(Toyooka S, Pass HI, Shivapurkar N, et al., Cancer Res 2001;61:5727-30; Tsou JA, Shen LY, Siegmund KD, et al., Lung Cancer 2005;47:193-204)。しかしながら、MPMとAdCaのメチル化状態のこれまでの検討により、その頻度に違いはあるが、候補マーカーの異常メチル化がある程度MPMとAdCaの両方で検出されることが示されている。さらに、MPMに特異的なメチル化マーカーがこれまで報告されたことはなかった。今回、本発明者らは、MPMとAdCaで全ゲノム領域のDNAメチル化プロフィールを解析し、特定の遺伝子座のDNAメチル化がMPMで頻繁に検出され、3つの遺伝子、すなわちTMEM30B、KAZALD1およびMAPK13遺伝子がMPMで特異的にメチル化されていたことを示した。特に、KAZALD1およびMAPK13遺伝子のDNAメチル化はヒト癌でこれまで報告されたことはなかったが、それらの異常なメチル化がMPMをAdCaから鑑別する有益なマーカーとして役立つはずである。
【0087】
まとめると、MPMとAdCaを全ゲノム領域のメチル化分析によって比較すると、これら2つの悪性腫瘍ではそれぞれ特徴的なDNAメチル化パターンを示し、そのパターンは、病態生理学的過程の違いから生じているようである。本発明者らは、今回、MPMの診断のための可能なマーカーを提案する。さらに、病理学的知見と関連するメチル化プロフィールによって分類されるMPMのサブセットを見出した。また、DNAメチル化が低頻度のMPM患者の予後が良好であることを見出した。これらの事実は、DNAメチル化異常がMPMの臨床上の特徴に影響を与えることを示している。したがって、プロモーターCpGアイランドのDNAメチル化およびそれと関連する遺伝子発現抑制がMPM発症に重要な役割を果たしている。
【産業上の利用可能性】
【0088】
本発明は、悪性胸膜中皮腫を肺癌から区別可能とする悪性胸膜中皮腫検出方法を提供するため、特にアスベストを原因とするような悪性胸膜中皮腫の確定診断のために医療上有用である。
【特許請求の範囲】
【請求項1】
悪性胸膜中皮腫を検出するための方法であって、被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を検出することを含み、該遺伝子がメチル化されているとき悪性胸膜中皮腫であると判定する、上記方法。
【請求項2】
悪性胸膜中皮腫に罹患した被験者の予後を判定する方法であって、該被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化の程度を測定することを含み、該遺伝子のメチル化程度が、予後不良被験者のメチル化程度と比べて低いとき、該疾患の予後が良好であると決定する、上記方法。
【請求項3】
前記TMEM30B遺伝子、KAZALD1遺伝子またはMAPK13遺伝子のプロモーターのCpG領域の塩基配列のメチル化を検出または測定する、請求項1または2に記載の方法。
【請求項4】
前記プロモーターのCpG領域の塩基配列が、配列番号78、79または80に示される塩基配列、あるいは、その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含み、かつ、プロモーター活性を有する塩基配列からなる、請求項3に記載の方法。
【請求項5】
前記メチル化をメチル化特異的PCR法によって検出することを含む、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
前記PCR法で使用するためのプライマーを、配列番号78、79または80に示される塩基配列中の、あるいは、その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含み、かつ、プロモーター活性を有する塩基配列中の、連続する17〜35塩基からなる配列を、メチル化特異的プライマーおよび/または非メチル化特異的プライマーとして選択する、請求項5に記載の方法。
【請求項7】
前記サンプルが血清、胸水、気管支洗浄液または患部組織である、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
請求項1〜7のいずれか1項に記載の方法で使用するためのキットであって、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を測定することが可能である、メチル化特異的PCR用の下記のプライマーセットを含むキット。
TMEM30B遺伝子のメチル化特異的プライマー
TTGGGAGGGTGTGGGTATTGTTC(配列番号66)
ACGCGCGAACTAACCGAAT(配列番号67)
CTGGGAGGGTGTGGGCACTGCTCG(配列番号81)
CGAGTAGTTTTCGGGTTTGC(配列番号95)
ACCGACGCGCGAACTAAC(配列番号96)
TMEM30B遺伝子の非メチル化特異的プライマー
GGAATTGGGAGGGTGTGGGTATTGTTT(配列番号68)
ACCTCCCTCTCCACACCAC(配列番号69)
GGAACTGGGAGGGTGTGGGCACTGCT(配列番号82)
TGAGTAGTTTTTGGGTTTGT(配列番号97)
ACCATAACACTCCAAATCATAACA(配列番号98)
KAZALD1遺伝子のメチル化特異的プライマー
GCGGGTCGTAGTATTTGTCG(配列番号70)
AACAAAACCAACCACCTTCG(配列番号71)
CGGGTCGTAGTATTTGTC(配列番号85)
GGTAAGTTTGCGGTCGTTTTC(配列番号99)
AACAAAACCAACCACCTTCG(配列番号100)
KAZALD1遺伝子の非メチル化特異的プライマー
TTATTTGTGGGTTGTAGTATTTGTT(配列番号72)
AAACCAACCACCTTCATCTCC(配列番号73)
TTATTCGCGGGTCGTAGTATTTGTC(配列番号86)
GTGGGTAAGTTTGTGGTTG(配列番号101)
AACAAAACCAACCACCTTCATCT(配列番号102)
MAPK13遺伝子のメチル化特異的プライマー
CGTGTTTTCGACGTACGTCGGTAGC(配列番号74)
ACCGCTAAATAACCCCGAAA(配列番号75)
CGTGTCCCCGACGCACGTCGGCAGC(配列番号89)
MAPK13遺伝子の非メチル化特異的プライマー
GGTTTTGTTGGGTAGTTTT(配列番号76)
TCCCACCTACCAACTCACC(配列番号77)
GGTCCCGCTGGGCAGCCC(配列番号90)
【請求項1】
悪性胸膜中皮腫を検出するための方法であって、被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を検出することを含み、該遺伝子がメチル化されているとき悪性胸膜中皮腫であると判定する、上記方法。
【請求項2】
悪性胸膜中皮腫に罹患した被験者の予後を判定する方法であって、該被験者からの生物学的サンプルにおいて、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化の程度を測定することを含み、該遺伝子のメチル化程度が、予後不良被験者のメチル化程度と比べて低いとき、該疾患の予後が良好であると決定する、上記方法。
【請求項3】
前記TMEM30B遺伝子、KAZALD1遺伝子またはMAPK13遺伝子のプロモーターのCpG領域の塩基配列のメチル化を検出または測定する、請求項1または2に記載の方法。
【請求項4】
前記プロモーターのCpG領域の塩基配列が、配列番号78、79または80に示される塩基配列、あるいは、その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含み、かつ、プロモーター活性を有する塩基配列からなる、請求項3に記載の方法。
【請求項5】
前記メチル化をメチル化特異的PCR法によって検出することを含む、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
前記PCR法で使用するためのプライマーを、配列番号78、79または80に示される塩基配列中の、あるいは、その塩基配列において1もしくは数個の塩基の欠失、置換または付加を含み、かつ、プロモーター活性を有する塩基配列中の、連続する17〜35塩基からなる配列を、メチル化特異的プライマーおよび/または非メチル化特異的プライマーとして選択する、請求項5に記載の方法。
【請求項7】
前記サンプルが血清、胸水、気管支洗浄液または患部組織である、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
請求項1〜7のいずれか1項に記載の方法で使用するためのキットであって、TMEM30B遺伝子、KAZALD1遺伝子およびMAPK13遺伝子からなる群から選択される少なくとも1つの遺伝子のメチル化を測定することが可能である、メチル化特異的PCR用の下記のプライマーセットを含むキット。
TMEM30B遺伝子のメチル化特異的プライマー
TTGGGAGGGTGTGGGTATTGTTC(配列番号66)
ACGCGCGAACTAACCGAAT(配列番号67)
CTGGGAGGGTGTGGGCACTGCTCG(配列番号81)
CGAGTAGTTTTCGGGTTTGC(配列番号95)
ACCGACGCGCGAACTAAC(配列番号96)
TMEM30B遺伝子の非メチル化特異的プライマー
GGAATTGGGAGGGTGTGGGTATTGTTT(配列番号68)
ACCTCCCTCTCCACACCAC(配列番号69)
GGAACTGGGAGGGTGTGGGCACTGCT(配列番号82)
TGAGTAGTTTTTGGGTTTGT(配列番号97)
ACCATAACACTCCAAATCATAACA(配列番号98)
KAZALD1遺伝子のメチル化特異的プライマー
GCGGGTCGTAGTATTTGTCG(配列番号70)
AACAAAACCAACCACCTTCG(配列番号71)
CGGGTCGTAGTATTTGTC(配列番号85)
GGTAAGTTTGCGGTCGTTTTC(配列番号99)
AACAAAACCAACCACCTTCG(配列番号100)
KAZALD1遺伝子の非メチル化特異的プライマー
TTATTTGTGGGTTGTAGTATTTGTT(配列番号72)
AAACCAACCACCTTCATCTCC(配列番号73)
TTATTCGCGGGTCGTAGTATTTGTC(配列番号86)
GTGGGTAAGTTTGTGGTTG(配列番号101)
AACAAAACCAACCACCTTCATCT(配列番号102)
MAPK13遺伝子のメチル化特異的プライマー
CGTGTTTTCGACGTACGTCGGTAGC(配列番号74)
ACCGCTAAATAACCCCGAAA(配列番号75)
CGTGTCCCCGACGCACGTCGGCAGC(配列番号89)
MAPK13遺伝子の非メチル化特異的プライマー
GGTTTTGTTGGGTAGTTTT(配列番号76)
TCCCACCTACCAACTCACC(配列番号77)
GGTCCCGCTGGGCAGCCC(配列番号90)
【図1A】

【図1B】

【図1C】
【図1D】
【図2】
【図3A】

【図3B】
【図3C】
【図3D】
【図4A】

【図4B】
【図4C】
【図4D】

【図1B】
【図1C】
【図1D】
【図2】
【図3A】
【図3B】
【図3C】
【図3D】
【図4A】
【図4B】
【図4C】
【図4D】
【公開番号】特開2010−279311(P2010−279311A)
【公開日】平成22年12月16日(2010.12.16)
【国際特許分類】
【出願番号】特願2009−136624(P2009−136624)
【出願日】平成21年6月5日(2009.6.5)
【出願人】(304031427)愛知県 (36)
【Fターム(参考)】
【公開日】平成22年12月16日(2010.12.16)
【国際特許分類】
【出願日】平成21年6月5日(2009.6.5)
【出願人】(304031427)愛知県 (36)
【Fターム(参考)】
[ Back to top ]