
成長因子複合体ならびに細胞の移動および増殖の調節
成長因子受容体に結合し活性化することができる、成長因子、例えばIGF−I、IGF−II、VEGFまたはPDGF、あるいは少なくとも該成長因子のドメインと、ビトロネクチンまたはフィブロネクチンのインテグリン受容体結合ドメインとを含んでなる単離タンパク質複合体が提供される。これらのタンパク質複合体は、オリゴタンパク質複合体の形態でも、単一の合成タンパク質の形態でもよく、単一の合成タンパク質の場合、成長因子およびビトロネクチン配列またはフィブロネクチン配列はリンカー配列により連結される。特定の形態においては、ビトロネクチン配列またはフィブロネクチン配列はヘパリン結合ドメインおよびポリ陰イオン性ドメインのうち少なくともいずれかを含まない。細胞の移動および増殖のうち少なくともいずれかを刺激または誘導するためのこれらのタンパク質複合体の使用も提供され、創傷治癒、組織工学、美容的および治療的処置、例えば皮膚置換および皮膚補充、ならびに上皮細胞の移動が必要とされる熱傷の治療における用途を有し得る。その他の実施形態では、本発明は、特に乳がんに関するがん細胞転移の抑制を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、成長因子受容体、例えば1型インスリン様成長因子受容体、ならびにビトロネクチンまたはフィブロネクチンの受容体であるインテグリンのいずれにも結合し活性化することができる各ドメインを有するタンパク質複合体に関する。特定の実施形態において、本発明は、インスリン様成長因子−I、インスリン様成長因子−II、血小板由来成長因子または血管内皮由来成長因子の受容体結合ドメイン、ならびにビトロネクチンまたはフィブロネクチンのインテグリン受容体結合ドメインを含んでなるキメラタンパク質に関する。特に本発明は、細胞移動を刺激するタンパク質複合体、ならびに細胞の移動および増殖のうち少なくともいずれかを促進または誘導する組成物および方法に関する。これらの組成物および方法は、創傷治癒、組織工学、美容上および治療上の処置(例えば皮膚置換および皮膚補充)ならびに上皮細胞の移動および増殖のうち少なくともいずれかが必要とされる熱傷の治療に用途を有し得る。その他の実施形態では、本発明は、特に乳がんに関連したがん細胞の転移の防止または抑制に関する本発明により提供される治療を提供する。本発明のキメラタンパク質は、インスリン様成長因子、ビトロネクチンおよびインスリン様成長因子結合タンパク質を含むタンパク質複合体の生物学的作用のアゴニストおよびアンタゴニストの作製のためにも有用であり得る。
【背景技術】
【0002】
インスリン様成長因子(IGF)であるIGF−IおよびIGF−IIは、広範囲の細胞プロセス、例えば過形成、DNA合成、分化、細胞周期の進行およびアポトーシスの抑制に関与する、細胞分裂促進性ペプチド成長因子である(非特許文献1〜3)。これらの作用は、IGFのチロシンキナーゼ結合型細胞表面受容体である、1型IGF受容体(IGF−IR)との結合により媒介される。また、IGFは、IGFBPと呼ばれる特定の結合タンパク質ファミリーにより厳密に調節されているが、IGFBPの主な役割は、遊離IGFを結合することによりIGFの半減期、特異性および活性を調節することである(非特許文献4)。
【0003】
近年、ビトロネクチン(VN)はIGF−IIと直接結合することが示された(非特許文献5)が、一方、IGF−Iは、特許文献1に記載されているように、ある種のIGFBPの存在下でVNと結合し得る。ECM組織化分子および接着分子であるVNがIGF−IIを結合する親和性は、生物学的関連を有する受容体であるIGF−IRに対するIGF−IIの親和性(非特許文献5)と同様である、という知見は、IGFの作用とECM中のVNとの間の特異的な物理的関連を明示している。さらに、VNに結合したIGF−IIは、in vitroでのヒトケラチノサイトにおける相乗的な機能的応答を刺激し得る(特許文献1)。
【0004】
VNは、血液中およびECM中で非常に豊富な糖タンパク質である。主に肝臓で合成されるが、しかし多数のその他のタイプの細胞によっても発現されるVNは、閉じた立体構造で血液中を循環し、そして開放または伸張した立体構造でECM中に沈着される(非特許文献6)。いずれの立体構造も、IGF−IIを結合すると考えられ(非特許文献5、特許文献1、非特許文献7)、かつ多様なその他のリガンド、例えばコラーゲン(非特許文献8)、グリコサミノグリカン(非特許文献9)、多数のその他のECMタンパク質、ならびに広範な種々のインテグリン、特にαVインテグリンも結合すると考えられる。実際、ビトロネクチンの主な役割は、RGD結合モチーフを介してこれらの細胞表面インテグリン受容体との接着性連結を提供する、ECM組織化分子としての役割である。VN受容体(αVインテグリン)は、増殖および侵潤に必要とされるアクチン細胞骨格の再編成を調節することが示されており、従って、VNの結合は細胞の接着および運動を調整する
(非特許文献10および11)。
【0005】
しかしながら、タンパク質複合体中に存在するIGFおよびVNのそれぞれの相対的寄与は、生物学的応答、例えば細胞の移動および増殖のうち少なくともいずれかの刺激という点において、依然として分かりにくい。なぜなら、IGF/IGFBPとVNとの間にタンパク質−タンパク質相互作用の部位があるからである。
【特許文献1】国際公開第02/24219号パンフレット
【非特許文献1】ケイス(Keiss )他、1994年、Hormone Research、第41巻、p.66
【非特許文献2】ウッドおよびイー(Wood & Yee)、2000年、J. Mammary Gland Biology and Neoplasia、第5巻、p.1
【非特許文献3】ジョンおよびクレモンス(Jones & Clemmons)、1995年、Endocrine Rev.、第16巻、p.3
【非特許文献4】クレモンス(Clemmons)、1998年、Mol. Cell. Endocrinol.、第140巻、p.19
【非特許文献5】アプトン(Upton )他、1999年、Endocrinology 、第140巻、p.2928‐31
【非特許文献6】シュバルツ(Schvartz)他、1999年、The International Journal of Biochemistry and Cell Biology、第31巻、p.531‐44
【非特許文献7】マクムーティ(McMurty )他、1996年、Endocrinology 、第150巻、p.149‐60
【非特許文献8】モーリス(Morris)他、1994年、Journal of Biological Chemistry 、第269巻、p.23845‐52
【非特許文献9】フランソワ(Francois)他、1999年、Journal of Biological Chemistry 、第274巻、p.37611‐19
【非特許文献10】デパスカール(DePasquale)、1998年、Histochemistry and Cell Biology 、第110巻、p.485‐94
【非特許文献11】ユアン(Huang )、2000年、Oncogene、第19巻、p.1915‐23
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、成長因子複合体ならびに細胞の移動および増殖を調節する方法を提供することである。
【課題を解決するための手段】
【0007】
本発明者等は、IGF−IIおよびVN、またはIGF−IおよびIGFBPおよびVNを含むタンパク質複合体が、IGF−I受容体(IGF−IR)およびVN−結合インテグリン受容体に結合し、かつ相乗的に同時活性化することにより、細胞の移動および増殖のうち少なくともいずれかを刺激する、ということを発見した。
【0008】
さらに、VNのポリ陰イオン性ドメインが同定されIGFまたはIGFBPの結合部位として提唱された。
したがって本発明は、広範には、成長因子ドメインの受容体結合ドメインと、インテグリン受容体に結合し得るビトロネクチンまたはフィブロネクチンのドメインとを含んでなる単離タンパク質複合体であって、成長因子およびインテグリン受容体を同時活性化することにより生物学的応答を引き出し得る単離タンパク質複合体に関する。
【0009】
第一の態様において、本発明は、単離タンパク質複合体であって、
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子
受容体に結合し得るドメイン、および
(ii)ビトロネクチン(VN)もしくはフィブロネクチン(FN)、または少なくともVNもしくはFNのインテグリン結合ドメイン
を含んでなる単離タンパク質複合体を提供する。
【0010】
第二の態様において、本発明は、合成キメラタンパク質の形態の単離タンパク質複合体であって、
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子受容体に結合し得るドメイン、および
(ii)ビトロネクチン(VN)もしくはフィブロネクチン(FN)、または少なくともVNもしくはFNのインテグリン結合ドメイン
のアミノ酸配列を含む単離タンパク質複合体を提供する。
【0011】
好ましくは、上記の態様によれば、成長因子はIGF−IまたはIGF−IIである。
さらに好ましくは、成長因子はIGF−Iである。
成長因子がIGF−Iである実施形態では、適切にはIGF−Iの前記少なくとも1つのドメインはIGF−Iの残基24を含む。
【0012】
成長因子がIGF−IIである実施形態では、適切にはIGF−IIの前記少なくとも1つのドメインはIGF−IIの残基27を含む。
代替的実施形態では、成長因子はVEGFまたはPDGFである。
【0013】
好ましくは、インテグリン受容体は、αVインテグリンである。
さらに好ましくは、インテグリン受容体は、αVβ3インテグリンまたはαVβ5インテグリンである。
【0014】
本発明のこの態様では、上記(i)および(ii)に対応するアミノ酸配列の、アミノ酸の欠失、付加、置換および突然変異のうち少なくともいずれかについても、その範囲内に含まれる。
【0015】
第三の態様において、本発明は、第二の態様の単離タンパク質複合体をコードする単離核酸を提供する。
第四の態様において、本発明は、発現ベクター中の1つまたは複数の調節配列に作動可能なように連結された第三の態様の単離核酸を含む遺伝子構築物を提供する。
【0016】
好ましくは、遺伝子構築物は発現構築物である。
第五の態様において、本発明は、第四の態様の遺伝子構築物を含む宿主細胞を提供する。
【0017】
第六の態様において、本発明は、第一の態様の単離タンパク質複合体または第二の態様の合成タンパク質と、薬学的に許容可能な担体、希釈剤または賦形剤とを含んでなる薬学的組成物を提供する。
【0018】
本発明のこの態様は、第五の態様の宿主細胞を含み、細胞が上記合成タンパク質を発現する薬学的組成物も包含する。
第七の態様において、本発明は、第二の態様の合成タンパク質に特異的な抗体を提供する。
【0019】
第八の態様において、本発明は、細胞移動の促進方法であって、成長因子受容体およびインテグリン受容体の両方に結合するために合成タンパク質を用いる工程を含む方法を提
供する。
【0020】
好ましくは、成長因子受容体はIGF−IRである。
好ましくは、インテグリン受容体はαVインテグリンである。
さらに好ましくは、インテグリン受容体は、αVβ3インテグリンまたはαVβ5インテグリンである。
【0021】
好ましい実施形態では、本発明のこの態様は、哺乳類、好ましくはヒトにおける創傷治癒を促すための、上皮細胞の移動および増殖のうち少なくともいずれかの促進または誘導に関する。
【0022】
好ましくは、上記合成タンパク質は、本発明の第一の態様によるものと同様である。
第九の態様において、本発明は、細胞の移動および増殖のうち少なくともいずれかを防止する方法であって、成長因子およびビトロネクチンもしくはフィブロネクチンを含んでなる複合体により成長因子受容体およびインテグリン受容体の両方の結合を防止、抑制、またはそうでなければ低減する工程を含む方法を提供する。
【0023】
好ましくは、成長因子受容体はIGF−IRである。
好ましくは、インテグリン受容体はαVインテグリンである。
さらに好ましくは、インテグリン受容体は、αVβ3インテグリンまたはαVβ5インテグリンである。
【0024】
好ましい実施形態では、本発明のこの態様は、哺乳類、好ましくはヒトにおける転移性がん細胞の移動および増殖のうち少なくともいずれかの防止または抑制に関する。
本発明のこの態様により意図される特定の例は、乳がん転移の防止または抑制である。
【0025】
また、第八および第九の態様の方法は、予防的および治療的処置方法を包含し得る、と理解される。
第十の態様において、本発明は、
(i)成長因子およびビトロネクチンまたはフィブロネクチンを含んでなるタンパク質複合体のアゴニストである分子、または
(ii)成長因子およびビトロネクチンまたはフィブロネクチンを含んでなるタンパク質複合体のアンタゴニストである分子
を作製するための、第一の態様の単離タンパク質複合体または第二の態様の合成タンパク質の用途を提供する。
【0026】
好ましい実施形態では、本発明は、以下の:
(i)IGF−II:VNタンパク質複合体またはIGF−I:IGFBP:VNタンパク質複合体のアゴニストである分子、または
(ii)IGF−II:VNタンパク質複合体またはIGF−I:IGFBP:VNタンパク質複合体のアンタゴニストである分子
を作製するための、第一の態様の合成タンパク質の用途を提供する。
【0027】
本発明のこの態様に従って作製されるアゴニストおよびアンタゴニストのうち少なくともいずれかは、創傷治癒、組織工学、皮膚再生、ならびにがん細胞の転移または皮膚の過増殖による障害(例えば瘢痕形成および乾癬)の防止、のうち少なくともいずれかを促すに際して特定の効能を有し得る。
【0028】
第十一の態様において、本発明は、第一または第二の態様の単離タンパク質複合体を含む生体適合材料を提供する。
特定の実施形態では、生体適合材料は、本発明の単離タンパク質複合体を、適切な含浸、被覆、またはその他の方法で含む外科用移植片、人工器官、足場材、創傷被覆材または熱傷被覆材等であり得る。
【発明を実施するための最良の形態】
【0029】
(発明の詳細な説明)
本発明は、IGF−IIおよびVNまたはIGF−IおよびIGFBPおよびVNを含んでなるタンパク質複合体が、応答性細胞により発現されるIGF−IR受容体およびVN結合インテグリン受容体に結合し、同受容体を介して細胞移動に生物学的作用を及ぼす、という発見から生じた。より具体的には、この二元的結合事象は、ケラチノサイトモデルおよび乳がん細胞モデルの両方において本発明者らにより示されたように、細胞の移動および増殖のうち少なくともいずれかを相乗作用的に刺激する。
【0030】
さらに、IGF−IIおよびIGFBPと相互作用するかまたは結合すると思われるVNのドメインは、成熟VNのアミノ酸53〜64に対応するポリ陰イオン性領域であるということが意外にも発見された。
【0031】
この発見により、本発明者等は、IGF−IRに結合し得るIGF−IまたはIGF−IIの少なくとも最小ドメインまたは最小領域をVNのインテグリン結合ドメインと組合せて含む単離タンパク質複合体を提供した。さらに具体的には、これらのドメインを含む単一で切れ目のないタンパク質が作製され得る。
【0032】
このようなタンパク質複合体は、複数のタンパク質を含むものであれ、単一の合成タンパク質の形態であれ、IGF−IRおよびVN結合インテグリン受容体に協調的に結合するかまたは同受容体を相互連結することにより、細胞の移動および増殖のうち少なくともいずれかならびに創傷治癒を促進するための有用な作用物質である、と予測される。同様に、IGF−IRおよびVN−結合インテグリン受容体の相互連結の防止は、がん細胞の転移を防止するために用いられ得る、ということが本発明者等により提唱される。この発見は、他の成長因子、例えばPDGFおよびVEGF(これらに限定されない)を含むタンパク質複合体に、そして他のインテグリン結合タンパク質、例えばフィブロネクチン(FN)にも拡張可能であり得る、ということも提唱される。
【0033】
本明細書全体を通して、別記しない限り、「〜を含む、含んでなる(comprise, comprises )」、および「含んでいる、含んでなる(comprising)」とは、提示された1または一群の完全体が、1つまたは複数の他の提示されていない1または一群の完全体を包含し得るよう、排他的というより包括的に用いられる。
【0034】
成長因子受容体結合ドメインおよびインテグリン結合ドメインの特定の状態においては、このようなドメインは、該ドメインのアミノ酸配列とともに、所望により他の付加的アミノ酸配列も含む。
【0035】
さらにまた、このようなドメインは、該ドメインのアミノ酸配列とともに、10個以下の、好ましくは5個以下の、さらに好ましくは4、3、2または1個以下の付加的アミノ酸「のみから本質的に構成される(consist essentially of)」、と理解されよう。
【0036】
さらにまた、このようなドメインは、任意の付加的アミノ酸の非存在下では、ドメインのアミノ酸配列「のみから構成される(consist of)」、と理解されよう。
本発明の目的のために、「単離(された)」とは、その天然状態から取り出されたか、または他の方法で人為的に操作された物質を意味する。単離物質は、その天然状態で該物質に通常随伴する構成成分を実質的または本質的に含有しないものでもよいし、あるいは
その天然状態で該物質に通常随伴する構成成分と一緒に人工状態で存在するよう操作されたものでもよい。単離物質は、天然の形態でも、化学合成形態でも、または組換え形態でもよい。
【0037】
本明細書中で用いる場合、「合成、合成の」とは、天然に存在せず、ヒトによる技術的介入により作製されることを意味する。合成のタンパク質および核酸に関しては、これは、当該技術分野で既知の組換え技術、化学合成技術またはコンビナトリアル技術により作製される分子を包含する。
【0038】
「タンパク質」とは、アミノ酸のポリマーを意味する。アミノ酸は、当該技術分野で既知であるような、天然または非天然のアミノ酸、D−またはL−アミノ酸でよい。「タンパク質」という用語には、当該技術分野で一般的に用いられるように、「糖タンパク質」、「リポタンパク質」等のような用語が挙げられ、かつ該用語を包含する。
【0039】
「ペプチド」とは、50個未満のアミノ酸を有するタンパク質である。
「ポリペプチド」とは、50個またはそれ以上のアミノ酸を有するタンパク質である。
本明細書中で上記したように、本発明は、特定の一態様において、単離タンパク質複合体を提供し、同単離タンパク質複合体は、
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子受容体に結合し得るドメイン、および
(ii)ビトロネクチンまたはフィブロネクチンの少なくとも1つのインテグリン結合ドメイン
を含んでなる。
【0040】
本明細書中で用いる場合、「成長因子」とは、in vitroおよびin vivoのうち少なくともいずれかにおいて細胞の増殖、分化、生存および移動のうち少なくともいずれかを調節し得る生物学的活性タンパク質である。
【0041】
好ましくは、成長因子は、IGF−I、IGF−II、VEGFおよびPDGFからなる群から選択される。
さらに好ましくは、成長因子は、IGF−IおよびIGF−IIから選択される。
【0042】
しかしながら本発明は、細胞の増殖、分化、生存および移動のうち少なくともいずれかを調節するその他の生物学的活性タンパク質、例えば上皮成長因子(EGF;ヘルディン(Heldin)他、1981, Science 4 1122-1123 )、繊維芽細胞成長因子(FGF;ナーコンブ(Nurcombe)他、2000, J. Biol. Chem. 275 30009-30018)、塩基性繊維芽細胞成長因子(bFGF;タラボレッティ(Taraboletti )他、1997, Cell Growth. Differ. 8 471-479)、オステオポンチン(ナム(Nam )他、2000, Endocrinol. 141 1100)、トロンボスポンジン−1(ナム(Nam )他著、2000(上記))、テネイシン−C(アライ(Arai)他、1996, J.Biol. Chem. 271 6099)、PAI−1(ナム(Nam )他、1997, Endocrinol. 138 2972)、プラスミノーゲン(キャンプベル(Campbell)他、1998, Am. J.Physiol.
275 E321 )、フィブリノーゲン(キャンプベル(Ca mpbell)他、1999, J. Biol. Chem
274 30215 )、フィブリン(キャンプベル(Campbell)他、1999(上記))またはトラ
ンスフェリン(バインジマー(Weinzimer )他、2001, J. Clin. Endocrinol. Metab. 86
1806 )も意図する。
【0043】
本発明の単離タンパク質複合体は、成長因子または成長因子の少なくとも1つのドメインであって同族の成長因子受容体と結合し得るドメインを含む。
これに関連して、「ドメイン」とは、同族の成長因子受容体に結合することが可能な、成長因子の少なくとも一部分または領域を意味する。典型的には、ただし限定されるもの
ではないが、同族の成長因子受容体は細胞により発現され、そして成長因子の上記少なくとも1つのドメインによる上記同族の成長因子受容体の結合または連結により、細胞の応答、例えば細胞の増殖、分化、生存および移動のうち少なくともいずれかが誘導される。
【0044】
特にIGF−Iに関しては、上記ドメインは適切にはアミノ酸残基24を含むが、アミノ酸残基24はロイシン残基ではない。
典型的には、上記残基はチロシンである。
【0045】
特にIGF−IIに関しては、上記ドメインは適切にはアミノ酸残基27を含むが、アミノ酸残基27はロイシン残基ではない。
典型的には、上記残基はチロシンである。
【0046】
特にIGF−Iに関しては、一実施形態では、上記ドメインはIGF−Iの残基1〜70のみから構成される。
別の実施形態では、上記ドメインはIGF−Iの残基4〜70のみから構成される。
【0047】
また、本発明の単離タンパク質複合体の別の構成成分は、ビトロネクチンまたはフィブロネクチンの少なくとも1つのインテグリン結合ドメインであることが理解されよう。
これは、αVインテグリンに結合し得るVNまたはFNの任意のドメインを含みそして包含する。
【0048】
さらに好ましくは、インテグリン受容体はαVβ3インテグリンまたはαVβ5インテグリンである。
本明細書中で以下にさらに詳細に記載されるように、本発明は、VN(そして同様にFN)のHBDは、単離タンパク質複合体の完全な生物学的活性に必要なわけではない、ということを示している。
【0049】
本発明の単離タンパク質複合体は、非共有結合により会合したオリゴタンパク質複合体でも、共有結合により(可逆的にまたは非可逆的に)架橋されたオリゴタンパク質複合体の形態でも、あるいは合成キメラタンパク質の形態でもよい、ということは上記から容易に理解されよう。
【0050】
したがって、特定の態様において、本発明は、
合成キメラタンパク質の形態の単離タンパク質複合体であって、
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子受容体に結合し得るドメイン、および
(ii)ビトロネクチン(VN)またはフィブロネクチン(FN)、あるいはVNまたはFNの少なくとも1つのインテグリン結合ドメイン
のアミノ酸配列を含んでなる単離タンパク質複合体を提供する。
【0051】
本明細書中で用いる場合、「キメラタンパク質」は、VNまたはFNのインテグリン受容体結合ドメイン、ならびに成長因子または成長因子の少なくとも1つの受容体結合ドメインに由来するアミノ酸の連続配列を含む。
【0052】
いかなる特定の理論にも縛られることは望ましくないが、合成キメラタンパク質は上記成長因子に対する同族の受容体ならびにVNまたはFNに対するインテグリン受容体を相互連結し、かつ同時活性化することによって、細胞移動を刺激、誘導、増強、さもなければ促進し得る、ということが提唱される。
【0053】
本発明のキメラタンパク質の利点は、化学合成または組換え手段により容易に生産され
ること、そして非共有結合によるオリゴタンパク質複合体に必要とされるタンパク質−タンパク質相互作用の保持には依存しないので、in vivoでより安定であると予測されることである。
【0054】
この点では、IGF−Iの受容体結合ドメインを含む単離タンパク質複合体はIGFBPも含むことになるにも関わらず、上記の作用様式に従えばIGFBPはIGF−I/VN合成キメラ中に存在しないことが好ましい、ということが提唱される。
【0055】
さらにまたVNに関して、本明細書中でさらに詳細に後述されるように、本発明者等は、IGF−IIまたはIGF−I/IGFBP複合体との相互作用に必要とされるのはおそらくVN(そして同様にFN)のポリ陰イオン性領域であると思われる、ということを示す。
【0056】
図7および図8を参照すると、ポリ陰イオン性領域は、成熟VN配列(配列番号2)の残基53〜64である。
上記の点から見て、本発明は、VNまたはFNのHBDおよびポリ陰イオン性領域のうち少なくともいずれかを含まない合成キメラタンパク質の実施形態を意図する。
【0057】
HBDおよびポリ陰イオン性領域のうち少なくともいずれかを含まないVNタンパク質およびそのアミノ酸配列に関しては、天然に存在するタンパク質、例えば54kDニワトリ卵黄VN(HBDを欠く)でもよいし、あるいはVNタンパク質またはアミノ酸配列を欠失、突然変異または切断により人為操作してHBDおよびポリ陰イオン性領域のうち少なくともいずれかが存在しないかまたは少なくとも実質的に機能を持たないようにしてもよい。
【0058】
タンパク質分解による消化および部位特異的突然変異誘発のような技法は、当該技術分野で既知であるように、この目的のために利用され得る。
特定の実施形態では、VNの上記少なくとも1つのインテグリン結合ドメインは、
(i)VNのアミノ酸残基1〜459;
(ii)VNのアミノ酸残基1〜379;
(iii)VNのアミノ酸残基1〜130;および
(iv)VNのアミノ酸残基1〜52
からなる群から選択されるアミノ酸配列を有する。
【0059】
含まれてもよい付加的アミノ酸配列は、
(v)VNのアミノ酸残基65〜459;
(vi)VNのアミノ酸残基347〜459;および
(vii)VNのアミノ酸残基347〜379
からなる群から選択されるアミノ酸配列を有する。
【0060】
上記の配列は、例えばVNのアミノ酸残基1〜130とVNのアミノ酸残基347〜459、またはVNのアミノ酸残基1〜52とVNのアミノ酸残基65〜459のように組合せて用いることができる。
【0061】
IGF−IおよびVNを含んでなるキメラタンパク質の具体例が図14に記述されているが、これらに限定されない。
さらにVEGFおよびVNまたはPDGFおよびVNを含んでなるキメラタンパク質の具体例が図15に記述されているが、これらに限定されない。
【0062】
好ましくは、キメラタンパク質はさらに、成長因子の配列とVNまたはFNのアミノ酸
配列との間に位置し、同配列と連続する「リンカー配列」を含む。
一実施形態では、上記リンカー配列は、1つまたは複数のグリシン残基および1つまたは複数のセリン残基を含んでなる。リンカー配列の具体例は、Gly4Ser(配列番号4);Gly4Ser3(配列番号5)および(Gly4Ser)3(配列番号6)から選択されるが、これらに限定されない。
【0063】
別の実施形態では、リンカー配列には、プラスミン切断認識部位、例えば以下の配列:
LeuIleLysMetLysPro(配列番号7)
が挙げられる。
【0064】
さらに別の実施形態では、リンカー配列には、コラゲナーゼ−3切断認識部位、例えば以下の配列:
GlnProGlnGlyLeuAlaLys(配列番号8)
が挙げられる。
【0065】
本発明は、本発明の合成キメラタンパク質の生物学的に活性な断片の用途、および本明細書中に例示した特定の成長因子受容体結合ドメインおよびインテグリン結合ドメインの生物学的に活性な断片の用途にも拡張する。
【0066】
一実施形態では、上記「生物学的に活性な断片」は、該断片の元のタンパク質の10%以上、好ましくは25%以上、さらに好ましくは50%以上、さらに好ましくは75、80、85、90または95%以上の生物学的活性を有する。
【0067】
別の実施形態では、上記「生物学的に活性な断片」は、該断片の元のタンパク質の10%以上、好ましくは25%以上、さらに好ましくは50%以上、さらに好ましくは75、80、85、90または95%以上の連続アミノ酸配列を有する。
【0068】
例えばHBDおよびポリ陰イオン性ドメインのうち少なくともいずれかを欠く、生物学的に活性なVN断片の具体例は、本明細書中で図14に提示されている。
本発明の変異体タンパク質複合体も意図される。
【0069】
典型的には、そしてタンパク質に関しては、「変異体」タンパク質は、異なるアミノ酸により置換された1つまたは複数のアミノ酸を有する。いくつかのアミノ酸は、おおむね類似の特性を有する他のアミノ酸を用いて、タンパク質の活性の性質を変えることなく変更可能である(保存的置換)、ということは当該技術分野でよく理解されている。
【0070】
基準となる配列、例えば成長因子、成長因子の受容体結合ドメイン、VNまたはFNのインテグリン結合ドメイン、IGFBPの1つまたは複数のアミノ酸残基、あるいは合成キメラタンパク質中に存在する1つまたは複数の対応する残基を、本発明の単離タンパク質複合体の生物学的活性を実質的に変えることなく、修飾し、または欠失させ、あるいは付加的配列を付加することができることは理解されよう。
【0071】
本発明により意図される成熟VN(配列番号2)における具体的な突然変異としては、(i)T50A;(ii)T57A;(iii)T50E;(iv)T57E;(v)S378E;(vi)S378A;および(v)S362E、が挙げられる。
【0072】
一実施形態では、タンパク質変異体は、基準となるアミノ酸配列と少なくとも70%、好ましくは少なくとも80%、さらに好ましくは少なくとも90%、95%、98%または99%の配列同一性を共有する。
【0073】
好ましくは、配列同一性は、基準となる配列の少なくとも60%に亘って、さらに好ましくは少なくとも75%に亘って、さらに好ましくは少なくとも90%に亘って、さらに好ましくは少なくとも95%、98%に亘って、あるいは実質的に全長に亘って測定される。
【0074】
配列同一性(%)を決定するために、コンピュータを用いたアルゴリズムの実行(インテリジェネティックス(Intelligenetics )によるGeneworks(登録商標)プログラム;米国ウィスコンシン州マディソン サイエンスドライブ575所在のウィスコンシン・ジェネティクス・ソフトウェア・パッケージ・リリース7.0(Wisconsin Genetics Software Package Release 7.0 )のGAP、BESTFIT、FASTAおよびTFASTA(参照により本明細書中で援用する))により、あるいは選択された種々の方法のいずれかにより作成された検討および最良のアラインメント(即ち比較ウインドウを上回る最高の相同性(%)を生じる)により、アミノ酸配列およびヌクレオチド配列のうち少なくともいずれかの最適アラインメントを実行することが可能である。例えばアルチュール(Altschule )他、1997, Nucl. Acids Res. 25 3389(この記載内容は、参照により本明細書中で援用される)に開示されているようなBLASTファミリーのプログラムも参照可能である。
【0075】
別の例では、「配列同一性」は、DNASISコンピュータプログラム(ウインドウズ用バージョン2.5;米国カリフォルニア州サウスサンフランシスコ所在の日立ソフトウェアエンジニアリング(Hitachi Software engineering Co., Ltd.)から入手可能)により算定される「マッチ率(%)」を意味すると理解することができる。
【0076】
配列分析の詳細な考察については、オースベル(Ausubel )他編、「分子生物学最新プロトコール(CURRENT PROTOCOLS IN MOLECULAR BIOLOGY)」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons Inc )、1995-1999 )の第19.3部に記載されている。
【0077】
本発明は、成長因子の受容体結合ドメイン、VNまたはFNのインテグリン結合ドメイン、あるいは前記ドメインを含んでなる単離タンパク質複合体の誘導体も意図する。
本明細書中で用いる場合、本発明の「誘導体」タンパク質は、例えば別の化学基の付加、共役または複合により、あるいは当該技術分野で既知の翻訳後修飾技法により変更されている。
【0078】
アミノ酸の「付加」には、ポリペプチドまたはその変異体と別のペプチドまたはタンパク質との融合が挙げられる。前記の別のタンパク質は、例えば、タンパク質の精製を助ける。例えばこれら別のタンパク質の例としては、ポリヒスチジンタグ、マルトース結合タンパク質、緑色蛍光タンパク質(GFP)、プロテインAまたはグルタチオンS−トランスフェラーゼ(GST)が挙げられる。
【0079】
本発明により意図されるその他の誘導体としては、側鎖に対する修飾、ペプチド、ポリペプチドまたはタンパク質合成の際の非天然アミノ酸およびそれらの誘導体のうち少なくともいずれかの組み入れ、ならびに架橋剤の使用および本発明のポリペプチド、断片および変異体に立体構造上の制約を与えるその他の方法が挙げられるが、これらに限定されない。本発明により意図される側鎖修飾の例としては、例えば無水酢酸を用いたアシル化;無水コハク酸および無水テトラヒドロフタル酸を用いたアミノ基のアシル化;メチルアセトイミデートを用いたアミド化;シアネートを用いたアミノ基のカルバモイル化;ピリドキサル−5−ホスフェートを用いたリシンのピリドキシル化とその後のNaBH4を用いた還元;アルデヒドとの反応による還元的アルキル化とその後のNaBH4を用いた還元;ならびに2,4,6−トリニトロベンゼンスルホン酸(TNBS)を用いたアミノ基の
トリニトロベンジル化など、アミノ基の修飾が挙げられる。
【0080】
カルボキシル基は、O−アシルイソ尿素の形成を介したカルボジイミド活性化とその後の誘導体化、例えば対応するアミドへの誘導体化により修飾され得る。
アルギニン残基のグアニジン基は、2,3−ブタンジオン、フェニルグリオキサルおよびグリオキサルのような試薬を用いた複素環式縮合産物の生成により修飾され得る。
【0081】
スルフヒドリル基は、例えばシステイン酸への過蟻酸酸化;4−クロロ水銀フェニルスルホン酸、4−クロロ水銀ベンゾエート、2−クロロ水銀−4−ニトロフェノール、塩化フェニル水銀およびその他の水銀剤を用いた水銀誘導体の形成;その他のチオール化合物との混合ジスルフィドの形成;マレイミド、無水マレイン酸またはその他の置換マレイミドとの反応;ヨード酢酸またはヨードアセトアミドを用いたカルボキシメチル化;ならびにアルカリ性pHでシアネートを用いたカルバモイル化などの方法により修飾され得る。
【0082】
トリプトファン残基は、例えば臭化2−ヒドロキシ−5−ニトロベンジルまたはハロゲン化スルホニルを用いたインドール環のアルキル化により、あるいはN−ブロモスクシンイミドを用いた酸化により修飾され得る。
【0083】
チロシン残基は、3−ニトロチロシン誘導体を形成するためのテトラニトロメタンを用いたニトロ化により修飾され得る。
ヒスチジン残基のイミダゾール環は、ジエチルピロカルボネートを用いたN−カルボエトキシル化により、あるいはヨード酢酸誘導体を用いたアルキル化により、修飾され得る。
【0084】
ペプチド合成中に非天然アミノ酸および誘導体を組み入れる例としては、4−アミノ酪酸、6−アミノヘキサン酸、4−アミノ−3−ヒドロキシ−5−フェニルペンタン酸、4−アミノ−3−ヒドロキシ−6−メチルヘプタン酸、t−ブチルグリシン、ノルロイシン、ノルバリン、フェニルグリシン、オルニチン、サルコシン、2−チエニルアラニンおよびアミノ酸のD異性体のうち少なくともいずれかの使用が挙げられるが、これらに限定されない。
【0085】
タンパク質の化学的誘導体化に適した方法の一例は、コリガン(Coligan )他編、「タンパク質科学の最新プロトコール(CURRENT PROTOCOLS IN PROTEIN SCIENCE)」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons )、1995-2001 )の第15章に提供されている。
【0086】
単離タンパク質複合体およびその個々の構成成分であるタンパク質(断片、変異体、誘導体および相同体を含む)は、当業者に既知の任意の適切な手法により調製され得る。
一実施形態では、本発明のタンパク質は化学合成により作製される。化学合成技法は当該技術分野で既知であるが、当業者は適切な方法の例について、コリガン(Coligan )他編、「タンパク質科学の最新プロトコール(CURRENT PROTOCOLS IN PROTEIN SCIENCE)」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons )、199 5-2001 )の第18章を参照してもよい。
【0087】
別の実施形態では、タンパク質は、組換えタンパク質として調製され得る。
組換えタンパク質の製造は当該技術分野で既知であるが、当業者は、例えばサムブルック(Sambrook)他、「分子クローニング.実験の手引き(MOLECULAR CLONING. A Laboratory Manual)」コールドスプリングハーバー出版(Cold Spring Harbor Press)、1989年(この記載内容は、参照により本明細書中で援用される)、特に第16章および第17章;オースベル(Ausubel )他編、「分子生物学最新プロトコール(CURRENT PROTOCOL
S IN MOLECULAR BIOLOGY)」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons Inc )、1995-1999 )(この記載内容は、参照により本明細書中で援用される)、特に第10章および第16章;ならびにコリガン(Coligan )他編、「タンパク質科学の最新プロトコール(CURRENT PROTOCOLS IN PROTEIN SCIENCE)」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons )、1995-2001 )(この記載内容は、参照により本明細書中で援用される)、特に第1章、第5章および第6章に記載されているような標準プロトコールを参照してもよい。
【0088】
一実施形態では、組換えタンパク質は、
(i)発現ベクター内の1つまたは複数の調節ヌクレオチド配列に作動可能なように連結された、前記タンパク質をコードする核酸を含む発現構築物を調製する工程、
(ii)発現構築物を用いて宿主細胞をトランスフェクションまたは形質転換する工程、および
(iii)前記宿主細胞中で組換えタンパク質を発現させる工程
を包含する方法により製造される。
【0089】
「発現ベクター」は、自己複製する染色体外ベクター、例えばプラスミド、または宿主ゲノム内に組み込まれるベクターのいずれでもよい。
「作動可能なように連結される」または「操作可能なように接続される」とは、前記1つまたは複数の調節ヌクレオチド配列が、本発明の組換え核酸に対して、該核酸の転写または該核酸によりコードされるタンパク質の翻訳を、開始し、調節し、またはそうでなければ制御するように配置される、ということを意味する。
【0090】
調節ヌクレオチド配列は一般に、発現のために用いられる宿主細胞に適したものとなろう。多数の種類の適切な発現ベクターおよび適切な調節配列が、種々の宿主細胞に関して当該技術分野で既知である。
【0091】
典型的には、前記1つまたは複数の調節ヌクレオチド配列としては、プロモーター配列、リーダー配列またはシグナル配列、リボソーム結合部位、転写開始配列および転写終結配列、翻訳開始配列および翻訳終結配列、スプライスドナー/アクセプター配列、ならびにエンハンサー配列またはアクチベーター配列が挙げられるが、これらに限定されない。
【0092】
構成的プロモーター(例えばCMV、RSV、アデノウイルス、SV40およびヒト伸長因子プロモーター)および誘導性/抑制性プロモーター(例えばtet抑制性プロモーター、およびIPTG誘導性、メタロチオニン誘導性またはエクジソン誘導性プロモーター)が当該技術分野で既知であり、そして本発明により意図される。また、プロモーターは、複数のプロモーターの要素を併合するハイブリッドプロモーターでもよいことは理解されよう。
【0093】
発現構築物は、本発明の組換えタンパク質が融合パートナーとの融合ポリペプチドとして発現されるよう、融合パートナー(典型的には発現ベクターにより提供される)も含み得る。融合パートナーの主な利点は、前記融合タンパク質の同定および精製のうち少なくともいずれかの助けになるということである。
【0094】
融合パートナーの既知の例としては、グルタチオン−S−トランスフェラーゼ(GST)、ヒトIgGのFc部分、マルトース結合タンパク質(MBP)およびヘキサヒスチジン(HIS6)があり、これらはアフィニティクロマトグラフィによる融合タンパク質の単離に特に有用であるが、これらに限定はされない。アフィニティクロマトグラフィによる融合タンパク質精製の目的のために、アフィニティクロマトグラフィのための適切な充填材は、それぞれグルタチオン共役樹脂、アミロース共役樹脂、およびニッケル共役樹脂
またはコバルト共役樹脂である。多数のこのような充填材が、例えば(HIS6)融合パートナーを用いるのに有用なQIAexpress(商標)システム(キアゲン社(Qiagen))、ならびにファルマシア(Pharmacia )のGST精製システムのように、「キット」の形態で入手可能である。
【0095】
場合によっては、融合パートナーがさらにプロテアーゼ切断部位、例えばXa因子による切断部位またはトロンビンによる切断部位を有し、関連プロテアーゼが本発明の融合タンパク質を部分消化することにより融合タンパク質から本発明の組換えポリペプチドを遊離させることが可能である。遊離したタンパク質は次に、その後のクロマトグラフィー分離により、融合パートナーから単離され得る。
【0096】
本発明の融合パートナーは、「エピトープタグ」もその範囲内に含む。「エピトープタグ」は特異的抗体が利用可能な通常は短いペプチド配列である。特異的モノクローナル抗体が容易に利用可能なエピトープタグの既知の例としては、c−myc、ヘマグルチニンおよびFLAGタグが挙げられる。
【0097】
発現のための適切な宿主細胞は原核生物でも真核生物でもよく、例えば大腸菌(例えばDH5α)、酵母細胞、バキュロウイルス発現系とともに利用されるSf9細胞、CHO細胞、COS、CV−1、NIH 3T3および293細胞であり得るが、しかしこれらに限定されない。
【0098】
発現構築物は、形質転換された宿主細胞に選択作用物質に対する耐性を付与する1つまたは複数の選択マーカーヌクレオチド配列をさらに含み得る。形質転換された細菌を選択するために有用な選択マーカーとしては、bla、kanRおよびtetRが挙げられる一方、形質転換された真核生物細胞は、ハイグロマイシン、G418およびピューロマイシンのようなマーカーにより選択され得るが、これらに限定はされない。
【0099】
宿主細胞中への遺伝物質の導入に関して、「形質転換する」および「トランスフェクションする」という用語が、宿主細胞中への遺伝物質の導入を表すために一般に用いられる。宿主細胞中に外来遺伝物質を導入するための多数の既知の方法が存在し、例としてはリン酸カルシウム沈殿法、電気穿孔法、リポフェクタミン、リポフェクチンおよびその他の親油性物質による送達、リン酸カルシウム沈殿法、DEAE−デキストラントランスフェクション、微粒子銃、マイクロインジェクション、ならびにプロトプラスト融合が挙げられるが、これらに限定されない。
【0100】
(単離核酸)
本発明は、本発明の合成キメラタンパク質(その変異体および相同体を含む)をコードする単離核酸を提供する。
【0101】
「核酸」という用語は、本明細書中で用いる場合、一本鎖または二本鎖のmRNA、RNA、cRNA、RNAiおよびDNA(cDNAおよびゲノムDNAを含む)、ならびにDNA−RNAハイブリッドを意味する。
【0102】
「ポリヌクレオチド」は、80またはそれ以上の連続ヌクレオチドを有する核酸であり、一方、「オリゴヌクレオチド」は、80未満の連続ヌクレオチドを有する。
「プローブ」は、例えばノーザンブロッティングまたはサザンブロッティングにおいて相補配列を検出する目的のために適切に標識された一本鎖または二本鎖のオリゴヌクレオチドまたはポリヌクレオチドであり得る。
【0103】
「プライマー」は、通常は一本鎖のオリゴヌクレオチドであり、好ましくは15〜50
連続ヌクレオチドを有し、相補的な核酸の「鋳型」とアニーリングして、DNAポリメラーゼ、例えばTaqポリメラーゼ、RNA依存性DNAポリメラーゼまたはSequenase(商標)の作用により鋳型依存性の様式で伸長可能である。
【0104】
本発明の合成核酸は、当該技術分野で知られているように、化学合成手法により、または核酸配列の増幅技術を利用する組換え方法により、あるいはそれらの組合せにより作製可能である。
【0105】
本発明に有用な化学合成プライマーおよびオリゴヌクレオチド、合成装置および関連技術は、一般にほとんどの実験室で利用可能であり、あるいは商業的供給元から購入可能である。
【0106】
適切な核酸増幅技法は当業者に既知であり、例としては、例えばオースベル(Ausubel )他の上記文献の第15章に記載されているようなポリメラーゼ連鎖反応(PCR)およびリガーゼ連鎖反応(LCR);例えば米国特許第5,422,252号に記載されているような鎖置換増幅(SDA);例えばリュー(Liu )他、1996, J. Am. Chem. Soc. 118 1587、国際出願国際公開第92/01813号パンフレットおよび国際出願国際公開第97/19193号パンフレットに記載されているようなローリングサークル複製(RCR);例えばスクナナン(Sooknanan )他、1994, Biotechniques 17 1077 に記載されているようなNASBA法(nucleic acid sequence-based amplification );ならびに例えばティアギ(Tyagi )他、1996, Proc. Natl. Acad. Sci. USA 93 5395に記載されているようなQ−βレプリカーゼ増幅が挙げられるが、これらに限定されない。
【0107】
好ましい核酸配列増幅技法は、PCRである。
本明細書中で用いる場合、「増幅産物」とは、核酸増幅技法により作製された核酸産物を指す。
【0108】
本発明の核酸を作製し、発現させるに際しては、本明細書中で例示された核酸が、同核酸によりコードされるアミノ酸配列を変えることなく容易に変更可能なように、コドン配列の冗長性の利点を利用することができることも理解されよう。
【0109】
特定の実施形態では、当該技術分野で既知であるように、組換え発現のために用いようとする宿主細胞の好ましい「コドン使用頻度」に従って核酸を最適化することができる。このことにより、選択的なコドン使用頻度がタンパク質の発現に影響を及ぼす特定の生物体またはその細胞中での最適な発現のために、核酸を効率的に調整することが可能である。
【0110】
したがって本発明は、本明細書中に例示された核酸と相同な合成核酸を包含する。
一実施形態では、核酸相同体は、本明細書中に記載された合成キメラタンパク質構築物のいずれか一つをコードする核酸と少なくとも70%、好ましくは少なくとも80%、さらに好ましくは少なくとも90%、さらに好ましくは少なくとも95%の配列同一性を共有する。
【0111】
好ましくは、配列同一性は、本発明のコード核酸の少なくとも70%に亘って、より好ましくは少なくとも80%、さらに好ましくは少なくとも90%、95%、または有益には実質的に全長に亘って測定される。
【0112】
別の実施形態では、核酸相同体は、ストリンジェンシーの高い条件下で、本明細書中に記載された合成キメラタンパク質構築物のいずれか一つをコードする核酸とハイブリダイズする。
【0113】
「ハイブリダイズ、ハイブリダイゼーション」とは、DNA−DNA、RNA−RNAまたはDNA−RNA二重鎖を生成する、少なくとも部分的に相補性のヌクレオチド配列の対合を意味するべく、本明細書中で用いられる。ハイブリダイズした配列は、当該技術分野で既知であるように、相補的なプリン間およびピリミジン間の塩基対合により生じる。
【0114】
この点では、修飾プリン(例えばイノシン、メチルイノシンおよびメチルアデノシン)および修飾ピリミジン(チオウリジンおよびメチルシトシン)も塩基対合に関与し得ることが理解されよう。
【0115】
「ストリンジェンシー」とは、本明細書中で用いる場合、ハイブリダイゼーション中の、温度およびイオン強度条件、ならびにある種の有機溶媒および洗剤のうち少なくともいずれかの有無を指す。ストリンジェンシーが高いほど、ハイブリダイズするヌクレオチド配列間で必要とされる相補性のレベルが高くなる。
【0116】
「ストリンジェントな条件」とは、相補的な塩基の頻度が高い核酸のみがハイブリダイズする条件を意味する。
本明細書中のストリンジェンシーの高い条件への言及には、
(i)42℃でのハイブリダイゼーションについては、少なくとも約31%(v/v)〜少なくとも約50%(v/v)のホルムアミドおよび少なくとも約0.01M〜少なくとも約0.15Mの塩、ならびに42℃での洗浄については少なくとも約0.01M〜少なくとも約0.15Mの塩;
(ii)65o Cでのハイブリダイゼーションについては、1%BSA、1mMのEDTA、0.5MのNaHPO4(pH7.2)、7%SDS、ならびに65℃を上回る温度で約1時間の洗浄については(a)0.1×SSC、0.1%SDS;または(b)0.5%BSA、1mMのEDTA、40mMのNaHPO4(pH7.2)、1%SDS;
(iii)68℃以上で約20分間の洗浄については、0.2×SSC、0.1%SDS
が含まれる。
【0117】
概して、洗浄は、Tm=69.3+0.41(G+C)%−12℃で実行される。一般に、ミスマッチの塩基数が1%増大するごとに二重鎖DNAのTmは約1℃低下する。
上記にもかかわらず、ストリンジェントな条件は、例えばオースベル(Ausubel )他の上記文献の第2.9章および第2.10章に、特に2.9.1〜2.9.20ページに記載されているように、当該技術分野で既知である。
【0118】
(抗体)
本発明は、本発明の合成キメラタンパク質(キメラタンパク質、またはその断片、変異体および誘導体のうち少なくともいずれかを含む)に対する抗体も意図する。本発明の抗体は、ポリクローナルでもモノクローナルでもよい。抗体の作製、精製および使用に適用可能な既知のプロトコールは、例えばコリガン(Coligan )他、「免疫学の最新プロトコール(CURRENT PROTOCOLS IN IMMUNOLOGY )」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons )、1991-1994 )の第2章に、ならびにハーロウとレーン(Harlow, E. & Lane, D. )著、「抗体:実験の手引き(Antibodies: A Laboratory Mannual)」、コールドスプリングハーバー所在のコールドスプリングハーバー研究所(Cold Spring Harbor Laboratory )、1988に見出され得る(これらの文献の記載内容は、参照により本明細書中で援用される)。
【0119】
一般に本発明の抗体は、本発明のポリペプチド、断片、変異体または誘導体と結合または共役する。例えば抗体は、ポリクローナル抗体を含み得る。このような抗体は、例えば、マウスまたはウサギなどの生産用生物種に本発明のポリペプチド、断片、変異体または誘導体を注射してポリクローナル抗血清を得ることにより調製され得る。ポリクローナル抗体の生産方法は、当業者に既知である。用いられ得るプロトコールの例は、例えばコリガン(Coligan )他、「免疫学の最新プロトコール(CURRENT PROTOCOLS IN IMMUNOLOGY )」(上記)に、ならびにハーロウとレーン(Harlow & Lane )、1988(上記)に記載されている。
【0120】
前記生産用生物種において得られるポリクローナル抗血清の代わりにモノクローナル抗体を作製してもよい。モノクローナル抗体は、例えばコーラーとミルスタイン(Koehler & Milstein)著、1975, Nature 256, 495 の論文(この記載内容は、参照により本明細書中で援用される)に記載されているような標準的方法を用いて、または例えばコリガン(Coligan )他、「免疫学の最新プロトコール(CURRENT PROTOCOLS IN IMMUNOLOGY )」(上記)に記載されているような近年の変法により、本発明のポリペプチド、断片、変異体または誘導体のうち1つまたは複数を接種された生産用生物種由来の脾臓その他の抗体産生細胞を不死化することにより作製することが可能である。
【0121】
本発明はまた、上記で言及されたポリクローナルまたはモノクローナル抗体のFc断片またはFab断片を含む抗体もその範囲内に含む。あるいは抗体は、本発明のBIXPタンパク質に対する一本鎖Fv抗体(scFvs)を含み得る。このようなscFvsは、例えば米国特許第5,091,513号、欧州特許第239,400号、またはウィンターとミルスタイン(Winter & Milstein )、1991, Nature 349, 293 の論文にそれぞれ記載されている方法に従って調製され得る(これらの文献の記載内容は、参照により本明細書中で援用される)。
【0122】
抗体または抗体断片に標識を会合させてもよい。
標識は、色原体、触媒、酵素、蛍光体、化学発光分子、ランタニドイオン、例えばユーロピウム(Eu34)、放射性同位体および直接目に見える標識を含む群から選択され得る。直接目に見える標識の場合、金属コロイドまたは非金属性粒子、染料粒子、酵素もしくは基質、有機ポリマー、ラテックス粒子、リポソームまたはシグナル生成物質を含有するその他のビヒクルなどが用いられ得る。
【0123】
標識として有用な多数の酵素が、米国特許第4,366,241号明細書、米国特許第4,843,000号明細書および米国特許第4,849,338号明細書(これらの記載内容はいずれも参照により本明細書中で援用される)に開示されている。本発明に有用な酵素標識としては、アルカリホスファターゼ、ホースラディッシュペルオキシダーゼ、ルシフェラーゼ、b−ガラクトシダーゼ、グルコースオキシダーゼ、リゾチーム、リンゴ酸デヒドロゲナーゼ等が挙げられる。酵素標識は、単独で用いてもよいし、または溶液中で二次酵素と組合せて用いてもよい。
【0124】
例としては、蛍光体は、フルオレセインイソチオシアネート(FITC)、オレゴングリーン、テトラメチルローダミンイソチオシアネート(TRITL)、アロフィコシアニン(APC)およびR−フィコエリトリン(RPE)であり得るが、しかしこれらに限定されない。
【0125】
(薬学的組成物)
本発明は、本発明の単離タンパク質複合体(その変異体および誘導体を含む)を含んでなる薬学的組成物も提供する。
【0126】
このような単離タンパク質複合体は、in vitroで形成された、または本発明の合成キメラタンパク質として形成された複数タンパク質の複合体を含む任意の形態であり得るが、これらに限定されない。
【0127】
本発明の薬学的組成物は、細胞移動、組織再生および創傷治癒を促進するか、そうでなければ助長するために用いられ得る。あるいは薬学的組成物は、二次部位への腫瘍細胞の移動を防止または抑制することにより腫瘍の転移を防止するために投与され得る。
【0128】
本組成物は、必要な場合、治療的または予防的処置に用いられ得る。例えば薬学的組成物は、皮膚の修復、創傷治癒、熱傷の治癒またはその他の皮膚科学的処置のための治療用または美容用調製物の形態で適用され得る。
【0129】
この点に関して、薬学的組成物は、該薬学的組成物を適切に含浸するか、被覆するか、または他の方法で含んでいる、生体適合材料、生体ポリマー、無機材料、例えばヒドロキシアパタイトまたはその誘導体、外科用移植片、人工器官、創傷被覆材または熱傷被覆材、圧定布、包帯等とともに、またはそれらの一構成成分として投与され得る。
【0130】
薬学的組成物は、適切な薬学的に許容可能な担体、希釈剤または賦形剤を含むのが適切である。
好ましくは、薬学的に許容可能な担体、希釈剤または賦形剤は、哺乳類に、さらに好ましくはヒトに投与するのに適している。
【0131】
「薬学的に許容可能な担体、希釈剤または賦形剤」とは、全身投与に安全に用いられ得る固体または液体の充填剤、希釈剤または封入物質を意味する。個々の投与経路に応じて、当該技術分野で既知の種々の担体が用いられ得る。これらの担体は、糖、デンプン、セルロースおよびその誘導体、麦芽、ゼラチン、タルク、硫酸カルシウム、植物油、合成油、ポリオール、アルギン酸、リン酸緩衝溶液、乳化剤、等張生理食塩水および塩、例えば無機酸塩(例えば塩酸塩、臭化物および硫酸塩)、有機酸、例えば酢酸、プロピオン酸およびマロン酸塩、ならびに発熱物質を含まない水、を含む群から選択され得る。
【0132】
薬学的に許容可能な担体、希釈剤および賦形剤について記載されている有用な参考文献は、「レミントンの薬剤学(Remington's Pharmaceutical Sciences )」、(米国ニュージャージー州所在のマック出版(Mack Publishing Co. )、1991)である(この文献の記載内容は、参照により本明細書中で援用される)。
【0133】
本発明の組成物を患者に提供するために、任意の安全な投与経路が用いられ得る。例えば、経口、直腸、非経口、舌下、口腔内、静脈内、関節内、筋肉内、皮内、皮下、吸入、眼内、腹腔内、脳室内、経皮投与等が用いられ得る。
【0134】
剤形としては、錠剤、分散液、懸濁液、注射液、溶液、シロップ、トローチ、カプセル、座薬、エアロゾル、経皮パッチ等が挙げられる。これらの剤形には、制御放出するために特別に設計された制御放出用デバイス、あるいは制御放出様式でも作用するように改良されたその他の型のインプラントを注入または植設することも含み得る。治療薬の制御放出は、疎水性ポリマー、例えばアクリル樹脂、蝋、高級脂肪族アルコール、ポリ乳酸およびポリグリコール酸、ならびにある種のセルロース誘導体、例えばヒドロキシプロピルメチルセルロースで治療薬を被覆することにより実行され得る。さらに制御放出は、その他のポリマーマトリックス、リポソームおよび微小球のうち少なくともいずれかを用いることにより実行され得る。
【0135】
上記の組成物は、薬剤処方物に適合する方法で、かつ薬学的に有効な量で投与されれば
よい。患者に投与される用量は、本発明に関しては、適切な期間に亘って、患者に有効な応答をもたらすのに十分なものであるべきである。投与されるべき作用物質の量は、治療を受ける対象者の年齢、性別、体重および全身健康状態など、対象者に応じて、かつ担当医の判断によって変わるであろう要因に応じて決めればよい。
【0136】
創傷治癒用の薬学的組成物に関しては、米国特許第5,936,064号明細書および国際公開第99/62536号パンフレット(これらの記載内容は、参照により本明細書中で援用される)を特に参照されたい。
【0137】
本発明の薬学的組成物は、例えばワクシニアなどのウイルスベクターのような発現ベクター、ならびに遺伝子療法に有用なウイルスベクターも含み得る。後者としては、例えばブラウンファルコ(Braun-Falco )他、1999, Gene Ther. 6 432に記載されているようなアデノウイルスおよびアデノ随伴ウイルス(AAV)、ブッチシャッカー(Buchshacher )他、2000, Blood 95 2499 に記載されているようなレトロウイルスおよびレンチウイルスベクター、ならびに単純ヘルペスウイルスおよびサイトメガロウイルス由来のベクターが挙げられる。内分泌の遺伝子療法に有用なウイルスベクターの概説は、ストーン(Stone )他、2000, J. Endocrinol. 164 103に提供されている。
【0138】
本発明は、米国特許第5,958,764号明細書に記載されているような上皮細胞の遺伝子発現を目的とする特別な発現ベクター、および米国特許第5,962,427号明細書に記載されているようなin vivoでの創傷治癒に適用するための特別な発現ベクターも利用し得る。
【0139】
(治療的使用)
本発明は、本発明の合成キメラタンパク質を含めた単離タンパク質複合体を用いた治療方法を提供する。これらの方法は特に、哺乳類、特にヒトの治療的および/または予防的処置を目的としている。
【0140】
しかしながら本発明による治療的使用は、家庭用動物およびペット、作業動物、例えばウマ、ラクダおよびグレイハウンド、家畜、実験室動物、ならびに異種移植のための細胞、器官および組織の供給源として用いられる動物のような哺乳類にも適用可能である。
【0141】
本発明は、本発明の合成キメラタンパク質を含めた単離タンパク質複合体が、皮膚の質または皮膚の外観を改善または強化するために投与される美容的処置の方法も意図する。
このような処置には、皮膚細胞の異常増殖に起因する皮膚障害、例えば乾癬および過形成性瘢痕形成の予防または治癒が含まれうる。
【0142】
あるいは、二次部位への腫瘍細胞の移動を阻止することにより腫瘍の転移が防止または抑制される処置方法が意図される。さらに、細胞の増殖を阻止することにより癌を治療する方法も意図される。
【0143】
特定の実施形態では、治療的および/または予防的処置において、本発明の合成キメラタンパク質を含めた単離タンパク質複合体は、同単離タンパク質複合体を適切に含浸するか、被覆するか、または他の方法で含んでいる、生体適合材料、生体ポリマー、無機材料、例えばフルオロヒドロキシアパタイト、外科用移植片、人工器官、創傷被覆材または熱傷被覆材、圧定布、包帯等とともに、またはそれらの一構成成分として利用され得る。
【0144】
このような方法は、本明細書中に上記したような薬学的組成物の投与を包含し、そして例えば米国特許第6,090,790号明細書に記載されているように特定組織部位への微細針による注射、米国特許第6,054,122号明細書に記載されているような、創
傷、熱傷または潰瘍に適用される局所用クリーム、ローションまたは封止包帯、あるいは国際公開第99/47070号パンフレットに記載されているような、組成物を放出するインプラントによるものであり得る。
【0145】
この点については、例えば米国特許第5,929,040号明細書および米国特許第5,962,427号明細書に記述されている方法に従って遺伝子療法も適用可能である。
例えば所望の成長因子発現を遺伝子工学的に操作することにより、皮膚代替物を作製する目的のために皮膚細胞を遺伝学的に改変し得る方法も存在する(サップ(Supp)他、2000, J. Invest. Dermatol. 114, 5 )。この分野の概説の一例は、ベバン(Bevan )他、Biotechnol. Gent. Eng. Rev. 16, 231 に提供されている。
【0146】
例えば国際公開第99/11789号パンフレットに記載されているように、トランスフェクションまたは形質転換した細胞をレシピエントに「植え付ける」ことも意図される。
【0147】
これらの方法を用いて、細胞移動を刺激することにより、創傷治癒および熱傷治癒、潰瘍などの皮膚病変の修復、組織置換および移植を助長または進行させることが可能であり、例えば自己由来の皮膚のin vitro培養、腎臓および肺などの内部器官の再上皮化、ならびに損傷神経組織の修復などにより実施される。
【0148】
皮膚の置換療法は当該技術分野で既知になってきており、該療法においては、例えばケーヒ(Kehe)他、1999, Arch. Dermatol. Res. 291, 600 に記載されているような共培養した上皮/ケラチノサイト細胞株を使用してもよいし、あるいは上皮細胞、真皮細胞およびケラチノサイト細胞のうち少なくともいずれか(通常は自己由来)のin vitro初代培養物を使用してもよい。これらの技法は、人工生体適合材料および合成ポリマーの「足場材」も利用し得る。
【0149】
当該技術分野の概説の例は、概して、タースキフとヴァシリエブ(Terskikh & Vasiliev )、1999, Int. Rev. Cytol. 188,41 およびイーグルスタインとファランガ(Eaglestein & Falanga)、1998, Cutis 62, 1 に提供されている。
【0150】
より具体的には、頭蓋顔面外科に有用な代替口腔粘膜の作製は、イズミ(Izumi )他、2000, J. Dent. Res. 79, 798 に記載されている。胎児ケラチノサイトおよび真皮繊維芽細胞を、ファウザ(Fauza )他、J. Pediatr. Surg. 33, 357 に記載されているように、皮膚病変を治療するための移植用の皮膚を生産するためにin vitroで増殖させてもよいし、一方、ヒアルロン酸由来の生体適合材料上でin vitroで培養された真皮および上皮の皮膚要素からの皮膚置換物は、熱傷の治療に潜在的に有用であることが示されている(ザッキ(Zacchi)他、1998, J. Biomed. Mater. Res. 40, 187)。
【0151】
ポリマーの足場も、例えばシェリダン(Sheridant )他、2000, J. Control Release 14, 91 およびファウザ(Fauza )他、1998、(上記)に記載されているように、皮膚の人工置換を容易にすることが意図されており、微小球も創傷および熱傷への皮膚細胞の送達のための作用物質として意図されている(ラフランスとアームストロング(LaFrance & Armstrong)、1999, Tissue Eng. 5, 153)。
【0152】
(アゴニストおよびアンタゴニストの作製)
本発明は、成長因子およびビトロネクチンまたはフィブロネクチンを含んでなる複合体、例えばIGF−II:VNまたはIGF−I:IGFBP:VN複合体のアゴニストまたはアンタゴニストを、同定し、スクリーニングし、設計し、またはそうでなければ作製するための、本発明の合成キメラタンパク質を含めた単離タンパク質複合体の使用を意図
する。このような作用物質は、「模倣物」であり得る。「模倣物」という用語は、本明細書中では、タンパク質またはペプチドの特定の機能性領域に類似するよう設計された分子を指すものとして用いられ、そして当該技術分野で既知であるように、該用語の範囲内には、「アゴニスト」、「類似体」および「アンタゴニスト」という用語が包含される。
【0153】
一実施形態では、IGF−II:VN複合体またはIGF−I:IGFBP:VN複合体によるIGF−IRおよびVN受容体の結合を模倣するアゴニストが作製される。このような分子は、創傷治癒、皮膚再生等に必要とされるような細胞移動の刺激剤としての有用性を有し得る。
【0154】
別の実施形態では、IGF:VN複合体またはIGF:IGFBP:VN複合体によるIGF−IRおよびVN受容体の結合を防止または抑制するアゴニストが作製される。このような分子は、細胞の移動および増殖のうち少なくともいずれかの阻害剤としての有用性を有することにより、有用な抗腫瘍剤を構成する可能性があり、そして皮膚細胞の異常増殖に起因する皮膚障害、例えば乾癬および過形成性瘢痕形成の治療においても有用性を有し得る。
【0155】
上記の模倣物、アゴニスト、アンタゴニストおよび類似体は、所望の生物学的活性および半減期を有するペプチド、ポリペプチドまたはその他の有機分子、好ましくは小有機分子であり得る。
【0156】
コンピュータ支援構造データベース検索は、模倣物を同定するための一手法として漸増的に利用されるようになっている。原則として模倣物を同定するのに適しているデータベース検索方法は、国際公開第94/18232号パンフレット(HIV抗原模倣物の作製に関する)、米国特許第5,752,019号明細書および国際公開第97/41526号パンフレット(EPO模倣物の同定に関する)に見出され得る(これらの文献の記載内容は、参照により本明細書中で援用される)。
【0157】
その他の方法としては、分子相互作用を同定する種々の生物物理学的技法が挙げられる。これらの技法により、候補分子が例えばIGF−IGFBP−VN複合体の形成に影響を及ぼすか否かに従って、候補分子をスクリーニングすることが可能になる。潜在的に有用な技法、例えば放射性リガンドの競合的結合アッセイ(関連する方法として、上記のアプトン(Upton )他、1999を参照)、分析的超遠心分離、微量熱量測定、表面プラズモン共鳴および光学バイオセンサを用いる方法に適用可能な方法は、コリガン(Coligan )他編「タンパク質科学の最新プロトコール(CURRENT PROTOCOLS IN PROTEIN SCIENCE)」、(ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons ) NY,
1997 )の第20章に提供されている(この文献の記載内容は、参照により本明細書中で援用される)。
【0158】
本発明をより理解し易く、そして実行し易くするために、当業者に以下の非限定な実施例を提供する。
【実施例1】
【0159】
[材料および方法]
(細胞培養)
10%FCSを含有するDMEM/Hams’F12(DMEM/F12)培地(1:1)(オーストラリア国ビクトリア州マルグレーブ(Mulgrave)所在のライフテクノロジー社(Life Technologies ))中で、ヒト乳腺がん細胞株MCF−7(ATCC#HTB−22)を増殖させた。培地を毎日交換し、細胞は80%コンフルエントの状態で0.25%トリプシン/0.5mMエチレンジアミン四酢酸(EDTA)溶液(英国ハンプシャ
ー所在のオキソイド社(Oxoid ))を用いて継代した。
【0160】
ヒト皮膚ケラチノサイト細胞株HaCATは、ノルベート・フューゼニヒ(Norbet Fusenig)教授(ハイデルベルク ノイエンハイマーフェルト(Neuenheimer Feld)内のドイツがん研究センター(German Cancer Research Center: DKFZ ))から入手した。10%FCSを含有するDMEM培地(ライフテクノロジー社)中で、HaCAT細胞株を増殖させた。培地を毎日交換し、細胞は80%コンフルエントの状態で0.25%トリプシン/0.5mM EDTA溶液(オキソイド社(Oxoid ))を用いて継代した。
【0161】
(VNおよびIGFBPへのIGFの予備結合)
細胞の機能を調べるin vitroアッセイのほとんどは、溶液中に外因性因子を加えるので、細胞は、アッセイの間中その物質を含有する溶液中に入っている。これは、細胞がin vivoで遭遇する環境ではない。むしろ、組織中の細胞は、細胞により合成されたECMによって支持され、取り囲まれており、該ECMの中にはホルモンおよびその他の因子が局在している。ECM分子への成長因子の結合を特定的に取り扱うこの試験では、24ウェルプレートの組織培養用プラスチックと、12.0μL孔のCostar
Transwells(商標)(米国ニューヨーク州ニューヨーク所在のコスター社(Costar))の下方チャンバおよび膜表面に、VN、IGFおよびIGFBPを予備結合する戦略を用いて、in vivo環境をより正確に反映することを試みた。
【0162】
300〜1000ngのVN(オーストラリア国ニューサウスウェルズ州アンナデール(Annandale )所在のプロメガ社(Promega ))を含有するDMEMまたはDMEM/F12培地300μLを24ウェル組織培養ディッシュに、またはTranswells(商標)の下方チャンバに添加し、37℃で2時間インキュベートした。結合していないVNを含有する培地を除去し、ウェルを0.5%ウシ血清アルブミン(RIA等級)(BSA)(シグマアルドリッチ社(Sigma Aldrich ))を含有する1mLのヘペス結合緩衝液(HBB)で洗浄した。次に1.0%BSAを含有するHBB 300μLをウェルに添加して37℃で30分間インキュベートし、組織培養ディッシュ内の非特異的結合部位をブロッキングした。次に、0.5%BSAを含有するHBB 1mLで、ウェルを再び洗浄した。次に0.5%BSAおよびIGF−IIまたはIGF−I+IGFBP(オーストラリア国サウスオーストラリア州アデレード(Adelaide)所在のグロペップ社(GroPep))を含有するHBB 300μLを添加し、プレートを再び2時間インキュベートした。非結合IGFおよびIGFBPを含有する溶液を除去し、ウェルをHBBで洗浄して、層流フード内で風乾した。
【0163】
(移動アッセイ)
リーブスレー(Leavesley )他、1993, Journal of Cell Biology 121: 163-70 に記載されているのと本質的に同様に、移動アッセイを実施した。無血清培地中で4時間のインキュベーションにより血清飢餓状態にした50,000個の細胞を、12.0μm孔Costar Transwells(商標)(12ウェルプレート型)の上部チャンバに播種した。5%CO2中で37℃にて5時間インキュベーションした後の、多孔性膜の下側表面に移動していた細胞を固定し、次に、0.1mMのホウ酸緩衝液(pH9)中のクリスタルバイオレットで染色した。10%酢酸中にクリスタルバイオレットを抽出し、分光分析によりこれらの抽出物の吸光度を測定することにより、結合した細胞の数を概算した。
【0164】
(統計学的分析)
先ず全データを陰性対照(−VN、−IGF、−IGFBP)に対する割合(%)として表すことにより、データを分析した。次に、等分散性のスチューデントt検定(両側検定)を用いて、VNのみの対照およびIGFのみの対照に対する有意性について、応答を
試験した。0.05未満のP値は、応答に有意差があることを示す。
【0165】
[結果]
(移動)
細胞移動は創傷治癒における重要な過程であり、VNおよびIGFはともに、細胞移動の媒介において役割を有することが立証されている。VNに結合したIGF−IIの、HaCATケラチノサイトの機能を変える能力について詳細に分析するために、Transwells(商標)を通り抜ける細胞移動を測定した。
【0166】
図1は、VNの存在下では、IGF−II誘導によるTranswells(商標)を通り抜けるHaCATヒトケラチノサイトの移動が、特に低濃度で増強されることを示す。各バーは3回の反復実験からのデータを表し、各処理は三連で試験した。
【0167】
乳癌細胞株MCF−7の移動も試験した。1μgのVNを12.0μmのTranswells(商標)の下方ウェルに予備結合させた場合、下方チャンバへの移動は5倍増加した。VNの非存在下でウェルに1〜100ngのIGF−IIを「予備結合」させると、移動が刺激されて2倍に増加した。しかしながら1〜100ngのIGF−IIを下方チャンバ中の1μgのVNに予備結合させた場合、細胞移動は8〜10倍に増加した(図2)。これらの応答は、IGF−II単独およびVN単独の作用より有意に高かった(p<0.01)。
【0168】
[L27]−IGF−IIは、IGF−IRに結合しないIGF−II類似体である。IGFは受容体IGF−IRを介して移動を媒介し、IGF−IRは信号を受けると考えられる。従って、VNに予備結合された[L27]−IGF−IIがTranswells(商標)を通り抜けるMCF−7細胞の移動を刺激する能力を調べるアッセイを実行した。このアッセイは、VN−[L27]−IGF−II複合体がVN単独の場合に得られるレベルを超えてMCF−7の移動を増強することはないということ、ならびに移動のレベルがVNに結合したIGF−IIに対する応答の場合に観察される移動のレベルより有意に低い(p<0.01)、ということを明示した(図3)。これらの結果は、VNに結合したIGF−IIから生じる移動の増強にIGF−IIとIGF−IRとの相互作用が関与することを示している。
【0169】
IGFBPは、細胞に対するIGF曝露の重要な調節物質である。IGFBPがここで観察されたVN:IGF−II複合体に対する移動応答に関与するか否かを確定するために、IGFBPにはほとんど結合しないが、IGF−IRに対する親和性は保持しているIGF−II類似体を用いてMCF−7細胞における移動アッセイを実行した。このIGF−II類似体des(1−6)−IGF−IIを用いたアッセイから、天然型のIGF−IIと比較した場合に移動応答に差がないことが明らかとなり、IGF−II:VN複合体がIGFBPとは独立に作用して細胞移動を増強することが示唆された(図4)。
【0170】
図5では、データから、αVインテグリン遮断抗体がVNとIGF−IIとの複合体に応答するMCF−7細胞の移動を実質的に低減させたことを示す。これらのデータは、VNに対するαVインテグリン受容体の結合および活性化が、IGF−II:VN複合体に応答した最適な細胞移動に必要であると思われることを示唆している。
【0171】
IGF−IRとほとんど結合しないIGF−I類似体L24−IGF−IについてMCF−7細胞の移動アッセイで調べた図6を参照すると、データから、
(1)L24−IGF−IおよびIGF−Iは、VNは存在するがIGFBP−5は存在しない条件下では、同じ作用を有すること;そして
(2)IGFBP−5の存在下では、IGF−IおよびVNが存在する場合は細胞移
動が増強されるが、L24−IGF−IおよびVNが存在する場合は増強されないこと
が実証される。
【0172】
上記のデータは、IGF−IRの活性化はIGF−I:IGFBP:VN複合体に応答して観察される細胞移動に必要であること、そしてさらにVNに対するインテグリン受容体の同時結合が必要であることを示唆している。
【0173】
この試験結果は、IGF−II:VN複合体およびIGF−I:IGFBP:VN複合体が細胞移動を有意に刺激する、ということをはじめて明示するものである。総合するとこれらのデータから、VN:IGF複合体が創傷治癒と機能的に関連しており、実際、乳がんの発症および進行における重要な因子であり得ることが示唆される。さらに、移動の増強には、IGF−IRおよびVN結合性インテグリン受容体の両方の受容体の活性化を必要とする。実際、VN:IGF複合体が移動を促進し、したがって乳がん細胞の転移を促進するのであれば、VN:IGF複合体の形成または成長因子受容体およびインテグリン受容体の同時活性化の阻害を目的とする薬剤は、非常に有効な治療薬であるということになる。
【実施例2】
【0174】
[方法および材料]
(卵黄ビトロネクチンの精製)
ナガノ(Nagano)他、1992, The Journal of Biological Chemistry 267: 24863-24870に記載された方法の変法を用いて、卵黄のビトロネクチン(VN)を精製した。この手法に用いられた溶液は全て、pH7.4であった。鶏卵(オーストラリア国クイーンズランド州トゥーウォン(Toowong )所在のコールズマイヤー(Coles-Myer)の農場から入手した卵黄を、2mMのフッ化フェニルメタンスルホニル(PMSF)を含有する等容積の冷リン酸緩衝生理食塩水(PBS)(0.16MのNaCl、10mMのリン酸ナトリウム)中に懸濁し、18,000g、4℃で20分間、遠心分離した。上清(卵黄プラズマ)を、5mMの2−β−メルカプトエタノールを含有する1mMのリン酸ナトリウムに対して4℃で一晩透析し、20,000g、4℃で20分間遠心分離した。上部の固体層(低密度リポタンパク質(LDL)分画)を回収し、15mlのPBS中に再懸濁した。
【0175】
3つのクロマトグラフィー技法、すなわち、ゲル濾過、Sepharose(商標)CL−6B(スウェーデン国ウプサラ所在のアマシャムバイオサイエンス社(Amersham Biosciences));ヒドロキシアパタイトHTP(米国カリフォルニア州リッチモンド所在のバイオラッド社(Bio-Rad ));およびイオン交換、Q‐Sepharose(登録商標)Fast Flow(アマシャムバイオサイエンス社)を用いて、LDL分画から卵黄VNを精製した。
【0176】
Sepharose(商標)CL−6Bカラム(カラム容量10ml、カラムサイズ:内径(ID)2.5cm×30cm)を、2段階、すなわち(i)2MのNaClを含有するPE緩衝液(5mM EDTA、10mM リン酸ナトリウム)および(ii)0.13MのNaClを有するPE緩衝液、を用いて平衡化した。50mlのLDL分画をPE緩衝液で1:1に希釈してSepharose(商標)CL−6Bカラムにかけた。同カラムから非結合画分を収集し、次に0.5MのNaClを含有する10mMのリン酸ナトリウムで予め平衡化したヒドロキシアパタイトマトリックスにバッチ式で適用した。該ヒドロキシアパタイトを平衡化緩衝液で洗浄し、その後10mMのリン酸ナトリウム20mlで洗浄した。次に該マトリックスをカラム(1.5cmID×8cm)に詰めて、200mMのリン酸ナトリウムでタンパク質を溶離させ、10×5ml画分を収集した。該溶離画分を、プレキャスト型の4〜20%勾配ポリアクリルアミドゲル(オーストラリア国ニューサウスウェールズ州フレンチフォレスト所在のグラディポア社(Gradipore ))
、SDS−PAGE(レムリ(Laemmli ), 1970)およびクーマシーブリリアントブルー(G−250、バイオラッド)染色を用いて、yVNの存在に関して分析した。バイオラッドの低分子量域マーカーを用いて、タンパク質の分子量を測定した。
【0177】
予測される分子量のyVN(54kDa)に相当する画分をプールし、10mMリン酸ナトリウムに対して4℃で一晩透析し、次に10mMリン酸ナトリウムで予め平衡化したQ Sepharose(登録商標)Fast Flowマトリックス(カラム容量5ml、カラムサイズ:ID 1cm×10cm)に適用した。該カラムを10mMのリン酸ナトリウム中の0.15MのNaClで洗浄し、10mMのリン酸ナトリウム中の0.25MのNaClでyVNを溶離させた。上記と同様に画分の分子量を測定した。
【0178】
(IGF:VN複合体の調製)
報告されているように(クリッカー(Kricker )他の上記文献、2003)、IGF:VN複合体を、96ウェルプレートおよびTranswells(商標)に予備結合させた。
【0179】
[結果]
図8は、完全長(75kDa)の血清VN(a)と短縮型(54kDa)の卵黄VN(b)との類似性を示す。特筆すべき主な類似性は、これらのタンパク質がともにRGD細胞結合部位およびポリ陰イオン性領域(IGF結合部位とされる)を有する、ということである。特筆すべき主な違いは、卵黄VNがヘパリン結合ドメインを欠くということである。
【0180】
図9は、溶離画分(レーンE)中に存在する主なタンパク質が、卵黄VNについて予測される大きさ(54kDa)である、ということを示している。このタンパク質がその後のアッセイに用いられたということも、特筆すべき重要なことである。
【0181】
図10は、IGFBP−3の存在下でVNが放射標識IGF−Iを結合する能力を実証している。したがってこの図から、54kDa卵黄VNが完全長の卵黄VNと同じレベルでIGF−I/IGFBP−3を結合する能力を有していることが示されている。このことは、IGF結合部位がヘパリン結合ドメイン(卵黄VN(54kDa)には欠けている)中には位置しないことを示唆しており、そしてポリ陰イオン性部位がIGF結合部位として提唱されることを支持している。
【0182】
図11は、IGF−I/IGFBP−3と複合体形成した場合のVN(血清VNまたは卵黄VN)の能力により、無処置群、IGF−I/IGFBP−3処置群およびVN単独処置群の対照を上回って細胞増殖(ミトコンドリアデヒドロゲナーゼ活性を評価するMMT技法により測定)が強化されることを示している。図11はまた、短縮型(54kDa)卵黄VNが、IGF−I/IGFBP−3と複合体形成した場合、完全長VNと同程度に細胞増殖を刺激し得る、ということも示している。
【0183】
図12は、IGF−I/IGFBP−3と複合体形成した場合のVN(血清VNまたは卵黄VN)の能力により、無処置群、IGF−I/IGFBP−3処置群およびVN単独処置群の対照を上回って細胞移動(Transwells(商標)移動アッセイによる)を強化し得ることを示している。したがって、短縮型(54kDa)卵黄VNが、IGF−I/IGFBP−3と複合体形成した場合、完全長VNと同程度に細胞増殖を刺激し得るという同じ結論が、図4と同様にこの図からも引き出され得る。総合すると、これらの図から、短縮型(54kDa)卵黄VNは、IGF−I/IGFBP−3と複合体形成した場合、完全長(75kDa)VNで観察されるのと同様のレベルに細胞の移動および増殖のいずれも刺激し得る、ということが示唆される。
【実施例3】
【0184】
VN:IGF−Iキメラタンパク質の形態の、本発明の合成キメラタンパク質の提唱例が、本明細書中に提供されている。
図14に種々に記述した提唱合成キメラタンパク質には、アミノ酸残基の改変を伴って、または伴わずに、IGF−Iと融合された、任意の完全長または短縮型のVNが含まれている。さらに本発明者等は、種々のペプチドリンカーを用いて、または用いずにVNおよびIGF−Iを融合することを提唱する。
【0185】
さらに本発明者等は、VNおよび成長因子、例えばVEGFおよびPDGFを含んでなるキメラタンパク質を意図し、その特定の実施形態を図15に記述する。
本明細書において用いられ、図13に示された成熟VN(配列番号2)およびIGF−I(配列番号3)に関する完全ペプチド配列は、NCBI(それぞれアクセス番号NP_0000629および1BQT)から入手した。VNに関して示した残基番号のアノテーションは成熟タンパク質のものであり、シグナルペプチドは除かれている。
【0186】
ビトロネクチンドメイン構造およびビトロネクチンリガンド結合部位に関しては、図7および8にそれぞれ記載されている。
(完全長および短縮型のVN)
細胞移動を調節し得る合成キメラタンパク質の一例は、完全長の成熟VNタンパク質およびIGF−Iタンパク質を含有する。
【0187】
A)VN(1・・・459):IGF−I(1・・・70)
細胞移動を調節し得る別の例は、残基380〜459(C末端80アミノ酸)の欠失を伴う成熟VNタンパク質を含有する。
【0188】
B)VN(1・・・379):IGF−I(1・・・70)
血清中の単量体VNは、2つの形態、すなわち、75kDaの一本鎖ポリペプチド、または内因性の切断を受けた二本鎖形態のVNで65kDaの大型断片がジスルフィド結合により10kDa小型断片に連結されて構成されているVN、として存在する。近年の研究から、これらの形態の間に機能的差異は認められないことが示されており、このことは、VN上のC末端80アミノ酸は機能的差異を付与しない、ということを示唆している(ギブソンとピーターソン(Gibson and Peterson ), 2001, Biochim Biophys Acta 1545,
289-304)。このことは、ブタVNがこのC末端領域を失う一方でその機能的活性は保持しているという知見(ヨネダ(Yoneda)他、1996, J Biochem (Tokyo) 120:954-60)により支持される。したがって、C末端80アミノ酸を取り除いた、VNの全ての機能的特性を依然として有するVNを含有してなる、より小型のキメラ分子を、本発明者等は提唱する。
【0189】
さらに別のキメラは、IGF−Iに連結されたVNのソマトメジンBドメインのみを含有する。この領域は、プラスミノーゲンアクチベーター−1(PAI−1)、ウロキナーゼプラスミノーゲンアクチベーター受容体(uPAR)およびインテグリン結合部位を含有する(シュバルツ(Schvarz )他、1999, Int J Biochem Cell Biol 31: 539-44)。
【0190】
このキメラは、ECM中の構成成分、例えばコラーゲンおよびグリコサミノグリカンと相互作用しないであろう。このキメラは、VN上の残基53〜459(接続領域、中央のβ−プロペラドメインおよびヘパリン結合ドメイン)の欠失を有している。
【0191】
C)VN(1・・・52):IGF−I(1・・・70)
VNの接続領域は、トロンビン−アンチトロンビン複合体、ならびにECM構成成分コラーゲンを結合する際に役割を演じると推測されている。ここで提唱されるキメラは、V
NのソマトメジンBドメイン、ならびにVN上の残基131〜459(中央のβ−プロペラドメインおよびヘパリン結合ドメイン)の欠失により創出される接続領域を含有する。
【0192】
D)VN(1・・・130):IGF−I(1・・・70)
さらなる例では、VN上の中央ドメインは最大であるが、しかし機能に関しては同タンパク質中であまりよく特性解析されていないドメインである。しかしながら、このドメイン中に観察されるβ−プロペラ構造はVNの多量体化に寄与し得る、と推測される(キュー(Xu)他、2001, Proteins 44: 312-20 )。
【0193】
本発明者等は、このドメインを欠失させて、VNの、ソマトメジンBドメイン内のリガンド結合領域、ポリ陰イオン性接続領域、およびポリ陽イオン性ヘパリン結合ドメイン(HBD)を保持し、しかしながら自己会合できない小さいキメラとすることを提唱する。この場合、VN上の残基131〜346(中央のβ−プロペラドメイン)の欠失を必要とする。
【0194】
E)VN(1・・・130、347〜459):IGF−I(1・・・70)
さらに別のキメラは、細胞外リガンドを結合し得ると考えられる最も小型のVNから構成される。該タンパク質は、VN上の中央ドメインおよびC末端80アミノ酸がいずれも除去されている。この場合、VN上の残基131〜346および380〜459(それぞれ中央のβ−プロペラドメインおよびC末端80アミノ酸)の欠失を要する。
【0195】
F)VN(1・・・130、347〜379):IGF−I(1・・・70)
キメラのさらなる例は、ヘパリン結合ドメインを伴わないC末端短縮型VNを含有する。したがって該タンパク質は、VNのソマトメジンBドメイン、接続領域および中央のβ−プロペラドメインを含有する。VNに関しては、二次ヘパリン結合部位と推定される部位がVNの中央のβ−プロペラドメイン内に提唱されているが、しかしギブソン(Gibson)他(ギブソン(Gibson)他、1999, J Biol Chem 274, 6432-42)は、該部位が機能性でないこと、そしてヘパリン結合ドメインはグリコサミノグリカン結合活性を完全するために寄与する、ということを実証した。したがって該キメラは、ヘパリンおよびヘパリン硫酸と相互作用しないであろう。このキメラは、VN上の残基347〜459(ヘパリン結合ドメイン)の欠失を有する。
【0196】
G)VN(1・・・346):IGF−I(1・・・70)
(VNおよびIGF−I上の残基の修飾)
VNは、カゼインキナーゼII(CK2)により残基T50およびT57がリン酸化されて、細胞の接着および伸展を促進し得る。VNのCK2−リン酸化類似体およびCK2−非リン酸化類似体(それぞれVN突然変異体(T50E、T57E)および(T50A、T57A)により模擬検討された)のいずれも、αvβ3およびαvβ5インテグリンに結合してERKシグナル伝達経路を活性化させるが、一方、αvβ3インテグリンに特異的に結合しているCK2−リン酸化VN類似体だけが、ホスファチジルイノシトール3キナーゼ(PI3−K)経路を活性化させた(セガー(Seger )他、1998, J Biol Chem 273: 24805-13; セガー(Seger )他、2001, J Biol Chem 276: 16998-7006 )。
【0197】
細胞の接着および伸展の増大をもたらすと思われるのは、このPI3−K経路の活性化である。したがって本発明者等は、αvβ3インテグリンとの結合後にPI3−K経路の活性化を促進または抑制するであろう突然変異を有するキメラを提唱する。したがってVN上のT50AおよびT57A置換を有するキメラ分子は、CK2−非リン酸化VNと類似し、ERK経路を介したシグナル伝達に限定される(H)が、一方、VN上のT50EおよびT57E置換を有する合成構築物は、CK2−リン酸化VNを模倣し、ERK経路およびPI3−K経路の両方を活性化して、細胞内シグナル伝達の変更をもたらし得る(
I)。
【0198】
H)VN(T50A、T57A):IGF−I
I)VN(T50E、T57E):IGF−I
VN上の残基S378にはcAMP依存性プロテインキナーゼ(PKA)リン酸化部位が存在する。この部位のリン酸化はPAI‐1のVNへの結合を低減し、したがってウロキナーゼ系におけるVNの役割を変える、ということが、PKAリン酸化VN類似体およびPKA非リン酸化VN類似体(それぞれVN突然変異体S378EおよびS378Aにより模擬検討された)を用いて実証されている(シュバルツ(Schvartz)他、2002, Arch
Biochem Biophys 397: 246-52)。
【0199】
したがって、キメラ(J)によるPAI−1結合を抑制するためのVN上のS378E突然変異、ならびにキメラタンパク質(K)内のPAI−1結合および安定化を促進するためのVN上のS378A突然変異、の両方を含有するキメラを本発明者等は提唱する。さらに、PAI−1のVNへの結合が、インテグリンを介した細胞移動を阻害すること(キョーラ(Kjoller )他、1997, Exp Cell Res 232: 420-9 )、ならびにuPARおよびインテグリンを介したVN上への細胞接着を阻害すること(デング(Deng)他、2001, J Cell Physiol 189: 23-33 )が示されているので、S378A突然変異は細胞移動を増強する可能性がある。興味深いことに、これらの知見は、プラスミノーゲン活性化の阻害剤としてのPAI−1の機能とは独立して観察された。
【0200】
J)VN(S378E):IGF−I
K)VN(S378A):IGF−I
ゲヒトマンとシャルティール(Gechtman and Shaltiel )、1997, Eur J Biochem 243:
493-501は、プロテインキナーゼC(PKC)がVNの残基S362をリン酸化し得ることを示した。このリン酸化は、VNのヘパリン結合ドメイン内に生じるプラスミンによる切断を低減する。したがってこの部位におけるプラスミン切断により、この領域内に結合するリガンドに対するVNの親和性が変化し、かつVNの半減期も変化する。したがって本発明者らは、リン酸化セリンを模倣し、その結果プラスミンによるキメラの切断を阻害するためのS362E置換の導入を提唱する。
【0201】
L)VN(S362E):IGF−I
IGFBPは、高い親和力で成長因子IGF−Iを結合するために、IGF−I上のN末端3残基を必要とすることが示されている(トーマス(Tomas )他、1991, J Endocrinol 128: 97-105)。
【0202】
したがって、N末端配列を介してVNに連結されたIGF−IがIGFBPを結合し得るとは考えられない。にもかかわらず、本発明者らは、IGFBPがIGF−Iと結合し、その結果VN:IGF−Iキメラタンパク質の生物学的活性を抑制する全ての機会を排除するために、N末端短縮型IGF−Iを含有するVN:IGF−Iキメラをさらに提唱する。この構築物は、IGF−I上の残基1〜3(IGFBP結合領域)の欠失を含む。
【0203】
M)VN:IGF−I(4・・・70)
VNのポリ陰イオン性領域はIGF−IIおよびIGFBPの結合に寄与することが示唆されている。したがって本発明者らは、VNからポリ陰イオン性ドメインを除去したさらに別のVN:IGF−Iキメラを提唱する。よってこのキメラは、IGF−IIまたはIGFBPに対して無力である。
【0204】
N)VN(1・・・52、65・・・459)IGF−I(1・・・70)
(VNとIGF−Iの融合)
VNおよびIGF−IのcDNAを、発現に先立って、ペプチドリンカー配列を挿入して、または挿入せずに、融合させてもよいことを提唱する。タンパク質を融合するための種々のリンカー配列が首尾よく用いられてきており、通常は、グリシンおよびセリン残基ならびに例えばトロンビン、コラゲナーゼまたはプラスミンなどのプロテアーゼの切断部位のうち少なくともいずれかを組合せて構成される。
【0205】
リンカー配列の例を以下に挙げるが、これらに限定されない。
(i)Gly4Ser(配列番号4);
(ii)Gly4Ser3(配列番号5);
(iii)(Gly4Ser)3(配列番号6);
(iv)LeuIleLysMetLysPro(配列番号7);および
(v)GlnProGlnGlyLeuAlaLys(配列番号8)
(VNと他のペプチド成長因子の融合)
細胞外マトリックスタンパク質VNを成長因子IGF−Iと融合することに加えて、VNと他のペプチド成長因子の融合も提唱する。具体的には、以下のキメラタンパク質の開発を提唱する(図16)。
【0206】
A)VN:PDGFα(1・・・210)(NCBIアクセス番号P04085)
B)VN:VEGF(1・・・102)(NCBIアクセス番号2VPFE)
それぞれの細胞表面受容体はインテグリンと、特にVN受容体であるαvβ3インテグリンと相互作用する、ということを本発明者らは提唱する。具体的には、PDGF受容体が、PDGFによる刺激後に、αvβ3インテグリンと免疫共沈降する、ということが示されている(シュネーラー(Schneller )他、EMBO J 16: 5600-7 )。
【0207】
2型VEGF受容体はその成長因子による刺激後にVN受容体と免疫共沈降する、ということも実証されている(ソルディ(Soldi )他、EMBO J 18: 882-92 )。
これらの成長因子に対する受容体がαvβ3インテグリンと相互作用するという知見は、これらの細胞表面受容体の細胞内シグナル伝達経路に対して調節および増強のうち少なくともいずれかをもたらすに際して、この相互作用が重要な役割を有することを示唆している。したがって、上記のキメラタンパク質により開始されるこれらの受容体の同時活性化および会合により、治療的用途に関連する顕著な生物学的応答を誘導し得る。
【0208】
本明細書全体を通して目的としたのは、本発明の好ましい実施形態を説明し、かつ本発明をいずれの一実施形態にも特定の特徴の集合体にも限定しないことであった。したがって、当業者には理解されることであるが、該開示にかんがみて、本発明の範囲を逸脱することなく、例示された特定の実施形態に種々の修正および変更をなすことができる。
【0209】
本明細書中で言及されたコンピュータプログラム、アルゴリズム、特許文献および科学文献は全て、参照により本明細書中で援用される。
【図面の簡単な説明】
【0210】
【図1】12μm孔のTranswells(商標)の上方チャンバ内に植え付けられたヒト皮膚ケラチノサイト細胞HaCATの、VNに予備結合されたIGF−II(黒色バー)またはVNの非存在下でディッシュに「結合された」IGF−II(灰色バー)で被覆された下方チャンバに応答した、下側表面への移動について示す図。各バーは、5時間インキュベーション後の下方膜上の細胞数の平均を表し、3回の反復実験(3連のウェル中で処理群についての分析を実施)から得ている。
【図2】12μm孔のTranswells(商標)の上方チャンバ内に植え付けられたヒト乳がん細胞MCF‐7の、VNに予備結合されたIGF−II(斜線バー)またはVNの非存在下でディッシュに「結合された」IGF−II(白色バー)で被覆された下方チャンバに応答した、下側表面への移動について示す図。各バーは、5時間インキュベーション後の下方膜上の細胞数の平均を表し、3回の反復実験(3連のウェル中で処理群についての分析を実施)から得ている。複合体の作用がVN単独の場合と有意に異なっているデータを星印で示す。
【図3】12μm孔のTranswells(商標)の上方チャンバ内に植え付けられたヒト乳がん細胞MCF‐7の、VN、VNに結合した天然型IGF−II(斜線バー)またはVNに結合したL27−IGF−II(黒色バー)で被覆されている下方チャンバへの移動について示す図。各データは、VNの非存在下で同一量のIGF−IIを含有する、VNを含まない対照(白色バー)と組み合わされている。各バーは、5時間インキュベーション後の下方膜上の細胞数の平均を表し、2回の反復実験(3連のウェル中で処理群についての分析を実施)から得ている。
【図4】12μm孔のTranswells(商標)の上方チャンバ内に植え付けられたヒト乳がん細胞MCF‐7の、VN単独、VNに結合した天然型IGF−II(斜線バー)またはVNに結合したDes(1−6)IGF−II(黒色バー)で予備結合されている下方チャンバへの移動について示す図。各データは、VNの非存在下で同一量のIGF−IIを含有する、VNを含まない対照(白色バー)と組み合わされている。各バーは、5時間インキュベーション後の下方膜上の細胞数の平均を表し、2回の反復実験(3連のウェル中で処理群についての分析を実施)から得ている。
【図5】ヒト乳がん細胞MCF‐7の、αVの機能を遮断するAbであるmAb2021Zの存在下で、IGF−IIに応答して、Transwells(商標)を通り抜ける移動について示す図。αV機能遮断Abで処置済みのMCF−7細胞を、VN+/−IGF−IIで被覆されているTranswells(商標)上に植え付け、5時間かけて移動させてこの多孔性膜を通過させた。次に、固定した細胞から染色剤を抽出して光学密度を読み取ることにより、膜を横断する細胞の数を測定した。次に、処理群をAbの存在下または非存在下におけるVN単独群の移動細胞数に対する割合(%)として表した。単回実験の4連処置からデータを合わせた。各バーはSEMである。星印は、非処理細胞またはAb処理細胞(Abを有する)の処理群間の有意差を示す(P<0.1)。
【図6】12μm孔のTranswells(商標)の上方チャンバ内に植え付けられたヒト乳がん細胞MCF‐7の、VN(白色バー);VN+IGFBP‐5(灰色バー);天然型IGF−I+VN(明色で塗りつぶされたバー)または天然型IGF−I+IGFBP‐5+VN(暗色で塗りつぶされたバー);L24−IGF−I+VN(左下がり斜線バー)またはL24−IGF−I+IGFBP‐5+VN(右下がり斜線バー)で被覆されている下方チャンバへの移動について示す図。各バーは、5時間インキュベーション後の下方膜上の細胞数の平均を表し、2回の反復実験(3連のウェル中で処理群についての分析を実施)から得ている。
【図7】ビトロネクチンのアミノ酸配列(配列番号1)を示す図。ビトロネクチン内の種々のドメインに関する残基の参照、ならびに残基の修飾部位、リガンド結合部位およびプロテアーゼ認識部位も示す。
【図8】(a)完全長VN(75kDa)および(b)卵黄VN(54kDa)の構造上の関係を、リガンド結合部位とともに示す図。哺乳類および鳥類の血清VNはともに同一のドメイン構造を有するが、アミノ酸配列には差が存在する。卵黄VN(54kDa)は、これらのタンパク質の短縮型である。用いた略号について以下に示す:Som B、ソマトメジンB;接続領域、接続ドメイン;ヘモペキシン、ヘモペキシン様リピート;HBD、ヘパリン結合ドメイン;PAI−1、プラスミノーゲン活性化抑制因子−1;uPAR、ウロキナーゼ型プラスミノーゲン活性化因子受容体;TAT、トロンビン−アンチトロンビンIII複合体;uPA、ウロキナーゼ型プラスミノーゲン活性化因子;−−−−、ポリ陰イオン性領域(塩基性領域);+++、ポリ陽イオン性領域(酸性領域)。
【図9】ニワトリ卵黄由来の54kDa卵黄VNの精製を示す図。Q−Sepharose(登録商標)マトリックスに装荷したタンパク質試料(レーンL)、および溶離された精製物(レーンE)のSDS−PAGE分析。レーンMは、分子量マーカー(バイオラッドの低分子量域マーカー、米国カリフォルニア州リッチモンド所在のバイオラッド)を示す。プレキャスト型の4〜20%勾配ポリアクリルアミドゲル(オーストラリア国ニューサウスウェールズ州フレンチフォレスト所在のグラディポア社(G radipore ))を用いて、タンパク質を分析した。
【図10】[125I]−IGF−I/IGFBP−3の、精製VNと結合する能力について評価する固体プレート結合アッセイを示す図。クリッカー(Kricker )他、2003 Endocrinology 144 2807-2815にすでに記載されているように、固体プレート結合アッセイを実行した。簡潔に述べれば、Immulon(登録商標)96ウェルプレートを精製VNで4℃にて一晩かけて予め被覆した。次に放射活性標識したIGF−I/IGFBP−3複合体を添加し、一晩VNに結合させた後、結合していない物質を除去した。ウェルに結合したVNに対する[125I]−IGF−I/IGFBP−3の結合を、γ計数器で測定した(n=18)。ヒトVN:ヒト血清から精製されたVN;卵黄VN75:75kDaの精製卵黄VN;卵黄VN54:54kDaの精製卵黄VN。
【図11】IGF:VN複合体に応答したHaCAT細胞の増殖に関する細胞増殖アッセイ(MTT)(48時間)を示す図。IGF:VN複合体でウェルを予め被覆し、HaCAT細胞を植え付け、48時間増殖させた。48時間後、MTT法(デニゾットおよびラング(Denizot & Lang)、1986 The Journal of Immunological Methods 89 271-277)を用いて、代謝活性により細胞増殖について複合体を評価した(n=3)。ヒトVN:ヒト血清から精製されたVN;卵黄VN75:75kDaの精製卵黄VN;卵黄VN54:54kDaの精製卵黄VN;IGF−I/BP3:インスリン様成長因子−Iおよびインスリン様成長因子結合タンパク質3。
【図12】IGF:VN複合体に応答したHaCATの移動に関するTranswells(商標)移動アッセイ(5時間)を示す図。上記と同様に(クリッカー(Kricker )他の上記文献、2003)、HaCAT細胞を、IGF−I:IGFBP−3:VN複合体で被覆したTranswells(商標)に植え付け、5時間移動させた。移動した細胞をクリスタルバイオレットで染色し、595nmで光学密度を読取った。各処理は、2連で実施した(n=2)。ヒトVN:ヒト血清から精製されたVN;卵黄VN75:75kDaの精製卵黄VN;卵黄VN54:54kDaの精製卵黄VN;IGF−I/BP3:インスリン様成長因子−Iおよびインスリン様成長因子結合タンパク質3。
【図13】(A)成熟ビトロネクチンタンパク質(配列番号2)、(B)IGF−I(配列番号3)および(C)好ましいリンカー配列(配列番号4〜8)のアミノ酸配列を示す図。
【図14】IGF−IおよびVNを含有するキメラタンパク質の実施形態(A)〜(N)のアミノ酸配列(配列番号9〜22)を示す図。
【図15】(A)PDGFおよびVNを含有するキメラタンパク質の実施形態のアミノ酸配列(配列番号23)、ならびに(B)VEGFおよびVNを含有するキメラタンパク質の実施形態のアミノ酸配列(配列番号24)を示す図。
【技術分野】
【0001】
本発明は、成長因子受容体、例えば1型インスリン様成長因子受容体、ならびにビトロネクチンまたはフィブロネクチンの受容体であるインテグリンのいずれにも結合し活性化することができる各ドメインを有するタンパク質複合体に関する。特定の実施形態において、本発明は、インスリン様成長因子−I、インスリン様成長因子−II、血小板由来成長因子または血管内皮由来成長因子の受容体結合ドメイン、ならびにビトロネクチンまたはフィブロネクチンのインテグリン受容体結合ドメインを含んでなるキメラタンパク質に関する。特に本発明は、細胞移動を刺激するタンパク質複合体、ならびに細胞の移動および増殖のうち少なくともいずれかを促進または誘導する組成物および方法に関する。これらの組成物および方法は、創傷治癒、組織工学、美容上および治療上の処置(例えば皮膚置換および皮膚補充)ならびに上皮細胞の移動および増殖のうち少なくともいずれかが必要とされる熱傷の治療に用途を有し得る。その他の実施形態では、本発明は、特に乳がんに関連したがん細胞の転移の防止または抑制に関する本発明により提供される治療を提供する。本発明のキメラタンパク質は、インスリン様成長因子、ビトロネクチンおよびインスリン様成長因子結合タンパク質を含むタンパク質複合体の生物学的作用のアゴニストおよびアンタゴニストの作製のためにも有用であり得る。
【背景技術】
【0002】
インスリン様成長因子(IGF)であるIGF−IおよびIGF−IIは、広範囲の細胞プロセス、例えば過形成、DNA合成、分化、細胞周期の進行およびアポトーシスの抑制に関与する、細胞分裂促進性ペプチド成長因子である(非特許文献1〜3)。これらの作用は、IGFのチロシンキナーゼ結合型細胞表面受容体である、1型IGF受容体(IGF−IR)との結合により媒介される。また、IGFは、IGFBPと呼ばれる特定の結合タンパク質ファミリーにより厳密に調節されているが、IGFBPの主な役割は、遊離IGFを結合することによりIGFの半減期、特異性および活性を調節することである(非特許文献4)。
【0003】
近年、ビトロネクチン(VN)はIGF−IIと直接結合することが示された(非特許文献5)が、一方、IGF−Iは、特許文献1に記載されているように、ある種のIGFBPの存在下でVNと結合し得る。ECM組織化分子および接着分子であるVNがIGF−IIを結合する親和性は、生物学的関連を有する受容体であるIGF−IRに対するIGF−IIの親和性(非特許文献5)と同様である、という知見は、IGFの作用とECM中のVNとの間の特異的な物理的関連を明示している。さらに、VNに結合したIGF−IIは、in vitroでのヒトケラチノサイトにおける相乗的な機能的応答を刺激し得る(特許文献1)。
【0004】
VNは、血液中およびECM中で非常に豊富な糖タンパク質である。主に肝臓で合成されるが、しかし多数のその他のタイプの細胞によっても発現されるVNは、閉じた立体構造で血液中を循環し、そして開放または伸張した立体構造でECM中に沈着される(非特許文献6)。いずれの立体構造も、IGF−IIを結合すると考えられ(非特許文献5、特許文献1、非特許文献7)、かつ多様なその他のリガンド、例えばコラーゲン(非特許文献8)、グリコサミノグリカン(非特許文献9)、多数のその他のECMタンパク質、ならびに広範な種々のインテグリン、特にαVインテグリンも結合すると考えられる。実際、ビトロネクチンの主な役割は、RGD結合モチーフを介してこれらの細胞表面インテグリン受容体との接着性連結を提供する、ECM組織化分子としての役割である。VN受容体(αVインテグリン)は、増殖および侵潤に必要とされるアクチン細胞骨格の再編成を調節することが示されており、従って、VNの結合は細胞の接着および運動を調整する
(非特許文献10および11)。
【0005】
しかしながら、タンパク質複合体中に存在するIGFおよびVNのそれぞれの相対的寄与は、生物学的応答、例えば細胞の移動および増殖のうち少なくともいずれかの刺激という点において、依然として分かりにくい。なぜなら、IGF/IGFBPとVNとの間にタンパク質−タンパク質相互作用の部位があるからである。
【特許文献1】国際公開第02/24219号パンフレット
【非特許文献1】ケイス(Keiss )他、1994年、Hormone Research、第41巻、p.66
【非特許文献2】ウッドおよびイー(Wood & Yee)、2000年、J. Mammary Gland Biology and Neoplasia、第5巻、p.1
【非特許文献3】ジョンおよびクレモンス(Jones & Clemmons)、1995年、Endocrine Rev.、第16巻、p.3
【非特許文献4】クレモンス(Clemmons)、1998年、Mol. Cell. Endocrinol.、第140巻、p.19
【非特許文献5】アプトン(Upton )他、1999年、Endocrinology 、第140巻、p.2928‐31
【非特許文献6】シュバルツ(Schvartz)他、1999年、The International Journal of Biochemistry and Cell Biology、第31巻、p.531‐44
【非特許文献7】マクムーティ(McMurty )他、1996年、Endocrinology 、第150巻、p.149‐60
【非特許文献8】モーリス(Morris)他、1994年、Journal of Biological Chemistry 、第269巻、p.23845‐52
【非特許文献9】フランソワ(Francois)他、1999年、Journal of Biological Chemistry 、第274巻、p.37611‐19
【非特許文献10】デパスカール(DePasquale)、1998年、Histochemistry and Cell Biology 、第110巻、p.485‐94
【非特許文献11】ユアン(Huang )、2000年、Oncogene、第19巻、p.1915‐23
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、成長因子複合体ならびに細胞の移動および増殖を調節する方法を提供することである。
【課題を解決するための手段】
【0007】
本発明者等は、IGF−IIおよびVN、またはIGF−IおよびIGFBPおよびVNを含むタンパク質複合体が、IGF−I受容体(IGF−IR)およびVN−結合インテグリン受容体に結合し、かつ相乗的に同時活性化することにより、細胞の移動および増殖のうち少なくともいずれかを刺激する、ということを発見した。
【0008】
さらに、VNのポリ陰イオン性ドメインが同定されIGFまたはIGFBPの結合部位として提唱された。
したがって本発明は、広範には、成長因子ドメインの受容体結合ドメインと、インテグリン受容体に結合し得るビトロネクチンまたはフィブロネクチンのドメインとを含んでなる単離タンパク質複合体であって、成長因子およびインテグリン受容体を同時活性化することにより生物学的応答を引き出し得る単離タンパク質複合体に関する。
【0009】
第一の態様において、本発明は、単離タンパク質複合体であって、
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子
受容体に結合し得るドメイン、および
(ii)ビトロネクチン(VN)もしくはフィブロネクチン(FN)、または少なくともVNもしくはFNのインテグリン結合ドメイン
を含んでなる単離タンパク質複合体を提供する。
【0010】
第二の態様において、本発明は、合成キメラタンパク質の形態の単離タンパク質複合体であって、
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子受容体に結合し得るドメイン、および
(ii)ビトロネクチン(VN)もしくはフィブロネクチン(FN)、または少なくともVNもしくはFNのインテグリン結合ドメイン
のアミノ酸配列を含む単離タンパク質複合体を提供する。
【0011】
好ましくは、上記の態様によれば、成長因子はIGF−IまたはIGF−IIである。
さらに好ましくは、成長因子はIGF−Iである。
成長因子がIGF−Iである実施形態では、適切にはIGF−Iの前記少なくとも1つのドメインはIGF−Iの残基24を含む。
【0012】
成長因子がIGF−IIである実施形態では、適切にはIGF−IIの前記少なくとも1つのドメインはIGF−IIの残基27を含む。
代替的実施形態では、成長因子はVEGFまたはPDGFである。
【0013】
好ましくは、インテグリン受容体は、αVインテグリンである。
さらに好ましくは、インテグリン受容体は、αVβ3インテグリンまたはαVβ5インテグリンである。
【0014】
本発明のこの態様では、上記(i)および(ii)に対応するアミノ酸配列の、アミノ酸の欠失、付加、置換および突然変異のうち少なくともいずれかについても、その範囲内に含まれる。
【0015】
第三の態様において、本発明は、第二の態様の単離タンパク質複合体をコードする単離核酸を提供する。
第四の態様において、本発明は、発現ベクター中の1つまたは複数の調節配列に作動可能なように連結された第三の態様の単離核酸を含む遺伝子構築物を提供する。
【0016】
好ましくは、遺伝子構築物は発現構築物である。
第五の態様において、本発明は、第四の態様の遺伝子構築物を含む宿主細胞を提供する。
【0017】
第六の態様において、本発明は、第一の態様の単離タンパク質複合体または第二の態様の合成タンパク質と、薬学的に許容可能な担体、希釈剤または賦形剤とを含んでなる薬学的組成物を提供する。
【0018】
本発明のこの態様は、第五の態様の宿主細胞を含み、細胞が上記合成タンパク質を発現する薬学的組成物も包含する。
第七の態様において、本発明は、第二の態様の合成タンパク質に特異的な抗体を提供する。
【0019】
第八の態様において、本発明は、細胞移動の促進方法であって、成長因子受容体およびインテグリン受容体の両方に結合するために合成タンパク質を用いる工程を含む方法を提
供する。
【0020】
好ましくは、成長因子受容体はIGF−IRである。
好ましくは、インテグリン受容体はαVインテグリンである。
さらに好ましくは、インテグリン受容体は、αVβ3インテグリンまたはαVβ5インテグリンである。
【0021】
好ましい実施形態では、本発明のこの態様は、哺乳類、好ましくはヒトにおける創傷治癒を促すための、上皮細胞の移動および増殖のうち少なくともいずれかの促進または誘導に関する。
【0022】
好ましくは、上記合成タンパク質は、本発明の第一の態様によるものと同様である。
第九の態様において、本発明は、細胞の移動および増殖のうち少なくともいずれかを防止する方法であって、成長因子およびビトロネクチンもしくはフィブロネクチンを含んでなる複合体により成長因子受容体およびインテグリン受容体の両方の結合を防止、抑制、またはそうでなければ低減する工程を含む方法を提供する。
【0023】
好ましくは、成長因子受容体はIGF−IRである。
好ましくは、インテグリン受容体はαVインテグリンである。
さらに好ましくは、インテグリン受容体は、αVβ3インテグリンまたはαVβ5インテグリンである。
【0024】
好ましい実施形態では、本発明のこの態様は、哺乳類、好ましくはヒトにおける転移性がん細胞の移動および増殖のうち少なくともいずれかの防止または抑制に関する。
本発明のこの態様により意図される特定の例は、乳がん転移の防止または抑制である。
【0025】
また、第八および第九の態様の方法は、予防的および治療的処置方法を包含し得る、と理解される。
第十の態様において、本発明は、
(i)成長因子およびビトロネクチンまたはフィブロネクチンを含んでなるタンパク質複合体のアゴニストである分子、または
(ii)成長因子およびビトロネクチンまたはフィブロネクチンを含んでなるタンパク質複合体のアンタゴニストである分子
を作製するための、第一の態様の単離タンパク質複合体または第二の態様の合成タンパク質の用途を提供する。
【0026】
好ましい実施形態では、本発明は、以下の:
(i)IGF−II:VNタンパク質複合体またはIGF−I:IGFBP:VNタンパク質複合体のアゴニストである分子、または
(ii)IGF−II:VNタンパク質複合体またはIGF−I:IGFBP:VNタンパク質複合体のアンタゴニストである分子
を作製するための、第一の態様の合成タンパク質の用途を提供する。
【0027】
本発明のこの態様に従って作製されるアゴニストおよびアンタゴニストのうち少なくともいずれかは、創傷治癒、組織工学、皮膚再生、ならびにがん細胞の転移または皮膚の過増殖による障害(例えば瘢痕形成および乾癬)の防止、のうち少なくともいずれかを促すに際して特定の効能を有し得る。
【0028】
第十一の態様において、本発明は、第一または第二の態様の単離タンパク質複合体を含む生体適合材料を提供する。
特定の実施形態では、生体適合材料は、本発明の単離タンパク質複合体を、適切な含浸、被覆、またはその他の方法で含む外科用移植片、人工器官、足場材、創傷被覆材または熱傷被覆材等であり得る。
【発明を実施するための最良の形態】
【0029】
(発明の詳細な説明)
本発明は、IGF−IIおよびVNまたはIGF−IおよびIGFBPおよびVNを含んでなるタンパク質複合体が、応答性細胞により発現されるIGF−IR受容体およびVN結合インテグリン受容体に結合し、同受容体を介して細胞移動に生物学的作用を及ぼす、という発見から生じた。より具体的には、この二元的結合事象は、ケラチノサイトモデルおよび乳がん細胞モデルの両方において本発明者らにより示されたように、細胞の移動および増殖のうち少なくともいずれかを相乗作用的に刺激する。
【0030】
さらに、IGF−IIおよびIGFBPと相互作用するかまたは結合すると思われるVNのドメインは、成熟VNのアミノ酸53〜64に対応するポリ陰イオン性領域であるということが意外にも発見された。
【0031】
この発見により、本発明者等は、IGF−IRに結合し得るIGF−IまたはIGF−IIの少なくとも最小ドメインまたは最小領域をVNのインテグリン結合ドメインと組合せて含む単離タンパク質複合体を提供した。さらに具体的には、これらのドメインを含む単一で切れ目のないタンパク質が作製され得る。
【0032】
このようなタンパク質複合体は、複数のタンパク質を含むものであれ、単一の合成タンパク質の形態であれ、IGF−IRおよびVN結合インテグリン受容体に協調的に結合するかまたは同受容体を相互連結することにより、細胞の移動および増殖のうち少なくともいずれかならびに創傷治癒を促進するための有用な作用物質である、と予測される。同様に、IGF−IRおよびVN−結合インテグリン受容体の相互連結の防止は、がん細胞の転移を防止するために用いられ得る、ということが本発明者等により提唱される。この発見は、他の成長因子、例えばPDGFおよびVEGF(これらに限定されない)を含むタンパク質複合体に、そして他のインテグリン結合タンパク質、例えばフィブロネクチン(FN)にも拡張可能であり得る、ということも提唱される。
【0033】
本明細書全体を通して、別記しない限り、「〜を含む、含んでなる(comprise, comprises )」、および「含んでいる、含んでなる(comprising)」とは、提示された1または一群の完全体が、1つまたは複数の他の提示されていない1または一群の完全体を包含し得るよう、排他的というより包括的に用いられる。
【0034】
成長因子受容体結合ドメインおよびインテグリン結合ドメインの特定の状態においては、このようなドメインは、該ドメインのアミノ酸配列とともに、所望により他の付加的アミノ酸配列も含む。
【0035】
さらにまた、このようなドメインは、該ドメインのアミノ酸配列とともに、10個以下の、好ましくは5個以下の、さらに好ましくは4、3、2または1個以下の付加的アミノ酸「のみから本質的に構成される(consist essentially of)」、と理解されよう。
【0036】
さらにまた、このようなドメインは、任意の付加的アミノ酸の非存在下では、ドメインのアミノ酸配列「のみから構成される(consist of)」、と理解されよう。
本発明の目的のために、「単離(された)」とは、その天然状態から取り出されたか、または他の方法で人為的に操作された物質を意味する。単離物質は、その天然状態で該物質に通常随伴する構成成分を実質的または本質的に含有しないものでもよいし、あるいは
その天然状態で該物質に通常随伴する構成成分と一緒に人工状態で存在するよう操作されたものでもよい。単離物質は、天然の形態でも、化学合成形態でも、または組換え形態でもよい。
【0037】
本明細書中で用いる場合、「合成、合成の」とは、天然に存在せず、ヒトによる技術的介入により作製されることを意味する。合成のタンパク質および核酸に関しては、これは、当該技術分野で既知の組換え技術、化学合成技術またはコンビナトリアル技術により作製される分子を包含する。
【0038】
「タンパク質」とは、アミノ酸のポリマーを意味する。アミノ酸は、当該技術分野で既知であるような、天然または非天然のアミノ酸、D−またはL−アミノ酸でよい。「タンパク質」という用語には、当該技術分野で一般的に用いられるように、「糖タンパク質」、「リポタンパク質」等のような用語が挙げられ、かつ該用語を包含する。
【0039】
「ペプチド」とは、50個未満のアミノ酸を有するタンパク質である。
「ポリペプチド」とは、50個またはそれ以上のアミノ酸を有するタンパク質である。
本明細書中で上記したように、本発明は、特定の一態様において、単離タンパク質複合体を提供し、同単離タンパク質複合体は、
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子受容体に結合し得るドメイン、および
(ii)ビトロネクチンまたはフィブロネクチンの少なくとも1つのインテグリン結合ドメイン
を含んでなる。
【0040】
本明細書中で用いる場合、「成長因子」とは、in vitroおよびin vivoのうち少なくともいずれかにおいて細胞の増殖、分化、生存および移動のうち少なくともいずれかを調節し得る生物学的活性タンパク質である。
【0041】
好ましくは、成長因子は、IGF−I、IGF−II、VEGFおよびPDGFからなる群から選択される。
さらに好ましくは、成長因子は、IGF−IおよびIGF−IIから選択される。
【0042】
しかしながら本発明は、細胞の増殖、分化、生存および移動のうち少なくともいずれかを調節するその他の生物学的活性タンパク質、例えば上皮成長因子(EGF;ヘルディン(Heldin)他、1981, Science 4 1122-1123 )、繊維芽細胞成長因子(FGF;ナーコンブ(Nurcombe)他、2000, J. Biol. Chem. 275 30009-30018)、塩基性繊維芽細胞成長因子(bFGF;タラボレッティ(Taraboletti )他、1997, Cell Growth. Differ. 8 471-479)、オステオポンチン(ナム(Nam )他、2000, Endocrinol. 141 1100)、トロンボスポンジン−1(ナム(Nam )他著、2000(上記))、テネイシン−C(アライ(Arai)他、1996, J.Biol. Chem. 271 6099)、PAI−1(ナム(Nam )他、1997, Endocrinol. 138 2972)、プラスミノーゲン(キャンプベル(Campbell)他、1998, Am. J.Physiol.
275 E321 )、フィブリノーゲン(キャンプベル(Ca mpbell)他、1999, J. Biol. Chem
274 30215 )、フィブリン(キャンプベル(Campbell)他、1999(上記))またはトラ
ンスフェリン(バインジマー(Weinzimer )他、2001, J. Clin. Endocrinol. Metab. 86
1806 )も意図する。
【0043】
本発明の単離タンパク質複合体は、成長因子または成長因子の少なくとも1つのドメインであって同族の成長因子受容体と結合し得るドメインを含む。
これに関連して、「ドメイン」とは、同族の成長因子受容体に結合することが可能な、成長因子の少なくとも一部分または領域を意味する。典型的には、ただし限定されるもの
ではないが、同族の成長因子受容体は細胞により発現され、そして成長因子の上記少なくとも1つのドメインによる上記同族の成長因子受容体の結合または連結により、細胞の応答、例えば細胞の増殖、分化、生存および移動のうち少なくともいずれかが誘導される。
【0044】
特にIGF−Iに関しては、上記ドメインは適切にはアミノ酸残基24を含むが、アミノ酸残基24はロイシン残基ではない。
典型的には、上記残基はチロシンである。
【0045】
特にIGF−IIに関しては、上記ドメインは適切にはアミノ酸残基27を含むが、アミノ酸残基27はロイシン残基ではない。
典型的には、上記残基はチロシンである。
【0046】
特にIGF−Iに関しては、一実施形態では、上記ドメインはIGF−Iの残基1〜70のみから構成される。
別の実施形態では、上記ドメインはIGF−Iの残基4〜70のみから構成される。
【0047】
また、本発明の単離タンパク質複合体の別の構成成分は、ビトロネクチンまたはフィブロネクチンの少なくとも1つのインテグリン結合ドメインであることが理解されよう。
これは、αVインテグリンに結合し得るVNまたはFNの任意のドメインを含みそして包含する。
【0048】
さらに好ましくは、インテグリン受容体はαVβ3インテグリンまたはαVβ5インテグリンである。
本明細書中で以下にさらに詳細に記載されるように、本発明は、VN(そして同様にFN)のHBDは、単離タンパク質複合体の完全な生物学的活性に必要なわけではない、ということを示している。
【0049】
本発明の単離タンパク質複合体は、非共有結合により会合したオリゴタンパク質複合体でも、共有結合により(可逆的にまたは非可逆的に)架橋されたオリゴタンパク質複合体の形態でも、あるいは合成キメラタンパク質の形態でもよい、ということは上記から容易に理解されよう。
【0050】
したがって、特定の態様において、本発明は、
合成キメラタンパク質の形態の単離タンパク質複合体であって、
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子受容体に結合し得るドメイン、および
(ii)ビトロネクチン(VN)またはフィブロネクチン(FN)、あるいはVNまたはFNの少なくとも1つのインテグリン結合ドメイン
のアミノ酸配列を含んでなる単離タンパク質複合体を提供する。
【0051】
本明細書中で用いる場合、「キメラタンパク質」は、VNまたはFNのインテグリン受容体結合ドメイン、ならびに成長因子または成長因子の少なくとも1つの受容体結合ドメインに由来するアミノ酸の連続配列を含む。
【0052】
いかなる特定の理論にも縛られることは望ましくないが、合成キメラタンパク質は上記成長因子に対する同族の受容体ならびにVNまたはFNに対するインテグリン受容体を相互連結し、かつ同時活性化することによって、細胞移動を刺激、誘導、増強、さもなければ促進し得る、ということが提唱される。
【0053】
本発明のキメラタンパク質の利点は、化学合成または組換え手段により容易に生産され
ること、そして非共有結合によるオリゴタンパク質複合体に必要とされるタンパク質−タンパク質相互作用の保持には依存しないので、in vivoでより安定であると予測されることである。
【0054】
この点では、IGF−Iの受容体結合ドメインを含む単離タンパク質複合体はIGFBPも含むことになるにも関わらず、上記の作用様式に従えばIGFBPはIGF−I/VN合成キメラ中に存在しないことが好ましい、ということが提唱される。
【0055】
さらにまたVNに関して、本明細書中でさらに詳細に後述されるように、本発明者等は、IGF−IIまたはIGF−I/IGFBP複合体との相互作用に必要とされるのはおそらくVN(そして同様にFN)のポリ陰イオン性領域であると思われる、ということを示す。
【0056】
図7および図8を参照すると、ポリ陰イオン性領域は、成熟VN配列(配列番号2)の残基53〜64である。
上記の点から見て、本発明は、VNまたはFNのHBDおよびポリ陰イオン性領域のうち少なくともいずれかを含まない合成キメラタンパク質の実施形態を意図する。
【0057】
HBDおよびポリ陰イオン性領域のうち少なくともいずれかを含まないVNタンパク質およびそのアミノ酸配列に関しては、天然に存在するタンパク質、例えば54kDニワトリ卵黄VN(HBDを欠く)でもよいし、あるいはVNタンパク質またはアミノ酸配列を欠失、突然変異または切断により人為操作してHBDおよびポリ陰イオン性領域のうち少なくともいずれかが存在しないかまたは少なくとも実質的に機能を持たないようにしてもよい。
【0058】
タンパク質分解による消化および部位特異的突然変異誘発のような技法は、当該技術分野で既知であるように、この目的のために利用され得る。
特定の実施形態では、VNの上記少なくとも1つのインテグリン結合ドメインは、
(i)VNのアミノ酸残基1〜459;
(ii)VNのアミノ酸残基1〜379;
(iii)VNのアミノ酸残基1〜130;および
(iv)VNのアミノ酸残基1〜52
からなる群から選択されるアミノ酸配列を有する。
【0059】
含まれてもよい付加的アミノ酸配列は、
(v)VNのアミノ酸残基65〜459;
(vi)VNのアミノ酸残基347〜459;および
(vii)VNのアミノ酸残基347〜379
からなる群から選択されるアミノ酸配列を有する。
【0060】
上記の配列は、例えばVNのアミノ酸残基1〜130とVNのアミノ酸残基347〜459、またはVNのアミノ酸残基1〜52とVNのアミノ酸残基65〜459のように組合せて用いることができる。
【0061】
IGF−IおよびVNを含んでなるキメラタンパク質の具体例が図14に記述されているが、これらに限定されない。
さらにVEGFおよびVNまたはPDGFおよびVNを含んでなるキメラタンパク質の具体例が図15に記述されているが、これらに限定されない。
【0062】
好ましくは、キメラタンパク質はさらに、成長因子の配列とVNまたはFNのアミノ酸
配列との間に位置し、同配列と連続する「リンカー配列」を含む。
一実施形態では、上記リンカー配列は、1つまたは複数のグリシン残基および1つまたは複数のセリン残基を含んでなる。リンカー配列の具体例は、Gly4Ser(配列番号4);Gly4Ser3(配列番号5)および(Gly4Ser)3(配列番号6)から選択されるが、これらに限定されない。
【0063】
別の実施形態では、リンカー配列には、プラスミン切断認識部位、例えば以下の配列:
LeuIleLysMetLysPro(配列番号7)
が挙げられる。
【0064】
さらに別の実施形態では、リンカー配列には、コラゲナーゼ−3切断認識部位、例えば以下の配列:
GlnProGlnGlyLeuAlaLys(配列番号8)
が挙げられる。
【0065】
本発明は、本発明の合成キメラタンパク質の生物学的に活性な断片の用途、および本明細書中に例示した特定の成長因子受容体結合ドメインおよびインテグリン結合ドメインの生物学的に活性な断片の用途にも拡張する。
【0066】
一実施形態では、上記「生物学的に活性な断片」は、該断片の元のタンパク質の10%以上、好ましくは25%以上、さらに好ましくは50%以上、さらに好ましくは75、80、85、90または95%以上の生物学的活性を有する。
【0067】
別の実施形態では、上記「生物学的に活性な断片」は、該断片の元のタンパク質の10%以上、好ましくは25%以上、さらに好ましくは50%以上、さらに好ましくは75、80、85、90または95%以上の連続アミノ酸配列を有する。
【0068】
例えばHBDおよびポリ陰イオン性ドメインのうち少なくともいずれかを欠く、生物学的に活性なVN断片の具体例は、本明細書中で図14に提示されている。
本発明の変異体タンパク質複合体も意図される。
【0069】
典型的には、そしてタンパク質に関しては、「変異体」タンパク質は、異なるアミノ酸により置換された1つまたは複数のアミノ酸を有する。いくつかのアミノ酸は、おおむね類似の特性を有する他のアミノ酸を用いて、タンパク質の活性の性質を変えることなく変更可能である(保存的置換)、ということは当該技術分野でよく理解されている。
【0070】
基準となる配列、例えば成長因子、成長因子の受容体結合ドメイン、VNまたはFNのインテグリン結合ドメイン、IGFBPの1つまたは複数のアミノ酸残基、あるいは合成キメラタンパク質中に存在する1つまたは複数の対応する残基を、本発明の単離タンパク質複合体の生物学的活性を実質的に変えることなく、修飾し、または欠失させ、あるいは付加的配列を付加することができることは理解されよう。
【0071】
本発明により意図される成熟VN(配列番号2)における具体的な突然変異としては、(i)T50A;(ii)T57A;(iii)T50E;(iv)T57E;(v)S378E;(vi)S378A;および(v)S362E、が挙げられる。
【0072】
一実施形態では、タンパク質変異体は、基準となるアミノ酸配列と少なくとも70%、好ましくは少なくとも80%、さらに好ましくは少なくとも90%、95%、98%または99%の配列同一性を共有する。
【0073】
好ましくは、配列同一性は、基準となる配列の少なくとも60%に亘って、さらに好ましくは少なくとも75%に亘って、さらに好ましくは少なくとも90%に亘って、さらに好ましくは少なくとも95%、98%に亘って、あるいは実質的に全長に亘って測定される。
【0074】
配列同一性(%)を決定するために、コンピュータを用いたアルゴリズムの実行(インテリジェネティックス(Intelligenetics )によるGeneworks(登録商標)プログラム;米国ウィスコンシン州マディソン サイエンスドライブ575所在のウィスコンシン・ジェネティクス・ソフトウェア・パッケージ・リリース7.0(Wisconsin Genetics Software Package Release 7.0 )のGAP、BESTFIT、FASTAおよびTFASTA(参照により本明細書中で援用する))により、あるいは選択された種々の方法のいずれかにより作成された検討および最良のアラインメント(即ち比較ウインドウを上回る最高の相同性(%)を生じる)により、アミノ酸配列およびヌクレオチド配列のうち少なくともいずれかの最適アラインメントを実行することが可能である。例えばアルチュール(Altschule )他、1997, Nucl. Acids Res. 25 3389(この記載内容は、参照により本明細書中で援用される)に開示されているようなBLASTファミリーのプログラムも参照可能である。
【0075】
別の例では、「配列同一性」は、DNASISコンピュータプログラム(ウインドウズ用バージョン2.5;米国カリフォルニア州サウスサンフランシスコ所在の日立ソフトウェアエンジニアリング(Hitachi Software engineering Co., Ltd.)から入手可能)により算定される「マッチ率(%)」を意味すると理解することができる。
【0076】
配列分析の詳細な考察については、オースベル(Ausubel )他編、「分子生物学最新プロトコール(CURRENT PROTOCOLS IN MOLECULAR BIOLOGY)」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons Inc )、1995-1999 )の第19.3部に記載されている。
【0077】
本発明は、成長因子の受容体結合ドメイン、VNまたはFNのインテグリン結合ドメイン、あるいは前記ドメインを含んでなる単離タンパク質複合体の誘導体も意図する。
本明細書中で用いる場合、本発明の「誘導体」タンパク質は、例えば別の化学基の付加、共役または複合により、あるいは当該技術分野で既知の翻訳後修飾技法により変更されている。
【0078】
アミノ酸の「付加」には、ポリペプチドまたはその変異体と別のペプチドまたはタンパク質との融合が挙げられる。前記の別のタンパク質は、例えば、タンパク質の精製を助ける。例えばこれら別のタンパク質の例としては、ポリヒスチジンタグ、マルトース結合タンパク質、緑色蛍光タンパク質(GFP)、プロテインAまたはグルタチオンS−トランスフェラーゼ(GST)が挙げられる。
【0079】
本発明により意図されるその他の誘導体としては、側鎖に対する修飾、ペプチド、ポリペプチドまたはタンパク質合成の際の非天然アミノ酸およびそれらの誘導体のうち少なくともいずれかの組み入れ、ならびに架橋剤の使用および本発明のポリペプチド、断片および変異体に立体構造上の制約を与えるその他の方法が挙げられるが、これらに限定されない。本発明により意図される側鎖修飾の例としては、例えば無水酢酸を用いたアシル化;無水コハク酸および無水テトラヒドロフタル酸を用いたアミノ基のアシル化;メチルアセトイミデートを用いたアミド化;シアネートを用いたアミノ基のカルバモイル化;ピリドキサル−5−ホスフェートを用いたリシンのピリドキシル化とその後のNaBH4を用いた還元;アルデヒドとの反応による還元的アルキル化とその後のNaBH4を用いた還元;ならびに2,4,6−トリニトロベンゼンスルホン酸(TNBS)を用いたアミノ基の
トリニトロベンジル化など、アミノ基の修飾が挙げられる。
【0080】
カルボキシル基は、O−アシルイソ尿素の形成を介したカルボジイミド活性化とその後の誘導体化、例えば対応するアミドへの誘導体化により修飾され得る。
アルギニン残基のグアニジン基は、2,3−ブタンジオン、フェニルグリオキサルおよびグリオキサルのような試薬を用いた複素環式縮合産物の生成により修飾され得る。
【0081】
スルフヒドリル基は、例えばシステイン酸への過蟻酸酸化;4−クロロ水銀フェニルスルホン酸、4−クロロ水銀ベンゾエート、2−クロロ水銀−4−ニトロフェノール、塩化フェニル水銀およびその他の水銀剤を用いた水銀誘導体の形成;その他のチオール化合物との混合ジスルフィドの形成;マレイミド、無水マレイン酸またはその他の置換マレイミドとの反応;ヨード酢酸またはヨードアセトアミドを用いたカルボキシメチル化;ならびにアルカリ性pHでシアネートを用いたカルバモイル化などの方法により修飾され得る。
【0082】
トリプトファン残基は、例えば臭化2−ヒドロキシ−5−ニトロベンジルまたはハロゲン化スルホニルを用いたインドール環のアルキル化により、あるいはN−ブロモスクシンイミドを用いた酸化により修飾され得る。
【0083】
チロシン残基は、3−ニトロチロシン誘導体を形成するためのテトラニトロメタンを用いたニトロ化により修飾され得る。
ヒスチジン残基のイミダゾール環は、ジエチルピロカルボネートを用いたN−カルボエトキシル化により、あるいはヨード酢酸誘導体を用いたアルキル化により、修飾され得る。
【0084】
ペプチド合成中に非天然アミノ酸および誘導体を組み入れる例としては、4−アミノ酪酸、6−アミノヘキサン酸、4−アミノ−3−ヒドロキシ−5−フェニルペンタン酸、4−アミノ−3−ヒドロキシ−6−メチルヘプタン酸、t−ブチルグリシン、ノルロイシン、ノルバリン、フェニルグリシン、オルニチン、サルコシン、2−チエニルアラニンおよびアミノ酸のD異性体のうち少なくともいずれかの使用が挙げられるが、これらに限定されない。
【0085】
タンパク質の化学的誘導体化に適した方法の一例は、コリガン(Coligan )他編、「タンパク質科学の最新プロトコール(CURRENT PROTOCOLS IN PROTEIN SCIENCE)」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons )、1995-2001 )の第15章に提供されている。
【0086】
単離タンパク質複合体およびその個々の構成成分であるタンパク質(断片、変異体、誘導体および相同体を含む)は、当業者に既知の任意の適切な手法により調製され得る。
一実施形態では、本発明のタンパク質は化学合成により作製される。化学合成技法は当該技術分野で既知であるが、当業者は適切な方法の例について、コリガン(Coligan )他編、「タンパク質科学の最新プロトコール(CURRENT PROTOCOLS IN PROTEIN SCIENCE)」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons )、199 5-2001 )の第18章を参照してもよい。
【0087】
別の実施形態では、タンパク質は、組換えタンパク質として調製され得る。
組換えタンパク質の製造は当該技術分野で既知であるが、当業者は、例えばサムブルック(Sambrook)他、「分子クローニング.実験の手引き(MOLECULAR CLONING. A Laboratory Manual)」コールドスプリングハーバー出版(Cold Spring Harbor Press)、1989年(この記載内容は、参照により本明細書中で援用される)、特に第16章および第17章;オースベル(Ausubel )他編、「分子生物学最新プロトコール(CURRENT PROTOCOL
S IN MOLECULAR BIOLOGY)」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons Inc )、1995-1999 )(この記載内容は、参照により本明細書中で援用される)、特に第10章および第16章;ならびにコリガン(Coligan )他編、「タンパク質科学の最新プロトコール(CURRENT PROTOCOLS IN PROTEIN SCIENCE)」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons )、1995-2001 )(この記載内容は、参照により本明細書中で援用される)、特に第1章、第5章および第6章に記載されているような標準プロトコールを参照してもよい。
【0088】
一実施形態では、組換えタンパク質は、
(i)発現ベクター内の1つまたは複数の調節ヌクレオチド配列に作動可能なように連結された、前記タンパク質をコードする核酸を含む発現構築物を調製する工程、
(ii)発現構築物を用いて宿主細胞をトランスフェクションまたは形質転換する工程、および
(iii)前記宿主細胞中で組換えタンパク質を発現させる工程
を包含する方法により製造される。
【0089】
「発現ベクター」は、自己複製する染色体外ベクター、例えばプラスミド、または宿主ゲノム内に組み込まれるベクターのいずれでもよい。
「作動可能なように連結される」または「操作可能なように接続される」とは、前記1つまたは複数の調節ヌクレオチド配列が、本発明の組換え核酸に対して、該核酸の転写または該核酸によりコードされるタンパク質の翻訳を、開始し、調節し、またはそうでなければ制御するように配置される、ということを意味する。
【0090】
調節ヌクレオチド配列は一般に、発現のために用いられる宿主細胞に適したものとなろう。多数の種類の適切な発現ベクターおよび適切な調節配列が、種々の宿主細胞に関して当該技術分野で既知である。
【0091】
典型的には、前記1つまたは複数の調節ヌクレオチド配列としては、プロモーター配列、リーダー配列またはシグナル配列、リボソーム結合部位、転写開始配列および転写終結配列、翻訳開始配列および翻訳終結配列、スプライスドナー/アクセプター配列、ならびにエンハンサー配列またはアクチベーター配列が挙げられるが、これらに限定されない。
【0092】
構成的プロモーター(例えばCMV、RSV、アデノウイルス、SV40およびヒト伸長因子プロモーター)および誘導性/抑制性プロモーター(例えばtet抑制性プロモーター、およびIPTG誘導性、メタロチオニン誘導性またはエクジソン誘導性プロモーター)が当該技術分野で既知であり、そして本発明により意図される。また、プロモーターは、複数のプロモーターの要素を併合するハイブリッドプロモーターでもよいことは理解されよう。
【0093】
発現構築物は、本発明の組換えタンパク質が融合パートナーとの融合ポリペプチドとして発現されるよう、融合パートナー(典型的には発現ベクターにより提供される)も含み得る。融合パートナーの主な利点は、前記融合タンパク質の同定および精製のうち少なくともいずれかの助けになるということである。
【0094】
融合パートナーの既知の例としては、グルタチオン−S−トランスフェラーゼ(GST)、ヒトIgGのFc部分、マルトース結合タンパク質(MBP)およびヘキサヒスチジン(HIS6)があり、これらはアフィニティクロマトグラフィによる融合タンパク質の単離に特に有用であるが、これらに限定はされない。アフィニティクロマトグラフィによる融合タンパク質精製の目的のために、アフィニティクロマトグラフィのための適切な充填材は、それぞれグルタチオン共役樹脂、アミロース共役樹脂、およびニッケル共役樹脂
またはコバルト共役樹脂である。多数のこのような充填材が、例えば(HIS6)融合パートナーを用いるのに有用なQIAexpress(商標)システム(キアゲン社(Qiagen))、ならびにファルマシア(Pharmacia )のGST精製システムのように、「キット」の形態で入手可能である。
【0095】
場合によっては、融合パートナーがさらにプロテアーゼ切断部位、例えばXa因子による切断部位またはトロンビンによる切断部位を有し、関連プロテアーゼが本発明の融合タンパク質を部分消化することにより融合タンパク質から本発明の組換えポリペプチドを遊離させることが可能である。遊離したタンパク質は次に、その後のクロマトグラフィー分離により、融合パートナーから単離され得る。
【0096】
本発明の融合パートナーは、「エピトープタグ」もその範囲内に含む。「エピトープタグ」は特異的抗体が利用可能な通常は短いペプチド配列である。特異的モノクローナル抗体が容易に利用可能なエピトープタグの既知の例としては、c−myc、ヘマグルチニンおよびFLAGタグが挙げられる。
【0097】
発現のための適切な宿主細胞は原核生物でも真核生物でもよく、例えば大腸菌(例えばDH5α)、酵母細胞、バキュロウイルス発現系とともに利用されるSf9細胞、CHO細胞、COS、CV−1、NIH 3T3および293細胞であり得るが、しかしこれらに限定されない。
【0098】
発現構築物は、形質転換された宿主細胞に選択作用物質に対する耐性を付与する1つまたは複数の選択マーカーヌクレオチド配列をさらに含み得る。形質転換された細菌を選択するために有用な選択マーカーとしては、bla、kanRおよびtetRが挙げられる一方、形質転換された真核生物細胞は、ハイグロマイシン、G418およびピューロマイシンのようなマーカーにより選択され得るが、これらに限定はされない。
【0099】
宿主細胞中への遺伝物質の導入に関して、「形質転換する」および「トランスフェクションする」という用語が、宿主細胞中への遺伝物質の導入を表すために一般に用いられる。宿主細胞中に外来遺伝物質を導入するための多数の既知の方法が存在し、例としてはリン酸カルシウム沈殿法、電気穿孔法、リポフェクタミン、リポフェクチンおよびその他の親油性物質による送達、リン酸カルシウム沈殿法、DEAE−デキストラントランスフェクション、微粒子銃、マイクロインジェクション、ならびにプロトプラスト融合が挙げられるが、これらに限定されない。
【0100】
(単離核酸)
本発明は、本発明の合成キメラタンパク質(その変異体および相同体を含む)をコードする単離核酸を提供する。
【0101】
「核酸」という用語は、本明細書中で用いる場合、一本鎖または二本鎖のmRNA、RNA、cRNA、RNAiおよびDNA(cDNAおよびゲノムDNAを含む)、ならびにDNA−RNAハイブリッドを意味する。
【0102】
「ポリヌクレオチド」は、80またはそれ以上の連続ヌクレオチドを有する核酸であり、一方、「オリゴヌクレオチド」は、80未満の連続ヌクレオチドを有する。
「プローブ」は、例えばノーザンブロッティングまたはサザンブロッティングにおいて相補配列を検出する目的のために適切に標識された一本鎖または二本鎖のオリゴヌクレオチドまたはポリヌクレオチドであり得る。
【0103】
「プライマー」は、通常は一本鎖のオリゴヌクレオチドであり、好ましくは15〜50
連続ヌクレオチドを有し、相補的な核酸の「鋳型」とアニーリングして、DNAポリメラーゼ、例えばTaqポリメラーゼ、RNA依存性DNAポリメラーゼまたはSequenase(商標)の作用により鋳型依存性の様式で伸長可能である。
【0104】
本発明の合成核酸は、当該技術分野で知られているように、化学合成手法により、または核酸配列の増幅技術を利用する組換え方法により、あるいはそれらの組合せにより作製可能である。
【0105】
本発明に有用な化学合成プライマーおよびオリゴヌクレオチド、合成装置および関連技術は、一般にほとんどの実験室で利用可能であり、あるいは商業的供給元から購入可能である。
【0106】
適切な核酸増幅技法は当業者に既知であり、例としては、例えばオースベル(Ausubel )他の上記文献の第15章に記載されているようなポリメラーゼ連鎖反応(PCR)およびリガーゼ連鎖反応(LCR);例えば米国特許第5,422,252号に記載されているような鎖置換増幅(SDA);例えばリュー(Liu )他、1996, J. Am. Chem. Soc. 118 1587、国際出願国際公開第92/01813号パンフレットおよび国際出願国際公開第97/19193号パンフレットに記載されているようなローリングサークル複製(RCR);例えばスクナナン(Sooknanan )他、1994, Biotechniques 17 1077 に記載されているようなNASBA法(nucleic acid sequence-based amplification );ならびに例えばティアギ(Tyagi )他、1996, Proc. Natl. Acad. Sci. USA 93 5395に記載されているようなQ−βレプリカーゼ増幅が挙げられるが、これらに限定されない。
【0107】
好ましい核酸配列増幅技法は、PCRである。
本明細書中で用いる場合、「増幅産物」とは、核酸増幅技法により作製された核酸産物を指す。
【0108】
本発明の核酸を作製し、発現させるに際しては、本明細書中で例示された核酸が、同核酸によりコードされるアミノ酸配列を変えることなく容易に変更可能なように、コドン配列の冗長性の利点を利用することができることも理解されよう。
【0109】
特定の実施形態では、当該技術分野で既知であるように、組換え発現のために用いようとする宿主細胞の好ましい「コドン使用頻度」に従って核酸を最適化することができる。このことにより、選択的なコドン使用頻度がタンパク質の発現に影響を及ぼす特定の生物体またはその細胞中での最適な発現のために、核酸を効率的に調整することが可能である。
【0110】
したがって本発明は、本明細書中に例示された核酸と相同な合成核酸を包含する。
一実施形態では、核酸相同体は、本明細書中に記載された合成キメラタンパク質構築物のいずれか一つをコードする核酸と少なくとも70%、好ましくは少なくとも80%、さらに好ましくは少なくとも90%、さらに好ましくは少なくとも95%の配列同一性を共有する。
【0111】
好ましくは、配列同一性は、本発明のコード核酸の少なくとも70%に亘って、より好ましくは少なくとも80%、さらに好ましくは少なくとも90%、95%、または有益には実質的に全長に亘って測定される。
【0112】
別の実施形態では、核酸相同体は、ストリンジェンシーの高い条件下で、本明細書中に記載された合成キメラタンパク質構築物のいずれか一つをコードする核酸とハイブリダイズする。
【0113】
「ハイブリダイズ、ハイブリダイゼーション」とは、DNA−DNA、RNA−RNAまたはDNA−RNA二重鎖を生成する、少なくとも部分的に相補性のヌクレオチド配列の対合を意味するべく、本明細書中で用いられる。ハイブリダイズした配列は、当該技術分野で既知であるように、相補的なプリン間およびピリミジン間の塩基対合により生じる。
【0114】
この点では、修飾プリン(例えばイノシン、メチルイノシンおよびメチルアデノシン)および修飾ピリミジン(チオウリジンおよびメチルシトシン)も塩基対合に関与し得ることが理解されよう。
【0115】
「ストリンジェンシー」とは、本明細書中で用いる場合、ハイブリダイゼーション中の、温度およびイオン強度条件、ならびにある種の有機溶媒および洗剤のうち少なくともいずれかの有無を指す。ストリンジェンシーが高いほど、ハイブリダイズするヌクレオチド配列間で必要とされる相補性のレベルが高くなる。
【0116】
「ストリンジェントな条件」とは、相補的な塩基の頻度が高い核酸のみがハイブリダイズする条件を意味する。
本明細書中のストリンジェンシーの高い条件への言及には、
(i)42℃でのハイブリダイゼーションについては、少なくとも約31%(v/v)〜少なくとも約50%(v/v)のホルムアミドおよび少なくとも約0.01M〜少なくとも約0.15Mの塩、ならびに42℃での洗浄については少なくとも約0.01M〜少なくとも約0.15Mの塩;
(ii)65o Cでのハイブリダイゼーションについては、1%BSA、1mMのEDTA、0.5MのNaHPO4(pH7.2)、7%SDS、ならびに65℃を上回る温度で約1時間の洗浄については(a)0.1×SSC、0.1%SDS;または(b)0.5%BSA、1mMのEDTA、40mMのNaHPO4(pH7.2)、1%SDS;
(iii)68℃以上で約20分間の洗浄については、0.2×SSC、0.1%SDS
が含まれる。
【0117】
概して、洗浄は、Tm=69.3+0.41(G+C)%−12℃で実行される。一般に、ミスマッチの塩基数が1%増大するごとに二重鎖DNAのTmは約1℃低下する。
上記にもかかわらず、ストリンジェントな条件は、例えばオースベル(Ausubel )他の上記文献の第2.9章および第2.10章に、特に2.9.1〜2.9.20ページに記載されているように、当該技術分野で既知である。
【0118】
(抗体)
本発明は、本発明の合成キメラタンパク質(キメラタンパク質、またはその断片、変異体および誘導体のうち少なくともいずれかを含む)に対する抗体も意図する。本発明の抗体は、ポリクローナルでもモノクローナルでもよい。抗体の作製、精製および使用に適用可能な既知のプロトコールは、例えばコリガン(Coligan )他、「免疫学の最新プロトコール(CURRENT PROTOCOLS IN IMMUNOLOGY )」、ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons )、1991-1994 )の第2章に、ならびにハーロウとレーン(Harlow, E. & Lane, D. )著、「抗体:実験の手引き(Antibodies: A Laboratory Mannual)」、コールドスプリングハーバー所在のコールドスプリングハーバー研究所(Cold Spring Harbor Laboratory )、1988に見出され得る(これらの文献の記載内容は、参照により本明細書中で援用される)。
【0119】
一般に本発明の抗体は、本発明のポリペプチド、断片、変異体または誘導体と結合または共役する。例えば抗体は、ポリクローナル抗体を含み得る。このような抗体は、例えば、マウスまたはウサギなどの生産用生物種に本発明のポリペプチド、断片、変異体または誘導体を注射してポリクローナル抗血清を得ることにより調製され得る。ポリクローナル抗体の生産方法は、当業者に既知である。用いられ得るプロトコールの例は、例えばコリガン(Coligan )他、「免疫学の最新プロトコール(CURRENT PROTOCOLS IN IMMUNOLOGY )」(上記)に、ならびにハーロウとレーン(Harlow & Lane )、1988(上記)に記載されている。
【0120】
前記生産用生物種において得られるポリクローナル抗血清の代わりにモノクローナル抗体を作製してもよい。モノクローナル抗体は、例えばコーラーとミルスタイン(Koehler & Milstein)著、1975, Nature 256, 495 の論文(この記載内容は、参照により本明細書中で援用される)に記載されているような標準的方法を用いて、または例えばコリガン(Coligan )他、「免疫学の最新プロトコール(CURRENT PROTOCOLS IN IMMUNOLOGY )」(上記)に記載されているような近年の変法により、本発明のポリペプチド、断片、変異体または誘導体のうち1つまたは複数を接種された生産用生物種由来の脾臓その他の抗体産生細胞を不死化することにより作製することが可能である。
【0121】
本発明はまた、上記で言及されたポリクローナルまたはモノクローナル抗体のFc断片またはFab断片を含む抗体もその範囲内に含む。あるいは抗体は、本発明のBIXPタンパク質に対する一本鎖Fv抗体(scFvs)を含み得る。このようなscFvsは、例えば米国特許第5,091,513号、欧州特許第239,400号、またはウィンターとミルスタイン(Winter & Milstein )、1991, Nature 349, 293 の論文にそれぞれ記載されている方法に従って調製され得る(これらの文献の記載内容は、参照により本明細書中で援用される)。
【0122】
抗体または抗体断片に標識を会合させてもよい。
標識は、色原体、触媒、酵素、蛍光体、化学発光分子、ランタニドイオン、例えばユーロピウム(Eu34)、放射性同位体および直接目に見える標識を含む群から選択され得る。直接目に見える標識の場合、金属コロイドまたは非金属性粒子、染料粒子、酵素もしくは基質、有機ポリマー、ラテックス粒子、リポソームまたはシグナル生成物質を含有するその他のビヒクルなどが用いられ得る。
【0123】
標識として有用な多数の酵素が、米国特許第4,366,241号明細書、米国特許第4,843,000号明細書および米国特許第4,849,338号明細書(これらの記載内容はいずれも参照により本明細書中で援用される)に開示されている。本発明に有用な酵素標識としては、アルカリホスファターゼ、ホースラディッシュペルオキシダーゼ、ルシフェラーゼ、b−ガラクトシダーゼ、グルコースオキシダーゼ、リゾチーム、リンゴ酸デヒドロゲナーゼ等が挙げられる。酵素標識は、単独で用いてもよいし、または溶液中で二次酵素と組合せて用いてもよい。
【0124】
例としては、蛍光体は、フルオレセインイソチオシアネート(FITC)、オレゴングリーン、テトラメチルローダミンイソチオシアネート(TRITL)、アロフィコシアニン(APC)およびR−フィコエリトリン(RPE)であり得るが、しかしこれらに限定されない。
【0125】
(薬学的組成物)
本発明は、本発明の単離タンパク質複合体(その変異体および誘導体を含む)を含んでなる薬学的組成物も提供する。
【0126】
このような単離タンパク質複合体は、in vitroで形成された、または本発明の合成キメラタンパク質として形成された複数タンパク質の複合体を含む任意の形態であり得るが、これらに限定されない。
【0127】
本発明の薬学的組成物は、細胞移動、組織再生および創傷治癒を促進するか、そうでなければ助長するために用いられ得る。あるいは薬学的組成物は、二次部位への腫瘍細胞の移動を防止または抑制することにより腫瘍の転移を防止するために投与され得る。
【0128】
本組成物は、必要な場合、治療的または予防的処置に用いられ得る。例えば薬学的組成物は、皮膚の修復、創傷治癒、熱傷の治癒またはその他の皮膚科学的処置のための治療用または美容用調製物の形態で適用され得る。
【0129】
この点に関して、薬学的組成物は、該薬学的組成物を適切に含浸するか、被覆するか、または他の方法で含んでいる、生体適合材料、生体ポリマー、無機材料、例えばヒドロキシアパタイトまたはその誘導体、外科用移植片、人工器官、創傷被覆材または熱傷被覆材、圧定布、包帯等とともに、またはそれらの一構成成分として投与され得る。
【0130】
薬学的組成物は、適切な薬学的に許容可能な担体、希釈剤または賦形剤を含むのが適切である。
好ましくは、薬学的に許容可能な担体、希釈剤または賦形剤は、哺乳類に、さらに好ましくはヒトに投与するのに適している。
【0131】
「薬学的に許容可能な担体、希釈剤または賦形剤」とは、全身投与に安全に用いられ得る固体または液体の充填剤、希釈剤または封入物質を意味する。個々の投与経路に応じて、当該技術分野で既知の種々の担体が用いられ得る。これらの担体は、糖、デンプン、セルロースおよびその誘導体、麦芽、ゼラチン、タルク、硫酸カルシウム、植物油、合成油、ポリオール、アルギン酸、リン酸緩衝溶液、乳化剤、等張生理食塩水および塩、例えば無機酸塩(例えば塩酸塩、臭化物および硫酸塩)、有機酸、例えば酢酸、プロピオン酸およびマロン酸塩、ならびに発熱物質を含まない水、を含む群から選択され得る。
【0132】
薬学的に許容可能な担体、希釈剤および賦形剤について記載されている有用な参考文献は、「レミントンの薬剤学(Remington's Pharmaceutical Sciences )」、(米国ニュージャージー州所在のマック出版(Mack Publishing Co. )、1991)である(この文献の記載内容は、参照により本明細書中で援用される)。
【0133】
本発明の組成物を患者に提供するために、任意の安全な投与経路が用いられ得る。例えば、経口、直腸、非経口、舌下、口腔内、静脈内、関節内、筋肉内、皮内、皮下、吸入、眼内、腹腔内、脳室内、経皮投与等が用いられ得る。
【0134】
剤形としては、錠剤、分散液、懸濁液、注射液、溶液、シロップ、トローチ、カプセル、座薬、エアロゾル、経皮パッチ等が挙げられる。これらの剤形には、制御放出するために特別に設計された制御放出用デバイス、あるいは制御放出様式でも作用するように改良されたその他の型のインプラントを注入または植設することも含み得る。治療薬の制御放出は、疎水性ポリマー、例えばアクリル樹脂、蝋、高級脂肪族アルコール、ポリ乳酸およびポリグリコール酸、ならびにある種のセルロース誘導体、例えばヒドロキシプロピルメチルセルロースで治療薬を被覆することにより実行され得る。さらに制御放出は、その他のポリマーマトリックス、リポソームおよび微小球のうち少なくともいずれかを用いることにより実行され得る。
【0135】
上記の組成物は、薬剤処方物に適合する方法で、かつ薬学的に有効な量で投与されれば
よい。患者に投与される用量は、本発明に関しては、適切な期間に亘って、患者に有効な応答をもたらすのに十分なものであるべきである。投与されるべき作用物質の量は、治療を受ける対象者の年齢、性別、体重および全身健康状態など、対象者に応じて、かつ担当医の判断によって変わるであろう要因に応じて決めればよい。
【0136】
創傷治癒用の薬学的組成物に関しては、米国特許第5,936,064号明細書および国際公開第99/62536号パンフレット(これらの記載内容は、参照により本明細書中で援用される)を特に参照されたい。
【0137】
本発明の薬学的組成物は、例えばワクシニアなどのウイルスベクターのような発現ベクター、ならびに遺伝子療法に有用なウイルスベクターも含み得る。後者としては、例えばブラウンファルコ(Braun-Falco )他、1999, Gene Ther. 6 432に記載されているようなアデノウイルスおよびアデノ随伴ウイルス(AAV)、ブッチシャッカー(Buchshacher )他、2000, Blood 95 2499 に記載されているようなレトロウイルスおよびレンチウイルスベクター、ならびに単純ヘルペスウイルスおよびサイトメガロウイルス由来のベクターが挙げられる。内分泌の遺伝子療法に有用なウイルスベクターの概説は、ストーン(Stone )他、2000, J. Endocrinol. 164 103に提供されている。
【0138】
本発明は、米国特許第5,958,764号明細書に記載されているような上皮細胞の遺伝子発現を目的とする特別な発現ベクター、および米国特許第5,962,427号明細書に記載されているようなin vivoでの創傷治癒に適用するための特別な発現ベクターも利用し得る。
【0139】
(治療的使用)
本発明は、本発明の合成キメラタンパク質を含めた単離タンパク質複合体を用いた治療方法を提供する。これらの方法は特に、哺乳類、特にヒトの治療的および/または予防的処置を目的としている。
【0140】
しかしながら本発明による治療的使用は、家庭用動物およびペット、作業動物、例えばウマ、ラクダおよびグレイハウンド、家畜、実験室動物、ならびに異種移植のための細胞、器官および組織の供給源として用いられる動物のような哺乳類にも適用可能である。
【0141】
本発明は、本発明の合成キメラタンパク質を含めた単離タンパク質複合体が、皮膚の質または皮膚の外観を改善または強化するために投与される美容的処置の方法も意図する。
このような処置には、皮膚細胞の異常増殖に起因する皮膚障害、例えば乾癬および過形成性瘢痕形成の予防または治癒が含まれうる。
【0142】
あるいは、二次部位への腫瘍細胞の移動を阻止することにより腫瘍の転移が防止または抑制される処置方法が意図される。さらに、細胞の増殖を阻止することにより癌を治療する方法も意図される。
【0143】
特定の実施形態では、治療的および/または予防的処置において、本発明の合成キメラタンパク質を含めた単離タンパク質複合体は、同単離タンパク質複合体を適切に含浸するか、被覆するか、または他の方法で含んでいる、生体適合材料、生体ポリマー、無機材料、例えばフルオロヒドロキシアパタイト、外科用移植片、人工器官、創傷被覆材または熱傷被覆材、圧定布、包帯等とともに、またはそれらの一構成成分として利用され得る。
【0144】
このような方法は、本明細書中に上記したような薬学的組成物の投与を包含し、そして例えば米国特許第6,090,790号明細書に記載されているように特定組織部位への微細針による注射、米国特許第6,054,122号明細書に記載されているような、創
傷、熱傷または潰瘍に適用される局所用クリーム、ローションまたは封止包帯、あるいは国際公開第99/47070号パンフレットに記載されているような、組成物を放出するインプラントによるものであり得る。
【0145】
この点については、例えば米国特許第5,929,040号明細書および米国特許第5,962,427号明細書に記述されている方法に従って遺伝子療法も適用可能である。
例えば所望の成長因子発現を遺伝子工学的に操作することにより、皮膚代替物を作製する目的のために皮膚細胞を遺伝学的に改変し得る方法も存在する(サップ(Supp)他、2000, J. Invest. Dermatol. 114, 5 )。この分野の概説の一例は、ベバン(Bevan )他、Biotechnol. Gent. Eng. Rev. 16, 231 に提供されている。
【0146】
例えば国際公開第99/11789号パンフレットに記載されているように、トランスフェクションまたは形質転換した細胞をレシピエントに「植え付ける」ことも意図される。
【0147】
これらの方法を用いて、細胞移動を刺激することにより、創傷治癒および熱傷治癒、潰瘍などの皮膚病変の修復、組織置換および移植を助長または進行させることが可能であり、例えば自己由来の皮膚のin vitro培養、腎臓および肺などの内部器官の再上皮化、ならびに損傷神経組織の修復などにより実施される。
【0148】
皮膚の置換療法は当該技術分野で既知になってきており、該療法においては、例えばケーヒ(Kehe)他、1999, Arch. Dermatol. Res. 291, 600 に記載されているような共培養した上皮/ケラチノサイト細胞株を使用してもよいし、あるいは上皮細胞、真皮細胞およびケラチノサイト細胞のうち少なくともいずれか(通常は自己由来)のin vitro初代培養物を使用してもよい。これらの技法は、人工生体適合材料および合成ポリマーの「足場材」も利用し得る。
【0149】
当該技術分野の概説の例は、概して、タースキフとヴァシリエブ(Terskikh & Vasiliev )、1999, Int. Rev. Cytol. 188,41 およびイーグルスタインとファランガ(Eaglestein & Falanga)、1998, Cutis 62, 1 に提供されている。
【0150】
より具体的には、頭蓋顔面外科に有用な代替口腔粘膜の作製は、イズミ(Izumi )他、2000, J. Dent. Res. 79, 798 に記載されている。胎児ケラチノサイトおよび真皮繊維芽細胞を、ファウザ(Fauza )他、J. Pediatr. Surg. 33, 357 に記載されているように、皮膚病変を治療するための移植用の皮膚を生産するためにin vitroで増殖させてもよいし、一方、ヒアルロン酸由来の生体適合材料上でin vitroで培養された真皮および上皮の皮膚要素からの皮膚置換物は、熱傷の治療に潜在的に有用であることが示されている(ザッキ(Zacchi)他、1998, J. Biomed. Mater. Res. 40, 187)。
【0151】
ポリマーの足場も、例えばシェリダン(Sheridant )他、2000, J. Control Release 14, 91 およびファウザ(Fauza )他、1998、(上記)に記載されているように、皮膚の人工置換を容易にすることが意図されており、微小球も創傷および熱傷への皮膚細胞の送達のための作用物質として意図されている(ラフランスとアームストロング(LaFrance & Armstrong)、1999, Tissue Eng. 5, 153)。
【0152】
(アゴニストおよびアンタゴニストの作製)
本発明は、成長因子およびビトロネクチンまたはフィブロネクチンを含んでなる複合体、例えばIGF−II:VNまたはIGF−I:IGFBP:VN複合体のアゴニストまたはアンタゴニストを、同定し、スクリーニングし、設計し、またはそうでなければ作製するための、本発明の合成キメラタンパク質を含めた単離タンパク質複合体の使用を意図
する。このような作用物質は、「模倣物」であり得る。「模倣物」という用語は、本明細書中では、タンパク質またはペプチドの特定の機能性領域に類似するよう設計された分子を指すものとして用いられ、そして当該技術分野で既知であるように、該用語の範囲内には、「アゴニスト」、「類似体」および「アンタゴニスト」という用語が包含される。
【0153】
一実施形態では、IGF−II:VN複合体またはIGF−I:IGFBP:VN複合体によるIGF−IRおよびVN受容体の結合を模倣するアゴニストが作製される。このような分子は、創傷治癒、皮膚再生等に必要とされるような細胞移動の刺激剤としての有用性を有し得る。
【0154】
別の実施形態では、IGF:VN複合体またはIGF:IGFBP:VN複合体によるIGF−IRおよびVN受容体の結合を防止または抑制するアゴニストが作製される。このような分子は、細胞の移動および増殖のうち少なくともいずれかの阻害剤としての有用性を有することにより、有用な抗腫瘍剤を構成する可能性があり、そして皮膚細胞の異常増殖に起因する皮膚障害、例えば乾癬および過形成性瘢痕形成の治療においても有用性を有し得る。
【0155】
上記の模倣物、アゴニスト、アンタゴニストおよび類似体は、所望の生物学的活性および半減期を有するペプチド、ポリペプチドまたはその他の有機分子、好ましくは小有機分子であり得る。
【0156】
コンピュータ支援構造データベース検索は、模倣物を同定するための一手法として漸増的に利用されるようになっている。原則として模倣物を同定するのに適しているデータベース検索方法は、国際公開第94/18232号パンフレット(HIV抗原模倣物の作製に関する)、米国特許第5,752,019号明細書および国際公開第97/41526号パンフレット(EPO模倣物の同定に関する)に見出され得る(これらの文献の記載内容は、参照により本明細書中で援用される)。
【0157】
その他の方法としては、分子相互作用を同定する種々の生物物理学的技法が挙げられる。これらの技法により、候補分子が例えばIGF−IGFBP−VN複合体の形成に影響を及ぼすか否かに従って、候補分子をスクリーニングすることが可能になる。潜在的に有用な技法、例えば放射性リガンドの競合的結合アッセイ(関連する方法として、上記のアプトン(Upton )他、1999を参照)、分析的超遠心分離、微量熱量測定、表面プラズモン共鳴および光学バイオセンサを用いる方法に適用可能な方法は、コリガン(Coligan )他編「タンパク質科学の最新プロトコール(CURRENT PROTOCOLS IN PROTEIN SCIENCE)」、(ニューヨーク所在のジョン・ワイリー・アンド・サンズ社(John Wiley & Sons ) NY,
1997 )の第20章に提供されている(この文献の記載内容は、参照により本明細書中で援用される)。
【0158】
本発明をより理解し易く、そして実行し易くするために、当業者に以下の非限定な実施例を提供する。
【実施例1】
【0159】
[材料および方法]
(細胞培養)
10%FCSを含有するDMEM/Hams’F12(DMEM/F12)培地(1:1)(オーストラリア国ビクトリア州マルグレーブ(Mulgrave)所在のライフテクノロジー社(Life Technologies ))中で、ヒト乳腺がん細胞株MCF−7(ATCC#HTB−22)を増殖させた。培地を毎日交換し、細胞は80%コンフルエントの状態で0.25%トリプシン/0.5mMエチレンジアミン四酢酸(EDTA)溶液(英国ハンプシャ
ー所在のオキソイド社(Oxoid ))を用いて継代した。
【0160】
ヒト皮膚ケラチノサイト細胞株HaCATは、ノルベート・フューゼニヒ(Norbet Fusenig)教授(ハイデルベルク ノイエンハイマーフェルト(Neuenheimer Feld)内のドイツがん研究センター(German Cancer Research Center: DKFZ ))から入手した。10%FCSを含有するDMEM培地(ライフテクノロジー社)中で、HaCAT細胞株を増殖させた。培地を毎日交換し、細胞は80%コンフルエントの状態で0.25%トリプシン/0.5mM EDTA溶液(オキソイド社(Oxoid ))を用いて継代した。
【0161】
(VNおよびIGFBPへのIGFの予備結合)
細胞の機能を調べるin vitroアッセイのほとんどは、溶液中に外因性因子を加えるので、細胞は、アッセイの間中その物質を含有する溶液中に入っている。これは、細胞がin vivoで遭遇する環境ではない。むしろ、組織中の細胞は、細胞により合成されたECMによって支持され、取り囲まれており、該ECMの中にはホルモンおよびその他の因子が局在している。ECM分子への成長因子の結合を特定的に取り扱うこの試験では、24ウェルプレートの組織培養用プラスチックと、12.0μL孔のCostar
Transwells(商標)(米国ニューヨーク州ニューヨーク所在のコスター社(Costar))の下方チャンバおよび膜表面に、VN、IGFおよびIGFBPを予備結合する戦略を用いて、in vivo環境をより正確に反映することを試みた。
【0162】
300〜1000ngのVN(オーストラリア国ニューサウスウェルズ州アンナデール(Annandale )所在のプロメガ社(Promega ))を含有するDMEMまたはDMEM/F12培地300μLを24ウェル組織培養ディッシュに、またはTranswells(商標)の下方チャンバに添加し、37℃で2時間インキュベートした。結合していないVNを含有する培地を除去し、ウェルを0.5%ウシ血清アルブミン(RIA等級)(BSA)(シグマアルドリッチ社(Sigma Aldrich ))を含有する1mLのヘペス結合緩衝液(HBB)で洗浄した。次に1.0%BSAを含有するHBB 300μLをウェルに添加して37℃で30分間インキュベートし、組織培養ディッシュ内の非特異的結合部位をブロッキングした。次に、0.5%BSAを含有するHBB 1mLで、ウェルを再び洗浄した。次に0.5%BSAおよびIGF−IIまたはIGF−I+IGFBP(オーストラリア国サウスオーストラリア州アデレード(Adelaide)所在のグロペップ社(GroPep))を含有するHBB 300μLを添加し、プレートを再び2時間インキュベートした。非結合IGFおよびIGFBPを含有する溶液を除去し、ウェルをHBBで洗浄して、層流フード内で風乾した。
【0163】
(移動アッセイ)
リーブスレー(Leavesley )他、1993, Journal of Cell Biology 121: 163-70 に記載されているのと本質的に同様に、移動アッセイを実施した。無血清培地中で4時間のインキュベーションにより血清飢餓状態にした50,000個の細胞を、12.0μm孔Costar Transwells(商標)(12ウェルプレート型)の上部チャンバに播種した。5%CO2中で37℃にて5時間インキュベーションした後の、多孔性膜の下側表面に移動していた細胞を固定し、次に、0.1mMのホウ酸緩衝液(pH9)中のクリスタルバイオレットで染色した。10%酢酸中にクリスタルバイオレットを抽出し、分光分析によりこれらの抽出物の吸光度を測定することにより、結合した細胞の数を概算した。
【0164】
(統計学的分析)
先ず全データを陰性対照(−VN、−IGF、−IGFBP)に対する割合(%)として表すことにより、データを分析した。次に、等分散性のスチューデントt検定(両側検定)を用いて、VNのみの対照およびIGFのみの対照に対する有意性について、応答を
試験した。0.05未満のP値は、応答に有意差があることを示す。
【0165】
[結果]
(移動)
細胞移動は創傷治癒における重要な過程であり、VNおよびIGFはともに、細胞移動の媒介において役割を有することが立証されている。VNに結合したIGF−IIの、HaCATケラチノサイトの機能を変える能力について詳細に分析するために、Transwells(商標)を通り抜ける細胞移動を測定した。
【0166】
図1は、VNの存在下では、IGF−II誘導によるTranswells(商標)を通り抜けるHaCATヒトケラチノサイトの移動が、特に低濃度で増強されることを示す。各バーは3回の反復実験からのデータを表し、各処理は三連で試験した。
【0167】
乳癌細胞株MCF−7の移動も試験した。1μgのVNを12.0μmのTranswells(商標)の下方ウェルに予備結合させた場合、下方チャンバへの移動は5倍増加した。VNの非存在下でウェルに1〜100ngのIGF−IIを「予備結合」させると、移動が刺激されて2倍に増加した。しかしながら1〜100ngのIGF−IIを下方チャンバ中の1μgのVNに予備結合させた場合、細胞移動は8〜10倍に増加した(図2)。これらの応答は、IGF−II単独およびVN単独の作用より有意に高かった(p<0.01)。
【0168】
[L27]−IGF−IIは、IGF−IRに結合しないIGF−II類似体である。IGFは受容体IGF−IRを介して移動を媒介し、IGF−IRは信号を受けると考えられる。従って、VNに予備結合された[L27]−IGF−IIがTranswells(商標)を通り抜けるMCF−7細胞の移動を刺激する能力を調べるアッセイを実行した。このアッセイは、VN−[L27]−IGF−II複合体がVN単独の場合に得られるレベルを超えてMCF−7の移動を増強することはないということ、ならびに移動のレベルがVNに結合したIGF−IIに対する応答の場合に観察される移動のレベルより有意に低い(p<0.01)、ということを明示した(図3)。これらの結果は、VNに結合したIGF−IIから生じる移動の増強にIGF−IIとIGF−IRとの相互作用が関与することを示している。
【0169】
IGFBPは、細胞に対するIGF曝露の重要な調節物質である。IGFBPがここで観察されたVN:IGF−II複合体に対する移動応答に関与するか否かを確定するために、IGFBPにはほとんど結合しないが、IGF−IRに対する親和性は保持しているIGF−II類似体を用いてMCF−7細胞における移動アッセイを実行した。このIGF−II類似体des(1−6)−IGF−IIを用いたアッセイから、天然型のIGF−IIと比較した場合に移動応答に差がないことが明らかとなり、IGF−II:VN複合体がIGFBPとは独立に作用して細胞移動を増強することが示唆された(図4)。
【0170】
図5では、データから、αVインテグリン遮断抗体がVNとIGF−IIとの複合体に応答するMCF−7細胞の移動を実質的に低減させたことを示す。これらのデータは、VNに対するαVインテグリン受容体の結合および活性化が、IGF−II:VN複合体に応答した最適な細胞移動に必要であると思われることを示唆している。
【0171】
IGF−IRとほとんど結合しないIGF−I類似体L24−IGF−IについてMCF−7細胞の移動アッセイで調べた図6を参照すると、データから、
(1)L24−IGF−IおよびIGF−Iは、VNは存在するがIGFBP−5は存在しない条件下では、同じ作用を有すること;そして
(2)IGFBP−5の存在下では、IGF−IおよびVNが存在する場合は細胞移
動が増強されるが、L24−IGF−IおよびVNが存在する場合は増強されないこと
が実証される。
【0172】
上記のデータは、IGF−IRの活性化はIGF−I:IGFBP:VN複合体に応答して観察される細胞移動に必要であること、そしてさらにVNに対するインテグリン受容体の同時結合が必要であることを示唆している。
【0173】
この試験結果は、IGF−II:VN複合体およびIGF−I:IGFBP:VN複合体が細胞移動を有意に刺激する、ということをはじめて明示するものである。総合するとこれらのデータから、VN:IGF複合体が創傷治癒と機能的に関連しており、実際、乳がんの発症および進行における重要な因子であり得ることが示唆される。さらに、移動の増強には、IGF−IRおよびVN結合性インテグリン受容体の両方の受容体の活性化を必要とする。実際、VN:IGF複合体が移動を促進し、したがって乳がん細胞の転移を促進するのであれば、VN:IGF複合体の形成または成長因子受容体およびインテグリン受容体の同時活性化の阻害を目的とする薬剤は、非常に有効な治療薬であるということになる。
【実施例2】
【0174】
[方法および材料]
(卵黄ビトロネクチンの精製)
ナガノ(Nagano)他、1992, The Journal of Biological Chemistry 267: 24863-24870に記載された方法の変法を用いて、卵黄のビトロネクチン(VN)を精製した。この手法に用いられた溶液は全て、pH7.4であった。鶏卵(オーストラリア国クイーンズランド州トゥーウォン(Toowong )所在のコールズマイヤー(Coles-Myer)の農場から入手した卵黄を、2mMのフッ化フェニルメタンスルホニル(PMSF)を含有する等容積の冷リン酸緩衝生理食塩水(PBS)(0.16MのNaCl、10mMのリン酸ナトリウム)中に懸濁し、18,000g、4℃で20分間、遠心分離した。上清(卵黄プラズマ)を、5mMの2−β−メルカプトエタノールを含有する1mMのリン酸ナトリウムに対して4℃で一晩透析し、20,000g、4℃で20分間遠心分離した。上部の固体層(低密度リポタンパク質(LDL)分画)を回収し、15mlのPBS中に再懸濁した。
【0175】
3つのクロマトグラフィー技法、すなわち、ゲル濾過、Sepharose(商標)CL−6B(スウェーデン国ウプサラ所在のアマシャムバイオサイエンス社(Amersham Biosciences));ヒドロキシアパタイトHTP(米国カリフォルニア州リッチモンド所在のバイオラッド社(Bio-Rad ));およびイオン交換、Q‐Sepharose(登録商標)Fast Flow(アマシャムバイオサイエンス社)を用いて、LDL分画から卵黄VNを精製した。
【0176】
Sepharose(商標)CL−6Bカラム(カラム容量10ml、カラムサイズ:内径(ID)2.5cm×30cm)を、2段階、すなわち(i)2MのNaClを含有するPE緩衝液(5mM EDTA、10mM リン酸ナトリウム)および(ii)0.13MのNaClを有するPE緩衝液、を用いて平衡化した。50mlのLDL分画をPE緩衝液で1:1に希釈してSepharose(商標)CL−6Bカラムにかけた。同カラムから非結合画分を収集し、次に0.5MのNaClを含有する10mMのリン酸ナトリウムで予め平衡化したヒドロキシアパタイトマトリックスにバッチ式で適用した。該ヒドロキシアパタイトを平衡化緩衝液で洗浄し、その後10mMのリン酸ナトリウム20mlで洗浄した。次に該マトリックスをカラム(1.5cmID×8cm)に詰めて、200mMのリン酸ナトリウムでタンパク質を溶離させ、10×5ml画分を収集した。該溶離画分を、プレキャスト型の4〜20%勾配ポリアクリルアミドゲル(オーストラリア国ニューサウスウェールズ州フレンチフォレスト所在のグラディポア社(Gradipore ))
、SDS−PAGE(レムリ(Laemmli ), 1970)およびクーマシーブリリアントブルー(G−250、バイオラッド)染色を用いて、yVNの存在に関して分析した。バイオラッドの低分子量域マーカーを用いて、タンパク質の分子量を測定した。
【0177】
予測される分子量のyVN(54kDa)に相当する画分をプールし、10mMリン酸ナトリウムに対して4℃で一晩透析し、次に10mMリン酸ナトリウムで予め平衡化したQ Sepharose(登録商標)Fast Flowマトリックス(カラム容量5ml、カラムサイズ:ID 1cm×10cm)に適用した。該カラムを10mMのリン酸ナトリウム中の0.15MのNaClで洗浄し、10mMのリン酸ナトリウム中の0.25MのNaClでyVNを溶離させた。上記と同様に画分の分子量を測定した。
【0178】
(IGF:VN複合体の調製)
報告されているように(クリッカー(Kricker )他の上記文献、2003)、IGF:VN複合体を、96ウェルプレートおよびTranswells(商標)に予備結合させた。
【0179】
[結果]
図8は、完全長(75kDa)の血清VN(a)と短縮型(54kDa)の卵黄VN(b)との類似性を示す。特筆すべき主な類似性は、これらのタンパク質がともにRGD細胞結合部位およびポリ陰イオン性領域(IGF結合部位とされる)を有する、ということである。特筆すべき主な違いは、卵黄VNがヘパリン結合ドメインを欠くということである。
【0180】
図9は、溶離画分(レーンE)中に存在する主なタンパク質が、卵黄VNについて予測される大きさ(54kDa)である、ということを示している。このタンパク質がその後のアッセイに用いられたということも、特筆すべき重要なことである。
【0181】
図10は、IGFBP−3の存在下でVNが放射標識IGF−Iを結合する能力を実証している。したがってこの図から、54kDa卵黄VNが完全長の卵黄VNと同じレベルでIGF−I/IGFBP−3を結合する能力を有していることが示されている。このことは、IGF結合部位がヘパリン結合ドメイン(卵黄VN(54kDa)には欠けている)中には位置しないことを示唆しており、そしてポリ陰イオン性部位がIGF結合部位として提唱されることを支持している。
【0182】
図11は、IGF−I/IGFBP−3と複合体形成した場合のVN(血清VNまたは卵黄VN)の能力により、無処置群、IGF−I/IGFBP−3処置群およびVN単独処置群の対照を上回って細胞増殖(ミトコンドリアデヒドロゲナーゼ活性を評価するMMT技法により測定)が強化されることを示している。図11はまた、短縮型(54kDa)卵黄VNが、IGF−I/IGFBP−3と複合体形成した場合、完全長VNと同程度に細胞増殖を刺激し得る、ということも示している。
【0183】
図12は、IGF−I/IGFBP−3と複合体形成した場合のVN(血清VNまたは卵黄VN)の能力により、無処置群、IGF−I/IGFBP−3処置群およびVN単独処置群の対照を上回って細胞移動(Transwells(商標)移動アッセイによる)を強化し得ることを示している。したがって、短縮型(54kDa)卵黄VNが、IGF−I/IGFBP−3と複合体形成した場合、完全長VNと同程度に細胞増殖を刺激し得るという同じ結論が、図4と同様にこの図からも引き出され得る。総合すると、これらの図から、短縮型(54kDa)卵黄VNは、IGF−I/IGFBP−3と複合体形成した場合、完全長(75kDa)VNで観察されるのと同様のレベルに細胞の移動および増殖のいずれも刺激し得る、ということが示唆される。
【実施例3】
【0184】
VN:IGF−Iキメラタンパク質の形態の、本発明の合成キメラタンパク質の提唱例が、本明細書中に提供されている。
図14に種々に記述した提唱合成キメラタンパク質には、アミノ酸残基の改変を伴って、または伴わずに、IGF−Iと融合された、任意の完全長または短縮型のVNが含まれている。さらに本発明者等は、種々のペプチドリンカーを用いて、または用いずにVNおよびIGF−Iを融合することを提唱する。
【0185】
さらに本発明者等は、VNおよび成長因子、例えばVEGFおよびPDGFを含んでなるキメラタンパク質を意図し、その特定の実施形態を図15に記述する。
本明細書において用いられ、図13に示された成熟VN(配列番号2)およびIGF−I(配列番号3)に関する完全ペプチド配列は、NCBI(それぞれアクセス番号NP_0000629および1BQT)から入手した。VNに関して示した残基番号のアノテーションは成熟タンパク質のものであり、シグナルペプチドは除かれている。
【0186】
ビトロネクチンドメイン構造およびビトロネクチンリガンド結合部位に関しては、図7および8にそれぞれ記載されている。
(完全長および短縮型のVN)
細胞移動を調節し得る合成キメラタンパク質の一例は、完全長の成熟VNタンパク質およびIGF−Iタンパク質を含有する。
【0187】
A)VN(1・・・459):IGF−I(1・・・70)
細胞移動を調節し得る別の例は、残基380〜459(C末端80アミノ酸)の欠失を伴う成熟VNタンパク質を含有する。
【0188】
B)VN(1・・・379):IGF−I(1・・・70)
血清中の単量体VNは、2つの形態、すなわち、75kDaの一本鎖ポリペプチド、または内因性の切断を受けた二本鎖形態のVNで65kDaの大型断片がジスルフィド結合により10kDa小型断片に連結されて構成されているVN、として存在する。近年の研究から、これらの形態の間に機能的差異は認められないことが示されており、このことは、VN上のC末端80アミノ酸は機能的差異を付与しない、ということを示唆している(ギブソンとピーターソン(Gibson and Peterson ), 2001, Biochim Biophys Acta 1545,
289-304)。このことは、ブタVNがこのC末端領域を失う一方でその機能的活性は保持しているという知見(ヨネダ(Yoneda)他、1996, J Biochem (Tokyo) 120:954-60)により支持される。したがって、C末端80アミノ酸を取り除いた、VNの全ての機能的特性を依然として有するVNを含有してなる、より小型のキメラ分子を、本発明者等は提唱する。
【0189】
さらに別のキメラは、IGF−Iに連結されたVNのソマトメジンBドメインのみを含有する。この領域は、プラスミノーゲンアクチベーター−1(PAI−1)、ウロキナーゼプラスミノーゲンアクチベーター受容体(uPAR)およびインテグリン結合部位を含有する(シュバルツ(Schvarz )他、1999, Int J Biochem Cell Biol 31: 539-44)。
【0190】
このキメラは、ECM中の構成成分、例えばコラーゲンおよびグリコサミノグリカンと相互作用しないであろう。このキメラは、VN上の残基53〜459(接続領域、中央のβ−プロペラドメインおよびヘパリン結合ドメイン)の欠失を有している。
【0191】
C)VN(1・・・52):IGF−I(1・・・70)
VNの接続領域は、トロンビン−アンチトロンビン複合体、ならびにECM構成成分コラーゲンを結合する際に役割を演じると推測されている。ここで提唱されるキメラは、V
NのソマトメジンBドメイン、ならびにVN上の残基131〜459(中央のβ−プロペラドメインおよびヘパリン結合ドメイン)の欠失により創出される接続領域を含有する。
【0192】
D)VN(1・・・130):IGF−I(1・・・70)
さらなる例では、VN上の中央ドメインは最大であるが、しかし機能に関しては同タンパク質中であまりよく特性解析されていないドメインである。しかしながら、このドメイン中に観察されるβ−プロペラ構造はVNの多量体化に寄与し得る、と推測される(キュー(Xu)他、2001, Proteins 44: 312-20 )。
【0193】
本発明者等は、このドメインを欠失させて、VNの、ソマトメジンBドメイン内のリガンド結合領域、ポリ陰イオン性接続領域、およびポリ陽イオン性ヘパリン結合ドメイン(HBD)を保持し、しかしながら自己会合できない小さいキメラとすることを提唱する。この場合、VN上の残基131〜346(中央のβ−プロペラドメイン)の欠失を必要とする。
【0194】
E)VN(1・・・130、347〜459):IGF−I(1・・・70)
さらに別のキメラは、細胞外リガンドを結合し得ると考えられる最も小型のVNから構成される。該タンパク質は、VN上の中央ドメインおよびC末端80アミノ酸がいずれも除去されている。この場合、VN上の残基131〜346および380〜459(それぞれ中央のβ−プロペラドメインおよびC末端80アミノ酸)の欠失を要する。
【0195】
F)VN(1・・・130、347〜379):IGF−I(1・・・70)
キメラのさらなる例は、ヘパリン結合ドメインを伴わないC末端短縮型VNを含有する。したがって該タンパク質は、VNのソマトメジンBドメイン、接続領域および中央のβ−プロペラドメインを含有する。VNに関しては、二次ヘパリン結合部位と推定される部位がVNの中央のβ−プロペラドメイン内に提唱されているが、しかしギブソン(Gibson)他(ギブソン(Gibson)他、1999, J Biol Chem 274, 6432-42)は、該部位が機能性でないこと、そしてヘパリン結合ドメインはグリコサミノグリカン結合活性を完全するために寄与する、ということを実証した。したがって該キメラは、ヘパリンおよびヘパリン硫酸と相互作用しないであろう。このキメラは、VN上の残基347〜459(ヘパリン結合ドメイン)の欠失を有する。
【0196】
G)VN(1・・・346):IGF−I(1・・・70)
(VNおよびIGF−I上の残基の修飾)
VNは、カゼインキナーゼII(CK2)により残基T50およびT57がリン酸化されて、細胞の接着および伸展を促進し得る。VNのCK2−リン酸化類似体およびCK2−非リン酸化類似体(それぞれVN突然変異体(T50E、T57E)および(T50A、T57A)により模擬検討された)のいずれも、αvβ3およびαvβ5インテグリンに結合してERKシグナル伝達経路を活性化させるが、一方、αvβ3インテグリンに特異的に結合しているCK2−リン酸化VN類似体だけが、ホスファチジルイノシトール3キナーゼ(PI3−K)経路を活性化させた(セガー(Seger )他、1998, J Biol Chem 273: 24805-13; セガー(Seger )他、2001, J Biol Chem 276: 16998-7006 )。
【0197】
細胞の接着および伸展の増大をもたらすと思われるのは、このPI3−K経路の活性化である。したがって本発明者等は、αvβ3インテグリンとの結合後にPI3−K経路の活性化を促進または抑制するであろう突然変異を有するキメラを提唱する。したがってVN上のT50AおよびT57A置換を有するキメラ分子は、CK2−非リン酸化VNと類似し、ERK経路を介したシグナル伝達に限定される(H)が、一方、VN上のT50EおよびT57E置換を有する合成構築物は、CK2−リン酸化VNを模倣し、ERK経路およびPI3−K経路の両方を活性化して、細胞内シグナル伝達の変更をもたらし得る(
I)。
【0198】
H)VN(T50A、T57A):IGF−I
I)VN(T50E、T57E):IGF−I
VN上の残基S378にはcAMP依存性プロテインキナーゼ(PKA)リン酸化部位が存在する。この部位のリン酸化はPAI‐1のVNへの結合を低減し、したがってウロキナーゼ系におけるVNの役割を変える、ということが、PKAリン酸化VN類似体およびPKA非リン酸化VN類似体(それぞれVN突然変異体S378EおよびS378Aにより模擬検討された)を用いて実証されている(シュバルツ(Schvartz)他、2002, Arch
Biochem Biophys 397: 246-52)。
【0199】
したがって、キメラ(J)によるPAI−1結合を抑制するためのVN上のS378E突然変異、ならびにキメラタンパク質(K)内のPAI−1結合および安定化を促進するためのVN上のS378A突然変異、の両方を含有するキメラを本発明者等は提唱する。さらに、PAI−1のVNへの結合が、インテグリンを介した細胞移動を阻害すること(キョーラ(Kjoller )他、1997, Exp Cell Res 232: 420-9 )、ならびにuPARおよびインテグリンを介したVN上への細胞接着を阻害すること(デング(Deng)他、2001, J Cell Physiol 189: 23-33 )が示されているので、S378A突然変異は細胞移動を増強する可能性がある。興味深いことに、これらの知見は、プラスミノーゲン活性化の阻害剤としてのPAI−1の機能とは独立して観察された。
【0200】
J)VN(S378E):IGF−I
K)VN(S378A):IGF−I
ゲヒトマンとシャルティール(Gechtman and Shaltiel )、1997, Eur J Biochem 243:
493-501は、プロテインキナーゼC(PKC)がVNの残基S362をリン酸化し得ることを示した。このリン酸化は、VNのヘパリン結合ドメイン内に生じるプラスミンによる切断を低減する。したがってこの部位におけるプラスミン切断により、この領域内に結合するリガンドに対するVNの親和性が変化し、かつVNの半減期も変化する。したがって本発明者らは、リン酸化セリンを模倣し、その結果プラスミンによるキメラの切断を阻害するためのS362E置換の導入を提唱する。
【0201】
L)VN(S362E):IGF−I
IGFBPは、高い親和力で成長因子IGF−Iを結合するために、IGF−I上のN末端3残基を必要とすることが示されている(トーマス(Tomas )他、1991, J Endocrinol 128: 97-105)。
【0202】
したがって、N末端配列を介してVNに連結されたIGF−IがIGFBPを結合し得るとは考えられない。にもかかわらず、本発明者らは、IGFBPがIGF−Iと結合し、その結果VN:IGF−Iキメラタンパク質の生物学的活性を抑制する全ての機会を排除するために、N末端短縮型IGF−Iを含有するVN:IGF−Iキメラをさらに提唱する。この構築物は、IGF−I上の残基1〜3(IGFBP結合領域)の欠失を含む。
【0203】
M)VN:IGF−I(4・・・70)
VNのポリ陰イオン性領域はIGF−IIおよびIGFBPの結合に寄与することが示唆されている。したがって本発明者らは、VNからポリ陰イオン性ドメインを除去したさらに別のVN:IGF−Iキメラを提唱する。よってこのキメラは、IGF−IIまたはIGFBPに対して無力である。
【0204】
N)VN(1・・・52、65・・・459)IGF−I(1・・・70)
(VNとIGF−Iの融合)
VNおよびIGF−IのcDNAを、発現に先立って、ペプチドリンカー配列を挿入して、または挿入せずに、融合させてもよいことを提唱する。タンパク質を融合するための種々のリンカー配列が首尾よく用いられてきており、通常は、グリシンおよびセリン残基ならびに例えばトロンビン、コラゲナーゼまたはプラスミンなどのプロテアーゼの切断部位のうち少なくともいずれかを組合せて構成される。
【0205】
リンカー配列の例を以下に挙げるが、これらに限定されない。
(i)Gly4Ser(配列番号4);
(ii)Gly4Ser3(配列番号5);
(iii)(Gly4Ser)3(配列番号6);
(iv)LeuIleLysMetLysPro(配列番号7);および
(v)GlnProGlnGlyLeuAlaLys(配列番号8)
(VNと他のペプチド成長因子の融合)
細胞外マトリックスタンパク質VNを成長因子IGF−Iと融合することに加えて、VNと他のペプチド成長因子の融合も提唱する。具体的には、以下のキメラタンパク質の開発を提唱する(図16)。
【0206】
A)VN:PDGFα(1・・・210)(NCBIアクセス番号P04085)
B)VN:VEGF(1・・・102)(NCBIアクセス番号2VPFE)
それぞれの細胞表面受容体はインテグリンと、特にVN受容体であるαvβ3インテグリンと相互作用する、ということを本発明者らは提唱する。具体的には、PDGF受容体が、PDGFによる刺激後に、αvβ3インテグリンと免疫共沈降する、ということが示されている(シュネーラー(Schneller )他、EMBO J 16: 5600-7 )。
【0207】
2型VEGF受容体はその成長因子による刺激後にVN受容体と免疫共沈降する、ということも実証されている(ソルディ(Soldi )他、EMBO J 18: 882-92 )。
これらの成長因子に対する受容体がαvβ3インテグリンと相互作用するという知見は、これらの細胞表面受容体の細胞内シグナル伝達経路に対して調節および増強のうち少なくともいずれかをもたらすに際して、この相互作用が重要な役割を有することを示唆している。したがって、上記のキメラタンパク質により開始されるこれらの受容体の同時活性化および会合により、治療的用途に関連する顕著な生物学的応答を誘導し得る。
【0208】
本明細書全体を通して目的としたのは、本発明の好ましい実施形態を説明し、かつ本発明をいずれの一実施形態にも特定の特徴の集合体にも限定しないことであった。したがって、当業者には理解されることであるが、該開示にかんがみて、本発明の範囲を逸脱することなく、例示された特定の実施形態に種々の修正および変更をなすことができる。
【0209】
本明細書中で言及されたコンピュータプログラム、アルゴリズム、特許文献および科学文献は全て、参照により本明細書中で援用される。
【図面の簡単な説明】
【0210】
【図1】12μm孔のTranswells(商標)の上方チャンバ内に植え付けられたヒト皮膚ケラチノサイト細胞HaCATの、VNに予備結合されたIGF−II(黒色バー)またはVNの非存在下でディッシュに「結合された」IGF−II(灰色バー)で被覆された下方チャンバに応答した、下側表面への移動について示す図。各バーは、5時間インキュベーション後の下方膜上の細胞数の平均を表し、3回の反復実験(3連のウェル中で処理群についての分析を実施)から得ている。
【図2】12μm孔のTranswells(商標)の上方チャンバ内に植え付けられたヒト乳がん細胞MCF‐7の、VNに予備結合されたIGF−II(斜線バー)またはVNの非存在下でディッシュに「結合された」IGF−II(白色バー)で被覆された下方チャンバに応答した、下側表面への移動について示す図。各バーは、5時間インキュベーション後の下方膜上の細胞数の平均を表し、3回の反復実験(3連のウェル中で処理群についての分析を実施)から得ている。複合体の作用がVN単独の場合と有意に異なっているデータを星印で示す。
【図3】12μm孔のTranswells(商標)の上方チャンバ内に植え付けられたヒト乳がん細胞MCF‐7の、VN、VNに結合した天然型IGF−II(斜線バー)またはVNに結合したL27−IGF−II(黒色バー)で被覆されている下方チャンバへの移動について示す図。各データは、VNの非存在下で同一量のIGF−IIを含有する、VNを含まない対照(白色バー)と組み合わされている。各バーは、5時間インキュベーション後の下方膜上の細胞数の平均を表し、2回の反復実験(3連のウェル中で処理群についての分析を実施)から得ている。
【図4】12μm孔のTranswells(商標)の上方チャンバ内に植え付けられたヒト乳がん細胞MCF‐7の、VN単独、VNに結合した天然型IGF−II(斜線バー)またはVNに結合したDes(1−6)IGF−II(黒色バー)で予備結合されている下方チャンバへの移動について示す図。各データは、VNの非存在下で同一量のIGF−IIを含有する、VNを含まない対照(白色バー)と組み合わされている。各バーは、5時間インキュベーション後の下方膜上の細胞数の平均を表し、2回の反復実験(3連のウェル中で処理群についての分析を実施)から得ている。
【図5】ヒト乳がん細胞MCF‐7の、αVの機能を遮断するAbであるmAb2021Zの存在下で、IGF−IIに応答して、Transwells(商標)を通り抜ける移動について示す図。αV機能遮断Abで処置済みのMCF−7細胞を、VN+/−IGF−IIで被覆されているTranswells(商標)上に植え付け、5時間かけて移動させてこの多孔性膜を通過させた。次に、固定した細胞から染色剤を抽出して光学密度を読み取ることにより、膜を横断する細胞の数を測定した。次に、処理群をAbの存在下または非存在下におけるVN単独群の移動細胞数に対する割合(%)として表した。単回実験の4連処置からデータを合わせた。各バーはSEMである。星印は、非処理細胞またはAb処理細胞(Abを有する)の処理群間の有意差を示す(P<0.1)。
【図6】12μm孔のTranswells(商標)の上方チャンバ内に植え付けられたヒト乳がん細胞MCF‐7の、VN(白色バー);VN+IGFBP‐5(灰色バー);天然型IGF−I+VN(明色で塗りつぶされたバー)または天然型IGF−I+IGFBP‐5+VN(暗色で塗りつぶされたバー);L24−IGF−I+VN(左下がり斜線バー)またはL24−IGF−I+IGFBP‐5+VN(右下がり斜線バー)で被覆されている下方チャンバへの移動について示す図。各バーは、5時間インキュベーション後の下方膜上の細胞数の平均を表し、2回の反復実験(3連のウェル中で処理群についての分析を実施)から得ている。
【図7】ビトロネクチンのアミノ酸配列(配列番号1)を示す図。ビトロネクチン内の種々のドメインに関する残基の参照、ならびに残基の修飾部位、リガンド結合部位およびプロテアーゼ認識部位も示す。
【図8】(a)完全長VN(75kDa)および(b)卵黄VN(54kDa)の構造上の関係を、リガンド結合部位とともに示す図。哺乳類および鳥類の血清VNはともに同一のドメイン構造を有するが、アミノ酸配列には差が存在する。卵黄VN(54kDa)は、これらのタンパク質の短縮型である。用いた略号について以下に示す:Som B、ソマトメジンB;接続領域、接続ドメイン;ヘモペキシン、ヘモペキシン様リピート;HBD、ヘパリン結合ドメイン;PAI−1、プラスミノーゲン活性化抑制因子−1;uPAR、ウロキナーゼ型プラスミノーゲン活性化因子受容体;TAT、トロンビン−アンチトロンビンIII複合体;uPA、ウロキナーゼ型プラスミノーゲン活性化因子;−−−−、ポリ陰イオン性領域(塩基性領域);+++、ポリ陽イオン性領域(酸性領域)。
【図9】ニワトリ卵黄由来の54kDa卵黄VNの精製を示す図。Q−Sepharose(登録商標)マトリックスに装荷したタンパク質試料(レーンL)、および溶離された精製物(レーンE)のSDS−PAGE分析。レーンMは、分子量マーカー(バイオラッドの低分子量域マーカー、米国カリフォルニア州リッチモンド所在のバイオラッド)を示す。プレキャスト型の4〜20%勾配ポリアクリルアミドゲル(オーストラリア国ニューサウスウェールズ州フレンチフォレスト所在のグラディポア社(G radipore ))を用いて、タンパク質を分析した。
【図10】[125I]−IGF−I/IGFBP−3の、精製VNと結合する能力について評価する固体プレート結合アッセイを示す図。クリッカー(Kricker )他、2003 Endocrinology 144 2807-2815にすでに記載されているように、固体プレート結合アッセイを実行した。簡潔に述べれば、Immulon(登録商標)96ウェルプレートを精製VNで4℃にて一晩かけて予め被覆した。次に放射活性標識したIGF−I/IGFBP−3複合体を添加し、一晩VNに結合させた後、結合していない物質を除去した。ウェルに結合したVNに対する[125I]−IGF−I/IGFBP−3の結合を、γ計数器で測定した(n=18)。ヒトVN:ヒト血清から精製されたVN;卵黄VN75:75kDaの精製卵黄VN;卵黄VN54:54kDaの精製卵黄VN。
【図11】IGF:VN複合体に応答したHaCAT細胞の増殖に関する細胞増殖アッセイ(MTT)(48時間)を示す図。IGF:VN複合体でウェルを予め被覆し、HaCAT細胞を植え付け、48時間増殖させた。48時間後、MTT法(デニゾットおよびラング(Denizot & Lang)、1986 The Journal of Immunological Methods 89 271-277)を用いて、代謝活性により細胞増殖について複合体を評価した(n=3)。ヒトVN:ヒト血清から精製されたVN;卵黄VN75:75kDaの精製卵黄VN;卵黄VN54:54kDaの精製卵黄VN;IGF−I/BP3:インスリン様成長因子−Iおよびインスリン様成長因子結合タンパク質3。
【図12】IGF:VN複合体に応答したHaCATの移動に関するTranswells(商標)移動アッセイ(5時間)を示す図。上記と同様に(クリッカー(Kricker )他の上記文献、2003)、HaCAT細胞を、IGF−I:IGFBP−3:VN複合体で被覆したTranswells(商標)に植え付け、5時間移動させた。移動した細胞をクリスタルバイオレットで染色し、595nmで光学密度を読取った。各処理は、2連で実施した(n=2)。ヒトVN:ヒト血清から精製されたVN;卵黄VN75:75kDaの精製卵黄VN;卵黄VN54:54kDaの精製卵黄VN;IGF−I/BP3:インスリン様成長因子−Iおよびインスリン様成長因子結合タンパク質3。
【図13】(A)成熟ビトロネクチンタンパク質(配列番号2)、(B)IGF−I(配列番号3)および(C)好ましいリンカー配列(配列番号4〜8)のアミノ酸配列を示す図。
【図14】IGF−IおよびVNを含有するキメラタンパク質の実施形態(A)〜(N)のアミノ酸配列(配列番号9〜22)を示す図。
【図15】(A)PDGFおよびVNを含有するキメラタンパク質の実施形態のアミノ酸配列(配列番号23)、ならびに(B)VEGFおよびVNを含有するキメラタンパク質の実施形態のアミノ酸配列(配列番号24)を示す図。
【特許請求の範囲】
【請求項1】
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子受容体に結合し得るドメイン、および
(ii)ヘパリン結合ドメイン(HBD)を含まないビトロネクチン(VN)またはフィブロネクチン(FN)
を含んでなる単離タンパク質複合体。
【請求項2】
前記成長因子はIGF−IまたはIGF−IIである、請求項1に記載の単離タンパク質複合体。
【請求項3】
前記IGF−IまたはIGF−IIの少なくとも1つのドメインはI型IGF受容体に結合し得る、請求項2に記載の単離タンパク質複合体。
【請求項4】
前記成長因子はIGF−Iであり、同族の成長因子受容体に結合し得るIGF−Iの少なくとも1つのドメインがIGF−Iの残基24を含む、請求項2に記載の単離タンパク質複合体。
【請求項5】
残基24はロイシンではない、請求項4に記載の単離タンパク質複合体。
【請求項6】
IGFBPをさらに含む、請求項4に記載の単離タンパク質複合体。
【請求項7】
IGFBPはIGFBP1、IGFBP2、IGFBP3、IGFBP4、IGFBP5およびIGFBP6からなる群から選択される、請求項6に記載の単離タンパク質複合体。
【請求項8】
IGFBPはIGFBP3およびIGFBP5から選択される、請求項7に記載の単離タンパク質複合体。
【請求項9】
成長因子はIGF−IIであり、同族の成長因子受容体に結合し得るIGF−IIの少なくとも1つのドメインがIGF−IIの残基27を含む、請求項2に記載の単離タンパク質複合体。
【請求項10】
残基27はロイシンではない、請求項9に記載の単離タンパク質複合体。
【請求項11】
HBDを含まないVNは天然に生じる種類のVNである、請求項1に記載の単離タンパク質複合体。
【請求項12】
HBDを含まないVNはVNの短縮型断片である、請求項1に記載の単離タンパク質複合体。
【請求項13】
HBDを含まないVNは、HBDが欠失または突然変異しているVNまたはVN断片である、請求項1に記載の単離タンパク質複合体。
【請求項14】
VNまたはFNは少なくとも1つのインテグリン結合ドメインを含む、請求項1に記載の単離タンパク質複合体。
【請求項15】
前記インテグリン結合ドメインはαVインテグリン結合ドメインである、請求項14に記載の単離タンパク質複合体。
【請求項16】
前記インテグリン結合ドメインはαVβ3インテグリン結合ドメインまたはαVβ5インテグリン結合ドメインである、請求項15に記載の単離タンパク質複合体。
【請求項17】
合成キメラタンパク質の形態の単離タンパク質複合体であって、
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子受容体に結合し得るドメイン、および
(ii)ビトロネクチン(VN)もしくはフィブロネクチン(FN)、またはVNもしくはFNの少なくとも1つのインテグリン結合ドメイン
のアミノ酸配列を含む、単離タンパク質複合体。
【請求項18】
前記インテグリン結合ドメインはαVインテグリン結合ドメインである、請求項17に記載の単離タンパク質複合体。
【請求項19】
前記インテグリン結合ドメインはαVβ3インテグリン結合ドメインまたはαVβ5インテグリン結合ドメインである、請求項18に記載の単離タンパク質複合体。
【請求項20】
ビトロネクチン(VN)もしくはフィブロネクチン(FN)、またはVNもしくはFNの前記少なくとも1つのインテグリン結合ドメインが、成熟VNタンパク質の残基53〜64に相当するポリ陰イオン性アミノ酸配列を含まないことを特徴とする、請求項17に記載の単離タンパク質複合体。
【請求項21】
ビトロネクチン(VN)もしくはフィブロネクチン(FN)、またはVNもしくはFNの前記少なくとも1つのインテグリン結合ドメインが、HBDを含まないことを特徴とする、請求項20に記載の単離タンパク質複合体。
【請求項22】
前記成長因子はIGF−I、IGF−II、VEGFまたはPDGFである、請求項17に記載の単離タンパク質複合体。
【請求項23】
前記成長因子はIGF−Iである、請求項22に記載の単離タンパク質複合体。
【請求項24】
IGFBPのアミノ酸配列を含まない、請求項23に記載の単離タンパク質複合体。
【請求項25】
同族の成長因子受容体に結合し得るIGF−Iの前記少なくとも1つのドメインはIGF−Iの残基24を含む、請求項23に記載の単離タンパク質複合体。
【請求項26】
ビトロネクチンのアミノ酸残基1〜52を含む、請求項17に記載の単離タンパク質複合体。
【請求項27】
ビトロネクチンのアミノ酸残基1〜130を含む、請求項17に記載の単離タンパク質複合体。
【請求項28】
ビトロネクチンのアミノ酸残基1〜346を含む、請求項17に記載の単離タンパク質複合体。
【請求項29】
ビトロネクチンのアミノ酸残基1〜379を含む、請求項17に記載の単離タンパク質複合体。
【請求項30】
ビトロネクチンのアミノ酸残基1〜459を含む、請求項17に記載の単離タンパク質複合体。
【請求項31】
ビトロネクチンのアミノ酸残基65〜459を含む、請求項26に記載の単離タンパク質複合体。
【請求項32】
ビトロネクチンのアミノ酸残基347〜379をさらに含む、請求項27に記載の単離タンパク質複合体。
【請求項33】
ビトロネクチンのアミノ酸残基347〜459をさらに含む、請求項27に記載の単離タンパク質複合体。
【請求項34】
IGF−Iのアミノ酸残基4〜70を含む、請求項23に記載の単離タンパク質複合体。
【請求項35】
IGF−Iのアミノ酸残基1〜70を含む、請求項23に記載の単離タンパク質複合体。
【請求項36】
少なくとも1つのリンカー配列をさらに含む、請求項17に記載の単離タンパク質複合体。
【請求項37】
前記リンカー配列はプロテアーゼ切断部位を含む、請求項36に記載の単離タンパク質複合体。
【請求項38】
前記リンカー配列は、
(i)Gly4Ser(配列番号4)、
(ii)Gly4Ser3(配列番号5)、
(iii)(Gly4Ser)3(配列番号6)、
(iv)LeuIleLysMetLysPro(配列番号7)、および
(v)GlnProGlnGlyLeuAlaLys(配列番号8)
からなる群から選択される、請求項36に記載の単離タンパク質複合体。
【請求項39】
VN中に1つまたは複数の突然変異を含む、請求項1または17に記載の単離タンパク質複合体。
【請求項40】
突然変異は、(i)T50A、(ii)T57A、(iii)T50E、(iv)T57E、(v)S378E、(vi)S378A、および(v)S362E、からなる群から選択される、請求項39に記載の単離タンパク質複合体。
【請求項41】
図14に記載されているアミノ酸配列を含む、請求項23に記載の単離タンパク質複合体。
【請求項42】
前記成長因子はVEGFであり、図15Aに記載されているアミノ酸配列を含む、請求項17に記載の単離タンパク質複合体。
【請求項43】
前記成長因子はPDGFであり、図15Bに記載されているアミノ酸配列を含む、請求項17に記載の単離タンパク質複合体。
【請求項44】
請求項1〜43のいずれか1項に記載の該単離タンパク質複合体をコードする単離核酸。
【請求項45】
ベクター中の1つまたは複数の調節ヌクレオチド配列に作動可能なように連結された請求項44に記載の単離核酸を含んでなる遺伝子構築物。
【請求項46】
前記単離核酸がプロモーターに作動可能なように連結された発現構築物である、請求項42に記載の遺伝子構築物。
【請求項47】
請求項46に記載の遺伝子構築物を含む宿主細胞。
【請求項48】
請求項1または請求項17に記載の単離タンパク質複合体ならびに薬学的に許容可能な担体、希釈剤または賦形剤を含んでなる薬学的組成物。
【請求項49】
請求項1または請求項17に記載の単離タンパク質複合体を、含浸、被覆、またはその他の方法で含む、外科用移植片、足場材、または人工器官。
【請求項50】
請求項1または請求項17に記載の単離タンパク質複合体を含む創傷被覆材または熱傷被覆材。
【請求項51】
細胞の移動および増殖のうち少なくともいずれかの促進方法であって、細胞により発現される成長因子受容体およびインテグリン受容体の両方に結合することにより前記細胞の移動および増殖のうち少なくともいずれかを誘導し、増強し、またはそうでなければ促進するために、請求項1または請求項17に記載の単離タンパク質複合体を用いる工程を含む方法。
【請求項52】
前記単離タンパク質複合体を動物に投与することによりin situで細胞の移動および増殖のうち少なくともいずれかを促進する、請求項51に記載の方法。
【請求項53】
上皮細胞の移動および増殖のうち少なくともいずれかを誘導し、増強し、またはそうでなければ促進することによって、in situでの創傷治癒を促すための、予防的または治療的に用いられる場合の請求項51に記載の方法。
【請求項54】
前記動物がヒトである請求項53に記載の方法。
【請求項55】
単離タンパク質複合体がin vitroで1つまたは複数の細胞または組織に投与される請求項51に記載の方法。
【請求項56】
細胞の移動および増殖のうち少なくともいずれかを防止する方法であって、成長因子およびビトロネクチンもしくはフィブロネクチンを含むタンパク質複合体により、成長因子受容体およびインテグリン受容体の両方の結合および活性化を防止、抑制、またはそうでなければ低減する工程を含む方法。
【請求項57】
動物におけるin situでの転移性がん細胞の移動および増殖のうち少なくともいずれかを防止または抑制するために、予防的または治療的に用いられる場合の請求項56に記載の方法。
【請求項58】
癌は乳がんである請求項57に記載の方法。
【請求項59】
動物におけるin situでの過形成性の瘢痕および乾癬のうち少なくともいずれかを防止または抑制するために、予防的または治療的に用いられる場合の請求項56に記載の方法。
【請求項60】
前記動物はヒトである請求項57または請求項59に記載の方法。
【請求項61】
成長因子およびビトロネクチンまたはフィブロネクチンを含むタンパク質複合体のアゴニストまたはアンタゴニストである分子を設計し、同定し、または作製するための請求項1または請求項17に記載の単離タンパク質複合体の使用。
【請求項62】
IGF−IIおよびビトロネクチン、またはIGF−I、IGFBPおよびビトロネクチンを含むタンパク質複合体のアゴニストを作製するための請求項61に記載の使用。
【請求項63】
IGF−IIおよびビトロネクチン、またはIGF−I、IGFBPおよびビトロネクチンを含むタンパク質複合体のアンタゴニストを作製するための請求項61に記載の使用。
【請求項1】
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子受容体に結合し得るドメイン、および
(ii)ヘパリン結合ドメイン(HBD)を含まないビトロネクチン(VN)またはフィブロネクチン(FN)
を含んでなる単離タンパク質複合体。
【請求項2】
前記成長因子はIGF−IまたはIGF−IIである、請求項1に記載の単離タンパク質複合体。
【請求項3】
前記IGF−IまたはIGF−IIの少なくとも1つのドメインはI型IGF受容体に結合し得る、請求項2に記載の単離タンパク質複合体。
【請求項4】
前記成長因子はIGF−Iであり、同族の成長因子受容体に結合し得るIGF−Iの少なくとも1つのドメインがIGF−Iの残基24を含む、請求項2に記載の単離タンパク質複合体。
【請求項5】
残基24はロイシンではない、請求項4に記載の単離タンパク質複合体。
【請求項6】
IGFBPをさらに含む、請求項4に記載の単離タンパク質複合体。
【請求項7】
IGFBPはIGFBP1、IGFBP2、IGFBP3、IGFBP4、IGFBP5およびIGFBP6からなる群から選択される、請求項6に記載の単離タンパク質複合体。
【請求項8】
IGFBPはIGFBP3およびIGFBP5から選択される、請求項7に記載の単離タンパク質複合体。
【請求項9】
成長因子はIGF−IIであり、同族の成長因子受容体に結合し得るIGF−IIの少なくとも1つのドメインがIGF−IIの残基27を含む、請求項2に記載の単離タンパク質複合体。
【請求項10】
残基27はロイシンではない、請求項9に記載の単離タンパク質複合体。
【請求項11】
HBDを含まないVNは天然に生じる種類のVNである、請求項1に記載の単離タンパク質複合体。
【請求項12】
HBDを含まないVNはVNの短縮型断片である、請求項1に記載の単離タンパク質複合体。
【請求項13】
HBDを含まないVNは、HBDが欠失または突然変異しているVNまたはVN断片である、請求項1に記載の単離タンパク質複合体。
【請求項14】
VNまたはFNは少なくとも1つのインテグリン結合ドメインを含む、請求項1に記載の単離タンパク質複合体。
【請求項15】
前記インテグリン結合ドメインはαVインテグリン結合ドメインである、請求項14に記載の単離タンパク質複合体。
【請求項16】
前記インテグリン結合ドメインはαVβ3インテグリン結合ドメインまたはαVβ5インテグリン結合ドメインである、請求項15に記載の単離タンパク質複合体。
【請求項17】
合成キメラタンパク質の形態の単離タンパク質複合体であって、
(i)成長因子、または成長因子の少なくとも1つのドメインであって同族の成長因子受容体に結合し得るドメイン、および
(ii)ビトロネクチン(VN)もしくはフィブロネクチン(FN)、またはVNもしくはFNの少なくとも1つのインテグリン結合ドメイン
のアミノ酸配列を含む、単離タンパク質複合体。
【請求項18】
前記インテグリン結合ドメインはαVインテグリン結合ドメインである、請求項17に記載の単離タンパク質複合体。
【請求項19】
前記インテグリン結合ドメインはαVβ3インテグリン結合ドメインまたはαVβ5インテグリン結合ドメインである、請求項18に記載の単離タンパク質複合体。
【請求項20】
ビトロネクチン(VN)もしくはフィブロネクチン(FN)、またはVNもしくはFNの前記少なくとも1つのインテグリン結合ドメインが、成熟VNタンパク質の残基53〜64に相当するポリ陰イオン性アミノ酸配列を含まないことを特徴とする、請求項17に記載の単離タンパク質複合体。
【請求項21】
ビトロネクチン(VN)もしくはフィブロネクチン(FN)、またはVNもしくはFNの前記少なくとも1つのインテグリン結合ドメインが、HBDを含まないことを特徴とする、請求項20に記載の単離タンパク質複合体。
【請求項22】
前記成長因子はIGF−I、IGF−II、VEGFまたはPDGFである、請求項17に記載の単離タンパク質複合体。
【請求項23】
前記成長因子はIGF−Iである、請求項22に記載の単離タンパク質複合体。
【請求項24】
IGFBPのアミノ酸配列を含まない、請求項23に記載の単離タンパク質複合体。
【請求項25】
同族の成長因子受容体に結合し得るIGF−Iの前記少なくとも1つのドメインはIGF−Iの残基24を含む、請求項23に記載の単離タンパク質複合体。
【請求項26】
ビトロネクチンのアミノ酸残基1〜52を含む、請求項17に記載の単離タンパク質複合体。
【請求項27】
ビトロネクチンのアミノ酸残基1〜130を含む、請求項17に記載の単離タンパク質複合体。
【請求項28】
ビトロネクチンのアミノ酸残基1〜346を含む、請求項17に記載の単離タンパク質複合体。
【請求項29】
ビトロネクチンのアミノ酸残基1〜379を含む、請求項17に記載の単離タンパク質複合体。
【請求項30】
ビトロネクチンのアミノ酸残基1〜459を含む、請求項17に記載の単離タンパク質複合体。
【請求項31】
ビトロネクチンのアミノ酸残基65〜459を含む、請求項26に記載の単離タンパク質複合体。
【請求項32】
ビトロネクチンのアミノ酸残基347〜379をさらに含む、請求項27に記載の単離タンパク質複合体。
【請求項33】
ビトロネクチンのアミノ酸残基347〜459をさらに含む、請求項27に記載の単離タンパク質複合体。
【請求項34】
IGF−Iのアミノ酸残基4〜70を含む、請求項23に記載の単離タンパク質複合体。
【請求項35】
IGF−Iのアミノ酸残基1〜70を含む、請求項23に記載の単離タンパク質複合体。
【請求項36】
少なくとも1つのリンカー配列をさらに含む、請求項17に記載の単離タンパク質複合体。
【請求項37】
前記リンカー配列はプロテアーゼ切断部位を含む、請求項36に記載の単離タンパク質複合体。
【請求項38】
前記リンカー配列は、
(i)Gly4Ser(配列番号4)、
(ii)Gly4Ser3(配列番号5)、
(iii)(Gly4Ser)3(配列番号6)、
(iv)LeuIleLysMetLysPro(配列番号7)、および
(v)GlnProGlnGlyLeuAlaLys(配列番号8)
からなる群から選択される、請求項36に記載の単離タンパク質複合体。
【請求項39】
VN中に1つまたは複数の突然変異を含む、請求項1または17に記載の単離タンパク質複合体。
【請求項40】
突然変異は、(i)T50A、(ii)T57A、(iii)T50E、(iv)T57E、(v)S378E、(vi)S378A、および(v)S362E、からなる群から選択される、請求項39に記載の単離タンパク質複合体。
【請求項41】
図14に記載されているアミノ酸配列を含む、請求項23に記載の単離タンパク質複合体。
【請求項42】
前記成長因子はVEGFであり、図15Aに記載されているアミノ酸配列を含む、請求項17に記載の単離タンパク質複合体。
【請求項43】
前記成長因子はPDGFであり、図15Bに記載されているアミノ酸配列を含む、請求項17に記載の単離タンパク質複合体。
【請求項44】
請求項1〜43のいずれか1項に記載の該単離タンパク質複合体をコードする単離核酸。
【請求項45】
ベクター中の1つまたは複数の調節ヌクレオチド配列に作動可能なように連結された請求項44に記載の単離核酸を含んでなる遺伝子構築物。
【請求項46】
前記単離核酸がプロモーターに作動可能なように連結された発現構築物である、請求項42に記載の遺伝子構築物。
【請求項47】
請求項46に記載の遺伝子構築物を含む宿主細胞。
【請求項48】
請求項1または請求項17に記載の単離タンパク質複合体ならびに薬学的に許容可能な担体、希釈剤または賦形剤を含んでなる薬学的組成物。
【請求項49】
請求項1または請求項17に記載の単離タンパク質複合体を、含浸、被覆、またはその他の方法で含む、外科用移植片、足場材、または人工器官。
【請求項50】
請求項1または請求項17に記載の単離タンパク質複合体を含む創傷被覆材または熱傷被覆材。
【請求項51】
細胞の移動および増殖のうち少なくともいずれかの促進方法であって、細胞により発現される成長因子受容体およびインテグリン受容体の両方に結合することにより前記細胞の移動および増殖のうち少なくともいずれかを誘導し、増強し、またはそうでなければ促進するために、請求項1または請求項17に記載の単離タンパク質複合体を用いる工程を含む方法。
【請求項52】
前記単離タンパク質複合体を動物に投与することによりin situで細胞の移動および増殖のうち少なくともいずれかを促進する、請求項51に記載の方法。
【請求項53】
上皮細胞の移動および増殖のうち少なくともいずれかを誘導し、増強し、またはそうでなければ促進することによって、in situでの創傷治癒を促すための、予防的または治療的に用いられる場合の請求項51に記載の方法。
【請求項54】
前記動物がヒトである請求項53に記載の方法。
【請求項55】
単離タンパク質複合体がin vitroで1つまたは複数の細胞または組織に投与される請求項51に記載の方法。
【請求項56】
細胞の移動および増殖のうち少なくともいずれかを防止する方法であって、成長因子およびビトロネクチンもしくはフィブロネクチンを含むタンパク質複合体により、成長因子受容体およびインテグリン受容体の両方の結合および活性化を防止、抑制、またはそうでなければ低減する工程を含む方法。
【請求項57】
動物におけるin situでの転移性がん細胞の移動および増殖のうち少なくともいずれかを防止または抑制するために、予防的または治療的に用いられる場合の請求項56に記載の方法。
【請求項58】
癌は乳がんである請求項57に記載の方法。
【請求項59】
動物におけるin situでの過形成性の瘢痕および乾癬のうち少なくともいずれかを防止または抑制するために、予防的または治療的に用いられる場合の請求項56に記載の方法。
【請求項60】
前記動物はヒトである請求項57または請求項59に記載の方法。
【請求項61】
成長因子およびビトロネクチンまたはフィブロネクチンを含むタンパク質複合体のアゴニストまたはアンタゴニストである分子を設計し、同定し、または作製するための請求項1または請求項17に記載の単離タンパク質複合体の使用。
【請求項62】
IGF−IIおよびビトロネクチン、またはIGF−I、IGFBPおよびビトロネクチンを含むタンパク質複合体のアゴニストを作製するための請求項61に記載の使用。
【請求項63】
IGF−IIおよびビトロネクチン、またはIGF−I、IGFBPおよびビトロネクチンを含むタンパク質複合体のアンタゴニストを作製するための請求項61に記載の使用。
【図1】


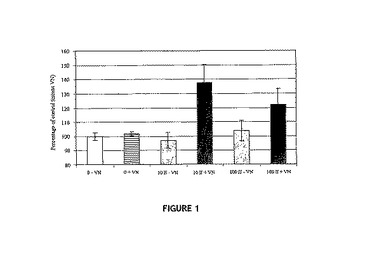
【図2】
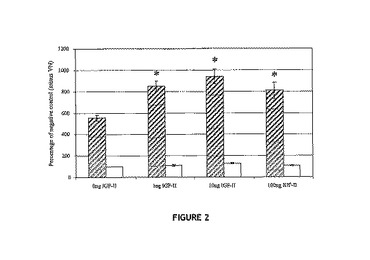
【図3】
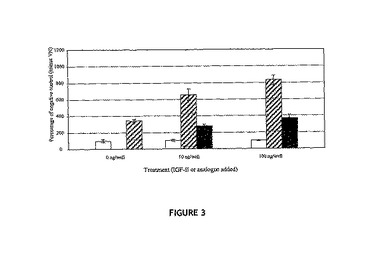
【図4】
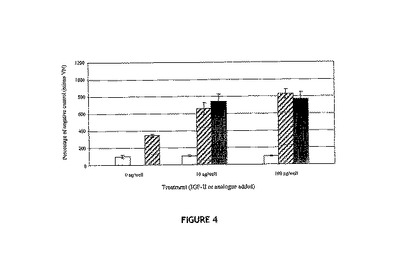
【図5】
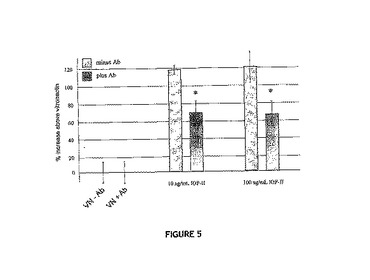
【図6】
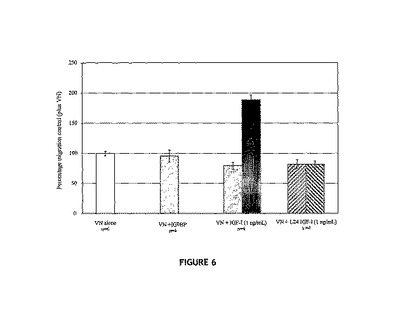
【図7】

【図8】
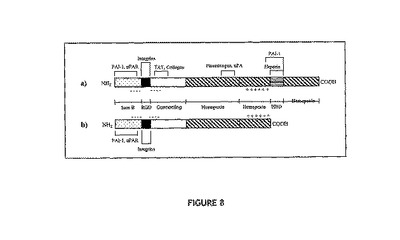
【図9】
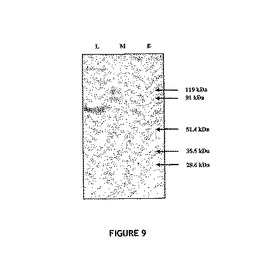
【図10】
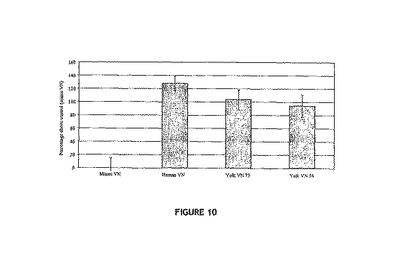
【図11】
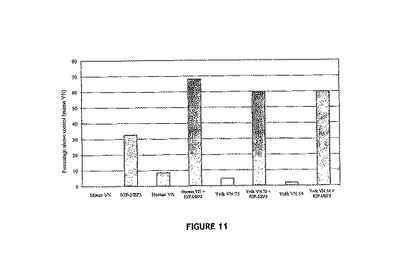
【図12】
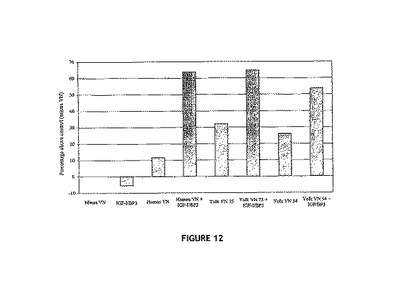
【図13】

【図14】




【図15】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公表番号】特表2007−523598(P2007−523598A)
【公表日】平成19年8月23日(2007.8.23)
【国際特許分類】
【出願番号】特願2006−501345(P2006−501345)
【出願日】平成16年2月5日(2004.2.5)
【国際出願番号】PCT/AU2004/000117
【国際公開番号】WO2004/069871
【国際公開日】平成16年8月19日(2004.8.19)
【出願人】(592253275)クイーンズランド ユニバーシティ オブ テクノロジー (13)
【Fターム(参考)】
【公表日】平成19年8月23日(2007.8.23)
【国際特許分類】
【出願日】平成16年2月5日(2004.2.5)
【国際出願番号】PCT/AU2004/000117
【国際公開番号】WO2004/069871
【国際公開日】平成16年8月19日(2004.8.19)
【出願人】(592253275)クイーンズランド ユニバーシティ オブ テクノロジー (13)
【Fターム(参考)】
[ Back to top ]