
抗原−ノロウイルスPドメインモノマーおよびダイマー、抗原−ノロウイルスP粒子分子、ならびにそれらの製造および使用法
Pドメインのみを有し、抗原−ノロウイルスPドメインモノマーと称される置換ノロウイルスカプシドタンパク質モノマーは、分子クローニングによって各Pドメインモノマー上に存在する3個の表面ループの1個以上に挿入された外来抗原を含む。抗原−Pドメインモノマーは、集合して、24個の抗原−Pドメインモノマーのコピーからなる、抗原−ノロウイルスP粒子と称される八面体形態を自然発生的に構築できる。各置換Pドメインモノマーは、各抗原−P粒子上の合計24〜72個の抗原コピーにつき、1〜3個の外来抗原のコピーを含む。抗原−P粒子は、外来ウイルス、例えばロタウイルスに感染した個体の診断、免疫および処置のための方法において有用であり、多くの感染症および非感染症に対する新規なワクチンの開発のための外来抗原提示用担体として利用できる。置換ノロウイルスP粒子は大腸菌および酵母において容易に製造でき、高度に安定であり、広範な物理化学的状態を許容する。修飾ノロウイルスPドメインモノマーは、表面ループに候補外来抗原を簡便に挿入するためのユーザーフレンドリーなクローニングカセットを提供するために、Pドメインモノマーの3個のループの1個以上に挿入された1個以上の制限認識部位を含む。P粒子−VP8キメラはまた、ロタウイルスおよびノロウイルス両方に対するデュアルワクチンとしても利用できる。

【発明の詳細な説明】
【技術分野】
【0001】
利害関係
本発明は、合衆国政府の支援の下になされ、National Institute of Health(NIH)、National Institute of Allergy and Infectious(認可番号R01 AI37093およびR01 AI055649)およびDepartment of Defense(認可番号PR033018)によってXi Jiangへと表彰された。この研究はUniversity of CincinnatiおよびCincinnati Children's Hospital Medical Center(Center for Clinical and Translational Science and TrainingからのT1助成金)によってMing Tanへも支援された。さらに、Ming TanへのNipert Foundationが出資する感染症助成金がこの研究に寄与した。合衆国政府は本発明についての一定の権利を保有しうる。
【0002】
発明の分野
本発明は、一般に、ノロウイルス(NOR)カプシドタンパク質のPドメインおよびNOR P粒子、抗原−ノロウイルスPドメインモノマー、抗原−ノロウイルスPドメインダイマーおよび抗原−ノロウイルスP粒子分子の製造、ならびに抗ウイルス剤、薬物送達系およびワクチン開発のためのそれらの使用に関する。
【背景技術】
【0003】
発明の背景
生体材料および生物工学は、現代医学の重要な部分となりつつある急速に成長している分野である。ウイルス性タンパク質ケージは、その万能性およびアレイを形成する傾向のため、提示系を構築するための理想的な基板である。遺伝子操作を通じて、自己集合性ウイルス構造タンパク質は、生体材料合成のための傑出したプラットフォームであり、そして設計されたパターンで外来分子を統合するためのスキャフォールドであり、これはワクチンまたは薬物として使用できる。
【0004】
抗原提示のためにウイルス粒子を用いる最近の例は、Manayani et alに記載の、炭疽菌(Bacillus anthracis)の180アミノ酸抗原挿入を有するフロックハウスウイルス(FHV)VLPのキメラである。組換えウイルス様粒子は、炭疽抗毒素として、かつ、炭疽菌ワクチンのための分子スキャフォールドとして機能し、炭疽抗毒素およびワクチンの機能を1個の化合物に組み合わせる。別の例は、Chatterji et al.に記載のCowpeaモザイクウイルス(CPMV)であり、これは、エピトープ提示ならびにペプチドおよびタンパク質の付着のためのマトリックスとしての両方のための、Cowpeaモザイクウイルスの使用を含む。全タンパク質は、正二十面体のCPMV粒子のコートタンパク質において遺伝的に操作されたリシンおよびシステイン残基と、化学的に架橋結合されている。また、ライム病菌(Borrelia burgdorferi)の表面抗原(OspA)を含むB型肝炎ウイルス(HBV)カプシド様粒子(CLP)が記載されている。
【0005】
ノロウイルス(NOR)は、以前は「ノーウォーク様ウイルス」(NLV)または小型球形ウイルスとして知られており、先進国および発展途上国の両方で生じる、伝染性急性胃腸炎の最も重要なウイルス性病原体である。これらの遺伝的に多様なウイルスは、2個の主要な遺伝子グループ(GIおよびGII)ならびに約30個の遺伝子型を含む。NORはカリシウイルス科(Caliciviridae)に属し、正二十面体の一本鎖のポジティブセンスRNAウイルスであり、そのカプシドは1個の主要構造タンパク質の180個のコピーからなる。
【0006】
過去、ヒトNORの生物学的特徴付けは、該ウイルスが細胞培養では効率的に増殖せず、ウイルス複製のための好適な動物モデルが構築されなかったために、妨害されていた。発生およびヒトボランティア試験から得られたヒト糞便サンプルが唯一のウイルス源であったが、糞便中のウイルス濃度は、通常の電子顕微鏡によるウイルス検出が不可能であったほど低い。しかし、近年の昆虫細胞中のバキュロウイルス(主として昆虫に感染する二本鎖DNAウイルス)によるNORカプシドタンパク質発現の成功が、NORの免疫学、疫学および発症機序の研究のための有用な別法を提供している。ウイルスカプシドタンパク質モノマーは、自己集合してウイルス様粒子(VLP)を作り出す。これらのVLPは形態学的および抗原的にヒト糞便において見られる真菌とは区別不可能であり、免疫学的アッセイの開発およびレセプター−ウイルス相互作用の研究に有用なツールを提供する。
【0007】
組換えNORカプシドタンパク質の原子構造は、それがT=3正二十面体を形成する90個のダイマーカプソメアに組織された180個のカプシドタンパク質モノマーを含むことを示している。低温電子顕微鏡およびX線結晶学からのデータは、該ウイルスカプシドタンパク質が2個の主要なドメイン、N末端シェル(S)ドメインおよびC末端突起(P)ドメインに折り畳まれることを示した。Sドメインは内部のシェルを形成し、一方でPドメインは、シェルから伸長もしくは突出するアーチ様構造を構築する。形態形成の研究により、Sドメインがカプシドの内部シェルの構築に必要な要素を含むが、Pドメインのダイマーサブユニット間の分子間接触がカプシドの安定性を増加することが示された。これら2個のドメインは、8〜10残基(アミノ酸)のヒンジを介して結合している。PドメインはさらにP1およびP2ドメインに分けられ、後者はカプシドの最も外側表面に位置する。SおよびP1ドメインとは異なって、P2ドメインは高い配列バリエーションを有する。Pドメインはウイルス粒子の最も外側表面に位置し、最も変化しうる配列を有するため、Pドメインが宿主相互作用、免疫認識、レセプター結合および免疫応答に関与すると考えられている。ヒンジを有する(しかしSドメインを欠く)単離されたPドメインが、ヒト組織−血液グループ抗原(HBGA)レセプターの構築を維持するダイマーをインビトロで形成することが示されている。
【0008】
HBGAは、複合グリカンと関連糖の異種グループである。NORは多様な、株特異的な手法で、ヒトHBGAをレセプターとして認識する。HBGAの中で、最も一般に遭遇する血液グループはABO(ABH)およびLewisである。ABHおよびLewis血液グループ系において抗原の形成に使用される生合成経路は、相互に関連している。
【0009】
ヒトHBGAは赤血球および血管内皮細胞を含む多くの細胞タイプ、ならびに消化器、尿生殖器および呼吸管の粘膜上皮に存在する。それらはまた、血液、唾液、消化器内容物および乳のような生物学的液体中で可溶形態でも存在しうる。HBGAは遺伝子的に制御され、ABO、Lewisおよび分泌遺伝子ファミリーとして知られる一組のグリコシルトランスフェラーゼを介して、単糖ユニットの段階的添加によって、一連の前駆体構造から合成される。
【0010】
ヒトHBGA系は高度に多形性であり、サイレントアレルを含む複数の遺伝子ファミリーによって制御される。細胞表面上でのHBGAのような多様化した分子の存在は、外部環境変化に対する宿主防御メカニズムの可能性を示す。それにもかかわらず、HBGAは多様な細菌性およびウイルス性病原体による感染と関連しており、ノロウイルスの「ドッキングステーション」を提供しうる。すなわち、HBGAは病原体にとっての認識標的であり、抗原とのレセプターリガンド結合を発現または形成する細胞への侵入を促進しうる。かかる相互作用の正確な性質は未だ知られていないが、生じる病原体と抗原構築との密接な関連は、感染プロセスにおける最初のステップとして、細胞への病原体の係留に重要な役割を果たしうる。
【0011】
NORによるヒトHBGAの認識は典型的なタンパク質−糖相互作用であり、ここで、ウイルスカプシドタンパク質の突起ドメインは、様々な株において広い多様性を有するHBGA抗原のオリゴ糖側鎖による界面を形成する。腸管内でのみ複製可能な病原体として、NORは、宿主防御系を克服するユニークな戦略を開発している。これは、なぜNOR関連疾患が世界中のあらゆる集団で一般的であり拡がっているかを説明する、それらの遺伝的および構造的バリエーションによって示されている。
【0012】
2003年12月2日に公開されたPCT公開公報US2003/101176(その全体について参照により本明細書に組み込む)は、複数の異なる組織−血液グループパターンの一つにおけるNOR株とABOおよびLewisHBGAの結合に関する。NORによるHBGAの認識は株特異的であり、多数の異なるHBGA結合パターンが同定されている。NORの多様性および宿主細胞表面における糖の多形性に基づいて、より多くの結合パターンが発見されうる。
【0013】
2006年12月28日に公開されたPCT公開公報US2006/138,514(その全体について参照により本明細書に組み込む)は、HBGAとの向上した結合親和性を示す、P粒子として知られる小さな粒子に関する。P粒子は、12個の同一のPドメインダイマーに組織される24個のPドメインモノマーによって構築されるT=1正二十面体である。12個(Pドメインダイマー)および24個(Pドメインモノマー)は、植物および動物ウイルスについて頻繁に生じる正二十面体シンメトリーについての完全単位数である。単離された、モノマーカプシドタンパク質のSドメインまたはヒンジを含まないPドメインは、12個のダイマーに組織される24個のPドメインモノマーからなる複合体である、T=1正二十面体P粒子を自然発生的に形成できる。P粒子は対応するHBGAと結合でき、NOR VLPとHBGAとの結合の強い遮断を示す。P粒子の自然発生的形成は、VA387、MOHおよびノーウォークウイルス(NV)を含む多様なNOR株で観察されている。NOR P粒子は、NOR感染の治療処置およびNOR感染に対するワクチンの作製において有用である。
【0014】
ロタウイルス(RV)およびNORはいずれも、疾患の大きな負担を招く世界中で一般的な病原体である。世界基準で、5歳未満の小児において毎年、全原因の胃腸炎が最大10億事象生じており、その13〜25%(〜1億3000万事象)がRVによって引き起こされる。RVは、小児の様々な下痢および脱水の主たる原因であり、毎年重度のRV胃腸炎によって35万〜60万人の5歳未満の小児が死亡している。それはまた、年間200万人の小児の入院に寄与しており、その費用は10億ドルを超えると予測される。他方、NORはあらゆる年齢の個体が罹患する急性胃腸炎の非細菌性流行の最も重要な原因である。NORは極めて接触感染性であり、急速に拡大して多様な状況で大流行を導く。近年の報告では、発展途上国においてNORは、毎年5歳未満の小児における1,091,000件の入院および218,000人の死亡を引き起こしていると予測された。米国において、食品媒介性病原体は、毎年推定7600万人に感染しており、325,000件の入院の原因である。NORは単独で、臨床治療およびリコールされた食品による損失のために、毎年3億5000万〜7億5000万ドルの損害を引き起こしている。
【0015】
2種の新たなRVワクチン(Rotarix(商標), GlaxoSmithKline および RotaTeq(登録商標), Merck)が近年導入されているが、これらのワクチンに関する十分に解決されていない次のような様々な問題が存在する:a)ワクチンにおいて見られるものよりも広い範囲のRV血清型にワクチン接種およびワクチン非接種小児が曝露された際の有効性;b)ワクチンが最も必要とされる貧困国にこれらのワクチンをどのように分配するかという問題を導く、ワクチンのコストおよび分配コスト;ならびにc)死亡率が最も高い発展途上国における保護のレベル。これらの弱毒化生ワクチンは、復帰し、最集合して、毒性株を生じる可能性がある。したがって、極めて有効なRV中和エピトープを含む、新世代のサブユニットワクチンが必要とされている。現在、NOR関連疾患に対する処置は存在しない。したがって、NORおよびRVに対して有効なワクチン、特に両方に対して保護ができる単一のワクチンの開発は、主要な臨床的必要を充足し、さらに本明細書に記載のP粒子ワクチンプラットフォームの顕著な販売潜在力を強調する。
【0016】
NOR感染の治療処置およびそれに対するワクチン開発における進歩にもかかわらず、他のウイルスタイプによって引き起こされる感染の同定の改善、他のウイルスタイプの治療処置、他のウイルスタイプに対するワクチン開発、ならびに特定の組織または臓器を標的とする改善された薬物送達系を開発する必要がなおも存在する。
【発明の概要】
【0017】
発明の概要
本発明は、NOR Pドメインモノマーの遠位部分が外来抗原、特に外来ウイルス性抗原を挿入できるペプチド鎖を含むという発見に関する。得られる抗原−Pドメインモノマーは、12個のダイマーに組織される24個の抗原−Pドメインモノマーからなる、典型的には八面体形態の、抗原−P粒子と称される、ナノ粒子に自然発生的に集合しうる。このP粒子は大腸菌において容易に製造され、極めて安定であり、免疫原性が高い。Pドメインモノマーあたり3個の表面ループが存在し、これは粒子あたり合計72個のループとなり、免疫増進のための外来抗原提示の潜在的部位である。抗原−P粒子は自然発生的に形成され、そしてPドメインモノマーの遠位部分で提示される外来抗原を有する。この抗原−P粒子形成プロセスは、抗原−Pダイマーの濃度に基づいて交換し、または動的に平衡化しうる。抗原−P粒子は、外来ウイルス性抗原に対する抗体を産生し、またかかる外来ウイルス感染のためのワクチンを作製するために有用である。
【0018】
本発明の第一の局面は、多様な感染性病原体の抗原を提示するためのプラットフォームとして使用するための修飾NOR P粒子に関し、したがってこれらの疾患に対する新たなワクチン戦略を提供する。このプラットフォームはまた、ワクチンの開発のため、あるいはがん、アレルギーおよび自己免疫疾患を含む非感染性疾患の処置のための、薬物の担体または薬物送達ビークルとしても有用である。多数の他のワクチンプラットフォームと比較して、本抗原−P粒子系は、単純な精製手法に伴う高収率および低製造コスト、ならびに高い抗原提示効率を含む多くの利点を有する。
【0019】
本発明の抗原−P粒子は、NORカプシドの突起(P)ドメインによって形成され、ダイマーに組織されるPドメインモノマーの24個のコピーを含む。抗原−P粒子の分子量およびサイズ(MW=約830kDaおよびΦ=20nm)は、サブユニットワクチンとして有効なサイズである。外来抗原は各Pドメインモノマーに存在する3個の表面ループの1個以上に挿入され、それによって各抗原−P粒子の表面で、外来抗原の72個までのコピーを提示する。1個の抗原−P粒子上の外来抗原の多重性は、外来抗原の抗原性および免疫原性を大きく向上させる。小さなポリペプチドから大きなタンパク質まで多様なタイプおよびサイズの外来抗原が3個の表面ループのいずれか1個以上に挿入されてよく、外来抗原の代表例としては次のものを含む:Hisタグ(7個のヒスチジン)、マウスサイトメガロウイルスのT細胞エピトープ(9アミノ酸)、シュードモナス属のEpi8抗原(10アミノ酸)、ロタウイルス(RV)のVP8(159アミノ酸)および緑色蛍光タンパク質(238アミノ酸)。
【0020】
NOR P粒子は、極めて安定であり、広範な物理化学的条件に耐性である。最も重要なことに、製造のために真核細胞系を必要とする多くの他の組換えサブユニットワクチンとは異なって、野生型NOR P粒子同様、抗原−P粒子は、大腸菌および酵母または他の細菌源において、極めて高い収率および単純な精製手段で容易に製造できる。
【0021】
本発明の第二の局面は、簡便なクローニングカセットの挿入によって、各Pドメインモノマー上に存在する表面ループの1個以上に候補外来抗原を挿入するためのユーザーフレンドリーな方法である。クローニングカセットは、典型的には、Pドメインベクターの1個以上のループに、制限部位または制限認識部位としても知られる特異的認識ヌクレオチド配列の1個以上、典型的には少なくとも1個の対を含む。制限部位は制限酵素によって認識される特異的なヌクレオチド配列である。該部位は一般にパリンドロームであり(制限酵素は通常ホモダイマーとして結合するため)、特定の制限酵素は典型的には、その認識部位内またはその近くのどこかの2個のヌクレオチド間の特定のDNA配列を切断する。例えば、一般的な制限酵素EcoRIは、パリンドローム配列GAATTCを認識し、DNA鎖の上下両方のGとAの間を切断し、各末端にオーバーハング(結合した相補鎖のないDNA鎖の末端部分)を残す。次いでこのオーバーハングまたは「付着末端」は、相補的オーバーハングを有するDNA片(例えばDNAの他のEcoRI切断断片)を加えまたはライゲート(DNAリガーゼを介して)するために使用できる。各Pドメインモノマー上に存在する表面ループの1個以上における制限部位を使用することで、表面ループのいずれか1個以上にクローニングカセットに含まれる外来遺伝子を置換でき、またPドメインモノマーの3個のループの異なる一つに、2個以上の異なる外来抗原または他のクローニングカセットを置換できる。
【0022】
本発明の別の局面は、12個のダイマーの対として、24個のPドメインモノマーの自然発生的集合による、抗原−P粒子の形成である。全てのPドメインモノマーが同一である場合、抗原−P粒子は同種P粒子と呼ばれる。
【0023】
本発明の別の局面は、2個以上の異なって修飾された抗原−Pドメインモノマー、または少なくとも1個の種々に修飾された抗原−Pドメインモノマーおよび野生型Pドメインモノマーを含む、異種抗原−P粒子の形成である。種々に修飾された抗原−Pドメインモノマーまたは野生型Pドメインモノマーは、広範な抗原−P粒子を得るために、任意の比(分子量または重量比)で使用できる。
【0024】
本発明の別の局面は、ウイルスに対する免疫保護を与えるための候補ワクチンとしての、P粒子内の各Pドメインモノマーの表面ループの少なくとも1個に挿入された特異的ウイルス性抗原、例えばロタウイルス(RV)抗原を有する、抗原−P粒子の使用である。
【0025】
本発明の別の局面は、ウイルス性疾患からヒトを保護する方法であって、滅菌、非毒性の薬学的に許容される担体中の、本明細書に記載のP粒子内のPドメインモノマーの表面ループの少なくとも1個に挿入された特異的ウイルス性抗原を有する抗原−P粒子の免疫量をヒトに投与することを含み、ここで前記抗原がウイルス性抗原である、方法に関する。
【0026】
本発明の別の局面は、ウイルス性疾患に対する個体の免疫応答を誘導する方法であって、本明細書に記載の抗原−P粒子および医薬担体を含む組成物の少なくとも1個の免疫学的有効量を個体に経口または非経口投与する工程を含み、ここで前記抗原がウイルス性抗原であり、量が個体の免疫応答を誘導するのに有効である、方法に関する。
【0027】
本発明の別の局面は、抗原−P粒子ベクター/ワクチンプラットフォームキットである。
【0028】
本発明の別の局面は、Pドメインモノマーの1個以上の完全に露出したループのクローニングカセットへの、固有または稀少な制限部位を含む制限部位の導入に関する。制限部位を含むクローニングカセットの挿入に関して、ループのクローニングカセットへのプロテアーゼ認識配列の挿入を避けることが推奨される。例えば、BgIII部位(AGATCT)は、典型的なトリプシン切断部位であるジアミノ酸制限部位をコードし、避けられるべきである。
【0029】
本発明の別の局面は、最適にある外来抗原を適合させるために、ループの露出を伸長するスペーサーまたはアームを加えることを含む、ループの1個以上の修飾に関する。
【0030】
本発明の別の局面は、RV VP8のような大きな抗原がより小さな外来抗原を有する隣のループを、このループにおける小さなエピトープの提示が困難となるように覆わないように、異なる抗原が複数挿入されている設計に関する。
【0031】
本発明の別の局面は、遠位表面の1個以上に挿入されたリガンドまたはシグナルペプチドを有する置換P粒子がもたらされる、Pドメインモノマーの表面ループの1個以上へのリガンドまたはシグナルペプチドの挿入に関する。このリガンドまたはシグナルペプチドは、次いで、特定の臓器または組織の対応するレセプターに置換P粒子を標的化し、それらの位置に移動させられる。例えば、ペプチドCNGRC(5アミノ酸)を有する置換P粒子は、CNGRCのレセプター(CD13)が高度に発現している腫瘍組織(すなわち癌)に移動できる。
【0032】
本発明の別の局面は、化学反応によって表面露出リシンおよびシステインを介してP粒子のループに挿入されている薬物のコンジュゲートを有する、置換P粒子モノマーならびにそのダイマーおよびP粒子に関する。この場合、置換P粒子は薬物の担体として使用される。
【0033】
本発明の別の局面は、表面ループの少なくとも1個に挿入されたリガンドまたはシグナルペプチドを有し、そして表面ループの少なくとも1個に挿入された薬物のコンジュゲートを有する、置換Pモノマーならびにそれから形成されたダイマーおよびP粒子に関する。
【0034】
本発明の別の局面は、修飾または抗原−Pドメインモノマーの修飾された遺伝子コードの遺伝子配列である。
【0035】
本発明の別の局面は、修飾または抗原−Pドメインモノマーの修飾された遺伝子コードを有する複製生物学的物体(例えば大腸菌)である。
【0036】
本発明の性質および利点は、本明細書に記載の、そして添付の特許請求の範囲によってより具体的に定義されている意図する新規構成、組合せおよび要素を示す下記図面および詳細な説明からより十分に理解されるが、先行技術によって除外されない限り、本発明の正確な態様における変化が特許請求の範囲に含まれることを意味すると理解される。
【図面の簡単な説明】
【0037】
【図1】P粒子−VP8キメラのCryo−EM構造を示す。(A)野生型P粒子。(B)P粒子−VP8キメラ。野生型P粒子と比較して、キメラは、中央にニックを伴う伸長された突起を示し、これはP2サブドメインと挿入されたVP8抗原との間の境界を示唆する。(A)および(B)における粒子の半径は、示したとおり異なるカラースキームによって示される。(C)ロタウイルス(Wa)VP8抗原の2つのコピーの結晶構造(模式図、緑および青)の、キメラの伸長された突起の密度マップ(透明なグレー)への重ね合わせであり、キメラP粒子上でのVP8抗原の露出が確認される。(D〜E)P粒子−VP8の突起を拡大した側面図(D)および上面図(E)。
【図2】NORカプシドおよびカプシドタンパク質(VP1)を示す。AおよびBは、ノーウォークウイルス様粒子(NOR VLP)の表面構造(A)および断面(B)を示す。CはVLPの基本単位であるVP1のダイマー(リボンモデル)を示す。各VP1は、N末端アーム(緑)、シェル(S)ドメイン(黄色)および突起(P)ドメインに分けられる。Pドメインはさらに、P1およびP2サブドメイン(それぞれ赤および青)に分けられ、P2はVLPの最も外側の表面に存在する。VP1の直線構造を同じカラーテーマでDに示す。
【0038】
【図3】NOR P粒子およびその表面ループの構造を示す。(A)はcryo−EMによって再構成されたノロウイルスP粒子の構造を示し、(B)は結晶構造(リボンモデル)において解明された、Pダイマーの最も外側の領域であるその突起の一つを示し、ダイマーの6個の表面ループを示している。赤−緑および青−黄色は、それぞれ2個のPドメインを示す。抗原挿入に好適な部位である各モノマーの3個の表面ループが示されている。2個のPドメインモノマーは安定なグローバルPダイマーを形成する。次いで、12個の同一のPダイマーが集合して、Pダイマーの濃度に依存して動力学的に交換および/または平衡するT=1正二十面体P粒子となる。Pドメイン間の分子内相互作用がP粒子形成の原動力である。末端結合システインは、分子内ジスルフィド結合を形成してP粒子を安定化できる。
【0039】
【図4】NOR Pダイマー−B−トリサッカライド複合体の結晶構造を示す。トリサッカライド(ウイルス性レセプター)は、リボンモデル中に存在するアーチ様Pダイマーの上部に、球で存在する。
【0040】
【図5】NOR P粒子が野生型NOR VLPと同様の免疫原性および免疫反応性を有することを示す。Aにおいて、NOR株VA387のP粒子によって誘導される過免疫抗体が、VLP誘導性抗体と同様の力価でVLPと反応した。VLP−(B)およびP粒子−(C)誘導性抗体は、NOR VLPとタイプA HBGAレセプターとの結合に対して同様の遮断効果を示した。
【0041】
【図6】cryo−EMによる小P粒子の3D構造再構成を示す。AおよびBは小P粒子の側面図および上面図を示し、Cは小P粒子のcryo−EMの密度マップへのPダイマーの結晶構造の重ね合わせを示す。
【0042】
【図7】P粒子形成および安定性の妥協なしに完全に露出される、P粒子のループ2での7xHisタグの露出を示す。(A)は7xHisタグを有するP粒子がTalon樹脂に良好に結合することを示す。GST−Pドメイン−Hisタグ融合タンパク質はトロンビンによって消化されて、P粒子−Hisタグキメラ(PP−Hisタグ)、GSTおよび他の共に精製されたタンパク質の混合物をもたらした。共に精製されたタンパク質と7xHis−P粒子(ロード前)をTalon樹脂と共にインキュベートした。7xHis−P粒子全ておよびわずかな共に精製されたタンパク質は、樹脂に結合した(フロースルー)。洗浄した後、結合した7xHis−P粒子をイミダゾールで樹脂から溶出した(溶出1および2)。マーカーは、上から下に113、92、50、35、29、21kDaを示すバンドを有する予め染色したタンパク質標準であった。(B)において、(A)の溶出のゲルろ過分析は、溶出した7xHis−P粒子の>98%がP粒子を形成したことを示した。
【0043】
【図8】露出したRV VP8を有するNOR P粒子を示す。SDS PAGEのパネルAは、アフィニティーカラム(レーン1)およびさらにP粒子(レーン2)およびPドメインダイマー(レーン3)フラクションでのゲルろ過によって精製された、組換えRV VP8−Pドメインキメラ(53.5kDa)を示す。パネルBおよびCは、VP8(パネルB、レーン4および5)またはNOR VLP(パネルC、レーン6および7)に対する抗体を用いた、VP8−P粒子キメラのウェスタン分析を示す。パネルDにおいて、アフィニティー精製VP8−Pドメインキメラ(レーン1)のゲルろ過は、該タンパク質の大部分がP粒子(〜1300kDa)を形成することを示す。パネルEは、被覆VP8−P粒子キメラ(赤、PP−hVP8C)、組換えVP8(青、hVP8C)およびP粒子のみ(緑、PP、コントロール)を検出するためにVP8に対する抗体を用いた、酵素免疫アッセイ(EIA)の結果を示す。レーンMは、上から113、92、52、35、29kDaのバンドで、予め染色したタンパク質マーカーを示す。
【0044】
【図9】ループ2上にクローニングカセットを有するP粒子ベクターを示す。P粒子の最も外側の結晶構造(リボンモデル)は、3個の表面ループの表示を有する図の下半分に示し、P粒子ベクターのクローニングカセットの詳細は、図の上半分に示す。
【0045】
【図10】RV VP8−P粒子キメラの製造を示す。ループ2にクローニングカセットを有するP粒子ベクター(A)は野生型NOR P粒子(B)を製造する。RV VP8コード配列(C)がP粒子ベクター(D)にクローン化されると、それはキメラの最も外側表面でのRV VP8の指標と共に、RV VP8−P粒子キメラ(E)を製造する。
【0046】
【図11】VP8−P粒子キメラによって誘導された抗体が、NORおよびRVの両方に反応性であり、NOR VLPとHBGAレセプターの結合の遮断を示した。グラフAは、それぞれP粒子およびRV VP8に対する抗体の反応性を示す。グラフBは、該抗体がNOR VLPとHBGAレセプターの結合を遮断することを示す。X軸は抗血清の希釈(倍率)を示す。
【0047】
【図12】P粒子ベクター、P粒子プラットフォームおよびキメラP粒子ワクチンの構築のフローチャートを示す。GST、PドメインおよびCは、グルタチオンS−トランスフェラーゼ(GST)遺伝子、NOR Pドメインおよびシステイン含有短ペプチドをそれぞれコードする配列を示す。両末端の太線は、pGEX−4T−1のプラスミドを示す。
【0048】
【図13】P粒子ベクターのデザインを示す。3個の表面ループの指標と共にP粒子の最も外側部分の結晶構造を、図の下側に示す。各表面ループで異なる制限酵素認識部位を含むクローニングカセットを有するP粒子ベクターの一つの代表的なデザイン選択肢を、図の上側部分に示す。
【図14】P粒子−Hisタグキメラの製造および分析を示す。(A)P粒子−Hisタグキメラの発現構築物。HisタグはN372とD374の間のループ2に挿入した。pGEX−4T−1は、GST遺伝子融合システムの発現ベクターである。丸で囲んだCは、システイン含有ペプチド(CDCRGDCFC)を示す。(B)結晶構造(模式図)においてP粒子の突起の遠位末端は、挿入されたHisタグが位置すると予測される、2個のN373の位置を示す(グレーおよびシアンのドットモデル)。(C)P粒子−Hisタグキメラの発現および精製。SDS PAGE分析は、GST−Pドメイン−Hisタグ融合タンパク質(GST融合体)が〜52kDaであることを示した。トロンビンによる溶液中での融合タンパク質の消化は、GST(〜27kDa)およびPドメイン−Hisタグキメラ(PD−Hisタグ)(〜35kDa、左パネル)をもたらした。PドメインHisタグキメラは、トロンビン消化によって精製樹脂からも放出されうる(右パネル)。Mは、上から下に113、92、50、35、29、21kDaを示すバンドで、予め染色したタンパク質マーカーである。(D)サイズ排除カラムSperdex 200を用いたトロンビン放出Pドメイン−Hisタグタンパク質のゲルろ過クロマトグラフィーの溶出曲線。3つの主要ピークはそれぞれ、空、P粒子−HisタグおよびPダイマー−Hisタグを示した。これらの3つのピークのサイズは、それぞれ青色デキストラン2000(〜2000kDa、空)、野生型P粒子(〜830kDa)および野生型Pダイマー(〜70kDa)でキャリブレートした。(E)ゲルろ過クロマトグラフィー(D)のフラクションをSDS PAGEで分析した。
【0049】
【図15】P粒子提示短ペプチドに対するマウスの反応を示す。(A)それぞれ等量のP粒子−Hisタグキメラ(PP−His、青)、Pダイマー−Hisタグキメラ(PD−His、紫)または野生型P粒子(WT PP、コントロール、黒)で免疫した後、EIAにおけるT. Maritimaの組換えHisタグ化αフコシダーゼに対するマウス血清の免疫反応性(n=5)。(B)(A)における血清の抗体力価を終点希釈アプローチによって測定した。(C)〜(F)P粒子の非露出短ペプチドに対するマウスの抗体反応。(C)埋め込みペプチドCDCRGDCFCを含むP粒子(PP−CDCRGDCFC)で免疫した後の、EIAにおける、CDCRGDCFCタグ化マルトース結合タンパク質(MBP)(to CDCRGDCFC、紫)および野生型P粒子(to WT PP、ポジティブコントロール、緑)に対するマウス血清の免疫反応性(n=5)。(A)(PP-His to His-tag、青)が比較のために含まれた場合の、P粒子−Hisタグで免疫した後の、T. MaritimaのHisタグ化αフコシダーゼに対するマウス血清の免疫反応。(D)終点希釈アプローチによって、(C)における血清の抗体力価を測定した。(E)埋め込みペプチドCNGRCを含むP粒子で免疫した後、CNGRCタグ化MBP(to CNGRC、紫)および野生型P粒子(to WT PP、ポジティブコントロール、緑)に対するマウス血清の免疫反応性。P粒子−Hisタグで免疫した後の、(A)(PP−His、青)のT. MaritimaのHisタグ化αフコシダーゼに対するマウス血清の免疫反応性を比較のために加えた。(F)終点希釈アプローチによって、(E)における血清の抗体力価を測定した。EIAアッセイのために、5ng/μlの異なる抗原[(A)および(B)についてHisタグ化αフコシダーゼ、(C)および(D)についてCDCRGDCFCタグ化MBP、(E)および(F)についてCNGRCタグ化MBP、ならびに(C)〜(F)について野生型P粒子]をマイクロタイタープレートに被覆した。指定の希釈での対応する血清を用いて、免疫反応性を測定した。*P<0.05。
【0050】
【図16】P粒子−VP8キメラの製造および分析を示す。(A)Pドメインコード化cDNA配列を含むベクターpGEX−4T−1に基づく、P粒子−VP8キメラの発現構築物。酵素部位Spe IおよびCla I/EcoRIを有するクローニングカセットのT368とL375との間のPドメインのループ2中に、ロタウイルス(Wa)VP8抗原を挿入した。丸で囲んだCはシステイン含有ペプチド(CDCRGDCFC)を示す。(B)P粒子−VP8キメラの発現および精製。SDS PAGE分析は、GST−P−VP8融合タンパク質(GST融合体)が〜80kDaであることを示した(左パネル)。トロンビンによる融合タンパク質の消化は、GST(〜27kDa)およびP−VP8キメラ(〜52kDa、中央パネル)をもたらした。遊離P−VP8キメラは、トロンビン消化によって精製ビーズからも放出されうる(右パネル)。レーンMは、上から下に113、92、50、35、29、21kDaを示すバンドで、予め染色したタンパク質マーカーであった。(C)サイズ排除カラムSuperdex 200によるトロンビン放出P−VP8タンパク質のゲルろ過クロマトグラフィーの溶出曲線。ピークフラクションのSDS PAGE分析を上に示す。それぞれ青色デキストラン2000(〜2000kDa、空)、野生型P粒子(〜830kDa)および野生型Pダイマー(〜70kDa)でカラムをキャリブレートした。空のそばの単一主要ピークは、ほぼ全てのP−VP8タンパク質がキメラP粒子を形成したことを示す。(D)P−VP8タンパク質(左パネル)は、ロタウイルスVP8(中央パネル)およびノロウイルスVLP(右パネル)に対する抗体と反応した。
【0051】
【図17】P粒子提示VP8に対するマウスの免疫応答を示す。等モル量のP粒子−VP8キメラおよび遊離VP8を用いて、アジュバントなしで鼻腔内(AおよびB、n=5〜7)またはフロイントアジュバントと共に皮下(C、n=6〜7)で、マウスを免疫した。遊離VP8およびGSTをEIAにおける抗体力価測定のための抗原として使用した。(AおよびB)WaのVP8(A)およびDS1(B)それぞれを含む遊離VP8抗原(遊離VP8)およびキメラP粒子(PP−VP8)で免疫した後の、VP8/GSTに対するマウス血清の抗体力価。(C)遊離VP8抗原(遊離VP8)、およびWa株のVP8を含むキメラP粒子(PP−VP8)で免疫した後の、VP8に対する抗体力価。共精製したGSTは内部コントロールとして利用した。**P<0.001。
【0052】
【図18】P粒子−VP8キメラでの免疫によって誘導されるマウス血清による、ロタウイルスの中和を示す。(A)アジュバントなしでP粒子−VP8(Wa、[P]8)キメラで鼻腔内免疫したマウス由来のマウス血清は、同じWa株(PP−VP8血清、青)の強い中和を示すが、遊離VP8で免疫したマウス由来の血清は顕著に低い中和を示す(VP8血清、黒)。抗原を投与されていないマウス由来の血清をネガティブコントロールとして用いた(コントロール血清、紫)。(B)P粒子VP8(DS1、[P]4)キメラで免疫したマウス由来の血清は、Waとの弱い交差中和を示し(PP−VP8血清、青)、一方でDS1の遊離VP8で免疫したマウス由来の血清(VP8血清)およびネガティブコントロール血清は、中和を示さなかった(黒および紫)。(C)遊離Wa VP8(VP8血清、黒)およびP粒子−VP8(Wa)キメラ(PP−VP8血清、青)でフロイントアジュバントと共に皮下免疫したマウス由来の血清のVP8に対する免疫反応性。抗原なしでの血清は、ネガティブコントロールとして用いる(コントロール血清、紫)。(D)P粒子−VP8キメラによるマウスの免疫によって誘導される(C)由来の血清(PP−VP8血清、青)は、遊離VP8による免疫によって誘導される血清(VP8血清、黒)のものよりも、顕著に高いロタウイルス(Wa)に対する中和力価を示した。抗原を投与されていないマウス由来のコントロール血清をネガティブコントロールとして用いた(コントロール血清、紫)。星印は2つの形態のVP8によって誘導される血清の中和間のP値を示す:*P<0.05;**P<0.005および***P<0.0005。
【0053】
【図19】P粒子−VP8キメラワクチンでマウスを免疫した後の、マウスロタウイルス感染に対する保護を示す。4種の異なるワクチンでワクチン接種後、マウスのロタウイルス排出(μg/ml)を測定し、マウスロタウイルス(EDIM)でチャレンジした。WT PPでは、野生型ノロウイルスP粒子(ベクターコントロール)をマウスにワクチン接種した(n=7)。PP−hVP8では、P粒子−VP8(Wa)キメラをマウスにワクチン接種した(n=5)。遊離mVP8では、遊離マウスVP8(EDIM)抗原をマウスにワクチン接種した(n=5)。PP−mVP8では、P粒子−VP8(EDIM)キメラをマウスにワクチン接種した(n=5)。データ計算および統計分析は表1に示す。
【0054】
【図20】P粒子−VP8キメラでの免疫によって誘導された抗体が、ノロウイルスVLPとHBGAレセプターの結合を遮断することを示す。(A)P粒子−VP8(Wa)キメラで免疫した後のマウス血清は、ノロウイルスP粒子と強く反応したが(PP−VP8)、遊離VP8誘導血清は、この反応性を示さなかった(遊離VP8)。(B)(A)由来のマウス血清はノロウイルスVLPとHBGAレセプターの結合を遮断したが(タイプA唾液、PP−VP8血清、青)、遊離VP8での免疫後に得られた血清は、この遮断を示さなかった(VP8血清、黒)。
【発明を実施するための形態】
【0055】
発明の詳細な説明
本明細書において使用するとき、用語「ノロウイルス」、「NOR」、「ノーウォーク様ウイルス」または「NLV」は、ノロウイルスファミリーの任意のウイルスを意味し、限定されないが次のものを含む:ノーウォークウイルス(NV)、MOH、Mexico、VA 207、VA 387、02−1419、C59、VA 115、Hawaii、Snow Mountain、Hillington、Toronto、Leeds、Amsterdam、Idaho Falls、Lordsdale、Grimsby、Southampton、Desert Shield、BirminghamおよびWhite Rivercap。
【0056】
本明細書において使用するとき、用語「Pドメインモノマー」および「P粒子」は、それぞれ野生型ノロウイルスPドメインモノマーおよび野生型ノロウイルスP粒子を意味する。
【0057】
本明細書において使用するとき、用語「修飾されたPドメインモノマー」および「修飾されたP粒子」は、分子クローニングによって各Pドメインモノマー上に存在する3個の表面ループの1個以上に、1個以上の制限部位を(典型的には対で)挿入し、それによって後で外来抗原、リガンド、薬物コンジュゲートまたはシグナルペプチドを挿入できるように表面ループにクローニングカセットを作成するように修飾されている、ノロウイルスPドメインモノマーおよびP粒子を(それぞれ)意味する。
【0058】
本明細書において使用するとき、用語「置換Pドメインモノマー」および「置換されたP粒子」は、分子クローニングによって各Pドメインモノマー上に存在する3個の表面ループの1個以上に挿入された少なくとも1個の外来抗原、リガンド、薬物コンジュゲートまたはシグナルペプチドを含む、ノロウイルスPドメインモノマーおよびP粒子を(それぞれ)意味する。
【0059】
本明細書において使用するとき、用語「抗原−Pドメインモノマー」または「抗原−ノロウイルスPドメインモノマー」は、外来抗原がPドメインモノマー上に存在する3個の表面ループの1個以上に挿入されている、置換Pドメインモノマーを意味する。
【0060】
本明細書において使用するとき、用語「抗原−P粒子」または「抗原−ノロウイルスP粒子」は、外来抗原が各Pドメインモノマー上に存在する3個の表面ループの1個以上に挿入されている、置換されたP粒子を意味する。
【0061】
4種の固有の非感染性NOR粒子、すなわちウイルス様粒子(VLP)、S粒子、P粒子および小P粒子は、多機能性を設計するための優れた候補を表すウイルス性タンパク質ケージであり、Tan et al., 2004, 「The P-domai of norovirus capsid protein forms dimer and binds to histo-blood group antigen receptors」, J Virol 78:6233-42およびTan et al., 2005, 「The p domain of norovirus capsid protein forms a subviral particle that binds to histo-blood group antigen receptors」, J Virol 79:14017-30に記載されている(それらの全体について、参照によりそれらの開示を本明細書に組み込む)。これらの粒子は、分子量420kDa(小P粒子)〜10.5mDa(VLP)を有し、サイズ14〜37nmを有する。NOR VLPは180個の完全カプシドタンパク質(VP1)から成り、3個のサブウイルス性粒子はVP1の一部から成る。シェル(S)ドメイン由来のS粒子、突起(P)ドメイン由来のP粒子および同じPドメイン由来であるがN末端にさらなるフラグタグを有する小P粒子。これら全ての粒子は、Tan et al., 2004, 上記; Tan et al., 2004, 「E. coli-expressed recombinant norovirus capsid proteins maintain authentic antigenicity and receptor binding capability」, J Med Virol 74:641-9; および Xia et al., 2007, 「Norovirus capsid protein expressed in yeast forms virus-like particles and stimulates systemic and mucosal immunity in mice following an oral administration of raw yeast extracts」, J Med Virol 79:74-83(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載のとおり、タンパク質がインビトロで発現されたとき自然発生的に集合する。
【0062】
野生型NORカプシドは、昆虫細胞において発現されたときウイルス様粒子(VLP)に自己集合する、55〜60kDaの単一の主要構造タンパク質(VP1)から成る(図2)。VLPは天然ウイルス粒子と形態学的および抗原的に同一であるが、感染性ではない。ノーウォークウイルスのX線結晶学的構造に基づくと、カプシドはVP1の90個のダイマーから成り、T=3正二十面体シンメトリーを示す。各VP1モノマー(530アミノ酸[aa])は、短いN末端領域(1〜49アミノ酸)、続いてシェル(S)ドメイン(50〜225アミノ酸)、ならびにP1(226〜278および406〜520アミノ酸)およびP2(279〜405アミノ酸)に分類できる突起(P)ドメインを含む。N末端/シェル(N/S)ドメインはカプシドの内部コアを形成し、VP1の最も保存的な部分であるが、Pドメインはカプシドの突出したアーチを形成し、より多様である。カプシドの表面に位置するP2サブドメインは、NOR株のゲノムにおいて最も高い多様性度を含む。それは株特異性、レセプター結合および潜在的中和抗体認識部位の決定基を含む。
【0063】
原子構造に従って、NORカプシドタンパク質のシェル(S)および突起(P1、P2)ドメイン(図2)は、8〜10アミノ酸ヒンジによって連結されている。P粒子のカプシドタンパク質(VP1)のレセプター結合領域は、Tan et al., 2004, 「The P domain of norovirus capsid protein forms dimer and binds to histo-blood group antigen receptors」, J Virol 78:6233-42(その全体について、参照によりその開示を本明細書に組み込む)に記載されているとおり、マッピングされている。
【0064】
NORは、感染のための宿主レセプターとしてヒト組織−血液グループ抗原(HBGA)を認識することが知られている。Sドメインの発現は宿主レセプターに結合する機能を有さない平滑粒子を形成するが、ヒンジを有するPドメインの発現は、レセプター結合機能を有するダイマーを形成するが、その結合親和性は低い。ヒンジなしのPドメインが発現されると、それはVLPのものと同様の顕著に高いレセプター結合親和性を有するサブウイルスP粒子(図1)を自然発生的に形成する。P粒子は12個のダイマーに組織される24個のPドメインモノマーからなる。P粒子は、対応するHBGAに結合でき、NOR VLPとHBGAレセプターの結合の強い遮断を示す。P粒子の自然発生的形成は、株VA387、MOHおよびノーウォークウイルス(NV)を含む多様なNOR株で観察されている。これらのP粒子は、電子顕微鏡(EM)下で中心に空洞を有するリング状のイメージを示し、分子量〜830kDaでゲルろ過で単一ピークを形成した。P粒子は極めて安定であり、Tan et al., 2005, 「The p domain of norovirus capsid protein forms a subviral particle that binds to histo-blood group antigen receptors」, J Virol 79:14017-30、およびTan et al., 2006, 「C-terminal arginine cluster is essential for receptor binding of norovirus capsid protein」, J Virol 80:7322-31(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載のとおり、大腸菌および酵母発現系の両方で、高収率で容易に製造できる。
【0065】
NOR P粒子は、ヒトNORの2つの主要な遺伝子群の異なる遺伝子群からのNOR P粒子の構築によって示されるように、NORに共通の特徴である。GI−1(ノーウォークウイルス)、GI−4(Koblenz433)、GI−8(Boxer)、GII−2(Melksham)、GII−3(Brattleboro321)、GII−4、GII−5(MOH)、GII−9(VA207)を含む異なるNOR遺伝子型を示す、40種類以上のNOR P粒子がこれまでに構築されている。世界中のNOR胃腸炎の65〜85%に関与する主たるNORタイプ(GII−4)について、25種類のP粒子が作成されている。これは、過去25年間で25種類の株が広まっていることを示す。
【0066】
P粒子の3D構造は、図1および3に示すとおり、7.6Åの分解能のcryo−EMによって再構成されている。P粒子は、12個のダイマーに組織される24個のPドメインモノマーを含む八面体シンメトリーである。P粒子の中心に空洞が存在する。Pダイマーの方向は、野生型カプシドのものと同様であり、P1サブドメインが空洞の中心に対して内側にあり、P2サブドメインがP粒子の最も外側の表面に対して外側にある。P粒子のcryo−EMの密度マップへのPダイマーの結晶構造の重ね合わせ(図4)は、HBGAレセプター結合界面がP粒子の最も外側の表面に位置することを示している。これは、HBGAレセプターとの強い結合機能を保持しているP粒子の観察と一致している。
【0067】
Pダイマーの原子構造は、プロトタイプノーウォークウイルスおよび主なGII−4株(VA387)においてX線結晶学によって解明されている(図4)。これらの結晶構造は、P粒子のcryo−EMの密度マップに良好にフィットする(図1)。3個の表面ループは、これらはアーチ様Pダイマーの上部(6個の表面ループ/Pダイマー、図3)またはNORカプシドおよびP粒子の最も外側の表面に対応する各Pモノマーの最も遠い末端で同定された。これらの表面ループは外来抗原提示のための優れた部位を提供し、詳細な結晶構造は、本明細書に開示するP粒子ワクチンプラットフォームの構造に基づく設計のための確かな基礎を提供する。
【0068】
HBGAレセプターとの結合について、P粒子はその外側表面に依存する。いずれか1個以上の制限酵素部位、抗原、リガンド、シグナルペプチドまたは薬物コンジュゲートが野生型Pドメインモノマーの表面ループに挿入されている場合、制限酵素部位、抗原、リガンド、シグナルペプチドまたは薬物コンジュゲートのサイズに部分的または完全に基づいて、それは形成されたP粒子の最も外側の部分を被覆し、1個以上の修飾もしくは置換されたループへのアクセスを隠すことができ、または修飾もしくは置換されたループのタンパク質構造を変化させて、形成されたP粒子がHBGAレセプターと結合するその能力を失わせることができる。我々は未だP粒子が最早HBGAに結合しないという前にはどの程度のループの変化が必要かということについての系統立てた分析は行っていない。しかし、我々は、制限部位をPドメインモノマーの中央ループ2に加えた場合に修飾されたP粒子がHBGAと結合しないことを見出している。
【0069】
ウイルスレセプター結合界面の同定は、図4に示すとおり、ノーウォークウイルスおよびVA387におけるA−またはB−トリサッカライド(ノロウイルスレセプター)との複合体におけるPダイマーの結晶化によって示されている。図示したとおり、図4は、B−トリサッカライド複合体(球体モデル)と関連したNOR Pダイマーの結晶構造(リボンモデル)を示す。レセプター結合界面はNORカプシドおよびP粒子の最も外側の表面と等しいアーチ様Pダイマーの上部に位置する。トリサッカライドとPダイマーのアミノ酸との間の大規模な水素結合ネットワークが観察される。ウイルスレセプターと相互作用していると予測されるこれらのアミノ酸の重要性は、Tan et al., 2008, 「Elucidation of strain-specific interaction of a GII-4 norovirus with HBGA receptors by site-directed mutagenesis study」,Virology 379:324-334(その全体について、参照によりその開示を本明細書に組み込む)に記載されているとおり、部位特異的突然変異誘発によって証明されている。
【0070】
4個のNOR粒子(VLP、S粒子、P粒子および小P粒子)のうち、P粒子は最も安定であり容易に製造できる。それは高度に免疫原性であり、広範な温度、pHならびに化学的および物理的条件に耐える。cryo−EMによるP粒子の3D構造(図1)およびPダイマーの原子構造(図2)は、Cao et al., 2007, 「Structural basis for the recognition of blood group trisaccharides by norovirus」, J Virol 81:5949-57、およびTan, 2008, 「Noroviral P-particle: Structure, function and applications in virus-host interaction」, Virology, Vol. 382, Issue 1, 5 December 2008, Pages 115-123(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載のとおり、P粒子の24個のモノマーまたは12個のPダイマーの各々の最も外側の表面に高度に露出している(図1、3および4)。構造に基づく配列アラインメントは、これらの露出ループが大きな配列の挿入に耐えることを示唆している。
【0071】
本発明の一つの局面において、タンパク質またはポリペプチド抗原性エピトープがこれらのループ、すなわちループ1(I293−H297)、ループ2(T371−D374)またはループ3(D391−N394)の一つで、あるいはその中に挿入されている場合、抗原−Pドメインモノマーまたはダイマーの自然発生的自己集合は、集合した抗原−P粒子の表面で24個の抗原エピトープを提示する。本発明のこの特徴は抗原提示、したがって下記の通りワクチンのための利点を提供する。Pダイマーの解明された分子構造は、したがって、かかる抗原挿入のための構造に基づく設計の確かな、詳細な情報を提供する。
【0072】
野生型Pドメインモノマーおよびダイマーのように、抗原−Pドメインモノマーがダイマーを、そしてダイマーが抗原−P粒子を形成するプロセスは、Pダイマーの濃度に基づいて動力学的に交換または平衡しうる。また野生型Pドメインモノマーおよびダイマーのように、システインが抗原−Pドメインモノマーの末端に結合しているとき、得られる抗原−P粒子は、システイン末端を有さないものよりもより安定となり、抗原−Pドメインモノマーの濃度に依存しない。各抗原−Pダイマーは、5本の折り畳み軸に沿った五角形を形成する、5個の周囲抗原−Pダイマーとの相互作用において同じ方向を有するべきである。結果として、抗原−P粒子の外側層はノロウイルスカプシドのアーチ構造と類似のP2サブドメインによって形成されるが、内側層はP2アーチに支えを提供するP1サブドメインによって作成される。他の正二十面体ウイルス粒子と同様に、抗原−P粒子の内側に空洞が存在すべきである。
【0073】
また野生型Pドメインモノマーおよびダイマーのように、本発明の抗原−Pドメインモノマーおよびダイマーは、インビトロで抗原−P粒子を自然発生的に形成することを防止するために、周知の方法で変性されていてもよい。同様に、本発明の抗原−P粒子は、それぞれの抗原−Pドメインモノマーおよびダイマーに自然発生的に分離しないように、変性されていてもよい。これは、所望により野生型Pドメインモノマーと共に、多様な抗原タイプの抗原−Pドメインモノマーの形成の、多様な分子割合へのインビトロでの混和または混合を可能として、次いで変性の影響が除かれたときに自然発生的に抗原−P粒子へと形成されうる。
【0074】
本発明の置換されたまたは抗原−P粒子は、安定であり、Tan et al., 2004, J Virol 78:6233-42およびTan et al., 2005, Virol 79:14017-30に記載のもののような周知の方法によって大腸菌および酵母において容易に製造できるため、野生型NOR P粒子のように、抗原ディスプレイのための魅力的な候補である。先行技術のManayani et al.のキメラFHV粒子およびChatterji et al.のキメラCPMV粒子はそれぞれ、昆虫および植物に感染するが(ヒトに対する影響は知られていない)、非感染性野生型NOR P粒子はヒトウイルスに由来し、候補ヒトワクチンとして同定されている。
【0075】
多くの固有の特徴のため、本発明の抗原−P粒子は、野生型NOR P粒子のように、ワクチンプラットフォームとして有用な商品となる。第一に、ベースのNOR P粒子はサブユニットワクチンにほぼ理想的なサイズ(〜840kDa、Φ=20nm)を有し、多価であり、完全サイズのウイルス様粒子(VLP)よりも容易に製造でき、単一ポリペプチドまたはタンパク質抗原よりも免疫原性である。第二に、抗原−P粒子は高収率かつ低コストで大腸菌で容易に製造でき、これは発展途上国にとって特に有用である。第三に、Pドメインモノマーの複数の(3個)表面ループおよび外来抗原の挿入のためのそれらの高い能力(少なくとも〜238アミノ酸まで)は、異なる複数の抗原の挿入を可能とする。したがって、得られたキメラ置換P粒子ワクチンは、多くの感染症および非感染症に対してさえも、広い適応を提供できる。第四に、外来抗原が挿入された少なくとも24個のコピーを含む均一なキメラ置換P粒子を作成するために、2〜3段階の単純な遺伝子操作のみが必要とされる。したがってこれらのワクチンは速やかに開発でき、インフルエンザウイルスのような急速に変化するウイルスファミリーに特に有用であり得る。第五に、制限部位を含む表面ループにクローニングカセットを有するPドメインモノマーベクターは、ユーザーフレンドリーなベクターであり、簡便なワクチンプラットフォームである。そして第六に、抗原−P粒子は、両方のヒトNORについての下痢および他の感染症に対する優れた候補ワクチンである。
【0076】
主たる感染症に加えて、本PドメインベクターおよびP粒子ワクチンプラットフォームは、抗原提示および/または薬物の担体もしくはがん、アレルギー、自己免疫疾患を含む非感染性疾患の処置のための薬物送達ビークルとしても有用であり得る。一つの態様において、薬物は、化学反応によって表面露出リシンおよびシステインを介してP粒子のループに挿入されていてよい。この場合、置換されたP粒子は、薬物の担体として用いられる。他の態様において、リガンドまたはシグナルペプチドは、表面ループの少なくとも一つに挿入されてよく、そして薬物のコンジュゲートも、表面ループの少なくとも一つに挿入されていてよい。これらの置換されたP粒子は、薬物に特定の組織または臓器を標的とさせる薬物送達系を提供する。
【0077】
本発明のP粒子は、さらに特異性を高め、当該技術分野の有用な進歩を利用するために、他の薬物送達アプローチと組み合わせてもよい。一つの例は、病原体感染または特定の化学的もしくは酵素的切断がないかぎり、そしてそれまで、生物学的に不活性であり、植栽棒哺乳類細胞の内部で活性薬物形態に変換される、プロドラッグを含む。かかる態様は、特定の微生物に感染した細胞または特定の腫瘍に罹患した細胞でのみプロドラッグが活性化される、プロドラッグ−P粒子の開発を特に含みうる。
【0078】
P粒子ベクターの固有の特徴のために、ワクチンプラットフォームは広い適応を有することができる。それは既知の保護エピトープを有する多くのワクチンを製造するために使用できる。したがって、P粒子ベクターワクチンプラットフォームの適応は、広範な臨床状態に潜在的に有益であり得る。さらに、このナノ粒子プラットフォームの柔軟性は、容易に拡張されうる。例えば、PドメインモノマーまたはP粒子の1個の表面ループが特定のレセプターを有するある組織または臓器にP粒子を誘導するシグナルペプチド(リガンド)を発現するが、ワクチンまたは薬物が効果を奏するように送達される第2の表面ループに結合されるように、粒子を開発できる。例えば、ペプチドCNGRC(5アミノ酸の鎖)で置換されたP粒子は、CNGRCのレセプター(CD13)が重度に発現されている腫瘍組織(すなわち癌腫)に移動できる。膜結合メタロプロテアーゼCD13のリガンドであるペプチド部分CNGRCは、CD13を発現する血管形成血管系の内皮細胞に結合でき、CD13を発現する内皮細胞におけるアポトーシスを誘導し、それによって腫瘍関連血管新生を阻害できる。
【0079】
さらに、2個以上のループが同一または異なる病原体由来の異なる抗原を発現するように、多価ワクチンが作製できる。置換されたP粒子は、野生型Pドメインモノマーおよび抗原−Pドメインモノマーを含む、異なる抗原−Pドメインモノマーの混合物から集合して、異種抗原−P粒子を形成できる。多様な外来抗原を同じループまたは3個のループの異なる一つに挿入して、広範な異種抗原−P粒子を提供できる。したがって、本P粒子およびPドメインモノマーベクタープラットフォームの市場価値は極めて高い。
【0080】
本発明の態様は、NORおよびロタウイルス(RV)に対するワクチンとして使用するための抗原−P粒子を含んでいてもよい。抗原−P粒子の一つの例は、RV VP8−P粒子キメラである。ロタウイルス(RV)およびNORは、疾患の大きな負担を生じる世界中で共通の病原体である。この抗原−P粒子は、毒性株を生じる復帰または再集合の可能性のない、単独ワクチンでNORおよびRVに対して有効なワクチンを提供する。
【0081】
1個以上のループの修飾の前に、P粒子形成を促進し安定化するためにPドメインのいずれかの末端にシステインを加え、そしてP粒子を不安定化しうるトリプシン処理に対する耐性を与えるためにPドメインのC末端に短いペプチド(CDCRGDCFC)を加えて、野生型Pドメインモノマーを製造できる。また、P粒子形成のために、PドメインモノマーのインタクトなC末端が必要であり、ワクチンプラットフォームとしてのP粒子形成および安定性のために、C末端でのアルギニンクラスターが重要である。
【0082】
小ペプチドタグ(DYKDDDDK、フラグタグ)がPドメインのC末端に結合したとき、NOR Pドメインモノマーはまた、小P粒子と呼ばれる別の複合体を形成した。この小P粒子は、6個のPダイマーに組織される12個のPモノマーを含む四面体シンメトリーを示す(図6)。小P粒子のcryo−EMの密度マップへのPダイマーの結晶構造の重ね合わせは、Pダイマーの方向がP粒子のものと同様であることを示した。インビトロでのHBGA結合アッセイは、小P粒子がNOR VLPおよびP粒子と同様の結合プロファイルを有することを示した。小P粒子の発見は、より小さなP粒子が特定の用途に必要である場合における、我々のP粒子系のさらなる候補を提供する。
【0083】
本発明のNORベースの抗原−P粒子は、各Pドメインダイマーまたはモノマーサブユニットの提示ループの1個以上において外来抗原を担持するように構築されうる。Hisタグ(7xヒスチジン)、マウスサイトメガロウイルス(MCMV)T細胞エピトープ(9アミノ酸)、シュードモナス属エピトープEpi8(14アミノ酸)およびRV表面抗原である2種のRV VP8コアタンパク質(各159アミノ酸)を含むがこれらに限定されない多様なポリペプチドおよび生物学的関連エピトープが、置換されたP粒子の形成および安定性に影響を与えることなく、図9に示すとおり、P粒子のループ1(I293−H297)、ループ2(T371−D374)またはループ3(D391−N394)のいずれか1つ以上に挿入されてよい。これはループ2について、図7のパネルBおよび図8のパネルDに示すとおり、ゲルろ過によって示される。Hisタグの露出はTalon樹脂との高い親和性によって示された(図7のパネルA)VP8の露出は、RV VP8に対する抗体を用いたEIA(図8のパネルE)およびcryo−EMを用いたVP8−P粒子の3D構造再構成(図10のパネルE)によって示されている。これらのデータは、P粒子が多様な外来抗原を提示するための有効なワクチンプラットフォームに発展しうるという我々の仮説を支持している。
【0084】
抗原−P粒子ベクターは、クローニングカセットと共に構築されてもよい。Pダイマーの結晶構造に基づいて、ループの先端を含むP粒子ベクターのループの一つ以上に制限部位を含むクローニングカセットを入れることができる。図9はP粒子ベクターのループ2の先端に入れたSpe I部位およびCla I部位を含むクローニングカセットを示す。クローニングカセットを含むこのP粒子ベクターは、図8および10に示すように、P粒子の表面へのRV VP8およびGFPの挿入成功に使用される。
【0085】
稀少および固有制限部位を含む制限部位、ならびにPドメインモノマーまたはP粒子の表面ループのいずれか一つ以上に制限部位を挿入する方法の例は、当業者に周知であり、Reed et al.の米国特許出願公開2004/0185556および2008/0085553(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載されている。これらの制限部位はDNA分子においてコードされ、典型的には二本鎖パリンドローム配列を含む。いくつかの制限酵素について、制限部位を提供するために4〜6ヌクレオチドで十分であるが、いくつかの制限酵素は、8個以上のヌクレオチドの制限部位を必要とする。例えば酵素EcoR1は下記6ヌクレオチド配列を認識する:5’G−A−A−T−T−C3’、ここで5’は「上流」末端と慣習により知られる分子の終点を示し、同様に3’は「下流」末端を示す。制限部位の相補鎖は、その逆平行鎖3’=G−A−A−T−T−C−5’である。
【0086】
NOR P粒子は大きな外来抗原挿入に耐えうる。例示のみを目的として、NOR P粒子のループ2に緑色蛍光タンパク質(GFP、238アミノ酸)を挿入したところ、得られたキメラ粒子はゲルろ過分析で測定したところ高度に安定である。GFPの露出は、GFP特異的抗体を用いたEIAおよび蛍光顕微鏡で示された(データは示さず)。
【0087】
ある方法の態様において、修飾されたNOR P粒子を用いて、NORおよび外来抗原両方に反応性の抗体を誘導する。具体的な、非限定的な例において、RV VP8−P粒子を用いて、RVおよびNORの両方に反応性の抗体を誘導する。VP8−P粒子キメラで免疫した哺乳類はNOR P粒子およびRV VP8の両方に反応性の抗体のレベルをもたらし、NOR VLPとHBGAレセプターの結合を遮断し、RVの複製を中和できる。
【0088】
NORの「中和アッセイ」およびRVチャレンジのマウスモデルが開発されている。NORは未だ細胞培養で培養することが難しく、NORの複製に有効な動物モデルが存在しない。NOR感染に対する抗体の潜在的保護を測定するため、NORとその宿主レセプターの結合の防止における抗体の能力を測定するためのHBGAレセプター遮断アッセイを開発した。このアッセイは、唾液またはオリゴ糖ベースのレセプター結合アッセイに基づいており、極めて感受性かつ特異性である。
【0089】
RV感染に対するワクチンの保護効能を測定するための成体マウスモデルも確立されており、RVワクチン開発を研究するために用いられている。ワクチン候補でマウスを免疫し、次いでマウスRV株でチャレンジした。免疫群におけるRV抗原の糞便排出量を、ワクチン効能の測定のためにコントロールまたは非免疫群における排出量と比較する。ワクチンに対する免疫応答は、抗体レベルおよびT細胞応答を測定することでも測定できる。
【0090】
本発明はまた、外来抗原の複数または複数−多価提示を有する抗原−P粒子の形成に関する。2個のループ(すなわちループ1および2、ループ2および3またはループ1および3)で2個のクローニングカセットを有する修飾されたP粒子ベクターは、多様な組合せで2個のカセットに外来抗原を挿入することによって構築でき、評価できる。3種の組合せの非限定的な例は、1)ループ1および2におけるHisタグおよびフラグタグ(YDKDDDDK);2)ループ1および2におけるHisタグおよびVP8抗原;ならびに3)ループ1および2における2個のVP8を含む。これらの態様は、同じ置換P粒子上で効率よく提示される2種の異なる小抗原、または1種の小抗原および1種の大抗原、または2種の大抗原を提供できる。キメラP粒子形成はゲルろ過によって測定され、挿入された抗原の露出は特異的抗体を用いたEIAによって試験できる。
【0091】
3個のループ(ループ1、2および3)全てで3個のクローニングカセットを有する修飾されたP粒子ベクターは、多様な組合せで3個のカセットに外来抗原を挿入して構築でき、評価できる。非限定的な例は、1)3個の7xHisタグ;ならびに2)7xHisタグ、mycタグ(EQKLISEEDL)およびフラグタグを含む。第一の組合せは、置換されたP粒子の各Pドメインモノマーの3個のループ全てで同じ小さなエピトープを重複して提示し、P粒子上で抗原の72個(3×24)コピーを与える。第二の組合せは、各Pドメインモノマーサブユニットで同時に提示される3種の異なる小エピトープを提供し、3種の異なる抗原に対する三価P粒子ワクチンをもたらす。
【0092】
したがって本発明は、簡便かつ多様な抗原挿入のためのP粒子ベクターを提供する。修飾されたP粒子の態様は、それぞれ3個の表面ループの各々で単一の固有のクローニングカセットを含む各々修飾されたPドメインモノマーである、3種のPドメインモノマーベクターを含みうる。修飾されたP粒子ベクターの別の態様は、3個のループのいずれか一つもしくはいずれか2つ(例えばループ1および2)または3個のループ全てにおいて2個のクローニングカセットを有するPドメインモノマーベクターを含む。修飾されたP粒子ベクターの別の態様は、各Pドメインモノマーの3個のループ全てにおいて3個のクローニングカセットを有していてよい。各クローニングカセットの酵素認識部位は、図13に示すとおり、目的の抗原のDNA配列のクローニングの必要に応じて修飾されてよい。これらのベクターは、1種類の抗原の単独提示または同一もしくは異なる抗原の複数提示のためものもである。異なるクローニングカセットを有するこれらのP粒子ベクターの構築は、当該技術分野で知られているとおり、部位特異的突然変異誘発によって達成されうる。
【0093】
変異誘発および組換えDNAクローニング手法。様々な修飾または置換されたP粒子ベクターの作製は、典型的には、NOR(特に、実施例のとおり、VA387株)のPドメインおよびシステインを有する短ペプチドをコードする配列を好適なプラスミド(特に、例示として、Amersham Biosciencesから入手可能なプラスミドpGEX−4T−1)中に含む、図12に示すような、野生型P粒子発現構築物で開始する。クローニングカセットは、Tan et al., 2003, 「Mutations 448 within the P2 domain of norovirus capsid affect binding to human histo-blood group 449 antigens: evidence for a binding pocket」, J Virol 77:12562-71、Tan et al., 2006, 「C-terminal arginine cluster is essential for receptor binding of norovirus capsid protein」, J Virol 80:7322-31、Tan et al., 2005, 「The p domain of norovirus capsid protein forms a subviral particle that binds to histo-blood group antigen receptors」, J Virol 79:14017-30およびTan et al., 2008, 「Elucidation of strain-specific interaction of a GII-4 norovirus with HBGA receptors by site-directed mutagenesis study」, Virology 379:324-334(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載のものを含む周知の方法による部位特異的突然変異誘発によって、一つずつ、図13に示すように、表面ループの特定部位でPドメイン配列に導入される。結晶学的研究による3個の表面ループの位置、組成および配列は、Bu et al., 2008, 「Structural basis for the receptor binding specificity of Norwalk virus」, J Virol 82:5340-7およびCao et al., 2007, 「Structural basis for the recognition of blood group trisaccharides by norovirus」, J Virol 81:5949-57(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載されている。
【0094】
構造および配列情報に基づいて、クローニングカセットとして特定の酵素認識部位が適切な位置に挿入されたものとして、特定のプライマーを設計し、合成できる。典型的には、表面ループの先端(最も露出した位置)での2個のアミノ酸がクローニングカセットの配列を挿入するために選択されてよく、これは2個のアミノ酸の置換または2個のアミノ酸の間に配列を挿入することによってよい。周知の通り、QuikChange Site-Directed Mutagenesis Kit(Stratagene)を用いてループにクローニングカセットを導入できる。特定の抗原についての正確な導入位置は、得られる抗原−P粒子キメラの安定性または効率を改善するために修飾されてもよい。より長い配列でのクローニングカセットの挿入のために、最適な挿入のために段階的部位特異的突然変異誘発を実施できる。次いで、部位特異的突然変異誘発の後得られた置換されたP粒子ベクターのプラスミドは、周知の方法によって、プラスミドDNA調製キット(Qiagen)を用いた大腸菌およびプラスミドDNA調製物へのプラスミド形質転換のための常套のクローニング手法によって増幅できる。周知の方法により配列決定することによって、挿入されたクローニングカセットの確認の後、抗原−P粒子ベクターは、抗原挿入の能力、キメラP粒子形成の安定性、および有用かつ効果的なワクチンとしての抗原提示の効率について、試験されてよい。
【0095】
適切なキメラP粒子形成は、抗原提示についてのP粒子ベクターの能力および効率を評価して決定できる。P粒子プラットフォームへの抗原またはエピトープの挿入後(すなわち、Pドメインモノマーに抗原またはエピトープの挿入後、ダイマー、次いで抗原−P粒子へのPドメインモノマーの集合)、得られたキメラタンパク質の各々は、サイズ排除カラム(例えば、Akta ファストパフォーマンス液体クロマトグラフィー(FPLC、HE healthcare))を用いたゲルろ過によって、標的抗原−P粒子キメラの形成について分析されてよい。正しく形成された標的キメラは、モノマーの分子量の〜24倍の予測分子量(MW)で単一のピークを形成する。例えば、HisタグP粒子は870kDaのMWを有し、VP8−P粒子は1240kDaのMW、そしてGFP−P粒子は1450kDaのMWを有する。P粒子による抗原の複数提示は、この計算に従う。予想されるMWを有するキメラP粒子の形成は、抗原挿入の成功の重要な指標である。キメラP粒子形成のさらなる証拠は、cryo−EMによるEM観察および3D再構成を含む(図10参照)。修飾された抗原−P粒子の安定な形成は、通常、抗原の優れた免疫原性を導き、対応する病原体に対する有望なワクチン候補であろう。
【0096】
キメラP粒子上の抗原露出の測定は、抗原提示についてのP粒子ベクターの能力および効率の別の重要な指標である。抗体特異的EIAおよびTalon樹脂結合アッセイのいずれかまたは両方が、この測定のために用いられる。抗体特異的EIAはHuang et al., 「Noroviruses bind to human ABO, Lewis, and secretor histo blood group antigens: identification of 4 distinct strain specific patterns,」 2003, J Infect Dis 188:19-31およびHuang et al., 「Norovirus and histo-blood group antigens: demonstration of a wide spectrum of 23 strain specificities and classification of two major binding groups among multiple 24 binding patterns,」 2005, J Virol 79:6714-22(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載されており、図8のパネルEおよび図11のパネルAに図示されている。精製された抗原−P粒子キメラはマイクロタイタープレートに被覆され、脱脂乳によって遮断されうる。露出している抗原/エピトープは、対応する抗体によって検出できる。シグナル強度は、二次抗体−HRPコンジュゲート、次いでHRP基質とインキュベート(inculcation)した後、吸光度(OD)によって記録されうる。抗原(例えばそれぞれHis、フラグおよびmycタグ)に対して特異的なモノクローナル抗体は、Cell Signalingから商業的に入手可能であるが、図8および11に示すとおり、RV VP8およびNOR P粒子に対する特異的抗体は本願出願人によって製造され、それから入手可能である。露出したHisタグの検出のために、別の強力な方法であるTalon樹脂結合アッセイを用いてもよく、ここでTalon樹脂(Clontech)はHisタグと特異的に結合できる(図7)。樹脂結合HisタグP粒子キメラはイミダゾール(Sigmaから入手可能)によって容易に放出されるため、このアッセイは、汚染タンパク質からHisタグP粒子キメラを精製するための単純なアプローチである(図7参照)。
【実施例】
【0097】
実験
キメラP粒子の調製。予め製造した[P]タイプ8株のヒトVP8を含むRV VP8−P粒子キメラ(図8、図10のパネルE)を用いる。マウスRVチャレンジ試験のためのマウスVP8を含むキメラワクチンのために、VP8−P粒子キメラのヒトVP8をマウス(EDIM)VP8で置換する。ヒトおよびマウスRVキメラP粒子はいずれも、確立された手法を用いて大腸菌で製造される。GSTアフィニティーカラム(Glutathione Sepharose 4 Fast Flow、HE Healthcare)による部分的精製の後、キメラP粒子はさらにサイズ排除カラムを用いたFPLC系(GE Healthcare)で精製される。得られたキメラP粒子調製物は、高い純度(〜95%)に達し、マウスを免疫する免疫源として使用される。図8に見られるとおり、SDS PAGEのパネルAは、アフィニティーカラム(レーン1)およびさらにP粒子(レーン2)およびPドメインダイマー(レーン3)フラクションでのゲルろ過(パネルD)によって精製された、組換えRV VP8−Pドメインキメラ(53.5kDa)を示す。パネルBおよびCは、VP8(パネルB、レーン4および5)またはNOR VLP(パネルC、レーン6および7)に対する抗体を用いた、VP8−P粒子キメラのウェスタン分析を示す。パネルDにおいて、アフィニティー精製VP8−Pドメインキメラ(レーン1)のゲルろ過は、該タンパク質の大部分がP粒子(〜1300kDa)を形成することを示す。パネルEは、被覆VP8−P粒子キメラ(PP−hVP8C)、組換えVP8(hVP8C)およびP粒子のみ(PP、コントロール)を検出するためにVP8に対する抗体を用いた、酵素免疫アッセイ(EIA)の結果を示す。レーンMは、上から113、92、52、35、29kDaのバンドで、予め染色したタンパク質マーカーを示す。図10は、RV VP8−P粒子キメラの製造を示す。ループ2にクローニングカセットを有するP粒子ベクター(A)は野生型NOR P粒子(B)を製造する。RV VP8コード配列(C)がP粒子ベクター(D)にクローン化されると、それはキメラの最も外側表面でのRV VP8の指標と共に、RV VP8−P粒子キメラ(E)を製造する。
【0098】
マウスの免疫。6週齢のメスBALB/cマウス(Charles River Labs)を高度精製RV VP8−P粒子キメラで、異なる用量で、アジュバント[LT(R192G)]ありまたはなしで、経口または鼻腔内で免疫する。野生型P粒子はコントロールとして含まれる。ワクチン接種の鼻腔内経路のために、体積50μl中、野生型P粒子およびVP8−P粒子キメラの3種の初期用量(5、15および45μg/マウス、3回用量)が投与される(鼻腔あたり25μl)。経口経路のために、1mlのシリンジを備えた20ゲージのステンレス鋼製給餌針を用いて胃内カニューレによって胃に直接、合計200μlの同量のワクチンを送達する。動物(12匹のマウス/群)を一般的外見および体重減少についてモニターして、ワクチンの有害な効果の可能性を評価する。回復期前および中の血清サンプルを回収し、抗原として組換えNOR VLPおよびRV VP8を用いたEIAによってNORおよびRVに対する抗体応答について試験する。NORおよびRVに対する分泌IgAのために、糞便サンプルを回収する。1つのマウス群(N=5)を、ネガティブコントロールとして、細菌発現グルタチオンSトランスフェラーゼ(GST)で免疫する。
【0099】
インビトロアッセイによる免疫応答の評価。異なる投与レジメンによるワクチンの免疫前後のマウス血清を回収し、Huang et al., 2003, J Infect Dis 188:19-31およびHuang et al., 2005, J Virol 79:6714-22に記載のように、タイプ特異的抗体検出EIAを用いてNORおよびRVに対するそれらの特異的抗体力価を測定する。糞便サンプル中の分泌IgAを、粘膜免疫の指標としてNORおよびRVに対する応答について試験する。RVについて、中和アッセイは、細胞培養中のRV複製の阻害のために血清サンプルで実施される。NORについて、NORレセプター遮断アッセイを実施して、潜在的「中和」活性として抗体によって、NOR VLPとそれらのHBGAレセプターとの結合の阻害を測定する。
【0100】
VA387は世界中のNOR関連胃腸炎の65〜85%に関与する主要なGII−4遺伝子型であるため、同じVA387 VLPまたはP粒子に対する同種抗体応答を用いる。NORレセプター結合のインビトロ遮断アッセイを用いて、NORに対する抗体の潜在的中和を評価する。VP8−P粒子ワクチンの異なる投与レジメンからのマウス血清を、HBGAレセプターとNOR VLPの結合に対する遮断効果について試験する。抗体の遮断効果を再度、同種株VA387とそれらに対するHBGA(A、B、H、Lewis BおよびLewis Y抗原)との結合について試験する。簡潔には、既知のHBGAまたは合成オリゴ糖で十分に特徴付けられた唾液をマイクロタイタープレートに被覆する。脱脂乳でブロックした後、30分間抗体と共にインキュベートしたNOR VLPをプレートに加える。HBGAレセプターと結合したVLPを、特異的VA387 VLPに対するモルモット過免疫血清、次いでHRP複合化ヤギ抗モルモットIgG(ICN、Aurora、OH)の添加によって検出する。シグナル強度(吸光度、OD)は波長450nmでELISAプレートリーダーによって読み取る。遮断効果は、抗体処理ありとなしとでの結合の比較によって決定する。
【0101】
プラーク減少を測定するための細胞培養を用いた中和アッセイを実施して、マウスにおいてVP8−P粒子キメラに対して誘導される抗体を試験する。[P]タイプのVP8−P粒子キメラと他のRVタイプの交差中和を、発明者の研究室で入手可能な様々なRV株を用いて試験する。組織培養順応Wa株、マウスRV EDIM株およびMA104ミドリザル腎臓細胞株が使用される。簡潔には、MA104細胞を6ウェルプレートで培養し、RV接種の連続希釈によって〜50プラーク/ウェルの形成についてRV力価を測定する。アッセイのために、RVは特定希釈の血清と60分間インキュベートし、6ウェルプレート中の細胞(MA104)に混合物を加える。2時間後プレートを洗浄し、次いで0.8%のアガロースを含む培地で覆う。37℃で4〜5日間インキュベートした後、各ウェルのプラーク数を計測する。処理していないウェルと比較して、血清を含むウェルでのプラーク数の減少は、血清中に存在する中和抗体の量を決定する。
【0102】
マウスRVチャレンジモデル。ワクチンの保護効能を試験するためのこのRVチャレンジモデルの手法は、Choi et al., 「Functional mapping of protective domains and epitopes in the rotavirus VP6 protein,」 2000, J Virol 74:11574-80; Choi et al., 「Functional mapping of protective epitopes within the rotavirus VP6 protein in mice belonging to different haplotypes,」 2003, Vaccine 21:761-7;およびMcNeal et al., 1999, J Virol 73:7565-73.41(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載されている。RV抗体を含まない6週齢のBALB/cマウス(Harlan-Sprague-Dawley、Indianapolis、Ind.)をVP8−P粒子キメラワクチンで経口または鼻腔内的に免疫する。最後の免疫の2〜4週間後、mice are 経口ly (via gavage、generally carried out by passing a feeding tube through the nose or mouth into the esophagus) challenged with murine RV EDIM strain at a dose of 100 50%排出用量と等しい4×104FFU(フコース形成単位)の用量でマウスRV EDIM株でマウスを経口的に(胃管栄養法で、一般的に鼻または口を介して食道へと給餌チューブを通して)チャレンジする。糞便中のRV抗原を測定するため、EDIMチャレンジの7日以上後に各マウスから2回糞便ペレットを採取し、1mlのアールバランス化食塩水中に保存する。分析までサンプルを凍結保存し、その時点でそれらをホモジナイズし、遠心分離してデブリを除去する。糞便サンプル中のRV抗原の量は、既報のとおりELISAで測定する。
【0103】
結果。結果は、VP8−P粒子が処置マウスにおいてRV−およびNOR−特異的抗体を誘導し、該抗体がNOR VLPとそのHBGAレセプターとの結合を遮断し、細胞培養中のRVの複製を中和し、そしてマウスチャレンジモデルにおいて免疫保護を提供することを示す。VP8−P粒子ワクチンは細胞傷害性または副作用を示さない。
【0104】
発現構築物。VA387ノロウイルス(GII.4)Pドメインコード配列およびシステイン含有ペプチドを含むpGEX−4T−1[グルタチオンSトランスフェラーゼ(GST) Gene fusion System、GE Healthcare life sciences]のP粒子発現ベクターを多様なキメラP粒子の構築のためのテンプレートとして用いた。P粒子Hisタグキメラを作製するために、Hisタグコード配列を部位特異的突然変異誘発によってN372とD374(図7および図14A)の間のループ2に挿入した(下記参照)。ヒトロタウイルスVP8抗原を含むキメラP粒子のために、まずループ2にSpeIおよびClaI/EcoRIの酵素部位を有するクローニングカセットを導入して、部位特異的突然変異誘発によってT369からD374の配列を置換した(図16A)。[P]8株(Wa、L65-L223、GenBank accession: VPXRWA)および[P]4株(DS1、L65-l223、GenBank accession: VPXRDS)のVP8抗原cDNA配列をカセットにクローン化した。P粒子にマウスロタウイルス(EDIM)VP8抗原(L65-L222、GenBank accession: AF039219)のcDNA配列をクローニングするため、XbaIおよびBgl II部位を有する別のクローニングカセットをN372とN373の間に挿入した。PダイマーHisタグキメラの作製のために、ヒンジを有するPドメインのN末端とHisタグを連結した。ペプチドCNGRCまたはCDCRGDCFCを含むキメラマルトース結合タンパク質(MBP)を作製するため、pGEX−4T−1のベクターにMBPのN末端と該ペプチドを融合した。pDEST17(Gateway、Invitrogen)(29)におけるHisタグ化サーモトガ・マリティマ(Thermotoga maritima)α−L−フコシダーゼ(GenBank accession: TM0306)の発現構築物は、Dr. HenrissatおよびDr. Bourne(Architecture et Fonction des Macromolecules Biologiques、UMR 6098、CNRSおよび Universites Aix-Marseille I and II、31 Chemin J. Aiguier、F-13402 Marseille、Cedex 20、France)から快く譲っていただいた。
【0105】
組換えタンパク質の発現および精製。0.25mMのイソプロピル−β−D−チオガラクトピラノシド(IPTG)の誘導により大腸菌(BL21)において、室温(〜25℃)で一夜、他に記載のとおりに組換えタンパク質を発現した。組換えGST融合タンパク質の精製は、Glutathione Sepharose 4 Fast Flow(GE Healthcare life Sciences)の樹脂を用いて、製造業者の指示書に従って実施した。ビーズ上または溶液中(リン酸緩衝化食塩水、PBS,pH7.4)のいずれかでトロンビン(GE Healthcare life Sciences)切断によって標的タンパク質からGSTを除去し、次いでゲルろ過クロマトグラフィーによってさらに精製した。Hisタグタンパク質の精製は、TALON His-Tag Purification Resins(Clontech、Mountain View、CA)を用いて、製造業者の指示書に従って実施した。250mMのイミダゾール(Sigma-Aldrich、St. Louis、MO)を含むPBSによって樹脂からHisタグタンパク質を溶出させた。樹脂精製タンパク質のさらなる精製は、ゲルろ過クロマトグラフィーによって実施した。
【0106】
ゲルろ過クロマトグラフィーは、既報の通りAKTA FPLC System(GE Healthcare life Sciences)で実施した。簡潔には、AKTA FPLC system(model 920、GE Healthcare life Sciences)で動力供給したサイズ排除カラムSuperdex 200(GE Healthcare life Sciences)にアフィニティーカラム精製タンパク質をロードした。溶出フラクションの分子量は、Gel Filtration Calibration Kits(GE Healthcare life Sciences)でキャリブレートした。あるいは、空ボリューム、キメラP粒子およびPダイマーのピークを、それぞれ青色デキストラン2000(〜2000kDa、GE Healthcare life Sciences)、野生型P粒子(830kDa)およびノロウイルスV387(GII.4)の野生型Pダイマー(69kDa)で決定できる。ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS PAGE)および/またはウェスタンブロット分析によって興味あるフラクションのタンパク質をさらに分析した。
【0107】
SDS PAGEおよびウェスタンブロット分析。10%ゲルを用いたSDS PAGEによって組換えタンパク質を分析した。他に記載のとおり、ノロウイルスVLP(VA387、GII.4、1:3000)またはロタウイルスVP8抗原(Wa、1:3000)に対する過免疫血清を用いたウェスタンブロット分析によって、組換えタンパク質の特異的組成を検出した。ブロットした膜を5%脱脂乳でブロックした。二次抗体−HRP(ホースラディッシュペルオキシダーゼ)コンジュゲート(1:5,000、ICN Pharmaceuticals、Costa Mesa、CA)を用い、そしてECL Eastern Blotting Detection Reagents (GE Healthcare life Sciences、Buckinghamshire、England)によってHRPを検出した。ECLシグナルはHyperfilm ECL(GE Healthcare life Sciences、Buckinghamshire、England)で捕捉した。
【0108】
既報の通り製造業者によって提供されたQuikChange Site-Directed Mutagenesis Kit(Stratagene、La Jolla、CA)の手法に従って部位特異的突然変異誘発を実施して、Hisタグおよび構成クローニングカセットをノロウイルスPドメインのループ2に挿入した。N372とD374との間のループ2(図7および図14A)に7個のヒスチジンの紐を挿入するためにプライマー対:caccactgacacaaaccaccaccaccatcatcaccacgatcttcaaactggcc/ggccagtttgaagatcgtggtgatgatggtggtggtggtttgt gtcagtggtgを用いた。さらに、ヒトロタウイルスVP8抗原のコード配列の挿入を促進するために、プライマー対:gttcaatacaccactagtacaaacatcgatatccttcaaactggc/ gccagtttgaaggatatcgatgtttgtactagtggtgtattgaacを用いて、ノロウイルスPドメインのループ2に3個の酵素部位(Spe IおよびCla I/EcoRI)を有するクローニングカセットを構築した。マウスロタウイルスVP8抗原のコード配列の挿入を補助するため、プライマー対:tacaccactgacavaaa ctc tagacacagatctaatgatcttcaaactgg/ccagtttgaagatcattagatctgtgtctagagtttgt gtcagtggtgtaを用いて、XbaIおよびBgl II部位を有する別のクローニングカセットを作製した。
【0109】
低温電子顕微鏡(cryo−EM)技術。
【0110】
Cryo−EMイメージ取得。我々の以前の研究に記載の野生型P粒子の構造再構成に用いた手法をここで採用した。簡潔には、精製P粒子−VP8(Wa)キメラのアリコート(3〜4μl)を液体窒素によって冷却した液体のエタン中、Quantifoil格子上でフラッシュ凍結した。サンプル格子を顕微鏡にのせ、200KVで操作するフィールド放出銃を備えたCM200低温顕微鏡を用いて、フィルム上で低線量イメージ(〜20e/A2)を記録した。公称倍率x50,000、デフォーカスレンジ2.0〜4.0μmでイメージを撮影した。顕微鏡写真を選択し、Nikon Super CoolScan 9000EDスキャナーをステップサイズ6.35μm/ピクセルで用いてデジタル化した。スキャンしたイメージをビニングして、さらなるイメージ処理および3D再構成のために、2.49Å/ピクセルでイメージの最終サンプリングを得た。
【0111】
Cryo−EMイメージ処理および3D再構成。EMANのboxerプログラムを用いてキメラP粒子のイメージを選択した。選択されたイメージを手作業で選別して、偽陽性を排除した。EMANのctfitプログラムを用いて、同じ顕微鏡写真に由来する粒子のセットに関連した、コントラスト−トランスファー関数(CTF)パラメーターを手作業で決定した。EMANのstartoctプログラムを用いて、キメラP粒子の初期モデルを作製した。次いで、EMANのrefineプログラムを用いて、新たなキメラ粒子の中心および方向を繰り返し決定し、収束するまでEMANのmake3dプログラムによって2Dイメージから3Dマップを再構成した。キメラP粒子の再構成によって八面体シンメトリーが与えられた。
【0112】
Cryo EMモデル評価および分析。UCSF Chimeraソフトウェアを用いて、ロタウイルスVP8抗原(Wa、2DWr、L65-L223)の結晶構造をP粒子−VP8(Wa)キメラの3D構造の伸長した突起に重ね合わせた。単純剛体運動がキメラP粒子の3D構造とX線構造の最良のマッチングの発見と考えられた。
【0113】
酵素免疫アッセイ(EIA)を用いて、P粒子−抗原キメラによって誘導されたマウス抗血清の免疫反応性および抗体力価を測定した。可変の抗血清のために異なる抗原を用いた:P粒子Hisタグキメラによって誘導された血清についてHisタグ化T. maritimaα−L−フコシダーゼ、非露出ペプチドCNGRCまたはCDCRGDCFCを含むP粒子によって誘導された血清についてMPB−CNGRCまたはMPB−CDCRGDCFC、P粒子−VP8キメラによって誘導された血清について遊離VP8、そして内部コントロールとしてP粒子−VP8キメラによって誘導された血清についてGST。抗原を96ウェルマイクロタイタープレート(Dynex Immulon; Dynatech、Franklin、Mass)に被覆した。5%脱脂乳でブロックした後、被覆した抗原と共に指定の希釈での血清をインキュベートした。他に記載のとおりに二次抗体−HRPコンジュゲートによって、結合した抗体を検出した。抗原に対する抗体の力価は、カットオフシグナル強度0.15での終点希釈として定義された。野生型P粒子またはPBSで免疫した動物由来の血清をネガティブコントロールとして用いた。
【0114】
HBGA結合および遮断アッセイ。基本的には他に記載のとおりに、唾液を利用した結合アッセイを実施した。簡潔には、既知のHBGA表現型を有する煮沸した唾液サンプルを1000倍に希釈し、96ウェルマイクロタイタープレート(Dynex Immulon; Dynatech、Franklin、MA)に被覆した。5%脱脂乳でブロックした後、VLPまたはノロウイルス(VA387、GII.4)のP粒子を加えた。我々自身が作製したウサギ抗VA387 VLP抗血清(1:3,300)を用いて結合したVLP/P粒子を検出し、次いでHRP複合化ヤギ抗ウサギIgG(ICN、Pharmaceuticals、Costa Mesa、CA)を加えた。VLPを被覆した唾液に加えた後、希釈した血清と共に30分間プレインキュベートして、ノロウイルスVLP−唾液結合に対するP粒子−VP8キメラによって誘導されたマウス血清の遮断効果を測定した。免疫した動物由来のマウス血清による遮断ありおよびなしで測定したODを比較することにより、遮断率を計算した。遊離VP8免疫動物由来の血清の遮断率をネガティブコントロールとして用いた。
【0115】
抗体応答のためのマウス免疫。6週齢のメスBALB/cマウス(Harlan-Sprague-Dawley、Indianapolis、IN)を精製キメラP粒子または遊離抗原で、用量5〜15μg/マウス、2週間間隔で3〜4回で免疫した。P粒子−またはPダイマー−提示Hisタグに対する免疫応答の比較のため、マウスに5μg/マウスの組換えP粒子−Hisタグキメラ、およびPダイマー−Hisタグまたは野生型P粒子を投与した(n=5)。非露出ペプチドに対する免疫応答の比較のため、マウスに非露出ペプチドCNGRCまたはCDCRGDCFCを含む組換えP粒子5μg/マウスを投与した。免疫源はフロイントアジュバントと共に皮下的に4回与えた。P粒子提示VP8および遊離VP8に対する免疫応答の比較のため、マウスに5μg/マウスの遊離VP8および15μg/マウスのP粒子−VP8キメラ(両抗原はVP8のモル量として同量である)を、アジュバントなしで鼻腔内またはフロイントアジュバントと共に皮下的に、3回投与で投与した(n=5〜7)。内部コントロールとして免疫源に等モル量のGSTを加えた。保護のためのマウスのP粒子−VP8ワクチンの免疫は下に記載した。免疫前および最終免疫の2週間後に、逆軌道毛細血管叢の穿刺によって血液を採取した。血清は、血液から4℃で一夜静置後遠心分離によって処理した。
【0116】
ロタウイルスプラークアッセイを実施して、細胞培養中のロタウイルス複製に対する、ロタウイルスVP8抗原を含むキメラP粒子によって誘導される抗血清の中和を測定した。MA104サル腎臓細胞で成長させた組織培養適応ロタウイルスWa株をこのアッセイに用いた。MA104細胞を6ウェルプレートで培養し、接種として〜50プラーク/ウェルのロタウイルス力価を用いた。アッセイのために、ロタウイルスを所定の希釈のマウス血清と共に60分間インキュベートした。次いで混合物を6ウェルプレート中のMA104細胞に加えた。2時間後、プレートを洗浄し、次いで5μg/mlのトリプシン(Invitrogen、Carlsbad、CA)および0.8%のアガロースを含む培地で被覆した。37℃で4〜5日インキュベーション後、各ウェルのプラーク数を計測した。無処理コントロールウェルのものと比較した抗血清処理したウェルにおけるプラーク数の減少によって、血清中の中和抗体の量を測定した。
【0117】
マウスロタウイルスチャレンジモデル。以前の研究に記載のロタウイルスチャレンジモデルに従って、P粒子−VP8ワクチンの保護効率を試験した。ロタウイルス抗体を含まない6週齢のBALB/cマウス(n=5〜7)(Harlan-Sprague-Dawley、Indianapolis、IN)をアジュバントなしで、マウスロタウイルス(EDIM)VP8抗原を含むキメラP粒子(15μg/マウス)で3回、鼻腔内免疫した。遊離マウスロタウイルスVP8抗原(5μg/マウス)およびヒトロタウイルス(Wa)VP8抗原を含むキメラP粒子(15μg/マウス)を比較のために含めた。さらに、野生型P粒子(ベクターコントロール)およびPBSをネガティブコントロールとして投与した。最後の免疫の2週間後、105の50%排出用量と等しい4×104FFU(フォーカス形成単位)の用量のマウスロタウイルスEDIM株で、経口胃管栄養法によってチャレンジした。糞便中のロタウイルス排出を測定するため、EDIMチャレンジ後6日間毎日各マウスから2個の糞便ペレットを採取し、1mlのアールバランス化食塩水中に保存した。分析までサンプルを凍結保存し、その時点でそれらをホモジナイズし、遠心分離してデブリを除去した。糞便サンプル中のロタウイルス抗原の量(μg/ml)は、既報のとおりELISAで測定した。
【0118】
統計的分析。GraphPad Prism version 5.00 for Windows(GraphPad Software、San Diego CA)およびMicrosoft Office Excel 2007を用いてグラフを作製した。GraphPad Prism version 5.00 for Windowsを用いたデータ群のt検定によってP値を決定した。
【0119】
結果
Hisタグを含むキメラP粒子の製造。我々の研究は、P粒子のループ2に小さなエピトープ、ポリヒスチジン(His)タグを挿入することで開始した(図3および14)。GST遺伝子融合系を用いた大腸菌におけるこのPタンパク質の発現および精製は、高収率(>10mg/リットル培養)で予測したサイズ(〜35kDa)を有するタンパク質が得られ、次いでトロンビンでGSTタグを除去するために(〜27kDa)タンパク質を消化した(図14C)。ゲルろ過クロマトグラフィー、次いでSDS PAGEおよびウェスタンブロット分析によってキメラPタンパク質のP粒子形成が示され、ここでキメラP粒子は〜840kDaの主要なピークを形成した(図14DおよびE、データは示さず)。キメラP粒子上での挿入したHisタグの露出は、Talon樹脂へのその特異的結合、続く250mMのイミダゾールによる溶出によって示され(図7A、溶出1および2)、これは高純度のキメラP粒子をもたらした。したがって、P粒子−Hisタグキメラはまた、アフィニティーTalon樹脂を用いて大腸菌からも精製できる(データは示さず)。
【0120】
P粒子担体によるHisタグの免疫増進。P粒子−Hisタグキメラで免疫した後マウスにおけるHisタグに対する免疫応答の試験によってこれを測定した(n=5)。Pダイマー提示HisタグよりもP粒子提示Hisタグで免疫したマウスにおいて、有為に高いHisタグに対する抗体力価が検出された(P<0.05、図15AおよびB)。PドメインのN末端(CNGRC)またはC末端(CDCRGDCFC)に融合した2種の短いペプチドは、P粒子提示Hisタグによって誘導されるものと比較して、有為に弱い免疫応答を誘導した(図15C〜F)。これらの2種のペプチドは、P粒子の内側に埋め込まれることが示されており、このことは、免疫増進のためにP粒子の表面上に挿入された抗原が適切に露出されることが重要であると示している。予想通り、全ての血清はノロウイルスP粒子プラットフォームと強く反応した(図15、データは示さず)。
【0121】
ロタウイルスVP8抗原を含むキメラP粒子の開発。我々は次に、159アミノ酸を含むロタウイルス(Wa)VP8抗原の挿入によって、より大きなポリペプチドを得るためのP粒子プラットフォームの能力を試験した。VP8挿入を促進するために、P粒子のループ2に3個の酵素部位(Spe IおよびCla I/EcoRI)を有するクローニングカセットを構築した(図16A)。大腸菌における構築物の発現によって、高収率(>15mg/リットル培養)でGST−P−VP8融合タンパク質(〜78kDa、図16Bの左パネル)が得られた。溶液中または精製ビーズ上のいずれかで、GST融合タンパク質のトロンビン消化によって、放出されたP−VP8キメラ(〜52kDa)が得られた(図16B、それぞれ中央および右パネル)。ゲルろ過クロマトグラフィーによってP−VP8キメラタンパク質が高い割合(>95%)でP粒子を形成することが示された(図16C)。ウェスタンブロット分析によって、P−VP8キメラタンパク質がノロウイルスVLP(VA387)およびロタウイルス(Wa)VP8に対する抗体両方と反応することが示された(図16D)。
【0122】
Cryo−EMおよび3次元イメージ再構成によって、P粒子−VP8キメラが野生型P粒子と同様の八面体シンメトリーのままであるが、キメラP粒子が顕著に大きいことが示された(図1AおよびB)。伸長した突起は、おそらく、Pダイマー由来の潜在的境界としてのニックによって認識されうる挿入されたVP8である。キメラの伸長した突起の密度マップにおける同じWa株(2)のロタウイルスVP8抗原の結晶構造の重ね合わせによって、伸長した突起が実際にVP8であることが確認された(図1C〜E)。
【0123】
VP8に対するP粒子増進免疫応答。次に我々は、マウスにおけるP粒子提示VP8抗原に対する免疫応答を試験した。等モル量のP粒子−VP8キメラおよび遊離VP8での免疫後、抗原として遊離VP8を用いたEIAによって得られたマウス血清を試験した。P粒子提示VP8に対する抗体力価は、アジュバントなしでの鼻腔内免疫による遊離VP8に対するものよりも有為に高かった(P<0.005)。同様の結果が、それぞれロタウイルスWaおよびDS−1のVP8抗原を含む2種のキメラP粒子とそれらの対応遊離VP8の比較において観察された(図17AおよびB)。これらの動物において遊離GSTに対してはわずかにのみ抗体力価が検出され、P粒子提示VP8の特異的免疫増進がさらに確認された。フロイントアジュバントと共に上記抗原で皮下免疫したマウスの免疫応答も試験したところ、遊離VP8とP粒子提示VP8の間でVP8に対する抗体力価の差はほとんど観察されなかった(図17C)。
【0124】
P粒子−VP8キメラはロタウイルスに対する中和抗体を誘導した。鼻腔内免疫を介したP粒子−VP8(Wa、[P]8)キメラによって誘導された過免疫抗血清は(図17A)、細胞培養中の同族ロタウイルス(Wa)複製を強く低減した。この中和力価は、遊離VP8での免疫によって誘導した中和力価よりも有為に高かった(P<0.0002)(図18A)。P粒子−VP8(DS−1、[P]4)キメラでの免疫後、Waに対する低レベルの交差中和も観察された(P<0.05、図18B)。反対に、DS−1の遊離VP8で免疫した動物由来の血清は、かかる交差中和を示さなかった。さらに、フロイントアジュバントと共に皮下免疫した動物由来の血清を用いた中和力価も測定した。驚くべきことに、EIAにおいて遊離VP8抗原と同様の免疫反応性を有する4対のマウス血清のうち(図18C)、P粒子−VP8(Wa)で免疫した動物由来の血清が全て、同じWa株の遊離VP8で免疫したマウス由来の血清のものよりも有為に高い中和力価を示した(P<0.005、図18D)。これらのデータは、キメラP粒子上のVP8がより良好に提示され、中和エピトープに必要な適切なコンホメーションを維持しうることを示した。
【0125】
P粒子−VP8キメラのワクチン接種はマウスロタウイルスチャレンジ後の低下した排出を引き起こした。次に我々は、マウスロタウイルスモデル(EDIM株)を用いてインビボで、P粒子−VP8キメラワクチンが保護を与えうるかを試験した。この目的で、EDIM VP8抗原を含むキメラP粒子を材料および方法に記載のとおりに構築し、マウスに投与した(N=5〜7)。同モル量の遊離EDIM VP8、ヒトVP8(Wa)を有するキメラP粒子または野生型P粒子で免疫した動物は、コントロールとして含めた。最後の免疫の2週間後、マウスEDIMロタウイルスで動物をチャレンジした。チャレンジの6日後に糞便サンプルを採取し、EIAで排出を検出した(表1、図19)。P粒子−VP8(EDIM)キメラをワクチン接種されたマウスは、最も少ない量のウイルス抗原を排出した。排出の減少は6日間で89%であった(P<0.05)。EDIMでのチャレンジの1日後では99.2%の減少であった。遊離マウスVP8で免疫したマウスは、次に少ない排出量であり、77%の平均保護であった(P<0.05、2〜5日)。ヒトロタウイルス(Wa)VP8を含むキメラP粒子ワクチンでの免疫は、23%減少と排出に対してわずかな効果を有し、これはEDIMに対する何らかの交差保護を示唆している。この群での6日目に、排出が顕著に減少した(63%、P<0.05)。
【0126】
【表1】
【0127】
ノロウイルスとHBGAの結合を遮断するP粒子−VP8キメラ誘導性抗体での免疫。また、P粒子−VP8(Wa)キメラでの免疫によって、マウスでの免疫応答におけるP粒子骨格の役割を試験した。予測したとおり、キメラ誘導性抗体は、EIAで測定したときノロウイルスVLPおよびP粒子と強く反応した(図20、データは示さず)。これらの血清はまた、HBGAレセプターとノロウイルスVLPの結合も遮断した(タイプA唾液、図20B)。ネガティブコントロールとして遊離VP8(Wa)で免疫した動物由来の血清はかかる遮断を示さなかった。この結果は、P粒子−VP8キメラがロタウイルスおよびノロウイルスの両方に対するデュアルワクチンであり得ることを示している。
【0128】
我々の以前の研究により、ノロウイルスP粒子が容易に製造でき、極めて安定で高度に免疫原性であり、これはノロウイルスに対するサブユニットワクチンとして使用できることが示された。我々はさらに、外来抗原の免疫増進のための新規ワクチンプラットフォームとしてP粒子が使用できることを示した。我々は、1)P粒子の表面ループがP粒子の形成および作製に影響を与えることなく外来抗原を挿入するための優れた部位であること、2)P粒子が少なくとも159アミノ酸までのサイズの外来抗原に耐えること、3)挿入抗原に対する増進された免疫応答がインビトロおよびインビボ両方での中和および保護実験によって示されることを示した。P粒子−VP8キメラはまた、ロタウイルスおよびノロウイルスの両方に対する有望なデュアルワクチンを提供した。したがって、複数多価外来挿入の能力を有するキメラ粒子およびその複数の表面ループを作製するための単純な手法は、P粒子を感染症および効果的なワクチンから利益を得るであろう他の状態の抗原提示のための魅力的なワクチンプラットフォームにする。
【0129】
本発明の第一次的な目標は、一般に免疫原性が低い小ポリペプチド抗原の免疫原性をP粒子が増進できるかを試験することである。我々は、モデルとしてHisタグを用いてこの問題を研究し、すばらしい結果を得た。2つの主要な要因、複数コピー数および挿入された抗原の表面露出が、観察された免疫増進に寄与している可能性がある。P粒子はPモノマーの24個のコピーからなり、このことはPダイマーと融合したHisタグとの比較において増進した免疫応答を説明できる(図15A)。抗原の表面露出の重要性は、PドメインのNまたはC末端と結合した2個の非露出ペプチドに対する低い免疫応答性によって示された(図15BおよびC)。したがって、P粒子はその大きなサイズ(830kDa)および適切な外来抗原の提示(これと異なるものは低い免疫原性を有する)によってアジュバントとして作用しうる。粒子あたりの抗原の増加した複数コピーは、増加した免疫応答を説明しうる別の特徴である。
【0130】
Hisタグペプチドに加えて、我々はP粒子に多数の他の小ペプチド、例えばマウスサイトメガロウイルスのT細胞エピトープ(9アミノ酸)、シュードモナス属Epi8エピトープ(14アミノ酸)およびインフルエンザAのM2細胞外エピトープ(23アミノ酸)の挿入に成功した(Tan and Jiang、未公表)。これらのデータは、P粒子が多様な小ペプチド抗原の免疫増進に容易に利用できることを示唆している。P粒子への様々なロタウイルスVP8の挿入成功は、広範な大型外来抗原のためのP粒子プラットフォームの適用を大きく拡張する。ロタウイルスVP8はウイルスカプシド上の突起タンパク質であり、ロタウイルス感染性に重要であると考えられている。また、それは中和抗体を誘導する2種の抗体の一つでもある。VP8タンパク質について多数の中和エピトープが同定されている。P粒子−VP8キメラの中和抗体誘導成功は、マウスにおけるロタウイルス排出に対して応答し、それに対する保護を提供する。このことは、これらのエピトープがP粒子担体上で提示されていることを示唆している。
【0131】
2種の近年導入されたロタウイルスワクチンは極めて有効であるが、新たな出現ウイルスのための新世代ワクチンが潜在的に必要とされうる。非感染性サブユニットワクチンは、現在のワクチンで心配されうる毒性株への復帰の危険がない。ロタウイルスのVLPワクチンの開発は数年来提案されている。ロタウイルスVLPワクチンは、バキュロウイルス宿主に複数のカプシドをコトランスフェクションする必要があるため、低い発現効率と高い製造コストの問題がある。反対に、P粒子−VP8キメラの作製は常套の大腸菌ベースのクローニングおよび発現手法のみを必要とし、これは効率がよくコストが低い。さらに、VP8についての交差中和エピトープが記載されている。我々は本稿において、P[4]ウイルス由来VP8を含むキメラP粒子でのマウスの免疫がP[8]ロタウイルスに対する交差中和を有することを示した。最小数のPタイプを含むカクテルワクチンもコスト的に有効であり得る。
【0132】
P粒子−BP8キメラによって誘導される抗体の、ノロウイルスVLPとHBGAの結合を遮断する能力は、予測外である。図1に示すとおり、P粒子のHBGA結合界面を含むPダイマーの遠位表面は、挿入されたVP8によってほとんど被覆されている。これはHBGAレセプターに対する該キメラの結合能の喪失を導く(データは示さず)。継続して遮断能が見られたことの一つの可能性は、挿入されたVP8抗原によって被覆されていても、P粒子−VP8キメラのHBGA結合界面のエピトープが抗体誘導のためになおもアクセス可能であるということである。あるいは、観察された糖遮断は、糖結合部位の近くでの抗体結合のためでありうる。どのようなメカニズムが関与するにせよ、キメラ誘導性抗体のノロウイルスVLPとHBGAとの結合を遮断する能力は、P粒子プラットフォームにさらなる価値を付け加える。ノロウイルスおよびロタウイルス両方に対するP粒子−VP8キメラのデュアルワクチンのコンセプトは、両方の感染の危険がある特定の集団に特に有用でありうる。
【0133】
ループ2のみがこの研究では試験されたが、他の2つのループにおける抗原挿入成功の可能性が予測され、我々の研究室で現在試験している。Pモノマーあたり3個の表面ループの利用可能性は、多目的ワクチン設計の機会を提供する。例えば、免疫応答を上昇させるため、同じエピトープまたは抗原を3個のループ全てに挿入して、粒子あたり72コピーの抗原に到達できる。それぞれの抗原のタンデム反復の挿入によって、より多いコピー数すらも作製できる。あるいは、異なる抗原を3個のループに挿入して、異なる病原体またはロタウイルスについて異なるVP8抗原に対する多価ワクチンを得ることができる。様々な目的のための機能的タグの挿入によって、さらなるワクチンテンプレートも作製できる。例えばHisタグの挿入は、精製手法をさらに単純にする。特定のリガンドまたはシグナル分子を用いて、宿主の特定の臓器、組織または細胞を標的とすることによって、免疫応答を刺激してもよい。
【0134】
P粒子プラットフォームのさらなる適用は、研究および診断的使用のための小ペプチドに対する抗体の製造用である。多様な疾患バイオマーカー(主としてペプチドエピトープ)が同定されており、これらのバイオマーカーに対する抗体が診断目的では重要である。小ペプチドは、DNAクローニングの単純な手法によってP粒子のループに容易に挿入できる。我々は、そのプロセスをさらに促進するクローニングカセットを含む簡便なP粒子ベクターを開発した。細菌における組換えキメラP粒子の発現の後、本研究で確立された手法にしたがって実験動物を免疫することによって、挿入したペプチド抗原に特異的な抗体を高力価で誘導できる。この方法による抗体製造は、ペプチド合成および免疫増進のためのキーホールリンペットヘモシニアン(KLH)のような巨大分子とペプチドとのコンジュゲートのようなコストのかかる工程を回避する。ノロウイルスPドメインは他のタンパク質のいずれとも相同性を有さない固有の配列を有しているため、他のタンパク質との交差反応性の心配にはおよばない。したがって、P粒子ワクチンプラットフォームは、生物医学的研究の多くの領域における抗体製造のための簡便なツールとして使用できる。
【0135】
本発明をその態様の解説によって説明し、その態様をかなり詳細に説明してきたが、いかなる方法においても、添付の特許請求の範囲がかかる詳細に限定されることを意図していない。さらなる利点および修飾が当業者に容易に理解される。したがって本発明は、そのより広い局面において、特定の詳細、代表的な系および方法、ならびに示し記載した実施例に限定されない。したがって、本発明の範囲または精神から離れることなく、かかる詳細から逸脱してもよい。
【技術分野】
【0001】
利害関係
本発明は、合衆国政府の支援の下になされ、National Institute of Health(NIH)、National Institute of Allergy and Infectious(認可番号R01 AI37093およびR01 AI055649)およびDepartment of Defense(認可番号PR033018)によってXi Jiangへと表彰された。この研究はUniversity of CincinnatiおよびCincinnati Children's Hospital Medical Center(Center for Clinical and Translational Science and TrainingからのT1助成金)によってMing Tanへも支援された。さらに、Ming TanへのNipert Foundationが出資する感染症助成金がこの研究に寄与した。合衆国政府は本発明についての一定の権利を保有しうる。
【0002】
発明の分野
本発明は、一般に、ノロウイルス(NOR)カプシドタンパク質のPドメインおよびNOR P粒子、抗原−ノロウイルスPドメインモノマー、抗原−ノロウイルスPドメインダイマーおよび抗原−ノロウイルスP粒子分子の製造、ならびに抗ウイルス剤、薬物送達系およびワクチン開発のためのそれらの使用に関する。
【背景技術】
【0003】
発明の背景
生体材料および生物工学は、現代医学の重要な部分となりつつある急速に成長している分野である。ウイルス性タンパク質ケージは、その万能性およびアレイを形成する傾向のため、提示系を構築するための理想的な基板である。遺伝子操作を通じて、自己集合性ウイルス構造タンパク質は、生体材料合成のための傑出したプラットフォームであり、そして設計されたパターンで外来分子を統合するためのスキャフォールドであり、これはワクチンまたは薬物として使用できる。
【0004】
抗原提示のためにウイルス粒子を用いる最近の例は、Manayani et alに記載の、炭疽菌(Bacillus anthracis)の180アミノ酸抗原挿入を有するフロックハウスウイルス(FHV)VLPのキメラである。組換えウイルス様粒子は、炭疽抗毒素として、かつ、炭疽菌ワクチンのための分子スキャフォールドとして機能し、炭疽抗毒素およびワクチンの機能を1個の化合物に組み合わせる。別の例は、Chatterji et al.に記載のCowpeaモザイクウイルス(CPMV)であり、これは、エピトープ提示ならびにペプチドおよびタンパク質の付着のためのマトリックスとしての両方のための、Cowpeaモザイクウイルスの使用を含む。全タンパク質は、正二十面体のCPMV粒子のコートタンパク質において遺伝的に操作されたリシンおよびシステイン残基と、化学的に架橋結合されている。また、ライム病菌(Borrelia burgdorferi)の表面抗原(OspA)を含むB型肝炎ウイルス(HBV)カプシド様粒子(CLP)が記載されている。
【0005】
ノロウイルス(NOR)は、以前は「ノーウォーク様ウイルス」(NLV)または小型球形ウイルスとして知られており、先進国および発展途上国の両方で生じる、伝染性急性胃腸炎の最も重要なウイルス性病原体である。これらの遺伝的に多様なウイルスは、2個の主要な遺伝子グループ(GIおよびGII)ならびに約30個の遺伝子型を含む。NORはカリシウイルス科(Caliciviridae)に属し、正二十面体の一本鎖のポジティブセンスRNAウイルスであり、そのカプシドは1個の主要構造タンパク質の180個のコピーからなる。
【0006】
過去、ヒトNORの生物学的特徴付けは、該ウイルスが細胞培養では効率的に増殖せず、ウイルス複製のための好適な動物モデルが構築されなかったために、妨害されていた。発生およびヒトボランティア試験から得られたヒト糞便サンプルが唯一のウイルス源であったが、糞便中のウイルス濃度は、通常の電子顕微鏡によるウイルス検出が不可能であったほど低い。しかし、近年の昆虫細胞中のバキュロウイルス(主として昆虫に感染する二本鎖DNAウイルス)によるNORカプシドタンパク質発現の成功が、NORの免疫学、疫学および発症機序の研究のための有用な別法を提供している。ウイルスカプシドタンパク質モノマーは、自己集合してウイルス様粒子(VLP)を作り出す。これらのVLPは形態学的および抗原的にヒト糞便において見られる真菌とは区別不可能であり、免疫学的アッセイの開発およびレセプター−ウイルス相互作用の研究に有用なツールを提供する。
【0007】
組換えNORカプシドタンパク質の原子構造は、それがT=3正二十面体を形成する90個のダイマーカプソメアに組織された180個のカプシドタンパク質モノマーを含むことを示している。低温電子顕微鏡およびX線結晶学からのデータは、該ウイルスカプシドタンパク質が2個の主要なドメイン、N末端シェル(S)ドメインおよびC末端突起(P)ドメインに折り畳まれることを示した。Sドメインは内部のシェルを形成し、一方でPドメインは、シェルから伸長もしくは突出するアーチ様構造を構築する。形態形成の研究により、Sドメインがカプシドの内部シェルの構築に必要な要素を含むが、Pドメインのダイマーサブユニット間の分子間接触がカプシドの安定性を増加することが示された。これら2個のドメインは、8〜10残基(アミノ酸)のヒンジを介して結合している。PドメインはさらにP1およびP2ドメインに分けられ、後者はカプシドの最も外側表面に位置する。SおよびP1ドメインとは異なって、P2ドメインは高い配列バリエーションを有する。Pドメインはウイルス粒子の最も外側表面に位置し、最も変化しうる配列を有するため、Pドメインが宿主相互作用、免疫認識、レセプター結合および免疫応答に関与すると考えられている。ヒンジを有する(しかしSドメインを欠く)単離されたPドメインが、ヒト組織−血液グループ抗原(HBGA)レセプターの構築を維持するダイマーをインビトロで形成することが示されている。
【0008】
HBGAは、複合グリカンと関連糖の異種グループである。NORは多様な、株特異的な手法で、ヒトHBGAをレセプターとして認識する。HBGAの中で、最も一般に遭遇する血液グループはABO(ABH)およびLewisである。ABHおよびLewis血液グループ系において抗原の形成に使用される生合成経路は、相互に関連している。
【0009】
ヒトHBGAは赤血球および血管内皮細胞を含む多くの細胞タイプ、ならびに消化器、尿生殖器および呼吸管の粘膜上皮に存在する。それらはまた、血液、唾液、消化器内容物および乳のような生物学的液体中で可溶形態でも存在しうる。HBGAは遺伝子的に制御され、ABO、Lewisおよび分泌遺伝子ファミリーとして知られる一組のグリコシルトランスフェラーゼを介して、単糖ユニットの段階的添加によって、一連の前駆体構造から合成される。
【0010】
ヒトHBGA系は高度に多形性であり、サイレントアレルを含む複数の遺伝子ファミリーによって制御される。細胞表面上でのHBGAのような多様化した分子の存在は、外部環境変化に対する宿主防御メカニズムの可能性を示す。それにもかかわらず、HBGAは多様な細菌性およびウイルス性病原体による感染と関連しており、ノロウイルスの「ドッキングステーション」を提供しうる。すなわち、HBGAは病原体にとっての認識標的であり、抗原とのレセプターリガンド結合を発現または形成する細胞への侵入を促進しうる。かかる相互作用の正確な性質は未だ知られていないが、生じる病原体と抗原構築との密接な関連は、感染プロセスにおける最初のステップとして、細胞への病原体の係留に重要な役割を果たしうる。
【0011】
NORによるヒトHBGAの認識は典型的なタンパク質−糖相互作用であり、ここで、ウイルスカプシドタンパク質の突起ドメインは、様々な株において広い多様性を有するHBGA抗原のオリゴ糖側鎖による界面を形成する。腸管内でのみ複製可能な病原体として、NORは、宿主防御系を克服するユニークな戦略を開発している。これは、なぜNOR関連疾患が世界中のあらゆる集団で一般的であり拡がっているかを説明する、それらの遺伝的および構造的バリエーションによって示されている。
【0012】
2003年12月2日に公開されたPCT公開公報US2003/101176(その全体について参照により本明細書に組み込む)は、複数の異なる組織−血液グループパターンの一つにおけるNOR株とABOおよびLewisHBGAの結合に関する。NORによるHBGAの認識は株特異的であり、多数の異なるHBGA結合パターンが同定されている。NORの多様性および宿主細胞表面における糖の多形性に基づいて、より多くの結合パターンが発見されうる。
【0013】
2006年12月28日に公開されたPCT公開公報US2006/138,514(その全体について参照により本明細書に組み込む)は、HBGAとの向上した結合親和性を示す、P粒子として知られる小さな粒子に関する。P粒子は、12個の同一のPドメインダイマーに組織される24個のPドメインモノマーによって構築されるT=1正二十面体である。12個(Pドメインダイマー)および24個(Pドメインモノマー)は、植物および動物ウイルスについて頻繁に生じる正二十面体シンメトリーについての完全単位数である。単離された、モノマーカプシドタンパク質のSドメインまたはヒンジを含まないPドメインは、12個のダイマーに組織される24個のPドメインモノマーからなる複合体である、T=1正二十面体P粒子を自然発生的に形成できる。P粒子は対応するHBGAと結合でき、NOR VLPとHBGAとの結合の強い遮断を示す。P粒子の自然発生的形成は、VA387、MOHおよびノーウォークウイルス(NV)を含む多様なNOR株で観察されている。NOR P粒子は、NOR感染の治療処置およびNOR感染に対するワクチンの作製において有用である。
【0014】
ロタウイルス(RV)およびNORはいずれも、疾患の大きな負担を招く世界中で一般的な病原体である。世界基準で、5歳未満の小児において毎年、全原因の胃腸炎が最大10億事象生じており、その13〜25%(〜1億3000万事象)がRVによって引き起こされる。RVは、小児の様々な下痢および脱水の主たる原因であり、毎年重度のRV胃腸炎によって35万〜60万人の5歳未満の小児が死亡している。それはまた、年間200万人の小児の入院に寄与しており、その費用は10億ドルを超えると予測される。他方、NORはあらゆる年齢の個体が罹患する急性胃腸炎の非細菌性流行の最も重要な原因である。NORは極めて接触感染性であり、急速に拡大して多様な状況で大流行を導く。近年の報告では、発展途上国においてNORは、毎年5歳未満の小児における1,091,000件の入院および218,000人の死亡を引き起こしていると予測された。米国において、食品媒介性病原体は、毎年推定7600万人に感染しており、325,000件の入院の原因である。NORは単独で、臨床治療およびリコールされた食品による損失のために、毎年3億5000万〜7億5000万ドルの損害を引き起こしている。
【0015】
2種の新たなRVワクチン(Rotarix(商標), GlaxoSmithKline および RotaTeq(登録商標), Merck)が近年導入されているが、これらのワクチンに関する十分に解決されていない次のような様々な問題が存在する:a)ワクチンにおいて見られるものよりも広い範囲のRV血清型にワクチン接種およびワクチン非接種小児が曝露された際の有効性;b)ワクチンが最も必要とされる貧困国にこれらのワクチンをどのように分配するかという問題を導く、ワクチンのコストおよび分配コスト;ならびにc)死亡率が最も高い発展途上国における保護のレベル。これらの弱毒化生ワクチンは、復帰し、最集合して、毒性株を生じる可能性がある。したがって、極めて有効なRV中和エピトープを含む、新世代のサブユニットワクチンが必要とされている。現在、NOR関連疾患に対する処置は存在しない。したがって、NORおよびRVに対して有効なワクチン、特に両方に対して保護ができる単一のワクチンの開発は、主要な臨床的必要を充足し、さらに本明細書に記載のP粒子ワクチンプラットフォームの顕著な販売潜在力を強調する。
【0016】
NOR感染の治療処置およびそれに対するワクチン開発における進歩にもかかわらず、他のウイルスタイプによって引き起こされる感染の同定の改善、他のウイルスタイプの治療処置、他のウイルスタイプに対するワクチン開発、ならびに特定の組織または臓器を標的とする改善された薬物送達系を開発する必要がなおも存在する。
【発明の概要】
【0017】
発明の概要
本発明は、NOR Pドメインモノマーの遠位部分が外来抗原、特に外来ウイルス性抗原を挿入できるペプチド鎖を含むという発見に関する。得られる抗原−Pドメインモノマーは、12個のダイマーに組織される24個の抗原−Pドメインモノマーからなる、典型的には八面体形態の、抗原−P粒子と称される、ナノ粒子に自然発生的に集合しうる。このP粒子は大腸菌において容易に製造され、極めて安定であり、免疫原性が高い。Pドメインモノマーあたり3個の表面ループが存在し、これは粒子あたり合計72個のループとなり、免疫増進のための外来抗原提示の潜在的部位である。抗原−P粒子は自然発生的に形成され、そしてPドメインモノマーの遠位部分で提示される外来抗原を有する。この抗原−P粒子形成プロセスは、抗原−Pダイマーの濃度に基づいて交換し、または動的に平衡化しうる。抗原−P粒子は、外来ウイルス性抗原に対する抗体を産生し、またかかる外来ウイルス感染のためのワクチンを作製するために有用である。
【0018】
本発明の第一の局面は、多様な感染性病原体の抗原を提示するためのプラットフォームとして使用するための修飾NOR P粒子に関し、したがってこれらの疾患に対する新たなワクチン戦略を提供する。このプラットフォームはまた、ワクチンの開発のため、あるいはがん、アレルギーおよび自己免疫疾患を含む非感染性疾患の処置のための、薬物の担体または薬物送達ビークルとしても有用である。多数の他のワクチンプラットフォームと比較して、本抗原−P粒子系は、単純な精製手法に伴う高収率および低製造コスト、ならびに高い抗原提示効率を含む多くの利点を有する。
【0019】
本発明の抗原−P粒子は、NORカプシドの突起(P)ドメインによって形成され、ダイマーに組織されるPドメインモノマーの24個のコピーを含む。抗原−P粒子の分子量およびサイズ(MW=約830kDaおよびΦ=20nm)は、サブユニットワクチンとして有効なサイズである。外来抗原は各Pドメインモノマーに存在する3個の表面ループの1個以上に挿入され、それによって各抗原−P粒子の表面で、外来抗原の72個までのコピーを提示する。1個の抗原−P粒子上の外来抗原の多重性は、外来抗原の抗原性および免疫原性を大きく向上させる。小さなポリペプチドから大きなタンパク質まで多様なタイプおよびサイズの外来抗原が3個の表面ループのいずれか1個以上に挿入されてよく、外来抗原の代表例としては次のものを含む:Hisタグ(7個のヒスチジン)、マウスサイトメガロウイルスのT細胞エピトープ(9アミノ酸)、シュードモナス属のEpi8抗原(10アミノ酸)、ロタウイルス(RV)のVP8(159アミノ酸)および緑色蛍光タンパク質(238アミノ酸)。
【0020】
NOR P粒子は、極めて安定であり、広範な物理化学的条件に耐性である。最も重要なことに、製造のために真核細胞系を必要とする多くの他の組換えサブユニットワクチンとは異なって、野生型NOR P粒子同様、抗原−P粒子は、大腸菌および酵母または他の細菌源において、極めて高い収率および単純な精製手段で容易に製造できる。
【0021】
本発明の第二の局面は、簡便なクローニングカセットの挿入によって、各Pドメインモノマー上に存在する表面ループの1個以上に候補外来抗原を挿入するためのユーザーフレンドリーな方法である。クローニングカセットは、典型的には、Pドメインベクターの1個以上のループに、制限部位または制限認識部位としても知られる特異的認識ヌクレオチド配列の1個以上、典型的には少なくとも1個の対を含む。制限部位は制限酵素によって認識される特異的なヌクレオチド配列である。該部位は一般にパリンドロームであり(制限酵素は通常ホモダイマーとして結合するため)、特定の制限酵素は典型的には、その認識部位内またはその近くのどこかの2個のヌクレオチド間の特定のDNA配列を切断する。例えば、一般的な制限酵素EcoRIは、パリンドローム配列GAATTCを認識し、DNA鎖の上下両方のGとAの間を切断し、各末端にオーバーハング(結合した相補鎖のないDNA鎖の末端部分)を残す。次いでこのオーバーハングまたは「付着末端」は、相補的オーバーハングを有するDNA片(例えばDNAの他のEcoRI切断断片)を加えまたはライゲート(DNAリガーゼを介して)するために使用できる。各Pドメインモノマー上に存在する表面ループの1個以上における制限部位を使用することで、表面ループのいずれか1個以上にクローニングカセットに含まれる外来遺伝子を置換でき、またPドメインモノマーの3個のループの異なる一つに、2個以上の異なる外来抗原または他のクローニングカセットを置換できる。
【0022】
本発明の別の局面は、12個のダイマーの対として、24個のPドメインモノマーの自然発生的集合による、抗原−P粒子の形成である。全てのPドメインモノマーが同一である場合、抗原−P粒子は同種P粒子と呼ばれる。
【0023】
本発明の別の局面は、2個以上の異なって修飾された抗原−Pドメインモノマー、または少なくとも1個の種々に修飾された抗原−Pドメインモノマーおよび野生型Pドメインモノマーを含む、異種抗原−P粒子の形成である。種々に修飾された抗原−Pドメインモノマーまたは野生型Pドメインモノマーは、広範な抗原−P粒子を得るために、任意の比(分子量または重量比)で使用できる。
【0024】
本発明の別の局面は、ウイルスに対する免疫保護を与えるための候補ワクチンとしての、P粒子内の各Pドメインモノマーの表面ループの少なくとも1個に挿入された特異的ウイルス性抗原、例えばロタウイルス(RV)抗原を有する、抗原−P粒子の使用である。
【0025】
本発明の別の局面は、ウイルス性疾患からヒトを保護する方法であって、滅菌、非毒性の薬学的に許容される担体中の、本明細書に記載のP粒子内のPドメインモノマーの表面ループの少なくとも1個に挿入された特異的ウイルス性抗原を有する抗原−P粒子の免疫量をヒトに投与することを含み、ここで前記抗原がウイルス性抗原である、方法に関する。
【0026】
本発明の別の局面は、ウイルス性疾患に対する個体の免疫応答を誘導する方法であって、本明細書に記載の抗原−P粒子および医薬担体を含む組成物の少なくとも1個の免疫学的有効量を個体に経口または非経口投与する工程を含み、ここで前記抗原がウイルス性抗原であり、量が個体の免疫応答を誘導するのに有効である、方法に関する。
【0027】
本発明の別の局面は、抗原−P粒子ベクター/ワクチンプラットフォームキットである。
【0028】
本発明の別の局面は、Pドメインモノマーの1個以上の完全に露出したループのクローニングカセットへの、固有または稀少な制限部位を含む制限部位の導入に関する。制限部位を含むクローニングカセットの挿入に関して、ループのクローニングカセットへのプロテアーゼ認識配列の挿入を避けることが推奨される。例えば、BgIII部位(AGATCT)は、典型的なトリプシン切断部位であるジアミノ酸制限部位をコードし、避けられるべきである。
【0029】
本発明の別の局面は、最適にある外来抗原を適合させるために、ループの露出を伸長するスペーサーまたはアームを加えることを含む、ループの1個以上の修飾に関する。
【0030】
本発明の別の局面は、RV VP8のような大きな抗原がより小さな外来抗原を有する隣のループを、このループにおける小さなエピトープの提示が困難となるように覆わないように、異なる抗原が複数挿入されている設計に関する。
【0031】
本発明の別の局面は、遠位表面の1個以上に挿入されたリガンドまたはシグナルペプチドを有する置換P粒子がもたらされる、Pドメインモノマーの表面ループの1個以上へのリガンドまたはシグナルペプチドの挿入に関する。このリガンドまたはシグナルペプチドは、次いで、特定の臓器または組織の対応するレセプターに置換P粒子を標的化し、それらの位置に移動させられる。例えば、ペプチドCNGRC(5アミノ酸)を有する置換P粒子は、CNGRCのレセプター(CD13)が高度に発現している腫瘍組織(すなわち癌)に移動できる。
【0032】
本発明の別の局面は、化学反応によって表面露出リシンおよびシステインを介してP粒子のループに挿入されている薬物のコンジュゲートを有する、置換P粒子モノマーならびにそのダイマーおよびP粒子に関する。この場合、置換P粒子は薬物の担体として使用される。
【0033】
本発明の別の局面は、表面ループの少なくとも1個に挿入されたリガンドまたはシグナルペプチドを有し、そして表面ループの少なくとも1個に挿入された薬物のコンジュゲートを有する、置換Pモノマーならびにそれから形成されたダイマーおよびP粒子に関する。
【0034】
本発明の別の局面は、修飾または抗原−Pドメインモノマーの修飾された遺伝子コードの遺伝子配列である。
【0035】
本発明の別の局面は、修飾または抗原−Pドメインモノマーの修飾された遺伝子コードを有する複製生物学的物体(例えば大腸菌)である。
【0036】
本発明の性質および利点は、本明細書に記載の、そして添付の特許請求の範囲によってより具体的に定義されている意図する新規構成、組合せおよび要素を示す下記図面および詳細な説明からより十分に理解されるが、先行技術によって除外されない限り、本発明の正確な態様における変化が特許請求の範囲に含まれることを意味すると理解される。
【図面の簡単な説明】
【0037】
【図1】P粒子−VP8キメラのCryo−EM構造を示す。(A)野生型P粒子。(B)P粒子−VP8キメラ。野生型P粒子と比較して、キメラは、中央にニックを伴う伸長された突起を示し、これはP2サブドメインと挿入されたVP8抗原との間の境界を示唆する。(A)および(B)における粒子の半径は、示したとおり異なるカラースキームによって示される。(C)ロタウイルス(Wa)VP8抗原の2つのコピーの結晶構造(模式図、緑および青)の、キメラの伸長された突起の密度マップ(透明なグレー)への重ね合わせであり、キメラP粒子上でのVP8抗原の露出が確認される。(D〜E)P粒子−VP8の突起を拡大した側面図(D)および上面図(E)。
【図2】NORカプシドおよびカプシドタンパク質(VP1)を示す。AおよびBは、ノーウォークウイルス様粒子(NOR VLP)の表面構造(A)および断面(B)を示す。CはVLPの基本単位であるVP1のダイマー(リボンモデル)を示す。各VP1は、N末端アーム(緑)、シェル(S)ドメイン(黄色)および突起(P)ドメインに分けられる。Pドメインはさらに、P1およびP2サブドメイン(それぞれ赤および青)に分けられ、P2はVLPの最も外側の表面に存在する。VP1の直線構造を同じカラーテーマでDに示す。
【0038】
【図3】NOR P粒子およびその表面ループの構造を示す。(A)はcryo−EMによって再構成されたノロウイルスP粒子の構造を示し、(B)は結晶構造(リボンモデル)において解明された、Pダイマーの最も外側の領域であるその突起の一つを示し、ダイマーの6個の表面ループを示している。赤−緑および青−黄色は、それぞれ2個のPドメインを示す。抗原挿入に好適な部位である各モノマーの3個の表面ループが示されている。2個のPドメインモノマーは安定なグローバルPダイマーを形成する。次いで、12個の同一のPダイマーが集合して、Pダイマーの濃度に依存して動力学的に交換および/または平衡するT=1正二十面体P粒子となる。Pドメイン間の分子内相互作用がP粒子形成の原動力である。末端結合システインは、分子内ジスルフィド結合を形成してP粒子を安定化できる。
【0039】
【図4】NOR Pダイマー−B−トリサッカライド複合体の結晶構造を示す。トリサッカライド(ウイルス性レセプター)は、リボンモデル中に存在するアーチ様Pダイマーの上部に、球で存在する。
【0040】
【図5】NOR P粒子が野生型NOR VLPと同様の免疫原性および免疫反応性を有することを示す。Aにおいて、NOR株VA387のP粒子によって誘導される過免疫抗体が、VLP誘導性抗体と同様の力価でVLPと反応した。VLP−(B)およびP粒子−(C)誘導性抗体は、NOR VLPとタイプA HBGAレセプターとの結合に対して同様の遮断効果を示した。
【0041】
【図6】cryo−EMによる小P粒子の3D構造再構成を示す。AおよびBは小P粒子の側面図および上面図を示し、Cは小P粒子のcryo−EMの密度マップへのPダイマーの結晶構造の重ね合わせを示す。
【0042】
【図7】P粒子形成および安定性の妥協なしに完全に露出される、P粒子のループ2での7xHisタグの露出を示す。(A)は7xHisタグを有するP粒子がTalon樹脂に良好に結合することを示す。GST−Pドメイン−Hisタグ融合タンパク質はトロンビンによって消化されて、P粒子−Hisタグキメラ(PP−Hisタグ)、GSTおよび他の共に精製されたタンパク質の混合物をもたらした。共に精製されたタンパク質と7xHis−P粒子(ロード前)をTalon樹脂と共にインキュベートした。7xHis−P粒子全ておよびわずかな共に精製されたタンパク質は、樹脂に結合した(フロースルー)。洗浄した後、結合した7xHis−P粒子をイミダゾールで樹脂から溶出した(溶出1および2)。マーカーは、上から下に113、92、50、35、29、21kDaを示すバンドを有する予め染色したタンパク質標準であった。(B)において、(A)の溶出のゲルろ過分析は、溶出した7xHis−P粒子の>98%がP粒子を形成したことを示した。
【0043】
【図8】露出したRV VP8を有するNOR P粒子を示す。SDS PAGEのパネルAは、アフィニティーカラム(レーン1)およびさらにP粒子(レーン2)およびPドメインダイマー(レーン3)フラクションでのゲルろ過によって精製された、組換えRV VP8−Pドメインキメラ(53.5kDa)を示す。パネルBおよびCは、VP8(パネルB、レーン4および5)またはNOR VLP(パネルC、レーン6および7)に対する抗体を用いた、VP8−P粒子キメラのウェスタン分析を示す。パネルDにおいて、アフィニティー精製VP8−Pドメインキメラ(レーン1)のゲルろ過は、該タンパク質の大部分がP粒子(〜1300kDa)を形成することを示す。パネルEは、被覆VP8−P粒子キメラ(赤、PP−hVP8C)、組換えVP8(青、hVP8C)およびP粒子のみ(緑、PP、コントロール)を検出するためにVP8に対する抗体を用いた、酵素免疫アッセイ(EIA)の結果を示す。レーンMは、上から113、92、52、35、29kDaのバンドで、予め染色したタンパク質マーカーを示す。
【0044】
【図9】ループ2上にクローニングカセットを有するP粒子ベクターを示す。P粒子の最も外側の結晶構造(リボンモデル)は、3個の表面ループの表示を有する図の下半分に示し、P粒子ベクターのクローニングカセットの詳細は、図の上半分に示す。
【0045】
【図10】RV VP8−P粒子キメラの製造を示す。ループ2にクローニングカセットを有するP粒子ベクター(A)は野生型NOR P粒子(B)を製造する。RV VP8コード配列(C)がP粒子ベクター(D)にクローン化されると、それはキメラの最も外側表面でのRV VP8の指標と共に、RV VP8−P粒子キメラ(E)を製造する。
【0046】
【図11】VP8−P粒子キメラによって誘導された抗体が、NORおよびRVの両方に反応性であり、NOR VLPとHBGAレセプターの結合の遮断を示した。グラフAは、それぞれP粒子およびRV VP8に対する抗体の反応性を示す。グラフBは、該抗体がNOR VLPとHBGAレセプターの結合を遮断することを示す。X軸は抗血清の希釈(倍率)を示す。
【0047】
【図12】P粒子ベクター、P粒子プラットフォームおよびキメラP粒子ワクチンの構築のフローチャートを示す。GST、PドメインおよびCは、グルタチオンS−トランスフェラーゼ(GST)遺伝子、NOR Pドメインおよびシステイン含有短ペプチドをそれぞれコードする配列を示す。両末端の太線は、pGEX−4T−1のプラスミドを示す。
【0048】
【図13】P粒子ベクターのデザインを示す。3個の表面ループの指標と共にP粒子の最も外側部分の結晶構造を、図の下側に示す。各表面ループで異なる制限酵素認識部位を含むクローニングカセットを有するP粒子ベクターの一つの代表的なデザイン選択肢を、図の上側部分に示す。
【図14】P粒子−Hisタグキメラの製造および分析を示す。(A)P粒子−Hisタグキメラの発現構築物。HisタグはN372とD374の間のループ2に挿入した。pGEX−4T−1は、GST遺伝子融合システムの発現ベクターである。丸で囲んだCは、システイン含有ペプチド(CDCRGDCFC)を示す。(B)結晶構造(模式図)においてP粒子の突起の遠位末端は、挿入されたHisタグが位置すると予測される、2個のN373の位置を示す(グレーおよびシアンのドットモデル)。(C)P粒子−Hisタグキメラの発現および精製。SDS PAGE分析は、GST−Pドメイン−Hisタグ融合タンパク質(GST融合体)が〜52kDaであることを示した。トロンビンによる溶液中での融合タンパク質の消化は、GST(〜27kDa)およびPドメイン−Hisタグキメラ(PD−Hisタグ)(〜35kDa、左パネル)をもたらした。PドメインHisタグキメラは、トロンビン消化によって精製樹脂からも放出されうる(右パネル)。Mは、上から下に113、92、50、35、29、21kDaを示すバンドで、予め染色したタンパク質マーカーである。(D)サイズ排除カラムSperdex 200を用いたトロンビン放出Pドメイン−Hisタグタンパク質のゲルろ過クロマトグラフィーの溶出曲線。3つの主要ピークはそれぞれ、空、P粒子−HisタグおよびPダイマー−Hisタグを示した。これらの3つのピークのサイズは、それぞれ青色デキストラン2000(〜2000kDa、空)、野生型P粒子(〜830kDa)および野生型Pダイマー(〜70kDa)でキャリブレートした。(E)ゲルろ過クロマトグラフィー(D)のフラクションをSDS PAGEで分析した。
【0049】
【図15】P粒子提示短ペプチドに対するマウスの反応を示す。(A)それぞれ等量のP粒子−Hisタグキメラ(PP−His、青)、Pダイマー−Hisタグキメラ(PD−His、紫)または野生型P粒子(WT PP、コントロール、黒)で免疫した後、EIAにおけるT. Maritimaの組換えHisタグ化αフコシダーゼに対するマウス血清の免疫反応性(n=5)。(B)(A)における血清の抗体力価を終点希釈アプローチによって測定した。(C)〜(F)P粒子の非露出短ペプチドに対するマウスの抗体反応。(C)埋め込みペプチドCDCRGDCFCを含むP粒子(PP−CDCRGDCFC)で免疫した後の、EIAにおける、CDCRGDCFCタグ化マルトース結合タンパク質(MBP)(to CDCRGDCFC、紫)および野生型P粒子(to WT PP、ポジティブコントロール、緑)に対するマウス血清の免疫反応性(n=5)。(A)(PP-His to His-tag、青)が比較のために含まれた場合の、P粒子−Hisタグで免疫した後の、T. MaritimaのHisタグ化αフコシダーゼに対するマウス血清の免疫反応。(D)終点希釈アプローチによって、(C)における血清の抗体力価を測定した。(E)埋め込みペプチドCNGRCを含むP粒子で免疫した後、CNGRCタグ化MBP(to CNGRC、紫)および野生型P粒子(to WT PP、ポジティブコントロール、緑)に対するマウス血清の免疫反応性。P粒子−Hisタグで免疫した後の、(A)(PP−His、青)のT. MaritimaのHisタグ化αフコシダーゼに対するマウス血清の免疫反応性を比較のために加えた。(F)終点希釈アプローチによって、(E)における血清の抗体力価を測定した。EIAアッセイのために、5ng/μlの異なる抗原[(A)および(B)についてHisタグ化αフコシダーゼ、(C)および(D)についてCDCRGDCFCタグ化MBP、(E)および(F)についてCNGRCタグ化MBP、ならびに(C)〜(F)について野生型P粒子]をマイクロタイタープレートに被覆した。指定の希釈での対応する血清を用いて、免疫反応性を測定した。*P<0.05。
【0050】
【図16】P粒子−VP8キメラの製造および分析を示す。(A)Pドメインコード化cDNA配列を含むベクターpGEX−4T−1に基づく、P粒子−VP8キメラの発現構築物。酵素部位Spe IおよびCla I/EcoRIを有するクローニングカセットのT368とL375との間のPドメインのループ2中に、ロタウイルス(Wa)VP8抗原を挿入した。丸で囲んだCはシステイン含有ペプチド(CDCRGDCFC)を示す。(B)P粒子−VP8キメラの発現および精製。SDS PAGE分析は、GST−P−VP8融合タンパク質(GST融合体)が〜80kDaであることを示した(左パネル)。トロンビンによる融合タンパク質の消化は、GST(〜27kDa)およびP−VP8キメラ(〜52kDa、中央パネル)をもたらした。遊離P−VP8キメラは、トロンビン消化によって精製ビーズからも放出されうる(右パネル)。レーンMは、上から下に113、92、50、35、29、21kDaを示すバンドで、予め染色したタンパク質マーカーであった。(C)サイズ排除カラムSuperdex 200によるトロンビン放出P−VP8タンパク質のゲルろ過クロマトグラフィーの溶出曲線。ピークフラクションのSDS PAGE分析を上に示す。それぞれ青色デキストラン2000(〜2000kDa、空)、野生型P粒子(〜830kDa)および野生型Pダイマー(〜70kDa)でカラムをキャリブレートした。空のそばの単一主要ピークは、ほぼ全てのP−VP8タンパク質がキメラP粒子を形成したことを示す。(D)P−VP8タンパク質(左パネル)は、ロタウイルスVP8(中央パネル)およびノロウイルスVLP(右パネル)に対する抗体と反応した。
【0051】
【図17】P粒子提示VP8に対するマウスの免疫応答を示す。等モル量のP粒子−VP8キメラおよび遊離VP8を用いて、アジュバントなしで鼻腔内(AおよびB、n=5〜7)またはフロイントアジュバントと共に皮下(C、n=6〜7)で、マウスを免疫した。遊離VP8およびGSTをEIAにおける抗体力価測定のための抗原として使用した。(AおよびB)WaのVP8(A)およびDS1(B)それぞれを含む遊離VP8抗原(遊離VP8)およびキメラP粒子(PP−VP8)で免疫した後の、VP8/GSTに対するマウス血清の抗体力価。(C)遊離VP8抗原(遊離VP8)、およびWa株のVP8を含むキメラP粒子(PP−VP8)で免疫した後の、VP8に対する抗体力価。共精製したGSTは内部コントロールとして利用した。**P<0.001。
【0052】
【図18】P粒子−VP8キメラでの免疫によって誘導されるマウス血清による、ロタウイルスの中和を示す。(A)アジュバントなしでP粒子−VP8(Wa、[P]8)キメラで鼻腔内免疫したマウス由来のマウス血清は、同じWa株(PP−VP8血清、青)の強い中和を示すが、遊離VP8で免疫したマウス由来の血清は顕著に低い中和を示す(VP8血清、黒)。抗原を投与されていないマウス由来の血清をネガティブコントロールとして用いた(コントロール血清、紫)。(B)P粒子VP8(DS1、[P]4)キメラで免疫したマウス由来の血清は、Waとの弱い交差中和を示し(PP−VP8血清、青)、一方でDS1の遊離VP8で免疫したマウス由来の血清(VP8血清)およびネガティブコントロール血清は、中和を示さなかった(黒および紫)。(C)遊離Wa VP8(VP8血清、黒)およびP粒子−VP8(Wa)キメラ(PP−VP8血清、青)でフロイントアジュバントと共に皮下免疫したマウス由来の血清のVP8に対する免疫反応性。抗原なしでの血清は、ネガティブコントロールとして用いる(コントロール血清、紫)。(D)P粒子−VP8キメラによるマウスの免疫によって誘導される(C)由来の血清(PP−VP8血清、青)は、遊離VP8による免疫によって誘導される血清(VP8血清、黒)のものよりも、顕著に高いロタウイルス(Wa)に対する中和力価を示した。抗原を投与されていないマウス由来のコントロール血清をネガティブコントロールとして用いた(コントロール血清、紫)。星印は2つの形態のVP8によって誘導される血清の中和間のP値を示す:*P<0.05;**P<0.005および***P<0.0005。
【0053】
【図19】P粒子−VP8キメラワクチンでマウスを免疫した後の、マウスロタウイルス感染に対する保護を示す。4種の異なるワクチンでワクチン接種後、マウスのロタウイルス排出(μg/ml)を測定し、マウスロタウイルス(EDIM)でチャレンジした。WT PPでは、野生型ノロウイルスP粒子(ベクターコントロール)をマウスにワクチン接種した(n=7)。PP−hVP8では、P粒子−VP8(Wa)キメラをマウスにワクチン接種した(n=5)。遊離mVP8では、遊離マウスVP8(EDIM)抗原をマウスにワクチン接種した(n=5)。PP−mVP8では、P粒子−VP8(EDIM)キメラをマウスにワクチン接種した(n=5)。データ計算および統計分析は表1に示す。
【0054】
【図20】P粒子−VP8キメラでの免疫によって誘導された抗体が、ノロウイルスVLPとHBGAレセプターの結合を遮断することを示す。(A)P粒子−VP8(Wa)キメラで免疫した後のマウス血清は、ノロウイルスP粒子と強く反応したが(PP−VP8)、遊離VP8誘導血清は、この反応性を示さなかった(遊離VP8)。(B)(A)由来のマウス血清はノロウイルスVLPとHBGAレセプターの結合を遮断したが(タイプA唾液、PP−VP8血清、青)、遊離VP8での免疫後に得られた血清は、この遮断を示さなかった(VP8血清、黒)。
【発明を実施するための形態】
【0055】
発明の詳細な説明
本明細書において使用するとき、用語「ノロウイルス」、「NOR」、「ノーウォーク様ウイルス」または「NLV」は、ノロウイルスファミリーの任意のウイルスを意味し、限定されないが次のものを含む:ノーウォークウイルス(NV)、MOH、Mexico、VA 207、VA 387、02−1419、C59、VA 115、Hawaii、Snow Mountain、Hillington、Toronto、Leeds、Amsterdam、Idaho Falls、Lordsdale、Grimsby、Southampton、Desert Shield、BirminghamおよびWhite Rivercap。
【0056】
本明細書において使用するとき、用語「Pドメインモノマー」および「P粒子」は、それぞれ野生型ノロウイルスPドメインモノマーおよび野生型ノロウイルスP粒子を意味する。
【0057】
本明細書において使用するとき、用語「修飾されたPドメインモノマー」および「修飾されたP粒子」は、分子クローニングによって各Pドメインモノマー上に存在する3個の表面ループの1個以上に、1個以上の制限部位を(典型的には対で)挿入し、それによって後で外来抗原、リガンド、薬物コンジュゲートまたはシグナルペプチドを挿入できるように表面ループにクローニングカセットを作成するように修飾されている、ノロウイルスPドメインモノマーおよびP粒子を(それぞれ)意味する。
【0058】
本明細書において使用するとき、用語「置換Pドメインモノマー」および「置換されたP粒子」は、分子クローニングによって各Pドメインモノマー上に存在する3個の表面ループの1個以上に挿入された少なくとも1個の外来抗原、リガンド、薬物コンジュゲートまたはシグナルペプチドを含む、ノロウイルスPドメインモノマーおよびP粒子を(それぞれ)意味する。
【0059】
本明細書において使用するとき、用語「抗原−Pドメインモノマー」または「抗原−ノロウイルスPドメインモノマー」は、外来抗原がPドメインモノマー上に存在する3個の表面ループの1個以上に挿入されている、置換Pドメインモノマーを意味する。
【0060】
本明細書において使用するとき、用語「抗原−P粒子」または「抗原−ノロウイルスP粒子」は、外来抗原が各Pドメインモノマー上に存在する3個の表面ループの1個以上に挿入されている、置換されたP粒子を意味する。
【0061】
4種の固有の非感染性NOR粒子、すなわちウイルス様粒子(VLP)、S粒子、P粒子および小P粒子は、多機能性を設計するための優れた候補を表すウイルス性タンパク質ケージであり、Tan et al., 2004, 「The P-domai of norovirus capsid protein forms dimer and binds to histo-blood group antigen receptors」, J Virol 78:6233-42およびTan et al., 2005, 「The p domain of norovirus capsid protein forms a subviral particle that binds to histo-blood group antigen receptors」, J Virol 79:14017-30に記載されている(それらの全体について、参照によりそれらの開示を本明細書に組み込む)。これらの粒子は、分子量420kDa(小P粒子)〜10.5mDa(VLP)を有し、サイズ14〜37nmを有する。NOR VLPは180個の完全カプシドタンパク質(VP1)から成り、3個のサブウイルス性粒子はVP1の一部から成る。シェル(S)ドメイン由来のS粒子、突起(P)ドメイン由来のP粒子および同じPドメイン由来であるがN末端にさらなるフラグタグを有する小P粒子。これら全ての粒子は、Tan et al., 2004, 上記; Tan et al., 2004, 「E. coli-expressed recombinant norovirus capsid proteins maintain authentic antigenicity and receptor binding capability」, J Med Virol 74:641-9; および Xia et al., 2007, 「Norovirus capsid protein expressed in yeast forms virus-like particles and stimulates systemic and mucosal immunity in mice following an oral administration of raw yeast extracts」, J Med Virol 79:74-83(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載のとおり、タンパク質がインビトロで発現されたとき自然発生的に集合する。
【0062】
野生型NORカプシドは、昆虫細胞において発現されたときウイルス様粒子(VLP)に自己集合する、55〜60kDaの単一の主要構造タンパク質(VP1)から成る(図2)。VLPは天然ウイルス粒子と形態学的および抗原的に同一であるが、感染性ではない。ノーウォークウイルスのX線結晶学的構造に基づくと、カプシドはVP1の90個のダイマーから成り、T=3正二十面体シンメトリーを示す。各VP1モノマー(530アミノ酸[aa])は、短いN末端領域(1〜49アミノ酸)、続いてシェル(S)ドメイン(50〜225アミノ酸)、ならびにP1(226〜278および406〜520アミノ酸)およびP2(279〜405アミノ酸)に分類できる突起(P)ドメインを含む。N末端/シェル(N/S)ドメインはカプシドの内部コアを形成し、VP1の最も保存的な部分であるが、Pドメインはカプシドの突出したアーチを形成し、より多様である。カプシドの表面に位置するP2サブドメインは、NOR株のゲノムにおいて最も高い多様性度を含む。それは株特異性、レセプター結合および潜在的中和抗体認識部位の決定基を含む。
【0063】
原子構造に従って、NORカプシドタンパク質のシェル(S)および突起(P1、P2)ドメイン(図2)は、8〜10アミノ酸ヒンジによって連結されている。P粒子のカプシドタンパク質(VP1)のレセプター結合領域は、Tan et al., 2004, 「The P domain of norovirus capsid protein forms dimer and binds to histo-blood group antigen receptors」, J Virol 78:6233-42(その全体について、参照によりその開示を本明細書に組み込む)に記載されているとおり、マッピングされている。
【0064】
NORは、感染のための宿主レセプターとしてヒト組織−血液グループ抗原(HBGA)を認識することが知られている。Sドメインの発現は宿主レセプターに結合する機能を有さない平滑粒子を形成するが、ヒンジを有するPドメインの発現は、レセプター結合機能を有するダイマーを形成するが、その結合親和性は低い。ヒンジなしのPドメインが発現されると、それはVLPのものと同様の顕著に高いレセプター結合親和性を有するサブウイルスP粒子(図1)を自然発生的に形成する。P粒子は12個のダイマーに組織される24個のPドメインモノマーからなる。P粒子は、対応するHBGAに結合でき、NOR VLPとHBGAレセプターの結合の強い遮断を示す。P粒子の自然発生的形成は、株VA387、MOHおよびノーウォークウイルス(NV)を含む多様なNOR株で観察されている。これらのP粒子は、電子顕微鏡(EM)下で中心に空洞を有するリング状のイメージを示し、分子量〜830kDaでゲルろ過で単一ピークを形成した。P粒子は極めて安定であり、Tan et al., 2005, 「The p domain of norovirus capsid protein forms a subviral particle that binds to histo-blood group antigen receptors」, J Virol 79:14017-30、およびTan et al., 2006, 「C-terminal arginine cluster is essential for receptor binding of norovirus capsid protein」, J Virol 80:7322-31(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載のとおり、大腸菌および酵母発現系の両方で、高収率で容易に製造できる。
【0065】
NOR P粒子は、ヒトNORの2つの主要な遺伝子群の異なる遺伝子群からのNOR P粒子の構築によって示されるように、NORに共通の特徴である。GI−1(ノーウォークウイルス)、GI−4(Koblenz433)、GI−8(Boxer)、GII−2(Melksham)、GII−3(Brattleboro321)、GII−4、GII−5(MOH)、GII−9(VA207)を含む異なるNOR遺伝子型を示す、40種類以上のNOR P粒子がこれまでに構築されている。世界中のNOR胃腸炎の65〜85%に関与する主たるNORタイプ(GII−4)について、25種類のP粒子が作成されている。これは、過去25年間で25種類の株が広まっていることを示す。
【0066】
P粒子の3D構造は、図1および3に示すとおり、7.6Åの分解能のcryo−EMによって再構成されている。P粒子は、12個のダイマーに組織される24個のPドメインモノマーを含む八面体シンメトリーである。P粒子の中心に空洞が存在する。Pダイマーの方向は、野生型カプシドのものと同様であり、P1サブドメインが空洞の中心に対して内側にあり、P2サブドメインがP粒子の最も外側の表面に対して外側にある。P粒子のcryo−EMの密度マップへのPダイマーの結晶構造の重ね合わせ(図4)は、HBGAレセプター結合界面がP粒子の最も外側の表面に位置することを示している。これは、HBGAレセプターとの強い結合機能を保持しているP粒子の観察と一致している。
【0067】
Pダイマーの原子構造は、プロトタイプノーウォークウイルスおよび主なGII−4株(VA387)においてX線結晶学によって解明されている(図4)。これらの結晶構造は、P粒子のcryo−EMの密度マップに良好にフィットする(図1)。3個の表面ループは、これらはアーチ様Pダイマーの上部(6個の表面ループ/Pダイマー、図3)またはNORカプシドおよびP粒子の最も外側の表面に対応する各Pモノマーの最も遠い末端で同定された。これらの表面ループは外来抗原提示のための優れた部位を提供し、詳細な結晶構造は、本明細書に開示するP粒子ワクチンプラットフォームの構造に基づく設計のための確かな基礎を提供する。
【0068】
HBGAレセプターとの結合について、P粒子はその外側表面に依存する。いずれか1個以上の制限酵素部位、抗原、リガンド、シグナルペプチドまたは薬物コンジュゲートが野生型Pドメインモノマーの表面ループに挿入されている場合、制限酵素部位、抗原、リガンド、シグナルペプチドまたは薬物コンジュゲートのサイズに部分的または完全に基づいて、それは形成されたP粒子の最も外側の部分を被覆し、1個以上の修飾もしくは置換されたループへのアクセスを隠すことができ、または修飾もしくは置換されたループのタンパク質構造を変化させて、形成されたP粒子がHBGAレセプターと結合するその能力を失わせることができる。我々は未だP粒子が最早HBGAに結合しないという前にはどの程度のループの変化が必要かということについての系統立てた分析は行っていない。しかし、我々は、制限部位をPドメインモノマーの中央ループ2に加えた場合に修飾されたP粒子がHBGAと結合しないことを見出している。
【0069】
ウイルスレセプター結合界面の同定は、図4に示すとおり、ノーウォークウイルスおよびVA387におけるA−またはB−トリサッカライド(ノロウイルスレセプター)との複合体におけるPダイマーの結晶化によって示されている。図示したとおり、図4は、B−トリサッカライド複合体(球体モデル)と関連したNOR Pダイマーの結晶構造(リボンモデル)を示す。レセプター結合界面はNORカプシドおよびP粒子の最も外側の表面と等しいアーチ様Pダイマーの上部に位置する。トリサッカライドとPダイマーのアミノ酸との間の大規模な水素結合ネットワークが観察される。ウイルスレセプターと相互作用していると予測されるこれらのアミノ酸の重要性は、Tan et al., 2008, 「Elucidation of strain-specific interaction of a GII-4 norovirus with HBGA receptors by site-directed mutagenesis study」,Virology 379:324-334(その全体について、参照によりその開示を本明細書に組み込む)に記載されているとおり、部位特異的突然変異誘発によって証明されている。
【0070】
4個のNOR粒子(VLP、S粒子、P粒子および小P粒子)のうち、P粒子は最も安定であり容易に製造できる。それは高度に免疫原性であり、広範な温度、pHならびに化学的および物理的条件に耐える。cryo−EMによるP粒子の3D構造(図1)およびPダイマーの原子構造(図2)は、Cao et al., 2007, 「Structural basis for the recognition of blood group trisaccharides by norovirus」, J Virol 81:5949-57、およびTan, 2008, 「Noroviral P-particle: Structure, function and applications in virus-host interaction」, Virology, Vol. 382, Issue 1, 5 December 2008, Pages 115-123(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載のとおり、P粒子の24個のモノマーまたは12個のPダイマーの各々の最も外側の表面に高度に露出している(図1、3および4)。構造に基づく配列アラインメントは、これらの露出ループが大きな配列の挿入に耐えることを示唆している。
【0071】
本発明の一つの局面において、タンパク質またはポリペプチド抗原性エピトープがこれらのループ、すなわちループ1(I293−H297)、ループ2(T371−D374)またはループ3(D391−N394)の一つで、あるいはその中に挿入されている場合、抗原−Pドメインモノマーまたはダイマーの自然発生的自己集合は、集合した抗原−P粒子の表面で24個の抗原エピトープを提示する。本発明のこの特徴は抗原提示、したがって下記の通りワクチンのための利点を提供する。Pダイマーの解明された分子構造は、したがって、かかる抗原挿入のための構造に基づく設計の確かな、詳細な情報を提供する。
【0072】
野生型Pドメインモノマーおよびダイマーのように、抗原−Pドメインモノマーがダイマーを、そしてダイマーが抗原−P粒子を形成するプロセスは、Pダイマーの濃度に基づいて動力学的に交換または平衡しうる。また野生型Pドメインモノマーおよびダイマーのように、システインが抗原−Pドメインモノマーの末端に結合しているとき、得られる抗原−P粒子は、システイン末端を有さないものよりもより安定となり、抗原−Pドメインモノマーの濃度に依存しない。各抗原−Pダイマーは、5本の折り畳み軸に沿った五角形を形成する、5個の周囲抗原−Pダイマーとの相互作用において同じ方向を有するべきである。結果として、抗原−P粒子の外側層はノロウイルスカプシドのアーチ構造と類似のP2サブドメインによって形成されるが、内側層はP2アーチに支えを提供するP1サブドメインによって作成される。他の正二十面体ウイルス粒子と同様に、抗原−P粒子の内側に空洞が存在すべきである。
【0073】
また野生型Pドメインモノマーおよびダイマーのように、本発明の抗原−Pドメインモノマーおよびダイマーは、インビトロで抗原−P粒子を自然発生的に形成することを防止するために、周知の方法で変性されていてもよい。同様に、本発明の抗原−P粒子は、それぞれの抗原−Pドメインモノマーおよびダイマーに自然発生的に分離しないように、変性されていてもよい。これは、所望により野生型Pドメインモノマーと共に、多様な抗原タイプの抗原−Pドメインモノマーの形成の、多様な分子割合へのインビトロでの混和または混合を可能として、次いで変性の影響が除かれたときに自然発生的に抗原−P粒子へと形成されうる。
【0074】
本発明の置換されたまたは抗原−P粒子は、安定であり、Tan et al., 2004, J Virol 78:6233-42およびTan et al., 2005, Virol 79:14017-30に記載のもののような周知の方法によって大腸菌および酵母において容易に製造できるため、野生型NOR P粒子のように、抗原ディスプレイのための魅力的な候補である。先行技術のManayani et al.のキメラFHV粒子およびChatterji et al.のキメラCPMV粒子はそれぞれ、昆虫および植物に感染するが(ヒトに対する影響は知られていない)、非感染性野生型NOR P粒子はヒトウイルスに由来し、候補ヒトワクチンとして同定されている。
【0075】
多くの固有の特徴のため、本発明の抗原−P粒子は、野生型NOR P粒子のように、ワクチンプラットフォームとして有用な商品となる。第一に、ベースのNOR P粒子はサブユニットワクチンにほぼ理想的なサイズ(〜840kDa、Φ=20nm)を有し、多価であり、完全サイズのウイルス様粒子(VLP)よりも容易に製造でき、単一ポリペプチドまたはタンパク質抗原よりも免疫原性である。第二に、抗原−P粒子は高収率かつ低コストで大腸菌で容易に製造でき、これは発展途上国にとって特に有用である。第三に、Pドメインモノマーの複数の(3個)表面ループおよび外来抗原の挿入のためのそれらの高い能力(少なくとも〜238アミノ酸まで)は、異なる複数の抗原の挿入を可能とする。したがって、得られたキメラ置換P粒子ワクチンは、多くの感染症および非感染症に対してさえも、広い適応を提供できる。第四に、外来抗原が挿入された少なくとも24個のコピーを含む均一なキメラ置換P粒子を作成するために、2〜3段階の単純な遺伝子操作のみが必要とされる。したがってこれらのワクチンは速やかに開発でき、インフルエンザウイルスのような急速に変化するウイルスファミリーに特に有用であり得る。第五に、制限部位を含む表面ループにクローニングカセットを有するPドメインモノマーベクターは、ユーザーフレンドリーなベクターであり、簡便なワクチンプラットフォームである。そして第六に、抗原−P粒子は、両方のヒトNORについての下痢および他の感染症に対する優れた候補ワクチンである。
【0076】
主たる感染症に加えて、本PドメインベクターおよびP粒子ワクチンプラットフォームは、抗原提示および/または薬物の担体もしくはがん、アレルギー、自己免疫疾患を含む非感染性疾患の処置のための薬物送達ビークルとしても有用であり得る。一つの態様において、薬物は、化学反応によって表面露出リシンおよびシステインを介してP粒子のループに挿入されていてよい。この場合、置換されたP粒子は、薬物の担体として用いられる。他の態様において、リガンドまたはシグナルペプチドは、表面ループの少なくとも一つに挿入されてよく、そして薬物のコンジュゲートも、表面ループの少なくとも一つに挿入されていてよい。これらの置換されたP粒子は、薬物に特定の組織または臓器を標的とさせる薬物送達系を提供する。
【0077】
本発明のP粒子は、さらに特異性を高め、当該技術分野の有用な進歩を利用するために、他の薬物送達アプローチと組み合わせてもよい。一つの例は、病原体感染または特定の化学的もしくは酵素的切断がないかぎり、そしてそれまで、生物学的に不活性であり、植栽棒哺乳類細胞の内部で活性薬物形態に変換される、プロドラッグを含む。かかる態様は、特定の微生物に感染した細胞または特定の腫瘍に罹患した細胞でのみプロドラッグが活性化される、プロドラッグ−P粒子の開発を特に含みうる。
【0078】
P粒子ベクターの固有の特徴のために、ワクチンプラットフォームは広い適応を有することができる。それは既知の保護エピトープを有する多くのワクチンを製造するために使用できる。したがって、P粒子ベクターワクチンプラットフォームの適応は、広範な臨床状態に潜在的に有益であり得る。さらに、このナノ粒子プラットフォームの柔軟性は、容易に拡張されうる。例えば、PドメインモノマーまたはP粒子の1個の表面ループが特定のレセプターを有するある組織または臓器にP粒子を誘導するシグナルペプチド(リガンド)を発現するが、ワクチンまたは薬物が効果を奏するように送達される第2の表面ループに結合されるように、粒子を開発できる。例えば、ペプチドCNGRC(5アミノ酸の鎖)で置換されたP粒子は、CNGRCのレセプター(CD13)が重度に発現されている腫瘍組織(すなわち癌腫)に移動できる。膜結合メタロプロテアーゼCD13のリガンドであるペプチド部分CNGRCは、CD13を発現する血管形成血管系の内皮細胞に結合でき、CD13を発現する内皮細胞におけるアポトーシスを誘導し、それによって腫瘍関連血管新生を阻害できる。
【0079】
さらに、2個以上のループが同一または異なる病原体由来の異なる抗原を発現するように、多価ワクチンが作製できる。置換されたP粒子は、野生型Pドメインモノマーおよび抗原−Pドメインモノマーを含む、異なる抗原−Pドメインモノマーの混合物から集合して、異種抗原−P粒子を形成できる。多様な外来抗原を同じループまたは3個のループの異なる一つに挿入して、広範な異種抗原−P粒子を提供できる。したがって、本P粒子およびPドメインモノマーベクタープラットフォームの市場価値は極めて高い。
【0080】
本発明の態様は、NORおよびロタウイルス(RV)に対するワクチンとして使用するための抗原−P粒子を含んでいてもよい。抗原−P粒子の一つの例は、RV VP8−P粒子キメラである。ロタウイルス(RV)およびNORは、疾患の大きな負担を生じる世界中で共通の病原体である。この抗原−P粒子は、毒性株を生じる復帰または再集合の可能性のない、単独ワクチンでNORおよびRVに対して有効なワクチンを提供する。
【0081】
1個以上のループの修飾の前に、P粒子形成を促進し安定化するためにPドメインのいずれかの末端にシステインを加え、そしてP粒子を不安定化しうるトリプシン処理に対する耐性を与えるためにPドメインのC末端に短いペプチド(CDCRGDCFC)を加えて、野生型Pドメインモノマーを製造できる。また、P粒子形成のために、PドメインモノマーのインタクトなC末端が必要であり、ワクチンプラットフォームとしてのP粒子形成および安定性のために、C末端でのアルギニンクラスターが重要である。
【0082】
小ペプチドタグ(DYKDDDDK、フラグタグ)がPドメインのC末端に結合したとき、NOR Pドメインモノマーはまた、小P粒子と呼ばれる別の複合体を形成した。この小P粒子は、6個のPダイマーに組織される12個のPモノマーを含む四面体シンメトリーを示す(図6)。小P粒子のcryo−EMの密度マップへのPダイマーの結晶構造の重ね合わせは、Pダイマーの方向がP粒子のものと同様であることを示した。インビトロでのHBGA結合アッセイは、小P粒子がNOR VLPおよびP粒子と同様の結合プロファイルを有することを示した。小P粒子の発見は、より小さなP粒子が特定の用途に必要である場合における、我々のP粒子系のさらなる候補を提供する。
【0083】
本発明のNORベースの抗原−P粒子は、各Pドメインダイマーまたはモノマーサブユニットの提示ループの1個以上において外来抗原を担持するように構築されうる。Hisタグ(7xヒスチジン)、マウスサイトメガロウイルス(MCMV)T細胞エピトープ(9アミノ酸)、シュードモナス属エピトープEpi8(14アミノ酸)およびRV表面抗原である2種のRV VP8コアタンパク質(各159アミノ酸)を含むがこれらに限定されない多様なポリペプチドおよび生物学的関連エピトープが、置換されたP粒子の形成および安定性に影響を与えることなく、図9に示すとおり、P粒子のループ1(I293−H297)、ループ2(T371−D374)またはループ3(D391−N394)のいずれか1つ以上に挿入されてよい。これはループ2について、図7のパネルBおよび図8のパネルDに示すとおり、ゲルろ過によって示される。Hisタグの露出はTalon樹脂との高い親和性によって示された(図7のパネルA)VP8の露出は、RV VP8に対する抗体を用いたEIA(図8のパネルE)およびcryo−EMを用いたVP8−P粒子の3D構造再構成(図10のパネルE)によって示されている。これらのデータは、P粒子が多様な外来抗原を提示するための有効なワクチンプラットフォームに発展しうるという我々の仮説を支持している。
【0084】
抗原−P粒子ベクターは、クローニングカセットと共に構築されてもよい。Pダイマーの結晶構造に基づいて、ループの先端を含むP粒子ベクターのループの一つ以上に制限部位を含むクローニングカセットを入れることができる。図9はP粒子ベクターのループ2の先端に入れたSpe I部位およびCla I部位を含むクローニングカセットを示す。クローニングカセットを含むこのP粒子ベクターは、図8および10に示すように、P粒子の表面へのRV VP8およびGFPの挿入成功に使用される。
【0085】
稀少および固有制限部位を含む制限部位、ならびにPドメインモノマーまたはP粒子の表面ループのいずれか一つ以上に制限部位を挿入する方法の例は、当業者に周知であり、Reed et al.の米国特許出願公開2004/0185556および2008/0085553(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載されている。これらの制限部位はDNA分子においてコードされ、典型的には二本鎖パリンドローム配列を含む。いくつかの制限酵素について、制限部位を提供するために4〜6ヌクレオチドで十分であるが、いくつかの制限酵素は、8個以上のヌクレオチドの制限部位を必要とする。例えば酵素EcoR1は下記6ヌクレオチド配列を認識する:5’G−A−A−T−T−C3’、ここで5’は「上流」末端と慣習により知られる分子の終点を示し、同様に3’は「下流」末端を示す。制限部位の相補鎖は、その逆平行鎖3’=G−A−A−T−T−C−5’である。
【0086】
NOR P粒子は大きな外来抗原挿入に耐えうる。例示のみを目的として、NOR P粒子のループ2に緑色蛍光タンパク質(GFP、238アミノ酸)を挿入したところ、得られたキメラ粒子はゲルろ過分析で測定したところ高度に安定である。GFPの露出は、GFP特異的抗体を用いたEIAおよび蛍光顕微鏡で示された(データは示さず)。
【0087】
ある方法の態様において、修飾されたNOR P粒子を用いて、NORおよび外来抗原両方に反応性の抗体を誘導する。具体的な、非限定的な例において、RV VP8−P粒子を用いて、RVおよびNORの両方に反応性の抗体を誘導する。VP8−P粒子キメラで免疫した哺乳類はNOR P粒子およびRV VP8の両方に反応性の抗体のレベルをもたらし、NOR VLPとHBGAレセプターの結合を遮断し、RVの複製を中和できる。
【0088】
NORの「中和アッセイ」およびRVチャレンジのマウスモデルが開発されている。NORは未だ細胞培養で培養することが難しく、NORの複製に有効な動物モデルが存在しない。NOR感染に対する抗体の潜在的保護を測定するため、NORとその宿主レセプターの結合の防止における抗体の能力を測定するためのHBGAレセプター遮断アッセイを開発した。このアッセイは、唾液またはオリゴ糖ベースのレセプター結合アッセイに基づいており、極めて感受性かつ特異性である。
【0089】
RV感染に対するワクチンの保護効能を測定するための成体マウスモデルも確立されており、RVワクチン開発を研究するために用いられている。ワクチン候補でマウスを免疫し、次いでマウスRV株でチャレンジした。免疫群におけるRV抗原の糞便排出量を、ワクチン効能の測定のためにコントロールまたは非免疫群における排出量と比較する。ワクチンに対する免疫応答は、抗体レベルおよびT細胞応答を測定することでも測定できる。
【0090】
本発明はまた、外来抗原の複数または複数−多価提示を有する抗原−P粒子の形成に関する。2個のループ(すなわちループ1および2、ループ2および3またはループ1および3)で2個のクローニングカセットを有する修飾されたP粒子ベクターは、多様な組合せで2個のカセットに外来抗原を挿入することによって構築でき、評価できる。3種の組合せの非限定的な例は、1)ループ1および2におけるHisタグおよびフラグタグ(YDKDDDDK);2)ループ1および2におけるHisタグおよびVP8抗原;ならびに3)ループ1および2における2個のVP8を含む。これらの態様は、同じ置換P粒子上で効率よく提示される2種の異なる小抗原、または1種の小抗原および1種の大抗原、または2種の大抗原を提供できる。キメラP粒子形成はゲルろ過によって測定され、挿入された抗原の露出は特異的抗体を用いたEIAによって試験できる。
【0091】
3個のループ(ループ1、2および3)全てで3個のクローニングカセットを有する修飾されたP粒子ベクターは、多様な組合せで3個のカセットに外来抗原を挿入して構築でき、評価できる。非限定的な例は、1)3個の7xHisタグ;ならびに2)7xHisタグ、mycタグ(EQKLISEEDL)およびフラグタグを含む。第一の組合せは、置換されたP粒子の各Pドメインモノマーの3個のループ全てで同じ小さなエピトープを重複して提示し、P粒子上で抗原の72個(3×24)コピーを与える。第二の組合せは、各Pドメインモノマーサブユニットで同時に提示される3種の異なる小エピトープを提供し、3種の異なる抗原に対する三価P粒子ワクチンをもたらす。
【0092】
したがって本発明は、簡便かつ多様な抗原挿入のためのP粒子ベクターを提供する。修飾されたP粒子の態様は、それぞれ3個の表面ループの各々で単一の固有のクローニングカセットを含む各々修飾されたPドメインモノマーである、3種のPドメインモノマーベクターを含みうる。修飾されたP粒子ベクターの別の態様は、3個のループのいずれか一つもしくはいずれか2つ(例えばループ1および2)または3個のループ全てにおいて2個のクローニングカセットを有するPドメインモノマーベクターを含む。修飾されたP粒子ベクターの別の態様は、各Pドメインモノマーの3個のループ全てにおいて3個のクローニングカセットを有していてよい。各クローニングカセットの酵素認識部位は、図13に示すとおり、目的の抗原のDNA配列のクローニングの必要に応じて修飾されてよい。これらのベクターは、1種類の抗原の単独提示または同一もしくは異なる抗原の複数提示のためものもである。異なるクローニングカセットを有するこれらのP粒子ベクターの構築は、当該技術分野で知られているとおり、部位特異的突然変異誘発によって達成されうる。
【0093】
変異誘発および組換えDNAクローニング手法。様々な修飾または置換されたP粒子ベクターの作製は、典型的には、NOR(特に、実施例のとおり、VA387株)のPドメインおよびシステインを有する短ペプチドをコードする配列を好適なプラスミド(特に、例示として、Amersham Biosciencesから入手可能なプラスミドpGEX−4T−1)中に含む、図12に示すような、野生型P粒子発現構築物で開始する。クローニングカセットは、Tan et al., 2003, 「Mutations 448 within the P2 domain of norovirus capsid affect binding to human histo-blood group 449 antigens: evidence for a binding pocket」, J Virol 77:12562-71、Tan et al., 2006, 「C-terminal arginine cluster is essential for receptor binding of norovirus capsid protein」, J Virol 80:7322-31、Tan et al., 2005, 「The p domain of norovirus capsid protein forms a subviral particle that binds to histo-blood group antigen receptors」, J Virol 79:14017-30およびTan et al., 2008, 「Elucidation of strain-specific interaction of a GII-4 norovirus with HBGA receptors by site-directed mutagenesis study」, Virology 379:324-334(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載のものを含む周知の方法による部位特異的突然変異誘発によって、一つずつ、図13に示すように、表面ループの特定部位でPドメイン配列に導入される。結晶学的研究による3個の表面ループの位置、組成および配列は、Bu et al., 2008, 「Structural basis for the receptor binding specificity of Norwalk virus」, J Virol 82:5340-7およびCao et al., 2007, 「Structural basis for the recognition of blood group trisaccharides by norovirus」, J Virol 81:5949-57(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載されている。
【0094】
構造および配列情報に基づいて、クローニングカセットとして特定の酵素認識部位が適切な位置に挿入されたものとして、特定のプライマーを設計し、合成できる。典型的には、表面ループの先端(最も露出した位置)での2個のアミノ酸がクローニングカセットの配列を挿入するために選択されてよく、これは2個のアミノ酸の置換または2個のアミノ酸の間に配列を挿入することによってよい。周知の通り、QuikChange Site-Directed Mutagenesis Kit(Stratagene)を用いてループにクローニングカセットを導入できる。特定の抗原についての正確な導入位置は、得られる抗原−P粒子キメラの安定性または効率を改善するために修飾されてもよい。より長い配列でのクローニングカセットの挿入のために、最適な挿入のために段階的部位特異的突然変異誘発を実施できる。次いで、部位特異的突然変異誘発の後得られた置換されたP粒子ベクターのプラスミドは、周知の方法によって、プラスミドDNA調製キット(Qiagen)を用いた大腸菌およびプラスミドDNA調製物へのプラスミド形質転換のための常套のクローニング手法によって増幅できる。周知の方法により配列決定することによって、挿入されたクローニングカセットの確認の後、抗原−P粒子ベクターは、抗原挿入の能力、キメラP粒子形成の安定性、および有用かつ効果的なワクチンとしての抗原提示の効率について、試験されてよい。
【0095】
適切なキメラP粒子形成は、抗原提示についてのP粒子ベクターの能力および効率を評価して決定できる。P粒子プラットフォームへの抗原またはエピトープの挿入後(すなわち、Pドメインモノマーに抗原またはエピトープの挿入後、ダイマー、次いで抗原−P粒子へのPドメインモノマーの集合)、得られたキメラタンパク質の各々は、サイズ排除カラム(例えば、Akta ファストパフォーマンス液体クロマトグラフィー(FPLC、HE healthcare))を用いたゲルろ過によって、標的抗原−P粒子キメラの形成について分析されてよい。正しく形成された標的キメラは、モノマーの分子量の〜24倍の予測分子量(MW)で単一のピークを形成する。例えば、HisタグP粒子は870kDaのMWを有し、VP8−P粒子は1240kDaのMW、そしてGFP−P粒子は1450kDaのMWを有する。P粒子による抗原の複数提示は、この計算に従う。予想されるMWを有するキメラP粒子の形成は、抗原挿入の成功の重要な指標である。キメラP粒子形成のさらなる証拠は、cryo−EMによるEM観察および3D再構成を含む(図10参照)。修飾された抗原−P粒子の安定な形成は、通常、抗原の優れた免疫原性を導き、対応する病原体に対する有望なワクチン候補であろう。
【0096】
キメラP粒子上の抗原露出の測定は、抗原提示についてのP粒子ベクターの能力および効率の別の重要な指標である。抗体特異的EIAおよびTalon樹脂結合アッセイのいずれかまたは両方が、この測定のために用いられる。抗体特異的EIAはHuang et al., 「Noroviruses bind to human ABO, Lewis, and secretor histo blood group antigens: identification of 4 distinct strain specific patterns,」 2003, J Infect Dis 188:19-31およびHuang et al., 「Norovirus and histo-blood group antigens: demonstration of a wide spectrum of 23 strain specificities and classification of two major binding groups among multiple 24 binding patterns,」 2005, J Virol 79:6714-22(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載されており、図8のパネルEおよび図11のパネルAに図示されている。精製された抗原−P粒子キメラはマイクロタイタープレートに被覆され、脱脂乳によって遮断されうる。露出している抗原/エピトープは、対応する抗体によって検出できる。シグナル強度は、二次抗体−HRPコンジュゲート、次いでHRP基質とインキュベート(inculcation)した後、吸光度(OD)によって記録されうる。抗原(例えばそれぞれHis、フラグおよびmycタグ)に対して特異的なモノクローナル抗体は、Cell Signalingから商業的に入手可能であるが、図8および11に示すとおり、RV VP8およびNOR P粒子に対する特異的抗体は本願出願人によって製造され、それから入手可能である。露出したHisタグの検出のために、別の強力な方法であるTalon樹脂結合アッセイを用いてもよく、ここでTalon樹脂(Clontech)はHisタグと特異的に結合できる(図7)。樹脂結合HisタグP粒子キメラはイミダゾール(Sigmaから入手可能)によって容易に放出されるため、このアッセイは、汚染タンパク質からHisタグP粒子キメラを精製するための単純なアプローチである(図7参照)。
【実施例】
【0097】
実験
キメラP粒子の調製。予め製造した[P]タイプ8株のヒトVP8を含むRV VP8−P粒子キメラ(図8、図10のパネルE)を用いる。マウスRVチャレンジ試験のためのマウスVP8を含むキメラワクチンのために、VP8−P粒子キメラのヒトVP8をマウス(EDIM)VP8で置換する。ヒトおよびマウスRVキメラP粒子はいずれも、確立された手法を用いて大腸菌で製造される。GSTアフィニティーカラム(Glutathione Sepharose 4 Fast Flow、HE Healthcare)による部分的精製の後、キメラP粒子はさらにサイズ排除カラムを用いたFPLC系(GE Healthcare)で精製される。得られたキメラP粒子調製物は、高い純度(〜95%)に達し、マウスを免疫する免疫源として使用される。図8に見られるとおり、SDS PAGEのパネルAは、アフィニティーカラム(レーン1)およびさらにP粒子(レーン2)およびPドメインダイマー(レーン3)フラクションでのゲルろ過(パネルD)によって精製された、組換えRV VP8−Pドメインキメラ(53.5kDa)を示す。パネルBおよびCは、VP8(パネルB、レーン4および5)またはNOR VLP(パネルC、レーン6および7)に対する抗体を用いた、VP8−P粒子キメラのウェスタン分析を示す。パネルDにおいて、アフィニティー精製VP8−Pドメインキメラ(レーン1)のゲルろ過は、該タンパク質の大部分がP粒子(〜1300kDa)を形成することを示す。パネルEは、被覆VP8−P粒子キメラ(PP−hVP8C)、組換えVP8(hVP8C)およびP粒子のみ(PP、コントロール)を検出するためにVP8に対する抗体を用いた、酵素免疫アッセイ(EIA)の結果を示す。レーンMは、上から113、92、52、35、29kDaのバンドで、予め染色したタンパク質マーカーを示す。図10は、RV VP8−P粒子キメラの製造を示す。ループ2にクローニングカセットを有するP粒子ベクター(A)は野生型NOR P粒子(B)を製造する。RV VP8コード配列(C)がP粒子ベクター(D)にクローン化されると、それはキメラの最も外側表面でのRV VP8の指標と共に、RV VP8−P粒子キメラ(E)を製造する。
【0098】
マウスの免疫。6週齢のメスBALB/cマウス(Charles River Labs)を高度精製RV VP8−P粒子キメラで、異なる用量で、アジュバント[LT(R192G)]ありまたはなしで、経口または鼻腔内で免疫する。野生型P粒子はコントロールとして含まれる。ワクチン接種の鼻腔内経路のために、体積50μl中、野生型P粒子およびVP8−P粒子キメラの3種の初期用量(5、15および45μg/マウス、3回用量)が投与される(鼻腔あたり25μl)。経口経路のために、1mlのシリンジを備えた20ゲージのステンレス鋼製給餌針を用いて胃内カニューレによって胃に直接、合計200μlの同量のワクチンを送達する。動物(12匹のマウス/群)を一般的外見および体重減少についてモニターして、ワクチンの有害な効果の可能性を評価する。回復期前および中の血清サンプルを回収し、抗原として組換えNOR VLPおよびRV VP8を用いたEIAによってNORおよびRVに対する抗体応答について試験する。NORおよびRVに対する分泌IgAのために、糞便サンプルを回収する。1つのマウス群(N=5)を、ネガティブコントロールとして、細菌発現グルタチオンSトランスフェラーゼ(GST)で免疫する。
【0099】
インビトロアッセイによる免疫応答の評価。異なる投与レジメンによるワクチンの免疫前後のマウス血清を回収し、Huang et al., 2003, J Infect Dis 188:19-31およびHuang et al., 2005, J Virol 79:6714-22に記載のように、タイプ特異的抗体検出EIAを用いてNORおよびRVに対するそれらの特異的抗体力価を測定する。糞便サンプル中の分泌IgAを、粘膜免疫の指標としてNORおよびRVに対する応答について試験する。RVについて、中和アッセイは、細胞培養中のRV複製の阻害のために血清サンプルで実施される。NORについて、NORレセプター遮断アッセイを実施して、潜在的「中和」活性として抗体によって、NOR VLPとそれらのHBGAレセプターとの結合の阻害を測定する。
【0100】
VA387は世界中のNOR関連胃腸炎の65〜85%に関与する主要なGII−4遺伝子型であるため、同じVA387 VLPまたはP粒子に対する同種抗体応答を用いる。NORレセプター結合のインビトロ遮断アッセイを用いて、NORに対する抗体の潜在的中和を評価する。VP8−P粒子ワクチンの異なる投与レジメンからのマウス血清を、HBGAレセプターとNOR VLPの結合に対する遮断効果について試験する。抗体の遮断効果を再度、同種株VA387とそれらに対するHBGA(A、B、H、Lewis BおよびLewis Y抗原)との結合について試験する。簡潔には、既知のHBGAまたは合成オリゴ糖で十分に特徴付けられた唾液をマイクロタイタープレートに被覆する。脱脂乳でブロックした後、30分間抗体と共にインキュベートしたNOR VLPをプレートに加える。HBGAレセプターと結合したVLPを、特異的VA387 VLPに対するモルモット過免疫血清、次いでHRP複合化ヤギ抗モルモットIgG(ICN、Aurora、OH)の添加によって検出する。シグナル強度(吸光度、OD)は波長450nmでELISAプレートリーダーによって読み取る。遮断効果は、抗体処理ありとなしとでの結合の比較によって決定する。
【0101】
プラーク減少を測定するための細胞培養を用いた中和アッセイを実施して、マウスにおいてVP8−P粒子キメラに対して誘導される抗体を試験する。[P]タイプのVP8−P粒子キメラと他のRVタイプの交差中和を、発明者の研究室で入手可能な様々なRV株を用いて試験する。組織培養順応Wa株、マウスRV EDIM株およびMA104ミドリザル腎臓細胞株が使用される。簡潔には、MA104細胞を6ウェルプレートで培養し、RV接種の連続希釈によって〜50プラーク/ウェルの形成についてRV力価を測定する。アッセイのために、RVは特定希釈の血清と60分間インキュベートし、6ウェルプレート中の細胞(MA104)に混合物を加える。2時間後プレートを洗浄し、次いで0.8%のアガロースを含む培地で覆う。37℃で4〜5日間インキュベートした後、各ウェルのプラーク数を計測する。処理していないウェルと比較して、血清を含むウェルでのプラーク数の減少は、血清中に存在する中和抗体の量を決定する。
【0102】
マウスRVチャレンジモデル。ワクチンの保護効能を試験するためのこのRVチャレンジモデルの手法は、Choi et al., 「Functional mapping of protective domains and epitopes in the rotavirus VP6 protein,」 2000, J Virol 74:11574-80; Choi et al., 「Functional mapping of protective epitopes within the rotavirus VP6 protein in mice belonging to different haplotypes,」 2003, Vaccine 21:761-7;およびMcNeal et al., 1999, J Virol 73:7565-73.41(それらの全体について、参照によりそれらの開示を本明細書に組み込む)に記載されている。RV抗体を含まない6週齢のBALB/cマウス(Harlan-Sprague-Dawley、Indianapolis、Ind.)をVP8−P粒子キメラワクチンで経口または鼻腔内的に免疫する。最後の免疫の2〜4週間後、mice are 経口ly (via gavage、generally carried out by passing a feeding tube through the nose or mouth into the esophagus) challenged with murine RV EDIM strain at a dose of 100 50%排出用量と等しい4×104FFU(フコース形成単位)の用量でマウスRV EDIM株でマウスを経口的に(胃管栄養法で、一般的に鼻または口を介して食道へと給餌チューブを通して)チャレンジする。糞便中のRV抗原を測定するため、EDIMチャレンジの7日以上後に各マウスから2回糞便ペレットを採取し、1mlのアールバランス化食塩水中に保存する。分析までサンプルを凍結保存し、その時点でそれらをホモジナイズし、遠心分離してデブリを除去する。糞便サンプル中のRV抗原の量は、既報のとおりELISAで測定する。
【0103】
結果。結果は、VP8−P粒子が処置マウスにおいてRV−およびNOR−特異的抗体を誘導し、該抗体がNOR VLPとそのHBGAレセプターとの結合を遮断し、細胞培養中のRVの複製を中和し、そしてマウスチャレンジモデルにおいて免疫保護を提供することを示す。VP8−P粒子ワクチンは細胞傷害性または副作用を示さない。
【0104】
発現構築物。VA387ノロウイルス(GII.4)Pドメインコード配列およびシステイン含有ペプチドを含むpGEX−4T−1[グルタチオンSトランスフェラーゼ(GST) Gene fusion System、GE Healthcare life sciences]のP粒子発現ベクターを多様なキメラP粒子の構築のためのテンプレートとして用いた。P粒子Hisタグキメラを作製するために、Hisタグコード配列を部位特異的突然変異誘発によってN372とD374(図7および図14A)の間のループ2に挿入した(下記参照)。ヒトロタウイルスVP8抗原を含むキメラP粒子のために、まずループ2にSpeIおよびClaI/EcoRIの酵素部位を有するクローニングカセットを導入して、部位特異的突然変異誘発によってT369からD374の配列を置換した(図16A)。[P]8株(Wa、L65-L223、GenBank accession: VPXRWA)および[P]4株(DS1、L65-l223、GenBank accession: VPXRDS)のVP8抗原cDNA配列をカセットにクローン化した。P粒子にマウスロタウイルス(EDIM)VP8抗原(L65-L222、GenBank accession: AF039219)のcDNA配列をクローニングするため、XbaIおよびBgl II部位を有する別のクローニングカセットをN372とN373の間に挿入した。PダイマーHisタグキメラの作製のために、ヒンジを有するPドメインのN末端とHisタグを連結した。ペプチドCNGRCまたはCDCRGDCFCを含むキメラマルトース結合タンパク質(MBP)を作製するため、pGEX−4T−1のベクターにMBPのN末端と該ペプチドを融合した。pDEST17(Gateway、Invitrogen)(29)におけるHisタグ化サーモトガ・マリティマ(Thermotoga maritima)α−L−フコシダーゼ(GenBank accession: TM0306)の発現構築物は、Dr. HenrissatおよびDr. Bourne(Architecture et Fonction des Macromolecules Biologiques、UMR 6098、CNRSおよび Universites Aix-Marseille I and II、31 Chemin J. Aiguier、F-13402 Marseille、Cedex 20、France)から快く譲っていただいた。
【0105】
組換えタンパク質の発現および精製。0.25mMのイソプロピル−β−D−チオガラクトピラノシド(IPTG)の誘導により大腸菌(BL21)において、室温(〜25℃)で一夜、他に記載のとおりに組換えタンパク質を発現した。組換えGST融合タンパク質の精製は、Glutathione Sepharose 4 Fast Flow(GE Healthcare life Sciences)の樹脂を用いて、製造業者の指示書に従って実施した。ビーズ上または溶液中(リン酸緩衝化食塩水、PBS,pH7.4)のいずれかでトロンビン(GE Healthcare life Sciences)切断によって標的タンパク質からGSTを除去し、次いでゲルろ過クロマトグラフィーによってさらに精製した。Hisタグタンパク質の精製は、TALON His-Tag Purification Resins(Clontech、Mountain View、CA)を用いて、製造業者の指示書に従って実施した。250mMのイミダゾール(Sigma-Aldrich、St. Louis、MO)を含むPBSによって樹脂からHisタグタンパク質を溶出させた。樹脂精製タンパク質のさらなる精製は、ゲルろ過クロマトグラフィーによって実施した。
【0106】
ゲルろ過クロマトグラフィーは、既報の通りAKTA FPLC System(GE Healthcare life Sciences)で実施した。簡潔には、AKTA FPLC system(model 920、GE Healthcare life Sciences)で動力供給したサイズ排除カラムSuperdex 200(GE Healthcare life Sciences)にアフィニティーカラム精製タンパク質をロードした。溶出フラクションの分子量は、Gel Filtration Calibration Kits(GE Healthcare life Sciences)でキャリブレートした。あるいは、空ボリューム、キメラP粒子およびPダイマーのピークを、それぞれ青色デキストラン2000(〜2000kDa、GE Healthcare life Sciences)、野生型P粒子(830kDa)およびノロウイルスV387(GII.4)の野生型Pダイマー(69kDa)で決定できる。ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS PAGE)および/またはウェスタンブロット分析によって興味あるフラクションのタンパク質をさらに分析した。
【0107】
SDS PAGEおよびウェスタンブロット分析。10%ゲルを用いたSDS PAGEによって組換えタンパク質を分析した。他に記載のとおり、ノロウイルスVLP(VA387、GII.4、1:3000)またはロタウイルスVP8抗原(Wa、1:3000)に対する過免疫血清を用いたウェスタンブロット分析によって、組換えタンパク質の特異的組成を検出した。ブロットした膜を5%脱脂乳でブロックした。二次抗体−HRP(ホースラディッシュペルオキシダーゼ)コンジュゲート(1:5,000、ICN Pharmaceuticals、Costa Mesa、CA)を用い、そしてECL Eastern Blotting Detection Reagents (GE Healthcare life Sciences、Buckinghamshire、England)によってHRPを検出した。ECLシグナルはHyperfilm ECL(GE Healthcare life Sciences、Buckinghamshire、England)で捕捉した。
【0108】
既報の通り製造業者によって提供されたQuikChange Site-Directed Mutagenesis Kit(Stratagene、La Jolla、CA)の手法に従って部位特異的突然変異誘発を実施して、Hisタグおよび構成クローニングカセットをノロウイルスPドメインのループ2に挿入した。N372とD374との間のループ2(図7および図14A)に7個のヒスチジンの紐を挿入するためにプライマー対:caccactgacacaaaccaccaccaccatcatcaccacgatcttcaaactggcc/ggccagtttgaagatcgtggtgatgatggtggtggtggtttgt gtcagtggtgを用いた。さらに、ヒトロタウイルスVP8抗原のコード配列の挿入を促進するために、プライマー対:gttcaatacaccactagtacaaacatcgatatccttcaaactggc/ gccagtttgaaggatatcgatgtttgtactagtggtgtattgaacを用いて、ノロウイルスPドメインのループ2に3個の酵素部位(Spe IおよびCla I/EcoRI)を有するクローニングカセットを構築した。マウスロタウイルスVP8抗原のコード配列の挿入を補助するため、プライマー対:tacaccactgacavaaa ctc tagacacagatctaatgatcttcaaactgg/ccagtttgaagatcattagatctgtgtctagagtttgt gtcagtggtgtaを用いて、XbaIおよびBgl II部位を有する別のクローニングカセットを作製した。
【0109】
低温電子顕微鏡(cryo−EM)技術。
【0110】
Cryo−EMイメージ取得。我々の以前の研究に記載の野生型P粒子の構造再構成に用いた手法をここで採用した。簡潔には、精製P粒子−VP8(Wa)キメラのアリコート(3〜4μl)を液体窒素によって冷却した液体のエタン中、Quantifoil格子上でフラッシュ凍結した。サンプル格子を顕微鏡にのせ、200KVで操作するフィールド放出銃を備えたCM200低温顕微鏡を用いて、フィルム上で低線量イメージ(〜20e/A2)を記録した。公称倍率x50,000、デフォーカスレンジ2.0〜4.0μmでイメージを撮影した。顕微鏡写真を選択し、Nikon Super CoolScan 9000EDスキャナーをステップサイズ6.35μm/ピクセルで用いてデジタル化した。スキャンしたイメージをビニングして、さらなるイメージ処理および3D再構成のために、2.49Å/ピクセルでイメージの最終サンプリングを得た。
【0111】
Cryo−EMイメージ処理および3D再構成。EMANのboxerプログラムを用いてキメラP粒子のイメージを選択した。選択されたイメージを手作業で選別して、偽陽性を排除した。EMANのctfitプログラムを用いて、同じ顕微鏡写真に由来する粒子のセットに関連した、コントラスト−トランスファー関数(CTF)パラメーターを手作業で決定した。EMANのstartoctプログラムを用いて、キメラP粒子の初期モデルを作製した。次いで、EMANのrefineプログラムを用いて、新たなキメラ粒子の中心および方向を繰り返し決定し、収束するまでEMANのmake3dプログラムによって2Dイメージから3Dマップを再構成した。キメラP粒子の再構成によって八面体シンメトリーが与えられた。
【0112】
Cryo EMモデル評価および分析。UCSF Chimeraソフトウェアを用いて、ロタウイルスVP8抗原(Wa、2DWr、L65-L223)の結晶構造をP粒子−VP8(Wa)キメラの3D構造の伸長した突起に重ね合わせた。単純剛体運動がキメラP粒子の3D構造とX線構造の最良のマッチングの発見と考えられた。
【0113】
酵素免疫アッセイ(EIA)を用いて、P粒子−抗原キメラによって誘導されたマウス抗血清の免疫反応性および抗体力価を測定した。可変の抗血清のために異なる抗原を用いた:P粒子Hisタグキメラによって誘導された血清についてHisタグ化T. maritimaα−L−フコシダーゼ、非露出ペプチドCNGRCまたはCDCRGDCFCを含むP粒子によって誘導された血清についてMPB−CNGRCまたはMPB−CDCRGDCFC、P粒子−VP8キメラによって誘導された血清について遊離VP8、そして内部コントロールとしてP粒子−VP8キメラによって誘導された血清についてGST。抗原を96ウェルマイクロタイタープレート(Dynex Immulon; Dynatech、Franklin、Mass)に被覆した。5%脱脂乳でブロックした後、被覆した抗原と共に指定の希釈での血清をインキュベートした。他に記載のとおりに二次抗体−HRPコンジュゲートによって、結合した抗体を検出した。抗原に対する抗体の力価は、カットオフシグナル強度0.15での終点希釈として定義された。野生型P粒子またはPBSで免疫した動物由来の血清をネガティブコントロールとして用いた。
【0114】
HBGA結合および遮断アッセイ。基本的には他に記載のとおりに、唾液を利用した結合アッセイを実施した。簡潔には、既知のHBGA表現型を有する煮沸した唾液サンプルを1000倍に希釈し、96ウェルマイクロタイタープレート(Dynex Immulon; Dynatech、Franklin、MA)に被覆した。5%脱脂乳でブロックした後、VLPまたはノロウイルス(VA387、GII.4)のP粒子を加えた。我々自身が作製したウサギ抗VA387 VLP抗血清(1:3,300)を用いて結合したVLP/P粒子を検出し、次いでHRP複合化ヤギ抗ウサギIgG(ICN、Pharmaceuticals、Costa Mesa、CA)を加えた。VLPを被覆した唾液に加えた後、希釈した血清と共に30分間プレインキュベートして、ノロウイルスVLP−唾液結合に対するP粒子−VP8キメラによって誘導されたマウス血清の遮断効果を測定した。免疫した動物由来のマウス血清による遮断ありおよびなしで測定したODを比較することにより、遮断率を計算した。遊離VP8免疫動物由来の血清の遮断率をネガティブコントロールとして用いた。
【0115】
抗体応答のためのマウス免疫。6週齢のメスBALB/cマウス(Harlan-Sprague-Dawley、Indianapolis、IN)を精製キメラP粒子または遊離抗原で、用量5〜15μg/マウス、2週間間隔で3〜4回で免疫した。P粒子−またはPダイマー−提示Hisタグに対する免疫応答の比較のため、マウスに5μg/マウスの組換えP粒子−Hisタグキメラ、およびPダイマー−Hisタグまたは野生型P粒子を投与した(n=5)。非露出ペプチドに対する免疫応答の比較のため、マウスに非露出ペプチドCNGRCまたはCDCRGDCFCを含む組換えP粒子5μg/マウスを投与した。免疫源はフロイントアジュバントと共に皮下的に4回与えた。P粒子提示VP8および遊離VP8に対する免疫応答の比較のため、マウスに5μg/マウスの遊離VP8および15μg/マウスのP粒子−VP8キメラ(両抗原はVP8のモル量として同量である)を、アジュバントなしで鼻腔内またはフロイントアジュバントと共に皮下的に、3回投与で投与した(n=5〜7)。内部コントロールとして免疫源に等モル量のGSTを加えた。保護のためのマウスのP粒子−VP8ワクチンの免疫は下に記載した。免疫前および最終免疫の2週間後に、逆軌道毛細血管叢の穿刺によって血液を採取した。血清は、血液から4℃で一夜静置後遠心分離によって処理した。
【0116】
ロタウイルスプラークアッセイを実施して、細胞培養中のロタウイルス複製に対する、ロタウイルスVP8抗原を含むキメラP粒子によって誘導される抗血清の中和を測定した。MA104サル腎臓細胞で成長させた組織培養適応ロタウイルスWa株をこのアッセイに用いた。MA104細胞を6ウェルプレートで培養し、接種として〜50プラーク/ウェルのロタウイルス力価を用いた。アッセイのために、ロタウイルスを所定の希釈のマウス血清と共に60分間インキュベートした。次いで混合物を6ウェルプレート中のMA104細胞に加えた。2時間後、プレートを洗浄し、次いで5μg/mlのトリプシン(Invitrogen、Carlsbad、CA)および0.8%のアガロースを含む培地で被覆した。37℃で4〜5日インキュベーション後、各ウェルのプラーク数を計測した。無処理コントロールウェルのものと比較した抗血清処理したウェルにおけるプラーク数の減少によって、血清中の中和抗体の量を測定した。
【0117】
マウスロタウイルスチャレンジモデル。以前の研究に記載のロタウイルスチャレンジモデルに従って、P粒子−VP8ワクチンの保護効率を試験した。ロタウイルス抗体を含まない6週齢のBALB/cマウス(n=5〜7)(Harlan-Sprague-Dawley、Indianapolis、IN)をアジュバントなしで、マウスロタウイルス(EDIM)VP8抗原を含むキメラP粒子(15μg/マウス)で3回、鼻腔内免疫した。遊離マウスロタウイルスVP8抗原(5μg/マウス)およびヒトロタウイルス(Wa)VP8抗原を含むキメラP粒子(15μg/マウス)を比較のために含めた。さらに、野生型P粒子(ベクターコントロール)およびPBSをネガティブコントロールとして投与した。最後の免疫の2週間後、105の50%排出用量と等しい4×104FFU(フォーカス形成単位)の用量のマウスロタウイルスEDIM株で、経口胃管栄養法によってチャレンジした。糞便中のロタウイルス排出を測定するため、EDIMチャレンジ後6日間毎日各マウスから2個の糞便ペレットを採取し、1mlのアールバランス化食塩水中に保存した。分析までサンプルを凍結保存し、その時点でそれらをホモジナイズし、遠心分離してデブリを除去した。糞便サンプル中のロタウイルス抗原の量(μg/ml)は、既報のとおりELISAで測定した。
【0118】
統計的分析。GraphPad Prism version 5.00 for Windows(GraphPad Software、San Diego CA)およびMicrosoft Office Excel 2007を用いてグラフを作製した。GraphPad Prism version 5.00 for Windowsを用いたデータ群のt検定によってP値を決定した。
【0119】
結果
Hisタグを含むキメラP粒子の製造。我々の研究は、P粒子のループ2に小さなエピトープ、ポリヒスチジン(His)タグを挿入することで開始した(図3および14)。GST遺伝子融合系を用いた大腸菌におけるこのPタンパク質の発現および精製は、高収率(>10mg/リットル培養)で予測したサイズ(〜35kDa)を有するタンパク質が得られ、次いでトロンビンでGSTタグを除去するために(〜27kDa)タンパク質を消化した(図14C)。ゲルろ過クロマトグラフィー、次いでSDS PAGEおよびウェスタンブロット分析によってキメラPタンパク質のP粒子形成が示され、ここでキメラP粒子は〜840kDaの主要なピークを形成した(図14DおよびE、データは示さず)。キメラP粒子上での挿入したHisタグの露出は、Talon樹脂へのその特異的結合、続く250mMのイミダゾールによる溶出によって示され(図7A、溶出1および2)、これは高純度のキメラP粒子をもたらした。したがって、P粒子−Hisタグキメラはまた、アフィニティーTalon樹脂を用いて大腸菌からも精製できる(データは示さず)。
【0120】
P粒子担体によるHisタグの免疫増進。P粒子−Hisタグキメラで免疫した後マウスにおけるHisタグに対する免疫応答の試験によってこれを測定した(n=5)。Pダイマー提示HisタグよりもP粒子提示Hisタグで免疫したマウスにおいて、有為に高いHisタグに対する抗体力価が検出された(P<0.05、図15AおよびB)。PドメインのN末端(CNGRC)またはC末端(CDCRGDCFC)に融合した2種の短いペプチドは、P粒子提示Hisタグによって誘導されるものと比較して、有為に弱い免疫応答を誘導した(図15C〜F)。これらの2種のペプチドは、P粒子の内側に埋め込まれることが示されており、このことは、免疫増進のためにP粒子の表面上に挿入された抗原が適切に露出されることが重要であると示している。予想通り、全ての血清はノロウイルスP粒子プラットフォームと強く反応した(図15、データは示さず)。
【0121】
ロタウイルスVP8抗原を含むキメラP粒子の開発。我々は次に、159アミノ酸を含むロタウイルス(Wa)VP8抗原の挿入によって、より大きなポリペプチドを得るためのP粒子プラットフォームの能力を試験した。VP8挿入を促進するために、P粒子のループ2に3個の酵素部位(Spe IおよびCla I/EcoRI)を有するクローニングカセットを構築した(図16A)。大腸菌における構築物の発現によって、高収率(>15mg/リットル培養)でGST−P−VP8融合タンパク質(〜78kDa、図16Bの左パネル)が得られた。溶液中または精製ビーズ上のいずれかで、GST融合タンパク質のトロンビン消化によって、放出されたP−VP8キメラ(〜52kDa)が得られた(図16B、それぞれ中央および右パネル)。ゲルろ過クロマトグラフィーによってP−VP8キメラタンパク質が高い割合(>95%)でP粒子を形成することが示された(図16C)。ウェスタンブロット分析によって、P−VP8キメラタンパク質がノロウイルスVLP(VA387)およびロタウイルス(Wa)VP8に対する抗体両方と反応することが示された(図16D)。
【0122】
Cryo−EMおよび3次元イメージ再構成によって、P粒子−VP8キメラが野生型P粒子と同様の八面体シンメトリーのままであるが、キメラP粒子が顕著に大きいことが示された(図1AおよびB)。伸長した突起は、おそらく、Pダイマー由来の潜在的境界としてのニックによって認識されうる挿入されたVP8である。キメラの伸長した突起の密度マップにおける同じWa株(2)のロタウイルスVP8抗原の結晶構造の重ね合わせによって、伸長した突起が実際にVP8であることが確認された(図1C〜E)。
【0123】
VP8に対するP粒子増進免疫応答。次に我々は、マウスにおけるP粒子提示VP8抗原に対する免疫応答を試験した。等モル量のP粒子−VP8キメラおよび遊離VP8での免疫後、抗原として遊離VP8を用いたEIAによって得られたマウス血清を試験した。P粒子提示VP8に対する抗体力価は、アジュバントなしでの鼻腔内免疫による遊離VP8に対するものよりも有為に高かった(P<0.005)。同様の結果が、それぞれロタウイルスWaおよびDS−1のVP8抗原を含む2種のキメラP粒子とそれらの対応遊離VP8の比較において観察された(図17AおよびB)。これらの動物において遊離GSTに対してはわずかにのみ抗体力価が検出され、P粒子提示VP8の特異的免疫増進がさらに確認された。フロイントアジュバントと共に上記抗原で皮下免疫したマウスの免疫応答も試験したところ、遊離VP8とP粒子提示VP8の間でVP8に対する抗体力価の差はほとんど観察されなかった(図17C)。
【0124】
P粒子−VP8キメラはロタウイルスに対する中和抗体を誘導した。鼻腔内免疫を介したP粒子−VP8(Wa、[P]8)キメラによって誘導された過免疫抗血清は(図17A)、細胞培養中の同族ロタウイルス(Wa)複製を強く低減した。この中和力価は、遊離VP8での免疫によって誘導した中和力価よりも有為に高かった(P<0.0002)(図18A)。P粒子−VP8(DS−1、[P]4)キメラでの免疫後、Waに対する低レベルの交差中和も観察された(P<0.05、図18B)。反対に、DS−1の遊離VP8で免疫した動物由来の血清は、かかる交差中和を示さなかった。さらに、フロイントアジュバントと共に皮下免疫した動物由来の血清を用いた中和力価も測定した。驚くべきことに、EIAにおいて遊離VP8抗原と同様の免疫反応性を有する4対のマウス血清のうち(図18C)、P粒子−VP8(Wa)で免疫した動物由来の血清が全て、同じWa株の遊離VP8で免疫したマウス由来の血清のものよりも有為に高い中和力価を示した(P<0.005、図18D)。これらのデータは、キメラP粒子上のVP8がより良好に提示され、中和エピトープに必要な適切なコンホメーションを維持しうることを示した。
【0125】
P粒子−VP8キメラのワクチン接種はマウスロタウイルスチャレンジ後の低下した排出を引き起こした。次に我々は、マウスロタウイルスモデル(EDIM株)を用いてインビボで、P粒子−VP8キメラワクチンが保護を与えうるかを試験した。この目的で、EDIM VP8抗原を含むキメラP粒子を材料および方法に記載のとおりに構築し、マウスに投与した(N=5〜7)。同モル量の遊離EDIM VP8、ヒトVP8(Wa)を有するキメラP粒子または野生型P粒子で免疫した動物は、コントロールとして含めた。最後の免疫の2週間後、マウスEDIMロタウイルスで動物をチャレンジした。チャレンジの6日後に糞便サンプルを採取し、EIAで排出を検出した(表1、図19)。P粒子−VP8(EDIM)キメラをワクチン接種されたマウスは、最も少ない量のウイルス抗原を排出した。排出の減少は6日間で89%であった(P<0.05)。EDIMでのチャレンジの1日後では99.2%の減少であった。遊離マウスVP8で免疫したマウスは、次に少ない排出量であり、77%の平均保護であった(P<0.05、2〜5日)。ヒトロタウイルス(Wa)VP8を含むキメラP粒子ワクチンでの免疫は、23%減少と排出に対してわずかな効果を有し、これはEDIMに対する何らかの交差保護を示唆している。この群での6日目に、排出が顕著に減少した(63%、P<0.05)。
【0126】
【表1】
【0127】
ノロウイルスとHBGAの結合を遮断するP粒子−VP8キメラ誘導性抗体での免疫。また、P粒子−VP8(Wa)キメラでの免疫によって、マウスでの免疫応答におけるP粒子骨格の役割を試験した。予測したとおり、キメラ誘導性抗体は、EIAで測定したときノロウイルスVLPおよびP粒子と強く反応した(図20、データは示さず)。これらの血清はまた、HBGAレセプターとノロウイルスVLPの結合も遮断した(タイプA唾液、図20B)。ネガティブコントロールとして遊離VP8(Wa)で免疫した動物由来の血清はかかる遮断を示さなかった。この結果は、P粒子−VP8キメラがロタウイルスおよびノロウイルスの両方に対するデュアルワクチンであり得ることを示している。
【0128】
我々の以前の研究により、ノロウイルスP粒子が容易に製造でき、極めて安定で高度に免疫原性であり、これはノロウイルスに対するサブユニットワクチンとして使用できることが示された。我々はさらに、外来抗原の免疫増進のための新規ワクチンプラットフォームとしてP粒子が使用できることを示した。我々は、1)P粒子の表面ループがP粒子の形成および作製に影響を与えることなく外来抗原を挿入するための優れた部位であること、2)P粒子が少なくとも159アミノ酸までのサイズの外来抗原に耐えること、3)挿入抗原に対する増進された免疫応答がインビトロおよびインビボ両方での中和および保護実験によって示されることを示した。P粒子−VP8キメラはまた、ロタウイルスおよびノロウイルスの両方に対する有望なデュアルワクチンを提供した。したがって、複数多価外来挿入の能力を有するキメラ粒子およびその複数の表面ループを作製するための単純な手法は、P粒子を感染症および効果的なワクチンから利益を得るであろう他の状態の抗原提示のための魅力的なワクチンプラットフォームにする。
【0129】
本発明の第一次的な目標は、一般に免疫原性が低い小ポリペプチド抗原の免疫原性をP粒子が増進できるかを試験することである。我々は、モデルとしてHisタグを用いてこの問題を研究し、すばらしい結果を得た。2つの主要な要因、複数コピー数および挿入された抗原の表面露出が、観察された免疫増進に寄与している可能性がある。P粒子はPモノマーの24個のコピーからなり、このことはPダイマーと融合したHisタグとの比較において増進した免疫応答を説明できる(図15A)。抗原の表面露出の重要性は、PドメインのNまたはC末端と結合した2個の非露出ペプチドに対する低い免疫応答性によって示された(図15BおよびC)。したがって、P粒子はその大きなサイズ(830kDa)および適切な外来抗原の提示(これと異なるものは低い免疫原性を有する)によってアジュバントとして作用しうる。粒子あたりの抗原の増加した複数コピーは、増加した免疫応答を説明しうる別の特徴である。
【0130】
Hisタグペプチドに加えて、我々はP粒子に多数の他の小ペプチド、例えばマウスサイトメガロウイルスのT細胞エピトープ(9アミノ酸)、シュードモナス属Epi8エピトープ(14アミノ酸)およびインフルエンザAのM2細胞外エピトープ(23アミノ酸)の挿入に成功した(Tan and Jiang、未公表)。これらのデータは、P粒子が多様な小ペプチド抗原の免疫増進に容易に利用できることを示唆している。P粒子への様々なロタウイルスVP8の挿入成功は、広範な大型外来抗原のためのP粒子プラットフォームの適用を大きく拡張する。ロタウイルスVP8はウイルスカプシド上の突起タンパク質であり、ロタウイルス感染性に重要であると考えられている。また、それは中和抗体を誘導する2種の抗体の一つでもある。VP8タンパク質について多数の中和エピトープが同定されている。P粒子−VP8キメラの中和抗体誘導成功は、マウスにおけるロタウイルス排出に対して応答し、それに対する保護を提供する。このことは、これらのエピトープがP粒子担体上で提示されていることを示唆している。
【0131】
2種の近年導入されたロタウイルスワクチンは極めて有効であるが、新たな出現ウイルスのための新世代ワクチンが潜在的に必要とされうる。非感染性サブユニットワクチンは、現在のワクチンで心配されうる毒性株への復帰の危険がない。ロタウイルスのVLPワクチンの開発は数年来提案されている。ロタウイルスVLPワクチンは、バキュロウイルス宿主に複数のカプシドをコトランスフェクションする必要があるため、低い発現効率と高い製造コストの問題がある。反対に、P粒子−VP8キメラの作製は常套の大腸菌ベースのクローニングおよび発現手法のみを必要とし、これは効率がよくコストが低い。さらに、VP8についての交差中和エピトープが記載されている。我々は本稿において、P[4]ウイルス由来VP8を含むキメラP粒子でのマウスの免疫がP[8]ロタウイルスに対する交差中和を有することを示した。最小数のPタイプを含むカクテルワクチンもコスト的に有効であり得る。
【0132】
P粒子−BP8キメラによって誘導される抗体の、ノロウイルスVLPとHBGAの結合を遮断する能力は、予測外である。図1に示すとおり、P粒子のHBGA結合界面を含むPダイマーの遠位表面は、挿入されたVP8によってほとんど被覆されている。これはHBGAレセプターに対する該キメラの結合能の喪失を導く(データは示さず)。継続して遮断能が見られたことの一つの可能性は、挿入されたVP8抗原によって被覆されていても、P粒子−VP8キメラのHBGA結合界面のエピトープが抗体誘導のためになおもアクセス可能であるということである。あるいは、観察された糖遮断は、糖結合部位の近くでの抗体結合のためでありうる。どのようなメカニズムが関与するにせよ、キメラ誘導性抗体のノロウイルスVLPとHBGAとの結合を遮断する能力は、P粒子プラットフォームにさらなる価値を付け加える。ノロウイルスおよびロタウイルス両方に対するP粒子−VP8キメラのデュアルワクチンのコンセプトは、両方の感染の危険がある特定の集団に特に有用でありうる。
【0133】
ループ2のみがこの研究では試験されたが、他の2つのループにおける抗原挿入成功の可能性が予測され、我々の研究室で現在試験している。Pモノマーあたり3個の表面ループの利用可能性は、多目的ワクチン設計の機会を提供する。例えば、免疫応答を上昇させるため、同じエピトープまたは抗原を3個のループ全てに挿入して、粒子あたり72コピーの抗原に到達できる。それぞれの抗原のタンデム反復の挿入によって、より多いコピー数すらも作製できる。あるいは、異なる抗原を3個のループに挿入して、異なる病原体またはロタウイルスについて異なるVP8抗原に対する多価ワクチンを得ることができる。様々な目的のための機能的タグの挿入によって、さらなるワクチンテンプレートも作製できる。例えばHisタグの挿入は、精製手法をさらに単純にする。特定のリガンドまたはシグナル分子を用いて、宿主の特定の臓器、組織または細胞を標的とすることによって、免疫応答を刺激してもよい。
【0134】
P粒子プラットフォームのさらなる適用は、研究および診断的使用のための小ペプチドに対する抗体の製造用である。多様な疾患バイオマーカー(主としてペプチドエピトープ)が同定されており、これらのバイオマーカーに対する抗体が診断目的では重要である。小ペプチドは、DNAクローニングの単純な手法によってP粒子のループに容易に挿入できる。我々は、そのプロセスをさらに促進するクローニングカセットを含む簡便なP粒子ベクターを開発した。細菌における組換えキメラP粒子の発現の後、本研究で確立された手法にしたがって実験動物を免疫することによって、挿入したペプチド抗原に特異的な抗体を高力価で誘導できる。この方法による抗体製造は、ペプチド合成および免疫増進のためのキーホールリンペットヘモシニアン(KLH)のような巨大分子とペプチドとのコンジュゲートのようなコストのかかる工程を回避する。ノロウイルスPドメインは他のタンパク質のいずれとも相同性を有さない固有の配列を有しているため、他のタンパク質との交差反応性の心配にはおよばない。したがって、P粒子ワクチンプラットフォームは、生物医学的研究の多くの領域における抗体製造のための簡便なツールとして使用できる。
【0135】
本発明をその態様の解説によって説明し、その態様をかなり詳細に説明してきたが、いかなる方法においても、添付の特許請求の範囲がかかる詳細に限定されることを意図していない。さらなる利点および修飾が当業者に容易に理解される。したがって本発明は、そのより広い局面において、特定の詳細、代表的な系および方法、ならびに示し記載した実施例に限定されない。したがって、本発明の範囲または精神から離れることなく、かかる詳細から逸脱してもよい。
【特許請求の範囲】
【請求項1】
少なくとも1個の表面ループを有するノロウイルス(NOR)Pドメインモノマーおよび該少なくとも1個の表面ループに挿入されたタンパク質またはポリペプチド抗原を含む、抗原−Pドメインモノマー。
【請求項2】
少なくとも1個の表面ループが、ループ1(I293−H297)、ループ2(T371−D374)およびループ3(D391−N394)の少なくとも1個を含む、請求項1の抗原−Pドメインモノマー。
【請求項3】
抗原がロタウイルス(RV)VP8抗原である、請求項1の抗原−Pドメインモノマー。
【請求項4】
タンパク質またはポリペプチド抗原が少なくとも2個の類似または異なる抗原を含む、請求項1の抗原−Pドメインモノマー。
【請求項5】
24個のモノマーからなる抗原−P粒子であって、P粒子が少なくとも1個の請求項1の抗原−Pドメインモノマー、および所望により1個以上の野生型NOR Pドメインモノマーを含み、ここで、抗原−P粒子が抗原単独と比較して抗原の免疫原性を向上しうる、抗原−P粒子。
【請求項6】
少なくとも1個の表面ループを有するNOR Pドメインモノマーを含むPドメインモノマーベクターであって、前記少なくとも1個の表面ループが少なくとも1個の制限部位の対を含む、Pドメインモノマーベクター。
【請求項7】
少なくとも1個の表面ループがループ1(I293−H297)、ループ2(T371−D374)およびループ3(D391−N394)の少なくとも1個を含む、請求項6のPドメインモノマーベクター。
【請求項8】
少なくとも1個の表面ループが、該少なくとも1個の表面ループの長さおよび露出を拡大するために、スペーサーを含む、請求項6のPドメインモノマー。
【請求項9】
哺乳動物にウイルス性抗原に対する免疫保護を与える方法であって、ワクチンとして請求項5の抗原−P粒子を哺乳動物に投与する段階を含む、方法。
【請求項10】
抗原がRV VP8抗原であり、P粒子−VP8キメラがロタウイルスおよびノロウイルス両方に対するデュアルワクチンとして作用する、請求項9の方法。
【請求項11】
少なくとも1個の表面ループが3個のループであり、RV VP8のような大きな抗原がより小さな外来抗原を有する隣のループを、小さなエピトープの露出が困難となるように覆わないように、モノマーの3個のループに異なる抗原が複数挿入されている、請求項1の抗原−Pドメインモノマー。
【請求項12】
Pドメインモノマーの1個以上の表面ループにリガンドまたはシグナルペプチドの挿入が存在することによって、遠位表面の1個以上に挿入されたリガンドまたはシグナルペプチドを有する置換P粒子の自然発生的形成がもたらされる、請求項1の抗原−Pドメインモノマー。
【請求項13】
挿入されたリガンドまたはシグナルペプチドによって、置換P粒子が特定の臓器または組織を標的として、それらの位置へと移動することが可能となる、請求項12の抗原−Pドメインモノマー。
【請求項14】
置換P粒子がPドメインモノマーの1個以上の表面ループに挿入されたペプチドCNGRC(5アミノ酸)を有する、請求項13の抗原−Pドメインモノマー。
【請求項15】
置換P粒子モノマーならびにそのダイマーおよびP粒子が、薬物の担体として使用するために化学反応によって、表面露出リシンおよびシステインを介してP粒子のループに挿入されている薬物のコンジュゲートを有する、請求項1の抗原−Pドメインモノマー。
【請求項16】
形成される置換PモノマーならびにそのダイマーおよびP粒子が、表面ループの少なくとも1個に挿入されたリガンドまたはシグナルペプチド、ならびに表面ループの少なくとも1個に挿入された薬物のコンジュゲートを有し、それによって疾患を有する特定の組織または臓器を標的とする薬物送達系を提供する、請求項1の抗原−Pドメインモノマー。
【請求項1】
少なくとも1個の表面ループを有するノロウイルス(NOR)Pドメインモノマーおよび該少なくとも1個の表面ループに挿入されたタンパク質またはポリペプチド抗原を含む、抗原−Pドメインモノマー。
【請求項2】
少なくとも1個の表面ループが、ループ1(I293−H297)、ループ2(T371−D374)およびループ3(D391−N394)の少なくとも1個を含む、請求項1の抗原−Pドメインモノマー。
【請求項3】
抗原がロタウイルス(RV)VP8抗原である、請求項1の抗原−Pドメインモノマー。
【請求項4】
タンパク質またはポリペプチド抗原が少なくとも2個の類似または異なる抗原を含む、請求項1の抗原−Pドメインモノマー。
【請求項5】
24個のモノマーからなる抗原−P粒子であって、P粒子が少なくとも1個の請求項1の抗原−Pドメインモノマー、および所望により1個以上の野生型NOR Pドメインモノマーを含み、ここで、抗原−P粒子が抗原単独と比較して抗原の免疫原性を向上しうる、抗原−P粒子。
【請求項6】
少なくとも1個の表面ループを有するNOR Pドメインモノマーを含むPドメインモノマーベクターであって、前記少なくとも1個の表面ループが少なくとも1個の制限部位の対を含む、Pドメインモノマーベクター。
【請求項7】
少なくとも1個の表面ループがループ1(I293−H297)、ループ2(T371−D374)およびループ3(D391−N394)の少なくとも1個を含む、請求項6のPドメインモノマーベクター。
【請求項8】
少なくとも1個の表面ループが、該少なくとも1個の表面ループの長さおよび露出を拡大するために、スペーサーを含む、請求項6のPドメインモノマー。
【請求項9】
哺乳動物にウイルス性抗原に対する免疫保護を与える方法であって、ワクチンとして請求項5の抗原−P粒子を哺乳動物に投与する段階を含む、方法。
【請求項10】
抗原がRV VP8抗原であり、P粒子−VP8キメラがロタウイルスおよびノロウイルス両方に対するデュアルワクチンとして作用する、請求項9の方法。
【請求項11】
少なくとも1個の表面ループが3個のループであり、RV VP8のような大きな抗原がより小さな外来抗原を有する隣のループを、小さなエピトープの露出が困難となるように覆わないように、モノマーの3個のループに異なる抗原が複数挿入されている、請求項1の抗原−Pドメインモノマー。
【請求項12】
Pドメインモノマーの1個以上の表面ループにリガンドまたはシグナルペプチドの挿入が存在することによって、遠位表面の1個以上に挿入されたリガンドまたはシグナルペプチドを有する置換P粒子の自然発生的形成がもたらされる、請求項1の抗原−Pドメインモノマー。
【請求項13】
挿入されたリガンドまたはシグナルペプチドによって、置換P粒子が特定の臓器または組織を標的として、それらの位置へと移動することが可能となる、請求項12の抗原−Pドメインモノマー。
【請求項14】
置換P粒子がPドメインモノマーの1個以上の表面ループに挿入されたペプチドCNGRC(5アミノ酸)を有する、請求項13の抗原−Pドメインモノマー。
【請求項15】
置換P粒子モノマーならびにそのダイマーおよびP粒子が、薬物の担体として使用するために化学反応によって、表面露出リシンおよびシステインを介してP粒子のループに挿入されている薬物のコンジュゲートを有する、請求項1の抗原−Pドメインモノマー。
【請求項16】
形成される置換PモノマーならびにそのダイマーおよびP粒子が、表面ループの少なくとも1個に挿入されたリガンドまたはシグナルペプチド、ならびに表面ループの少なくとも1個に挿入された薬物のコンジュゲートを有し、それによって疾患を有する特定の組織または臓器を標的とする薬物送達系を提供する、請求項1の抗原−Pドメインモノマー。
【図1】

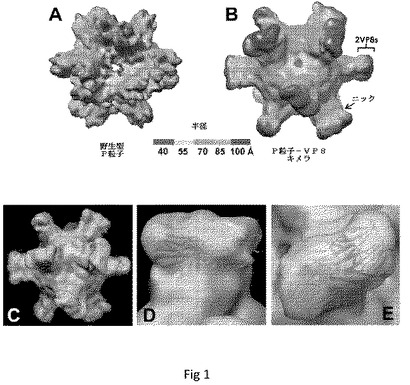
【図2】
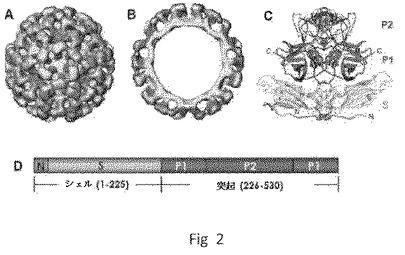
【図3】
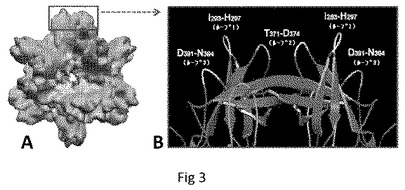
【図4】
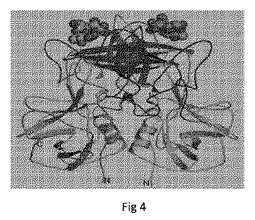
【図5】
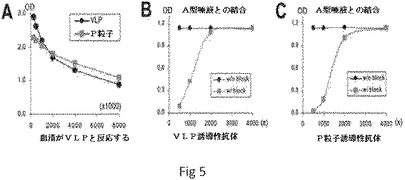
【図6】
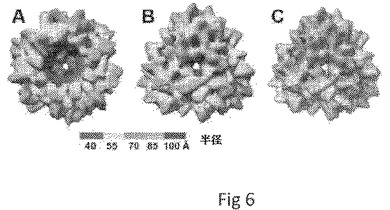
【図7】
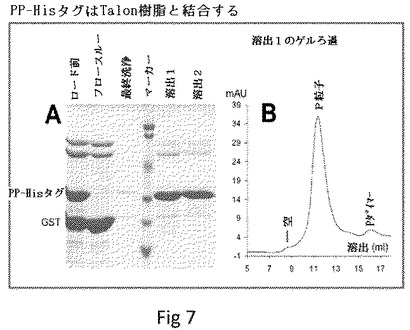
【図8】
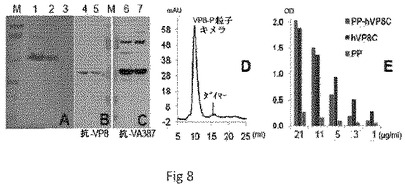
【図9】
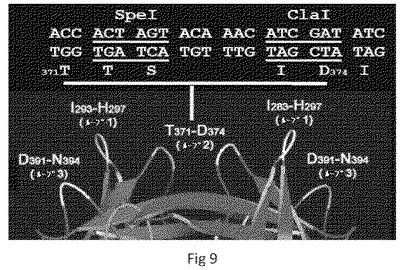
【図10】
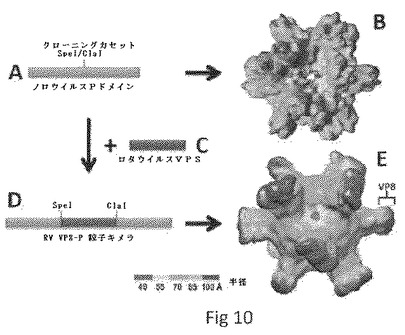
【図11】
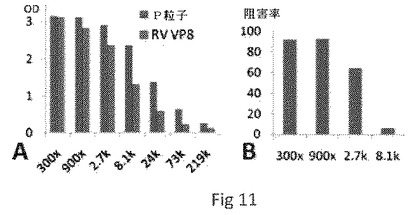
【図12】
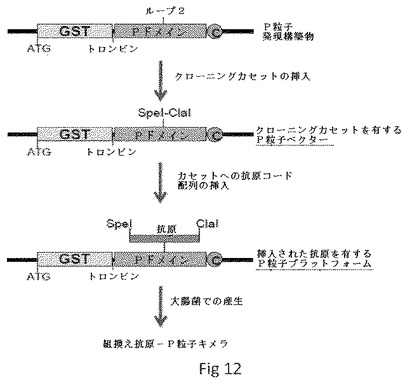
【図13】
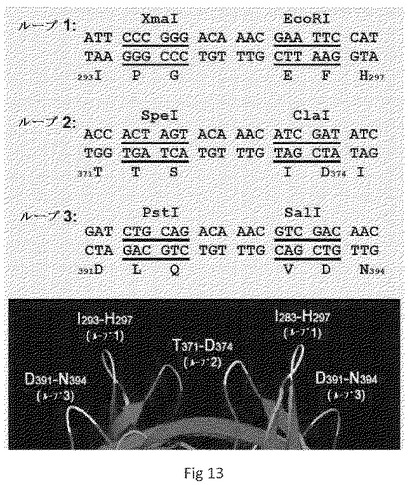
【図14】
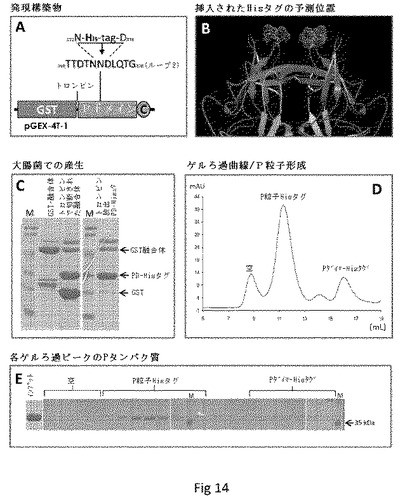
【図15】
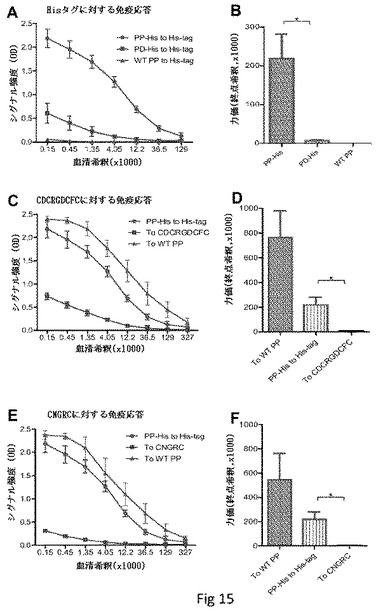
【図16】
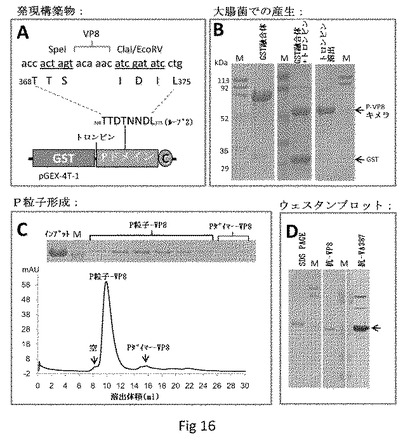
【図17】
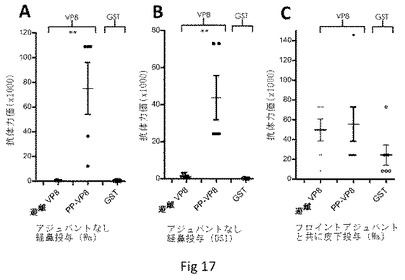
【図18】
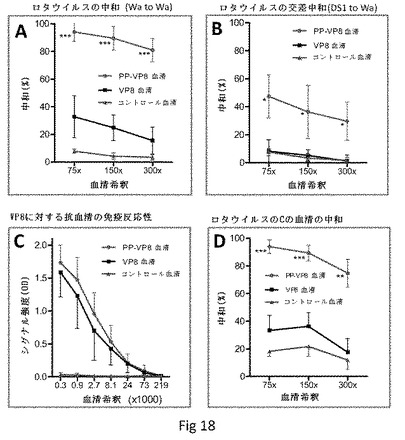
【図19】
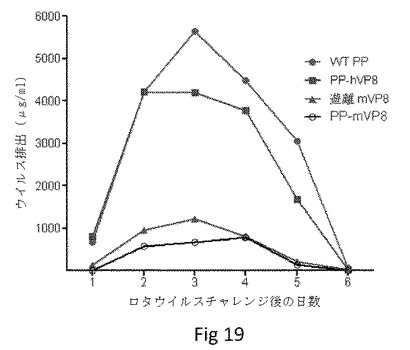
【図20】
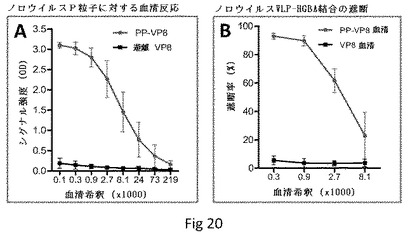
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【公表番号】特表2012−529298(P2012−529298A)
【公表日】平成24年11月22日(2012.11.22)
【国際特許分類】
【出願番号】特願2012−515109(P2012−515109)
【出願日】平成22年6月9日(2010.6.9)
【国際出願番号】PCT/US2010/038008
【国際公開番号】WO2010/144602
【国際公開日】平成22年12月16日(2010.12.16)
【出願人】(500469235)チルドレンズ ホスピタル メディカル センター (40)
【Fターム(参考)】
【公表日】平成24年11月22日(2012.11.22)
【国際特許分類】
【出願日】平成22年6月9日(2010.6.9)
【国際出願番号】PCT/US2010/038008
【国際公開番号】WO2010/144602
【国際公開日】平成22年12月16日(2010.12.16)
【出願人】(500469235)チルドレンズ ホスピタル メディカル センター (40)
【Fターム(参考)】
[ Back to top ]