
抗炎症化合物及びその利用
【課題】炎症性疾患を治療するための、抗炎症化合物、その薬学的組成物、及び、その利用法を提供する。抗炎症化合物を同定する方法と、細胞中のNF-κB依存性標的遺伝子発現を抑制する方法についても提供する。
【解決手段】構造:Xa-Xb,(ただし式中、Xaは、6乃至15個のアミノ酸残基を含む膜移行ドメインであり、そしてXbはNEMO結合配列である)を含む抗炎症化合物。NEMOがIκB-キナーゼ-β(IKKβ)と相互作用するのを、NEMO結合ドメイン(NBD)で遮断することによって、サイトカインの伝達するNF-κB活性化を、選択的に抑制する。
【解決手段】構造:Xa-Xb,(ただし式中、Xaは、6乃至15個のアミノ酸残基を含む膜移行ドメインであり、そしてXbはNEMO結合配列である)を含む抗炎症化合物。NEMOがIκB-キナーゼ-β(IKKβ)と相互作用するのを、NEMO結合ドメイン(NBD)で遮断することによって、サイトカインの伝達するNF-κB活性化を、選択的に抑制する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、NEMOがNEMO結合ドメイン(NBD)でIκB-キナーゼ-β(IKKβ)と相互作用するのを遮断することで、サイトカイン媒介性NF-κB活性化を選択的に抑制する組成物及び方法に関するものである。IKKβとNEMOとの相互作用を遮断すると、IKKβキナーゼ活性化が抑制され、ひいてはIκBのリン酸化が減少する。IκBのリン酸化は、サイトカイン媒介性NF-κB活性化において必須なステップである。
【背景技術】
【0002】
関連出願
本出願は、2000年5月2日出願の米国暫定特許出願第60/201,261号及び2000年8月22日出願の米国特許出願第09/643,260号の優先権を主張するものである。これら各関連出願の内容全体を、言及をもってここに編入することとする。
【0003】
米国政府による助成
本研究は、米国国立保健研究所から助成金による支援を受けた(AI33443)。
【0004】
発明の背景
NF-κBは、炎症誘発性応答に関与する遺伝子の誘導を担う細胞外シグナルを伝達する転写因子である(Baltimore ら (1998)、米国特許第5,804,374号)。NF-κBは、刺激を受けていない細胞の大半の細胞質に、IκBとして公知の複数の阻害性タンパク質の一つとの非共有結合的相互作用を通じて繋ぎ止められている(May & Ghosh, (1997) Semin. Cancer. Biol. 8, 63-73; May & Ghosh, (1998) Immunol. Today 19, 80-88; Ghosh et al., (1998) Annu. Rev. Immunol. 16, 225-260)。TNFαなどの炎症誘発性応答に関連した細胞刺激を受けるとキナーゼ群が活性化するが、活性化したキナーゼはIκBをリン酸化して、NF-κBを活性化する。IκBをリン酸化するキナーゼはIκB-キナーゼ(IKK)と呼ばれる。
【0005】
リン酸化したIκBはユビキチン化及び分解へと向かう。IκBが分解してNF-κBから解離すると、NF-κB上に核内局在化シグナルが現れ、活性NF-κBの核内移行が起きて、NF-κB応答性遺伝子が上方調節される(May & Ghosh, (1997) Semin. Cancer. Biol. 8, 63-73; May & Ghosh, (1998) Immunol. Today 19, 80-88; Ghosh et al., (1998) Annu. Rev. Immunol. 16, 225-260; Siebenlist et al., (1994) Annu. Rev. Cell Biol. 12, 405-455)。従ってIκBのリン酸化は、NF-κBが媒介する炎症誘発性応答の調節における重要な一ステップなのである。
【0006】
IκBをリン酸化するキナーゼ群の同定及び特徴付けが行われたことで、NF-κB活性化に関与するシグナル伝達経路の解明が進んだ。これまでのところ、いくつか異なるサブタイプのIKKが同定されている。IKKαは当初、ヒーラ細胞でTNFα刺激で誘導されるIκBキナーゼとして同定された(DiDonato et al., (1997) Nature 388, 548-554)。IKKαに相同なIκB-キナーゼがもう一つ同定され、IKKβと呼ばれており、TNFα刺激後に誘導される主要なIκB-キナーゼであることが判明している(Takeda et al., (1999) Science 284, 313-316; Hu et al., (1999) Science 284, 316-320; Li et al., (1999) Science 284, 321-325; Pot et al., (2000) 米国特許第6,030,834号;Woronicz & Goeddel (1999) 米国特許第5,939,302号)。IKKα及びIKKβの全体的なホモロジーは52%であり、キナーゼドメインにおけるホモロジーは65%である(Zandi et al., (1997) Cell 91, 243-252)。
【0007】
IκBプロテインキナーゼ(IKK)はIκBを特定のセリン残基でリン酸化する。例えば、これらはIκBαの32位及び36位のセリンを特異的にリン酸化する(Traenckner et al., (1995) EMBO J. 14, 2876-2883; DiDonato et al., (1996) Mol. Cell. Biol. 16, 1295-1304)。両方の部位でリン酸化することが、IκBαを効率的に分解に向かわせるのに必要である。さらに、IKKα及びIKKβの活性化は通常、NF-κB活性化物質に対する応答として起きるため、触媒として不活性な変異型IKKα及びIKKβを用いると、TNFα及びIL-1などのサイトカインによるNF-κB刺激を遮断することができる(Regnier et al., (997) Cell 90, 373-383; Delhase et al., (1999) Science 284, 309-313)。従って、IκBプロテインキナーゼはNF-κB活性化プロセスの調節に必須なのである。
【0008】
IKKα及びIKKβは、カルボキシル近位側のへリックス−ループ−へリックス(H-L-H)ドメインから、ロイシン・ジッパードメインによって隔てられたアミノ末端のセリン−スレオニンキナーゼドメインを含有する異なる構造モチーフを有する。このような構造上の特徴は他のキナーゼとは異なり、この非触媒ドメインが、タンパク質対タンパク質の相互作用に関与していると考えられる。従って、IKKに結合するタンパク質は、NF-κBの活性を調節したり(Marcu et al., (1999)米国特許第5,972,655号)、そしておそらくはNF-κBの誘導などの下流の事象も調節することができると思われる。例えば、NEMO(NF-κB必須調節因子)は、IKKに結合してキナーゼ活性を促進することが明らかにされたタンパク質である(Yamaoke et al., (1998) Cell 93, 1231-1240; Rothwarf et al., (1998) Nature 395, 287-300; Mercurio et al., (1999) Mol. Cell. Biol. 19, 1526-1538; Haraj & Sun, (1999) J. Biol. Chem. 274, 22911-22914; Jin & Jeang, (1999) J. Biomed. Sci. 6, 115-120)。
【0009】
炎症は、傷害に対する血管化生体組織の反応であると定義されている。従って、炎症は、物理的、化学的もしくは生物学的作用物質が原因で起きた傷害又は異常な刺激に対する、損傷した血管及び隣接組織の細胞学的及び化学的反応が混合した基本的、定型的な現象である。通常、炎症の結果、体液及び血球が損傷部位に蓄積するが、炎症は一般には治癒の一過程でなる。しかし、炎症はときに、炎症の正常な進行機能の不全が原因で有害ともなる。炎症性疾患は、炎症に関する、炎症を特徴とする、炎症を起こす、炎症の結果である、又は、炎症の影響を受けるようになる、ものである。炎症性疾患又は異常の例には、限定はしないが、喘息、肺炎症、結核、らい、サルコイドーシス、及び珪肺症などの慢性肉芽腫性疾患、腎炎、アミロイド症、リウマチ性関節炎、強直性脊椎炎、慢性気管支炎、強皮症、狼瘡、多発性筋炎、虫垂炎、炎症性腸疾患、潰瘍、シェーグレン症候群、ライター症候群、乾癬、骨盤内炎症性疾患、眼窩内炎症性疾患、血栓性疾患、及び、アトピー性皮膚炎及び接触性皮膚炎を含め、ウルシツタ、花粉、昆虫による刺し傷及び特定の食物などの環境刺激に対する不適切なアレルギー反応、がある。
【0010】
炎症性疾患は世界的な問題となっている。疾患の負荷研究により、結核は世界中で死亡原因の上位10位以内に入っていることが再確認された。喘息には成人人口の5%、そして小児人口の10乃至15%が罹患している(Armetti and Nicosia (1999) Boll Chim. Farm. 138(11):599)。喘息は、広汎なしかし様々な空気流閉塞を伴う慢性炎症性疾患である。
【0011】
敗血症も炎症性疾患の一つであり、様々な膿形成性等の病原性微生物や、又はそれらの毒素が被験体の血中又は組織中に存在することが原因で起きる。敗血症の特徴は、感染時の細菌生成物に対する全身の炎症性応答である。発熱などの敗血症の症状は、少なくとも部分的には、感染性物質に対する身体の炎症性応答が起こすものである。
従って、炎症性疾患を治療するのに有用な化合物への大きな需要が依然、あるのである。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】米国特許第5,804,374号
【特許文献2】米国特許第6,030,834号
【特許文献3】米国特許第5,972,655号
【発明の概要】
【発明が解決しようとする課題】
【0013】
本発明は、炎症性疾患を治療するための抗炎症化合物、その薬学的組成物、及びその利用法を提供するものである。本発明は、少なくとも部分的に、IκB-キナーゼ-α(IKKα)及びIκB-キナーゼ-β(IKKβ)上のNEMO結合ドメイン(NBD)の同定に基づく。
【0014】
従って、ある態様では、本発明は、NEMO結合ドメイン(NBD)を含む抗炎症化合物を提供する。
【0015】
一実施例では、本発明は、NEMO結合ドメインと、少なくとも一つの膜移行ドメインとの融合を含む抗炎症化合物を提供する。ある好適な実施例では、前記膜移行ドメインは、本発明の抗炎症化合物のインビボでの膜移行を促進するものである。前記膜移行ドメインは、例えば、アンテナペディア(antennapedia)ホメオドメインの第三へリックス又はHIV-1 Tatたんぱくであってもよい。一実施例では、前記NEMO結合ドメインは、SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19に記載した配列を有するポリペプチドである。
【0016】
別の実施例では、本発明は、(a)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列を含む、又は、から成るペプチド;(b)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列のうちの少なくとも三つのアミノ酸から成るフラグメント;(c)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列の保存的アミノ酸置換を含むペプチド;及び(d)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列の天然型アミノ酸配列バリアント、を含む抗炎症化合物を提供する。
【0017】
別の態様では、本発明は、例えば一種以上の薬学的に許容可能な担体を含む薬学的組成物など、本発明の抗炎症化合物を含む薬学的組成物を提供する。
【0018】
さらに別の態様では、本発明は、喘息、肺炎症又は癌など、被験体の炎症性疾患を治療する方法を特徴とする。本方法は、本発明の一種以上の抗炎症化合物を治療上有効量、被験体に投与するステップを含む。機序の限定を受けることを意図する訳ではないが、本発明の抗炎症化合物は、急性及び慢性炎症の部位に白血球が動員されるのを遮断したり、白血球上のE-セレクチンの発現を下方調節したり、又は、破骨細胞分化を遮断することにより、(直接的又は間接的に)作用すると考えられる。
【0019】
別の態様では、本発明は、細胞におけるE-セレクチンなどのNF-κB依存性標的遺伝子の発現を抑制する方法を提供する。本方法は、本発明の抗炎症化合物に細胞を接触させることにより、細胞内のNF-κB依存性標的遺伝子の発現を抑制するステップを含む。さらに別の態様では、本発明は、有効量の本発明の抗炎症化合物に細胞を接触させて、細胞内のNF-κB誘導を抑制することにより、細胞内のNF-κB誘導(例えばIKKα及び/又はIKKβ依存性誘導)を抑制する方法を提供する。本発明の一実施例では、このような方法は、少なくとも一つの膜移行ドメインを含有する抗炎症化合物を利用するものである。本発明のさらに別の具体的な実施例では、このような方法で用いられる抗炎症化合物は、SEQ ID NO:2、4、5、6、11、12、16、17、又は18の配列を含むアミノ酸配列を含有する。
【0020】
別の態様では、本発明は、抗炎症化合物を同定する方法を提供する。本方法は、NEMO及びNF-κBを発現する細胞をテスト化合物に暴露するステップと;前記テスト化合物が、当該細胞によるNF-κBの活性化を調節するかどうかを調べることで、抗炎症化合物を同定するステップと、を含む。
【0021】
別の態様では、本発明は、NEMOを発現する細胞をテスト化合物に暴露するステップと、前記テスト化合物がNEMOの活性を調節するかどうかを調べることで、NEMOの活性を調節する化合物などの抗炎症化合物を同定するステップと、により、抗炎症化合物を同定する方法を提供する。
【0022】
本発明の抗炎症化合物の具体的な利点の一つは、これらは、IKKを通じたNF-κB誘導を遮断しながらも、NF-κBの基礎活性を抑制しないことである。
【0023】
本発明の他の特徴及び利点は、以下の詳細な説明及び請求の範囲から明白となるであろう。
【0024】
I.概略の解説
本発明は、炎症性疾患を治療するための抗炎症化合物、その薬学的組成物、及びその利用法を提供するものである。本発明は、少なくとも部分的に、IκB-キナーゼ-α(IKKα)及びIκB-キナーゼ-β(IKKβ)上のNEMO結合ドメイン(NBD)の同定に基づく。
【0025】
機序に限定されることを意図する訳ではないが、本発明の抗炎症化合物は、NEMOとIKK(例えばIKKβ又はIKKα)のNEMO結合ドメイン(NBD)での相互作用を遮断して、IκBのリン酸化、分解、及びその後のNF-κBからの解離を抑制することにより、作用すると考えられる。この抑制の結果、炎症誘発性応答に関係したNF-κB活性化が遮断される。
【0026】
さらに本発明は、抗炎症化合物をスクリーニングし、同定する方法も提供する。
【0027】
II.定義
他に特に定義しない限り、ここで用いた全ての技術用語及び科学用語は、本発明が関係する分野の当業者が通常理解するのと同じ意味を有するものである。
【0028】
ここで用いる用語「結合」とは、限定はしないが、酵素対基質、抗体対抗原、DNA鎖対それらの相補鎖など、分子間相互の付着を言う。結合が起きるのは、分子表面の部分同士の間で形状及び化学的性質が「相補的」になっているからである。よくある喩えは、酵素がどのように基質の周りに嵌め合うかを記述するのに用いられる「鍵穴と鍵」である。
【0029】
用語「融合ペプチド」又は「融合ポリペプチド」又は「融合たんぱく」には、二つの異なるアミノ酸配列を組み合わせて得られるペプチド、ポリペプチド又はタンパク質が含まれる。典型的には、ある一個のペプチド、ポリペプチド又はタンパク質の部分的な配列が、別の異種のペプチド、ポリペプチド又はタンパク質に、当業で公知の技術を用いて連結されている。
【0030】
「保存的アミノ酸置換」とは、アミノ酸残基が、同様な側鎖を有するアミノ酸残基に置換されることを言う。塩基性側鎖(例えばリシン、アルギニン、ヒスチジン)、酸性側鎖(例えばアスパラギン酸、グルタミン酸)、無電荷の極性側鎖(例えばグリシン、アスパラギン、グルタミン、セリン、スレオニン、チロシン、システイン)、非極性側鎖(例えばアラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン)、β-分岐側鎖(例えばスレオニン、バリン、イソロイシン)及び芳香族側鎖(例えばチロシン、フェニルアラニン、トリプトファン、ヒスチジン)を含め、同様な側鎖を有するアミノ酸残基の仲間は当業で公知である。本発明の抗炎症化合物で行うことのできる保存的置換の非限定的な例には、D型フェニルアラニンのD型チロシン、D型ピリジルアラニン又はD型ホモフェニルアラニンとの置換、D型ロイシンのD型バリン又は他の脂肪族側鎖を有する天然もしくは非天然アミノ酸との置換、及び/又は、D型バリンのD型ロイシン又は脂肪族側鎖を有する他の天然もしくは非天然アミノ酸との置換、がある。
【0031】
ここで用いる用語「ドメイン」とは、限定はしないが、例えば疎水性、極性、球状及び螺旋状のドメイン又は性質など、共通の物理化学的特徴を持つ分子又は構造の一部分を言う。結合ドメインの具体的な例には、限定はしないが、DNA結合ドメイン及びATP結合ドメイン、がある。
【0032】
ここで用いる用語「膜移行ドメイン」とは、細胞膜を透過できて、付着させたペプチドをインビボで細胞内に輸送するのに利用されるペプチドを言う。膜移行ドメインには、限定はしないが、アンテナペディアホメオドメインたんぱくの第三へリックス及びHIV-1たんぱくTatがある。他の膜移行ドメインも当業で公知であり、例えばDerossi et al., (1994) J. Biol. Chem. 269, 10444-40450; Lindgren et al., (2000) Trends Pharmacol. Sci 21, 99-103; Ho et al., Cancer Research 61, 474-477 (2001); 米国特許第5,888,762号;米国特許第6,015,787号;米国特許第5,846,743号;米国特許第5,747,641号;米国特許第5,804,604号;及び公開済みPCT出願WO98/52614号、WO00/29427号、及びWO99/29721号に解説されたものがある。前出の各参考文献の内容全体を言及をもってここに編入することとする。
【0033】
ここで用いる用語「IκB」(IカッパB)とは、抑制たんぱくのファミリーのうちでNF-κB誘導の調節で働く構造上関連した仲間のいずれか一つを言う。
【0034】
ここで用いる用語「IκB-キナーゼ」又は「IκBプロテインキナーゼ」又は「IκB-キナーゼ複合体」又は「IκBプロテインキナーゼ複合体」又は「IKK」とは、IκBをリン酸化するキナーゼを言う。
【0035】
ここで用いる用語「IKKα」とは、IκB-キナーゼ複合体のαサブユニットを言う。ここで用いる用語「IKKβ」とは、IκB-キナーゼ複合体のβサブユニットを言う。
【0036】
ここで用いる用語「NEMO」(NF-κB必須調節因子)、「IKKγ」又は「IKKAP」とは、IKKに結合してキナーゼ活性を促進するタンパク質を言う。
【0037】
ここで用いる用語「NEMO結合ドメイン」又は「NBD」には、NEMOが通常(IKKα又はIKKβなどの)IKKと相互作用する領域にあってNEMOに結合できるあらゆるドメインが含まれる。NEMO結合ドメインには、NEMOとの相互作用に重要な、例えば、野生型IKKβのα2領域(残基737−742)、又は野生型IKKαの相当する6個のアミノ酸配列(残基738−743)がある。野生型IKKβNBDの核酸配列及び対応するアミノ酸配列を、それぞれSEQ ID NO:1(GenBank受託番号AR067807;ヌクレオチド2203−2235)及びSEQ ID NO:2に公開する。
【0038】
ここで用いる場合の用語「類似体」、「誘導体」及び「ミメティック」には、あるペプチド構造の化学構造を模倣すると共に、このペプチド構造の機能的性質を維持した分子が包含されるものと、意図されている。ペプチド類似体、誘導体及びミメティックをデザインする方法は当業で公知である。例えば、Farmer, P. S. in Drug Design (E. J. Ariens, ed.) Academic Press, New York, 1980, vol. 10, pp. 119-143; Ball. J. B. and Alewood, P. E. (1990) J. Mol. Recognition 3:55; Morgan, B. A. and Gainor, J. A. (1989) Ann. Rep. Med. Chem. 24:243; adn Freidinger, R. M. (1989) Trends Pharmacol. Sci. 10:270を参照されたい。さらにSawyer, T. K. (1995) "Peptidomimetic Design and Chemical Approaches to Peptide Metabolism" in Taylor, M. D. and Amidon, G. L. (eds.) Peptide-Based Drug Design: Controlling Transport and Metabolism, Chapter 17; Smith, A. B. 3rd, et al. (1995) J. Am. Chem. Soc. 117:11113-11123; Smith, A. B. 3rd, et al. (1994) J. Am. Chem. Soc. 116:9947-9962; and Hirschman, R., et al. (1993) J. Am. Chem. Soc. 115:12550-12568も参照されたい。
【0039】
ここで用いる「化合物X(例えばペプチド又はアミノ酸)の誘導体」とは、当該化合物上の一つ以上の反応基を置換基で誘導変化させてある形のXを言う。ペプチド誘導体の例には、アミノ酸側鎖、ペプチド骨格、又はアミノ末端又はカルボキシ末端が誘導変化させてある(例えばメチル化アミド結合を持つペプチド化合物など)ペプチドがある。ここで用いる化合物Xの「類似体」とは、Xの機能的活性に必要なXの化学構造は残っているが、Xとは異なる化学構造もいくつか含んでいるような化合物を言う。天然型ペプチドの類似体の一例は、一つ以上の非天然発生型のアミノ酸を含有するペプチドである。ここで用いる、化合物Xの「ミメティック」とは、Xの機能的活性に必要なXの化学構造が、Xのコンホメーションを模倣した他の化学構造に置換されているような化合物を言う。ペプチドミメティックの例には、ペプチド骨格が一つ以上のベンゾジアゼピン分子に置換されたようなペプチド化合物がある(例えばJames, G. L. et al. (1993) Science 260:1937-1942)。
【0040】
用語ミメティック、特にペプチドミメティックには、同配体が含まれるものと、意図されている。ここで用いる「同配体」という用語には、第一の構造の立体的コンホメーションが、第二の構造に特異的な結合部位に適合しているために、第二の化学構造に置換することのできる化学構造が含まれるものと、意図されている。この用語には特に、当業者に公知のペプチド骨格修飾(即ちアミド結合ミメティック)が含まれる。このような修飾には、アミド窒素の修飾、α位炭素の修飾、アミドカルボニルの修飾、アミド結合の完全な置換、伸長、欠失、又は骨格架橋、がある。ψ[CH2S]、ψ[CH2NH]、ψ[CSNH2]、ψ[NHCO]、ψ[COCH2]、及びψ[(E)又は(Z)CH=CH]を含め、いくつかのペプチド骨格修飾が公知である。上で用いた命名法において、ψはアミド結合がないことを示す。アミド基を置換する構造を括弧内に明示した。
【0041】
他の可能な修飾には、N-アルキル(又はアリール)置換(ψ[CONR])、又はラクタム又は他の環式構造を構築するための骨格の架橋がある。本発明の抗炎症化合物の他の誘導体には、C末端ヒドロキシメチル誘導体、O-修飾誘導体(例えばC末端ヒドロキシメチルベンジルエーテル)、アルキルアミド及びヒドラジドなどの置換アミドを含むN末端修飾誘導体や、C末端フェニルアラニン残基をフェネチルアミド類似体に(例えばトリペプチドVal-Phe-Pheの類似体としてのVal-Phe-フェネチルアミドなど)置換してある抗炎症化合物、がある。
【0042】
ここで用いる用語「野生型」とは、天然発生種の仲間の大半に特徴的であり、変異種の遺伝子型及び表現型とは対照的な遺伝子型及び表現型を言う。
【0043】
III.具体的な実施例
A.抗炎症化合物
本発明は、NEMO結合ドメイン(NBD)を含む抗炎症化合物を提供するものである。NEMOが通常、IKK(例えばIKKα又はIKKβ)と相互作用する領域で、NEMOに結合することのできるドメインを含む分子であれば、本発明の抗炎症化合物を調製するのに利用できよう。このような分子の例には、D型及び/又はL型立体配置のアミノ酸を含むペプチド;ペプチド化合物の誘導体、類似体、及びミメティック;抗体(例えばポリクローナル、モノクローナル、ヒト化、抗イディオタイプ、キメラ、及び一本鎖抗体や、Fab、F(ab')2、Fab発現ライブラリフラグメント、及び抗体のエピトープ結合フラグメントなど);及び小型の有機及び無機分子(例えばコンビナトリアル・ライブラリ及び天然生成物ライブラリから得られる分子など)、がある。
【0044】
ある好適な実施例では、本発明の抗炎症化合物は、NEMO結合ドメインと、本発明の抗炎症化合物のインビボでの膜移行を促進する少なくとも一つの膜移行ドメインとの融合を含んで成る。
【0045】
本発明の抗炎症化合物を、IKKα又はIKKβのNBDの野生型アミノ酸配列(SEQ ID NO:2)に基づいて設計してもよい。NEMOに結合できれば、IKKα又はIKKβのNBDの野生型アミノ酸配列のいかなるフラグメントを用いても、本発明の抗炎症化合物を調製できよう。(ここに解説した方法を用いた)これらの野生型配列の点変異、挿入、又は欠失を利用して、更なる抗炎症化合物を作製してもよい。SEQ ID NO:2に記載したペプチドの737位、740位及び742位に保存的アミノ酸置換を含有するペプチドは、特に有用な本発明の抗炎症化合物である(ペプチドがNEMOに結合する能力に有意な影響を及ぼさない保存的置換の例については表1を参照されたい)。さらに、NEMOに結合する能力の残った、IKKβ遺伝子の天然発生型対立遺伝子バリアントを用いても抗炎症化合物を調製できよう。
【0046】
実施例の一つでは、本発明の抗炎症化合物は、(a)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列を含む、又は、から成るペプチド;(b)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列のうちの少なくとも三つのアミノ酸から成るペプチドフラグメント;(c)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列に保存的アミノ酸置換を含有するペプチド;及び(d)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列の天然型アミノ酸配列バリアント、を含む。
【0047】
さらに本発明の抗炎症化合物には、特異抗体又はT細胞抗原受容体(Harlow & Lane, (1988) Antibodies - A Laboratory Manual, Cold Spring Harbor Laboratory Pressを参照されたい)のような体細胞組換えペプチド受容体などのNEMO特異的受容体や、一種、二種及び三種ハイブリッドスクリーンなどの検定で同定された他の天然細胞内結合物質や、以下に解説するものなどの化学的ライブラリのスクリーンで同定された非天然細胞内結合物質を含めてもよい。
【0048】
本発明の抗炎症化合物は、NEMOを下方調節することができる。下方調節とは、ここでは、NEMO、そのリガンド又は活性化物質の活性化、機能又は合成の減少であると、定義しておく。さらにこれには、NEMO遺伝子、そのタンパク質生成物、リガンド又は活性化物質の分解の増加も包含されると、定義しておく。下方調節は、例えばNEMOのIKK(例えばIKKβ又はIKKα)への結合を不安定にしたり;又はIκBのリン酸化を遮断してこのタンパク質のその後の分解を起こさせるなど、多数の方法で行うことができる。
【0049】
IKKβによるIκBのリン酸化の結果、Iκbのユビキチン化及び分解が起き、さらにはIκBの解離が起きて、NF-κBの核内移行が可能となり、炎症応答に重要な遺伝子が上方調節される。従って、本発明の抗炎症化合物を用いれば、NF-κB機能を下方調節できよう。さらにNF-κBの下方調節は、NBD又はNEMO自体を狙ったポリクローナルもしくはモノクローナル抗体又はそのフラグメントを含む抗炎症化合物を利用しても、達成できるであろう。さらに本発明は、NBD又はNEMO自体に対して充分な親和性で結合して、IKKβとのNEMO相互作用を遮断するなどに必要な三次元構造を有するような小分子も包含する。IκBの分解を減少させ、NF-κBの活性化を減少させるIKKβ遮断法により、これらの小分子は、炎症を治療又は予防する治療薬として有用なものとなる。
【0050】
ある実施例では、本発明は、式:
Xa-Xb、
の抗炎症化合物を提供するが、ただし式中、Xa は、6乃至15個のアミノ酸残基を含む膜移行ドメインであり、そしてXb はNEMO結合配列である。前記化合物には、選択に応じて、N末端、C末端又は両者に、修飾基を含めてもよい。
【0051】
Xb は、6乃至9個のアミノ酸残基を含むNEMO結合配列である。ある実施例では、Xbは、以下の構造:
(Y)n-X1-X2-X3-X4-X5-X6-(A)m
から成り、ただし式中、n 及びm はそれぞれ個別に、0 又は1であり、そしてA 及びYはそれぞれ1乃至約3個のアミノ酸残基を含む。nが1であるとき、Yは配列TAであると好ましい。mが1であるとき、Aは配列QTEであると好ましい。X1 はL、A、I 又はノルロイシン(Nle)である。X2 はD、E、N、Q、ホモセリン(Hser)又は2-ケトプロピルアラニン(2-ケトプロピ-A)である。X3 は W、F Y、4-ビフェニル-アラニン (Bpa)、ホモフェニルアラニン (Hphe)、2-ナフチルアラニン (2-Nal)、1-ナフチルアラニン (1-Nal)、又はシクロキセキシル(原語cycloxexyl)-アラニン (Cha)である。X4 は S、A、E、L、T、ノルロイシン (Nle)、又はホモセリン (Hser)である。X5 はW、H、ホモフェニルアラニン (Hphe)、2-ナフチルアラニン (2-Nal)、1-ナフチルアラニン (1-Nal)、O-ベンジルセリン (SeroBn)、又は3-ピリジルアラニン (3-Pal)である。そしてX6 はL、A、I、又はノルロイシン (Nle)である。
【0052】
好ましくは、Xb は、TALDWSWLQTE (SEQ ID NO:28); LDWSWLQTE (SEQ ID NO:29); TALDWSWL (SEQ ID NO:30); ALDWSWLQTE (SEQ ID NO:31); LDWSWLQTE (SEQ ID NO:32); LDWSWL (SEQ ID NO:33); TALDWSWLQT (SEQ ID NO:34); TALDWSWLQ (SEQ ID NO:35); ALDWSWLQT (SEQ ID NO:36); LDWSWLQ (SEQ ID NO:37); LDWSWLQT (SEQ ID NO:38); ADWSWL (SEQ ID NO:39); LDWSWA (SEQ ID NO:40); ADWSWA (SEQ ID NO:41); LDFSWL (SEQ ID NO:42); LDYSWL (SEQ ID NO:43); LDWAWL (SEQ ID NO:44); LDWEWL (SEQ ID NO:45); TAADWSWLQTE (SEQ ID NO:46); ADWSWLQTE (SEQ ID NO:47); TAADWSWL (SEQ ID NO:48); AADWSWLQTE (SEQ ID NO:49); ADWSWLQTE (SEQ ID NO:50); ADWSWL (SEQ ID NO:51); TAADWSWLQT (SEQ ID NO:52); TAADWSWLQ (SEQ ID NO:53); AADWSWLQT (SEQ ID NO:54); ADWSWLQ (SEQ ID NO:55); ADWSWLQT (SEQ ID NO:56); ALDWSWAQTE (SEQ ID NO:57); LDWSWAQTE (SEQ ID NO:58); TALDWSWA (SEQ ID NO:59); ALDWSWAQTE (SEQ ID NO:60); LDWSWAQTE (SEQ ID NO:61); LDWSWA (SEQ ID NO:62); TALDWSWAQT (SEQ ID NO:63); TALDWSWAQ (SEQ ID NO:64); ALDWSWAQT (SEQ ID NO:65); LDWSWAQ (SEQ ID NO:66); LDWSWAQT (SEQ ID NO:67); TAADWSWAQTE (SEQ ID NO:68); ADWSWAQTE (SEQ ID NO:69); TAADWSWA (SEQ ID NO:70); AADWSWAQTE (SEQ ID NO:71); ADWSWAQTE (SEQ ID NO:72); ADWSWA (SEQ ID NO:73); TAADWSWAQT (SEQ ID NO:74); TAADWSWAQ (SEQ ID NO:75); AADWSWAQT (SEQ ID NO:76); ADWSWAQ (SEQ ID NO:77); ADWSWAQT (SEQ ID NO:78); TALDFSWLQTE (SEQ ID NO:79); LDFSWLQTE (SEQ ID NO:80); TALDFSWL (SEQ ID NO:81); ALDFSWLQTE (SEQ ID NO:82); LDFSWLQTE (SEQ ID NO:83); LDFSWL (SEQ ID NO:84); TALDFSWLQT (SEQ ID NO:85); TALDFSWLQ (SEQ ID NO:86); ALDFSWLQT (SEQ ID NO:87); LDFSWLQ (SEQ ID NO:88); LDFSWLQT (SEQ ID NO:89); TALDYSWLQTE (SEQ ID NO:90); LDYSWLQTE (SEQ ID NO:91); TALDYSWL (SEQ ID NO:92); ALDYSWLQTE (SEQ ID NO:93); LDYSWLQTE (SEQ ID NO:94); LDYSWL (SEQ ID NO:95); TALDYSWLQT (SEQ ID NO:96); TALDYSWLQ (SEQ ID NO:97); ALDYSWLQT (SEQ ID NO:98); LDYSWLQ (SEQ ID NO:99); LDYSWLQT (SEQ ID NO:100); TALDWAWLQTE (SEQ ID NO:101); LDWAWLQTE (SEQ ID NO:102); TALDWAWL (SEQ ID NO:103); ALDWAWLQTE (SEQ ID NO:104); LDWAWLQTE (SEQ ID NO:105); LDWAWL (SEQ ID NO:106); TALDWAWLQT (SEQ ID NO:107); TALDWAWLQ (SEQ ID NO:108); ALDWAWLQT (SEQ ID NO:109); LDWAWLQ (SEQ ID NO:110); LDWAWLQT (SEQ ID NO:111); TALDWEWLQTE (SEQ ID NO:112); LDWEWLQTE (SEQ ID NO:113); TALDWEWL (SEQ ID NO:114); ALDWEWLQTE (SEQ ID NO:115); LDWEWLQTE (SEQ ID NO:116); LDWEWL (SEQ ID NO:117); TALDWEWLQT (SEQ ID NO:118); TALDWEWLQ (SEQ ID NO:119); ALDWEWLQT (SEQ ID NO:120); LDWEWLQ (SEQ ID NO:121); 及び LDWEWLQT (SEQ ID NO:122)からなる群より選択される配列であるとよい。
【0053】
Xa は、6乃至15個のアミノ酸残基、好ましくは6乃至12個、又は6乃至10個のアミノ酸残基から成る膜移行ドメインである。好ましくは、Xa は、少なくとも5個の塩基性アミノ酸残基、好ましくは L-アルギニン、D-アルギニン、L-リシン及びD-リシンから個別に選択される少なくとも5個のアミノ酸残基を含む膜移行ドメインであるとよい。適した膜移行ドメインには、ここに開示したものがある。
【0054】
実施例の一つでは、Xa はアミノ酸配列RRMKWKK (SEQ ID NO:123); YGRKKRRQRRR (SEQ ID NO:124); ygrkkrrqrrr (SEQ ID NO:125); YARKARRQARR (SEQ ID NO:126); yarkarrqarr (SEQ ID NO:127); YARAARRAARR (SEQ ID NO:128); yaraarraarr (SEQ ID NO:129); rrmkwkk (SEQ ID NO:130); (R)y 及び (r)y、(ただしこの場合y は6乃至11である)から選択される。小文字はD型アミノ酸残基を指し、大文字はL型アミノ酸残基を指す。
【0055】
適したペプチドXa-Xb の例には、以下の配列:RRMKWKKTALDWSWLQTE (SEQ ID NO:131); rrmkwkkTALDWSWLQTE (SEQ ID NO:132); YGRKKRRQRRRTALDWSWLQTE (SEQ ID NO:133); ygrkkrrqrrrTALDWSWLQTE (SEQ ID NO:134); rrrrrrrTALDWSWLQTE (SEQ ID NO:135); RRRRRRRTALDWSWLQTE (SEQ ID NO:136); YARKARRQARRTALDWSWLQTE (SEQ ID NO:137); yarkarrqarrTALDWSWLQTE (SEQ ID NO:138); YARAARRAARRTALDWSWLQTE (SEQ ID NO:139); yaraarraarrTALDWSWLQTE (SEQ ID NO:140) YGRKKRRQRRRLDWSWL (SEQ ID NO:141); ygrkkrrqrrrLDWSWL (SEQ ID NO:142); RRMKWKKLDWSWL (SEQ ID NO:143); rrmkwkkLDWSWL (SEQ ID NO:144); rrrrrrrLDWSWL (SEQ ID NO:145); YARAARRAARRLDWSWL (SEQ ID NO:146); yaraarraarrLDWSWL (SEQ ID NO:147); 及びRRRRRRRLDWSWL (SEQ ID NO:148)を有するものがある。
【0056】
本発明の抗炎症化合物には、選択に応じ、C末端、N末端又は両方に付着した修飾基を含めてもよい。例えば、C末端に付着させることのできる適した修飾基には、置換及び非置換アミノ基、例えば-NH2、-NH(アルキル) 及び-N(アルキル)2 基;及びアルコキシ基、例えば直線状、分岐式又は環式のC1-C6-アルコキシ基、など、がある。好適なC末端修飾基は、-NH2 基である。N末端に付着させることのできる適した修飾基には、アシル基、例えばアセチル基;及びアルキル基、好ましくはC1-C6-アルキル基、より好ましくはメチル、がある。
【0057】
本発明の抗炎症化合物においては、膜移行ドメインであるXaが当該化合物のアミノ末端にあって、NEMO結合ドメインであるXbが当該化合物のカルボキシル末端にあってもよい (Xa- Xb)。反対に、本発明の抗炎症化合物において、膜移行ドメインであるXaが当該化合物のカルボキシル末端にあって、NEMO結合配列であるXbが当該化合物のアミノ末端にあってもよい(Xb- Xa)。
【0058】
本発明の具体的な抗炎症化合物には、以下に挙げるものが含まれる。
【0059】
化合物番号
1 H-RRMKWKKTALDWSWLQTE-NH2
2 H-YGRKKRRQRRRTALDWSWLQTE-NH2
3 H-rrrrrrrTALDWSWLQTE-NH2
4 H-YARKARRQARRTALDWSWLQTE-NH2
5 H-YARAARRAARRTALDWSWLQTE-NH2
6 H-RRMKWKKLDWSWL-NH2
7 H-rrmkwkkLDWSWL-NH2
8 H-rrrrrrrLDWSWL-NH2
9 H-YARAARRAARRLDWSWL-NH2
10 H-yaraarraarrLDWSWL-NH2
11 H-YGRKKRRQRRRLDWSWL- NH2
【0060】
B.スクリーニング検定
加えて本発明は、抗炎症化合物を同定するスクリーニング法も提供する。本抗炎症化合物は、IKKβ分子のNBDで相互作用することにより、この分子の機能を遮断したり、合成を妨げたり、又はその生物学的安定性を低下させるであろう。生物学的安定性は、当該分子の合成とその分解との間の時間の測定値である。例えば、あるタンパク質、ペプチド又はペプチドミメティック(Kauvar, Nature Biotech. (1996) 14, 709)治療薬の安定性は、その配列を変更して酵素分解をより受けやすくするようにすれば、短縮することができる。
【0061】
さらに本発明は、IKKβ機能を失活させる、又は、IKKβ機能のアンタゴニストとして働く化合物をスクリーニングする方法も包含する。このような化合物は、IKKβもしくはNF-κBタンパク質レベルの変化に関連した病的状態の調節に有用であろう。
【0062】
さらに本発明は、例えば、IKKβと、この分子のNBDで相互作用する化合物、又は、NEMOと相互作用することでNEMOとIKKβとの相互作用を遮断する化合物など、本発明のタンパク質の結合相手を単離及び同定する方法も提供する。本発明のタンパク質と、候補結合相手又は細胞抽出物もしくは画分との混合は、候補結合相手と本発明のタンパク質との結合が可能なような条件下で行う。混合後、本発明のタンパク質と結合したペプチド、ポリペプチド、タンパク質又は他の分子を、混合物から分離する。次に、本発明のタンパク質に結合した結合相手を取り外し、さらに解析することもできる。結合相手を同定及び単離するには、当該タンパク質全体、例えば全IKKβペプチドを利用することもできる。反対に、当該タンパク質の一フラグメントを利用してもよい。例えば、NBDを含むペプチドフラグメントを利用して、IKKβとNEMOとの相互作用を遮断することもできる。
【0063】
細胞抽出物を得るには多種の方法を利用できる。細胞は、物理的又は化学的破砕法のいずれを利用して破砕してもよい。物理的破砕法の例には、音波破砕法及び機械的せん断法がある。化学的溶解法の例には、限定はしないが、界面活性剤による溶解法及び酵素溶解法がある。当業者であれば、本方法で利用する抽出物を得るのに、細胞抽出物を調製する方法を容易に適合できよう。
【0064】
細胞抽出物を調製したら、その抽出物を、IKKβ又はNEMOに、当該タンパク質と結合相手との結合が可能な条件下で混合する。多種の条件を利用できるが、最も好ましい条件は、ヒト細胞の細胞質で見られる条件に近いものである。浸透圧モル濃度、pH、温度、及び、用いる細胞抽出物の濃度などの特徴は、当該タンパク質と結合相手との結合が至適になるよう、変更することができる。
【0065】
適当な条件下で混合した後、結合した複合体を混合液から分離する。多種の技術を利用して混合液を分離できる。例えば、本発明のタンパク質に特異的な抗体を用いれば、結合相手との複合体を免疫沈降させることができる。代わりに、クロマトグラフィー及び濃度−沈渣遠心分離法などの標準的な化学的分離技術も利用できる。
【0066】
抽出物中に見られる結合しなかった細胞構成成分を除去した後、従来の方法を利用して結合相手を複合体から解離させることができる。例えば、混合液の塩濃度又はpHを変化させても、解離が可能である。
【0067】
混合抽出物から、結合済みの結合相手との組を分離する手助けとして、タンパク質を固体の支持体上に固定してもよい。例えば、タンパク質をニトロセルロース・マトリックス又はアクリル製ビーズに付着させてもよい。タンパク質を固体の支持体上に付着させると、抽出物中に見られる他の構成成分からペプチド−結合相手の組を分離するのに便利である。同定される結合相手は、単一のタンパク質であるかも、又は二種以上のタンパク質からなる複合体であるかも知れない。代わりに、結合相手は、Takayama et al., (1997) Methods Mol. Biol. 69, 171-814 又はSauder et al., (1996) J. Gen. Virol. 77, 991-996の手法に基づくファー−ウェスタン検定を用いて同定でき、又は、エピトープ・タグを付けたタンパク質又はGST融合タンパク質を用いて同定できよう。
【0068】
選択によっては、本発明のペプチドをコードする核酸分子を酵母二種ハイブリッド系で利用してもよい。酵母二種ハイブリッド系は、他のタンパク質結合相手を同定するのに用いられてきており、ここに解説した核酸分子を利用できるよう、容易に適合が可能である(例えばストラータジーン社Hybrizap(R)、二種ハイブリッド系を参照されたい)。
【0069】
本発明の別の実施例は、NEMO又はIKKβの少なくとも一つの活性を調節する作用物質を同定する方法を提供するものである。このような方法又は検定では、所望の活性を観察又は検出するいかなる手段を用いてもよい。
【0070】
あるフォーマットでは、テスト対象の作用物質に暴露した細胞集団と、暴露していないコントロール細胞集団との間で、本発明のタンパク質の相対量を検定してもよい。このフォーマットでは、特異抗体などのプローブを用いて、異なる細胞集団中のタンパク質の発現の差を観察する。細胞株又は集団を、テスト対象の作用物質に、適当な条件下及び時間、暴露する。細胞ライセートは、暴露した細胞株又は集団と、コントロールの非暴露細胞株又は集団とから調製してもよい。次に、この細胞ライセートをプローブで解析する。
【0071】
抗体プローブの作製は、当該ペプチドを用いた適した免疫処置プロトコールで適切なほ乳類ホストを免疫処置することで行う。NBDを含むペプチド又はタンパク質は、充分な長さの物であり、免疫原性を高めるのに必要であれば、適した担体に共役させておく。BSA、KLH、又は他の担体たんぱくなどの担体を用いて免疫原性共役体を調製する方法は当業で公知である。場合によっては、カルボジイミド試薬などを用いた直接共役法が効果的であろう。別の場合には、ピアース・ケミカル社が市販するものなどの結合試薬が、ハプテンを利用するのに好ましいであろう。ハプテンペプチドは、アミノ末端又はカルボキシ末端のいずれかでシステイン残基で伸長させたり、又はシステイン残基を散在させるなどして担体への結合を促進することもできる。免疫原の投与は多くの場合、当業で広く公知であるように、適した期間にわたって、そして適したアジュバントを用いて、注射で行われる。免疫処置期間中は、抗体価を測定して、抗体形成が充分であるかを調べる。この方法で生成させたポリクローナル抗血清は用途によっては充分かも知れないが、薬学的組成物のためには、モノクローナル製剤の利用が好ましい。所望のモノクローナル抗体を分泌する不死化細胞系は、公知のように、Kohler & Milstein, (1992) Biotechnology 24, 524-526の標準的方法や、リンパ球又は脾細胞の不死化を行う改良法を利用して、調製できる。所望の抗体を分泌する不死化細胞系は、抗原を当該ペプチドハプテン、ペプチド又はタンパク質とするイムノアッセイでスクリーニングする。
【0072】
所望の抗体を分泌する適切な不死化細胞培養株を同定したら、この細胞をインビトロで培養しても、又は腹水液中で生成させてもよい。所望のモノクローナル抗体は、この培養上清又は腹水上清から、回収できよう。免疫学的に有意な部分を含有するモノクローナルもしくはポリクローナル抗血清のフラグメントをアンタゴニストとして用いてもよいが、インタクト抗体を用いてもよい。F(ab')2フラグメントのFab、Fab'など、免疫学的に反応性のフラグメントを利用するのが、特に治療上は好ましいことが多い。なぜならこれらのフラグメントは、免疫グロブリン全体よりも免疫原性が一般に低いからである。
【0073】
さらに抗体又はフラグメントは、組換え法により、現在の技術を用いても作製できる。さらに、当該タンパク質の所望の領域に特異的に結合する抗体領域を、複数の種を由来とするキメラとして作製することもできる。上記の方法で検定される作用物質は、無作為に選択しても、又は合理的に選択又は設計してもよい。ここで用いる場合の、作用物質を無作為に選択するとは、本発明のタンパク質のみや、それが結合する基質、結合相手、等々との結合に関与する特定の配列を考慮することなく、作用物質を無作為に選択することを言う。無作為に選択される作用物質の一例は、化学的ライブラリ又はペプチド・コンビナトリアルライブラリ、又は生物の成長ブロスの利用である。
【0074】
ここで用いる場合、ある作用物質が合理的に選択又は設計されるとは、標的部位の配列、及び/又は、作用物質の作用に関連したそのコンホメーション、を考慮に入れた非無作為的基礎に基づいて作用物質が選択されることを言う。IKKβ上のNBDか、又は、NEMO上のIKKβ結合ドメイン、を含むペプチド配列を利用すれば、作用物質を合理的に選択又は合理的に設計することができる。例えば、合理的に選択されたペプチド物質は、SEQ ID NO:2のアミノ酸配列とアミノ酸配列が同一になったペプチドであったり、又は、その保存的置換を持つペプチドであるかも知れない。
【0075】
本発明の化合物は、例としては、ペプチド、小分子、ビタミン誘導体や、糖質であってもよい。本発明の化合物の構造上の性質には何ら制限はないことは、当業者であれば容易に認識できよう。
【0076】
本発明のペプチド化合物は、当業で公知のように、標準的な固相(又は液相)ペプチド合成法を利用して調製できる。加えて、これらのペプチドをコードするDNAを、市販のオリゴヌクレオチド合成装置を用いて合成したり、標準的な組換え産生系を用いて組換えにより産生させてもよい。遺伝子にコードされていないアミノ酸を含めねばならない場合、固相ペプチド合成を利用した生成が必要となる。
【0077】
さらに本発明は、NBD及びその保存的ヌクレオチド置換を有するペプチドを、好ましくは単離された形でコードする、単離された核酸分子を提供するものである。保存的ヌクレオチド置換には、大半のアミノ酸が二つ以上のコドンを有することが原因で、ある特定のアミノ酸の暗号化に影響を与えないようなヌクレオチド置換がある(King & Stansfield (編集者)、A Dictionary of Genetics, Oxford University Press, 1997 、19ページを参照されたい)。従って保存的ヌクレオチド置換には、サイレント変異や、使用コドンの差も含まれる。例えば、本発明には、SEQ ID NO:2に記載のペプチドをコードする、SEQ ID NO:1に記載の核酸分子や、その保存的ヌクレオチド置換が含まれる。さらに本発明は、SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、及び19に記載のペプチドをコードする核酸及びその保存的ヌクレオチド置換も包含する。SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、及び19に記載のペプチドをコードするヌクレオチド配列には複数の順列が可能であるため、これらのペプチドをコードする核酸はいかなるものも、本発明の包含するところである。
【0078】
本発明の包含する核酸の具体的な例には、限定はしないが、以下のものが含まれる。(1)SEQ ID NO:2のペプチドのアミノ酸は、核酸配列TTAGATTGGTCTTGGTTA(SEQ ID NO:24)又はTTGGACTGGTCCTGGCTA(SEQ ID NO:25)にコードされていてもよい;そして(2)SEQ ID NO:15のペプチドのアミノ酸は、核酸配列TTAGATTGGTCTTATCTG(SEQ ID NO:26)又はCTTGACTGGTCATACTTA(SEQ ID NO:27)にコードされていてもよい。
【0079】
ここで用いる場合の、核酸分子が「単離されている」とは、核酸の源にある他のポリペプチドをコードする混入核酸から当該核酸分子が略、分離されていることを言う。翻訳中のタンパク質配列にアミノ酸の欠失、追加、又は変更が起きて、核酸自体の一次構造に対する修飾が、ペプチドの活性を破壊せずに起きることもある。このような置換又は他の変化の結果生ずるのは、本発明が考察する範囲に包含される核酸のコードするアミノ酸配列を有するペプチドである。
【0080】
本発明の化合物の他の分類は、本発明のタンパク質の重要な部分と免疫反応性である抗体である。抗体物質は、抗体に狙わせようとするタンパク質部分を抗原性領域として含有するペプチドで、適したほ乳類被験体を免疫処置して、得る。
【0081】
C.高スループット検定
導入−高スループット・スクリーニングの力を、NEMOと相互作用できる新規な抗炎症化合物の検索に用いる。高スループット・スクリーニングに関する概略的な情報については、例えばCost-Effective Strategies for Automated and Accelerated High-Throughput Screening, IBCS Biomedical Library Series, IBC United States Conferences, 1996; Devlin (編集者)、High Throughput Screening, Marcel Dekker 1998; 米国特許第5,763,263号を参照されたい。高スループット検定では、一種以上の異なる検定技術を利用する。
【0082】
免疫診断及びイムノアッセイ − 生物体液などの複合混合液中に通常低濃度で存在する特定の生化学的物質を測定するのに用いられている一群の技術があるが、このような技術は適宜調製及び選択された抗体がそれらに相補の抗原に対して示す特異性及び高親和性を利用するものである。測定対象となる物質は、必須条件として、免疫原性高分子又はハプテン小分子のいずれかの抗原性でなくてはならない。各試料に、既知の、限られた量の特異抗体を加え、結合:遊離比としてしばしば表される、この抗体と結合した抗原の比率を、放射性同位元素(ラジオイムノアッセイ)、蛍光分子(蛍光イムノアッセイ)、安定な遊離ラジカル(スピンイムノアッセイ)、酵素(酵素イムノアッセイ)等の容易に識別可能な標識で標識した形の抗原を指標として利用して、推定する。
【0083】
酵素結合免疫吸着検定(ELISA);ラジオイムノアッセイ(RIA);蛍光イムノアッセイ(FIA);化学発光イムノアッセイ(CLIA);及びコロイド状金粒子を用いた抗体の標識付け(イムノゴールド法)を含め、抗体には様々な態様で標識を付けることができる。
【0084】
一般的な検定・フォーマットには、サンドウィッチ検定、競合的又は競合検定、ラテックス凝集反応検定、ホモジェネアス(homogeneous)検定、微量定量プレートフォーマット及び微小粒子ベースの検定、がある。
【0085】
酵素結合免疫吸着検定(ELISA) − ELISAは、放射性化学物質の危険性及び蛍光検出系の費用を避けた免疫化学技術である。代わりに、この検定では酵素を指標として用いる。ELISAは、定量的イムノアッセイの一つの形であり、抗体(又は抗原)を不溶性の担体表面に結合させたものの利用に基づく。この担体表面を利用して、テスト溶液中の関連抗原(又は抗体)を「捕獲」するのである。こうして、前もって当該抗原(又は抗体)に共有結合させてある適当な酵素の活性を測定することで、抗原−抗体の複合体を検出する。
【0086】
ELISA技術に関する情報については、例えばCrowther, ELISA: Theory and Practise (Methods in Molecular Biology, Vol. 42), Humana Press, 1995; Challacombe & Kemeny, ELISA and Other Solid Phase Immunoassays: Theoretical and Practival Aspects, John Wiley, 1998; Kemeny, A Practical Guide to ELISA, Pergamon Press, 1991; Ishikawa, Ultrasensitive and Rapid Enzyme Immunoassay (Laboratory Techniques in Biochemistry and Molecular Biology, Vol. 27), Elsevier, 1991を参照されたい。
【0087】
酵素に関する比色検定 − 比色法とは、試薬と、標準量及びテスト量の両方の化合物とを反応させたときの色を、しばしば比色計を用いて比較することで、ある化合物の濃度又は量を判定するようなあらゆる定量的化学解析法を言う。比色計は、色の輝度又は色の輝度の違いを、視覚的又は光電学的に測定する装置である。
【0088】
ベータ−ガラクトシダーゼ酵素活性の標準的比色検定は、当業者に公知である(例えばNorton et al., (1985) Mol. Cell. Biol. 5, 281-290)。比色検定は、全細胞ライセートで、O-ニトロフェニル-ベータ-D-ガラクトピラノシド(ONPG、シグマ社製)を標準的比色ベータ-ガラクトシダーゼ検定(Sambrood et al., (1989) Molecular Cloning - A Laboratory Manual, Cold Spring Harbor Press)の基質として用いて行うことができる。自動化比色検定も、米国特許第5,733,720号が解説するようにベータ-ガラクトシダーゼ活性の検出に利用できる。
【0089】
免疫蛍光検定 − 免疫蛍光法又は免疫蛍光顕微鏡法は、抗原又は抗体を蛍光染料に結合させて蛍光性にした後、組織切片又はスメア中の相補の抗体又は抗原と反応させる技術である。次に、抗原又は抗体の位置を、この蛍光に紫外線を当てて顕微鏡で観察することで、判定することができる。
【0090】
免疫蛍光技術に関する概略的情報については、例えばKnapp et al., (1978) Immunofluorescence and Related Staining Techniques, Elsevier; Allan, (1999) Protein Localization by Fluorescent Microscopy: A Practical Approach (The Practical Approach Series, Vol. 218) Oxford University Press; Beutner, (1983) Defined Immunofluorescence and Related Cytochemical Methods, New York Academy of Sciences; Caul, (1993) Immunofluorescence Antigen Detection Techniques in Diagnostic Microbiology, Cambridge University Pressを参照されたい。本発明に応用できる免疫蛍光技術の詳細な解説については、米国特許第5,912,176号、第5,869,264号、第5,866,319号及び第5,861,259号を参照されたい。
【0091】
バイオチップ − 本発明のペプチドをアレイ又はマイクロアレイに利用して、本発明の核酸又は対応するタンパク質と相互作用する作用物質の高スループットスクリーニングを行うこともできる。
【0092】
「アレイ」又は「マイクロアレイ」とは、一般に、オリゴヌクレオチドとしても知られる定義済みの核酸断片で各位置又はプローブ・セルが占められているような格子系を言う。アレイ自体が、数千のプローブ・セルを多様な格子状に有する高密度核酸及びペプチドマイクロアレイである「チップ」又は「バイオチップ」と呼ばれることがある。
【0093】
典型的な分子検出チップには、認識部位、結合部位又はハイブリダイゼーション部位のアレイが上に並んだ基板がある。各部位は、所定の構造を有する分子と結合又はハイブリダイズする各分子受容基を有する。アレイ又はチップの表面を形成するのに利用できる固体の支持基板には、ガラス、ポリスチレン、ポリイミド、二酸化珪素及び窒化珪素などの有機及び無機の基板がある。プローブを電極に直接付着させる場合、プローブと共役体を形成できる物質で、電極表面を作製しなければならない。
【0094】
アレイを作製したら、試料溶液を分子検出チップに付着させると、試料中の分子が一箇所以上の部位で結合又はハイブリダイズする。結合が起きた部位を検出し、こうして試料中の一つ以上の構造を導き出す。標識群の試料を検出する方法は伝統的な検出法であり、その中には放射性標識、蛍光標識及びビオチン標識があるが、電気的シグナル伝達を含め、他の方法も利用できる。
【0095】
本発明の方法は、試料の高スループットが必要な場合には特に便利であろう。具体的には、本発明は、リガンドスクリーニングの場合や、複合体混合物の組成を決定する場合に、有用である。
【0096】
ポリペプチドは、生物学において構造及び機能間の関係を調べるための代表的な系である。二十種類の天然型アミノ酸が縮合してポリマー分子を形成すると、幅広い三次元立体配置を形成するが、その形状の各々は特定のアミノ酸配列及び溶媒条件の所産である。例えば、アミノ酸5個分の長さのポリマーを、二十種類の天然型アミノ酸を用いて形成するときに考えられるポリペプチドの立体配置の数は300万を超える。典型的なタンパク質は、100を越えるアミノ酸長である。
【0097】
典型的な用途としては、特徴付けようとする一種以上の物質を含有する複合溶液を、ポリペプチドを含んで成るポリマーアレイに接触させる。本発明のポリペプチドは、標準的固相技術を用いるなど、当業で公知の伝統的方法で調製できる。この標準的方法には、排他的固相合成、部分的固相合成法、画分凝縮法、伝統的溶液合成、及び組換えDNA技術がある(Merrifield, (1963) Am. Chem. Soc. 85, 2149-2152を参照されたい)。
【0098】
ある好適な実施例では、アレイのポリペプチド又はタンパク質を他の共受容基に結合させて、アレイ上でヘテロ二重鎖を形成させてもよい。さらに別の実施例では、アレイのポリペプチド又はタンパク質をペプチド又は小分子に結合させることもできる。
【0099】
D.本発明の抗炎症化合物の用途
本発明の抗炎症化合物(例えばNEMOの発現を調節する化合物、又は、NEMOの少なくとも一つの活性のアゴニスト又はアンタゴニストなどの化合物、など)を用いて、被験体の炎症を調節したり、炎症性疾患を治療又は診断することができよう。本方法には、本発明の抗炎症化合物を、炎症性疾患を治療するのに有効量、被験体に投与するステップが含まれる。
【0100】
ここで用いる「炎症性疾患」には、炎症を特徴とする、炎症で引き起こされる、炎症の結果起きる、又は炎症の影響を受けるようになる、疾患又は異常が含まれるものと、意図されている。炎症性疾患は、NEMO又はIKKβ機能及び活性に関連した、及び/又は、NF-κB媒介性プロセスに関連した生物学的及び病的プロセスが原因であっても、又は、関連していてもよい。炎症性疾患又は異常の例には、限定はしないが、急性及び慢性の炎症性異常、例えば喘息、乾癬、リウマチ性関節炎、変形性関節炎、乾癬性関節炎、炎症性腸疾患(クローン病、潰瘍性大腸炎)、敗血症、脈管炎、及び滑液包炎;自己免疫疾患、例えば狼瘡、多発性筋痛、リウマチ、強皮症、ウェグナー肉芽腫症、一過性動脈炎、クリオグロブリン血症、及び多発性硬化症;移植片拒絶;骨粗鬆症;実質性腫瘍を含む癌(例えば肺、CNS、結腸、腎臓、及び膵臓);アルツハイマー病;アテローム性硬化症;ウィルス(例えばHIV又はインフルエンザ)感染;慢性ウィルス(例えばエプスタイン−バー、サイトメガロウィルス、単純疱疹ウィルス)感染;及び毛細血管拡張性運動失調、がある。
【0101】
病的プロセスとは、有害な作用を生むような種類の生物学的プロセスを言う。例えば、NF-κBの発現調節不全には、特定の病的プロセスが根底にある炎症誘発性プロセスが伴う。ここで用いる場合の、ある抗炎症化合物が病的プロセスを調節する、とは、当該化合物が、このプロセスの程度又は重篤度を低下させることを言う。例えば、NEMO又はIKKβの発現又は少なくとも一つの活性を何らかの態様で低下、促進又は調節するような抗炎症化合物を投与すると、炎症誘発性の応答が妨げられたり、病的プロセスが調節されるであろう。
【0102】
従って、NF-κB炎症性成分を伴う疾患を治療するのに、本発明の抗炎症化合物を用いてもよい。このような疾患には、限定はしないが、骨粗鬆症、リウマチ性関節炎、アテローム性硬化症、喘息(Ray & Cohn, (1999) J. Clin. Invest. 104, 985-993; Christman et al., (2000) Chest 117, 1482-1487)及びアルツハイマー病、がある。NF-κB炎症性成分を伴う疾患のレビューについては、Epstein, (1997) New Eng. J. Med. 336, 1066-1071; Lee et al., (1998) J. Clin. Pharmacol. 38, 981-993; Brand et al., (1997) Exp. Physiol. 82, 297-304を参照されたい。
【0103】
本発明の抗炎症化合物が、治療にとって有用と思われる、炎症誘発性応答を伴う病的プロセスには、限定はしないが、喘息、アレルギー、例えばアレルギー性鼻炎、ユティカリア(uticaria)、アナフィラキシー、薬物過敏症、食物過敏症、等々;皮膚の炎症、例えば皮膚炎、湿疹、乾癬、接触性皮膚炎、日焼け、加齢、等々;関節炎、例えば変形性関節炎、乾癬性関節炎、狼瘡、脊椎炎、等々、がある。抗炎症化合物は、慢性閉塞性肺疾患及び慢性炎症性長疾患の治療にも有用である。さらに本発明の抗炎症化合物は、移植や癌治療での免疫抑制を含め、コルチコステロイドを用いるあらゆる用途で、コルチコステロイドの代わりに用いてもよい。
【0104】
ここで用いる用語「被験体」には、温血動物、好ましくはヒトを含むほ乳類、が含まれる。ある好適な実施例では、被験体は霊長類である。さらにより好適な実施例では、前記霊長類はヒトである。
【0105】
ここで用いる用語、被験体に「投与する」には、(ここに解説するような)製剤の形の抗炎症化合物などの抗炎症化合物を、被験体に、非経口又は経口経路による送達、筋肉内注射、皮下/皮内注射、静脈内注射、口腔内投与、経皮送達や、直腸、結腸、膣、鼻孔内もしくは呼吸器官経路(吸入など)による投与を含め、被験体の所望の位置に化合物を送達するのに適した何らかの経路で、投薬する、送達又は投与することが、含まれる。
【0106】
ここで用いる用語「有効量」は、被験体において、炎症性異常を治療するのに充分であるなど、所望の結果を達成するのに必要な投薬量及び時間で、有効となる量を包含する。ここで定義するような、本発明の抗炎症化合物の有効量は、被験体の疾患状態、年齢及び体重や、化合物が被験体の所望の応答を引き出す能力などの因子に応じて様々であろう。投薬計画は最適な治療応答が得られるよう、調節してもよい。有効量はまた、化合物の毒性もしくは有害作用(副作用など)が、治療上有益な効果を下回るような量である。
【0107】
本発明の抗炎症化合物の治療上有効量(即ち有効投薬量)は、約0.001乃至30mg/kg体重の範囲内であろうが、好ましくは約0.01乃至25mg/kg体重、より好ましくは約0.1乃至20mg/kg体重、そしてさらにより好ましくは約1乃至10mg/kg、2乃至9mg/kg、3乃至8mg/kg、4乃至7mg/kg、又は5乃至6mg/kg体重であるとよい。当業者であれば、限定はしないが、疾患又は異常の重篤度、以前の治療歴、被験体の全身の健康及び/又は年齢、及び他の罹患状態を含め、特定の因子が、被験体を効果的に治療するのに必要な投薬量を左右する場合があることは、理解されよう。さらに、治療上有効量の本発明の抗炎症化合物を用いた被験体の治療には、一回の治療を含めることができるが、好ましくは連続した治療が含まれる。一実施例では、約0.1乃至20mg/kg体重の範囲の本発明の抗炎症化合物で、一週間当たり1回、約1乃至10週間、好ましくは2乃至8週間、より好ましくは約3乃至7週間、そしてさらにより好ましくは約4、5、又は6週間、被験体を治療する。さらに、治療に用いる本発明の抗炎症化合物の有効投薬量を、特定の治療経過の間、増減させてもよいことは理解されよう。
【0108】
本発明の抗炎症化合物を単独で提供してもよいが、ある特定の病的プロセスを調節する他の作用薬と組み合わせて提供してもよい。例えば、本発明の抗炎症化合物を、他の公知の抗炎症剤と組み合わせて投与することもできる。本発明の方法で用いても良い公知の抗炎症剤は、Harrison's Principles of Internal Medicine, Thirteenth Edition, Eds. T.R. Harrison et al. McGraw-Hill N.Y., NY; 及び the Physicians Desk Reference 50th Edition 1997, Oradell New Jersey, Medical Economics Co.,(これら文献の内容全体を、言及をもってここに編入することを明示しておく)に見ることができる。本発明の抗炎症化合物及び付加的な抗炎症剤は、同じ薬学的組成物として被験体に投与しても、又は、異なる薬学的組成物として(同時又は異なる時期に)被験体に投与してもよい。
【0109】
さらに本発明は、細胞内でIκBが関与するシグナル伝達を調節する方法も提供する。本方法は、抗炎症化合物に細胞を接触させるなどにより、IKKβ活性を調節するステップを含む。この抗炎症化合物は、例えば、NEMOがNBDでIKKβと相互作用するのを抑制することで、IKKβ機能を抑制するものであってもよい。細胞は培養物内にあっても、インシツー、即ち天然ホスト内にあってもよい。
【0110】
診断用に利用する場合、本発明の抗炎症化合物に、蛍光性、放射性、化学発光性等の容易に検出できる分子で標識を付けてもよい。標識は、抗炎症化合物に直接又は間接的に共役させてよい。
【0111】
E.製剤
さらに本発明は、本発明の抗炎症化合物を、薬学的に許容可能な担体と一緒に含んで成る薬学的組成物を包含する。薬学的に許容可能な担体は、水や、ピーナッツ油、大豆油、鉱油、ごま油、等々の石油、動物性油脂、植物性油脂、又は合成のものも含む油脂類など、無菌の液体であってよい。当該薬学的組成物を静脈内投与する場合には、水が好適な担体である。生理食塩水や、水性デキストロース及びグリセロール溶液が、特に注射用溶液の場合には、液体の担体として、利用できる。適した薬剤用担体は、Gennaro et al., (1995) Remington's Pharmaceutical Sciences, Mack Publishing Companyに解説されている。本発明の組成物には、薬理学的に活性な物質に加え、活性化合物を加工して、作用部位への送達に薬学的に利用できる製剤にするのに便利な添加剤及び佐剤を含む薬学的に許容可能な担体を含有させてもよい。非経口投与用に適した製剤には、水溶性の塩など、水溶性の形の活性化合物の水溶液がある。加えて、活性化合物の懸濁液を、適宜油性の注射用懸濁液として、投与してもよい。適した親油性溶媒又は賦形剤には、脂肪油、例えばごま油、又は合成の脂肪酸エステル、例えばオレイン酸エチル又はトリグリセリド、がある。水性の注射用懸濁液には、カルボキシメチルセルロースナトリウム、ソルビトール、及びデキストランを含め、懸濁液の粘性を増す物質を含めてもよい。選択に応じ、懸濁液にはさらに安定剤も含めてもよい。さらに、細胞への送達用に、リポソームを用いて当該作用薬を被包してもよい。
【0112】
本発明に基づく全身投与用の製剤は、腸内用、非経口用又は局所用投与に向けて、調合してもよい。実際、有効成分の全身への投与を行うために、三種類の製剤すべてを同時に用いてもよい。
【0113】
経口投与に適した製剤には、被覆錠剤、エリキシル、懸濁液、シロップ又は吸入剤や、これらの制御放出型を含め、硬質もしくは軟質ゼラチンカプセル、丸剤、錠剤、がある。
【0114】
さらに、少なくとも数週間から一ヶ月又はそれ以上の期間にわたって被験体への抗炎症化合物の送達を持続させる薬学的組成物に、本発明の抗炎症化合物を組み込んでもよい。このような製剤は、各内容を言及をもってここに編入することとする米国特許第5,968,895号及び第6,180,608号B1に解説されている。
【0115】
本発明の抗炎症化合物を、非経口、皮下、静脈内、筋肉内、腹腔内、経皮又は口腔内経路で投与してもよい。あるいは、又は同時に、投与を経口経路で行ったり、又は、吸入もしくは肺への直接の潅注で行ってもよい。投与する投薬量は、レシピエントの年齢、健康、及び体重、あれば併用療法の種類、治療の頻度、及び、所望の効果の性質、に左右されるであろう。
【0116】
ここで解説した治療法で用いる抗炎症化合物は、治療しようとする状態、部位特異的治療の必要性、投与する薬物の量、等の考慮点などの考慮点に応じて、全身又は局所的に投与してよい。
【0117】
局所投与を利用してもよい。溶液、懸濁液、ゲル、軟膏(ointment)又は軟膏剤(salve)などの通常の局所用製剤を用いてもよい。このような局所用製剤の製剤は、例えばレミントンズ・ファーマシューティカル・サイエンセズなどに例示された薬剤調剤業でよく解説されている。局所用投与の場合、これらの化合物を粉末又はスプレー、特にエーロゾル型として投与してもよいであろう。全身投与用に適合させた薬学的組成物に有効成分を含めて投与してもよい。公知のように、薬物を全身投与する場合、それを粉末、丸剤、錠剤に調製しても、又は、経口投与に向けたシロップ又はエリキシルとして調製してもよい。静脈内、腹腔内又は病変部内投与の場合、注射で投与できる溶液又は懸濁液として、化合物を調製することになるであろう。場合によっては、これらの化合物を座薬型に調製したり、又は、皮下又は筋肉内注射用のデポジット(deposit)の長時間放出製剤として調製すると、便利であろう。ある好適な実施例では、本発明の抗炎症化合物を吸入で投与してもよい。吸入治療の場合、当該化合物は、定量吸入器による投与に便利な溶液か、又は、乾燥粉末吸入器に適した形であってもよい。
【0118】
有効量とは、目的たんぱくの活性を調節したり、又はレベルを変化させるような量である。有効量は条件に応じて様々であり、場合によっては、治療しようとする状態の重篤度や、治療に対する患者の感受性によっても、様々であろう。従って、通常の実験を通じて、その場その場で有効量を決めるとよいであろう。しかしながら、本発明に基づいて腫瘍を治療する場合、重量で0.001乃至5パーセント、好ましくは約0.01乃至1パーセントを含有する製剤が、多くの場合に治療上有効量となるであろうことが予測できる。全身投与の場合、一日当たり体重1kg当たり0.01乃至100mg、より好ましくは1kg当たり約0.1乃至10mgの量が、多くの場合に治療効果を生じることとなろう。
【0119】
本発明の方法を実施する際、本発明の化合物を単独で用いても、又は併用してもよく、あるいは他の治療的又は診断的作用薬と併用してもよい。いくつかの好適な実施例では、本発明の化合物を、広く認められた医療行為に基づいてこれらの状態に典型的に処方されている他の化合物と同時に投与してもよい。本発明の化合物を、通常は哺乳動物で、好ましくはヒトで、インビボで利用することもできる。
【0120】
さらに別の実施例では、標的たんぱくの機能又は調節を、治療上の利益に向けて変更するタンパク質を含め、化学的部分に本発明の抗炎症化合物を結合させてもよい。これらのタンパク質には、炎症性異常の治療にとってさらに治療上有益となると思われる他のサイトカイン阻害剤及び成長因子阻害剤が組み合わされていてもよい。加えて、本発明の抗炎症化合物に、当業で公知の何らかの架橋試薬を用いたリン酸化を行って、ビオチニレート、チオエート、アセチレート、イオジネートに共役させてもよい。
【0121】
F.分子生物学、微生物学及び組換えDNA技術
本発明によれば、上で解説したか、又は、下記の実施例で論じるように、従来の分子生物学、微生物学及び組換えDNA技術を利用することになるであろう。このような技術は文献に十二分に説明されている。例えばSambrook et al., (1989) Molecular Cloning - A Laboratory Manual, Cold Spring Harbor Press; Glover, (1985) DNA Cloning: A Practical Approach; Gait, (1984) Oligonucleotide Synthesis; Harlow & Lane, (1988) Antibodies - A Laboratory Manual, Cold Spring Harbor Press; Roe et al., (1996) DNA Isolation and Sequencing: Essential Techniques, John Wiley; 及びAusubel et. al., (1995) Current Protocols in Molecular Biology, Greene Publishingを参照されたい。
【0122】
G.アンチセンスRNA
アンチセンス分子は、特定のmRNAに相補なヌクレオチド配列を持つRNA又は一本鎖DNA分子である。研究室で調製したアンチセンス分子を、研究対象の遺伝子が転写した正常mRNAを含有する細胞に注射すると、このアンチセンス分子はこのmRNAと塩基対を形成することで、mRNAのタンパク質への翻訳を妨げることができる。その結果できる二本鎖RNA又はRNA/DNAは、このような分子に特異的に結合する酵素で消化される。従って、mRNAの消去が起きて遺伝子産物の翻訳が遮断される。このように、アンチセンス分子は、有害たんぱくの産生を遮断する医療において有用である。アンチセンスRNAを作製し、利用する方法は当業者に公知である(例えばLichtenstein & Nellen (編集者), Antisense Technology: A Practical Approach, Oxford University Press, 1997; Agrawal & Crooke, Antisense Research and Application (Handbook of Experimental Pharmacology, Vol. 131), Springer Verlag, 1998; Gibson, Antisense and Ribozyme Methodology: Laboratory Companion, Chapman & Hall, 1997; Mol & Van Der Krol, Antisense Nucleic Acids and Proteins, Marcel Dekker; Weiss, Antisense Oligodeoxynucleotides and Antisense RNA: Novel Pharmacological and Therapeutic Agents, CRC Press, 1997; Stanley et al., (1993) Antisense Research and Applications, CRC Press; Stein & Krieg, (1998) Applied Antisense Oligonucleotide Technologyを参照されたい)。
【0123】
本発明によるアンチセンス分子及びリボ酵素は、核酸分子の合成について当業で公知のいかなる方法で調製してもよい。これらの中には、固相ホスホールアミジト化学合成など、オリゴヌクレオチドを化学合成する技術がある。代わりに、DNA配列のインビトロ及びインビボ転写でもRNA分子を作製できよう。このようなDNA配列を、T7又はSP6などの適したRNAポリメラーゼプロモータを持つ多種のベクタに組み込んでもよい。代わりに、アンチセンスRNAを構成的又は誘導的に合成するこれらのcDNAコンストラクトを細胞株、細胞、又は組織内に導入することもできる。RNA分子は、細胞内での安定性や半減期を向上させるよう、修飾してもよい。可能な修飾には、限定はしないが、当該分子の5’末端及び/又は3’末端へのフランキング配列の追加、又は、当該分子の骨格内にホスホジエステラーゼ結合の代わりにホスホロチオエート又は2'O-メチルを用いる、などがある。この発想を進展させて、イオシン、ケオシン、及びワイブトシンなどの非伝統的な塩基を含めたり、内因性エンドヌクレアーゼが容易には認識できない、アセチル修飾型、メチル修飾型、チオ修飾型、等の修飾型アデニン、シチジン、グアニン、チミン、及びウリジンを含めてもよい。
【0124】
H.融合タンパク質
融合タンパク質は、二つの遺伝子の融合の結果生じる発現産物である。このようなタンパク質は、例えば組換えDNA発現研究で生じたり、又は、腫瘍遺伝子がgagに融合したいくつかのウィルス性腫瘍遺伝子で天然に生じることもある。
【0125】
時には、細菌系で発現させるために、細菌性プロモータの制御下に真核生物のクローン遺伝子を配置する必要性から、融合タンパク質が生ずることもある。こうして、細菌系の配列は、真核生物たんぱくに連結した形で発現することが多い。融合タンパク質は、異種遺伝子産物の構造解析や、精製、機能、及び発現解析に用いられる。
【0126】
融合タンパク質は、目的の核酸を組換えDNA技術によりレシピエント・プラスミド内に挿入し、プラスミド遺伝子の停止コドンに替えたときに生じさせることのできるハイブリッドたんぱく分子である。融合されたタンパク質は、プラスミドたんぱく配列の一部分を持つアミノ末端で始まり、目的のタンパク質で終わる。
【0127】
融合タンパク質の作製は当業者に公知である(例えば米国特許第5,908,756号、第5,907,085号、第5,906,819号、第5,905,146号、第5,895,813、第5,891,643号、第5,891,628号、第5,891,432号、第5,889,169号、第5,889,150号、第5,888,981号、第5,888,773号、第5,886,150号、第5,886,149号、第5,885,833号、第5,885,803号、第5,885,779号、第5,885,580号、第5,883,124号、第5,882,941号、第5,882,894号、第5,882,864号、第5,879,917号、第5,879,893号、第5,876,972号、第5,874,304号、及び第5,874,290号を参照されたい)。臨床及び研究医療で用いられている特定の種類の融合分子に関連する構造、性質、用途、及び問題の概略的レビューについては、例えばChamow et al., (1999) Antibody Fusion Proteins, John Wileyを参照されたい。
【0128】
I.ペプチドミメティック
さらに本発明は、IKKβ上のNBDの三次元構造を模倣して、NEMOに結合することで、NEMOがこのNBDで結合するのを遮断するペプチドミメティックなどの、ペプチドミメティックも包含する。このようなペプチドミメティックは、例えばより生産コストが低い、化学的安定性が高い、薬理学的性質(半減期、吸収、効能及び効験)が向上している、特異性が変更されている(例えば幅広い生物活性など)、抗原性が低い、等々を含め、天然型のペプチドに比べて有意な長所を有していることがある。
【0129】
ある一つの形としては、ミメティックは、タンパク質の二次構造の要素を模倣したペプチド含有分子である。例えばJohnson et al., (1993) Peptide Turn Mimetics in Biotechnology and Pharmacy, Pezzuto et al., (Editors) Chapman & Hallを参照されたい。ペプチドミメティックを利用する根拠は、タンパク質のペプチド骨格は、主に、抗体対抗原などの分子間相互作用を促進するような方向にアミノ酸側鎖を方向付けるよう、並んでいることである。ペプチドミメティックでも、天然分子に類似の分子間相互作用が可能であると予測できる。別の形では、ペプチド類似体は、テンプレートペプチドのものに類似の性質を持つ非ペプチド化合物として、製薬業で広く用いられている。これらの種類の非ペプチド化合物も「ペプチドミメティック」又は「ペプチドミメティック」と呼ばれており( 言及をもってここに編入するFauchere, (1986) Adv. Drug Res. 15, 29-69; Veber & Freidinger, (1985) Trends Neurosci. 8, 392-396; and Evans et al., (1987) J. Med. Chem. 30, 1229-1239)、一般にコンピュータによる分子モデリングで開発されている。
【0130】
治療上有用なペプチドに構造上類似のペプチドミメティックを用いれば、同等の治療効果又は予防効果が得られるであろう。一般に、ペプチドミメティックは、NBDなどのパラダイムポリペプチド(即ち、生化学的性質又は薬理活性を有するポリペプチド)に構造上類似であるが、一箇所以上のペプチド結合が、選択に応じて-CH2NH-、-CH2S-、-CH2-CH2-、-CH=CH- (cis 及びtrans)、-COCH2-、-CH(OH)CH2-、及び-CH2SO-からなる群より選択される結合に、当業で公知の方法であり、さらに以下の文献:Weinstein, (1983) Chemistry and Biochemistry of Amino Acids, Peptides and Proteins, Marcel Dekker; Morley, (1980) Trends Pharmacol. Sci. 1, 463-468 (概略的レビュー); Hudson et al., (1979) Int. J. Pept. Protein Res. 14, 177-185 (-CH2NH-、CH2CH2-); Spatola et al., (1986) Life Sci. 38, 1243-1249 (-CH2-S); Hann, (1982) J. Chem. Soc. Perkin Trans. 1, 307-314 (-CH-CH-, cis 及びtrans); Almquist et al., (1980) J. Med. Chem. 23, 1392-1398 (-COCH2-); Jennings-White et al., (1982) Tetrahedron Lett. 23, 2533 (-COCH2-); 米国特許出願第4,424,207号 (-CH(OH)CH2-); Holladay et al., (1983) Tetrahedron Lett. 24, 4401-4404 (-C(OH)CH2-); 及び Hruby, (1982) Life Sci. 31, 189-199 (-CH2-S-)(これらの文献各々を、言及をもってここに編入する)に解説された方法で、置換されている。
【0131】
ペプチドミメティックの標識付けには、通常、定量的構造−活性データ及び/又は分子モデリングで予測された、ペプチドミメティック上の非干渉位置に、直接的に又はスペーサ(例えばアミド基)を介して、一つ以上の標識を共有結合させる方法がある。このような非干渉位置とは、一般に、当該ペプチドミメティックが結合して治療効果を生ずる相手である高分子と直接接触しない(例えばNBD-NEMO複合体上の接点ではない)位置である。ペプチドミメティックの誘導体化(標識付けなど)は、当該ペプチドミメティックの所望の生物活性又は薬理活性に大きく干渉するものであってはならない。NBDペプチドミメティックは、有機成分でアミノ酸を置換する方法を通じ、構造ベースの薬物デザインにより構築することができる(例えば Hughes, (1980) Philos. Trans. R. Soc. Lond. 290, 387-394; Hodgson, (1991) Biotechnol. 9, 19-21; Suckling, (1991) Sci. Prog. 75, 323-359を参照されたい)。
【0132】
コンビナトリアル化学法を用いて薬物ライブラリを作製すると、ペプチドミメティックの利用を向上させることができる。ペプチドミメティックの設計には、NBD(例えばIKKβ上のNBD)のNEMOへの結合を増加又は減少させるアミノ酸変異の特定が役立つであろう。例として、このような変異を表1で特定した。利用の可能な方法には、酵母二種ハイブリッド法(Chien et al., (1991) Proc. Natl. Acad. Sci. USA 88, 9578-9582を参照されたい)及びファージ展示法の利用がある。二種ハイブリッド法では、タンパク質対タンパク質の相互作用を酵母で検出する (Fields et al., (1989) Nature 340, 245-246)。ファージ展示法では、固定したタンパク質と、ラムダ及びM13などのファージの表面上に発現させたタンパク質との間の相互作用を検出する(Amberg et al., (1993) Strategies 6, 2-4; Hogrefe et al., (1993) Gene 128, 119-126)。これらの方法により、タンパク質対タンパク質の相互作用による正及び負の淘汰や、このような相互作用を決定する配列の同定が可能である。
【0133】
ペプチド合成及びペプチドミメティックに関する概略的情報については、例えば、ここに言及をもって各全文を編入することとするJones, (1992) Amino Acid and Peptide Synthesis, Oxford University Press; Jung, (1997) Combinatorial Peptide and Nonpeptide Libraries: A Handbook, John Wiley; and Bodanszky et al., (1993) Peptide Chemistry: A Practical Textbook, 2nd Revised Edition, Springer Verlagを参照されたい。
【0134】
J.トランスジェニック動物
トランスジェニック動物は、組換え、外因性もしくはクローン形成した遺伝物質を実験的に移入させた、遺伝子改変動物である。このような遺伝物質はしばしば導入遺伝子と呼ばれる。導入遺伝子の核酸配列は、この核酸配列が存在しない、もしくは、通常見られないようなゲノム上の遺伝子座や、又は、この導入遺伝子にとって正常な遺伝子座のいずれにも、組み込まれることがある。導入遺伝子は、標的動物と同じ種のゲノム由来の核酸配列や、又は、異なる種のゲノム由来の核酸配列から成る場合がある。
【0135】
「生殖細胞系トランスジェニック動物」という用語は、遺伝子改変又は遺伝情報が、生殖系の細胞に導入されたことで、このトランスジェニック動物に、遺伝情報を子孫に伝える能力がもたらされたようなトランスジェニック動物を言う。実際にこのような子孫が、その改変又は遺伝情報の一部又は全部を有していれば、彼らもトランスジェニック動物である。
【0136】
改変又は遺伝情報は、レシピエントが属する動物種にとって異種であっても、特定の個体レシピエントにとってのみ、異種であっても、又は、レシピエントが既に持つ遺伝情報であってもよい。最後者の場合、改変したもしくは導入された遺伝子に、天然遺伝子とは異なる発現をさせてもよい。
【0137】
トランスジェニック動物は、トランスフェクション、電気穿孔法、顕微注射、胚性幹細胞への遺伝子ターゲティング、及び組換えウィルス及びレトロウィルス感染法を含め、多種の様々な方法で作製することができる(例えば米国特許第4,736,866号、米国特許第5,602,307号、Mullins et al., (1993) Hypertension 22, 630-633; Brenin et al., (1997) Surg. Oncol. 6, 99-110; Tuan, (1997) Recombinant Gene Expression Protocols, Methods in Molecular Biology No. 62, Humana Pressを参照されたい)。
【0138】
数多くの組換えもしくはトランスジェニックマウスが作製されてきており、その中には、活性化腫瘍遺伝子配列を発現するもの(米国特許第 4,736,866号、);シミアンSV40 T-抗原を発現するもの(米国特許第5,728,915号);インターフェロン調節因子1(IRF-1)の発現を欠失させたもの(米国特許第5,731,490号);ドーパミン作動性機能不全を呈するもの(米国特許第5,723,719号);血圧制御に関与する少なくとも一つのヒト遺伝子を発現するもの(米国特許第5,731,489号);天然型アルツハイマー病における状態に、より大きな類似性を呈するもの(米国特許第5,720,936号);細胞接着を媒介する能力を低下させたもの(米国特許第5,602,307号);ウシ成長ホルモン遺伝子を持つもの(Clutter et al., (1996) Genetics 143, 1753-1760) 又は、完全にヒト抗体応答を生じられるもの(Zou et al., (1993) Science 262, 1271-1274)、がある。
【0139】
マウス及びラットは、依然、大半のトランスジェニック実験で選択される動物であるが、場合によっては、別の動物種を用いるのが好ましいか、又は必要なことがある。トランスジェニック法は、ヒツジ、ヤギ、ブタ、イヌ、ネコ、サル、チンパンジー、ハムスター、ウサギ、ウシ及びモルモットを含め、多種のマウス以外の動物で利用されて、成功が収められてきた(Kim et al., (1997) Mol. Reprod. Dev. 46, 515-526; Houdebine, (1995) Reprod. Nutr. Dev. 35, 609-617; Petters, (1994) Reprod. Fertil. Dev. 6, 643-645; Schnieke et al., (1997) Science 278, 2130-2133; Amoah, (1997) J. Animal Science 75, 578-585を参照されたい)。
【0140】
核酸断片を組換えコンピテントほ乳類細胞へ導入する方法は、複数の核酸分子の同時形質転換にとって有利な方法であれば、いかなるものでもよい。トランスジェニック動物を作製する手法の詳細は、米国特許第5,489,743号及び米国特許第5,602,307号の開示を含め、当業者が容易に利用できるものである。
【0141】
本発明は、NBDをコードする遺伝子を発現するトランスジェニック動物と、この野生型遺伝子に比較したときの保存的及び非保存的アミノ酸置換を生ずるようなこの遺伝子の変異を、包含するものである。
さらに本発明を以下の実施例で解説することとするが、同実施例を限定的なものと捉えられてはならない。本出願を通じて引用された全参考文献、特許及び公開済み特許出願の内容や、図面及び配列表を、言及をもってここに編入することとする。
【図面の簡単な説明】
【0142】
【図1A】図1は、NEMOがIKKβのCOOH末端と相互作用することを示した実験結果を示す。図1A:GST単独又はGST-NEMOを、グルタチオン-アガロースを用いて細菌ライセートから沈降させ、SDS-PAGE(10%)で分離し、このゲルをクーマシー・ブルーで染色した(左側パネル)。等量のGST又はGST-NEMOを次のGSTプル−ダウン実験で用いた。右側パネルに表したスキームは、NEMO相互作用領域を調べるのに用いた、IKKβのCOOH及びNH2末端切断型変異体を表す。
【図1B】IKKβ変異体をクローンし、インビトロ翻訳で発現させ(インプット;左側パネル)て、GSTプル−ダウンに用いた(右側パネル)。
【図1C】野生型IKKβ及びIKKβ-(644-756)をインビトロ翻訳させ(左側パネル)、GSTプル−ダウン解析に用いた(左側パネル)。
【図1D】ヒーラ細胞を、pBIIX-ルシフェラーゼレポータプラスミドと一緒に、ベクタのみか、又は、次第に濃度を上げた(0.25、0.5、1.0μg/ml)、xpressタグ付きIKKβ-(644-756)コンストラクトで、一時的にトランスフェクトした。48時間後、細胞をTNFα(10ng/ml)又はIL-1β(10ng/ml)で4時間処置し、その後NF-κB活性を測定した。ライセートの一部を抗xpress(挿入図)を用いてウェスタンブロット解析したところ、発現したたんぱくも次第に高レベルになっていることが分かる。
【図2A】図2は、NEMOの一番目のαへリックス領域がIKKβへの結合に必要であることを示した実験結果を表す。図2A:残基V644からS756までのCOOH末端のみから成る切断型のIKKβをGSTに融合し(GST-644-756)、細菌で発現させた。グルタチオンアガロースで沈殿させた後、GST単独及びGST-(644-756)をSDS-PAGE(10%)で分離し、そのゲルをクーマシー・ブルーで染色した(左側パネル)。等量の各タンパク質を次のGSTプル−ダウン解析に用いた。NEMOの様々なNH2及びCOOH末端切断型を構築し、[35S]-メチオニンで標識し、インビトロプル−ダウンに用いた(右側パネル)。GST-(644-756)と相互作用した変異体を(+)で示す。変異体のいずれも、GST単独とは相互作用しなかった。
【図2B】野生型NEMOと、一番目のαへリックス領域を欠失させた欠失変異体(del.αH)とをインビトロで翻訳し(左側パネル:インプット)、上記のタンパク質を用いたGSTプル−ダウンに利用した(A:左側)。
【図2C】ヒーラ細胞に、pcDNA-3(ベクタ)又は次第に濃度を高くしたdel.αH(0.25、0.5、1.0μg/ml)のいずれかと一緒に、pBIIx-ルシフェラーゼを48時間、トランスフェクトし、次に、TNFα(10ng/ml)で4時間処置した。次に細胞を溶解させ、NF-κB活性をルシフェラーゼ検定で測定した。
【図3A】図3は、NEMOとの相互作用や、機能的キナーゼ活性には、IKKβ COOH-末端のIKKα-相同領域が必要であることを示した実験結果を示す。図3A:最末端のCOOh末端(1-733)、セリンなしの領域(1-707)、セリンリッチなドメイン(1-662)及びα1領域(1-644)を順に省略した切断変異型のIKKβをインビトロ翻訳で発現及び標識付けし、GST-NEMOによるGSTプル−ダウンに用いた(図1A)。変異体のいずれも、GST単独とは相互作用しなかった。
【図3B】IKKβ及びIKKαの最末端COOH末端の配列アラインメント。α2領域及びグルタミン酸リッチな領域を示し、アミノ酸が6個同一になっている部分は網がけした。
【図3C】野生型IKKβ及び切断変異型(1-733及び1-744)を[35S]-メチオニンで標識し(インプット)、GST単独又はGST-NEMOによるインビトロプル−ダウンに用いた。
【図3D】ヒーラ細胞に、1μg/ウェルの提示したFLAGでタグを付けたコンストラクトで48時間、トランスフェクトした後、抗FLAGを用いて免疫沈降させた。[32P]で標識したγATPを含有するキナーゼバッファ中でこの免疫沈降物を15分間、30℃でインキュベートし、1%トリトン-100を含有する溶解バッファで洗浄した。その結果得られた複合体を、SDS-PAGE(10%)で分離し、オートラジオグラフィで観察した(上側パネル)。下側のパネルは同一試料の免疫ブロットであり、各レーンにトランスフェクトしたタンパク質が等量、現れている。
【図3E】ヒーラ細胞に、提示したコンストラクト1μg/ml又は空のベクター(pcDNA-3)を、pBIIx-ルシフェラーゼと一緒に48時間かけてトランスフェクトした。NF-κB活性はルシフェラーゼ検定で調べた。(F)FLAGでタグを付けた形のIKKβ(野生型)又はIKKβ(1-733)を48時間かけてトランスフェクトしたヒーラ細胞を、処置しない(-)か、又はTNFα(10ng/ml)で7分間、処置した(+)。溶解させ、抗FLAGを用いて免疫沈降させた後、免疫複合体キナーゼ検定(上側パネル)を行った。同一試料を免疫沈降させ、抗FLAGで免疫ブロットした(下側パネル)。
【図4A】図4は、IKKβ及びIKKαとのNEMOの結合の結果、NEMO結合ドメイン(NBD)は6個のCOOH末端アミノ酸であることが判明した実験結果を示す。図4A:提示するように、合計DNA濃度が2μg/ウェルになるようにベクタ単独、FLAGでタグを付けたIKKα又はIKkβ(1μg/ウェル)又はxpressでタグを付けたNEMO(1μg・ウェル)を一時的にトランスフェクトしたCOS細胞。溶解後、免疫沈降(IP)を抗FLAG(M2)を用いて行い、沈降物の含有量を、抗FLAG(M2)又は抗xpressを用いた免疫ブロット(IB)で観察した。IP前のライセートの一部を抗xpressで免疫ブロットして、同等のレベルのNEMO発現がトランスフェクト後の細胞にあることを確認した。
【図4B】野生型IKKα及びIKKα-(1-737)を発現させ、標識し(インプット)、GST又はGST-NEMOを用いたGSTプル−ダウンに用いた。
【図4C】ヒトIKKiをコードする完全長cDNAを、RT-PCRにより、ヒーラ細胞mRNAからExpandTMロングテンプレートPCRシステム(ベーリンガー・マンハイム社製)、正方向プライマー(5’-CTAGTCGAATTCACCATGCAGAGCACAGCCAATTAC) (SEQ ID NO: 22) 及び逆方向プライマー(3’-CTAGTCTCTAGATTAGACATCAGGAGGTGCTGG) (SEQ ID NO: 23)を用いて得て、pcDNA-3のEcoRI 及びXbaI 部位にクローンした。GSTプル−ダウン解析は、[35S]-メチオニンで標識したIKKα、IKKβ及びIKKiを用いて行った。
【図4D】NBDを欠失した欠失変異型のIKKβ(del.NBD)を[35S]-メチオニンで標識し て(インプット)、GSTプルダウン解析に用いた。
【図4E】ヒトIKKβのCOOH末端(N721-S756)のフォーシャー−プリスカ(Fauchere-Pliska)疎水性プロットを、MacVector□(バージョン6.5.3)ソフトウェアを用いて作製した。NBD(L737-L742)は四角で囲んである。
【図4F】COS細胞に、48時間かけて、合計2μgのDNA/ウェルになるよう、ベクタのみ、ベクタにNEMO-FLAGを加えたもの、又は、NEMO-FLAGに、提示するようにNBD内に点変異を含有するxpressでタグを付けた形のIKKβ-(1-744)を加えたもの、でトランスフェクトした。溶解させ、抗FLAG(M2)を用いて免疫沈降させた後、抗FLAG又は抗xpressで免疫ブロット解析を行った。IP前のタンパク質発現レベルは、抗xpressで免疫ブロットして調べた(下側パネル)。
【図4G】ヒーラ細胞を、48時間かけて、pBIIX-ルシフェラーゼと一緒に、提示したコンストラクトで一時的にトランスフェクトして、ライセート中のNF-κB活性をルシフェラーゼ検定で測定した。
【図5A】図5は、IKKβ NBDを包含する細胞透過性ペプチドがIKKβ/NEMO相互作用、TNFαで誘導されるNF-κB活性化及びNF-κB依存性遺伝子発現を抑制することを示した実験結果を示す。図5A:GST-プル−ダウン解析を、GST-NEMOとインビトロで翻訳されたIKKβの組(上側パネル)か、又はGST-IKKβ-(644-756)とインビトロで翻訳されたNEMOの組(下側パネル)を用いて行った。この検定は、次第に濃度を上昇させた(125、250、500又は1000μM)の変異型(MUT)もしくは野生型(WT)NBDペプチドの不在下(ペプチドなし)又は存在下で行った。
【図5B】ヒーラ細胞はいずれかのペプチド(200μM)と一緒に、指定した時間、インキュベートした。溶解後、IKK複合体を、抗NEMOを用いて免疫沈降させ、その結果得られた免疫ブロットを抗IKKβでプローブした。
【図5C】ヒーラ細胞を、48時間かけて、pBIIX-ルシフェラーゼでトランスフェクトした後、変異型もしくは野生型NBDペプチド(それぞれ100及び200μM)の不在下(コントロール)又は存在下で、2時間、インキュベートした。その後、細胞をTNFα(10ng/ml)で提示したように処置する(左側パネル)か、又は、その後4時間、未処置(右側パネル)のままにしてから、NF-κβ活性化をルシフェラーゼ検定で測定した。
【図5D】ヒーラ細胞を3時間、次第に濃度を上昇させた(50、100又は200μM)の各ペプチドと一緒にインキュベートし、その後15分間、TNFα(10ng/ml)で、提示したように処置した(+)。溶解後、核抽出液を作製し、各試料からの10μgのタンパク質を、[32P]標識付κB部位プローブを用いたEMSAに利用した。
【図5E】始原HUVECを2時間、野生型(左側)又は変異型(右側)NBDペプチド(100μM)と一緒にプレインキュベートした後、TNFα(10ng/ml)でその後6時間、刺激した。コントロール細胞にはペプチドは与えなかった。細胞を抗E-セレクチン(H4/18)又は非結合性コントロール抗体(K16/16)で染色し、FACS(ベクトン・ディッキンソン社製、FACSort)で発現を測定した。輪郭は、同じ条件下でTNFα及びコントロール抗体の不在下(網掛け部分)及び存在下(実線)でのE-セレクチン染色を示す(断続線、TNFαなし;点線、TNFαあり)。
【図6A】図6は、野生型NBDペプチドが、NF-κBが誘導する遺伝子発現及び実験的に誘導した炎症を抑制することを示した実験結果を示す。図6A:マウスに賦形剤(VEH)、デキサメタゾン(DEX)又はNBDペプチドを局所的に処置して、PMA誘発性の耳水腫を起こして、実施例8で説明したように測定した。データは、耳の厚さの平均差±SD(未処置のコントロール[-]及び賦形剤[VEH]の両方に比較したときに*=p<0.05)で表されている。
【図6】ザイモサン(ZYM)でマウスに誘発した腹膜炎に及ぼす、デキサメタゾン(DEX)の作用に比較したときのNBDペプチドの作用を、実施例8で解説したように調べた。コントロールマウスには、リン酸緩衝生理食塩水(PBS)を注射した。
【図7】図7は、破骨細胞分化は野生型により用量依存的に抑制されたが、変異型NBDペプチドでは抑制されなかったことを示した実験結果を示す。データは、三重式試料±SDの平均値で表されている。
【図8A】図8は、ヒトIKKβのNEMO結合ドメイン(NBD)内のD738の変異解析結果を示す。図8A:IKKβの738位のアスパラギン酸残基をアラニン、アスパラギン又はグルタミン酸に、PCR変異誘発法を用いて置換した。
【図8B】図8Aで示したIKKβ(D738)変異体にインビトロ転写及び翻訳で35S-メチオニンで標識した後、前に解説されたようにGST-NEMOを用いたGSTプル−ダウン解析に用いた。
【図8C】Fugene6トランスフェクション法を利用して、上述のpcDNA-3、IKKβ又はD738変異体のいずれかと一緒にpBIIx-ルシフェラーゼを持つNF-κB依存性レポータ・コンストラクトで、ヒーラ細胞を一時的にトランスフェクトした。48時間後、細胞を溶解させ、ルシフェラーゼ活性を前に解説された通りに調べた。
【図9A】図9は、ヒトIKKβのNBD内のW739及びW741の変異解析結果を示す。図9A:IKKβの739位及び741位のトリプトファン残基を、アラニン、フェニルアラニン、チロシン又はアルギニンに、PCR変異誘発法を用いて置換した。
【図9B】FLAGでタグを付けたNEMOと一緒に、ベクタのみ(pcDNA-3.1-xpress)、IKKβ、W739A、W739F又はW739YでCOS細胞を一時的にトランスフェクトした。48時間後、細胞を溶解させ、抗FLAG(M2)共役アガロースビーズを利用して複合体を免疫沈降(IP)させた。免疫沈降前に、各ライセートの一部(5%)を解析用にとっておいた(pre-IP)。試料中のタンパク質をSDS-PAGE (10%)で分離し、FLAG(M2)又はxpressを認識する抗体を用いた免疫ブロッティング(IB)で解析した。上側二つのパネルは、xpressのタグが付いたIKKβを示し、下側のパネルはFLAGのタグが付いたNEMOを表す。
【図9C】COS細胞に図示のプラスミドでトランスフェクトした後、Bで解説したように免疫沈降及び免疫ブロット解析を行った。
【図9D】図9Cと同じく図示のプラスミドと一緒にpBIIx-ルシフェラーゼでヒーラ細胞を一時的にトランスフェクトし、48時間後にライセート中のルシフェラーゼ活性を調べた。
【図10A】図10は、ヒトIKKβのNBD内のS740の変異解析結果を示す。IKKβの740位のセリン残基をアラニン又はグルタミン酸に、PCR変異誘発法を用いて置換した。
【図10B】COS細胞に図示のプラスミドで一時的にトランスフェクトし、図2Bで解説したように免疫沈降及び免疫ブロット解析を行った。
【図10C】IKKβ-FLAG又はS740E-FLAGのいずれかでヒーラ細胞を48時間かけて一時的にトランスフェクトした後、TNFα(1μg/ml)で図示の時間、処置した。溶解後、抗FLAG(M2)共役アガロースビーズを用いて複合体を沈降させ、前に解説されたように、GST-IκBα(1-90)を基質として用いて免疫-複合体キナーゼ検定を行った。
【図11A】図11は、IKKα NBDの変異解析結果を示す。図11A:IKKαのNBDを含む各残基(L738乃至L743)を、PCR変異誘発法によりアラニンに置換した。COS細胞を、ベクタのみ(pcDNA-3.1-xpress)又はxpressでタグを付けた形のIKKα及びNBD変異体のいずれかと一緒にNEMO-FLAGで図示のように一時的にトランスフェクトした。このIKkα-NEMO複合体の免疫沈降及び免疫ブロット解析を図2Bで解説したように行った。
【図11B】ヒーラ細胞を、図示のプラスミドと一緒にpBIIx-ルシフェラーゼで一時的にトランスフェクトし、48時間後に、ライセート中のルシフェラーゼ活性を調べた。
【図12A】図12は、IKKβ NBDを包含するペプチドは、IKKαのNEMOとの相互作用を妨げることを実証した実験結果を示す。図12A:NBD野生型及びスクランブル型コントロールペプチドの配列。この野生型ペプチドは、IKKβの残基734から744に対応する。
【図12B】GST-NEMOと、インビトロで転写・翻訳したIKKα(上側パネル)及びIKKβ(中央のパネル)とを用いて、賦形剤(2%DMSO)、スクランブル型もしくは野生型NBDペプチド(各ペプチドを500及び1000μM)の存在下又は不在下でGSTプル−ダウン解析を行った。下側のパネルは、クーマシー・ブルー染色したゲルを示し、いずれのペプチドも、沈降に用いたグルタチオン-アガロースビーズとのGST-NEMOの相互作用に影響を与えなかったことを実証している。
【図12C】ある範囲の濃度の野生型NBDペプチドの存在下でGST-NEMOを用いてIKKα及びIKKβのGSTプル−ダウンを行った後に得たオートラジオグラフィのバンドの比重解析。挿入図は一実験例を示す。データは、ピクセル密度をコントロール(ペプチドなし)に対するパーセンテージで表しており、平均値±sd(n=11)で表している。解析はNIH-イメージソフトウェアを用いて行った。
【発明を実施するための形態】
【実施例】
【0143】
実施例1: IKKβ上のNEMO結合ドメインの同定
我々は、IKKβ上のNEMO相互作用ドメインを同定するために、インビトロ・プルダウン検定(Zhong et al., (1997) Cell 89, 413-424)を、NH2末端でグルタチオンS-トランスフェラーゼに融合させた完全長NEMO(GST-NEMO;図1A)の細菌発現版を用いて行った。IKKβの様々な機能的ドメイン(触媒ドメイン、ロイシンジッパー及びへリックス-ループ-へリックス;図1A)を欠失した多様な切断変異体が構築された。
【0144】
IKKα及びIKKβの完全長cDNAクローンのサブクローニング及び変異誘発はすべて、クローンPfu DNA-ポリメラーゼ(ストラータジーン社製)を用いたPCRで行った。野生型及び変異型IKKβcDNAを、pcDNA-3 又は pcDNA-3.1-xpress (インビトロジェン社製)のKpnI 及び NotI 制限部位に挿入し、 IKKαcDNAはすべて、同じベクタのEcoRI 及びXhoI 部位に挿入した。FLAGで標識した形の両キナーゼは、pFLAG-CMV-2 (シグマ社製)にサブクローンして構築した。GST-IKKβ-(644-756)については、このPCR断片を、 pGEX-4T1 (ファルマシア社製)のEcoRI 及びXhoI部位に挿入した。ヒトNEMOをコードする完全長cDNAは、逆転写酵素(RT)-PCRにより、ヒーラ細胞mRNAから、ExpandTM ロング・テンプレートPCRシステム(ベーリンガーマンハイム社製)及びプライマペア(5'-ATAGACGAATTCAATAGGCACCTCTGGAAG) (SEQ ID NO: 20) 及び(3'-TAGGACCTCGAGCTACTCAATGCACTCCATG) (SEQ ID NO: 21)を用いて得た。その結果得られたPCR断片を、pcDNA-3 又はpcDNA-s.1-xpressのEcoRI 及びXhoI 部位に挿入した。その後のNEMO変異体はすべて、PCRにより、Pfu DNA-ポリメラーゼを用いて構築した。GST-NEMO は、この完全長cDNAをpGEX-4T1のEcoRI 及び XhoI 部位にサブクローンして、構築した。
【0145】
これらの変異体には、インビトロ翻訳で [35S]-メチオニンの標識を付け、(インプット;図1B)、GST のみか、又はGST-NEMOと混合し、グルタチオン−アガロースを用いて沈降させた。変異体のいずれも、GSTのみとは相互作用しなかったが、一方、野生型と、IKKβのNH2末端切断型の三つすべて(307-756、458-756及び486-756)が、GST-NEMOと相互作用した(図1B(右側パネル)。対照的に、COOH-末端切断型変異体(1-456、1-605又は1-644)のいずれも、GST-NEMOで沈降しなかった。これらの結果は、NEMOが、へリックス-ループ-へリックス(HLH)ドメインから遠いIKKβのCOOH末端にある領域と、相互作用することを示している。次に、IKKβのアミノ酸644(HLHの直後)からCOOH末端(残基756)までの領域のみから成る変異体を構築した。図1Cに示すように、この変異体はGSTでは沈降しなかったが、GST-NEMOとは相互作用したことから、この領域が、これら二つの分子間の相互作用を媒介していることが裏付けられた。
【0146】
次に、IL-1β及びTNFαで誘導されるNF-κB活性化に対するIKKβ-(644-756)の作用を、NF-κB依存性レポータプラスミド(pBIIX-ルシフェラーゼ)と一緒に前記変異体でヒーラ細胞を一時的にトランスフェクトして調べた(Kopp & Ghosh, (1994) Science 265, 956-959)。トランスフェクション実験では、ヒーラ細胞及びCOS細胞を24ウェル(1×105細胞/ウェル)プレート又は6ウェル(5×105細胞/ウェル)プレートのいずれかに接種して、24時間成長させてから、Fugene6(ロシュ社製)でメーカのプロトコルに従ってDNAをトランスフェクトした。24ウェルプレート及び6ウェルプレートの細胞に、それぞれ合計1μg又は2μgのDNAを与えた。48時間後に細胞を TNT (200 mM NaCl、20 mM トリス-pH 8.0、1% トリトン-100)で溶解させ、そのライセートを用いて、免疫沈降又はルシフェラーゼ検定(プリメージ・ルシフェラーゼ検定系)を行った。
【0147】
図1は、これらのサイトカインにより誘導されるNF-κBの活性化を、IKKβ-(644-756)が用量依存的に抑制したことを示す。これらの結果では、コアIκB-キナーゼ複合体から内因性NEMOを滴定することにより、IKKβ-(644-756)がドミナントネガティブとして作用することを示す。NEMOによる調節タンパクの動員がないと、IL-1β及びTNFα誘導シグナルに対してIKKβは不応性となり、起きるはずのIKKβ活性化が起きない。構造的にはNEMOは、コイルド・コイル及びロイシン・ジッパーモチーフからなる二つの主要な部分を含有する長いαらせん領域と、COOH末端亜鉛フィンガードメインとから成る(図2A)(Mercurio et al., (1999) Mol. Cell. Biol. 19, 1526-1538; Yamaoka et al., (1998) Cell 93, 1231-1240; Rothwarf et al., (1998) Nature 395, 297-300)。NEMOがIKKβと相互作用するのに必要なその領域を特定しようとするこれまでの研究は、互いに矛盾する結果を出していた(Harhaj et al., (1999) J. Biol. Chem. 274, 15297-15300)。この疑問に対処するために、IKKβ-(644-756)(図2A)及びNEMOの多様な[35S]-メチオニン標識付き切断型変異体(図2A)のGST-融合タンパクを用いたGST-プル-ダウン検定を行った。図2A(右側パネル)は、これらの実験結果を要約したものであり、IKKβ-(644-756) はNEMO-(1-196)、-(1-302)及び-(44-419) とは相互作用するが、NEMO-(197-419) もしくは-(86-419)とはしないことが実証された。同一の結果が、FLAGでタグしたIKKβ及びこのNEMO変異体を一時的にトランスフェクトしたCOS細胞又はHEK293細胞のライセートを用いた免疫沈降実験でも得られた(データは図示せず)。
【0148】
すべての免疫沈降で、6ウェルプレートで成長させたヒーラ細胞又はCOS細胞は500μlのTNTで溶解させた。FLAGでタグを付けたタンパク質は、20 μlの抗FLAG (M2)-共役アガロースビーズ(シグマ社製)を用いて、4℃で2時間かけて、トランスフェクトした細胞のライセートから沈降させた。内因性IKKβ又はNEMOの免疫沈降は、1 μgの特異ウサギポリクローナル抗体(サンタクルズ社製)と、20μlのプロテインAセファロース(アマーシャム−ファルマシア社製)を用いて行った。免疫ブロットについては、沈降物をTNTで三回、PBSで二回、洗浄してから、SDS-試料バッファ中に懸濁させた。タンパク質をSDS-PAGE(10%)で分離し、PVDFメンブレンに写し取り、高画質化学発光法(アマーシャム−ファルマシア社製)で観察した。
【0149】
これらの結果から、NEMOの一番目のαへリックスを含む一領域である、残基44から86の間に相互作用ドメインがあることが確定された。こうして、一番目のコイルド・コイル−ドメインまでのαへリックスを欠失させた変異体(残基T50−L93;del.αH)を作製した。この変異体はIKKβ-(644-756)とは相互作用しなかった(図2B)。さらに、pBIIX-ルシフェラーゼを用いたトランスフェクション実験では、del.αHが、TNFαで誘導されるNF-κB活性を抑制したことが実証され(図2C)、IKKβとは相互作用できないNEMOのCOOH末端が、NF-κBのドミナントネガティブ抑制因子であることが裏付けられた(Mercurio et al., (1999) Mol. Cell. Biol. 19, 1526-1538; Rothwarf et al., (1998) Nature 395, 297-300)。まとめると、図1及び2は、IKKβ及びNEMOの間の相互作用は、IKKβのCOOH末端と、NEMOの一番目のαへリックス領域とを通じて起きることを示している。これらの発見から、NEMOのNH2末端がNEMOをIKK-複合体に繋ぎ止め、いくつかのタンパク質対タンパク質相互作用ドメインを含有するこの分子のその他の部分は剥き出しのままであり、IKK機能の上流の調節因子との相互作用に使われるのだ、ということが分かる。
【0150】
実施例2: NBDでの相互作用を通じたIKKβ機能のNEMO調節
IKKβのNEMO相互作用ドメインを十二分に特徴付けるために、残基V644からS756までの間の切断型変異体(図3A)をさらに構築した。HLHの直後から、662位のシステインまでのアミノ酸配列はIKKα(図3Aではα1と示した)と72%の同一性を示す。この後の、E707までの領域は、かつて、自己リン酸化のターゲットであり、炎症誘発性サイトカインによる刺激後のIKKβ活性の下方調節に機能すると報告されたことのあるセリンリッチなドメインである(Delhase et al., (1999) Science 284, 309-313)。この後に続く配列は733位まで、全くセリン残基を含まない。これらの領域のそれぞれを配列から削除した変異体を[35S]-メチオニンで標識し、上述したGST-プル-ダウン検定に用いた。図3Aは、これらの実験結果を要約したものであり、このIKKβ変異体のいずれもGST-NEMOで沈降しなかったことから、その相互作用ドメインは、残基F734及びS756の間の最先端のCOOH末端にあることが分かる。
【0151】
IKKβのこの短い部分を、IKKαの相当する領域に比較すると、驚くべき二つの構造上の特徴があることが分かる(図3B)。第一に、IKKβのF734からT744までの配列(図3Bのα2)は、IKKαの同等の配列(IKKβのL737からL742及びIKKαのL738からL743)と同一である。第二に、IKKβのこの配列は、IKKαのCOOH末端残基(E745)よりも12個のアミノ酸分、長く延びているが、この12個のアミノ酸は酸性の高い領域を含んで成り、そのうちの残基5個はグルタミン酸である(図3B)。IKKβのα2領域と、IKKαの最先端COOH末端との間に著しい類似性があることを、NEMOはIKKαとはインビトロで相互作用しないという以前の報告 (Mercurio et al., (1999) Mol. Cell. Biol. 19, 1526-1538; Yamaoka et al., (1998) Cell 93, 1231-1240; Rothwarf et al., (1998) Nature 395, 297-300)と合わせて考えると、NEMO相互作用ドメインは、IKKβのグルタミン酸リッチな部分(E745からS756)であろうという仮説が導かれた。
【0152】
この仮説を調べるために、この領域を削除した切断変異体を作製し(1-744;図3C)、GST-NEMOと相互作用するその能力について調査した。この変異体は、野生型IKKβと同程度に、GST-NEMOと結合した(図3C)。これらの結果は、一時的にトランスフェクトしたCOS細胞のライセートから、エピトープでタグを付けたNEMO及びIKKβ-(1-744)を同時免疫沈降させることにより、確認した。これらの発見は、IKKβのNEMO相互作用ドメインがこのCOOH末端のα2領域にあることを実証するものである。
【0153】
次にIKKβ COOH末端切断変異体を用いて、IKKβの基礎活性及び誘導後活性に対するNEMO結合の作用を調べた。図3Dは、IKKβをV644で切断して、セリンリッチな領域(図3Aを参照されたい)を除去したところ、基礎自己リン酸化反応が完全に失われたことを示している。対照的に、このセリンリッチな領域(1-733)を含有する変異体は、野生型IKKβよりもはるかに高いレベルの自己リン酸化を呈した(図3D)。興味深いことに、NEMO結合α2領域を含有するIKKβ-(1-744)の自己リン酸化レベルは、この野生型キナーゼで観察されるものと同一であった。これらの変異が基礎キナーゼ活性に及ぼす影響を調べるために、変異体をヒーラ細胞に一時的にトランスフェクトし、NF-κB活性を、実施例1で解説したようにルシフェラーゼ検定で調べた。図3Dに示すその結果は、IKKβ-(1-644)はNF-κB活性を誘導しなかったが、IKKβ-(1-733)は野生型(図3E)よりも高い活性化を引き起こしたことを示す。さらに、IKKβ-(1-744)が誘導するNF-κB活性は、野生型IKKβが誘導するそれと同一であった。
【0154】
これらの結果は、IKKβの基礎自己リン酸化及びキナーゼ活性は、NEMOがこのキナーゼと結合する能力に依存していることを、実証している。これらの観察の解釈の一つは、NEMOがホスファターゼをIKK-複合体に動員して、このCOOH末端のセリンリッチな領域を狙わせることで、基礎IKKβ機能を調節しているのではないか、という解釈であろう。従って、NEMOを結合させられないとホスファターゼを動員できず、この領域のリン酸化が増加することになる。
【0155】
α2領域の消失が、IKKβの触媒活性に及ぼす作用を直接調べようと、免疫−複合体キナーゼ検定を、トランスフェクト後のヒーラ細胞から採ったライセートで行った(図3F)。免疫−複合体キナーゼ検定については、沈降物をTNTで洗浄してから、さらにキナーゼバッファ (20 mM HEPES pH 7.5、20 mM MgCl2、1 mM EDTA、2 mM NaF、2 mM β-グリセロホスフェート、1 mM DTT、10μM ATP)で洗浄した。次に、沈降物を、15分間、30℃で、 GST-IκBα-(1-90)及び10μCi [32P]-γ-標識ATP (アマーシャム・ファルマシア社製)を含有する20μlのキナーゼバッファ中でインキュベートした。基質をグルタチオン−アガロース(アマーシャム・ファルマシア社製)を用いて沈降させ、SDS-PAGE (10%)で分離した。キナーゼ活性はオートラジオグラフィで調べた。オートラジオグラフィでは、リン酸化したタンパク質がキナーゼ複合体と結合して見えたが、それは、免疫沈降させた複合体を、GST−基質沈降の前に取り除かなかったからである。IKKβ(野生型)の活性は、非処置細胞では低かったが、TNFαで処置後は著しく上昇した。図3Eに示したデータと合致して、IKKβ-(1-733)の基礎活性は野生型よりも著しく高かったが、この活性は、TNFαで処置してもそれ以上、上昇しなかった(図3F)。さらに、IKKβ-(1-744)の基礎触媒活性及びTNFαによる誘導後の触媒活性は、IKKβ(WT)の活性と同一であった。リン酸化GST-IκBαに加え、自己リン酸化IKKβも検出された(図3F、上のバンド)。TNFα処置後、IKKβ(WT)及びIKKβ-(1-744)は急速に自己リン酸化したが、既に高い値となっているIKKβ-(1-733)の基礎リン酸化は僅かに上昇したのみだった(図3F)。以前の研究では、自己リン酸化は、このタンパク質にコンホメーション上の変化を起こすことで、TNFαが誘導するIKKβ活性を下方調節するよう働くと示されている (Delhase et al., (1999) Science 284, 309-313)。まとめると、これらの発見(図3D−F)は、NEMOが存在しないとき、IKKβは自己リン酸化して基礎活性となり、TNFαで誘導されるシグナルに対して不応性となることを実証するものであり、NEMOが、IKKβ活性の活性化だけでなく、下方調節にも基本的な役割を果たしていることを示している。
【0156】
ある一つのリン酸化タンパク質を表す別のバンドが、TNFαで誘導したIKKβ(WT)細胞及びIKKβ-(1-744)トランスフェクト細胞から採った試料中にのみ、現れた(図3F)。このタンパク質の分子量(49kD)から、これが、沈降した複合体と結合した内因性NEMOであることが明確に分かる。このことは、IKKβ-(1-733)トランスフェクト細胞のいずれの沈降物(+/-TNFα)にも、このバンドがないことが裏付けとなっている。この沈降複合体をSDSで解離させ、特異的抗NEMO抗体を用いて[32P]標識付きNEMOを再沈降させることで、このタンパク質をリン酸化NEMOと同定した。従って、NEMOの誘導性リン酸化は、IKK複合体の更なる活性調節段階であるのかも知れない。
【0157】
実施例3: IKKA上のNBDの同定
IKKβのα2領域はIKKαのCOOH末端に酷似しているため(図3B)、IKKαがNEMOと相互作用する能力を調べた。
【0158】
xpressでタグを付けたNEMOと、FLAGでタグを付けた形のIKKαもしくはIKKβのいずれかとを一緒に一時的にトランスフェクトしたCOS細胞のライセートの免疫沈降を抗FLAG を用いて実施例1で解説したように行った。図4Aは、NEMOが、IKKβ及びIKKαの両方と同等に良好に相互作用したことを示す。この実験では、IKKαとの相互作用が直接的なものではなく、代わりに、内因性IKKβ、FLAG-IKKα及びxpress-NEMOを含有する複合体が形成されたことが原因である知れない可能性がある。従って、GST-プル-ダウン検定を、GST-NEMOと、[35S]-メチオニン標識した形の、野生型IKKαか、もしくは8個のCOOH末端アミノ酸を欠失させた切断型IKKα変異体(1-737):図4B)とを用いて、行った。上に記載した発見と合致して(図4A)、しかし以前の報告(Mercurio et al., (1999) Mol. Cell. Biol. 19, 1526-1538; Yamaoka et al., (1998) Cell 93, 1231-1240; Rothwarf et al., (1998) Nature 395, 297-300)とは対照的に、野生型IKKαはNEMOとインビトロでも相互作用したが、切断型変異体はしなかった(図4B)。これらの結果は、IKKαがNEMOと相互作用することを実証するだけでなく、IKKαとIKKβのα2領域に共通の、6個のアミノ酸を含有するCOOH末端領域を介してこの相互作用が行われていることも、示している(図3B)。遺伝子ターゲティング研究では、TNFαによるIKKα及びIKKβの活性化に、奥深い違いがあることが実証されている (Woronicz et al., (1997) Science 278, 866-869; Zandi et al., (1997) Cell 91, 243-252; Mercurio et al., (1997) Science 278, 860-866; DiDonato et al., (1997) Nature 388, 548-554; R gnier et al., (1997) Cell 90, 373-383)。
【0159】
本発見は、この違いの根底にあるのが、NEMOの動員の違いではないことを示唆している (Delhase et al., (1999) Science 284, 309-313; Takeda et al., (1999) Science 284, 313-316; Hu et al., (1999) Science 284, 316-20; Li et al., (1999) Science 284, 321-325; Li et al., (1999) J. Exp. Med. 189, 1839-1845; Li et al., (1999) Genes Dev. 13, 1322-1328; Tanaka et al., (1999) Immunity 10, 421-429)。反対に、この違いは、各キナーゼが、NEMO結合シグナリング成分を活性化応答に向けて、おそらくは個々のキナーゼの本来の調節的特徴の違いを通じて、動員する能力に、あるのではないかと思われる。
【0160】
この短いCOOH末端配列が、IKKのNEMO相互作用ドメインを構成していることの更なる証拠は、最近解説されたIKK関連キナーゼIKKi (Shimada et al., (1999) Int. Immunol. 11, 1357-1362)がNEMOと相互作用する能力を我々が調べたときに、得られた。IKKα及びIKKβの配列を比較すると(Shimada et al., (1999) Int. Immunol. 11, 1357-1362; Woronicz et al., (1997) Science 278, 866-869; Zandi et al., (1997) Cell 91, 243-52; Mercurio et al., (1997) Science 278, 860-866; DiDonato et al., (1997) Nature 388, 548-554; Regnier et al., (1997) Cell 90, 373-383)、IKKiは、そのCOOH末端にα2領域を含有していないことが分かり(Shimada et al., (1999) Int. Immunol. 11, 1357-1362)、これがNEMO結合ドメインであるということと一致して、我々は、IKKiが、GST-NEMOとは相互作用しないことをプル-ダウン検定で見いだした(図4C)。この発見は、NEMOはIKKiの機能的活性には必要ではないことを示唆しており、これは、IKKiがTNFαまたはIL-1βの誘導するシグナルのいずれにも応答できないことでも、裏付けられている(Shimada et al., (1999) Int. Immunol. 11, 1357-1362)。
【0161】
実施例4: NBD中のアミノ酸残基の変異
(NEMO結合ドメイン(NBD)と指定する)IKKβのα2領域と、IKKαの同等の6個のアミノ酸配列とが、NEMOとの相互作用にとって重要であると確信したところで、L737からL742までの6個のアミノ酸を欠失させたIKKβの欠失変異体(del.NBD)を構築した。この欠失変異体はGST-NEMOと結合しなかった(図4D)。NBDの予測上の構造的及び生化学的特徴を、周囲の残基との関連で調べたところ、これが、IKKβ COOH末端の親水性領域内で、柔軟性のない疎水性の「ポケット」を構成していることが示唆された(図4E)。このことは、NBDが、結合したNEMOの一番目のαへリックス領域内に埋没してしまい(図2)、水性の環境にこれが曝されるのが妨げられて、強力な分子間相互作用を維持する、というモデルを示唆している。この相互作用が実際、この疎水性部分の機能であるかどうかは、まだ調べなければならないが、W739又はW741のいずれかをアラニンに置換すると、NEMOのIKKβとの結合が妨げられることを我々は見いだした(図4F)。従って、これらの疎水性トリプトファン残基のそれぞれが、機能的NBDを維持するのに重要なのである。加えて、D737がアラニンに変異しても、NEMOの相互作用が妨げられたことから、この位置に負電荷の残基があることがNEMO機能にとって必要であることも、分かった。これらの変異とは対照的に、L737、S740又はL742をアラニンに置換しても、NEMO結合には影響を与えなかった。これらのアラニン置換がIKKβ機能に及ぼす作用を調べるために、ヒーラ細胞に、前記点変異体のそれぞれを、pBIIX-ルシフェラーゼレポータと一緒に同時トランスフェクトした。IKKβの基礎活性は結合NEMOのないときの方が高いという観察と合致して、NEMOに結合しないIKKβ-(1-733)変異体(図3E)(D738A、W739A及びW741A)は、野生型IKKβ又はIKKβ-(1-744)(図4G)よりも、より高いレベルにNF-κBを活性化した。対照的に、NEMO結合を損なわなかった置換を含有する変異体 (L737A、S740A及び L742A) は、コントロールと同じレベルまで、NF-κBを誘導した。これらの結果から、NEMOは、内因性IKKβ活性の下方調節に重要な役割を果たしていることが分かる。
【0162】
実施例3で解説したGSTプルダウン検定を利用して、NBDでの変異についてさらに解析を行い(表1を参照されたい)、NEMOのIKKβへの結合に及ぼすその影響ついて調べた。
【0163】
【表1】

【0164】
実施例5: NBDと相互作用してNEMO結合を遮断する作用薬
NBDは比較的に小さいため、コアIKK複合体の破壊を狙いとした化合物の開発にとって、魅力的なターゲットとなっている。IKKβNBDに延びる細胞透過性ペプチドを設計し、それらがIKKβ-NEMO相互作用を解離させる能力を調べることで、この方法の今日性を調査した。
【0165】
この研究で用いた二つのNBDペプチドの配列は[DRQIKIWFQNRRMKWKK]TALDWSWLQTE (野生型) (SEQ ID NO:18) 及び [DRQIKIWFQNRRMKWKK]TALDASALQTE (変異型) (SEQ ID NO:19)であった。アンテナペディア・ホメオドメイン(antennapedia homeodomain)配列 (Derossi et al., (1994) J. Biol. Chem. 269, 10444-10450; 米国特許第5,888,762号;米国特許第6,015,787号;米国特許第6,080,724号)を括弧内に入れ、WからAへの変異位置に下線を加えた。両ペプチドともDMSOで溶解させて、ストック濃度20 mMとした。すべての実験で、DMSO単独のコントロールは、ペプチドなしコントロールから差がなかった。
【0166】
野生型NBDペプチドは、IKKβのT735からE745までの領域を、膜移行を媒介することが示されているアンテナペディア・ホメオドメインの第三へリックス(Derossi et al., (1994) J. Biol. Chem. 269, 10444-10450)を由来とする配列に融合させたものから構成した。変異体は同一であるが、例外としてNBDのトリプトファン残基(W739及びW741)をアラニンに変異させてあった。図5Aは、野生型のNBDが、GST-NEMOによる[35S]-標識付IKKβのプルダウン及びGST-IKKβ-(644-756)による[35S]-標識付NEMOのプルダウンを、インビトロで用量依存的に抑制し、変異型は抑制しなかったことを示す。このNBDペプチドが細胞内へ進入してIKKβ-NEMO相互作用を抑制する能力について調べようと、ヒーラ細胞をこれらペプチドと一緒に様々な時間、インキュベートし、抗NEMOを用いてそのIKK複合体を免疫沈降させた。インビトロでのデータ(図5A)と合致して、野生型は内因性IKK複合体の形成を損なったが、変異型は損なわなかった(図5B)。
【0167】
実施例6: NEMO機能を遮断する作用薬
次に、シグナルにより誘導されるNF-κBの活性化に及ぼすこのNBDペプチドの作用を調べた。ヒーラ細胞にpBIIX-ルシフェラーゼレポータをトランスフェクトした後、細胞を野生型又は変異型ペプチドと一緒にプレインキュベートし、TNFαで処置し、NF-κB活性化をルシフェラーゼレポータ検定で測定した。図5C(左側パネル)に示すように、野生型NBDペプチドは、TNFαで誘導されるNF-κB活性化を抑制したが、変異型では何の作用もなかった。興味深いことに、基礎NF-κB活性は、野生型ペプチドで処置すると上昇した(図5C;右側パネル)、これは、前の変異解析(図3E−F及び4G)の結果と一致する発見である。このことは、NEMOを除去すると、IKKの基礎内因性活性が上昇するが、TNFαに対するその応答性は喪失させることを、示している。電気泳動度シフトアッセイ(EMSA)を用いた別の解析でも、野生型NBDペプチドだけが、TNFαで誘導される活性化やNF-κBの核内移行を抑制したことを実証できた(図5D)。まとめると、これらの結果は、インタクトNBDペプチドが細胞内へ送達されると、IKKβ-NEMO間の相互作用が損なわれて、活性化NF-κBEからの炎症誘発性シグナルが妨げられることを実証するものである。対照的に、NEMO相互作用を維持するのに重要なトリプトファン残基に変異を持つペプチドによる伝達では、何の作用もないのである。
【0168】
実施例7: E-セレクチンを下方調節できる作用薬
炎症性応答の開始及び維持に関与するタンパク質の多くにとって、それらの遺伝子発現が誘導されるには、NF-κBが活性化することが必要である(Ghosh et al., (1998) Annu. Rev. Immunol. 16, 225-260; May & Ghosh, (1998) Immunol. Today 19, 80-88)。このようなタンパク質の一つであるE-セレクチンはIL-1又はTNFαなどの炎症誘発性刺激による活性化後の血管内皮細胞の管腔表面に発現する白血球接着分子である (Pober et al., (1986) J. Immunol. 436, 1680-1687; Bevilacqua et al., (1987) Proc. Natl. Acad. Sci. USA 84, 9238-9242; Collins et al., (1995) FASEB J. 9, 899-909)。E-セレクチン及び他のNF-κB依存性接着分子の発現は、急性及び慢性炎症の部位に白血球が捕獲及び動員されるのに、重要である。前記NBDペプチドの抗炎症能を評価するために、始原ヒト臍静脈内皮細胞(HUVEC)を野生型及び変異型ペプチドで予備処置し、E-セレクチンの発現をTNFαで誘導した。基礎NF-κB活性化に及ぼす作用と一致して(図5C)、野生型NBDペプチドは低レベルのE-セレクチン発現を誘導した(図5E)。しかし、TNFαで処置後は、この野生型ではE-セレクチンの発現が著しく減少したが、変異型では減少しなかった(図5E)。野生型NBDペプチドによる抑制により、TNFαのない状態でこのペプチドが誘導するレベルまで、発現が減少した。
【0169】
本発明の重要性は二つのレベルで見ることができる。第一に、出願人は、NEMOがIKKと結合するのに必要な構造上の要件を特定し、IKKβとの結合は、NBD内の三つのアミノ酸(D738、W739及びW741)に依存していることを見いだした。さらに、NEMOは、IKKβの活性化で機能するだけでなく、IKK複合体の本来の基礎活性を抑制する上でも重要な役割を有するのである。二番目のレベルの重要性は、NBDをターゲットとする薬物の臨床上の明かな用途である。出願人は、NBDを包含する細胞透過性ペプチドは、TNFαで誘導されるNF-αB活性化を抑制できるだけでなく、始原ヒト内皮細胞で、NF-κB依存性標的遺伝子であるE-セレクチンの発現を減少させることも、実証した。NBDは僅かに6アミノ酸長であり、従ってコアIKK複合体を損なうペプチドミメティック化合物を設計する方法は当業者の技術範囲内にある。複合体を破壊するこの作用は、IKKの基礎活性の上昇にあるため、NBDをターゲティングする化合物で処置すると、NF-κBの活性を完全に遮断する投薬をしたときに起きるであろう、肝細胞アポトーシスを原因とするものなど、毒性の問題を避けることができる。このように、NBDの同定は、IKK複合体に活性化シグナルが達しないようにし、と同時に低レベルのNF-κB活性は維持しつつも、毒性の副作用が生じる可能性を避けた新規な抗炎症薬物を開発する手段である。
【0170】
実施例8: インビボにおける炎症性応答の、NBDペプチドを媒介とする抑制
急性炎症反応の二つの異なるモデルを用いて、動物における炎症性応答を抑制する能力について、当該NBDペプチドを調べた。一番目のモデルでは、ホルボール-12-ミリステート-13-アセテート(PMA)を用いてマウスの耳に水腫を起こさせ、NBDペプチドを局所投与してその作用を測定した。PMAを用いた耳水腫は、以前に解説された(Chang et al., (1987) Eur. J. Pharmacol. 142, 197-205)通りに、年齢及び性別を同じにしたマウスの複式のグループで起こした。20μlのPMA (5μg/耳)エタノール 溶液の投与の30分前及び30分後に、20μlのNBDペプチド(200 μg/耳)、デキサメタゾン (40μg/耳)又は賦形剤(DMSO:エタノール;25:75 v/v) をマウスの右耳に局所投与した。PMA投与後6時間で、耳の膨潤をマイクロゲージを用いて測定し、処置(右)側耳及び非処置(左)側耳の間の厚さの違いの平均値として表した。このデータの統計学的解析は、スチューデントのt検定を用いて行った。p<0.05の値を統計上有意とみなした。
【0171】
図6Aは、野生型ペプチドが、PMAで誘導される耳の肥厚を著しく減少させ(77±3%抑制;p<0.05)、デキサメタゾンで観察されるレベル(82±9%抑制;p<0.05)まで下げたことを示す。対照的に、等量の変異型で観察される作用は、有意ではなかった(p= 0.09)。いずれのペプチドも、PMAなしで投与したときには何ら作用はなかった(図示せず)。
【0172】
二番目のモデルでは、ザイモサンを単独か、又は、デキサメタゾン又はNBDペプチドと組み合わせて腹腔内(i.p.)注射して、マウスに腹膜炎を起こさせた。ザイモサンで起こした腹膜炎では、前に解説された(Getting et al., (1998) Immunology 95, 625-630)ように、年齢及び性別を合わせた複式グループのマウス (C57BL/6NCR)で、腹腔滲出液を測定し、炎症性細胞の採集を行った。一部分の動物には、1mlのザイモサン(1mg/ml)と、デキサメタゾン(100mg/ml)又はNBDペプチド(200mg/ml)のいずれかとを同時に注射した。炎症性滲出液中のNOX(硝酸及び亜硝酸化合物)の濃度を、メーカのプロトコルに従って比色検定キット(アレキシス・コーポレーション社製)を用いて測定した。
【0173】
図6Bに示すように、ザイモサンの注射により、炎症性滲出液が蓄積し、多核白血球(PMN)が、これらの動物の腹膜に移動した。マウスを野生型NBDペプチド又はデキサメタゾンで処置した場合、滲出液形成及びPMN蓄積が著しく減少したが、変異型では何ら影響はなかった。
【0174】
多様なインビボ研究で、滲出液形成及び白血球の炎症部位への移行に、NOが役割を果たしていることが実証されている(Ialenti et al., (1992) Eur. J. Pharmacol. 211, 177-182; Ialenti et al., (1993) Br. J. Pharmacol. 110, 701-706; Iuvone et al., (1998) Br. J. Pharmacol. 123, 1325-1330)。従って、ザイモサンで処置したマウスの腹腔滲出液における、NBDペプチドのNOX蓄積に対する作用を調べた。図6C(下側パネル)は、デキサメタゾン及び野生型ペプチドがそれぞれ86±7%及び66±4%、NOXを減少させたが、変異型では何の作用もなかったことを示す。これらの結果は、滲出液形成及び細胞蓄積の減少が、NF-κB活性化の抑制と、NO形成の減少とに密接に相関していることを実証したこれまでの研究と合致している(D’Acquisto et al., (1999) Eur. J. Pharmacol. 369, 223-236; D’Acquisto et al., (1999) Naunyn-Schmeideberg’s Arch. Pharmacol. 360, 670-675)。従って、野生型NBDペプチドは、実験動物モデルでは、炎症の有効な抑制物質である。
【0175】
実施例9: NBDペプチドによる破骨細胞分化の抑制
骨の形態発生及びリモデリングのプロセスには、骨芽細胞による骨マトリックスの合成と、破骨細胞による骨再吸収との間にバランスが維持されることが必要である(Suda et al., (1992) Endocr. Rev. 13, 66-80; Suda et al., (1999) Endocr. Rev. 20, 345-357)。骨再吸収中の破骨細胞は、骨髄前駆細胞と、コロニー刺激因子(CSF-1)、インターロイキン-1(IL-1)、腫瘍壊死因子-α(TNF-α)、IL-6及びIL-11を含め、様々な段階で破骨細胞分化に影響を与える(Suda et al., (1992) Endocr. Rev. 13, 66-80; Suda et al., (1999) Endocr. Rev. 20, 345-357)多様な可溶性因子とから分化する多核巨細胞である。破骨細胞発生にとって重要な因子の一つは、RANKL(NF-κBリガンドの受容体活性化物質)という名称の最近解説された分子であり、この分子は、ODF (破骨細胞分化因子)、OPGL(オステオプロテゲリンリガンド)及びTRANCE (TNF-関連活性化により誘導されるサイトカイン)(Kong et al., (1999) Nature, 397, 315-323; Lacey et al., (1998) Cell 93, 165-176; Suda et al., (1999) Endocr. Rev. 20, 345-357; Wong et al., (1999) J. Leukoc. Biol. 65, 715-724; Yasuda et al., (1998) Proc. Natl. Acad. Sci. USA 95, 3597-3602)としても公知である。RANKLの受容体は、RANK(NF-κBの受容体活性化物質)という名称のTNF受容体ファミリーの仲間であり (Anderson et al., (1997) Nature 390, 175-179; Dougall et al., (1999) Genes Dev. 13, 2412-2424)、RANKLが結合すると、NF-κB活性化が誘導される(Anderson et al., (1997) Nature 390, 175-179; Darnay et al., (1998) J. Biol. Chem. 273, 20551-20555; Darnay et al., (1999) J. Biol. Chem. 274, 7724-31; Suda et al., (1999) Endocr. Rev. 20, 345-357; Wong et al., (1998) J. Biol. Chem. 273, 28355-28359)。さらに、破骨細胞の分化はNF-κB活性化にも依存しており、遺伝子ターゲティング研究では、成熟破骨細胞は、p50及びp52NF-κBサブユニット欠損マウスでは発生できないことが実証されている(Franzoso et al., (1997) Genes Dev. 11, 3482-3496)。
【0176】
骨粗鬆症は、破骨細胞に依存する骨再吸収が媒介する広汎な骨量減少を特徴とする重篤な衰弱性疾患である(Suda et al., (1992) Endocr. Rev. 13, 66-80; Suda et al., (1999) Endocr. Rev. 20, 345-357)。従って、NF-κBの活性化を破骨細胞前駆細胞で選択的に抑制すると、破骨細胞分化が妨げられて、骨粗鬆症の治療のための治療上有効な薬物の基礎となる可能性がある。よって、本NBDペプチドが破骨細胞分化に及ぼす作用を、以前に解説されたインビトロモデル(Jimi et al., (1999) Exp. Cell Res. 247, 84-93)を利用して調べた。48ウェル組織培養プレートに接種したマウス骨髄細胞を、ヒトマクロファージコロニー刺激因子 (M-CSF; 20 ng/ml)及びヒトRANKL(100 ng/ml)と一緒に、6日間、様々な濃度(6.25、12.5及び25 mM)の変異型もしくは野生型NBDペプチドの不在下又は存在下でインキュベートした。その後これらの細胞を固定し、破骨細胞表現型マーカである酒石酸塩耐性酸性ホスファターゼ(TRAP)で染色し、4つ以上の核を含有するTRAP陽性多核性細胞を破骨細胞として計数した。三重複試料を計数し、結果を平均値±SDで計算した。図7に示すように、野生型ペプチドは破骨細胞分化を用量依存的に抑制したが、変異型ペプチドはしなかった。
【0177】
このデータは、NF-κB活性化を抑制する細胞透過性NBDペプチドで、コアIKK複合体を破壊すると、RANKLで誘導される破骨細胞分化が妨げられることを実証しており、NBDを特異的に狙う薬物は、骨粗鬆症の治療にとって有効であろうことを示唆している。これらのインビトロ研究の延長として、同じペプチドを、インビボで骨粗鬆症に及ぼす作用について解析することもできる。重篤な骨粗鬆症を呈する卵巣切除マウス(チャールズ・リバー・ラブズ)を、このNBDペプチドで処置し、処置期間中の骨密度に対する作用を調べる。
【0178】
実施例10: NF-κBが媒介する他の疾患に対するNBDペプチドの作用
加えて、NBDペプチドが喘息に及ぼす作用を調べることも可能である。細気管支上皮細胞、T細胞及び細気管支マクロファージでのNF-κB活性化が、喘息患者及び喘息のモデル動物の気道で観察されている(Ray & Cohn, (1999) J. Clin. Invest. 104, 985-993; Christman et al., (2000) Chest 117, 1482-1487)。さらに、喘息を誘発する物質の多くはNF-κB活性化を引き起こし、喘息に関与するタンパク質(例えば白血球接着分子、多種のケモカイン、誘導性酸化窒素シンターゼなど)をコードする遺伝子の多くがNF-κB依存性である。樹立された喘息のマウスモデル(Kleeberger et al., (1990) Am. J. Physiol. 258, 313-320)を用いて、NBDペプチドを噴霧投与したときの、喘息に関連するこれらの状態の進行への作用を調べることもできる。同様な態様で、敗血症性ショックに対するNBDペプチドの作用も測定することができる。敗血症性ショックには、リポ多糖体(LPS)などの細菌性エンドトキシンが誘導する多くのNF-κB依存性遺伝子(即ちTNF、IL-1)の発現が関与している。LPSは、グラム陰性細菌の細胞壁の主要な構成成分を成し、免疫原性が高く、内因性発熱物質IL-1及びTNFの産生を刺激する(Sell et al., (1996) Immunology, Immunopathology & Immunity, Appleton & Lange)。敗血症性ショックに対するNBDペプチドの作用を調べるには、マウスにNBDペプチド及びLPSを注射し、動物の生存率を評価する。
【0179】
実施例11: NEMOとの相互作用におけるNBD内の各アミノ酸の相対的寄与及び重要性 上述の実施例で示したように、IKKα及びIKKβのNEMO結合ドメイン(NBD)は、両キナーゼのC末端最先端にある6個の保存されたアミノ酸(IKKβのL737 から L742及びIKKαのL738からL743)から成る。上記のNBD中の各アミノ酸がNEMOとの相互作用に対して持つ相対的寄与及び重要性をより明確に理解しようと、この実験を行った。各残基を多様な保存的アミノ酸及び非保存的アミノ酸に置換して、IKKβNBDの広汎な変異解析を行った。
【0180】
ロイシン残基(L737又はL742)又はセリン740のいずれを置換しても、NEMOのIKKβとの結合には影響を与えなかったことは、これらの残基のいずれも、この相互作用を維持するのに重要な役割は果たしていないと判断した。これらのアミノ酸に複数の変異があると結合に影響するかを調べるために、L737 及びS740 か、S740及びL742のいずれかをアラニンに置換した(それぞれLS及びSLと命名)変異体を構築した。GSTプル−ダウン及びCOS細胞トランスフェクション−免疫沈降−イムノブロット解析を行ったところ、LS及びSL変異体の両方とも、野生型IKKβと同程度に、NEMOと結合することが判明したため、これらの残基がこの相互作用に大きくは寄与していない証拠が新たに得られた。さらに、LS及びSLの両方とも、一時的トランスフェクション検定でNF-κB依存性ルシフェラーゼレポータ・コンストラクト(pBIIx-ルシフェラーゼ)の活性化で測定すると、IKKβだけでなくNF-κBも活性化する。両ロイシン残基(LL)の二重アラニン変異体や、三重変異体(LSL)を作れば、NEMO結合に対するこれらの残基の重要性に関する前記データを裏付けるのに役立つであろう。
【0181】
NEMO結合又はNF-κB活性化のいずれにも、上記の変異は作用を及ぼさないのと対照的に、NBD内のアスパラギン酸残基(D738)をアラニンに置換すると、IKKβのNEMOへの結合が妨げられた。さらに、この置換により、IKKβの基礎NF-κB活性化能力が2乃至3倍に上昇した。これらの結果は、IKK複合体の基礎活性を維持するのに、NEMO結合が一定の役割を果たしていることを実証するものである。興味深いことに、ヒーラ細胞を細胞透過性NBDペプチドで処置しても、基礎NF-κB活性はある程度、上昇したことから、NEMO結合が失われると基礎IKK活性が上昇することが、さらに裏付けられた。
【0182】
NBD内の738位の残基の性質を調べるために、アスパラギン酸をアスパラギン(D738N)又はグルタミン酸(D738E;図8A)のいずれかに置換した。これらの保存的置換は、この位置にある残基の形状(N)又は形状及び電荷(E)のいずれかを維持するものである。図8Bに示すように、いずれの置換も、IKKβのNEMO結合能には影響を与えなかったが、アラニン置換では結合が妨げられた。これらのデータは、IKKβ及びNEMO間の相互作用にとって重要なのは、この位置にあるアミノ酸側鎖の電荷ではなく形状(特に二番目の炭素の存在)であることを、実証している。ここに開示した前述の観察と一致して、いずれの変異も、IKKβの基礎活性には影響を及ぼさなかったが、アラニンでの置換では、活性が上昇した(図8C)。
【0183】
上述したように、NBD内のトリプトファン残基(W739及びW741)は両方とも、NEMOとの相互作用を維持するのに重要である。これらの位置にある残基の芳香族の構造を維持するような保存的置換の影響を、フェニルアラニン(F)又はチロシン(Y)のいずれかにトリプトファンを置換して、調べた(図9A)。さらに、両トリプトファンをアルギニンに変異させたが、これは、最も普通に天然で起きるトリプトファン変異である、コーディングコドン内に一箇所の塩基の変化だけを要する非保存的置換である。図9Bに示すように、W739F及びW739Y変異体の両方とも、IKKβと同程度にNEMOと結合したが、W739Rは結合しなかった(図9C)。アラニン置換の作用(図9B)と合わせると、これらの発見は、この位置にある残基の芳香族性が、NBDの機能にとって重要であることを示している。W739と同様、W742をフェニルアラニン(W742F)に置換すると、NEMOとの結合に影響は出なかったが、アルギニン(W742R)に変異させると、結合が妨げられた(図9D)と判断した。W739とは対照的に、チロシン(W742Y)に置換すると、NEMOとの結合が妨げられたことから、この位置にあるアミノ酸側鎖に水酸基部分が存在すれば、NEMOの結合を妨げるのに充分であることが実証された。この発見は、NBD内のある残基が、たとえそれが人工的に挿入されたアミノ酸であっても、リン酸化すれば、IKKβのNEMOとの結合が妨げられることを示唆していることになろう。
【0184】
実施例12:致死量リポ多糖体マウスモデルにおける抗炎症化合物の評価
この実験では、本発明の抗炎症化合物が、致死量のリポ多糖体(LPS)を与えたマウスを救助する能力を評価した。LPSは、敗血症で見られる、死亡を含む多くの応答を誘発する細菌生成物である。このモデルでは、サルモネラ−チフィムリウム(Salmonella typhimurium)LPSのリン酸緩衝血清(PBS)溶液30mg/kg(600μg/20gマウス)を、オスのC57BL/6マウスに静注投与した。この用量は、これを投与したマウスの100%で致死量であることをコントロール実験で確定された量である。マウスは、このLPS注射直後と、LPS注射から24時間後に、テストペプチドを静注(1%ジメチルスルホキシドのPBS溶液に溶かして)して処置した。LPS投与後最長で8日間、一日当たり2回、マウスを観察し、マウスの生存期間及び生存数を記録した。
【0185】
この研究結果を表2に示すが、同表では、各ペプチド及び用量について、投薬したマウスの合計数及び5日後の生存マウス数を示す。
【0186】
【表2】
【0187】
表2に示した結果は、このモデルでは、化合物2、3、及び5−11を5mg/kgの用量で静脈投与したときに、致死量のLPS投与からの有意な保護があったことを実証している。また化合物10は1mg/kg静注の用量でも有意な保護をもたらした。
【0188】
実施例13: コンカナバリンA誘発性肝炎におけるペプチドの評価
この実験では、本発明の抗炎症化合物が、コンカナバリンAで誘発した肝炎からマウスを救助する能力。
【0189】
コンカナバリンAは糖質に結合する種類のタンパク質であるレクチンである。あるタンパク質の一部が糖質である場合、このレクチンはそのタンパク質に結合する。コンカナバリンAは、細胞表面にあるタンパク質に結合することで、Tリンパ球を含む数多くの細胞を刺激する。これらのTリンパ球は、コンカナバリンAによる刺激で放出される他の媒介物質と協働して、やはりコンカナバリンAを結合させた肝細胞を攻撃して、肝細胞を死滅させる。Tリンパ球のこのような関与により、このモデルは、ヒトのウィルス感染に似たものとなっている。しかしながら、この急性モデルでは、TNFα応答もその一部を成す。
【0190】
体重18g乃至22gの40匹のオスのC57BL/6マウスを、下に示すようにそれぞれ8匹のマウスから成る5つの処置群に分けた。
【0191】
【表3】
【0192】
マウスに拘束具を着け、テストペプチド又は賦形剤を1%DMSOのPBS溶液に加えたものを尾に静脈注射(i.v.)した。これらマウスには、すぐに、15mg/kgのコンカナバリンAの無菌PBS溶液を静脈注射した。注射量は5ml/kg(100μI/20gマウス)であり、コンカナバリンA濃度は3.0mg/mlであった。翌朝(18乃至24時間後)、これらのマウスをCO2吸入で安楽死させ、心臓穿刺で血液を採集した。次に血清を分離し、AST及びALTについて分析した。
【0193】
この研究の結果を表4に示す。ALT値及びAST値はシグマ−フランケル単位/ml、平均値±SEMで表してある。
【0194】
【表4】
【0195】
上記の結果は、化合物10が、5mg/kgの用量で、コンカナバリンAで誘発させた肝損傷から保護できることを示す。肉眼病理でもこの結論が裏付けられ、肝臓はコンカナバリンAで損傷したが、この損傷は、5mg/kgの用量の化合物10で妨げられたことが示唆された。
【0196】
当業者であれば、ごく通常の実験を行うのみで、ここに解説した本発明の具体的実施例と同等の例を数多く認識され、又は確証できることであろう。このような同等物は、以下の請求の範囲の包含するところと、意図されている。
【技術分野】
【0001】
本発明は、NEMOがNEMO結合ドメイン(NBD)でIκB-キナーゼ-β(IKKβ)と相互作用するのを遮断することで、サイトカイン媒介性NF-κB活性化を選択的に抑制する組成物及び方法に関するものである。IKKβとNEMOとの相互作用を遮断すると、IKKβキナーゼ活性化が抑制され、ひいてはIκBのリン酸化が減少する。IκBのリン酸化は、サイトカイン媒介性NF-κB活性化において必須なステップである。
【背景技術】
【0002】
関連出願
本出願は、2000年5月2日出願の米国暫定特許出願第60/201,261号及び2000年8月22日出願の米国特許出願第09/643,260号の優先権を主張するものである。これら各関連出願の内容全体を、言及をもってここに編入することとする。
【0003】
米国政府による助成
本研究は、米国国立保健研究所から助成金による支援を受けた(AI33443)。
【0004】
発明の背景
NF-κBは、炎症誘発性応答に関与する遺伝子の誘導を担う細胞外シグナルを伝達する転写因子である(Baltimore ら (1998)、米国特許第5,804,374号)。NF-κBは、刺激を受けていない細胞の大半の細胞質に、IκBとして公知の複数の阻害性タンパク質の一つとの非共有結合的相互作用を通じて繋ぎ止められている(May & Ghosh, (1997) Semin. Cancer. Biol. 8, 63-73; May & Ghosh, (1998) Immunol. Today 19, 80-88; Ghosh et al., (1998) Annu. Rev. Immunol. 16, 225-260)。TNFαなどの炎症誘発性応答に関連した細胞刺激を受けるとキナーゼ群が活性化するが、活性化したキナーゼはIκBをリン酸化して、NF-κBを活性化する。IκBをリン酸化するキナーゼはIκB-キナーゼ(IKK)と呼ばれる。
【0005】
リン酸化したIκBはユビキチン化及び分解へと向かう。IκBが分解してNF-κBから解離すると、NF-κB上に核内局在化シグナルが現れ、活性NF-κBの核内移行が起きて、NF-κB応答性遺伝子が上方調節される(May & Ghosh, (1997) Semin. Cancer. Biol. 8, 63-73; May & Ghosh, (1998) Immunol. Today 19, 80-88; Ghosh et al., (1998) Annu. Rev. Immunol. 16, 225-260; Siebenlist et al., (1994) Annu. Rev. Cell Biol. 12, 405-455)。従ってIκBのリン酸化は、NF-κBが媒介する炎症誘発性応答の調節における重要な一ステップなのである。
【0006】
IκBをリン酸化するキナーゼ群の同定及び特徴付けが行われたことで、NF-κB活性化に関与するシグナル伝達経路の解明が進んだ。これまでのところ、いくつか異なるサブタイプのIKKが同定されている。IKKαは当初、ヒーラ細胞でTNFα刺激で誘導されるIκBキナーゼとして同定された(DiDonato et al., (1997) Nature 388, 548-554)。IKKαに相同なIκB-キナーゼがもう一つ同定され、IKKβと呼ばれており、TNFα刺激後に誘導される主要なIκB-キナーゼであることが判明している(Takeda et al., (1999) Science 284, 313-316; Hu et al., (1999) Science 284, 316-320; Li et al., (1999) Science 284, 321-325; Pot et al., (2000) 米国特許第6,030,834号;Woronicz & Goeddel (1999) 米国特許第5,939,302号)。IKKα及びIKKβの全体的なホモロジーは52%であり、キナーゼドメインにおけるホモロジーは65%である(Zandi et al., (1997) Cell 91, 243-252)。
【0007】
IκBプロテインキナーゼ(IKK)はIκBを特定のセリン残基でリン酸化する。例えば、これらはIκBαの32位及び36位のセリンを特異的にリン酸化する(Traenckner et al., (1995) EMBO J. 14, 2876-2883; DiDonato et al., (1996) Mol. Cell. Biol. 16, 1295-1304)。両方の部位でリン酸化することが、IκBαを効率的に分解に向かわせるのに必要である。さらに、IKKα及びIKKβの活性化は通常、NF-κB活性化物質に対する応答として起きるため、触媒として不活性な変異型IKKα及びIKKβを用いると、TNFα及びIL-1などのサイトカインによるNF-κB刺激を遮断することができる(Regnier et al., (997) Cell 90, 373-383; Delhase et al., (1999) Science 284, 309-313)。従って、IκBプロテインキナーゼはNF-κB活性化プロセスの調節に必須なのである。
【0008】
IKKα及びIKKβは、カルボキシル近位側のへリックス−ループ−へリックス(H-L-H)ドメインから、ロイシン・ジッパードメインによって隔てられたアミノ末端のセリン−スレオニンキナーゼドメインを含有する異なる構造モチーフを有する。このような構造上の特徴は他のキナーゼとは異なり、この非触媒ドメインが、タンパク質対タンパク質の相互作用に関与していると考えられる。従って、IKKに結合するタンパク質は、NF-κBの活性を調節したり(Marcu et al., (1999)米国特許第5,972,655号)、そしておそらくはNF-κBの誘導などの下流の事象も調節することができると思われる。例えば、NEMO(NF-κB必須調節因子)は、IKKに結合してキナーゼ活性を促進することが明らかにされたタンパク質である(Yamaoke et al., (1998) Cell 93, 1231-1240; Rothwarf et al., (1998) Nature 395, 287-300; Mercurio et al., (1999) Mol. Cell. Biol. 19, 1526-1538; Haraj & Sun, (1999) J. Biol. Chem. 274, 22911-22914; Jin & Jeang, (1999) J. Biomed. Sci. 6, 115-120)。
【0009】
炎症は、傷害に対する血管化生体組織の反応であると定義されている。従って、炎症は、物理的、化学的もしくは生物学的作用物質が原因で起きた傷害又は異常な刺激に対する、損傷した血管及び隣接組織の細胞学的及び化学的反応が混合した基本的、定型的な現象である。通常、炎症の結果、体液及び血球が損傷部位に蓄積するが、炎症は一般には治癒の一過程でなる。しかし、炎症はときに、炎症の正常な進行機能の不全が原因で有害ともなる。炎症性疾患は、炎症に関する、炎症を特徴とする、炎症を起こす、炎症の結果である、又は、炎症の影響を受けるようになる、ものである。炎症性疾患又は異常の例には、限定はしないが、喘息、肺炎症、結核、らい、サルコイドーシス、及び珪肺症などの慢性肉芽腫性疾患、腎炎、アミロイド症、リウマチ性関節炎、強直性脊椎炎、慢性気管支炎、強皮症、狼瘡、多発性筋炎、虫垂炎、炎症性腸疾患、潰瘍、シェーグレン症候群、ライター症候群、乾癬、骨盤内炎症性疾患、眼窩内炎症性疾患、血栓性疾患、及び、アトピー性皮膚炎及び接触性皮膚炎を含め、ウルシツタ、花粉、昆虫による刺し傷及び特定の食物などの環境刺激に対する不適切なアレルギー反応、がある。
【0010】
炎症性疾患は世界的な問題となっている。疾患の負荷研究により、結核は世界中で死亡原因の上位10位以内に入っていることが再確認された。喘息には成人人口の5%、そして小児人口の10乃至15%が罹患している(Armetti and Nicosia (1999) Boll Chim. Farm. 138(11):599)。喘息は、広汎なしかし様々な空気流閉塞を伴う慢性炎症性疾患である。
【0011】
敗血症も炎症性疾患の一つであり、様々な膿形成性等の病原性微生物や、又はそれらの毒素が被験体の血中又は組織中に存在することが原因で起きる。敗血症の特徴は、感染時の細菌生成物に対する全身の炎症性応答である。発熱などの敗血症の症状は、少なくとも部分的には、感染性物質に対する身体の炎症性応答が起こすものである。
従って、炎症性疾患を治療するのに有用な化合物への大きな需要が依然、あるのである。
【先行技術文献】
【特許文献】
【0012】
【特許文献1】米国特許第5,804,374号
【特許文献2】米国特許第6,030,834号
【特許文献3】米国特許第5,972,655号
【発明の概要】
【発明が解決しようとする課題】
【0013】
本発明は、炎症性疾患を治療するための抗炎症化合物、その薬学的組成物、及びその利用法を提供するものである。本発明は、少なくとも部分的に、IκB-キナーゼ-α(IKKα)及びIκB-キナーゼ-β(IKKβ)上のNEMO結合ドメイン(NBD)の同定に基づく。
【0014】
従って、ある態様では、本発明は、NEMO結合ドメイン(NBD)を含む抗炎症化合物を提供する。
【0015】
一実施例では、本発明は、NEMO結合ドメインと、少なくとも一つの膜移行ドメインとの融合を含む抗炎症化合物を提供する。ある好適な実施例では、前記膜移行ドメインは、本発明の抗炎症化合物のインビボでの膜移行を促進するものである。前記膜移行ドメインは、例えば、アンテナペディア(antennapedia)ホメオドメインの第三へリックス又はHIV-1 Tatたんぱくであってもよい。一実施例では、前記NEMO結合ドメインは、SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19に記載した配列を有するポリペプチドである。
【0016】
別の実施例では、本発明は、(a)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列を含む、又は、から成るペプチド;(b)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列のうちの少なくとも三つのアミノ酸から成るフラグメント;(c)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列の保存的アミノ酸置換を含むペプチド;及び(d)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列の天然型アミノ酸配列バリアント、を含む抗炎症化合物を提供する。
【0017】
別の態様では、本発明は、例えば一種以上の薬学的に許容可能な担体を含む薬学的組成物など、本発明の抗炎症化合物を含む薬学的組成物を提供する。
【0018】
さらに別の態様では、本発明は、喘息、肺炎症又は癌など、被験体の炎症性疾患を治療する方法を特徴とする。本方法は、本発明の一種以上の抗炎症化合物を治療上有効量、被験体に投与するステップを含む。機序の限定を受けることを意図する訳ではないが、本発明の抗炎症化合物は、急性及び慢性炎症の部位に白血球が動員されるのを遮断したり、白血球上のE-セレクチンの発現を下方調節したり、又は、破骨細胞分化を遮断することにより、(直接的又は間接的に)作用すると考えられる。
【0019】
別の態様では、本発明は、細胞におけるE-セレクチンなどのNF-κB依存性標的遺伝子の発現を抑制する方法を提供する。本方法は、本発明の抗炎症化合物に細胞を接触させることにより、細胞内のNF-κB依存性標的遺伝子の発現を抑制するステップを含む。さらに別の態様では、本発明は、有効量の本発明の抗炎症化合物に細胞を接触させて、細胞内のNF-κB誘導を抑制することにより、細胞内のNF-κB誘導(例えばIKKα及び/又はIKKβ依存性誘導)を抑制する方法を提供する。本発明の一実施例では、このような方法は、少なくとも一つの膜移行ドメインを含有する抗炎症化合物を利用するものである。本発明のさらに別の具体的な実施例では、このような方法で用いられる抗炎症化合物は、SEQ ID NO:2、4、5、6、11、12、16、17、又は18の配列を含むアミノ酸配列を含有する。
【0020】
別の態様では、本発明は、抗炎症化合物を同定する方法を提供する。本方法は、NEMO及びNF-κBを発現する細胞をテスト化合物に暴露するステップと;前記テスト化合物が、当該細胞によるNF-κBの活性化を調節するかどうかを調べることで、抗炎症化合物を同定するステップと、を含む。
【0021】
別の態様では、本発明は、NEMOを発現する細胞をテスト化合物に暴露するステップと、前記テスト化合物がNEMOの活性を調節するかどうかを調べることで、NEMOの活性を調節する化合物などの抗炎症化合物を同定するステップと、により、抗炎症化合物を同定する方法を提供する。
【0022】
本発明の抗炎症化合物の具体的な利点の一つは、これらは、IKKを通じたNF-κB誘導を遮断しながらも、NF-κBの基礎活性を抑制しないことである。
【0023】
本発明の他の特徴及び利点は、以下の詳細な説明及び請求の範囲から明白となるであろう。
【0024】
I.概略の解説
本発明は、炎症性疾患を治療するための抗炎症化合物、その薬学的組成物、及びその利用法を提供するものである。本発明は、少なくとも部分的に、IκB-キナーゼ-α(IKKα)及びIκB-キナーゼ-β(IKKβ)上のNEMO結合ドメイン(NBD)の同定に基づく。
【0025】
機序に限定されることを意図する訳ではないが、本発明の抗炎症化合物は、NEMOとIKK(例えばIKKβ又はIKKα)のNEMO結合ドメイン(NBD)での相互作用を遮断して、IκBのリン酸化、分解、及びその後のNF-κBからの解離を抑制することにより、作用すると考えられる。この抑制の結果、炎症誘発性応答に関係したNF-κB活性化が遮断される。
【0026】
さらに本発明は、抗炎症化合物をスクリーニングし、同定する方法も提供する。
【0027】
II.定義
他に特に定義しない限り、ここで用いた全ての技術用語及び科学用語は、本発明が関係する分野の当業者が通常理解するのと同じ意味を有するものである。
【0028】
ここで用いる用語「結合」とは、限定はしないが、酵素対基質、抗体対抗原、DNA鎖対それらの相補鎖など、分子間相互の付着を言う。結合が起きるのは、分子表面の部分同士の間で形状及び化学的性質が「相補的」になっているからである。よくある喩えは、酵素がどのように基質の周りに嵌め合うかを記述するのに用いられる「鍵穴と鍵」である。
【0029】
用語「融合ペプチド」又は「融合ポリペプチド」又は「融合たんぱく」には、二つの異なるアミノ酸配列を組み合わせて得られるペプチド、ポリペプチド又はタンパク質が含まれる。典型的には、ある一個のペプチド、ポリペプチド又はタンパク質の部分的な配列が、別の異種のペプチド、ポリペプチド又はタンパク質に、当業で公知の技術を用いて連結されている。
【0030】
「保存的アミノ酸置換」とは、アミノ酸残基が、同様な側鎖を有するアミノ酸残基に置換されることを言う。塩基性側鎖(例えばリシン、アルギニン、ヒスチジン)、酸性側鎖(例えばアスパラギン酸、グルタミン酸)、無電荷の極性側鎖(例えばグリシン、アスパラギン、グルタミン、セリン、スレオニン、チロシン、システイン)、非極性側鎖(例えばアラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン)、β-分岐側鎖(例えばスレオニン、バリン、イソロイシン)及び芳香族側鎖(例えばチロシン、フェニルアラニン、トリプトファン、ヒスチジン)を含め、同様な側鎖を有するアミノ酸残基の仲間は当業で公知である。本発明の抗炎症化合物で行うことのできる保存的置換の非限定的な例には、D型フェニルアラニンのD型チロシン、D型ピリジルアラニン又はD型ホモフェニルアラニンとの置換、D型ロイシンのD型バリン又は他の脂肪族側鎖を有する天然もしくは非天然アミノ酸との置換、及び/又は、D型バリンのD型ロイシン又は脂肪族側鎖を有する他の天然もしくは非天然アミノ酸との置換、がある。
【0031】
ここで用いる用語「ドメイン」とは、限定はしないが、例えば疎水性、極性、球状及び螺旋状のドメイン又は性質など、共通の物理化学的特徴を持つ分子又は構造の一部分を言う。結合ドメインの具体的な例には、限定はしないが、DNA結合ドメイン及びATP結合ドメイン、がある。
【0032】
ここで用いる用語「膜移行ドメイン」とは、細胞膜を透過できて、付着させたペプチドをインビボで細胞内に輸送するのに利用されるペプチドを言う。膜移行ドメインには、限定はしないが、アンテナペディアホメオドメインたんぱくの第三へリックス及びHIV-1たんぱくTatがある。他の膜移行ドメインも当業で公知であり、例えばDerossi et al., (1994) J. Biol. Chem. 269, 10444-40450; Lindgren et al., (2000) Trends Pharmacol. Sci 21, 99-103; Ho et al., Cancer Research 61, 474-477 (2001); 米国特許第5,888,762号;米国特許第6,015,787号;米国特許第5,846,743号;米国特許第5,747,641号;米国特許第5,804,604号;及び公開済みPCT出願WO98/52614号、WO00/29427号、及びWO99/29721号に解説されたものがある。前出の各参考文献の内容全体を言及をもってここに編入することとする。
【0033】
ここで用いる用語「IκB」(IカッパB)とは、抑制たんぱくのファミリーのうちでNF-κB誘導の調節で働く構造上関連した仲間のいずれか一つを言う。
【0034】
ここで用いる用語「IκB-キナーゼ」又は「IκBプロテインキナーゼ」又は「IκB-キナーゼ複合体」又は「IκBプロテインキナーゼ複合体」又は「IKK」とは、IκBをリン酸化するキナーゼを言う。
【0035】
ここで用いる用語「IKKα」とは、IκB-キナーゼ複合体のαサブユニットを言う。ここで用いる用語「IKKβ」とは、IκB-キナーゼ複合体のβサブユニットを言う。
【0036】
ここで用いる用語「NEMO」(NF-κB必須調節因子)、「IKKγ」又は「IKKAP」とは、IKKに結合してキナーゼ活性を促進するタンパク質を言う。
【0037】
ここで用いる用語「NEMO結合ドメイン」又は「NBD」には、NEMOが通常(IKKα又はIKKβなどの)IKKと相互作用する領域にあってNEMOに結合できるあらゆるドメインが含まれる。NEMO結合ドメインには、NEMOとの相互作用に重要な、例えば、野生型IKKβのα2領域(残基737−742)、又は野生型IKKαの相当する6個のアミノ酸配列(残基738−743)がある。野生型IKKβNBDの核酸配列及び対応するアミノ酸配列を、それぞれSEQ ID NO:1(GenBank受託番号AR067807;ヌクレオチド2203−2235)及びSEQ ID NO:2に公開する。
【0038】
ここで用いる場合の用語「類似体」、「誘導体」及び「ミメティック」には、あるペプチド構造の化学構造を模倣すると共に、このペプチド構造の機能的性質を維持した分子が包含されるものと、意図されている。ペプチド類似体、誘導体及びミメティックをデザインする方法は当業で公知である。例えば、Farmer, P. S. in Drug Design (E. J. Ariens, ed.) Academic Press, New York, 1980, vol. 10, pp. 119-143; Ball. J. B. and Alewood, P. E. (1990) J. Mol. Recognition 3:55; Morgan, B. A. and Gainor, J. A. (1989) Ann. Rep. Med. Chem. 24:243; adn Freidinger, R. M. (1989) Trends Pharmacol. Sci. 10:270を参照されたい。さらにSawyer, T. K. (1995) "Peptidomimetic Design and Chemical Approaches to Peptide Metabolism" in Taylor, M. D. and Amidon, G. L. (eds.) Peptide-Based Drug Design: Controlling Transport and Metabolism, Chapter 17; Smith, A. B. 3rd, et al. (1995) J. Am. Chem. Soc. 117:11113-11123; Smith, A. B. 3rd, et al. (1994) J. Am. Chem. Soc. 116:9947-9962; and Hirschman, R., et al. (1993) J. Am. Chem. Soc. 115:12550-12568も参照されたい。
【0039】
ここで用いる「化合物X(例えばペプチド又はアミノ酸)の誘導体」とは、当該化合物上の一つ以上の反応基を置換基で誘導変化させてある形のXを言う。ペプチド誘導体の例には、アミノ酸側鎖、ペプチド骨格、又はアミノ末端又はカルボキシ末端が誘導変化させてある(例えばメチル化アミド結合を持つペプチド化合物など)ペプチドがある。ここで用いる化合物Xの「類似体」とは、Xの機能的活性に必要なXの化学構造は残っているが、Xとは異なる化学構造もいくつか含んでいるような化合物を言う。天然型ペプチドの類似体の一例は、一つ以上の非天然発生型のアミノ酸を含有するペプチドである。ここで用いる、化合物Xの「ミメティック」とは、Xの機能的活性に必要なXの化学構造が、Xのコンホメーションを模倣した他の化学構造に置換されているような化合物を言う。ペプチドミメティックの例には、ペプチド骨格が一つ以上のベンゾジアゼピン分子に置換されたようなペプチド化合物がある(例えばJames, G. L. et al. (1993) Science 260:1937-1942)。
【0040】
用語ミメティック、特にペプチドミメティックには、同配体が含まれるものと、意図されている。ここで用いる「同配体」という用語には、第一の構造の立体的コンホメーションが、第二の構造に特異的な結合部位に適合しているために、第二の化学構造に置換することのできる化学構造が含まれるものと、意図されている。この用語には特に、当業者に公知のペプチド骨格修飾(即ちアミド結合ミメティック)が含まれる。このような修飾には、アミド窒素の修飾、α位炭素の修飾、アミドカルボニルの修飾、アミド結合の完全な置換、伸長、欠失、又は骨格架橋、がある。ψ[CH2S]、ψ[CH2NH]、ψ[CSNH2]、ψ[NHCO]、ψ[COCH2]、及びψ[(E)又は(Z)CH=CH]を含め、いくつかのペプチド骨格修飾が公知である。上で用いた命名法において、ψはアミド結合がないことを示す。アミド基を置換する構造を括弧内に明示した。
【0041】
他の可能な修飾には、N-アルキル(又はアリール)置換(ψ[CONR])、又はラクタム又は他の環式構造を構築するための骨格の架橋がある。本発明の抗炎症化合物の他の誘導体には、C末端ヒドロキシメチル誘導体、O-修飾誘導体(例えばC末端ヒドロキシメチルベンジルエーテル)、アルキルアミド及びヒドラジドなどの置換アミドを含むN末端修飾誘導体や、C末端フェニルアラニン残基をフェネチルアミド類似体に(例えばトリペプチドVal-Phe-Pheの類似体としてのVal-Phe-フェネチルアミドなど)置換してある抗炎症化合物、がある。
【0042】
ここで用いる用語「野生型」とは、天然発生種の仲間の大半に特徴的であり、変異種の遺伝子型及び表現型とは対照的な遺伝子型及び表現型を言う。
【0043】
III.具体的な実施例
A.抗炎症化合物
本発明は、NEMO結合ドメイン(NBD)を含む抗炎症化合物を提供するものである。NEMOが通常、IKK(例えばIKKα又はIKKβ)と相互作用する領域で、NEMOに結合することのできるドメインを含む分子であれば、本発明の抗炎症化合物を調製するのに利用できよう。このような分子の例には、D型及び/又はL型立体配置のアミノ酸を含むペプチド;ペプチド化合物の誘導体、類似体、及びミメティック;抗体(例えばポリクローナル、モノクローナル、ヒト化、抗イディオタイプ、キメラ、及び一本鎖抗体や、Fab、F(ab')2、Fab発現ライブラリフラグメント、及び抗体のエピトープ結合フラグメントなど);及び小型の有機及び無機分子(例えばコンビナトリアル・ライブラリ及び天然生成物ライブラリから得られる分子など)、がある。
【0044】
ある好適な実施例では、本発明の抗炎症化合物は、NEMO結合ドメインと、本発明の抗炎症化合物のインビボでの膜移行を促進する少なくとも一つの膜移行ドメインとの融合を含んで成る。
【0045】
本発明の抗炎症化合物を、IKKα又はIKKβのNBDの野生型アミノ酸配列(SEQ ID NO:2)に基づいて設計してもよい。NEMOに結合できれば、IKKα又はIKKβのNBDの野生型アミノ酸配列のいかなるフラグメントを用いても、本発明の抗炎症化合物を調製できよう。(ここに解説した方法を用いた)これらの野生型配列の点変異、挿入、又は欠失を利用して、更なる抗炎症化合物を作製してもよい。SEQ ID NO:2に記載したペプチドの737位、740位及び742位に保存的アミノ酸置換を含有するペプチドは、特に有用な本発明の抗炎症化合物である(ペプチドがNEMOに結合する能力に有意な影響を及ぼさない保存的置換の例については表1を参照されたい)。さらに、NEMOに結合する能力の残った、IKKβ遺伝子の天然発生型対立遺伝子バリアントを用いても抗炎症化合物を調製できよう。
【0046】
実施例の一つでは、本発明の抗炎症化合物は、(a)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列を含む、又は、から成るペプチド;(b)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列のうちの少なくとも三つのアミノ酸から成るペプチドフラグメント;(c)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列に保存的アミノ酸置換を含有するペプチド;及び(d)SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、又は19のアミノ酸配列の天然型アミノ酸配列バリアント、を含む。
【0047】
さらに本発明の抗炎症化合物には、特異抗体又はT細胞抗原受容体(Harlow & Lane, (1988) Antibodies - A Laboratory Manual, Cold Spring Harbor Laboratory Pressを参照されたい)のような体細胞組換えペプチド受容体などのNEMO特異的受容体や、一種、二種及び三種ハイブリッドスクリーンなどの検定で同定された他の天然細胞内結合物質や、以下に解説するものなどの化学的ライブラリのスクリーンで同定された非天然細胞内結合物質を含めてもよい。
【0048】
本発明の抗炎症化合物は、NEMOを下方調節することができる。下方調節とは、ここでは、NEMO、そのリガンド又は活性化物質の活性化、機能又は合成の減少であると、定義しておく。さらにこれには、NEMO遺伝子、そのタンパク質生成物、リガンド又は活性化物質の分解の増加も包含されると、定義しておく。下方調節は、例えばNEMOのIKK(例えばIKKβ又はIKKα)への結合を不安定にしたり;又はIκBのリン酸化を遮断してこのタンパク質のその後の分解を起こさせるなど、多数の方法で行うことができる。
【0049】
IKKβによるIκBのリン酸化の結果、Iκbのユビキチン化及び分解が起き、さらにはIκBの解離が起きて、NF-κBの核内移行が可能となり、炎症応答に重要な遺伝子が上方調節される。従って、本発明の抗炎症化合物を用いれば、NF-κB機能を下方調節できよう。さらにNF-κBの下方調節は、NBD又はNEMO自体を狙ったポリクローナルもしくはモノクローナル抗体又はそのフラグメントを含む抗炎症化合物を利用しても、達成できるであろう。さらに本発明は、NBD又はNEMO自体に対して充分な親和性で結合して、IKKβとのNEMO相互作用を遮断するなどに必要な三次元構造を有するような小分子も包含する。IκBの分解を減少させ、NF-κBの活性化を減少させるIKKβ遮断法により、これらの小分子は、炎症を治療又は予防する治療薬として有用なものとなる。
【0050】
ある実施例では、本発明は、式:
Xa-Xb、
の抗炎症化合物を提供するが、ただし式中、Xa は、6乃至15個のアミノ酸残基を含む膜移行ドメインであり、そしてXb はNEMO結合配列である。前記化合物には、選択に応じて、N末端、C末端又は両者に、修飾基を含めてもよい。
【0051】
Xb は、6乃至9個のアミノ酸残基を含むNEMO結合配列である。ある実施例では、Xbは、以下の構造:
(Y)n-X1-X2-X3-X4-X5-X6-(A)m
から成り、ただし式中、n 及びm はそれぞれ個別に、0 又は1であり、そしてA 及びYはそれぞれ1乃至約3個のアミノ酸残基を含む。nが1であるとき、Yは配列TAであると好ましい。mが1であるとき、Aは配列QTEであると好ましい。X1 はL、A、I 又はノルロイシン(Nle)である。X2 はD、E、N、Q、ホモセリン(Hser)又は2-ケトプロピルアラニン(2-ケトプロピ-A)である。X3 は W、F Y、4-ビフェニル-アラニン (Bpa)、ホモフェニルアラニン (Hphe)、2-ナフチルアラニン (2-Nal)、1-ナフチルアラニン (1-Nal)、又はシクロキセキシル(原語cycloxexyl)-アラニン (Cha)である。X4 は S、A、E、L、T、ノルロイシン (Nle)、又はホモセリン (Hser)である。X5 はW、H、ホモフェニルアラニン (Hphe)、2-ナフチルアラニン (2-Nal)、1-ナフチルアラニン (1-Nal)、O-ベンジルセリン (SeroBn)、又は3-ピリジルアラニン (3-Pal)である。そしてX6 はL、A、I、又はノルロイシン (Nle)である。
【0052】
好ましくは、Xb は、TALDWSWLQTE (SEQ ID NO:28); LDWSWLQTE (SEQ ID NO:29); TALDWSWL (SEQ ID NO:30); ALDWSWLQTE (SEQ ID NO:31); LDWSWLQTE (SEQ ID NO:32); LDWSWL (SEQ ID NO:33); TALDWSWLQT (SEQ ID NO:34); TALDWSWLQ (SEQ ID NO:35); ALDWSWLQT (SEQ ID NO:36); LDWSWLQ (SEQ ID NO:37); LDWSWLQT (SEQ ID NO:38); ADWSWL (SEQ ID NO:39); LDWSWA (SEQ ID NO:40); ADWSWA (SEQ ID NO:41); LDFSWL (SEQ ID NO:42); LDYSWL (SEQ ID NO:43); LDWAWL (SEQ ID NO:44); LDWEWL (SEQ ID NO:45); TAADWSWLQTE (SEQ ID NO:46); ADWSWLQTE (SEQ ID NO:47); TAADWSWL (SEQ ID NO:48); AADWSWLQTE (SEQ ID NO:49); ADWSWLQTE (SEQ ID NO:50); ADWSWL (SEQ ID NO:51); TAADWSWLQT (SEQ ID NO:52); TAADWSWLQ (SEQ ID NO:53); AADWSWLQT (SEQ ID NO:54); ADWSWLQ (SEQ ID NO:55); ADWSWLQT (SEQ ID NO:56); ALDWSWAQTE (SEQ ID NO:57); LDWSWAQTE (SEQ ID NO:58); TALDWSWA (SEQ ID NO:59); ALDWSWAQTE (SEQ ID NO:60); LDWSWAQTE (SEQ ID NO:61); LDWSWA (SEQ ID NO:62); TALDWSWAQT (SEQ ID NO:63); TALDWSWAQ (SEQ ID NO:64); ALDWSWAQT (SEQ ID NO:65); LDWSWAQ (SEQ ID NO:66); LDWSWAQT (SEQ ID NO:67); TAADWSWAQTE (SEQ ID NO:68); ADWSWAQTE (SEQ ID NO:69); TAADWSWA (SEQ ID NO:70); AADWSWAQTE (SEQ ID NO:71); ADWSWAQTE (SEQ ID NO:72); ADWSWA (SEQ ID NO:73); TAADWSWAQT (SEQ ID NO:74); TAADWSWAQ (SEQ ID NO:75); AADWSWAQT (SEQ ID NO:76); ADWSWAQ (SEQ ID NO:77); ADWSWAQT (SEQ ID NO:78); TALDFSWLQTE (SEQ ID NO:79); LDFSWLQTE (SEQ ID NO:80); TALDFSWL (SEQ ID NO:81); ALDFSWLQTE (SEQ ID NO:82); LDFSWLQTE (SEQ ID NO:83); LDFSWL (SEQ ID NO:84); TALDFSWLQT (SEQ ID NO:85); TALDFSWLQ (SEQ ID NO:86); ALDFSWLQT (SEQ ID NO:87); LDFSWLQ (SEQ ID NO:88); LDFSWLQT (SEQ ID NO:89); TALDYSWLQTE (SEQ ID NO:90); LDYSWLQTE (SEQ ID NO:91); TALDYSWL (SEQ ID NO:92); ALDYSWLQTE (SEQ ID NO:93); LDYSWLQTE (SEQ ID NO:94); LDYSWL (SEQ ID NO:95); TALDYSWLQT (SEQ ID NO:96); TALDYSWLQ (SEQ ID NO:97); ALDYSWLQT (SEQ ID NO:98); LDYSWLQ (SEQ ID NO:99); LDYSWLQT (SEQ ID NO:100); TALDWAWLQTE (SEQ ID NO:101); LDWAWLQTE (SEQ ID NO:102); TALDWAWL (SEQ ID NO:103); ALDWAWLQTE (SEQ ID NO:104); LDWAWLQTE (SEQ ID NO:105); LDWAWL (SEQ ID NO:106); TALDWAWLQT (SEQ ID NO:107); TALDWAWLQ (SEQ ID NO:108); ALDWAWLQT (SEQ ID NO:109); LDWAWLQ (SEQ ID NO:110); LDWAWLQT (SEQ ID NO:111); TALDWEWLQTE (SEQ ID NO:112); LDWEWLQTE (SEQ ID NO:113); TALDWEWL (SEQ ID NO:114); ALDWEWLQTE (SEQ ID NO:115); LDWEWLQTE (SEQ ID NO:116); LDWEWL (SEQ ID NO:117); TALDWEWLQT (SEQ ID NO:118); TALDWEWLQ (SEQ ID NO:119); ALDWEWLQT (SEQ ID NO:120); LDWEWLQ (SEQ ID NO:121); 及び LDWEWLQT (SEQ ID NO:122)からなる群より選択される配列であるとよい。
【0053】
Xa は、6乃至15個のアミノ酸残基、好ましくは6乃至12個、又は6乃至10個のアミノ酸残基から成る膜移行ドメインである。好ましくは、Xa は、少なくとも5個の塩基性アミノ酸残基、好ましくは L-アルギニン、D-アルギニン、L-リシン及びD-リシンから個別に選択される少なくとも5個のアミノ酸残基を含む膜移行ドメインであるとよい。適した膜移行ドメインには、ここに開示したものがある。
【0054】
実施例の一つでは、Xa はアミノ酸配列RRMKWKK (SEQ ID NO:123); YGRKKRRQRRR (SEQ ID NO:124); ygrkkrrqrrr (SEQ ID NO:125); YARKARRQARR (SEQ ID NO:126); yarkarrqarr (SEQ ID NO:127); YARAARRAARR (SEQ ID NO:128); yaraarraarr (SEQ ID NO:129); rrmkwkk (SEQ ID NO:130); (R)y 及び (r)y、(ただしこの場合y は6乃至11である)から選択される。小文字はD型アミノ酸残基を指し、大文字はL型アミノ酸残基を指す。
【0055】
適したペプチドXa-Xb の例には、以下の配列:RRMKWKKTALDWSWLQTE (SEQ ID NO:131); rrmkwkkTALDWSWLQTE (SEQ ID NO:132); YGRKKRRQRRRTALDWSWLQTE (SEQ ID NO:133); ygrkkrrqrrrTALDWSWLQTE (SEQ ID NO:134); rrrrrrrTALDWSWLQTE (SEQ ID NO:135); RRRRRRRTALDWSWLQTE (SEQ ID NO:136); YARKARRQARRTALDWSWLQTE (SEQ ID NO:137); yarkarrqarrTALDWSWLQTE (SEQ ID NO:138); YARAARRAARRTALDWSWLQTE (SEQ ID NO:139); yaraarraarrTALDWSWLQTE (SEQ ID NO:140) YGRKKRRQRRRLDWSWL (SEQ ID NO:141); ygrkkrrqrrrLDWSWL (SEQ ID NO:142); RRMKWKKLDWSWL (SEQ ID NO:143); rrmkwkkLDWSWL (SEQ ID NO:144); rrrrrrrLDWSWL (SEQ ID NO:145); YARAARRAARRLDWSWL (SEQ ID NO:146); yaraarraarrLDWSWL (SEQ ID NO:147); 及びRRRRRRRLDWSWL (SEQ ID NO:148)を有するものがある。
【0056】
本発明の抗炎症化合物には、選択に応じ、C末端、N末端又は両方に付着した修飾基を含めてもよい。例えば、C末端に付着させることのできる適した修飾基には、置換及び非置換アミノ基、例えば-NH2、-NH(アルキル) 及び-N(アルキル)2 基;及びアルコキシ基、例えば直線状、分岐式又は環式のC1-C6-アルコキシ基、など、がある。好適なC末端修飾基は、-NH2 基である。N末端に付着させることのできる適した修飾基には、アシル基、例えばアセチル基;及びアルキル基、好ましくはC1-C6-アルキル基、より好ましくはメチル、がある。
【0057】
本発明の抗炎症化合物においては、膜移行ドメインであるXaが当該化合物のアミノ末端にあって、NEMO結合ドメインであるXbが当該化合物のカルボキシル末端にあってもよい (Xa- Xb)。反対に、本発明の抗炎症化合物において、膜移行ドメインであるXaが当該化合物のカルボキシル末端にあって、NEMO結合配列であるXbが当該化合物のアミノ末端にあってもよい(Xb- Xa)。
【0058】
本発明の具体的な抗炎症化合物には、以下に挙げるものが含まれる。
【0059】
化合物番号
1 H-RRMKWKKTALDWSWLQTE-NH2
2 H-YGRKKRRQRRRTALDWSWLQTE-NH2
3 H-rrrrrrrTALDWSWLQTE-NH2
4 H-YARKARRQARRTALDWSWLQTE-NH2
5 H-YARAARRAARRTALDWSWLQTE-NH2
6 H-RRMKWKKLDWSWL-NH2
7 H-rrmkwkkLDWSWL-NH2
8 H-rrrrrrrLDWSWL-NH2
9 H-YARAARRAARRLDWSWL-NH2
10 H-yaraarraarrLDWSWL-NH2
11 H-YGRKKRRQRRRLDWSWL- NH2
【0060】
B.スクリーニング検定
加えて本発明は、抗炎症化合物を同定するスクリーニング法も提供する。本抗炎症化合物は、IKKβ分子のNBDで相互作用することにより、この分子の機能を遮断したり、合成を妨げたり、又はその生物学的安定性を低下させるであろう。生物学的安定性は、当該分子の合成とその分解との間の時間の測定値である。例えば、あるタンパク質、ペプチド又はペプチドミメティック(Kauvar, Nature Biotech. (1996) 14, 709)治療薬の安定性は、その配列を変更して酵素分解をより受けやすくするようにすれば、短縮することができる。
【0061】
さらに本発明は、IKKβ機能を失活させる、又は、IKKβ機能のアンタゴニストとして働く化合物をスクリーニングする方法も包含する。このような化合物は、IKKβもしくはNF-κBタンパク質レベルの変化に関連した病的状態の調節に有用であろう。
【0062】
さらに本発明は、例えば、IKKβと、この分子のNBDで相互作用する化合物、又は、NEMOと相互作用することでNEMOとIKKβとの相互作用を遮断する化合物など、本発明のタンパク質の結合相手を単離及び同定する方法も提供する。本発明のタンパク質と、候補結合相手又は細胞抽出物もしくは画分との混合は、候補結合相手と本発明のタンパク質との結合が可能なような条件下で行う。混合後、本発明のタンパク質と結合したペプチド、ポリペプチド、タンパク質又は他の分子を、混合物から分離する。次に、本発明のタンパク質に結合した結合相手を取り外し、さらに解析することもできる。結合相手を同定及び単離するには、当該タンパク質全体、例えば全IKKβペプチドを利用することもできる。反対に、当該タンパク質の一フラグメントを利用してもよい。例えば、NBDを含むペプチドフラグメントを利用して、IKKβとNEMOとの相互作用を遮断することもできる。
【0063】
細胞抽出物を得るには多種の方法を利用できる。細胞は、物理的又は化学的破砕法のいずれを利用して破砕してもよい。物理的破砕法の例には、音波破砕法及び機械的せん断法がある。化学的溶解法の例には、限定はしないが、界面活性剤による溶解法及び酵素溶解法がある。当業者であれば、本方法で利用する抽出物を得るのに、細胞抽出物を調製する方法を容易に適合できよう。
【0064】
細胞抽出物を調製したら、その抽出物を、IKKβ又はNEMOに、当該タンパク質と結合相手との結合が可能な条件下で混合する。多種の条件を利用できるが、最も好ましい条件は、ヒト細胞の細胞質で見られる条件に近いものである。浸透圧モル濃度、pH、温度、及び、用いる細胞抽出物の濃度などの特徴は、当該タンパク質と結合相手との結合が至適になるよう、変更することができる。
【0065】
適当な条件下で混合した後、結合した複合体を混合液から分離する。多種の技術を利用して混合液を分離できる。例えば、本発明のタンパク質に特異的な抗体を用いれば、結合相手との複合体を免疫沈降させることができる。代わりに、クロマトグラフィー及び濃度−沈渣遠心分離法などの標準的な化学的分離技術も利用できる。
【0066】
抽出物中に見られる結合しなかった細胞構成成分を除去した後、従来の方法を利用して結合相手を複合体から解離させることができる。例えば、混合液の塩濃度又はpHを変化させても、解離が可能である。
【0067】
混合抽出物から、結合済みの結合相手との組を分離する手助けとして、タンパク質を固体の支持体上に固定してもよい。例えば、タンパク質をニトロセルロース・マトリックス又はアクリル製ビーズに付着させてもよい。タンパク質を固体の支持体上に付着させると、抽出物中に見られる他の構成成分からペプチド−結合相手の組を分離するのに便利である。同定される結合相手は、単一のタンパク質であるかも、又は二種以上のタンパク質からなる複合体であるかも知れない。代わりに、結合相手は、Takayama et al., (1997) Methods Mol. Biol. 69, 171-814 又はSauder et al., (1996) J. Gen. Virol. 77, 991-996の手法に基づくファー−ウェスタン検定を用いて同定でき、又は、エピトープ・タグを付けたタンパク質又はGST融合タンパク質を用いて同定できよう。
【0068】
選択によっては、本発明のペプチドをコードする核酸分子を酵母二種ハイブリッド系で利用してもよい。酵母二種ハイブリッド系は、他のタンパク質結合相手を同定するのに用いられてきており、ここに解説した核酸分子を利用できるよう、容易に適合が可能である(例えばストラータジーン社Hybrizap(R)、二種ハイブリッド系を参照されたい)。
【0069】
本発明の別の実施例は、NEMO又はIKKβの少なくとも一つの活性を調節する作用物質を同定する方法を提供するものである。このような方法又は検定では、所望の活性を観察又は検出するいかなる手段を用いてもよい。
【0070】
あるフォーマットでは、テスト対象の作用物質に暴露した細胞集団と、暴露していないコントロール細胞集団との間で、本発明のタンパク質の相対量を検定してもよい。このフォーマットでは、特異抗体などのプローブを用いて、異なる細胞集団中のタンパク質の発現の差を観察する。細胞株又は集団を、テスト対象の作用物質に、適当な条件下及び時間、暴露する。細胞ライセートは、暴露した細胞株又は集団と、コントロールの非暴露細胞株又は集団とから調製してもよい。次に、この細胞ライセートをプローブで解析する。
【0071】
抗体プローブの作製は、当該ペプチドを用いた適した免疫処置プロトコールで適切なほ乳類ホストを免疫処置することで行う。NBDを含むペプチド又はタンパク質は、充分な長さの物であり、免疫原性を高めるのに必要であれば、適した担体に共役させておく。BSA、KLH、又は他の担体たんぱくなどの担体を用いて免疫原性共役体を調製する方法は当業で公知である。場合によっては、カルボジイミド試薬などを用いた直接共役法が効果的であろう。別の場合には、ピアース・ケミカル社が市販するものなどの結合試薬が、ハプテンを利用するのに好ましいであろう。ハプテンペプチドは、アミノ末端又はカルボキシ末端のいずれかでシステイン残基で伸長させたり、又はシステイン残基を散在させるなどして担体への結合を促進することもできる。免疫原の投与は多くの場合、当業で広く公知であるように、適した期間にわたって、そして適したアジュバントを用いて、注射で行われる。免疫処置期間中は、抗体価を測定して、抗体形成が充分であるかを調べる。この方法で生成させたポリクローナル抗血清は用途によっては充分かも知れないが、薬学的組成物のためには、モノクローナル製剤の利用が好ましい。所望のモノクローナル抗体を分泌する不死化細胞系は、公知のように、Kohler & Milstein, (1992) Biotechnology 24, 524-526の標準的方法や、リンパ球又は脾細胞の不死化を行う改良法を利用して、調製できる。所望の抗体を分泌する不死化細胞系は、抗原を当該ペプチドハプテン、ペプチド又はタンパク質とするイムノアッセイでスクリーニングする。
【0072】
所望の抗体を分泌する適切な不死化細胞培養株を同定したら、この細胞をインビトロで培養しても、又は腹水液中で生成させてもよい。所望のモノクローナル抗体は、この培養上清又は腹水上清から、回収できよう。免疫学的に有意な部分を含有するモノクローナルもしくはポリクローナル抗血清のフラグメントをアンタゴニストとして用いてもよいが、インタクト抗体を用いてもよい。F(ab')2フラグメントのFab、Fab'など、免疫学的に反応性のフラグメントを利用するのが、特に治療上は好ましいことが多い。なぜならこれらのフラグメントは、免疫グロブリン全体よりも免疫原性が一般に低いからである。
【0073】
さらに抗体又はフラグメントは、組換え法により、現在の技術を用いても作製できる。さらに、当該タンパク質の所望の領域に特異的に結合する抗体領域を、複数の種を由来とするキメラとして作製することもできる。上記の方法で検定される作用物質は、無作為に選択しても、又は合理的に選択又は設計してもよい。ここで用いる場合の、作用物質を無作為に選択するとは、本発明のタンパク質のみや、それが結合する基質、結合相手、等々との結合に関与する特定の配列を考慮することなく、作用物質を無作為に選択することを言う。無作為に選択される作用物質の一例は、化学的ライブラリ又はペプチド・コンビナトリアルライブラリ、又は生物の成長ブロスの利用である。
【0074】
ここで用いる場合、ある作用物質が合理的に選択又は設計されるとは、標的部位の配列、及び/又は、作用物質の作用に関連したそのコンホメーション、を考慮に入れた非無作為的基礎に基づいて作用物質が選択されることを言う。IKKβ上のNBDか、又は、NEMO上のIKKβ結合ドメイン、を含むペプチド配列を利用すれば、作用物質を合理的に選択又は合理的に設計することができる。例えば、合理的に選択されたペプチド物質は、SEQ ID NO:2のアミノ酸配列とアミノ酸配列が同一になったペプチドであったり、又は、その保存的置換を持つペプチドであるかも知れない。
【0075】
本発明の化合物は、例としては、ペプチド、小分子、ビタミン誘導体や、糖質であってもよい。本発明の化合物の構造上の性質には何ら制限はないことは、当業者であれば容易に認識できよう。
【0076】
本発明のペプチド化合物は、当業で公知のように、標準的な固相(又は液相)ペプチド合成法を利用して調製できる。加えて、これらのペプチドをコードするDNAを、市販のオリゴヌクレオチド合成装置を用いて合成したり、標準的な組換え産生系を用いて組換えにより産生させてもよい。遺伝子にコードされていないアミノ酸を含めねばならない場合、固相ペプチド合成を利用した生成が必要となる。
【0077】
さらに本発明は、NBD及びその保存的ヌクレオチド置換を有するペプチドを、好ましくは単離された形でコードする、単離された核酸分子を提供するものである。保存的ヌクレオチド置換には、大半のアミノ酸が二つ以上のコドンを有することが原因で、ある特定のアミノ酸の暗号化に影響を与えないようなヌクレオチド置換がある(King & Stansfield (編集者)、A Dictionary of Genetics, Oxford University Press, 1997 、19ページを参照されたい)。従って保存的ヌクレオチド置換には、サイレント変異や、使用コドンの差も含まれる。例えば、本発明には、SEQ ID NO:2に記載のペプチドをコードする、SEQ ID NO:1に記載の核酸分子や、その保存的ヌクレオチド置換が含まれる。さらに本発明は、SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、及び19に記載のペプチドをコードする核酸及びその保存的ヌクレオチド置換も包含する。SEQ ID NO:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、及び19に記載のペプチドをコードするヌクレオチド配列には複数の順列が可能であるため、これらのペプチドをコードする核酸はいかなるものも、本発明の包含するところである。
【0078】
本発明の包含する核酸の具体的な例には、限定はしないが、以下のものが含まれる。(1)SEQ ID NO:2のペプチドのアミノ酸は、核酸配列TTAGATTGGTCTTGGTTA(SEQ ID NO:24)又はTTGGACTGGTCCTGGCTA(SEQ ID NO:25)にコードされていてもよい;そして(2)SEQ ID NO:15のペプチドのアミノ酸は、核酸配列TTAGATTGGTCTTATCTG(SEQ ID NO:26)又はCTTGACTGGTCATACTTA(SEQ ID NO:27)にコードされていてもよい。
【0079】
ここで用いる場合の、核酸分子が「単離されている」とは、核酸の源にある他のポリペプチドをコードする混入核酸から当該核酸分子が略、分離されていることを言う。翻訳中のタンパク質配列にアミノ酸の欠失、追加、又は変更が起きて、核酸自体の一次構造に対する修飾が、ペプチドの活性を破壊せずに起きることもある。このような置換又は他の変化の結果生ずるのは、本発明が考察する範囲に包含される核酸のコードするアミノ酸配列を有するペプチドである。
【0080】
本発明の化合物の他の分類は、本発明のタンパク質の重要な部分と免疫反応性である抗体である。抗体物質は、抗体に狙わせようとするタンパク質部分を抗原性領域として含有するペプチドで、適したほ乳類被験体を免疫処置して、得る。
【0081】
C.高スループット検定
導入−高スループット・スクリーニングの力を、NEMOと相互作用できる新規な抗炎症化合物の検索に用いる。高スループット・スクリーニングに関する概略的な情報については、例えばCost-Effective Strategies for Automated and Accelerated High-Throughput Screening, IBCS Biomedical Library Series, IBC United States Conferences, 1996; Devlin (編集者)、High Throughput Screening, Marcel Dekker 1998; 米国特許第5,763,263号を参照されたい。高スループット検定では、一種以上の異なる検定技術を利用する。
【0082】
免疫診断及びイムノアッセイ − 生物体液などの複合混合液中に通常低濃度で存在する特定の生化学的物質を測定するのに用いられている一群の技術があるが、このような技術は適宜調製及び選択された抗体がそれらに相補の抗原に対して示す特異性及び高親和性を利用するものである。測定対象となる物質は、必須条件として、免疫原性高分子又はハプテン小分子のいずれかの抗原性でなくてはならない。各試料に、既知の、限られた量の特異抗体を加え、結合:遊離比としてしばしば表される、この抗体と結合した抗原の比率を、放射性同位元素(ラジオイムノアッセイ)、蛍光分子(蛍光イムノアッセイ)、安定な遊離ラジカル(スピンイムノアッセイ)、酵素(酵素イムノアッセイ)等の容易に識別可能な標識で標識した形の抗原を指標として利用して、推定する。
【0083】
酵素結合免疫吸着検定(ELISA);ラジオイムノアッセイ(RIA);蛍光イムノアッセイ(FIA);化学発光イムノアッセイ(CLIA);及びコロイド状金粒子を用いた抗体の標識付け(イムノゴールド法)を含め、抗体には様々な態様で標識を付けることができる。
【0084】
一般的な検定・フォーマットには、サンドウィッチ検定、競合的又は競合検定、ラテックス凝集反応検定、ホモジェネアス(homogeneous)検定、微量定量プレートフォーマット及び微小粒子ベースの検定、がある。
【0085】
酵素結合免疫吸着検定(ELISA) − ELISAは、放射性化学物質の危険性及び蛍光検出系の費用を避けた免疫化学技術である。代わりに、この検定では酵素を指標として用いる。ELISAは、定量的イムノアッセイの一つの形であり、抗体(又は抗原)を不溶性の担体表面に結合させたものの利用に基づく。この担体表面を利用して、テスト溶液中の関連抗原(又は抗体)を「捕獲」するのである。こうして、前もって当該抗原(又は抗体)に共有結合させてある適当な酵素の活性を測定することで、抗原−抗体の複合体を検出する。
【0086】
ELISA技術に関する情報については、例えばCrowther, ELISA: Theory and Practise (Methods in Molecular Biology, Vol. 42), Humana Press, 1995; Challacombe & Kemeny, ELISA and Other Solid Phase Immunoassays: Theoretical and Practival Aspects, John Wiley, 1998; Kemeny, A Practical Guide to ELISA, Pergamon Press, 1991; Ishikawa, Ultrasensitive and Rapid Enzyme Immunoassay (Laboratory Techniques in Biochemistry and Molecular Biology, Vol. 27), Elsevier, 1991を参照されたい。
【0087】
酵素に関する比色検定 − 比色法とは、試薬と、標準量及びテスト量の両方の化合物とを反応させたときの色を、しばしば比色計を用いて比較することで、ある化合物の濃度又は量を判定するようなあらゆる定量的化学解析法を言う。比色計は、色の輝度又は色の輝度の違いを、視覚的又は光電学的に測定する装置である。
【0088】
ベータ−ガラクトシダーゼ酵素活性の標準的比色検定は、当業者に公知である(例えばNorton et al., (1985) Mol. Cell. Biol. 5, 281-290)。比色検定は、全細胞ライセートで、O-ニトロフェニル-ベータ-D-ガラクトピラノシド(ONPG、シグマ社製)を標準的比色ベータ-ガラクトシダーゼ検定(Sambrood et al., (1989) Molecular Cloning - A Laboratory Manual, Cold Spring Harbor Press)の基質として用いて行うことができる。自動化比色検定も、米国特許第5,733,720号が解説するようにベータ-ガラクトシダーゼ活性の検出に利用できる。
【0089】
免疫蛍光検定 − 免疫蛍光法又は免疫蛍光顕微鏡法は、抗原又は抗体を蛍光染料に結合させて蛍光性にした後、組織切片又はスメア中の相補の抗体又は抗原と反応させる技術である。次に、抗原又は抗体の位置を、この蛍光に紫外線を当てて顕微鏡で観察することで、判定することができる。
【0090】
免疫蛍光技術に関する概略的情報については、例えばKnapp et al., (1978) Immunofluorescence and Related Staining Techniques, Elsevier; Allan, (1999) Protein Localization by Fluorescent Microscopy: A Practical Approach (The Practical Approach Series, Vol. 218) Oxford University Press; Beutner, (1983) Defined Immunofluorescence and Related Cytochemical Methods, New York Academy of Sciences; Caul, (1993) Immunofluorescence Antigen Detection Techniques in Diagnostic Microbiology, Cambridge University Pressを参照されたい。本発明に応用できる免疫蛍光技術の詳細な解説については、米国特許第5,912,176号、第5,869,264号、第5,866,319号及び第5,861,259号を参照されたい。
【0091】
バイオチップ − 本発明のペプチドをアレイ又はマイクロアレイに利用して、本発明の核酸又は対応するタンパク質と相互作用する作用物質の高スループットスクリーニングを行うこともできる。
【0092】
「アレイ」又は「マイクロアレイ」とは、一般に、オリゴヌクレオチドとしても知られる定義済みの核酸断片で各位置又はプローブ・セルが占められているような格子系を言う。アレイ自体が、数千のプローブ・セルを多様な格子状に有する高密度核酸及びペプチドマイクロアレイである「チップ」又は「バイオチップ」と呼ばれることがある。
【0093】
典型的な分子検出チップには、認識部位、結合部位又はハイブリダイゼーション部位のアレイが上に並んだ基板がある。各部位は、所定の構造を有する分子と結合又はハイブリダイズする各分子受容基を有する。アレイ又はチップの表面を形成するのに利用できる固体の支持基板には、ガラス、ポリスチレン、ポリイミド、二酸化珪素及び窒化珪素などの有機及び無機の基板がある。プローブを電極に直接付着させる場合、プローブと共役体を形成できる物質で、電極表面を作製しなければならない。
【0094】
アレイを作製したら、試料溶液を分子検出チップに付着させると、試料中の分子が一箇所以上の部位で結合又はハイブリダイズする。結合が起きた部位を検出し、こうして試料中の一つ以上の構造を導き出す。標識群の試料を検出する方法は伝統的な検出法であり、その中には放射性標識、蛍光標識及びビオチン標識があるが、電気的シグナル伝達を含め、他の方法も利用できる。
【0095】
本発明の方法は、試料の高スループットが必要な場合には特に便利であろう。具体的には、本発明は、リガンドスクリーニングの場合や、複合体混合物の組成を決定する場合に、有用である。
【0096】
ポリペプチドは、生物学において構造及び機能間の関係を調べるための代表的な系である。二十種類の天然型アミノ酸が縮合してポリマー分子を形成すると、幅広い三次元立体配置を形成するが、その形状の各々は特定のアミノ酸配列及び溶媒条件の所産である。例えば、アミノ酸5個分の長さのポリマーを、二十種類の天然型アミノ酸を用いて形成するときに考えられるポリペプチドの立体配置の数は300万を超える。典型的なタンパク質は、100を越えるアミノ酸長である。
【0097】
典型的な用途としては、特徴付けようとする一種以上の物質を含有する複合溶液を、ポリペプチドを含んで成るポリマーアレイに接触させる。本発明のポリペプチドは、標準的固相技術を用いるなど、当業で公知の伝統的方法で調製できる。この標準的方法には、排他的固相合成、部分的固相合成法、画分凝縮法、伝統的溶液合成、及び組換えDNA技術がある(Merrifield, (1963) Am. Chem. Soc. 85, 2149-2152を参照されたい)。
【0098】
ある好適な実施例では、アレイのポリペプチド又はタンパク質を他の共受容基に結合させて、アレイ上でヘテロ二重鎖を形成させてもよい。さらに別の実施例では、アレイのポリペプチド又はタンパク質をペプチド又は小分子に結合させることもできる。
【0099】
D.本発明の抗炎症化合物の用途
本発明の抗炎症化合物(例えばNEMOの発現を調節する化合物、又は、NEMOの少なくとも一つの活性のアゴニスト又はアンタゴニストなどの化合物、など)を用いて、被験体の炎症を調節したり、炎症性疾患を治療又は診断することができよう。本方法には、本発明の抗炎症化合物を、炎症性疾患を治療するのに有効量、被験体に投与するステップが含まれる。
【0100】
ここで用いる「炎症性疾患」には、炎症を特徴とする、炎症で引き起こされる、炎症の結果起きる、又は炎症の影響を受けるようになる、疾患又は異常が含まれるものと、意図されている。炎症性疾患は、NEMO又はIKKβ機能及び活性に関連した、及び/又は、NF-κB媒介性プロセスに関連した生物学的及び病的プロセスが原因であっても、又は、関連していてもよい。炎症性疾患又は異常の例には、限定はしないが、急性及び慢性の炎症性異常、例えば喘息、乾癬、リウマチ性関節炎、変形性関節炎、乾癬性関節炎、炎症性腸疾患(クローン病、潰瘍性大腸炎)、敗血症、脈管炎、及び滑液包炎;自己免疫疾患、例えば狼瘡、多発性筋痛、リウマチ、強皮症、ウェグナー肉芽腫症、一過性動脈炎、クリオグロブリン血症、及び多発性硬化症;移植片拒絶;骨粗鬆症;実質性腫瘍を含む癌(例えば肺、CNS、結腸、腎臓、及び膵臓);アルツハイマー病;アテローム性硬化症;ウィルス(例えばHIV又はインフルエンザ)感染;慢性ウィルス(例えばエプスタイン−バー、サイトメガロウィルス、単純疱疹ウィルス)感染;及び毛細血管拡張性運動失調、がある。
【0101】
病的プロセスとは、有害な作用を生むような種類の生物学的プロセスを言う。例えば、NF-κBの発現調節不全には、特定の病的プロセスが根底にある炎症誘発性プロセスが伴う。ここで用いる場合の、ある抗炎症化合物が病的プロセスを調節する、とは、当該化合物が、このプロセスの程度又は重篤度を低下させることを言う。例えば、NEMO又はIKKβの発現又は少なくとも一つの活性を何らかの態様で低下、促進又は調節するような抗炎症化合物を投与すると、炎症誘発性の応答が妨げられたり、病的プロセスが調節されるであろう。
【0102】
従って、NF-κB炎症性成分を伴う疾患を治療するのに、本発明の抗炎症化合物を用いてもよい。このような疾患には、限定はしないが、骨粗鬆症、リウマチ性関節炎、アテローム性硬化症、喘息(Ray & Cohn, (1999) J. Clin. Invest. 104, 985-993; Christman et al., (2000) Chest 117, 1482-1487)及びアルツハイマー病、がある。NF-κB炎症性成分を伴う疾患のレビューについては、Epstein, (1997) New Eng. J. Med. 336, 1066-1071; Lee et al., (1998) J. Clin. Pharmacol. 38, 981-993; Brand et al., (1997) Exp. Physiol. 82, 297-304を参照されたい。
【0103】
本発明の抗炎症化合物が、治療にとって有用と思われる、炎症誘発性応答を伴う病的プロセスには、限定はしないが、喘息、アレルギー、例えばアレルギー性鼻炎、ユティカリア(uticaria)、アナフィラキシー、薬物過敏症、食物過敏症、等々;皮膚の炎症、例えば皮膚炎、湿疹、乾癬、接触性皮膚炎、日焼け、加齢、等々;関節炎、例えば変形性関節炎、乾癬性関節炎、狼瘡、脊椎炎、等々、がある。抗炎症化合物は、慢性閉塞性肺疾患及び慢性炎症性長疾患の治療にも有用である。さらに本発明の抗炎症化合物は、移植や癌治療での免疫抑制を含め、コルチコステロイドを用いるあらゆる用途で、コルチコステロイドの代わりに用いてもよい。
【0104】
ここで用いる用語「被験体」には、温血動物、好ましくはヒトを含むほ乳類、が含まれる。ある好適な実施例では、被験体は霊長類である。さらにより好適な実施例では、前記霊長類はヒトである。
【0105】
ここで用いる用語、被験体に「投与する」には、(ここに解説するような)製剤の形の抗炎症化合物などの抗炎症化合物を、被験体に、非経口又は経口経路による送達、筋肉内注射、皮下/皮内注射、静脈内注射、口腔内投与、経皮送達や、直腸、結腸、膣、鼻孔内もしくは呼吸器官経路(吸入など)による投与を含め、被験体の所望の位置に化合物を送達するのに適した何らかの経路で、投薬する、送達又は投与することが、含まれる。
【0106】
ここで用いる用語「有効量」は、被験体において、炎症性異常を治療するのに充分であるなど、所望の結果を達成するのに必要な投薬量及び時間で、有効となる量を包含する。ここで定義するような、本発明の抗炎症化合物の有効量は、被験体の疾患状態、年齢及び体重や、化合物が被験体の所望の応答を引き出す能力などの因子に応じて様々であろう。投薬計画は最適な治療応答が得られるよう、調節してもよい。有効量はまた、化合物の毒性もしくは有害作用(副作用など)が、治療上有益な効果を下回るような量である。
【0107】
本発明の抗炎症化合物の治療上有効量(即ち有効投薬量)は、約0.001乃至30mg/kg体重の範囲内であろうが、好ましくは約0.01乃至25mg/kg体重、より好ましくは約0.1乃至20mg/kg体重、そしてさらにより好ましくは約1乃至10mg/kg、2乃至9mg/kg、3乃至8mg/kg、4乃至7mg/kg、又は5乃至6mg/kg体重であるとよい。当業者であれば、限定はしないが、疾患又は異常の重篤度、以前の治療歴、被験体の全身の健康及び/又は年齢、及び他の罹患状態を含め、特定の因子が、被験体を効果的に治療するのに必要な投薬量を左右する場合があることは、理解されよう。さらに、治療上有効量の本発明の抗炎症化合物を用いた被験体の治療には、一回の治療を含めることができるが、好ましくは連続した治療が含まれる。一実施例では、約0.1乃至20mg/kg体重の範囲の本発明の抗炎症化合物で、一週間当たり1回、約1乃至10週間、好ましくは2乃至8週間、より好ましくは約3乃至7週間、そしてさらにより好ましくは約4、5、又は6週間、被験体を治療する。さらに、治療に用いる本発明の抗炎症化合物の有効投薬量を、特定の治療経過の間、増減させてもよいことは理解されよう。
【0108】
本発明の抗炎症化合物を単独で提供してもよいが、ある特定の病的プロセスを調節する他の作用薬と組み合わせて提供してもよい。例えば、本発明の抗炎症化合物を、他の公知の抗炎症剤と組み合わせて投与することもできる。本発明の方法で用いても良い公知の抗炎症剤は、Harrison's Principles of Internal Medicine, Thirteenth Edition, Eds. T.R. Harrison et al. McGraw-Hill N.Y., NY; 及び the Physicians Desk Reference 50th Edition 1997, Oradell New Jersey, Medical Economics Co.,(これら文献の内容全体を、言及をもってここに編入することを明示しておく)に見ることができる。本発明の抗炎症化合物及び付加的な抗炎症剤は、同じ薬学的組成物として被験体に投与しても、又は、異なる薬学的組成物として(同時又は異なる時期に)被験体に投与してもよい。
【0109】
さらに本発明は、細胞内でIκBが関与するシグナル伝達を調節する方法も提供する。本方法は、抗炎症化合物に細胞を接触させるなどにより、IKKβ活性を調節するステップを含む。この抗炎症化合物は、例えば、NEMOがNBDでIKKβと相互作用するのを抑制することで、IKKβ機能を抑制するものであってもよい。細胞は培養物内にあっても、インシツー、即ち天然ホスト内にあってもよい。
【0110】
診断用に利用する場合、本発明の抗炎症化合物に、蛍光性、放射性、化学発光性等の容易に検出できる分子で標識を付けてもよい。標識は、抗炎症化合物に直接又は間接的に共役させてよい。
【0111】
E.製剤
さらに本発明は、本発明の抗炎症化合物を、薬学的に許容可能な担体と一緒に含んで成る薬学的組成物を包含する。薬学的に許容可能な担体は、水や、ピーナッツ油、大豆油、鉱油、ごま油、等々の石油、動物性油脂、植物性油脂、又は合成のものも含む油脂類など、無菌の液体であってよい。当該薬学的組成物を静脈内投与する場合には、水が好適な担体である。生理食塩水や、水性デキストロース及びグリセロール溶液が、特に注射用溶液の場合には、液体の担体として、利用できる。適した薬剤用担体は、Gennaro et al., (1995) Remington's Pharmaceutical Sciences, Mack Publishing Companyに解説されている。本発明の組成物には、薬理学的に活性な物質に加え、活性化合物を加工して、作用部位への送達に薬学的に利用できる製剤にするのに便利な添加剤及び佐剤を含む薬学的に許容可能な担体を含有させてもよい。非経口投与用に適した製剤には、水溶性の塩など、水溶性の形の活性化合物の水溶液がある。加えて、活性化合物の懸濁液を、適宜油性の注射用懸濁液として、投与してもよい。適した親油性溶媒又は賦形剤には、脂肪油、例えばごま油、又は合成の脂肪酸エステル、例えばオレイン酸エチル又はトリグリセリド、がある。水性の注射用懸濁液には、カルボキシメチルセルロースナトリウム、ソルビトール、及びデキストランを含め、懸濁液の粘性を増す物質を含めてもよい。選択に応じ、懸濁液にはさらに安定剤も含めてもよい。さらに、細胞への送達用に、リポソームを用いて当該作用薬を被包してもよい。
【0112】
本発明に基づく全身投与用の製剤は、腸内用、非経口用又は局所用投与に向けて、調合してもよい。実際、有効成分の全身への投与を行うために、三種類の製剤すべてを同時に用いてもよい。
【0113】
経口投与に適した製剤には、被覆錠剤、エリキシル、懸濁液、シロップ又は吸入剤や、これらの制御放出型を含め、硬質もしくは軟質ゼラチンカプセル、丸剤、錠剤、がある。
【0114】
さらに、少なくとも数週間から一ヶ月又はそれ以上の期間にわたって被験体への抗炎症化合物の送達を持続させる薬学的組成物に、本発明の抗炎症化合物を組み込んでもよい。このような製剤は、各内容を言及をもってここに編入することとする米国特許第5,968,895号及び第6,180,608号B1に解説されている。
【0115】
本発明の抗炎症化合物を、非経口、皮下、静脈内、筋肉内、腹腔内、経皮又は口腔内経路で投与してもよい。あるいは、又は同時に、投与を経口経路で行ったり、又は、吸入もしくは肺への直接の潅注で行ってもよい。投与する投薬量は、レシピエントの年齢、健康、及び体重、あれば併用療法の種類、治療の頻度、及び、所望の効果の性質、に左右されるであろう。
【0116】
ここで解説した治療法で用いる抗炎症化合物は、治療しようとする状態、部位特異的治療の必要性、投与する薬物の量、等の考慮点などの考慮点に応じて、全身又は局所的に投与してよい。
【0117】
局所投与を利用してもよい。溶液、懸濁液、ゲル、軟膏(ointment)又は軟膏剤(salve)などの通常の局所用製剤を用いてもよい。このような局所用製剤の製剤は、例えばレミントンズ・ファーマシューティカル・サイエンセズなどに例示された薬剤調剤業でよく解説されている。局所用投与の場合、これらの化合物を粉末又はスプレー、特にエーロゾル型として投与してもよいであろう。全身投与用に適合させた薬学的組成物に有効成分を含めて投与してもよい。公知のように、薬物を全身投与する場合、それを粉末、丸剤、錠剤に調製しても、又は、経口投与に向けたシロップ又はエリキシルとして調製してもよい。静脈内、腹腔内又は病変部内投与の場合、注射で投与できる溶液又は懸濁液として、化合物を調製することになるであろう。場合によっては、これらの化合物を座薬型に調製したり、又は、皮下又は筋肉内注射用のデポジット(deposit)の長時間放出製剤として調製すると、便利であろう。ある好適な実施例では、本発明の抗炎症化合物を吸入で投与してもよい。吸入治療の場合、当該化合物は、定量吸入器による投与に便利な溶液か、又は、乾燥粉末吸入器に適した形であってもよい。
【0118】
有効量とは、目的たんぱくの活性を調節したり、又はレベルを変化させるような量である。有効量は条件に応じて様々であり、場合によっては、治療しようとする状態の重篤度や、治療に対する患者の感受性によっても、様々であろう。従って、通常の実験を通じて、その場その場で有効量を決めるとよいであろう。しかしながら、本発明に基づいて腫瘍を治療する場合、重量で0.001乃至5パーセント、好ましくは約0.01乃至1パーセントを含有する製剤が、多くの場合に治療上有効量となるであろうことが予測できる。全身投与の場合、一日当たり体重1kg当たり0.01乃至100mg、より好ましくは1kg当たり約0.1乃至10mgの量が、多くの場合に治療効果を生じることとなろう。
【0119】
本発明の方法を実施する際、本発明の化合物を単独で用いても、又は併用してもよく、あるいは他の治療的又は診断的作用薬と併用してもよい。いくつかの好適な実施例では、本発明の化合物を、広く認められた医療行為に基づいてこれらの状態に典型的に処方されている他の化合物と同時に投与してもよい。本発明の化合物を、通常は哺乳動物で、好ましくはヒトで、インビボで利用することもできる。
【0120】
さらに別の実施例では、標的たんぱくの機能又は調節を、治療上の利益に向けて変更するタンパク質を含め、化学的部分に本発明の抗炎症化合物を結合させてもよい。これらのタンパク質には、炎症性異常の治療にとってさらに治療上有益となると思われる他のサイトカイン阻害剤及び成長因子阻害剤が組み合わされていてもよい。加えて、本発明の抗炎症化合物に、当業で公知の何らかの架橋試薬を用いたリン酸化を行って、ビオチニレート、チオエート、アセチレート、イオジネートに共役させてもよい。
【0121】
F.分子生物学、微生物学及び組換えDNA技術
本発明によれば、上で解説したか、又は、下記の実施例で論じるように、従来の分子生物学、微生物学及び組換えDNA技術を利用することになるであろう。このような技術は文献に十二分に説明されている。例えばSambrook et al., (1989) Molecular Cloning - A Laboratory Manual, Cold Spring Harbor Press; Glover, (1985) DNA Cloning: A Practical Approach; Gait, (1984) Oligonucleotide Synthesis; Harlow & Lane, (1988) Antibodies - A Laboratory Manual, Cold Spring Harbor Press; Roe et al., (1996) DNA Isolation and Sequencing: Essential Techniques, John Wiley; 及びAusubel et. al., (1995) Current Protocols in Molecular Biology, Greene Publishingを参照されたい。
【0122】
G.アンチセンスRNA
アンチセンス分子は、特定のmRNAに相補なヌクレオチド配列を持つRNA又は一本鎖DNA分子である。研究室で調製したアンチセンス分子を、研究対象の遺伝子が転写した正常mRNAを含有する細胞に注射すると、このアンチセンス分子はこのmRNAと塩基対を形成することで、mRNAのタンパク質への翻訳を妨げることができる。その結果できる二本鎖RNA又はRNA/DNAは、このような分子に特異的に結合する酵素で消化される。従って、mRNAの消去が起きて遺伝子産物の翻訳が遮断される。このように、アンチセンス分子は、有害たんぱくの産生を遮断する医療において有用である。アンチセンスRNAを作製し、利用する方法は当業者に公知である(例えばLichtenstein & Nellen (編集者), Antisense Technology: A Practical Approach, Oxford University Press, 1997; Agrawal & Crooke, Antisense Research and Application (Handbook of Experimental Pharmacology, Vol. 131), Springer Verlag, 1998; Gibson, Antisense and Ribozyme Methodology: Laboratory Companion, Chapman & Hall, 1997; Mol & Van Der Krol, Antisense Nucleic Acids and Proteins, Marcel Dekker; Weiss, Antisense Oligodeoxynucleotides and Antisense RNA: Novel Pharmacological and Therapeutic Agents, CRC Press, 1997; Stanley et al., (1993) Antisense Research and Applications, CRC Press; Stein & Krieg, (1998) Applied Antisense Oligonucleotide Technologyを参照されたい)。
【0123】
本発明によるアンチセンス分子及びリボ酵素は、核酸分子の合成について当業で公知のいかなる方法で調製してもよい。これらの中には、固相ホスホールアミジト化学合成など、オリゴヌクレオチドを化学合成する技術がある。代わりに、DNA配列のインビトロ及びインビボ転写でもRNA分子を作製できよう。このようなDNA配列を、T7又はSP6などの適したRNAポリメラーゼプロモータを持つ多種のベクタに組み込んでもよい。代わりに、アンチセンスRNAを構成的又は誘導的に合成するこれらのcDNAコンストラクトを細胞株、細胞、又は組織内に導入することもできる。RNA分子は、細胞内での安定性や半減期を向上させるよう、修飾してもよい。可能な修飾には、限定はしないが、当該分子の5’末端及び/又は3’末端へのフランキング配列の追加、又は、当該分子の骨格内にホスホジエステラーゼ結合の代わりにホスホロチオエート又は2'O-メチルを用いる、などがある。この発想を進展させて、イオシン、ケオシン、及びワイブトシンなどの非伝統的な塩基を含めたり、内因性エンドヌクレアーゼが容易には認識できない、アセチル修飾型、メチル修飾型、チオ修飾型、等の修飾型アデニン、シチジン、グアニン、チミン、及びウリジンを含めてもよい。
【0124】
H.融合タンパク質
融合タンパク質は、二つの遺伝子の融合の結果生じる発現産物である。このようなタンパク質は、例えば組換えDNA発現研究で生じたり、又は、腫瘍遺伝子がgagに融合したいくつかのウィルス性腫瘍遺伝子で天然に生じることもある。
【0125】
時には、細菌系で発現させるために、細菌性プロモータの制御下に真核生物のクローン遺伝子を配置する必要性から、融合タンパク質が生ずることもある。こうして、細菌系の配列は、真核生物たんぱくに連結した形で発現することが多い。融合タンパク質は、異種遺伝子産物の構造解析や、精製、機能、及び発現解析に用いられる。
【0126】
融合タンパク質は、目的の核酸を組換えDNA技術によりレシピエント・プラスミド内に挿入し、プラスミド遺伝子の停止コドンに替えたときに生じさせることのできるハイブリッドたんぱく分子である。融合されたタンパク質は、プラスミドたんぱく配列の一部分を持つアミノ末端で始まり、目的のタンパク質で終わる。
【0127】
融合タンパク質の作製は当業者に公知である(例えば米国特許第5,908,756号、第5,907,085号、第5,906,819号、第5,905,146号、第5,895,813、第5,891,643号、第5,891,628号、第5,891,432号、第5,889,169号、第5,889,150号、第5,888,981号、第5,888,773号、第5,886,150号、第5,886,149号、第5,885,833号、第5,885,803号、第5,885,779号、第5,885,580号、第5,883,124号、第5,882,941号、第5,882,894号、第5,882,864号、第5,879,917号、第5,879,893号、第5,876,972号、第5,874,304号、及び第5,874,290号を参照されたい)。臨床及び研究医療で用いられている特定の種類の融合分子に関連する構造、性質、用途、及び問題の概略的レビューについては、例えばChamow et al., (1999) Antibody Fusion Proteins, John Wileyを参照されたい。
【0128】
I.ペプチドミメティック
さらに本発明は、IKKβ上のNBDの三次元構造を模倣して、NEMOに結合することで、NEMOがこのNBDで結合するのを遮断するペプチドミメティックなどの、ペプチドミメティックも包含する。このようなペプチドミメティックは、例えばより生産コストが低い、化学的安定性が高い、薬理学的性質(半減期、吸収、効能及び効験)が向上している、特異性が変更されている(例えば幅広い生物活性など)、抗原性が低い、等々を含め、天然型のペプチドに比べて有意な長所を有していることがある。
【0129】
ある一つの形としては、ミメティックは、タンパク質の二次構造の要素を模倣したペプチド含有分子である。例えばJohnson et al., (1993) Peptide Turn Mimetics in Biotechnology and Pharmacy, Pezzuto et al., (Editors) Chapman & Hallを参照されたい。ペプチドミメティックを利用する根拠は、タンパク質のペプチド骨格は、主に、抗体対抗原などの分子間相互作用を促進するような方向にアミノ酸側鎖を方向付けるよう、並んでいることである。ペプチドミメティックでも、天然分子に類似の分子間相互作用が可能であると予測できる。別の形では、ペプチド類似体は、テンプレートペプチドのものに類似の性質を持つ非ペプチド化合物として、製薬業で広く用いられている。これらの種類の非ペプチド化合物も「ペプチドミメティック」又は「ペプチドミメティック」と呼ばれており( 言及をもってここに編入するFauchere, (1986) Adv. Drug Res. 15, 29-69; Veber & Freidinger, (1985) Trends Neurosci. 8, 392-396; and Evans et al., (1987) J. Med. Chem. 30, 1229-1239)、一般にコンピュータによる分子モデリングで開発されている。
【0130】
治療上有用なペプチドに構造上類似のペプチドミメティックを用いれば、同等の治療効果又は予防効果が得られるであろう。一般に、ペプチドミメティックは、NBDなどのパラダイムポリペプチド(即ち、生化学的性質又は薬理活性を有するポリペプチド)に構造上類似であるが、一箇所以上のペプチド結合が、選択に応じて-CH2NH-、-CH2S-、-CH2-CH2-、-CH=CH- (cis 及びtrans)、-COCH2-、-CH(OH)CH2-、及び-CH2SO-からなる群より選択される結合に、当業で公知の方法であり、さらに以下の文献:Weinstein, (1983) Chemistry and Biochemistry of Amino Acids, Peptides and Proteins, Marcel Dekker; Morley, (1980) Trends Pharmacol. Sci. 1, 463-468 (概略的レビュー); Hudson et al., (1979) Int. J. Pept. Protein Res. 14, 177-185 (-CH2NH-、CH2CH2-); Spatola et al., (1986) Life Sci. 38, 1243-1249 (-CH2-S); Hann, (1982) J. Chem. Soc. Perkin Trans. 1, 307-314 (-CH-CH-, cis 及びtrans); Almquist et al., (1980) J. Med. Chem. 23, 1392-1398 (-COCH2-); Jennings-White et al., (1982) Tetrahedron Lett. 23, 2533 (-COCH2-); 米国特許出願第4,424,207号 (-CH(OH)CH2-); Holladay et al., (1983) Tetrahedron Lett. 24, 4401-4404 (-C(OH)CH2-); 及び Hruby, (1982) Life Sci. 31, 189-199 (-CH2-S-)(これらの文献各々を、言及をもってここに編入する)に解説された方法で、置換されている。
【0131】
ペプチドミメティックの標識付けには、通常、定量的構造−活性データ及び/又は分子モデリングで予測された、ペプチドミメティック上の非干渉位置に、直接的に又はスペーサ(例えばアミド基)を介して、一つ以上の標識を共有結合させる方法がある。このような非干渉位置とは、一般に、当該ペプチドミメティックが結合して治療効果を生ずる相手である高分子と直接接触しない(例えばNBD-NEMO複合体上の接点ではない)位置である。ペプチドミメティックの誘導体化(標識付けなど)は、当該ペプチドミメティックの所望の生物活性又は薬理活性に大きく干渉するものであってはならない。NBDペプチドミメティックは、有機成分でアミノ酸を置換する方法を通じ、構造ベースの薬物デザインにより構築することができる(例えば Hughes, (1980) Philos. Trans. R. Soc. Lond. 290, 387-394; Hodgson, (1991) Biotechnol. 9, 19-21; Suckling, (1991) Sci. Prog. 75, 323-359を参照されたい)。
【0132】
コンビナトリアル化学法を用いて薬物ライブラリを作製すると、ペプチドミメティックの利用を向上させることができる。ペプチドミメティックの設計には、NBD(例えばIKKβ上のNBD)のNEMOへの結合を増加又は減少させるアミノ酸変異の特定が役立つであろう。例として、このような変異を表1で特定した。利用の可能な方法には、酵母二種ハイブリッド法(Chien et al., (1991) Proc. Natl. Acad. Sci. USA 88, 9578-9582を参照されたい)及びファージ展示法の利用がある。二種ハイブリッド法では、タンパク質対タンパク質の相互作用を酵母で検出する (Fields et al., (1989) Nature 340, 245-246)。ファージ展示法では、固定したタンパク質と、ラムダ及びM13などのファージの表面上に発現させたタンパク質との間の相互作用を検出する(Amberg et al., (1993) Strategies 6, 2-4; Hogrefe et al., (1993) Gene 128, 119-126)。これらの方法により、タンパク質対タンパク質の相互作用による正及び負の淘汰や、このような相互作用を決定する配列の同定が可能である。
【0133】
ペプチド合成及びペプチドミメティックに関する概略的情報については、例えば、ここに言及をもって各全文を編入することとするJones, (1992) Amino Acid and Peptide Synthesis, Oxford University Press; Jung, (1997) Combinatorial Peptide and Nonpeptide Libraries: A Handbook, John Wiley; and Bodanszky et al., (1993) Peptide Chemistry: A Practical Textbook, 2nd Revised Edition, Springer Verlagを参照されたい。
【0134】
J.トランスジェニック動物
トランスジェニック動物は、組換え、外因性もしくはクローン形成した遺伝物質を実験的に移入させた、遺伝子改変動物である。このような遺伝物質はしばしば導入遺伝子と呼ばれる。導入遺伝子の核酸配列は、この核酸配列が存在しない、もしくは、通常見られないようなゲノム上の遺伝子座や、又は、この導入遺伝子にとって正常な遺伝子座のいずれにも、組み込まれることがある。導入遺伝子は、標的動物と同じ種のゲノム由来の核酸配列や、又は、異なる種のゲノム由来の核酸配列から成る場合がある。
【0135】
「生殖細胞系トランスジェニック動物」という用語は、遺伝子改変又は遺伝情報が、生殖系の細胞に導入されたことで、このトランスジェニック動物に、遺伝情報を子孫に伝える能力がもたらされたようなトランスジェニック動物を言う。実際にこのような子孫が、その改変又は遺伝情報の一部又は全部を有していれば、彼らもトランスジェニック動物である。
【0136】
改変又は遺伝情報は、レシピエントが属する動物種にとって異種であっても、特定の個体レシピエントにとってのみ、異種であっても、又は、レシピエントが既に持つ遺伝情報であってもよい。最後者の場合、改変したもしくは導入された遺伝子に、天然遺伝子とは異なる発現をさせてもよい。
【0137】
トランスジェニック動物は、トランスフェクション、電気穿孔法、顕微注射、胚性幹細胞への遺伝子ターゲティング、及び組換えウィルス及びレトロウィルス感染法を含め、多種の様々な方法で作製することができる(例えば米国特許第4,736,866号、米国特許第5,602,307号、Mullins et al., (1993) Hypertension 22, 630-633; Brenin et al., (1997) Surg. Oncol. 6, 99-110; Tuan, (1997) Recombinant Gene Expression Protocols, Methods in Molecular Biology No. 62, Humana Pressを参照されたい)。
【0138】
数多くの組換えもしくはトランスジェニックマウスが作製されてきており、その中には、活性化腫瘍遺伝子配列を発現するもの(米国特許第 4,736,866号、);シミアンSV40 T-抗原を発現するもの(米国特許第5,728,915号);インターフェロン調節因子1(IRF-1)の発現を欠失させたもの(米国特許第5,731,490号);ドーパミン作動性機能不全を呈するもの(米国特許第5,723,719号);血圧制御に関与する少なくとも一つのヒト遺伝子を発現するもの(米国特許第5,731,489号);天然型アルツハイマー病における状態に、より大きな類似性を呈するもの(米国特許第5,720,936号);細胞接着を媒介する能力を低下させたもの(米国特許第5,602,307号);ウシ成長ホルモン遺伝子を持つもの(Clutter et al., (1996) Genetics 143, 1753-1760) 又は、完全にヒト抗体応答を生じられるもの(Zou et al., (1993) Science 262, 1271-1274)、がある。
【0139】
マウス及びラットは、依然、大半のトランスジェニック実験で選択される動物であるが、場合によっては、別の動物種を用いるのが好ましいか、又は必要なことがある。トランスジェニック法は、ヒツジ、ヤギ、ブタ、イヌ、ネコ、サル、チンパンジー、ハムスター、ウサギ、ウシ及びモルモットを含め、多種のマウス以外の動物で利用されて、成功が収められてきた(Kim et al., (1997) Mol. Reprod. Dev. 46, 515-526; Houdebine, (1995) Reprod. Nutr. Dev. 35, 609-617; Petters, (1994) Reprod. Fertil. Dev. 6, 643-645; Schnieke et al., (1997) Science 278, 2130-2133; Amoah, (1997) J. Animal Science 75, 578-585を参照されたい)。
【0140】
核酸断片を組換えコンピテントほ乳類細胞へ導入する方法は、複数の核酸分子の同時形質転換にとって有利な方法であれば、いかなるものでもよい。トランスジェニック動物を作製する手法の詳細は、米国特許第5,489,743号及び米国特許第5,602,307号の開示を含め、当業者が容易に利用できるものである。
【0141】
本発明は、NBDをコードする遺伝子を発現するトランスジェニック動物と、この野生型遺伝子に比較したときの保存的及び非保存的アミノ酸置換を生ずるようなこの遺伝子の変異を、包含するものである。
さらに本発明を以下の実施例で解説することとするが、同実施例を限定的なものと捉えられてはならない。本出願を通じて引用された全参考文献、特許及び公開済み特許出願の内容や、図面及び配列表を、言及をもってここに編入することとする。
【図面の簡単な説明】
【0142】
【図1A】図1は、NEMOがIKKβのCOOH末端と相互作用することを示した実験結果を示す。図1A:GST単独又はGST-NEMOを、グルタチオン-アガロースを用いて細菌ライセートから沈降させ、SDS-PAGE(10%)で分離し、このゲルをクーマシー・ブルーで染色した(左側パネル)。等量のGST又はGST-NEMOを次のGSTプル−ダウン実験で用いた。右側パネルに表したスキームは、NEMO相互作用領域を調べるのに用いた、IKKβのCOOH及びNH2末端切断型変異体を表す。
【図1B】IKKβ変異体をクローンし、インビトロ翻訳で発現させ(インプット;左側パネル)て、GSTプル−ダウンに用いた(右側パネル)。
【図1C】野生型IKKβ及びIKKβ-(644-756)をインビトロ翻訳させ(左側パネル)、GSTプル−ダウン解析に用いた(左側パネル)。
【図1D】ヒーラ細胞を、pBIIX-ルシフェラーゼレポータプラスミドと一緒に、ベクタのみか、又は、次第に濃度を上げた(0.25、0.5、1.0μg/ml)、xpressタグ付きIKKβ-(644-756)コンストラクトで、一時的にトランスフェクトした。48時間後、細胞をTNFα(10ng/ml)又はIL-1β(10ng/ml)で4時間処置し、その後NF-κB活性を測定した。ライセートの一部を抗xpress(挿入図)を用いてウェスタンブロット解析したところ、発現したたんぱくも次第に高レベルになっていることが分かる。
【図2A】図2は、NEMOの一番目のαへリックス領域がIKKβへの結合に必要であることを示した実験結果を表す。図2A:残基V644からS756までのCOOH末端のみから成る切断型のIKKβをGSTに融合し(GST-644-756)、細菌で発現させた。グルタチオンアガロースで沈殿させた後、GST単独及びGST-(644-756)をSDS-PAGE(10%)で分離し、そのゲルをクーマシー・ブルーで染色した(左側パネル)。等量の各タンパク質を次のGSTプル−ダウン解析に用いた。NEMOの様々なNH2及びCOOH末端切断型を構築し、[35S]-メチオニンで標識し、インビトロプル−ダウンに用いた(右側パネル)。GST-(644-756)と相互作用した変異体を(+)で示す。変異体のいずれも、GST単独とは相互作用しなかった。
【図2B】野生型NEMOと、一番目のαへリックス領域を欠失させた欠失変異体(del.αH)とをインビトロで翻訳し(左側パネル:インプット)、上記のタンパク質を用いたGSTプル−ダウンに利用した(A:左側)。
【図2C】ヒーラ細胞に、pcDNA-3(ベクタ)又は次第に濃度を高くしたdel.αH(0.25、0.5、1.0μg/ml)のいずれかと一緒に、pBIIx-ルシフェラーゼを48時間、トランスフェクトし、次に、TNFα(10ng/ml)で4時間処置した。次に細胞を溶解させ、NF-κB活性をルシフェラーゼ検定で測定した。
【図3A】図3は、NEMOとの相互作用や、機能的キナーゼ活性には、IKKβ COOH-末端のIKKα-相同領域が必要であることを示した実験結果を示す。図3A:最末端のCOOh末端(1-733)、セリンなしの領域(1-707)、セリンリッチなドメイン(1-662)及びα1領域(1-644)を順に省略した切断変異型のIKKβをインビトロ翻訳で発現及び標識付けし、GST-NEMOによるGSTプル−ダウンに用いた(図1A)。変異体のいずれも、GST単独とは相互作用しなかった。
【図3B】IKKβ及びIKKαの最末端COOH末端の配列アラインメント。α2領域及びグルタミン酸リッチな領域を示し、アミノ酸が6個同一になっている部分は網がけした。
【図3C】野生型IKKβ及び切断変異型(1-733及び1-744)を[35S]-メチオニンで標識し(インプット)、GST単独又はGST-NEMOによるインビトロプル−ダウンに用いた。
【図3D】ヒーラ細胞に、1μg/ウェルの提示したFLAGでタグを付けたコンストラクトで48時間、トランスフェクトした後、抗FLAGを用いて免疫沈降させた。[32P]で標識したγATPを含有するキナーゼバッファ中でこの免疫沈降物を15分間、30℃でインキュベートし、1%トリトン-100を含有する溶解バッファで洗浄した。その結果得られた複合体を、SDS-PAGE(10%)で分離し、オートラジオグラフィで観察した(上側パネル)。下側のパネルは同一試料の免疫ブロットであり、各レーンにトランスフェクトしたタンパク質が等量、現れている。
【図3E】ヒーラ細胞に、提示したコンストラクト1μg/ml又は空のベクター(pcDNA-3)を、pBIIx-ルシフェラーゼと一緒に48時間かけてトランスフェクトした。NF-κB活性はルシフェラーゼ検定で調べた。(F)FLAGでタグを付けた形のIKKβ(野生型)又はIKKβ(1-733)を48時間かけてトランスフェクトしたヒーラ細胞を、処置しない(-)か、又はTNFα(10ng/ml)で7分間、処置した(+)。溶解させ、抗FLAGを用いて免疫沈降させた後、免疫複合体キナーゼ検定(上側パネル)を行った。同一試料を免疫沈降させ、抗FLAGで免疫ブロットした(下側パネル)。
【図4A】図4は、IKKβ及びIKKαとのNEMOの結合の結果、NEMO結合ドメイン(NBD)は6個のCOOH末端アミノ酸であることが判明した実験結果を示す。図4A:提示するように、合計DNA濃度が2μg/ウェルになるようにベクタ単独、FLAGでタグを付けたIKKα又はIKkβ(1μg/ウェル)又はxpressでタグを付けたNEMO(1μg・ウェル)を一時的にトランスフェクトしたCOS細胞。溶解後、免疫沈降(IP)を抗FLAG(M2)を用いて行い、沈降物の含有量を、抗FLAG(M2)又は抗xpressを用いた免疫ブロット(IB)で観察した。IP前のライセートの一部を抗xpressで免疫ブロットして、同等のレベルのNEMO発現がトランスフェクト後の細胞にあることを確認した。
【図4B】野生型IKKα及びIKKα-(1-737)を発現させ、標識し(インプット)、GST又はGST-NEMOを用いたGSTプル−ダウンに用いた。
【図4C】ヒトIKKiをコードする完全長cDNAを、RT-PCRにより、ヒーラ細胞mRNAからExpandTMロングテンプレートPCRシステム(ベーリンガー・マンハイム社製)、正方向プライマー(5’-CTAGTCGAATTCACCATGCAGAGCACAGCCAATTAC) (SEQ ID NO: 22) 及び逆方向プライマー(3’-CTAGTCTCTAGATTAGACATCAGGAGGTGCTGG) (SEQ ID NO: 23)を用いて得て、pcDNA-3のEcoRI 及びXbaI 部位にクローンした。GSTプル−ダウン解析は、[35S]-メチオニンで標識したIKKα、IKKβ及びIKKiを用いて行った。
【図4D】NBDを欠失した欠失変異型のIKKβ(del.NBD)を[35S]-メチオニンで標識し て(インプット)、GSTプルダウン解析に用いた。
【図4E】ヒトIKKβのCOOH末端(N721-S756)のフォーシャー−プリスカ(Fauchere-Pliska)疎水性プロットを、MacVector□(バージョン6.5.3)ソフトウェアを用いて作製した。NBD(L737-L742)は四角で囲んである。
【図4F】COS細胞に、48時間かけて、合計2μgのDNA/ウェルになるよう、ベクタのみ、ベクタにNEMO-FLAGを加えたもの、又は、NEMO-FLAGに、提示するようにNBD内に点変異を含有するxpressでタグを付けた形のIKKβ-(1-744)を加えたもの、でトランスフェクトした。溶解させ、抗FLAG(M2)を用いて免疫沈降させた後、抗FLAG又は抗xpressで免疫ブロット解析を行った。IP前のタンパク質発現レベルは、抗xpressで免疫ブロットして調べた(下側パネル)。
【図4G】ヒーラ細胞を、48時間かけて、pBIIX-ルシフェラーゼと一緒に、提示したコンストラクトで一時的にトランスフェクトして、ライセート中のNF-κB活性をルシフェラーゼ検定で測定した。
【図5A】図5は、IKKβ NBDを包含する細胞透過性ペプチドがIKKβ/NEMO相互作用、TNFαで誘導されるNF-κB活性化及びNF-κB依存性遺伝子発現を抑制することを示した実験結果を示す。図5A:GST-プル−ダウン解析を、GST-NEMOとインビトロで翻訳されたIKKβの組(上側パネル)か、又はGST-IKKβ-(644-756)とインビトロで翻訳されたNEMOの組(下側パネル)を用いて行った。この検定は、次第に濃度を上昇させた(125、250、500又は1000μM)の変異型(MUT)もしくは野生型(WT)NBDペプチドの不在下(ペプチドなし)又は存在下で行った。
【図5B】ヒーラ細胞はいずれかのペプチド(200μM)と一緒に、指定した時間、インキュベートした。溶解後、IKK複合体を、抗NEMOを用いて免疫沈降させ、その結果得られた免疫ブロットを抗IKKβでプローブした。
【図5C】ヒーラ細胞を、48時間かけて、pBIIX-ルシフェラーゼでトランスフェクトした後、変異型もしくは野生型NBDペプチド(それぞれ100及び200μM)の不在下(コントロール)又は存在下で、2時間、インキュベートした。その後、細胞をTNFα(10ng/ml)で提示したように処置する(左側パネル)か、又は、その後4時間、未処置(右側パネル)のままにしてから、NF-κβ活性化をルシフェラーゼ検定で測定した。
【図5D】ヒーラ細胞を3時間、次第に濃度を上昇させた(50、100又は200μM)の各ペプチドと一緒にインキュベートし、その後15分間、TNFα(10ng/ml)で、提示したように処置した(+)。溶解後、核抽出液を作製し、各試料からの10μgのタンパク質を、[32P]標識付κB部位プローブを用いたEMSAに利用した。
【図5E】始原HUVECを2時間、野生型(左側)又は変異型(右側)NBDペプチド(100μM)と一緒にプレインキュベートした後、TNFα(10ng/ml)でその後6時間、刺激した。コントロール細胞にはペプチドは与えなかった。細胞を抗E-セレクチン(H4/18)又は非結合性コントロール抗体(K16/16)で染色し、FACS(ベクトン・ディッキンソン社製、FACSort)で発現を測定した。輪郭は、同じ条件下でTNFα及びコントロール抗体の不在下(網掛け部分)及び存在下(実線)でのE-セレクチン染色を示す(断続線、TNFαなし;点線、TNFαあり)。
【図6A】図6は、野生型NBDペプチドが、NF-κBが誘導する遺伝子発現及び実験的に誘導した炎症を抑制することを示した実験結果を示す。図6A:マウスに賦形剤(VEH)、デキサメタゾン(DEX)又はNBDペプチドを局所的に処置して、PMA誘発性の耳水腫を起こして、実施例8で説明したように測定した。データは、耳の厚さの平均差±SD(未処置のコントロール[-]及び賦形剤[VEH]の両方に比較したときに*=p<0.05)で表されている。
【図6】ザイモサン(ZYM)でマウスに誘発した腹膜炎に及ぼす、デキサメタゾン(DEX)の作用に比較したときのNBDペプチドの作用を、実施例8で解説したように調べた。コントロールマウスには、リン酸緩衝生理食塩水(PBS)を注射した。
【図7】図7は、破骨細胞分化は野生型により用量依存的に抑制されたが、変異型NBDペプチドでは抑制されなかったことを示した実験結果を示す。データは、三重式試料±SDの平均値で表されている。
【図8A】図8は、ヒトIKKβのNEMO結合ドメイン(NBD)内のD738の変異解析結果を示す。図8A:IKKβの738位のアスパラギン酸残基をアラニン、アスパラギン又はグルタミン酸に、PCR変異誘発法を用いて置換した。
【図8B】図8Aで示したIKKβ(D738)変異体にインビトロ転写及び翻訳で35S-メチオニンで標識した後、前に解説されたようにGST-NEMOを用いたGSTプル−ダウン解析に用いた。
【図8C】Fugene6トランスフェクション法を利用して、上述のpcDNA-3、IKKβ又はD738変異体のいずれかと一緒にpBIIx-ルシフェラーゼを持つNF-κB依存性レポータ・コンストラクトで、ヒーラ細胞を一時的にトランスフェクトした。48時間後、細胞を溶解させ、ルシフェラーゼ活性を前に解説された通りに調べた。
【図9A】図9は、ヒトIKKβのNBD内のW739及びW741の変異解析結果を示す。図9A:IKKβの739位及び741位のトリプトファン残基を、アラニン、フェニルアラニン、チロシン又はアルギニンに、PCR変異誘発法を用いて置換した。
【図9B】FLAGでタグを付けたNEMOと一緒に、ベクタのみ(pcDNA-3.1-xpress)、IKKβ、W739A、W739F又はW739YでCOS細胞を一時的にトランスフェクトした。48時間後、細胞を溶解させ、抗FLAG(M2)共役アガロースビーズを利用して複合体を免疫沈降(IP)させた。免疫沈降前に、各ライセートの一部(5%)を解析用にとっておいた(pre-IP)。試料中のタンパク質をSDS-PAGE (10%)で分離し、FLAG(M2)又はxpressを認識する抗体を用いた免疫ブロッティング(IB)で解析した。上側二つのパネルは、xpressのタグが付いたIKKβを示し、下側のパネルはFLAGのタグが付いたNEMOを表す。
【図9C】COS細胞に図示のプラスミドでトランスフェクトした後、Bで解説したように免疫沈降及び免疫ブロット解析を行った。
【図9D】図9Cと同じく図示のプラスミドと一緒にpBIIx-ルシフェラーゼでヒーラ細胞を一時的にトランスフェクトし、48時間後にライセート中のルシフェラーゼ活性を調べた。
【図10A】図10は、ヒトIKKβのNBD内のS740の変異解析結果を示す。IKKβの740位のセリン残基をアラニン又はグルタミン酸に、PCR変異誘発法を用いて置換した。
【図10B】COS細胞に図示のプラスミドで一時的にトランスフェクトし、図2Bで解説したように免疫沈降及び免疫ブロット解析を行った。
【図10C】IKKβ-FLAG又はS740E-FLAGのいずれかでヒーラ細胞を48時間かけて一時的にトランスフェクトした後、TNFα(1μg/ml)で図示の時間、処置した。溶解後、抗FLAG(M2)共役アガロースビーズを用いて複合体を沈降させ、前に解説されたように、GST-IκBα(1-90)を基質として用いて免疫-複合体キナーゼ検定を行った。
【図11A】図11は、IKKα NBDの変異解析結果を示す。図11A:IKKαのNBDを含む各残基(L738乃至L743)を、PCR変異誘発法によりアラニンに置換した。COS細胞を、ベクタのみ(pcDNA-3.1-xpress)又はxpressでタグを付けた形のIKKα及びNBD変異体のいずれかと一緒にNEMO-FLAGで図示のように一時的にトランスフェクトした。このIKkα-NEMO複合体の免疫沈降及び免疫ブロット解析を図2Bで解説したように行った。
【図11B】ヒーラ細胞を、図示のプラスミドと一緒にpBIIx-ルシフェラーゼで一時的にトランスフェクトし、48時間後に、ライセート中のルシフェラーゼ活性を調べた。
【図12A】図12は、IKKβ NBDを包含するペプチドは、IKKαのNEMOとの相互作用を妨げることを実証した実験結果を示す。図12A:NBD野生型及びスクランブル型コントロールペプチドの配列。この野生型ペプチドは、IKKβの残基734から744に対応する。
【図12B】GST-NEMOと、インビトロで転写・翻訳したIKKα(上側パネル)及びIKKβ(中央のパネル)とを用いて、賦形剤(2%DMSO)、スクランブル型もしくは野生型NBDペプチド(各ペプチドを500及び1000μM)の存在下又は不在下でGSTプル−ダウン解析を行った。下側のパネルは、クーマシー・ブルー染色したゲルを示し、いずれのペプチドも、沈降に用いたグルタチオン-アガロースビーズとのGST-NEMOの相互作用に影響を与えなかったことを実証している。
【図12C】ある範囲の濃度の野生型NBDペプチドの存在下でGST-NEMOを用いてIKKα及びIKKβのGSTプル−ダウンを行った後に得たオートラジオグラフィのバンドの比重解析。挿入図は一実験例を示す。データは、ピクセル密度をコントロール(ペプチドなし)に対するパーセンテージで表しており、平均値±sd(n=11)で表している。解析はNIH-イメージソフトウェアを用いて行った。
【発明を実施するための形態】
【実施例】
【0143】
実施例1: IKKβ上のNEMO結合ドメインの同定
我々は、IKKβ上のNEMO相互作用ドメインを同定するために、インビトロ・プルダウン検定(Zhong et al., (1997) Cell 89, 413-424)を、NH2末端でグルタチオンS-トランスフェラーゼに融合させた完全長NEMO(GST-NEMO;図1A)の細菌発現版を用いて行った。IKKβの様々な機能的ドメイン(触媒ドメイン、ロイシンジッパー及びへリックス-ループ-へリックス;図1A)を欠失した多様な切断変異体が構築された。
【0144】
IKKα及びIKKβの完全長cDNAクローンのサブクローニング及び変異誘発はすべて、クローンPfu DNA-ポリメラーゼ(ストラータジーン社製)を用いたPCRで行った。野生型及び変異型IKKβcDNAを、pcDNA-3 又は pcDNA-3.1-xpress (インビトロジェン社製)のKpnI 及び NotI 制限部位に挿入し、 IKKαcDNAはすべて、同じベクタのEcoRI 及びXhoI 部位に挿入した。FLAGで標識した形の両キナーゼは、pFLAG-CMV-2 (シグマ社製)にサブクローンして構築した。GST-IKKβ-(644-756)については、このPCR断片を、 pGEX-4T1 (ファルマシア社製)のEcoRI 及びXhoI部位に挿入した。ヒトNEMOをコードする完全長cDNAは、逆転写酵素(RT)-PCRにより、ヒーラ細胞mRNAから、ExpandTM ロング・テンプレートPCRシステム(ベーリンガーマンハイム社製)及びプライマペア(5'-ATAGACGAATTCAATAGGCACCTCTGGAAG) (SEQ ID NO: 20) 及び(3'-TAGGACCTCGAGCTACTCAATGCACTCCATG) (SEQ ID NO: 21)を用いて得た。その結果得られたPCR断片を、pcDNA-3 又はpcDNA-s.1-xpressのEcoRI 及びXhoI 部位に挿入した。その後のNEMO変異体はすべて、PCRにより、Pfu DNA-ポリメラーゼを用いて構築した。GST-NEMO は、この完全長cDNAをpGEX-4T1のEcoRI 及び XhoI 部位にサブクローンして、構築した。
【0145】
これらの変異体には、インビトロ翻訳で [35S]-メチオニンの標識を付け、(インプット;図1B)、GST のみか、又はGST-NEMOと混合し、グルタチオン−アガロースを用いて沈降させた。変異体のいずれも、GSTのみとは相互作用しなかったが、一方、野生型と、IKKβのNH2末端切断型の三つすべて(307-756、458-756及び486-756)が、GST-NEMOと相互作用した(図1B(右側パネル)。対照的に、COOH-末端切断型変異体(1-456、1-605又は1-644)のいずれも、GST-NEMOで沈降しなかった。これらの結果は、NEMOが、へリックス-ループ-へリックス(HLH)ドメインから遠いIKKβのCOOH末端にある領域と、相互作用することを示している。次に、IKKβのアミノ酸644(HLHの直後)からCOOH末端(残基756)までの領域のみから成る変異体を構築した。図1Cに示すように、この変異体はGSTでは沈降しなかったが、GST-NEMOとは相互作用したことから、この領域が、これら二つの分子間の相互作用を媒介していることが裏付けられた。
【0146】
次に、IL-1β及びTNFαで誘導されるNF-κB活性化に対するIKKβ-(644-756)の作用を、NF-κB依存性レポータプラスミド(pBIIX-ルシフェラーゼ)と一緒に前記変異体でヒーラ細胞を一時的にトランスフェクトして調べた(Kopp & Ghosh, (1994) Science 265, 956-959)。トランスフェクション実験では、ヒーラ細胞及びCOS細胞を24ウェル(1×105細胞/ウェル)プレート又は6ウェル(5×105細胞/ウェル)プレートのいずれかに接種して、24時間成長させてから、Fugene6(ロシュ社製)でメーカのプロトコルに従ってDNAをトランスフェクトした。24ウェルプレート及び6ウェルプレートの細胞に、それぞれ合計1μg又は2μgのDNAを与えた。48時間後に細胞を TNT (200 mM NaCl、20 mM トリス-pH 8.0、1% トリトン-100)で溶解させ、そのライセートを用いて、免疫沈降又はルシフェラーゼ検定(プリメージ・ルシフェラーゼ検定系)を行った。
【0147】
図1は、これらのサイトカインにより誘導されるNF-κBの活性化を、IKKβ-(644-756)が用量依存的に抑制したことを示す。これらの結果では、コアIκB-キナーゼ複合体から内因性NEMOを滴定することにより、IKKβ-(644-756)がドミナントネガティブとして作用することを示す。NEMOによる調節タンパクの動員がないと、IL-1β及びTNFα誘導シグナルに対してIKKβは不応性となり、起きるはずのIKKβ活性化が起きない。構造的にはNEMOは、コイルド・コイル及びロイシン・ジッパーモチーフからなる二つの主要な部分を含有する長いαらせん領域と、COOH末端亜鉛フィンガードメインとから成る(図2A)(Mercurio et al., (1999) Mol. Cell. Biol. 19, 1526-1538; Yamaoka et al., (1998) Cell 93, 1231-1240; Rothwarf et al., (1998) Nature 395, 297-300)。NEMOがIKKβと相互作用するのに必要なその領域を特定しようとするこれまでの研究は、互いに矛盾する結果を出していた(Harhaj et al., (1999) J. Biol. Chem. 274, 15297-15300)。この疑問に対処するために、IKKβ-(644-756)(図2A)及びNEMOの多様な[35S]-メチオニン標識付き切断型変異体(図2A)のGST-融合タンパクを用いたGST-プル-ダウン検定を行った。図2A(右側パネル)は、これらの実験結果を要約したものであり、IKKβ-(644-756) はNEMO-(1-196)、-(1-302)及び-(44-419) とは相互作用するが、NEMO-(197-419) もしくは-(86-419)とはしないことが実証された。同一の結果が、FLAGでタグしたIKKβ及びこのNEMO変異体を一時的にトランスフェクトしたCOS細胞又はHEK293細胞のライセートを用いた免疫沈降実験でも得られた(データは図示せず)。
【0148】
すべての免疫沈降で、6ウェルプレートで成長させたヒーラ細胞又はCOS細胞は500μlのTNTで溶解させた。FLAGでタグを付けたタンパク質は、20 μlの抗FLAG (M2)-共役アガロースビーズ(シグマ社製)を用いて、4℃で2時間かけて、トランスフェクトした細胞のライセートから沈降させた。内因性IKKβ又はNEMOの免疫沈降は、1 μgの特異ウサギポリクローナル抗体(サンタクルズ社製)と、20μlのプロテインAセファロース(アマーシャム−ファルマシア社製)を用いて行った。免疫ブロットについては、沈降物をTNTで三回、PBSで二回、洗浄してから、SDS-試料バッファ中に懸濁させた。タンパク質をSDS-PAGE(10%)で分離し、PVDFメンブレンに写し取り、高画質化学発光法(アマーシャム−ファルマシア社製)で観察した。
【0149】
これらの結果から、NEMOの一番目のαへリックスを含む一領域である、残基44から86の間に相互作用ドメインがあることが確定された。こうして、一番目のコイルド・コイル−ドメインまでのαへリックスを欠失させた変異体(残基T50−L93;del.αH)を作製した。この変異体はIKKβ-(644-756)とは相互作用しなかった(図2B)。さらに、pBIIX-ルシフェラーゼを用いたトランスフェクション実験では、del.αHが、TNFαで誘導されるNF-κB活性を抑制したことが実証され(図2C)、IKKβとは相互作用できないNEMOのCOOH末端が、NF-κBのドミナントネガティブ抑制因子であることが裏付けられた(Mercurio et al., (1999) Mol. Cell. Biol. 19, 1526-1538; Rothwarf et al., (1998) Nature 395, 297-300)。まとめると、図1及び2は、IKKβ及びNEMOの間の相互作用は、IKKβのCOOH末端と、NEMOの一番目のαへリックス領域とを通じて起きることを示している。これらの発見から、NEMOのNH2末端がNEMOをIKK-複合体に繋ぎ止め、いくつかのタンパク質対タンパク質相互作用ドメインを含有するこの分子のその他の部分は剥き出しのままであり、IKK機能の上流の調節因子との相互作用に使われるのだ、ということが分かる。
【0150】
実施例2: NBDでの相互作用を通じたIKKβ機能のNEMO調節
IKKβのNEMO相互作用ドメインを十二分に特徴付けるために、残基V644からS756までの間の切断型変異体(図3A)をさらに構築した。HLHの直後から、662位のシステインまでのアミノ酸配列はIKKα(図3Aではα1と示した)と72%の同一性を示す。この後の、E707までの領域は、かつて、自己リン酸化のターゲットであり、炎症誘発性サイトカインによる刺激後のIKKβ活性の下方調節に機能すると報告されたことのあるセリンリッチなドメインである(Delhase et al., (1999) Science 284, 309-313)。この後に続く配列は733位まで、全くセリン残基を含まない。これらの領域のそれぞれを配列から削除した変異体を[35S]-メチオニンで標識し、上述したGST-プル-ダウン検定に用いた。図3Aは、これらの実験結果を要約したものであり、このIKKβ変異体のいずれもGST-NEMOで沈降しなかったことから、その相互作用ドメインは、残基F734及びS756の間の最先端のCOOH末端にあることが分かる。
【0151】
IKKβのこの短い部分を、IKKαの相当する領域に比較すると、驚くべき二つの構造上の特徴があることが分かる(図3B)。第一に、IKKβのF734からT744までの配列(図3Bのα2)は、IKKαの同等の配列(IKKβのL737からL742及びIKKαのL738からL743)と同一である。第二に、IKKβのこの配列は、IKKαのCOOH末端残基(E745)よりも12個のアミノ酸分、長く延びているが、この12個のアミノ酸は酸性の高い領域を含んで成り、そのうちの残基5個はグルタミン酸である(図3B)。IKKβのα2領域と、IKKαの最先端COOH末端との間に著しい類似性があることを、NEMOはIKKαとはインビトロで相互作用しないという以前の報告 (Mercurio et al., (1999) Mol. Cell. Biol. 19, 1526-1538; Yamaoka et al., (1998) Cell 93, 1231-1240; Rothwarf et al., (1998) Nature 395, 297-300)と合わせて考えると、NEMO相互作用ドメインは、IKKβのグルタミン酸リッチな部分(E745からS756)であろうという仮説が導かれた。
【0152】
この仮説を調べるために、この領域を削除した切断変異体を作製し(1-744;図3C)、GST-NEMOと相互作用するその能力について調査した。この変異体は、野生型IKKβと同程度に、GST-NEMOと結合した(図3C)。これらの結果は、一時的にトランスフェクトしたCOS細胞のライセートから、エピトープでタグを付けたNEMO及びIKKβ-(1-744)を同時免疫沈降させることにより、確認した。これらの発見は、IKKβのNEMO相互作用ドメインがこのCOOH末端のα2領域にあることを実証するものである。
【0153】
次にIKKβ COOH末端切断変異体を用いて、IKKβの基礎活性及び誘導後活性に対するNEMO結合の作用を調べた。図3Dは、IKKβをV644で切断して、セリンリッチな領域(図3Aを参照されたい)を除去したところ、基礎自己リン酸化反応が完全に失われたことを示している。対照的に、このセリンリッチな領域(1-733)を含有する変異体は、野生型IKKβよりもはるかに高いレベルの自己リン酸化を呈した(図3D)。興味深いことに、NEMO結合α2領域を含有するIKKβ-(1-744)の自己リン酸化レベルは、この野生型キナーゼで観察されるものと同一であった。これらの変異が基礎キナーゼ活性に及ぼす影響を調べるために、変異体をヒーラ細胞に一時的にトランスフェクトし、NF-κB活性を、実施例1で解説したようにルシフェラーゼ検定で調べた。図3Dに示すその結果は、IKKβ-(1-644)はNF-κB活性を誘導しなかったが、IKKβ-(1-733)は野生型(図3E)よりも高い活性化を引き起こしたことを示す。さらに、IKKβ-(1-744)が誘導するNF-κB活性は、野生型IKKβが誘導するそれと同一であった。
【0154】
これらの結果は、IKKβの基礎自己リン酸化及びキナーゼ活性は、NEMOがこのキナーゼと結合する能力に依存していることを、実証している。これらの観察の解釈の一つは、NEMOがホスファターゼをIKK-複合体に動員して、このCOOH末端のセリンリッチな領域を狙わせることで、基礎IKKβ機能を調節しているのではないか、という解釈であろう。従って、NEMOを結合させられないとホスファターゼを動員できず、この領域のリン酸化が増加することになる。
【0155】
α2領域の消失が、IKKβの触媒活性に及ぼす作用を直接調べようと、免疫−複合体キナーゼ検定を、トランスフェクト後のヒーラ細胞から採ったライセートで行った(図3F)。免疫−複合体キナーゼ検定については、沈降物をTNTで洗浄してから、さらにキナーゼバッファ (20 mM HEPES pH 7.5、20 mM MgCl2、1 mM EDTA、2 mM NaF、2 mM β-グリセロホスフェート、1 mM DTT、10μM ATP)で洗浄した。次に、沈降物を、15分間、30℃で、 GST-IκBα-(1-90)及び10μCi [32P]-γ-標識ATP (アマーシャム・ファルマシア社製)を含有する20μlのキナーゼバッファ中でインキュベートした。基質をグルタチオン−アガロース(アマーシャム・ファルマシア社製)を用いて沈降させ、SDS-PAGE (10%)で分離した。キナーゼ活性はオートラジオグラフィで調べた。オートラジオグラフィでは、リン酸化したタンパク質がキナーゼ複合体と結合して見えたが、それは、免疫沈降させた複合体を、GST−基質沈降の前に取り除かなかったからである。IKKβ(野生型)の活性は、非処置細胞では低かったが、TNFαで処置後は著しく上昇した。図3Eに示したデータと合致して、IKKβ-(1-733)の基礎活性は野生型よりも著しく高かったが、この活性は、TNFαで処置してもそれ以上、上昇しなかった(図3F)。さらに、IKKβ-(1-744)の基礎触媒活性及びTNFαによる誘導後の触媒活性は、IKKβ(WT)の活性と同一であった。リン酸化GST-IκBαに加え、自己リン酸化IKKβも検出された(図3F、上のバンド)。TNFα処置後、IKKβ(WT)及びIKKβ-(1-744)は急速に自己リン酸化したが、既に高い値となっているIKKβ-(1-733)の基礎リン酸化は僅かに上昇したのみだった(図3F)。以前の研究では、自己リン酸化は、このタンパク質にコンホメーション上の変化を起こすことで、TNFαが誘導するIKKβ活性を下方調節するよう働くと示されている (Delhase et al., (1999) Science 284, 309-313)。まとめると、これらの発見(図3D−F)は、NEMOが存在しないとき、IKKβは自己リン酸化して基礎活性となり、TNFαで誘導されるシグナルに対して不応性となることを実証するものであり、NEMOが、IKKβ活性の活性化だけでなく、下方調節にも基本的な役割を果たしていることを示している。
【0156】
ある一つのリン酸化タンパク質を表す別のバンドが、TNFαで誘導したIKKβ(WT)細胞及びIKKβ-(1-744)トランスフェクト細胞から採った試料中にのみ、現れた(図3F)。このタンパク質の分子量(49kD)から、これが、沈降した複合体と結合した内因性NEMOであることが明確に分かる。このことは、IKKβ-(1-733)トランスフェクト細胞のいずれの沈降物(+/-TNFα)にも、このバンドがないことが裏付けとなっている。この沈降複合体をSDSで解離させ、特異的抗NEMO抗体を用いて[32P]標識付きNEMOを再沈降させることで、このタンパク質をリン酸化NEMOと同定した。従って、NEMOの誘導性リン酸化は、IKK複合体の更なる活性調節段階であるのかも知れない。
【0157】
実施例3: IKKA上のNBDの同定
IKKβのα2領域はIKKαのCOOH末端に酷似しているため(図3B)、IKKαがNEMOと相互作用する能力を調べた。
【0158】
xpressでタグを付けたNEMOと、FLAGでタグを付けた形のIKKαもしくはIKKβのいずれかとを一緒に一時的にトランスフェクトしたCOS細胞のライセートの免疫沈降を抗FLAG を用いて実施例1で解説したように行った。図4Aは、NEMOが、IKKβ及びIKKαの両方と同等に良好に相互作用したことを示す。この実験では、IKKαとの相互作用が直接的なものではなく、代わりに、内因性IKKβ、FLAG-IKKα及びxpress-NEMOを含有する複合体が形成されたことが原因である知れない可能性がある。従って、GST-プル-ダウン検定を、GST-NEMOと、[35S]-メチオニン標識した形の、野生型IKKαか、もしくは8個のCOOH末端アミノ酸を欠失させた切断型IKKα変異体(1-737):図4B)とを用いて、行った。上に記載した発見と合致して(図4A)、しかし以前の報告(Mercurio et al., (1999) Mol. Cell. Biol. 19, 1526-1538; Yamaoka et al., (1998) Cell 93, 1231-1240; Rothwarf et al., (1998) Nature 395, 297-300)とは対照的に、野生型IKKαはNEMOとインビトロでも相互作用したが、切断型変異体はしなかった(図4B)。これらの結果は、IKKαがNEMOと相互作用することを実証するだけでなく、IKKαとIKKβのα2領域に共通の、6個のアミノ酸を含有するCOOH末端領域を介してこの相互作用が行われていることも、示している(図3B)。遺伝子ターゲティング研究では、TNFαによるIKKα及びIKKβの活性化に、奥深い違いがあることが実証されている (Woronicz et al., (1997) Science 278, 866-869; Zandi et al., (1997) Cell 91, 243-252; Mercurio et al., (1997) Science 278, 860-866; DiDonato et al., (1997) Nature 388, 548-554; R gnier et al., (1997) Cell 90, 373-383)。
【0159】
本発見は、この違いの根底にあるのが、NEMOの動員の違いではないことを示唆している (Delhase et al., (1999) Science 284, 309-313; Takeda et al., (1999) Science 284, 313-316; Hu et al., (1999) Science 284, 316-20; Li et al., (1999) Science 284, 321-325; Li et al., (1999) J. Exp. Med. 189, 1839-1845; Li et al., (1999) Genes Dev. 13, 1322-1328; Tanaka et al., (1999) Immunity 10, 421-429)。反対に、この違いは、各キナーゼが、NEMO結合シグナリング成分を活性化応答に向けて、おそらくは個々のキナーゼの本来の調節的特徴の違いを通じて、動員する能力に、あるのではないかと思われる。
【0160】
この短いCOOH末端配列が、IKKのNEMO相互作用ドメインを構成していることの更なる証拠は、最近解説されたIKK関連キナーゼIKKi (Shimada et al., (1999) Int. Immunol. 11, 1357-1362)がNEMOと相互作用する能力を我々が調べたときに、得られた。IKKα及びIKKβの配列を比較すると(Shimada et al., (1999) Int. Immunol. 11, 1357-1362; Woronicz et al., (1997) Science 278, 866-869; Zandi et al., (1997) Cell 91, 243-52; Mercurio et al., (1997) Science 278, 860-866; DiDonato et al., (1997) Nature 388, 548-554; Regnier et al., (1997) Cell 90, 373-383)、IKKiは、そのCOOH末端にα2領域を含有していないことが分かり(Shimada et al., (1999) Int. Immunol. 11, 1357-1362)、これがNEMO結合ドメインであるということと一致して、我々は、IKKiが、GST-NEMOとは相互作用しないことをプル-ダウン検定で見いだした(図4C)。この発見は、NEMOはIKKiの機能的活性には必要ではないことを示唆しており、これは、IKKiがTNFαまたはIL-1βの誘導するシグナルのいずれにも応答できないことでも、裏付けられている(Shimada et al., (1999) Int. Immunol. 11, 1357-1362)。
【0161】
実施例4: NBD中のアミノ酸残基の変異
(NEMO結合ドメイン(NBD)と指定する)IKKβのα2領域と、IKKαの同等の6個のアミノ酸配列とが、NEMOとの相互作用にとって重要であると確信したところで、L737からL742までの6個のアミノ酸を欠失させたIKKβの欠失変異体(del.NBD)を構築した。この欠失変異体はGST-NEMOと結合しなかった(図4D)。NBDの予測上の構造的及び生化学的特徴を、周囲の残基との関連で調べたところ、これが、IKKβ COOH末端の親水性領域内で、柔軟性のない疎水性の「ポケット」を構成していることが示唆された(図4E)。このことは、NBDが、結合したNEMOの一番目のαへリックス領域内に埋没してしまい(図2)、水性の環境にこれが曝されるのが妨げられて、強力な分子間相互作用を維持する、というモデルを示唆している。この相互作用が実際、この疎水性部分の機能であるかどうかは、まだ調べなければならないが、W739又はW741のいずれかをアラニンに置換すると、NEMOのIKKβとの結合が妨げられることを我々は見いだした(図4F)。従って、これらの疎水性トリプトファン残基のそれぞれが、機能的NBDを維持するのに重要なのである。加えて、D737がアラニンに変異しても、NEMOの相互作用が妨げられたことから、この位置に負電荷の残基があることがNEMO機能にとって必要であることも、分かった。これらの変異とは対照的に、L737、S740又はL742をアラニンに置換しても、NEMO結合には影響を与えなかった。これらのアラニン置換がIKKβ機能に及ぼす作用を調べるために、ヒーラ細胞に、前記点変異体のそれぞれを、pBIIX-ルシフェラーゼレポータと一緒に同時トランスフェクトした。IKKβの基礎活性は結合NEMOのないときの方が高いという観察と合致して、NEMOに結合しないIKKβ-(1-733)変異体(図3E)(D738A、W739A及びW741A)は、野生型IKKβ又はIKKβ-(1-744)(図4G)よりも、より高いレベルにNF-κBを活性化した。対照的に、NEMO結合を損なわなかった置換を含有する変異体 (L737A、S740A及び L742A) は、コントロールと同じレベルまで、NF-κBを誘導した。これらの結果から、NEMOは、内因性IKKβ活性の下方調節に重要な役割を果たしていることが分かる。
【0162】
実施例3で解説したGSTプルダウン検定を利用して、NBDでの変異についてさらに解析を行い(表1を参照されたい)、NEMOのIKKβへの結合に及ぼすその影響ついて調べた。
【0163】
【表1】
【0164】
実施例5: NBDと相互作用してNEMO結合を遮断する作用薬
NBDは比較的に小さいため、コアIKK複合体の破壊を狙いとした化合物の開発にとって、魅力的なターゲットとなっている。IKKβNBDに延びる細胞透過性ペプチドを設計し、それらがIKKβ-NEMO相互作用を解離させる能力を調べることで、この方法の今日性を調査した。
【0165】
この研究で用いた二つのNBDペプチドの配列は[DRQIKIWFQNRRMKWKK]TALDWSWLQTE (野生型) (SEQ ID NO:18) 及び [DRQIKIWFQNRRMKWKK]TALDASALQTE (変異型) (SEQ ID NO:19)であった。アンテナペディア・ホメオドメイン(antennapedia homeodomain)配列 (Derossi et al., (1994) J. Biol. Chem. 269, 10444-10450; 米国特許第5,888,762号;米国特許第6,015,787号;米国特許第6,080,724号)を括弧内に入れ、WからAへの変異位置に下線を加えた。両ペプチドともDMSOで溶解させて、ストック濃度20 mMとした。すべての実験で、DMSO単独のコントロールは、ペプチドなしコントロールから差がなかった。
【0166】
野生型NBDペプチドは、IKKβのT735からE745までの領域を、膜移行を媒介することが示されているアンテナペディア・ホメオドメインの第三へリックス(Derossi et al., (1994) J. Biol. Chem. 269, 10444-10450)を由来とする配列に融合させたものから構成した。変異体は同一であるが、例外としてNBDのトリプトファン残基(W739及びW741)をアラニンに変異させてあった。図5Aは、野生型のNBDが、GST-NEMOによる[35S]-標識付IKKβのプルダウン及びGST-IKKβ-(644-756)による[35S]-標識付NEMOのプルダウンを、インビトロで用量依存的に抑制し、変異型は抑制しなかったことを示す。このNBDペプチドが細胞内へ進入してIKKβ-NEMO相互作用を抑制する能力について調べようと、ヒーラ細胞をこれらペプチドと一緒に様々な時間、インキュベートし、抗NEMOを用いてそのIKK複合体を免疫沈降させた。インビトロでのデータ(図5A)と合致して、野生型は内因性IKK複合体の形成を損なったが、変異型は損なわなかった(図5B)。
【0167】
実施例6: NEMO機能を遮断する作用薬
次に、シグナルにより誘導されるNF-κBの活性化に及ぼすこのNBDペプチドの作用を調べた。ヒーラ細胞にpBIIX-ルシフェラーゼレポータをトランスフェクトした後、細胞を野生型又は変異型ペプチドと一緒にプレインキュベートし、TNFαで処置し、NF-κB活性化をルシフェラーゼレポータ検定で測定した。図5C(左側パネル)に示すように、野生型NBDペプチドは、TNFαで誘導されるNF-κB活性化を抑制したが、変異型では何の作用もなかった。興味深いことに、基礎NF-κB活性は、野生型ペプチドで処置すると上昇した(図5C;右側パネル)、これは、前の変異解析(図3E−F及び4G)の結果と一致する発見である。このことは、NEMOを除去すると、IKKの基礎内因性活性が上昇するが、TNFαに対するその応答性は喪失させることを、示している。電気泳動度シフトアッセイ(EMSA)を用いた別の解析でも、野生型NBDペプチドだけが、TNFαで誘導される活性化やNF-κBの核内移行を抑制したことを実証できた(図5D)。まとめると、これらの結果は、インタクトNBDペプチドが細胞内へ送達されると、IKKβ-NEMO間の相互作用が損なわれて、活性化NF-κBEからの炎症誘発性シグナルが妨げられることを実証するものである。対照的に、NEMO相互作用を維持するのに重要なトリプトファン残基に変異を持つペプチドによる伝達では、何の作用もないのである。
【0168】
実施例7: E-セレクチンを下方調節できる作用薬
炎症性応答の開始及び維持に関与するタンパク質の多くにとって、それらの遺伝子発現が誘導されるには、NF-κBが活性化することが必要である(Ghosh et al., (1998) Annu. Rev. Immunol. 16, 225-260; May & Ghosh, (1998) Immunol. Today 19, 80-88)。このようなタンパク質の一つであるE-セレクチンはIL-1又はTNFαなどの炎症誘発性刺激による活性化後の血管内皮細胞の管腔表面に発現する白血球接着分子である (Pober et al., (1986) J. Immunol. 436, 1680-1687; Bevilacqua et al., (1987) Proc. Natl. Acad. Sci. USA 84, 9238-9242; Collins et al., (1995) FASEB J. 9, 899-909)。E-セレクチン及び他のNF-κB依存性接着分子の発現は、急性及び慢性炎症の部位に白血球が捕獲及び動員されるのに、重要である。前記NBDペプチドの抗炎症能を評価するために、始原ヒト臍静脈内皮細胞(HUVEC)を野生型及び変異型ペプチドで予備処置し、E-セレクチンの発現をTNFαで誘導した。基礎NF-κB活性化に及ぼす作用と一致して(図5C)、野生型NBDペプチドは低レベルのE-セレクチン発現を誘導した(図5E)。しかし、TNFαで処置後は、この野生型ではE-セレクチンの発現が著しく減少したが、変異型では減少しなかった(図5E)。野生型NBDペプチドによる抑制により、TNFαのない状態でこのペプチドが誘導するレベルまで、発現が減少した。
【0169】
本発明の重要性は二つのレベルで見ることができる。第一に、出願人は、NEMOがIKKと結合するのに必要な構造上の要件を特定し、IKKβとの結合は、NBD内の三つのアミノ酸(D738、W739及びW741)に依存していることを見いだした。さらに、NEMOは、IKKβの活性化で機能するだけでなく、IKK複合体の本来の基礎活性を抑制する上でも重要な役割を有するのである。二番目のレベルの重要性は、NBDをターゲットとする薬物の臨床上の明かな用途である。出願人は、NBDを包含する細胞透過性ペプチドは、TNFαで誘導されるNF-αB活性化を抑制できるだけでなく、始原ヒト内皮細胞で、NF-κB依存性標的遺伝子であるE-セレクチンの発現を減少させることも、実証した。NBDは僅かに6アミノ酸長であり、従ってコアIKK複合体を損なうペプチドミメティック化合物を設計する方法は当業者の技術範囲内にある。複合体を破壊するこの作用は、IKKの基礎活性の上昇にあるため、NBDをターゲティングする化合物で処置すると、NF-κBの活性を完全に遮断する投薬をしたときに起きるであろう、肝細胞アポトーシスを原因とするものなど、毒性の問題を避けることができる。このように、NBDの同定は、IKK複合体に活性化シグナルが達しないようにし、と同時に低レベルのNF-κB活性は維持しつつも、毒性の副作用が生じる可能性を避けた新規な抗炎症薬物を開発する手段である。
【0170】
実施例8: インビボにおける炎症性応答の、NBDペプチドを媒介とする抑制
急性炎症反応の二つの異なるモデルを用いて、動物における炎症性応答を抑制する能力について、当該NBDペプチドを調べた。一番目のモデルでは、ホルボール-12-ミリステート-13-アセテート(PMA)を用いてマウスの耳に水腫を起こさせ、NBDペプチドを局所投与してその作用を測定した。PMAを用いた耳水腫は、以前に解説された(Chang et al., (1987) Eur. J. Pharmacol. 142, 197-205)通りに、年齢及び性別を同じにしたマウスの複式のグループで起こした。20μlのPMA (5μg/耳)エタノール 溶液の投与の30分前及び30分後に、20μlのNBDペプチド(200 μg/耳)、デキサメタゾン (40μg/耳)又は賦形剤(DMSO:エタノール;25:75 v/v) をマウスの右耳に局所投与した。PMA投与後6時間で、耳の膨潤をマイクロゲージを用いて測定し、処置(右)側耳及び非処置(左)側耳の間の厚さの違いの平均値として表した。このデータの統計学的解析は、スチューデントのt検定を用いて行った。p<0.05の値を統計上有意とみなした。
【0171】
図6Aは、野生型ペプチドが、PMAで誘導される耳の肥厚を著しく減少させ(77±3%抑制;p<0.05)、デキサメタゾンで観察されるレベル(82±9%抑制;p<0.05)まで下げたことを示す。対照的に、等量の変異型で観察される作用は、有意ではなかった(p= 0.09)。いずれのペプチドも、PMAなしで投与したときには何ら作用はなかった(図示せず)。
【0172】
二番目のモデルでは、ザイモサンを単独か、又は、デキサメタゾン又はNBDペプチドと組み合わせて腹腔内(i.p.)注射して、マウスに腹膜炎を起こさせた。ザイモサンで起こした腹膜炎では、前に解説された(Getting et al., (1998) Immunology 95, 625-630)ように、年齢及び性別を合わせた複式グループのマウス (C57BL/6NCR)で、腹腔滲出液を測定し、炎症性細胞の採集を行った。一部分の動物には、1mlのザイモサン(1mg/ml)と、デキサメタゾン(100mg/ml)又はNBDペプチド(200mg/ml)のいずれかとを同時に注射した。炎症性滲出液中のNOX(硝酸及び亜硝酸化合物)の濃度を、メーカのプロトコルに従って比色検定キット(アレキシス・コーポレーション社製)を用いて測定した。
【0173】
図6Bに示すように、ザイモサンの注射により、炎症性滲出液が蓄積し、多核白血球(PMN)が、これらの動物の腹膜に移動した。マウスを野生型NBDペプチド又はデキサメタゾンで処置した場合、滲出液形成及びPMN蓄積が著しく減少したが、変異型では何ら影響はなかった。
【0174】
多様なインビボ研究で、滲出液形成及び白血球の炎症部位への移行に、NOが役割を果たしていることが実証されている(Ialenti et al., (1992) Eur. J. Pharmacol. 211, 177-182; Ialenti et al., (1993) Br. J. Pharmacol. 110, 701-706; Iuvone et al., (1998) Br. J. Pharmacol. 123, 1325-1330)。従って、ザイモサンで処置したマウスの腹腔滲出液における、NBDペプチドのNOX蓄積に対する作用を調べた。図6C(下側パネル)は、デキサメタゾン及び野生型ペプチドがそれぞれ86±7%及び66±4%、NOXを減少させたが、変異型では何の作用もなかったことを示す。これらの結果は、滲出液形成及び細胞蓄積の減少が、NF-κB活性化の抑制と、NO形成の減少とに密接に相関していることを実証したこれまでの研究と合致している(D’Acquisto et al., (1999) Eur. J. Pharmacol. 369, 223-236; D’Acquisto et al., (1999) Naunyn-Schmeideberg’s Arch. Pharmacol. 360, 670-675)。従って、野生型NBDペプチドは、実験動物モデルでは、炎症の有効な抑制物質である。
【0175】
実施例9: NBDペプチドによる破骨細胞分化の抑制
骨の形態発生及びリモデリングのプロセスには、骨芽細胞による骨マトリックスの合成と、破骨細胞による骨再吸収との間にバランスが維持されることが必要である(Suda et al., (1992) Endocr. Rev. 13, 66-80; Suda et al., (1999) Endocr. Rev. 20, 345-357)。骨再吸収中の破骨細胞は、骨髄前駆細胞と、コロニー刺激因子(CSF-1)、インターロイキン-1(IL-1)、腫瘍壊死因子-α(TNF-α)、IL-6及びIL-11を含め、様々な段階で破骨細胞分化に影響を与える(Suda et al., (1992) Endocr. Rev. 13, 66-80; Suda et al., (1999) Endocr. Rev. 20, 345-357)多様な可溶性因子とから分化する多核巨細胞である。破骨細胞発生にとって重要な因子の一つは、RANKL(NF-κBリガンドの受容体活性化物質)という名称の最近解説された分子であり、この分子は、ODF (破骨細胞分化因子)、OPGL(オステオプロテゲリンリガンド)及びTRANCE (TNF-関連活性化により誘導されるサイトカイン)(Kong et al., (1999) Nature, 397, 315-323; Lacey et al., (1998) Cell 93, 165-176; Suda et al., (1999) Endocr. Rev. 20, 345-357; Wong et al., (1999) J. Leukoc. Biol. 65, 715-724; Yasuda et al., (1998) Proc. Natl. Acad. Sci. USA 95, 3597-3602)としても公知である。RANKLの受容体は、RANK(NF-κBの受容体活性化物質)という名称のTNF受容体ファミリーの仲間であり (Anderson et al., (1997) Nature 390, 175-179; Dougall et al., (1999) Genes Dev. 13, 2412-2424)、RANKLが結合すると、NF-κB活性化が誘導される(Anderson et al., (1997) Nature 390, 175-179; Darnay et al., (1998) J. Biol. Chem. 273, 20551-20555; Darnay et al., (1999) J. Biol. Chem. 274, 7724-31; Suda et al., (1999) Endocr. Rev. 20, 345-357; Wong et al., (1998) J. Biol. Chem. 273, 28355-28359)。さらに、破骨細胞の分化はNF-κB活性化にも依存しており、遺伝子ターゲティング研究では、成熟破骨細胞は、p50及びp52NF-κBサブユニット欠損マウスでは発生できないことが実証されている(Franzoso et al., (1997) Genes Dev. 11, 3482-3496)。
【0176】
骨粗鬆症は、破骨細胞に依存する骨再吸収が媒介する広汎な骨量減少を特徴とする重篤な衰弱性疾患である(Suda et al., (1992) Endocr. Rev. 13, 66-80; Suda et al., (1999) Endocr. Rev. 20, 345-357)。従って、NF-κBの活性化を破骨細胞前駆細胞で選択的に抑制すると、破骨細胞分化が妨げられて、骨粗鬆症の治療のための治療上有効な薬物の基礎となる可能性がある。よって、本NBDペプチドが破骨細胞分化に及ぼす作用を、以前に解説されたインビトロモデル(Jimi et al., (1999) Exp. Cell Res. 247, 84-93)を利用して調べた。48ウェル組織培養プレートに接種したマウス骨髄細胞を、ヒトマクロファージコロニー刺激因子 (M-CSF; 20 ng/ml)及びヒトRANKL(100 ng/ml)と一緒に、6日間、様々な濃度(6.25、12.5及び25 mM)の変異型もしくは野生型NBDペプチドの不在下又は存在下でインキュベートした。その後これらの細胞を固定し、破骨細胞表現型マーカである酒石酸塩耐性酸性ホスファターゼ(TRAP)で染色し、4つ以上の核を含有するTRAP陽性多核性細胞を破骨細胞として計数した。三重複試料を計数し、結果を平均値±SDで計算した。図7に示すように、野生型ペプチドは破骨細胞分化を用量依存的に抑制したが、変異型ペプチドはしなかった。
【0177】
このデータは、NF-κB活性化を抑制する細胞透過性NBDペプチドで、コアIKK複合体を破壊すると、RANKLで誘導される破骨細胞分化が妨げられることを実証しており、NBDを特異的に狙う薬物は、骨粗鬆症の治療にとって有効であろうことを示唆している。これらのインビトロ研究の延長として、同じペプチドを、インビボで骨粗鬆症に及ぼす作用について解析することもできる。重篤な骨粗鬆症を呈する卵巣切除マウス(チャールズ・リバー・ラブズ)を、このNBDペプチドで処置し、処置期間中の骨密度に対する作用を調べる。
【0178】
実施例10: NF-κBが媒介する他の疾患に対するNBDペプチドの作用
加えて、NBDペプチドが喘息に及ぼす作用を調べることも可能である。細気管支上皮細胞、T細胞及び細気管支マクロファージでのNF-κB活性化が、喘息患者及び喘息のモデル動物の気道で観察されている(Ray & Cohn, (1999) J. Clin. Invest. 104, 985-993; Christman et al., (2000) Chest 117, 1482-1487)。さらに、喘息を誘発する物質の多くはNF-κB活性化を引き起こし、喘息に関与するタンパク質(例えば白血球接着分子、多種のケモカイン、誘導性酸化窒素シンターゼなど)をコードする遺伝子の多くがNF-κB依存性である。樹立された喘息のマウスモデル(Kleeberger et al., (1990) Am. J. Physiol. 258, 313-320)を用いて、NBDペプチドを噴霧投与したときの、喘息に関連するこれらの状態の進行への作用を調べることもできる。同様な態様で、敗血症性ショックに対するNBDペプチドの作用も測定することができる。敗血症性ショックには、リポ多糖体(LPS)などの細菌性エンドトキシンが誘導する多くのNF-κB依存性遺伝子(即ちTNF、IL-1)の発現が関与している。LPSは、グラム陰性細菌の細胞壁の主要な構成成分を成し、免疫原性が高く、内因性発熱物質IL-1及びTNFの産生を刺激する(Sell et al., (1996) Immunology, Immunopathology & Immunity, Appleton & Lange)。敗血症性ショックに対するNBDペプチドの作用を調べるには、マウスにNBDペプチド及びLPSを注射し、動物の生存率を評価する。
【0179】
実施例11: NEMOとの相互作用におけるNBD内の各アミノ酸の相対的寄与及び重要性 上述の実施例で示したように、IKKα及びIKKβのNEMO結合ドメイン(NBD)は、両キナーゼのC末端最先端にある6個の保存されたアミノ酸(IKKβのL737 から L742及びIKKαのL738からL743)から成る。上記のNBD中の各アミノ酸がNEMOとの相互作用に対して持つ相対的寄与及び重要性をより明確に理解しようと、この実験を行った。各残基を多様な保存的アミノ酸及び非保存的アミノ酸に置換して、IKKβNBDの広汎な変異解析を行った。
【0180】
ロイシン残基(L737又はL742)又はセリン740のいずれを置換しても、NEMOのIKKβとの結合には影響を与えなかったことは、これらの残基のいずれも、この相互作用を維持するのに重要な役割は果たしていないと判断した。これらのアミノ酸に複数の変異があると結合に影響するかを調べるために、L737 及びS740 か、S740及びL742のいずれかをアラニンに置換した(それぞれLS及びSLと命名)変異体を構築した。GSTプル−ダウン及びCOS細胞トランスフェクション−免疫沈降−イムノブロット解析を行ったところ、LS及びSL変異体の両方とも、野生型IKKβと同程度に、NEMOと結合することが判明したため、これらの残基がこの相互作用に大きくは寄与していない証拠が新たに得られた。さらに、LS及びSLの両方とも、一時的トランスフェクション検定でNF-κB依存性ルシフェラーゼレポータ・コンストラクト(pBIIx-ルシフェラーゼ)の活性化で測定すると、IKKβだけでなくNF-κBも活性化する。両ロイシン残基(LL)の二重アラニン変異体や、三重変異体(LSL)を作れば、NEMO結合に対するこれらの残基の重要性に関する前記データを裏付けるのに役立つであろう。
【0181】
NEMO結合又はNF-κB活性化のいずれにも、上記の変異は作用を及ぼさないのと対照的に、NBD内のアスパラギン酸残基(D738)をアラニンに置換すると、IKKβのNEMOへの結合が妨げられた。さらに、この置換により、IKKβの基礎NF-κB活性化能力が2乃至3倍に上昇した。これらの結果は、IKK複合体の基礎活性を維持するのに、NEMO結合が一定の役割を果たしていることを実証するものである。興味深いことに、ヒーラ細胞を細胞透過性NBDペプチドで処置しても、基礎NF-κB活性はある程度、上昇したことから、NEMO結合が失われると基礎IKK活性が上昇することが、さらに裏付けられた。
【0182】
NBD内の738位の残基の性質を調べるために、アスパラギン酸をアスパラギン(D738N)又はグルタミン酸(D738E;図8A)のいずれかに置換した。これらの保存的置換は、この位置にある残基の形状(N)又は形状及び電荷(E)のいずれかを維持するものである。図8Bに示すように、いずれの置換も、IKKβのNEMO結合能には影響を与えなかったが、アラニン置換では結合が妨げられた。これらのデータは、IKKβ及びNEMO間の相互作用にとって重要なのは、この位置にあるアミノ酸側鎖の電荷ではなく形状(特に二番目の炭素の存在)であることを、実証している。ここに開示した前述の観察と一致して、いずれの変異も、IKKβの基礎活性には影響を及ぼさなかったが、アラニンでの置換では、活性が上昇した(図8C)。
【0183】
上述したように、NBD内のトリプトファン残基(W739及びW741)は両方とも、NEMOとの相互作用を維持するのに重要である。これらの位置にある残基の芳香族の構造を維持するような保存的置換の影響を、フェニルアラニン(F)又はチロシン(Y)のいずれかにトリプトファンを置換して、調べた(図9A)。さらに、両トリプトファンをアルギニンに変異させたが、これは、最も普通に天然で起きるトリプトファン変異である、コーディングコドン内に一箇所の塩基の変化だけを要する非保存的置換である。図9Bに示すように、W739F及びW739Y変異体の両方とも、IKKβと同程度にNEMOと結合したが、W739Rは結合しなかった(図9C)。アラニン置換の作用(図9B)と合わせると、これらの発見は、この位置にある残基の芳香族性が、NBDの機能にとって重要であることを示している。W739と同様、W742をフェニルアラニン(W742F)に置換すると、NEMOとの結合に影響は出なかったが、アルギニン(W742R)に変異させると、結合が妨げられた(図9D)と判断した。W739とは対照的に、チロシン(W742Y)に置換すると、NEMOとの結合が妨げられたことから、この位置にあるアミノ酸側鎖に水酸基部分が存在すれば、NEMOの結合を妨げるのに充分であることが実証された。この発見は、NBD内のある残基が、たとえそれが人工的に挿入されたアミノ酸であっても、リン酸化すれば、IKKβのNEMOとの結合が妨げられることを示唆していることになろう。
【0184】
実施例12:致死量リポ多糖体マウスモデルにおける抗炎症化合物の評価
この実験では、本発明の抗炎症化合物が、致死量のリポ多糖体(LPS)を与えたマウスを救助する能力を評価した。LPSは、敗血症で見られる、死亡を含む多くの応答を誘発する細菌生成物である。このモデルでは、サルモネラ−チフィムリウム(Salmonella typhimurium)LPSのリン酸緩衝血清(PBS)溶液30mg/kg(600μg/20gマウス)を、オスのC57BL/6マウスに静注投与した。この用量は、これを投与したマウスの100%で致死量であることをコントロール実験で確定された量である。マウスは、このLPS注射直後と、LPS注射から24時間後に、テストペプチドを静注(1%ジメチルスルホキシドのPBS溶液に溶かして)して処置した。LPS投与後最長で8日間、一日当たり2回、マウスを観察し、マウスの生存期間及び生存数を記録した。
【0185】
この研究結果を表2に示すが、同表では、各ペプチド及び用量について、投薬したマウスの合計数及び5日後の生存マウス数を示す。
【0186】
【表2】
【0187】
表2に示した結果は、このモデルでは、化合物2、3、及び5−11を5mg/kgの用量で静脈投与したときに、致死量のLPS投与からの有意な保護があったことを実証している。また化合物10は1mg/kg静注の用量でも有意な保護をもたらした。
【0188】
実施例13: コンカナバリンA誘発性肝炎におけるペプチドの評価
この実験では、本発明の抗炎症化合物が、コンカナバリンAで誘発した肝炎からマウスを救助する能力。
【0189】
コンカナバリンAは糖質に結合する種類のタンパク質であるレクチンである。あるタンパク質の一部が糖質である場合、このレクチンはそのタンパク質に結合する。コンカナバリンAは、細胞表面にあるタンパク質に結合することで、Tリンパ球を含む数多くの細胞を刺激する。これらのTリンパ球は、コンカナバリンAによる刺激で放出される他の媒介物質と協働して、やはりコンカナバリンAを結合させた肝細胞を攻撃して、肝細胞を死滅させる。Tリンパ球のこのような関与により、このモデルは、ヒトのウィルス感染に似たものとなっている。しかしながら、この急性モデルでは、TNFα応答もその一部を成す。
【0190】
体重18g乃至22gの40匹のオスのC57BL/6マウスを、下に示すようにそれぞれ8匹のマウスから成る5つの処置群に分けた。
【0191】
【表3】
【0192】
マウスに拘束具を着け、テストペプチド又は賦形剤を1%DMSOのPBS溶液に加えたものを尾に静脈注射(i.v.)した。これらマウスには、すぐに、15mg/kgのコンカナバリンAの無菌PBS溶液を静脈注射した。注射量は5ml/kg(100μI/20gマウス)であり、コンカナバリンA濃度は3.0mg/mlであった。翌朝(18乃至24時間後)、これらのマウスをCO2吸入で安楽死させ、心臓穿刺で血液を採集した。次に血清を分離し、AST及びALTについて分析した。
【0193】
この研究の結果を表4に示す。ALT値及びAST値はシグマ−フランケル単位/ml、平均値±SEMで表してある。
【0194】
【表4】
【0195】
上記の結果は、化合物10が、5mg/kgの用量で、コンカナバリンAで誘発させた肝損傷から保護できることを示す。肉眼病理でもこの結論が裏付けられ、肝臓はコンカナバリンAで損傷したが、この損傷は、5mg/kgの用量の化合物10で妨げられたことが示唆された。
【0196】
当業者であれば、ごく通常の実験を行うのみで、ここに解説した本発明の具体的実施例と同等の例を数多く認識され、又は確証できることであろう。このような同等物は、以下の請求の範囲の包含するところと、意図されている。
【特許請求の範囲】
【請求項1】
構造:
Xa-Xb,
(ただし式中、Xa は、6乃至15個のアミノ酸残基を含む膜移行ドメインであり、そしてXb はNEMO結合配列である)を含む抗炎症化合物。
【請求項2】
修飾基をさらに含む、請求項1に記載の抗炎症化合物。
【請求項3】
Xb が以下の構造:
(Y)n-X1-X2-X3-X4-X5-X6-(A)m
(但し式中、
n及び m はそれぞれ個別に0 又は1であり;
A及びYはそれぞれ1乃至約3個のアミノ酸残基を含み;
X1 はL、A、I 又はノルロイシン (Nle)であり;
X2 はD、E、N、Q、ホモセリン (Hser)又は2-ケトプロピルアラニン (2-ケトプロピ-A)であり;
X3 はW、F Y、4-ビフェニル-アラニン (Bpa)、ホモフェニルアラニン (Hphe)、2-ナフチルアラニン (2-Nal)、1-ナフチルアラニン (1-Nal)、又はシクロキセキシル(原語cycloxexyl)-アラニン (Cha)であり;
X4 はS、A、E、L、T、ノルロイシン (Nle)、又はホモセリン (Hser)であり;
X5 はW、H、ホモフェニルアラニン (Hphe)、2-ナフチルアラニン (2-Nal)、1-ナフチルアラニン (1-Nal)、O-ベンジルセリン (SeroBn)、又は3-ピリジルアラニン (3-Pal)であり;そして
X6 はL、A、I、又はノルロイシン (Nle)である)
から成る、請求項1に記載の抗炎症化合物。
【請求項4】
nが1であり、そしてYが配列TAである、請求項1に記載の抗炎症化合物。
【請求項5】
mが1であり、そしてAが配列QTEである、請求項1に記載の抗炎症化合物。
【請求項6】
Xb が、TALDWSWLQTE; LDWSWLQTE; TALDWSWL; ALDWSWLQTE; LDWSWLQTE; LDWSWL; TALDWSWLQT; TALDWSWLQ; ALDWSWLQT; LDWSWLQ; LDWSWLQT; ADWSWL; LDWSWA; ADWSWA; LDFSWL; LDYSWL; LDWAWL; LDWEWL; TAADWSWLQTE; ADWSWLQTE; TAADWSWL; AADWSWLQTE; ADWSWLQTE; ADWSWL; TAADWSWLQT; TAADWSWLQ; AADWSWLQT; ADWSWLQ; ADWSWLQT; ALDWSWAQTE; LDWSWAQTE; TALDWSWA; ALDWSWAQTE; LDWSWAQTE; LDWSWA; TALDWSWAQT; TALDWSWAQ; ALDWSWAQT; LDWSWAQ; LDWSWAQT; TAADWSWAQTE; ADWSWAQTE; TAADWSWA; AADWSWAQTE; ADWSWAQTE; ADWSWA; TAADWSWAQT; TAADWSWAQ; AADWSWAQT; ADWSWAQ; ADWSWAQT; TALDFSWLQTE; LDFSWLQTE; TALDFSWL; ALDFSWLQTE; LDFSWLQTE; LDFSWL; TALDFSWLQT; TALDFSWLQ; ALDFSWLQT; LDFSWLQ; LDFSWLQT; TALDYSWLQTE; LDYSWLQTE; TALDYSWL; ALDYSWLQTE; LDYSWLQTE; LDYSWL; TALDYSWLQT; TALDYSWLQ; ALDYSWLQT; LDYSWLQ; LDYSWLQT; TALDWAWLQTE; LDWAWLQTE; TALDWAWL; ALDWAWLQTE; LDWAWLQTE; LDWAWL; TALDWAWLQT; TALDWAWLQ; ALDWAWLQT; LDWAWLQ; LDWAWLQT; TALDWEWLQTE; LDWEWLQTE; TALDWEWL; ALDWEWLQTE; LDWEWLQTE; LDWEWL; TALDWEWLQT; TALDWEWLQ; ALDWEWLQT; LDWEWLQ; 及びLDWEWLQTからなる群より選択される配列である、請求項1に記載の抗炎症化合物。
【請求項7】
Xa が6乃至12個のアミノ酸残基から成る、請求項1に記載の抗炎症化合物。
【請求項8】
Xa が6乃至10個のアミノ酸残基から成る、請求項1に記載の抗炎症化合物。
【請求項9】
Xa が、少なくとも5個の塩基性アミノ酸残基を含む、請求項1に記載の抗炎症化合物。
【請求項10】
Xa がL-アルギニン、D-アルギニン、L-リシン及びD-リシンから個別に選択される少なくとも5個のアミノ酸残基を含む、請求項1に記載の抗炎症化合物。
【請求項11】
Xa がRRMKWKK; YGRKKRRQRRR; ygrkkrrqrrr; YARKARRQARR; yarkarrqarr; YARAARRAARR; yaraarraarr; rrmkwkk、RRRRRR、RRRRRRR、RRRRRRRR、RRRRRRRRR、RRRRRRRRRR、RRRRRRRRRRR、rrrrrr、rrrrrrr、rrrrrrrr、rrrrrrrrr、rrrrrrrrrr、及び rrrrrrrrrrrからなる群より選択される、請求項1に記載の抗炎症化合物。
【請求項12】
RRMKWKKTALDWSWLQTE; rrmkwkkTALDWSWLQTE; YGRKKRRQRRRTALDWSWLQTE; ygrkkrrqrrrTALDWSWLQTE; rrrrrrrTALDWSWLQTE; RRRRRRRTALDWSWLQTE; YARKARRQARRTALDWSWLQTE; yarkarrqarrTALDWSWLQTE YARAARRAARRTALDWSWLQTE; yaraarraarrTALDWSWLQTE YGRKKRRQRRRLDWSWL; ygrkkrrqrrrLDWSWL; RRMKWKKLDWSWL; rrmkwkkLDWSWL; rrrrrrrLDWSWL; YARAARRAARRLDWSWL; yaraarraarrLDWSWL; 及びRRRRRRRLDWSWLからなる群より選択されるアミノ酸配列を含む抗炎症化合物。
【請求項13】
H-RRMKWKKTALDWSWLQTE-NH2;
H-YGRKKRRQRRRTALDWSWLQTE-NH2;
H-rrrrrrrTALDWSWLQTE-NH2;
H-YARKARRQARRTALDWSWLQTE-NH2;
H-YARAARRAARRTALDWSWLQTE-NH2;
H-RRMKWKKLDWSWL-NH2;
H-rrmkwkkLDWSWL-NH2;
H-rrrrrrrLDWSWL-NH2;
H-YARAARRAARRLDWSWL-NH2;
H-yaraarraarrLDWSWL-NH2; 及び
H-YGRKKRRQRRRLDWSWL- NH2
からなる群より選択される構造を有する抗炎症化合物。
【請求項1】
構造:
Xa-Xb,
(ただし式中、Xa は、6乃至15個のアミノ酸残基を含む膜移行ドメインであり、そしてXb はNEMO結合配列である)を含む抗炎症化合物。
【請求項2】
修飾基をさらに含む、請求項1に記載の抗炎症化合物。
【請求項3】
Xb が以下の構造:
(Y)n-X1-X2-X3-X4-X5-X6-(A)m
(但し式中、
n及び m はそれぞれ個別に0 又は1であり;
A及びYはそれぞれ1乃至約3個のアミノ酸残基を含み;
X1 はL、A、I 又はノルロイシン (Nle)であり;
X2 はD、E、N、Q、ホモセリン (Hser)又は2-ケトプロピルアラニン (2-ケトプロピ-A)であり;
X3 はW、F Y、4-ビフェニル-アラニン (Bpa)、ホモフェニルアラニン (Hphe)、2-ナフチルアラニン (2-Nal)、1-ナフチルアラニン (1-Nal)、又はシクロキセキシル(原語cycloxexyl)-アラニン (Cha)であり;
X4 はS、A、E、L、T、ノルロイシン (Nle)、又はホモセリン (Hser)であり;
X5 はW、H、ホモフェニルアラニン (Hphe)、2-ナフチルアラニン (2-Nal)、1-ナフチルアラニン (1-Nal)、O-ベンジルセリン (SeroBn)、又は3-ピリジルアラニン (3-Pal)であり;そして
X6 はL、A、I、又はノルロイシン (Nle)である)
から成る、請求項1に記載の抗炎症化合物。
【請求項4】
nが1であり、そしてYが配列TAである、請求項1に記載の抗炎症化合物。
【請求項5】
mが1であり、そしてAが配列QTEである、請求項1に記載の抗炎症化合物。
【請求項6】
Xb が、TALDWSWLQTE; LDWSWLQTE; TALDWSWL; ALDWSWLQTE; LDWSWLQTE; LDWSWL; TALDWSWLQT; TALDWSWLQ; ALDWSWLQT; LDWSWLQ; LDWSWLQT; ADWSWL; LDWSWA; ADWSWA; LDFSWL; LDYSWL; LDWAWL; LDWEWL; TAADWSWLQTE; ADWSWLQTE; TAADWSWL; AADWSWLQTE; ADWSWLQTE; ADWSWL; TAADWSWLQT; TAADWSWLQ; AADWSWLQT; ADWSWLQ; ADWSWLQT; ALDWSWAQTE; LDWSWAQTE; TALDWSWA; ALDWSWAQTE; LDWSWAQTE; LDWSWA; TALDWSWAQT; TALDWSWAQ; ALDWSWAQT; LDWSWAQ; LDWSWAQT; TAADWSWAQTE; ADWSWAQTE; TAADWSWA; AADWSWAQTE; ADWSWAQTE; ADWSWA; TAADWSWAQT; TAADWSWAQ; AADWSWAQT; ADWSWAQ; ADWSWAQT; TALDFSWLQTE; LDFSWLQTE; TALDFSWL; ALDFSWLQTE; LDFSWLQTE; LDFSWL; TALDFSWLQT; TALDFSWLQ; ALDFSWLQT; LDFSWLQ; LDFSWLQT; TALDYSWLQTE; LDYSWLQTE; TALDYSWL; ALDYSWLQTE; LDYSWLQTE; LDYSWL; TALDYSWLQT; TALDYSWLQ; ALDYSWLQT; LDYSWLQ; LDYSWLQT; TALDWAWLQTE; LDWAWLQTE; TALDWAWL; ALDWAWLQTE; LDWAWLQTE; LDWAWL; TALDWAWLQT; TALDWAWLQ; ALDWAWLQT; LDWAWLQ; LDWAWLQT; TALDWEWLQTE; LDWEWLQTE; TALDWEWL; ALDWEWLQTE; LDWEWLQTE; LDWEWL; TALDWEWLQT; TALDWEWLQ; ALDWEWLQT; LDWEWLQ; 及びLDWEWLQTからなる群より選択される配列である、請求項1に記載の抗炎症化合物。
【請求項7】
Xa が6乃至12個のアミノ酸残基から成る、請求項1に記載の抗炎症化合物。
【請求項8】
Xa が6乃至10個のアミノ酸残基から成る、請求項1に記載の抗炎症化合物。
【請求項9】
Xa が、少なくとも5個の塩基性アミノ酸残基を含む、請求項1に記載の抗炎症化合物。
【請求項10】
Xa がL-アルギニン、D-アルギニン、L-リシン及びD-リシンから個別に選択される少なくとも5個のアミノ酸残基を含む、請求項1に記載の抗炎症化合物。
【請求項11】
Xa がRRMKWKK; YGRKKRRQRRR; ygrkkrrqrrr; YARKARRQARR; yarkarrqarr; YARAARRAARR; yaraarraarr; rrmkwkk、RRRRRR、RRRRRRR、RRRRRRRR、RRRRRRRRR、RRRRRRRRRR、RRRRRRRRRRR、rrrrrr、rrrrrrr、rrrrrrrr、rrrrrrrrr、rrrrrrrrrr、及び rrrrrrrrrrrからなる群より選択される、請求項1に記載の抗炎症化合物。
【請求項12】
RRMKWKKTALDWSWLQTE; rrmkwkkTALDWSWLQTE; YGRKKRRQRRRTALDWSWLQTE; ygrkkrrqrrrTALDWSWLQTE; rrrrrrrTALDWSWLQTE; RRRRRRRTALDWSWLQTE; YARKARRQARRTALDWSWLQTE; yarkarrqarrTALDWSWLQTE YARAARRAARRTALDWSWLQTE; yaraarraarrTALDWSWLQTE YGRKKRRQRRRLDWSWL; ygrkkrrqrrrLDWSWL; RRMKWKKLDWSWL; rrmkwkkLDWSWL; rrrrrrrLDWSWL; YARAARRAARRLDWSWL; yaraarraarrLDWSWL; 及びRRRRRRRLDWSWLからなる群より選択されるアミノ酸配列を含む抗炎症化合物。
【請求項13】
H-RRMKWKKTALDWSWLQTE-NH2;
H-YGRKKRRQRRRTALDWSWLQTE-NH2;
H-rrrrrrrTALDWSWLQTE-NH2;
H-YARKARRQARRTALDWSWLQTE-NH2;
H-YARAARRAARRTALDWSWLQTE-NH2;
H-RRMKWKKLDWSWL-NH2;
H-rrmkwkkLDWSWL-NH2;
H-rrrrrrrLDWSWL-NH2;
H-YARAARRAARRLDWSWL-NH2;
H-yaraarraarrLDWSWL-NH2; 及び
H-YGRKKRRQRRRLDWSWL- NH2
からなる群より選択される構造を有する抗炎症化合物。
【図1A】

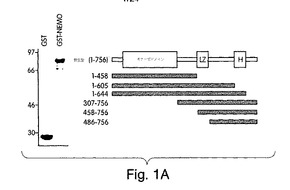
【図1B】
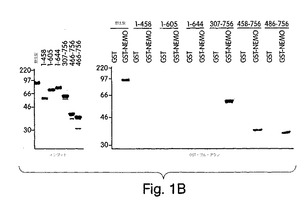
【図1C】
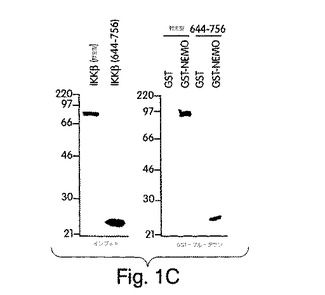
【図1D】
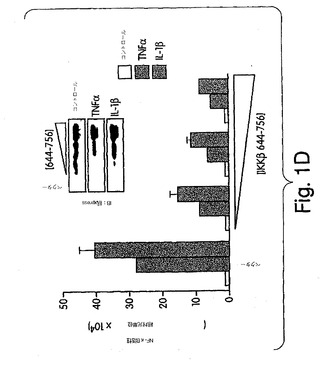
【図2A】
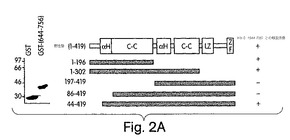
【図2B】
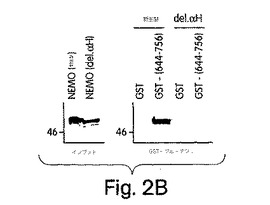
【図2C】
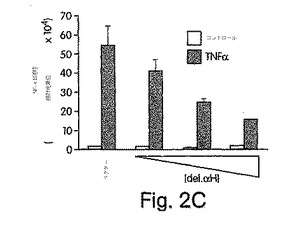
【図3A】
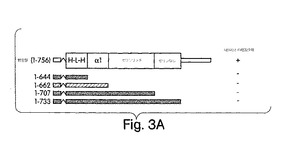
【図3B】
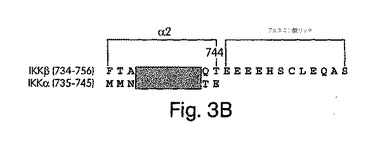
【図3C】
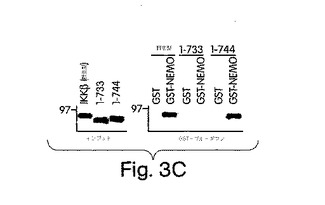
【図3D】
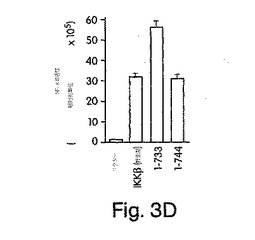
【図4A】
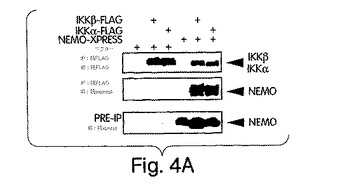
【図4B】
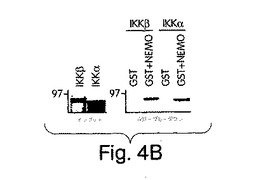
【図4C】
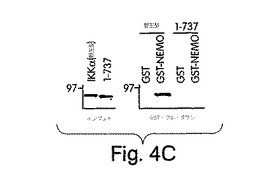
【図4D】
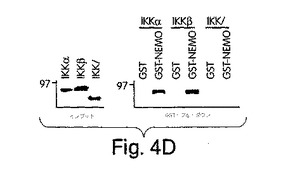
【図4E】
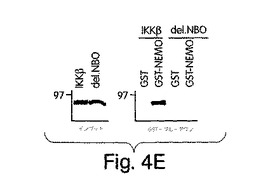
【図4F】
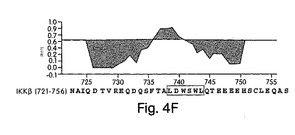
【図4G】
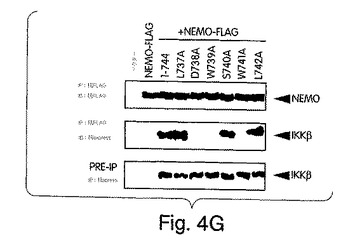
【図4H】
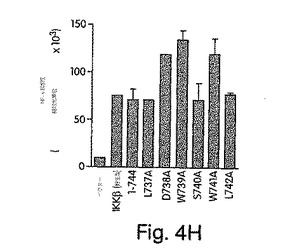
【図5A】
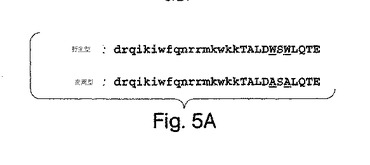
【図5B】
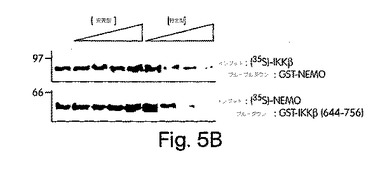
【図5C】
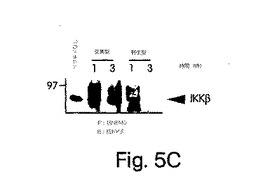
【図5D】
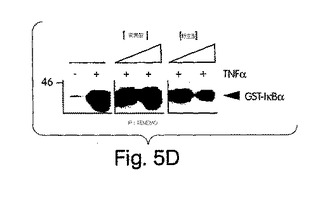
【図5E】
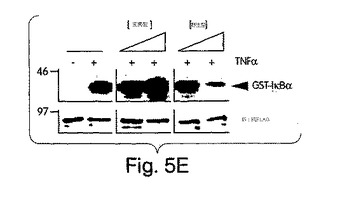
【図5F】
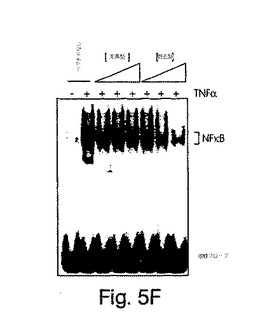
【図5G】
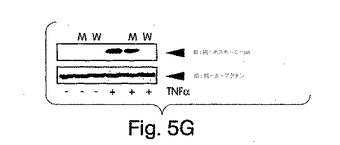
【図5H】
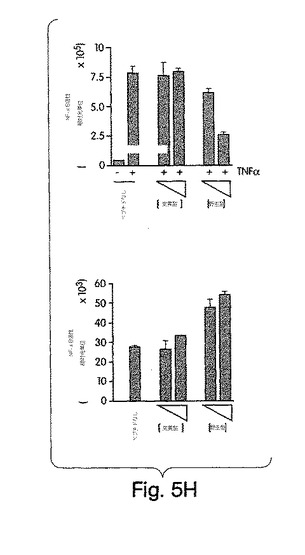
【図6A】
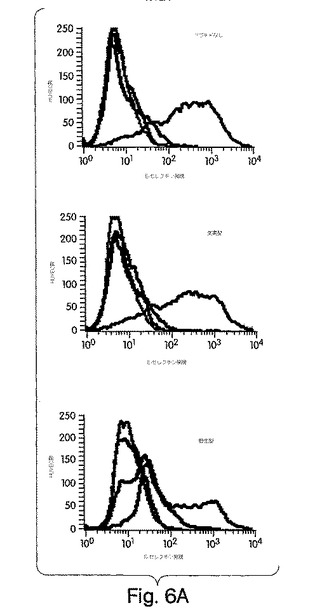
【図6B】
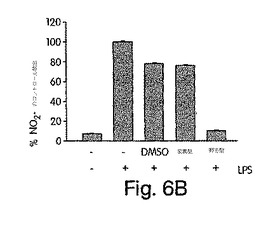
【図6C】
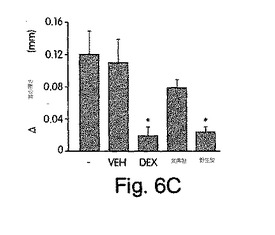
【図6D】
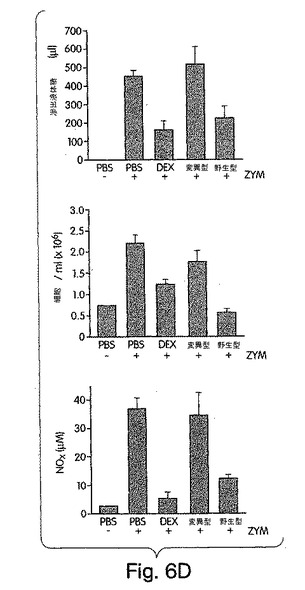
【図7】
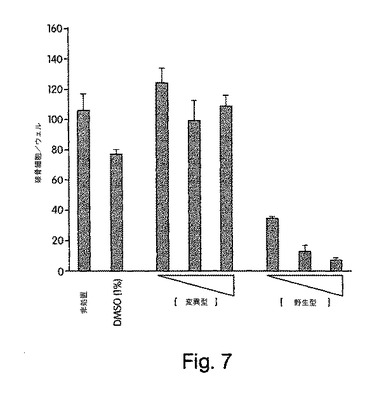
【図8A】
【図8B】
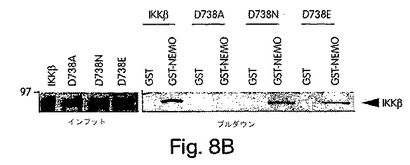
【図8C】
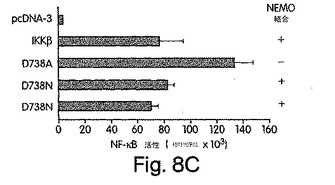
【図9A】
【図9B】
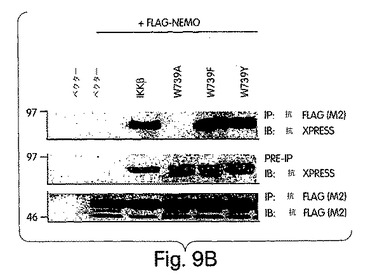
【図9C】
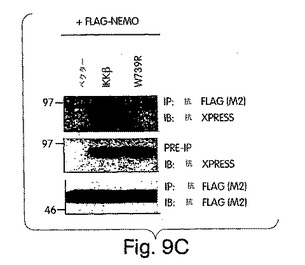
【図9D】
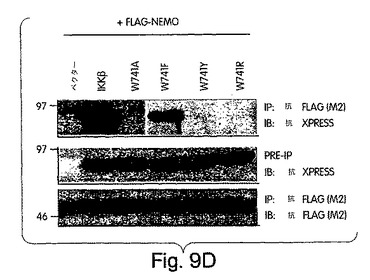
【図9E】
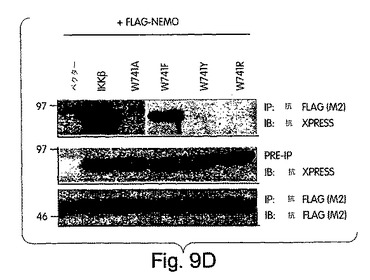
【図9F】
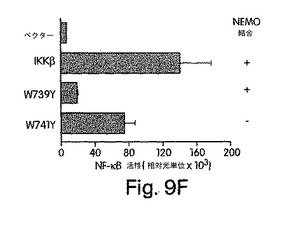
【図10A】
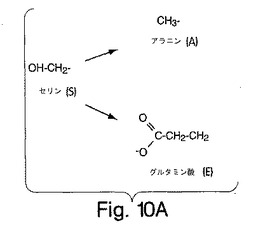
【図10B】
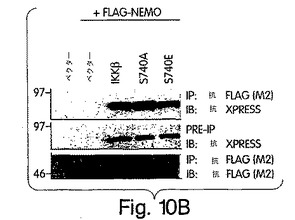
【図10C】
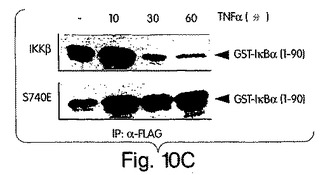
【図11A】
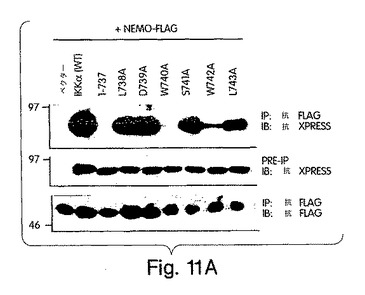
【図11B】
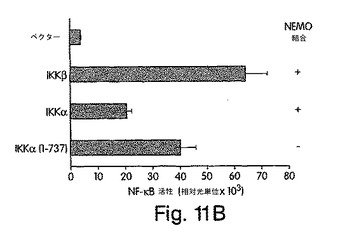
【図12A】
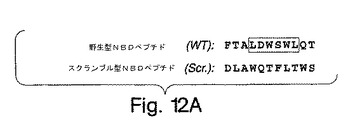
【図12B】
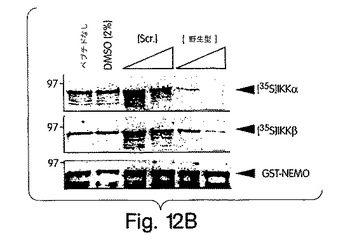
【図12C】
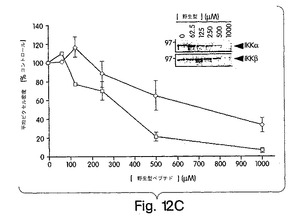
【図1B】
【図1C】
【図1D】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【図3D】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【図4G】
【図4H】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図5G】
【図5H】
【図6A】
【図6B】
【図6C】
【図6D】
【図7】
【図8A】
【図8B】
【図8C】
【図9A】
【図9B】
【図9C】
【図9D】
【図9E】
【図9F】
【図10A】
【図10B】
【図10C】
【図11A】
【図11B】
【図12A】
【図12B】
【図12C】
【公開番号】特開2013−56876(P2013−56876A)
【公開日】平成25年3月28日(2013.3.28)
【国際特許分類】
【出願番号】特願2011−270735(P2011−270735)
【出願日】平成23年12月9日(2011.12.9)
【分割の表示】特願2001−580978(P2001−580978)の分割
【原出願日】平成13年5月2日(2001.5.2)
【出願人】(399043691)プリーシス・ファーマシューティカルズ・インコーポレイテッド (14)
【氏名又は名称原語表記】PRAECIS PHARMACEUTICALS,INC.
【出願人】(500114601)イエール ユニバーシティー (4)
【Fターム(参考)】
【公開日】平成25年3月28日(2013.3.28)
【国際特許分類】
【出願日】平成23年12月9日(2011.12.9)
【分割の表示】特願2001−580978(P2001−580978)の分割
【原出願日】平成13年5月2日(2001.5.2)
【出願人】(399043691)プリーシス・ファーマシューティカルズ・インコーポレイテッド (14)
【氏名又は名称原語表記】PRAECIS PHARMACEUTICALS,INC.
【出願人】(500114601)イエール ユニバーシティー (4)
【Fターム(参考)】
[ Back to top ]