
抗体結合タンパク質およびその製造方法
【課題】 ヒト抗体に対する結合親和性が向上したタンパク質、およびその製造方法を提供すること。
【解決の手段】 ヒトFcγRI細胞外領域の全体配列または概ね全体配列を含んだポリペプチドを複数連結したタンパク質、前記タンパク質をコードするポリヌクレオチド、前記ポリヌクレオチドを含む発現ベクター、および前記ベクターで宿主を形質転換して得られる形質転換体を用いることで、ヒト抗体に対する結合親和性が向上したタンパク質を製造することができた。
【解決の手段】 ヒトFcγRI細胞外領域の全体配列または概ね全体配列を含んだポリペプチドを複数連結したタンパク質、前記タンパク質をコードするポリヌクレオチド、前記ポリヌクレオチドを含む発現ベクター、および前記ベクターで宿主を形質転換して得られる形質転換体を用いることで、ヒト抗体に対する結合親和性が向上したタンパク質を製造することができた。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒト抗体に対する結合親和性の高いタンパク質、および遺伝子工学的手法を用いた前記タンパク質の製造方法に関する。
【背景技術】
【0002】
Fcレセプターとは免疫グロブリン分子のFc領域に結合する一群のタンパク質分子である。個々の分子種は免疫グロブリンスーパーファミリーに属するFc認識ドメインによって、単一の、あるいは同じグループの免疫グロブリンイソタイプをFcレセプター上の結合ドメインによって認識する。これによって一定の免疫応答においてどのアクセサリー細胞が動員されるかが決まってくる(非特許文献1)。Fcレセプターは、さらにサブタイプに分類することができ、IgG(免疫グロブリンG)に対するレセプターはFcγRI、FcγRIIa、FcγRIIb、FcγRIIIの存在が報告されている(非特許文献1)。なかでも、FcγRIとIgGの結合親和性は高く、その平衡解離定数(KD)は10−8M以下である(非特許文献2)。FcγRIはIgG(免疫グロブリンG)に対するレセプターであり、単球とマクロファージ上に構成的に発現され、好中球および好酸球上においては誘導的に発現される。FcγRIは、細胞外領域、細胞膜貫通領域、細胞質内領域に区分され、IgGとの結合は、IgG分子Fc領域とFcγRIの細胞外領域で起こり、その後細胞質へとシグナルが伝達される。FcγRIはIgGとの結合に直接関わるα鎖と、γ鎖の2種類のサブユニットによって構成されており、γ鎖は細胞膜と細胞外領域の境界にあるアミノ酸システインを介した共有結合によりホモダイマーを形成している(非特許文献1)。
【0003】
近年になり、Fcレセプターの予想外の免疫抑制的な生物学的特性は、特に、自己免疫疾患または自己免疫症候群、移植物の拒絶および悪性リンパ増殖の領域において医薬として注目を浴びつつある(非特許文献2)。
【0004】
ヒトFcレセプターFcγRIα鎖のアミノ酸配列および遺伝子塩基配列は非特許文献3により明らかにされ、その後、遺伝子組換え技術により、大腸菌(特許文献1)あるいは動物細胞を利用した発現が報告されている(非特許文献4)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特表2002−531086号公報
【非特許文献】
【0006】
【非特許文献1】J.V.Ravetch等,Annu.Rev.Immunol.,9,457,1991
【非特許文献2】Toshiyuki Takai,Jpn.J.Clin.Immunol.,28,318,2005
【非特許文献3】J.M.Allen等,Science,243,378,1989
【非特許文献4】A.Paetz等,Biochem.Biophys.Res.Commun.,338,1811,2005
【非特許文献5】Yasukawa K.等、J.Biochem.,108,673,1990
【発明の概要】
【発明が解決しようとする課題】
【0007】
前述した通り、ヒトFcレセプターFcγRIのIgG分子Fc領域に対する結合親和性は平衡解離定数(KD)として10−8M以下と極めて高いものの、血液中に含まれるIgG分子を捕捉・定量するためには、分子あたりの抗体結合性をさらに向上させる必要があった。そこで本発明は、ヒトIgG抗体に対する結合親和性が向上したタンパク質、および前記タンパク質の製造方法を提供することを目的としている。
【課題を解決するための手段】
【0008】
上記課題を鑑みてなされた本発明は、以下の発明を包含する:
(1)配列番号1に記載のアミノ酸配列からなるヒトFcレセプターFcγRIのうち、細胞外領域の全体領域または概ね全体領域を含むポリペプチドを複数連結した、タンパク質。
【0009】
(2)配列番号4または5に記載のアミノ酸配列からなる、タンパク質。
【0010】
(3)(1)または(2)に記載のタンパク質をコードする、ポリヌクレオチド。
【0011】
(4)(3)に記載のポリヌクレオチドを含む、発現ベクター。
【0012】
(5)(4)に記載のベクターで宿主を形質転換して得られる、形質転換体。
【0013】
(6)(5)に記載の形質転換体を用いた、タンパク質の製造方法。
【0014】
以下、本発明について詳細に説明する。
【0015】
1.本発明のタンパク質
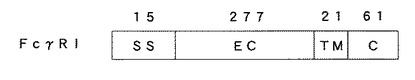
ヒトFcレセプターFcγRIタンパク質は図1に示すように、N末端側から15アミノ酸のシグナルペプチド領域(SS、配列番号1に記載のアミノ酸配列のうち−15番目から−1番目までの領域)、277アミノ酸の細胞外領域(EC、配列番号1に記載のアミノ酸配列のうち1番目から277番目までの領域)、21アミノ酸の細胞膜貫通領域(TM、配列番号1に記載のアミノ酸配列のうち278番目から298番目までの領域)、61アミノ酸の細胞内領域(C、配列番号1に記載のアミノ酸配列のうち299番目から359番目までの領域)からなる。このうち、IgG分子Fc領域と結合親和性を有するのは細胞外領域であり、当該領域でIgG抗体を直接捕捉している。
【0016】
本発明のタンパク質における、複数連結するポリペプチドは、配列番号1に記載のアミノ酸配列からなるヒトFcγRIのうち、細胞外領域の全体配列(1番目から277番目までの領域)、または細胞外領域の概ね全体配列を含んでいればよく、細胞外領域のN末端側にあるシグナル配列の全体または一部を含んでもよいし、細胞外領域のC末端側にある細胞膜貫通領域の一部を含んでもよい。なお、細胞外領域の概ね全体配列とは配列番号1に記載のアミノ酸配列からなるヒトFcγRIのうち、細胞外領域の全体配列(1番目から277番目までの領域)のN末端側および/またはC末端側にある1から10アミノ酸を削除した配列のことをいう。
【0017】
本発明のタンパク質における、ヒトFcγRI細胞外領域の全体配列または概ね全体配列を含んだポリペプチドの連結数は、タンパク質の分子量が概ね10万を超えると取り扱いが困難となることから、2つまたは3つが好ましい。また、ヒトFcγRI細胞外領域の全体配列または概ね全体配列を含んだポリペプチドを連結する際には、直接前記ポリペプチド同士を連結させてもよいし、IgG分子Fc領域に対する結合親和性、および菌体外に分泌可能なだけの可溶性を有する範囲で適宜リンカーペプチドを挿入してもよい。
【0018】
本発明のタンパク質の一態様として、N末端側から15アミノ酸のシグナルペプチド領域、1つ目の細胞外領域(配列番号1に記載のアミノ酸配列のうち1番目から274番目までの配列)、連結領域(2アミノ酸)、2つ目の細胞外領域(配列番号1に記載のアミノ酸配列のうち6番目から274番目までのアミノ酸)からなる配列番号4に記載のタンパク質をあげることができる。
【0019】
本発明のタンパク質の別の態様として、N末端側から15アミノ酸のシグナルペプチド領域、1つ目の細胞外領域(配列番号1に記載のアミノ酸配列のうち1番目から274番目までのアミノ酸)、連結領域(2アミノ酸)、2つ目の細胞外領域(配列番号1に記載のアミノ酸配列のうち6番目から274番目までのアミノ酸)、連結領域(2塩基)、3つ目の細胞外領域(2つ目の細胞外領域と同じアミノ酸)からなる配列番号5に記載のタンパク質をあげることができる。
【0020】
2.本発明のタンパク質をコードするポリヌクレオチド
本発明のタンパク質をコードするポリヌクレオチドは、配列番号1に記載のアミノ酸配列からなるヒトFcγRIのうち、細胞外領域の全体配列または概ね全体配列を含むポリペプチドを複数連結したタンパク質をコードするポリヌクレオチドのことをいう。なお、遺伝子組換え技術を用いて本発明のタンパク質を製造する場合は、前記タンパク質のアミノ酸配列からヌクレオチド配列に変換する際に、組換え対象宿主におけるコドンの使用頻度を考慮のうえ変換するのが好ましい。一例としてBrevibacillus choshinensisを宿主とする場合は、セリン(Ser)ではTCA、ロイシン(Leu)ではCTA、アルギニン(Arg)ではCGG/AGA/AGG、イソロイシン(Ile)ではATAへの変換をそれぞれ避けて変換するのが好ましい(特願2008−046438号参照)。
【0021】
本発明のタンパク質をコードするポリヌクレオチドの一態様として、配列番号4に記載のアミノ酸配列からヌクレオチド配列に変換した配列番号2に記載のポリヌクレオチド、および配列番号5に記載のアミノ酸配列からヌクレオチド配列に変換した配列番号3に記載のポリヌクレオチドをあげることができる。
【0022】
3.本発明のタンパク質製造方法
本発明のタンパク質の製造方法としては、配列番号1に記載のアミノ酸配列からなるヒトFcγRIから適当な試薬を用いて細胞外領域の全体配列または概ね全体配列を含んだポリペプチドを調製し、前記ペプチド同士を直接またはリンカーペプチドを介したペプチド結合により製造する方法、および本発明のタンパク質をコードするポリヌクレオチド配列を設計し、前記ポリヌクレオチドから遺伝子工学的手法を用いて製造する方法があげられるが、後者の製造方法が、本発明のタンパク質を簡便かつ大量に生産できる点で好ましい。以下、後者の製造方法について詳細に説明する。
【0023】
本発明のタンパク質を遺伝子工学的手法を用いて製造するために必要な、前記タンパク質をコードするポリヌクレオチドの取得方法としては、
(1)目的とする本発明のタンパク質のアミノ酸配列からヌクレオチド配列に変換し、前記ヌクレオチド配列を含むポリヌクレオチドを人工的に合成して取得する方法、
(2)ヒトFcγRI細胞外領域(配列番号1に記載のヌクレオチド配列のうち58番目から888番目までの配列)の全体配列または概ね全体配列を含むポリヌクレオチドを、直接人工的に、またはヒトFcγRIのcDNA等からPCR法といったDNA増幅法を用いて調製し、調製した前記ポリヌクレオチドを適当な方法で連結し取得する方法、
が例示できる。
【0024】
(1)の方法で取得する場合において、アミノ酸配列からヌクレオチド配列に変換する際は、前述したように、組換え対象宿主におけるコドンの使用頻度を考慮して変換するのが好ましい。(2)の方法で取得する場合において、配列番号1に記載のヒトFcγRIのヌクレオチド配列から直接人工的にヒトFcγRI細胞外領域の全体配列または概ね全体配列を含むポリヌクレオチドを調製する際は、組換え対象宿主におけるコドンの使用頻度を考慮のうえ、使用頻度が低いコドン、いわゆるレアコドン(rare codon)に該当する場合は、使用頻度の高いコドンにあらかじめ変換後、調製するのが好ましい(特願2008−046438号参照)。
【0025】
また、(1)、(2)いずれの方法で本発明のタンパク質をコードするポリヌクレオチドを取得する場合においても、ヒトFcγRI細胞外領域の全体配列または概ね全体配列を含んだポリペプチドをコードするポリヌクレオチド同士を、または前記ポリヌクレオチドに発現ベクターを連結させるための制限酵素認識配列、もしくは発現したヒトFcγRIの簡便な精製・定量を可能にするためのタグ配列を付加してもよい。前記付加する制限酵素認識配列としては、ベクターおよび連結するDNA断片のポリヌクレオチド配列を考慮のうえ適切な配列を選択すればよい。前記付加するタグ配列としては、遺伝子工学で多用される、ポリヒスチジンタグやc−mycタグといったオリゴペプチドをコードするヌクレオチド配列が例示できる。また、タグ配列を付加する箇所は前記ポリヌクレオチドの5’末端側に付加してもよいし、3’末端側に付加してもよい。
【0026】
本発明のタンパク質を遺伝子工学的手法を用いて製造するために使用する発現ベクターとしては、本発明のタンパク質をコードするポリヌクレオチド、前記ポリヌクレオチドを発現させるためのポリヌクレオチドおよび宿主中で発現ベクターを複製するための複製起点を有し、かつ、選定された宿主を形質転換できるものであれば、適宜選択し使用できるが、通常は取り扱いの容易な発現プラスミドを用いる。本発明のタンパク質をコードするポリヌクレオチドを発現させるためのポリヌクレオチドとしては、乳糖プロモータ系、トリプトファンプロモータ系、GAL4プロモータ系、SV40プロモータ系、アデノウイルスプロモータ系に由来するポリヌクレオチドが例示でき、宿主との関係において適宜選定すればよいが、宿主がCOS細胞やCHO細胞といった細胞の場合は、プロモーター系としてSV40プロモータ系に由来するポリヌクレオチドが好ましい。また、前記発現ベクターを用いて形質転換する操作にあたり、発現ベクターが導入されなかった宿主と導入された宿主との選別を可能にするために、発現ベクター中にアンピシリンといった薬剤に対する耐性を宿主に付与するためのポリヌクレオチド(薬剤耐性遺伝子)を含んでいるのが好ましい。
【0027】
本発明のタンパク質を遺伝子工学的手法を用いて製造するために使用する宿主としては、安定的にタンパク質を発現可能なCOS細胞やCHO細胞(チャイニーズハムスターの卵巣細胞)を例示することができる。また、前記細胞に、本発明のタンパク質をコードするポリヌクレオチドを含んだ発現プラスミドを導入し、さらに特定の薬剤に対する耐性も付与した細胞について、薬剤に対する耐性の強さから前記細胞のスクリーニングを行なうことで、本発明のタンパク質を高発現する細胞を得ることができる。前記スクリーニングにおいて、薬剤としてはメソトレキセート(MTX)、耐性濃度としては数μM以上がそれぞれ好ましい。MTXに対する耐性の付与方法は、本発明のタンパク質をコードするポリヌクレオチドを含む発現プラスミドにジヒドロ葉酸レダクターゼ遺伝子(dhfr)が含まれていればよい。
【0028】
本発明のタンパク質はヒトFcγRIの細胞外領域から構成されているため、生産に用いた培養液または培養上清から、通常の生理活性タンパク質回収法によって分離精製することができる。分離精製方法としては、市販のHPLCやカラムクロマトグラフィーを例示することができる。また、培養液中から前記タンパク質を生産する際に、培養液中の他のタンパク質の減少、例えば、宿主が細胞の場合は、培養細胞を無血清培地で培養して前記タンパク質を発現させることにより分離回収の効率を高めることができる。
【発明の効果】
【0029】
本発明のタンパク質は、配列番号1に記載のアミノ酸配列からなるヒトFcレセプターFcγRIのうちIgG抗体を直接捕捉する機能を有する細胞外領域の全体領域または概ね全体領域を含んだポリペプチドを、前記捕捉する機能を有したまま複数連結していることを特徴としており、配列番号1に記載のヒトFcγRI、およびヒトFcγRIの細胞外領域が1つのみからなるタンパク質と比較し、IgG抗体に対する親和性が向上している。このことは、個体発生および免疫機構の研究、および前記成果に基づく治療診断薬等の開発に大きな意義を持つ。また、抗FcγRI抗体を作製するための免疫源、およびFcγRIの免疫化学測定方法の標準物質として用いることもできる。
【0030】
また、本発明のタンパク質を適切なクロマトグラフィー担体(例えば、His・Bind Resinゲル(Novagen社製)、HisTrap NHS−Activated HP(GEヘルスケアバイオサイエンス社製)、トレシル基活性化ダイナビーズ(Dynal社製))に固定化することで、ヒトモノクローナル抗体といった抗体を精製するための吸着剤として用いることもできる。
【0031】
さらに、本発明のタンパク質をコードしたポリヌクレオチドを含む発現ベクター、および前記ベクターで宿主を形質転換して得られる形質転換体を用いて、本発明のタンパク質を製造することで、IgG抗体に対する結合親和性が向上したタンパク質を簡便かつ大量に製造することができる。
【図面の簡単な説明】
【0032】
【図1】ヒトFcレセプターFcγRIの構造を示す図。SSはシグナルペプチド、ECは細胞外領域、TMは細胞膜貫通領域、Cは細胞質内領域をそれぞれコードする領域を示す。図の上部の数字は、それぞれの領域のアミノ酸数を示す。
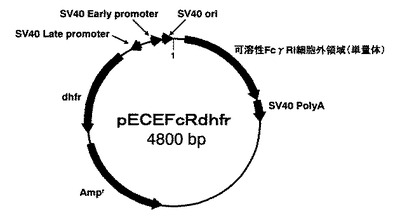
【図2】発現プラスミドpECEFcRdhfrの構造を示す図。
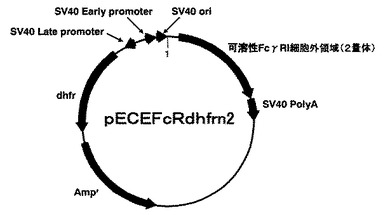
【図3】発現プラスミドpECEFcRdhfrn2の構造を示す図。
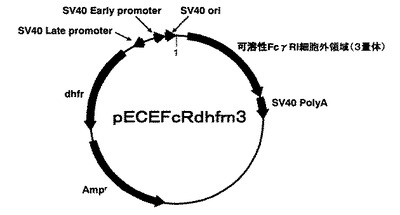
【図4】発現プラスミドpECEFcRdhfrn3の構造を示す図。
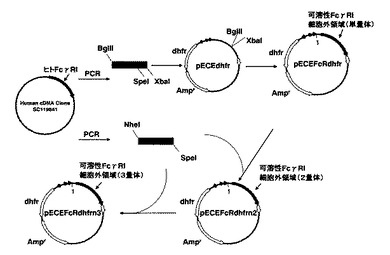
【図5】発現プラスミドの作製概略を示す図。
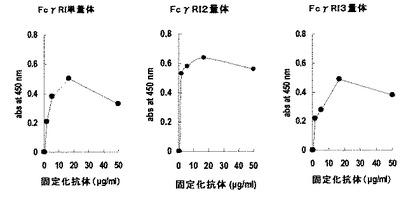
【図6】形質転換したCOS7培養上清の抗体結合活性を示す図。横軸は抗体固定化濃度、縦軸は450nmにおける吸光度を示す。
【実施例】
【0033】
以下、本発明をさらに詳細に説明するために実施例を示すが、本発明は実施例に限定されるものではない。
【0034】
実施例1 発現プラスミドの作製(その1)
ヒトFcレセプターFcγRI細胞外領域からなるタンパク質を発現可能なプラスミドの作製を、以下の方法で行なった。
(1)ヒトFcγRIをコードするポリヌクレオチドを以下の方法で調製した。
(1−1)Human cDNA clone SC119841プラスミド(Origene社製)をテンプレートとし、配列番号6(5’−GA[AGATCT]ATGTGGTTCTTGACAACTCTGCTCC−3’:角かっこ部分は制限酵素BglIIサイト)と配列番号7(5’−CG[TCTAGA]TCAGTGGTGGTGGTGGTGGTG[ACTAGT]GACAGGAGTTGGTAACTGGAGGCC−3’:角かっこ部分はそれぞれ制限酵素XbaI、SpeIサイト)のオリゴヌクレオチドをPCRプライマーとして使用した。なお、ヒトFcγRIの調製および定量を行なうために、発現させるタンパク質のC末端側にポリヒスチジンタグが付加されるようにPCRプライマーを設計した。
(1−2)PCR反応を、94℃・5分の熱処理後、94℃・30秒間の第一ステップ、65℃・30秒間の第二ステップ、72℃・1分間の第三ステップを25サイクル行ない、最後に、72℃・7分の条件で行なった。反応液組成を表1に示す。
【0035】
【表1】

【0036】
(1−3)PCR反応終了後、0.9%のアガロース電気泳動にて、設計通りのサイズに相当するバンドを確認した。
(1−4)目的産物に相当するバンドをアガロースゲルから抽出(QIAquick Gel extraction kit(商品名):キアゲン社製)し、ヒトFcγRIをコードするポリヌクレオチドを調製した。
(2)(1)で調製したポリヌクレオチドを制限酵素BglIIとXbaIにより消化し、これらの制限酵素により事前に消化したプラスミドpECEdhfr(非特許文献5)とライゲーションし(Ligation Kit Ver.2:タカラバイオ社製)、50μg/mLの抗生物質カルベシニリンを添加したLB寒天培地を用いて大腸菌JM109株を形質転換した。
(3)(2)の形質転換体を50μg/mLの抗生物質カルベシニリンを含むLB培地により培養(37℃、18時間)し、QIAprep Spin Miniprep Kit(商品名、キアゲン社製)によりプラスミドを抽出した。
(4)抽出プラスミドを制限酵素BglIIとXbaIにより消化し、0.9%のアガロース電気泳動に供した。
【0037】
結果、インサートサイズから設計通りであることを確認し、これをpECEFcRdhfrとした。図2にpECEFcRdhfrの構造概略を示す。
【0038】
実施例2 発現プラスミドの作製(その2)
実施例1で作製した発現プラスミドpECEFcRdhfr(図2)から、以下の方法で、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質を発現可能なプラスミドを作製した。
(1)ヒトFcγRI細胞外領域の概ね全体配列を機能的に連結するため、Human cDNA clone SC119841プラスミド(Origene社製)をテンプレートとし、配列番号8(5’−CG[GCTAGC]GCAGTGATCACTTTGCAGCCTCCATTG−3’:角かっこ部分は制限酵素NheIサイト)と配列番号9(5’−GG[ACTAGT]GACAGGAGTTGGTAACTGGAGGC−3’:角かっこ部分は、制限酵素SpeIサイト)のオリゴヌクレオチドをPCRプライマーとして使用しPCR反応によりポリヌクレオチドを増幅した。なお、ヒトFcγRIの調製および定量を行なうために、発現させるタンパク質のC末端側にポリヒスチジンタグが付加されるようにPCRプライマーを設計した。また、PCR反応の反応条件は実施例1と同様にして行なった。
(2)(1)で調製したポリヌクレオチドを、制限酵素NheIとSpeIにより消化し、事前に制限酵素SpeIにより消化したpECEFcRdhfr(図2)に挿入した。
(3)実施例1と同様の方法で大腸菌JM109株に形質転換し、前記形質転換された大腸菌株からプラスミドを調製することで、ヒトFcγRI細胞外領域の概ね全体配列を含むポリヌクレオチドを2つ連結した発現プラスミドpECFFcRdhfrn2を作製した。図3にpECEFcRdhfrn2の構造概略を示す。
(4)さらに(1)で調製したポリヌクレオチドを、事前に制限酵素SpeIで消化したpECEFcRdhfrn2(図3)に挿入することにより、ヒトFcγRI細胞外領域の概ね全体配列を含むポリヌクレオチドを3つ連結した発現プラスミドpECEFcEdhfrn3を作製した。図4にpECEFcEdhfrn3の構造概略を示す。また、pECEFcRdhfrn3までの作製手順を図5に示す。
【0039】
なお、本実施例と同様な方法で、ヒトFcレセプターFcγRI細胞外領域の概ね全体配列を含むポリヌクレオチドを4つ以上連結した発現プラスミドの作製を試みたものの、前記プラスミドを作製することはできなかった。
【0040】
実施例3 発現プラスミドの塩基配列確認
実施例1および2で作製したpECEFcRdhfr(図2)、pECEFcRdhfrn2(図3)、pECEFcRdhfrn3(図4)に挿入されている、ヒトFcγRIをコードするヌクレオチド配列をチェーンターミネータ法に基づくBig Dye Terminator v3.1 Cycle Sequencing kit(商品名、PEアプライドバイオシステム社製)を用いてサイクルシークエンス反応に供し、全自動DNAシークエンサーABI Prism 310 DNA analyzer(PEアプライドバイオシステム社製)において解析した。なお、シークエンス用プライマーとして、配列番号10(5’−AGCTGTATGGGGTCACTTCG−3’)と配列番号11(5’−TTTTTCCACTGGAATTCTAACC−3’)と配列番号12(5’−AGTGGGGATGTCACAGATGC−3’)と配列番号13(5’−AGGAACACATCCTCTGAATACC−3’)に示すオリゴヌクレオチドを使用した。
【0041】
解析の結果、pECEFcRdhfr、pECEFcRdhfrn2、pECEFcRdhfrn3に挿入されている、ヒトFcγRIをコードするヌクレオチド配列はいずれも設計通りであることを確認した。ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを2つ連結したタンパク質をコードするヌクレオチド配列を配列番号2(pECEFcRdhfrn2)に、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを3つ連結したタンパク質をコードするヌクレオチド配列を配列番号3(pECEFcRdhfrn3)に、それぞれ示す。
【0042】
実施例4 発現したタンパク質の抗体結合活性の確認
実施例1および2で作製したpECEFcRdhfr、pECEFcRdhfrn2およびpECEFcRdhfrn3プラスミドをリポフェクトアミン(インビトロジェン社製)によりCOS7細胞にそれぞれ導入し、10% FCSを含むD−MEM培地(GIBCO社製)で培養することで(37℃、3日間)、一過的に細胞外へタンパク質を発現させ、培養後、培養液を回収した。回収した培養上清の抗体結合活性を以下の方法でELISA法により評価した。
(1)96穴のELISAプレート(Nunc社製)に100μg/mLから段階的に希釈したガンマグロブリン製剤(化学及血清療法研究所製)を各ウェルに100μLずつ添加し、4℃で18時間静置することによりガンマグロブリンを固相に固定化した。
(2)TBS緩衝液(0.2%(w/v)Tween 20、150mM NaClを含むTris−HCl緩衝液(pH8.0))で洗浄後、Starting Block Blocking Buffers(PIERCE社製)によりブロッキング操作を行なった。
(3)TBS緩衝液で洗浄後、調製した培養上清を100μL添加し、(1)で固定化したヒトガンマグロブリンと反応させた(30℃、2時間)。
(4)(3)の反応終了後、TBS緩衝液で洗浄し、His−probe(H−15)HRP抗体(Santa Cruz Biotechnology社製)を添加した。
(5)(4)の反応終了後、TBS緩衝液で洗浄し、TMB Peroxidase Substrate(KPL社製)を添加し450nmの吸光度を測定した。
【0043】
評価結果を図6に示す。図6の通り、pECEFcRdhfr(図6に記載の単量体)、pECEFcRdhfrn2(図6に記載の2量体)、pECEFcRdhfrn3(図6に記載の3量体)により形質転換されたCOS7細胞の培養上清にいずれも抗体結合活性が検出された。すなわち、pECEFcRdhfrn2、pECEFcRdhfrn3に挿入された、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質をコードするポリヌクレオチドにより、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質を得ることができ、かつ、当該タンパク質がヒト抗体への結合活性を保持していることを示している。なお、対照として、pECEFcRdhfrにより形質転換したCOS7培養液を使用した。
【0044】
実施例5 発現したタンパク質の調製
以下の方法で、実施例4で得られた培養上清から、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質を分離精製した。
(1)実施例4で得られた、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質が含まれた培養上清約1Lを、20mM酢酸緩衝液(pH5.2)で透析し脱塩処理を施した。
(2)20mM酢酸緩衝液(pH5.2)で平衡化した300mLのStreamline SPゲル(商品名、GEヘルスケアバイオサイエンス社製)を備えた吸着流動床システム(GEヘルスケアバイオサイエンス社製)に添加し、同緩衝液で洗浄後、10%グリセロール、1M NaClを含む20mMリン酸緩衝液(pH7.4)により溶出し、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチド複数連結したタンパク質が含まれた濃縮画分を調製した。
(3)(2)で得られた濃縮画分を、20mM イミダゾール、0.5M NaClを含む20mM リン酸緩衝液(pH7.4)に透析し、同緩衝液で平衡化したHisTrap HSカラム(商品名、GEヘルスケアバイオサイエンス社製)に添加し、洗浄後、500mM イミダゾール、0.5M NaClを含む20mM リン酸緩衝液(pH7.4)で溶出することで、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質精製標品約500μg得た。なお、タンパク質濃度の定量は、ウシガンマグロブリンを標準タンパク質としてブラッドフォード法に基づくプロテインアッセイキット(Bio−Rad社製)を用いて行なった。なお、対照として、pECEFcRdhfrにより形質転換したCOS7細胞より調製した培養液から、上記方法と同様の方法でヒトFcγRI細胞外領域を1つのみ含むタンパク質を調製している。
【0045】
実施例6 発現したタンパク質の表面プラズモン共鳴解析
実施例5で作製した精製タンパク質の抗体結合特性を表面プラズモン共鳴解析装置BIAcore T100(商品名、GEヘルスケアバイオサイエンス社製)を用いて解析した。
(1)ヒト抗体のチップ固定
ヒト抗体IgG1(コスモバイオ社製)を、ランニング緩衝液HBS−EPを用いて終濃度約5μg/mLとなるように希釈した。次いで、BIAcore T100の取扱説明書に従いセンサーチップCM5(GEヘルスケアバイオサイエンス社製)に390RU固定した。
(2)精製FcγRIのヒト抗体IgG1の結合解析
抗体との結合解析を行なうために、ランニング緩衝液で任意に希釈した、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質を、BIAcore T100に供した。抗体との結合性(RU)は希釈したタンパク質濃度に従い変化した。この結果を用いて、上記解析装置の専用ソフトであるBIAcore T100 Evaluation Softwareにより前記タンパク質のヒト抗体への反応特性を解析した。
【0046】
解析結果を表2に示す。表2の通り、今回作製したヒトFcγRI細胞外領域の概ね全体配列を複数連結したタンパク質は、細胞外領域を1つのみ含むヒトFcγRI(表2に記載の単量体)と比較し、ヒト抗体に対する結合親和性が向上しており、平衡解離定数の比較で、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを2つ連結したタンパク質(表2に記載の2量体)で単量体の約16倍、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを3つ連結したタンパク質(表2に記載の3量体)で単量体の約85倍、それぞれ向上していた。
【0047】
【表2】
【技術分野】
【0001】
本発明は、ヒト抗体に対する結合親和性の高いタンパク質、および遺伝子工学的手法を用いた前記タンパク質の製造方法に関する。
【背景技術】
【0002】
Fcレセプターとは免疫グロブリン分子のFc領域に結合する一群のタンパク質分子である。個々の分子種は免疫グロブリンスーパーファミリーに属するFc認識ドメインによって、単一の、あるいは同じグループの免疫グロブリンイソタイプをFcレセプター上の結合ドメインによって認識する。これによって一定の免疫応答においてどのアクセサリー細胞が動員されるかが決まってくる(非特許文献1)。Fcレセプターは、さらにサブタイプに分類することができ、IgG(免疫グロブリンG)に対するレセプターはFcγRI、FcγRIIa、FcγRIIb、FcγRIIIの存在が報告されている(非特許文献1)。なかでも、FcγRIとIgGの結合親和性は高く、その平衡解離定数(KD)は10−8M以下である(非特許文献2)。FcγRIはIgG(免疫グロブリンG)に対するレセプターであり、単球とマクロファージ上に構成的に発現され、好中球および好酸球上においては誘導的に発現される。FcγRIは、細胞外領域、細胞膜貫通領域、細胞質内領域に区分され、IgGとの結合は、IgG分子Fc領域とFcγRIの細胞外領域で起こり、その後細胞質へとシグナルが伝達される。FcγRIはIgGとの結合に直接関わるα鎖と、γ鎖の2種類のサブユニットによって構成されており、γ鎖は細胞膜と細胞外領域の境界にあるアミノ酸システインを介した共有結合によりホモダイマーを形成している(非特許文献1)。
【0003】
近年になり、Fcレセプターの予想外の免疫抑制的な生物学的特性は、特に、自己免疫疾患または自己免疫症候群、移植物の拒絶および悪性リンパ増殖の領域において医薬として注目を浴びつつある(非特許文献2)。
【0004】
ヒトFcレセプターFcγRIα鎖のアミノ酸配列および遺伝子塩基配列は非特許文献3により明らかにされ、その後、遺伝子組換え技術により、大腸菌(特許文献1)あるいは動物細胞を利用した発現が報告されている(非特許文献4)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特表2002−531086号公報
【非特許文献】
【0006】
【非特許文献1】J.V.Ravetch等,Annu.Rev.Immunol.,9,457,1991
【非特許文献2】Toshiyuki Takai,Jpn.J.Clin.Immunol.,28,318,2005
【非特許文献3】J.M.Allen等,Science,243,378,1989
【非特許文献4】A.Paetz等,Biochem.Biophys.Res.Commun.,338,1811,2005
【非特許文献5】Yasukawa K.等、J.Biochem.,108,673,1990
【発明の概要】
【発明が解決しようとする課題】
【0007】
前述した通り、ヒトFcレセプターFcγRIのIgG分子Fc領域に対する結合親和性は平衡解離定数(KD)として10−8M以下と極めて高いものの、血液中に含まれるIgG分子を捕捉・定量するためには、分子あたりの抗体結合性をさらに向上させる必要があった。そこで本発明は、ヒトIgG抗体に対する結合親和性が向上したタンパク質、および前記タンパク質の製造方法を提供することを目的としている。
【課題を解決するための手段】
【0008】
上記課題を鑑みてなされた本発明は、以下の発明を包含する:
(1)配列番号1に記載のアミノ酸配列からなるヒトFcレセプターFcγRIのうち、細胞外領域の全体領域または概ね全体領域を含むポリペプチドを複数連結した、タンパク質。
【0009】
(2)配列番号4または5に記載のアミノ酸配列からなる、タンパク質。
【0010】
(3)(1)または(2)に記載のタンパク質をコードする、ポリヌクレオチド。
【0011】
(4)(3)に記載のポリヌクレオチドを含む、発現ベクター。
【0012】
(5)(4)に記載のベクターで宿主を形質転換して得られる、形質転換体。
【0013】
(6)(5)に記載の形質転換体を用いた、タンパク質の製造方法。
【0014】
以下、本発明について詳細に説明する。
【0015】
1.本発明のタンパク質
ヒトFcレセプターFcγRIタンパク質は図1に示すように、N末端側から15アミノ酸のシグナルペプチド領域(SS、配列番号1に記載のアミノ酸配列のうち−15番目から−1番目までの領域)、277アミノ酸の細胞外領域(EC、配列番号1に記載のアミノ酸配列のうち1番目から277番目までの領域)、21アミノ酸の細胞膜貫通領域(TM、配列番号1に記載のアミノ酸配列のうち278番目から298番目までの領域)、61アミノ酸の細胞内領域(C、配列番号1に記載のアミノ酸配列のうち299番目から359番目までの領域)からなる。このうち、IgG分子Fc領域と結合親和性を有するのは細胞外領域であり、当該領域でIgG抗体を直接捕捉している。
【0016】
本発明のタンパク質における、複数連結するポリペプチドは、配列番号1に記載のアミノ酸配列からなるヒトFcγRIのうち、細胞外領域の全体配列(1番目から277番目までの領域)、または細胞外領域の概ね全体配列を含んでいればよく、細胞外領域のN末端側にあるシグナル配列の全体または一部を含んでもよいし、細胞外領域のC末端側にある細胞膜貫通領域の一部を含んでもよい。なお、細胞外領域の概ね全体配列とは配列番号1に記載のアミノ酸配列からなるヒトFcγRIのうち、細胞外領域の全体配列(1番目から277番目までの領域)のN末端側および/またはC末端側にある1から10アミノ酸を削除した配列のことをいう。
【0017】
本発明のタンパク質における、ヒトFcγRI細胞外領域の全体配列または概ね全体配列を含んだポリペプチドの連結数は、タンパク質の分子量が概ね10万を超えると取り扱いが困難となることから、2つまたは3つが好ましい。また、ヒトFcγRI細胞外領域の全体配列または概ね全体配列を含んだポリペプチドを連結する際には、直接前記ポリペプチド同士を連結させてもよいし、IgG分子Fc領域に対する結合親和性、および菌体外に分泌可能なだけの可溶性を有する範囲で適宜リンカーペプチドを挿入してもよい。
【0018】
本発明のタンパク質の一態様として、N末端側から15アミノ酸のシグナルペプチド領域、1つ目の細胞外領域(配列番号1に記載のアミノ酸配列のうち1番目から274番目までの配列)、連結領域(2アミノ酸)、2つ目の細胞外領域(配列番号1に記載のアミノ酸配列のうち6番目から274番目までのアミノ酸)からなる配列番号4に記載のタンパク質をあげることができる。
【0019】
本発明のタンパク質の別の態様として、N末端側から15アミノ酸のシグナルペプチド領域、1つ目の細胞外領域(配列番号1に記載のアミノ酸配列のうち1番目から274番目までのアミノ酸)、連結領域(2アミノ酸)、2つ目の細胞外領域(配列番号1に記載のアミノ酸配列のうち6番目から274番目までのアミノ酸)、連結領域(2塩基)、3つ目の細胞外領域(2つ目の細胞外領域と同じアミノ酸)からなる配列番号5に記載のタンパク質をあげることができる。
【0020】
2.本発明のタンパク質をコードするポリヌクレオチド
本発明のタンパク質をコードするポリヌクレオチドは、配列番号1に記載のアミノ酸配列からなるヒトFcγRIのうち、細胞外領域の全体配列または概ね全体配列を含むポリペプチドを複数連結したタンパク質をコードするポリヌクレオチドのことをいう。なお、遺伝子組換え技術を用いて本発明のタンパク質を製造する場合は、前記タンパク質のアミノ酸配列からヌクレオチド配列に変換する際に、組換え対象宿主におけるコドンの使用頻度を考慮のうえ変換するのが好ましい。一例としてBrevibacillus choshinensisを宿主とする場合は、セリン(Ser)ではTCA、ロイシン(Leu)ではCTA、アルギニン(Arg)ではCGG/AGA/AGG、イソロイシン(Ile)ではATAへの変換をそれぞれ避けて変換するのが好ましい(特願2008−046438号参照)。
【0021】
本発明のタンパク質をコードするポリヌクレオチドの一態様として、配列番号4に記載のアミノ酸配列からヌクレオチド配列に変換した配列番号2に記載のポリヌクレオチド、および配列番号5に記載のアミノ酸配列からヌクレオチド配列に変換した配列番号3に記載のポリヌクレオチドをあげることができる。
【0022】
3.本発明のタンパク質製造方法
本発明のタンパク質の製造方法としては、配列番号1に記載のアミノ酸配列からなるヒトFcγRIから適当な試薬を用いて細胞外領域の全体配列または概ね全体配列を含んだポリペプチドを調製し、前記ペプチド同士を直接またはリンカーペプチドを介したペプチド結合により製造する方法、および本発明のタンパク質をコードするポリヌクレオチド配列を設計し、前記ポリヌクレオチドから遺伝子工学的手法を用いて製造する方法があげられるが、後者の製造方法が、本発明のタンパク質を簡便かつ大量に生産できる点で好ましい。以下、後者の製造方法について詳細に説明する。
【0023】
本発明のタンパク質を遺伝子工学的手法を用いて製造するために必要な、前記タンパク質をコードするポリヌクレオチドの取得方法としては、
(1)目的とする本発明のタンパク質のアミノ酸配列からヌクレオチド配列に変換し、前記ヌクレオチド配列を含むポリヌクレオチドを人工的に合成して取得する方法、
(2)ヒトFcγRI細胞外領域(配列番号1に記載のヌクレオチド配列のうち58番目から888番目までの配列)の全体配列または概ね全体配列を含むポリヌクレオチドを、直接人工的に、またはヒトFcγRIのcDNA等からPCR法といったDNA増幅法を用いて調製し、調製した前記ポリヌクレオチドを適当な方法で連結し取得する方法、
が例示できる。
【0024】
(1)の方法で取得する場合において、アミノ酸配列からヌクレオチド配列に変換する際は、前述したように、組換え対象宿主におけるコドンの使用頻度を考慮して変換するのが好ましい。(2)の方法で取得する場合において、配列番号1に記載のヒトFcγRIのヌクレオチド配列から直接人工的にヒトFcγRI細胞外領域の全体配列または概ね全体配列を含むポリヌクレオチドを調製する際は、組換え対象宿主におけるコドンの使用頻度を考慮のうえ、使用頻度が低いコドン、いわゆるレアコドン(rare codon)に該当する場合は、使用頻度の高いコドンにあらかじめ変換後、調製するのが好ましい(特願2008−046438号参照)。
【0025】
また、(1)、(2)いずれの方法で本発明のタンパク質をコードするポリヌクレオチドを取得する場合においても、ヒトFcγRI細胞外領域の全体配列または概ね全体配列を含んだポリペプチドをコードするポリヌクレオチド同士を、または前記ポリヌクレオチドに発現ベクターを連結させるための制限酵素認識配列、もしくは発現したヒトFcγRIの簡便な精製・定量を可能にするためのタグ配列を付加してもよい。前記付加する制限酵素認識配列としては、ベクターおよび連結するDNA断片のポリヌクレオチド配列を考慮のうえ適切な配列を選択すればよい。前記付加するタグ配列としては、遺伝子工学で多用される、ポリヒスチジンタグやc−mycタグといったオリゴペプチドをコードするヌクレオチド配列が例示できる。また、タグ配列を付加する箇所は前記ポリヌクレオチドの5’末端側に付加してもよいし、3’末端側に付加してもよい。
【0026】
本発明のタンパク質を遺伝子工学的手法を用いて製造するために使用する発現ベクターとしては、本発明のタンパク質をコードするポリヌクレオチド、前記ポリヌクレオチドを発現させるためのポリヌクレオチドおよび宿主中で発現ベクターを複製するための複製起点を有し、かつ、選定された宿主を形質転換できるものであれば、適宜選択し使用できるが、通常は取り扱いの容易な発現プラスミドを用いる。本発明のタンパク質をコードするポリヌクレオチドを発現させるためのポリヌクレオチドとしては、乳糖プロモータ系、トリプトファンプロモータ系、GAL4プロモータ系、SV40プロモータ系、アデノウイルスプロモータ系に由来するポリヌクレオチドが例示でき、宿主との関係において適宜選定すればよいが、宿主がCOS細胞やCHO細胞といった細胞の場合は、プロモーター系としてSV40プロモータ系に由来するポリヌクレオチドが好ましい。また、前記発現ベクターを用いて形質転換する操作にあたり、発現ベクターが導入されなかった宿主と導入された宿主との選別を可能にするために、発現ベクター中にアンピシリンといった薬剤に対する耐性を宿主に付与するためのポリヌクレオチド(薬剤耐性遺伝子)を含んでいるのが好ましい。
【0027】
本発明のタンパク質を遺伝子工学的手法を用いて製造するために使用する宿主としては、安定的にタンパク質を発現可能なCOS細胞やCHO細胞(チャイニーズハムスターの卵巣細胞)を例示することができる。また、前記細胞に、本発明のタンパク質をコードするポリヌクレオチドを含んだ発現プラスミドを導入し、さらに特定の薬剤に対する耐性も付与した細胞について、薬剤に対する耐性の強さから前記細胞のスクリーニングを行なうことで、本発明のタンパク質を高発現する細胞を得ることができる。前記スクリーニングにおいて、薬剤としてはメソトレキセート(MTX)、耐性濃度としては数μM以上がそれぞれ好ましい。MTXに対する耐性の付与方法は、本発明のタンパク質をコードするポリヌクレオチドを含む発現プラスミドにジヒドロ葉酸レダクターゼ遺伝子(dhfr)が含まれていればよい。
【0028】
本発明のタンパク質はヒトFcγRIの細胞外領域から構成されているため、生産に用いた培養液または培養上清から、通常の生理活性タンパク質回収法によって分離精製することができる。分離精製方法としては、市販のHPLCやカラムクロマトグラフィーを例示することができる。また、培養液中から前記タンパク質を生産する際に、培養液中の他のタンパク質の減少、例えば、宿主が細胞の場合は、培養細胞を無血清培地で培養して前記タンパク質を発現させることにより分離回収の効率を高めることができる。
【発明の効果】
【0029】
本発明のタンパク質は、配列番号1に記載のアミノ酸配列からなるヒトFcレセプターFcγRIのうちIgG抗体を直接捕捉する機能を有する細胞外領域の全体領域または概ね全体領域を含んだポリペプチドを、前記捕捉する機能を有したまま複数連結していることを特徴としており、配列番号1に記載のヒトFcγRI、およびヒトFcγRIの細胞外領域が1つのみからなるタンパク質と比較し、IgG抗体に対する親和性が向上している。このことは、個体発生および免疫機構の研究、および前記成果に基づく治療診断薬等の開発に大きな意義を持つ。また、抗FcγRI抗体を作製するための免疫源、およびFcγRIの免疫化学測定方法の標準物質として用いることもできる。
【0030】
また、本発明のタンパク質を適切なクロマトグラフィー担体(例えば、His・Bind Resinゲル(Novagen社製)、HisTrap NHS−Activated HP(GEヘルスケアバイオサイエンス社製)、トレシル基活性化ダイナビーズ(Dynal社製))に固定化することで、ヒトモノクローナル抗体といった抗体を精製するための吸着剤として用いることもできる。
【0031】
さらに、本発明のタンパク質をコードしたポリヌクレオチドを含む発現ベクター、および前記ベクターで宿主を形質転換して得られる形質転換体を用いて、本発明のタンパク質を製造することで、IgG抗体に対する結合親和性が向上したタンパク質を簡便かつ大量に製造することができる。
【図面の簡単な説明】
【0032】
【図1】ヒトFcレセプターFcγRIの構造を示す図。SSはシグナルペプチド、ECは細胞外領域、TMは細胞膜貫通領域、Cは細胞質内領域をそれぞれコードする領域を示す。図の上部の数字は、それぞれの領域のアミノ酸数を示す。
【図2】発現プラスミドpECEFcRdhfrの構造を示す図。
【図3】発現プラスミドpECEFcRdhfrn2の構造を示す図。
【図4】発現プラスミドpECEFcRdhfrn3の構造を示す図。
【図5】発現プラスミドの作製概略を示す図。
【図6】形質転換したCOS7培養上清の抗体結合活性を示す図。横軸は抗体固定化濃度、縦軸は450nmにおける吸光度を示す。
【実施例】
【0033】
以下、本発明をさらに詳細に説明するために実施例を示すが、本発明は実施例に限定されるものではない。
【0034】
実施例1 発現プラスミドの作製(その1)
ヒトFcレセプターFcγRI細胞外領域からなるタンパク質を発現可能なプラスミドの作製を、以下の方法で行なった。
(1)ヒトFcγRIをコードするポリヌクレオチドを以下の方法で調製した。
(1−1)Human cDNA clone SC119841プラスミド(Origene社製)をテンプレートとし、配列番号6(5’−GA[AGATCT]ATGTGGTTCTTGACAACTCTGCTCC−3’:角かっこ部分は制限酵素BglIIサイト)と配列番号7(5’−CG[TCTAGA]TCAGTGGTGGTGGTGGTGGTG[ACTAGT]GACAGGAGTTGGTAACTGGAGGCC−3’:角かっこ部分はそれぞれ制限酵素XbaI、SpeIサイト)のオリゴヌクレオチドをPCRプライマーとして使用した。なお、ヒトFcγRIの調製および定量を行なうために、発現させるタンパク質のC末端側にポリヒスチジンタグが付加されるようにPCRプライマーを設計した。
(1−2)PCR反応を、94℃・5分の熱処理後、94℃・30秒間の第一ステップ、65℃・30秒間の第二ステップ、72℃・1分間の第三ステップを25サイクル行ない、最後に、72℃・7分の条件で行なった。反応液組成を表1に示す。
【0035】
【表1】
【0036】
(1−3)PCR反応終了後、0.9%のアガロース電気泳動にて、設計通りのサイズに相当するバンドを確認した。
(1−4)目的産物に相当するバンドをアガロースゲルから抽出(QIAquick Gel extraction kit(商品名):キアゲン社製)し、ヒトFcγRIをコードするポリヌクレオチドを調製した。
(2)(1)で調製したポリヌクレオチドを制限酵素BglIIとXbaIにより消化し、これらの制限酵素により事前に消化したプラスミドpECEdhfr(非特許文献5)とライゲーションし(Ligation Kit Ver.2:タカラバイオ社製)、50μg/mLの抗生物質カルベシニリンを添加したLB寒天培地を用いて大腸菌JM109株を形質転換した。
(3)(2)の形質転換体を50μg/mLの抗生物質カルベシニリンを含むLB培地により培養(37℃、18時間)し、QIAprep Spin Miniprep Kit(商品名、キアゲン社製)によりプラスミドを抽出した。
(4)抽出プラスミドを制限酵素BglIIとXbaIにより消化し、0.9%のアガロース電気泳動に供した。
【0037】
結果、インサートサイズから設計通りであることを確認し、これをpECEFcRdhfrとした。図2にpECEFcRdhfrの構造概略を示す。
【0038】
実施例2 発現プラスミドの作製(その2)
実施例1で作製した発現プラスミドpECEFcRdhfr(図2)から、以下の方法で、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質を発現可能なプラスミドを作製した。
(1)ヒトFcγRI細胞外領域の概ね全体配列を機能的に連結するため、Human cDNA clone SC119841プラスミド(Origene社製)をテンプレートとし、配列番号8(5’−CG[GCTAGC]GCAGTGATCACTTTGCAGCCTCCATTG−3’:角かっこ部分は制限酵素NheIサイト)と配列番号9(5’−GG[ACTAGT]GACAGGAGTTGGTAACTGGAGGC−3’:角かっこ部分は、制限酵素SpeIサイト)のオリゴヌクレオチドをPCRプライマーとして使用しPCR反応によりポリヌクレオチドを増幅した。なお、ヒトFcγRIの調製および定量を行なうために、発現させるタンパク質のC末端側にポリヒスチジンタグが付加されるようにPCRプライマーを設計した。また、PCR反応の反応条件は実施例1と同様にして行なった。
(2)(1)で調製したポリヌクレオチドを、制限酵素NheIとSpeIにより消化し、事前に制限酵素SpeIにより消化したpECEFcRdhfr(図2)に挿入した。
(3)実施例1と同様の方法で大腸菌JM109株に形質転換し、前記形質転換された大腸菌株からプラスミドを調製することで、ヒトFcγRI細胞外領域の概ね全体配列を含むポリヌクレオチドを2つ連結した発現プラスミドpECFFcRdhfrn2を作製した。図3にpECEFcRdhfrn2の構造概略を示す。
(4)さらに(1)で調製したポリヌクレオチドを、事前に制限酵素SpeIで消化したpECEFcRdhfrn2(図3)に挿入することにより、ヒトFcγRI細胞外領域の概ね全体配列を含むポリヌクレオチドを3つ連結した発現プラスミドpECEFcEdhfrn3を作製した。図4にpECEFcEdhfrn3の構造概略を示す。また、pECEFcRdhfrn3までの作製手順を図5に示す。
【0039】
なお、本実施例と同様な方法で、ヒトFcレセプターFcγRI細胞外領域の概ね全体配列を含むポリヌクレオチドを4つ以上連結した発現プラスミドの作製を試みたものの、前記プラスミドを作製することはできなかった。
【0040】
実施例3 発現プラスミドの塩基配列確認
実施例1および2で作製したpECEFcRdhfr(図2)、pECEFcRdhfrn2(図3)、pECEFcRdhfrn3(図4)に挿入されている、ヒトFcγRIをコードするヌクレオチド配列をチェーンターミネータ法に基づくBig Dye Terminator v3.1 Cycle Sequencing kit(商品名、PEアプライドバイオシステム社製)を用いてサイクルシークエンス反応に供し、全自動DNAシークエンサーABI Prism 310 DNA analyzer(PEアプライドバイオシステム社製)において解析した。なお、シークエンス用プライマーとして、配列番号10(5’−AGCTGTATGGGGTCACTTCG−3’)と配列番号11(5’−TTTTTCCACTGGAATTCTAACC−3’)と配列番号12(5’−AGTGGGGATGTCACAGATGC−3’)と配列番号13(5’−AGGAACACATCCTCTGAATACC−3’)に示すオリゴヌクレオチドを使用した。
【0041】
解析の結果、pECEFcRdhfr、pECEFcRdhfrn2、pECEFcRdhfrn3に挿入されている、ヒトFcγRIをコードするヌクレオチド配列はいずれも設計通りであることを確認した。ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを2つ連結したタンパク質をコードするヌクレオチド配列を配列番号2(pECEFcRdhfrn2)に、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを3つ連結したタンパク質をコードするヌクレオチド配列を配列番号3(pECEFcRdhfrn3)に、それぞれ示す。
【0042】
実施例4 発現したタンパク質の抗体結合活性の確認
実施例1および2で作製したpECEFcRdhfr、pECEFcRdhfrn2およびpECEFcRdhfrn3プラスミドをリポフェクトアミン(インビトロジェン社製)によりCOS7細胞にそれぞれ導入し、10% FCSを含むD−MEM培地(GIBCO社製)で培養することで(37℃、3日間)、一過的に細胞外へタンパク質を発現させ、培養後、培養液を回収した。回収した培養上清の抗体結合活性を以下の方法でELISA法により評価した。
(1)96穴のELISAプレート(Nunc社製)に100μg/mLから段階的に希釈したガンマグロブリン製剤(化学及血清療法研究所製)を各ウェルに100μLずつ添加し、4℃で18時間静置することによりガンマグロブリンを固相に固定化した。
(2)TBS緩衝液(0.2%(w/v)Tween 20、150mM NaClを含むTris−HCl緩衝液(pH8.0))で洗浄後、Starting Block Blocking Buffers(PIERCE社製)によりブロッキング操作を行なった。
(3)TBS緩衝液で洗浄後、調製した培養上清を100μL添加し、(1)で固定化したヒトガンマグロブリンと反応させた(30℃、2時間)。
(4)(3)の反応終了後、TBS緩衝液で洗浄し、His−probe(H−15)HRP抗体(Santa Cruz Biotechnology社製)を添加した。
(5)(4)の反応終了後、TBS緩衝液で洗浄し、TMB Peroxidase Substrate(KPL社製)を添加し450nmの吸光度を測定した。
【0043】
評価結果を図6に示す。図6の通り、pECEFcRdhfr(図6に記載の単量体)、pECEFcRdhfrn2(図6に記載の2量体)、pECEFcRdhfrn3(図6に記載の3量体)により形質転換されたCOS7細胞の培養上清にいずれも抗体結合活性が検出された。すなわち、pECEFcRdhfrn2、pECEFcRdhfrn3に挿入された、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質をコードするポリヌクレオチドにより、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質を得ることができ、かつ、当該タンパク質がヒト抗体への結合活性を保持していることを示している。なお、対照として、pECEFcRdhfrにより形質転換したCOS7培養液を使用した。
【0044】
実施例5 発現したタンパク質の調製
以下の方法で、実施例4で得られた培養上清から、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質を分離精製した。
(1)実施例4で得られた、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質が含まれた培養上清約1Lを、20mM酢酸緩衝液(pH5.2)で透析し脱塩処理を施した。
(2)20mM酢酸緩衝液(pH5.2)で平衡化した300mLのStreamline SPゲル(商品名、GEヘルスケアバイオサイエンス社製)を備えた吸着流動床システム(GEヘルスケアバイオサイエンス社製)に添加し、同緩衝液で洗浄後、10%グリセロール、1M NaClを含む20mMリン酸緩衝液(pH7.4)により溶出し、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチド複数連結したタンパク質が含まれた濃縮画分を調製した。
(3)(2)で得られた濃縮画分を、20mM イミダゾール、0.5M NaClを含む20mM リン酸緩衝液(pH7.4)に透析し、同緩衝液で平衡化したHisTrap HSカラム(商品名、GEヘルスケアバイオサイエンス社製)に添加し、洗浄後、500mM イミダゾール、0.5M NaClを含む20mM リン酸緩衝液(pH7.4)で溶出することで、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質精製標品約500μg得た。なお、タンパク質濃度の定量は、ウシガンマグロブリンを標準タンパク質としてブラッドフォード法に基づくプロテインアッセイキット(Bio−Rad社製)を用いて行なった。なお、対照として、pECEFcRdhfrにより形質転換したCOS7細胞より調製した培養液から、上記方法と同様の方法でヒトFcγRI細胞外領域を1つのみ含むタンパク質を調製している。
【0045】
実施例6 発現したタンパク質の表面プラズモン共鳴解析
実施例5で作製した精製タンパク質の抗体結合特性を表面プラズモン共鳴解析装置BIAcore T100(商品名、GEヘルスケアバイオサイエンス社製)を用いて解析した。
(1)ヒト抗体のチップ固定
ヒト抗体IgG1(コスモバイオ社製)を、ランニング緩衝液HBS−EPを用いて終濃度約5μg/mLとなるように希釈した。次いで、BIAcore T100の取扱説明書に従いセンサーチップCM5(GEヘルスケアバイオサイエンス社製)に390RU固定した。
(2)精製FcγRIのヒト抗体IgG1の結合解析
抗体との結合解析を行なうために、ランニング緩衝液で任意に希釈した、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを複数連結したタンパク質を、BIAcore T100に供した。抗体との結合性(RU)は希釈したタンパク質濃度に従い変化した。この結果を用いて、上記解析装置の専用ソフトであるBIAcore T100 Evaluation Softwareにより前記タンパク質のヒト抗体への反応特性を解析した。
【0046】
解析結果を表2に示す。表2の通り、今回作製したヒトFcγRI細胞外領域の概ね全体配列を複数連結したタンパク質は、細胞外領域を1つのみ含むヒトFcγRI(表2に記載の単量体)と比較し、ヒト抗体に対する結合親和性が向上しており、平衡解離定数の比較で、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを2つ連結したタンパク質(表2に記載の2量体)で単量体の約16倍、ヒトFcγRI細胞外領域の概ね全体配列を含むポリペプチドを3つ連結したタンパク質(表2に記載の3量体)で単量体の約85倍、それぞれ向上していた。
【0047】
【表2】
【特許請求の範囲】
【請求項1】
配列番号1に記載のアミノ酸配列からなるヒトFcレセプターFcγRIのうち、細胞外領域の全体領域または概ね全体領域を含むポリペプチドを複数連結した、タンパク質。
【請求項2】
配列番号4または5に記載のアミノ酸配列からなる、タンパク質。
【請求項3】
請求項1または2に記載のタンパク質をコードする、ポリヌクレオチド。
【請求項4】
請求項3に記載のポリヌクレオチドを含む、発現ベクター。
【請求項5】
請求項4に記載のベクターで宿主を形質転換して得られる、形質転換体。
【請求項6】
請求項5に記載の形質転換体を用いた、タンパク質の製造方法。
【請求項1】
配列番号1に記載のアミノ酸配列からなるヒトFcレセプターFcγRIのうち、細胞外領域の全体領域または概ね全体領域を含むポリペプチドを複数連結した、タンパク質。
【請求項2】
配列番号4または5に記載のアミノ酸配列からなる、タンパク質。
【請求項3】
請求項1または2に記載のタンパク質をコードする、ポリヌクレオチド。
【請求項4】
請求項3に記載のポリヌクレオチドを含む、発現ベクター。
【請求項5】
請求項4に記載のベクターで宿主を形質転換して得られる、形質転換体。
【請求項6】
請求項5に記載の形質転換体を用いた、タンパク質の製造方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2011−115083(P2011−115083A)
【公開日】平成23年6月16日(2011.6.16)
【国際特許分類】
【出願番号】特願2009−274353(P2009−274353)
【出願日】平成21年12月2日(2009.12.2)
【出願人】(000003300)東ソー株式会社 (1,901)
【Fターム(参考)】
【公開日】平成23年6月16日(2011.6.16)
【国際特許分類】
【出願日】平成21年12月2日(2009.12.2)
【出願人】(000003300)東ソー株式会社 (1,901)
【Fターム(参考)】
[ Back to top ]