
截形グリア細胞系由来神経栄養因子
【課題】ドーパミン作動性細胞によるドーパミン取り込みを促進し、神経細胞の生存を促進する截形グリア細胞系由来神経栄養因子(截形GDNF)タンパク質と呼ぶ新規タンパク質を提供する。
【解決手段】截形グリア細胞系由来神経栄養因子(GDNF)タンパク質産物並びにその付加、置換及び内部欠失変異体及び誘導体。截形GDNFタンパク質産物をコードするポリヌクレオチド。宿主細胞を適切な栄養培地で培養し、場合により、前記細胞又は前記栄養培地から前記截形GDNFを単離する截形GDNFタンパク質の製造方法。截形GDNFタンパク質産物を含む医薬組成物。
【解決手段】截形グリア細胞系由来神経栄養因子(GDNF)タンパク質産物並びにその付加、置換及び内部欠失変異体及び誘導体。截形GDNFタンパク質産物をコードするポリヌクレオチド。宿主細胞を適切な栄養培地で培養し、場合により、前記細胞又は前記栄養培地から前記截形GDNFを単離する截形GDNFタンパク質の製造方法。截形GDNFタンパク質産物を含む医薬組成物。
【発明の詳細な説明】
【技術分野】
【0001】
一般には、本発明はドーパミン作動性ニューロンによるドーパミンの取り込みを促進し、パーキンソン病で死滅するニューロンの生存を助長する能力をもつ本明細書中でグリア細胞系由来神経栄養因子(グリア由来神経栄養因子又はGDNFとも呼ぶ)と呼ぶタンパク質に関する。本発明はより詳細には新規截形(truncated)GDNFタンパク質に関する。
【背景技術】
【0002】
神経栄養因子は神経系又は神経系により刺激される非神経組織に存在するタンパク質であり、神経及び/又はグリア細胞のの生存を促進し、その表現型分化を維持する機能をもつ(Varonら,Ann.Rev.Neuroscience 1:327,1979; Thoenenら,Science 229:238,1985)。この生理的役割により、神経栄養因子は種々の神経変性疾患で生じる神経細胞の変性及び分化機能の低下を治療するのに有用である。
【0003】
特定神経栄養因子が神経損傷の治療に潜在的に有用であるためには、損傷神経細胞の種類がその因子に応答性でなければならない。一般に、種々の神経栄養因子は全く異なる種類の神経細胞に作用する。従って、種々の形態の疾病又は傷害で生じ得る損傷ニューロン種の各々を治療できるように種々の異なる神経栄養因子を利用できることが有利である。
【0004】
神経栄養因子は種々の無関係の傷害から応答性ニューロンを保護することができる。例えば、神経成長因子(NGF)はその軸索突起の切断により生じる死(Richら,J.Neurocytol 16:261,1987; Ottoら,J.Neurosci.83:156,1987)、胚発生中の個体発生死(Hamburgerら,J.Neurosci.4:767,1984)及びタキソール又はシスプラチンの投与により生じる損傷(Apfelら,Ann.Neurol.29:87,1991)から感覚ニューロンの有意部分を救済する。この明白な保護の一般性に鑑み、ある神経栄養因子が実験的損傷から応答性ニューロンを保護するならば、その因果関係は不明であるとしても、このような神経栄養因子は患者にこれらのニューロンの損傷を引き起こす疾病を治療するのに有用であり得るという概念に至った。
【0005】
所与の神経栄養因子は適正なニューロン特異性をもつ以外に、医薬治療として使用するのに十分な量で入手できなければならない。神経栄養因子は一般に組織中に少量しか存在しない(例えばHoferとBarde,Nature 331:261,1988; Linら,Science 246:1023,1989)ので、医薬的量の神経栄養因子を動物組織から直接調製するのは困難である。代替手段として、所望のタンパク質を製造するために組換え発現系を使用することが望ましい。
【0006】
Linらは黒質ドーパミン作動性ニューロンの胚前駆体に及ぼす神経栄養活性について生物サンプルをスクリーニングする方法を過去に記載している(参考資料としてその開示内容を本明細書の一部とする1994年5月23日付け米国特許出願第08/182,183号とその親出願、1992年9月17日付けPCT/US92/07888(WO93/06116)、及びヨーロッパ特許出願第92921022.7号(公開EP第610254号)参照)。パーキンソン病は線条体を刺激する中脳内のドーパミン作動性ニューロンの変性により特徴付けられるで、このバイオアッセイはパーキンソン病の治療に使用可能な神経栄養因子を同定するのに有用である(Friedmanら,Neuro.Sci.Lett.79:65−72,1987)。 Linらは更に、このような材料の1例であるグリオブラストーマ細胞系B49からの条件付け培地から精製した新規神経栄養因子の特徴付けについても記載している(Schubertら,Nature 249:224−27,1974)。この細胞系からの条件付け培地はドーパミン作動性神経栄養活性を含むことが過去に報告されている(Bohnら,Soc.Neurosci.Abs.15:277,1989)。Linらの開示以前に、グリア細胞系由来神経栄養因子(GDNF)は独立した生物活性物質として同定されておらず、即ち実質的に純粋なタンパク質として単離されていなかった。更に、LinらはGDNFをコードするヒト遺伝子のクローニング方法、GDNFをコードするヒト遺伝子の核酸配列及びGDNFタンパク質のアミノ酸配列を記載している。GDNF遺伝子は発現ベクターにサブクローニングされ、ベクターを使用して生物学的に活性なGDNFを発現させている。GDNFタンパク質はジスルフィド結合により結合した22kDaの2個の134アミノ酸サブユニットから構成されるホモダイマーである。パーキンソン病等の神経損傷及び神経関連疾患の予防及び治療にGDNFを使用することも記載されている。
【0007】
GDNF治療は1種以上の神経細胞の生存及び/又は適正な機能を損なう症状に起因する神経損傷の治療に役立つ。このような神経損傷は多種多様の原因により生じると考えられる。神経損傷は、(1)傷害部位の近傍の軸索突起及び/又は神経細胞体の変性を生じる物理的傷害、(2)神経系の一部への血流の一時的又は永久的停止(例えば卒中)、(3)化学療法剤(例えば癌治療剤であるシスプラチンやエイズ治療剤であるジデオキシシチジン(ddC))等の神経毒素への意図的又は偶発的暴露、(4)糖尿病や腎不全等の慢性代謝病、あるいは(5)特定ニューロンポピュレーションの変性に起因するパーキンソン病、アルツハイマー病及び筋萎縮性軸索硬化症(ALS)等の神経変性病により、1種以上の神経細胞に生じると考えられる。
【0008】
GDNF治療はパーキンソン病等の黒質のドーパミン作動性ニューロンの変性を伴う神経変性症状の治療に特に有用であると考えられる。現在行われているパーキンソン病の唯一の治療は、線条体中のドーパミン濃度の増加を目的とした一時的軽減に過ぎない。GDNF治療は、線条体中のドーパミン作動性神経末端のドーパミン作動性神経伝達を増加する(その結果として症状を緩和する)だけでなく、変性プロセスの進行を遅らせ、あるいは停止させ、損傷黒質経路を修復し、その機能を回復する効果があると予想される。GDNFはヒト患者でドーパミン作動性神経細胞の他の形態の損傷又は機能不全を治療するにも有用であると考えられる。このような損傷又は機能不全は、精神分裂病や他の形態の精神病で生じ得る。このような症状に現在行われている唯一の治療は対症療法であり、ドーパミンレセプター又はドーパミン取り込み部位に作用する薬剤が必要であり、これらのレセプターを含むニューロンポピュレーションを刺激するドーパミン作動性ニューロンの機能不全が疾病プロセスに関係があるらしいという見地に立っている。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】米国特許出願第08/182,183号
【特許文献2】国際公開第93/06116号
【特許文献3】欧州特許出願公開第610254号
【非特許文献】
【0010】
【非特許文献1】Varonら,Ann.Rev.Neuroscience 1:327,1979; Thoenenら,Science 229:238,1985
【非特許文献2】Richら,J.Neurocytol 16:261,1987; Ottoら,J.Neurosci.83:156,1987
【非特許文献3】Hamburgerら,J.Neurosci.4:767,1984
【非特許文献4】Apfelら,Ann.Neurol.29:87,1991
【非特許文献5】HoferとBarde,Nature 331:261,1988; Linら,Science 246:1023,1989
【非特許文献6】Friedmanら,Neuro.Sci.Lett.79:65−72,1987
【非特許文献7】Schubertら,Nature 249:224−27,1974
【非特許文献8】Bohnら,Soc.Neurosci.Abs.15:277,1989
【発明の概要】
【0011】
1側面において、本発明は新規截形グリア細胞系由来神経栄養因子(GDNF)タンパク質産物を提供する。1態様では、組換え遺伝子工学技術により截形GDNFタンパク質を製造する。別の態様では、化学的方法又は組換え技術と化学的方法の組み合わせにより截形GDNFタンパク質を合成する。
【0012】
本発明の截形GDNFタンパク質産物はアミノ酸配列X−[Cys41−Cys133]−Yにより表されるタンパク質を含む。成熟GDNFタンパク質と比較し易くするために、図1のアミノ酸残基ナンバリング図(配列番号2)を使用する。
[Cys41−Cys133]は図1に示すCys41〜Cys133のアミノ酸配列(配列番号2)を表す。YはCys133のカルボキシ末端基又はIle134のカルボキシ末端アミノ酸残基を表す。XはCys41のメチオニン化もしくは非メチオニン化アミン基又は以下の群:
【0013】
【表1】

から選択されるアミノ末端アミノ酸残基を表す。前記截形GDNFタンパク質産物はX−[Cys41−Cys133]−Yにより表されるようなアミノ酸配列をもつ截形GDNFタンパク質産物とその変異体及び誘導体を含むと考えられる。従って、本発明の截形GDNFタンパク質産物はX−[Cys41−Cys133]−Yにより表されるアミノ酸配列の付加、置換及び内部欠失変異体及び誘導体も含む。截形GDNFタンパク質産物は更に、截形GDNFタンパク質のメチオニン化又は非メチオニン化形態とグリコシル化又は非グリコシル化形態も含む。
【0014】
本発明の截形GDNFタンパク質の例としては、メチオニン化又は非メチオニン化形態の[Arg16−Ile134]、[Asn22−Ile134]、[Pro23−Ile134]、[Ser26−Ile134]、[Arg32−Ile134]、[Gly33−Ile134]、[Lys37−Ile134]及び[Asn38−Ile134]截形GDNFタンパク質とその変異体及び誘導体が挙げられる。本発明の好ましい截形GDNFタンパク質としては、メチオニン化又は非メチオニン化形態の[Lys37−Ile134]及び[Asn38−Ile134]截形GDNFタンパク質とその変異体及び誘導体が挙げられる。置換変異体の例は[Asn22ΔSer22−Ile134]及び[Pro23−Lys37ΔAsn37−Ile134]截形GDNFタンパク質である。付加変異体の例はSer−[Pro23−Ile134]截形GDNFタンパク質である。
【0015】
本発明の別の側面では、截形GDNFタンパク質はグリコシル化形態で製造してもよいし、非グリコシル化形態でもよい。截形GDNFタンパク質の誘導体は、一般に水溶性ポリマーに截形GDNFタンパク質が結合している。例えば、截形GDNFタンパク質を1個以上のポリエチレングリコール分子に結合すると、水性環境での截形GDNFタンパク質産物の沈殿を減らすことができる。
【0016】
本発明の更に別の側面は、截形GDNFタンパク質をコードする種々のポリヌクレオチドに関する。これらの核酸配列は一般に真核又は原核宿主細胞で截形GDNFを発現させるのに使用され、発現産物又はその誘導体はドーパミン作動性細胞によるドーパミン取り込みを増加することができる。ポリヌクレオチドは細胞治療又は遺伝子治療用にも使用できる。適切な核酸配列としては、図面に具体的に示す配列や、付加的縮重配列(additional degenerate sequences)及び天然対立遺伝子変異配列が挙げられる。
【0017】
本発明の別の側面は、増幅及び/又は発現調節配列に作動的に連結した截形GDNFタンパク質をコードするポリヌクレオチドを含むベクターに関する。原核及び真核宿主細胞の両者を前記ベクターで安定的に形質転換及びトランスフェクトし、截形グリア由来神経栄養因子を発現させることができる。本発明は更に、前記形質転換又はトランスフェクトした宿主細胞を適切な栄養培地で培養し、場合により、前記細胞により発現される截形GDNFを宿主細胞及び/又は栄養培地から単離することを特徴とする、截形GDNFタンパク質の組換え製造にも関する。本発明は更に、遺伝子治療又は細胞治療における截形
GDNFをコードするポリヌクレオチドと該ポリヌクレオチドを含むベクターの使用にも関する。
【0018】
別の側面では、本発明は成熟GDNFタンパク質と該タンパク質から誘導される1種以上の截形GDNFタンパク質の混合物を含む組換え製造GDNF組成物にも関し、成熟GDNFタンパク質は約44kDaの分子量をもち、截形GDNFタンパク質は約36〜40kDaの分子量をもつ。GDNF組成物は約36kDaの分子量をもつ第1の種と約40kDaの分子量をもつ第2の種との少なくとも2種の截形GDNF種を含むものでもよい。約40kDaの分子量をもつ截形GDNF種は約22kDaの分子量をもつGDNFモノマーと約18kDaの分子量をもつ截形GDNFモノマーのヘテロダイマーである。前記混合物から截形GDNF種の1種以上を治療用に単離することも考えられる。
【0019】
本発明の別の側面は、截形GDNFタンパク質産物を含む医薬組成物に関する。一般に、截形GDNFタンパク質産物は医薬的に許容可能なビヒクルと組み合わせて調剤する。製造、貯蔵、取り扱い、送達及び/又は効力を助長するために種々の他の配合剤を使用してもよい。本発明の別の側面では、截形GDNFタンパク質産物はドーパミン作動性ニューロンのドーパミン取り込み及び生存を強化する。従って、截形GDNFタンパク質産物は、パーキンソン病等の傷害又は疾病により生じる神経系の損傷の治療に特に適している。
【0020】
本発明の他の側面及び利点は、本発明の実施を詳細に説明する以下の記載から当業者に容易に理解されよう。
【0021】
発明の詳細な説明
ヒトグリア細胞系由来神経栄養因子(hGDNF)は前駆物質として合成され、プロセシングにより134アミノ酸の成熟タンパク質として分泌される。成熟ヒトGDNFは図1に示すアミノ酸配列(配列番号2)をもつことが認められている。
【0022】
本発明は、その生物活性を維持しながら成熟GDNFタンパク質の寸法を小さくできるという予想外の発見に基づく(本明細書では「切断」もしくは「截形」タンパク質又は截形GDNFタンパク質とも言う)。切断タンパク質はチャイニーズハムスター卵巣(CHO)細胞でGDNFの組換え製造中に最初に発見された。要約すると、組換えヒトGDNF(rhGDNF)は次のように作製した。成熟ヒトGDNFタンパク質の完全オープンリーディングフレームをコードする核酸配列を発現プラスミドにクローニングした。(DNA配列決定によりGenBank中のhGDNF配列の等価物として)核酸配列が正しいことを確認し、成熟ヒトGDNFの公表配列(Linら,Science 260,1130−1132,1993)に等しいアミノ酸配列に翻訳した。プラスミドDNAを直鎖化し、リン酸カルシウム沈殿法を用いてジヒドロ葉酸レダクターゼ欠損CHO細胞(CHOd−細胞)にトランスフェクトした。トランスフェクトした細胞を選択培地で培養し、選択プロセスに耐えて生存したコロニーを選択し、hGDNF発現を個々に分析した。
【0023】
個々のクローンから無血清条件付け培地を集め、hGDNFに特異的な抗血清を用いてウェスタンブロット分析した。抗血清は、大腸菌で発現させた組換えhGDNFで免疫したウサギから誘導したウサギポリクローナル抗体から構成した。還元条件下でこれらのサンプル中に存在するhGDNFは、約22kDa及び18kDaの見かけの分子量をもつ2つの主バンドに分解した。各バンドは夫々約22+22.5kDa及び18+17.5kDaのごく近接する二重項(簡単にするために、以下の文中ではこれらの二重項を22kDa及び18kDaバンド又は種と呼ぶ)から構成されていた。
【0024】
GDNFは約20〜22kDaの分子量をもつ成熟GDNFタンパク質の2つの同一のサブユニットから構成されるジスルフィド結合ホモダイマーとして存在することが過去に報告されている。非還元条件下でGDNFを分析すると、32〜42kDa(Linら,Science 260,1130−1132,1993)又は33〜45kDa(Linら,J.Neurochem.63(2),758−768,1994)の幅広のバンドが確認されたと報告されている。レンジの存在は成熟モノマー上のグリコシル化の不均一に起因すると解釈され、脱グリコシル化実験で更に確証されている。
【0025】
本発明の22kDaバンドは文献に報告されている成熟GDNFタンパク質に対応するが、18kDaバンドは従来報告されていない。22kDa及び18kDaタンパク質の相対量は個々のクローンから採取したサンプルにより相違した。更に、同一クローンからの複数の回収物で2つのバンドの比が異なることが判明した。更に、CHOで発現されたGDNFタンパク質を貯蔵すると、多くの場合には18kDaバンドの存在が増加すると同時に22kDaバンドが減少することが判明した。
【0026】
形質転換CHOd−細胞からの条件付け培地を非還元条件下でウェスタンブロットにより分析すると、36、40及び44kDaの見かけの分子量をもつ3つの明確に分解したバンドが観察された。この知見も従来の報告に反するものであった。これらのバンドの相対強度は一定していないが、サンプルの各々に存在する22及び18kDaモノマーの比とよく相関していた。モノクローナル抗血清により更に分析すると、非還元ゲル中の3つのバンドは2種のモノマーから構成される3種の可能な二量体に対応することが判明した。最大の44kDaタンパク質は従来報告されているような2個の22kDa成熟GDNFタンパク質の二量体である。中間の40kDaタンパク質は、一方の成熟タンパク質の分子量が18kDa形態まで減少した二量体から構成される。最小の36kDa二量体は2個の18kDaタンパク質を含むと思われ、即ち22kDa形態の両者の分子量が減少している。このデータは、新規形態のGDNFモノマーの存在だけでなく、二量体構造における切断GDNFタンパク質の存在をも立証した最初のものである。貯蔵すると、サンプルのモノマー組成は切断形態及び対応する二量体種の組成に向かって変化することも判明し、即ち36kDaタンパク質の量が増加することが判明した。
【0027】
次に、従来報告されている成熟GDNFタンパク質と比較してタンパク質のどの部分が除去又は変異して分子量が減少するのかを調べる研究を行った。まず最初に分子量の減少はグリコシル化の変化には起因しないことが分かった。
【0028】
GDNFは2つの潜在的N結合グリコシル化部位を含み、グリコシル化されると報告されている。しかし、切断タンパク質は単に成熟GDNFの非グリコシル化又は低グリコシル化形態ではない。これは、サンプルをN−グリカナーゼ、O−グリカナーゼ及びノイラミニダーゼで処理する脱グリコシル化実験で実証された。還元ゲル上で18kDaタンパク質はN−グリカナーゼ消化により13.5kDaバンドまで還元され、N結合糖の4.5kDaの等価物の存在を示した。ノイラミニダーゼ及びO−グリカナーゼで処理すると、18kDaバンドは17kDaまで僅かにシフトした。従って、タンパク質にO結合糖が存在すると予想された。成熟22kDaバンドはグリコシル化されると報告されており、同様にN−グリカナーゼにより18kDaまで(即ち同様に4.5kDaだけ)還元された。これは、ゲル上の22kDaバンドに特異的なモノクローナル抗体を使用することにより更に確認された。非還元二量体のグリカナーゼ消化パターンはより複雑であったが、解明可能であり、3形態の初期帰属に一致した。
【0029】
その結果、タンパク質の分子量の4.5kDaの減少はグリコシル化の変化ではなく約30〜35アミノ酸残基の欠失に起因すると考えられた。この欠失は以下の理由で成熟GDNFタンパク質のアミノ末端に最も生じ易いと予想された。成熟GDNFは合計7個のシスチンを含む。欠失がカルボキシル末端から開始したならば、7個のシスチンのうちの2〜4個が失われ、その結果、不活性タンパク質が生じ易いと思われる。しかし、主に切断形態から構成される試料をバイオアッセイにかけ、そのドーパミン作動性ニューロン神経栄養活性を測定した処、この試料は成熟形GDNFを高比率で含む試料と同等の活性を示した。
【0030】
次に、精製タンパク質のアミノ酸配列分析により切断部位を決定した。Applied Biosystems 494Aタンパク質シーケンサーを10サイクル使用して製造業者の指示に従ってサンプルを配列決定した。アミノ酸配列分析技術及び手順は当業者に公知であるが、タンパク質の配列決定に関する詳細な説明は、参考資料としてその開示内容を本明細書の一部とするFaussetら,Electrophoresis 12:22−27,1991及び1990年8月24日付け米国特許出願第576,316号(1990年10月4日付けヨーロッパ特許出願第90310899号、公開EP第423980号、発明の名称「幹細胞因子」)に記載されている。分析の結果、切断タンパク質のアミノ末端は「RGQRGK」即ちArg−Gly−Gln−Arg−Gly−Lysであることが判明した。即ち、条件付け培地中の成熟タンパク質から最初から31個のアミノ酸が除去されていた。アミノ酸Arg32から始まる切断タンパク質の残りのアミノ酸配列は、この欠失以外は図1に示す成熟GDNFアミノ酸配列(配列番号2)に一致していた。
【0031】
[Arg32−Ile134]截形GDNFタンパク質は、ドーパミン作動性ニューロンアッセイで定量的に活性であることが判明した。ドーパミン作動性神経栄養活性アッセイは、パーキンソン病の治療に有益であると考えられる神経栄養因子を同定するために使用されている。このアッセイは従来記載されているアッセイ(参考資料としてその開示内容を本明細書の一部とするFriedmanら,Neuro.Sci.Lett.79:65−72,1987)を基礎とし、Linら(1994年5月23日付け米国特許出願第08/182,183号とその親出願、1992年9月17日付けPCT/US92/07888(WO93/06116)及びヨーロッパ特許出願第92921022.7号(公開EP第610254号))に記載されているように改変できる。アッセイの詳細な説明は後記実施例5に記載する。
【0032】
その後の精製操作と、アミノ酸配列決定の結果、成熟GDNFのN末端から最初から36個のアミノ酸残基が除去された別のタンパク質即ちKNRG(C)VL−−のN末端配列をもつ[Lys37−Ile134]截形GDNFタンパク質が発見された。この場合も、切断タンパク質の残りのアミノ酸残基はこの欠失以外は成熟ヒトGDNFアミノ酸配列と一致していた。
[Lys37−Ile134]截形GDNFタンパク質もドーパミン取り込みバイオアッセイで分析した。この截形GDNFタンパク質は、大腸菌で発現させた精製組換え成熟GDNFと同様に約50pg/mlのED50で活性であることが判明した。
【0033】
更に、細菌で発現させた成熟GDNFを截形型に改変できることも分かった。(Linらの上記米国特許出願第08/182,183号に記載されているように)形質転換大腸菌で発現させた成熟GDNFをCHO由来条件付け培地と共にインキュベートした。組換え大腸菌GDNFは還元ゲル上で17kDaの見かけの分子量をもつ。材料をCHO細胞条件付け培地と混合し、4℃で5日間インキュベートした処、タンパク質は12.5kDaまで完全に切断された。この切断は1時間又は24時間のインキュベーションでは不完全であり、このような条件下で時間依存性プロセスであることが示唆された。
0.1%ウシ胎児血清を含む培地と共に組換え大腸菌GDNFを一晩インキュベートしただけでは切断形態が生成されないことも判明した。従って、切断プロセスが生じるには培養に生きた細胞の存在が必要であると思われる。従って、切断現象は組織によってはin vivoでも生じると考えられる。
【0034】
更に、(同様にLinらの上記米国特許出願第08/182,183号に記載されているような)ペジル化GDNF等の成熟大腸菌発現hGDNFの誘導体をCHO由来条件付け培地の存在下で截形型に加工できることが判明した。成熟GDNFをアミノ末端でペジル化すると、その循環クリアランス時間を増すことができる。ペジル化はタンパク質の寸法を増加し、修飾成熟
GDNFは還元条件下で約45kDaで移動する。ペジル化大腸菌GDNFをCHO細胞(未トランスフェクト)条件付け培地と共にインキュベートすると、非ペジル化成熟形態と同様に12.5kDaバンドが生じた。どちらの場合も12.5kDa種は非還元ゲル上でジスルフィド結合二量体として存在していた。N末端ペジル化成熟タンパク質からこの切断形が生成されたことから、タンパク質のN末端に切断現象が生じたのは切断プロセス中にペジル化残基が失われたためであると考えられる。
【0035】
これらの知見と、in vivoでも切断現象が生じると予想されることから、GDNFタンパク質の截形型は生理的条件下のhGDNFの最終天然プロセシング形態であると考えられる。従って、治療用截形GDNFタンパク質又はその誘導体を製造すると有利であると思われた。例えば、直接発現させるか又は合成した截形GDNFタンパク質(例えば[Arg32−Ile134]截形GDNFタンパク質)は上記タンパク分解活性に耐性であると予想される。更に、ペジル化[Arg32−Ile134]截形GDNFタンパク質等の截形GDNF誘導体を製造することが所望される場合には、得られる誘導体は成熟GDNF誘導体で観察される特異的切断に非感受性であるという利点をもつと予想される。
【0036】
截形GDNFタンパク産物には付加的利点も予想できる。第1に、[Arg32−Ile134]截形GDNFタンパク質等の截形タンパク質のpIは約10から約8.0〜8.5まで低下するであろう。従って、タンパク質は極めて低塩基性となり、その結果、鞘内注射部位等の投与部位におけるレセプター結合の改善や細胞毒性の低下等の有益な効果が得られる。第2に、成熟GDNFアミノ酸配列の最初から26個のアミノ酸には、Arg−Asn−Arg(アミノ酸14〜16)とGlu−Asn−Ser(アミノ酸24〜26)の2つのアミド分解部位が存在する。截形GDNFタンパク質にはこれらの部位の一方又は両方が存在しないので、タンパク質の安定性が増すと予想される。
【0037】
截形GDNFタンパク質産物
基本態様では、本発明の截形GDNFタンパク質は成熟GDNFタンパク質と比較し易くするために図1のアミノ酸残基ナンバリング図を用いた下記アミノ酸配列:
X−[Cys41−Cys133]−Y
(式中、[Cys41−Cys133]は図1に示すCys41〜Cys133のアミノ酸配列(配列番号1)を表し、YはCys133のカルボキシ末端基又はIle134のカルボキシ末端アミノ酸残基を表し、XはCys41のメチオニン化もしくは非メチオニン化アミン基又は以下の群:
【0038】
【表2】
から選択されるアミノ末端アミノ酸残基を表す)により表すことができる。
【0039】
本明細書で使用する「截形GDNFタンパク質産物」なる用語は、生物学的に活性な合成又は組換え截形GDNFタンパク質、成熟GDNFから産生される截形GDNFタンパク質、生物学的に活性な截形GDNF変異体(挿入、置換及び欠失変異体を含む)、及びその化学修飾誘導体を含む。配列番号2に示すアミノ酸配列をもつヒトGDNFタンパク質に実質的に相同な截形GDNFタンパク質も含まれる。
【0040】
本明細書で使用する「生物学的に活性」なる用語は、截形GDNFタンパク質が配列番号2に示すアミノ酸配列をもつGDNFタンパク質と必ずしも同一程度まで全く同一の性質を示す必要はないが、これに類似の神経栄養性を示すことを意味する。目的とする特定神経栄養性の選択は、截形GDNFタンパク質産物を投与する用途に依存する。截形GDNFタンパク質産物は生物学的に活性であり、後記実施例に記載するような典型的バイオアッセイ等でドーパミン取り込み及びチロシンヒドロキシラーゼ(TH)発現を評価すると、成熟GDNFタンパク質に類似のドーパミン作動性ニューロン生存特性を示す。
【0041】
本明細書で使用する「実質的に相同」なる用語は、配列番号2に示すアミノ酸配列をもつヒトGDNFタンパク質に対して好ましくは>70%、より好ましくは>80%、更に好ましくは>90%又は更には>95%の相同度を意味する。本明細書に記載する相同百分率は、参考資料としてその開示内容を本明細書の一部とするDayhoff,Atlas of Protein Sequence and Structure Vol.5,p.124(1972),National Biochemical Research Foundation,Washington,D.C.に記載されているように、整列を助長するのに100アミノ酸長の4個のギャップを導入すればよい場合に、比較する配列中の同一のアミノ酸残基であって整列する2種の配列の小さいほうに存在するアミノ酸残基の百分率として計算される。更に、配列番号2のGDNFに対する抗体との交差反応性により単離できるか、又は配列番号1のGDNFをコードする遺伝子もしくは該遺伝子のセグメントとのハイブリダイゼーションによりその遺伝子を単離できるならば、任意の截形GDNFタンパク質を実質的に相同とみなす。
【0042】
本明細書の記載から当業者に容易に理解されるように、実質的に相同なタンパク質はX−[Cys41−Cys133]−Yにより表される截形GDNFタンパク質のアミノ酸残基の1個以上の欠失、付加又は置換を含む。このような変異体の作製については以下に詳述する。本発明は明らかに「截形」GDNFタンパク質に関するものであるので、アミノ末端付加変異体はメチオニン残基又は非GDNFアミノ酸残基もしくは配列を含むと考えられるが、成熟GDNFタンパク質を再構成するようなアミノ酸残基の付加は含まないことも理解されよう。天然対立遺伝子突然変異体又は変異体に基づく截形GDNFタンパク質も本発明の範囲に含まれる。変異体截形GDNFタンパク質の製造については以下に詳述する。
【0043】
Linら(上記米国特許出願第08/182,183号)は、成熟GDNFのカルボキシル末端から夫々6番目及び5番目の残基であるLys−Arg残基のタンパク分解プロセシングによる成熟GDNFのカルボキシル末端截形を記載している(即ち図1のアミノ酸残基ナンバリング(配列番号1又は配列番号2でも同じ)によるとLys129−Arg130)。このような截形は成熟GDNFタンパク質から2個のシステイン残基を除去すると思われる。その結果、タンパク質の不適正な折り畳みを生じ易く、不活性なタンパク質を形成する恐れがある。これに対して本発明のX−[Cys41−Cys133]−Y截形GDNFタンパク質産物はCys131及びCys133残基を維持しており、ドーパミン取り込みアッセイにより活性タンパク質であると判定される。
【0044】
本発明の1態様では、好ましい截形GDNFタンパク質産物は1個以上のアミド分解部位を欠失している。このようなアミド分解部位の欠失は精製タンパク質の生化学的安定性を増すと共に、予想される分解物を減らし、貯蔵安定性の高いタンパク質をもたらすと考えられる。截形GDNFタンパク質産物の1例は、成熟タンパク質のアミド分解をもたらすような部位を欠失する[Ser26−Ile134]截形GDNFタンパク質である。あるいは、[Arg16−Ile134]截形GDNFタンパク質は成熟タンパク質に存在する少なくとも最初のアミド分解部位を欠失している。
【0045】
本発明の好ましい截形GDNFタンパク質産物は[Arg32−Ile134]截形GDNFタンパク質である。この截形
GDNFタンパク質は成熟タンパク質のタンパク分解切断部位又はその近傍部位を欠失している。従って、この截形GDNFタンパク質はin vivoでも生じると思われるプロセシング現象に耐性であると予想される。本発明の好ましい別の截形GDNFタンパク質産物は[Lys37−Ile134]截形GDNFタンパク質である。この截形は、夫々N及びC末端からGly40及びIle134までの残基を除去する他の截形と同様に、截形タンパク質のpIを更に低下させると思われる。本発明の最も好ましい截形GDNFタンパク質産物は成熟GDNFタンパク質に存在する全システイン残基を維持しながら、発現及び製造中又はin vivo投与後の截形GDNFタンパク質の迅速なタンパク分解プロセシングに関与する部位を欠失している。これらの好ましいタンパク質としては、[Arg32−Ile134]、[Gly33−Ile134]、[Gln34−Ile134]、[Arg35−Ile134]、[Gly36−Ile134]、[Lys37−Ile134]、[Asn38−Ile134]及び[Arg39−Ile134]截形GDNFタンパク質産物が挙げられる。
【0046】
Linら(上記米国特許出願第07/855,413号)により成熟GDNFについて従来記載されている結果と同様に、本発明の截形GDNFタンパク質は黒質ドーパミン作動性ニューロンの胚前駆体によるドーパミン取り込みを増加できることが判明した。截形GDNFタンパク質のバイオアッセイについては、後記実施例4に詳述する。
【0047】
新規截形GDNFタンパク質は一般に、他の(非GDNF)タンパク性材料を実質的に含まない截形GDNFタンパク質を形成するように単離精製される。好ましくは、截形GDNFタンパク質産物は、截形GDNFタンパク質産物の製造に使用する製造技術に起因し得る他のタンパク質を約80%除去している。より好ましくは、截形GDNFタンパク質産物は他のタンパク質を約90%除去し、特に好ましくは他のタンパク質を約95%除去し、最も好ましくは他のタンパク質を約>98%除去している。更に、本発明は均一な截形GDNFタンパク質を製造するためのポリヌクレオチド配列を提供するという独自の利点を提供する。例えば、[Arg32−Ile134]截形GDNFタンパク質をコードするポリヌクレオチド配列を使用すると、大腸菌及び他の適当な発現系で截形GDNFタンパク質を組換え製造することができる。換言するならば、新規ポリヌクレオチドはタンパク分解プロセシングに非感受性であるか又はこのようなプロセシングもしくは上述したような他の生化学プロセシング効果に対する感受性の低い截形GDNFタンパク質を製造することができる。こうして、新規ポリヌクレオチドにより単一種の截形GDNFタンパク質を容易に製造及び/又は単離することができ、従って、截形GDNFタンパク質及び/又はその産物は上記ヘテロ及びホモダイマーの混合物を全く又は少量しか含まない。他方、最終截形GDNFタンパク質産物は、追って詳述するように投与前に他の因子、化学組成物及び/又は適切な医薬配合剤と配合できることが理解されよう。
【0048】
本発明の1側面では、截形GDNFタンパク質は高純度で比較的多量のタンパク質を獲得できるという理由から組換え技術により製造すると有利である。組換え截形GDNFタンパク質形態は該タンパク質のグリコシル化及び非グリコシル化形態を含み、細菌、哺乳動物又は昆虫細胞系で発現される。あるいは、截形GDNFタンパク質は化学的に合成することもできる。本発明の好ましい製造方法については追って詳述する。
【0049】
截形GDNF変異体及び誘導体
A.截形GDNF変異体
本発明の別の側面は截形GDNFタンパク質の変異体に関する。本明細書で使用する「截形GDNFタンパク質産物」なる用語は、天然GDNFのアミノ酸配列の残基からアミノ酸が欠失(「欠失変異体」)、挿入(「付加変異体」)又は置換(「置換変異体」)している変異体タンパク質を含む。このような変異体は、タンパク質をコードするDNAに適当なヌクレオチド変異を導入するか又は所望のタンパク質のin vitro化学合成により作製される。最終タンパク質がGDNF生物活性をもつのであれば、欠失、挿入及び置換の多数の組み合わせが可能であることが当業者に理解されよう。
【0050】
1個以上の選択されたアミノ酸残基の置換、挿入又は欠失のための突然変異誘発技術は当業者に周知である(例えば参考資料としてその開示内容を本明細書の一部とする米国特許第4,518,584号参照)。アミノ酸配列変異体の構築には、突然変異部位の位置と突然変異の種類という2つの主な変更が存在する。截形GDNF変異体に設計に際して、突然変異部位の位置と突然変異の種類は修飾しようとする生化学特性により異なる。突然変異部位は、例えば(1)まず同類アミノ酸選択で置換した後、得られた結果に応じてよりラジカルな選択で置換するか、(2)標的アミノ酸残基を欠失させるか、(3)位置決定した部位の隣接位置にアミノ酸残基を挿入することにより、個々に又は連続して修飾することができる。
【0051】
アミノ酸配列欠失は一般に約1〜30アミノ酸残基、より通常は約1〜10残基、典型的には約1〜5残基である。例えば、Cys41のN末端に配置されたアミノ酸残基の「X」部分の欠失は約1〜30残基でもよいが、[Cys41−Cys133]のシステイン残基はタンパク質折り畳みを破壊しないように、一般にその位置に応じて約1〜5残基である。截形GDNFタンパク質における欠失はトランスフォーミング成長因子−β(TGF−β)ファミリーメンバーに対して相同性の低い領域に形成できる。他のTGF−βファミリー配列に対して実質的に相同な領域で截形GDNFタンパク質から欠失させると、生物活性がより有意に変化する傾向がある。総欠失及び/又は連続欠失の数は、影響を受ける領域、例えばシステイン架橋における截形GDNFタンパク質の三次構造を維持するように選択される。
【0052】
アミノ酸配列付加としては、1残基〜100残基以上の長さのアミノ及び/又はカルボキシル末端融合と、単一又は多重アミノ酸残基の内部配列内挿入が挙げられる。内部付加は一般に約1〜10アミノ酸残基、より一般には約1〜5アミノ酸残基、通常は約1〜3アミノ酸残基であり得る。上述のように、本発明のアミノ末端付加変異体はメチオニン(例えば細菌組換え細胞培養におけるGDNFの直接発現のアーチファクト)又は非GDNFアミノ酸残基もしくは配列の付加を含むと考えられる。アミノ末端付加変異体は、成熟GDNFタンパク質の再構成をもたらすようなアミノ酸残基の付加を含まない。末端挿入の別の例として、組換え宿主細胞からのタンパク質の分泌を助長するように異種N末端シグナル配列をN末端と融合してもよい。このようなシグナル配列は一般に、所期宿主細胞種から得られ、従ってこのような細胞種と相同である。挿入又は付加変異体は、他の神経栄養因子の配列に由来するアミノ酸配列を含んでいてもよい。
【0053】
別の群の変異体はアミノ酸置換変異体である。これらの変異体は截形GDNFタンパク質から少なくとも1個のアミノ酸残基が除去され、その代わりに別の残基が挿入されている。例えば、Met残基を更に除去し易くするために天然Asn22をSerに置換した図5参照。X−[Cys41−Cys133]−Yアミノ酸配列式と截形GDNFタンパク質産物の本発明の定義を使用すると、このような截形GDNFタンパク質は置換変異体Met−[Asn22ΔSer22−Ile134]截形GDNFタンパク質又は付加変異体Met−Ser−[Pro23−Ile134]截形GDNFタンパク質と呼ぶことができる。置換変異体は、アミノ酸置換を生じるか否かに拘わらず種ポピュレーションの天然ヌクレオチド配列変異を特徴とする対立遺伝子変異体でもよい。
【0054】
截形GDNFタンパク質の配列の具体的な突然変異としては、グリコシル化部位(例えばセリン、スレオニン又はアスパラギン)の修飾が挙げられる。グリコシル化又は単に部分的なグリコシル化の不在は、任意のアスパラギン結合グリコシル化認識部位又はO結合炭水化物の付加により修飾されたタンパク質の任意部位におけるアミノ酸置換又は欠失に起因すると考えられる。アスパラギン結合グリコシル化認識部位は適当な細胞グリコシル化酵素により特異的に認識されるトリペプチド配列を含む。これらのトリペプチド配列はAsn−Xaa−Thr又はAsn−Xaa−Serであり、ここでXaaはPro以外の任意のアミノ酸でよい。グリコシル化認識部位の第1又は第3番目のアミノ酸位置の一方又は両方に種々のアミノ酸置換又は欠失(及び/又は第2位のアミノ酸欠失)が存在すると、修飾トリペプチド配列にグリコシル化は生じない。従って、適当に改変されたヌクレオチド配列が発現されると、前記部位がグリコシル化されていない変異体を生じる。あるいは、截形GDNFタンパク質にグリコシル化部位を付加するように配列を修飾してもよい。
【0055】
突然変異誘発に用いる截形GDNFアミノ酸残基又は領域の指定方法の1例はCunninghamとWells(Science,244: 1081−1085,1988)により記載されている「アラニン走査突然変異誘発」と呼ばれる方法である。この方法では、標的残基のアミノ酸残基又は基(例えばArg、Asp、His、Lys及びGlu等の帯電残基)を指定し、中性又は負電荷をもつアミノ酸(最も好ましくはアラニン又はポリアラニン)で置換し、アミノ酸を細胞の内側又は外側の周囲水性環境と相互作用させる。次に、置換部位に付加又は代替残基を導入することにより、置換に機能的感受性を示す領域を限定する。こうしてアミノ酸配列修飾を導入する部位を予め決定し、所与の部位での突然変異の性能を最適化するために、アラニン走査又はランダム突然変異誘発を実施し、所望の活性及び活性度の最適組み合わせを求めて変異体をスクリーニングする。
【0056】
置換突然変異誘発に最も有利な部位としては、種々の種からのGDNFタンパク質に存在するアミノ酸が側鎖の大きさ、電荷及び/又は疎水性の点で実質的に相違する部位が挙げられる。他の有用な部位としては、種々の種から得られるGDNF様タンパク質の特定残基が同一である部位が挙げられる。このような位置は一般にタンパク質の生物活性に重要である。まず最初に、これらの部位を比較的同類的に置換して修飾する。このような同類置換を表1の好ましい置換の欄に示す。このような置換により生物活性が変化するならば、より実質的な変異(典型的な置換)を導入及び/又は他の付加/欠失を実施し、得られた産物をスクリーニングする。
【0057】
【表3】
【0058】
アミノ酸配列に穏当な改変(及びコーディング核酸配列の対応する修飾)を加えると、後記実施例に記載する截形GDNFタンパク質に類似の機能的及び化学的特性をもつ截形GDNFタンパク質を生じると予想される。他方、截形GDNFタンパク質の機能的及び/又は化学的特性の実質的変更は、(a)置換領域のポリペプチド主鎖の構造(例えばシート又は螺旋構造等)、(b)標的部位のタンパク質の電荷もしくは疎水性、又は(c)側鎖の大きさのいずれかを維持する効果の点で有意に異なる置換を選択することにより達せられる。天然残基は共通の側鎖性質に基づいて次のように分類される。
1)疎水性:ノルロイシン、Met、Ala、Val、Leu、Ile、
2)中性親水性:Cys、Ser、Thr、
3)酸性:Asp、Glu、
4)塩基性:Asn、Gln、His、Lys、Arg、
5)側鎖配向に影響する残基:Gly、Pro、及び
6)芳香族:Trp、Tyr、Phe。
【0059】
穏当でない置換は、これらの分類の1つのメンバーを別のメンバーに置換するものと言える。このような置換残基は、他のTGF−βタンパク質と相同の截形GDNFタンパク質の領域に導入してもよいし、タンパク質の非相同領域に導入してもよい。
【0060】
B.截形GDNF誘導体
本明細書の開示に基づき、当業者は截形GDNF又は截形
GDNF変異体の化学修飾誘導体を作製できる。截形GDNFタンパク質の誘導体化に最も適した化学種としては水溶性ポリマーが挙げられる。水溶性ポリマーはこれと結合したタンパク質が生理的環境等の水性環境で沈殿しないという理由で望ましい。ポリマーは治療用製品又は組成物の製造に医薬的に許容可能であることが好ましい。当業者はポリマー/タンパク質結合体を治療に使用するか否か、治療用の場合には所望の投与量、循環時間、耐タンパク分解性及び他の要因に基づいて所望のポリマーを選択することができよう。誘導体化の効果は、所望形態(即ち浸透圧ポンプ、より好ましくは注射もしくは注入、又は他の経口、肺もしくは他の送達経路用剤形)で誘導体を投与し、その効果を判定することにより確認することができる。
【0061】
適切な水溶性ポリマーの非限定的な例としては、ポリエチレングリコール(PEG)、エチレングリコール/プロピレングリコールのコポリマー、モノメトキシポリエチレングリコール、カルボキシメチルセルロース、デキストラン、ポリビニルアルコール、ポリビニルピロリドン、ポリ−1,3−ジオキソラン、ポリ−1,3,6−トリオキサン、エチレン/無水マレイン酸コポリマー、ポリアミノ酸(ホモポリマー又はランダムポリマー)、ポリ(n−ビニルピロリドン)ポリエチレングリコール、プロピレングリコールホモポリマー、ポリプロピレンオキシド/エチレンオキシドコポリマー、ポリオキシエチル化ポリオール(例えばグリセロール)、ポリエチレングリコールプロピオンアルデヒド及びその混合物が挙げられる。本明細書で使用するポリエチレングリコールとは、モノ(C1〜C10)アルコキシ又はアリールオキシポリエチレングリコールのように、他のタンパク質を誘導体化するために使用されているPEG形態の任意のものを意味する。ポリエチレングリコールプロピオンアルデヒドはその水中安定性により製造が有利であると思われる。ポリマーは任意の分子量でよく、枝分かれしていてもいなくてもよい。
【0062】
本発明は特に、少なくとも1個のPEG分子に結合した截形GDNFタンパク質を含む截形GDNFタンパク質産物に関する。別の側面では、本発明はアシル又はアルキル結合を介して少なくとも1個のPEG分子に結合した截形GDNFタンパク質に関する。
【0063】
ペジル化は当該技術分野で公知のペジル化反応の任意のものにより実施することができる。例えば、Focus on Growth Factors 3(2):4−10(1992); EP第0154316号;EP第0401384号;及びMalikら,Exp.Hematol.20:1028−1035(1992)(塩化トレシルを使用したGM−CSFのペジル化の報告)参照。ペジル化は反応性水溶性ポリマーとのアシル化反応又はアルキル化反応を介して実施するのが好ましい。これらの好ましい誘導体化手段については以下に詳述する。アシル化反応では、単一反応性エステル基をもつポリマーを選択するのが好ましい。還元アルキル化反応では、単一反応性アルデヒド基をもつポリマーを選択するのが好ましい。更に、単一反応基(例えばアシル化には活性エステル、アルキル化にはアルデヒド)をもつように選択ポリマーを修飾し、重合度を調節できるようにしてもよい。天然グリコシル残基は通常は哺乳動物組換え発現系により簡便に製造されるので、一般に、水溶性ポリマーは天然グリコシル残基からは選択しない。
【0064】
アシル化
本発明では、アシル化によるペジル化は一般にポリエチレングリコールの活性エステル誘導体を截形GDNFタンパク質と反応させることにより行われる。ペジル化工程を実施するためには任意の公知又は今後発見される反応性PEG分子を使用することができる。好ましい活性化PEGエステルはN−ヒドロキシスクシンイミド(「NHS」)にエステル化したPEGである。本明細書で使用する「アシル化」とは、非限定的な例として截形GDNFタンパク質とPEG:アミド、カルバメート、ウレタン等の水溶性ポリマーの結合を意味する。Bioconjugate Chem.5:133−140(1994)参照。反応条件はペジル化技術で公知の条件又は今後開発される条件の任意のものから選択することができるが、修飾しようとする截形GDNFタンパク質を不活化するような温度、溶媒及びpHレベル等の反応条件への暴露は回避又は制限すべきである。
【0065】
アシル化によるペジル化の結果、一般にリジンε−アミノ基がアシル結合基を介してペジル化されたポリペジル化截形GDNFタンパク質が得られる。アミドによる結合が好ましい。また、得られる生成物が実質的に単独で(例えば≧95%)モノ、ジ又はトリペジル化されていることも好ましい。他方、使用する特定反応条件に依存する量でペジル化度の高い結合物が若干形成される場合もある。所望により、特に透析、塩析、限外濾過、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー及び電気泳動等の標準精製技術により混合物から高純度ペジル化結合体を製造することもできる。
【0066】
アルキル化
本発明では、アルキル化によるペジル化は一般に還元剤の存在下でPEGの末端アルデヒド誘導体を截形GDNFタンパク質と反応させることにより実施される。アルキル化によるペジル化もポリペジル化截形GDNFタンパク質を生成することができる。更に、実質的にタンパク質のN末端のα−アミノ基のみでペジル化を助長するように反応条件を操作することもできる(即ちモノペジル化種)。モノペジル化又はポリペジル化のいずれの場合もPEG基は−CH2−NH基を介してタンパク質に結合しているのが好ましい。特に−CH2−基に関して、この型の結合を本明細書では「アルキル」結合と呼ぶ。
【0067】
特定タンパク質で誘導体化に利用可能な種々の型の第1アミノ基(N末端に対するリジン)の反応性の差を利用する還元アルキル化により、選択的N末端化学修飾を実施することができる。適当な反応条件下では、カルボニル基含有ポリマーによりN末端でタンパク質の実質的に選択的な誘導体化が達せられる。例えば、リジン残基のε−アミノ基とタンパク質のN末端残基のα−アミノ基のpKa差を利用できるようなpHで反応を実施することにより、タンパク質を選択的にN末端ペジル化することができる。このような選択的誘導体化により、水溶性ポリマーとタンパク質の結合が制御され、ポリマーとの結合は主にタンパク質のN末端で行われ、リジン側鎖アミノ基等の他の反応基の有意修飾は生じない。還元アルキル化を使用する場合、水溶性ポリマーはタンパク質との結合に関わる反応性アルデヒドをただ1つだけもつことが好ましい。例えば、単一反応性アルデヒドを含むポリエチレングリコールプロピオンアルデヒドを使用する。
【0068】
本発明は、PEG基がアシル又はアルキル基を介して結合したペジル化截形GDNFタンパク質を含む。上述のように、このような截形GDNFタンパク質産物はモノペジル化物でもポリペジル化物(例えば2〜6個、好ましくは2〜5個のPEG基を含む)でもよい。PEG基は一般にアミノ酸のα又はε−アミノ基でタンパク質に結合しているが、適切な反応条件下でPEG基に結合するように十分反応性の基であるならば、タンパク質の任意の反応基にPEG基を結合できるとも考えられる。例えば、遊離アミノ基又はカルボキシル基等の反応基を介してポリエチレングリコールをタンパク質に共有結合させてもよい。反応基は活性化PEG分子を結合可能な基である。遊離アミノ基をもつアミノ酸残基としては、リジン残基とN末端アミノ酸残基が挙げられる。遊離カルボキシル基をもつアミノ酸残基としては、アスパラギン酸残基、グルタミン酸残基及びC末端アミノ酸残基が挙げられる。スルフヒドリル基もPEG分子を結合する反応基として使用することができる。治療目的では、例えばN末端又はリジン基等のアミノ基に結合するのが一般には好ましい。レセプター結合が所望される場合にはレセプター結合に重要な残基に結合するのは避けるべきである。
【0069】
1側面では、本発明はポリマー分子が実質的に単独位置のみ(即ち≧95%)に結合しているモノポリマー/截形GDNFタンパク質結合体の実質的に均一な調製物を提供する。より詳細には、PEGを使用する場合には、本発明は場合により抗原結合基を欠失し、截形GDNFタンパク質と直接結合したPEG分子をもつペジル化截形GDNFタンパク質も提供する。
【0070】
更に、グリコシル化、非グリコシル化又は脱グリコシル化截形GDNFタンパク質を使用して誘導体を製造することもできる。一般には非グリコシル化截形GDNFタンパク質を使用する。例えば、原核生物で発現させた[Arg32−Ile134]截形GDNFタンパク質を化学誘導体化し、アシル又はアルキル基を介して結合した1個又は複数(例えば2〜4個)のPEG分子を含むようにすることができる。
【0071】
一般に、化学誘導体化は生物学的に活性な物質を活性化ポリマー分子と反応させるために使用するのに適した任意の条件下で実施することができる。ペジル化截形GDNFタンパク質の製造方法は一般に、(a)截形GDNFタンパク質が1個以上のPEG基と結合するような条件下で截形GDNFタンパク質をポリエチレングリコール(例えばPEGの反応性エステル又はアルデヒド誘導体)と反応させる段階と、(b)反応物を得る段階を含む。一般に、アシル化反応に最適な反応条件は公知パラメーターと所望の結果に基づいて個々に決定される。例えば、PEG対タンパク質の比が大きいほど、ポリペジル化物の百分率が高い。(過剰の未反応タンパク質又はポリマーが存在しないという意味での反応効率に関する)最適比は所望の誘導体化度(例えばモノ、ジ、トリ等)、選択するポリマーの分子量、ポリマーが枝分かれしているか否か、使用する反応条件等の因子により決定することができる。
【0072】
モノポリマー/截形GDNFタンパク質結合体の実質的に均一なポピュレーションを製造するための還元アルキル化は、一般に(a)截形GDNFタンパク質のアミノ末端でα−アミノ基の選択的修飾を可能にするのに適したpHで還元条件下に截形GDNFタンパク質を反応性PEG分子と反応させる段階と、(b)反応物を得る段階を含む。
【0073】
モノポリマー/截形GDNFタンパク質結合体の実質的に均一なポピュレーションでは、還元アルキル化反応条件は截形GDNFタンパク質のN末端に水溶性ポリマー部分を選択的に結合できるような条件である。このような反応条件は、一般にリジンアミノ基とN末端のαアミノ基の間にpKa差を提供する(pKaはアミノ基の50%がプロトン化され、50%がプロトン化されないpHである)。pHは使用するポリマーとタンパク質の比にも影響を与える。一般に、pHが低いほど、高いポリマー対タンパク質比が所望される(即ちN末端αアミノ基が低反応性であるほど、最適条件に達するために多量のポリマーが必要となる)。pHが高い場合には、ポリマー対タンパク質比を大きくする必要はない(即ち高反応性基を利用できるので、必要なポリマー分子数は少なくて済む)。本発明の目的では、pHは一般に3〜9、好ましくは3〜6の範囲である。
【0074】
ポリマーの分子量も考慮すべきである。一般に、ポリマーの分子量が大きいほど、タンパク質に結合できるポリマー分子の数は少ない。また、これらのパラメーターを最適化する際にはポリマーの分枝度も考慮すべきである。一般に、分子量が大きいほど(又は分枝度が高いほど)ポリマー対タンパク質比は高い。一般に、本発明で意図するペジル化反応では、好ましい平均分子量は約2kDa〜約100kDaである(「約」なる用語はポリエチレングリコールの調製物中に指定分子量よりも高い分子や低い分子があることを示す)。好ましい平均分子量は約5kDa〜約50kDa、特に好ましくは約12kDa〜約25kDaである。水溶性ポリマーと截形GDNFタンパク質の比は一般に1:1〜100:1、好ましくは(ポリペジル化では)1:1〜20:1、及び(モノペジル化では)1:1〜5:1である。
【0075】
上記条件を使用すると、還元アルキル化はアミノ末端にα−アミノ基をもつ任意の截形GDNFタンパク質にポリマーを選択的に結合することができ、モノポリマー/截形GDNFタンパク質結合体の実質的に均一な調製物を提供する。本明細書で使用する「モノポリマー/截形GDNFタンパク質結合体」なる用語は截形GDNFタンパク質に結合した単一ポリマー分子を含む誘導体を意味する。モノポリマー/截形GDNFタンパク質結合体は、リジンアミノ側鎖でなくN末端に配置されたポリマー分子をもつことが好ましい。調製物は好ましくは>90%モノポリマー/截形GDNFタンパク質結合体、より好ましくは>95%モノポリマー/截形GDNFタンパク質結合体であり、検出可能なタンパク質の残余は未反応である(即ちポリマー部分をもたないタンパク質)。
【0076】
還元アルキル化では、還元剤は水溶液中で安定であるべきであり、初期還元アルキル化プロセスで形成されるシッフ塩基のみを還元できることが好ましい。還元剤の例はホウ水素化ナトリウム、シアノホウ水素化ナトリウム、ジメチルアミンボラン、トリメチルアミンボラン及びピリジンボランから構成される群から選択することができる。特に好ましい還元剤はシアノホウ水素化ナトリウムである。溶媒、反応時間、温度等の他の反応パラメーターと生成物の精製手段は水溶性ポリマーによるタンパク質の誘導体化に関する一般に入手可能な情報に基づいて個々に決定することができる。
【0077】
アシル化及び/又はアルキル化方法によりポリマー/タンパク質結合体の混合物を製造するように選択してもよく、本発明は、混合物に配合するモノポリマー/タンパク質結合体の割合を選択できるという利点がある。従って、所望により様々な数(即ちジ、トリ、テトラ等)のポリマー分子を結合したタンパク質の混合物を製造し、本発明の方法を使用して製造したモノポリマー/タンパク質結合体材料と組み合わせ、所定の割合のモノポリマー/タンパク質結合体を含む混合物を取得できる。
【0078】
截形GDNFタンパク質をコードするポリヌクレオチド
本発明は更に截形GDNFタンパク質をコードする新規ポリヌクレオチドを提供する。ハイブリダイゼーションプローブ又は増幅プライマーとして使用する場合、核酸配列は実質的に他の全核酸配列を含まない。組換えタンパク質発現で使用するには、融合タンパク質が所望される場合を除き、核酸配列は一般に他のタンパク質をコードする核酸配列を実質的に含まない。本明細書の記載に基づき、汎用コドン表を使用すると、当業者は截形GDNFタンパク質のアミノ酸配列をコードする全核酸配列を容易に決定することができる。本発明で好ましい核酸配列としては、[Arg16−Ile134]、[Ser26−Ile134]、[Arg32−Ile134]及び[Lys37−Ile134]截形GDNFタンパク質をコードするポリヌクレオチドが挙げられる。種々のポリヌクレオチドの例は図5、6及び7に示すものや、截形GDNFタンパク質をコードする図1、3及び4の部分である。截形GDNFタンパク質をコードする新規ポリヌクレオチドが人工又は天然を問わず変異体截形GDNFタンパク質をコードする核酸配列を含むことも当業者に理解されよう。
【0079】
以下の記載に従って組換え発現技術を実施すると、これらのポリヌクレオチドを製造し、種々の截形GDNFタンパク質を発現させることができる。例えば、截形GDNFタンパク質をコードする核酸配列を適当なベクターに挿入することにより、当業者は大量の所望のヌクレオチド配列を容易に製造することができる。その後、この配列を使用して検出プローブ又は増幅プライマーを作製することができる。あるいは、截形GDNFタンパク質をコードするポリヌクレオチドを発現ベクターに挿入することもできる。発現ベクターを適当な宿主に導入することにより、所望の截形GDNFタンパク質を大量に製造することができる。
【0080】
以下に詳述するように、核酸配列の増殖及び/又は截形GDNFタンパク質の製造には多数の宿主/ベクター系を利用できる。非限定的な例としては、プラスミド、ウイルス及び挿入ベクターと、原核及び真核宿主が挙げられる。当業者は異種DNAを増殖又は発現することが可能な宿主/ベクター系を改変して本発明の配列を産生又は発現させることができる。
【0081】
このような組換え技術により、本発明の截形GDNFタンパク質は商用量で容易に製造される。更に、本明細書の開示に鑑み、新規核酸配列は図面に具体的に示す截形GDNFタンパク質をコードする縮重核酸配列、このような截形GDNFタンパク質の変異体、及び好ましくはストリンジェントハイブリダイゼーション条件下でこれらの核酸配列の相補体とハイブリダイズする核酸配列(Maniatisら,Molecular Cloning (A Laboratory Manual); Cold Spring Harbor Laboratory,387〜389頁,1982参照)を含むことが当業者に理解されよう。ストリンジェントハイブリダイゼーション条件の例は、62〜67℃で4×SSC中でハイブリダイゼーション後、62〜67℃で0.1×SSCで約1時間の洗浄である。他のストリンジェントハイブリダイゼーション条件は40〜45℃で45〜55%ホルムアミド、4×SSC中でのハイブリダイゼーションである。リラックスハイブリダイゼーション条件下で截形GDNFタンパク質の相補的配列とハイブリダイズし、本発明の截形GDNFタンパク質をコードするDNA配列も本発明に含まれる。このようなリラックスストリンジェンシーハイブリダイゼーション条件の例は45〜55℃で4×SSC又は40〜45℃で30〜40%ホルムアミドを用いたハイブリダイゼーションである。
【0082】
本発明は、截形GDNFタンパク質をコードするDNA配列と共にベクターDNAを含む組換えDNA構築物も提供する。このようなDNA構築物において、(シグナルペプチドの有無に拘わらず)截形GDNFタンパク質をコードする核酸配列は選択された宿主で截形GDNFタンパク質の複製及び/又は発現を誘導することが可能な適切な発現制御又は調節配列と作動的に結合している。
【0083】
截形GDNFタンパク質の組換え発現
截形GDNFをコードするポリヌクレオチドの作製
截形GDNFをコードする核酸配列又は成熟GDNF出発材料は、非限定的な例として化学的合成、cDNA又はゲノムライブラリースクリーニング、発現ライブラリースクリーニング及び/又はcDNAのPCR増幅等の種々の方法で容易に得ることができる。これらの方法及びこのような核酸配列を単離するために有用な他の方法は、例えばSambrookら(Molecular Cloning: A Laboratory Manual,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY,1989)、Ausubelら編(Current Protocols in Molecular Biology,Current Protocols Press,1994)及びBergerとKimmel(Methods in Enzymology: Guide to Molecular Cloning Techniques,vol.152,Academic Press,Inc.,San Diego,CA,1987)により記載されている。GDNFをコードする好ましい核酸配列は哺乳動物配列である。
【0084】
截形GDNFタンパク質をコードする核酸配列の化学的合成は、例えばEngelsら(Angew.Chem.Intl.Ed.,28:716−734,1989)により記載されているような当該技術分野で周知の方法を使用しても実施することができる。これらの方法のうちでは、特にホスホトリエステル、ホスホラミダイト及びH−ホスホネートによる核酸配列合成法が挙げられる。截形GDNFタンパク質をコードする核酸配列は数百塩基対(bp)又はヌクレオチド長である。例えば約100ヌクレオチド長を上回る大きい核酸配列を数個のフラグメントとして合成することができる。その後、フラグメントを相互に連結すると、截形GDNFタンパク質をコードする核酸配列を形成することができる。好ましい方法の1例は、標準ホスホラミダイト化学を使用したポリマーによる合成である。
【0085】
あるいは、適当なcDNAライブラリー(即ちタンパク質を発現すると考えられる1種以上の組織材料から調製したライブラリー)又はゲノムライブラリー(全ゲノムDNAから調製したライブラリー)をスクリーニングすることにより適切な核酸配列を得ることもできる。cDNAライブラリーの材料は一般には妥当な量のGDNFを発現すると考えられる任意の種からの組織である。ゲノムライブラリーの材料はGDNF又はGDNF同族体をコードする遺伝子をもつと考えられる任意の哺乳動物又は他の種からの任意の組織であり得る。ライブラリーに存在するGDNF又はGDNF同族体cDNA又は遺伝子と選択的にハイブリダイズする1種以上のプローブ(クローニングしようとするGDNF又はGDNF同族体cDNA又は遺伝子に対して許容可能な相同レベルをもつオリゴヌクレオチド、cDNA又はゲノムDNAフラグメント)を使用し、GDNF cDNA/遺伝子の存在についてライブラリーをスクリーニングすることができる。このようなライブラリースクリーニングに一般に使用するプローブは通常はライブラリーの調製に用いた種と同一又は類似の種からのGDNF DNA配列の小領域をコードする。あるいは、プローブは本明細書に記載するような縮重配列でもよい。
【0086】
ライブラリースクリーニングは一般に、プローブ又はプライマーに対して有意相同レベルをもつクローンとの結合を可能にし且つ非特異的結合を阻止するストリンジェンシー条件下でライブラリー中のクローンにオリゴヌクレオチドプローブ又はcDNAをアニールすることにより実施される。典型的なハイブリダイゼーション及び洗浄ストリンジェンシー条件はcDNA又はオリゴヌクレオチドプローブの寸法(即ちヌクレオチド長数)や、プローブが縮重配列であるか否かにも依存する。ハイブリダイゼーション溶液を設計するときはクローンの獲得確率(即ちcDNAライブラリーをスクリーニングするのか又はゲノムライブラリーか、cDNAライブラリーであるならば、目的cDNAが高レベルで存在する確率)も考慮する。
【0087】
DNAフラグメント(例えばcDNA)をプローブとして使用する場合には、典型的なハイブリダイゼーション条件は、上記Ausubelら編に記載されているような条件を含む。ハイブリダイゼーション後、プローブ寸法、クローンに対するプローブの予想される相同性、スクリーニングするライブラリーの種類、スクリーニングするクローン数等の数種の因子に依存して適当なストリンジェンシーでライブラリーを含むブロットを洗浄する。(通常はイオン強度が低く、比較的高温で使用される)ストリンジェント洗浄溶液の例を以下に挙げる。このようなストリンジェント洗浄液の1例は55〜65℃で0.015M NaCl,0.005Mクエン酸ナトリウム及び0.1%SDSである。別のストリンジェント緩衝液の例は約40〜50℃で1mM Na2EDTA,40mM NaHPO4,pH7.2及び1%SDSである。他のストリンジェント洗浄液の例は約50〜65℃で0.2×SSC及び0.1%SDSである。
【0088】
更に、オリゴヌクレオチドプローブを使用してcDNA又はゲノムライブラリーをスクリーニングする場合に典型的なストリンジェントな洗浄条件のプロトコルも存在する。例えば、第1のプロトコルではプローブの長さに応じて約35〜62℃の温度で0.05%ピロリン酸ナトリウムを含む6×SSCを使用する。例えば、14塩基プローブは35〜40℃、17塩基プローブは45〜50℃、20塩基プローブは52〜57℃、23塩基プローブは57〜63℃で洗浄する。非特異的なバックグラウンド結合が高いと思われる場合には温度を2〜3℃上げてもよい。第2のプロトコルではテトラメチルアンモニウムクロリド(TMAC)を洗浄に使用する。このようなストリンジェント洗浄溶液の1例は3M TMAC,50mM Tris−HCl,pH8.0及び0.2%SDSである。
【0089】
GDNFタンパク質をコードする核酸配列を得るために利用可能な別の方法はポリメラーゼ連鎖反応(PCR)である。この方法では、GDNFを発現する組織からポリ(A)+RNA又は全RNAを抽出する。次に逆転写酵素を使用してRNAからcDNAを調製する。次に、一般にはGDNF cDNAの2つの別個の領域に相補的な2種のプライマー(オリゴヌクレオチド)をTaqポリメラーゼ等のポリメラーゼと共にcDNAに加えると、ポリメラーゼは2種のプライマーの間のcDNA領域を増幅する。
【0090】
所望の截形GDNFタンパク質をコードする核酸配列を作製するのに選択した方法がオリゴヌクレオチドプライマー又はプローブの使用を必要とする場合(例えばPCR、cDNA又はゲノムライブラリースクリーニング)には、プローブ又はプライマーとして選択するオリゴヌクレオチド配列は、ライブラリースクリーニング又はPCR増幅中に生じる非特異的結合の量を最小限にするように十分な長さと十分明確な配列をもつべきである。プローブ又はプライマーの実際の配列は、通常は同一遺伝子又は別の生物からの類似遺伝子に由来する保存又は高相同性配列又は領域に基づく。場合により、プローブ又はプライマーは完全又は部分縮重配列でもよく、即ち異なるコドンを使用しながら全て同一のアミノ酸配列をコードするプローブ/プライマー混合物でもよい。縮重プローブを作製する代わりに、種によって異なるコドン位置の一部又は全部にイノシンを配置してもよい。オリゴヌクレオチドプローブ又はプライマーは上記のようなDNAの化学合成法により作製できる。
【0091】
GDNFをコードするこれらの核酸配列に基づく截形GDNFタンパク質とその突然変異体又は変異体配列も本発明の範囲に含まれる。上述のように、突然変異体又は変異体配列は野生型配列に比較して1個以上のヌクレオチド置換、欠失及び/又は挿入を含み、その結果として野生型アミノ酸配列に比較してアミノ酸配列変異を発現する配列である。場合によっては、自然対立遺伝子変異の存在により、天然GDNFアミノ酸突然変異体又は変異体が存在することもある。このような天然突然変異体又は変異体に基づく截形GDNFタンパク質も本発明の範囲に含まれる。合成突然変異体配列の作製は当該技術分野で周知であり、例えばWellsら(Gene,34:315,1985)
やSambrookら,前出に記載されている。
【0092】
ベクター
截形GDNFタンパク質をコードするcDNA又はゲノム
DNAをベクターに挿入して更にクローニング(DNAの増幅)又は発現させる。利用可能なベクターは市販されており、又はベクターを特別に構築してもよい。適当なベクターの選択又は構築は、1)DNA増幅に使用するのか、DNA発現に使用するのか、2)ベクターに挿入するDNAの寸法、及び3)ベクターで形質転換する宿主細胞(例えば哺乳動物、昆虫、酵母、真菌、植物又は細菌細胞)に依存する。各ベクターはその機能(DNAの増幅又はDNAの発現)と目的宿主細胞との適合性に依存して種々の成分を含む。ベクター成分の非限定的な例としては一般に、シグナル配列、複製起点、1個以上の選択又はマーカー遺伝子、エンハンサーエレメント、プロモーター、転写終結配列等の1種以上が挙げられる。これらの成分は天然材料から得てもよいし、公知手順で合成してもよい。本発明のベクターは選択された宿主細胞による截形GDNFタンパク質の発現を誘導、制御又は他の方法で司ることが可能な下記発現制御又は調節配列の1種以上に作動的に連結した目的截形GDNFタンパク質をコードする核酸配列を含む。
【0093】
シグナル配列
シグナル配列はベクターの成分でもよいし、ベクターに挿入するGDNF DNAの一部でもよい。天然GDNF DNAは、タンパク質の翻訳後プロセシング中に切断されて成熟GDNFタンパク質を形成するタンパク質のアミノ末端のシグナル配列をコードする。天然シグナル配列及び他のプレ−プロ配列をもつ截形GDNFポリヌクレオチドと、天然シグナル配列を欠失し、異種シグナル配列で置換した截形GDNFポリヌクレオチドが本発明の範囲に含まれる。選択する異種シグナル配列は宿主細胞により認識及びプロセシングされる配列、即ちシグナルペプチダーゼにより切断される配列とすべきである。天然GDNFシグナル配列を認識及びプロセシングしない原核宿主細胞では、シグナル配列を例えばアルカリホスファターゼ、ペニシリナーゼ又は熱安定性エントロトキシンIIリーダーの群から選択される原核シグナル配列で置換する。酵母分泌では、天然GDNFシグナル配列を酵母インベルターゼ、α因子又は酸ホスファターゼリーダーで置換すればよい。哺乳動物細胞発現では、天然シグナル配列でよいが、他の哺乳動物シグナル配列も利用できる。
【0094】
複製起点
発現及びクローニングベクターは一般に、1種以上の選択された宿主細胞でベクターを複製させる核酸配列を含む。クローニングベクターでは、この配列は一般にはベクターを宿主染色体DNAから独立して複製させる配列であり、複製起点又は自律複製配列を含む。このような配列は種々の細菌、酵母及びウイルスで周知である。プラスミドpBR322からの複製起点は殆どのグラム陰性菌に利用でき、哺乳動物細胞でのクローニングベクターには種々の起点(例えばSV40、ポリオーマ、アデノウイルス、VSV又はBPV)が有用である。一般に、哺乳動物発現ベクターには複製起点成分は不要である(例えばSV40起点を使用することが多いが、これは単に初期プロモーターを含むという理由に過ぎない)。
【0095】
選択遺伝子
発現及びクローニングベクターは一般に選択遺伝子を含む。この遺伝子は選択培地で培養する場合に形質転換宿主細胞の生存又は増殖に必要な「マーカー」タンパク質をコードする。ベクターで形質転換されなかった宿主細胞は選択遺伝子を含まないので、培地中で生存しない。典型的な選択遺伝子は、(a)抗生物質又は他の毒素(例えばアンピシリン、ネオマイシン、メトトレキセート又はテトラサイクリン)に対する耐性を与え、(b)栄養要求欠損を補充し、又は(c)培地から得られない必須栄養を供給するタンパク質をコードする。
【0096】
発現される遺伝子を増幅するために他の選択遺伝子を使用してもよい。増幅は、増殖に必須のタンパク質の産生に大いに必要とされる遺伝子が組換え細胞の連続世代の染色体内でタンデムに反復するプロセスである。哺乳動物細胞に利用可能な選択マーカーの例としては、ジヒドロ葉酸レダクターゼ(DHFR)とチミジンキナーゼが挙げられる。形質転換哺乳動物細胞を選択圧下におき、形質転換細胞のみを特別に改変してベクター中に存在するマーカーにより生存できるようにする。選択圧は、培地中の選択物質の濃度が連続的に変化して選択遺伝子と截形GDNFをコードするDNAの両者を増幅させるような条件下で形質転換細胞を培養することにより加えられる。その結果、増幅DNAから多量の截形GDNFが合成される。
【0097】
例えば、DHFRの競合的アンタゴニストであるメトトレキセートを含む培地で全形質転換細胞を培養することにより、
DHFR選択遺伝子で形質転換した細胞をまず同定する。野生型DHFRを使用する場合に適当な宿主細胞は、DHFR活性を欠損するチャイニーズハムスター卵巣細胞系である(例えばUrlaubとChasin,Proc.Natl.Acad.Sci.,USA 77(7):4216−4220(1980)参照)。次に、形質転換細胞を高レベルのメトトレキセートに暴露する。その結果、DHFR遺伝子の多重コピーが合成され、それと同時に発現ベクター中に存在する他のDNA(例えば截形GDNFタンパク質をコードするDNA)の多重コピーも合成される。
【0098】
プロモーター
本発明の発現及びクローニングベクターは一般に、宿主生物により認識され、截形GDNFタンパク質をコードする核酸配列に作動的に連結されたプロモーターを含む。プロモーターは構造遺伝子の開始コドンの上流(5’)に配置された非翻訳配列(一般に約100〜1000bp以内)であり、截形GDNFをコードする配列等の特定核酸配列の転写及び翻訳を制御する。プロモーターは従来、誘導的プロモーターと構成的プロモーターの2種類に分類されている。誘導的プロモーターは栄養又は温度変化の有無等の培養条件の何らかの変化に応答してその制御下のDNAから高レベルの転写を開始する。種々の潜在宿主細胞により認識される多数のプロモーターが周知である。これらのプロモーターは、制限酵素消化によりソースDNAからプロモーターを除去し、所望のプロモーター配列をベクターに挿入することにより、截形GDNFをコードするDNAに作動的に連結される。天然GDNFプロモーター配列を使用して截形GDNF DNAの直接増幅及び/又は発現を誘導することもできる。他方、天然プロモーターよりも多量の発現タンパク質の転写と高収率を可能にし、使用に選択した宿主細胞系に適合可能である場合には、異種プロモーターが好ましい。
【0099】
原核宿主で使用するのに利用可能なプロモーターとしては、β−ラクタマーゼ及びラクトースプロモーター系、アルカリホスファターゼ、トリプトファン(trp)プロモーター系、及びハイブリッドプロモーター(例えばtacプロモーター)が挙げられる。他の公知細菌プロモーターも利用できる。これらのプロモーターのヌクレオチド配列は公表されているので、当業者は任意の必要な制限部位を提供するのに必要なリンカー又はアダプターを使用して所望のDNA配列に連結することができる。
【0100】
酵母宿主で使用するのに利用可能なプロモーター配列も当該技術分野で周知である。酵母エンハンサーを酵母プロモーターと併用すると有利である。哺乳動物細胞で使用するのに適したプロモーターは周知であり、例えばポリオーマウイルス、鶏痘ウイルス、アデノウイルス(例えばアデノウイルス2)、ウシパピローマウイルス、トリサルコーマウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルス及び最も好ましい例としてサルウイルス40(SV40)等のウイルスのゲノムから得られるものが挙げられる。他の利用可能な哺乳動物プロモーターとしては、異種哺乳動物プロモーター(例えば熱ショックプロモーター及びアクチンプロモーター)が挙げられる。CHO細胞でGDNFタンパク質を産生させるのに本発明で使用したプロモーターはSRαである。Takebeら,Mol.Cell,Biol.8(1):466−472(1988)参照。適切な発現ベクターの1例は、追って詳述するpDSRα2である。
【0101】
エンハンサーエレメント
本発明の截形GDNFタンパク質をコードするDNA配列の高等真核生物による転写を増加するために、ベクターにエンハンサー配列を挿入してもよい。エンハンサーは通常は約10〜300bp長のDNAのシス作用エレメントであり、その転写を増加するようにプロモーターに作用する。エンハンサーは向きや位置から比較的独立している。エンハンサーは転写単位の5’及び3’に検出されている。哺乳動物遺伝子から入手可能な数種のエンハンサー配列が知られている(例えばグロブリン、エラスターゼ、アルブミン、α−フェトプロテイン及びインスリン)。しかし、一般にはウイルスからのエンハンサーを使用する。SV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、ポリオーマエンハンサー及びアデノウイルスエンハンサーが真核プロモーターの活性化を亢進するエレメントの例である。エンハンサーは截形GDNF DNAの5’又は3’位でベクターにスプライスしてもよいが、一般にはプロモーターから5’位に配置する。
【0102】
転写終結
真核宿主細胞(酵母、真菌、昆虫、植物、動物、ヒト又は他の多細胞生物からの有核細胞)で使用する発現ベクターは更に、転写の終結とmRNAの安定化に必要な配列も含む。このような配列は通常は真核DNA又はcDNAの5’及び場合により3’非翻訳領域から得られる。これらの領域は截形GDNFをコードするmRNAの非翻訳部分でポリアデニル化フラグメントとして転写されたヌクレオチドセグメントを含む。
【0103】
所望の截形GDNFコーディング配列と共に上記成分の1種以上を含む適切なベクターの構築は標準連結技術により実施される。単離プラスミド又はDNAフラグメントを切断し、順化し、必要なプラスミドを生成するために所望の順序に再連結する。正しい配列が構築されたことを確認するためには、連結混合物を使用して大腸菌を形質転換し、形質転換に成功した細胞を上記のようなアンピシリン又はテトラサイクリン耐性等の公知技術により選択すればよい。次いで形質転換細胞からプラスミドを調製し、制限エンドヌクレアーゼにより分析し、及び/又は配列決定し、所望の構築物の存在を確認する。
【0104】
哺乳動物細胞で截形GDNFをコードするDNAの一過性発現を可能にするベクターを使用してもよい。一般に、一過性発現は宿主細胞で効率的に複製することが可能な発現ベクターを使用し、宿主細胞に発現ベクターの多数のコピーを蓄積させ、その結果、発現ベクターによりコードされる所望のタンパク質を高レベルで合成させる。適切な発現ベクターと宿主細胞を含む一過性発現系は、クローン化DNAによりコードされるタンパク質の簡便な正の同定と、所望の生物学的又は生理的性質に関する前記タンパク質の迅速なスクリーニングを可能にする。従って、一過性発現系はタンパク質の変異体を同定するのに特に有用である。
【0105】
宿主細胞の選択及び形質転換
組換え截形GDNFタンパク質を発現させるのに使用する核酸配列で形質転換した宿主細胞(例えば細菌、哺乳動物、昆虫、酵母又は植物細胞)も本発明により提供される。形質転換した宿主細胞を、核酸配列の発現を可能にする適当な条件下で培養する。適切な宿主細胞と形質転換、培養、増幅、スクリーニング並びに生成物産生及び精製方法は当該技術分野で周知である。例えばGethingとSambrook,Nature 293:620−625(1981); Kaufmanら,Mol.Cell.Biol.,5(7):1750−1759(1985); 又はHoweleyら,米国特許第4,419,446号参照。截形GDNFは、成熟GDNFの発現に関するLinらの記載(米国特許出願第07/855,413号、国際出願第PCT/US92/07888号、WO93/06116)に従って大腸菌で発現させることができる。他の典型的な材料及び方法については以下に詳述する。形質転換した宿主細胞を適切な培地で培養した後、発現された因子を場合により当業者に公知の適当な手段により培地から(又は細胞内で発現される場合には細胞から)回収、単離及び精製する。
【0106】
本発明のベクターをクローニング及び発現させるのに適した宿主細胞は上述のような原核、酵母又は高級真核細胞である。原核宿主細胞の非限定的な例としては、グラム陰性又はグラム陽性生物等の真正細菌が挙げられ、例えば大腸菌、バシラス属(例えば枯草菌)、シュードモナス種(例えばP.aeruginosa)、Salmonella typhimurium又はSerratia marcescansである。あるいは、PCR又は他の核酸ポリメラーゼ反応等のin vitroクローニング法も利用できる。
【0107】
原核宿主細胞に加え、糸状菌類や酵母等の真核微生物も截形GDNFタンパク質の発現宿主として利用できる。下級真核宿主微生物のうちではSaccharomyces cerevisiae(俗名パン酵母)が最もよく使用されるが、多数の他の属、種及び株も周知であり、一般に入手可能である。
【0108】
グリコシル化截形GDNFタンパク質の発現に適した宿主細胞は多細胞生物から得られる。このような宿主細胞は複雑なプロセシング及びグリコシル化活性が可能である。脊椎動物細胞であるか、植物及び昆虫細胞を含む無脊椎動物細胞であるかを問わず、原則として任意の高級真核細胞培養物を使用できる。脊椎動物細胞の培養(組織培養)増殖は周知方法であるので、一般には脊椎動物細胞を使用する。有用な哺乳動物宿主細胞系の非限定的な例としては、SV40により形質転換したサル腎CV1系(COS−7)、ヒト胎児腎系(293又は懸濁培養増殖のためにサブクローニングした293細胞)、乳児ハムスター腎細胞、及びチャイニーズハムスター卵巣細胞が挙げられる。他の利用可能な哺乳動物細胞系の非限定的な例としては、HeLa、マウスL−929細胞、Swiss、Balb−cもしくはNIHマウスに由来する3T3系、BHK又はHaKハムスター細胞系が挙げられる。
【0109】
本発明に利用可能な宿主細胞として細菌細胞も有用である。例えば、種々の大腸菌株(例えばHB101、DH5α、DH10及びMC1061)がバイオテクノロジー分野で宿主細胞として周知である。ストレプトミセス種等の種々の株も使用できる。截形GDNFタンパク質を製造するのに本発明で好適な宿主細胞は細菌細胞(例えば大腸菌)と哺乳動物細胞(例えばチャイニーズハムスター卵巣細胞、COS細胞等)である。
【0110】
宿主細胞を上記発現又はクローニングベクターでトランスフェクト、好ましくは形質転換し、慣用栄養培地で培養する。培地はプロモーターの誘導、形質転換細胞の選択又は所望の配列をコードする遺伝子の増幅に適するように改変してもよい。トランスフェクション及び形質転換は、利用する宿主細胞に適するように選択された当業者に周知の標準技術を使用して実施される。例えば、細胞壁をもたない哺乳動物細胞では、リン酸カルシウム沈殿法を使用できる。エレクトロポレーション、マイクロインジェクション及び他の公知技術も使用できる。
【0111】
宿主細胞の培養
本発明の截形GDNFタンパク質を製造するために使用する形質転換細胞は適切な培地で培養する。培地は必要に応じてホルモン及び/又は他の成長因子(例えばインスリン、トランスフェリン又は上皮成長因子)、塩類(例えば塩化ナトリウム、カルシウム、マグネシウム及びリン酸塩)、緩衝液(例えば
HEPES)、ヌクレオシド(例えばアデノシン及びチミジン)、抗生物質(例えばゲンタマイシン)、微量元素(通常はマイクロモル範囲の最終濃度で存在する無機化合物として定義される)、及びグルコース又は他のエネルギー源を補充してもよい。当業者に理解される通り、他の補充剤も適当な濃度で加えてもよい。選択した宿主細胞で使用するのに適した温度、pH等の培養条件も当業者に周知である。
【0112】
截形GDNFタンパク質は相同組換え、即ちGDNFをコードするDNAを既に含んでいる細胞に導入した制御エレメントを使用する組換え製造方法により製造することも可能である。相同組換えは、転写活性遺伝子に突然変異を誘導又は修正するように遺伝子をターゲティングするために本来開発された技術である(Kucherlapati,Prog.in Nucl.Acid Res.and Mol.Biol.36:301(1989))。基本技術は哺乳動物ゲノムの特定領域に特定突然変異を導入する方法(Thomasら,Cell.44:419−428,1986; ThomasとCapecchi,Cell.51:503−512,1987; Doetschmanら,Proc.Natl.Acad.Sci.85:8583−8587,1988)として又は欠損遺伝子内の特定突然変異を修正するため(Doetschmanら,Nature.330:576−578,1987)に開発された。典型的な相同組換え技術は、参考資料としてその開示内容を本明細書の一部とする米国特許第5,272,071号(EP第91903051号、EP公開第505500号、PCT/US90/07642、国際公開WO91/09955)に記載されている。
【0113】
相同組換えにより、ゲノムに挿入しようとするDNA配列をターゲティングDNAに結合することにより目的遺伝子の特定領域に導くことができる。ターゲティングDNAはゲノムDNAのある領域に相補的(相同)なDNAである。ゲノムの特定領域に相補的なターゲティングDNAの小断片をDNA複製プロセス中に親鎖と接触させる。細胞に挿入されたDNAは、共通相同領域を介して内因DNAの他の断片とハイブリダイズし、従って、これと組換えするという一般性質をもつ。突然変異又は異なるDNA配列を含むオリゴヌクレオチドにこの相補鎖を結合するならば、この相補鎖は組換えの結果として新しく合成された鎖にも組み込まれる。プルーフリーディング機能の結果として、新しいDNA配列を鋳型として使用することができる。従って、転写されたDNAはゲノムに組み込まれる。
【0114】
GDNFの核酸配列等の特定遺伝子の配列が分かっている場合には、遺伝子の選択領域に相補的なDNAの断片であるプレ−プロ配列又は発現制御配列を合成することができ、あるいは目的領域の境界となる特定認識部位で天然DNAを適当に制限消化するなどの方法でこのような配列が得られる。この断片は細胞に挿入するとターゲティング配列として機能し、ゲノムの内側でその相同領域とハイブリダイズする。DNA複製中にこのハイブリダイゼーションが生じるならば、このDNA断片とこれに結合した任意の付加的配列は岡崎フラグメントとして作用し、新しく合成されたDNA娘鎖にバックステッチされる。
【0115】
本発明では、GDNFタンパク質の発現に相互作用すると考えられるDNAの領域がターゲティングDNAのこれらの断片に結合している。例えば、所望の截形GDNFをコードする
DNAの転写に作用するに十分近傍の位置及び向きで所期宿主細胞のゲノムにプロモーター/エンハンサーエレメント、サプレッサー、又は外因転写調節エレメントを挿入する。制御エレメントは截形GDNFをコードせず、宿主細胞ゲノムに存在するDNAの一部を制御する。従って、截形GDNFタンパク質の発現は、截形GDNF遺伝子自体をコードするDNAのトランスフェクションによって達せられるのではなく、截形GDNFタンパク質の転写のための認識可能なシグナルを内因遺伝子配列に提供するDNA調節セグメントと結合したターゲティングDNA(目的の内因遺伝子に対して相同の領域を含む)の使用によって達せられる。
【0116】
本発明によると、相同組換え法を使用して通常は転写に不活性なGDNF遺伝子を含む細胞を改変し、GDNFを発現する細胞を作製することもできる。その後、GDNFタンパク質をプロセシングすると、截形GDNFタンパク質を形成することができる。
【0117】
截形GDNF医薬組成物
截形GDNFタンパク質産物医薬組成物は一般に、1種以上の医薬的及び生理的に許容可能な配合剤と混合した治療上有効な量の截形GDNFタンパク質産物を含む。利用可能な配合剤の非限定的な例としては、酸化防止剤、防腐剤、着色剤、矯味矯臭剤、希釈剤、乳化剤、懸濁剤、溶剤、充填剤、増量剤、緩衝液、送達ビヒクル、希釈液、賦形剤及び/又は医薬アジュバントが挙げられる。例えば、利用可能なビヒクルとしては注射用水、生理的食品水、又は人工脳髄液(CSF)が挙げられ、場合により、非経口投与用組成物に通常用いられる他の材料を補充してもよい。天然緩衝食品水又は血清アルブミンと混合した食品水も典型的ビヒクルである。
【0118】
ビヒクル中の主溶媒は水性でも非水性でもよい。更に、ビヒクルは製剤のpH、オスモル濃度、粘度、透明度、色、滅菌度、安定性、溶解速度又は臭いを変更又は維持するために他の医薬的に許容可能な賦形剤を含んでいてもよい。また、ビヒクルは截形GDNFタンパク質産物の安定性、溶解速度又は放出速度を変更又は維持するため、あるいは截形GDNFタンパク質産物の血液脳関門吸収又は通過を助長するために更に別の医薬的に許容可能な賦形剤を含んでいてもよい。このような賦形剤は単位剤形もしくは多剤形の非経口投与製剤又は移植ポンプからの連続もしくは周期的注入によるCSFへの直接注入用製剤を調剤するために通常慣用的に使用されている物質である。
【0119】
治療組成物を調剤したら、溶液、懸濁液、ゲル、エマルション、固体又は脱水もしくは凍結乾燥粉末として滅菌バイアルに保存することができる。このような製剤は即使用可能な形態で保存してもよいし、例えば投与前に再構成が必要な凍結乾燥形態で保存してもよい。
【0120】
最適医薬製剤は投与経路と所望の投与量に依存して当業者により決定される。例えば、参考資料としてその開示内容を本明細書の一部とするRemington’s Pharmaceutical Science,第18版(1990,Mack Publishing Co.,Easton,PA 18042)1435〜1712頁参照。組成物は更に、ポリ乳酸、ポリグルコール酸等のポリマー化合物の粒状調製物を含んでいてもよいし、リポソームに封入してもよい。ヒアルロン酸も使用でき、これは循環持続時間を促進する効果があると考えられる。このような組成物は本発明のタンパク質及び誘導体の物理的状態、安定性、in vivo放出速度、及びin vivoクリアランス速度を変更できる。
【0121】
非経口徐放製剤、吸入ミスト、経口活性製剤又は座薬等の他の有効な投与形態も考えられる。本発明の截形GDNFタンパク質産物医薬組成物は例えば脳髄内注射等の非経口投与用に調製される。このような非経口投与治療組成物は一般に医薬的に許容可能なビヒクル中に截形GDNFタンパク質産物を含む非経口的に許容可能なパイロジェンフリー水溶液の形態である。好ましいビヒクルの1例は生理的食塩水である。
【0122】
截形GDNFタンパク質産物を含む所定の製剤は経口投与することも考えられる。このように投与する截形GDNFタンパク質産物はカプセル封入することができ、固体剤形の配合に慣用的に使用されているキャリヤーの存在下又は不在下で調剤できる。カプセルは生体利用性が最大となり且つ全身循環前の分解が最小となる時点で胃腸管に製剤の活性部分を放出するように設計することができる。截形GDNFタンパク質産物の吸収を助長するために付加的な賦形剤を加えてもよい。希釈剤、矯味矯臭剤、低融点ろう、植物油、潤滑剤、懸濁剤、錠剤崩壊剤及び結合剤も使用してもよい。
【0123】
截形GDNFタンパク質産物の投与
截形GDNFタンパク質産物は皮下、筋肉内、静脈内、経肺、経皮、鞘内又は大脳内経路により非経口投与することができる。血液脳関門を通過しないタンパク質成長因子は大脳内に直接投与するか、又は関門間を輸送する他のエレメントと共に投与すればよい。截形GDNFタンパク質産物は脳髄内又は脳もしくは脊髄くも膜下腔に投与するのが好ましい。截形GDNFタンパク質産物は脳実質に直接大脳内投与してもよい。生分解性ポリマーマトリックスに埋封した神経栄養因子を含む徐放インプラントを脳に移植し、截形GDNFタンパク質産物を送達することもできる。截形GDNFタンパク質産物は、血液脳関門を通過するように化学的に修飾又はパッケージした形態で大脳外に投与してもよいし、截形GDNFタンパク質産物の関門通過を促進することが可能な1種以上の物質と併用投与してもよい。例えば、NGFとモノクローナル抗トランスフェリンレセプター抗体の結合体はトランスフェリンレセプターとの結合により脳に輸送されることが示されている。所望用量の截形GDNFタンパク質産物を投与するためには、毎日又はより低頻度で繰り返し注射してもよいし、一定又はプログラム可能な流量の移植ポンプから截形GDNFタンパク質産物を連続又は周期的に注入してもよい。投与頻度は調剤する截形GDNFタンパク質産物の薬物動態パラメーターと、投与経路によって異なる。
【0124】
投与方法に関係なく、比用量は一般に体重又は体表面積に従って計算される。脳疾患では、比用量は一般に患者の概算大脳重量に従って計算され、大脳重量も体重又は体表面積に基づいて推定できる。上記製剤の各々を利用する治療に適した用量を決定するために必要な更に精密な計算は、特に本明細書に開示する用量情報及びアッセイを参考にして当業者により日常的方法で実施される。用量が適当であるか否かは、適当な用量応答データと組み合わせて使用量を決定する確立アッセイを使用して確認することができる。特定症状の治療方法に用いる最終投与計画は、薬剤作用に影響する種々の因子(例えば患者の年齢、症状、体重、性別及び食事、感染の重篤度、投与時間並びに他の臨床因子)を考慮して主治医により決定される。
【0125】
本発明の截形GDNFタンパク質産物は更に、神経疾患の治療で単独又は他の成長因子と組み合わせて使用できる。例えば、截形GDNFタンパク質産物は所定形態の神経疾患の治療に神経成長因子と併用できる。更に、化学組成物を含む他の因子又は他の分子を截形GDNFタンパク質産物と併用してもよい。パーキンソン病の治療では、截形GDNFタンパク質産物を単独又はレボドパと併用投与することが考えられ、截形GDNFは内因ドーパミンの産生と高濃度のドーパミンのニューロン取り込みを強化すると思われる。
【0126】
上述のように、所定のニューロン細胞ポピュレーション又はある種の傷害もしくは疾病を治療するためには、付加的な神経栄養又はニューロン栄養因子が有用又は必要であるとも考えられる。截形GDNFと併用できる他の因子の非限定的な例としては、マイトジェン(例えばインスリン、インスリン様成長因子、上皮成長因子、血管作用成長因子、下垂体アデニル酸シクラーゼ活性化ポリペプチド、インターフェロン及びソマトスタチン)、神経栄養因子(例えば脳由来神経栄養因子、ニューロトロフィン−3、ニューロトロフィン−4/5、ニューロトロフィン−6、インスリン様成長因子、繊毛神経栄養因子、酸性及び塩基性フィブロブラスト成長因子、フィブロブラスト成長因子−5、トランスフォーミング成長因子−β、コカインアンフェタミン調節転写産物(CART)並びに成熟GDNF)、及び他の成長因子(例えば上皮成長因子、白血病阻害因子、インターロイキン、インターフェロン及びコロニー刺激因子)並びにこれらの因子の機能的等価物である分子及び材料が挙げられる。
【0127】
治療によっては截形GDNFタンパク質産物の連続投与又は持続送達が有利であると考えられる。連続投与は注入ポンプを使用するなど機械的手段により達成できるが、他の連続又は準連続投与方法も実施できると考えられる。例えば、化学誘導体化によりタンパク質の持続放出形態が得られ、所定の投与計画に基づいて予測可能な量で血流中の連続的存在を可能にする。従って、截形GDNFタンパク質産物はこのような連続投与を実現するように誘導体化された截形GDNFタンパク質を含む。
【0128】
截形GDNFタンパク質細胞治療、例えば截形GDNFタンパク質を産生する細胞の大脳内移植も考えられる。本発明のこの態様では、生物活性形態の截形GDNFタンパク質を合成及び分泌することが可能な細胞を患者に移植することが考えられる。このような截形GDNFタンパク質産生細胞は、通常は神経栄養因子を産生しないが、截形GDNFを産生するように改変された細胞や、截形GDNFタンパク質の発現及び分泌に適したポリヌクレオチドで形質転換することによりそのGDNF産生能を増加した細胞であり得る。外来種のGDNFの投与により患者に免疫反応が生じる可能性を最小限にするためには、細胞はヒト起源であり且つヒト截形GDNFタンパク質を産生することが好ましい。
【0129】
細胞の脳組織浸潤を避けるために移植細胞をカプセル封入してもよい。截形GDNFタンパク質産物を放出できるが、患者の免疫系又は周囲組織からの他の有害因子による細胞の破壊を阻止する生体適合性の半透ポリマーエンクロージャー又は膜にヒト又は非ヒト動物細胞を封入して患者に移植することができる。あるいは、截形GDNFを産生するようにex vivo形質転換した患者自身の細胞をカプセル封入せずに患者に直接移植してもよい。
【0130】
生きた細胞の膜封入方法は当業者に周知であり、封入細胞の作製と患者へのその移植は実施可能である。例えば、参考資料としてその開示内容を本明細書の一部とする米国特許第4,892,538号、5,011,472号及び5,106,627号参照。生きた細胞を封入するシステムは、特に参考資料としてその開示内容を本明細書の一部とするAebischerらのPCT出願WO91/10425にも記載されている。
更に、参考資料としてその開示内容を本明細書の一部とするAebischerらのPCT出願WO91/10470; Winnら,Exper.Neurol.,113:322−329,1991; Aebischerら,Exper.Neurol.,111:269−275,1991; Trescoら,ASAIO,38:17−23,1992も参照されたい。
【0131】
截形GDNFタンパク質をコードする核酸配列を患者に直接導入する截形GDNFタンパク質in vivo遺伝子治療も考えられる。例えば、アデノ随伴ウイルスベクター等の適当な送達ベクターを介して又は介さずに核酸構築物の局所注射により、截形GDNFタンパク質をコードする核酸配列を標的細胞に導入する。別のウイルスベクターの非限定的な例としては、レトロウイルス、アデノウイルス、単純ヘルペスウイルス及びパピローマウイルスベクターが挙げられる。所望の核酸構築物もしくは所望の核酸配列を含む他の適当な送達ベクターの局所注射、リポソームによる導入、直接注射(裸のDNA)、レセプターによる導入(リガンド−DNA複合体)、又は微粒子ボンバードメント(遺伝子銃)により、in vivo物理的導入が可能である。
【0132】
本明細書に記載する截形GDNFタンパク質産物製剤は獣医学及びヒト適用に使用することができ、「患者」なる用語は限定的に解釈すべきでないことに留意されたい。獣医学適用の場合では、用量範囲は上記と同一にすべきである。
【0133】
本発明の截形GDNFタンパク質を更に特性決定する手段として、X−[Cys41−Cys133]−Yアミノ酸配列内のエピトープ等の截形GDNFタンパク質に結合する抗体を製造することができる。当業者は周知の公表手順を使用し、本発明のアミノ酸配列によりコードされる種々のタンパク質を特異的に認識してこれと結合するモノクローナル及びポリクローナル抗体又は組換え抗体を取得することができる。その後、このような抗体を使用して截形GDNFタンパク質を精製及び特性決定することができる。あるいは、これらの抗体はその抗原であるタンパク質の治療阻害剤としても使用できる。
【0134】
本発明の他の側面及び利点は、下記実施例から理解されよう。実施例1は哺乳動物細胞系における成熟GDNFの発現と截形GDNFタンパク質の作製に関する。実施例2は細菌細胞系における成熟GDNFの発現に関する。実施例3は細菌細胞系における種々の截形GDNFタンパク質の発現に関する。実施例4はドーパミン作動性ニューロン神経栄養活性アッセイで成熟GDNFタンパク質と截形GDNFタンパク質の生物活性を比較する。
【図面の簡単な説明】
【0135】
【図1】成熟ヒトグリア細胞系由来神経栄養因子(hGDNF)をコードする核酸配列(配列番号1)を示す。成熟ヒトGDNFタンパク質のアミノ酸配列(配列番号2)も示す。
【図2】組換え截形GDNFタンパク質の発現のために利用したプラスミド構築図を示す。
【図3A】GDNF及び截形GDNFポリヌクレオチドをコードする別の核酸配列(配列番号39)の制限地図を示す。
【図3B】GDNF及び截形GDNFポリヌクレオチドをコードする別の核酸配列(配列番号39)の制限地図を示す。
【図4】GDNF及び截形GDNFポリヌクレオチドをコードする更に別の核酸配列(配列番号40)の制限地図を示す。
【図5】[Pro23−Lys37ΔAsn37−Ile134]截形GDNFタンパク質置換変異体(配列番号42)をコードする核酸配列(配列番号41)を示す。このタンパク質はMet−Ser−[Pro23−Lys37ΔAsn37−Ile134]截形GDNFタンパク質付加/置換変異体と表すこともできる。
【図6】[Arg32−Ile134]截形GDNFタンパク質(配列番号44)をコードする核酸配列(配列番号43)を示す。
【図7】[Gly33−Ile134]截形GDNFタンパク質(配列番号46)をコードする核酸配列(配列番号45)を示す。
【図8】数種の截形GDNFタンパク質の例であるMet−[Arg32−Ile134](配列番号48)、Met−[Gly33−Ile134](配列番号49)及びMet−Ser−[Pro23−Lys37ΔAsn37−Ile134](配列番号50)と成熟hGDNFのアミノ酸配列(配列番号47)の比較を示す。
【0136】
実施例
【実施例1】
【0137】
CHO細胞における成熟ヒトGDNFの発現及びCHO細胞由来截形GDNFタンパク質の精製
材料
下記材料を使用してジヒドロ葉酸レダクターゼ欠損CHO細胞(例えばUrlaubとChasin,Proc.Natl.Acad.Sci.,USA 77(7):4216−4220(1980)に記載されているようなCHOd−細胞)でヒトGDNFを発現させる。
【0138】
CHOd−培地は、ダルベッコ改変イーグル培地(DMEM)−高グルコース(Gibco/BRL)、5%ウシ胎児血清
(HyClone)、MEM非必須アミノ酸(1%)(Gibco/BRL)、ヒポキサンチン/チミジン(1%)(Gibco/BRL)、及びグルタミン/ペニシリン/ストレプトマイシン(1%)(Irvine Scientific)を含むものとした。
【0139】
選択培地は、DMEM(高グルコース)、5%透析ウシ胎児血清(HyClone)、MEM非必須アミノ酸、及びグルタミン/ペニシリン/ストレプトマイシンを含むものとした。
【0140】
2×HEPES緩衝食塩水(HBS)は、280mM NaCl、10mM KCl、1.5mM Na2HPO4、12mMデキストロース、及び50mM HEPESを含むものとした。
【0141】
Tris緩衝食塩水+Tween(TBST)は、137
mM NaCl、20mM Tris/HCl,pH7.5、及び0.1% Tween−20を含むものとした。方法
トランスフェクション及び選択
CHOd−細胞(20継代)をCHOd−増殖培地中細胞8×105個/皿の密度で60mm組織培養皿(Falcon)に接種した。翌日、トランスフェクションから約3時間前に細胞上の培地を新鮮な培地に交換した。
【0142】
周知技術を使用して適当なGDNF cDNAを含むプラスミド構築物を調製した。例えば、参考資料としてその開示内容を本明細書の一部とする本願出願人と同一名義の1990年3月29日付け同時係属米国特許出願第501,904号(1990年5月18日付けヨーロッパ特許出願第90305433号、公開EP第398753号及びWO90/14363(1990)も参照)に記載の方法に実質的に従ってプラスミド構築物pDSRα2を調製した。ベクターの構造編成を示すプラスミド地図の例を図2に示す。図1、3及び4に示す配列等の成熟GDNFタンパク質をコードする種々の核酸配列を使用してもよいことが当業者に理解されよう。
【0143】
pcDNA3に基づく発現ベクター(Invitrogen,San Diego,CA)から制限酵素消化により、ヒトGDNFコーディング配列とコンセンサスKozak配列CCACC(ATG)を含むHindIII−Xbal DNAフラグメントを回収した。HindIII/Xbalで切断したpDSRα2にDNAフラグメントを直接クローニングした。得られたプラスミドをpSW5と命名した。トランスフェクションの前にpSW5のプラスミドDNAをPuvI部位で直鎖化した。
【0144】
pDSRα2(図2)はプラスミドpCD(OkayamaとBerg,Mol.Cell Biol.3:280−289,1983)の誘導体であり、(1)SV40ポリアデニル化シグナルをウシ濾胞刺激ホルモンのα−サブユニットであるα−bFSHからのシグナルで置換し(Goodwinら,Nucleic Acids Res.11:6873−6882,1983)、(ii)形質転換細胞の選択及び増殖を可能にするように発現カセットの下流にマウスジヒドロ葉酸レダクターゼミニ遺伝子(Gasserら,Proc.Natl.Acad.Sci.79:6522−6526,1982)を挿入し、(iii)ヒトT細胞I型白血病ウイルス(HTLV−I)の長い末端反復配列(LTR)の「Rエレメント」と「U5」配列の一部を含む267bpフラグメントを従来の記載(Takebeら,Mol.Cell Biol.8:466−472,1988)通りにSV40プロモーターとスプライスシグナルの間にクローニング及び挿入したという主に3点を改変した。
【0145】
最終濃度3.0μg/皿のGDNF−プラスミドDNA、7.0μg/皿のマウス腎ゲノムキャリヤーDNA(Clontech)、25μl/皿の2.5M CaCl2及び最終容量250μl/皿までの滅菌蒸留水を含むDNA溶液を調製した。pDSRα2ベクターDNA又はキャリヤーDNA単独を含むDNA溶液も夫々正負対照として同様に調製した。溶液に気泡を通しながら、DNA溶液を等容量の2×HEPES緩衝食塩水に滴下した。DNA/HBS溶液を室温で30分間インキュベートした。
【0146】
CHOd−細胞培養物から培地を除去し、DNA溶液500μl/皿を加えた。皿を室温で30分間インキュベートした後、CHOd−培地(5.0ml)を各皿に加えた。次に、皿を37℃で一晩インキュベートした。
【0147】
翌日に培地を新鮮なCHOd−培地に交換した。その翌日に細胞がコンフルエントに達した時点で培養物をトリプシン処理し、1×60mm皿対8×100mm皿の比で100mm皿(Falcon)に再プレーティングした。細胞を選択培地で再プレーティングした。2〜3日おきに培養物に新鮮な培地を再補充した。
【0148】
15日後に、トランスフェクトした細胞のコロニーをガラスクローニングシリンダーで単離し、トリプシン処理し、24穴皿(Falcon)に再プレーティングした。GDNF/pSW5をトランスフェクトした細胞から合計40個のコロニーを単離した。皿に残っている細胞をトリプシン処理し、プールし、2個の100mm皿に再プレーティングした(各DNA構築物毎に1個のプール)。
【0149】
トランスフェクトした細胞のスクリーニング
24穴及びプール培養物をコンフルエントまで増殖させ、その時点で増殖培地を除去し、無血清培地(400μl/ウェル即ち4ml/皿)に交換した。細胞を48時間インキュベートし、条件付け培地を回収した。条件付け培地サンプルのGDNFタンパク質発現をウェスタンブロットにより分析した。条件付け培地のアリコート(20μl又は40μl)を電気泳動サンプル緩衝液(β−メルカプトエタノールを含むか又は含まない)で希釈した。β−メルカプトエタノールを含むサンプルを3分間煮沸した(還元条件)。還元及び非還元サンプルの両者を16%Tris−グリシンゲル(Novex)で試験した。ゲルをニトロセルロースフィルター(Schleicher and Schuell BA−83、0.2μ)にエレクトロブロットした。ブロットをTBSTで濯いだ後、TBST中5%ドライミルク(Carnation)のブロッキング溶液中で室温で30分間インキュベートした。次にブロットをGDNF抗血清(大腸菌由来GDNFに対するウサギポリクローナル抗血清、5%ドライミルク/TBST中1:1000)で室温で1時間処理した。次にブロットをTBSTで濯ぎ、1×10分間及び2×5分間1%ドライミルク/TBSTで洗浄した。次に抗ウサギIg−ウマ西洋ワサビペルオキシダーゼ結合二次抗体(1%ドライミルク/TBST中1:15,000)で20分間処理した。ブロットを濯ぎ、TBSTで1×20分間及び2×10分間洗浄した後、ECL試薬(Amersham)で1分間処理し、Hyperfilm−ECL(Amersham)を感光した。
【0150】
下記プロセスは、CHOで発現させたGDNFとCHOに由来する切断GDNFホモダイマーを条件付け培地1リットルから精製するプロセスである。CHO培地には有意なプロテアーゼ作用があり、残基31で鎖を切断するので、この手順では精製中にプロテアーゼ阻害剤を使用してもよい。
【0151】
ステップ1.ビーズクロマトグラフィー:
1/50容量の1M MES,pH6.0を加えることにより、無血清条件付け培地を20mM 2−[N−モルホリノ]エタンスルホネート(MES),pH6.0にした。20mM MES,pH6.0で平衡化したSP Sepharose Big Bead樹脂(Pharmacia)25mlを加え、4℃で1時間撹拌した。樹脂を沈殿させて条件付け培地をデカントすることにより樹脂を集めた。デカントした培地を焼結ディスクフィルターで濾過し、未沈殿樹脂を回収した。沈殿した樹脂と濾過により回収した樹脂を再懸濁し、直径2.5cmのカラムに注入し、3カラム容量の0.15M NaCl,20mM MES,pH6.0(A緩衝液)で洗浄した。280nmの吸光度をモニターしながら0.2カラム容量/分の流速でA緩衝液から1.0M NaCl,20mM MES,pH6.0(B緩衝液)への300nlのグラジエントでタンパク質を溶離した。1.1カラム容量を含むフラクションを集めた。フラクション中のGDNFの存在をウェスタンブロット分析により検出した。GDNFを含むフラクションを更に精製するためにプールした。GDNFは0.3〜0.6M NaClで溶出した。
【0152】
ステップ2.HPLC C4クロマトグラフィー:
ステップ1からのプールを0.1%(v/v)トリフルオロ酢酸(TFA)とし、0.45μフィルターで減圧濾過し、
10%アセトニトリル、0.1%TFA水溶液(A緩衝液)で条件付けしたVydac C4カラム(0.46×25cm)にアプライした。280nmの吸光度を測定しながらA緩衝液から90%アセトニトリル、0.1%TFA水溶液(B緩衝液)への2%/分の直線勾配で50分間かけてタンパク質を溶離した。1mlフラクションを集め、GDNFの存在をウェスタンブロット分析により検出した。GDNFは45%〜55%アセトニトリルで溶出した。フラクションを減圧乾涸した。
【0153】
ステップ3.高性能Sクロマトグラフィー:
ステップ2からのGDNFを含むフラクションを1mlの0.15M NaCl,10mM Tris,pH8.0に再可溶化し、0.75×7.5cm TSK−Gel 5WP高性能Sカラム(Toso Haas)にアプライした。0.15M NaCl,10mM Tris,pH8.0(A緩衝液)から1.0M NaCl,10mM Tris,pH8.0(B緩衝液)への0.4%/分の直線勾配で流速1ml/分で50分間かけて溶離した。1分毎にフラクションを集め、280nmの吸光度を測定した。35%B緩衝液で勾配を10分間かけて6.5%/分まで変化させた。フラクションのウェスタンブロット分析は、4個の主GDNF成分を示した。これらの成分のうちの3個は0.4%/分勾配中に溶出し、4番目の成分は6.5%/分勾配中に溶出した。同様の成分から適当なプールを調製し、配列決定した。配列決定分析の結果、約29〜36kDプールは[Arg32−Ile134]截形GDNFタンパク質であることが確認された。約38〜40kDの成分は[Arg32−Ile134]截形GDNF/成熟GDNFヘテロダイマーであることが確認された。最後に、勾配の最後の部分で単離された約41〜44kDの成分は成熟GDNFホモダイマーであることが配列決定により確認された。
【実施例2】
【0154】
大腸菌における成熟ヒトGDNFの産生
成熟ヒトGDNFの細菌発現は、Linらの方法(参考資料としてその開示内容を本明細書の一部とする1994年5月23日付け米国特許出願第08/182,183号とその親出願、1992年9月17日付けPCT/US92/07888(WO93/06116)及びヨーロッパ特許出願第92921022.7号(公開EP第610254号))により実施できる。本発明の記載に基づき、大腸菌及び他の細菌での適切な発現に種々の材料及び方法を容易に使用又は応用できることが当業者に理解されよう。例えば、図1、3及び4に示すような代替ポリヌクレオチドを発現プロセスで使用してもよい。
【0155】
発現された成熟GDNFの再生及び精製
形質転換細胞を(特に指定しない限り)5℃で次のように処理した。最終容量200mlの5mM EDTAを含む25mM Tris,pH8.5に細胞ペースト(30g)を懸濁し、15%(w/v)の最終細胞スラリーを得た。Biospecハンドヘルド低剪断ホモジナイザーを使用して細胞を十分に分散させた。スラリーを14,500psiのマイクロ流動化装置に2回通し、細胞を破壊して封入体を放出させた。得られたホモジネートを次に16,000×gで30分間遠心分離した。上記と同様にBiospecホモジナイザーを使用し、遠心分離により得られた封入体のペレットを最終容量240mlまでの冷水に再懸濁して洗浄し、スラリーを形成した。このスラリーのサンプルをGDNF発現レベルのHPLC分析用に取り分けた。残りのスラリーを16,000×gで30分間遠心分離した。上清を捨て、遠心びんに少量の冷水を加えて静かに渦形成し、封入体ペレットの上にまばらに形成された膜層を除去した。Biospecホモジナイザーで十分な容量の冷水を使用してGDNF2mg/mlの濃度にペレットを再懸濁した。次に最終封入体懸濁液(25ml)と180mM システインHCl及び50mM Tris HCl,pH8.7を含む8MグアニジンHCl(25ml)を混合することにより、封入体を可溶化した。可溶化混合物を25℃で60〜90分間撹拌した後、20mM Tris HCl,pH8.75と0.2MグアニジンHClを含む2M尿素(450ml,5℃)に混合しながら注入した。再生混合物を5℃で72時間ゆっくりと撹拌した。
【0156】
再生したGDNFを次のように部分精製した。5℃の20
mM酢酸ナトリウム緩衝液(250ml,pH5)を迅速に撹拌しながら再生混合物に加え、pHを氷酢酸で5に調整した。生じた沈殿を5℃で13,600×gで45分間遠心分離して除去した。この遠心分離からの上清を次の精製段階のロード溶液として使用し、SP−大ビーズ樹脂(Pharmacia)によるカチオン交換クロマトグラフィーにかけた。カラムは、20mM酢酸ナトリウム(pH5)を平衡、濯ぎ及び溶離緩衝系として使用して5℃で操作した。樹脂ベッド(5ml)を5カラム容量(CV)の0.2N NaOHで処理した後、酢酸緩衝液(5CV)で平衡化した。ロード溶液(190ml)を0.5CV/分でカラムにアプライした後、同一流速で10CVの酢酸緩衝液で濯いだ。次に0.1CV/分の流速で酢酸緩衝液中NaCl0.3Mから0.9Mの20CV直線勾配でGDNFを樹脂から溶離した。カラム溶出液の280nmの吸光度をモニターし、フラクションとして集めてSDS−PAGEによりアッセイした。10%ピーク高さのGDNFピークの前縁から10%ピーク高さのピークの後縁までのGDNFを含むフラクションをプールした。このプール中のタンパク質は完全にGDNFであり、使用した産生株に応じて改変GDNF形として32%〜12%の汚染を含んでいた。次にプールをPBS又は他の組成の緩衝液で透析し、場合により限外濾過により25mg/mlまで濃縮した。この手順により精製した野生型及び類似形態のGDNFを逆相HPLC、カチオン交換HPLC、質量スペクトル分析及びエンドトキシンレベルにより特性決定し、調製物の純度を対応する産生株と比較した。
【実施例3】
【0157】
大腸菌における截形GDNFの組換え製造
Linらに記載されている方法(上記1994年5月23日付け米国特許出願第08/182,183号)に実質的に従って典型的なGDNFタンパク質を製造した。上述したような代替細菌発現材料及び方法を使用してもよい。夫々図5、6及び7に示すような[Pro23−Ile134]、[Arg32−Ile134]及び[Gly33−Ile134]截形GDNFタンパク質を大腸菌で発現させた。これらの典型的截形GDNFタンパク質をコードするポリヌクレオチドは図5、6及び7に示すように構築したが、図1、3及び4に示すような対応するポリヌクレオチドを使用してもよい。これらのポリヌクレオチドは、参考資料としてその開示内容を本明細書の一部とするPCR Technology,Principles and Applications for DNA Amplification,Henry A.Erlich編,Stockton Press,NY,1989(第6章、Using PCR to Engineer DNA)に記載されているような標準PCR手順により構築した。
【実施例4】
【0158】
ドーパミン作動性ニューロン神経栄養活性のバイオアッセイ
実施例3の大腸菌で発現させた[Pro23−Ile134]、[Arg32−Ile134]、[Gly33−Ile134]及び[Lys37−Ile134]截形GDNFタンパク質と実施例1のCHO由来[Arg32−Ile134]截形GDNFタンパク質が黒質ドーパミン作動性ニューロンによるドーパミン取り込みを促進する能力を定量的に評価した。
【0159】
材料
截形GDNFタンパク質の存在下におけるドーパミン作動性ニューロンの生存を評価するアッセイでは、次の材料を使用する。
【0160】
細胞培地
高グルコースダルベッコ改変イーグル培地(DMEM、カタログ番号#11965−092)、ハムF12培地(F12、#11765−021)、重炭酸ナトリウムを含まないライボビッツL15培地(#41300−039)、B27培地補充剤(#17504−010)、ペニシリン/ストレプトマイシン(#15070−014)、L−グルタミン(#25030−016)、ダルベッコのリン酸緩衝食塩水(D−PBS、#14190−052)、カルシウム及びマグネシウム塩を含むハンク平衡塩類溶液(HBSS、#24020−026)、N−2−ヒドロキシエチルピペラジン−N’−2−エタンスルホン酸(HEPES、#15630−015)、マウスラミニン(#23017−015)及びウシ血清アルブミンフラクションV(#110−18−017)は全てGIBCO,Grand Island,NYから入手した。熱不活化ウマ血清はHyClone,Logan,Utahから入手した。コンアルブミン(C−7786)、臭化水素酸ポリ−L−オルニチン(P−3655)、ウシインスリン(I−5500)、ヒトトランスフェリン(T−2252)、プトレッシン(P−6024)、プロゲステロン(P−6149)、亜セレン酸ナトリウム(S−9133)、メトリザミド(N−3383)は全てSigma Chemical Company,Saint−Louis,MOから入手した。パパイン、デオキシリボヌクレアーゼI(DNアーゼ)及びオボアルブミン(パパイン解離系)はWorthington Biochemicals,Freehold,NJから入手した。
【0161】
Falcon滅菌96穴マイクロプレート(#3072)、組織培養プラスチック容器及びポリプロピレン遠心分離管はBecton−Dickinson,Oxnard,CAから入手した。Nunc Lab−Tek組織培養チャンバーカバーガラス(#136439)はBaxter,Irvine,CAから入手し、20μm(#460)ナイロンメッシュはTetko,Elmsford,NYから入手し、4”解剖鉗子と4”鉗子鋏はRoboz Surgical,Washington,DCから入手した。
【0162】
抗体及び関連試薬
ポリクローナルウサギ抗チロシンヒドロキシラーゼ抗体(TE101)はEugene Tech,Ridgefield Park,NJから入手し、ポリクローナルウサギ抗ニューロン特異的エノラーゼ抗体(NSE,AB951)はChemicon,Temecula,CAから入手し、ビオチン化ヤギ抗ウサギIgG及びペルオキシダーゼ結合アビジン/ビオチン複合体(ABC Elite;VectastainキットPK−6100)はVector Laboratories,Burlingame,CAから入手した。3’,3’−ジアミノベンジジンはCappel Laboratories,West Chester,PAから入手した。PBS中のSuperblockブロッキング緩衝液(#37515)はPierce Chemical Company,Rockford,ILから入手した。Triton X−100(X−100)、Nonidet P−40(N6507)及び過酸化水素(30%v/v;H1009)はSigmaから入手した。
【0163】
GBR−12909ドーパミン取り込み阻害剤(D−052)はRBI,Natick,MAから入手した。3H−ドーパミン(トリチウム化ドーパミン、NE−131;21Ci/mmol)はNew England Nuclear,Boston,MAから入手した。Optiphase SupermixシンチレーションカクテルはWallac,Turku,フィンランドから入手した。透明ViewPlate−96マイクロプレート(#6005182)はPackard Instruments Corporation,Meriden,CTから入手した。他の全試薬は特に指定しない限り、Sigma Chemical Companyから入手した。
【0164】
培地の調製
DMEMとF12培地の1:1混合物として基本培地を調製し、50倍濃縮ストック溶液としてB27培地補充剤を補充した。最終濃度約2mMのL−グルタミン、約100IU/lのペニシリン及び約100mg/lのストレプトマイシンを加えた。熱不活化ウマ血清を最終濃度約15%まで加えた。混合後、pHを約7.3に調整し、培地を4℃に維持した。培地は、実験内変動を最小限にするように使用直前に新たに調製した。タンパク質吸着を最小限にするように常にプラスチックピペット及び容器を使用した。
【0165】
培養基質
基質ニューロンの最適結合と神経突起生長を助長するために、マイクロタイタープレート表面(培養基質)を次のようにポリ−L−オルニチンとラミニンで順次被覆して修飾した。プレート表面を0.1M硼酸(pH8.4)中0.1mg/ml滅菌ポリ−L−オルニチン溶液で室温で少なくとも1時間完全に被覆した後、Super−Q水で滅菌洗浄した。次に水洗物を吸引し、PBS中1.0μg/mlマウスラミニン溶液を加え、37℃で2時間インキュベートした。これらの手順は、結果の再現性を確保するためにプレートの使用直前に実施した。
【0166】
胎児ラット黒質培養物の調製
胎児ラット脳を黒質ドーパミン作動性ニューロン源として使用した。胎生15日の予定妊娠Sprague−Dawleyラットを使用した。各実験毎に最大36匹の胎児(約3リットル)を処理した。妊娠したラットをCO2に暴露して殺し、その腹腔を解剖鋏で切り開き、子宮から胎児を取り出した。次に胎児の脳を切開し、血液と髄膜を除去し、確定解剖目標(AltmanとBayer,Atlas of Prenatal Rat Brain Development,CRC Press,Boca Raton,FL,1995)を使用して黒質を含む腹側被蓋領域を解剖した。組織を氷冷D−PBSに集め、解離培地(HBSS中120単位パパイン及び2000単位DNアーゼ)10mlに移した後、約200rpmに設定した回転プラットフォーム震盪機で約37℃で45分間インキュベートした。次に火仕上げパスツールピペットで細胞を潰して分散させ、20μm Nitexメッシュで篩別して未解離組織を捨て、IEC臨床遠心機を使用して200×gで5分間遠心分離した。得られた細胞ペレットを、オボアルブミンと約500単位のDNアーゼを含むHBSSに再懸濁し、4%オボアルブミン溶液(HBSS中)に重層し、500×gで約10分間遠心分離した。最終ペレットを完全培地(上記参照)に再懸濁し、約28,000細胞/mlに調整し、ポリオルニチンとラミニンで予め被覆しておいた96穴マイクロプレートの6mmウェルに分注(90μl)した。すぐに細胞の結合が生じ、プレーティング効率は約75%であった。
【0167】
ドーパミン作動性ニューロンの免疫組織化学
Louisら(J.Pharmacol.Exp.Therap.,262:1274−1283,1992; Science,259:689−692,1993)により記載されている間接イムノペルオキシダーゼ法を下記のように若干改変して使用し、黒質の培養物中のドーパミン作動性ニューロンを特性決定した。培養物をD−PBS,pH7.4中4%パラホルムアルデヒドで室温で約30分間固定した後、D−PBS(6mmウェル当たり200μl)で3回洗浄した。次に、固定した培養物を1%NP−40を含むPBS中Superblockブロッキング緩衝液中でインキュベートし、抗体の浸透を増加した。次に、一次ウサギ抗チロシンヒドロキシラーゼ抗体を同一緩衝液に約1:2000に希釈して加え、回転震盪機で37℃で1時間インキュベートした。D−PBSで3回洗浄後、ヤギ抗ウサギビオチン化IgGを約1:500に希釈して使用して結合抗体を検出し、これらの二次抗体を細胞と共に37℃で約1時間インキュベートした。次に細胞をD−PBSで3回洗浄し、二次抗体を1:500に希釈したアビジン−ビオチン−ペルオキシダーゼ複合体で検出し、細胞を37℃で約45分間インキュベートした。D−PBSで更に3回洗浄後、0.04%3’,3’−ジアミノベンジジン−(HCl)4、0.06%NiCl2及び0.02%過酸化水素を含む0.1M Tris−HCl,pH7.4の溶液中で培養物を5〜20分間反応させた。
【0168】
ニューロン生存の測定
黒質培養物を固定し、前記のように免疫染色処理した後、200倍明視野顕微鏡で試験した。チロシンヒドロキシラーゼで染色するニューロンの数を96穴マイクロプレートの6mmウェル全体で数えた。生存可能なニューロンは定形細胞体をもち、主軸索様突起と数個の樹状突起があるとみなした。不整形の空胞のある核周囲部細胞体や分裂した神経突起をもつなど変性の徴候を示すニューロンは計数から除いた(但し、変性ニューロンの大部分は培養基質から分離していた)。ドーパミン作動性ニューロン細胞数は、TH陽性ニューロン/6mmウェル又は対照ドーパミン作動性ニューロン密度に対する倍数として表した。
【0169】
ドーパミン取り込みの測定
透明ViewPlate−96マイクロプレートで調製しておいた15日齢胎児ラット黒質ニューロンの培養物でドーパミン取り込みを測定した。約120mM NaCl、4.7mM KCl、1.8mM CaCl2、1.2mM MgSO4、32mM NaHPO4、1.3mM EDTA及び5.6mM D−グルコースを含む改変クレブス−リンゲル溶液、pH7.4から構成される予熱取り込み緩衝液(約100μl)で培養物を洗浄した。取り込み緩衝液には、ドーパミンの酸化を阻止するために1mMアスコルビン酸と50μMパルジリンも加えた。次に細胞を取り込み緩衝液中で37℃で約10分間プレインキュベートした。次に、トリチウム化したドーパミン(3H−DA、21Ci/mmol)を取り込み緩衝液75μl中約50nMの濃度で黒質培養物に加え、培養物を37℃で約60分間インキュベートした。ドーパミン取り込み阻害剤GBR−12909(1μM)を含む取り込み緩衝液と共に培養物をインキュベートすることにより、非特異的ドーパミン取り込みを測定した。非特異的取り込みは総取り込みの約1%未満であった。インキュベーション培地を吸引して取り込みアッセイを停止した後、氷冷取り込み緩衝液(約120μl)で3回迅速に洗浄した。次に、Optiphase Supermixシンチレーションカクテル(200μl)を加えて細胞を溶解させ、Wallac MicrobetaPlus 96穴マイクロプレートカウンターを使用してシンチレーションスペクトル分析により放射能を測定した(即ち培養物中に保持されたトリチウムのシンチレーション計数によりドーパミン取り込みを分析する)。結果をdpm/6mmウェル又は対照培養物に対する倍数として表す。
【0170】
アッセイ
ドーパミン作動性ニューロン生存及び形態学的発生
ドーパミン作動性ニューロンを多く含む15日齢胎児(E15)ラット黒質の培養物を使用して、截形GDNFタンパク質がドーパミン作動性ニューロンの生存に及ぼす効果を立証した。ポリオルニチンとラミニンで被覆した96穴マイクロプレートで種々の濃度(約1pg/ml〜約10ng/ml)の下記タンパク質、即ち大腸菌で発現させた成熟hGDNF、大腸菌で発現させた[Pro23−Ile134]、[Arg32−Ile134]、[Gly33−Ile134]及び[Lys37−Ile134]截形Glyタンパク質、CHO細胞で発現させた成熟hGDNF、並びにCHO細胞由来[Arg32−Ile134]截形GDNFタンパク質の存在下又は単独で培養物を6日間まで増殖させた。培地は15%熱不活化ウマ血清(E15培養物)又は2.5%熱不活化ウマ血清、D−グルコース、HEPES、インスリン及びトランスフェリン(P6培養物)を補充したDMEM/F12から構成した。ドーパミン生合成の律速酵素であるチロシンヒドロキシラーゼ(TH)の免疫染色をドーパミン作動性ニューロンのマーカーとして使用した。菱脳中のノルアドレナリン作動性ニューロンもTHに陽性染色するので、中脳の脳質に限定した領域を解剖し、ノルアドレナリン作動性細胞体を含む尾側の領域を避けるように十分注意した。6日後に、E15培養物は主に(上記)ニューロン特異的エノラーゼ免疫染色により確認される約70%のニューロンと、30%の(平坦で暗い外観をもつ)非ニューロン細胞から構成され、ドーパミン作動性ニューロンはニューロンポピュレーションの約10〜15%であった。
【0171】
6日後に培養物をパラホルムアルデヒドで固定し、これらの培養物中のドーパミン作動性ニューロンを識別するマーカーであるチロシンヒドロキシラーゼで免疫染色した。6mmウェル中に存在する全チロシンヒドロキシラーゼ陽性ニューロンを明視野顕微鏡で計数した。各実験条件毎に3〜6個の異なるウェルを分析した。結果を対照培養物中に存在するチロシンヒドロキシラーゼ陽性ニューロンの数の百分率として表した。
【0172】
CHO細胞で発現させたGDNF又は大腸菌で発現させたGDNFの2種のGDNF1.0ng/mlで処理したE15黒質の培養物は未処理対照培養物よりも夫々約38%及び27%多くのTH免疫反応性ニューロンを含んでおり、どちらのGDNF種もドーパミン作動性ニューロンの生存を促進することを示した。1.0ng/mlの截形GDNFタンパク質で処理したE15黒質の培養物は、in vitroで6日後に培養物中のTH陽性ニューロン数に同様の増加を示し、CHO細胞由来[Arg32−Ile134]截形GDNFタンパク質では42%、大腸菌で発現させた[Arg32−Ile134]及び[Gly33−Ile134]截形GDNFタンパク質では夫々26%及び17%増加した。
【0173】
対照培養物と成熟及び截形GDNFタンパク質で処理した培養物を比較すると更に、全GDNFタンパク質がドーパミン作動性ニューロンの形態学的分化に顕著な効果をもつことも判明した。[Arg32−Ile134]及び[Gly33−Ile134]截形GDNFタンパク質の効果は夫々の対応する成熟GDNFタンパク質と同一であった。全GDNF処理培養物中のTH免疫反応性ニューロンは、対照培養物中のTH陽性ニューロンに比較して有意に複雑で広範囲の神経突起分枝をもち、神経突起分枝度が高く、細胞体の総寸法が大きかった。
【0174】
ドーパミン取り込み
ドーパミン取り込みは高親和性ドーパミン再取り込みトランスポーター部位の数及び活性の尺度であり、ドーパミン作動性ニューロンの機能的分化を表す。成熟GDNF又は截形GDNFタンパク質の存在下又は不在下でin vitroで6日後にE15ラット黒質の培養物でドーパミン取り込みを測定した。これらの培養物においてドーパミン取り込みはドーパミン作動性ニューロンの特徴的薬理プロフィルをもち、即ちドーパミン作動性ニューロンに特異的なドーパミン輸送阻害剤(ID50=20nM)である1.0μM GBR−12909によりほぼ完全にブロックされた(>98%)。従って、ノルエピネフリン輸送体を介してドーパミンを取り込むことができるが、GBR−12909阻害に非感受性の汚染性ノルアドレナリン作動性ニューロンの存在はドーパミン取り込み測定値に影響していないと考えられる。CHO細胞で発現させた成熟GDNFとCHO由来[Arg32−Ile134]截形GDNFタンパク質の効果は同一であり、約65%増加し、ED50は約20pg/mlであった。図5に示すような大腸菌で発現させた[Pro23−Lys37ΔAsn37−Ile134]截形Dタンパク質は65%の増加を示し、ED50は約40pg/mlであった。大腸菌で発現させた成熟タンパク質と大腸菌で発現させた[Arg32−Ile134]、[Gly33−Ile134]及び[Lys37−Ile134]截形GDNFタンパク質のドーパミン取り込み効果は同一であり、約50%増加し、ED50は約50pg/mlであった。
【0175】
以上の結果は、截形GDNFタンパク質が黒質ドーパミン作動性ニューロンの強力な生存促進及び分化誘導因子として作用することを示す。従って、截形GDNFタンパク質は情動の鋭さの低下、随意及び不随意筋運動の減退、筋硬直及び振戦を特徴とする神経障害であるパーキンソン病の治療に特に有利であると考えられる。このような症状は黒質内のドーパミン産生ニューロンの進行性変性に起因すると考えられる。これらのニューロン(「ドーパミン作動性ニューロン」)の変性は線条体と呼ばれる脳の隣接領域でドーパミンの低下をもたらす。
【0176】
図面の簡単な説明
本発明の多数の特徴及び利点は図面を参考に容易に理解されよう。
【0177】
図1は成熟ヒトグリア細胞系由来神経栄養因子(hGDNF)をコードする核酸配列(配列番号1)を示す。成熟ヒトGDNFタンパク質のアミノ酸配列(配列番号2)も示す。
【0178】
図2は組換え截形GDNFタンパク質の発現のために利用したプラスミド構築図を示す。
【0179】
図3はGDNF及び截形GDNFポリヌクレオチドをコードする別の核酸配列(配列番号39)の制限地図を示す。
【0180】
図4はGDNF及び截形GDNFポリヌクレオチドをコードする更に別の核酸配列(配列番号40)の制限地図を示す。
【0181】
図5は[Pro23−Lys37ΔAsn37−Ile134]截形GDNFタンパク質置換変異体(配列番号42)をコードする核酸配列(配列番号41)を示す。このタンパク質はMet−Ser−[Pro23−Lys37ΔAsn37−Ile134]截形GDNFタンパク質付加/置換変異体と表すこともできる。
【0182】
図6は[Arg32−Ile134]截形GDNFタンパク質(配列番号44)をコードする核酸配列(配列番号43)を示す。
【0183】
図7は[Gly33−Ile134]截形GDNFタンパク質(配列番号46)をコードする核酸配列(配列番号45)を示す。
【0184】
図8は数種の截形GDNFタンパク質の例であるMet−[Arg32−Ile134](配列番号48)、Met−[Gly33−Ile134](配列番号49)及びMet−Ser−[Pro23−Lys37ΔAsn37−Ile134](配列番号50)と成熟hGDNFのアミノ酸配列(配列番号47)の比較を示す。
【技術分野】
【0001】
一般には、本発明はドーパミン作動性ニューロンによるドーパミンの取り込みを促進し、パーキンソン病で死滅するニューロンの生存を助長する能力をもつ本明細書中でグリア細胞系由来神経栄養因子(グリア由来神経栄養因子又はGDNFとも呼ぶ)と呼ぶタンパク質に関する。本発明はより詳細には新規截形(truncated)GDNFタンパク質に関する。
【背景技術】
【0002】
神経栄養因子は神経系又は神経系により刺激される非神経組織に存在するタンパク質であり、神経及び/又はグリア細胞のの生存を促進し、その表現型分化を維持する機能をもつ(Varonら,Ann.Rev.Neuroscience 1:327,1979; Thoenenら,Science 229:238,1985)。この生理的役割により、神経栄養因子は種々の神経変性疾患で生じる神経細胞の変性及び分化機能の低下を治療するのに有用である。
【0003】
特定神経栄養因子が神経損傷の治療に潜在的に有用であるためには、損傷神経細胞の種類がその因子に応答性でなければならない。一般に、種々の神経栄養因子は全く異なる種類の神経細胞に作用する。従って、種々の形態の疾病又は傷害で生じ得る損傷ニューロン種の各々を治療できるように種々の異なる神経栄養因子を利用できることが有利である。
【0004】
神経栄養因子は種々の無関係の傷害から応答性ニューロンを保護することができる。例えば、神経成長因子(NGF)はその軸索突起の切断により生じる死(Richら,J.Neurocytol 16:261,1987; Ottoら,J.Neurosci.83:156,1987)、胚発生中の個体発生死(Hamburgerら,J.Neurosci.4:767,1984)及びタキソール又はシスプラチンの投与により生じる損傷(Apfelら,Ann.Neurol.29:87,1991)から感覚ニューロンの有意部分を救済する。この明白な保護の一般性に鑑み、ある神経栄養因子が実験的損傷から応答性ニューロンを保護するならば、その因果関係は不明であるとしても、このような神経栄養因子は患者にこれらのニューロンの損傷を引き起こす疾病を治療するのに有用であり得るという概念に至った。
【0005】
所与の神経栄養因子は適正なニューロン特異性をもつ以外に、医薬治療として使用するのに十分な量で入手できなければならない。神経栄養因子は一般に組織中に少量しか存在しない(例えばHoferとBarde,Nature 331:261,1988; Linら,Science 246:1023,1989)ので、医薬的量の神経栄養因子を動物組織から直接調製するのは困難である。代替手段として、所望のタンパク質を製造するために組換え発現系を使用することが望ましい。
【0006】
Linらは黒質ドーパミン作動性ニューロンの胚前駆体に及ぼす神経栄養活性について生物サンプルをスクリーニングする方法を過去に記載している(参考資料としてその開示内容を本明細書の一部とする1994年5月23日付け米国特許出願第08/182,183号とその親出願、1992年9月17日付けPCT/US92/07888(WO93/06116)、及びヨーロッパ特許出願第92921022.7号(公開EP第610254号)参照)。パーキンソン病は線条体を刺激する中脳内のドーパミン作動性ニューロンの変性により特徴付けられるで、このバイオアッセイはパーキンソン病の治療に使用可能な神経栄養因子を同定するのに有用である(Friedmanら,Neuro.Sci.Lett.79:65−72,1987)。 Linらは更に、このような材料の1例であるグリオブラストーマ細胞系B49からの条件付け培地から精製した新規神経栄養因子の特徴付けについても記載している(Schubertら,Nature 249:224−27,1974)。この細胞系からの条件付け培地はドーパミン作動性神経栄養活性を含むことが過去に報告されている(Bohnら,Soc.Neurosci.Abs.15:277,1989)。Linらの開示以前に、グリア細胞系由来神経栄養因子(GDNF)は独立した生物活性物質として同定されておらず、即ち実質的に純粋なタンパク質として単離されていなかった。更に、LinらはGDNFをコードするヒト遺伝子のクローニング方法、GDNFをコードするヒト遺伝子の核酸配列及びGDNFタンパク質のアミノ酸配列を記載している。GDNF遺伝子は発現ベクターにサブクローニングされ、ベクターを使用して生物学的に活性なGDNFを発現させている。GDNFタンパク質はジスルフィド結合により結合した22kDaの2個の134アミノ酸サブユニットから構成されるホモダイマーである。パーキンソン病等の神経損傷及び神経関連疾患の予防及び治療にGDNFを使用することも記載されている。
【0007】
GDNF治療は1種以上の神経細胞の生存及び/又は適正な機能を損なう症状に起因する神経損傷の治療に役立つ。このような神経損傷は多種多様の原因により生じると考えられる。神経損傷は、(1)傷害部位の近傍の軸索突起及び/又は神経細胞体の変性を生じる物理的傷害、(2)神経系の一部への血流の一時的又は永久的停止(例えば卒中)、(3)化学療法剤(例えば癌治療剤であるシスプラチンやエイズ治療剤であるジデオキシシチジン(ddC))等の神経毒素への意図的又は偶発的暴露、(4)糖尿病や腎不全等の慢性代謝病、あるいは(5)特定ニューロンポピュレーションの変性に起因するパーキンソン病、アルツハイマー病及び筋萎縮性軸索硬化症(ALS)等の神経変性病により、1種以上の神経細胞に生じると考えられる。
【0008】
GDNF治療はパーキンソン病等の黒質のドーパミン作動性ニューロンの変性を伴う神経変性症状の治療に特に有用であると考えられる。現在行われているパーキンソン病の唯一の治療は、線条体中のドーパミン濃度の増加を目的とした一時的軽減に過ぎない。GDNF治療は、線条体中のドーパミン作動性神経末端のドーパミン作動性神経伝達を増加する(その結果として症状を緩和する)だけでなく、変性プロセスの進行を遅らせ、あるいは停止させ、損傷黒質経路を修復し、その機能を回復する効果があると予想される。GDNFはヒト患者でドーパミン作動性神経細胞の他の形態の損傷又は機能不全を治療するにも有用であると考えられる。このような損傷又は機能不全は、精神分裂病や他の形態の精神病で生じ得る。このような症状に現在行われている唯一の治療は対症療法であり、ドーパミンレセプター又はドーパミン取り込み部位に作用する薬剤が必要であり、これらのレセプターを含むニューロンポピュレーションを刺激するドーパミン作動性ニューロンの機能不全が疾病プロセスに関係があるらしいという見地に立っている。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】米国特許出願第08/182,183号
【特許文献2】国際公開第93/06116号
【特許文献3】欧州特許出願公開第610254号
【非特許文献】
【0010】
【非特許文献1】Varonら,Ann.Rev.Neuroscience 1:327,1979; Thoenenら,Science 229:238,1985
【非特許文献2】Richら,J.Neurocytol 16:261,1987; Ottoら,J.Neurosci.83:156,1987
【非特許文献3】Hamburgerら,J.Neurosci.4:767,1984
【非特許文献4】Apfelら,Ann.Neurol.29:87,1991
【非特許文献5】HoferとBarde,Nature 331:261,1988; Linら,Science 246:1023,1989
【非特許文献6】Friedmanら,Neuro.Sci.Lett.79:65−72,1987
【非特許文献7】Schubertら,Nature 249:224−27,1974
【非特許文献8】Bohnら,Soc.Neurosci.Abs.15:277,1989
【発明の概要】
【0011】
1側面において、本発明は新規截形グリア細胞系由来神経栄養因子(GDNF)タンパク質産物を提供する。1態様では、組換え遺伝子工学技術により截形GDNFタンパク質を製造する。別の態様では、化学的方法又は組換え技術と化学的方法の組み合わせにより截形GDNFタンパク質を合成する。
【0012】
本発明の截形GDNFタンパク質産物はアミノ酸配列X−[Cys41−Cys133]−Yにより表されるタンパク質を含む。成熟GDNFタンパク質と比較し易くするために、図1のアミノ酸残基ナンバリング図(配列番号2)を使用する。
[Cys41−Cys133]は図1に示すCys41〜Cys133のアミノ酸配列(配列番号2)を表す。YはCys133のカルボキシ末端基又はIle134のカルボキシ末端アミノ酸残基を表す。XはCys41のメチオニン化もしくは非メチオニン化アミン基又は以下の群:
【0013】
【表1】
から選択されるアミノ末端アミノ酸残基を表す。前記截形GDNFタンパク質産物はX−[Cys41−Cys133]−Yにより表されるようなアミノ酸配列をもつ截形GDNFタンパク質産物とその変異体及び誘導体を含むと考えられる。従って、本発明の截形GDNFタンパク質産物はX−[Cys41−Cys133]−Yにより表されるアミノ酸配列の付加、置換及び内部欠失変異体及び誘導体も含む。截形GDNFタンパク質産物は更に、截形GDNFタンパク質のメチオニン化又は非メチオニン化形態とグリコシル化又は非グリコシル化形態も含む。
【0014】
本発明の截形GDNFタンパク質の例としては、メチオニン化又は非メチオニン化形態の[Arg16−Ile134]、[Asn22−Ile134]、[Pro23−Ile134]、[Ser26−Ile134]、[Arg32−Ile134]、[Gly33−Ile134]、[Lys37−Ile134]及び[Asn38−Ile134]截形GDNFタンパク質とその変異体及び誘導体が挙げられる。本発明の好ましい截形GDNFタンパク質としては、メチオニン化又は非メチオニン化形態の[Lys37−Ile134]及び[Asn38−Ile134]截形GDNFタンパク質とその変異体及び誘導体が挙げられる。置換変異体の例は[Asn22ΔSer22−Ile134]及び[Pro23−Lys37ΔAsn37−Ile134]截形GDNFタンパク質である。付加変異体の例はSer−[Pro23−Ile134]截形GDNFタンパク質である。
【0015】
本発明の別の側面では、截形GDNFタンパク質はグリコシル化形態で製造してもよいし、非グリコシル化形態でもよい。截形GDNFタンパク質の誘導体は、一般に水溶性ポリマーに截形GDNFタンパク質が結合している。例えば、截形GDNFタンパク質を1個以上のポリエチレングリコール分子に結合すると、水性環境での截形GDNFタンパク質産物の沈殿を減らすことができる。
【0016】
本発明の更に別の側面は、截形GDNFタンパク質をコードする種々のポリヌクレオチドに関する。これらの核酸配列は一般に真核又は原核宿主細胞で截形GDNFを発現させるのに使用され、発現産物又はその誘導体はドーパミン作動性細胞によるドーパミン取り込みを増加することができる。ポリヌクレオチドは細胞治療又は遺伝子治療用にも使用できる。適切な核酸配列としては、図面に具体的に示す配列や、付加的縮重配列(additional degenerate sequences)及び天然対立遺伝子変異配列が挙げられる。
【0017】
本発明の別の側面は、増幅及び/又は発現調節配列に作動的に連結した截形GDNFタンパク質をコードするポリヌクレオチドを含むベクターに関する。原核及び真核宿主細胞の両者を前記ベクターで安定的に形質転換及びトランスフェクトし、截形グリア由来神経栄養因子を発現させることができる。本発明は更に、前記形質転換又はトランスフェクトした宿主細胞を適切な栄養培地で培養し、場合により、前記細胞により発現される截形GDNFを宿主細胞及び/又は栄養培地から単離することを特徴とする、截形GDNFタンパク質の組換え製造にも関する。本発明は更に、遺伝子治療又は細胞治療における截形
GDNFをコードするポリヌクレオチドと該ポリヌクレオチドを含むベクターの使用にも関する。
【0018】
別の側面では、本発明は成熟GDNFタンパク質と該タンパク質から誘導される1種以上の截形GDNFタンパク質の混合物を含む組換え製造GDNF組成物にも関し、成熟GDNFタンパク質は約44kDaの分子量をもち、截形GDNFタンパク質は約36〜40kDaの分子量をもつ。GDNF組成物は約36kDaの分子量をもつ第1の種と約40kDaの分子量をもつ第2の種との少なくとも2種の截形GDNF種を含むものでもよい。約40kDaの分子量をもつ截形GDNF種は約22kDaの分子量をもつGDNFモノマーと約18kDaの分子量をもつ截形GDNFモノマーのヘテロダイマーである。前記混合物から截形GDNF種の1種以上を治療用に単離することも考えられる。
【0019】
本発明の別の側面は、截形GDNFタンパク質産物を含む医薬組成物に関する。一般に、截形GDNFタンパク質産物は医薬的に許容可能なビヒクルと組み合わせて調剤する。製造、貯蔵、取り扱い、送達及び/又は効力を助長するために種々の他の配合剤を使用してもよい。本発明の別の側面では、截形GDNFタンパク質産物はドーパミン作動性ニューロンのドーパミン取り込み及び生存を強化する。従って、截形GDNFタンパク質産物は、パーキンソン病等の傷害又は疾病により生じる神経系の損傷の治療に特に適している。
【0020】
本発明の他の側面及び利点は、本発明の実施を詳細に説明する以下の記載から当業者に容易に理解されよう。
【0021】
発明の詳細な説明
ヒトグリア細胞系由来神経栄養因子(hGDNF)は前駆物質として合成され、プロセシングにより134アミノ酸の成熟タンパク質として分泌される。成熟ヒトGDNFは図1に示すアミノ酸配列(配列番号2)をもつことが認められている。
【0022】
本発明は、その生物活性を維持しながら成熟GDNFタンパク質の寸法を小さくできるという予想外の発見に基づく(本明細書では「切断」もしくは「截形」タンパク質又は截形GDNFタンパク質とも言う)。切断タンパク質はチャイニーズハムスター卵巣(CHO)細胞でGDNFの組換え製造中に最初に発見された。要約すると、組換えヒトGDNF(rhGDNF)は次のように作製した。成熟ヒトGDNFタンパク質の完全オープンリーディングフレームをコードする核酸配列を発現プラスミドにクローニングした。(DNA配列決定によりGenBank中のhGDNF配列の等価物として)核酸配列が正しいことを確認し、成熟ヒトGDNFの公表配列(Linら,Science 260,1130−1132,1993)に等しいアミノ酸配列に翻訳した。プラスミドDNAを直鎖化し、リン酸カルシウム沈殿法を用いてジヒドロ葉酸レダクターゼ欠損CHO細胞(CHOd−細胞)にトランスフェクトした。トランスフェクトした細胞を選択培地で培養し、選択プロセスに耐えて生存したコロニーを選択し、hGDNF発現を個々に分析した。
【0023】
個々のクローンから無血清条件付け培地を集め、hGDNFに特異的な抗血清を用いてウェスタンブロット分析した。抗血清は、大腸菌で発現させた組換えhGDNFで免疫したウサギから誘導したウサギポリクローナル抗体から構成した。還元条件下でこれらのサンプル中に存在するhGDNFは、約22kDa及び18kDaの見かけの分子量をもつ2つの主バンドに分解した。各バンドは夫々約22+22.5kDa及び18+17.5kDaのごく近接する二重項(簡単にするために、以下の文中ではこれらの二重項を22kDa及び18kDaバンド又は種と呼ぶ)から構成されていた。
【0024】
GDNFは約20〜22kDaの分子量をもつ成熟GDNFタンパク質の2つの同一のサブユニットから構成されるジスルフィド結合ホモダイマーとして存在することが過去に報告されている。非還元条件下でGDNFを分析すると、32〜42kDa(Linら,Science 260,1130−1132,1993)又は33〜45kDa(Linら,J.Neurochem.63(2),758−768,1994)の幅広のバンドが確認されたと報告されている。レンジの存在は成熟モノマー上のグリコシル化の不均一に起因すると解釈され、脱グリコシル化実験で更に確証されている。
【0025】
本発明の22kDaバンドは文献に報告されている成熟GDNFタンパク質に対応するが、18kDaバンドは従来報告されていない。22kDa及び18kDaタンパク質の相対量は個々のクローンから採取したサンプルにより相違した。更に、同一クローンからの複数の回収物で2つのバンドの比が異なることが判明した。更に、CHOで発現されたGDNFタンパク質を貯蔵すると、多くの場合には18kDaバンドの存在が増加すると同時に22kDaバンドが減少することが判明した。
【0026】
形質転換CHOd−細胞からの条件付け培地を非還元条件下でウェスタンブロットにより分析すると、36、40及び44kDaの見かけの分子量をもつ3つの明確に分解したバンドが観察された。この知見も従来の報告に反するものであった。これらのバンドの相対強度は一定していないが、サンプルの各々に存在する22及び18kDaモノマーの比とよく相関していた。モノクローナル抗血清により更に分析すると、非還元ゲル中の3つのバンドは2種のモノマーから構成される3種の可能な二量体に対応することが判明した。最大の44kDaタンパク質は従来報告されているような2個の22kDa成熟GDNFタンパク質の二量体である。中間の40kDaタンパク質は、一方の成熟タンパク質の分子量が18kDa形態まで減少した二量体から構成される。最小の36kDa二量体は2個の18kDaタンパク質を含むと思われ、即ち22kDa形態の両者の分子量が減少している。このデータは、新規形態のGDNFモノマーの存在だけでなく、二量体構造における切断GDNFタンパク質の存在をも立証した最初のものである。貯蔵すると、サンプルのモノマー組成は切断形態及び対応する二量体種の組成に向かって変化することも判明し、即ち36kDaタンパク質の量が増加することが判明した。
【0027】
次に、従来報告されている成熟GDNFタンパク質と比較してタンパク質のどの部分が除去又は変異して分子量が減少するのかを調べる研究を行った。まず最初に分子量の減少はグリコシル化の変化には起因しないことが分かった。
【0028】
GDNFは2つの潜在的N結合グリコシル化部位を含み、グリコシル化されると報告されている。しかし、切断タンパク質は単に成熟GDNFの非グリコシル化又は低グリコシル化形態ではない。これは、サンプルをN−グリカナーゼ、O−グリカナーゼ及びノイラミニダーゼで処理する脱グリコシル化実験で実証された。還元ゲル上で18kDaタンパク質はN−グリカナーゼ消化により13.5kDaバンドまで還元され、N結合糖の4.5kDaの等価物の存在を示した。ノイラミニダーゼ及びO−グリカナーゼで処理すると、18kDaバンドは17kDaまで僅かにシフトした。従って、タンパク質にO結合糖が存在すると予想された。成熟22kDaバンドはグリコシル化されると報告されており、同様にN−グリカナーゼにより18kDaまで(即ち同様に4.5kDaだけ)還元された。これは、ゲル上の22kDaバンドに特異的なモノクローナル抗体を使用することにより更に確認された。非還元二量体のグリカナーゼ消化パターンはより複雑であったが、解明可能であり、3形態の初期帰属に一致した。
【0029】
その結果、タンパク質の分子量の4.5kDaの減少はグリコシル化の変化ではなく約30〜35アミノ酸残基の欠失に起因すると考えられた。この欠失は以下の理由で成熟GDNFタンパク質のアミノ末端に最も生じ易いと予想された。成熟GDNFは合計7個のシスチンを含む。欠失がカルボキシル末端から開始したならば、7個のシスチンのうちの2〜4個が失われ、その結果、不活性タンパク質が生じ易いと思われる。しかし、主に切断形態から構成される試料をバイオアッセイにかけ、そのドーパミン作動性ニューロン神経栄養活性を測定した処、この試料は成熟形GDNFを高比率で含む試料と同等の活性を示した。
【0030】
次に、精製タンパク質のアミノ酸配列分析により切断部位を決定した。Applied Biosystems 494Aタンパク質シーケンサーを10サイクル使用して製造業者の指示に従ってサンプルを配列決定した。アミノ酸配列分析技術及び手順は当業者に公知であるが、タンパク質の配列決定に関する詳細な説明は、参考資料としてその開示内容を本明細書の一部とするFaussetら,Electrophoresis 12:22−27,1991及び1990年8月24日付け米国特許出願第576,316号(1990年10月4日付けヨーロッパ特許出願第90310899号、公開EP第423980号、発明の名称「幹細胞因子」)に記載されている。分析の結果、切断タンパク質のアミノ末端は「RGQRGK」即ちArg−Gly−Gln−Arg−Gly−Lysであることが判明した。即ち、条件付け培地中の成熟タンパク質から最初から31個のアミノ酸が除去されていた。アミノ酸Arg32から始まる切断タンパク質の残りのアミノ酸配列は、この欠失以外は図1に示す成熟GDNFアミノ酸配列(配列番号2)に一致していた。
【0031】
[Arg32−Ile134]截形GDNFタンパク質は、ドーパミン作動性ニューロンアッセイで定量的に活性であることが判明した。ドーパミン作動性神経栄養活性アッセイは、パーキンソン病の治療に有益であると考えられる神経栄養因子を同定するために使用されている。このアッセイは従来記載されているアッセイ(参考資料としてその開示内容を本明細書の一部とするFriedmanら,Neuro.Sci.Lett.79:65−72,1987)を基礎とし、Linら(1994年5月23日付け米国特許出願第08/182,183号とその親出願、1992年9月17日付けPCT/US92/07888(WO93/06116)及びヨーロッパ特許出願第92921022.7号(公開EP第610254号))に記載されているように改変できる。アッセイの詳細な説明は後記実施例5に記載する。
【0032】
その後の精製操作と、アミノ酸配列決定の結果、成熟GDNFのN末端から最初から36個のアミノ酸残基が除去された別のタンパク質即ちKNRG(C)VL−−のN末端配列をもつ[Lys37−Ile134]截形GDNFタンパク質が発見された。この場合も、切断タンパク質の残りのアミノ酸残基はこの欠失以外は成熟ヒトGDNFアミノ酸配列と一致していた。
[Lys37−Ile134]截形GDNFタンパク質もドーパミン取り込みバイオアッセイで分析した。この截形GDNFタンパク質は、大腸菌で発現させた精製組換え成熟GDNFと同様に約50pg/mlのED50で活性であることが判明した。
【0033】
更に、細菌で発現させた成熟GDNFを截形型に改変できることも分かった。(Linらの上記米国特許出願第08/182,183号に記載されているように)形質転換大腸菌で発現させた成熟GDNFをCHO由来条件付け培地と共にインキュベートした。組換え大腸菌GDNFは還元ゲル上で17kDaの見かけの分子量をもつ。材料をCHO細胞条件付け培地と混合し、4℃で5日間インキュベートした処、タンパク質は12.5kDaまで完全に切断された。この切断は1時間又は24時間のインキュベーションでは不完全であり、このような条件下で時間依存性プロセスであることが示唆された。
0.1%ウシ胎児血清を含む培地と共に組換え大腸菌GDNFを一晩インキュベートしただけでは切断形態が生成されないことも判明した。従って、切断プロセスが生じるには培養に生きた細胞の存在が必要であると思われる。従って、切断現象は組織によってはin vivoでも生じると考えられる。
【0034】
更に、(同様にLinらの上記米国特許出願第08/182,183号に記載されているような)ペジル化GDNF等の成熟大腸菌発現hGDNFの誘導体をCHO由来条件付け培地の存在下で截形型に加工できることが判明した。成熟GDNFをアミノ末端でペジル化すると、その循環クリアランス時間を増すことができる。ペジル化はタンパク質の寸法を増加し、修飾成熟
GDNFは還元条件下で約45kDaで移動する。ペジル化大腸菌GDNFをCHO細胞(未トランスフェクト)条件付け培地と共にインキュベートすると、非ペジル化成熟形態と同様に12.5kDaバンドが生じた。どちらの場合も12.5kDa種は非還元ゲル上でジスルフィド結合二量体として存在していた。N末端ペジル化成熟タンパク質からこの切断形が生成されたことから、タンパク質のN末端に切断現象が生じたのは切断プロセス中にペジル化残基が失われたためであると考えられる。
【0035】
これらの知見と、in vivoでも切断現象が生じると予想されることから、GDNFタンパク質の截形型は生理的条件下のhGDNFの最終天然プロセシング形態であると考えられる。従って、治療用截形GDNFタンパク質又はその誘導体を製造すると有利であると思われた。例えば、直接発現させるか又は合成した截形GDNFタンパク質(例えば[Arg32−Ile134]截形GDNFタンパク質)は上記タンパク分解活性に耐性であると予想される。更に、ペジル化[Arg32−Ile134]截形GDNFタンパク質等の截形GDNF誘導体を製造することが所望される場合には、得られる誘導体は成熟GDNF誘導体で観察される特異的切断に非感受性であるという利点をもつと予想される。
【0036】
截形GDNFタンパク産物には付加的利点も予想できる。第1に、[Arg32−Ile134]截形GDNFタンパク質等の截形タンパク質のpIは約10から約8.0〜8.5まで低下するであろう。従って、タンパク質は極めて低塩基性となり、その結果、鞘内注射部位等の投与部位におけるレセプター結合の改善や細胞毒性の低下等の有益な効果が得られる。第2に、成熟GDNFアミノ酸配列の最初から26個のアミノ酸には、Arg−Asn−Arg(アミノ酸14〜16)とGlu−Asn−Ser(アミノ酸24〜26)の2つのアミド分解部位が存在する。截形GDNFタンパク質にはこれらの部位の一方又は両方が存在しないので、タンパク質の安定性が増すと予想される。
【0037】
截形GDNFタンパク質産物
基本態様では、本発明の截形GDNFタンパク質は成熟GDNFタンパク質と比較し易くするために図1のアミノ酸残基ナンバリング図を用いた下記アミノ酸配列:
X−[Cys41−Cys133]−Y
(式中、[Cys41−Cys133]は図1に示すCys41〜Cys133のアミノ酸配列(配列番号1)を表し、YはCys133のカルボキシ末端基又はIle134のカルボキシ末端アミノ酸残基を表し、XはCys41のメチオニン化もしくは非メチオニン化アミン基又は以下の群:
【0038】
【表2】
から選択されるアミノ末端アミノ酸残基を表す)により表すことができる。
【0039】
本明細書で使用する「截形GDNFタンパク質産物」なる用語は、生物学的に活性な合成又は組換え截形GDNFタンパク質、成熟GDNFから産生される截形GDNFタンパク質、生物学的に活性な截形GDNF変異体(挿入、置換及び欠失変異体を含む)、及びその化学修飾誘導体を含む。配列番号2に示すアミノ酸配列をもつヒトGDNFタンパク質に実質的に相同な截形GDNFタンパク質も含まれる。
【0040】
本明細書で使用する「生物学的に活性」なる用語は、截形GDNFタンパク質が配列番号2に示すアミノ酸配列をもつGDNFタンパク質と必ずしも同一程度まで全く同一の性質を示す必要はないが、これに類似の神経栄養性を示すことを意味する。目的とする特定神経栄養性の選択は、截形GDNFタンパク質産物を投与する用途に依存する。截形GDNFタンパク質産物は生物学的に活性であり、後記実施例に記載するような典型的バイオアッセイ等でドーパミン取り込み及びチロシンヒドロキシラーゼ(TH)発現を評価すると、成熟GDNFタンパク質に類似のドーパミン作動性ニューロン生存特性を示す。
【0041】
本明細書で使用する「実質的に相同」なる用語は、配列番号2に示すアミノ酸配列をもつヒトGDNFタンパク質に対して好ましくは>70%、より好ましくは>80%、更に好ましくは>90%又は更には>95%の相同度を意味する。本明細書に記載する相同百分率は、参考資料としてその開示内容を本明細書の一部とするDayhoff,Atlas of Protein Sequence and Structure Vol.5,p.124(1972),National Biochemical Research Foundation,Washington,D.C.に記載されているように、整列を助長するのに100アミノ酸長の4個のギャップを導入すればよい場合に、比較する配列中の同一のアミノ酸残基であって整列する2種の配列の小さいほうに存在するアミノ酸残基の百分率として計算される。更に、配列番号2のGDNFに対する抗体との交差反応性により単離できるか、又は配列番号1のGDNFをコードする遺伝子もしくは該遺伝子のセグメントとのハイブリダイゼーションによりその遺伝子を単離できるならば、任意の截形GDNFタンパク質を実質的に相同とみなす。
【0042】
本明細書の記載から当業者に容易に理解されるように、実質的に相同なタンパク質はX−[Cys41−Cys133]−Yにより表される截形GDNFタンパク質のアミノ酸残基の1個以上の欠失、付加又は置換を含む。このような変異体の作製については以下に詳述する。本発明は明らかに「截形」GDNFタンパク質に関するものであるので、アミノ末端付加変異体はメチオニン残基又は非GDNFアミノ酸残基もしくは配列を含むと考えられるが、成熟GDNFタンパク質を再構成するようなアミノ酸残基の付加は含まないことも理解されよう。天然対立遺伝子突然変異体又は変異体に基づく截形GDNFタンパク質も本発明の範囲に含まれる。変異体截形GDNFタンパク質の製造については以下に詳述する。
【0043】
Linら(上記米国特許出願第08/182,183号)は、成熟GDNFのカルボキシル末端から夫々6番目及び5番目の残基であるLys−Arg残基のタンパク分解プロセシングによる成熟GDNFのカルボキシル末端截形を記載している(即ち図1のアミノ酸残基ナンバリング(配列番号1又は配列番号2でも同じ)によるとLys129−Arg130)。このような截形は成熟GDNFタンパク質から2個のシステイン残基を除去すると思われる。その結果、タンパク質の不適正な折り畳みを生じ易く、不活性なタンパク質を形成する恐れがある。これに対して本発明のX−[Cys41−Cys133]−Y截形GDNFタンパク質産物はCys131及びCys133残基を維持しており、ドーパミン取り込みアッセイにより活性タンパク質であると判定される。
【0044】
本発明の1態様では、好ましい截形GDNFタンパク質産物は1個以上のアミド分解部位を欠失している。このようなアミド分解部位の欠失は精製タンパク質の生化学的安定性を増すと共に、予想される分解物を減らし、貯蔵安定性の高いタンパク質をもたらすと考えられる。截形GDNFタンパク質産物の1例は、成熟タンパク質のアミド分解をもたらすような部位を欠失する[Ser26−Ile134]截形GDNFタンパク質である。あるいは、[Arg16−Ile134]截形GDNFタンパク質は成熟タンパク質に存在する少なくとも最初のアミド分解部位を欠失している。
【0045】
本発明の好ましい截形GDNFタンパク質産物は[Arg32−Ile134]截形GDNFタンパク質である。この截形
GDNFタンパク質は成熟タンパク質のタンパク分解切断部位又はその近傍部位を欠失している。従って、この截形GDNFタンパク質はin vivoでも生じると思われるプロセシング現象に耐性であると予想される。本発明の好ましい別の截形GDNFタンパク質産物は[Lys37−Ile134]截形GDNFタンパク質である。この截形は、夫々N及びC末端からGly40及びIle134までの残基を除去する他の截形と同様に、截形タンパク質のpIを更に低下させると思われる。本発明の最も好ましい截形GDNFタンパク質産物は成熟GDNFタンパク質に存在する全システイン残基を維持しながら、発現及び製造中又はin vivo投与後の截形GDNFタンパク質の迅速なタンパク分解プロセシングに関与する部位を欠失している。これらの好ましいタンパク質としては、[Arg32−Ile134]、[Gly33−Ile134]、[Gln34−Ile134]、[Arg35−Ile134]、[Gly36−Ile134]、[Lys37−Ile134]、[Asn38−Ile134]及び[Arg39−Ile134]截形GDNFタンパク質産物が挙げられる。
【0046】
Linら(上記米国特許出願第07/855,413号)により成熟GDNFについて従来記載されている結果と同様に、本発明の截形GDNFタンパク質は黒質ドーパミン作動性ニューロンの胚前駆体によるドーパミン取り込みを増加できることが判明した。截形GDNFタンパク質のバイオアッセイについては、後記実施例4に詳述する。
【0047】
新規截形GDNFタンパク質は一般に、他の(非GDNF)タンパク性材料を実質的に含まない截形GDNFタンパク質を形成するように単離精製される。好ましくは、截形GDNFタンパク質産物は、截形GDNFタンパク質産物の製造に使用する製造技術に起因し得る他のタンパク質を約80%除去している。より好ましくは、截形GDNFタンパク質産物は他のタンパク質を約90%除去し、特に好ましくは他のタンパク質を約95%除去し、最も好ましくは他のタンパク質を約>98%除去している。更に、本発明は均一な截形GDNFタンパク質を製造するためのポリヌクレオチド配列を提供するという独自の利点を提供する。例えば、[Arg32−Ile134]截形GDNFタンパク質をコードするポリヌクレオチド配列を使用すると、大腸菌及び他の適当な発現系で截形GDNFタンパク質を組換え製造することができる。換言するならば、新規ポリヌクレオチドはタンパク分解プロセシングに非感受性であるか又はこのようなプロセシングもしくは上述したような他の生化学プロセシング効果に対する感受性の低い截形GDNFタンパク質を製造することができる。こうして、新規ポリヌクレオチドにより単一種の截形GDNFタンパク質を容易に製造及び/又は単離することができ、従って、截形GDNFタンパク質及び/又はその産物は上記ヘテロ及びホモダイマーの混合物を全く又は少量しか含まない。他方、最終截形GDNFタンパク質産物は、追って詳述するように投与前に他の因子、化学組成物及び/又は適切な医薬配合剤と配合できることが理解されよう。
【0048】
本発明の1側面では、截形GDNFタンパク質は高純度で比較的多量のタンパク質を獲得できるという理由から組換え技術により製造すると有利である。組換え截形GDNFタンパク質形態は該タンパク質のグリコシル化及び非グリコシル化形態を含み、細菌、哺乳動物又は昆虫細胞系で発現される。あるいは、截形GDNFタンパク質は化学的に合成することもできる。本発明の好ましい製造方法については追って詳述する。
【0049】
截形GDNF変異体及び誘導体
A.截形GDNF変異体
本発明の別の側面は截形GDNFタンパク質の変異体に関する。本明細書で使用する「截形GDNFタンパク質産物」なる用語は、天然GDNFのアミノ酸配列の残基からアミノ酸が欠失(「欠失変異体」)、挿入(「付加変異体」)又は置換(「置換変異体」)している変異体タンパク質を含む。このような変異体は、タンパク質をコードするDNAに適当なヌクレオチド変異を導入するか又は所望のタンパク質のin vitro化学合成により作製される。最終タンパク質がGDNF生物活性をもつのであれば、欠失、挿入及び置換の多数の組み合わせが可能であることが当業者に理解されよう。
【0050】
1個以上の選択されたアミノ酸残基の置換、挿入又は欠失のための突然変異誘発技術は当業者に周知である(例えば参考資料としてその開示内容を本明細書の一部とする米国特許第4,518,584号参照)。アミノ酸配列変異体の構築には、突然変異部位の位置と突然変異の種類という2つの主な変更が存在する。截形GDNF変異体に設計に際して、突然変異部位の位置と突然変異の種類は修飾しようとする生化学特性により異なる。突然変異部位は、例えば(1)まず同類アミノ酸選択で置換した後、得られた結果に応じてよりラジカルな選択で置換するか、(2)標的アミノ酸残基を欠失させるか、(3)位置決定した部位の隣接位置にアミノ酸残基を挿入することにより、個々に又は連続して修飾することができる。
【0051】
アミノ酸配列欠失は一般に約1〜30アミノ酸残基、より通常は約1〜10残基、典型的には約1〜5残基である。例えば、Cys41のN末端に配置されたアミノ酸残基の「X」部分の欠失は約1〜30残基でもよいが、[Cys41−Cys133]のシステイン残基はタンパク質折り畳みを破壊しないように、一般にその位置に応じて約1〜5残基である。截形GDNFタンパク質における欠失はトランスフォーミング成長因子−β(TGF−β)ファミリーメンバーに対して相同性の低い領域に形成できる。他のTGF−βファミリー配列に対して実質的に相同な領域で截形GDNFタンパク質から欠失させると、生物活性がより有意に変化する傾向がある。総欠失及び/又は連続欠失の数は、影響を受ける領域、例えばシステイン架橋における截形GDNFタンパク質の三次構造を維持するように選択される。
【0052】
アミノ酸配列付加としては、1残基〜100残基以上の長さのアミノ及び/又はカルボキシル末端融合と、単一又は多重アミノ酸残基の内部配列内挿入が挙げられる。内部付加は一般に約1〜10アミノ酸残基、より一般には約1〜5アミノ酸残基、通常は約1〜3アミノ酸残基であり得る。上述のように、本発明のアミノ末端付加変異体はメチオニン(例えば細菌組換え細胞培養におけるGDNFの直接発現のアーチファクト)又は非GDNFアミノ酸残基もしくは配列の付加を含むと考えられる。アミノ末端付加変異体は、成熟GDNFタンパク質の再構成をもたらすようなアミノ酸残基の付加を含まない。末端挿入の別の例として、組換え宿主細胞からのタンパク質の分泌を助長するように異種N末端シグナル配列をN末端と融合してもよい。このようなシグナル配列は一般に、所期宿主細胞種から得られ、従ってこのような細胞種と相同である。挿入又は付加変異体は、他の神経栄養因子の配列に由来するアミノ酸配列を含んでいてもよい。
【0053】
別の群の変異体はアミノ酸置換変異体である。これらの変異体は截形GDNFタンパク質から少なくとも1個のアミノ酸残基が除去され、その代わりに別の残基が挿入されている。例えば、Met残基を更に除去し易くするために天然Asn22をSerに置換した図5参照。X−[Cys41−Cys133]−Yアミノ酸配列式と截形GDNFタンパク質産物の本発明の定義を使用すると、このような截形GDNFタンパク質は置換変異体Met−[Asn22ΔSer22−Ile134]截形GDNFタンパク質又は付加変異体Met−Ser−[Pro23−Ile134]截形GDNFタンパク質と呼ぶことができる。置換変異体は、アミノ酸置換を生じるか否かに拘わらず種ポピュレーションの天然ヌクレオチド配列変異を特徴とする対立遺伝子変異体でもよい。
【0054】
截形GDNFタンパク質の配列の具体的な突然変異としては、グリコシル化部位(例えばセリン、スレオニン又はアスパラギン)の修飾が挙げられる。グリコシル化又は単に部分的なグリコシル化の不在は、任意のアスパラギン結合グリコシル化認識部位又はO結合炭水化物の付加により修飾されたタンパク質の任意部位におけるアミノ酸置換又は欠失に起因すると考えられる。アスパラギン結合グリコシル化認識部位は適当な細胞グリコシル化酵素により特異的に認識されるトリペプチド配列を含む。これらのトリペプチド配列はAsn−Xaa−Thr又はAsn−Xaa−Serであり、ここでXaaはPro以外の任意のアミノ酸でよい。グリコシル化認識部位の第1又は第3番目のアミノ酸位置の一方又は両方に種々のアミノ酸置換又は欠失(及び/又は第2位のアミノ酸欠失)が存在すると、修飾トリペプチド配列にグリコシル化は生じない。従って、適当に改変されたヌクレオチド配列が発現されると、前記部位がグリコシル化されていない変異体を生じる。あるいは、截形GDNFタンパク質にグリコシル化部位を付加するように配列を修飾してもよい。
【0055】
突然変異誘発に用いる截形GDNFアミノ酸残基又は領域の指定方法の1例はCunninghamとWells(Science,244: 1081−1085,1988)により記載されている「アラニン走査突然変異誘発」と呼ばれる方法である。この方法では、標的残基のアミノ酸残基又は基(例えばArg、Asp、His、Lys及びGlu等の帯電残基)を指定し、中性又は負電荷をもつアミノ酸(最も好ましくはアラニン又はポリアラニン)で置換し、アミノ酸を細胞の内側又は外側の周囲水性環境と相互作用させる。次に、置換部位に付加又は代替残基を導入することにより、置換に機能的感受性を示す領域を限定する。こうしてアミノ酸配列修飾を導入する部位を予め決定し、所与の部位での突然変異の性能を最適化するために、アラニン走査又はランダム突然変異誘発を実施し、所望の活性及び活性度の最適組み合わせを求めて変異体をスクリーニングする。
【0056】
置換突然変異誘発に最も有利な部位としては、種々の種からのGDNFタンパク質に存在するアミノ酸が側鎖の大きさ、電荷及び/又は疎水性の点で実質的に相違する部位が挙げられる。他の有用な部位としては、種々の種から得られるGDNF様タンパク質の特定残基が同一である部位が挙げられる。このような位置は一般にタンパク質の生物活性に重要である。まず最初に、これらの部位を比較的同類的に置換して修飾する。このような同類置換を表1の好ましい置換の欄に示す。このような置換により生物活性が変化するならば、より実質的な変異(典型的な置換)を導入及び/又は他の付加/欠失を実施し、得られた産物をスクリーニングする。
【0057】
【表3】
【0058】
アミノ酸配列に穏当な改変(及びコーディング核酸配列の対応する修飾)を加えると、後記実施例に記載する截形GDNFタンパク質に類似の機能的及び化学的特性をもつ截形GDNFタンパク質を生じると予想される。他方、截形GDNFタンパク質の機能的及び/又は化学的特性の実質的変更は、(a)置換領域のポリペプチド主鎖の構造(例えばシート又は螺旋構造等)、(b)標的部位のタンパク質の電荷もしくは疎水性、又は(c)側鎖の大きさのいずれかを維持する効果の点で有意に異なる置換を選択することにより達せられる。天然残基は共通の側鎖性質に基づいて次のように分類される。
1)疎水性:ノルロイシン、Met、Ala、Val、Leu、Ile、
2)中性親水性:Cys、Ser、Thr、
3)酸性:Asp、Glu、
4)塩基性:Asn、Gln、His、Lys、Arg、
5)側鎖配向に影響する残基:Gly、Pro、及び
6)芳香族:Trp、Tyr、Phe。
【0059】
穏当でない置換は、これらの分類の1つのメンバーを別のメンバーに置換するものと言える。このような置換残基は、他のTGF−βタンパク質と相同の截形GDNFタンパク質の領域に導入してもよいし、タンパク質の非相同領域に導入してもよい。
【0060】
B.截形GDNF誘導体
本明細書の開示に基づき、当業者は截形GDNF又は截形
GDNF変異体の化学修飾誘導体を作製できる。截形GDNFタンパク質の誘導体化に最も適した化学種としては水溶性ポリマーが挙げられる。水溶性ポリマーはこれと結合したタンパク質が生理的環境等の水性環境で沈殿しないという理由で望ましい。ポリマーは治療用製品又は組成物の製造に医薬的に許容可能であることが好ましい。当業者はポリマー/タンパク質結合体を治療に使用するか否か、治療用の場合には所望の投与量、循環時間、耐タンパク分解性及び他の要因に基づいて所望のポリマーを選択することができよう。誘導体化の効果は、所望形態(即ち浸透圧ポンプ、より好ましくは注射もしくは注入、又は他の経口、肺もしくは他の送達経路用剤形)で誘導体を投与し、その効果を判定することにより確認することができる。
【0061】
適切な水溶性ポリマーの非限定的な例としては、ポリエチレングリコール(PEG)、エチレングリコール/プロピレングリコールのコポリマー、モノメトキシポリエチレングリコール、カルボキシメチルセルロース、デキストラン、ポリビニルアルコール、ポリビニルピロリドン、ポリ−1,3−ジオキソラン、ポリ−1,3,6−トリオキサン、エチレン/無水マレイン酸コポリマー、ポリアミノ酸(ホモポリマー又はランダムポリマー)、ポリ(n−ビニルピロリドン)ポリエチレングリコール、プロピレングリコールホモポリマー、ポリプロピレンオキシド/エチレンオキシドコポリマー、ポリオキシエチル化ポリオール(例えばグリセロール)、ポリエチレングリコールプロピオンアルデヒド及びその混合物が挙げられる。本明細書で使用するポリエチレングリコールとは、モノ(C1〜C10)アルコキシ又はアリールオキシポリエチレングリコールのように、他のタンパク質を誘導体化するために使用されているPEG形態の任意のものを意味する。ポリエチレングリコールプロピオンアルデヒドはその水中安定性により製造が有利であると思われる。ポリマーは任意の分子量でよく、枝分かれしていてもいなくてもよい。
【0062】
本発明は特に、少なくとも1個のPEG分子に結合した截形GDNFタンパク質を含む截形GDNFタンパク質産物に関する。別の側面では、本発明はアシル又はアルキル結合を介して少なくとも1個のPEG分子に結合した截形GDNFタンパク質に関する。
【0063】
ペジル化は当該技術分野で公知のペジル化反応の任意のものにより実施することができる。例えば、Focus on Growth Factors 3(2):4−10(1992); EP第0154316号;EP第0401384号;及びMalikら,Exp.Hematol.20:1028−1035(1992)(塩化トレシルを使用したGM−CSFのペジル化の報告)参照。ペジル化は反応性水溶性ポリマーとのアシル化反応又はアルキル化反応を介して実施するのが好ましい。これらの好ましい誘導体化手段については以下に詳述する。アシル化反応では、単一反応性エステル基をもつポリマーを選択するのが好ましい。還元アルキル化反応では、単一反応性アルデヒド基をもつポリマーを選択するのが好ましい。更に、単一反応基(例えばアシル化には活性エステル、アルキル化にはアルデヒド)をもつように選択ポリマーを修飾し、重合度を調節できるようにしてもよい。天然グリコシル残基は通常は哺乳動物組換え発現系により簡便に製造されるので、一般に、水溶性ポリマーは天然グリコシル残基からは選択しない。
【0064】
アシル化
本発明では、アシル化によるペジル化は一般にポリエチレングリコールの活性エステル誘導体を截形GDNFタンパク質と反応させることにより行われる。ペジル化工程を実施するためには任意の公知又は今後発見される反応性PEG分子を使用することができる。好ましい活性化PEGエステルはN−ヒドロキシスクシンイミド(「NHS」)にエステル化したPEGである。本明細書で使用する「アシル化」とは、非限定的な例として截形GDNFタンパク質とPEG:アミド、カルバメート、ウレタン等の水溶性ポリマーの結合を意味する。Bioconjugate Chem.5:133−140(1994)参照。反応条件はペジル化技術で公知の条件又は今後開発される条件の任意のものから選択することができるが、修飾しようとする截形GDNFタンパク質を不活化するような温度、溶媒及びpHレベル等の反応条件への暴露は回避又は制限すべきである。
【0065】
アシル化によるペジル化の結果、一般にリジンε−アミノ基がアシル結合基を介してペジル化されたポリペジル化截形GDNFタンパク質が得られる。アミドによる結合が好ましい。また、得られる生成物が実質的に単独で(例えば≧95%)モノ、ジ又はトリペジル化されていることも好ましい。他方、使用する特定反応条件に依存する量でペジル化度の高い結合物が若干形成される場合もある。所望により、特に透析、塩析、限外濾過、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー及び電気泳動等の標準精製技術により混合物から高純度ペジル化結合体を製造することもできる。
【0066】
アルキル化
本発明では、アルキル化によるペジル化は一般に還元剤の存在下でPEGの末端アルデヒド誘導体を截形GDNFタンパク質と反応させることにより実施される。アルキル化によるペジル化もポリペジル化截形GDNFタンパク質を生成することができる。更に、実質的にタンパク質のN末端のα−アミノ基のみでペジル化を助長するように反応条件を操作することもできる(即ちモノペジル化種)。モノペジル化又はポリペジル化のいずれの場合もPEG基は−CH2−NH基を介してタンパク質に結合しているのが好ましい。特に−CH2−基に関して、この型の結合を本明細書では「アルキル」結合と呼ぶ。
【0067】
特定タンパク質で誘導体化に利用可能な種々の型の第1アミノ基(N末端に対するリジン)の反応性の差を利用する還元アルキル化により、選択的N末端化学修飾を実施することができる。適当な反応条件下では、カルボニル基含有ポリマーによりN末端でタンパク質の実質的に選択的な誘導体化が達せられる。例えば、リジン残基のε−アミノ基とタンパク質のN末端残基のα−アミノ基のpKa差を利用できるようなpHで反応を実施することにより、タンパク質を選択的にN末端ペジル化することができる。このような選択的誘導体化により、水溶性ポリマーとタンパク質の結合が制御され、ポリマーとの結合は主にタンパク質のN末端で行われ、リジン側鎖アミノ基等の他の反応基の有意修飾は生じない。還元アルキル化を使用する場合、水溶性ポリマーはタンパク質との結合に関わる反応性アルデヒドをただ1つだけもつことが好ましい。例えば、単一反応性アルデヒドを含むポリエチレングリコールプロピオンアルデヒドを使用する。
【0068】
本発明は、PEG基がアシル又はアルキル基を介して結合したペジル化截形GDNFタンパク質を含む。上述のように、このような截形GDNFタンパク質産物はモノペジル化物でもポリペジル化物(例えば2〜6個、好ましくは2〜5個のPEG基を含む)でもよい。PEG基は一般にアミノ酸のα又はε−アミノ基でタンパク質に結合しているが、適切な反応条件下でPEG基に結合するように十分反応性の基であるならば、タンパク質の任意の反応基にPEG基を結合できるとも考えられる。例えば、遊離アミノ基又はカルボキシル基等の反応基を介してポリエチレングリコールをタンパク質に共有結合させてもよい。反応基は活性化PEG分子を結合可能な基である。遊離アミノ基をもつアミノ酸残基としては、リジン残基とN末端アミノ酸残基が挙げられる。遊離カルボキシル基をもつアミノ酸残基としては、アスパラギン酸残基、グルタミン酸残基及びC末端アミノ酸残基が挙げられる。スルフヒドリル基もPEG分子を結合する反応基として使用することができる。治療目的では、例えばN末端又はリジン基等のアミノ基に結合するのが一般には好ましい。レセプター結合が所望される場合にはレセプター結合に重要な残基に結合するのは避けるべきである。
【0069】
1側面では、本発明はポリマー分子が実質的に単独位置のみ(即ち≧95%)に結合しているモノポリマー/截形GDNFタンパク質結合体の実質的に均一な調製物を提供する。より詳細には、PEGを使用する場合には、本発明は場合により抗原結合基を欠失し、截形GDNFタンパク質と直接結合したPEG分子をもつペジル化截形GDNFタンパク質も提供する。
【0070】
更に、グリコシル化、非グリコシル化又は脱グリコシル化截形GDNFタンパク質を使用して誘導体を製造することもできる。一般には非グリコシル化截形GDNFタンパク質を使用する。例えば、原核生物で発現させた[Arg32−Ile134]截形GDNFタンパク質を化学誘導体化し、アシル又はアルキル基を介して結合した1個又は複数(例えば2〜4個)のPEG分子を含むようにすることができる。
【0071】
一般に、化学誘導体化は生物学的に活性な物質を活性化ポリマー分子と反応させるために使用するのに適した任意の条件下で実施することができる。ペジル化截形GDNFタンパク質の製造方法は一般に、(a)截形GDNFタンパク質が1個以上のPEG基と結合するような条件下で截形GDNFタンパク質をポリエチレングリコール(例えばPEGの反応性エステル又はアルデヒド誘導体)と反応させる段階と、(b)反応物を得る段階を含む。一般に、アシル化反応に最適な反応条件は公知パラメーターと所望の結果に基づいて個々に決定される。例えば、PEG対タンパク質の比が大きいほど、ポリペジル化物の百分率が高い。(過剰の未反応タンパク質又はポリマーが存在しないという意味での反応効率に関する)最適比は所望の誘導体化度(例えばモノ、ジ、トリ等)、選択するポリマーの分子量、ポリマーが枝分かれしているか否か、使用する反応条件等の因子により決定することができる。
【0072】
モノポリマー/截形GDNFタンパク質結合体の実質的に均一なポピュレーションを製造するための還元アルキル化は、一般に(a)截形GDNFタンパク質のアミノ末端でα−アミノ基の選択的修飾を可能にするのに適したpHで還元条件下に截形GDNFタンパク質を反応性PEG分子と反応させる段階と、(b)反応物を得る段階を含む。
【0073】
モノポリマー/截形GDNFタンパク質結合体の実質的に均一なポピュレーションでは、還元アルキル化反応条件は截形GDNFタンパク質のN末端に水溶性ポリマー部分を選択的に結合できるような条件である。このような反応条件は、一般にリジンアミノ基とN末端のαアミノ基の間にpKa差を提供する(pKaはアミノ基の50%がプロトン化され、50%がプロトン化されないpHである)。pHは使用するポリマーとタンパク質の比にも影響を与える。一般に、pHが低いほど、高いポリマー対タンパク質比が所望される(即ちN末端αアミノ基が低反応性であるほど、最適条件に達するために多量のポリマーが必要となる)。pHが高い場合には、ポリマー対タンパク質比を大きくする必要はない(即ち高反応性基を利用できるので、必要なポリマー分子数は少なくて済む)。本発明の目的では、pHは一般に3〜9、好ましくは3〜6の範囲である。
【0074】
ポリマーの分子量も考慮すべきである。一般に、ポリマーの分子量が大きいほど、タンパク質に結合できるポリマー分子の数は少ない。また、これらのパラメーターを最適化する際にはポリマーの分枝度も考慮すべきである。一般に、分子量が大きいほど(又は分枝度が高いほど)ポリマー対タンパク質比は高い。一般に、本発明で意図するペジル化反応では、好ましい平均分子量は約2kDa〜約100kDaである(「約」なる用語はポリエチレングリコールの調製物中に指定分子量よりも高い分子や低い分子があることを示す)。好ましい平均分子量は約5kDa〜約50kDa、特に好ましくは約12kDa〜約25kDaである。水溶性ポリマーと截形GDNFタンパク質の比は一般に1:1〜100:1、好ましくは(ポリペジル化では)1:1〜20:1、及び(モノペジル化では)1:1〜5:1である。
【0075】
上記条件を使用すると、還元アルキル化はアミノ末端にα−アミノ基をもつ任意の截形GDNFタンパク質にポリマーを選択的に結合することができ、モノポリマー/截形GDNFタンパク質結合体の実質的に均一な調製物を提供する。本明細書で使用する「モノポリマー/截形GDNFタンパク質結合体」なる用語は截形GDNFタンパク質に結合した単一ポリマー分子を含む誘導体を意味する。モノポリマー/截形GDNFタンパク質結合体は、リジンアミノ側鎖でなくN末端に配置されたポリマー分子をもつことが好ましい。調製物は好ましくは>90%モノポリマー/截形GDNFタンパク質結合体、より好ましくは>95%モノポリマー/截形GDNFタンパク質結合体であり、検出可能なタンパク質の残余は未反応である(即ちポリマー部分をもたないタンパク質)。
【0076】
還元アルキル化では、還元剤は水溶液中で安定であるべきであり、初期還元アルキル化プロセスで形成されるシッフ塩基のみを還元できることが好ましい。還元剤の例はホウ水素化ナトリウム、シアノホウ水素化ナトリウム、ジメチルアミンボラン、トリメチルアミンボラン及びピリジンボランから構成される群から選択することができる。特に好ましい還元剤はシアノホウ水素化ナトリウムである。溶媒、反応時間、温度等の他の反応パラメーターと生成物の精製手段は水溶性ポリマーによるタンパク質の誘導体化に関する一般に入手可能な情報に基づいて個々に決定することができる。
【0077】
アシル化及び/又はアルキル化方法によりポリマー/タンパク質結合体の混合物を製造するように選択してもよく、本発明は、混合物に配合するモノポリマー/タンパク質結合体の割合を選択できるという利点がある。従って、所望により様々な数(即ちジ、トリ、テトラ等)のポリマー分子を結合したタンパク質の混合物を製造し、本発明の方法を使用して製造したモノポリマー/タンパク質結合体材料と組み合わせ、所定の割合のモノポリマー/タンパク質結合体を含む混合物を取得できる。
【0078】
截形GDNFタンパク質をコードするポリヌクレオチド
本発明は更に截形GDNFタンパク質をコードする新規ポリヌクレオチドを提供する。ハイブリダイゼーションプローブ又は増幅プライマーとして使用する場合、核酸配列は実質的に他の全核酸配列を含まない。組換えタンパク質発現で使用するには、融合タンパク質が所望される場合を除き、核酸配列は一般に他のタンパク質をコードする核酸配列を実質的に含まない。本明細書の記載に基づき、汎用コドン表を使用すると、当業者は截形GDNFタンパク質のアミノ酸配列をコードする全核酸配列を容易に決定することができる。本発明で好ましい核酸配列としては、[Arg16−Ile134]、[Ser26−Ile134]、[Arg32−Ile134]及び[Lys37−Ile134]截形GDNFタンパク質をコードするポリヌクレオチドが挙げられる。種々のポリヌクレオチドの例は図5、6及び7に示すものや、截形GDNFタンパク質をコードする図1、3及び4の部分である。截形GDNFタンパク質をコードする新規ポリヌクレオチドが人工又は天然を問わず変異体截形GDNFタンパク質をコードする核酸配列を含むことも当業者に理解されよう。
【0079】
以下の記載に従って組換え発現技術を実施すると、これらのポリヌクレオチドを製造し、種々の截形GDNFタンパク質を発現させることができる。例えば、截形GDNFタンパク質をコードする核酸配列を適当なベクターに挿入することにより、当業者は大量の所望のヌクレオチド配列を容易に製造することができる。その後、この配列を使用して検出プローブ又は増幅プライマーを作製することができる。あるいは、截形GDNFタンパク質をコードするポリヌクレオチドを発現ベクターに挿入することもできる。発現ベクターを適当な宿主に導入することにより、所望の截形GDNFタンパク質を大量に製造することができる。
【0080】
以下に詳述するように、核酸配列の増殖及び/又は截形GDNFタンパク質の製造には多数の宿主/ベクター系を利用できる。非限定的な例としては、プラスミド、ウイルス及び挿入ベクターと、原核及び真核宿主が挙げられる。当業者は異種DNAを増殖又は発現することが可能な宿主/ベクター系を改変して本発明の配列を産生又は発現させることができる。
【0081】
このような組換え技術により、本発明の截形GDNFタンパク質は商用量で容易に製造される。更に、本明細書の開示に鑑み、新規核酸配列は図面に具体的に示す截形GDNFタンパク質をコードする縮重核酸配列、このような截形GDNFタンパク質の変異体、及び好ましくはストリンジェントハイブリダイゼーション条件下でこれらの核酸配列の相補体とハイブリダイズする核酸配列(Maniatisら,Molecular Cloning (A Laboratory Manual); Cold Spring Harbor Laboratory,387〜389頁,1982参照)を含むことが当業者に理解されよう。ストリンジェントハイブリダイゼーション条件の例は、62〜67℃で4×SSC中でハイブリダイゼーション後、62〜67℃で0.1×SSCで約1時間の洗浄である。他のストリンジェントハイブリダイゼーション条件は40〜45℃で45〜55%ホルムアミド、4×SSC中でのハイブリダイゼーションである。リラックスハイブリダイゼーション条件下で截形GDNFタンパク質の相補的配列とハイブリダイズし、本発明の截形GDNFタンパク質をコードするDNA配列も本発明に含まれる。このようなリラックスストリンジェンシーハイブリダイゼーション条件の例は45〜55℃で4×SSC又は40〜45℃で30〜40%ホルムアミドを用いたハイブリダイゼーションである。
【0082】
本発明は、截形GDNFタンパク質をコードするDNA配列と共にベクターDNAを含む組換えDNA構築物も提供する。このようなDNA構築物において、(シグナルペプチドの有無に拘わらず)截形GDNFタンパク質をコードする核酸配列は選択された宿主で截形GDNFタンパク質の複製及び/又は発現を誘導することが可能な適切な発現制御又は調節配列と作動的に結合している。
【0083】
截形GDNFタンパク質の組換え発現
截形GDNFをコードするポリヌクレオチドの作製
截形GDNFをコードする核酸配列又は成熟GDNF出発材料は、非限定的な例として化学的合成、cDNA又はゲノムライブラリースクリーニング、発現ライブラリースクリーニング及び/又はcDNAのPCR増幅等の種々の方法で容易に得ることができる。これらの方法及びこのような核酸配列を単離するために有用な他の方法は、例えばSambrookら(Molecular Cloning: A Laboratory Manual,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY,1989)、Ausubelら編(Current Protocols in Molecular Biology,Current Protocols Press,1994)及びBergerとKimmel(Methods in Enzymology: Guide to Molecular Cloning Techniques,vol.152,Academic Press,Inc.,San Diego,CA,1987)により記載されている。GDNFをコードする好ましい核酸配列は哺乳動物配列である。
【0084】
截形GDNFタンパク質をコードする核酸配列の化学的合成は、例えばEngelsら(Angew.Chem.Intl.Ed.,28:716−734,1989)により記載されているような当該技術分野で周知の方法を使用しても実施することができる。これらの方法のうちでは、特にホスホトリエステル、ホスホラミダイト及びH−ホスホネートによる核酸配列合成法が挙げられる。截形GDNFタンパク質をコードする核酸配列は数百塩基対(bp)又はヌクレオチド長である。例えば約100ヌクレオチド長を上回る大きい核酸配列を数個のフラグメントとして合成することができる。その後、フラグメントを相互に連結すると、截形GDNFタンパク質をコードする核酸配列を形成することができる。好ましい方法の1例は、標準ホスホラミダイト化学を使用したポリマーによる合成である。
【0085】
あるいは、適当なcDNAライブラリー(即ちタンパク質を発現すると考えられる1種以上の組織材料から調製したライブラリー)又はゲノムライブラリー(全ゲノムDNAから調製したライブラリー)をスクリーニングすることにより適切な核酸配列を得ることもできる。cDNAライブラリーの材料は一般には妥当な量のGDNFを発現すると考えられる任意の種からの組織である。ゲノムライブラリーの材料はGDNF又はGDNF同族体をコードする遺伝子をもつと考えられる任意の哺乳動物又は他の種からの任意の組織であり得る。ライブラリーに存在するGDNF又はGDNF同族体cDNA又は遺伝子と選択的にハイブリダイズする1種以上のプローブ(クローニングしようとするGDNF又はGDNF同族体cDNA又は遺伝子に対して許容可能な相同レベルをもつオリゴヌクレオチド、cDNA又はゲノムDNAフラグメント)を使用し、GDNF cDNA/遺伝子の存在についてライブラリーをスクリーニングすることができる。このようなライブラリースクリーニングに一般に使用するプローブは通常はライブラリーの調製に用いた種と同一又は類似の種からのGDNF DNA配列の小領域をコードする。あるいは、プローブは本明細書に記載するような縮重配列でもよい。
【0086】
ライブラリースクリーニングは一般に、プローブ又はプライマーに対して有意相同レベルをもつクローンとの結合を可能にし且つ非特異的結合を阻止するストリンジェンシー条件下でライブラリー中のクローンにオリゴヌクレオチドプローブ又はcDNAをアニールすることにより実施される。典型的なハイブリダイゼーション及び洗浄ストリンジェンシー条件はcDNA又はオリゴヌクレオチドプローブの寸法(即ちヌクレオチド長数)や、プローブが縮重配列であるか否かにも依存する。ハイブリダイゼーション溶液を設計するときはクローンの獲得確率(即ちcDNAライブラリーをスクリーニングするのか又はゲノムライブラリーか、cDNAライブラリーであるならば、目的cDNAが高レベルで存在する確率)も考慮する。
【0087】
DNAフラグメント(例えばcDNA)をプローブとして使用する場合には、典型的なハイブリダイゼーション条件は、上記Ausubelら編に記載されているような条件を含む。ハイブリダイゼーション後、プローブ寸法、クローンに対するプローブの予想される相同性、スクリーニングするライブラリーの種類、スクリーニングするクローン数等の数種の因子に依存して適当なストリンジェンシーでライブラリーを含むブロットを洗浄する。(通常はイオン強度が低く、比較的高温で使用される)ストリンジェント洗浄溶液の例を以下に挙げる。このようなストリンジェント洗浄液の1例は55〜65℃で0.015M NaCl,0.005Mクエン酸ナトリウム及び0.1%SDSである。別のストリンジェント緩衝液の例は約40〜50℃で1mM Na2EDTA,40mM NaHPO4,pH7.2及び1%SDSである。他のストリンジェント洗浄液の例は約50〜65℃で0.2×SSC及び0.1%SDSである。
【0088】
更に、オリゴヌクレオチドプローブを使用してcDNA又はゲノムライブラリーをスクリーニングする場合に典型的なストリンジェントな洗浄条件のプロトコルも存在する。例えば、第1のプロトコルではプローブの長さに応じて約35〜62℃の温度で0.05%ピロリン酸ナトリウムを含む6×SSCを使用する。例えば、14塩基プローブは35〜40℃、17塩基プローブは45〜50℃、20塩基プローブは52〜57℃、23塩基プローブは57〜63℃で洗浄する。非特異的なバックグラウンド結合が高いと思われる場合には温度を2〜3℃上げてもよい。第2のプロトコルではテトラメチルアンモニウムクロリド(TMAC)を洗浄に使用する。このようなストリンジェント洗浄溶液の1例は3M TMAC,50mM Tris−HCl,pH8.0及び0.2%SDSである。
【0089】
GDNFタンパク質をコードする核酸配列を得るために利用可能な別の方法はポリメラーゼ連鎖反応(PCR)である。この方法では、GDNFを発現する組織からポリ(A)+RNA又は全RNAを抽出する。次に逆転写酵素を使用してRNAからcDNAを調製する。次に、一般にはGDNF cDNAの2つの別個の領域に相補的な2種のプライマー(オリゴヌクレオチド)をTaqポリメラーゼ等のポリメラーゼと共にcDNAに加えると、ポリメラーゼは2種のプライマーの間のcDNA領域を増幅する。
【0090】
所望の截形GDNFタンパク質をコードする核酸配列を作製するのに選択した方法がオリゴヌクレオチドプライマー又はプローブの使用を必要とする場合(例えばPCR、cDNA又はゲノムライブラリースクリーニング)には、プローブ又はプライマーとして選択するオリゴヌクレオチド配列は、ライブラリースクリーニング又はPCR増幅中に生じる非特異的結合の量を最小限にするように十分な長さと十分明確な配列をもつべきである。プローブ又はプライマーの実際の配列は、通常は同一遺伝子又は別の生物からの類似遺伝子に由来する保存又は高相同性配列又は領域に基づく。場合により、プローブ又はプライマーは完全又は部分縮重配列でもよく、即ち異なるコドンを使用しながら全て同一のアミノ酸配列をコードするプローブ/プライマー混合物でもよい。縮重プローブを作製する代わりに、種によって異なるコドン位置の一部又は全部にイノシンを配置してもよい。オリゴヌクレオチドプローブ又はプライマーは上記のようなDNAの化学合成法により作製できる。
【0091】
GDNFをコードするこれらの核酸配列に基づく截形GDNFタンパク質とその突然変異体又は変異体配列も本発明の範囲に含まれる。上述のように、突然変異体又は変異体配列は野生型配列に比較して1個以上のヌクレオチド置換、欠失及び/又は挿入を含み、その結果として野生型アミノ酸配列に比較してアミノ酸配列変異を発現する配列である。場合によっては、自然対立遺伝子変異の存在により、天然GDNFアミノ酸突然変異体又は変異体が存在することもある。このような天然突然変異体又は変異体に基づく截形GDNFタンパク質も本発明の範囲に含まれる。合成突然変異体配列の作製は当該技術分野で周知であり、例えばWellsら(Gene,34:315,1985)
やSambrookら,前出に記載されている。
【0092】
ベクター
截形GDNFタンパク質をコードするcDNA又はゲノム
DNAをベクターに挿入して更にクローニング(DNAの増幅)又は発現させる。利用可能なベクターは市販されており、又はベクターを特別に構築してもよい。適当なベクターの選択又は構築は、1)DNA増幅に使用するのか、DNA発現に使用するのか、2)ベクターに挿入するDNAの寸法、及び3)ベクターで形質転換する宿主細胞(例えば哺乳動物、昆虫、酵母、真菌、植物又は細菌細胞)に依存する。各ベクターはその機能(DNAの増幅又はDNAの発現)と目的宿主細胞との適合性に依存して種々の成分を含む。ベクター成分の非限定的な例としては一般に、シグナル配列、複製起点、1個以上の選択又はマーカー遺伝子、エンハンサーエレメント、プロモーター、転写終結配列等の1種以上が挙げられる。これらの成分は天然材料から得てもよいし、公知手順で合成してもよい。本発明のベクターは選択された宿主細胞による截形GDNFタンパク質の発現を誘導、制御又は他の方法で司ることが可能な下記発現制御又は調節配列の1種以上に作動的に連結した目的截形GDNFタンパク質をコードする核酸配列を含む。
【0093】
シグナル配列
シグナル配列はベクターの成分でもよいし、ベクターに挿入するGDNF DNAの一部でもよい。天然GDNF DNAは、タンパク質の翻訳後プロセシング中に切断されて成熟GDNFタンパク質を形成するタンパク質のアミノ末端のシグナル配列をコードする。天然シグナル配列及び他のプレ−プロ配列をもつ截形GDNFポリヌクレオチドと、天然シグナル配列を欠失し、異種シグナル配列で置換した截形GDNFポリヌクレオチドが本発明の範囲に含まれる。選択する異種シグナル配列は宿主細胞により認識及びプロセシングされる配列、即ちシグナルペプチダーゼにより切断される配列とすべきである。天然GDNFシグナル配列を認識及びプロセシングしない原核宿主細胞では、シグナル配列を例えばアルカリホスファターゼ、ペニシリナーゼ又は熱安定性エントロトキシンIIリーダーの群から選択される原核シグナル配列で置換する。酵母分泌では、天然GDNFシグナル配列を酵母インベルターゼ、α因子又は酸ホスファターゼリーダーで置換すればよい。哺乳動物細胞発現では、天然シグナル配列でよいが、他の哺乳動物シグナル配列も利用できる。
【0094】
複製起点
発現及びクローニングベクターは一般に、1種以上の選択された宿主細胞でベクターを複製させる核酸配列を含む。クローニングベクターでは、この配列は一般にはベクターを宿主染色体DNAから独立して複製させる配列であり、複製起点又は自律複製配列を含む。このような配列は種々の細菌、酵母及びウイルスで周知である。プラスミドpBR322からの複製起点は殆どのグラム陰性菌に利用でき、哺乳動物細胞でのクローニングベクターには種々の起点(例えばSV40、ポリオーマ、アデノウイルス、VSV又はBPV)が有用である。一般に、哺乳動物発現ベクターには複製起点成分は不要である(例えばSV40起点を使用することが多いが、これは単に初期プロモーターを含むという理由に過ぎない)。
【0095】
選択遺伝子
発現及びクローニングベクターは一般に選択遺伝子を含む。この遺伝子は選択培地で培養する場合に形質転換宿主細胞の生存又は増殖に必要な「マーカー」タンパク質をコードする。ベクターで形質転換されなかった宿主細胞は選択遺伝子を含まないので、培地中で生存しない。典型的な選択遺伝子は、(a)抗生物質又は他の毒素(例えばアンピシリン、ネオマイシン、メトトレキセート又はテトラサイクリン)に対する耐性を与え、(b)栄養要求欠損を補充し、又は(c)培地から得られない必須栄養を供給するタンパク質をコードする。
【0096】
発現される遺伝子を増幅するために他の選択遺伝子を使用してもよい。増幅は、増殖に必須のタンパク質の産生に大いに必要とされる遺伝子が組換え細胞の連続世代の染色体内でタンデムに反復するプロセスである。哺乳動物細胞に利用可能な選択マーカーの例としては、ジヒドロ葉酸レダクターゼ(DHFR)とチミジンキナーゼが挙げられる。形質転換哺乳動物細胞を選択圧下におき、形質転換細胞のみを特別に改変してベクター中に存在するマーカーにより生存できるようにする。選択圧は、培地中の選択物質の濃度が連続的に変化して選択遺伝子と截形GDNFをコードするDNAの両者を増幅させるような条件下で形質転換細胞を培養することにより加えられる。その結果、増幅DNAから多量の截形GDNFが合成される。
【0097】
例えば、DHFRの競合的アンタゴニストであるメトトレキセートを含む培地で全形質転換細胞を培養することにより、
DHFR選択遺伝子で形質転換した細胞をまず同定する。野生型DHFRを使用する場合に適当な宿主細胞は、DHFR活性を欠損するチャイニーズハムスター卵巣細胞系である(例えばUrlaubとChasin,Proc.Natl.Acad.Sci.,USA 77(7):4216−4220(1980)参照)。次に、形質転換細胞を高レベルのメトトレキセートに暴露する。その結果、DHFR遺伝子の多重コピーが合成され、それと同時に発現ベクター中に存在する他のDNA(例えば截形GDNFタンパク質をコードするDNA)の多重コピーも合成される。
【0098】
プロモーター
本発明の発現及びクローニングベクターは一般に、宿主生物により認識され、截形GDNFタンパク質をコードする核酸配列に作動的に連結されたプロモーターを含む。プロモーターは構造遺伝子の開始コドンの上流(5’)に配置された非翻訳配列(一般に約100〜1000bp以内)であり、截形GDNFをコードする配列等の特定核酸配列の転写及び翻訳を制御する。プロモーターは従来、誘導的プロモーターと構成的プロモーターの2種類に分類されている。誘導的プロモーターは栄養又は温度変化の有無等の培養条件の何らかの変化に応答してその制御下のDNAから高レベルの転写を開始する。種々の潜在宿主細胞により認識される多数のプロモーターが周知である。これらのプロモーターは、制限酵素消化によりソースDNAからプロモーターを除去し、所望のプロモーター配列をベクターに挿入することにより、截形GDNFをコードするDNAに作動的に連結される。天然GDNFプロモーター配列を使用して截形GDNF DNAの直接増幅及び/又は発現を誘導することもできる。他方、天然プロモーターよりも多量の発現タンパク質の転写と高収率を可能にし、使用に選択した宿主細胞系に適合可能である場合には、異種プロモーターが好ましい。
【0099】
原核宿主で使用するのに利用可能なプロモーターとしては、β−ラクタマーゼ及びラクトースプロモーター系、アルカリホスファターゼ、トリプトファン(trp)プロモーター系、及びハイブリッドプロモーター(例えばtacプロモーター)が挙げられる。他の公知細菌プロモーターも利用できる。これらのプロモーターのヌクレオチド配列は公表されているので、当業者は任意の必要な制限部位を提供するのに必要なリンカー又はアダプターを使用して所望のDNA配列に連結することができる。
【0100】
酵母宿主で使用するのに利用可能なプロモーター配列も当該技術分野で周知である。酵母エンハンサーを酵母プロモーターと併用すると有利である。哺乳動物細胞で使用するのに適したプロモーターは周知であり、例えばポリオーマウイルス、鶏痘ウイルス、アデノウイルス(例えばアデノウイルス2)、ウシパピローマウイルス、トリサルコーマウイルス、サイトメガロウイルス、レトロウイルス、B型肝炎ウイルス及び最も好ましい例としてサルウイルス40(SV40)等のウイルスのゲノムから得られるものが挙げられる。他の利用可能な哺乳動物プロモーターとしては、異種哺乳動物プロモーター(例えば熱ショックプロモーター及びアクチンプロモーター)が挙げられる。CHO細胞でGDNFタンパク質を産生させるのに本発明で使用したプロモーターはSRαである。Takebeら,Mol.Cell,Biol.8(1):466−472(1988)参照。適切な発現ベクターの1例は、追って詳述するpDSRα2である。
【0101】
エンハンサーエレメント
本発明の截形GDNFタンパク質をコードするDNA配列の高等真核生物による転写を増加するために、ベクターにエンハンサー配列を挿入してもよい。エンハンサーは通常は約10〜300bp長のDNAのシス作用エレメントであり、その転写を増加するようにプロモーターに作用する。エンハンサーは向きや位置から比較的独立している。エンハンサーは転写単位の5’及び3’に検出されている。哺乳動物遺伝子から入手可能な数種のエンハンサー配列が知られている(例えばグロブリン、エラスターゼ、アルブミン、α−フェトプロテイン及びインスリン)。しかし、一般にはウイルスからのエンハンサーを使用する。SV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、ポリオーマエンハンサー及びアデノウイルスエンハンサーが真核プロモーターの活性化を亢進するエレメントの例である。エンハンサーは截形GDNF DNAの5’又は3’位でベクターにスプライスしてもよいが、一般にはプロモーターから5’位に配置する。
【0102】
転写終結
真核宿主細胞(酵母、真菌、昆虫、植物、動物、ヒト又は他の多細胞生物からの有核細胞)で使用する発現ベクターは更に、転写の終結とmRNAの安定化に必要な配列も含む。このような配列は通常は真核DNA又はcDNAの5’及び場合により3’非翻訳領域から得られる。これらの領域は截形GDNFをコードするmRNAの非翻訳部分でポリアデニル化フラグメントとして転写されたヌクレオチドセグメントを含む。
【0103】
所望の截形GDNFコーディング配列と共に上記成分の1種以上を含む適切なベクターの構築は標準連結技術により実施される。単離プラスミド又はDNAフラグメントを切断し、順化し、必要なプラスミドを生成するために所望の順序に再連結する。正しい配列が構築されたことを確認するためには、連結混合物を使用して大腸菌を形質転換し、形質転換に成功した細胞を上記のようなアンピシリン又はテトラサイクリン耐性等の公知技術により選択すればよい。次いで形質転換細胞からプラスミドを調製し、制限エンドヌクレアーゼにより分析し、及び/又は配列決定し、所望の構築物の存在を確認する。
【0104】
哺乳動物細胞で截形GDNFをコードするDNAの一過性発現を可能にするベクターを使用してもよい。一般に、一過性発現は宿主細胞で効率的に複製することが可能な発現ベクターを使用し、宿主細胞に発現ベクターの多数のコピーを蓄積させ、その結果、発現ベクターによりコードされる所望のタンパク質を高レベルで合成させる。適切な発現ベクターと宿主細胞を含む一過性発現系は、クローン化DNAによりコードされるタンパク質の簡便な正の同定と、所望の生物学的又は生理的性質に関する前記タンパク質の迅速なスクリーニングを可能にする。従って、一過性発現系はタンパク質の変異体を同定するのに特に有用である。
【0105】
宿主細胞の選択及び形質転換
組換え截形GDNFタンパク質を発現させるのに使用する核酸配列で形質転換した宿主細胞(例えば細菌、哺乳動物、昆虫、酵母又は植物細胞)も本発明により提供される。形質転換した宿主細胞を、核酸配列の発現を可能にする適当な条件下で培養する。適切な宿主細胞と形質転換、培養、増幅、スクリーニング並びに生成物産生及び精製方法は当該技術分野で周知である。例えばGethingとSambrook,Nature 293:620−625(1981); Kaufmanら,Mol.Cell.Biol.,5(7):1750−1759(1985); 又はHoweleyら,米国特許第4,419,446号参照。截形GDNFは、成熟GDNFの発現に関するLinらの記載(米国特許出願第07/855,413号、国際出願第PCT/US92/07888号、WO93/06116)に従って大腸菌で発現させることができる。他の典型的な材料及び方法については以下に詳述する。形質転換した宿主細胞を適切な培地で培養した後、発現された因子を場合により当業者に公知の適当な手段により培地から(又は細胞内で発現される場合には細胞から)回収、単離及び精製する。
【0106】
本発明のベクターをクローニング及び発現させるのに適した宿主細胞は上述のような原核、酵母又は高級真核細胞である。原核宿主細胞の非限定的な例としては、グラム陰性又はグラム陽性生物等の真正細菌が挙げられ、例えば大腸菌、バシラス属(例えば枯草菌)、シュードモナス種(例えばP.aeruginosa)、Salmonella typhimurium又はSerratia marcescansである。あるいは、PCR又は他の核酸ポリメラーゼ反応等のin vitroクローニング法も利用できる。
【0107】
原核宿主細胞に加え、糸状菌類や酵母等の真核微生物も截形GDNFタンパク質の発現宿主として利用できる。下級真核宿主微生物のうちではSaccharomyces cerevisiae(俗名パン酵母)が最もよく使用されるが、多数の他の属、種及び株も周知であり、一般に入手可能である。
【0108】
グリコシル化截形GDNFタンパク質の発現に適した宿主細胞は多細胞生物から得られる。このような宿主細胞は複雑なプロセシング及びグリコシル化活性が可能である。脊椎動物細胞であるか、植物及び昆虫細胞を含む無脊椎動物細胞であるかを問わず、原則として任意の高級真核細胞培養物を使用できる。脊椎動物細胞の培養(組織培養)増殖は周知方法であるので、一般には脊椎動物細胞を使用する。有用な哺乳動物宿主細胞系の非限定的な例としては、SV40により形質転換したサル腎CV1系(COS−7)、ヒト胎児腎系(293又は懸濁培養増殖のためにサブクローニングした293細胞)、乳児ハムスター腎細胞、及びチャイニーズハムスター卵巣細胞が挙げられる。他の利用可能な哺乳動物細胞系の非限定的な例としては、HeLa、マウスL−929細胞、Swiss、Balb−cもしくはNIHマウスに由来する3T3系、BHK又はHaKハムスター細胞系が挙げられる。
【0109】
本発明に利用可能な宿主細胞として細菌細胞も有用である。例えば、種々の大腸菌株(例えばHB101、DH5α、DH10及びMC1061)がバイオテクノロジー分野で宿主細胞として周知である。ストレプトミセス種等の種々の株も使用できる。截形GDNFタンパク質を製造するのに本発明で好適な宿主細胞は細菌細胞(例えば大腸菌)と哺乳動物細胞(例えばチャイニーズハムスター卵巣細胞、COS細胞等)である。
【0110】
宿主細胞を上記発現又はクローニングベクターでトランスフェクト、好ましくは形質転換し、慣用栄養培地で培養する。培地はプロモーターの誘導、形質転換細胞の選択又は所望の配列をコードする遺伝子の増幅に適するように改変してもよい。トランスフェクション及び形質転換は、利用する宿主細胞に適するように選択された当業者に周知の標準技術を使用して実施される。例えば、細胞壁をもたない哺乳動物細胞では、リン酸カルシウム沈殿法を使用できる。エレクトロポレーション、マイクロインジェクション及び他の公知技術も使用できる。
【0111】
宿主細胞の培養
本発明の截形GDNFタンパク質を製造するために使用する形質転換細胞は適切な培地で培養する。培地は必要に応じてホルモン及び/又は他の成長因子(例えばインスリン、トランスフェリン又は上皮成長因子)、塩類(例えば塩化ナトリウム、カルシウム、マグネシウム及びリン酸塩)、緩衝液(例えば
HEPES)、ヌクレオシド(例えばアデノシン及びチミジン)、抗生物質(例えばゲンタマイシン)、微量元素(通常はマイクロモル範囲の最終濃度で存在する無機化合物として定義される)、及びグルコース又は他のエネルギー源を補充してもよい。当業者に理解される通り、他の補充剤も適当な濃度で加えてもよい。選択した宿主細胞で使用するのに適した温度、pH等の培養条件も当業者に周知である。
【0112】
截形GDNFタンパク質は相同組換え、即ちGDNFをコードするDNAを既に含んでいる細胞に導入した制御エレメントを使用する組換え製造方法により製造することも可能である。相同組換えは、転写活性遺伝子に突然変異を誘導又は修正するように遺伝子をターゲティングするために本来開発された技術である(Kucherlapati,Prog.in Nucl.Acid Res.and Mol.Biol.36:301(1989))。基本技術は哺乳動物ゲノムの特定領域に特定突然変異を導入する方法(Thomasら,Cell.44:419−428,1986; ThomasとCapecchi,Cell.51:503−512,1987; Doetschmanら,Proc.Natl.Acad.Sci.85:8583−8587,1988)として又は欠損遺伝子内の特定突然変異を修正するため(Doetschmanら,Nature.330:576−578,1987)に開発された。典型的な相同組換え技術は、参考資料としてその開示内容を本明細書の一部とする米国特許第5,272,071号(EP第91903051号、EP公開第505500号、PCT/US90/07642、国際公開WO91/09955)に記載されている。
【0113】
相同組換えにより、ゲノムに挿入しようとするDNA配列をターゲティングDNAに結合することにより目的遺伝子の特定領域に導くことができる。ターゲティングDNAはゲノムDNAのある領域に相補的(相同)なDNAである。ゲノムの特定領域に相補的なターゲティングDNAの小断片をDNA複製プロセス中に親鎖と接触させる。細胞に挿入されたDNAは、共通相同領域を介して内因DNAの他の断片とハイブリダイズし、従って、これと組換えするという一般性質をもつ。突然変異又は異なるDNA配列を含むオリゴヌクレオチドにこの相補鎖を結合するならば、この相補鎖は組換えの結果として新しく合成された鎖にも組み込まれる。プルーフリーディング機能の結果として、新しいDNA配列を鋳型として使用することができる。従って、転写されたDNAはゲノムに組み込まれる。
【0114】
GDNFの核酸配列等の特定遺伝子の配列が分かっている場合には、遺伝子の選択領域に相補的なDNAの断片であるプレ−プロ配列又は発現制御配列を合成することができ、あるいは目的領域の境界となる特定認識部位で天然DNAを適当に制限消化するなどの方法でこのような配列が得られる。この断片は細胞に挿入するとターゲティング配列として機能し、ゲノムの内側でその相同領域とハイブリダイズする。DNA複製中にこのハイブリダイゼーションが生じるならば、このDNA断片とこれに結合した任意の付加的配列は岡崎フラグメントとして作用し、新しく合成されたDNA娘鎖にバックステッチされる。
【0115】
本発明では、GDNFタンパク質の発現に相互作用すると考えられるDNAの領域がターゲティングDNAのこれらの断片に結合している。例えば、所望の截形GDNFをコードする
DNAの転写に作用するに十分近傍の位置及び向きで所期宿主細胞のゲノムにプロモーター/エンハンサーエレメント、サプレッサー、又は外因転写調節エレメントを挿入する。制御エレメントは截形GDNFをコードせず、宿主細胞ゲノムに存在するDNAの一部を制御する。従って、截形GDNFタンパク質の発現は、截形GDNF遺伝子自体をコードするDNAのトランスフェクションによって達せられるのではなく、截形GDNFタンパク質の転写のための認識可能なシグナルを内因遺伝子配列に提供するDNA調節セグメントと結合したターゲティングDNA(目的の内因遺伝子に対して相同の領域を含む)の使用によって達せられる。
【0116】
本発明によると、相同組換え法を使用して通常は転写に不活性なGDNF遺伝子を含む細胞を改変し、GDNFを発現する細胞を作製することもできる。その後、GDNFタンパク質をプロセシングすると、截形GDNFタンパク質を形成することができる。
【0117】
截形GDNF医薬組成物
截形GDNFタンパク質産物医薬組成物は一般に、1種以上の医薬的及び生理的に許容可能な配合剤と混合した治療上有効な量の截形GDNFタンパク質産物を含む。利用可能な配合剤の非限定的な例としては、酸化防止剤、防腐剤、着色剤、矯味矯臭剤、希釈剤、乳化剤、懸濁剤、溶剤、充填剤、増量剤、緩衝液、送達ビヒクル、希釈液、賦形剤及び/又は医薬アジュバントが挙げられる。例えば、利用可能なビヒクルとしては注射用水、生理的食品水、又は人工脳髄液(CSF)が挙げられ、場合により、非経口投与用組成物に通常用いられる他の材料を補充してもよい。天然緩衝食品水又は血清アルブミンと混合した食品水も典型的ビヒクルである。
【0118】
ビヒクル中の主溶媒は水性でも非水性でもよい。更に、ビヒクルは製剤のpH、オスモル濃度、粘度、透明度、色、滅菌度、安定性、溶解速度又は臭いを変更又は維持するために他の医薬的に許容可能な賦形剤を含んでいてもよい。また、ビヒクルは截形GDNFタンパク質産物の安定性、溶解速度又は放出速度を変更又は維持するため、あるいは截形GDNFタンパク質産物の血液脳関門吸収又は通過を助長するために更に別の医薬的に許容可能な賦形剤を含んでいてもよい。このような賦形剤は単位剤形もしくは多剤形の非経口投与製剤又は移植ポンプからの連続もしくは周期的注入によるCSFへの直接注入用製剤を調剤するために通常慣用的に使用されている物質である。
【0119】
治療組成物を調剤したら、溶液、懸濁液、ゲル、エマルション、固体又は脱水もしくは凍結乾燥粉末として滅菌バイアルに保存することができる。このような製剤は即使用可能な形態で保存してもよいし、例えば投与前に再構成が必要な凍結乾燥形態で保存してもよい。
【0120】
最適医薬製剤は投与経路と所望の投与量に依存して当業者により決定される。例えば、参考資料としてその開示内容を本明細書の一部とするRemington’s Pharmaceutical Science,第18版(1990,Mack Publishing Co.,Easton,PA 18042)1435〜1712頁参照。組成物は更に、ポリ乳酸、ポリグルコール酸等のポリマー化合物の粒状調製物を含んでいてもよいし、リポソームに封入してもよい。ヒアルロン酸も使用でき、これは循環持続時間を促進する効果があると考えられる。このような組成物は本発明のタンパク質及び誘導体の物理的状態、安定性、in vivo放出速度、及びin vivoクリアランス速度を変更できる。
【0121】
非経口徐放製剤、吸入ミスト、経口活性製剤又は座薬等の他の有効な投与形態も考えられる。本発明の截形GDNFタンパク質産物医薬組成物は例えば脳髄内注射等の非経口投与用に調製される。このような非経口投与治療組成物は一般に医薬的に許容可能なビヒクル中に截形GDNFタンパク質産物を含む非経口的に許容可能なパイロジェンフリー水溶液の形態である。好ましいビヒクルの1例は生理的食塩水である。
【0122】
截形GDNFタンパク質産物を含む所定の製剤は経口投与することも考えられる。このように投与する截形GDNFタンパク質産物はカプセル封入することができ、固体剤形の配合に慣用的に使用されているキャリヤーの存在下又は不在下で調剤できる。カプセルは生体利用性が最大となり且つ全身循環前の分解が最小となる時点で胃腸管に製剤の活性部分を放出するように設計することができる。截形GDNFタンパク質産物の吸収を助長するために付加的な賦形剤を加えてもよい。希釈剤、矯味矯臭剤、低融点ろう、植物油、潤滑剤、懸濁剤、錠剤崩壊剤及び結合剤も使用してもよい。
【0123】
截形GDNFタンパク質産物の投与
截形GDNFタンパク質産物は皮下、筋肉内、静脈内、経肺、経皮、鞘内又は大脳内経路により非経口投与することができる。血液脳関門を通過しないタンパク質成長因子は大脳内に直接投与するか、又は関門間を輸送する他のエレメントと共に投与すればよい。截形GDNFタンパク質産物は脳髄内又は脳もしくは脊髄くも膜下腔に投与するのが好ましい。截形GDNFタンパク質産物は脳実質に直接大脳内投与してもよい。生分解性ポリマーマトリックスに埋封した神経栄養因子を含む徐放インプラントを脳に移植し、截形GDNFタンパク質産物を送達することもできる。截形GDNFタンパク質産物は、血液脳関門を通過するように化学的に修飾又はパッケージした形態で大脳外に投与してもよいし、截形GDNFタンパク質産物の関門通過を促進することが可能な1種以上の物質と併用投与してもよい。例えば、NGFとモノクローナル抗トランスフェリンレセプター抗体の結合体はトランスフェリンレセプターとの結合により脳に輸送されることが示されている。所望用量の截形GDNFタンパク質産物を投与するためには、毎日又はより低頻度で繰り返し注射してもよいし、一定又はプログラム可能な流量の移植ポンプから截形GDNFタンパク質産物を連続又は周期的に注入してもよい。投与頻度は調剤する截形GDNFタンパク質産物の薬物動態パラメーターと、投与経路によって異なる。
【0124】
投与方法に関係なく、比用量は一般に体重又は体表面積に従って計算される。脳疾患では、比用量は一般に患者の概算大脳重量に従って計算され、大脳重量も体重又は体表面積に基づいて推定できる。上記製剤の各々を利用する治療に適した用量を決定するために必要な更に精密な計算は、特に本明細書に開示する用量情報及びアッセイを参考にして当業者により日常的方法で実施される。用量が適当であるか否かは、適当な用量応答データと組み合わせて使用量を決定する確立アッセイを使用して確認することができる。特定症状の治療方法に用いる最終投与計画は、薬剤作用に影響する種々の因子(例えば患者の年齢、症状、体重、性別及び食事、感染の重篤度、投与時間並びに他の臨床因子)を考慮して主治医により決定される。
【0125】
本発明の截形GDNFタンパク質産物は更に、神経疾患の治療で単独又は他の成長因子と組み合わせて使用できる。例えば、截形GDNFタンパク質産物は所定形態の神経疾患の治療に神経成長因子と併用できる。更に、化学組成物を含む他の因子又は他の分子を截形GDNFタンパク質産物と併用してもよい。パーキンソン病の治療では、截形GDNFタンパク質産物を単独又はレボドパと併用投与することが考えられ、截形GDNFは内因ドーパミンの産生と高濃度のドーパミンのニューロン取り込みを強化すると思われる。
【0126】
上述のように、所定のニューロン細胞ポピュレーション又はある種の傷害もしくは疾病を治療するためには、付加的な神経栄養又はニューロン栄養因子が有用又は必要であるとも考えられる。截形GDNFと併用できる他の因子の非限定的な例としては、マイトジェン(例えばインスリン、インスリン様成長因子、上皮成長因子、血管作用成長因子、下垂体アデニル酸シクラーゼ活性化ポリペプチド、インターフェロン及びソマトスタチン)、神経栄養因子(例えば脳由来神経栄養因子、ニューロトロフィン−3、ニューロトロフィン−4/5、ニューロトロフィン−6、インスリン様成長因子、繊毛神経栄養因子、酸性及び塩基性フィブロブラスト成長因子、フィブロブラスト成長因子−5、トランスフォーミング成長因子−β、コカインアンフェタミン調節転写産物(CART)並びに成熟GDNF)、及び他の成長因子(例えば上皮成長因子、白血病阻害因子、インターロイキン、インターフェロン及びコロニー刺激因子)並びにこれらの因子の機能的等価物である分子及び材料が挙げられる。
【0127】
治療によっては截形GDNFタンパク質産物の連続投与又は持続送達が有利であると考えられる。連続投与は注入ポンプを使用するなど機械的手段により達成できるが、他の連続又は準連続投与方法も実施できると考えられる。例えば、化学誘導体化によりタンパク質の持続放出形態が得られ、所定の投与計画に基づいて予測可能な量で血流中の連続的存在を可能にする。従って、截形GDNFタンパク質産物はこのような連続投与を実現するように誘導体化された截形GDNFタンパク質を含む。
【0128】
截形GDNFタンパク質細胞治療、例えば截形GDNFタンパク質を産生する細胞の大脳内移植も考えられる。本発明のこの態様では、生物活性形態の截形GDNFタンパク質を合成及び分泌することが可能な細胞を患者に移植することが考えられる。このような截形GDNFタンパク質産生細胞は、通常は神経栄養因子を産生しないが、截形GDNFを産生するように改変された細胞や、截形GDNFタンパク質の発現及び分泌に適したポリヌクレオチドで形質転換することによりそのGDNF産生能を増加した細胞であり得る。外来種のGDNFの投与により患者に免疫反応が生じる可能性を最小限にするためには、細胞はヒト起源であり且つヒト截形GDNFタンパク質を産生することが好ましい。
【0129】
細胞の脳組織浸潤を避けるために移植細胞をカプセル封入してもよい。截形GDNFタンパク質産物を放出できるが、患者の免疫系又は周囲組織からの他の有害因子による細胞の破壊を阻止する生体適合性の半透ポリマーエンクロージャー又は膜にヒト又は非ヒト動物細胞を封入して患者に移植することができる。あるいは、截形GDNFを産生するようにex vivo形質転換した患者自身の細胞をカプセル封入せずに患者に直接移植してもよい。
【0130】
生きた細胞の膜封入方法は当業者に周知であり、封入細胞の作製と患者へのその移植は実施可能である。例えば、参考資料としてその開示内容を本明細書の一部とする米国特許第4,892,538号、5,011,472号及び5,106,627号参照。生きた細胞を封入するシステムは、特に参考資料としてその開示内容を本明細書の一部とするAebischerらのPCT出願WO91/10425にも記載されている。
更に、参考資料としてその開示内容を本明細書の一部とするAebischerらのPCT出願WO91/10470; Winnら,Exper.Neurol.,113:322−329,1991; Aebischerら,Exper.Neurol.,111:269−275,1991; Trescoら,ASAIO,38:17−23,1992も参照されたい。
【0131】
截形GDNFタンパク質をコードする核酸配列を患者に直接導入する截形GDNFタンパク質in vivo遺伝子治療も考えられる。例えば、アデノ随伴ウイルスベクター等の適当な送達ベクターを介して又は介さずに核酸構築物の局所注射により、截形GDNFタンパク質をコードする核酸配列を標的細胞に導入する。別のウイルスベクターの非限定的な例としては、レトロウイルス、アデノウイルス、単純ヘルペスウイルス及びパピローマウイルスベクターが挙げられる。所望の核酸構築物もしくは所望の核酸配列を含む他の適当な送達ベクターの局所注射、リポソームによる導入、直接注射(裸のDNA)、レセプターによる導入(リガンド−DNA複合体)、又は微粒子ボンバードメント(遺伝子銃)により、in vivo物理的導入が可能である。
【0132】
本明細書に記載する截形GDNFタンパク質産物製剤は獣医学及びヒト適用に使用することができ、「患者」なる用語は限定的に解釈すべきでないことに留意されたい。獣医学適用の場合では、用量範囲は上記と同一にすべきである。
【0133】
本発明の截形GDNFタンパク質を更に特性決定する手段として、X−[Cys41−Cys133]−Yアミノ酸配列内のエピトープ等の截形GDNFタンパク質に結合する抗体を製造することができる。当業者は周知の公表手順を使用し、本発明のアミノ酸配列によりコードされる種々のタンパク質を特異的に認識してこれと結合するモノクローナル及びポリクローナル抗体又は組換え抗体を取得することができる。その後、このような抗体を使用して截形GDNFタンパク質を精製及び特性決定することができる。あるいは、これらの抗体はその抗原であるタンパク質の治療阻害剤としても使用できる。
【0134】
本発明の他の側面及び利点は、下記実施例から理解されよう。実施例1は哺乳動物細胞系における成熟GDNFの発現と截形GDNFタンパク質の作製に関する。実施例2は細菌細胞系における成熟GDNFの発現に関する。実施例3は細菌細胞系における種々の截形GDNFタンパク質の発現に関する。実施例4はドーパミン作動性ニューロン神経栄養活性アッセイで成熟GDNFタンパク質と截形GDNFタンパク質の生物活性を比較する。
【図面の簡単な説明】
【0135】
【図1】成熟ヒトグリア細胞系由来神経栄養因子(hGDNF)をコードする核酸配列(配列番号1)を示す。成熟ヒトGDNFタンパク質のアミノ酸配列(配列番号2)も示す。
【図2】組換え截形GDNFタンパク質の発現のために利用したプラスミド構築図を示す。
【図3A】GDNF及び截形GDNFポリヌクレオチドをコードする別の核酸配列(配列番号39)の制限地図を示す。
【図3B】GDNF及び截形GDNFポリヌクレオチドをコードする別の核酸配列(配列番号39)の制限地図を示す。
【図4】GDNF及び截形GDNFポリヌクレオチドをコードする更に別の核酸配列(配列番号40)の制限地図を示す。
【図5】[Pro23−Lys37ΔAsn37−Ile134]截形GDNFタンパク質置換変異体(配列番号42)をコードする核酸配列(配列番号41)を示す。このタンパク質はMet−Ser−[Pro23−Lys37ΔAsn37−Ile134]截形GDNFタンパク質付加/置換変異体と表すこともできる。
【図6】[Arg32−Ile134]截形GDNFタンパク質(配列番号44)をコードする核酸配列(配列番号43)を示す。
【図7】[Gly33−Ile134]截形GDNFタンパク質(配列番号46)をコードする核酸配列(配列番号45)を示す。
【図8】数種の截形GDNFタンパク質の例であるMet−[Arg32−Ile134](配列番号48)、Met−[Gly33−Ile134](配列番号49)及びMet−Ser−[Pro23−Lys37ΔAsn37−Ile134](配列番号50)と成熟hGDNFのアミノ酸配列(配列番号47)の比較を示す。
【0136】
実施例
【実施例1】
【0137】
CHO細胞における成熟ヒトGDNFの発現及びCHO細胞由来截形GDNFタンパク質の精製
材料
下記材料を使用してジヒドロ葉酸レダクターゼ欠損CHO細胞(例えばUrlaubとChasin,Proc.Natl.Acad.Sci.,USA 77(7):4216−4220(1980)に記載されているようなCHOd−細胞)でヒトGDNFを発現させる。
【0138】
CHOd−培地は、ダルベッコ改変イーグル培地(DMEM)−高グルコース(Gibco/BRL)、5%ウシ胎児血清
(HyClone)、MEM非必須アミノ酸(1%)(Gibco/BRL)、ヒポキサンチン/チミジン(1%)(Gibco/BRL)、及びグルタミン/ペニシリン/ストレプトマイシン(1%)(Irvine Scientific)を含むものとした。
【0139】
選択培地は、DMEM(高グルコース)、5%透析ウシ胎児血清(HyClone)、MEM非必須アミノ酸、及びグルタミン/ペニシリン/ストレプトマイシンを含むものとした。
【0140】
2×HEPES緩衝食塩水(HBS)は、280mM NaCl、10mM KCl、1.5mM Na2HPO4、12mMデキストロース、及び50mM HEPESを含むものとした。
【0141】
Tris緩衝食塩水+Tween(TBST)は、137
mM NaCl、20mM Tris/HCl,pH7.5、及び0.1% Tween−20を含むものとした。方法
トランスフェクション及び選択
CHOd−細胞(20継代)をCHOd−増殖培地中細胞8×105個/皿の密度で60mm組織培養皿(Falcon)に接種した。翌日、トランスフェクションから約3時間前に細胞上の培地を新鮮な培地に交換した。
【0142】
周知技術を使用して適当なGDNF cDNAを含むプラスミド構築物を調製した。例えば、参考資料としてその開示内容を本明細書の一部とする本願出願人と同一名義の1990年3月29日付け同時係属米国特許出願第501,904号(1990年5月18日付けヨーロッパ特許出願第90305433号、公開EP第398753号及びWO90/14363(1990)も参照)に記載の方法に実質的に従ってプラスミド構築物pDSRα2を調製した。ベクターの構造編成を示すプラスミド地図の例を図2に示す。図1、3及び4に示す配列等の成熟GDNFタンパク質をコードする種々の核酸配列を使用してもよいことが当業者に理解されよう。
【0143】
pcDNA3に基づく発現ベクター(Invitrogen,San Diego,CA)から制限酵素消化により、ヒトGDNFコーディング配列とコンセンサスKozak配列CCACC(ATG)を含むHindIII−Xbal DNAフラグメントを回収した。HindIII/Xbalで切断したpDSRα2にDNAフラグメントを直接クローニングした。得られたプラスミドをpSW5と命名した。トランスフェクションの前にpSW5のプラスミドDNAをPuvI部位で直鎖化した。
【0144】
pDSRα2(図2)はプラスミドpCD(OkayamaとBerg,Mol.Cell Biol.3:280−289,1983)の誘導体であり、(1)SV40ポリアデニル化シグナルをウシ濾胞刺激ホルモンのα−サブユニットであるα−bFSHからのシグナルで置換し(Goodwinら,Nucleic Acids Res.11:6873−6882,1983)、(ii)形質転換細胞の選択及び増殖を可能にするように発現カセットの下流にマウスジヒドロ葉酸レダクターゼミニ遺伝子(Gasserら,Proc.Natl.Acad.Sci.79:6522−6526,1982)を挿入し、(iii)ヒトT細胞I型白血病ウイルス(HTLV−I)の長い末端反復配列(LTR)の「Rエレメント」と「U5」配列の一部を含む267bpフラグメントを従来の記載(Takebeら,Mol.Cell Biol.8:466−472,1988)通りにSV40プロモーターとスプライスシグナルの間にクローニング及び挿入したという主に3点を改変した。
【0145】
最終濃度3.0μg/皿のGDNF−プラスミドDNA、7.0μg/皿のマウス腎ゲノムキャリヤーDNA(Clontech)、25μl/皿の2.5M CaCl2及び最終容量250μl/皿までの滅菌蒸留水を含むDNA溶液を調製した。pDSRα2ベクターDNA又はキャリヤーDNA単独を含むDNA溶液も夫々正負対照として同様に調製した。溶液に気泡を通しながら、DNA溶液を等容量の2×HEPES緩衝食塩水に滴下した。DNA/HBS溶液を室温で30分間インキュベートした。
【0146】
CHOd−細胞培養物から培地を除去し、DNA溶液500μl/皿を加えた。皿を室温で30分間インキュベートした後、CHOd−培地(5.0ml)を各皿に加えた。次に、皿を37℃で一晩インキュベートした。
【0147】
翌日に培地を新鮮なCHOd−培地に交換した。その翌日に細胞がコンフルエントに達した時点で培養物をトリプシン処理し、1×60mm皿対8×100mm皿の比で100mm皿(Falcon)に再プレーティングした。細胞を選択培地で再プレーティングした。2〜3日おきに培養物に新鮮な培地を再補充した。
【0148】
15日後に、トランスフェクトした細胞のコロニーをガラスクローニングシリンダーで単離し、トリプシン処理し、24穴皿(Falcon)に再プレーティングした。GDNF/pSW5をトランスフェクトした細胞から合計40個のコロニーを単離した。皿に残っている細胞をトリプシン処理し、プールし、2個の100mm皿に再プレーティングした(各DNA構築物毎に1個のプール)。
【0149】
トランスフェクトした細胞のスクリーニング
24穴及びプール培養物をコンフルエントまで増殖させ、その時点で増殖培地を除去し、無血清培地(400μl/ウェル即ち4ml/皿)に交換した。細胞を48時間インキュベートし、条件付け培地を回収した。条件付け培地サンプルのGDNFタンパク質発現をウェスタンブロットにより分析した。条件付け培地のアリコート(20μl又は40μl)を電気泳動サンプル緩衝液(β−メルカプトエタノールを含むか又は含まない)で希釈した。β−メルカプトエタノールを含むサンプルを3分間煮沸した(還元条件)。還元及び非還元サンプルの両者を16%Tris−グリシンゲル(Novex)で試験した。ゲルをニトロセルロースフィルター(Schleicher and Schuell BA−83、0.2μ)にエレクトロブロットした。ブロットをTBSTで濯いだ後、TBST中5%ドライミルク(Carnation)のブロッキング溶液中で室温で30分間インキュベートした。次にブロットをGDNF抗血清(大腸菌由来GDNFに対するウサギポリクローナル抗血清、5%ドライミルク/TBST中1:1000)で室温で1時間処理した。次にブロットをTBSTで濯ぎ、1×10分間及び2×5分間1%ドライミルク/TBSTで洗浄した。次に抗ウサギIg−ウマ西洋ワサビペルオキシダーゼ結合二次抗体(1%ドライミルク/TBST中1:15,000)で20分間処理した。ブロットを濯ぎ、TBSTで1×20分間及び2×10分間洗浄した後、ECL試薬(Amersham)で1分間処理し、Hyperfilm−ECL(Amersham)を感光した。
【0150】
下記プロセスは、CHOで発現させたGDNFとCHOに由来する切断GDNFホモダイマーを条件付け培地1リットルから精製するプロセスである。CHO培地には有意なプロテアーゼ作用があり、残基31で鎖を切断するので、この手順では精製中にプロテアーゼ阻害剤を使用してもよい。
【0151】
ステップ1.ビーズクロマトグラフィー:
1/50容量の1M MES,pH6.0を加えることにより、無血清条件付け培地を20mM 2−[N−モルホリノ]エタンスルホネート(MES),pH6.0にした。20mM MES,pH6.0で平衡化したSP Sepharose Big Bead樹脂(Pharmacia)25mlを加え、4℃で1時間撹拌した。樹脂を沈殿させて条件付け培地をデカントすることにより樹脂を集めた。デカントした培地を焼結ディスクフィルターで濾過し、未沈殿樹脂を回収した。沈殿した樹脂と濾過により回収した樹脂を再懸濁し、直径2.5cmのカラムに注入し、3カラム容量の0.15M NaCl,20mM MES,pH6.0(A緩衝液)で洗浄した。280nmの吸光度をモニターしながら0.2カラム容量/分の流速でA緩衝液から1.0M NaCl,20mM MES,pH6.0(B緩衝液)への300nlのグラジエントでタンパク質を溶離した。1.1カラム容量を含むフラクションを集めた。フラクション中のGDNFの存在をウェスタンブロット分析により検出した。GDNFを含むフラクションを更に精製するためにプールした。GDNFは0.3〜0.6M NaClで溶出した。
【0152】
ステップ2.HPLC C4クロマトグラフィー:
ステップ1からのプールを0.1%(v/v)トリフルオロ酢酸(TFA)とし、0.45μフィルターで減圧濾過し、
10%アセトニトリル、0.1%TFA水溶液(A緩衝液)で条件付けしたVydac C4カラム(0.46×25cm)にアプライした。280nmの吸光度を測定しながらA緩衝液から90%アセトニトリル、0.1%TFA水溶液(B緩衝液)への2%/分の直線勾配で50分間かけてタンパク質を溶離した。1mlフラクションを集め、GDNFの存在をウェスタンブロット分析により検出した。GDNFは45%〜55%アセトニトリルで溶出した。フラクションを減圧乾涸した。
【0153】
ステップ3.高性能Sクロマトグラフィー:
ステップ2からのGDNFを含むフラクションを1mlの0.15M NaCl,10mM Tris,pH8.0に再可溶化し、0.75×7.5cm TSK−Gel 5WP高性能Sカラム(Toso Haas)にアプライした。0.15M NaCl,10mM Tris,pH8.0(A緩衝液)から1.0M NaCl,10mM Tris,pH8.0(B緩衝液)への0.4%/分の直線勾配で流速1ml/分で50分間かけて溶離した。1分毎にフラクションを集め、280nmの吸光度を測定した。35%B緩衝液で勾配を10分間かけて6.5%/分まで変化させた。フラクションのウェスタンブロット分析は、4個の主GDNF成分を示した。これらの成分のうちの3個は0.4%/分勾配中に溶出し、4番目の成分は6.5%/分勾配中に溶出した。同様の成分から適当なプールを調製し、配列決定した。配列決定分析の結果、約29〜36kDプールは[Arg32−Ile134]截形GDNFタンパク質であることが確認された。約38〜40kDの成分は[Arg32−Ile134]截形GDNF/成熟GDNFヘテロダイマーであることが確認された。最後に、勾配の最後の部分で単離された約41〜44kDの成分は成熟GDNFホモダイマーであることが配列決定により確認された。
【実施例2】
【0154】
大腸菌における成熟ヒトGDNFの産生
成熟ヒトGDNFの細菌発現は、Linらの方法(参考資料としてその開示内容を本明細書の一部とする1994年5月23日付け米国特許出願第08/182,183号とその親出願、1992年9月17日付けPCT/US92/07888(WO93/06116)及びヨーロッパ特許出願第92921022.7号(公開EP第610254号))により実施できる。本発明の記載に基づき、大腸菌及び他の細菌での適切な発現に種々の材料及び方法を容易に使用又は応用できることが当業者に理解されよう。例えば、図1、3及び4に示すような代替ポリヌクレオチドを発現プロセスで使用してもよい。
【0155】
発現された成熟GDNFの再生及び精製
形質転換細胞を(特に指定しない限り)5℃で次のように処理した。最終容量200mlの5mM EDTAを含む25mM Tris,pH8.5に細胞ペースト(30g)を懸濁し、15%(w/v)の最終細胞スラリーを得た。Biospecハンドヘルド低剪断ホモジナイザーを使用して細胞を十分に分散させた。スラリーを14,500psiのマイクロ流動化装置に2回通し、細胞を破壊して封入体を放出させた。得られたホモジネートを次に16,000×gで30分間遠心分離した。上記と同様にBiospecホモジナイザーを使用し、遠心分離により得られた封入体のペレットを最終容量240mlまでの冷水に再懸濁して洗浄し、スラリーを形成した。このスラリーのサンプルをGDNF発現レベルのHPLC分析用に取り分けた。残りのスラリーを16,000×gで30分間遠心分離した。上清を捨て、遠心びんに少量の冷水を加えて静かに渦形成し、封入体ペレットの上にまばらに形成された膜層を除去した。Biospecホモジナイザーで十分な容量の冷水を使用してGDNF2mg/mlの濃度にペレットを再懸濁した。次に最終封入体懸濁液(25ml)と180mM システインHCl及び50mM Tris HCl,pH8.7を含む8MグアニジンHCl(25ml)を混合することにより、封入体を可溶化した。可溶化混合物を25℃で60〜90分間撹拌した後、20mM Tris HCl,pH8.75と0.2MグアニジンHClを含む2M尿素(450ml,5℃)に混合しながら注入した。再生混合物を5℃で72時間ゆっくりと撹拌した。
【0156】
再生したGDNFを次のように部分精製した。5℃の20
mM酢酸ナトリウム緩衝液(250ml,pH5)を迅速に撹拌しながら再生混合物に加え、pHを氷酢酸で5に調整した。生じた沈殿を5℃で13,600×gで45分間遠心分離して除去した。この遠心分離からの上清を次の精製段階のロード溶液として使用し、SP−大ビーズ樹脂(Pharmacia)によるカチオン交換クロマトグラフィーにかけた。カラムは、20mM酢酸ナトリウム(pH5)を平衡、濯ぎ及び溶離緩衝系として使用して5℃で操作した。樹脂ベッド(5ml)を5カラム容量(CV)の0.2N NaOHで処理した後、酢酸緩衝液(5CV)で平衡化した。ロード溶液(190ml)を0.5CV/分でカラムにアプライした後、同一流速で10CVの酢酸緩衝液で濯いだ。次に0.1CV/分の流速で酢酸緩衝液中NaCl0.3Mから0.9Mの20CV直線勾配でGDNFを樹脂から溶離した。カラム溶出液の280nmの吸光度をモニターし、フラクションとして集めてSDS−PAGEによりアッセイした。10%ピーク高さのGDNFピークの前縁から10%ピーク高さのピークの後縁までのGDNFを含むフラクションをプールした。このプール中のタンパク質は完全にGDNFであり、使用した産生株に応じて改変GDNF形として32%〜12%の汚染を含んでいた。次にプールをPBS又は他の組成の緩衝液で透析し、場合により限外濾過により25mg/mlまで濃縮した。この手順により精製した野生型及び類似形態のGDNFを逆相HPLC、カチオン交換HPLC、質量スペクトル分析及びエンドトキシンレベルにより特性決定し、調製物の純度を対応する産生株と比較した。
【実施例3】
【0157】
大腸菌における截形GDNFの組換え製造
Linらに記載されている方法(上記1994年5月23日付け米国特許出願第08/182,183号)に実質的に従って典型的なGDNFタンパク質を製造した。上述したような代替細菌発現材料及び方法を使用してもよい。夫々図5、6及び7に示すような[Pro23−Ile134]、[Arg32−Ile134]及び[Gly33−Ile134]截形GDNFタンパク質を大腸菌で発現させた。これらの典型的截形GDNFタンパク質をコードするポリヌクレオチドは図5、6及び7に示すように構築したが、図1、3及び4に示すような対応するポリヌクレオチドを使用してもよい。これらのポリヌクレオチドは、参考資料としてその開示内容を本明細書の一部とするPCR Technology,Principles and Applications for DNA Amplification,Henry A.Erlich編,Stockton Press,NY,1989(第6章、Using PCR to Engineer DNA)に記載されているような標準PCR手順により構築した。
【実施例4】
【0158】
ドーパミン作動性ニューロン神経栄養活性のバイオアッセイ
実施例3の大腸菌で発現させた[Pro23−Ile134]、[Arg32−Ile134]、[Gly33−Ile134]及び[Lys37−Ile134]截形GDNFタンパク質と実施例1のCHO由来[Arg32−Ile134]截形GDNFタンパク質が黒質ドーパミン作動性ニューロンによるドーパミン取り込みを促進する能力を定量的に評価した。
【0159】
材料
截形GDNFタンパク質の存在下におけるドーパミン作動性ニューロンの生存を評価するアッセイでは、次の材料を使用する。
【0160】
細胞培地
高グルコースダルベッコ改変イーグル培地(DMEM、カタログ番号#11965−092)、ハムF12培地(F12、#11765−021)、重炭酸ナトリウムを含まないライボビッツL15培地(#41300−039)、B27培地補充剤(#17504−010)、ペニシリン/ストレプトマイシン(#15070−014)、L−グルタミン(#25030−016)、ダルベッコのリン酸緩衝食塩水(D−PBS、#14190−052)、カルシウム及びマグネシウム塩を含むハンク平衡塩類溶液(HBSS、#24020−026)、N−2−ヒドロキシエチルピペラジン−N’−2−エタンスルホン酸(HEPES、#15630−015)、マウスラミニン(#23017−015)及びウシ血清アルブミンフラクションV(#110−18−017)は全てGIBCO,Grand Island,NYから入手した。熱不活化ウマ血清はHyClone,Logan,Utahから入手した。コンアルブミン(C−7786)、臭化水素酸ポリ−L−オルニチン(P−3655)、ウシインスリン(I−5500)、ヒトトランスフェリン(T−2252)、プトレッシン(P−6024)、プロゲステロン(P−6149)、亜セレン酸ナトリウム(S−9133)、メトリザミド(N−3383)は全てSigma Chemical Company,Saint−Louis,MOから入手した。パパイン、デオキシリボヌクレアーゼI(DNアーゼ)及びオボアルブミン(パパイン解離系)はWorthington Biochemicals,Freehold,NJから入手した。
【0161】
Falcon滅菌96穴マイクロプレート(#3072)、組織培養プラスチック容器及びポリプロピレン遠心分離管はBecton−Dickinson,Oxnard,CAから入手した。Nunc Lab−Tek組織培養チャンバーカバーガラス(#136439)はBaxter,Irvine,CAから入手し、20μm(#460)ナイロンメッシュはTetko,Elmsford,NYから入手し、4”解剖鉗子と4”鉗子鋏はRoboz Surgical,Washington,DCから入手した。
【0162】
抗体及び関連試薬
ポリクローナルウサギ抗チロシンヒドロキシラーゼ抗体(TE101)はEugene Tech,Ridgefield Park,NJから入手し、ポリクローナルウサギ抗ニューロン特異的エノラーゼ抗体(NSE,AB951)はChemicon,Temecula,CAから入手し、ビオチン化ヤギ抗ウサギIgG及びペルオキシダーゼ結合アビジン/ビオチン複合体(ABC Elite;VectastainキットPK−6100)はVector Laboratories,Burlingame,CAから入手した。3’,3’−ジアミノベンジジンはCappel Laboratories,West Chester,PAから入手した。PBS中のSuperblockブロッキング緩衝液(#37515)はPierce Chemical Company,Rockford,ILから入手した。Triton X−100(X−100)、Nonidet P−40(N6507)及び過酸化水素(30%v/v;H1009)はSigmaから入手した。
【0163】
GBR−12909ドーパミン取り込み阻害剤(D−052)はRBI,Natick,MAから入手した。3H−ドーパミン(トリチウム化ドーパミン、NE−131;21Ci/mmol)はNew England Nuclear,Boston,MAから入手した。Optiphase SupermixシンチレーションカクテルはWallac,Turku,フィンランドから入手した。透明ViewPlate−96マイクロプレート(#6005182)はPackard Instruments Corporation,Meriden,CTから入手した。他の全試薬は特に指定しない限り、Sigma Chemical Companyから入手した。
【0164】
培地の調製
DMEMとF12培地の1:1混合物として基本培地を調製し、50倍濃縮ストック溶液としてB27培地補充剤を補充した。最終濃度約2mMのL−グルタミン、約100IU/lのペニシリン及び約100mg/lのストレプトマイシンを加えた。熱不活化ウマ血清を最終濃度約15%まで加えた。混合後、pHを約7.3に調整し、培地を4℃に維持した。培地は、実験内変動を最小限にするように使用直前に新たに調製した。タンパク質吸着を最小限にするように常にプラスチックピペット及び容器を使用した。
【0165】
培養基質
基質ニューロンの最適結合と神経突起生長を助長するために、マイクロタイタープレート表面(培養基質)を次のようにポリ−L−オルニチンとラミニンで順次被覆して修飾した。プレート表面を0.1M硼酸(pH8.4)中0.1mg/ml滅菌ポリ−L−オルニチン溶液で室温で少なくとも1時間完全に被覆した後、Super−Q水で滅菌洗浄した。次に水洗物を吸引し、PBS中1.0μg/mlマウスラミニン溶液を加え、37℃で2時間インキュベートした。これらの手順は、結果の再現性を確保するためにプレートの使用直前に実施した。
【0166】
胎児ラット黒質培養物の調製
胎児ラット脳を黒質ドーパミン作動性ニューロン源として使用した。胎生15日の予定妊娠Sprague−Dawleyラットを使用した。各実験毎に最大36匹の胎児(約3リットル)を処理した。妊娠したラットをCO2に暴露して殺し、その腹腔を解剖鋏で切り開き、子宮から胎児を取り出した。次に胎児の脳を切開し、血液と髄膜を除去し、確定解剖目標(AltmanとBayer,Atlas of Prenatal Rat Brain Development,CRC Press,Boca Raton,FL,1995)を使用して黒質を含む腹側被蓋領域を解剖した。組織を氷冷D−PBSに集め、解離培地(HBSS中120単位パパイン及び2000単位DNアーゼ)10mlに移した後、約200rpmに設定した回転プラットフォーム震盪機で約37℃で45分間インキュベートした。次に火仕上げパスツールピペットで細胞を潰して分散させ、20μm Nitexメッシュで篩別して未解離組織を捨て、IEC臨床遠心機を使用して200×gで5分間遠心分離した。得られた細胞ペレットを、オボアルブミンと約500単位のDNアーゼを含むHBSSに再懸濁し、4%オボアルブミン溶液(HBSS中)に重層し、500×gで約10分間遠心分離した。最終ペレットを完全培地(上記参照)に再懸濁し、約28,000細胞/mlに調整し、ポリオルニチンとラミニンで予め被覆しておいた96穴マイクロプレートの6mmウェルに分注(90μl)した。すぐに細胞の結合が生じ、プレーティング効率は約75%であった。
【0167】
ドーパミン作動性ニューロンの免疫組織化学
Louisら(J.Pharmacol.Exp.Therap.,262:1274−1283,1992; Science,259:689−692,1993)により記載されている間接イムノペルオキシダーゼ法を下記のように若干改変して使用し、黒質の培養物中のドーパミン作動性ニューロンを特性決定した。培養物をD−PBS,pH7.4中4%パラホルムアルデヒドで室温で約30分間固定した後、D−PBS(6mmウェル当たり200μl)で3回洗浄した。次に、固定した培養物を1%NP−40を含むPBS中Superblockブロッキング緩衝液中でインキュベートし、抗体の浸透を増加した。次に、一次ウサギ抗チロシンヒドロキシラーゼ抗体を同一緩衝液に約1:2000に希釈して加え、回転震盪機で37℃で1時間インキュベートした。D−PBSで3回洗浄後、ヤギ抗ウサギビオチン化IgGを約1:500に希釈して使用して結合抗体を検出し、これらの二次抗体を細胞と共に37℃で約1時間インキュベートした。次に細胞をD−PBSで3回洗浄し、二次抗体を1:500に希釈したアビジン−ビオチン−ペルオキシダーゼ複合体で検出し、細胞を37℃で約45分間インキュベートした。D−PBSで更に3回洗浄後、0.04%3’,3’−ジアミノベンジジン−(HCl)4、0.06%NiCl2及び0.02%過酸化水素を含む0.1M Tris−HCl,pH7.4の溶液中で培養物を5〜20分間反応させた。
【0168】
ニューロン生存の測定
黒質培養物を固定し、前記のように免疫染色処理した後、200倍明視野顕微鏡で試験した。チロシンヒドロキシラーゼで染色するニューロンの数を96穴マイクロプレートの6mmウェル全体で数えた。生存可能なニューロンは定形細胞体をもち、主軸索様突起と数個の樹状突起があるとみなした。不整形の空胞のある核周囲部細胞体や分裂した神経突起をもつなど変性の徴候を示すニューロンは計数から除いた(但し、変性ニューロンの大部分は培養基質から分離していた)。ドーパミン作動性ニューロン細胞数は、TH陽性ニューロン/6mmウェル又は対照ドーパミン作動性ニューロン密度に対する倍数として表した。
【0169】
ドーパミン取り込みの測定
透明ViewPlate−96マイクロプレートで調製しておいた15日齢胎児ラット黒質ニューロンの培養物でドーパミン取り込みを測定した。約120mM NaCl、4.7mM KCl、1.8mM CaCl2、1.2mM MgSO4、32mM NaHPO4、1.3mM EDTA及び5.6mM D−グルコースを含む改変クレブス−リンゲル溶液、pH7.4から構成される予熱取り込み緩衝液(約100μl)で培養物を洗浄した。取り込み緩衝液には、ドーパミンの酸化を阻止するために1mMアスコルビン酸と50μMパルジリンも加えた。次に細胞を取り込み緩衝液中で37℃で約10分間プレインキュベートした。次に、トリチウム化したドーパミン(3H−DA、21Ci/mmol)を取り込み緩衝液75μl中約50nMの濃度で黒質培養物に加え、培養物を37℃で約60分間インキュベートした。ドーパミン取り込み阻害剤GBR−12909(1μM)を含む取り込み緩衝液と共に培養物をインキュベートすることにより、非特異的ドーパミン取り込みを測定した。非特異的取り込みは総取り込みの約1%未満であった。インキュベーション培地を吸引して取り込みアッセイを停止した後、氷冷取り込み緩衝液(約120μl)で3回迅速に洗浄した。次に、Optiphase Supermixシンチレーションカクテル(200μl)を加えて細胞を溶解させ、Wallac MicrobetaPlus 96穴マイクロプレートカウンターを使用してシンチレーションスペクトル分析により放射能を測定した(即ち培養物中に保持されたトリチウムのシンチレーション計数によりドーパミン取り込みを分析する)。結果をdpm/6mmウェル又は対照培養物に対する倍数として表す。
【0170】
アッセイ
ドーパミン作動性ニューロン生存及び形態学的発生
ドーパミン作動性ニューロンを多く含む15日齢胎児(E15)ラット黒質の培養物を使用して、截形GDNFタンパク質がドーパミン作動性ニューロンの生存に及ぼす効果を立証した。ポリオルニチンとラミニンで被覆した96穴マイクロプレートで種々の濃度(約1pg/ml〜約10ng/ml)の下記タンパク質、即ち大腸菌で発現させた成熟hGDNF、大腸菌で発現させた[Pro23−Ile134]、[Arg32−Ile134]、[Gly33−Ile134]及び[Lys37−Ile134]截形Glyタンパク質、CHO細胞で発現させた成熟hGDNF、並びにCHO細胞由来[Arg32−Ile134]截形GDNFタンパク質の存在下又は単独で培養物を6日間まで増殖させた。培地は15%熱不活化ウマ血清(E15培養物)又は2.5%熱不活化ウマ血清、D−グルコース、HEPES、インスリン及びトランスフェリン(P6培養物)を補充したDMEM/F12から構成した。ドーパミン生合成の律速酵素であるチロシンヒドロキシラーゼ(TH)の免疫染色をドーパミン作動性ニューロンのマーカーとして使用した。菱脳中のノルアドレナリン作動性ニューロンもTHに陽性染色するので、中脳の脳質に限定した領域を解剖し、ノルアドレナリン作動性細胞体を含む尾側の領域を避けるように十分注意した。6日後に、E15培養物は主に(上記)ニューロン特異的エノラーゼ免疫染色により確認される約70%のニューロンと、30%の(平坦で暗い外観をもつ)非ニューロン細胞から構成され、ドーパミン作動性ニューロンはニューロンポピュレーションの約10〜15%であった。
【0171】
6日後に培養物をパラホルムアルデヒドで固定し、これらの培養物中のドーパミン作動性ニューロンを識別するマーカーであるチロシンヒドロキシラーゼで免疫染色した。6mmウェル中に存在する全チロシンヒドロキシラーゼ陽性ニューロンを明視野顕微鏡で計数した。各実験条件毎に3〜6個の異なるウェルを分析した。結果を対照培養物中に存在するチロシンヒドロキシラーゼ陽性ニューロンの数の百分率として表した。
【0172】
CHO細胞で発現させたGDNF又は大腸菌で発現させたGDNFの2種のGDNF1.0ng/mlで処理したE15黒質の培養物は未処理対照培養物よりも夫々約38%及び27%多くのTH免疫反応性ニューロンを含んでおり、どちらのGDNF種もドーパミン作動性ニューロンの生存を促進することを示した。1.0ng/mlの截形GDNFタンパク質で処理したE15黒質の培養物は、in vitroで6日後に培養物中のTH陽性ニューロン数に同様の増加を示し、CHO細胞由来[Arg32−Ile134]截形GDNFタンパク質では42%、大腸菌で発現させた[Arg32−Ile134]及び[Gly33−Ile134]截形GDNFタンパク質では夫々26%及び17%増加した。
【0173】
対照培養物と成熟及び截形GDNFタンパク質で処理した培養物を比較すると更に、全GDNFタンパク質がドーパミン作動性ニューロンの形態学的分化に顕著な効果をもつことも判明した。[Arg32−Ile134]及び[Gly33−Ile134]截形GDNFタンパク質の効果は夫々の対応する成熟GDNFタンパク質と同一であった。全GDNF処理培養物中のTH免疫反応性ニューロンは、対照培養物中のTH陽性ニューロンに比較して有意に複雑で広範囲の神経突起分枝をもち、神経突起分枝度が高く、細胞体の総寸法が大きかった。
【0174】
ドーパミン取り込み
ドーパミン取り込みは高親和性ドーパミン再取り込みトランスポーター部位の数及び活性の尺度であり、ドーパミン作動性ニューロンの機能的分化を表す。成熟GDNF又は截形GDNFタンパク質の存在下又は不在下でin vitroで6日後にE15ラット黒質の培養物でドーパミン取り込みを測定した。これらの培養物においてドーパミン取り込みはドーパミン作動性ニューロンの特徴的薬理プロフィルをもち、即ちドーパミン作動性ニューロンに特異的なドーパミン輸送阻害剤(ID50=20nM)である1.0μM GBR−12909によりほぼ完全にブロックされた(>98%)。従って、ノルエピネフリン輸送体を介してドーパミンを取り込むことができるが、GBR−12909阻害に非感受性の汚染性ノルアドレナリン作動性ニューロンの存在はドーパミン取り込み測定値に影響していないと考えられる。CHO細胞で発現させた成熟GDNFとCHO由来[Arg32−Ile134]截形GDNFタンパク質の効果は同一であり、約65%増加し、ED50は約20pg/mlであった。図5に示すような大腸菌で発現させた[Pro23−Lys37ΔAsn37−Ile134]截形Dタンパク質は65%の増加を示し、ED50は約40pg/mlであった。大腸菌で発現させた成熟タンパク質と大腸菌で発現させた[Arg32−Ile134]、[Gly33−Ile134]及び[Lys37−Ile134]截形GDNFタンパク質のドーパミン取り込み効果は同一であり、約50%増加し、ED50は約50pg/mlであった。
【0175】
以上の結果は、截形GDNFタンパク質が黒質ドーパミン作動性ニューロンの強力な生存促進及び分化誘導因子として作用することを示す。従って、截形GDNFタンパク質は情動の鋭さの低下、随意及び不随意筋運動の減退、筋硬直及び振戦を特徴とする神経障害であるパーキンソン病の治療に特に有利であると考えられる。このような症状は黒質内のドーパミン産生ニューロンの進行性変性に起因すると考えられる。これらのニューロン(「ドーパミン作動性ニューロン」)の変性は線条体と呼ばれる脳の隣接領域でドーパミンの低下をもたらす。
【0176】
図面の簡単な説明
本発明の多数の特徴及び利点は図面を参考に容易に理解されよう。
【0177】
図1は成熟ヒトグリア細胞系由来神経栄養因子(hGDNF)をコードする核酸配列(配列番号1)を示す。成熟ヒトGDNFタンパク質のアミノ酸配列(配列番号2)も示す。
【0178】
図2は組換え截形GDNFタンパク質の発現のために利用したプラスミド構築図を示す。
【0179】
図3はGDNF及び截形GDNFポリヌクレオチドをコードする別の核酸配列(配列番号39)の制限地図を示す。
【0180】
図4はGDNF及び截形GDNFポリヌクレオチドをコードする更に別の核酸配列(配列番号40)の制限地図を示す。
【0181】
図5は[Pro23−Lys37ΔAsn37−Ile134]截形GDNFタンパク質置換変異体(配列番号42)をコードする核酸配列(配列番号41)を示す。このタンパク質はMet−Ser−[Pro23−Lys37ΔAsn37−Ile134]截形GDNFタンパク質付加/置換変異体と表すこともできる。
【0182】
図6は[Arg32−Ile134]截形GDNFタンパク質(配列番号44)をコードする核酸配列(配列番号43)を示す。
【0183】
図7は[Gly33−Ile134]截形GDNFタンパク質(配列番号46)をコードする核酸配列(配列番号45)を示す。
【0184】
図8は数種の截形GDNFタンパク質の例であるMet−[Arg32−Ile134](配列番号48)、Met−[Gly33−Ile134](配列番号49)及びMet−Ser−[Pro23−Lys37ΔAsn37−Ile134](配列番号50)と成熟hGDNFのアミノ酸配列(配列番号47)の比較を示す。
【特許請求の範囲】
【請求項1】
アミノ酸配列:
X−[Cys41−Cys133]−Y
(式中、[Cys41−Cys133]は図1(配列番号2)に示すCys41〜Cys133のアミノ酸配列を表し、YはCys133のカルボキシ末端基又はIle134のカルボキシ末端アミノ酸残基を表し、XはCys41のメチオニン化もしくは非メチオニン化アミン基又は以下の群:
【表1】
から選択されるアミノ末端アミノ酸残基を表す)をもつ截形グリア細胞系由来神経栄養因子(GDNF)タンパク質産物並びにその付加、置換及び内部欠失変異体及び誘導体。
【請求項2】
X=RQAAA ANPENSRGKG RRGQRGKNRG(配列番号24)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項3】
X=NPENSRGKG RRGQRGKNRG(配列番号18)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項4】
X=PENSRGKG RRGQRGKNRG(配列番号17)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項5】
X=SRGKG RRGQRGKNRG(配列番号14)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項6】
X=RGQRGKNRG(配列番号8)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項7】
X=GQRGKNRG(配列番号7)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項8】
X=KNRG(配列番号3)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項9】
X=NRGである請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項10】
前記アミノ酸配列がグリコシル化されている請求項1から9のいずれか一項に記載の截形GDNFタンパク質産物。
【請求項11】
前記アミノ酸配列がグリコシル化されていない請求項1から9のいずれか一項に記載の截形GDNFタンパク質産物。
【請求項12】
前記誘導体が水溶性ポリマーに結合したX−[Cys41−Cys133]−Yアミノ酸配列である請求項1に記載の截形GDNFタンパク質産物。
【請求項13】
請求項1に記載の截形GDNFタンパク質産物をコードするポリヌクレオチド。
【請求項14】
図1に示す配列の一部を含む請求項13に記載のポリヌクレオチド。
【請求項15】
図3に示す配列の一部を含む請求項13に記載のポリヌクレオチド。
【請求項16】
図4に示す配列の一部を含む請求項13に記載のポリヌクレオチド。
【請求項17】
図5に示す配列を含む請求項13に記載のポリヌクレオチド。
【請求項18】
図6に示す配列を含む請求項13に記載のポリヌクレオチド。
【請求項19】
図7に示す配列を含む請求項13に記載のポリヌクレオチド。
【請求項20】
発現制御配列に作動的に連結した請求項13に記載のポリヌクレオチドを含むベクター。
【請求項21】
請求項13に記載のポリヌクレオチドで形質転換又はトランスフェクトされた原核又は真核宿主細胞。
【請求項22】
截形GDNFタンパク質の製造方法であって、請求項
21に記載の宿主細胞を適切な栄養培地で培養し、場合により、前記細胞又は前記栄養培地から前記截形GDNFを単離することを特徴とする前記方法。
【請求項23】
前記宿主細胞が大腸菌である請求項22に記載の截形
GDNFタンパク質の製造方法。
【請求項24】
前記宿主細胞がチャイニーズハムスター卵巣細胞である請求項22に記載の截形GDNFタンパク質の製造方法。
【請求項25】
截形グリア細胞系由来神経栄養因子(GDNF)タンパク質の製造方法であって、
(a)請求項20に記載のベクターで形質転換又はトランスフェクトされた原核又は真核宿主細胞を培養する段階と、
(b)前記宿主細胞が截形GDNFタンパク質を発現できるような条件下に前記宿主細胞を維持する段階と、
(c)場合により、前記宿主細胞により発現された截形GDNF
タンパク質を単離する段階を含む前記方法。
【請求項26】
請求項13に記載の外因ポリヌクレオチドを含む原核又は真核宿主細胞の組換え発現産物である截形GDNFタンパク質。
【請求項27】
医薬的に許容可能なビヒクルと共に請求項1に記載の截形GDNFタンパク質産物を含む医薬組成物。
【請求項28】
医薬的に許容可能なビヒクルと共に請求項22に記載の方法により製造された截形GDNFタンパク質を含む医薬組成物。
【請求項29】
医薬的に許容可能なビヒクルと共に請求項25に記載の方法により製造された截形GDNFタンパク質を含む医薬組成物。
【請求項30】
請求項27に記載の医薬組成物を患者に投与することを特徴とするパーキンソン病の治療方法。
【請求項31】
請求項13に記載のポリヌクレオチド配列を患者に投与し、前記截形GDNFタンパク質をin vivo産生させることを特徴とするパーキンソン病の治療方法。
【請求項32】
請求項13に記載のポリヌクレオチド配列で形質転換された細胞を患者に移植し、前記截形GDNFタンパク質をin vivo産生させることを特徴とするパーキンソン病の治療方法。
【請求項33】
成熟GDNFタンパク質と1種以上の截形GDNFタンパク質を含むグリア細胞系由来神経栄養因子(GDNF)組成物であって、前記成熟GDNFタンパク質が約44kDaの分子量をもち、前記截形GDNFタンパク質が約29〜40kDa
の分子量をもつ前記GDNF組成物。
【請求項34】
少なくとも2種の截形GDNFタンパク質を含み、第1種が約36kDaの分子量をもち、第2種が約40kDaの分子量をもつ請求項33に記載のGDNF組成物。
【請求項35】
約40kDaの分子量をもつ前記第2種の截形GDNFが約22kDaの分子量をもつ成熟GDNFモノマーと約18kDaの分子量をもつ截形GDNFモノマーのヘテロダイマーである請求項34に記載のGDNF組成物。
【請求項36】
請求項33に記載のGDNF組成物から単離され、約29
〜40kDaの分子量をもつ截形GDNFタンパク質。
【請求項37】
請求項33に記載のGDNF組成物から単離され、約29
〜36kDaの分子量をもつ截形GDNFタンパク質。
【請求項38】
組換えにより改変された細菌又は哺乳動物細胞により発現される成熟GDNFタンパク質から誘導される截形GDNFタンパク質であって、アミノ酸配列:
X−[Cys41−Cys133]−Y
(式中、[Cys41−Cys133]は図1(配列番号2)に示す
Cys41〜Cys133のアミノ酸配列を表し、YはCys133のカ
ルボキシ末端基又はIle134のカルボキシ末端アミノ酸
残基を表し、XはCys41のアミン基又は以下の群:
【表2】
から選択されるアミノ末端アミノ酸残基を表す)をもつ截形
GDNFタンパク質並びにその付加、置換及び内部欠失変異体。
【請求項39】
Xが
【表3】
から構成される群から選択される請求項38に記載の截形
GDNFタンパク質及びその変異体。
【請求項40】
成熟GDNFタンパク質が組換えにより改変された細菌細胞により発現され、前記截形GDNFタンパク質がin vitro又はin vivo産生される請求項38に記載の截形GDNFタンパク質。
【請求項41】
治療上有効な量の請求項1に記載の截形GDNFタンパク質産物を1種以上の医薬的に許容可能なビヒクルと混合してなる医薬組成物の製造方法。
【請求項42】
疾病又は傷害に起因する神経系の損傷を治療するための請求項1に記載の截形GDNFタンパク質産物の使用。
【請求項43】
パーキンソン病の治療のための請求項42に記載の截形GDNFタンパク質産物の使用。
【請求項44】
疾病又は傷害に起因する神経系の損傷を治療するための医薬組成物の製造のための請求項1に記載の截形GDNFタンパク質産物の使用。
【請求項1】
アミノ酸配列:
X−[Cys41−Cys133]−Y
(式中、[Cys41−Cys133]は図1(配列番号2)に示すCys41〜Cys133のアミノ酸配列を表し、YはCys133のカルボキシ末端基又はIle134のカルボキシ末端アミノ酸残基を表し、XはCys41のメチオニン化もしくは非メチオニン化アミン基又は以下の群:
【表1】
から選択されるアミノ末端アミノ酸残基を表す)をもつ截形グリア細胞系由来神経栄養因子(GDNF)タンパク質産物並びにその付加、置換及び内部欠失変異体及び誘導体。
【請求項2】
X=RQAAA ANPENSRGKG RRGQRGKNRG(配列番号24)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項3】
X=NPENSRGKG RRGQRGKNRG(配列番号18)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項4】
X=PENSRGKG RRGQRGKNRG(配列番号17)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項5】
X=SRGKG RRGQRGKNRG(配列番号14)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項6】
X=RGQRGKNRG(配列番号8)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項7】
X=GQRGKNRG(配列番号7)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項8】
X=KNRG(配列番号3)である請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項9】
X=NRGである請求項1に記載の截形GDNFタンパク質産物又はその変異体。
【請求項10】
前記アミノ酸配列がグリコシル化されている請求項1から9のいずれか一項に記載の截形GDNFタンパク質産物。
【請求項11】
前記アミノ酸配列がグリコシル化されていない請求項1から9のいずれか一項に記載の截形GDNFタンパク質産物。
【請求項12】
前記誘導体が水溶性ポリマーに結合したX−[Cys41−Cys133]−Yアミノ酸配列である請求項1に記載の截形GDNFタンパク質産物。
【請求項13】
請求項1に記載の截形GDNFタンパク質産物をコードするポリヌクレオチド。
【請求項14】
図1に示す配列の一部を含む請求項13に記載のポリヌクレオチド。
【請求項15】
図3に示す配列の一部を含む請求項13に記載のポリヌクレオチド。
【請求項16】
図4に示す配列の一部を含む請求項13に記載のポリヌクレオチド。
【請求項17】
図5に示す配列を含む請求項13に記載のポリヌクレオチド。
【請求項18】
図6に示す配列を含む請求項13に記載のポリヌクレオチド。
【請求項19】
図7に示す配列を含む請求項13に記載のポリヌクレオチド。
【請求項20】
発現制御配列に作動的に連結した請求項13に記載のポリヌクレオチドを含むベクター。
【請求項21】
請求項13に記載のポリヌクレオチドで形質転換又はトランスフェクトされた原核又は真核宿主細胞。
【請求項22】
截形GDNFタンパク質の製造方法であって、請求項
21に記載の宿主細胞を適切な栄養培地で培養し、場合により、前記細胞又は前記栄養培地から前記截形GDNFを単離することを特徴とする前記方法。
【請求項23】
前記宿主細胞が大腸菌である請求項22に記載の截形
GDNFタンパク質の製造方法。
【請求項24】
前記宿主細胞がチャイニーズハムスター卵巣細胞である請求項22に記載の截形GDNFタンパク質の製造方法。
【請求項25】
截形グリア細胞系由来神経栄養因子(GDNF)タンパク質の製造方法であって、
(a)請求項20に記載のベクターで形質転換又はトランスフェクトされた原核又は真核宿主細胞を培養する段階と、
(b)前記宿主細胞が截形GDNFタンパク質を発現できるような条件下に前記宿主細胞を維持する段階と、
(c)場合により、前記宿主細胞により発現された截形GDNF
タンパク質を単離する段階を含む前記方法。
【請求項26】
請求項13に記載の外因ポリヌクレオチドを含む原核又は真核宿主細胞の組換え発現産物である截形GDNFタンパク質。
【請求項27】
医薬的に許容可能なビヒクルと共に請求項1に記載の截形GDNFタンパク質産物を含む医薬組成物。
【請求項28】
医薬的に許容可能なビヒクルと共に請求項22に記載の方法により製造された截形GDNFタンパク質を含む医薬組成物。
【請求項29】
医薬的に許容可能なビヒクルと共に請求項25に記載の方法により製造された截形GDNFタンパク質を含む医薬組成物。
【請求項30】
請求項27に記載の医薬組成物を患者に投与することを特徴とするパーキンソン病の治療方法。
【請求項31】
請求項13に記載のポリヌクレオチド配列を患者に投与し、前記截形GDNFタンパク質をin vivo産生させることを特徴とするパーキンソン病の治療方法。
【請求項32】
請求項13に記載のポリヌクレオチド配列で形質転換された細胞を患者に移植し、前記截形GDNFタンパク質をin vivo産生させることを特徴とするパーキンソン病の治療方法。
【請求項33】
成熟GDNFタンパク質と1種以上の截形GDNFタンパク質を含むグリア細胞系由来神経栄養因子(GDNF)組成物であって、前記成熟GDNFタンパク質が約44kDaの分子量をもち、前記截形GDNFタンパク質が約29〜40kDa
の分子量をもつ前記GDNF組成物。
【請求項34】
少なくとも2種の截形GDNFタンパク質を含み、第1種が約36kDaの分子量をもち、第2種が約40kDaの分子量をもつ請求項33に記載のGDNF組成物。
【請求項35】
約40kDaの分子量をもつ前記第2種の截形GDNFが約22kDaの分子量をもつ成熟GDNFモノマーと約18kDaの分子量をもつ截形GDNFモノマーのヘテロダイマーである請求項34に記載のGDNF組成物。
【請求項36】
請求項33に記載のGDNF組成物から単離され、約29
〜40kDaの分子量をもつ截形GDNFタンパク質。
【請求項37】
請求項33に記載のGDNF組成物から単離され、約29
〜36kDaの分子量をもつ截形GDNFタンパク質。
【請求項38】
組換えにより改変された細菌又は哺乳動物細胞により発現される成熟GDNFタンパク質から誘導される截形GDNFタンパク質であって、アミノ酸配列:
X−[Cys41−Cys133]−Y
(式中、[Cys41−Cys133]は図1(配列番号2)に示す
Cys41〜Cys133のアミノ酸配列を表し、YはCys133のカ
ルボキシ末端基又はIle134のカルボキシ末端アミノ酸
残基を表し、XはCys41のアミン基又は以下の群:
【表2】
から選択されるアミノ末端アミノ酸残基を表す)をもつ截形
GDNFタンパク質並びにその付加、置換及び内部欠失変異体。
【請求項39】
Xが
【表3】
から構成される群から選択される請求項38に記載の截形
GDNFタンパク質及びその変異体。
【請求項40】
成熟GDNFタンパク質が組換えにより改変された細菌細胞により発現され、前記截形GDNFタンパク質がin vitro又はin vivo産生される請求項38に記載の截形GDNFタンパク質。
【請求項41】
治療上有効な量の請求項1に記載の截形GDNFタンパク質産物を1種以上の医薬的に許容可能なビヒクルと混合してなる医薬組成物の製造方法。
【請求項42】
疾病又は傷害に起因する神経系の損傷を治療するための請求項1に記載の截形GDNFタンパク質産物の使用。
【請求項43】
パーキンソン病の治療のための請求項42に記載の截形GDNFタンパク質産物の使用。
【請求項44】
疾病又は傷害に起因する神経系の損傷を治療するための医薬組成物の製造のための請求項1に記載の截形GDNFタンパク質産物の使用。
【図1】


【図2】
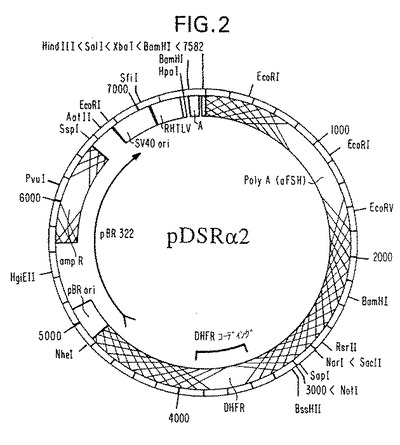
【図3A】
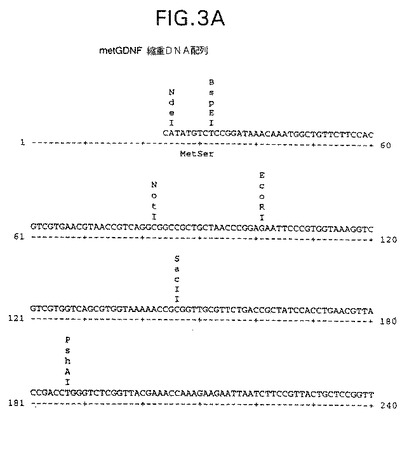
【図3B】

【図4】

【図5】
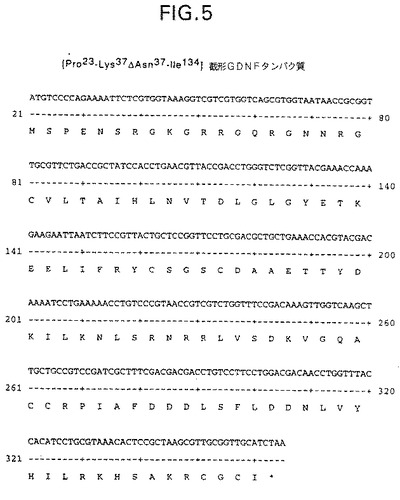
【図6】

【図7】

【図8】

【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2010−279360(P2010−279360A)
【公開日】平成22年12月16日(2010.12.16)
【国際特許分類】
【外国語出願】
【出願番号】特願2010−140665(P2010−140665)
【出願日】平成22年6月21日(2010.6.21)
【分割の表示】特願2007−229870(P2007−229870)の分割
【原出願日】平成8年9月16日(1996.9.16)
【出願人】(500049716)アムジエン・インコーポレーテツド (242)
【Fターム(参考)】
【公開日】平成22年12月16日(2010.12.16)
【国際特許分類】
【出願番号】特願2010−140665(P2010−140665)
【出願日】平成22年6月21日(2010.6.21)
【分割の表示】特願2007−229870(P2007−229870)の分割
【原出願日】平成8年9月16日(1996.9.16)
【出願人】(500049716)アムジエン・インコーポレーテツド (242)
【Fターム(参考)】
[ Back to top ]