
抗体を表面に提示したバキュロウイルス
【課題】抗原に対する特異性および親和性に優れた抗体を表面に提示できるバキュロウイルスを提供すること。
【解決手段】本発明は、抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルスを提供し、このバキュロウイルスは、バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質として発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現し、表面に提示された抗体またはそのFabフラグメントが抗原と結合し得る。
【解決手段】本発明は、抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルスを提供し、このバキュロウイルスは、バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質として発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現し、表面に提示された抗体またはそのFabフラグメントが抗原と結合し得る。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、抗体を表面に提示した組換えバキュロウイルスに関する。
【背景技術】
【0002】
抗体作製の手段として、一般に、ファージディスプレイ法が用いられている。ファージディスプレイ法では、大腸菌を宿主として繊維状ファージを構成するタンパク質の遺伝子に各々連結させた抗体断片の遺伝子群を発現させて、その表面にさまざまな抗体断片が提示されたファージのライブラリーを作製し、そしてその中から特定の抗原に対する特異性または親和性を有する抗体断片が提示されたファージを選択する。抗体断片を発現したファージは、その抗体断片の遺伝子をコードしたDNAを有するので、特定の抗原に結合するファージを回収すれば、目的の抗体の遺伝子を単離することができる。得られた抗体遺伝子を動物細胞などにおいて発現させることにより、抗体タンパク質の生産が可能となる。
【0003】
ファージディスプレイ法は、動物を免疫する必要がないこと、ヒト由来の抗体のファージディスプレイライブラリーを用いればヒト型の抗体を取得できることなどの利点を有する。しかしながら、ファージディスプレイ法は、大腸菌を宿主とするため、高分子量のタンパク質ほど提示効率が悪い。このため、抗体の可変部(VHとVL)をペプチドリンカーで接続した一本鎖抗体(scFv)を提示する場合が多く、プロテアーゼによる分解または抗原との親和性の低下が避けられない。また、固相に固定した抗原に対して特異的に結合するファージを選択および回収するバイオパンニングによる選択効率が悪いなどの欠点を有している。
【0004】
昆虫に感染して病気を起こすウイルスであるバキュロウイルスは、環状の二本鎖DNAを遺伝子としてもつエンベロープウイルスで、鱗翅目、膜翅目および双翅目などの昆虫に感受性を示す。核多角体病ウイルス(NPV)は、このようなバキュロウイルスの一種であり、感染した昆虫細胞の核内に多角体(ポリヒドラ)と呼ばれる封入体を大量に生成する一群のウイルスである。多角体は、分子量31kDaのポリヘドリンタンパク質から構成され、感染後期に大量に生成されて、その中に多数のウイルス粒子を埋め込んでいる。多角体はウイルスが自然界で生存するためには必須であるが、ウイルスの増殖には必要ないので、ポリヘドリン遺伝子の代わりに発現を意図する外来遺伝子を挿入してもウイルスは全く支障なく感染し増殖する。このような性質を利用して、昆虫細胞−バキュロウイルス系は、ポリヘドリンプロモーターの下流に外来遺伝子を挿入した組換えバキュロウイルスを培養昆虫細胞に感染させ、ウイルス感染した細胞に外来タンパク質を生産させる組換えタンパク質発現系として利用されている。
【0005】
バキュロウイルスの1種であるオートグラファ・カリフォルニカ核多角体病ウイルス(Autographa californica nucleopolyhedrovirus, AcNPV)のエンベロープを構成する糖タンパク質gp64をコードする遺伝子の5’末端側に連結させた外来タンパク質の遺伝子を発現させることにより、そのタンパク質は、gp64と融合した形でウイルスの表面に提示されることが報告されている(非特許文献1)。非特許文献1に記載の方法は、昆虫細胞を宿主として用いるため、哺乳動物細胞により発現されるような、複雑な構造を有するタンパク質または翻訳後修飾されたタンパク質を、本来の機能を保持した状態でウイルスの表面に提示し得る。
【特許文献1】特開2006−087427号公報
【非特許文献1】Grabherr, R.ら, Trends Biotechnol., 19, 231-236 (2001)
【非特許文献2】Miyashita, H.ら, J. Mol. Biol., 267, 1247-1257 (1997)
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明は、抗原に対する特異性および親和性に優れた抗体を表面に提示できるバキュロウイルスを提供することを目的とする。
【課題を解決するための手段】
【0007】
本発明は、抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルス(以下、単に「出芽型組換えバキュロウイルス」ともいう)を提供し、このバキュロウイルスは、バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質として発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現し、表面に提示された該抗体またはそのFabフラグメントが抗原と結合し得る。
【0008】
本発明はまた、抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルスを作製する方法を提供し、この方法は、
バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質を発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現する、組換えバキュロウイルスを作製する工程
を含む。
【0009】
1つの実施態様では、上記組換えバキュロウイルスを作製する工程が、上記バキュロウイルスエンベロープ構成糖タンパク質をコードする遺伝子の5’末端側に連結した、該糖タンパク質の分泌シグナル配列および上記抗体またはそのFabフラグメントを構成する一方のペプチド鎖をコードする遺伝子を含み、かつ分泌シグナル配列および該抗体またはそのFabフラグメントの他方のペプチド鎖をコードする遺伝子を含むトランスファーベクターを構築する工程;および
該トランスファーベクターを介して目的遺伝子を昆虫細胞に導入する工程
を含む。
【0010】
別の実施態様では、上記抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルスを作製する方法は、
前記組換えバキュロウィルスを昆虫細胞に感染させる工程;および
前記感染させた昆虫細胞を培養してバキュロウイルスを出芽させる工程
をさらに含む。
【0011】
本発明はまた、抗体またはそのFabフラグメントを表面に提示する出芽型組換えバキュロウイルスディスプレイライブラリーを作製する方法を提供し、この方法は、
異なる抗体またはそのFabフラグメントを発現する少なくとも2種の組換えバキュロウイルスを作製する工程であって、該組換えバキュロウイルスが各々、バキュロウイルスエンベロープ構成糖タンパク質と抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質を発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現する、工程;
該少なくとも2種の組換えバキュロウィルスを昆虫細胞に感染させる工程;および
該感染させた昆虫細胞を培養して、バキュロウイルスを出芽させる工程
を含む。
【0012】
本発明はまた、上記方法により作製されたバキュロウイルスディスプレイライブラリーを提供する。
【0013】
本発明はさらに、抗体またはそのFabフラグメントをスクリーニングする方法を提供し、この方法は、
上記ライブラリーに抗原を結合させる工程;および
該抗原が結合した該ライブラリーの抗体またはそのFabフラグメントを検出する工程
を含む。
【発明の効果】
【0014】
本発明によれば、抗原に対する特異性および親和性に優れた抗体を表面に提示できる出芽型バキュロウイルスが得られる。このバキュロウイルスは、抗体分子のH鎖とL鎖とが本来の形で会合したFabフラグメントを表面に提示できる。
【発明を実施するための最良の形態】
【0015】
本発明の出芽型組換えバキュロウイルスは、バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質として発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現し得る。便宜上、糖タンパク質と連結されるペプチド鎖を鎖Aとし、他方のペプチド鎖を鎖Bという。鎖AがH鎖である場合、鎖BはL鎖であり、その逆であってもよい。
【0016】
本発明において、出芽型組換えバキュロウイルスの表面に提示される抗体は、任意の抗体であり得る。本発明によって出芽型組換えバキュロウイルスの表面に提示される抗体は、その抗原と結合可能な任意の形態をとり得、代表的には、Fabフラグメントの形態となり得る。抗体のH鎖のFdフラグメントとL鎖とがジスルフィド結合したFab形態を構成するように、その抗体のH鎖およびL鎖が選択される。H鎖は、少なくともFab形成に必要な領域を含む一部分、例えば、Fdフラグメントおよびこれを含むフラグメントであり得、または全長であってもよい。Fdフラグメントが好適に用いられ得る。L鎖は、全長、またはH鎖とFabを形成するのに必要な領域を含む任意の長さのフラグメントであり得る。全長のL鎖が好適に用いられ得る。H鎖およびL鎖は、Fabの形成に影響しない限り、変異が導入されていてもよい。
【0017】
バキュロウイルスとしては、オートグラファ・カリフォルニカ核多角体病ウイルス(AcNPV)およびボンビックス・モリ(Bombyx mori)核多角体病ウイルス(BmNPV)が用いられ得る。
【0018】
宿主としては、バキュロウイルスの宿主となる昆虫細胞であれば特に制限なく利用できる。宿主は、トランスファーベクターの移入により組換えバキュロウイルスを生成し得、そして感染によりバキュロウイルスを増殖し得る。AcNPVの宿主としては、ハスモンヨトウの近縁種のスポドプテラ・フルギペルダ(Spodoptera frugiperda)由来のSf細胞などが挙げられる。細胞株Sf9およびSf21などは、スポドプテラ・フルギペルダ幼虫の卵巣組織から確立しており、インビトロジェン株式会社、メルク株式会社、もしくは日本ベクトンディッキンソン株式会社、または寄託機関からも入手可能である。また、イラクサギンウワバ(Trichoplusia ni)由来のBTI-TN-5B1-4細胞(High FiveTM)も利用することができ、本細胞株はインビトロジェン株式会社から入手可能である。BmNPVの宿主としてはBmN4細胞などが挙げられ、日本農産工業株式会社から入手可能である。
【0019】
本発明の出芽型組換えバキュロウイルスの作製は、当業者が通常用いる方法に従って行われ得る。以下、簡単に説明する。
【0020】
発現させるべきペプチド鎖(鎖Aまたは鎖B、あるいはその両方)の遺伝子をトランスファーベクターに挿入して組換えトランスファーベクターを構築する。例えば、抗体産生細胞のmRNAから逆転写酵素により調製されたcDNAライブラリーを鋳型にして、PCRにより、上記で説明したようなFabフラグメントを形成する組合せとなるペプチド鎖をコードする抗体遺伝子の組合せ、例えば、Fdフラグメント遺伝子およびL鎖遺伝子、または、H鎖遺伝子およびL鎖遺伝子をクローニングする。この際、PCRに用いるプライマーには、遺伝子増幅後にベクターに組込めるように制限酵素認識配列のオリゴヌクレオチドを付加しておくことが好ましい。増幅した抗体遺伝子およびベクターを制限酵素で消化した後、抗体遺伝子をベクターに挿入する。鎖Aをコードする遺伝子は、バキュロウイルスエンベロープ構成糖タンパク質(例えば、gp64)をコードする遺伝子の5’末端側に配置され得る。トランスファーベクターの全体の大きさは一般的には数kb〜10kb程度であり、そのうちの約3kbはプラスミド由来の骨格であり、アンピシリンなどの抗生物質耐性遺伝子および細菌のDNA複製起点を含んでいる。トランスファーベクターには、挿入遺伝子を発現させるためのプロモーターを含むことが好ましい。プロモーターとしては、ポリヘドリンプロモーター(PPH)、p10遺伝子のプロモーター(Pp10)、キャプシド遺伝子のプロモーターなどが挙げられる。これらのベクターはさらに、分泌のためのシグナル配列を含み得る。分泌シグナル配列としては、ショウジョウバエ(Drosophila)由来のBiP分泌シグナル配列、ミツバチメリチン分泌シグナル配列などが挙げられる。バキュロウイルスエンベロープ構成糖タンパク質と連結する鎖Aの場合は、この糖タンパク質の分泌シグナル配列が好ましく用いられ、その場合、鎖Aをコードする遺伝子は、この分泌シグナル配列と、糖タンパク質をコードする構造遺伝子との間に配置され得る。トランスファーベクターは、目的遺伝子とポリヘドリン遺伝子との間で相同組換えを生じさせるために、ポリヘドリン遺伝子の5’領域および3’領域をそれぞれ数kbずつ含んでいてもよいが、これは必ずしも必要ではない。
【0021】
以下の実施例では、鎖Aおよび鎖Bをコードする両方の遺伝子を挿入したトランスファーベクターを用いているが、鎖Aおよび鎖Bのそれぞれを単独で挿入している複数のトランスファーベクターを用いてもよい。トランスファーベクターの種類は特に限定されない。トランスファーベクターの具体例としては、AcNPV系トランスファーベクターとしては、pVL1392/1393、pAcG1、pAcG2T、pAcG3X、pAcGHLT-A/B/C、pAcGP67-A/B/C、pAcHLT-A/B/C、pAcMP2/3、pAcSG2、pAcSecG2T、pAcUW21、pAcAB3/4、pAcDB3、pAcUW51、pBACシリーズ、pFastBacTMシリーズ(pFastBacTM 1、pFastBacTM Dualなど)、pDESTTM8/10/20、pBlueBacシリーズ、pMelBac A/B/Cなどが挙げられる(以上、日本ベクトンディッキンソン株式会社、メルク株式会社、インビトロジェン株式会社などから入手可能)。
【0022】
鎖Aおよび鎖Bをコードする両方の遺伝子の発現を可能にするトランスファーベクターとしては、pFastBacTM Dual(インビトロジェン株式会社)が好ましく用いられ得る。pFastBacTM Dualは、2つのプロモーターPPHおよびPp10が向かい合って互いに反対の方向に配置されており、これらのプロモーターからそれぞれの挿入遺伝子を発現させ得る。PPHおよびPp10のプロモーターの下流に配置される遺伝子は特に制限されず、鎖Aまたは鎖B、あるいはH鎖またはL鎖のどちらの遺伝子も、いずれかのプロモーターの下流に配置され得る。このトランスファーベクターにおいても、鎖Aをコードする遺伝子は、バキュロウイルスエンベロープ構成糖タンパク質(例えば、gp64)をコードする遺伝子の5’末端側に配置され得る。これらの鎖をコードする遺伝子は、上述したような分泌シグナル配列と連結され得、特に、鎖Aをコードする遺伝子は、好ましくは、連結される糖タンパク質の分泌シグナル配列とこの糖タンパク質をコードする構造遺伝子との間に配置され得る。
【0023】
次に、上記のトランスファーベクターを介して目的遺伝子を昆虫細胞に導入する。この導入された昆虫細胞内での相同組換えが利用され得る。相同組換えを利用する方法の場合、上記の組換えトランスファーベクターをバキュロウイルスゲノムDNAと混合し、宿主として用いる培養細胞に移入するか、あるいは予めバキュロウイルスに感染させた宿主として用いる培養細胞に上記のトランスファーベクターを移入し、組換えトランスファーベクターとウイルスゲノムDNAとの間に相同組換えを起こさせ、組換えバキュロウイルスを作製し得る。また、組換えバキュロウイルスを作製するために、昆虫細胞での相同組換えを利用せずに、大腸菌内で組換えバキュロウイルスDNAを作製し、これを昆虫細胞にトランスフェクトする方法も使用され得る。このような方法としては、例えば、Bac-to-Bac(登録商標)バキュロウイルス発現システム(インビトロジェン株式会社)の使用が挙げられる。Bac-to-Bacバキュロウイルス発現システムを用いる方法では、トランスファーベクターを直接昆虫細胞に移入するのではなく、まずpFastBacTMベクターのようなトランスファーベクターで大腸菌(例えば、DH10Bacコンピテント大腸菌(インビトロジェン株式会社);これは、lacZ-mini-attTn7融合を含むバクミドを含む)を形質転換し、この大腸菌が形成した組換えバクミドDNAを分離し、その後このDNAを昆虫細胞にトランスフェクトして目的遺伝子の導入された組換えバキュロウイルスを作製する。宿主として用いる培養細胞としては、上記の宿主が挙げられる。通常、昆虫培養細胞(例えば、Sf9細胞)が用いられ得る。培養条件は、当業者により適宜決定されるが、Sf9細胞を用いた場合は、10%ウシ胎児血清を含む培地または市販の無血清培地などで、27℃前後で培養することが好ましい。
【0024】
このようにして作製された組換えバキュロウイルスは、常法、例えばプラークアッセイなどによって精製することができる。なお、このようにして作製された組換えウイルスは、核多角体病ウイルスのポリヘドリンの遺伝子領域に外来のDNAが置換または挿入されており多角体を形成することができないため、非組換えウイルスと容易に区別することが可能である。
【0025】
さらに、上記組換えバキュロウイルスを宿主(例えば、Sf9およびSf21などの培養細胞)に感染させて、一定時間(例えば、48〜120時間)インキュベートすることにより、組換えバキュロウイルスを増殖させることもできる。増殖した組換えバキュロウイルスは、感染に際して出芽し得、さらに出芽型バキュロウイルスとして細胞外に放出され得る。組換えバキュロウイルスは1種類のみ感染させてもよいし、2種類以上の組換えバキュロウイルスを組み合わせて感染させることもできる。
【0026】
出芽型バキュロウイルスの回収は、例えば、以下のように行われ得る。感染細胞の培養液を500〜2,000gで遠心分離して、出芽型バキュロウイルスを含む上清を回収し得る。さらに、この上清を約30,000〜50,000gで遠心分離して出芽型バキュロウイルスを含む沈殿物を得ることもできる。
【0027】
本発明の出芽型組換えバキュロウイルスは、鎖Aおよび鎖Bで構成される抗体をその表面に提示し得る。本発明によれば、組換えバキュロウイルスは、その出芽に際して、鎖Aと鎖Bとの間のジスルフィド結合により形成されたFabフラグメントまたはそのようなFab形態を含む抗体を、感染した昆虫細胞の表面に提示し得る。このFabフラグメントまたはそのようなFab形態を含む抗体は、組換えバキュロウイルスが昆虫細胞から出芽する際に、出芽型バキュロウイルスの表面に提示され得る。このFabフラグメントまたはそのようなFab形態を含む抗体は、その抗原と結合可能な形態である。
【0028】
したがって、本発明によれば、特定の抗原と反応する抗体をスクリーニングまたは同定するために、細胞外に放出された出芽型組換えバキュロウイルスまたは出芽型組換えバキュロウイルスが感染した昆虫細胞が用いられ得る。例えば、蛍光標識した抗原をウイルス感染細胞の表面に発現したFabフラグメントと結合させ、フローサイトメーターを用いて強い蛍光を発する細胞をソーティングすることにより、目的の抗体を発現したバキュロウイルスを選択し得る。これにより、バイオパンニングにおいて問題となる非特異吸着を回避でき、そして簡便かつハイスループットな選択が可能となると期待される。
【0029】
さらに、本発明によれば、ゲノム由来の抗体遺伝子ライブラリーまたは変異体遺伝子プールから、Fabフラグメントを網羅的に表面提示したバキュロウイルスディスプレイライブラリーを作製することもできる。バキュロウイルスディスプレイライブラリーは、上記と同様にして、異なるFabフラグメントを発現する少なくとも2種の組換えバキュロウイルスを作製し、これらのウイルスを昆虫細胞に感染させ、感染した昆虫細胞を培養してバキュロウイルスを出芽させることにより作製され得る。鎖Aおよび鎖Bをコードする両方の遺伝子の発現を可能にするトランスファーベクター(例えば、pFastBacTM Dual)を異なるFabフラグメントを発現するようにそれぞれ構築し、これらの構築された複数のトランスファーベクターを用いて種々の組換えバキュロウイルスを作製し、これらの組換えバキュロウイルスを昆虫細胞に感染させることが好ましい。バキュロウイルスディスプレイライブラリーを用いて抗原を結合させ、抗原が結合したFabフラグメントを検出することによって、抗原が結合する抗体を同定することができる。検出を容易にするために、抗原は、蛍光などにより標識され得る。さらに、この目的とするFabを提示したバキュロウイルスを選別することにより、目的抗体の遺伝子も取得することができる。取得した目的の抗体遺伝子は哺乳動物細胞などに導入して抗体を産生できる。
【0030】
本発明の出芽型組換えバキュロウイルスが提示するFabフラグメントは、抗体のH鎖とL鎖が本来の形で会合しているため、抗体の可変部をペプチドリンカーで接続したscFvに比べて、抗原に対する特異性と親和性に優れているのみならず安定性も高い。したがって、本発明は、タンパク質の網羅的な定量解析を目的とした抗体マイクロアレイの構築、および治療用抗体医薬の開発などにおける新規抗体の創出手段として有用であり得る。
【実施例】
【0031】
以下の実施例では、クロラムフェニコールモノエステルの加水分解反応を触媒するマウス由来の触媒抗体6D9(非特許文献2)のFabフラグメントの発現を検討した。6D9のFabフラグメントを構成するFdフラグメント(またはL鎖(Lc))をgp64との融合タンパク質として発現し、一方Lc(またはFd)を分泌タンパク質として発現するような組換えAcNPVの作製を試みた。
【0032】
以下の実施例で使用したプラスミドpXIHA-6D9 HcおよびpXIHA-6D9 Lcは、FdフラグメントおよびL鎖をコードするDNA配列を含むプラスミドpARA-6D9Fabから構築したプラスミドである。pARA-6D9Fabの構築に関しては、非特許文献2に記載されている。このプラスミドpARA-6D9Fabは、大阪府立大学先端科学研究所の藤井郁雄教授から分与された。pXIHA-6D9 HcおよびpXIHA-6D9 Lcは、プラスミドpARA-6D9Fabから特許文献1に記載の方法に従って調製した(pARA-6D9Fabは、特許文献1では「pARA7Fab」と記載のプラスミドに相当する)。
【0033】
(実施例1:Fabフラグメント表面提示ベクターの構築)
(1−1.pBACsurf-1への6D9のFdフラグメント遺伝子の挿入)
pARA-6D9Fabを鋳型とし、上流部分に制限酵素KpnIの認識塩基配列(プライマーの4位〜9位に相当、そして10位にフレームシフトを防ぐためにアデニンを挿入)を有するプライマー1(配列番号1)および下流部分に制限酵素SmaIの認識配列(プライマーの4位〜9位、そして10位にフレームシフトを防ぐためにアデニンを挿入)を有するプライマー2(配列番号2)を用いて、6D9のFdフラグメントの遺伝子をPCRにより増幅した。増幅したFdフラグメント遺伝子およびプラスミドpBACsurf-1(メルク株式会社)を、いずれもKpnIおよびSmaIで消化し、次いでFdフラグメント遺伝子をpBACsurf-1と連結し、pBACsurf-1-Fdを得た。
【0034】
(1−2.pBACsurf-1への6D9のL鎖遺伝子の挿入)
pARA-6D9Fabを鋳型とし、上流部分に制限酵素KpnIの認識配列(プライマーの4位〜9位に相当、そして10位にフレームシフトを防ぐためにアデニンを挿入)を有するプライマー3(配列番号3)および下流部分に制限酵素SmaIの認識配列(プライマーの4位〜9位に相当、そして10位にフレームシフトを防ぐためにアデニンを挿入)を有するプライマー4(配列番号4)を用いて、6D9のL鎖の遺伝子をPCRにより増幅した。増幅したL鎖遺伝子およびpBACsurf-1を、いずれもKpnIおよびSmaIで消化し、次いでL鎖遺伝子をpBACsurf-1と連結し、pBACsurf-1-Lcを得た。
【0035】
(1−3.pFastBacTM Dualのポリヘドリンプロモーター(PPH)の下流へのFdフラグメント遺伝子の挿入)
pXIHA-6D9 Hcを鋳型とし、上流部分に制限酵素EcoRIの認識配列(プライマーの4位〜9位に相当)および引き続きDrosophila由来のBiP分泌シグナル配列の一部(プライマーの10位〜39位に相当)を有するプライマー5(配列番号5)および下流部分に制限酵素HindIIIの認識配列(プライマーの4位〜9位に相当)を有するプライマー6(配列番号6)を用いてPCRを行った。PCR増幅で得られたBiP分泌シグナル配列が連結された6D9のFdフラグメントの遺伝子およびプラスミドpFastBacTM Dual(インビトロジェン株式会社)を、いずれもEcoRIおよびHindIIIで消化し、次いでBiP分泌シグナル配列が連結されたFdフラグメント遺伝子をpFastBac Dualと連結し、pFastBac -/Fdを得た。
【0036】
(1−4.pFastBacTM Dualのp10プロモーター(Pp10)の下流へのFdフラグメント遺伝子の挿入)
pXIHA-6D9 Hcを鋳型とし、上流部分に制限酵素SmaIの認識配列(プライマーの4位〜9位に相当)および引き続きDrosophila由来のBiP分泌シグナル配列の一部(プライマーの10位〜33位に相当)を有するプライマー7(配列番号7)および下流部分に制限酵素KpnIの認識配列(プライマーの4位〜9位に相当)を有するプライマー8(配列番号7)を用いてPCRを行った。PCR増幅で得られたBiP分泌シグナル配列が連結された6D9のFdフラグメントの遺伝子およびプラスミドpFastBacTM Dualを、いずれもSmaIおよびKpnIで消化し、次いでBiP分泌シグナル配列が連結されたFdフラグメント遺伝子をpFastBac Dualと連結し、pFastBac Fd/-を得た。
【0037】
(1−5.pFastBac DualのPPHの下流へのL鎖遺伝子の挿入)
pXIHA-6D9 Lcを鋳型とし、上記のプライマー5および下流部分に制限酵素HindIIIの認識配列(プライマーの4位〜9位に相当)を有するプライマー9(配列番号9)を用いてPCRを行った。PCR増幅で得られたBiP分泌シグナル配列が連結された6D9のL鎖の遺伝子およびプラスミドpFastBacTM Dualを、いずれもEcoRIおよびHindIIIで消化し、次いでBiP分泌シグナル配列が連結されたL鎖遺伝子をpFastBac Dualと連結し、pFastBac -/Lcを得た。
【0038】
(1−6.pFastBacTM DualのPp10の下流へのL鎖遺伝子の挿入)
pXIHA-6D9 Lcを鋳型とし、上記のプライマー7および下流部分に制限酵素KpnIの認識配列(プライマーの4位〜9位に相当)を有するプライマー10(配列番号10)を用いてPCRを行った。PCR増幅で得られたBiP分泌シグナル配列が連結された6D9のL鎖の遺伝子およびプラスミドpFastBacTM Dualを、いずれもSmaIおよびKpnIで消化し、次いでBiP分泌シグナル配列が連結されたL鎖遺伝子をpFastBac Dualと連結し、pFastBac Lc/-を得た。
【0039】
(1−7.pFastBac -/FdへのL鎖遺伝子およびgp64遺伝子のハイブリッド遺伝子の挿入)
上記実施例1−2で得られたpBACsurf-1-Lcを鋳型とし、上流部分に制限酵素NheIの認識配列(プライマーの4位〜9位に相当)および引き続きgp64の分泌シグナル配列の一部(プライマーの10位〜37位に相当)を有するプライマー11(配列番号11)および下流部分に制限酵素NheIの認識配列(プライマーの4位〜9位に相当)を有するプライマー12(配列番号12)を用いて、gp64の分泌シグナル配列、6D9のL鎖の遺伝子、およびgp64遺伝子からなるハイブリッド遺伝子をPCRにより増幅した。増幅したハイブリッド遺伝子および上記実施例1−3で得られたpFastBac -/Fdを、いずれもNheIで消化し、次いでハイブリッド遺伝子をpFastBac -/Fdと連結し、そして正しい方向にハイブリッド遺伝子が挿入されたpFastBac Lc-gp64/Fdを回収した。
【0040】
(1−8.pFastBac Fd/-へのL鎖遺伝子およびgp64遺伝子のハイブリッド遺伝子の挿入)
上記実施例1−2で得られたpBACsurf-1-Lcを鋳型とし、上流部分に制限酵素EcoRIの認識配列(プライマーの4位〜9位に相当)および引き続きgp64の分泌シグナル配列の一部(プライマーの10位〜34位に相当)を有するプライマー13(配列番号13)および下流部分に制限酵素HindIIIの認識配列(プライマーの4位〜9位に相当)を有するプライマー14(配列番号14)を用いて、gp64の分泌シグナル配列、6D9のL鎖の遺伝子、およびgp64遺伝子からなるハイブリッド遺伝子をPCRにより増幅した。増幅したハイブリッド遺伝子および上記実施例1−4で得られたpFastBac Fd/-を、いずれもEcoRIおよびHindIIIで消化し、次いでハイブリッド遺伝子をpFastBac Fd/-と連結し、pFastBac Fd/Lc-gp64を得た。
【0041】
(1−9.pFastBac -/LcへのFdフラグメント遺伝子およびgp64遺伝子のハイブリッド遺伝子の挿入)
上記実施例1−1で得られたpBACsurf-1-Fdを鋳型とし、プライマー11およびプライマー12を用いて、gp64の分泌シグナル配列、6D9のFdフラグメントの遺伝子、およびgp64遺伝子からなるハイブリッド遺伝子をPCRにより増幅した。増幅したハイブリッド遺伝子および上記実施例1−5で得られたpFastBac -/Lcをいずれも、NheIで消化し、次いでハイブリッド遺伝子をpFastBac -/Lcと連結し、そして正しい方向にハイブリッド遺伝子が挿入されたpFastBac Fd-gp64/Lcを回収した。
【0042】
(1−10.pFastBac Lc/-へのFdフラグメント遺伝子およびgp64遺伝子のハイブリッド遺伝子の挿入)
上記実施例1−1で得られたpBACsurf-1-Fdを鋳型とし、プライマー13およびプライマー14を用いて、gp64の分泌シグナル配列、6D9のFdフラグメントの遺伝子、およびgp64遺伝子からなるハイブリッド遺伝子をPCRにより増幅した。増幅したハイブリッド遺伝子および上記実施例1−6で得られたpFastBac Lc/-を、いずれもEcoRIおよびHindIIIで消化し、次いでハイブリッド遺伝子をpFastBac Lc/-と連結し、pFastBac Lc/Fd-gp64を得た。
【0043】
(実施例2:組換えバキュロウイルスの作製)
大腸菌内で組換えバキュロウイルスDNAを作製する方法(Bac-to-Bacバキュロウイルス発現システム(インビトロジェン株式会社)を使用し、基本的にマニュアルに記載の方法にしたがって、組換えバキュロウイルスを作製した。
【0044】
上記実施例1で作製した4種類のベクターのpFastBac Lc-gp64/Fd、pFastBac Fd/Lc-gp64、pFastBac Fd-gp64/Lc、およびpFastBac Lc/Fd-gp64のそれぞれ1ngを、DH10Bacコンピテント大腸菌(インビトロジェン株式会社)100μlに対して加え、大腸菌を形質転換した。その後、S.O.C.培地(2%トリプトン、0.5%酵母エキス、10mM NaCl、2.5mM KCl、10mM MgCl2、10mM MgSO4、20mM グルコース)4mlを加え、試験管にて37℃で2〜3時間振とう培養を行った。細胞懸濁培地100μlをLB寒天培地(1%トリプトン、0.5%酵母エキス、0.5% NaCl、2%寒天、50μg/mlカナマイシン、7μg/mlゲンタマイシン、10μg/mlテトラサイクリン、100μg/ml Bluo-gal、40μg/ml IPTG)に塗布して24時間インキュベートした。生成した白色のコロニーを採取し、S.N.A.P.TMミディプレップキット(インビトロジェン株式会社)を用いて組換えバクミドDNAを分離した。直径35mmのペトリディッシュまたは6ウェルプレートに昆虫細胞Sf9(インビトロジェン株式会社)を2×106細胞/cm3の細胞密度で2mlのグレース培地(インビトロジェン株式会社)に播種し、27℃にて24時間培養した。この培養液に、上記組換えバクミドDNA1μgに対してトランスフェクション試薬FuGENE 6(ロシュ・ダイアグノスティックス)を3μlの割合で含む混合液を添加して、昆虫細胞Sf9をトランスフェクトし、27℃にて72時間インキュベートした。これにより、出芽型組換えバキュロウイルスが昆虫細胞から生じた。インキュベート後、出芽型組換えバキュロウイルスを含む培養上清を2000rpmにて5分間の遠心分離により回収し、10%ウシ胎児血清を含むTNM-FH培地(シグマアルドリッチジャパン)を用いて培養しているSf9細胞に添加し、27℃にて72時間インキュベートした。再度この操作を繰り返し、組換えバキュロウイルスを増殖させた。
【0045】
(実施例3:組換えバキュロウイルスの抗体表面提示の解析)
上記実施例2の出芽型組換えバキュロウイルスを含む培養上清を、抗原としてクロラムフェニコールモノエステルの加水分解反応の遷移状態アナログをハプテン基として結合したウシ血清アルブミン、および検出抗体としてヤギ抗マウスκ鎖-HRP(Exalpha Biologicals)を用いる酵素免疫測定法(ELISA)によって分析した。405nmでの吸光度を測定した。
【0046】
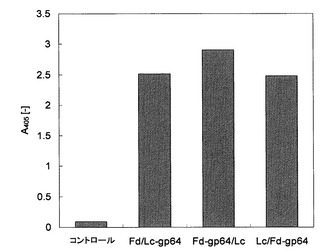
図1は、pFastBac Fd/Lc-gp64、pFastBac Fd-gp64/Lc、およびpFastBac Lc/Fd-gp64のそれぞれを含む培養上清をELISAにより分析した結果を示す。図1において、コントロールは、比較のためにウイルス未感染のSf9の培養上清を用いた場合の結果である。図1から明らかなように、上記のいずれのベクターを用いた場合もウイルス未感染のSf9の培養上清と比べて、高い吸光度が得られた。
【0047】
また、上記実施例2の出芽型組換えバキュロウイルスを含む培養上清をSf9の細胞懸濁培地2mlに添加し、そして27℃にて24時間インキュベートして、組換えバキュロウイルスをSf9に感染させた。この細胞懸濁培地に、FITC標識したヤギ抗マウスIgGであるFITC-F(ab’)2ヤギ抗マウスIgG(インビトロジェン株式会社)5μlを加えて30分間インキュベートし、引き続き細胞をリン酸緩衝液で洗浄し、そしてフローサイトメーター(FACSCalibur、日本ベクトン・ディッキンソン株式会社)を用いて解析した。
【0048】
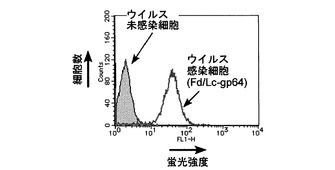
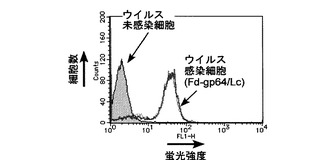
pFastBac Fd/Lc-gp64およびpFastBac Fd-gp64/Lcを用いて作製した組換えバキュロウイルスの場合のフローサイトメーター解析結果を、それぞれ図2および3に示す。これらの図に見られるように、これら2種類の組換えバキュロウイルスに感染した細胞は、ウイルス未感染のSf9と比較して、高い蛍光強度を示した。
【0049】
これらの結果は、作製した組換えバキュロウイルスの感染によって、昆虫細胞Sf9の表面にマウスの抗体分子が発現および提示されることを示している。組換えバキュロウイルスが出芽する際に、抗体分子は、出芽型バキュロウイルスの表面に提示され得る。これらの結果から、作製した組換えバキュロウイルスは、出芽の際に抗原結合活性を有するFabフラグメントを表面に提示したと考えられる。
【0050】
この結果はまた、蛍光標識した抗原をウイルス感染細胞の表面に発現したFabフラグメントと結合させ、そしてフローサイトメーターを用いて強い蛍光を発する細胞をソーティングすることにより、目的の抗体が発現されたバキュロウイルスを選択できることを示唆する。
【産業上の利用可能性】
【0051】
本発明の出芽型組換えバキュロウイルスを用いることにより、抗原に対する特異性および親和性に優れ、そして安定性も高い抗体を作製することができる。このような抗体作製技術は、タンパク質の網羅的な定量解析を目的とした抗体マイクロアレイの構築や治療用抗体医薬の開発などにおいて、新規抗体の創出手段として有用であると考えられる。
【図面の簡単な説明】
【0052】
【図1】pFastBac Fd/Lc-gp64、pFastBac Fd-gp64/Lc、およびpFastBac Lc/Fd-gp64のそれぞれを含む培養上清をELISAにより分析した結果を示すグラフである。
【図2】pFastBac Fd/Lc-gp64を用いて作製した組換えバキュロウイルスを感染させたSf9および未感染Sf9のフローサイトメーターによる解析結果を示す図である。
【図3】pFastBac Fd-gp64/Lcを用いて作製した組換えバキュロウイルスを感染させたSf9および未感染Sf9のフローサイトメーターによる解析結果を示す図である。
【技術分野】
【0001】
本発明は、抗体を表面に提示した組換えバキュロウイルスに関する。
【背景技術】
【0002】
抗体作製の手段として、一般に、ファージディスプレイ法が用いられている。ファージディスプレイ法では、大腸菌を宿主として繊維状ファージを構成するタンパク質の遺伝子に各々連結させた抗体断片の遺伝子群を発現させて、その表面にさまざまな抗体断片が提示されたファージのライブラリーを作製し、そしてその中から特定の抗原に対する特異性または親和性を有する抗体断片が提示されたファージを選択する。抗体断片を発現したファージは、その抗体断片の遺伝子をコードしたDNAを有するので、特定の抗原に結合するファージを回収すれば、目的の抗体の遺伝子を単離することができる。得られた抗体遺伝子を動物細胞などにおいて発現させることにより、抗体タンパク質の生産が可能となる。
【0003】
ファージディスプレイ法は、動物を免疫する必要がないこと、ヒト由来の抗体のファージディスプレイライブラリーを用いればヒト型の抗体を取得できることなどの利点を有する。しかしながら、ファージディスプレイ法は、大腸菌を宿主とするため、高分子量のタンパク質ほど提示効率が悪い。このため、抗体の可変部(VHとVL)をペプチドリンカーで接続した一本鎖抗体(scFv)を提示する場合が多く、プロテアーゼによる分解または抗原との親和性の低下が避けられない。また、固相に固定した抗原に対して特異的に結合するファージを選択および回収するバイオパンニングによる選択効率が悪いなどの欠点を有している。
【0004】
昆虫に感染して病気を起こすウイルスであるバキュロウイルスは、環状の二本鎖DNAを遺伝子としてもつエンベロープウイルスで、鱗翅目、膜翅目および双翅目などの昆虫に感受性を示す。核多角体病ウイルス(NPV)は、このようなバキュロウイルスの一種であり、感染した昆虫細胞の核内に多角体(ポリヒドラ)と呼ばれる封入体を大量に生成する一群のウイルスである。多角体は、分子量31kDaのポリヘドリンタンパク質から構成され、感染後期に大量に生成されて、その中に多数のウイルス粒子を埋め込んでいる。多角体はウイルスが自然界で生存するためには必須であるが、ウイルスの増殖には必要ないので、ポリヘドリン遺伝子の代わりに発現を意図する外来遺伝子を挿入してもウイルスは全く支障なく感染し増殖する。このような性質を利用して、昆虫細胞−バキュロウイルス系は、ポリヘドリンプロモーターの下流に外来遺伝子を挿入した組換えバキュロウイルスを培養昆虫細胞に感染させ、ウイルス感染した細胞に外来タンパク質を生産させる組換えタンパク質発現系として利用されている。
【0005】
バキュロウイルスの1種であるオートグラファ・カリフォルニカ核多角体病ウイルス(Autographa californica nucleopolyhedrovirus, AcNPV)のエンベロープを構成する糖タンパク質gp64をコードする遺伝子の5’末端側に連結させた外来タンパク質の遺伝子を発現させることにより、そのタンパク質は、gp64と融合した形でウイルスの表面に提示されることが報告されている(非特許文献1)。非特許文献1に記載の方法は、昆虫細胞を宿主として用いるため、哺乳動物細胞により発現されるような、複雑な構造を有するタンパク質または翻訳後修飾されたタンパク質を、本来の機能を保持した状態でウイルスの表面に提示し得る。
【特許文献1】特開2006−087427号公報
【非特許文献1】Grabherr, R.ら, Trends Biotechnol., 19, 231-236 (2001)
【非特許文献2】Miyashita, H.ら, J. Mol. Biol., 267, 1247-1257 (1997)
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明は、抗原に対する特異性および親和性に優れた抗体を表面に提示できるバキュロウイルスを提供することを目的とする。
【課題を解決するための手段】
【0007】
本発明は、抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルス(以下、単に「出芽型組換えバキュロウイルス」ともいう)を提供し、このバキュロウイルスは、バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質として発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現し、表面に提示された該抗体またはそのFabフラグメントが抗原と結合し得る。
【0008】
本発明はまた、抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルスを作製する方法を提供し、この方法は、
バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質を発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現する、組換えバキュロウイルスを作製する工程
を含む。
【0009】
1つの実施態様では、上記組換えバキュロウイルスを作製する工程が、上記バキュロウイルスエンベロープ構成糖タンパク質をコードする遺伝子の5’末端側に連結した、該糖タンパク質の分泌シグナル配列および上記抗体またはそのFabフラグメントを構成する一方のペプチド鎖をコードする遺伝子を含み、かつ分泌シグナル配列および該抗体またはそのFabフラグメントの他方のペプチド鎖をコードする遺伝子を含むトランスファーベクターを構築する工程;および
該トランスファーベクターを介して目的遺伝子を昆虫細胞に導入する工程
を含む。
【0010】
別の実施態様では、上記抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルスを作製する方法は、
前記組換えバキュロウィルスを昆虫細胞に感染させる工程;および
前記感染させた昆虫細胞を培養してバキュロウイルスを出芽させる工程
をさらに含む。
【0011】
本発明はまた、抗体またはそのFabフラグメントを表面に提示する出芽型組換えバキュロウイルスディスプレイライブラリーを作製する方法を提供し、この方法は、
異なる抗体またはそのFabフラグメントを発現する少なくとも2種の組換えバキュロウイルスを作製する工程であって、該組換えバキュロウイルスが各々、バキュロウイルスエンベロープ構成糖タンパク質と抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質を発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現する、工程;
該少なくとも2種の組換えバキュロウィルスを昆虫細胞に感染させる工程;および
該感染させた昆虫細胞を培養して、バキュロウイルスを出芽させる工程
を含む。
【0012】
本発明はまた、上記方法により作製されたバキュロウイルスディスプレイライブラリーを提供する。
【0013】
本発明はさらに、抗体またはそのFabフラグメントをスクリーニングする方法を提供し、この方法は、
上記ライブラリーに抗原を結合させる工程;および
該抗原が結合した該ライブラリーの抗体またはそのFabフラグメントを検出する工程
を含む。
【発明の効果】
【0014】
本発明によれば、抗原に対する特異性および親和性に優れた抗体を表面に提示できる出芽型バキュロウイルスが得られる。このバキュロウイルスは、抗体分子のH鎖とL鎖とが本来の形で会合したFabフラグメントを表面に提示できる。
【発明を実施するための最良の形態】
【0015】
本発明の出芽型組換えバキュロウイルスは、バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質として発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現し得る。便宜上、糖タンパク質と連結されるペプチド鎖を鎖Aとし、他方のペプチド鎖を鎖Bという。鎖AがH鎖である場合、鎖BはL鎖であり、その逆であってもよい。
【0016】
本発明において、出芽型組換えバキュロウイルスの表面に提示される抗体は、任意の抗体であり得る。本発明によって出芽型組換えバキュロウイルスの表面に提示される抗体は、その抗原と結合可能な任意の形態をとり得、代表的には、Fabフラグメントの形態となり得る。抗体のH鎖のFdフラグメントとL鎖とがジスルフィド結合したFab形態を構成するように、その抗体のH鎖およびL鎖が選択される。H鎖は、少なくともFab形成に必要な領域を含む一部分、例えば、Fdフラグメントおよびこれを含むフラグメントであり得、または全長であってもよい。Fdフラグメントが好適に用いられ得る。L鎖は、全長、またはH鎖とFabを形成するのに必要な領域を含む任意の長さのフラグメントであり得る。全長のL鎖が好適に用いられ得る。H鎖およびL鎖は、Fabの形成に影響しない限り、変異が導入されていてもよい。
【0017】
バキュロウイルスとしては、オートグラファ・カリフォルニカ核多角体病ウイルス(AcNPV)およびボンビックス・モリ(Bombyx mori)核多角体病ウイルス(BmNPV)が用いられ得る。
【0018】
宿主としては、バキュロウイルスの宿主となる昆虫細胞であれば特に制限なく利用できる。宿主は、トランスファーベクターの移入により組換えバキュロウイルスを生成し得、そして感染によりバキュロウイルスを増殖し得る。AcNPVの宿主としては、ハスモンヨトウの近縁種のスポドプテラ・フルギペルダ(Spodoptera frugiperda)由来のSf細胞などが挙げられる。細胞株Sf9およびSf21などは、スポドプテラ・フルギペルダ幼虫の卵巣組織から確立しており、インビトロジェン株式会社、メルク株式会社、もしくは日本ベクトンディッキンソン株式会社、または寄託機関からも入手可能である。また、イラクサギンウワバ(Trichoplusia ni)由来のBTI-TN-5B1-4細胞(High FiveTM)も利用することができ、本細胞株はインビトロジェン株式会社から入手可能である。BmNPVの宿主としてはBmN4細胞などが挙げられ、日本農産工業株式会社から入手可能である。
【0019】
本発明の出芽型組換えバキュロウイルスの作製は、当業者が通常用いる方法に従って行われ得る。以下、簡単に説明する。
【0020】
発現させるべきペプチド鎖(鎖Aまたは鎖B、あるいはその両方)の遺伝子をトランスファーベクターに挿入して組換えトランスファーベクターを構築する。例えば、抗体産生細胞のmRNAから逆転写酵素により調製されたcDNAライブラリーを鋳型にして、PCRにより、上記で説明したようなFabフラグメントを形成する組合せとなるペプチド鎖をコードする抗体遺伝子の組合せ、例えば、Fdフラグメント遺伝子およびL鎖遺伝子、または、H鎖遺伝子およびL鎖遺伝子をクローニングする。この際、PCRに用いるプライマーには、遺伝子増幅後にベクターに組込めるように制限酵素認識配列のオリゴヌクレオチドを付加しておくことが好ましい。増幅した抗体遺伝子およびベクターを制限酵素で消化した後、抗体遺伝子をベクターに挿入する。鎖Aをコードする遺伝子は、バキュロウイルスエンベロープ構成糖タンパク質(例えば、gp64)をコードする遺伝子の5’末端側に配置され得る。トランスファーベクターの全体の大きさは一般的には数kb〜10kb程度であり、そのうちの約3kbはプラスミド由来の骨格であり、アンピシリンなどの抗生物質耐性遺伝子および細菌のDNA複製起点を含んでいる。トランスファーベクターには、挿入遺伝子を発現させるためのプロモーターを含むことが好ましい。プロモーターとしては、ポリヘドリンプロモーター(PPH)、p10遺伝子のプロモーター(Pp10)、キャプシド遺伝子のプロモーターなどが挙げられる。これらのベクターはさらに、分泌のためのシグナル配列を含み得る。分泌シグナル配列としては、ショウジョウバエ(Drosophila)由来のBiP分泌シグナル配列、ミツバチメリチン分泌シグナル配列などが挙げられる。バキュロウイルスエンベロープ構成糖タンパク質と連結する鎖Aの場合は、この糖タンパク質の分泌シグナル配列が好ましく用いられ、その場合、鎖Aをコードする遺伝子は、この分泌シグナル配列と、糖タンパク質をコードする構造遺伝子との間に配置され得る。トランスファーベクターは、目的遺伝子とポリヘドリン遺伝子との間で相同組換えを生じさせるために、ポリヘドリン遺伝子の5’領域および3’領域をそれぞれ数kbずつ含んでいてもよいが、これは必ずしも必要ではない。
【0021】
以下の実施例では、鎖Aおよび鎖Bをコードする両方の遺伝子を挿入したトランスファーベクターを用いているが、鎖Aおよび鎖Bのそれぞれを単独で挿入している複数のトランスファーベクターを用いてもよい。トランスファーベクターの種類は特に限定されない。トランスファーベクターの具体例としては、AcNPV系トランスファーベクターとしては、pVL1392/1393、pAcG1、pAcG2T、pAcG3X、pAcGHLT-A/B/C、pAcGP67-A/B/C、pAcHLT-A/B/C、pAcMP2/3、pAcSG2、pAcSecG2T、pAcUW21、pAcAB3/4、pAcDB3、pAcUW51、pBACシリーズ、pFastBacTMシリーズ(pFastBacTM 1、pFastBacTM Dualなど)、pDESTTM8/10/20、pBlueBacシリーズ、pMelBac A/B/Cなどが挙げられる(以上、日本ベクトンディッキンソン株式会社、メルク株式会社、インビトロジェン株式会社などから入手可能)。
【0022】
鎖Aおよび鎖Bをコードする両方の遺伝子の発現を可能にするトランスファーベクターとしては、pFastBacTM Dual(インビトロジェン株式会社)が好ましく用いられ得る。pFastBacTM Dualは、2つのプロモーターPPHおよびPp10が向かい合って互いに反対の方向に配置されており、これらのプロモーターからそれぞれの挿入遺伝子を発現させ得る。PPHおよびPp10のプロモーターの下流に配置される遺伝子は特に制限されず、鎖Aまたは鎖B、あるいはH鎖またはL鎖のどちらの遺伝子も、いずれかのプロモーターの下流に配置され得る。このトランスファーベクターにおいても、鎖Aをコードする遺伝子は、バキュロウイルスエンベロープ構成糖タンパク質(例えば、gp64)をコードする遺伝子の5’末端側に配置され得る。これらの鎖をコードする遺伝子は、上述したような分泌シグナル配列と連結され得、特に、鎖Aをコードする遺伝子は、好ましくは、連結される糖タンパク質の分泌シグナル配列とこの糖タンパク質をコードする構造遺伝子との間に配置され得る。
【0023】
次に、上記のトランスファーベクターを介して目的遺伝子を昆虫細胞に導入する。この導入された昆虫細胞内での相同組換えが利用され得る。相同組換えを利用する方法の場合、上記の組換えトランスファーベクターをバキュロウイルスゲノムDNAと混合し、宿主として用いる培養細胞に移入するか、あるいは予めバキュロウイルスに感染させた宿主として用いる培養細胞に上記のトランスファーベクターを移入し、組換えトランスファーベクターとウイルスゲノムDNAとの間に相同組換えを起こさせ、組換えバキュロウイルスを作製し得る。また、組換えバキュロウイルスを作製するために、昆虫細胞での相同組換えを利用せずに、大腸菌内で組換えバキュロウイルスDNAを作製し、これを昆虫細胞にトランスフェクトする方法も使用され得る。このような方法としては、例えば、Bac-to-Bac(登録商標)バキュロウイルス発現システム(インビトロジェン株式会社)の使用が挙げられる。Bac-to-Bacバキュロウイルス発現システムを用いる方法では、トランスファーベクターを直接昆虫細胞に移入するのではなく、まずpFastBacTMベクターのようなトランスファーベクターで大腸菌(例えば、DH10Bacコンピテント大腸菌(インビトロジェン株式会社);これは、lacZ-mini-attTn7融合を含むバクミドを含む)を形質転換し、この大腸菌が形成した組換えバクミドDNAを分離し、その後このDNAを昆虫細胞にトランスフェクトして目的遺伝子の導入された組換えバキュロウイルスを作製する。宿主として用いる培養細胞としては、上記の宿主が挙げられる。通常、昆虫培養細胞(例えば、Sf9細胞)が用いられ得る。培養条件は、当業者により適宜決定されるが、Sf9細胞を用いた場合は、10%ウシ胎児血清を含む培地または市販の無血清培地などで、27℃前後で培養することが好ましい。
【0024】
このようにして作製された組換えバキュロウイルスは、常法、例えばプラークアッセイなどによって精製することができる。なお、このようにして作製された組換えウイルスは、核多角体病ウイルスのポリヘドリンの遺伝子領域に外来のDNAが置換または挿入されており多角体を形成することができないため、非組換えウイルスと容易に区別することが可能である。
【0025】
さらに、上記組換えバキュロウイルスを宿主(例えば、Sf9およびSf21などの培養細胞)に感染させて、一定時間(例えば、48〜120時間)インキュベートすることにより、組換えバキュロウイルスを増殖させることもできる。増殖した組換えバキュロウイルスは、感染に際して出芽し得、さらに出芽型バキュロウイルスとして細胞外に放出され得る。組換えバキュロウイルスは1種類のみ感染させてもよいし、2種類以上の組換えバキュロウイルスを組み合わせて感染させることもできる。
【0026】
出芽型バキュロウイルスの回収は、例えば、以下のように行われ得る。感染細胞の培養液を500〜2,000gで遠心分離して、出芽型バキュロウイルスを含む上清を回収し得る。さらに、この上清を約30,000〜50,000gで遠心分離して出芽型バキュロウイルスを含む沈殿物を得ることもできる。
【0027】
本発明の出芽型組換えバキュロウイルスは、鎖Aおよび鎖Bで構成される抗体をその表面に提示し得る。本発明によれば、組換えバキュロウイルスは、その出芽に際して、鎖Aと鎖Bとの間のジスルフィド結合により形成されたFabフラグメントまたはそのようなFab形態を含む抗体を、感染した昆虫細胞の表面に提示し得る。このFabフラグメントまたはそのようなFab形態を含む抗体は、組換えバキュロウイルスが昆虫細胞から出芽する際に、出芽型バキュロウイルスの表面に提示され得る。このFabフラグメントまたはそのようなFab形態を含む抗体は、その抗原と結合可能な形態である。
【0028】
したがって、本発明によれば、特定の抗原と反応する抗体をスクリーニングまたは同定するために、細胞外に放出された出芽型組換えバキュロウイルスまたは出芽型組換えバキュロウイルスが感染した昆虫細胞が用いられ得る。例えば、蛍光標識した抗原をウイルス感染細胞の表面に発現したFabフラグメントと結合させ、フローサイトメーターを用いて強い蛍光を発する細胞をソーティングすることにより、目的の抗体を発現したバキュロウイルスを選択し得る。これにより、バイオパンニングにおいて問題となる非特異吸着を回避でき、そして簡便かつハイスループットな選択が可能となると期待される。
【0029】
さらに、本発明によれば、ゲノム由来の抗体遺伝子ライブラリーまたは変異体遺伝子プールから、Fabフラグメントを網羅的に表面提示したバキュロウイルスディスプレイライブラリーを作製することもできる。バキュロウイルスディスプレイライブラリーは、上記と同様にして、異なるFabフラグメントを発現する少なくとも2種の組換えバキュロウイルスを作製し、これらのウイルスを昆虫細胞に感染させ、感染した昆虫細胞を培養してバキュロウイルスを出芽させることにより作製され得る。鎖Aおよび鎖Bをコードする両方の遺伝子の発現を可能にするトランスファーベクター(例えば、pFastBacTM Dual)を異なるFabフラグメントを発現するようにそれぞれ構築し、これらの構築された複数のトランスファーベクターを用いて種々の組換えバキュロウイルスを作製し、これらの組換えバキュロウイルスを昆虫細胞に感染させることが好ましい。バキュロウイルスディスプレイライブラリーを用いて抗原を結合させ、抗原が結合したFabフラグメントを検出することによって、抗原が結合する抗体を同定することができる。検出を容易にするために、抗原は、蛍光などにより標識され得る。さらに、この目的とするFabを提示したバキュロウイルスを選別することにより、目的抗体の遺伝子も取得することができる。取得した目的の抗体遺伝子は哺乳動物細胞などに導入して抗体を産生できる。
【0030】
本発明の出芽型組換えバキュロウイルスが提示するFabフラグメントは、抗体のH鎖とL鎖が本来の形で会合しているため、抗体の可変部をペプチドリンカーで接続したscFvに比べて、抗原に対する特異性と親和性に優れているのみならず安定性も高い。したがって、本発明は、タンパク質の網羅的な定量解析を目的とした抗体マイクロアレイの構築、および治療用抗体医薬の開発などにおける新規抗体の創出手段として有用であり得る。
【実施例】
【0031】
以下の実施例では、クロラムフェニコールモノエステルの加水分解反応を触媒するマウス由来の触媒抗体6D9(非特許文献2)のFabフラグメントの発現を検討した。6D9のFabフラグメントを構成するFdフラグメント(またはL鎖(Lc))をgp64との融合タンパク質として発現し、一方Lc(またはFd)を分泌タンパク質として発現するような組換えAcNPVの作製を試みた。
【0032】
以下の実施例で使用したプラスミドpXIHA-6D9 HcおよびpXIHA-6D9 Lcは、FdフラグメントおよびL鎖をコードするDNA配列を含むプラスミドpARA-6D9Fabから構築したプラスミドである。pARA-6D9Fabの構築に関しては、非特許文献2に記載されている。このプラスミドpARA-6D9Fabは、大阪府立大学先端科学研究所の藤井郁雄教授から分与された。pXIHA-6D9 HcおよびpXIHA-6D9 Lcは、プラスミドpARA-6D9Fabから特許文献1に記載の方法に従って調製した(pARA-6D9Fabは、特許文献1では「pARA7Fab」と記載のプラスミドに相当する)。
【0033】
(実施例1:Fabフラグメント表面提示ベクターの構築)
(1−1.pBACsurf-1への6D9のFdフラグメント遺伝子の挿入)
pARA-6D9Fabを鋳型とし、上流部分に制限酵素KpnIの認識塩基配列(プライマーの4位〜9位に相当、そして10位にフレームシフトを防ぐためにアデニンを挿入)を有するプライマー1(配列番号1)および下流部分に制限酵素SmaIの認識配列(プライマーの4位〜9位、そして10位にフレームシフトを防ぐためにアデニンを挿入)を有するプライマー2(配列番号2)を用いて、6D9のFdフラグメントの遺伝子をPCRにより増幅した。増幅したFdフラグメント遺伝子およびプラスミドpBACsurf-1(メルク株式会社)を、いずれもKpnIおよびSmaIで消化し、次いでFdフラグメント遺伝子をpBACsurf-1と連結し、pBACsurf-1-Fdを得た。
【0034】
(1−2.pBACsurf-1への6D9のL鎖遺伝子の挿入)
pARA-6D9Fabを鋳型とし、上流部分に制限酵素KpnIの認識配列(プライマーの4位〜9位に相当、そして10位にフレームシフトを防ぐためにアデニンを挿入)を有するプライマー3(配列番号3)および下流部分に制限酵素SmaIの認識配列(プライマーの4位〜9位に相当、そして10位にフレームシフトを防ぐためにアデニンを挿入)を有するプライマー4(配列番号4)を用いて、6D9のL鎖の遺伝子をPCRにより増幅した。増幅したL鎖遺伝子およびpBACsurf-1を、いずれもKpnIおよびSmaIで消化し、次いでL鎖遺伝子をpBACsurf-1と連結し、pBACsurf-1-Lcを得た。
【0035】
(1−3.pFastBacTM Dualのポリヘドリンプロモーター(PPH)の下流へのFdフラグメント遺伝子の挿入)
pXIHA-6D9 Hcを鋳型とし、上流部分に制限酵素EcoRIの認識配列(プライマーの4位〜9位に相当)および引き続きDrosophila由来のBiP分泌シグナル配列の一部(プライマーの10位〜39位に相当)を有するプライマー5(配列番号5)および下流部分に制限酵素HindIIIの認識配列(プライマーの4位〜9位に相当)を有するプライマー6(配列番号6)を用いてPCRを行った。PCR増幅で得られたBiP分泌シグナル配列が連結された6D9のFdフラグメントの遺伝子およびプラスミドpFastBacTM Dual(インビトロジェン株式会社)を、いずれもEcoRIおよびHindIIIで消化し、次いでBiP分泌シグナル配列が連結されたFdフラグメント遺伝子をpFastBac Dualと連結し、pFastBac -/Fdを得た。
【0036】
(1−4.pFastBacTM Dualのp10プロモーター(Pp10)の下流へのFdフラグメント遺伝子の挿入)
pXIHA-6D9 Hcを鋳型とし、上流部分に制限酵素SmaIの認識配列(プライマーの4位〜9位に相当)および引き続きDrosophila由来のBiP分泌シグナル配列の一部(プライマーの10位〜33位に相当)を有するプライマー7(配列番号7)および下流部分に制限酵素KpnIの認識配列(プライマーの4位〜9位に相当)を有するプライマー8(配列番号7)を用いてPCRを行った。PCR増幅で得られたBiP分泌シグナル配列が連結された6D9のFdフラグメントの遺伝子およびプラスミドpFastBacTM Dualを、いずれもSmaIおよびKpnIで消化し、次いでBiP分泌シグナル配列が連結されたFdフラグメント遺伝子をpFastBac Dualと連結し、pFastBac Fd/-を得た。
【0037】
(1−5.pFastBac DualのPPHの下流へのL鎖遺伝子の挿入)
pXIHA-6D9 Lcを鋳型とし、上記のプライマー5および下流部分に制限酵素HindIIIの認識配列(プライマーの4位〜9位に相当)を有するプライマー9(配列番号9)を用いてPCRを行った。PCR増幅で得られたBiP分泌シグナル配列が連結された6D9のL鎖の遺伝子およびプラスミドpFastBacTM Dualを、いずれもEcoRIおよびHindIIIで消化し、次いでBiP分泌シグナル配列が連結されたL鎖遺伝子をpFastBac Dualと連結し、pFastBac -/Lcを得た。
【0038】
(1−6.pFastBacTM DualのPp10の下流へのL鎖遺伝子の挿入)
pXIHA-6D9 Lcを鋳型とし、上記のプライマー7および下流部分に制限酵素KpnIの認識配列(プライマーの4位〜9位に相当)を有するプライマー10(配列番号10)を用いてPCRを行った。PCR増幅で得られたBiP分泌シグナル配列が連結された6D9のL鎖の遺伝子およびプラスミドpFastBacTM Dualを、いずれもSmaIおよびKpnIで消化し、次いでBiP分泌シグナル配列が連結されたL鎖遺伝子をpFastBac Dualと連結し、pFastBac Lc/-を得た。
【0039】
(1−7.pFastBac -/FdへのL鎖遺伝子およびgp64遺伝子のハイブリッド遺伝子の挿入)
上記実施例1−2で得られたpBACsurf-1-Lcを鋳型とし、上流部分に制限酵素NheIの認識配列(プライマーの4位〜9位に相当)および引き続きgp64の分泌シグナル配列の一部(プライマーの10位〜37位に相当)を有するプライマー11(配列番号11)および下流部分に制限酵素NheIの認識配列(プライマーの4位〜9位に相当)を有するプライマー12(配列番号12)を用いて、gp64の分泌シグナル配列、6D9のL鎖の遺伝子、およびgp64遺伝子からなるハイブリッド遺伝子をPCRにより増幅した。増幅したハイブリッド遺伝子および上記実施例1−3で得られたpFastBac -/Fdを、いずれもNheIで消化し、次いでハイブリッド遺伝子をpFastBac -/Fdと連結し、そして正しい方向にハイブリッド遺伝子が挿入されたpFastBac Lc-gp64/Fdを回収した。
【0040】
(1−8.pFastBac Fd/-へのL鎖遺伝子およびgp64遺伝子のハイブリッド遺伝子の挿入)
上記実施例1−2で得られたpBACsurf-1-Lcを鋳型とし、上流部分に制限酵素EcoRIの認識配列(プライマーの4位〜9位に相当)および引き続きgp64の分泌シグナル配列の一部(プライマーの10位〜34位に相当)を有するプライマー13(配列番号13)および下流部分に制限酵素HindIIIの認識配列(プライマーの4位〜9位に相当)を有するプライマー14(配列番号14)を用いて、gp64の分泌シグナル配列、6D9のL鎖の遺伝子、およびgp64遺伝子からなるハイブリッド遺伝子をPCRにより増幅した。増幅したハイブリッド遺伝子および上記実施例1−4で得られたpFastBac Fd/-を、いずれもEcoRIおよびHindIIIで消化し、次いでハイブリッド遺伝子をpFastBac Fd/-と連結し、pFastBac Fd/Lc-gp64を得た。
【0041】
(1−9.pFastBac -/LcへのFdフラグメント遺伝子およびgp64遺伝子のハイブリッド遺伝子の挿入)
上記実施例1−1で得られたpBACsurf-1-Fdを鋳型とし、プライマー11およびプライマー12を用いて、gp64の分泌シグナル配列、6D9のFdフラグメントの遺伝子、およびgp64遺伝子からなるハイブリッド遺伝子をPCRにより増幅した。増幅したハイブリッド遺伝子および上記実施例1−5で得られたpFastBac -/Lcをいずれも、NheIで消化し、次いでハイブリッド遺伝子をpFastBac -/Lcと連結し、そして正しい方向にハイブリッド遺伝子が挿入されたpFastBac Fd-gp64/Lcを回収した。
【0042】
(1−10.pFastBac Lc/-へのFdフラグメント遺伝子およびgp64遺伝子のハイブリッド遺伝子の挿入)
上記実施例1−1で得られたpBACsurf-1-Fdを鋳型とし、プライマー13およびプライマー14を用いて、gp64の分泌シグナル配列、6D9のFdフラグメントの遺伝子、およびgp64遺伝子からなるハイブリッド遺伝子をPCRにより増幅した。増幅したハイブリッド遺伝子および上記実施例1−6で得られたpFastBac Lc/-を、いずれもEcoRIおよびHindIIIで消化し、次いでハイブリッド遺伝子をpFastBac Lc/-と連結し、pFastBac Lc/Fd-gp64を得た。
【0043】
(実施例2:組換えバキュロウイルスの作製)
大腸菌内で組換えバキュロウイルスDNAを作製する方法(Bac-to-Bacバキュロウイルス発現システム(インビトロジェン株式会社)を使用し、基本的にマニュアルに記載の方法にしたがって、組換えバキュロウイルスを作製した。
【0044】
上記実施例1で作製した4種類のベクターのpFastBac Lc-gp64/Fd、pFastBac Fd/Lc-gp64、pFastBac Fd-gp64/Lc、およびpFastBac Lc/Fd-gp64のそれぞれ1ngを、DH10Bacコンピテント大腸菌(インビトロジェン株式会社)100μlに対して加え、大腸菌を形質転換した。その後、S.O.C.培地(2%トリプトン、0.5%酵母エキス、10mM NaCl、2.5mM KCl、10mM MgCl2、10mM MgSO4、20mM グルコース)4mlを加え、試験管にて37℃で2〜3時間振とう培養を行った。細胞懸濁培地100μlをLB寒天培地(1%トリプトン、0.5%酵母エキス、0.5% NaCl、2%寒天、50μg/mlカナマイシン、7μg/mlゲンタマイシン、10μg/mlテトラサイクリン、100μg/ml Bluo-gal、40μg/ml IPTG)に塗布して24時間インキュベートした。生成した白色のコロニーを採取し、S.N.A.P.TMミディプレップキット(インビトロジェン株式会社)を用いて組換えバクミドDNAを分離した。直径35mmのペトリディッシュまたは6ウェルプレートに昆虫細胞Sf9(インビトロジェン株式会社)を2×106細胞/cm3の細胞密度で2mlのグレース培地(インビトロジェン株式会社)に播種し、27℃にて24時間培養した。この培養液に、上記組換えバクミドDNA1μgに対してトランスフェクション試薬FuGENE 6(ロシュ・ダイアグノスティックス)を3μlの割合で含む混合液を添加して、昆虫細胞Sf9をトランスフェクトし、27℃にて72時間インキュベートした。これにより、出芽型組換えバキュロウイルスが昆虫細胞から生じた。インキュベート後、出芽型組換えバキュロウイルスを含む培養上清を2000rpmにて5分間の遠心分離により回収し、10%ウシ胎児血清を含むTNM-FH培地(シグマアルドリッチジャパン)を用いて培養しているSf9細胞に添加し、27℃にて72時間インキュベートした。再度この操作を繰り返し、組換えバキュロウイルスを増殖させた。
【0045】
(実施例3:組換えバキュロウイルスの抗体表面提示の解析)
上記実施例2の出芽型組換えバキュロウイルスを含む培養上清を、抗原としてクロラムフェニコールモノエステルの加水分解反応の遷移状態アナログをハプテン基として結合したウシ血清アルブミン、および検出抗体としてヤギ抗マウスκ鎖-HRP(Exalpha Biologicals)を用いる酵素免疫測定法(ELISA)によって分析した。405nmでの吸光度を測定した。
【0046】
図1は、pFastBac Fd/Lc-gp64、pFastBac Fd-gp64/Lc、およびpFastBac Lc/Fd-gp64のそれぞれを含む培養上清をELISAにより分析した結果を示す。図1において、コントロールは、比較のためにウイルス未感染のSf9の培養上清を用いた場合の結果である。図1から明らかなように、上記のいずれのベクターを用いた場合もウイルス未感染のSf9の培養上清と比べて、高い吸光度が得られた。
【0047】
また、上記実施例2の出芽型組換えバキュロウイルスを含む培養上清をSf9の細胞懸濁培地2mlに添加し、そして27℃にて24時間インキュベートして、組換えバキュロウイルスをSf9に感染させた。この細胞懸濁培地に、FITC標識したヤギ抗マウスIgGであるFITC-F(ab’)2ヤギ抗マウスIgG(インビトロジェン株式会社)5μlを加えて30分間インキュベートし、引き続き細胞をリン酸緩衝液で洗浄し、そしてフローサイトメーター(FACSCalibur、日本ベクトン・ディッキンソン株式会社)を用いて解析した。
【0048】
pFastBac Fd/Lc-gp64およびpFastBac Fd-gp64/Lcを用いて作製した組換えバキュロウイルスの場合のフローサイトメーター解析結果を、それぞれ図2および3に示す。これらの図に見られるように、これら2種類の組換えバキュロウイルスに感染した細胞は、ウイルス未感染のSf9と比較して、高い蛍光強度を示した。
【0049】
これらの結果は、作製した組換えバキュロウイルスの感染によって、昆虫細胞Sf9の表面にマウスの抗体分子が発現および提示されることを示している。組換えバキュロウイルスが出芽する際に、抗体分子は、出芽型バキュロウイルスの表面に提示され得る。これらの結果から、作製した組換えバキュロウイルスは、出芽の際に抗原結合活性を有するFabフラグメントを表面に提示したと考えられる。
【0050】
この結果はまた、蛍光標識した抗原をウイルス感染細胞の表面に発現したFabフラグメントと結合させ、そしてフローサイトメーターを用いて強い蛍光を発する細胞をソーティングすることにより、目的の抗体が発現されたバキュロウイルスを選択できることを示唆する。
【産業上の利用可能性】
【0051】
本発明の出芽型組換えバキュロウイルスを用いることにより、抗原に対する特異性および親和性に優れ、そして安定性も高い抗体を作製することができる。このような抗体作製技術は、タンパク質の網羅的な定量解析を目的とした抗体マイクロアレイの構築や治療用抗体医薬の開発などにおいて、新規抗体の創出手段として有用であると考えられる。
【図面の簡単な説明】
【0052】
【図1】pFastBac Fd/Lc-gp64、pFastBac Fd-gp64/Lc、およびpFastBac Lc/Fd-gp64のそれぞれを含む培養上清をELISAにより分析した結果を示すグラフである。
【図2】pFastBac Fd/Lc-gp64を用いて作製した組換えバキュロウイルスを感染させたSf9および未感染Sf9のフローサイトメーターによる解析結果を示す図である。
【図3】pFastBac Fd-gp64/Lcを用いて作製した組換えバキュロウイルスを感染させたSf9および未感染Sf9のフローサイトメーターによる解析結果を示す図である。
【特許請求の範囲】
【請求項1】
抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルスであって、バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質として発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現し、表面に提示された該抗体またはそのFabフラグメントが抗原と結合し得る、出芽型組換えバキュロウイルス。
【請求項2】
抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルスを作製する方法であって、
バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質を発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現する、組換えバキュロウイルスを作製する工程
を含む、方法。
【請求項3】
前記組換えバキュロウイルスを作製する工程が、前記バキュロウイルスエンベロープ構成糖タンパク質をコードする遺伝子の5’末端側に連結した、該糖タンパク質の分泌シグナル配列および前記抗体またはそのFabフラグメントを構成する一方のペプチド鎖をコードする遺伝子を含み、かつ分泌シグナル配列および該抗体またはそのFabフラグメントの他方のペプチド鎖をコードする遺伝子を含むトランスファーベクターを構築する工程;および
該トランスファーベクターを介して目的遺伝子を昆虫細胞に導入する工程
を含む、請求項2に記載の方法。
【請求項4】
前記組換えバキュロウィルスを昆虫細胞に感染させる工程;および
前記感染させた昆虫細胞を培養してバキュロウイルスを出芽させる工程
をさらに含む、請求項2または3に記載の方法。
【請求項5】
抗体またはそのFabフラグメントを表面に提示する出芽型組換えバキュロウイルスディスプレイライブラリーを作製する方法であって、
異なる抗体またはそのFabフラグメントを発現する少なくとも2種の組換えバキュロウイルスを作製する工程であって、該組換えバキュロウイルスが各々、バキュロウイルスエンベロープ構成糖タンパク質と抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質を発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現する、工程;
該少なくとも2種の組換えバキュロウィルスを昆虫細胞に感染させる工程;および
該感染させた昆虫細胞を培養して、バキュロウイルスを出芽させる工程
を含む、方法。
【請求項6】
請求項5に記載の方法により作製されたバキュロウイルスディスプレイライブラリー。
【請求項7】
抗体またはそのFabフラグメントをスクリーニングする方法であって、
請求項6に記載のライブラリーに抗原を結合させる工程;および
該抗原が結合した該ライブラリーの抗体またはそのFabフラグメントを検出する工程
を含む、方法。
【請求項1】
抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルスであって、バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質として発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現し、表面に提示された該抗体またはそのFabフラグメントが抗原と結合し得る、出芽型組換えバキュロウイルス。
【請求項2】
抗体またはそのFabフラグメントを表面に提示し得る出芽型組換えバキュロウイルスを作製する方法であって、
バキュロウイルスエンベロープ構成糖タンパク質と該抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質を発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現する、組換えバキュロウイルスを作製する工程
を含む、方法。
【請求項3】
前記組換えバキュロウイルスを作製する工程が、前記バキュロウイルスエンベロープ構成糖タンパク質をコードする遺伝子の5’末端側に連結した、該糖タンパク質の分泌シグナル配列および前記抗体またはそのFabフラグメントを構成する一方のペプチド鎖をコードする遺伝子を含み、かつ分泌シグナル配列および該抗体またはそのFabフラグメントの他方のペプチド鎖をコードする遺伝子を含むトランスファーベクターを構築する工程;および
該トランスファーベクターを介して目的遺伝子を昆虫細胞に導入する工程
を含む、請求項2に記載の方法。
【請求項4】
前記組換えバキュロウィルスを昆虫細胞に感染させる工程;および
前記感染させた昆虫細胞を培養してバキュロウイルスを出芽させる工程
をさらに含む、請求項2または3に記載の方法。
【請求項5】
抗体またはそのFabフラグメントを表面に提示する出芽型組換えバキュロウイルスディスプレイライブラリーを作製する方法であって、
異なる抗体またはそのFabフラグメントを発現する少なくとも2種の組換えバキュロウイルスを作製する工程であって、該組換えバキュロウイルスが各々、バキュロウイルスエンベロープ構成糖タンパク質と抗体またはそのFabフラグメントを構成する一方のペプチド鎖との融合タンパク質を発現し、かつ該抗体またはそのFabフラグメントを構成する他方のペプチド鎖を分泌タンパク質として発現する、工程;
該少なくとも2種の組換えバキュロウィルスを昆虫細胞に感染させる工程;および
該感染させた昆虫細胞を培養して、バキュロウイルスを出芽させる工程
を含む、方法。
【請求項6】
請求項5に記載の方法により作製されたバキュロウイルスディスプレイライブラリー。
【請求項7】
抗体またはそのFabフラグメントをスクリーニングする方法であって、
請求項6に記載のライブラリーに抗原を結合させる工程;および
該抗原が結合した該ライブラリーの抗体またはそのFabフラグメントを検出する工程
を含む、方法。
【図1】


【図2】
【図3】
【図2】
【図3】
【公開番号】特開2008−194014(P2008−194014A)
【公開日】平成20年8月28日(2008.8.28)
【国際特許分類】
【出願番号】特願2007−35538(P2007−35538)
【出願日】平成19年2月15日(2007.2.15)
【出願人】(502059825)Bio−energy株式会社 (16)
【出願人】(504150450)国立大学法人神戸大学 (421)
【Fターム(参考)】
【公開日】平成20年8月28日(2008.8.28)
【国際特許分類】
【出願日】平成19年2月15日(2007.2.15)
【出願人】(502059825)Bio−energy株式会社 (16)
【出願人】(504150450)国立大学法人神戸大学 (421)
【Fターム(参考)】
[ Back to top ]