
抗ウイルス薬および癌に対する医薬を製造するためのパラポックスウイルスovis株の使用
【課題】感染性または非感染性の免疫不全のための免疫治療剤の提供。
【解決手段】ヒトおよび動物におけるウイルス感染および癌に対する医薬の製造のための、パラポックスウイルスovis株NZ2、NZ−7、NZ−10およびorf−11の1つに分類上属するウイルスの使用。
【解決手段】ヒトおよび動物におけるウイルス感染および癌に対する医薬の製造のための、パラポックスウイルスovis株NZ2、NZ−7、NZ−10およびorf−11の1つに分類上属するウイルスの使用。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、感染性または非感染性の免疫不全のための免疫治療剤としての、パラポックスウイルスovis株の使用、および腫瘍性疾患およびウイルス感染ならびにそのような感染を伴う疾患を治療するためのパラポックスウイルスovis株の使用、さらにヒトおよび動物において使用するための医薬の製造のためのパラポックスウイルスovis株の使用に関する。
【0002】
本発明はさらに、パラポックスウイルスovis株、およびそれから調製される医薬形態の、ストレス(例えば手術)後の感染性疾患を予防または軽減するためのストレス後感染防御における免疫治療剤または免疫予防剤としての使用;手術または介入前(例えば、プロテーゼの移植前または歯科的介入の前)に投与することによる感染性疾患の予防または軽減のための感染予防におけるそれらの使用、(例えば気管の、パピローマウイルス感染の、ヘルペスウイルスによる感染の、HIV感染の、および肝炎ウイルスによる感染のような内蔵のウイルス感染の)急性および慢性のウイルス感染の感染後防御または治療におけるそれらの使用、創傷治癒過程を促進するための創傷治癒におけるそれらの使用、および治癒がごくわずかまたは全く治癒しない創傷の治癒の支援のためのそれらの使用(例えば、脚の潰瘍)、多発性硬化症、喘息、いぼおよびその他の皮膚の新形成等の疾患に対するそれらの使用、様々なアレルギー疾患に対するそれらの使用、全身的なアレルギーの発症を阻止するためのそれらの使用、および局所的なアレルギーに対するそれらの使用、および例えば老人患者における健康状態の改善のためのそれらの使用に関し、本発明において用いられるパラポックスウイルスovis株はNZ2、NZ−7、NZ−10およびorf−11である。
【0003】
本発明はさらに、特定の細胞、例えばWI−38, MRC−5もしくはVero細胞、で継代および/または適合させることによって得られたこれらの株の子孫、またはこれらの株もしくは子孫から得られたウイルスの一部もしくは断片を使用することも可能である。一部とは、適当な系、例えば培養線維芽細胞において、適当なベクター、例えばワクシニアの助けかりて発現するゲノムまたはサブゲノム断片であると理解される。断片とは、例えば超音波処理により物理的に破壊された粒子の、例えばクロマトグラフィー等の生化学的な精製によって得られる一部分であると理解される。
【0004】
本発明はさらに、医薬および医薬調製物を製造するためのパラポックスウイルスovisの該株の使用に関する。さらに、本発明は、抗ウイルス治療または癌治療のための医薬および医薬調製物を製造するための、他の治療法と組み合わせた、パラポックスウイルスovisの該株の使用に関する。
【背景技術】
【0005】
潜在的で慢性的な持続的ウイルス感染は、免疫抑制によって活性化または再活性化し得ること、また逆に、潜在的であるウイルス(例えば潜在的なヘルペスウイルス感染は免疫抑制に関連して再発する:ストレスまたはコルチゾン投与に関連する陰唇疱疹)によって誘発される急性疾患を免疫系が抑制することが知られている。さらに、慢性的に持続するおよび潜在的なウイルス感染は、低分子量を有する従来の抗ウイルス物質を用いて治療することは困難であるかまたは不可能でさえある。
【0006】
その理由は、そのような感染に関連するウイルスの酵素活性の不在(例えば、最初にヌクレオシドインヒビターをウイルスの核酸に取り込んでこのインヒビターが例えばウイルスDNAの鎖を終止させるようにするウイルスポリメラーゼの不在;例えば、抗ウイルス化合物を最初にリン酸化してこの化合物が活性になるようにする必要のある何らかのウイルスチミジンキナーゼ活性の不在)、または宿主の免疫系による、感染したまたは変性した細胞の、例えば癌細胞の、またはウイルス抗原の、何らかの認識の欠如であり得る。
【0007】
さらに、臨床上治りにくいウイルス感染に関連して、別のウイルスによる重感染が、その臨床上治りにくいウイルスに対する抗ウイルス効果をもたらし得ることが知られている1)。著者1)は、T細胞、ナチュラルキラー細胞およびマクロファージよって分泌される、IFN−γおよびTNF−αなどのインターフェロンに対するこの効果の関連を実証することができた。
【0008】
クラスI制限的細胞傷害性T細胞がHBVトランスジェニックマウスにおける肝細胞HBV遺伝子発現を阻害することが可能であること、肝細胞をなんら破壊することなくこの過程が起こること、およびこの過程がTNF−αおよびIFN−γによって発揮されることを実証した別の初期の研究2)がこの著者によって得られた結果によって確認された。
【0009】
「パラ特異的免疫」を誘導するための物質、即ち「パラ免疫誘導物質」と呼ばれる物質は、獣医の現場において、比較的長い間、治療的、感染後防御的および予防的に用いられてきた。パラ特異的免疫誘導物質は、例えば、化学的に不活化されたパラポックスウイルスovis、D1701株(DE3504940)からなる。BAYPAIV1UN(登録商標)はこのウイルス(パラポックスウイルスovis、D1701株)をもとに調製された製品である。
【0010】
動物において、不活化されたパラポックスウイルスは、非常に多岐にわたる病原体によって引き起こされた感染に対する非特異的な保護を誘導する。この保護は、内因的な防御システムの一部を構成する様々なメカニズムによって媒介されると考えられる。
【0011】
これらのメカニズムには、感染の誘導、ナチュラルキラー細胞の活性化、「コロニー刺激活性」(CSA)の誘導およびリンパ球増殖の刺激が含まれる。作用機序の初期の研究は、インターロイキン2およびインターフェロンαの刺激効果を実証した3)。
【発明の概要】
【発明が解決しようとする課題】
【0012】
この背景に基づき、本発明の目的は、パラポックスウイルスovisD1701株によるパラ特異的免疫反応の上記の一般化された誘導を質的に増大させるようにパラポックスウイルスovisの優れた効果の治療上の有用性を更に改善し、より優れた抗ウイルスまたは抗腫瘍効果を低用量を用いて達成することができるように改善することである。またこの治療効果は副作用が少ないと期待される。
【0013】
したがって、本発明の目的は、パラポックスウイルスの免疫学的効果を改善することである。この目的は、慣用的に用いられているD1701株の代わりにパラポックスウイルスovisの上記の株を使用することにより達成される。
【課題を解決するための手段】
【0014】
本発明は、ヒトおよび動物におけるウイルス感染および癌に対する医薬の製造のための、分類上パラポックスウイルスovis株NZ2、NZ−7、NZ−10またはorf−11の1つに属するウイルスの使用に関する。
【0015】
本発明はさらに、ヒトおよび動物におけるウイルス感染および癌に対する医薬の製造のための、本発明の株の子孫の使用であって、該子孫が、適当な細胞系、例えばWI−38およびMRC−5、Vero細胞等のヒト細胞、BK−Kl3A47/RegまたはMDBK等のウシ細胞およびMDOK等のヒツジ細胞に継代または適合させることによって得られる、使用、さらに該株およびそれらの継代および適合化した変異体の部分または断片の使用であって、該一部が培養線維芽細胞等の適当な系においてワクシニアウイルス等の適当なベクターの助けをかりて発現するゲノムまたはサブゲノム断片であると理解され、断片が、発現したまたは物理的に破壊したウイルス粒子のクロマトグラフィー等の生化学的精製によって得られる一部分であると理解される、使用、およびさらにパラポックスウイルスovisの1つの株の使用であって、自己免疫疾患のためのおよび気道および内蔵の急性および慢性ウイルス感染のための免疫治療剤または免疫予防剤としての医薬および医薬粗製物の製造のための、使用、およびさらに該株およびそれから得られた誘導体の1つの使用であって、ストレス感染後防御および感染後の感染性疾患を予防または軽減するための、さらに手術および歯科的介入のなかでの感染予防に関連する医薬および医薬粗製物の製造のための、使用、およびさらに該株およびそれから得られた誘導体の1つの使用であって、例えば気道の、パピローマウイルス感染の、ヘルペス属のウイルスによる感染の、HIV感染の、または内蔵のウイルス感染、例えば肝炎ウイルスによる感染等の急性および慢性ウイルス感染の感染後防御または治療において使用するための、使用、およびさらに該株およびそれから得られた誘導体の1つの使用であって、創傷治癒過程を促進するために、および治癒がごくわずかまたは全く治癒しない創傷の治癒を支援するために創傷に対して、および脚の潰瘍に対して使用するための、使用、およびさらに該株およびそれから得られた誘導体の1つの使用であって、アレルギー疾患、乾癬、神経皮膚炎および他の自己免疫疾患、例えばループスエリテマトーデス等、の様々な疾患に対して使用するための、および例えば高齢の患者において、健康状態を改善するための、使用、およびさらに該株およびそれから得られた誘導体の1つの使用であって、内蔵の、例えばクーロン病、皮膚の、血液の、中枢神経系および、目を含むその付加的な構造の、癌を含む炎症性、変性および増殖性疾患に対して使用するための医薬および医薬粗製物の製造のための、使用、およびさらにヒトおよび動物における抗ウイルス治療または癌治療のための医薬および医薬調製物の製造のための他の治療法と組合せての、該株およびそれから得られた誘導体の1つの使用に関する。
【0016】
好ましくは、本発明は、経口投与のためのおよび/または経口投与のための耐胃液製剤における、他の治療法と組み合わせた、医薬および医薬粗製物の製造のためのパラポックスウイルスovisの1つの株の使用に関する。
【0017】
本明細書に例示的に記載されたパラポックスウイルスovis NZ−2は、European Collection of Cell Cultures, Centre for Applied Microbiology and Research, Porton Down, Salisbury, Wiltshire, SP4 0JG, United Kingdomにおいて2001年7月10日に寄託された。受託番号はECACC−01071006である。
【図面の簡単な説明】
【0018】
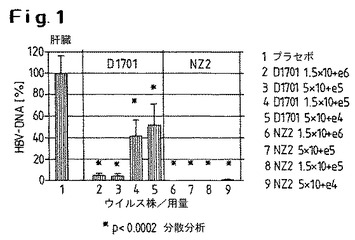
【図1】D1701株またはNZ2株によるHBV−トランスジェニックマウスの処置の結果を示す。
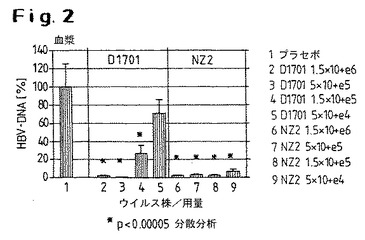
【図2】血漿において得られたD1701株またはNZ2株を用いたHBV−トランスジェニックマウスの処置の結果を示す。
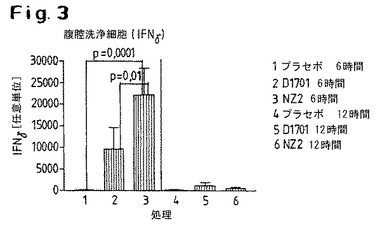
【図3】D1701株またはNZ2株による処置後に誘導されたγインターフェロンの発現レベルを示す。
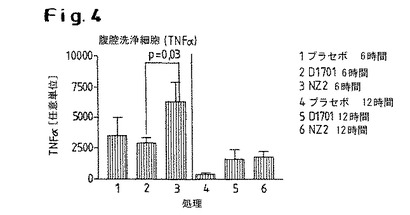
【図4】D1701株またはNZ2株による処置後に誘導されたTNFαの発現レベルを示す。
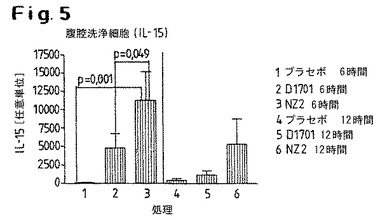
【図5】D1701株またはNZ2株による処置後に誘導されたIL−15の発現レベルを示す。
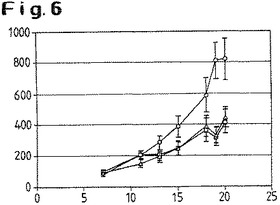
【図6】腫瘍を有するヌードマウスにおけるD1701株またはNZ2株による処置後の腫瘍の平均の大きさを示す(mg)。○:グループ1(プラセボ(PBS))、△:グループ2(D1701株)、□:グループ3(NZ2株)。
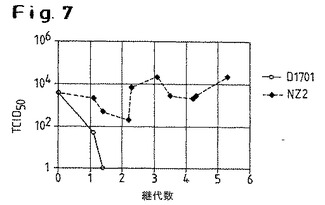
【図7】NZ2株およびD1701株のヒトMRC−5細胞における継代能力を示す。ヒトMRC−5細胞をNZ2株またはD1701株に感染させ、得られた細胞上清中のウイルス力価(TCID50)を継代数に対してプロットした。
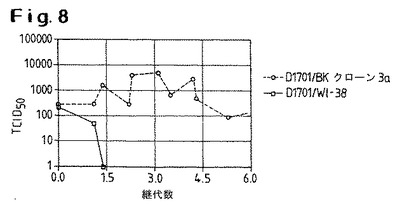
【図8】ウシBKクローン3a細胞およびヒトWI−38細胞でのD1701株の継代において得られた、継代数に対してプロットされたウイルス力価を示す。
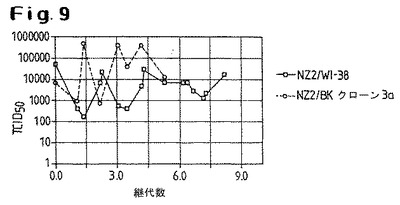
【図9】ウシBKクローン3a細胞およびヒトWI−38細胞でのNZ2株の継代において得られた、継代数に対してプロットされたウイルス力価を示す。
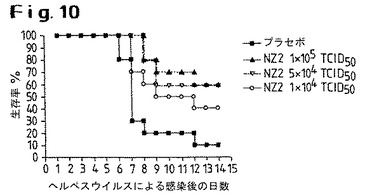
【図10】ヘルペスウイルス感染後の生存率に対する、NZ2株の用量依存的効果を示す。
【発明を実施するための形態】
【0019】
一例として以下のものを発見した:
実施例1.B型肝炎ウイルスのおける治療活性の実証−トランスジェニックマウス
B型肝炎ウイルス−トランスジェニックマウス(HBV株1.3X-tg)を「概念実証」実験に用いた。1群あたり8〜10週齢の雄性マウスを7匹用いた。実験開始から1日目および4日目に0.15mLの体積で種々の用量を腹腔内投与した。
【0020】
投与については、個々の投与群について、以下の希釈率の各ウイルス株を調製した:
第1用量 1.5×106TCID50
第2用量 5×105TCID50
第3用量 1.5×105TCID50
第4用量 5×104TCID50
滅菌した、ピロゲンフリーのPBSをプラセボ対照として用いた。
ウイルスは以下のようにして濃縮した:500mLのパラポックスウイルスovisNZ2株(力価、約2×105TCID50/mL)およびパラポックスウイルスovisD1701株(力価1×107TCID50/mL)から得られた上清を同じウイルス力価に調整した。Beckman超遠心(SW28ローター、28000RPM、4℃にて3時間)をこの目的に使用した。超遠心の後、適当な体積の希釈用培地を用いてペレットを1×107TCID50/mLの力価に調整した。
【0021】
逆滴定を確認するために、対応する用量のアリコートを用い、これらの用量の力価を確認した。確認用のアリコートを除去した後、対応する用量を56℃で1時間不活化した。
希釈用培地:
10mLのEMEM 10×
2.7mLの2g/L炭酸水素塩1mLの1%グルタミン
86.3mLの2回蒸留水
【0022】
5日目に、動物を安楽死させ肝臓と血液を採取した。各動物の肝臓約20mgをQIAamp組織キット(Qiagen、Hilden)を用いて処理し、DNAの濃度を分光計で測定し、1%アガロースゲルでの電気泳動によりその完全性をチェックした。ドットブロットハイブリダイゼーション法で、このDNAをHBV特異的プローブとハイブリダイズさせた。RNAによって生じるシグナルを排除するためにDNAは予めRNAseA(Qiagen、Hilden)で処理しておいた。200μLのDNA(10μg)をナイロンメンブレン(Boehringer Mannheim)に載せ、それぞれのケースにつき3分間ソークI(0.5N NaOH;1MNaCl)で4回処理し、ソークII(3M NaCl;0.5Mtris−HCI、pH7.4)で2回処理した;次いでDNAを120℃で30分間焼いた後、プローブを含まない標準ハイブリダイゼーションバッファー(5×SSC、N−ラウリルサルコシン、0.1%w/v;SDS、0.02%;ブロッキング試薬、1×および100μgの新鮮な精子DNA/mL)を用いて60℃で30分間プレハイブリダイズさせた。この工程の後、全HBVゲノムを含むDNA(20〜40ng/mLハイブリダイゼーション緩衝液)をランダムオリゴヌクレオチドプライム化プローブとハイブリダイズさせた。その後、フィルターを、4×SSC/0.1%SDS、2×SSC/0.1%SDSおよび1×SSC/0.1%SDS中で64℃にて10分間洗浄した。
【0023】
免疫学的検出は、製造者の説明書にしたがって、CDP−StarRシステム(Boehringer Mannheim)を用いて行った。評価のためにLumilmagerR(Boehringer Mannheim)を使用した。血液中のHBV特異的DNAを定量的PCRにより定量した。まず、EDTA血を遠心することにより血漿を単離した。DNAをHighPure 16 System Viral Nucleic Acid Kit (Boehringer Mannheim)および定量的PCRを用いて精製し、ABI PRISM 7700 Sequence Detection System (PE Applied Biosystems)によってHBV特異的シグナルについて試験した。以下のプライマーとプローブを用いた:
ayw-570f (センス) 5'- CTGTACCAAACCTTCGGACGG - 3'
ayw-670r (アンチセンス) 5'- AGGAGAAACGGGCTGAGGC - 3'
プローブ:
ayw-613t 5'- CCATCATCCTGGGCTTTCGGAAAATT - 3'。
【0024】
DNAを50μLの反応液 (この反応液には、1.4mMの各dNTP、4.75mMのMgCl2、15pmolの各プライマーおよびプローブ、5μLの10倍量のPCR緩衝液[すべてのPCR試薬はTagManコア試薬キット(Perkin Elmer/Roche Molecular Systems Inc.)から入手した]および1.25UのTaq DNAポリメラーゼおよび0.25UのAmp Eraseを含有する)中で増幅させた。最初の変性工程(95℃にて10分間)の後、試料を40サイクルの変性(95℃、30秒)およびアニーリング/伸張 (56℃、1分間)に付した。生成物をABI PRISM(登録商標)7700 Sequence Detection System スタンダードソフトウエアを用いて解析した。
【0025】
組織化学的解析は、B型肝炎ウイルスコア抗原に対して検出された抗体(Dako)を用いて行った。このため、1つの肝葉の一部を4%ホルムアルデヒドで一晩固定化し、パラフィンに包埋し、切片(5μm)を作成した。パラフィンを除去し再水和した後、内因性のペルオキシダーゼ活性を3%H2O2を用いて20分間クエンチした。非特異的結合を通常のヒツジ血清を用いてブロックした。次いで、この切片を1:500に希釈した抗体と室温で30分間インキュベーションした。その後の工程はすべてVectastain ABCキット(Vector Laboratories)を用いて説明書にしたがって行った。
【0026】
この免疫反応を、3,3'−ジアミノベンジジンテトラクロリドおよび過酸化水素を用いて可視化した。この切片をヘマトキシリン/エオシンで対比染色した。
【0027】
結果を、分散および多重比較(post hoc comparison)により統計的に解析した。
【0028】
結論として、意外にも、NZ2株を用いた場合、既知のパラポックスovisD1701株で得られた効果と比較して抗ウイルス効果の増幅がみられた。このことにより、はじめて、パラポックスウイルスovisを用いて、これまでに知られているパラ免疫誘導物質で達成された効果の強度とは有意に異なる強度を有する免疫系の複合的能力(complex capacity)を誘導することが可能になる。
【0029】
意外にも、以下の結果が得られた:
肝臓:D1701で処置した動物と比較して、HBV特異的DNAの有意に大きな減少がNZ2で処置した動物において得られた。NZ2の抗ウイルス活性は、D1701の活性よりもさらに強力である:用量が最大の群では、NZ2の投与によってHBV特異的DNAが、D1701を同量投与した場合よりも、45倍以上も効果的に減少するのに対し、用量が最低の群では57倍も効果的に減少する。血漿では、NZ2の投与によりHBV特異的DNAが最低用量の群において、D1701を同量の投与した群よりも10倍以上効果的に減少する。このことは、パラポックスウイルスovisNZ2株の治療効果は、D1701株の効果よりも有意に優れていることを示している。
【0030】
図では、プラセボ群(100%として示した)と比較して、HBV特異的シグナルの減少が示されている。
図1は、D1701株またはNZ2株によるHBV−トランスジェニックマウスの処置の結果を示す。プラセボ群に対して、いずれの染色もおよびすべてのケースで肝臓においてHBV特異的DNAが有意に減少するが、NZ2株を用いた場合その減少は大きくなる。最低のNZ2用量ではいずれも、HBV特異的DNAの減少は、等価なD1701投与群の場合において有意に大きい。
図2は、血漿において得られたD1701株またはNZ2株を用いたHBV−トランスジェニックマウスの処置の結果を示す。いずれの染色およびすべての投与群において、プラセボ群と比較して、HBV特異的DNAは血漿において有意に減少するが、NZ2株を用いる場合その減少は大きくなる。NZ2の最低投与量とは対照的に、D1701株の最低投与量はもはや有意の抗ウイルス効果を有しない。
【0031】
実施例2.サイトカインの誘導
7〜8週齢の雌性Balb/cマウスを無菌条件下で飼育し、実験に用いた。マウスを各群6匹の群に無作為にグループ分けした。以下の処置スキームを適用した:
グループ1: プラセボ
グループ2: パラポックスウイルスovisD1701株;5×104TCID50/用量
グループ3: パラポックスウイルスovisNZ2株;5×104TCID50/用量
グループ4: プラセボ
グループ5: パラポックスウイルスovisD1701株;5×104TCID50/用量
グループ6: パラポックスウイルスovisNZ2株;5×104TCID50/用量
【0032】
投与した体積は10mL/kgであり、投与は腹腔内投与で行った。
【0033】
グループ1〜3の動物を投与から6時間後に屠殺し、グループ4〜6の動物は投与から12時間後に屠殺した:腹腔細胞を氷冷PBSを用いた洗浄により得、門脈および腸間膜リンパ節を単離した。腹腔細胞を遠心工程により濃縮した(Eppendorf 卓上遠心器で、室温にて3000 rpmで5分間)後0.2mLの溶解緩衝液(溶解溶液:25 mM クエン酸ナトリウム、4Mグアニジウムイソチオシアネート、0.5%N−ラウリルサルコシン)中に溶解し、ショック凍結し、RNAを調製するときまで−75℃で保存した。
【0034】
全RNAを酸性フェノール/クロロホルム抽出により調製した。このため、溶解緩衝液中で凍結した試料を室温で融解し、伸張のため以下の溶液で処理した:溶解緩衝液1/10体積の2Mクエン酸ナトリウム(pH4.0)、溶解緩衝液1体積の1水飽和フェノールおよび溶解緩衝液1/5体積のクロロホルム/イソアミールアルコール(24:1)。これらの試薬を攪拌機で10秒間混合した後、氷上で10分間保って温度を一定にした。15365gで4℃にて30分間遠心することによりこの相を分離した。その後、水相を新しい容器に移し、この相に存在するRNAを単離するために、Rnaid(登録商標)plusキット(DIANOVA)から8mLのRNA−MATRIXを加えた。および全体を室温で15分間インキュベーションした。得られたRNA/RNA−MATRIX複合体を7000gでの遠心によりペレット化し、上清を捨てた。次いでペレットを、それぞれのケースについて2回250mLのRNA−WASH(DIANOVA)で洗浄した後、最後に洗浄した後、真空遠心で乾燥させた。20−30mLのRNAse不含蒸留水を添加することによってRNAを溶出し、全体を55℃に15分間加熱した。室温にて7000gで1分間遠心した後、RNA溶液を新しい容器に移すことによりマトリックスを分離した。
【0035】
RNAの量をゲル電気泳動によりチェックした。RNAは−70℃で保存した。
【0036】
オリゴ(dT)プライマーを出発分子として使用してmRNAを逆転写することによりcDNAを合成した。以下の成分を合成混合物に含有させた:200ng−2μgの全RNA、2μLのM−MLV逆転写酵素(200U/μL) (GIBCOBRL)、8μLの適当な5×RT緩衝液(GIBCOBRL)、1μLのDTT(0.1M)(GIBCOBRL)、4μLのdNTP(2.5mM)(SIGMA)、2μLのオリゴ(dT)12-18プライマー(100μg/mL)(PROMEGA)、1μLのヒト胎盤インヒビター(10000U/mL)(GIBCOBRL)および水を加えて合計40μLにする。混合物を室温に10分間保った後、37℃で45分間インキュベーションした;次いで、95℃に3分間加熱し氷上で急速に冷却した。この方法により合成したcDNAを−20℃で保存した。
【0037】
cDNAの量を「ハウスキーピング」遺伝子(β-アクチン)を用いて標準化した。定量的PCRをABI PRISM 7700 Sequence Detection System (PE Applied Biosystems)を用いて行った。以下のプライマーを用いた:
β−アクチン センス: 5'-TGG AAT CCT GTG GCA TCC ATG AAA C-3'
アンチセンス:5'-TAA AAC GCA GCT CAG TAA CAG TCC G-3'
IFN−γ センス: 5'-AGCGGC TGA CTG AAC TCA GAT TGT AG-3'
アンチセンス:5'-GTC ACA GTT TTC AGC TGT ATA GGG-3'
IFN−α センス: 5'-GGC AGG TCT ACT TTG GAG TCA TTG C-3'
アンチセンス:5'-ACA TTC GAG GCT CCA GTG AAT TCG G-3'
IL−15 センス: 5'-GCC AAC TGG ATA GAT GTA AGA TAT GAC CT-3'
アンチセンス:5'-CGT GTT GAT GAA CAT TTG GAC AAT GCG TAT-3'
【0038】
DNAは、25μLの体積の反応液(この反応液には、1.4mMの各dNTP、4mMのMgCl2、0.3μmolの各プライマーおよびプローブ、2.5μLのSYBRグリーンを含む10倍PCR緩衝液[すべてのPCR試薬はSYBRグリーンPCRコア試薬キット(PerkinElmer/RocheMolecularSystemsInc.)から入手した]および1.25UのTaqDNAポリメラーゼおよび0.25UのAmpEraseを含有する)中で増幅させた。最初の変性工程(95℃にて10分間)の後、試料を40サイクルの変性(95℃、30秒)およびアニーリング/伸張(60℃,1.30分間)に付した。生成物をABIPRISM(登録商標)7700 Sequence Detection System スタンダードソフトウエアを用いて解析した。結果を、分散および多重比較により統計的に解析した。
【0039】
意外にも以下の結果が得られた:
1.D1701株またはNZ2株による処置後、γインターフェロンの発現が投与後6時間および12時間後の両方で誘導されている(図3)。NZ2株のケースでは、この誘導がプラセボおよびD1701株の両方に関して有意に高い。D1701の投与後に得られたγ−インターフェロン発現の程度は、プラセボ対照とは有意な差異はない。この図は、腹腔洗浄により得られた細胞において測定された値を示している。
【0040】
2.D1701株による処置後、TNFαの発現が投与後12時間で誘導されるのに対し、NZ2株での処置後では6時間後および12時間後の両方で誘導されている(6時間の値と比較して12時間後では減少を既に観察することができる:図4)。NZ2株のケースでは、投与後6時間でのこの誘導は、D1701株のケースで観察される値よりも有意に高い。この図は、腹腔洗浄により得られた細胞において測定された値を示している。
【0041】
3.D1701株またはNZ2株での処置後、6時間および12時間後の両方でIL−15の発現が誘導される(図5)。NZ2株の場合、この誘導は、投与後6時間でD1701株またはプラセボのケースよりも有意に高い。D1701の投与後に観察されるIL−15発現の程度は、プラセボ対照で観察される程度よりも有意に高い。この図は、腹腔洗浄により得られた細胞において測定された値を示している。
【0042】
実施例3.腫瘍を有するヌードマウスにおける治療効果の実証
インキュベーターにて、MDA−MB231細胞(ATCC#HTB26)を、完全培地(88 5 DMEM、10%FBS、1%ペニシリン/ストレプトマイシン、1%L−グルタミン(いずれもGibco Life Technologies))中、37℃にて5%CO2の存在下で培養した。移植の日に、細胞を約70%コンフルエントであった。この細胞をトリプシン処理し、HBSSで洗浄し、細胞数を計測し、予め冷却したPBSを用いて2.5×107細胞/mLに調整した。雌性の NCrヌードマウス(taconic)を用いた。このマウスは8〜10週齢であり、体重は約22gであった。操作はすべて無菌状態で行った。5×106細胞を合計0.2mlの体積で脇腹の領域に皮下注射した。この後、マウスを、腫瘍重量が平均約80mgに達するまでさらに7日間飼育した。腫瘍を測定してマウスを各ケースにつき10匹の群に無作為にグループ分けした。各群に以下のものを投与した:
グループ1: プラセボ(PBS)
グループ2: パラポックスウイルスovis、D1701株
グループ3: パラポックスウイルスovis、NZ2株
【0043】
D1701を2.5×105TCID50の用量で投与し、NZ2は1×105TCID50の用量を投与した。各群においてこれらの用量を3日おきに4回投与した。腫瘍を1週間に2回測定した。Studentテストを用いて有意度を求めた。
【0044】
図6は、グループ1〜3の実験期間(日)中の腫瘍の平均の大きさを示す(mg)(記号:グループ1○、グループ2△;グループ3□)。驚くべきことに、腫瘍に対する活性(活性が対照群と比較して有意(p<0.05)であった)がこの実験系でみとめられた。NZ2株はこの点について、D1701株よりも効力が顕著であることがみとめられた。同等の効果を達成するために、NZ2はD1701と比較して約半分しか必要としなかった。
【0045】
この知見は、腫瘍を有するヌードマウスにおける、本発明のウイルス調製物の治療効果を明白に裏付けるものである。ヌードマウスは免疫不全であり、機能性のT細胞は有していない。この実験系において、腫瘍に対する活性は、ナチュラルキラー(NK)細胞、他の細胞およびサイトカイン/ケモカインの直接の効果に起因するものと推定される。NZ2の優れた効果は、完全なおよびインタクトな免疫系の場合ではより一層明白な証拠となるであろう。
【0046】
実施例4.NZ2とD1701とのその他の生物学的な差異
D1701株とは対照的に、パラポックスウイルスNZ2はヒト細胞系において継続的に継代することができることが分かった。このことは、複製の挙動および/またはウイルスレセプターにおけるNZ2とD1701との間の本質的な違いを示している。
【0047】
ヒト細胞系への適合化は、ヒト細胞系におけるウイルス株の製造のための重要な前提条件である。
4.a)ヒトMRC−5細胞に対する継続的に継代される能力
図7は、NZ2およびD1701株をヒト二倍体細胞系MRC−5に適合させる試みを示す。MRC−5は生物学的に活性な化合物およびワクチンの製造に適している。両株について同じ初発力価を用い、対応する培養細胞上清をMRC−5細胞において5回継代させた。図は、各ケースにおいて感染させたMRC−5細胞から得られた上清の試料の力価をTCID50で示し、継代数に対してプロットした。D1701ではなくNZ2株のみが、ウシ培養腎細胞に対して逆滴定可能であった(BK クローン3a)。これは、感染および複製挙動におけるこの2つの株の間の根本的な生物学的違いの存在を示している。同時に、この結果は、NZ2がD1701よりも広範なスペクトルで複製され得ること、それによりヒト細胞をベースとした製造方法の可能性がもたらされることを示している。
【0048】
4.b)NZ2およびD1701のWI−38細胞およびBKクローン3a細胞おける継代能力
NZ2とD1701株の間の根本的な生物学的違いは、図8および図9に示された実験によってさらに明白に実証される。図8は、ウシBKクローン3a細胞およびヒトWI−38細胞でのD1701の継代において得られた、継代数に対してプロットされたウイルス力価を示す。D1701はBKクローン3a細胞では継続的に継代することができるが、ヒトWI−38細胞ではできないことがみとめられる。
【0049】
この状況は、NZ2株については異なる。この株は、BKクローン3a細胞とWI−38細胞の両方について数日間に渡って継続的に継代することができる(図9)。
【0050】
これらの結果はまた、NZ2およびD1701の感染および複製挙動における明白な違いを示している。
【0051】
4.c)WI−38での継代後のヘルペスウイルス投与試験におけるNZ2の用量依存的効果
WI−38細胞において継代させたNZ2株の免疫刺激特性を調べるため、マウスに対してヘルペスウイルス投与試験を行った。各ケース10匹の実験動物からなる3つの群に対し、それぞれ、1×104TCID50、5×104TCID50および1×105TCID50で処置し、対照群にはプラセボを投与した。図10は、ヘルペスウイルスによる感染後の時間経過に対する4つの実験群の生存率を示す。意外にも、NZ2は、WI−38で継代させた結果、免疫刺激特性は失われなかった。
【0052】
この実験例における実験結果はすべて、NZ2とD1701の感染および複製挙動における根本的な違いを示している。意外にも、D1701の場合とは対照的に、免疫調節物質として用いるためのNZ2の調製物は産生細胞系としてウシ腎細胞を用いる必要はない。
【0053】
潜在的で慢性的に持続するウイルス感染に対するTh1免疫反応の影響の知られている状況4)、5)、また増殖性の疾患例えば癌89、およびパラポックスウイルスovisNZ2株の免疫調節特性がパラポックスウイルスD1701株よりも優れているという事実に基づけは、パラポックスウイルスovisNZ2株、または上記の株の1つを基礎とする免疫調節物質の、ヒトおよび動物における、単独療法または生物学的に活性な物質、例えば抗ウイルス薬、低分子量化合物との組合せとしての使用は、B型肝炎ウイルス、またはC型肝炎ウイルスまたは肝炎の原因ウイルスの群に由来するその他の病原体のいずれかによる感染、さらに、内臓の他のウイルス感染、並びに様々なタイプの単純ヘルペスウイルス(HSV)、様々なタイプのヒトパピローマウイルス(HPV)、ヒト免疫不全ウイルス(HIV)、水疱瘡ウイルス、ヒトサイトメガロウイルス(HCMV)による、他の疾患を同様に伴う感染および動物における対応するウイルス性疾患の抗ウイルス治療が可能であり治療上有用である。
【0054】
さらに、示された作用機序に基づけば、上記のパラポックスウイルスovis株の上記の株は、特に、以下の予防的または治療的処置を行うことについて、ある程度の成功の見込みをもって使用することができる:
この治療法による処置を暴露後迅速に行う場合、ヘルペスウイルス感染に関する再発の防止、感染後防御、即ち、ウイルス感染の確立の阻止(例えば、HIV)7)。この作用機序に基づいて癌の処置も同様に可能である8)、9)。
【0055】
臨床上の問題の性質によって、パラポックスウイルスをベースとする治療剤を全身的、即ち、例えば筋肉内、皮下、腹腔内、静脈内または経口的に、あるいは局所的に投与する。これに関連して、パラポックスウイルスは、精製された凍結乾燥状態で存在するかおよび/または投与の直前に適当な溶媒に懸濁させる、あるいは別の適当な製剤中に存在させるか耐胃液投与剤型または別の経口投与剤型中に存在させることができる。
【0056】
適当な調製物はまた、特定の細胞、例えばWI−38、MRC−5またはVero細胞で継代および/または適合させることによって得られるNZ2の子孫、および上記のその他の株またはNZ2の一部もしくは断片、およびそれらの上記の株またはそれらの子孫から調製することもできる。一部とは、適当な系、例えば培養線維芽細胞、において適当なベクター、例えばワクシニア、の助けをかりて発現するゲノムまたはサブゲノム断片であると理解される。断片とは、例えば超音波処理によって物理的に破壊した粒子の生化学的精製、例えばクロマトグラフィー、によって得られる一部分であると理解される。
【0057】
これに関連して、その臨床上の問題が必要とする条件を満たす時間的な計画にしたがって、いくつかの投与または長期間の処置が必要になるかもしれない。
【0058】
即ち、例えば以下の計画に従う使用が癌治療の場合には特に期待できることが分かった(但し、これらに何ら限定されない):
各ケースにつき106〜107TCID50(組織培養細胞感染量)の筋肉内投与を3日毎に4週間、次いで2週間の休止;各ケースについて106〜107TCID50の筋肉内投与を再開し、3日毎に4週間行った後、2週間の休止;各ケースについて106〜107TCID50の筋肉内投与を再開し、3日毎に4週間行った後、2週間の休止;重篤度および治療の成功度によって、これらのサイクルをさらに増やしてもよい;あるいは、製剤を少なくとも3ヶ月間4日〜5日毎に投与するという計画を示すこともできる。
【0059】
例えば、慢性のウイルス感染の場合、3日毎に合計5回106〜107TCID50を腹部に皮下投与するか、三角筋もしくは大腿四頭筋の領域に筋肉内投与する。その疾患の必要によってはこの計画を分けることが可能である。風邪の予防については、製剤をうがい用として用い、病原菌の危険が存在する限りは、毎日繰り返す。
【0060】
口の部分の外科的介入(例えば、歯の手術)後の感染を防ぐために、その介入の前夜に、本製剤で1〜2分間うがいを行う。
【0061】
参考文献:
1. Guidotti, L.G., Borrow, P., Hobbs, M.V., Matzke, B., Gresser, I., Oldstone, M.B.A., and Chisari, F.V. (1996): Viral cross talk: Intracellular inactivation of the hepatitis B virus during an unrelated viral infection of the liver. Proc. Natl. Acad. Sci. USA. 93:4589-4594.
2. Guidotti, L.G., Ando, K., Hobbs, M.V., Ishikawa, T., Runkel, L., Schreiber, R.D., and Chisari, F.V. (1994): Cytotoxic T lymphocytes inhibit hepatitis B virus gene expression by a noncytolytic mechanism in transgenic mice. Proc. Natl. Acad. Sci. USA. 91:3764-3768.
3. Steinmassl,G., G.Wolf (1990): Bildung von Interleukin 2 and Interferondurch mononukleare Leukozyten des Schweines nach in vitro-Stimulation mit verschiedenen Viruspraparaten (Formation of interleukin 2 and interferon by pig mononuclear leukocytes following in-vitro stimulation with various virus preparations). J.Vet.Med.B37,5,321-331.
4. P. Lucin, S. Jonjic, M. Messerle, B. Polic, H. Hengel, U.H. Koszinowski (1994): Late-Phase inhibition of murine cytomegalovirus replication by synergistic action of interferon gamma and tumor necrosis factor alpha. J. Gen. Viro175-101-110;P.M.
5. Smith, R.M. Wolcott, R. Chervenak, S.R. Jennings (1994): Control of acute cutaneous herpes-simplex virus-Infection - T-cell mediated viral clearance is dependent upon interferon gamma. Virology 202 (1):76-88].
6. Y. Kawanashi, N. Hayashi, K. Katayama, K. ueda, T. Takehara, E. Miyoshi, E. Mita, A. Kasahara, H. Fusamoto, T. Kamada (1995): Tumor necrosis factor alpha and interferon gamma inhibit synergistically viral replication in hepatitis B virus replicating cells. J. Medical Virology 47 (3):272-277.
7. Dhawan, S., L.M. Wahl, A. Heredia, Y.H. Zhang, J.S. Epstein, M.S. Meltzer, I.K. Hewlett (1995): Interferon gamma inhibits HIV-induced invasiveness of Monocytes. J. Leukocyte Biology, 58 (6):713-716.
8. J.F. Bromberg, C.M. Horvath, Z.L. Wen, R.D. Schreiber, J.E.Darnell (1996): Transcriptionally active statl is required for the antiproliferative effects of both interferon alpha and interferon gamma. PNAS 93(15):7673-7678.
9. M.Klouche, H.Kirchner, F.Holzel (1994): Antiproliferative effects of inter-feron gamma in combination with alpha-difluoromethylornithine on human carcinoma cell cultures. J.Cancer Research and Clinical Oncology 120(12):706].
【技術分野】
【0001】
本発明は、感染性または非感染性の免疫不全のための免疫治療剤としての、パラポックスウイルスovis株の使用、および腫瘍性疾患およびウイルス感染ならびにそのような感染を伴う疾患を治療するためのパラポックスウイルスovis株の使用、さらにヒトおよび動物において使用するための医薬の製造のためのパラポックスウイルスovis株の使用に関する。
【0002】
本発明はさらに、パラポックスウイルスovis株、およびそれから調製される医薬形態の、ストレス(例えば手術)後の感染性疾患を予防または軽減するためのストレス後感染防御における免疫治療剤または免疫予防剤としての使用;手術または介入前(例えば、プロテーゼの移植前または歯科的介入の前)に投与することによる感染性疾患の予防または軽減のための感染予防におけるそれらの使用、(例えば気管の、パピローマウイルス感染の、ヘルペスウイルスによる感染の、HIV感染の、および肝炎ウイルスによる感染のような内蔵のウイルス感染の)急性および慢性のウイルス感染の感染後防御または治療におけるそれらの使用、創傷治癒過程を促進するための創傷治癒におけるそれらの使用、および治癒がごくわずかまたは全く治癒しない創傷の治癒の支援のためのそれらの使用(例えば、脚の潰瘍)、多発性硬化症、喘息、いぼおよびその他の皮膚の新形成等の疾患に対するそれらの使用、様々なアレルギー疾患に対するそれらの使用、全身的なアレルギーの発症を阻止するためのそれらの使用、および局所的なアレルギーに対するそれらの使用、および例えば老人患者における健康状態の改善のためのそれらの使用に関し、本発明において用いられるパラポックスウイルスovis株はNZ2、NZ−7、NZ−10およびorf−11である。
【0003】
本発明はさらに、特定の細胞、例えばWI−38, MRC−5もしくはVero細胞、で継代および/または適合させることによって得られたこれらの株の子孫、またはこれらの株もしくは子孫から得られたウイルスの一部もしくは断片を使用することも可能である。一部とは、適当な系、例えば培養線維芽細胞において、適当なベクター、例えばワクシニアの助けかりて発現するゲノムまたはサブゲノム断片であると理解される。断片とは、例えば超音波処理により物理的に破壊された粒子の、例えばクロマトグラフィー等の生化学的な精製によって得られる一部分であると理解される。
【0004】
本発明はさらに、医薬および医薬調製物を製造するためのパラポックスウイルスovisの該株の使用に関する。さらに、本発明は、抗ウイルス治療または癌治療のための医薬および医薬調製物を製造するための、他の治療法と組み合わせた、パラポックスウイルスovisの該株の使用に関する。
【背景技術】
【0005】
潜在的で慢性的な持続的ウイルス感染は、免疫抑制によって活性化または再活性化し得ること、また逆に、潜在的であるウイルス(例えば潜在的なヘルペスウイルス感染は免疫抑制に関連して再発する:ストレスまたはコルチゾン投与に関連する陰唇疱疹)によって誘発される急性疾患を免疫系が抑制することが知られている。さらに、慢性的に持続するおよび潜在的なウイルス感染は、低分子量を有する従来の抗ウイルス物質を用いて治療することは困難であるかまたは不可能でさえある。
【0006】
その理由は、そのような感染に関連するウイルスの酵素活性の不在(例えば、最初にヌクレオシドインヒビターをウイルスの核酸に取り込んでこのインヒビターが例えばウイルスDNAの鎖を終止させるようにするウイルスポリメラーゼの不在;例えば、抗ウイルス化合物を最初にリン酸化してこの化合物が活性になるようにする必要のある何らかのウイルスチミジンキナーゼ活性の不在)、または宿主の免疫系による、感染したまたは変性した細胞の、例えば癌細胞の、またはウイルス抗原の、何らかの認識の欠如であり得る。
【0007】
さらに、臨床上治りにくいウイルス感染に関連して、別のウイルスによる重感染が、その臨床上治りにくいウイルスに対する抗ウイルス効果をもたらし得ることが知られている1)。著者1)は、T細胞、ナチュラルキラー細胞およびマクロファージよって分泌される、IFN−γおよびTNF−αなどのインターフェロンに対するこの効果の関連を実証することができた。
【0008】
クラスI制限的細胞傷害性T細胞がHBVトランスジェニックマウスにおける肝細胞HBV遺伝子発現を阻害することが可能であること、肝細胞をなんら破壊することなくこの過程が起こること、およびこの過程がTNF−αおよびIFN−γによって発揮されることを実証した別の初期の研究2)がこの著者によって得られた結果によって確認された。
【0009】
「パラ特異的免疫」を誘導するための物質、即ち「パラ免疫誘導物質」と呼ばれる物質は、獣医の現場において、比較的長い間、治療的、感染後防御的および予防的に用いられてきた。パラ特異的免疫誘導物質は、例えば、化学的に不活化されたパラポックスウイルスovis、D1701株(DE3504940)からなる。BAYPAIV1UN(登録商標)はこのウイルス(パラポックスウイルスovis、D1701株)をもとに調製された製品である。
【0010】
動物において、不活化されたパラポックスウイルスは、非常に多岐にわたる病原体によって引き起こされた感染に対する非特異的な保護を誘導する。この保護は、内因的な防御システムの一部を構成する様々なメカニズムによって媒介されると考えられる。
【0011】
これらのメカニズムには、感染の誘導、ナチュラルキラー細胞の活性化、「コロニー刺激活性」(CSA)の誘導およびリンパ球増殖の刺激が含まれる。作用機序の初期の研究は、インターロイキン2およびインターフェロンαの刺激効果を実証した3)。
【発明の概要】
【発明が解決しようとする課題】
【0012】
この背景に基づき、本発明の目的は、パラポックスウイルスovisD1701株によるパラ特異的免疫反応の上記の一般化された誘導を質的に増大させるようにパラポックスウイルスovisの優れた効果の治療上の有用性を更に改善し、より優れた抗ウイルスまたは抗腫瘍効果を低用量を用いて達成することができるように改善することである。またこの治療効果は副作用が少ないと期待される。
【0013】
したがって、本発明の目的は、パラポックスウイルスの免疫学的効果を改善することである。この目的は、慣用的に用いられているD1701株の代わりにパラポックスウイルスovisの上記の株を使用することにより達成される。
【課題を解決するための手段】
【0014】
本発明は、ヒトおよび動物におけるウイルス感染および癌に対する医薬の製造のための、分類上パラポックスウイルスovis株NZ2、NZ−7、NZ−10またはorf−11の1つに属するウイルスの使用に関する。
【0015】
本発明はさらに、ヒトおよび動物におけるウイルス感染および癌に対する医薬の製造のための、本発明の株の子孫の使用であって、該子孫が、適当な細胞系、例えばWI−38およびMRC−5、Vero細胞等のヒト細胞、BK−Kl3A47/RegまたはMDBK等のウシ細胞およびMDOK等のヒツジ細胞に継代または適合させることによって得られる、使用、さらに該株およびそれらの継代および適合化した変異体の部分または断片の使用であって、該一部が培養線維芽細胞等の適当な系においてワクシニアウイルス等の適当なベクターの助けをかりて発現するゲノムまたはサブゲノム断片であると理解され、断片が、発現したまたは物理的に破壊したウイルス粒子のクロマトグラフィー等の生化学的精製によって得られる一部分であると理解される、使用、およびさらにパラポックスウイルスovisの1つの株の使用であって、自己免疫疾患のためのおよび気道および内蔵の急性および慢性ウイルス感染のための免疫治療剤または免疫予防剤としての医薬および医薬粗製物の製造のための、使用、およびさらに該株およびそれから得られた誘導体の1つの使用であって、ストレス感染後防御および感染後の感染性疾患を予防または軽減するための、さらに手術および歯科的介入のなかでの感染予防に関連する医薬および医薬粗製物の製造のための、使用、およびさらに該株およびそれから得られた誘導体の1つの使用であって、例えば気道の、パピローマウイルス感染の、ヘルペス属のウイルスによる感染の、HIV感染の、または内蔵のウイルス感染、例えば肝炎ウイルスによる感染等の急性および慢性ウイルス感染の感染後防御または治療において使用するための、使用、およびさらに該株およびそれから得られた誘導体の1つの使用であって、創傷治癒過程を促進するために、および治癒がごくわずかまたは全く治癒しない創傷の治癒を支援するために創傷に対して、および脚の潰瘍に対して使用するための、使用、およびさらに該株およびそれから得られた誘導体の1つの使用であって、アレルギー疾患、乾癬、神経皮膚炎および他の自己免疫疾患、例えばループスエリテマトーデス等、の様々な疾患に対して使用するための、および例えば高齢の患者において、健康状態を改善するための、使用、およびさらに該株およびそれから得られた誘導体の1つの使用であって、内蔵の、例えばクーロン病、皮膚の、血液の、中枢神経系および、目を含むその付加的な構造の、癌を含む炎症性、変性および増殖性疾患に対して使用するための医薬および医薬粗製物の製造のための、使用、およびさらにヒトおよび動物における抗ウイルス治療または癌治療のための医薬および医薬調製物の製造のための他の治療法と組合せての、該株およびそれから得られた誘導体の1つの使用に関する。
【0016】
好ましくは、本発明は、経口投与のためのおよび/または経口投与のための耐胃液製剤における、他の治療法と組み合わせた、医薬および医薬粗製物の製造のためのパラポックスウイルスovisの1つの株の使用に関する。
【0017】
本明細書に例示的に記載されたパラポックスウイルスovis NZ−2は、European Collection of Cell Cultures, Centre for Applied Microbiology and Research, Porton Down, Salisbury, Wiltshire, SP4 0JG, United Kingdomにおいて2001年7月10日に寄託された。受託番号はECACC−01071006である。
【図面の簡単な説明】
【0018】
【図1】D1701株またはNZ2株によるHBV−トランスジェニックマウスの処置の結果を示す。
【図2】血漿において得られたD1701株またはNZ2株を用いたHBV−トランスジェニックマウスの処置の結果を示す。
【図3】D1701株またはNZ2株による処置後に誘導されたγインターフェロンの発現レベルを示す。
【図4】D1701株またはNZ2株による処置後に誘導されたTNFαの発現レベルを示す。
【図5】D1701株またはNZ2株による処置後に誘導されたIL−15の発現レベルを示す。
【図6】腫瘍を有するヌードマウスにおけるD1701株またはNZ2株による処置後の腫瘍の平均の大きさを示す(mg)。○:グループ1(プラセボ(PBS))、△:グループ2(D1701株)、□:グループ3(NZ2株)。
【図7】NZ2株およびD1701株のヒトMRC−5細胞における継代能力を示す。ヒトMRC−5細胞をNZ2株またはD1701株に感染させ、得られた細胞上清中のウイルス力価(TCID50)を継代数に対してプロットした。
【図8】ウシBKクローン3a細胞およびヒトWI−38細胞でのD1701株の継代において得られた、継代数に対してプロットされたウイルス力価を示す。
【図9】ウシBKクローン3a細胞およびヒトWI−38細胞でのNZ2株の継代において得られた、継代数に対してプロットされたウイルス力価を示す。
【図10】ヘルペスウイルス感染後の生存率に対する、NZ2株の用量依存的効果を示す。
【発明を実施するための形態】
【0019】
一例として以下のものを発見した:
実施例1.B型肝炎ウイルスのおける治療活性の実証−トランスジェニックマウス
B型肝炎ウイルス−トランスジェニックマウス(HBV株1.3X-tg)を「概念実証」実験に用いた。1群あたり8〜10週齢の雄性マウスを7匹用いた。実験開始から1日目および4日目に0.15mLの体積で種々の用量を腹腔内投与した。
【0020】
投与については、個々の投与群について、以下の希釈率の各ウイルス株を調製した:
第1用量 1.5×106TCID50
第2用量 5×105TCID50
第3用量 1.5×105TCID50
第4用量 5×104TCID50
滅菌した、ピロゲンフリーのPBSをプラセボ対照として用いた。
ウイルスは以下のようにして濃縮した:500mLのパラポックスウイルスovisNZ2株(力価、約2×105TCID50/mL)およびパラポックスウイルスovisD1701株(力価1×107TCID50/mL)から得られた上清を同じウイルス力価に調整した。Beckman超遠心(SW28ローター、28000RPM、4℃にて3時間)をこの目的に使用した。超遠心の後、適当な体積の希釈用培地を用いてペレットを1×107TCID50/mLの力価に調整した。
【0021】
逆滴定を確認するために、対応する用量のアリコートを用い、これらの用量の力価を確認した。確認用のアリコートを除去した後、対応する用量を56℃で1時間不活化した。
希釈用培地:
10mLのEMEM 10×
2.7mLの2g/L炭酸水素塩1mLの1%グルタミン
86.3mLの2回蒸留水
【0022】
5日目に、動物を安楽死させ肝臓と血液を採取した。各動物の肝臓約20mgをQIAamp組織キット(Qiagen、Hilden)を用いて処理し、DNAの濃度を分光計で測定し、1%アガロースゲルでの電気泳動によりその完全性をチェックした。ドットブロットハイブリダイゼーション法で、このDNAをHBV特異的プローブとハイブリダイズさせた。RNAによって生じるシグナルを排除するためにDNAは予めRNAseA(Qiagen、Hilden)で処理しておいた。200μLのDNA(10μg)をナイロンメンブレン(Boehringer Mannheim)に載せ、それぞれのケースにつき3分間ソークI(0.5N NaOH;1MNaCl)で4回処理し、ソークII(3M NaCl;0.5Mtris−HCI、pH7.4)で2回処理した;次いでDNAを120℃で30分間焼いた後、プローブを含まない標準ハイブリダイゼーションバッファー(5×SSC、N−ラウリルサルコシン、0.1%w/v;SDS、0.02%;ブロッキング試薬、1×および100μgの新鮮な精子DNA/mL)を用いて60℃で30分間プレハイブリダイズさせた。この工程の後、全HBVゲノムを含むDNA(20〜40ng/mLハイブリダイゼーション緩衝液)をランダムオリゴヌクレオチドプライム化プローブとハイブリダイズさせた。その後、フィルターを、4×SSC/0.1%SDS、2×SSC/0.1%SDSおよび1×SSC/0.1%SDS中で64℃にて10分間洗浄した。
【0023】
免疫学的検出は、製造者の説明書にしたがって、CDP−StarRシステム(Boehringer Mannheim)を用いて行った。評価のためにLumilmagerR(Boehringer Mannheim)を使用した。血液中のHBV特異的DNAを定量的PCRにより定量した。まず、EDTA血を遠心することにより血漿を単離した。DNAをHighPure 16 System Viral Nucleic Acid Kit (Boehringer Mannheim)および定量的PCRを用いて精製し、ABI PRISM 7700 Sequence Detection System (PE Applied Biosystems)によってHBV特異的シグナルについて試験した。以下のプライマーとプローブを用いた:
ayw-570f (センス) 5'- CTGTACCAAACCTTCGGACGG - 3'
ayw-670r (アンチセンス) 5'- AGGAGAAACGGGCTGAGGC - 3'
プローブ:
ayw-613t 5'- CCATCATCCTGGGCTTTCGGAAAATT - 3'。
【0024】
DNAを50μLの反応液 (この反応液には、1.4mMの各dNTP、4.75mMのMgCl2、15pmolの各プライマーおよびプローブ、5μLの10倍量のPCR緩衝液[すべてのPCR試薬はTagManコア試薬キット(Perkin Elmer/Roche Molecular Systems Inc.)から入手した]および1.25UのTaq DNAポリメラーゼおよび0.25UのAmp Eraseを含有する)中で増幅させた。最初の変性工程(95℃にて10分間)の後、試料を40サイクルの変性(95℃、30秒)およびアニーリング/伸張 (56℃、1分間)に付した。生成物をABI PRISM(登録商標)7700 Sequence Detection System スタンダードソフトウエアを用いて解析した。
【0025】
組織化学的解析は、B型肝炎ウイルスコア抗原に対して検出された抗体(Dako)を用いて行った。このため、1つの肝葉の一部を4%ホルムアルデヒドで一晩固定化し、パラフィンに包埋し、切片(5μm)を作成した。パラフィンを除去し再水和した後、内因性のペルオキシダーゼ活性を3%H2O2を用いて20分間クエンチした。非特異的結合を通常のヒツジ血清を用いてブロックした。次いで、この切片を1:500に希釈した抗体と室温で30分間インキュベーションした。その後の工程はすべてVectastain ABCキット(Vector Laboratories)を用いて説明書にしたがって行った。
【0026】
この免疫反応を、3,3'−ジアミノベンジジンテトラクロリドおよび過酸化水素を用いて可視化した。この切片をヘマトキシリン/エオシンで対比染色した。
【0027】
結果を、分散および多重比較(post hoc comparison)により統計的に解析した。
【0028】
結論として、意外にも、NZ2株を用いた場合、既知のパラポックスovisD1701株で得られた効果と比較して抗ウイルス効果の増幅がみられた。このことにより、はじめて、パラポックスウイルスovisを用いて、これまでに知られているパラ免疫誘導物質で達成された効果の強度とは有意に異なる強度を有する免疫系の複合的能力(complex capacity)を誘導することが可能になる。
【0029】
意外にも、以下の結果が得られた:
肝臓:D1701で処置した動物と比較して、HBV特異的DNAの有意に大きな減少がNZ2で処置した動物において得られた。NZ2の抗ウイルス活性は、D1701の活性よりもさらに強力である:用量が最大の群では、NZ2の投与によってHBV特異的DNAが、D1701を同量投与した場合よりも、45倍以上も効果的に減少するのに対し、用量が最低の群では57倍も効果的に減少する。血漿では、NZ2の投与によりHBV特異的DNAが最低用量の群において、D1701を同量の投与した群よりも10倍以上効果的に減少する。このことは、パラポックスウイルスovisNZ2株の治療効果は、D1701株の効果よりも有意に優れていることを示している。
【0030】
図では、プラセボ群(100%として示した)と比較して、HBV特異的シグナルの減少が示されている。
図1は、D1701株またはNZ2株によるHBV−トランスジェニックマウスの処置の結果を示す。プラセボ群に対して、いずれの染色もおよびすべてのケースで肝臓においてHBV特異的DNAが有意に減少するが、NZ2株を用いた場合その減少は大きくなる。最低のNZ2用量ではいずれも、HBV特異的DNAの減少は、等価なD1701投与群の場合において有意に大きい。
図2は、血漿において得られたD1701株またはNZ2株を用いたHBV−トランスジェニックマウスの処置の結果を示す。いずれの染色およびすべての投与群において、プラセボ群と比較して、HBV特異的DNAは血漿において有意に減少するが、NZ2株を用いる場合その減少は大きくなる。NZ2の最低投与量とは対照的に、D1701株の最低投与量はもはや有意の抗ウイルス効果を有しない。
【0031】
実施例2.サイトカインの誘導
7〜8週齢の雌性Balb/cマウスを無菌条件下で飼育し、実験に用いた。マウスを各群6匹の群に無作為にグループ分けした。以下の処置スキームを適用した:
グループ1: プラセボ
グループ2: パラポックスウイルスovisD1701株;5×104TCID50/用量
グループ3: パラポックスウイルスovisNZ2株;5×104TCID50/用量
グループ4: プラセボ
グループ5: パラポックスウイルスovisD1701株;5×104TCID50/用量
グループ6: パラポックスウイルスovisNZ2株;5×104TCID50/用量
【0032】
投与した体積は10mL/kgであり、投与は腹腔内投与で行った。
【0033】
グループ1〜3の動物を投与から6時間後に屠殺し、グループ4〜6の動物は投与から12時間後に屠殺した:腹腔細胞を氷冷PBSを用いた洗浄により得、門脈および腸間膜リンパ節を単離した。腹腔細胞を遠心工程により濃縮した(Eppendorf 卓上遠心器で、室温にて3000 rpmで5分間)後0.2mLの溶解緩衝液(溶解溶液:25 mM クエン酸ナトリウム、4Mグアニジウムイソチオシアネート、0.5%N−ラウリルサルコシン)中に溶解し、ショック凍結し、RNAを調製するときまで−75℃で保存した。
【0034】
全RNAを酸性フェノール/クロロホルム抽出により調製した。このため、溶解緩衝液中で凍結した試料を室温で融解し、伸張のため以下の溶液で処理した:溶解緩衝液1/10体積の2Mクエン酸ナトリウム(pH4.0)、溶解緩衝液1体積の1水飽和フェノールおよび溶解緩衝液1/5体積のクロロホルム/イソアミールアルコール(24:1)。これらの試薬を攪拌機で10秒間混合した後、氷上で10分間保って温度を一定にした。15365gで4℃にて30分間遠心することによりこの相を分離した。その後、水相を新しい容器に移し、この相に存在するRNAを単離するために、Rnaid(登録商標)plusキット(DIANOVA)から8mLのRNA−MATRIXを加えた。および全体を室温で15分間インキュベーションした。得られたRNA/RNA−MATRIX複合体を7000gでの遠心によりペレット化し、上清を捨てた。次いでペレットを、それぞれのケースについて2回250mLのRNA−WASH(DIANOVA)で洗浄した後、最後に洗浄した後、真空遠心で乾燥させた。20−30mLのRNAse不含蒸留水を添加することによってRNAを溶出し、全体を55℃に15分間加熱した。室温にて7000gで1分間遠心した後、RNA溶液を新しい容器に移すことによりマトリックスを分離した。
【0035】
RNAの量をゲル電気泳動によりチェックした。RNAは−70℃で保存した。
【0036】
オリゴ(dT)プライマーを出発分子として使用してmRNAを逆転写することによりcDNAを合成した。以下の成分を合成混合物に含有させた:200ng−2μgの全RNA、2μLのM−MLV逆転写酵素(200U/μL) (GIBCOBRL)、8μLの適当な5×RT緩衝液(GIBCOBRL)、1μLのDTT(0.1M)(GIBCOBRL)、4μLのdNTP(2.5mM)(SIGMA)、2μLのオリゴ(dT)12-18プライマー(100μg/mL)(PROMEGA)、1μLのヒト胎盤インヒビター(10000U/mL)(GIBCOBRL)および水を加えて合計40μLにする。混合物を室温に10分間保った後、37℃で45分間インキュベーションした;次いで、95℃に3分間加熱し氷上で急速に冷却した。この方法により合成したcDNAを−20℃で保存した。
【0037】
cDNAの量を「ハウスキーピング」遺伝子(β-アクチン)を用いて標準化した。定量的PCRをABI PRISM 7700 Sequence Detection System (PE Applied Biosystems)を用いて行った。以下のプライマーを用いた:
β−アクチン センス: 5'-TGG AAT CCT GTG GCA TCC ATG AAA C-3'
アンチセンス:5'-TAA AAC GCA GCT CAG TAA CAG TCC G-3'
IFN−γ センス: 5'-AGCGGC TGA CTG AAC TCA GAT TGT AG-3'
アンチセンス:5'-GTC ACA GTT TTC AGC TGT ATA GGG-3'
IFN−α センス: 5'-GGC AGG TCT ACT TTG GAG TCA TTG C-3'
アンチセンス:5'-ACA TTC GAG GCT CCA GTG AAT TCG G-3'
IL−15 センス: 5'-GCC AAC TGG ATA GAT GTA AGA TAT GAC CT-3'
アンチセンス:5'-CGT GTT GAT GAA CAT TTG GAC AAT GCG TAT-3'
【0038】
DNAは、25μLの体積の反応液(この反応液には、1.4mMの各dNTP、4mMのMgCl2、0.3μmolの各プライマーおよびプローブ、2.5μLのSYBRグリーンを含む10倍PCR緩衝液[すべてのPCR試薬はSYBRグリーンPCRコア試薬キット(PerkinElmer/RocheMolecularSystemsInc.)から入手した]および1.25UのTaqDNAポリメラーゼおよび0.25UのAmpEraseを含有する)中で増幅させた。最初の変性工程(95℃にて10分間)の後、試料を40サイクルの変性(95℃、30秒)およびアニーリング/伸張(60℃,1.30分間)に付した。生成物をABIPRISM(登録商標)7700 Sequence Detection System スタンダードソフトウエアを用いて解析した。結果を、分散および多重比較により統計的に解析した。
【0039】
意外にも以下の結果が得られた:
1.D1701株またはNZ2株による処置後、γインターフェロンの発現が投与後6時間および12時間後の両方で誘導されている(図3)。NZ2株のケースでは、この誘導がプラセボおよびD1701株の両方に関して有意に高い。D1701の投与後に得られたγ−インターフェロン発現の程度は、プラセボ対照とは有意な差異はない。この図は、腹腔洗浄により得られた細胞において測定された値を示している。
【0040】
2.D1701株による処置後、TNFαの発現が投与後12時間で誘導されるのに対し、NZ2株での処置後では6時間後および12時間後の両方で誘導されている(6時間の値と比較して12時間後では減少を既に観察することができる:図4)。NZ2株のケースでは、投与後6時間でのこの誘導は、D1701株のケースで観察される値よりも有意に高い。この図は、腹腔洗浄により得られた細胞において測定された値を示している。
【0041】
3.D1701株またはNZ2株での処置後、6時間および12時間後の両方でIL−15の発現が誘導される(図5)。NZ2株の場合、この誘導は、投与後6時間でD1701株またはプラセボのケースよりも有意に高い。D1701の投与後に観察されるIL−15発現の程度は、プラセボ対照で観察される程度よりも有意に高い。この図は、腹腔洗浄により得られた細胞において測定された値を示している。
【0042】
実施例3.腫瘍を有するヌードマウスにおける治療効果の実証
インキュベーターにて、MDA−MB231細胞(ATCC#HTB26)を、完全培地(88 5 DMEM、10%FBS、1%ペニシリン/ストレプトマイシン、1%L−グルタミン(いずれもGibco Life Technologies))中、37℃にて5%CO2の存在下で培養した。移植の日に、細胞を約70%コンフルエントであった。この細胞をトリプシン処理し、HBSSで洗浄し、細胞数を計測し、予め冷却したPBSを用いて2.5×107細胞/mLに調整した。雌性の NCrヌードマウス(taconic)を用いた。このマウスは8〜10週齢であり、体重は約22gであった。操作はすべて無菌状態で行った。5×106細胞を合計0.2mlの体積で脇腹の領域に皮下注射した。この後、マウスを、腫瘍重量が平均約80mgに達するまでさらに7日間飼育した。腫瘍を測定してマウスを各ケースにつき10匹の群に無作為にグループ分けした。各群に以下のものを投与した:
グループ1: プラセボ(PBS)
グループ2: パラポックスウイルスovis、D1701株
グループ3: パラポックスウイルスovis、NZ2株
【0043】
D1701を2.5×105TCID50の用量で投与し、NZ2は1×105TCID50の用量を投与した。各群においてこれらの用量を3日おきに4回投与した。腫瘍を1週間に2回測定した。Studentテストを用いて有意度を求めた。
【0044】
図6は、グループ1〜3の実験期間(日)中の腫瘍の平均の大きさを示す(mg)(記号:グループ1○、グループ2△;グループ3□)。驚くべきことに、腫瘍に対する活性(活性が対照群と比較して有意(p<0.05)であった)がこの実験系でみとめられた。NZ2株はこの点について、D1701株よりも効力が顕著であることがみとめられた。同等の効果を達成するために、NZ2はD1701と比較して約半分しか必要としなかった。
【0045】
この知見は、腫瘍を有するヌードマウスにおける、本発明のウイルス調製物の治療効果を明白に裏付けるものである。ヌードマウスは免疫不全であり、機能性のT細胞は有していない。この実験系において、腫瘍に対する活性は、ナチュラルキラー(NK)細胞、他の細胞およびサイトカイン/ケモカインの直接の効果に起因するものと推定される。NZ2の優れた効果は、完全なおよびインタクトな免疫系の場合ではより一層明白な証拠となるであろう。
【0046】
実施例4.NZ2とD1701とのその他の生物学的な差異
D1701株とは対照的に、パラポックスウイルスNZ2はヒト細胞系において継続的に継代することができることが分かった。このことは、複製の挙動および/またはウイルスレセプターにおけるNZ2とD1701との間の本質的な違いを示している。
【0047】
ヒト細胞系への適合化は、ヒト細胞系におけるウイルス株の製造のための重要な前提条件である。
4.a)ヒトMRC−5細胞に対する継続的に継代される能力
図7は、NZ2およびD1701株をヒト二倍体細胞系MRC−5に適合させる試みを示す。MRC−5は生物学的に活性な化合物およびワクチンの製造に適している。両株について同じ初発力価を用い、対応する培養細胞上清をMRC−5細胞において5回継代させた。図は、各ケースにおいて感染させたMRC−5細胞から得られた上清の試料の力価をTCID50で示し、継代数に対してプロットした。D1701ではなくNZ2株のみが、ウシ培養腎細胞に対して逆滴定可能であった(BK クローン3a)。これは、感染および複製挙動におけるこの2つの株の間の根本的な生物学的違いの存在を示している。同時に、この結果は、NZ2がD1701よりも広範なスペクトルで複製され得ること、それによりヒト細胞をベースとした製造方法の可能性がもたらされることを示している。
【0048】
4.b)NZ2およびD1701のWI−38細胞およびBKクローン3a細胞おける継代能力
NZ2とD1701株の間の根本的な生物学的違いは、図8および図9に示された実験によってさらに明白に実証される。図8は、ウシBKクローン3a細胞およびヒトWI−38細胞でのD1701の継代において得られた、継代数に対してプロットされたウイルス力価を示す。D1701はBKクローン3a細胞では継続的に継代することができるが、ヒトWI−38細胞ではできないことがみとめられる。
【0049】
この状況は、NZ2株については異なる。この株は、BKクローン3a細胞とWI−38細胞の両方について数日間に渡って継続的に継代することができる(図9)。
【0050】
これらの結果はまた、NZ2およびD1701の感染および複製挙動における明白な違いを示している。
【0051】
4.c)WI−38での継代後のヘルペスウイルス投与試験におけるNZ2の用量依存的効果
WI−38細胞において継代させたNZ2株の免疫刺激特性を調べるため、マウスに対してヘルペスウイルス投与試験を行った。各ケース10匹の実験動物からなる3つの群に対し、それぞれ、1×104TCID50、5×104TCID50および1×105TCID50で処置し、対照群にはプラセボを投与した。図10は、ヘルペスウイルスによる感染後の時間経過に対する4つの実験群の生存率を示す。意外にも、NZ2は、WI−38で継代させた結果、免疫刺激特性は失われなかった。
【0052】
この実験例における実験結果はすべて、NZ2とD1701の感染および複製挙動における根本的な違いを示している。意外にも、D1701の場合とは対照的に、免疫調節物質として用いるためのNZ2の調製物は産生細胞系としてウシ腎細胞を用いる必要はない。
【0053】
潜在的で慢性的に持続するウイルス感染に対するTh1免疫反応の影響の知られている状況4)、5)、また増殖性の疾患例えば癌89、およびパラポックスウイルスovisNZ2株の免疫調節特性がパラポックスウイルスD1701株よりも優れているという事実に基づけは、パラポックスウイルスovisNZ2株、または上記の株の1つを基礎とする免疫調節物質の、ヒトおよび動物における、単独療法または生物学的に活性な物質、例えば抗ウイルス薬、低分子量化合物との組合せとしての使用は、B型肝炎ウイルス、またはC型肝炎ウイルスまたは肝炎の原因ウイルスの群に由来するその他の病原体のいずれかによる感染、さらに、内臓の他のウイルス感染、並びに様々なタイプの単純ヘルペスウイルス(HSV)、様々なタイプのヒトパピローマウイルス(HPV)、ヒト免疫不全ウイルス(HIV)、水疱瘡ウイルス、ヒトサイトメガロウイルス(HCMV)による、他の疾患を同様に伴う感染および動物における対応するウイルス性疾患の抗ウイルス治療が可能であり治療上有用である。
【0054】
さらに、示された作用機序に基づけば、上記のパラポックスウイルスovis株の上記の株は、特に、以下の予防的または治療的処置を行うことについて、ある程度の成功の見込みをもって使用することができる:
この治療法による処置を暴露後迅速に行う場合、ヘルペスウイルス感染に関する再発の防止、感染後防御、即ち、ウイルス感染の確立の阻止(例えば、HIV)7)。この作用機序に基づいて癌の処置も同様に可能である8)、9)。
【0055】
臨床上の問題の性質によって、パラポックスウイルスをベースとする治療剤を全身的、即ち、例えば筋肉内、皮下、腹腔内、静脈内または経口的に、あるいは局所的に投与する。これに関連して、パラポックスウイルスは、精製された凍結乾燥状態で存在するかおよび/または投与の直前に適当な溶媒に懸濁させる、あるいは別の適当な製剤中に存在させるか耐胃液投与剤型または別の経口投与剤型中に存在させることができる。
【0056】
適当な調製物はまた、特定の細胞、例えばWI−38、MRC−5またはVero細胞で継代および/または適合させることによって得られるNZ2の子孫、および上記のその他の株またはNZ2の一部もしくは断片、およびそれらの上記の株またはそれらの子孫から調製することもできる。一部とは、適当な系、例えば培養線維芽細胞、において適当なベクター、例えばワクシニア、の助けをかりて発現するゲノムまたはサブゲノム断片であると理解される。断片とは、例えば超音波処理によって物理的に破壊した粒子の生化学的精製、例えばクロマトグラフィー、によって得られる一部分であると理解される。
【0057】
これに関連して、その臨床上の問題が必要とする条件を満たす時間的な計画にしたがって、いくつかの投与または長期間の処置が必要になるかもしれない。
【0058】
即ち、例えば以下の計画に従う使用が癌治療の場合には特に期待できることが分かった(但し、これらに何ら限定されない):
各ケースにつき106〜107TCID50(組織培養細胞感染量)の筋肉内投与を3日毎に4週間、次いで2週間の休止;各ケースについて106〜107TCID50の筋肉内投与を再開し、3日毎に4週間行った後、2週間の休止;各ケースについて106〜107TCID50の筋肉内投与を再開し、3日毎に4週間行った後、2週間の休止;重篤度および治療の成功度によって、これらのサイクルをさらに増やしてもよい;あるいは、製剤を少なくとも3ヶ月間4日〜5日毎に投与するという計画を示すこともできる。
【0059】
例えば、慢性のウイルス感染の場合、3日毎に合計5回106〜107TCID50を腹部に皮下投与するか、三角筋もしくは大腿四頭筋の領域に筋肉内投与する。その疾患の必要によってはこの計画を分けることが可能である。風邪の予防については、製剤をうがい用として用い、病原菌の危険が存在する限りは、毎日繰り返す。
【0060】
口の部分の外科的介入(例えば、歯の手術)後の感染を防ぐために、その介入の前夜に、本製剤で1〜2分間うがいを行う。
【0061】
参考文献:
1. Guidotti, L.G., Borrow, P., Hobbs, M.V., Matzke, B., Gresser, I., Oldstone, M.B.A., and Chisari, F.V. (1996): Viral cross talk: Intracellular inactivation of the hepatitis B virus during an unrelated viral infection of the liver. Proc. Natl. Acad. Sci. USA. 93:4589-4594.
2. Guidotti, L.G., Ando, K., Hobbs, M.V., Ishikawa, T., Runkel, L., Schreiber, R.D., and Chisari, F.V. (1994): Cytotoxic T lymphocytes inhibit hepatitis B virus gene expression by a noncytolytic mechanism in transgenic mice. Proc. Natl. Acad. Sci. USA. 91:3764-3768.
3. Steinmassl,G., G.Wolf (1990): Bildung von Interleukin 2 and Interferondurch mononukleare Leukozyten des Schweines nach in vitro-Stimulation mit verschiedenen Viruspraparaten (Formation of interleukin 2 and interferon by pig mononuclear leukocytes following in-vitro stimulation with various virus preparations). J.Vet.Med.B37,5,321-331.
4. P. Lucin, S. Jonjic, M. Messerle, B. Polic, H. Hengel, U.H. Koszinowski (1994): Late-Phase inhibition of murine cytomegalovirus replication by synergistic action of interferon gamma and tumor necrosis factor alpha. J. Gen. Viro175-101-110;P.M.
5. Smith, R.M. Wolcott, R. Chervenak, S.R. Jennings (1994): Control of acute cutaneous herpes-simplex virus-Infection - T-cell mediated viral clearance is dependent upon interferon gamma. Virology 202 (1):76-88].
6. Y. Kawanashi, N. Hayashi, K. Katayama, K. ueda, T. Takehara, E. Miyoshi, E. Mita, A. Kasahara, H. Fusamoto, T. Kamada (1995): Tumor necrosis factor alpha and interferon gamma inhibit synergistically viral replication in hepatitis B virus replicating cells. J. Medical Virology 47 (3):272-277.
7. Dhawan, S., L.M. Wahl, A. Heredia, Y.H. Zhang, J.S. Epstein, M.S. Meltzer, I.K. Hewlett (1995): Interferon gamma inhibits HIV-induced invasiveness of Monocytes. J. Leukocyte Biology, 58 (6):713-716.
8. J.F. Bromberg, C.M. Horvath, Z.L. Wen, R.D. Schreiber, J.E.Darnell (1996): Transcriptionally active statl is required for the antiproliferative effects of both interferon alpha and interferon gamma. PNAS 93(15):7673-7678.
9. M.Klouche, H.Kirchner, F.Holzel (1994): Antiproliferative effects of inter-feron gamma in combination with alpha-difluoromethylornithine on human carcinoma cell cultures. J.Cancer Research and Clinical Oncology 120(12):706].
【特許請求の範囲】
【請求項1】
ヒトおよび動物におけるウイルス感染および癌に対する医薬の製造のための、パラポックスウイルスovis株NZ2、NZ−7、NZ−10およびorf−11の1つに分類上属するウイルスの使用。
【請求項1】
ヒトおよび動物におけるウイルス感染および癌に対する医薬の製造のための、パラポックスウイルスovis株NZ2、NZ−7、NZ−10およびorf−11の1つに分類上属するウイルスの使用。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2012−121902(P2012−121902A)
【公開日】平成24年6月28日(2012.6.28)
【国際特許分類】
【出願番号】特願2012−21508(P2012−21508)
【出願日】平成24年2月3日(2012.2.3)
【分割の表示】特願2002−508456(P2002−508456)の分割
【原出願日】平成13年7月11日(2001.7.11)
【出願人】(506207901)アイキュリス・ゲゼルシャフト・ミット・ベシュレンクテル・ハフツング・ウント・コムパニー・コマンディットゲゼルシャフト (30)
【氏名又は名称原語表記】AiCuris GmbH & Co. KG
【Fターム(参考)】
【公開日】平成24年6月28日(2012.6.28)
【国際特許分類】
【出願日】平成24年2月3日(2012.2.3)
【分割の表示】特願2002−508456(P2002−508456)の分割
【原出願日】平成13年7月11日(2001.7.11)
【出願人】(506207901)アイキュリス・ゲゼルシャフト・ミット・ベシュレンクテル・ハフツング・ウント・コムパニー・コマンディットゲゼルシャフト (30)
【氏名又は名称原語表記】AiCuris GmbH & Co. KG
【Fターム(参考)】
[ Back to top ]