
感染因子を含まないエンベロープウイルスベースVLPを単離するための改良法
感染因子を含まず、感染因子を不活化する前の対応するVLPと実質的に同程度に免疫原性である、エンベロープウイルスベースのウイルス様粒子(VLP)が記載される。エンベロープウイルスベースのVLP調製物における感染因子を不活化する改良法であって、VLPの免疫原性に有害な影響を及ぼさない方法もまた記載される。上記方法は、(a)エンベロープウイルスベースのウイルス様粒子調製物を作製するのに用いられる宿主細胞、または宿主細胞の成分から、エンベロープウイルスベースのウイルス様粒子調製物を単離するステップと、(b)エンベロープウイルスベースのウイルス様粒子調製物における実質的にすべての感染因子を不活化するのに十分な線量の電磁放射を、調製物へと適用するステップとを含む。

【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、感染因子(infectious agent)を含まない、エンベロープウイルスベースのウイルス様粒子(VLP)を単離する分野に関する。好ましい例において、該分野は、エンベロープウイルスベースのVLPの免疫原性に有害な影響を及ぼさない、感染因子を不活化する方法を包含する。特定の実施形態では、エンベロープウイルスベースのVLPを、昆虫細胞ベースの発現系において作製する。
【背景技術】
【0002】
(発明の背景)
アセンブリーは、宿主細胞内で見出されるアクセサリー因子を必要とすることが多いため、エンベロープウイルスベースのVLPを作製するには、宿主細胞の発現系内におけるVLPの発現およびアセンブリーを必要とすることが典型的である。宿主細胞内における生物学的成分の発現には、感染因子による汚染の危険性が付きまとう。タンパク質および多糖など、比較的小さな生物学的成分については、そのような作用物質を除去するのに、フィルターによる滅菌が一般に用いられるが、UVによる不活化または化学的な不活化など、他の方法もまた用いられている(単独で、またはフィルターベースの方法と組み合わせて用いられる)。例えば、オーエスキー病ウイルスの糖タンパク質D遺伝子を発現させるのに用いるときのバキュロウイルスを不活化するには、光化学的不活性化が用いられており、該発現するタンパク質に対する抗体の結合の低下は何ら観察されていない(非特許文献1を参照されたい)。しかし、VLPなど、比較的大きな生物学的成分については、VLPはサイズが感染因子(例えば、ウイルス)に近接し、濾過によりそのような感染因子から分離することが不可能なことが多いので、フィルターによる滅菌は適用可能でないことが多い。したがって、VLPを調製するときの感染因子を不活化するには、他の方法を用いる必要がある。ブタパルボウイルス(PPV:非エンベロープまたはカプシドウイルス)VLPを生成させるのに用いるときのバキュロウイルスを不活化するには、そのような他の方法が試みられている。Ruedaらは、バキュロウイルスに対する低温殺菌、化学的不活化、および、洗浄剤による不活化を比較し、PPV VLPの免疫原性に対して影響を及ぼさないことを見出した(例えば、非特許文献2を参照されたい)。
【先行技術文献】
【非特許文献】
【0003】
【非特許文献1】S.A. Weightmanら、Journal of Virological Methods(1999年)81巻:179〜182頁
【非特許文献2】P. Ruedaら、Vaccine(2001年)19巻:726〜734頁
【発明の概要】
【発明が解決しようとする課題】
【0004】
しかし、感染因子を不活化する典型的な方法をエンベロープウイルスベースのVLPに適用すると、該エンベロープウイルスベースのVLPの免疫原性が顕著に低下することが、思いがけず判明した。したがって、エンベロープウイルスベースのVLPに対して用いることができ、該VLPの免疫原性に有害な影響を及ぼすことなく感染因子を不活化する不活化法が必要とされている。
【課題を解決するための手段】
【0005】
(概要)
本発明の好ましい実施形態は、エンベロープウイルスベースのVLPの免疫原性に有害な影響を及ぼさない、本明細書で開示される、感染因子を不活化するための各種の方法および組成物と、不活化を受けていないVLP調製物と実質的に同じ免疫原性を有する、感染因子を含まないエンベロープウイルスベースのVLP調製物を含む組成物とを提供することにより、この必要を満たす。このような好ましい実施形態は、試験された電磁放射ベースの不活化法(単独の不活化法、または該電磁放射に反応性の化学物質を伴う不活化法)が、他の純粋に化学物質ベースの不活化法と比較して、エンベロープウイルスベースのVLPの免疫原性を結果として低下させないという驚くべき観察に基づく。
【0006】
本発明の態様は、感染因子を実質的に含まないエンベロープウイルスベースのウイルス様粒子調製物を単離する方法であって、(a)該エンベロープウイルスベースのウイルス様粒子調製物を作製するのに用いられる宿主細胞、または該宿主細胞の成分から、該エンベロープウイルスベースのウイルス様粒子調製物を単離するステップと、(b)該エンベロープウイルスベースのウイルス様粒子調製物における実質的にすべての感染因子を不活化するのに十分な線量の電磁放射を、該調製物へと適用するステップとを含み、ステップ(b)の後の該エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の該エンベロープウイルスベースのウイルス様粒子調製物と実質的に同じ免疫原性を有する方法を包含する。ある実施形態では、分離するステップ(a)が、少なくとも1つの遠心分離ステップを含む。前出の実施形態と組み合わせ得る別の実施形態では、分離するステップ(a)がまた、少なくとも1つの濾過ステップも包含し、該少なくとも1つの濾過ステップを、ノーマルフロー濾過、限外濾過、またはタンジェンシャルフロー(tangential flow)濾過からさらに選択することができる。前出の実施形態と組み合わせ得る別の実施形態では、分離するステップ(a)がまた、クロマトグラフィーステップも包含し、該少なくとも1つのクロマトグラフィーステップを、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー、疎水性相互作用クロマトグラフィー、混合方式クロマトグラフィー、逆相クロマトグラフィー、およびサイズ除外クロマトグラフィーからなる群よりさらに選択することができる。
【0007】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、電磁放射を、可視光、x線放射、紫外線放射、およびガンマ放射からなる群より選択することができ、該紫外線放射を、UV−A、UV−B、およびUV−Cからなる群よりさらに選択する場合もあり、該紫外線放射の波長が、320nm〜400nmの場合もある。
【0008】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、宿主細胞が、昆虫細胞または哺乳動物細胞である。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、宿主細胞が昆虫細胞であり、該昆虫細胞に、エンベロープウイルスベースのウイルス様粒子のうちの少なくとも1つの成分を発現するバキュロウイルス発現ベクターを感染させる。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、宿主細胞を、Bombyx mori宿主細胞、Spodoptera frugiperda宿主細胞、Choristoneura fumiferana宿主細胞、Heliothis virescens宿主細胞、Heliothis zea宿主細胞、Helicoverpa zea宿主細胞、Helicoverpa virescens宿主細胞、Orgyia pseudotsugata宿主細胞、Lymantria dispar宿主細胞、Plutella xylostella宿主細胞、Malacostoma disstria宿主細胞、Trichoplusia ni宿主細胞、Pieris rapae宿主細胞、Mamestra configurata宿主細胞、Mamestra brassica宿主細胞、およびHyalophora cecropia宿主細胞からなる群より選択する。宿主細胞が哺乳動物細胞である、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該哺乳動物細胞を、MRC−5細胞、Vero細胞、PER.C6(TM)細胞、チャイニーズハムスター卵巣細胞、およびHEK293細胞から選択する。昆虫細胞を宿主細胞として包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、電磁放射線量が、エンベロープウイルスベースのウイルス様粒子調製物におけるバキュロウイルスを不活化させるのに十分である。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、宿主細胞が哺乳動物細胞であり、前記哺乳動物細胞に、エンベロープウイルスベースのウイルス様粒子の少なくとも1つの成分を発現する、アデノウイルス発現ベクター、アデノ関連ウイルス発現ベクター、アルファウイルス発現ベクター、ヘルペスウイルス発現ベクター、ポックスウイルス発現ベクター、またはレトロウイルス発現ベクターを感染させる。アデノウイルス発現ベクター、アデノ関連ウイルス発現ベクター、アルファウイルス発現ベクター、ヘルペスウイルス発現ベクター、ポックスウイルス発現ベクター、またはレトロウイルス発現ベクターを包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、電磁放射線量が、エンベロープウイルスベースのウイルス様粒子調製物における、アデノウイルス、アデノ関連ウイルス、アルファウイルス、ヘルペスウイルス、ポックスウイルス、またはレトロウイルスを適宜不活化させるのに十分である。
【0009】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、適用するステップ(b)の前に、エンベロープウイルスベースのウイルス様粒子調製物にDNAインターカレート化合物を添加することができ、該DNAインターカレート化合物は、場合によって、光反応性のこともあり、ソラーレン、イソソラーレン、ならびにこれらの誘導体および類似体からなる群より選択することもある。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、電磁放射を、紫外線放射および可視光からなる群より選択することができる。
【0010】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該方法が、(c)エンベロープウイルスベースのウイルス様粒子調製物にアジュバントを添加するステップをさらに含み得る。
【0011】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、分離するステップ(a)の前に、(i)gagポリペプチドと、天然では脂質ラフトと会合しない抗原に連結した脂質ラフト会合ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記gagポリペプチドと、前記抗原に連結した脂質ラフト会合ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、宿主細胞内においてエンベロープウイルスベースのウイルス様粒子を作製する。
【0012】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、分離するステップ(a)の前に、(i)RSウイルスMポリペプチドと、RSウイルスFポリペプチドとを発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記RSウイルスMポリペプチドと、前記RSウイルスFポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、宿主細胞内においてエンベロープウイルスベースのウイルス様粒子を作製する。前出の実施形態は、場合によって、前記1または複数の発現ベクターから、RSウイルスGポリペプチドをさらに発現させることがある。
【0013】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、分離するステップ(a)の前に、(i)レンチウイルスおよびアルファレトロウイルスからなる群より選択されるレトロウイルスのgagポリペプチドと、RSウイルスFポリペプチドとを発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記レトロウイルスのgagポリペプチドと、前記RSウイルスFポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、宿主細胞内においてエンベロープウイルスベースのウイルス様粒子を作製する。前出の実施形態は、場合によって、前記1または複数の発現ベクターから、RSウイルスGポリペプチドをさらに発現させることがある。
【0014】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、分離するステップ(a)の前に、(i)gagポリペプチドと、インフルエンザ赤血球凝集素ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記gagポリペプチドと、前記インフルエンザ赤血球凝集素ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、宿主細胞内においてエンベロープウイルスベースのウイルス様粒子を作製する。
【0015】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、分離するステップ(a)の前に、(i)インフルエンザM1ポリペプチドと、赤血球凝集素ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記インフルエンザM1ポリペプチドと、前記赤血球凝集素ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、宿主細胞内においてエンベロープウイルスベースのウイルス様粒子を作製する。赤血球凝集素ポリペプチドを包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該1または複数の発現ベクターが、ノイラミニダーゼポリペプチドをさらに発現し得る。1または複数の発現ベクターを包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該1または複数の発現ベクターが、ウイルスベクター(複数可)であり得、該ウイルスベクター(複数可)を、バキュロウイルス、アルファウイルス、アデノ関連ウイルス、アデノウイルス、ヘルペスウイルス、ポックスウイルス、およびレトロウイルスからなる群よりさらに選択することができる。
【0016】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、ウイルスベクターを用いて、宿主細胞内において、エンベロープウイルスベースのウイルス様粒子の少なくとも1つの成分を発現させ、また、場合によって、該感染因子が、該ウイルスベクターを含む。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、調製物が、1ミリリットル当たり20個未満の感染因子;1ミリリットル当たり15個未満の感染因子;1ミリリットル当たり10個未満の感染因子;1ミリリットル当たり8個未満の感染因子;1ミリリットル当たり6個未満の感染因子;または1ミリリットル当たり5個未満の感染因子を含む。
【0017】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、ステップ(b)の後のエンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも50パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも60パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも70パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも80パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも85パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも90パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも95パーセントの免疫原性を有する。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、感染因子を含まないようにされた後のエンベロープウイルスベースのウイルス様粒子調製物は、感染因子を含まないようにされる前のエンベロープウイルスベースのVLP調製物より大きな免疫原性を有するか、または、これと比較して免疫原性が増大する。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含み、免疫原性を予測する(predictor)赤血球凝集アッセイを用いて、該エンベロープウイルスベースのウイルス様粒子の物理的および生化学的な完全性を測定することもでき、または、エンベロープウイルスベースのウイルス様粒子により免疫化した動物の血清におけるHAI活性を、免疫原性の尺度として決定することもできる。免疫原性を特定する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、動物にエンベロープウイルスベースのウイルス様粒子(RSウイルス(RSV)ポリペプチドを包含し得る)をワクチン接種し、ELISAアッセイもしくはウイルス中和アッセイにより、またはウェスタンブロットにより抗体力価を測定するか、あるいは増殖アッセイ、ELISPOTアッセイ、またはサイトカイン放出アッセイによりT細胞反応を測定することにより、該免疫原性を直接測定することができる。
【0018】
本発明の別の態様は、感染因子を実質的に含まないエンベロープウイルスベースのウイルス様粒子を含む、エンベロープウイルスベースのウイルス様粒子調製物であって、該エンベロープウイルスベースのウイルス様粒子が、感染因子を実質的に含まないわけではないエンベロープウイルスベースのウイルス様粒子と実質的に同じ免疫原性を有する調製物を包含する。
【0019】
ある実施形態では、エンベロープウイルスベースのウイルス様粒子が、昆虫または哺乳動物グリコシル化をさらに含む。前出の実施形態と組み合わせ得る別の実施形態では、昆虫グリコシル化を、Bombyx mori; Spodoptera frugiperda; Choristoneura fumiferana; Heliothis virescens; Heliothis zea; Helicoverpa zea; Helicoverpa virescens; Orgyia pseudotsugata; Lymantria dispar; Plutella xylostella; Malacostoma disstria; Trichoplusia ni; Pieris rapae; Mamestra configurata; Mamestra brassica;およびHyalophora cecropiaからなる群よりさらに選択することができる。前出の実施形態と組み合わせ得る別の実施形態では、哺乳動物グリコシル化を、ヒトグリコシル化(PER.C6(TM)細胞、MRC−5細胞、およびHEK293細胞によりもたらされるグリコシル化を含めた)、サルグリコシル化(Vero細胞によりもたらされるグリコシル化を含めた)、また、げっ歯動物グリコシル化(チャイニーズハムスター卵巣細胞によりもたらされるグリコシル化を含めた)からなる群よりさらに選択することができる。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、共有結合した光化学作用物質、タンパク質サブユニットの三次構造または四次構造においてUV照射もしくはガンマ照射により誘導される変化、ガンマ照射により誘導される化学結合の開裂、またはUV照射もしくはガンマ照射により誘導される、脂質の酸化、タンパク質の架橋形成、アミノ酸の酸化、およびアミノ酸の修飾からなる群より選択される化学修飾から選択される1または複数の欠損をさらに示す。好ましい実施形態では、エンベロープウイルスベースのウイルス様粒子が、該欠損のすべてを示す。特定の実施形態では、このような欠損の検出を、免疫原性の低下が見られないことを介して推測することもでき、または、適切な技法(例えば、ウイルス様粒子を含むポリペプチド内における共有結合の変化に関して欠損が見られないことを確認する場合の質量分析、また、ウイルス様粒子を含むポリペプチドの三次構造または四次構造内において、欠損が見られないことを確認する場合の分析的HPLC)の適用を介して推測することもできる。
【0020】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、gagポリペプチドと、非ウイルス性脂質ラフト会合ポリペプチド、または天然では脂質ラフトと会合しない、連結を形成するための抗原に連結した脂質ラフト会合ポリペプチドとを含み、場合によって、該非ウイルス性脂質ラフト会合ポリペプチドを、GPIアンカーポリペプチド、ミリストイル化配列ポリペプチド、パルミトイル化配列ポリペプチド、二重アセチル化配列ポリペプチド、シグナル伝達ポリペプチド、および膜輸送ポリペプチドからなる群よりさらに選択することもでき、GPIアンカーポリペプチド、ミリストイル化配列ポリペプチド、パルミトイル化配列ポリペプチド、二重アセチル化配列ポリペプチド、カベオリン(cavelin)ポリペプチド、フロチリンポリペプチド、シンタクシン−1ポリペプチド、シンタクシン−4ポリペプチド、シナプシンIポリペプチド、アデューシンポリペプチド、VAMP2ポリペプチド、VAMP/シナプトブレビンポリペプチド、シナプトブレビンIIポリペプチド、SNAREポリペプチド、SNAP−25ポリペプチド、SNAP−23ポリペプチド、シナプトタグミンIポリペプチド、およびシナプトタグミンIIポリペプチドからなる群よりさらに選択することもできる。ウイルス性脂質ラフト会合ポリペプチドは、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、融合タンパク質ポリペプチド、糖タンパク質ポリペプチド、およびエンベロープタンパク質ポリペプチドからなる群よりさらに選択することができる。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、膜結合エンベロープタンパク質ポリペプチドをさらに含む。
【0021】
連結を包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該連結を、共有結合、イオン性相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、および抗体−抗原相互作用からなる群よりさらに選択することができ、該共有結合は、場合によって、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、およびジスルフィド結合からなる群よりさらに選択することもできる。脂質ラフト会合ポリペプチドを包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該脂質ラフト会合ポリペプチドが、内在性膜タンパク質である。抗原を包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該抗原を、タンパク質、ポリペプチド、グリコポリペプチド、リポポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性または非ペプチド性模倣体、低分子、脂質、糖脂質、および炭水化物からなる群よりさらに選択することができる。
【0022】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチド、RSウイルスMポリペプチド、RSウイルスGポリペプチド、および/またはRSウイルスFポリペプチドをさらに含む。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、gagポリペプチドおよび赤血球凝集素ポリペプチド;レンチウイルスおよびアルファレトロウイルスからなる群より選択されるレトロウイルスのgagポリペプチド、ならびにRSウイルスFポリペプチド(また、場合によって、Gポリペプチド);またはインフルエンザM1ポリペプチドおよび赤血球凝集素ポリペプチドを含む。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、ノイラミニダーゼポリペプチドをさらに包含する。
【0023】
前出の実施形態のいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子調製物が、前記ウイルス様粒子と会合するアジュバントを包含する。アジュバントを包含する特定の実施形態では、処方ステップにおいて、該アジュバントを、エンベロープウイルスベースのウイルス様粒子と混合することができる。アジュバントは、ウイルス様粒子の内側または外側に位置する場合もあり、ウイルス様粒子に組み込まれる場合もある。前出の実施形態のいずれかと組み合わせ得るさらに別の実施形態では、アジュバントを、前記ウイルス様粒子と共有結合させて、共有結合を形成させることができる。
【0024】
本発明の別の態様は、免疫系の疾患または症状を処置または予防する方法であって、免疫原性量の、前出の実施形態のうちのいずれかによるエンベロープウイルスベースのウイルス様粒子調製物、または前出の方法およびその実施形態のうちのいずれかを用いて単離したエンベロープウイルスベースのウイルス様粒子調製物を、被験体に投与するステップを含む方法を包含する。一実施形態では、被験体がヒトである。前出の実施形態と組み合わせ得る別の実施形態では、投与するステップにより、被験体において、防御的免疫化反応が誘導される。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、投与するステップを、皮下(subcutaneous)送達、皮内送達、皮下(subdermal)送達、筋肉内送達、経口(peroral)送達、経口(oral)送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群よりさらに選択することができる。
【0025】
本発明の別の態様は、免疫原性量の、前出の実施形態のうちのいずれかによるエンベロープウイルスベースのウイルス様粒子調製物、または前出の方法およびその実施形態のうちのいずれかを用いて単離したエンベロープウイルスベースのウイルス様粒子調製物を含む医薬組成物を包含する。
【0026】
前出の態様およびそれらの実施形態は、本明細書で開示される実施形態のうちのいずれかとさらに組み合わせることができる。前出の実施形態、および/または本明細書で開示されるさらなる実施形態のうちのいずれかと共に包含され得る、本発明のさらなる態様は、本明細書の全体において見出すことができる。
【図面の簡単な説明】
【0027】
【図1】図1は、HAおよびNAを含有するキメラVLPの分析を示す図である。組換えバキュロウイルス発現細胞の上清は、細胞性細片を除去され、PR/8 H1N1 VLPは、ショ糖クッションによって100,000×gで遠心分離され、再懸濁され、不連続ショ糖密度勾配で再度遠心分離された。(A)勾配全体でのPR/8 H1N1 VLP赤血球凝集活性。(B)PR/8 H1N1 VLPのSDS−PAGE分析、GagおよびHAの共移動を示している。(C)A/Russia/77(H1N1)に特異的な抗体を使用する勾配画分4〜16のH1N1特異的ウェスタンブロット。(D)ピーク勾配画分由来のネガティブ染色試料の電子顕微鏡写真。(E)分子量を増加させてHA産物の電気泳動移動度を減少させるために、無関係な配列でHA遺伝子がそのアミノ末端で伸長された精製PR/8 H1N1 VLPのSDS−PAGE。(F)A/Solomon Islands/3/2006(H1N1)を示す精製VLPのSDS−PAGE、Gag対HAの比を示す。
【図2】図2は、不連続ショ糖勾配で遠心分離したH5N1 VLPでの赤血球凝集活性およびノイラミニダーゼ活性の検査を示すグラフである。H5N1 VLPは、図1でH1N1 VLPについて示した通り調製されたが、ショ糖勾配画分は、赤血球凝集活性およびノイラミニダーゼ活性の両方についてアッセイされた。NA活性は蛍光基質2’−(4−メチルウンベリフェリル)−α−D−N−アセチルノイラミン酸を使用して測定された。(A)は、Vietnam H5N1 VLPについての活性を示す。(B)はIndonesia H5N1 VLPについての活性を示す。
【図3】図3は、VLP免疫原性およびチャレンジ防御を示すグラフである。16匹のマウス3群は、キメラH1N1 VLP(A/PR/8/34を示す)で筋肉内または腹腔内免疫化を介して初回および追加免疫化を受けた。すべてのVLP処方物は、1用量あたりHAおよそ0.7μgを含有した。追加の4週間後に動物は10LD50のA/PR/8/34(H1N1)でチャレンジされた。(A)追加後2週間でのA/PR/8/34(H1N1)に特異的なHAI活性。(B)追加後2週間でのA/PR/8/34に特異的なIgG1およびIgG2A応答の定量。(C)H1N1チャレンジに続く重量減少によって示されるチャレンジ防御。
【図4】図4は、VLPおよび生きているウイルスがHAIアッセイにおいて同様に挙動することを示すグラフである。PR/8(H1N1)VLPで免疫化したマウス由来の血清を生きているPR/8(H1N1)ウイルスの4HAユニットまたは対応するVLPの4HAユニットのいずれかを使用してHAI活性について検査した。HK/68(H3N2)VLPで免疫化したマウス由来の血清を同様にHK/68ウイルスおよびVLPに対して検査した。データは、HAIアッセイにおいてウイルスとVLPとの間での同様の挙動を示した。
【図5】図5は、VLP免疫化フェレットにおける追加後免疫応答を示すグラフである。フェレットは初回および追加免疫化をそれぞれ0日目および28日目にH1N1、H5N1および裸のVLPで受けた。H1N1およびH5N1 VLP用量は、1用量あたりHAおよそ5μgを含有した。28日目および42日目の血清試料を(A)A/PR/8/34(H1N1)特異的HAI活性および(B)A/Vietnam/1203/04(H5N1)特異的マイクロ中和活性について分析した。
【図6】図6は、H1N1、H5N1および裸のVLPで免疫化したフェレットでのチャレンジ後の体重減少および生存を示すグラフである。追加免疫化を受けた2週間後にVLPワクチン接種フェレットは、1×106TCID50のA/Vietnam/1203/04(H5N1)でチャレンジされた。(A)すべての群についての平均体重減少データ(生存データをグラフ説明に示す)。(B)B群中のH1N1ワクチン接種動物についての個々の体重減少データ。
【図7】図7は、H1N1、H5N1および裸のVLPで免疫化されたフェレットでのチャレンジ後の鼻洗浄液ウイルス力価を示すグラフである。追加免疫化を受けた2週間後にVLPワクチン接種フェレットは、1×106TCID50のA/Vietnam/1203/04(H5N1)でチャレンジされた。(A)チャレンジ後3日目(45日目)の鼻洗浄液ウイルス力価。(B)チャレンジ後5日目(47日目)の鼻洗浄液ウイルス力価。
【発明を実施するための形態】
【0028】
(好ましい実施形態の詳細な説明)
本発明の好ましい実施形態には、感染因子を不活化する方法に供されたエンベロープウイルスベースのVLP調製物であって、そのような不活化法に供されていないVLPと実質的に同じ免疫原性をVLPが保持することを可能とする調製物と;エンベロープウイルスベースのVLP調製物における感染因子を不活化するこのような方法と;このような調製物をワクチン組成物へとさらに加工する方法と、このようなワクチン組成物を用いる方法とが含まれるが、これらに限定されない。
【0029】
理論に制約されることを望まないが、本発明の特定の好ましい実施形態は、以下の実施例に示すように、PPV VLPなど、他の非エンベロープウイルスベースのVLPとは異なり、エンベロープウイルスベースのVLPが、感染因子を不活化するのに用いられる標準的な不活化法に対して感受性であるという驚くべき発見に基づいている。しかし、同じく驚くべきことに、UV−A照射+光化学作用物質、UV−C照射、およびガンマ照射など、電磁ベースの不活化システムは、汚染性のエンベロープウイルスを不活化するのに効果的である一方で、該エンベロープウイルスベースのVLPの免疫原性を維持し、このために、このような不活化法は、ワクチンにおいて用いられるエンベロープウイルスベースのVLPを調製するのに理想的となっている。
【0030】
エンベロープウイルスベースのVLPを作製する好ましい方法は、好ましくはポリペプチド抗原の共発現(coexpression)を含めた、昆虫細胞内における発現を介する。バキュロウイルス発現系内における各種のレトロウイルスから得ることができるgag VLPの収量が大きい(23、24、46、49、52〜58)ため、gagポリペプチドを用いてVLPを作製することがさらにより好ましい。天然のレトロウイルスアセンブリー過程において、gagポリペプチドはC末端伸長を本質的に包含する。すなわち、天然において、機能的なgagタンパク質は、リボソームのフレームシフトに起因して、レトロウイルスプロテアーゼ活性、逆転写酵素活性、およびインテグラーゼ活性を含有する長いC末端伸長を有する。ラウス肉腫ウイルスのgag(59)およびMLVのgag(60)の両方について、人工的な伸長を伴う機能的なgagタンパク質の作製が達成されている。gagのC末端操作がこのように柔軟であることから、他の抗原および免疫刺激性タンパク質の配列など、他のポリペプチドを組み込むのに重要な部位が得られる。エンベロープウイルスベースのVLPを作製する別の好ましい方法は、インフルエンザHAタンパク質およびインフルエンザM1タンパク質(また、場合によって、インフルエンザNAタンパク質)の共発現を介する。昆虫細胞内におけるこれら2つのタンパク質の共発現は、エンベロープウイルスベースのVLPを作製する有効な方法であることが示されている(111〜112)。
【0031】
エンベロープウイルスベースのVLPの好ましい例には、(i)gagポリペプチドと、抗原に連結した脂質ラフト会合ポリペプチドとを含むVLP;(ii)RSウイルスMポリペプチドと、RSウイルスFポリペプチドと(また、場合によって、RSウイルスGポリペプチドと)を含むVLP;(iii)レンチウイルスおよびアルファレトロウイルスからなる群より選択されるレトロウイルスのgagポリペプチドと、RSウイルスFポリペプチドとを含むVLP;(iv)gagポリペプチドと、インフルエンザ赤血球凝集素ポリペプチドと(また、場合によって、ノイラミニダーゼポリペプチドと)を含むVLP;ならびに(v)インフルエンザM1ポリペプチドと、赤血球凝集素ポリペプチドと(また、場合によって、ノイラミニダーゼポリペプチドと)を含むVLPが含まれる。
【0032】
1つのウイルスに由来するコア粒子と、別のウイルスに由来する表面抗原とを含有するキメラVLPの作製は、シュードタイピング(pseudotyping)と呼ばれる。おそらく、インフルエンザHAタンパク質およびインフルエンザNAタンパク質が、脂質ラフトドメイン内に濃縮される(61、62)一方、出芽過程では、ミリストイル化した(myristolated)gagタンパク質もまた、脂質ラフトドメインの内部表面において濃縮される(63)ため、gagポリペプチドが効率的にシュードタイピングされるのは、これらのタンパク質を伴うときである。
【0033】
本明細書で記載される本発明の実施形態は、キメラVLPを形成するための基盤として、天然では脂質ラフトと会合しない抗原に連結した脂質ラフト会合ポリペプチドを包含するVLPプラットフォームに適合する。
【0034】
開示される方法およびプロトコールを実施するには、別段に示さない限り、当業者の能力の範囲内にある、化学、分子生物学、微生物学、組換えDNA、および免疫学の従来の技法を用いる。このような技法は、文献において説明されている。例えば、J. Sambrook、E. F. Fritsch、およびT. Maniatis、1989年、「Molecular Cloning: A Laboratory Manual」、第2版、1〜3巻、Cold Spring Harbor Laboratory Press社; Ausubel, F. M.ら(1995年、および定期的な補遺;「Current Protocols in Molecular Biology」、9、13、および16章、ニューヨーク州、ニューヨーク、John Wiley & Sons社); B. Roe、J. Crabtree、およびA. Kahn、1996年、「DNA Isolation and Sequencing: Essential Techniques」、John Wiley & Sons社; J. M. PolakおよびJames O’D. McGee、1990年、「In Situ Hybridization: Principles and Practice」、Oxford University Press社; M. J. Gait(編)、1984年、「Oligonucleotide Synthesis: A Practical Approach」、Irl Press社;ならびにD. M. J. LilleyおよびJ. E. Dahlberg、1992年、「Methods of Enzymology: DNA Structure Part A: Synthesis and Physical Analysis of DNA Methods in Enzymology」、Academic Press社を参照されたい。
【0035】
(定義)
本明細書で用いられる「エンベロープウイルスベースのVLP」とは、エンベロープウイルスに由来する1または複数の成分を用いて形成されるウイルス様粒子を指す。好ましい例には、gagポリペプチドを用いて作製したVLPと、インフルエンザM1ポリペプチドおよび/または赤血球凝集素ポリペプチド(また、場合によって、ノイラミニダーゼポリペプチド)を用いて作製したVLPと、レンチウイルスのgagポリペプチドおよびアルファレトロウイルスのgagポリペプチドからなる群ならびにRSウイルス(RSV)Fポリペプチド(および、場合によって、RSウイルスGポリペプチド)を用いて作製したVLPと、RSウイルス(RSV)Mおよび/またはFポリペプチド(および、場合によって、RSウイルスGポリペプチド)を用いて作製したVLPとが含まれるが、これらに限定されない。
【0036】
さらなる例には、エンベロープウイルスベースのVLPを形成するのに用い得る、エボラウイルスおよびマールブルクウイルスなどのフィロウイルス(例えば、細胞内においてフィロウイルスに由来するウイルス性GPおよびウイルス性VP40を共発現させると、脂質ラフト内におけるこれら2つのウイルス性タンパク質の会合により、VLPが生成される(米国特許公開第20060099225号を参照されたい));SARSウイルスなどのコロナウイルス(例えば、コロナウイルスのVLP形成には、Eタンパク質およびMタンパク質で十分である(Fischerら、J. Virol.(1998年)、72巻:7885〜7894頁;およびVennemaら、EMBO J.(1996年)、15巻:2020〜2028頁を参照されたい));RSウイルス(RSV)など、パラミクソウイルス科のウイルス(例えば、RSVのMタンパク質を発現させると、VLPが生成される(例えば、米国特許公開第20080233150号を参照されたい));ならびに西ナイルウイルスなどのフラビウイルス科(例えば、バキュロウイルス発現系内で西ナイルウイルスのprM遺伝子およびE遺伝子を含む構築物を発現させると、VLPが生成される(例えば、米国特許公開第20080233150号を参照されたい))が含まれる。
【0037】
本明細書で用いられる「感染因子を含まない」とは、感染可能な活性作用物質の不在を指す。このような試料は、不活性であり、感染不能な作用物質を含有し得る。例を目的として述べると、バキュロウイルスがもはや感染不能であるように処理された、バキュロウイルスを含有する試料は、不活化されたバキュロウイルスをなおも含有するが、感染因子を含まない。さらに、試料は、感染可能な活性作用物質を絶対的に含まないことが必要なわけではなく、これをヒト用または動物用のワクチンとして意図される目的に適宜用い得る(すなわち、ヒト用または動物用のワクチン内における感染因子の許容レベルを統轄する任意の米国連邦規則を適宜満たす)程度に十分に活性作用物質を含まないことだけが必要である。特定の実施形態では、感染因子を含まないようにされた後のエンベロープウイルスベースのVLP調製物の用量当たりの感染単位またはml当たりの感染単位は、感染因子を含まないようにされる前のエンベロープウイルスベースのVLP調製物の少なくとも10分の1、少なくとも100分の1、少なくとも1000分の1、または少なくとも10,000分の1に低下する。例示を目的として述べると、感染因子には、1または複数の宿主細胞内において、VLPを含むポリペプチド(複数可)を発現させるのに用いるベクター(複数可)、および外部の細菌性汚染物質、真菌性汚染物質、またはウイルス性汚染物質、さらにまた、内因性の病原体(例えば、宿主ゲノム内ではサイレントである(silent)ことが典型的である、レトロウイルス性エレメントまたはレトロトランスポゾン性エレメントの再活性化など、供給源物質または宿主細胞に由来する)が含まれ得る。
【0038】
本明細書で用いられる「実質的に同じ免疫原性」とは、感染因子を含まないようにされる前の調製物と比較した、感染因子を含まないようにされた後のエンベロープウイルスベースのVLP調製物の免疫原性を指す。特定の実施形態では、感染因子を含まないようにされた後のエンベロープウイルスベースのVLP調製物は、感染因子を含まないようにされる前のエンベロープウイルスベースのVLP調製物の少なくとも50パーセント、少なくとも60パーセント、少なくとも70パーセント、少なくとも80パーセント、少なくとも85パーセント、少なくとも90パーセント、または少なくとも95パーセントの免疫原性を有する。特定の実施形態では、感染因子を含まないようにされた後のエンベロープウイルスベースのVLP調製物は、感染因子を含まないようにされる前のエンベロープウイルスベースのVLP調製物より大きな免疫原性を有するか、または、これと比較して免疫原性が増大する。免疫原性の好ましい尺度は、接種後に得られる、VLP組成物に対する抗体力価である。インフルエンザワクチンの場合、免疫原性の好ましい尺度は、以下の実施例によるHAI活性である。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含み、免疫原性を予測する赤血球凝集アッセイを用いて、該エンベロープウイルスベースのウイルス様粒子の物理的および生化学的な完全性を測定することもでき、または、エンベロープウイルスベースのウイルス様粒子により免疫化した動物の血清におけるHAI活性を、免疫原性の尺度として決定することもできる。免疫原性を特定する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、動物にエンベロープウイルスベースのウイルス様粒子(RSウイルス(RSV)ポリペプチドを包含し得る)をワクチン接種し、ELISAアッセイもしくはウイルス中和化アッセイにより、またはウェスタンブロットにより抗体力価を測定するか、あるいは増殖アッセイ、ELISPOTアッセイ、またはサイトカイン放出アッセイによりT細胞反応を測定することにより、該免疫原性を直接測定することができる。
【0039】
「昆虫グリコシル化」とは、昆虫により、また、昆虫細胞ベースの発現系によりもたらされるグリコシル化パターンを指す。このようなグリコシル化パターンには、天然においてもたらされるグリコシル化、および哺乳動物のグリコシル化酵素を包含するように改変された昆虫細胞が、天然において、哺乳動物または哺乳動物細胞ベースの発現系によりもたらされるグリコシル化パターンではなく、「哺乳動物様の」グリコシル化をもたらすだけである限りにおいて、このような改変昆虫細胞によりもたらされるグリコシル化パターンも含まれ得る。昆虫グリコシル化パターンの好ましい例には、
【0040】
【化1】
など、バキュロウイルス発現系および関連のウイルス発現系に適合する昆虫細胞が含まれる。
【0041】
「哺乳動物グリコシル化」とは、哺乳動物により、また、哺乳動物細胞ベースの発現系によりもたらされるグリコシル化パターンを指す。このようなグリコシル化パターンには、天然においてもたらされるグリコシル化、およびこのような細胞では見出されないか、または発現することが典型的ではないグリコシル化酵素を包含するように改変された哺乳動物細胞が、天然において、哺乳動物以外または哺乳動物以外の細胞ベースの発現系によりもたらされるグリコシル化パターンではなく、哺乳動物グリコシル化または非天然グリコシル化をもたらすだけである限りにおいて、このような改変哺乳動物細胞によりもたらされるグリコシル化パターンも含まれ得る。昆虫グリコシル化パターンの好ましい例には、ヒト細胞(PER.C6(TM)細胞、MRC−5細胞、およびHEK293細胞によりもたらされるグリコシル化を含めた)、サル細胞(Vero細胞によりもたらされるグリコシル化を含めた)、また、げっ歯動物細胞(チャイニーズハムスター卵巣細胞によりもたらされるグリコシル化を含めた)など、公知のウイルス発現系に適合する哺乳動物細胞が含まれる。
【0042】
本明細書で用いられる「Gagポリペプチド」とは、本明細書で記載するウイルス様粒子の形成の一因となる、レトロウイルス由来の構造ポリペプチドである。gagポリペプチドは、エンベロープウイルスベースのVLPを形成するのに好ましい手段である。一部の実施形態では、RNAをパッケージングする傾向、または粒子形成および出芽の効率など、特定の特徴に影響を及ぼす目的で、gagポリペプチドを意図的に変異させることができる。レトロウイルスのゲノムは、3つの主要な遺伝子産物をコードする:構造タンパク質をコードするgag遺伝子の産物;逆転写酵素および関連のタンパク質分解性ポリペプチド、ヌクレアーゼおよびインテグラーゼに関連する機能をコードするpol遺伝子の産物;envの産物(それによりコードされる糖タンパク質の膜タンパク質が、感染細胞の表面上において、また、放出された成熟ウイルス粒子の表面上においても検出される)。すべてのレトロウイルスのgag遺伝子は、全体的な構造類似性を有し、レトロウイルスの各群内においても、アミノ酸レベルで保存されている。gag遺伝子は、逆転写酵素を除くコアタンパク質をもたらす。
【0043】
MLVの場合、Gagの前駆体であるポリタンパク質は、Pr65Gagであり、該前駆体上におけるその順序がNH2−p15−pp12−p30−p10−COOHである4つのタンパク質へと切断される。これらの切断は、ウイルス性プロテアーゼを介し、また、ウイルスに応じて、ウイルス放出の前または後において生じ得る。MLV Gagタンパク質は、グリコシル化形態および非グリコシル化形態で存在する。グリコシル化形態はgPr80Gagから切り出され、gPr80Gagは、非グリコシル化Pr65Gagに対応するAUGコドンの上流に位置する、インフレームの、異なる開始コドンから合成される。グリコシル化Gagを合成しない、MLVの欠失変異体がなおも感染性であり、また、非グリコシル化Gagがなおもウイルス様粒子を形成し得ることから、グリコシル化イベントの重要性について、疑問が投げかけられる。ウイルスがコードするプロテアーゼにより、HIV−1 Gagの前駆体であるpr55Gagが翻訳後に切断されることから、N−ミリストイル化されて内部でリン酸化されたp17マトリックスタンパク質(p17MA)、リン酸化されたp24カプシドタンパク質(p24CA)、ならびにヌクレオカプシドタンパク質であるp15(p15NC)がもたらされ、p15は、さらにp9およびp6へと切断される。
【0044】
構造面において、原型的なGagポリタンパク質は、レトロウイルスのgag遺伝子において常に同じ順序で生じる3つの主要なタンパク質:マトリックスタンパク質(MA)(マトリックスという名称を共有するが、MAとは異なるタンパク質である、インフルエンザマトリックスタンパク質M1と混同しないようにされたい)、カプシドタンパク質(CA)、およびヌクレオカプシドタンパク質(NC)に分けられる。成熟タンパク質へのGagポリタンパク質のプロセシングは、レトロウイルスがコードするプロテアーゼにより触媒され、新たに出芽するウイルス粒子が成熟するときに生じる。機能面において、Gagポリタンパク質は、3つのドメイン:細胞膜をGagポリタンパク質の標的とする膜結合ドメインと;Gagの重合化を促進する相互作用ドメインと;宿主細胞からの新生ビリオンの放出を促進する後期ドメインとに分けられる。アセンブリーを媒介するGagタンパク質の形態は、ポリタンパク質である。したがって、アセンブリードメインは、後期に形成される任意の切断産物内にもれなく収まる必要はない。こうして、本明細書で包含されるGagポリタンパク質は、VLPの形成および放出に重要な機能的エレメントを包含する。これらの重要な機能的エレメントに関する最新技術の進歩は大きい。例えば、
【0045】
【化2】
を参照されたい。
【0046】
本発明の特定のVLPにおいて用いられるgagポリペプチドは、VLPを形成するための機能的エレメントを最小限において包含するだろう。gagポリペプチドは、場合によって、そのコード配列をgagポリペプチドのコード配列にスプライシングすることによって作製し得る、1または複数のさらなるポリペプチドを包含し得る。gagポリペプチド内へのさらなるポリペプチドの挿入に好ましい部位は、C末端である。
【0047】
Gagポリペプチドに好ましいレトロウイルスの供給源には、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス(トリ白血病ウイルス、またはラウス肉腫ウイルスなど)、ベータレトロウイルス(マウス乳癌ウイルス、ヤーグジークテヒツジレトロウイルス、およびメーソン−ファイザーサルウイルスなど)、ガンマレトロウイルス(マウス白血病ウイルス、ネコ白血病ウイルス、細網内皮症ウイルス、およびテナガザル白血病ウイルスなど)、デルタレトロウイルス(ヒトTリンパ向性ウイルス、およびウシ白血病ウイルスなど)、イプシロンレトロウイルス(ウォールアイ皮膚肉腫ウイルスなど)、またはレンチウイルス(1型ヒト免疫不全ウイルス、HIV−2、サル免疫不全ウイルス、ネコ免疫不全ウイルス、ウマ感染性貧血ウイルス、およびヤギ関節炎脳炎ウイルス)が含まれる。
【0048】
本明細書で用いられる「脂質ラフト」とは、ウイルス粒子のアセンブリー過程においてgagポリペプチドが濃縮される、細胞膜のマイクロドメインを指す。
【0049】
本明細書で用いられる「脂質ラフト会合ポリペプチド」とは、脂質ラフトと直接的または間接的に会合する任意のポリペプチドを指す。本発明で用いられる具体的な脂質ラフト会合ポリペプチドは、キメラウイルス様粒子の所望の使用に依存する。
【0050】
脂質ラフト会合ポリペプチドは、内在性膜タンパク質、膜との会合を引き起こすタンパク質修飾により脂質ラフトと直接的に会合するタンパク質、または脂質ラフト会合ポリペプチドにより脂質ラフトと間接的に会合するポリペプチドであり得る。
【0051】
脂質アンカーを有する多くのタンパク質が、脂質ラフトと会合する。ポリペプチドを脂質ラフトへと連結する脂質アンカーには、GPIアンカー、ミリストイル化、パルミトイル化、および二重アセチル化が含まれる。
【0052】
多くの異なる種類のポリペプチドが、脂質ラフトと会合する。脂質ラフトは、シグナル伝達、膜輸送、ウイルスの侵入、ウイルスのアセンブリー、また、アセンブルした粒子の出芽を含めた、多くの生物学的活性のためのプラットフォームとして機能し、したがって、これらの過程に関与する各種のポリペプチドと会合する。
【0053】
シグナル伝達カスケードに関与する各種のポリペプチドは、シグナル伝達プラットフォームとして機能する脂質ラフトと会合する。シグナル伝達プラットフォームとして機能する脂質ラフトの1つの種類は、カベオラと呼ばれている。それは、カベオリンファミリーに由来するポリペプチド(例えば、カベオリンおよび/またはフロチリン(flottillin))を含有する、細胞膜のフラスコ型の陥入部である。
【0054】
膜輸送ポリペプチドは、膜輸送プラットフォームとして機能する脂質ラフトと会合する。例には、シンタクシン−1タンパク質、シンタクシン−4タンパク質、シナプシンIタンパク質、アデューシンタンパク質、VAMP2タンパク質、VAMP/シナプトブレビンタンパク質、シナプトブレビンIIタンパク質、SNAREタンパク質、SNAP−25タンパク質、SNAP−23タンパク質、シナプトタグミンIタンパク質、およびシナプトタグミンIIタンパク質など、エンドサイトーシスおよびエクソサイトーシスに関与するタンパク質が含まれる。
【0055】
ウイルス受容体、ウイルス受容体−共受容体複合体、ウイルス侵入過程の調節を補助する他の任意の成分は、ウイルス侵入に特化した膜輸送プラットフォームとして機能する脂質ラフトと会合する。脂質ラフトと会合するウイルス受容体の例には、崩壊促進因子(DAFまたはCD55)、多くのエンテロウイルスの受容体であるGPIアンカー膜糖タンパク質;ガングリオシド、Hsc70タンパク質、アルファ2−ベータ1インテグリンおよびアルファ5−ベータ2インテグリンを含めた複数の成分を含有する複合体である、A群ロタウイルスの受容体;HIVウイルス、MLVウイルス、麻疹ウイルス、およびEbolaウイルスなど、複数のエンベロープウイルスの糖タンパク質;ならびにCD5ポリペプチド、CCR5ポリペプチド、およびnefポリペプチドなど、HIVの侵入に関与するポリペプチドが含まれる。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. And Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。
【0056】
ウイルス粒子のアセンブリーに関与するポリペプチドは、ウイルスアセンブリープラットフォームとして機能する脂質ラフトと会合する。このようなポリペプチドの例には、HAインフルエンザエンベロープ糖タンパク質およびNAインフルエンザエンベロープ糖タンパク質、麻疹ウイルスに由来するHタンパク質および成熟F1−F2融合タンパク質、ならびにHIVウイルスに由来するgp160、gp41、およびPr55gagが含まれる。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. & Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。
【0057】
アセンブルしたウイルスの出芽に関与するポリペプチドは、ウイルス出芽プラットフォームとして機能する脂質ラフトと会合する。宿主細胞からのHIV−1の出芽は、膜ラフト内で生じることを示唆するデータが存在する。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. And Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。ウイルスの出芽に関与するポリペプチドについての一般的な情報は、「Fields Virology」(第4版)、2001年において見出すことができる。
【0058】
好ましい脂質ラフト会合ポリペプチドには、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、融合タンパク質ポリペプチド、糖タンパク質ポリペプチド、およびエンベロープタンパク質ポリペプチドなどのウイルスポリペプチドが含まれる。これらのポリペプチドの各々は、任意の種類のウイルスに由来し得るが、特定の実施形態は、HIV−1ウイルスに由来するエンベロープタンパク質、RSウイルスまたは麻疹ウイルスに由来する融合タンパク質、RSウイルス、単純ヘルペスウイルス、またはエボラウイルスに由来する糖タンパク質、また、麻疹ウイルスに由来する赤血球凝集素タンパク質を包含する。
【0059】
好ましい非ウイルス性病原体の脂質ラフト会合ポリペプチドは、Plasmodium falciparum、Plasmodium malariae、Plasmodium ovale、およびPlasmodium vivaxなどのPlasmodium属; Toxoplasma gondii; Trypanosoma brucei、Trypanosoma cruzi; Schistosoma haematobium、Schistosoma mansoni、Schistosoma japonicum; Leishmania donovani; Giardia intestinalis; Cryptosporidium parvumなどが含まれるがこれらに限定されない病原性の原虫、蠕虫、また、他の真核微生物の病原体から得ることができる。このような非ウイルス性脂質ラフト会合ポリペプチドは、それ自体が抗原として作用するので、天然では脂質ラフトと会合しない抗原に連結することなく用いることができる。
【0060】
本明細書で用いられる「インフルエンザM1ポリペプチド」は、ウイルスエンベロープ内におけるウイルス被覆の形成を媒介する、インフルエンザウイルスタンパク質に由来する。該M1タンパク質は、ウイルスの出芽を駆動し、また、ウイルス粒子の主要なタンパク質成分でもあり、ウイルス粒子内では、ウイルスエンベロープおよび内在性膜タンパク質と、ゲノムリボ核タンパク質との間の中間層を形成する。M1ポリペプチドはまた、正に帯電したアミノ酸に富む、M1ポリペプチド内における結合モチーフ(RKLKR)に依存する非特異的な様式で、ウイルスRNAに結合するとも考えられている。
【0061】
ウイルス性脂質ラフト会合ポリペプチドの好ましい例は、赤血球凝集素ポリペプチドである。本明細書で用いられる「赤血球凝集素ポリペプチド」は、感染される細胞に対するウイルスの結合を媒介するインフルエンザウイルスタンパク質に由来する。赤血球凝集素ポリペプチドはまた、同等の麻疹ウイルスタンパク質にも由来し得る。このタンパク質は、単一の膜貫通ドメインにより、インフルエンザウイルスの表面へとアンカリングすることが見出される抗原性の糖タンパク質である。インフルエンザ赤血球凝集素のうち、H1〜H16と名付けられた、少なくとも16のサブタイプが同定されている。H1、H2、およびH3は、ヒトインフルエンザウイルスにおいて見出される。H5またはH7の赤血球凝集素を有する、高病原性鳥類インフルエンザウイルスがヒトに感染する割合は低いことが判明している。鳥類ウイルス株のH5型赤血球凝集素内における単一のアミノ酸変化がヒト患者において見出されており、これが受容体の特異性を変化させることを可能にし、H5赤血球凝集素が鳥類H5N1ウイルスの受容体特異性を顕著に変化させることによって、これらのウイルスに、ヒト受容体に結合する能力がもたらされると報告されている(109および110)。この知見は、通常ヒトに感染しないH5N1ウイルスが変異し、ヒト細胞に効率的に感染できるようになることを説明する。
【0062】
赤血球凝集素は、ホモ三量体の内在性膜ポリペプチドである。天然では、その膜貫通ドメインが、脂質ラフトドメインと会合し、これにより、gagポリペプチドと会合することが可能となり、VLP内へと組み込まれる。それは、円筒様の形状をしており、約135Åの長さである。HAを構成する3つの同一の単量体は、中央のコイルドコイルと、VLPの表面上に露出されるシアル酸結合部位を含有する球形の頭状部分とを形成する。HAの単量体は、グリコシル化され、2つのより小型のポリペプチド:HA1サブユニットおよびHA2サブユニットへと切断される、単一のポリペプチド前駆体として合成される。HA2サブユニットは、膜へとアンカリングされる、三量体のコイルドコイルを形成し、HA1サブユニットは、球形の頭状部分を形成する。
【0063】
本発明の特定のVLPにおいて脂質ラフト会合ポリペプチドとして用いられる赤血球凝集素ポリペプチドは、最小限において、膜アンカードメインを包含するものとする。赤血球凝集素ポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、H1、H2、H3、H5、H7、およびH9赤血球凝集素に由来することが好ましい。加えて、赤血球凝集素ポリペプチドは、異なるインフルエンザ赤血球凝集素のキメラでもあり得る。赤血球凝集素ポリペプチドは、1または複数のさらなるポリペプチドのコード配列を、赤血球凝集素ポリペプチドのコード配列にスプライシングすることにより作製し得る、天然では脂質ラフトと会合しない、1または複数のさらなる抗原を包含することが好ましい。赤血球凝集素ポリペプチド内へのさらなるポリペプチドの挿入に好ましい部位は、N末端である。
【0064】
本発明の特定のVLPにおいて抗原として用いられる赤血球凝集素ポリペプチドは、最小限において、膜アンカードメイン、また、赤血球凝集素に由来する少なくとも1つのエピトープを包含するものとする。赤血球凝集素ポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、H1、H2、H3、H5、H7、およびH9赤血球凝集素に由来することが好ましい。加えて、赤血球凝集素ポリペプチドは、異なるインフルエンザ赤血球凝集素のキメラでもあり得る。赤血球凝集素ポリペプチドは、場合によって、そのコード配列を赤血球凝集素ポリペプチドのコード配列にスプライシングすることにより作製し得る、1または複数のさらなるポリペプチドを包含し得る。赤血球凝集素ポリペプチド内へのさらなるポリペプチドの挿入に好ましい部位は、N末端である。
【0065】
ウイルス性脂質ラフト会合ポリペプチドの好ましい別の例は、ノイラミニダーゼポリペプチドである。本明細書で用いられる「ノイラミニダーゼポリペプチド」は、糖タンパク質から末端のシアル酸残基を切断することにより、細胞からのインフルエンザウイルスの放出を媒介する、インフルエンザウイルスタンパク質に由来する。ノイラミニダーゼ糖タンパク質は、ウイルス表面上において発現する。ノイラミニダーゼタンパク質は四量体であり、ベータ−ピンホイール構造を有する球形の頭状部分、細いストーク状領域、また、単一の膜貫通ドメインによりウイルス膜内に該タンパク質をアンカリングする小さな疎水性領域からなる共通構造を共有する。シアル酸残基を切断するための活性部位は、すべてのインフルエンザAウイルス内に保存されている、15の荷電アミノ酸により形成される、各サブユニット表面上のポケットを包含する。インフルエンザノイラミニダーゼのうち、N1〜N9と名付けられた、少なくとも9つのサブタイプが同定されている。
【0066】
本発明の特定のVLPにおいて用いられるノイラミニダーゼポリペプチドは、最小限において、膜アンカードメインを包含するものとする。機能的領域に関する最新技術は極めて高度である。例えば、Vargheseら、Nature、303巻、35〜40頁、1983年; Colmanら、Nature、303巻、41〜44頁、1983年; Lentzら、Biochem、26巻、5321〜5385頁、1987年; Websterら、Virol.、135巻、30〜42頁、1984年を参照されたい。ノイラミニダーゼポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、N1ノイラミニダーゼおよびN2ノイラミニダーゼに由来することが好ましい。加えて、ノイラミニダーゼポリペプチドは、異なるインフルエンザノイラミニダーゼのキメラでもあり得る。ノイラミニダーゼポリペプチドは、1または複数のさらなるポリペプチドのコード配列を赤血球凝集素ポリペプチドにスプライシングすることにより作製し得る、天然では脂質ラフトと会合しない1または複数のさらなる抗原を包含することが好ましい。ノイラミニダーゼポリペプチドコード配列内へのさらなるポリペプチドの挿入に好ましい部位は、C末端である。
【0067】
本発明の特定のVLPにおいて抗原として用いられるノイラミニダーゼポリペプチドは、最小限において、膜アンカードメイン、また、少なくとも1つのシアル酸残基切断活性を包含するものとする。機能的領域に関する最新技術は極めて高度である。例えば、Vargheseら、Nature、303巻、35〜40頁、1983年; Colmanら、Nature、303巻、41〜44頁、1983年; Lentzら、Biochem、26巻、5321〜5385頁、1987年; Websterら、Virol.、135巻、30〜42頁、1984年を参照されたい。ノイラミニダーゼポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、N1ノイラミニダーゼおよびN2ノイラミニダーゼに由来することが好ましい。加えて、ノイラミニダーゼポリペプチドは、異なるインフルエンザノイラミニダーゼのキメラでもあり得る。ノイラミニダーゼポリペプチドは、場合によって、そのコード配列をノイラミニダーゼポリペプチドのコード配列にスプライシングすることにより作製し得る、1または複数のさらなるポリペプチドを包含し得る。ノイラミニダーゼポリペプチド内へのさらなるポリペプチドの挿入に好ましい部位は、C末端である。
【0068】
脂質ラフト会合ポリペプチドの好ましい別の例は、ファシクリンI(FasI)と称する、昆虫由来の接着タンパク質である。本明細書で用いられる「ファシクリンIポリペプチド」は、胚の発生に関与する昆虫タンパク質に由来する。この非ウイルス性タンパク質は、昆虫細胞内のバキュロウイルス発現系において発現させることができ、これにより、FasIの脂質ラフト会合がもたらされる(J. Virol.、77巻、6265〜6273頁、2003年)。したがって、ファシクリンIポリペプチドに異種抗原を結合させて、gagと共に共発現させると、VLP内にキメラ分子が組み込まれる。本発明のVLPにおいて用いられるファシクリンIポリペプチドは、最小限において、膜アンカードメインを包含するものとする。
【0069】
脂質ラフト会合ポリペプチドの好ましい別の例は、G糖タンパク質と称する、RSVに由来するウイルス性付着タンパク質である。本明細書で用いられる「Gグリコポリペプチド」は、RSV G糖タンパク質に由来する。近年のデータは、脂質ラフトドメインが、インフルエンザウイルスにとって重要であるのと同様に、RSV粒子の出芽にも重要であることを示している(Virol、327巻、175〜185頁、2004年; Arch. Virol.、149巻、199〜210頁、2004年; Virol.、300巻、244〜254頁、2002年)。RSVに由来するG糖タンパク質は、ウイルス性付着タンパク質であり、およびRSV感染に対する防御抗原として機能する、32.5kdの内在性膜タンパク質である。インフルエンザウイルスに由来する赤血球凝集素と同様、その抗原性は、それに付着した、任意の非脂質ラフト抗原の抗原性を増強し得る。天然において、RSVは、ノイラミニダーゼ活性を有するタンパク質を発現しないので、gagおよびRSV GからなるVLPは、効率的な作製および放出に、NAの存在を必要としない可能性が高い。したがって、gag(アルファレトロウイルスのgagなど)およびGグリコポリペプチドをコードする発現ベクターを開発すれば、結果として、膜内に組み込まれたGグリコポリペプチドを含有するVLPが作製される。非脂質ラフト外来抗原を付着させる形での、Gグリコポリペプチドに対する任意の改変は、結果として、該外来抗原に対して顕著な免疫反応を誘導することが可能なキメラVLPをもたらす。
【0070】
VLPが、その文脈により、エンベロープベースのウイルスに基づかないか、または本明細書で開示される特定のエンベロープベースのウイルスの特定の成分に基づくウイルス様粒子を指す場合を除き、用語「エンベロープウイルスベースのウイルス様粒子」と「VLP」とは、本明細書の全体において互換的に用いられる。
【0071】
(抗原)
本発明の特定の態様は、エンベロープウイルスベースのVLP調製物と会合するさらなる抗原を包含する。このようなさらなる抗原は、同じ組成物中に組み込むことができ、さらにまた、VLPと共有結合させることもでき、非共有結合させることもできる。好ましい実施形態では、gagポリペプチド、インフルエンザM1ポリペプチド、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、および/または他の脂質ラフト会合ポリペプチドが、天然では脂質ラフトと会合しないであろう抗原を含有する、エンベロープウイルスベースのVLPを形成するのに容易に適合可能なプラットフォームである。本節では、開示されるVLPと共に用いるのに好ましい抗原について記載する。
【0072】
(抗原と脂質ラフト会合ポリペプチドとの連結)
天然では脂質ラフトと会合しない抗原、または天然では細胞膜と会合しない抗原を含有するVLPを形成する手段として、gagポリペプチド、インフルエンザM1ポリペプチド、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、および/または別の脂質ラフト会合ポリペプチドと、該抗原との連結を形成することができる。単一の抗原または複数の抗原に脂質ラフト会合ポリペプチドを連結することにより、VLPの免疫原性を増大させるか、各種の病原体に免疫原性を付与するか、または、特定の病原体の各種の株に免疫原性を付与することができる。
【0073】
抗原と脂質ラフト会合ポリペプチドとの間の連結は、該抗原を結果としてVLP内へと組み込むのに十分な任意の種類の連結であり得る。結合は、共有結合、イオン性相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、または抗体−抗原相互作用であり得る。好ましい実施形態では、結合は、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、またはジスルフィド結合などの共有結合である。
【0074】
抗原は、脂質ラフト会合ポリペプチドに対する既存の結合を伴なって組換えにより作製することもでき、単離物質として作製し、次いで、後に、脂質ラフト会合ポリペプチドに連結することもできる。
【0075】
(抗原の種類)
本明細書で用いられる抗原は、免疫反応を誘発することが可能であり、また、天然では脂質ラフトと会合しない任意の物質であり得る。抗原には、タンパク質、ポリペプチド(活性タンパク質、また、タンパク質内における個々のポリペプチドエピトープを含めた)、グリコポリペプチド、リポポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性および非ペプチド性模倣体、ならびに他の分子、低分子、脂質、糖脂質、ならびに炭水化物が含まれるが、これらに限定されない。天然において、抗原が脂質ラフトと直接的または間接的に会合しない場合、脂質ラフト会合ポリペプチドに連結しない限り、それをVLP内へと組み込むことは期待されない。抗原は、疾患または障害に関与する任意の抗原、例えば、微生物抗原(例えば、ウイルス抗原、細菌抗原、真菌抗原、原虫抗原、蠕虫抗原、酵母抗原など)、腫瘍抗原、アレルゲンなどであり得る。
【0076】
(抗原の供給源)
本明細書で記載される抗原は、化学的または酵素的に合成することもでき、組換えにより作製することもでき、天然の供給源から単離することもでき、前出の組合せでもあり得る。抗原は、精製することもでき、部分精製することもでき、粗抽出物でもあり得る。
【0077】
ポリペプチド抗原は、液体クロマトグラフィー(例えば、高速液体クロマトグラフィー、高速タンパク質液体クロマトグラフィーなど)、サイズ除外クロマトグラフィー、ゲル電気泳動(一次元ゲル電気泳動、二次元ゲル電気泳動を含めた)、アフィニティークロマトグラフィー、または他の精製法が含まれるがこれらに限定されない、当技術分野で公知の標準的なタンパク質精製法を用いて、天然の供給源から単離することができる。多くの実施形態では、抗原が、例えば、約50%〜約75%純粋、約75%〜約85%純粋、約85%〜約90%純粋、約90%〜約95%純粋、約95%〜約98%純粋、約98%〜約99%純粋、または99%超純粋の精製抗原である。
【0078】
固相ペプチド合成法を用いることができるが、このような技法は、当業者に公知である。Jones、「The Chemical Synthesis of Peptides」(オックスフォード、Clarendon Press社)(1994年)を参照されたい。一般に、このような方法では、活性化させた単量体のユニットを、伸長するペプチド鎖を結合させた固相へと連鎖的に付加することにより、ペプチドを作製する。
【0079】
ポリペプチドを作製するには、十分に確立された組換えDNA法を、脂質ラフト会合ポリペプチドと同じベクター内において用いることもでき、この場合、例えば、ポリペプチドをコードするヌクレオチド配列を含む発現構築物は、適切な宿主細胞(例えば、インビトロの細胞培養物における単細胞実体として増殖させる真核宿主細胞、例えば、酵母細胞、昆虫細胞、哺乳動物細胞など)または、原核細胞(例えば、インビトロの細胞培養物中)中に導入され、これにより遺伝的に改変された宿主細胞を作製し、適切な培養条件下において、該遺伝的に改変された宿主細胞によりタンパク質を作製する。
【0080】
(ウイルス抗原)
適切なウイルス抗原には、以下の群:Retroviridae科(例えば、HIV−1などのヒト免疫不全ウイルス(また、HTLV−III、LAV、もしくはHTLV−III/LAV、またはHIV−IIIとも称する));また、HIV−LPなど、他の単離物; Picomaviridae科(例えば、ポリオウイルス、A型肝炎ウイルス;エンテロウイルス、ヒトコクサッキーウイルス、ライノウイルス、エコーウイルス); Calciviridae科(例えば、ノーウォークウイルスおよび関連ウイルスを含めた、胃腸炎を引き起こすウイルス株); Togaviridae科(例えば、ウマ脳炎ウイルス、風疹ウイルス); Flaviridae科(例えば、デング熱ウイルス、脳炎ウイルス、黄熱病ウイルス); Coronoviridae科(例えば、コロナウイルス); Rhabdoviradae科(例えば、水疱性口内炎ウイルス、狂犬病ウイルス); Coronaviridae科(例えば、コロナウイルス); Rhabdoviridae科(例えば、水疱性口内炎ウイルス、狂犬病ウイルス); Filoviridae科(例えば、エボラウイルス); Paramyxoviridae科(例えば、パラインフルエンザウイルス、ムンプスウイルス、麻疹ウイルス、RSウイルス); Orthomyxoviridae科(例えば、インフルエンザウイルス); Bungaviridae科(例えば、ハンタンウイルス、ブンガウイルス、フレボウイルス、およびナイロウイルス); Arena viridae科(出血熱ウイルス); Reoviridae科(例えば、レオウイルス、オルビウイルス、およびロタウイルス); Bimaviridae科; Hepadnaviridae科(B型肝炎ウイルス); Parvovirida科(パルボウイルス); Papovavirida(パピローマウイルス、ポリオーマウイルス); Adenoviridae科(大半のアデノウイルス); Herpesviridae科(1型および2型の単純ヘルペスウイルス(HSV)、水痘帯状疱疹ウイルス、サイトメガロウイルス(CMV)、ヘルペスウイルス); Poxyiridae科(痘瘡ウイルス、ワクシニアウイルス、ポックスウイルス);およびIridoviridae科(例えば、アフリカブタコレラウイルス);ならびに未分類のウイルス(例えば、海綿状脳症の病因因子、デルタ肝炎の因子(B型肝炎ウイルスの欠損サテライトウイルスと思われる)、非A型肝炎、非B型肝炎の因子(クラス1=内部感染による;クラス2=非経口感染による(すなわち、C型肝炎));およびアストロウイルス)のうちの1または複数のウイルスと関連する(例えば、これらにより合成される)抗原が含まれる。
【0081】
(ノロウイルス(Norvirus)抗原)
本明細書で開示されるVLPには、ノロウイルス科に由来する各種の抗原が含まれ得ることが好ましい。「ノーウォーク様ウイルス」とも呼ばれるノロウイルスは、Caliciviridaeウイルス科内の4つの属のうちの1つを占める。ノロウイルス属内には、遺伝子群Iおよび遺伝子群IIと名付けられた、2つの主要な遺伝子群が存在する。遺伝子群Iのノロウイルス株には、ノーウォークウイルス、サウサンプトンウイルス、デザートシールドウイルス、およびチバウイルスが含まれる。遺伝子群IIのノロウイルス株には、ヒューストンウイルス、ハワイウイルス、ローズデールウイルス、グリムズビーウイルス、メキシコウイルス、およびスノーマウンテンエージェントが含まれる(Parker, T.D.ら、J. Virol.(2005年)、79巻(12号):7402〜9頁; Hale, A.D.ら、J Clin. Micro.(2000年)、38巻(4号):1656〜1660頁)。ノーウォークウイルス(NV)は、世界中における、大半の急性ウイルス性胃腸炎の流行的発生の一因となる、ヒトカリチウイルス群の原型株である。ノーウォークウイルスのカプシドタンパク質は、2つのドメイン:殻ドメイン(S)および突出部ドメイン(P)を有する。Pドメイン(アミノ酸226〜530;ノーウォーク株の番号付け)は、2つのサブドメインであるP1およびP2に分けられる。P2ドメインは、P1ドメイン内における127アミノ酸の挿入(アミノ酸279〜405)であり、フォールディングされた単量体の最遠位面に位置する。P2ドメインは、ノロウイルス株のうちで最も保存の程度が低いVP1領域であり、P2内の超可変領域は、受容体の結合および免疫反応性において重要な役割を果たすと考えられている。Pドメインが外部に位置することを踏まえるなら、それは、本明細書で開示されるVLPワクチンのための抗原として用いるのに好ましい抗原、またはポリペプチドエピトープの供給源である。P2ドメインは、遺伝子群Iまたは遺伝子群IIのノロウイルス株に好ましい抗原である。広範にわたるノロウイルス株を通じて保存されるP2ドメインのうちのある領域内に存在するエピトープとして近年になって同定されたmAb 61.21エピトープ、ならびにmAb 54.6エピトープ(Lochridge, V.P.ら、J Gen. Virol.(2005年)、86巻:2799〜2806頁)がさらにより好ましい。
【0082】
(インフルエンザ抗原)
本明細書で開示されるVLPは、赤血球凝集素およびノイラミニダーゼ以外に、またはこれらに加えて、インフルエンザに由来する各種の抗原を包含し得る。さらなる好ましいインフルエンザ抗原は、M2ポリペプチドである。インフルエンザウイルスのM2ポリペプチドは、スプライシングイベント後において、RNAセグメント7(マトリックスセグメント)によりコードされる、97アミノ酸の小型のクラスIII内在性膜タンパク質である(80、81)。ウイルス粒子上に存在するM2は極めてわずかであり、感染細胞上においてより豊富に見出すことができる。M2は、ウイルスの侵入に必要な、プロトン選択性イオンチャネルとして用いられる(82、83)。感染中または従来のワクチン接種中の免疫原性は最小限であることから、その保存が説明されるが、代替的なフォーマットで存在する場合は、より免疫原性であり防御性である(84〜86)。これは、インビボにおけるM2モノクローナル抗体の受動伝達により、ウイルスクリアランスが加速化され、結果として防御がもたらされるという観察(87)と符合する。M2の外部ドメインエピトープを、融合タンパク質として、HBVのコア粒子に連結すると、非経口接種および鼻腔内接種のいずれによっても、マウスにおいて防御性であり、3回の繰り返しコピーを該コアタンパク質のN末端へと融合させると、最も免疫原性となる(88〜90)。これは、エピトープ密度が増大すると免疫原性が増大することを示す他のキャリア−ハプテンデータ(91)と符合する。
【0083】
M2ワクチンを鼻腔内送達する場合、良好な防御を達成するにはアジュバントが必要とされ、LTR192G(88、90)およびCTA1−DD(89)により良好な結果が達成されている。ペプチドはまた、KLH、またはN. meningitidesの外膜タンパク質複合体、またはヒトパピローマウイルスのVLPなどのキャリアに化学的に結合体化することもでき、マウスおよび他の動物におけるワクチンとして防御的である(92、93)。
【0084】
M2タンパク質は、高度に保存的ではあるが、配列の相違を全く示さないわけではない。一般的な株であるA/PR/8/34(H1N1)およびA/Aichi/68(H3N2)のM2外部ドメインエピトープは、A/Hong Kong/156/97(H5N1)を除き、今日配列決定されている他のすべてのヒト株と、免疫学的に交差反応性であることが示された(92)。インフルエンザデータベースの配列の検討からも、A/Vietnam/1203/04など、他のより近年の病原性H5N1ヒト単離物のM2配列内において、同様の相違が示されている。この知見は、M2エピトープを組み込む、有効なH5特異的汎流行ワクチンが、ヒトH1単離物およびH3単離物中において現在広まっているM2配列ではなく、病原性の鳥類株に固有のM2配列を反映する必要があることを裏付ける。
【0085】
インフルエンザウイルスに由来するさらなるタンパク質(HA、NA、およびM2以外のタンパク質)も、共発現により、またはさらなる抗原の全部もしくは一部を、gagポリペプチドもしくはHAポリペプチドに連結することにより、VLPワクチン内へと組み込むことができる。これらのさらなる抗原には、PB2、PB1、PA、核タンパク質、マトリックス(M1)、NS1、およびNS2が含まれる。これら後者の抗原は、一般に、中和抗体反応の標的ではなく、T細胞により認識される重要なエピトープを含有し得る。このようなエピトープに対してVLPワクチンにより誘導されるT細胞反応が、防御的免疫性を促進するのに有益であることが判明し得る。
【0086】
(他の病原性抗原)
適切な細菌性抗原には、例えば、病原性Pasteurella属種、Staphylococci属種、およびStreptococcus属種などの病原性グラム陽性菌;また、Neisseria属、Escherichia属、Bordetella属、Campylobacter属、Legionella属、Pseudomonas属、Shigella属、Vibrio属、Yersinia属、Salmonella属、Haemophilus属、Brucella属、Francisella属、およびBacterioides属のグラム陰性菌などのグラム陰性菌を含めた、各種の病原性細菌のうちのいずれかと関連する(例えば、これらにより合成され、また、これらに内因性である)抗原が含まれる。例えば、Schaechter, M、H. Medoff、D. Schlesinger、「Mechanisms of Microbial Disease」、ボルチモア、Williams and Wilkins社(1989年)を参照されたい。
【0087】
感染性の病原性真菌と関連する(例えば、これらにより合成され、また、これらに内因性である)適切な抗原には、Cryptococcus neoformans、Histoplasma capsulatum、Coccidioides immitis、Blastomyces dermatitidis、およびCandida albicans、Candida glabrata、Aspergillus fumigata、Aspergillus flavus、およびSporothrix schenckiiが含まれるがこれらに限定されない感染性真菌と関連する抗原が含まれる。
【0088】
病原性原虫、蠕虫、および他の真核微生物病原体と関連する(例えば、これらにより合成され、また、これらに内因性である)適切な抗原には、Plasmodium falciparum、Plasmodium malariae、Plasmodium ovale、およびPlasmodium vivaxなどのPlasmodium属; Toxoplasma gondii; Trypanosoma brucei、Trypanosoma cruzi; Schistosoma haematobium、Schistosoma mansoni、Schistosoma japonicum; Leishmania donovani; Giardia intestinalis; Cryptosporidium parvum等が含まれるがこれらに限定されない原虫、蠕虫、および他の真核微生物病原体と関連する抗原が含まれる。
【0089】
適切な抗原には、Helicobacter pyloris、Borelia burgdorferi、Legionella pneumophila、Mycobacteria属種(例えば、M. tuberculosis、M. avium、M. intracellulare、M. kansaii、M. gordonae)、Staphylococcus aureus、Neisseria gonorrhoeae、Neisseria meningitidis、Listeria monocytogenes、Chlamydia trachomatis、Streptococcus pyogenes(Streptococcus属のA群)、Streptococcus agalactiae(Streptococcus属のB群)、Streptococcus属(viridans群)、Streptococcus faecalis、Streptococcus bovis、Streptococcus属(嫌気性の種)、Streptococcus pneumoniae、病原性のCampylobacter属種、Enterococcus属種、Haemophilus influenzae、Bacillus anthracis、Corynebacterium diphtheriae、Corynebacterium(corynebacterium)属種、Erysipelothrix rhusiopathiae、Clostridium perfringens、Clostridium tetani、Enterobacter aerogenes、Klebsiella pneumoniae、Pasturella multocida、Bacteroides属種、Fusobacterium nucleatum、Streptobacillus moniliformis、Treponema pallidium、Treponema pertenue、Leptospira属、Rickettsia属、およびActinomyces israeliなどの病原性微生物と関連する(例えば、これらにより合成され、また、これらに内因性である)抗原が含まれる。病原性E. coli株の非限定的な例は、ATCC第31618号、同第23505号、同第43886号、同第43892号、同第35401号、同第43896号、同第33985号、同第31619号、および同第31617号である。
【0090】
細胞内病原体と関連する各種のポリペプチドまたは他の抗原のうちのいずれかを、VLP内に組み込むことができる。細胞内病原体と関連するポリペプチドエピトープおよびペプチドエピトープとは、その断片が、MHCクラスI分子と併せて、CD8+リンパ球の表面上におけるT細胞抗原受容体により認識される、例えば、これにより結合されるように、感染細胞の表面上に提示される、細胞内病原体と関連する(例えば、これによりコードされる)任意のポリペプチドである。細胞内病原体と関連するポリペプチドエピトープおよびペプチドエピトープは当技術分野において公知であり、これらには、ヒト免疫不全ウイルス、例えば、HIV gp120またはその抗原性断片と関連する抗原;サイトメガロウイルス抗原; Mycobacterium属抗原(例えば、Mycobacterium avium、Mycobacterium tuberculosisなど);Pneumocystic carinii(PCP)抗原;41−3、AMA−1、CSP、PFEMP−1、GBP−130、MSP−1、PFS−16、SERPなど、Plasmodium falciparumまたは他の任意のマラリア種と関連する抗原が含まれるがこれらに限定されない、マラリア抗原;真菌抗原;酵母抗原(例えば、Candida属種抗原); Toxoplasma gondii、Toxoplasma encephalitis、または他のToxoplasma属種と関連する抗原が含まれるがこれらに限定されないトキソプラズマ抗原;エプスタイン−バーウイルス(EBV)による抗原; Plasmodium属抗原(例えば、gp190/MSP1など)などが含まれるが、これらに限定されない。
【0091】
好ましいVLPワクチンは、Bacillus anthracisに対するものであり得る。Bacillus anthracisは、好気性または通性嫌気性でグラム陽性の非運動性桿菌であり、幅1.0μm、長さ3.0〜5.0μmである。B. anthracisは、悪条件下でも耐性の高い内性胞子を形成し、これは、感染した動物がかつて死亡した場所の土壌で見出すことができる。本明細書で開示されるVLPワクチン内で用いるのに好ましい抗原は、哺乳動物細胞上の受容体に結合し、B. anthracisが疾患を引き起こす能力にとって極めて重要な83kDaのタンパク質である防御抗原(PA)である。より好ましい抗原は、被験体においてBacillus anthracisに対する防御的免疫性を誘導するのに用い得る、Bacillus anthracis PAのC末端における140アミノ酸の断片である。炭疽菌に対するVLPワクチン内において用いられる他の例示的な抗原は、炭疽菌胞子に由来する抗原(例えば、BclA)、該細菌の栄養期に由来する抗原(例えば、細胞壁抗原、被膜抗原(例えば、ポリ−ガンマ−D−グルタミン酸、またはPGA)、分泌抗原(例えば、防御抗原、致死性因子、または浮腫因子などの外毒素))である。VLPワクチンで用いるのに好ましい別の抗原は、B. anthracisに固有のアントロースを含有する四糖である(Daubenspeck J.M.ら、J. Biol. Chem.(2004年)、279巻:30945頁)。四糖は、脂質ラフト会合ポリペプチドに連結することができ、これにより、VLPワクチンとの抗原の会合が可能となる。
【0092】
(腫瘍関連抗原)
公知の各種腫瘍特異抗原または腫瘍関連抗原(TAA)のうちのいずれかを、VLPに組み込むことができる。全TAAを用い得るが、その必要はない。代わりに、TAAの一部、例えば、エピトープを用いることができる。VLP内で用い得る腫瘍関連抗原(またはそれらのエピトープを含有する断片)には、MAGE−2、MAGE−3、MUC−1、MUC−2、HER−2、高分子量黒色腫関連抗原であるMAA、GD2、癌胎児性抗原(CEA)、TAG−72、卵巣関連抗原であるOV−TL3およびMOV18、TUAN、アルファ−フェトプロテイン(AFP)、OFP、CA−125、CA−50、CA−19−9、腎腫瘍関連抗原であるG250、EGP−40(EpCAMとしても知られる)、S100(悪性黒色腫関連抗原)、p53、およびp21rasが含まれるがこれらに限定されない。前出のうちのいずれかを含めた、任意のTAA(またはそれらのエピトープ)の合成類似体を用いることができる。さらに、1または複数のTAA(またはそれらのエピトープ)の組合せを、組成物中に組み込むことができる。
【0093】
(アレルゲン)
一態様では、VLPワクチンの一部である抗原が、各種のアレルゲンのうちのいずれかであり得る。アレルゲンベースのワクチンを用い、被験体におけるアレルゲンに対する忍容性を誘導することができる。チロシンとの共沈殿(co−precipitation)を伴うアレルゲンワクチンの例は、米国特許第3,792,159号、同第4,070,455号、および同第6,440,426号において見出すことができる。
【0094】
各種のアレルゲンのうちのいずれかをVLP内に組み込むことができる。アレルゲンには、環境空中アレルゲン;ブタクサ/花粉症などの植物の花粉;雑草花粉アレルゲン;芝草花粉アレルゲン;ジョンソングラス;樹木花粉アレルゲン;ライグラス;イエダニアレルゲン(例えば、Der p I、Der f Iなど)などのクモ形類動物アレルゲン;コナダニ(storage mite)アレルゲン;スギ花粉/花粉症;カビ胞子アレルゲン;動物アレルゲン(例えば、イヌ、モルモット、ハムスター、アレチネズミ、ラット、マウスなどによるアレルゲン);食物アレルゲン(例えば、甲殻類;ラッカセイなどの堅果;柑橘類によるアレルゲン);昆虫アレルゲン;毒(膜翅目、スズメバチ(yellow jacket)、ミツバチ、カリバチ、スズメバチ(hornet)、ヒアリ);ゴキブリ、ノミ、蚊などによる他の環境昆虫アレルゲン;連鎖球菌抗原などの細菌アレルゲン; Ascaris属抗原などの寄生虫アレルゲン;ウイルス抗原;真菌胞子;薬物アレルゲン;抗生剤;ペニシリンおよび関連化合物;他の抗生剤;ホルモン(インスリン)、酵素(ストレプトキナーゼ)などの全タンパク質;不完全抗原またはハプテンとして作用することが可能なすべての薬物およびそれらの代謝物;ハプテンとして作用し、アレルゲンとして機能することが可能な、産業用化学物質および代謝物(例えば、酸無水物(トリメリト酸無水物など)およびイソシアン酸塩(ジイソシアン酸トルエンなど));小麦粉などの職業アレルゲン(例えば、パン屋喘息(Baker’s asthma)を引き起こすアレルゲン)、トウゴマ、コーヒー豆、また、上記で記載した産業用化学物質;ノミアレルゲン;ヒト以外の動物におけるヒトタンパク質が含まれるが、これらに限定されない。
【0095】
アレルゲンには、細胞、細胞抽出物、タンパク質、ポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性および非ペプチド性模倣体、ならびに他の分子、低分子、脂質、糖脂質、ならびに炭水化物が含まれるが、これらに限定されない。
【0096】
特定の天然の動物アレルゲンおよび植物アレルゲンの例には、以下の属:Canine属(Canis familiaris); Dermatophagoides属(例えば、Dermatophagoides farinae); Felis属(Felis domesticus); Ambrosia属(Ambrosia artemiisfolia); Lolium属(例えば、Lolium perenneまたはLolium multiflorum); Cryptomeria属(Cryptomeria japonica); Altemaria属(Altemaria altemata); Alder属; Alnus属(Alnus gultinoas); Betula属(Betula verrucosa); Quercus属(Quercus alba); Olea属(Olea europa); Artemisia属(Artemisia vulgaris); Plantago属(例えば、Plantago lanceolata); Parietaria属(例えば、Parietaria officinalisまたはParietaria judaica); Blattella属(例えば、Blattella germanica); Apis属(例えば、Apis multiflorum); Cupressus属(例えば、Cupressus sempervirens、Cupressus arizonica、およびCupressus macrocarpa); Juniperus属(例えば、Juniperus sabinoides、Juniperus virginiana、Juniperus communis、およびJuniperus ashei); Thuya属(例えば、Thuya orientalis); Chamaecyparis属(例えば、Chamaecyparis obtusa); Periplaneta属(例えば、Periplaneta americana); Agropyron属(例えば、Agropyron repens); Secale属(例えば、Secale cereale); Triticum属(例えば、Triticum aestivum); Dactylis属(例えば、Dactylis glomerata); Festuca属(例えば、Festuca elatior); Poa属(例えば、PoapratensisまたはPoa compressa); Avena属(例えば、Avena sativa); Holcus属(例えば、Holcus lanatus); Anthoxanthum属(例えば、Anthoxanthum odoratum); Arrhenatherum属(例えば、Arrhenatherun elatius); Agrostis属(例えば、Agrostis alba); Phleum属(例えば、Phleum pratense); Phalaris属(例えば、Phalaris arundinacea); Paspalum属(例えば、Paspalum notatum); Sorghum属(例えば、Sorghum halepensis); ならびにBromus属(例えば、Bromus inermis)に特異的なタンパク質が含まれるが、これらに限定されない。
【0097】
(エンベロープウイルスベースのVLPを作製するのに好ましい方法)
エンベロープウイルスベースのVLPは、当業者に適用可能な任意の方法により作製することができる。エンベロープウイルスベースのVLPは、VLPを形成する一因となる1または複数のポリペプチドを包含することが典型的である。加えて、エンベロープウイルスベースのVLPは、膜(脂質ラフトを含めた)会合ポリペプチドなど、1または複数のさらなるポリペプチドを包含することにより、(VLPを形成する一因となる1または複数のポリペプチドの一部として、天然において、または人工的に存在する抗原以外のさらなる)抗原をもたらすことができる。好ましい実施形態では、ポリペプチドを、任意の適用可能なタンパク質発現系、好ましくは、哺乳動物細胞による発現系および昆虫細胞による発現系など、細胞膜内に脂質ラフトドメインを包含する細胞ベースの系において共発現させることができる。
【0098】
組換えによるVLP用ポリペプチドの発現は、該ポリペプチドのうちの1または複数をコードするポリヌクレオチドを含有する発現ベクターを伴う。ポリペプチドのうちの1または複数をコードするポリヌクレオチドが得られたら、当技術分野で周知の技法を用いる組換えDNA法により、該ポリペプチドを作製するためのベクターを作製することができる。このようにして、本明細書では、VLPポリペプチドをコードするヌクレオチド配列のうちのいずれかを含有するポリヌクレオチドを発現することによりタンパク質を調製する方法が記載される。当業者に周知の方法を用いて、VLPポリペプチドをコードする配列と、適切な転写制御シグナルおよび翻訳制御シグナルとを含有する発現ベクターを構築することができる。例えば、これらの方法には、インビトロにおける組換えDNA法、合成法、また、インビボにおける遺伝子組換えが含まれる。したがって、本発明は、すべてが1または複数のプロモーターに作動可能に連結した、gagポリペプチドと、抗原に連結した脂質ラフト会合ポリペプチドとをコードするヌクレオチド配列を含む複製可能なベクターを提供する。
【0099】
従来の技法により、発現ベクターを宿主細胞へと導入することができ、次いで、従来の技法により、トランスフェクトされた細胞を培養して、VLPポリペプチド(複数可)を作製することができる。したがって、本発明は、異種プロモーターに作動可能に連結したVLPポリペプチドのうちの1または複数をコードするポリヌクレオチドを含有する宿主細胞を包含する。VLPを作製するのに好ましい実施形態では、以下で詳述する通り、gagポリペプチドと、抗原に連結した脂質ラフト会合ポリペプチドとの両方をコードするベクターを、宿主細胞内で共発現させて、VLPを作製することができる。
【0100】
各種の宿主発現ベクター系を用いて、VLPポリペプチドを発現させることができる。このような宿主発現系は、好ましくは共発現によりVLPを生成させる目的で、VLPポリペプチドを作製し得るビヒクルを表す。広範囲にわたる宿主を、適切な発現ベクター構築物において用いることができ、脂質ラフトベースのアセンブリーに依拠する場合、好ましい宿主発現系は、VLPのアセンブリーに適する脂質ラフトを有する宿主である。これらには、VLPポリペプチドコード配列を含有する組換えバクテリオファージDNA発現ベクター、プラスミドDNA発現ベクター、もしくはコスミドDNA発現ベクターにより形質転換した細菌(例えば、E. coli、B. subtilis);VLPポリペプチドコード配列を含有する組換え酵母発現ベクターにより形質転換した酵母(例えば、Saccharomyces属、Pichia属);VLPポリペプチドのコード配列を含有する組換えウイルス発現ベクター(例えば、バキュロウイルス)を感染させた昆虫細胞系;VLPポリペプチドのコード配列を含有する組換えウイルス発現ベクター(例えば、カリフラワーモザイクウイルス、CaMV;タバコモザイクウイルス、TMV)を感染させた植物細胞系、もしくはVLPポリペプチドのコード配列を含有する組換えプラスミド発現ベクター(例えば、Tiプラスミド)により形質転換した植物細胞系;または、哺乳動物細胞のゲノムに由来するプロモーター(例えば、メタロチオネインプロモーター)、もしくは哺乳動物ウイルスに由来するプロモーター(例えば、アデノウイルス後期プロモーター;ワクシニアウイルス7.5Kプロモーター)を含有する組換え発現構築物を保有する、哺乳動物細胞系(例えば、COS細胞、CHO細胞、BHK細胞、293細胞、3T3細胞)などの微生物が含まれるがこれらに限定されない。それらのいずれもがVLPのアセンブリーに適する脂質ラフトを有するため、VLPポリペプチドを発現させるには、哺乳動物細胞を用いることが好ましく、また、昆虫細胞を用いることがより好ましい。例えば、ヒトサイトメガロウイルスに由来する主要前初期(intermediate early)遺伝子プロモーターエレメントなどのベクターと組合わされる、MRC−5細胞、Vero細胞、PER.C6(TM)細胞、チャイニーズハムスター卵巣細胞(CHO)、およびHEK293細胞などの哺乳動物細胞は、VLPポリペプチドの有効な発現系である(Foeckingら、Gene、45巻:101頁(1986年); Cockettら、Bio/Technology、8巻:2頁(1990年))。
【0101】
昆虫系では、Autographa californica核多角体病ウイルス(AcNPV)を、外来遺伝子を発現するためのベクターとして用いることができる。該ウイルスは、Spodoptera frugiperda細胞内で増殖する。VLPポリペプチドのコード配列(複数可)は、ウイルスの非必須領域(例えば、ポリヘドリン遺伝子領域)内へと個別にクローニングすることができ、また、AcNPVプロモーター(例えば、ポリヘドリンプロモーター)の制御下に置くことができる。
【0102】
哺乳動物の宿主細胞では、多くのウイルスベースの発現系を用いることができる。発現ベクターとしてアデノウイルスを用いる場合は、対象のVLPポリペプチド配列(複数可)を、アデノウイルス転写/翻訳制御複合体、例えば、後期プロモーターおよび三連リーダー(tripartite leader)配列にライゲーションすることができる。次いで、インビトロまたはインビボにおける組換えにより、このキメラ遺伝子を、アデノウイルスゲノム内に挿入することができる。ウイルスゲノムの非必須領域(例えば、E1領域またはE3領域)内に挿入する結果、感染宿主内において生存可能であり、また、VLPポリペプチド(複数可)を発現することが可能な組換えウイルスがもたらされる(例えば、LoganおよびShenk、Proc. Natl. Acad. Sci. USA、81巻:355〜359頁(1984年)を参照されたい)。挿入されたVLPポリペプチドのコード配列(複数可)を効率的に翻訳するには、特定の開始シグナルもまた必要とされ得る。これらのシグナルには、ATG開始コドン、および隣接配列が含まれる。さらに、全挿入配列の翻訳を確保するには、該開始コドンが、所望されるコード配列のリーディングフレームと同フレームでなければならない。これらの外因性翻訳制御シグナルおよび開始コドンは、由来が多様であることが可能であり、天然および合成のいずれもが可能である。発現の効率は、適切な転写エンハンサーエレメント、転写ターミネーターなどを組み込むことにより、増強することができる(Bittnerら、Methods in Enzymol.、153巻:51〜544頁(1987年)を参照されたい)。1つの例は、Stratagene社製のAdEASY−XL(TM)システムなど、アデノウイルスベースのベクター系において用いられる、ヒトCMV前初期プロモーターである。
【0103】
加えて、挿入した配列の発現を調節するか、または該遺伝子産物を、所望される特定の形で修飾およびプロセシングする宿主細胞株も選択することができる。タンパク質産物に対するこのような修飾(例えば、グリコシル化)およびプロセシング(例えば、切断または膜への輸送)は、VLPを作製するのに、またはVLPポリペプチド、もしくはアジュバントなどのさらなるポリペプチド、もしくはさらなる抗原が機能するのに重要であり得る。種々の宿主細胞は、タンパク質および遺伝子産物の翻訳後プロセシングおよび修飾について特徴的で特異的な機構を有する。発現する外来タンパク質の適正な修飾およびプロセシングを確保するために、適切な細胞株または宿主系を選択することができる。この目的で、一次転写物を適正にプロセシングし、遺伝子産物を適正にグリコシル化およびリン酸化するための細胞機構を保有する真核宿主細胞を用いることができる。
【0104】
宿主細胞には、gagポリペプチドをコードする第1のベクターと、ウイルス性膜抗原、または抗原に連結した脂質ラフト会合ポリペプチドをコードする第2のベクターという、本発明の2つの発現ベクターを共トランスフェクト(co−transfect)することができる。2つのベクターは、各VLPポリペプチドの同等の発現を可能とする、同一の選択マーカーを含有し得る。代替的に、gagポリペプチドと、抗原に連結した脂質ラフト会合ポリペプチドとの両方をコードし、また、これらの両方を発現することが可能な単一のベクターを用いることもできる。
【0105】
宿主細胞によりVLPが生成されたら、ポリペプチドの精製についての当技術分野で公知の任意の方法により、例えば、クロマトグラフィー(例えば、イオン交換クロマトグラフィー、特に、該ポリペプチドに付加した任意のアフィニティー精製タグによるアフィニティークロマトグラフィー、およびサイズ除外クロマトグラフィー)、遠心分離、溶解度差(differential solubility)により、またはタンパク質もしくは他の高分子を精製するための他の任意の標準的な技法によりこれを精製することができる。加えて、VLPポリペプチドを、本明細書で記載されるか、またはこれ以外に当技術分野で公知の異種ポリペプチド配列に融合させ、VLPの精製を容易にすることもできる。精製後、VLPポリペプチドに対する共有結合を介して、または他の非共有結合機構により、さらなる抗原またはアジュバントなどのさらなるエレメントを、VLPへと物理的に連結することができる。哺乳動物細胞および昆虫細胞など、脂質ラフトドメインを有する宿主細胞内においてVLPポリペプチドを共発現させる好ましい実施形態では、VLPが自己アセンブルして放出され、これにより、上記の方法のうちのいずれかによるVLPの精製が可能となる。VLPの好ましい実施形態には、例えば、同種ウイルスのタンパク質から操作されたVLP、例えば、インフルエンザウイルスに由来するM1、HA、また、場合によってNAから構築されたVLP、また、異種ウイルスのタンパク質から操作されたVLP、例えば、MLVもしくはHIVまたは他のレトロウイルスに由来するGagタンパク質を、異なるウイルスに由来する抗原、例えば、インフルエンザHAおよびNAと共に操作することにより形成したVLPが含まれる。
【0106】
(GagベースのVLPを作製する好ましい方法)
当業者に適用可能な任意の方法により、VLPを容易にアッセンブルすることができ、この結果、gagポリペプチドと、天然では脂質ラフトと会合しない抗原に連結した脂質ラフト会合ポリペプチドとを包含するVLPがアッセンブルされることが好ましい。好ましい実施形態では、任意の適用可能なタンパク質発現系、好ましくは、哺乳動物細胞による発現系および昆虫細胞による発現系など、脂質内に脂質ラフトドメインを包含する細胞ベースの発現系内において、ポリペプチドを共発現させることができる。
【0107】
gagポリペプチドを用いて形成されるVLP発現の例が数多く公表されており、これにより、VLPを作製するのに適用可能な発現系の範囲が示されている。複数のレトロウイルスによる研究は、他のウイルス成分の不在下で発現したGagポリペプチドが、細胞表面におけるVLPの形成および出芽に十分であることを裏付けている(WillsおよびCraven、AIDS、5巻、639〜654頁、1991年; Zhouら、3. Virol.、68巻、2556〜2569頁、1994年; Morikawaら、Virology、183巻、288〜297頁、1991年; Royerら、Virology、184巻、417〜422頁、1991年; Gheysenら、Cell、59巻、103〜112頁、1989年; Hughesら、Virology、193巻、242〜255頁、1993年; Yamshchikovら、Virology、214巻、50〜58頁、1995年)。バキュロウイルスベクターを用いる昆虫細胞内において、Gag前駆体が発現するとVLPが形成されることについては、複数の研究グループにより裏付けられている(Delchambreら、EMBO J.、8巻、2653〜2660頁、1989年; Luoら、Virology、179巻、874〜880頁、1990年; Royer ら、Virology、184巻、417〜422頁、1991年; Morikawa ら、Virology、183巻、288〜297頁、1991年; Zhouら、J. Virol.、68巻、2556〜2569頁、1994年; Gheysenら、Cell、59巻、103〜112頁、1989年; Hughesら、Virology、193巻、242〜255頁、1993年; Yamshchikovら、Virology、214巻、50〜58頁、1995年)。これらのVLPは、未成熟レンチウイルス粒子に類似しており、効率的にアッセンブルされて昆虫細胞の細胞膜から出芽により放出される。
【0108】
Gag前駆体のアミノ末端領域は、ウイルスアセンブリーに必要とされる、細胞表面への輸送および膜結合のためのターゲッティングシグナルであることが報告されている(Yuら、J. Virol.、66巻、4966〜4971頁、1992年; an, Xら、J. Virol.、67巻、6387〜6394頁、1993年; Zhouら、J. Virol.、68巻、2556〜2569頁、1994年; LeeおよびLinial、J. Virol.、68巻、6644〜6654頁、1994年; Dorfmanら、J. Virol.、68巻、1689〜1696頁、1994年; Fackeら、J. Virol.、67巻、4972〜4980頁、1993年)。ワクシニアウイルスによる発現系を用いて、Gag構造タンパク質、ならびにEnv糖タンパク質であるgp120およびgp41を含有する、組換えHIVベースのVLPのアセンブリーが報告されている(Haffarら、J. Virol.、66巻、4279〜4287頁、1992年)。
【0109】
(エンベロープウイルスベースのVLP調製物における感染因子を不活化するのに好ましい方法)
電子放射は、エンベロープウイルスベースのVLPの免疫原性を実質的に低下させることなく、感染因子を不活化することが可能なので、不活化の好ましい方法は、電子放射を介する方法である。電磁放射で好ましい3つの方式のすべて(すなわち、光反応性化合物を伴うUV照射、UV照射単独、およびガンマ照射)には、血液、食品、ワクチンなど、多種多様な試料における病原体を不活化するのに用いられてきた長い歴史があるため、不活化用電磁放射を適用するための多種多様な装置が市販されており、これらは、本明細書で開示される方法を実施するのにほとんど〜まったく改変せずに用いることができる。さらに、当技術分野では、波長および線量の最適化がルーチン化しており、したがって、当業者の能力範囲内で容易に実施される。
【0110】
(光反応性化合物を伴うUV照射)
電磁放射による例示的な不活化法は、光反応性化合物、好ましくは、感染因子中のポリヌクレオチドと反応する光反応性化合物の存在下における、UV−A照射などの紫外線照射の組合せである。
【0111】
好ましい光反応性化合物には、アクチノマイシン、アントラサイクリノン、アントラマイシン、ベンゾジピロン、フルオレン、フルオレノン、フロクマリン、イソアロキサジン、マイトマイシン、monostral fast blue、norphillin A、フェナントリジン、フェナザチオニウム塩、フェナジン、フェノチアジン、フェニルアジド、キノリン、およびチオキサンテノン(thiaxanthenones)が含まれる。好ましい種は、2つの主要なカテゴリーのうちの1つに属するフロクマリンである。第1のカテゴリーは、ソラーレン[7H−フロ(3,2−g)−(1)−ベンゾピラン−7−オン、または6−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、直鎖型であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の6位に結合している。第2のカテゴリーは、イソソラーレン[2H−フロ(2,3−h)−(1)−ベンゾピラン−2−オン、または4−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、角度型(angular)であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の8位に結合している。3、4、5、8、4’、または5’位において直鎖型フロクマリンを置換することによりソラーレン誘導体を作製し得る一方、3、4、5、6、4’、または5位において角度型フロクマリンを置換することによりイソソラーレン誘導体を作製し得る。ソラーレンは、二本鎖核酸の塩基対間にインターカレートすることができ、長波長の紫外光(UVA)を吸収すると、ピリミジン塩基に対する共有結合付加体を形成する。例えば、G. D. Ciminoら、Ann. Rev. Biochem.、54巻:1151頁(1985年); Hearstら、Quart. Rev. Biophys.、17巻:1頁(1984頁)を参照されたい。
【0112】
好ましいUV(または場合によって、可視光)放射の波長は、適切な反応および/または光付加体が生成される波長に依存し、これは、光反応性化学物質の化学反応に依存する。例示を目的として述べると、多くのソラーレンには、320〜380nmの波長にあるUV放射が最も有効であり、330〜360nmが最大の有効性を示す。同様のUV−A波長はまた、419nmなどの可視光と組み合わせても用い得る光反応性化合物であるリボフラビンと合わせても、病原体を不活化するのに極めて有効である。
【0113】
(UV照射単独)
光反応性化合物の存在下におけるUV照射に加え、感染因子は、UV照射単独でも不活化することができる。好ましい実施形態では、放射は、波長が、約180〜320nm、または約225〜290nm、または約254nm(すなわち、ポリヌクレオチドの高い吸光度ピークを有し、タンパク質吸収は低いスペクトル領域)であるUV−C放射である。UV−C放射は、エンベロープを形成する脂質二重層、また、エンベロープ内のタンパク質など、本明細書で開示されるエンベロープウイルスベースのVLPの成分に対して、安定性および免疫原性の両面で有害性が低い一方で、感染因子を不活化するのに十分なエネルギーを保持するので好ましい。しかし、例えば、UV−AおよびUV−Bなど、他の種類のUV放射もまた用いることができる。
【0114】
(ガンマ照射)
本明細書で開示される方法を実施して組成物を作製するには、ガンマ照射(すなわち、イオン化放射)もまた用いることができる。この好ましい実施形態では、10〜60kGyのガンマ照射線量が、病原体の不活化に有効である。ガンマ照射は、感染因子のゲノムをコードするポリヌクレオチド内において鎖の切断を導入することにより、感染因子を直接的に不活化することもでき、または、該ポリヌクレオチドを攻撃するフリーラジカルを生成することにより、感染因子を間接的に不活化することもできる。ガンマ照射と共にフリーラジカルスカベンジャー(free radical scavenger)および低温を用い、ラジカルを介してエンベロープVLPの脂質成分およびタンパク質成分にもたらされる損傷を抑制することができる。
【0115】
(エンベロープウイルスベースのVLPを用いる好ましい方法)
(処方物)
本明細書で記載されるエンベロープウイルスベースのVLPの好ましい使用は、ワクチン調製物としての使用である。典型的に、このようなワクチンは、液体溶液または懸濁物の注射剤として調製されるが、注射前に液体中に溶解させるか、または懸濁させるのに適する固体形態もまた調製することができる。このような調製物はまた、乳化させることもでき、乾燥粉末として作製することもできる。免疫原性の有効成分は、薬学的に許容され、また、有効成分に適合する、賦形剤と混合することが多い。適切な賦形剤は、例えば、水、生理食塩液、デキストロース、グリセロール、エタノールなどであり、また、これらの組合せである。加えて、所望の場合、ワクチンは、保湿剤もしくは乳化剤、pH緩衝剤、または該ワクチンの有効性を増強するアジュバントなどの補助物質を含有し得る。
【0116】
ワクチンは、注射、例えば、皮下(subcutaneously)注射、皮内注射、皮下(subdermally)注射、または筋肉内注射を介する、従来の非経口投与が可能である。他の投与方式に適する、さらなる処方物には坐剤が含まれ、場合によっては、経口処方物、鼻腔内処方物、口腔内処方物、舌下処方物、腹腔内処方物、膣内処方物、経肛門処方物、および頭蓋内処方物も含まれる。坐剤の場合、従来の結合剤およびキャリアには、例えば、ポリアルキレングリコール(polyalkalene glycol)またはトリグリセリドが含まれる場合があり、このような坐剤は、0.5%〜10%、好ましくは1〜2%の範囲で有効成分を含有する混合物から形成することができる。特定の実施形態では、脂肪酸グリセリドまたはココアバターの混合物などの低温溶融ワックスをまず溶融させ、本明細書で記載されるエンベロープウイルスベースのVLPを、例えば、撹拌することにより、均一に分散させる。次いで、溶融した均一の混合物を、好都合なサイズの型に流し込み、冷却して固形化させる。
【0117】
鼻腔内送達に適する処方物には、液体および乾燥粉末が含まれる。処方物は、例えば、医薬グレードのマンニトール、ラクトース、スクロース、トレハロース、およびキトサンなど、通常用いられる賦形剤を包含する。液体処方物または粉末処方物において、キトサンなどの粘膜付着剤を用いて、粘膜絨毛による、鼻腔内投与された処方物のクリアランスを遅延させることができる。マンニトールおよびスクロースなどの糖を、液体処方物では安定化剤として用いることができ、また、乾燥粉末処方物では安定化剤およびバルキング剤として用いることができる。加えて、モノホスホリルリピドA(MPL)などのアジュバントを、液体処方物および乾燥粉末処方物の両方において、免疫刺激性アジュバントとして用いることもできる。
【0118】
経口送達に適する処方物には、液体、固体、半固体、ゲル、錠剤、カプセル、ロゼンジなどが含まれる。経口送達に適する処方物には、錠剤、ロゼンジ、カプセル、ゲル、液体、食品、飲料、栄養補助食品などが含まれる。処方物は、例えば、医薬グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなど、通常用いられる賦形剤を包含する。他のエンベロープウイルスベースのVLPワクチン組成物は、溶液、懸濁物、丸薬、持続放出処方物、または粉末の形態を取る場合があり、10〜95%、好ましくは25〜70%の有効成分を含有する場合がある。経口処方物の場合、コレラ毒素が興味深い処方物パートナー(また、可能な結合パートナー)である。
【0119】
膣内投与用に処方する場合、エンベロープウイルスベースのVLPワクチンは、ペッサリー、タンポン、クリーム、ジェル、ペースト、フォーム、またはスプレーの形態であり得る。前出の処方物のうちのいずれもが、エンベロープウイルスベースのVLPに加えて、キャリアなど、当技術分野で適切であることが公知の薬剤を含有し得る。
【0120】
一部の実施形態では、エンベロープウイルスベースのVLPワクチンを、全身送達用に処方する場合もあり、局所送達用に処方する場合もある。このような処方物は、当技術分野において周知である。非経口ビヒクルには、塩化ナトリウム溶液、リンゲルデキストロース液、デキストロースおよび塩化ナトリウムによる溶液、乳酸加リンゲル液、または固定油が含まれる。静脈内ビヒクルには、流体および栄養物質補給剤、電解質補給剤(リンゲルデキストロース液に基づく補給剤など)などが含まれる。全身投与経路および局所投与経路には、例えば、皮内経路、局所適用経路、静脈内経路、筋肉内経路などが含まれる。
【0121】
エンベロープウイルスベースのVLPは、中性処方物または塩ベースの処方物を含めたワクチンへと処方することができる。薬学的に許容される塩には、酸添加塩(ペプチドの遊離アミノ基と共に形成され、また、例えば、塩酸もしくはリン酸などの無機酸、または酢酸、シュウ酸、酒石酸、マンデル酸などの有機酸と共に形成される)が含まれる。遊離カルボキシル基と共に形成される塩はまた、例えば、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、または水酸化鉄などの無機塩基、また、イソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、プロカインなどの有機塩基に由来し得る。
【0122】
ワクチンは、投薬処方物に適合した形で、また、治療的に有効であり免疫原性となる量で投与することができる。投与量は、例えば、個体の免疫系が免疫反応を起こす能力、また、所望される防御の程度を含め、処置される被験体に依存する。適切な用量範囲は、1回のワクチン接種当たりの有効成分が数百マイクログラムのオーダーであり、好ましい範囲は、約0.5μg〜1000μgの範囲、好ましくは1μg〜500μgの範囲、またとりわけ、約10μg〜100μgの範囲など、約0.1μg〜2000μgの範囲である(1〜10mgの範囲にある高量も意図される)ことが好ましい。初回投与および追加投与に適するレジメンもまた可変的であるが、初回投与にその後の接種または他の投与が続く形が典型的である。
【0123】
適用方式は多様に変化させることができる。従来のワクチン投与方法のうちのいずれもが適用可能である。これらには、固体の生理学的に許容されるベース上における経口適用、または注射など、生理学的に許容される分散剤による非経口適用が含まれる。ワクチンの用量は、投与経路に依存し、ワクチン接種されるヒトの年齢、および抗原処方物により変化する。
【0124】
ワクチン処方物の一部は、それだけでワクチンとして十分に免疫原性であるが、他の処方物では、該ワクチンがアジュバント物質をさらに包含すると、免疫反応が増強される場合もある。
【0125】
とりわけ、鼻腔内ベース、経口ベース、または肺内ベースの送達処方物の場合、送達および免疫原性を改善するには、粘膜付着を改善する送達剤もまた用いることができる。このような化合物の1つである、キチンのN−脱アセチル化形態であるキトサンは、多くの医薬処方物で用いられる(32)。粘膜絨毛によるクリアランスを遅延させ、粘膜による抗原の取込みおよびプロセシングのためのより多くの時間を可能にする能力のために、キトサンは、鼻腔内におけるワクチン送達にとって魅力的な粘膜付着剤である(33、34)。加えて、それは、接着結合を一過性に開口させ、これにより、NALTへの抗原の経上皮輸送が増強され得る。近年のヒト試験では、キトサンを伴うが、さらなるアジュバントは伴わない、三価の不活化インフルエンザワクチンを鼻腔内投与したところ、筋肉内接種後に得られたセロコンバージョンおよびHI力価をごくわずかに下回るセロコンバージョンおよびHI力価がもたらされた(33)。
【0126】
キトサンはまた、遺伝的操作により解毒したE. coli易熱性エンテロトキシンの変異体であるLTK63など、鼻腔内において良好に機能するアジュバントと共に処方することもできる。これは、キトサンにより付与される送達および付着の利益の上に、免疫刺激効果を付加し、その結果、粘膜反応および全身反応の増強をもたらす(35)。
【0127】
最後に、キトサン処方物はまた、ワクチンの安定性を改善し、粘膜絨毛によるクリアランスにおいて液体処方物を上回る遅延を結果としてもたらすことが示されている乾燥粉末フォーマットでも調製することができることに注意すべきである(42)。これは、キトサンと共に処方された、鼻腔内用乾燥粉末によるジフテリア毒素ワクチンを伴う、近年のヒト臨床試験において見られ、ここで、鼻腔内経路は、従来の筋肉内経路と同程度に有効であり、分泌性IgA反応の利益が付加された(43)。該ワクチンはまた、忍容性も極めて良好であった。キトサンおよびMPLを含有する炭疽菌用の鼻腔内乾燥粉末ワクチンは、ウサギにおいて、筋肉内接種より強力な反応を誘導し、また、エアゾールによる胞子チャレンジ(challenge)に対しても防御的であった(44)。
【0128】
鼻腔内ワクチンは、下気道に対してより良好に作用する非経口投与ワクチンに対して、上気道および下気道に作用し得るので、好ましい処方物を表す。これは、アレルゲンベースのワクチンには忍容性を誘導し、また、病原体ベースのワクチンには免疫性を誘導する点で有益であり得る。
【0129】
鼻腔内ワクチンは、上気道および下気道の両方において防御をもたらすのに加え、針による接種の煩雑さを回避し、また、粒子抗原および/または可溶性抗原が、鼻咽頭関連リンパ組織(NALT)と相互作用することを介して、粘膜および全身における体液性反応および細胞性反応の両方を誘導する手段をもたらす(16〜19)。歴史的に、鼻腔内経路は、非経口接種ほど有効ではなかったが、新規の送達処方物であるエンベロープウイルスベースのVLPおよびアジュバントを用いることにより、この枠組みが変わりつつある。実際、鼻腔粘膜内にはシアル酸を含有する受容体が豊富であるため、鼻腔内送達には、機能性の赤血球凝集素ポリペプチドを含有するエンベロープウイルスベースのVLPがとりわけよく適する場合があり、その結果、HA抗原の結合が増強され、粘膜絨毛によるクリアランスが低減される可能性がある。
【0130】
(アジュバント)
ワクチンに対するアジュバント効果を達成する各種の方法が公知であり、本明細書で開示されるエンベロープウイルスベースのVLPと共に用いることができる。一般的な原理および方法は、「The Theory and Practical Application of Adjuvants」、1995年、Duncan E. S. Stewart−Tull(編)、John Wiley & Sons社、ISBN 0−471−95170−6において詳述されており、また、「Vaccines: New Generation Immunological Adjuvants」、1995年、Gregoriadis Gら(編)、ニューヨーク、Plenum Press社、ISBN 0−306−45283−9においても詳述されている。
【0131】
一部の実施形態では、エンベロープウイルスベースのVLPワクチンが、重量ベースの比率約10:1〜約1010:1のエンベロープウイルスベースのVLP:アジュバント、例えば、約10:1〜約100:1、約100:1〜約103:1、約103:1〜約104:1、約104:1〜約105:1、約105:1〜約106:1、約106:1〜約107:1、約107:1〜約108:1、約108:1〜約109:1、または約109:1〜約1010:1のエンベロープウイルスベースのVLP:アジュバントで、少なくとも1つのアジュバントと混合したエンベロープウイルスベースのVLPを包含する。当業者は、アジュバントに関する情報、また、最適の比率を決定する日常的実験により、適切な比率を容易に決定することができる。
【0132】
アジュバントの好ましい例は、エンベロープウイルスベースのVLPのポリペプチド成分と共発現させるか、または該ポリペプチド成分と融合させて、キメラポリペプチドを作製することにより、本明細書で記載されるエンベロープウイルスベースのVLPに容易に添加し得るポリペプチドアジュバントである。鞭毛の主要なタンパク質構成要素である細菌性フラジェリンは、それが、toll様受容体であるTLR5を介する先天性免疫系により認識されるために、アジュバントタンパク質としてますます注目を集めている、好ましいアジュバントである(65)。TLR5を介するフラジェリンシグナル伝達は、DCの成熟化および移動の他、マクロファージ、好中球、および腸上皮細胞の活性化も誘導することにより、先天性免疫機能および後天性免疫機能の両方に影響を及ぼし、その結果、炎症促進(proinflammatory)メディエーターを生成させる(66〜72)。
【0133】
TLR5は、フラジェリン単量体内における保存的構造(これはフラジェリン単量体タンパク質に固有であり、また、鞭毛機能に必要とされる)を認識し、このことは免疫学的圧力に応答したその変異を妨げる(73)。該受容体は、100fMの濃度に対して感受性であるが、完全な線維は認識しない。鞭毛が単量体へと解体されることが、結合および刺激に必要とされる。
【0134】
アジュバントとしてのフラジェリンは、非経口投与または鼻腔内投与された異種抗原に対する防御反応を誘導する強力な活性を有し(66、74〜77)、また、DNAワクチンに対するアジュバント効果もまた報告されている(78)。マウスまたはサルにおいてフラジェリンを用いると、Th2バイアス(これは、インフルエンザなどの呼吸器ウイルスには適切である)は観察されるが、IgE誘導の証拠は観察されていない。加えて、サルにおける鼻腔内投与または全身投与後における、局所炎症反応も全身炎症反応も報告されていない(74)。TLR5を介して、MyD88依存的な形でなされるフラジェリンシグナル伝達、また、TLRを介する、他のすべてのMyD88依存的シグナルは、Th1バイアスを結果としてもたらすことが示されている(67、79)ため、フラジェリンを使用した後で誘発される反応がTh2特徴を示すことは、ある意味で驚くべきことである。重要なことに、フラジェリンに対して予め存在する抗体は、アジュバントの有効性に対してさほどの影響を及ぼさない(74)ことから、フラジェリンは、複数回使用のためのアジュバントとして魅力的となっている。
【0135】
多くの近年における鼻腔内ワクチン試験における共通の主題は、ワクチンの有効性を改善するアジュバントおよび/または送達系の使用である。このような一研究では、遺伝的に解毒されたE. coli易熱性エンテロトキシンアジュバント(LT R192G)を含有するインフルエンザH3ワクチンが、H5チャレンジに対する異種サブタイプ的な防御を結果としてもたらしたが、これは、鼻腔内送達後に限られた。防御は、交差中和抗体の誘導に基づき、これにより、新たなワクチンの開発における鼻腔内経路についての重要な示唆が示された(22)。
【0136】
サイトカイン、コロニー刺激因子(例えば、GM−CSF、CSFなど);腫瘍壊死因子;インターロイキン2、インターロイキン7、インターロイキン12、インターフェロン、および他の同様の増殖因子はまた、アジュバントとして用いることができ、これらは
エンベロープウイルスベースのVLPのポリペプチド成分と混合または融合させることにより、該ワクチン内に容易に組み込み得るので、これらもまた好ましい。
【0137】
一部の実施形態では、本明細書で開示されるエンベロープウイルスベースのVLPワクチン組成物が、5’−TCG−3’配列を含む、核酸のTLR9リガンド;イミダゾキノリンのTLR7リガンド;置換グアニンのTLR7/8リガンド;ロキソリビン、7−デアザデオキシグアノシン、7−チア−8−オキソデオキシグアノシン、イミキモド(R−837)、およびレシキモド(R−848)など、他のTLR7リガンドなど、Toll様受容体を介して作用する他のアジュバントを包含し得る。
【0138】
特定のアジュバントは、樹状細胞などのAPCによるワクチン分子の取込みを促進し、これらを活性化する。非限定的な例は、免疫標的化アジュバント;毒素、サイトカイン、およびマイコバクテリア誘導体などの免疫調節アジュバント;油処方物;ポリマー;ミセル形成アジュバント;サポニン;免疫刺激複合体マトリックス(ISCOMマトリックス);粒子;DDA;アルミニウムアジュバント;DNAアジュバント;MPL;および封入アジュバントからなる群より選択される。
【0139】
アジュバントのさらなる例には、作用物質、例えば、緩衝生理食塩液中0.05〜0.1パーセントの溶液として一般的に用いられるアルミニウム塩、例えば、水酸化物またはリン酸塩(ミョウバン)(例えば、Nicklas(1992年)、Res. Immunol.、143巻:489〜493頁を参照されたい)、0.25パーセントの溶液として用いられる糖の合成ポリマー(例えば、Carbopol(登録商標))との混合物、それぞれ、70℃〜101℃の範囲の温度で30秒間〜2分間にわたる熱処理によるワクチン中のタンパク質の凝集物が含まれ、また、架橋形成剤による凝集物も可能である。ペプシン処理した、アルブミンに対する抗体(Fab断片)での再活性化による凝集物、C. parvumなどの細菌細胞またはグラム陰性菌の内毒素もしくはリポ多糖成分との混合物、モノオレイン酸マンニド(Aracel A)などの生理学的に許容される油ビヒクルにおけるエマルジョン、または保護置換物として用いられる、ペルフルオロカーボン(Fluosol−DA)の20パーセント溶液によるエマルジョンもまた用いることができる。スクアレンおよびIFAなどの油との混合物もまた好ましい。
【0140】
DDA(ジメチルジオクタデシルアンモニウムブロミド)は、アジュバントの興味深い候補物質であるが、フロイントの完全アジュバントおよびフロイントの不完全アジュバントの他、QuilAおよびQS21などのキラヤサポニンもまた興味深い。さらなる可能性には、モノホスホリルリピドA(MPL(登録商標))、ムラミルジペプチド(MDP)、およびトレオニルムラミルジペプチド(tMDP)など、リポ多糖のポリ[ジ(カルボキシレートフェノキシ)ホスファゼン](poly[di(earboxylatophenoxy)phosphazene)(PCPP)誘導体が含まれる。例えば、アルミニウム塩と併せた、モノホスホリルリピドA、好ましくは、3−脱−O−アシル化モノホスホリルリピドAの組合せを含めた、主にTh1型の反応をもたらすリポ多糖ベースのアジュバントが好ましい。MPL(登録商標)アジュバントは、GlaxoSmithKline社から市販されている(例えば、米国特許第4,436,727号;同第4,877,611号;同第4,866,034号;および同第4,912,094号を参照されたい)。
【0141】
リポソーム処方物もまた、アジュバント効果を付与することが知られており、したがって、リポソームアジュバントは、エンベロープウイルスベースのVLPと合わされる、好ましい例である。
【0142】
免疫刺激複合体マトリックス型(ISCOM(登録商標)マトリックス)アジュバントは、とりわけ、この型のアジュバントが、APCによるMHCクラスII発現を上方制御することが可能であることが示されているため、本発明に従う好ましい選択である。ISCOMマトリックスは、Quillaja saponariaに由来する(場合によって分画された)サポニン(トリテルペノイド)、コレステロール、およびリン脂質からなる。VLP中のものなどの免疫原性タンパク質と混合する場合、結果として得られる粒子状処方物は、サポニンが60〜70%w/wを構成することが可能であり、コレステロールおよびリン脂質が10〜15%w/wを構成することが可能であり、タンパク質が10〜15%w/wを構成することが可能な、ISCOM粒子として公知の処方物である。免疫刺激複合体の組成および使用に関する詳細は、例えば、アジュバントを扱う上述の教科書中に見出し得るが、また、Morein Bら、1995年、Clin. Immunother.、3巻:461〜475頁の他、Barr I GおよびMitchell G F、1996年、Immunol. and Cell Biol.、74巻:8〜25頁も、完全な免疫刺激複合体を調製するのに有益な教示を提供している。
【0143】
本明細書で開示されるエンベロープウイルスベースのVLPワクチンとアジュバントの組合せにおいて用い得るサポニンには、それらがISCOM形態にある場合であれ、そうでない場合であれ、米国特許第5,057,540号;および「Saponins as vaccine adjuvants」、Kensil, C. R.、Crit Rev Ther Drug Carrier Syst、1996年、12巻(1〜2号):1〜55頁;およびEP0362279B1で記載される、Quil Aと称する、Quillaja Saponaria Molinaの樹皮に由来するサポニン、また、それらの画分が含まれる。Quil Aの特に好ましい画分は、QS21、QS7、およびQS17である。
【0144】
β−エスシンは、本発明のアジュバント組成物中で用いるのに好ましい、別の溶血性サポニンである。エスシンは、「メルクインデックス」(第12版:エントリー3737)において、ラテン名: Aesculus hippocastanumであるトチノキの種子内で発生するサポニンの混合物として記載されている。クロマトグラフィーおよび精製(Fiedler、Arzneimittel−Forsch.、4巻、213頁(1953年))による、また、イオン交換樹脂(Erbringら、米国特許第3,238,190号)によるその単離が記載されている。エスシンの画分は精製されており、生物学的に活性であることが示されている(Yoshikawa Mら(Chem Pharm Bull(東京)、1996年8月、44巻(8号):1454〜1464頁))。β−エスシン(escin)はまた、エスシン(aescin)としても公知である。
【0145】
本発明において用いるのに好ましい別の溶血性サポニンは、ジギトニンである。ジギトニンは、「メルクインデックス」(第12版:エントリー3204)において、Digitalis purpureaの種子に由来し、また、Gisvoldら、J.Am.Pharm.Assoc.、1934年、23巻、664頁;およびRuhenstroth−Bauer、Physiol.Chem.、1955年、301巻、621頁において記載される手順により精製されるサポニンとして記載されている。その使用は、コレステロールを判定するための臨床試薬として記載されている。
【0146】
アジュバント効果を達成する別の興味深い(したがって、好ましい)可能性は、Gosselinら、1992年において記載される技法を用いることである。略述すると、本発明の抗原などの関与抗原の提示は、該抗原を、単球/マクロファージ上におけるFC受容体に対する抗体(または抗原に結合する抗体断片)に結合体化することにより増強することができる。とりわけ、抗原と抗FCRIとの結合体は、ワクチン接種のための免疫原性を増強することが示されている。該抗体は、生成後において、または生成の一部として(エンベロープウイルスベースのVLPのポリペプチド成分のうちのいずれか1つに対する融合体としての発現によることを含む)、エンベロープウイルスベースのVLPに結合体化することができる。
【0147】
他の可能性は、標的化物質および免疫調節物質(すなわち、サイトカイン)の使用を伴う。加えて、ポリI:Cなど、合成のサイトカイン誘導物質もまた用いることができる。
【0148】
適切なマイコバクテリア誘導体は、ムラミルジペプチド、フロイントの完全アジュバント、RIBI(モンタナ州、ハミルトン、RiBi ImmunoChem Reserch社製)、ならびに、TDMおよびTDEなど、トレハロースのジエステルからなる群より選択することができる。
【0149】
適切な免疫標的化アジュバントの例には、CD40リガンドおよびCD40抗体、またはこれらの特異的結合断片(上記の議論を参照されたい)、マンノース、Fab断片、ならびにCTLA−4が含まれる。
【0150】
適切なポリマーアジュバントの例には、炭水化物、例えば、デキストラン、PEG、デンプン、マンナン、およびマンノース;可塑性ポリマー;ならびにラテックスビーズなどのラテックスが含まれる。
【0151】
免疫反応を調節するさらに別の興味深い方法は、「仮想リンパ節」(VLN)(ImmunoTherapy, Inc., 360 Lexington Avenue, New York, N.Y. 10017−6501により開発された、特許品の医療デバイス)内に免疫原(場合によって、アジュバント、ならびに薬学的に許容されるキャリアおよびビヒクルと併せて)を組み込むことである。VLN(細長型の管状デバイス)は、リンパ節の構造および機能を模倣する。VLNを皮下に挿入することにより、サイトカインおよびケモカインの急増による無菌的炎症部位が創出される。T細胞およびB細胞の他、APCも、迅速に危険シグナルに応答し、炎症部位へと至り、VLNの多孔性マトリックスの内部に蓄積される。VLNを用いると、抗原に対する免疫反応を誘発するのに必要とされる抗原用量が低下し、VLNを用いるワクチン接種により付与される免疫的防御が、Ribiをアジュバントとして用いる従来の免疫化を凌駕することが示されている。この技法は、Gelber Cら、1998年、「Elicitation of Robust Cellular and Humoral Immune Responses to Small Amounts of Immunogens Using a Novel Medical Device Designated the Virtual Lymph Node」、1998年10月12〜15日、カリフォルニア州、アプトス、シースケープリゾート、「From the Laboratory to the Clinic, Book of Abstracts」において略述されている。
【0152】
エンベロープウイルスベースのVLPワクチンと共に、オリゴヌクレオチドをアジュバントとして用いることができ、少なくとも3つの、またはより好ましくは、少なくとも6つ以上のヌクレオチドにより隔てられた2つ以上のジヌクレオチドCpGモチーフを含有することが好ましい。CpGを含有するオリゴヌクレオチド(ここで、CpGジヌクレオチドは、メチル化されていない)は、主にTh1反応を誘導する。このようなオリゴヌクレオチドは周知であり、例えば、WO96/02555、WO99/33488、ならびに米国特許第6,008,200号および同第5,856,462号において記載されている。
【0153】
このようなオリゴヌクレオチドによるアジュバントは、デオキシヌクレオチドであり得る。好ましい実施形態では、該オリゴヌクレオチド内のヌクレオチド骨格は、ホスホロジチオエート結合であるか、またはより好ましくは、ホスホロチオエート結合であるが、骨格結合が混合したオリゴヌクレオチドを含め、ホスホジエステル骨格、また、PNAなど他のヌクレオチド骨格も本発明の範囲内にある。ホスホロチオエートオリゴヌクレオチドまたはホスホロジチオエートオリゴヌクレオチドを作製する方法は、米国特許第5,666,153、米国特許第5,278,302号、およびW095/26204において記載されている。
【0154】
好ましいオリゴヌクレオチドの例は、以下の配列を有する。配列は、ホスホロチオエート修飾されたヌクレオチド骨格を含有することが好ましい。
【0155】
【化3】
代替的な好ましいCpGオリゴヌクレオチドには、これらに対する重要でない欠失または付加を伴う上記の配列が含まれる。アジュバントとしてのCpGオリゴヌクレオチドは、当技術分野で公知である任意の方法(例えば、EP468520)により合成することができる。自動合成器を用いてこのようなオリゴヌクレオチドを合成し得ることが好ましい。このようなオリゴヌクレオチドによるアジュバントは、10〜50塩基の長さであり得る。別のアジュバント系は、CpGを含有するオリゴヌクレオチドと、サポニン誘導体との組合せを伴い、特に、CpGとQS21との組合せが、WO00/09159において開示されている。
【0156】
多くの単相または複相のエマルジョン系が記載されている。エマルジョンが、エンベロープウイルスベースのVLPの構造を破壊しないように、当業者は、エンベロープウイルスベースのVLPと共に用いられるこのようなエマルジョン系を容易に適合させることができる。水中油エマルジョンによるアジュバントは、それ自体で、アジュバント組成物として有用であることが示唆されており(EPO399843B)、また、水中油エマルジョンと他の活性薬剤との組合せも、ワクチン用のアジュバントとして記載されている(WO95/17210;WO98/56414;WO99/12565;WO99/11241)。油中水エマルジョン(米国特許第5,422,109号;EP 0480982B2)、および水中油中水エマルジョン(米国特許第5,424,067;EP0480981B)など、他の油エマルジョンによるアジュバントが記載されている。
【0157】
本明細書で記載されるエンベロープウイルスベースのVLPワクチンと共に用いられる油エマルジョンによるアジュバントは、天然の場合もあり、合成の場合もあり、また、鉱油性の場合もあり、有機油性の場合もある。鉱油および有機油の例は、当業者に容易に明らかである。
【0158】
任意の水中油組成物が、ヒトへの投与に適するためには、エマルジョン系の油相が、代謝性油を包含することが好ましい。代謝性油という用語の意味は、当技術分野において周知である。代謝性とは、「代謝により変換可能であること」と定義することができる(「Dorland’s Illustrated Medical Dictionary」、W.B. Sanders Company社、第25版(1974年))。油は、レシピエントに毒性でなく、代謝により変換可能な、任意の植物油、魚油、動物油、または合成油であることが可能である。堅果油(ラッカセイ油など)、種油、穀類油が、植物油の一般的な供給源である。合成油もまた本発明の一部であり、これらには、NEOBEE(登録商標)および他の油など、市販の油が含まれ得る。スクアレン(2,6,10,15,19,23−ヘキサメチル−2,6,10,14,18,22−テトラコサヘキサエン)は、サメの肝油中に高量で見出され、また、オリーブ油、コムギ胚種油、コメぬか油、および酵母中に低量で見出される不飽和油であり、本発明で用いるのに特に好ましい油である。スクアレンは、コレステロールの生合成における中間体であるという事実により、代謝性油である(「メルクインデックス」、第10版、エントリー8619)。
【0159】
特に好ましい油エマルジョンは水中油エマルジョンであり、特に、水中スクアレンエマルジョンである。
【0160】
加えて、本発明の最も好ましい油エマルジョンによるアジュバントには、抗酸化剤が含まれ、油であるα−トコフェロール(ビタミンE;EP0382271B1)が好ましい。
【0161】
WO95/17210およびWO99/11241は、場合によって、免疫刺激剤であるQS21および/または3D−MPLと共に処方される、スクアレン、α−トコフェロール、TWEEN 80に基づくエマルジョンアジュバントを開示している。WO99/12565は、油相へのステロールの添加による、これらのスクアレンエマルジョンに対する改善を開示している。加えて、エマルジョンを安定化させるために、トリカプリリン(C27H50O6)などのトリグリセリドを、油相へと添加することもできる(WO98/56414)。
【0162】
安定的な水中油エマルジョン中で見出される油滴のサイズは、光子相関分光分析により測定したときに、1ミクロン未満であることが好ましく、実質的に30〜600nmの範囲内、好ましくは、実質的に直径約30〜500nm、また、最も好ましくは、実質的に直径150〜500nm、また、特に直径約150nmであり得る。この点で、油滴数の80%が好ましい範囲内にあるべきであり、より好ましくは油滴数の90%超、また、最も好ましくは油滴数の95%超が規定のサイズ範囲内にある。本発明の油エマルジョン中に存在する成分の量は、従来、スクアレンなどの油が2〜10%の範囲内であり;また、存在する場合、アルファトコフェロールが2〜10%であり、ポリオキシエチレンソルビタンモノオレエートなどの界面活性剤が0.3〜3%である。油:アルファトコフェロールの比率は、より安定的なエマルジョンがもたらされるため、1以下であることが好ましい。Span 85もまた、約1%のレベルで存在し得る。一部の場合には、本明細書で開示されるエンベロープウイルスベースのVLPワクチンが、安定化剤をさらに含有することが有利であり得る。
【0163】
水中油エマルジョンを作製する方法は、当業者に周知である。一般に、該方法は、油相を、PBS/TWEEN80(登録商標)溶液などの界面活性剤と混合し、その後、ホモジナイザーを用いるホモジナイゼーションを行うステップを包含するが、該混合物に、シリンジ針内を2回流過させるステップを含む方法が、少量の液体をホモジナイズするのには適切であることが、当業者には明らかである。同様に、当業者は、Microfluidizer(M110S型マイクロ流体作製器;最大入力圧6バールで2分間にわたり、最大50回の流過(約850バールの出力圧))において乳化過程を適合させることで、より少量または多量のエマルジョンを作製することができる。この適合は、必要とされる直径の油滴を有する調製物が達成されるまでの、結果として得られるエマルジョンの測定を含む日常的な実験により達成し得る。
【0164】
本明細書で開示されるエンベロープウイルスベースのVLPによるワクチン調製物は、該ワクチンを、鼻腔内投与、筋肉内投与、腹腔内投与、皮内投与、経皮投与、静脈内投与、または皮下投与を介して投与することにより、ウイルス感染を受けやすいか、またはこれを患う、哺乳動物または鳥類を保護または処置するのに用いることができる。ワクチン調製物を全身投与する方法には、従来のシリンジおよび針、または固体ワクチンを弾道送達(ballistic delivery)するためにデザインされたデバイス(WO99/27961)、または針なしの高圧液体ジェットデバイス(米国特許第4,596,556号;米国特許第5,993,412号)、または経皮パッチ(WO97/48440;WO98/28037)が含まれ得る。エンベロープウイルスベースのVLPワクチンはまた、皮膚に適用することもできる(経皮(transdermalまたはtranscutaneous)送達;WO98/20734;WO98/28037)。したがって、本明細書で開示されるエンベロープウイルスベースのVLPワクチンは、エンベロープウイルスベースのVLPワクチンまたはアジュバント組成物を予め充填した全身投与用の送達デバイスを包含する。したがって、個体、好ましくは哺乳動物または鳥類において免疫反応を誘導する方法であって、本明細書で記載されるエンベロープウイルスベースのVLP組成物のうちのいずれかを含み、また、場合によって、アジュバントおよび/またはキャリアを包含するワクチンを該個体へと投与するステップを含み、該ワクチンが、非経口経路または全身経路により投与される方法が提供される。
【0165】
本発明のワクチン調製物は、該ワクチンを、経口/経消化管経路、または経鼻経路などの粘膜経路を介して投与することにより、ウイルス感染を受けやすいか、またはこれを患う、哺乳動物または鳥類を保護または処置するのに用い得ることが好ましい。代替的な粘膜経路は、膣内経路および直腸内経路である。好ましい経粘膜経路投与は、経鼻経路を介し、鼻腔内ワクチン接種と称する。免疫化される個体の鼻咽頭内へと、ワクチンの液滴形態、スプレー形態、または乾燥粉末形態を投与することを含め、鼻腔内ワクチン接種の方法は、当技術分野において周知である。したがって、噴霧化またはエアゾール化されたワクチン処方物が、本明細書で開示されるエンベロープウイルスベースのVLPワクチンの好ましい形態である。経口投与用の胃液耐性のカプセルおよび顆粒などの腸溶処方物、直腸内投与または膣内投与用の坐剤もまた、本明細書で開示されるエンベロープウイルスベースのVLPワクチンの処方物である。
【0166】
本明細書で開示される、好ましいエンベロープウイルスベースのVLPによるワクチン組成物は、経粘膜ワクチン接種により全身ワクチン接種を代替するヒトにおける適用に適する経粘膜ワクチンのクラスの一例である。
【0167】
エンベロープウイルスベースのVLPワクチンはまた、経口経路を介して投与することもできる。このような場合、薬学的に許容される賦形剤にはまた、アルカリ性緩衝液、または腸溶性のカプセルもしくはマイクロ顆粒も含まれ得る。エンベロープウイルスベースのVLPワクチンはまた、膣内経路によっても投与することができる。このような場合、薬学的に許容される賦形剤には、乳化剤、CARBOPOL(登録商標)などのポリマー、また、膣内クリームおよび膣内坐剤の他の公知の安定化剤も含まれ得る。エンベロープウイルスベースのVLPワクチンはまた、直腸内経路によっても投与することができる。このような場合、賦形剤には、直腸内坐剤を形成するために当技術分野で公知のワックスおよびポリマーも含まれ得る。
【0168】
代替的に、エンベロープウイルスベースのVLPによるワクチン処方物は、キトサン(上記で記載した)または他のポリカチオンポリマー、ポリラクチド粒子およびポリラクチド−コグリコリド粒子、ポリ−N−アセチルグルコサミンベースのポリマーマトリックス、多糖または化学修飾した多糖からなる粒子、リポソームおよび脂質ベースの粒子、グリセロールモノエステルからなる粒子などからなるワクチンビヒクルと組み合わせることができる。サポニンはまた、コレステロールの存在下において処方して、リポソームまたはISCOMなどの粒子構造を形成することもできる。さらに、サポニンは、非粒子状の溶液もしくは懸濁物中、または小ラメラ(paucilamelar)リポソームもしくはISCOMなどの粒子状構造内において、ポリオキシエチレンエーテルまたはポリオキシエチレンエステルと併せて処方することもできる。
【0169】
本明細書で記載されるエンベロープウイルスベースのVLPを用いる医薬組成物およびワクチン組成物中で用いられる、さらなる例示的なアジュバントには、SAF(米国、カリフォルニア州、Chiron社製)、MF−59(Chiron社製、例えば、Granoffら(1997年)、Infect Immun.、65巻(5号):1710〜1715頁を参照されたい)、SBASシリーズのアジュバント(例えば、SB−AS2(SmithKline Beecham社製のアジュバントシステム♯2;MPLおよびQS21を含有する水中油エマルジョン);SBAS−4(SmithKline Beecham社製のアジュバントシステム♯4;ミョウバンおよびMPLを含有する)、ベルギー、リクセンサルト、SmithKline Beecham社から市販されている)、Detox(Enhanzyn(登録商標))(GlaxoSmithKline社製)、RC−512、RC−522、RC−527、RC−529、RC−544、およびRC−560(GlaxoSmithKline社製)、ならびに、係属中の米国特許出願第08/853,826号、および同第09/074,720号において記載されるものなどの、他のアミノアルキルグルコサミド4−ホスフェート(AGP)が含まれる。
【0170】
アジュバントの他の例には、HunterによるTiterMax(登録商標)アジュバント(ジョージア州、ノークロス、CytRx社製);Gerbuアジュバント(ドイツ、ガイベルク、Gerbu Biotechnik GmbH社製);ニトロセルロース(NilssonおよびLarsson(1992年)、Res. Immunol.、143巻:553〜557頁);Seppic ISAシリーズのMontamideアジュバント(例えば、ISA−51、ISA−57、ISA−720、ISA−151など;フランス、パリ、Seppic社製)など、鉱油エマルジョン、非鉱油エマルジョン、油中水エマルジョン、または水中油エマルジョンを含めたミョウバン(例えば、水酸化アルミニウム、リン酸アルミニウム)エマルジョンベースの処方物;およびPROVAX(登録商標)(IDEC Pharmaceuticals社製);OM−174(脂質Aと関連するグルコサミンジサッカリド); Leishmania属の伸長因子;CRL 1005など、ミセルを形成する非イオン性のブロックコポリマー;ならびにSyntexアジュバント処方物が含まれるがこれらに限定されない。例えば、O’Haganら(2001年)、Biomol Eng.、18巻(3号):69〜85頁;および「Vaccine Adjuvants: Preparation Methods and Research Protocols」、D. O’Hagan編(2000年)、Humana Press社を参照されたい。
【0171】
他の好ましいアジュバントには、一般式
HO(CH 2CH2O)n−A−R, (I)
[式中、nは1〜50であり、Aは結合または−−C(O)−−であり、RはC1〜50アルキル、またはフェニルC1〜50アルキルである]のアジュバント分子が含まれる。
【0172】
本発明の一実施形態は、一般式(I)[式中、nは1〜50、好ましくは4〜24、最も好ましくは9であり;R成分はC1〜50、好ましくはC4〜C20アルキル、また最も好ましくはC12アルキルであり;また、Aは結合である]のポリオキシエチレンエーテルを含むワクチン処方物からなる。ポリオキシエチレンエーテルの濃度は、0.1〜20%、好ましくは0.1〜10%、また最も好ましくは0.1〜1%の範囲であるべきである。好ましいポリオキシエチレンエーテルは、以下の群:ポリオキシエチレン−9−ラウリルエーテル、ポリオキシエチレン−9−ステアリルエーテル(polyoxyethylene−9−steoryl ether)、ポリオキシエチレン−8−ステアリルエーテル、ポリオキシエチレン−4−ラウリルエーテル、ポリオキシエチレン−35−ラウリルエーテル、およびポリオキシエチレン−23−ラウリルエーテルから選択される。ポリオキシエチレンラウリルエーテルなどのポリオキシエチレンエーテルは、「メルクインデックス」(第12版;エントリー7717)で記載されている。これらのアジュバント分子は、WO99/52549で記載されている。
【0173】
所望の場合、上記の一般式(I)に従うポリオキシエチレンエーテルを、別のアジュバントと組み合わせることができる。例えば、好ましいアジュバントの組合せは、上で記載したCpGとの組合せであることが好ましい。
【0174】
本明細書で開示されるエンベロープウイルスベースのVLPワクチンと共に用いるのに適する、薬学的に許容される賦形剤のさらなる例には、水、リン酸緩衝生理食塩液、等張性緩衝液が含まれる。
【0175】
本発明は、以下の非限定的な実施例を参照することにより、よりよく理解される。本明細書で記載される通り、本発明は、天然では脂質ラフトと会合しない抗原に連結した、任意の種類の脂質ラフト会合ポリペプチドを組み込む、キメラエンベロープウイルスベースのVLPを包含する。以下の実施例では、本発明の代表的な実施形態である、インフルエンザ抗原を伴う、キメラエンベロープウイルスベースのVLPが記載される。
【実施例】
【0176】
(実施例1−インフルエンザ偽型Gag VLPの作製)
MLV Gag遺伝子ならびに種々のインフルエンザAサブタイプのHAおよびNA遺伝子を、下に記載の通り個々にpFastBac1バキュロウイルス導入ベクターのポリヘドリンプロモーターの後ろにクローニングした。「三重遺伝子」発現ベクターを構築するために1つのHAおよび1つのNAベクターの転写単位全体をSnaB IおよびHpa Iでの切断によって切り出し、これらの平滑末端断片をGag遺伝子導入ベクター中のGag転写単位のいずれかの側の唯一のSnaB IおよびHpa Iクローニング部位にそれぞれ導入した。これは、ヘッドトゥーテール(head−to−tail)様式で配置された3種の別々の転写単位(HA、Gag、NA)を含有する単一のプラスミドを生じた。次いで種々のインフルエンザAサブタイプを示すpFB−HA−pGag−pNA3重導入ベクターをキット製造者(Invitrogen、Carlsbad、CA)によって記載された通りバキュロウイルスゲノムへの組み換えのためにDH10Bac細胞中に形質転換した。
【0177】
(MLV GagならびにインフルエンザHAおよびNA遺伝子)
マウス白血病ウイルスのGag遺伝子はプラスミドpAMS(ATCC、Manassas、VA)から以下のプライマー:5’CACCATGGGCCAGACTGTTACC3’(配列番号6)および5’CTACTAGTCATCTAGGGTCAGGAG3’(配列番号7)を使用するPCRによって得た。最初のプライマーの5’末端のCACC伸長は、Gateway cloning technology(Invitrogen)を用いるベクターpENTR D−TOPOへの一方向性の挿入を促進し、プラスミドpENTR−Gagを生じる。Gagコード配列をDNA配列決定によって確認し、次いでpFastbac1に以下の通り導入した:pENTR−GagをAsc Iで切断し、末端をKlenow DNAポリメラーゼでの処理によって平滑化し、その後DNAをNot Iで切断した。最初にpFastbac1をSph Iで切断し、末端を平滑化し、次いでNot Iで切断した後に、得られたGag断片をpFastbac1ベクターDNAにライゲーションした。
【0178】
インフルエンザA/PR/8/34(H1N1)およびA/Hong Kong/68(H3N2)のHAおよびNA遺伝子をRT−PCRによってクローニングした。ウイルスRNAをQIAmp MinElute Virus Spin Kit(Qiagen、Valencia、CA)を使用して卵培養ウイルスから調製し、第1鎖cDNA反応をAccuscript High Fidelity 1st Strand cDNA Synthesis kit(Stratagene、La Jolla、CA)を使用して実施した。第1鎖cDNA合成に続いて、HAおよびNA断片を増幅するために標準的PCR反応を使用した。PR/8 H1遺伝子のためのプライマーは以下の通り:5’CACCATGAAGGCAAACCTACTGGTCC3’(配列番号8)および5’TCAGATGCATATTCTGCACTGC3’(配列番号9)。PR/8 N1遺伝子のためのプライマーは以下の通り:5’CACCATGAATCCAAATCAGAAAATAATAACCATTCC3’(配列番号10)および5’CTACTTGTCAATGGTGAATGGCAAC3’(配列番号11)。A/Hong Kong/68 H3遺伝子のためのプライマーは以下の通り:5’CACCATGAAGACCATCATTGCTTTGAGC3’(配列番号12)および5’TCAAATGCAAATGTTGCACCTAATGTTGCC3’(配列番号13)。A/Hong Kong/68 N2遺伝子のためのプライマーは以下の通り:5’ATATAGGCGCGCCACCATGAATCCAAATCAAAAGATAATAACAATTGGC3’(配列番号14)および5’ATATAGCGGCCGCTTATATAGGCATGAAATTGATGTTCGC3’(配列番号15)。A/PR/8/34(H1N1)のHAおよびNA遺伝子ならびにA/Hong Kong/68(H3N2)のHA遺伝子を最初にpENTR D−TOPOにクローニングし、配列決定によって確認し、次いで上に記載の通りpFastbac1に導入した。A/Hong Kong/68(H3N2)のNA遺伝子をAscIおよびNot IでPCR断片末端を整えた後にBssHII−NotI切断pFastbac1に直接クローニングした。候補クローンを配列決定によって確認した。
【0179】
A/Vietnam/1203/04およびA/Indonesia/5/05のH5およびN1遺伝子を含有しているプラスミドクローンはDr.Ruben Donis(Branch Chief、Molecular Virology and Vaccines、CDC、Atlanta、GA USA)から得た。H5クローンは、成熟切断部位に多塩基領域の欠失を含んでいた。提供されたプラスミドを配列確認用のpENTR D−TOPOへの挿入のための「Vietnam」および「Indonesia」H5およびN1 PCR断片を作製するための鋳型として使用した。H5プライマーは、以下の通り:5’CACCATGGAGAAAATAGTGCTTC3’(配列番号16)および5’TTAAATGCAAATTCTGCATTGTAACG3’(配列番号17)。N1プライマーは以下の通り:5’CACCATGAATCCAAATCAGAAGATAATAACC3’(配列番号18)および5’CTACTTGTCAATGGTGAATGGC3’(配列番号19)。配列確認後、個々のH5およびN1遺伝子は、上に記載の通りpFastbac1に導入した。
【0180】
A/Wisconsin/67/2005(H3N2)のHAおよびNA遺伝子をRT−PCRによって上に記載の通りウイルスRNAからクローニングした。H3遺伝子のためのプライマーは以下の通り:5’ATATAGGCGCGCCACCATGAAGACTATCATTGCTTTGAGC3’(配列番号20)および5’ATATAGCGGCCGCTCAAATGCAAATGTTGCACCTAATGTTGCC3’(配列番号21)。N2遺伝子のためのプライマーは以下の通り:5’ATATAGGCGCGCCACCATGAATCCAAATCAAAAGATAATAACGATTGGC3’(配列番号22)および5’ATATAGCGGCCGCTTATATAGGCATGAGATTGATGTCCG3’(配列番号23)。H3およびN2遺伝子断片をそれぞれAscIおよびNotIで整え、BssHII−NotI切断pFastbac1にクローニングした。
【0181】
A/Solomon Islands/3/2006のH1およびN1遺伝子をコードするカスタム合成遺伝子をGeneArt(Regensburg、ドイツ)から得て、昆虫細胞での発現のためにコドンを最適化した。各断片は、NotI−KpnI−切断pFastbac1への直接クローニング用にNotIおよびKpnI部位を5’および3’末端にそれぞれ含有した。
【0182】
組換えDNAの使用を含む作業の実施において、研究者らは組換えDNA分子に関与する研究のためのガイドラインを遵守した;注、Federal Register、1994年7月5日、第59巻、第127号。
【0183】
(VLPの精製)
昆虫細胞Sf9をSF900−II培地で培養し、1mlあたり細胞5×105個で200mlスピナーフラスコに播種した。細胞を27℃で1mlあたり細胞2×106個の密度まで培養し、その時点でVLPコード組換えバキュロウイルスの第2継代接種材料をおよそ0.1から1.0の感染多重度で加えた。細胞生存率が20%またはそれより低くまで低下したときに培養液を回収した。培地を2,000rpm、15分間遠心分離することによって細胞細片を除去し、その後32ml一定分量をトリス緩衝生理食塩水(TBS)、pH7.4中の30%ショ糖のクッション4ml上に重層した。VLPをショ糖クッションによって25,000rpm、1時間、10℃でBeckman SW28ローター(100,000×g)で遠心分離した。培地200ml由来のVLPを合計6mlのTBSに再懸濁し、次いで単一の20〜60%不連続ショ糖勾配に重層し、25,000rpm、1時間、10℃で遠心分離した。ショ糖勾配を底から1.5ml画分へと分取し、赤血球凝集アッセイおよびノイラミニダーゼアッセイ(下を参照されたい)ならびにSDS−PAGEおよびウェスタンブロットによって分析した。ショ糖勾配精製したVLPをショ糖存在下、4℃で保存し、少なくとも6カ月間安定(HA活性に関して)であることを見出した。VLPを播種の前にショ糖溶液から遠心分離し、トリス緩衝生理食塩水に再懸濁した。本明細書において報告する動物実験に利用したVLPワクチンは凍結されていないが、しかしH1N1 VLPの少なくとも1回の凍結融解をHA活性の測定可能な損失を伴わずに実施できることが測定された。
【0184】
(赤血球凝集アッセイ)
赤血球凝集アッセイを0.5%ヒヨコ赤血球を使用して、動物インフルエンザ診断および調査についてのWHO手順書(WHO Manual on Animal Influenza Diagnosis and Surveillance)に記載の通り実施した[41]。
【0185】
(ノイラミニダーゼアッセイ)
ノイラミニダーゼ活性を蛍光基質2’−(4−メチルウンベリフェリル)−α−D−N−アセチルノイラミン酸(MUN)を使用してVLP調製物およびショ糖勾配画分中で検出した。PBS中のVLP含有試料の漸増希釈物(50μl)を黒色平底96ウエルプレートに入れ、PBS 50μlを各試料ウエルに添加した。NA活性を35mM MES pH6.5、4mM CaCl2、150mM NaCl、300μM MUNの50μlの添加によって検出した。プレートを37℃で1時間インキュベートし、次いで83%エタノール中0.14N NaOHの1ウエルあたり100μlでの添加によって停止させた。プレートを励起波長および発光波長それぞれ365nmおよび455nmを使用して蛍光光度計で読み取った。すべての蛍光データは、基質を含有しているがVLPを含有していない対照ウエルから得たバックグラウンド値に対して補正した。補正した蛍光値を次いでアッセイの線形範囲にあるデータ点を決定するために希釈係数で割った。
【0186】
A/PR/8/34(H1N1)を示すHA−Gag−NA VLPを上に記載の「三重遺伝子」組換えバキュロウイルスを感染させたSf9細胞の培地から不連続ショ糖密度勾配での遠心分離によって精製した。赤血球凝集アッセイによる勾配画分の分析は、勾配の中央の顕著な可視バンドに関連したHA活性の強いピーク(図1A)を明らかにした。このピークは、SDS−PAGEゲル上の顕著な65kd産物とも一致があり、GagおよびHAの共移動(図1B)、ならびにHAおよびNAについてのウェスタンブロットシグナル(図1C)と一致した。最後に電子顕微鏡によるピーク画分の直接分析は、出芽エンジンとしてのレトロウイルスGagタンパク質の使用から予測される通りインフルエンザよりもガンマレトロウイルスを連想させる多量の100nm球状粒子を示した(図1D)。これらの粒子はインフルエンザウイルスおよびインフルエンザマトリックスベースのVLPとは形態学的に異なると考えられる一方で、プールしたピーク画分での赤血球凝集活性が、勾配精製した卵培養A/PR/8/34ウイルスについて観察されたものと同様である総VLPタンパク質1mg当たり2〜3×105HA単位の範囲内で測定され、これらの粒子への機能的HAスパイクの顕著な組み込みを実証していることに注目することは重要である。
【0187】
PR/8 H1N1 VLP中のGag対HAの比は、Gagより遅く移動する改変HA産物を作製するようにHAコード配列がそのアミノ末端で伸長された改変PR/8 H1N1 VLP発現ベクターを使用しておよそ3〜4:1と測定された(図1E)。PR/8 H1N1 VLPに関しておよそ3〜4:1のGag対HAの比は、本発明者らが作製し、A/Wisconsin/67/2005(H3N2)、A/Solomon Islands/3/2006(H1N1)およびB/Malaysia/2506/2004などの現在のヒトインフルエンザ株を示す他のVLPと一致している。これら後者のVLPは、SDSゲル上でGagよりも天然で遅い移動度を示すHA分子を含有し、同様に3〜4:1のGag対HAの比を示す(図1F、Solomon Islands H1N1 VLP)。
【0188】
HAに加えて、顕著なノイラミニダーゼ生物活性もショ糖勾配でのVLPピークにおいて見出されており、これらのデータをH5N1 VLPについて図2に示す。A/Vietnam/1204/2004およびA/Indonesia/5/2005株を示すH5N1 VLPを、上に記載のVLPと同様のやり方で同様の収率および赤血球凝集特異的活性を伴って作製した。NA活性が、HA活性ピークの重い側にしばしば観察されるHA活性の肩と重なって勾配中深くに広がる傾向があることに注目することは興味深い。このパターンは、今日までに検査したすべてのサブタイプ(H1N1、H3N2およびH5N1)について観察されており、VLP集団におけるHA:NA比が連続的である可能性と一致している(実施例7を参照されたい)。
【0189】
(実施例2−マウスにおけるH1N1 VLP 免疫原性および防御)
実施例2は、H1N1 VLPでのマウスの免疫化を実証する。
【0190】
(マウス免疫化およびチャレンジ)
6〜8週齢、雌性Balb/cマウスをHAおよそ0.7〜1.0μgを含有するトリス緩衝生理食塩水(TBS)中のVLP処方物で腹腔内接種(100μl)または筋肉内接種(30μl)を用いて免疫化した。初期実験において、モノホスホリルリピドA(解毒化脂質A、Avanti Polar Lipids、Alabaster、AL)20μgをアジュバントとして添加した。初回および追加免疫化は、4週間間隔を置いて離し、各免疫化に続いて2週間、側方顔面静脈から血液試料を採取した。免疫化マウスおよび対照マウスをPBS 50μl中の卵培養A/PR/8/34の10LD50で鼻腔内でチャレンジし、体重減少および病的状態について毎日モニターした。瀕死であるかまたは刺激に反応しないことが見出された動物は、CO2吸入で安楽死させた。動物を使用する研究を実施する際に研究者らは、National Research Councilの実験動物資源局(Institute of Laboratory Animal Resources)の実験動物の管理と使用に関する委員会(Committee on Care and Use of Laboratory Animals)によって作成された「実験動物の管理と使用のための指針」を遵守した[40]。
【0191】
(赤血球凝集抑制アッセイ)
赤血球凝集抑制(HAI)アッセイは、0.5%ヒヨコ赤血球を使用して動物インフルエンザ診断および調査についてのWHO手順書に記載の通り実施した[41]。H5N1特異的HAIアッセイに関して、1.0%ウマ赤血球も使用し、沈降時間を1時間に延ばした。H5N1 HAIアッセイはまた、精製H5N1 VLPも感染性ウイルスの代替の凝集作用物質として使用した(実施例4を参照されたい)。
【0192】
ショ糖勾配画分中のPR/8 H1N1 VLPをHA活性に基づいてプールし、ショ糖から沈降させて取り出し、アジュバントとしてのモノホスホリルリピドA(MPL)を伴うかまたは伴わない腹腔内または筋肉内経路を用いるマウスの免疫化のためにPBS中に再懸濁した。1群あたり16匹の動物が、免疫化あたりHAおよそ0.7μgを含有し、4週間隔てた初回および追加免疫化を受け、無処置動物16匹は対照とした。図3Aは、追加後2週間の免疫化マウスにおける強いA/PR/8/34特異的HAI活性を示し、これらの応答はほとんどIgG2aアイソタイプのものであった(図3B)。追加後およそ4週間で各群を10LD50のマウス適合A/PR/8/34(H1N1)でチャレンジした。H1N1特異的液性応答から予測される通り、免疫化したすべての動物は病的状態または体重減少の徴候無くH1N1チャレンジを生き残ったが、H1N1チャレンジした無処置動物8匹すべては8匹中7匹の死を伴って強い病的状態および体重減少を示した(図3C)。この初回試験での免疫応答の強さから、腹腔内経路投与をさらなる実験から除外した。
【0193】
(実施例3−マウスにおけるH3N2およびH5N1 VLP免疫原性)
実施例3は、H3N2またはH5N1 VLPでのマウスの免疫化を実証する。マウスの免疫化およびチャレンジの方法ならびにHAIアッセイの方法は上の実施例2に記載の通り実施した。
【0194】
(インフルエンザ特異的抗体ELISA)
卵培養インフルエンザウイルス(A/PR/8/34(H1N1)またはA/Aichi/68(H3N2))を尿膜腔液から、TBS中の30%ショ糖クッションによってMLS 50ローター、36,000RPM(100,000×g)、1時間、10℃での遠心分離で取り出した。ウイルスペレットをPBSに再懸濁し、タンパク質含有量をBCAアッセイ(Pierce Biotechnology、Rockford、IL)によって定量し、1mlあたり5μgに調整し、平底ELISAプレートを1ウエルあたり100μl、1晩、4℃でコートするために使用した。翌日プレートを0.05%Tween20を含むPBS(PBS−T)で洗浄し、StartingBlock(PBS)(Pierce Biotechnology)1ウエルあたり350μlで30分間ブロックした。血清試料をStartingBlock T20(PBS)(Pierce Biotechnology)で1:500に希釈して最上列に添加し、各カラムで下がる毎に連続的に3倍希釈し、室温で3時間または4℃で1晩インキュベートした。プレートをPBS−Tで3回洗浄し、StartingBlock T20(PBS)中に1:1000希釈したヤギ抗マウスIgG−HRP結合体(Southern Biotech、Birmingham、AL)100μlを各ウエルに添加した。プレートをさらに1.5時間室温でインキュベートし、PBS−Tで3回洗浄し、次いでABTS基質(Pierce Biotechnology)100μlを各ウエルに添加した。プレートを室温、45分後に405nmで読み取った。ELISA力価を、バックグラウンド吸光度値の1.75倍の吸光度値を生じた最も高い血清希釈の逆数を取ることによって算出した。このカットオフレベルは、すべての無処置血清試料対照からのすべての疑陽性を排除した。
【0195】
H5N1特異的ELISAを、プレートを組換えH5 Vietnam抗原(Protein Sciences Inc.、Meriden、CT)またはスプリットH5N1ワクチン処方物1mlあたり3μg(Dr.Sally Mossman、GlaxoSmithKline、Rixensart、Belgiumからの寄贈)でコートした以外は同様に実施した。
【0196】
また、IgG1およびIgG2a特異的ELISAを、総IgGではなくIgG1およびIgG2aに特異的な2次抗体を使用したこと以外は同様のやり方で実施した。定量のために、特異的2次抗体との反応性についての標準曲線を作成するために一連のIgG1およびIgG2a濃度標準を一連のウエルにコートした。
【0197】
(結果)
A/Hong Kong/68(H3N2)を示すVLPを実施例1に記載の通り作製した。H3N2およびH5N1 VLP(PR/8 H1N1 VLPと共に)の免疫原性を表1に示す通りマウスの免疫化試験において評価した(MPLアジュバントの存在下で1用量あたりHAおよそ1μgを含有する筋肉内初回および追加免疫化がすべての動物において強いHAI活性を誘導した)。興味深いことに、相当な交差クレードHAI活性がIndonesia H5N1免疫化群とVietnam H5N1免疫化群との間でウマRBC HAIアッセイを使用して観察され[43]、H5N1チャレンジに対する顕著な交差クレード防御を誘導する可能性を示唆した。
【0198】
【表1】
表2は、この同じ実験(A/PR/8/34(H1N1)、A/Aichi(H3N2)および組換えサブユニットH5 HA抗原(Vietnam)への強いサブタイプ特異的応答がそれぞれのワクチンについて観察された)からの抗体応答のELISA分析の結果を示す。重要なことにH5N1 VLPは、共通N1抗原の理由によって予測される(2つのN1抗原は約70年間のドリフトによって分離されているが(16.1%のアミノ酸配列相違(アラインメントデータ未記載)))A/PR/8/34(H1N1)に対する弱いELISA活性を誘導した。表2はまた、群2のH1N1 VLP免疫化マウスを8週間休ませ、次いでMPLアジュバントと共におよび共にではなくH3N2 VLP(初回および追加)で再免疫化した再免疫化実験の結果を示す。重要なことに先行する2回のH1N1 VLP免疫化後のH3N2応答は、無処置マウスで誘導されたものと同等であった(群3)。これらのデータは、動物が第2のVLPサブタイプで再免疫化された場合にGagおよび共通昆虫細胞膜抗原などの異なるVLPサブタイプ間で共有する抗原に対する既存の免疫応答が強力なインフルエンザ特異的応答の誘発を抑制しないことを示す。再免疫化データは、ワクチン性能がMPL添加の有無において同一であったことから観察された免疫応答にMPLアジュバントがほとんど寄与しないことも示す。したがってMPLは続く実験から除いた。
【0199】
【表2】
(実施例4−インフルエンザ−偽型VLPはHAIアッセイにおいて生きているウイルスを代替できる)
表1に示すH5N1 HAIアッセイは、適切なH5N1ウイルス株を入手できないことから生きているウイルスの代替としてH5N1 IndonesiaおよびVietnam VLPを使用して実施した。HAIアッセイを実施例2に記載の通り実施した。インフルエンザ偽型Gag VLPが、H1N1およびH3N2サブタイプの両方について図4に示す通りHAIアッセイにおいて生きているウイルスの性能をかなりの程度まで模倣できることに注目することは重要である。この実験においてH1N1およびH3N2 VLP免疫化マウス由来の免疫血清を生きているウイルスおよび対応するVLPの両方を使用するHAIアッセイについて検査し、アッセイ方法間で顕著に類似する力価を明らかにした。これらの結果は、HAIアッセイにおける偽型VLPとウイルスとの間での同様の性能を実証するだけでなく、生きているインフルエンザウイルスをHA活性および密度(ワクチン性能に関して重要であると考えられる)に関して模倣する偽型VLPの能力についての追加的証拠を提供する。
【0200】
(実施例5−フェレットにおける高病原性H5N1チャレンジ)
マウスにおけるVLPの免疫原性に基づいて本発明者らは、H5N1およびH1N1 VLPワクチンを使用して、HPAI H5N1チャレンジに対して誘導され得る防御の程度を決定するためのフェレットの免疫化およびチャレンジ試験を実施した。
【0201】
(フェレットワクチン接種およびチャレンジ)
32匹の雄性フェレット、8〜16週齢(Triple、F Farms、Sayer、PA)(現在広まっているインフルエンザウイルスに関して赤血球凝集抑制により血清学的に陰性)を、下に記載のVLPワクチン処方物で0日目および28日目にワクチン接種した。フェレットVLPワクチンは、生理食塩水中にアジュバントを含まずに処方され、1用量あたりHAおよそ5μgを含有した。血液試料(前大静脈を用いて)を0日目、28日目および42日目に採取した。体重測定は、チャレンジの1週間前から研究終了(63日目)まで通して毎日行った。温度をIPTT−300埋め込み型トランスポンダーをDAS−7000携帯型プローブ(BioMedic Data Systems、Seafort、DE)と共に用いて測定した。各免疫化群の8匹のフェレットの内7匹をチャレンジのために無作為選択した。動物を筋肉内注射を用いてテラゾール(Telezol)(16mg/kg)で麻酔し、PBS 0.6ml中のA/Vietnam/1203/04ウイルスの106TCID50を鼻腔内に接種した。チャレンジウイルスは、Dr.Alexander Klimov(CDC、Atlanta、GA、USA)によって提供された。フェレットを体温および体重の変化についてモニターし、疾患の臨床徴候について観察した。神経学的機能障害を示したかまたは瀕死になったフェレットは、安楽死させた。動物実験は、実験動物の管理および使用に関する指針(Guide for the Care and Use of Laboratory Animals)[40]に従って、国際実験動物管理評価公認協会(Association for Assessment and Accreditation of Laboratory Animal Care International)公認動物機関のBattelle施設内動物管理使用委員会(Battelle’s Institutional Animal Care and Use Committee)の指導の下で実施した。
【0202】
(フェレット鼻洗浄液中のウイルスの定量)
ウイルス流出(shedding)をチャレンジ後3日目および5日目に麻酔したフェレットから採取した鼻洗浄液中で測定した。合計1mlでの鼻洗浄用に軟性カテーテルを繋いだ1mLシリンジを使用してPBS 500μLで各鼻腔を素早く洗い流した。鼻洗浄液を滅菌標本カップに集め、クライオバイアル(cryovial)に移し、TCID50での計数まで−70℃保存した。鼻洗浄液中のウイルスをメイディン・ダービー・イヌ腎臓(MDCK)細胞での組織培養感染用量中央値(median tissue culture infectious dose)によって定量した。ウイルスの連続希釈物をEMEM(2mM グルタミン、1mMピルビン酸ナトリウムおよび1%ペニシリン−ストレプトマイシンで増量)中、96ウエル形式の細胞単層に5連で入れた。単層を96時間、37℃、5%CO2でインキュベートした。細胞変性効果(CPE)を示したウエルを陽性と記録し、データをSpearman Karber法[42]を使用して分析し、TCID50/mLとして報告した。
【0203】
(マイクロ中和アッセイ)
血清中和抗体を標準的マイクロ中和アッセイを使用して測定した。熱不活化、連続希釈検査血清300μLを、ウイルス600TCID50を含有する等量のEMEM(2mM グルタミン、1mMピルビン酸ナトリウムおよび1%ペニシリン−ストレプトマイシンで増量)と混合した。37℃、1時間のインキュベーションに続いて、各希釈物100μLを5連で96ウエル形式中のMDCK細胞に入れた。37℃、5%CO2での4日間のインキュベーションに続いて、CPEを含有するウエルを陰性と記録した。データをSpearman Karber法を使用して分析し、中和用量中央値(ND50)として報告した。
【0204】
32匹のフェレットを無作為に4群(1群あたり動物8匹)にし、gagだけのVLP、H1N1(PR/8/34)VLP、Indonesia H5N1 VLP およびVietnam H5N1 VLPでそれぞれ免疫化した。VLPをMPLアジュバントを含まない生理食塩水中に処方し、初回および追加免疫化は、4週間離した。免疫化はHAおよそ5μgを含み、筋肉内に投与した。追加免疫化を施した後、各群の動物8匹の内7匹を無作為に選択し、続くチャレンジのために封じ込め施設に移し、順化させた。図5は、H1N1への(図5A、実施例2に記載の通り実施したHAIアッセイ)およびVietnam H5N1への(図5B、マイクロ中和アッセイ)免疫応答を示す(ここで、強いH1N1およびH5N1特異的免疫応答が単回免疫化に続いてのそれぞれのワクチンによって誘発された)。H1N1特異的応答は、追加免疫化後にさらに増大しなかったが、H5N1マイクロ中和力価は、2回目の免疫化後にわずかに増大した。驚くべきことに低レベルH5N1中和力価が追加免疫化後にH1N1免疫化動物7匹の内の5匹で検出された。H5N1中和活性に加えて、IndonesiaおよびVietnam株の両方に特異的な強いH5N1 HAI活性が、IndonesiaおよびVietnam VLPを使用するウマRBC HAIアッセイを使用して両方のH5N1ワクチン接種群において検出された。顕著な交差クレードHAI活性が両方のH5N1ワクチン接種群において検出されたこれらのデータを表3に示す。
【0205】
【表3】
追加の2週間後、すべてのフェレットを106 TCID50のHPAI A/Vietnam/1203/04(H5N1)でチャレンジし、体重減少/生存およびウイルス力価データをそれぞれ図6および7に示す。チャレンジ前のH5N1中和力価から予測される通り、すべてのIndonesiaおよびVietnam H5N1 VLPワクチン接種動物が病的状態および体重減少をほとんど示さずにチャレンジを生き残ったが、すべてのGag VLPワクチン接種対照は強い病的状態を示し、動物7匹のうち6匹が安楽死を必要とした(図6A)。対照群でのこのレベルの病的状態および生存率(7匹の内1匹)は、この同じ用量のA/Vietnam/1203/04(H5N1)チャレンジウイルスについて以前報告された[44]ものと類似していた。驚くべきことに、H1N1ワクチン接種動物7匹のうち6匹に中程度の病的状態が観察されただけであり、この群のフェレットで実験の生活期間において安楽死を必要としたものは無かった。H1N1ワクチン接種群の動物について個々の体重減少データを図6Bに示す。H1N1ワクチン群の動物1匹(フェレット449)は経時的に顕著な体重減少を示したが、食欲不振(持続的な症状であるだけである)を伴なう許容できる敏捷性および活動レベルのため安楽死させなかった。フェレット449における体重減少は、コホートの平均重量を減少させ、チャレンジ後7日目以降の高い標準偏差を生じさせた。
【0206】
生存、活動度および体重減少データは、3および5日目の鼻洗浄液からのチャレンジ後のウイルス単離データと明らかに一致していた(図7AおよびB)。ここで、H5N1 VLPワクチン接種動物は、チャレンジ後に顕著な病的状態を示さず、チャレンジ後3および5日目の鼻洗浄液中に検出可能なウイルスも示さなかった。対照的にGag VLPワクチン接種フェレットにおいて観察された強度の病的状態および死亡率は、同様のチャレンジ用量を受けた無処置動物について以前報告されたのと同様のレベルでの、チャレンジ後3および5日後両日における高いウイルスレベルと関連していた[44、45]。H1N1 VLPワクチン接種フェレットにおいて観察された中程度の病的状態と一致して、チャレンジ後のウイルス力価は、対照群と比較して3日目で減少し、5日目には1匹を除くすべての動物において存在しなかった。5日目に鼻洗浄液ウイルス力価を有したH1N1ワクチン接種動物1匹は、より強い病的状態および体重減少を示したのと同じ動物(449)であった。H1N1群中の残りの動物6匹におけるウイルスの迅速なクリアランスは、軽減された病的状態およびより少ない体重減少と関連していた。
【0207】
H1N1 VLPワクチン接種フェレットにおけるHPAI H5N1チャレンジに対する部分的な防御に関する機序は公知ではないが、A/PR/8/34(H1N1)とA/Vietnam/1203/04(H5N1)とで共有するN1抗原に関連する可能性がある。表2のマウス免疫原性研究は、両方のH5N1 VLPワクチンによる弱いH1N1 ELISA反応性の誘発を実証する(実施例3を参照されたい)。同様に表4は、H1N1ワクチンによって誘発された部分的な防御と一致して、H1N1 VLPワクチンがフェレットにおいて卵培養スプリットH5N1ウイルス調製物に対する弱いELISA反応性を誘発したこと示す。これらのELISAデータは、H5N1チャレンジに対するH1N1媒介防御でのN1応答の役割を確認するものではないが、それらは、この可能性と一致する。
【0208】
【表4】
(実施例6−感染性バキュロウイルスはVLP免疫原性に寄与しない)
生きているバキュロウイルス粒子が顕著な先天免疫刺激およびアジュバント効果を示すこと[46〜48]、およびこれらの特性が、除去または不活化されなければVLPなどの昆虫細胞由来産物の免疫原性において役割を演じ得ることが実証されている。この現象を調査するために本発明者らは、ウイルス、A/Wisconsin/67/2005(H3N2)を示すVLPを精製するために使用したショ糖密度勾配においてバキュロウイルス粒子の力価を計り、同じ画分中の1mlあたり粒子およそ2×1012個のVLP濃度と比較して1mlあたりおよそ2×108pfuのバキュロウイルス力価を見出した。VLP粒子定量はVLPのタンパク質濃度を測定し、推定されるVLP分子量1.2×108ダルトン(ガンマレトロウイルス粒子1個あたりGagの複製物1500個およびGag対HAの比4:1に基づく)を使用することによって概算した。バキュロウイルスを超える大過剰量のVLPにも関わらず感染性バキュロウイルスがVLP調製物中に存在し、免疫原性へのその寄与は調査に重要であった。
【0209】
(VLP調製物中のバキュロウイルスの不活化)
Wisconsin/67(H3N2)VLP調製物中の感染性バキュロウイルスを、ソラーレン誘導体4’−アミノメチル−4,5’,8−トリメチルソラーレン塩酸塩(AMT)(Sigma−Aldrich、St.Lous、MO)存在下で長波長UV照射によって、またはベータ−プロピオラクトン(BPL)での処理によって不活化した。VLPを含有するプールしたショ糖勾配画分をUVまたはBPL処理のいずれかに供し、その後VLPを不活化溶液から100,000×g、トリス緩衝生理食塩水、pH7.4中の30%ショ糖クッションによる1時間、10℃での遠心分離によって回収した。UV不活化は1mlあたりAMT30μgの添加によって実施し、VLP懸濁物の1ml一定分量を6ウエル滅菌組織培養プレートの個々のウエルに添加した。培養プレート(蓋なし)を予め長波長照射用に再構成したCL−1000UV crosslinker(UVP、Upland、CA)に置き、365nm照射2.0〜2.5ジュールに、0.25ジュール毎に穏やかに撹拌しながら供した。BPL不活化についてはBPLを最終濃度0.2%で添加し、試料を室温で3時間インキュベートした。
【0210】
Wisconsin H3N2 VLPの3つの調製物を長波長UV/ソラーレン処理、ベータ−プロピオラクトン処理またはモック不活化にそれぞれ供し、ワクチン調製物をバキュロウイルス感染性について力価測定した。両方の不活化処理は、Sf9細胞でのエンドポイント力価測定によって測定した通り、ワクチン用量あたり10個未満の感染性バキュロウイルス粒子まで昆虫細胞感染力の低減をもたらした一方で、モック不活化VLP調製物は、ワクチン用量あたり105から106個の間の感染性バキュロウイルス粒子を含有していた。追加的に、UV/ソラーレン処理VLPは、HA活性の損失を示さなかったが、BPL処理VLPは、HA抗原の直接アルキル化および/またはVLPの完全性の破壊を示すHA活性の16〜20の低下を受けた。上の処理VLPから調製した筋肉内ワクチン用量は、初回免疫化用にはHAおよそ1μgを、追加免疫化用にはHA 0.5μgを含有した。表5は、UV/ソラーレン処理を用いるバキュロウイルス不活化後にVLP免疫原性の損失が無いことを実証する、3つのワクチン調製物についての免疫原性データを示し、これはHA活性の完全な保持と一致する。これらのデータは、観察された偽型VLP免疫原性のために感染性バキュロウイルスの潜在的アジュバント特性が必要ではないことを実証している。対照的にBPL処理VLPは、大きく減少した免疫原性を示し、BPL処理の結果としてのHA活性における顕著な減少と一致した。
【0211】
【表5】
(実施例7−考察)
本発明者らは、マウスおよびフェレットにおいて強い免疫原性および防御を示すキメラインフルエンザVLPの作製のためにMLV Gag遺伝子産物の堅調な粒子出芽特性を使用した。Gag媒介粒子出芽は、会合の膜部位へのこれらの分子の標的化を補助する翻訳後ミリスチル化に依存し[31、32]、Gag媒介粒子出芽がGagとカベオリンとの会合を介して脂質ラフトドメインを通って優先的に生じることの証拠が蓄積している[33]。この工程は、昆虫細胞および哺乳動物細胞の両方において効率的に生じ、Gagと組み込まれる抗原の細胞質側末端との間での特異的相互作用を必要することなく出芽粒子への脂質ラフト標的化内在性膜抗原の組み込みを可能にする。インフルエンザに加えて、このVLPプラットフォームは、パラミクソウイルス抗原の組み込みに同様に適用できるはずであり、このことは、複数の呼吸器感染疾患に対するキメラVLPワクチンファミリーの開発を可能にする。
【0212】
本発明者らは、昆虫細胞におけるキメラ粒子のスケールアップ(scaling up)作製、HAおよびNA含有量および生物学的活性についての粒子の特徴付けならびに、強い免疫原性および防御効果を示すマウスおよびフェレットにおける多数のワクチン研究の開始によるインフルエンザHAのGag粒子への組み込みの初期の知見に基づいている。H1N1、H3N2およびH5N1サブタイプを示すキメラインフルエンザVLPは、すべてバキュロウイルス昆虫細胞系において同様の効率で産生され、すべてがウイルスまたはVLPタンパク質1mgあたりのHA単位に関して、ショ糖勾配精製された生きているインフルエンザウイルスで観察されたのと同様の赤血球凝集特異的活性を示した。赤血球凝集抑制アッセイにおいてVLPが、生きているインフルエンザウイルスを容易に代替できるという事実に加えて、これらの観察は、キメラVLP表面上のHA抗原提示および密度が生きているインフルエンザで見られるものをかなりの程度まで模倣するという説得力のある証拠を提供する。本発明者らは、インフルエンザA以外のウイルスに向けたこの系の柔軟性を示すインフルエンザBウイルス抗原含有VLPの作製も実証している(データ未記載)。
【0213】
3種すべての遺伝子(HA、GagおよびNA)の1種のバキュロウイルスベクターへの組み込みは、VLP作製を大きく簡素化し、産物収量は昆虫細胞における感染多重度に決定的には依存しない。今日までに作製されたすべてのキメラインフルエンザA VLPは、同様の収率およびショ糖勾配バンドパターンを示す。電子顕微鏡によりこれらのVLPは、より多形性である生きているインフルエンザまたはインフルエンザマトリックスベースのVLP粒子とは著しく異なる均一な100nm球状粒子であると考えられる。VLPサイズのこの均一性は、この技術がヒトでの評価用の臨床グレード材料の作製のためにさらにスケールアップされることから重要である。キメラVLPが大きさにおいて均一である場合、図2に示す通り(ここでは、HAおよびNA活性の両方が単一のショ糖勾配全体で測定される)、HAおよびNAの相対的含有量は連続的であると考えられる。Gagとの予測されない物理的相互作用を伴なう、HAおよびNAの脂質ラフト中での標的化および選別は、無作為ではない可能性があり、これは、HAおよびNAの相対含有量での多様性を示し得るVLPを生じる。これは、HAおよびNA抗原がVLPの表面上に多様な状況で提示されることで免疫学的利益を有する場合がある。バキュロウイルスgp64エンベロープ糖タンパク質は脂質ラフトタンパク質ではなく、VLP中に顕著な程度にまで蓄積するとは予測されないことに注目することも重要である[49]。
【0214】
H1N1、H3N2およびH5N1サブタイプを示すキメラVLPは、マウスおよびフェレットにおいて添加アジュバントの非存在下で強く免疫原性であり、HAI、中和およびELISAアッセイにおいて測定される通りそれらそれぞれのウイルスに対する強い応答をもたらす。これらの強い応答は、マウスでの相同チャレンジ(homologous challenge)およびフェレットでの交差クレードH5N1チャレンジに対する病的状態についての証拠を示さない強固な防御を生じる。フェレットでのH5N1に対するH5N1交差クレード防御は新たなものではないが、本発明者らが、Indonesia H5N1またはVietnam H5N1をワクチン接種した動物においてVietnamウイルスの標準的1×106TCID50チャレンジ用量を使用したチャレンジ後の鼻洗浄液中にいかなるウイルスも検出できなかったことは注目に値する。すべての先行するフェレットH5N1ワクチン試験は、ホモサブタイプのH5N1チャレンジ後に上気道における顕著な量のウイルス複製を報告している[45、50、51]。
【0215】
偽型VLP媒介防御の機構の理解は、さらなる研究を必要とするが、ワクチン有効性を示すための有力な要因はこれらのワクチンによって誘発される免疫応答全体の強さである。VLPの強い免疫原性は、本質的に任意の細胞(抗原提示細胞を含む)への効率的な結合を促進できる(これは増強された抗原提示をもたらす)、VLP上の機能性HAスパイクの存在に部分的による可能性がある。細胞結合能力についての証拠は、ヒヨコ赤血球を使用する赤血球凝集アッセイおよび赤血球凝集抑制アッセイの両方において観察されたVLPの強いHA活性である。本発明者らは、種々のサブタイプのVLPを用いてヒトRBCに向けた強いHA活性も実証した(データ未記載)。
【0216】
HAによって媒介される直接細胞結合能力に加えて、生きているバキュロウイルス粒子は短期間先天免疫を刺激でき、他の抗原と混合された場合にアジュバントとして作用できることが実証されていることから、バキュロウイルスベクター由来VLPワクチンの免疫原性はバキュロウイルス粒子を混入させることによって増強され得ることが示唆されている[46〜48]。バキュロウイルス粒子バンドは、本発明者らの手法においてはショ糖勾配でVLPよりもわずかに低く、プラークアッセイによるバキュロウイルスの定量およびタンパク質濃度によるVLPの定量は、VLP調製物においてバキュロウイルスをおよそ4桁超えるVLPの存在量を明らかにした。しかし、本発明者らは本明細書において記載する実験のほとんどにおいて使用したVLP調製物からバキュロウイルスを除外しなかった。しかし表5は、混入バキュロウイルス粒子を長波長UV/ソラーレン処理またはベータ−プロピオラクトン(BPL)処理のいずれかによって不活化したVLP免疫原性実験の結果を示している。不活化の両方法が核酸を広く標的化する一方で、本発明者らは、BPL処理後にVLP関連HA活性の顕著な減少を観察したが、UV/ソラーレン処理後にHA活性の減少は観察しなかった。ワクチン接種後のHAI力価は、昆虫細胞上での力価測定によって測定されたバキュロウイルス感染性の欠如にもかかわらず、UV/ソラーレン処理VLPが無処理VLPと比較して免疫原性の減少を被らなかったことを明らかにした。したがって混入している生きたバキュロウイルスは、これらのVLPに関連する免疫原性にほとんど影響を有さないと考えられる。興味深いことにBLP処理VLPは、VLP関連HA活性の16〜20分の1への減少によって明らかである通りおそらくVLPの直接アルキル化によってそれらの免疫原性をほとんど失っている。予測外に、混入バキュロウイルス粒子のUV/ソラーレン不活化は、VLP免疫原性に効果を有さず、したがってVLP作製における不活化の理想的方法である。
【0217】
VLP調製物中の低レベルのバキュロウイルス混入の問題は、HA偽型バキュロウイルス粒子が、観察されたインフルエンザ特異的免疫原性にどの程度寄与できるかについての追加的疑問を提起した。なぜ可能性が低いのかについては、いくつかの原因がある。第1にバキュロウイルス出芽ビリオン(BV)の会合および出芽は、脂質ラフトドメインを標的化しない[49]膜結合バキュロウイルスgp64に依存する[54]。第2に、ポリヘドリンプロモーターが活性化するとBV産生は遮断されることから、大部分のBV産生は一時的に大部分のGag−VLP会合から外れると予測され、Gag−HA−NA VLP会合をバキュロウイルス感染周期のかなり遅い段階で駆動する。第3に、BVとVLPとの会合の相対位置および時期に幾分かの重複があると仮定すると、BVおよびVLPの相対量における大きな相違が考慮されなければならない。上に記載の通り本発明者らの算定は、混入しているバキュロウイルス粒子の104倍過剰であるVLPを示す。そのような比は、使用された投与量レベルにおいて混入HA偽型BVがワクチン効果に顕著に寄与する可能性を下げる。
【0218】
要約すると本発明者らは、MLV gagならびに種々のインフルエンザAサブタイプ由来のインフルエンザHAおよびNAからなる偽型VLPの作製およびマウスおよびフェレットでの防御免疫原性を記載する。これらのVLPは、株にかかわらず迅速かつ一定して産生され、強い免疫原性を示す。VLP調製物由来のバキュロウイルス粒子を不活化する2つの異なる方法についての本発明者らの研究は、UV/ソラーレン処理による不活化がVLPにその完全なレベルの免疫原性を保持させるという驚くべき発見をもたらした。これらのVLPの免疫原性を既存のおよび提案されたインフルエンザワクチンと比較するため実験、ならびにこのプラットフォームの有用性を他の呼吸器ウイルスに拡張するための実験は、進行中である。
【0219】
(実施例8−UV−A、UV−Cおよび可視光によるウイルス不活化方法)
以下の実施例において本発明者らは、UV−A/リボフラビン、UV−C照射単独および可視光/リボフラビンによるウイルス不活化の好ましい方法を記載する。ウイルス不活化は、バイオリアクター回収物(bioreactor offloads)、遠心分離または濾過によるバイオリアクター回収後の精製開始材料、中間精製画分(連続流式勾配遠心分離画分、クロマトグラフィー素通り画分またはクロマトグラフィー溶出画分)および/または最終精製産物を含むエンベロープVLPの作製において生成された任意の画分について実施され得る。
【0220】
ウイルス不活化研究用に、エンベロープVLP含有試料を適切な放射−透明用具(保存袋またはチューブなど)に入れ、ウイルス不活化をこれだけに限らないが、1)単一のまたは複数の放射源を対象の試料を囲む所定の距離だが任意の配置に置く配置;2)所定の試料流路を提供するためにチューブを任意の商業的に入手できる放射源に繋いだ配置(試料は蠕動ポンプの手段によって流路を流れる)または;3)上に記載の流路をエンペロープVLP精製のための標準的装置操作の一部として(例えばタンジェンシャルフロー濾過またはクロマトグラフィー分離中、インラインで)物理的に統合した配置が挙げられ得るデバイスを使用して実施した。
【0221】
UV−A/リボフラビンを使用する病原体不活化のために、エンベロープVLP試料をリボフラビン濃度10〜100μMに調整し、265〜370nmの間の照射波長を使用するエネルギー量0〜50J/cm2に供する。UV−Cを使用する病原体不活化は、VLP試料を254nmの照射波長を使用するエネルギー量0〜2J/cm2で処理することによって実施する。可視光/リボフラビンを使用する病原体不活化のために、エンベロープVLP試料をリボブラビン濃度100〜500μMに調整し、次いで400〜500nmの間の照射波長を使用するエネルギー量0〜500J/cm2に供する。上の研究においてpH、温度および遊離ラジカルスカベンジャーの効果などの試料条件を不活化条件を最適化するために評価する。代表的な試料を種々の量の照射に曝露し、ウイルス不活化(例えばバキュロウイルス不活化)を標準的感染性アッセイによって評価する。エンベロープVLPへの最小の損傷を示すために、分析試験のパネルを処理後のエンベロープVLPの同一性、安定性および免疫原性を示すために実施する。適切なウイルス不活化条件が定められれば、移転可能な製造工程のより代表的である材料のレベルにおいて、選択されたウイルス不活化方法の効果を確実にするために、様々な深度の試料においてスケールアップ研究を実施する。
【0222】
(実施例9−ガンマ照射によるウイルス不活化方法)
ガンマ照射を使用する病原体不活化のために、エンベロープVLP試料(液体試料および凍結試料の両方として)に、遊離ラジカルスカベンジャーの存在下または非存在下で照射する。代表的な試料を0から60kGyに増大させるエネルギー量に供し、ウイルス不活化(例えばバキュロウイルス不活化)を標準的感染性アッセイによって評価する。エンベロープVLPへの最小の損傷を実証するために、分析試験のパネルを処理後のエンベロープVLPの同一性、安定性および免疫原性を示すために実施する。適切なウイルス不活化条件が定められれば、移転可能な製造工程のより代表的である材料のレベルにおいて、選択されたウイルス不活化方法の効果を確実にするために、様々な深度の試料においてスケールアップ研究を実施する。
【0223】
(さらなる参考文献)
【0224】
【数1】
【0225】
【数2】
【0226】
【数3】
【0227】
【数4】
【0228】
【数5】
【0229】
【数6】
【0230】
【数7】
【0231】
【数8】
【技術分野】
【0001】
(発明の分野)
本発明は、感染因子(infectious agent)を含まない、エンベロープウイルスベースのウイルス様粒子(VLP)を単離する分野に関する。好ましい例において、該分野は、エンベロープウイルスベースのVLPの免疫原性に有害な影響を及ぼさない、感染因子を不活化する方法を包含する。特定の実施形態では、エンベロープウイルスベースのVLPを、昆虫細胞ベースの発現系において作製する。
【背景技術】
【0002】
(発明の背景)
アセンブリーは、宿主細胞内で見出されるアクセサリー因子を必要とすることが多いため、エンベロープウイルスベースのVLPを作製するには、宿主細胞の発現系内におけるVLPの発現およびアセンブリーを必要とすることが典型的である。宿主細胞内における生物学的成分の発現には、感染因子による汚染の危険性が付きまとう。タンパク質および多糖など、比較的小さな生物学的成分については、そのような作用物質を除去するのに、フィルターによる滅菌が一般に用いられるが、UVによる不活化または化学的な不活化など、他の方法もまた用いられている(単独で、またはフィルターベースの方法と組み合わせて用いられる)。例えば、オーエスキー病ウイルスの糖タンパク質D遺伝子を発現させるのに用いるときのバキュロウイルスを不活化するには、光化学的不活性化が用いられており、該発現するタンパク質に対する抗体の結合の低下は何ら観察されていない(非特許文献1を参照されたい)。しかし、VLPなど、比較的大きな生物学的成分については、VLPはサイズが感染因子(例えば、ウイルス)に近接し、濾過によりそのような感染因子から分離することが不可能なことが多いので、フィルターによる滅菌は適用可能でないことが多い。したがって、VLPを調製するときの感染因子を不活化するには、他の方法を用いる必要がある。ブタパルボウイルス(PPV:非エンベロープまたはカプシドウイルス)VLPを生成させるのに用いるときのバキュロウイルスを不活化するには、そのような他の方法が試みられている。Ruedaらは、バキュロウイルスに対する低温殺菌、化学的不活化、および、洗浄剤による不活化を比較し、PPV VLPの免疫原性に対して影響を及ぼさないことを見出した(例えば、非特許文献2を参照されたい)。
【先行技術文献】
【非特許文献】
【0003】
【非特許文献1】S.A. Weightmanら、Journal of Virological Methods(1999年)81巻:179〜182頁
【非特許文献2】P. Ruedaら、Vaccine(2001年)19巻:726〜734頁
【発明の概要】
【発明が解決しようとする課題】
【0004】
しかし、感染因子を不活化する典型的な方法をエンベロープウイルスベースのVLPに適用すると、該エンベロープウイルスベースのVLPの免疫原性が顕著に低下することが、思いがけず判明した。したがって、エンベロープウイルスベースのVLPに対して用いることができ、該VLPの免疫原性に有害な影響を及ぼすことなく感染因子を不活化する不活化法が必要とされている。
【課題を解決するための手段】
【0005】
(概要)
本発明の好ましい実施形態は、エンベロープウイルスベースのVLPの免疫原性に有害な影響を及ぼさない、本明細書で開示される、感染因子を不活化するための各種の方法および組成物と、不活化を受けていないVLP調製物と実質的に同じ免疫原性を有する、感染因子を含まないエンベロープウイルスベースのVLP調製物を含む組成物とを提供することにより、この必要を満たす。このような好ましい実施形態は、試験された電磁放射ベースの不活化法(単独の不活化法、または該電磁放射に反応性の化学物質を伴う不活化法)が、他の純粋に化学物質ベースの不活化法と比較して、エンベロープウイルスベースのVLPの免疫原性を結果として低下させないという驚くべき観察に基づく。
【0006】
本発明の態様は、感染因子を実質的に含まないエンベロープウイルスベースのウイルス様粒子調製物を単離する方法であって、(a)該エンベロープウイルスベースのウイルス様粒子調製物を作製するのに用いられる宿主細胞、または該宿主細胞の成分から、該エンベロープウイルスベースのウイルス様粒子調製物を単離するステップと、(b)該エンベロープウイルスベースのウイルス様粒子調製物における実質的にすべての感染因子を不活化するのに十分な線量の電磁放射を、該調製物へと適用するステップとを含み、ステップ(b)の後の該エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の該エンベロープウイルスベースのウイルス様粒子調製物と実質的に同じ免疫原性を有する方法を包含する。ある実施形態では、分離するステップ(a)が、少なくとも1つの遠心分離ステップを含む。前出の実施形態と組み合わせ得る別の実施形態では、分離するステップ(a)がまた、少なくとも1つの濾過ステップも包含し、該少なくとも1つの濾過ステップを、ノーマルフロー濾過、限外濾過、またはタンジェンシャルフロー(tangential flow)濾過からさらに選択することができる。前出の実施形態と組み合わせ得る別の実施形態では、分離するステップ(a)がまた、クロマトグラフィーステップも包含し、該少なくとも1つのクロマトグラフィーステップを、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー、疎水性相互作用クロマトグラフィー、混合方式クロマトグラフィー、逆相クロマトグラフィー、およびサイズ除外クロマトグラフィーからなる群よりさらに選択することができる。
【0007】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、電磁放射を、可視光、x線放射、紫外線放射、およびガンマ放射からなる群より選択することができ、該紫外線放射を、UV−A、UV−B、およびUV−Cからなる群よりさらに選択する場合もあり、該紫外線放射の波長が、320nm〜400nmの場合もある。
【0008】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、宿主細胞が、昆虫細胞または哺乳動物細胞である。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、宿主細胞が昆虫細胞であり、該昆虫細胞に、エンベロープウイルスベースのウイルス様粒子のうちの少なくとも1つの成分を発現するバキュロウイルス発現ベクターを感染させる。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、宿主細胞を、Bombyx mori宿主細胞、Spodoptera frugiperda宿主細胞、Choristoneura fumiferana宿主細胞、Heliothis virescens宿主細胞、Heliothis zea宿主細胞、Helicoverpa zea宿主細胞、Helicoverpa virescens宿主細胞、Orgyia pseudotsugata宿主細胞、Lymantria dispar宿主細胞、Plutella xylostella宿主細胞、Malacostoma disstria宿主細胞、Trichoplusia ni宿主細胞、Pieris rapae宿主細胞、Mamestra configurata宿主細胞、Mamestra brassica宿主細胞、およびHyalophora cecropia宿主細胞からなる群より選択する。宿主細胞が哺乳動物細胞である、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該哺乳動物細胞を、MRC−5細胞、Vero細胞、PER.C6(TM)細胞、チャイニーズハムスター卵巣細胞、およびHEK293細胞から選択する。昆虫細胞を宿主細胞として包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、電磁放射線量が、エンベロープウイルスベースのウイルス様粒子調製物におけるバキュロウイルスを不活化させるのに十分である。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、宿主細胞が哺乳動物細胞であり、前記哺乳動物細胞に、エンベロープウイルスベースのウイルス様粒子の少なくとも1つの成分を発現する、アデノウイルス発現ベクター、アデノ関連ウイルス発現ベクター、アルファウイルス発現ベクター、ヘルペスウイルス発現ベクター、ポックスウイルス発現ベクター、またはレトロウイルス発現ベクターを感染させる。アデノウイルス発現ベクター、アデノ関連ウイルス発現ベクター、アルファウイルス発現ベクター、ヘルペスウイルス発現ベクター、ポックスウイルス発現ベクター、またはレトロウイルス発現ベクターを包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、電磁放射線量が、エンベロープウイルスベースのウイルス様粒子調製物における、アデノウイルス、アデノ関連ウイルス、アルファウイルス、ヘルペスウイルス、ポックスウイルス、またはレトロウイルスを適宜不活化させるのに十分である。
【0009】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、適用するステップ(b)の前に、エンベロープウイルスベースのウイルス様粒子調製物にDNAインターカレート化合物を添加することができ、該DNAインターカレート化合物は、場合によって、光反応性のこともあり、ソラーレン、イソソラーレン、ならびにこれらの誘導体および類似体からなる群より選択することもある。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、電磁放射を、紫外線放射および可視光からなる群より選択することができる。
【0010】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該方法が、(c)エンベロープウイルスベースのウイルス様粒子調製物にアジュバントを添加するステップをさらに含み得る。
【0011】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、分離するステップ(a)の前に、(i)gagポリペプチドと、天然では脂質ラフトと会合しない抗原に連結した脂質ラフト会合ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記gagポリペプチドと、前記抗原に連結した脂質ラフト会合ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、宿主細胞内においてエンベロープウイルスベースのウイルス様粒子を作製する。
【0012】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、分離するステップ(a)の前に、(i)RSウイルスMポリペプチドと、RSウイルスFポリペプチドとを発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記RSウイルスMポリペプチドと、前記RSウイルスFポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、宿主細胞内においてエンベロープウイルスベースのウイルス様粒子を作製する。前出の実施形態は、場合によって、前記1または複数の発現ベクターから、RSウイルスGポリペプチドをさらに発現させることがある。
【0013】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、分離するステップ(a)の前に、(i)レンチウイルスおよびアルファレトロウイルスからなる群より選択されるレトロウイルスのgagポリペプチドと、RSウイルスFポリペプチドとを発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記レトロウイルスのgagポリペプチドと、前記RSウイルスFポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、宿主細胞内においてエンベロープウイルスベースのウイルス様粒子を作製する。前出の実施形態は、場合によって、前記1または複数の発現ベクターから、RSウイルスGポリペプチドをさらに発現させることがある。
【0014】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、分離するステップ(a)の前に、(i)gagポリペプチドと、インフルエンザ赤血球凝集素ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記gagポリペプチドと、前記インフルエンザ赤血球凝集素ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、宿主細胞内においてエンベロープウイルスベースのウイルス様粒子を作製する。
【0015】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、分離するステップ(a)の前に、(i)インフルエンザM1ポリペプチドと、赤血球凝集素ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記インフルエンザM1ポリペプチドと、前記赤血球凝集素ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、宿主細胞内においてエンベロープウイルスベースのウイルス様粒子を作製する。赤血球凝集素ポリペプチドを包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該1または複数の発現ベクターが、ノイラミニダーゼポリペプチドをさらに発現し得る。1または複数の発現ベクターを包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該1または複数の発現ベクターが、ウイルスベクター(複数可)であり得、該ウイルスベクター(複数可)を、バキュロウイルス、アルファウイルス、アデノ関連ウイルス、アデノウイルス、ヘルペスウイルス、ポックスウイルス、およびレトロウイルスからなる群よりさらに選択することができる。
【0016】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、ウイルスベクターを用いて、宿主細胞内において、エンベロープウイルスベースのウイルス様粒子の少なくとも1つの成分を発現させ、また、場合によって、該感染因子が、該ウイルスベクターを含む。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、調製物が、1ミリリットル当たり20個未満の感染因子;1ミリリットル当たり15個未満の感染因子;1ミリリットル当たり10個未満の感染因子;1ミリリットル当たり8個未満の感染因子;1ミリリットル当たり6個未満の感染因子;または1ミリリットル当たり5個未満の感染因子を含む。
【0017】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、ステップ(b)の後のエンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも50パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも60パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも70パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも80パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも85パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも90パーセントの免疫原性、ステップ(b)の前のエンベロープウイルスベースのウイルス様粒子調製物の少なくとも95パーセントの免疫原性を有する。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、感染因子を含まないようにされた後のエンベロープウイルスベースのウイルス様粒子調製物は、感染因子を含まないようにされる前のエンベロープウイルスベースのVLP調製物より大きな免疫原性を有するか、または、これと比較して免疫原性が増大する。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含み、免疫原性を予測する(predictor)赤血球凝集アッセイを用いて、該エンベロープウイルスベースのウイルス様粒子の物理的および生化学的な完全性を測定することもでき、または、エンベロープウイルスベースのウイルス様粒子により免疫化した動物の血清におけるHAI活性を、免疫原性の尺度として決定することもできる。免疫原性を特定する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、動物にエンベロープウイルスベースのウイルス様粒子(RSウイルス(RSV)ポリペプチドを包含し得る)をワクチン接種し、ELISAアッセイもしくはウイルス中和アッセイにより、またはウェスタンブロットにより抗体力価を測定するか、あるいは増殖アッセイ、ELISPOTアッセイ、またはサイトカイン放出アッセイによりT細胞反応を測定することにより、該免疫原性を直接測定することができる。
【0018】
本発明の別の態様は、感染因子を実質的に含まないエンベロープウイルスベースのウイルス様粒子を含む、エンベロープウイルスベースのウイルス様粒子調製物であって、該エンベロープウイルスベースのウイルス様粒子が、感染因子を実質的に含まないわけではないエンベロープウイルスベースのウイルス様粒子と実質的に同じ免疫原性を有する調製物を包含する。
【0019】
ある実施形態では、エンベロープウイルスベースのウイルス様粒子が、昆虫または哺乳動物グリコシル化をさらに含む。前出の実施形態と組み合わせ得る別の実施形態では、昆虫グリコシル化を、Bombyx mori; Spodoptera frugiperda; Choristoneura fumiferana; Heliothis virescens; Heliothis zea; Helicoverpa zea; Helicoverpa virescens; Orgyia pseudotsugata; Lymantria dispar; Plutella xylostella; Malacostoma disstria; Trichoplusia ni; Pieris rapae; Mamestra configurata; Mamestra brassica;およびHyalophora cecropiaからなる群よりさらに選択することができる。前出の実施形態と組み合わせ得る別の実施形態では、哺乳動物グリコシル化を、ヒトグリコシル化(PER.C6(TM)細胞、MRC−5細胞、およびHEK293細胞によりもたらされるグリコシル化を含めた)、サルグリコシル化(Vero細胞によりもたらされるグリコシル化を含めた)、また、げっ歯動物グリコシル化(チャイニーズハムスター卵巣細胞によりもたらされるグリコシル化を含めた)からなる群よりさらに選択することができる。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、共有結合した光化学作用物質、タンパク質サブユニットの三次構造または四次構造においてUV照射もしくはガンマ照射により誘導される変化、ガンマ照射により誘導される化学結合の開裂、またはUV照射もしくはガンマ照射により誘導される、脂質の酸化、タンパク質の架橋形成、アミノ酸の酸化、およびアミノ酸の修飾からなる群より選択される化学修飾から選択される1または複数の欠損をさらに示す。好ましい実施形態では、エンベロープウイルスベースのウイルス様粒子が、該欠損のすべてを示す。特定の実施形態では、このような欠損の検出を、免疫原性の低下が見られないことを介して推測することもでき、または、適切な技法(例えば、ウイルス様粒子を含むポリペプチド内における共有結合の変化に関して欠損が見られないことを確認する場合の質量分析、また、ウイルス様粒子を含むポリペプチドの三次構造または四次構造内において、欠損が見られないことを確認する場合の分析的HPLC)の適用を介して推測することもできる。
【0020】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、gagポリペプチドと、非ウイルス性脂質ラフト会合ポリペプチド、または天然では脂質ラフトと会合しない、連結を形成するための抗原に連結した脂質ラフト会合ポリペプチドとを含み、場合によって、該非ウイルス性脂質ラフト会合ポリペプチドを、GPIアンカーポリペプチド、ミリストイル化配列ポリペプチド、パルミトイル化配列ポリペプチド、二重アセチル化配列ポリペプチド、シグナル伝達ポリペプチド、および膜輸送ポリペプチドからなる群よりさらに選択することもでき、GPIアンカーポリペプチド、ミリストイル化配列ポリペプチド、パルミトイル化配列ポリペプチド、二重アセチル化配列ポリペプチド、カベオリン(cavelin)ポリペプチド、フロチリンポリペプチド、シンタクシン−1ポリペプチド、シンタクシン−4ポリペプチド、シナプシンIポリペプチド、アデューシンポリペプチド、VAMP2ポリペプチド、VAMP/シナプトブレビンポリペプチド、シナプトブレビンIIポリペプチド、SNAREポリペプチド、SNAP−25ポリペプチド、SNAP−23ポリペプチド、シナプトタグミンIポリペプチド、およびシナプトタグミンIIポリペプチドからなる群よりさらに選択することもできる。ウイルス性脂質ラフト会合ポリペプチドは、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、融合タンパク質ポリペプチド、糖タンパク質ポリペプチド、およびエンベロープタンパク質ポリペプチドからなる群よりさらに選択することができる。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、膜結合エンベロープタンパク質ポリペプチドをさらに含む。
【0021】
連結を包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該連結を、共有結合、イオン性相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、および抗体−抗原相互作用からなる群よりさらに選択することができ、該共有結合は、場合によって、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、およびジスルフィド結合からなる群よりさらに選択することもできる。脂質ラフト会合ポリペプチドを包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該脂質ラフト会合ポリペプチドが、内在性膜タンパク質である。抗原を包含する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、該抗原を、タンパク質、ポリペプチド、グリコポリペプチド、リポポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性または非ペプチド性模倣体、低分子、脂質、糖脂質、および炭水化物からなる群よりさらに選択することができる。
【0022】
前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチド、RSウイルスMポリペプチド、RSウイルスGポリペプチド、および/またはRSウイルスFポリペプチドをさらに含む。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、gagポリペプチドおよび赤血球凝集素ポリペプチド;レンチウイルスおよびアルファレトロウイルスからなる群より選択されるレトロウイルスのgagポリペプチド、ならびにRSウイルスFポリペプチド(また、場合によって、Gポリペプチド);またはインフルエンザM1ポリペプチドおよび赤血球凝集素ポリペプチドを含む。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、ノイラミニダーゼポリペプチドをさらに包含する。
【0023】
前出の実施形態のいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子調製物が、前記ウイルス様粒子と会合するアジュバントを包含する。アジュバントを包含する特定の実施形態では、処方ステップにおいて、該アジュバントを、エンベロープウイルスベースのウイルス様粒子と混合することができる。アジュバントは、ウイルス様粒子の内側または外側に位置する場合もあり、ウイルス様粒子に組み込まれる場合もある。前出の実施形態のいずれかと組み合わせ得るさらに別の実施形態では、アジュバントを、前記ウイルス様粒子と共有結合させて、共有結合を形成させることができる。
【0024】
本発明の別の態様は、免疫系の疾患または症状を処置または予防する方法であって、免疫原性量の、前出の実施形態のうちのいずれかによるエンベロープウイルスベースのウイルス様粒子調製物、または前出の方法およびその実施形態のうちのいずれかを用いて単離したエンベロープウイルスベースのウイルス様粒子調製物を、被験体に投与するステップを含む方法を包含する。一実施形態では、被験体がヒトである。前出の実施形態と組み合わせ得る別の実施形態では、投与するステップにより、被験体において、防御的免疫化反応が誘導される。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、投与するステップを、皮下(subcutaneous)送達、皮内送達、皮下(subdermal)送達、筋肉内送達、経口(peroral)送達、経口(oral)送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群よりさらに選択することができる。
【0025】
本発明の別の態様は、免疫原性量の、前出の実施形態のうちのいずれかによるエンベロープウイルスベースのウイルス様粒子調製物、または前出の方法およびその実施形態のうちのいずれかを用いて単離したエンベロープウイルスベースのウイルス様粒子調製物を含む医薬組成物を包含する。
【0026】
前出の態様およびそれらの実施形態は、本明細書で開示される実施形態のうちのいずれかとさらに組み合わせることができる。前出の実施形態、および/または本明細書で開示されるさらなる実施形態のうちのいずれかと共に包含され得る、本発明のさらなる態様は、本明細書の全体において見出すことができる。
【図面の簡単な説明】
【0027】
【図1】図1は、HAおよびNAを含有するキメラVLPの分析を示す図である。組換えバキュロウイルス発現細胞の上清は、細胞性細片を除去され、PR/8 H1N1 VLPは、ショ糖クッションによって100,000×gで遠心分離され、再懸濁され、不連続ショ糖密度勾配で再度遠心分離された。(A)勾配全体でのPR/8 H1N1 VLP赤血球凝集活性。(B)PR/8 H1N1 VLPのSDS−PAGE分析、GagおよびHAの共移動を示している。(C)A/Russia/77(H1N1)に特異的な抗体を使用する勾配画分4〜16のH1N1特異的ウェスタンブロット。(D)ピーク勾配画分由来のネガティブ染色試料の電子顕微鏡写真。(E)分子量を増加させてHA産物の電気泳動移動度を減少させるために、無関係な配列でHA遺伝子がそのアミノ末端で伸長された精製PR/8 H1N1 VLPのSDS−PAGE。(F)A/Solomon Islands/3/2006(H1N1)を示す精製VLPのSDS−PAGE、Gag対HAの比を示す。
【図2】図2は、不連続ショ糖勾配で遠心分離したH5N1 VLPでの赤血球凝集活性およびノイラミニダーゼ活性の検査を示すグラフである。H5N1 VLPは、図1でH1N1 VLPについて示した通り調製されたが、ショ糖勾配画分は、赤血球凝集活性およびノイラミニダーゼ活性の両方についてアッセイされた。NA活性は蛍光基質2’−(4−メチルウンベリフェリル)−α−D−N−アセチルノイラミン酸を使用して測定された。(A)は、Vietnam H5N1 VLPについての活性を示す。(B)はIndonesia H5N1 VLPについての活性を示す。
【図3】図3は、VLP免疫原性およびチャレンジ防御を示すグラフである。16匹のマウス3群は、キメラH1N1 VLP(A/PR/8/34を示す)で筋肉内または腹腔内免疫化を介して初回および追加免疫化を受けた。すべてのVLP処方物は、1用量あたりHAおよそ0.7μgを含有した。追加の4週間後に動物は10LD50のA/PR/8/34(H1N1)でチャレンジされた。(A)追加後2週間でのA/PR/8/34(H1N1)に特異的なHAI活性。(B)追加後2週間でのA/PR/8/34に特異的なIgG1およびIgG2A応答の定量。(C)H1N1チャレンジに続く重量減少によって示されるチャレンジ防御。
【図4】図4は、VLPおよび生きているウイルスがHAIアッセイにおいて同様に挙動することを示すグラフである。PR/8(H1N1)VLPで免疫化したマウス由来の血清を生きているPR/8(H1N1)ウイルスの4HAユニットまたは対応するVLPの4HAユニットのいずれかを使用してHAI活性について検査した。HK/68(H3N2)VLPで免疫化したマウス由来の血清を同様にHK/68ウイルスおよびVLPに対して検査した。データは、HAIアッセイにおいてウイルスとVLPとの間での同様の挙動を示した。
【図5】図5は、VLP免疫化フェレットにおける追加後免疫応答を示すグラフである。フェレットは初回および追加免疫化をそれぞれ0日目および28日目にH1N1、H5N1および裸のVLPで受けた。H1N1およびH5N1 VLP用量は、1用量あたりHAおよそ5μgを含有した。28日目および42日目の血清試料を(A)A/PR/8/34(H1N1)特異的HAI活性および(B)A/Vietnam/1203/04(H5N1)特異的マイクロ中和活性について分析した。
【図6】図6は、H1N1、H5N1および裸のVLPで免疫化したフェレットでのチャレンジ後の体重減少および生存を示すグラフである。追加免疫化を受けた2週間後にVLPワクチン接種フェレットは、1×106TCID50のA/Vietnam/1203/04(H5N1)でチャレンジされた。(A)すべての群についての平均体重減少データ(生存データをグラフ説明に示す)。(B)B群中のH1N1ワクチン接種動物についての個々の体重減少データ。
【図7】図7は、H1N1、H5N1および裸のVLPで免疫化されたフェレットでのチャレンジ後の鼻洗浄液ウイルス力価を示すグラフである。追加免疫化を受けた2週間後にVLPワクチン接種フェレットは、1×106TCID50のA/Vietnam/1203/04(H5N1)でチャレンジされた。(A)チャレンジ後3日目(45日目)の鼻洗浄液ウイルス力価。(B)チャレンジ後5日目(47日目)の鼻洗浄液ウイルス力価。
【発明を実施するための形態】
【0028】
(好ましい実施形態の詳細な説明)
本発明の好ましい実施形態には、感染因子を不活化する方法に供されたエンベロープウイルスベースのVLP調製物であって、そのような不活化法に供されていないVLPと実質的に同じ免疫原性をVLPが保持することを可能とする調製物と;エンベロープウイルスベースのVLP調製物における感染因子を不活化するこのような方法と;このような調製物をワクチン組成物へとさらに加工する方法と、このようなワクチン組成物を用いる方法とが含まれるが、これらに限定されない。
【0029】
理論に制約されることを望まないが、本発明の特定の好ましい実施形態は、以下の実施例に示すように、PPV VLPなど、他の非エンベロープウイルスベースのVLPとは異なり、エンベロープウイルスベースのVLPが、感染因子を不活化するのに用いられる標準的な不活化法に対して感受性であるという驚くべき発見に基づいている。しかし、同じく驚くべきことに、UV−A照射+光化学作用物質、UV−C照射、およびガンマ照射など、電磁ベースの不活化システムは、汚染性のエンベロープウイルスを不活化するのに効果的である一方で、該エンベロープウイルスベースのVLPの免疫原性を維持し、このために、このような不活化法は、ワクチンにおいて用いられるエンベロープウイルスベースのVLPを調製するのに理想的となっている。
【0030】
エンベロープウイルスベースのVLPを作製する好ましい方法は、好ましくはポリペプチド抗原の共発現(coexpression)を含めた、昆虫細胞内における発現を介する。バキュロウイルス発現系内における各種のレトロウイルスから得ることができるgag VLPの収量が大きい(23、24、46、49、52〜58)ため、gagポリペプチドを用いてVLPを作製することがさらにより好ましい。天然のレトロウイルスアセンブリー過程において、gagポリペプチドはC末端伸長を本質的に包含する。すなわち、天然において、機能的なgagタンパク質は、リボソームのフレームシフトに起因して、レトロウイルスプロテアーゼ活性、逆転写酵素活性、およびインテグラーゼ活性を含有する長いC末端伸長を有する。ラウス肉腫ウイルスのgag(59)およびMLVのgag(60)の両方について、人工的な伸長を伴う機能的なgagタンパク質の作製が達成されている。gagのC末端操作がこのように柔軟であることから、他の抗原および免疫刺激性タンパク質の配列など、他のポリペプチドを組み込むのに重要な部位が得られる。エンベロープウイルスベースのVLPを作製する別の好ましい方法は、インフルエンザHAタンパク質およびインフルエンザM1タンパク質(また、場合によって、インフルエンザNAタンパク質)の共発現を介する。昆虫細胞内におけるこれら2つのタンパク質の共発現は、エンベロープウイルスベースのVLPを作製する有効な方法であることが示されている(111〜112)。
【0031】
エンベロープウイルスベースのVLPの好ましい例には、(i)gagポリペプチドと、抗原に連結した脂質ラフト会合ポリペプチドとを含むVLP;(ii)RSウイルスMポリペプチドと、RSウイルスFポリペプチドと(また、場合によって、RSウイルスGポリペプチドと)を含むVLP;(iii)レンチウイルスおよびアルファレトロウイルスからなる群より選択されるレトロウイルスのgagポリペプチドと、RSウイルスFポリペプチドとを含むVLP;(iv)gagポリペプチドと、インフルエンザ赤血球凝集素ポリペプチドと(また、場合によって、ノイラミニダーゼポリペプチドと)を含むVLP;ならびに(v)インフルエンザM1ポリペプチドと、赤血球凝集素ポリペプチドと(また、場合によって、ノイラミニダーゼポリペプチドと)を含むVLPが含まれる。
【0032】
1つのウイルスに由来するコア粒子と、別のウイルスに由来する表面抗原とを含有するキメラVLPの作製は、シュードタイピング(pseudotyping)と呼ばれる。おそらく、インフルエンザHAタンパク質およびインフルエンザNAタンパク質が、脂質ラフトドメイン内に濃縮される(61、62)一方、出芽過程では、ミリストイル化した(myristolated)gagタンパク質もまた、脂質ラフトドメインの内部表面において濃縮される(63)ため、gagポリペプチドが効率的にシュードタイピングされるのは、これらのタンパク質を伴うときである。
【0033】
本明細書で記載される本発明の実施形態は、キメラVLPを形成するための基盤として、天然では脂質ラフトと会合しない抗原に連結した脂質ラフト会合ポリペプチドを包含するVLPプラットフォームに適合する。
【0034】
開示される方法およびプロトコールを実施するには、別段に示さない限り、当業者の能力の範囲内にある、化学、分子生物学、微生物学、組換えDNA、および免疫学の従来の技法を用いる。このような技法は、文献において説明されている。例えば、J. Sambrook、E. F. Fritsch、およびT. Maniatis、1989年、「Molecular Cloning: A Laboratory Manual」、第2版、1〜3巻、Cold Spring Harbor Laboratory Press社; Ausubel, F. M.ら(1995年、および定期的な補遺;「Current Protocols in Molecular Biology」、9、13、および16章、ニューヨーク州、ニューヨーク、John Wiley & Sons社); B. Roe、J. Crabtree、およびA. Kahn、1996年、「DNA Isolation and Sequencing: Essential Techniques」、John Wiley & Sons社; J. M. PolakおよびJames O’D. McGee、1990年、「In Situ Hybridization: Principles and Practice」、Oxford University Press社; M. J. Gait(編)、1984年、「Oligonucleotide Synthesis: A Practical Approach」、Irl Press社;ならびにD. M. J. LilleyおよびJ. E. Dahlberg、1992年、「Methods of Enzymology: DNA Structure Part A: Synthesis and Physical Analysis of DNA Methods in Enzymology」、Academic Press社を参照されたい。
【0035】
(定義)
本明細書で用いられる「エンベロープウイルスベースのVLP」とは、エンベロープウイルスに由来する1または複数の成分を用いて形成されるウイルス様粒子を指す。好ましい例には、gagポリペプチドを用いて作製したVLPと、インフルエンザM1ポリペプチドおよび/または赤血球凝集素ポリペプチド(また、場合によって、ノイラミニダーゼポリペプチド)を用いて作製したVLPと、レンチウイルスのgagポリペプチドおよびアルファレトロウイルスのgagポリペプチドからなる群ならびにRSウイルス(RSV)Fポリペプチド(および、場合によって、RSウイルスGポリペプチド)を用いて作製したVLPと、RSウイルス(RSV)Mおよび/またはFポリペプチド(および、場合によって、RSウイルスGポリペプチド)を用いて作製したVLPとが含まれるが、これらに限定されない。
【0036】
さらなる例には、エンベロープウイルスベースのVLPを形成するのに用い得る、エボラウイルスおよびマールブルクウイルスなどのフィロウイルス(例えば、細胞内においてフィロウイルスに由来するウイルス性GPおよびウイルス性VP40を共発現させると、脂質ラフト内におけるこれら2つのウイルス性タンパク質の会合により、VLPが生成される(米国特許公開第20060099225号を参照されたい));SARSウイルスなどのコロナウイルス(例えば、コロナウイルスのVLP形成には、Eタンパク質およびMタンパク質で十分である(Fischerら、J. Virol.(1998年)、72巻:7885〜7894頁;およびVennemaら、EMBO J.(1996年)、15巻:2020〜2028頁を参照されたい));RSウイルス(RSV)など、パラミクソウイルス科のウイルス(例えば、RSVのMタンパク質を発現させると、VLPが生成される(例えば、米国特許公開第20080233150号を参照されたい));ならびに西ナイルウイルスなどのフラビウイルス科(例えば、バキュロウイルス発現系内で西ナイルウイルスのprM遺伝子およびE遺伝子を含む構築物を発現させると、VLPが生成される(例えば、米国特許公開第20080233150号を参照されたい))が含まれる。
【0037】
本明細書で用いられる「感染因子を含まない」とは、感染可能な活性作用物質の不在を指す。このような試料は、不活性であり、感染不能な作用物質を含有し得る。例を目的として述べると、バキュロウイルスがもはや感染不能であるように処理された、バキュロウイルスを含有する試料は、不活化されたバキュロウイルスをなおも含有するが、感染因子を含まない。さらに、試料は、感染可能な活性作用物質を絶対的に含まないことが必要なわけではなく、これをヒト用または動物用のワクチンとして意図される目的に適宜用い得る(すなわち、ヒト用または動物用のワクチン内における感染因子の許容レベルを統轄する任意の米国連邦規則を適宜満たす)程度に十分に活性作用物質を含まないことだけが必要である。特定の実施形態では、感染因子を含まないようにされた後のエンベロープウイルスベースのVLP調製物の用量当たりの感染単位またはml当たりの感染単位は、感染因子を含まないようにされる前のエンベロープウイルスベースのVLP調製物の少なくとも10分の1、少なくとも100分の1、少なくとも1000分の1、または少なくとも10,000分の1に低下する。例示を目的として述べると、感染因子には、1または複数の宿主細胞内において、VLPを含むポリペプチド(複数可)を発現させるのに用いるベクター(複数可)、および外部の細菌性汚染物質、真菌性汚染物質、またはウイルス性汚染物質、さらにまた、内因性の病原体(例えば、宿主ゲノム内ではサイレントである(silent)ことが典型的である、レトロウイルス性エレメントまたはレトロトランスポゾン性エレメントの再活性化など、供給源物質または宿主細胞に由来する)が含まれ得る。
【0038】
本明細書で用いられる「実質的に同じ免疫原性」とは、感染因子を含まないようにされる前の調製物と比較した、感染因子を含まないようにされた後のエンベロープウイルスベースのVLP調製物の免疫原性を指す。特定の実施形態では、感染因子を含まないようにされた後のエンベロープウイルスベースのVLP調製物は、感染因子を含まないようにされる前のエンベロープウイルスベースのVLP調製物の少なくとも50パーセント、少なくとも60パーセント、少なくとも70パーセント、少なくとも80パーセント、少なくとも85パーセント、少なくとも90パーセント、または少なくとも95パーセントの免疫原性を有する。特定の実施形態では、感染因子を含まないようにされた後のエンベロープウイルスベースのVLP調製物は、感染因子を含まないようにされる前のエンベロープウイルスベースのVLP調製物より大きな免疫原性を有するか、または、これと比較して免疫原性が増大する。免疫原性の好ましい尺度は、接種後に得られる、VLP組成物に対する抗体力価である。インフルエンザワクチンの場合、免疫原性の好ましい尺度は、以下の実施例によるHAI活性である。前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、エンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含み、免疫原性を予測する赤血球凝集アッセイを用いて、該エンベロープウイルスベースのウイルス様粒子の物理的および生化学的な完全性を測定することもでき、または、エンベロープウイルスベースのウイルス様粒子により免疫化した動物の血清におけるHAI活性を、免疫原性の尺度として決定することもできる。免疫原性を特定する、前出の実施形態のうちのいずれかと組み合わせ得るさらに別の実施形態では、動物にエンベロープウイルスベースのウイルス様粒子(RSウイルス(RSV)ポリペプチドを包含し得る)をワクチン接種し、ELISAアッセイもしくはウイルス中和化アッセイにより、またはウェスタンブロットにより抗体力価を測定するか、あるいは増殖アッセイ、ELISPOTアッセイ、またはサイトカイン放出アッセイによりT細胞反応を測定することにより、該免疫原性を直接測定することができる。
【0039】
「昆虫グリコシル化」とは、昆虫により、また、昆虫細胞ベースの発現系によりもたらされるグリコシル化パターンを指す。このようなグリコシル化パターンには、天然においてもたらされるグリコシル化、および哺乳動物のグリコシル化酵素を包含するように改変された昆虫細胞が、天然において、哺乳動物または哺乳動物細胞ベースの発現系によりもたらされるグリコシル化パターンではなく、「哺乳動物様の」グリコシル化をもたらすだけである限りにおいて、このような改変昆虫細胞によりもたらされるグリコシル化パターンも含まれ得る。昆虫グリコシル化パターンの好ましい例には、
【0040】
【化1】
など、バキュロウイルス発現系および関連のウイルス発現系に適合する昆虫細胞が含まれる。
【0041】
「哺乳動物グリコシル化」とは、哺乳動物により、また、哺乳動物細胞ベースの発現系によりもたらされるグリコシル化パターンを指す。このようなグリコシル化パターンには、天然においてもたらされるグリコシル化、およびこのような細胞では見出されないか、または発現することが典型的ではないグリコシル化酵素を包含するように改変された哺乳動物細胞が、天然において、哺乳動物以外または哺乳動物以外の細胞ベースの発現系によりもたらされるグリコシル化パターンではなく、哺乳動物グリコシル化または非天然グリコシル化をもたらすだけである限りにおいて、このような改変哺乳動物細胞によりもたらされるグリコシル化パターンも含まれ得る。昆虫グリコシル化パターンの好ましい例には、ヒト細胞(PER.C6(TM)細胞、MRC−5細胞、およびHEK293細胞によりもたらされるグリコシル化を含めた)、サル細胞(Vero細胞によりもたらされるグリコシル化を含めた)、また、げっ歯動物細胞(チャイニーズハムスター卵巣細胞によりもたらされるグリコシル化を含めた)など、公知のウイルス発現系に適合する哺乳動物細胞が含まれる。
【0042】
本明細書で用いられる「Gagポリペプチド」とは、本明細書で記載するウイルス様粒子の形成の一因となる、レトロウイルス由来の構造ポリペプチドである。gagポリペプチドは、エンベロープウイルスベースのVLPを形成するのに好ましい手段である。一部の実施形態では、RNAをパッケージングする傾向、または粒子形成および出芽の効率など、特定の特徴に影響を及ぼす目的で、gagポリペプチドを意図的に変異させることができる。レトロウイルスのゲノムは、3つの主要な遺伝子産物をコードする:構造タンパク質をコードするgag遺伝子の産物;逆転写酵素および関連のタンパク質分解性ポリペプチド、ヌクレアーゼおよびインテグラーゼに関連する機能をコードするpol遺伝子の産物;envの産物(それによりコードされる糖タンパク質の膜タンパク質が、感染細胞の表面上において、また、放出された成熟ウイルス粒子の表面上においても検出される)。すべてのレトロウイルスのgag遺伝子は、全体的な構造類似性を有し、レトロウイルスの各群内においても、アミノ酸レベルで保存されている。gag遺伝子は、逆転写酵素を除くコアタンパク質をもたらす。
【0043】
MLVの場合、Gagの前駆体であるポリタンパク質は、Pr65Gagであり、該前駆体上におけるその順序がNH2−p15−pp12−p30−p10−COOHである4つのタンパク質へと切断される。これらの切断は、ウイルス性プロテアーゼを介し、また、ウイルスに応じて、ウイルス放出の前または後において生じ得る。MLV Gagタンパク質は、グリコシル化形態および非グリコシル化形態で存在する。グリコシル化形態はgPr80Gagから切り出され、gPr80Gagは、非グリコシル化Pr65Gagに対応するAUGコドンの上流に位置する、インフレームの、異なる開始コドンから合成される。グリコシル化Gagを合成しない、MLVの欠失変異体がなおも感染性であり、また、非グリコシル化Gagがなおもウイルス様粒子を形成し得ることから、グリコシル化イベントの重要性について、疑問が投げかけられる。ウイルスがコードするプロテアーゼにより、HIV−1 Gagの前駆体であるpr55Gagが翻訳後に切断されることから、N−ミリストイル化されて内部でリン酸化されたp17マトリックスタンパク質(p17MA)、リン酸化されたp24カプシドタンパク質(p24CA)、ならびにヌクレオカプシドタンパク質であるp15(p15NC)がもたらされ、p15は、さらにp9およびp6へと切断される。
【0044】
構造面において、原型的なGagポリタンパク質は、レトロウイルスのgag遺伝子において常に同じ順序で生じる3つの主要なタンパク質:マトリックスタンパク質(MA)(マトリックスという名称を共有するが、MAとは異なるタンパク質である、インフルエンザマトリックスタンパク質M1と混同しないようにされたい)、カプシドタンパク質(CA)、およびヌクレオカプシドタンパク質(NC)に分けられる。成熟タンパク質へのGagポリタンパク質のプロセシングは、レトロウイルスがコードするプロテアーゼにより触媒され、新たに出芽するウイルス粒子が成熟するときに生じる。機能面において、Gagポリタンパク質は、3つのドメイン:細胞膜をGagポリタンパク質の標的とする膜結合ドメインと;Gagの重合化を促進する相互作用ドメインと;宿主細胞からの新生ビリオンの放出を促進する後期ドメインとに分けられる。アセンブリーを媒介するGagタンパク質の形態は、ポリタンパク質である。したがって、アセンブリードメインは、後期に形成される任意の切断産物内にもれなく収まる必要はない。こうして、本明細書で包含されるGagポリタンパク質は、VLPの形成および放出に重要な機能的エレメントを包含する。これらの重要な機能的エレメントに関する最新技術の進歩は大きい。例えば、
【0045】
【化2】
を参照されたい。
【0046】
本発明の特定のVLPにおいて用いられるgagポリペプチドは、VLPを形成するための機能的エレメントを最小限において包含するだろう。gagポリペプチドは、場合によって、そのコード配列をgagポリペプチドのコード配列にスプライシングすることによって作製し得る、1または複数のさらなるポリペプチドを包含し得る。gagポリペプチド内へのさらなるポリペプチドの挿入に好ましい部位は、C末端である。
【0047】
Gagポリペプチドに好ましいレトロウイルスの供給源には、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス(トリ白血病ウイルス、またはラウス肉腫ウイルスなど)、ベータレトロウイルス(マウス乳癌ウイルス、ヤーグジークテヒツジレトロウイルス、およびメーソン−ファイザーサルウイルスなど)、ガンマレトロウイルス(マウス白血病ウイルス、ネコ白血病ウイルス、細網内皮症ウイルス、およびテナガザル白血病ウイルスなど)、デルタレトロウイルス(ヒトTリンパ向性ウイルス、およびウシ白血病ウイルスなど)、イプシロンレトロウイルス(ウォールアイ皮膚肉腫ウイルスなど)、またはレンチウイルス(1型ヒト免疫不全ウイルス、HIV−2、サル免疫不全ウイルス、ネコ免疫不全ウイルス、ウマ感染性貧血ウイルス、およびヤギ関節炎脳炎ウイルス)が含まれる。
【0048】
本明細書で用いられる「脂質ラフト」とは、ウイルス粒子のアセンブリー過程においてgagポリペプチドが濃縮される、細胞膜のマイクロドメインを指す。
【0049】
本明細書で用いられる「脂質ラフト会合ポリペプチド」とは、脂質ラフトと直接的または間接的に会合する任意のポリペプチドを指す。本発明で用いられる具体的な脂質ラフト会合ポリペプチドは、キメラウイルス様粒子の所望の使用に依存する。
【0050】
脂質ラフト会合ポリペプチドは、内在性膜タンパク質、膜との会合を引き起こすタンパク質修飾により脂質ラフトと直接的に会合するタンパク質、または脂質ラフト会合ポリペプチドにより脂質ラフトと間接的に会合するポリペプチドであり得る。
【0051】
脂質アンカーを有する多くのタンパク質が、脂質ラフトと会合する。ポリペプチドを脂質ラフトへと連結する脂質アンカーには、GPIアンカー、ミリストイル化、パルミトイル化、および二重アセチル化が含まれる。
【0052】
多くの異なる種類のポリペプチドが、脂質ラフトと会合する。脂質ラフトは、シグナル伝達、膜輸送、ウイルスの侵入、ウイルスのアセンブリー、また、アセンブルした粒子の出芽を含めた、多くの生物学的活性のためのプラットフォームとして機能し、したがって、これらの過程に関与する各種のポリペプチドと会合する。
【0053】
シグナル伝達カスケードに関与する各種のポリペプチドは、シグナル伝達プラットフォームとして機能する脂質ラフトと会合する。シグナル伝達プラットフォームとして機能する脂質ラフトの1つの種類は、カベオラと呼ばれている。それは、カベオリンファミリーに由来するポリペプチド(例えば、カベオリンおよび/またはフロチリン(flottillin))を含有する、細胞膜のフラスコ型の陥入部である。
【0054】
膜輸送ポリペプチドは、膜輸送プラットフォームとして機能する脂質ラフトと会合する。例には、シンタクシン−1タンパク質、シンタクシン−4タンパク質、シナプシンIタンパク質、アデューシンタンパク質、VAMP2タンパク質、VAMP/シナプトブレビンタンパク質、シナプトブレビンIIタンパク質、SNAREタンパク質、SNAP−25タンパク質、SNAP−23タンパク質、シナプトタグミンIタンパク質、およびシナプトタグミンIIタンパク質など、エンドサイトーシスおよびエクソサイトーシスに関与するタンパク質が含まれる。
【0055】
ウイルス受容体、ウイルス受容体−共受容体複合体、ウイルス侵入過程の調節を補助する他の任意の成分は、ウイルス侵入に特化した膜輸送プラットフォームとして機能する脂質ラフトと会合する。脂質ラフトと会合するウイルス受容体の例には、崩壊促進因子(DAFまたはCD55)、多くのエンテロウイルスの受容体であるGPIアンカー膜糖タンパク質;ガングリオシド、Hsc70タンパク質、アルファ2−ベータ1インテグリンおよびアルファ5−ベータ2インテグリンを含めた複数の成分を含有する複合体である、A群ロタウイルスの受容体;HIVウイルス、MLVウイルス、麻疹ウイルス、およびEbolaウイルスなど、複数のエンベロープウイルスの糖タンパク質;ならびにCD5ポリペプチド、CCR5ポリペプチド、およびnefポリペプチドなど、HIVの侵入に関与するポリペプチドが含まれる。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. And Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。
【0056】
ウイルス粒子のアセンブリーに関与するポリペプチドは、ウイルスアセンブリープラットフォームとして機能する脂質ラフトと会合する。このようなポリペプチドの例には、HAインフルエンザエンベロープ糖タンパク質およびNAインフルエンザエンベロープ糖タンパク質、麻疹ウイルスに由来するHタンパク質および成熟F1−F2融合タンパク質、ならびにHIVウイルスに由来するgp160、gp41、およびPr55gagが含まれる。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. & Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。
【0057】
アセンブルしたウイルスの出芽に関与するポリペプチドは、ウイルス出芽プラットフォームとして機能する脂質ラフトと会合する。宿主細胞からのHIV−1の出芽は、膜ラフト内で生じることを示唆するデータが存在する。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. And Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。ウイルスの出芽に関与するポリペプチドについての一般的な情報は、「Fields Virology」(第4版)、2001年において見出すことができる。
【0058】
好ましい脂質ラフト会合ポリペプチドには、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、融合タンパク質ポリペプチド、糖タンパク質ポリペプチド、およびエンベロープタンパク質ポリペプチドなどのウイルスポリペプチドが含まれる。これらのポリペプチドの各々は、任意の種類のウイルスに由来し得るが、特定の実施形態は、HIV−1ウイルスに由来するエンベロープタンパク質、RSウイルスまたは麻疹ウイルスに由来する融合タンパク質、RSウイルス、単純ヘルペスウイルス、またはエボラウイルスに由来する糖タンパク質、また、麻疹ウイルスに由来する赤血球凝集素タンパク質を包含する。
【0059】
好ましい非ウイルス性病原体の脂質ラフト会合ポリペプチドは、Plasmodium falciparum、Plasmodium malariae、Plasmodium ovale、およびPlasmodium vivaxなどのPlasmodium属; Toxoplasma gondii; Trypanosoma brucei、Trypanosoma cruzi; Schistosoma haematobium、Schistosoma mansoni、Schistosoma japonicum; Leishmania donovani; Giardia intestinalis; Cryptosporidium parvumなどが含まれるがこれらに限定されない病原性の原虫、蠕虫、また、他の真核微生物の病原体から得ることができる。このような非ウイルス性脂質ラフト会合ポリペプチドは、それ自体が抗原として作用するので、天然では脂質ラフトと会合しない抗原に連結することなく用いることができる。
【0060】
本明細書で用いられる「インフルエンザM1ポリペプチド」は、ウイルスエンベロープ内におけるウイルス被覆の形成を媒介する、インフルエンザウイルスタンパク質に由来する。該M1タンパク質は、ウイルスの出芽を駆動し、また、ウイルス粒子の主要なタンパク質成分でもあり、ウイルス粒子内では、ウイルスエンベロープおよび内在性膜タンパク質と、ゲノムリボ核タンパク質との間の中間層を形成する。M1ポリペプチドはまた、正に帯電したアミノ酸に富む、M1ポリペプチド内における結合モチーフ(RKLKR)に依存する非特異的な様式で、ウイルスRNAに結合するとも考えられている。
【0061】
ウイルス性脂質ラフト会合ポリペプチドの好ましい例は、赤血球凝集素ポリペプチドである。本明細書で用いられる「赤血球凝集素ポリペプチド」は、感染される細胞に対するウイルスの結合を媒介するインフルエンザウイルスタンパク質に由来する。赤血球凝集素ポリペプチドはまた、同等の麻疹ウイルスタンパク質にも由来し得る。このタンパク質は、単一の膜貫通ドメインにより、インフルエンザウイルスの表面へとアンカリングすることが見出される抗原性の糖タンパク質である。インフルエンザ赤血球凝集素のうち、H1〜H16と名付けられた、少なくとも16のサブタイプが同定されている。H1、H2、およびH3は、ヒトインフルエンザウイルスにおいて見出される。H5またはH7の赤血球凝集素を有する、高病原性鳥類インフルエンザウイルスがヒトに感染する割合は低いことが判明している。鳥類ウイルス株のH5型赤血球凝集素内における単一のアミノ酸変化がヒト患者において見出されており、これが受容体の特異性を変化させることを可能にし、H5赤血球凝集素が鳥類H5N1ウイルスの受容体特異性を顕著に変化させることによって、これらのウイルスに、ヒト受容体に結合する能力がもたらされると報告されている(109および110)。この知見は、通常ヒトに感染しないH5N1ウイルスが変異し、ヒト細胞に効率的に感染できるようになることを説明する。
【0062】
赤血球凝集素は、ホモ三量体の内在性膜ポリペプチドである。天然では、その膜貫通ドメインが、脂質ラフトドメインと会合し、これにより、gagポリペプチドと会合することが可能となり、VLP内へと組み込まれる。それは、円筒様の形状をしており、約135Åの長さである。HAを構成する3つの同一の単量体は、中央のコイルドコイルと、VLPの表面上に露出されるシアル酸結合部位を含有する球形の頭状部分とを形成する。HAの単量体は、グリコシル化され、2つのより小型のポリペプチド:HA1サブユニットおよびHA2サブユニットへと切断される、単一のポリペプチド前駆体として合成される。HA2サブユニットは、膜へとアンカリングされる、三量体のコイルドコイルを形成し、HA1サブユニットは、球形の頭状部分を形成する。
【0063】
本発明の特定のVLPにおいて脂質ラフト会合ポリペプチドとして用いられる赤血球凝集素ポリペプチドは、最小限において、膜アンカードメインを包含するものとする。赤血球凝集素ポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、H1、H2、H3、H5、H7、およびH9赤血球凝集素に由来することが好ましい。加えて、赤血球凝集素ポリペプチドは、異なるインフルエンザ赤血球凝集素のキメラでもあり得る。赤血球凝集素ポリペプチドは、1または複数のさらなるポリペプチドのコード配列を、赤血球凝集素ポリペプチドのコード配列にスプライシングすることにより作製し得る、天然では脂質ラフトと会合しない、1または複数のさらなる抗原を包含することが好ましい。赤血球凝集素ポリペプチド内へのさらなるポリペプチドの挿入に好ましい部位は、N末端である。
【0064】
本発明の特定のVLPにおいて抗原として用いられる赤血球凝集素ポリペプチドは、最小限において、膜アンカードメイン、また、赤血球凝集素に由来する少なくとも1つのエピトープを包含するものとする。赤血球凝集素ポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、H1、H2、H3、H5、H7、およびH9赤血球凝集素に由来することが好ましい。加えて、赤血球凝集素ポリペプチドは、異なるインフルエンザ赤血球凝集素のキメラでもあり得る。赤血球凝集素ポリペプチドは、場合によって、そのコード配列を赤血球凝集素ポリペプチドのコード配列にスプライシングすることにより作製し得る、1または複数のさらなるポリペプチドを包含し得る。赤血球凝集素ポリペプチド内へのさらなるポリペプチドの挿入に好ましい部位は、N末端である。
【0065】
ウイルス性脂質ラフト会合ポリペプチドの好ましい別の例は、ノイラミニダーゼポリペプチドである。本明細書で用いられる「ノイラミニダーゼポリペプチド」は、糖タンパク質から末端のシアル酸残基を切断することにより、細胞からのインフルエンザウイルスの放出を媒介する、インフルエンザウイルスタンパク質に由来する。ノイラミニダーゼ糖タンパク質は、ウイルス表面上において発現する。ノイラミニダーゼタンパク質は四量体であり、ベータ−ピンホイール構造を有する球形の頭状部分、細いストーク状領域、また、単一の膜貫通ドメインによりウイルス膜内に該タンパク質をアンカリングする小さな疎水性領域からなる共通構造を共有する。シアル酸残基を切断するための活性部位は、すべてのインフルエンザAウイルス内に保存されている、15の荷電アミノ酸により形成される、各サブユニット表面上のポケットを包含する。インフルエンザノイラミニダーゼのうち、N1〜N9と名付けられた、少なくとも9つのサブタイプが同定されている。
【0066】
本発明の特定のVLPにおいて用いられるノイラミニダーゼポリペプチドは、最小限において、膜アンカードメインを包含するものとする。機能的領域に関する最新技術は極めて高度である。例えば、Vargheseら、Nature、303巻、35〜40頁、1983年; Colmanら、Nature、303巻、41〜44頁、1983年; Lentzら、Biochem、26巻、5321〜5385頁、1987年; Websterら、Virol.、135巻、30〜42頁、1984年を参照されたい。ノイラミニダーゼポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、N1ノイラミニダーゼおよびN2ノイラミニダーゼに由来することが好ましい。加えて、ノイラミニダーゼポリペプチドは、異なるインフルエンザノイラミニダーゼのキメラでもあり得る。ノイラミニダーゼポリペプチドは、1または複数のさらなるポリペプチドのコード配列を赤血球凝集素ポリペプチドにスプライシングすることにより作製し得る、天然では脂質ラフトと会合しない1または複数のさらなる抗原を包含することが好ましい。ノイラミニダーゼポリペプチドコード配列内へのさらなるポリペプチドの挿入に好ましい部位は、C末端である。
【0067】
本発明の特定のVLPにおいて抗原として用いられるノイラミニダーゼポリペプチドは、最小限において、膜アンカードメイン、また、少なくとも1つのシアル酸残基切断活性を包含するものとする。機能的領域に関する最新技術は極めて高度である。例えば、Vargheseら、Nature、303巻、35〜40頁、1983年; Colmanら、Nature、303巻、41〜44頁、1983年; Lentzら、Biochem、26巻、5321〜5385頁、1987年; Websterら、Virol.、135巻、30〜42頁、1984年を参照されたい。ノイラミニダーゼポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、N1ノイラミニダーゼおよびN2ノイラミニダーゼに由来することが好ましい。加えて、ノイラミニダーゼポリペプチドは、異なるインフルエンザノイラミニダーゼのキメラでもあり得る。ノイラミニダーゼポリペプチドは、場合によって、そのコード配列をノイラミニダーゼポリペプチドのコード配列にスプライシングすることにより作製し得る、1または複数のさらなるポリペプチドを包含し得る。ノイラミニダーゼポリペプチド内へのさらなるポリペプチドの挿入に好ましい部位は、C末端である。
【0068】
脂質ラフト会合ポリペプチドの好ましい別の例は、ファシクリンI(FasI)と称する、昆虫由来の接着タンパク質である。本明細書で用いられる「ファシクリンIポリペプチド」は、胚の発生に関与する昆虫タンパク質に由来する。この非ウイルス性タンパク質は、昆虫細胞内のバキュロウイルス発現系において発現させることができ、これにより、FasIの脂質ラフト会合がもたらされる(J. Virol.、77巻、6265〜6273頁、2003年)。したがって、ファシクリンIポリペプチドに異種抗原を結合させて、gagと共に共発現させると、VLP内にキメラ分子が組み込まれる。本発明のVLPにおいて用いられるファシクリンIポリペプチドは、最小限において、膜アンカードメインを包含するものとする。
【0069】
脂質ラフト会合ポリペプチドの好ましい別の例は、G糖タンパク質と称する、RSVに由来するウイルス性付着タンパク質である。本明細書で用いられる「Gグリコポリペプチド」は、RSV G糖タンパク質に由来する。近年のデータは、脂質ラフトドメインが、インフルエンザウイルスにとって重要であるのと同様に、RSV粒子の出芽にも重要であることを示している(Virol、327巻、175〜185頁、2004年; Arch. Virol.、149巻、199〜210頁、2004年; Virol.、300巻、244〜254頁、2002年)。RSVに由来するG糖タンパク質は、ウイルス性付着タンパク質であり、およびRSV感染に対する防御抗原として機能する、32.5kdの内在性膜タンパク質である。インフルエンザウイルスに由来する赤血球凝集素と同様、その抗原性は、それに付着した、任意の非脂質ラフト抗原の抗原性を増強し得る。天然において、RSVは、ノイラミニダーゼ活性を有するタンパク質を発現しないので、gagおよびRSV GからなるVLPは、効率的な作製および放出に、NAの存在を必要としない可能性が高い。したがって、gag(アルファレトロウイルスのgagなど)およびGグリコポリペプチドをコードする発現ベクターを開発すれば、結果として、膜内に組み込まれたGグリコポリペプチドを含有するVLPが作製される。非脂質ラフト外来抗原を付着させる形での、Gグリコポリペプチドに対する任意の改変は、結果として、該外来抗原に対して顕著な免疫反応を誘導することが可能なキメラVLPをもたらす。
【0070】
VLPが、その文脈により、エンベロープベースのウイルスに基づかないか、または本明細書で開示される特定のエンベロープベースのウイルスの特定の成分に基づくウイルス様粒子を指す場合を除き、用語「エンベロープウイルスベースのウイルス様粒子」と「VLP」とは、本明細書の全体において互換的に用いられる。
【0071】
(抗原)
本発明の特定の態様は、エンベロープウイルスベースのVLP調製物と会合するさらなる抗原を包含する。このようなさらなる抗原は、同じ組成物中に組み込むことができ、さらにまた、VLPと共有結合させることもでき、非共有結合させることもできる。好ましい実施形態では、gagポリペプチド、インフルエンザM1ポリペプチド、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、および/または他の脂質ラフト会合ポリペプチドが、天然では脂質ラフトと会合しないであろう抗原を含有する、エンベロープウイルスベースのVLPを形成するのに容易に適合可能なプラットフォームである。本節では、開示されるVLPと共に用いるのに好ましい抗原について記載する。
【0072】
(抗原と脂質ラフト会合ポリペプチドとの連結)
天然では脂質ラフトと会合しない抗原、または天然では細胞膜と会合しない抗原を含有するVLPを形成する手段として、gagポリペプチド、インフルエンザM1ポリペプチド、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、および/または別の脂質ラフト会合ポリペプチドと、該抗原との連結を形成することができる。単一の抗原または複数の抗原に脂質ラフト会合ポリペプチドを連結することにより、VLPの免疫原性を増大させるか、各種の病原体に免疫原性を付与するか、または、特定の病原体の各種の株に免疫原性を付与することができる。
【0073】
抗原と脂質ラフト会合ポリペプチドとの間の連結は、該抗原を結果としてVLP内へと組み込むのに十分な任意の種類の連結であり得る。結合は、共有結合、イオン性相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、または抗体−抗原相互作用であり得る。好ましい実施形態では、結合は、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、またはジスルフィド結合などの共有結合である。
【0074】
抗原は、脂質ラフト会合ポリペプチドに対する既存の結合を伴なって組換えにより作製することもでき、単離物質として作製し、次いで、後に、脂質ラフト会合ポリペプチドに連結することもできる。
【0075】
(抗原の種類)
本明細書で用いられる抗原は、免疫反応を誘発することが可能であり、また、天然では脂質ラフトと会合しない任意の物質であり得る。抗原には、タンパク質、ポリペプチド(活性タンパク質、また、タンパク質内における個々のポリペプチドエピトープを含めた)、グリコポリペプチド、リポポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性および非ペプチド性模倣体、ならびに他の分子、低分子、脂質、糖脂質、ならびに炭水化物が含まれるが、これらに限定されない。天然において、抗原が脂質ラフトと直接的または間接的に会合しない場合、脂質ラフト会合ポリペプチドに連結しない限り、それをVLP内へと組み込むことは期待されない。抗原は、疾患または障害に関与する任意の抗原、例えば、微生物抗原(例えば、ウイルス抗原、細菌抗原、真菌抗原、原虫抗原、蠕虫抗原、酵母抗原など)、腫瘍抗原、アレルゲンなどであり得る。
【0076】
(抗原の供給源)
本明細書で記載される抗原は、化学的または酵素的に合成することもでき、組換えにより作製することもでき、天然の供給源から単離することもでき、前出の組合せでもあり得る。抗原は、精製することもでき、部分精製することもでき、粗抽出物でもあり得る。
【0077】
ポリペプチド抗原は、液体クロマトグラフィー(例えば、高速液体クロマトグラフィー、高速タンパク質液体クロマトグラフィーなど)、サイズ除外クロマトグラフィー、ゲル電気泳動(一次元ゲル電気泳動、二次元ゲル電気泳動を含めた)、アフィニティークロマトグラフィー、または他の精製法が含まれるがこれらに限定されない、当技術分野で公知の標準的なタンパク質精製法を用いて、天然の供給源から単離することができる。多くの実施形態では、抗原が、例えば、約50%〜約75%純粋、約75%〜約85%純粋、約85%〜約90%純粋、約90%〜約95%純粋、約95%〜約98%純粋、約98%〜約99%純粋、または99%超純粋の精製抗原である。
【0078】
固相ペプチド合成法を用いることができるが、このような技法は、当業者に公知である。Jones、「The Chemical Synthesis of Peptides」(オックスフォード、Clarendon Press社)(1994年)を参照されたい。一般に、このような方法では、活性化させた単量体のユニットを、伸長するペプチド鎖を結合させた固相へと連鎖的に付加することにより、ペプチドを作製する。
【0079】
ポリペプチドを作製するには、十分に確立された組換えDNA法を、脂質ラフト会合ポリペプチドと同じベクター内において用いることもでき、この場合、例えば、ポリペプチドをコードするヌクレオチド配列を含む発現構築物は、適切な宿主細胞(例えば、インビトロの細胞培養物における単細胞実体として増殖させる真核宿主細胞、例えば、酵母細胞、昆虫細胞、哺乳動物細胞など)または、原核細胞(例えば、インビトロの細胞培養物中)中に導入され、これにより遺伝的に改変された宿主細胞を作製し、適切な培養条件下において、該遺伝的に改変された宿主細胞によりタンパク質を作製する。
【0080】
(ウイルス抗原)
適切なウイルス抗原には、以下の群:Retroviridae科(例えば、HIV−1などのヒト免疫不全ウイルス(また、HTLV−III、LAV、もしくはHTLV−III/LAV、またはHIV−IIIとも称する));また、HIV−LPなど、他の単離物; Picomaviridae科(例えば、ポリオウイルス、A型肝炎ウイルス;エンテロウイルス、ヒトコクサッキーウイルス、ライノウイルス、エコーウイルス); Calciviridae科(例えば、ノーウォークウイルスおよび関連ウイルスを含めた、胃腸炎を引き起こすウイルス株); Togaviridae科(例えば、ウマ脳炎ウイルス、風疹ウイルス); Flaviridae科(例えば、デング熱ウイルス、脳炎ウイルス、黄熱病ウイルス); Coronoviridae科(例えば、コロナウイルス); Rhabdoviradae科(例えば、水疱性口内炎ウイルス、狂犬病ウイルス); Coronaviridae科(例えば、コロナウイルス); Rhabdoviridae科(例えば、水疱性口内炎ウイルス、狂犬病ウイルス); Filoviridae科(例えば、エボラウイルス); Paramyxoviridae科(例えば、パラインフルエンザウイルス、ムンプスウイルス、麻疹ウイルス、RSウイルス); Orthomyxoviridae科(例えば、インフルエンザウイルス); Bungaviridae科(例えば、ハンタンウイルス、ブンガウイルス、フレボウイルス、およびナイロウイルス); Arena viridae科(出血熱ウイルス); Reoviridae科(例えば、レオウイルス、オルビウイルス、およびロタウイルス); Bimaviridae科; Hepadnaviridae科(B型肝炎ウイルス); Parvovirida科(パルボウイルス); Papovavirida(パピローマウイルス、ポリオーマウイルス); Adenoviridae科(大半のアデノウイルス); Herpesviridae科(1型および2型の単純ヘルペスウイルス(HSV)、水痘帯状疱疹ウイルス、サイトメガロウイルス(CMV)、ヘルペスウイルス); Poxyiridae科(痘瘡ウイルス、ワクシニアウイルス、ポックスウイルス);およびIridoviridae科(例えば、アフリカブタコレラウイルス);ならびに未分類のウイルス(例えば、海綿状脳症の病因因子、デルタ肝炎の因子(B型肝炎ウイルスの欠損サテライトウイルスと思われる)、非A型肝炎、非B型肝炎の因子(クラス1=内部感染による;クラス2=非経口感染による(すなわち、C型肝炎));およびアストロウイルス)のうちの1または複数のウイルスと関連する(例えば、これらにより合成される)抗原が含まれる。
【0081】
(ノロウイルス(Norvirus)抗原)
本明細書で開示されるVLPには、ノロウイルス科に由来する各種の抗原が含まれ得ることが好ましい。「ノーウォーク様ウイルス」とも呼ばれるノロウイルスは、Caliciviridaeウイルス科内の4つの属のうちの1つを占める。ノロウイルス属内には、遺伝子群Iおよび遺伝子群IIと名付けられた、2つの主要な遺伝子群が存在する。遺伝子群Iのノロウイルス株には、ノーウォークウイルス、サウサンプトンウイルス、デザートシールドウイルス、およびチバウイルスが含まれる。遺伝子群IIのノロウイルス株には、ヒューストンウイルス、ハワイウイルス、ローズデールウイルス、グリムズビーウイルス、メキシコウイルス、およびスノーマウンテンエージェントが含まれる(Parker, T.D.ら、J. Virol.(2005年)、79巻(12号):7402〜9頁; Hale, A.D.ら、J Clin. Micro.(2000年)、38巻(4号):1656〜1660頁)。ノーウォークウイルス(NV)は、世界中における、大半の急性ウイルス性胃腸炎の流行的発生の一因となる、ヒトカリチウイルス群の原型株である。ノーウォークウイルスのカプシドタンパク質は、2つのドメイン:殻ドメイン(S)および突出部ドメイン(P)を有する。Pドメイン(アミノ酸226〜530;ノーウォーク株の番号付け)は、2つのサブドメインであるP1およびP2に分けられる。P2ドメインは、P1ドメイン内における127アミノ酸の挿入(アミノ酸279〜405)であり、フォールディングされた単量体の最遠位面に位置する。P2ドメインは、ノロウイルス株のうちで最も保存の程度が低いVP1領域であり、P2内の超可変領域は、受容体の結合および免疫反応性において重要な役割を果たすと考えられている。Pドメインが外部に位置することを踏まえるなら、それは、本明細書で開示されるVLPワクチンのための抗原として用いるのに好ましい抗原、またはポリペプチドエピトープの供給源である。P2ドメインは、遺伝子群Iまたは遺伝子群IIのノロウイルス株に好ましい抗原である。広範にわたるノロウイルス株を通じて保存されるP2ドメインのうちのある領域内に存在するエピトープとして近年になって同定されたmAb 61.21エピトープ、ならびにmAb 54.6エピトープ(Lochridge, V.P.ら、J Gen. Virol.(2005年)、86巻:2799〜2806頁)がさらにより好ましい。
【0082】
(インフルエンザ抗原)
本明細書で開示されるVLPは、赤血球凝集素およびノイラミニダーゼ以外に、またはこれらに加えて、インフルエンザに由来する各種の抗原を包含し得る。さらなる好ましいインフルエンザ抗原は、M2ポリペプチドである。インフルエンザウイルスのM2ポリペプチドは、スプライシングイベント後において、RNAセグメント7(マトリックスセグメント)によりコードされる、97アミノ酸の小型のクラスIII内在性膜タンパク質である(80、81)。ウイルス粒子上に存在するM2は極めてわずかであり、感染細胞上においてより豊富に見出すことができる。M2は、ウイルスの侵入に必要な、プロトン選択性イオンチャネルとして用いられる(82、83)。感染中または従来のワクチン接種中の免疫原性は最小限であることから、その保存が説明されるが、代替的なフォーマットで存在する場合は、より免疫原性であり防御性である(84〜86)。これは、インビボにおけるM2モノクローナル抗体の受動伝達により、ウイルスクリアランスが加速化され、結果として防御がもたらされるという観察(87)と符合する。M2の外部ドメインエピトープを、融合タンパク質として、HBVのコア粒子に連結すると、非経口接種および鼻腔内接種のいずれによっても、マウスにおいて防御性であり、3回の繰り返しコピーを該コアタンパク質のN末端へと融合させると、最も免疫原性となる(88〜90)。これは、エピトープ密度が増大すると免疫原性が増大することを示す他のキャリア−ハプテンデータ(91)と符合する。
【0083】
M2ワクチンを鼻腔内送達する場合、良好な防御を達成するにはアジュバントが必要とされ、LTR192G(88、90)およびCTA1−DD(89)により良好な結果が達成されている。ペプチドはまた、KLH、またはN. meningitidesの外膜タンパク質複合体、またはヒトパピローマウイルスのVLPなどのキャリアに化学的に結合体化することもでき、マウスおよび他の動物におけるワクチンとして防御的である(92、93)。
【0084】
M2タンパク質は、高度に保存的ではあるが、配列の相違を全く示さないわけではない。一般的な株であるA/PR/8/34(H1N1)およびA/Aichi/68(H3N2)のM2外部ドメインエピトープは、A/Hong Kong/156/97(H5N1)を除き、今日配列決定されている他のすべてのヒト株と、免疫学的に交差反応性であることが示された(92)。インフルエンザデータベースの配列の検討からも、A/Vietnam/1203/04など、他のより近年の病原性H5N1ヒト単離物のM2配列内において、同様の相違が示されている。この知見は、M2エピトープを組み込む、有効なH5特異的汎流行ワクチンが、ヒトH1単離物およびH3単離物中において現在広まっているM2配列ではなく、病原性の鳥類株に固有のM2配列を反映する必要があることを裏付ける。
【0085】
インフルエンザウイルスに由来するさらなるタンパク質(HA、NA、およびM2以外のタンパク質)も、共発現により、またはさらなる抗原の全部もしくは一部を、gagポリペプチドもしくはHAポリペプチドに連結することにより、VLPワクチン内へと組み込むことができる。これらのさらなる抗原には、PB2、PB1、PA、核タンパク質、マトリックス(M1)、NS1、およびNS2が含まれる。これら後者の抗原は、一般に、中和抗体反応の標的ではなく、T細胞により認識される重要なエピトープを含有し得る。このようなエピトープに対してVLPワクチンにより誘導されるT細胞反応が、防御的免疫性を促進するのに有益であることが判明し得る。
【0086】
(他の病原性抗原)
適切な細菌性抗原には、例えば、病原性Pasteurella属種、Staphylococci属種、およびStreptococcus属種などの病原性グラム陽性菌;また、Neisseria属、Escherichia属、Bordetella属、Campylobacter属、Legionella属、Pseudomonas属、Shigella属、Vibrio属、Yersinia属、Salmonella属、Haemophilus属、Brucella属、Francisella属、およびBacterioides属のグラム陰性菌などのグラム陰性菌を含めた、各種の病原性細菌のうちのいずれかと関連する(例えば、これらにより合成され、また、これらに内因性である)抗原が含まれる。例えば、Schaechter, M、H. Medoff、D. Schlesinger、「Mechanisms of Microbial Disease」、ボルチモア、Williams and Wilkins社(1989年)を参照されたい。
【0087】
感染性の病原性真菌と関連する(例えば、これらにより合成され、また、これらに内因性である)適切な抗原には、Cryptococcus neoformans、Histoplasma capsulatum、Coccidioides immitis、Blastomyces dermatitidis、およびCandida albicans、Candida glabrata、Aspergillus fumigata、Aspergillus flavus、およびSporothrix schenckiiが含まれるがこれらに限定されない感染性真菌と関連する抗原が含まれる。
【0088】
病原性原虫、蠕虫、および他の真核微生物病原体と関連する(例えば、これらにより合成され、また、これらに内因性である)適切な抗原には、Plasmodium falciparum、Plasmodium malariae、Plasmodium ovale、およびPlasmodium vivaxなどのPlasmodium属; Toxoplasma gondii; Trypanosoma brucei、Trypanosoma cruzi; Schistosoma haematobium、Schistosoma mansoni、Schistosoma japonicum; Leishmania donovani; Giardia intestinalis; Cryptosporidium parvum等が含まれるがこれらに限定されない原虫、蠕虫、および他の真核微生物病原体と関連する抗原が含まれる。
【0089】
適切な抗原には、Helicobacter pyloris、Borelia burgdorferi、Legionella pneumophila、Mycobacteria属種(例えば、M. tuberculosis、M. avium、M. intracellulare、M. kansaii、M. gordonae)、Staphylococcus aureus、Neisseria gonorrhoeae、Neisseria meningitidis、Listeria monocytogenes、Chlamydia trachomatis、Streptococcus pyogenes(Streptococcus属のA群)、Streptococcus agalactiae(Streptococcus属のB群)、Streptococcus属(viridans群)、Streptococcus faecalis、Streptococcus bovis、Streptococcus属(嫌気性の種)、Streptococcus pneumoniae、病原性のCampylobacter属種、Enterococcus属種、Haemophilus influenzae、Bacillus anthracis、Corynebacterium diphtheriae、Corynebacterium(corynebacterium)属種、Erysipelothrix rhusiopathiae、Clostridium perfringens、Clostridium tetani、Enterobacter aerogenes、Klebsiella pneumoniae、Pasturella multocida、Bacteroides属種、Fusobacterium nucleatum、Streptobacillus moniliformis、Treponema pallidium、Treponema pertenue、Leptospira属、Rickettsia属、およびActinomyces israeliなどの病原性微生物と関連する(例えば、これらにより合成され、また、これらに内因性である)抗原が含まれる。病原性E. coli株の非限定的な例は、ATCC第31618号、同第23505号、同第43886号、同第43892号、同第35401号、同第43896号、同第33985号、同第31619号、および同第31617号である。
【0090】
細胞内病原体と関連する各種のポリペプチドまたは他の抗原のうちのいずれかを、VLP内に組み込むことができる。細胞内病原体と関連するポリペプチドエピトープおよびペプチドエピトープとは、その断片が、MHCクラスI分子と併せて、CD8+リンパ球の表面上におけるT細胞抗原受容体により認識される、例えば、これにより結合されるように、感染細胞の表面上に提示される、細胞内病原体と関連する(例えば、これによりコードされる)任意のポリペプチドである。細胞内病原体と関連するポリペプチドエピトープおよびペプチドエピトープは当技術分野において公知であり、これらには、ヒト免疫不全ウイルス、例えば、HIV gp120またはその抗原性断片と関連する抗原;サイトメガロウイルス抗原; Mycobacterium属抗原(例えば、Mycobacterium avium、Mycobacterium tuberculosisなど);Pneumocystic carinii(PCP)抗原;41−3、AMA−1、CSP、PFEMP−1、GBP−130、MSP−1、PFS−16、SERPなど、Plasmodium falciparumまたは他の任意のマラリア種と関連する抗原が含まれるがこれらに限定されない、マラリア抗原;真菌抗原;酵母抗原(例えば、Candida属種抗原); Toxoplasma gondii、Toxoplasma encephalitis、または他のToxoplasma属種と関連する抗原が含まれるがこれらに限定されないトキソプラズマ抗原;エプスタイン−バーウイルス(EBV)による抗原; Plasmodium属抗原(例えば、gp190/MSP1など)などが含まれるが、これらに限定されない。
【0091】
好ましいVLPワクチンは、Bacillus anthracisに対するものであり得る。Bacillus anthracisは、好気性または通性嫌気性でグラム陽性の非運動性桿菌であり、幅1.0μm、長さ3.0〜5.0μmである。B. anthracisは、悪条件下でも耐性の高い内性胞子を形成し、これは、感染した動物がかつて死亡した場所の土壌で見出すことができる。本明細書で開示されるVLPワクチン内で用いるのに好ましい抗原は、哺乳動物細胞上の受容体に結合し、B. anthracisが疾患を引き起こす能力にとって極めて重要な83kDaのタンパク質である防御抗原(PA)である。より好ましい抗原は、被験体においてBacillus anthracisに対する防御的免疫性を誘導するのに用い得る、Bacillus anthracis PAのC末端における140アミノ酸の断片である。炭疽菌に対するVLPワクチン内において用いられる他の例示的な抗原は、炭疽菌胞子に由来する抗原(例えば、BclA)、該細菌の栄養期に由来する抗原(例えば、細胞壁抗原、被膜抗原(例えば、ポリ−ガンマ−D−グルタミン酸、またはPGA)、分泌抗原(例えば、防御抗原、致死性因子、または浮腫因子などの外毒素))である。VLPワクチンで用いるのに好ましい別の抗原は、B. anthracisに固有のアントロースを含有する四糖である(Daubenspeck J.M.ら、J. Biol. Chem.(2004年)、279巻:30945頁)。四糖は、脂質ラフト会合ポリペプチドに連結することができ、これにより、VLPワクチンとの抗原の会合が可能となる。
【0092】
(腫瘍関連抗原)
公知の各種腫瘍特異抗原または腫瘍関連抗原(TAA)のうちのいずれかを、VLPに組み込むことができる。全TAAを用い得るが、その必要はない。代わりに、TAAの一部、例えば、エピトープを用いることができる。VLP内で用い得る腫瘍関連抗原(またはそれらのエピトープを含有する断片)には、MAGE−2、MAGE−3、MUC−1、MUC−2、HER−2、高分子量黒色腫関連抗原であるMAA、GD2、癌胎児性抗原(CEA)、TAG−72、卵巣関連抗原であるOV−TL3およびMOV18、TUAN、アルファ−フェトプロテイン(AFP)、OFP、CA−125、CA−50、CA−19−9、腎腫瘍関連抗原であるG250、EGP−40(EpCAMとしても知られる)、S100(悪性黒色腫関連抗原)、p53、およびp21rasが含まれるがこれらに限定されない。前出のうちのいずれかを含めた、任意のTAA(またはそれらのエピトープ)の合成類似体を用いることができる。さらに、1または複数のTAA(またはそれらのエピトープ)の組合せを、組成物中に組み込むことができる。
【0093】
(アレルゲン)
一態様では、VLPワクチンの一部である抗原が、各種のアレルゲンのうちのいずれかであり得る。アレルゲンベースのワクチンを用い、被験体におけるアレルゲンに対する忍容性を誘導することができる。チロシンとの共沈殿(co−precipitation)を伴うアレルゲンワクチンの例は、米国特許第3,792,159号、同第4,070,455号、および同第6,440,426号において見出すことができる。
【0094】
各種のアレルゲンのうちのいずれかをVLP内に組み込むことができる。アレルゲンには、環境空中アレルゲン;ブタクサ/花粉症などの植物の花粉;雑草花粉アレルゲン;芝草花粉アレルゲン;ジョンソングラス;樹木花粉アレルゲン;ライグラス;イエダニアレルゲン(例えば、Der p I、Der f Iなど)などのクモ形類動物アレルゲン;コナダニ(storage mite)アレルゲン;スギ花粉/花粉症;カビ胞子アレルゲン;動物アレルゲン(例えば、イヌ、モルモット、ハムスター、アレチネズミ、ラット、マウスなどによるアレルゲン);食物アレルゲン(例えば、甲殻類;ラッカセイなどの堅果;柑橘類によるアレルゲン);昆虫アレルゲン;毒(膜翅目、スズメバチ(yellow jacket)、ミツバチ、カリバチ、スズメバチ(hornet)、ヒアリ);ゴキブリ、ノミ、蚊などによる他の環境昆虫アレルゲン;連鎖球菌抗原などの細菌アレルゲン; Ascaris属抗原などの寄生虫アレルゲン;ウイルス抗原;真菌胞子;薬物アレルゲン;抗生剤;ペニシリンおよび関連化合物;他の抗生剤;ホルモン(インスリン)、酵素(ストレプトキナーゼ)などの全タンパク質;不完全抗原またはハプテンとして作用することが可能なすべての薬物およびそれらの代謝物;ハプテンとして作用し、アレルゲンとして機能することが可能な、産業用化学物質および代謝物(例えば、酸無水物(トリメリト酸無水物など)およびイソシアン酸塩(ジイソシアン酸トルエンなど));小麦粉などの職業アレルゲン(例えば、パン屋喘息(Baker’s asthma)を引き起こすアレルゲン)、トウゴマ、コーヒー豆、また、上記で記載した産業用化学物質;ノミアレルゲン;ヒト以外の動物におけるヒトタンパク質が含まれるが、これらに限定されない。
【0095】
アレルゲンには、細胞、細胞抽出物、タンパク質、ポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性および非ペプチド性模倣体、ならびに他の分子、低分子、脂質、糖脂質、ならびに炭水化物が含まれるが、これらに限定されない。
【0096】
特定の天然の動物アレルゲンおよび植物アレルゲンの例には、以下の属:Canine属(Canis familiaris); Dermatophagoides属(例えば、Dermatophagoides farinae); Felis属(Felis domesticus); Ambrosia属(Ambrosia artemiisfolia); Lolium属(例えば、Lolium perenneまたはLolium multiflorum); Cryptomeria属(Cryptomeria japonica); Altemaria属(Altemaria altemata); Alder属; Alnus属(Alnus gultinoas); Betula属(Betula verrucosa); Quercus属(Quercus alba); Olea属(Olea europa); Artemisia属(Artemisia vulgaris); Plantago属(例えば、Plantago lanceolata); Parietaria属(例えば、Parietaria officinalisまたはParietaria judaica); Blattella属(例えば、Blattella germanica); Apis属(例えば、Apis multiflorum); Cupressus属(例えば、Cupressus sempervirens、Cupressus arizonica、およびCupressus macrocarpa); Juniperus属(例えば、Juniperus sabinoides、Juniperus virginiana、Juniperus communis、およびJuniperus ashei); Thuya属(例えば、Thuya orientalis); Chamaecyparis属(例えば、Chamaecyparis obtusa); Periplaneta属(例えば、Periplaneta americana); Agropyron属(例えば、Agropyron repens); Secale属(例えば、Secale cereale); Triticum属(例えば、Triticum aestivum); Dactylis属(例えば、Dactylis glomerata); Festuca属(例えば、Festuca elatior); Poa属(例えば、PoapratensisまたはPoa compressa); Avena属(例えば、Avena sativa); Holcus属(例えば、Holcus lanatus); Anthoxanthum属(例えば、Anthoxanthum odoratum); Arrhenatherum属(例えば、Arrhenatherun elatius); Agrostis属(例えば、Agrostis alba); Phleum属(例えば、Phleum pratense); Phalaris属(例えば、Phalaris arundinacea); Paspalum属(例えば、Paspalum notatum); Sorghum属(例えば、Sorghum halepensis); ならびにBromus属(例えば、Bromus inermis)に特異的なタンパク質が含まれるが、これらに限定されない。
【0097】
(エンベロープウイルスベースのVLPを作製するのに好ましい方法)
エンベロープウイルスベースのVLPは、当業者に適用可能な任意の方法により作製することができる。エンベロープウイルスベースのVLPは、VLPを形成する一因となる1または複数のポリペプチドを包含することが典型的である。加えて、エンベロープウイルスベースのVLPは、膜(脂質ラフトを含めた)会合ポリペプチドなど、1または複数のさらなるポリペプチドを包含することにより、(VLPを形成する一因となる1または複数のポリペプチドの一部として、天然において、または人工的に存在する抗原以外のさらなる)抗原をもたらすことができる。好ましい実施形態では、ポリペプチドを、任意の適用可能なタンパク質発現系、好ましくは、哺乳動物細胞による発現系および昆虫細胞による発現系など、細胞膜内に脂質ラフトドメインを包含する細胞ベースの系において共発現させることができる。
【0098】
組換えによるVLP用ポリペプチドの発現は、該ポリペプチドのうちの1または複数をコードするポリヌクレオチドを含有する発現ベクターを伴う。ポリペプチドのうちの1または複数をコードするポリヌクレオチドが得られたら、当技術分野で周知の技法を用いる組換えDNA法により、該ポリペプチドを作製するためのベクターを作製することができる。このようにして、本明細書では、VLPポリペプチドをコードするヌクレオチド配列のうちのいずれかを含有するポリヌクレオチドを発現することによりタンパク質を調製する方法が記載される。当業者に周知の方法を用いて、VLPポリペプチドをコードする配列と、適切な転写制御シグナルおよび翻訳制御シグナルとを含有する発現ベクターを構築することができる。例えば、これらの方法には、インビトロにおける組換えDNA法、合成法、また、インビボにおける遺伝子組換えが含まれる。したがって、本発明は、すべてが1または複数のプロモーターに作動可能に連結した、gagポリペプチドと、抗原に連結した脂質ラフト会合ポリペプチドとをコードするヌクレオチド配列を含む複製可能なベクターを提供する。
【0099】
従来の技法により、発現ベクターを宿主細胞へと導入することができ、次いで、従来の技法により、トランスフェクトされた細胞を培養して、VLPポリペプチド(複数可)を作製することができる。したがって、本発明は、異種プロモーターに作動可能に連結したVLPポリペプチドのうちの1または複数をコードするポリヌクレオチドを含有する宿主細胞を包含する。VLPを作製するのに好ましい実施形態では、以下で詳述する通り、gagポリペプチドと、抗原に連結した脂質ラフト会合ポリペプチドとの両方をコードするベクターを、宿主細胞内で共発現させて、VLPを作製することができる。
【0100】
各種の宿主発現ベクター系を用いて、VLPポリペプチドを発現させることができる。このような宿主発現系は、好ましくは共発現によりVLPを生成させる目的で、VLPポリペプチドを作製し得るビヒクルを表す。広範囲にわたる宿主を、適切な発現ベクター構築物において用いることができ、脂質ラフトベースのアセンブリーに依拠する場合、好ましい宿主発現系は、VLPのアセンブリーに適する脂質ラフトを有する宿主である。これらには、VLPポリペプチドコード配列を含有する組換えバクテリオファージDNA発現ベクター、プラスミドDNA発現ベクター、もしくはコスミドDNA発現ベクターにより形質転換した細菌(例えば、E. coli、B. subtilis);VLPポリペプチドコード配列を含有する組換え酵母発現ベクターにより形質転換した酵母(例えば、Saccharomyces属、Pichia属);VLPポリペプチドのコード配列を含有する組換えウイルス発現ベクター(例えば、バキュロウイルス)を感染させた昆虫細胞系;VLPポリペプチドのコード配列を含有する組換えウイルス発現ベクター(例えば、カリフラワーモザイクウイルス、CaMV;タバコモザイクウイルス、TMV)を感染させた植物細胞系、もしくはVLPポリペプチドのコード配列を含有する組換えプラスミド発現ベクター(例えば、Tiプラスミド)により形質転換した植物細胞系;または、哺乳動物細胞のゲノムに由来するプロモーター(例えば、メタロチオネインプロモーター)、もしくは哺乳動物ウイルスに由来するプロモーター(例えば、アデノウイルス後期プロモーター;ワクシニアウイルス7.5Kプロモーター)を含有する組換え発現構築物を保有する、哺乳動物細胞系(例えば、COS細胞、CHO細胞、BHK細胞、293細胞、3T3細胞)などの微生物が含まれるがこれらに限定されない。それらのいずれもがVLPのアセンブリーに適する脂質ラフトを有するため、VLPポリペプチドを発現させるには、哺乳動物細胞を用いることが好ましく、また、昆虫細胞を用いることがより好ましい。例えば、ヒトサイトメガロウイルスに由来する主要前初期(intermediate early)遺伝子プロモーターエレメントなどのベクターと組合わされる、MRC−5細胞、Vero細胞、PER.C6(TM)細胞、チャイニーズハムスター卵巣細胞(CHO)、およびHEK293細胞などの哺乳動物細胞は、VLPポリペプチドの有効な発現系である(Foeckingら、Gene、45巻:101頁(1986年); Cockettら、Bio/Technology、8巻:2頁(1990年))。
【0101】
昆虫系では、Autographa californica核多角体病ウイルス(AcNPV)を、外来遺伝子を発現するためのベクターとして用いることができる。該ウイルスは、Spodoptera frugiperda細胞内で増殖する。VLPポリペプチドのコード配列(複数可)は、ウイルスの非必須領域(例えば、ポリヘドリン遺伝子領域)内へと個別にクローニングすることができ、また、AcNPVプロモーター(例えば、ポリヘドリンプロモーター)の制御下に置くことができる。
【0102】
哺乳動物の宿主細胞では、多くのウイルスベースの発現系を用いることができる。発現ベクターとしてアデノウイルスを用いる場合は、対象のVLPポリペプチド配列(複数可)を、アデノウイルス転写/翻訳制御複合体、例えば、後期プロモーターおよび三連リーダー(tripartite leader)配列にライゲーションすることができる。次いで、インビトロまたはインビボにおける組換えにより、このキメラ遺伝子を、アデノウイルスゲノム内に挿入することができる。ウイルスゲノムの非必須領域(例えば、E1領域またはE3領域)内に挿入する結果、感染宿主内において生存可能であり、また、VLPポリペプチド(複数可)を発現することが可能な組換えウイルスがもたらされる(例えば、LoganおよびShenk、Proc. Natl. Acad. Sci. USA、81巻:355〜359頁(1984年)を参照されたい)。挿入されたVLPポリペプチドのコード配列(複数可)を効率的に翻訳するには、特定の開始シグナルもまた必要とされ得る。これらのシグナルには、ATG開始コドン、および隣接配列が含まれる。さらに、全挿入配列の翻訳を確保するには、該開始コドンが、所望されるコード配列のリーディングフレームと同フレームでなければならない。これらの外因性翻訳制御シグナルおよび開始コドンは、由来が多様であることが可能であり、天然および合成のいずれもが可能である。発現の効率は、適切な転写エンハンサーエレメント、転写ターミネーターなどを組み込むことにより、増強することができる(Bittnerら、Methods in Enzymol.、153巻:51〜544頁(1987年)を参照されたい)。1つの例は、Stratagene社製のAdEASY−XL(TM)システムなど、アデノウイルスベースのベクター系において用いられる、ヒトCMV前初期プロモーターである。
【0103】
加えて、挿入した配列の発現を調節するか、または該遺伝子産物を、所望される特定の形で修飾およびプロセシングする宿主細胞株も選択することができる。タンパク質産物に対するこのような修飾(例えば、グリコシル化)およびプロセシング(例えば、切断または膜への輸送)は、VLPを作製するのに、またはVLPポリペプチド、もしくはアジュバントなどのさらなるポリペプチド、もしくはさらなる抗原が機能するのに重要であり得る。種々の宿主細胞は、タンパク質および遺伝子産物の翻訳後プロセシングおよび修飾について特徴的で特異的な機構を有する。発現する外来タンパク質の適正な修飾およびプロセシングを確保するために、適切な細胞株または宿主系を選択することができる。この目的で、一次転写物を適正にプロセシングし、遺伝子産物を適正にグリコシル化およびリン酸化するための細胞機構を保有する真核宿主細胞を用いることができる。
【0104】
宿主細胞には、gagポリペプチドをコードする第1のベクターと、ウイルス性膜抗原、または抗原に連結した脂質ラフト会合ポリペプチドをコードする第2のベクターという、本発明の2つの発現ベクターを共トランスフェクト(co−transfect)することができる。2つのベクターは、各VLPポリペプチドの同等の発現を可能とする、同一の選択マーカーを含有し得る。代替的に、gagポリペプチドと、抗原に連結した脂質ラフト会合ポリペプチドとの両方をコードし、また、これらの両方を発現することが可能な単一のベクターを用いることもできる。
【0105】
宿主細胞によりVLPが生成されたら、ポリペプチドの精製についての当技術分野で公知の任意の方法により、例えば、クロマトグラフィー(例えば、イオン交換クロマトグラフィー、特に、該ポリペプチドに付加した任意のアフィニティー精製タグによるアフィニティークロマトグラフィー、およびサイズ除外クロマトグラフィー)、遠心分離、溶解度差(differential solubility)により、またはタンパク質もしくは他の高分子を精製するための他の任意の標準的な技法によりこれを精製することができる。加えて、VLPポリペプチドを、本明細書で記載されるか、またはこれ以外に当技術分野で公知の異種ポリペプチド配列に融合させ、VLPの精製を容易にすることもできる。精製後、VLPポリペプチドに対する共有結合を介して、または他の非共有結合機構により、さらなる抗原またはアジュバントなどのさらなるエレメントを、VLPへと物理的に連結することができる。哺乳動物細胞および昆虫細胞など、脂質ラフトドメインを有する宿主細胞内においてVLPポリペプチドを共発現させる好ましい実施形態では、VLPが自己アセンブルして放出され、これにより、上記の方法のうちのいずれかによるVLPの精製が可能となる。VLPの好ましい実施形態には、例えば、同種ウイルスのタンパク質から操作されたVLP、例えば、インフルエンザウイルスに由来するM1、HA、また、場合によってNAから構築されたVLP、また、異種ウイルスのタンパク質から操作されたVLP、例えば、MLVもしくはHIVまたは他のレトロウイルスに由来するGagタンパク質を、異なるウイルスに由来する抗原、例えば、インフルエンザHAおよびNAと共に操作することにより形成したVLPが含まれる。
【0106】
(GagベースのVLPを作製する好ましい方法)
当業者に適用可能な任意の方法により、VLPを容易にアッセンブルすることができ、この結果、gagポリペプチドと、天然では脂質ラフトと会合しない抗原に連結した脂質ラフト会合ポリペプチドとを包含するVLPがアッセンブルされることが好ましい。好ましい実施形態では、任意の適用可能なタンパク質発現系、好ましくは、哺乳動物細胞による発現系および昆虫細胞による発現系など、脂質内に脂質ラフトドメインを包含する細胞ベースの発現系内において、ポリペプチドを共発現させることができる。
【0107】
gagポリペプチドを用いて形成されるVLP発現の例が数多く公表されており、これにより、VLPを作製するのに適用可能な発現系の範囲が示されている。複数のレトロウイルスによる研究は、他のウイルス成分の不在下で発現したGagポリペプチドが、細胞表面におけるVLPの形成および出芽に十分であることを裏付けている(WillsおよびCraven、AIDS、5巻、639〜654頁、1991年; Zhouら、3. Virol.、68巻、2556〜2569頁、1994年; Morikawaら、Virology、183巻、288〜297頁、1991年; Royerら、Virology、184巻、417〜422頁、1991年; Gheysenら、Cell、59巻、103〜112頁、1989年; Hughesら、Virology、193巻、242〜255頁、1993年; Yamshchikovら、Virology、214巻、50〜58頁、1995年)。バキュロウイルスベクターを用いる昆虫細胞内において、Gag前駆体が発現するとVLPが形成されることについては、複数の研究グループにより裏付けられている(Delchambreら、EMBO J.、8巻、2653〜2660頁、1989年; Luoら、Virology、179巻、874〜880頁、1990年; Royer ら、Virology、184巻、417〜422頁、1991年; Morikawa ら、Virology、183巻、288〜297頁、1991年; Zhouら、J. Virol.、68巻、2556〜2569頁、1994年; Gheysenら、Cell、59巻、103〜112頁、1989年; Hughesら、Virology、193巻、242〜255頁、1993年; Yamshchikovら、Virology、214巻、50〜58頁、1995年)。これらのVLPは、未成熟レンチウイルス粒子に類似しており、効率的にアッセンブルされて昆虫細胞の細胞膜から出芽により放出される。
【0108】
Gag前駆体のアミノ末端領域は、ウイルスアセンブリーに必要とされる、細胞表面への輸送および膜結合のためのターゲッティングシグナルであることが報告されている(Yuら、J. Virol.、66巻、4966〜4971頁、1992年; an, Xら、J. Virol.、67巻、6387〜6394頁、1993年; Zhouら、J. Virol.、68巻、2556〜2569頁、1994年; LeeおよびLinial、J. Virol.、68巻、6644〜6654頁、1994年; Dorfmanら、J. Virol.、68巻、1689〜1696頁、1994年; Fackeら、J. Virol.、67巻、4972〜4980頁、1993年)。ワクシニアウイルスによる発現系を用いて、Gag構造タンパク質、ならびにEnv糖タンパク質であるgp120およびgp41を含有する、組換えHIVベースのVLPのアセンブリーが報告されている(Haffarら、J. Virol.、66巻、4279〜4287頁、1992年)。
【0109】
(エンベロープウイルスベースのVLP調製物における感染因子を不活化するのに好ましい方法)
電子放射は、エンベロープウイルスベースのVLPの免疫原性を実質的に低下させることなく、感染因子を不活化することが可能なので、不活化の好ましい方法は、電子放射を介する方法である。電磁放射で好ましい3つの方式のすべて(すなわち、光反応性化合物を伴うUV照射、UV照射単独、およびガンマ照射)には、血液、食品、ワクチンなど、多種多様な試料における病原体を不活化するのに用いられてきた長い歴史があるため、不活化用電磁放射を適用するための多種多様な装置が市販されており、これらは、本明細書で開示される方法を実施するのにほとんど〜まったく改変せずに用いることができる。さらに、当技術分野では、波長および線量の最適化がルーチン化しており、したがって、当業者の能力範囲内で容易に実施される。
【0110】
(光反応性化合物を伴うUV照射)
電磁放射による例示的な不活化法は、光反応性化合物、好ましくは、感染因子中のポリヌクレオチドと反応する光反応性化合物の存在下における、UV−A照射などの紫外線照射の組合せである。
【0111】
好ましい光反応性化合物には、アクチノマイシン、アントラサイクリノン、アントラマイシン、ベンゾジピロン、フルオレン、フルオレノン、フロクマリン、イソアロキサジン、マイトマイシン、monostral fast blue、norphillin A、フェナントリジン、フェナザチオニウム塩、フェナジン、フェノチアジン、フェニルアジド、キノリン、およびチオキサンテノン(thiaxanthenones)が含まれる。好ましい種は、2つの主要なカテゴリーのうちの1つに属するフロクマリンである。第1のカテゴリーは、ソラーレン[7H−フロ(3,2−g)−(1)−ベンゾピラン−7−オン、または6−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、直鎖型であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の6位に結合している。第2のカテゴリーは、イソソラーレン[2H−フロ(2,3−h)−(1)−ベンゾピラン−2−オン、または4−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、角度型(angular)であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の8位に結合している。3、4、5、8、4’、または5’位において直鎖型フロクマリンを置換することによりソラーレン誘導体を作製し得る一方、3、4、5、6、4’、または5位において角度型フロクマリンを置換することによりイソソラーレン誘導体を作製し得る。ソラーレンは、二本鎖核酸の塩基対間にインターカレートすることができ、長波長の紫外光(UVA)を吸収すると、ピリミジン塩基に対する共有結合付加体を形成する。例えば、G. D. Ciminoら、Ann. Rev. Biochem.、54巻:1151頁(1985年); Hearstら、Quart. Rev. Biophys.、17巻:1頁(1984頁)を参照されたい。
【0112】
好ましいUV(または場合によって、可視光)放射の波長は、適切な反応および/または光付加体が生成される波長に依存し、これは、光反応性化学物質の化学反応に依存する。例示を目的として述べると、多くのソラーレンには、320〜380nmの波長にあるUV放射が最も有効であり、330〜360nmが最大の有効性を示す。同様のUV−A波長はまた、419nmなどの可視光と組み合わせても用い得る光反応性化合物であるリボフラビンと合わせても、病原体を不活化するのに極めて有効である。
【0113】
(UV照射単独)
光反応性化合物の存在下におけるUV照射に加え、感染因子は、UV照射単独でも不活化することができる。好ましい実施形態では、放射は、波長が、約180〜320nm、または約225〜290nm、または約254nm(すなわち、ポリヌクレオチドの高い吸光度ピークを有し、タンパク質吸収は低いスペクトル領域)であるUV−C放射である。UV−C放射は、エンベロープを形成する脂質二重層、また、エンベロープ内のタンパク質など、本明細書で開示されるエンベロープウイルスベースのVLPの成分に対して、安定性および免疫原性の両面で有害性が低い一方で、感染因子を不活化するのに十分なエネルギーを保持するので好ましい。しかし、例えば、UV−AおよびUV−Bなど、他の種類のUV放射もまた用いることができる。
【0114】
(ガンマ照射)
本明細書で開示される方法を実施して組成物を作製するには、ガンマ照射(すなわち、イオン化放射)もまた用いることができる。この好ましい実施形態では、10〜60kGyのガンマ照射線量が、病原体の不活化に有効である。ガンマ照射は、感染因子のゲノムをコードするポリヌクレオチド内において鎖の切断を導入することにより、感染因子を直接的に不活化することもでき、または、該ポリヌクレオチドを攻撃するフリーラジカルを生成することにより、感染因子を間接的に不活化することもできる。ガンマ照射と共にフリーラジカルスカベンジャー(free radical scavenger)および低温を用い、ラジカルを介してエンベロープVLPの脂質成分およびタンパク質成分にもたらされる損傷を抑制することができる。
【0115】
(エンベロープウイルスベースのVLPを用いる好ましい方法)
(処方物)
本明細書で記載されるエンベロープウイルスベースのVLPの好ましい使用は、ワクチン調製物としての使用である。典型的に、このようなワクチンは、液体溶液または懸濁物の注射剤として調製されるが、注射前に液体中に溶解させるか、または懸濁させるのに適する固体形態もまた調製することができる。このような調製物はまた、乳化させることもでき、乾燥粉末として作製することもできる。免疫原性の有効成分は、薬学的に許容され、また、有効成分に適合する、賦形剤と混合することが多い。適切な賦形剤は、例えば、水、生理食塩液、デキストロース、グリセロール、エタノールなどであり、また、これらの組合せである。加えて、所望の場合、ワクチンは、保湿剤もしくは乳化剤、pH緩衝剤、または該ワクチンの有効性を増強するアジュバントなどの補助物質を含有し得る。
【0116】
ワクチンは、注射、例えば、皮下(subcutaneously)注射、皮内注射、皮下(subdermally)注射、または筋肉内注射を介する、従来の非経口投与が可能である。他の投与方式に適する、さらなる処方物には坐剤が含まれ、場合によっては、経口処方物、鼻腔内処方物、口腔内処方物、舌下処方物、腹腔内処方物、膣内処方物、経肛門処方物、および頭蓋内処方物も含まれる。坐剤の場合、従来の結合剤およびキャリアには、例えば、ポリアルキレングリコール(polyalkalene glycol)またはトリグリセリドが含まれる場合があり、このような坐剤は、0.5%〜10%、好ましくは1〜2%の範囲で有効成分を含有する混合物から形成することができる。特定の実施形態では、脂肪酸グリセリドまたはココアバターの混合物などの低温溶融ワックスをまず溶融させ、本明細書で記載されるエンベロープウイルスベースのVLPを、例えば、撹拌することにより、均一に分散させる。次いで、溶融した均一の混合物を、好都合なサイズの型に流し込み、冷却して固形化させる。
【0117】
鼻腔内送達に適する処方物には、液体および乾燥粉末が含まれる。処方物は、例えば、医薬グレードのマンニトール、ラクトース、スクロース、トレハロース、およびキトサンなど、通常用いられる賦形剤を包含する。液体処方物または粉末処方物において、キトサンなどの粘膜付着剤を用いて、粘膜絨毛による、鼻腔内投与された処方物のクリアランスを遅延させることができる。マンニトールおよびスクロースなどの糖を、液体処方物では安定化剤として用いることができ、また、乾燥粉末処方物では安定化剤およびバルキング剤として用いることができる。加えて、モノホスホリルリピドA(MPL)などのアジュバントを、液体処方物および乾燥粉末処方物の両方において、免疫刺激性アジュバントとして用いることもできる。
【0118】
経口送達に適する処方物には、液体、固体、半固体、ゲル、錠剤、カプセル、ロゼンジなどが含まれる。経口送達に適する処方物には、錠剤、ロゼンジ、カプセル、ゲル、液体、食品、飲料、栄養補助食品などが含まれる。処方物は、例えば、医薬グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなど、通常用いられる賦形剤を包含する。他のエンベロープウイルスベースのVLPワクチン組成物は、溶液、懸濁物、丸薬、持続放出処方物、または粉末の形態を取る場合があり、10〜95%、好ましくは25〜70%の有効成分を含有する場合がある。経口処方物の場合、コレラ毒素が興味深い処方物パートナー(また、可能な結合パートナー)である。
【0119】
膣内投与用に処方する場合、エンベロープウイルスベースのVLPワクチンは、ペッサリー、タンポン、クリーム、ジェル、ペースト、フォーム、またはスプレーの形態であり得る。前出の処方物のうちのいずれもが、エンベロープウイルスベースのVLPに加えて、キャリアなど、当技術分野で適切であることが公知の薬剤を含有し得る。
【0120】
一部の実施形態では、エンベロープウイルスベースのVLPワクチンを、全身送達用に処方する場合もあり、局所送達用に処方する場合もある。このような処方物は、当技術分野において周知である。非経口ビヒクルには、塩化ナトリウム溶液、リンゲルデキストロース液、デキストロースおよび塩化ナトリウムによる溶液、乳酸加リンゲル液、または固定油が含まれる。静脈内ビヒクルには、流体および栄養物質補給剤、電解質補給剤(リンゲルデキストロース液に基づく補給剤など)などが含まれる。全身投与経路および局所投与経路には、例えば、皮内経路、局所適用経路、静脈内経路、筋肉内経路などが含まれる。
【0121】
エンベロープウイルスベースのVLPは、中性処方物または塩ベースの処方物を含めたワクチンへと処方することができる。薬学的に許容される塩には、酸添加塩(ペプチドの遊離アミノ基と共に形成され、また、例えば、塩酸もしくはリン酸などの無機酸、または酢酸、シュウ酸、酒石酸、マンデル酸などの有機酸と共に形成される)が含まれる。遊離カルボキシル基と共に形成される塩はまた、例えば、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、または水酸化鉄などの無機塩基、また、イソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、プロカインなどの有機塩基に由来し得る。
【0122】
ワクチンは、投薬処方物に適合した形で、また、治療的に有効であり免疫原性となる量で投与することができる。投与量は、例えば、個体の免疫系が免疫反応を起こす能力、また、所望される防御の程度を含め、処置される被験体に依存する。適切な用量範囲は、1回のワクチン接種当たりの有効成分が数百マイクログラムのオーダーであり、好ましい範囲は、約0.5μg〜1000μgの範囲、好ましくは1μg〜500μgの範囲、またとりわけ、約10μg〜100μgの範囲など、約0.1μg〜2000μgの範囲である(1〜10mgの範囲にある高量も意図される)ことが好ましい。初回投与および追加投与に適するレジメンもまた可変的であるが、初回投与にその後の接種または他の投与が続く形が典型的である。
【0123】
適用方式は多様に変化させることができる。従来のワクチン投与方法のうちのいずれもが適用可能である。これらには、固体の生理学的に許容されるベース上における経口適用、または注射など、生理学的に許容される分散剤による非経口適用が含まれる。ワクチンの用量は、投与経路に依存し、ワクチン接種されるヒトの年齢、および抗原処方物により変化する。
【0124】
ワクチン処方物の一部は、それだけでワクチンとして十分に免疫原性であるが、他の処方物では、該ワクチンがアジュバント物質をさらに包含すると、免疫反応が増強される場合もある。
【0125】
とりわけ、鼻腔内ベース、経口ベース、または肺内ベースの送達処方物の場合、送達および免疫原性を改善するには、粘膜付着を改善する送達剤もまた用いることができる。このような化合物の1つである、キチンのN−脱アセチル化形態であるキトサンは、多くの医薬処方物で用いられる(32)。粘膜絨毛によるクリアランスを遅延させ、粘膜による抗原の取込みおよびプロセシングのためのより多くの時間を可能にする能力のために、キトサンは、鼻腔内におけるワクチン送達にとって魅力的な粘膜付着剤である(33、34)。加えて、それは、接着結合を一過性に開口させ、これにより、NALTへの抗原の経上皮輸送が増強され得る。近年のヒト試験では、キトサンを伴うが、さらなるアジュバントは伴わない、三価の不活化インフルエンザワクチンを鼻腔内投与したところ、筋肉内接種後に得られたセロコンバージョンおよびHI力価をごくわずかに下回るセロコンバージョンおよびHI力価がもたらされた(33)。
【0126】
キトサンはまた、遺伝的操作により解毒したE. coli易熱性エンテロトキシンの変異体であるLTK63など、鼻腔内において良好に機能するアジュバントと共に処方することもできる。これは、キトサンにより付与される送達および付着の利益の上に、免疫刺激効果を付加し、その結果、粘膜反応および全身反応の増強をもたらす(35)。
【0127】
最後に、キトサン処方物はまた、ワクチンの安定性を改善し、粘膜絨毛によるクリアランスにおいて液体処方物を上回る遅延を結果としてもたらすことが示されている乾燥粉末フォーマットでも調製することができることに注意すべきである(42)。これは、キトサンと共に処方された、鼻腔内用乾燥粉末によるジフテリア毒素ワクチンを伴う、近年のヒト臨床試験において見られ、ここで、鼻腔内経路は、従来の筋肉内経路と同程度に有効であり、分泌性IgA反応の利益が付加された(43)。該ワクチンはまた、忍容性も極めて良好であった。キトサンおよびMPLを含有する炭疽菌用の鼻腔内乾燥粉末ワクチンは、ウサギにおいて、筋肉内接種より強力な反応を誘導し、また、エアゾールによる胞子チャレンジ(challenge)に対しても防御的であった(44)。
【0128】
鼻腔内ワクチンは、下気道に対してより良好に作用する非経口投与ワクチンに対して、上気道および下気道に作用し得るので、好ましい処方物を表す。これは、アレルゲンベースのワクチンには忍容性を誘導し、また、病原体ベースのワクチンには免疫性を誘導する点で有益であり得る。
【0129】
鼻腔内ワクチンは、上気道および下気道の両方において防御をもたらすのに加え、針による接種の煩雑さを回避し、また、粒子抗原および/または可溶性抗原が、鼻咽頭関連リンパ組織(NALT)と相互作用することを介して、粘膜および全身における体液性反応および細胞性反応の両方を誘導する手段をもたらす(16〜19)。歴史的に、鼻腔内経路は、非経口接種ほど有効ではなかったが、新規の送達処方物であるエンベロープウイルスベースのVLPおよびアジュバントを用いることにより、この枠組みが変わりつつある。実際、鼻腔粘膜内にはシアル酸を含有する受容体が豊富であるため、鼻腔内送達には、機能性の赤血球凝集素ポリペプチドを含有するエンベロープウイルスベースのVLPがとりわけよく適する場合があり、その結果、HA抗原の結合が増強され、粘膜絨毛によるクリアランスが低減される可能性がある。
【0130】
(アジュバント)
ワクチンに対するアジュバント効果を達成する各種の方法が公知であり、本明細書で開示されるエンベロープウイルスベースのVLPと共に用いることができる。一般的な原理および方法は、「The Theory and Practical Application of Adjuvants」、1995年、Duncan E. S. Stewart−Tull(編)、John Wiley & Sons社、ISBN 0−471−95170−6において詳述されており、また、「Vaccines: New Generation Immunological Adjuvants」、1995年、Gregoriadis Gら(編)、ニューヨーク、Plenum Press社、ISBN 0−306−45283−9においても詳述されている。
【0131】
一部の実施形態では、エンベロープウイルスベースのVLPワクチンが、重量ベースの比率約10:1〜約1010:1のエンベロープウイルスベースのVLP:アジュバント、例えば、約10:1〜約100:1、約100:1〜約103:1、約103:1〜約104:1、約104:1〜約105:1、約105:1〜約106:1、約106:1〜約107:1、約107:1〜約108:1、約108:1〜約109:1、または約109:1〜約1010:1のエンベロープウイルスベースのVLP:アジュバントで、少なくとも1つのアジュバントと混合したエンベロープウイルスベースのVLPを包含する。当業者は、アジュバントに関する情報、また、最適の比率を決定する日常的実験により、適切な比率を容易に決定することができる。
【0132】
アジュバントの好ましい例は、エンベロープウイルスベースのVLPのポリペプチド成分と共発現させるか、または該ポリペプチド成分と融合させて、キメラポリペプチドを作製することにより、本明細書で記載されるエンベロープウイルスベースのVLPに容易に添加し得るポリペプチドアジュバントである。鞭毛の主要なタンパク質構成要素である細菌性フラジェリンは、それが、toll様受容体であるTLR5を介する先天性免疫系により認識されるために、アジュバントタンパク質としてますます注目を集めている、好ましいアジュバントである(65)。TLR5を介するフラジェリンシグナル伝達は、DCの成熟化および移動の他、マクロファージ、好中球、および腸上皮細胞の活性化も誘導することにより、先天性免疫機能および後天性免疫機能の両方に影響を及ぼし、その結果、炎症促進(proinflammatory)メディエーターを生成させる(66〜72)。
【0133】
TLR5は、フラジェリン単量体内における保存的構造(これはフラジェリン単量体タンパク質に固有であり、また、鞭毛機能に必要とされる)を認識し、このことは免疫学的圧力に応答したその変異を妨げる(73)。該受容体は、100fMの濃度に対して感受性であるが、完全な線維は認識しない。鞭毛が単量体へと解体されることが、結合および刺激に必要とされる。
【0134】
アジュバントとしてのフラジェリンは、非経口投与または鼻腔内投与された異種抗原に対する防御反応を誘導する強力な活性を有し(66、74〜77)、また、DNAワクチンに対するアジュバント効果もまた報告されている(78)。マウスまたはサルにおいてフラジェリンを用いると、Th2バイアス(これは、インフルエンザなどの呼吸器ウイルスには適切である)は観察されるが、IgE誘導の証拠は観察されていない。加えて、サルにおける鼻腔内投与または全身投与後における、局所炎症反応も全身炎症反応も報告されていない(74)。TLR5を介して、MyD88依存的な形でなされるフラジェリンシグナル伝達、また、TLRを介する、他のすべてのMyD88依存的シグナルは、Th1バイアスを結果としてもたらすことが示されている(67、79)ため、フラジェリンを使用した後で誘発される反応がTh2特徴を示すことは、ある意味で驚くべきことである。重要なことに、フラジェリンに対して予め存在する抗体は、アジュバントの有効性に対してさほどの影響を及ぼさない(74)ことから、フラジェリンは、複数回使用のためのアジュバントとして魅力的となっている。
【0135】
多くの近年における鼻腔内ワクチン試験における共通の主題は、ワクチンの有効性を改善するアジュバントおよび/または送達系の使用である。このような一研究では、遺伝的に解毒されたE. coli易熱性エンテロトキシンアジュバント(LT R192G)を含有するインフルエンザH3ワクチンが、H5チャレンジに対する異種サブタイプ的な防御を結果としてもたらしたが、これは、鼻腔内送達後に限られた。防御は、交差中和抗体の誘導に基づき、これにより、新たなワクチンの開発における鼻腔内経路についての重要な示唆が示された(22)。
【0136】
サイトカイン、コロニー刺激因子(例えば、GM−CSF、CSFなど);腫瘍壊死因子;インターロイキン2、インターロイキン7、インターロイキン12、インターフェロン、および他の同様の増殖因子はまた、アジュバントとして用いることができ、これらは
エンベロープウイルスベースのVLPのポリペプチド成分と混合または融合させることにより、該ワクチン内に容易に組み込み得るので、これらもまた好ましい。
【0137】
一部の実施形態では、本明細書で開示されるエンベロープウイルスベースのVLPワクチン組成物が、5’−TCG−3’配列を含む、核酸のTLR9リガンド;イミダゾキノリンのTLR7リガンド;置換グアニンのTLR7/8リガンド;ロキソリビン、7−デアザデオキシグアノシン、7−チア−8−オキソデオキシグアノシン、イミキモド(R−837)、およびレシキモド(R−848)など、他のTLR7リガンドなど、Toll様受容体を介して作用する他のアジュバントを包含し得る。
【0138】
特定のアジュバントは、樹状細胞などのAPCによるワクチン分子の取込みを促進し、これらを活性化する。非限定的な例は、免疫標的化アジュバント;毒素、サイトカイン、およびマイコバクテリア誘導体などの免疫調節アジュバント;油処方物;ポリマー;ミセル形成アジュバント;サポニン;免疫刺激複合体マトリックス(ISCOMマトリックス);粒子;DDA;アルミニウムアジュバント;DNAアジュバント;MPL;および封入アジュバントからなる群より選択される。
【0139】
アジュバントのさらなる例には、作用物質、例えば、緩衝生理食塩液中0.05〜0.1パーセントの溶液として一般的に用いられるアルミニウム塩、例えば、水酸化物またはリン酸塩(ミョウバン)(例えば、Nicklas(1992年)、Res. Immunol.、143巻:489〜493頁を参照されたい)、0.25パーセントの溶液として用いられる糖の合成ポリマー(例えば、Carbopol(登録商標))との混合物、それぞれ、70℃〜101℃の範囲の温度で30秒間〜2分間にわたる熱処理によるワクチン中のタンパク質の凝集物が含まれ、また、架橋形成剤による凝集物も可能である。ペプシン処理した、アルブミンに対する抗体(Fab断片)での再活性化による凝集物、C. parvumなどの細菌細胞またはグラム陰性菌の内毒素もしくはリポ多糖成分との混合物、モノオレイン酸マンニド(Aracel A)などの生理学的に許容される油ビヒクルにおけるエマルジョン、または保護置換物として用いられる、ペルフルオロカーボン(Fluosol−DA)の20パーセント溶液によるエマルジョンもまた用いることができる。スクアレンおよびIFAなどの油との混合物もまた好ましい。
【0140】
DDA(ジメチルジオクタデシルアンモニウムブロミド)は、アジュバントの興味深い候補物質であるが、フロイントの完全アジュバントおよびフロイントの不完全アジュバントの他、QuilAおよびQS21などのキラヤサポニンもまた興味深い。さらなる可能性には、モノホスホリルリピドA(MPL(登録商標))、ムラミルジペプチド(MDP)、およびトレオニルムラミルジペプチド(tMDP)など、リポ多糖のポリ[ジ(カルボキシレートフェノキシ)ホスファゼン](poly[di(earboxylatophenoxy)phosphazene)(PCPP)誘導体が含まれる。例えば、アルミニウム塩と併せた、モノホスホリルリピドA、好ましくは、3−脱−O−アシル化モノホスホリルリピドAの組合せを含めた、主にTh1型の反応をもたらすリポ多糖ベースのアジュバントが好ましい。MPL(登録商標)アジュバントは、GlaxoSmithKline社から市販されている(例えば、米国特許第4,436,727号;同第4,877,611号;同第4,866,034号;および同第4,912,094号を参照されたい)。
【0141】
リポソーム処方物もまた、アジュバント効果を付与することが知られており、したがって、リポソームアジュバントは、エンベロープウイルスベースのVLPと合わされる、好ましい例である。
【0142】
免疫刺激複合体マトリックス型(ISCOM(登録商標)マトリックス)アジュバントは、とりわけ、この型のアジュバントが、APCによるMHCクラスII発現を上方制御することが可能であることが示されているため、本発明に従う好ましい選択である。ISCOMマトリックスは、Quillaja saponariaに由来する(場合によって分画された)サポニン(トリテルペノイド)、コレステロール、およびリン脂質からなる。VLP中のものなどの免疫原性タンパク質と混合する場合、結果として得られる粒子状処方物は、サポニンが60〜70%w/wを構成することが可能であり、コレステロールおよびリン脂質が10〜15%w/wを構成することが可能であり、タンパク質が10〜15%w/wを構成することが可能な、ISCOM粒子として公知の処方物である。免疫刺激複合体の組成および使用に関する詳細は、例えば、アジュバントを扱う上述の教科書中に見出し得るが、また、Morein Bら、1995年、Clin. Immunother.、3巻:461〜475頁の他、Barr I GおよびMitchell G F、1996年、Immunol. and Cell Biol.、74巻:8〜25頁も、完全な免疫刺激複合体を調製するのに有益な教示を提供している。
【0143】
本明細書で開示されるエンベロープウイルスベースのVLPワクチンとアジュバントの組合せにおいて用い得るサポニンには、それらがISCOM形態にある場合であれ、そうでない場合であれ、米国特許第5,057,540号;および「Saponins as vaccine adjuvants」、Kensil, C. R.、Crit Rev Ther Drug Carrier Syst、1996年、12巻(1〜2号):1〜55頁;およびEP0362279B1で記載される、Quil Aと称する、Quillaja Saponaria Molinaの樹皮に由来するサポニン、また、それらの画分が含まれる。Quil Aの特に好ましい画分は、QS21、QS7、およびQS17である。
【0144】
β−エスシンは、本発明のアジュバント組成物中で用いるのに好ましい、別の溶血性サポニンである。エスシンは、「メルクインデックス」(第12版:エントリー3737)において、ラテン名: Aesculus hippocastanumであるトチノキの種子内で発生するサポニンの混合物として記載されている。クロマトグラフィーおよび精製(Fiedler、Arzneimittel−Forsch.、4巻、213頁(1953年))による、また、イオン交換樹脂(Erbringら、米国特許第3,238,190号)によるその単離が記載されている。エスシンの画分は精製されており、生物学的に活性であることが示されている(Yoshikawa Mら(Chem Pharm Bull(東京)、1996年8月、44巻(8号):1454〜1464頁))。β−エスシン(escin)はまた、エスシン(aescin)としても公知である。
【0145】
本発明において用いるのに好ましい別の溶血性サポニンは、ジギトニンである。ジギトニンは、「メルクインデックス」(第12版:エントリー3204)において、Digitalis purpureaの種子に由来し、また、Gisvoldら、J.Am.Pharm.Assoc.、1934年、23巻、664頁;およびRuhenstroth−Bauer、Physiol.Chem.、1955年、301巻、621頁において記載される手順により精製されるサポニンとして記載されている。その使用は、コレステロールを判定するための臨床試薬として記載されている。
【0146】
アジュバント効果を達成する別の興味深い(したがって、好ましい)可能性は、Gosselinら、1992年において記載される技法を用いることである。略述すると、本発明の抗原などの関与抗原の提示は、該抗原を、単球/マクロファージ上におけるFC受容体に対する抗体(または抗原に結合する抗体断片)に結合体化することにより増強することができる。とりわけ、抗原と抗FCRIとの結合体は、ワクチン接種のための免疫原性を増強することが示されている。該抗体は、生成後において、または生成の一部として(エンベロープウイルスベースのVLPのポリペプチド成分のうちのいずれか1つに対する融合体としての発現によることを含む)、エンベロープウイルスベースのVLPに結合体化することができる。
【0147】
他の可能性は、標的化物質および免疫調節物質(すなわち、サイトカイン)の使用を伴う。加えて、ポリI:Cなど、合成のサイトカイン誘導物質もまた用いることができる。
【0148】
適切なマイコバクテリア誘導体は、ムラミルジペプチド、フロイントの完全アジュバント、RIBI(モンタナ州、ハミルトン、RiBi ImmunoChem Reserch社製)、ならびに、TDMおよびTDEなど、トレハロースのジエステルからなる群より選択することができる。
【0149】
適切な免疫標的化アジュバントの例には、CD40リガンドおよびCD40抗体、またはこれらの特異的結合断片(上記の議論を参照されたい)、マンノース、Fab断片、ならびにCTLA−4が含まれる。
【0150】
適切なポリマーアジュバントの例には、炭水化物、例えば、デキストラン、PEG、デンプン、マンナン、およびマンノース;可塑性ポリマー;ならびにラテックスビーズなどのラテックスが含まれる。
【0151】
免疫反応を調節するさらに別の興味深い方法は、「仮想リンパ節」(VLN)(ImmunoTherapy, Inc., 360 Lexington Avenue, New York, N.Y. 10017−6501により開発された、特許品の医療デバイス)内に免疫原(場合によって、アジュバント、ならびに薬学的に許容されるキャリアおよびビヒクルと併せて)を組み込むことである。VLN(細長型の管状デバイス)は、リンパ節の構造および機能を模倣する。VLNを皮下に挿入することにより、サイトカインおよびケモカインの急増による無菌的炎症部位が創出される。T細胞およびB細胞の他、APCも、迅速に危険シグナルに応答し、炎症部位へと至り、VLNの多孔性マトリックスの内部に蓄積される。VLNを用いると、抗原に対する免疫反応を誘発するのに必要とされる抗原用量が低下し、VLNを用いるワクチン接種により付与される免疫的防御が、Ribiをアジュバントとして用いる従来の免疫化を凌駕することが示されている。この技法は、Gelber Cら、1998年、「Elicitation of Robust Cellular and Humoral Immune Responses to Small Amounts of Immunogens Using a Novel Medical Device Designated the Virtual Lymph Node」、1998年10月12〜15日、カリフォルニア州、アプトス、シースケープリゾート、「From the Laboratory to the Clinic, Book of Abstracts」において略述されている。
【0152】
エンベロープウイルスベースのVLPワクチンと共に、オリゴヌクレオチドをアジュバントとして用いることができ、少なくとも3つの、またはより好ましくは、少なくとも6つ以上のヌクレオチドにより隔てられた2つ以上のジヌクレオチドCpGモチーフを含有することが好ましい。CpGを含有するオリゴヌクレオチド(ここで、CpGジヌクレオチドは、メチル化されていない)は、主にTh1反応を誘導する。このようなオリゴヌクレオチドは周知であり、例えば、WO96/02555、WO99/33488、ならびに米国特許第6,008,200号および同第5,856,462号において記載されている。
【0153】
このようなオリゴヌクレオチドによるアジュバントは、デオキシヌクレオチドであり得る。好ましい実施形態では、該オリゴヌクレオチド内のヌクレオチド骨格は、ホスホロジチオエート結合であるか、またはより好ましくは、ホスホロチオエート結合であるが、骨格結合が混合したオリゴヌクレオチドを含め、ホスホジエステル骨格、また、PNAなど他のヌクレオチド骨格も本発明の範囲内にある。ホスホロチオエートオリゴヌクレオチドまたはホスホロジチオエートオリゴヌクレオチドを作製する方法は、米国特許第5,666,153、米国特許第5,278,302号、およびW095/26204において記載されている。
【0154】
好ましいオリゴヌクレオチドの例は、以下の配列を有する。配列は、ホスホロチオエート修飾されたヌクレオチド骨格を含有することが好ましい。
【0155】
【化3】
代替的な好ましいCpGオリゴヌクレオチドには、これらに対する重要でない欠失または付加を伴う上記の配列が含まれる。アジュバントとしてのCpGオリゴヌクレオチドは、当技術分野で公知である任意の方法(例えば、EP468520)により合成することができる。自動合成器を用いてこのようなオリゴヌクレオチドを合成し得ることが好ましい。このようなオリゴヌクレオチドによるアジュバントは、10〜50塩基の長さであり得る。別のアジュバント系は、CpGを含有するオリゴヌクレオチドと、サポニン誘導体との組合せを伴い、特に、CpGとQS21との組合せが、WO00/09159において開示されている。
【0156】
多くの単相または複相のエマルジョン系が記載されている。エマルジョンが、エンベロープウイルスベースのVLPの構造を破壊しないように、当業者は、エンベロープウイルスベースのVLPと共に用いられるこのようなエマルジョン系を容易に適合させることができる。水中油エマルジョンによるアジュバントは、それ自体で、アジュバント組成物として有用であることが示唆されており(EPO399843B)、また、水中油エマルジョンと他の活性薬剤との組合せも、ワクチン用のアジュバントとして記載されている(WO95/17210;WO98/56414;WO99/12565;WO99/11241)。油中水エマルジョン(米国特許第5,422,109号;EP 0480982B2)、および水中油中水エマルジョン(米国特許第5,424,067;EP0480981B)など、他の油エマルジョンによるアジュバントが記載されている。
【0157】
本明細書で記載されるエンベロープウイルスベースのVLPワクチンと共に用いられる油エマルジョンによるアジュバントは、天然の場合もあり、合成の場合もあり、また、鉱油性の場合もあり、有機油性の場合もある。鉱油および有機油の例は、当業者に容易に明らかである。
【0158】
任意の水中油組成物が、ヒトへの投与に適するためには、エマルジョン系の油相が、代謝性油を包含することが好ましい。代謝性油という用語の意味は、当技術分野において周知である。代謝性とは、「代謝により変換可能であること」と定義することができる(「Dorland’s Illustrated Medical Dictionary」、W.B. Sanders Company社、第25版(1974年))。油は、レシピエントに毒性でなく、代謝により変換可能な、任意の植物油、魚油、動物油、または合成油であることが可能である。堅果油(ラッカセイ油など)、種油、穀類油が、植物油の一般的な供給源である。合成油もまた本発明の一部であり、これらには、NEOBEE(登録商標)および他の油など、市販の油が含まれ得る。スクアレン(2,6,10,15,19,23−ヘキサメチル−2,6,10,14,18,22−テトラコサヘキサエン)は、サメの肝油中に高量で見出され、また、オリーブ油、コムギ胚種油、コメぬか油、および酵母中に低量で見出される不飽和油であり、本発明で用いるのに特に好ましい油である。スクアレンは、コレステロールの生合成における中間体であるという事実により、代謝性油である(「メルクインデックス」、第10版、エントリー8619)。
【0159】
特に好ましい油エマルジョンは水中油エマルジョンであり、特に、水中スクアレンエマルジョンである。
【0160】
加えて、本発明の最も好ましい油エマルジョンによるアジュバントには、抗酸化剤が含まれ、油であるα−トコフェロール(ビタミンE;EP0382271B1)が好ましい。
【0161】
WO95/17210およびWO99/11241は、場合によって、免疫刺激剤であるQS21および/または3D−MPLと共に処方される、スクアレン、α−トコフェロール、TWEEN 80に基づくエマルジョンアジュバントを開示している。WO99/12565は、油相へのステロールの添加による、これらのスクアレンエマルジョンに対する改善を開示している。加えて、エマルジョンを安定化させるために、トリカプリリン(C27H50O6)などのトリグリセリドを、油相へと添加することもできる(WO98/56414)。
【0162】
安定的な水中油エマルジョン中で見出される油滴のサイズは、光子相関分光分析により測定したときに、1ミクロン未満であることが好ましく、実質的に30〜600nmの範囲内、好ましくは、実質的に直径約30〜500nm、また、最も好ましくは、実質的に直径150〜500nm、また、特に直径約150nmであり得る。この点で、油滴数の80%が好ましい範囲内にあるべきであり、より好ましくは油滴数の90%超、また、最も好ましくは油滴数の95%超が規定のサイズ範囲内にある。本発明の油エマルジョン中に存在する成分の量は、従来、スクアレンなどの油が2〜10%の範囲内であり;また、存在する場合、アルファトコフェロールが2〜10%であり、ポリオキシエチレンソルビタンモノオレエートなどの界面活性剤が0.3〜3%である。油:アルファトコフェロールの比率は、より安定的なエマルジョンがもたらされるため、1以下であることが好ましい。Span 85もまた、約1%のレベルで存在し得る。一部の場合には、本明細書で開示されるエンベロープウイルスベースのVLPワクチンが、安定化剤をさらに含有することが有利であり得る。
【0163】
水中油エマルジョンを作製する方法は、当業者に周知である。一般に、該方法は、油相を、PBS/TWEEN80(登録商標)溶液などの界面活性剤と混合し、その後、ホモジナイザーを用いるホモジナイゼーションを行うステップを包含するが、該混合物に、シリンジ針内を2回流過させるステップを含む方法が、少量の液体をホモジナイズするのには適切であることが、当業者には明らかである。同様に、当業者は、Microfluidizer(M110S型マイクロ流体作製器;最大入力圧6バールで2分間にわたり、最大50回の流過(約850バールの出力圧))において乳化過程を適合させることで、より少量または多量のエマルジョンを作製することができる。この適合は、必要とされる直径の油滴を有する調製物が達成されるまでの、結果として得られるエマルジョンの測定を含む日常的な実験により達成し得る。
【0164】
本明細書で開示されるエンベロープウイルスベースのVLPによるワクチン調製物は、該ワクチンを、鼻腔内投与、筋肉内投与、腹腔内投与、皮内投与、経皮投与、静脈内投与、または皮下投与を介して投与することにより、ウイルス感染を受けやすいか、またはこれを患う、哺乳動物または鳥類を保護または処置するのに用いることができる。ワクチン調製物を全身投与する方法には、従来のシリンジおよび針、または固体ワクチンを弾道送達(ballistic delivery)するためにデザインされたデバイス(WO99/27961)、または針なしの高圧液体ジェットデバイス(米国特許第4,596,556号;米国特許第5,993,412号)、または経皮パッチ(WO97/48440;WO98/28037)が含まれ得る。エンベロープウイルスベースのVLPワクチンはまた、皮膚に適用することもできる(経皮(transdermalまたはtranscutaneous)送達;WO98/20734;WO98/28037)。したがって、本明細書で開示されるエンベロープウイルスベースのVLPワクチンは、エンベロープウイルスベースのVLPワクチンまたはアジュバント組成物を予め充填した全身投与用の送達デバイスを包含する。したがって、個体、好ましくは哺乳動物または鳥類において免疫反応を誘導する方法であって、本明細書で記載されるエンベロープウイルスベースのVLP組成物のうちのいずれかを含み、また、場合によって、アジュバントおよび/またはキャリアを包含するワクチンを該個体へと投与するステップを含み、該ワクチンが、非経口経路または全身経路により投与される方法が提供される。
【0165】
本発明のワクチン調製物は、該ワクチンを、経口/経消化管経路、または経鼻経路などの粘膜経路を介して投与することにより、ウイルス感染を受けやすいか、またはこれを患う、哺乳動物または鳥類を保護または処置するのに用い得ることが好ましい。代替的な粘膜経路は、膣内経路および直腸内経路である。好ましい経粘膜経路投与は、経鼻経路を介し、鼻腔内ワクチン接種と称する。免疫化される個体の鼻咽頭内へと、ワクチンの液滴形態、スプレー形態、または乾燥粉末形態を投与することを含め、鼻腔内ワクチン接種の方法は、当技術分野において周知である。したがって、噴霧化またはエアゾール化されたワクチン処方物が、本明細書で開示されるエンベロープウイルスベースのVLPワクチンの好ましい形態である。経口投与用の胃液耐性のカプセルおよび顆粒などの腸溶処方物、直腸内投与または膣内投与用の坐剤もまた、本明細書で開示されるエンベロープウイルスベースのVLPワクチンの処方物である。
【0166】
本明細書で開示される、好ましいエンベロープウイルスベースのVLPによるワクチン組成物は、経粘膜ワクチン接種により全身ワクチン接種を代替するヒトにおける適用に適する経粘膜ワクチンのクラスの一例である。
【0167】
エンベロープウイルスベースのVLPワクチンはまた、経口経路を介して投与することもできる。このような場合、薬学的に許容される賦形剤にはまた、アルカリ性緩衝液、または腸溶性のカプセルもしくはマイクロ顆粒も含まれ得る。エンベロープウイルスベースのVLPワクチンはまた、膣内経路によっても投与することができる。このような場合、薬学的に許容される賦形剤には、乳化剤、CARBOPOL(登録商標)などのポリマー、また、膣内クリームおよび膣内坐剤の他の公知の安定化剤も含まれ得る。エンベロープウイルスベースのVLPワクチンはまた、直腸内経路によっても投与することができる。このような場合、賦形剤には、直腸内坐剤を形成するために当技術分野で公知のワックスおよびポリマーも含まれ得る。
【0168】
代替的に、エンベロープウイルスベースのVLPによるワクチン処方物は、キトサン(上記で記載した)または他のポリカチオンポリマー、ポリラクチド粒子およびポリラクチド−コグリコリド粒子、ポリ−N−アセチルグルコサミンベースのポリマーマトリックス、多糖または化学修飾した多糖からなる粒子、リポソームおよび脂質ベースの粒子、グリセロールモノエステルからなる粒子などからなるワクチンビヒクルと組み合わせることができる。サポニンはまた、コレステロールの存在下において処方して、リポソームまたはISCOMなどの粒子構造を形成することもできる。さらに、サポニンは、非粒子状の溶液もしくは懸濁物中、または小ラメラ(paucilamelar)リポソームもしくはISCOMなどの粒子状構造内において、ポリオキシエチレンエーテルまたはポリオキシエチレンエステルと併せて処方することもできる。
【0169】
本明細書で記載されるエンベロープウイルスベースのVLPを用いる医薬組成物およびワクチン組成物中で用いられる、さらなる例示的なアジュバントには、SAF(米国、カリフォルニア州、Chiron社製)、MF−59(Chiron社製、例えば、Granoffら(1997年)、Infect Immun.、65巻(5号):1710〜1715頁を参照されたい)、SBASシリーズのアジュバント(例えば、SB−AS2(SmithKline Beecham社製のアジュバントシステム♯2;MPLおよびQS21を含有する水中油エマルジョン);SBAS−4(SmithKline Beecham社製のアジュバントシステム♯4;ミョウバンおよびMPLを含有する)、ベルギー、リクセンサルト、SmithKline Beecham社から市販されている)、Detox(Enhanzyn(登録商標))(GlaxoSmithKline社製)、RC−512、RC−522、RC−527、RC−529、RC−544、およびRC−560(GlaxoSmithKline社製)、ならびに、係属中の米国特許出願第08/853,826号、および同第09/074,720号において記載されるものなどの、他のアミノアルキルグルコサミド4−ホスフェート(AGP)が含まれる。
【0170】
アジュバントの他の例には、HunterによるTiterMax(登録商標)アジュバント(ジョージア州、ノークロス、CytRx社製);Gerbuアジュバント(ドイツ、ガイベルク、Gerbu Biotechnik GmbH社製);ニトロセルロース(NilssonおよびLarsson(1992年)、Res. Immunol.、143巻:553〜557頁);Seppic ISAシリーズのMontamideアジュバント(例えば、ISA−51、ISA−57、ISA−720、ISA−151など;フランス、パリ、Seppic社製)など、鉱油エマルジョン、非鉱油エマルジョン、油中水エマルジョン、または水中油エマルジョンを含めたミョウバン(例えば、水酸化アルミニウム、リン酸アルミニウム)エマルジョンベースの処方物;およびPROVAX(登録商標)(IDEC Pharmaceuticals社製);OM−174(脂質Aと関連するグルコサミンジサッカリド); Leishmania属の伸長因子;CRL 1005など、ミセルを形成する非イオン性のブロックコポリマー;ならびにSyntexアジュバント処方物が含まれるがこれらに限定されない。例えば、O’Haganら(2001年)、Biomol Eng.、18巻(3号):69〜85頁;および「Vaccine Adjuvants: Preparation Methods and Research Protocols」、D. O’Hagan編(2000年)、Humana Press社を参照されたい。
【0171】
他の好ましいアジュバントには、一般式
HO(CH 2CH2O)n−A−R, (I)
[式中、nは1〜50であり、Aは結合または−−C(O)−−であり、RはC1〜50アルキル、またはフェニルC1〜50アルキルである]のアジュバント分子が含まれる。
【0172】
本発明の一実施形態は、一般式(I)[式中、nは1〜50、好ましくは4〜24、最も好ましくは9であり;R成分はC1〜50、好ましくはC4〜C20アルキル、また最も好ましくはC12アルキルであり;また、Aは結合である]のポリオキシエチレンエーテルを含むワクチン処方物からなる。ポリオキシエチレンエーテルの濃度は、0.1〜20%、好ましくは0.1〜10%、また最も好ましくは0.1〜1%の範囲であるべきである。好ましいポリオキシエチレンエーテルは、以下の群:ポリオキシエチレン−9−ラウリルエーテル、ポリオキシエチレン−9−ステアリルエーテル(polyoxyethylene−9−steoryl ether)、ポリオキシエチレン−8−ステアリルエーテル、ポリオキシエチレン−4−ラウリルエーテル、ポリオキシエチレン−35−ラウリルエーテル、およびポリオキシエチレン−23−ラウリルエーテルから選択される。ポリオキシエチレンラウリルエーテルなどのポリオキシエチレンエーテルは、「メルクインデックス」(第12版;エントリー7717)で記載されている。これらのアジュバント分子は、WO99/52549で記載されている。
【0173】
所望の場合、上記の一般式(I)に従うポリオキシエチレンエーテルを、別のアジュバントと組み合わせることができる。例えば、好ましいアジュバントの組合せは、上で記載したCpGとの組合せであることが好ましい。
【0174】
本明細書で開示されるエンベロープウイルスベースのVLPワクチンと共に用いるのに適する、薬学的に許容される賦形剤のさらなる例には、水、リン酸緩衝生理食塩液、等張性緩衝液が含まれる。
【0175】
本発明は、以下の非限定的な実施例を参照することにより、よりよく理解される。本明細書で記載される通り、本発明は、天然では脂質ラフトと会合しない抗原に連結した、任意の種類の脂質ラフト会合ポリペプチドを組み込む、キメラエンベロープウイルスベースのVLPを包含する。以下の実施例では、本発明の代表的な実施形態である、インフルエンザ抗原を伴う、キメラエンベロープウイルスベースのVLPが記載される。
【実施例】
【0176】
(実施例1−インフルエンザ偽型Gag VLPの作製)
MLV Gag遺伝子ならびに種々のインフルエンザAサブタイプのHAおよびNA遺伝子を、下に記載の通り個々にpFastBac1バキュロウイルス導入ベクターのポリヘドリンプロモーターの後ろにクローニングした。「三重遺伝子」発現ベクターを構築するために1つのHAおよび1つのNAベクターの転写単位全体をSnaB IおよびHpa Iでの切断によって切り出し、これらの平滑末端断片をGag遺伝子導入ベクター中のGag転写単位のいずれかの側の唯一のSnaB IおよびHpa Iクローニング部位にそれぞれ導入した。これは、ヘッドトゥーテール(head−to−tail)様式で配置された3種の別々の転写単位(HA、Gag、NA)を含有する単一のプラスミドを生じた。次いで種々のインフルエンザAサブタイプを示すpFB−HA−pGag−pNA3重導入ベクターをキット製造者(Invitrogen、Carlsbad、CA)によって記載された通りバキュロウイルスゲノムへの組み換えのためにDH10Bac細胞中に形質転換した。
【0177】
(MLV GagならびにインフルエンザHAおよびNA遺伝子)
マウス白血病ウイルスのGag遺伝子はプラスミドpAMS(ATCC、Manassas、VA)から以下のプライマー:5’CACCATGGGCCAGACTGTTACC3’(配列番号6)および5’CTACTAGTCATCTAGGGTCAGGAG3’(配列番号7)を使用するPCRによって得た。最初のプライマーの5’末端のCACC伸長は、Gateway cloning technology(Invitrogen)を用いるベクターpENTR D−TOPOへの一方向性の挿入を促進し、プラスミドpENTR−Gagを生じる。Gagコード配列をDNA配列決定によって確認し、次いでpFastbac1に以下の通り導入した:pENTR−GagをAsc Iで切断し、末端をKlenow DNAポリメラーゼでの処理によって平滑化し、その後DNAをNot Iで切断した。最初にpFastbac1をSph Iで切断し、末端を平滑化し、次いでNot Iで切断した後に、得られたGag断片をpFastbac1ベクターDNAにライゲーションした。
【0178】
インフルエンザA/PR/8/34(H1N1)およびA/Hong Kong/68(H3N2)のHAおよびNA遺伝子をRT−PCRによってクローニングした。ウイルスRNAをQIAmp MinElute Virus Spin Kit(Qiagen、Valencia、CA)を使用して卵培養ウイルスから調製し、第1鎖cDNA反応をAccuscript High Fidelity 1st Strand cDNA Synthesis kit(Stratagene、La Jolla、CA)を使用して実施した。第1鎖cDNA合成に続いて、HAおよびNA断片を増幅するために標準的PCR反応を使用した。PR/8 H1遺伝子のためのプライマーは以下の通り:5’CACCATGAAGGCAAACCTACTGGTCC3’(配列番号8)および5’TCAGATGCATATTCTGCACTGC3’(配列番号9)。PR/8 N1遺伝子のためのプライマーは以下の通り:5’CACCATGAATCCAAATCAGAAAATAATAACCATTCC3’(配列番号10)および5’CTACTTGTCAATGGTGAATGGCAAC3’(配列番号11)。A/Hong Kong/68 H3遺伝子のためのプライマーは以下の通り:5’CACCATGAAGACCATCATTGCTTTGAGC3’(配列番号12)および5’TCAAATGCAAATGTTGCACCTAATGTTGCC3’(配列番号13)。A/Hong Kong/68 N2遺伝子のためのプライマーは以下の通り:5’ATATAGGCGCGCCACCATGAATCCAAATCAAAAGATAATAACAATTGGC3’(配列番号14)および5’ATATAGCGGCCGCTTATATAGGCATGAAATTGATGTTCGC3’(配列番号15)。A/PR/8/34(H1N1)のHAおよびNA遺伝子ならびにA/Hong Kong/68(H3N2)のHA遺伝子を最初にpENTR D−TOPOにクローニングし、配列決定によって確認し、次いで上に記載の通りpFastbac1に導入した。A/Hong Kong/68(H3N2)のNA遺伝子をAscIおよびNot IでPCR断片末端を整えた後にBssHII−NotI切断pFastbac1に直接クローニングした。候補クローンを配列決定によって確認した。
【0179】
A/Vietnam/1203/04およびA/Indonesia/5/05のH5およびN1遺伝子を含有しているプラスミドクローンはDr.Ruben Donis(Branch Chief、Molecular Virology and Vaccines、CDC、Atlanta、GA USA)から得た。H5クローンは、成熟切断部位に多塩基領域の欠失を含んでいた。提供されたプラスミドを配列確認用のpENTR D−TOPOへの挿入のための「Vietnam」および「Indonesia」H5およびN1 PCR断片を作製するための鋳型として使用した。H5プライマーは、以下の通り:5’CACCATGGAGAAAATAGTGCTTC3’(配列番号16)および5’TTAAATGCAAATTCTGCATTGTAACG3’(配列番号17)。N1プライマーは以下の通り:5’CACCATGAATCCAAATCAGAAGATAATAACC3’(配列番号18)および5’CTACTTGTCAATGGTGAATGGC3’(配列番号19)。配列確認後、個々のH5およびN1遺伝子は、上に記載の通りpFastbac1に導入した。
【0180】
A/Wisconsin/67/2005(H3N2)のHAおよびNA遺伝子をRT−PCRによって上に記載の通りウイルスRNAからクローニングした。H3遺伝子のためのプライマーは以下の通り:5’ATATAGGCGCGCCACCATGAAGACTATCATTGCTTTGAGC3’(配列番号20)および5’ATATAGCGGCCGCTCAAATGCAAATGTTGCACCTAATGTTGCC3’(配列番号21)。N2遺伝子のためのプライマーは以下の通り:5’ATATAGGCGCGCCACCATGAATCCAAATCAAAAGATAATAACGATTGGC3’(配列番号22)および5’ATATAGCGGCCGCTTATATAGGCATGAGATTGATGTCCG3’(配列番号23)。H3およびN2遺伝子断片をそれぞれAscIおよびNotIで整え、BssHII−NotI切断pFastbac1にクローニングした。
【0181】
A/Solomon Islands/3/2006のH1およびN1遺伝子をコードするカスタム合成遺伝子をGeneArt(Regensburg、ドイツ)から得て、昆虫細胞での発現のためにコドンを最適化した。各断片は、NotI−KpnI−切断pFastbac1への直接クローニング用にNotIおよびKpnI部位を5’および3’末端にそれぞれ含有した。
【0182】
組換えDNAの使用を含む作業の実施において、研究者らは組換えDNA分子に関与する研究のためのガイドラインを遵守した;注、Federal Register、1994年7月5日、第59巻、第127号。
【0183】
(VLPの精製)
昆虫細胞Sf9をSF900−II培地で培養し、1mlあたり細胞5×105個で200mlスピナーフラスコに播種した。細胞を27℃で1mlあたり細胞2×106個の密度まで培養し、その時点でVLPコード組換えバキュロウイルスの第2継代接種材料をおよそ0.1から1.0の感染多重度で加えた。細胞生存率が20%またはそれより低くまで低下したときに培養液を回収した。培地を2,000rpm、15分間遠心分離することによって細胞細片を除去し、その後32ml一定分量をトリス緩衝生理食塩水(TBS)、pH7.4中の30%ショ糖のクッション4ml上に重層した。VLPをショ糖クッションによって25,000rpm、1時間、10℃でBeckman SW28ローター(100,000×g)で遠心分離した。培地200ml由来のVLPを合計6mlのTBSに再懸濁し、次いで単一の20〜60%不連続ショ糖勾配に重層し、25,000rpm、1時間、10℃で遠心分離した。ショ糖勾配を底から1.5ml画分へと分取し、赤血球凝集アッセイおよびノイラミニダーゼアッセイ(下を参照されたい)ならびにSDS−PAGEおよびウェスタンブロットによって分析した。ショ糖勾配精製したVLPをショ糖存在下、4℃で保存し、少なくとも6カ月間安定(HA活性に関して)であることを見出した。VLPを播種の前にショ糖溶液から遠心分離し、トリス緩衝生理食塩水に再懸濁した。本明細書において報告する動物実験に利用したVLPワクチンは凍結されていないが、しかしH1N1 VLPの少なくとも1回の凍結融解をHA活性の測定可能な損失を伴わずに実施できることが測定された。
【0184】
(赤血球凝集アッセイ)
赤血球凝集アッセイを0.5%ヒヨコ赤血球を使用して、動物インフルエンザ診断および調査についてのWHO手順書(WHO Manual on Animal Influenza Diagnosis and Surveillance)に記載の通り実施した[41]。
【0185】
(ノイラミニダーゼアッセイ)
ノイラミニダーゼ活性を蛍光基質2’−(4−メチルウンベリフェリル)−α−D−N−アセチルノイラミン酸(MUN)を使用してVLP調製物およびショ糖勾配画分中で検出した。PBS中のVLP含有試料の漸増希釈物(50μl)を黒色平底96ウエルプレートに入れ、PBS 50μlを各試料ウエルに添加した。NA活性を35mM MES pH6.5、4mM CaCl2、150mM NaCl、300μM MUNの50μlの添加によって検出した。プレートを37℃で1時間インキュベートし、次いで83%エタノール中0.14N NaOHの1ウエルあたり100μlでの添加によって停止させた。プレートを励起波長および発光波長それぞれ365nmおよび455nmを使用して蛍光光度計で読み取った。すべての蛍光データは、基質を含有しているがVLPを含有していない対照ウエルから得たバックグラウンド値に対して補正した。補正した蛍光値を次いでアッセイの線形範囲にあるデータ点を決定するために希釈係数で割った。
【0186】
A/PR/8/34(H1N1)を示すHA−Gag−NA VLPを上に記載の「三重遺伝子」組換えバキュロウイルスを感染させたSf9細胞の培地から不連続ショ糖密度勾配での遠心分離によって精製した。赤血球凝集アッセイによる勾配画分の分析は、勾配の中央の顕著な可視バンドに関連したHA活性の強いピーク(図1A)を明らかにした。このピークは、SDS−PAGEゲル上の顕著な65kd産物とも一致があり、GagおよびHAの共移動(図1B)、ならびにHAおよびNAについてのウェスタンブロットシグナル(図1C)と一致した。最後に電子顕微鏡によるピーク画分の直接分析は、出芽エンジンとしてのレトロウイルスGagタンパク質の使用から予測される通りインフルエンザよりもガンマレトロウイルスを連想させる多量の100nm球状粒子を示した(図1D)。これらの粒子はインフルエンザウイルスおよびインフルエンザマトリックスベースのVLPとは形態学的に異なると考えられる一方で、プールしたピーク画分での赤血球凝集活性が、勾配精製した卵培養A/PR/8/34ウイルスについて観察されたものと同様である総VLPタンパク質1mg当たり2〜3×105HA単位の範囲内で測定され、これらの粒子への機能的HAスパイクの顕著な組み込みを実証していることに注目することは重要である。
【0187】
PR/8 H1N1 VLP中のGag対HAの比は、Gagより遅く移動する改変HA産物を作製するようにHAコード配列がそのアミノ末端で伸長された改変PR/8 H1N1 VLP発現ベクターを使用しておよそ3〜4:1と測定された(図1E)。PR/8 H1N1 VLPに関しておよそ3〜4:1のGag対HAの比は、本発明者らが作製し、A/Wisconsin/67/2005(H3N2)、A/Solomon Islands/3/2006(H1N1)およびB/Malaysia/2506/2004などの現在のヒトインフルエンザ株を示す他のVLPと一致している。これら後者のVLPは、SDSゲル上でGagよりも天然で遅い移動度を示すHA分子を含有し、同様に3〜4:1のGag対HAの比を示す(図1F、Solomon Islands H1N1 VLP)。
【0188】
HAに加えて、顕著なノイラミニダーゼ生物活性もショ糖勾配でのVLPピークにおいて見出されており、これらのデータをH5N1 VLPについて図2に示す。A/Vietnam/1204/2004およびA/Indonesia/5/2005株を示すH5N1 VLPを、上に記載のVLPと同様のやり方で同様の収率および赤血球凝集特異的活性を伴って作製した。NA活性が、HA活性ピークの重い側にしばしば観察されるHA活性の肩と重なって勾配中深くに広がる傾向があることに注目することは興味深い。このパターンは、今日までに検査したすべてのサブタイプ(H1N1、H3N2およびH5N1)について観察されており、VLP集団におけるHA:NA比が連続的である可能性と一致している(実施例7を参照されたい)。
【0189】
(実施例2−マウスにおけるH1N1 VLP 免疫原性および防御)
実施例2は、H1N1 VLPでのマウスの免疫化を実証する。
【0190】
(マウス免疫化およびチャレンジ)
6〜8週齢、雌性Balb/cマウスをHAおよそ0.7〜1.0μgを含有するトリス緩衝生理食塩水(TBS)中のVLP処方物で腹腔内接種(100μl)または筋肉内接種(30μl)を用いて免疫化した。初期実験において、モノホスホリルリピドA(解毒化脂質A、Avanti Polar Lipids、Alabaster、AL)20μgをアジュバントとして添加した。初回および追加免疫化は、4週間間隔を置いて離し、各免疫化に続いて2週間、側方顔面静脈から血液試料を採取した。免疫化マウスおよび対照マウスをPBS 50μl中の卵培養A/PR/8/34の10LD50で鼻腔内でチャレンジし、体重減少および病的状態について毎日モニターした。瀕死であるかまたは刺激に反応しないことが見出された動物は、CO2吸入で安楽死させた。動物を使用する研究を実施する際に研究者らは、National Research Councilの実験動物資源局(Institute of Laboratory Animal Resources)の実験動物の管理と使用に関する委員会(Committee on Care and Use of Laboratory Animals)によって作成された「実験動物の管理と使用のための指針」を遵守した[40]。
【0191】
(赤血球凝集抑制アッセイ)
赤血球凝集抑制(HAI)アッセイは、0.5%ヒヨコ赤血球を使用して動物インフルエンザ診断および調査についてのWHO手順書に記載の通り実施した[41]。H5N1特異的HAIアッセイに関して、1.0%ウマ赤血球も使用し、沈降時間を1時間に延ばした。H5N1 HAIアッセイはまた、精製H5N1 VLPも感染性ウイルスの代替の凝集作用物質として使用した(実施例4を参照されたい)。
【0192】
ショ糖勾配画分中のPR/8 H1N1 VLPをHA活性に基づいてプールし、ショ糖から沈降させて取り出し、アジュバントとしてのモノホスホリルリピドA(MPL)を伴うかまたは伴わない腹腔内または筋肉内経路を用いるマウスの免疫化のためにPBS中に再懸濁した。1群あたり16匹の動物が、免疫化あたりHAおよそ0.7μgを含有し、4週間隔てた初回および追加免疫化を受け、無処置動物16匹は対照とした。図3Aは、追加後2週間の免疫化マウスにおける強いA/PR/8/34特異的HAI活性を示し、これらの応答はほとんどIgG2aアイソタイプのものであった(図3B)。追加後およそ4週間で各群を10LD50のマウス適合A/PR/8/34(H1N1)でチャレンジした。H1N1特異的液性応答から予測される通り、免疫化したすべての動物は病的状態または体重減少の徴候無くH1N1チャレンジを生き残ったが、H1N1チャレンジした無処置動物8匹すべては8匹中7匹の死を伴って強い病的状態および体重減少を示した(図3C)。この初回試験での免疫応答の強さから、腹腔内経路投与をさらなる実験から除外した。
【0193】
(実施例3−マウスにおけるH3N2およびH5N1 VLP免疫原性)
実施例3は、H3N2またはH5N1 VLPでのマウスの免疫化を実証する。マウスの免疫化およびチャレンジの方法ならびにHAIアッセイの方法は上の実施例2に記載の通り実施した。
【0194】
(インフルエンザ特異的抗体ELISA)
卵培養インフルエンザウイルス(A/PR/8/34(H1N1)またはA/Aichi/68(H3N2))を尿膜腔液から、TBS中の30%ショ糖クッションによってMLS 50ローター、36,000RPM(100,000×g)、1時間、10℃での遠心分離で取り出した。ウイルスペレットをPBSに再懸濁し、タンパク質含有量をBCAアッセイ(Pierce Biotechnology、Rockford、IL)によって定量し、1mlあたり5μgに調整し、平底ELISAプレートを1ウエルあたり100μl、1晩、4℃でコートするために使用した。翌日プレートを0.05%Tween20を含むPBS(PBS−T)で洗浄し、StartingBlock(PBS)(Pierce Biotechnology)1ウエルあたり350μlで30分間ブロックした。血清試料をStartingBlock T20(PBS)(Pierce Biotechnology)で1:500に希釈して最上列に添加し、各カラムで下がる毎に連続的に3倍希釈し、室温で3時間または4℃で1晩インキュベートした。プレートをPBS−Tで3回洗浄し、StartingBlock T20(PBS)中に1:1000希釈したヤギ抗マウスIgG−HRP結合体(Southern Biotech、Birmingham、AL)100μlを各ウエルに添加した。プレートをさらに1.5時間室温でインキュベートし、PBS−Tで3回洗浄し、次いでABTS基質(Pierce Biotechnology)100μlを各ウエルに添加した。プレートを室温、45分後に405nmで読み取った。ELISA力価を、バックグラウンド吸光度値の1.75倍の吸光度値を生じた最も高い血清希釈の逆数を取ることによって算出した。このカットオフレベルは、すべての無処置血清試料対照からのすべての疑陽性を排除した。
【0195】
H5N1特異的ELISAを、プレートを組換えH5 Vietnam抗原(Protein Sciences Inc.、Meriden、CT)またはスプリットH5N1ワクチン処方物1mlあたり3μg(Dr.Sally Mossman、GlaxoSmithKline、Rixensart、Belgiumからの寄贈)でコートした以外は同様に実施した。
【0196】
また、IgG1およびIgG2a特異的ELISAを、総IgGではなくIgG1およびIgG2aに特異的な2次抗体を使用したこと以外は同様のやり方で実施した。定量のために、特異的2次抗体との反応性についての標準曲線を作成するために一連のIgG1およびIgG2a濃度標準を一連のウエルにコートした。
【0197】
(結果)
A/Hong Kong/68(H3N2)を示すVLPを実施例1に記載の通り作製した。H3N2およびH5N1 VLP(PR/8 H1N1 VLPと共に)の免疫原性を表1に示す通りマウスの免疫化試験において評価した(MPLアジュバントの存在下で1用量あたりHAおよそ1μgを含有する筋肉内初回および追加免疫化がすべての動物において強いHAI活性を誘導した)。興味深いことに、相当な交差クレードHAI活性がIndonesia H5N1免疫化群とVietnam H5N1免疫化群との間でウマRBC HAIアッセイを使用して観察され[43]、H5N1チャレンジに対する顕著な交差クレード防御を誘導する可能性を示唆した。
【0198】
【表1】
表2は、この同じ実験(A/PR/8/34(H1N1)、A/Aichi(H3N2)および組換えサブユニットH5 HA抗原(Vietnam)への強いサブタイプ特異的応答がそれぞれのワクチンについて観察された)からの抗体応答のELISA分析の結果を示す。重要なことにH5N1 VLPは、共通N1抗原の理由によって予測される(2つのN1抗原は約70年間のドリフトによって分離されているが(16.1%のアミノ酸配列相違(アラインメントデータ未記載)))A/PR/8/34(H1N1)に対する弱いELISA活性を誘導した。表2はまた、群2のH1N1 VLP免疫化マウスを8週間休ませ、次いでMPLアジュバントと共におよび共にではなくH3N2 VLP(初回および追加)で再免疫化した再免疫化実験の結果を示す。重要なことに先行する2回のH1N1 VLP免疫化後のH3N2応答は、無処置マウスで誘導されたものと同等であった(群3)。これらのデータは、動物が第2のVLPサブタイプで再免疫化された場合にGagおよび共通昆虫細胞膜抗原などの異なるVLPサブタイプ間で共有する抗原に対する既存の免疫応答が強力なインフルエンザ特異的応答の誘発を抑制しないことを示す。再免疫化データは、ワクチン性能がMPL添加の有無において同一であったことから観察された免疫応答にMPLアジュバントがほとんど寄与しないことも示す。したがってMPLは続く実験から除いた。
【0199】
【表2】
(実施例4−インフルエンザ−偽型VLPはHAIアッセイにおいて生きているウイルスを代替できる)
表1に示すH5N1 HAIアッセイは、適切なH5N1ウイルス株を入手できないことから生きているウイルスの代替としてH5N1 IndonesiaおよびVietnam VLPを使用して実施した。HAIアッセイを実施例2に記載の通り実施した。インフルエンザ偽型Gag VLPが、H1N1およびH3N2サブタイプの両方について図4に示す通りHAIアッセイにおいて生きているウイルスの性能をかなりの程度まで模倣できることに注目することは重要である。この実験においてH1N1およびH3N2 VLP免疫化マウス由来の免疫血清を生きているウイルスおよび対応するVLPの両方を使用するHAIアッセイについて検査し、アッセイ方法間で顕著に類似する力価を明らかにした。これらの結果は、HAIアッセイにおける偽型VLPとウイルスとの間での同様の性能を実証するだけでなく、生きているインフルエンザウイルスをHA活性および密度(ワクチン性能に関して重要であると考えられる)に関して模倣する偽型VLPの能力についての追加的証拠を提供する。
【0200】
(実施例5−フェレットにおける高病原性H5N1チャレンジ)
マウスにおけるVLPの免疫原性に基づいて本発明者らは、H5N1およびH1N1 VLPワクチンを使用して、HPAI H5N1チャレンジに対して誘導され得る防御の程度を決定するためのフェレットの免疫化およびチャレンジ試験を実施した。
【0201】
(フェレットワクチン接種およびチャレンジ)
32匹の雄性フェレット、8〜16週齢(Triple、F Farms、Sayer、PA)(現在広まっているインフルエンザウイルスに関して赤血球凝集抑制により血清学的に陰性)を、下に記載のVLPワクチン処方物で0日目および28日目にワクチン接種した。フェレットVLPワクチンは、生理食塩水中にアジュバントを含まずに処方され、1用量あたりHAおよそ5μgを含有した。血液試料(前大静脈を用いて)を0日目、28日目および42日目に採取した。体重測定は、チャレンジの1週間前から研究終了(63日目)まで通して毎日行った。温度をIPTT−300埋め込み型トランスポンダーをDAS−7000携帯型プローブ(BioMedic Data Systems、Seafort、DE)と共に用いて測定した。各免疫化群の8匹のフェレットの内7匹をチャレンジのために無作為選択した。動物を筋肉内注射を用いてテラゾール(Telezol)(16mg/kg)で麻酔し、PBS 0.6ml中のA/Vietnam/1203/04ウイルスの106TCID50を鼻腔内に接種した。チャレンジウイルスは、Dr.Alexander Klimov(CDC、Atlanta、GA、USA)によって提供された。フェレットを体温および体重の変化についてモニターし、疾患の臨床徴候について観察した。神経学的機能障害を示したかまたは瀕死になったフェレットは、安楽死させた。動物実験は、実験動物の管理および使用に関する指針(Guide for the Care and Use of Laboratory Animals)[40]に従って、国際実験動物管理評価公認協会(Association for Assessment and Accreditation of Laboratory Animal Care International)公認動物機関のBattelle施設内動物管理使用委員会(Battelle’s Institutional Animal Care and Use Committee)の指導の下で実施した。
【0202】
(フェレット鼻洗浄液中のウイルスの定量)
ウイルス流出(shedding)をチャレンジ後3日目および5日目に麻酔したフェレットから採取した鼻洗浄液中で測定した。合計1mlでの鼻洗浄用に軟性カテーテルを繋いだ1mLシリンジを使用してPBS 500μLで各鼻腔を素早く洗い流した。鼻洗浄液を滅菌標本カップに集め、クライオバイアル(cryovial)に移し、TCID50での計数まで−70℃保存した。鼻洗浄液中のウイルスをメイディン・ダービー・イヌ腎臓(MDCK)細胞での組織培養感染用量中央値(median tissue culture infectious dose)によって定量した。ウイルスの連続希釈物をEMEM(2mM グルタミン、1mMピルビン酸ナトリウムおよび1%ペニシリン−ストレプトマイシンで増量)中、96ウエル形式の細胞単層に5連で入れた。単層を96時間、37℃、5%CO2でインキュベートした。細胞変性効果(CPE)を示したウエルを陽性と記録し、データをSpearman Karber法[42]を使用して分析し、TCID50/mLとして報告した。
【0203】
(マイクロ中和アッセイ)
血清中和抗体を標準的マイクロ中和アッセイを使用して測定した。熱不活化、連続希釈検査血清300μLを、ウイルス600TCID50を含有する等量のEMEM(2mM グルタミン、1mMピルビン酸ナトリウムおよび1%ペニシリン−ストレプトマイシンで増量)と混合した。37℃、1時間のインキュベーションに続いて、各希釈物100μLを5連で96ウエル形式中のMDCK細胞に入れた。37℃、5%CO2での4日間のインキュベーションに続いて、CPEを含有するウエルを陰性と記録した。データをSpearman Karber法を使用して分析し、中和用量中央値(ND50)として報告した。
【0204】
32匹のフェレットを無作為に4群(1群あたり動物8匹)にし、gagだけのVLP、H1N1(PR/8/34)VLP、Indonesia H5N1 VLP およびVietnam H5N1 VLPでそれぞれ免疫化した。VLPをMPLアジュバントを含まない生理食塩水中に処方し、初回および追加免疫化は、4週間離した。免疫化はHAおよそ5μgを含み、筋肉内に投与した。追加免疫化を施した後、各群の動物8匹の内7匹を無作為に選択し、続くチャレンジのために封じ込め施設に移し、順化させた。図5は、H1N1への(図5A、実施例2に記載の通り実施したHAIアッセイ)およびVietnam H5N1への(図5B、マイクロ中和アッセイ)免疫応答を示す(ここで、強いH1N1およびH5N1特異的免疫応答が単回免疫化に続いてのそれぞれのワクチンによって誘発された)。H1N1特異的応答は、追加免疫化後にさらに増大しなかったが、H5N1マイクロ中和力価は、2回目の免疫化後にわずかに増大した。驚くべきことに低レベルH5N1中和力価が追加免疫化後にH1N1免疫化動物7匹の内の5匹で検出された。H5N1中和活性に加えて、IndonesiaおよびVietnam株の両方に特異的な強いH5N1 HAI活性が、IndonesiaおよびVietnam VLPを使用するウマRBC HAIアッセイを使用して両方のH5N1ワクチン接種群において検出された。顕著な交差クレードHAI活性が両方のH5N1ワクチン接種群において検出されたこれらのデータを表3に示す。
【0205】
【表3】
追加の2週間後、すべてのフェレットを106 TCID50のHPAI A/Vietnam/1203/04(H5N1)でチャレンジし、体重減少/生存およびウイルス力価データをそれぞれ図6および7に示す。チャレンジ前のH5N1中和力価から予測される通り、すべてのIndonesiaおよびVietnam H5N1 VLPワクチン接種動物が病的状態および体重減少をほとんど示さずにチャレンジを生き残ったが、すべてのGag VLPワクチン接種対照は強い病的状態を示し、動物7匹のうち6匹が安楽死を必要とした(図6A)。対照群でのこのレベルの病的状態および生存率(7匹の内1匹)は、この同じ用量のA/Vietnam/1203/04(H5N1)チャレンジウイルスについて以前報告された[44]ものと類似していた。驚くべきことに、H1N1ワクチン接種動物7匹のうち6匹に中程度の病的状態が観察されただけであり、この群のフェレットで実験の生活期間において安楽死を必要としたものは無かった。H1N1ワクチン接種群の動物について個々の体重減少データを図6Bに示す。H1N1ワクチン群の動物1匹(フェレット449)は経時的に顕著な体重減少を示したが、食欲不振(持続的な症状であるだけである)を伴なう許容できる敏捷性および活動レベルのため安楽死させなかった。フェレット449における体重減少は、コホートの平均重量を減少させ、チャレンジ後7日目以降の高い標準偏差を生じさせた。
【0206】
生存、活動度および体重減少データは、3および5日目の鼻洗浄液からのチャレンジ後のウイルス単離データと明らかに一致していた(図7AおよびB)。ここで、H5N1 VLPワクチン接種動物は、チャレンジ後に顕著な病的状態を示さず、チャレンジ後3および5日目の鼻洗浄液中に検出可能なウイルスも示さなかった。対照的にGag VLPワクチン接種フェレットにおいて観察された強度の病的状態および死亡率は、同様のチャレンジ用量を受けた無処置動物について以前報告されたのと同様のレベルでの、チャレンジ後3および5日後両日における高いウイルスレベルと関連していた[44、45]。H1N1 VLPワクチン接種フェレットにおいて観察された中程度の病的状態と一致して、チャレンジ後のウイルス力価は、対照群と比較して3日目で減少し、5日目には1匹を除くすべての動物において存在しなかった。5日目に鼻洗浄液ウイルス力価を有したH1N1ワクチン接種動物1匹は、より強い病的状態および体重減少を示したのと同じ動物(449)であった。H1N1群中の残りの動物6匹におけるウイルスの迅速なクリアランスは、軽減された病的状態およびより少ない体重減少と関連していた。
【0207】
H1N1 VLPワクチン接種フェレットにおけるHPAI H5N1チャレンジに対する部分的な防御に関する機序は公知ではないが、A/PR/8/34(H1N1)とA/Vietnam/1203/04(H5N1)とで共有するN1抗原に関連する可能性がある。表2のマウス免疫原性研究は、両方のH5N1 VLPワクチンによる弱いH1N1 ELISA反応性の誘発を実証する(実施例3を参照されたい)。同様に表4は、H1N1ワクチンによって誘発された部分的な防御と一致して、H1N1 VLPワクチンがフェレットにおいて卵培養スプリットH5N1ウイルス調製物に対する弱いELISA反応性を誘発したこと示す。これらのELISAデータは、H5N1チャレンジに対するH1N1媒介防御でのN1応答の役割を確認するものではないが、それらは、この可能性と一致する。
【0208】
【表4】
(実施例6−感染性バキュロウイルスはVLP免疫原性に寄与しない)
生きているバキュロウイルス粒子が顕著な先天免疫刺激およびアジュバント効果を示すこと[46〜48]、およびこれらの特性が、除去または不活化されなければVLPなどの昆虫細胞由来産物の免疫原性において役割を演じ得ることが実証されている。この現象を調査するために本発明者らは、ウイルス、A/Wisconsin/67/2005(H3N2)を示すVLPを精製するために使用したショ糖密度勾配においてバキュロウイルス粒子の力価を計り、同じ画分中の1mlあたり粒子およそ2×1012個のVLP濃度と比較して1mlあたりおよそ2×108pfuのバキュロウイルス力価を見出した。VLP粒子定量はVLPのタンパク質濃度を測定し、推定されるVLP分子量1.2×108ダルトン(ガンマレトロウイルス粒子1個あたりGagの複製物1500個およびGag対HAの比4:1に基づく)を使用することによって概算した。バキュロウイルスを超える大過剰量のVLPにも関わらず感染性バキュロウイルスがVLP調製物中に存在し、免疫原性へのその寄与は調査に重要であった。
【0209】
(VLP調製物中のバキュロウイルスの不活化)
Wisconsin/67(H3N2)VLP調製物中の感染性バキュロウイルスを、ソラーレン誘導体4’−アミノメチル−4,5’,8−トリメチルソラーレン塩酸塩(AMT)(Sigma−Aldrich、St.Lous、MO)存在下で長波長UV照射によって、またはベータ−プロピオラクトン(BPL)での処理によって不活化した。VLPを含有するプールしたショ糖勾配画分をUVまたはBPL処理のいずれかに供し、その後VLPを不活化溶液から100,000×g、トリス緩衝生理食塩水、pH7.4中の30%ショ糖クッションによる1時間、10℃での遠心分離によって回収した。UV不活化は1mlあたりAMT30μgの添加によって実施し、VLP懸濁物の1ml一定分量を6ウエル滅菌組織培養プレートの個々のウエルに添加した。培養プレート(蓋なし)を予め長波長照射用に再構成したCL−1000UV crosslinker(UVP、Upland、CA)に置き、365nm照射2.0〜2.5ジュールに、0.25ジュール毎に穏やかに撹拌しながら供した。BPL不活化についてはBPLを最終濃度0.2%で添加し、試料を室温で3時間インキュベートした。
【0210】
Wisconsin H3N2 VLPの3つの調製物を長波長UV/ソラーレン処理、ベータ−プロピオラクトン処理またはモック不活化にそれぞれ供し、ワクチン調製物をバキュロウイルス感染性について力価測定した。両方の不活化処理は、Sf9細胞でのエンドポイント力価測定によって測定した通り、ワクチン用量あたり10個未満の感染性バキュロウイルス粒子まで昆虫細胞感染力の低減をもたらした一方で、モック不活化VLP調製物は、ワクチン用量あたり105から106個の間の感染性バキュロウイルス粒子を含有していた。追加的に、UV/ソラーレン処理VLPは、HA活性の損失を示さなかったが、BPL処理VLPは、HA抗原の直接アルキル化および/またはVLPの完全性の破壊を示すHA活性の16〜20の低下を受けた。上の処理VLPから調製した筋肉内ワクチン用量は、初回免疫化用にはHAおよそ1μgを、追加免疫化用にはHA 0.5μgを含有した。表5は、UV/ソラーレン処理を用いるバキュロウイルス不活化後にVLP免疫原性の損失が無いことを実証する、3つのワクチン調製物についての免疫原性データを示し、これはHA活性の完全な保持と一致する。これらのデータは、観察された偽型VLP免疫原性のために感染性バキュロウイルスの潜在的アジュバント特性が必要ではないことを実証している。対照的にBPL処理VLPは、大きく減少した免疫原性を示し、BPL処理の結果としてのHA活性における顕著な減少と一致した。
【0211】
【表5】
(実施例7−考察)
本発明者らは、マウスおよびフェレットにおいて強い免疫原性および防御を示すキメラインフルエンザVLPの作製のためにMLV Gag遺伝子産物の堅調な粒子出芽特性を使用した。Gag媒介粒子出芽は、会合の膜部位へのこれらの分子の標的化を補助する翻訳後ミリスチル化に依存し[31、32]、Gag媒介粒子出芽がGagとカベオリンとの会合を介して脂質ラフトドメインを通って優先的に生じることの証拠が蓄積している[33]。この工程は、昆虫細胞および哺乳動物細胞の両方において効率的に生じ、Gagと組み込まれる抗原の細胞質側末端との間での特異的相互作用を必要することなく出芽粒子への脂質ラフト標的化内在性膜抗原の組み込みを可能にする。インフルエンザに加えて、このVLPプラットフォームは、パラミクソウイルス抗原の組み込みに同様に適用できるはずであり、このことは、複数の呼吸器感染疾患に対するキメラVLPワクチンファミリーの開発を可能にする。
【0212】
本発明者らは、昆虫細胞におけるキメラ粒子のスケールアップ(scaling up)作製、HAおよびNA含有量および生物学的活性についての粒子の特徴付けならびに、強い免疫原性および防御効果を示すマウスおよびフェレットにおける多数のワクチン研究の開始によるインフルエンザHAのGag粒子への組み込みの初期の知見に基づいている。H1N1、H3N2およびH5N1サブタイプを示すキメラインフルエンザVLPは、すべてバキュロウイルス昆虫細胞系において同様の効率で産生され、すべてがウイルスまたはVLPタンパク質1mgあたりのHA単位に関して、ショ糖勾配精製された生きているインフルエンザウイルスで観察されたのと同様の赤血球凝集特異的活性を示した。赤血球凝集抑制アッセイにおいてVLPが、生きているインフルエンザウイルスを容易に代替できるという事実に加えて、これらの観察は、キメラVLP表面上のHA抗原提示および密度が生きているインフルエンザで見られるものをかなりの程度まで模倣するという説得力のある証拠を提供する。本発明者らは、インフルエンザA以外のウイルスに向けたこの系の柔軟性を示すインフルエンザBウイルス抗原含有VLPの作製も実証している(データ未記載)。
【0213】
3種すべての遺伝子(HA、GagおよびNA)の1種のバキュロウイルスベクターへの組み込みは、VLP作製を大きく簡素化し、産物収量は昆虫細胞における感染多重度に決定的には依存しない。今日までに作製されたすべてのキメラインフルエンザA VLPは、同様の収率およびショ糖勾配バンドパターンを示す。電子顕微鏡によりこれらのVLPは、より多形性である生きているインフルエンザまたはインフルエンザマトリックスベースのVLP粒子とは著しく異なる均一な100nm球状粒子であると考えられる。VLPサイズのこの均一性は、この技術がヒトでの評価用の臨床グレード材料の作製のためにさらにスケールアップされることから重要である。キメラVLPが大きさにおいて均一である場合、図2に示す通り(ここでは、HAおよびNA活性の両方が単一のショ糖勾配全体で測定される)、HAおよびNAの相対的含有量は連続的であると考えられる。Gagとの予測されない物理的相互作用を伴なう、HAおよびNAの脂質ラフト中での標的化および選別は、無作為ではない可能性があり、これは、HAおよびNAの相対含有量での多様性を示し得るVLPを生じる。これは、HAおよびNA抗原がVLPの表面上に多様な状況で提示されることで免疫学的利益を有する場合がある。バキュロウイルスgp64エンベロープ糖タンパク質は脂質ラフトタンパク質ではなく、VLP中に顕著な程度にまで蓄積するとは予測されないことに注目することも重要である[49]。
【0214】
H1N1、H3N2およびH5N1サブタイプを示すキメラVLPは、マウスおよびフェレットにおいて添加アジュバントの非存在下で強く免疫原性であり、HAI、中和およびELISAアッセイにおいて測定される通りそれらそれぞれのウイルスに対する強い応答をもたらす。これらの強い応答は、マウスでの相同チャレンジ(homologous challenge)およびフェレットでの交差クレードH5N1チャレンジに対する病的状態についての証拠を示さない強固な防御を生じる。フェレットでのH5N1に対するH5N1交差クレード防御は新たなものではないが、本発明者らが、Indonesia H5N1またはVietnam H5N1をワクチン接種した動物においてVietnamウイルスの標準的1×106TCID50チャレンジ用量を使用したチャレンジ後の鼻洗浄液中にいかなるウイルスも検出できなかったことは注目に値する。すべての先行するフェレットH5N1ワクチン試験は、ホモサブタイプのH5N1チャレンジ後に上気道における顕著な量のウイルス複製を報告している[45、50、51]。
【0215】
偽型VLP媒介防御の機構の理解は、さらなる研究を必要とするが、ワクチン有効性を示すための有力な要因はこれらのワクチンによって誘発される免疫応答全体の強さである。VLPの強い免疫原性は、本質的に任意の細胞(抗原提示細胞を含む)への効率的な結合を促進できる(これは増強された抗原提示をもたらす)、VLP上の機能性HAスパイクの存在に部分的による可能性がある。細胞結合能力についての証拠は、ヒヨコ赤血球を使用する赤血球凝集アッセイおよび赤血球凝集抑制アッセイの両方において観察されたVLPの強いHA活性である。本発明者らは、種々のサブタイプのVLPを用いてヒトRBCに向けた強いHA活性も実証した(データ未記載)。
【0216】
HAによって媒介される直接細胞結合能力に加えて、生きているバキュロウイルス粒子は短期間先天免疫を刺激でき、他の抗原と混合された場合にアジュバントとして作用できることが実証されていることから、バキュロウイルスベクター由来VLPワクチンの免疫原性はバキュロウイルス粒子を混入させることによって増強され得ることが示唆されている[46〜48]。バキュロウイルス粒子バンドは、本発明者らの手法においてはショ糖勾配でVLPよりもわずかに低く、プラークアッセイによるバキュロウイルスの定量およびタンパク質濃度によるVLPの定量は、VLP調製物においてバキュロウイルスをおよそ4桁超えるVLPの存在量を明らかにした。しかし、本発明者らは本明細書において記載する実験のほとんどにおいて使用したVLP調製物からバキュロウイルスを除外しなかった。しかし表5は、混入バキュロウイルス粒子を長波長UV/ソラーレン処理またはベータ−プロピオラクトン(BPL)処理のいずれかによって不活化したVLP免疫原性実験の結果を示している。不活化の両方法が核酸を広く標的化する一方で、本発明者らは、BPL処理後にVLP関連HA活性の顕著な減少を観察したが、UV/ソラーレン処理後にHA活性の減少は観察しなかった。ワクチン接種後のHAI力価は、昆虫細胞上での力価測定によって測定されたバキュロウイルス感染性の欠如にもかかわらず、UV/ソラーレン処理VLPが無処理VLPと比較して免疫原性の減少を被らなかったことを明らかにした。したがって混入している生きたバキュロウイルスは、これらのVLPに関連する免疫原性にほとんど影響を有さないと考えられる。興味深いことにBLP処理VLPは、VLP関連HA活性の16〜20分の1への減少によって明らかである通りおそらくVLPの直接アルキル化によってそれらの免疫原性をほとんど失っている。予測外に、混入バキュロウイルス粒子のUV/ソラーレン不活化は、VLP免疫原性に効果を有さず、したがってVLP作製における不活化の理想的方法である。
【0217】
VLP調製物中の低レベルのバキュロウイルス混入の問題は、HA偽型バキュロウイルス粒子が、観察されたインフルエンザ特異的免疫原性にどの程度寄与できるかについての追加的疑問を提起した。なぜ可能性が低いのかについては、いくつかの原因がある。第1にバキュロウイルス出芽ビリオン(BV)の会合および出芽は、脂質ラフトドメインを標的化しない[49]膜結合バキュロウイルスgp64に依存する[54]。第2に、ポリヘドリンプロモーターが活性化するとBV産生は遮断されることから、大部分のBV産生は一時的に大部分のGag−VLP会合から外れると予測され、Gag−HA−NA VLP会合をバキュロウイルス感染周期のかなり遅い段階で駆動する。第3に、BVとVLPとの会合の相対位置および時期に幾分かの重複があると仮定すると、BVおよびVLPの相対量における大きな相違が考慮されなければならない。上に記載の通り本発明者らの算定は、混入しているバキュロウイルス粒子の104倍過剰であるVLPを示す。そのような比は、使用された投与量レベルにおいて混入HA偽型BVがワクチン効果に顕著に寄与する可能性を下げる。
【0218】
要約すると本発明者らは、MLV gagならびに種々のインフルエンザAサブタイプ由来のインフルエンザHAおよびNAからなる偽型VLPの作製およびマウスおよびフェレットでの防御免疫原性を記載する。これらのVLPは、株にかかわらず迅速かつ一定して産生され、強い免疫原性を示す。VLP調製物由来のバキュロウイルス粒子を不活化する2つの異なる方法についての本発明者らの研究は、UV/ソラーレン処理による不活化がVLPにその完全なレベルの免疫原性を保持させるという驚くべき発見をもたらした。これらのVLPの免疫原性を既存のおよび提案されたインフルエンザワクチンと比較するため実験、ならびにこのプラットフォームの有用性を他の呼吸器ウイルスに拡張するための実験は、進行中である。
【0219】
(実施例8−UV−A、UV−Cおよび可視光によるウイルス不活化方法)
以下の実施例において本発明者らは、UV−A/リボフラビン、UV−C照射単独および可視光/リボフラビンによるウイルス不活化の好ましい方法を記載する。ウイルス不活化は、バイオリアクター回収物(bioreactor offloads)、遠心分離または濾過によるバイオリアクター回収後の精製開始材料、中間精製画分(連続流式勾配遠心分離画分、クロマトグラフィー素通り画分またはクロマトグラフィー溶出画分)および/または最終精製産物を含むエンベロープVLPの作製において生成された任意の画分について実施され得る。
【0220】
ウイルス不活化研究用に、エンベロープVLP含有試料を適切な放射−透明用具(保存袋またはチューブなど)に入れ、ウイルス不活化をこれだけに限らないが、1)単一のまたは複数の放射源を対象の試料を囲む所定の距離だが任意の配置に置く配置;2)所定の試料流路を提供するためにチューブを任意の商業的に入手できる放射源に繋いだ配置(試料は蠕動ポンプの手段によって流路を流れる)または;3)上に記載の流路をエンペロープVLP精製のための標準的装置操作の一部として(例えばタンジェンシャルフロー濾過またはクロマトグラフィー分離中、インラインで)物理的に統合した配置が挙げられ得るデバイスを使用して実施した。
【0221】
UV−A/リボフラビンを使用する病原体不活化のために、エンベロープVLP試料をリボフラビン濃度10〜100μMに調整し、265〜370nmの間の照射波長を使用するエネルギー量0〜50J/cm2に供する。UV−Cを使用する病原体不活化は、VLP試料を254nmの照射波長を使用するエネルギー量0〜2J/cm2で処理することによって実施する。可視光/リボフラビンを使用する病原体不活化のために、エンベロープVLP試料をリボブラビン濃度100〜500μMに調整し、次いで400〜500nmの間の照射波長を使用するエネルギー量0〜500J/cm2に供する。上の研究においてpH、温度および遊離ラジカルスカベンジャーの効果などの試料条件を不活化条件を最適化するために評価する。代表的な試料を種々の量の照射に曝露し、ウイルス不活化(例えばバキュロウイルス不活化)を標準的感染性アッセイによって評価する。エンベロープVLPへの最小の損傷を示すために、分析試験のパネルを処理後のエンベロープVLPの同一性、安定性および免疫原性を示すために実施する。適切なウイルス不活化条件が定められれば、移転可能な製造工程のより代表的である材料のレベルにおいて、選択されたウイルス不活化方法の効果を確実にするために、様々な深度の試料においてスケールアップ研究を実施する。
【0222】
(実施例9−ガンマ照射によるウイルス不活化方法)
ガンマ照射を使用する病原体不活化のために、エンベロープVLP試料(液体試料および凍結試料の両方として)に、遊離ラジカルスカベンジャーの存在下または非存在下で照射する。代表的な試料を0から60kGyに増大させるエネルギー量に供し、ウイルス不活化(例えばバキュロウイルス不活化)を標準的感染性アッセイによって評価する。エンベロープVLPへの最小の損傷を実証するために、分析試験のパネルを処理後のエンベロープVLPの同一性、安定性および免疫原性を示すために実施する。適切なウイルス不活化条件が定められれば、移転可能な製造工程のより代表的である材料のレベルにおいて、選択されたウイルス不活化方法の効果を確実にするために、様々な深度の試料においてスケールアップ研究を実施する。
【0223】
(さらなる参考文献)
【0224】
【数1】
【0225】
【数2】
【0226】
【数3】
【0227】
【数4】
【0228】
【数5】
【0229】
【数6】
【0230】
【数7】
【0231】
【数8】
【特許請求の範囲】
【請求項1】
感染因子を実質的に含まないエンベロープウイルスベースのウイルス様粒子調製物を単離する方法であって、
(a)前記エンベロープウイルスベースのウイルス様粒子調製物を作製するのに用いられる宿主細胞、または前記宿主細胞の成分から、前記エンベロープウイルスベースのウイルス様粒子調製物を単離するステップと、
(b)前記エンベロープウイルスベースのウイルス様粒子調製物における実質的にすべての感染因子を不活化するのに十分な線量の電磁放射を、前記調製物へと適用するステップと
を含み、ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物は、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物と実質的に同じ免疫原性を有する、方法。
【請求項2】
前記分離するステップ(a)が、少なくとも1つのクロマトグラフィーステップを含む、請求項1に記載の方法。
【請求項3】
前記少なくとも1つのクロマトグラフィーステップが、イオン交換クロマトグラフィー、アフィニティー−クロマトグラフィー、疎水性相互作用クロマトグラフィー、混合方式クロマトグラフィー、逆相クロマトグラフィー、およびサイズ除外クロマトグラフィーからなる群より選択される、請求項2に記載の方法。
【請求項4】
前記分離するステップ(a)が、少なくとも1つの濾過ステップを含む、請求項1から3のうちのいずれかに記載の方法。
【請求項5】
前記少なくとも1つの濾過ステップが、ノーマルフロー濾過、限外濾過、またはタンジェンシャルフロー濾過である、請求項4に記載の方法。
【請求項6】
前記分離するステップ(a)が、少なくとも1つの遠心分離ステップを含む、請求項1から5のうちのいずれかに記載の方法。
【請求項7】
前記電磁放射が、可視光、x線放射、紫外線放射、およびガンマ放射からなる群より選択される、請求項1から6のうちのいずれかに記載の方法。
【請求項8】
前記紫外線放射が、UV−A、UV−B、およびUV−Cからなる群より選択される、請求項7に記載の方法。
【請求項9】
前記紫外線放射の波長が、320nm〜400nmである、請求項7から8のうちのいずれかに記載の方法。
【請求項10】
前記宿主細胞が、昆虫細胞または哺乳動物細胞である、請求項1から9のうちのいずれかに記載の方法。
【請求項11】
前記宿主細胞が昆虫細胞であり、前記昆虫細胞は、前記エンベロープウイルスベースのウイルス様粒子の少なくとも1つの成分を発現するバキュロウイルス発現ベクターで感染させられる、請求項1から10のうちのいずれかに記載の方法。
【請求項12】
前記宿主細胞が、Bombyx mori宿主細胞、Spodoptera frugiperda宿主細胞、Choristoneura fumiferana宿主細胞、Heliothis virescens宿主細胞、Heliothis zea宿主細胞、Helicoverpa zea宿主細胞、Helicoverpa virescens宿主細胞、Orgyia pseudotsugata宿主細胞、Lymantria dispar宿主細胞、Plutella xylostella宿主細胞、Malacostoma disstria宿主細胞、Trichoplusia ni宿主細胞、Pieris rapae宿主細胞、Mamestra configurata宿主細胞、Mamestra brassica宿主細胞、およびHyalophora cecropia宿主細胞からなる群より選択される、請求項1から11のうちのいずれかに記載の方法。
【請求項13】
前記電磁放射線量が、前記エンベロープウイルスベースのウイルス様粒子調製物におけるバキュロウイルスを不活化させるのに十分である、請求項11から12のうちのいずれかに記載の方法。
【請求項14】
前記宿主細胞が哺乳動物細胞であり、前記哺乳動物細胞は、前記エンベロープウイルスベースのウイルス様粒子の少なくとも1つの成分を発現する、アデノウイルス発現ベクター、アデノ関連ウイルス発現ベクター、アルファウイルス発現ベクター、ヘルペスウイルス発現ベクター、ポックスウイルス発現ベクター、またはレトロウイルス発現ベクターで感染させられる、請求項1から10のうちのいずれかに記載の方法。
【請求項15】
前記適用するステップ(b)の前に、前記エンベロープウイルスベースのウイルス様粒子調製物にDNAインターカレート化合物を添加するステップをさらに含む、請求項1から14のうちのいずれかに記載の方法。
【請求項16】
前記DNAインターカレート化合物が、光反応性である、請求項15に記載の方法。
【請求項17】
前記DNAインターカレート化合物が、ソラーレン、イソソラーレン、ならびにこれらの誘導体および類似体からなる群より選択される、請求項15に記載の方法。
【請求項18】
前記電磁放射が、紫外線放射および可視光からなる群より選択される、請求項1から17のうちのいずれかに記載の方法。
【請求項19】
(c)前記エンベロープウイルスベースのウイルス様粒子調製物にアジュバントを添加するステップをさらに含む、請求項1から1718のうちのいずれかに記載の方法。
【請求項20】
前記分離するステップ(a)の前に、(i)gagポリペプチドと、天然では脂質ラフトと会合しない抗原に連結した脂質ラフト会合ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記gagポリペプチドと、前記抗原に連結した脂質ラフト会合ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、前記宿主細胞内において前記エンベロープウイルスベースのウイルス様粒子を作製する、請求項1から19のうちのいずれかに記載の方法。
【請求項21】
前記分離するステップ(a)の前に、(i)RSウイルスMポリペプチドと、RSウイルスFポリペプチドとを発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記RSウイルスMポリペプチドと、前記RSウイルスFポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、前記宿主細胞内において前記エンベロープウイルスベースのウイルス様粒子を作製する、請求項1から19のうちのいずれかに記載の方法。
【請求項22】
前記1または複数の発現ベクターが、RSウイルスGポリペプチドをさらに発現する、請求項21に記載の方法。
【請求項23】
前記分離するステップ(a)の前に、(i)gagポリペプチドと、インフルエンザ赤血球凝集素ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記gagポリペプチドと、前記インフルエンザ赤血球凝集素ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、前記宿主細胞内において前記エンベロープウイルスベースのウイルス様粒子を作製する、請求項1から19のうちのいずれかに記載の方法。
【請求項24】
前記分離するステップ(a)の前に、(i)インフルエンザM1ポリペプチドと、赤血球凝集素ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記インフルエンザM1ポリペプチドと、前記赤血球凝集素ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、前記宿主細胞内において前記エンベロープウイルスベースのウイルス様粒子を作製する、請求項1から19のうちのいずれかに記載の方法。
【請求項25】
前記1または複数の発現ベクターが、ノイラミニダーゼポリペプチドをさらに発現する、請求項23から24のうちのいずれかに記載の方法。
【請求項26】
前記1または複数の発現ベクターが、ウイルスベクターである、請求項20から25のうちのいずれかに記載の方法。
【請求項27】
前記ウイルスベクターが、バキュロウイルス、アデノウイルス、アデノ関連ウイルス、アルファウイルス、ヘルペスウイルス、ポックスウイルス、およびレトロウイルスからなる群より選択される、請求項26に記載の方法。
【請求項28】
ウイルスベクターを用いて、前記宿主細胞内において、前記エンベロープウイルスベースのウイルス様粒子の少なくとも1つの成分を発現させる、請求項1から27のうちのいずれかに記載の方法。
【請求項29】
前記感染因子が、前記ウイルスベクターを含む、請求項28に記載の方法。
【請求項30】
前記調製物が、1ミリリットル当たり20個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項31】
前記調製物が、1ミリリットル当たり15個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項32】
前記調製物が、1ミリリットル当たり10個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項33】
前記調製物が、1ミリリットル当たり8個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項34】
前記調製物が、1ミリリットル当たり6個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項35】
前記調製物が、1ミリリットル当たり5個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項36】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも50パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項37】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも60パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項38】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも70パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項39】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも80パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項40】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも85パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項41】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも90パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項42】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも95パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項43】
前記エンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含み、前記免疫原性が、赤血球凝集抑制アッセイを用いて測定される、請求項1から42のうちのいずれかに記載の方法。
【請求項44】
感染因子を実質的に含まないエンベロープウイルスベースのウイルス様粒子を含む、エンベロープウイルスベースのウイルス様粒子調製物であって、前記エンベロープウイルスベースのウイルス様粒子が、感染因子を実質的に含まないわけではないエンベロープウイルスベースのウイルス様粒子と実質的に同じ免疫原性を有する、調製物。
【請求項45】
前記エンベロープウイルスベースのウイルス様粒子が、昆虫または哺乳動物グリコシル化をさらに含む、請求項44に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項46】
前記昆虫グリコシル化が、Autographa californica;Bombyx mori;Spodoptera frugiperda;Choristoneura fumiferana;Heliothis virescens;Heliothis zea;Helicoverpa zea;Helicoverpa virescens;Orgyia pseudotsugata;Lymantria dispar;Plutella xylostella;Malacostoma disstria;Trichoplusia ni;Pieris rapae;Mamestra configurata;Mamestra brassica;およびHyalophora cecropiaからなる群より選択される、請求項44または45に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項47】
前記エンベロープウイルスベースのウイルス様粒子が、共有結合した光化学作用物質、タンパク質サブユニットの三次構造もしくは四次構造においてUV照射もしくはガンマ照射により誘導される変化、ガンマ照射により誘導される化学結合の開裂、またはUV照射もしくはガンマ照射により誘導される、脂質の酸化、タンパク質の架橋形成、アミノ酸の酸化、およびアミノ酸の修飾からなる群より選択される化学修飾から選択される1または複数の欠損をさらに示す、請求項44から46のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項48】
前記エンベロープウイルスベースのウイルス様粒子が、以下の欠損:共有結合した光化学作用物質、タンパク質サブユニットの三次構造もしくは四次構造においてUV照射もしくはガンマ照射により誘導される変化、ガンマ照射により誘導される化学結合の開裂、またはUV照射もしくはガンマ照射により誘導される、脂質の酸化、タンパク質の架橋形成、アミノ酸の酸化、およびアミノ酸の修飾からなる群より選択される化学修飾のいずれもを含まない、請求項47に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項49】
前記エンベロープウイルスベースのウイルス様粒子が、
(a)gagポリペプチドと、
(b)非ウイルス性脂質ラフト会合ポリペプチド、または連結を形成するための抗原に連結した脂質ラフト会合ポリペプチドであって、前記抗原は天然では脂質ラフトと会合しない、ポリペプチドと
を含む、請求項44から48のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項50】
前記非ウイルス性脂質ラフト会合ポリペプチドが、GPIアンカーポリペプチド、ミリストイル化配列ポリペプチド、パルミトイル化配列ポリペプチド、二重アセチル化配列ポリペプチド、シグナル伝達ポリペプチド、および膜輸送ポリペプチドからなる群より選択される、請求項49に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項51】
前記非ウイルス性脂質ラフト会合ポリペプチドが、GPIアンカーポリペプチド、ミリストイル化配列ポリペプチド、パルミトイル化配列ポリペプチド、二重アセチル化配列ポリペプチド、カベオリンポリペプチド、フロチリンポリペプチド、シンタクシン−1ポリペプチド、シンタクシン−4ポリペプチド、シナプシンIポリペプチド、アデューシンポリペプチド、VAMP2ポリペプチド、VAMP/シナプトブレビンポリペプチド、シナプトブレビンIIポリペプチド、SNAREポリペプチド、SNAP−25ポリペプチド、SNAP−23ポリペプチド、シナプトタグミンIポリペプチド、およびシナプトタグミンIIポリペプチドからなる群より選択される、請求項49に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項52】
前記ウイルス性脂質ラフト会合ポリペプチドが、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、融合タンパク質ポリペプチド、糖タンパク質ポリペプチド、およびエンベロープタンパク質ポリペプチドからなる群より選択される、請求項49に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項53】
前記連結が、共有結合、イオン性相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、および抗体−抗原相互作用からなる群より選択される、請求項49から52のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項54】
前記連結が、共有結合である、請求項49から52のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項55】
前記共有結合が、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、およびジスルフィド結合からなる群より選択される、請求項54に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項56】
前記脂質ラフト会合ポリペプチドが、内在性膜タンパク質である、請求項49から55のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項57】
前記抗原が、タンパク質、ポリペプチド、グリコポリペプチド、リポポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性または非ペプチド性模倣体、低分子、脂質、糖脂質、および炭水化物からなる群より選択される、請求項49から56のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項58】
赤血球凝集素ポリペプチドをさらに含む、請求項44から57のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項59】
RSウイルスMポリペプチドをさらに含む、請求項44から58のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項60】
RSウイルスGポリペプチドをさらに含む、請求項44から59のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項61】
RSウイルスFポリペプチドをさらに含む、請求項44から60のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項62】
前記エンベロープウイルスベースのウイルス様粒子が、
(a)gagポリペプチドと、
(b)赤血球凝集素ポリペプチドと
を含む、請求項44から47のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項63】
前記エンベロープウイルスベースのウイルス様粒子が、
(a)インフルエンザM1ポリペプチドと、
(b)赤血球凝集素ポリペプチドと
を含む、請求項44から47のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項64】
ノイラミニダーゼポリペプチドをさらに含む、請求項44から63に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項65】
前記ウイルス様粒子と会合するアジュバントをさらに含む、請求項44から64に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項66】
前記アジュバントが、前記ウイルス様粒子の内側に位置する、請求項65に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項67】
前記アジュバントが、前記ウイルス様粒子の外側に位置する、請求項65に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項68】
前記アジュバントを、前記gagポリペプチドと共有結合させて、共有結合を形成させる、請求項65から67のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項69】
前記アジュバントを、前記脂質ラフト会合ポリペプチドと共有結合させて、共有結合を形成させる、請求項65から67のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項70】
前記アジュバントが、フラジェリンのアジュバント活性断片を含む、請求項65から69のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項71】
免疫系の疾患または症状を処置または予防する方法であって、免疫原性量の、請求項44から70のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物、または請求項1から43のうちのいずれかに記載の方法を用いて単離したエンベロープウイルスベースのウイルス様粒子調製物を投与するステップを含む、方法。
【請求項72】
前記投与するステップにより、被験体において、防御的免疫化反応が誘導される、請求項71に記載の方法。
【請求項73】
前記投与するステップが、皮下送達、皮内送達、皮下送達、筋肉内送達、経口送達、経口送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群より選択される、請求項71から72のうちのいずれかに記載の方法。
【請求項74】
免疫原性量の、請求項44から70のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物、または請求項1から43のうちのいずれかに記載の方法を用いて単離したエンベロープウイルスベースのウイルス様粒子調製物と、薬学的に許容されるキャリアとを含む、医薬組成物。
【請求項1】
感染因子を実質的に含まないエンベロープウイルスベースのウイルス様粒子調製物を単離する方法であって、
(a)前記エンベロープウイルスベースのウイルス様粒子調製物を作製するのに用いられる宿主細胞、または前記宿主細胞の成分から、前記エンベロープウイルスベースのウイルス様粒子調製物を単離するステップと、
(b)前記エンベロープウイルスベースのウイルス様粒子調製物における実質的にすべての感染因子を不活化するのに十分な線量の電磁放射を、前記調製物へと適用するステップと
を含み、ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物は、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物と実質的に同じ免疫原性を有する、方法。
【請求項2】
前記分離するステップ(a)が、少なくとも1つのクロマトグラフィーステップを含む、請求項1に記載の方法。
【請求項3】
前記少なくとも1つのクロマトグラフィーステップが、イオン交換クロマトグラフィー、アフィニティー−クロマトグラフィー、疎水性相互作用クロマトグラフィー、混合方式クロマトグラフィー、逆相クロマトグラフィー、およびサイズ除外クロマトグラフィーからなる群より選択される、請求項2に記載の方法。
【請求項4】
前記分離するステップ(a)が、少なくとも1つの濾過ステップを含む、請求項1から3のうちのいずれかに記載の方法。
【請求項5】
前記少なくとも1つの濾過ステップが、ノーマルフロー濾過、限外濾過、またはタンジェンシャルフロー濾過である、請求項4に記載の方法。
【請求項6】
前記分離するステップ(a)が、少なくとも1つの遠心分離ステップを含む、請求項1から5のうちのいずれかに記載の方法。
【請求項7】
前記電磁放射が、可視光、x線放射、紫外線放射、およびガンマ放射からなる群より選択される、請求項1から6のうちのいずれかに記載の方法。
【請求項8】
前記紫外線放射が、UV−A、UV−B、およびUV−Cからなる群より選択される、請求項7に記載の方法。
【請求項9】
前記紫外線放射の波長が、320nm〜400nmである、請求項7から8のうちのいずれかに記載の方法。
【請求項10】
前記宿主細胞が、昆虫細胞または哺乳動物細胞である、請求項1から9のうちのいずれかに記載の方法。
【請求項11】
前記宿主細胞が昆虫細胞であり、前記昆虫細胞は、前記エンベロープウイルスベースのウイルス様粒子の少なくとも1つの成分を発現するバキュロウイルス発現ベクターで感染させられる、請求項1から10のうちのいずれかに記載の方法。
【請求項12】
前記宿主細胞が、Bombyx mori宿主細胞、Spodoptera frugiperda宿主細胞、Choristoneura fumiferana宿主細胞、Heliothis virescens宿主細胞、Heliothis zea宿主細胞、Helicoverpa zea宿主細胞、Helicoverpa virescens宿主細胞、Orgyia pseudotsugata宿主細胞、Lymantria dispar宿主細胞、Plutella xylostella宿主細胞、Malacostoma disstria宿主細胞、Trichoplusia ni宿主細胞、Pieris rapae宿主細胞、Mamestra configurata宿主細胞、Mamestra brassica宿主細胞、およびHyalophora cecropia宿主細胞からなる群より選択される、請求項1から11のうちのいずれかに記載の方法。
【請求項13】
前記電磁放射線量が、前記エンベロープウイルスベースのウイルス様粒子調製物におけるバキュロウイルスを不活化させるのに十分である、請求項11から12のうちのいずれかに記載の方法。
【請求項14】
前記宿主細胞が哺乳動物細胞であり、前記哺乳動物細胞は、前記エンベロープウイルスベースのウイルス様粒子の少なくとも1つの成分を発現する、アデノウイルス発現ベクター、アデノ関連ウイルス発現ベクター、アルファウイルス発現ベクター、ヘルペスウイルス発現ベクター、ポックスウイルス発現ベクター、またはレトロウイルス発現ベクターで感染させられる、請求項1から10のうちのいずれかに記載の方法。
【請求項15】
前記適用するステップ(b)の前に、前記エンベロープウイルスベースのウイルス様粒子調製物にDNAインターカレート化合物を添加するステップをさらに含む、請求項1から14のうちのいずれかに記載の方法。
【請求項16】
前記DNAインターカレート化合物が、光反応性である、請求項15に記載の方法。
【請求項17】
前記DNAインターカレート化合物が、ソラーレン、イソソラーレン、ならびにこれらの誘導体および類似体からなる群より選択される、請求項15に記載の方法。
【請求項18】
前記電磁放射が、紫外線放射および可視光からなる群より選択される、請求項1から17のうちのいずれかに記載の方法。
【請求項19】
(c)前記エンベロープウイルスベースのウイルス様粒子調製物にアジュバントを添加するステップをさらに含む、請求項1から1718のうちのいずれかに記載の方法。
【請求項20】
前記分離するステップ(a)の前に、(i)gagポリペプチドと、天然では脂質ラフトと会合しない抗原に連結した脂質ラフト会合ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記gagポリペプチドと、前記抗原に連結した脂質ラフト会合ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、前記宿主細胞内において前記エンベロープウイルスベースのウイルス様粒子を作製する、請求項1から19のうちのいずれかに記載の方法。
【請求項21】
前記分離するステップ(a)の前に、(i)RSウイルスMポリペプチドと、RSウイルスFポリペプチドとを発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記RSウイルスMポリペプチドと、前記RSウイルスFポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、前記宿主細胞内において前記エンベロープウイルスベースのウイルス様粒子を作製する、請求項1から19のうちのいずれかに記載の方法。
【請求項22】
前記1または複数の発現ベクターが、RSウイルスGポリペプチドをさらに発現する、請求項21に記載の方法。
【請求項23】
前記分離するステップ(a)の前に、(i)gagポリペプチドと、インフルエンザ赤血球凝集素ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記gagポリペプチドと、前記インフルエンザ赤血球凝集素ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、前記宿主細胞内において前記エンベロープウイルスベースのウイルス様粒子を作製する、請求項1から19のうちのいずれかに記載の方法。
【請求項24】
前記分離するステップ(a)の前に、(i)インフルエンザM1ポリペプチドと、赤血球凝集素ポリペプチドとを併せて発現する、1または複数の発現ベクターを供給するステップと;(ii)前記1または複数の発現ベクターを細胞内へと導入するステップと;(iii)前記インフルエンザM1ポリペプチドと、前記赤血球凝集素ポリペプチドとを発現させて、前記ウイルス様粒子を作製するステップとにより、前記宿主細胞内において前記エンベロープウイルスベースのウイルス様粒子を作製する、請求項1から19のうちのいずれかに記載の方法。
【請求項25】
前記1または複数の発現ベクターが、ノイラミニダーゼポリペプチドをさらに発現する、請求項23から24のうちのいずれかに記載の方法。
【請求項26】
前記1または複数の発現ベクターが、ウイルスベクターである、請求項20から25のうちのいずれかに記載の方法。
【請求項27】
前記ウイルスベクターが、バキュロウイルス、アデノウイルス、アデノ関連ウイルス、アルファウイルス、ヘルペスウイルス、ポックスウイルス、およびレトロウイルスからなる群より選択される、請求項26に記載の方法。
【請求項28】
ウイルスベクターを用いて、前記宿主細胞内において、前記エンベロープウイルスベースのウイルス様粒子の少なくとも1つの成分を発現させる、請求項1から27のうちのいずれかに記載の方法。
【請求項29】
前記感染因子が、前記ウイルスベクターを含む、請求項28に記載の方法。
【請求項30】
前記調製物が、1ミリリットル当たり20個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項31】
前記調製物が、1ミリリットル当たり15個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項32】
前記調製物が、1ミリリットル当たり10個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項33】
前記調製物が、1ミリリットル当たり8個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項34】
前記調製物が、1ミリリットル当たり6個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項35】
前記調製物が、1ミリリットル当たり5個未満の感染因子を含む、請求項1から29のうちのいずれかに記載の方法。
【請求項36】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも50パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項37】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも60パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項38】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも70パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項39】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも80パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項40】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも85パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項41】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも90パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項42】
ステップ(b)の後の前記エンベロープウイルスベースのウイルス様粒子調製物が、ステップ(b)の前の前記エンベロープウイルスベースのウイルス様粒子調製物の少なくとも95パーセントの免疫原性を有する、請求項1から35のうちのいずれかに記載の方法。
【請求項43】
前記エンベロープウイルスベースのウイルス様粒子が、赤血球凝集素ポリペプチドを含み、前記免疫原性が、赤血球凝集抑制アッセイを用いて測定される、請求項1から42のうちのいずれかに記載の方法。
【請求項44】
感染因子を実質的に含まないエンベロープウイルスベースのウイルス様粒子を含む、エンベロープウイルスベースのウイルス様粒子調製物であって、前記エンベロープウイルスベースのウイルス様粒子が、感染因子を実質的に含まないわけではないエンベロープウイルスベースのウイルス様粒子と実質的に同じ免疫原性を有する、調製物。
【請求項45】
前記エンベロープウイルスベースのウイルス様粒子が、昆虫または哺乳動物グリコシル化をさらに含む、請求項44に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項46】
前記昆虫グリコシル化が、Autographa californica;Bombyx mori;Spodoptera frugiperda;Choristoneura fumiferana;Heliothis virescens;Heliothis zea;Helicoverpa zea;Helicoverpa virescens;Orgyia pseudotsugata;Lymantria dispar;Plutella xylostella;Malacostoma disstria;Trichoplusia ni;Pieris rapae;Mamestra configurata;Mamestra brassica;およびHyalophora cecropiaからなる群より選択される、請求項44または45に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項47】
前記エンベロープウイルスベースのウイルス様粒子が、共有結合した光化学作用物質、タンパク質サブユニットの三次構造もしくは四次構造においてUV照射もしくはガンマ照射により誘導される変化、ガンマ照射により誘導される化学結合の開裂、またはUV照射もしくはガンマ照射により誘導される、脂質の酸化、タンパク質の架橋形成、アミノ酸の酸化、およびアミノ酸の修飾からなる群より選択される化学修飾から選択される1または複数の欠損をさらに示す、請求項44から46のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項48】
前記エンベロープウイルスベースのウイルス様粒子が、以下の欠損:共有結合した光化学作用物質、タンパク質サブユニットの三次構造もしくは四次構造においてUV照射もしくはガンマ照射により誘導される変化、ガンマ照射により誘導される化学結合の開裂、またはUV照射もしくはガンマ照射により誘導される、脂質の酸化、タンパク質の架橋形成、アミノ酸の酸化、およびアミノ酸の修飾からなる群より選択される化学修飾のいずれもを含まない、請求項47に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項49】
前記エンベロープウイルスベースのウイルス様粒子が、
(a)gagポリペプチドと、
(b)非ウイルス性脂質ラフト会合ポリペプチド、または連結を形成するための抗原に連結した脂質ラフト会合ポリペプチドであって、前記抗原は天然では脂質ラフトと会合しない、ポリペプチドと
を含む、請求項44から48のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項50】
前記非ウイルス性脂質ラフト会合ポリペプチドが、GPIアンカーポリペプチド、ミリストイル化配列ポリペプチド、パルミトイル化配列ポリペプチド、二重アセチル化配列ポリペプチド、シグナル伝達ポリペプチド、および膜輸送ポリペプチドからなる群より選択される、請求項49に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項51】
前記非ウイルス性脂質ラフト会合ポリペプチドが、GPIアンカーポリペプチド、ミリストイル化配列ポリペプチド、パルミトイル化配列ポリペプチド、二重アセチル化配列ポリペプチド、カベオリンポリペプチド、フロチリンポリペプチド、シンタクシン−1ポリペプチド、シンタクシン−4ポリペプチド、シナプシンIポリペプチド、アデューシンポリペプチド、VAMP2ポリペプチド、VAMP/シナプトブレビンポリペプチド、シナプトブレビンIIポリペプチド、SNAREポリペプチド、SNAP−25ポリペプチド、SNAP−23ポリペプチド、シナプトタグミンIポリペプチド、およびシナプトタグミンIIポリペプチドからなる群より選択される、請求項49に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項52】
前記ウイルス性脂質ラフト会合ポリペプチドが、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、融合タンパク質ポリペプチド、糖タンパク質ポリペプチド、およびエンベロープタンパク質ポリペプチドからなる群より選択される、請求項49に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項53】
前記連結が、共有結合、イオン性相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、および抗体−抗原相互作用からなる群より選択される、請求項49から52のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項54】
前記連結が、共有結合である、請求項49から52のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項55】
前記共有結合が、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、およびジスルフィド結合からなる群より選択される、請求項54に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項56】
前記脂質ラフト会合ポリペプチドが、内在性膜タンパク質である、請求項49から55のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項57】
前記抗原が、タンパク質、ポリペプチド、グリコポリペプチド、リポポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性または非ペプチド性模倣体、低分子、脂質、糖脂質、および炭水化物からなる群より選択される、請求項49から56のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項58】
赤血球凝集素ポリペプチドをさらに含む、請求項44から57のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項59】
RSウイルスMポリペプチドをさらに含む、請求項44から58のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項60】
RSウイルスGポリペプチドをさらに含む、請求項44から59のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項61】
RSウイルスFポリペプチドをさらに含む、請求項44から60のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項62】
前記エンベロープウイルスベースのウイルス様粒子が、
(a)gagポリペプチドと、
(b)赤血球凝集素ポリペプチドと
を含む、請求項44から47のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項63】
前記エンベロープウイルスベースのウイルス様粒子が、
(a)インフルエンザM1ポリペプチドと、
(b)赤血球凝集素ポリペプチドと
を含む、請求項44から47のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項64】
ノイラミニダーゼポリペプチドをさらに含む、請求項44から63に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項65】
前記ウイルス様粒子と会合するアジュバントをさらに含む、請求項44から64に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項66】
前記アジュバントが、前記ウイルス様粒子の内側に位置する、請求項65に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項67】
前記アジュバントが、前記ウイルス様粒子の外側に位置する、請求項65に記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項68】
前記アジュバントを、前記gagポリペプチドと共有結合させて、共有結合を形成させる、請求項65から67のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項69】
前記アジュバントを、前記脂質ラフト会合ポリペプチドと共有結合させて、共有結合を形成させる、請求項65から67のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項70】
前記アジュバントが、フラジェリンのアジュバント活性断片を含む、請求項65から69のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物。
【請求項71】
免疫系の疾患または症状を処置または予防する方法であって、免疫原性量の、請求項44から70のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物、または請求項1から43のうちのいずれかに記載の方法を用いて単離したエンベロープウイルスベースのウイルス様粒子調製物を投与するステップを含む、方法。
【請求項72】
前記投与するステップにより、被験体において、防御的免疫化反応が誘導される、請求項71に記載の方法。
【請求項73】
前記投与するステップが、皮下送達、皮内送達、皮下送達、筋肉内送達、経口送達、経口送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群より選択される、請求項71から72のうちのいずれかに記載の方法。
【請求項74】
免疫原性量の、請求項44から70のうちのいずれかに記載のエンベロープウイルスベースのウイルス様粒子調製物、または請求項1から43のうちのいずれかに記載の方法を用いて単離したエンベロープウイルスベースのウイルス様粒子調製物と、薬学的に許容されるキャリアとを含む、医薬組成物。
【図2】

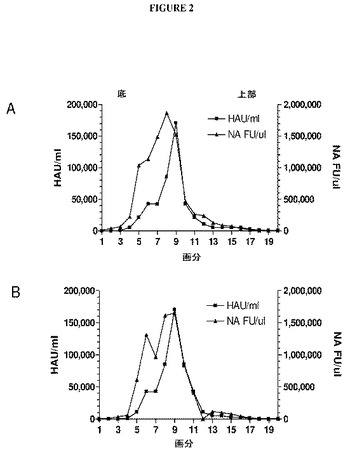
【図3】
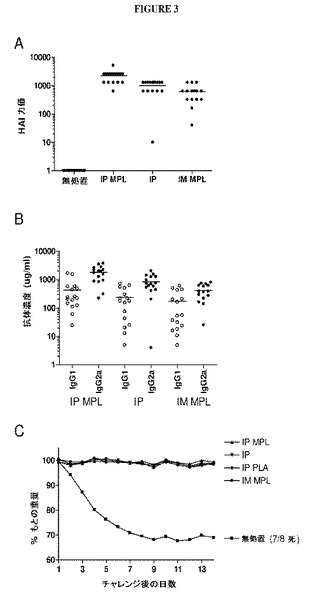
【図4】
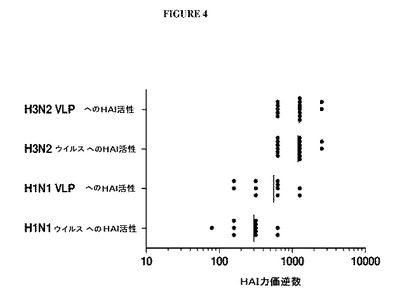
【図5】
【図6】
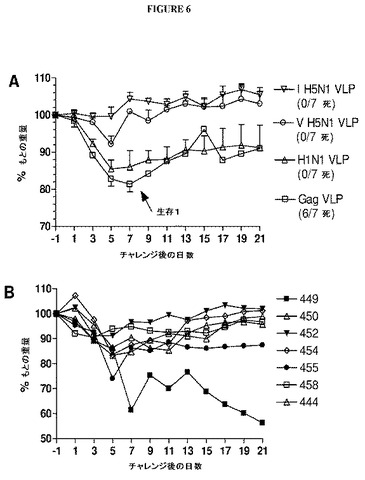
【図7】
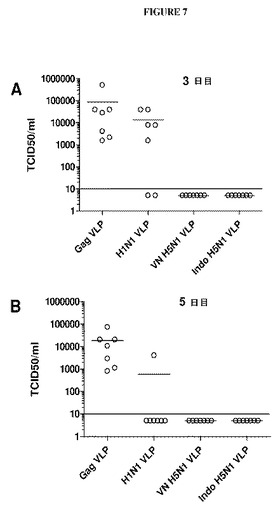
【図1】
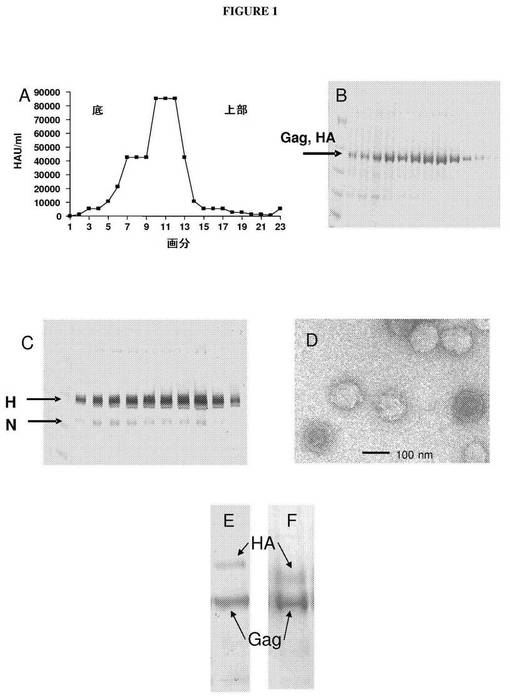
【図3】
【図4】
【図5】
【図6】
【図7】
【図1】
【公表番号】特表2012−507985(P2012−507985A)
【公表日】平成24年4月5日(2012.4.5)
【国際特許分類】
【出願番号】特願2011−534885(P2011−534885)
【出願日】平成21年11月3日(2009.11.3)
【国際出願番号】PCT/US2009/063128
【国際公開番号】WO2010/062757
【国際公開日】平成22年6月3日(2010.6.3)
【出願人】(509024190)リゴサイト ファーマシューティカルズ インコーポレイテッド (6)
【氏名又は名称原語表記】LIGOCYTE PHARMACEUTICALS,INC.
【住所又は居所原語表記】2155 Analysis Drive,Bozeman,MT 59718−6831,United States of America
【Fターム(参考)】
【公表日】平成24年4月5日(2012.4.5)
【国際特許分類】
【出願日】平成21年11月3日(2009.11.3)
【国際出願番号】PCT/US2009/063128
【国際公開番号】WO2010/062757
【国際公開日】平成22年6月3日(2010.6.3)
【出願人】(509024190)リゴサイト ファーマシューティカルズ インコーポレイテッド (6)
【氏名又は名称原語表記】LIGOCYTE PHARMACEUTICALS,INC.
【住所又は居所原語表記】2155 Analysis Drive,Bozeman,MT 59718−6831,United States of America
【Fターム(参考)】
[ Back to top ]