
所望のポリヌクレオチドの発現を達成するためのセンス構築物の使用と組合わせた内因的遺伝子のダウンレギュレーションを達成するためのsiRNAの使用
本発明は、宿主における内因的遺伝子の発現をノックアウトまたはノックダウンするための内因的遺伝子をターゲットとするsiRNAの組合わせ使用、およびポリヌクレオチドによりコードされるタンパク質を宿主において発現させるために、宿主に送出ベヒクル/発現ベクター中の遺伝子をコードするポリヌクレオチドを送出すことに関する。

【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、内因的遺伝子の発現をノックダウンまたはノックアウトするための内因的遺伝子をターゲットとするsiRNAの組合わせ使用、およびその遺伝子を発現させるための外因的遺伝子をコードするポリヌクレオチドの使用に関する。
【背景技術】
【0002】
比較的最近において、タンパク質の発現を阻害するために二本鎖RNA (“dsRNA”) を使用できることを研究者らは観察した。遺伝子を阻害する能力は人間の疾患を治療する広い可能性を有し、そして多数の研究者らおよび商業的エンティティーは現在この技術に基づく療法の開発にかなりの財源を投じている。
【0003】
遺伝子サイレンシングを誘導する二本鎖RNAは少なくとも3つのレベルで発生することができる: (i) 転写不活性化、これはRNAガイドDNAまたはヒストンメチル化を意味する; (ii) mRNA分解を誘導するsiRNA; および (iii) 転写アテニュエーションを誘導するmRNA。
【0004】
哺乳動物細胞においてサイレンシング (RNAインターフェレンス、またはRNAi) を誘導するRNAの主要な機構はmRNA分解であることが一般に考えられる。哺乳動物細胞においてRNAiを使用する初期の試みはdsRNAの長い鎖の使用に集中した。しかしながら、RNAiを誘導するこれらの試みは、部分的にインターフェロンの応答の誘導のために、成功が制限され、これは一般にターゲット特異的と反対に、タンパク質合成の阻害を生ずる。こうして、長いdsRNAは哺乳動物系におけるRNAiのための価値のあるオプションではない。
【0005】
より最近において、短い (18〜30 bp) RNAデュプレックスを培養において哺乳動物細胞中に導入するとき、ターゲットmRNAの配列特異的阻害はインターフェロンの応答を誘導しないで実現できることが示された。これらの短いdsRNAのあるもの、小さい阻害性RNAs (“siRNAs”) と呼ぶ、は1モル以下の濃度において触媒的に作用して細胞中のターゲットmRNAの95%より多くを切り離すことができる。
【0006】
siRNA活性についての機構の説明、ならびにその用途のいくつかは、下記の文献に記載されている: Provost他 (2002) Ribonuclease Activity and RNA Binding of Recombinant Human Dicer、EMBO J. 21 (21): 5864-5874; Tabara他 (2002) The dsRNA Binding Protein RDE-4 Interacts with RDE-1、DCR-1 and a DexH-box Helicase to Direct RNA in C. elegans、Cell 109 (7): 861-71; Ketting他 (2002) Dicer Functions in RNA Interference and in Synthesis of Small RNA Involved in Developmental Timing in C. elegans; Martinez他、Single-Stranded Antisense siRNAs Guide Target RNA Cleavage in RNAi、Cell 110 (5): 563; Hutvagner & Zamore (2002) A microRNA in a multiple-turnover RNAi enzyme complex、Science 297: 2056。
【0007】
機械的見解から、植物および無脊椎動物の細胞中への長い二本鎖RNAの導入は、ダイサー (Dicer) として知られているIII型エンドヌクレアーゼによりsiRNAに破壊される。Sharp、RNA interference - 2001、Genes Div. 2001、15: 485。ダイサー、リボヌクレアーゼ-III様酵素はdsRNAをプロセシングして、19〜23塩基対の短い干渉性RNAにし、これらのRNAは特徴ある2塩基の3’ オーバーハングを有する。Bernstein、Caudy、Hammond & Hannon (2001) Role for a bidentate Ribonuclease in the initiation step of RNA interference、Nature 409: 363。
【0008】
次いでsiRNAはRNA誘導サイレンシング複合体 (RISC) 中に組込まれ、ここで1または2以上のヘリカーゼはsiRNAデュプレックスを巻戻し、相補的アンチセンス鎖がターゲットを認識できるようにする。Nykanen、Halve & Zamore (2001) ATP requirements and small interfering RNA structure in the RNA interference pathway、Cell 107: 309。適当なターゲットmRNAに結合すると、RISC内の1または2以上のリボヌクレアーゼはターゲットを切り離してサイレンシングを誘導する。Elbashir、Lendeckel & Tuschl (2001) RNA interference is mediated by 21 - and 22 - nucleotide RNAs、Genes Dev. 15: 188、図1。
【0009】
インターフェレンス効果は長く持続することができ、そして多数回の細胞分裂後に検出することができる。その上、RNAiは配列特異性を示す。Kisielow M.他(2002) Isoform-specific knockdown and expression of adaptor protein ShcA using small interferon RNA、J. Biochem. 363: 1-5。こうして、RNAi機構は転写の1つの型を特異的にノックダウンできるが、密接に関係するmRNAに影響を与えない。siRNAはこれらの性質を有するために、遺伝子発現を阻害し、遺伝子機能および薬物ターゲット確認を研究する潜在的に価値のある道具である。その上、siRNAは下記に対する治療剤として潜在的に有効である: (1) 遺伝子の過剰発現または異常発現により引き起こされる疾患; および (2) 突然変異を含有する遺伝子の発現により発生する疾患。
【0010】
有望なsiRNA従属的サイレンシングは多数の因子に依存する。RNAiにおける最も議論を呼びそうな問題の1つは、siRNA設計の必要性の問題、すなわち、使用するsiRNAの配列を考慮することである。C. elegansおよび植物における初期の研究は長いdsRNAの導入により設計の問題を回避した (例えば、下記の文献を参照のこと、Fire A. 他 (1998) Nature 391: 806-811) 。この原始生物において、長いdsRNA分子はダイサーによりsiRNAに切断され、こうして潜在的に全体の転写物をカバーできるデュプレックスの多様な集団を発生させた。
【0011】
これらの分子のいくつかの小部分は非機能的である (すなわち、サイレンシングをほとんど、あるいはまったく誘導しない) が、1または2以上は高度に機能的である可能性を有し、これにより問題の遺伝子をサイレンシングし、siRNA設計の必要性を軽減する。不都合なことには、インターフェロン応答のために、この同一のアプローチは哺乳動物系に利用不可能である。この効果はダイサー切断をバイパスし、siRNAを直接導入することによって回避できるが、この計画は選択したsiRNA配列が非機能的または半機能的であるという危険を伴う。
【0012】
siRNA設計はRNAiのきわめて重要な要素ではないという見解を多数の研究が示してきている。他方において、この分野における他の研究は、siRNAの設計に注意を払うことによってRNAiをいっそう効率的とすることができる可能性を探求し始めた。
【0013】
種々の疾患または障害を治療するために、ある種のタンパク質をアップレギュレートすることは望ましいが、これは必要であるすべてのことではないことがある。例えば、内因的タンパク質または異なるタンパク質の発現をノックダウンまたはノックアウトするために、siRNAの組合わせ使用は必要であることがある。本発明はこの要求を満足させ、そして癌、特に多発性骨髄腫を治療する方法を提供する。
【0014】
多発性骨髄腫を含む癌は、アポトーシスを誘導する能力から利益を受ける疾患である。多発性骨髄腫の慣用療法は、化学療法、幹細胞移植、幹細胞移植を伴う高投与量の化学療法、およびサルベージ療法を含む。化学療法は、Thalomid(商標)(タリドミド) 、ボルテゾミド、Aredia(商標)(パミドロネート) 、ステロイド、およびZometa(商標)(ゾレドロン酸) を使用する治療を含む。しかしながら多数の化学療法剤は毒性であり、非癌細胞、例えば骨髄、胃腸の内層、および毛胞を活発に分割する。したがって、化学療法は血球数の減少、悪心、嘔吐、下痢、および毛髪喪失を生ずることがある。
【0015】
典型的には、慣用化学療法または標準的投与量の化学療法は、多発性骨髄腫の患者のための一次的または初期の治療である。高投与量の化学療法および幹細胞移植のための調整において、患者はまた化学療法を受けることがある。移植前に存在する腫瘍を減少させるために、誘導療法 (幹細胞移植前の慣用化学療法) を使用できる。ある種の化学療法剤は他の薬剤よりも誘導療法のために適当である。なぜなら、それらは骨髄細胞に対する毒性が低く、骨髄からの幹細胞の収量を大きくするからである。誘導療法のために適当な化学療法剤の例は次の通りである: デキサメタゾン、タリドミド/デキサメタゾン、VAD (ビンクリスチン、Adriamycin(商標)(ドキソルビシン) およびデキサメタゾンの組合わせ) 、およびDVd (ペギル化リポソームドキソルビシン (Doxil(商標)、Caelyx(商標)) 、ビンクリスチンおよび還元スケジュールデキサメタゾンの組合わせ) 。
【0016】
多発性骨髄腫の標準的治療は、メルファランとプレドニソン (コルチコステロイド薬剤) との組合わせであり、50%の応答比率を達成する。不都合なことには、メルファランはアルキル化剤であり、誘導療法にそれほど適当ではない。コルチコステロイド (ことにデキサメタゾン) は、特により高齢の患者および化学療法に耐えることができない患者において、多発性骨髄腫の療法に時々単独で使用される。また、デキサメタゾンは、単独でまたは他の薬剤との組合わせで、誘導療法において使用される。VADは誘導療法において最も普通に使用されるが、最近誘導療法に有効であることが示された。ボルテゾミドは最近多発性骨髄腫の治療のために承認されたが、非常に毒性である。しかしながら、現存する治療剤はいずれも十分な治癒の可能性を提供しない。こうして、癌および多発性骨髄腫のための適当な治療がなお要求されている。本発明はこの要求を満足する。
【発明の概要】
【0017】
本発明は、宿主における内因的遺伝子の発現をノックアウトまたはノックダウンするための内因的遺伝子をターゲットとするsiRNAの組合わせ使用、および送出ビヒクル/発現ベクター中の遺伝子をコードするポリヌクレオチドを送出して、ポリヌクレオチドによりコードされるタンパク質を宿主において発現させることに関する。正常 (非不完全) タンパク質をコードするポリヌクレオチド (またはタンパク質それ自体) を宿主に投与し、発現させ (ポリヌクレオチドの場合において) 、こうして正常タンパク質がその必要な機能を実行できるようにする。siRNAは好ましくは遺伝子のある領域をターゲットするように設計し、それが不完全なタンパク質の内因的発現をノックダウンまたはノックアウトするが、同時に正常タンパク質をコードする投与したポリヌクレオチドの外因的発現に影響を与えないようにする。
【0018】
本発明は、eIF5A1の3’ 末端をターゲットとするeIF5A1 siRNAの複合体と、突然変異体eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターとを含んでなる組成物であって、突然変異体eIF5A1はハイプシン化 (hypusinate) されることができず、かつsiRNAおよび発現ベクターはポリエチレンイミンに対して複合化して複合体を形成している組成物を提供する。
【0019】
本発明は、ターゲット遺伝子をターゲットとして被検体におけるターゲット遺伝子の内因的発現を抑制するsiRNAと、被検体において発現されることができるターゲットタンパク質をコードするポリヌクレオチドとを含んでなる組成物を提供する。ある種の態様において、ポリヌクレオチドはRNAI抵抗性プラスミド中に存在する (siRNAにより抑制されないであろう) 。siRNAおよびプラスミドはポリエチレンイミンに対して複合化して複合体を形成している。
【0020】
ある種の態様において、siRNAは図25に示す配列を有し、ここで突然変異体eIF5A1をコードするポリヌクレオチドはeIF5A1K50Rである。発現ベクターは突然変異体eIF5A1をコードするポリヌクレオチドと、被検体におけるポリヌクレオチドの発現を提供するように作用可能に連鎖されたプロモーターとを含んでなる。プロモーターは好ましくは組織特異的または系統的である。例えば、癌を治療するために組成物を使用する場合、プロモーターは癌が存在する組織に対して組織特異的であることが好ましい。例えば、多発性骨髄腫を治療するために、B細胞特異的プロモーター、例えばB29を使用することが好ましい。ある種の態様において、発現ベクターはpCpGプラスミドを含んでなる。
【0021】
ある種の態様において、eIF5A1 siRNAと突然変異体eIF5A1ポリヌクレオチドを含んでなる発現ベクターは独立してポリエチレンイミン、例えばin vivo JetPEITMに対して複合化している。他の態様において、eIF5A1 siRNAと突然変異体eIF5A1ポリヌクレオチドを含んでなる発現ベクターは一緒になってポリエチレンイミンに対して複合化している。
【0022】
さらに、本発明は、eIF5A1の3’ 末端をターゲットとするeIF5A1 siRNAと、突然変異体eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターとを含んでなる組成物であって、突然変異体eIF5A1はハイプシン化されることができず、かつsiRNAおよび発現ベクターは癌を治療するために被検体に送出される組成物を提供する。癌は多発性骨髄腫を包含する任意の癌であることができる。
【0023】
さらに、本発明は、本発明の組成物を被検体 (哺乳動物およびヒトを包含するが、これらに限定されない) に投与することを含んでなる癌を治療する方法を提供する。
【0024】
組成物は任意の許容される経路、例えば静脈内、腹腔内または腫瘍内であるが、これらに限定されない経路で投与できる。siRNAおよび発現ベクターは異なる時間に異なる経路を経て投与できるか、あるいは同時に同一経路を経て一緒に投与できる。例えば、限定されないが、siRNAはそのままでまたは担体、例えばin vivo JetPEITMに対して複合化して静脈内に送出すことができ、そして発現ベクターは腫瘍内に投与ことができるか、あるいはsiRNAおよび発現ベクターの両方は静脈内または腫瘍内に投与することができる、およびその他。
【0025】
本発明は、癌細胞の成長を阻害しおよび/または癌細胞を殺す方法を提供する。また、本発明は、癌細胞の転移能力を阻害または遅延させる方法を提供する。癌成長の阻害は、腫瘍サイズの減少、腫瘍成長の低下を包含し、そしてまた腫瘍の完全な寛解を包含することができる。癌は任意の癌または腫瘍であることができ、結腸癌、結腸直腸癌、膀胱癌、頸部腺癌および肺癌を包含するが、これらに限定されない。好ましくは、癌は多発性骨髄腫である。
【0026】
好ましい態様において、elF-5Aはハイプシン化されることができない突然変異体である。典型的な突然変異体は本明細書において説明されている。
elF-5AまたはelF-5Aをコードするポリヌクレオチド (elF-5Aを発現させるために) を被検体に提供することに加えて、siRNAを提供してelF-5Aの内因的発現をノックアウトまたはノックダウンする。
【0027】
また、本発明は、多発性骨髄腫の患者における多発性骨髄腫細胞を殺す薬剤を製造するために、elF5A、eIF5A1をコードするポリヌクレオチドおよびeIF5A1に対するsiRNAを使用することに関する。好ましくは、突然変異体elF-5Aをコードするポリヌクレオチドはハイプシン化されることができない。
【0028】
また、本発明は、鎌状白血球貧血を治療する方法を提供する。 鎌状白血球貧血を患っている患者に、健康なヘモグロビン遺伝子 (例えば、HBB) をコードするポリヌクレオチドを投与する。また、不完全なヘモグロビン遺伝子をコードする遺伝子 (例えば、突然変異体HbSをコードする遺伝子) をターゲットとするsiRNAを患者に投与して、不完全なタンパク質の発現をノックダウンまたはノックアウトする。さらに、本発明は、鎌状白血球貧血の治療において普通に使用される薬剤または治療を施すことを含むことができる。
【図面の簡単な説明】
【0029】
【図1】図1は、ヒトeIF-5A1のアミノ酸配列を提供し、種々の重要部位を示す。
【図2】図2は、K50およびK67におけるeIF-5A1の突然変異がトランスフェクトされたタンパク質の蓄積を増加させることを示す。実施例1参照。
【図3】図3は、K47、K50およびK67におけるeIF5A1の突然変異がトランスフェクトされたタンパク質の蓄積を増加させることを示す。実施例2参照。
【0030】
【図4】図4は、KAS細胞中にトランスフェクトされたとき、K50およびK67におけるeIF5A1の突然変異がアポトーシスを誘導させることを示す。実施例3参照。
【図5】図5は、KAS細胞中にトランスフェクトされたとき、K50およびK67におけるeIF5A1の突然変異がアポトーシスを誘導させることを示す。実施例4参照。
【図6】図6は、KAS細胞中にトランスフェクトされたとき、K50およびK67におけるeIF5A1の突然変異がアポトーシスを誘導させることを示す。実施例5参照。
【0031】
【図7A】図7Aは、siRNAを使用するトランスフェクションおよび、KAS細胞におけるアポトーシス生ずる、eIF-5A1を発現するように修飾されたアデノウイルスを使用する治療を示す。実施例6A参照。
【図7B】図7Bは、eIF5A1 siRNA (ターゲット#1 (配列番号1)) (図25に示すsiRNA構築物の配列) を使用する前処理が内因的eIF5A1の発現を減少させたが、アデノウイルスが発現したRNAi抵抗性eIF5A1K50Aの蓄積を可能とすることを示す。実施例6B参照。
【0032】
【図7C】図7Cは、アデノウイルス感染前のターゲット#1に対するeIF5A1 siRNAを使用する前処理がヒト多発性骨髄腫細胞におけるリン酸化NF-kBの発現を減少させることを示す。実施例6C参照。
【図7D】図7Dは、アデノウイルス感染前のターゲット#1に対するeIF5A1 siRNAを使用する前処理がヒト多発性骨髄腫細胞におけるリン酸化NF-kB及びICAM-1の発現を減少させることを示す。実施例6D参照。
【図7E】図7Eは、ヒト多発性骨髄腫細胞におけるelF5AのsiRNA仲介抑制がNF-kB DNA結合活性のLPS仲介誘導を抑制することを示す。elF5A siRNAによるNFkB活性の阻害は、elF5AK50Rの過剰発現と組合わせたとき、アポトーシスの誘導を増加するその能力を説明する。なぜなら、NF-kBは多数の前生存率および抗アポトーシス経路を調節するからである。
【0033】
【図7F】図7Fは、siRNAを使用するKAS細胞の前処理がIL-6の存在下のeIF5A1K50R遺伝子送出によるアポトーシスを増加させることを示す。実施例6E参照。
【0034】
【図8】図8は、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の成長を遅延させることを示す。示したデータは各グループにおけるすべてのマウスについての腫瘍体積である。実施例7参照。
【図9】図9は、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の成長を遅延させることを示す。示したデータは平均腫瘍体積/グループ±標準誤差である。実施例7参照。
【0035】
【図10】図10は、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の重量を減少させることを示す。実施例7参照。
【図11】図11は、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の成長を遅延させ、腫瘍を収縮させることを示す。実施例8参照。
【図12】図12は、eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の腫瘍内 (i.t.) 投与が多発性骨髄腫皮下腫瘍の腫瘍収縮を生じさせることを示す。実施例9参照。
【0036】
【図13A】図13Aは、eIF5A1プラスミドおよびeIF5A1 siRNAを使用する治療が多発性骨髄腫皮下腫瘍の成長を遅延させ、腫瘍を収縮させることを示す。
【図13B】図13Bは、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が腫瘍の収縮を生じさせることを示す。
【図13C】図13Cは、eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の腫瘍内 (i.t.) 投与が多発性骨髄腫皮下腫瘍の腫瘍収縮を生じさせることを示す。
【0037】
【図14】図14は、eIF5A1プラスミドおよびeIF5A1 siRNAの静脈内共投与が多発性骨髄腫皮下腫瘍の成長を遅延させることを示す。実施例10参照。
【図15】図15は、eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の静脈内 (i.v.) または腹腔内 (i.p.) 投与が多発性骨髄腫皮下腫瘍の成長を遅延させることを示す。実施例11参照。
【図16】図16は、eIF5A1プラスミドおよびeIF5A1 siRNAを使用する治療が多発性骨髄腫皮下腫瘍の成長を遅延させることを示す。
【0038】
【図17】図17は、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の成長を遅延させ、腫瘍を収縮させることを示す。実施例12参照。
【図18】図18は、eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の腫瘍内 (i.t.) 投与が多発性骨髄腫皮下腫瘍の腫瘍収縮を生じさせることを示す。実施例13参照。
【図19】図19は、EF1またはB29プロモーターのいずれかにより推進されたeIF5A1K50Rプラスミド、およびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の増殖を遅延させ、腫瘍を収縮させることを示す (KAS-SQ-5) 。実施例14参照。
【0039】
【図20】図20は、eIF5A1 siRNAの共投与がEF1またはB29プロモーターいずれかにより推進されるeIF5A1K50Rプラスミドの多発性骨髄腫皮下腫瘍に対する抗腫瘍作用を増加させ、存在する腫瘍を減少させることを示す (KAS-SQ-5) 。実施例15参照。
【図21】図21は、eIF5A1 siRNAが肺腺癌細胞におけるAd-elF5Aによる感染から生ずるアポトーシス誘導を相乗的に増加させる。実施例16参照。
【図22】図22はpExp5Aの地図を示し、その構築は実施例17に記載する。
【0040】
【図23】図23はpExp5A (3371 bp) の予測される配列を示す。実施例17参照。
【図24】図24は、種々の細胞系統におけるeIF5A1K50Rの再発現を示す。実施例18参照。
【図25】図25は、ターゲット配列および好ましいeIF5A1 siRNAの配列を示す。
【図26】図26は、多発性骨髄腫における効能研究の結果を提供する。実施例21参照。
【図27】図27は、多発性骨髄腫における効能研究の結果を提供する。実施例21参照。
【0041】
【図28】図28は、eIF5A1K50R cDNAの配列を提供する。
【図29】図29は、ヒトeIF5A1K50Rに対するヒトeIF-5A1のアラインメントを提供する。
【図30】図30は、HA-eIF5A1K50R発現に対するDNA: siRNA比の効果を示す。実施例23参照。
【図31】図31は、ナノ粒子トランスフェクションより誘導されるアポトーシスに対するDNA: siRNA比の効果を示す。実施例24参照。
【0042】
【図32】図32は、eIF5A1K50RプラスミドおよびeIF5A1 siRNA (siSTABLEまたは非siSTABLE) を含有するPEI複合体 (N/P = 6または8) の投与が多発性骨髄腫皮下腫瘍の成長を阻害し、腫瘍収縮を生じさせることを示す。実施例25参照。
【図33】図33は、JET PEITMナノ粒子が腫瘍組織により効果的に吸収されること、およびナノがプラスミドおよびsiRNAを同一細胞に送出していることを示す。実施例26参照。
【発明を実施するための形態】
【0043】
発明の詳細な説明
本発明は、被検体における内因的遺伝子の発現をノックアウトまたはノックダウンするための内因的遺伝子をターゲットとするsiRNAと、ポリヌクレオチドによりコードされるタンパク質の宿主における発現を提供するために被検体に提供される送出ビヒクル/発現ベクター中の遺伝子をコードするポリヌクレオチドとの組合わせた使用に関する。
【0044】
この組成物は不完全なまたは突然変異体のタンパク質の存在により引き起こされる疾患または症状を有する被検体の治療において有効である、すなわち、被検体において生産されるタンパク質がその必要な機能を実行できないか、あるいはその構造が不完全なために代謝経路または生体分子の相互作用を失う場合において有効である。siRNAは不完全なタンパク質をコードする遺伝子をターゲットとし、その不完全なタンパク質の発現をノックダウンまたはノックアウトするように設計されている。正常 (不完全でない) タンパク質をコードするポリヌクレオチドを被検体に投与し、その被検体において発現させて、正常タンパク質がその必要な機能を実行できるようにする。
【0045】
他の態様において、所望のタンパク質をコードするポリヌクレオチドを投与する代わりに、タンパク質を被検体に投与する。用語「タンパク質」、「ペプチド」および「ポリペプチド」は本明細書中で互換的に使用される。
【0046】
siRNAは遺伝子のある領域をターゲットとし、不完全なタンパク質の内因的発現をノックダウンまたはノックアウトするが、同時に正常タンパク質をコードする投与したポリヌクレオチドの外因的発現に影響を与えないことが好ましい。例えば、siRNAは3’ UTRをターゲットとし、投与したセンス構築物 (タンパク質をコードするポリヌクレオチド) の外因的発現に影響を与えないようにすることができる。不完全な遺伝子の外因的発現をノックダウンまたはノックアウトすることによって、外因的ポリヌクレオチドから発現された正常タンパク質と競合する不完全なタンパク質はほとんど、あるいはまったく存在しないであろう。
【0047】
この適用が有効である疾患の状態の1例は鎌状白血球貧血に関する。鎌状白血球貧血は、ヘモグロビン、すなわち、体全体を通じて酸素の運搬を促進する赤血球 (RBCs) 中に見出されるタンパク質である。
【0048】
ある人が突然変異体ヘモグロビン (Hbs) の発現を生ずる2つの異常な遺伝子 (各親から1つ) を受け継ぐとき、鎌状白血球貧血が起こる。突然変異体ヘモグロビンはRBCsの形状を変化させる。正常ヘモグロビン (ヘモグロビンAまたはHbA) を有する赤血球は血流中を通して容易に動き、体のすべての細胞に酸素を送出す。赤血球は非常に細い血管でさえ「押分けて進む」ことができる。ヘモグロビンの異常形態 (HbS) は一緒に凝集し、赤血球を粘着性、剛性、より脆弱とし、そしてそれらを湾曲した、鎌形にする傾向がある。
【0049】
数百のHBB遺伝子変異型は知られているが、鎌状白血球貧血は最も普通にヘモグロビン変異型HbSにより引き起こされる。この変異型において、疎水性アミノ酸バリンはHBBポリペプチド鎖の第6アミノ酸位置に親水性グルタミン酸の位置を取る。この置換はタンパク質構造の外側に疎水性スポットをつくり、これは隣接するヘモグロビン分子のβ鎖の疎水性領域に粘着する。HbS分子の剛性線維へのこの凝集 (重合) は、赤血球の「鎌形化」を引き起こす。
【0050】
赤血球が体全体を通じて種々の組織に運んだ酸素を赤血球が放出した後、重合は起こる。いったん赤血球は肺に戻り、ここでヘモグロビンは酸素に結合することができ、HbS分子の長い線維は解重合または分解して単一分子になる。重合と解重合との間のサイクリングは赤血球の膜を剛性にさせる。これらの赤血球が酸素を運んでないときの、赤血球の剛性およびそれらの変形した形状は小さい血管を遮断することがある。この遮断は短期間の疼痛発現を引き起こし、器官を損傷させることがある。
【0051】
鎌状白血球貧血は、常染色体劣性遺伝障害である。疾患を発現させるために、人は2コピーのHbS変異型または1コピーのHbSおよび1コピーの他の変異型を受け継がなくてはならない。キャリヤーは1コピーの正常HBB遺伝子 (HbA) および1コピーのHbSを有し、鎌状白血球の特徴を有するとして記載され、疾患の症状を発現しない。
【0052】
こうして、本発明の1つの態様は鎌状白血球貧血の患者を治療する方法を提供する。HBB遺伝子をターゲットとするsiRNAを患者に投与する。siRNAはヘモグロビンのHbs変異型の発現をノックダウンし、好ましくはノックアウトするように設計する。正常ヘモグロビンをコードするポリヌクレオチドは被検体に提供して、被検体が正常ヘモグロビンを発現するようにする。また、siRNAは正常ヘモグロビンをコードする外因的ポリヌクレオチドの発現を妨害しないように、設計される。こうして、被検体はもはや変異型ヘモグロビンを作らず (または実質的により少なくする) 、その代わりに正常な健康的ヘモグロビンを作り、正常に機能する、いっそう正常な赤血球を生ずる。
【0053】
本発明は、また、翻訳後修飾がタンパク質に対して起こる状況において有効であり、この翻訳後修飾は疾患の状態を引き起こすか、あるいはその状態に導く。siRNAを使用して内因的タンパク質の発現をノックダウンするので、翻訳後修飾はほとんど、あるいはまったく利用不可能である。次いで、タンパク質をコードするポリヌクレオチドを外因的発現のために患者に提供する。タンパク質は修飾されるので、翻訳後に修飾されることはできない。次いでこのタンパク質は適当な使用のために体に利用可能であるが、翻訳後に修飾されることはできないので、疾患の状態に導かないであろう。
【0054】
当業者は異なる翻訳後修飾を理解するであろう。例えば、翻訳後、アミノ酸の翻訳後修飾は、アミノ酸の化学的特質 (例えば、シトルリン化) を変化させるか、あるいは構造的変化、例えばジサルファイド架橋の形成を行うことによって、他の生化学的官能基、アセテート、ホスフェート、種々の脂質および炭水化物をアミノ酸に結合させることによって、タンパク質機能の範囲を拡大する。また、酵素はタンパク質のアミノ末端からアミノ酸を除去するか、あるいはタンパク質鎖を中央において切断することができる。
【0055】
例えば、ペプチドホルモンのインスリンはジサルファイド結合が形成された後2回切断され、そしてポリペプチドは鎖の中央から除去される; 生ずるタンパク質はジサルファイド結合により結合された2つのポリペプチド鎖から成る。また、最も新生のポリペプチドはアミノ酸メチオニンで開始する。なぜなら、mRNA上の「開始」コドンもまたこのアミノ酸をコードするからである。このアミノ酸は通常翻訳後修飾の間に除去される。リン酸化のような、他の修飾はタンパク質の挙動をコントロールする、例えば、酵素を活性化または不活性するための普通の機構の一部分である。他の翻訳後修飾は、デオキシハイプシンシンターゼ (DHS) による真核開始因子5A (elF5A) のハイプシン化を包含する。
【0056】
こうして、本発明は、タンパク質をコードするポリヌクレオチドを患者に投与し、患者において発現させる、被検体における遺伝子の発現を変更する方法を提供する。タンパク質は正常/野生型タンパク質または突然変異したタンパク質であることができる。対応する内因的遺伝子の発現は、被検体に投与されたsiRNAで抑制される。
【0057】
この方法は、さらに、ポリヌクレオチドが被検体において発現されてターゲットタンパク質を産生する、ターゲットタンパク質をコードするポリヌクレオチドを含んでなる構築物を提供することを含んでなる。内因的遺伝子が不完全なタンパク質を発現するある種の態様において、正常/健康なタンパク質をコードするようにポリヌクレオチドを設計する。siRNAを投与して不完全な内因的タンパク質の発現を抑制する。内因的遺伝子が正常の健康なタンパク質を発現するある種の態様において、正常/健康なまたは非突然変異体のタンパク質を使用するとき起こるような、転写後修飾されることができない突然変異体タンパク質をコードするように、ポリヌクレオチドを設計する。siRNAを投与して内因的タンパク質の発現を抑制して、このタンパク質が転写後修飾に利用されることが少ないようにする。
【0058】
ある種の態様において、内因的ポリヌクレオチドの発現に影響を与えないように、遺伝子の領域をターゲットするようにsiRNAを選択または設計する。例えば、siRNAは3’ UTRまたは3’ 末端をターゲットとすることができる。
【0059】
siRNAは裸siRNAまたは血清のために安定化した裸siRNAとして患者に送出すことができる。siRNAは全身的に、すなわち、IPまたはIV注射することができる。選択的に、siRNAは体の所望の区域に局所的に注射しまたは送出すことができる。ある種の態様において、siRNAは送出ビヒクルで投与することができ、送出ビヒクルの例はデンドリマー、リポソームまたはポリマーであるが、これらに限定されない。
【0060】
所望のタンパク質をコードするポリヌクレオチドは、ヌクレオチドの発現を提供または可能とする任意の送出手段により、投与することができる。用語「ポリヌクレオチド」または「ヌクレオチド」は本明細書中で互換的に使用される。送出は任意のウイルスまたは非ウイルス機構により実施することができ、このような機構はプラスミド、発現ベクター、ウイルス構築物、アデノウイルス構築物、デンドリマー、リポソームまたはポリマーであるが、これらに限定されない。
【0061】
ある種の態様において、CpGジヌクレオチドが減少した発現プラスミドを使用してポリヌクレオチドを発現させる。ポリヌクレオチドの発現を促進できる任意のプロモーターを使用することができ、これは療法に所望の適用に基づいて選択することができる。例えば、多発性骨髄腫を殺すために、B細胞に対して特異的なプロモーター、例えばヒトB29プロモーター/エンハンサーは望ましい。他の態様において、プロモーターは他の組織特異的プロモーターであることができるか、あるいは系統的プロモーターであることができる。
【0062】
ターゲットタンパク質をコードするポリヌクレオチドは、IVまたは皮下注射、または他の任意の生物学的に適当な送出機構により送出すことができる。
選択的に、ポリヌクレオチドはリポソームにより送出すことができるか、あるいはDNA (またはプラスミドまたは発現ベクター) をターゲット腫瘍または癌細胞に送出す任意の他の適当な「担体」または「ビヒクル」により送出すことができる。例えば、合成DNA送出系の概観についての下記を参照のこと: Luo、Dan 他、Nature Biotechnology、Vol. 18、January 2000、pp. 33-37。
【0063】
こうして、ナノメートルサイズのビヒクル、例えばリポソーム、デンドリマーまたは同様な非毒性ナノ粒子を経てヌクレオチド/プラスミド/発現ベクターを送出すことが好ましいことがある。ビヒクルは、ヌクレオチド/プラスミド/発現ベクターを早期のクリアランスから保護するか、あるいは有効量のヌクレオチド/プラスミド/発現ベクターを被検体、腫瘍または癌細胞に送出す間免疫応答の発生からヌクレオチド/プラスミド/発現ベクターを保護することが好ましい。典型的なビヒクルは、ヌクレオチド/プラスミド/発現ベクターに関連する簡単なナノ粒子から、いっそう複雑なペギミル化ビヒクル、例えば、特異的細胞レセプターをターゲットとするその表面に取り付けられたリガンドを有するペギミル化リポソームまでの範囲である。
【0064】
リポソームおよびペギミル化リポソームはこの分野において知られている。慣用リポソームにおいて、送出すべき分子 (すなわち、小さい薬剤、タンパク質、ヌクレオチドまたはプラスミド) をリポソームの中央キャビティイ内に含有させる。当業者は認識するように、ターゲットとした「ステルス」、および分子送出に有効なカチオン性リポソームがまた存在する。例えば、下記を参照のこと: Hortobagyi、Gabriel N 他、J. Clinical Oncology、Vol. 19、Issue 14 (July) 2001: 3422-3433およびYu、Wei 他、Nucleic Acids Research、2004、32 (5): c48。リポソームは静脈内に注射することができ、修飾してそれらの表面をいっそう親水性とすることができる (ポリエチレングリコールを2層に添加することによって (「ペギミル化」) 、これは血流中のそれらの循環時間を増加させる) 。
【0065】
これらは「ステルス」リポソームとして知られており、親水性 (水溶性) 抗癌剤、例えばドキソルビジンおよびミトキサントロンのための担体として特に有効である。ターゲット細胞、例えば、腫瘍細胞に対する薬剤担持リポソームの特異的結合特性を増加させるために、特異的分子、例えば、抗体、タンパク質、ペプチドおよびその他をリポソーム表面上に取り付けることができる。例えば、癌細胞上に存在するレセプターに対する抗体を使用して、癌細胞に対してリポソームをターゲッティングすることができる。多発性骨髄腫をターゲッティングする場合、例えば、葉酸塩、IL-6またはトランスフェリンを使用してリポソームを多発性骨髄腫細胞にターゲッティングすることができる。
【0066】
また、デンドリマーはこの分野において知られており、そして好ましい送出ビヒクルを提供する。例えば、下記を参照のこと: Marjoros、Istvan J 他、”PAMAM Dendrimer - Based Multifunctional Conjugate for Cancer Therapy: Synthesis, Characterization, and Functionality”、Biomacromolecules、Vol. 7、No. 2、2006: 572-579、および Marjoros、Istvan J 他、J. Med. Chem. 2005、48: 5892-5899、デンドリマーの論考について。
【0067】
好ましい態様において、送出ビヒクルはポリエチレンイミンナノ粒子を含んでなる。典型的なポリエチレンイミンナノ粒子はin vivo-jetPEITM (現在Polyplus Transfection, Inc. により製造されている) である。in vivo-jetPEITMはカチオン性ポリマートランスフェクション剤であり、DNAおよびsiRNA送出剤として有効である。in vivo-jetPEITM (Polyplus Transfection製) は、動物において信頼性ある核酸送出を提供する線形ポリエチレンイミン試薬である。
【0068】
それは下記において使用される: 遺伝子治療 (Ohana 他、2004、Gene Ther. Mol. Bio. 8: 181-192; Vernejoul 他、2002、Cancer Research 62: 6124-31) 、RNAインターフェレンス (Urbain - Klein 他、2004、Gene Therapy 23: 1-6; Grezelinski 他、2006、Human Gene Therapy 17: 751-66) 、および遺伝ワクチン接種 (Garzon 他、2005、Vaccine 23: 1384-92) 。in vivo JET-PEIは現在ヒト臨床実験において癌遺伝子治療のための送出ビヒクルとして使用されている (Lemkine 他、2002、Mol. Cell. Neurosci. 19: 165-174) 。
【0069】
in vivo-jetPEITMは核酸を大よそ50 nmのナノ粒子に縮合させ、これらのナノ粒子は数時間安定である。この独特の保護機構の結果として、注射後の血球凝集は他の療法と比較して減少し、組織内拡散の制限、赤血球凝集および微小塞栓が防止される。これらのナノ粒子は十分に小さく、組織中に拡散し、エンドサイトーシスにより細胞の中に入る。in vivo-jetPEITMは、核酸のエンドソームからの放出および核膜を横切る移動を促進する。
【0070】
好ましい態様において、siRNAおよびポリヌクレオチドを含んでなるベクター/プラスミドの両方はin vivo-jetPEITM複合体を経て被検体に投与される。siRNAおよびポリヌクレオチドを含んでなるベクター/プラスミドは、ポリマー複合体、例えば、ポリエチレンイミンまたはin vivo-jetPEITM複合体を経て一緒に複合化することができるか、あるいはポリマーに対して別々に複合化することができる。例えば、siRNAおよびポリヌクレオチドを含んでなるベクター/プラスミドを被検体に別々に投与する (時間および/または送出部位の意味において別々に) 場合、異なる担体に対して複合化されたsiRNAおよびポリヌクレオチドを使用することが好ましい。投与を同時にかつ同一部位において実施する場合、siRNAおよびポリヌクレオチドを一緒に複合化することが好ましいことがある。
【0071】
他の態様において、プラスミドまたはベクターを投与して被検体において発現させるポリヌクレオチドを送出す代わりに、タンパク質それ自体を被検体に送出す。タンパク質は分離するか、あるいは合成することができる。
【0072】
本発明の1つの態様は、哺乳動物およびヒトを包含する被検体において癌を治療する方法を提供する。癌の治療は下記を包含するが、これらに限定されない: 癌細胞におけるアポトーシス、癌細胞を殺すこと、癌細胞の数の減少および腫瘍の体積/重量の減少。この方法は、eIF5A1 siRNAと、突然変異体eIF5A1をコードするポリヌクレオチドとを含んでなる組成物を投与することを含んでなる。組成物およびeIF5A1 siRNA、および突然変異体eIF5A1をコードするポリヌクレオチドは、本明細書において後述する。
【0073】
すべての細胞は、真核性開始因子5A (“elF-5A”) (または本明細書中で「因子5A」と呼ぶ) を産生する。哺乳動物細胞はelF-5Aの2つのアイソタイプ (eIF-5A1およびeIF-5A2) を産生する。eIF-5A1はアポトーシスを行う細胞中でアップレギュレートされるので、eIF-5A1はアポトーシス特異的elF-5Aと呼ばれてきている。ヒトeIF-5A1は受け入れ番号NM 001970を有し、図1に示されている。eIF-5A1は、アポトーシスに必要なタンパク質をコードするmRNA類の核サブセットの往復の原因となると考えられる。下記の文献を参照のこと: Liu & Tartakoff (1997) Supplement Society for Cell、8、462a、Abstract No. 2476、37th American Society for Cell Biology Annual Meeting, and Rosorius 他 (1999) J. Cell Science、112、2369-2380。
【0074】
両方の因子5Aは、デオキシハイプシンシンターゼ (“DHS”) により転写後修飾される。DHSはelF-5A類をハイプシン化する。ハイプシンは、ユニークアミノ酸であり、すべての検査した真核生物および古細菌中に見出されるが、真正細菌において見出されず、elF-5Aは唯一の既知のハイプシン含有タンパク質である。Park (1988) J. Biol. Chem. 263、7447-7449; Schumann & Klink (1989) System. Appl. Microbiol. 11、103-107; Bartig 他 (1990) System. Appl. Microbiol. 13、112-116; Gordon 他 (1987a) J. Biol. Chem. 262、16585-16589。
【0075】
ハイプシン化elF-5Aは2つの翻訳後の段階において形成される: 第1段階はジオキシハイプシン残基の形成であり、この形成はスペルミジンの4-アミノブチル部分が前駆体elF-5Aの特異的リシンへ転移することによってなされ、これはデオキシハイプシンシンターゼにより触媒される。第2段階は、この4-アミノブチル部分がデオキシハイプシンヒドロキシラーゼによりヒドロキシル化されてハイプシンを形成することを含む。
【0076】
elF-5Aのアミノ酸配列は種間で十分に保存され、elF-5A中のハイプシン残基を取り囲むアミノ酸配列の厳格な保存が存在し、これはこの修飾が生存に重要であることがあることを示唆する。Park 他 (1993) Biofactors 4、95-104。この仮定は酵母菌において今日までに発見されたelF-5Aの両方のイソ型の不活性化、またはDHS遺伝子 (これはそれらの活性化の第1段階を触媒する) の不活性化が細胞分裂をブロックするという観察によりさらに支持される。Schnier 他 (1991) Mol. Cell. Biol. 11、3105-3114; Sasaki 他 (1996) FEBS Lett. 384、151-154; Park 他 (1998) J. Biol. Chem. 273、1677-1683。
【0077】
しかしながら、酵母菌におけるelF-5Aタンパク質の消耗は全タンパク質合成をわずかに減少しただけであり、elF-5Aがタンパク質の全体的合成よりもむしろmRNA類の特異的サブセットの翻訳に必要とされるであろうことを示唆する。Kang 他 (1993) 、“Effect of initiation factor elF-5A depletion on cell proliferation and protein synthesis”、Tuite M (編者) 、Protein Synthesis and Targeting in Yeast、NATO Series H。elF-5Aを結合するリガンドは高度に保存されたモチーフを共有するという最近の発見は、elF-5Aの重要性をまた支持する。Xu & Chen (2001) J. Biol. Chem. 276、2555-2561。さらに、変性されたelF-5Aのハイプシン残基はRNAに特異的に結合する配列にとって必須であることが発見され、そして結合はリボヌクレアーゼからの保護を提供しなかった。
【0078】
本発明者らは、elF-5Aをコードするポリヌクレオチドを細胞に投与するとき、それらの細胞のアポトーシスが増加することを示した。次いで癌細胞中で発現されるeIF-5A1ポリヌクレオチドを投与することによって、癌細胞がアポトーシスするようにできることを本発明者らは示した。下記の同時係属出願を参照のこと: 10/200,148; 11/287,460; 11/293,391および11/637,835、それらのすべてはそれらの全体において引用することによって本明細書の一部とされる。
【0079】
さらに、本発明は、細胞が因子5Aのハイプシン化形態の構築物を有するとき、細胞は生存モードに入り、通常オーバータイムであるので、アポトーシスを行わない。顕著には、癌細胞において、有意な量のハイプシン化因子5Aが存在し、こうして細胞はアポトーシスに入らない (そして死なない) 。こうして、癌細胞を殺すことによって癌を治療する (癌細胞をアポトーシス経路に入れる) ために、eIF-5A1をコードするポリヌクレオチドを被検体に、または癌細胞または腫瘍に投与してeIF-5A1の発現を増加させ、これは引き続いて癌細胞のアポトーシスを引き起こし、究極的に細胞を死亡させ、腫瘍を収縮させる。しかしながら、eIF-5A1タンパク質をコードするポリヌクレオチドのみを提供してeIF-5A1の遺伝子発現をアップレギュレートし、また、eIF-5A1の内因的発現をノックダウンするためにsiRNAを使用しない場合、綱引きが存在する: アポトーシス経路に細胞を向けるeIF-5A1の発現は、生存経路に細胞を向けるハイプシン化因子5Aの存在と競合する。
【0080】
本発明はこの綱引きを排除し、eIF-5A1の発現のみを増加させることを超えた改良を提供する。被検体または細胞に投与されたポリヌクレオチドは突然変異されるので、生ずる発現されたタンパク質はハイプシン化されることができない。さらに、因子5Aの内因的発現はelF-5AをターゲットとするsiRNAによりをノックアウト/ノックダウンされるので、ハイプシン化されるelF-5Aはほとんどあるいはまったく存在しない。こうして、細胞中にハイプシン化されたelF-5Aはまったく (または実質的にほとんど) 存在しないので、それらは生存モードに押しやられない。
【0081】
突然変異したeIF-5A1をコードするポリヌクレオチドは好ましくは突然変異されるので、ハイプシン化されることができず、こうして細胞を生存モードに推進するために利用されない。例えば、1つの態様において、elF-5Aをコードするポリヌクレオチドは突然変異されるので、通常DHSによりハイプシン化される、位置50におけるリシン (K) はアラニン (A) (これはハイプシン化されることができない) に変化される。この突然変異体をK50Aと表示する。
【0082】
他の態様において、位置67におけるリシンはアルギニン (R) に変化される。この突然変異体を (K67R) と表示する。他の態様において、位置67におけるリシンはアラニン (A) に変化され、(K67A) と表示する。他の態様において、位置50におけるリシンはアルギニン (K50R) に変化され、他の態様は位置47におけるリシン (K) がアルギニン (K47R) に変化される突然変異体を提供する。
【0083】
他の態様において、二重突然変異体を使用する。1つの二重突然変異体は、位置50におけるリシン (K) がアルギニン (R) に変化されており、そして位置67におけるリシン (K) がアルギニン (R) に変化されている突然変異体である。この二重突然変異体をK50R/K67Rと呼ぶ。この二重突然変異体は同様にハイプシン化されることができないが、アミノ酸変化は単一突然変異体 (K50R) 程度に多くeIF-5A1の三次元構造を変更しない。こうして二重突然変異体は、三次元形状に非常に類似し、野生型としてフォールディングし、こうして単一突然変異体よりもいっそう安定であるタンパク質を提供する。二重突然変異体は、いっそう安定であるので、体中でより長く存在して、より長い療法的利益を提供する。こうして、体は因子5Aを有し、因子5Aは正常の細胞機能を必要とするが、ハイプシン化されることができないので、細胞は細胞の生存モードに拘束されず、アポトーシスを回避する。
【0084】
他の二重突然変異体は、位置47におけるリシン (K) がアルギニン (R) に変化されており、そして位置50におけるリシンがアルギニン (R) に変化されている突然変異体である。この突然変異体をK47R/K50Rと呼ぶ。本発明は、位置50におけるリシン (L) がアラニン (A) に変化されており、そして位置67におけるリシンがアラニン (A) に変化されている他の二重突然変異体を提供する。この突然変異体をK50A/K67Aと呼ぶ。
【0085】
体は正常の細胞生存および健康な細胞増殖のために因子5Aを必要とするので、siRNAが全身的に送出される場合、siRNAをもつ被検体において発現を完全に遮断しないことが好ましい。発現の遮断において良好でないsiRNAを使用する (すなわち、発現を停止または減少させるが、発現を完全には遮断しない) か、あるいは、選択的に、投与/治療の養生法を利用して発現レベルをバランスさせて健康な細胞の正常な成長または機能化を可能とするが、また癌細胞をアポトーシスさせることによって、elF-5A発現のコントロールを達成することができる。
【0086】
選択的に、局所的siRNA送出を利用できる。siRNAを局所的に癌細胞または腫瘍に送出す場合、発現をノックアウトすることが好ましい。発現をノックアウトすることよって、因子5Aはハイプシン化できる付近に存在せず、こうして細胞を生存モードにロックするハイプシン化elF-5Aは存在しない。siRNAは局所的に癌または腫瘍に送出されるので、細胞増殖に利用可能なelF-5Aを用意する必要がない。
【0087】
ある種の態様において、内因的遺伝子はeIF5A1である。eIF5A1をターゲットとするsiRNAを被検体に投与して内因的eIF-5A1の発現を抑制する。ある種の態様において、siRNAは配列番号1または配列2を含んでなるか、あるいは内因的eIF-5A1の発現を抑制するeIF5A1をターゲットとする任意のsiRNAである。ある種の態様において、eIF5A1はヒトeIF-5A1 (図1に示す) であり、そして被検体はヒトである。eIF5A1をターゲットとする他のsiRNAは既知であり、そして下記の同時係属出願に記載されている: 11/134,445; 11/287,460; 11/184,982; 11/293,391; 11/725,520; 11/725,470; 11/637,835。他の態様において、被検体は哺乳動物であり、そしてeIF5A1は哺乳動物に対して特異的である。例えば、被検体はイヌであり、そしてeIF5A1はイヌeIF5A1である。
【0088】
ある種の態様において、siRNAは図25に示すsiRNA構築物から本質的に成る。例えば、siRNAはeIF5A1をターゲットとする核酸を含有するが、またオーバーハング、例えばUまたはT核酸を含有するか、あるいはまたタグ、例えばhisタグ (しばしばHAタグと呼ばれる - これはin vivo研究においてしばしば使用される) を含有する。5’ または3’ 末端 (または、例えば、図25に示す核酸の連続的ひも内にさえ) に結合した分子または追加の核酸は、siRNA構築物がターゲット遺伝子の発現を減少させることができるかぎり、「から本質的に成る」の範囲内に含まれかつ入る。好ましくは、外因的ポリヌクレオチドの発現を生じないように、siRNAはeIF5A1遺伝子の領域をターゲットとする。例えば、eIF5A1 siRNAは3’ UTRまたは3’ 末端をターゲットとする。図25に示すsiRNAは典型的なeIF5A1 siRNAである。
【0089】
ポリヌクレオチドはeIF5A1をコードし、ここでポリヌクレオチドはeIF5A1変異型をコードするように突然変異されている。変異型eIF5A1が翻訳後に修飾されることができない (ハイプシン化されることができない) ように、突然変異したeIF5A1を設計する。典型的な突然変異体は本明細書において上に論じられている。
【0090】
充実性腫瘍を包含する癌の場合において、siRNAを腫瘍に直接送出すことは望ましいことがある。siRNAは時間ならびにポリヌクレオチドからの送出部位に関して別々に投与するか、あるいは同一時間におよび/または同一送出部位に一緒に投与することができる。当業者は理解するように、siRNA投与のタイミングは内因的タンパク質が翻訳されているときであり、そしてそれが既に作られた後ではない。
【0091】
本発明者らは、eIF5A1が正常組織に対して無毒であることを初期に示した (係属中の出願番号11/293,391、出願2005年11月28日、これは引用することによって本明細書の一部とされる) が、送出複合体 (elF5Aポリヌクレオチド/プラスミド/発現ベクターの直接投与に比較して) は好ましいことがある。好ましい送出系は有効量のeIF5A1を被検体、腫瘍または癌細胞のグループに提供し、ならびに好ましくは腫瘍または癌細胞のグループにターゲテッド送出を提供する。こうして、ある種の態様において、ナノメートルサイズのビヒクル、例えば、リポソーム、デンドリマーまたは同様な非毒性ナノ粒子、例えばポリエチレンイミンポリマー (例えば、in vivo JetPEITM複合体) によってeIF5A1ヌクレオチド/プラスミド/発現ベクターを送出すことが好ましい。
【0092】
また、eIF5A1タンパク質を腫瘍の部位に直接送出すことができる。当業者はeIF5A1タンパク質送出のための投与量および治療養生法の長さを決定できるであろう。
eIF5A1によるアポトーシス誘導の分子基準を下に論ずる。
【0093】
死レセプターシグナリング
Ad-eIF5A1 (野生型eIF5A1をもつアデノウイルス) またはAd-eIF5A1 (K50A) を使用する癌細胞の治療はカスパーゼ8、これは死レセプターにより開始される - リガンド結合性、およびカスパーゼ3、エクシキュウーショナーカスパーゼの活性化を誘導する。これらはeIF5A1の間接的作用であるようであり、そしてカスパーゼ8およびカスパーゼ3がeIF5A1 (K50A) (これはハイプシン化されることができない) を使用する治療後にまた活性化されるという事実は、この作用がリシン50 eIF5A1に帰することを示す。また、Ad-eIF5A1を使用する治療は、TNFR1のアップレギュレーションで前に示したように、死レセプターをアップレギュレートするように見える。
【0094】
ミトコンドリア経路
アポトーシスのミトコンドリア経路におけるリシン50 eIF5A1の直接的または間接的掛かり合いは、カスパーゼ9がeIF5A1またはeIF5A1 (K50A) による癌細胞の治療により活性化されるという発見を含む多数の観察により支持される。その上、p53はミトコンドリアアポトーシス経路の活性化においてある役割を演じ、eIF5A1により調節されるように思われる。
【0095】
例えば、アクチノマイシンDを使用する癌細胞の治療はp53をアップレギュレートし、そしてp53のこのアップレギュレーションはeIF5A1 siRNAにより開始される。これと一致して、Ad-eIF5A1を使用する癌細胞の治療は、p53 mRNAをアップレギュレートする。また、eIF5A1を使用する癌細胞の治療は、細胞質ゾルからミトコンドリアへのBaxの移動を誘導し、ミトコンドリア膜の喪失およびミトコンドリア空間内から細胞質ゾル中へのシトクロムCの放出を保証する。さらに、この治療はすべて前アポトーシスである切断したBcl2、BimおよびスプライスドBimをアップレギュレートする。
【0096】
MAPKシグナリング
さらに、本発明は、MAPKシグナリングにおけるeIF5A1の掛かり合いがアポトーシスに関係するという証拠を獲得した。例えば、Ad-eIF5A1を使用する癌細胞の治療は、P-JNKをアップレギュレートし、これは引き続いて抗アポトーシスBcl2を阻害する。さらに、Ad-eIF5A1およびAd-eIF5A1 (K50A) の両方はP-p38の形成を誘導し、これは引き続いて下記を包含する種々の前アポトーシス剤を衝撃することによってアポトーシスを開始することができる: TNFR1およびTNF; FASおよびFASL; カスパーゼ8; Bid; シトクロムCおよびカスパーゼ3。
【0097】
NF-κBシグナリング
NF-κBシグナリングが骨髄腫の増殖を支持するという証拠が存在する。例えば、骨髄ストロマ細胞に対する骨髄腫細胞の接着はIL-6のNF-κB依存的転写アップレギュレーションを誘導し、ここでIL-6は多発性骨髄腫における増殖因子および抗アポトーシス因子の両方である [Chauhan 他 (1996) Blood 87、1104] 。さらに、骨髄腫細胞が分泌するTNF-αは骨髄ストロマ細胞中のNF-κB活性化し、これによりIL-6の転写および分泌をアップレギュレートする。
【0098】
また、TNF-αは骨髄腫細胞中のNF-κBを活性化し、骨髄腫細胞および骨髄ストロマ細胞の両方上の細胞内接着分子-1 (ICAM-1; CD 54) および脈管細胞接着分子-1 (VCAM-1; CD 106) をアップレギュレートする [Hideshima 他 (2001) Oncogene 20、4519] 。これは引き続いて骨髄腫細胞と骨髄ストロマ細胞との関連を増強する [Hideshima 他 (2001) Oncogene 20、4519] 。逆に、これらの作用はTNF-α誘導NF-κB活性化により開始される [Hideshima 他 (2001) Oncogene 20、4519] 。事実、NF-κBは骨髄腫細胞中のTNF-α誘導アポトーシスに対する保護を仲介することが期待される [Hideshima 他 (2002) JBC 277、16639] 。これらおよび他の観察は、NF-κBシグナリングが多発性骨髄腫療法のための魅力的ターゲットありうるという見解を促進する。
【0099】
本発明者らは、eIF5A1 siRNAがヒト骨髄腫細胞におけるNF-κBの活性化およびICAM-1の形成の両方を阻害することを示した。これらの観察はeIF5A1がNF-κBの活性化においてある役割を演ずることを示し、そしてNF-κBの活性化の保証的作用が特質において前生存的である限り、我々はこの活性化がハイプシン化eIF5A1により直接的または間接的に仲介されることを予測する。
【0100】
IL-1
骨髄腫細胞による前炎症性サイトカインIL-1の過剰生産は、骨組織の悪化に導く多発性骨髄腫の特徴的特性である。eIF5A1 siRNAは、マウスにおけるLPS挑戦より誘導されるIL-1の過剰生産を劇的に減少することが示された。
【0101】
本発明の1つの態様は、多発性骨髄腫を治療する方法を提供する。多発性骨髄腫 (“MM”) は、骨髄中の悪性プラズマ細胞の発現および骨溶解性病変の存在により特徴づけられる進行性致死的疾患である。多発性骨髄腫はプラズマ細胞の不治であるが、治療可能な癌である。プラズマ細胞は免疫系の重要な部分であり、感染および疾患に対する戦いを促進する免疫グロブリン (抗体) を産生する。多発性骨髄腫は、骨髄中の異常なプラズマ細胞の極端な数、および無傷のモノクローナル免疫グロブリン (IgG、IgA、IgDまたはIgE; 「M-タンパク質」) またはベンス・ジョーンズタンパク質 (遊離モノクローナル軽鎖) の過剰生産により特徴づけられる。低カルシウム血症、貧血、腎臓障害、細菌感染に対する感受性の増加、および正常免疫グロブリンの産生悪化は多発性骨髄腫の普通の臨床的症状の発現である。また、多発性骨髄腫は、通常骨盤、肋骨および頭蓋における、広汎性骨粗しょう症によりしばしば特徴づけられる。
【0102】
本発明は、多発性骨髄腫において見出される同時のフィードバックループのために、多発性骨髄腫の治療に十分に適していると思われる。例えば、多発性骨髄腫は骨髄中でIL-1を低い濃度で産生する。IL-1は引き続いてストロマ細胞を刺激してIL-6を産生させ、次いでIL-6は多発性骨髄腫の増殖を刺激するようになる。本発明者らが以前に示したように (係属中の出願11/725,539および11/184,982参照) 、eIF-5A1に対して向けられたsiRNAは前炎症性サイトカイン、例えばIL-1; TNF-αおよびIL-8の発現を阻害する。こうして、siRNAはelF-5Aの発現をノックダウンしてハイプシン化の利用可能性を少なくするばかりではなく、またそれはIL-1/IL-6フィードバックループをカットオフまたは減少させるであろう。
【0103】
ヒトelF5AをターゲットとするsiRNAを使用して腫瘍中の内因的ハイプシン化elF5Aのレベルを抑制したが、ハイプシン化できないelF5Aの突然変異体 (elF5AK50R) を発現するRNAi抵抗性プラスミドを使用して非修飾elF5Aのレベルをin vivo上昇させた。elF5A siRNAを含有するPEIナノ複合体の腫瘍内注射は、対照siRNAを含有する複合体に対して80% (***p = 0.0003) より多くMM腫瘍増殖を阻害し、ハイプシン化elF5Aレベルの抑制が抗腫瘍作用を有することを示す。elF5AK50R発現プラスミドを含有するPEI複合体は同様な作用を有し、対照プラスミドを含有する複合体に対して70% (**p = 0.001) より多く腫瘍増殖を阻害した。
【0104】
こうして、MM腫瘍増殖は増殖促進ハイプシン化elF5Aの抑制により阻害できるか、あるいはelF5Aの前アポトーシス非ハイプシン化形態のレベルの増加により阻害できる。elF5A siRNAおよびRNAi抵抗性elF5AK50Rプラスミドの両方を含有する複合体の腫瘍内送出は腫瘍増殖に対して相乗作用を有し、そして有意に腫瘍を有意に収縮させ、94% (***p = 0.0002) だけ腫瘍増殖を阻害した。また、elF5A siRNA/elF5AK50R PEI複合体の静脈内送出は腫瘍増殖を95% (**p = 0.002) だけ効果的に減少させ、治療の全身的送出が実行可能であることを示した。
【0105】
elF5A siRNA/elF5AK50R pDNA PEI複合体の局所的送出および全身的送出の両方は、多発性骨髄腫において有意な抗腫瘍応答を生じた。
【0106】
さらに、本発明は、多発性骨髄腫を包含する癌の治療において有効な組成物を提供する。好ましい態様において、組成物はハイプシン化できない点突然変異化eIF5A1をコードするプラスミドDNAと、内因的ヒトeIF5A1を選択的に抑制するが、プラスミドによりコードされる点突然変異化eIF5A1に対して作用しないeIF5A1 siRNAとの複合体である。eIF5A1 siRNAおよび突然変異体eIF5A1をコードするポリヌクレオチドは前述した。プラスミドDNAおよびsiRNAの両方はPEI (ポリエチレンイミン) ナノ粒子に対して複合化されていることが好ましい。それらは別々に複合化でき、別々にまたは一緒に投与することができるか、あるいは一緒に複合化することができる。DNAおよびRNAはPEI上の正に帯電したアミノ基に結合し、ナノ粒子が細胞中に吸収されるとき放出される。PEI-核酸複合体は分裂している細胞および分裂していない細胞の両方に効率よく吸収されることが証明された。
【0107】
プラスミドDNAはeIF5A1 (K50A) のようにハイプシン化されることができず、したがって、強くアポトーシス性であるeIF5A1 (K50R) をコードすることが好ましい。eIF5A1 (K50R) の発現はB細胞特異的プロモーターにより調節されることが好ましい。
【0108】
eIF5A1 siRNAは内因的ヒトeIF5A1の3’ 末端に対して特異的であることが好ましく、トランスeIF5A1 (K50R) の発現に対して影響を与えない。典型的な好ましいeIF5A1 siRNAは、図25に示すsiRNAを含んでなり、から本質的に成るか、あるいはから成る。eIF5A1 siRNAを包含するための根本的理由は次の通りである: (1) ほとんどすべてがハイプシン化され、それゆえ前生存形態である、内因的eIF5A1を消耗するため; (2) NF-κBの活性化を阻害し、これによりIL-6の産生および細胞内接着分子の形成を減少させるため; および (3) IL-1の形成を阻害するため。eIF5A1 siRNAはeIF5A1 (K50R) と相乗的に作用して、骨髄腫細胞におけるアポトーシスを誘導する。上記 (2) および (3) が前生存的事象である限り、それらはハイプシン化eIF5A1により仲介されることが期待され、それゆえハイプシン化できないeIF5A1 (K50R) により影響を受けない。このアプローチは非ハイプシン化elF5Aのより大きいプールを生じ、多発性骨髄腫細胞を包含する癌細胞をアポトーシスに導き、健康な細胞にほとんど影響を与えない。
【0109】
好ましい組成物を本明細書中でSNS01と呼ぶ。SNS01は、安全性を増強させるためにB細胞由来の細胞 (骨髄腫細胞を包含する) への発現を制限するプロモーターにより推進されるelF5AK50RをコードするRNAi抵抗性プラスミドDNAと、ヌクレアーゼ耐性を増強させるためにdTdT 3’ オーバーハングをもつヒトelF5AをターゲットとするsiRNAの両方を含有する複合体であり、siRNAおよびプラスミドはin vivo JetPEITMに対して複合化されている。
【実施例】
【0110】
実施例1: eIF-5A1の野生型および変異型を使用するHeLaS3細胞のトランスフェクション
下記を包含するHAタッグドeIF5A1変異型を発現するプラスミドとともにリポフェクタミン (Lipofectamine) 2000を使用して、HeLa S3細胞をトランスフェクトした: 野生型eIF5A1 (WT) 、eIF5A1K50R (K50R) 、eIF5A1K67R (K67R) 、eIF5A1K67A (K67A) 、eIF5A1K47R/K50R (K4750R) 、eIF5A1K50R/K67R (K5067R) 、またはeIF5A1K50A/K67A (K5067A) 。LacZを発現するプラスミドを対照として使用した。トランスフェクション後24および48時間 (A) または28および52時間 (B) において、細胞ライゼイトを収集し、SDS-PAGEにより分画した。
【0111】
トランスフェクトしたeIF5A1の発現レベルをHAに対する抗体で検出した。結果: 推定上のユビキン化部位のリシンにおけるeIF5A1の突然変異化 (K67R) は、eIF5A1トランスジーンの蓄積を野生型 (A) より増加させた。また、ハイプシン化のために要求されるリシンにおけるeIF5A1の突然変異化 (K50R) は、eIF5A1トランスジーンの蓄積を野生型eIF5A1 (B) より増加させた。eIF5A1の二重突然変異化形態 (K50A/K67A) は、非突然変異化野生型eIF5A1トランスジーン (A + B) と比較したとき、特に十分に発現された。図2参照。
【0112】
実施例2: eIF-5A1の野生型および変異型を使用するKAS細胞のトランスフェクション
下記を包含するHAタッグドeIF5A1変異型を発現するプラスミドとともにPAMAMデンドリマー (FMD44) を使用して、KAS細胞をトランスフェクトした: 野生型eIF5A1 (5A1) 、eIF5A1K67A (K67A) 、eIF5A1K50A/K67A (K50A K67A) 、eIF5A1K50R (K50R) 、eIF5A1K47R (K47R) 、eIF5A1K67R (K67R) 、eIF5A1K47R/K50R (K47R K50R) 、またはeIF5A1K50R/K67R (K50R K67R) 。LacZを発現するプラスミドを対照として使用した。トランスフェクション後48時間において、細胞ライゼイトを収集し、SDS-PAGEにより分画した。トランスフェクトしたeIF5A1の発現レベルをHAに対する抗体で検出した。同じ負荷を、アクチンに対する抗体を用いて評価した。
【0113】
結果: 推定上のユビキン化部位のリシンにおけるeIF5A1の突然変異化 (K67AまたはK67R) は、eIF5A1トランスジーンの蓄積を野生型より増加させた。また、ハイプシン化のために要求されるリシンにおけるeIF5A1の突然変異化 (K50R) またはアセチル化部位におけるeIF5A1の突然変異化 (K47R) は、eIF5A1トランスジーンの蓄積を野生型eIF5A1より増加させた。eIF5A1の二重突然変異化 (K50A/K67A) は、非突然変異化野生型eIF5A1トランスジーンと比較したとき、より高いレベルで発現された。図3参照。
【0114】
実施例3: PAMAMデンドリマーを使用するKAS細胞のトランスフェクション
下記を包含するHAタッグドeIF5A1変異型を発現するプラスミドとともにPAMAMデンドリマー (FMD45-2) を使用して、KAS細胞をトランスフェクトした: eIF5A1K50R (K50R) 、eIF5A1K50A/K67A (K50A K67A) 、またはeIF5A1K50R/K67R (K50R K67R) 。LacZを発現するプラスミドを対照として使用した。トランスフェクション後72時間において、細胞をアネキシン (Annexin) /PIで染色し、FACSにより分析した。
【0115】
アネキシンVで陽性に染色され、PI (ヨウ化プロピジウム) で陰性に染色された細胞はアポトーシスの初期段階 (Ann+/PI-) にあると考慮し、そしてアネキシンVおよびPIの両方で陽性に染色された細胞はアポトーシスの最終段階 (Ann+/PI+) にあると考慮した。結果: ハイプシン化部位 (K50R) または推定上のユビキン化部位 (K67R) のリシンにおけるeIF5A1の突然変異化、ならびに二重突然変異体 (K50A/K67A) は、LacZ対照のレベルよりも有意に高いKAS細胞のアポトーシスを生じた。図4参照。
【0116】
実施例4: eIF-5A1およびeIF-5A1変異型を発現するプラスミドを使用するKAS細胞のトランスフェクション
下記を包含するHAタッグドeIF5A1変異型を発現するプラスミドとともにリポフェクタミン2000を使用して、KAS細胞をトランスフェクトした: eIF5A1K50A (K50A) 、eIF5A1K50R (K50R) 、eIF5A1K67R (K67R) 、eIF5A1K50A/K67A (K50A K67A) 、またはeIF5A1K50R/K67R (K50R K67R) 。LacZを発現するプラスミドを対照として使用した。
【0117】
トランスフェクション後72時間において、細胞をアネキシン/PIで染色し、FACSにより分析した。アネキシンVで陽性に染色され、PI (ヨウ化プロピジウム) で陰性に染色された細胞はアポトーシスの初期段階 (Ann+/PI-) にあると考慮し、そしてアネキシンVおよびPIの両方で陽性に染色された細胞はアポトーシスの最終段階 (Ann+/PI+) にあると考慮した。結果: ハイプシン化部位 (K50R) または推定上のユビキン化部位 (K67R) のリシンにおけるeIF5A1の突然変異化、ならびに二重突然変異体 (K50A/K67A) は、LacZ対照のレベルよりも有意に高いKAS細胞のアポトーシスを生じた。図5参照。
【0118】
実施例5: KAS細胞の治療に突然変異化eIF-5A1を使用するアポトーシスの発生
HAタッグドeIF5A1変異型eIF5A1K50R (K50R) またはeIF5A1K50A/K67A (K50A/K67A) を発現するプラスミドとともにリポフェクタミン2000を使用して、KAS細胞をトランスフェクトした。LacZを発現するプラスミドを対照として使用した。トランスフェクション後72時間において、細胞をアネキシン/PIで染色し、FACSにより分析した。アネキシンVで陽性に染色され、PI (ヨウ化プロピジウム) で陰性に染色された細胞はアポトーシスの初期段階 (Ann+/PI-) にあると考慮し、そしてアネキシンVおよびPIの両方で陽性に染色された細胞はアポトーシスの最終段階 (Ann+/PI+) にあると考慮した。結果: ハイプシン化部位 (K50R) のリシンにおけるeIF5A1の突然変異化またはハイプシン化部位および推定上のユビキン化部位 (K50A/K67A) の両方におけるeIF5A1の突然変異化は、LacZ対照のレベルよりも有意に高いKAS細胞のアポトーシスを生じた。図6参照。
【0119】
実施例6A: 多発性骨髄腫細胞のsiRNA/アデノウイルス仲介死滅
KAS (ヒト多発性骨髄腫) をS10培地 [4 ng/ml IL-6、10% 胎仔ウシ血清 (FBS) およびペニシリン/ストレプトマイシン (P/S) を含むRPMI 1640] 中で維持した。リポフェクタミン2000 (Invitrogen) を使用して58.7 ピコモルのsiRNAでKAS細胞をトランスフェクトした。偽トランスフェクトした細胞をsiRNAの非存在にリポフェクタミン2000で治療した。トランスフェクションを無抗体S10培地中で実施した。
【0120】
a) ヒトeIF5A1をターゲットとするsiRNA:
eIF5A1 siRNAターゲット#1 (このsiRNAはヒトeIF5A1のこの領域をターゲットとする: 5’ - AAGCTGGACTCCTCCTACACA - 3’ (配列番号_) 。このsiRNA配列は図25に示されており、本明細書においてしばしばh5A1と呼ぶ) 。
eIF5A1 siRNAターゲット#2 eIF5A1 (このsiRNAはヒトeIF5A1のこの領域をターゲットとする: 5’ - AAAGGAATGACTTCCAGCTGA - 3’ (配列番号_) 。このsiRNA配列は本明細書においてしばしばh5A1-ALTと呼ぶ) 。
b) 対照siRNA: 対照siRNAは下記の配列を有した: センス鎖、5’ - ACACAUCCUCCUCAGGUCGdTdT - 3’; およびアンチセンス鎖、3’ - dTdTUGUGUAGGAGGAGUCCAGC - 5’。
【0121】
使用した他の対照は、好ましくないオフ・ターゲッティング作用を制限するために検査したミクロアレイを有したので、ダーマコン (Dharmacon) からの非ターゲッティング確認siRNAを包含する。例えば、NF-κBを研究するin vitro研究のために、使用した対照はダーマコンの非ターゲッティングsiRNA (配列D-001700-01) であり、そしてin vivo研究のために、使用した対照はダーマコンの非ターゲッティングsiRNA (配列D-001810-01) であった。
【0122】
トランスフェクション後4時間に、細胞をペレット化し、1 mlのS10培地中に再懸濁させた。初期のsiRNAトランスフェクション後72時間に、トランスフェクトされたKAS細胞を数計測し、24ウェルのプレート中に300,000細胞/ウェルで播種し、そして同一siRNAで2回目のトランスフェクションを実施した。
【0123】
トランスフェクション後4時間に、細胞をペレット化し、3000 ifuのAd-LacZ (β-ガラクトシダーゼを発現するアデノウイルス) またはAd-5A1M (ヒトeIF5A1K50Rを発現するアデノウイルス) を含有するS10培地の1 ml中 (IL-6を含まない) に再懸濁させた。
72時間後に細胞を収集し、アネキシンV - FTTCおよびPI (BD Bioscience) で染色することにより、次いでFACS分析することにより、アポトーシスを分析した。
【0124】
a) 初期アポトーシスをアネキシン - FTTCで陽性に、PIで陰性に染色される細胞として定義した (Ann +/PI -) 。
b) 全アポトーシスを初期アポトーシス (Ann +/PI -) または後期アポトーシス/壊死 (Ann +/PI +) における細胞の合計として定義した。
【0125】
5A1 siRNAターゲッティング#1はヒトeIF5A1の3’ UTRをターゲットとし、したがってアデノウイルスからのeIF5A1の発現に影響を与えないであろう。5A1 siRNAターゲッティング#2はヒトeIF5A1のオープンリーディングフレーム内をターゲットとし、それゆえそれはアデノウイルスからのeIF5A1の発現を潜在的に妨害できるであろう。
結果: siRNAで治療し、eIF5A1K50R変異型を発現するアデノウイルスを感染させた細胞は、非治療細胞およびsiRNAのみで治療した細胞よりも大きい数の、アポトーシスを行う。図7参照。
【0126】
実施例6B: eIF5A1ターゲット#1に対するeIF5A1 siRNAの前処理 (図25に示す) は内因的eIF-5A1の発現を減少させたが、アデノウイルスにより発現されるRNAi抵抗性eIF5A1K50Rの蓄積を可能とする。
対照siRNA (C) または2つのsiRNAターゲッティングeIF5A1 (#1および#2) の1つとともにリポフェクタミン2000を使用して、KAS細胞をトランスフェクトした。eIF5A1 siRNA #1はeIF5A1の3’ UTRをターゲットとし、したがってそれはeIF5A1のオープンリーディングフレームのみを含有するので、アデノウイルスからのeIF5A1の発現を妨害しない。siRNAの配列を図25に示す。eIF5A1 #2 siRNAはeIF5A1のオープンリーディングフレームをターゲットとし、したがって内因的および外因的に発現されたeIF5A1の両方の発現に影響を与えるであろう。
【0127】
初期トランスフェクション時間後72時間に、細胞を第2回目に同一siRNAでトランスフェクトした。4時間後、トランスフェクション複合体を細胞から取出し、Ad-LacZ (L) またはAd-eIF5A1K50R (5M) のいずれかを含有する成長培地 (-) IL-6と置換した。72時間後、細胞ライゼイトを収集し、elF5Aに対する抗体およびアクチンを使用するウェスタンブロットにより分析した。図7B参照。ウイルスが発現したeIF5A1の蓄積が観察され (レーン1対レーン2) 、そしてeIF5A1 siRNAターゲッティング #1および #2により発現されたelF5A発現の減少は明瞭に見ることができる (レーン5および7対レーン3) 。期待するように、eIF5A1 siRNA #1はウイルスが発現したeIF5A1K50Rの蓄積に影響を与えない (レーン6対レーン4) が、eIF5A1 siRNA #2はウイルスが発現したトランスジーンの蓄積に適度にのみ影響を与える (レーン8対レーン4) 。
【0128】
実施例6C: アデノウイルス感染前のターゲット#1に対するeIF5A1 siRNAの前処理はヒト多発性骨髄腫細胞におけるリン酸化NF-κBの発現を減少させる。
対照siRNA (hC) またはeIF5A1をターゲットとするsiRNA (#1) とともにリポフェクタミン2000を使用して、KAS細胞をトランスフェクトした。eIF5A1 siRNA #1はeIF5A1の3’ UTRをターゲットとし、したがってeIF5A1のオープンリーディングフレームのみを含有するので、アデノウイルスからのeIF5A1の発現を妨害しない。初期トランスフェクション時間後72時間に、細胞を第2回目に同一siRNAでトランスフェクトした。
【0129】
4時間後、トランスフェクション複合体を細胞から取出し、Ad-LacZ (L) またはAd-eIF5A1K50R (5M) のいずれかを含有する成長培地 (+) IL-6と置換した。24時間後、細胞ライゼイトを収集し、ホスホ-NF-κB p65 (Ser 536) に対する抗体およびelF5Aを使用するウェスタンブロットにより分析した。期待するように、eIF5A1 siRNA #1はウイルスが発現したeIF5A1K50Rの蓄積に影響を与えない。セリン536におけるNF-κB p65のリン酸化は、活性化、核局在化、タンパク質・タンパク質相互作用および転写活性を調節する。図7C参照。
【0130】
実施例6D: アデノウイルス感染前のeIF5A1 siRNA #1を使用する前処理はヒト多発性骨髄腫細胞におけるリン酸化NF-κBおよびICAM-1の発現を減少させる。
対照siRNA (C) またはeIF5A1をターゲットとする2つのsiRNA (#1および#2) の1つともにリポフェクタミン2000を使用して、KAS細胞をトランスフェクトした。初期トランスフェクション時間後72時間に、細胞を第2回目に同一siRNAでトランスフェクトした。4時間後、トランスフェクション複合体を細胞から取出し、成長培地 (+) IL-6と置換した。2回目のトランスフェクション後24時間に、細胞を40 ng/mlのTNF-αで刺激し、細胞ライゼイトを0、4または24時間に収集し、ホスホ-NF-κB p65 (Ser 536) に対する抗体、ICAM-1およびアクチンを使用するウェスタンブロットにより分析した。
【0131】
両方のeIF5A1特異的siRNAを使用するトランスフェクション後、TNF-α誘導NF-κB p65リン酸化 (Ser 536) およびICAM-1の発現の減少が観察された。セリン536におけるNF-κB p65のリン酸化は、活性化、核局在化、タンパク質・タンパク質相互作用および転写活性を調節する。ICAM-1は、多発性骨髄腫の病原性に関係すると考えられる、細胞間接着表面の糖タンパク質である。図7D参照。
【0132】
実施例6E: siRNAを使用するKAS細胞の前処理は、IL-6の存在下のeIF5A1K50R遺伝子送出によるアポトーシスを増加させる。
リポフェクタミン2000を使用して対照siRNA (hcon) またはヒトeIF5A1 siRNA (h5A1) でKAS細胞をトランスフェクトした。72時間後、細胞をsiRNAで再トランスフェクトした。siRNAトランスフェクション培地を除去した後4時間に、空ベクター (mcs) またはeIF5A1K50R (K50R) プラスミドのPEI複合体を細胞に添加した。研究を通じて使用した成長培地はIL-6を含有した。72時間後、細胞をアネキシン/PIで染色し、FACS分析すると、アポトーシスが測定された。図7F参照。
【0133】
実施例7: eIF-5A1プラスミドおよびeIF5A1 siRNAの共投与は多発性骨髄腫皮下腫瘍の増殖を遅延させる (図8〜10) 。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。6匹の3〜5週齢のSCID/CB 17マウスの側腹部に、200 μlのPBS中の1000万のKAS-6/lの骨髄腫細胞を注射し、腫瘍が4 mm3の最大サイズに到達したとき、治療を開始した。
【0134】
対照マウスにpCpG-mcs (空ベクター) および対照siRNAを含有するPEI複合体を2回/週腫瘍内に注射した (対照グループは3匹のマウスから構成した: C-1、C-2およびC-3) 。治療したマウスにRNAi抵抗性プラスミドpCpG-eIF5A1 K50RおよびeIF5A1 siRNAを含有するPEI複合体を2回/週腫瘍内に注射した (治療したグループは3匹のマウスから構成した: 5A-1、5A-2および5A-3) 。注射は腫瘍内の多数の部位に実施して還流を防止し、遅い注射速度を使用して吸収を増加させた。図8中のデータはグループ中のすべてのマウスについての腫瘍体積を示す。図9に示すデータは平均腫瘍体積±標準誤差である。星印は統計的有意性を意味する (* = p < 0.025; n = 3) 。
【0135】
図10は、eIF-5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の重量を減少させることを示す。示すデータは平均腫瘍体積±標準誤差である。星印は統計的有意性を意味する (* = p < 0.05; n = 3) 。
2×0.1 mlのJET-PEI (PolyPlus) をin vivo試験において使用した。N/P比は8であった。PEI/DNA/siRNA複合体を5%のグルコース中で0.1 mlの全体積で形成した。複合体を形成するプロトコルは次の通りであった:
【0136】
1. 成分を温室にする。無菌を保持する。
2. 20μgのプラスミドDNA (2 mg/mlにおいて約10μl) および10μgのsiRNA (1 mg/mlにおいて10μl) を25μlの全体積に希釈する。無菌水を使用して差を構成する。
3. 25μlの10%グルコースの添加によりDNA溶液の体積を50μlの5%グルコースに調節する。おだやかに渦形成し、短時間遠心する。
【0137】
4. 4.8μlのin vivo JET-PEIを25μlの10%グルコースの全体積に希釈する。50μlの無菌水で体積を5%グルコースの最終濃度に調節する。おだやかに渦形成し、短時間遠心する。
5. 直ちに50μlの希釈PEIを50μlの希釈DNAに添加する (順序を逆にしない) 。短時間渦形成し、直ちに回転する。
6. 注射前に15分間インキュベートする。複合体は6時間安定である。
【0138】
無CpGクローニングベクターおよびpCpGプラスミドを入手した (InvivoGenから) 。これらのプラスミドはCpGジヌクレオチドを完全に欠如し、pCpGと命名する。これらのプラスミドはin vitroおよびin vivoにおいて高いレベルのトランスジーン発現を生じ、CMVに基づくプラスミドと対照的に、in vivoにおいて持続した発現を可能とする。pCpGプラスミドは、自然にCpGジヌクレオチドを欠如し、すべてのCpGを除去するように変性されているか、あるいは完全に合成された要素、例えば、選択可能なマーカーまたはリポーターをコードする遺伝子を含有する。
【0139】
遺伝暗号を形成する16ジヌクレオチドの間で、CGは非必須の唯一のジヌクレオチドでありかつ置換できるという事実により、これらの新しいアレレの合成は可能である。8コドンは5つの異なるアミノ酸をコードするCGを含有する。野生型と同一にとどまり、こうしてそれらの野生型対応物と同程度に活性であるアミノ酸配列を有するタンパク質を新たなアレレを生成する同じアミノ酸をコードする2つのコドンを少なくとも選択することによって、すべての8コドンは置換できる。これらの新しいアレレはpMODと命名するプラスミドにおいて入手可能であり、このプラスミドからそれらを容易に切除することができる。
【0140】
pCpGプラスミドはin vivoにおいて長く持続する発現を可能とし、TLR9を発現する細胞系統を使用する、in vivoおよびin vitroにおける遺伝子発現に対するCpGの作用、ならびに生得的および後天的免疫系に対するそれらの作用を研究するための価値のある道具を表す。
【0141】
空ベクター、pCpG-mcs (Invitrogen) は、発現された遺伝子産物をもたず、多重クローニング部位のみをもち、対照ベクターとして使用されたベクターである。HAタッグドeIF5A1K50R cDNAを、LacZ遺伝子が除去されている、pCpG-LacZベクター (Invitrogen) のNcoIおよびNheI部位中にサブクローニングして、治療ベクターpCpG-eIF-5A1 (K50R) を発生させた。DNAはキット (Endo-Free Qiagen) を使用して調製した。エンドドキシンレベルを測定し、これは < 0.03 EU/μgである; DNAは水中で2 mg/mlであるべきである。
【0142】
実験において使用した対照siRNAは、ダーマコン (Dharmacon) からのミクロアレイ確認非ターゲッティング対照siRNAであった (D-00180-01) 。血清中の分解を防止するために修飾した (siSTABLE) siRNAを得た。
【0143】
実験において使用したeIF5A1 siRNAは、ヒトeIF5A1の3’ UTRに対して設計した。eIF5A1 siRNAとマウスeIF5A1との間に類似性は存在せず、したがってsiRNAはヒト (マウスではない) eIF5A1のみを抑制すべきである。また、siRNAはeIF5A2 (ヒトまたはマウス) に対して類似性をもたない。血清中の分解を防止するために修飾した (siSTABLE) siRNAを得た。eIF5A1 siRNAは下記のターゲット配列を有する:
5’ GCU GGA CUC CUC CUA CAC A (UU) 3’
【0144】
SiSTABLE siRNAを水中に1 mg/mlで溶解した (- 20℃においてアリコートで貯蔵した) 。
デジタルキャリパーを使用して、長さ (l) および幅 (w) の腫瘍寸法を2〜3回/週測定した。下記の方程式を使用して、腫瘍体積を計算した:
l = 長さ; 最小寸法
w = 幅; 最大寸法
腫瘍体積 (mm3) = l2×w×0.5
【0145】
統計的解析
スチューデントのt検定を統計的解析に使用した。有意性は95% (p < 0.05) を超える信頼水準であると考える。
【0146】
実施例8: eIF5A1プラスミドおよびeIF5A1 siRNAの共投与は多発性骨髄腫皮下腫瘍の増殖を遅延し、腫瘍を収縮させる。
他の研究において、再びSCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。対照マウスにpCpG-mcs (空ベクター) および対照siRNAを含有するPEI複合体を2回/週腫瘍内に注射した (対照グループG-1、G-2およびG-3) 。治療したマウスにRNAi抵抗性プラスミドpCpG-eIF5A1K50R (20μgのプラスミドDNA) およびeIF5A1 siRNA (10μgのsiRNA) を含有するPEI複合体を2回/週腫瘍内に注射した (治療したグループG-4、G-5およびG-6) 。図11に示すデータは平均腫瘍体積±標準誤差である。星印は統計的有意性を意味する (* = p < 0.025; n = 3) 。21日間にわたって6回の注射を実施した (赤色矢印) 。
【0147】
実施例9: eIF5A1 siRNAを静脈内 (i.v.) に投与しかつPEI/eIF5A1K50Rプラスミドを腫瘍内 (i.t.) に投与すると、多発性骨髄腫皮下腫瘍の腫瘍収縮を生ずる (グループ2B) 。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、50μgの対照siRNA (対照グループ) またはヒトeIF5A1 siRNA (治療したグループ) の初期尾注射で治療を開始した。続いて、対照マウスにpCpG-mcsを含有するPEI複合体を2回/週腫瘍内に注射することにより治療した (空ベクター; 対照グループ; G-1、G-2およびG-3) 。続いて、治療したマウスにRNAi抵抗性プラスミドpCpG-eIF5A1K50R (20μgのプラスミドDNA) を含有するPEI複合体を2回/週腫瘍内に注射することにより治療した (治療したグループ; G-4、G-5およびG-6) 。
【0148】
対照マウスに1回/週のi.v. 注射により対照siRNAを投与し続けた (対照グループR-1、R-2およびR-3) 。治療したマウスに1回/週のi.v. 注射によりヒトeIF5A1 siRNA (20μg) を投与し続けた (治療したグループR-4、R-5およびR-6) 。図12に示すデータは各グループにおけるすべてのマウスについての腫瘍体積である。PEI/DNAの6回の腫瘍内注射 (赤色矢印) および4回のi.v. 注射 (青色矢印) を21日間の期間にわたって実施した。
【0149】
図13は実施例8および9からの結果のオーバーレイを提供する。SCIDマウスにKAS細胞を皮下注射した。図13に示すデータはそれぞれのグループにおけるマウスの平均腫瘍体積±標準誤差である。星印は治療したグループと対照グループとの間の統計的有意性を意味する (** = p < 0.01; *** = p < 0.001; n = 3) 。
【0150】
PEI複合体を形成するプロトコル:
1. 成分を温室にする。無菌を保持する。
2. プラスミドDNAまたはプラスミドDNA + siRNAを全体積25μlに希釈する。無菌水を使用して体積を調節する。
a) プラスミドDNAのみの複合体について:
20μgのプラスミドDNA (約2 mg/mlにおいて10μl) を全体積25μlに希釈する。無菌水を使用して差を構成する。
【0151】
b) プラスミドDNA + siRNAの複合体について:
20μgのプラスミドDNA (約2 mg/mlにおいて10μl) および10μgのsiRNA (1 mg/mlにおいて10μl) を全体積25μlに希釈する。無菌水を使用して差を構成する。
3. DNA溶液の体積を25μlの10%グルコースの添加により50μlの5%グルコースに調節する。おだやかに渦形成し、短時間遠心する。
【0152】
4. in vivo JETPEIを全体積25μlの10%グルコースに希釈する。
a) プラスミドDNAのみの複合体について:
3.2μlのin vivo JETPEIを全体積25μlの10%グルコースに希釈する。体積を無菌水で50μlに調節して5%グルコースの最終濃度にする。おだやかに渦形成し、短時間遠心する。
b) プラスミドDNA + siRNAの複合体について:
4.8μlのin vivo JETPEIを全体積25μlの10%グルコースに希釈する。体積を無菌水で50μlに調節して5%グルコースの最終濃度にする。おだやかに渦形成し、短時間遠心する。
【0153】
5. 直ちに50μlの希釈PEIを50μlの希釈DNAに添加する (順序を逆にしない) 。短時間渦形成し、直ちに回転する。
6. 注射前に15分間インキュベートする。複合体は6時間安定である。
【0154】
siRNAの尾静脈注射に関して、初期siRNA注射は50マイクログラムであった。siRNAをPBS中で0.4 mg/mlに希釈した。125μl/マウス (50μg) を尾静脈に注射した。引き続いて血清安定化siRNAを20μg/マウスで2回/週注射した。siRNAをPBS中で0.4 mg/mlに希釈した。50μl/マウス (20μg) を尾静脈に注射した。
【0155】
図13Bは、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が腫瘍を収縮させることを示す。SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。マウスにRNAi抵抗性プラスミドpCpG-eIF5A1K50RおよびeIF5A1 siRNAを含有するPEI複合体を2回/週腫瘍内に注射した (治療したグループ; G-4、G-5およびG-6) 。21日間にわたって6回の注射を実施した。治療の開始後42日に、マウスを殺し、腫瘍部位下の皮膚を開き、腫瘍増殖の証拠を検査した。グループ2Aの治療マウスのいずれにおいても、腫瘍増殖は観察されなかった。
【0156】
図13Cは、eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の腫瘍内 (i.t.) 投与が多発性骨髄腫皮下腫瘍の腫瘍収縮を生じさせることを示す。SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、50μgのヒトeIF5A1 siRNAの初期注射で治療を開始した (治療したグループ) 。引き続いてマウスにRNAi抵抗性プラスミドpCpG-eIF5A1K50Rを含有するPEI複合体を2回/週腫瘍内に注射して治療した (治療したグループ; R-4、R-5およびR-6) 。マウスにヒトeIF5A1 siRNAを1回/週注射により投与し続けた。治療開始後21日に治療を終えた。治療の開始後42日に、マウスを殺し、腫瘍部位下の皮膚を開き、腫瘍増殖の証拠を検査した。治療グループの1匹のマウスにおいて腫瘍増殖の証拠は存在しなかった (グループ2B) 。
【0157】
実施例10: eIF5A1プラスミドおよびeIF5A1 siRNAの静脈内共投与は多発性骨髄腫皮下腫瘍の増殖を遅延する。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、50μgの対照siRNA (対照グループ) またはヒトeIF5A1 siRNA (治療したグループ) の初期注射で治療を開始した。引き続いてマウスにpCpG-mcsを含有するPEI複合体 (空ベクター; 対照グループA1、A2およびA3) またはRNAi抵抗性プラスミドpCpG-eIF5A1K50Rを含有するPEI複合体 (治療したグループ; A4、A5およびA6) を2回/週静脈内注射 (赤色矢印) または腹腔内注射 (緑色矢印) により治療した。マウスに対照siRNA (対照グループA1、A2およびA3) またはヒトeIF5A1 siRNA (治療したグループ; A4、A5およびA6) をi.v. 注射 (青色矢印) により1回/週投与し続けた。示したデータは各グループにおけるすべてのマウスについての腫瘍体積である。図14に示すデータは各グループにおけるすべてのマウスについての腫瘍体積である。
【0158】
実施例11: eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の静脈内 (i.v.) または腹腔内 (i.p.) 投与は多発性骨髄腫皮下腫瘍の増殖を遅延させる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、50μgの対照siRNA (対照グループ) またはヒトeIF5A1 siRNA (治療したグループ) の初期注射で治療を開始した。引き続いて対照マウスをpCpG-mcsを含有するPEI複合体で静脈内または腹腔内注射により1回/週治療した (空ベクター; 対照グループは3匹のマウスであった: B1、B2およびB3) 。
【0159】
引き続いて治療したマウスをRNAi抵抗性プラスミドpCpG-eIF5A1K50Rを含有するPEI複合体で静脈内または腹腔内注射により1回/週治療した (治療したグループ: B4、B5およびB6) 。マウスに対照siRNA (対照グループ: B1、B2およびB3) またはヒトeIF5A1 siRNA (治療したグループは3匹のマウスであった: B4、B5およびB6) をi.v. 注射により1回/週投与し続けた。50μgの初期siRNA注射により、実験を開始した (図15中のグラフ上の第2日) 。引き続いて20μgのsiRNAを1回/週注射した。siRNAは裸で、すなわち、送出ビヒクルを使用しないで投与した。PEI複合体は20μgのプラスミドDNAを含有した。初期PEIをi.p. 注射し、引き続いてi.v. 注射した。図15に示すデータは、各グループにおけるすべてのマウスについての腫瘍体積である。
【0160】
図16は実施例10および11のオーバーレイを提供する。SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。マウスの1組に対照siRNA (対照; グループA) またはeIF5A1 siRNA (治療した; グループA) を1回/週i.v. 注射し、そしてpCpG-mcsを含有するPEI複合体 (対照; グループA) またはRNAi抵抗性プラスミドpCpG-eIF5A1K50Rを含有するPEI複合体 (治療した; グループA) を1回/週i.v. またはi.p. 注射した。
【0161】
マウスの第2組にpCpG-mcs (空ベクター) および対照siRNAを含有するPEI複合体 (対照; グループB) またはRNAi抵抗性プラスミドpCpG-eIF5A1K50RおよびeIF5A1を含有するPEI複合体 (治療した; グループB) を2回/週i.v. またはi.p. 注射した。示すデータは各グループにおけるマウスについての平均腫瘍体積±標準誤差である。星印は治療したグループと対照グループとの間の統計的有意性を意味する (* = p < 0.05; *** = p < 0.001; n = 3) 。
PEI複合体およびsiRNAを調製するプロトコルは前の実施例に記載されている通りである。
【0162】
実施例12: eIF5A1プラスミドおよびeIF5A1 siRNAの共投与は多発性骨髄腫皮下腫瘍の成長を遅延させ、腫瘍を収縮させる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。対照マウスにpCpG-mcs (空ベクター) および対照siRNAを含有するPEI複合体を2回/週腫瘍内注射した (対照グループは3匹のマウスを有した: 対照1、対照2および対照3) 。治療したマウスにRNAi抵抗性プラスミドpCpG-eIF5A1K50RおよびeIF5A1 siRNAを含有するPEI複合体を2回/週腫瘍内注射した (治療したグループは4匹のマウスを有した: 5A-1、5A-2、5A-3および5A-4) 。PEI複合体の腫瘍内注射は20μgのプラスミドDNAおよび10μgのsiRNAの両方を含有した。図17に示すデータは各グループにおけるすべてのマウスについての腫瘍体積である。
【0163】
実施例13: eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の腫瘍内 (i.t.) 投与は多発性骨髄腫皮下腫瘍の腫瘍収縮を生じさせる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、50μgの対照siRNA (対照グループは3匹のマウスを有した: 対照1、対照2および対照3) またはヒトeIF5A1 siRNA (治療したグループは3匹のマウスを有した: 5A-1、5A-2および 5A-3) の初期注射で治療を開始した。引き続いて対照マウスをpCpG-mcs (20μg) を含有するPEI複合体の2回/週の腫瘍内注射により治療した (対照グループ1〜3) 。
【0164】
引き続いて治療したマウスをRNAi抵抗性プラスミドpCpG-eIF5A1K50R (20μg) を含有するPEI複合体の2回/週の腫瘍内注射により治療した (5A-1、5A-2および5A-3) 。対照マウスに対照siRNA (20μg) を尾静脈 i.v. 注射により2回/週投与し続けた。治療したマウスにヒトeIF5A1 siRNA (20μg) を尾静脈 i.v. 注射により2回/週投与し続けた。腫瘍内注射48時間前に注射した。siRNAは裸で、すなわち、送出ビヒクルを使用しないで投与した。図18に示すデータは各グループにおけるすべてのマウスについての腫瘍体積である。
【0165】
実施例14: EF1またはB29プロモーターのいずれかにより推進されたeIF5A1K50Rプラスミド、およびeIF5A1 siRNAの共投与は多発性骨髄腫皮下腫瘍の増殖を遅延させ、腫瘍を収縮させる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。マウスに対照ベクター (G1およびG2) またはB29プロモーター (G3およびG4) もしくはEF1プロモーター (G5およびG6) により推進されるeIF5A1プラスミドまたは対照siRNA (G1、G3、G5) またはh5A1 siRNA (G2、G4、G6) を含有するPEI複合体を2回/週腫瘍内注射した。
【0166】
示すデータは各グループの平均腫瘍体積±標準誤差である。注: B29プロモーターはB細胞特異的プロモーターであることを意図した。しかしながら、この研究において使用したB29プロモーター/mCMVエンハンサーはin vitro KAS細胞中でHA-eIF5A1K50Rの高い発現を推進させることが発見されたが、B細胞特異的であるように思われない (CMVエンハンサーのためであろう) 。図19参照。
【0167】
実施例15: eIF5A1 siRNAの共投与はEF1またはB29プロモーターいずれかにより推進されるeIF5A1K50Rプラスミドの多発性骨髄腫皮下腫瘍に対する抗腫瘍作用を増加させ、存在する腫瘍を減少させる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。マウスに対照ベクター (G1およびG2) またはB29プロモーター (G3およびG4) もしくはEF1プロモーター (G5およびG6) により推進されるeIF5A1プラスミドまたは対照siRNA (G1、G3、G5) またはh5A1 siRNA (G2、G4、G6) を含有するPEI複合体を2回/週腫瘍内注射した。治療開始後24日にマウスを殺し、皮下腫瘍を取出し、秤量した。示すデータはすべてのグループの平均腫瘍体積±標準誤差である。図20参照。
【0168】
実施例16: eIF5A1 siRNAは肺腺癌細胞におけるAd-elF5Aによる感染から生ずるアポトーシス誘導を相乗的に増加させる。
A549細胞にAd-LacZまたはAd-elF5Aを感染させた。トランスフェクション培地を細胞に添加し、直ちにウイルスを添加することによって、細胞を対照siRNAまたはsiRNAターゲッティングヒトeIF5A1 (h5A1) でトランスフェクトした。siRNAでトランスフェクトし、ウイルスで感染させた4時間後、培地を新鮮な培地と置換し、細胞を72時間インキュベートした後、アネキシン/PIで標識化してアポトーシス細胞を検出した。注: この細胞系統におけるelF5Aの過剰発現は限定的量のDHSおよびDOHHの理由から非ハイプシン化elF5Aが蓄積し、したがってelF5AK50Rの過剰発現として同一前アポトーシス作用が生ずる。これらのデータが示すように、同時のハイプシン化elF5Aの抑制および非ハイプシン化elF5Aの過剰発現により生じたアポトーシスにおける相乗効果はその上非骨髄腫腫瘍型において観察される。図21参照。
【0169】
実施例17: pExp5Aの構築
pExp5Aは、主としてB細胞系統の細胞においてヒトeIF5A1K50Rの発現を推進するように設計された、CpGジヌクレオチドが減少した発現プラスミドである。このベクターはpCpG-LacZ、すなわち、CpGジヌクレオチドを完全に欠如するプラスミド (Invitrogen) である。大腸菌 (E. coli) における複製および選択に必要なすべての要素はCpGジヌクレオチドを含まない。eIF5A1K50RのB細胞特異的発現を推進するために、本来のCMVエンハンサー/プロモーターおよびCpG-LacZベクターからのLacZ遺伝子は、それぞれ、ヒト最小B細胞特異的プロモーター (B29/CD79b; Invitrogen) およびヒトeIF5A1K50Rと置換されてきている。
【0170】
B29プロモーターの活性を増大しかつ非B細胞における発現を減少させるために、B29 DHS4.4 3’ エンハンサーがeIF5A1発現カセットの下流のプラスミド中に導入されてきている (Malone 他、2006、B29 gene silencing in pituitary cells is regulated by its 3’ enhancer. J. Mol. Biol. 362: 173-183) 。B29最小プロモーター、eIF5A1K50RおよびB29 DHS4.4 3’ エンハンサーの組込みは、32 CpGジヌクレオチドをベクター中に導入した。
【0171】
大腸菌 (E. coli) における発現のための要素
複製起点: 大腸菌 (E. coli) R6Kγori.
* R6Kγ複製起点が存在するために、pCpGプラスミドはpir突然変異体遺伝子を発現する大腸菌 (E. coli) においてのみ発現されることができる。それらは標準的大腸菌 (E. coli) 株において複製しないであろう。したがって、pCpGプラスミドは大腸菌 (E. coli) GT115株、すなわち、Dcmメチル化を欠如するpir突然変異体 (Invivogen) を提供する。
細菌プロモーター: EM2K、細菌EM7プロモーターの無CpGバージョン。
選択可能なマーカー: ZeocinTM 抵抗性遺伝子; CpGをもたない合成アレレ。
【0172】
哺乳動物細胞における発現のための要素
哺乳動物プロモーター: B細胞における組織特異的発現のためのヒト-167 bp最小B29 (CD79b) プロモーター。また、合成イントロン (I 140) が5’ UTRに存在する。
ポリアデニル化シグナル: 後期SV40ポリアデニル化シグナルの無CpGジヌクレオチドバージョン。
3’ エンハンサー: ヒトB29 DHS4.4 3’ エンハンサー。
MAR: 細菌の転写単位と哺乳動物の転写単位との間に、2つの無CpGマトリックス結合領域 (MAR) が存在する。1つのMARはヒトIFN-β遺伝子の5’ 領域に由来し、そして1つはβ-グロビン遺伝子の5’ 領域に由来する。
pExp5A (3371 bp) の予測される配列は図23に示されている。
【0173】
eIF5A1K50Rのアミノ酸配列
* K50R突然変異に下線が引かれている
【0174】
【化1】
【0175】
pExp5Aの構築 - 構築の概略:
工程1: B29 DHS4.4 3’ エンハンサーをクローニングし、pGEM T easy (Promega) 中にサブクローニングする -- pGEM-4.4 enh #8をつくる。
工程2: 最小B29プロモーターをpCpG-LacZ (Invivogen) 中にサブクローニングする - B29-5#3をつくる。
工程3: HA-eIF5A1K50RをB29-5#3ベクター中にサブクローニングする - B29-5#3-eIF5A1K50Rをつくる。
【0176】
工程4: pCpG-mcs (Invitrogen) 中に新しい多重クローニング部位をつくる - pCpG-Linker4をつくる。
工程5: B29 DHS4.4 3’ エンハンサーをpCpG-Linker4中にサブクローニングする - pCpG-DHS4.4をつくる。
工程6: B29プロモーター + HA-eIF5A1K50R + SV40 pA発現カセットをpCpG-DHS4.4中にサブクローニングしてpExp-5をつくる。
工程7: HA-eIF5A1K50RをもたないpExp-5中にHA-eIF5A1K50Rと置き換えて最終のベクターpExp5Aをつくる。
【0177】
構築の詳細:
工程1: B29 DHS4.4 3’ エンハンサーのクローニングおよびpGEM T easy (Promega) 中へのサブクローニング -- pGEM-4.4 enh #8をつくる。
【0178】
下記のプライマーを使用してKAS細胞 (多発性骨髄腫細胞系統) から単離したゲノムDNAからPCRにより、B29 DHS4.4 3’ エンハンサーをクローニングした: 前方向5’ - CAGCAAGGGAGCACCTATG - 3’ および逆方向5’ - GTTGCAGTGAGCGGAGATG - 3’ 。ヒトCD79B/GH-1遺伝子間領域 (Accession AB062674) の配列を使用してプライマーを設計した。生ずる608 bpのPCRフラグメントをpGEM(商標)Teasyクローニングベクター (Promega) 中にサブクローニングし、配列決定した。Komatsu 他 2002 Novel regulatory regions found downstream of the rat B29/Ig-b gene. Eur. J. Biochem. 269: 1227-1236。
【0179】
pGEM-4.4 enh #8中のB29 DS4.4 3’ エンハンサーPCRフラグメントの配列 (297 bp)
【0180】
【化2】
【0181】
pGEM-4.4 enh #8中のB29 DHS4.4 3’ エンハンサーPCRフラグメントの配列 (297 bp) とヒトCD79B/GH-1遺伝子間領域 (Accession AB062674) の配列とのアラインメント。
【0182】
【化3】
【0183】
工程2: pCpG-LacZ (Invivogen) 中への最小B29プロモーターのサブクローニング -- B29-5 #3をつくる。
下記のプライマーを使用して全長のヒトB29プロモーターを有する商用プラスミド (pDrive-hB29; Invivogen) から、最小-167ヒトB29プロモーターを増幅した: 前方向5’ - CCAACTAGTGCGACCGCCAAACCTTAGC - 3’ ; 逆方向5’ - CAAAAGCTTGACAACGTCCGAGGCTCCTTGG - 3’ 。生ずるPCRフラグメントをSpeIおよびHindIIIで消化し、pCpG-LacZベクター (Invivogen) のSpeIおよびHindIII部位中に結合してB29-5 #3をつくった。
【0184】
B29-5 #3中のB29最小プロモーターPCRフラグメントの配列 (188 bp)
【0185】
【化4】
【0186】
B29-5 #3中のB29最小プロモーターPCRフラグメント (B29 min) とpDrive-hB29からの全長ヒトB29プロモーターとのアラインメント
【0187】
【化5】
【0188】
工程3: B29-5 #3ベクター中のHA-eIF5A1K50Rのサブクローニング -- pB29-eIF5A1K50R_7をつくる。
DNAテンプレートとしてpHM6-eIF5A1K50Rおよび下記のプライマーを使用してPCRにより、HA-eIF5A1K50Rを増幅した: 前方向5’ - CGCCATGGACATGTACCCTTACGACGTCCCAGACTACGCTGCAGATGATTTGGACTTCGAG - 3’ および逆方向5’ - CGCGCTAGCCAGTTATTTTGCCATCGCC - 3’ 。生ずるPCRフラグメントをNcoIおよびNheIで消化し、B29-5 #3のNcoIおよびNheI部位中にサブクローニングしてLacZを置換した。
【0189】
pB29-eIF5A1K50R_7中のHA-eIF5A1K50RPCRフラグメントの配列
【0190】
【化6】
【0191】
pB29-eIF5A1K50R_7中のHA-eIF5A1K50R PCRフラグメントの翻訳
HAエピトープ
eIF5A1K50R
K50R
【0192】
【化7】
【0193】
pB29-eIF5A1K50R_7中のHA-eIF5A1K50R PCRフラグメントとヒトeIF5A1 (Accession # NP 001961) とのアラインメント
【0194】
【化8】
【0195】
工程4: pCpG-mcs (Invivogen) 中に新しい多重クローニング部位をつくる -- pCpG-Linker4をつくる。
pCpGクローニングベクター、pCpG-mcs G2 (Invivogen) をEcoRIで消化して、mCMVエンハンサー、hEFIプロモーター、合成イントロン、多重クローニング部位およびSV40ポリアデニル化シグナルを含有する哺乳動物発現カセットを除去した。次いでEcoRI消化pCpG-mcs G2ベクターをEcoRI粘着末端をもつ合成リンカーに結合して、新しい多重クローニング部位をもつ無プロモーターベクター (pCpG-Linker4) をつくる。
【0196】
【化9】
【0197】
pCpG-Linker4中の新しい多重クローニング部位を取り囲む領域の配列
【0198】
【化10】
【0199】
工程5: pCpG-Linker4中のB29 DS4.4 3’ エンハンサーのサブクローニング - pCpG-DHS4.4をつくる。
テンプレートとしてpGEM-4.4 enh #8を使用しそして下記のプライマーを使用してPCRにより、B29 DHS4.4 3’ エンハンサーを増幅した: 前方向5’ - GAAGCGGCCGCACCACCCTGGGCCAGGCTGG - 3’ および逆方向5’ -CCACGCGTAGAGGTGTTAAAAAGTCTTTAGGTAAAG - 3’ 。生ずるPCRフラグメントをNotIおよびMluIで消化し、pCpG-Linker4中の新しい多重クローニング部位中のNotIおよびMluI部位中に結合し、pCpG-DHS4.4をつくる。
【0200】
【化11】
【0201】
工程6: B29プロモーター + HA-eIF5A1K50R + SV40 pA発現カセットpCpG-DHS4.4へのサブクローニング -- pExp-5をつくる。
下記のプライマーを使用してPCRにより、pB29-eIF5A1K50R_7 (工程3) から最小B29プロモーター、合成イントロン、HA-eIF5A1K50RおよびSV40 pAを含有するB29-eIF5A1発現カセットを増幅した: 前方向5’ - GTTATCGATACTAGTGCGACCGCCAAACC - 3’ および逆方向5’ - CAAGCGGCCGCCATACCACATTTGTAGAGGTTTTAC - 3’ 。生ずるPCRフラグメントをClaIおよびNotIで消化し、pCpG-DHS4.4の多重クローニング部位中のClaIおよびNotI部位中にサブクローニングしてpExp-5をつくった。
【0202】
工程7: pExp-5中のHA-eIF5A1K50Rを非HA-eIF5A1K50Rで置換して最終ベクター、pExp5Aをつくる。
pExp-5プラスミドをNcoIおよびNheIで消化して、HA-eIF5A1K50Rを除去した。下記のプライマーを使用してPCRにより、pHM6-eIF5A1K50Rから非HAタッグドeIF5A1K50R PCRフラグメントを増幅した: 前方向5’ - CACCATGGCAGATGATTTGGACTTC - 3’ および逆方向5’ - CGCGCTAGCCAGTTATTTTGCCATCGCC - 3’ 。生ずるPCR生成物をNcoIおよびNheIで消化し、B29-5 #3のNcoIおよびNheI部位に結合してB29- K50Rを発生させた。B29-K50RをNcoIおよびNheIで消化し、470 bpのeIF5A1K50Rフラグメントをゲル精製し、NcoI/NheI消化pExp-5に結合させて最終空ベクター、pExp5Aを発生させた。
【0203】
実施例18: pExp5Aの試験
リポフェクタミン2000を使用して種々の細胞系統をプラスミドでトランスフェクトし、トランスフェクション後24時間に抗HA抗体 (Roche) を使用するウェスタンブロットにより、HA-eIF5A1K50Rの発現を決定した。試験した異なる細胞系統は次の通りであった: P3X63Ag8.653 (マウスBリンパ芽球 - 骨髄腫) 、KAS (ヒト骨髄腫) 、HepG2 (ヒト肝臓肝細胞性癌腫) 、T24 (ヒト膀胱癌腫) 、HT-29 (ヒト結腸直腸腺癌) 、HEK-293 (ヒト胚腎細胞) 、PC3 (ヒト前立腺腺癌) 、HeLa (ヒト頸部腺癌) およびA549 (肺癌腫) 。図24参照。
【0204】
pExp-5プラスミドは、構成的EF1プロモーター (CpG-eIF5A1K50R) を有するプラスミドに匹敵するレベルで、ヒトおよびマウスの両方の骨髄腫細胞系統においてHA-eIF5A1K50Rを発現する。しかしながら、pExp-5により推進されたHA-eIF5A1K50Rの発現は、構成的プロモーターによる発現と比較して、非B細胞系統において限定される。1つの例外はHEK-293細胞、ヒト胚腎臓細胞系統においてであり、ここでpExp-5のトランスフェクション後に高いレベルのHA-eIF5A1K50Rの発現が観察され - これは細胞系統の胚の特質のためであることがある; この時点において、pExp-5が成人腎細胞において発現するかどうかは我々は知らない。
【0205】
毒性研究および臨床的実験において使用する最終プラスミドは、HA-eIF5A1K50Rが非HAタッグドeIF5A1K50R (pExp5A) で置換されているpExp-5のバージョンであろう。pExp-5は最小ヒトB29プロモーター/エンハンサーの制御下にHAタッグドeIF5A1K50Rを含有する; HA-eIF5A1K50Rの発現を構成的発現をもつプラスミドにより推進されるものならびに全長B29プロモーターを含有するプラスミドと比較した。
【0206】
実施例19: in vivo JETPEITMナノ粒子の形成
20 gのマウスに1.5 mg/kg (0.1 ml) -- 1.5 mg/kg = 1.0 mg pExp5A/kg + 0.5 mg h5A/kg -- DNA: siRNA比 = 2:1の投与量で注射するために、in vivo JETPEITMナノ粒子の複合体を形成することをこの実施例において説明する。
【0207】
プラスミドDNAおよびsiRNAを全体積25 mlに希釈する。無菌水を使用して体積を調節する。* 20 mgのプラスミドDNA (10 ml、2 mg/ml) および10 mgのsiRNA (10 ml、1 mg/ml) を全体積25 mlに希釈する。無菌水を使用して差を構成する。25 mlの10%グルコース (PEIキットとともに準備されている) の添加により、DNA溶液の体積を50 mlの5%グルコースに調節する。おだやかに渦形成し、短時間遠心する。in vivo JETPEITMを全体積25 mlの水中に希釈する。* 3.6 mlのin vivo JETPEITMを全体積25 mlの水中に希釈する。最終体積を10%グルコースで50 mlに調節して、5%グルコースの最終濃度にする。おだやかに渦形成し、短時間遠心する。直ちに50 mlの希薄PEIを50 mlの希薄DNA に添加する (順序を逆にしない) 。短時間渦形成し、直ちに回転する。
【0208】
形成後、複合体は8〜10時間安定である。複合体のN/P比は6であるべきである。N/P比はin vivo jetPEIの正に帯電した窒素残基の数/DNAおよびsiRNAの負に帯電したリン酸塩残基の数の比である。DNAおよびsiRNAは同一数のリン酸塩残基の数/gを含有する。したがって、N/P比は複合体中のイオン釣合いの測度である。複合体のN/P比を増加させると、複合体の毒性が増加する。in vivo JET-PEIは150 mM溶液 (窒素残基として表す) として提供されるが、DNAは1 mg中に3モルのアニオンリン酸塩を含有する。最終体積のDNAの最終濃度は0.5 mg/mlを超えるべきではない。DNAは高品質であるべきであり、水中で調製される。in vivo jetPEIおよび10%グルコースを使用前に室温にする。
【0209】
実施例20: マウスにおける静脈内SNS01およびSNS-EF1/UUを使用する投与量範囲の発見および反復投与の研究
SNS01は本発明の1つの態様である - それは多発性骨髄腫の治療をターゲットとする癌療法の生物学である。SNS01は3つの成分から構成されている: elF5Aの前アポトーシス突然変異体を発現するDNAベクター (図22参照); 癌細胞の増殖/抗アポトーシスを促進する自然elF5AをターゲットとするsiRNA (図25中の配列参照); および送出ビヒクルとして作用するポリエチレンイミンと呼ばれる合成ポリマー (in vivo jetPEI; Polyplus Transfection Inc.) 。
【0210】
研究の目的は、最大許容投与量およびマウスへの静脈内SNS01の治療投与量の長期投与の可能性を決定することであった。2つの別々の研究を実施した。最大許容投与量の研究 (研究ID: MTD) は8日の研究であり、マウスに増加する量のSNS01 (2.2 mg/kgから3.7 mg/kgまで) を静脈内に2回投与し、臨床的徴候、体重、器官重量および肝臓酵素を監視して毒性を評価した。9週の反復投与研究 (研究ID: EX6) は、SNS-EF1/UUならびにその成分の2回/週の治療投与量 (1.5 mg/kg) の長期投与後に、毒性を評価するように設計された研究であった。
【0211】
SNS-EF1/UUはSNS01の前臨床的バージョンであり、SNS01複合体におけるようにB細胞特異的プロモーターよりむしろ構成的プロモーター (すべての組織においてすべての時間において発現するプロモーター) によりelF5AK50Rの発現が推進されることにおいて主として異なる。SNS01におけるB細胞特異的B29プロモーターの使用は、骨髄腫細胞を包含する、B細胞由来の細胞に前アポトーシスelF5A突然変異体の発現を限定することによって、療法の安全性を増強するように設計されている。また、EX6研究は、マウス組織においてelF5Aを抑制する毒性作用が存在するかどうかを決定するためにマウス特異的elF5A siRNAを投与したマウスのグループを含んだ。反復投与研究における毒性は、臨床的徴候、体重、血行性、肝臓酵素血行性ならびに組織病理学を監視することによって評価した。
【0212】
【表1】
【0213】
【表2】
【0214】
前臨床実験において、SNS01は0.75 mg/kg〜1.5 mg/kgの投与量で治療的であることが示された (研究EX9) 。8日の投与量範囲発見研究 (研究ID: MTD) において、治療的範囲よりも有意に高い投与量を試験して投与量領域に上限を決定した。試験物品の2回/週の静脈内投与量は2.2 mg/kgおよび2.9 mg/kgのより低い投与量で十分に許容されたが、1匹のマウスは2.9 mg/kgにおいて病的状態に到達し、安楽死させた。3.3 mg/kgまたはそれ以上投与量は、ほぼ20〜25%の生存率を生じた。したがって、最大許容投与量は2.2 mg/kg〜2.9 mg/kgであり、0.75 mg/kg〜1.5 mg/kgの治療範囲より十分に上である。
【0215】
9週の反復投与研究 (研究ID: EX6) において、マウスに治療的投与量 (1.5 mg/kg) のSNS-EF1/UUを2回/週尾静脈注射し、試験物品に関係する毒性作用は研究期間にわたって観察されなかった。また、DNAおよびsiRNAを反復して別々に試験し、マウスは両方を十分に許容した。ヒトelF5A siRNAはマウスにおいて活性ではないので、マウスelF5A特異的siRNAをまたこの研究に含めた。マウスelF5A siRNAの慢性投与に関係する毒性作用は、9週の期間に観察されなかった。これらの結果が示すように、治療的投与量のSNS01およびSNS-EF1/UUは、長期間にわたって投与したときでさえ、マウスに対して無毒である。
【0216】
【表3】
【0217】
試験システムおよび研究設計
この研究のすべての面はウォータールー・アニマル・ケアー・コミッティー大学 (the University of Waterloo Animal Care Committee) (カナダオンタリオ州ウォータールー) (設立:Canadian Council on Animal Care and the Province of Ontario Animals for Research Actによる) が設立したガイドラインに従って実施した。
【0218】
CD-1およびBALB/cマウスはチャールス・リバー・ラボラトリーズ (Charles River Laboratories) (カナダケベック州) から入手した。両方の研究のマウスに、尾静脈静脈内注射により試験物品を2回/週与えた。遅い注射 (約2〜3分) を使用して、0.2 mlより大きい体積を送出した。
8日の研究のマウスは研究の開始時にほぼ6〜9週齢であった。9週の反復投与研究のためのマウスは研究の開始時にほぼ5〜6週齢であった。
【0219】
【表4】
【0220】
【表5】
【0221】
8日最大許容投与量の研究 (MTD)
2投与量の8日研究は、SNS01の最大許容投与量を決定するように設計された投与量範囲発見研究であった。投与量範囲は2.2 mg/kg〜3.7 mg/kgであり、0.75 mg/kg〜1.5 mg/kgの治療投与量範囲よりかなり上である。SNS01の最低投与量 (2.2 mg/kg) において、わずかにしわの寄った毛皮を示し、1時間以内に消散する活性の減少を除外して、毒性の臨床的徴候は観察されなかった。第2回の2.2 mg/kgのSNS01の注射後、毒性の臨床的徴候は観察されなかった。すべてのマウスは研究を通じて彼らの体重を維持した。器官の巨視的変化は観察されなかった。器官重量/体重の比は、肝臓重量/体重の比の適度の増加を除外して、対照グループから変化しなかった。しかしながら、この比の増加はいかなるより高い投与レベルのグループにおいても観察されなかったので、試験物品に関係することはありそうもない。
【0222】
5匹のマウスのうちの4匹は2.9 mg/kgのSNS01を許容し、毒性の臨床的徴候はなかった。しかしながら1匹のマウスは注射の1時間以内に痙攣および軽症の呼吸窮迫を経験し、ヒト的に安楽死させなければならなかった。残りのマウスにおいて2回目のSNS01の注射後、毒性の臨床的徴候はなかった。マウスは研究を通じて彼らの体重を維持し、器官における巨視的変化または器官重量/体重における変化は観察されなかった。2.9 mg/kgのSNS01の2回の投与後、ALTの血清レベルのわずかの減少が存在した。
【0223】
期待されるように、3.3 mg/kgまたはそれ以上におけるSNS01の投与は十分に許容されず、両方のグループにおいて、1匹を除くすべてのマウスはヒト的に安楽死させなくてはならなかった。病的状態のためにマウスをヒト的に安楽死させなくてはならなかったすべての場合において、臨床的徴候は注射の1時間以内に現れ、高い投与量のPEIを使用した他の報告された研究と一致した。3.3 mg/kgおよび3.7 mg/kgの生存するマウスは注射後4時間以内に回復し、2回目の注射を投与しなかったが、研究を通じて彼らの重量を維持した。したがって、SNS01の最大許容投与量は2.2 mg/kg〜2.9 mg/kgであるように思われる。
【0224】
9週の反復投与量の研究 (EX6)
9週の反復投与量の研究の目的は、SNS-EF1/UU、すなわち、SNS01の開発間に前臨床的研究に使用された複合体の治療投与量 (1.5 mg/kg) の臨床的投与の安全性を評価することであった。SNS-EF1/UUはSNS01と有意に異ならず、主要な差は材料が研究等級であること、およびすべての細胞型において活性である構成的ヒトPEIプロモーターによりelF5AK50R発現が推進されることである。SNS01はB細胞特異的プロモーターを使用してelF5AK50R発現を推進させるが、この安全性の研究における構成的プロモーターの使用は非B細胞組織における突然変異体elF5AK50Rの蓄積から生ずる毒性の評価可能とする。
【0225】
9週の反復投与量の研究の他の面は、SNS-EF1/UUの個々の成分の安全性を試験するグループを含めることであった。DNAグループ (Ex6-G3) にelF5Aプラスミドおよび非ターゲッティング対照siRNAを含有する複合体を投与したが、siRNAグループ (Ex6-G4) にヒトelF5A (h5A1) および非発現プラスミドを含有する複合体を投与した。試験物品SNS-EF1/UUは内因的マウスelF5Aの発現に影響を与えないヒトelF5A siRNAを含有するが、この研究の他の特徴は非発現プラスミドとマウスelF5Aを効率よくターゲットとするsiRNAを含有するPEI複合体を投与したグループ (Ex6-G6) を含めたことであった。このグループは活性elF5A siRNAの慢性投与の安全性の評価を可能とした。
【0226】
すべての動物は予定した犠牲日まで生存した。毒性の臨床的徴候は9週の研究過程にわたっていずれのグループにおいても観察されず、すべてのグループにおけるマウスは研究期間の間に重量を増加し続けた。赤血球および白血球を治療開始後3および6週に計測し、すべての投与グループにおいて正常であった。血清肝臓酵素レベルを犠牲後に測定し、すべてのマウスについて正常範囲内であった。器官の巨視的外観の変化はいずれのグループにおいても観察されなかった。主要な器官の組織病理学的分析を独立病理学者が実施し、試験物品に帰する毒性は明らかにされなかった。
【0227】
マウスは治療投与量のSNS-EF1/UUの慢性投与を十分に許容し、悪影響は観察されなかった。さらに、マウス特異的elF5A siRNAの慢性投与は毒性作用を明らかにせず、ヒトelF5A siRNAを含有するPEI複合体の投与はヒトにとって安全であろうことが示された。
【0228】
実施例21: 多発性骨髄腫腫瘍を有するマウスに静脈内SNS-B29/UUおよびSNS01投与する治療効能研究
SNS01は上に記載した通りである。試験物品SNS-B29/UUはSNS01の前臨床的バージョンである。SNS-B29/UUはSNS01と非常にわずかに異なり、主要な差は成分がGLP等級よりむしろ研究等級であることである。報告されたこの研究の目的は、SNS-B29/UUの最小作用投与量を決定すること、およびSNS01を含んでなるGLP等級材料が前臨床的研究において使用した研究等級材料と同様に働くことを確認することであった。反復投与の腫瘍研究 (研究ID: EX9) は5週の研究であり、マウスにおける皮下腫瘍増殖を阻害する順次減少する投与量のSNS-B29/UUの能力を評価して、SNS01の最適な治療投与量を決定した。また、臨床的徴候、体重および器官重量を監視することによって、治療した動物の毒性徴候を評価した。
【0229】
【表6】
【0230】
【表7】
【0231】
皮下ヒト多発性骨髄腫腫瘍を有するSCIDマウスにおいて、SNS-B29/UUの治療範囲を決定した。0.15 mg/kg〜1.5 mg/kgのSNS-B29/UUの投与量を試験した。2回/週の腫瘍体積の測定および犠牲後の腫瘍組織の切除および秤量により、試験物品の抗腫瘍効能を決定した。0.75 mg/kg〜1.5 mg/kgのSNS-B29/UUの投与量は腫瘍を有意に収縮させ、SNS-B29/UUの治療範囲が0.75 mg/kg〜1.5 mg/kgの範囲にあることを示す。また、皮下腫瘍増殖の効率的阻害は0.38 mg/kg SNS-B29/UUにおいて観察されたが、腫瘍収縮は観察されなかった。腫瘍増殖のある程度の阻害は0.15 mg/kg程度に低い投与量においてさえ観察され、広い治療範囲を示した。図26および27参照。
【0232】
GLP等級の成分を使用して製造されたSNS01の効能をSNS-B29/UUと比較し、腫瘍増殖の阻害において匹敵する効能を有することが発見された。腫瘍を有するSCIDマウスはSNS01およびSNS-B29/UUを使用する治療に十分に許容性であり、マウスは研究を通じて重量を増加し続けた。
【0233】
【表8】
【0234】
試験システムおよび研究設計
雌C.B.17/IcrHsd-Prkdc (SCID) マウスをハーラン (Harlan) (米国インジアナ州インジアナポリス) から入手した。5〜6週齢のマウスの右側腹部に10×106の生存可能なKAS-6/1 (ヒト多発性骨髄腫) 細胞を注射して、皮下腫瘍を確立した。腫瘍がほぼ20〜40 mm3のサイズ (腫瘍細胞注射後ほぼ4週) に到達したとき、SNS-B29/UUの治療を開始した。腫瘍がほぼ130 mm3のサイズの到達したとき (腫瘍細胞注射後ほぼ6週) 、SNS01を使用する治療を開始した。マウスに尾静脈の静脈内により試験物品を2回/週投与した。
【0235】
【表9】
【0236】
反復投与腫瘍研究
SNS-B29/UUの最小有効治療投与量を決定し、そして研究等級試験物品SNS-B29/UUにより証明される腫瘍阻害活性をGLP等級SNS01試験物品が保持することを決定するように、反復投与腫瘍研究を設計した。二次的目的は、治療したマウスを臨床的徴候、体重および器官重量について治療したマウスを監視することによって、治療の毒性作用を評価することであった。デジタルキャリパーを使用して腫瘍体積を2回/週測定して、試験物品の治療抗腫瘍活性をモニターした。殺した後、腫瘍を摘出し、そして秤量した。
【0237】
すべてのマウスは予定した犠牲日まで生存した。非発現性プラスミドおよび非ターゲッティングsiRNAを含有するPEIナノ複合体で治療した対照マウスは犠牲時間に284 mm3の平均腫瘍体積を有したが、1.5 mg/kgのSNS-B29/UUで治療したマウスはわずかに13 mm3の平均腫瘍体積を有し、腫瘍増殖が95% (*p = 0.026) 減少した。しかしながら、1.5 mg/kgのSNS-B29/UUで治療したマウスから腫瘍を切除しようと試みたとき、マウスのいずれにも腫瘍の証拠は発見されなかった。SNS-B29/UUの投与量を半分の0.75 mg/kgに減少しても、腫瘍体積および重量はなおそれぞれ91% (*p = 0.03) および87% (*p = 0.04) 減少し、1匹のマウスにおいて、腫瘍は完全に消失した。したがって、SNS-B29/UUの2回/週注射のための最適な治療投与量は0.75 mg/kg〜1.5 mg/kgの間であるように思われる。0.15 mg/kg程度に低い2回/週SNS-B29/UUの投与量は最終腫瘍体積を60%減少させ、SNS-B29/UUが広い投与量範囲にわたって効力のある抗腫瘍活性を有することを示す。
【0238】
腫瘍増殖を阻害することに加えて、0.75 mg/kgおよび1.5 mg/kgのSNS-B29/UUおよびSNS01を使用する治療は腫瘍体積を有意に減少させ、この治療が、多分腫瘍中のアポトーシスの誘導により、腫瘍退縮を誘導できることを示す。0.75 mg/kgおよび1.5 mg/kgの投与量レベルのSNS-B29/UUで治療した腫瘍担持マウスにおける腫瘍体積の変化の百分率は、それぞれ、-244%および-245%であった。対照マウスの腫瘍は、同一時間の間にサイズが2000%より大きく増加した。
【0239】
また、SNS01の2回/週の注射は多発性骨髄腫腫瘍を有意に収縮させた。1.5 mg/kgのSNS01で治療したマウスの腫瘍体積の変化の百分率は-349%であり、SNS01がちょうどSNS-B29/UUのように有効であることを示した。GLP等級の材料の使用は、事実、生物学的活性を増加させることができる。なぜなら、SNS01を使用する治療はわずかに25日間の治療後腫瘍体積を349%減少させたが、SNS-B29/UUは35日間の治療後腫瘍体積を245%減少させたからである。さらに、SNS01で治療した腫瘍は非常に大きく (約130 mm3) 、SNS01を使用する治療が十分に確立された腫瘍に対して有効であることを示す。
【0240】
すべてのマウスは治療を十分に許容し、毒性の臨床的徴候は観察されなかった。すべてのグループにおけるマウスは、研究を通じて重量を増加し続けた。器官の巨視的外観は壊死において観察されず、器官重量/体重が有意に変化した。
【0241】
したがって、SNS01 (およびその前臨床的バージョンSNS-B29/UU) はSCIDマウスに十分に許容され、2回/週の静脈内注射により送出すとき、ヒト多発性骨髄腫腫瘍の皮下的治療において極めて有効である。試験したSNS-B29/UUのすべての投与量は腫瘍増殖の阻害において有効であるが、1.5 mg/kgの最高投与量は治療を受けたすべてのマウスにおいて腫瘍を首尾よく排除した。
【0242】
実施例22: プラスミドDNAおよびsiRNAポリエチレンイミン (JetPEI) 複合体の生物分布
緑色蛍光タンパク質 (“GFP”) GFP発現構築物を使用して、PEI複合体により送出されたプラスミドDNAの局在化を決定した。2つのプロモーターを使用してGFP発現を推進した: EF1: 偏在性プロモーター (EF1::GFP) またはB29: B細胞特異的プロモーター (B29::GFP) 。20μgのGFPプラスミドDNAおよび10μgの蛍光標識化 (DY547) h5A1 siRNAを含有するPEI複合体を6のN/P比で製造した。BALB/Cマウスに5%グルコースまたはPEI複合体を連続2日間静脈内注射した。最初の注射後72時間に、マウスを安楽死させ、マウスの器官を収集し、共焦点顕微鏡検査によりGFP発現およびDY547-siRNAを分析した。
【0243】
骨髄: ほとんどの場合において、DY547-siRNAの証拠は存在したが、GFP発現の証拠は存在しなかった。器官収集のタイミングはGFPのピーク発現と一致せず、GFPシグナルのクエンチングは存在しうるか、あるいはGFPは発現されることはないであろう。しかしながら、ある場合において同一骨髄細胞に共局在化したGFPおよびDY547が観察された。したがって、これはPEIナノ粒子が静脈内注射により与えられたとき生きている動物中の骨髄細胞をトランスフェクトできるという証拠を提供する。
【0244】
肺: ほとんどの場合において、DY547-siRNAの証拠は存在したが、GFP発現の証拠は存在しなかった。器官収集のタイミングはGFPのピーク発現と一致せず、GFPシグナルのクエンチングは存在しうるか、あるいはGFPは発現されることはないであろう。
脾臓: またDY547-siRNAの存在に陽性である細胞中に共局在化するGFP発現の証拠 (PEIプロモーターより推進されるとき) が見られた。B29プロモーターにより推進されるとき、GFPの発現は脾細胞において非常に低い。これが示すように、PEIナノ粒子は脾臓の細胞をトランスフェクトするようである。
【0245】
腎臓: GFPまたはDY547は観察されず、ナノ粒子が腎臓に入らないことを示した。
肝臓: ほとんどの場合においてDY547-siRNAの証拠は存在したが、GFP発現は存在しなかった。これはPEIナノ粒子が肝臓の細胞をトランスフェクトしている証拠を提供する。
心臓: 心臓組織中のEF1::GFPおよびDY547-siRNAの共局在化が見られ、こうしてPEIナノ粒子がこの器官をトランスフェクトできること示す。GFPはB29プロモーターとともに観察されなかった。
【0246】
実施例23: HA-elF5AK50R発現に対するDNA: siRNA比の効果
B29-HA-elF5AK50R (B細胞特異的プロモーターにより推進されるプラスミド) およびh5A1 siRNAを含有するナノ粒子でKAS細胞をトランスフェクトした。異なる比でpExp5Aおよびh5A1 siRNAを含有するJET PEITMナノ粒子を作り、室温において4時間インキュベートした後、KAS細胞に添加した。トランスフェクション後4時間に、ナノ粒子を含有する培地を新鮮な培地と交換した。24時間後、細胞ライゼイトを収集し、HAに対する抗体とともにウェスタンブロット分析のために使用した。DNA: siRNAの比を2:1の標準的比から変化させた。HA-elF5AK50R蓄積は1:0、3:1および2:1においてピークとなった。図30参照。
【0247】
実施例24: ナノ粒子のトランスフェクションにより誘導されるアポトーシスに対するDNA: siRNAの比の効果
pExp5Aおよびh5A1 siRNAを異なる比で含有するナノ粒子を作り、室温において4時間インキュベートした後、KAS細胞に添加した。トランスフェクション4時間後、ナノ粒子を含有する培地を新鮮な培地で置換した。48時間後、細胞を収集し、アンキシンV/PIで標識化し、FACSにより分析した。アポトーシスの誘導は、2:1の標準的DNA: siRNA比のナノ粒子でトランスフェクトした細胞において最高であった。図31参照。
【0248】
実施例25: elF5AK50RプラスミドおよびeIF5A1 siRNA (siSTABLEまたは非siSTABLE) を含有するPEI複合体 (N/P = 6または8) の投与は多発性骨髄腫皮下腫瘍の増殖を阻害し、腫瘍を収縮させる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。マウスに下記を2回/週静脈内注射した: (G1) 20 mgのpCpG-mcs (空ベクター) および10 mgの対照siRNAをN/P = 8で含有するPEI複合体 (中程度の投与量); (G5) 20 mgのRNAi抵抗性プラスミドpCpG-elF5AK50Rおよび10 mgのsiSTABLE h5A1 siRNAをN/P = 8で含有するPEI複合体 (中程度の投与量、siSTABLE); (G8) 20 mgのRNAi抵抗性プラスミドpCpG-elF5AK50Rおよび10 mgのh5A1 siRNAをN/P = 6で含有するPEI複合体 (中程度の投与量、N/P = 6) 。示したデータは各グループにおけるマウスについての個々の腫瘍体積である。最初の治療後第40日に、最終の注射を実施した。図32参照。
【0249】
実施例26: 腫瘍組織はJET PEITMナノ粒子を効率よく吸収し、ナノ粒子はプラスミドおよびsiRNAを同一細胞に送出す。
pExp-GFP (B細胞特異的プロモーターの制御下のGFP) およびDY547-siRNA (蛍光標識化siRNA) を含有するナノ粒子を注射した後48時間に、腫瘍を切断した。共焦点顕微鏡検査によりGFPおよびDY547の共局在化発現が観察され、腫瘍組織はナノ粒子を効率よく吸収し、ナノ粒子はプラスミドおよびsiRNAを同一細胞に送出していることを示される。図33参照。
【技術分野】
【0001】
発明の分野
本発明は、内因的遺伝子の発現をノックダウンまたはノックアウトするための内因的遺伝子をターゲットとするsiRNAの組合わせ使用、およびその遺伝子を発現させるための外因的遺伝子をコードするポリヌクレオチドの使用に関する。
【背景技術】
【0002】
比較的最近において、タンパク質の発現を阻害するために二本鎖RNA (“dsRNA”) を使用できることを研究者らは観察した。遺伝子を阻害する能力は人間の疾患を治療する広い可能性を有し、そして多数の研究者らおよび商業的エンティティーは現在この技術に基づく療法の開発にかなりの財源を投じている。
【0003】
遺伝子サイレンシングを誘導する二本鎖RNAは少なくとも3つのレベルで発生することができる: (i) 転写不活性化、これはRNAガイドDNAまたはヒストンメチル化を意味する; (ii) mRNA分解を誘導するsiRNA; および (iii) 転写アテニュエーションを誘導するmRNA。
【0004】
哺乳動物細胞においてサイレンシング (RNAインターフェレンス、またはRNAi) を誘導するRNAの主要な機構はmRNA分解であることが一般に考えられる。哺乳動物細胞においてRNAiを使用する初期の試みはdsRNAの長い鎖の使用に集中した。しかしながら、RNAiを誘導するこれらの試みは、部分的にインターフェロンの応答の誘導のために、成功が制限され、これは一般にターゲット特異的と反対に、タンパク質合成の阻害を生ずる。こうして、長いdsRNAは哺乳動物系におけるRNAiのための価値のあるオプションではない。
【0005】
より最近において、短い (18〜30 bp) RNAデュプレックスを培養において哺乳動物細胞中に導入するとき、ターゲットmRNAの配列特異的阻害はインターフェロンの応答を誘導しないで実現できることが示された。これらの短いdsRNAのあるもの、小さい阻害性RNAs (“siRNAs”) と呼ぶ、は1モル以下の濃度において触媒的に作用して細胞中のターゲットmRNAの95%より多くを切り離すことができる。
【0006】
siRNA活性についての機構の説明、ならびにその用途のいくつかは、下記の文献に記載されている: Provost他 (2002) Ribonuclease Activity and RNA Binding of Recombinant Human Dicer、EMBO J. 21 (21): 5864-5874; Tabara他 (2002) The dsRNA Binding Protein RDE-4 Interacts with RDE-1、DCR-1 and a DexH-box Helicase to Direct RNA in C. elegans、Cell 109 (7): 861-71; Ketting他 (2002) Dicer Functions in RNA Interference and in Synthesis of Small RNA Involved in Developmental Timing in C. elegans; Martinez他、Single-Stranded Antisense siRNAs Guide Target RNA Cleavage in RNAi、Cell 110 (5): 563; Hutvagner & Zamore (2002) A microRNA in a multiple-turnover RNAi enzyme complex、Science 297: 2056。
【0007】
機械的見解から、植物および無脊椎動物の細胞中への長い二本鎖RNAの導入は、ダイサー (Dicer) として知られているIII型エンドヌクレアーゼによりsiRNAに破壊される。Sharp、RNA interference - 2001、Genes Div. 2001、15: 485。ダイサー、リボヌクレアーゼ-III様酵素はdsRNAをプロセシングして、19〜23塩基対の短い干渉性RNAにし、これらのRNAは特徴ある2塩基の3’ オーバーハングを有する。Bernstein、Caudy、Hammond & Hannon (2001) Role for a bidentate Ribonuclease in the initiation step of RNA interference、Nature 409: 363。
【0008】
次いでsiRNAはRNA誘導サイレンシング複合体 (RISC) 中に組込まれ、ここで1または2以上のヘリカーゼはsiRNAデュプレックスを巻戻し、相補的アンチセンス鎖がターゲットを認識できるようにする。Nykanen、Halve & Zamore (2001) ATP requirements and small interfering RNA structure in the RNA interference pathway、Cell 107: 309。適当なターゲットmRNAに結合すると、RISC内の1または2以上のリボヌクレアーゼはターゲットを切り離してサイレンシングを誘導する。Elbashir、Lendeckel & Tuschl (2001) RNA interference is mediated by 21 - and 22 - nucleotide RNAs、Genes Dev. 15: 188、図1。
【0009】
インターフェレンス効果は長く持続することができ、そして多数回の細胞分裂後に検出することができる。その上、RNAiは配列特異性を示す。Kisielow M.他(2002) Isoform-specific knockdown and expression of adaptor protein ShcA using small interferon RNA、J. Biochem. 363: 1-5。こうして、RNAi機構は転写の1つの型を特異的にノックダウンできるが、密接に関係するmRNAに影響を与えない。siRNAはこれらの性質を有するために、遺伝子発現を阻害し、遺伝子機能および薬物ターゲット確認を研究する潜在的に価値のある道具である。その上、siRNAは下記に対する治療剤として潜在的に有効である: (1) 遺伝子の過剰発現または異常発現により引き起こされる疾患; および (2) 突然変異を含有する遺伝子の発現により発生する疾患。
【0010】
有望なsiRNA従属的サイレンシングは多数の因子に依存する。RNAiにおける最も議論を呼びそうな問題の1つは、siRNA設計の必要性の問題、すなわち、使用するsiRNAの配列を考慮することである。C. elegansおよび植物における初期の研究は長いdsRNAの導入により設計の問題を回避した (例えば、下記の文献を参照のこと、Fire A. 他 (1998) Nature 391: 806-811) 。この原始生物において、長いdsRNA分子はダイサーによりsiRNAに切断され、こうして潜在的に全体の転写物をカバーできるデュプレックスの多様な集団を発生させた。
【0011】
これらの分子のいくつかの小部分は非機能的である (すなわち、サイレンシングをほとんど、あるいはまったく誘導しない) が、1または2以上は高度に機能的である可能性を有し、これにより問題の遺伝子をサイレンシングし、siRNA設計の必要性を軽減する。不都合なことには、インターフェロン応答のために、この同一のアプローチは哺乳動物系に利用不可能である。この効果はダイサー切断をバイパスし、siRNAを直接導入することによって回避できるが、この計画は選択したsiRNA配列が非機能的または半機能的であるという危険を伴う。
【0012】
siRNA設計はRNAiのきわめて重要な要素ではないという見解を多数の研究が示してきている。他方において、この分野における他の研究は、siRNAの設計に注意を払うことによってRNAiをいっそう効率的とすることができる可能性を探求し始めた。
【0013】
種々の疾患または障害を治療するために、ある種のタンパク質をアップレギュレートすることは望ましいが、これは必要であるすべてのことではないことがある。例えば、内因的タンパク質または異なるタンパク質の発現をノックダウンまたはノックアウトするために、siRNAの組合わせ使用は必要であることがある。本発明はこの要求を満足させ、そして癌、特に多発性骨髄腫を治療する方法を提供する。
【0014】
多発性骨髄腫を含む癌は、アポトーシスを誘導する能力から利益を受ける疾患である。多発性骨髄腫の慣用療法は、化学療法、幹細胞移植、幹細胞移植を伴う高投与量の化学療法、およびサルベージ療法を含む。化学療法は、Thalomid(商標)(タリドミド) 、ボルテゾミド、Aredia(商標)(パミドロネート) 、ステロイド、およびZometa(商標)(ゾレドロン酸) を使用する治療を含む。しかしながら多数の化学療法剤は毒性であり、非癌細胞、例えば骨髄、胃腸の内層、および毛胞を活発に分割する。したがって、化学療法は血球数の減少、悪心、嘔吐、下痢、および毛髪喪失を生ずることがある。
【0015】
典型的には、慣用化学療法または標準的投与量の化学療法は、多発性骨髄腫の患者のための一次的または初期の治療である。高投与量の化学療法および幹細胞移植のための調整において、患者はまた化学療法を受けることがある。移植前に存在する腫瘍を減少させるために、誘導療法 (幹細胞移植前の慣用化学療法) を使用できる。ある種の化学療法剤は他の薬剤よりも誘導療法のために適当である。なぜなら、それらは骨髄細胞に対する毒性が低く、骨髄からの幹細胞の収量を大きくするからである。誘導療法のために適当な化学療法剤の例は次の通りである: デキサメタゾン、タリドミド/デキサメタゾン、VAD (ビンクリスチン、Adriamycin(商標)(ドキソルビシン) およびデキサメタゾンの組合わせ) 、およびDVd (ペギル化リポソームドキソルビシン (Doxil(商標)、Caelyx(商標)) 、ビンクリスチンおよび還元スケジュールデキサメタゾンの組合わせ) 。
【0016】
多発性骨髄腫の標準的治療は、メルファランとプレドニソン (コルチコステロイド薬剤) との組合わせであり、50%の応答比率を達成する。不都合なことには、メルファランはアルキル化剤であり、誘導療法にそれほど適当ではない。コルチコステロイド (ことにデキサメタゾン) は、特により高齢の患者および化学療法に耐えることができない患者において、多発性骨髄腫の療法に時々単独で使用される。また、デキサメタゾンは、単独でまたは他の薬剤との組合わせで、誘導療法において使用される。VADは誘導療法において最も普通に使用されるが、最近誘導療法に有効であることが示された。ボルテゾミドは最近多発性骨髄腫の治療のために承認されたが、非常に毒性である。しかしながら、現存する治療剤はいずれも十分な治癒の可能性を提供しない。こうして、癌および多発性骨髄腫のための適当な治療がなお要求されている。本発明はこの要求を満足する。
【発明の概要】
【0017】
本発明は、宿主における内因的遺伝子の発現をノックアウトまたはノックダウンするための内因的遺伝子をターゲットとするsiRNAの組合わせ使用、および送出ビヒクル/発現ベクター中の遺伝子をコードするポリヌクレオチドを送出して、ポリヌクレオチドによりコードされるタンパク質を宿主において発現させることに関する。正常 (非不完全) タンパク質をコードするポリヌクレオチド (またはタンパク質それ自体) を宿主に投与し、発現させ (ポリヌクレオチドの場合において) 、こうして正常タンパク質がその必要な機能を実行できるようにする。siRNAは好ましくは遺伝子のある領域をターゲットするように設計し、それが不完全なタンパク質の内因的発現をノックダウンまたはノックアウトするが、同時に正常タンパク質をコードする投与したポリヌクレオチドの外因的発現に影響を与えないようにする。
【0018】
本発明は、eIF5A1の3’ 末端をターゲットとするeIF5A1 siRNAの複合体と、突然変異体eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターとを含んでなる組成物であって、突然変異体eIF5A1はハイプシン化 (hypusinate) されることができず、かつsiRNAおよび発現ベクターはポリエチレンイミンに対して複合化して複合体を形成している組成物を提供する。
【0019】
本発明は、ターゲット遺伝子をターゲットとして被検体におけるターゲット遺伝子の内因的発現を抑制するsiRNAと、被検体において発現されることができるターゲットタンパク質をコードするポリヌクレオチドとを含んでなる組成物を提供する。ある種の態様において、ポリヌクレオチドはRNAI抵抗性プラスミド中に存在する (siRNAにより抑制されないであろう) 。siRNAおよびプラスミドはポリエチレンイミンに対して複合化して複合体を形成している。
【0020】
ある種の態様において、siRNAは図25に示す配列を有し、ここで突然変異体eIF5A1をコードするポリヌクレオチドはeIF5A1K50Rである。発現ベクターは突然変異体eIF5A1をコードするポリヌクレオチドと、被検体におけるポリヌクレオチドの発現を提供するように作用可能に連鎖されたプロモーターとを含んでなる。プロモーターは好ましくは組織特異的または系統的である。例えば、癌を治療するために組成物を使用する場合、プロモーターは癌が存在する組織に対して組織特異的であることが好ましい。例えば、多発性骨髄腫を治療するために、B細胞特異的プロモーター、例えばB29を使用することが好ましい。ある種の態様において、発現ベクターはpCpGプラスミドを含んでなる。
【0021】
ある種の態様において、eIF5A1 siRNAと突然変異体eIF5A1ポリヌクレオチドを含んでなる発現ベクターは独立してポリエチレンイミン、例えばin vivo JetPEITMに対して複合化している。他の態様において、eIF5A1 siRNAと突然変異体eIF5A1ポリヌクレオチドを含んでなる発現ベクターは一緒になってポリエチレンイミンに対して複合化している。
【0022】
さらに、本発明は、eIF5A1の3’ 末端をターゲットとするeIF5A1 siRNAと、突然変異体eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターとを含んでなる組成物であって、突然変異体eIF5A1はハイプシン化されることができず、かつsiRNAおよび発現ベクターは癌を治療するために被検体に送出される組成物を提供する。癌は多発性骨髄腫を包含する任意の癌であることができる。
【0023】
さらに、本発明は、本発明の組成物を被検体 (哺乳動物およびヒトを包含するが、これらに限定されない) に投与することを含んでなる癌を治療する方法を提供する。
【0024】
組成物は任意の許容される経路、例えば静脈内、腹腔内または腫瘍内であるが、これらに限定されない経路で投与できる。siRNAおよび発現ベクターは異なる時間に異なる経路を経て投与できるか、あるいは同時に同一経路を経て一緒に投与できる。例えば、限定されないが、siRNAはそのままでまたは担体、例えばin vivo JetPEITMに対して複合化して静脈内に送出すことができ、そして発現ベクターは腫瘍内に投与ことができるか、あるいはsiRNAおよび発現ベクターの両方は静脈内または腫瘍内に投与することができる、およびその他。
【0025】
本発明は、癌細胞の成長を阻害しおよび/または癌細胞を殺す方法を提供する。また、本発明は、癌細胞の転移能力を阻害または遅延させる方法を提供する。癌成長の阻害は、腫瘍サイズの減少、腫瘍成長の低下を包含し、そしてまた腫瘍の完全な寛解を包含することができる。癌は任意の癌または腫瘍であることができ、結腸癌、結腸直腸癌、膀胱癌、頸部腺癌および肺癌を包含するが、これらに限定されない。好ましくは、癌は多発性骨髄腫である。
【0026】
好ましい態様において、elF-5Aはハイプシン化されることができない突然変異体である。典型的な突然変異体は本明細書において説明されている。
elF-5AまたはelF-5Aをコードするポリヌクレオチド (elF-5Aを発現させるために) を被検体に提供することに加えて、siRNAを提供してelF-5Aの内因的発現をノックアウトまたはノックダウンする。
【0027】
また、本発明は、多発性骨髄腫の患者における多発性骨髄腫細胞を殺す薬剤を製造するために、elF5A、eIF5A1をコードするポリヌクレオチドおよびeIF5A1に対するsiRNAを使用することに関する。好ましくは、突然変異体elF-5Aをコードするポリヌクレオチドはハイプシン化されることができない。
【0028】
また、本発明は、鎌状白血球貧血を治療する方法を提供する。 鎌状白血球貧血を患っている患者に、健康なヘモグロビン遺伝子 (例えば、HBB) をコードするポリヌクレオチドを投与する。また、不完全なヘモグロビン遺伝子をコードする遺伝子 (例えば、突然変異体HbSをコードする遺伝子) をターゲットとするsiRNAを患者に投与して、不完全なタンパク質の発現をノックダウンまたはノックアウトする。さらに、本発明は、鎌状白血球貧血の治療において普通に使用される薬剤または治療を施すことを含むことができる。
【図面の簡単な説明】
【0029】
【図1】図1は、ヒトeIF-5A1のアミノ酸配列を提供し、種々の重要部位を示す。
【図2】図2は、K50およびK67におけるeIF-5A1の突然変異がトランスフェクトされたタンパク質の蓄積を増加させることを示す。実施例1参照。
【図3】図3は、K47、K50およびK67におけるeIF5A1の突然変異がトランスフェクトされたタンパク質の蓄積を増加させることを示す。実施例2参照。
【0030】
【図4】図4は、KAS細胞中にトランスフェクトされたとき、K50およびK67におけるeIF5A1の突然変異がアポトーシスを誘導させることを示す。実施例3参照。
【図5】図5は、KAS細胞中にトランスフェクトされたとき、K50およびK67におけるeIF5A1の突然変異がアポトーシスを誘導させることを示す。実施例4参照。
【図6】図6は、KAS細胞中にトランスフェクトされたとき、K50およびK67におけるeIF5A1の突然変異がアポトーシスを誘導させることを示す。実施例5参照。
【0031】
【図7A】図7Aは、siRNAを使用するトランスフェクションおよび、KAS細胞におけるアポトーシス生ずる、eIF-5A1を発現するように修飾されたアデノウイルスを使用する治療を示す。実施例6A参照。
【図7B】図7Bは、eIF5A1 siRNA (ターゲット#1 (配列番号1)) (図25に示すsiRNA構築物の配列) を使用する前処理が内因的eIF5A1の発現を減少させたが、アデノウイルスが発現したRNAi抵抗性eIF5A1K50Aの蓄積を可能とすることを示す。実施例6B参照。
【0032】
【図7C】図7Cは、アデノウイルス感染前のターゲット#1に対するeIF5A1 siRNAを使用する前処理がヒト多発性骨髄腫細胞におけるリン酸化NF-kBの発現を減少させることを示す。実施例6C参照。
【図7D】図7Dは、アデノウイルス感染前のターゲット#1に対するeIF5A1 siRNAを使用する前処理がヒト多発性骨髄腫細胞におけるリン酸化NF-kB及びICAM-1の発現を減少させることを示す。実施例6D参照。
【図7E】図7Eは、ヒト多発性骨髄腫細胞におけるelF5AのsiRNA仲介抑制がNF-kB DNA結合活性のLPS仲介誘導を抑制することを示す。elF5A siRNAによるNFkB活性の阻害は、elF5AK50Rの過剰発現と組合わせたとき、アポトーシスの誘導を増加するその能力を説明する。なぜなら、NF-kBは多数の前生存率および抗アポトーシス経路を調節するからである。
【0033】
【図7F】図7Fは、siRNAを使用するKAS細胞の前処理がIL-6の存在下のeIF5A1K50R遺伝子送出によるアポトーシスを増加させることを示す。実施例6E参照。
【0034】
【図8】図8は、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の成長を遅延させることを示す。示したデータは各グループにおけるすべてのマウスについての腫瘍体積である。実施例7参照。
【図9】図9は、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の成長を遅延させることを示す。示したデータは平均腫瘍体積/グループ±標準誤差である。実施例7参照。
【0035】
【図10】図10は、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の重量を減少させることを示す。実施例7参照。
【図11】図11は、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の成長を遅延させ、腫瘍を収縮させることを示す。実施例8参照。
【図12】図12は、eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の腫瘍内 (i.t.) 投与が多発性骨髄腫皮下腫瘍の腫瘍収縮を生じさせることを示す。実施例9参照。
【0036】
【図13A】図13Aは、eIF5A1プラスミドおよびeIF5A1 siRNAを使用する治療が多発性骨髄腫皮下腫瘍の成長を遅延させ、腫瘍を収縮させることを示す。
【図13B】図13Bは、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が腫瘍の収縮を生じさせることを示す。
【図13C】図13Cは、eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の腫瘍内 (i.t.) 投与が多発性骨髄腫皮下腫瘍の腫瘍収縮を生じさせることを示す。
【0037】
【図14】図14は、eIF5A1プラスミドおよびeIF5A1 siRNAの静脈内共投与が多発性骨髄腫皮下腫瘍の成長を遅延させることを示す。実施例10参照。
【図15】図15は、eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の静脈内 (i.v.) または腹腔内 (i.p.) 投与が多発性骨髄腫皮下腫瘍の成長を遅延させることを示す。実施例11参照。
【図16】図16は、eIF5A1プラスミドおよびeIF5A1 siRNAを使用する治療が多発性骨髄腫皮下腫瘍の成長を遅延させることを示す。
【0038】
【図17】図17は、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の成長を遅延させ、腫瘍を収縮させることを示す。実施例12参照。
【図18】図18は、eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の腫瘍内 (i.t.) 投与が多発性骨髄腫皮下腫瘍の腫瘍収縮を生じさせることを示す。実施例13参照。
【図19】図19は、EF1またはB29プロモーターのいずれかにより推進されたeIF5A1K50Rプラスミド、およびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の増殖を遅延させ、腫瘍を収縮させることを示す (KAS-SQ-5) 。実施例14参照。
【0039】
【図20】図20は、eIF5A1 siRNAの共投与がEF1またはB29プロモーターいずれかにより推進されるeIF5A1K50Rプラスミドの多発性骨髄腫皮下腫瘍に対する抗腫瘍作用を増加させ、存在する腫瘍を減少させることを示す (KAS-SQ-5) 。実施例15参照。
【図21】図21は、eIF5A1 siRNAが肺腺癌細胞におけるAd-elF5Aによる感染から生ずるアポトーシス誘導を相乗的に増加させる。実施例16参照。
【図22】図22はpExp5Aの地図を示し、その構築は実施例17に記載する。
【0040】
【図23】図23はpExp5A (3371 bp) の予測される配列を示す。実施例17参照。
【図24】図24は、種々の細胞系統におけるeIF5A1K50Rの再発現を示す。実施例18参照。
【図25】図25は、ターゲット配列および好ましいeIF5A1 siRNAの配列を示す。
【図26】図26は、多発性骨髄腫における効能研究の結果を提供する。実施例21参照。
【図27】図27は、多発性骨髄腫における効能研究の結果を提供する。実施例21参照。
【0041】
【図28】図28は、eIF5A1K50R cDNAの配列を提供する。
【図29】図29は、ヒトeIF5A1K50Rに対するヒトeIF-5A1のアラインメントを提供する。
【図30】図30は、HA-eIF5A1K50R発現に対するDNA: siRNA比の効果を示す。実施例23参照。
【図31】図31は、ナノ粒子トランスフェクションより誘導されるアポトーシスに対するDNA: siRNA比の効果を示す。実施例24参照。
【0042】
【図32】図32は、eIF5A1K50RプラスミドおよびeIF5A1 siRNA (siSTABLEまたは非siSTABLE) を含有するPEI複合体 (N/P = 6または8) の投与が多発性骨髄腫皮下腫瘍の成長を阻害し、腫瘍収縮を生じさせることを示す。実施例25参照。
【図33】図33は、JET PEITMナノ粒子が腫瘍組織により効果的に吸収されること、およびナノがプラスミドおよびsiRNAを同一細胞に送出していることを示す。実施例26参照。
【発明を実施するための形態】
【0043】
発明の詳細な説明
本発明は、被検体における内因的遺伝子の発現をノックアウトまたはノックダウンするための内因的遺伝子をターゲットとするsiRNAと、ポリヌクレオチドによりコードされるタンパク質の宿主における発現を提供するために被検体に提供される送出ビヒクル/発現ベクター中の遺伝子をコードするポリヌクレオチドとの組合わせた使用に関する。
【0044】
この組成物は不完全なまたは突然変異体のタンパク質の存在により引き起こされる疾患または症状を有する被検体の治療において有効である、すなわち、被検体において生産されるタンパク質がその必要な機能を実行できないか、あるいはその構造が不完全なために代謝経路または生体分子の相互作用を失う場合において有効である。siRNAは不完全なタンパク質をコードする遺伝子をターゲットとし、その不完全なタンパク質の発現をノックダウンまたはノックアウトするように設計されている。正常 (不完全でない) タンパク質をコードするポリヌクレオチドを被検体に投与し、その被検体において発現させて、正常タンパク質がその必要な機能を実行できるようにする。
【0045】
他の態様において、所望のタンパク質をコードするポリヌクレオチドを投与する代わりに、タンパク質を被検体に投与する。用語「タンパク質」、「ペプチド」および「ポリペプチド」は本明細書中で互換的に使用される。
【0046】
siRNAは遺伝子のある領域をターゲットとし、不完全なタンパク質の内因的発現をノックダウンまたはノックアウトするが、同時に正常タンパク質をコードする投与したポリヌクレオチドの外因的発現に影響を与えないことが好ましい。例えば、siRNAは3’ UTRをターゲットとし、投与したセンス構築物 (タンパク質をコードするポリヌクレオチド) の外因的発現に影響を与えないようにすることができる。不完全な遺伝子の外因的発現をノックダウンまたはノックアウトすることによって、外因的ポリヌクレオチドから発現された正常タンパク質と競合する不完全なタンパク質はほとんど、あるいはまったく存在しないであろう。
【0047】
この適用が有効である疾患の状態の1例は鎌状白血球貧血に関する。鎌状白血球貧血は、ヘモグロビン、すなわち、体全体を通じて酸素の運搬を促進する赤血球 (RBCs) 中に見出されるタンパク質である。
【0048】
ある人が突然変異体ヘモグロビン (Hbs) の発現を生ずる2つの異常な遺伝子 (各親から1つ) を受け継ぐとき、鎌状白血球貧血が起こる。突然変異体ヘモグロビンはRBCsの形状を変化させる。正常ヘモグロビン (ヘモグロビンAまたはHbA) を有する赤血球は血流中を通して容易に動き、体のすべての細胞に酸素を送出す。赤血球は非常に細い血管でさえ「押分けて進む」ことができる。ヘモグロビンの異常形態 (HbS) は一緒に凝集し、赤血球を粘着性、剛性、より脆弱とし、そしてそれらを湾曲した、鎌形にする傾向がある。
【0049】
数百のHBB遺伝子変異型は知られているが、鎌状白血球貧血は最も普通にヘモグロビン変異型HbSにより引き起こされる。この変異型において、疎水性アミノ酸バリンはHBBポリペプチド鎖の第6アミノ酸位置に親水性グルタミン酸の位置を取る。この置換はタンパク質構造の外側に疎水性スポットをつくり、これは隣接するヘモグロビン分子のβ鎖の疎水性領域に粘着する。HbS分子の剛性線維へのこの凝集 (重合) は、赤血球の「鎌形化」を引き起こす。
【0050】
赤血球が体全体を通じて種々の組織に運んだ酸素を赤血球が放出した後、重合は起こる。いったん赤血球は肺に戻り、ここでヘモグロビンは酸素に結合することができ、HbS分子の長い線維は解重合または分解して単一分子になる。重合と解重合との間のサイクリングは赤血球の膜を剛性にさせる。これらの赤血球が酸素を運んでないときの、赤血球の剛性およびそれらの変形した形状は小さい血管を遮断することがある。この遮断は短期間の疼痛発現を引き起こし、器官を損傷させることがある。
【0051】
鎌状白血球貧血は、常染色体劣性遺伝障害である。疾患を発現させるために、人は2コピーのHbS変異型または1コピーのHbSおよび1コピーの他の変異型を受け継がなくてはならない。キャリヤーは1コピーの正常HBB遺伝子 (HbA) および1コピーのHbSを有し、鎌状白血球の特徴を有するとして記載され、疾患の症状を発現しない。
【0052】
こうして、本発明の1つの態様は鎌状白血球貧血の患者を治療する方法を提供する。HBB遺伝子をターゲットとするsiRNAを患者に投与する。siRNAはヘモグロビンのHbs変異型の発現をノックダウンし、好ましくはノックアウトするように設計する。正常ヘモグロビンをコードするポリヌクレオチドは被検体に提供して、被検体が正常ヘモグロビンを発現するようにする。また、siRNAは正常ヘモグロビンをコードする外因的ポリヌクレオチドの発現を妨害しないように、設計される。こうして、被検体はもはや変異型ヘモグロビンを作らず (または実質的により少なくする) 、その代わりに正常な健康的ヘモグロビンを作り、正常に機能する、いっそう正常な赤血球を生ずる。
【0053】
本発明は、また、翻訳後修飾がタンパク質に対して起こる状況において有効であり、この翻訳後修飾は疾患の状態を引き起こすか、あるいはその状態に導く。siRNAを使用して内因的タンパク質の発現をノックダウンするので、翻訳後修飾はほとんど、あるいはまったく利用不可能である。次いで、タンパク質をコードするポリヌクレオチドを外因的発現のために患者に提供する。タンパク質は修飾されるので、翻訳後に修飾されることはできない。次いでこのタンパク質は適当な使用のために体に利用可能であるが、翻訳後に修飾されることはできないので、疾患の状態に導かないであろう。
【0054】
当業者は異なる翻訳後修飾を理解するであろう。例えば、翻訳後、アミノ酸の翻訳後修飾は、アミノ酸の化学的特質 (例えば、シトルリン化) を変化させるか、あるいは構造的変化、例えばジサルファイド架橋の形成を行うことによって、他の生化学的官能基、アセテート、ホスフェート、種々の脂質および炭水化物をアミノ酸に結合させることによって、タンパク質機能の範囲を拡大する。また、酵素はタンパク質のアミノ末端からアミノ酸を除去するか、あるいはタンパク質鎖を中央において切断することができる。
【0055】
例えば、ペプチドホルモンのインスリンはジサルファイド結合が形成された後2回切断され、そしてポリペプチドは鎖の中央から除去される; 生ずるタンパク質はジサルファイド結合により結合された2つのポリペプチド鎖から成る。また、最も新生のポリペプチドはアミノ酸メチオニンで開始する。なぜなら、mRNA上の「開始」コドンもまたこのアミノ酸をコードするからである。このアミノ酸は通常翻訳後修飾の間に除去される。リン酸化のような、他の修飾はタンパク質の挙動をコントロールする、例えば、酵素を活性化または不活性するための普通の機構の一部分である。他の翻訳後修飾は、デオキシハイプシンシンターゼ (DHS) による真核開始因子5A (elF5A) のハイプシン化を包含する。
【0056】
こうして、本発明は、タンパク質をコードするポリヌクレオチドを患者に投与し、患者において発現させる、被検体における遺伝子の発現を変更する方法を提供する。タンパク質は正常/野生型タンパク質または突然変異したタンパク質であることができる。対応する内因的遺伝子の発現は、被検体に投与されたsiRNAで抑制される。
【0057】
この方法は、さらに、ポリヌクレオチドが被検体において発現されてターゲットタンパク質を産生する、ターゲットタンパク質をコードするポリヌクレオチドを含んでなる構築物を提供することを含んでなる。内因的遺伝子が不完全なタンパク質を発現するある種の態様において、正常/健康なタンパク質をコードするようにポリヌクレオチドを設計する。siRNAを投与して不完全な内因的タンパク質の発現を抑制する。内因的遺伝子が正常の健康なタンパク質を発現するある種の態様において、正常/健康なまたは非突然変異体のタンパク質を使用するとき起こるような、転写後修飾されることができない突然変異体タンパク質をコードするように、ポリヌクレオチドを設計する。siRNAを投与して内因的タンパク質の発現を抑制して、このタンパク質が転写後修飾に利用されることが少ないようにする。
【0058】
ある種の態様において、内因的ポリヌクレオチドの発現に影響を与えないように、遺伝子の領域をターゲットするようにsiRNAを選択または設計する。例えば、siRNAは3’ UTRまたは3’ 末端をターゲットとすることができる。
【0059】
siRNAは裸siRNAまたは血清のために安定化した裸siRNAとして患者に送出すことができる。siRNAは全身的に、すなわち、IPまたはIV注射することができる。選択的に、siRNAは体の所望の区域に局所的に注射しまたは送出すことができる。ある種の態様において、siRNAは送出ビヒクルで投与することができ、送出ビヒクルの例はデンドリマー、リポソームまたはポリマーであるが、これらに限定されない。
【0060】
所望のタンパク質をコードするポリヌクレオチドは、ヌクレオチドの発現を提供または可能とする任意の送出手段により、投与することができる。用語「ポリヌクレオチド」または「ヌクレオチド」は本明細書中で互換的に使用される。送出は任意のウイルスまたは非ウイルス機構により実施することができ、このような機構はプラスミド、発現ベクター、ウイルス構築物、アデノウイルス構築物、デンドリマー、リポソームまたはポリマーであるが、これらに限定されない。
【0061】
ある種の態様において、CpGジヌクレオチドが減少した発現プラスミドを使用してポリヌクレオチドを発現させる。ポリヌクレオチドの発現を促進できる任意のプロモーターを使用することができ、これは療法に所望の適用に基づいて選択することができる。例えば、多発性骨髄腫を殺すために、B細胞に対して特異的なプロモーター、例えばヒトB29プロモーター/エンハンサーは望ましい。他の態様において、プロモーターは他の組織特異的プロモーターであることができるか、あるいは系統的プロモーターであることができる。
【0062】
ターゲットタンパク質をコードするポリヌクレオチドは、IVまたは皮下注射、または他の任意の生物学的に適当な送出機構により送出すことができる。
選択的に、ポリヌクレオチドはリポソームにより送出すことができるか、あるいはDNA (またはプラスミドまたは発現ベクター) をターゲット腫瘍または癌細胞に送出す任意の他の適当な「担体」または「ビヒクル」により送出すことができる。例えば、合成DNA送出系の概観についての下記を参照のこと: Luo、Dan 他、Nature Biotechnology、Vol. 18、January 2000、pp. 33-37。
【0063】
こうして、ナノメートルサイズのビヒクル、例えばリポソーム、デンドリマーまたは同様な非毒性ナノ粒子を経てヌクレオチド/プラスミド/発現ベクターを送出すことが好ましいことがある。ビヒクルは、ヌクレオチド/プラスミド/発現ベクターを早期のクリアランスから保護するか、あるいは有効量のヌクレオチド/プラスミド/発現ベクターを被検体、腫瘍または癌細胞に送出す間免疫応答の発生からヌクレオチド/プラスミド/発現ベクターを保護することが好ましい。典型的なビヒクルは、ヌクレオチド/プラスミド/発現ベクターに関連する簡単なナノ粒子から、いっそう複雑なペギミル化ビヒクル、例えば、特異的細胞レセプターをターゲットとするその表面に取り付けられたリガンドを有するペギミル化リポソームまでの範囲である。
【0064】
リポソームおよびペギミル化リポソームはこの分野において知られている。慣用リポソームにおいて、送出すべき分子 (すなわち、小さい薬剤、タンパク質、ヌクレオチドまたはプラスミド) をリポソームの中央キャビティイ内に含有させる。当業者は認識するように、ターゲットとした「ステルス」、および分子送出に有効なカチオン性リポソームがまた存在する。例えば、下記を参照のこと: Hortobagyi、Gabriel N 他、J. Clinical Oncology、Vol. 19、Issue 14 (July) 2001: 3422-3433およびYu、Wei 他、Nucleic Acids Research、2004、32 (5): c48。リポソームは静脈内に注射することができ、修飾してそれらの表面をいっそう親水性とすることができる (ポリエチレングリコールを2層に添加することによって (「ペギミル化」) 、これは血流中のそれらの循環時間を増加させる) 。
【0065】
これらは「ステルス」リポソームとして知られており、親水性 (水溶性) 抗癌剤、例えばドキソルビジンおよびミトキサントロンのための担体として特に有効である。ターゲット細胞、例えば、腫瘍細胞に対する薬剤担持リポソームの特異的結合特性を増加させるために、特異的分子、例えば、抗体、タンパク質、ペプチドおよびその他をリポソーム表面上に取り付けることができる。例えば、癌細胞上に存在するレセプターに対する抗体を使用して、癌細胞に対してリポソームをターゲッティングすることができる。多発性骨髄腫をターゲッティングする場合、例えば、葉酸塩、IL-6またはトランスフェリンを使用してリポソームを多発性骨髄腫細胞にターゲッティングすることができる。
【0066】
また、デンドリマーはこの分野において知られており、そして好ましい送出ビヒクルを提供する。例えば、下記を参照のこと: Marjoros、Istvan J 他、”PAMAM Dendrimer - Based Multifunctional Conjugate for Cancer Therapy: Synthesis, Characterization, and Functionality”、Biomacromolecules、Vol. 7、No. 2、2006: 572-579、および Marjoros、Istvan J 他、J. Med. Chem. 2005、48: 5892-5899、デンドリマーの論考について。
【0067】
好ましい態様において、送出ビヒクルはポリエチレンイミンナノ粒子を含んでなる。典型的なポリエチレンイミンナノ粒子はin vivo-jetPEITM (現在Polyplus Transfection, Inc. により製造されている) である。in vivo-jetPEITMはカチオン性ポリマートランスフェクション剤であり、DNAおよびsiRNA送出剤として有効である。in vivo-jetPEITM (Polyplus Transfection製) は、動物において信頼性ある核酸送出を提供する線形ポリエチレンイミン試薬である。
【0068】
それは下記において使用される: 遺伝子治療 (Ohana 他、2004、Gene Ther. Mol. Bio. 8: 181-192; Vernejoul 他、2002、Cancer Research 62: 6124-31) 、RNAインターフェレンス (Urbain - Klein 他、2004、Gene Therapy 23: 1-6; Grezelinski 他、2006、Human Gene Therapy 17: 751-66) 、および遺伝ワクチン接種 (Garzon 他、2005、Vaccine 23: 1384-92) 。in vivo JET-PEIは現在ヒト臨床実験において癌遺伝子治療のための送出ビヒクルとして使用されている (Lemkine 他、2002、Mol. Cell. Neurosci. 19: 165-174) 。
【0069】
in vivo-jetPEITMは核酸を大よそ50 nmのナノ粒子に縮合させ、これらのナノ粒子は数時間安定である。この独特の保護機構の結果として、注射後の血球凝集は他の療法と比較して減少し、組織内拡散の制限、赤血球凝集および微小塞栓が防止される。これらのナノ粒子は十分に小さく、組織中に拡散し、エンドサイトーシスにより細胞の中に入る。in vivo-jetPEITMは、核酸のエンドソームからの放出および核膜を横切る移動を促進する。
【0070】
好ましい態様において、siRNAおよびポリヌクレオチドを含んでなるベクター/プラスミドの両方はin vivo-jetPEITM複合体を経て被検体に投与される。siRNAおよびポリヌクレオチドを含んでなるベクター/プラスミドは、ポリマー複合体、例えば、ポリエチレンイミンまたはin vivo-jetPEITM複合体を経て一緒に複合化することができるか、あるいはポリマーに対して別々に複合化することができる。例えば、siRNAおよびポリヌクレオチドを含んでなるベクター/プラスミドを被検体に別々に投与する (時間および/または送出部位の意味において別々に) 場合、異なる担体に対して複合化されたsiRNAおよびポリヌクレオチドを使用することが好ましい。投与を同時にかつ同一部位において実施する場合、siRNAおよびポリヌクレオチドを一緒に複合化することが好ましいことがある。
【0071】
他の態様において、プラスミドまたはベクターを投与して被検体において発現させるポリヌクレオチドを送出す代わりに、タンパク質それ自体を被検体に送出す。タンパク質は分離するか、あるいは合成することができる。
【0072】
本発明の1つの態様は、哺乳動物およびヒトを包含する被検体において癌を治療する方法を提供する。癌の治療は下記を包含するが、これらに限定されない: 癌細胞におけるアポトーシス、癌細胞を殺すこと、癌細胞の数の減少および腫瘍の体積/重量の減少。この方法は、eIF5A1 siRNAと、突然変異体eIF5A1をコードするポリヌクレオチドとを含んでなる組成物を投与することを含んでなる。組成物およびeIF5A1 siRNA、および突然変異体eIF5A1をコードするポリヌクレオチドは、本明細書において後述する。
【0073】
すべての細胞は、真核性開始因子5A (“elF-5A”) (または本明細書中で「因子5A」と呼ぶ) を産生する。哺乳動物細胞はelF-5Aの2つのアイソタイプ (eIF-5A1およびeIF-5A2) を産生する。eIF-5A1はアポトーシスを行う細胞中でアップレギュレートされるので、eIF-5A1はアポトーシス特異的elF-5Aと呼ばれてきている。ヒトeIF-5A1は受け入れ番号NM 001970を有し、図1に示されている。eIF-5A1は、アポトーシスに必要なタンパク質をコードするmRNA類の核サブセットの往復の原因となると考えられる。下記の文献を参照のこと: Liu & Tartakoff (1997) Supplement Society for Cell、8、462a、Abstract No. 2476、37th American Society for Cell Biology Annual Meeting, and Rosorius 他 (1999) J. Cell Science、112、2369-2380。
【0074】
両方の因子5Aは、デオキシハイプシンシンターゼ (“DHS”) により転写後修飾される。DHSはelF-5A類をハイプシン化する。ハイプシンは、ユニークアミノ酸であり、すべての検査した真核生物および古細菌中に見出されるが、真正細菌において見出されず、elF-5Aは唯一の既知のハイプシン含有タンパク質である。Park (1988) J. Biol. Chem. 263、7447-7449; Schumann & Klink (1989) System. Appl. Microbiol. 11、103-107; Bartig 他 (1990) System. Appl. Microbiol. 13、112-116; Gordon 他 (1987a) J. Biol. Chem. 262、16585-16589。
【0075】
ハイプシン化elF-5Aは2つの翻訳後の段階において形成される: 第1段階はジオキシハイプシン残基の形成であり、この形成はスペルミジンの4-アミノブチル部分が前駆体elF-5Aの特異的リシンへ転移することによってなされ、これはデオキシハイプシンシンターゼにより触媒される。第2段階は、この4-アミノブチル部分がデオキシハイプシンヒドロキシラーゼによりヒドロキシル化されてハイプシンを形成することを含む。
【0076】
elF-5Aのアミノ酸配列は種間で十分に保存され、elF-5A中のハイプシン残基を取り囲むアミノ酸配列の厳格な保存が存在し、これはこの修飾が生存に重要であることがあることを示唆する。Park 他 (1993) Biofactors 4、95-104。この仮定は酵母菌において今日までに発見されたelF-5Aの両方のイソ型の不活性化、またはDHS遺伝子 (これはそれらの活性化の第1段階を触媒する) の不活性化が細胞分裂をブロックするという観察によりさらに支持される。Schnier 他 (1991) Mol. Cell. Biol. 11、3105-3114; Sasaki 他 (1996) FEBS Lett. 384、151-154; Park 他 (1998) J. Biol. Chem. 273、1677-1683。
【0077】
しかしながら、酵母菌におけるelF-5Aタンパク質の消耗は全タンパク質合成をわずかに減少しただけであり、elF-5Aがタンパク質の全体的合成よりもむしろmRNA類の特異的サブセットの翻訳に必要とされるであろうことを示唆する。Kang 他 (1993) 、“Effect of initiation factor elF-5A depletion on cell proliferation and protein synthesis”、Tuite M (編者) 、Protein Synthesis and Targeting in Yeast、NATO Series H。elF-5Aを結合するリガンドは高度に保存されたモチーフを共有するという最近の発見は、elF-5Aの重要性をまた支持する。Xu & Chen (2001) J. Biol. Chem. 276、2555-2561。さらに、変性されたelF-5Aのハイプシン残基はRNAに特異的に結合する配列にとって必須であることが発見され、そして結合はリボヌクレアーゼからの保護を提供しなかった。
【0078】
本発明者らは、elF-5Aをコードするポリヌクレオチドを細胞に投与するとき、それらの細胞のアポトーシスが増加することを示した。次いで癌細胞中で発現されるeIF-5A1ポリヌクレオチドを投与することによって、癌細胞がアポトーシスするようにできることを本発明者らは示した。下記の同時係属出願を参照のこと: 10/200,148; 11/287,460; 11/293,391および11/637,835、それらのすべてはそれらの全体において引用することによって本明細書の一部とされる。
【0079】
さらに、本発明は、細胞が因子5Aのハイプシン化形態の構築物を有するとき、細胞は生存モードに入り、通常オーバータイムであるので、アポトーシスを行わない。顕著には、癌細胞において、有意な量のハイプシン化因子5Aが存在し、こうして細胞はアポトーシスに入らない (そして死なない) 。こうして、癌細胞を殺すことによって癌を治療する (癌細胞をアポトーシス経路に入れる) ために、eIF-5A1をコードするポリヌクレオチドを被検体に、または癌細胞または腫瘍に投与してeIF-5A1の発現を増加させ、これは引き続いて癌細胞のアポトーシスを引き起こし、究極的に細胞を死亡させ、腫瘍を収縮させる。しかしながら、eIF-5A1タンパク質をコードするポリヌクレオチドのみを提供してeIF-5A1の遺伝子発現をアップレギュレートし、また、eIF-5A1の内因的発現をノックダウンするためにsiRNAを使用しない場合、綱引きが存在する: アポトーシス経路に細胞を向けるeIF-5A1の発現は、生存経路に細胞を向けるハイプシン化因子5Aの存在と競合する。
【0080】
本発明はこの綱引きを排除し、eIF-5A1の発現のみを増加させることを超えた改良を提供する。被検体または細胞に投与されたポリヌクレオチドは突然変異されるので、生ずる発現されたタンパク質はハイプシン化されることができない。さらに、因子5Aの内因的発現はelF-5AをターゲットとするsiRNAによりをノックアウト/ノックダウンされるので、ハイプシン化されるelF-5Aはほとんどあるいはまったく存在しない。こうして、細胞中にハイプシン化されたelF-5Aはまったく (または実質的にほとんど) 存在しないので、それらは生存モードに押しやられない。
【0081】
突然変異したeIF-5A1をコードするポリヌクレオチドは好ましくは突然変異されるので、ハイプシン化されることができず、こうして細胞を生存モードに推進するために利用されない。例えば、1つの態様において、elF-5Aをコードするポリヌクレオチドは突然変異されるので、通常DHSによりハイプシン化される、位置50におけるリシン (K) はアラニン (A) (これはハイプシン化されることができない) に変化される。この突然変異体をK50Aと表示する。
【0082】
他の態様において、位置67におけるリシンはアルギニン (R) に変化される。この突然変異体を (K67R) と表示する。他の態様において、位置67におけるリシンはアラニン (A) に変化され、(K67A) と表示する。他の態様において、位置50におけるリシンはアルギニン (K50R) に変化され、他の態様は位置47におけるリシン (K) がアルギニン (K47R) に変化される突然変異体を提供する。
【0083】
他の態様において、二重突然変異体を使用する。1つの二重突然変異体は、位置50におけるリシン (K) がアルギニン (R) に変化されており、そして位置67におけるリシン (K) がアルギニン (R) に変化されている突然変異体である。この二重突然変異体をK50R/K67Rと呼ぶ。この二重突然変異体は同様にハイプシン化されることができないが、アミノ酸変化は単一突然変異体 (K50R) 程度に多くeIF-5A1の三次元構造を変更しない。こうして二重突然変異体は、三次元形状に非常に類似し、野生型としてフォールディングし、こうして単一突然変異体よりもいっそう安定であるタンパク質を提供する。二重突然変異体は、いっそう安定であるので、体中でより長く存在して、より長い療法的利益を提供する。こうして、体は因子5Aを有し、因子5Aは正常の細胞機能を必要とするが、ハイプシン化されることができないので、細胞は細胞の生存モードに拘束されず、アポトーシスを回避する。
【0084】
他の二重突然変異体は、位置47におけるリシン (K) がアルギニン (R) に変化されており、そして位置50におけるリシンがアルギニン (R) に変化されている突然変異体である。この突然変異体をK47R/K50Rと呼ぶ。本発明は、位置50におけるリシン (L) がアラニン (A) に変化されており、そして位置67におけるリシンがアラニン (A) に変化されている他の二重突然変異体を提供する。この突然変異体をK50A/K67Aと呼ぶ。
【0085】
体は正常の細胞生存および健康な細胞増殖のために因子5Aを必要とするので、siRNAが全身的に送出される場合、siRNAをもつ被検体において発現を完全に遮断しないことが好ましい。発現の遮断において良好でないsiRNAを使用する (すなわち、発現を停止または減少させるが、発現を完全には遮断しない) か、あるいは、選択的に、投与/治療の養生法を利用して発現レベルをバランスさせて健康な細胞の正常な成長または機能化を可能とするが、また癌細胞をアポトーシスさせることによって、elF-5A発現のコントロールを達成することができる。
【0086】
選択的に、局所的siRNA送出を利用できる。siRNAを局所的に癌細胞または腫瘍に送出す場合、発現をノックアウトすることが好ましい。発現をノックアウトすることよって、因子5Aはハイプシン化できる付近に存在せず、こうして細胞を生存モードにロックするハイプシン化elF-5Aは存在しない。siRNAは局所的に癌または腫瘍に送出されるので、細胞増殖に利用可能なelF-5Aを用意する必要がない。
【0087】
ある種の態様において、内因的遺伝子はeIF5A1である。eIF5A1をターゲットとするsiRNAを被検体に投与して内因的eIF-5A1の発現を抑制する。ある種の態様において、siRNAは配列番号1または配列2を含んでなるか、あるいは内因的eIF-5A1の発現を抑制するeIF5A1をターゲットとする任意のsiRNAである。ある種の態様において、eIF5A1はヒトeIF-5A1 (図1に示す) であり、そして被検体はヒトである。eIF5A1をターゲットとする他のsiRNAは既知であり、そして下記の同時係属出願に記載されている: 11/134,445; 11/287,460; 11/184,982; 11/293,391; 11/725,520; 11/725,470; 11/637,835。他の態様において、被検体は哺乳動物であり、そしてeIF5A1は哺乳動物に対して特異的である。例えば、被検体はイヌであり、そしてeIF5A1はイヌeIF5A1である。
【0088】
ある種の態様において、siRNAは図25に示すsiRNA構築物から本質的に成る。例えば、siRNAはeIF5A1をターゲットとする核酸を含有するが、またオーバーハング、例えばUまたはT核酸を含有するか、あるいはまたタグ、例えばhisタグ (しばしばHAタグと呼ばれる - これはin vivo研究においてしばしば使用される) を含有する。5’ または3’ 末端 (または、例えば、図25に示す核酸の連続的ひも内にさえ) に結合した分子または追加の核酸は、siRNA構築物がターゲット遺伝子の発現を減少させることができるかぎり、「から本質的に成る」の範囲内に含まれかつ入る。好ましくは、外因的ポリヌクレオチドの発現を生じないように、siRNAはeIF5A1遺伝子の領域をターゲットとする。例えば、eIF5A1 siRNAは3’ UTRまたは3’ 末端をターゲットとする。図25に示すsiRNAは典型的なeIF5A1 siRNAである。
【0089】
ポリヌクレオチドはeIF5A1をコードし、ここでポリヌクレオチドはeIF5A1変異型をコードするように突然変異されている。変異型eIF5A1が翻訳後に修飾されることができない (ハイプシン化されることができない) ように、突然変異したeIF5A1を設計する。典型的な突然変異体は本明細書において上に論じられている。
【0090】
充実性腫瘍を包含する癌の場合において、siRNAを腫瘍に直接送出すことは望ましいことがある。siRNAは時間ならびにポリヌクレオチドからの送出部位に関して別々に投与するか、あるいは同一時間におよび/または同一送出部位に一緒に投与することができる。当業者は理解するように、siRNA投与のタイミングは内因的タンパク質が翻訳されているときであり、そしてそれが既に作られた後ではない。
【0091】
本発明者らは、eIF5A1が正常組織に対して無毒であることを初期に示した (係属中の出願番号11/293,391、出願2005年11月28日、これは引用することによって本明細書の一部とされる) が、送出複合体 (elF5Aポリヌクレオチド/プラスミド/発現ベクターの直接投与に比較して) は好ましいことがある。好ましい送出系は有効量のeIF5A1を被検体、腫瘍または癌細胞のグループに提供し、ならびに好ましくは腫瘍または癌細胞のグループにターゲテッド送出を提供する。こうして、ある種の態様において、ナノメートルサイズのビヒクル、例えば、リポソーム、デンドリマーまたは同様な非毒性ナノ粒子、例えばポリエチレンイミンポリマー (例えば、in vivo JetPEITM複合体) によってeIF5A1ヌクレオチド/プラスミド/発現ベクターを送出すことが好ましい。
【0092】
また、eIF5A1タンパク質を腫瘍の部位に直接送出すことができる。当業者はeIF5A1タンパク質送出のための投与量および治療養生法の長さを決定できるであろう。
eIF5A1によるアポトーシス誘導の分子基準を下に論ずる。
【0093】
死レセプターシグナリング
Ad-eIF5A1 (野生型eIF5A1をもつアデノウイルス) またはAd-eIF5A1 (K50A) を使用する癌細胞の治療はカスパーゼ8、これは死レセプターにより開始される - リガンド結合性、およびカスパーゼ3、エクシキュウーショナーカスパーゼの活性化を誘導する。これらはeIF5A1の間接的作用であるようであり、そしてカスパーゼ8およびカスパーゼ3がeIF5A1 (K50A) (これはハイプシン化されることができない) を使用する治療後にまた活性化されるという事実は、この作用がリシン50 eIF5A1に帰することを示す。また、Ad-eIF5A1を使用する治療は、TNFR1のアップレギュレーションで前に示したように、死レセプターをアップレギュレートするように見える。
【0094】
ミトコンドリア経路
アポトーシスのミトコンドリア経路におけるリシン50 eIF5A1の直接的または間接的掛かり合いは、カスパーゼ9がeIF5A1またはeIF5A1 (K50A) による癌細胞の治療により活性化されるという発見を含む多数の観察により支持される。その上、p53はミトコンドリアアポトーシス経路の活性化においてある役割を演じ、eIF5A1により調節されるように思われる。
【0095】
例えば、アクチノマイシンDを使用する癌細胞の治療はp53をアップレギュレートし、そしてp53のこのアップレギュレーションはeIF5A1 siRNAにより開始される。これと一致して、Ad-eIF5A1を使用する癌細胞の治療は、p53 mRNAをアップレギュレートする。また、eIF5A1を使用する癌細胞の治療は、細胞質ゾルからミトコンドリアへのBaxの移動を誘導し、ミトコンドリア膜の喪失およびミトコンドリア空間内から細胞質ゾル中へのシトクロムCの放出を保証する。さらに、この治療はすべて前アポトーシスである切断したBcl2、BimおよびスプライスドBimをアップレギュレートする。
【0096】
MAPKシグナリング
さらに、本発明は、MAPKシグナリングにおけるeIF5A1の掛かり合いがアポトーシスに関係するという証拠を獲得した。例えば、Ad-eIF5A1を使用する癌細胞の治療は、P-JNKをアップレギュレートし、これは引き続いて抗アポトーシスBcl2を阻害する。さらに、Ad-eIF5A1およびAd-eIF5A1 (K50A) の両方はP-p38の形成を誘導し、これは引き続いて下記を包含する種々の前アポトーシス剤を衝撃することによってアポトーシスを開始することができる: TNFR1およびTNF; FASおよびFASL; カスパーゼ8; Bid; シトクロムCおよびカスパーゼ3。
【0097】
NF-κBシグナリング
NF-κBシグナリングが骨髄腫の増殖を支持するという証拠が存在する。例えば、骨髄ストロマ細胞に対する骨髄腫細胞の接着はIL-6のNF-κB依存的転写アップレギュレーションを誘導し、ここでIL-6は多発性骨髄腫における増殖因子および抗アポトーシス因子の両方である [Chauhan 他 (1996) Blood 87、1104] 。さらに、骨髄腫細胞が分泌するTNF-αは骨髄ストロマ細胞中のNF-κB活性化し、これによりIL-6の転写および分泌をアップレギュレートする。
【0098】
また、TNF-αは骨髄腫細胞中のNF-κBを活性化し、骨髄腫細胞および骨髄ストロマ細胞の両方上の細胞内接着分子-1 (ICAM-1; CD 54) および脈管細胞接着分子-1 (VCAM-1; CD 106) をアップレギュレートする [Hideshima 他 (2001) Oncogene 20、4519] 。これは引き続いて骨髄腫細胞と骨髄ストロマ細胞との関連を増強する [Hideshima 他 (2001) Oncogene 20、4519] 。逆に、これらの作用はTNF-α誘導NF-κB活性化により開始される [Hideshima 他 (2001) Oncogene 20、4519] 。事実、NF-κBは骨髄腫細胞中のTNF-α誘導アポトーシスに対する保護を仲介することが期待される [Hideshima 他 (2002) JBC 277、16639] 。これらおよび他の観察は、NF-κBシグナリングが多発性骨髄腫療法のための魅力的ターゲットありうるという見解を促進する。
【0099】
本発明者らは、eIF5A1 siRNAがヒト骨髄腫細胞におけるNF-κBの活性化およびICAM-1の形成の両方を阻害することを示した。これらの観察はeIF5A1がNF-κBの活性化においてある役割を演ずることを示し、そしてNF-κBの活性化の保証的作用が特質において前生存的である限り、我々はこの活性化がハイプシン化eIF5A1により直接的または間接的に仲介されることを予測する。
【0100】
IL-1
骨髄腫細胞による前炎症性サイトカインIL-1の過剰生産は、骨組織の悪化に導く多発性骨髄腫の特徴的特性である。eIF5A1 siRNAは、マウスにおけるLPS挑戦より誘導されるIL-1の過剰生産を劇的に減少することが示された。
【0101】
本発明の1つの態様は、多発性骨髄腫を治療する方法を提供する。多発性骨髄腫 (“MM”) は、骨髄中の悪性プラズマ細胞の発現および骨溶解性病変の存在により特徴づけられる進行性致死的疾患である。多発性骨髄腫はプラズマ細胞の不治であるが、治療可能な癌である。プラズマ細胞は免疫系の重要な部分であり、感染および疾患に対する戦いを促進する免疫グロブリン (抗体) を産生する。多発性骨髄腫は、骨髄中の異常なプラズマ細胞の極端な数、および無傷のモノクローナル免疫グロブリン (IgG、IgA、IgDまたはIgE; 「M-タンパク質」) またはベンス・ジョーンズタンパク質 (遊離モノクローナル軽鎖) の過剰生産により特徴づけられる。低カルシウム血症、貧血、腎臓障害、細菌感染に対する感受性の増加、および正常免疫グロブリンの産生悪化は多発性骨髄腫の普通の臨床的症状の発現である。また、多発性骨髄腫は、通常骨盤、肋骨および頭蓋における、広汎性骨粗しょう症によりしばしば特徴づけられる。
【0102】
本発明は、多発性骨髄腫において見出される同時のフィードバックループのために、多発性骨髄腫の治療に十分に適していると思われる。例えば、多発性骨髄腫は骨髄中でIL-1を低い濃度で産生する。IL-1は引き続いてストロマ細胞を刺激してIL-6を産生させ、次いでIL-6は多発性骨髄腫の増殖を刺激するようになる。本発明者らが以前に示したように (係属中の出願11/725,539および11/184,982参照) 、eIF-5A1に対して向けられたsiRNAは前炎症性サイトカイン、例えばIL-1; TNF-αおよびIL-8の発現を阻害する。こうして、siRNAはelF-5Aの発現をノックダウンしてハイプシン化の利用可能性を少なくするばかりではなく、またそれはIL-1/IL-6フィードバックループをカットオフまたは減少させるであろう。
【0103】
ヒトelF5AをターゲットとするsiRNAを使用して腫瘍中の内因的ハイプシン化elF5Aのレベルを抑制したが、ハイプシン化できないelF5Aの突然変異体 (elF5AK50R) を発現するRNAi抵抗性プラスミドを使用して非修飾elF5Aのレベルをin vivo上昇させた。elF5A siRNAを含有するPEIナノ複合体の腫瘍内注射は、対照siRNAを含有する複合体に対して80% (***p = 0.0003) より多くMM腫瘍増殖を阻害し、ハイプシン化elF5Aレベルの抑制が抗腫瘍作用を有することを示す。elF5AK50R発現プラスミドを含有するPEI複合体は同様な作用を有し、対照プラスミドを含有する複合体に対して70% (**p = 0.001) より多く腫瘍増殖を阻害した。
【0104】
こうして、MM腫瘍増殖は増殖促進ハイプシン化elF5Aの抑制により阻害できるか、あるいはelF5Aの前アポトーシス非ハイプシン化形態のレベルの増加により阻害できる。elF5A siRNAおよびRNAi抵抗性elF5AK50Rプラスミドの両方を含有する複合体の腫瘍内送出は腫瘍増殖に対して相乗作用を有し、そして有意に腫瘍を有意に収縮させ、94% (***p = 0.0002) だけ腫瘍増殖を阻害した。また、elF5A siRNA/elF5AK50R PEI複合体の静脈内送出は腫瘍増殖を95% (**p = 0.002) だけ効果的に減少させ、治療の全身的送出が実行可能であることを示した。
【0105】
elF5A siRNA/elF5AK50R pDNA PEI複合体の局所的送出および全身的送出の両方は、多発性骨髄腫において有意な抗腫瘍応答を生じた。
【0106】
さらに、本発明は、多発性骨髄腫を包含する癌の治療において有効な組成物を提供する。好ましい態様において、組成物はハイプシン化できない点突然変異化eIF5A1をコードするプラスミドDNAと、内因的ヒトeIF5A1を選択的に抑制するが、プラスミドによりコードされる点突然変異化eIF5A1に対して作用しないeIF5A1 siRNAとの複合体である。eIF5A1 siRNAおよび突然変異体eIF5A1をコードするポリヌクレオチドは前述した。プラスミドDNAおよびsiRNAの両方はPEI (ポリエチレンイミン) ナノ粒子に対して複合化されていることが好ましい。それらは別々に複合化でき、別々にまたは一緒に投与することができるか、あるいは一緒に複合化することができる。DNAおよびRNAはPEI上の正に帯電したアミノ基に結合し、ナノ粒子が細胞中に吸収されるとき放出される。PEI-核酸複合体は分裂している細胞および分裂していない細胞の両方に効率よく吸収されることが証明された。
【0107】
プラスミドDNAはeIF5A1 (K50A) のようにハイプシン化されることができず、したがって、強くアポトーシス性であるeIF5A1 (K50R) をコードすることが好ましい。eIF5A1 (K50R) の発現はB細胞特異的プロモーターにより調節されることが好ましい。
【0108】
eIF5A1 siRNAは内因的ヒトeIF5A1の3’ 末端に対して特異的であることが好ましく、トランスeIF5A1 (K50R) の発現に対して影響を与えない。典型的な好ましいeIF5A1 siRNAは、図25に示すsiRNAを含んでなり、から本質的に成るか、あるいはから成る。eIF5A1 siRNAを包含するための根本的理由は次の通りである: (1) ほとんどすべてがハイプシン化され、それゆえ前生存形態である、内因的eIF5A1を消耗するため; (2) NF-κBの活性化を阻害し、これによりIL-6の産生および細胞内接着分子の形成を減少させるため; および (3) IL-1の形成を阻害するため。eIF5A1 siRNAはeIF5A1 (K50R) と相乗的に作用して、骨髄腫細胞におけるアポトーシスを誘導する。上記 (2) および (3) が前生存的事象である限り、それらはハイプシン化eIF5A1により仲介されることが期待され、それゆえハイプシン化できないeIF5A1 (K50R) により影響を受けない。このアプローチは非ハイプシン化elF5Aのより大きいプールを生じ、多発性骨髄腫細胞を包含する癌細胞をアポトーシスに導き、健康な細胞にほとんど影響を与えない。
【0109】
好ましい組成物を本明細書中でSNS01と呼ぶ。SNS01は、安全性を増強させるためにB細胞由来の細胞 (骨髄腫細胞を包含する) への発現を制限するプロモーターにより推進されるelF5AK50RをコードするRNAi抵抗性プラスミドDNAと、ヌクレアーゼ耐性を増強させるためにdTdT 3’ オーバーハングをもつヒトelF5AをターゲットとするsiRNAの両方を含有する複合体であり、siRNAおよびプラスミドはin vivo JetPEITMに対して複合化されている。
【実施例】
【0110】
実施例1: eIF-5A1の野生型および変異型を使用するHeLaS3細胞のトランスフェクション
下記を包含するHAタッグドeIF5A1変異型を発現するプラスミドとともにリポフェクタミン (Lipofectamine) 2000を使用して、HeLa S3細胞をトランスフェクトした: 野生型eIF5A1 (WT) 、eIF5A1K50R (K50R) 、eIF5A1K67R (K67R) 、eIF5A1K67A (K67A) 、eIF5A1K47R/K50R (K4750R) 、eIF5A1K50R/K67R (K5067R) 、またはeIF5A1K50A/K67A (K5067A) 。LacZを発現するプラスミドを対照として使用した。トランスフェクション後24および48時間 (A) または28および52時間 (B) において、細胞ライゼイトを収集し、SDS-PAGEにより分画した。
【0111】
トランスフェクトしたeIF5A1の発現レベルをHAに対する抗体で検出した。結果: 推定上のユビキン化部位のリシンにおけるeIF5A1の突然変異化 (K67R) は、eIF5A1トランスジーンの蓄積を野生型 (A) より増加させた。また、ハイプシン化のために要求されるリシンにおけるeIF5A1の突然変異化 (K50R) は、eIF5A1トランスジーンの蓄積を野生型eIF5A1 (B) より増加させた。eIF5A1の二重突然変異化形態 (K50A/K67A) は、非突然変異化野生型eIF5A1トランスジーン (A + B) と比較したとき、特に十分に発現された。図2参照。
【0112】
実施例2: eIF-5A1の野生型および変異型を使用するKAS細胞のトランスフェクション
下記を包含するHAタッグドeIF5A1変異型を発現するプラスミドとともにPAMAMデンドリマー (FMD44) を使用して、KAS細胞をトランスフェクトした: 野生型eIF5A1 (5A1) 、eIF5A1K67A (K67A) 、eIF5A1K50A/K67A (K50A K67A) 、eIF5A1K50R (K50R) 、eIF5A1K47R (K47R) 、eIF5A1K67R (K67R) 、eIF5A1K47R/K50R (K47R K50R) 、またはeIF5A1K50R/K67R (K50R K67R) 。LacZを発現するプラスミドを対照として使用した。トランスフェクション後48時間において、細胞ライゼイトを収集し、SDS-PAGEにより分画した。トランスフェクトしたeIF5A1の発現レベルをHAに対する抗体で検出した。同じ負荷を、アクチンに対する抗体を用いて評価した。
【0113】
結果: 推定上のユビキン化部位のリシンにおけるeIF5A1の突然変異化 (K67AまたはK67R) は、eIF5A1トランスジーンの蓄積を野生型より増加させた。また、ハイプシン化のために要求されるリシンにおけるeIF5A1の突然変異化 (K50R) またはアセチル化部位におけるeIF5A1の突然変異化 (K47R) は、eIF5A1トランスジーンの蓄積を野生型eIF5A1より増加させた。eIF5A1の二重突然変異化 (K50A/K67A) は、非突然変異化野生型eIF5A1トランスジーンと比較したとき、より高いレベルで発現された。図3参照。
【0114】
実施例3: PAMAMデンドリマーを使用するKAS細胞のトランスフェクション
下記を包含するHAタッグドeIF5A1変異型を発現するプラスミドとともにPAMAMデンドリマー (FMD45-2) を使用して、KAS細胞をトランスフェクトした: eIF5A1K50R (K50R) 、eIF5A1K50A/K67A (K50A K67A) 、またはeIF5A1K50R/K67R (K50R K67R) 。LacZを発現するプラスミドを対照として使用した。トランスフェクション後72時間において、細胞をアネキシン (Annexin) /PIで染色し、FACSにより分析した。
【0115】
アネキシンVで陽性に染色され、PI (ヨウ化プロピジウム) で陰性に染色された細胞はアポトーシスの初期段階 (Ann+/PI-) にあると考慮し、そしてアネキシンVおよびPIの両方で陽性に染色された細胞はアポトーシスの最終段階 (Ann+/PI+) にあると考慮した。結果: ハイプシン化部位 (K50R) または推定上のユビキン化部位 (K67R) のリシンにおけるeIF5A1の突然変異化、ならびに二重突然変異体 (K50A/K67A) は、LacZ対照のレベルよりも有意に高いKAS細胞のアポトーシスを生じた。図4参照。
【0116】
実施例4: eIF-5A1およびeIF-5A1変異型を発現するプラスミドを使用するKAS細胞のトランスフェクション
下記を包含するHAタッグドeIF5A1変異型を発現するプラスミドとともにリポフェクタミン2000を使用して、KAS細胞をトランスフェクトした: eIF5A1K50A (K50A) 、eIF5A1K50R (K50R) 、eIF5A1K67R (K67R) 、eIF5A1K50A/K67A (K50A K67A) 、またはeIF5A1K50R/K67R (K50R K67R) 。LacZを発現するプラスミドを対照として使用した。
【0117】
トランスフェクション後72時間において、細胞をアネキシン/PIで染色し、FACSにより分析した。アネキシンVで陽性に染色され、PI (ヨウ化プロピジウム) で陰性に染色された細胞はアポトーシスの初期段階 (Ann+/PI-) にあると考慮し、そしてアネキシンVおよびPIの両方で陽性に染色された細胞はアポトーシスの最終段階 (Ann+/PI+) にあると考慮した。結果: ハイプシン化部位 (K50R) または推定上のユビキン化部位 (K67R) のリシンにおけるeIF5A1の突然変異化、ならびに二重突然変異体 (K50A/K67A) は、LacZ対照のレベルよりも有意に高いKAS細胞のアポトーシスを生じた。図5参照。
【0118】
実施例5: KAS細胞の治療に突然変異化eIF-5A1を使用するアポトーシスの発生
HAタッグドeIF5A1変異型eIF5A1K50R (K50R) またはeIF5A1K50A/K67A (K50A/K67A) を発現するプラスミドとともにリポフェクタミン2000を使用して、KAS細胞をトランスフェクトした。LacZを発現するプラスミドを対照として使用した。トランスフェクション後72時間において、細胞をアネキシン/PIで染色し、FACSにより分析した。アネキシンVで陽性に染色され、PI (ヨウ化プロピジウム) で陰性に染色された細胞はアポトーシスの初期段階 (Ann+/PI-) にあると考慮し、そしてアネキシンVおよびPIの両方で陽性に染色された細胞はアポトーシスの最終段階 (Ann+/PI+) にあると考慮した。結果: ハイプシン化部位 (K50R) のリシンにおけるeIF5A1の突然変異化またはハイプシン化部位および推定上のユビキン化部位 (K50A/K67A) の両方におけるeIF5A1の突然変異化は、LacZ対照のレベルよりも有意に高いKAS細胞のアポトーシスを生じた。図6参照。
【0119】
実施例6A: 多発性骨髄腫細胞のsiRNA/アデノウイルス仲介死滅
KAS (ヒト多発性骨髄腫) をS10培地 [4 ng/ml IL-6、10% 胎仔ウシ血清 (FBS) およびペニシリン/ストレプトマイシン (P/S) を含むRPMI 1640] 中で維持した。リポフェクタミン2000 (Invitrogen) を使用して58.7 ピコモルのsiRNAでKAS細胞をトランスフェクトした。偽トランスフェクトした細胞をsiRNAの非存在にリポフェクタミン2000で治療した。トランスフェクションを無抗体S10培地中で実施した。
【0120】
a) ヒトeIF5A1をターゲットとするsiRNA:
eIF5A1 siRNAターゲット#1 (このsiRNAはヒトeIF5A1のこの領域をターゲットとする: 5’ - AAGCTGGACTCCTCCTACACA - 3’ (配列番号_) 。このsiRNA配列は図25に示されており、本明細書においてしばしばh5A1と呼ぶ) 。
eIF5A1 siRNAターゲット#2 eIF5A1 (このsiRNAはヒトeIF5A1のこの領域をターゲットとする: 5’ - AAAGGAATGACTTCCAGCTGA - 3’ (配列番号_) 。このsiRNA配列は本明細書においてしばしばh5A1-ALTと呼ぶ) 。
b) 対照siRNA: 対照siRNAは下記の配列を有した: センス鎖、5’ - ACACAUCCUCCUCAGGUCGdTdT - 3’; およびアンチセンス鎖、3’ - dTdTUGUGUAGGAGGAGUCCAGC - 5’。
【0121】
使用した他の対照は、好ましくないオフ・ターゲッティング作用を制限するために検査したミクロアレイを有したので、ダーマコン (Dharmacon) からの非ターゲッティング確認siRNAを包含する。例えば、NF-κBを研究するin vitro研究のために、使用した対照はダーマコンの非ターゲッティングsiRNA (配列D-001700-01) であり、そしてin vivo研究のために、使用した対照はダーマコンの非ターゲッティングsiRNA (配列D-001810-01) であった。
【0122】
トランスフェクション後4時間に、細胞をペレット化し、1 mlのS10培地中に再懸濁させた。初期のsiRNAトランスフェクション後72時間に、トランスフェクトされたKAS細胞を数計測し、24ウェルのプレート中に300,000細胞/ウェルで播種し、そして同一siRNAで2回目のトランスフェクションを実施した。
【0123】
トランスフェクション後4時間に、細胞をペレット化し、3000 ifuのAd-LacZ (β-ガラクトシダーゼを発現するアデノウイルス) またはAd-5A1M (ヒトeIF5A1K50Rを発現するアデノウイルス) を含有するS10培地の1 ml中 (IL-6を含まない) に再懸濁させた。
72時間後に細胞を収集し、アネキシンV - FTTCおよびPI (BD Bioscience) で染色することにより、次いでFACS分析することにより、アポトーシスを分析した。
【0124】
a) 初期アポトーシスをアネキシン - FTTCで陽性に、PIで陰性に染色される細胞として定義した (Ann +/PI -) 。
b) 全アポトーシスを初期アポトーシス (Ann +/PI -) または後期アポトーシス/壊死 (Ann +/PI +) における細胞の合計として定義した。
【0125】
5A1 siRNAターゲッティング#1はヒトeIF5A1の3’ UTRをターゲットとし、したがってアデノウイルスからのeIF5A1の発現に影響を与えないであろう。5A1 siRNAターゲッティング#2はヒトeIF5A1のオープンリーディングフレーム内をターゲットとし、それゆえそれはアデノウイルスからのeIF5A1の発現を潜在的に妨害できるであろう。
結果: siRNAで治療し、eIF5A1K50R変異型を発現するアデノウイルスを感染させた細胞は、非治療細胞およびsiRNAのみで治療した細胞よりも大きい数の、アポトーシスを行う。図7参照。
【0126】
実施例6B: eIF5A1ターゲット#1に対するeIF5A1 siRNAの前処理 (図25に示す) は内因的eIF-5A1の発現を減少させたが、アデノウイルスにより発現されるRNAi抵抗性eIF5A1K50Rの蓄積を可能とする。
対照siRNA (C) または2つのsiRNAターゲッティングeIF5A1 (#1および#2) の1つとともにリポフェクタミン2000を使用して、KAS細胞をトランスフェクトした。eIF5A1 siRNA #1はeIF5A1の3’ UTRをターゲットとし、したがってそれはeIF5A1のオープンリーディングフレームのみを含有するので、アデノウイルスからのeIF5A1の発現を妨害しない。siRNAの配列を図25に示す。eIF5A1 #2 siRNAはeIF5A1のオープンリーディングフレームをターゲットとし、したがって内因的および外因的に発現されたeIF5A1の両方の発現に影響を与えるであろう。
【0127】
初期トランスフェクション時間後72時間に、細胞を第2回目に同一siRNAでトランスフェクトした。4時間後、トランスフェクション複合体を細胞から取出し、Ad-LacZ (L) またはAd-eIF5A1K50R (5M) のいずれかを含有する成長培地 (-) IL-6と置換した。72時間後、細胞ライゼイトを収集し、elF5Aに対する抗体およびアクチンを使用するウェスタンブロットにより分析した。図7B参照。ウイルスが発現したeIF5A1の蓄積が観察され (レーン1対レーン2) 、そしてeIF5A1 siRNAターゲッティング #1および #2により発現されたelF5A発現の減少は明瞭に見ることができる (レーン5および7対レーン3) 。期待するように、eIF5A1 siRNA #1はウイルスが発現したeIF5A1K50Rの蓄積に影響を与えない (レーン6対レーン4) が、eIF5A1 siRNA #2はウイルスが発現したトランスジーンの蓄積に適度にのみ影響を与える (レーン8対レーン4) 。
【0128】
実施例6C: アデノウイルス感染前のターゲット#1に対するeIF5A1 siRNAの前処理はヒト多発性骨髄腫細胞におけるリン酸化NF-κBの発現を減少させる。
対照siRNA (hC) またはeIF5A1をターゲットとするsiRNA (#1) とともにリポフェクタミン2000を使用して、KAS細胞をトランスフェクトした。eIF5A1 siRNA #1はeIF5A1の3’ UTRをターゲットとし、したがってeIF5A1のオープンリーディングフレームのみを含有するので、アデノウイルスからのeIF5A1の発現を妨害しない。初期トランスフェクション時間後72時間に、細胞を第2回目に同一siRNAでトランスフェクトした。
【0129】
4時間後、トランスフェクション複合体を細胞から取出し、Ad-LacZ (L) またはAd-eIF5A1K50R (5M) のいずれかを含有する成長培地 (+) IL-6と置換した。24時間後、細胞ライゼイトを収集し、ホスホ-NF-κB p65 (Ser 536) に対する抗体およびelF5Aを使用するウェスタンブロットにより分析した。期待するように、eIF5A1 siRNA #1はウイルスが発現したeIF5A1K50Rの蓄積に影響を与えない。セリン536におけるNF-κB p65のリン酸化は、活性化、核局在化、タンパク質・タンパク質相互作用および転写活性を調節する。図7C参照。
【0130】
実施例6D: アデノウイルス感染前のeIF5A1 siRNA #1を使用する前処理はヒト多発性骨髄腫細胞におけるリン酸化NF-κBおよびICAM-1の発現を減少させる。
対照siRNA (C) またはeIF5A1をターゲットとする2つのsiRNA (#1および#2) の1つともにリポフェクタミン2000を使用して、KAS細胞をトランスフェクトした。初期トランスフェクション時間後72時間に、細胞を第2回目に同一siRNAでトランスフェクトした。4時間後、トランスフェクション複合体を細胞から取出し、成長培地 (+) IL-6と置換した。2回目のトランスフェクション後24時間に、細胞を40 ng/mlのTNF-αで刺激し、細胞ライゼイトを0、4または24時間に収集し、ホスホ-NF-κB p65 (Ser 536) に対する抗体、ICAM-1およびアクチンを使用するウェスタンブロットにより分析した。
【0131】
両方のeIF5A1特異的siRNAを使用するトランスフェクション後、TNF-α誘導NF-κB p65リン酸化 (Ser 536) およびICAM-1の発現の減少が観察された。セリン536におけるNF-κB p65のリン酸化は、活性化、核局在化、タンパク質・タンパク質相互作用および転写活性を調節する。ICAM-1は、多発性骨髄腫の病原性に関係すると考えられる、細胞間接着表面の糖タンパク質である。図7D参照。
【0132】
実施例6E: siRNAを使用するKAS細胞の前処理は、IL-6の存在下のeIF5A1K50R遺伝子送出によるアポトーシスを増加させる。
リポフェクタミン2000を使用して対照siRNA (hcon) またはヒトeIF5A1 siRNA (h5A1) でKAS細胞をトランスフェクトした。72時間後、細胞をsiRNAで再トランスフェクトした。siRNAトランスフェクション培地を除去した後4時間に、空ベクター (mcs) またはeIF5A1K50R (K50R) プラスミドのPEI複合体を細胞に添加した。研究を通じて使用した成長培地はIL-6を含有した。72時間後、細胞をアネキシン/PIで染色し、FACS分析すると、アポトーシスが測定された。図7F参照。
【0133】
実施例7: eIF-5A1プラスミドおよびeIF5A1 siRNAの共投与は多発性骨髄腫皮下腫瘍の増殖を遅延させる (図8〜10) 。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。6匹の3〜5週齢のSCID/CB 17マウスの側腹部に、200 μlのPBS中の1000万のKAS-6/lの骨髄腫細胞を注射し、腫瘍が4 mm3の最大サイズに到達したとき、治療を開始した。
【0134】
対照マウスにpCpG-mcs (空ベクター) および対照siRNAを含有するPEI複合体を2回/週腫瘍内に注射した (対照グループは3匹のマウスから構成した: C-1、C-2およびC-3) 。治療したマウスにRNAi抵抗性プラスミドpCpG-eIF5A1 K50RおよびeIF5A1 siRNAを含有するPEI複合体を2回/週腫瘍内に注射した (治療したグループは3匹のマウスから構成した: 5A-1、5A-2および5A-3) 。注射は腫瘍内の多数の部位に実施して還流を防止し、遅い注射速度を使用して吸収を増加させた。図8中のデータはグループ中のすべてのマウスについての腫瘍体積を示す。図9に示すデータは平均腫瘍体積±標準誤差である。星印は統計的有意性を意味する (* = p < 0.025; n = 3) 。
【0135】
図10は、eIF-5A1プラスミドおよびeIF5A1 siRNAの共投与が多発性骨髄腫皮下腫瘍の重量を減少させることを示す。示すデータは平均腫瘍体積±標準誤差である。星印は統計的有意性を意味する (* = p < 0.05; n = 3) 。
2×0.1 mlのJET-PEI (PolyPlus) をin vivo試験において使用した。N/P比は8であった。PEI/DNA/siRNA複合体を5%のグルコース中で0.1 mlの全体積で形成した。複合体を形成するプロトコルは次の通りであった:
【0136】
1. 成分を温室にする。無菌を保持する。
2. 20μgのプラスミドDNA (2 mg/mlにおいて約10μl) および10μgのsiRNA (1 mg/mlにおいて10μl) を25μlの全体積に希釈する。無菌水を使用して差を構成する。
3. 25μlの10%グルコースの添加によりDNA溶液の体積を50μlの5%グルコースに調節する。おだやかに渦形成し、短時間遠心する。
【0137】
4. 4.8μlのin vivo JET-PEIを25μlの10%グルコースの全体積に希釈する。50μlの無菌水で体積を5%グルコースの最終濃度に調節する。おだやかに渦形成し、短時間遠心する。
5. 直ちに50μlの希釈PEIを50μlの希釈DNAに添加する (順序を逆にしない) 。短時間渦形成し、直ちに回転する。
6. 注射前に15分間インキュベートする。複合体は6時間安定である。
【0138】
無CpGクローニングベクターおよびpCpGプラスミドを入手した (InvivoGenから) 。これらのプラスミドはCpGジヌクレオチドを完全に欠如し、pCpGと命名する。これらのプラスミドはin vitroおよびin vivoにおいて高いレベルのトランスジーン発現を生じ、CMVに基づくプラスミドと対照的に、in vivoにおいて持続した発現を可能とする。pCpGプラスミドは、自然にCpGジヌクレオチドを欠如し、すべてのCpGを除去するように変性されているか、あるいは完全に合成された要素、例えば、選択可能なマーカーまたはリポーターをコードする遺伝子を含有する。
【0139】
遺伝暗号を形成する16ジヌクレオチドの間で、CGは非必須の唯一のジヌクレオチドでありかつ置換できるという事実により、これらの新しいアレレの合成は可能である。8コドンは5つの異なるアミノ酸をコードするCGを含有する。野生型と同一にとどまり、こうしてそれらの野生型対応物と同程度に活性であるアミノ酸配列を有するタンパク質を新たなアレレを生成する同じアミノ酸をコードする2つのコドンを少なくとも選択することによって、すべての8コドンは置換できる。これらの新しいアレレはpMODと命名するプラスミドにおいて入手可能であり、このプラスミドからそれらを容易に切除することができる。
【0140】
pCpGプラスミドはin vivoにおいて長く持続する発現を可能とし、TLR9を発現する細胞系統を使用する、in vivoおよびin vitroにおける遺伝子発現に対するCpGの作用、ならびに生得的および後天的免疫系に対するそれらの作用を研究するための価値のある道具を表す。
【0141】
空ベクター、pCpG-mcs (Invitrogen) は、発現された遺伝子産物をもたず、多重クローニング部位のみをもち、対照ベクターとして使用されたベクターである。HAタッグドeIF5A1K50R cDNAを、LacZ遺伝子が除去されている、pCpG-LacZベクター (Invitrogen) のNcoIおよびNheI部位中にサブクローニングして、治療ベクターpCpG-eIF-5A1 (K50R) を発生させた。DNAはキット (Endo-Free Qiagen) を使用して調製した。エンドドキシンレベルを測定し、これは < 0.03 EU/μgである; DNAは水中で2 mg/mlであるべきである。
【0142】
実験において使用した対照siRNAは、ダーマコン (Dharmacon) からのミクロアレイ確認非ターゲッティング対照siRNAであった (D-00180-01) 。血清中の分解を防止するために修飾した (siSTABLE) siRNAを得た。
【0143】
実験において使用したeIF5A1 siRNAは、ヒトeIF5A1の3’ UTRに対して設計した。eIF5A1 siRNAとマウスeIF5A1との間に類似性は存在せず、したがってsiRNAはヒト (マウスではない) eIF5A1のみを抑制すべきである。また、siRNAはeIF5A2 (ヒトまたはマウス) に対して類似性をもたない。血清中の分解を防止するために修飾した (siSTABLE) siRNAを得た。eIF5A1 siRNAは下記のターゲット配列を有する:
5’ GCU GGA CUC CUC CUA CAC A (UU) 3’
【0144】
SiSTABLE siRNAを水中に1 mg/mlで溶解した (- 20℃においてアリコートで貯蔵した) 。
デジタルキャリパーを使用して、長さ (l) および幅 (w) の腫瘍寸法を2〜3回/週測定した。下記の方程式を使用して、腫瘍体積を計算した:
l = 長さ; 最小寸法
w = 幅; 最大寸法
腫瘍体積 (mm3) = l2×w×0.5
【0145】
統計的解析
スチューデントのt検定を統計的解析に使用した。有意性は95% (p < 0.05) を超える信頼水準であると考える。
【0146】
実施例8: eIF5A1プラスミドおよびeIF5A1 siRNAの共投与は多発性骨髄腫皮下腫瘍の増殖を遅延し、腫瘍を収縮させる。
他の研究において、再びSCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。対照マウスにpCpG-mcs (空ベクター) および対照siRNAを含有するPEI複合体を2回/週腫瘍内に注射した (対照グループG-1、G-2およびG-3) 。治療したマウスにRNAi抵抗性プラスミドpCpG-eIF5A1K50R (20μgのプラスミドDNA) およびeIF5A1 siRNA (10μgのsiRNA) を含有するPEI複合体を2回/週腫瘍内に注射した (治療したグループG-4、G-5およびG-6) 。図11に示すデータは平均腫瘍体積±標準誤差である。星印は統計的有意性を意味する (* = p < 0.025; n = 3) 。21日間にわたって6回の注射を実施した (赤色矢印) 。
【0147】
実施例9: eIF5A1 siRNAを静脈内 (i.v.) に投与しかつPEI/eIF5A1K50Rプラスミドを腫瘍内 (i.t.) に投与すると、多発性骨髄腫皮下腫瘍の腫瘍収縮を生ずる (グループ2B) 。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、50μgの対照siRNA (対照グループ) またはヒトeIF5A1 siRNA (治療したグループ) の初期尾注射で治療を開始した。続いて、対照マウスにpCpG-mcsを含有するPEI複合体を2回/週腫瘍内に注射することにより治療した (空ベクター; 対照グループ; G-1、G-2およびG-3) 。続いて、治療したマウスにRNAi抵抗性プラスミドpCpG-eIF5A1K50R (20μgのプラスミドDNA) を含有するPEI複合体を2回/週腫瘍内に注射することにより治療した (治療したグループ; G-4、G-5およびG-6) 。
【0148】
対照マウスに1回/週のi.v. 注射により対照siRNAを投与し続けた (対照グループR-1、R-2およびR-3) 。治療したマウスに1回/週のi.v. 注射によりヒトeIF5A1 siRNA (20μg) を投与し続けた (治療したグループR-4、R-5およびR-6) 。図12に示すデータは各グループにおけるすべてのマウスについての腫瘍体積である。PEI/DNAの6回の腫瘍内注射 (赤色矢印) および4回のi.v. 注射 (青色矢印) を21日間の期間にわたって実施した。
【0149】
図13は実施例8および9からの結果のオーバーレイを提供する。SCIDマウスにKAS細胞を皮下注射した。図13に示すデータはそれぞれのグループにおけるマウスの平均腫瘍体積±標準誤差である。星印は治療したグループと対照グループとの間の統計的有意性を意味する (** = p < 0.01; *** = p < 0.001; n = 3) 。
【0150】
PEI複合体を形成するプロトコル:
1. 成分を温室にする。無菌を保持する。
2. プラスミドDNAまたはプラスミドDNA + siRNAを全体積25μlに希釈する。無菌水を使用して体積を調節する。
a) プラスミドDNAのみの複合体について:
20μgのプラスミドDNA (約2 mg/mlにおいて10μl) を全体積25μlに希釈する。無菌水を使用して差を構成する。
【0151】
b) プラスミドDNA + siRNAの複合体について:
20μgのプラスミドDNA (約2 mg/mlにおいて10μl) および10μgのsiRNA (1 mg/mlにおいて10μl) を全体積25μlに希釈する。無菌水を使用して差を構成する。
3. DNA溶液の体積を25μlの10%グルコースの添加により50μlの5%グルコースに調節する。おだやかに渦形成し、短時間遠心する。
【0152】
4. in vivo JETPEIを全体積25μlの10%グルコースに希釈する。
a) プラスミドDNAのみの複合体について:
3.2μlのin vivo JETPEIを全体積25μlの10%グルコースに希釈する。体積を無菌水で50μlに調節して5%グルコースの最終濃度にする。おだやかに渦形成し、短時間遠心する。
b) プラスミドDNA + siRNAの複合体について:
4.8μlのin vivo JETPEIを全体積25μlの10%グルコースに希釈する。体積を無菌水で50μlに調節して5%グルコースの最終濃度にする。おだやかに渦形成し、短時間遠心する。
【0153】
5. 直ちに50μlの希釈PEIを50μlの希釈DNAに添加する (順序を逆にしない) 。短時間渦形成し、直ちに回転する。
6. 注射前に15分間インキュベートする。複合体は6時間安定である。
【0154】
siRNAの尾静脈注射に関して、初期siRNA注射は50マイクログラムであった。siRNAをPBS中で0.4 mg/mlに希釈した。125μl/マウス (50μg) を尾静脈に注射した。引き続いて血清安定化siRNAを20μg/マウスで2回/週注射した。siRNAをPBS中で0.4 mg/mlに希釈した。50μl/マウス (20μg) を尾静脈に注射した。
【0155】
図13Bは、eIF5A1プラスミドおよびeIF5A1 siRNAの共投与が腫瘍を収縮させることを示す。SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。マウスにRNAi抵抗性プラスミドpCpG-eIF5A1K50RおよびeIF5A1 siRNAを含有するPEI複合体を2回/週腫瘍内に注射した (治療したグループ; G-4、G-5およびG-6) 。21日間にわたって6回の注射を実施した。治療の開始後42日に、マウスを殺し、腫瘍部位下の皮膚を開き、腫瘍増殖の証拠を検査した。グループ2Aの治療マウスのいずれにおいても、腫瘍増殖は観察されなかった。
【0156】
図13Cは、eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の腫瘍内 (i.t.) 投与が多発性骨髄腫皮下腫瘍の腫瘍収縮を生じさせることを示す。SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、50μgのヒトeIF5A1 siRNAの初期注射で治療を開始した (治療したグループ) 。引き続いてマウスにRNAi抵抗性プラスミドpCpG-eIF5A1K50Rを含有するPEI複合体を2回/週腫瘍内に注射して治療した (治療したグループ; R-4、R-5およびR-6) 。マウスにヒトeIF5A1 siRNAを1回/週注射により投与し続けた。治療開始後21日に治療を終えた。治療の開始後42日に、マウスを殺し、腫瘍部位下の皮膚を開き、腫瘍増殖の証拠を検査した。治療グループの1匹のマウスにおいて腫瘍増殖の証拠は存在しなかった (グループ2B) 。
【0157】
実施例10: eIF5A1プラスミドおよびeIF5A1 siRNAの静脈内共投与は多発性骨髄腫皮下腫瘍の増殖を遅延する。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、50μgの対照siRNA (対照グループ) またはヒトeIF5A1 siRNA (治療したグループ) の初期注射で治療を開始した。引き続いてマウスにpCpG-mcsを含有するPEI複合体 (空ベクター; 対照グループA1、A2およびA3) またはRNAi抵抗性プラスミドpCpG-eIF5A1K50Rを含有するPEI複合体 (治療したグループ; A4、A5およびA6) を2回/週静脈内注射 (赤色矢印) または腹腔内注射 (緑色矢印) により治療した。マウスに対照siRNA (対照グループA1、A2およびA3) またはヒトeIF5A1 siRNA (治療したグループ; A4、A5およびA6) をi.v. 注射 (青色矢印) により1回/週投与し続けた。示したデータは各グループにおけるすべてのマウスについての腫瘍体積である。図14に示すデータは各グループにおけるすべてのマウスについての腫瘍体積である。
【0158】
実施例11: eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の静脈内 (i.v.) または腹腔内 (i.p.) 投与は多発性骨髄腫皮下腫瘍の増殖を遅延させる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、50μgの対照siRNA (対照グループ) またはヒトeIF5A1 siRNA (治療したグループ) の初期注射で治療を開始した。引き続いて対照マウスをpCpG-mcsを含有するPEI複合体で静脈内または腹腔内注射により1回/週治療した (空ベクター; 対照グループは3匹のマウスであった: B1、B2およびB3) 。
【0159】
引き続いて治療したマウスをRNAi抵抗性プラスミドpCpG-eIF5A1K50Rを含有するPEI複合体で静脈内または腹腔内注射により1回/週治療した (治療したグループ: B4、B5およびB6) 。マウスに対照siRNA (対照グループ: B1、B2およびB3) またはヒトeIF5A1 siRNA (治療したグループは3匹のマウスであった: B4、B5およびB6) をi.v. 注射により1回/週投与し続けた。50μgの初期siRNA注射により、実験を開始した (図15中のグラフ上の第2日) 。引き続いて20μgのsiRNAを1回/週注射した。siRNAは裸で、すなわち、送出ビヒクルを使用しないで投与した。PEI複合体は20μgのプラスミドDNAを含有した。初期PEIをi.p. 注射し、引き続いてi.v. 注射した。図15に示すデータは、各グループにおけるすべてのマウスについての腫瘍体積である。
【0160】
図16は実施例10および11のオーバーレイを提供する。SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。マウスの1組に対照siRNA (対照; グループA) またはeIF5A1 siRNA (治療した; グループA) を1回/週i.v. 注射し、そしてpCpG-mcsを含有するPEI複合体 (対照; グループA) またはRNAi抵抗性プラスミドpCpG-eIF5A1K50Rを含有するPEI複合体 (治療した; グループA) を1回/週i.v. またはi.p. 注射した。
【0161】
マウスの第2組にpCpG-mcs (空ベクター) および対照siRNAを含有するPEI複合体 (対照; グループB) またはRNAi抵抗性プラスミドpCpG-eIF5A1K50RおよびeIF5A1を含有するPEI複合体 (治療した; グループB) を2回/週i.v. またはi.p. 注射した。示すデータは各グループにおけるマウスについての平均腫瘍体積±標準誤差である。星印は治療したグループと対照グループとの間の統計的有意性を意味する (* = p < 0.05; *** = p < 0.001; n = 3) 。
PEI複合体およびsiRNAを調製するプロトコルは前の実施例に記載されている通りである。
【0162】
実施例12: eIF5A1プラスミドおよびeIF5A1 siRNAの共投与は多発性骨髄腫皮下腫瘍の成長を遅延させ、腫瘍を収縮させる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。対照マウスにpCpG-mcs (空ベクター) および対照siRNAを含有するPEI複合体を2回/週腫瘍内注射した (対照グループは3匹のマウスを有した: 対照1、対照2および対照3) 。治療したマウスにRNAi抵抗性プラスミドpCpG-eIF5A1K50RおよびeIF5A1 siRNAを含有するPEI複合体を2回/週腫瘍内注射した (治療したグループは4匹のマウスを有した: 5A-1、5A-2、5A-3および5A-4) 。PEI複合体の腫瘍内注射は20μgのプラスミドDNAおよび10μgのsiRNAの両方を含有した。図17に示すデータは各グループにおけるすべてのマウスについての腫瘍体積である。
【0163】
実施例13: eIF5A1 siRNA の静脈内 (i.v.) およびPEI/eIF5A1K50Rプラスミド複合体の腫瘍内 (i.t.) 投与は多発性骨髄腫皮下腫瘍の腫瘍収縮を生じさせる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、50μgの対照siRNA (対照グループは3匹のマウスを有した: 対照1、対照2および対照3) またはヒトeIF5A1 siRNA (治療したグループは3匹のマウスを有した: 5A-1、5A-2および 5A-3) の初期注射で治療を開始した。引き続いて対照マウスをpCpG-mcs (20μg) を含有するPEI複合体の2回/週の腫瘍内注射により治療した (対照グループ1〜3) 。
【0164】
引き続いて治療したマウスをRNAi抵抗性プラスミドpCpG-eIF5A1K50R (20μg) を含有するPEI複合体の2回/週の腫瘍内注射により治療した (5A-1、5A-2および5A-3) 。対照マウスに対照siRNA (20μg) を尾静脈 i.v. 注射により2回/週投与し続けた。治療したマウスにヒトeIF5A1 siRNA (20μg) を尾静脈 i.v. 注射により2回/週投与し続けた。腫瘍内注射48時間前に注射した。siRNAは裸で、すなわち、送出ビヒクルを使用しないで投与した。図18に示すデータは各グループにおけるすべてのマウスについての腫瘍体積である。
【0165】
実施例14: EF1またはB29プロモーターのいずれかにより推進されたeIF5A1K50Rプラスミド、およびeIF5A1 siRNAの共投与は多発性骨髄腫皮下腫瘍の増殖を遅延させ、腫瘍を収縮させる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。マウスに対照ベクター (G1およびG2) またはB29プロモーター (G3およびG4) もしくはEF1プロモーター (G5およびG6) により推進されるeIF5A1プラスミドまたは対照siRNA (G1、G3、G5) またはh5A1 siRNA (G2、G4、G6) を含有するPEI複合体を2回/週腫瘍内注射した。
【0166】
示すデータは各グループの平均腫瘍体積±標準誤差である。注: B29プロモーターはB細胞特異的プロモーターであることを意図した。しかしながら、この研究において使用したB29プロモーター/mCMVエンハンサーはin vitro KAS細胞中でHA-eIF5A1K50Rの高い発現を推進させることが発見されたが、B細胞特異的であるように思われない (CMVエンハンサーのためであろう) 。図19参照。
【0167】
実施例15: eIF5A1 siRNAの共投与はEF1またはB29プロモーターいずれかにより推進されるeIF5A1K50Rプラスミドの多発性骨髄腫皮下腫瘍に対する抗腫瘍作用を増加させ、存在する腫瘍を減少させる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。マウスに対照ベクター (G1およびG2) またはB29プロモーター (G3およびG4) もしくはEF1プロモーター (G5およびG6) により推進されるeIF5A1プラスミドまたは対照siRNA (G1、G3、G5) またはh5A1 siRNA (G2、G4、G6) を含有するPEI複合体を2回/週腫瘍内注射した。治療開始後24日にマウスを殺し、皮下腫瘍を取出し、秤量した。示すデータはすべてのグループの平均腫瘍体積±標準誤差である。図20参照。
【0168】
実施例16: eIF5A1 siRNAは肺腺癌細胞におけるAd-elF5Aによる感染から生ずるアポトーシス誘導を相乗的に増加させる。
A549細胞にAd-LacZまたはAd-elF5Aを感染させた。トランスフェクション培地を細胞に添加し、直ちにウイルスを添加することによって、細胞を対照siRNAまたはsiRNAターゲッティングヒトeIF5A1 (h5A1) でトランスフェクトした。siRNAでトランスフェクトし、ウイルスで感染させた4時間後、培地を新鮮な培地と置換し、細胞を72時間インキュベートした後、アネキシン/PIで標識化してアポトーシス細胞を検出した。注: この細胞系統におけるelF5Aの過剰発現は限定的量のDHSおよびDOHHの理由から非ハイプシン化elF5Aが蓄積し、したがってelF5AK50Rの過剰発現として同一前アポトーシス作用が生ずる。これらのデータが示すように、同時のハイプシン化elF5Aの抑制および非ハイプシン化elF5Aの過剰発現により生じたアポトーシスにおける相乗効果はその上非骨髄腫腫瘍型において観察される。図21参照。
【0169】
実施例17: pExp5Aの構築
pExp5Aは、主としてB細胞系統の細胞においてヒトeIF5A1K50Rの発現を推進するように設計された、CpGジヌクレオチドが減少した発現プラスミドである。このベクターはpCpG-LacZ、すなわち、CpGジヌクレオチドを完全に欠如するプラスミド (Invitrogen) である。大腸菌 (E. coli) における複製および選択に必要なすべての要素はCpGジヌクレオチドを含まない。eIF5A1K50RのB細胞特異的発現を推進するために、本来のCMVエンハンサー/プロモーターおよびCpG-LacZベクターからのLacZ遺伝子は、それぞれ、ヒト最小B細胞特異的プロモーター (B29/CD79b; Invitrogen) およびヒトeIF5A1K50Rと置換されてきている。
【0170】
B29プロモーターの活性を増大しかつ非B細胞における発現を減少させるために、B29 DHS4.4 3’ エンハンサーがeIF5A1発現カセットの下流のプラスミド中に導入されてきている (Malone 他、2006、B29 gene silencing in pituitary cells is regulated by its 3’ enhancer. J. Mol. Biol. 362: 173-183) 。B29最小プロモーター、eIF5A1K50RおよびB29 DHS4.4 3’ エンハンサーの組込みは、32 CpGジヌクレオチドをベクター中に導入した。
【0171】
大腸菌 (E. coli) における発現のための要素
複製起点: 大腸菌 (E. coli) R6Kγori.
* R6Kγ複製起点が存在するために、pCpGプラスミドはpir突然変異体遺伝子を発現する大腸菌 (E. coli) においてのみ発現されることができる。それらは標準的大腸菌 (E. coli) 株において複製しないであろう。したがって、pCpGプラスミドは大腸菌 (E. coli) GT115株、すなわち、Dcmメチル化を欠如するpir突然変異体 (Invivogen) を提供する。
細菌プロモーター: EM2K、細菌EM7プロモーターの無CpGバージョン。
選択可能なマーカー: ZeocinTM 抵抗性遺伝子; CpGをもたない合成アレレ。
【0172】
哺乳動物細胞における発現のための要素
哺乳動物プロモーター: B細胞における組織特異的発現のためのヒト-167 bp最小B29 (CD79b) プロモーター。また、合成イントロン (I 140) が5’ UTRに存在する。
ポリアデニル化シグナル: 後期SV40ポリアデニル化シグナルの無CpGジヌクレオチドバージョン。
3’ エンハンサー: ヒトB29 DHS4.4 3’ エンハンサー。
MAR: 細菌の転写単位と哺乳動物の転写単位との間に、2つの無CpGマトリックス結合領域 (MAR) が存在する。1つのMARはヒトIFN-β遺伝子の5’ 領域に由来し、そして1つはβ-グロビン遺伝子の5’ 領域に由来する。
pExp5A (3371 bp) の予測される配列は図23に示されている。
【0173】
eIF5A1K50Rのアミノ酸配列
* K50R突然変異に下線が引かれている
【0174】
【化1】
【0175】
pExp5Aの構築 - 構築の概略:
工程1: B29 DHS4.4 3’ エンハンサーをクローニングし、pGEM T easy (Promega) 中にサブクローニングする -- pGEM-4.4 enh #8をつくる。
工程2: 最小B29プロモーターをpCpG-LacZ (Invivogen) 中にサブクローニングする - B29-5#3をつくる。
工程3: HA-eIF5A1K50RをB29-5#3ベクター中にサブクローニングする - B29-5#3-eIF5A1K50Rをつくる。
【0176】
工程4: pCpG-mcs (Invitrogen) 中に新しい多重クローニング部位をつくる - pCpG-Linker4をつくる。
工程5: B29 DHS4.4 3’ エンハンサーをpCpG-Linker4中にサブクローニングする - pCpG-DHS4.4をつくる。
工程6: B29プロモーター + HA-eIF5A1K50R + SV40 pA発現カセットをpCpG-DHS4.4中にサブクローニングしてpExp-5をつくる。
工程7: HA-eIF5A1K50RをもたないpExp-5中にHA-eIF5A1K50Rと置き換えて最終のベクターpExp5Aをつくる。
【0177】
構築の詳細:
工程1: B29 DHS4.4 3’ エンハンサーのクローニングおよびpGEM T easy (Promega) 中へのサブクローニング -- pGEM-4.4 enh #8をつくる。
【0178】
下記のプライマーを使用してKAS細胞 (多発性骨髄腫細胞系統) から単離したゲノムDNAからPCRにより、B29 DHS4.4 3’ エンハンサーをクローニングした: 前方向5’ - CAGCAAGGGAGCACCTATG - 3’ および逆方向5’ - GTTGCAGTGAGCGGAGATG - 3’ 。ヒトCD79B/GH-1遺伝子間領域 (Accession AB062674) の配列を使用してプライマーを設計した。生ずる608 bpのPCRフラグメントをpGEM(商標)Teasyクローニングベクター (Promega) 中にサブクローニングし、配列決定した。Komatsu 他 2002 Novel regulatory regions found downstream of the rat B29/Ig-b gene. Eur. J. Biochem. 269: 1227-1236。
【0179】
pGEM-4.4 enh #8中のB29 DS4.4 3’ エンハンサーPCRフラグメントの配列 (297 bp)
【0180】
【化2】
【0181】
pGEM-4.4 enh #8中のB29 DHS4.4 3’ エンハンサーPCRフラグメントの配列 (297 bp) とヒトCD79B/GH-1遺伝子間領域 (Accession AB062674) の配列とのアラインメント。
【0182】
【化3】
【0183】
工程2: pCpG-LacZ (Invivogen) 中への最小B29プロモーターのサブクローニング -- B29-5 #3をつくる。
下記のプライマーを使用して全長のヒトB29プロモーターを有する商用プラスミド (pDrive-hB29; Invivogen) から、最小-167ヒトB29プロモーターを増幅した: 前方向5’ - CCAACTAGTGCGACCGCCAAACCTTAGC - 3’ ; 逆方向5’ - CAAAAGCTTGACAACGTCCGAGGCTCCTTGG - 3’ 。生ずるPCRフラグメントをSpeIおよびHindIIIで消化し、pCpG-LacZベクター (Invivogen) のSpeIおよびHindIII部位中に結合してB29-5 #3をつくった。
【0184】
B29-5 #3中のB29最小プロモーターPCRフラグメントの配列 (188 bp)
【0185】
【化4】
【0186】
B29-5 #3中のB29最小プロモーターPCRフラグメント (B29 min) とpDrive-hB29からの全長ヒトB29プロモーターとのアラインメント
【0187】
【化5】
【0188】
工程3: B29-5 #3ベクター中のHA-eIF5A1K50Rのサブクローニング -- pB29-eIF5A1K50R_7をつくる。
DNAテンプレートとしてpHM6-eIF5A1K50Rおよび下記のプライマーを使用してPCRにより、HA-eIF5A1K50Rを増幅した: 前方向5’ - CGCCATGGACATGTACCCTTACGACGTCCCAGACTACGCTGCAGATGATTTGGACTTCGAG - 3’ および逆方向5’ - CGCGCTAGCCAGTTATTTTGCCATCGCC - 3’ 。生ずるPCRフラグメントをNcoIおよびNheIで消化し、B29-5 #3のNcoIおよびNheI部位中にサブクローニングしてLacZを置換した。
【0189】
pB29-eIF5A1K50R_7中のHA-eIF5A1K50RPCRフラグメントの配列
【0190】
【化6】
【0191】
pB29-eIF5A1K50R_7中のHA-eIF5A1K50R PCRフラグメントの翻訳
HAエピトープ
eIF5A1K50R
K50R
【0192】
【化7】
【0193】
pB29-eIF5A1K50R_7中のHA-eIF5A1K50R PCRフラグメントとヒトeIF5A1 (Accession # NP 001961) とのアラインメント
【0194】
【化8】
【0195】
工程4: pCpG-mcs (Invivogen) 中に新しい多重クローニング部位をつくる -- pCpG-Linker4をつくる。
pCpGクローニングベクター、pCpG-mcs G2 (Invivogen) をEcoRIで消化して、mCMVエンハンサー、hEFIプロモーター、合成イントロン、多重クローニング部位およびSV40ポリアデニル化シグナルを含有する哺乳動物発現カセットを除去した。次いでEcoRI消化pCpG-mcs G2ベクターをEcoRI粘着末端をもつ合成リンカーに結合して、新しい多重クローニング部位をもつ無プロモーターベクター (pCpG-Linker4) をつくる。
【0196】
【化9】
【0197】
pCpG-Linker4中の新しい多重クローニング部位を取り囲む領域の配列
【0198】
【化10】
【0199】
工程5: pCpG-Linker4中のB29 DS4.4 3’ エンハンサーのサブクローニング - pCpG-DHS4.4をつくる。
テンプレートとしてpGEM-4.4 enh #8を使用しそして下記のプライマーを使用してPCRにより、B29 DHS4.4 3’ エンハンサーを増幅した: 前方向5’ - GAAGCGGCCGCACCACCCTGGGCCAGGCTGG - 3’ および逆方向5’ -CCACGCGTAGAGGTGTTAAAAAGTCTTTAGGTAAAG - 3’ 。生ずるPCRフラグメントをNotIおよびMluIで消化し、pCpG-Linker4中の新しい多重クローニング部位中のNotIおよびMluI部位中に結合し、pCpG-DHS4.4をつくる。
【0200】
【化11】
【0201】
工程6: B29プロモーター + HA-eIF5A1K50R + SV40 pA発現カセットpCpG-DHS4.4へのサブクローニング -- pExp-5をつくる。
下記のプライマーを使用してPCRにより、pB29-eIF5A1K50R_7 (工程3) から最小B29プロモーター、合成イントロン、HA-eIF5A1K50RおよびSV40 pAを含有するB29-eIF5A1発現カセットを増幅した: 前方向5’ - GTTATCGATACTAGTGCGACCGCCAAACC - 3’ および逆方向5’ - CAAGCGGCCGCCATACCACATTTGTAGAGGTTTTAC - 3’ 。生ずるPCRフラグメントをClaIおよびNotIで消化し、pCpG-DHS4.4の多重クローニング部位中のClaIおよびNotI部位中にサブクローニングしてpExp-5をつくった。
【0202】
工程7: pExp-5中のHA-eIF5A1K50Rを非HA-eIF5A1K50Rで置換して最終ベクター、pExp5Aをつくる。
pExp-5プラスミドをNcoIおよびNheIで消化して、HA-eIF5A1K50Rを除去した。下記のプライマーを使用してPCRにより、pHM6-eIF5A1K50Rから非HAタッグドeIF5A1K50R PCRフラグメントを増幅した: 前方向5’ - CACCATGGCAGATGATTTGGACTTC - 3’ および逆方向5’ - CGCGCTAGCCAGTTATTTTGCCATCGCC - 3’ 。生ずるPCR生成物をNcoIおよびNheIで消化し、B29-5 #3のNcoIおよびNheI部位に結合してB29- K50Rを発生させた。B29-K50RをNcoIおよびNheIで消化し、470 bpのeIF5A1K50Rフラグメントをゲル精製し、NcoI/NheI消化pExp-5に結合させて最終空ベクター、pExp5Aを発生させた。
【0203】
実施例18: pExp5Aの試験
リポフェクタミン2000を使用して種々の細胞系統をプラスミドでトランスフェクトし、トランスフェクション後24時間に抗HA抗体 (Roche) を使用するウェスタンブロットにより、HA-eIF5A1K50Rの発現を決定した。試験した異なる細胞系統は次の通りであった: P3X63Ag8.653 (マウスBリンパ芽球 - 骨髄腫) 、KAS (ヒト骨髄腫) 、HepG2 (ヒト肝臓肝細胞性癌腫) 、T24 (ヒト膀胱癌腫) 、HT-29 (ヒト結腸直腸腺癌) 、HEK-293 (ヒト胚腎細胞) 、PC3 (ヒト前立腺腺癌) 、HeLa (ヒト頸部腺癌) およびA549 (肺癌腫) 。図24参照。
【0204】
pExp-5プラスミドは、構成的EF1プロモーター (CpG-eIF5A1K50R) を有するプラスミドに匹敵するレベルで、ヒトおよびマウスの両方の骨髄腫細胞系統においてHA-eIF5A1K50Rを発現する。しかしながら、pExp-5により推進されたHA-eIF5A1K50Rの発現は、構成的プロモーターによる発現と比較して、非B細胞系統において限定される。1つの例外はHEK-293細胞、ヒト胚腎臓細胞系統においてであり、ここでpExp-5のトランスフェクション後に高いレベルのHA-eIF5A1K50Rの発現が観察され - これは細胞系統の胚の特質のためであることがある; この時点において、pExp-5が成人腎細胞において発現するかどうかは我々は知らない。
【0205】
毒性研究および臨床的実験において使用する最終プラスミドは、HA-eIF5A1K50Rが非HAタッグドeIF5A1K50R (pExp5A) で置換されているpExp-5のバージョンであろう。pExp-5は最小ヒトB29プロモーター/エンハンサーの制御下にHAタッグドeIF5A1K50Rを含有する; HA-eIF5A1K50Rの発現を構成的発現をもつプラスミドにより推進されるものならびに全長B29プロモーターを含有するプラスミドと比較した。
【0206】
実施例19: in vivo JETPEITMナノ粒子の形成
20 gのマウスに1.5 mg/kg (0.1 ml) -- 1.5 mg/kg = 1.0 mg pExp5A/kg + 0.5 mg h5A/kg -- DNA: siRNA比 = 2:1の投与量で注射するために、in vivo JETPEITMナノ粒子の複合体を形成することをこの実施例において説明する。
【0207】
プラスミドDNAおよびsiRNAを全体積25 mlに希釈する。無菌水を使用して体積を調節する。* 20 mgのプラスミドDNA (10 ml、2 mg/ml) および10 mgのsiRNA (10 ml、1 mg/ml) を全体積25 mlに希釈する。無菌水を使用して差を構成する。25 mlの10%グルコース (PEIキットとともに準備されている) の添加により、DNA溶液の体積を50 mlの5%グルコースに調節する。おだやかに渦形成し、短時間遠心する。in vivo JETPEITMを全体積25 mlの水中に希釈する。* 3.6 mlのin vivo JETPEITMを全体積25 mlの水中に希釈する。最終体積を10%グルコースで50 mlに調節して、5%グルコースの最終濃度にする。おだやかに渦形成し、短時間遠心する。直ちに50 mlの希薄PEIを50 mlの希薄DNA に添加する (順序を逆にしない) 。短時間渦形成し、直ちに回転する。
【0208】
形成後、複合体は8〜10時間安定である。複合体のN/P比は6であるべきである。N/P比はin vivo jetPEIの正に帯電した窒素残基の数/DNAおよびsiRNAの負に帯電したリン酸塩残基の数の比である。DNAおよびsiRNAは同一数のリン酸塩残基の数/gを含有する。したがって、N/P比は複合体中のイオン釣合いの測度である。複合体のN/P比を増加させると、複合体の毒性が増加する。in vivo JET-PEIは150 mM溶液 (窒素残基として表す) として提供されるが、DNAは1 mg中に3モルのアニオンリン酸塩を含有する。最終体積のDNAの最終濃度は0.5 mg/mlを超えるべきではない。DNAは高品質であるべきであり、水中で調製される。in vivo jetPEIおよび10%グルコースを使用前に室温にする。
【0209】
実施例20: マウスにおける静脈内SNS01およびSNS-EF1/UUを使用する投与量範囲の発見および反復投与の研究
SNS01は本発明の1つの態様である - それは多発性骨髄腫の治療をターゲットとする癌療法の生物学である。SNS01は3つの成分から構成されている: elF5Aの前アポトーシス突然変異体を発現するDNAベクター (図22参照); 癌細胞の増殖/抗アポトーシスを促進する自然elF5AをターゲットとするsiRNA (図25中の配列参照); および送出ビヒクルとして作用するポリエチレンイミンと呼ばれる合成ポリマー (in vivo jetPEI; Polyplus Transfection Inc.) 。
【0210】
研究の目的は、最大許容投与量およびマウスへの静脈内SNS01の治療投与量の長期投与の可能性を決定することであった。2つの別々の研究を実施した。最大許容投与量の研究 (研究ID: MTD) は8日の研究であり、マウスに増加する量のSNS01 (2.2 mg/kgから3.7 mg/kgまで) を静脈内に2回投与し、臨床的徴候、体重、器官重量および肝臓酵素を監視して毒性を評価した。9週の反復投与研究 (研究ID: EX6) は、SNS-EF1/UUならびにその成分の2回/週の治療投与量 (1.5 mg/kg) の長期投与後に、毒性を評価するように設計された研究であった。
【0211】
SNS-EF1/UUはSNS01の前臨床的バージョンであり、SNS01複合体におけるようにB細胞特異的プロモーターよりむしろ構成的プロモーター (すべての組織においてすべての時間において発現するプロモーター) によりelF5AK50Rの発現が推進されることにおいて主として異なる。SNS01におけるB細胞特異的B29プロモーターの使用は、骨髄腫細胞を包含する、B細胞由来の細胞に前アポトーシスelF5A突然変異体の発現を限定することによって、療法の安全性を増強するように設計されている。また、EX6研究は、マウス組織においてelF5Aを抑制する毒性作用が存在するかどうかを決定するためにマウス特異的elF5A siRNAを投与したマウスのグループを含んだ。反復投与研究における毒性は、臨床的徴候、体重、血行性、肝臓酵素血行性ならびに組織病理学を監視することによって評価した。
【0212】
【表1】
【0213】
【表2】
【0214】
前臨床実験において、SNS01は0.75 mg/kg〜1.5 mg/kgの投与量で治療的であることが示された (研究EX9) 。8日の投与量範囲発見研究 (研究ID: MTD) において、治療的範囲よりも有意に高い投与量を試験して投与量領域に上限を決定した。試験物品の2回/週の静脈内投与量は2.2 mg/kgおよび2.9 mg/kgのより低い投与量で十分に許容されたが、1匹のマウスは2.9 mg/kgにおいて病的状態に到達し、安楽死させた。3.3 mg/kgまたはそれ以上投与量は、ほぼ20〜25%の生存率を生じた。したがって、最大許容投与量は2.2 mg/kg〜2.9 mg/kgであり、0.75 mg/kg〜1.5 mg/kgの治療範囲より十分に上である。
【0215】
9週の反復投与研究 (研究ID: EX6) において、マウスに治療的投与量 (1.5 mg/kg) のSNS-EF1/UUを2回/週尾静脈注射し、試験物品に関係する毒性作用は研究期間にわたって観察されなかった。また、DNAおよびsiRNAを反復して別々に試験し、マウスは両方を十分に許容した。ヒトelF5A siRNAはマウスにおいて活性ではないので、マウスelF5A特異的siRNAをまたこの研究に含めた。マウスelF5A siRNAの慢性投与に関係する毒性作用は、9週の期間に観察されなかった。これらの結果が示すように、治療的投与量のSNS01およびSNS-EF1/UUは、長期間にわたって投与したときでさえ、マウスに対して無毒である。
【0216】
【表3】
【0217】
試験システムおよび研究設計
この研究のすべての面はウォータールー・アニマル・ケアー・コミッティー大学 (the University of Waterloo Animal Care Committee) (カナダオンタリオ州ウォータールー) (設立:Canadian Council on Animal Care and the Province of Ontario Animals for Research Actによる) が設立したガイドラインに従って実施した。
【0218】
CD-1およびBALB/cマウスはチャールス・リバー・ラボラトリーズ (Charles River Laboratories) (カナダケベック州) から入手した。両方の研究のマウスに、尾静脈静脈内注射により試験物品を2回/週与えた。遅い注射 (約2〜3分) を使用して、0.2 mlより大きい体積を送出した。
8日の研究のマウスは研究の開始時にほぼ6〜9週齢であった。9週の反復投与研究のためのマウスは研究の開始時にほぼ5〜6週齢であった。
【0219】
【表4】
【0220】
【表5】
【0221】
8日最大許容投与量の研究 (MTD)
2投与量の8日研究は、SNS01の最大許容投与量を決定するように設計された投与量範囲発見研究であった。投与量範囲は2.2 mg/kg〜3.7 mg/kgであり、0.75 mg/kg〜1.5 mg/kgの治療投与量範囲よりかなり上である。SNS01の最低投与量 (2.2 mg/kg) において、わずかにしわの寄った毛皮を示し、1時間以内に消散する活性の減少を除外して、毒性の臨床的徴候は観察されなかった。第2回の2.2 mg/kgのSNS01の注射後、毒性の臨床的徴候は観察されなかった。すべてのマウスは研究を通じて彼らの体重を維持した。器官の巨視的変化は観察されなかった。器官重量/体重の比は、肝臓重量/体重の比の適度の増加を除外して、対照グループから変化しなかった。しかしながら、この比の増加はいかなるより高い投与レベルのグループにおいても観察されなかったので、試験物品に関係することはありそうもない。
【0222】
5匹のマウスのうちの4匹は2.9 mg/kgのSNS01を許容し、毒性の臨床的徴候はなかった。しかしながら1匹のマウスは注射の1時間以内に痙攣および軽症の呼吸窮迫を経験し、ヒト的に安楽死させなければならなかった。残りのマウスにおいて2回目のSNS01の注射後、毒性の臨床的徴候はなかった。マウスは研究を通じて彼らの体重を維持し、器官における巨視的変化または器官重量/体重における変化は観察されなかった。2.9 mg/kgのSNS01の2回の投与後、ALTの血清レベルのわずかの減少が存在した。
【0223】
期待されるように、3.3 mg/kgまたはそれ以上におけるSNS01の投与は十分に許容されず、両方のグループにおいて、1匹を除くすべてのマウスはヒト的に安楽死させなくてはならなかった。病的状態のためにマウスをヒト的に安楽死させなくてはならなかったすべての場合において、臨床的徴候は注射の1時間以内に現れ、高い投与量のPEIを使用した他の報告された研究と一致した。3.3 mg/kgおよび3.7 mg/kgの生存するマウスは注射後4時間以内に回復し、2回目の注射を投与しなかったが、研究を通じて彼らの重量を維持した。したがって、SNS01の最大許容投与量は2.2 mg/kg〜2.9 mg/kgであるように思われる。
【0224】
9週の反復投与量の研究 (EX6)
9週の反復投与量の研究の目的は、SNS-EF1/UU、すなわち、SNS01の開発間に前臨床的研究に使用された複合体の治療投与量 (1.5 mg/kg) の臨床的投与の安全性を評価することであった。SNS-EF1/UUはSNS01と有意に異ならず、主要な差は材料が研究等級であること、およびすべての細胞型において活性である構成的ヒトPEIプロモーターによりelF5AK50R発現が推進されることである。SNS01はB細胞特異的プロモーターを使用してelF5AK50R発現を推進させるが、この安全性の研究における構成的プロモーターの使用は非B細胞組織における突然変異体elF5AK50Rの蓄積から生ずる毒性の評価可能とする。
【0225】
9週の反復投与量の研究の他の面は、SNS-EF1/UUの個々の成分の安全性を試験するグループを含めることであった。DNAグループ (Ex6-G3) にelF5Aプラスミドおよび非ターゲッティング対照siRNAを含有する複合体を投与したが、siRNAグループ (Ex6-G4) にヒトelF5A (h5A1) および非発現プラスミドを含有する複合体を投与した。試験物品SNS-EF1/UUは内因的マウスelF5Aの発現に影響を与えないヒトelF5A siRNAを含有するが、この研究の他の特徴は非発現プラスミドとマウスelF5Aを効率よくターゲットとするsiRNAを含有するPEI複合体を投与したグループ (Ex6-G6) を含めたことであった。このグループは活性elF5A siRNAの慢性投与の安全性の評価を可能とした。
【0226】
すべての動物は予定した犠牲日まで生存した。毒性の臨床的徴候は9週の研究過程にわたっていずれのグループにおいても観察されず、すべてのグループにおけるマウスは研究期間の間に重量を増加し続けた。赤血球および白血球を治療開始後3および6週に計測し、すべての投与グループにおいて正常であった。血清肝臓酵素レベルを犠牲後に測定し、すべてのマウスについて正常範囲内であった。器官の巨視的外観の変化はいずれのグループにおいても観察されなかった。主要な器官の組織病理学的分析を独立病理学者が実施し、試験物品に帰する毒性は明らかにされなかった。
【0227】
マウスは治療投与量のSNS-EF1/UUの慢性投与を十分に許容し、悪影響は観察されなかった。さらに、マウス特異的elF5A siRNAの慢性投与は毒性作用を明らかにせず、ヒトelF5A siRNAを含有するPEI複合体の投与はヒトにとって安全であろうことが示された。
【0228】
実施例21: 多発性骨髄腫腫瘍を有するマウスに静脈内SNS-B29/UUおよびSNS01投与する治療効能研究
SNS01は上に記載した通りである。試験物品SNS-B29/UUはSNS01の前臨床的バージョンである。SNS-B29/UUはSNS01と非常にわずかに異なり、主要な差は成分がGLP等級よりむしろ研究等級であることである。報告されたこの研究の目的は、SNS-B29/UUの最小作用投与量を決定すること、およびSNS01を含んでなるGLP等級材料が前臨床的研究において使用した研究等級材料と同様に働くことを確認することであった。反復投与の腫瘍研究 (研究ID: EX9) は5週の研究であり、マウスにおける皮下腫瘍増殖を阻害する順次減少する投与量のSNS-B29/UUの能力を評価して、SNS01の最適な治療投与量を決定した。また、臨床的徴候、体重および器官重量を監視することによって、治療した動物の毒性徴候を評価した。
【0229】
【表6】
【0230】
【表7】
【0231】
皮下ヒト多発性骨髄腫腫瘍を有するSCIDマウスにおいて、SNS-B29/UUの治療範囲を決定した。0.15 mg/kg〜1.5 mg/kgのSNS-B29/UUの投与量を試験した。2回/週の腫瘍体積の測定および犠牲後の腫瘍組織の切除および秤量により、試験物品の抗腫瘍効能を決定した。0.75 mg/kg〜1.5 mg/kgのSNS-B29/UUの投与量は腫瘍を有意に収縮させ、SNS-B29/UUの治療範囲が0.75 mg/kg〜1.5 mg/kgの範囲にあることを示す。また、皮下腫瘍増殖の効率的阻害は0.38 mg/kg SNS-B29/UUにおいて観察されたが、腫瘍収縮は観察されなかった。腫瘍増殖のある程度の阻害は0.15 mg/kg程度に低い投与量においてさえ観察され、広い治療範囲を示した。図26および27参照。
【0232】
GLP等級の成分を使用して製造されたSNS01の効能をSNS-B29/UUと比較し、腫瘍増殖の阻害において匹敵する効能を有することが発見された。腫瘍を有するSCIDマウスはSNS01およびSNS-B29/UUを使用する治療に十分に許容性であり、マウスは研究を通じて重量を増加し続けた。
【0233】
【表8】
【0234】
試験システムおよび研究設計
雌C.B.17/IcrHsd-Prkdc (SCID) マウスをハーラン (Harlan) (米国インジアナ州インジアナポリス) から入手した。5〜6週齢のマウスの右側腹部に10×106の生存可能なKAS-6/1 (ヒト多発性骨髄腫) 細胞を注射して、皮下腫瘍を確立した。腫瘍がほぼ20〜40 mm3のサイズ (腫瘍細胞注射後ほぼ4週) に到達したとき、SNS-B29/UUの治療を開始した。腫瘍がほぼ130 mm3のサイズの到達したとき (腫瘍細胞注射後ほぼ6週) 、SNS01を使用する治療を開始した。マウスに尾静脈の静脈内により試験物品を2回/週投与した。
【0235】
【表9】
【0236】
反復投与腫瘍研究
SNS-B29/UUの最小有効治療投与量を決定し、そして研究等級試験物品SNS-B29/UUにより証明される腫瘍阻害活性をGLP等級SNS01試験物品が保持することを決定するように、反復投与腫瘍研究を設計した。二次的目的は、治療したマウスを臨床的徴候、体重および器官重量について治療したマウスを監視することによって、治療の毒性作用を評価することであった。デジタルキャリパーを使用して腫瘍体積を2回/週測定して、試験物品の治療抗腫瘍活性をモニターした。殺した後、腫瘍を摘出し、そして秤量した。
【0237】
すべてのマウスは予定した犠牲日まで生存した。非発現性プラスミドおよび非ターゲッティングsiRNAを含有するPEIナノ複合体で治療した対照マウスは犠牲時間に284 mm3の平均腫瘍体積を有したが、1.5 mg/kgのSNS-B29/UUで治療したマウスはわずかに13 mm3の平均腫瘍体積を有し、腫瘍増殖が95% (*p = 0.026) 減少した。しかしながら、1.5 mg/kgのSNS-B29/UUで治療したマウスから腫瘍を切除しようと試みたとき、マウスのいずれにも腫瘍の証拠は発見されなかった。SNS-B29/UUの投与量を半分の0.75 mg/kgに減少しても、腫瘍体積および重量はなおそれぞれ91% (*p = 0.03) および87% (*p = 0.04) 減少し、1匹のマウスにおいて、腫瘍は完全に消失した。したがって、SNS-B29/UUの2回/週注射のための最適な治療投与量は0.75 mg/kg〜1.5 mg/kgの間であるように思われる。0.15 mg/kg程度に低い2回/週SNS-B29/UUの投与量は最終腫瘍体積を60%減少させ、SNS-B29/UUが広い投与量範囲にわたって効力のある抗腫瘍活性を有することを示す。
【0238】
腫瘍増殖を阻害することに加えて、0.75 mg/kgおよび1.5 mg/kgのSNS-B29/UUおよびSNS01を使用する治療は腫瘍体積を有意に減少させ、この治療が、多分腫瘍中のアポトーシスの誘導により、腫瘍退縮を誘導できることを示す。0.75 mg/kgおよび1.5 mg/kgの投与量レベルのSNS-B29/UUで治療した腫瘍担持マウスにおける腫瘍体積の変化の百分率は、それぞれ、-244%および-245%であった。対照マウスの腫瘍は、同一時間の間にサイズが2000%より大きく増加した。
【0239】
また、SNS01の2回/週の注射は多発性骨髄腫腫瘍を有意に収縮させた。1.5 mg/kgのSNS01で治療したマウスの腫瘍体積の変化の百分率は-349%であり、SNS01がちょうどSNS-B29/UUのように有効であることを示した。GLP等級の材料の使用は、事実、生物学的活性を増加させることができる。なぜなら、SNS01を使用する治療はわずかに25日間の治療後腫瘍体積を349%減少させたが、SNS-B29/UUは35日間の治療後腫瘍体積を245%減少させたからである。さらに、SNS01で治療した腫瘍は非常に大きく (約130 mm3) 、SNS01を使用する治療が十分に確立された腫瘍に対して有効であることを示す。
【0240】
すべてのマウスは治療を十分に許容し、毒性の臨床的徴候は観察されなかった。すべてのグループにおけるマウスは、研究を通じて重量を増加し続けた。器官の巨視的外観は壊死において観察されず、器官重量/体重が有意に変化した。
【0241】
したがって、SNS01 (およびその前臨床的バージョンSNS-B29/UU) はSCIDマウスに十分に許容され、2回/週の静脈内注射により送出すとき、ヒト多発性骨髄腫腫瘍の皮下的治療において極めて有効である。試験したSNS-B29/UUのすべての投与量は腫瘍増殖の阻害において有効であるが、1.5 mg/kgの最高投与量は治療を受けたすべてのマウスにおいて腫瘍を首尾よく排除した。
【0242】
実施例22: プラスミドDNAおよびsiRNAポリエチレンイミン (JetPEI) 複合体の生物分布
緑色蛍光タンパク質 (“GFP”) GFP発現構築物を使用して、PEI複合体により送出されたプラスミドDNAの局在化を決定した。2つのプロモーターを使用してGFP発現を推進した: EF1: 偏在性プロモーター (EF1::GFP) またはB29: B細胞特異的プロモーター (B29::GFP) 。20μgのGFPプラスミドDNAおよび10μgの蛍光標識化 (DY547) h5A1 siRNAを含有するPEI複合体を6のN/P比で製造した。BALB/Cマウスに5%グルコースまたはPEI複合体を連続2日間静脈内注射した。最初の注射後72時間に、マウスを安楽死させ、マウスの器官を収集し、共焦点顕微鏡検査によりGFP発現およびDY547-siRNAを分析した。
【0243】
骨髄: ほとんどの場合において、DY547-siRNAの証拠は存在したが、GFP発現の証拠は存在しなかった。器官収集のタイミングはGFPのピーク発現と一致せず、GFPシグナルのクエンチングは存在しうるか、あるいはGFPは発現されることはないであろう。しかしながら、ある場合において同一骨髄細胞に共局在化したGFPおよびDY547が観察された。したがって、これはPEIナノ粒子が静脈内注射により与えられたとき生きている動物中の骨髄細胞をトランスフェクトできるという証拠を提供する。
【0244】
肺: ほとんどの場合において、DY547-siRNAの証拠は存在したが、GFP発現の証拠は存在しなかった。器官収集のタイミングはGFPのピーク発現と一致せず、GFPシグナルのクエンチングは存在しうるか、あるいはGFPは発現されることはないであろう。
脾臓: またDY547-siRNAの存在に陽性である細胞中に共局在化するGFP発現の証拠 (PEIプロモーターより推進されるとき) が見られた。B29プロモーターにより推進されるとき、GFPの発現は脾細胞において非常に低い。これが示すように、PEIナノ粒子は脾臓の細胞をトランスフェクトするようである。
【0245】
腎臓: GFPまたはDY547は観察されず、ナノ粒子が腎臓に入らないことを示した。
肝臓: ほとんどの場合においてDY547-siRNAの証拠は存在したが、GFP発現は存在しなかった。これはPEIナノ粒子が肝臓の細胞をトランスフェクトしている証拠を提供する。
心臓: 心臓組織中のEF1::GFPおよびDY547-siRNAの共局在化が見られ、こうしてPEIナノ粒子がこの器官をトランスフェクトできること示す。GFPはB29プロモーターとともに観察されなかった。
【0246】
実施例23: HA-elF5AK50R発現に対するDNA: siRNA比の効果
B29-HA-elF5AK50R (B細胞特異的プロモーターにより推進されるプラスミド) およびh5A1 siRNAを含有するナノ粒子でKAS細胞をトランスフェクトした。異なる比でpExp5Aおよびh5A1 siRNAを含有するJET PEITMナノ粒子を作り、室温において4時間インキュベートした後、KAS細胞に添加した。トランスフェクション後4時間に、ナノ粒子を含有する培地を新鮮な培地と交換した。24時間後、細胞ライゼイトを収集し、HAに対する抗体とともにウェスタンブロット分析のために使用した。DNA: siRNAの比を2:1の標準的比から変化させた。HA-elF5AK50R蓄積は1:0、3:1および2:1においてピークとなった。図30参照。
【0247】
実施例24: ナノ粒子のトランスフェクションにより誘導されるアポトーシスに対するDNA: siRNAの比の効果
pExp5Aおよびh5A1 siRNAを異なる比で含有するナノ粒子を作り、室温において4時間インキュベートした後、KAS細胞に添加した。トランスフェクション4時間後、ナノ粒子を含有する培地を新鮮な培地で置換した。48時間後、細胞を収集し、アンキシンV/PIで標識化し、FACSにより分析した。アポトーシスの誘導は、2:1の標準的DNA: siRNA比のナノ粒子でトランスフェクトした細胞において最高であった。図31参照。
【0248】
実施例25: elF5AK50RプラスミドおよびeIF5A1 siRNA (siSTABLEまたは非siSTABLE) を含有するPEI複合体 (N/P = 6または8) の投与は多発性骨髄腫皮下腫瘍の増殖を阻害し、腫瘍を収縮させる。
SCIDマウスにKAS細胞を皮下注射した。明白な腫瘍が観察されたとき、治療を開始した。マウスに下記を2回/週静脈内注射した: (G1) 20 mgのpCpG-mcs (空ベクター) および10 mgの対照siRNAをN/P = 8で含有するPEI複合体 (中程度の投与量); (G5) 20 mgのRNAi抵抗性プラスミドpCpG-elF5AK50Rおよび10 mgのsiSTABLE h5A1 siRNAをN/P = 8で含有するPEI複合体 (中程度の投与量、siSTABLE); (G8) 20 mgのRNAi抵抗性プラスミドpCpG-elF5AK50Rおよび10 mgのh5A1 siRNAをN/P = 6で含有するPEI複合体 (中程度の投与量、N/P = 6) 。示したデータは各グループにおけるマウスについての個々の腫瘍体積である。最初の治療後第40日に、最終の注射を実施した。図32参照。
【0249】
実施例26: 腫瘍組織はJET PEITMナノ粒子を効率よく吸収し、ナノ粒子はプラスミドおよびsiRNAを同一細胞に送出す。
pExp-GFP (B細胞特異的プロモーターの制御下のGFP) およびDY547-siRNA (蛍光標識化siRNA) を含有するナノ粒子を注射した後48時間に、腫瘍を切断した。共焦点顕微鏡検査によりGFPおよびDY547の共局在化発現が観察され、腫瘍組織はナノ粒子を効率よく吸収し、ナノ粒子はプラスミドおよびsiRNAを同一細胞に送出していることを示される。図33参照。
【特許請求の範囲】
【請求項1】
eIF5A1の3’ 末端をターゲットとするeIF5A1 siRNAの複合体と、突然変異体eIF5A1をコードするポリヌクレオチドとを含んでなる発現ベクターとを含んでなる組成物であって、突然変異体eIF5A1はハイプシン化されることができず、かつsiRNAおよび発現ベクターはポリエチレンイミンに対して複合化されて複合体を形成している組成物。
【請求項2】
ターゲット遺伝子をターゲットとして被検体におけるターゲット遺伝子の内因的発現を抑制するsiRNAと、被検体においてRNAi抵抗性プラスミド中で発現されることができるターゲットタンパク質をコードするポリヌクレオチドとを含んでなる組成物であって、siRNAおよびプラスミドはポリエチレンイミンに対して複合化されて複合体を形成している組成物。
【請求項3】
siRNAが図25に示す配列を有し、そして突然変異体eIF5A1をコードするポリヌクレオチドがeIF5A1K50Rである、請求項1の組成物。
【請求項4】
組織特異的プロモーターを含んでなる請求項3の組成物。
【請求項5】
B細胞特異的プロモーターを含んでなる請求項4の組成物。
【請求項6】
B細胞プロモーターがB29である請求項5の組成物。
【請求項7】
発現ベクターがpCpGを含んでなる請求項3の組成物。
【請求項8】
eIF5A1 siRNAと突然変異体eIF5A1ポリヌクレオチドを含んでなる発現ベクターが独立してポリエチレンイミンに対して複合化している、請求項1の組成物。
【請求項9】
eIF5A1 siRNAと突然変異体eIF5A1ポリヌクレオチドを含んでなる発現ベクターが一緒になってポリエチレンイミンに対して複合化している、請求項1の組成物。
【請求項10】
eIF5A1の3’ 末端をターゲットとするeIF5A1 siRNAと、突然変異体eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターとを含んでなる組成物であって、突然変異体eIF5A1がハイプシン化されることができず、かつsiRNAおよび発現ベクターが癌を治療するために被検体に送出される組成物。
【請求項11】
癌が多発性骨髄腫である、請求項10の組成物。
【請求項12】
請求項10の組成物を被検体に投与することを含んでなる癌を治療する方法。
【請求項13】
請求項1の組成物を被検体に投与することを含んでなる癌を治療する方法。
【請求項14】
組成物を静脈内、腹腔内または腫瘍内に投与する、請求項12の方法。
【請求項15】
eIF5A1の3’ 末端をターゲットとするsiRNAと、突然変異体eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターとを異なる経路を経て送出す、請求項13の方法。
【請求項16】
組成物を注射により約0.15 mg/kg〜約1.5 mg/kgの投与量で毎週2回投与する、請求項12の方法。
【請求項17】
組成物を注射により約0.75 mg/kg〜約1.5 mg/kgの投与量で毎週2回投与する、請求項12の方法。
【請求項1】
eIF5A1の3’ 末端をターゲットとするeIF5A1 siRNAの複合体と、突然変異体eIF5A1をコードするポリヌクレオチドとを含んでなる発現ベクターとを含んでなる組成物であって、突然変異体eIF5A1はハイプシン化されることができず、かつsiRNAおよび発現ベクターはポリエチレンイミンに対して複合化されて複合体を形成している組成物。
【請求項2】
ターゲット遺伝子をターゲットとして被検体におけるターゲット遺伝子の内因的発現を抑制するsiRNAと、被検体においてRNAi抵抗性プラスミド中で発現されることができるターゲットタンパク質をコードするポリヌクレオチドとを含んでなる組成物であって、siRNAおよびプラスミドはポリエチレンイミンに対して複合化されて複合体を形成している組成物。
【請求項3】
siRNAが図25に示す配列を有し、そして突然変異体eIF5A1をコードするポリヌクレオチドがeIF5A1K50Rである、請求項1の組成物。
【請求項4】
組織特異的プロモーターを含んでなる請求項3の組成物。
【請求項5】
B細胞特異的プロモーターを含んでなる請求項4の組成物。
【請求項6】
B細胞プロモーターがB29である請求項5の組成物。
【請求項7】
発現ベクターがpCpGを含んでなる請求項3の組成物。
【請求項8】
eIF5A1 siRNAと突然変異体eIF5A1ポリヌクレオチドを含んでなる発現ベクターが独立してポリエチレンイミンに対して複合化している、請求項1の組成物。
【請求項9】
eIF5A1 siRNAと突然変異体eIF5A1ポリヌクレオチドを含んでなる発現ベクターが一緒になってポリエチレンイミンに対して複合化している、請求項1の組成物。
【請求項10】
eIF5A1の3’ 末端をターゲットとするeIF5A1 siRNAと、突然変異体eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターとを含んでなる組成物であって、突然変異体eIF5A1がハイプシン化されることができず、かつsiRNAおよび発現ベクターが癌を治療するために被検体に送出される組成物。
【請求項11】
癌が多発性骨髄腫である、請求項10の組成物。
【請求項12】
請求項10の組成物を被検体に投与することを含んでなる癌を治療する方法。
【請求項13】
請求項1の組成物を被検体に投与することを含んでなる癌を治療する方法。
【請求項14】
組成物を静脈内、腹腔内または腫瘍内に投与する、請求項12の方法。
【請求項15】
eIF5A1の3’ 末端をターゲットとするsiRNAと、突然変異体eIF5A1をコードするポリヌクレオチドを含んでなる発現ベクターとを異なる経路を経て送出す、請求項13の方法。
【請求項16】
組成物を注射により約0.15 mg/kg〜約1.5 mg/kgの投与量で毎週2回投与する、請求項12の方法。
【請求項17】
組成物を注射により約0.75 mg/kg〜約1.5 mg/kgの投与量で毎週2回投与する、請求項12の方法。
【図1】


【図2】
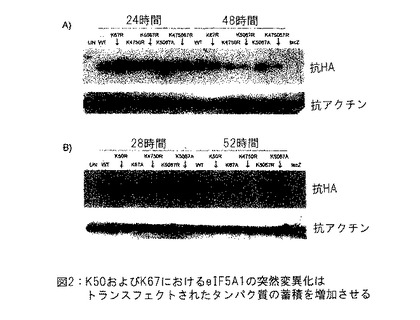
【図3】

【図4】
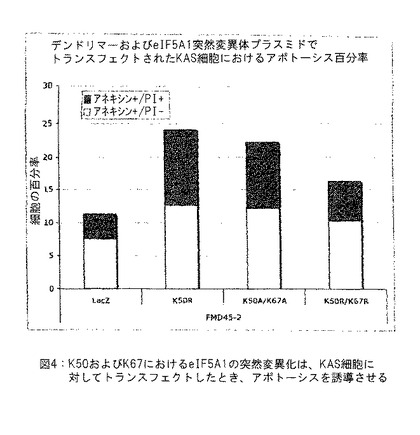
【図5】

【図6】

【図7A】

【図7B】

【図7C】

【図7D】
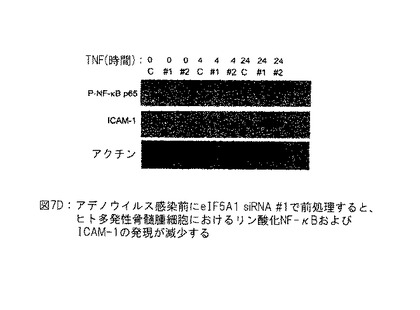
【図7E】
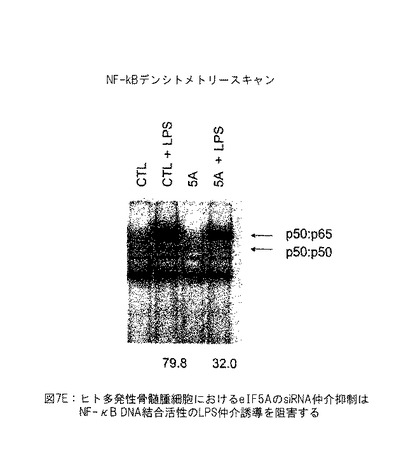
【図7F】
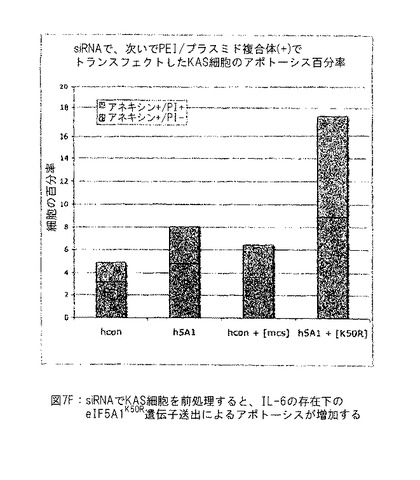
【図8】
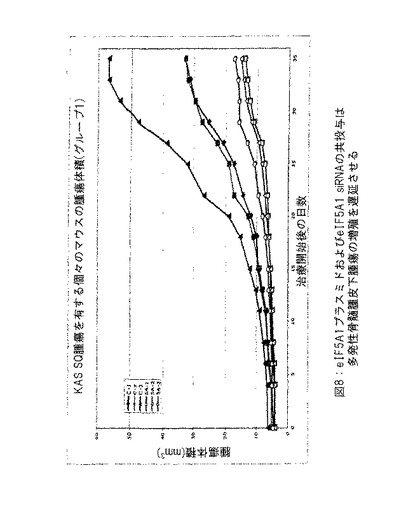
【図9】
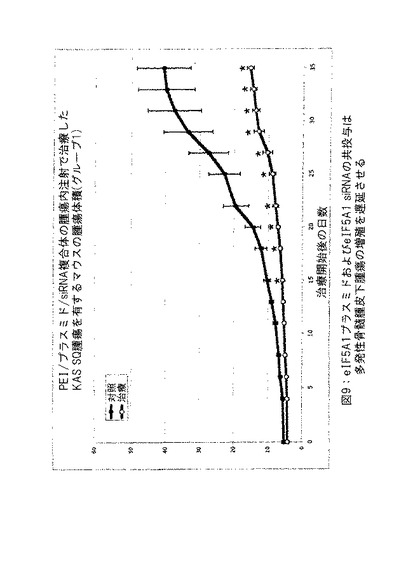
【図10】

【図11】
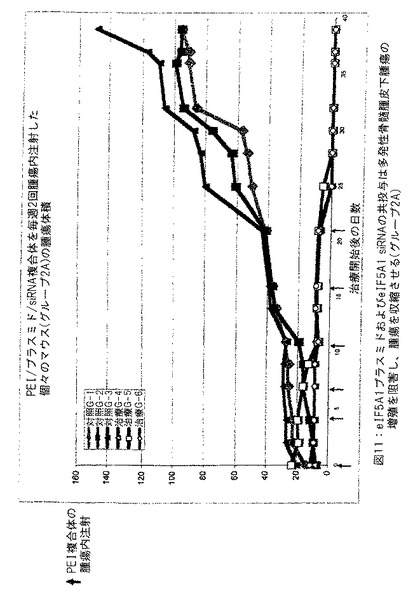
【図12】

【図13A】
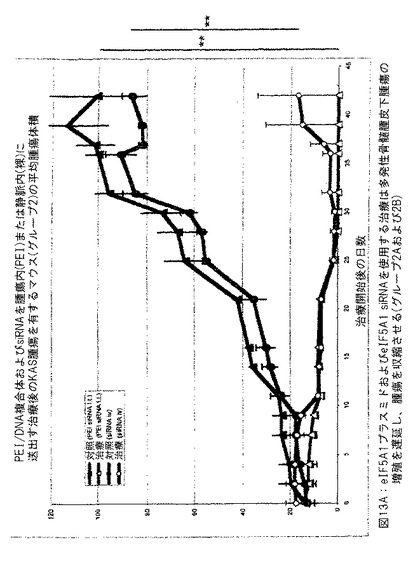
【図13B】
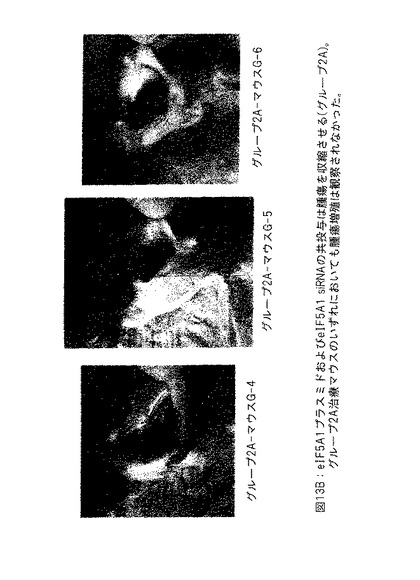
【図13C】
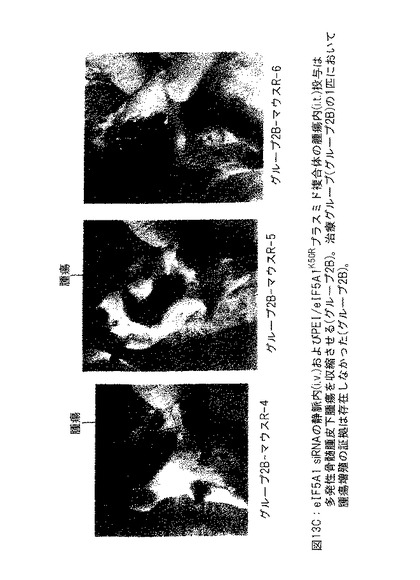
【図14】

【図15】
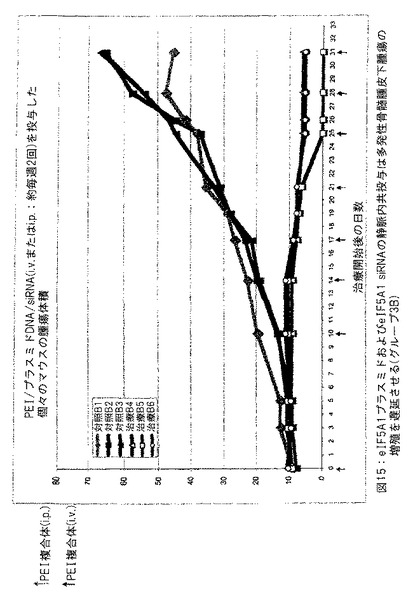
【図16】

【図17】
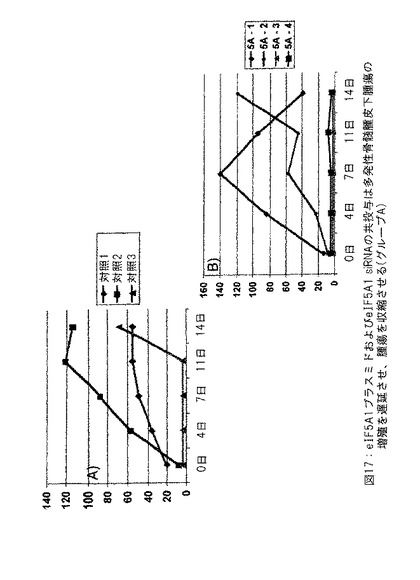
【図18】
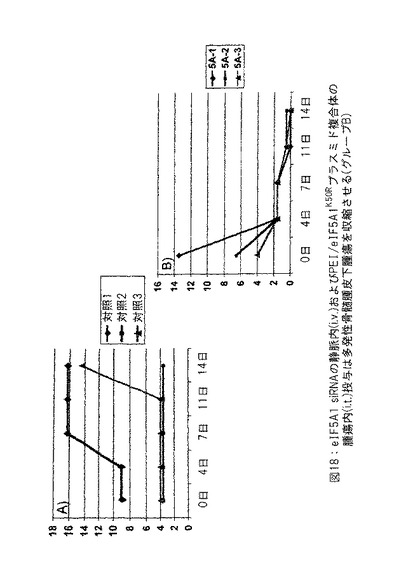
【図19】

【図20】
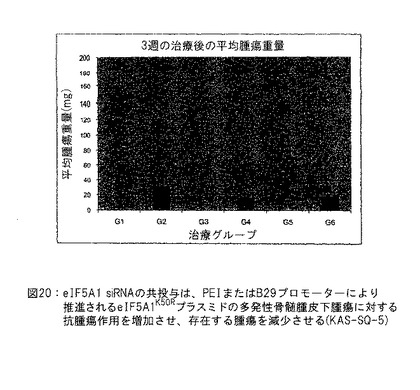
【図21】
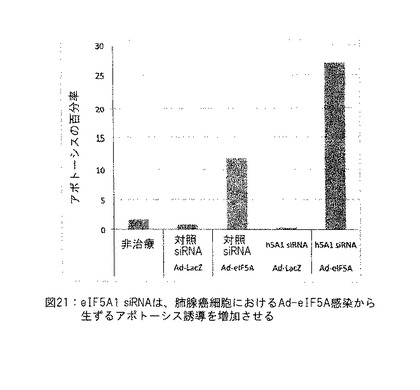
【図22】

【図23】

【図24】
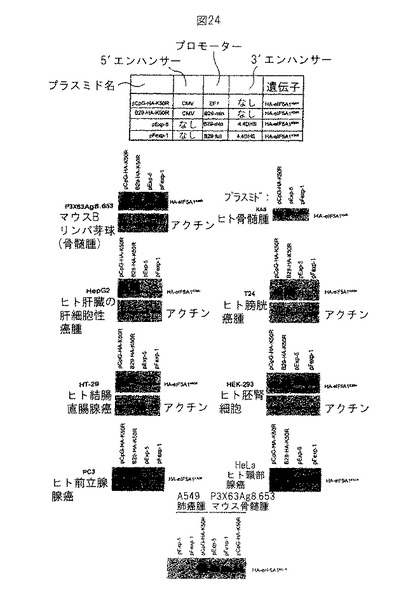
【図25】

【図26】

【図27】
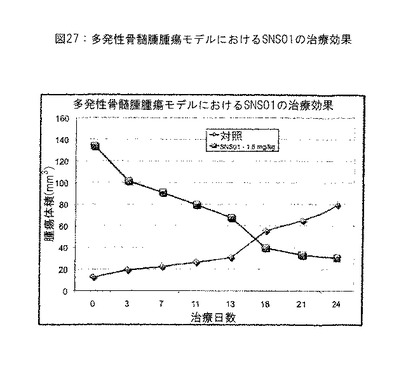
【図28】

【図29】

【図30】

【図31】
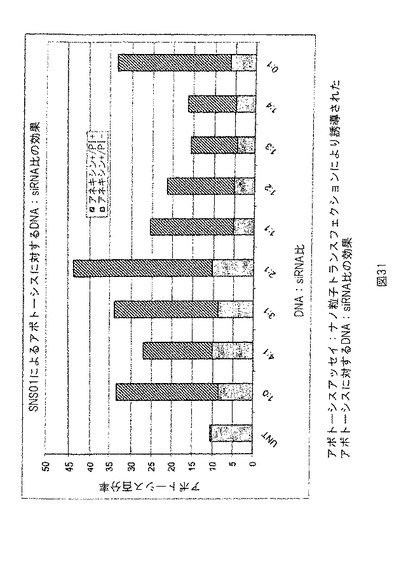
【図32】

【図33】
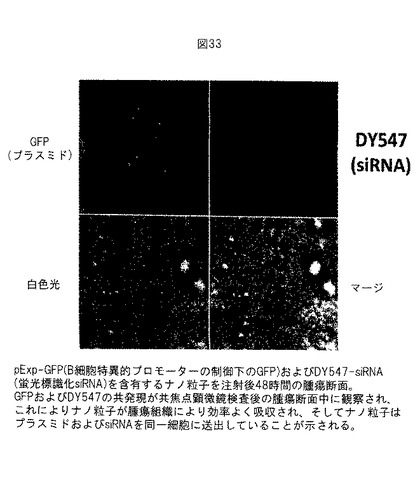
【図2】
【図3】
【図4】
【図5】
【図6】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13A】
【図13B】
【図13C】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【公表番号】特表2011−516035(P2011−516035A)
【公表日】平成23年5月26日(2011.5.26)
【国際特許分類】
【出願番号】特願2010−549939(P2010−549939)
【出願日】平成21年3月9日(2009.3.9)
【国際出願番号】PCT/US2009/036583
【国際公開番号】WO2009/114487
【国際公開日】平成21年9月17日(2009.9.17)
【出願人】(502458132)セネスコ テクノロジーズ,インコーポレイティド (16)
【Fターム(参考)】
【公表日】平成23年5月26日(2011.5.26)
【国際特許分類】
【出願日】平成21年3月9日(2009.3.9)
【国際出願番号】PCT/US2009/036583
【国際公開番号】WO2009/114487
【国際公開日】平成21年9月17日(2009.9.17)
【出願人】(502458132)セネスコ テクノロジーズ,インコーポレイティド (16)
【Fターム(参考)】
[ Back to top ]