
抗体模倣物および他の結合タンパク質のためのタンパク質骨格
【課題】関心対象のどのような化合物とも結合するように導出(evolve)しうる新規のタンパク質のファミリーを提供する。
【解決手段】免疫グロブリンの折畳みを含み、骨格として使用できるタンパク質をコードする核酸、ならびに診断方法におけるこのようなタンパク質の使用法、および化合物に結合する新規の分子種およびそれらのリガンドを展開する方法におけるこのようなタンパク質の使用法。
【解決手段】免疫グロブリンの折畳みを含み、骨格として使用できるタンパク質をコードする核酸、ならびに診断方法におけるこのようなタンパク質の使用法、および化合物に結合する新規の分子種およびそれらのリガンドを展開する方法におけるこのようなタンパク質の使用法。
【発明の詳細な説明】
【技術分野】
【0001】
発明の背景
本発明は、例えば新規の結合特性を持つ生成物の生産に有用なタンパク質骨格に関する。
【背景技術】
【0002】
一般にタンパク質骨格と呼ばれる比較的限定された三次元構造を持つタンパク質は、操作された生成物を設計するための試薬として用いることができる。これらの骨格は、特定の、もしくは無作為な配列の変異を受け入れる1つまたは複数の領域を典型的に含んでおり、このような配列の無作為化は、タンパク質ライブラリーを生産し、そのライブラリーから求める生成物を選択するためにしばしば行われる。このような骨格が有用な1つの特定の分野は、抗体設計の分野である。
【0003】
試薬または薬剤を得るために哺乳類の免疫系を操作する多くの方法が以前より試みられてきた。これらには、特定の抗原に反応するポリクローナル抗体の混合物を得るために関心対象の抗原を動物に注射する段階、ハイブリドーマ培養細胞にモノクローナル抗体を生産させる段階(KoehlerおよびMilstein、Nature、第256巻、第495頁、1975年(非特許文献1))、新規の認識特性または最適化された認識特性を得るために現存のモノクローナル抗体を改変する段階、望ましい結合特性を持つ新規の抗体断片を作製する段階、ならびに単鎖抗体(抗体分子の重鎖および軽鎖の可変領域を可動性のペプチドリンカーと連結することにより作製される)を無作為化してファージディスプレイ法で抗原との結合により選択する段階(Clacksonら、Nature、第352巻、第624頁、1991年(非特許文献2))が含まれる。
【0004】
さらに、新規の結合特性を持つタンパク質を得るために、免疫グロブリン以外のいくつかのタンパク質骨格が提案されている。例えば「ミニボディ」骨格は、免疫グロブリンの折り畳みに関連しており、モノクローナル抗体の重鎖の可変ドメインから3つのβ鎖を除去することで設計されている(Tramontanoら、J. Mol. Recognit.、第7巻、第9頁、1994年(非特許文献3))。このタンパク質は61残基を含んでおり、これを使用して2つの超可変ループを生じうる。これらの2つのループは無作為化され、抗原との結合により生成物が選択されるが、これまでのところ、溶解性の問題により、この枠組みには幾らかの限定された効用しかないようである。ループを示すのに使用される別の枠組みは、テンダミスタット(tendamistat)であり、74残基で、サンドイッチ状の6本のβシート鎖が2つのジスルフィド結合によって結合している(McConnellおよびHoess、J. Mol. Biol.、第250巻、第460頁、1995年(非特許文献4))。この骨格には3つのループが含まれているが、これまでのところ、これらのループのうちの2つについてのみ、無作為化の可能性が試されている。
【0005】
他のタンパク質も枠組みとして試験されてきており、αヘリックスの表面に無作為化された残基(Nordら、Nat. Biotechnol.、第15巻、第772頁、1997年(非特許文献5)、Nordら、Protein Eng.、第8巻、第601頁、1995年(非特許文献6))、αヘリックス束中のαヘリックス間にあるループ(KuおよびSchultz、Proc. Natl. Acad. Sci. USA、第92巻、第6552頁、1995年(非特許文献7))、および小さいプロテアーゼ阻害剤のようにジスルフィド架橋で束ねられたループを提示するために使用されている(Marklandら、Biochemistry、第35巻、第8045頁、1996年(非特許文献8);Marklandら、Biochemistry、第35巻、第8058頁、1996年(非特許文献9);RottgenおよびCollins、Gene、第164巻、第243頁、1995年(非特許文献10);Wangら、J. Biol. Chem.、第270巻、第12250頁、1995年(非特許文献11))。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】KoehlerおよびMilstein、Nature、第256巻、第495頁、1975年
【非特許文献2】Clacksonら、Nature、第352巻、第624頁、1991年
【非特許文献3】Tramontanoら、J. Mol. Recognit.、第7巻、第9頁、1994年
【非特許文献4】McConnellおよびHoess、J. Mol. Biol.、第250巻、第460頁、1995年
【非特許文献5】Nordら、Nat. Biotechnol.、第15巻、第772頁、1997年
【非特許文献6】Nordら、Protein Eng.、第8巻、第601頁、1995年
【非特許文献7】KuおよびSchultz、Proc. Natl. Acad. Sci. USA、第92巻、第6552頁、1995年
【非特許文献8】Marklandら、Biochemistry、第35巻、第8045頁、1996年
【非特許文献9】Marklandら、Biochemistry、第35巻、第8058頁、1996年
【非特許文献10】RottgenおよびCollins、Gene、第164巻、第243頁、1995年
【非特許文献11】Wangら、J. Biol. Chem.、第270巻、第12250頁、1995年
【発明の概要】
【0007】
本発明は、関心対象のどのような化合物とも結合するように導出(evolve)しうる新規のタンパク質のファミリーを提供する。フィブロネクチン型III(Fn3)もしくはFn3様ドメイン由来の骨格を一般に使用したこれらのタンパク質は、特徴的な天然型抗体または操作された抗体(すなわち、ポリクローナル抗体、モノクローナル抗体、もしくは単鎖の抗体)の様式で機能し、さらに構造的な利点を持つ。特に、これらの抗体模倣物の構造は、標準的には抗体の構造および機能の喪失につながるような条件下でさえも、最適な折り畳み、安定性、および溶解性を持つように設計されている。
【0008】
これらの抗体模倣物は、実質的に関心対象の任意の化合物(例えば任意のタンパク質)とも結合しうるタンパク質を設計する目的で使用することができる。例えば、本明細書に述べられた10Fn3に基づく分子を、抗体の可変領域の相補性決定領域(CDR)と位置および構造において類似した無作為化されたFn3ループを1つまたは複数有する集団を形成するために、および/または、Fn3の他の3つの可溶性露出ループを無作為化するために、定方向展開(directed evolution)に供する骨格として使用することができる。このような定方向展開の方法により、関心対象の抗原に高い親和性を持つ抗体様分子が生産される。さらに、本明細書に述べられた骨格を規定された露出ループ(例えば、前もって無作為化され、抗原との結合に基づいて選択されたループ)を示し、このように導入されたループと結合する分子の展開(evolution)の方向付けをするためにも使用することができる。この型の選択は、個々の任意のCDR様ループ、または非線形エピトープに結合した2つもしくは3つ全部のCDR様ループを認識する認識分子を同定するために行ってもよい。
【0009】
したがって、第1の局面では、本発明は無作為化されたまたは変異した骨格タンパク質に関する。特に、本発明は免疫グロブリン様の折畳みを持つドメインを含む非抗体タンパク質に関するが、その非抗体タンパク質は変異したアミノ酸配列を持つことにより基準タンパク質から誘導され、この非抗体タンパク質は基準タンパク質が非抗体タンパク質ほど強く結合しない化合物に対して少なくとも1μMの強さのKdで結合する。
【0010】
さらに、本発明は免疫グロブリン様の折畳みをもつドメインを含む骨格タンパク質から誘導される非抗体タンパク質に関し、誘導体タンパク質中のドメインのアミノ酸配列は、骨格タンパク質中のドメインのアミノ酸配列と50%以上同一である。
【0011】
別の態様では、本発明は少なくとも1つの無作為化ループを持つフィブロネクチン3型ドメインを含むタンパク質に関するが、このタンパク質はFn3ドメインが対応する天然に存在するFn3ドメインの結合しない化合物に結合できるということにより特徴づけられる。
【0012】
種々の好ましい態様では、本発明のこれらの任意のタンパク質は、少なくとも500 nMのKd、好ましくは少なくとも100 nMまたは10 nMのKd、およびより好ましくは少なくとも1 nM、500 pM、100 pM、または20 pMの強さのKdでその標的化合物に結合する。タンパク質は好ましくは1個、2個、または3個の変異ループを含み、少なくとも1個のループ、および好ましくは2個または3個全てのループが化合物に対するタンパク質の結合に貢献する。さらに、基準タンパク質は好ましくはジスルフィド結合を持たず、誘導体タンパク質は少なくとも1つのジスルフィド結合を持つ可能性がある。
【0013】
特定の態様に関して、免疫グロブリン様の折畳みをもつドメインは、好ましくは10 kD未満または7.5 kDを越える分子量、およびより好ましくは7.5〜10 kDの間の分子量を持つ。本発明のタンパク質は生理的条件下で単量体であるか、例えば二量体のような多量体である。他の好ましい態様では、本発明の変異タンパク質を誘導するために用いた基準タンパク質は、天然に存在する哺乳類タンパク質(例えば、ヒトのタンパク質)である;免疫グロブリン様の折畳みをもつドメインは、変異しており、基準タンパク質の免疫グロブリン様の折畳みと比較して最高50%、および好ましくは最高34%の変異したアミノ酸を含む。さらに、免疫グロブリン様の折畳みをもつドメインは、約50〜150アミノ酸を含み、より好ましくは約50アミノ酸を含む。
【0014】
本発明の誘導体タンパク質は、好ましいタンパク質、フィブロネクチンまたはフィブロネクチン二量体、テナシン、N-カドヘリン、E-カドヘリン、ICAM、タイチン、GCSF受容体、サイトカイン受容体、グリコシダーゼ阻害剤、抗生物質色素蛋白質、ミエリン膜接着分子P0、CD8、CD4、CD2、クラスI MHC、T細胞抗原受容体、CD1、C2、およびVCAM-1のI-setドメイン、ミオシン結合タンパク質CのI-set免疫グロブリンドメイン、ミオシン結合タンパク質HのI-set免疫グロブリンドメイン、テロキンのI-set免疫グロブリンドメイン、NCAM、トウィチン、ニューログリアン、成長ホルモン受容体、エリスロポエチン受容体、プロラクチン受容体、インターフェロンγ受容体、βガラクトシダーゼ/グルクロニダーゼ、βグルクロニダーゼ、トランスグルタミナーゼ、T細胞抗原受容体、スーパーオキシド・ジスムターゼ、組織因子ドメイン、シトクロームF,緑色蛍光タンパク質、GroEL、およびタウマチンを含むがこれらに限定されることのない任意の適当な基準タンパク質から誘導され得る。
【0015】
Fn3ドメインを含むタンパク質の別の好ましい態様では、フィブロネクチン3型ドメインは哺乳類(例えばヒト)フィブロネクチン3型ドメインである;このタンパク質はフィブロネクチン3型ドメインの10番目のモジュール(10Fn3)を含む。そのようなタンパク質では、化合物の結合は、好ましくは1個、2個、または3個の10Fn3ループによって媒介される。別の好ましい態様では、10Fn3の第2の(DE)ループが天然に存在する分子に比較して伸長しているか、10Fn3がインテグリン結合モチーフを欠いている場合がある。これらの分子では、インテグリン結合モチーフは、アミノ酸配列によって置換されている可能性があり、ここでは極性アミノ酸-中性アミノ酸-酸性アミノ酸の配列(N末端からC末端の方向)がインテグリン結合モチーフを置き換えている;または、1つの好ましい配列は、セリン-グリシン-グルタミン酸である。別の好ましい態様では、フィブロネクチン3型ドメインを含む本発明のタンパク質は、ジスルフィド結合を欠いている。
【0016】
本発明の任意のタンパク質(例えば、フィブロネクチン3型ドメインを含むタンパク質)は、融合タンパク質の一部として製剤化されていてもよい。化合物との結合または化合物との結合の選択に融合タンパク質が使用される場合は、融合タンパク質はそれ自身は関心対象の化合物に結合しない異種のタンパク質を含む。異種タンパク質は、例えば、抗体または抗体ドメイン(たとえば免疫グロブリンFcドメイン)、補体タンパク質、毒素タンパク質,またはアルブミンタンパク質でよい。また、本発明の任意のタンパク質(たとえば、フィブロネクチン3型ドメインタンパク質)は、核酸(たとえば、RNA)と共有結合していてもよく、核酸はタンパク質をコードしていてもよい。さらに、このタンパク質は多量体でもよく、または特にインテグリン結合モチーフを欠失する場合には、生理学的に許容される担体として製剤化されていてもよい。
【0017】
本発明はさらに、βシート配列に少なくとも1つの変異を持つフィブロネクチン3型ドメインを含むタンパク質の特徴を含んでいる。また、これらのタンパク質は、対応する天然型のフィブロネクチンとは結合しないまたはそれほどは強く結合しない化合物と結合することができることにより特徴付けられる。
【0018】
本発明の任意のタンパク質は、固相支持体(例えば、ビーズまたはチップ)上に固定することができ、これらの骨格は固相支持体上でアレイを含む任意の形に配置できる。
【0019】
関連する局面では、本発明はさらに本発明の任意のタンパク質をコードする核酸を特徴とする。好ましい態様では、核酸はDNAまたはRNAである。
【0020】
別の関連する局面では、本発明はまた、フィブロネクチン3型ドメインのインテグリン結合ドメインの除去を伴う、フィブロネクチン3型ドメインを含み、哺乳類に対して薬学的に許容される、タンパク質の生成方法も特徴とする。本方法は、上述の任意のフィブロネクチン3型ドメイン含有タンパク質に適用でき、ヒトの治療的応用のためのタンパク質の生成に特に有用である。本発明は、インテグリン結合ドメインを持たない、そのようなフィブロネクチン3型ドメイン含有タンパク質も特徴とする。
【0021】
別の関連する局面では、本発明は、関心対象の化合物に結合する誘導非抗体タンパク質を得る方法を特徴とする。そのような方法の1つは:(a) 免疫グロブリン様の折畳みを含む非抗体骨格タンパク質を提供する段階であって、骨格タンパク質は1μMほどの強いKdで化合物には結合しない;(b) 非抗体骨格タンパク質の変異した誘導体を作製し、それによって変異したタンパク質のライブラリーを作製する段階;(c) ライブラリーに化合物を接触させる段階;(d) ライブラリーから少なくとも1μMのKdの強さで化合物に結合する少なくとも1つの誘導体タンパク質を選択する段階;および(e) 選択的に、繰り返しの段階(b)の非抗体骨格タンパク質をその前の段階(d)で置き換えて段階(b)〜(d)を繰り返す段階を含む。このテクニックは、本発明の任意のタンパク質(例えば、フィブロネクチン3型ドメインを含む任意のタンパク質)についても行なうことができる。
【0022】
さらに別の関連する局面において、本発明は関心対象の化合物と結合しうる無作為化または変異した本発明のタンパク質を得る、もしくは導出するために、または無作為化または変異したモチーフを含む特定のタンパク質と結合しうる化合物(例えば、タンパク質)を得る、もしくは導出するために使用されうるスクリーニング法を特徴としている。さらに、本発明は、関心対象の化合物またはタンパク質のいずれかを得るために、2つの方法を任意の順番で結合したスクリーニング法を特徴としている。
【0023】
特に、関心対象の無作為化または変異したタンパク質を単離または同定するのに有用な最初のスクリーニング法は、(a)関心対象の化合物に候補タンパク質を接触させる段階であって、候補タンパク質は免疫グロブリン様の折畳みをもつドメインを含む誘導非抗体タンパク質であり、非抗体タンパク質は変異したアミノ酸配列を持つことにより基準タンパク質から誘導され、非抗体タンパク質は基準タンパク質がそれほどは強く結合しない化合物に対して少なくとも1μMのKdの強さで結合し、接触が化合物とタンパク質の複合体の形成を可能にする条件下で行われる段階;および(b) 複合体からタンパク質に結合する化合物を得る段階を含んでいる。この一般的なテクニックは、少なくとも1つの無作為化または変異したループを持つフィブロネクチン3型ドメインタンパク質で実行してもよい。
【0024】
第2のスクリーニング方法は、本発明のタンパク質に結合する化合物を単離または同定するためのものである。この方法は、免疫グロブリン様の折畳みをもつドメインを含み、変異したアミノ酸配列を持つことで基準タンパク質から誘導される非抗体タンパク質から始めるが、この非抗体タンパク質は基準タンパク質がそれほど強く結合しない化合物に対して少なくとも1μMの強さのKdで結合する。この誘導体タンパク質に候補化合物を接触させるが、接触は化合物とタンパク質の複合体の形成が可能な条件下で行われ、誘導体タンパク質に結合する化合物を複合体から得る。やはり、この一般的テクニックは、例えば、変異したフィブロネクチン3型ドメインを持つタンパク質のような、本発明の任意のタンパク質を用いて実行できる。
【0025】
また、本発明は、本発明のタンパク質(たとえば、フィブロネクチン3型骨格タンパク質およびその誘導体)を用いる診断方法を特徴とする。そのような診断方法は、試料(例えば、生物試料)に対して行ない、試料中で1つの分析物を検出したり、または多くの異なる分析物を同時に検出することができる。本方法では、本明細書中に記述される任意の骨格分子を利用できる。好ましくは、本方法は、(a)化合物分析物に結合する本発明のタンパク質を、試料に接触させる段階であって、化合物-タンパク質複合体の形成を可能にする条件下で接触が行われる段階;および (b) 複合体を検出し、それにより試料中の化合物を検出する段階、を含む。また、この方法は試料中の化合物レベルを検出するのみならず定量するためにも使用できる。
【0026】
本明細書に記載される任意の選択または診断方法の好ましい態様では、本発明のタンパク質は標的化合物に対して少なくとも1μMまたは500 nMの強さのKd、好ましくは少なくとも100 nMまたは10 nMの強さのKd、およびより好ましくは少なくとも1 nM、500 pM、100 pM、または20 pMの強さのKdですらその標的化合物に結合する。タンパク質は好ましくは1個、2個、または3個の変異ループを含み、少なくとも1個のループ、および好ましくは2個または3個全てのループが化合物に対するタンパク質の結合に貢献する。さらに、基準タンパク質は好ましくはジスルフィド結合を持たず、誘導体タンパク質は少なくとも1つのジスルフィド結合を持つ可能性がある。
【0027】
本方法の特定の態様に関して、免疫グロブリン様の折畳みをもつドメインは、好ましくは10 kD未満または7.5 kDを越える分子量、およびより好ましくは7.5〜10 kDの間の分子量を持つ。本発明のタンパク質は生理的条件下で単量体であるか、例えば二量体のような多量体である。他の好ましい態様では、本発明の変異タンパク質を誘導するために用いた基準タンパク質は、天然に存在する哺乳類タンパク質(例えば、ヒトのタンパク質)である;免疫グロブリン様の折畳みをもつドメインは変異しており、基準タンパク質の免疫グロブリン様の折畳みと比較して最高50%、および好ましくは最高34%の変異したアミノ酸を含む。さらに、免疫グロブリン様の折畳みをもつドメインは、約50〜150アミノ酸を含み、より好ましくは約50アミノ酸を含む。
【0028】
本発明の方法で使用される誘導体タンパク質は、好ましいタンパク質、フィブロネクチンまたはフィブロネクチン二量体、テナシン、N-カドヘリン、E-カドヘリン、ICAM、タイチン、GCSF受容体、サイトカイン受容体、グリコシダーゼ阻害剤、抗生物質色素蛋白質、ミエリン膜接着分子PO、CD8、CD4、CD2、クラスI MHC、T細胞抗原受容体、CD1、C2、およびVCAM-1のI-setドメイン、ミオシン結合タンパク質CのI-set免疫グロブリンドメイン、ミオシン結合タンパク質HのI-set免疫グロブリンドメイン、テロキンのI-set免疫グロブリンドメイン、NCAM、トウィチン、ニューログリアン、成長ホルモン受容体、エリスロポエチン受容体、プロラクチン受容体、インターフェロンγ受容体、βガラクトシダーゼ/グルクロニダーゼ、βグルクロニダーゼ、トランスグルタミナーゼ、T細胞抗原受容体、スーパーオキシド・ジスムターゼ、組織因子ドメイン、シトクロームF,緑色蛍光タンパク質、GroEL、およびタウマチンを含むがこれらに限定されることのない任意の適当な基準タンパク質から誘導され得る。
【0029】
また、本明細書に記述される選択方法の段階は、サイクルの間にさらなる変異または無作為化を行なって繰り返すことができる。例えば、変異または無作為化したフィブロネクチン3型ドメインを持つタンパク質に関する方法では、段階(b)で得られたタンパク質のフィブロネクチン3型ドメインの少なくとも1個のループを変異させ、さらに無作為化されたタンパク質を用いて段階(a)および(b)を繰り返すか、段階(b)で得られた化合物を修飾し、さらに修飾された化合物を用いて段階(a)および(b)を繰り返すことができる。これらの方法では、化合物は好ましくはタンパク質であり、フィブロネクチン3型ドメインは好ましくは哺乳類(例えばヒト)フィブロネクチン3型ドメインである。別の好ましい態様では、このタンパク質はフィブロネクチン3型ドメインの10番目のモジュール(10Fn3)を含み、結合は1個、2個、または3個の10Fn3ループによって媒介される。また、10Fn3の第2の(DE)ループが天然に存在する分子と比較して伸長しているか、10Fn3がインテグリン結合モチーフを欠いている場合がある。やはり上述のように、インテグリン結合モチーフは置換されている場合があり、ここでは、極性アミノ酸-中性アミノ酸-酸性アミノ酸の配列(N末端からC末端の方向)がインテグリン結合モチーフを置き換えている;または、1つの好ましい配列は、セリン-グリシン-グルタミン酸である。
【0030】
本明細書に記述される選択および診断方法は、本発明の任意のタンパク質を用いて実行できる(例えば、フィブロネクチン3型ドメインを含むタンパク質)。また、これらの任意のタンパク質は、それ自身は関心対象の化合物に結合しない異種のタンパク質(例えば、抗体または抗体ドメイン(免疫グロブリンFcドメインを含む)、または補体タンパク質、毒素タンパク質、またはアルブミンタンパク質)との融合タンパク質の一部として製剤化されていてもよい。また、選択および診断方法は、核酸(たとえば、RNAまたはタンパク質をコードする任意の核酸)と共有結合していてもよい。さらに、単量体または二量体のような多量体としてのこれらのタンパク質(例えば、フィブロネクチンドメインを含むタンパク質)を用いて、選択および診断方法を実行することができる。
【0031】
好ましくは、選択および診断方法は、固相支持体上への結合した標的の固定化を含んでいる。好ましい固相支持体には、カラム(例えば、アガロースカラムのような親和性カラム)またはマイクロチップ、またはビーズが含まれる。または、タンパク質(例えばFn3ドメインを含むタンパク質)を固定化し、これに1個または複数の潜在的結合標的を接触させることもできる。
【0032】
診断方法では、化合物はしばしばタンパク質であるが、試料中の任意の他の分析物でもよい。検出は、X線撮影、蛍光検出、質量分光、または表面プラズモン共鳴を含むが、これらに限定されない、任意の標準的な手法で行われる。
【0033】
最後の局面では、本発明は少なくとも1μMの強さのKdで腫瘍壊死因子α(TNF-α)に結合する非抗体タンパク質で、TNF-α受容体(例えば、天然に存在するTNF-α受容体、例えば哺乳類またはヒトのTNF-α受容体)とは20%未満の同一な配列を持つタンパク質の特徴を含んでいる。
【0034】
好ましい態様では、このタンパク質は、変異したフィブロネクチン3型ドメインを含み、タンパク質はDE、BC、およびFGループに変異を持つ。好ましくは変異したFGループは野生型のFGループと同じ長さである。別の好ましい態様では、タンパク質は、約50〜150アミノ酸、および好ましくは約50アミノ酸からなる、免疫グロブリン様の折畳み(好ましくは10 kD未満、7.5 kDを越える、または7.5〜10 kDの間の分子量を持つ)を含む。
【0035】
本発明のTNF-α結合体は、少なくとも1μMの強さ、好ましくは少なくとも500 nM、100 nM、または10 nMの強さのKd、およびより好ましくは少なくとも1 nMまたは500 pMの強さ、および最も好ましくは100 pMまたは20 pMの強さのKdでTNF-αに結合する。これらのタンパク質は好ましくは1個、2個、または3個の変異ループを含み、少なくとも1個のループ、および好ましくは2個または3個全てのループが、TNF-αに対するこの非抗体タンパク質の結合に貢献する。別の好ましい態様では、この非抗体タンパク質は少なくとも1つのジスルフィド結合を持ち、非抗体タンパク質は生理的条件下で単量体または二量体である。
【0036】
TNF-α結合体は、固相支持体、(例えば、ビーズまたはチップ)上に固定することができ、アレイの一部となっていてもよい。また、TNF-α結合体には、異種タンパク質(例えば、TNF-α、免疫グロブリンFcドメイン、補体タンパク質、またはアルブミンタンパク質に結合しない抗体または抗体ドメインである異種タンパク質)が結合していてもよい。
【0037】
必要ならば、タンパク質は変異フィブロネクチン3型ドメイン(例えば、フィブロネクチン3型ドメインの変異した10番目のモジュール(10Fn3)のようなヒトフィブロネクチン3型ドメイン由来のもの)を含むことができる。また、タンパク質は10Fn3インテグリン結合モチーフを欠如していてもよい。TNF-α結合体は、好ましくは10Fn3のループ(例えば、ループ配列PW(A/G))に非天然配列を含み、10Fn3のβシートに非天然配列を含む可能性がある。本発明の特に好ましいTNF-α結合体は、図25(配列番号:34〜140)に示されている。
【0038】
また、関連する局面では、本発明は本発明の任意のTNF-α結合タンパク質、および図25(配列番号:34〜140)の任意のアミノ酸配列を含む任意のタンパク質上のループ構造をコードする核酸の特徴を含んでいる。
【0039】
本明細書では「非抗体タンパク質」は、天然でも哺乳類の免疫後でも哺乳類のB細胞によって産生されないタンパク質を意味する。この用語はまた、長さが100アミノ酸以上、好ましくは80アミノ酸以上、および最も好ましくは50アミノ酸以上の抗体断片も含まない。
【0040】
「免疫グロブリン様の折畳み」は、逆方向に平行の2層のβシートを含む約80〜150アミノ酸残基のタンパク質ドメインであって、2つのβシートの平らな疎水性の面が互いに押し付けられているタンパク質ドメインを意味する。本発明のタンパク質は、共有結合または非共有結合で、より大きな構造に会合したいくつかの免疫グロブリン様の折畳みを含む可能性がある。
【0041】
「骨格」は、結合のような特定の好ましい性質を持つタンパク質枠組みを選択または設計するために用いられるタンパク質を意味する。骨格からタンパク質を設計するときには、枠組みの望ましい特性に重要なアミノ酸残基は保持され、他の残基を変化させることができる。そのような骨格は、異なる性質を持つタンパク質誘導体の間で変化するアミノ酸残基を50%未満持ち、そのような誘導体の間で一定の残基を50%以上持つ。最も一般的には、これらの一定の残基は、その性質に関係なく、全ての変化するドメインに、同一の全体的な3次元折畳みを与える。
【0042】
「フィブロネクチン3型ドメイン」は互いに充填し合ってタンパク質のコアを形成する2個のβシート間に散在する7個もしくは8個のβ鎖を持つドメインであって、かつβ鎖を互いに連結して、溶媒に露出されたループをさらに含むドメインを意味する。βシートサンドイッチの両端には、このようなループが少なくとも3個存在し、この端はβ鎖の方向と垂直なタンパク質の境界である。好ましくは、フィブロネクチン3型ドメインは、RCSB (Research Collaboratory for Structural Bioinformatics) プロテインデータベース(Protein Data Base)より入手可能であって、「1ttg」(ID=「1ttg」(1つのttg))として引用される10Fn3ドメインの構造をコードする配列とアミノ酸レベルで少なくとも30%の同一性を呈し、および好ましくはアミノ酸レベルで少なくとも50%の同一性を呈する配列を含んでいる。この定義において引用される配列の同一性は、モレキュラー・シミュレーション(Molecular Simulation)(San Diego、CA)より入手可能なホモロジー・プログラムによって決定される。本発明はさらに、10Fn3関連分子の重合体であって、単量体構造の使用の延長に当たり、ポリタンパク質のサブユニットが配列において同一であるかもしくはないものを含む。
【0043】
「天然に存在する」とは、生きている生物によってコードされる任意のタンパク質を意味する。
【0044】
「無作為化された」または「変異した」とは、鋳型配列に比べて1個または複数のアミノ酸変異を含むことを意味する。「無作為化する」または「変異させる」とは、そのようなアミノ酸変異を配列中の導入する過程を意味する。無作為化または変異は、一般的には核酸コード配列の意図的な、盲目の、または自発的な変異の変化によって形成され、例えば、PCR、変異性PCR、またはDNA化学合成のような任意の手法によって作製される。「対応する非変異タンパク質」とは、導入されたアミノ酸変異以外は、配列の同一なタンパク質を意味する。
【0045】
「タンパク質」とは、長さ、翻訳後修飾、もしくは機能にかかわらず、2個またはそれ以上のアミノ酸の任意の配列を意味する。「タンパク質」および「ペプチド」は、本明細書において互換的に用いられる。
【0046】
「RNA」とは、二つまたはそれ以上の共有結合した、天然または改変されたリボヌクレオチドの配列を意味する。この用語に含まれる改変されたRNAの一つの例は、ホスホロチオエートRNAである。
【0047】
「DNA」とは、二つまたはそれ以上の共有結合した、天然または改変されたデオキシリボヌクレオチドの配列を意味する。
【0048】
「核酸」とは、任意の2つまたはそれ以上の共有結合したヌクレオチドまたはヌクレオチド類似体もしくは誘導体を意味する。本明細書において用いられるように、この用語にはDNA、RNA、およびPNAが含まれるが、これらに限定されることはない。
【0049】
「薬学的に許容される」とは、重大な医学的に有害な結果なしに、動物(例えば哺乳類)に投与しうる化合物またはタンパク質を意味する。
【0050】
「生理学的に許容される担体」とは、処理された宿主に対して重大な有害な影響を持たず、投与される化合物の治療上の性質を維持する担体を意味する。一つの例示的な生理学的に許容される担体は生理食塩水である。他の生理学的に許容される担体およびその処方は当業者には自明であって、例えば「レミントンの薬剤科学(Remington's Pharmaceutical Sciences)」、(第18版)、A. Gennaro編、1990年、Mack Publishing Company、Easton、PA、に述べられており、参照として本明細書に組み入れられている。
【0051】
「融合タンパク質」とは、(i)本発明の骨格タンパク質が(ii)第2の異なる(異種)タンパク質に結合したタンパク質を意味する。「融合タンパク質」はすべてアミノ酸から構成されるが、「核酸-タンパク質融合体」および「RNA-タンパク質融合体」はどちらもアミノ酸のひと配列(タンパク質成分)に結合した核酸のひと配列(核酸またはRNA成分)を含むという点で、「融合タンパク質」は「核酸-タンパク質融合体」および「RNA-タンパク質融合体」とは異なっている。
【0052】
「選択」とは、ある分子を集団の他の分子から実質的に分離することを意味する。本明細書において用いられるように、「選択」段階は、選択段階後には集団中の望ましい分子が望ましくない分子に比べて少なくとも2倍、好ましくは少なくとも30倍、より好ましくは少なくとも100倍、最も好ましくは少なくとも1000倍に濃縮されることを提供する。選択段階は何回繰り返してもよく、所与の方法中で異なる型の選択段階が組み合わされていてもよい。
【0053】
本明細書において用いられる「結合パートナー」とは、所望の関心対象の化合物(例えば、タンパク質)のある部分に対する特異的、共有結合、または非共有結合親和性を有するいかなる分子も意味する。結合パートナーの例には、抗原/抗体対、タンパク質/阻害剤対、受容体/リガンド対(例えば、ホルモン受容体/ペプチドホルモン対のような細胞表面受容体/リガンド対)、酵素/基質対(例えば、キナーゼ/基質対)、レクチン/炭化水素対、オリゴマーまたはヘテロオリゴマータンパク質凝集体、DNA結合タンパク質/DNA結合部位、RNA/タンパク質対、および核酸二本鎖、ヘテロ二本鎖、または連結鎖のメンバー、並びに、他の分子(例えば、化合物またはタンパク質)のいかなる部分とも1つもしくはそれ以上の共有結合または非共有結合を形成することができる任意の分子が含まれるが、これらに限定されることはない。
【0054】
「固相支持体」とは、それに対して直接または間接的に(例えば、他の抗体またはプロテインAのような他の結合パートナー中間体を通じて)抗体模倣体もしくは親和性複合体が結合してもよい、またはその中に抗体模倣体もしくは親和性複合体が埋め込まれてもよい(例えば、受容体またはチャンネルを通じて)、任意のカラム(もしくはカラム材料)、ビーズ、試験管、マイクロタイター皿、固体粒子(例えば、アガロースもしくはセファロース)、マイクロチップ(例えば、シリコン、シリコンガラス、もしくは金チップ)、またはメンブレン(例えば、無機メンブレン、ニトロセルロース、またはリポソームもしくは小胞のメンブレン)を意味するがこれらに限定されることはない。
【0055】
本発明によって、多くの利点が提供される。例えば、以下により詳細に述べられているように、本抗体模倣物は還元条件下での安定性および高濃度での溶解性など、改良された生物物理学的性質を呈する。さらに、これらの分子は大腸菌(E. coli)などの原核生物系、酵母などの真核生物系、およびウサギ網状赤血球溶解物系などのインビトロ翻訳系において容易に発現し、折り畳まれる。さらに、これらの分子は、RNA-タンパク質融合技術を用いたインビトロ選択(RobertsおよびSzostak、Proc. Natl. Acad. Sci. USA、第94巻、第12297頁、1997年;Szostakら、米国特許出願第09/007,005号および米国特許出願第09/247,190号;Szostakら、国際公開公報第98/31700号)、ファージ・ディスプレイ(例えば、SmithおよびPetrenko、Chem. Rev.、第97巻、第317頁、1997年を参照)、および酵母ディスプレイ系(例えば、BoderおよびWittrup、Nature Biotech.、第15巻、第553頁、1997年を参照)を含む選択を多回数繰り返すことを含む親和性成熟(affinity maturation)技術にきわめて適している。
【0056】
本発明の他の特徴および利点は、以下の詳細な説明および特許請求の範囲から明らかになると思われる。
【0057】
詳細な説明
本明細書に記載される新規の抗体模倣物は、抗体に由来する断片、および例えば前掲の枠組みのような非抗体の枠組みのどちらよりも優れているように設計されている。
【0058】
これらの抗体模倣物が抗体断片に勝っている主要な利点は構造である。これらの抗体模倣体は、安定性で、可溶性の構造の骨格全体に由来している。例えば、Fn3骨格は人体で見いだされる。結果として、これらの作製は、抗体の天然の折り畳みの一部を除去することを含み、無傷の抗体における可変ドメインおよび定常ドメイン間の界面のような疎水性環境に埋まっているアミノ酸残基がしばしばさらされている抗体断片よりも、これらは優れた折り畳みおよび熱安定性の性質を呈する。このような疎水性残基の溶媒への曝露は、抗体断片の凝集の可能性を増加させる。
【0059】
さらに、本明細書に記載の骨格は、特定の条件下で抗体断片の正しい折り畳みを妨害もしくは阻害すると報告されているジスルフィド結合を持たない。本骨格の自然な折り畳み安定性はジスルフィド結合に依存していないため、ジスルフィド結合が減少すると分解してしまう抗体およびそれらの断片とは異なり、還元条件下でも安定である。
【0060】
さらに、これらの骨格は、抗体分子の機能的な利点を提供する。特に、10Fn3モジュールは免疫グロブリンではないという事実にかかわらず、その全体的な折り畳みはIgGの重鎖の可変領域に近く(図2)、CDRに類似した3つのフィブロネクチン・ループを天然抗体と同様な相対的配向性で示すことが可能にされている。この構造のために、本抗体模倣物は、天然と類似した抗原結合の性質および抗体への親和性を持ち、インビボにおける抗体の親和性成熟化の過程に類似したインビトロでループを無作為化して混合する戦略をとることができる可能性を有する。
【0061】
例示的な骨格(例えばフィブロネクチンに基づく骨格)、ならびにその標的リガンドと同様に新規の結合タンパク質を同定し、選択し、および導出するための使用が以下に述べられている。これらの実施例は本発明を例示する目的で提供されており、本発明を限定するためのものではない。
【0062】
10Fn3の構造モチーフ
本発明の好ましい抗体模倣物は、哺乳類の血液および構造タンパク質で見出された共通ドメインであるフィブロネクチン3型モジュール(Fn3)の構造に基づいている。このドメインはタンパク質配列データベースの中に400回よりも多く出現し、フィブロネクチン、テンシン、細胞内の細胞骨格タンパク質、および原核生物の酵素を含むこれまでにシークエンシングされたタンパク質の2%に出現すると見積もられている(BorkおよびDoolittle、Proc. Natl. Acad. Sci. USA、第89巻、第8990頁、1992年;Borkら、Nature Biotech.、第15巻、第553頁、1997年;Meinkeら、J. Bacteriol.、第175巻、第1910頁、1993年;Watanabeら、J. Biol. Chem.、第265巻、第15659頁、1990年)。特定の骨格は94個のアミノ酸残基を含むヒトFn3の10番目のモジュール(10Fn3)である。このドメインの全体的な折り畳みは、最も小さな機能的抗体断片であり、ラクダおよびラマのIgG(図1、図2)において抗原認識単位の全体を含む重鎖の可変領域のものと密接に関連している。ラクダおよびラマのドメインと10Fn3ドメインの間の主要な差異は、(i)10Fn3のほうがβ鎖が少ないこと(7対9)、および(ii)ラクダおよびラマのドメインでは、互いに充填し合う2個のβシートがジスルフィド架橋により連結されているが、10Fn3ではそうでないことである。
【0063】
IgGの重鎖にある抗原結合ループに対応する10Fn3の3個のループは、アミノ酸残基21〜31位(BC)、51〜56位(DE)、76〜88位(FG)の間を通っている(図3)。BCのループの長さおよびDEのループの長さはそれぞれ10残基および6残基であり、抗体の重鎖で見出される対応する抗原認識ループの狭い範囲内、すなわちそれぞれ7〜10残基および4〜8残基に位置する。したがって、無作為化され、抗原との高親和性によって選択される場合には、これらの2個のループは抗体中の対応するループと同等に抗原と接触できる。
【0064】
対照的に、10Fn3のFGループは長さが12残基で、一方で抗体の重鎖にある対応するループは4残基〜28残基の範囲である。それゆえ、抗原との結合を最適化するために、10Fn3のFGループの長さは、好ましくは4残基〜28残基のCDR3の範囲を覆うように、長さおよび配列が無作為化され、抗原結合において可能な最も高い可動性および親和性を獲得する。実際に、一般的には、インビトロもしくはインビボの親和性成熟化において、(以下により詳細に述べるように)抗体模倣物のCDR様ループの配列と同様に長さも無作為化してもよい。
【0065】
10番目のヒトフィブロネクチン3型ドメインである10Fn3は、低温においてさえ急速に再び折り畳まれる。その背骨のコンフォメーションは、5℃において1秒間以内に回復する。10Fn3の熱力学的安定性は高く(ΔGU=24kJ/mol=5.7kcal/mol)、110℃の高い融解温度と相関する。
【0066】
10Fn3の生理学的な役割の1つは、体液中に可溶型としておよび細胞外マトリックス中で不溶型として存在する糖タンパク質であるフィブロネクチンのサブユニットである(Dickinsonら、J. Mol. Biol.、第236巻、第1079頁、1994年)。220〜250kDのフィブロネクチンの単量体は、12個の1型モジュール、2個の2型モジュール、および17個のフィブロネクチン3型モジュールを含んでいる(PottsおよびCampbell、Curr. Opin. Cell Biol.、第6巻、第648頁、1994年)。インテグリン、ヘパリン、およびコンドロイチン硫酸塩とフィブロネクチンとの結合には異なる3型モジュールが含まれている。10Fn3は、その曝露されたループの1つにあるインテグリン結合Arg-Gly-Asp(RGD)モチーフを介して細胞接着を媒介することが見出されている。フィブリノーゲン、フォン・ビルブラント因子、およびビトロネクチンのような他のタンパク質でも、類似のRGDモチーフがインテグリン結合に含まれることが示されている(Hynesら、Cell、第69巻、第11頁、1992年)。10Fn3については、他のマトリックスもしくは細胞結合的な役割は述べられていない。
【0067】
RGDを含む短いペプチドに比べて10Fn3の接着活性がわずかに強いという知見は、10Fn3の細胞結合活性が10Fn3の構造全体に分散しているというよりはRGDペプチドに局在しているという結果(Baronら、Biochemistry、第31巻、第2068頁、1992年)と一致する。RGDモチーフを持たない10Fn3は他の血漿タンパク質もしくは細胞外マトリックスと結合しそうもないという事実から、10Fn3は抗体に換わる有用な骨格である。さらに、血流の天然フィブリノーゲン中に10Fn3が存在することは、元々の生物において10Fn3自体が免疫原生ではないとおもわれることを示唆している。
【0068】
さらに、本発明者らは、無作為化に耐性を持ち、抗体模倣物の多様なプールの生産を助ける曝露されたループ配列が10Fn3の枠組みにあることを決定した。この決定は、10Fn3の配列の可動性を試すことで行われた。特に、関連タンパク質の配列はもちろん、他の源から得られたフィブロネクチンの配列とヒト10Fn3の配列を整列化させ(図4)、この整列化の結果をヒト10Fn3ドメインの三次元構造上に位置づけた(図5)。この整列化により、保存された残基の大部分はβシートサンドイッチのコアに見出され、著しく変異しやすい残基は両βシートの溶媒に接触しうる表面上、および抗体模倣物の親和性成熟化において超可変ループとして働き、溶媒と接触しうる3つのループ上でN末端およびC末端を含むβシートの端に沿って局在することが判明した。これらの結果を考慮すると、これら3つのループの無作為化には、10Fn3の枠組み自体の全体的な折り畳みもしくは安定性に反する効果はないように思われる。
【0069】
この解析は、ヒト10Fn3の配列では最小限でもアミノ酸1〜9位、44〜50位、61〜54位、82〜94位(βシートの端);19位、21位、31〜46位(偶数)、79〜65位(奇数)(両βシートの溶媒に接触しうる表面);21〜31位、51〜56位、76〜88位(CDR様の溶媒に接触しうるループ);ならびに14〜16位および36〜45位(溶媒に接触しうる他のループおよびβターン)は、新規のもしくは改良された化合物結合タンパク質を導出するために無作為化されてもよいことを示している。さらに、以上に討議されたように、溶媒に曝露された1つもしくは複数のループの長さにおける変化も、このような定方向展開法に含まれる。
【0070】
または、βシート配列の変化を利用して新規のタンパク質を導出してもよい。これらの変異は骨格を変化させ、それにより間接的に1つまたは複数のループ構造を変化させる。この方法を用いた場合は、配列に変異を過剰に行うことなく、むしろほとんど変異を導入してはいけない。この方法では、βシート配列に、好ましくは3-20の間以下の変異が導入されるべきである。
【0071】
配列の変異は、例えばTaqポリメラーゼによる変異導入(TindallおよびKunkel, Biochemistry 27:6008 (1988))、断片の組換え、またはその組み合せを含む任意の手法で行なえる。同様に、例えば、CDR様ループの長さおよび配列の変化、または選択されたループに見つかった有利な枠組みの変異に基づく構造の再設計による、ライブラリーの構造の多様性の増加を用いて、抗体模倣物骨格をさらに改良することができる。
【0072】
抗体模倣体融合体
本明細書に述べられた抗体模倣物は、他のタンパク質ドメインと融合していてもよい。例えばこれらの模倣物は、IgG(Fc)の定常領域と10Fn3モジュール、好ましくは10Fn3のC末端のような抗体模倣体を融合させることで、ヒト免疫応答に組み込まれていてもよい。このような10Fn3-Fc融合分子などにおけるFcは、免疫応答の補体構成要素を活性化し、抗体模倣物の治療上の価値を増加させる。同様に、10Fn3のような抗体模倣体とC1qのような補体タンパク質との融合は、細胞を標的化するために用いることができ、10Fn3の抗体模倣体と毒素の融合は、特定の抗原を運搬する細胞を特異的に破壊するために用いることができる。さらに、いかなる型の抗体骨格(例えば10Fn3)であっても、アルブミンと融合させて血流中での半減期および組織への浸透を増加させることができる。これらの任意の融合体を、例えば一般的に入手可能な遺伝子配列を用いて構築した組換え融合遺伝子から融合タンパク質を発現させるなどの常法により生産することができる。
【0073】
骨格の多量体
単量体に加えて、本明細書に述べられている任意の骨格構築物を、結合価を増加させて、それにより抗原との結合活性を増加させるために、抗体模倣物の二量体または多量体として生産してもよい。このような多量体は、共有結合により生産されてもよい。例えば、個々の10Fn3モジュールは天然の8Fn3-9Fn3-10Fn3のC末端からN末端の結合を模倣したり、または定常領域を介して結合した抗体の二量体を模倣することによって結合させてもよい。10Fn3-Fc構築物を、10Fn3-Fc::Fc-10Fn3という一般的な図式の二量体を設計するために利用してもよい。Fc::Fcの界面に操作された結合は共有結合であっても、非共有結合であってもよい。さらに、このようなより高いオーダーの構造を作製するために、10Fn3ハイブリッドのようなハイブリッドにおいてFc以外の相手との二量体化もしくは多量体化を利用することができる。
【0074】
特定の例としては、共有結合した多量体は、多量体をコードする融合遺伝子を構築する段階によって、または単量体の配列中にシステイン残基のコドンを操作し、発現した生成物間でジスルフィド結合の形成を生じさせる段階によって作製されてもよい。非共有結合した多量体もまた、様々な技術により生産することができる。これらには、正電荷および/または負電荷を帯びた残基に対応するコドンを単量体の配列に導入し、発現した生成物間(および、その結果として単量体間)において、これらの残基間で相互作用を生じさせておくことが含まれる。本方法は、例えばフィブロネクチンの負電荷を帯びた残基のように、単量体サブユニット中に天然に存在する荷電残基の利点を用いることで単純化することができる。非共有結合した抗体模倣物を生産する別の方法は、相互作用することが知られているタンパク質またはタンパク質ドメインのコード配列を単量体遺伝子(例えば、アミノ末端もしくはカルボキシル末端)に導入することである。このようなタンパク質もしくはタンパク質ドメインは、コイルドコイルモチーフ、ロイシンジッパーモチーフ、および二量体もしくはより高いオーダーの多量体を直接形成することが知られている任意の多数のタンパク質サブユニット(またはその断片)を含んでいる。
【0075】
フィブロネクチン様分子
10Fn3は抗体模倣物の生産に好ましい骨格を表すにもかかわらず、本明細書に記載の分子においては、他の分子が10Fn3を置換していてもよい。これらに限定されることはないが、ヒト以外の動物および原核生物から得られる関連したFn3モジュール、ならびにヒトフィブロネクチンモジュール1Fn3-9Fn3および11Fn3-17Fn3を含む。さらに、テネイシンおよびアンジュリンのような10Fn3と配列に相同性を持つ他のタンパク質から得られたFn3モジュールを使用してもよい。免疫グロブリン様の折畳みを持つ(ただしVHドメインに関連しない配列を持つ)他の代表的な骨格は図21に示されており、N-カドヘリン、ICAM-2、タイチン、GCSF受容体、サイトカイン受容体、グリコシダーゼ阻害剤、E-カドヘリン、および抗生物質色素蛋白質が含まれる。関連する構造を持つ別のドメインは、ミエリン膜接着分子PO、CD8、CD4、CD2、クラスI MHC、T細胞抗原受容体、CD1、C2、およびVCAM-1のI-setドメイン、ミオシン結合タンパク質CのI-set免疫グロブリンドメイン、ミオシン結合タンパク質HのI-set免疫グロブリンドメイン、テロキンのI-set免疫グロブリンドメイン、テリキン、NCAM、トウィチン、ニューログリアン、成長ホルモン受容体、エリスロポエチン受容体、プロラクチン受容体、GC-SF受容体、インターフェロンγ受容体、βガラクトシダーゼ/グルクロニダーゼ、βグルクロニダーゼ、トランスグルタミナーゼに由来する可能性がある。または、免疫グロブリン様の折畳みを1つまたは複数含む任意の他のタンパク質が利用できる。そのようなタンパク質は、例えば、プログラムSCOP(Murzinら、J. Mol. Biol. 247:536 (1995); Lo Conteら、Nucleic Acids Res, 25:257 (2000))を用いて同定できる。
【0076】
一般に、VHドメインに構造的関連性を示す任意の分子(例えば、上述のコンピュータプログラムで同定される)は、抗体模倣物として使用できる。そのような分子は、フィブロネクチンのように、分子のN末端に3つのループ、C末端に3つのループを含んでおり、その各々を無作為化して多様なライブラリーを作製できる;または、そのような表面の無作為化可能なループが抗原結合に参加できるほど空間的に近接しているかぎり、より多くのループを持つより大きなドメインも使用できる。図22は、互いに近接した3個を越える数のループをもつ有用なドメインの例を示す。これらの例には、各々無作為化できるループを4つ持つT細胞抗原受容体およびスーパーオキシド・ジスムターゼ;および3つの無作為化可能なループが2つのドメインの一部である(ループの合計数が6となる)ような2つの類似したドメインを3セットずつ持つ、Fn3二量体、組織因子ドメイン、およびサイトカイン受容体ドメインが含まれる。
【0077】
または、空間的に充分近接した可変ループを持つ任意のタンパク質を用いて、候補結合タンパク質の生産ができる。例えば、免疫グロブリン様の折畳みには構造的に関連していなくても、空間的に関連した、溶媒が接することのできるループを持つ大きなタンパク質が使用できる。例には、シトクロームF、緑色蛍光タンパク質、GroEL、およびタウマチンが含まれるが、これらに限定されない(図23)。これらのタンパク質のディスプレイするループを無作為化して、本明細書に記載するように無作為化したライブラリーから優れた結合をする分子を選択できる。そのサイズのために、抗体-抗原相互作用よりも、はるかに大きな抗原結合表面を示す分子が得られる可能性がある。このタイプの他の有用な折畳みは、数多くのループ、特に平行するβシートまたはいくつかのαへリックスの間に位置するループを持つ候補タンパク質を見るためにプログラムSCOP(Murzinら、J. Mol. Biol. 247:536 (1995))を用いても同定できる。
【0078】
異なる生物および親タンパク質から得られるモジュールは、異なる適用に適切である可能性がある。例えば、抗体模倣物の設計では、治療薬を意図する生物に天然に存在するフィブロネクチンまたはフィブロネクチン様分子から、そのタンパク質を作製するのが最も望ましいことがある。反対に、診断薬または研究用試薬のようなインビトロ適用に使用する抗体模倣物は、起源の生物の重要度は低いまたは関係ない。
【0079】
これらの任意の分子について、本明細書に記述する任意の方法によりライブラリーを作製し、結合するタンパク質を選択するために使用できる。
【0080】
骨格に基づく結合タンパク質の定方向展開
本明細書に記載の抗体模倣物は、新規または改良された結合タンパク質を導出するために、いかなる技術において使用してもよい。一つの特定の例として、結合の標的をカラム樹脂またはマイクロタイタープレートのウェルのような固相支持体に固定化し、候補となる骨格に基づく結合タンパク質のライブラリーに標的を接触させる。このようなライブラリーは、10Fn3のCDR様ループの配列および/または長さを無作為化することで野生型10Fn3の骨格から構築された10Fn3クローンのような抗体模倣体クローンから成っていてもよい。望ましい場合には、このライブラリーは、例えばショスタック(Szostak)ら、米国特許出願第09/007,005号および同第09/247,190号;ショスタック(Szostak)ら、国際公開公報第98/31700号;ならびにロバーツ(Roberts)およびショスタック(Szostak)、Proc. Natl. Acad. Sci. USA、1997年、第94巻、第12297〜12302頁に記載の技術によって生産されたRNA-タンパク質融合体ライブラリーであってもよい。またはDNA-タンパク質ライブラリーであってもよい(例えば、Lohse、「DNA-タンパク質融合体およびそれらの使用法(DNA-Protein Fusions and Uses Thereof)」、米国特許出願第60/110,549号、米国特許出願第09/459,190号、および国際公開公報第00/32823号)。この融合体ライブラリーを固定された標的と共にインキュベートし、支持体を洗浄して非特異的な結合物を除去し、高ストリンジェントな条件下における最も強い結合物を溶出してPCRにかけて配列情報を回収するか、または配列にさらなる変異を加えたりもしくは加えなかったりして選択段階を繰り返すのに使用することができる結合物の新規のライブラリーを作製する。抗原に対して十分な親和性を持つ結合物を得られるまでには、選択ラウンドが多数回行われる可能性がある。
【0081】
一つの特定の例においては、10Fn3の骨格を選択の標的として使用することができる。例えば、10残基のループ中に存在する特異的なペプチド配列に結合するタンパク質が必要な場合は、このループの1つが10の長さで所望の配列に設定された単一の10Fn3クローンを構築する。新規のクローンをインビボで発現させて精製し、次に支持抗体に固定化する。適切な骨格に基づいたRNA-タンパク質融合体ライブラリーを支持体と相互作用させ、その後洗浄して、所望の分子を溶出し、上述したように再度選択する。
【0082】
同様に、本明細書に記載の骨格(例えば10Fn3の骨格)は、例えば10Fn3のループにおいて、骨格によって提示されたペプチド配列と相互作用する天然タンパク質を見出すためにも使用することができる。上述したように10Fn3タンパク質のような骨格タンパク質を固定化し、提示されたループへの結合物に対してRNA-タンパク質融合体ライブラリーをスクリーニングする。選択ラウンドを複数回行うことで結合物を濃縮し、DNAのシークエンシングにより同定される。
【0083】
さらに上述の方法において、RNAタンパク質ライブラリーを定方向展開のための例示的なライブラリーとして示したが、いかなる型の骨格に基づくライブラリーも本発明の選択方法において使用することができる。
【0084】
使用
本明細書に記載の抗体模倣物は、関心対象の任意の抗原とも結合するように導出することができる。これらのタンパク質は天然抗体に比べて熱力学的に優れた性質を持ち、インビトロで急速に導出することができる。したがって、これらの抗体模倣物は、研究、治療、診断の分野を含む、抗体が使用されるすべての分野においても抗体の代わりに用いることができる。さらに、これらの骨格は抗体に比べて可溶性でかつ安定性の性質を持つので、本明細書に述べられた抗体模倣物は抗体分子が破壊されたり、または不活性化されたりするような条件下でも使用することができる。最後に、本発明の骨格は事実上いかなる化合物とも結合するように導出することができるので、これらの分子は完全に新規であり、研究、治療、および診断の分野における使用も見出されている結合タンパク質を提供する。
【0085】
実験結果
上述された例示的な骨格分子は、例えば以下のような選択手順で生産され、かつ検査された。
【0086】
ライブラリーの構築
複合体のライブラリーは、CDR様ループに対応する無作為化された部位を各々1個含む3つの断片から構築した。無作為化された残基は、図18に下線をつけた配列で示されているが、10Fn3 BCループ(ラマVHのCDR-H1に対応)の残基23〜29;10Fn3 DEループ(ラマVHのCDR-H2に対応)の残基52〜55;および10Fn3 FGループ(ラマVHのCDR-H3に対応)の残基78〜87である。断片は、その中にあるCDR-H様ループの名前に基づいて、BC、DE、およびFGと名付けた。各々の断片には、10Fn3および無作為化された配列に加えて、His6ドメインのN末端またはFLAGペプチドタグのC末端をコードする伸長部分が含まれた。各DNA断片には、2つの断片間(すなわち、BCおよびDE断片間もしくはDEおよびFG断片間)の各接合部にEar IタイプIIS制限エンドヌクレアーゼの認識配列が含まれた。この制限酵素は、すべての外来非10Fn3配列を除去し、隣接する断片を一緒にスプライシングさせた。さらに変異および選択ラウンドの間に、3つの10Fn3断片を組換えのように混合させる。
【0087】
野生型のヒト10Fn3遺伝子は、プライマー

を用いて、ヒト肝臓ライブラリー(Maxim Biotech、カリフォルニア州南サンフランシスコ)からクローニングした。10Fn3クローンのPCRで得られた野生型セグメントおよびオリゴヌクレオチド合成で得られた無作為化セグメントから、3つの異なるライブラリーが構築された。BCr-DEr-FGrライブラリーは、BC、DE、およびFGループの選択された残基を無作為化することによって得られた;BCr-DEwt-FGrライブラリーは、BCおよびFGループの選択された残基を無作為化し、DEループ配列を野生型のまま残すことによって得られた;BCwt-DEwt-FGrライブラリーは、FGループのみの選択された残基を無作為化することによって得られた。
【0088】
BCr、DE rおよびFGr断片は、合成された。各断片は2つの重複するオリゴヌクレオチドから構成され、最初にアニーリングさせた後、断片の2本鎖DNAの形状を形成するように伸長させた。3つの断片を構築および加工するのに使用されるオリゴヌクレオチドを以下の表に示す:「先端(top)」および「末尾(bottom)」の種は、10Fn3コード配列全体を含むオリゴヌクレオチドである。これらのオリゴヌクレオチドの呼称において、「N」は、A、T、C、またはGを示し、「S」は、CおよびGを示す。
T7Tmv(インビトロ翻訳に必要なT7プロモーターおよびTMV非翻訳領域を導入する):
Unispl-s(Robertsら、1997、上記、ピューロマイシン含有リンカーにmRNAを連結するために用いられるスプリントオリゴヌクレオチド)
A18---2PEG(DNA-ピュロマイシンリンカー):
5’-(A)18(PEG)2CCPur(配列番号:14)
【0089】
オリゴヌクレオチド対BCTopおよびBCBot-flag8を用いて無作為化BCループを含む断片が構築された;対DETopおよびDE Bot-flag8を用いて無作為化DEループを含む断片が構築された;対BCTopおよびDE3-flag8を用いてBCwt-DE wt断片をPCR増幅した;および対FGTopおよびFG Bot-flag8を用いて無作為化FGループを含む断片が作製された。オリゴヌクレオチド対(各500pmol)を100μlの10mM Tris 7.5、50mM NaCl中で85℃にて10分間アニーリングさせ、次にゆっくりと(0.5〜1時間)室温に冷却した。1本鎖の突出部分にアニーリングした断片を、アニーリングしたオリゴ各100μlアリコートに対して100Uのクレノー(New England Biolabs、Beverly、MA)ならびに838.5μlのH2O、9μlの1M Tris 7.5、5μlの1M MgCl2、20μlの10mM dNTP、および7.5μlの1M DTTで調製された緩衝液を用いて伸長させた。伸長反応は、25℃にて1時間行った。
【0090】
無作為配列によって導入される停止コドンの頻度を下げるために、無作為化された残基は、(NNS)nによってコードされたが、Nは任意のヌクレオチド、SはCおよびGの等モル混合物を表す;3つの停止コドンのうちの1つのみ(TAG)がNNSの制限に従う。遺伝子断片は、10Fn3をコードする配列に加えて、5’タバコモザイクウイルス(TMV)非翻訳領域およびT7プロモーター、ならびに5’ヘキサヒスチジンタンパク質精製タグおよび3’ FLAGエピトープ精製タグをコードする配列も含んでいた。また、3つの断片の組み立てを容易にするために、上述のように、Ear I制限エンドヌクレアーゼ認識部位が、隣接する断片の重複部分に組み込まれた。
【0091】
次に、ショスタック(Szostak)ら、米国特許出願第09/007,005号および同第09/247,190号;ショスタック(Szostak)ら、国際公開公報第98/31700号;ならびにロバーツ(Roberts)およびショスタック(Szostak)、Proc. Natl. Acad. Sci. USA、1997年、第94巻、第12297〜12302頁によって開発された技術を用いて、各2本鎖断片をRNA-タンパク質融合体(PROfusion(商標))に形質転換した。簡単に言えば、アンビオンインビトロ転写キット(Ambion in vitro transcription kit)、T7-MEGAshortscript(商標)(Ambion、Austin、TX)を用いて断片を転写し、得られたmRNAをゲル精製し、かつDNAリガーゼ(Promega、ウィスコンシン州マジソン)を用いて5’リン酸化DNA-ピューロマイシンリンカー(好ましくは5’ dA18PEG2dCdCPur)に連結した。ショスタック(Szostak)(上記)に記述されるようにして、mRNAはDNAスプリントオリゴヌクレオチド
を用いてDNAリンカーと整列化させた。アンビオンウサギ網状赤血球溶解物に基づく翻訳キット(Ambion rabbit reticulocyte lysate-based translation kit)を用いて、35Sメチオニンの存在下で、mRNA-DNA-ピューロマイシン分子を翻訳した。得られたmRNA-DNA-ピューロマイシン-融合体タンパク質をオリゴ(dT)セルロース(タイプ7、Amersham Pharmacia、ニュージャージー州ピスカタウェイ)を用いて精製し、逆転写酵素(Superscript(商標)II、Gibco, Life Technologies、メリーランド州ロックビル)および上述したRTプライマー(Unisplint-SもしくはflagASA)を用いて、製造者の指示(好ましくは、70℃で2分間アニーリングおよび42℃で40分間反応)に従ってDNAの相補鎖を合成した。
【0092】
各断片について得られたアニーリングcDNAを持つRNAタンパク質融合体は、そのペプチド精製タグに適した樹脂、すなわちHis6タグにはNi-NTAアガロース(Qiagen、カリフォルニア州バレンシア)およびFLAGタグにはM2抗Flagアガロース(Sigma、ミズーリ州セントルイス)を用いて、製造者に推奨された方法に従って精製した。KOH溶出によって回収された断片のコードする遺伝情報を、ファルマシアレディートゥゴーPCRビーズ(Pharmacia Ready-to-Go PCR Beads)、10pmolの5'および3' PCRプライマーを用いて、以下のPCRプログラム(Pharmacia、Piscataway、NJ)に従ってPCRにより増幅した:1段階目は95℃で3分間、2段階目は95℃で30秒間、58/62℃で30秒間、72℃で1分間を必要に応じて20/25/30サイクル、3段階目は72℃で5分間、4段階目は最後まで4℃(通常、25サイクル)。
【0093】
得られたDNAを、1μgのDNAにつき5〜6 UのEar I(ニュー・イングランド・バイオラボ(New England Biolabs))で切断した。反応は、T4 DNAリガーゼ緩衝液(New England Biolabs)中で37℃にて1時間行い、続いて選択的に、Ear Iを失活させるために70℃にて15分間インキュベートした。無作為化されたループを持つ10Fn3遺伝子の全長を形成させるために、等量のBC断片、DE断片、およびFG断片を混合して連結した。連結には10Uの新鮮なEar I(New England Biolabs)および20UのT4 DNAリガーゼ(Promega、ウィスコンシン州マディソン)が必要であり、37℃にて1時間かかった。Ear Iおよびリガーゼは、65℃で15分間インキュベートして失活させた。
【0094】
3つの異なるライブラリーBCwt-DEwt-FGr、BCr-DEwt-FGr、および、BCr-DEr-FGrは、上述のようにして構築された。各々が10個の無作為化された残基を持つFGループの形を含んでいた。最初のライブラリーのBCおよびDEループは、野生型の10Fn3配列を持っていた;7個の無作為化された残基を持つBCループおよび野生型のDEループが第2のライブラリーを構成していた;7つの無作為化された残基を持つBCループおよび4つの無作為化された残基を持つDEループが第3のライブラリーを構成していた。これら3つのライブラリーの各々におけるFGループの複雑性は1013だった;さらなる2つの無作為化されたループを合わせると、潜在的な複雑性は研究室で標本抽出するには高すぎるほどになった。これらのライブラリーの組み合せは1012のユニークなクローンをもつマスターライブラリーを提供していた。
【0095】
元の無作為化されたBCr-DEr-FGrライブラリーから無作為に選んだ76個のクローンの配列は、無作為化されたループに何のパターンも示さなかった(データは示さず);ライブラリーのアミノ酸頻度は、各残基をコードするコドンの数に比例しており、各位置あたり1%(グルタミン酸、メチオニン、トリプトファン)と14%(プロリン)の間だった。反対に、保存されたβシート枠組中の残基が野生型のまま保存される平均確率は、99%だった。
【0096】
等モルの3つのライブラリーは(各々DNA 2 pmol)、選択工程を単純化するために混合され、1つのマスターライブラリーとし、標的との結合それ自体によって、特定の誘発に最も適したライブラリーを選択することにした。ラウンド3から10の親和性による精製ではM2-セファロース(以下参照)が使用されたこと以外、ショスタック(Szostak)ら、米国特許出願第09/007,005号および同第09/247,190号;ショスタック(Szostak)ら、国際公開公報第98/31700号;ならびにロバーツ(Roberts)およびショスタック(Szostak)、Proc. Natl. Acad. Sci. USA、1997年、第94巻、第12297〜12302頁に述べられた一般的な方法に従って、マスターライブラリーからRNA-タンパク質融合体を得た(図8)。
【0097】
融合体の選択
RNA-タンパク質融合体の形態のマスターライブラリーが、TNF-αとの結合を選択するために供された(Pepro Tech、ニュージャージー州ロッキーヒル)。最初に2つの方法が行われた:1つでは標的をアガロースカラムに固定化し、1つでは標的をBIACOREチップに固定化した。最初に、アガロースカラムへのバックグラウンドの結合体を最小限にするために条件の大幅な最適化を行い、50mM HEPES pH 7.4、0.02% Triton、100μg/ml変性されたサケ精子DNAという好ましい緩衝液の条件を得た。この緩衝液中では、10Fn3-RNA融合体のTNF-α-セファロースへの非特異的結合は0.3%であった。10Fn3-RNA/cDNAライブラリーのTNF-α-セファロースへの非特異的結合バックグラウンドは0.1%であることが見出された。
【0098】
TNF-α-セファロース上での各々の選択ラウンドにおいて、残存している非特異的な結合体のいかなるものも除去するために、最初にライブラリーを非誘導体化されたセファロースと共に1時間プレインキュベートし、この前除去により得られた流出液をTNF-α-セファロースと共にさらに1時間インキュベートした。TNF-α-セファロースは3〜30分間洗浄した。
【0099】
各選択後に、0.3M NaOHまたは0.1M KOHを用いて複合体のcDNA成分を固相支持体から溶出させ、PCRにより増幅させた。所望の大きさのDNAのバンドは、選択ラウンドを複数回行っても残留していた(図9)。2つの他の選択法でも同様な結果が観察されたが、アガロースカラム選択で得られたデータのみを図9に示す。
【0100】
この選択の最初の7ラウンドにおいては、Fn3-RNA/cDNA分子の標的への結合は低いままであり;対照的に、選択の異なる段階においてDNAプールから遊離タンパク質が翻訳された場合には、カラムに結合した種の割合はラウンド間で有意に増加した(図10)。
【0101】
後の選択では、選択の最初の8ラウンド(R1〜R8)で選択された融合体プールは、バックグラウンドに近いレベル(<0.25%)でTNF-α-セファロースに結合した(図13)。選択の第9ラウンド(R9)後、TNF-α-セファロースへの融合体の結合は、0.7%と急上昇し、第10ラウンド(R10)後は、結合はさらに7%に増加した(図13)。選択は、10 mL中10 mg TNF/gセファロースでエポキシ活性化セファロース(商標)6B(Amersham Pharmacia)に固定化されたTNF-αを用いて行われた。使用前にTNF-α誘導化セファロースは、結合緩衝液(50 mM HEPES, pH 7.4, 0.02% Triton, 0.1 mg/mL変性されたサケ精子DNA(Ambion))中で、4℃で一晩ブロックされた。
【0102】
上述のように、10Fn3に基づくマスターライブラリーを転写し、ピュロマイシンを持つリンカーに連結し、5-10μL/300μL 35S-メチオニンの存在下でmRNA-タンパク質ライブラリーに翻訳し、オリゴ(dT)セルロース上で親和性により精製し、DNA/mRNA-タンパク質ライブラリーに逆転写し、M2-セファロース上で親和性により精製した(3〜10ラウンド)。DNA-mRNA-タンパク質融合体ライブラリー分子4 x 1012の異なる配列20コピーに当たる40 pmolを回収し、第1ラウンド(R1)の選択を行なった。
【0103】
選択の第1段階では、40 pmolのDNA-mRNA-タンパク質ライブラリーを、TNF-αの非存在下で誘導化したエポキシセファロース30μLを含む300μLの結合緩衝液中で、回転させながら4℃で1時間インキュベートした。第2ラウンドでは、24 pmolのライブラリーを添加し、残りの8ラウンドでは、0.1〜2 pmolのライブラリーを添加した。上清はMico Bio-Spin(登録商標)クロマトグラフィーカラム(BIO-RAD、カリフォルニア州ハーキュリーズ)で微量遠心して回収し、300μLの結合緩衝液中で30μLのTNF-α-セファロース(6μM)と4℃で1時間インキュベートした(7〜10ラウンドでは、結合緩衝液にはさらに1 mg/mLのBSAが含まれていた)。TNF-α-セファロースはスピンカラムで回収し、3 x 300μLの結合緩衝液で洗い、100μLの0.1 M KOHで溶出し、最後に1μLの1 M Tris 8.0, 8μLの1 M HClで中和した。ライブラリー、溶出前後のTNF-α-セファロース、洗浄液、および溶出液のサンプルは、シンチレーションカウンターでサンプル中の35S-メチオニンを計数して定量した。選択の次のラウンドは、PCR増幅によって新しいDNA/mRNA-タンパク質プールを形成することから始まり、これをPCR産物から転写、翻訳、および逆転写した。
【0104】
第9ラウンドおよび第10ラウンド後の溶出から得られたDNAプールをTOPO(商標)TA(登録商標), pCR2.1クローニングベクター(Invitrogen、カリフォルニア州カールズバッド)にクローニングし、大腸菌を形質転換した。30〜100クローンを選択し、プラスミドミニプレップ(Qiagen)に増殖させた。R9から38クローン、R10から29クローンを無作為に選択し、配列決定した(DNAシーケンシングコアファシリティー、マサチューセッツ総合病院、分子生物学部門、マサチューセッツ州ボストン)。得られたタンパク質配列の整列化には、プログラムClustalW60を使用した。
【0105】
TNF-α結合クローンのアミノ酸残基配列
R9およびR10から得られた61クローンのうちの38は、ユニークなアミノ酸配列を持っており、これは驚くべき多様性である。おそらくTNF-αに対する優れた結合のために複数回単離された10のクローンは、表1に示されている(全体の配列は図25)。
【0106】
得られたプールから無作為に選択された61のクローンのうち、1つ(クローンT09.08、配列は示さず)のみがBCwt-DEwt-FGrライブラリー由来で、別の6個がBCr-DEwt-FGrライブラリー由来だった。残りの54個(ウィナーの88%)がBCr-DEr-FGrライブラリーから選択されたということは、いくつかのループが標的に協力してTNF-αに結合することの重要性を示す。
【0107】
選択されたループ配列の最も一般的なモチーフはPWA(S/T)で、これは61のクローンのうちの33のDEループに見られた;より緩く定義されたPW(A/G)の配列は41/61のクローンに見られた。抗体VHドメインの類似したCDR-H2ループは一般的に抗原結合には小さな貢献しかしないので、特定のDE配列に対するそのような強い選択は、驚くべきことである。一方、DEループの長さが短いために、ライブラリーには可能なテトラペプチド配列がそれぞれ107コピー存在すると期待され、そのため選択された性質に対するDEループの任意の貢献を最適化するのが容易になる。他のFn3ドメインの調査(Dickensonら、J. Mol. Biol. 236:1079-1092 (1994))は、10Fn3の残基52に相当する位置には、野生型のグリシンと同様の頻度でプロリンが見られることを示す;同様に、アラニン、グリシン、および野生型リジンは、すべて10Fn3位置54に相当する位置によく見られる。その結果、位置52および54で選択される残基は、少なくとも好ましい生物物理学的性質を持つ点で一貫していると思われる。対照的に、10Fn3残基53に相当する位置ではトリプトファンは見られず、トリプトファン53はTNF-αとの結合への貢献のためというような、本発明に特異的な理由で選択された可能性がある。これは他の抗原に対する後の選択でもこのモチーフが見られないこととも一致しており、やはりPWA/Gモチーフは10Fn3ドメインの安定性または溶解性というよりも、TNF-αへの結合に直接貢献している可能性が高い。ループDEでPWA/Gが好まれることは、選択時にBCr-DEr-FGrライブラリーが好まれる別の理由を示唆する:BCr-DEr-FGrライブラリーのみが無作為化されたDEループを含み、標的との結合にPWA/G配列が重要ならば他の2つのライブラリーとの競争に勝つと考えられる。
【0108】
BCループで最も頻繁に選択された配列はNRSGLQS (12/61)(配列番号:31)であり、FGループで最も頻繁に選択された配列はAQTGHHLHDK (6/61)(配列番号:32)だった。NRSGLQS BCループとAQTGHHLHDK FGループは同一の分子には見つかれなかったが、3つの無作為化されたループのうちの2つに、最も頻繁に単離された配列を含むクローンが2つ見つかった。これらのクローン
は、調べたクローンのうちでTNF-αからの解離定数が最も低い4つのうちの2つだった(表1)。
【0109】
プルーフリード活性を持たないTaqポリメラーゼを使用したため、PCRのラウンドごとに、10Fn3のCDR様ループとβシート折畳みの両方に、1塩基対につき0.01%の割合、すなわちPCRの1ラウンドにつき10Fn3遺伝子につき3%、および選択1ラウンドにつき約75%の割合で、さらなる無作為化変異が導入された。したがって、野生型のまま保存された残基、および非野生型の安定した配列として保存された残基は、これらの配列はその優れた性質のために選択されたことを意味すると考えられる。変異したループでは、オリゴヌクレオチド合成によって導入された変異かPCR変異導入による変異かを区別するのは不可能だが、βシートの折畳みでは、選択された変異の大部分はTaqエラーに起因する。選択されたクローンはタンパク質の折畳みにいくつかの保存された変化を示すが、これは意図的に無作為化したものではない。図18は無作為化されなかったが、それでも選択中に変異した10Fn3βシートにおける残基を示す。この変異導入は、61クローンのうちの26〜28の頻度で起こった;これらの変異は野生型10Fn3配列の下に矢印で示されており選択された残基を示す文字が付けられている。特に、61クローンのうちの28ではロイシン18からアルギニンまたはグルタミンへ変異しており、26クローンではスレオニン56からイソロイシンに変異している。図19はこれらの折畳みの変異の位置を示す。位置56はループDEの幹にあり、したがってこのループの立体配座および標的結合特性に影響すると期待される。TNF-α結合ループからの位置18の距離は、この変異の選択的優位性は、ループBCの立体配座への間接的影響、またはタンパク質の安定性への影響に起因する可能性を示唆する(図19)。これは、頻繁に見られる野生型からの変異L18RおよびT56Iを含むクローンT10.06の位置18を野生型のロイシンに復帰させた変異導入の実験でも支持されている。この変化により、変異体のKdは約10倍に上昇した。T10.06(L18)タンパク質のTNF-αに対する弱くなった結合は、位置18の残基は、おそらくループBCへβ鎖を通して伝えられる小さな構造変化によって、CDR様ループによる標的への結合に影響することを示唆する。
【0110】
選択されたTNF-α結合プールの親和性と特異性
9回および10回目の選択後のTNF-αの遊離タンパク質プールの見かけの平均Kd値は、区別不可能(それぞれ4nMおよび6 nM;表1)だった;この親和性の類似は最後のラウンドの選択での濃縮が比較的低レベル(10倍)だったこと、および2つのプールの配列組成が類似していることと一貫している。さらに4回の選択を行なった後の遊離タンパク質プールの見かけの平均Kdは3 nMで、やはりR9およびR10プールと区別不能だった。
【0111】
10回目の選択後に選択されたプールの結合特異性を評価するために、我々はセファロースに固定された3つのサイトカイン、選択の標的であるTNF-α、および無関係のIL-1αおよびIL-13に対する2つの異なる遊離タンパク質プールの結合を比較した。第1のプールは最初の選択前(R0)の無作為化DNAライブラリーから、第2のプールは10回目の選択後(R10)のライブラリーから転写および翻訳した。
【0112】
これらの実験を実行するために、10回目の選択後の溶出液のPCR産物を35Sメチオニンの存在下でインビトロで転写および翻訳したが、mRNA-タンパク質融合体の形成はさせなかった。TNF-α-セファロース、IL-1α-セファロース、IL-13-セファロースに、それぞれ約10μM、30μM、および50μMで結合した、および未誘導化セファロースに結合した、得られた遊離タンパク質の画分は、TNF-α-セファロースに対するDNA/mRNAタンパク質融合体の結合について上述の手順を用いて、比較した(図20)。各標的に結合した選択されたプールの量は、洗浄したビーズのシンチレーション計測で測定した。
【0113】
図20は、R0のTNF-α、IL-1α、およびIL-13に対する結合は類似していたが(それぞれ2%、4%、および3%)、10回の選択度には標的TNF-αには32%、IL-1αには3%、およびIL-13には1%結合することを示す。TNF-αに対する絶対的および相対的なタンパク質結合の増加は、10Fn3骨格および標的特異的結合体を選択するためのDNA/mRNAタンパク質融合体に基づく選択システムの能力を示す。
【0114】
結合の特異性をさらに調べるために、クローンT09.12をマイクロアレイ形式(以下に一般的に説明されている)に固定し、可溶性TNF-αに対する結合を調べた。このクローンに対するTNF-αの特異的結合は、蛍光検出(図24A)および質量分光(図24B)で検出した。質量分光については、干渉する可能性のある様々なタンパク質を含む代表的な複雑な生体液である、牛胎児血清の存在下で結合測定が行われた。蛍光検出(図24A)については、野生型10Fn3およびT09.12変異体のRNA-10Fn3融合体の混合物(表1)を、融合分子のRNA部分に相補的なオリゴヌクレオチドが24の反復フィーチャーで600ミクロンのピッチで固定されたDNAマイクロアレイにハイブリダイズさせた。洗浄によりハイブリダイズしなかった融合体を除去した後、その表面をビオチン-TNF-α(TBS、0.02% Tween-20、0.2% BSA中2.6μg/mL)に暴露し、洗浄し、空気乾燥した。キャプチャーされたビオチン-TNF-αは、ScanArray 5000システム(GSI Lumonics)を用いて、Cy3標識の抗ビオチンモノクローナル抗体(Sigma)により、検出した。質量分光検出については、T09.12変異体のRNA-10Fn3融合体(図24B)および野生型10Fn3(図24C)をRNase Aで処理してタンパク質およびDNAリンカーの融合体を作製した。得られたDNA結合タンパク質をDNAリンカーに相補的なオリゴヌクレオチドを固定したガラスカバースリップアレイにハイブリダイズさせた(図24Bおよび24C;融合体は図24Dには使用されていない)。洗浄後、上述の表面をTNF-α(90% v/v PBS/10%牛胎児血清中1.5 mg/mL)に暴露した。乾燥したチップにMALDIマトリックスをスポットし、Voyager DE MALDI-TOF質量分光計(PerSeptive Biosystems)で分析した。モノマーのTNF-αに期待される分子質量に対応する17.4 kDのシグナルが、T09.12タンパク質を含む200μmフィーチャー上に検出されたが(図24A)、野生型10Fn3(図24B)およびDNAタンパク質融合体を含まないフィーチャー(図24C)には検出されなかった。
【0115】
選択されたTNF-α結合クローンのKd
9回および10回目の選択で作製された2つのプールに複数回現れる全てのクローン、およびBCwt-DEwt-FGrライブラリーに由来する唯一のクローン(クローンT09.08)について、解離定数が決定された。
【0116】
これらの結合定数を決定するために、ピエス(Pierce)(イリノイ州ロックフォード)のNHS-LC-LCビオチン試薬を用いてビオチン化TNF-αを調製した。MALDI-TOF質量分光を用いて、モノマーのTNF-αの80%以上、したがって、三量体の99%以上がビオチン化されていると見積もられた。
【0117】
R9およびR10プール(および以下に説明されるR14およびM12プール)ならびにこれらの2つのプールに由来する解析されたクローンについて、200μL 10 mM HEPES, pH 7.4, 150 mM NaCl, 1% BSA, 0.02% Triton中で、0.25 nMのインビトロ翻訳35S-メチオニン標識遊離タンパク質を、ビオチン化TNF-αと17 pMおよび23 nMの間の濃度で室温で1時間インキュベートした。その後、96ウェルのイージータイター(Easy-Titer)(商標)ELIFAシステム(Pierce)を用いて各サンプルをあらかじめぬらしたSAM2R ビオチンキャプチャーメンブレン(Promega、ウィスコンシン州マジソン)にかけた。減圧下で、各スポットを200μLのHBS pH 7.4, 1% BSA, 0.05% Tritonで洗った。次に、メンブレン全体を緩衝液ですすいで空気乾燥した。メンブレンをストレージ・ホスホ・スクリーン(Storage Phosphor Screen)(Molecular Dynamics、カリフォルニア州サニーベール)に一晩露出し、得られた各々のスポットの強度を、STORM 860ホスホイメージャーを用いてImageQuaNT濃度計プログラム(Molecular Dynamics)をにより定量した。結合のKdは平衡状態を得られた結合曲線に当てはめて決定された(KaleidaGraph、Synergy Software)。実験誤差は2〜4の独立した実験から見積もられた。
【0118】
これらの実験では、Kdは1〜24 nMの狭い範囲に入っていた(表1)。各々2つのループに最も多く単離される配列を含むT09.12およびT10.06クローンは、それぞれ4および2 nMという低いKdを持っていた。それに対して、クローンT09.07およびT10.15のような頻度のより低いループを持ついくつかのクローンも、同様に強い結合を示した。
【0119】
遊離タンパク質と、同じ配列に由来するcDNA/mRNA-タンパク質複合体のTNF-α結合の比較の標本を見ると、2つの解離定数は互いの実験誤差範囲内であり、これはタンパク質自身の標的結合特性を選択するために、cDNA/mRNA-タンパク質複合体を使用することが可能になるというシステムの性質を示す。
【0120】
TNF-α結合クローンの厳密度の高い選択
重複したクローンが単離されたにも関わらず、9回および10回の選択後のTNF-α結合プールには、数多くの異なるクローン、すなわち、標本にした61クローンでは38の異なる配列が、含まれていた。したがって、これらのクローンの中で優れたTNF-α結合を示すサブセットを回収するために、より厳密な結合要件を用いたさらなる選択が行われた。従って、さらに4回の選択(R11-R14)が標的濃度のコントロールが容易な溶液中で行われた。TNF-αの濃度は0.5 nMに限定され、DNA/mRNA-10Fn3プールの濃度は0.1 nMに限定された。さらに、ストレプトアビジンコートの常磁性ビーズに結合した10Fn3/ TNF-α複合体の洗浄の長さと温度も、引き上げられた。
【0121】
具体的には、以下のように選択が行われた。ラウンド11-13では、上述のように構築された0.1 nM DNA/mRNA-10Fn3融合体ライブラリーを、結合緩衝液であらかじめブロックした100μLのダイナビーズ(Dynabeads)(登録商標) M-280(ストレプトアビジンコートの常磁性ビーズ、Dynal、ニューヨーク州レークサクセス)と4℃で1時間回転させて、あらかじめきれいにした。得られた融合体混合物を、上述の結合緩衝液300 μL中で0.5 nMビオチン化TNF-αと混合し、複合体を4℃で1時間インキュベートした。次に、結合緩衝液中で一晩ブロックした1.3 g/cm3で再懸濁したダイナビーズ(登録商標) M-280ストレプトアビジン100μlを混合物に添加し、回転させながらさらに45分間4℃でインキュベートした。Dynal MPC-Sラック上で常磁性ビーズを上清から分離し、上清を除去し、結合緩衝液を用いてビーズをR 11およびR12の場合は1分間、15分間、および30分間洗浄し、R13〜R14の場合は1分間洗浄後、9回の10分間洗浄を行なった。100μl 0.1 M KOHで2回洗うことによって、洗浄したDNA/mRNA-10Fn3:TNF-αビオチン:ストレプトアビジンビーズ複合体からDNAを溶出し、上述のように、カラムを用いた選択によって次の世代のDNA/mRNA-10Fn3融合ライブラリーを作製した。ラウンド14は、選択がさらに150 mM NaClを加えた条件で30分間行われたという点でR11〜R13とは異なっていた。温度を上昇させた以外は、R14とR13の洗浄手順は同じだった。
【0122】
さらに4回の選択後(R14)に溶出されたDNA由来の無作為に選択した22のクローンは、15の異なるループ構造を持っていた(表2;全体の配列は図25)。上述のようにR10で単離されていたクローンT10.06は、8回選択されたが、R9プールで単離されたR09.31を含めた残りの配列は、各々1回単離された。9回目および10回目の単離株と同様に、R14クローンはDEループ中PWA/G配列に偏っており(22クローン中18)、4つの新しい非野生型DE配列も観察された。
【0123】
R14遊離タンパク質プールの見かけの平均Kd値3 nMは9回および10回のプール(それぞれ4および6 nM)と類似していたが、R14プールから単離されたいくつかのクローンのKd値は、R9およびR10プールで観察された最低値よりも1桁低かった(表2)。TNF-αに最も強く結合したクローンT14.07およびT14.25のKdは90 pmolだった。したがって、最後の4回の選択に使用した条件は、ナノモル以下のKdを持つ10Fn3分子に有利になるために充分厳密でありながら、そのような分子を排除してしまうほど厳密ではなかった。
【0124】
突然変異誘発による親和性成熟
上述のように、本明細書に記述される選択では、さらにライブラリーの多様性を増やすために、選択段階の全てまたは一部の後に変異誘発を行なうこともできる。1つの平行選択戦略では、ラウンドの間のDNA増幅に変異性PCRを組み込んだ(CadwellおよびJoyce, PCR Methods Appl. 2:28 (1992))。このテクニックは、上述のR8後に溶出される多様なDNAプールから始められた。このプールを変異性PCRで増幅したが、プールを7つの等量の部分に分け、標的頻度0.8%, 1.6%, 2.4%, 3.2%, 4.0%, 4.8%, および5.6%で変異誘発した。この7つのPCR反応を合わせ、混合物からcDNA/RNA-タンパク質融合体を作製し、溶液中で選択ラウンドを行なった。第2回の変異誘発ラウンドM10の前に、3つの別の反応0.8%, 1.6%, および2.4%で変異性PCRを行なった。2つの残るラウンドM11およびM12は、標準的なTaq PCRを用いて行なった。変異誘発以外は、M9-M12の選択条件はR11-R14と同じだった。調べた20のM12クローンは、前の2つの選択プロトコールを用いて選択したクローンよりも強い結合を示した(表3;全配列は図25)。TNF-αへの最も強い結合はM12.04で、観察されたKdは20 pMだった。これらの結果は、選択後期における低レベルの無作為の変異導入が、選択された抗体模倣物の結合親和性(20 pM対90 pM)、および選択できる速度(12回対14回)の両方を改善し得ることを示す。さらに、この変異導入の手法における強い結合体の頻度は約5%であるのに対し、他の選択では約3%だった。
【0125】
Fn結合体の優位性
選択単位として共有結合のmRNA-タンパク質融合体を用いておこなったTNF-αに結合できる10Fn3の選択は、20 pMという低い解離定数を持つ分子が勝ち残った。これらのKd値は、他の抗体模倣物骨格および選択方法を用いる他の選択の基準と比較して勝るものである。したがって、10Fn3に基づく骨格および共有結合のmRNA-タンパク質融合体に基づくインビトロの選択方法は、広範囲の抗原に対する抗体模倣物の開発に利用できる可能性がある。さらに、本明細書に記述されるナノモルレベル以下のTNF-α結合10Fn3変異体は、治療、研究、診断のための薬剤となる可能性がある。さらに、このインビトロ選択方法は自動化できるので、そのような骨格および選択方法の組み合せは、ゲノムスケールでの用途がある。
【0126】
本選択の成功に寄与した1つの要素は、10Fn3の3つのCDR様ループの全ての無作為化である。1つまたは2つの無作為化ループのみを含む類似のライブラリーは、3つの無作為化CDR様ループを持つライブラリーよりも、強い結合体を含む可能性が低い。
【0127】
上記に報告された選択では、無作為化ループは対応する野生型の10Fn3ループの長さのままである。ライブラリーの多様性をさらに高めるために、ループの長さおよびその配列も変化させ、ライブラリーの構築に使用した野生型骨格に10Fn3βシート中の好ましい変異を組み込み、抗体の模倣という点ではさらにうまく構造の選択ができる無作為化βシート骨格を持つライブラリーを作製できる。
【0128】
本明細書に記述した選択と類似したものは、他の結合種標的(例えばIL-1またはIL-13)を用いても実行できる。
【0129】
動物実験
野生型10Fn3は、インテグリン結合性のトリペプチドモチーフであるアルギニン78-グリシン79-アスパラギン酸80(「RGDモチーフ」)をFGループの先端に含んでいる。このトリペプチドに基づくインテグリンとの結合および潜在性の炎症応答をインビボで回避するために、密接に関連した野生型11Fn3ドメインで見出される配列で、活性を持たない配列であるセリン78-グリシン79-グルタミン酸80(「SGE変異体」)を含む10Fn3の変異体を生産した。このSGE変異体は、N末端にHis6タグを付けて大腸菌中で遊離タンパク質として発現し、金属キレートカラムに続いてサイズ排除カラム上で均質性によって精製される。
【0130】
特に、His6-10Fn3(SGE)をコードするDNA配列をpET9a発現ベクターにクローニングし、BL21 DE3 pLysS細胞を形質転換した。次に、50μg/mlのカナマイシンを含むLB培地中で37℃にて振盪しながらA260=1.0になるまで培養物を増殖させ、その後0.4mM IPTGにより誘導された。誘導された培養物を同じ条件下でさらに一晩(14〜18時間)インキュベートし;標準の低速度遠心分離により細菌を回収した。元の培養容量の1/50倍の溶解緩衝液(50mM Tris 8.0、0.5M NaCl、5% グリセロール、0.05% Triton X-100、および1mM PMSF)に細胞沈殿を再懸濁し、得られたペーストをMicrofluidics Corporation Microfluidizer M110-EHに3回通すことにより細胞を溶解させた。溶解物を遠心分離して清澄化し、上清を0.45μmのフィルターに通して濾過し、続いて0.2μmのフィルターに通して濾過した。清澄化された溶解物の100mlを5mlのタロンコバルトカラム(クロンテック(Clontech)、パロアルト(Palo Alto)、CA)に載せ、70mlの溶解緩衝液で洗浄し、0mM〜30mM イミダゾールの直線勾配を含む溶解緩衝液で溶出した。すべての段階を通じて、カラムを通す流速は1ml/分であった。溶出したタンパク質を15,000〜20,000のPEGに対して透析し(分子量カットオフ値、3,500)、10倍に濃縮した。得られた試料を緩衝液1(グリセロールを含まない溶解緩衝液)中で透析し、緩衝液1で平衡化した16×60mmのセファクリル100のサイズ排除カラム上に1回に5mlずつ載せた。カラムに0.8ml/分で緩衝液1を流し、所望の分子量のタンパク質を含む全画分をプールして、上述のように10倍に濃縮し、次にPBS中で透析した。得られた試料でエンドトキシンスクリーニングおよび動物実験を行った(Toxikon(MA))。
【0131】
これまでに検査された試料中のエンドトキシンの水準は、検査法の検出水準以下であった。予備的な毒性学の研究において、このタンパク質を2匹のマウスに治療量の100倍と見積もられる2.6mg/マウスを注射した。動物は研究の2週間のあいだ、はっきりとした悪い影響なしに生存した。これらの安全な結果は、静脈内薬に取り込まれた10Fn3の使用を支持する。
【0132】
インビボでの使用するための他の構築物
8kDの10Fn3ドメインの半減期を延ばすために、天然抗体を模倣するより大きい分子も構築されている。この10Fn3-Fc分子は、宿主のIgGの定常領域にある-CH1-CH2-CH3(図11)もしくは-CH2-CH3ドメインを含んでおり、これらの構築物中では、10Fn3ドメインはIgGのVHドメインの代わりにN末端に結合されている(図11および図12)。このような抗体様構築物は、天然の免疫応答を利用できる可能性と同様に、タンパク質の薬物動態を改善させるべきである。
【0133】
10Fn3-CH1-CH2-CH3クローンのマウス型を構築するために、最初にマウス肝脾cDNAライブラリー(クロンテック(Clontech))から-CH1-CH2-CH3領域を増幅し、次にpET25ベクターに連結した。クローニングに使用したプライマーは5' Fc Nestおよび3' 5 Fc Nestであり、回収したインサートの末端に適切な制限酵素部位を結合するのに使用したプライマーは、5' Fc HIIIおよび3' Fc Nheであった。
【0134】
このクローンからCH1領域を除去し、より短いFc部分を作製して10Fn3-CH2-CH3クローンとするために、さらなるPCRを用いた。各クローンの5'末端に10Fn3をコードする配列を連結した。同一のマウス脾臓cDNAライブラリーからクローニングされた野生型10Fn3でも、または分子の変異もしくは無作為化により得られた改変10Fn3でも使用することができる。マウス野生型10Fn3のクローニングに使用したオリゴヌクレオチドは、以下であった:
Mo 5PCR-NdeI:
Mo5PCR-His-NdeI(His6精製タグを有する、代わりのN末端用):
【0135】
ヒトオリゴヌクレオチド配列において、同一の方法を用いることにより、ヒトの等価なクローンが構築される。
【0136】
タンパク質チップ適用における抗体模倣体の骨格
本明細書に記載の任意の抗体模倣体は、マイクロチップのような固体支持体上で免疫化してもよい。10Fn3骨格のような本明細書の骨格がタンパク質チップ適用に適しているのは、 (1) ベンチまたは自動化装置で迅速に選択できる多くの結合基を支持する能力があり、かつ (2) 生物物理学的特性が優れているためである。
【0137】
10Fn3の用途の広い結合特性は、Fn3免疫グロブリン様、βサンドイッチ型折畳みによって示されるループに関連する。上述のように、これらのループは抗体の可変ドメインの相補性決定領域に類似しており、抗体ループと同様な様式で共同して抗原に結合できる。本発明者らの系では、多様なmRNA-10Fn3融合体のライブラリーを作製するために、10Fn3ループのBC(例えば21残基〜30残基)、DE(例えば51残基〜56残基)、およびFG(例えば76残基〜87残基)は、配列、長さ、または配列と長さの両方が、無作為化されている。そのようなライブラリーにおける結合物は、高親和性の結合物の小さな集団が得られるまで、固定化またはタグ付き標的に対する親和性に基づいて濃縮される。また、誤りがちの(error-prone)PCRおよび組換えを用いて、選択された結合物の親和性の成熟を促進することができる。迅速で効率の良い選択および親和性成熟プロトコールにより、多数の標的に対する結合物を短時間で選択することができる。
【0138】
タンパク質チップ上に固定される結合物の骨格として、10Fn3ドメインは、抗体断片および単鎖抗体よりも小さく、扱いやすいという利点を持つ。例えば、安定性および溶解度に大きな差があり、構造的に必須なジスルフィド結合を保つために酸化環境を必要とする単鎖骨格または単離された抗体可変ドメインとは異なり、10Fn3は非常に安定で、その融解温度は110℃、溶解度は>16 mg/mLである。また、10Fn3骨格はジスルフィド結合も遊離のシステインも含まない;したがって、その環境の酸化還元電位には影響されない。10Fn3のもう1つの利点は、抗原結合ループおよびN末端が、βサンドイッチの、C末端とは反対側の端にあるということである;そのため、10Fn3骨格がC末端でチップに結合すると、抗原結合ループが、分析する溶液に最大限で接触可能になる。10Fn3はわずか94のアミノ酸残基からなる単一ドメインであるため、約250残基の単鎖抗体よりも高密度でチップ表面に固定することが可能である。また、10Fn3骨格は、このドメインの溶解度が高いことからもわかるように、親水性であるため、チップ表面に対する望まれない結合を最小化する。
【0139】
10Fn3骨格の安定性と、ライブラリー作製および結合物選択のための適合性は、テネイシン、Nカドヘリン、Eカドヘリン、ICAM、タイチン、GCSF-R、サイトカイン受容体、グリコシダーゼ阻害剤、および抗生物質色素タンパク質のドメインのような、免疫グロブリン様の折畳みを持つ、タンパク質ドメインの10Fn3様の大きなクラスにも共通している可能性がある。このようなドメインに共通の重要な特徴は、2つのβシートが提供する安定した枠組みであり、このβシートは互いに圧縮され、シートの1つの縁につき、少なくとも3つの溶媒が接触可能であるループによって結合する。そのようなループを無作為化して、枠組みの構造を破壊することなく、潜在的な結合物のライブラリーを作製することができる(上記)。さらに10Fn3と同様に、これらのループのいずれか(または他のタンパク質由来の類似のループ)は、単独でまたは他のループと組み合わせて、固体支持表面上に対して固定化させてもよい。
【0140】
Fn3を基にした抗体模倣体の固定化
チップ表面にFn3を基にした抗体模倣体のような抗体模倣体を固定するために、いくつかの典型的な手法が利用できる。例えば、このような抗体模倣体はRNA-タンパク質融合体として、融合体のRNA部分をワトソン・クリックハイブリダイゼーションによって、チップ表面に固定化したベースの相補的DNAに固定することができる(例えば、Adressable Protein Assays、米国特許出願第60/080,686号;米国特許出願第09/282,734号、および国際公開公報第99/51773号;およびMethods for Encoding and Sorting In Vitro Translated Protein、米国特許出願第60/151,261号および米国特許出願第09/648,040号に記述)。または、抗体模倣体は、チップ表面に直接遊離のタンパク質として固定できる。チップ表面に抗体模倣体を沈着させるためには、手作業およびロボット装置が使用できる。アレイ形式で高密度に抗体模倣体を沈着させるためには、スポットロボットが使用できる(例えば、Luekingら、Anal Biochem. 1999 May 15; 270(1):103-11の方法による)。抗体模倣体をチップ表面に結合させるためには、別の方法も利用できる。「酵素学の方法(Methods in Enzymology )」(M. MosbachおよびB. Danielsson編)、第135巻および136巻、Academic Press、Orlando、Florida、1987;ニルソン(Nilsson)ら、Protein Expr. Purif. 1997 Oct;11(1):1〜16、およびその参考文献に記述されるものを含め、いくつかの標準的な固定化手法が使用できる。抗体模倣体の方向性を持った固定化は、チップに結合した抗体模倣体の結合容量を増加させる際に役立つ。方向性を持った結合のための典型的な手法は、リュー(Lu)ら、The Analyst (1996)、vol. 121、p29R-32R;およびターコバ(Turkova)、J Chromatogr B Biomed Sci App. 1999 Feb 5;722(1-2):11-31に記述されている。さらに、抗体模倣体をチップ表面に結合させるための本明細書に記述されるいかなる方法も、抗体模倣体をビーズ、または他の支持体に固定するために使用できる。
【0141】
標的タンパク質の捕捉と検出
骨格結合物の選択された集団を用いて、例えば、生物試料のような試料中で、分析物標的の検出および/または定量ができる。この種の診断解析を行なうためには、関心対象の標的に対する選択された骨格結合体を適当な支持体上に固定して、多機能タンパク質チップを作製する。次に、チップに試料を載せ、試料中の成分で結合物に結合するものを、固定化した結合物の標的特異性に基づいて同定する。この手法を用いると、試料中の1つまたは複数の成分が、同時に同定または定量できる(例えば、試料のプロファイリングのための手段として)。
【0142】
標的の検出方法は、結合したタンパク質標的レベルの測定を可能にするものであり、X線撮影、蛍光スキャニング、質量分光(MS)、および表面プラズモン共鳴(SPR)を含むがこれらに限定されるものではない。リン光画像化システム(Molecular Dynamics、Sunnyvale、CA)を用いたオートラジオグラフィも、例えば35Sメチオニンで放射標識した標的タンパク質の検出と定量に使用できる。レーザースキャナーを用いた蛍光スキャニング(以下を参照のこと)は、蛍光標識した標的の検出と定量に使用できる。または、蛍光スキャニングを用いて、それ自身が標的タンパク質に結合する蛍光標識リガンドを検出することもできる(例えば、蛍光標識した標的特異的抗体、または標的-ビオチンに結合する、蛍光標識したストレプトアビジン、以下に記述)。
【0143】
質量分光は、結合した標的をその分子量によって検出および同定するために使用できる。結合した標的タンパク質の脱離は、下記のようにして、チップ表面から直接レーザーを用いて行なうことができる。質量の検出では、分子量に基づいて、リン酸化またはグリコシル化のような翻訳後修飾を含む標的の修飾も決定することができる。表面プラズモン共鳴は、骨格結合物が適当な金表面(例えば、スウェーデンのBiacoreより入手可能)に固定されている場合に結合したタンパク質標的の定量に使用することができる。
【0144】
以下に、該結合物(この場合、タンパク質TNF-αに特異的なFn結合物)の選択方法、および選択された集団を用いたチップ上での検出を例示する。本実施例は、本発明を説明するためのものであり、制限するためのものではない。
【0145】
10Fn3骨格に基づくTNF-α結合物の選択
チップ上で骨格を選択するための1つの典型例では、無作為化ループBC、DE、およびFGを持つヒト10Fn3変異体のライブラリーを用いて、TNF-αに対して10Fn3に基づく選択が行われた。ライブラリーは、1つの無作為化ループを含め、ヒト10Fn3の約3分の1をコードするヌクレオチド配列をそれぞれ含む、3つのDNA断片から構築された。上記のループ残基をコードするDNA配列は、関心対象の残基のコドンが (NNS)nで置換されたように、オリゴヌクレオチド合成によって再構築された。式中、Nは4つのデオキシリボヌクレオチド(A, C, GまたはT)のいずれかであり、SはCまたはGを表す。各断片のC末端は、FLAG精製タグの配列を含んでいた。
【0146】
ショスタック(Szostak)ら(RobertsおよびSzostak、Proc. Natl. Acad. Sci. USA 94:12297, 1997; Szostakら、米国特許出願第09/007,005号、および米国特許出願第09/247,190号;Szostakら、国際公開公報第98/31700号)に記述されるように、クレノー酵素で伸長後、各DNAを転写し、ピューロマイシン含有DNAリンカーと連結し、インビトロで翻訳し、mRNA-ペプチド融合体を作製し、これをDNA-mRNA-ペプチド融合体に逆転写した。FLAGタグの付いたペプチドをM2アガロースに結合することにより、フレームシフトまたは余分の終止コドンを含むものから、全長の融合分子が分離された;精製した全長の融合体に結合したDNAをPCRにより増幅した後、3つのDNA断片をEar I 制限エンドヌクレアーゼにより切断し、連結して全長の鋳型を形成させた。鋳型を転写し、ピューロマイシン含有DNAリンカーに連結し、翻訳して10Fn3-RNA/cDNAライブラリーを作製した。その後、これを逆転写してDNA-mRNA-ペプチド融合ライブラリーを作製し、後に選択に使用した。
【0147】
TNF-α結合物の選択は、50 mM HEPES、pH 7.4、0.02% Triton-X、0.1 mg/mLサケ精子DNA中で行われた。PROfusion(商標)ライブラリーは、セファロース固定TNF-αとインキュベートした;洗浄後、最も強い結合物に伴うDNAを0.1 M KOHで溶出し、PCR増幅、転写、連結、翻訳、逆転写し、次の選択ラウンドの開始材料とした。
【0148】
そのような選択を10ラウンド行なった(図13に示す);見かけの平均Kdが120 nMでTNF-αセファロースに結合するPROfusion(商標)プールが得られた。解析されたプールの特定のクローン成分は、50 nM〜500 nMの範囲でTNF-α結合を示した。
【0149】
固定化、標的タンパク質の捕捉、およびMALDI-TOF検出
Fn3融合体をチップ表面に固定するための第1段階として、自動DNA合成装置(PE BioSystems Expedite 8909)で、固相支持体ホスホラミダイド法でオリゴヌクレオチド捕捉プローブを調製した。全ての試薬はグレン・リサーチ(Glen Research)から入手した。合成は、ジスルフィド結合を含む固相支持体から開始して、最終的に3'末端のチオール官能性が得られた。添加される最初の4つの単量体は、ヘキサエチレンオキシドユニットで、20のT単量体が続いた。5'末端のDMT基は除去されなかった。捕捉プローブは固相支持体から開裂し、水酸化アンモニウムで脱保護し、真空遠心分離で乾燥するまで濃縮し、トリエチルアンモニウム酢酸緩衝液中のアセトニトリル勾配を用いて、逆相HPLCで精製した。HPLCの適当な画分を回収し、真空遠心分離で乾燥するまで蒸発させ、30分間の80% AcOH処理によって5'末端のDMT基を除去した。酸は気化により除去し、次にオリゴヌクレオチドは100 mM DTTで30分間処理してジスルフィド結合を開裂させた。DTTはEtOAcで繰り返し抽出して除去した。オリゴヌクレオチドは残りの水相からエタノール沈殿させ、逆相HPLCによって純度を確認した。
【0150】
3'チオールの捕捉プローブは脱気した1×PBS緩衝液中で250μMに調節して、少量の水を含む、アルゴンをフラッシュした(argon-flushed)チェンバー中で、単一の液滴(75μL)として9×9 mmの金をコートしたチップ(Biacore)に塗布した。室温で18時間後、捕捉プローブ溶液を除去し、機能化したチップを穏やかに撹拌しながら50 mLの1×PBS緩衝液で洗浄し(各15分間、2回)、同様にして50 mLの水ですすいだ(各15分間、2回)。残りの液体を注意深く除去して、機能化したチップは直ちに使用するか、またはアルゴン下で4℃で保存した。
【0151】
ラウンド10のTNF-α選択(上記)で得られた10Fn3融合体プールの約1 pmolを数時間リボヌクレアーゼ処理し、70μL中で5×SSCに調節し、上述の機能化した金チップに単一の液滴として塗布した。50μL容量のガスケット装置を使って、機能化チップで融合混合液を密封し、装置を4℃で連続的に回転させた。18時間後に、装置を解体し、金チップを穏やかに撹拌しながら50 mLの5×SSCで10分間洗浄した。チップ表面から過剰な液体を注意深く除去し、チップを4℃で10分間、ブロッキング溶液(1×TBS + 0.02% トゥイーン20 + 0.25% BSA)で不動態化した。過剰な液体を注意深く除去し、同一組成のブロッキング溶液中に500μg/mL TNF-αを含む溶液を単一の液滴としてチップに塗布し、ピペットマンで液滴を時々混合しながら4℃で2時間インキュベートした。結合溶液を除去した後、チップを穏やかに撹拌しながら4℃で5分間洗浄(50 mL 1×TBS + 0.02%トゥイーン20)し、室温で乾燥させた。第2のチップは、ハイブリダイゼーション混合液に融合体が添加されなかったこと以外は、上記と同様に調製した。
【0152】
次に、MALDI-TOFマトリックス(1:1エタノール/10% 蟻酸水溶液中、15 mg/mL 3,5-ジメトキシ-4-ヒドロキシケイ皮酸)を、高精度3軸ロボット(MicroGrid, BioRobotics)を用いて、金チップに均一に塗布した。マトリックスを384ウェルのマイクロタイタープレートからチップに移すのに16ピンのツールが使用され、600ミクロンのピッチを有する直径200ミクロンの形質が作られた。MALDI-TOF質量分析器(Voyager DE, PerSeptive Biosystems)の機器設定は以下の通りであった:加速電圧=25k、グリッド電圧=92%、ガイドワイヤ電圧=0.05%、遅延=200オン、レーザー出力=2400、低質量ゲート=1500、陰イオン=オフ。金チップはチップのレベルをステージのレベルと同じに保つように、改変されたMALDI試料ステージ上に1つずつ置かれ、適切なフライト距離を可能にした。装置のビデオモニターおよび運動制御システムを用いて、レーザービームが個々のマトリックス形質に向かうようにした。
【0153】
図14および図15は、それぞれ10Fn3融合体チップおよび非融合体チップの質量スペクトルを示す。いずれも、スペクトルを集めるために少数の200ミクロンの形質が分析されたが、図15のほうが、かなり多くのデータ収集を要した。17.4 kDaのシグナルはTNF-α単量体に相当する。
【0154】
固定化、標的タンパク質の捕捉、および蛍光検出
洗浄済の1×3インチの顕微鏡スライドグラス(Goldseal, #3010)を、ナノストップ(Nanostrip)(Cyantek)で15分間、10% NaOH水溶液、70℃で3分間、および1% HCl水溶液で1分間、各試薬の後に脱イオン水で十分にすすぎながら処理した。その後、スライドは無水硫酸カルシウム上で真空乾燥器中で数時間乾燥させた。95%アセトン/5%水中の1%アミノプロピルトリメトキシシランを調製し、20分間加水分解させた。スライドグラスは、穏やかに撹拌しながら加水分解したシラン溶液に5分間浸した。毎回新しい95%アセトン/5%水を用いて、スライドを5分間穏やかに撹拌しながら10回洗浄し、過剰なシランを除去した。その後、スライドを110℃で20分間加熱することにより、保存処理した。シラン処理したスライドは、新しく調製した90% DMF/10%ピリジン中の0.2%フェニレン1,4-ジイソチオシアネート溶液に、穏やかに撹拌しながら2時間浸した。その後、90% DMF/10%ピリジン、メタノール、およびアセトンで順次洗浄した。空気乾燥後、機能化したスライドは、真空乾燥器中で無水硫酸カルシウム上で0℃で保存した。市販のアミン反応性スライド(3-D Link, Surmodics)を用いて、同様な結果が得られた。
【0155】
オリゴヌクレオチド捕捉プローブは、従来のホスホラミダイド化学を用いて、自動DNA合成装置(PE BioSystems Expedite 8909)で調製された。全ての試薬はグレン・リサーチ(Glen Research)から入手した。合成は、保護されているアミノ官能性を直交して持つ固相支持体から開始し、それにより3'末端のアミンが最後の脱保護段階まで曝露されないようにした。添加される最初の4つの単量体は、ヘキサエチレンオキシドユニットで、標準的なA, G, CおよびT単量体が続いた。すべての捕捉オリゴ配列は、固相支持体から開裂され、水酸化アンモニウムで脱保護し、乾燥するまで濃縮し、エタノール沈殿し、かつトリエチルアンモニウム酢酸緩衝液中のアセトニトリル勾配を用いて、逆相HPLCで精製した。HPLCの適当な画分を回収し、真空遠心分離で乾燥するまで蒸発させ、水の一部とともに蒸発させた。
【0156】
精製したアミン標識捕捉オリゴは、10%グリセロールを含む50 mM炭酸ナトリウム緩衝液(pH 9.0)中で、250μMの濃度に調節した。プローブを3軸ロボット(MicroGrid, BioRobotics)を用いて、アミン反応性ガラス表面上の5×5×6アレイパターン中の規定の位置にスポットした。16ピンのツールを用いて、384ウェルのマイクロタイタープレートから液体を移し、600ミクロンのピッチを有する200ミクロンの形質を作製した。24の形質の各サブグリッドは、単一の捕捉プローブ(すなわち、24の重複するスポット)に相当する。アレイは湿度が飽和した環境中で、室温で12時間〜18時間インキュベートした。接着反応は、チップを2%水酸化アンモニウム水溶液中に穏やかに撹拌しながら5分間浸すことで停止させ、蒸留水ですすいだ(各5分3回)。最後にアレイを室温で30分間10×PBSに浸漬してから、蒸留水で5分すすいだ。
【0157】
10Fn3 mRNAと共に特異的かつ熱力学的に等エネルギーの配列が、自己集合および10Fn3タンパク質固定のための捕捉ポイントとして役立つことが明らかにされた。ソフトウェアプログラムHybSimulator v4.0 (Advanced Gene Computing Technology, Inc.)は、潜在的な捕捉プローブの同定および解析を容易にした。6つの固有の捕捉プローブが選択され、チップ上にプリントされ、そのうちの3つは10Fn3融合体プールのmRNAの共通領域(CP3', CP5'およびCPflag)に相補的である。残りの3つの配列(CPneg1, CPneg2およびCPneg3)は相補的ではなく、部分的に陰性対照として働く。上述のように、捕捉プローブは各々3'アミノ末端および4つのヘキサエチレンオキシドスペーサーユニットを持っている。以下は、使用した捕捉プローブの配列リストである(5'→3'):
ラウンド10のTNF-α選択由来の約1 pmolの10Fn3融合体プールを、総容量350μL中での0.02%トゥイーン20および2 mMバナジルリボヌクレオチド複合体を含む、5×SSCに調節した。400μLガスケット装置下で全体量をマイクロアレイに添加し、この集合を室温で18時間、連続的に回転させた。ハイブリダイゼーション後、スライドを500 mLの5×SSC、2.5×SSC、および1×SSCで順次5分ずつ洗浄した。液体の痕跡は遠心分離で除去し、スライドを空気乾燥させた。
【0158】
組換えヒトTNF-α(500μg、凍結乾燥、PreproTech由来)を230μLの1×PBSに入れ、Microdialyzerユニット(3,500 MWCO, Pierce)中で、700 mLの撹拌した1×PBSに対して4℃で18時間透析した。透析したTNF-αをEZ-Link NHS-LC-LCビオチン化試薬(20μg、Pierce)で0℃で2時間処理し、Microdialyzerユニット(3,500 MWCO, Pierce)中で、700 mLの撹拌した1×PBSに対して4℃で18時間再び透析した。得られた結合体をMALDI-TOF質量分析器によって分析し、単一のビオチン部分でほぼ完全に機能化されることが見出された。
【0159】
以下の過程は、各々4℃で連続回転または混合しながら行なった。タンパク質マイクロアレイ表面は、0.02%トゥイーン20および0.2% BSAを含む1×TBS(200μL)による60分間の処理によって、不動態化した。ビオチン化TNF-α(不動態化緩衝液中100 nM濃度に作製)を、マイクロアレイに120分間接触させた。マイクロアレイは0.02%トゥイーン20を含む1×TBSで洗浄した(50 mL3回、各5分間)。蛍光標識したストレプトアビジン(Molecular Probes由来の2.5μg/mL アレクサ 546-ストレプトアビジン結合体、不動態化緩衝液中に作製)をマイクロアレイに60分間接触させた。マイクロアレイを0.02%トゥイーン20を含む1×TBSで洗浄し(50 mL2回、各5分間)、続いて1×TBSで3分間すすいだ。液体の痕跡は遠心分離で除去し、スライドを室温で空気乾燥させた。
【0160】
蛍光レーザースキャニングは、GSI Lumonics ScanArray 5000システムで、10μMピクセルの解像度ならびにアレクサ 546色素用の予め設定された励起波長および発光波長を用いて行なった。リン光画像(phosphorimage)分析は、分子動態学ストーム(storm)システムを用いて行なった。マイクロアレイとリン光体保存スクリーンを直接接触させて、曝露時間は48時間であった。リン光画像スキャニングは50ミクロンの解像度の設定で行われ、データはImageQuant v.4.3ソフトウェアで抽出した。
【0161】
図16および図17は、同一アレイのリン光画像および蛍光スキャンである。リン光画像は、35Sメチオニンシグナルに基づいて、10Fn3融合体がどこにハイブリダイズしたかを示す。蛍光スキャンは、標識TNF-αがどこに結合したかを示す。
【0162】
他の態様
他の態様は、特許請求の範囲内である。
【0163】
本明細書で言及される全ての刊行物、特許、および特許出願は、参照として本明細書に組み入れられる。
【図面の簡単な説明】
【0164】
【図1】ラクダ(濃青)およびラマ(淡青)より得られた抗体の重鎖における可変領域の構造間の比較を2つの配向からそれぞれ示す写真である。
【図2】ラクダ抗体の重鎖の可変領域(濃青)、ラマ抗体の重鎖における可変領域(淡青)、および10番目のフィブロネクチン3型モジュール(10Fn3)(黄色)の構造間の比較を示す写真である。
【図3】10番目のフィブロネクチン3型モジュール(10Fn3)を示す写真であり、IgG重鎖における抗原結合ループに対応するループを赤色で強調した。
【図4】フィブロネクチン3型タンパク質ドメインおよび関連タンパク質ドメイン間の配列の整列化を図示するグラフである。
【図5】10Fn3ドメイン、ならびにフィブロネクチン、テネイシン、コラーゲン、およびアンジュリンを含む15個の関連タンパク質間の構造の類似性を示す写真である。この写真では、領域は以下のように分類されている:定常領域は濃青、保存領域は淡青、中性領域は白、可変領域は赤、およびRGDインテグリン結合領域(可変)は黄。
【図6】9番目および10番目のフィブロネクチン3型モジュールの空間充填モデルを2つの異なる配向からそれぞれ示す写真である。2つのモジュールおよびインテグリン結合ループ(RGD)が分類されている。この図では、青が正電荷を帯びた残基を示し、赤が負電荷を帯びた残基を示し、白が無電荷の残基を示している。
【図7】7〜10番目のフィブロネクチン3型モジュールの空間充填モデルを3つの異なる配向からそれぞれ示す写真である。4つのモジュールが分類されている。この図では、青が正電荷を帯びた残基を示し、赤が負電荷を帯びた残基を示し、白が無電荷の残基を示している。
【図8】フィブロネクチン3型ドメインを含むRNA-タンパク質融合体の異なる塩条件下における構造を図示する写真である。
【図9】PCRシグナル解析によって測定されたフィブロネクチン3型ドメインを含むRNA-タンパク質融合体の選択を図示する一連の写真である。
【図10】RNA-タンパク質融合体および遊離タンパク質の選択間における比較、ならびに本明細書に記載の選択間において、TNF-αとの結合%での増加を図示するグラフである。
【図11】IgG、10Fn3、Fn-CH1-CH2-CH3、およびFn-CH2-CH3を示す一連の概略図である(左上から時計回りに)。
【図12】IgG(X線結晶学)および10Fn3(NMRおよびX線結晶学)の既知の三次元構造に基づいたFn-CH1-CH2-CH3の分子モデルを示す写真である。
【図13】TNF-α結合物の10Fn3に基づく核酸-タンパク質融合体の例示的な選択の時間経過を示すグラフである。TNF-α-セファロースに結合した、核酸-タンパク質融合体のプール(白抜きのひし形)および遊離のタンパク質のプール(白抜きの丸)の割合、ならびに未誘導化セファロースに結合した遊離タンパク質のプール(黒丸)の割合が示されている。
【図14】TNF-αFn結合物によるTNF-α結合を示すグラフである。特に、これらの図はそれぞれ10Fn3融合体チップおよび非融合体チップから得られた質量スペクトルデータを示す。
【図15】TNF-αFn結合物によるTNF-α結合を示すグラフである。特に、これらの図はそれぞれ10Fn3融合体チップおよび非融合体チップから得られた質量スペクトルデータを示す。
【図16】それぞれ10Fn3アレイのリン光画像および蛍光スキャンであり、TNF-α結合を示す。
【図17】それぞれ10Fn3アレイのリン光画像および蛍光スキャンであり、TNF-α結合を示す。
【図18】ラマVHドメインおよび野生型ヒト10Fn3ドメインの一次配列の整列化を示すグラフである。2つの配列の間の相同残基が示されている。選択されたクローンの約45%で変異している無作為化ループの外の10Fn3残基は、野生型10Fn3配列の下の矢印で示されており、選択された残基を示す文字が付けられている。
【図19】ラマVHドメインおよび野生型ヒト10Fn3ドメインの模式図である。変異した枠組みの残基が示されている。
【図20】元のライブラリー(R0)およびTNF-αを用いた10回の選択後(R10)から翻訳された遊離タンパク質プールの結合効率および特異性を示すグラフである。未誘導化セファロース、TNF-α-セファロース、IL-1α-セファロース、およびIL-13-セファロースに対するタンパク質プールの結合が比較されている。
【図21】最大3個のループのディスプレイのための一連のIgG様骨格である。
【図22】最大4個、または6個のループのディスプレイのための一連のIgG様骨格である。
【図23】ループ構造のディスプレイのための、IgGに関連のない一連の骨格である。
【図24】固相表面に固定化された模倣物による標的(TNF-α)の特異的キャプチャを示す写真およびグラフである。
【図25−1】本発明の典型的なTNF-α結合体(配列番号:33〜140)を示すグラフである。
【図25−2】本発明の典型的なTNF-α結合体(配列番号:33〜140)を示すグラフである。
【図25−3】本発明の典型的なTNF-α結合体(配列番号:33〜140)を示すグラフである。
【図25−4】本発明の典型的なTNF-α結合体(配列番号:33〜140)を示すグラフである。
【技術分野】
【0001】
発明の背景
本発明は、例えば新規の結合特性を持つ生成物の生産に有用なタンパク質骨格に関する。
【背景技術】
【0002】
一般にタンパク質骨格と呼ばれる比較的限定された三次元構造を持つタンパク質は、操作された生成物を設計するための試薬として用いることができる。これらの骨格は、特定の、もしくは無作為な配列の変異を受け入れる1つまたは複数の領域を典型的に含んでおり、このような配列の無作為化は、タンパク質ライブラリーを生産し、そのライブラリーから求める生成物を選択するためにしばしば行われる。このような骨格が有用な1つの特定の分野は、抗体設計の分野である。
【0003】
試薬または薬剤を得るために哺乳類の免疫系を操作する多くの方法が以前より試みられてきた。これらには、特定の抗原に反応するポリクローナル抗体の混合物を得るために関心対象の抗原を動物に注射する段階、ハイブリドーマ培養細胞にモノクローナル抗体を生産させる段階(KoehlerおよびMilstein、Nature、第256巻、第495頁、1975年(非特許文献1))、新規の認識特性または最適化された認識特性を得るために現存のモノクローナル抗体を改変する段階、望ましい結合特性を持つ新規の抗体断片を作製する段階、ならびに単鎖抗体(抗体分子の重鎖および軽鎖の可変領域を可動性のペプチドリンカーと連結することにより作製される)を無作為化してファージディスプレイ法で抗原との結合により選択する段階(Clacksonら、Nature、第352巻、第624頁、1991年(非特許文献2))が含まれる。
【0004】
さらに、新規の結合特性を持つタンパク質を得るために、免疫グロブリン以外のいくつかのタンパク質骨格が提案されている。例えば「ミニボディ」骨格は、免疫グロブリンの折り畳みに関連しており、モノクローナル抗体の重鎖の可変ドメインから3つのβ鎖を除去することで設計されている(Tramontanoら、J. Mol. Recognit.、第7巻、第9頁、1994年(非特許文献3))。このタンパク質は61残基を含んでおり、これを使用して2つの超可変ループを生じうる。これらの2つのループは無作為化され、抗原との結合により生成物が選択されるが、これまでのところ、溶解性の問題により、この枠組みには幾らかの限定された効用しかないようである。ループを示すのに使用される別の枠組みは、テンダミスタット(tendamistat)であり、74残基で、サンドイッチ状の6本のβシート鎖が2つのジスルフィド結合によって結合している(McConnellおよびHoess、J. Mol. Biol.、第250巻、第460頁、1995年(非特許文献4))。この骨格には3つのループが含まれているが、これまでのところ、これらのループのうちの2つについてのみ、無作為化の可能性が試されている。
【0005】
他のタンパク質も枠組みとして試験されてきており、αヘリックスの表面に無作為化された残基(Nordら、Nat. Biotechnol.、第15巻、第772頁、1997年(非特許文献5)、Nordら、Protein Eng.、第8巻、第601頁、1995年(非特許文献6))、αヘリックス束中のαヘリックス間にあるループ(KuおよびSchultz、Proc. Natl. Acad. Sci. USA、第92巻、第6552頁、1995年(非特許文献7))、および小さいプロテアーゼ阻害剤のようにジスルフィド架橋で束ねられたループを提示するために使用されている(Marklandら、Biochemistry、第35巻、第8045頁、1996年(非特許文献8);Marklandら、Biochemistry、第35巻、第8058頁、1996年(非特許文献9);RottgenおよびCollins、Gene、第164巻、第243頁、1995年(非特許文献10);Wangら、J. Biol. Chem.、第270巻、第12250頁、1995年(非特許文献11))。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】KoehlerおよびMilstein、Nature、第256巻、第495頁、1975年
【非特許文献2】Clacksonら、Nature、第352巻、第624頁、1991年
【非特許文献3】Tramontanoら、J. Mol. Recognit.、第7巻、第9頁、1994年
【非特許文献4】McConnellおよびHoess、J. Mol. Biol.、第250巻、第460頁、1995年
【非特許文献5】Nordら、Nat. Biotechnol.、第15巻、第772頁、1997年
【非特許文献6】Nordら、Protein Eng.、第8巻、第601頁、1995年
【非特許文献7】KuおよびSchultz、Proc. Natl. Acad. Sci. USA、第92巻、第6552頁、1995年
【非特許文献8】Marklandら、Biochemistry、第35巻、第8045頁、1996年
【非特許文献9】Marklandら、Biochemistry、第35巻、第8058頁、1996年
【非特許文献10】RottgenおよびCollins、Gene、第164巻、第243頁、1995年
【非特許文献11】Wangら、J. Biol. Chem.、第270巻、第12250頁、1995年
【発明の概要】
【0007】
本発明は、関心対象のどのような化合物とも結合するように導出(evolve)しうる新規のタンパク質のファミリーを提供する。フィブロネクチン型III(Fn3)もしくはFn3様ドメイン由来の骨格を一般に使用したこれらのタンパク質は、特徴的な天然型抗体または操作された抗体(すなわち、ポリクローナル抗体、モノクローナル抗体、もしくは単鎖の抗体)の様式で機能し、さらに構造的な利点を持つ。特に、これらの抗体模倣物の構造は、標準的には抗体の構造および機能の喪失につながるような条件下でさえも、最適な折り畳み、安定性、および溶解性を持つように設計されている。
【0008】
これらの抗体模倣物は、実質的に関心対象の任意の化合物(例えば任意のタンパク質)とも結合しうるタンパク質を設計する目的で使用することができる。例えば、本明細書に述べられた10Fn3に基づく分子を、抗体の可変領域の相補性決定領域(CDR)と位置および構造において類似した無作為化されたFn3ループを1つまたは複数有する集団を形成するために、および/または、Fn3の他の3つの可溶性露出ループを無作為化するために、定方向展開(directed evolution)に供する骨格として使用することができる。このような定方向展開の方法により、関心対象の抗原に高い親和性を持つ抗体様分子が生産される。さらに、本明細書に述べられた骨格を規定された露出ループ(例えば、前もって無作為化され、抗原との結合に基づいて選択されたループ)を示し、このように導入されたループと結合する分子の展開(evolution)の方向付けをするためにも使用することができる。この型の選択は、個々の任意のCDR様ループ、または非線形エピトープに結合した2つもしくは3つ全部のCDR様ループを認識する認識分子を同定するために行ってもよい。
【0009】
したがって、第1の局面では、本発明は無作為化されたまたは変異した骨格タンパク質に関する。特に、本発明は免疫グロブリン様の折畳みを持つドメインを含む非抗体タンパク質に関するが、その非抗体タンパク質は変異したアミノ酸配列を持つことにより基準タンパク質から誘導され、この非抗体タンパク質は基準タンパク質が非抗体タンパク質ほど強く結合しない化合物に対して少なくとも1μMの強さのKdで結合する。
【0010】
さらに、本発明は免疫グロブリン様の折畳みをもつドメインを含む骨格タンパク質から誘導される非抗体タンパク質に関し、誘導体タンパク質中のドメインのアミノ酸配列は、骨格タンパク質中のドメインのアミノ酸配列と50%以上同一である。
【0011】
別の態様では、本発明は少なくとも1つの無作為化ループを持つフィブロネクチン3型ドメインを含むタンパク質に関するが、このタンパク質はFn3ドメインが対応する天然に存在するFn3ドメインの結合しない化合物に結合できるということにより特徴づけられる。
【0012】
種々の好ましい態様では、本発明のこれらの任意のタンパク質は、少なくとも500 nMのKd、好ましくは少なくとも100 nMまたは10 nMのKd、およびより好ましくは少なくとも1 nM、500 pM、100 pM、または20 pMの強さのKdでその標的化合物に結合する。タンパク質は好ましくは1個、2個、または3個の変異ループを含み、少なくとも1個のループ、および好ましくは2個または3個全てのループが化合物に対するタンパク質の結合に貢献する。さらに、基準タンパク質は好ましくはジスルフィド結合を持たず、誘導体タンパク質は少なくとも1つのジスルフィド結合を持つ可能性がある。
【0013】
特定の態様に関して、免疫グロブリン様の折畳みをもつドメインは、好ましくは10 kD未満または7.5 kDを越える分子量、およびより好ましくは7.5〜10 kDの間の分子量を持つ。本発明のタンパク質は生理的条件下で単量体であるか、例えば二量体のような多量体である。他の好ましい態様では、本発明の変異タンパク質を誘導するために用いた基準タンパク質は、天然に存在する哺乳類タンパク質(例えば、ヒトのタンパク質)である;免疫グロブリン様の折畳みをもつドメインは、変異しており、基準タンパク質の免疫グロブリン様の折畳みと比較して最高50%、および好ましくは最高34%の変異したアミノ酸を含む。さらに、免疫グロブリン様の折畳みをもつドメインは、約50〜150アミノ酸を含み、より好ましくは約50アミノ酸を含む。
【0014】
本発明の誘導体タンパク質は、好ましいタンパク質、フィブロネクチンまたはフィブロネクチン二量体、テナシン、N-カドヘリン、E-カドヘリン、ICAM、タイチン、GCSF受容体、サイトカイン受容体、グリコシダーゼ阻害剤、抗生物質色素蛋白質、ミエリン膜接着分子P0、CD8、CD4、CD2、クラスI MHC、T細胞抗原受容体、CD1、C2、およびVCAM-1のI-setドメイン、ミオシン結合タンパク質CのI-set免疫グロブリンドメイン、ミオシン結合タンパク質HのI-set免疫グロブリンドメイン、テロキンのI-set免疫グロブリンドメイン、NCAM、トウィチン、ニューログリアン、成長ホルモン受容体、エリスロポエチン受容体、プロラクチン受容体、インターフェロンγ受容体、βガラクトシダーゼ/グルクロニダーゼ、βグルクロニダーゼ、トランスグルタミナーゼ、T細胞抗原受容体、スーパーオキシド・ジスムターゼ、組織因子ドメイン、シトクロームF,緑色蛍光タンパク質、GroEL、およびタウマチンを含むがこれらに限定されることのない任意の適当な基準タンパク質から誘導され得る。
【0015】
Fn3ドメインを含むタンパク質の別の好ましい態様では、フィブロネクチン3型ドメインは哺乳類(例えばヒト)フィブロネクチン3型ドメインである;このタンパク質はフィブロネクチン3型ドメインの10番目のモジュール(10Fn3)を含む。そのようなタンパク質では、化合物の結合は、好ましくは1個、2個、または3個の10Fn3ループによって媒介される。別の好ましい態様では、10Fn3の第2の(DE)ループが天然に存在する分子に比較して伸長しているか、10Fn3がインテグリン結合モチーフを欠いている場合がある。これらの分子では、インテグリン結合モチーフは、アミノ酸配列によって置換されている可能性があり、ここでは極性アミノ酸-中性アミノ酸-酸性アミノ酸の配列(N末端からC末端の方向)がインテグリン結合モチーフを置き換えている;または、1つの好ましい配列は、セリン-グリシン-グルタミン酸である。別の好ましい態様では、フィブロネクチン3型ドメインを含む本発明のタンパク質は、ジスルフィド結合を欠いている。
【0016】
本発明の任意のタンパク質(例えば、フィブロネクチン3型ドメインを含むタンパク質)は、融合タンパク質の一部として製剤化されていてもよい。化合物との結合または化合物との結合の選択に融合タンパク質が使用される場合は、融合タンパク質はそれ自身は関心対象の化合物に結合しない異種のタンパク質を含む。異種タンパク質は、例えば、抗体または抗体ドメイン(たとえば免疫グロブリンFcドメイン)、補体タンパク質、毒素タンパク質,またはアルブミンタンパク質でよい。また、本発明の任意のタンパク質(たとえば、フィブロネクチン3型ドメインタンパク質)は、核酸(たとえば、RNA)と共有結合していてもよく、核酸はタンパク質をコードしていてもよい。さらに、このタンパク質は多量体でもよく、または特にインテグリン結合モチーフを欠失する場合には、生理学的に許容される担体として製剤化されていてもよい。
【0017】
本発明はさらに、βシート配列に少なくとも1つの変異を持つフィブロネクチン3型ドメインを含むタンパク質の特徴を含んでいる。また、これらのタンパク質は、対応する天然型のフィブロネクチンとは結合しないまたはそれほどは強く結合しない化合物と結合することができることにより特徴付けられる。
【0018】
本発明の任意のタンパク質は、固相支持体(例えば、ビーズまたはチップ)上に固定することができ、これらの骨格は固相支持体上でアレイを含む任意の形に配置できる。
【0019】
関連する局面では、本発明はさらに本発明の任意のタンパク質をコードする核酸を特徴とする。好ましい態様では、核酸はDNAまたはRNAである。
【0020】
別の関連する局面では、本発明はまた、フィブロネクチン3型ドメインのインテグリン結合ドメインの除去を伴う、フィブロネクチン3型ドメインを含み、哺乳類に対して薬学的に許容される、タンパク質の生成方法も特徴とする。本方法は、上述の任意のフィブロネクチン3型ドメイン含有タンパク質に適用でき、ヒトの治療的応用のためのタンパク質の生成に特に有用である。本発明は、インテグリン結合ドメインを持たない、そのようなフィブロネクチン3型ドメイン含有タンパク質も特徴とする。
【0021】
別の関連する局面では、本発明は、関心対象の化合物に結合する誘導非抗体タンパク質を得る方法を特徴とする。そのような方法の1つは:(a) 免疫グロブリン様の折畳みを含む非抗体骨格タンパク質を提供する段階であって、骨格タンパク質は1μMほどの強いKdで化合物には結合しない;(b) 非抗体骨格タンパク質の変異した誘導体を作製し、それによって変異したタンパク質のライブラリーを作製する段階;(c) ライブラリーに化合物を接触させる段階;(d) ライブラリーから少なくとも1μMのKdの強さで化合物に結合する少なくとも1つの誘導体タンパク質を選択する段階;および(e) 選択的に、繰り返しの段階(b)の非抗体骨格タンパク質をその前の段階(d)で置き換えて段階(b)〜(d)を繰り返す段階を含む。このテクニックは、本発明の任意のタンパク質(例えば、フィブロネクチン3型ドメインを含む任意のタンパク質)についても行なうことができる。
【0022】
さらに別の関連する局面において、本発明は関心対象の化合物と結合しうる無作為化または変異した本発明のタンパク質を得る、もしくは導出するために、または無作為化または変異したモチーフを含む特定のタンパク質と結合しうる化合物(例えば、タンパク質)を得る、もしくは導出するために使用されうるスクリーニング法を特徴としている。さらに、本発明は、関心対象の化合物またはタンパク質のいずれかを得るために、2つの方法を任意の順番で結合したスクリーニング法を特徴としている。
【0023】
特に、関心対象の無作為化または変異したタンパク質を単離または同定するのに有用な最初のスクリーニング法は、(a)関心対象の化合物に候補タンパク質を接触させる段階であって、候補タンパク質は免疫グロブリン様の折畳みをもつドメインを含む誘導非抗体タンパク質であり、非抗体タンパク質は変異したアミノ酸配列を持つことにより基準タンパク質から誘導され、非抗体タンパク質は基準タンパク質がそれほどは強く結合しない化合物に対して少なくとも1μMのKdの強さで結合し、接触が化合物とタンパク質の複合体の形成を可能にする条件下で行われる段階;および(b) 複合体からタンパク質に結合する化合物を得る段階を含んでいる。この一般的なテクニックは、少なくとも1つの無作為化または変異したループを持つフィブロネクチン3型ドメインタンパク質で実行してもよい。
【0024】
第2のスクリーニング方法は、本発明のタンパク質に結合する化合物を単離または同定するためのものである。この方法は、免疫グロブリン様の折畳みをもつドメインを含み、変異したアミノ酸配列を持つことで基準タンパク質から誘導される非抗体タンパク質から始めるが、この非抗体タンパク質は基準タンパク質がそれほど強く結合しない化合物に対して少なくとも1μMの強さのKdで結合する。この誘導体タンパク質に候補化合物を接触させるが、接触は化合物とタンパク質の複合体の形成が可能な条件下で行われ、誘導体タンパク質に結合する化合物を複合体から得る。やはり、この一般的テクニックは、例えば、変異したフィブロネクチン3型ドメインを持つタンパク質のような、本発明の任意のタンパク質を用いて実行できる。
【0025】
また、本発明は、本発明のタンパク質(たとえば、フィブロネクチン3型骨格タンパク質およびその誘導体)を用いる診断方法を特徴とする。そのような診断方法は、試料(例えば、生物試料)に対して行ない、試料中で1つの分析物を検出したり、または多くの異なる分析物を同時に検出することができる。本方法では、本明細書中に記述される任意の骨格分子を利用できる。好ましくは、本方法は、(a)化合物分析物に結合する本発明のタンパク質を、試料に接触させる段階であって、化合物-タンパク質複合体の形成を可能にする条件下で接触が行われる段階;および (b) 複合体を検出し、それにより試料中の化合物を検出する段階、を含む。また、この方法は試料中の化合物レベルを検出するのみならず定量するためにも使用できる。
【0026】
本明細書に記載される任意の選択または診断方法の好ましい態様では、本発明のタンパク質は標的化合物に対して少なくとも1μMまたは500 nMの強さのKd、好ましくは少なくとも100 nMまたは10 nMの強さのKd、およびより好ましくは少なくとも1 nM、500 pM、100 pM、または20 pMの強さのKdですらその標的化合物に結合する。タンパク質は好ましくは1個、2個、または3個の変異ループを含み、少なくとも1個のループ、および好ましくは2個または3個全てのループが化合物に対するタンパク質の結合に貢献する。さらに、基準タンパク質は好ましくはジスルフィド結合を持たず、誘導体タンパク質は少なくとも1つのジスルフィド結合を持つ可能性がある。
【0027】
本方法の特定の態様に関して、免疫グロブリン様の折畳みをもつドメインは、好ましくは10 kD未満または7.5 kDを越える分子量、およびより好ましくは7.5〜10 kDの間の分子量を持つ。本発明のタンパク質は生理的条件下で単量体であるか、例えば二量体のような多量体である。他の好ましい態様では、本発明の変異タンパク質を誘導するために用いた基準タンパク質は、天然に存在する哺乳類タンパク質(例えば、ヒトのタンパク質)である;免疫グロブリン様の折畳みをもつドメインは変異しており、基準タンパク質の免疫グロブリン様の折畳みと比較して最高50%、および好ましくは最高34%の変異したアミノ酸を含む。さらに、免疫グロブリン様の折畳みをもつドメインは、約50〜150アミノ酸を含み、より好ましくは約50アミノ酸を含む。
【0028】
本発明の方法で使用される誘導体タンパク質は、好ましいタンパク質、フィブロネクチンまたはフィブロネクチン二量体、テナシン、N-カドヘリン、E-カドヘリン、ICAM、タイチン、GCSF受容体、サイトカイン受容体、グリコシダーゼ阻害剤、抗生物質色素蛋白質、ミエリン膜接着分子PO、CD8、CD4、CD2、クラスI MHC、T細胞抗原受容体、CD1、C2、およびVCAM-1のI-setドメイン、ミオシン結合タンパク質CのI-set免疫グロブリンドメイン、ミオシン結合タンパク質HのI-set免疫グロブリンドメイン、テロキンのI-set免疫グロブリンドメイン、NCAM、トウィチン、ニューログリアン、成長ホルモン受容体、エリスロポエチン受容体、プロラクチン受容体、インターフェロンγ受容体、βガラクトシダーゼ/グルクロニダーゼ、βグルクロニダーゼ、トランスグルタミナーゼ、T細胞抗原受容体、スーパーオキシド・ジスムターゼ、組織因子ドメイン、シトクロームF,緑色蛍光タンパク質、GroEL、およびタウマチンを含むがこれらに限定されることのない任意の適当な基準タンパク質から誘導され得る。
【0029】
また、本明細書に記述される選択方法の段階は、サイクルの間にさらなる変異または無作為化を行なって繰り返すことができる。例えば、変異または無作為化したフィブロネクチン3型ドメインを持つタンパク質に関する方法では、段階(b)で得られたタンパク質のフィブロネクチン3型ドメインの少なくとも1個のループを変異させ、さらに無作為化されたタンパク質を用いて段階(a)および(b)を繰り返すか、段階(b)で得られた化合物を修飾し、さらに修飾された化合物を用いて段階(a)および(b)を繰り返すことができる。これらの方法では、化合物は好ましくはタンパク質であり、フィブロネクチン3型ドメインは好ましくは哺乳類(例えばヒト)フィブロネクチン3型ドメインである。別の好ましい態様では、このタンパク質はフィブロネクチン3型ドメインの10番目のモジュール(10Fn3)を含み、結合は1個、2個、または3個の10Fn3ループによって媒介される。また、10Fn3の第2の(DE)ループが天然に存在する分子と比較して伸長しているか、10Fn3がインテグリン結合モチーフを欠いている場合がある。やはり上述のように、インテグリン結合モチーフは置換されている場合があり、ここでは、極性アミノ酸-中性アミノ酸-酸性アミノ酸の配列(N末端からC末端の方向)がインテグリン結合モチーフを置き換えている;または、1つの好ましい配列は、セリン-グリシン-グルタミン酸である。
【0030】
本明細書に記述される選択および診断方法は、本発明の任意のタンパク質を用いて実行できる(例えば、フィブロネクチン3型ドメインを含むタンパク質)。また、これらの任意のタンパク質は、それ自身は関心対象の化合物に結合しない異種のタンパク質(例えば、抗体または抗体ドメイン(免疫グロブリンFcドメインを含む)、または補体タンパク質、毒素タンパク質、またはアルブミンタンパク質)との融合タンパク質の一部として製剤化されていてもよい。また、選択および診断方法は、核酸(たとえば、RNAまたはタンパク質をコードする任意の核酸)と共有結合していてもよい。さらに、単量体または二量体のような多量体としてのこれらのタンパク質(例えば、フィブロネクチンドメインを含むタンパク質)を用いて、選択および診断方法を実行することができる。
【0031】
好ましくは、選択および診断方法は、固相支持体上への結合した標的の固定化を含んでいる。好ましい固相支持体には、カラム(例えば、アガロースカラムのような親和性カラム)またはマイクロチップ、またはビーズが含まれる。または、タンパク質(例えばFn3ドメインを含むタンパク質)を固定化し、これに1個または複数の潜在的結合標的を接触させることもできる。
【0032】
診断方法では、化合物はしばしばタンパク質であるが、試料中の任意の他の分析物でもよい。検出は、X線撮影、蛍光検出、質量分光、または表面プラズモン共鳴を含むが、これらに限定されない、任意の標準的な手法で行われる。
【0033】
最後の局面では、本発明は少なくとも1μMの強さのKdで腫瘍壊死因子α(TNF-α)に結合する非抗体タンパク質で、TNF-α受容体(例えば、天然に存在するTNF-α受容体、例えば哺乳類またはヒトのTNF-α受容体)とは20%未満の同一な配列を持つタンパク質の特徴を含んでいる。
【0034】
好ましい態様では、このタンパク質は、変異したフィブロネクチン3型ドメインを含み、タンパク質はDE、BC、およびFGループに変異を持つ。好ましくは変異したFGループは野生型のFGループと同じ長さである。別の好ましい態様では、タンパク質は、約50〜150アミノ酸、および好ましくは約50アミノ酸からなる、免疫グロブリン様の折畳み(好ましくは10 kD未満、7.5 kDを越える、または7.5〜10 kDの間の分子量を持つ)を含む。
【0035】
本発明のTNF-α結合体は、少なくとも1μMの強さ、好ましくは少なくとも500 nM、100 nM、または10 nMの強さのKd、およびより好ましくは少なくとも1 nMまたは500 pMの強さ、および最も好ましくは100 pMまたは20 pMの強さのKdでTNF-αに結合する。これらのタンパク質は好ましくは1個、2個、または3個の変異ループを含み、少なくとも1個のループ、および好ましくは2個または3個全てのループが、TNF-αに対するこの非抗体タンパク質の結合に貢献する。別の好ましい態様では、この非抗体タンパク質は少なくとも1つのジスルフィド結合を持ち、非抗体タンパク質は生理的条件下で単量体または二量体である。
【0036】
TNF-α結合体は、固相支持体、(例えば、ビーズまたはチップ)上に固定することができ、アレイの一部となっていてもよい。また、TNF-α結合体には、異種タンパク質(例えば、TNF-α、免疫グロブリンFcドメイン、補体タンパク質、またはアルブミンタンパク質に結合しない抗体または抗体ドメインである異種タンパク質)が結合していてもよい。
【0037】
必要ならば、タンパク質は変異フィブロネクチン3型ドメイン(例えば、フィブロネクチン3型ドメインの変異した10番目のモジュール(10Fn3)のようなヒトフィブロネクチン3型ドメイン由来のもの)を含むことができる。また、タンパク質は10Fn3インテグリン結合モチーフを欠如していてもよい。TNF-α結合体は、好ましくは10Fn3のループ(例えば、ループ配列PW(A/G))に非天然配列を含み、10Fn3のβシートに非天然配列を含む可能性がある。本発明の特に好ましいTNF-α結合体は、図25(配列番号:34〜140)に示されている。
【0038】
また、関連する局面では、本発明は本発明の任意のTNF-α結合タンパク質、および図25(配列番号:34〜140)の任意のアミノ酸配列を含む任意のタンパク質上のループ構造をコードする核酸の特徴を含んでいる。
【0039】
本明細書では「非抗体タンパク質」は、天然でも哺乳類の免疫後でも哺乳類のB細胞によって産生されないタンパク質を意味する。この用語はまた、長さが100アミノ酸以上、好ましくは80アミノ酸以上、および最も好ましくは50アミノ酸以上の抗体断片も含まない。
【0040】
「免疫グロブリン様の折畳み」は、逆方向に平行の2層のβシートを含む約80〜150アミノ酸残基のタンパク質ドメインであって、2つのβシートの平らな疎水性の面が互いに押し付けられているタンパク質ドメインを意味する。本発明のタンパク質は、共有結合または非共有結合で、より大きな構造に会合したいくつかの免疫グロブリン様の折畳みを含む可能性がある。
【0041】
「骨格」は、結合のような特定の好ましい性質を持つタンパク質枠組みを選択または設計するために用いられるタンパク質を意味する。骨格からタンパク質を設計するときには、枠組みの望ましい特性に重要なアミノ酸残基は保持され、他の残基を変化させることができる。そのような骨格は、異なる性質を持つタンパク質誘導体の間で変化するアミノ酸残基を50%未満持ち、そのような誘導体の間で一定の残基を50%以上持つ。最も一般的には、これらの一定の残基は、その性質に関係なく、全ての変化するドメインに、同一の全体的な3次元折畳みを与える。
【0042】
「フィブロネクチン3型ドメイン」は互いに充填し合ってタンパク質のコアを形成する2個のβシート間に散在する7個もしくは8個のβ鎖を持つドメインであって、かつβ鎖を互いに連結して、溶媒に露出されたループをさらに含むドメインを意味する。βシートサンドイッチの両端には、このようなループが少なくとも3個存在し、この端はβ鎖の方向と垂直なタンパク質の境界である。好ましくは、フィブロネクチン3型ドメインは、RCSB (Research Collaboratory for Structural Bioinformatics) プロテインデータベース(Protein Data Base)より入手可能であって、「1ttg」(ID=「1ttg」(1つのttg))として引用される10Fn3ドメインの構造をコードする配列とアミノ酸レベルで少なくとも30%の同一性を呈し、および好ましくはアミノ酸レベルで少なくとも50%の同一性を呈する配列を含んでいる。この定義において引用される配列の同一性は、モレキュラー・シミュレーション(Molecular Simulation)(San Diego、CA)より入手可能なホモロジー・プログラムによって決定される。本発明はさらに、10Fn3関連分子の重合体であって、単量体構造の使用の延長に当たり、ポリタンパク質のサブユニットが配列において同一であるかもしくはないものを含む。
【0043】
「天然に存在する」とは、生きている生物によってコードされる任意のタンパク質を意味する。
【0044】
「無作為化された」または「変異した」とは、鋳型配列に比べて1個または複数のアミノ酸変異を含むことを意味する。「無作為化する」または「変異させる」とは、そのようなアミノ酸変異を配列中の導入する過程を意味する。無作為化または変異は、一般的には核酸コード配列の意図的な、盲目の、または自発的な変異の変化によって形成され、例えば、PCR、変異性PCR、またはDNA化学合成のような任意の手法によって作製される。「対応する非変異タンパク質」とは、導入されたアミノ酸変異以外は、配列の同一なタンパク質を意味する。
【0045】
「タンパク質」とは、長さ、翻訳後修飾、もしくは機能にかかわらず、2個またはそれ以上のアミノ酸の任意の配列を意味する。「タンパク質」および「ペプチド」は、本明細書において互換的に用いられる。
【0046】
「RNA」とは、二つまたはそれ以上の共有結合した、天然または改変されたリボヌクレオチドの配列を意味する。この用語に含まれる改変されたRNAの一つの例は、ホスホロチオエートRNAである。
【0047】
「DNA」とは、二つまたはそれ以上の共有結合した、天然または改変されたデオキシリボヌクレオチドの配列を意味する。
【0048】
「核酸」とは、任意の2つまたはそれ以上の共有結合したヌクレオチドまたはヌクレオチド類似体もしくは誘導体を意味する。本明細書において用いられるように、この用語にはDNA、RNA、およびPNAが含まれるが、これらに限定されることはない。
【0049】
「薬学的に許容される」とは、重大な医学的に有害な結果なしに、動物(例えば哺乳類)に投与しうる化合物またはタンパク質を意味する。
【0050】
「生理学的に許容される担体」とは、処理された宿主に対して重大な有害な影響を持たず、投与される化合物の治療上の性質を維持する担体を意味する。一つの例示的な生理学的に許容される担体は生理食塩水である。他の生理学的に許容される担体およびその処方は当業者には自明であって、例えば「レミントンの薬剤科学(Remington's Pharmaceutical Sciences)」、(第18版)、A. Gennaro編、1990年、Mack Publishing Company、Easton、PA、に述べられており、参照として本明細書に組み入れられている。
【0051】
「融合タンパク質」とは、(i)本発明の骨格タンパク質が(ii)第2の異なる(異種)タンパク質に結合したタンパク質を意味する。「融合タンパク質」はすべてアミノ酸から構成されるが、「核酸-タンパク質融合体」および「RNA-タンパク質融合体」はどちらもアミノ酸のひと配列(タンパク質成分)に結合した核酸のひと配列(核酸またはRNA成分)を含むという点で、「融合タンパク質」は「核酸-タンパク質融合体」および「RNA-タンパク質融合体」とは異なっている。
【0052】
「選択」とは、ある分子を集団の他の分子から実質的に分離することを意味する。本明細書において用いられるように、「選択」段階は、選択段階後には集団中の望ましい分子が望ましくない分子に比べて少なくとも2倍、好ましくは少なくとも30倍、より好ましくは少なくとも100倍、最も好ましくは少なくとも1000倍に濃縮されることを提供する。選択段階は何回繰り返してもよく、所与の方法中で異なる型の選択段階が組み合わされていてもよい。
【0053】
本明細書において用いられる「結合パートナー」とは、所望の関心対象の化合物(例えば、タンパク質)のある部分に対する特異的、共有結合、または非共有結合親和性を有するいかなる分子も意味する。結合パートナーの例には、抗原/抗体対、タンパク質/阻害剤対、受容体/リガンド対(例えば、ホルモン受容体/ペプチドホルモン対のような細胞表面受容体/リガンド対)、酵素/基質対(例えば、キナーゼ/基質対)、レクチン/炭化水素対、オリゴマーまたはヘテロオリゴマータンパク質凝集体、DNA結合タンパク質/DNA結合部位、RNA/タンパク質対、および核酸二本鎖、ヘテロ二本鎖、または連結鎖のメンバー、並びに、他の分子(例えば、化合物またはタンパク質)のいかなる部分とも1つもしくはそれ以上の共有結合または非共有結合を形成することができる任意の分子が含まれるが、これらに限定されることはない。
【0054】
「固相支持体」とは、それに対して直接または間接的に(例えば、他の抗体またはプロテインAのような他の結合パートナー中間体を通じて)抗体模倣体もしくは親和性複合体が結合してもよい、またはその中に抗体模倣体もしくは親和性複合体が埋め込まれてもよい(例えば、受容体またはチャンネルを通じて)、任意のカラム(もしくはカラム材料)、ビーズ、試験管、マイクロタイター皿、固体粒子(例えば、アガロースもしくはセファロース)、マイクロチップ(例えば、シリコン、シリコンガラス、もしくは金チップ)、またはメンブレン(例えば、無機メンブレン、ニトロセルロース、またはリポソームもしくは小胞のメンブレン)を意味するがこれらに限定されることはない。
【0055】
本発明によって、多くの利点が提供される。例えば、以下により詳細に述べられているように、本抗体模倣物は還元条件下での安定性および高濃度での溶解性など、改良された生物物理学的性質を呈する。さらに、これらの分子は大腸菌(E. coli)などの原核生物系、酵母などの真核生物系、およびウサギ網状赤血球溶解物系などのインビトロ翻訳系において容易に発現し、折り畳まれる。さらに、これらの分子は、RNA-タンパク質融合技術を用いたインビトロ選択(RobertsおよびSzostak、Proc. Natl. Acad. Sci. USA、第94巻、第12297頁、1997年;Szostakら、米国特許出願第09/007,005号および米国特許出願第09/247,190号;Szostakら、国際公開公報第98/31700号)、ファージ・ディスプレイ(例えば、SmithおよびPetrenko、Chem. Rev.、第97巻、第317頁、1997年を参照)、および酵母ディスプレイ系(例えば、BoderおよびWittrup、Nature Biotech.、第15巻、第553頁、1997年を参照)を含む選択を多回数繰り返すことを含む親和性成熟(affinity maturation)技術にきわめて適している。
【0056】
本発明の他の特徴および利点は、以下の詳細な説明および特許請求の範囲から明らかになると思われる。
【0057】
詳細な説明
本明細書に記載される新規の抗体模倣物は、抗体に由来する断片、および例えば前掲の枠組みのような非抗体の枠組みのどちらよりも優れているように設計されている。
【0058】
これらの抗体模倣物が抗体断片に勝っている主要な利点は構造である。これらの抗体模倣体は、安定性で、可溶性の構造の骨格全体に由来している。例えば、Fn3骨格は人体で見いだされる。結果として、これらの作製は、抗体の天然の折り畳みの一部を除去することを含み、無傷の抗体における可変ドメインおよび定常ドメイン間の界面のような疎水性環境に埋まっているアミノ酸残基がしばしばさらされている抗体断片よりも、これらは優れた折り畳みおよび熱安定性の性質を呈する。このような疎水性残基の溶媒への曝露は、抗体断片の凝集の可能性を増加させる。
【0059】
さらに、本明細書に記載の骨格は、特定の条件下で抗体断片の正しい折り畳みを妨害もしくは阻害すると報告されているジスルフィド結合を持たない。本骨格の自然な折り畳み安定性はジスルフィド結合に依存していないため、ジスルフィド結合が減少すると分解してしまう抗体およびそれらの断片とは異なり、還元条件下でも安定である。
【0060】
さらに、これらの骨格は、抗体分子の機能的な利点を提供する。特に、10Fn3モジュールは免疫グロブリンではないという事実にかかわらず、その全体的な折り畳みはIgGの重鎖の可変領域に近く(図2)、CDRに類似した3つのフィブロネクチン・ループを天然抗体と同様な相対的配向性で示すことが可能にされている。この構造のために、本抗体模倣物は、天然と類似した抗原結合の性質および抗体への親和性を持ち、インビボにおける抗体の親和性成熟化の過程に類似したインビトロでループを無作為化して混合する戦略をとることができる可能性を有する。
【0061】
例示的な骨格(例えばフィブロネクチンに基づく骨格)、ならびにその標的リガンドと同様に新規の結合タンパク質を同定し、選択し、および導出するための使用が以下に述べられている。これらの実施例は本発明を例示する目的で提供されており、本発明を限定するためのものではない。
【0062】
10Fn3の構造モチーフ
本発明の好ましい抗体模倣物は、哺乳類の血液および構造タンパク質で見出された共通ドメインであるフィブロネクチン3型モジュール(Fn3)の構造に基づいている。このドメインはタンパク質配列データベースの中に400回よりも多く出現し、フィブロネクチン、テンシン、細胞内の細胞骨格タンパク質、および原核生物の酵素を含むこれまでにシークエンシングされたタンパク質の2%に出現すると見積もられている(BorkおよびDoolittle、Proc. Natl. Acad. Sci. USA、第89巻、第8990頁、1992年;Borkら、Nature Biotech.、第15巻、第553頁、1997年;Meinkeら、J. Bacteriol.、第175巻、第1910頁、1993年;Watanabeら、J. Biol. Chem.、第265巻、第15659頁、1990年)。特定の骨格は94個のアミノ酸残基を含むヒトFn3の10番目のモジュール(10Fn3)である。このドメインの全体的な折り畳みは、最も小さな機能的抗体断片であり、ラクダおよびラマのIgG(図1、図2)において抗原認識単位の全体を含む重鎖の可変領域のものと密接に関連している。ラクダおよびラマのドメインと10Fn3ドメインの間の主要な差異は、(i)10Fn3のほうがβ鎖が少ないこと(7対9)、および(ii)ラクダおよびラマのドメインでは、互いに充填し合う2個のβシートがジスルフィド架橋により連結されているが、10Fn3ではそうでないことである。
【0063】
IgGの重鎖にある抗原結合ループに対応する10Fn3の3個のループは、アミノ酸残基21〜31位(BC)、51〜56位(DE)、76〜88位(FG)の間を通っている(図3)。BCのループの長さおよびDEのループの長さはそれぞれ10残基および6残基であり、抗体の重鎖で見出される対応する抗原認識ループの狭い範囲内、すなわちそれぞれ7〜10残基および4〜8残基に位置する。したがって、無作為化され、抗原との高親和性によって選択される場合には、これらの2個のループは抗体中の対応するループと同等に抗原と接触できる。
【0064】
対照的に、10Fn3のFGループは長さが12残基で、一方で抗体の重鎖にある対応するループは4残基〜28残基の範囲である。それゆえ、抗原との結合を最適化するために、10Fn3のFGループの長さは、好ましくは4残基〜28残基のCDR3の範囲を覆うように、長さおよび配列が無作為化され、抗原結合において可能な最も高い可動性および親和性を獲得する。実際に、一般的には、インビトロもしくはインビボの親和性成熟化において、(以下により詳細に述べるように)抗体模倣物のCDR様ループの配列と同様に長さも無作為化してもよい。
【0065】
10番目のヒトフィブロネクチン3型ドメインである10Fn3は、低温においてさえ急速に再び折り畳まれる。その背骨のコンフォメーションは、5℃において1秒間以内に回復する。10Fn3の熱力学的安定性は高く(ΔGU=24kJ/mol=5.7kcal/mol)、110℃の高い融解温度と相関する。
【0066】
10Fn3の生理学的な役割の1つは、体液中に可溶型としておよび細胞外マトリックス中で不溶型として存在する糖タンパク質であるフィブロネクチンのサブユニットである(Dickinsonら、J. Mol. Biol.、第236巻、第1079頁、1994年)。220〜250kDのフィブロネクチンの単量体は、12個の1型モジュール、2個の2型モジュール、および17個のフィブロネクチン3型モジュールを含んでいる(PottsおよびCampbell、Curr. Opin. Cell Biol.、第6巻、第648頁、1994年)。インテグリン、ヘパリン、およびコンドロイチン硫酸塩とフィブロネクチンとの結合には異なる3型モジュールが含まれている。10Fn3は、その曝露されたループの1つにあるインテグリン結合Arg-Gly-Asp(RGD)モチーフを介して細胞接着を媒介することが見出されている。フィブリノーゲン、フォン・ビルブラント因子、およびビトロネクチンのような他のタンパク質でも、類似のRGDモチーフがインテグリン結合に含まれることが示されている(Hynesら、Cell、第69巻、第11頁、1992年)。10Fn3については、他のマトリックスもしくは細胞結合的な役割は述べられていない。
【0067】
RGDを含む短いペプチドに比べて10Fn3の接着活性がわずかに強いという知見は、10Fn3の細胞結合活性が10Fn3の構造全体に分散しているというよりはRGDペプチドに局在しているという結果(Baronら、Biochemistry、第31巻、第2068頁、1992年)と一致する。RGDモチーフを持たない10Fn3は他の血漿タンパク質もしくは細胞外マトリックスと結合しそうもないという事実から、10Fn3は抗体に換わる有用な骨格である。さらに、血流の天然フィブリノーゲン中に10Fn3が存在することは、元々の生物において10Fn3自体が免疫原生ではないとおもわれることを示唆している。
【0068】
さらに、本発明者らは、無作為化に耐性を持ち、抗体模倣物の多様なプールの生産を助ける曝露されたループ配列が10Fn3の枠組みにあることを決定した。この決定は、10Fn3の配列の可動性を試すことで行われた。特に、関連タンパク質の配列はもちろん、他の源から得られたフィブロネクチンの配列とヒト10Fn3の配列を整列化させ(図4)、この整列化の結果をヒト10Fn3ドメインの三次元構造上に位置づけた(図5)。この整列化により、保存された残基の大部分はβシートサンドイッチのコアに見出され、著しく変異しやすい残基は両βシートの溶媒に接触しうる表面上、および抗体模倣物の親和性成熟化において超可変ループとして働き、溶媒と接触しうる3つのループ上でN末端およびC末端を含むβシートの端に沿って局在することが判明した。これらの結果を考慮すると、これら3つのループの無作為化には、10Fn3の枠組み自体の全体的な折り畳みもしくは安定性に反する効果はないように思われる。
【0069】
この解析は、ヒト10Fn3の配列では最小限でもアミノ酸1〜9位、44〜50位、61〜54位、82〜94位(βシートの端);19位、21位、31〜46位(偶数)、79〜65位(奇数)(両βシートの溶媒に接触しうる表面);21〜31位、51〜56位、76〜88位(CDR様の溶媒に接触しうるループ);ならびに14〜16位および36〜45位(溶媒に接触しうる他のループおよびβターン)は、新規のもしくは改良された化合物結合タンパク質を導出するために無作為化されてもよいことを示している。さらに、以上に討議されたように、溶媒に曝露された1つもしくは複数のループの長さにおける変化も、このような定方向展開法に含まれる。
【0070】
または、βシート配列の変化を利用して新規のタンパク質を導出してもよい。これらの変異は骨格を変化させ、それにより間接的に1つまたは複数のループ構造を変化させる。この方法を用いた場合は、配列に変異を過剰に行うことなく、むしろほとんど変異を導入してはいけない。この方法では、βシート配列に、好ましくは3-20の間以下の変異が導入されるべきである。
【0071】
配列の変異は、例えばTaqポリメラーゼによる変異導入(TindallおよびKunkel, Biochemistry 27:6008 (1988))、断片の組換え、またはその組み合せを含む任意の手法で行なえる。同様に、例えば、CDR様ループの長さおよび配列の変化、または選択されたループに見つかった有利な枠組みの変異に基づく構造の再設計による、ライブラリーの構造の多様性の増加を用いて、抗体模倣物骨格をさらに改良することができる。
【0072】
抗体模倣体融合体
本明細書に述べられた抗体模倣物は、他のタンパク質ドメインと融合していてもよい。例えばこれらの模倣物は、IgG(Fc)の定常領域と10Fn3モジュール、好ましくは10Fn3のC末端のような抗体模倣体を融合させることで、ヒト免疫応答に組み込まれていてもよい。このような10Fn3-Fc融合分子などにおけるFcは、免疫応答の補体構成要素を活性化し、抗体模倣物の治療上の価値を増加させる。同様に、10Fn3のような抗体模倣体とC1qのような補体タンパク質との融合は、細胞を標的化するために用いることができ、10Fn3の抗体模倣体と毒素の融合は、特定の抗原を運搬する細胞を特異的に破壊するために用いることができる。さらに、いかなる型の抗体骨格(例えば10Fn3)であっても、アルブミンと融合させて血流中での半減期および組織への浸透を増加させることができる。これらの任意の融合体を、例えば一般的に入手可能な遺伝子配列を用いて構築した組換え融合遺伝子から融合タンパク質を発現させるなどの常法により生産することができる。
【0073】
骨格の多量体
単量体に加えて、本明細書に述べられている任意の骨格構築物を、結合価を増加させて、それにより抗原との結合活性を増加させるために、抗体模倣物の二量体または多量体として生産してもよい。このような多量体は、共有結合により生産されてもよい。例えば、個々の10Fn3モジュールは天然の8Fn3-9Fn3-10Fn3のC末端からN末端の結合を模倣したり、または定常領域を介して結合した抗体の二量体を模倣することによって結合させてもよい。10Fn3-Fc構築物を、10Fn3-Fc::Fc-10Fn3という一般的な図式の二量体を設計するために利用してもよい。Fc::Fcの界面に操作された結合は共有結合であっても、非共有結合であってもよい。さらに、このようなより高いオーダーの構造を作製するために、10Fn3ハイブリッドのようなハイブリッドにおいてFc以外の相手との二量体化もしくは多量体化を利用することができる。
【0074】
特定の例としては、共有結合した多量体は、多量体をコードする融合遺伝子を構築する段階によって、または単量体の配列中にシステイン残基のコドンを操作し、発現した生成物間でジスルフィド結合の形成を生じさせる段階によって作製されてもよい。非共有結合した多量体もまた、様々な技術により生産することができる。これらには、正電荷および/または負電荷を帯びた残基に対応するコドンを単量体の配列に導入し、発現した生成物間(および、その結果として単量体間)において、これらの残基間で相互作用を生じさせておくことが含まれる。本方法は、例えばフィブロネクチンの負電荷を帯びた残基のように、単量体サブユニット中に天然に存在する荷電残基の利点を用いることで単純化することができる。非共有結合した抗体模倣物を生産する別の方法は、相互作用することが知られているタンパク質またはタンパク質ドメインのコード配列を単量体遺伝子(例えば、アミノ末端もしくはカルボキシル末端)に導入することである。このようなタンパク質もしくはタンパク質ドメインは、コイルドコイルモチーフ、ロイシンジッパーモチーフ、および二量体もしくはより高いオーダーの多量体を直接形成することが知られている任意の多数のタンパク質サブユニット(またはその断片)を含んでいる。
【0075】
フィブロネクチン様分子
10Fn3は抗体模倣物の生産に好ましい骨格を表すにもかかわらず、本明細書に記載の分子においては、他の分子が10Fn3を置換していてもよい。これらに限定されることはないが、ヒト以外の動物および原核生物から得られる関連したFn3モジュール、ならびにヒトフィブロネクチンモジュール1Fn3-9Fn3および11Fn3-17Fn3を含む。さらに、テネイシンおよびアンジュリンのような10Fn3と配列に相同性を持つ他のタンパク質から得られたFn3モジュールを使用してもよい。免疫グロブリン様の折畳みを持つ(ただしVHドメインに関連しない配列を持つ)他の代表的な骨格は図21に示されており、N-カドヘリン、ICAM-2、タイチン、GCSF受容体、サイトカイン受容体、グリコシダーゼ阻害剤、E-カドヘリン、および抗生物質色素蛋白質が含まれる。関連する構造を持つ別のドメインは、ミエリン膜接着分子PO、CD8、CD4、CD2、クラスI MHC、T細胞抗原受容体、CD1、C2、およびVCAM-1のI-setドメイン、ミオシン結合タンパク質CのI-set免疫グロブリンドメイン、ミオシン結合タンパク質HのI-set免疫グロブリンドメイン、テロキンのI-set免疫グロブリンドメイン、テリキン、NCAM、トウィチン、ニューログリアン、成長ホルモン受容体、エリスロポエチン受容体、プロラクチン受容体、GC-SF受容体、インターフェロンγ受容体、βガラクトシダーゼ/グルクロニダーゼ、βグルクロニダーゼ、トランスグルタミナーゼに由来する可能性がある。または、免疫グロブリン様の折畳みを1つまたは複数含む任意の他のタンパク質が利用できる。そのようなタンパク質は、例えば、プログラムSCOP(Murzinら、J. Mol. Biol. 247:536 (1995); Lo Conteら、Nucleic Acids Res, 25:257 (2000))を用いて同定できる。
【0076】
一般に、VHドメインに構造的関連性を示す任意の分子(例えば、上述のコンピュータプログラムで同定される)は、抗体模倣物として使用できる。そのような分子は、フィブロネクチンのように、分子のN末端に3つのループ、C末端に3つのループを含んでおり、その各々を無作為化して多様なライブラリーを作製できる;または、そのような表面の無作為化可能なループが抗原結合に参加できるほど空間的に近接しているかぎり、より多くのループを持つより大きなドメインも使用できる。図22は、互いに近接した3個を越える数のループをもつ有用なドメインの例を示す。これらの例には、各々無作為化できるループを4つ持つT細胞抗原受容体およびスーパーオキシド・ジスムターゼ;および3つの無作為化可能なループが2つのドメインの一部である(ループの合計数が6となる)ような2つの類似したドメインを3セットずつ持つ、Fn3二量体、組織因子ドメイン、およびサイトカイン受容体ドメインが含まれる。
【0077】
または、空間的に充分近接した可変ループを持つ任意のタンパク質を用いて、候補結合タンパク質の生産ができる。例えば、免疫グロブリン様の折畳みには構造的に関連していなくても、空間的に関連した、溶媒が接することのできるループを持つ大きなタンパク質が使用できる。例には、シトクロームF、緑色蛍光タンパク質、GroEL、およびタウマチンが含まれるが、これらに限定されない(図23)。これらのタンパク質のディスプレイするループを無作為化して、本明細書に記載するように無作為化したライブラリーから優れた結合をする分子を選択できる。そのサイズのために、抗体-抗原相互作用よりも、はるかに大きな抗原結合表面を示す分子が得られる可能性がある。このタイプの他の有用な折畳みは、数多くのループ、特に平行するβシートまたはいくつかのαへリックスの間に位置するループを持つ候補タンパク質を見るためにプログラムSCOP(Murzinら、J. Mol. Biol. 247:536 (1995))を用いても同定できる。
【0078】
異なる生物および親タンパク質から得られるモジュールは、異なる適用に適切である可能性がある。例えば、抗体模倣物の設計では、治療薬を意図する生物に天然に存在するフィブロネクチンまたはフィブロネクチン様分子から、そのタンパク質を作製するのが最も望ましいことがある。反対に、診断薬または研究用試薬のようなインビトロ適用に使用する抗体模倣物は、起源の生物の重要度は低いまたは関係ない。
【0079】
これらの任意の分子について、本明細書に記述する任意の方法によりライブラリーを作製し、結合するタンパク質を選択するために使用できる。
【0080】
骨格に基づく結合タンパク質の定方向展開
本明細書に記載の抗体模倣物は、新規または改良された結合タンパク質を導出するために、いかなる技術において使用してもよい。一つの特定の例として、結合の標的をカラム樹脂またはマイクロタイタープレートのウェルのような固相支持体に固定化し、候補となる骨格に基づく結合タンパク質のライブラリーに標的を接触させる。このようなライブラリーは、10Fn3のCDR様ループの配列および/または長さを無作為化することで野生型10Fn3の骨格から構築された10Fn3クローンのような抗体模倣体クローンから成っていてもよい。望ましい場合には、このライブラリーは、例えばショスタック(Szostak)ら、米国特許出願第09/007,005号および同第09/247,190号;ショスタック(Szostak)ら、国際公開公報第98/31700号;ならびにロバーツ(Roberts)およびショスタック(Szostak)、Proc. Natl. Acad. Sci. USA、1997年、第94巻、第12297〜12302頁に記載の技術によって生産されたRNA-タンパク質融合体ライブラリーであってもよい。またはDNA-タンパク質ライブラリーであってもよい(例えば、Lohse、「DNA-タンパク質融合体およびそれらの使用法(DNA-Protein Fusions and Uses Thereof)」、米国特許出願第60/110,549号、米国特許出願第09/459,190号、および国際公開公報第00/32823号)。この融合体ライブラリーを固定された標的と共にインキュベートし、支持体を洗浄して非特異的な結合物を除去し、高ストリンジェントな条件下における最も強い結合物を溶出してPCRにかけて配列情報を回収するか、または配列にさらなる変異を加えたりもしくは加えなかったりして選択段階を繰り返すのに使用することができる結合物の新規のライブラリーを作製する。抗原に対して十分な親和性を持つ結合物を得られるまでには、選択ラウンドが多数回行われる可能性がある。
【0081】
一つの特定の例においては、10Fn3の骨格を選択の標的として使用することができる。例えば、10残基のループ中に存在する特異的なペプチド配列に結合するタンパク質が必要な場合は、このループの1つが10の長さで所望の配列に設定された単一の10Fn3クローンを構築する。新規のクローンをインビボで発現させて精製し、次に支持抗体に固定化する。適切な骨格に基づいたRNA-タンパク質融合体ライブラリーを支持体と相互作用させ、その後洗浄して、所望の分子を溶出し、上述したように再度選択する。
【0082】
同様に、本明細書に記載の骨格(例えば10Fn3の骨格)は、例えば10Fn3のループにおいて、骨格によって提示されたペプチド配列と相互作用する天然タンパク質を見出すためにも使用することができる。上述したように10Fn3タンパク質のような骨格タンパク質を固定化し、提示されたループへの結合物に対してRNA-タンパク質融合体ライブラリーをスクリーニングする。選択ラウンドを複数回行うことで結合物を濃縮し、DNAのシークエンシングにより同定される。
【0083】
さらに上述の方法において、RNAタンパク質ライブラリーを定方向展開のための例示的なライブラリーとして示したが、いかなる型の骨格に基づくライブラリーも本発明の選択方法において使用することができる。
【0084】
使用
本明細書に記載の抗体模倣物は、関心対象の任意の抗原とも結合するように導出することができる。これらのタンパク質は天然抗体に比べて熱力学的に優れた性質を持ち、インビトロで急速に導出することができる。したがって、これらの抗体模倣物は、研究、治療、診断の分野を含む、抗体が使用されるすべての分野においても抗体の代わりに用いることができる。さらに、これらの骨格は抗体に比べて可溶性でかつ安定性の性質を持つので、本明細書に述べられた抗体模倣物は抗体分子が破壊されたり、または不活性化されたりするような条件下でも使用することができる。最後に、本発明の骨格は事実上いかなる化合物とも結合するように導出することができるので、これらの分子は完全に新規であり、研究、治療、および診断の分野における使用も見出されている結合タンパク質を提供する。
【0085】
実験結果
上述された例示的な骨格分子は、例えば以下のような選択手順で生産され、かつ検査された。
【0086】
ライブラリーの構築
複合体のライブラリーは、CDR様ループに対応する無作為化された部位を各々1個含む3つの断片から構築した。無作為化された残基は、図18に下線をつけた配列で示されているが、10Fn3 BCループ(ラマVHのCDR-H1に対応)の残基23〜29;10Fn3 DEループ(ラマVHのCDR-H2に対応)の残基52〜55;および10Fn3 FGループ(ラマVHのCDR-H3に対応)の残基78〜87である。断片は、その中にあるCDR-H様ループの名前に基づいて、BC、DE、およびFGと名付けた。各々の断片には、10Fn3および無作為化された配列に加えて、His6ドメインのN末端またはFLAGペプチドタグのC末端をコードする伸長部分が含まれた。各DNA断片には、2つの断片間(すなわち、BCおよびDE断片間もしくはDEおよびFG断片間)の各接合部にEar IタイプIIS制限エンドヌクレアーゼの認識配列が含まれた。この制限酵素は、すべての外来非10Fn3配列を除去し、隣接する断片を一緒にスプライシングさせた。さらに変異および選択ラウンドの間に、3つの10Fn3断片を組換えのように混合させる。
【0087】
野生型のヒト10Fn3遺伝子は、プライマー
を用いて、ヒト肝臓ライブラリー(Maxim Biotech、カリフォルニア州南サンフランシスコ)からクローニングした。10Fn3クローンのPCRで得られた野生型セグメントおよびオリゴヌクレオチド合成で得られた無作為化セグメントから、3つの異なるライブラリーが構築された。BCr-DEr-FGrライブラリーは、BC、DE、およびFGループの選択された残基を無作為化することによって得られた;BCr-DEwt-FGrライブラリーは、BCおよびFGループの選択された残基を無作為化し、DEループ配列を野生型のまま残すことによって得られた;BCwt-DEwt-FGrライブラリーは、FGループのみの選択された残基を無作為化することによって得られた。
【0088】
BCr、DE rおよびFGr断片は、合成された。各断片は2つの重複するオリゴヌクレオチドから構成され、最初にアニーリングさせた後、断片の2本鎖DNAの形状を形成するように伸長させた。3つの断片を構築および加工するのに使用されるオリゴヌクレオチドを以下の表に示す:「先端(top)」および「末尾(bottom)」の種は、10Fn3コード配列全体を含むオリゴヌクレオチドである。これらのオリゴヌクレオチドの呼称において、「N」は、A、T、C、またはGを示し、「S」は、CおよびGを示す。
T7Tmv(インビトロ翻訳に必要なT7プロモーターおよびTMV非翻訳領域を導入する):
Unispl-s(Robertsら、1997、上記、ピューロマイシン含有リンカーにmRNAを連結するために用いられるスプリントオリゴヌクレオチド)
A18---2PEG(DNA-ピュロマイシンリンカー):
5’-(A)18(PEG)2CCPur(配列番号:14)
【0089】
オリゴヌクレオチド対BCTopおよびBCBot-flag8を用いて無作為化BCループを含む断片が構築された;対DETopおよびDE Bot-flag8を用いて無作為化DEループを含む断片が構築された;対BCTopおよびDE3-flag8を用いてBCwt-DE wt断片をPCR増幅した;および対FGTopおよびFG Bot-flag8を用いて無作為化FGループを含む断片が作製された。オリゴヌクレオチド対(各500pmol)を100μlの10mM Tris 7.5、50mM NaCl中で85℃にて10分間アニーリングさせ、次にゆっくりと(0.5〜1時間)室温に冷却した。1本鎖の突出部分にアニーリングした断片を、アニーリングしたオリゴ各100μlアリコートに対して100Uのクレノー(New England Biolabs、Beverly、MA)ならびに838.5μlのH2O、9μlの1M Tris 7.5、5μlの1M MgCl2、20μlの10mM dNTP、および7.5μlの1M DTTで調製された緩衝液を用いて伸長させた。伸長反応は、25℃にて1時間行った。
【0090】
無作為配列によって導入される停止コドンの頻度を下げるために、無作為化された残基は、(NNS)nによってコードされたが、Nは任意のヌクレオチド、SはCおよびGの等モル混合物を表す;3つの停止コドンのうちの1つのみ(TAG)がNNSの制限に従う。遺伝子断片は、10Fn3をコードする配列に加えて、5’タバコモザイクウイルス(TMV)非翻訳領域およびT7プロモーター、ならびに5’ヘキサヒスチジンタンパク質精製タグおよび3’ FLAGエピトープ精製タグをコードする配列も含んでいた。また、3つの断片の組み立てを容易にするために、上述のように、Ear I制限エンドヌクレアーゼ認識部位が、隣接する断片の重複部分に組み込まれた。
【0091】
次に、ショスタック(Szostak)ら、米国特許出願第09/007,005号および同第09/247,190号;ショスタック(Szostak)ら、国際公開公報第98/31700号;ならびにロバーツ(Roberts)およびショスタック(Szostak)、Proc. Natl. Acad. Sci. USA、1997年、第94巻、第12297〜12302頁によって開発された技術を用いて、各2本鎖断片をRNA-タンパク質融合体(PROfusion(商標))に形質転換した。簡単に言えば、アンビオンインビトロ転写キット(Ambion in vitro transcription kit)、T7-MEGAshortscript(商標)(Ambion、Austin、TX)を用いて断片を転写し、得られたmRNAをゲル精製し、かつDNAリガーゼ(Promega、ウィスコンシン州マジソン)を用いて5’リン酸化DNA-ピューロマイシンリンカー(好ましくは5’ dA18PEG2dCdCPur)に連結した。ショスタック(Szostak)(上記)に記述されるようにして、mRNAはDNAスプリントオリゴヌクレオチド
を用いてDNAリンカーと整列化させた。アンビオンウサギ網状赤血球溶解物に基づく翻訳キット(Ambion rabbit reticulocyte lysate-based translation kit)を用いて、35Sメチオニンの存在下で、mRNA-DNA-ピューロマイシン分子を翻訳した。得られたmRNA-DNA-ピューロマイシン-融合体タンパク質をオリゴ(dT)セルロース(タイプ7、Amersham Pharmacia、ニュージャージー州ピスカタウェイ)を用いて精製し、逆転写酵素(Superscript(商標)II、Gibco, Life Technologies、メリーランド州ロックビル)および上述したRTプライマー(Unisplint-SもしくはflagASA)を用いて、製造者の指示(好ましくは、70℃で2分間アニーリングおよび42℃で40分間反応)に従ってDNAの相補鎖を合成した。
【0092】
各断片について得られたアニーリングcDNAを持つRNAタンパク質融合体は、そのペプチド精製タグに適した樹脂、すなわちHis6タグにはNi-NTAアガロース(Qiagen、カリフォルニア州バレンシア)およびFLAGタグにはM2抗Flagアガロース(Sigma、ミズーリ州セントルイス)を用いて、製造者に推奨された方法に従って精製した。KOH溶出によって回収された断片のコードする遺伝情報を、ファルマシアレディートゥゴーPCRビーズ(Pharmacia Ready-to-Go PCR Beads)、10pmolの5'および3' PCRプライマーを用いて、以下のPCRプログラム(Pharmacia、Piscataway、NJ)に従ってPCRにより増幅した:1段階目は95℃で3分間、2段階目は95℃で30秒間、58/62℃で30秒間、72℃で1分間を必要に応じて20/25/30サイクル、3段階目は72℃で5分間、4段階目は最後まで4℃(通常、25サイクル)。
【0093】
得られたDNAを、1μgのDNAにつき5〜6 UのEar I(ニュー・イングランド・バイオラボ(New England Biolabs))で切断した。反応は、T4 DNAリガーゼ緩衝液(New England Biolabs)中で37℃にて1時間行い、続いて選択的に、Ear Iを失活させるために70℃にて15分間インキュベートした。無作為化されたループを持つ10Fn3遺伝子の全長を形成させるために、等量のBC断片、DE断片、およびFG断片を混合して連結した。連結には10Uの新鮮なEar I(New England Biolabs)および20UのT4 DNAリガーゼ(Promega、ウィスコンシン州マディソン)が必要であり、37℃にて1時間かかった。Ear Iおよびリガーゼは、65℃で15分間インキュベートして失活させた。
【0094】
3つの異なるライブラリーBCwt-DEwt-FGr、BCr-DEwt-FGr、および、BCr-DEr-FGrは、上述のようにして構築された。各々が10個の無作為化された残基を持つFGループの形を含んでいた。最初のライブラリーのBCおよびDEループは、野生型の10Fn3配列を持っていた;7個の無作為化された残基を持つBCループおよび野生型のDEループが第2のライブラリーを構成していた;7つの無作為化された残基を持つBCループおよび4つの無作為化された残基を持つDEループが第3のライブラリーを構成していた。これら3つのライブラリーの各々におけるFGループの複雑性は1013だった;さらなる2つの無作為化されたループを合わせると、潜在的な複雑性は研究室で標本抽出するには高すぎるほどになった。これらのライブラリーの組み合せは1012のユニークなクローンをもつマスターライブラリーを提供していた。
【0095】
元の無作為化されたBCr-DEr-FGrライブラリーから無作為に選んだ76個のクローンの配列は、無作為化されたループに何のパターンも示さなかった(データは示さず);ライブラリーのアミノ酸頻度は、各残基をコードするコドンの数に比例しており、各位置あたり1%(グルタミン酸、メチオニン、トリプトファン)と14%(プロリン)の間だった。反対に、保存されたβシート枠組中の残基が野生型のまま保存される平均確率は、99%だった。
【0096】
等モルの3つのライブラリーは(各々DNA 2 pmol)、選択工程を単純化するために混合され、1つのマスターライブラリーとし、標的との結合それ自体によって、特定の誘発に最も適したライブラリーを選択することにした。ラウンド3から10の親和性による精製ではM2-セファロース(以下参照)が使用されたこと以外、ショスタック(Szostak)ら、米国特許出願第09/007,005号および同第09/247,190号;ショスタック(Szostak)ら、国際公開公報第98/31700号;ならびにロバーツ(Roberts)およびショスタック(Szostak)、Proc. Natl. Acad. Sci. USA、1997年、第94巻、第12297〜12302頁に述べられた一般的な方法に従って、マスターライブラリーからRNA-タンパク質融合体を得た(図8)。
【0097】
融合体の選択
RNA-タンパク質融合体の形態のマスターライブラリーが、TNF-αとの結合を選択するために供された(Pepro Tech、ニュージャージー州ロッキーヒル)。最初に2つの方法が行われた:1つでは標的をアガロースカラムに固定化し、1つでは標的をBIACOREチップに固定化した。最初に、アガロースカラムへのバックグラウンドの結合体を最小限にするために条件の大幅な最適化を行い、50mM HEPES pH 7.4、0.02% Triton、100μg/ml変性されたサケ精子DNAという好ましい緩衝液の条件を得た。この緩衝液中では、10Fn3-RNA融合体のTNF-α-セファロースへの非特異的結合は0.3%であった。10Fn3-RNA/cDNAライブラリーのTNF-α-セファロースへの非特異的結合バックグラウンドは0.1%であることが見出された。
【0098】
TNF-α-セファロース上での各々の選択ラウンドにおいて、残存している非特異的な結合体のいかなるものも除去するために、最初にライブラリーを非誘導体化されたセファロースと共に1時間プレインキュベートし、この前除去により得られた流出液をTNF-α-セファロースと共にさらに1時間インキュベートした。TNF-α-セファロースは3〜30分間洗浄した。
【0099】
各選択後に、0.3M NaOHまたは0.1M KOHを用いて複合体のcDNA成分を固相支持体から溶出させ、PCRにより増幅させた。所望の大きさのDNAのバンドは、選択ラウンドを複数回行っても残留していた(図9)。2つの他の選択法でも同様な結果が観察されたが、アガロースカラム選択で得られたデータのみを図9に示す。
【0100】
この選択の最初の7ラウンドにおいては、Fn3-RNA/cDNA分子の標的への結合は低いままであり;対照的に、選択の異なる段階においてDNAプールから遊離タンパク質が翻訳された場合には、カラムに結合した種の割合はラウンド間で有意に増加した(図10)。
【0101】
後の選択では、選択の最初の8ラウンド(R1〜R8)で選択された融合体プールは、バックグラウンドに近いレベル(<0.25%)でTNF-α-セファロースに結合した(図13)。選択の第9ラウンド(R9)後、TNF-α-セファロースへの融合体の結合は、0.7%と急上昇し、第10ラウンド(R10)後は、結合はさらに7%に増加した(図13)。選択は、10 mL中10 mg TNF/gセファロースでエポキシ活性化セファロース(商標)6B(Amersham Pharmacia)に固定化されたTNF-αを用いて行われた。使用前にTNF-α誘導化セファロースは、結合緩衝液(50 mM HEPES, pH 7.4, 0.02% Triton, 0.1 mg/mL変性されたサケ精子DNA(Ambion))中で、4℃で一晩ブロックされた。
【0102】
上述のように、10Fn3に基づくマスターライブラリーを転写し、ピュロマイシンを持つリンカーに連結し、5-10μL/300μL 35S-メチオニンの存在下でmRNA-タンパク質ライブラリーに翻訳し、オリゴ(dT)セルロース上で親和性により精製し、DNA/mRNA-タンパク質ライブラリーに逆転写し、M2-セファロース上で親和性により精製した(3〜10ラウンド)。DNA-mRNA-タンパク質融合体ライブラリー分子4 x 1012の異なる配列20コピーに当たる40 pmolを回収し、第1ラウンド(R1)の選択を行なった。
【0103】
選択の第1段階では、40 pmolのDNA-mRNA-タンパク質ライブラリーを、TNF-αの非存在下で誘導化したエポキシセファロース30μLを含む300μLの結合緩衝液中で、回転させながら4℃で1時間インキュベートした。第2ラウンドでは、24 pmolのライブラリーを添加し、残りの8ラウンドでは、0.1〜2 pmolのライブラリーを添加した。上清はMico Bio-Spin(登録商標)クロマトグラフィーカラム(BIO-RAD、カリフォルニア州ハーキュリーズ)で微量遠心して回収し、300μLの結合緩衝液中で30μLのTNF-α-セファロース(6μM)と4℃で1時間インキュベートした(7〜10ラウンドでは、結合緩衝液にはさらに1 mg/mLのBSAが含まれていた)。TNF-α-セファロースはスピンカラムで回収し、3 x 300μLの結合緩衝液で洗い、100μLの0.1 M KOHで溶出し、最後に1μLの1 M Tris 8.0, 8μLの1 M HClで中和した。ライブラリー、溶出前後のTNF-α-セファロース、洗浄液、および溶出液のサンプルは、シンチレーションカウンターでサンプル中の35S-メチオニンを計数して定量した。選択の次のラウンドは、PCR増幅によって新しいDNA/mRNA-タンパク質プールを形成することから始まり、これをPCR産物から転写、翻訳、および逆転写した。
【0104】
第9ラウンドおよび第10ラウンド後の溶出から得られたDNAプールをTOPO(商標)TA(登録商標), pCR2.1クローニングベクター(Invitrogen、カリフォルニア州カールズバッド)にクローニングし、大腸菌を形質転換した。30〜100クローンを選択し、プラスミドミニプレップ(Qiagen)に増殖させた。R9から38クローン、R10から29クローンを無作為に選択し、配列決定した(DNAシーケンシングコアファシリティー、マサチューセッツ総合病院、分子生物学部門、マサチューセッツ州ボストン)。得られたタンパク質配列の整列化には、プログラムClustalW60を使用した。
【0105】
TNF-α結合クローンのアミノ酸残基配列
R9およびR10から得られた61クローンのうちの38は、ユニークなアミノ酸配列を持っており、これは驚くべき多様性である。おそらくTNF-αに対する優れた結合のために複数回単離された10のクローンは、表1に示されている(全体の配列は図25)。
【0106】
得られたプールから無作為に選択された61のクローンのうち、1つ(クローンT09.08、配列は示さず)のみがBCwt-DEwt-FGrライブラリー由来で、別の6個がBCr-DEwt-FGrライブラリー由来だった。残りの54個(ウィナーの88%)がBCr-DEr-FGrライブラリーから選択されたということは、いくつかのループが標的に協力してTNF-αに結合することの重要性を示す。
【0107】
選択されたループ配列の最も一般的なモチーフはPWA(S/T)で、これは61のクローンのうちの33のDEループに見られた;より緩く定義されたPW(A/G)の配列は41/61のクローンに見られた。抗体VHドメインの類似したCDR-H2ループは一般的に抗原結合には小さな貢献しかしないので、特定のDE配列に対するそのような強い選択は、驚くべきことである。一方、DEループの長さが短いために、ライブラリーには可能なテトラペプチド配列がそれぞれ107コピー存在すると期待され、そのため選択された性質に対するDEループの任意の貢献を最適化するのが容易になる。他のFn3ドメインの調査(Dickensonら、J. Mol. Biol. 236:1079-1092 (1994))は、10Fn3の残基52に相当する位置には、野生型のグリシンと同様の頻度でプロリンが見られることを示す;同様に、アラニン、グリシン、および野生型リジンは、すべて10Fn3位置54に相当する位置によく見られる。その結果、位置52および54で選択される残基は、少なくとも好ましい生物物理学的性質を持つ点で一貫していると思われる。対照的に、10Fn3残基53に相当する位置ではトリプトファンは見られず、トリプトファン53はTNF-αとの結合への貢献のためというような、本発明に特異的な理由で選択された可能性がある。これは他の抗原に対する後の選択でもこのモチーフが見られないこととも一致しており、やはりPWA/Gモチーフは10Fn3ドメインの安定性または溶解性というよりも、TNF-αへの結合に直接貢献している可能性が高い。ループDEでPWA/Gが好まれることは、選択時にBCr-DEr-FGrライブラリーが好まれる別の理由を示唆する:BCr-DEr-FGrライブラリーのみが無作為化されたDEループを含み、標的との結合にPWA/G配列が重要ならば他の2つのライブラリーとの競争に勝つと考えられる。
【0108】
BCループで最も頻繁に選択された配列はNRSGLQS (12/61)(配列番号:31)であり、FGループで最も頻繁に選択された配列はAQTGHHLHDK (6/61)(配列番号:32)だった。NRSGLQS BCループとAQTGHHLHDK FGループは同一の分子には見つかれなかったが、3つの無作為化されたループのうちの2つに、最も頻繁に単離された配列を含むクローンが2つ見つかった。これらのクローン
は、調べたクローンのうちでTNF-αからの解離定数が最も低い4つのうちの2つだった(表1)。
【0109】
プルーフリード活性を持たないTaqポリメラーゼを使用したため、PCRのラウンドごとに、10Fn3のCDR様ループとβシート折畳みの両方に、1塩基対につき0.01%の割合、すなわちPCRの1ラウンドにつき10Fn3遺伝子につき3%、および選択1ラウンドにつき約75%の割合で、さらなる無作為化変異が導入された。したがって、野生型のまま保存された残基、および非野生型の安定した配列として保存された残基は、これらの配列はその優れた性質のために選択されたことを意味すると考えられる。変異したループでは、オリゴヌクレオチド合成によって導入された変異かPCR変異導入による変異かを区別するのは不可能だが、βシートの折畳みでは、選択された変異の大部分はTaqエラーに起因する。選択されたクローンはタンパク質の折畳みにいくつかの保存された変化を示すが、これは意図的に無作為化したものではない。図18は無作為化されなかったが、それでも選択中に変異した10Fn3βシートにおける残基を示す。この変異導入は、61クローンのうちの26〜28の頻度で起こった;これらの変異は野生型10Fn3配列の下に矢印で示されており選択された残基を示す文字が付けられている。特に、61クローンのうちの28ではロイシン18からアルギニンまたはグルタミンへ変異しており、26クローンではスレオニン56からイソロイシンに変異している。図19はこれらの折畳みの変異の位置を示す。位置56はループDEの幹にあり、したがってこのループの立体配座および標的結合特性に影響すると期待される。TNF-α結合ループからの位置18の距離は、この変異の選択的優位性は、ループBCの立体配座への間接的影響、またはタンパク質の安定性への影響に起因する可能性を示唆する(図19)。これは、頻繁に見られる野生型からの変異L18RおよびT56Iを含むクローンT10.06の位置18を野生型のロイシンに復帰させた変異導入の実験でも支持されている。この変化により、変異体のKdは約10倍に上昇した。T10.06(L18)タンパク質のTNF-αに対する弱くなった結合は、位置18の残基は、おそらくループBCへβ鎖を通して伝えられる小さな構造変化によって、CDR様ループによる標的への結合に影響することを示唆する。
【0110】
選択されたTNF-α結合プールの親和性と特異性
9回および10回目の選択後のTNF-αの遊離タンパク質プールの見かけの平均Kd値は、区別不可能(それぞれ4nMおよび6 nM;表1)だった;この親和性の類似は最後のラウンドの選択での濃縮が比較的低レベル(10倍)だったこと、および2つのプールの配列組成が類似していることと一貫している。さらに4回の選択を行なった後の遊離タンパク質プールの見かけの平均Kdは3 nMで、やはりR9およびR10プールと区別不能だった。
【0111】
10回目の選択後に選択されたプールの結合特異性を評価するために、我々はセファロースに固定された3つのサイトカイン、選択の標的であるTNF-α、および無関係のIL-1αおよびIL-13に対する2つの異なる遊離タンパク質プールの結合を比較した。第1のプールは最初の選択前(R0)の無作為化DNAライブラリーから、第2のプールは10回目の選択後(R10)のライブラリーから転写および翻訳した。
【0112】
これらの実験を実行するために、10回目の選択後の溶出液のPCR産物を35Sメチオニンの存在下でインビトロで転写および翻訳したが、mRNA-タンパク質融合体の形成はさせなかった。TNF-α-セファロース、IL-1α-セファロース、IL-13-セファロースに、それぞれ約10μM、30μM、および50μMで結合した、および未誘導化セファロースに結合した、得られた遊離タンパク質の画分は、TNF-α-セファロースに対するDNA/mRNAタンパク質融合体の結合について上述の手順を用いて、比較した(図20)。各標的に結合した選択されたプールの量は、洗浄したビーズのシンチレーション計測で測定した。
【0113】
図20は、R0のTNF-α、IL-1α、およびIL-13に対する結合は類似していたが(それぞれ2%、4%、および3%)、10回の選択度には標的TNF-αには32%、IL-1αには3%、およびIL-13には1%結合することを示す。TNF-αに対する絶対的および相対的なタンパク質結合の増加は、10Fn3骨格および標的特異的結合体を選択するためのDNA/mRNAタンパク質融合体に基づく選択システムの能力を示す。
【0114】
結合の特異性をさらに調べるために、クローンT09.12をマイクロアレイ形式(以下に一般的に説明されている)に固定し、可溶性TNF-αに対する結合を調べた。このクローンに対するTNF-αの特異的結合は、蛍光検出(図24A)および質量分光(図24B)で検出した。質量分光については、干渉する可能性のある様々なタンパク質を含む代表的な複雑な生体液である、牛胎児血清の存在下で結合測定が行われた。蛍光検出(図24A)については、野生型10Fn3およびT09.12変異体のRNA-10Fn3融合体の混合物(表1)を、融合分子のRNA部分に相補的なオリゴヌクレオチドが24の反復フィーチャーで600ミクロンのピッチで固定されたDNAマイクロアレイにハイブリダイズさせた。洗浄によりハイブリダイズしなかった融合体を除去した後、その表面をビオチン-TNF-α(TBS、0.02% Tween-20、0.2% BSA中2.6μg/mL)に暴露し、洗浄し、空気乾燥した。キャプチャーされたビオチン-TNF-αは、ScanArray 5000システム(GSI Lumonics)を用いて、Cy3標識の抗ビオチンモノクローナル抗体(Sigma)により、検出した。質量分光検出については、T09.12変異体のRNA-10Fn3融合体(図24B)および野生型10Fn3(図24C)をRNase Aで処理してタンパク質およびDNAリンカーの融合体を作製した。得られたDNA結合タンパク質をDNAリンカーに相補的なオリゴヌクレオチドを固定したガラスカバースリップアレイにハイブリダイズさせた(図24Bおよび24C;融合体は図24Dには使用されていない)。洗浄後、上述の表面をTNF-α(90% v/v PBS/10%牛胎児血清中1.5 mg/mL)に暴露した。乾燥したチップにMALDIマトリックスをスポットし、Voyager DE MALDI-TOF質量分光計(PerSeptive Biosystems)で分析した。モノマーのTNF-αに期待される分子質量に対応する17.4 kDのシグナルが、T09.12タンパク質を含む200μmフィーチャー上に検出されたが(図24A)、野生型10Fn3(図24B)およびDNAタンパク質融合体を含まないフィーチャー(図24C)には検出されなかった。
【0115】
選択されたTNF-α結合クローンのKd
9回および10回目の選択で作製された2つのプールに複数回現れる全てのクローン、およびBCwt-DEwt-FGrライブラリーに由来する唯一のクローン(クローンT09.08)について、解離定数が決定された。
【0116】
これらの結合定数を決定するために、ピエス(Pierce)(イリノイ州ロックフォード)のNHS-LC-LCビオチン試薬を用いてビオチン化TNF-αを調製した。MALDI-TOF質量分光を用いて、モノマーのTNF-αの80%以上、したがって、三量体の99%以上がビオチン化されていると見積もられた。
【0117】
R9およびR10プール(および以下に説明されるR14およびM12プール)ならびにこれらの2つのプールに由来する解析されたクローンについて、200μL 10 mM HEPES, pH 7.4, 150 mM NaCl, 1% BSA, 0.02% Triton中で、0.25 nMのインビトロ翻訳35S-メチオニン標識遊離タンパク質を、ビオチン化TNF-αと17 pMおよび23 nMの間の濃度で室温で1時間インキュベートした。その後、96ウェルのイージータイター(Easy-Titer)(商標)ELIFAシステム(Pierce)を用いて各サンプルをあらかじめぬらしたSAM2R ビオチンキャプチャーメンブレン(Promega、ウィスコンシン州マジソン)にかけた。減圧下で、各スポットを200μLのHBS pH 7.4, 1% BSA, 0.05% Tritonで洗った。次に、メンブレン全体を緩衝液ですすいで空気乾燥した。メンブレンをストレージ・ホスホ・スクリーン(Storage Phosphor Screen)(Molecular Dynamics、カリフォルニア州サニーベール)に一晩露出し、得られた各々のスポットの強度を、STORM 860ホスホイメージャーを用いてImageQuaNT濃度計プログラム(Molecular Dynamics)をにより定量した。結合のKdは平衡状態を得られた結合曲線に当てはめて決定された(KaleidaGraph、Synergy Software)。実験誤差は2〜4の独立した実験から見積もられた。
【0118】
これらの実験では、Kdは1〜24 nMの狭い範囲に入っていた(表1)。各々2つのループに最も多く単離される配列を含むT09.12およびT10.06クローンは、それぞれ4および2 nMという低いKdを持っていた。それに対して、クローンT09.07およびT10.15のような頻度のより低いループを持ついくつかのクローンも、同様に強い結合を示した。
【0119】
遊離タンパク質と、同じ配列に由来するcDNA/mRNA-タンパク質複合体のTNF-α結合の比較の標本を見ると、2つの解離定数は互いの実験誤差範囲内であり、これはタンパク質自身の標的結合特性を選択するために、cDNA/mRNA-タンパク質複合体を使用することが可能になるというシステムの性質を示す。
【0120】
TNF-α結合クローンの厳密度の高い選択
重複したクローンが単離されたにも関わらず、9回および10回の選択後のTNF-α結合プールには、数多くの異なるクローン、すなわち、標本にした61クローンでは38の異なる配列が、含まれていた。したがって、これらのクローンの中で優れたTNF-α結合を示すサブセットを回収するために、より厳密な結合要件を用いたさらなる選択が行われた。従って、さらに4回の選択(R11-R14)が標的濃度のコントロールが容易な溶液中で行われた。TNF-αの濃度は0.5 nMに限定され、DNA/mRNA-10Fn3プールの濃度は0.1 nMに限定された。さらに、ストレプトアビジンコートの常磁性ビーズに結合した10Fn3/ TNF-α複合体の洗浄の長さと温度も、引き上げられた。
【0121】
具体的には、以下のように選択が行われた。ラウンド11-13では、上述のように構築された0.1 nM DNA/mRNA-10Fn3融合体ライブラリーを、結合緩衝液であらかじめブロックした100μLのダイナビーズ(Dynabeads)(登録商標) M-280(ストレプトアビジンコートの常磁性ビーズ、Dynal、ニューヨーク州レークサクセス)と4℃で1時間回転させて、あらかじめきれいにした。得られた融合体混合物を、上述の結合緩衝液300 μL中で0.5 nMビオチン化TNF-αと混合し、複合体を4℃で1時間インキュベートした。次に、結合緩衝液中で一晩ブロックした1.3 g/cm3で再懸濁したダイナビーズ(登録商標) M-280ストレプトアビジン100μlを混合物に添加し、回転させながらさらに45分間4℃でインキュベートした。Dynal MPC-Sラック上で常磁性ビーズを上清から分離し、上清を除去し、結合緩衝液を用いてビーズをR 11およびR12の場合は1分間、15分間、および30分間洗浄し、R13〜R14の場合は1分間洗浄後、9回の10分間洗浄を行なった。100μl 0.1 M KOHで2回洗うことによって、洗浄したDNA/mRNA-10Fn3:TNF-αビオチン:ストレプトアビジンビーズ複合体からDNAを溶出し、上述のように、カラムを用いた選択によって次の世代のDNA/mRNA-10Fn3融合ライブラリーを作製した。ラウンド14は、選択がさらに150 mM NaClを加えた条件で30分間行われたという点でR11〜R13とは異なっていた。温度を上昇させた以外は、R14とR13の洗浄手順は同じだった。
【0122】
さらに4回の選択後(R14)に溶出されたDNA由来の無作為に選択した22のクローンは、15の異なるループ構造を持っていた(表2;全体の配列は図25)。上述のようにR10で単離されていたクローンT10.06は、8回選択されたが、R9プールで単離されたR09.31を含めた残りの配列は、各々1回単離された。9回目および10回目の単離株と同様に、R14クローンはDEループ中PWA/G配列に偏っており(22クローン中18)、4つの新しい非野生型DE配列も観察された。
【0123】
R14遊離タンパク質プールの見かけの平均Kd値3 nMは9回および10回のプール(それぞれ4および6 nM)と類似していたが、R14プールから単離されたいくつかのクローンのKd値は、R9およびR10プールで観察された最低値よりも1桁低かった(表2)。TNF-αに最も強く結合したクローンT14.07およびT14.25のKdは90 pmolだった。したがって、最後の4回の選択に使用した条件は、ナノモル以下のKdを持つ10Fn3分子に有利になるために充分厳密でありながら、そのような分子を排除してしまうほど厳密ではなかった。
【0124】
突然変異誘発による親和性成熟
上述のように、本明細書に記述される選択では、さらにライブラリーの多様性を増やすために、選択段階の全てまたは一部の後に変異誘発を行なうこともできる。1つの平行選択戦略では、ラウンドの間のDNA増幅に変異性PCRを組み込んだ(CadwellおよびJoyce, PCR Methods Appl. 2:28 (1992))。このテクニックは、上述のR8後に溶出される多様なDNAプールから始められた。このプールを変異性PCRで増幅したが、プールを7つの等量の部分に分け、標的頻度0.8%, 1.6%, 2.4%, 3.2%, 4.0%, 4.8%, および5.6%で変異誘発した。この7つのPCR反応を合わせ、混合物からcDNA/RNA-タンパク質融合体を作製し、溶液中で選択ラウンドを行なった。第2回の変異誘発ラウンドM10の前に、3つの別の反応0.8%, 1.6%, および2.4%で変異性PCRを行なった。2つの残るラウンドM11およびM12は、標準的なTaq PCRを用いて行なった。変異誘発以外は、M9-M12の選択条件はR11-R14と同じだった。調べた20のM12クローンは、前の2つの選択プロトコールを用いて選択したクローンよりも強い結合を示した(表3;全配列は図25)。TNF-αへの最も強い結合はM12.04で、観察されたKdは20 pMだった。これらの結果は、選択後期における低レベルの無作為の変異導入が、選択された抗体模倣物の結合親和性(20 pM対90 pM)、および選択できる速度(12回対14回)の両方を改善し得ることを示す。さらに、この変異導入の手法における強い結合体の頻度は約5%であるのに対し、他の選択では約3%だった。
【0125】
Fn結合体の優位性
選択単位として共有結合のmRNA-タンパク質融合体を用いておこなったTNF-αに結合できる10Fn3の選択は、20 pMという低い解離定数を持つ分子が勝ち残った。これらのKd値は、他の抗体模倣物骨格および選択方法を用いる他の選択の基準と比較して勝るものである。したがって、10Fn3に基づく骨格および共有結合のmRNA-タンパク質融合体に基づくインビトロの選択方法は、広範囲の抗原に対する抗体模倣物の開発に利用できる可能性がある。さらに、本明細書に記述されるナノモルレベル以下のTNF-α結合10Fn3変異体は、治療、研究、診断のための薬剤となる可能性がある。さらに、このインビトロ選択方法は自動化できるので、そのような骨格および選択方法の組み合せは、ゲノムスケールでの用途がある。
【0126】
本選択の成功に寄与した1つの要素は、10Fn3の3つのCDR様ループの全ての無作為化である。1つまたは2つの無作為化ループのみを含む類似のライブラリーは、3つの無作為化CDR様ループを持つライブラリーよりも、強い結合体を含む可能性が低い。
【0127】
上記に報告された選択では、無作為化ループは対応する野生型の10Fn3ループの長さのままである。ライブラリーの多様性をさらに高めるために、ループの長さおよびその配列も変化させ、ライブラリーの構築に使用した野生型骨格に10Fn3βシート中の好ましい変異を組み込み、抗体の模倣という点ではさらにうまく構造の選択ができる無作為化βシート骨格を持つライブラリーを作製できる。
【0128】
本明細書に記述した選択と類似したものは、他の結合種標的(例えばIL-1またはIL-13)を用いても実行できる。
【0129】
動物実験
野生型10Fn3は、インテグリン結合性のトリペプチドモチーフであるアルギニン78-グリシン79-アスパラギン酸80(「RGDモチーフ」)をFGループの先端に含んでいる。このトリペプチドに基づくインテグリンとの結合および潜在性の炎症応答をインビボで回避するために、密接に関連した野生型11Fn3ドメインで見出される配列で、活性を持たない配列であるセリン78-グリシン79-グルタミン酸80(「SGE変異体」)を含む10Fn3の変異体を生産した。このSGE変異体は、N末端にHis6タグを付けて大腸菌中で遊離タンパク質として発現し、金属キレートカラムに続いてサイズ排除カラム上で均質性によって精製される。
【0130】
特に、His6-10Fn3(SGE)をコードするDNA配列をpET9a発現ベクターにクローニングし、BL21 DE3 pLysS細胞を形質転換した。次に、50μg/mlのカナマイシンを含むLB培地中で37℃にて振盪しながらA260=1.0になるまで培養物を増殖させ、その後0.4mM IPTGにより誘導された。誘導された培養物を同じ条件下でさらに一晩(14〜18時間)インキュベートし;標準の低速度遠心分離により細菌を回収した。元の培養容量の1/50倍の溶解緩衝液(50mM Tris 8.0、0.5M NaCl、5% グリセロール、0.05% Triton X-100、および1mM PMSF)に細胞沈殿を再懸濁し、得られたペーストをMicrofluidics Corporation Microfluidizer M110-EHに3回通すことにより細胞を溶解させた。溶解物を遠心分離して清澄化し、上清を0.45μmのフィルターに通して濾過し、続いて0.2μmのフィルターに通して濾過した。清澄化された溶解物の100mlを5mlのタロンコバルトカラム(クロンテック(Clontech)、パロアルト(Palo Alto)、CA)に載せ、70mlの溶解緩衝液で洗浄し、0mM〜30mM イミダゾールの直線勾配を含む溶解緩衝液で溶出した。すべての段階を通じて、カラムを通す流速は1ml/分であった。溶出したタンパク質を15,000〜20,000のPEGに対して透析し(分子量カットオフ値、3,500)、10倍に濃縮した。得られた試料を緩衝液1(グリセロールを含まない溶解緩衝液)中で透析し、緩衝液1で平衡化した16×60mmのセファクリル100のサイズ排除カラム上に1回に5mlずつ載せた。カラムに0.8ml/分で緩衝液1を流し、所望の分子量のタンパク質を含む全画分をプールして、上述のように10倍に濃縮し、次にPBS中で透析した。得られた試料でエンドトキシンスクリーニングおよび動物実験を行った(Toxikon(MA))。
【0131】
これまでに検査された試料中のエンドトキシンの水準は、検査法の検出水準以下であった。予備的な毒性学の研究において、このタンパク質を2匹のマウスに治療量の100倍と見積もられる2.6mg/マウスを注射した。動物は研究の2週間のあいだ、はっきりとした悪い影響なしに生存した。これらの安全な結果は、静脈内薬に取り込まれた10Fn3の使用を支持する。
【0132】
インビボでの使用するための他の構築物
8kDの10Fn3ドメインの半減期を延ばすために、天然抗体を模倣するより大きい分子も構築されている。この10Fn3-Fc分子は、宿主のIgGの定常領域にある-CH1-CH2-CH3(図11)もしくは-CH2-CH3ドメインを含んでおり、これらの構築物中では、10Fn3ドメインはIgGのVHドメインの代わりにN末端に結合されている(図11および図12)。このような抗体様構築物は、天然の免疫応答を利用できる可能性と同様に、タンパク質の薬物動態を改善させるべきである。
【0133】
10Fn3-CH1-CH2-CH3クローンのマウス型を構築するために、最初にマウス肝脾cDNAライブラリー(クロンテック(Clontech))から-CH1-CH2-CH3領域を増幅し、次にpET25ベクターに連結した。クローニングに使用したプライマーは5' Fc Nestおよび3' 5 Fc Nestであり、回収したインサートの末端に適切な制限酵素部位を結合するのに使用したプライマーは、5' Fc HIIIおよび3' Fc Nheであった。
【0134】
このクローンからCH1領域を除去し、より短いFc部分を作製して10Fn3-CH2-CH3クローンとするために、さらなるPCRを用いた。各クローンの5'末端に10Fn3をコードする配列を連結した。同一のマウス脾臓cDNAライブラリーからクローニングされた野生型10Fn3でも、または分子の変異もしくは無作為化により得られた改変10Fn3でも使用することができる。マウス野生型10Fn3のクローニングに使用したオリゴヌクレオチドは、以下であった:
Mo 5PCR-NdeI:
Mo5PCR-His-NdeI(His6精製タグを有する、代わりのN末端用):
【0135】
ヒトオリゴヌクレオチド配列において、同一の方法を用いることにより、ヒトの等価なクローンが構築される。
【0136】
タンパク質チップ適用における抗体模倣体の骨格
本明細書に記載の任意の抗体模倣体は、マイクロチップのような固体支持体上で免疫化してもよい。10Fn3骨格のような本明細書の骨格がタンパク質チップ適用に適しているのは、 (1) ベンチまたは自動化装置で迅速に選択できる多くの結合基を支持する能力があり、かつ (2) 生物物理学的特性が優れているためである。
【0137】
10Fn3の用途の広い結合特性は、Fn3免疫グロブリン様、βサンドイッチ型折畳みによって示されるループに関連する。上述のように、これらのループは抗体の可変ドメインの相補性決定領域に類似しており、抗体ループと同様な様式で共同して抗原に結合できる。本発明者らの系では、多様なmRNA-10Fn3融合体のライブラリーを作製するために、10Fn3ループのBC(例えば21残基〜30残基)、DE(例えば51残基〜56残基)、およびFG(例えば76残基〜87残基)は、配列、長さ、または配列と長さの両方が、無作為化されている。そのようなライブラリーにおける結合物は、高親和性の結合物の小さな集団が得られるまで、固定化またはタグ付き標的に対する親和性に基づいて濃縮される。また、誤りがちの(error-prone)PCRおよび組換えを用いて、選択された結合物の親和性の成熟を促進することができる。迅速で効率の良い選択および親和性成熟プロトコールにより、多数の標的に対する結合物を短時間で選択することができる。
【0138】
タンパク質チップ上に固定される結合物の骨格として、10Fn3ドメインは、抗体断片および単鎖抗体よりも小さく、扱いやすいという利点を持つ。例えば、安定性および溶解度に大きな差があり、構造的に必須なジスルフィド結合を保つために酸化環境を必要とする単鎖骨格または単離された抗体可変ドメインとは異なり、10Fn3は非常に安定で、その融解温度は110℃、溶解度は>16 mg/mLである。また、10Fn3骨格はジスルフィド結合も遊離のシステインも含まない;したがって、その環境の酸化還元電位には影響されない。10Fn3のもう1つの利点は、抗原結合ループおよびN末端が、βサンドイッチの、C末端とは反対側の端にあるということである;そのため、10Fn3骨格がC末端でチップに結合すると、抗原結合ループが、分析する溶液に最大限で接触可能になる。10Fn3はわずか94のアミノ酸残基からなる単一ドメインであるため、約250残基の単鎖抗体よりも高密度でチップ表面に固定することが可能である。また、10Fn3骨格は、このドメインの溶解度が高いことからもわかるように、親水性であるため、チップ表面に対する望まれない結合を最小化する。
【0139】
10Fn3骨格の安定性と、ライブラリー作製および結合物選択のための適合性は、テネイシン、Nカドヘリン、Eカドヘリン、ICAM、タイチン、GCSF-R、サイトカイン受容体、グリコシダーゼ阻害剤、および抗生物質色素タンパク質のドメインのような、免疫グロブリン様の折畳みを持つ、タンパク質ドメインの10Fn3様の大きなクラスにも共通している可能性がある。このようなドメインに共通の重要な特徴は、2つのβシートが提供する安定した枠組みであり、このβシートは互いに圧縮され、シートの1つの縁につき、少なくとも3つの溶媒が接触可能であるループによって結合する。そのようなループを無作為化して、枠組みの構造を破壊することなく、潜在的な結合物のライブラリーを作製することができる(上記)。さらに10Fn3と同様に、これらのループのいずれか(または他のタンパク質由来の類似のループ)は、単独でまたは他のループと組み合わせて、固体支持表面上に対して固定化させてもよい。
【0140】
Fn3を基にした抗体模倣体の固定化
チップ表面にFn3を基にした抗体模倣体のような抗体模倣体を固定するために、いくつかの典型的な手法が利用できる。例えば、このような抗体模倣体はRNA-タンパク質融合体として、融合体のRNA部分をワトソン・クリックハイブリダイゼーションによって、チップ表面に固定化したベースの相補的DNAに固定することができる(例えば、Adressable Protein Assays、米国特許出願第60/080,686号;米国特許出願第09/282,734号、および国際公開公報第99/51773号;およびMethods for Encoding and Sorting In Vitro Translated Protein、米国特許出願第60/151,261号および米国特許出願第09/648,040号に記述)。または、抗体模倣体は、チップ表面に直接遊離のタンパク質として固定できる。チップ表面に抗体模倣体を沈着させるためには、手作業およびロボット装置が使用できる。アレイ形式で高密度に抗体模倣体を沈着させるためには、スポットロボットが使用できる(例えば、Luekingら、Anal Biochem. 1999 May 15; 270(1):103-11の方法による)。抗体模倣体をチップ表面に結合させるためには、別の方法も利用できる。「酵素学の方法(Methods in Enzymology )」(M. MosbachおよびB. Danielsson編)、第135巻および136巻、Academic Press、Orlando、Florida、1987;ニルソン(Nilsson)ら、Protein Expr. Purif. 1997 Oct;11(1):1〜16、およびその参考文献に記述されるものを含め、いくつかの標準的な固定化手法が使用できる。抗体模倣体の方向性を持った固定化は、チップに結合した抗体模倣体の結合容量を増加させる際に役立つ。方向性を持った結合のための典型的な手法は、リュー(Lu)ら、The Analyst (1996)、vol. 121、p29R-32R;およびターコバ(Turkova)、J Chromatogr B Biomed Sci App. 1999 Feb 5;722(1-2):11-31に記述されている。さらに、抗体模倣体をチップ表面に結合させるための本明細書に記述されるいかなる方法も、抗体模倣体をビーズ、または他の支持体に固定するために使用できる。
【0141】
標的タンパク質の捕捉と検出
骨格結合物の選択された集団を用いて、例えば、生物試料のような試料中で、分析物標的の検出および/または定量ができる。この種の診断解析を行なうためには、関心対象の標的に対する選択された骨格結合体を適当な支持体上に固定して、多機能タンパク質チップを作製する。次に、チップに試料を載せ、試料中の成分で結合物に結合するものを、固定化した結合物の標的特異性に基づいて同定する。この手法を用いると、試料中の1つまたは複数の成分が、同時に同定または定量できる(例えば、試料のプロファイリングのための手段として)。
【0142】
標的の検出方法は、結合したタンパク質標的レベルの測定を可能にするものであり、X線撮影、蛍光スキャニング、質量分光(MS)、および表面プラズモン共鳴(SPR)を含むがこれらに限定されるものではない。リン光画像化システム(Molecular Dynamics、Sunnyvale、CA)を用いたオートラジオグラフィも、例えば35Sメチオニンで放射標識した標的タンパク質の検出と定量に使用できる。レーザースキャナーを用いた蛍光スキャニング(以下を参照のこと)は、蛍光標識した標的の検出と定量に使用できる。または、蛍光スキャニングを用いて、それ自身が標的タンパク質に結合する蛍光標識リガンドを検出することもできる(例えば、蛍光標識した標的特異的抗体、または標的-ビオチンに結合する、蛍光標識したストレプトアビジン、以下に記述)。
【0143】
質量分光は、結合した標的をその分子量によって検出および同定するために使用できる。結合した標的タンパク質の脱離は、下記のようにして、チップ表面から直接レーザーを用いて行なうことができる。質量の検出では、分子量に基づいて、リン酸化またはグリコシル化のような翻訳後修飾を含む標的の修飾も決定することができる。表面プラズモン共鳴は、骨格結合物が適当な金表面(例えば、スウェーデンのBiacoreより入手可能)に固定されている場合に結合したタンパク質標的の定量に使用することができる。
【0144】
以下に、該結合物(この場合、タンパク質TNF-αに特異的なFn結合物)の選択方法、および選択された集団を用いたチップ上での検出を例示する。本実施例は、本発明を説明するためのものであり、制限するためのものではない。
【0145】
10Fn3骨格に基づくTNF-α結合物の選択
チップ上で骨格を選択するための1つの典型例では、無作為化ループBC、DE、およびFGを持つヒト10Fn3変異体のライブラリーを用いて、TNF-αに対して10Fn3に基づく選択が行われた。ライブラリーは、1つの無作為化ループを含め、ヒト10Fn3の約3分の1をコードするヌクレオチド配列をそれぞれ含む、3つのDNA断片から構築された。上記のループ残基をコードするDNA配列は、関心対象の残基のコドンが (NNS)nで置換されたように、オリゴヌクレオチド合成によって再構築された。式中、Nは4つのデオキシリボヌクレオチド(A, C, GまたはT)のいずれかであり、SはCまたはGを表す。各断片のC末端は、FLAG精製タグの配列を含んでいた。
【0146】
ショスタック(Szostak)ら(RobertsおよびSzostak、Proc. Natl. Acad. Sci. USA 94:12297, 1997; Szostakら、米国特許出願第09/007,005号、および米国特許出願第09/247,190号;Szostakら、国際公開公報第98/31700号)に記述されるように、クレノー酵素で伸長後、各DNAを転写し、ピューロマイシン含有DNAリンカーと連結し、インビトロで翻訳し、mRNA-ペプチド融合体を作製し、これをDNA-mRNA-ペプチド融合体に逆転写した。FLAGタグの付いたペプチドをM2アガロースに結合することにより、フレームシフトまたは余分の終止コドンを含むものから、全長の融合分子が分離された;精製した全長の融合体に結合したDNAをPCRにより増幅した後、3つのDNA断片をEar I 制限エンドヌクレアーゼにより切断し、連結して全長の鋳型を形成させた。鋳型を転写し、ピューロマイシン含有DNAリンカーに連結し、翻訳して10Fn3-RNA/cDNAライブラリーを作製した。その後、これを逆転写してDNA-mRNA-ペプチド融合ライブラリーを作製し、後に選択に使用した。
【0147】
TNF-α結合物の選択は、50 mM HEPES、pH 7.4、0.02% Triton-X、0.1 mg/mLサケ精子DNA中で行われた。PROfusion(商標)ライブラリーは、セファロース固定TNF-αとインキュベートした;洗浄後、最も強い結合物に伴うDNAを0.1 M KOHで溶出し、PCR増幅、転写、連結、翻訳、逆転写し、次の選択ラウンドの開始材料とした。
【0148】
そのような選択を10ラウンド行なった(図13に示す);見かけの平均Kdが120 nMでTNF-αセファロースに結合するPROfusion(商標)プールが得られた。解析されたプールの特定のクローン成分は、50 nM〜500 nMの範囲でTNF-α結合を示した。
【0149】
固定化、標的タンパク質の捕捉、およびMALDI-TOF検出
Fn3融合体をチップ表面に固定するための第1段階として、自動DNA合成装置(PE BioSystems Expedite 8909)で、固相支持体ホスホラミダイド法でオリゴヌクレオチド捕捉プローブを調製した。全ての試薬はグレン・リサーチ(Glen Research)から入手した。合成は、ジスルフィド結合を含む固相支持体から開始して、最終的に3'末端のチオール官能性が得られた。添加される最初の4つの単量体は、ヘキサエチレンオキシドユニットで、20のT単量体が続いた。5'末端のDMT基は除去されなかった。捕捉プローブは固相支持体から開裂し、水酸化アンモニウムで脱保護し、真空遠心分離で乾燥するまで濃縮し、トリエチルアンモニウム酢酸緩衝液中のアセトニトリル勾配を用いて、逆相HPLCで精製した。HPLCの適当な画分を回収し、真空遠心分離で乾燥するまで蒸発させ、30分間の80% AcOH処理によって5'末端のDMT基を除去した。酸は気化により除去し、次にオリゴヌクレオチドは100 mM DTTで30分間処理してジスルフィド結合を開裂させた。DTTはEtOAcで繰り返し抽出して除去した。オリゴヌクレオチドは残りの水相からエタノール沈殿させ、逆相HPLCによって純度を確認した。
【0150】
3'チオールの捕捉プローブは脱気した1×PBS緩衝液中で250μMに調節して、少量の水を含む、アルゴンをフラッシュした(argon-flushed)チェンバー中で、単一の液滴(75μL)として9×9 mmの金をコートしたチップ(Biacore)に塗布した。室温で18時間後、捕捉プローブ溶液を除去し、機能化したチップを穏やかに撹拌しながら50 mLの1×PBS緩衝液で洗浄し(各15分間、2回)、同様にして50 mLの水ですすいだ(各15分間、2回)。残りの液体を注意深く除去して、機能化したチップは直ちに使用するか、またはアルゴン下で4℃で保存した。
【0151】
ラウンド10のTNF-α選択(上記)で得られた10Fn3融合体プールの約1 pmolを数時間リボヌクレアーゼ処理し、70μL中で5×SSCに調節し、上述の機能化した金チップに単一の液滴として塗布した。50μL容量のガスケット装置を使って、機能化チップで融合混合液を密封し、装置を4℃で連続的に回転させた。18時間後に、装置を解体し、金チップを穏やかに撹拌しながら50 mLの5×SSCで10分間洗浄した。チップ表面から過剰な液体を注意深く除去し、チップを4℃で10分間、ブロッキング溶液(1×TBS + 0.02% トゥイーン20 + 0.25% BSA)で不動態化した。過剰な液体を注意深く除去し、同一組成のブロッキング溶液中に500μg/mL TNF-αを含む溶液を単一の液滴としてチップに塗布し、ピペットマンで液滴を時々混合しながら4℃で2時間インキュベートした。結合溶液を除去した後、チップを穏やかに撹拌しながら4℃で5分間洗浄(50 mL 1×TBS + 0.02%トゥイーン20)し、室温で乾燥させた。第2のチップは、ハイブリダイゼーション混合液に融合体が添加されなかったこと以外は、上記と同様に調製した。
【0152】
次に、MALDI-TOFマトリックス(1:1エタノール/10% 蟻酸水溶液中、15 mg/mL 3,5-ジメトキシ-4-ヒドロキシケイ皮酸)を、高精度3軸ロボット(MicroGrid, BioRobotics)を用いて、金チップに均一に塗布した。マトリックスを384ウェルのマイクロタイタープレートからチップに移すのに16ピンのツールが使用され、600ミクロンのピッチを有する直径200ミクロンの形質が作られた。MALDI-TOF質量分析器(Voyager DE, PerSeptive Biosystems)の機器設定は以下の通りであった:加速電圧=25k、グリッド電圧=92%、ガイドワイヤ電圧=0.05%、遅延=200オン、レーザー出力=2400、低質量ゲート=1500、陰イオン=オフ。金チップはチップのレベルをステージのレベルと同じに保つように、改変されたMALDI試料ステージ上に1つずつ置かれ、適切なフライト距離を可能にした。装置のビデオモニターおよび運動制御システムを用いて、レーザービームが個々のマトリックス形質に向かうようにした。
【0153】
図14および図15は、それぞれ10Fn3融合体チップおよび非融合体チップの質量スペクトルを示す。いずれも、スペクトルを集めるために少数の200ミクロンの形質が分析されたが、図15のほうが、かなり多くのデータ収集を要した。17.4 kDaのシグナルはTNF-α単量体に相当する。
【0154】
固定化、標的タンパク質の捕捉、および蛍光検出
洗浄済の1×3インチの顕微鏡スライドグラス(Goldseal, #3010)を、ナノストップ(Nanostrip)(Cyantek)で15分間、10% NaOH水溶液、70℃で3分間、および1% HCl水溶液で1分間、各試薬の後に脱イオン水で十分にすすぎながら処理した。その後、スライドは無水硫酸カルシウム上で真空乾燥器中で数時間乾燥させた。95%アセトン/5%水中の1%アミノプロピルトリメトキシシランを調製し、20分間加水分解させた。スライドグラスは、穏やかに撹拌しながら加水分解したシラン溶液に5分間浸した。毎回新しい95%アセトン/5%水を用いて、スライドを5分間穏やかに撹拌しながら10回洗浄し、過剰なシランを除去した。その後、スライドを110℃で20分間加熱することにより、保存処理した。シラン処理したスライドは、新しく調製した90% DMF/10%ピリジン中の0.2%フェニレン1,4-ジイソチオシアネート溶液に、穏やかに撹拌しながら2時間浸した。その後、90% DMF/10%ピリジン、メタノール、およびアセトンで順次洗浄した。空気乾燥後、機能化したスライドは、真空乾燥器中で無水硫酸カルシウム上で0℃で保存した。市販のアミン反応性スライド(3-D Link, Surmodics)を用いて、同様な結果が得られた。
【0155】
オリゴヌクレオチド捕捉プローブは、従来のホスホラミダイド化学を用いて、自動DNA合成装置(PE BioSystems Expedite 8909)で調製された。全ての試薬はグレン・リサーチ(Glen Research)から入手した。合成は、保護されているアミノ官能性を直交して持つ固相支持体から開始し、それにより3'末端のアミンが最後の脱保護段階まで曝露されないようにした。添加される最初の4つの単量体は、ヘキサエチレンオキシドユニットで、標準的なA, G, CおよびT単量体が続いた。すべての捕捉オリゴ配列は、固相支持体から開裂され、水酸化アンモニウムで脱保護し、乾燥するまで濃縮し、エタノール沈殿し、かつトリエチルアンモニウム酢酸緩衝液中のアセトニトリル勾配を用いて、逆相HPLCで精製した。HPLCの適当な画分を回収し、真空遠心分離で乾燥するまで蒸発させ、水の一部とともに蒸発させた。
【0156】
精製したアミン標識捕捉オリゴは、10%グリセロールを含む50 mM炭酸ナトリウム緩衝液(pH 9.0)中で、250μMの濃度に調節した。プローブを3軸ロボット(MicroGrid, BioRobotics)を用いて、アミン反応性ガラス表面上の5×5×6アレイパターン中の規定の位置にスポットした。16ピンのツールを用いて、384ウェルのマイクロタイタープレートから液体を移し、600ミクロンのピッチを有する200ミクロンの形質を作製した。24の形質の各サブグリッドは、単一の捕捉プローブ(すなわち、24の重複するスポット)に相当する。アレイは湿度が飽和した環境中で、室温で12時間〜18時間インキュベートした。接着反応は、チップを2%水酸化アンモニウム水溶液中に穏やかに撹拌しながら5分間浸すことで停止させ、蒸留水ですすいだ(各5分3回)。最後にアレイを室温で30分間10×PBSに浸漬してから、蒸留水で5分すすいだ。
【0157】
10Fn3 mRNAと共に特異的かつ熱力学的に等エネルギーの配列が、自己集合および10Fn3タンパク質固定のための捕捉ポイントとして役立つことが明らかにされた。ソフトウェアプログラムHybSimulator v4.0 (Advanced Gene Computing Technology, Inc.)は、潜在的な捕捉プローブの同定および解析を容易にした。6つの固有の捕捉プローブが選択され、チップ上にプリントされ、そのうちの3つは10Fn3融合体プールのmRNAの共通領域(CP3', CP5'およびCPflag)に相補的である。残りの3つの配列(CPneg1, CPneg2およびCPneg3)は相補的ではなく、部分的に陰性対照として働く。上述のように、捕捉プローブは各々3'アミノ末端および4つのヘキサエチレンオキシドスペーサーユニットを持っている。以下は、使用した捕捉プローブの配列リストである(5'→3'):
ラウンド10のTNF-α選択由来の約1 pmolの10Fn3融合体プールを、総容量350μL中での0.02%トゥイーン20および2 mMバナジルリボヌクレオチド複合体を含む、5×SSCに調節した。400μLガスケット装置下で全体量をマイクロアレイに添加し、この集合を室温で18時間、連続的に回転させた。ハイブリダイゼーション後、スライドを500 mLの5×SSC、2.5×SSC、および1×SSCで順次5分ずつ洗浄した。液体の痕跡は遠心分離で除去し、スライドを空気乾燥させた。
【0158】
組換えヒトTNF-α(500μg、凍結乾燥、PreproTech由来)を230μLの1×PBSに入れ、Microdialyzerユニット(3,500 MWCO, Pierce)中で、700 mLの撹拌した1×PBSに対して4℃で18時間透析した。透析したTNF-αをEZ-Link NHS-LC-LCビオチン化試薬(20μg、Pierce)で0℃で2時間処理し、Microdialyzerユニット(3,500 MWCO, Pierce)中で、700 mLの撹拌した1×PBSに対して4℃で18時間再び透析した。得られた結合体をMALDI-TOF質量分析器によって分析し、単一のビオチン部分でほぼ完全に機能化されることが見出された。
【0159】
以下の過程は、各々4℃で連続回転または混合しながら行なった。タンパク質マイクロアレイ表面は、0.02%トゥイーン20および0.2% BSAを含む1×TBS(200μL)による60分間の処理によって、不動態化した。ビオチン化TNF-α(不動態化緩衝液中100 nM濃度に作製)を、マイクロアレイに120分間接触させた。マイクロアレイは0.02%トゥイーン20を含む1×TBSで洗浄した(50 mL3回、各5分間)。蛍光標識したストレプトアビジン(Molecular Probes由来の2.5μg/mL アレクサ 546-ストレプトアビジン結合体、不動態化緩衝液中に作製)をマイクロアレイに60分間接触させた。マイクロアレイを0.02%トゥイーン20を含む1×TBSで洗浄し(50 mL2回、各5分間)、続いて1×TBSで3分間すすいだ。液体の痕跡は遠心分離で除去し、スライドを室温で空気乾燥させた。
【0160】
蛍光レーザースキャニングは、GSI Lumonics ScanArray 5000システムで、10μMピクセルの解像度ならびにアレクサ 546色素用の予め設定された励起波長および発光波長を用いて行なった。リン光画像(phosphorimage)分析は、分子動態学ストーム(storm)システムを用いて行なった。マイクロアレイとリン光体保存スクリーンを直接接触させて、曝露時間は48時間であった。リン光画像スキャニングは50ミクロンの解像度の設定で行われ、データはImageQuant v.4.3ソフトウェアで抽出した。
【0161】
図16および図17は、同一アレイのリン光画像および蛍光スキャンである。リン光画像は、35Sメチオニンシグナルに基づいて、10Fn3融合体がどこにハイブリダイズしたかを示す。蛍光スキャンは、標識TNF-αがどこに結合したかを示す。
【0162】
他の態様
他の態様は、特許請求の範囲内である。
【0163】
本明細書で言及される全ての刊行物、特許、および特許出願は、参照として本明細書に組み入れられる。
【図面の簡単な説明】
【0164】
【図1】ラクダ(濃青)およびラマ(淡青)より得られた抗体の重鎖における可変領域の構造間の比較を2つの配向からそれぞれ示す写真である。
【図2】ラクダ抗体の重鎖の可変領域(濃青)、ラマ抗体の重鎖における可変領域(淡青)、および10番目のフィブロネクチン3型モジュール(10Fn3)(黄色)の構造間の比較を示す写真である。
【図3】10番目のフィブロネクチン3型モジュール(10Fn3)を示す写真であり、IgG重鎖における抗原結合ループに対応するループを赤色で強調した。
【図4】フィブロネクチン3型タンパク質ドメインおよび関連タンパク質ドメイン間の配列の整列化を図示するグラフである。
【図5】10Fn3ドメイン、ならびにフィブロネクチン、テネイシン、コラーゲン、およびアンジュリンを含む15個の関連タンパク質間の構造の類似性を示す写真である。この写真では、領域は以下のように分類されている:定常領域は濃青、保存領域は淡青、中性領域は白、可変領域は赤、およびRGDインテグリン結合領域(可変)は黄。
【図6】9番目および10番目のフィブロネクチン3型モジュールの空間充填モデルを2つの異なる配向からそれぞれ示す写真である。2つのモジュールおよびインテグリン結合ループ(RGD)が分類されている。この図では、青が正電荷を帯びた残基を示し、赤が負電荷を帯びた残基を示し、白が無電荷の残基を示している。
【図7】7〜10番目のフィブロネクチン3型モジュールの空間充填モデルを3つの異なる配向からそれぞれ示す写真である。4つのモジュールが分類されている。この図では、青が正電荷を帯びた残基を示し、赤が負電荷を帯びた残基を示し、白が無電荷の残基を示している。
【図8】フィブロネクチン3型ドメインを含むRNA-タンパク質融合体の異なる塩条件下における構造を図示する写真である。
【図9】PCRシグナル解析によって測定されたフィブロネクチン3型ドメインを含むRNA-タンパク質融合体の選択を図示する一連の写真である。
【図10】RNA-タンパク質融合体および遊離タンパク質の選択間における比較、ならびに本明細書に記載の選択間において、TNF-αとの結合%での増加を図示するグラフである。
【図11】IgG、10Fn3、Fn-CH1-CH2-CH3、およびFn-CH2-CH3を示す一連の概略図である(左上から時計回りに)。
【図12】IgG(X線結晶学)および10Fn3(NMRおよびX線結晶学)の既知の三次元構造に基づいたFn-CH1-CH2-CH3の分子モデルを示す写真である。
【図13】TNF-α結合物の10Fn3に基づく核酸-タンパク質融合体の例示的な選択の時間経過を示すグラフである。TNF-α-セファロースに結合した、核酸-タンパク質融合体のプール(白抜きのひし形)および遊離のタンパク質のプール(白抜きの丸)の割合、ならびに未誘導化セファロースに結合した遊離タンパク質のプール(黒丸)の割合が示されている。
【図14】TNF-αFn結合物によるTNF-α結合を示すグラフである。特に、これらの図はそれぞれ10Fn3融合体チップおよび非融合体チップから得られた質量スペクトルデータを示す。
【図15】TNF-αFn結合物によるTNF-α結合を示すグラフである。特に、これらの図はそれぞれ10Fn3融合体チップおよび非融合体チップから得られた質量スペクトルデータを示す。
【図16】それぞれ10Fn3アレイのリン光画像および蛍光スキャンであり、TNF-α結合を示す。
【図17】それぞれ10Fn3アレイのリン光画像および蛍光スキャンであり、TNF-α結合を示す。
【図18】ラマVHドメインおよび野生型ヒト10Fn3ドメインの一次配列の整列化を示すグラフである。2つの配列の間の相同残基が示されている。選択されたクローンの約45%で変異している無作為化ループの外の10Fn3残基は、野生型10Fn3配列の下の矢印で示されており、選択された残基を示す文字が付けられている。
【図19】ラマVHドメインおよび野生型ヒト10Fn3ドメインの模式図である。変異した枠組みの残基が示されている。
【図20】元のライブラリー(R0)およびTNF-αを用いた10回の選択後(R10)から翻訳された遊離タンパク質プールの結合効率および特異性を示すグラフである。未誘導化セファロース、TNF-α-セファロース、IL-1α-セファロース、およびIL-13-セファロースに対するタンパク質プールの結合が比較されている。
【図21】最大3個のループのディスプレイのための一連のIgG様骨格である。
【図22】最大4個、または6個のループのディスプレイのための一連のIgG様骨格である。
【図23】ループ構造のディスプレイのための、IgGに関連のない一連の骨格である。
【図24】固相表面に固定化された模倣物による標的(TNF-α)の特異的キャプチャを示す写真およびグラフである。
【図25−1】本発明の典型的なTNF-α結合体(配列番号:33〜140)を示すグラフである。
【図25−2】本発明の典型的なTNF-α結合体(配列番号:33〜140)を示すグラフである。
【図25−3】本発明の典型的なTNF-α結合体(配列番号:33〜140)を示すグラフである。
【図25−4】本発明の典型的なTNF-α結合体(配列番号:33〜140)を示すグラフである。
【特許請求の範囲】
【請求項1】
免疫グロブリン様の折畳みをもつドメインを含む非抗体誘導体タンパク質であって、変異したアミノ酸配列を持つことにより基準タンパク質から誘導され、該基準タンパク質がそれほど強く結合しない化合物に対して少なくとも10nMの強さのKdで結合する、非抗体誘導体タンパク質。
【請求項2】
少なくとも1 nMの強さ、少なくとも500 pMの強さ、少なくとも100 pMの強さ、または少なくとも20 pMの強さのKdで結合する、請求項1記載の誘導体タンパク質。
【請求項3】
1、2、または3個の変異したループを含み、該ループの少なくとも1個が化合物に対する誘導体タンパク質の結合に寄与する、請求項1記載の誘導体タンパク質。
【請求項4】
変異ループのうちの少なくとも2つ、または3つの変異ループが化合物に対する誘導体タンパク質の結合に寄与する、請求項3記載の誘導体タンパク質。
【請求項5】
基準タンパク質がジスルフィド結合を持たない、請求項1記載の誘導体タンパク質。
【請求項6】
誘導体タンパク質が少なくとも1つのジスルフィド結合を持つ、請求項1記載の誘導体タンパク質。
【請求項7】
免疫グロブリン様折畳みを持つドメインが、10 kD未満、7.5 kDを越える、または7.5〜10 kDの間の分子量を持つ、請求項1記載の誘導体タンパク質。
【請求項8】
生理的条件下でモノマーまたはダイマーである、請求項1記載の誘導体タンパク質。
【請求項9】
基準タンパク質が天然に存在する哺乳類タンパク質である、請求項1記載の誘導体タンパク質。
【請求項10】
免疫グロブリン様折畳みを持つドメインが、変異しており、基準タンパク質の免疫グロブリン様折畳みと比較して、34%までの変異アミノ酸を含む、請求項1記載の誘導体タンパク質。
【請求項11】
免疫グロブリン様折畳みを持つドメインが、約50〜150アミノ酸、好ましくは約50アミノ酸からなる、請求項1記載の誘導体タンパク質。
【請求項12】
固相支持体上に固定された、請求項1記載の誘導体タンパク質。
【請求項13】
固相支持体上に固定されたアレイの一部である、請求項12記載の誘導体タンパク質。
【請求項14】
固相支持体がチップまたはビーズである、請求項12記載の誘導体タンパク質。
【請求項15】
該化合物に結合できない異種タンパク質に結合した、請求項1記載の誘導体タンパク質。
【請求項16】
異種タンパク質が抗体または抗体ドメインであり、免疫グロブリンFcドメインを含むまたは補体タンパク質、毒素タンパク質、およびアルブミンタンパク質からなる群より選択される、請求項15記載の誘導体タンパク質。
【請求項17】
核酸に共有結合している、請求項1記載の誘導体タンパク質。
【請求項18】
核酸がそのタンパク質をコードしている、請求項1記載の誘導体タンパク質。
【請求項19】
フィブロネクチンまたはフィブロネクチン二量体、テナシン、N-カドヘリン、E-カドヘリン、ICAM、タイチン、GCSF受容体、サイトカイン受容体、グリコシダーゼ阻害剤、抗生物質色素蛋白質、ミエリン膜接着分子PO、CD8、CD4、CD2、クラスI MHC、T細胞抗原受容体、VCAM-1のCD1、C2およびI-setドメイン、ミオシン結合タンパク質CのI-set免疫グロブリンドメイン、ミオシン結合タンパク質HのI-set免疫グロブリンドメイン、テロキンのI-set免疫グロブリンドメイン、NCAM、トウィチン、ニューログリアン、成長ホルモン受容体、エリスロポエチン受容体、プロラクチン受容体、インターフェロンγ受容体、βガラクトシダーゼ/グルクロニダーゼ、βグルクロニダーゼ、トランスグルタミナーゼ、T細胞抗原受容体、スーパーオキシド・ジスムターゼ、組織因子ドメイン、シトクロームF,緑色蛍光タンパク質、GroEL、またはタウマチンから誘導される、請求項1記載の誘導体タンパク質。
【請求項20】
請求項1記載の誘導体タンパク質をコードする核酸。
【請求項21】
以下の段階を含む、化合物に結合する誘導非抗体タンパク質を得る方法:
(a) 免疫グロブリン様の折畳みを含む非抗体骨格タンパク質を提供する段階であって、該骨格タンパク質が10 nMの強さのKdで化合物に結合しない段階;
(b) 該非抗体骨格タンパク質の変異した誘導体を作製し、それによって変異タンパク質のライブラリーを構築する段階;
(c) 該化合物を該ライブラリーに接触させる段階;
(d) 少なくとも10 nMの強さのKdで該化合物に結合する少なくとも1つの誘導体タンパク質を該ライブラリーから選択する段階;および
(d) 選択的に、段階(b)の非抗体骨格タンパク質をその前の段階(d)の産物で置き換えて段階(b)〜(d)を繰り返す段階。
【請求項22】
以下の段階を含む、化合物に結合する非抗体タンパク質を得る方法:
(a) 候補タンパク質を該化合物に接触させる段階であって、該候補タンパク質は免疫グロブリン様の折畳みを持つドメインを含む誘導非抗体タンパク質であって、該非抗体タンパク質は変異したアミノ酸配列を持つことにより基準タンパク質から誘導され、該非抗体タンパク質は該基準タンパク質がそれほど強く結合しない化合物に少なくとも10 nMの強さのKdで化合物に結合し、該接触が化合物-タンパク質複合体の形成が可能な条件下で行われる段階;および
(b) 該複合体から該化合物に結合する該誘導体タンパク質を得る段階。
【請求項23】
以下の段階を含む、非抗体タンパク質に結合する化合物を得る方法であって、非抗体タンパク質は免疫グロブリン様の折畳みを持つドメインを含み、変異したアミノ酸配列を持つことで基準タンパク質から誘導され、該非抗体タンパク質は該基準タンパク質がそれほど強く結合しない化合物に少なくとも10 nMの強さのKdで化合物に結合する方法:
(a) 候補化合物を該誘導体タンパク質に接触させる段階であって、接触は化合物-タンパク質複合体の形成は可能な条件下で行われる;および
(b) 該複合体から、該誘導体タンパク質に結合する該化合物を得る段階。
【請求項24】
以下の段階を含む、試料中の化合物を検出する方法:
(a)免疫グロブリン様の折畳みを持つドメインを含む非抗体タンパク質を該試料に接触させる段階であって、該非抗体タンパク質は変異したアミノ酸配列を持つことによって基準タンパク質から誘導され、該非抗体タンパク質は該基準タンパク質がそれほど強く結合しない化合物に少なくとも10 nMの強さのKdで化合物に結合し、該接触が化合物-タンパク質複合体の形成が可能な条件下で行われる段階;および
(b) 該複合体を検出することによって該試料中の該化合物を検出する段階。
【請求項25】
腫瘍壊死因子α(TNF-α)受容体に対する同一性が20%未満の配列を持つ、少なくとも1μMの強さのKdでTNF-αに結合する非抗体タンパク質。
【請求項26】
DE、BCおよびFGループに変異を持つ、変異したフィブロネクチン3型ドメインを含む、請求項25記載の非抗体タンパク質。
【請求項27】
FGループが野生型のFGループと同じ長さである、請求項26記載の非抗体タンパク質。
【請求項28】
免疫グロブリン様の折畳みを含む、請求項25記載の非抗体タンパク質。
【請求項29】
免疫グロブリン様の折畳みが10 kD未満、7.5 kDを越える、または7.5〜10 kDの間の分子量を持つ、請求項28記載の非抗体タンパク質。
【請求項30】
免疫グロブリン様の折畳みが、約50〜150アミノ酸、および好ましくは約50アミノ酸からなる、請求項29記載の非抗体タンパク質。
【請求項31】
少なくとも500 nM、100 nM、10 nM、1 nM、500 pM、100 pM、または20 pMの強さのKdでTNF-αに結合する、請求項25記載の非抗体タンパク質。
【請求項32】
1個、2個、または3個の変異ループを含む、請求項25記載の非抗体タンパク質。
【請求項33】
少なくとも1個、および好ましくは2個または3個すべての変異ループがTNF-αに対する非抗体タンパク質の結合に寄与する、請求項32記載の非抗体タンパク質。
【請求項34】
少なくとも1つのジスルフィド結合を持つ、請求項25記載の非抗体タンパク質。
【請求項35】
生理的条件下で単量体または二量体である、請求項25記載の非抗体タンパク質。
【請求項36】
固相支持体上に固定されている、請求項25記載の非抗体タンパク質。
【請求項37】
固相支持体がチップまたはビーズである、請求項36記載の非抗体タンパク質。
【請求項38】
固相支持体上に固定されたアレイの一部である、請求項36記載の非抗体タンパク質。
【請求項39】
異種タンパク質に結合している、請求項25記載の非抗体タンパク質。
【請求項40】
異種タンパク質がTNF-α、免疫グロブリンFcドメイン、補体タンパク質、またはアルブミンタンパク質に結合しない抗体または抗体ドメインである、請求項39記載の非抗体タンパク質。
【請求項41】
変異したフィブロネクチン3型ドメインおよび好ましくは変異したヒトフィブロネクチン3型ドメインを含む、請求項25記載の非抗体タンパク質。
【請求項42】
変異したフィブロネクチン3型ドメインが、フィブロネクチン3型ドメインの変異した10番目のモジュール(10Fn3)である、請求項41記載の非抗体タンパク質。
【請求項43】
10Fn3インテグリン結合モチーフを持たない、請求項42記載の非抗体タンパク質。
【請求項44】
10Fn3のループ中に非天然の配列および好ましくはループ配列PW(A/G)を含み、および/または10Fn3のβシート中に非天然の配列を含む、請求項42記載の非抗体タンパク質。
【請求項45】
図25(配列番号:34〜140)の配列の任意の1つを含む、請求項25記載の非抗体タンパク質。
【請求項46】
請求項25記載の非抗体タンパク質をコードする核酸。
【請求項47】
ループが図25(配列番号:34〜140)のアミノ酸配列の任意の1つを含む、タンパク質のループ構造。
【請求項1】
免疫グロブリン様の折畳みをもつドメインを含む非抗体誘導体タンパク質であって、変異したアミノ酸配列を持つことにより基準タンパク質から誘導され、該基準タンパク質がそれほど強く結合しない化合物に対して少なくとも10nMの強さのKdで結合する、非抗体誘導体タンパク質。
【請求項2】
少なくとも1 nMの強さ、少なくとも500 pMの強さ、少なくとも100 pMの強さ、または少なくとも20 pMの強さのKdで結合する、請求項1記載の誘導体タンパク質。
【請求項3】
1、2、または3個の変異したループを含み、該ループの少なくとも1個が化合物に対する誘導体タンパク質の結合に寄与する、請求項1記載の誘導体タンパク質。
【請求項4】
変異ループのうちの少なくとも2つ、または3つの変異ループが化合物に対する誘導体タンパク質の結合に寄与する、請求項3記載の誘導体タンパク質。
【請求項5】
基準タンパク質がジスルフィド結合を持たない、請求項1記載の誘導体タンパク質。
【請求項6】
誘導体タンパク質が少なくとも1つのジスルフィド結合を持つ、請求項1記載の誘導体タンパク質。
【請求項7】
免疫グロブリン様折畳みを持つドメインが、10 kD未満、7.5 kDを越える、または7.5〜10 kDの間の分子量を持つ、請求項1記載の誘導体タンパク質。
【請求項8】
生理的条件下でモノマーまたはダイマーである、請求項1記載の誘導体タンパク質。
【請求項9】
基準タンパク質が天然に存在する哺乳類タンパク質である、請求項1記載の誘導体タンパク質。
【請求項10】
免疫グロブリン様折畳みを持つドメインが、変異しており、基準タンパク質の免疫グロブリン様折畳みと比較して、34%までの変異アミノ酸を含む、請求項1記載の誘導体タンパク質。
【請求項11】
免疫グロブリン様折畳みを持つドメインが、約50〜150アミノ酸、好ましくは約50アミノ酸からなる、請求項1記載の誘導体タンパク質。
【請求項12】
固相支持体上に固定された、請求項1記載の誘導体タンパク質。
【請求項13】
固相支持体上に固定されたアレイの一部である、請求項12記載の誘導体タンパク質。
【請求項14】
固相支持体がチップまたはビーズである、請求項12記載の誘導体タンパク質。
【請求項15】
該化合物に結合できない異種タンパク質に結合した、請求項1記載の誘導体タンパク質。
【請求項16】
異種タンパク質が抗体または抗体ドメインであり、免疫グロブリンFcドメインを含むまたは補体タンパク質、毒素タンパク質、およびアルブミンタンパク質からなる群より選択される、請求項15記載の誘導体タンパク質。
【請求項17】
核酸に共有結合している、請求項1記載の誘導体タンパク質。
【請求項18】
核酸がそのタンパク質をコードしている、請求項1記載の誘導体タンパク質。
【請求項19】
フィブロネクチンまたはフィブロネクチン二量体、テナシン、N-カドヘリン、E-カドヘリン、ICAM、タイチン、GCSF受容体、サイトカイン受容体、グリコシダーゼ阻害剤、抗生物質色素蛋白質、ミエリン膜接着分子PO、CD8、CD4、CD2、クラスI MHC、T細胞抗原受容体、VCAM-1のCD1、C2およびI-setドメイン、ミオシン結合タンパク質CのI-set免疫グロブリンドメイン、ミオシン結合タンパク質HのI-set免疫グロブリンドメイン、テロキンのI-set免疫グロブリンドメイン、NCAM、トウィチン、ニューログリアン、成長ホルモン受容体、エリスロポエチン受容体、プロラクチン受容体、インターフェロンγ受容体、βガラクトシダーゼ/グルクロニダーゼ、βグルクロニダーゼ、トランスグルタミナーゼ、T細胞抗原受容体、スーパーオキシド・ジスムターゼ、組織因子ドメイン、シトクロームF,緑色蛍光タンパク質、GroEL、またはタウマチンから誘導される、請求項1記載の誘導体タンパク質。
【請求項20】
請求項1記載の誘導体タンパク質をコードする核酸。
【請求項21】
以下の段階を含む、化合物に結合する誘導非抗体タンパク質を得る方法:
(a) 免疫グロブリン様の折畳みを含む非抗体骨格タンパク質を提供する段階であって、該骨格タンパク質が10 nMの強さのKdで化合物に結合しない段階;
(b) 該非抗体骨格タンパク質の変異した誘導体を作製し、それによって変異タンパク質のライブラリーを構築する段階;
(c) 該化合物を該ライブラリーに接触させる段階;
(d) 少なくとも10 nMの強さのKdで該化合物に結合する少なくとも1つの誘導体タンパク質を該ライブラリーから選択する段階;および
(d) 選択的に、段階(b)の非抗体骨格タンパク質をその前の段階(d)の産物で置き換えて段階(b)〜(d)を繰り返す段階。
【請求項22】
以下の段階を含む、化合物に結合する非抗体タンパク質を得る方法:
(a) 候補タンパク質を該化合物に接触させる段階であって、該候補タンパク質は免疫グロブリン様の折畳みを持つドメインを含む誘導非抗体タンパク質であって、該非抗体タンパク質は変異したアミノ酸配列を持つことにより基準タンパク質から誘導され、該非抗体タンパク質は該基準タンパク質がそれほど強く結合しない化合物に少なくとも10 nMの強さのKdで化合物に結合し、該接触が化合物-タンパク質複合体の形成が可能な条件下で行われる段階;および
(b) 該複合体から該化合物に結合する該誘導体タンパク質を得る段階。
【請求項23】
以下の段階を含む、非抗体タンパク質に結合する化合物を得る方法であって、非抗体タンパク質は免疫グロブリン様の折畳みを持つドメインを含み、変異したアミノ酸配列を持つことで基準タンパク質から誘導され、該非抗体タンパク質は該基準タンパク質がそれほど強く結合しない化合物に少なくとも10 nMの強さのKdで化合物に結合する方法:
(a) 候補化合物を該誘導体タンパク質に接触させる段階であって、接触は化合物-タンパク質複合体の形成は可能な条件下で行われる;および
(b) 該複合体から、該誘導体タンパク質に結合する該化合物を得る段階。
【請求項24】
以下の段階を含む、試料中の化合物を検出する方法:
(a)免疫グロブリン様の折畳みを持つドメインを含む非抗体タンパク質を該試料に接触させる段階であって、該非抗体タンパク質は変異したアミノ酸配列を持つことによって基準タンパク質から誘導され、該非抗体タンパク質は該基準タンパク質がそれほど強く結合しない化合物に少なくとも10 nMの強さのKdで化合物に結合し、該接触が化合物-タンパク質複合体の形成が可能な条件下で行われる段階;および
(b) 該複合体を検出することによって該試料中の該化合物を検出する段階。
【請求項25】
腫瘍壊死因子α(TNF-α)受容体に対する同一性が20%未満の配列を持つ、少なくとも1μMの強さのKdでTNF-αに結合する非抗体タンパク質。
【請求項26】
DE、BCおよびFGループに変異を持つ、変異したフィブロネクチン3型ドメインを含む、請求項25記載の非抗体タンパク質。
【請求項27】
FGループが野生型のFGループと同じ長さである、請求項26記載の非抗体タンパク質。
【請求項28】
免疫グロブリン様の折畳みを含む、請求項25記載の非抗体タンパク質。
【請求項29】
免疫グロブリン様の折畳みが10 kD未満、7.5 kDを越える、または7.5〜10 kDの間の分子量を持つ、請求項28記載の非抗体タンパク質。
【請求項30】
免疫グロブリン様の折畳みが、約50〜150アミノ酸、および好ましくは約50アミノ酸からなる、請求項29記載の非抗体タンパク質。
【請求項31】
少なくとも500 nM、100 nM、10 nM、1 nM、500 pM、100 pM、または20 pMの強さのKdでTNF-αに結合する、請求項25記載の非抗体タンパク質。
【請求項32】
1個、2個、または3個の変異ループを含む、請求項25記載の非抗体タンパク質。
【請求項33】
少なくとも1個、および好ましくは2個または3個すべての変異ループがTNF-αに対する非抗体タンパク質の結合に寄与する、請求項32記載の非抗体タンパク質。
【請求項34】
少なくとも1つのジスルフィド結合を持つ、請求項25記載の非抗体タンパク質。
【請求項35】
生理的条件下で単量体または二量体である、請求項25記載の非抗体タンパク質。
【請求項36】
固相支持体上に固定されている、請求項25記載の非抗体タンパク質。
【請求項37】
固相支持体がチップまたはビーズである、請求項36記載の非抗体タンパク質。
【請求項38】
固相支持体上に固定されたアレイの一部である、請求項36記載の非抗体タンパク質。
【請求項39】
異種タンパク質に結合している、請求項25記載の非抗体タンパク質。
【請求項40】
異種タンパク質がTNF-α、免疫グロブリンFcドメイン、補体タンパク質、またはアルブミンタンパク質に結合しない抗体または抗体ドメインである、請求項39記載の非抗体タンパク質。
【請求項41】
変異したフィブロネクチン3型ドメインおよび好ましくは変異したヒトフィブロネクチン3型ドメインを含む、請求項25記載の非抗体タンパク質。
【請求項42】
変異したフィブロネクチン3型ドメインが、フィブロネクチン3型ドメインの変異した10番目のモジュール(10Fn3)である、請求項41記載の非抗体タンパク質。
【請求項43】
10Fn3インテグリン結合モチーフを持たない、請求項42記載の非抗体タンパク質。
【請求項44】
10Fn3のループ中に非天然の配列および好ましくはループ配列PW(A/G)を含み、および/または10Fn3のβシート中に非天然の配列を含む、請求項42記載の非抗体タンパク質。
【請求項45】
図25(配列番号:34〜140)の配列の任意の1つを含む、請求項25記載の非抗体タンパク質。
【請求項46】
請求項25記載の非抗体タンパク質をコードする核酸。
【請求項47】
ループが図25(配列番号:34〜140)のアミノ酸配列の任意の1つを含む、タンパク質のループ構造。
【図1】


【図2】

【図3】

【図4】

【図5】
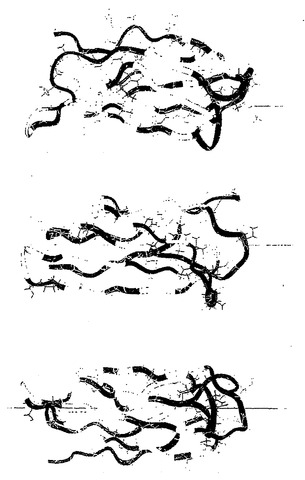
【図6】
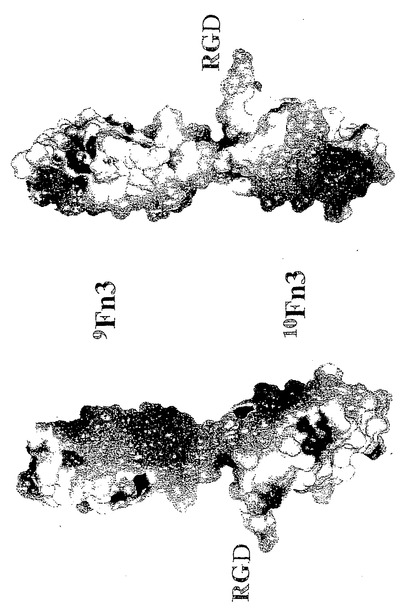
【図7】

【図8】
【図9】
【図10】
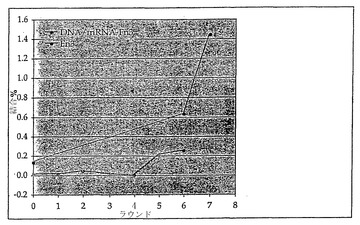
【図11】
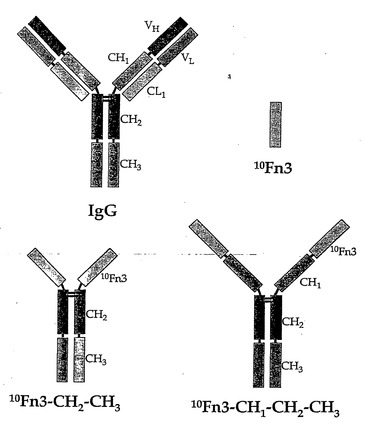
【図12】
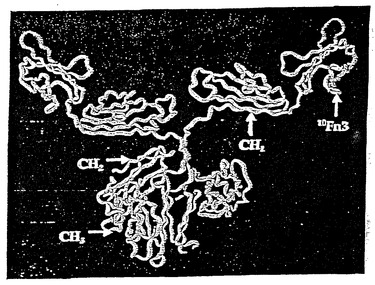
【図13】
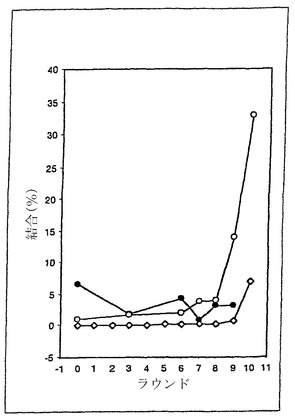
【図14】
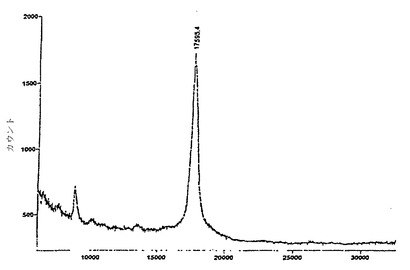
【図15】
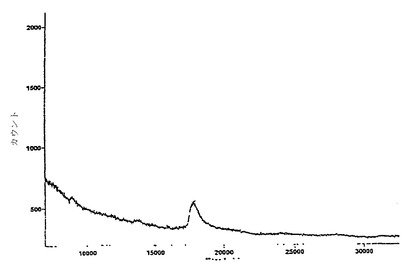
【図16】
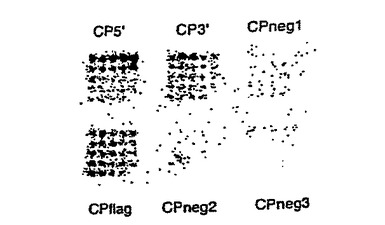
【図17】
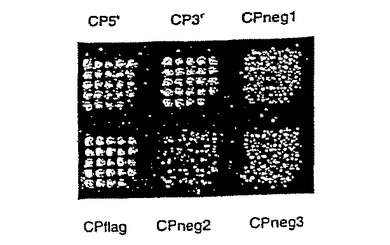
【図18】
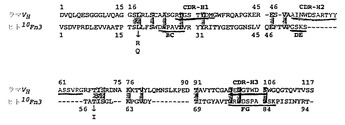
【図19】
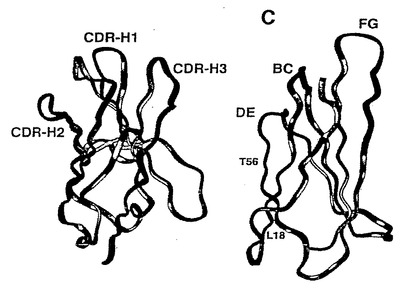
【図20】
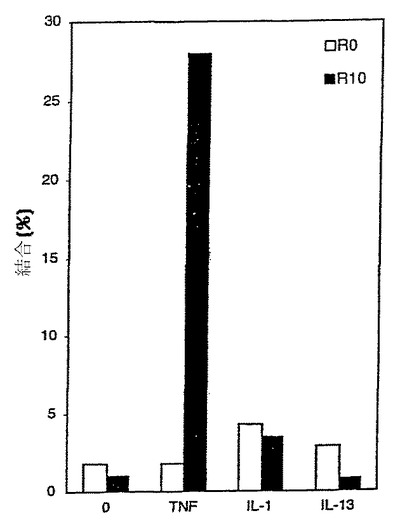
【図21】

【図22】

【図23】

【図24】
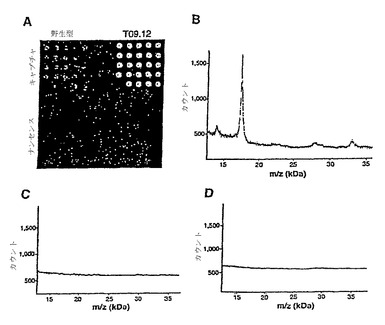
【図25−1】

【図25−2】

【図25−3】

【図25−4】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25−1】
【図25−2】
【図25−3】
【図25−4】
【公開番号】特開2009−183292(P2009−183292A)
【公開日】平成21年8月20日(2009.8.20)
【国際特許分類】
【出願番号】特願2009−78046(P2009−78046)
【出願日】平成21年3月27日(2009.3.27)
【分割の表示】特願2002−536306(P2002−536306)の分割
【原出願日】平成13年10月16日(2001.10.16)
【出願人】(509054371)ブリストル−マイヤーズ スクウィブ カンパニー (10)
【Fターム(参考)】
【公開日】平成21年8月20日(2009.8.20)
【国際特許分類】
【出願日】平成21年3月27日(2009.3.27)
【分割の表示】特願2002−536306(P2002−536306)の分割
【原出願日】平成13年10月16日(2001.10.16)
【出願人】(509054371)ブリストル−マイヤーズ スクウィブ カンパニー (10)
【Fターム(参考)】
[ Back to top ]