
抗体を産生する方法
免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を産生するための方法であって、該断片は免疫グロブリン重および軽鎖の可変ドメインの少なくとも機能的な部分を具備している方法が記載される。前記方法は、重および軽鎖を2つの別々の宿主細胞で産生させることと、免疫グロブリンの分子または断片をエクスビボでリフォールディングさせることとを備える。
【発明の詳細な説明】
【発明の開示】
【0001】
[発明の分野]
本発明は、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を産生するための方法に関する。本方法において、免疫グロブリンの軽および重鎖の配列は、同じ細胞培養物中の別々(separate)の宿主細胞中で発現させることができ、次に免疫グロブリンまたは免疫グロブリン断片のエクスビボでの会合がなされる。
【0002】
[発明の背景]
モノクローナル抗体を、ハイブリドーマ技術によって又は組換え型の発現によって産生することができる。組換え型の発現によって、抗体の設計および産生を改善することに関してさらなるオプションが提供される。様々な形式の抗体または抗体断片を、組換え型で発現させることができる。そして、親和性、特異性、および糖鎖形成などの特性を、遺伝子工学で変更して、抗体または抗体断片の特性を改善することができる。組換え型で発現させた抗体の生産性は、ハイブリドーマから得られる収量(yield)を一般的には超え、組換え型で産生された抗体では生産コストが低くなる。
【0003】
US6331415(Cabillyら)は、重および軽鎖が単一細胞中で単一のベクターから又は2つの別々のベクターから同時に発現される免疫グロブリンの組換え型を産生する方法を記載している。
【0004】
Wibbenmeyerら(1999, Biochim Biophys Acta 1430(2):191-202)およびLeeおよびKwak(2003, J. Biotechnology 101:189-198)は、大腸菌(E. coli)の別々の培養物中で発現させたプラスミドを用いて、別々に産生した重および軽鎖からのモノクローナル抗体の産生を記載している。H-およびL-鎖を、封入体として発現し、細胞溶解物から精製し、インビトロでリフォールディングした。
【0005】
US5643745は、2つの核を含んでいる糸状菌を培養することによってヘテロダイマーを産生する方法を記載しており、そのうちの最初のものは修飾されて第1のサブユニットのヘテロダイマーを発現し、そのうちの第2のものは修飾されて第2のサブユニットを発現する。
【0006】
1つの細胞発現系が便利である。というのも、抗体は、精製後に構造的に使える状態となるからである。しかしながら、他の因子は、制御することが困難であろう(例えば、軽および重鎖を、至適な収量を得るために所望の比率で発現させること)。他方で、グラム陰性の大腸菌中での発現はエンドトキシンの混入を生じる可能性があり、完全長の免疫グロブリンの発現は問題がある。
【0007】
従って、特定の要求に適し、上記の課題を解決する、組換え型抗体を産生するための代替法が、必要とされる。
【0008】
[発明の概要]
本発明は、免疫グロブリン重および軽鎖の可変ドメインの少なくとも機能的な部分を具備している、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を産生するための方法を提供し、該方法は重および軽鎖を、真核生物細胞およびグラム陽性菌からなる群から選択される2つの別々の宿主細胞で産生させること;及び免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片をエクスビボでリフォールディングさせることを備える。
【0009】
従って、本発明は、免疫グロブリン分子または免疫学的に機能的なその断片を産生する方法を提供し、該方法は以下を備える:
(a)第1の宿主細胞を、少なくとも免疫グロブリン重鎖の可変ドメインを具備している第1のポリペプチドをコード化しているヌクレオチド配列を具備している第1の核酸で形質転換すること;
(b)第2の宿主細胞を、少なくとも免疫グロブリン軽鎖の可変ドメインを具備している第2のポリペプチドをコード化しているヌクレオチド配列を具備している第2の核酸で形質転換すること;
(c)第1および第2の核酸配列を発現させること;
(d)第1および第2のポリペプチドを精製すること;および
(e)第1および第2のポリペプチドがリフォールディングして、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を形成することを認容すること;
ここで、第1および第2の宿主細胞は、別々に真核生物細胞およびグラム陽性菌からなる群から選択される。
【0010】
前記免疫グロブリン分子は、IgA、IgD、IgE、IgG、およびIgM免疫グロブリンなどから選択することができる。前記免疫学的に機能的な断片は、Fab断片、Fab’断片、Fab'-SH断片、F(ab’)2断片、Fv断片、VHH断片、ドメイン抗体、ディアボディー(diabody)、および多重特異性の抗体または抗体断片などから選択することができる。前記真核生物細胞は、哺乳類細胞、昆虫細胞、植物細胞、および真菌細胞などから選択することができる。前記第1および第2の宿主細胞は、例えば、COS細胞、BHK細胞、HEK293細胞、DUKX細胞、Saccharomyces spp細胞、Kluyveromyces spp細胞、Aspergillus spp細胞、Neurospora spp細胞、Fusarium spp細胞、Trichoderma spp細胞、およびLepidoptera spp細胞からなる群から別々に選択することができる。別の側面において、前記第1および第2の宿主細胞は、同じ細胞タイプの細胞であるか又は異なる細胞タイプの細胞である。
【0011】
一側面において、前記第1および第2の宿主細胞は、同じ培養で成長させられる。別の側面において、前記第1および第2の宿主細胞は、別々の培養で成長させられる。別の側面において、精製する工程は、Obelix陽イオン交換カラムを用いる精製を備えてもよい。一側面において、前記第1の宿主細胞は免疫グロブリン軽鎖をコード化している核酸を発現しなく、第2の宿主細胞は免疫グロブリン重鎖をコード化している核酸を発現しない。別の側面において、前記第1および第2の核酸は、1以上のモノクローナル抗体産生細胞から由来する。モノクローナル抗体産生細胞は、例えば、ハイブリドーマ、ポリドーマ(polydoma)および不死化させたB細胞から選択することができる。
【0012】
一側面において、リフォールディングは、前記第1および第2のポリペプチドを次の(a)および(b)から選択される条件下で混合することを備える:(a)第1と第2のポリペプチドとの比が約1:1、温度は約室温、およびpHは約7;並びに(b)第1と第2のポリペプチドとの比が約1:1、温度は約5゜C、およびpHは約8.0〜8.5の範囲。別の更なる側面において、前記第1および第2のポリペプチドは、約0.5M L-アルギニン-HCl、約0.9mM酸化型グルタチオン(GSSG)、および約2mM EDTAを含んでいる溶液中で混合される。
【0013】
また、本発明は、免疫グロブリン分子または免疫学的に機能的なその断片を産生する方法を提供し、該方法は以下を備える:
(a)第1の宿主細胞を、少なくとも免疫グロブリン重鎖の可変ドメインを具備している第1のポリペプチドをコード化しているヌクレオチド配列を具備している第1の核酸で形質転換すること;
(b)第2の宿主細胞を、少なくとも免疫グロブリン軽鎖の可変ドメインを具備している第2のポリペプチドをコード化しているヌクレオチド配列を具備している第2の核酸で形質転換すること;
(c)第1および第2の核酸配列を発現させること;
(d)第1および第2のポリペプチドの混合物を含んでいる溶液を透析すること;および
(e)第1および第2のポリペプチドがリフォールディングして、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を形成することを認容すること;
ここで、第1および第2の宿主細胞は、別々に真核生物細胞およびグラム陽性菌からなる群から選択される。
【0014】
一側面において、第1および第2の宿主細胞は、同じ培養で成長させられる(その溶液は、第1および第2の宿主細胞が成長させられた培養培地である)。別の側面において、前記溶液中の第1および第2のポリペプチドの相対量(relative amount)は、約1:2〜約2:1の範囲である。
【0015】
また、本発明は、抗体を精製するための方法を提供し、該方法は抗体を含んでいる溶液をObelix陽イオン交換カラムに適用すること、および抗体を精製することを備える。一側面において、前記方法は、次の工程のうち少なくとも1つを備える:(a)濾過した細胞培養(任意でpH調製される)を前記カラムに適用すること;(b)溶媒を溶出緩衝液に添加すること;および(c)塩グラジエントを増加させることによって抗体を溶出すること。特定の側面において、工程(c)は、工程(b)の前に実施される。代替的な溶出戦略には、約6.0のpHを有し、塩およびグリセロールを含んでいる溶出緩衝液(例えば、約30mMクエン酸, 約25mM NaCl, 約30%グリセロール、約6,0のpH)、約7.5〜8.5のpHを有している溶出緩衝液(例えば、Tris-緩衝剤)、約pH6.0から約6〜約9の範囲のpH(例えば、pH7.5〜8.5)までのpHグラジエント、および約6,5〜約7.0で0〜約1Mの塩(例えば、NaCl)でのグラジエント溶出の使用が含まれるが、これらに限定されない。
【0016】
また、本発明は、免疫グロブリン分子または免疫グロブリン重および軽鎖の少なくとも可変性ドメインを含んでいる免疫学的に機能的な免疫グロブリン断片を産生する方法を提供し、該方法は以下の工程を備える:
(a)重および軽鎖を、真核生物細胞およびグラム陽性細菌からなる群から選択される2つの別々の宿主細胞で独立に産生すること;
(b)重および軽鎖を精製すること;および
(c)免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片をインビトロでリフォールディングさせること。
【0017】
これらの及び他の側面は、以下により完全に記載される。
【0018】
[発明の記載]
本発明は、次の発見に基づくものである;その発見とは、免疫グロブリン分子の別々のポリペプチド鎖又はその断片が、別々の宿主細胞で独立に産生され、その後にリフォールドされて最終的な分子が形成されることである。宿主細胞を、同じ培地で共培養するか又は別々に培養することができる。発現後、鎖内および鎖間のジスルフィド形成の双方を、生体外の適切な反応条件下で行い、これによって正確に会合し、フォールドした抗体の形成が許容される。
【0019】
免疫グロブリン重(HC)および軽(LC)鎖(又は同じものの断片)を含んでいる及びコード化している宿主細胞は、それぞれ同じ、同一の細胞タイプのもの(例えば、同じ哺乳類細胞株からのもの)である。或いは、それぞれHCおよびLCを発現している宿主細胞は、異なる起源(例えば、哺乳類宿主細胞において発現されているHC、およびグラム陽性菌において発現されているLC)のものである。さらにまた、重および軽鎖をコード化しているDNA配列は、1つのハイブリドーマ細胞株から又は2以上のモノクローナル抗体産生性ハイブリドーマから取得しえる。
【0020】
上記のとおり、重および軽鎖を発現している宿主細胞は、それぞれ別々の培養および個々の培地、または同じ培地の共培養で成長させることができる。別々の宿主細胞で組換え型を産生させた後、HCおよびLC鎖を、当該技術分野において既知の方法によって又は本明細書中に記載された方法によって精製することができる。例えば、例は、高い伝導度を伴うタンパク質を結合することができる高度に疎水性の樹脂(Obelix陽イオン交換カラム, アマシャムから利用可能、カタログNo.11-0010)を用いる精製を記載している。
【0021】
完全な免疫グロブリン分子または機能的な免疫グロブリン断片を供給するための残りの最終工程は、重および軽鎖のジスルフィド結合形成による再会合が関与する(本発明において、リフォールディングと称される)。リフォールディング(再生とも称される)を、Jin-Lian Xingなど(2004; World J Gastroenterol 10(14):2029-2033)およびLeeおよびKwak(2003; Journal of Biotechnology 101:189-198)に記載のとおりに実施することができる。特定の態様において、リフォールディングは、重および軽鎖の混合物(又はその断片)の透析によって達成される(混合物中の重鎖および軽鎖の量は1:2〜2:1の範囲である)。更なる一態様において、前記範囲は約1:1である。前記態様において、前記宿主細胞は、同じ培養培地に含有される。培地中のHCおよびLC(又はその断片)の自己会合物(self-assemble)、および機能的な免疫グロブリンまたは断片は、前記培地から収穫することができる。HCおよびLCの混合物を含んでいる培養培地の透析工程には、リフォールディング処理を任意で含んでもよい。
【0022】
一態様において、HC鎖を発現する宿主細胞は、LC鎖をコード化しているDNAを含有または発現しない及び/又はLC鎖を発現する宿主細胞はHC鎖をコード化しているDNAを含有または発現しない。
【0023】
本発明の方法によって、グラム陰性菌(例えば、大腸菌)中で免疫グロブリンを発現させることと比較して、幾つかの利点が提供される。前記利点には、次のものが含まれる:
(i)エンドトキシンが存在しない、
(ii)高収量のタンパク質が取得される(というのも、封入体からタンパク質をリフォールディングさせる必要がないからである)、
(iii)完全長の免疫グロブリンを産生することができる、および
(iV)抗体の糖鎖形成パターンを、宿主となる生体に依存して変化させることができる。
【0024】
項目(i)に関して、本明細書中で使用されるエンドトキシンは、腸内細菌性のリポ多糖の毒性活性を意味し、グラム陰性細菌の外膜に見出されるものである。
【0025】
項目(ii)および(v)に関して、グラム陰性菌(例えば、大腸菌)は、大量のタンパク質が所望される場合、産生宿主細胞として適切ではない。大量の所望のタンパク質を大腸菌で生産した結果、しばしば封入体の形成を生じ、引続いてリフォールディングがなされる。対照的に、グラム陽性細菌は外膜を有さないが、タンパク質を細胞質から細胞外間隙へと直接的に分泌させるグリカン層を有する。相対的に単純な搬出機構によって、組換えタンパク質の分泌が高収量で促進される。
【0026】
項目(iii)に関して、完全長の免疫グロブリン分子はサイズが大きいので、大腸菌で取得することが困難である。リフォールディングにおける最近の報告に関して、完全なIgG分子が、大腸菌中で産生された(Simmons et al 2002 J. Immunol. Methods 263:133-147を参照されたい)。
【0027】
項目(iv)関して、薬学的な適用に関して開発された大抵のタンパク質は、真核生物性の宿主細胞中で産生された場合に、該タンパク質のポリペプチドバックボーンに付着したオリゴ糖を有する。一般的には、係る糖タンパク質の糖鎖は、アスパラギン残基のアミド基にN-グリコシド結合で又はセリンまたはスレオニン残基のヒドロキシル基にO-グリコシド結合で付着されてもよい。糖鎖形成は、タンパク質の適切な機能にしばしば必要とされ、適切なフォールディング、機能、および安定性を保証する。原核生物は、タンパク質の翻訳後修飾を実施する能力を欠いている。従って、タンパク質の糖鎖形成は、係る系で取得することができない。真菌類および酵母細胞を操作して、適切な糖鎖形成パターンを有するタンパク質を産生させることができる(Ballew and Gerngross 2004 Expert Opin. Biol. Ther. 4:623-626)。
【0028】
本発明によると、上記の効果は、重および軽鎖を、真核生物細胞およびグラム陽性菌からなる群から選択される2つの別々の宿主細胞中で独立に産生することによって提供される。本発明において、「独立に(independently)」の用語は、次の事項を意味する;その事項とは、個々の重鎖(HC)および軽鎖(LC)(又はその断片)の産生を、宿主細胞培養の異なる宿主細胞、異なる培養培地、異なる発現ベクター、および/または異なる物理的な条件(例えば、温度、レドックス条件、pH)などの使用によって、独立にコントロールする又は制御することができることである。HCおよびLC鎖(又はその断片)の産生後に、完全長の抗体または抗体断片へのエクスビボでのリフォールディングは、培養培地中で直接的に達成することができる(2つの別々のHCおよびLC鎖を発現している宿主細胞が、それぞれ同じ細胞培養中に存在する場合);或いは、本明細書中の他の箇所で記載した、LCおよびHCまたは断片の1以上の連続的(joint)または別個の精製工程、HCおよび/またはLC鎖溶液を濃縮するための及び/又は緩衝液を交換するための透析、及び特定のリフォールディング緩衝液への移動又は該緩衝液での希釈後に、達成することができる。リフォールディング条件を、当該技術分野における既知の方法にしたがって、各抗体又は抗体断片に関して選択する又は至適化することができる。典型的には、リフォールディングは、約+4゜C〜約+40゜Cまたは約+4゜C〜約室温の範囲の温度で、約5〜約9または約5.5〜約8.5のpHで得ることができる。リフォールディングを至適化するために使用しえる例示的な緩衝液には、リン酸、クエン酸-リン酸、酢酸(acetate)、およびトリス、同様にCO2でpHが制御される細胞の培養培地が含まれる。特定のリフォールディング条件は、例1に記載される。他の例示的なリフォールディング条件には、約1:1のHC:LC(またはHC:LC断片)比、約室温の温度、および中性のpHが含まれる。別の例示的なリフォールディング条件には、約1:1のHC:LC(またはHC:LC断片)比、約5゜Cの温度、約0.1 M Tris-HCl緩衝剤、約0.5MのL-アルギニン-HCl、酸化還元系として約0.9mMの酸化型グルタチオン(GSSG)、約2mMのEDTA、約8.0〜8.5のpHが含まれる。一側面において、リフォールディング溶液は、pHが約7.4で約100mMの尿素を含んでいる20mM Tris-HCl緩衝液に対して、平衡化した透析緩衝液における伝導度が約3.0〜3.5mSの範囲の値にまで減少するまで透析される。
【0029】
抗体
「抗体」および「免疫グロブリン分子」の用語は、本明細書中で互換的に使用され、モノクローナル抗体を意味する。重鎖中の定常ドメインの種類に応じて、抗体は、5つの主要なクラス:IgA、IgD、IgE、IgG及びIgMの1つに割り当てられる。これらのうち幾つかは、IgG1、IgG2、IgG3、IgG4などのように、サブクラス又はアイソタイプにさらに分けられる。免疫グロブリンの異なるクラスに対応する重鎖定常ドメインは、それぞれ「アルファ」、「デルタ」、「イプシロン」、「ガンマ」及び「ミュー」と称される。免疫グロブリンの異なるクラスのサブユニット構造及び三次元立体配置は周知である。IgGおよび/またはIgM(一般に生理学的な/臨床的な状況で一般的に使用され、研究室で容易に作出できる)は、本発明で使用する抗体の例示的なクラスである。
【0030】
本発明による、「IgG」分子の免疫グロブリンは、完全な抗体であってもよいし、機能的な免疫グロブリン断片であってもよい。モノクローナル抗体の断片または誘導体を作出する方法を、以下に開示する。
【0031】
当業者は所望の免疫グロブリン分子をどのように提供するかについて知っており、更なる詳細は以下で記載される。前記免疫グロブリン分子は、例えば、IgA、IgD、IgE、IgG、またはIgMの免疫グロブリンであってもよい。
【0032】
本明細書中で記載される「重鎖(HC)」または「軽鎖(LC)」は、完全長抗体の重および軽鎖または抗体断片の重鎖および軽鎖を具備する。
【0033】
本明細書中に使用される、「抗体断片(antibody fragment)」または「免疫グロブリン分子断片(immunoglobulin molecule fragment)」は、完全長抗体の一部を具備し、抗原と結合する能力がある。典型的には、抗体断片は、HCおよびLCの少なくとも1,2,3,4,5つの又は全ての抗体CDR領域、又は全体の可変重鎖(VH)または可変軽鎖(VL)部分を具備する。例示的な抗体断片は、Fab, F(ab)2, F(ab’)2, Fd, scFv, dsFv断片, VHH断片, ドメイン抗体(VHおよび/またはVL)、同様に、2以上の完全長抗体の抗原結合部位を含んでいる多重特異性(二重特異的なものを含む)抗体構築物が含まれるが、これらに限定されない。
【0034】
「免疫原(immunogen)」の用語は、免疫寛容ではなく、体液性の抗体および/または細胞性の免疫応答を誘導する能力がある物質である。「免疫原」の用語は、「抗原(antigen)」と時々互換的に使用されるが、その用語は免疫応答を刺激する及びその産物(例えば、抗体)と反応する能力があることを特定する。対照的に、「抗原」は、抗体と反応する物質を意味させるために確保(reserved)される。主要な免疫原は、微生物から遊離しているか又はそれに付着している、タンパク質およびリポ多糖である。
【0035】
本明細書中で使用される「エクスビボ(ex vivo)」の用語は、任意の身体の外側(即ち、インビボで生じないプロセス)および任意の生細胞の外側(即ち、細胞内で生じないプロセス)を意味する。
【0036】
「免疫原性(immunogenic)」の用語は、体液性の抗体および/または細胞性の免疫応答を誘導する能力を意味する。
【0037】
抗体産生
本発明は、重(H)および軽(L)鎖の少なくとも可変ドメインが別々の宿主細胞中で産生され、自由選択で単離され、そしてインビトロでリフォールドされる免疫グロブリン分子または断片の組換え型の産生に関する。本発明に適用するためのVHおよびVL配列は、当該技術分野において既知の様々な技術の何れかによって産生される抗体から取得することができる。典型的には、それらは、非ヒト動物(好ましくは、マウス)を所望の抗原または免疫原を含んでいる免疫原で免疫することによって提供される。或いは、抗体は、例えば、Wardら(Nature 341 (1989) 544)に開示されているように、免疫グロブリンのコンビナトリアルライブラリーを選択することによって提供しえる。
【0038】
ヒト以外の哺乳動物を抗原で免疫化する工程は、マウスでの抗体の産生を刺激するための、当該技術で周知の任意の様式で実施することができる(例えば、 E. Harlow and D. Lane, Antibodies: A Laboratory Manual., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1988)を参照されたい)。好適な態様において、非ヒト動物は、げっ歯類(例えば、マウス、ラットなど)、ウシ、ブタ、ウマ、ウサギ、ヤギ、ヒツジなどの哺乳動物である。また、ヒト以外の哺乳動物を、XenomouseTM(Abgenix)又はHuMAb−MouseTM(Medarex)などの、「ヒト」抗体を産生するように、遺伝的に改変し、又は操作することができる。典型的には、免疫原は、任意で完全フロイントアジュバントなどのアジュバントとともに、緩衝液中に懸濁又は溶解される。免疫原の量、緩衝液の種類及びアジュバントの量を決定する方法は、当業者に周知であり、いかなる意味においても、本発明を限定するものではない。これらのパラメータは、異なる免疫原については異なる場合があり得るが、容易に解明される。
【0039】
同様に、抗体の産生を刺激するのに十分な免疫化の位置(location)及び頻度も、当該技術において周知である。典型的な免疫化プロトコールでは、1日目に、ヒト以外の動物に抗原を腹腔内注射し、約1週後に再度抗原を腹腔内注射する。この後、必要に応じて、不完全フロイントアジュバントなどのアジュバントとともに、20日目前後に、抗原のリコール注射(recall injections)を行う。リコール注射は、静脈内に行われ、連続数日間、反復することができる。その後、通例、アジュバントなしに、静脈内又は腹腔内の何れかで、40日目にブースター注射を行う。このプロトコールによって、約40日後に、抗原特異的抗体産生B細胞が産生される。免疫化に使用される抗原に対して誘導される抗体を発現するB細胞の産生をもたらす限り、他のプロトコールも使用することができる。
【0040】
別の態様では、非免疫のヒト以外の哺乳動物からリンパ球を単離し、インビトロで成長させ、次いで、細胞培養において免疫原に曝露する。次いで、リンパ球を採集し、以下に記載された融合工程を実施する。
【0041】
モノクローナル抗体の場合、次の工程は、免疫されたヒト以外の哺乳動物から脾細胞を単離した後、これらの脾細胞を不死化された細胞と融合して、抗体を産生するハイブリドーマを形成させる。ヒト以外の哺乳動物からの脾細胞の単離は当該技術において周知であり、典型的には、麻酔されたヒト以外の哺乳動物から脾臓を取り出し、脾臓を小片に切断し、脾臓カプセル(splenic capsule)から細胞ろ過器のナイロンメッシュを通して、脾細胞を適切な緩衝液中に絞り取り、単一細胞の懸濁液を得る。細胞を洗浄し、遠心し、赤血球を溶解する緩衝液中に再懸濁する。この溶液を再び遠心し、最後に、ペレット中に残存するリンパ球を新鮮な緩衝液に再懸濁する。
【0042】
一度単離し、単一の細胞懸濁液中に存在させ、リンパ球を不死化細胞株と融合する。これは、典型的には、マウスのミエローマ細胞株であるが、ハイブリドーマの作製に有用な他の多くの不死化細胞株も、当該技術において公知である。好ましいマウスのミエローマ株には、「Salk Institute Cell Distribution Center, San Diego, Calif. U.S.A」から入手可能なMOPC−21及びMPC−11マウス腫瘍、並びに「American Type Culture Collection, Rockville, Maryland U.S.A」から入手可能なX63、Ag8653及びSP−2細胞から得られるマウスのミエローマ株が含まれるが、これらに限定されるものではない。融合は、ポリエチレングリコールなどを用いて行われる。次いで、得られたハイブリドーマを、融合されていない親ミエローマ細胞の増殖又は生存を阻害する1以上の物質を含有する選択培地中で増殖させる。例えば、親ミエローマ細胞が酵素ヒポキサンチングアニンホスホリボシル転移酵素(HGPRT又はHPRT)を欠如していれば、ハイブリドーマに対する培地は、典型的には、HGPRT欠損細胞の増殖を抑える物質であるヒポキサンチン、アミノプテリン及びチミジン(HAT培地)を含むであろう。
【0043】
ハイブリドーマは、典型的には、マクロファージの支持細胞上で増殖される。マクロファージは、脾細胞を単離するために使用されるヒト以外の哺乳動物の同腹仔から得ることが好ましく、典型的には、ハイブリドーマを播種する数日前に、不完全フロイントアジュバントなどで刺激される。融合法は、文献〔Goding, "Monoclonal Antibodies: Principles and Practice," pp. 59-103 (Academic Press, 1986)〕中に記載されている。
【0044】
前記細胞は、コロニー形成と抗体産生に十分な時間、選択培地中で増殖させる。これは、通常、7日から14日の間である。次いで、このハイブリドーマコロニーを、免疫原/抗原と結合する抗体の産生についてアッセイする。その中でハイブリドーマが増殖されるウェルに適合できる任意のアッセイを使用し得るが、このアッセイは、典型的には、比色分析のELISAタイプのアッセイである。他のアッセイには、免疫沈降及びラジオイムノアッセイが含まれる。一以上の別個のコロニーが存在するかどうかを決定するために、所望の抗体の産生に関して陽性のウェルを調べる。二以上のコロニーが存在すれば、細胞を再度クローニングし、単一の細胞のみが、所望の抗体を産生するコロニーを確実に生じるように増殖させ得る。モノクローナル抗体が唯一つ検出され、産生されるように、典型的には、単一の明白なコロニーを有する陽性細胞を再クローニングし、再アッセイする。
【0045】
モノクローナル抗体を産生することが確認されているハイブリドーマは、次にDMEM又はRPMI−1640などの、適切な培地中で、大量に増殖される。或いは、ハイブリドーマ細胞は、動物内の腹水癌として、インビボで増殖させることができる。
【0046】
所望のモノクローナル抗体を産生するまで十分に増殖させた後、モノクローナル抗体を含有する増殖培地(又は腹水液)を細胞から分離し、その中に存在するモノクローナル抗体を精製する。典型的には、精製は、ゲル電気泳動、透析、プロテインA若しくはプロテインG−セファロース、又はアガロース又はセファロースビーズなどの固相支持体に連結された抗マウスIgを用いたクロマトグラフィーによって達成される(全て、例えば、「Antibody Purification Handbook, Amersham Biosciences, publication No. 18-1037-46, Edition AC」に記載されており、その開示内容は、参照により、本明細書に援用される)。結合された抗体は、典型的には、低pH緩衝液(pH3.0以下のグリシン又は酢酸緩衝液)を使用することによって、プロテインAまたはプロテインGカラムから溶出され、抗体含有画分を直ちに中和する。必要に応じて、これらの画分をプールし、透析し、濃縮する。
【0047】
本発明の特定の側面として、Obelix陽イオン交換体を抗体の精製に使用することができる。Obelix陽イオン交換体は、高伝導度で及びpI(抗体に関する)よりも高pHで抗体と結合する。これによって、精製の性能(capability)が影響される。精製を、プロピレンジオールなどを添加することによって更に変化させて、疎水性相互作用を本陽イオン交換体カラム上で利用することができる。
【0048】
本発明の方法に使用されるモノクローナル抗体をコードするDNAは、従来の手順を用いて(例えば、マウス抗体の重鎖及び軽鎖をコードする遺伝子に特異的に結合することができるオリゴヌクレオチドプローブを使用することによって)、容易に単離され、配列決定される。DNAを単離したら、DNAを発現ベクター中に配置することができ、次いで、発現ベクターを宿主細胞(トランスフェクションが行われていない状態で、免疫グロブリンタンパク質を産生しない、細菌細胞、サルのCOS細胞、チャイニーズハムスター卵巣(CHO)細胞又は骨髄腫細胞など)中にトランスフェクションして、組換え宿主細胞中でモノクローナル抗体の合成を得る。抗体をコード化しているDNAの細菌中での組換え発現は、当該技術で周知である〔例えば、Skerra et al., Curr. Opinion in Immunol., 5, pp. 256 (1993);およびPluckthun, Immunol. Revs., 130, pp. 151 (1992)を参照〕。例えば、所望の抗原と結合する抗体をコード化しているDNAは、ハイブリドーマから単離され、適切な宿主へのトランスフェクションのために適切な発現ベクターに配置される。次いで、抗体、又はそれらの断片(ヒト化された様式のモノクローナル抗体、抗体の活性断片、又は抗体の抗原認識部分を含むキメラ抗体など)の発明にしたがって組換え産生するために、宿主を使用する。
【0049】
免疫グロブリンポリペプチドをコード化しているDNA配列は、通常は組換え型ベクターに挿入される(これは、組換えDNA処置に都合よく供される、任意のベクターでよい);また、ベクターの選択は、それが導入される宿主細胞にしばしば依存する。ベクター成分には、1以上の次の因子、即ち:プロモーター、シグナル配列、複製開始点、1以上の選択マーカー、および転写ターミネーター配列が一般的に含まれるが、これらに限定されない。従って、前記ベクターは自律的に複製するベクター、即ち、染色体外の実体(extrachromosomal entity)として存在し、その複製が染色体複製に非依存性であるベクターである(例えば、プラスミド)。或いは、前記ベクターは次のようなベクター、即ち、宿主細胞に導入された際に、前記宿主細胞ゲノムに統合され、それが統合された染色体と共に複製されるベクターであってもよい。
【0050】
前記ベクターは、好ましくは発現ベクター中の免疫グロブリンポリペプチドをコード化しているDNA配列が、前記DNAの転写に必要とされる付加的なセグメントに動作可能に連結される、発現ベクターである。通常は、前記発現ベクターは、プラスミドまたはウイルスのDNAに由来するものである、或いは両方のエレメントを含有していてもよい。「動作可能に連結(operably linked)」の用語は、前記セグメントが、それらがそれらの意図する目的に関して協調して機能するように配置されることを示す(例えば、転写は、プロモーターで開始され、ポリペプチドをコード化しているDNA配列を通して進行する)。
【0051】
ポリペプチドの発現に使用するための発現ベクターは、クローン化された遺伝子またはcDNAの転写を方向付ける能力を有するプロモーターを具備する。前記プロモーターは、選択された宿主細胞において転写活性を呈する、任意のDNA配列でよく、前記宿主細胞に対して相同性(homologous)の又は異種性(heterologous)のタンパク質をコード化している遺伝子に由来するものであってもよい。
【0052】
グラム陽性細菌においてポリペプチドバリアントをコード化しているcDNAの転写を駆動させるための適切なプロモーターの例は、AmyP(Kruger et al 2002 Nature Biotechnol 20:702-706)、NisA(de Ruyter et al 1996 J. Bacteriol 178:3434-3439)、Spac(Yansura and Henner 1984 Proc. Natl. Acad. Sci. USA 81:439-443)、XylA(Eichenbaum et al 1998 Appl. Environ. Microbiol. 63:3451-3457)、LacA(Platteeuw et al 1994 Appl. Environ. Microbiol. 60:587-593)、UW85 upp(Dunn and Handelsman 1999 Gene 226:297-305)である。
【0053】
哺乳類細胞におけるポリペプチドバリアント(polypeptide variant)をコード化しているDNAの転写を方向付けるための適切なプロモーターの例は、SV40プロモーター(Subramani et al., Mol.Cell. Biol. 1 (1981), 854-864)、MT-1(メタロチオネイン遺伝子)プロモーター(Palmiter et al., Science 222 (1983), 809 - 814 )、CMVプロモーター(Boshart et al.,Cell. 41:521-530, 1985)、またはアデノウイルス2主要後期プロモーター(Kaufman and Sharp, Mol.Cell. Biol, 2:1304-1319, 1982)である。
【0054】
昆虫細胞における使用に関して適切なプロモーターは、ポリヘドリンプロモーター(US 4,745,051; Vasuvedan et al., FEBS Lett. 311, (1992) 7 - 11)、P10プロモーター(J.M. Vlak et al., J. Gen. Virology 69, 1988, pp. 765-776)、Autographa californica多核体病ウイルス塩基性タンパク質プロモーター(EP 397 485)、バキュロウイルス前初期遺伝子1プロモーター(US 5,155,037; US 5,162,222)、またはバキュロウイルス39K遅延性-初期遺伝子プロモーター(US 5,155,037; US 5,162,222)である。
【0055】
酵母宿主細胞における使用に関して適切なプロモーターの例には、酵母の解糖遺伝子(glycolytic genes)からのプロモーター(Hitzeman et al., J. Biol. Chem. 255 (1980), 12073 - 12080; Alber and Kawasaki, J. Mol. Appl. Gen. 1 (1982), 419 - 434)またはアルコール脱水素酵素遺伝子(Young et al., in Genetic Engineering of Microorganisms for Chemicals (Hollaender et al, eds.), Plenum Press, New York, 1982)、またはTPI1(US 4,599,311)またはADH2-4c (Russell et al., Nature 304 (1983), 652 - 654)プロモーターが含まれる。
【0056】
糸状菌(filamentous fungus)宿主細胞における使用に関して適切なプロモーターの例は、例えば、ADH3プロモーター(McKnight et al., The EMBO J. 4 (1985), 2093 - 2099 )またはtpiAプロモーターである。他の有用なプロモーターの例は、A.oryzae TAKAアミラーゼ、Rhizomucor mieheiアスパラギン酸プロテイナーゼ、A.niger中性α-アミラーゼ、A.niger酸安定性α-アミラーゼ(acid stable α-amylase)、A.niger、またはA.awamoriグルコアミラーゼ(gluA)、Rhizomucor mieheiリパーゼ、A.oryzaeアルカリ性プロテアーゼ、A.oryzae三炭糖リン酸塩イソメラーゼ、またはA.nidulansアセトアミダーゼをコード化している遺伝子に由来するものである。好適なものは、TAKA-アミラーゼおよびgluAプロモーターである。適切なプロモーターは、例えば、EP 238023およびEP 383779に記載されている。
【0057】
また、前記ヒトポリペプチドをコード化しているDNA配列は、必要な場合には、適切なターミネーター〔例えば、ヒト成長ホルモンターミネーター(Palmiter et al., Science 222, 1983, pp. 809-814)またはTPI1(Alber and Kawasaki, J. Mol. Appl. Gen. 1, 1982, pp. 419-434)またはADH3(McKnight et al., The EMBO J. 4, 1985, pp. 2093-2099)ターミネーター〕に動作可能に連結されてもよい。また、発現ベクターは、前記プロモーターから下流で且つポリペプチド配列自身に関する挿入部位から上流に位置するRNAスプライス部位のセットを含んでいてもよい。好適なRNAスプライス部位は、アデノウイルスおよび/または免疫グロブリン遺伝子から取得し得る。発現ベクターに含まれるものは、挿入部位の下流に位置する、ポリアデニル化シグナルである。特に好適なポリアデニル化シグナルには、SV40の早期の若しくは遅発型のポリアデニル化シグナル(Kaufman and Sharp, ibid.)、アデノウイルス5Elb領域、ヒト成長ホルモン遺伝子ターミネーターのポリアデニル化シグナル(DeNoto et al. Nucl. Acids Res. 9:3719-3730, 1981)、または前記遺伝子のポリアデニル化シグナルが含まれる。また、発現ベクターは、非コードの(noncoding)ウイルス性のリーダー配列〔例えば、アデノウイルス2 トリパタイト(tripartite)リーダー(前記プロモーターおよびRNAスプライス部位の間に位置する〕;およびエンハンサー配列(例えばSV40エンハンサー)を含む。
【0058】
本発明のポリペプチドを宿主細胞の分泌経路(secretory pathway)へと方向付ける(direct)ために、分泌性シグナル配列(リーダー配列、プレプロ配列、またはプレ配列としても知られる)を組換えベクターに提供してもよい。分泌性シグナル配列は、正しい読み枠で、前記ポリペプチドをコード化しているDNA配列と連結される。分泌性シグナル配列は、前記ペプチドをコード化しているDNA配列に対して、5'に通常配置される。分泌性シグナル配列は、前記タンパク質と通常関連するものであってもよい又は別の分泌タンパク質をコード化している遺伝子からのものであってもよい。グラム陽性細菌において、L.amylovorusのα-アミラーゼ遺伝子のシグナル配列が、使用された(Kruger et al 2002 Nature Biotechnol 20:702-706)。
【0059】
酵母細胞からの分泌に関して、分泌性シグナル配列は、発現したポリペプチドの前記細胞の分泌経路への効率的な方向付けを保証する任意のシグナルペプチドをコード化してもよい。前記シグナルペプチドは、天然の(naturally occurring)シグナルペプチド、又はその機能的な部分であってもよい、又は合成ペプチドであってもよい。適切なシグナルペプチドは、α-因子シグナルペプチド(cf. US 4,870,008)、マウス唾液アミラーゼのシグナルペプチド(cf. O. Hagenbuchle et al., Nature 289, 1981, pp. 643-646)、修飾したカルボキシペプチダーゼシグナルペプチド(cf. L.A. Valls et al., Cell 48, 1987, pp. 887-897)、酵母BAR1シグナルペプチド(cf. WO 87/02670)、または酵母アスパラギン酸プロテアーゼ3(YAP3)シグナルペプチド(cf. M. Egel-Mitani et al., Yeast 6, 1990, pp. 127-137)であることが見出されている。
【0060】
酵母における効率的な分泌に関して、リーダーペプチドをコード化している配列が、シグナル配列の下流およびポリペプチドをコード化しているDNA配列の上流に挿入されてもよい。リーダーペプチドの機能は、発現されたペプチドを小胞体からゴルジ体へと、更に培養培地への分泌に関連する分泌小胞(secretory vesicle)へと方向付けることを可能にすることである(即ち、ポリペプチドの、細胞壁を横切った輸送又は少なくとも細胞膜を介した酵母細胞の細胞膜周辺腔への輸送)。前記リーダーペプチドは、酵母α-因子リーダーであってもよい(その使用は、例えば、US 4,546,082、US 4,870,008、EP 16 201、EP 123 294、EP 123 544およびEP 163 529に記載されている)。或いは、前記リーダーペプチドは合成プロペプチドであってもよく、つまり自然界でみつけることができないリーダーペプチドであってもよい。合成リーダーペプチドは、例えば、WO 89/02463またはWO 92/11378に記載されたように構築し得る。
【0061】
糸状菌における使用に関して、前記シグナルペプチドは、コウジカビ属の種(Aspergillus sp)のアミラーゼまたはグルコアミラーゼをコード化している遺伝子、Rhizomucor mieheiのリパーゼまたはプロテアーゼまたはHumicola lanuginosaのリパーゼをコード化している遺伝子から由来するものであってもよい。前記シグナルペプチドは、好ましくはA.oryzae TAKAアミラーゼ、A.niger中性アルファ-アミラーゼ、A.niger酸安定性アミラーゼ、またはA.nigerグルコアミラーゼをコード化している遺伝子から由来するものである。適切なシグナルペプチドは、例えば、EP 238023およびEP 215594に開示されている。
【0062】
昆虫細胞における使用に関して、前記シグナルペプチドは、昆虫遺伝子(cf. WO 90/05783)、例えば、lepidopteran Manduca sextaの脂質動員ホルモン前駆物質シグナルペプチド(cf. US 5,023,328)から由来するものであってもよい。
【0063】
それぞれ、ポリペプチドをコード化しているDNA配列、プロモーターおよび自由選択のターミネーター、および/または分泌性シグナル配列を連結するために及びそれらを適切なベクター(このベクターは複製に必要な情報を含有している)挿入するために使用される処置は、当業者に周知である(例えば、Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, New York, 1989を参照されたい)。
【0064】
哺乳類細胞をトランスフェクションする及び前記細胞に導入したDNA配列を発現させる方法は、例えば、文献〔Kaufman and Sharp, J. Mol. Biol. 159 (1982), 601 - 621; Southern and Berg, J. Mol. Appl. Genet. 1 (1982), 327 - 341; Loyter et al., Proc. Natl. Acad. Sci. USA 79 (1982), 422 - 426; Wigler et al., Cell 14 (1978), 725; Corsaro and Pearson, Somatic Cell Genetics 7 (1981), 603, Graham and van der Eb, Virology 52 (1973), 456; およびNeumann et al., EMBO J. 1 (1982), 841 - 845〕に記載されている。
【0065】
クローン化したDNA配列は、培養した哺乳類細胞に、例えば、リン酸カルシウムに媒介されるトランスフェクション(Wigler et al., Cell 14:725-732, 1978; Corsaro and Pearson, Somatic Cell Genetics 7:603-616, 1981; Graham and Van der Eb, Virology 52d:456-467, 1973)または電気穿孔法(Neumann et al., EMBO J. 1:841-845, 1982)により導入される。外来性(exogenous)のDNAを発現する細胞を同定する及び選択するため、一般的に選択可能な表現型(選択可能なマーカー)が、所望の遺伝子またはcDNAと共に導入される。好適な選択可能なマーカーには、薬物(例えば、ネオマイシン、ハイグロマイシン、およびメトトレキセート)に対する耐性を与える遺伝子が含まれる。前記選択可能なマーカーは、増幅可能な選択可能なマーカーであってもよい。好適な増幅可能な選択可能なマーカーは、ジヒドロ葉酸レダクターゼ(DHFR)配列である。選択可能なマーカーはThilly(Mammalian Cell Technology, Butterworth Publishers, Stoneham, MA、本明細書中に参照により援用される)により論評されている。当業者は、適切な選択可能なマーカーを容易に選択することができる。
【0066】
選択可能なマーカーは、細胞中に、別々のプラスミド上で、所望の遺伝子と同時に導入し得る、又はそれらを同じプラスミド上で導入し得る。同じプラスミド上で、選択可能なマーカーおよび所望の遺伝子が、異なるプロモーターまたは同じプロモーターの制御下で存在し得る;この後者の配置はジシストロン性メッセージ(a dicistronic message)を産生する。このタイプの構築物は既知である(例えば、LevinsonおよびSimonsen、U.S.4,713,339)。「キャリアDNA(carrier DNA)」として知られる付加的なDNAを、前記細胞に導入される混合物に付加することも有利であろう。
【0067】
細胞が前記DNAを受け取った後、それらは適切な成長培地において成長し(典型的には1〜2日)、所望の遺伝子の発現が開始される。本明細書中に使用される「適切な成長培地(appropriate growth medium)」の用語は、細胞の成長に及び所望のポリペプチドバリアントの発現に要求される、栄養物および他の成分を含有している培地を意味する。培地は、一般に炭素源、窒素源、必須アミノ酸類、必須な糖類、ビタミン類、塩類、リン脂質類、タンパク質、および成長因子を含む。ガンマ-カルボキシル化されたタンパク質の産生に関して、培地はビタミンK(好ましくは、約0.1μg/mlから約5μg/mlの濃度で)を含有する。次に薬物選択が、安定な様式で選択可能なマーカーを発現している細胞の成長に関する選択に適用される。増幅可能で選択可能なマーカーでトランスフェクションされた細胞に関して、薬物濃度を増加して、クローン化された配列のコピー数の増加に関して選択し得る。これによって発現レベルが増加される。安定的にトランスフェクションされた細胞のクローンは、次に所望のポリペプチドバリアントの発現に関して選抜される。
【0068】
前記免疫グロブリンポリペプチドをコード化しているDNA配列が導入された宿主細胞は、必要に応じて、翻訳後修飾されたポリペプチドを産生する能力を有する任意の細胞でありえ、該宿主細胞には、酵母、真菌類、および高等真核細胞が含まれる。翻訳後修飾は、原核生物の発現系では得られない。
【0069】
本発明の一態様において、真核生物細胞は、哺乳類細胞、昆虫細胞、植物細胞、および真菌細胞(酵母細胞を含む)から選択される。
【0070】
原核細胞の例は、大腸菌(Cabilly et al US 6331415)などのグラム陰性細胞またはBacilli、Clostridia、Staphylococci、Lactobailli、またはLactococci(de Vos et al 1997 Curr. Opin. Biotechnol. 8:547-553)などのグラム陽性細菌であってもよい。組換えタンパク質をグラム陽性細菌において発現させる例示的な方法は、US5821088に記載される。
【0071】
本発明における使用に関する哺乳類細胞株は、COS−1(ATCC CRL 1650)、乳児ハムスター腎臓(BHK)およびHEK293(ATCC CRL 1573; Graham et al., J. Gen. Virol. 36:59-72, 1977)細胞株である。好適なBHK株化細胞は、tk- ts13 BHK株化細胞(Waechter and Baserga, Proc. Natl. Acad. Sci. USA 79:1106-1110, 1982, 本明細書中に参照により援用される)であり、以下でBHK570細胞と称する。BHK570株化細胞は、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection, 12301 Parklawn Dr., Rockville, Md. 20852)にATCC受託番号CRL10314で寄託されている。また、tk- ts13 BHK細胞株も、ATCCから受託番号CRL1632で利用可能である。加えて、本発明に使用し得る、いくつかの他の細胞株には、Rat Hep I(ラット ヘパトーマ;ATCC CRL 1600)、Rat HepII(ラット ヘパトーマ;ATCC CRL 1548)、TCMK(ATCC CCL 139)、ヒト肺(Human lung)(ATCC HB 8065)、NCTC1469(ATCC CCL 9.1)、CHO(ATCC CCL 61)、およびDUKX細胞(Urlaub and Chasin, Proc. Natl. Acad. Sci. USA 77:4216-4220, 1980)が含まれる。
【0072】
適切な酵母細胞の例には、Saccharomyces spp.またはSchizosaccharomyces spp.の細胞が、特にSaccharomyces cerevisiaeまたはSaccharomyces kluyveriの株が含まれる。酵母細胞を異種性のDNAで形質転換する及びそれからの異種性のポリペプチドを産生させるための方法は、例えば、US 4,599,311、US 4,931,373、US 4,870,008、5,037,743、および US 4,845,075に記載され、これら全ては参照によって援用される。形質転換された細胞は、選択可能なマーカー(一般に薬剤抵抗性または特定の栄養分(例えば、ロイシン)の非存在下で成長する能力)により決定される表現型により選択される。酵母における使用に関して好適なベクターは、US 4,931,373に開示されるPOT1ベクターである。ポリペプチドをコード化しているDNA配列は、シグナル配列および任意でリーダー配列(例えば、上記のような)により先行されていてもよい。適切な酵母細胞の更なる例は、Kluyveromyces(例えば、K.lactis)、hansenula(例えば、H.polymorpha)、またはPichia(例えば、P.pastoris)の株である(Gleeson et al., J. Gen. Microbiol. 132, 1986, pp. 3459-3465; US4882279を参照されたい)。
【0073】
他の真菌細胞の例は、糸状菌、例えば、コウジカビ属 spp.(Aspergillus spp.)、パンカビ属 spp.(Neurospora spp.)、フザリウム spp.(Fusarium spp.)、またはトリコデルマ属 spp.(Trichoderma spp.)の細胞、特にA.oryzae、A.nidulans、およびA.nigerの株である。コウジカビ属 spp.のタンパク質の発現に関する使用は、例えば、EP 272 277、EP 238 023、EP 184 438に記載されている。F.oxysporumの形質転換は、例えば、文献(Malardier et al., 1989 (Gene 78: 147-156))に記載のとおり実施し得る。トリコデルマ属 spp.の形質転換は、例えば、EP 244 234に記載のとおり実施し得る。
【0074】
糸状菌が宿主細胞として使用される場合、それを本発明のDNA構築物で、前記DNA構築物を宿主染色体に統合させて組換え宿主細胞(recombinant host cell)を得ることにより形質転換し得る。この統合は一般的には有利であると考えられる、というのも前記DNA配列が前記細胞において安定的に維持される可能性が高いからである。前記DNA配列の宿主染色体への統合は、従来の方法(例えば、相同性の又は異種性の組換えによる)にしたがって実施し得る。
【0075】
昆虫細胞の形質転換及びその中での異種性のポリペプチドの産生は、US 4,745,051;US 4,879,236;US 5,155,037;5,162,222;EP 397,485に記載のとおり実施し得る(これらの文献の全ては本明細書中に参照によって援用される)。宿主として使用される昆虫株化細胞は、鱗翅目株化細胞(例えば、Spodoptera frugiperda細胞またはTrichoplusia ni細胞)であってもよい(cf. US 5,077,214)。培養条件は、例えば、WO 89/01029またはWO 89/01028に、または上述の文献の何れかに記載されたとおりであってもよい。
【0076】
上記の形質転換された又はトランスフェクションした宿主細胞は、次に適切な栄養培地中で、免疫グロブリンポリペプチドの発現を許容する条件下で培養され、その後、生じたペプチドの全部または一部を培養物から回収し得る。前記細胞を培養するために使用される培地は、前記宿主細胞を成長させるために適切な任意の従来培地、例えば、適切なサプリメントを含有する最小の又は複合的(complex)な培地であってもよい。適切な培地は、商業上の提供者から購入される又は公開されたレシピ(例えば、アメリカン・タイプ・カルチャー・コレクションのカタログ中の)にしたがって調製し得る。前記細胞により産生されたポリペプチドは、次に培養培地(culture medium)から従来の処理により回収または精製される〔この処理には宿主細胞を培地から遠心分離または濾過により分離すること、上清もしくは濾過物のタンパク性成分を塩(例えば、硫酸アンモニウム)で沈殿させ、様々なクロマトグラフィー処理(例えば、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、アフィニティークロマトグラフィー)により精製すること、等が含まれる〕、この処理は問題のポリペプチドのタイプに依存して行われる。クロマトグラフィー処理において、ポリペプチドはカラムから溶液中に溶出される。一側面において、前記ポリペプチドは、培養培地からの精製の前または後に透析されて、ポリペプチドが所望の溶液中に得られる。
【0077】
モノクローナル抗体の断片及び誘導体
抗体断片の例には、Fab、Fab'、Fab'−SH,F(ab')2及びFv断片;二重特異性抗体;連続するアミノ残基の一つの連続配列からなる一次構造を有するポリペプチド(本明細書において、「一本鎖抗体断片」又は「一本鎖ポリペプチド」と称される。)である任意の抗体断片〔(1)一本鎖Fv(scFv)分子、(2)軽鎖可変ドメインを一つだけ含有する一本鎖ポリペプチド、又は軽鎖可変ドメインの3つのCDRを含有し、付随する重鎖部分を伴わないそれらの断片、及び(3)重鎖可変領域を一つだけ含有する一本鎖ポリペプチド、又は重鎖可変ドメインの3つのCDRを含有し、付随する軽鎖部分を伴わないそれらの断片などが含まれるが、これらに限定されない。〕;並びに抗体断片から形成される多重特異性抗体(例えば、二重特異性抗体)が含まれる。「免疫反応性断片(Immunoreactive fragments)」は、無傷(intact)の抗体の一部(一般的には、抗原結合部位又は可変領域)を含む。本発明によって産生された免疫学的に機能的な免疫グロブリン断片は、1以上の抗体の少なくとも重鎖(VH鎖)の可変ドメインおよび少なくとも軽鎖(VL鎖)の可変ドメインを具備してもよい。
【0078】
本発明の方法を産生のために使用することができる、2つの例示的な抗原結合免疫グロブリン断片は、FabおよびFvである。小さいFv(即ち、可変性の断片)は、VLおよびVHのみから構成される。Fvの組換え型バージョンは、単鎖可変断片(ScFv)と称される。ScFvにおける2つの断片は、可動性のペプチドリンカー〔通常では15アミノ酸リンカーが(G4S)3の配列で使用される〕と人工的に連結され、単一のポリペプチド鎖として発現される。前記リンカーによって、VHおよびVLの会合が許容されて、抗原結合部位が形成される。2つのポリペプチドからなる大きなFab(抗原結合断片)のうち、一方は軽鎖可変部および定常ドメイン VL−CLを含み、他方は可変ドメインおよび1つの定常ドメインを含んでいる短縮型(truncated)の重鎖 VH−CH1を含む。無傷のIgG免疫グロブリンのように、2つの鎖は共にジスルフィド結合で連結される。ScFv抗体は2つのジスルフィド結合を含有するが、他方でFabsは5つのジスルフィド結合を含有する(安定的なフォールディングのために全てが形成されなければならない)。
【0079】
ScFvおよびFab断片の発現は、真核生物、酵母、真菌、および細菌を含む様々な宿主細胞において実施される。グラム陰性細菌において、システインチオールからジスルフィドへの酸化は、タンパク質が細胞質の高度に還元的な環境からペリプラズム中のより酸化的な環境へと輸送された後にのみ通常は発生する。ペリプラズムへと発現されたタンパク質は、浸透圧ショックによって又は全細胞ライセートから回収することができる。抗体を37゜Cで長期にわたり高レベルで発現させることによって、大腸菌の外膜は透過性となり、タンパク質を培養培地から回収することができる。しかしながら、外膜における脂質の存在によって、エンドトキシン汚染に関する問題が生じる。グラム陽性細菌および酵母は外膜を有さず、タンパク質を細胞質から細胞外空間へと直接的に分泌させることができる。結果的に、エンドトキシンは存在せず、相対的に単純な輸送機構によって、組換え型タンパク質の高収量での分泌が促進される。
【0080】
本発明の抗体の断片および誘導体は、当該技術で公知の技術によって作出することができる。例えば、Fab又はF(ab')2断片は、従来の技術に従って、前記単離された抗体をプロテアーゼ消化することによって作出することができる。或いは、本発明の抗体を産生しているハイブリドーマのDNAは、本発明の断片をコードするように修飾しえる。次いで、修飾されたDNAは発現ベクター中に挿入され、適切な細胞を形質転換し又はトランスフェクションするために使用され、次いで、前記細胞が所望の断片を発現する。
【0081】
他のタイプの断片には、重鎖抗体(VHH)断片の重鎖の可変ドメイン〔例えば、Muyldermans, Reviews in Mol. Biotechnol. 2001, 74:277-302 およびSpinelli et al., Biochemistry. 2000;39(6):1217-22に記載されたもの〕および抗体の最小の機能的な結合単位であるドメイン抗体(dAbs)が含まれ、これはヒト抗体の重(VH)または軽(VL)鎖の何れかの可変領域に対応する。ドメイン抗体及びそれらを調製するための例示的な方法は、例えば、米国特許第6,696,245号, WO05/035572, WO04/101790, WO04/081026, WO04/058821, WO04/003019およびWO03/002609に記載される。
【0082】
多重特異性の抗体または抗体断片は、1、2(二重特異性)、又は3つの特異性を含む任意の適切な数の特異性を有することが可能である。例えば、二重特異性抗体は、ハイブリドーマの融合またはFab’断片の連結〔例えば、Songsivilai & Lachmann Clin. Exp. Immunol. 79: 315-321 (1990)およびKostelny et al. J. Immunol. 148:1547-1553 (1992)を参照されたい〕を含む様々な既知の方法によって産生される。また、三重特異性の抗体を調製する方法は、当該技術において既知である〔例えば、Tutt et al. J. Immunol. 147: 60 (1991)を参照されたい〕。
【0083】
別の態様において、本発明の抗体を産生するハイブリドーマのDNAは、例えば、相同な非ヒト配列に代えて、ヒト重鎖及び軽鎖定常ドメインに対するコード配列を置換することによって〔例えば、Morrison et al., Proc. Natl. Acad. Sci. U.S.A., 81, pp. 6851 (1984)〕、又は、非免疫グロブリンポリペプチドに対するコード配列の全部又は一部を、免疫グロブリンコード配列に共有結合することによって、発現ベクター中に挿入する前に修飾することができる。このように、元の抗体の結合特異性を有する「キメラ(chimeric)」又は「ハイブリッド(hybrid)」抗体が調製される。典型的には、このような非免疫グロブリンポリペプチドは、本発明の抗体の定常ドメインの代用とされる。
【0084】
好ましい実施形態において、本発明の抗体はモノクローナル抗体である。本発明の抗体の「ヒト化された」形態は、特異的なキメラ免疫グロブリン、免疫グロブリン鎖、又は、マウス免疫グロブリンに由来する最小配列を含有する、それらの断片〔Fv、Fab、Fab'、F(ab')2又は抗体の他の抗原結合配列など〕である。通例、ヒト化抗体は、レシピエントの相補性決定領域(CDR)由来の残基が、元の抗体の望ましい特異性、親和性及び能力を維持しつつ、元の抗体(ドナー抗体)のCDR由来の残基によって置換されている、ヒト免疫グロブリン(レシピエント抗体)である。一部の事例では、ヒト免疫グロブリンのFvフレームワーク残基は、対応する非ヒト残基によって置換されることができる。さらに、ヒト化抗体は、レシピエント抗体又は移入されたCDR若しくはフレームワーク配列中の何れにも存在しない残基を含むことができる。これらの修飾は、抗体性能をさらに改良し、最適化するために施される。一般に、ヒト化抗体は、少なくとも一つ、典型的には二つの可変ドメインの実質的に全てを含むであろう。ヒト化抗体中では、元の抗体(original antibody)のCDR領域に対応する全て又は実質的に全てのCDR領域及び全て又は実質的に全てのフレームワーク領域(FR領域)は、ヒト免疫グロブリンコンセンサス配列のものである。ヒト化抗体は、最適には、通例ヒト免疫グロブリンの免疫グロブリン定常領域(Fc)の少なくとも一部も含むであろう。更なる詳細に関しては、文献〔Jones et al., Nature, 321, pp. 522 (1986); Reichmann et al., Nature, 332, pp. 323 (1988);およびPresta, Curr. Op. Struct. Biol., 2, pp. 593 (1992)〕を参照のこと。
【0085】
本発明に使用する抗体をヒト化する方法は、当該技術において周知である。一般的に、本発明のヒト化抗体は、元の抗体からヒト化抗体中に導入されたアミノ酸残基を一又は複数有する。これらのマウス又は他の非ヒトアミノ酸残基は、しばしば、「移入(import)」残基と称され、通例、「移入」可変ドメインから採取される。ヒト化は、本質的に、Winter及び共同研究者の方法に従って、実施することができる〔Jones et al., Nature, 321, pp. 522 (1986); Riechmann et al., Nature, 332, pp. 323 (1988); Verhoeyen et al., Science, 239, pp. 1534 (1988)〕。従って、このような「ヒト化」抗体は、元の抗体の対応する配列によって、無傷のヒト可変ドメインが僅かに置換されているキメラ抗体である(Cabillyらに対する米国特許第4,816,567号)。実際、本発明のヒト化抗体は、典型的には、幾つかのCDR残基と、おそらくは幾つかのFR残基とが、元の抗体中の類似の部位から得られる残基によって置換されているヒト抗体である。
【0086】
ヒト化抗体の作出に使用すべきヒト可変ドメインの選択(軽鎖及び重鎖ドメインの両方)は、抗原性を減少させるのに極めて重要である。いわゆる「ベストフィット(best-fit)」法にしたがって、本発明の抗体の可変ドメインの配列は、公知のヒト可変ドメイン配列の全ライブラリーに対してスクリーニングされる。次いで、マウスの配列に最も近いヒト配列が、ヒト化抗体に対するヒトフレームワーク(FR)として受容される〔Sims et al., J. Immunol., 151, pp. 2296 (1993); Chothia and Lesk, J. Mol. Biol., 196, pp. 901 (1987)〕。別の方法は、軽鎖又は重鎖の特定のサブグループの全てのヒト抗体のコンセンサス配列から得られるフレームワークを使用する。同じフレームワークを、数個の異なるヒト化抗体に対して使用することができる〔Carter et al., Proc. Natl. Acad. Sci. U.S.A., 89, pp. 4285 (1992); Presta et al., J. Immunol., 51, pp. 1993)〕。
【0087】
別の方法は、免疫化に使用されるマウスとして、XenoMouse(登録商標)(Abgenix, Fremont, CA)を用いて「ヒト化」モノクローナル抗体を作出することである。XenoMouseは、機能的ヒト免疫グロブリン遺伝子によって、マウスの免疫グロブリン遺伝子が置換されているマウス宿主である。このため、本マウスによって産生される抗体、又は本マウスのB細胞から作製されるハイブリドーマ中で産生される抗体は、すでにヒト化されている。XenoMouseは、米国特許第6,162,963号に記載されている。HuMAb−MouseTM(Medarex)を用いて、類似の方法を行うことが可能である。
【0088】
ヒト抗体は、例えば、ヒト抗体レパートリーを発現するように操作された他のトランスジェニック動物を免疫化のために使用することによる〔Jakobovitz et al., Nature 362 (1993) 255〕、又はファージディスプレイ法を使用して抗体レパートリーを選択することによるなどの、様々な他の技術に従って作製することもできる。このような技術は当業者に公知であり、本願に開示されているモノクローナル抗体から出発して、実施することができる。
【0089】
本発明の抗体は、「キメラ」抗体(免疫グロブリン)に誘導化することも可能であり(キメラ抗体においては、重鎖及び/又は軽鎖の一部は元の抗体中の対応する配列と同一又は相同であるが、鎖の残りは別の種に由来し、又は別の抗体クラス若しくはサブクラスに属する抗体中の対応する配列と同一若しくは相同である)、所望の生物活性を発揮する限りにおいて、係る抗体の断片に誘導化することも可能である〔Cabilly et al., supra; Morrison et al., Proc. Natl. Acad. Sci. U.S.A., 81, pp. 6851 (1984)〕。
【0090】
以下の例は、さらに本発明の特定の側面を説明するが、本発明を限定しない。
【0091】
[例]
例1
抗体の軽および重鎖の分離およびリフォールディング(再生)
ヒト組織因子(TF)を認識するヒト抗体TF36F40およびヒト成長ホルモン(hGH)を認識するマウス抗体hGH3を、以下のとおり使用した。各抗体を、6M塩化グアニジウム(guanidinium chloride)および50mMリン酸ナトリウムpH8.4を用いて変性させた。ジスルフィド結合を、ジチオスレイトール(DTT)を終濃度100mMまで添加することによって還元し、SDS-PAGEおよびウエスタンブロッティングを還元または非還元の両方の条件下でサンプルを分析する前に室温で30分間インキュベートした。ペルオキシダーゼ結合型ヤギ-抗-ヒトIgG重鎖およびヤギ-抗-ヒトカッパー軽鎖を、それぞれTF36F40重鎖(HC)および軽鎖(LC)を検出するために適用した。ペルオキシダーゼ結合型ヤギ-抗-マウスIgG重鎖およびヤギ-抗-マウスカッパー軽鎖を、それぞれhGH3の重および軽鎖を検出するために適用した。
【0092】
各抗体の軽および重鎖を、Superose12カラム(Amersham Bioscience)を用いたサイズ排除クロマトグラフィー(SEC)で引続いて分離した。洗浄および平衡化の条件は、製造者により提供されている条件をもちいた。タンパク質を、6 M塩化グアニジウム, 50mMリン酸ナトリウムpH 8.4および50mM DTTを用いて、前記カラムから溶出した。フラクションを、SDS-PAGEおよびウエスタンブロッティングで上記のとおり分析した(図2および3)。
【0093】
hGH3抗体をリフォールディングさせるために、軽および重鎖のフラクションを等モルの比率で混合し、100mM Tris HCl(pH7.5)および10mM EDTAに対して1時間、37゜Cで透析した。リフォールディングさせた抗体を無菌0.2μmフィルターを通す前に、全長IgGをリン酸緩衝生理食塩水(PBS)の存在下でサイズ排除クロマトグラフィーで単離した。サンプルを、使用するまで4゜Cで保存した。
【0094】
代替的な実験において、hGH3抗体をリフォールディングさせるために、軽および重鎖フラクションを等モルの比率で混合し、以下の緩衝液に対して透析した:
1) 4Mグアニジウム塩酸, 50mM Na リン酸, 5 mM DTT, pH8.4で90min、室温;
2) 2Mグアニジウム塩酸, 50mM Na リン酸, 5 mM DTT, pH8.4で90min、室温;
3) 1Mグアニジウム塩酸, 50mM Na リン酸, 5 mM DTT, pH8.4で90min、室温;
4) 0.5Mグアニジウム塩酸, 50mM Na リン酸, 5 mM DTT, pH8.4で90min、室温;および
5) 50mM Na リン酸塩, 5mM DTT, pH8.4で72hrs, 室温で処理して、DTTおよびジスルフィド架橋の酸化を認容させる。
【0095】
最終的な容量は、2mlであった。500ulを、14kDaのカットオフ値を有するセントリコン フィルターで限外濾過によって濃縮した。両方の溶液を、SDS−PAGEで引続いて分析した(図4)。
【0096】
例2
ELISA
インビトロで会合させたHuTF36F40およびhGH3抗体は、標準的な手順を用いたELISAでアッセイされた。簡単に説明すると、ELISAマイクロタイタープレート(Maxisorp, Nunc, Denmark)を、1μg/mlのヒトhGHまたはヒトTF(TF1−119)を1xPBS中に含有するもので一晩、4゜Cでコートした。インビトロで会合させたhGH3またはHuTF36F40抗体サンプルが添加され、hGHまたはヒトTFへの結合が、それぞれペルオキシダーゼ結合型ヤギ-抗-ヒトまたはマウスIgG(Sigma)を用いて検出された。
【0097】
例3
表面プラズモン共鳴
Biacore3000光学的バイオセンサーを用いて、hGHまたはヒトTFに対するインビトロで会合させた抗体の親和性が評価された。親和性を決定するために、約10000RUの抗原はセンサー表面にEDC/NHSカップリング化学で固定化された。その後、抗体は、約5μl/minの流速で約3minフローセル(flow cell)へと注入され、その各自の抗原(hGHまたはヒトTF)と会合された(hGHまたはヒトTF)。会合フェーズに続き、表面は、ランニング緩衝剤(HBS-EP, pH 7.4, 0.005%の洗浄剤P20を含有している)で5μl/minの流速で2min洗浄された。センサーグラムのデータは、Bia評価ソフトウェア3.0を用いて分析された。
【0098】
例4
トータルRNAのハイブリドーマ細胞からの単離
組織因子に対する抗体を分泌している、4x106のハイブリドーマ細胞(TF−36F40)が、QiagenのRNeasy Mini Kitを用いたトータルRNAの単離に使用された。前記細胞は、5min、1000rpmで沈殿され、10μl/mlのベータ-メルカプトエタノールを含有している350μlのRLT緩衝剤の添加によって破壊された。そのライセートは、QiagenのQIAshredderカラムに移され、最高速度で2min遠心分離された。フロースルーは、1容量の70%エタノールと混合された。RNeasyスピンカラムごとに、700μlまでサンプルが適用され、14000rpmで遠心分離され、フロースルーが廃棄された。700μlのRW1緩衝剤が、カラムごとに適用され、カラムを洗浄するために15s間、14000rpmで遠心された。前記カラムは、500μlのRPE緩衝剤で2回洗浄され、14000rpmで15s遠心された。前記カラムを乾燥させるため、付加的に2min、14000rpmで遠心された。前記カラムは、新しい収集チュウブに移される。RNAは50μlのヌクレアーゼ フリーの水で溶出され、そして1min、14000rpmで遠心された。RNA濃度が、OD=260nmでの吸光度によって測定された。RNAは、必要とされるまで-80゜Cで保存された。
【0099】
例5
cDNA合成
1μgのRNAを、SMART RACE cDNA Amplification Kit(Clontech)を用いた第1鎖のcDNA合成に使用した。5'-RACE-Ready cDNAの調製に関して、単離されたRNAを含有している反応混合物〔上記のとおり、バックプライマー5'-CDSプライマーバック(backprimer 5'-CDS primer back)、およびSMART II Aオリゴ〕を、調製し、72゜Cで約2minインキュベートし、次に氷上で約2min冷却し、1xFirst-Strand緩衝剤、DTT(20mM)、dNTP(10mM)およびPowerScript逆転写酵素を添加した。反応混合物を、42゜Cで1.5時間インキュベートし、トリシン−EDTA緩衝剤を添加し、72゜Cで7minインキュベーションした。サンプルは-20゜Cで保存することができる。
【0100】
例6
ヒト軽鎖(LC)およびヒト重鎖(HC)の増幅およびクローニング
1xAdvantage HF 2 PCR緩衝剤、dNTP(10mM)および1xAdvantage HF 2ポリメラーゼ混合物を含有しているPCR(ポリメラーゼ連鎖反応)の反応混合物は、上記のとおり作出されたcDNAからのVLおよびVH双方の可変領域の別々の増幅に関して用意される。
【0101】
LCの増幅に関して、以下のプライマーが使用される:
UPM(ユニバーサル プライマー 混合物):
5'-CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT-3'(配列番号1)
5'-CTAATACGACTCACTATAGGG-3'(配列番号2)
P1:
5'-CGCGGCTAGCACACTCTCCCCTGTTGAAGCTC-3'(配列番号3)
HCの増幅に関して、以下のプライマーが使用される:
UPM(ユニバーサル プライマー 混合物):
5'-CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT-3'(配列番号1)
5'-CTAATACGACTCACTATAGGG-3'(配列番号2)
P2:
5'-TCATTTACCCGGGGACAGGGAGA-3'(配列番号4)
PCRは、3ラウンド実施された。ラウンド1: PCRは、94゜Cで5sおよび72゜Cで3minを5サイクル実施された。ラウンド2: PCRは、94゜Cで5s、70゜Cで10s、および72゜Cで1minを5サイクル実施された。ラウンド3: PCRは、94゜Cで5s、68゜Cで10s、および72゜Cで1minを28サイクル実施された。
【0102】
前記PCR産物が1%アガロースゲル上での電気泳動法によって分析され、DNAがQIAEX11アガロースゲル抽出キット(Qiagenより)を用いてゲルから精製された。
【0103】
精製されたPCR産物は、TOPO TAクローニングキット(インビトロゲンより)を用いてPCR4-TOPOへと導入され、TOP10コンピテント細胞の形質転換のために使用された。
【0104】
コロニーの適切な量は、Taqポリメラーゼ、1xTaqポリメラーゼ緩衝剤、dNTP(10mM)および以下のプライマーおよびPCRプログラムを用いて、コロニーPCRによって分析された:
M13フォワード:
5'-GTAAAACGACGGCCAG-3'(配列番号5)
M13リバース:
5'-CAGGAAACAGCTATGAC-3'(配列番号6)
PCRプログラム:
25サイクルを、94゜Cで30s、55゜Cで30s、および72゜Cで1min行った。
【0105】
各々、LCおよびHCの挿入物を具備しているクローンからのプラスミドDNAが、抽出され、上記のプライマーM13フォワードおよびM13リバースを用いて配列決定された。TF−36F40抗体遺伝子の生じた配列は、それぞれ配列番号7および8としてリストされる。また、コード化された軽鎖および重鎖配列が、それぞれ配列番号9および10に記載される。
【0106】
例7
抗体遺伝子の哺乳類発現ベクターへの導入
ヒトTF−36F40のそれぞれLCおよびHC配列を開示している配列データ(配列番号7および8)に基づいて、プライマーがそれぞれLCおよびHC遺伝子の増幅に関して設計された。抗体領域をPCRでフォーマットして、コザック配列、リーダー配列およびユニークな制限酵素部位を含ませた。この事項は、5'PCRプライマーを、HindIII部位、コザック配列を導入し、可変軽鎖領域のリーダー配列の5'端に相同性であるように、デザインすることによって達成された。3'プライマーは、それぞれ、定常領域の軽鎖および重鎖領域のC末端の3'末端と相同性であり、3'端にXbaI部位が導入された。前記HC領域は類似の様式で作製された。但し、NotIおよびBamHI部位が、各々5'および3'端にHindIIIおよびXbaIの代わりに導入された。DNA断片は、それぞれHindIII+XbaIおよびNotI+BamHIで消化され、そしてアンピシリン抵抗性をコード化しているベータラクタマーゼ遺伝子および大腸菌複製開始点(pUC)を含んでいる市販の真核生物性発現ベクター〔InvitrogenのpCDNA3(+)〕へと結合された。結合されたプラスミドを使用して、大腸菌が形質転換された。プラスミドDNAが、これらのアンピシリン耐性の細菌集団から調製され、哺乳類細胞へのトランスフェクションに使用された。
【0107】
例8
哺乳類細胞のトランスフェクション
上記で記載したクローン化したDNAsは、製造者の推奨する方法にしたがいLipofectamine TM 2000(Cat. No.11668-019, Invitrogen)を用いて、哺乳類細胞へと導入された。
【0108】
例9
哺乳類細胞での抗体LCおよびHCの発現
細胞を、製造者のプロトコール(Cat.No.11668-019, Invitrogen)に記載のとおり成長させる。細胞は、上清の収穫および続く精製およびエクソビボでの会合の前に、6日成長させられる。
【0109】
LCおよびHCを発現している細胞の共培養は、それぞれ次の様式で実施された、即ち:HEK293細胞が、それぞれLCおよびHCベクター構築物でトランスフェクションされ、2つの培養物を混合する前に16時間37゜Cで成長させた。6日後、混合した培養物からの上清が、例2および3に記載のとおりに分析された。抗体のエクスビボでの会合を至適化するために、宿主細胞培養の宿主細胞、発現ベクター、培養培地および/または異なる生物物理学的な条件(例えば、温度、レドックス条件、pH)などのパラメータを変動させた。LCおよびHCの濃度およびLC:HC比は、これらのパラメータを変化させることによって至適化することができる。1実験において、次の条件を使用した、即ち:HEK293細胞を、抗体LCおよびHCを発現するpCDNA3(+)に準拠したベクター構築物でトランスフェクションし、25μg/mlジェネテシン(GIBCO cat. No. 10131-019)を含有しているOptimum1培地(GIBCO cat. no. 51985-026)を添加したGlutamax-I中で、37゜C で200rpmで成長させた。
【0110】
例10
グラム陽性細菌中での抗体LCおよびHCの形質転換および発現
ヒトTF-36F40のそれぞれLCおよびHC配列を開示している配列データ(配列番号7および8)に基づいて、プライマーはそれぞれLCおよびHC遺伝子の増幅に関して設計された。抗体領域は、PCRで整えられて、HCに関してNotI/BamHI部位およびLCに関してHindIII/XbaI部位を含むが、天然のシグナルペプチドは含まない。
【0111】
HCおよびLCのDNA断片が、消化され、グラム陽性菌(Streptococcus gordonii)SPEX発現ベクター(Myscofski et al 2000 Protein Expression Purif)の修正バージョンへと別々に連結された(このベクター中で、発現はP2プロモーターで駆動される)。
【0112】
上記のクローン化したDNAは、グラム陽性細菌へと、以前に記載されたとおり導入された(Myscofski et al 2000 Protein Expression Purif.)。細胞は、上清の収穫および引続く精製およびエクスビボ会合の前に、ブレイン―ハートインフュージョンブロス(BHI, Difco)中で、一晩、37゜Cで成長された。
【0113】
LCおよびHCを発現している細胞の共培養は、上記と同じ様式で実施された。
【0114】
例11
抗体断片の精製
LCは上清からプロテインLクロマトグラフィーを用いて精製され、HCは上清からプロテインAで精製された(当該技術において、既知の処理にしたがって)。或いは、LCは、マトリックスにEDC/NHSカップリング化学を用いて固定化された、VLCLに対する抗体を用いて上清から精製された;および/または、HCは、VHCH1-3に対する抗体を用いて上清から精製された。
【0115】
例12
抗体断片のインビトロ会合
TF36F40のLCおよびHCの、完全長TF抗体へのインビトロ会合は、例1に記載のとおり実施された。前記抗体は、BCAタンパク質アッセイ(Pierce, Rockford, IL)を用いて定量され、例2および3に記載のとおりアッセイされた。
【0116】
例13
Obelix陽イオン交換体による精製
Obelix陽イオン交換体は、Amersham(Cat. No.11-0010)から市販されている。
【0117】
使用するためには、濾過した細胞培養のpHは、約3.0〜9.0(好ましくは、約pH7.0)の範囲内である。前記サンプルは、約5〜40(好ましくは、約10)mg/ml 充填カラム材料の範囲でカラムに適用される。
【0118】
非結合性の物質は、緩衝液A(緩衝液Aは、好適な範囲のpHを有し、所望の抗体の結合が許容されるために、十分に低い伝導率を有する緩衝液と規定される)を1〜5のCV(カラム容量)で適用することによってカラムから洗浄除去された。
【0119】
引き続いて、前記抗体は、伝導率を増加させること、それにより静電反発力を増加させることを介してカラムから溶出された。これは低から高い塩濃度での段階的グラジエントまたは連続的グラジエントによって達成することができる。好ましくは、NaClが塩として選択されるが、他のタイプの塩も使用することができる。
【0120】
プロパンジオールを、溶出緩衝液に添加することができる。この態様において、陽イオン交換体を、疎水性相互作用クロマトグラフィーカラムとして使用することができる。高い塩析効果を有する塩(例えば、酢酸ナトリウム)を添加することによって、陽イオン交換体を疎水性相互作用クロマトグラフィーカラムに変化させることができる。従って、パラメータ(例えば、塩、pH、伝導率、および疎水性)を変化させることによって、前記カラムを同じ操作の間に2様式以上で機能させることができる。前記クロマトグラフィー手順は、典型的には4〜25゜Cで実施される。
【0121】
1実験において、次の条件を使用した:抗体(Ab)の適用物を、培養物中のpHをpH5,5に調整し、0,2μmフィルターで濾過することによって調製された。EDTAおよびベンザミジン-HClを、1mMまで添加した。
【0122】
カラム材料(column material)を、30mMクエン酸緩衝液pH6,0で平衡化した。そして、適用物を、室温で20cv/h、約5mg Ab/ml カラム材料の条件で適用した。
【0123】
適用後に、カラムを、平衡化緩衝液で洗浄した。30mMクエン酸、25mM NaCl、30%グリセロールpH6,0での洗浄を、平衡化緩衝液での洗浄に続いて実施した。
【0124】
溶出は、異なる様式で実施することができる。1実験において、溶出を、Tris-緩衝液pH7,5〜8,5で実施した(図5および6)。別の実験において、0〜1M NaClの塩をpH6,5〜7で適用するグラジエント溶出を使用した。
【0125】
Abの溶出後、前記カラムは、1M NaOHで再生された。
【0126】
* * *
公報、特許出願及び特許を含む、本明細書に引用された全ての参考文献は、各参考文献が個別的且つ具体的に参照により援用され、その全体が本明細書に記載されている場合と同じ程度に、参照により本明細書に援用される。
【0127】
本明細書で使用されている全ての見出し及び小見出しは、便宜的に使用されているものにすぎず、いかなる意味においても、本発明を限定するものと解釈してはならない。
【0128】
本明細書に別段の記載がなく、又は明確に反対の記載がなければ、考えられる全ての変形における上記事項のあらゆる組み合わせが、本発明によって包含される。
【0129】
本発明を記載する際に使用される「a」及び「an」及び「the」という用語及び類似の指示記号(referents)は、本明細書に別段の記載がなく、又は明確に反対の記載がなければ、単数及び複数の両方を包含するものと解釈しなければならない。
【0130】
本明細書において値の範囲を引用することは、当該範囲に含まれる各個別の値を個別的に表すための簡略法としての役割を果たすことを意図しているにすぎず、本明細書中に別段の記載がなければ、各個別の値は、本明細書に個別的に記載されている場合と同様に、本明細書中に取り込まれる。別段の記載がなければ、本明細書に記載されている全ての値は、対応する近似値(approximate values)を代表している(例えば、具体的な因子又は測定に関して記載されている全ての例示的な値は、適宜、「約」と修飾される、対応する近似的な測定値も表すと考えることができる)。
【0131】
本明細書に別段の記載がなく、又は明確に反対の記載がなければ、本明細書に記載されている全ての方法は、任意の適切な順序で実施することができる。
【0132】
本明細書中で記載される、任意の及び全ての例又は例示的な用語(「例えば」など)は、単に本発明をよりよく説明することを意図しており、他で指摘しない限り本発明の範囲を限定しない。明示の記載がなければ、本明細書中の言語(language)は、何らかの事項が本発明の実施に不可欠であることを示唆するものと解釈すべきでない。
【0133】
本明細書での特許文書(patent documents)の引用(citation)および援用(incorporation)は、便宜上の目的でのみなされ、係る特許文書の有効性(validity)、特許性(patentability)、および/または強制力(enforceability)の如何なる見解(view)をも反映するものではない。
【0134】
本明細書に別段の記載がなく、又は明確に反対の記載がなければ、ある事項に関して「含む(comprising)」、「有する(having)」、「含む(including)」又は「含有する(containing)」などの用語を用いて、本発明の任意の側面又は態様を本明細書で記載することは、当該事項「からなる(consists of)」、「〜から本質的になる(consists essentially of)」、「実質的に〜を含む(substantially comprises)」本発明の類似の側面又は態様に対する支持を与えることを意図するものである(例えば、特定の事項を含むと本明細書に記載されている組成物は、本明細書に別段の記載がなく、又は明確に反対の記載がなければ、その事項からなる組成物も記載するものとして理解すべきである)。
【0135】
本発明は、本明細書に提示されている前記側面又は特許請求の範囲に記載されている主題のあらゆる修飾及び均等物を、適用可能な法律によって許容される最大限度まで含む。
【図面の簡単な説明】
【0136】
【図1】図1は、抗体の重および軽鎖の発現と、引続く完全長の機能的な抗体へのエクスビボ会合とを示す図である。
【図2】図2は、例1に記載のような変性および還元後の、hGH3抗体のHPLCクロマトグラフィーを示す図である。
【図3】図3は、変性および還元したhGH3抗体(例1に記載のような)のSDS−PAGEおよびクーマシー染色を示す〔HPLC精製前(レーン2)およびHPLCフラクション5(レーン3), フラクション6(レーン4), フラクション7(レーン5), フラクション8(レーン6)〕。レーン1は、Mark12マーカーを示す。
【図4】図4は、Mark12マーカー(レーン1)およびhGH3抗体の変性、リフォールディング、およびサイズ-排除クロマトグラフィー後のサンプル(レーン2)のSDS-PAGE分析を示す。
【図5】図5は、Obelix陽イオン交換体によるIgG精製のクロマトグラムを示す。
【図6】図6は、Obelix陽イオン交換体でのIgGの精製から取得されたフラクションの非還元SDS-PAGE分析を示す。レーン1:Mark12マーカー;レーン2:IgG標準;レーン3:適用物;レーン4:通過物(Run through);レーン5:フラクションC7;レーン6:フラクションD7;レーン7:フラクションE1;レーン8:フラクションE2, レーン9:フラクションE3;レーン 10:フラクションE4。
【発明の開示】
【0001】
[発明の分野]
本発明は、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を産生するための方法に関する。本方法において、免疫グロブリンの軽および重鎖の配列は、同じ細胞培養物中の別々(separate)の宿主細胞中で発現させることができ、次に免疫グロブリンまたは免疫グロブリン断片のエクスビボでの会合がなされる。
【0002】
[発明の背景]
モノクローナル抗体を、ハイブリドーマ技術によって又は組換え型の発現によって産生することができる。組換え型の発現によって、抗体の設計および産生を改善することに関してさらなるオプションが提供される。様々な形式の抗体または抗体断片を、組換え型で発現させることができる。そして、親和性、特異性、および糖鎖形成などの特性を、遺伝子工学で変更して、抗体または抗体断片の特性を改善することができる。組換え型で発現させた抗体の生産性は、ハイブリドーマから得られる収量(yield)を一般的には超え、組換え型で産生された抗体では生産コストが低くなる。
【0003】
US6331415(Cabillyら)は、重および軽鎖が単一細胞中で単一のベクターから又は2つの別々のベクターから同時に発現される免疫グロブリンの組換え型を産生する方法を記載している。
【0004】
Wibbenmeyerら(1999, Biochim Biophys Acta 1430(2):191-202)およびLeeおよびKwak(2003, J. Biotechnology 101:189-198)は、大腸菌(E. coli)の別々の培養物中で発現させたプラスミドを用いて、別々に産生した重および軽鎖からのモノクローナル抗体の産生を記載している。H-およびL-鎖を、封入体として発現し、細胞溶解物から精製し、インビトロでリフォールディングした。
【0005】
US5643745は、2つの核を含んでいる糸状菌を培養することによってヘテロダイマーを産生する方法を記載しており、そのうちの最初のものは修飾されて第1のサブユニットのヘテロダイマーを発現し、そのうちの第2のものは修飾されて第2のサブユニットを発現する。
【0006】
1つの細胞発現系が便利である。というのも、抗体は、精製後に構造的に使える状態となるからである。しかしながら、他の因子は、制御することが困難であろう(例えば、軽および重鎖を、至適な収量を得るために所望の比率で発現させること)。他方で、グラム陰性の大腸菌中での発現はエンドトキシンの混入を生じる可能性があり、完全長の免疫グロブリンの発現は問題がある。
【0007】
従って、特定の要求に適し、上記の課題を解決する、組換え型抗体を産生するための代替法が、必要とされる。
【0008】
[発明の概要]
本発明は、免疫グロブリン重および軽鎖の可変ドメインの少なくとも機能的な部分を具備している、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を産生するための方法を提供し、該方法は重および軽鎖を、真核生物細胞およびグラム陽性菌からなる群から選択される2つの別々の宿主細胞で産生させること;及び免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片をエクスビボでリフォールディングさせることを備える。
【0009】
従って、本発明は、免疫グロブリン分子または免疫学的に機能的なその断片を産生する方法を提供し、該方法は以下を備える:
(a)第1の宿主細胞を、少なくとも免疫グロブリン重鎖の可変ドメインを具備している第1のポリペプチドをコード化しているヌクレオチド配列を具備している第1の核酸で形質転換すること;
(b)第2の宿主細胞を、少なくとも免疫グロブリン軽鎖の可変ドメインを具備している第2のポリペプチドをコード化しているヌクレオチド配列を具備している第2の核酸で形質転換すること;
(c)第1および第2の核酸配列を発現させること;
(d)第1および第2のポリペプチドを精製すること;および
(e)第1および第2のポリペプチドがリフォールディングして、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を形成することを認容すること;
ここで、第1および第2の宿主細胞は、別々に真核生物細胞およびグラム陽性菌からなる群から選択される。
【0010】
前記免疫グロブリン分子は、IgA、IgD、IgE、IgG、およびIgM免疫グロブリンなどから選択することができる。前記免疫学的に機能的な断片は、Fab断片、Fab’断片、Fab'-SH断片、F(ab’)2断片、Fv断片、VHH断片、ドメイン抗体、ディアボディー(diabody)、および多重特異性の抗体または抗体断片などから選択することができる。前記真核生物細胞は、哺乳類細胞、昆虫細胞、植物細胞、および真菌細胞などから選択することができる。前記第1および第2の宿主細胞は、例えば、COS細胞、BHK細胞、HEK293細胞、DUKX細胞、Saccharomyces spp細胞、Kluyveromyces spp細胞、Aspergillus spp細胞、Neurospora spp細胞、Fusarium spp細胞、Trichoderma spp細胞、およびLepidoptera spp細胞からなる群から別々に選択することができる。別の側面において、前記第1および第2の宿主細胞は、同じ細胞タイプの細胞であるか又は異なる細胞タイプの細胞である。
【0011】
一側面において、前記第1および第2の宿主細胞は、同じ培養で成長させられる。別の側面において、前記第1および第2の宿主細胞は、別々の培養で成長させられる。別の側面において、精製する工程は、Obelix陽イオン交換カラムを用いる精製を備えてもよい。一側面において、前記第1の宿主細胞は免疫グロブリン軽鎖をコード化している核酸を発現しなく、第2の宿主細胞は免疫グロブリン重鎖をコード化している核酸を発現しない。別の側面において、前記第1および第2の核酸は、1以上のモノクローナル抗体産生細胞から由来する。モノクローナル抗体産生細胞は、例えば、ハイブリドーマ、ポリドーマ(polydoma)および不死化させたB細胞から選択することができる。
【0012】
一側面において、リフォールディングは、前記第1および第2のポリペプチドを次の(a)および(b)から選択される条件下で混合することを備える:(a)第1と第2のポリペプチドとの比が約1:1、温度は約室温、およびpHは約7;並びに(b)第1と第2のポリペプチドとの比が約1:1、温度は約5゜C、およびpHは約8.0〜8.5の範囲。別の更なる側面において、前記第1および第2のポリペプチドは、約0.5M L-アルギニン-HCl、約0.9mM酸化型グルタチオン(GSSG)、および約2mM EDTAを含んでいる溶液中で混合される。
【0013】
また、本発明は、免疫グロブリン分子または免疫学的に機能的なその断片を産生する方法を提供し、該方法は以下を備える:
(a)第1の宿主細胞を、少なくとも免疫グロブリン重鎖の可変ドメインを具備している第1のポリペプチドをコード化しているヌクレオチド配列を具備している第1の核酸で形質転換すること;
(b)第2の宿主細胞を、少なくとも免疫グロブリン軽鎖の可変ドメインを具備している第2のポリペプチドをコード化しているヌクレオチド配列を具備している第2の核酸で形質転換すること;
(c)第1および第2の核酸配列を発現させること;
(d)第1および第2のポリペプチドの混合物を含んでいる溶液を透析すること;および
(e)第1および第2のポリペプチドがリフォールディングして、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を形成することを認容すること;
ここで、第1および第2の宿主細胞は、別々に真核生物細胞およびグラム陽性菌からなる群から選択される。
【0014】
一側面において、第1および第2の宿主細胞は、同じ培養で成長させられる(その溶液は、第1および第2の宿主細胞が成長させられた培養培地である)。別の側面において、前記溶液中の第1および第2のポリペプチドの相対量(relative amount)は、約1:2〜約2:1の範囲である。
【0015】
また、本発明は、抗体を精製するための方法を提供し、該方法は抗体を含んでいる溶液をObelix陽イオン交換カラムに適用すること、および抗体を精製することを備える。一側面において、前記方法は、次の工程のうち少なくとも1つを備える:(a)濾過した細胞培養(任意でpH調製される)を前記カラムに適用すること;(b)溶媒を溶出緩衝液に添加すること;および(c)塩グラジエントを増加させることによって抗体を溶出すること。特定の側面において、工程(c)は、工程(b)の前に実施される。代替的な溶出戦略には、約6.0のpHを有し、塩およびグリセロールを含んでいる溶出緩衝液(例えば、約30mMクエン酸, 約25mM NaCl, 約30%グリセロール、約6,0のpH)、約7.5〜8.5のpHを有している溶出緩衝液(例えば、Tris-緩衝剤)、約pH6.0から約6〜約9の範囲のpH(例えば、pH7.5〜8.5)までのpHグラジエント、および約6,5〜約7.0で0〜約1Mの塩(例えば、NaCl)でのグラジエント溶出の使用が含まれるが、これらに限定されない。
【0016】
また、本発明は、免疫グロブリン分子または免疫グロブリン重および軽鎖の少なくとも可変性ドメインを含んでいる免疫学的に機能的な免疫グロブリン断片を産生する方法を提供し、該方法は以下の工程を備える:
(a)重および軽鎖を、真核生物細胞およびグラム陽性細菌からなる群から選択される2つの別々の宿主細胞で独立に産生すること;
(b)重および軽鎖を精製すること;および
(c)免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片をインビトロでリフォールディングさせること。
【0017】
これらの及び他の側面は、以下により完全に記載される。
【0018】
[発明の記載]
本発明は、次の発見に基づくものである;その発見とは、免疫グロブリン分子の別々のポリペプチド鎖又はその断片が、別々の宿主細胞で独立に産生され、その後にリフォールドされて最終的な分子が形成されることである。宿主細胞を、同じ培地で共培養するか又は別々に培養することができる。発現後、鎖内および鎖間のジスルフィド形成の双方を、生体外の適切な反応条件下で行い、これによって正確に会合し、フォールドした抗体の形成が許容される。
【0019】
免疫グロブリン重(HC)および軽(LC)鎖(又は同じものの断片)を含んでいる及びコード化している宿主細胞は、それぞれ同じ、同一の細胞タイプのもの(例えば、同じ哺乳類細胞株からのもの)である。或いは、それぞれHCおよびLCを発現している宿主細胞は、異なる起源(例えば、哺乳類宿主細胞において発現されているHC、およびグラム陽性菌において発現されているLC)のものである。さらにまた、重および軽鎖をコード化しているDNA配列は、1つのハイブリドーマ細胞株から又は2以上のモノクローナル抗体産生性ハイブリドーマから取得しえる。
【0020】
上記のとおり、重および軽鎖を発現している宿主細胞は、それぞれ別々の培養および個々の培地、または同じ培地の共培養で成長させることができる。別々の宿主細胞で組換え型を産生させた後、HCおよびLC鎖を、当該技術分野において既知の方法によって又は本明細書中に記載された方法によって精製することができる。例えば、例は、高い伝導度を伴うタンパク質を結合することができる高度に疎水性の樹脂(Obelix陽イオン交換カラム, アマシャムから利用可能、カタログNo.11-0010)を用いる精製を記載している。
【0021】
完全な免疫グロブリン分子または機能的な免疫グロブリン断片を供給するための残りの最終工程は、重および軽鎖のジスルフィド結合形成による再会合が関与する(本発明において、リフォールディングと称される)。リフォールディング(再生とも称される)を、Jin-Lian Xingなど(2004; World J Gastroenterol 10(14):2029-2033)およびLeeおよびKwak(2003; Journal of Biotechnology 101:189-198)に記載のとおりに実施することができる。特定の態様において、リフォールディングは、重および軽鎖の混合物(又はその断片)の透析によって達成される(混合物中の重鎖および軽鎖の量は1:2〜2:1の範囲である)。更なる一態様において、前記範囲は約1:1である。前記態様において、前記宿主細胞は、同じ培養培地に含有される。培地中のHCおよびLC(又はその断片)の自己会合物(self-assemble)、および機能的な免疫グロブリンまたは断片は、前記培地から収穫することができる。HCおよびLCの混合物を含んでいる培養培地の透析工程には、リフォールディング処理を任意で含んでもよい。
【0022】
一態様において、HC鎖を発現する宿主細胞は、LC鎖をコード化しているDNAを含有または発現しない及び/又はLC鎖を発現する宿主細胞はHC鎖をコード化しているDNAを含有または発現しない。
【0023】
本発明の方法によって、グラム陰性菌(例えば、大腸菌)中で免疫グロブリンを発現させることと比較して、幾つかの利点が提供される。前記利点には、次のものが含まれる:
(i)エンドトキシンが存在しない、
(ii)高収量のタンパク質が取得される(というのも、封入体からタンパク質をリフォールディングさせる必要がないからである)、
(iii)完全長の免疫グロブリンを産生することができる、および
(iV)抗体の糖鎖形成パターンを、宿主となる生体に依存して変化させることができる。
【0024】
項目(i)に関して、本明細書中で使用されるエンドトキシンは、腸内細菌性のリポ多糖の毒性活性を意味し、グラム陰性細菌の外膜に見出されるものである。
【0025】
項目(ii)および(v)に関して、グラム陰性菌(例えば、大腸菌)は、大量のタンパク質が所望される場合、産生宿主細胞として適切ではない。大量の所望のタンパク質を大腸菌で生産した結果、しばしば封入体の形成を生じ、引続いてリフォールディングがなされる。対照的に、グラム陽性細菌は外膜を有さないが、タンパク質を細胞質から細胞外間隙へと直接的に分泌させるグリカン層を有する。相対的に単純な搬出機構によって、組換えタンパク質の分泌が高収量で促進される。
【0026】
項目(iii)に関して、完全長の免疫グロブリン分子はサイズが大きいので、大腸菌で取得することが困難である。リフォールディングにおける最近の報告に関して、完全なIgG分子が、大腸菌中で産生された(Simmons et al 2002 J. Immunol. Methods 263:133-147を参照されたい)。
【0027】
項目(iv)関して、薬学的な適用に関して開発された大抵のタンパク質は、真核生物性の宿主細胞中で産生された場合に、該タンパク質のポリペプチドバックボーンに付着したオリゴ糖を有する。一般的には、係る糖タンパク質の糖鎖は、アスパラギン残基のアミド基にN-グリコシド結合で又はセリンまたはスレオニン残基のヒドロキシル基にO-グリコシド結合で付着されてもよい。糖鎖形成は、タンパク質の適切な機能にしばしば必要とされ、適切なフォールディング、機能、および安定性を保証する。原核生物は、タンパク質の翻訳後修飾を実施する能力を欠いている。従って、タンパク質の糖鎖形成は、係る系で取得することができない。真菌類および酵母細胞を操作して、適切な糖鎖形成パターンを有するタンパク質を産生させることができる(Ballew and Gerngross 2004 Expert Opin. Biol. Ther. 4:623-626)。
【0028】
本発明によると、上記の効果は、重および軽鎖を、真核生物細胞およびグラム陽性菌からなる群から選択される2つの別々の宿主細胞中で独立に産生することによって提供される。本発明において、「独立に(independently)」の用語は、次の事項を意味する;その事項とは、個々の重鎖(HC)および軽鎖(LC)(又はその断片)の産生を、宿主細胞培養の異なる宿主細胞、異なる培養培地、異なる発現ベクター、および/または異なる物理的な条件(例えば、温度、レドックス条件、pH)などの使用によって、独立にコントロールする又は制御することができることである。HCおよびLC鎖(又はその断片)の産生後に、完全長の抗体または抗体断片へのエクスビボでのリフォールディングは、培養培地中で直接的に達成することができる(2つの別々のHCおよびLC鎖を発現している宿主細胞が、それぞれ同じ細胞培養中に存在する場合);或いは、本明細書中の他の箇所で記載した、LCおよびHCまたは断片の1以上の連続的(joint)または別個の精製工程、HCおよび/またはLC鎖溶液を濃縮するための及び/又は緩衝液を交換するための透析、及び特定のリフォールディング緩衝液への移動又は該緩衝液での希釈後に、達成することができる。リフォールディング条件を、当該技術分野における既知の方法にしたがって、各抗体又は抗体断片に関して選択する又は至適化することができる。典型的には、リフォールディングは、約+4゜C〜約+40゜Cまたは約+4゜C〜約室温の範囲の温度で、約5〜約9または約5.5〜約8.5のpHで得ることができる。リフォールディングを至適化するために使用しえる例示的な緩衝液には、リン酸、クエン酸-リン酸、酢酸(acetate)、およびトリス、同様にCO2でpHが制御される細胞の培養培地が含まれる。特定のリフォールディング条件は、例1に記載される。他の例示的なリフォールディング条件には、約1:1のHC:LC(またはHC:LC断片)比、約室温の温度、および中性のpHが含まれる。別の例示的なリフォールディング条件には、約1:1のHC:LC(またはHC:LC断片)比、約5゜Cの温度、約0.1 M Tris-HCl緩衝剤、約0.5MのL-アルギニン-HCl、酸化還元系として約0.9mMの酸化型グルタチオン(GSSG)、約2mMのEDTA、約8.0〜8.5のpHが含まれる。一側面において、リフォールディング溶液は、pHが約7.4で約100mMの尿素を含んでいる20mM Tris-HCl緩衝液に対して、平衡化した透析緩衝液における伝導度が約3.0〜3.5mSの範囲の値にまで減少するまで透析される。
【0029】
抗体
「抗体」および「免疫グロブリン分子」の用語は、本明細書中で互換的に使用され、モノクローナル抗体を意味する。重鎖中の定常ドメインの種類に応じて、抗体は、5つの主要なクラス:IgA、IgD、IgE、IgG及びIgMの1つに割り当てられる。これらのうち幾つかは、IgG1、IgG2、IgG3、IgG4などのように、サブクラス又はアイソタイプにさらに分けられる。免疫グロブリンの異なるクラスに対応する重鎖定常ドメインは、それぞれ「アルファ」、「デルタ」、「イプシロン」、「ガンマ」及び「ミュー」と称される。免疫グロブリンの異なるクラスのサブユニット構造及び三次元立体配置は周知である。IgGおよび/またはIgM(一般に生理学的な/臨床的な状況で一般的に使用され、研究室で容易に作出できる)は、本発明で使用する抗体の例示的なクラスである。
【0030】
本発明による、「IgG」分子の免疫グロブリンは、完全な抗体であってもよいし、機能的な免疫グロブリン断片であってもよい。モノクローナル抗体の断片または誘導体を作出する方法を、以下に開示する。
【0031】
当業者は所望の免疫グロブリン分子をどのように提供するかについて知っており、更なる詳細は以下で記載される。前記免疫グロブリン分子は、例えば、IgA、IgD、IgE、IgG、またはIgMの免疫グロブリンであってもよい。
【0032】
本明細書中で記載される「重鎖(HC)」または「軽鎖(LC)」は、完全長抗体の重および軽鎖または抗体断片の重鎖および軽鎖を具備する。
【0033】
本明細書中に使用される、「抗体断片(antibody fragment)」または「免疫グロブリン分子断片(immunoglobulin molecule fragment)」は、完全長抗体の一部を具備し、抗原と結合する能力がある。典型的には、抗体断片は、HCおよびLCの少なくとも1,2,3,4,5つの又は全ての抗体CDR領域、又は全体の可変重鎖(VH)または可変軽鎖(VL)部分を具備する。例示的な抗体断片は、Fab, F(ab)2, F(ab’)2, Fd, scFv, dsFv断片, VHH断片, ドメイン抗体(VHおよび/またはVL)、同様に、2以上の完全長抗体の抗原結合部位を含んでいる多重特異性(二重特異的なものを含む)抗体構築物が含まれるが、これらに限定されない。
【0034】
「免疫原(immunogen)」の用語は、免疫寛容ではなく、体液性の抗体および/または細胞性の免疫応答を誘導する能力がある物質である。「免疫原」の用語は、「抗原(antigen)」と時々互換的に使用されるが、その用語は免疫応答を刺激する及びその産物(例えば、抗体)と反応する能力があることを特定する。対照的に、「抗原」は、抗体と反応する物質を意味させるために確保(reserved)される。主要な免疫原は、微生物から遊離しているか又はそれに付着している、タンパク質およびリポ多糖である。
【0035】
本明細書中で使用される「エクスビボ(ex vivo)」の用語は、任意の身体の外側(即ち、インビボで生じないプロセス)および任意の生細胞の外側(即ち、細胞内で生じないプロセス)を意味する。
【0036】
「免疫原性(immunogenic)」の用語は、体液性の抗体および/または細胞性の免疫応答を誘導する能力を意味する。
【0037】
抗体産生
本発明は、重(H)および軽(L)鎖の少なくとも可変ドメインが別々の宿主細胞中で産生され、自由選択で単離され、そしてインビトロでリフォールドされる免疫グロブリン分子または断片の組換え型の産生に関する。本発明に適用するためのVHおよびVL配列は、当該技術分野において既知の様々な技術の何れかによって産生される抗体から取得することができる。典型的には、それらは、非ヒト動物(好ましくは、マウス)を所望の抗原または免疫原を含んでいる免疫原で免疫することによって提供される。或いは、抗体は、例えば、Wardら(Nature 341 (1989) 544)に開示されているように、免疫グロブリンのコンビナトリアルライブラリーを選択することによって提供しえる。
【0038】
ヒト以外の哺乳動物を抗原で免疫化する工程は、マウスでの抗体の産生を刺激するための、当該技術で周知の任意の様式で実施することができる(例えば、 E. Harlow and D. Lane, Antibodies: A Laboratory Manual., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1988)を参照されたい)。好適な態様において、非ヒト動物は、げっ歯類(例えば、マウス、ラットなど)、ウシ、ブタ、ウマ、ウサギ、ヤギ、ヒツジなどの哺乳動物である。また、ヒト以外の哺乳動物を、XenomouseTM(Abgenix)又はHuMAb−MouseTM(Medarex)などの、「ヒト」抗体を産生するように、遺伝的に改変し、又は操作することができる。典型的には、免疫原は、任意で完全フロイントアジュバントなどのアジュバントとともに、緩衝液中に懸濁又は溶解される。免疫原の量、緩衝液の種類及びアジュバントの量を決定する方法は、当業者に周知であり、いかなる意味においても、本発明を限定するものではない。これらのパラメータは、異なる免疫原については異なる場合があり得るが、容易に解明される。
【0039】
同様に、抗体の産生を刺激するのに十分な免疫化の位置(location)及び頻度も、当該技術において周知である。典型的な免疫化プロトコールでは、1日目に、ヒト以外の動物に抗原を腹腔内注射し、約1週後に再度抗原を腹腔内注射する。この後、必要に応じて、不完全フロイントアジュバントなどのアジュバントとともに、20日目前後に、抗原のリコール注射(recall injections)を行う。リコール注射は、静脈内に行われ、連続数日間、反復することができる。その後、通例、アジュバントなしに、静脈内又は腹腔内の何れかで、40日目にブースター注射を行う。このプロトコールによって、約40日後に、抗原特異的抗体産生B細胞が産生される。免疫化に使用される抗原に対して誘導される抗体を発現するB細胞の産生をもたらす限り、他のプロトコールも使用することができる。
【0040】
別の態様では、非免疫のヒト以外の哺乳動物からリンパ球を単離し、インビトロで成長させ、次いで、細胞培養において免疫原に曝露する。次いで、リンパ球を採集し、以下に記載された融合工程を実施する。
【0041】
モノクローナル抗体の場合、次の工程は、免疫されたヒト以外の哺乳動物から脾細胞を単離した後、これらの脾細胞を不死化された細胞と融合して、抗体を産生するハイブリドーマを形成させる。ヒト以外の哺乳動物からの脾細胞の単離は当該技術において周知であり、典型的には、麻酔されたヒト以外の哺乳動物から脾臓を取り出し、脾臓を小片に切断し、脾臓カプセル(splenic capsule)から細胞ろ過器のナイロンメッシュを通して、脾細胞を適切な緩衝液中に絞り取り、単一細胞の懸濁液を得る。細胞を洗浄し、遠心し、赤血球を溶解する緩衝液中に再懸濁する。この溶液を再び遠心し、最後に、ペレット中に残存するリンパ球を新鮮な緩衝液に再懸濁する。
【0042】
一度単離し、単一の細胞懸濁液中に存在させ、リンパ球を不死化細胞株と融合する。これは、典型的には、マウスのミエローマ細胞株であるが、ハイブリドーマの作製に有用な他の多くの不死化細胞株も、当該技術において公知である。好ましいマウスのミエローマ株には、「Salk Institute Cell Distribution Center, San Diego, Calif. U.S.A」から入手可能なMOPC−21及びMPC−11マウス腫瘍、並びに「American Type Culture Collection, Rockville, Maryland U.S.A」から入手可能なX63、Ag8653及びSP−2細胞から得られるマウスのミエローマ株が含まれるが、これらに限定されるものではない。融合は、ポリエチレングリコールなどを用いて行われる。次いで、得られたハイブリドーマを、融合されていない親ミエローマ細胞の増殖又は生存を阻害する1以上の物質を含有する選択培地中で増殖させる。例えば、親ミエローマ細胞が酵素ヒポキサンチングアニンホスホリボシル転移酵素(HGPRT又はHPRT)を欠如していれば、ハイブリドーマに対する培地は、典型的には、HGPRT欠損細胞の増殖を抑える物質であるヒポキサンチン、アミノプテリン及びチミジン(HAT培地)を含むであろう。
【0043】
ハイブリドーマは、典型的には、マクロファージの支持細胞上で増殖される。マクロファージは、脾細胞を単離するために使用されるヒト以外の哺乳動物の同腹仔から得ることが好ましく、典型的には、ハイブリドーマを播種する数日前に、不完全フロイントアジュバントなどで刺激される。融合法は、文献〔Goding, "Monoclonal Antibodies: Principles and Practice," pp. 59-103 (Academic Press, 1986)〕中に記載されている。
【0044】
前記細胞は、コロニー形成と抗体産生に十分な時間、選択培地中で増殖させる。これは、通常、7日から14日の間である。次いで、このハイブリドーマコロニーを、免疫原/抗原と結合する抗体の産生についてアッセイする。その中でハイブリドーマが増殖されるウェルに適合できる任意のアッセイを使用し得るが、このアッセイは、典型的には、比色分析のELISAタイプのアッセイである。他のアッセイには、免疫沈降及びラジオイムノアッセイが含まれる。一以上の別個のコロニーが存在するかどうかを決定するために、所望の抗体の産生に関して陽性のウェルを調べる。二以上のコロニーが存在すれば、細胞を再度クローニングし、単一の細胞のみが、所望の抗体を産生するコロニーを確実に生じるように増殖させ得る。モノクローナル抗体が唯一つ検出され、産生されるように、典型的には、単一の明白なコロニーを有する陽性細胞を再クローニングし、再アッセイする。
【0045】
モノクローナル抗体を産生することが確認されているハイブリドーマは、次にDMEM又はRPMI−1640などの、適切な培地中で、大量に増殖される。或いは、ハイブリドーマ細胞は、動物内の腹水癌として、インビボで増殖させることができる。
【0046】
所望のモノクローナル抗体を産生するまで十分に増殖させた後、モノクローナル抗体を含有する増殖培地(又は腹水液)を細胞から分離し、その中に存在するモノクローナル抗体を精製する。典型的には、精製は、ゲル電気泳動、透析、プロテインA若しくはプロテインG−セファロース、又はアガロース又はセファロースビーズなどの固相支持体に連結された抗マウスIgを用いたクロマトグラフィーによって達成される(全て、例えば、「Antibody Purification Handbook, Amersham Biosciences, publication No. 18-1037-46, Edition AC」に記載されており、その開示内容は、参照により、本明細書に援用される)。結合された抗体は、典型的には、低pH緩衝液(pH3.0以下のグリシン又は酢酸緩衝液)を使用することによって、プロテインAまたはプロテインGカラムから溶出され、抗体含有画分を直ちに中和する。必要に応じて、これらの画分をプールし、透析し、濃縮する。
【0047】
本発明の特定の側面として、Obelix陽イオン交換体を抗体の精製に使用することができる。Obelix陽イオン交換体は、高伝導度で及びpI(抗体に関する)よりも高pHで抗体と結合する。これによって、精製の性能(capability)が影響される。精製を、プロピレンジオールなどを添加することによって更に変化させて、疎水性相互作用を本陽イオン交換体カラム上で利用することができる。
【0048】
本発明の方法に使用されるモノクローナル抗体をコードするDNAは、従来の手順を用いて(例えば、マウス抗体の重鎖及び軽鎖をコードする遺伝子に特異的に結合することができるオリゴヌクレオチドプローブを使用することによって)、容易に単離され、配列決定される。DNAを単離したら、DNAを発現ベクター中に配置することができ、次いで、発現ベクターを宿主細胞(トランスフェクションが行われていない状態で、免疫グロブリンタンパク質を産生しない、細菌細胞、サルのCOS細胞、チャイニーズハムスター卵巣(CHO)細胞又は骨髄腫細胞など)中にトランスフェクションして、組換え宿主細胞中でモノクローナル抗体の合成を得る。抗体をコード化しているDNAの細菌中での組換え発現は、当該技術で周知である〔例えば、Skerra et al., Curr. Opinion in Immunol., 5, pp. 256 (1993);およびPluckthun, Immunol. Revs., 130, pp. 151 (1992)を参照〕。例えば、所望の抗原と結合する抗体をコード化しているDNAは、ハイブリドーマから単離され、適切な宿主へのトランスフェクションのために適切な発現ベクターに配置される。次いで、抗体、又はそれらの断片(ヒト化された様式のモノクローナル抗体、抗体の活性断片、又は抗体の抗原認識部分を含むキメラ抗体など)の発明にしたがって組換え産生するために、宿主を使用する。
【0049】
免疫グロブリンポリペプチドをコード化しているDNA配列は、通常は組換え型ベクターに挿入される(これは、組換えDNA処置に都合よく供される、任意のベクターでよい);また、ベクターの選択は、それが導入される宿主細胞にしばしば依存する。ベクター成分には、1以上の次の因子、即ち:プロモーター、シグナル配列、複製開始点、1以上の選択マーカー、および転写ターミネーター配列が一般的に含まれるが、これらに限定されない。従って、前記ベクターは自律的に複製するベクター、即ち、染色体外の実体(extrachromosomal entity)として存在し、その複製が染色体複製に非依存性であるベクターである(例えば、プラスミド)。或いは、前記ベクターは次のようなベクター、即ち、宿主細胞に導入された際に、前記宿主細胞ゲノムに統合され、それが統合された染色体と共に複製されるベクターであってもよい。
【0050】
前記ベクターは、好ましくは発現ベクター中の免疫グロブリンポリペプチドをコード化しているDNA配列が、前記DNAの転写に必要とされる付加的なセグメントに動作可能に連結される、発現ベクターである。通常は、前記発現ベクターは、プラスミドまたはウイルスのDNAに由来するものである、或いは両方のエレメントを含有していてもよい。「動作可能に連結(operably linked)」の用語は、前記セグメントが、それらがそれらの意図する目的に関して協調して機能するように配置されることを示す(例えば、転写は、プロモーターで開始され、ポリペプチドをコード化しているDNA配列を通して進行する)。
【0051】
ポリペプチドの発現に使用するための発現ベクターは、クローン化された遺伝子またはcDNAの転写を方向付ける能力を有するプロモーターを具備する。前記プロモーターは、選択された宿主細胞において転写活性を呈する、任意のDNA配列でよく、前記宿主細胞に対して相同性(homologous)の又は異種性(heterologous)のタンパク質をコード化している遺伝子に由来するものであってもよい。
【0052】
グラム陽性細菌においてポリペプチドバリアントをコード化しているcDNAの転写を駆動させるための適切なプロモーターの例は、AmyP(Kruger et al 2002 Nature Biotechnol 20:702-706)、NisA(de Ruyter et al 1996 J. Bacteriol 178:3434-3439)、Spac(Yansura and Henner 1984 Proc. Natl. Acad. Sci. USA 81:439-443)、XylA(Eichenbaum et al 1998 Appl. Environ. Microbiol. 63:3451-3457)、LacA(Platteeuw et al 1994 Appl. Environ. Microbiol. 60:587-593)、UW85 upp(Dunn and Handelsman 1999 Gene 226:297-305)である。
【0053】
哺乳類細胞におけるポリペプチドバリアント(polypeptide variant)をコード化しているDNAの転写を方向付けるための適切なプロモーターの例は、SV40プロモーター(Subramani et al., Mol.Cell. Biol. 1 (1981), 854-864)、MT-1(メタロチオネイン遺伝子)プロモーター(Palmiter et al., Science 222 (1983), 809 - 814 )、CMVプロモーター(Boshart et al.,Cell. 41:521-530, 1985)、またはアデノウイルス2主要後期プロモーター(Kaufman and Sharp, Mol.Cell. Biol, 2:1304-1319, 1982)である。
【0054】
昆虫細胞における使用に関して適切なプロモーターは、ポリヘドリンプロモーター(US 4,745,051; Vasuvedan et al., FEBS Lett. 311, (1992) 7 - 11)、P10プロモーター(J.M. Vlak et al., J. Gen. Virology 69, 1988, pp. 765-776)、Autographa californica多核体病ウイルス塩基性タンパク質プロモーター(EP 397 485)、バキュロウイルス前初期遺伝子1プロモーター(US 5,155,037; US 5,162,222)、またはバキュロウイルス39K遅延性-初期遺伝子プロモーター(US 5,155,037; US 5,162,222)である。
【0055】
酵母宿主細胞における使用に関して適切なプロモーターの例には、酵母の解糖遺伝子(glycolytic genes)からのプロモーター(Hitzeman et al., J. Biol. Chem. 255 (1980), 12073 - 12080; Alber and Kawasaki, J. Mol. Appl. Gen. 1 (1982), 419 - 434)またはアルコール脱水素酵素遺伝子(Young et al., in Genetic Engineering of Microorganisms for Chemicals (Hollaender et al, eds.), Plenum Press, New York, 1982)、またはTPI1(US 4,599,311)またはADH2-4c (Russell et al., Nature 304 (1983), 652 - 654)プロモーターが含まれる。
【0056】
糸状菌(filamentous fungus)宿主細胞における使用に関して適切なプロモーターの例は、例えば、ADH3プロモーター(McKnight et al., The EMBO J. 4 (1985), 2093 - 2099 )またはtpiAプロモーターである。他の有用なプロモーターの例は、A.oryzae TAKAアミラーゼ、Rhizomucor mieheiアスパラギン酸プロテイナーゼ、A.niger中性α-アミラーゼ、A.niger酸安定性α-アミラーゼ(acid stable α-amylase)、A.niger、またはA.awamoriグルコアミラーゼ(gluA)、Rhizomucor mieheiリパーゼ、A.oryzaeアルカリ性プロテアーゼ、A.oryzae三炭糖リン酸塩イソメラーゼ、またはA.nidulansアセトアミダーゼをコード化している遺伝子に由来するものである。好適なものは、TAKA-アミラーゼおよびgluAプロモーターである。適切なプロモーターは、例えば、EP 238023およびEP 383779に記載されている。
【0057】
また、前記ヒトポリペプチドをコード化しているDNA配列は、必要な場合には、適切なターミネーター〔例えば、ヒト成長ホルモンターミネーター(Palmiter et al., Science 222, 1983, pp. 809-814)またはTPI1(Alber and Kawasaki, J. Mol. Appl. Gen. 1, 1982, pp. 419-434)またはADH3(McKnight et al., The EMBO J. 4, 1985, pp. 2093-2099)ターミネーター〕に動作可能に連結されてもよい。また、発現ベクターは、前記プロモーターから下流で且つポリペプチド配列自身に関する挿入部位から上流に位置するRNAスプライス部位のセットを含んでいてもよい。好適なRNAスプライス部位は、アデノウイルスおよび/または免疫グロブリン遺伝子から取得し得る。発現ベクターに含まれるものは、挿入部位の下流に位置する、ポリアデニル化シグナルである。特に好適なポリアデニル化シグナルには、SV40の早期の若しくは遅発型のポリアデニル化シグナル(Kaufman and Sharp, ibid.)、アデノウイルス5Elb領域、ヒト成長ホルモン遺伝子ターミネーターのポリアデニル化シグナル(DeNoto et al. Nucl. Acids Res. 9:3719-3730, 1981)、または前記遺伝子のポリアデニル化シグナルが含まれる。また、発現ベクターは、非コードの(noncoding)ウイルス性のリーダー配列〔例えば、アデノウイルス2 トリパタイト(tripartite)リーダー(前記プロモーターおよびRNAスプライス部位の間に位置する〕;およびエンハンサー配列(例えばSV40エンハンサー)を含む。
【0058】
本発明のポリペプチドを宿主細胞の分泌経路(secretory pathway)へと方向付ける(direct)ために、分泌性シグナル配列(リーダー配列、プレプロ配列、またはプレ配列としても知られる)を組換えベクターに提供してもよい。分泌性シグナル配列は、正しい読み枠で、前記ポリペプチドをコード化しているDNA配列と連結される。分泌性シグナル配列は、前記ペプチドをコード化しているDNA配列に対して、5'に通常配置される。分泌性シグナル配列は、前記タンパク質と通常関連するものであってもよい又は別の分泌タンパク質をコード化している遺伝子からのものであってもよい。グラム陽性細菌において、L.amylovorusのα-アミラーゼ遺伝子のシグナル配列が、使用された(Kruger et al 2002 Nature Biotechnol 20:702-706)。
【0059】
酵母細胞からの分泌に関して、分泌性シグナル配列は、発現したポリペプチドの前記細胞の分泌経路への効率的な方向付けを保証する任意のシグナルペプチドをコード化してもよい。前記シグナルペプチドは、天然の(naturally occurring)シグナルペプチド、又はその機能的な部分であってもよい、又は合成ペプチドであってもよい。適切なシグナルペプチドは、α-因子シグナルペプチド(cf. US 4,870,008)、マウス唾液アミラーゼのシグナルペプチド(cf. O. Hagenbuchle et al., Nature 289, 1981, pp. 643-646)、修飾したカルボキシペプチダーゼシグナルペプチド(cf. L.A. Valls et al., Cell 48, 1987, pp. 887-897)、酵母BAR1シグナルペプチド(cf. WO 87/02670)、または酵母アスパラギン酸プロテアーゼ3(YAP3)シグナルペプチド(cf. M. Egel-Mitani et al., Yeast 6, 1990, pp. 127-137)であることが見出されている。
【0060】
酵母における効率的な分泌に関して、リーダーペプチドをコード化している配列が、シグナル配列の下流およびポリペプチドをコード化しているDNA配列の上流に挿入されてもよい。リーダーペプチドの機能は、発現されたペプチドを小胞体からゴルジ体へと、更に培養培地への分泌に関連する分泌小胞(secretory vesicle)へと方向付けることを可能にすることである(即ち、ポリペプチドの、細胞壁を横切った輸送又は少なくとも細胞膜を介した酵母細胞の細胞膜周辺腔への輸送)。前記リーダーペプチドは、酵母α-因子リーダーであってもよい(その使用は、例えば、US 4,546,082、US 4,870,008、EP 16 201、EP 123 294、EP 123 544およびEP 163 529に記載されている)。或いは、前記リーダーペプチドは合成プロペプチドであってもよく、つまり自然界でみつけることができないリーダーペプチドであってもよい。合成リーダーペプチドは、例えば、WO 89/02463またはWO 92/11378に記載されたように構築し得る。
【0061】
糸状菌における使用に関して、前記シグナルペプチドは、コウジカビ属の種(Aspergillus sp)のアミラーゼまたはグルコアミラーゼをコード化している遺伝子、Rhizomucor mieheiのリパーゼまたはプロテアーゼまたはHumicola lanuginosaのリパーゼをコード化している遺伝子から由来するものであってもよい。前記シグナルペプチドは、好ましくはA.oryzae TAKAアミラーゼ、A.niger中性アルファ-アミラーゼ、A.niger酸安定性アミラーゼ、またはA.nigerグルコアミラーゼをコード化している遺伝子から由来するものである。適切なシグナルペプチドは、例えば、EP 238023およびEP 215594に開示されている。
【0062】
昆虫細胞における使用に関して、前記シグナルペプチドは、昆虫遺伝子(cf. WO 90/05783)、例えば、lepidopteran Manduca sextaの脂質動員ホルモン前駆物質シグナルペプチド(cf. US 5,023,328)から由来するものであってもよい。
【0063】
それぞれ、ポリペプチドをコード化しているDNA配列、プロモーターおよび自由選択のターミネーター、および/または分泌性シグナル配列を連結するために及びそれらを適切なベクター(このベクターは複製に必要な情報を含有している)挿入するために使用される処置は、当業者に周知である(例えば、Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, New York, 1989を参照されたい)。
【0064】
哺乳類細胞をトランスフェクションする及び前記細胞に導入したDNA配列を発現させる方法は、例えば、文献〔Kaufman and Sharp, J. Mol. Biol. 159 (1982), 601 - 621; Southern and Berg, J. Mol. Appl. Genet. 1 (1982), 327 - 341; Loyter et al., Proc. Natl. Acad. Sci. USA 79 (1982), 422 - 426; Wigler et al., Cell 14 (1978), 725; Corsaro and Pearson, Somatic Cell Genetics 7 (1981), 603, Graham and van der Eb, Virology 52 (1973), 456; およびNeumann et al., EMBO J. 1 (1982), 841 - 845〕に記載されている。
【0065】
クローン化したDNA配列は、培養した哺乳類細胞に、例えば、リン酸カルシウムに媒介されるトランスフェクション(Wigler et al., Cell 14:725-732, 1978; Corsaro and Pearson, Somatic Cell Genetics 7:603-616, 1981; Graham and Van der Eb, Virology 52d:456-467, 1973)または電気穿孔法(Neumann et al., EMBO J. 1:841-845, 1982)により導入される。外来性(exogenous)のDNAを発現する細胞を同定する及び選択するため、一般的に選択可能な表現型(選択可能なマーカー)が、所望の遺伝子またはcDNAと共に導入される。好適な選択可能なマーカーには、薬物(例えば、ネオマイシン、ハイグロマイシン、およびメトトレキセート)に対する耐性を与える遺伝子が含まれる。前記選択可能なマーカーは、増幅可能な選択可能なマーカーであってもよい。好適な増幅可能な選択可能なマーカーは、ジヒドロ葉酸レダクターゼ(DHFR)配列である。選択可能なマーカーはThilly(Mammalian Cell Technology, Butterworth Publishers, Stoneham, MA、本明細書中に参照により援用される)により論評されている。当業者は、適切な選択可能なマーカーを容易に選択することができる。
【0066】
選択可能なマーカーは、細胞中に、別々のプラスミド上で、所望の遺伝子と同時に導入し得る、又はそれらを同じプラスミド上で導入し得る。同じプラスミド上で、選択可能なマーカーおよび所望の遺伝子が、異なるプロモーターまたは同じプロモーターの制御下で存在し得る;この後者の配置はジシストロン性メッセージ(a dicistronic message)を産生する。このタイプの構築物は既知である(例えば、LevinsonおよびSimonsen、U.S.4,713,339)。「キャリアDNA(carrier DNA)」として知られる付加的なDNAを、前記細胞に導入される混合物に付加することも有利であろう。
【0067】
細胞が前記DNAを受け取った後、それらは適切な成長培地において成長し(典型的には1〜2日)、所望の遺伝子の発現が開始される。本明細書中に使用される「適切な成長培地(appropriate growth medium)」の用語は、細胞の成長に及び所望のポリペプチドバリアントの発現に要求される、栄養物および他の成分を含有している培地を意味する。培地は、一般に炭素源、窒素源、必須アミノ酸類、必須な糖類、ビタミン類、塩類、リン脂質類、タンパク質、および成長因子を含む。ガンマ-カルボキシル化されたタンパク質の産生に関して、培地はビタミンK(好ましくは、約0.1μg/mlから約5μg/mlの濃度で)を含有する。次に薬物選択が、安定な様式で選択可能なマーカーを発現している細胞の成長に関する選択に適用される。増幅可能で選択可能なマーカーでトランスフェクションされた細胞に関して、薬物濃度を増加して、クローン化された配列のコピー数の増加に関して選択し得る。これによって発現レベルが増加される。安定的にトランスフェクションされた細胞のクローンは、次に所望のポリペプチドバリアントの発現に関して選抜される。
【0068】
前記免疫グロブリンポリペプチドをコード化しているDNA配列が導入された宿主細胞は、必要に応じて、翻訳後修飾されたポリペプチドを産生する能力を有する任意の細胞でありえ、該宿主細胞には、酵母、真菌類、および高等真核細胞が含まれる。翻訳後修飾は、原核生物の発現系では得られない。
【0069】
本発明の一態様において、真核生物細胞は、哺乳類細胞、昆虫細胞、植物細胞、および真菌細胞(酵母細胞を含む)から選択される。
【0070】
原核細胞の例は、大腸菌(Cabilly et al US 6331415)などのグラム陰性細胞またはBacilli、Clostridia、Staphylococci、Lactobailli、またはLactococci(de Vos et al 1997 Curr. Opin. Biotechnol. 8:547-553)などのグラム陽性細菌であってもよい。組換えタンパク質をグラム陽性細菌において発現させる例示的な方法は、US5821088に記載される。
【0071】
本発明における使用に関する哺乳類細胞株は、COS−1(ATCC CRL 1650)、乳児ハムスター腎臓(BHK)およびHEK293(ATCC CRL 1573; Graham et al., J. Gen. Virol. 36:59-72, 1977)細胞株である。好適なBHK株化細胞は、tk- ts13 BHK株化細胞(Waechter and Baserga, Proc. Natl. Acad. Sci. USA 79:1106-1110, 1982, 本明細書中に参照により援用される)であり、以下でBHK570細胞と称する。BHK570株化細胞は、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection, 12301 Parklawn Dr., Rockville, Md. 20852)にATCC受託番号CRL10314で寄託されている。また、tk- ts13 BHK細胞株も、ATCCから受託番号CRL1632で利用可能である。加えて、本発明に使用し得る、いくつかの他の細胞株には、Rat Hep I(ラット ヘパトーマ;ATCC CRL 1600)、Rat HepII(ラット ヘパトーマ;ATCC CRL 1548)、TCMK(ATCC CCL 139)、ヒト肺(Human lung)(ATCC HB 8065)、NCTC1469(ATCC CCL 9.1)、CHO(ATCC CCL 61)、およびDUKX細胞(Urlaub and Chasin, Proc. Natl. Acad. Sci. USA 77:4216-4220, 1980)が含まれる。
【0072】
適切な酵母細胞の例には、Saccharomyces spp.またはSchizosaccharomyces spp.の細胞が、特にSaccharomyces cerevisiaeまたはSaccharomyces kluyveriの株が含まれる。酵母細胞を異種性のDNAで形質転換する及びそれからの異種性のポリペプチドを産生させるための方法は、例えば、US 4,599,311、US 4,931,373、US 4,870,008、5,037,743、および US 4,845,075に記載され、これら全ては参照によって援用される。形質転換された細胞は、選択可能なマーカー(一般に薬剤抵抗性または特定の栄養分(例えば、ロイシン)の非存在下で成長する能力)により決定される表現型により選択される。酵母における使用に関して好適なベクターは、US 4,931,373に開示されるPOT1ベクターである。ポリペプチドをコード化しているDNA配列は、シグナル配列および任意でリーダー配列(例えば、上記のような)により先行されていてもよい。適切な酵母細胞の更なる例は、Kluyveromyces(例えば、K.lactis)、hansenula(例えば、H.polymorpha)、またはPichia(例えば、P.pastoris)の株である(Gleeson et al., J. Gen. Microbiol. 132, 1986, pp. 3459-3465; US4882279を参照されたい)。
【0073】
他の真菌細胞の例は、糸状菌、例えば、コウジカビ属 spp.(Aspergillus spp.)、パンカビ属 spp.(Neurospora spp.)、フザリウム spp.(Fusarium spp.)、またはトリコデルマ属 spp.(Trichoderma spp.)の細胞、特にA.oryzae、A.nidulans、およびA.nigerの株である。コウジカビ属 spp.のタンパク質の発現に関する使用は、例えば、EP 272 277、EP 238 023、EP 184 438に記載されている。F.oxysporumの形質転換は、例えば、文献(Malardier et al., 1989 (Gene 78: 147-156))に記載のとおり実施し得る。トリコデルマ属 spp.の形質転換は、例えば、EP 244 234に記載のとおり実施し得る。
【0074】
糸状菌が宿主細胞として使用される場合、それを本発明のDNA構築物で、前記DNA構築物を宿主染色体に統合させて組換え宿主細胞(recombinant host cell)を得ることにより形質転換し得る。この統合は一般的には有利であると考えられる、というのも前記DNA配列が前記細胞において安定的に維持される可能性が高いからである。前記DNA配列の宿主染色体への統合は、従来の方法(例えば、相同性の又は異種性の組換えによる)にしたがって実施し得る。
【0075】
昆虫細胞の形質転換及びその中での異種性のポリペプチドの産生は、US 4,745,051;US 4,879,236;US 5,155,037;5,162,222;EP 397,485に記載のとおり実施し得る(これらの文献の全ては本明細書中に参照によって援用される)。宿主として使用される昆虫株化細胞は、鱗翅目株化細胞(例えば、Spodoptera frugiperda細胞またはTrichoplusia ni細胞)であってもよい(cf. US 5,077,214)。培養条件は、例えば、WO 89/01029またはWO 89/01028に、または上述の文献の何れかに記載されたとおりであってもよい。
【0076】
上記の形質転換された又はトランスフェクションした宿主細胞は、次に適切な栄養培地中で、免疫グロブリンポリペプチドの発現を許容する条件下で培養され、その後、生じたペプチドの全部または一部を培養物から回収し得る。前記細胞を培養するために使用される培地は、前記宿主細胞を成長させるために適切な任意の従来培地、例えば、適切なサプリメントを含有する最小の又は複合的(complex)な培地であってもよい。適切な培地は、商業上の提供者から購入される又は公開されたレシピ(例えば、アメリカン・タイプ・カルチャー・コレクションのカタログ中の)にしたがって調製し得る。前記細胞により産生されたポリペプチドは、次に培養培地(culture medium)から従来の処理により回収または精製される〔この処理には宿主細胞を培地から遠心分離または濾過により分離すること、上清もしくは濾過物のタンパク性成分を塩(例えば、硫酸アンモニウム)で沈殿させ、様々なクロマトグラフィー処理(例えば、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、アフィニティークロマトグラフィー)により精製すること、等が含まれる〕、この処理は問題のポリペプチドのタイプに依存して行われる。クロマトグラフィー処理において、ポリペプチドはカラムから溶液中に溶出される。一側面において、前記ポリペプチドは、培養培地からの精製の前または後に透析されて、ポリペプチドが所望の溶液中に得られる。
【0077】
モノクローナル抗体の断片及び誘導体
抗体断片の例には、Fab、Fab'、Fab'−SH,F(ab')2及びFv断片;二重特異性抗体;連続するアミノ残基の一つの連続配列からなる一次構造を有するポリペプチド(本明細書において、「一本鎖抗体断片」又は「一本鎖ポリペプチド」と称される。)である任意の抗体断片〔(1)一本鎖Fv(scFv)分子、(2)軽鎖可変ドメインを一つだけ含有する一本鎖ポリペプチド、又は軽鎖可変ドメインの3つのCDRを含有し、付随する重鎖部分を伴わないそれらの断片、及び(3)重鎖可変領域を一つだけ含有する一本鎖ポリペプチド、又は重鎖可変ドメインの3つのCDRを含有し、付随する軽鎖部分を伴わないそれらの断片などが含まれるが、これらに限定されない。〕;並びに抗体断片から形成される多重特異性抗体(例えば、二重特異性抗体)が含まれる。「免疫反応性断片(Immunoreactive fragments)」は、無傷(intact)の抗体の一部(一般的には、抗原結合部位又は可変領域)を含む。本発明によって産生された免疫学的に機能的な免疫グロブリン断片は、1以上の抗体の少なくとも重鎖(VH鎖)の可変ドメインおよび少なくとも軽鎖(VL鎖)の可変ドメインを具備してもよい。
【0078】
本発明の方法を産生のために使用することができる、2つの例示的な抗原結合免疫グロブリン断片は、FabおよびFvである。小さいFv(即ち、可変性の断片)は、VLおよびVHのみから構成される。Fvの組換え型バージョンは、単鎖可変断片(ScFv)と称される。ScFvにおける2つの断片は、可動性のペプチドリンカー〔通常では15アミノ酸リンカーが(G4S)3の配列で使用される〕と人工的に連結され、単一のポリペプチド鎖として発現される。前記リンカーによって、VHおよびVLの会合が許容されて、抗原結合部位が形成される。2つのポリペプチドからなる大きなFab(抗原結合断片)のうち、一方は軽鎖可変部および定常ドメイン VL−CLを含み、他方は可変ドメインおよび1つの定常ドメインを含んでいる短縮型(truncated)の重鎖 VH−CH1を含む。無傷のIgG免疫グロブリンのように、2つの鎖は共にジスルフィド結合で連結される。ScFv抗体は2つのジスルフィド結合を含有するが、他方でFabsは5つのジスルフィド結合を含有する(安定的なフォールディングのために全てが形成されなければならない)。
【0079】
ScFvおよびFab断片の発現は、真核生物、酵母、真菌、および細菌を含む様々な宿主細胞において実施される。グラム陰性細菌において、システインチオールからジスルフィドへの酸化は、タンパク質が細胞質の高度に還元的な環境からペリプラズム中のより酸化的な環境へと輸送された後にのみ通常は発生する。ペリプラズムへと発現されたタンパク質は、浸透圧ショックによって又は全細胞ライセートから回収することができる。抗体を37゜Cで長期にわたり高レベルで発現させることによって、大腸菌の外膜は透過性となり、タンパク質を培養培地から回収することができる。しかしながら、外膜における脂質の存在によって、エンドトキシン汚染に関する問題が生じる。グラム陽性細菌および酵母は外膜を有さず、タンパク質を細胞質から細胞外空間へと直接的に分泌させることができる。結果的に、エンドトキシンは存在せず、相対的に単純な輸送機構によって、組換え型タンパク質の高収量での分泌が促進される。
【0080】
本発明の抗体の断片および誘導体は、当該技術で公知の技術によって作出することができる。例えば、Fab又はF(ab')2断片は、従来の技術に従って、前記単離された抗体をプロテアーゼ消化することによって作出することができる。或いは、本発明の抗体を産生しているハイブリドーマのDNAは、本発明の断片をコードするように修飾しえる。次いで、修飾されたDNAは発現ベクター中に挿入され、適切な細胞を形質転換し又はトランスフェクションするために使用され、次いで、前記細胞が所望の断片を発現する。
【0081】
他のタイプの断片には、重鎖抗体(VHH)断片の重鎖の可変ドメイン〔例えば、Muyldermans, Reviews in Mol. Biotechnol. 2001, 74:277-302 およびSpinelli et al., Biochemistry. 2000;39(6):1217-22に記載されたもの〕および抗体の最小の機能的な結合単位であるドメイン抗体(dAbs)が含まれ、これはヒト抗体の重(VH)または軽(VL)鎖の何れかの可変領域に対応する。ドメイン抗体及びそれらを調製するための例示的な方法は、例えば、米国特許第6,696,245号, WO05/035572, WO04/101790, WO04/081026, WO04/058821, WO04/003019およびWO03/002609に記載される。
【0082】
多重特異性の抗体または抗体断片は、1、2(二重特異性)、又は3つの特異性を含む任意の適切な数の特異性を有することが可能である。例えば、二重特異性抗体は、ハイブリドーマの融合またはFab’断片の連結〔例えば、Songsivilai & Lachmann Clin. Exp. Immunol. 79: 315-321 (1990)およびKostelny et al. J. Immunol. 148:1547-1553 (1992)を参照されたい〕を含む様々な既知の方法によって産生される。また、三重特異性の抗体を調製する方法は、当該技術において既知である〔例えば、Tutt et al. J. Immunol. 147: 60 (1991)を参照されたい〕。
【0083】
別の態様において、本発明の抗体を産生するハイブリドーマのDNAは、例えば、相同な非ヒト配列に代えて、ヒト重鎖及び軽鎖定常ドメインに対するコード配列を置換することによって〔例えば、Morrison et al., Proc. Natl. Acad. Sci. U.S.A., 81, pp. 6851 (1984)〕、又は、非免疫グロブリンポリペプチドに対するコード配列の全部又は一部を、免疫グロブリンコード配列に共有結合することによって、発現ベクター中に挿入する前に修飾することができる。このように、元の抗体の結合特異性を有する「キメラ(chimeric)」又は「ハイブリッド(hybrid)」抗体が調製される。典型的には、このような非免疫グロブリンポリペプチドは、本発明の抗体の定常ドメインの代用とされる。
【0084】
好ましい実施形態において、本発明の抗体はモノクローナル抗体である。本発明の抗体の「ヒト化された」形態は、特異的なキメラ免疫グロブリン、免疫グロブリン鎖、又は、マウス免疫グロブリンに由来する最小配列を含有する、それらの断片〔Fv、Fab、Fab'、F(ab')2又は抗体の他の抗原結合配列など〕である。通例、ヒト化抗体は、レシピエントの相補性決定領域(CDR)由来の残基が、元の抗体の望ましい特異性、親和性及び能力を維持しつつ、元の抗体(ドナー抗体)のCDR由来の残基によって置換されている、ヒト免疫グロブリン(レシピエント抗体)である。一部の事例では、ヒト免疫グロブリンのFvフレームワーク残基は、対応する非ヒト残基によって置換されることができる。さらに、ヒト化抗体は、レシピエント抗体又は移入されたCDR若しくはフレームワーク配列中の何れにも存在しない残基を含むことができる。これらの修飾は、抗体性能をさらに改良し、最適化するために施される。一般に、ヒト化抗体は、少なくとも一つ、典型的には二つの可変ドメインの実質的に全てを含むであろう。ヒト化抗体中では、元の抗体(original antibody)のCDR領域に対応する全て又は実質的に全てのCDR領域及び全て又は実質的に全てのフレームワーク領域(FR領域)は、ヒト免疫グロブリンコンセンサス配列のものである。ヒト化抗体は、最適には、通例ヒト免疫グロブリンの免疫グロブリン定常領域(Fc)の少なくとも一部も含むであろう。更なる詳細に関しては、文献〔Jones et al., Nature, 321, pp. 522 (1986); Reichmann et al., Nature, 332, pp. 323 (1988);およびPresta, Curr. Op. Struct. Biol., 2, pp. 593 (1992)〕を参照のこと。
【0085】
本発明に使用する抗体をヒト化する方法は、当該技術において周知である。一般的に、本発明のヒト化抗体は、元の抗体からヒト化抗体中に導入されたアミノ酸残基を一又は複数有する。これらのマウス又は他の非ヒトアミノ酸残基は、しばしば、「移入(import)」残基と称され、通例、「移入」可変ドメインから採取される。ヒト化は、本質的に、Winter及び共同研究者の方法に従って、実施することができる〔Jones et al., Nature, 321, pp. 522 (1986); Riechmann et al., Nature, 332, pp. 323 (1988); Verhoeyen et al., Science, 239, pp. 1534 (1988)〕。従って、このような「ヒト化」抗体は、元の抗体の対応する配列によって、無傷のヒト可変ドメインが僅かに置換されているキメラ抗体である(Cabillyらに対する米国特許第4,816,567号)。実際、本発明のヒト化抗体は、典型的には、幾つかのCDR残基と、おそらくは幾つかのFR残基とが、元の抗体中の類似の部位から得られる残基によって置換されているヒト抗体である。
【0086】
ヒト化抗体の作出に使用すべきヒト可変ドメインの選択(軽鎖及び重鎖ドメインの両方)は、抗原性を減少させるのに極めて重要である。いわゆる「ベストフィット(best-fit)」法にしたがって、本発明の抗体の可変ドメインの配列は、公知のヒト可変ドメイン配列の全ライブラリーに対してスクリーニングされる。次いで、マウスの配列に最も近いヒト配列が、ヒト化抗体に対するヒトフレームワーク(FR)として受容される〔Sims et al., J. Immunol., 151, pp. 2296 (1993); Chothia and Lesk, J. Mol. Biol., 196, pp. 901 (1987)〕。別の方法は、軽鎖又は重鎖の特定のサブグループの全てのヒト抗体のコンセンサス配列から得られるフレームワークを使用する。同じフレームワークを、数個の異なるヒト化抗体に対して使用することができる〔Carter et al., Proc. Natl. Acad. Sci. U.S.A., 89, pp. 4285 (1992); Presta et al., J. Immunol., 51, pp. 1993)〕。
【0087】
別の方法は、免疫化に使用されるマウスとして、XenoMouse(登録商標)(Abgenix, Fremont, CA)を用いて「ヒト化」モノクローナル抗体を作出することである。XenoMouseは、機能的ヒト免疫グロブリン遺伝子によって、マウスの免疫グロブリン遺伝子が置換されているマウス宿主である。このため、本マウスによって産生される抗体、又は本マウスのB細胞から作製されるハイブリドーマ中で産生される抗体は、すでにヒト化されている。XenoMouseは、米国特許第6,162,963号に記載されている。HuMAb−MouseTM(Medarex)を用いて、類似の方法を行うことが可能である。
【0088】
ヒト抗体は、例えば、ヒト抗体レパートリーを発現するように操作された他のトランスジェニック動物を免疫化のために使用することによる〔Jakobovitz et al., Nature 362 (1993) 255〕、又はファージディスプレイ法を使用して抗体レパートリーを選択することによるなどの、様々な他の技術に従って作製することもできる。このような技術は当業者に公知であり、本願に開示されているモノクローナル抗体から出発して、実施することができる。
【0089】
本発明の抗体は、「キメラ」抗体(免疫グロブリン)に誘導化することも可能であり(キメラ抗体においては、重鎖及び/又は軽鎖の一部は元の抗体中の対応する配列と同一又は相同であるが、鎖の残りは別の種に由来し、又は別の抗体クラス若しくはサブクラスに属する抗体中の対応する配列と同一若しくは相同である)、所望の生物活性を発揮する限りにおいて、係る抗体の断片に誘導化することも可能である〔Cabilly et al., supra; Morrison et al., Proc. Natl. Acad. Sci. U.S.A., 81, pp. 6851 (1984)〕。
【0090】
以下の例は、さらに本発明の特定の側面を説明するが、本発明を限定しない。
【0091】
[例]
例1
抗体の軽および重鎖の分離およびリフォールディング(再生)
ヒト組織因子(TF)を認識するヒト抗体TF36F40およびヒト成長ホルモン(hGH)を認識するマウス抗体hGH3を、以下のとおり使用した。各抗体を、6M塩化グアニジウム(guanidinium chloride)および50mMリン酸ナトリウムpH8.4を用いて変性させた。ジスルフィド結合を、ジチオスレイトール(DTT)を終濃度100mMまで添加することによって還元し、SDS-PAGEおよびウエスタンブロッティングを還元または非還元の両方の条件下でサンプルを分析する前に室温で30分間インキュベートした。ペルオキシダーゼ結合型ヤギ-抗-ヒトIgG重鎖およびヤギ-抗-ヒトカッパー軽鎖を、それぞれTF36F40重鎖(HC)および軽鎖(LC)を検出するために適用した。ペルオキシダーゼ結合型ヤギ-抗-マウスIgG重鎖およびヤギ-抗-マウスカッパー軽鎖を、それぞれhGH3の重および軽鎖を検出するために適用した。
【0092】
各抗体の軽および重鎖を、Superose12カラム(Amersham Bioscience)を用いたサイズ排除クロマトグラフィー(SEC)で引続いて分離した。洗浄および平衡化の条件は、製造者により提供されている条件をもちいた。タンパク質を、6 M塩化グアニジウム, 50mMリン酸ナトリウムpH 8.4および50mM DTTを用いて、前記カラムから溶出した。フラクションを、SDS-PAGEおよびウエスタンブロッティングで上記のとおり分析した(図2および3)。
【0093】
hGH3抗体をリフォールディングさせるために、軽および重鎖のフラクションを等モルの比率で混合し、100mM Tris HCl(pH7.5)および10mM EDTAに対して1時間、37゜Cで透析した。リフォールディングさせた抗体を無菌0.2μmフィルターを通す前に、全長IgGをリン酸緩衝生理食塩水(PBS)の存在下でサイズ排除クロマトグラフィーで単離した。サンプルを、使用するまで4゜Cで保存した。
【0094】
代替的な実験において、hGH3抗体をリフォールディングさせるために、軽および重鎖フラクションを等モルの比率で混合し、以下の緩衝液に対して透析した:
1) 4Mグアニジウム塩酸, 50mM Na リン酸, 5 mM DTT, pH8.4で90min、室温;
2) 2Mグアニジウム塩酸, 50mM Na リン酸, 5 mM DTT, pH8.4で90min、室温;
3) 1Mグアニジウム塩酸, 50mM Na リン酸, 5 mM DTT, pH8.4で90min、室温;
4) 0.5Mグアニジウム塩酸, 50mM Na リン酸, 5 mM DTT, pH8.4で90min、室温;および
5) 50mM Na リン酸塩, 5mM DTT, pH8.4で72hrs, 室温で処理して、DTTおよびジスルフィド架橋の酸化を認容させる。
【0095】
最終的な容量は、2mlであった。500ulを、14kDaのカットオフ値を有するセントリコン フィルターで限外濾過によって濃縮した。両方の溶液を、SDS−PAGEで引続いて分析した(図4)。
【0096】
例2
ELISA
インビトロで会合させたHuTF36F40およびhGH3抗体は、標準的な手順を用いたELISAでアッセイされた。簡単に説明すると、ELISAマイクロタイタープレート(Maxisorp, Nunc, Denmark)を、1μg/mlのヒトhGHまたはヒトTF(TF1−119)を1xPBS中に含有するもので一晩、4゜Cでコートした。インビトロで会合させたhGH3またはHuTF36F40抗体サンプルが添加され、hGHまたはヒトTFへの結合が、それぞれペルオキシダーゼ結合型ヤギ-抗-ヒトまたはマウスIgG(Sigma)を用いて検出された。
【0097】
例3
表面プラズモン共鳴
Biacore3000光学的バイオセンサーを用いて、hGHまたはヒトTFに対するインビトロで会合させた抗体の親和性が評価された。親和性を決定するために、約10000RUの抗原はセンサー表面にEDC/NHSカップリング化学で固定化された。その後、抗体は、約5μl/minの流速で約3minフローセル(flow cell)へと注入され、その各自の抗原(hGHまたはヒトTF)と会合された(hGHまたはヒトTF)。会合フェーズに続き、表面は、ランニング緩衝剤(HBS-EP, pH 7.4, 0.005%の洗浄剤P20を含有している)で5μl/minの流速で2min洗浄された。センサーグラムのデータは、Bia評価ソフトウェア3.0を用いて分析された。
【0098】
例4
トータルRNAのハイブリドーマ細胞からの単離
組織因子に対する抗体を分泌している、4x106のハイブリドーマ細胞(TF−36F40)が、QiagenのRNeasy Mini Kitを用いたトータルRNAの単離に使用された。前記細胞は、5min、1000rpmで沈殿され、10μl/mlのベータ-メルカプトエタノールを含有している350μlのRLT緩衝剤の添加によって破壊された。そのライセートは、QiagenのQIAshredderカラムに移され、最高速度で2min遠心分離された。フロースルーは、1容量の70%エタノールと混合された。RNeasyスピンカラムごとに、700μlまでサンプルが適用され、14000rpmで遠心分離され、フロースルーが廃棄された。700μlのRW1緩衝剤が、カラムごとに適用され、カラムを洗浄するために15s間、14000rpmで遠心された。前記カラムは、500μlのRPE緩衝剤で2回洗浄され、14000rpmで15s遠心された。前記カラムを乾燥させるため、付加的に2min、14000rpmで遠心された。前記カラムは、新しい収集チュウブに移される。RNAは50μlのヌクレアーゼ フリーの水で溶出され、そして1min、14000rpmで遠心された。RNA濃度が、OD=260nmでの吸光度によって測定された。RNAは、必要とされるまで-80゜Cで保存された。
【0099】
例5
cDNA合成
1μgのRNAを、SMART RACE cDNA Amplification Kit(Clontech)を用いた第1鎖のcDNA合成に使用した。5'-RACE-Ready cDNAの調製に関して、単離されたRNAを含有している反応混合物〔上記のとおり、バックプライマー5'-CDSプライマーバック(backprimer 5'-CDS primer back)、およびSMART II Aオリゴ〕を、調製し、72゜Cで約2minインキュベートし、次に氷上で約2min冷却し、1xFirst-Strand緩衝剤、DTT(20mM)、dNTP(10mM)およびPowerScript逆転写酵素を添加した。反応混合物を、42゜Cで1.5時間インキュベートし、トリシン−EDTA緩衝剤を添加し、72゜Cで7minインキュベーションした。サンプルは-20゜Cで保存することができる。
【0100】
例6
ヒト軽鎖(LC)およびヒト重鎖(HC)の増幅およびクローニング
1xAdvantage HF 2 PCR緩衝剤、dNTP(10mM)および1xAdvantage HF 2ポリメラーゼ混合物を含有しているPCR(ポリメラーゼ連鎖反応)の反応混合物は、上記のとおり作出されたcDNAからのVLおよびVH双方の可変領域の別々の増幅に関して用意される。
【0101】
LCの増幅に関して、以下のプライマーが使用される:
UPM(ユニバーサル プライマー 混合物):
5'-CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT-3'(配列番号1)
5'-CTAATACGACTCACTATAGGG-3'(配列番号2)
P1:
5'-CGCGGCTAGCACACTCTCCCCTGTTGAAGCTC-3'(配列番号3)
HCの増幅に関して、以下のプライマーが使用される:
UPM(ユニバーサル プライマー 混合物):
5'-CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT-3'(配列番号1)
5'-CTAATACGACTCACTATAGGG-3'(配列番号2)
P2:
5'-TCATTTACCCGGGGACAGGGAGA-3'(配列番号4)
PCRは、3ラウンド実施された。ラウンド1: PCRは、94゜Cで5sおよび72゜Cで3minを5サイクル実施された。ラウンド2: PCRは、94゜Cで5s、70゜Cで10s、および72゜Cで1minを5サイクル実施された。ラウンド3: PCRは、94゜Cで5s、68゜Cで10s、および72゜Cで1minを28サイクル実施された。
【0102】
前記PCR産物が1%アガロースゲル上での電気泳動法によって分析され、DNAがQIAEX11アガロースゲル抽出キット(Qiagenより)を用いてゲルから精製された。
【0103】
精製されたPCR産物は、TOPO TAクローニングキット(インビトロゲンより)を用いてPCR4-TOPOへと導入され、TOP10コンピテント細胞の形質転換のために使用された。
【0104】
コロニーの適切な量は、Taqポリメラーゼ、1xTaqポリメラーゼ緩衝剤、dNTP(10mM)および以下のプライマーおよびPCRプログラムを用いて、コロニーPCRによって分析された:
M13フォワード:
5'-GTAAAACGACGGCCAG-3'(配列番号5)
M13リバース:
5'-CAGGAAACAGCTATGAC-3'(配列番号6)
PCRプログラム:
25サイクルを、94゜Cで30s、55゜Cで30s、および72゜Cで1min行った。
【0105】
各々、LCおよびHCの挿入物を具備しているクローンからのプラスミドDNAが、抽出され、上記のプライマーM13フォワードおよびM13リバースを用いて配列決定された。TF−36F40抗体遺伝子の生じた配列は、それぞれ配列番号7および8としてリストされる。また、コード化された軽鎖および重鎖配列が、それぞれ配列番号9および10に記載される。
【0106】
例7
抗体遺伝子の哺乳類発現ベクターへの導入
ヒトTF−36F40のそれぞれLCおよびHC配列を開示している配列データ(配列番号7および8)に基づいて、プライマーがそれぞれLCおよびHC遺伝子の増幅に関して設計された。抗体領域をPCRでフォーマットして、コザック配列、リーダー配列およびユニークな制限酵素部位を含ませた。この事項は、5'PCRプライマーを、HindIII部位、コザック配列を導入し、可変軽鎖領域のリーダー配列の5'端に相同性であるように、デザインすることによって達成された。3'プライマーは、それぞれ、定常領域の軽鎖および重鎖領域のC末端の3'末端と相同性であり、3'端にXbaI部位が導入された。前記HC領域は類似の様式で作製された。但し、NotIおよびBamHI部位が、各々5'および3'端にHindIIIおよびXbaIの代わりに導入された。DNA断片は、それぞれHindIII+XbaIおよびNotI+BamHIで消化され、そしてアンピシリン抵抗性をコード化しているベータラクタマーゼ遺伝子および大腸菌複製開始点(pUC)を含んでいる市販の真核生物性発現ベクター〔InvitrogenのpCDNA3(+)〕へと結合された。結合されたプラスミドを使用して、大腸菌が形質転換された。プラスミドDNAが、これらのアンピシリン耐性の細菌集団から調製され、哺乳類細胞へのトランスフェクションに使用された。
【0107】
例8
哺乳類細胞のトランスフェクション
上記で記載したクローン化したDNAsは、製造者の推奨する方法にしたがいLipofectamine TM 2000(Cat. No.11668-019, Invitrogen)を用いて、哺乳類細胞へと導入された。
【0108】
例9
哺乳類細胞での抗体LCおよびHCの発現
細胞を、製造者のプロトコール(Cat.No.11668-019, Invitrogen)に記載のとおり成長させる。細胞は、上清の収穫および続く精製およびエクソビボでの会合の前に、6日成長させられる。
【0109】
LCおよびHCを発現している細胞の共培養は、それぞれ次の様式で実施された、即ち:HEK293細胞が、それぞれLCおよびHCベクター構築物でトランスフェクションされ、2つの培養物を混合する前に16時間37゜Cで成長させた。6日後、混合した培養物からの上清が、例2および3に記載のとおりに分析された。抗体のエクスビボでの会合を至適化するために、宿主細胞培養の宿主細胞、発現ベクター、培養培地および/または異なる生物物理学的な条件(例えば、温度、レドックス条件、pH)などのパラメータを変動させた。LCおよびHCの濃度およびLC:HC比は、これらのパラメータを変化させることによって至適化することができる。1実験において、次の条件を使用した、即ち:HEK293細胞を、抗体LCおよびHCを発現するpCDNA3(+)に準拠したベクター構築物でトランスフェクションし、25μg/mlジェネテシン(GIBCO cat. No. 10131-019)を含有しているOptimum1培地(GIBCO cat. no. 51985-026)を添加したGlutamax-I中で、37゜C で200rpmで成長させた。
【0110】
例10
グラム陽性細菌中での抗体LCおよびHCの形質転換および発現
ヒトTF-36F40のそれぞれLCおよびHC配列を開示している配列データ(配列番号7および8)に基づいて、プライマーはそれぞれLCおよびHC遺伝子の増幅に関して設計された。抗体領域は、PCRで整えられて、HCに関してNotI/BamHI部位およびLCに関してHindIII/XbaI部位を含むが、天然のシグナルペプチドは含まない。
【0111】
HCおよびLCのDNA断片が、消化され、グラム陽性菌(Streptococcus gordonii)SPEX発現ベクター(Myscofski et al 2000 Protein Expression Purif)の修正バージョンへと別々に連結された(このベクター中で、発現はP2プロモーターで駆動される)。
【0112】
上記のクローン化したDNAは、グラム陽性細菌へと、以前に記載されたとおり導入された(Myscofski et al 2000 Protein Expression Purif.)。細胞は、上清の収穫および引続く精製およびエクスビボ会合の前に、ブレイン―ハートインフュージョンブロス(BHI, Difco)中で、一晩、37゜Cで成長された。
【0113】
LCおよびHCを発現している細胞の共培養は、上記と同じ様式で実施された。
【0114】
例11
抗体断片の精製
LCは上清からプロテインLクロマトグラフィーを用いて精製され、HCは上清からプロテインAで精製された(当該技術において、既知の処理にしたがって)。或いは、LCは、マトリックスにEDC/NHSカップリング化学を用いて固定化された、VLCLに対する抗体を用いて上清から精製された;および/または、HCは、VHCH1-3に対する抗体を用いて上清から精製された。
【0115】
例12
抗体断片のインビトロ会合
TF36F40のLCおよびHCの、完全長TF抗体へのインビトロ会合は、例1に記載のとおり実施された。前記抗体は、BCAタンパク質アッセイ(Pierce, Rockford, IL)を用いて定量され、例2および3に記載のとおりアッセイされた。
【0116】
例13
Obelix陽イオン交換体による精製
Obelix陽イオン交換体は、Amersham(Cat. No.11-0010)から市販されている。
【0117】
使用するためには、濾過した細胞培養のpHは、約3.0〜9.0(好ましくは、約pH7.0)の範囲内である。前記サンプルは、約5〜40(好ましくは、約10)mg/ml 充填カラム材料の範囲でカラムに適用される。
【0118】
非結合性の物質は、緩衝液A(緩衝液Aは、好適な範囲のpHを有し、所望の抗体の結合が許容されるために、十分に低い伝導率を有する緩衝液と規定される)を1〜5のCV(カラム容量)で適用することによってカラムから洗浄除去された。
【0119】
引き続いて、前記抗体は、伝導率を増加させること、それにより静電反発力を増加させることを介してカラムから溶出された。これは低から高い塩濃度での段階的グラジエントまたは連続的グラジエントによって達成することができる。好ましくは、NaClが塩として選択されるが、他のタイプの塩も使用することができる。
【0120】
プロパンジオールを、溶出緩衝液に添加することができる。この態様において、陽イオン交換体を、疎水性相互作用クロマトグラフィーカラムとして使用することができる。高い塩析効果を有する塩(例えば、酢酸ナトリウム)を添加することによって、陽イオン交換体を疎水性相互作用クロマトグラフィーカラムに変化させることができる。従って、パラメータ(例えば、塩、pH、伝導率、および疎水性)を変化させることによって、前記カラムを同じ操作の間に2様式以上で機能させることができる。前記クロマトグラフィー手順は、典型的には4〜25゜Cで実施される。
【0121】
1実験において、次の条件を使用した:抗体(Ab)の適用物を、培養物中のpHをpH5,5に調整し、0,2μmフィルターで濾過することによって調製された。EDTAおよびベンザミジン-HClを、1mMまで添加した。
【0122】
カラム材料(column material)を、30mMクエン酸緩衝液pH6,0で平衡化した。そして、適用物を、室温で20cv/h、約5mg Ab/ml カラム材料の条件で適用した。
【0123】
適用後に、カラムを、平衡化緩衝液で洗浄した。30mMクエン酸、25mM NaCl、30%グリセロールpH6,0での洗浄を、平衡化緩衝液での洗浄に続いて実施した。
【0124】
溶出は、異なる様式で実施することができる。1実験において、溶出を、Tris-緩衝液pH7,5〜8,5で実施した(図5および6)。別の実験において、0〜1M NaClの塩をpH6,5〜7で適用するグラジエント溶出を使用した。
【0125】
Abの溶出後、前記カラムは、1M NaOHで再生された。
【0126】
* * *
公報、特許出願及び特許を含む、本明細書に引用された全ての参考文献は、各参考文献が個別的且つ具体的に参照により援用され、その全体が本明細書に記載されている場合と同じ程度に、参照により本明細書に援用される。
【0127】
本明細書で使用されている全ての見出し及び小見出しは、便宜的に使用されているものにすぎず、いかなる意味においても、本発明を限定するものと解釈してはならない。
【0128】
本明細書に別段の記載がなく、又は明確に反対の記載がなければ、考えられる全ての変形における上記事項のあらゆる組み合わせが、本発明によって包含される。
【0129】
本発明を記載する際に使用される「a」及び「an」及び「the」という用語及び類似の指示記号(referents)は、本明細書に別段の記載がなく、又は明確に反対の記載がなければ、単数及び複数の両方を包含するものと解釈しなければならない。
【0130】
本明細書において値の範囲を引用することは、当該範囲に含まれる各個別の値を個別的に表すための簡略法としての役割を果たすことを意図しているにすぎず、本明細書中に別段の記載がなければ、各個別の値は、本明細書に個別的に記載されている場合と同様に、本明細書中に取り込まれる。別段の記載がなければ、本明細書に記載されている全ての値は、対応する近似値(approximate values)を代表している(例えば、具体的な因子又は測定に関して記載されている全ての例示的な値は、適宜、「約」と修飾される、対応する近似的な測定値も表すと考えることができる)。
【0131】
本明細書に別段の記載がなく、又は明確に反対の記載がなければ、本明細書に記載されている全ての方法は、任意の適切な順序で実施することができる。
【0132】
本明細書中で記載される、任意の及び全ての例又は例示的な用語(「例えば」など)は、単に本発明をよりよく説明することを意図しており、他で指摘しない限り本発明の範囲を限定しない。明示の記載がなければ、本明細書中の言語(language)は、何らかの事項が本発明の実施に不可欠であることを示唆するものと解釈すべきでない。
【0133】
本明細書での特許文書(patent documents)の引用(citation)および援用(incorporation)は、便宜上の目的でのみなされ、係る特許文書の有効性(validity)、特許性(patentability)、および/または強制力(enforceability)の如何なる見解(view)をも反映するものではない。
【0134】
本明細書に別段の記載がなく、又は明確に反対の記載がなければ、ある事項に関して「含む(comprising)」、「有する(having)」、「含む(including)」又は「含有する(containing)」などの用語を用いて、本発明の任意の側面又は態様を本明細書で記載することは、当該事項「からなる(consists of)」、「〜から本質的になる(consists essentially of)」、「実質的に〜を含む(substantially comprises)」本発明の類似の側面又は態様に対する支持を与えることを意図するものである(例えば、特定の事項を含むと本明細書に記載されている組成物は、本明細書に別段の記載がなく、又は明確に反対の記載がなければ、その事項からなる組成物も記載するものとして理解すべきである)。
【0135】
本発明は、本明細書に提示されている前記側面又は特許請求の範囲に記載されている主題のあらゆる修飾及び均等物を、適用可能な法律によって許容される最大限度まで含む。
【図面の簡単な説明】
【0136】
【図1】図1は、抗体の重および軽鎖の発現と、引続く完全長の機能的な抗体へのエクスビボ会合とを示す図である。
【図2】図2は、例1に記載のような変性および還元後の、hGH3抗体のHPLCクロマトグラフィーを示す図である。
【図3】図3は、変性および還元したhGH3抗体(例1に記載のような)のSDS−PAGEおよびクーマシー染色を示す〔HPLC精製前(レーン2)およびHPLCフラクション5(レーン3), フラクション6(レーン4), フラクション7(レーン5), フラクション8(レーン6)〕。レーン1は、Mark12マーカーを示す。
【図4】図4は、Mark12マーカー(レーン1)およびhGH3抗体の変性、リフォールディング、およびサイズ-排除クロマトグラフィー後のサンプル(レーン2)のSDS-PAGE分析を示す。
【図5】図5は、Obelix陽イオン交換体によるIgG精製のクロマトグラムを示す。
【図6】図6は、Obelix陽イオン交換体でのIgGの精製から取得されたフラクションの非還元SDS-PAGE分析を示す。レーン1:Mark12マーカー;レーン2:IgG標準;レーン3:適用物;レーン4:通過物(Run through);レーン5:フラクションC7;レーン6:フラクションD7;レーン7:フラクションE1;レーン8:フラクションE2, レーン9:フラクションE3;レーン 10:フラクションE4。
【特許請求の範囲】
【請求項1】
免疫グロブリン分子または免疫学的に機能的なその断片を産生する方法であって:
(a)第1の宿主細胞を、少なくとも免疫グロブリン重鎖の可変ドメインを具備している第1のポリペプチドをコード化しているヌクレオチド配列を具備している第1の核酸で形質転換すること;
(b)第2の宿主細胞を、少なくとも免疫グロブリン軽鎖の可変ドメインを具備している第2のポリペプチドをコード化しているヌクレオチド配列を具備している第2の核酸で形質転換すること;
(c)第1および第2の核酸配列を発現させること;
(d)第1および第2のポリペプチドを精製すること;および
(e)第1および第2のポリペプチドがリフォールディングして、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を形成することを認容すること;
を備え、
第1および第2の宿主細胞は、別々に真核生物細胞およびグラム陽性菌からなる群から選択される方法。
【請求項2】
請求項1に記載の方法であって、前記第1の宿主細胞は免疫グロブリン軽鎖をコード化している核酸を発現しなく、第2の宿主細胞は免疫グロブリン重鎖をコード化している核酸を発現しない方法。
【請求項3】
請求項1〜2の何れか1項に記載の方法であって、前記免疫グロブリン分子は、IgA、IgD、IgE、IgG、およびIgM免疫グロブリンからなる群から選択される方法。
【請求項4】
請求項1〜3の何れか1項に記載の方法であって、前記免疫学的に機能的な断片は、Fab断片、Fab'断片、Fab'-SH断片、F(ab')2断片、Fv断片、VHH断片、ドメイン抗体、ディアボディー(diabody)、および多重特異性の抗体または抗体断片からなる群から選択される方法。
【請求項5】
請求項1〜4の何れか1項に記載の方法であって、前記第1および第2の宿主細胞は、同じ培養で成長させられる方法。
【請求項6】
請求項1〜4の何れか1項に記載の方法であって、前記第1および第2の宿主細胞は、別々の培養で成長させられる方法。
【請求項7】
請求項1〜6の何れか1項に記載の方法であって、前記精製することが、Obelix陽イオン交換カラムを用いる精製を備える方法。
【請求項8】
請求項1〜7の何れか1項に記載の方法であって、前記真核生物細胞は、哺乳類細胞、昆虫細胞、植物細胞、および真菌細胞からなる群から選択される方法。
【請求項9】
請求項1〜8の何れか1項に記載の方法であって、前記第1および第2の宿主細胞は、COS細胞、BHK細胞、HEK293細胞、DUKX細胞、Saccharomyces spp細胞、Kluyveromyces spp細胞、Aspergillus spp細胞、Neurospora spp細胞、Fusarium spp細胞、Trichoderma spp細胞、およびLepidoptera spp細胞からなる群から別々に選択される方法。
【請求項10】
請求項1〜9の何れか1項に記載の方法であって、前記第1および第2の宿主細胞は、同じ細胞タイプの細胞である方法。
【請求項11】
請求項1〜9の何れか1項に記載の方法であって、前記第1および第2の宿主細胞は、異なる細胞タイプの細胞である方法。
【請求項12】
請求項1〜11の何れか1項に記載の方法であって、前記第1および第2の核酸は、1以上のモノクローナル抗体産生細胞から由来する方法。
【請求項13】
請求項12に記載の方法であって、前記モノクローナル抗体産生細胞は、ハイブリドーマ、ポリドーマ(polydoma)および不死化させたB細胞からなる群から選択される方法。
【請求項14】
請求項1〜13の何れか1項に記載の方法であって、工程(e)は、前記第1および第2のポリペプチドを以下から選択される条件下で混合することを備える方法:
(a)第1および第2のポリペプチドの比は約1:1、温度は約室温、およびpHは約7;および
(b)第1および第2のポリペプチドの比は約1:1、温度は約5゜C、およびpHは約8.0〜8.5の範囲。
【請求項15】
請求項14に記載の方法であって、前記第1および第2のポリペプチドは、約0.5MのL-アルギニン-HCl、約0.9mMの酸化型グルタチオン(GSSG)、および約2mMのEDTAを含んでいる溶液中で混合される方法。
【請求項16】
免疫グロブリン分子または免疫学的に機能的なその断片を産生する方法であって:
(a)第1の宿主細胞を、少なくとも免疫グロブリン重鎖の可変ドメインを具備している第1のポリペプチドをコード化しているヌクレオチド配列を具備している第1の核酸で形質転換すること;
(b)第2の宿主細胞を、少なくとも免疫グロブリン軽鎖の可変ドメインを具備している第2のポリペプチドをコード化しているヌクレオチド配列を具備している第2の核酸で形質転換すること;
(c)第1および第2の核酸配列を発現させること;
(d)第1および第2のポリペプチドの混合物を含んでいる溶液を透析すること;および
(e)第1および第2のポリペプチドがリフォールディングして、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を形成することを認容すること;
を備え、
第1および第2の宿主細胞は、別々に真核生物細胞およびグラム陽性菌からなる群から選択される方法。
【請求項17】
請求項16に記載の方法であって、前記第1および第2の宿主細胞は同じ培養で成長させられ、その溶液は前記第1および第2の宿主細胞が成長させられた培養培地である方法。
【請求項18】
請求項16および17の何れか1項に記載の方法であって、前記溶液中の第1および第2のポリペプチドの相対量は、約1:2〜約2:1の範囲である方法。
【請求項19】
抗体を精製する方法であって、抗体を含んでいる溶液をObelix陽イオン交換カラムに適用すること、および精製された抗体を溶出することを備える方法。
【請求項20】
請求項19に記載の方法であって、以下の工程のうち少なくとも1つを備える方法:
(a)濾過した細胞培養を前記カラムに適用すること(ここで濾過した細胞培養は任意でpHが調製される);
(b)溶媒を溶出緩衝液に添加すること;および
(c)抗体を塩グラジエントを増加することによって溶出させること。
【請求項21】
請求項16に記載の方法であって、工程(c)は工程(b)の前に実施される方法。
【請求項22】
免疫グロブリン分子または免疫グロブリン重および軽鎖の少なくとも可変性ドメインを含んでいる免疫学的に機能的な免疫グロブリン断片を産生するための方法であって、以下の工程を備える方法:
(a)重および軽鎖を、真核生物細胞およびグラム陽性細菌からなる群から選択される2つの別々の宿主細胞で独立に産生すること;
(b)重および軽鎖を精製すること;および
(c)免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片をインビトロでリフォールディングさせること。
【請求項1】
免疫グロブリン分子または免疫学的に機能的なその断片を産生する方法であって:
(a)第1の宿主細胞を、少なくとも免疫グロブリン重鎖の可変ドメインを具備している第1のポリペプチドをコード化しているヌクレオチド配列を具備している第1の核酸で形質転換すること;
(b)第2の宿主細胞を、少なくとも免疫グロブリン軽鎖の可変ドメインを具備している第2のポリペプチドをコード化しているヌクレオチド配列を具備している第2の核酸で形質転換すること;
(c)第1および第2の核酸配列を発現させること;
(d)第1および第2のポリペプチドを精製すること;および
(e)第1および第2のポリペプチドがリフォールディングして、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を形成することを認容すること;
を備え、
第1および第2の宿主細胞は、別々に真核生物細胞およびグラム陽性菌からなる群から選択される方法。
【請求項2】
請求項1に記載の方法であって、前記第1の宿主細胞は免疫グロブリン軽鎖をコード化している核酸を発現しなく、第2の宿主細胞は免疫グロブリン重鎖をコード化している核酸を発現しない方法。
【請求項3】
請求項1〜2の何れか1項に記載の方法であって、前記免疫グロブリン分子は、IgA、IgD、IgE、IgG、およびIgM免疫グロブリンからなる群から選択される方法。
【請求項4】
請求項1〜3の何れか1項に記載の方法であって、前記免疫学的に機能的な断片は、Fab断片、Fab'断片、Fab'-SH断片、F(ab')2断片、Fv断片、VHH断片、ドメイン抗体、ディアボディー(diabody)、および多重特異性の抗体または抗体断片からなる群から選択される方法。
【請求項5】
請求項1〜4の何れか1項に記載の方法であって、前記第1および第2の宿主細胞は、同じ培養で成長させられる方法。
【請求項6】
請求項1〜4の何れか1項に記載の方法であって、前記第1および第2の宿主細胞は、別々の培養で成長させられる方法。
【請求項7】
請求項1〜6の何れか1項に記載の方法であって、前記精製することが、Obelix陽イオン交換カラムを用いる精製を備える方法。
【請求項8】
請求項1〜7の何れか1項に記載の方法であって、前記真核生物細胞は、哺乳類細胞、昆虫細胞、植物細胞、および真菌細胞からなる群から選択される方法。
【請求項9】
請求項1〜8の何れか1項に記載の方法であって、前記第1および第2の宿主細胞は、COS細胞、BHK細胞、HEK293細胞、DUKX細胞、Saccharomyces spp細胞、Kluyveromyces spp細胞、Aspergillus spp細胞、Neurospora spp細胞、Fusarium spp細胞、Trichoderma spp細胞、およびLepidoptera spp細胞からなる群から別々に選択される方法。
【請求項10】
請求項1〜9の何れか1項に記載の方法であって、前記第1および第2の宿主細胞は、同じ細胞タイプの細胞である方法。
【請求項11】
請求項1〜9の何れか1項に記載の方法であって、前記第1および第2の宿主細胞は、異なる細胞タイプの細胞である方法。
【請求項12】
請求項1〜11の何れか1項に記載の方法であって、前記第1および第2の核酸は、1以上のモノクローナル抗体産生細胞から由来する方法。
【請求項13】
請求項12に記載の方法であって、前記モノクローナル抗体産生細胞は、ハイブリドーマ、ポリドーマ(polydoma)および不死化させたB細胞からなる群から選択される方法。
【請求項14】
請求項1〜13の何れか1項に記載の方法であって、工程(e)は、前記第1および第2のポリペプチドを以下から選択される条件下で混合することを備える方法:
(a)第1および第2のポリペプチドの比は約1:1、温度は約室温、およびpHは約7;および
(b)第1および第2のポリペプチドの比は約1:1、温度は約5゜C、およびpHは約8.0〜8.5の範囲。
【請求項15】
請求項14に記載の方法であって、前記第1および第2のポリペプチドは、約0.5MのL-アルギニン-HCl、約0.9mMの酸化型グルタチオン(GSSG)、および約2mMのEDTAを含んでいる溶液中で混合される方法。
【請求項16】
免疫グロブリン分子または免疫学的に機能的なその断片を産生する方法であって:
(a)第1の宿主細胞を、少なくとも免疫グロブリン重鎖の可変ドメインを具備している第1のポリペプチドをコード化しているヌクレオチド配列を具備している第1の核酸で形質転換すること;
(b)第2の宿主細胞を、少なくとも免疫グロブリン軽鎖の可変ドメインを具備している第2のポリペプチドをコード化しているヌクレオチド配列を具備している第2の核酸で形質転換すること;
(c)第1および第2の核酸配列を発現させること;
(d)第1および第2のポリペプチドの混合物を含んでいる溶液を透析すること;および
(e)第1および第2のポリペプチドがリフォールディングして、免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片を形成することを認容すること;
を備え、
第1および第2の宿主細胞は、別々に真核生物細胞およびグラム陽性菌からなる群から選択される方法。
【請求項17】
請求項16に記載の方法であって、前記第1および第2の宿主細胞は同じ培養で成長させられ、その溶液は前記第1および第2の宿主細胞が成長させられた培養培地である方法。
【請求項18】
請求項16および17の何れか1項に記載の方法であって、前記溶液中の第1および第2のポリペプチドの相対量は、約1:2〜約2:1の範囲である方法。
【請求項19】
抗体を精製する方法であって、抗体を含んでいる溶液をObelix陽イオン交換カラムに適用すること、および精製された抗体を溶出することを備える方法。
【請求項20】
請求項19に記載の方法であって、以下の工程のうち少なくとも1つを備える方法:
(a)濾過した細胞培養を前記カラムに適用すること(ここで濾過した細胞培養は任意でpHが調製される);
(b)溶媒を溶出緩衝液に添加すること;および
(c)抗体を塩グラジエントを増加することによって溶出させること。
【請求項21】
請求項16に記載の方法であって、工程(c)は工程(b)の前に実施される方法。
【請求項22】
免疫グロブリン分子または免疫グロブリン重および軽鎖の少なくとも可変性ドメインを含んでいる免疫学的に機能的な免疫グロブリン断片を産生するための方法であって、以下の工程を備える方法:
(a)重および軽鎖を、真核生物細胞およびグラム陽性細菌からなる群から選択される2つの別々の宿主細胞で独立に産生すること;
(b)重および軽鎖を精製すること;および
(c)免疫グロブリン分子または免疫学的に機能的な免疫グロブリン断片をインビトロでリフォールディングさせること。
【図1】


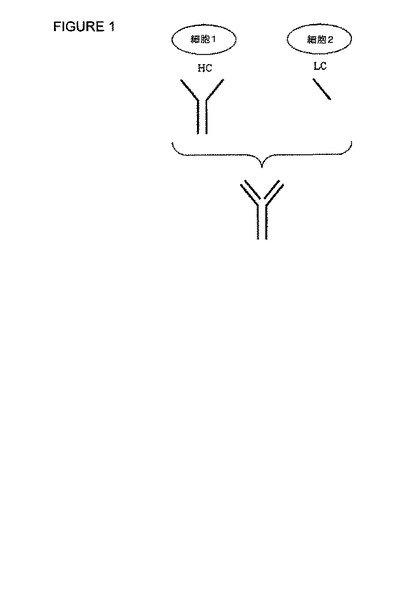
【図2】
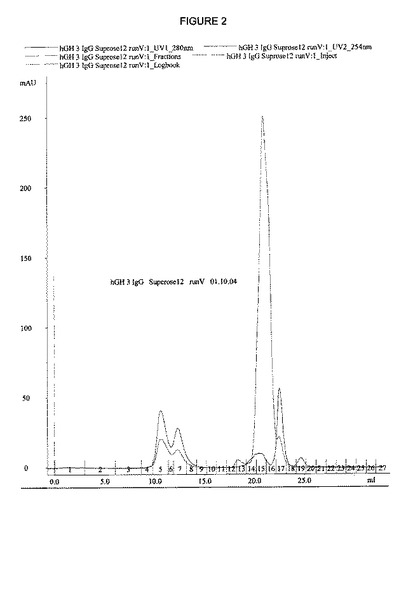
【図3】
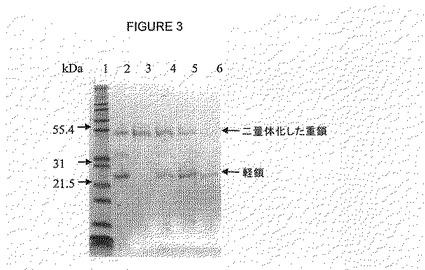
【図4】
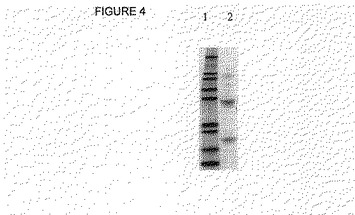
【図5】
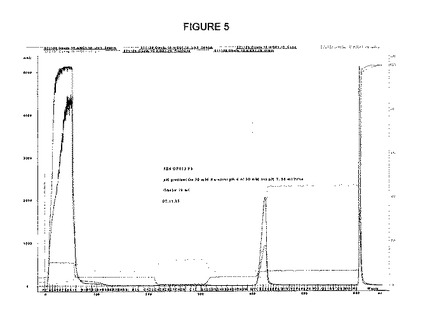
【図6】
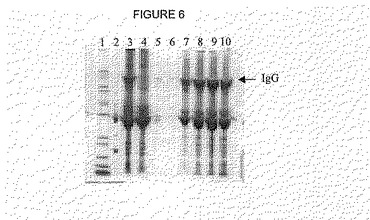
【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2008−521420(P2008−521420A)
【公表日】平成20年6月26日(2008.6.26)
【国際特許分類】
【出願番号】特願2007−543846(P2007−543846)
【出願日】平成17年11月30日(2005.11.30)
【国際出願番号】PCT/EP2005/056351
【国際公開番号】WO2006/058890
【国際公開日】平成18年6月8日(2006.6.8)
【出願人】(391032071)ノボ ノルディスク アクティーゼルスカブ (148)
【氏名又は名称原語表記】NOVO NORDISK AKTIE SELSXAB
【Fターム(参考)】
【公表日】平成20年6月26日(2008.6.26)
【国際特許分類】
【出願日】平成17年11月30日(2005.11.30)
【国際出願番号】PCT/EP2005/056351
【国際公開番号】WO2006/058890
【国際公開日】平成18年6月8日(2006.6.8)
【出願人】(391032071)ノボ ノルディスク アクティーゼルスカブ (148)
【氏名又は名称原語表記】NOVO NORDISK AKTIE SELSXAB
【Fターム(参考)】
[ Back to top ]