
悪性腫瘍細胞抑制タンパク質、悪性腫瘍細胞抑制遺伝子、悪性腫瘍細胞抑制ウイルスベクター及びそれを用いたキット
【課題】 悪性腫瘍細胞において過剰発現し、抗悪性腫瘍作用を示すウイルスベクター及びそれを用いたキットを提供する。
【解決手段】 以下の(a)又は(b)の悪性腫瘍細胞抑制タンパク質。(a)ヒト由来の癌細胞の増殖,転移,浸潤を抑制する特定のアミノ酸配列を有するタンパク質(b)上記タンパク質(a)のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を有するタンパク質
【解決手段】 以下の(a)又は(b)の悪性腫瘍細胞抑制タンパク質。(a)ヒト由来の癌細胞の増殖,転移,浸潤を抑制する特定のアミノ酸配列を有するタンパク質(b)上記タンパク質(a)のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を有するタンパク質
【発明の詳細な説明】
【技術分野】
【0001】
本発明は悪性腫瘍細胞抑制タンパク質、悪性腫瘍細胞抑制遺伝子、及び悪性腫瘍細胞抑制組換えベクターに関する。より詳細には、本発明は、悪性腫瘍細胞特異的に抗腫瘍作用を示すウイルスベクター及びそれを用いたキットに関する。
【背景技術】
【0002】
これまで、ヒトゲノムおよびマウスゲノムの解明の一環として、ヒトおよびマウスcDNAの解明が行われてきた。マウスゲノムはほとんどヒトゲノムそのものを反映しており、マウスで単離された遺伝子は直接ヒト疾患の研究に用いることができ、ヒト疾患の研究モデルとしてマウスが活用されている。特に、マウスcDNAマイクロアレイを用いた異なる細胞間の遺伝子発現差の網羅的解析技術の開発や、種々のヒト癌とその対応する器官の正常組織との間の遺伝子発現差の検討が行われている。その結果、種々の新規癌関連遺伝子の同定がなされてきた。例えば、種々の癌抑制遺伝子が同定・解析されて、前臨床試験(動物試験)で癌の抑制、治療の標的としてその有用性が示されている(P53,p16waf-1,Rb,PTEN,TSLC-1,KAI-1等の遺伝子)。
ところで、全長cDNAの網羅的な遺伝子配列決定により、様々な遺伝子配列そのものは大量に決定されている。しかし、それらのほとんどの遺伝子が生体内で、どのような働きがあるかはいまだ決定できていないのが現状である(例えば、非特許文献1、2参照。)。
現在、癌の治療の1つとして、遺伝子治療が行われている。しかし、遺伝子治療では、安全性のために非増殖性ウイルスベクターを用いて遺伝子導入するため、標的細胞の範囲にしか適用できず、また、導入効率が低いために満足のいく治療効果が得られていない。
また、癌化した細胞又は不死化した細胞株においてテロメラーゼの活性が増大している頻度が高く、生殖系列の細胞、血球系細胞、上皮系幹細胞等以外の正常な体細胞ではテロメラーゼの活性はほとんど検出されないことが知られている。
【0003】
【非特許文献1】Okazaki,Y.,Hayashizaki,Y. et al.: Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs Nature 420(6915),563-573(2002)
【非特許文献2】Strausberg,R.L. et al.; Generation and initial analysis of more than 15,000 full-length human and mouse cDNA sequences Proc.Natl.Acad.Sci.U.S.A. 99(26),16899-16903(2002)
【発明の開示】
【発明が解決しようとする課題】
【0004】
本発明の目的は、悪性腫瘍細胞において特異的に増殖・発現し、抗腫瘍作用を示すウイルスベクター及びそれを用いたキットを提供することにある。
【課題を解決するための手段】
【0005】
本発明者らは、癌細胞の増殖および浸潤や転移を抑制する遺伝子を標的として網羅的発現解析を行った結果、網羅的にcDNA配列決定された遺伝子の中から、悪性腫瘍抑制遺伝子SVS-1(マウスSVS-1(mSVS-1)およびヒトSVS-1(hSVS-1)遺伝子)を発見し、前記遺伝子が悪性腫瘍細胞の増殖を抑制する機能を有することを見出した。本発明はこの知見に基づき完成するに至った。
すなわち、本発明は、
(1)下記(a)又は(b)の悪性腫瘍細胞抑制タンパク質、
(a)配列番号1に示すアミノ酸配列を有するタンパク質
(b)配列番号1に示すアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を有するタンパク質
(2)下記(c)又は(d)の悪性腫瘍細胞抑制遺伝子、
(c)配列番号2に示す塩基配列からなるDNA
(d)配列番号2に示す塩基酸配列において1若しくは数個の塩基が欠失、置換、付加及び/又は挿入された塩基配列からなるDNA
(3) 転写因子が結合するプロモーター配列の下流に(1)項に記載の悪性腫瘍細胞抑制タンパク質コードする遺伝子を含む組換えベクター、
(4) テロメラーゼプロモーターの下流に(3)項記載の転写因子をコードする遺伝子を含むことを特徴とする組換えアデノウイルスベクター、
(5) 前記(3)項に記載の組換えベクター及び(4)項に記載のアデノウイルスベクターを含む悪性腫瘍細胞抑制ウイルスベクターキット、
(6) テロメラーゼプロモーターの下流に(1)項に記載の悪性腫瘍細胞抑制タンパク質をコードする遺伝子を含む組換えアデノウイルスベクター、及び
(7) テロメラーゼプロモーターの下流に転写因子をコードする遺伝子を含み、かつ前記転写因子が結合するプロモーター配列の下流に(1)項に記載の悪性腫瘍細胞抑制タンパク質コードする遺伝子を含む組換えアデノウイルスベクター
を提供するものである。
【発明の効果】
【0006】
本発明の配列番号1で表される配列を有するタンパク質は、悪性腫瘍細胞抑制機能を有する。
本発明の悪性腫瘍細胞抑制遺伝子は、癌治療用医薬組成物(ウイルスベクター、創薬等)の開発に貢献できる。
本発明のウイルスベクター及びそれを用いたキットは、悪性腫瘍細胞特異的抑制機能を有する。
【発明を実施するための最良の形態】
【0007】
以下、本発明を詳細に説明する。
まず、本発明の遺伝子等の悪性腫瘍細胞抑制遺伝子の取得について説明する。
悪性腫瘍細胞抑制遺伝子のスクリーニングはマウスcDNAマイクロアレイ法等により行なうことができる。
ここで、マウスゲノムはほとんどヒトゲノムそのものを反映しており、マウスで単離された遺伝子は直接ヒト疾患の研究に用いることができる(例えば、Okazaki,Y.,Hayashizaki,Y. et al.: Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs Nature 420(6915),563-573(2002)参照。)。
cDNAマイクロアレイ法については、例えば、監修 林崎良英 編集 岡崎康司「必ずデータが出る DNAマイクロアレイ実践マニュアル」(羊土社)、Mark Schena編、加藤郁之進監訳「DNAマイクロアレイ」(発売所丸善)等に記載の方法に準じて行なうことができる。
例えば、マウス正常せんい穿細胞NIH3T3と活性化癌遺伝子v-K-rasによるトランスフォーマント(TF、癌化)Ki-3T3細胞との間の遺伝子の発現差を20,000個(20K)のマウスcDNAを搭載したマイクロアレイを用いて解析できる。実施例の項で後述するように、親株NIH3T3に比較して癌化細胞Ki-3T3で2倍以上に高発現している遺伝子が433個、1/2以下に発現が抑制されている遺伝子が313個同定することができる。
さらに上記発現が低下している遺伝子の翻訳領域についてNCBI Conserved Domain Search(http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml)等の機能ドメイン検索により前記遺伝子がコードするタンパク質の機能を推定することができる。これにより癌抑制活性を示すことが予想される遺伝子を複数同定することができる。
その中の一つとして、本発明の悪性腫瘍細胞抑制タンパク質(配列番号1;以下、単にSVS-1タンパク質ということもある。)をコードする配列番号2で表される遺伝子(以下、単にSVS-1遺伝子という。)を取得することができる。
図1は、悪性腫瘍細胞抑制タンパク質のアミノ酸配列(配列番号1)を示す図である。図2は、悪性腫瘍細胞抑制遺伝子の塩基配列(配列番号2)と図1で示したアミノ酸配列(配列番号1)との相関を示す図である。
【0008】
次に、前記悪性腫瘍細胞抑制遺伝子を悪性腫瘍細胞に導入するためのベクターの構築について説明する。
本発明のウイルスベクターは、多くの癌細胞がテロメラーゼ活性を有することから、テロメラーゼのプロモーターを導入した遺伝子を有することにより、悪性腫瘍細胞を特異的に死滅させることを特徴とする。
本発明は、前記SVS-1遺伝子安定導入腫瘍細胞を樹立するための組換えベクター、及び融合転写因子(Gal4VP2)をコード(配列番号3)するアデノウイルスベクターを含む悪性腫瘍細胞抑制ウイルスベクターキットとすることができる。前記キットにおいて、前記SVS-1遺伝子安定導入腫瘍細胞を樹立するための組換えベクターと融合転写因子(Gal4VP2)をコードするアデノウイルスベクターとが混合されていてもいなくてもよい。前記キットが組成物である場合には、組成比は特に制限はないが、1:1が好ましい。
前記SVS-1遺伝子安定導入腫瘍細胞を樹立するための組換えベクターは、融合転写因子(Gal4VP2)が結合するプロモーター配列(G5E4T)の下流に前記悪性腫瘍細胞抑制タンパク質(配列番号1)をコードする遺伝子を含む組換えベクターであることが好ましい。
用いる宿主ベクターはプラスミドベクターpG5(カリフォルニア大学ロサンゼルス校医学部泌尿器科(University of California at Los Angels, Department of Urology School of Medicine) L Wu教授らから譲り受けたベクター。例えば、L. Zhang et al., Molec. Therapy 5(3):223-232,2002.参照。)等が好ましい。
図3は、プロモーター配列(G5E4T)(配列番号4)を示す図である。図3から明らかなようにプロモーター配列(G5E4T)はG5DNA結合部位及びE4TATAプロモーターを有する。
前記悪性腫瘍細胞抑制タンパク質(配列番号1)をコードする遺伝子にFlagタグ遺伝子(配列番号5)を直結させること(配列番号6)で、抗FLAG抗体により前記悪性腫瘍細胞抑制タンパク質の発現を確認することができる。
また、G418耐性遺伝子を有するベクター(例えば、pcDNA3.1v5his(Neo)ベクター(インビトロジェン社製))を共導入することで、SVS-1遺伝子安定導入腫瘍細胞クローンをG418薬剤選択することができる。
【0009】
上記安定導入腫瘍細胞においてSVS-1遺伝子を誘導的に発現させるための融合転写因子(Gal4VP2)発現アデノウイルスベクター(以下単にAd-Gal4VP2ウイルスということもある。)は、M.Lyer et al.,Proc. Nat. Acad. Sci.98:14595-14600,2002.、L. Zhang et al., Molec. Therapy 5(3):223-232,2002.、JY Adams et al.,Nature Medicine 8(8):891-896,2002.、L Wu et al.,Trends in Molec. Medicine 9(10):421-429,2003.等に記載の手順に準じて製造できる。
具体的には、まず、アデノウイルスベクターキット(AdEasyキット(ストラタジーン社製))中のpShuttle-CMVのCMVプロモーターの下流の制限酵素XhoI-XbaI部位にGal4VP2遺伝子(配列番号3)を導入する。そして、これをコンピテント大腸菌BJ5183-AD1(ストラタジーン社製)へトランスフォーメーションすることにより、コンピテント大腸菌内に導入されているアデノウィルスゲノムとpShuttle-CMV-Gal4VP2が組み換えを起こしGal4VP2を構成的に発現するアデノウィルスベクターDNAを大腸菌内に構築することができる。このDNAを大腸菌から調製し、アデノウイルスのE1A遺伝子を発現するヒト腎細胞株HEK293に導入することにより目的のGal4VP2発現アデノウイルスベクターを調製することができる。
【0010】
特に、癌特異的発現のためのhTERTプロモーターを含有するGal4VP2発現アデノウイルスベクターも同様に作製することができる。
具体的には、pcDNA3-Gal4VP2から制限酵素Hind III, Xba IによりGal4VP2(配列番号3)を切り出し、シャトル・プラスミド(pShuttle)の同制限酵素サイトHind III, Xba Iで切断したベクターに挿入し、pShuttle-Gal4VP2を得る。次ぎにpGL-basic(プロメガ社製)の制限酵素Mlu I-Bgl II部位にhTERTプロモーターを入れたベクターpGL3328(金沢大学医学部 京 哲博士から譲り受けたベクター。例えば、Takakura et al:Cloning of human telomerase catalytic subunit(hTERT)gene promoter and identification of proximal core promoter sequences essential for transcriptional activation in immortalized and cancer cells. Cancer Res.59:551-557,1999参照。)をPvu II,Bgl IIで切断しhTERTプロモーターのコア部分を切り出し、これを平滑末端化し、hTERTを得る。pShuttle-Gal4VP2をBgl IIで消化し、平滑末端化する。これに前記hTERTを挿入しライゲーションするとpShuttle-hTERT-Gal4VP2が調製される。ただし、hTERTの向きが両方向性のものがあるので、制限酵素処理断片の生じ方によって目的の方向にhTERTが挿入されたDNAのみを選択することができる。このようにして、pShuttle-hTERT-Gal4VP2を作製することができる。そして、これをコンピテント大腸菌BJ5183-AD1(ストラタジーン社製)へ導入し、トランスフォーメーションし、コンピテント大腸菌内に予め入っているアデノウィルスゲノムと導入したpShuttle-hTERT-Gal4VP2との組み換えを起こさせGal4VP2を癌特異的に発現するアデノウィルスベクターDNAを大腸菌内に構築することができる。
このDNAを大腸菌から調製し、ヒト腎細胞株HEK293に導入することにより癌特異的発現のためのhTERTプロモーターを含有するGal4VP2発現アデノウイルスベクターを調製することができる。
【0011】
前記融合転写因子(Gal4VP2)は転写因子Gal4のDNA結合ドメインとHSVウイルスの転写因子VP16の転写活性化ドメインVP2とを融合した複合転写因子であり、人工的なプロモーターG5E4T特異的に結合し、その下流の遺伝子を誘導的に発現させること、及び目的遺伝子を「2段階の転写活性化(TSTA法,Two Step Transciption Activation)」を行うことにより転写効率を飛躍的に向上させることができるという観点から融合させたものである。
また、本発明の融合転写因子(Gal4VP2)発現アデノウイルスベクターは、ヒトテロメラーゼのプロモーターを融合転写因子(Gal4VP2)遺伝子の上流に有するのは、テロメラーゼ活性を有する悪性腫瘍細胞内で転写を促進することができるからである。ヒトテロメラーゼのプロモーターであれば種類などは限定されないが、その中でもhTERTが好ましい。
【0012】
本発明は、テロメラーゼプロモーターの下流に前記悪性腫瘍細胞抑制タンパク質(配列番号1)コードする遺伝子を含む組換えアデノウイルスベクターとすることもできる。
より好ましくは、テロメラーゼプロモーターの下流に転写因子をコードする遺伝子を含み、かつ前記転写因子が結合するプロモーター配列の下流に前記悪性腫瘍細胞抑制タンパク質コードする遺伝子を含む組換えアデノウイルスベクターである。
本発明において用いられるウイルスは特に限定されないが、安全性等の点からアデノウイルスが好ましい。また、アデノウイルスの中でも、使用の簡便さ等の点からタイプ5のアデノウイルスが特に好ましい。
【0013】
本発明のウイルスベクターは、以下の理由で副作用が生じる可能性は極めて低いと考えられ、非常に安全な製剤であるということができる。(1)正常の体細胞ではテロメラーゼ活性がほとんどなく、また、造血細胞等の浮遊細胞には本発明のウイルスベクターは感染しにくい。(2)本発明のウイルスベクターは転写因子Gal4VP2により誘導されるプロモーター配列(G5E4T)の下流遺伝子の発現を著しく増強することを特徴とし、通常の遺伝子治療で用いられている非誘導性ウイルスよりも低い濃度で使用することができる。(3)本発明のウイルスベクターが過剰に投与された場合であっても、生体内の通常の免疫作用によって抗ウイルス作用が働く。
このことから、本発明の悪性腫瘍細胞抑制遺伝子が、悪性腫瘍細胞特異的抑制に有用であることが分かる。
【0014】
次に、非増殖型SVS-1遺伝子組換えアデノウイルスの制限増殖型アデノウイルスとの共感染によるSVS-1遺伝子の発現と抗悪性腫瘍性の増強について説明する。
非増殖型組換えアデノウイルスの発現する遺伝子は制限増殖型(癌細胞でのみ増殖する)アデノウイルスの共感染によって非増殖型のウイルスも癌細胞では共に増殖し遺伝子も増幅され、その結果、組換え遺伝子の発現は大幅に増強されること、細胞効果も著しく増強されることが示されている(例えば、Sunamura M, Oonuma M, Motoi F, Abe, H, Saitoh Y, Hoshida T, Ottomo S, Hori A and Matsuno S:Gene therapy for pancreatic cancer targeting the genomic alterations of tumor suppressor genes using replication-selective oncolytic adenovirus. Hum Cell. 15:138-150,2002.、Nagayama Y, Nakao K, Mizuguchi H, Hayakawa T and Niwa M:Enhanced antitumor effect of combined replicative adenovirus and nonreplicative adenovirus expressing interleukin-12 in an immunocompetent mouse model. Gene Ther. 10:1400-1403,2003.等参照。)。
すなわち、本発明のウイルスベクターとAxE1AdBなどの制限増殖型のアデノウイルスとの共感染によってSVS-1遺伝子の悪性腫瘍内での発現は著しく増強され、従って抗腫瘍性も著しく増大するものと予想される。
【0015】
次に本発明のウイルスベクターの使用について説明する。
本発明のウイルスベクターは、抗悪性腫瘍細胞ベクターとして使用することができる。例えば、単なる癌の治療だけでなく、手術後の再発予防、転移の防止及び/又は予防等にも使用できる。
本発明のウイルスベクターを適用する癌の種類としては、限定されるものではなく、あらゆる種類の癌に用いることができる。特に、例えば、胃、大腸、肺、肝、前立腺、膵、食道、膀胱、胆嚢・胆管、乳房、子宮、甲状腺、卵巣等における固形癌に有効である。
【0016】
本発明のウイルスベクターは、そのまま患部に適用することもできるし、あらゆる公知の方法、例えば、静脈、筋肉、腹腔内又は皮下といった注射、鼻腔、口腔又は肺からの吸入、経口投与、坐剤、外用剤等により生体(対象となる細胞や臓器)に導入することもできる。
生体、即ち悪性腫瘍細胞に導入、即ち感染した本発明のウイルスベクターは、該細胞内で増殖し、該細胞を死滅させることができる。そうすることによって、癌を治療したり、悪性腫瘍細胞の増殖を抑制したり、転移を防いだりすることができる。
また、例えば凍結などの方法により扱いやすくした後、そのまま若しくは賦形剤、増量剤、結合剤、滑沢剤等公知の薬学的に許容される担体、公知の添加剤(緩衝剤、等張化剤、キレート剤、着色剤、保存剤、香料、風味剤、甘味剤等が含まれる。)などと混合して医薬組成物として調整することができる。
本発明のウイルスベクターは、錠剤、カプセル剤、散剤、穎粒剤、丸剤、液剤、シロップ剤等の経口投与剤、注射剤、外用剤、坐剤、点眼剤等の非経口投与剤などの形態に応じて、経口投与又は非経口投与することができる。好ましくは、筋肉、腹腔等への局部注射、静脈への注射等が例示される。
【0017】
投与量は、有効成分の種類、投与経路、投与対象、患者の年齢、体重、性別、症状その他の条件により適宜選択されるが、一日投与量として、通常有効成分である本発明のウイルスベクターの量を1x107〜1x1012PFU程度、好ましくは1x108〜1x1011PFU程度とするのがよく、1日1回投与することもでき、数回に分けて投与することもできる。
また、本発明のウイルスベクターを使用する際には、公知の免疫抑制剤等を用いることにより、生体の免疫を抑制し、該ウイルスが感染し易くすることもできる。
更に、本発明のウイルスベクターは、従来の遺伝子治療で用いられている例えばp53遺伝子を含むような非増殖性ウイルス、公知の抗癌剤及び放射線からなる群から選ばれる少なくとも1種の抗癌剤を併用することもできる。
【0018】
次に本発明のタンパク質に特異的に結合する抗体について説明する。
本発明のタンパク質と特異的に結合する抗体の調製方法としては、通常用いられる公知の方法を用いることができ、抗原として用いられるポリペプチドについても、公知の方法に従って抗原性が高くエピトープ(抗原決定基)として適した配列を選択して用いることができる。エピトープの選択方法としては、例えばEpitope Adviser(富士通九州ンステムエンジニアリング社製)等の市販のソフトウェアを用いることができる。
【0019】
上記の抗原として用いるポリぺプチドは、本発明のSVS-1タンパク質そのものを用いることができる。抗原となるポリペプチドは、公知の方法に従って適当な溶液等に調製して、哺乳動物、例えばウサギ、マウス、ラット等に免疫を行えばよいが、安定的な免疫を行ったり抗体価を高めるために抗原ペプチドを適当なキャリアタンパク質とのコンジュゲートにして用いたり、アジュバント等を加えて免疫を行うのが好ましい。
【0020】
免疫に際しての抗原の投与経路は特に限定されず、例えば皮下、腹腔内、静脈内、あるいは筋肉内等のいずれの経路を用いてもよい。具体的には、例えばBALB/cマウスに抗原ポリペプチドを数日〜数週間おきに数回接種する方法等が用いられる。また、抗原の摂取量としては、抗原がポリベプチドの場合0.3〜0.5mg/1回程度が好ましいが、免疫する動物種によっては適宜調節される。
【0021】
免疫後、適宜試験的に採血を行って固相酵素免疫検定法(以下、これを「ELISA法」と称することがある)やウエスタンブロッティング等の方法で抗体価の上昇を確認し、十分に抗体価の上昇した動物から採血を行う。これに抗体の調製に用いられる適当な処理を行えばポリクローナル抗体を得ることができる。具体的には、例えば、公知の方法に従い血清から抗体成分を精製した精製抗体を取得する方法等が挙げられる。抗体成分の精製は、遠析、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等の方法を用いることができる。
また、該動物の脾臓細胞とミエローマ細胞とを用いて公知の方法に従って融合させたハイブリドーマを用いる(Milstein,et al.,Nature,256,495(1975))ことによりモノクローナル抗体を作製することもできる。
また、本発明の悪性腫瘍細胞抑制タンパク質としてヒト由来のものを取得した場合には、かかるポリペプチド、あるいはその部分ペプチドを抗原として、ヒト末梢血リンパ球を移植したSevere combined immune deficiency(SCID)マウスに上記した方法と同様にして免疫し、該免疫動物の抗体産生細胞とヒトのミエローマ細胞とのハイブリドーマを作製することによってもヒト型抗体を作製することができる(Mosier,D.E.,et al.Nature,335,256-259(1988):Duchosal,M.A.,etal.,Nature,355,258-262(1992))。
【0022】
また、取得したヒト型抗体を産生するハイブリドーマからRNAを抽出し、目的のヒト型抗体をコードする遺伝子をクローニングして、この遺伝子を適当なベクターに挿入し、これを適当な宿主に導入して発現させることにより、さらに大量にヒト型抗体を作製することができる。ここで、抗原との結合性の低い抗体は、それ自体既知の進化工学的手法を用いることによりさらに結合性の高い抗体として取得することもできる。一価性抗体等の部分フラグメントは、例えばパパイン等を用いてFab部分とFc部分を切断し、アフィニティカラム等を用いてFab部分を回収することによって作製することができる。
かくして得られる本発明の悪性腫瘍細胞抑制タンパク質と特異的に結合する抗体は、本発明の悪性腫瘍細胞抑制タンパク質に特異的に結合することによって該タンパク質の検出方法に用いることができる。このとき、任意の2次抗体検出により検出定量できる。
【実施例】
【0023】
次に、実施例に基づいて本発明をより詳細に説明するが、本発明はこれらにより限定されるものではない。
【0024】
(1)マウスcDNAマイクロアレイによる悪性腫瘍細胞抑制遺伝子のスクリーニング
まず、マウス正常せんい穿細胞NIH3T3と活性化癌遺伝子v-K-rasによるトランスフォーマントKi-3T3細胞との間の遺伝子の発現差を20,000個(20K)のマウスcDNAを搭載したマイクロアレイを用いて解析した。その結果、親株NIH3T3に比較して癌化細胞Ki-3T3で2倍以上に高発現している遺伝子が433個、1/2以下に発現が抑制されている遺伝子が313個同定された。
(2)機能ドメイン検索
上記発現低下している遺伝子の中から、それらの遺伝子のコードするタンパクの機能ドメイン検索から癌抑制活性を示すことが予想された遺伝子をいくつか同定し、その中の一つとして得たSVS-1遺伝子(配列番号2)のタンパク質への翻訳領域について、前記NCBI Conserved Domain Search(http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml)のドメイン検索を行なった。
ドメイン検索の結果得られた予測されるドメインを下記表1に示す。
【0025】
【表1】

【0026】
表1から明らかなように、SVS-1タンパク質は、公知の癌抑制遺伝子drsに存在するsushiドメインを有していたこと、sushi(CCP,SCR)ドメイン、AMOPドメイン、膜貫通領域等が存在することから膜タンパク質であり細胞接着等に関与することが推測でき、また、細胞膜に存在し、細胞接着に関わるタンパクには癌抑制効果を示す遺伝子が多く見つかっていることから悪性腫瘍細胞抑制機能があることが推定された。「(6)悪性腫瘍細胞抑制試験」で後述するように、SVS-1タンパク質が細胞膜に局在し、細胞接着等に関与するタンパク質であることを示したことからもこの推定は支持される。
(3)SVS-1遺伝子安定導入株(HT/SVS-1及びHeLa/SVS-1)の樹立
まず、プラスミドベクターpG5(カリフォルニア大学ロサンゼルス校医学部泌尿器科 L Wu教授らから譲り受けたベクター、例えば、L. Zhang et al., Molec. Therapy 5(3): 223-232,2002.参照。)の融合転写因子(Gal4VP2)によって認識され結合されるDNA上のプロモーター配列(G5E4T)(配列番号4)の下流の制限酵素Xba I部位に、Flagタグ遺伝子(配列番号5)を直結させたSVS-1遺伝子(配列番号6)を挿入しプラスミドベクターpG5-Flag-svs1を構築した。
つぎに、癌細胞HT1080細胞(せんい肉腫)とHeLa細胞(子宮頚部癌)にそれぞれ、リポフェクション法により、前記pG5-Flag-svs1ベクターとG418耐性遺伝子を有するpcDNA3.1v5his(Neo)ベクター(インビトロジェン社製)を共導入し、2週間のG418薬剤選択の後、SVS-1遺伝子安定導入腫瘍細胞クローンを得た。なお、HT1080細胞に関しては、4クローン得られたうちの1クローン、HeLa細胞に関しては、24クローン得られたうちの2クローンが発現誘導する株であった。
【0027】
(4)融合転写因子発現ベクターの構築
上記安定導入腫瘍細胞においてSVS-1遺伝子を誘導的に発現させるための融合転写因子(Gal4VP2)発現アデノウイルスベクター(Ad-Gal4VP2ウイルス)を、M.Lyer et al.,Proc. Nat. Acad. Sci.98:14595-14600,2002.、L. Zhang et al., Molec. Therapy 5(3): 223-232,2002、JY Adams et al.,Nature Medicine 8(8):891-896,2002.、L Wu et al.,Trends in Molec. Medicine 9(10):421-429,2003.等に記載の手順に準じて製造した。
具体的には、pcDNA3-Gal4VP2からHind III, Xba IによりGal4VP2(配列番号3)を切り出し、pShuttleの同制限酵素サイトHind III,Xba Iで切断したベクターに挿入した(pShuttel-Gal4VP2)。次ぎにpGL-basic(プロメガ社製)の制限酵素Mlu I-Bgl II部位にhTERTプロモーターを入れたベクターpGL3328(金沢大学医学部 京 哲博士から譲り受けた。)をPvu II,Bgl IIで切断しhTERTプロモーターのコア部分を切り出し、これを平滑末端化しておいた(hTERT)。pShuttle-Gal4VP2をBgl IIで消化し、平滑末端化した。これに前記hTERTを挿入しライゲーションするとpShuttle-hTERT-Gal4VP2が出来上がった。ただし、hTERTの向きが両方向性のものがあるので、制限酵素処理断片の生じ方によって目的の方向にhTERTが挿入されたDNAのみを選択した。かくしてpShuttle-hTERT-Gal4VP2を作製した(収量10μg)。そして、これをコンピテント大腸菌BJ5183-AD1(ストラタジーン社製)(1x1010細胞)へ導入し、LB寒天培地で37℃で一晩培養し、トランスフォーメーションした。コンピテント大腸菌内に予め入っているアデノウィルスゲノムと導入したpShuttle-hTERT-Gal4VP2との組み換えを起こさせGal4VP2を癌特異的に発現するアデノウィルスベクターDNAを大腸菌内に構築した。
このDNAを大腸菌から調製し、ヒト腎細胞株HEK293に導入することにより癌特異的発現のためのhTERTプロモーターを含有するGal4VP2発現アデノウイルスベクターを調製した(収量1x1013PFU)。
(5)SVS-1遺伝子誘導的発現の解析
(5−1)前記SVS-1導入株(HT/SVS-1又はHeLa/SVS-1)に前記Ad-Gal4VP2ウイルスを感染させ、誘導的にSVS-1遺伝子を発現させた。誘導発現培養条件を表2に示す。
【0028】
【表2】
【0029】
なお、moiは感染多重度、FBSはウシ胎仔血清である。
(5−2)発現を確認するため、モック(Mock;コントロールとして空ウイルスを感染させた細胞)との比較により、Ad-Gal4VP2ウイルスによる発現誘導を抗FLAG抗体を用いた免疫蛍光法により顕微鏡観察した。
まず、癌細胞HT1080細胞について観察すると(倍率200倍)、ヘキスト染色法(生きたままの細胞の核を染色する方法)によれば、モック、Ad-Gal4VP2ウイルス感染細胞いずれも染色させた。しかし、抗FLAG抗体を用いた免疫蛍光法により観察(倍率200倍)すると、モック細胞は蛍光がない一方、Ad-Gal4VP2ウイルス感染細胞は蛍光付きで観察された。
次に、HeLa細胞について観察すると(倍率400倍)、ヘキスト染色法(生きたままの細胞の核を染色する方法)によれば、モック、Ad-Gal4VP2ウイルス感染細胞いずれも染色させた。しかし、抗FLAG抗体を用いた免疫蛍光法により観察(倍率400倍)すると、モック細胞は蛍光がない一方、Ad-Gal4VP2ウイルス感染細胞は蛍光付きで観察された。
よって、Ad-Gal4VP2ウイルス感染細胞ではFLAGタグ付きのSVS-1タンパク質が誘導的に発現されていることが分かる。
【0030】
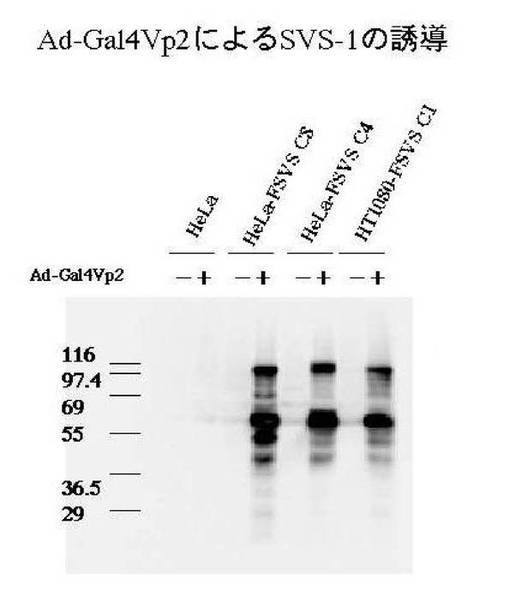
(5−3)Ad-Gal4VP2ウイルスにより誘導発現されることをFLAG抗体(αFLAG)を用いてウエスタンブロット法により解析を行った。結果を図3に示す。
図3から明らかなようにAd-Gal4VP2ウイルスによる感染細胞のみに約100kDaの完全長のSVS-1タンパクに由来するバンドと約60kDaのプロセシングを受けた断片のバンドが見られ、Ad-Gal4VP2ウイルスにより感染されなければ発現しないことがわかる。
【0031】
(6)悪性腫瘍細胞抑制試験
まず、SVS-1遺伝子を発現させることにより細胞骨格を破壊し球形化する試験をした。
(a)マウス正常せんい芽細胞NIH3T3に前記SVS-1のcDNAをトランスフェクトした時に細胞骨格のうち、ミクロフィラメント(アクチン繊維)が消失し、細胞が球形化した。
(b)位相差、及びSVS-1タンパクをオレンジ蛍光標識した顕微鏡観察したところ(倍率400%)、HeLa細胞にSVS-1を誘導発現した時は殆ど球状化し、凝集体(aggregation)を形成していた。対照実験として行ったAd/GFP(Green Fluorescent Protein)を感染・発現した細胞(位相差、及び緑蛍光による顕微鏡観察(倍率400%))はそのような像はみられなかった。
以上(a)、(b)の結果からSVS-1遺伝子を悪性腫瘍細胞において誘導発現することにより細胞骨格を破壊することができることがわかる。
【0032】
また、SVS-1タンパク質を誘導発現することにより悪性腫瘍細胞の浸潤能や転移能を低下させることが推定される。
すなわち、共焦点顕微鏡像観察(倍率800%)により、TritonX100により細胞膜の透過性を上げた細胞ではSVS-1タンパクは細胞内構造体(小胞体)と細胞膜上に、TritonX100未処理細胞(抗体が細胞内に侵入できず細胞表明の外にある抗原だけを染色する)では細胞表面の外側に発現している事が明瞭に示された事、また、上記(a)、(b)で示したように細胞形態を球状化し、細胞骨格を破壊する事から生体内での悪性腫瘍細胞の浸潤性や転移性を低下させることが推定される。上記推定は癌転移抑制遺伝子として同定された「TSLC-1」遺伝子の細胞内分布や凝集体の形成など、SVS-1と共通する性質であることから推定したものである(国立がんセンターと東京大学のグループによる研究、J.Biol.Chem.277:31014-31019,2002など)。
【0033】
以上のように、本発明の配列番号1で表される配列を有するタンパク質が、誘導発現されることにより悪性腫瘍細胞を抑制することがわかる。
また、癌細胞HT1080細胞(せんい肉腫)とHeLa細胞(子宮頚部癌)を用いた試験結果であることから、動物体内においてSVS-1遺伝子を悪性腫瘍細胞特異的に誘導発現することができ、悪性腫瘍細胞の細胞骨格を破壊することができることは明らかである。
【図面の簡単な説明】
【0034】
【図1】図1は、悪性腫瘍細胞抑制タンパク質のアミノ酸配列(配列番号1)を示す図である。
【図2】図2は、悪性腫瘍細胞抑制遺伝子の塩基配列(配列番号2)と図1で示したアミノ酸配列(配列番号1)との相関を示す図である。
【図3】図3は、G5E4T配列(配列番号4)を示す図である。
【図4】図4は、FLAG抗体(αFLAG)を用いたウエスタンブロットを示す図である。
【技術分野】
【0001】
本発明は悪性腫瘍細胞抑制タンパク質、悪性腫瘍細胞抑制遺伝子、及び悪性腫瘍細胞抑制組換えベクターに関する。より詳細には、本発明は、悪性腫瘍細胞特異的に抗腫瘍作用を示すウイルスベクター及びそれを用いたキットに関する。
【背景技術】
【0002】
これまで、ヒトゲノムおよびマウスゲノムの解明の一環として、ヒトおよびマウスcDNAの解明が行われてきた。マウスゲノムはほとんどヒトゲノムそのものを反映しており、マウスで単離された遺伝子は直接ヒト疾患の研究に用いることができ、ヒト疾患の研究モデルとしてマウスが活用されている。特に、マウスcDNAマイクロアレイを用いた異なる細胞間の遺伝子発現差の網羅的解析技術の開発や、種々のヒト癌とその対応する器官の正常組織との間の遺伝子発現差の検討が行われている。その結果、種々の新規癌関連遺伝子の同定がなされてきた。例えば、種々の癌抑制遺伝子が同定・解析されて、前臨床試験(動物試験)で癌の抑制、治療の標的としてその有用性が示されている(P53,p16waf-1,Rb,PTEN,TSLC-1,KAI-1等の遺伝子)。
ところで、全長cDNAの網羅的な遺伝子配列決定により、様々な遺伝子配列そのものは大量に決定されている。しかし、それらのほとんどの遺伝子が生体内で、どのような働きがあるかはいまだ決定できていないのが現状である(例えば、非特許文献1、2参照。)。
現在、癌の治療の1つとして、遺伝子治療が行われている。しかし、遺伝子治療では、安全性のために非増殖性ウイルスベクターを用いて遺伝子導入するため、標的細胞の範囲にしか適用できず、また、導入効率が低いために満足のいく治療効果が得られていない。
また、癌化した細胞又は不死化した細胞株においてテロメラーゼの活性が増大している頻度が高く、生殖系列の細胞、血球系細胞、上皮系幹細胞等以外の正常な体細胞ではテロメラーゼの活性はほとんど検出されないことが知られている。
【0003】
【非特許文献1】Okazaki,Y.,Hayashizaki,Y. et al.: Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs Nature 420(6915),563-573(2002)
【非特許文献2】Strausberg,R.L. et al.; Generation and initial analysis of more than 15,000 full-length human and mouse cDNA sequences Proc.Natl.Acad.Sci.U.S.A. 99(26),16899-16903(2002)
【発明の開示】
【発明が解決しようとする課題】
【0004】
本発明の目的は、悪性腫瘍細胞において特異的に増殖・発現し、抗腫瘍作用を示すウイルスベクター及びそれを用いたキットを提供することにある。
【課題を解決するための手段】
【0005】
本発明者らは、癌細胞の増殖および浸潤や転移を抑制する遺伝子を標的として網羅的発現解析を行った結果、網羅的にcDNA配列決定された遺伝子の中から、悪性腫瘍抑制遺伝子SVS-1(マウスSVS-1(mSVS-1)およびヒトSVS-1(hSVS-1)遺伝子)を発見し、前記遺伝子が悪性腫瘍細胞の増殖を抑制する機能を有することを見出した。本発明はこの知見に基づき完成するに至った。
すなわち、本発明は、
(1)下記(a)又は(b)の悪性腫瘍細胞抑制タンパク質、
(a)配列番号1に示すアミノ酸配列を有するタンパク質
(b)配列番号1に示すアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を有するタンパク質
(2)下記(c)又は(d)の悪性腫瘍細胞抑制遺伝子、
(c)配列番号2に示す塩基配列からなるDNA
(d)配列番号2に示す塩基酸配列において1若しくは数個の塩基が欠失、置換、付加及び/又は挿入された塩基配列からなるDNA
(3) 転写因子が結合するプロモーター配列の下流に(1)項に記載の悪性腫瘍細胞抑制タンパク質コードする遺伝子を含む組換えベクター、
(4) テロメラーゼプロモーターの下流に(3)項記載の転写因子をコードする遺伝子を含むことを特徴とする組換えアデノウイルスベクター、
(5) 前記(3)項に記載の組換えベクター及び(4)項に記載のアデノウイルスベクターを含む悪性腫瘍細胞抑制ウイルスベクターキット、
(6) テロメラーゼプロモーターの下流に(1)項に記載の悪性腫瘍細胞抑制タンパク質をコードする遺伝子を含む組換えアデノウイルスベクター、及び
(7) テロメラーゼプロモーターの下流に転写因子をコードする遺伝子を含み、かつ前記転写因子が結合するプロモーター配列の下流に(1)項に記載の悪性腫瘍細胞抑制タンパク質コードする遺伝子を含む組換えアデノウイルスベクター
を提供するものである。
【発明の効果】
【0006】
本発明の配列番号1で表される配列を有するタンパク質は、悪性腫瘍細胞抑制機能を有する。
本発明の悪性腫瘍細胞抑制遺伝子は、癌治療用医薬組成物(ウイルスベクター、創薬等)の開発に貢献できる。
本発明のウイルスベクター及びそれを用いたキットは、悪性腫瘍細胞特異的抑制機能を有する。
【発明を実施するための最良の形態】
【0007】
以下、本発明を詳細に説明する。
まず、本発明の遺伝子等の悪性腫瘍細胞抑制遺伝子の取得について説明する。
悪性腫瘍細胞抑制遺伝子のスクリーニングはマウスcDNAマイクロアレイ法等により行なうことができる。
ここで、マウスゲノムはほとんどヒトゲノムそのものを反映しており、マウスで単離された遺伝子は直接ヒト疾患の研究に用いることができる(例えば、Okazaki,Y.,Hayashizaki,Y. et al.: Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs Nature 420(6915),563-573(2002)参照。)。
cDNAマイクロアレイ法については、例えば、監修 林崎良英 編集 岡崎康司「必ずデータが出る DNAマイクロアレイ実践マニュアル」(羊土社)、Mark Schena編、加藤郁之進監訳「DNAマイクロアレイ」(発売所丸善)等に記載の方法に準じて行なうことができる。
例えば、マウス正常せんい穿細胞NIH3T3と活性化癌遺伝子v-K-rasによるトランスフォーマント(TF、癌化)Ki-3T3細胞との間の遺伝子の発現差を20,000個(20K)のマウスcDNAを搭載したマイクロアレイを用いて解析できる。実施例の項で後述するように、親株NIH3T3に比較して癌化細胞Ki-3T3で2倍以上に高発現している遺伝子が433個、1/2以下に発現が抑制されている遺伝子が313個同定することができる。
さらに上記発現が低下している遺伝子の翻訳領域についてNCBI Conserved Domain Search(http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml)等の機能ドメイン検索により前記遺伝子がコードするタンパク質の機能を推定することができる。これにより癌抑制活性を示すことが予想される遺伝子を複数同定することができる。
その中の一つとして、本発明の悪性腫瘍細胞抑制タンパク質(配列番号1;以下、単にSVS-1タンパク質ということもある。)をコードする配列番号2で表される遺伝子(以下、単にSVS-1遺伝子という。)を取得することができる。
図1は、悪性腫瘍細胞抑制タンパク質のアミノ酸配列(配列番号1)を示す図である。図2は、悪性腫瘍細胞抑制遺伝子の塩基配列(配列番号2)と図1で示したアミノ酸配列(配列番号1)との相関を示す図である。
【0008】
次に、前記悪性腫瘍細胞抑制遺伝子を悪性腫瘍細胞に導入するためのベクターの構築について説明する。
本発明のウイルスベクターは、多くの癌細胞がテロメラーゼ活性を有することから、テロメラーゼのプロモーターを導入した遺伝子を有することにより、悪性腫瘍細胞を特異的に死滅させることを特徴とする。
本発明は、前記SVS-1遺伝子安定導入腫瘍細胞を樹立するための組換えベクター、及び融合転写因子(Gal4VP2)をコード(配列番号3)するアデノウイルスベクターを含む悪性腫瘍細胞抑制ウイルスベクターキットとすることができる。前記キットにおいて、前記SVS-1遺伝子安定導入腫瘍細胞を樹立するための組換えベクターと融合転写因子(Gal4VP2)をコードするアデノウイルスベクターとが混合されていてもいなくてもよい。前記キットが組成物である場合には、組成比は特に制限はないが、1:1が好ましい。
前記SVS-1遺伝子安定導入腫瘍細胞を樹立するための組換えベクターは、融合転写因子(Gal4VP2)が結合するプロモーター配列(G5E4T)の下流に前記悪性腫瘍細胞抑制タンパク質(配列番号1)をコードする遺伝子を含む組換えベクターであることが好ましい。
用いる宿主ベクターはプラスミドベクターpG5(カリフォルニア大学ロサンゼルス校医学部泌尿器科(University of California at Los Angels, Department of Urology School of Medicine) L Wu教授らから譲り受けたベクター。例えば、L. Zhang et al., Molec. Therapy 5(3):223-232,2002.参照。)等が好ましい。
図3は、プロモーター配列(G5E4T)(配列番号4)を示す図である。図3から明らかなようにプロモーター配列(G5E4T)はG5DNA結合部位及びE4TATAプロモーターを有する。
前記悪性腫瘍細胞抑制タンパク質(配列番号1)をコードする遺伝子にFlagタグ遺伝子(配列番号5)を直結させること(配列番号6)で、抗FLAG抗体により前記悪性腫瘍細胞抑制タンパク質の発現を確認することができる。
また、G418耐性遺伝子を有するベクター(例えば、pcDNA3.1v5his(Neo)ベクター(インビトロジェン社製))を共導入することで、SVS-1遺伝子安定導入腫瘍細胞クローンをG418薬剤選択することができる。
【0009】
上記安定導入腫瘍細胞においてSVS-1遺伝子を誘導的に発現させるための融合転写因子(Gal4VP2)発現アデノウイルスベクター(以下単にAd-Gal4VP2ウイルスということもある。)は、M.Lyer et al.,Proc. Nat. Acad. Sci.98:14595-14600,2002.、L. Zhang et al., Molec. Therapy 5(3):223-232,2002.、JY Adams et al.,Nature Medicine 8(8):891-896,2002.、L Wu et al.,Trends in Molec. Medicine 9(10):421-429,2003.等に記載の手順に準じて製造できる。
具体的には、まず、アデノウイルスベクターキット(AdEasyキット(ストラタジーン社製))中のpShuttle-CMVのCMVプロモーターの下流の制限酵素XhoI-XbaI部位にGal4VP2遺伝子(配列番号3)を導入する。そして、これをコンピテント大腸菌BJ5183-AD1(ストラタジーン社製)へトランスフォーメーションすることにより、コンピテント大腸菌内に導入されているアデノウィルスゲノムとpShuttle-CMV-Gal4VP2が組み換えを起こしGal4VP2を構成的に発現するアデノウィルスベクターDNAを大腸菌内に構築することができる。このDNAを大腸菌から調製し、アデノウイルスのE1A遺伝子を発現するヒト腎細胞株HEK293に導入することにより目的のGal4VP2発現アデノウイルスベクターを調製することができる。
【0010】
特に、癌特異的発現のためのhTERTプロモーターを含有するGal4VP2発現アデノウイルスベクターも同様に作製することができる。
具体的には、pcDNA3-Gal4VP2から制限酵素Hind III, Xba IによりGal4VP2(配列番号3)を切り出し、シャトル・プラスミド(pShuttle)の同制限酵素サイトHind III, Xba Iで切断したベクターに挿入し、pShuttle-Gal4VP2を得る。次ぎにpGL-basic(プロメガ社製)の制限酵素Mlu I-Bgl II部位にhTERTプロモーターを入れたベクターpGL3328(金沢大学医学部 京 哲博士から譲り受けたベクター。例えば、Takakura et al:Cloning of human telomerase catalytic subunit(hTERT)gene promoter and identification of proximal core promoter sequences essential for transcriptional activation in immortalized and cancer cells. Cancer Res.59:551-557,1999参照。)をPvu II,Bgl IIで切断しhTERTプロモーターのコア部分を切り出し、これを平滑末端化し、hTERTを得る。pShuttle-Gal4VP2をBgl IIで消化し、平滑末端化する。これに前記hTERTを挿入しライゲーションするとpShuttle-hTERT-Gal4VP2が調製される。ただし、hTERTの向きが両方向性のものがあるので、制限酵素処理断片の生じ方によって目的の方向にhTERTが挿入されたDNAのみを選択することができる。このようにして、pShuttle-hTERT-Gal4VP2を作製することができる。そして、これをコンピテント大腸菌BJ5183-AD1(ストラタジーン社製)へ導入し、トランスフォーメーションし、コンピテント大腸菌内に予め入っているアデノウィルスゲノムと導入したpShuttle-hTERT-Gal4VP2との組み換えを起こさせGal4VP2を癌特異的に発現するアデノウィルスベクターDNAを大腸菌内に構築することができる。
このDNAを大腸菌から調製し、ヒト腎細胞株HEK293に導入することにより癌特異的発現のためのhTERTプロモーターを含有するGal4VP2発現アデノウイルスベクターを調製することができる。
【0011】
前記融合転写因子(Gal4VP2)は転写因子Gal4のDNA結合ドメインとHSVウイルスの転写因子VP16の転写活性化ドメインVP2とを融合した複合転写因子であり、人工的なプロモーターG5E4T特異的に結合し、その下流の遺伝子を誘導的に発現させること、及び目的遺伝子を「2段階の転写活性化(TSTA法,Two Step Transciption Activation)」を行うことにより転写効率を飛躍的に向上させることができるという観点から融合させたものである。
また、本発明の融合転写因子(Gal4VP2)発現アデノウイルスベクターは、ヒトテロメラーゼのプロモーターを融合転写因子(Gal4VP2)遺伝子の上流に有するのは、テロメラーゼ活性を有する悪性腫瘍細胞内で転写を促進することができるからである。ヒトテロメラーゼのプロモーターであれば種類などは限定されないが、その中でもhTERTが好ましい。
【0012】
本発明は、テロメラーゼプロモーターの下流に前記悪性腫瘍細胞抑制タンパク質(配列番号1)コードする遺伝子を含む組換えアデノウイルスベクターとすることもできる。
より好ましくは、テロメラーゼプロモーターの下流に転写因子をコードする遺伝子を含み、かつ前記転写因子が結合するプロモーター配列の下流に前記悪性腫瘍細胞抑制タンパク質コードする遺伝子を含む組換えアデノウイルスベクターである。
本発明において用いられるウイルスは特に限定されないが、安全性等の点からアデノウイルスが好ましい。また、アデノウイルスの中でも、使用の簡便さ等の点からタイプ5のアデノウイルスが特に好ましい。
【0013】
本発明のウイルスベクターは、以下の理由で副作用が生じる可能性は極めて低いと考えられ、非常に安全な製剤であるということができる。(1)正常の体細胞ではテロメラーゼ活性がほとんどなく、また、造血細胞等の浮遊細胞には本発明のウイルスベクターは感染しにくい。(2)本発明のウイルスベクターは転写因子Gal4VP2により誘導されるプロモーター配列(G5E4T)の下流遺伝子の発現を著しく増強することを特徴とし、通常の遺伝子治療で用いられている非誘導性ウイルスよりも低い濃度で使用することができる。(3)本発明のウイルスベクターが過剰に投与された場合であっても、生体内の通常の免疫作用によって抗ウイルス作用が働く。
このことから、本発明の悪性腫瘍細胞抑制遺伝子が、悪性腫瘍細胞特異的抑制に有用であることが分かる。
【0014】
次に、非増殖型SVS-1遺伝子組換えアデノウイルスの制限増殖型アデノウイルスとの共感染によるSVS-1遺伝子の発現と抗悪性腫瘍性の増強について説明する。
非増殖型組換えアデノウイルスの発現する遺伝子は制限増殖型(癌細胞でのみ増殖する)アデノウイルスの共感染によって非増殖型のウイルスも癌細胞では共に増殖し遺伝子も増幅され、その結果、組換え遺伝子の発現は大幅に増強されること、細胞効果も著しく増強されることが示されている(例えば、Sunamura M, Oonuma M, Motoi F, Abe, H, Saitoh Y, Hoshida T, Ottomo S, Hori A and Matsuno S:Gene therapy for pancreatic cancer targeting the genomic alterations of tumor suppressor genes using replication-selective oncolytic adenovirus. Hum Cell. 15:138-150,2002.、Nagayama Y, Nakao K, Mizuguchi H, Hayakawa T and Niwa M:Enhanced antitumor effect of combined replicative adenovirus and nonreplicative adenovirus expressing interleukin-12 in an immunocompetent mouse model. Gene Ther. 10:1400-1403,2003.等参照。)。
すなわち、本発明のウイルスベクターとAxE1AdBなどの制限増殖型のアデノウイルスとの共感染によってSVS-1遺伝子の悪性腫瘍内での発現は著しく増強され、従って抗腫瘍性も著しく増大するものと予想される。
【0015】
次に本発明のウイルスベクターの使用について説明する。
本発明のウイルスベクターは、抗悪性腫瘍細胞ベクターとして使用することができる。例えば、単なる癌の治療だけでなく、手術後の再発予防、転移の防止及び/又は予防等にも使用できる。
本発明のウイルスベクターを適用する癌の種類としては、限定されるものではなく、あらゆる種類の癌に用いることができる。特に、例えば、胃、大腸、肺、肝、前立腺、膵、食道、膀胱、胆嚢・胆管、乳房、子宮、甲状腺、卵巣等における固形癌に有効である。
【0016】
本発明のウイルスベクターは、そのまま患部に適用することもできるし、あらゆる公知の方法、例えば、静脈、筋肉、腹腔内又は皮下といった注射、鼻腔、口腔又は肺からの吸入、経口投与、坐剤、外用剤等により生体(対象となる細胞や臓器)に導入することもできる。
生体、即ち悪性腫瘍細胞に導入、即ち感染した本発明のウイルスベクターは、該細胞内で増殖し、該細胞を死滅させることができる。そうすることによって、癌を治療したり、悪性腫瘍細胞の増殖を抑制したり、転移を防いだりすることができる。
また、例えば凍結などの方法により扱いやすくした後、そのまま若しくは賦形剤、増量剤、結合剤、滑沢剤等公知の薬学的に許容される担体、公知の添加剤(緩衝剤、等張化剤、キレート剤、着色剤、保存剤、香料、風味剤、甘味剤等が含まれる。)などと混合して医薬組成物として調整することができる。
本発明のウイルスベクターは、錠剤、カプセル剤、散剤、穎粒剤、丸剤、液剤、シロップ剤等の経口投与剤、注射剤、外用剤、坐剤、点眼剤等の非経口投与剤などの形態に応じて、経口投与又は非経口投与することができる。好ましくは、筋肉、腹腔等への局部注射、静脈への注射等が例示される。
【0017】
投与量は、有効成分の種類、投与経路、投与対象、患者の年齢、体重、性別、症状その他の条件により適宜選択されるが、一日投与量として、通常有効成分である本発明のウイルスベクターの量を1x107〜1x1012PFU程度、好ましくは1x108〜1x1011PFU程度とするのがよく、1日1回投与することもでき、数回に分けて投与することもできる。
また、本発明のウイルスベクターを使用する際には、公知の免疫抑制剤等を用いることにより、生体の免疫を抑制し、該ウイルスが感染し易くすることもできる。
更に、本発明のウイルスベクターは、従来の遺伝子治療で用いられている例えばp53遺伝子を含むような非増殖性ウイルス、公知の抗癌剤及び放射線からなる群から選ばれる少なくとも1種の抗癌剤を併用することもできる。
【0018】
次に本発明のタンパク質に特異的に結合する抗体について説明する。
本発明のタンパク質と特異的に結合する抗体の調製方法としては、通常用いられる公知の方法を用いることができ、抗原として用いられるポリペプチドについても、公知の方法に従って抗原性が高くエピトープ(抗原決定基)として適した配列を選択して用いることができる。エピトープの選択方法としては、例えばEpitope Adviser(富士通九州ンステムエンジニアリング社製)等の市販のソフトウェアを用いることができる。
【0019】
上記の抗原として用いるポリぺプチドは、本発明のSVS-1タンパク質そのものを用いることができる。抗原となるポリペプチドは、公知の方法に従って適当な溶液等に調製して、哺乳動物、例えばウサギ、マウス、ラット等に免疫を行えばよいが、安定的な免疫を行ったり抗体価を高めるために抗原ペプチドを適当なキャリアタンパク質とのコンジュゲートにして用いたり、アジュバント等を加えて免疫を行うのが好ましい。
【0020】
免疫に際しての抗原の投与経路は特に限定されず、例えば皮下、腹腔内、静脈内、あるいは筋肉内等のいずれの経路を用いてもよい。具体的には、例えばBALB/cマウスに抗原ポリペプチドを数日〜数週間おきに数回接種する方法等が用いられる。また、抗原の摂取量としては、抗原がポリベプチドの場合0.3〜0.5mg/1回程度が好ましいが、免疫する動物種によっては適宜調節される。
【0021】
免疫後、適宜試験的に採血を行って固相酵素免疫検定法(以下、これを「ELISA法」と称することがある)やウエスタンブロッティング等の方法で抗体価の上昇を確認し、十分に抗体価の上昇した動物から採血を行う。これに抗体の調製に用いられる適当な処理を行えばポリクローナル抗体を得ることができる。具体的には、例えば、公知の方法に従い血清から抗体成分を精製した精製抗体を取得する方法等が挙げられる。抗体成分の精製は、遠析、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等の方法を用いることができる。
また、該動物の脾臓細胞とミエローマ細胞とを用いて公知の方法に従って融合させたハイブリドーマを用いる(Milstein,et al.,Nature,256,495(1975))ことによりモノクローナル抗体を作製することもできる。
また、本発明の悪性腫瘍細胞抑制タンパク質としてヒト由来のものを取得した場合には、かかるポリペプチド、あるいはその部分ペプチドを抗原として、ヒト末梢血リンパ球を移植したSevere combined immune deficiency(SCID)マウスに上記した方法と同様にして免疫し、該免疫動物の抗体産生細胞とヒトのミエローマ細胞とのハイブリドーマを作製することによってもヒト型抗体を作製することができる(Mosier,D.E.,et al.Nature,335,256-259(1988):Duchosal,M.A.,etal.,Nature,355,258-262(1992))。
【0022】
また、取得したヒト型抗体を産生するハイブリドーマからRNAを抽出し、目的のヒト型抗体をコードする遺伝子をクローニングして、この遺伝子を適当なベクターに挿入し、これを適当な宿主に導入して発現させることにより、さらに大量にヒト型抗体を作製することができる。ここで、抗原との結合性の低い抗体は、それ自体既知の進化工学的手法を用いることによりさらに結合性の高い抗体として取得することもできる。一価性抗体等の部分フラグメントは、例えばパパイン等を用いてFab部分とFc部分を切断し、アフィニティカラム等を用いてFab部分を回収することによって作製することができる。
かくして得られる本発明の悪性腫瘍細胞抑制タンパク質と特異的に結合する抗体は、本発明の悪性腫瘍細胞抑制タンパク質に特異的に結合することによって該タンパク質の検出方法に用いることができる。このとき、任意の2次抗体検出により検出定量できる。
【実施例】
【0023】
次に、実施例に基づいて本発明をより詳細に説明するが、本発明はこれらにより限定されるものではない。
【0024】
(1)マウスcDNAマイクロアレイによる悪性腫瘍細胞抑制遺伝子のスクリーニング
まず、マウス正常せんい穿細胞NIH3T3と活性化癌遺伝子v-K-rasによるトランスフォーマントKi-3T3細胞との間の遺伝子の発現差を20,000個(20K)のマウスcDNAを搭載したマイクロアレイを用いて解析した。その結果、親株NIH3T3に比較して癌化細胞Ki-3T3で2倍以上に高発現している遺伝子が433個、1/2以下に発現が抑制されている遺伝子が313個同定された。
(2)機能ドメイン検索
上記発現低下している遺伝子の中から、それらの遺伝子のコードするタンパクの機能ドメイン検索から癌抑制活性を示すことが予想された遺伝子をいくつか同定し、その中の一つとして得たSVS-1遺伝子(配列番号2)のタンパク質への翻訳領域について、前記NCBI Conserved Domain Search(http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml)のドメイン検索を行なった。
ドメイン検索の結果得られた予測されるドメインを下記表1に示す。
【0025】
【表1】
【0026】
表1から明らかなように、SVS-1タンパク質は、公知の癌抑制遺伝子drsに存在するsushiドメインを有していたこと、sushi(CCP,SCR)ドメイン、AMOPドメイン、膜貫通領域等が存在することから膜タンパク質であり細胞接着等に関与することが推測でき、また、細胞膜に存在し、細胞接着に関わるタンパクには癌抑制効果を示す遺伝子が多く見つかっていることから悪性腫瘍細胞抑制機能があることが推定された。「(6)悪性腫瘍細胞抑制試験」で後述するように、SVS-1タンパク質が細胞膜に局在し、細胞接着等に関与するタンパク質であることを示したことからもこの推定は支持される。
(3)SVS-1遺伝子安定導入株(HT/SVS-1及びHeLa/SVS-1)の樹立
まず、プラスミドベクターpG5(カリフォルニア大学ロサンゼルス校医学部泌尿器科 L Wu教授らから譲り受けたベクター、例えば、L. Zhang et al., Molec. Therapy 5(3): 223-232,2002.参照。)の融合転写因子(Gal4VP2)によって認識され結合されるDNA上のプロモーター配列(G5E4T)(配列番号4)の下流の制限酵素Xba I部位に、Flagタグ遺伝子(配列番号5)を直結させたSVS-1遺伝子(配列番号6)を挿入しプラスミドベクターpG5-Flag-svs1を構築した。
つぎに、癌細胞HT1080細胞(せんい肉腫)とHeLa細胞(子宮頚部癌)にそれぞれ、リポフェクション法により、前記pG5-Flag-svs1ベクターとG418耐性遺伝子を有するpcDNA3.1v5his(Neo)ベクター(インビトロジェン社製)を共導入し、2週間のG418薬剤選択の後、SVS-1遺伝子安定導入腫瘍細胞クローンを得た。なお、HT1080細胞に関しては、4クローン得られたうちの1クローン、HeLa細胞に関しては、24クローン得られたうちの2クローンが発現誘導する株であった。
【0027】
(4)融合転写因子発現ベクターの構築
上記安定導入腫瘍細胞においてSVS-1遺伝子を誘導的に発現させるための融合転写因子(Gal4VP2)発現アデノウイルスベクター(Ad-Gal4VP2ウイルス)を、M.Lyer et al.,Proc. Nat. Acad. Sci.98:14595-14600,2002.、L. Zhang et al., Molec. Therapy 5(3): 223-232,2002、JY Adams et al.,Nature Medicine 8(8):891-896,2002.、L Wu et al.,Trends in Molec. Medicine 9(10):421-429,2003.等に記載の手順に準じて製造した。
具体的には、pcDNA3-Gal4VP2からHind III, Xba IによりGal4VP2(配列番号3)を切り出し、pShuttleの同制限酵素サイトHind III,Xba Iで切断したベクターに挿入した(pShuttel-Gal4VP2)。次ぎにpGL-basic(プロメガ社製)の制限酵素Mlu I-Bgl II部位にhTERTプロモーターを入れたベクターpGL3328(金沢大学医学部 京 哲博士から譲り受けた。)をPvu II,Bgl IIで切断しhTERTプロモーターのコア部分を切り出し、これを平滑末端化しておいた(hTERT)。pShuttle-Gal4VP2をBgl IIで消化し、平滑末端化した。これに前記hTERTを挿入しライゲーションするとpShuttle-hTERT-Gal4VP2が出来上がった。ただし、hTERTの向きが両方向性のものがあるので、制限酵素処理断片の生じ方によって目的の方向にhTERTが挿入されたDNAのみを選択した。かくしてpShuttle-hTERT-Gal4VP2を作製した(収量10μg)。そして、これをコンピテント大腸菌BJ5183-AD1(ストラタジーン社製)(1x1010細胞)へ導入し、LB寒天培地で37℃で一晩培養し、トランスフォーメーションした。コンピテント大腸菌内に予め入っているアデノウィルスゲノムと導入したpShuttle-hTERT-Gal4VP2との組み換えを起こさせGal4VP2を癌特異的に発現するアデノウィルスベクターDNAを大腸菌内に構築した。
このDNAを大腸菌から調製し、ヒト腎細胞株HEK293に導入することにより癌特異的発現のためのhTERTプロモーターを含有するGal4VP2発現アデノウイルスベクターを調製した(収量1x1013PFU)。
(5)SVS-1遺伝子誘導的発現の解析
(5−1)前記SVS-1導入株(HT/SVS-1又はHeLa/SVS-1)に前記Ad-Gal4VP2ウイルスを感染させ、誘導的にSVS-1遺伝子を発現させた。誘導発現培養条件を表2に示す。
【0028】
【表2】
【0029】
なお、moiは感染多重度、FBSはウシ胎仔血清である。
(5−2)発現を確認するため、モック(Mock;コントロールとして空ウイルスを感染させた細胞)との比較により、Ad-Gal4VP2ウイルスによる発現誘導を抗FLAG抗体を用いた免疫蛍光法により顕微鏡観察した。
まず、癌細胞HT1080細胞について観察すると(倍率200倍)、ヘキスト染色法(生きたままの細胞の核を染色する方法)によれば、モック、Ad-Gal4VP2ウイルス感染細胞いずれも染色させた。しかし、抗FLAG抗体を用いた免疫蛍光法により観察(倍率200倍)すると、モック細胞は蛍光がない一方、Ad-Gal4VP2ウイルス感染細胞は蛍光付きで観察された。
次に、HeLa細胞について観察すると(倍率400倍)、ヘキスト染色法(生きたままの細胞の核を染色する方法)によれば、モック、Ad-Gal4VP2ウイルス感染細胞いずれも染色させた。しかし、抗FLAG抗体を用いた免疫蛍光法により観察(倍率400倍)すると、モック細胞は蛍光がない一方、Ad-Gal4VP2ウイルス感染細胞は蛍光付きで観察された。
よって、Ad-Gal4VP2ウイルス感染細胞ではFLAGタグ付きのSVS-1タンパク質が誘導的に発現されていることが分かる。
【0030】
(5−3)Ad-Gal4VP2ウイルスにより誘導発現されることをFLAG抗体(αFLAG)を用いてウエスタンブロット法により解析を行った。結果を図3に示す。
図3から明らかなようにAd-Gal4VP2ウイルスによる感染細胞のみに約100kDaの完全長のSVS-1タンパクに由来するバンドと約60kDaのプロセシングを受けた断片のバンドが見られ、Ad-Gal4VP2ウイルスにより感染されなければ発現しないことがわかる。
【0031】
(6)悪性腫瘍細胞抑制試験
まず、SVS-1遺伝子を発現させることにより細胞骨格を破壊し球形化する試験をした。
(a)マウス正常せんい芽細胞NIH3T3に前記SVS-1のcDNAをトランスフェクトした時に細胞骨格のうち、ミクロフィラメント(アクチン繊維)が消失し、細胞が球形化した。
(b)位相差、及びSVS-1タンパクをオレンジ蛍光標識した顕微鏡観察したところ(倍率400%)、HeLa細胞にSVS-1を誘導発現した時は殆ど球状化し、凝集体(aggregation)を形成していた。対照実験として行ったAd/GFP(Green Fluorescent Protein)を感染・発現した細胞(位相差、及び緑蛍光による顕微鏡観察(倍率400%))はそのような像はみられなかった。
以上(a)、(b)の結果からSVS-1遺伝子を悪性腫瘍細胞において誘導発現することにより細胞骨格を破壊することができることがわかる。
【0032】
また、SVS-1タンパク質を誘導発現することにより悪性腫瘍細胞の浸潤能や転移能を低下させることが推定される。
すなわち、共焦点顕微鏡像観察(倍率800%)により、TritonX100により細胞膜の透過性を上げた細胞ではSVS-1タンパクは細胞内構造体(小胞体)と細胞膜上に、TritonX100未処理細胞(抗体が細胞内に侵入できず細胞表明の外にある抗原だけを染色する)では細胞表面の外側に発現している事が明瞭に示された事、また、上記(a)、(b)で示したように細胞形態を球状化し、細胞骨格を破壊する事から生体内での悪性腫瘍細胞の浸潤性や転移性を低下させることが推定される。上記推定は癌転移抑制遺伝子として同定された「TSLC-1」遺伝子の細胞内分布や凝集体の形成など、SVS-1と共通する性質であることから推定したものである(国立がんセンターと東京大学のグループによる研究、J.Biol.Chem.277:31014-31019,2002など)。
【0033】
以上のように、本発明の配列番号1で表される配列を有するタンパク質が、誘導発現されることにより悪性腫瘍細胞を抑制することがわかる。
また、癌細胞HT1080細胞(せんい肉腫)とHeLa細胞(子宮頚部癌)を用いた試験結果であることから、動物体内においてSVS-1遺伝子を悪性腫瘍細胞特異的に誘導発現することができ、悪性腫瘍細胞の細胞骨格を破壊することができることは明らかである。
【図面の簡単な説明】
【0034】
【図1】図1は、悪性腫瘍細胞抑制タンパク質のアミノ酸配列(配列番号1)を示す図である。
【図2】図2は、悪性腫瘍細胞抑制遺伝子の塩基配列(配列番号2)と図1で示したアミノ酸配列(配列番号1)との相関を示す図である。
【図3】図3は、G5E4T配列(配列番号4)を示す図である。
【図4】図4は、FLAG抗体(αFLAG)を用いたウエスタンブロットを示す図である。
【特許請求の範囲】
【請求項1】
下記(a)又は(b)の悪性腫瘍細胞抑制タンパク質。
(a)配列番号1に示すアミノ酸配列を有するタンパク質
(b)配列番号1に示すアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を有するタンパク質
【請求項2】
下記(c)又は(d)の悪性腫瘍細胞抑制遺伝子。
(c)配列番号2に示す塩基配列からなるDNA
(d)配列番号2に示す塩基酸配列において1若しくは数個の塩基が欠失、置換、付加及び/又は挿入された塩基配列からなるDNA
【請求項3】
転写因子が結合するプロモーター配列の下流に請求項1に記載の悪性腫瘍細胞抑制タンパク質コードする遺伝子を含む組換えベクター。
【請求項4】
テロメラーゼプロモーターの下流に請求項3記載の転写因子をコードする遺伝子を含むことを特徴とする組換えアデノウイルスベクター。
【請求項5】
請求項3に記載の組換えベクター及び請求項4に記載のアデノウイルスベクターを含む悪性腫瘍細胞抑制ウイルスベクターキット。
【請求項6】
テロメラーゼプロモーターの下流に請求項1に記載の悪性腫瘍細胞抑制タンパク質をコードする遺伝子を含む組換えアデノウイルスベクター。
【請求項7】
テロメラーゼプロモーターの下流に転写因子をコードする遺伝子を含み、かつ前記転写因子が結合するプロモーター配列の下流に請求項1に記載の悪性腫瘍細胞抑制タンパク質コードする遺伝子を含む組換えアデノウイルスベクター。
【請求項1】
下記(a)又は(b)の悪性腫瘍細胞抑制タンパク質。
(a)配列番号1に示すアミノ酸配列を有するタンパク質
(b)配列番号1に示すアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を有するタンパク質
【請求項2】
下記(c)又は(d)の悪性腫瘍細胞抑制遺伝子。
(c)配列番号2に示す塩基配列からなるDNA
(d)配列番号2に示す塩基酸配列において1若しくは数個の塩基が欠失、置換、付加及び/又は挿入された塩基配列からなるDNA
【請求項3】
転写因子が結合するプロモーター配列の下流に請求項1に記載の悪性腫瘍細胞抑制タンパク質コードする遺伝子を含む組換えベクター。
【請求項4】
テロメラーゼプロモーターの下流に請求項3記載の転写因子をコードする遺伝子を含むことを特徴とする組換えアデノウイルスベクター。
【請求項5】
請求項3に記載の組換えベクター及び請求項4に記載のアデノウイルスベクターを含む悪性腫瘍細胞抑制ウイルスベクターキット。
【請求項6】
テロメラーゼプロモーターの下流に請求項1に記載の悪性腫瘍細胞抑制タンパク質をコードする遺伝子を含む組換えアデノウイルスベクター。
【請求項7】
テロメラーゼプロモーターの下流に転写因子をコードする遺伝子を含み、かつ前記転写因子が結合するプロモーター配列の下流に請求項1に記載の悪性腫瘍細胞抑制タンパク質コードする遺伝子を含む組換えアデノウイルスベクター。
【図3】


【図1】

【図2】

【図4】
【図1】
【図2】
【図4】
【公開番号】特開2007−6779(P2007−6779A)
【公開日】平成19年1月18日(2007.1.18)
【国際特許分類】
【出願番号】特願2005−191789(P2005−191789)
【出願日】平成17年6月30日(2005.6.30)
【出願人】(800000080)タマティーエルオー株式会社 (255)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【出願人】(598123138)学校法人 創価大学 (49)
【Fターム(参考)】
【公開日】平成19年1月18日(2007.1.18)
【国際特許分類】
【出願日】平成17年6月30日(2005.6.30)
【出願人】(800000080)タマティーエルオー株式会社 (255)
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【出願人】(598123138)学校法人 創価大学 (49)
【Fターム(参考)】
[ Back to top ]