
抗生物質の相乗効果の同定法と応用法
本発明は、βラクタム系抗生物質と任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、GlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との抗菌効果のある組み合わせを、それを必要とするヒトまたは動物に投与する段階を含む、ヒトおよび動物における細菌感染症の治療に有用な薬学的組成物を提供する。以下の段階を含む抗生物質の相乗剤を発見する方法がさらに提供される:a)細胞内における遺伝子産物の活性または量を低減するように、遺伝子産物をコードする核酸に対するアンチセンス核酸を細胞内で発現し、それによって抗生物質に感作された細胞を産生する段階;b)抗生物質に対する細胞の感作を特徴付け、アンチセンス遺伝子の非存在下において必要とされる濃度の5分の1またはそれ未満で抗生物質の効力をもたらす抗生物質と遺伝子との対を選択する段階;c)選択された相乗的遺伝子に対応する遺伝子産物を阻害する化学化合物をスクリーニングする段階;およびd)阻害が細菌中で起こるように、選択された相乗的遺伝子に対応する遺伝子産物を阻害する化学的類似体を選択または作成する段階。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、概して細菌性疾患に関し、より具体的には、単独投与される化合物のいずれよりも極めて低い化合物の用量で細菌感染症の治療制御を可能にする化合物の組み合わせ、特に、βラクタム系抗生物質と組み合わせたMurBの阻害剤に関する。
【背景技術】
【0002】
背景情報
利用可能な別個のクラスの抗微生物化合物はほとんどなく、抗生物質耐性菌を伴う感染症を含む、細菌感染症の単独および併用薬物治療の範囲を限定する。したがって、抗菌化学療法研究は、新たな抗菌薬の開発のための新規標的の発見に重点的に取り組んできた。新たな抗菌化合物の発見への代替的アプローチは、抗生物質の相乗剤の発見である。抗微生物療法における相乗効果は周知であり、組み合わせて使用される抗生物質の超相加的活性を説明するために使用される。例えば、細菌感染症の治療において、ペニシリンまたはアンピシリンとストレプトマイシンまたはゲンタマイシン等の組み合わせは、腸球菌感染症に対して超相加的効果を有することが示されている。同様に、カルベニシリンまたはチカルシリンをゲンタマイシンまたはトブラマイシン等のアミノグリコシド剤と組み合わせると、緑膿菌(Pseudomonas aeruginosa)感染症の治療において相乗効果を呈する。ストレプトマイシンをテトラサイクリンと共に使用する併用療法は、いずれかの単独の薬剤よりもブルセラ症の治療において効果的であり、クロラムフェニコールにスルホンアミドを加えた混合物は、インフルエンザ菌(Haemophilus influenzae)による髄膜炎に対してより効果的である。
【0003】
抗菌剤の効力を予測するために使用される一方法は、Scribnerら(1982,Antimicrobial Agents and Chemotherapy 21(6):939‐943)(非特許文献1)によって、およびGoodman&Gilman(1980,The Pharmacological Basis of Therapeutics,Sixth Edition,pp.1097‐1098)(非特許文献2)において説明されており、チェッカーボードアッセイと称される。このアッセイは、培養液中に、個別のおよび両方を組み合わせた抗生物質の連続2倍希釈液を伴い、続いて試験される微生物がこれに接種される。インキュベーション後、個別にまたは組み合わせて使用された各薬物の最小発育阻止濃度(minimum inhibitory concentration;MIC)が決定される(注記:MICは、培地中における増殖を阻害する薬物の最低濃度である)。相乗効果は、組み合わせて使用した場合、各薬物のMICの減少によって示される。拮抗作用は、組み合わせて使用した場合、いずれかまたは両方の薬物におけるMICの増加によって示される。
【0004】
上記の例において、相乗効果は、抗生物質対の経験的試験によって発見される。相乗剤は、代謝的解毒によって抗生物質を不活性化する病原体の能力を阻害する化合物、抗生物質を搬出する細胞ポンプを阻害する化合物、および抗生物質の最小発育阻止濃度(MIC)を別の手法で減少させる化合物を含んでもよい。アモキシシリンおよびクラブラン酸を含有する組み合わせの開発は、そのような取り組みを説明する最たる例である。アモキシシリンはアミノペニシリンであり、ベータラクタマーゼ産生細菌によって分解される。クラブラン酸は、ベータラクタマーゼの活性を阻害するが、それ自身の抗菌活性を欠くことが発見された(Reading C&Cole M,1977.A beta lactamase inhibiting beta‐lactam from Streptomyces clavuligenus;Antimicrob Agents Chemother,11,p‐852‐857(非特許文献3);Reading C,Farmer T&Cole,M 1983,The Beta lactamase stability of Amoxycillin with beta lactamase inhibiting Clavulanic acid,J.Antimicrob Chemother.,11,p‐27‐32(非特許文献4);Todd P A&Benfield P 1990.Amoxycillin/Clavulanic acid,an up‐date of its antibacterial activity,pharmacokinetic properties&Therapeutic use,Drugs 39,p‐264‐307(非特許文献5))。
【0005】
クラブラン酸の場合、ベータラクタム系抗生物質に対する耐性を与える上での、ベータラクタマーゼの重要性が十分に理解された。この理解により、ベータラクタマーゼ阻害剤の標的開発が可能となった。
【0006】
任意の特定の抗生物質の有効性を限定する上で重要なその他のプロセスもあり得る。異なるプロセスを理解することは、クラブラン酸の場合とほぼ同様に、相乗剤開発のための標的の同定をもたらし得る。したがって、標的相乗剤開発が可能となるように、抗生物質活性を限定するプロセスを見いだす一般的手法を有することは、有用であると考えられる。
【0007】
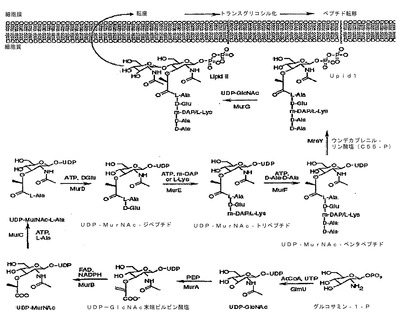
細菌細胞壁生合成は細菌における必須プロセスであり、該プロセスにおいては、細菌の内部でペプチドグリカンが産生され、外膜へ移送され、架橋され合って細胞壁を形成する。ペニシリンおよびセファロスポリンを含む重要な治療抗生物質は、ペニシリン結合タンパク質によって触媒される架橋プロセスを阻害する。図1は、該プロセスの図である。特に関心が寄せられているのは、MurA酵素(UDP‐N‐アセチルグルコサミンエノールピルビル転移酵素、EC2.5.1.7としても公知である)によって触媒される段階であり、これはmurA遺伝子によってコードされ、ホスホマイシンによって阻害される。MurB酵素によって触媒され、murB遺伝子(UDP‐N‐アセチルエノールピルビルグルコサミン還元酵素、EC1.1.1.158)によってコードされる次の段階にも関心が寄せられている。
【0008】
細胞膜の外部で起こるプロセス(トランスグリコシル化およびペプチド転移)により、ペプチドグリカンモノマーが架橋される。これらの反応は、ペニシリンおよびセファロスポリン系抗生物質の標的部位であるペニシリン結合タンパク質(PBP)によって触媒される。細菌中には複数のPBPがあり、それぞれの種が独自の分類を有している。PBPは、細胞1個あたり数百から千または二千の少数のコピーにおいて存在し、低分子量(低Mr)PBP、高MrPBP、およびβラクタマーゼを含む。これら3つのクラスはすべて、それらの触媒機能において重要な活性部位セリン残基を有する。ペニシリンおよびセファロスポリンは、PBPを不活性化する自殺基質として作用することによって細胞壁生合成の最終段階を阻害する。βラクタマーゼは、ペニシリンおよびセファロスポリンに作用する活性部位セリンを有するPBPに構造的に関係している。しかしながら、βラクタマーゼの場合、ペニシリンまたはセファロスポリンとの相互作用によって抗生物質の加水分解をもたらし、PBPと相互に作用するそれらの能力を失わせてしまう。
【0009】
文献中には、ホスホマイシンの、セファロスポリンおよびペニシリンとの組み合わせに関する多くの研究がある。Alvarezら(1985年)(非特許文献6)は、ホスホマイシン‐セファマンドールを伴う株の66%およびホスホマイシン‐メチシリンを伴う株の46%において、相乗効果が観察されたことを報告した。この研究において報告された相乗効果は、両方の薬物の最小発育阻止濃度(MIC)が4分の1減少した場合に定義された。Utsuiら(1986年)(非特許文献7)は、インビトロおよびインビボにおけるセフメタゾールおよびセフォタキシム単独の場合、ならびにホスホマイシンと組み合わせた場合の、黄色ブドウ球菌(Staphylococcus aureus)のメチシリンおよびセフェム耐性株に対する抗菌活性について報告した。この研究は、両方の薬物のMICにおける4分の1の低減を相乗効果の尺度として示すものであった。この研究はまた、併用治療における細菌の死滅加速という、別の形態の相乗効果も実証した。
【0010】
Utsuiらの研究は、ホスホマイシンによって誘導された特異的なPBPプロファイルの検出を含むものであった。
【0011】
いくつかのPBPのレベルは、ホスホマイシンに対する曝露時に著しく低減された。セファロスポリンはPBPを不活性化することによって作用するため、研究者らは、PBPの低減は、阻害すべき標的タンパク質数を低減させることにより、セファロスポリンをより効果的にするであろうとの結論を下した。研究者らはさらに、ペプチドグリカンモノマーがPBP転写の誘導因子であると推測した。ホスホマイシンによるペプチドグリカン生合成の阻害は、ペプチドグリカンのレベルを低減させ、それによってPBPの転写を阻害すると考えられた。
【0012】
ペプチドグリカン生合成酵素とベータラクタマーゼ阻害剤との相互作用についての遺伝学的研究は、メチシリン耐性黄色ブドウ球菌におけるmurEまたはmurFへの干渉が、ベータラクタム系オキサシリンに対する感受性を増大させたことを実証している(Sieradzki et al.,1997(非特許文献8);Sobral et al.,2003(非特許文献9);Gardete et al.,2004(非特許文献10))。これらの研究が含意するのは、ベータラクタム耐性の発現にはMurEおよびMurF活性が必要であるということである。したがって、MurEまたはMurFの阻害剤が、オキサシリンおよびその他のベータラクタム系抗生物質に対するメチシリン耐性黄色ブドウ球菌の感受性も増大させ得ることは当然である。
【0013】
これらの研究のいずれも、ペプチドグリカン生合成酵素の化学阻害剤が、ベータラクタム系抗生物質に対する非耐性細菌の感受性を増大させるとは予期していない。
【0014】
アンチセンスは、細菌中において新たな抗生物質の標的を同定する際に有用である。そのような標的を発見するため、およびアンチセンスクローンを感作細胞として使用してアンチセンス標的の阻害剤を検出するための、アンチセンスの使用を実証する多くの出版物がある。
【0015】
これらの発明の背後にある概念は、(1)アンチセンス発現は、細菌に「必須の要素」に特異的に関連する遺伝子を区別し、それによって抗生物質発見に使用するための標的のセットを定義することができること、および(2)アンチセンス発現は、アンチセンスによって標的とされるmRNAに対応するタンパク質を特異的に阻害する化合物に対して感作された細胞をもたらし得ることである。このアンチセンスの後者の使用は、化学ライブラリまたは天然産物コレクションから標的特異的阻害剤を見いだす際における感作細胞の使用をもたらし得る。
【0016】
これらの参考文献のいずれも、相乗剤発見のための標的を同定するためのアンチセンスの使用については記述していない。
【0017】
抗生物質の対の体系的な試験によって、病原体による抗生物質代謝を減少させる化合物の標的研究によって、または、特定の抗生物質を用いる化学物質または抽出物のランダムまたは合理的スクリーニングによって、抗生物質の有用な相乗的組み合わせを発見することができる。
【0018】
アンチセンスは、必須遺伝子を発見するため、および、新規抗生物質化合物の発見用のツールを開発するために使用されている。
【0019】
【非特許文献1】Scribnerら(1982,Antimicrobial Agents and Chemotherapy 21(6):939‐943)
【非特許文献2】Goodman&Gilman(1980,The Pharmacological Basis of Therapeutics,Sixth Edition,pp.1097‐1098)
【非特許文献3】Reading C&Cole M,1977.A beta lactamase inhibiting beta‐lactam from Streptomyces clavuligenus;Antimicrob Agents Chemother,11,p‐852‐857
【非特許文献4】Reading C,Farmer T&Cole,M 1983,The Beta lactamase stability of Amoxycillin with beta lactamase inhibiting Clavulanic acid,J.Antimicrob Chemother.,11,p‐27‐32
【非特許文献5】Todd P A&Benfield P 1990.Amoxycillin/Clavulanic acid,an up‐date of its antibacterial activity,pharmacokinetic properties&Therapeutic use,Drugs 39,p‐264‐307
【非特許文献6】Alvarezら(1985年)
【非特許文献7】Utsuiら(1986年)
【非特許文献8】Sieradzki et al.,1997
【非特許文献9】Sobral et al.,2003
【非特許文献10】Gardete et al.,2004
【発明の開示】
【0020】
発明の概要
概して、本発明は、抗生物質の相乗的遺伝子を発見する方法であって、
a.細胞内における遺伝子産物の活性または量を低減するように、遺伝子産物をコードする核酸に対するアンチセンス核酸を細胞内で発現し、それによって抗生物質に感作された細胞を産生する段階、および
b.抗生物質に対する細胞の感作を特徴付け、アンチセンス遺伝子の非存在下において必要とされる濃度の5分の1またはそれ未満で抗生物質の効力をもたらす抗生物質と遺伝子との対を選択する段階
を含む、方法を提供する。
【0021】
本発明は、抗生物質の相乗剤を発見する方法であって、
a.選択された相乗的遺伝子に対応する遺伝子産物を阻害する化学化合物をスクリーニングする段階、
b.阻害が細菌中で起こるように、選択された相乗的遺伝子に対応する遺伝子産物を阻害する化学的類似体を選択または作成する段階、および
c.相乗的阻害剤を抗生物質と組み合わせ、それぞれの種々の濃度における組み合わせの抗微生物活性を決定する段階
を含む、方法も提供する。
【0022】
一つの態様において、本発明は、ベータラクタム系抗生物質の抗菌能を増大させ、それによって動物における細菌感染症を予防または治療するための方法において有用性を有する方法を提供する。
【0023】
別の態様において、本発明は、MurBの阻害剤と細胞壁阻害抗生物質、特にベータラクタム系抗生物質との組み合わせを含む。
【0024】
微生物遺伝子断片を同定するための改良法によって、アンチセンス配向で異所的に発現した場合、関心対象の微生物遺伝子の転写後発現阻害をもたらし、それによって微生物中において観察および計測され得る表現型変化を引き起こすことができる。首尾よく同定されると、そのようなアンチセンス配向断片の条件発現を使用して、細胞内における特定の遺伝子産物の濃度を低下させ、それによって該細胞を、それ自体が遺伝子産物の特異的阻害剤である第2の物質に過剰感作させることができる。観察された表現型が、繁殖する能力の損失を構成する場合、この過感受性は、抗微生物活性を有する化合物の作用機序を試験するためのアッセイの基礎となり得る。そのようなアッセイにより、繁殖の損失を引き起こす能力が、関心対象の標的遺伝子産物およびそれらの化合物に対する特定の効果と関連付けられている抗微生物化合物と、観察される関連がない化合物とを区別することができるであろう。該改良法は、最も適切なアンチセンスDNA断片を明確に同定することにより、そのようなアッセイの開発を加速すると考えられる。
【0025】
2つの先行特許、米国特許第6228579号および米国特許第6924101号では、微生物ゲノムDNAのDNA断片を、微生物中においてアンチセンス配向で発現した場合に繁殖を低減または妨害するそれらの能力を調査したプロセスを記述している。手短に述べると、ゲノムDNAを断片化し、その断片をプラスミド上の誘導性プロモータの後にクローニングし、該プラスミドを関心対象の微生物に導入し、その後、クローニングされた断片の発現の依存的存在または非存在をもたらす誘導因子化合物または刺激剤の存在下および非存在下で、繁殖を計測および比較する。
【0026】
断片は、センス配向またはアンチセンス配向のいずれかで、プロモータに対してクローニングされ得る。断片がアンチセンス配向である場合、微生物の繁殖の低減または妨害を引き起こすそれらの断片は、該断片の由来源である遺伝子の機能に特異的な機序によって該低減または妨害を行うことが想定され、この遺伝子が、コードする細胞機能が正常な繁殖のために必要とされる遺伝子であることが示唆される。
【0027】
しかしながら、標的遺伝子の特異的阻害に至るアンチセンス断片を同定するための改良法を使用することにより、本発明者らは、上記の記述が真実ではないことを示した。実際に、本発明者らは、上記のようなプロモータから発現する断片は、示されている遺伝子および遺伝子産物に特異的でない多数の手段によって繁殖を低減し得ると認識している。見せかけのセンスまたはアンチセンス配向のいずれかで発現される断片は、1)細胞中における別の無関係な機能に対してアプタマーまたはその他の種類の阻害剤として作用し得るRNA種、2)元の遺伝子のものとは別のリーディングフレームにおいて発現し得、非特異的な毒性を有するペプチドを産生するRNA種をもたらし得る。このため、上記の特許において記述されているようなアンチセンスベースのスクリーニングは偽陽性をもたらし得る、すなわち、繁殖に必須である機能をコードしない遺伝子が、その遺伝子の増殖阻害アンチセンス断片のスクリーニングにおいて見かけ上のみ、そのようなものとして同定され得る。これは、繁殖等に必須なものとして、およびそのような可能性のある抗生物質の開発標的としての、遺伝子の誤同定につながる場合がある。
【0028】
生物のゲノム全体ではなく、1つの遺伝子または関心対象の遺伝子に焦点を当てる。これは、標的特異的表現型効果を引き起こす断片のスクリーニングにおいて、該遺伝子または関心対象の遺伝子のはるかに優れた描写を可能にする。特定の遺伝子または関心対象の遺伝子に対するアンチセンス断片を見出すために、ゲノムDNA全体を使用することにより、結果としてスクリーニングが長くなる。
【0029】
表現型変化を引き起こすアンチセンス断片を同定するための発見プロセスへの検証段階の組み込みは、該アンチセンス断片の由来源である遺伝子から発現した無傷のmRNAの特異的低減に伴う。
【0030】
発明の詳細な説明
「ベータラクタム」ベータラクタム(βラクタム)は、3つの炭素原子および1つの窒素原子からなる、ヘテロ原子環状構造を有するラクタムである。ベータラクタム環は、ペニシリン等、いくつかの抗生物質の一部であることから、それらはベータラクタム系抗生物質とも称される。これらの抗生物質は、細菌細胞壁合成を阻害することによって働く。ベータラクタム系抗生物質クラスの例としては、ペニシリン、セファロスポリン、およびカルバペネムが含まれる。

【0031】
「抗生物質」は、感染症を制御するために使用される化合物である。抗生物質の例としては、以下が含まれる。
細胞壁阻害剤
ベータラクタム(ペニシリンG、ペニシリンV、ナフシリン、メチシリン、オキサシリン、クロキサシリン、ジクロキサシリン、フルクロキサシリン、アンピシリン、アモキサシリン、カルベニシリン、チカルシリン、アゾシリン、メズロシリン、ピペラシリン、セファロスポリン、セファロチン、セファゾリン、セファレキシン、セフロキシム、セファマンドール、セフォキシチン、セファクロル、モキサラクタム、セフォペラゾン、セフタジジム、セフトリアキソン、クラブラン酸、スルバクタム、イミペネム、アズトレオナム、セフォキシチン、セフォゾリン、セファクロル)
ペプチドグリカン生合成の阻害剤は、以下を含むがこれらに限定されない。
1.GlmU、MurAの阻害剤(ホスホマイシン)
2.MurBの阻害剤(ジヒドロピロロン)
3.ウンデカプレニルピロリン酸合成酵素の阻害剤(ヒダントイン、スルホンアミド)
4.LpxCの阻害剤(UDP‐3‐O‐アシルN‐アセチルグルコサミン脱アセチル化酵素)
リポペプチド(ダプトマイシン)
イソプレノイド生合成阻害剤(バシトラシン、ホスミドマイシン)
脂肪酸生合成阻害剤(セルレニン、トリクロサン、イソニアジド)
タンパク質合成阻害剤
1.アミノグリコシド(ストレプトマイシン、ゲンタマイシン、およびカナマイシン)、
2.テトラサイクリン
3.クロラムフェニコール、
4.マクロライド(エリスロマイシン、アジスロマイシン、クラリスロマイシン)
5.リンコサミド(リンコマイシンおよびクリンダマイシン)
6.オキサゾリジノン(リネゾリド)
ムピロシン等のtRNA合成酵素阻害剤
1.MetRS阻害剤(カテコール、プロリン、キノロン)
2.ペプチド脱ホルミル酵素阻害剤(ヒドロキサマート)
リファンピシン等のRNA合成阻害剤
キノロン等のDNAジャイレース阻害剤(ナリジクス酸、シプロフロキサシン)
抗葉酸剤(トリメトプリムおよびスルファメトキサゾール)
【0032】
「相乗効果」は、≦0.5のフラクショナル阻止濃度(fractional inhibitory concentration)指数(FICI)として定義され、ここでFICIは、2つの化合物を組み合わせた場合の個々の構成要素のフラクショナル阻止濃度(FIC)の和として定義され、FICは組み合わせた場合の化合物の最小発育阻止濃度(MIC)を化合物単独のMICで割った比率として定義される。
【0033】
「アンチセンス相乗効果」は、誘導因子の存在下における化合物の50%発育阻止濃度(IC50)を、誘導因子の非存在下におけるIC50で割った比率≧5として定義される。
【0034】
本発明者らのアンチセンスへの関心は、新たな抗生物質候補の作用機序を追跡する際に使用するための、炭疽菌の特定のアンチセンス株の開発にあった。本発明者らの標的の1つはMurB、つまりペプチドグリカン生合成の第2の段階である(図1)。すべての病原菌はMurBを必要とするが、現在MurBを阻害することによって作用する抗生物質はない。
【0035】
その機能的murB遺伝子(murB‐2)に対する誘導性アンチセンスを有する炭疽菌株の作成は、誘導因子の存在下において増殖減弱された株をもたらした。選択された株は、murB遺伝子の種々の部分に対応するアンチセンス挿入断片を有することが決定された。続いて、増殖減弱および抗生物質のパネルに対する感受性の誘導因子依存性の観点から、それらの株を特徴付けた。予想に反して、株のいくつかは抗生物質に対する感受性において相違を示さなかった。したがって、抗生物質発見のためのアンチセンスベースのツールを作成する際における重大な付加的段階は、ある特定の抗生物質に対する感受性増大の実証である。本発明者らは、他者らによって作成されたアンチセンス株の多くは、阻害剤に対して過感受性でないため、スクリーニングまたは作用機構決定において有用ではないのではないかとの疑念を抱いている。
【0036】
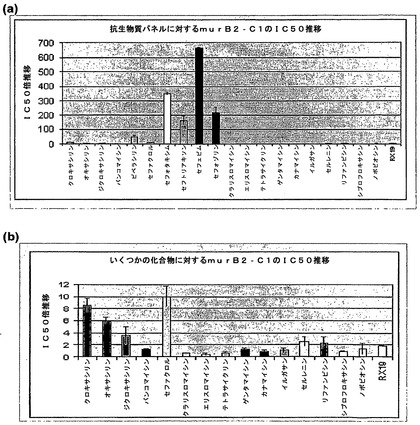
確認されたmurBアンチセンス株の、細胞壁阻害抗生物質について、予想外且つ劇的なアンチセンス相乗効果が観察された。4つの場合(セフェピム、セフトリアキソン、セフォキシチン、またはセフォタキシム)において、murBアンチセンス発現は、非誘導条件にある細胞に対して、100〜600倍低い濃度の抗生物質に対して感受性のある細胞をもたらした(図3)。その他の細胞壁阻害抗生物質に対する感受性は、増大されたが程度の低いものであった。タンパク質合成、DNA合成、またはRNA合成等、その他の経路を標的とする抗生物質に対する感受性は、増大されなかった。その他の遺伝子を標的とするアンチセンス株は、試験したいかなる抗生物質に対しても感受性におけるこのような異例の増大がなかったことから、該効果はペプチドグリカン生合成経路内の遺伝子に特異的であると考えられる。
【0037】
図4は、抗生物質のパネルに対するmurAアンチセンス株の感受性を示す。murBアンチセンス株に観察されたように、murAアンチセンス発現は、細胞壁阻害抗生物質に対する感受性は何倍にもなったが、その他の経路を標的とする抗生物質に対してはそうでない細胞をもたらした。ペプチドグリカン生合成におけるその他の遺伝子を標的とするアンチセンス株にも同様の結果が得られた。
【0038】
標的間における限られた相乗的な相互作用については前例があるが、MurBアンチセンス株とセファロスポリンとの間における相乗効果の大きさについては前例がない。本発明者らには、アンチセンスの代わりに標的阻害剤を使用して相乗効果が達成され得ることを示す、セフォタキシムと組み合わせたいくつかの弱MurB阻害剤についての限定的なデータがある(図5)。本発明者らはまた、この現象が炭疽菌に限定されるものでなく、黄色ブドウ球菌中におけるmurBアンチセンス発現が、ベータラクタム系抗生物質に対して細胞を過剰感作させることも示した(図6)。最後に、本発明者らは、抗生物質化学療法における重要なパラメータである「死滅時間(Time‐to‐kill)」の観点から、この相乗効果を特徴付けるプロセスにある。本発明者らは、セファロスポリンとMurB阻害との組み合わせが、セファロスポリン単独での殺菌結果にかかる時間に対して、殺菌効果にかかる時間を短くすると期待している。
【0039】
本発明者らはさらに、MurB阻害が、そうしなければそのような感染症の治療時に効果のないメチシリンおよびその他のペニシリンに対して、メチシリン耐性細菌を再感作すると期待している。
【0040】
炭疽菌におけるMurBおよびセファロスポリンに関するこの観察により、本発明者らは、この方法論を用いて発見され得る抗生物質と遺伝子標的とのその他の組み合わせを期待している。本発明者らは、これらの観察は、その他の重要な病原体、特に黄色ブドウ球菌に移し変えることができると期待している。図6は、murBに対する誘導性アンチセンスで操作された黄色ブドウ球菌の株におけるセフォキシチンおよびジクロキサシリンの効能の増大を示す。抗生物質のパネルに対する黄色ブドウ球菌株の感受性は、ベータラクタム系抗生物質の誘導因子の存在下において増大したが、その他の作用機構の抗生物質では増大しなかった。
【0041】
本発明者らは、抗生物質と遺伝子との、その他の予想外の相乗的対を明らかにする能力も期待している。これらの対は、その他の抗生物質に対する相乗剤の開発のための標的部位の発見を可能にし得る。菌類および哺乳動物細胞を含むその他の生物におけるアンチセンスは、抗癌剤、向精神薬等を含む多くのクラスの薬物の有効性を増強するための、相乗的標的部位および最終的には相乗化合物の発見のために、同様の様式で活用され得る。
【0042】
したがって、任意の所与の抗生物質と任意の特定の標的の阻害剤との間における相乗的な相互作用の潜在性を決定できることが有用であると考えられる。そのような決定は、所与の抗生物質の効能を増大させる化学化合物のスクリーニングの構成を可能にすると考えられる。誘導性アンチセンスは、相乗剤の発見および開発用の標的を決定するためのシステムの基礎となり得る。アンチセンスのこの使用は、遺伝子の必要性を発見する際におけるその使用と対照をなす。遺伝子の必要性を発見するためのアンチセンスの使用については他者らによって記述されているが、アンチセンスのこの使用は、必須遺伝子、または必須ではないが抗生物質の効能を何らかの手法で限定する遺伝子のいずれかの検出をもたらし得る。
【0043】
薬物の相乗的組み合わせの使用は、使用される用量の低下または化学療法の時間が短くなることによる薬物の副作用低下、感染症のより迅速な治癒によって入院が短くなること、制御される病原体のスペクトルの増大、抗生物質に対する耐性発現の発生率の減少を含む、従来の単一化合物化学療法よりも多くの利点を有し得る。
【0044】
抗生物質の相乗剤発見のための関心対象の遺伝子を決定するためのアンチセンスツールの重要性は、それが、抗生物質の作用機序に関する特定の知識を持たずに、および相乗的標的用に特定の化合物を使用することなく、新規スクリーニングを開発するための経験的方法を提供することである。このアンチセンスの使用は、遺伝子と任意の所与の病原体に対する抗生物質感受性との間の予想外の関係を明らかにすることができる。図7は、抗生物質のための相乗的標的の発見についてのプロセスの流れを示す。
【0045】
本発明者らは、MurBとベータラクタム系抗生物質との間の相乗的な相互作用を発見した。この発見は、治療法の投薬量を低減すること、治療時間を減少させること、または病原体制御のスペクトルを増大させることによって、抗感染症化学療法を増強することができる、MurB阻害剤とベータラクタムとの組み合わせの開発を予期するものである。
【0046】
本発明者らは、特定の遺伝子のための誘導性アンチセンスを使用し、誘導因子の存在下および非存在下における生物の抗生物質感受性を比較して、任意の所与の抗生物質の相乗的標的を発見するための一般法を開発した。
【0047】
本発明者らは、薬物と相乗作用のある標的を発見し、それによって薬効を増大させることを可能にするための、アンチセンスの一般的使用を予期している。この使用は、抗ウイルス剤、抗真菌剤、抗寄生虫剤、抗腫瘍剤、殺虫剤、除草剤、および化学療法によって処置が施されるその他の生物系にまでおよび得る。
【0048】
実施例
以下の例は、本発明を例示することを意図し、限定することを意図しない。
【0049】
以下は、本発明の方法を表す説明的例である。
1)ゲノムDNAをテンプレートとして使用して、関心対象の遺伝子(例えば、オペロン)をPCR増幅する。調査する遺伝子のいずれかの末端の約200bpを含むように注意する。
2)該PCR断片を超音波処理によって断片化し、結果として生じた断片を平滑末端化し、約350〜400bpより大きくないようにサイズ選択し、次いで本発明者らの発現プラスミドであるpBAX‐2内の誘導性プロモータの直ぐ下流のポリリンカーにクローニングする。このライブラリの作成は、通常、大腸菌(E.coli)において為される。
3)続いて、ライブラリのDNAを、関心対象の遺伝子が由来する生物、例えば、炭疽菌または黄色ブドウ球菌に形質転換する。ピッキングによって個々のコロニーを収集し、プラスミドの存在を選択する成長培地中で、96ウェル培養皿において培養する。
4)細胞が増殖した後、誘導化合物キシロースが含まれるかまたは含まれないかのいずれかの元の成長培地の固体版に、該細胞をレプリカプレーティングする(例えば、ピンツールを使用して)。
5)これらのレプリカプレートを一晩増殖させ、続いて、プラスとマイナスのキシロース対として比較し、キシロースプレート上においてのみ著しい増殖感受性を示すコロニーを、さらなる研究のために非キシロースプレートから選択する。
6)キシロース成長感受性株のpBAX‐2プラスミドの挿入断片を、DNA配列決定法によって分析する。これにより、PCR増幅した関心対象の遺伝子において断片が何処に由来するかを同定し、該関心対象の遺伝子および誘導性プロモータの両方に対してセンスまたはアンチセンス配向であるかを確定する。
7)センスおよびアンチセンスの相対数を記録する。本発明者らは、遺伝子が必須である場合、キシロース増殖感受性株のプールにおいて、アンチセンス配向断片を主に得ることを見出した。遺伝子が必須でない場合、スクリーニングされる同等数のコロニーにおいて増殖感受性が見られない傾向があるか、または、より同等な数のアンチセンスおよびセンスが見られる。キシローススクリーニングを適用する前に形質転換細胞を無作為に選択する場合、得られるアンチセンス対センスの数は、ほぼ等しいままであることを見出すであろうことに留意されたい。
8)さらなる分析のために、多数のアンチセンスクローンを選択する。これらは、関心対象の遺伝子における相対的位置(本発明者らのmRNAレベルを分析する現在の能力では、さらなる3’断片に制限される)および相対的サイズについて選択する。多くの場合、断片が大きくなるにつれて、非特異的な増殖感受性を引き起こす可能性が大きくなる。
9)続いて、これらのアンチセンスクローンに対して2つの研究を行う。アンチセンス断片の発現の誘導を受けて、関心対象の遺伝子からの転写物の内因性レベルを計測する。関心対象の遺伝子に対応し、対照遺伝子のものには対応しないメッセージにおける特異的な分解または低減は、アンチセンスが、関心対象の遺伝子の特異的な転写後発現阻害剤として働いていることの指標である。
10)もう1つのアッセイは、アンチセンス断片の発現の誘導を受けて、一般的な増殖効果とは異なる特異的な表現型変化を計測することである。本発明者らのDHFRアンチセンスの場合、試験はアンチセンスがチミジンの栄養要求性を引き起こし得るか否かを見るものであった。チミジンの生合成はDHFRによって合成されるテトラヒドロ葉酸塩の大部分を要するものであり、栄養要求性はDHFR突然変異の古典的効果である。もう1つの行うべきことは、特定の抗生物質等、関心対象の遺伝子の細胞産物に特異的なアンチセンスの誘導による、阻害化合物に対する過感受性を計測することである。アンチセンスは、関心対象の遺伝子に特異的でなく、無関係な細胞標的に特異的である、阻害剤または抗生物質に対する過感受性を引き起こさないはずである。
【0050】
上記の例を参照して本発明を説明したが、本発明の精神および範囲には、改変および変形が包含されることが理解されるであろう。したがって、本発明は、特許請求の範囲によってのみ限定される。
【図面の簡単な説明】
【0051】
【図1】細菌中におけるペプチドグリカンモノマー単位の生合成経路を示す。
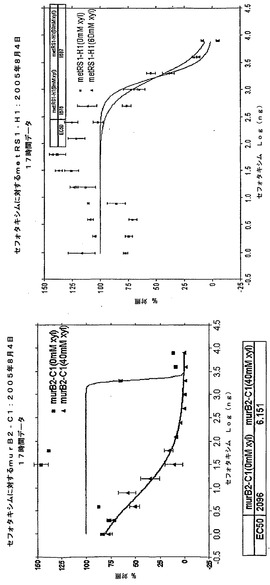
【図2】阻止濃度下のアンチセンス誘導因子の存在下および非存在下における、セフォタキシムへのmurB2アンチセンスクローンの応答を示す。左パネル:murB2アンチセンスクローン。上部線:誘導因子なし。下部線:+40mMキシロース。右パネル:metRSアンチセンスクローン。EC50+キシロース/EC50(−キシロース)の比率により、「IC50推移」が求められる。この例において推移を計算すると、(2096/6.1)=343である。
【図3】(a)セファロスポリン、ならびに(b)ベータラクタム系クロキサシリン、オキサシリン、およびジクロキサシリンを含むその他の抗生物質に対する、炭疽菌(B.anthracis)murB‐2アンチセンス株の感受性を示す。
【図4】種々の抗生物質に対する炭疽菌murAlアンチセンス株の感受性を示す。
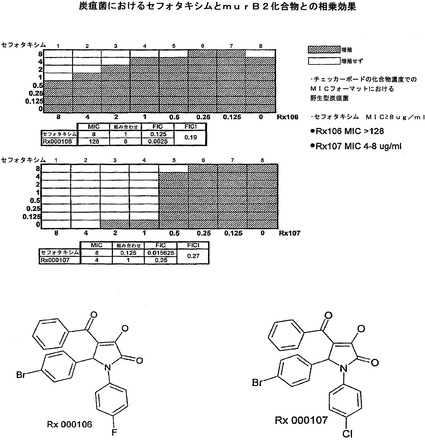
【図5】セフォタキシムおよびジヒドロピロロン(dihydropyrrolone)MurB阻害剤Rx000106またはRx000107を使用した、野生型炭疽菌におけるFICI実験を示す。チェッカーボード実験においては、化合物のフラクショナル阻止濃度(FIC)は、単独および組み合わせた化合物の最小発育阻止濃度(MIC)の比率として計算される。FIC指数(FICI)は、2つの化合物のFICの和である。
【図6】murBアンチセンスを用いて操作された炭疽菌を示す。
【図7】相乗的標的および相乗剤の発見のためのプロセスを示す。
【図8】標的配列の表である。
【技術分野】
【0001】
発明の分野
本発明は、概して細菌性疾患に関し、より具体的には、単独投与される化合物のいずれよりも極めて低い化合物の用量で細菌感染症の治療制御を可能にする化合物の組み合わせ、特に、βラクタム系抗生物質と組み合わせたMurBの阻害剤に関する。
【背景技術】
【0002】
背景情報
利用可能な別個のクラスの抗微生物化合物はほとんどなく、抗生物質耐性菌を伴う感染症を含む、細菌感染症の単独および併用薬物治療の範囲を限定する。したがって、抗菌化学療法研究は、新たな抗菌薬の開発のための新規標的の発見に重点的に取り組んできた。新たな抗菌化合物の発見への代替的アプローチは、抗生物質の相乗剤の発見である。抗微生物療法における相乗効果は周知であり、組み合わせて使用される抗生物質の超相加的活性を説明するために使用される。例えば、細菌感染症の治療において、ペニシリンまたはアンピシリンとストレプトマイシンまたはゲンタマイシン等の組み合わせは、腸球菌感染症に対して超相加的効果を有することが示されている。同様に、カルベニシリンまたはチカルシリンをゲンタマイシンまたはトブラマイシン等のアミノグリコシド剤と組み合わせると、緑膿菌(Pseudomonas aeruginosa)感染症の治療において相乗効果を呈する。ストレプトマイシンをテトラサイクリンと共に使用する併用療法は、いずれかの単独の薬剤よりもブルセラ症の治療において効果的であり、クロラムフェニコールにスルホンアミドを加えた混合物は、インフルエンザ菌(Haemophilus influenzae)による髄膜炎に対してより効果的である。
【0003】
抗菌剤の効力を予測するために使用される一方法は、Scribnerら(1982,Antimicrobial Agents and Chemotherapy 21(6):939‐943)(非特許文献1)によって、およびGoodman&Gilman(1980,The Pharmacological Basis of Therapeutics,Sixth Edition,pp.1097‐1098)(非特許文献2)において説明されており、チェッカーボードアッセイと称される。このアッセイは、培養液中に、個別のおよび両方を組み合わせた抗生物質の連続2倍希釈液を伴い、続いて試験される微生物がこれに接種される。インキュベーション後、個別にまたは組み合わせて使用された各薬物の最小発育阻止濃度(minimum inhibitory concentration;MIC)が決定される(注記:MICは、培地中における増殖を阻害する薬物の最低濃度である)。相乗効果は、組み合わせて使用した場合、各薬物のMICの減少によって示される。拮抗作用は、組み合わせて使用した場合、いずれかまたは両方の薬物におけるMICの増加によって示される。
【0004】
上記の例において、相乗効果は、抗生物質対の経験的試験によって発見される。相乗剤は、代謝的解毒によって抗生物質を不活性化する病原体の能力を阻害する化合物、抗生物質を搬出する細胞ポンプを阻害する化合物、および抗生物質の最小発育阻止濃度(MIC)を別の手法で減少させる化合物を含んでもよい。アモキシシリンおよびクラブラン酸を含有する組み合わせの開発は、そのような取り組みを説明する最たる例である。アモキシシリンはアミノペニシリンであり、ベータラクタマーゼ産生細菌によって分解される。クラブラン酸は、ベータラクタマーゼの活性を阻害するが、それ自身の抗菌活性を欠くことが発見された(Reading C&Cole M,1977.A beta lactamase inhibiting beta‐lactam from Streptomyces clavuligenus;Antimicrob Agents Chemother,11,p‐852‐857(非特許文献3);Reading C,Farmer T&Cole,M 1983,The Beta lactamase stability of Amoxycillin with beta lactamase inhibiting Clavulanic acid,J.Antimicrob Chemother.,11,p‐27‐32(非特許文献4);Todd P A&Benfield P 1990.Amoxycillin/Clavulanic acid,an up‐date of its antibacterial activity,pharmacokinetic properties&Therapeutic use,Drugs 39,p‐264‐307(非特許文献5))。
【0005】
クラブラン酸の場合、ベータラクタム系抗生物質に対する耐性を与える上での、ベータラクタマーゼの重要性が十分に理解された。この理解により、ベータラクタマーゼ阻害剤の標的開発が可能となった。
【0006】
任意の特定の抗生物質の有効性を限定する上で重要なその他のプロセスもあり得る。異なるプロセスを理解することは、クラブラン酸の場合とほぼ同様に、相乗剤開発のための標的の同定をもたらし得る。したがって、標的相乗剤開発が可能となるように、抗生物質活性を限定するプロセスを見いだす一般的手法を有することは、有用であると考えられる。
【0007】
細菌細胞壁生合成は細菌における必須プロセスであり、該プロセスにおいては、細菌の内部でペプチドグリカンが産生され、外膜へ移送され、架橋され合って細胞壁を形成する。ペニシリンおよびセファロスポリンを含む重要な治療抗生物質は、ペニシリン結合タンパク質によって触媒される架橋プロセスを阻害する。図1は、該プロセスの図である。特に関心が寄せられているのは、MurA酵素(UDP‐N‐アセチルグルコサミンエノールピルビル転移酵素、EC2.5.1.7としても公知である)によって触媒される段階であり、これはmurA遺伝子によってコードされ、ホスホマイシンによって阻害される。MurB酵素によって触媒され、murB遺伝子(UDP‐N‐アセチルエノールピルビルグルコサミン還元酵素、EC1.1.1.158)によってコードされる次の段階にも関心が寄せられている。
【0008】
細胞膜の外部で起こるプロセス(トランスグリコシル化およびペプチド転移)により、ペプチドグリカンモノマーが架橋される。これらの反応は、ペニシリンおよびセファロスポリン系抗生物質の標的部位であるペニシリン結合タンパク質(PBP)によって触媒される。細菌中には複数のPBPがあり、それぞれの種が独自の分類を有している。PBPは、細胞1個あたり数百から千または二千の少数のコピーにおいて存在し、低分子量(低Mr)PBP、高MrPBP、およびβラクタマーゼを含む。これら3つのクラスはすべて、それらの触媒機能において重要な活性部位セリン残基を有する。ペニシリンおよびセファロスポリンは、PBPを不活性化する自殺基質として作用することによって細胞壁生合成の最終段階を阻害する。βラクタマーゼは、ペニシリンおよびセファロスポリンに作用する活性部位セリンを有するPBPに構造的に関係している。しかしながら、βラクタマーゼの場合、ペニシリンまたはセファロスポリンとの相互作用によって抗生物質の加水分解をもたらし、PBPと相互に作用するそれらの能力を失わせてしまう。
【0009】
文献中には、ホスホマイシンの、セファロスポリンおよびペニシリンとの組み合わせに関する多くの研究がある。Alvarezら(1985年)(非特許文献6)は、ホスホマイシン‐セファマンドールを伴う株の66%およびホスホマイシン‐メチシリンを伴う株の46%において、相乗効果が観察されたことを報告した。この研究において報告された相乗効果は、両方の薬物の最小発育阻止濃度(MIC)が4分の1減少した場合に定義された。Utsuiら(1986年)(非特許文献7)は、インビトロおよびインビボにおけるセフメタゾールおよびセフォタキシム単独の場合、ならびにホスホマイシンと組み合わせた場合の、黄色ブドウ球菌(Staphylococcus aureus)のメチシリンおよびセフェム耐性株に対する抗菌活性について報告した。この研究は、両方の薬物のMICにおける4分の1の低減を相乗効果の尺度として示すものであった。この研究はまた、併用治療における細菌の死滅加速という、別の形態の相乗効果も実証した。
【0010】
Utsuiらの研究は、ホスホマイシンによって誘導された特異的なPBPプロファイルの検出を含むものであった。
【0011】
いくつかのPBPのレベルは、ホスホマイシンに対する曝露時に著しく低減された。セファロスポリンはPBPを不活性化することによって作用するため、研究者らは、PBPの低減は、阻害すべき標的タンパク質数を低減させることにより、セファロスポリンをより効果的にするであろうとの結論を下した。研究者らはさらに、ペプチドグリカンモノマーがPBP転写の誘導因子であると推測した。ホスホマイシンによるペプチドグリカン生合成の阻害は、ペプチドグリカンのレベルを低減させ、それによってPBPの転写を阻害すると考えられた。
【0012】
ペプチドグリカン生合成酵素とベータラクタマーゼ阻害剤との相互作用についての遺伝学的研究は、メチシリン耐性黄色ブドウ球菌におけるmurEまたはmurFへの干渉が、ベータラクタム系オキサシリンに対する感受性を増大させたことを実証している(Sieradzki et al.,1997(非特許文献8);Sobral et al.,2003(非特許文献9);Gardete et al.,2004(非特許文献10))。これらの研究が含意するのは、ベータラクタム耐性の発現にはMurEおよびMurF活性が必要であるということである。したがって、MurEまたはMurFの阻害剤が、オキサシリンおよびその他のベータラクタム系抗生物質に対するメチシリン耐性黄色ブドウ球菌の感受性も増大させ得ることは当然である。
【0013】
これらの研究のいずれも、ペプチドグリカン生合成酵素の化学阻害剤が、ベータラクタム系抗生物質に対する非耐性細菌の感受性を増大させるとは予期していない。
【0014】
アンチセンスは、細菌中において新たな抗生物質の標的を同定する際に有用である。そのような標的を発見するため、およびアンチセンスクローンを感作細胞として使用してアンチセンス標的の阻害剤を検出するための、アンチセンスの使用を実証する多くの出版物がある。
【0015】
これらの発明の背後にある概念は、(1)アンチセンス発現は、細菌に「必須の要素」に特異的に関連する遺伝子を区別し、それによって抗生物質発見に使用するための標的のセットを定義することができること、および(2)アンチセンス発現は、アンチセンスによって標的とされるmRNAに対応するタンパク質を特異的に阻害する化合物に対して感作された細胞をもたらし得ることである。このアンチセンスの後者の使用は、化学ライブラリまたは天然産物コレクションから標的特異的阻害剤を見いだす際における感作細胞の使用をもたらし得る。
【0016】
これらの参考文献のいずれも、相乗剤発見のための標的を同定するためのアンチセンスの使用については記述していない。
【0017】
抗生物質の対の体系的な試験によって、病原体による抗生物質代謝を減少させる化合物の標的研究によって、または、特定の抗生物質を用いる化学物質または抽出物のランダムまたは合理的スクリーニングによって、抗生物質の有用な相乗的組み合わせを発見することができる。
【0018】
アンチセンスは、必須遺伝子を発見するため、および、新規抗生物質化合物の発見用のツールを開発するために使用されている。
【0019】
【非特許文献1】Scribnerら(1982,Antimicrobial Agents and Chemotherapy 21(6):939‐943)
【非特許文献2】Goodman&Gilman(1980,The Pharmacological Basis of Therapeutics,Sixth Edition,pp.1097‐1098)
【非特許文献3】Reading C&Cole M,1977.A beta lactamase inhibiting beta‐lactam from Streptomyces clavuligenus;Antimicrob Agents Chemother,11,p‐852‐857
【非特許文献4】Reading C,Farmer T&Cole,M 1983,The Beta lactamase stability of Amoxycillin with beta lactamase inhibiting Clavulanic acid,J.Antimicrob Chemother.,11,p‐27‐32
【非特許文献5】Todd P A&Benfield P 1990.Amoxycillin/Clavulanic acid,an up‐date of its antibacterial activity,pharmacokinetic properties&Therapeutic use,Drugs 39,p‐264‐307
【非特許文献6】Alvarezら(1985年)
【非特許文献7】Utsuiら(1986年)
【非特許文献8】Sieradzki et al.,1997
【非特許文献9】Sobral et al.,2003
【非特許文献10】Gardete et al.,2004
【発明の開示】
【0020】
発明の概要
概して、本発明は、抗生物質の相乗的遺伝子を発見する方法であって、
a.細胞内における遺伝子産物の活性または量を低減するように、遺伝子産物をコードする核酸に対するアンチセンス核酸を細胞内で発現し、それによって抗生物質に感作された細胞を産生する段階、および
b.抗生物質に対する細胞の感作を特徴付け、アンチセンス遺伝子の非存在下において必要とされる濃度の5分の1またはそれ未満で抗生物質の効力をもたらす抗生物質と遺伝子との対を選択する段階
を含む、方法を提供する。
【0021】
本発明は、抗生物質の相乗剤を発見する方法であって、
a.選択された相乗的遺伝子に対応する遺伝子産物を阻害する化学化合物をスクリーニングする段階、
b.阻害が細菌中で起こるように、選択された相乗的遺伝子に対応する遺伝子産物を阻害する化学的類似体を選択または作成する段階、および
c.相乗的阻害剤を抗生物質と組み合わせ、それぞれの種々の濃度における組み合わせの抗微生物活性を決定する段階
を含む、方法も提供する。
【0022】
一つの態様において、本発明は、ベータラクタム系抗生物質の抗菌能を増大させ、それによって動物における細菌感染症を予防または治療するための方法において有用性を有する方法を提供する。
【0023】
別の態様において、本発明は、MurBの阻害剤と細胞壁阻害抗生物質、特にベータラクタム系抗生物質との組み合わせを含む。
【0024】
微生物遺伝子断片を同定するための改良法によって、アンチセンス配向で異所的に発現した場合、関心対象の微生物遺伝子の転写後発現阻害をもたらし、それによって微生物中において観察および計測され得る表現型変化を引き起こすことができる。首尾よく同定されると、そのようなアンチセンス配向断片の条件発現を使用して、細胞内における特定の遺伝子産物の濃度を低下させ、それによって該細胞を、それ自体が遺伝子産物の特異的阻害剤である第2の物質に過剰感作させることができる。観察された表現型が、繁殖する能力の損失を構成する場合、この過感受性は、抗微生物活性を有する化合物の作用機序を試験するためのアッセイの基礎となり得る。そのようなアッセイにより、繁殖の損失を引き起こす能力が、関心対象の標的遺伝子産物およびそれらの化合物に対する特定の効果と関連付けられている抗微生物化合物と、観察される関連がない化合物とを区別することができるであろう。該改良法は、最も適切なアンチセンスDNA断片を明確に同定することにより、そのようなアッセイの開発を加速すると考えられる。
【0025】
2つの先行特許、米国特許第6228579号および米国特許第6924101号では、微生物ゲノムDNAのDNA断片を、微生物中においてアンチセンス配向で発現した場合に繁殖を低減または妨害するそれらの能力を調査したプロセスを記述している。手短に述べると、ゲノムDNAを断片化し、その断片をプラスミド上の誘導性プロモータの後にクローニングし、該プラスミドを関心対象の微生物に導入し、その後、クローニングされた断片の発現の依存的存在または非存在をもたらす誘導因子化合物または刺激剤の存在下および非存在下で、繁殖を計測および比較する。
【0026】
断片は、センス配向またはアンチセンス配向のいずれかで、プロモータに対してクローニングされ得る。断片がアンチセンス配向である場合、微生物の繁殖の低減または妨害を引き起こすそれらの断片は、該断片の由来源である遺伝子の機能に特異的な機序によって該低減または妨害を行うことが想定され、この遺伝子が、コードする細胞機能が正常な繁殖のために必要とされる遺伝子であることが示唆される。
【0027】
しかしながら、標的遺伝子の特異的阻害に至るアンチセンス断片を同定するための改良法を使用することにより、本発明者らは、上記の記述が真実ではないことを示した。実際に、本発明者らは、上記のようなプロモータから発現する断片は、示されている遺伝子および遺伝子産物に特異的でない多数の手段によって繁殖を低減し得ると認識している。見せかけのセンスまたはアンチセンス配向のいずれかで発現される断片は、1)細胞中における別の無関係な機能に対してアプタマーまたはその他の種類の阻害剤として作用し得るRNA種、2)元の遺伝子のものとは別のリーディングフレームにおいて発現し得、非特異的な毒性を有するペプチドを産生するRNA種をもたらし得る。このため、上記の特許において記述されているようなアンチセンスベースのスクリーニングは偽陽性をもたらし得る、すなわち、繁殖に必須である機能をコードしない遺伝子が、その遺伝子の増殖阻害アンチセンス断片のスクリーニングにおいて見かけ上のみ、そのようなものとして同定され得る。これは、繁殖等に必須なものとして、およびそのような可能性のある抗生物質の開発標的としての、遺伝子の誤同定につながる場合がある。
【0028】
生物のゲノム全体ではなく、1つの遺伝子または関心対象の遺伝子に焦点を当てる。これは、標的特異的表現型効果を引き起こす断片のスクリーニングにおいて、該遺伝子または関心対象の遺伝子のはるかに優れた描写を可能にする。特定の遺伝子または関心対象の遺伝子に対するアンチセンス断片を見出すために、ゲノムDNA全体を使用することにより、結果としてスクリーニングが長くなる。
【0029】
表現型変化を引き起こすアンチセンス断片を同定するための発見プロセスへの検証段階の組み込みは、該アンチセンス断片の由来源である遺伝子から発現した無傷のmRNAの特異的低減に伴う。
【0030】
発明の詳細な説明
「ベータラクタム」ベータラクタム(βラクタム)は、3つの炭素原子および1つの窒素原子からなる、ヘテロ原子環状構造を有するラクタムである。ベータラクタム環は、ペニシリン等、いくつかの抗生物質の一部であることから、それらはベータラクタム系抗生物質とも称される。これらの抗生物質は、細菌細胞壁合成を阻害することによって働く。ベータラクタム系抗生物質クラスの例としては、ペニシリン、セファロスポリン、およびカルバペネムが含まれる。
【0031】
「抗生物質」は、感染症を制御するために使用される化合物である。抗生物質の例としては、以下が含まれる。
細胞壁阻害剤
ベータラクタム(ペニシリンG、ペニシリンV、ナフシリン、メチシリン、オキサシリン、クロキサシリン、ジクロキサシリン、フルクロキサシリン、アンピシリン、アモキサシリン、カルベニシリン、チカルシリン、アゾシリン、メズロシリン、ピペラシリン、セファロスポリン、セファロチン、セファゾリン、セファレキシン、セフロキシム、セファマンドール、セフォキシチン、セファクロル、モキサラクタム、セフォペラゾン、セフタジジム、セフトリアキソン、クラブラン酸、スルバクタム、イミペネム、アズトレオナム、セフォキシチン、セフォゾリン、セファクロル)
ペプチドグリカン生合成の阻害剤は、以下を含むがこれらに限定されない。
1.GlmU、MurAの阻害剤(ホスホマイシン)
2.MurBの阻害剤(ジヒドロピロロン)
3.ウンデカプレニルピロリン酸合成酵素の阻害剤(ヒダントイン、スルホンアミド)
4.LpxCの阻害剤(UDP‐3‐O‐アシルN‐アセチルグルコサミン脱アセチル化酵素)
リポペプチド(ダプトマイシン)
イソプレノイド生合成阻害剤(バシトラシン、ホスミドマイシン)
脂肪酸生合成阻害剤(セルレニン、トリクロサン、イソニアジド)
タンパク質合成阻害剤
1.アミノグリコシド(ストレプトマイシン、ゲンタマイシン、およびカナマイシン)、
2.テトラサイクリン
3.クロラムフェニコール、
4.マクロライド(エリスロマイシン、アジスロマイシン、クラリスロマイシン)
5.リンコサミド(リンコマイシンおよびクリンダマイシン)
6.オキサゾリジノン(リネゾリド)
ムピロシン等のtRNA合成酵素阻害剤
1.MetRS阻害剤(カテコール、プロリン、キノロン)
2.ペプチド脱ホルミル酵素阻害剤(ヒドロキサマート)
リファンピシン等のRNA合成阻害剤
キノロン等のDNAジャイレース阻害剤(ナリジクス酸、シプロフロキサシン)
抗葉酸剤(トリメトプリムおよびスルファメトキサゾール)
【0032】
「相乗効果」は、≦0.5のフラクショナル阻止濃度(fractional inhibitory concentration)指数(FICI)として定義され、ここでFICIは、2つの化合物を組み合わせた場合の個々の構成要素のフラクショナル阻止濃度(FIC)の和として定義され、FICは組み合わせた場合の化合物の最小発育阻止濃度(MIC)を化合物単独のMICで割った比率として定義される。
【0033】
「アンチセンス相乗効果」は、誘導因子の存在下における化合物の50%発育阻止濃度(IC50)を、誘導因子の非存在下におけるIC50で割った比率≧5として定義される。
【0034】
本発明者らのアンチセンスへの関心は、新たな抗生物質候補の作用機序を追跡する際に使用するための、炭疽菌の特定のアンチセンス株の開発にあった。本発明者らの標的の1つはMurB、つまりペプチドグリカン生合成の第2の段階である(図1)。すべての病原菌はMurBを必要とするが、現在MurBを阻害することによって作用する抗生物質はない。
【0035】
その機能的murB遺伝子(murB‐2)に対する誘導性アンチセンスを有する炭疽菌株の作成は、誘導因子の存在下において増殖減弱された株をもたらした。選択された株は、murB遺伝子の種々の部分に対応するアンチセンス挿入断片を有することが決定された。続いて、増殖減弱および抗生物質のパネルに対する感受性の誘導因子依存性の観点から、それらの株を特徴付けた。予想に反して、株のいくつかは抗生物質に対する感受性において相違を示さなかった。したがって、抗生物質発見のためのアンチセンスベースのツールを作成する際における重大な付加的段階は、ある特定の抗生物質に対する感受性増大の実証である。本発明者らは、他者らによって作成されたアンチセンス株の多くは、阻害剤に対して過感受性でないため、スクリーニングまたは作用機構決定において有用ではないのではないかとの疑念を抱いている。
【0036】
確認されたmurBアンチセンス株の、細胞壁阻害抗生物質について、予想外且つ劇的なアンチセンス相乗効果が観察された。4つの場合(セフェピム、セフトリアキソン、セフォキシチン、またはセフォタキシム)において、murBアンチセンス発現は、非誘導条件にある細胞に対して、100〜600倍低い濃度の抗生物質に対して感受性のある細胞をもたらした(図3)。その他の細胞壁阻害抗生物質に対する感受性は、増大されたが程度の低いものであった。タンパク質合成、DNA合成、またはRNA合成等、その他の経路を標的とする抗生物質に対する感受性は、増大されなかった。その他の遺伝子を標的とするアンチセンス株は、試験したいかなる抗生物質に対しても感受性におけるこのような異例の増大がなかったことから、該効果はペプチドグリカン生合成経路内の遺伝子に特異的であると考えられる。
【0037】
図4は、抗生物質のパネルに対するmurAアンチセンス株の感受性を示す。murBアンチセンス株に観察されたように、murAアンチセンス発現は、細胞壁阻害抗生物質に対する感受性は何倍にもなったが、その他の経路を標的とする抗生物質に対してはそうでない細胞をもたらした。ペプチドグリカン生合成におけるその他の遺伝子を標的とするアンチセンス株にも同様の結果が得られた。
【0038】
標的間における限られた相乗的な相互作用については前例があるが、MurBアンチセンス株とセファロスポリンとの間における相乗効果の大きさについては前例がない。本発明者らには、アンチセンスの代わりに標的阻害剤を使用して相乗効果が達成され得ることを示す、セフォタキシムと組み合わせたいくつかの弱MurB阻害剤についての限定的なデータがある(図5)。本発明者らはまた、この現象が炭疽菌に限定されるものでなく、黄色ブドウ球菌中におけるmurBアンチセンス発現が、ベータラクタム系抗生物質に対して細胞を過剰感作させることも示した(図6)。最後に、本発明者らは、抗生物質化学療法における重要なパラメータである「死滅時間(Time‐to‐kill)」の観点から、この相乗効果を特徴付けるプロセスにある。本発明者らは、セファロスポリンとMurB阻害との組み合わせが、セファロスポリン単独での殺菌結果にかかる時間に対して、殺菌効果にかかる時間を短くすると期待している。
【0039】
本発明者らはさらに、MurB阻害が、そうしなければそのような感染症の治療時に効果のないメチシリンおよびその他のペニシリンに対して、メチシリン耐性細菌を再感作すると期待している。
【0040】
炭疽菌におけるMurBおよびセファロスポリンに関するこの観察により、本発明者らは、この方法論を用いて発見され得る抗生物質と遺伝子標的とのその他の組み合わせを期待している。本発明者らは、これらの観察は、その他の重要な病原体、特に黄色ブドウ球菌に移し変えることができると期待している。図6は、murBに対する誘導性アンチセンスで操作された黄色ブドウ球菌の株におけるセフォキシチンおよびジクロキサシリンの効能の増大を示す。抗生物質のパネルに対する黄色ブドウ球菌株の感受性は、ベータラクタム系抗生物質の誘導因子の存在下において増大したが、その他の作用機構の抗生物質では増大しなかった。
【0041】
本発明者らは、抗生物質と遺伝子との、その他の予想外の相乗的対を明らかにする能力も期待している。これらの対は、その他の抗生物質に対する相乗剤の開発のための標的部位の発見を可能にし得る。菌類および哺乳動物細胞を含むその他の生物におけるアンチセンスは、抗癌剤、向精神薬等を含む多くのクラスの薬物の有効性を増強するための、相乗的標的部位および最終的には相乗化合物の発見のために、同様の様式で活用され得る。
【0042】
したがって、任意の所与の抗生物質と任意の特定の標的の阻害剤との間における相乗的な相互作用の潜在性を決定できることが有用であると考えられる。そのような決定は、所与の抗生物質の効能を増大させる化学化合物のスクリーニングの構成を可能にすると考えられる。誘導性アンチセンスは、相乗剤の発見および開発用の標的を決定するためのシステムの基礎となり得る。アンチセンスのこの使用は、遺伝子の必要性を発見する際におけるその使用と対照をなす。遺伝子の必要性を発見するためのアンチセンスの使用については他者らによって記述されているが、アンチセンスのこの使用は、必須遺伝子、または必須ではないが抗生物質の効能を何らかの手法で限定する遺伝子のいずれかの検出をもたらし得る。
【0043】
薬物の相乗的組み合わせの使用は、使用される用量の低下または化学療法の時間が短くなることによる薬物の副作用低下、感染症のより迅速な治癒によって入院が短くなること、制御される病原体のスペクトルの増大、抗生物質に対する耐性発現の発生率の減少を含む、従来の単一化合物化学療法よりも多くの利点を有し得る。
【0044】
抗生物質の相乗剤発見のための関心対象の遺伝子を決定するためのアンチセンスツールの重要性は、それが、抗生物質の作用機序に関する特定の知識を持たずに、および相乗的標的用に特定の化合物を使用することなく、新規スクリーニングを開発するための経験的方法を提供することである。このアンチセンスの使用は、遺伝子と任意の所与の病原体に対する抗生物質感受性との間の予想外の関係を明らかにすることができる。図7は、抗生物質のための相乗的標的の発見についてのプロセスの流れを示す。
【0045】
本発明者らは、MurBとベータラクタム系抗生物質との間の相乗的な相互作用を発見した。この発見は、治療法の投薬量を低減すること、治療時間を減少させること、または病原体制御のスペクトルを増大させることによって、抗感染症化学療法を増強することができる、MurB阻害剤とベータラクタムとの組み合わせの開発を予期するものである。
【0046】
本発明者らは、特定の遺伝子のための誘導性アンチセンスを使用し、誘導因子の存在下および非存在下における生物の抗生物質感受性を比較して、任意の所与の抗生物質の相乗的標的を発見するための一般法を開発した。
【0047】
本発明者らは、薬物と相乗作用のある標的を発見し、それによって薬効を増大させることを可能にするための、アンチセンスの一般的使用を予期している。この使用は、抗ウイルス剤、抗真菌剤、抗寄生虫剤、抗腫瘍剤、殺虫剤、除草剤、および化学療法によって処置が施されるその他の生物系にまでおよび得る。
【0048】
実施例
以下の例は、本発明を例示することを意図し、限定することを意図しない。
【0049】
以下は、本発明の方法を表す説明的例である。
1)ゲノムDNAをテンプレートとして使用して、関心対象の遺伝子(例えば、オペロン)をPCR増幅する。調査する遺伝子のいずれかの末端の約200bpを含むように注意する。
2)該PCR断片を超音波処理によって断片化し、結果として生じた断片を平滑末端化し、約350〜400bpより大きくないようにサイズ選択し、次いで本発明者らの発現プラスミドであるpBAX‐2内の誘導性プロモータの直ぐ下流のポリリンカーにクローニングする。このライブラリの作成は、通常、大腸菌(E.coli)において為される。
3)続いて、ライブラリのDNAを、関心対象の遺伝子が由来する生物、例えば、炭疽菌または黄色ブドウ球菌に形質転換する。ピッキングによって個々のコロニーを収集し、プラスミドの存在を選択する成長培地中で、96ウェル培養皿において培養する。
4)細胞が増殖した後、誘導化合物キシロースが含まれるかまたは含まれないかのいずれかの元の成長培地の固体版に、該細胞をレプリカプレーティングする(例えば、ピンツールを使用して)。
5)これらのレプリカプレートを一晩増殖させ、続いて、プラスとマイナスのキシロース対として比較し、キシロースプレート上においてのみ著しい増殖感受性を示すコロニーを、さらなる研究のために非キシロースプレートから選択する。
6)キシロース成長感受性株のpBAX‐2プラスミドの挿入断片を、DNA配列決定法によって分析する。これにより、PCR増幅した関心対象の遺伝子において断片が何処に由来するかを同定し、該関心対象の遺伝子および誘導性プロモータの両方に対してセンスまたはアンチセンス配向であるかを確定する。
7)センスおよびアンチセンスの相対数を記録する。本発明者らは、遺伝子が必須である場合、キシロース増殖感受性株のプールにおいて、アンチセンス配向断片を主に得ることを見出した。遺伝子が必須でない場合、スクリーニングされる同等数のコロニーにおいて増殖感受性が見られない傾向があるか、または、より同等な数のアンチセンスおよびセンスが見られる。キシローススクリーニングを適用する前に形質転換細胞を無作為に選択する場合、得られるアンチセンス対センスの数は、ほぼ等しいままであることを見出すであろうことに留意されたい。
8)さらなる分析のために、多数のアンチセンスクローンを選択する。これらは、関心対象の遺伝子における相対的位置(本発明者らのmRNAレベルを分析する現在の能力では、さらなる3’断片に制限される)および相対的サイズについて選択する。多くの場合、断片が大きくなるにつれて、非特異的な増殖感受性を引き起こす可能性が大きくなる。
9)続いて、これらのアンチセンスクローンに対して2つの研究を行う。アンチセンス断片の発現の誘導を受けて、関心対象の遺伝子からの転写物の内因性レベルを計測する。関心対象の遺伝子に対応し、対照遺伝子のものには対応しないメッセージにおける特異的な分解または低減は、アンチセンスが、関心対象の遺伝子の特異的な転写後発現阻害剤として働いていることの指標である。
10)もう1つのアッセイは、アンチセンス断片の発現の誘導を受けて、一般的な増殖効果とは異なる特異的な表現型変化を計測することである。本発明者らのDHFRアンチセンスの場合、試験はアンチセンスがチミジンの栄養要求性を引き起こし得るか否かを見るものであった。チミジンの生合成はDHFRによって合成されるテトラヒドロ葉酸塩の大部分を要するものであり、栄養要求性はDHFR突然変異の古典的効果である。もう1つの行うべきことは、特定の抗生物質等、関心対象の遺伝子の細胞産物に特異的なアンチセンスの誘導による、阻害化合物に対する過感受性を計測することである。アンチセンスは、関心対象の遺伝子に特異的でなく、無関係な細胞標的に特異的である、阻害剤または抗生物質に対する過感受性を引き起こさないはずである。
【0050】
上記の例を参照して本発明を説明したが、本発明の精神および範囲には、改変および変形が包含されることが理解されるであろう。したがって、本発明は、特許請求の範囲によってのみ限定される。
【図面の簡単な説明】
【0051】
【図1】細菌中におけるペプチドグリカンモノマー単位の生合成経路を示す。
【図2】阻止濃度下のアンチセンス誘導因子の存在下および非存在下における、セフォタキシムへのmurB2アンチセンスクローンの応答を示す。左パネル:murB2アンチセンスクローン。上部線:誘導因子なし。下部線:+40mMキシロース。右パネル:metRSアンチセンスクローン。EC50+キシロース/EC50(−キシロース)の比率により、「IC50推移」が求められる。この例において推移を計算すると、(2096/6.1)=343である。
【図3】(a)セファロスポリン、ならびに(b)ベータラクタム系クロキサシリン、オキサシリン、およびジクロキサシリンを含むその他の抗生物質に対する、炭疽菌(B.anthracis)murB‐2アンチセンス株の感受性を示す。
【図4】種々の抗生物質に対する炭疽菌murAlアンチセンス株の感受性を示す。
【図5】セフォタキシムおよびジヒドロピロロン(dihydropyrrolone)MurB阻害剤Rx000106またはRx000107を使用した、野生型炭疽菌におけるFICI実験を示す。チェッカーボード実験においては、化合物のフラクショナル阻止濃度(FIC)は、単独および組み合わせた化合物の最小発育阻止濃度(MIC)の比率として計算される。FIC指数(FICI)は、2つの化合物のFICの和である。
【図6】murBアンチセンスを用いて操作された炭疽菌を示す。
【図7】相乗的標的および相乗剤の発見のためのプロセスを示す。
【図8】標的配列の表である。
【特許請求の範囲】
【請求項1】
βラクタム系抗生物質と細菌性MurB酵素の阻害剤との抗菌効果のある組み合わせを、それを必要とするヒトまたは動物に投与する段階を含む、ヒトおよび動物における細菌感染症の治療に有用な薬学的組成物。
【請求項2】
βラクタム系抗生物質と任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、GlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との抗菌効果のある組み合わせを、それを必要とするヒトまたは動物に投与する段階を含む、ヒトおよび動物における細菌感染症の治療に有用な薬学的組成物。
【請求項3】
βラクタム系抗生物質に対して<0.2のフラクショナル阻止濃度(fractional inhibitory concentration)指数(FICI)をもたらす、βラクタムとMurB酵素の阻害剤との組み合わせ。
【請求項4】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、セファロスポリンとMurB阻害剤との組み合わせ。
【請求項5】
抗グラム陽性細菌剤としての、セファロスポリンとMurB阻害剤との組み合わせ。
【請求項6】
炭疽病の治療用の、セファロスポリンとMurB阻害剤との組み合わせ。
【請求項7】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、セファロスポリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項8】
抗グラム陽性剤に対する抗菌剤としての、セファロスポリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項9】
炭疽病の治療用の広域スペクトルの抗菌剤としての、セファロスポリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項10】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、カルバペネムとMurB阻害剤との組み合わせ。
【請求項11】
抗グラム陽性剤としての、カルバペネムとMurB阻害剤との組み合わせ。
【請求項12】
炭疽病の治療用の、カルバペネムとMurB阻害剤との組み合わせ。
【請求項13】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、カルバペネムと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項14】
抗グラム陽性剤に対する広域スペクトルの抗菌剤としての、カルバペネムと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項15】
炭疽病の治療用の広域スペクトルの抗菌剤としての、カルバペネムと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項16】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、ペニシリンとMurB阻害剤との組み合わせ。
【請求項17】
抗グラム陽性剤としての、ペニシリンとMurB阻害剤との組み合わせ。
【請求項18】
炭疽病の治療用の、ペニシリンとMurB阻害剤との組み合わせ。
【請求項19】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、ペニシリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項20】
抗グラム陽性剤に対する広域スペクトルの抗菌剤としての、ペニシリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項21】
炭疽病の治療用の広域スペクトルの抗菌剤としての、ペニシリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項22】
メチシリン耐性感染症の治療用の、ペニシリンとMurB阻害剤との組み合わせ。
【請求項23】
インビトロでFICI≦0.3をもたらす、ペニシリンとMurB阻害剤との組み合わせ。
【請求項24】
インビトロでFICI≦0.3をもたらす、ムピロシンとrelA阻害剤との組み合わせ。
【請求項25】
インビトロでFICI≦0.3をもたらす、MetRS阻害剤とrelA阻害剤との組み合わせ。
【請求項26】
以下の段階を含む、抗生物質の相乗剤を発見する方法:
a.細胞内における遺伝子産物の活性または量を低減するように、遺伝子産物をコードする核酸に対するアンチセンス核酸を細胞内で発現し、それによって抗生物質に感作された細胞を産生する段階、
b.抗生物質に対する細胞の感作を特徴付け、アンチセンス遺伝子の非存在下において必要とされる濃度の5分の1またはそれ未満で抗生物質の効力をもたらす抗生物質と遺伝子との対を選択する段階、
c.選択された相乗的遺伝子に対応する遺伝子産物を阻害する化学化合物をスクリーニングする段階、
d.阻害が細菌中で起こるように、選択された相乗的遺伝子に対応する遺伝子産物を阻害する化学的類似体を選択または作成する段階。
【請求項27】
アンチセンス核酸は、誘導性プロモータから転写される、請求項1記載の方法。
【請求項28】
アンチセンス核酸を亜致死レベルに誘導する濃度の誘導因子に、細胞を接触させる段階をさらに含む、請求項2記載の方法。
【請求項29】
亜致死濃度の誘導因子は、抗生物質に対する感受性が少なくとも5倍に増大されている、請求項3記載の方法。
【請求項30】
遺伝子産物はポリペプチドである、請求項1記載の方法。
【請求項31】
遺伝子産物はRNAである、請求項1記載の方法。
【請求項32】
細胞は、細菌細胞、真菌細胞、植物細胞、および動物細胞からなる群より選択される、請求項1記載の方法。
【請求項33】
細胞は、黄色ブドウ球菌(Staphylococcus aureus)、緑膿菌(Pseudomonas aeruginosa)、エンテロバクター・クロアカエ(Enterobacter cloacae)、ヘリコバクター・ピロリ(Helicobacter pylori)、淋菌(Neisseria gonorrhoeae)、大便連鎖球菌(Enterococcus faecalis)、肺炎連鎖球菌(Streptococcus pneumoniae)、インフルエンザ菌(Haemophilus influenzae)、ネズミチフス菌(Salmonella typhimurium)、サッカロマイセス・セレビシエ(Saccharomyces cerevisae)、カンジダ・アルビカンス(Candida albicans)、クリプトコックス・ネオフォルマンス(Cryptococcus neoformans)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、肺炎桿菌(Klebsiella pneumonia)、チフス菌(Salmonella typhi)、パラチフス菌(Salmonella paratyphi)、サルモネラ・コレラスイス(Salmonella cholerasuis)、表皮ブドウ球菌(Staphylococcus epidermidis)、ヒト型結核菌(Mycobacterium tuberculosis)、らい菌(Mycobacterium leprae)、梅毒トレポネーマ(Treponema pallidum)、炭疽菌(Bacillus anthracis)、ペスト菌(Yersinia pestis)、ボツリヌス菌(Clostridium botulinum)、カンピロバクター・ジェジュニ(Campylobacter jejuni)、クラミジア・トラコマチス(Chlamydia trachomatus)、およびクラミジア・ニューモニエ(Chlamydia pneumoniae)、または上記の種のいずれかの属に含まれる任意の種からなる群より選択される生物に由来する、請求項1記載の方法。
【請求項34】
細胞は炭疽菌細胞である、請求項1記載の方法。
【請求項35】
候補化合物は天然産物である、請求項1記載の方法。
【請求項36】
以下の段階を含む、微生物の繁殖の阻害に使用するために候補化合物をスクリーニングする方法:(a)第1の微生物において繁殖に必要とされる遺伝子または遺伝子産物を同定する段階、(b)第2の微生物において該遺伝子または遺伝子産物のホモログを同定する段階、(c)該第2の微生物において該ホモログの活性を阻害する阻害性核酸配列を同定する段階、(d)繁殖阻害量の該阻害性核酸に該第2の微生物を接触させ、それによって該第2の微生物を感作する段階、(e)段階(d)の感作された微生物を候補化合物に接触させる段階、および(f)該候補化合物が、該感作された微生物ほど感作されていない微生物の繁殖を阻害するより大きな程度で、該候補化合物が該感作された微生物の繁殖を阻害するか否かを決定する段階。
【請求項37】
MurBの阻害剤を得るために化合物ライブラリまたは天然産物ライブラリをスクリーニングし、その後、抗生物質を用いて化合物の相乗活性を試験することによる、相乗剤としての化合物の同定法。
【請求項38】
任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤を得るために化合物ライブラリまたは天然産物ライブラリをスクリーニングし、その後、抗生物質を用いて化合物の相乗活性を試験することによる、相乗剤としての化合物の同定法。
【請求項39】
濃度20%または抗生物質のMIC未満の抗生物質の存在下において、細菌培養物を使用して化合物ライブラリまたは天然産物ライブラリから化合物を選択すること、MIC下レベルの抗生物質の存在下においてのみ細菌を死滅させる化合物をスコアリングすること、および選択された化合物のMurB阻害をインビトロ酵素アッセイにおいて試験することによる、MurB阻害剤としての化合物の同定法。
【請求項40】
濃度20%または抗生物質のMIC未満の抗生物質の存在下において、細菌培養物を使用して化合物ライブラリまたは天然産物ライブラリから化合物を選択すること、MIC下レベルの抗生物質の存在下においてのみ細菌を死滅させる化合物をスコアリングすること、および選択された化合物の、任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害をインビトロ酵素アッセイにおいて試験することによる、任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤としての化合物の同定法。
【請求項41】
βラクタムの抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項42】
細菌細胞壁に対する作用機序を有する化合物を伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項43】
細胞壁阻害抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項44】
DNAジャイレースに対する作用機序を有する化合物を伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項45】
タンパク質合成に対する作用機序を有する化合物を伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項46】
ダプトマイシンを伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項47】
バンコマイシンを伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項48】
シプロフロキサシンを伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項49】
トリメトプリムを伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項50】
細菌細胞壁に対する作用機序を有する化合物を伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項51】
抗生物質と遺伝子標的との相乗的組み合わせを同定するプロセス:
a.細菌遺伝子を長さ200塩基以下のランダムな断片に分割する段階、
b.誘導性プロモータおよび選択可能なマーカーを含有する適切なプラスミドに、断片をライゲーションする段階、
c.誘導因子の存在下において増殖阻害を呈する形質転換細胞を選択する段階、
d.誘導因子の非存在下において選択されたクローンの抗生物質感受性と比較して、1つまたは複数の抗生物質に対する感作をもたらす誘導因子の用量を決定する段階、
e.抗生物質の種々のクラスおよび抗生物質の作用機序の範囲を示す抗生物質のパネルに対する感受性について、誘導因子の存在下におけるクローンをプロファイリングする段階。
【請求項52】
所与の抗生物質の相乗剤を同定するプロセス:
a.(27)において同定された遺伝子の産物に基づいて、インビトロ酵素アッセイを使用して化合物ライブラリをスクリーニングする段階、
b.阻害活性を示す化合物を同定する段階、
c.細菌中の遺伝子産物を阻害する(b)の化学的類似体を作成する段階、
d.化学的類似体を抗生物質と組み合わせて、相乗抗菌活性について試験する段階。
【請求項53】
抗生物質の相乗剤を同定するプロセス:
a.最小発育阻止濃度の8分の1の抗生物質を含有する培地の存在下および非存在下において増殖する細菌を使用して、化合物ライブラリをスクリーニングする段階、
b.抗生物質の最小発育阻止濃度の8分の1の抗生物質の存在下においてのみ阻害活性を示す化合物を同定する段階。
【請求項54】
微生物遺伝子または関心対象の遺伝子であるDNA断片を生成、検出、および使用するための方法であって、ある特定の標的微生物内においてアンチセンス配向で異所的に発現した場合、微生物内の関心対象の遺伝子に特異的な転写後発現阻害を引き起こすことができる、方法。
【請求項55】
転写後発現阻害は、アンチセンス配向のDNA断片における発現レベルの増加または減少を受けて直接的に強度が増加または減少され得る、請求項54記載の方法。
【請求項56】
アンチセンス配向のDNA断片の発現レベルは、実験的に調節され得る、請求項55記載の方法。
【請求項57】
転写後発現阻害の強度の増加または減少は、標的微生物において観察できる表現型変化の計測可能な強度に、直接的且つ依存的な増加または減少をもたらす、請求項55記載の方法。
【請求項58】
DNA断片を画定するためのスクリーニングは、アンチセンス配向のDNA断片の高レベル発現により引き起こされるがアンチセンス配向のDNA断片の低レベル発現からは明白でない、標的微生物における計測可能な表現型変化の検出に依存する、請求項54〜56のいずれか一項記載の方法。
【請求項59】
アンチセンス配向断片の発現によって関心対象の遺伝子に引き起こされる表現型変化は、関心対象の遺伝子の特異的な転写後発現阻害に伴う、請求項58記載の方法。
【請求項60】
関心対象の遺伝子の転写後発現阻害は、RT‐PCR法、ノーザンブロット法、マイクロアレイ法、またはその他適切な方法による関心対象の遺伝子のmRNAの定量化によって計測され、機能的および/または遺伝的に無関係な対照遺伝子の、同様に計測されたmRNAレベルと比較される、請求項59記載の方法。
【請求項61】
アンチセンス配向断片は、炭疽菌遺伝子murB‐2の断片であり、SEQ ID:Ba‐murB2‐C1に明記される配列を含む、請求項59記載の方法。
【請求項62】
アンチセンス配向断片は、炭疽菌遺伝子murB‐2の断片であり、SEQ ID:Ba‐murB2‐H1に明記される配列を含む、請求項59記載の方法。
【請求項63】
アンチセンス配向断片は、炭疽菌遺伝子murB‐2の断片であり、SEQ ID:Ba‐murB2‐D1に明記される配列を含む、請求項59記載の方法。
【請求項64】
アンチセンス配向断片は、炭疽菌遺伝子murB‐2の断片であり、SEQ ID:Ba‐murB2‐D2に明記される配列を含む、請求項59記載の方法。
【請求項65】
アンチセンス配向断片は、炭疽菌遺伝子metSの断片であり、SEQ ID:Ba‐metRS‐H1に明記される配列を含む、請求項59記載の方法。
【請求項66】
アンチセンス配向断片は、炭疽菌遺伝子metSの断片であり、SEQ ID:Ba‐metRS‐H2に明記される配列を含む、請求項59記載の方法。
【請求項67】
アンチセンス配向断片は、炭疽菌遺伝子metSの断片であり、SEQ ID:Ba‐metRS‐H6に明記される配列を含む、請求項59記載の方法。
【請求項68】
アンチセンス配向断片は、炭疽菌遺伝子metSの断片であり、SEQ ID:Ba‐metRS‐E4に明記される配列を含む、請求項59記載の方法。
【請求項69】
アンチセンス配向断片は、炭疽菌遺伝子uppSの断片であり、SEQ ID:Ba‐uppS‐UG9に明記される配列を含む、請求項59記載の方法。
【請求項70】
アンチセンス配向断片は、炭疽菌遺伝子uppSの断片であり、SEQ ID:Ba‐uppS‐UA3に明記される配列を含む、請求項59記載の方法。
【請求項71】
アンチセンス配向断片は、炭疽菌遺伝子dfrAの断片であり、SEQ ID:Ba‐dfrA‐2G1に明記される配列を含む、請求項59記載の方法。
【請求項72】
アンチセンス配向断片は、炭疽菌遺伝子dfrAの断片であり、SEQ ID:Ba‐dfrA‐2G6に明記される配列を含む、請求項59記載の方法。
【請求項73】
アンチセンス配向断片は、黄色ブドウ球菌遺伝子murBの断片であり、SEQ ID:Sa‐murB‐E9に明記される配列を含む、請求項59記載の方法。
【請求項74】
アンチセンス配向断片は、黄色ブドウ球菌遺伝子murBの断片であり、SEQ ID:Sa‐murB‐F7に明記される配列を含む、請求項59記載の方法。
【請求項75】
アンチセンス配向断片は、黄色ブドウ球菌遺伝子murBの断片であり、SEQ ID:Sa‐murB‐B9に明記される配列を含む、請求項59記載の方法。
【請求項1】
βラクタム系抗生物質と細菌性MurB酵素の阻害剤との抗菌効果のある組み合わせを、それを必要とするヒトまたは動物に投与する段階を含む、ヒトおよび動物における細菌感染症の治療に有用な薬学的組成物。
【請求項2】
βラクタム系抗生物質と任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、GlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との抗菌効果のある組み合わせを、それを必要とするヒトまたは動物に投与する段階を含む、ヒトおよび動物における細菌感染症の治療に有用な薬学的組成物。
【請求項3】
βラクタム系抗生物質に対して<0.2のフラクショナル阻止濃度(fractional inhibitory concentration)指数(FICI)をもたらす、βラクタムとMurB酵素の阻害剤との組み合わせ。
【請求項4】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、セファロスポリンとMurB阻害剤との組み合わせ。
【請求項5】
抗グラム陽性細菌剤としての、セファロスポリンとMurB阻害剤との組み合わせ。
【請求項6】
炭疽病の治療用の、セファロスポリンとMurB阻害剤との組み合わせ。
【請求項7】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、セファロスポリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項8】
抗グラム陽性剤に対する抗菌剤としての、セファロスポリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項9】
炭疽病の治療用の広域スペクトルの抗菌剤としての、セファロスポリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項10】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、カルバペネムとMurB阻害剤との組み合わせ。
【請求項11】
抗グラム陽性剤としての、カルバペネムとMurB阻害剤との組み合わせ。
【請求項12】
炭疽病の治療用の、カルバペネムとMurB阻害剤との組み合わせ。
【請求項13】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、カルバペネムと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項14】
抗グラム陽性剤に対する広域スペクトルの抗菌剤としての、カルバペネムと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項15】
炭疽病の治療用の広域スペクトルの抗菌剤としての、カルバペネムと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項16】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、ペニシリンとMurB阻害剤との組み合わせ。
【請求項17】
抗グラム陽性剤としての、ペニシリンとMurB阻害剤との組み合わせ。
【請求項18】
炭疽病の治療用の、ペニシリンとMurB阻害剤との組み合わせ。
【請求項19】
グラム陽性およびグラム陰性病原菌に対する広域スペクトルの抗菌剤としての、ペニシリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項20】
抗グラム陽性剤に対する広域スペクトルの抗菌剤としての、ペニシリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項21】
炭疽病の治療用の広域スペクトルの抗菌剤としての、ペニシリンと任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤との組み合わせ。
【請求項22】
メチシリン耐性感染症の治療用の、ペニシリンとMurB阻害剤との組み合わせ。
【請求項23】
インビトロでFICI≦0.3をもたらす、ペニシリンとMurB阻害剤との組み合わせ。
【請求項24】
インビトロでFICI≦0.3をもたらす、ムピロシンとrelA阻害剤との組み合わせ。
【請求項25】
インビトロでFICI≦0.3をもたらす、MetRS阻害剤とrelA阻害剤との組み合わせ。
【請求項26】
以下の段階を含む、抗生物質の相乗剤を発見する方法:
a.細胞内における遺伝子産物の活性または量を低減するように、遺伝子産物をコードする核酸に対するアンチセンス核酸を細胞内で発現し、それによって抗生物質に感作された細胞を産生する段階、
b.抗生物質に対する細胞の感作を特徴付け、アンチセンス遺伝子の非存在下において必要とされる濃度の5分の1またはそれ未満で抗生物質の効力をもたらす抗生物質と遺伝子との対を選択する段階、
c.選択された相乗的遺伝子に対応する遺伝子産物を阻害する化学化合物をスクリーニングする段階、
d.阻害が細菌中で起こるように、選択された相乗的遺伝子に対応する遺伝子産物を阻害する化学的類似体を選択または作成する段階。
【請求項27】
アンチセンス核酸は、誘導性プロモータから転写される、請求項1記載の方法。
【請求項28】
アンチセンス核酸を亜致死レベルに誘導する濃度の誘導因子に、細胞を接触させる段階をさらに含む、請求項2記載の方法。
【請求項29】
亜致死濃度の誘導因子は、抗生物質に対する感受性が少なくとも5倍に増大されている、請求項3記載の方法。
【請求項30】
遺伝子産物はポリペプチドである、請求項1記載の方法。
【請求項31】
遺伝子産物はRNAである、請求項1記載の方法。
【請求項32】
細胞は、細菌細胞、真菌細胞、植物細胞、および動物細胞からなる群より選択される、請求項1記載の方法。
【請求項33】
細胞は、黄色ブドウ球菌(Staphylococcus aureus)、緑膿菌(Pseudomonas aeruginosa)、エンテロバクター・クロアカエ(Enterobacter cloacae)、ヘリコバクター・ピロリ(Helicobacter pylori)、淋菌(Neisseria gonorrhoeae)、大便連鎖球菌(Enterococcus faecalis)、肺炎連鎖球菌(Streptococcus pneumoniae)、インフルエンザ菌(Haemophilus influenzae)、ネズミチフス菌(Salmonella typhimurium)、サッカロマイセス・セレビシエ(Saccharomyces cerevisae)、カンジダ・アルビカンス(Candida albicans)、クリプトコックス・ネオフォルマンス(Cryptococcus neoformans)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、肺炎桿菌(Klebsiella pneumonia)、チフス菌(Salmonella typhi)、パラチフス菌(Salmonella paratyphi)、サルモネラ・コレラスイス(Salmonella cholerasuis)、表皮ブドウ球菌(Staphylococcus epidermidis)、ヒト型結核菌(Mycobacterium tuberculosis)、らい菌(Mycobacterium leprae)、梅毒トレポネーマ(Treponema pallidum)、炭疽菌(Bacillus anthracis)、ペスト菌(Yersinia pestis)、ボツリヌス菌(Clostridium botulinum)、カンピロバクター・ジェジュニ(Campylobacter jejuni)、クラミジア・トラコマチス(Chlamydia trachomatus)、およびクラミジア・ニューモニエ(Chlamydia pneumoniae)、または上記の種のいずれかの属に含まれる任意の種からなる群より選択される生物に由来する、請求項1記載の方法。
【請求項34】
細胞は炭疽菌細胞である、請求項1記載の方法。
【請求項35】
候補化合物は天然産物である、請求項1記載の方法。
【請求項36】
以下の段階を含む、微生物の繁殖の阻害に使用するために候補化合物をスクリーニングする方法:(a)第1の微生物において繁殖に必要とされる遺伝子または遺伝子産物を同定する段階、(b)第2の微生物において該遺伝子または遺伝子産物のホモログを同定する段階、(c)該第2の微生物において該ホモログの活性を阻害する阻害性核酸配列を同定する段階、(d)繁殖阻害量の該阻害性核酸に該第2の微生物を接触させ、それによって該第2の微生物を感作する段階、(e)段階(d)の感作された微生物を候補化合物に接触させる段階、および(f)該候補化合物が、該感作された微生物ほど感作されていない微生物の繁殖を阻害するより大きな程度で、該候補化合物が該感作された微生物の繁殖を阻害するか否かを決定する段階。
【請求項37】
MurBの阻害剤を得るために化合物ライブラリまたは天然産物ライブラリをスクリーニングし、その後、抗生物質を用いて化合物の相乗活性を試験することによる、相乗剤としての化合物の同定法。
【請求項38】
任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤を得るために化合物ライブラリまたは天然産物ライブラリをスクリーニングし、その後、抗生物質を用いて化合物の相乗活性を試験することによる、相乗剤としての化合物の同定法。
【請求項39】
濃度20%または抗生物質のMIC未満の抗生物質の存在下において、細菌培養物を使用して化合物ライブラリまたは天然産物ライブラリから化合物を選択すること、MIC下レベルの抗生物質の存在下においてのみ細菌を死滅させる化合物をスコアリングすること、および選択された化合物のMurB阻害をインビトロ酵素アッセイにおいて試験することによる、MurB阻害剤としての化合物の同定法。
【請求項40】
濃度20%または抗生物質のMIC未満の抗生物質の存在下において、細菌培養物を使用して化合物ライブラリまたは天然産物ライブラリから化合物を選択すること、MIC下レベルの抗生物質の存在下においてのみ細菌を死滅させる化合物をスコアリングすること、および選択された化合物の、任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害をインビトロ酵素アッセイにおいて試験することによる、任意の細菌性ペプチドグリカン生合成酵素、特にGlmU、MurA、MurB、MurC、MurD、MurE、MurF、MurG、MraY、およびUppSの阻害剤としての化合物の同定法。
【請求項41】
βラクタムの抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項42】
細菌細胞壁に対する作用機序を有する化合物を伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項43】
細胞壁阻害抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項44】
DNAジャイレースに対する作用機序を有する化合物を伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項45】
タンパク質合成に対する作用機序を有する化合物を伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項46】
ダプトマイシンを伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項47】
バンコマイシンを伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項48】
シプロフロキサシンを伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項49】
トリメトプリムを伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項50】
細菌細胞壁に対する作用機序を有する化合物を伴う抗感染症療法用の相乗化合物を同定するための、アンチセンス遺伝子の使用。
【請求項51】
抗生物質と遺伝子標的との相乗的組み合わせを同定するプロセス:
a.細菌遺伝子を長さ200塩基以下のランダムな断片に分割する段階、
b.誘導性プロモータおよび選択可能なマーカーを含有する適切なプラスミドに、断片をライゲーションする段階、
c.誘導因子の存在下において増殖阻害を呈する形質転換細胞を選択する段階、
d.誘導因子の非存在下において選択されたクローンの抗生物質感受性と比較して、1つまたは複数の抗生物質に対する感作をもたらす誘導因子の用量を決定する段階、
e.抗生物質の種々のクラスおよび抗生物質の作用機序の範囲を示す抗生物質のパネルに対する感受性について、誘導因子の存在下におけるクローンをプロファイリングする段階。
【請求項52】
所与の抗生物質の相乗剤を同定するプロセス:
a.(27)において同定された遺伝子の産物に基づいて、インビトロ酵素アッセイを使用して化合物ライブラリをスクリーニングする段階、
b.阻害活性を示す化合物を同定する段階、
c.細菌中の遺伝子産物を阻害する(b)の化学的類似体を作成する段階、
d.化学的類似体を抗生物質と組み合わせて、相乗抗菌活性について試験する段階。
【請求項53】
抗生物質の相乗剤を同定するプロセス:
a.最小発育阻止濃度の8分の1の抗生物質を含有する培地の存在下および非存在下において増殖する細菌を使用して、化合物ライブラリをスクリーニングする段階、
b.抗生物質の最小発育阻止濃度の8分の1の抗生物質の存在下においてのみ阻害活性を示す化合物を同定する段階。
【請求項54】
微生物遺伝子または関心対象の遺伝子であるDNA断片を生成、検出、および使用するための方法であって、ある特定の標的微生物内においてアンチセンス配向で異所的に発現した場合、微生物内の関心対象の遺伝子に特異的な転写後発現阻害を引き起こすことができる、方法。
【請求項55】
転写後発現阻害は、アンチセンス配向のDNA断片における発現レベルの増加または減少を受けて直接的に強度が増加または減少され得る、請求項54記載の方法。
【請求項56】
アンチセンス配向のDNA断片の発現レベルは、実験的に調節され得る、請求項55記載の方法。
【請求項57】
転写後発現阻害の強度の増加または減少は、標的微生物において観察できる表現型変化の計測可能な強度に、直接的且つ依存的な増加または減少をもたらす、請求項55記載の方法。
【請求項58】
DNA断片を画定するためのスクリーニングは、アンチセンス配向のDNA断片の高レベル発現により引き起こされるがアンチセンス配向のDNA断片の低レベル発現からは明白でない、標的微生物における計測可能な表現型変化の検出に依存する、請求項54〜56のいずれか一項記載の方法。
【請求項59】
アンチセンス配向断片の発現によって関心対象の遺伝子に引き起こされる表現型変化は、関心対象の遺伝子の特異的な転写後発現阻害に伴う、請求項58記載の方法。
【請求項60】
関心対象の遺伝子の転写後発現阻害は、RT‐PCR法、ノーザンブロット法、マイクロアレイ法、またはその他適切な方法による関心対象の遺伝子のmRNAの定量化によって計測され、機能的および/または遺伝的に無関係な対照遺伝子の、同様に計測されたmRNAレベルと比較される、請求項59記載の方法。
【請求項61】
アンチセンス配向断片は、炭疽菌遺伝子murB‐2の断片であり、SEQ ID:Ba‐murB2‐C1に明記される配列を含む、請求項59記載の方法。
【請求項62】
アンチセンス配向断片は、炭疽菌遺伝子murB‐2の断片であり、SEQ ID:Ba‐murB2‐H1に明記される配列を含む、請求項59記載の方法。
【請求項63】
アンチセンス配向断片は、炭疽菌遺伝子murB‐2の断片であり、SEQ ID:Ba‐murB2‐D1に明記される配列を含む、請求項59記載の方法。
【請求項64】
アンチセンス配向断片は、炭疽菌遺伝子murB‐2の断片であり、SEQ ID:Ba‐murB2‐D2に明記される配列を含む、請求項59記載の方法。
【請求項65】
アンチセンス配向断片は、炭疽菌遺伝子metSの断片であり、SEQ ID:Ba‐metRS‐H1に明記される配列を含む、請求項59記載の方法。
【請求項66】
アンチセンス配向断片は、炭疽菌遺伝子metSの断片であり、SEQ ID:Ba‐metRS‐H2に明記される配列を含む、請求項59記載の方法。
【請求項67】
アンチセンス配向断片は、炭疽菌遺伝子metSの断片であり、SEQ ID:Ba‐metRS‐H6に明記される配列を含む、請求項59記載の方法。
【請求項68】
アンチセンス配向断片は、炭疽菌遺伝子metSの断片であり、SEQ ID:Ba‐metRS‐E4に明記される配列を含む、請求項59記載の方法。
【請求項69】
アンチセンス配向断片は、炭疽菌遺伝子uppSの断片であり、SEQ ID:Ba‐uppS‐UG9に明記される配列を含む、請求項59記載の方法。
【請求項70】
アンチセンス配向断片は、炭疽菌遺伝子uppSの断片であり、SEQ ID:Ba‐uppS‐UA3に明記される配列を含む、請求項59記載の方法。
【請求項71】
アンチセンス配向断片は、炭疽菌遺伝子dfrAの断片であり、SEQ ID:Ba‐dfrA‐2G1に明記される配列を含む、請求項59記載の方法。
【請求項72】
アンチセンス配向断片は、炭疽菌遺伝子dfrAの断片であり、SEQ ID:Ba‐dfrA‐2G6に明記される配列を含む、請求項59記載の方法。
【請求項73】
アンチセンス配向断片は、黄色ブドウ球菌遺伝子murBの断片であり、SEQ ID:Sa‐murB‐E9に明記される配列を含む、請求項59記載の方法。
【請求項74】
アンチセンス配向断片は、黄色ブドウ球菌遺伝子murBの断片であり、SEQ ID:Sa‐murB‐F7に明記される配列を含む、請求項59記載の方法。
【請求項75】
アンチセンス配向断片は、黄色ブドウ球菌遺伝子murBの断片であり、SEQ ID:Sa‐murB‐B9に明記される配列を含む、請求項59記載の方法。
【図1】

【図2】
【図3】
【図4】

【図5】
【図6】
【図7】
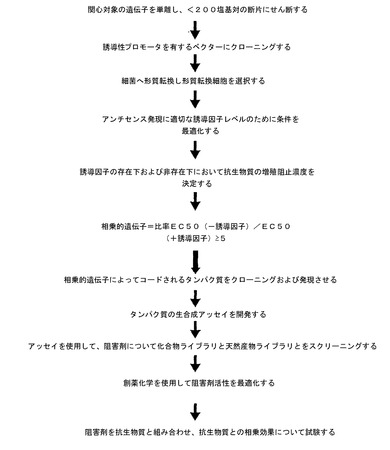
【図8】
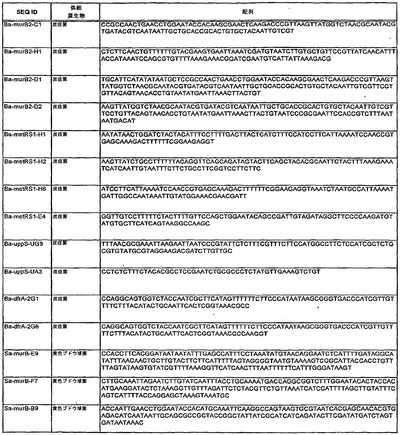
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2009−523120(P2009−523120A)
【公表日】平成21年6月18日(2009.6.18)
【国際特許分類】
【出願番号】特願2008−544559(P2008−544559)
【出願日】平成18年12月8日(2006.12.8)
【国際出願番号】PCT/US2006/047057
【国際公開番号】WO2007/070430
【国際公開日】平成19年6月21日(2007.6.21)
【出願人】(508171871)アールエックス3 ファーマシューティカルズ インコーポレイテッド (1)
【Fターム(参考)】
【公表日】平成21年6月18日(2009.6.18)
【国際特許分類】
【出願日】平成18年12月8日(2006.12.8)
【国際出願番号】PCT/US2006/047057
【国際公開番号】WO2007/070430
【国際公開日】平成19年6月21日(2007.6.21)
【出願人】(508171871)アールエックス3 ファーマシューティカルズ インコーポレイテッド (1)
【Fターム(参考)】
[ Back to top ]