
抗生物質フリープラスミド
本発明は、抗生物質選別圧を用いないでグラム陰性細菌プラスミドを維持する方法を提供する。さらにまた、本発明は、製造したドラッグレスプラスミド(異種遺伝子を含むドラッグレスプラスミドを含む)に関する。本発明はまた、異種遺伝子を含むドラッグレスプラスミドを含む処方物および/または組成物、前記ドラッグレスプラスミドを用いて発現させたタンパク質または免疫原を含む処方物および/または組成物、およびそのような処方物および/または組成物を宿主に投与する方法を提供する。本発明は、前記ドラッグレスプラスミドを含むグラム陰性細菌に関する。
【発明の詳細な説明】
【技術分野】
【0001】
参照による一体化
本出願は、米国仮特許出願61/180,755号(2009年5月22日出願)の利益を主張する。
本発明は、抗生物質選別圧を用いることなくグラム陰性細菌でプラスミドを維持および製造する方法に関する。さらにまた、本発明は、製造したドラッグレスプラスミド、並びに前記ドラッグレスプラスミドを含む処方物および/または組成物、および前記ドラッグレスプラスミドを用いて製造したタンパク質または免疫原を含む処方物および/または組成物、および前記処方物および/または組成物を宿主に投与する方法に関する。本発明は、前記ドラッグレスプラスミドを含むグラム陰性細菌に関する。
【背景技術】
【0002】
今日まで、医薬として使用するために安全であり強力かつ効率的であると考えられるプラスミドDNAベクターは存在しない。誘導されたプラスミド中に抗生物質耐性遺伝子が存在することが、新しい遺伝子療法およびDNAワクチン利用における短所の1つである。
プラスミドは、染色体DNAから分離し、染色体DNAとは別個に複製することができる染色体外DNA分子である(G. Lipps, ed. (2008). Plasmids: Current Research and Future Trends. Caister Academic Press. ISBN978-1-904455-35-6)。天然では、プラスミドは細菌中に通常存在するが、時に真核細胞生物で見出される。それらは、適切な宿主内で自律的に複製することができる、伝達可能な遺伝的エレメント(またはレプリコン)と考えられる。プラスミドは裸のDNAであり、新しい宿主に伝達されるために遺伝物質を包み込むために必要な遺伝子をコードしない。したがって、宿主から宿主へのプラスミドの伝達には、接合または遺伝的エレメントの意図的取り込みを可能にする形質転換による宿主遺伝子発現の変化によって直接的で機械的な伝達が要求される(G. Lipps, 2008)。
遺伝子療法およびワクチン免疫を目的とするプラスミドDNA(pDNA)の使用は、ヒトおよび動物のヘルスケアで顕著な潜在能力を有する新規な技術である(J. Mairhofer et al. (2008) Biotechnol J, 3:83-89)。さらにまた、プラスミドは遺伝学および生物工学研究室で重要なツールとして供される(そのような研究室では、プラスミドは個々の遺伝子の増幅または発現に通常的に用いられる)(David W Russell, Joseph Sambrook (2001), Molecular cloning: a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory)。
【0003】
プラスミドは、細菌集団内で水平的遺伝子伝達のためのメカニズムを提供し、典型的にはある環境状況下で選別優位性を提供する。例えば、プラスミドは、天然には存在しない抗生物質に対する耐性を提供する遺伝子を保持することができる。抗生物質の他にまた別のマーカーも存在する。例えば、プラスミドによって産生されるタンパク質は毒素として作用することができる(前記毒素はまたある環境状況下で選別優位性を提供する)。プラスミドはまた、窒素元素を固定する能力または扱いにくい有機化合物を分解する能力をもつ細菌を提供することができる(前記能力はまたある環境状況下で選別優位性を提供する)(G. Lipps, 2008)。
抗生物質耐性遺伝子を用いる選別の場合、細胞を該当する抗生物質に対して耐性にするタンパク質を生成する遺伝子が挿入されたプラスミドが調製される。次に、このプラスミドは形質転換と称されるプロセスによって細菌に挿入される。続いて、この細菌は該当する抗生物質に曝露される。前記プラスミドは細菌を前記抗生物質に対して耐性にするので、前記プラスミドのコピーを取り込んだ細菌をのみがこの抗生物質から生き残る。
問題の遺伝子は抗生物質耐性マーカーを含むプラスミドを用いてデリバ−される。そのような遺伝子は、典型的にはマルチクローニング部位(MCS(またはポリリンカー))に挿入される。抗生物質耐性遺伝子が発現され、発現されたタンパク質は当該抗生物質を分解する。このようにして、抗生物質は、改変された細菌のみを選別するフィルターとして機能する。ここでこれらの細菌を大量に増殖させ採集して溶解し(しばしばアルカリ溶解方法が用いられる)、問題のプラスミドを単離することができる。
【0004】
医薬におけるこの抗生物質の成功は今では問題となってしまった。多くの細菌(感染症の病原体を含む)がすでに耐性を有し、もはや特定の抗生物質で制御することができない。抗生物質はヒトおよび動物の医薬であまりにも頻繁に用いられた。さらにまた、抗生物質が成果の強化物質として動物飼料に長期間にわたって添加されたという事実ははるかに重大である。この慣行は今ではほとんどが禁止されているが、ばらまかれた抗生物質は、対応する耐性遺伝子をもつ細菌に生存優位性を与えた。さらにまた、細菌中の耐性遺伝子は、可動性DNAユニット上にしばしば配置され、前記ユニットは異なる種間で交換され得る。
このような背景に対して、細菌はマーカー遺伝子を同化し、最終的に、これまで処方されてきた抗生物質が効かない病原体を生じる懸念がある。環境中の微生物(例えば病原体)への遺伝子の伝達も起こり得よう(D.B. Murphy, S.L. Epstein, Guidance for Industry: Guidance for humann somatic cell therapy and gene therapy, Food and Drug Administration, Rockville 1998)。別の安全性に関する懸念は、ヒトの染色体への抗生物質耐性遺伝子の組み込みの可能性である(H.A. Smith, D.M. Klinman (2001) Curr Opin Biotechnol, 12:299-203)。
さらにまた、これらの遺伝子の構成的発現は細菌宿主細胞に不必要な代謝負荷を負わせるので、そのような遺伝子は、プラスミドの製造過程に相当な影響を及ぼす可能性がある(R.M. Cranenburgh et al. (2001) Nucleic Acids Res, 29, e26;A. Rozkov et al. (2006) Enzyme Microb Technol 39:47-50)。これらの遺伝子を除去することによりプラスミドのサイズを縮小することは、pDNAの安定性および発酵プロセスによって得られる収量の改善をもたらすであろう(M.A. Smith et al. (1998) Can J Microbiol, 44:351-355)。
【0005】
したがって、公衆/消費者の潜在的受容リスクのために、規制当局の従来の推奨にしたがい、最終生成物(商品)(裸のDNAワクチン)で抗生物質耐性遺伝子を使用しないことが当分野で絶対に必要である。食品医薬局(FDA)および国連世界保健機関(WHO)は、DNAワクチンの品質を担保し、さらに感染症を予防するために抗生物質耐性マーカーの使用を規制している。同様に、EUデリバレートレリース・ディレクティブ(2002年から発効)は、“ヒトの健康または環境に有害な影響を与えるおそれがある遺伝的改変生物における抗生物質耐性マーカーの使用の段階的廃止”を要求している。
慣習的マーカーの短所は実際的研究においてすら明白になりつつある。例えば、バクトフェクション技術の産業利用のために抗生物質フリーデリバリー系をもつことが所望される。バクトフェクション技術は、侵入細菌を用いる真核細胞へのプラスミドDNAのデリバリーである。さらにまた、発酵過程中の不要な代謝負荷を軽減する技術的な要求が存在する(前記はより高いODおよびDNAプラスミドのより高い収量をもたらすであろう)。
患者の腸内細菌に抗生物質耐性遺伝子をまき散らすという懸念に対して、また別の選別方法が設計された。前記には、栄養要求性補完、リプレッサー滴定、タンパク質系解毒薬/毒素選別系、およびRNA系選別性マーカーの使用が含まれる(以下を参照されたい:J.A. Williams et al. Plasmid DNA vaccine vector design: Impact on efficacy, safety and upstream production, Biotechnol Adv (2009). Doi:10.1016/j.biotechadv.1009.02.003)。
【0006】
R.M. Cranenburghらは、複雑な培地中で組換えプラスミドの抗生物質フリー選別および安定的維持を促進する2つの新規な大腸菌(Escherichia coli)株(DH1lacdapDおよびDH1lacP2dapD)の構築を報告した。前記の株は、必須の染色体遺伝子(dapD)をlacオペレーター/プロモーターの制御下に含んでいた(R.M.Cranenburgh et al. 2001)。IPTG(dapDの発現を誘導する)またはDAPが補充されなければ、これらの細胞は溶解するが、しかしながらこれらの株がlacオペレーターを含むマルチコピープラスミドで形質転換されると、前記オペレーターはLacIリプレッサーを競合的に滴定してlacプロモーターからdapDを発現させる。したがって、リプレッサー滴定選別によって任意の培地で増殖する形質転換体の能力によって、形質転換体を簡単に単離および増殖させることができる。抗生物質耐性遺伝子または他のタンパク質発現配列はプラスミドで要求されず、プラスミドの選別に抗生物質は不要である。
最近Mairhoferらは、いずれの選別マーカーおよび他の付加配列もプラスミドで使用することなくプラスミドを選別および維持するために供される細菌宿主株の設計を探索した。プラスミドの複製阻害因子RNA IがRNA-RNAアンチセンス反応によって増殖に必須の遺伝子の翻訳を抑圧することができるように、いくつかの細菌株を改変した(J. Mairhofer et al. (2008) Biotechnol J 3:83-89)。必須遺伝子(murA)をリプレッサータンパク質(tetR)がその発現を妨害するように改変した(J. Mairhofer et. al., 2008)。プラスミド(したがってRNA I)が存在する場合のみ、tetRが弱まりmurAが発現される。前記著者らは、多様な改変大腸菌株によって市場で入手可能な種々のプラスミドを選別できることを報告した。彼らはさらに、いずれの選別マーカーも含まないミニマリスト的プラスミドを設計した。
本出願におけるいずれの文書の引用または認定も、そのような文書が本発明の先行技術として利用可能であることを容認するものではない。
【発明の概要】
【0007】
抗生物質フリープラスミドが、前記抗生物質フリープラスミドを含むグラム陰性細菌と同様に提供される。免疫原またはタンパク質をコードする異種遺伝子を含む抗生物質フリープラスミドを含む組成物および前記抗生物質フリープラスミドを用いて発現された免疫原またはタンパク質を含む組成物もまた提供される。細菌染色体の非必須領域に1つまたは2つ以上の異種ポリヌクレオチドを含むように、グラム陰性細菌が操作される。前記抗生物質フリープラスミドはリプレッサーをコードするポリヌクレオチドを含み、前記リプレッサーは細菌の染色体上の異種ポリヌクレオチドの発現を調節する。前記抗生物質フリープラスミドは、さらに免疫原またはタンパク質をコードする1つまたは2つ以上のポリヌクレオチドを含むことができる。
本発明の方法は、抗生物質フリープラスミドを製造する方法および前記抗生物質フリープラスミドを用いて哺乳動物細胞に外来遺伝子を導入するための方法を含む。
これらの実施態様および他の実施態様は、以下の詳細な説明によって開示されまたは以下の詳細な説明から明白であり、さらにそれら実施態様は以下の詳細な説明に包含されている。
例示の手段により提供する以下の詳細な説明(記載した具体的実施態様に本発明を限定しようとするものではない)は、付随の図面(参照により本明細書に含まれる)と一緒にして理解されよう。
【図面の簡単な説明】
【0008】
【図1】ドラッグレス概念の第一の構成要素を示す。前記は、ショ糖を含む培地にプレートしたとき形質転換細菌の急速な死をもたらす、グラム陰性細菌のレバンスクラーゼをコードするsacB遺伝子のクローニングである。
【図2】ドラッグレスまたは抗生物質フリー概念の第二および第三の構成成分を示す。前記はλファージの免疫領域である。ここで、cIはcIリプレッサーをコードし、croはCroタンパク質をコードし、Nは転写抗ターミネーターをコードし、cIIはcIアクチベーターをコードし、cIIIはcIIIプロテアーゼ阻害因子をコードし、OL1、2および3並びにOR1、2および3はオペレーターであり、PLおよびPRは右側および左側プロモーターであり、PREはリプレッサー確立のためのプロモーターであり、PRMはリプレッサー維持のためのプロモーターである。
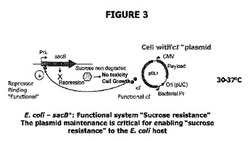
【図3】“ドラッグレス”概念の理論的大要を示す。ここで、cIリプレッサー活性(pDL1(DrugLess Plasmid)プラスミドから発現される)は、宿主細胞染色体上に存在するλPrプロモーター下に配置された有毒sacB遺伝子生成物の転写を阻害し、ショ糖存在下の宿主大腸菌細胞の生存能力は、プラスミド由来のλcIリプレッサータンパク質の十分な発現レベルによって担保される。
【図4】不活化された系の“ショ糖感受性”を示す。この系では、42℃の温度への切換えは、cIリプレッサーのλPrプロモーターとの結合活性を阻害し、宿主細胞をショ糖に感受性にする。
【図5】宿主細胞株と抗生物質フリーpDL1プラスミドの合体(pDL1による形質転換から生じる)およびショ糖を含む培地での増殖(ドラッグレスプラスミドの産生をもたらす)を示す。
【図6】A−Cは実験計画を示す。図6Aは大要を提供する。図6Bは、ΔpurN(またはΔedA)親株はカナマイシン耐性であり、30℃−37℃でインキュベートしたときそのショ糖感受性を付与するSacB+を構成的に発現することを示す。ΔpurN(またはΔedA)親株をpPB829またはpPB838プラスミドのどちらかで形質転換したとき、これらの細胞は、30℃−37℃でインキュベートしたときショ糖の存在下で生存し増殖する能力を獲得する。図6Cは、pPB829またはpPB838プラスミド(cIプラスミド)のどちらかで形質転換したΔpurN(またはΔedA)親株は、カナマイシン、ショ糖またはクロラムフェニコールを含むLB培地で同様に増殖できることを示す。ショ糖存在下でプレートしたとき、30−37℃から42℃への温度の切換えは細胞死を引き起こす。
【図7】クロラムフェニコールアセチルトランスフェラーゼ(CatR)をコードする遺伝子で目印を付された、λcI遺伝子を含む両プラスミド(pPB829およびpPB838)を示す。pPB838はpMCS5の誘導体で、前記誘導体ではクロラムフェニコールアセチルトランスフェラーゼ(cat)がアンピシリン耐性遺伝子と入れ代わっている。このプラスミドは、カナマイシン遺伝子の弱いプロモータ(P1)の制御下に配置されたcI遺伝子を含む。pPB829はcat遺伝子を含むpVR1012プラスミド誘導体である。このプラスミドはcI遺伝子を含み、前記は弱いプロモーター(P1)の制御下に配置されている。ΔpurN(またはΔedA)親大腸菌株に導入されたとき、これらのプラスミドはショ糖の存在下での細胞の増殖を可能にする。クロラムフェニコールは概念の明白な立証を可能にする。
【図8】A−Eは宿主株の操作を示す。図8Aおよび8BはλPr::sacBΩkanカセットの作製を示す。前記カセットは、それぞれedAまたはpurN遺伝子の対立遺伝子置換によって大腸菌染色体に挿入される。図8Cは、ダブルsacBカセット大腸菌株を作製するために、ΔpurNΩλPr::sacB Km大腸菌宿主株にedA遺伝子を対立遺伝子置換するために用いられる完全なsacBカセット(ΔpurNΩλPr::sacBcat)の、PCRおよび結合PCRによる操作を示す。
【図9】A、BおよびCは、宿主株の操作、すなわち大腸菌のedAおよびpurN欠失のデモンストレーションのためにPCRの使用を示す。図9Aは、2つのPCR生成物(それぞれ151および729bp)を部分として示す。それら生成物を精製し、さらにPB1186およびPB1189プライマー並びにPhusion DNAポリメラーゼとともに第二のPCR工程で鋳型として用いる。図9BはPCRチェックを示し、λPr::sacB+Ωkanカセットの大腸菌染色体への導入を示す。図9Cは、λPr::sacB+ΩkanカセットがedAおよび/またはpurN遺伝子のいずれかにおける対立遺伝子置換によって遺伝子座に正確に挿入されることを示す。図9Dは、二重sacBカセット宿主株の操作、すなわち対立遺伝子置換による大腸菌のedAおよびpurNの欠失のデモンストレーションのためにPCRの使用を示す。PCRチェックは、λPr::sacB+ΩkanおよびλPr::sacB+ΩCatカセットの両方が、シークェンシングによって確認されるように、大腸菌の染色体の特定の遺伝子座に正確に導入されることを示す。図9Dに示すように、レーン1はBJ5183wt、レーン2はBJ5138ΔpurN、レーン3はBJ5138ΔedA、レーン4はBJ5183ΔpurNΔedAで希釈せずに1μL、レーン5はBJ5183ΔpurNΔedAで1/10希釈、レーン6は水で0bp(コントロール)、AはBJ5183wt、BはBJ5138ΔpurN、CはBJ5138ΔedA、DはBJ5183ΔpurNΔedAで希釈せずに3μL、EはBJ5183ΔpurNΔedAで希釈せずに1μL、FはBJ5183ΔpurNΔedAで1/10希釈、GはBJ5183ΔpurNΔedAで1/50希釈、Hは水で0bp(コントロール)である。
【図10】Aは、宿主株(ΔpurNλPr::sacB+Ωkan)はショ糖に対して高度に感受性であること(増殖しない)、pPB829またはpPB838プラスミドで形質転換した細胞はショ糖の存在下において30℃で良好に増殖したこと、各cIプラスミドから合成されたcI遺伝子生成物によってsacB遺伝子発現は完全に抑圧されたこと、およびクロラムフェニコールの存在下で増殖する同様な細胞を用いて平行して実施したコントロール実験によって明確に示されるようにプラスミドの維持は100%有効であったことを示す。42℃への温度上昇は、cI遺伝子生成物の不活化およびショ糖またはクロラムフェニコールのどちらかを含むLB寒天にプレートされた細胞の死をもたらす。この厳しいプラスミド維持実験は、ショ糖またはクロラムフェニコールの存在下で細胞を2回続けて継代して増殖させた後実施された。図10B−Cは、二重sacBカセット大腸菌株(ΔpurNλPr::sacB+Ωkan ΔedAλPr::sacB+ΩCat)は、30℃および42℃でインキュベートしたときショ糖の存在下で高度に感受性である(増殖しない)が、一方、ショ糖上にプレートされたとき1つのSacBカセットをもつ大腸菌株とともにいくつかの偶発的変異体が出現することを示している。図10Cはまた、1つのSacBカセットをもつ大腸菌株と比較した二重sacBカセット大腸菌株が要求する最低のショ糖濃度を示す(10%から最終の2%まで)。
【図11】増殖細胞中でのpPB829およびpPB838プラスミドの安定性を示す。
【図12】Aは、λPr::sacB 1つのsacBカセットでは30℃で偶発的変異が発生しないことを示す。図12B−Cは、2つのsacBカセット(1つではなくて)の存在は、30℃から37℃の範囲の温度ではショ糖感受性についてより強力な厳格性を付与することを示している(それでもなお2%ショ糖が最適である)。大腸菌染色体に1つのsacBカセットが存在する場合、同様な温度で2%から4%の範囲のショ糖濃度でより低い厳格性が存在する。この平板培養アッセイで確認されたように、2つのsacBカセットをもつ大腸菌株のショ糖に対する変異率は、37℃でインキュベートしたとき最低ショ糖濃度(2%)では検出不能であったが、1つのsacBカセット大腸菌株の変異率は3.8 x 10-6から5 x 10-5の範囲であった。別個の実験セットによって、2つのsacBカセットの大腸菌株における変異率は、2%ショ糖の存在下においては37℃でインキュベートしたときほぼ5 x 10-10であることが示された。
【図13】A−Bは、クロラムフェニコール(Cat)の目印を有するcIプラスミドで形質転換した細胞を選別圧力として抗生物質を使用することなく寒天プレート上でスクリーニングすることができる実験手順を示す。
【図14】A−Cは、cIプラスミドのみで(pPB829およびpPB838)形質転換した細胞の選別に対する、選別圧力として抗生物質を使用しない前記実験手順の有効性を示す。
【図15】A−Bは、1コピーのsacBカセットを含む増殖中の大腸菌細胞でのpPB829およびpPB838プラスミドの安定性および産生を示す。C−Eは、1つのsacBカセットおよび2つのsacBカセットをもつ大腸菌株(それぞれΔpurNλPr::sacB+ΩkanおよびΔpurNλPr::sacB+Ωkan ΔedAλPr::sacB+Ωcat)におけるcatフリーpPB885プラスミドの安定性および産生を示す。図15Fは、CHO細胞における抗生物質フリープラスミド(pPB896/選別圧としてショ糖)および抗生物質プラスミド(pCG105/選別圧としてCat)のトランスフェクション有効性(GFP発現)を示す。これら2つの選別圧間(ショ糖対抗生物質としてのCat)において発現GFPタンパク質に関する明白な相違は認められない。
【図16】pCRblunt+PB1184-1185プラスミドマップを示す。
【図17】pPB828の制限マップおよび特徴を示す。
【図18】pPB829の制限マップおよび特徴を示す。
【図19】pPB838のプラスミドマップおよび特徴を示す。
【図20】pPB844のプラスミドマップおよび特徴を示す。
【図21】pPB845のプラスミドマップおよび特徴を示す。
【図22】pPB846のプラスミドマップおよび特徴を示す。
【図23】pPB847のプラスミドマップおよび特徴を示す。
【図24】pPB885のプラスミドマップを示す。pPB885の特徴物マップは以下を示す:2つのCDS:cIリプレッサー:324−1037;AP(R):2321−2990(相補性)、本来の位置(補完物1626..2483);2つの複製起点:pUC起点:1500−2173(相補性)、本来の位置(補完物805..1478);f1起点:2991−3301(本来の位置2615−3053)。
【図25】pPB896のプラスミドマップを示す。pPB896の特徴物マップは以下を示す:3つのCDS:GFP:1675−2397(本来の位置1676…2398);cIリプレッサー:4089−4802;Cm(R):4828−5484(相補性)、補完物(950−1606);1つのイントロン:イントロンA:805−1625(本来の位置805−1625);1つの真核細胞プロモーター:CMVプロモーター:1−683;1つの複製起点:3133−3640(本来の位置2425..2932);1つのターミネーター:BGH転写ターミネーター/PolyA:2405−2951(本来の位置1697..2243);1つの5'UTR:CMV IE5'UTR:684−804(本来の位置684..804)。
【図26】cIリプレッサータンパク質の配列アラインメントおよび配列同一性のパーセンテージを示す。
【図27】sacBタンパク質の配列アラインメントおよび配列同一性のパーセンテージを示す。
【図28】ポリヌクレオチドおよびタンパク質に割り当てた配列番号を示す表である。
【図29】cIリプレッサータンパク質をコードするポリヌクレオチドの配列アラインメントおよびヌクレオチドレベルにおける配列同一性のパーセンテージを示す。
【図30】sacBタンパク質をコードするポリヌクレオチドの配列アラインメントおよびヌクレオチドレベルにおける配列同一性のパーセンテージを示す。
【図31】P1プロモーター、cIの天然のプロモーター、λPrプロモーター、λPrプロモーター+5'UTR+sacB遺伝子、P1プロモーター+cI遺伝子、およびcIの天然のプロモーター+cI遺伝子の配列を示す。
【発明を実施するための形態】
【0009】
詳細な説明
本開示および特に特許請求の範囲では、例えば“comprises”、“comprised”および“comprising”などのような用語は、米国特許法でそれらに与えられた意味を有し得る。例えば、それら用語は“includes”、“included”および“including”などを意味することができ、さらに、例えば“consisting essentially of”および“consists essentially of”のような用語は、米国特許法でそれらに与えられた意味を有し得る。例えば、それら用語は明示的に列挙されていない成分も許容するが、しかし従来技術で見出されている成分、または当該発明の基本的なまたは新規な特徴に影響を及ぼす成分は除かれる。
特段の説明がなければ、本明細書で用いられる全ての技術用語および学術用語は、本開示が属する分野の業者のある者が一般的に理解する意味と同じ意味を有する。単数用語の“a”、“an”および“the”は、文脈が明らかにそうでないことを示していないかぎり、対応する複数形を含む。同様に、“or”という語は、文脈が明らかにそうでないことを示していないかぎり、“and”を含むことが意図される。
【0010】
“動物”は哺乳動物、鳥類などが意図される。動物または宿主には、哺乳動物およびヒトが含まれる。動物は、ウマ科の動物(例えばウマ)、イヌ科の動物(例えばイヌ、オオカミ、キツネ、コヨーテ、ジャッカル)、ネコ科の動物(例えばライオン、トラ、イエネコ、野生のネコ、他の大型のネコおよび他のネコ科の動物(チーターおよびオオヤマネコを含む))、ヒツジ類(例えばヒツジ)、ウシ科の動物(例えばウシ)、ブタ類(例えばブタ)、鳥類(例えばニワトリ、アヒル、カモ、シチメンチョウ、ウズラ、キジ、オウム、ウソ、タカ、カラス、ダチョウ、エミュおよびヒクイドリ)、霊長類(例えばキツネザル、メガネザル、サル、テナガザル、ヒヒ)および魚類が含まれる。“動物”という用語はまた、全ての発育期(胚および胎児期を含む)の個々の動物を含む。
“タンパク質”、“ペプチド”、“ポリペプチド”および“ポリペプチドフラグメント”という用語は本明細書で互換的に用いられ、任意の長さのアミノ酸残基ポリマーを指す。ポリマーは線状でも分枝していてもよく、改変アミノ酸またはアミノ酸アナローグを含むことができ、さらにアミノ酸以外の化学的部分によって中断されていてもよい。前記用語はまた、天然に改変されているか、または介入(例えばジスルフィド結合形成、グリコシル化、リピド化、アセチル化、リン酸化)または他の任意の操作もしくは修飾(例えば標識または生物活性成分との結合)によって改変されているアミノ酸ポリマーを包含する。
【0011】
“核酸”または“ポリヌクレオチド”という用語は互換的に用いられ、線状もしくは分枝状、一本鎖状もしくは二本鎖状、または前記の混合であるRNAまたはDNAを指す。前記用語はまた、RNA/DNAハイブリッドを包含する。以下はポリヌクレオチドの非限定的な例である:遺伝子または遺伝子フラグメント、エキソン、イントロン、mRNA、tRNA、rRNA、リボザイム、cDNA、組換えポリヌクレオチド、分枝ポリヌクレオチド、プラスミド、ベクター、任意の配列の単離DNA、任意の配列の単離RNA、核酸プローブおよびプライマー。ポリヌクレオチドは、改変ヌクレオチド、例えばメチル化ヌクレオチドおよびヌクレオチドアナローグ、ウラシル、他の糖および連結基(例えばフルオロリボースおよびチオレート)、並びにヌクレオチド分枝を含むことができる。ヌクレオチドの配列は、重合の後で例えば標識成分との結合によってさらに改変することができる。本定義に含まれる他のタイプの改変は、キャップ、天然に存在する1つまたは2つ以上のヌクレオチドのアナローグによる置換、ポリヌクレオチドをタンパク質、金属イオン、標識成分、他のポリヌクレオチドまたは固相に結合させるための手段の導入である。ポリヌクレオチドは、化学的合成によって入手するか、または微生物から誘導することができる。
“ドラッグレスプラスミド”または“抗生物質フリープラスミド”という用語は互換的に用いられ、抗生物質選別遺伝子を含まないDNAプラスミドを指す。
“遺伝子”という用語は広範囲に用いられ、生物学的機能と結びついた任意のポリヌクレオチドセグメントを指す。したがって、遺伝子には、ゲノム配列の場合のイントロンおよびエキソン、cDNAの場合のコード配列および/または前記配列のための調節配列が含まれる。例えば、遺伝子はまた、mRNAまたは機能的RNAを発現するか、または具体的なタンパク質をコードする核酸フラグメントを指し、前記には調節配列が含まれる。
【0012】
本明細書に開示する主題は、抗生物質耐性遺伝子の使用を回避することによって細菌プラスミドDNAベクターを作製し、安全なワクチンおよび免疫原性組成物を製造する新規なアプローチに関する。
本明細書に開示する主題は、グラム陰性宿主細菌細胞内で多数のプラスミドコピーを維持するための新規な概念(3つの構成成分に基づく)を提示する。第一の構成成分はグラム陰性宿主細菌であり、前記細菌は規定の培養条件下で当該細菌にとって有毒な生成物を発現し、前記有毒遺伝子は当該細菌染色体の非必須領域に挿入されている。第二の構成成分は、当該グラム陰性細菌の染色体上の遺伝子の存在であり、前記遺伝子は、厳密に制御され得る構成的プロモーターの制御下で有毒生成物をコードする。第三の構成成分は、前記宿主染色体上の有毒遺伝子に作動できるように連結されたプロモーターを調節するプラスミド由来の特異的なリプレッサーの発現である。
本発明で意図されるグラム陰性細菌には、アビバクテリウム(Avibacterium)、ブルセラ(Brucella)、大腸菌(Escherichia coli)、ヘモフィルス(Haemophilus)(例えばヘモフィルス・スイス(Haemophilus suis))、サルモネラ(Salmonella)(例えばサルモネラ・エンテリディス(Salmonella enteridis)、ネズミチフス菌(Salmonella typhimurium)、サルモネラ・インファンチス(Salmonella infantis))、シゲラ(Shigella)、パスツレラ(Pasteurella)およびリメイレラ(Rimeirella)が含まれるが、ただしこれらに限定されない。
ある実施態様では、有毒遺伝子は、レバンスクラーゼをコードするsacB構造遺伝子である。別の実施態様では、枯草菌(Bacillus subtilis)のレバンスクラーゼをコードする枯草菌のsacB構造遺伝子である。その天然の環境でのsacBの発現はグラム陽性細菌にとっては無害であるが、グラム陰性細菌で発現される場合、ショ糖含有培地にグラム陰性細菌がプレートされたとき形質転換細菌の急速な死を引き起こす。
【0013】
ある特徴では、本発明はsacBタンパク質(レバンスクラーゼ)を提供する。別の特徴では、本発明は、配列番号:4、60、62、64、66、68、70、72または74に示す配列を有するsacBタンパク質およびその変種またはフラグメントを提供する。さらに別の特徴では、本発明は、配列番号:4、60、62、64、66、68、70、72または74に示す配列を有するポリペプチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するsacBタンパク質を提供する。さらに別の特徴では、本発明は、上記で認定したsacBタンパク質(配列番号:4、60、62、64、66、68、70、72または74)のフラグメントおよび変種を提供する(前記は周知の分子生物学的技術を用いて当業者により容易に調製され得る)。変種は、本発明のポリペプチド(特に配列番号:4、60、62、64、66、68、70、72または74に示すアミノ酸配列)と少なくとも約75%、80%、85%、90%、95%、96%、97%、98%または99%のアミノ酸配列同一性を有する相同なポリペプチドである。
【0014】
別の特徴では、本発明は、sacBタンパク質(レバンスクラーゼ)をコードするポリヌクレオチド(例えばsacB遺伝子)、例えば配列番号:4、60、62、64、66、68、70、72または74に示す配列を有するsacBタンパク質をコードするポリヌクレオチドを提供する。さらに別の特徴では、本発明は、配列番号:4、60、62、64、66、68、70、72または74に示す配列を有するポリペプチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するsacBタンパク質、または保存的変種、対立遺伝子変種、ホモローグ、またはこれらポリペプチドの1つの少なくとも8つまたは10の連続するアミノ酸を含むフラグメント、またはこれらポリペプチドの組み合わせをコードするポリヌクレオチドを提供する。別の特徴では、本発明は、配列番号:3、61、63、65、67、69、71または73に示すヌクレオチド配列を有するポリヌクレオチドまたはその変種を提供する。さらに別の特徴では、本発明は、配列番号:3、61、63、65、67、69、71または73に示す配列を有するポリヌクレオチドの1つと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するポリヌクレオチドまたはその変種を提供する。
【0015】
本発明で利用できる他の有毒な遺伝子にはリステリア(Listeria)またはスタフィロコッカス(Staphylococcus)遺伝子が含まれるが、ただしこれらに限定されない。有毒な遺伝子は、有毒な遺伝子生成物(有毒タンパク質またはRNA)として発現されるDNAであってもよいが、またそのもの自体が有毒であるものおよび単独で有毒であるものであってもよい。そのような有毒遺伝子生成物の例は当分野では周知であり、制限エンドヌクレアーゼ(例えばDpnI)、CcdA/CcdB(S. Maki et al. 1996, J Mol Biol 256:473-482)、および抑圧機能の非存在下で宿主を殺す遺伝子(例えばkicB)が含まれるが、ただしこれらに限定されない。また別には、有毒遺伝子はin vitroで選別可能であり得る(例えば制限部位)。
本明細書で用いられるように、“ホモローグ”という用語にはオルトローグ、アナローグおよびパラローグが含まれる。野生型ポリペプチドのアナローグ、オルトローグおよびパラローグは、翻訳後修飾により、アミノ酸配列の相違により、またはその両方により野生型ポリペプチドと相違し得る。特に、本発明のホモローグは、一般的には野生型ポリペプチドまたはポリヌクレオチド配列の全体または部分に関して少なくとも80−85%、85−90%、90−95%または95%、96%、97%、98%または99%の配列同一性を示し、かつ類似の機能を示すであろう。
【0016】
“変種”は実質的に類似する配列を意味することが意図される。ポリヌクレオチドについては、変種は、天然のポリヌクレオチド内の1つまたは2つ以上の部位の1つまたは2つ以上の欠失および/または付加、および/または天然のポリヌクレオチド内の1つまたは2つ以上の部位の1つまたは2つ以上のヌクレオチドの置換を含む。本明細書で用いられるように、“天然の”ポリヌクレオチドまたはポリペプチドは、それぞれ天然に存在するヌクレオチド配列またはアミノ酸配列を含む。本発明の特定のポリヌクレオチド(すなわち参照ポリヌクレオチド)の変種はまた、変種ポリヌクレオチドによってコードされるポリペプチドおよび参照ポリヌクレオチドによってコードされるポリペプチド間のパーセント配列同一性を比較することによって評価することができる。“変種”タンパク質は、天然のタンパク質内の1つまたは2つ以上の部位の1つまたは2つ以上のアミノ酸の欠失または付加、および/または天然のタンパク質内の1つまたは2つ以上の部位の1つまたは2つ以上のアミノ酸の置換によって天然のタンパク質から誘導されたタンパク質を意味する。本発明に包含される変種タンパク質は生物学的に活性を有する。すなわちそれらは免疫応答を誘引する能力を有する。
【0017】
変種には対立遺伝子変種が含まれる。“対立遺伝子変種”という用語は、タンパク質のアミノ酸配列に変化をもたらし、さらに天然の集団(例えばウイルス種またはウイルス変種)内に存在する多形性を含むポリヌクレオチドまたはポリペプチドを指す。そのような天然の対立遺伝子変種は、典型的にはポリヌクレオチドまたはポリペプチドで1−5%の変動を生じる。対立遺伝子変種は、多数の異なる種の中の問題の核酸配列をシークェンシングすることによって識別することができる。前記は、そのような種の中で同じ遺伝子座を識別するハイブリダイゼーションプローブを用いることによって容易に実施することができる。そのような核酸の多様性およびその結果生じたアミノ酸多形性または変種(それらは天然の対立遺伝子変種の結果であり、問題の遺伝子の機能的活性を変化させない)の任意のものおよび全てが本発明の範囲内に含まれる。
“保存的変種”という用語は、アミノ酸残基の生物学的に類似する別の残基による置換、またはコードされるアミノ酸残基が変化しないかまたは生物学的に類似する別の残基となるような核酸配列内のヌクレオチドの置換を指す。これに関して特に好ましい置換は、上記に記載したように一般的に性質が保存的であろう。
【0018】
開示のポリヌクレオチドには、遺伝暗号の結果として、例えば特定の宿主のための最適なコドン使用頻度の結果として縮退性である配列が含まれる。本明細書で用いられるように、“最適化”とは、ある種においてその発現を高めるために一般的に操作されるポリヌクレオチドを指す。本発明のポリペプチドをコードする最適化ポリヌクレオチドを提供するために、ポリペプチドのDNA配列は、1)個別の種において高度に発現される遺伝子の優先的コドンを含むように;2)前記種で実質的に見出されるヌクレオチド塩基組成のA+TまたはG+C含量と類似する含量を含むように;3)前記種の開始配列を形成するように;または4)脱安定化、不適切なポリアデニル化、RNAの分解および停止を引き起こす配列、または二次構造ヘアピンまたはRNAスプライス部位を形成する配列を除去するように改変できる。前記の種における本発明のポリペプチド(例えばsacBタンパク質またはcIリプレッサータンパク質)の増加は、真核生物および原核生物または個別の種のコドン使用頻度の頻度分布を利用することによって達成できる。“優先的コドン使用頻度”という用語は、あるアミノ酸を指定するためにヌクレオチドコドンの使用において個々の宿主細胞によって示される優先性を指す。20の天然のアミノ酸が存在し、そのほとんどが2つ以上のコドンによって指定される。したがって、ヌクレオチド配列によってコードされるポリペプチドのアミノ酸配列が機能的に変化しないかぎり、全ての縮退ヌクレオチド配列が本発明に含まれる。
【0019】
配列に関して“同一性”とは、2つの配列の短い方のヌクレオチドまたはアミノ酸の数で割った、同一ヌクレオチドまたはアミノ酸を有する位置の数を意味し、この場合、2つの配列のアラインメントはWilburとLipmannのアルゴリズム(Wilbur & Lipman)にしたがって決定できる。RNA配列がDNA配列と類似している、またはある程度の配列同一性もしくは配列相同性を有するというとき、DNA配列中のチミジン(T)はRNA配列中のウラシル(U)と等価であると考えられる。したがって、DNA配列中のチミジン(T)をRNA配列中のウラシル(U)と等価であると考えることによって、RNA配列は本発明の範囲内にあり、DNA配列から誘導することができる。2つのアミノ酸配列の配列同一性または配列類似性、または2つのヌクレオチド配列間の配列同一性は、Vector NTIソフトウェアパッケージ(Invitrogen, 1600 Faraday Ave., Carlsbad, CA)を用いて決定できる。
【0020】
別の実施態様では、本発明の有毒な遺伝子に作動できるように連結されるプロモーターは、λファージ由来のプロモーターである。λファージは、大腸菌内で生活する溶原性ファージである。ファージがその宿主細胞内に入ると、ファージは自身を宿主のDNAに組み入れる。この状態でλはプロファージと呼ばれ、宿主に大きな害を引き起こすことなく宿主ゲノム内に留まる。このようにして、プロファージは宿主の細胞分割のたびに二倍になる。この状態で発現されるプロファージのDNAは、宿主細胞のストレスの徴候に注意するタンパク質をコードする。ストレスは、飢餓、毒物(抗生物質のような)および宿主を損傷または破壊することができる他の因子の結果であり得る。ストレス状態が検出されたとき、プロファージは再び活性化し、宿主細胞のDNAからそれ自身を切り出し、その溶菌周期に入る。再活性化されたファージは宿主DNAから離れ、そのタンパク質工場を初期化して新ファージの多数のコピーを生産する。新ファージの構築によって宿主の全資源が枯渇したとき、細胞が溶解し(細胞膜が破壊され)、新ファージが放出される。
【0021】
ラムダリプレッサー遺伝子系は以下から成る(染色体の左から右へ):cI遺伝子、OR3、OR2、OR1、cro遺伝子。cI遺伝子はλリプレッサー(“cIリプレッサータンパク質”)をコードする。cIリプレッサータンパク質をコードするゲノム領域は免疫領域として知られている。cIリプレッサータンパク質は、遺伝子転写の正および負の両方の調節因子である。前記は、λファージがその宿主細菌の染色体上で“休眠状態”に留まることを可能にする。バクテリオファージλの溶原性状態は、調節タンパク質cIとOR(右オペレーター)およびOL(左オペレーター)(それぞれλPlおよびλPrプロモーターに由来し、溶菌状態に必要なタンパク質の転写を妨げる)が結合することによって維持される。溶原性λファージを保持する細菌細胞はλファージによる更なる感染に対して免疫である。cIリプレッサータンパク質はさらに別のファージ粒子の感染による溶菌の進行を阻害する。
この実施態様のある特徴では、プロモーターは配列番号:5に示す配列を有するポリヌクレオチドを含む。別の特徴では、プロモーターは、配列番号:5に示す配列を有するポリヌクレオチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するポリヌクレオチドを含む。
【0022】
ある実施態様では、本発明のリプレッサータンパク質は、cIリプレッサータンパク質、例えば温度感受性のcI857リプレッサーである。溶原としてcI857を保持するλファージは39℃より低い温度で増殖するが、続いて温度上昇によって溶菌性増殖を誘発するであろう。30℃では、cIリプレッサータンパク質は活性であり、感染ファージの右および左オペレーターと結合する。これによっていずれのファージタンパク質の転写も妨害され、したがって溶菌が妨げられる。しかしながら42℃では、cIリプレッサーは不活化され、プロモーターオペレーターと結合できない。
ある特徴では、本発明はcIリプレッサータンパク質を提供する。別の特徴では、本発明は、配列番号:2、44、46、48、50、52、54、56または58に示す配列を有するcIリプレッサータンパク質およびその変種またはフラグメントを提供する。さらに別の特徴では、本発明は、配列番号:2、44、46、48、50、52、54、56または58に示す配列を有するポリペプチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するcIリプレッサータンパク質を提供する。さらに別の特徴では、本発明は上記で認定したcIリプレッサータンパク質(配列番号:2、44、46、48、50、52、54、56または58)のフラグメントおよび変種を提供する(前記は周知の分子生物学的技術を用いて当業者によって容易に調製され得る)。変種は、本発明のポリペプチド、特に、配列番号:2、44、46、48、50、52、54、56または58に示すアミノ酸配列と少なくとも約75%、80%、85%、90%、95%、96%、97%、98%または99%の同一性を有する相同なポリペプチドである。
【0023】
別の特徴では、本発明は、cIリプレッサータンパク質をコードするポリヌクレオチド(例えばcI遺伝子)、例えば配列番号:2、44、46、48、50、52、54、56または58に示す配列を有するcIリプレッサータンパク質をコードするポリヌクレオチドを提供する。さらに別の特徴では、本発明は、配列番号:2、44、46、48、50、52、54、56または58に示す配列を有するポリペプチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するcIリプレッサータンパク質、または保存的変種、対立遺伝子変種、ホモローグ、またはこれらポリペプチドの1つの少なくとも8つまたは10の連続するアミノ酸を含むフラグメント、またはこれらポリペプチドの組み合わせをコードするポリヌクレオチドを提供する。別の特徴では、本発明は、配列番号:1、45、47、49、51、53、55、57または59に示すヌクレオチド配列を有するポリヌクレオチドまたはその変種を提供する。さらに別の特徴では、本発明は、配列番号:1、45、47、49、51、53、55、57または59に示す配列を有するポリヌクレオチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するポリヌクレオチド、またはその変種を提供する。
【0024】
ある実施態様では、cI遺伝子を駆動するプロモーターは天然のcI 遺伝子プロモーターである。別の実施態様では、cI遺伝子を駆動するプロモーターはカナマイシン遺伝子の弱いプロモーター(P1)である。別の実施態様では、プロモーターは配列番号:29、83または30に示す配列を有するポリヌクレオチドを含む。さらに別の実施態様では、プロモーターは、配列番号:29、83または30に示す配列を有するポリヌクレオチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するポリヌクレオチドを含む。
本発明の方法で利用できる他のプロモーター/リプレッサー対には、CarA-CarSリプレッサー-アンチリプレッサー対により制御されるcarBプロモーター、成長ホルモン遺伝子プロモーターおよびリプレッサー、並びにLacリプレッサー(lacl)およびPtrc-2プロモーターが含まれるが、ただしこれらに限定されない。
【0025】
変異導入のための基質として機能する変異プロモーター/リプレッサー対は、現在入手可能であり当分野で報告されているようなものから選択することができる。変異リプレッサーとそれらのオペレーターとの結合に関する研究(以下を参照されたい:Nelson and Sauer, 1985, Cell 42:549;Nelson and Sauer, 1986, J Mol Biol 192:27;およびGussin et al. 1983, Lambda II, (Hendrix, Roberts, Stahl and Weisberg, eds) Cold Spring Harbor Press, Cold Spring Harbor, NY, p.93-123)は、減速されるが停止されることはない転写速度を提供する既知の変異プロモーターおよび/または変異リプレッサー配列の選別を可能にする。この選別プロモーター領域を異種発現プラスミドに取り込み、続いて通常の位置特異的変異導入によって前記領域を改変する(例えば以下を参照されたい:Morinaga et al. 1984, Biotechnology, 2:636)。また別には、リプレッサーコード配列を類似の態様で変更する。生じた変異リプレッサー/プロモーター対を、位置特異的変異導入に付されていない野生型プラスミドと発現を比較することによって、異種タンパク質の分泌を促進するその能力について解析する。
本明細書に開示する主題はドラッグレスまたは抗生物質フリーの概念を提供する。本概念では、プラスミドから発現されるリプレッサー活性(例えばcIリプレッサー)は、細胞宿主染色体上に存在するλファージプロモーターの下に配置された有毒遺伝子生成物(例えばsacB遺伝子生成物)の転写を阻害し、さらに基質(例えばショ糖)の存在下における宿主細胞の生存活性は、プラスミド由来リプレッサータンパク質(例えばλcIリプレッサータンパク質)がプラスミドから十分なレベルで発現されることによって担保される。抗生物質フリープラスミドを含む宿主株のショ糖存在下での増殖は、効率的な系のDNAプラスミドの維持および産生を担保する。
プラスミドという用語は、本発明のポリヌクレオチドおよび所望の宿主または標的の細胞における前記ポリヌクレオチドのin vivo発現に必要なエレメントを含む任意のDNA転写ユニットをカバーし、さらに前記ユニットに関して、スーパーコイルまたは非スーパーコイルプラスミド、環状プラスミドが、線状形と同様に本発明の範囲内に含まれることが特記される。
【0026】
本発明のある実施態様は、1つまたは2つ以上のプロモーターに作動できるように連結された1つまたは2つ以上のリプレッサー遺伝子を含むドラッグレスプラスミドに関する。ある特徴では、ドラッグレスプラスミドはcIリプレッサータンパク質をコードするポリヌクレオチドを含む。別の特徴では、ドラッグレスプラスミドはcIリプレッサータンパク質をコードするポリヌクレオチドに作動できるように連結されたプロモーターを含む。プロモーターは、天然のcI遺伝子プロモーターまたはカナマイシン遺伝子の弱いプロモーターであり得る。別の特徴では、ドラッグレスプラスミドはさらに、免疫原またはタンパク質をコードする異種ポリヌクレオチドを含む。別の特徴では、ドラッグレスプラスミドは、免疫原またはタンパク質をコードする異種ポリヌクレオチドに作動できるように連結されたプロモーターを含む。当分野で公知の任意のプロモーターを、本発明のドラッグレスプラスミドで用いることができる。前記には、細菌性、酵母、菌類、昆虫、哺乳動物および植物のプロモーターが含まれる。プロモーターは、ヒトまたはネズミ起源の、または場合によって別の起源(例えばラットまたはモルモット)を有する極初期サイトメガロウイルスプロモーター(CMV-IE)、スーパープロモーター(M. Ni et al. 1995, Plant J 7:661-676)であるが、ただし前記に限定されない。CMV-IEプロモーターは実際のプロモーター部分を含むことができるが、前記はエンハンサー部分と結合していてもいなくてもよい。EP-A-260148、EP-A-323597、米国特許5,168,062号、5,385,839号および4,968,615号をPCT出願WO87/03905と同様に参照することができる。グラム陰性細菌に由来する他のプロモーターもまた適切である。前記には、アビバクテリウム、ブルセラ、大腸菌、ヘモフィルス(例えばヘモフィルス・スイス)、サルモネラ(例えばサルモネラ・エンテリディス、ネズミチフス菌、サルモネラ・インファンチス)、シゲラ、パスツレラおよびリメイレラが含まれるが、ただしこれらに限定されない。
【0027】
プラスミドは他の発現制御エレメントを含むことができる。安定化配列、例えばイントロン配列を取り込むことは有利である。イントロン配列は、例えばhCMV-IEの最初のイントロン(PCT出願WO1,989/01036)、ウサギβグロビン遺伝子のイントロンII(van Ooyen et al. 1979)である。別の実施態様では、プラスミドは3'UTRを含むことができる。3'UTRは、アグロバクテリウムノパリンシンターゼ(Nos)3'UTRであるが、ただし前記に限定されない。
プラスミドおよびポックスウイルス以外のウイルスベクターのためのポリアデニル化(ポリA)シグナルに関しては、ウシ成長ホルモン(BGH)遺伝子のポリ(A)シグナル(US 5,122,458参照)、またはウサギβグロビン遺伝子ポリ(A)シグナル、またはSV40ウイルスのポリ(A)シグナルが有用であり得る。
“宿主細胞”は、外因性ポリヌクレオチド(例えば組換えプラスミドまたはベクター)の投与によって既に遺伝的に改変されているかまたは遺伝的に改変することができる原核細胞または真核細胞を意味する。遺伝的に改変された細胞というとき、前記用語は最初に改変された細胞およびその子孫の両方を指す。
【0028】
本発明の別の実施態様は、ドラッグレスプラスミドを含むグラム陰性細菌宿主株に関する。この実施態様のある特徴は、ドラッグレスプラスミドおよび宿主染色体の1つまたは2つ以上の非必須領域に挿入された異種ポリヌクレオチドを含むグラム陰性細菌宿主株に関し、この場合、前記異種ポリヌクレオチドは、リプレッサーにより厳密に調節され得るプロモーターに作動できるように連結されている。この実施態様の別の特徴では、宿主染色体に挿入された異種ポリヌクレオチドはsacB遺伝子である。この実施態様の別の特徴では、sacB遺伝子は、プロモーター(例えばλファージの右のプロモーター)に作動できるように連結される。sacB遺伝子およびプロモーターはマルチコピー(例えば2または3コピー)として宿主染色体に挿入できることは理解されよう。宿主株の非必須領域は、大腸菌染色体のdeAまたはpurN遺伝子であり得る。deAまたはpurN遺伝子は非必須であり、これら遺伝子の欠失は細菌の増殖速度に影響を与えない(J. Kim et al. Biochemistry. 46,44:12501-12511)。ある特徴では、sacB遺伝子およびプロモーターのカセットの1コピーが、deA遺伝子座またはpurN遺伝子座のどちらかに対立遺伝子置換によって挿入される。別の特徴では、sacB遺伝子およびプロモーターのカセットの2コピーが、deA遺伝子座またはpurN遺伝子座のどちらかに挿入される。さらに別の特徴では、sacB遺伝子およびプロモーターのカセットの2コピーが宿主染色体に挿入され、この場合sacB遺伝子およびプロモーターカセットの1つのコピーがdeA遺伝子座に挿入され、他方のコピーがpurN遺伝子座に挿入される。sacB遺伝子およびプロモーターのカセットは、プロモーターに作動できるように連結されたsacB遺伝子を含むポリヌクレオチドを指す。ある特徴では、sacB遺伝子およびプロモーターのカセットは、sacB遺伝子およびλファージの右のプロモーターを含む。別の特徴では、sacB遺伝子およびプロモーターのカセットは、配列番号:75に示す配列を有するポリヌクレオチドを含む。
【0029】
本明細書に開示する主題はさらに、免疫原またはタンパク質をコードする異種遺伝子を含むドラッグレスプラスミドを含むワクチンもしくは組成物、またはドラッグレスプラスミドを用いて発現させた免疫原またはタンパク質を含むワクチンもしくは組成物に関する。前記ワクチンまたは組成物はさらに医薬的に許容できる担体を含むことができる。
ある実施態様では、免疫原は、ネコの病原体、例えばネコヘルペスウイルス(FHV)、ネコカリシウイルス(FCV)、ネコ白血病ウイルス(FeLV)、ネコ免疫不全ウイルス(FIV)(ただしこれらに限定されない)、狂犬病ウイルスなどおよび前記の組み合わせから選択される。
別の実施態様では、免疫原は、イヌの病原体(狂犬病ウイルス、イヌヘルペスウイルス(CHV)、イヌパルボプイルス(CPV)、イヌコロナウイルス、レプトスピラ・カニコラ(Leptospira canicola)を含むが、ただしこれらに限定されない)、レプトスピラ・イクテロヘモラージアエ(L. ichterohaemorragiae)、レプトスピラ・グリッポチフォサ(L. grippotyphosa)、ボレリア・バーグドルフェリ(Borrelia burgdorferi)、ボルデテラ・ブロンキセプチカ(Bordetella bronchiseptica)など、および前記の組み合わせから選択される。
さらに別の実施態様では、免疫原は、ウマの病原体、例えばウマヘルペスウイルス(1型または4型)、ウマインフルエンザウイルス、破傷風、西ナイルウイルス、ストレプトコッカス・エクィ(Streptococcus equi)、ロドコッカス・エクィ(Rhodococcus equi)など、または前記の組合せから選択される。
【0030】
さらに別の実施態様では、免疫原は、ウシの病原体、例えば狂犬病ウイルス、ウシロタウイルス、ウシパラインフルエンザウイルス3型(bPIV-3)、ウシコロナウイルス、ウシウイルス性下痢ウイルス(BVDV)、口蹄疫ウイルス(FMDV)、ウシ呼吸器合胞体ウイルス(BRSV)、伝染性ウシ鼻気管炎ウイルス(IBR)、大腸菌、パスツレラ・マルトシダ(Pasteurella multocida)、パスツレラ・ヘモリチカ(P. haemolytica)、サルモネラ、クリプトスポリジウム(Cryptosporidium)など、および前記の組合せから選択される。
さらに別の実施態様では、免疫原は、ブタの病原体(例えばブタインフルエンザウイルス(SIV)、ブタシルコウイルス2型(PCV-2)、ブタ生殖器呼吸器症候群ウイルス(PRRS)、仮性狂犬病ウイルス(PRV)、ブタパルボウイルス(PPV)を含むが、ただしこれらに限定されない)、FMDV、マイコプラズマ・ヒオプニューモニアエ(Mycoplasma hyopneumoniae)、エリシペロトリクス・リューシオパシアエ(Erysipelothrix rhusiopathiae)、パスツレラ・マルトシダ、ボルデテラ・ブロンキセプチカ、大腸菌、ブルータングウイルス、アフリカウマ病ウイルス、リフトバレー熱、ニパ(Nipah)ウイルスなど、および前記の組合せから選択される。
【0031】
“抗原”または“免役原”という用語は互換的に用いられ、宿主動物で特異的な免疫応答をを誘発する物質を意味する。抗原は、免疫原性特性を有する挿入物を含む組換えベクター;宿主動物に提示されたとき免疫応答を誘発することができるDNAの断片またはフラグメント;タンパク質、ポリペプチド、ペプチド、エピトープ、ハプテンまたは前記の任意の組合せを含むことができる。また別には、免疫原または抗原は毒素または抗毒素を含むことができる。
本明細書で用いられるように、“医薬的に許容できる担体”および“医薬的に許容できるベヒクル”は相互に用いることができ、副作用を示さずに宿主に注射できる、ワクチン抗原収容のための液体ベヒクルを指す。当分野で公知の医薬的に許容できる適切な担体には、無菌的な水、食塩水、グルコース、デキストロースまたは緩衝溶液が含まれるが、ただしこれらに限定されない。担体は補助剤を含むことができ、前記には、希釈剤、安定化剤(すなわち糖類およびアミノ酸)、保存料、湿潤剤、乳化剤、pH緩衝剤、粘性強化添加剤、着色剤などが含まれるが、ただしこれらに限定されない。
【0032】
プラスミドに有益である(ただし絶対的に適切というわけではないが)第四アンモニウム塩を含む陽イオン性脂質は、有利には以下の式を有するものである:
【0033】
【化1】

【0034】
式中、R1は、12から18の炭素原子を有する飽和または不飽和の直鎖脂肪族ラジカルであり、R2は、2または3個の炭素原子を含む別の脂肪族ラジカルであり、Xは、アミンまたはヒドロキシル基(例えばDMRIE)である。別の実施態様では、陽イオン性脂質は、中性脂質(例えばDOPE)と結合することができる。
これらの陽イオン性脂質でとりわけ好ましいものはDNRIE(N-(2-ヒドロキシエチル)-N,N-ジメチル-2,3-ビス(テトラデシルオキシ)-1-プロパンアンモニウム(WO96/34109)であり、有利には中性脂質(有利にはDOPE(ジオレオイル-ホスファチジル-エタノールアミン))(Behr, 1994)と結合してDMRIE-DOPEを生成する。
有利には、アジュバントを含むプラスミド混合物は即席に、有利には調製物の投与と同時にまたは調製物の投与前近くに(例えば投与前近くまたは投与の前に)形成される。プラスミド-アジュバント混合物は、有利には投与前に十分な時間、例えば投与前に約10から約60分、例えば投与前にほぼ30分を与えて混合物が複合体を生じるように形成される。
DOPEが存在するときは、DMRIE:DOPE分子比は有利には約95:約5から約5:95、より有利には約1:約1、例えば1:1である。
DMRIEまたはDMRIE-DOPEアジュバント:プラスミド重量比は、約50:約1から約1:約10、例えば約10:約1から約1:約5、有利には約1:約1から約1:約2、例えば1:1から1:2である。
【0035】
別の実施態様では、医薬的にまたは獣医的に許容できる担体、賦形剤またはベヒクルは油中水エマルジョンであり得る。適切な油中水エマルジョンの例には、安定で4℃で液状である、油を基剤とする油中水ワクチンエマルジョンが含まれる。前記エマルジョンは、6から50v/v%、好ましくは12から25v/v%の抗原含有水相、50から94v/v%の油相(非代謝性油(例えばパラフィン油のような鉱物油)および/または代謝性油(例えば植物油)または脂肪酸、ポリオールまたはアルコールエステルを全部としてまたは部分として含む)、0.2から20p/v%、好ましくは3から8p/v%の界面活性剤(後者は全部としてまたは部分として含まれる)、または混合物としてポリグリセロールエステル(前記ポリグリセロールエステルは、好ましくはポリグリセロール(ポリ)リシンオレエート)またはポリオキシエチレンリシン油または他の水素添加ポリオキシエチレンリシン油を含む。油中水エマルジョンで用いることができる界面活性剤の例には、エトキシル化ソルビタンエステル(例えばポリオキシエチレン(20)ソルビタンモノオレエート(Tween80(登録商標))(AppliChem社(Cheshire, CT)から入手できる)およびソルビタンエステル(例えばソルビタンモノオレエート(Span 80(登録商標))(Sigma Aldrich(St. Louis, MO)から入手できる))が含まれる。さらに、油中水エマルジョンに関しては、米国特許6,919,084号(例えばその実施例8)を参照されたい(前記文献は参照により本明細書に含まれる)。いくつかの実施態様では、抗原含有水相は、1つまたは2つ以上の緩衝剤を含む食塩水溶液を含む。適切な緩衝溶液の例はリン酸緩衝食塩水である。有利な実施態様では、油中水エマルジョンは、水/油/水(W/O/W)三重エマルジョンである(例えば米国特許6,358,500号を参照されたい(前記文献は参照により本明細書に含まれる))。他の適切なエマルジョンの例は米国特許7,371,395号に記載されている(前記文献は参照により本明細書に含まれる)。
【0036】
ワクチンまたは組成物は、受容動物の齢、性別、体重、種および状態のような要件、および投与ルートを考慮しながら、医学または獣医学分野の業者によく知られた用量または技術によって投与することができる。投与ルートは、経皮、経粘膜投与(例えば経口、鼻内、肛門、膣投与)または非経口ルート(皮内、筋肉内、皮下、静脈内または腹腔内投与)であり得る。ワクチンまたは組成物は単独で投与できるが、また他の処置または治療と一緒にまたは連続して投与できる。投与形態には懸濁物、シロップまたはエリキシル、および非経口、皮下、皮内、筋肉内または静脈内投与用調製物(例えば注射可能投与物)、例えば無菌的懸濁物またはエマルジョンが含まれ得る。ワクチンまたは組成物は、スプレーとして投与するか、または食べ物および/または水に混合するか、または適切な担体、希釈剤または賦形剤(例えば無菌的な水、生理学的食塩水、グルコースなど)との混合物としてデリバ−することができる。組成物は、補助物質、例えば湿潤化もしくは乳化剤、pH緩衝剤、アジュバント、ゲル化もしくは粘性強化用添加剤、保存料、香料、着色剤などを、所望される投与経路および調製物に応じて含むことができる。標準的な薬学テキスト、例えば“Remington's Pharmaceutical Sciences”(1990)を参考にして、煩雑な実験を実施することなく適切な調製物を製造することができる。
【0037】
本明細書に開示する主題はさらに、以下の工程を含むドラッグプラスミドの製造方法に関する:1)宿主染色体の1つまたは2つ以上の非必須領域に対立遺伝子置換によって挿入された異種ポリヌクレオチドを含むグラム陰性細菌宿主株を操作する工程;2)cIリプレッサータンパク質をコードするポリヌクレオチドを含むDNAプラスミドを構築する工程;3)cIリプレッサータンパク質をコードする遺伝子を含むDNAプラスミドで細菌宿主株を形質転換する工程;4)形質転換細菌宿主株をショ糖の存在下において30℃から42℃の範囲の温度で増殖させる工程;および5)DNAプラスミドを回収する工程。
この実施態様のある特徴では、DNAプラスミドはさらに免疫原またはタンパク質をコードする異種ポリヌクレオチドを含み、このプラスミドでは異種ポリヌクレオチドは作動できるようにプロモーターに連結される。前記プロモーターは原核細胞または真核細胞で機能するプロモーター、例えばCMVプロモーターでもよい。
本明細書に開示する主題はさらに、以下の工程を含む、ドラッグレスプラスミドを用いてタンパク質または免疫原を製造する方法に関する:1)宿主染色体の1つまたは2つ以上の非必須領域に対立遺伝子置換によって挿入された異種ポリヌクレオチドを含むグラム陰性細菌宿主株を操作する工程;2)cIリプレッサータンパク質をコードするポリヌクレオチドおよび免疫原またはタンパク質をコードする遺伝子を含むDNAプラスミドを構築する工程;3)cIリプレッサータンパク質をコードするポリヌクレオチドおよび免疫原またはタンパク質をコードする遺伝子を含むDNAプラスミドで細菌宿主株を形質転換する工程;4)形質転換細菌宿主株をショ糖の存在下において30℃から42℃の範囲の温度で増殖させる工程;および5)免疫原またはタンパク質を回収する工程。
【0038】
この実施態様のある特徴では、免疫原またはタンパク質をコードする遺伝子は原核細胞で機能するプロモーターに作動できるように連結される。プロモーターはグラム陰性細菌由来のプロモーターでもよく、アビバクテリウム、ブルセラ、大腸菌、ヘモフィルス(例えばヘモフィルス・スイス)、サルモネラ(例えばサルモネラ・エンテリディス、ネズミチフス菌、サルモネラ・インファンチス)、シゲラ、パスツレラおよびリメイレラから単離されたプロモーターが含まれるが、ただしこれらに限定されない。
この実施態様のある特徴では、宿主染色体に挿入される異種ポリヌクレオチドはsacBタンパク質をコードする。別の特徴では、ショ糖濃度は、0%から約20%、約1%から約20%、約1%から約15%、約1%から約10%の範囲であり得る。さらに別の特徴では、ショ糖濃度は、約1%から約10%、約1%から約2%、約2%から約3%、約3%から約4%、約4%から約5%、約5%から約6%、約6%から約7%、約7%から約8%、約8%から約9%または約9%から約10%であり得る。さらに別の特徴では、ショ糖濃度は、約1%、約2%、約3%、約4%、約5%、約6%、約7%、約8%、約9%または約10%であり得る。
この実施態様のある特徴では、温度は約30℃から約42℃の範囲であり得る。別の特徴では、温度は、約30℃から約41℃、約30℃から約39℃、約30℃から約38℃、約30℃から約37℃の範囲であり得る。さらに別の特徴では、温度は、約30℃、約31℃、約33℃、約34℃、約35℃、約36℃、約37℃、約38℃、約39℃、約40℃または約41℃であり得る。
【0039】
“回収”という用語は、培養液から採集、抽出、採集または精製することを含むが、ただし前記に限定されない。
“単離された”生物学的成分(例えばポリヌクレオチド、DNAプラスミド、タンパク質または細胞小器官)とは、前記成分が通常存在する生物の細胞内の他の生物学的成分、例えば他の染色体性および染色体外DNAおよびRNA、タンパク質並びに細胞小器官から実質的に分離または精製されている成分を指す。“単離されている”ポリヌクレオチド(DNAプラスミドを含む)およびタンパク質には、標準的な精製方法によって精製されたポリヌクレオチドおよびタンパク質が含まれる(例えば以下を参照されたい:David W. Russell, Joseph Sambrook (2001), Molecular Cloning: a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory)。前記用語はまた、化学的合成と同様に組み換え技術によって調製されたポリヌクレオチドおよびタンパク質を包含する。
本明細書で用いられる“精製された”という用語は完全に純粋であることを要求せず、むしろ相対的用語として意図される。したがって、例えば、精製されたポリペプチドまたはポリヌクレオチド調製物は、当該ポリペプチドまたはポリヌクレオチドがその天然の環境で存在するよりも濃縮されている調製物である。すなわち、当該ポリペプチドまたはポリヌクレオチドは細胞性成分から分離されてある。“実質的に精製された”とは、細胞性成分または物質の少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、または少なくとも98%、またはそれ以上が取り除かれているようなものが意図される。同様に、ポリペプチドまたはポリヌクレオチドは部分的に精製されてあってもよい。“部分的に精製”とは、細胞性成分または物質の60%未満が除去されることを意味する。
【0040】
本発明はさらに免疫または防御応答を動物で誘発する方法を提供する。前記方法は、本発明の免疫学的組成物、ワクチンまたは組成物を動物に投与する工程を含む。誘引される免疫応答は、特に抗体および/または細胞性免疫応答、および特にガンマインターフェロン応答である。
特に、本発明は、病原微生物による動物の感染(例えばウイルス、細菌、菌類または原生動物寄生による感染)に対して免疫するか、または前記を予防するか、または前記によって引き起こされる症状を緩和するための方法を提供する。本発明の方法は脊椎動物で有用であり、前記脊椎動物にはヒト、イヌ科の動物(例えばイヌ)、ネコ科の動物(例えばネコ)、ウマ科の動物(例えばウマ)、ウシ科の動物(例えばウシ)およびブタのような動物(例えばブタ)だけでなく鳥類(ニワトリ、シチメンチョウ、アヒル、カモ、ウズラ、キジ、オウム、ウソ、タカ、カラスおよび走鳥類(ダチョウ、エミュおよびヒクイドリ)を含むが、ただしこれらに限定されない)が含まれるが、ただしこれらに限定されない。本発明の方法はまた魚類にDNAワクチンを提供するために有用である。
本発明の具体的な特徴では、これらの方法は、本発明にしたがって製造したワクチン組成物を投与することによって分娩前に妊娠雌動物をワクチン接種することから成る。これらの方法はさらに、ワクチン接種プロトコルにより誘引される防御抗体の誘導およびワクチン接種妊娠雌動物からそれらの子孫への前記防御抗体の伝達を含む。そのような母性抗体の伝達は続いて当該子孫を病気から防御する。
【0041】
本発明にしたがって製造したワクチン組成物の投与量は、ワクチン接種される動物の種、品種、齢、サイズ、ワクチン接種歴および健康状態に左右されるであろう。抗原濃度、追加のワクチン成分および投与経路(すなわち皮下、皮内、経口、筋肉内または静脈内投与)のような他の要件もまた有効な投与量に影響を与えるであろう。ワクチン投与量は、ワクチンの抗原濃度、投与経路、並びにワクチン接種される動物の齢および状態に基づいて容易に決定することができる。各抗原バッチを個々に計測することができる。また別には、MPD(最少防御用量)試験および他のスクリーニング方法と同様に種々の投与量による秩序だった免疫原性試験を用いて、煩雑な実験を実施することなく本発明のワクチン組成物のための有効な投与量を決定することができる。以下に提供する実施例から、本明細書に記載したワクチン組成物の使用に適切な投与量および体積は極めて明白であろう。もっとも重要な要素は、当該投与量によって自然感染に対して少なくとも部分的な防御効果が提供されることである(前記は自然感染に付随する死亡率および罹患率の減少によって立証される)。適切な体積は、当業者によって同様に容易に確認される。例えば、鳥類の種では、1用量の体積は約0.1mLから約0.5mL、有利には約0.3mLから約0.5mLであり得る。ネコ科、イヌ科およびウマ科の種では、1用量の体積は約0.2mLから約3.0mL、有利には約0.3mLから約2.0mL、より有利には約0.5mLから約1.0mLであり得る。ウシ類およびブタ類の種では、前記用量体積は約0.2mLから約5.0mL、有利には約0.3mLから約3.0mL、より有利には0.5mLから約2.0mLであり得る。
【0042】
早期に免疫応答を強化するために、または最後の投与から長期間が経過したときには、好ましくは、周期的な期間間隔での反復ワクチン接種を実施することができる。本発明のある実施態様得は、ワクチン組成物は非経口注射として(すなわち皮下、皮内または筋肉内に)投与される。組成物は1回の投与として投与されるか、または別の実施態様では、約2から約6週間、好ましくは約2から約5週間の間隔で与えられる約2回から約5回の反復投与で与えることが。しかしながら、当業者は、投与回数およびワクチン接種の間の間隔は、多数の要件(ワクチン接種動物の齢、動物の状態、免疫経路、1回あたりに利用可能な抗原量などを含むが、ただしこれらに限定されない)に左右されることを承知しているであろう。最初のワクチン接種の場合、前記期間は一般的に1週間よりも長く、好ましくは約2から約5週間であろう。以前にワクチン接種された動物の場合、好ましくは、約1年間隔のブースターワクチン接種(妊娠前および妊娠中)が実施される。
本発明はまた、無針注入器(例えばPigjet(登録商標)、Avijet(登録商標)、Dermojet(登録商標)またはBiojector(登録商標)(Bioject, Oregon, USA))を用いてワクチン組成物を投与すること意図する。当業者は、煩雑な実験を実施することなく、複数の要件(例えばワクチン接種される動物の種、動物の齢および体重など)に関して要求されるように前記注入器の詳細を調節することができる。
本発明はさらに、活性成分(例えば免疫原または医薬組成物)を含む第一のバイアルおよび第二のバイアルに本発明にしたがって作製した希釈剤を含むキットに関する。前記免疫原は凍結乾燥形、乾燥形であっても、または本明細書に記載した水溶液中に存在してもよい。
本発明は、以下の非限定的な実施例の手段によってこれからさらに説明されるであろう。
【実施例1】
【0043】
抗生物質フリープラスミド維持の概念
以下の実施例は、グラム陰性宿主細胞内に多数のプラスミドコピーを維持するための、3つの成分に基づく新規な概念を示す:
1.所定の培養条件下でグラム陰性細菌にとって有毒な生成物を発現するグラム陰性細菌(前記有毒な遺伝子は、細菌宿主染色体の1つまたは2つ以上の非必須領域に挿入される);
2.厳密に調節することができる構成的プロモーターの制御下にある有毒生成物をコードする遺伝子が細菌染色体に存在すること;および
3.宿主染色体上で有毒遺伝子に作動できるように連結されたプロモーターを調節する特異的リプレッサーがプラスミドから発現されること。
この設計では、グラム陰性細菌宿主が所定の培養条件下(例えばショ糖の存在下)にあるときは、前記宿主の増殖を制御するのはプラスミドの存在である。
第一の成分では、グラム陰性細菌は形質転換され、レバンスクラーゼをコードするsacB遺伝子を発現する(図1参照)。第二および第三の成分は、cIリプレッサー遺伝子生成物および右λプロモーターに関する。λバクテリオファージのcI遺伝子はλリプレッサーをコードする。
cIリプレッサータンパク質をコードするゲノム領域は免疫領域として知られている。免疫領域は図2に示されている。cI遺伝子生成物を発現するプラスミド系は、λプロモーターの制御下に配置されたsacB遺伝子の転写を阻害するはずである。そのような抑圧はショ糖に対する耐性を付与し、プラスミドの維持および宿主細胞の増殖の両方を可能にするはずである(図1参照)。
図3は“ドラッグレス”の概念を示す。この概念では、cIリプレッサー活性(プラスミドから発現される)が、細胞宿主染色体に存在するλPrプロモーター下に配置されたsacB遺伝子生成物の転写を阻害し、さらにショ糖の存在下における宿主大腸菌の生存活性は、プラスミドから発現されるλcIリプレッサータンパク質の十分なレベルによって担保される。cIリプレッサーとその特異的プロモーターとの結合は、30℃から37℃の温度で最適である。
特定の事例では(例えば温度感受性cI857リプレッサーを用いるとき)、cIリプレッサーは40℃より高い温度ではプロモーターと結合しないであろう。したがって、42℃への切換えは、cIリプレッサーのλPrプロモーターとの結合活性を阻害し、結果として宿主細胞をショ糖(より具体的にはレバン副生成物)に感受性にするはずである。ショ糖の存在下における42℃でのインキュベーションは完全系における機能性の有効性を立証するためのad hoc条件であり、この系の完全な機能性を支持する“負の証明”を構成する(図4および図6C参照)。実際、Catまたはショ糖のどちらかの存在下で増殖するpPB829またはpPB838のどちらかで形質転換した親細胞λPr::sacBpurNΩkanは、LBショ糖寒天に播種し42℃でインキュベートしたとき生存しなかった。このことは、親細胞の生存活性はcI遺伝子を含むプラスミドの維持に依存することを示している。
【実施例2】
【0044】
λプロモーター/レバンスクラーゼ遺伝子を保持する大腸菌宿主株の作製
λPr::sacBpurNΩkanカセットを含む2つの操作宿主株を調製した。この場合、カセット(ラムダプロモーター(λPr)の制御下にsacB遺伝子を含む)にカナマイシンで目印を付し、続いてedAまたはpurN遺伝子の対立遺伝子置換によって大腸菌染色体に導入する。これらの遺伝子は必須ではなく、前記の欠失は増殖速度に影響を与えない。形質転換細胞はショ糖に対して高度に感受性になった。
図8A−Bは、大腸菌染色体にそれぞれedAまたはpurN遺伝子の対立遺伝子置換によって挿入されるべきλPr::sacBpurNΩkanカセットが、結合PCRによって融合される前にPCR増幅された6つの別個の成分を含むことを示す。
図8Dは、λPr::sacBpurNΩkanカセットが、edAまたはpurN遺伝子の対立遺伝子置換によって大腸菌の染色体に挿入されたことを示す。このカセット挿入は、線状DNA鋳型(染色体遺伝子座の上流および下流の相同領域によってフランキングされた欠失遺伝子を含む)で大腸菌を形質転換することによって実施し、さらに組換え熟練大腸菌株(例えばrecD, recB recC sbcA変異体)に伝達した。親株の操作のための大腸菌株はStratageneのBJ5183株(ref# 200154)で、遺伝子型はendA1 sbcBC recBC galK met thi-1 bioT hsdR(Strr)であった。提示したように、この大腸菌野生型の最初の表現型はKanS、CatSおよびSucroseRであった。この最新の表現型は、抗生物質フリープラスミドの機能性の提示に関する更なる実験に用いられた親株のものであった。
図8Cは、Prλ::sacBΩkan大腸菌染色体にedA遺伝子の対立遺伝子置換によって挿入されるべきλPr::sacBΩcatカセットが、結合PCRによって融合される前にPCR増幅された6つの別個の成分を含んでいたことを示す。対立遺伝子交換によりedA遺伝子の欠失を標的とする新規なλPr::sacBΩcatカセットの目的は、2つのsacBカセットを発現する二重欠失変異体ΔedAΔpurNを操作することであった。
図8Eは、親株をcIプラスミド(pPB829またはpPB838)で形質転換したとき、細胞がカナマイシン耐性(KanR)、クロラムフェニコール耐性(CatR)およびショ糖耐性(SucroseR)になったことを示す(親株+cIプラスミド[pPB829またはpPB838])。図は、概念を立証するために、sacBがラムダプロモーターの制御下に配置され、構築物がedAまたはpurN遺伝子の対立遺伝子置換によって大腸菌の染色体に導入されたときに、操作された宿主株(ΔpurNλPr::sacB+Ωkan)または(ΔedAλPr::sacB+Ωkan)およびドラッグプラスミドが同時に存在することを示している。形質転換細胞は高度にショ糖感受性になった。cIリプレッサーを含むクロラムフェニコールの目印をもつプラスミド(pPB838またはpPB829)がΔpurNλPr::sacB+ΩkanまたはΔedAλPr::sacB+Ωkan株に導入されたとき、形質転換細胞はカナマイシン、クロラムフェニコール、およびショ糖耐性になった。
【0045】
λプロモーター(λPr)が、purN欠失のためにPB1232およびPB1233を用いて増幅された。
PB1232プライマー(配列番号:6):
(CCGAACAACGCGTGGTTATCGACACCGCAAGGGATAAATATCTAACACCG) および
PB1232プライマー(配列番号:7)
(CAAACTTTTTGATGTTCATATCCATCTGATCCTCTTCAAAAGGCCACCTG)
λプロモーター(λPr)が、edA欠失のためにPB1234およびPB1233を用いて増幅された。
PB1234プライマー(配列番号:8):
(GACGACAAATTTGTAATCAGGCGAGAGCACCGCAAGGGATAAATATCTAACACCG)。
λプロモーター(λPr)(配列番号:5)の増幅はDNA鋳型としてpLDR8プラスミド(ATCC#77357)を用いて実施した。sacB遺伝子(配列番号:3)は、λPr::sacBΩkanカセットの操作のために結合PCRを用いたときは、PB1192(配列番号:9)(ATGGATATGAACATCAAAAAGTTTGC)およびPB1193(配列番号:10)(AAACAAATAGGGGTTCCGCGCACATTTATTTGTTAACTGTTAATTGTCCTTG)プライマーを用い、DNA鋳型としてはpNB350(Merialの特許物)を用いて増幅した(図8A−B参照)。λPr::sacBΩcatカセットの操作のためには、リバースプライマーPB1320(配列番号:80)(GCCGATCAACGTCTCATTTTCGCCGTTAACAGATCTTTATTTGTTAACTGTTAATTGTCCTTG)がPB1193の代わりに用いられた。blaプロモーターは、PB1194(配列番号:11)(ATGTGCGCGGAACCCCTATTTG)およびPB1195(配列番号:12)(GACGTTTCCCGTTGAATATGGCTCATACTCTTCCTTTTTCAATATTATTGAAGC)プライマーを用い、DNA鋳型としてはpCMVβを用いて増幅された。カナマイシン耐性遺伝子は、PB1196(配列番号:13)(ATGAGCCATATTCAACGGGAAACG)およびPB1197(配列番号:14)(GAAAAACGCCAGCGGCAGAGCTGGCGCTTAGAAAAACTCATCGAGCATCAAATG)プライマーを用い、DNA鋳型としてはプラスミドpLL14(カナマイシン遺伝子の弱いプロモーターを含む、Merialの特許物)を用いて増幅された。その天然のcatプロモーターの制御下に配置されたクロラムフェニコール(cat)耐性遺伝子は、PB1321(配列番号:81)((AGATCTGTTAACGGCGAAAATGAG)およびPB1322(配列番号:82)(AAAACGCTACAAAAATGCCCGATCCTTTACGCCCCGCCCTGCCACTCATCGC)プライマーを用い、DNA鋳型としてはpPB829を用いて増幅された。purN遺伝子の5'フランキング領域は、PB1237(配列番号:15)(TTTGCGGCCGCTGGTGGTGGTCGCCATGTGCGTTAATGACC)およびPB1199(配列番号:16)(TATTCGATAACCACGCGTTGTTCGG)を用い、DNA鋳型としては大腸菌SCS1株のゲノムDNAを用いて増幅された。purN遺伝子の3'フランキング領域は、PB1200(配列番号:17)(GCGCCAGCTCTGCCGCTGGCGTTTTTC)およびPB1238(配列番号:18)(TTTGGATCCGCTGGTGGATATCATCAAGGCAGTAACGCAGAATG)プライマーを用い、DNA鋳型としては大腸菌SCS1株のゲノムDNAを用いて増幅された。edA遺伝子の5'フランキング領域は、PB1235(配列番号:19)(TTTGCGGCCGCTGGTGGTTGAGAACCAGGTGATTGAAGCGCC)およびPB1209(配列番号:20)(CTCTCGCCTGATTACAAATTTGTCGTC)プライマーを用い、DNA鋳型としてはSCS1のゲノムDNAを用いて増幅された。edA遺伝子の3'フランキング領域は、以下のプライマーPB1210(配列番号:21)(AGGATCGGGCATTTTTGTAGCGT)およびPB1236(配列番号:22)(TTTCTAGAGCTGGTGGCGACTACCGTGAATCCTGGCAACC)を用い、DNA鋳型としてはプラスミドpLDR8を用いて増幅された。
【0046】
大腸菌染色体に伝達されるべきλPr::sacBΩkanカセットまたはλPr::sacBΩcatのこれら6つの個々のPCR生成物を結合PCRに続いて融合させた(図8A、B、C)これら6つの個々の増幅フラグメントをPCR clean-upキット(Geneclean turbo kit; MP Biomedicals, CA, USA)を用いて精製し、さらに6つ全てのフラグメントを用いプライマーは使用せずに第2ラウンドのPCRをセットした。PCRの条件は以下のとおりであった:PCR混合物(最終25μL)、1μLの各PCRフラグメント、2μLのdNTP(各々2.5mM、5μLの5x PCR緩衝液、11.5μLの滅菌蒸留水、0.5μLのPhusionDNAポリメラーゼ(Finenzymes, Finland);PCRサイクル:98℃で30秒、(98℃で10秒、60℃で30秒、72℃で5分)を15サイクル、および72℃で10分。第三ラウンドのPCR増幅は、1μLの精製した第2ラウンドの生成物で、purN(対立遺伝子置換)のためにはPB1235およびPB1236を用い、edA(対立遺伝子置換)のためにはPB1237およびPB1238を用いて実施した。PCR条件は以下のとおりであった:PCR混合物(最終50μL)、1μLの第二のPCRフラグメント、1μLのdNTP(各々2.5mM、10μLの5x PCR緩衝液、37μLの滅菌蒸留水、0.25μLのフォーワードプライマー、0.25μLのリバースプライマー、0.5μLのPhusionDNAポリメラーゼ(Finenzymes, Finland);PCRサイクル:98℃で30秒、(98℃で15秒、60℃で30秒、72℃で4分)を35サイクル、および72℃で10分。最終的なPCR結合アンプリコン(purN対立遺伝子置換については4007bp、およびedA対立遺伝子置換については3407bp)は、アガロースゲルでチェックし、大腸菌の形質転換前に精製した。各結合PCRアンプリコンは制限分析によってチェックした。
標的遺伝子座(edAまたはpurN)の境界のDNA配列に相同な2つの長い領域によってフランキングされた、λPrプロモーター(配列番号:75)の制御下に配置されたsacB遺伝子およびblaプロモーターの制御下に配置されたKan耐性遺伝子をコードする線状DNAフラグメント(λPr::sacBΩkan)が、エレクトロポレーションによって大腸菌の染色体に組み込まれた。ほぼ300の形質転換候補体(ΔpurNλPr::sacBΩkanまたはΔedAλPr::sacBΩkanシングル変異体)が、選別圧としてカナマイシンを含むLB寒天プレートで得られた。20個のコロニーをランダムに採取し、カナマイシン含有プレートでストリーキングすることにより精製し、PCRによってedAまたはpurN遺伝子座へのλPr::sacBΩkanカセットの挿入を立証した(図9B参照)。PB1196およびPB1197を用いたPCRを実施し、染色体へのKanamycinの挿入をチェックした。PB1192およびPB1193を用いてPCRを実施し、染色体にsacB遺伝子が存在することを立証した。PB1204(配列番号:23)(GTGGTGCTTATTTCCGGCAACGG)およびPB1205(配列番号:24)(CCAGCCACGCGGCGTTTTCGTGC)を用いてPCRを実施し、染色体にpurN遺伝子が存在しないことを立証した。PB1214(配列番号:25)(GACCACCGGCCCGGTTGTACCGG)およびPB1215(配列番号:26)(CGGACCCGCGATCGCCTGCAGG)を用いてPCRを実施し、edA遺伝子が存在しないことを立証した(図9B参照)。PB1213(配列番号:27)(GGTGGATGGCGTCCATTTCTGTGC)およびPB1196プライマーを用いてPCRを実施し、edA遺伝子座の右側に正しくカセットが挿入されていることを立証した。PB1212(配列番号:28)(CAAAAGTGTTAAGCGGTAACCTG)およびPB1233プライマーを用いてPCRを実施し、edA遺伝子座の左に正しくカセットが挿入されていることを立証した。PB1203およびPB1196プライマーを用いてPCRを実施し、purN遺伝子座の右に正しくカセットが挿入されていることを立証した。PB1202およびPB1233プライマーを用いてPCRを実施し、purN遺伝子座の左に正しくカセットが挿入されていることを立証した(図9C参照)。全てのPCRチェックによって正しい遺伝子座への組込みが確認された。さらにまた、sacB遺伝子の機能性もまた図10Bに示すように確認された。この操作した親株ΔpurNλPr::sacBΩkanはショ糖を含むLB寒天プレートでは増殖できなかった(図10)。
標的遺伝子座(edA)の境界のDNA配列に相同な2つの長い領域によってフランキングされた、sacB遺伝子(λPrプロモーターの制御下に配置されている)およびCat耐性遺伝子(その天然のcatプロモーターの制御下に配置されている)をコードする線状DNAフラグメント(λPr::sacBΩcat)が、エレクトロポレーションによってΔpurNλPr::sacBΩkan大腸菌株の染色体に組み込まれた。ほぼ200の形質転換候補体(ΔpurNλPr::sacBΩkanΔedAλPr::sacBΩcatダブル変異体)が、選別圧としてクロラムフェニコールを含むLB寒天プレートで得られた。20個のコロニーをランダムに採取し、クロラムフェニコール上でストリーキングすることにより精製し、プライマーセットPB1212およびPB1213を用いたPCRによって、edA遺伝子座へのλPr::sacBΩcatカセットの挿入を立証した(図9C参照)。PCR生成物はシークェンシングによってチェックし、λPr::sacBΩcatとの同一性を立証した。
宿主株の操作は図8Dに示されている。図8Eは、宿主株(1つのsacBカセット[λPr::sacBΩkan])およびドラッグレスプラスミドの同時存在を示す。最初の大腸菌の遺伝子型はKanS、CatS、SucroseRである。大腸菌ΔpurN Prλ::sacBΩkanの染色体へのカセットの組込みは、新規な遺伝子型KanR、CatS、SucroseSを生じた。cIリプレッサーを保持するプラスミドの導入によって、最終的な遺伝子型KanR、CatR、SucroseRが生成された。
【実施例3】
【0047】
cIリプレッサー発現カセットを保持するプラスミドの作製(pPB838-pPB844からpPB847-pPB885-pPB896)
プラスミド構築の要旨
pPB828から誘導したプラスミド、例えばpPB829およびpPB844-pPB847を作製した。前記は、それ自身のプロモーターまたはカナマイシン遺伝子の弱いプロモーター(P1)のどちらかの制御下にあるλファージのcI ORFを含む。カナマイシン遺伝子の弱いプロモーター(P1)の制御下に配置されたcI遺伝子を含むpPB383プラスミドを中間プラスミドの市販ベクターpMCS5を用いて作製した(pMCS5ではアンピシリン耐性遺伝子がクロラムフェニコールアセチルトランスフェラーゼ遺伝子(cat)に置換されていた)(図19)。
pPB844およびpPB845プラスミドはcI遺伝子(P1プロモーターの制御下に配置されている)を含む。前記cI遺伝子は、cat遺伝子を含むpVR1012誘導体(pVR1020または1012プラスミド、VICAL Inc.(Luke et al. 1997;Hartikka et al. 1996;例えば米国特許5,846,946号および6,451,769号参照))であるpPB828でクローニングされた。pPB844およびpPB845プラスミドは、cI遺伝子の当該プラスミド中での向きが異なっている(図20−21)。
pPB846およびpPB847プラスミドはcI遺伝子(それ自身の(天然の)プロモーター(配列番号:30)の制御下に配置されている)を含む(前記cI遺伝子は、cat遺伝子を含むpVR1012誘導体であるpPB828でクローニングされた)。これらのプラスミドは、cI遺伝子の当該プラスミド中での向きが異なっている(図22−23)。
pPB885はpPB838プラスミド(cI遺伝子はカナマイシン遺伝子の弱いプロモーター(P1)の制御下に配置されている)から調製され、前記では、クロラムフェニコールアセチルトランスフェラーゼ遺伝子(cat)はAvrII制限消化によって除去された(図24)。
pPB829から誘導されたプラスミド、例えばpPB896は、カナマイシン遺伝子の弱いプロモーター(P1)の制御下に配置されたcI遺伝子を含む。pPB896プラスミドは、真核細胞CMVプロモーターの制御下に配置されたGFPマーカー遺伝子を含む(図25)。
これらのプラスミドをドラッグレス系で異種ポリヌクレオチド、対象の遺伝子(トランスジーン)のクローニングのためにベクターとして用いた。各プラスミドは抗生物質耐性遺伝子(cat)を含み、前記は、固有の制限酵素によって切り出して除去し、適切な大腸菌sacB+株でショ糖の存在下において抗生物質選別圧(例えばクロラムフェニコール)によることなくプラスミドの増殖を可能にすることができる。
これらのプラスミドの抗生物質耐性遺伝子(cat)は概念の証明のためにのみ用いられた。最終的なドラッグレスプラスミドではこのcat遺伝子は最終的に除去される(図13−15および実施例4を参照)。
プラスミドpLDR8(ATCC No#77357)由来のcI遺伝子はリプレッサータンパク質cI857変異体を生成する(NCBI: AB248924, Cloning vector pND707, C.A. Love et al. Gene 1996, 17; 176(1-2):49-53)。
【0048】
A.弱いプロモーターP1の制御下に配置されたcIリプレッサーORFを含むプラスミドpPB829の構築
pPB828プラスミド:(pVR1012系プラスミド+Cat遺伝子)(図17)
pPB828誘導プラスミド、例えばpPB829を作製した。前記はカナマイシン遺伝子の弱いプロモーター(P1)の制御下にあるλファージのcI ORFおよび固有のEcoRI消化部位によって切り出すことができるCat遺伝子を含む。クロラムフェニコールアセチルトランスフェラーゼ(cat)遺伝子を含み、pPB828プラスミドを生じる、pVR1012系発現ベクターを作製した。pPB627(Merialの特許物質)由来のcat遺伝子はpNB335(Merialの特許物質)の誘導体であった(pNB335はそれ自体最初のプラスミドpSW23T(クローニングプラスミドGenBankアクセッション番号AY733066)の誘導体である)。cat遺伝子に対応するDNAフラグメント(798bp)(DNAについては配列番号:31、タンパク質については配列番号:32)は、プライマーPB1184(配列番号:33)(GAATTCCGGTCCGGGCGAAAATGAGACGTTGATCGGC3)(EcoRI部位(下線部)を含む)およびPB1185(配列番号:34)(CCTAGGCTGTGTTAATTAAGGCGCGCCGAATTCCGGTCCGTTACGCCCCGCCCTGCCACTCATCGC)(EcoRI部位(下線部)を含む)を用い、鋳型のpNB350およびPhusionTM High-Fidelity DNAポリメラーゼ(Finnzymes, 02150 Espoo, Finland)を用いてPCRによって入手した。得られたPCR生成物(832bp)をpCRblunt Topoベクター(Invitrogen, CA, USA)に連結し、プラスミドpCRblunt/PB1184-1185(4351bp)を得た(図16)。pCRblunt/PB1184-1185の実体は制限分析(PvuII消化)によって確認した。Cat遺伝子に対応するDNAフラグメントは、pCRblunt/PB1184-1185プラスミドのEcl136IIおよびEcoRVによる酵素消化によって入手した。897bpフラグメントをpVR1012系プラスミド(先にMscIおよびStuIで消化しておいた(3303bp))に連結してプラスミドpPB828を作製した(図17)。pPB828の実体は制限分析によって確認した(挿入物のサイズ決定のためにはPvuIIおよびNcoI消化、および遺伝子catが良好に除去されていることを立証するためにはEcoRI消化)。
pPB829プラスミド:(pPB828プラスミド+P1::cI遺伝子)(図18)
P1プロモーター(カナマイシン耐性遺伝子の弱いプロモーター)の制御下にあるcI遺伝子を含む構築物は融合PCRを用いて入手した。弱いプロモーターP1に対応するDNAフラグメントは、プライマーPB1186(配列番号:35)(TCATACCAGGCCTAGGTGATACGCCTATTTTTATAGGTTAATG)およびPB1187(配列番号:36)(AACACCCCTTGTATTACTGTTTATG)を用い、鋳型としてpLL14(Merialの特許出願US2005/0164946を参照されたい)、さらにPhusion DNAポリメラーゼを用いたPCRによって入手した。cI遺伝子(配列番号:1)に対応するDNAフラグメントは、プライマーPB1188(配列番号:37)(AATACAAGGGGTGTTATGAGCACAAAAAAGAAACCATTAACAC)(5'末端のP1配列の相補性領域(下線部)を含む)およびPB1189(配列番号:38)(CCGGAATTCGGCGCGTCAGCCAAACGTCTCTTCAGGCCACTG)を用い、鋳型としてpLDR8(ATCC#77357)、さらにPhusion DNAポリメラーゼを用いたPCRによって入手した。2つのPCR生成物(それぞれ151および729bp)を精製し、第二のPCR工程(PB1186およびPB1189プライマーおよびPhusion DNAポリメラーゼ(Finnzymes, Finland)を使用)で鋳型として用いた(図24参照)。2つのPCR生成物(それぞれ151および729bp)を精製し、AscIおよびAvrIIで消化したpPB828と連結して、プラスミドpPB829(4970bp)を作製した(図9A)。連結は、Clontech(Takara Bio Europe, St-Germain-en-Laye, France)の“In-Fusion PCR Cloning Kit”(Cat No. 740590.250)を用いて実施した。HindIII消化分析に続いてクローンpPB829を選別した。cI遺伝子およびP1プロモーターの配列の完全性は、プラスミドpPB829のシークェンシングによって確認した。
pPB896プラスミド:(pPB829プラスミド+P1::cI遺伝子+GFP)(図25)
GFPマーカー遺伝子は、真核細胞CMVプロモーター(配列番号:43)の制御下に配置した。GFP遺伝子は、pCG105プラスミド(真核細胞CMVプロモーターの制御下に配置されたGFP遺伝子を有するpPB828誘導プラスミド、Merialの特許物)のNotI/BglIIの両方による制限消化により調製した。この消化GFP遺伝子を、先にNotI/BglIIで切断しておいたpPB829プラスミドでクローニングしpPB896プラスミドを生成した。
【0049】
B.中間プラスミド(pPB838)およびP1プロモーター(カナマイシン遺伝子の弱いプロモーター)の制御下にあるcI遺伝子を含むプラスミドpPB844-pPB845の構築
pPB837.1プラスミド:(pMCS5プラスミド+Cat遺伝子)
Cat遺伝子に対応するDNAフラグメントは、プライマーPB1182(配列番号:39)(AGATCTGTTAACGGCGAAAATGAGACGTTGATCGGC)(5'末端にBglII部位を含む)およびPB1183(配列番号:40)(GTCGACGTTAACTTACGCCCCGCCCTGCCACTCATCGC)(5'末端にSalI部位を含む)、鋳型としてpPB791(プラスミドpNB350の誘導体)、およびPhusion DNAポリメラーゼを用いてPCRによって入手した。
PCRプライマーはpNB350のcat配列を土台にして設計した。3つの別個のPCR生成物(779bp)をプールし、pCRIIベクターに連結してプラスミドpCRII+PB1182-1183(4734bp)を入手した。pCRII+PB1182-1183のクローンをSalI/BglII消化に続いて選別した。DNAフラグメントA(2950bp)を得るために、pMCS5プラスミド(MoBiTec, Germany)をSalIおよびBglIIで消化した。pCRII+PB1182-1183をSalIおよびBglIIで消化し、フラグメントSalI-BglIIをアガロースゲルから単離した(797bp:フラグメントB)。フラグメントAおよびBを連結してプラスミドpPB837.1(3747bp)を生成した。
pPB837.2:blaプロモーターをもたない(pMCS5+Cat)
blaプロモーター領域を含まないプラスミドpPB837.2を作製した。プラスミドpPB837.1をNaeIおよびXmnIで消化し、3300bpのフラグメントNaeI-XmnIを単離し、アガロースゲルで精製した(フラグメントA)。このフラグメントAを連結してpPB837.2(3300bp)を得た。BglI消化に続いてクローンpPB837.2を選別した。
pPB838プラスミド(blaプロモーターのないpMCS5+Cat)+P1::cI(図22)
P1プロモーター(カナマイシン耐性遺伝子の弱いプロモーター)の制御下にあるcI遺伝子を含む構築物は融合PCRを用いて入手した。弱いプロモーターP1に対応するDNAフラグメントは、プライマーPB1186およびPB1187、鋳型としてPLL14、およびPhusion DNAポリメラーゼを用いてPCRにより入手した。cI遺伝子に対応するDNAフラグメントは、プライマーPB1188およびPB1189、鋳型としてpLDR8(ATCC#77357)、およびPhusion DNAポリメラーゼ(Finnzymes. Finland)を用いてPCRによって入手した。2つのPCR生成物(それぞれ151および729bp)を精製し、第二のPCR工程でPB1186およびPB1189プライマー並びにPhusion DNAポリメラーゼとともに鋳型として用いた。2つのPCR生成物(880bp)をプールし、pCRIIbluntベクターに連結してプラスミドpCRblunt+PB1186-1189(4400bp)を得た。クローンpCRblunt+PB1186-1189をEcoRV/SpeI消化に続いて選別した。
EcoRV-SpeIフラグメントA(936bp)を得るために、pCRblunt+PB1186-1189をEcoRVおよびSpeIで消化した。pPB837.2をEcoRVおよびSpeIで消化し、フラグメントEcoRV-SpeIをアガロースゲルから単離した(3464p:フラグメントB)。フラグメントAおよびBを連結してプラスミドpPB838(4216bp)を生成した。HindIII消化に続いてクローンpPB838を選別した。cI遺伝子およびP1プロモーターの配列の完全性はシークェンシングによって確認した。
pPB885プラスミド(blaプロモーターのないpMCS5+Cat)+P1::cI(図24)
このプラスミドはpPB838プラスミドから誘導した(前記では、クロラムフェニコールアセチルトランスフェラーゼ遺伝子(cat)はAvrII制限消化によって除去された)(図24)。pPB885プラスミドは、カナマイシン耐性遺伝子の弱いプロモーター(P1)の制御下にcI遺伝子を含む。pPB885プラスミドは、ΔpurNλPr::sacBΩkan大腸菌宿主株の形質転換に続いて、実施例4(下記に示す)に記載するように10%ショ糖補充LB寒天プレート上での直接選別によって単離された。
プラスミドpPB844-pPB845(図23−24)
P1プロモーターの制御下のcI遺伝子に対応するDNAフラグメントは、プライマーPB1186およびPB1263(配列番号:41)(TCAGCCAAACGTCTCTTCAGGCCAC)、鋳型としてpPB838、並びにPhusion DNAポリメラーゼを用いてPCRによって入手した。4つの別個のPCR生成物PB1186-PB1263(864bp)をプールし、pCRbluntベクターに連結してプラスミドpCRblunt+PB1186-1263(4383bp)を入手した。pCRblunt+PB1186-1263のクローンをHindII消化に続いて選別した。
pCRblunt+PB1186-1263プラスミドをEcoRIで消化し、末端をクレノーポリメラーゼで充填して平滑端フラグメント(929bp:フラグメントA)を生成した。pPB828プラスミドをXmnIで消化し、得られたXmnI-XmnIフラグメント(フラグメントB:4135bp)を精製した。フラグメントAおよびBを連結してプラスミドpPB844またはpPB845を生成した。候補物をBamHI/HindIII消化によってスクリーニングした。pPB844プラスミド(図20参照)では、cI遺伝子は、ベクターのMCS(マルチクローニング部位)でクローニングされるべき対象のトランスジーンとCMVプロモーター(配列番号:43)の制御下で同じ向きに存在する。pPB845プラスミド(図21参照)では、cIは反対方向に向いている。
【0050】
C.その天然のプロモーターをもつcI遺伝子プラスミドpPB846-pPB847の入手
それ自身のプロモーター(cIの天然のプロモーター:配列番号:30)の制御下のcI遺伝子に対応するDNAフラグメントは、プライマーPB1266(配列番号:42)(GCTGACTCATACCAGGCACGCACGGTGTTAGATATTTATCCC)およびPB1263、鋳型としてpLDR8、並びにPhusion DNAポリメラーゼを用いてPCRによって入手した。
pPB846-pPB847プラスミド
4つの別個のPCR生成物PB1266-PB1263(777bp)をプールし、pCRbluntベクターに連結してプラスミドpCRblunt+PB1266-1263(4296bp)を入手した。pCRblunt+PB1266-1263のクローンをHindII消化に続いて選別した。
pCRblunt+PB1266-1263プラスミドをEcoRIで消化し、末端をクレノーポリメラーゼで充填して平滑端フラグメント(842bp:フラグメントA)を生成した。pPB828プラスミドをXmnIで消化し、得られたXmnI-XmnIフラグメント(フラグメントB:4135bp)を精製した。フラグメントAおよびBを連結してプラスミドpPB846またはpPB847を生成した。候補物をBamHI/HindIII消化によってスクリーニングした。pPB846プラスミド(図22参照)では、cI遺伝子は、ベクターのMCS部位でクローニングされるべき対象のトランスジーンとCMVプロモーターの制御下で同じ向きに存在する。pPB847プラスミド(図23参照)では、cI遺伝子は反対方向に向いている。
D.構築の結果
図7は、ドラッグレスプラスミドの調製に用いられたcIリプレッサーを保持するpPB838およびpPB829の構築を示す。両プラスミドはクロラムフェニコール(CatR)遺伝子で目印を付されていた。これらのプラスミドのCat遺伝子は概念の証明のためにのみ要求されたが、最終的には除去することができる(図13−15および実施例4を参照されたい)。
【実施例4】
【0051】
大腸菌宿主細胞のcIプラスミドによる形質転換およびショ糖寒天プレートでの直接選別
クロラムフェニコール(Cat)の目印をもつcIプラスミドによって形質転換した細胞を選別圧として抗生物質を使用することなく寒天プレートでスクリーニングできる実験的方法は有効であることが示された(図13−15参照)。大腸菌λPr::sacB+ΔpurNΩkanのコンピテント細胞を1μgのpPB838またはpPB829プラスミドで形質転換し、LB液体ブロスにて30℃で3時間インキュベートした。希釈した(10-3から10-4)形質転換細胞(pPB838またはpPB829プラスミドで)の100μLアリコットを10%のショ糖を含むLB寒天プレートに播種した。10-3から10-4の範囲のこれら希釈物は本物の形質転換細胞(例えばcIプラスミドを有する)の選別に適切であるように思われる。なぜならば、ショ糖含有LB寒天プレートで増殖するショ糖耐性細胞の98−100%がCatRであったからである。このことは、これら形質転換細胞はcIプラスミド(pPB838またはpPB829)を有することを示唆している。
図13Aは、大腸菌λPr::sacB+ΔpurNΩkanのコンピテント細胞が、1μgのpPB838またはpPB829プラスミドで形質転換され、ドットスポット(各ドットに4μL)の前にLB液体ブロスにて30℃で3時間インキュベートされたことを示す。図は、クロラムフェニコール(15μg/mL)またはショ糖(10%)を含むLB寒天プレートでのLog希釈(未希釈から6Log希釈の範囲)を示す。大腸菌λPr::sacB+の非形質転換コンピテント親株細胞を同様な条件下での標準細胞増殖として用いた。期待したとおり、標準細胞はcatおよびショ糖のような選別プレートでは増殖しなかった。それにもかかわらず、長期間(3日間)のインキュベーションのために、いくつかの偶発的ショ糖耐性変異体がこの条件下で出現した。このレベルの偶発的変異体は、本物の形質転換細胞のスクリーニングについて考慮されるべきベースラインを規定する。この実験条件下では、2Log差がcatおよびショ糖で増殖する形質転換細胞で観察された。前記各規定のコンピテント細胞は、一時的に細胞壁の脆弱を示す。形質転換細胞をエレクトロポレーション後にLB液体培地でインキュベーションすることによって、エレクトロポレーションに続く前述の細胞壁の脆弱並びに細胞壁および細胞膜の誘発孔の復活が可能になる。コンピテント細胞は、λPr::sacB+ΔpurNΩkanカセットの挿入のために遺伝的にショ糖に感受性であるので、ショ糖の存在下での形質転換細胞の回復は、Catでの細胞の回復と比較して2Log低下した。これらの観察は、100μLの形質転換細胞を選別プレートに播種することによって確認された(図13B参照)。
図13Bは、catおよびショ糖上での形質転換有効率を決定し、さらにLBショ糖寒天プレートで増殖できる本物の形質転換細胞のスクリーニングの成功のために考慮すべき偶発的変異体出現率のベースラインをより厳密に規定するために、100μLの形質転換細胞および非形質転換標準細胞を適切なLog希釈を用いてcatまたはショ糖含有LB寒天プレートに播種したことを示す。以前のドットスポット細胞実験(図13A参照)と一致して、偶発的ショ糖耐性変異体は、形質転換細胞の10-3希釈をショ糖含有LB寒天プレートに播種したときは検出できなかった。以前に示したように(図13A参照)、この条件下ではcatおよびショ糖上での選別で形質転換効率に2Log差が存在した。10-3から10-4の範囲の希釈は、ショ糖含有LB寒天に播種したとき、本物の形質転換細胞(例えばcIプラスミドを有する)の選別に適切であるように思われた。
図14A−Cは、選別圧として抗生物質を使用することなく、プラスミド(pPB829およびpPB838)で形質転換した細胞を選別する実験的方法の有効性を示す。
図14Aは、cIプラスミドを有する本物の形質転換細胞のスクリーニングの有効性を立証するためにレプリカプレートアッセイを用いた実験の概要を示す。図13Bに示したものと同様に、コンピテント親細胞λPr::sacB+ΔpurNΩkanをcIプラスミド(pPB829およびpPB838)で形質転換し、30℃で3日間インキュベートする前に適切なLog希釈物を選別(Catまたはショ糖)LB寒天プレートに播種した。その後、20から50の範囲のCFU(コロニー形成単位)を示したプレートのレプリカを、30℃でさらに3日間インキュベートする前にCatまたはショ糖またはその組合せを含む別の新しいLB寒天プレートで作成した。理論的には、Cat耐性コロニーはcIプラスミドを増殖させる細胞と一致するはずである。
図14Bは、CatR CFUは、レプリカプレートアッセイ後にショ糖LB寒天プレートおよびCat/ショ糖LB寒天プレートで得られた数と同一であることを確認した実験結果を示す。図14B(パネルB)に示すように、CatR CFUは、i)全てがショ糖耐性、ii)全てがCatおよびショ糖の組合せに耐性であり、全ての親株は形質転換後にcIプラスミドを保持することを示している。パネルCは、ショ糖上で直接選別した10-3および10-4希釈の形質転換細胞はまた、ショ糖耐性であり、さらにCatおよびショ糖の組合せに耐性であることを示し、これらショ糖耐性CFUはまたcIプラスミドを含む形質転換細胞と一致することを示す。実際、非形質転換親細胞をショ糖含有LB寒天プレートに播種したとき、ショ糖耐性CFUは、ショ糖およびCatの組合せ上で複製させたとき期待したように増殖しなかった(クロラムフェニコール耐性はpPB829またはpPB838プラスミドのどちらかによって持ち込まれた表現型の属性であるからである)。
図14Cは、レプリカプレートアッセイ(図14B参照)から得られた統計学的データ、より具体的には、抗生物質を使用することなく選別されたcIプラスミド保有コロニーのパーセンテージを示す。この実験条件下では、cIプラスミド保有コロニーのパーセンテージは98%から100%の範囲である。
【実施例5】
【0052】
cIプラスミドを維持できることを示す、ショ糖が存在するまたは存在しない種々の培養条件
大腸菌の染色体に挿入されたλPr::sacB+カセットの機能性をチェックするために、 sacBを発現する大腸菌細胞、ΔpurNλPr::sacB+Ωkanをショ糖の存在下または非存在下で増殖させた。
図4はこの不活化系の“ショ糖感受性”を示す(前記不活化系では、42℃への切換えは、cIリプレッサーのλPrプロモーターとの結合活性を阻害し、宿主細胞をショ糖感受性にする)。cIリプレッサータンパク質(プラスミドから発現される)は、宿主染色体に位置するλPrプロモーターの制御下に置かれた有毒なsacB遺伝子生成物の転写を阻害する。ショ糖の存在下での宿主大腸菌細胞の生存活性は、プラスミドから発現されるλcIリプレッサータンパク質の十分なレベルによって担保される。図5は、ショ糖存在下での宿主細胞と抗生物質フリーpDL1プラスミドとの組合せによるドラッグレスプラスミド(pDL:DrugLess Plasmid)の生成を示す。概念の証明のための実験計画はさらに図6A、6Bおよび6Cに示される。
ショ糖の存在下または非存在下で増殖させたときのsacB発現宿主細胞、ΔpurNΩλPr::sacB+Ωkanの生理学的特徴を表1に示す。
表1:ショ糖の存在下および非存在下で増殖させたときのsacB+宿主細胞(ΔpurNΩλPr::sacB+)の生理学的特徴
【0053】
ショ糖の存在下または非存在下で増殖させたときの、pDL1プラスミド保持および非保持sacB発現宿主細胞(λPr::sacB+)の生理学的特徴を表2に示す。
表2:ショ糖の存在下および非存在下で増殖させたときのpDL1プラスミド保持および非保持(λPr::sacB+)宿主細胞の生理学的特徴
図10−11は、遺伝的に操作した宿主株は30−37℃でショ糖に対して高度に耐性であるが、42℃では増殖は認められないことが確認された結果を提供する。図10は、pPB829プラスミドまたはpPB838プラスミドを保持する大腸菌宿主株(ΔpurNλPr::sacB+Ωkan)の2回目の継代はショ糖の存在下において32℃で良好に増殖したことを示す。sacB遺伝子発現は、両方のcIプラスミドから合成されたcI遺伝子生成物によって完全に抑圧された。プラスミドの維持は100%有効で(CatRのコントロールに対して)、細胞はショ糖の存在下で生存できた。42℃では、cI遺伝子生成物は不活化された。プラスミドは維持されず、細胞はショ糖の存在下で死滅した。このことは、ショ糖の存在下における細胞の生存活性は機能的cIプラスミドの存在に帰せられることを示している。
【実施例6】
【0054】
抗生物質選別圧を使用せずに5継代にわたってcIプラスミドが安定であることを示す培養条件およびショ糖耐性の偶発的変異体の欠如
ドラッグレスプラスミドを生成したら、安定性およびショ糖またはCat含有新鮮ブロス培養液中での数代にわたるそれらの強固さについて試験した。
図11は、増殖細胞中でのpPB829およびpPB838プラスミドの安定性を示す。前記安定性には、クロラムフェニコールまたはショ糖による選別中に明白な遺伝的再編成が存在しないことが含まれる。ショ糖およびクロラムフェニコールの存在下で増殖する培養に匹敵し得るプラスミドの収量および維持が5連続継代の後でも認められた。抗生物質系および非抗生物質系は、プラスミドの維持および増殖細胞からの選別に関しては少なくとも等価であった。
図12は、細胞を30℃で増殖させたときは、親大腸菌に挿入されたλPr::sacBカセットで偶発的変異は発生しないことを示す。この観察は、細胞をショ糖の存在下で増殖させたとき、そのような大腸菌の遺伝的背景におけるcIプラスミドの安定性をはっきりと示している(図10参照)。この変異発生率実験は、親株(purNが欠失しsacB遺伝子を発現する)を用いて実施した。30℃では18時間インキュベートした後で変異は発生しないことが示された。染色体にsacB+カセットを保持する親細胞のクローン培養をショ糖含有LBで増殖させた。細胞を1.0のODで採集し、100μLアリコットをショ糖含有LB寒天プレートに播種した。sacB遺伝子の存在は細胞死を引き起こすので、偶発的変異発生率は、30℃での一晩インキュベーション後のショ糖耐性コロニーのスコアによって判定した。sacB+カセットを保持するがcIプラスミド(pPB829およびpPB838)で形質転換されていない標準細胞はショ糖耐性となり、スコア判定のために、ショ糖補充LB寒天プレートに播種(100μL)する前にOD1.0の培養を5Log希釈することが必要であった。一晩(18時間インキュベーション)培養の後で親細胞培養(ΔpurNλPr::sacBΩkan)に関してショ糖耐性コロニーは検出されず、一方、pPB829およびpPB838プラスミドで形質転換したΔpurNλPr::sacBΩkanの5Log希釈は増殖した。細胞を37℃でインキュベートしたとき、36時間がいくつかのショ糖耐性コロニー(ΔpurNλPr::sacBΩkan)の検出に必要であった。
37℃でインキュベートしたときには、偶発的変異発生は、36時間後に1.3 x 105細胞当たり1変異発生率で生じた。ショ糖の存在下で増殖している細胞でのcIプラスミドの維持は30℃での一晩インキュベーションで有効であるが、一方、親株の増殖は検出されなかった。
【実施例7】
【0055】
前述の1つのsacBカセット保有大腸菌宿主株(ΔpurNλPr::sacBΩkan)に第二のsacBカセット(ΔedAλPr::sacBΩcat)を付加することによる、ショ糖に対する偶発的変異発生率低下のための親大腸菌宿主株の改良
図8Cおよび図9Dの両図は、2つのsacBカセット(ΔpurNλPr::sacBΩkan ΔedAλPr::sacBΩcat)を保有する大腸菌の新規な親宿主株の操作態様を示す。
図10B−Cは、二重sacBカセット大腸菌株(ΔpurNλPr::sacB+ΩkanΔedAλPr::sacB+ΩCat)は、30℃および42℃でインキュベートしたとき、ショ糖の存在下で高度に感受性である(増殖しない)が、一方、1つのsacBカセットを有する大腸菌株に関しては10%ショ糖に播種したときでさえ、いくつかの偶発的変異体(ΔpurNλPr::sacBΩkan)が出現したことを示す。
図10Cはまた、1つのsacBカセットをもつ大腸菌(最終ショ糖濃度10%)と比較して二重sacBカセット大腸菌株が要求する最低ショ糖濃度(最終2%)を示す。さらにまた、低いショ糖%および最高温度(例えば42℃)で示されるように、2つのsacBカセットをもつ大腸菌株はショ糖に対してはるかに感受性のようである。このことは、約37℃の温度および約2%の(またはわずかに低い)ショ糖濃度が、増殖細胞でcI遺伝子リプレッサーを保持するプラスミドを維持するための最適条件と一致し得ることを示唆する。
図12B−Cは、2つの(1つではなく)sacBカセットの存在は、30℃から37℃の範囲の温度でショ糖感受性に対する強固さの向上を付与することを示す(しかもなお2%ショ糖が最適である)。2つのsacBカセットを含むプラスミドで形質転換した大腸菌細胞、BJ5183ΔpurNΩλPr::sacBKmΔedaΩλPr::sacBCatクローン#1およびBJ5183ΔpurNΩλPr::sacBKmΔedaΩλPr::sacBCatクローン#2、並びに1つのsacBカセットを含むプラスミドで形質転換した大腸菌細胞、BJ5183ΔpurNΩλPr::sacBKmクローン#16およびBJ5183ΔedaΩλPr::sacBKmクローン#11を約1.0のOD600まで増殖させた。各培養の(10-5希釈)約100μLを、0%、2%および4%のショ糖を含むLB/寒天プレートにCFUスコア判定のために播種した。このプレートを30℃から37℃でインキュベートした。37℃でのインキュベーションは、2%および4%のショ糖の存在下における二重sacBカセット発現親株の強固さを判定するために用いた。図12B−Cおよび下記の表3−7は、大腸菌染色体中の1つのsacBカセットの存在は、同様の温度(例えば30℃および37℃)での2%から4%の範囲のショ糖濃度では強固さは低下することを示す。このプレートアッセイで確認されたように、2つのsacBカセットをもつ大腸菌株に関するショ糖に対する変異発生率は、37℃でインキュベートしたとき最低ショ糖濃度(2%)では検出できなかったが、1つのsacBカセットをもつ大腸菌株に関する変異発生率は、3.8 10-6から5 10-5の範囲であった。別個の実験セットによって、2つのsacBカセットをもつ大腸菌株の変異発生率は、2%のショ糖の存在下において37℃でインキュベートしたときほぼ5.10-10であることが示された。
【0056】
表3
表4
表5
表6
表7
1*:BJ5183ΔpurNΩλPr::sacBKmΔedaΩλPr::sacBCatクローン#1
2*:BJ5183ΔpurNΩλPr::sacBKmΔedaΩλPr::sacBCatクローン#2
3*:BJ5183ΔpurNΩλPr::sacBKmクローン#16
4*:BJ5183ΔedaΩλPr::sacBKmクローン#11
【実施例8】
【0057】
抗生物質選別圧の非存在下で得られるプラスミドの維持および収量(cIプラスミドは宿主株内において高コピー数で維持される)
sacBを発現しcIプラスミド(pPB829またはpPB838)を保持する大腸菌宿主細胞を抗生物質選別圧の非存在下(例えばショ糖の存在下)で、またはコントロールとしてCatの存在下で増殖させた。
図11では、プラスミド収量は2および5継代で決定した。30℃で維持した培養からプラスミドを抽出し、ゲル電気泳動に付した。HindII制限消化は、増殖中およびCatまたはショ糖による選別中にpPB829またはpPB838で明白な遺伝的再編成は発生しなかったことを示している。さらにまた、Catまたはショ糖選別圧下でのcIプラスミド選別中に、同様なcIプラスミド維持が認められた。
図15A−Bは、培養におけるcIプラスミド(pPB829およびpPB838、sacBカセットの1コピーを含む)の安定性および維持の有効性を示す。前記安定性には、クロラムフェニコールまたはショ糖による選別後に遺伝的再編成が発生しないことが含まれる。ショ糖またはクロラムフェニコールの存在下で増殖させたcIプラスミドの効率的なプラスミド維持および安定性が観察された。さらにまた、プラスミド収量は、pPB829またはpPB838で形質転換した親宿主株を選別圧としてショ糖の存在下において30℃で増殖させたとき、選別圧としてCatの存在下で培養したときよりもはるかに高かった(約700/細胞)。プラスミドの維持および収量は極めて効率的であり、コピー数は、Catの存在下で培養した細胞と比較して2倍多かった。宿主細胞をショ糖の存在下で培養したときのこのcIプラスミド収量の向上は、cI発現を制御する弱いプロモーター(P1)によるものであった。実際、カナマイシン遺伝子の弱いプロモーター(P1)の制御下でのcIリプレッサーの発現は、λPrプロモーターの制御下に置かれたsacB遺伝子の毒性をより良好に無効にするためにプラスミド収量に対して正の作用を有していた。
以前に示したように(図11A参照)、抗生物質系および非抗生物質系は、プラスミドの維持およびcIプラスミド保持増殖細胞の選別に関しては等価である。図15Bは、プラスミド収量(コピー数)は、pPB829またはpPB838で形質転換した親宿主株を選別圧としてショ糖の存在下で増殖させたとき、選別圧としてCatの存在下で培養したときよりもはるかに高かった(約700/細胞)ことを示している。プラスミドの維持および収量は極めて効率的であり、コピー数は、抗生物質選別、例えばCatの存在下で培養した細胞と比較して2倍多かった
【実施例9】
【0058】
cIプラスミドに抗生物質遺伝子が存在しない場合のプラスミドの維持および収量:1つのsacBカセットおよび2つのsacBカセットをもつ大腸菌株間の比較解析
1つのsacBカセットおよび2つのsacBカセットをもつ大腸菌株(それぞれΔpurNλPr::sacB+ΩkanおよびΔpurNλPr::sacB+ΩkanΔedAλPr::sacB+Ωcat)の両方におけるcatフリーpPB885プラスミドの安定性および収量を示す。ΔpurNΩλPr::sacBKm(1つのsacBカセット)およびΔpurNΩλPr::sacBKmΔedaΩλPr::sacBcat(2つのsacBカセット)の両親株をcIcatフリーpPB885プラスミドで形質転換し、ショ糖プレートへの播種とその後の培養の前に3時間インキュベートした。プラスミド収量は、2つのsacBカセットの大腸菌株および1つのsacBカセットの大腸菌株に対してそれぞれ5%および10%ショ糖補充LBで3継代して決定した。37℃で維持した培養からpPB885を抽出し、ゲル電気泳動に付した。PvuII制限消化は、増殖およびショ糖選別の間にどちらのpPB885でも明白な遺伝的再編成は発生しないことを示した(図15D参照)。このゲルが示すように、10%および5%のどちらのショ糖選別後でもそれぞれ1つのsacBカセットおよび2つのsacBカセットをもつ両大腸菌で、明白な遺伝的再編成は生じなかった。
図15Eは、1つのsacBカセットおよび2つのsacBカセットをもつ両大腸菌株(それぞれΔpurNλPr::sacB+ΩkanおよびΔpurNλPr::sacB+ΩkanΔedAλPr::sacB+Ωcat)でのcatフリーpPB885プラスミドの収量を示す。興味深いことに、プラスミド収量(コピー数)は、1つのsacBカセットをもつ大腸菌株から得られたプラスミド収量(ほぼ142/細胞)と比較して2つのsacBカセットをもつ大腸菌株ではるかに高かった(ほぼ278/細胞).この実験で得られたプラスミド収量は以前の比較実験で得られたプラスミド収量より低いように思われる(図15B参照)。これは主として以下の事実によるものである:i)pPB885による形質転換後に、最良のクローンを同定するためにより多くのクローンを評価する必要があること、およびii)さらにcatフリーpPB885プラスミドで形質転換した2つのsacBカセットをもつ大腸菌株の増殖条件は最適ではなかったということ。実際、我々は、このもっとも新しい株の最適増殖条件は約37℃で約2%(またはわずかに低い)ショ糖であることを示した。しかしながら、この実験結果は、最適で強固な条件下では、プラスミド収量は、1つのsacBカセットをもつ大腸菌株と比較して2つのsacBカセットをもつ大腸菌株でより多量であり得ることを示唆している。
【実施例10】
【0059】
CHO細胞への抗生物質フリーcIプラスミドのトランスフェクション
図15Eおよび下記の表8は、抗生物質フリープラスミド(pPB896/選別圧としてショ糖)および抗生物質プラスミド(pCG105/選別圧としてCat)のCHO細胞へのプラスミドトランスフェクション効率(GFP発現)を示す。発現されるGFPに関してこれら2つの選別圧(ショ糖対抗生物質としてCat)間で明白な差異は認められない。pCG105プラスミド(CMVプロモーターの制御下に配置されたGFP遺伝子を有するpPB828誘導プラスミド(Merialの特許物))およびpPB896(pPB829プラスミド+P1::cI遺伝子+GFP)プラスミドをCHO細胞のトランスフェクションに用いて、従来の抗生物質プラスミドに対して抗生物質フリープラスミド概念の機能性を評価した。この実験は、1つのsacBカセットをもつ大腸菌宿主株の培養から単離したプラスミドを用いて実施した。pCG105は選別圧としてカナマイシンを用いて大腸菌で増殖させ、一方、pPB896は選別圧として10%ショ糖を用いて大腸菌で増殖させた。DNAプラスミドを抽出し、同じ濃度で投与した。
図15は、リポフェクタミン2000を用いたCHO-K1細胞の一過性トランスフェクション後の、pCG105およびpPB896によりコードされるGFPタンパク質のin vitro発現を示す。直径6cmのプレート中の90%コンフルエンシーのCHO-K1細胞に、各々5μgのプラスミドおよび10μLのリポフェクタミンを用い製造業者の指示にしたがいながらトランスフェクションを実施した。トランスフェクション後に、1%のSVFを含むMEM-glutamax培養液で細胞を培養した。培養を採集し、蛍光顕微鏡(図15)およびフローサイトメトリー(FACS Calibur(Becton Dickinson))(表8)を用いて解析した。
図15は、トランスフェクションを実施した両方のCHO-K1細胞で同様なGFPエピフルオレセンスが可視化されたことを示す。クロラムフェニコールおよびショ糖の選別圧下でpCG105およびpPB896プラスミドは比較可能な効率でCHO-K1細胞にトランスフェクトできた。
表8は、pCG105およびpPB896プラスミドのCHOトランスフェクション効率を示す。この表に示すように、CHO細胞にpCG105(59%のGFP発現)およびpPB896(48%のGFP発現)をトランスフェクトしたとき、比較可能なGFPタンパク質発現CHO細胞の%が測定された。これらの結果は、CHOのトランスフェクションおよび最終的にはDNAワクチンでの応用のためにショ糖系抗生物質フリープラスミドを使用することの有効性を立証した。
表8
これらの実験は、染色体上に存在する有毒遺伝子の制御はプラスミド上に存在するリプレッサーにより達成できることを示す。この系は、抗生物質の選別圧の非存在下で完全に機能を発揮する。この系は宿主細胞の5継代にわたって安定である。この系は、宿主細胞が細胞当たり高いプラスミドコピー数を達成することを可能にする。この系は、最少合成培養液と完ぺきに適合し得る。
これまで本発明の好ましい実施態様を詳細に説明してきたが、上記で規定した本発明は、その多くの変型が本発明の範囲を逸脱することなく可能であるので、上述の記載に示す具体的な内容に限定されないことは理解されよう。
本明細書に引用または参照した全ての文書(“本明細書引用文書”)、および本明細書引用文書中に引用または参照された全ての文書は、製造業者の指示、説明書、製品明細書および本明細書に記載したまたは参照により含まれる任意の文書中の任意の製品のプロダクトシートとともに、参照により本明細書に含まれ、本発明の実施で利用することができる。
【技術分野】
【0001】
参照による一体化
本出願は、米国仮特許出願61/180,755号(2009年5月22日出願)の利益を主張する。
本発明は、抗生物質選別圧を用いることなくグラム陰性細菌でプラスミドを維持および製造する方法に関する。さらにまた、本発明は、製造したドラッグレスプラスミド、並びに前記ドラッグレスプラスミドを含む処方物および/または組成物、および前記ドラッグレスプラスミドを用いて製造したタンパク質または免疫原を含む処方物および/または組成物、および前記処方物および/または組成物を宿主に投与する方法に関する。本発明は、前記ドラッグレスプラスミドを含むグラム陰性細菌に関する。
【背景技術】
【0002】
今日まで、医薬として使用するために安全であり強力かつ効率的であると考えられるプラスミドDNAベクターは存在しない。誘導されたプラスミド中に抗生物質耐性遺伝子が存在することが、新しい遺伝子療法およびDNAワクチン利用における短所の1つである。
プラスミドは、染色体DNAから分離し、染色体DNAとは別個に複製することができる染色体外DNA分子である(G. Lipps, ed. (2008). Plasmids: Current Research and Future Trends. Caister Academic Press. ISBN978-1-904455-35-6)。天然では、プラスミドは細菌中に通常存在するが、時に真核細胞生物で見出される。それらは、適切な宿主内で自律的に複製することができる、伝達可能な遺伝的エレメント(またはレプリコン)と考えられる。プラスミドは裸のDNAであり、新しい宿主に伝達されるために遺伝物質を包み込むために必要な遺伝子をコードしない。したがって、宿主から宿主へのプラスミドの伝達には、接合または遺伝的エレメントの意図的取り込みを可能にする形質転換による宿主遺伝子発現の変化によって直接的で機械的な伝達が要求される(G. Lipps, 2008)。
遺伝子療法およびワクチン免疫を目的とするプラスミドDNA(pDNA)の使用は、ヒトおよび動物のヘルスケアで顕著な潜在能力を有する新規な技術である(J. Mairhofer et al. (2008) Biotechnol J, 3:83-89)。さらにまた、プラスミドは遺伝学および生物工学研究室で重要なツールとして供される(そのような研究室では、プラスミドは個々の遺伝子の増幅または発現に通常的に用いられる)(David W Russell, Joseph Sambrook (2001), Molecular cloning: a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory)。
【0003】
プラスミドは、細菌集団内で水平的遺伝子伝達のためのメカニズムを提供し、典型的にはある環境状況下で選別優位性を提供する。例えば、プラスミドは、天然には存在しない抗生物質に対する耐性を提供する遺伝子を保持することができる。抗生物質の他にまた別のマーカーも存在する。例えば、プラスミドによって産生されるタンパク質は毒素として作用することができる(前記毒素はまたある環境状況下で選別優位性を提供する)。プラスミドはまた、窒素元素を固定する能力または扱いにくい有機化合物を分解する能力をもつ細菌を提供することができる(前記能力はまたある環境状況下で選別優位性を提供する)(G. Lipps, 2008)。
抗生物質耐性遺伝子を用いる選別の場合、細胞を該当する抗生物質に対して耐性にするタンパク質を生成する遺伝子が挿入されたプラスミドが調製される。次に、このプラスミドは形質転換と称されるプロセスによって細菌に挿入される。続いて、この細菌は該当する抗生物質に曝露される。前記プラスミドは細菌を前記抗生物質に対して耐性にするので、前記プラスミドのコピーを取り込んだ細菌をのみがこの抗生物質から生き残る。
問題の遺伝子は抗生物質耐性マーカーを含むプラスミドを用いてデリバ−される。そのような遺伝子は、典型的にはマルチクローニング部位(MCS(またはポリリンカー))に挿入される。抗生物質耐性遺伝子が発現され、発現されたタンパク質は当該抗生物質を分解する。このようにして、抗生物質は、改変された細菌のみを選別するフィルターとして機能する。ここでこれらの細菌を大量に増殖させ採集して溶解し(しばしばアルカリ溶解方法が用いられる)、問題のプラスミドを単離することができる。
【0004】
医薬におけるこの抗生物質の成功は今では問題となってしまった。多くの細菌(感染症の病原体を含む)がすでに耐性を有し、もはや特定の抗生物質で制御することができない。抗生物質はヒトおよび動物の医薬であまりにも頻繁に用いられた。さらにまた、抗生物質が成果の強化物質として動物飼料に長期間にわたって添加されたという事実ははるかに重大である。この慣行は今ではほとんどが禁止されているが、ばらまかれた抗生物質は、対応する耐性遺伝子をもつ細菌に生存優位性を与えた。さらにまた、細菌中の耐性遺伝子は、可動性DNAユニット上にしばしば配置され、前記ユニットは異なる種間で交換され得る。
このような背景に対して、細菌はマーカー遺伝子を同化し、最終的に、これまで処方されてきた抗生物質が効かない病原体を生じる懸念がある。環境中の微生物(例えば病原体)への遺伝子の伝達も起こり得よう(D.B. Murphy, S.L. Epstein, Guidance for Industry: Guidance for humann somatic cell therapy and gene therapy, Food and Drug Administration, Rockville 1998)。別の安全性に関する懸念は、ヒトの染色体への抗生物質耐性遺伝子の組み込みの可能性である(H.A. Smith, D.M. Klinman (2001) Curr Opin Biotechnol, 12:299-203)。
さらにまた、これらの遺伝子の構成的発現は細菌宿主細胞に不必要な代謝負荷を負わせるので、そのような遺伝子は、プラスミドの製造過程に相当な影響を及ぼす可能性がある(R.M. Cranenburgh et al. (2001) Nucleic Acids Res, 29, e26;A. Rozkov et al. (2006) Enzyme Microb Technol 39:47-50)。これらの遺伝子を除去することによりプラスミドのサイズを縮小することは、pDNAの安定性および発酵プロセスによって得られる収量の改善をもたらすであろう(M.A. Smith et al. (1998) Can J Microbiol, 44:351-355)。
【0005】
したがって、公衆/消費者の潜在的受容リスクのために、規制当局の従来の推奨にしたがい、最終生成物(商品)(裸のDNAワクチン)で抗生物質耐性遺伝子を使用しないことが当分野で絶対に必要である。食品医薬局(FDA)および国連世界保健機関(WHO)は、DNAワクチンの品質を担保し、さらに感染症を予防するために抗生物質耐性マーカーの使用を規制している。同様に、EUデリバレートレリース・ディレクティブ(2002年から発効)は、“ヒトの健康または環境に有害な影響を与えるおそれがある遺伝的改変生物における抗生物質耐性マーカーの使用の段階的廃止”を要求している。
慣習的マーカーの短所は実際的研究においてすら明白になりつつある。例えば、バクトフェクション技術の産業利用のために抗生物質フリーデリバリー系をもつことが所望される。バクトフェクション技術は、侵入細菌を用いる真核細胞へのプラスミドDNAのデリバリーである。さらにまた、発酵過程中の不要な代謝負荷を軽減する技術的な要求が存在する(前記はより高いODおよびDNAプラスミドのより高い収量をもたらすであろう)。
患者の腸内細菌に抗生物質耐性遺伝子をまき散らすという懸念に対して、また別の選別方法が設計された。前記には、栄養要求性補完、リプレッサー滴定、タンパク質系解毒薬/毒素選別系、およびRNA系選別性マーカーの使用が含まれる(以下を参照されたい:J.A. Williams et al. Plasmid DNA vaccine vector design: Impact on efficacy, safety and upstream production, Biotechnol Adv (2009). Doi:10.1016/j.biotechadv.1009.02.003)。
【0006】
R.M. Cranenburghらは、複雑な培地中で組換えプラスミドの抗生物質フリー選別および安定的維持を促進する2つの新規な大腸菌(Escherichia coli)株(DH1lacdapDおよびDH1lacP2dapD)の構築を報告した。前記の株は、必須の染色体遺伝子(dapD)をlacオペレーター/プロモーターの制御下に含んでいた(R.M.Cranenburgh et al. 2001)。IPTG(dapDの発現を誘導する)またはDAPが補充されなければ、これらの細胞は溶解するが、しかしながらこれらの株がlacオペレーターを含むマルチコピープラスミドで形質転換されると、前記オペレーターはLacIリプレッサーを競合的に滴定してlacプロモーターからdapDを発現させる。したがって、リプレッサー滴定選別によって任意の培地で増殖する形質転換体の能力によって、形質転換体を簡単に単離および増殖させることができる。抗生物質耐性遺伝子または他のタンパク質発現配列はプラスミドで要求されず、プラスミドの選別に抗生物質は不要である。
最近Mairhoferらは、いずれの選別マーカーおよび他の付加配列もプラスミドで使用することなくプラスミドを選別および維持するために供される細菌宿主株の設計を探索した。プラスミドの複製阻害因子RNA IがRNA-RNAアンチセンス反応によって増殖に必須の遺伝子の翻訳を抑圧することができるように、いくつかの細菌株を改変した(J. Mairhofer et al. (2008) Biotechnol J 3:83-89)。必須遺伝子(murA)をリプレッサータンパク質(tetR)がその発現を妨害するように改変した(J. Mairhofer et. al., 2008)。プラスミド(したがってRNA I)が存在する場合のみ、tetRが弱まりmurAが発現される。前記著者らは、多様な改変大腸菌株によって市場で入手可能な種々のプラスミドを選別できることを報告した。彼らはさらに、いずれの選別マーカーも含まないミニマリスト的プラスミドを設計した。
本出願におけるいずれの文書の引用または認定も、そのような文書が本発明の先行技術として利用可能であることを容認するものではない。
【発明の概要】
【0007】
抗生物質フリープラスミドが、前記抗生物質フリープラスミドを含むグラム陰性細菌と同様に提供される。免疫原またはタンパク質をコードする異種遺伝子を含む抗生物質フリープラスミドを含む組成物および前記抗生物質フリープラスミドを用いて発現された免疫原またはタンパク質を含む組成物もまた提供される。細菌染色体の非必須領域に1つまたは2つ以上の異種ポリヌクレオチドを含むように、グラム陰性細菌が操作される。前記抗生物質フリープラスミドはリプレッサーをコードするポリヌクレオチドを含み、前記リプレッサーは細菌の染色体上の異種ポリヌクレオチドの発現を調節する。前記抗生物質フリープラスミドは、さらに免疫原またはタンパク質をコードする1つまたは2つ以上のポリヌクレオチドを含むことができる。
本発明の方法は、抗生物質フリープラスミドを製造する方法および前記抗生物質フリープラスミドを用いて哺乳動物細胞に外来遺伝子を導入するための方法を含む。
これらの実施態様および他の実施態様は、以下の詳細な説明によって開示されまたは以下の詳細な説明から明白であり、さらにそれら実施態様は以下の詳細な説明に包含されている。
例示の手段により提供する以下の詳細な説明(記載した具体的実施態様に本発明を限定しようとするものではない)は、付随の図面(参照により本明細書に含まれる)と一緒にして理解されよう。
【図面の簡単な説明】
【0008】
【図1】ドラッグレス概念の第一の構成要素を示す。前記は、ショ糖を含む培地にプレートしたとき形質転換細菌の急速な死をもたらす、グラム陰性細菌のレバンスクラーゼをコードするsacB遺伝子のクローニングである。
【図2】ドラッグレスまたは抗生物質フリー概念の第二および第三の構成成分を示す。前記はλファージの免疫領域である。ここで、cIはcIリプレッサーをコードし、croはCroタンパク質をコードし、Nは転写抗ターミネーターをコードし、cIIはcIアクチベーターをコードし、cIIIはcIIIプロテアーゼ阻害因子をコードし、OL1、2および3並びにOR1、2および3はオペレーターであり、PLおよびPRは右側および左側プロモーターであり、PREはリプレッサー確立のためのプロモーターであり、PRMはリプレッサー維持のためのプロモーターである。
【図3】“ドラッグレス”概念の理論的大要を示す。ここで、cIリプレッサー活性(pDL1(DrugLess Plasmid)プラスミドから発現される)は、宿主細胞染色体上に存在するλPrプロモーター下に配置された有毒sacB遺伝子生成物の転写を阻害し、ショ糖存在下の宿主大腸菌細胞の生存能力は、プラスミド由来のλcIリプレッサータンパク質の十分な発現レベルによって担保される。
【図4】不活化された系の“ショ糖感受性”を示す。この系では、42℃の温度への切換えは、cIリプレッサーのλPrプロモーターとの結合活性を阻害し、宿主細胞をショ糖に感受性にする。
【図5】宿主細胞株と抗生物質フリーpDL1プラスミドの合体(pDL1による形質転換から生じる)およびショ糖を含む培地での増殖(ドラッグレスプラスミドの産生をもたらす)を示す。
【図6】A−Cは実験計画を示す。図6Aは大要を提供する。図6Bは、ΔpurN(またはΔedA)親株はカナマイシン耐性であり、30℃−37℃でインキュベートしたときそのショ糖感受性を付与するSacB+を構成的に発現することを示す。ΔpurN(またはΔedA)親株をpPB829またはpPB838プラスミドのどちらかで形質転換したとき、これらの細胞は、30℃−37℃でインキュベートしたときショ糖の存在下で生存し増殖する能力を獲得する。図6Cは、pPB829またはpPB838プラスミド(cIプラスミド)のどちらかで形質転換したΔpurN(またはΔedA)親株は、カナマイシン、ショ糖またはクロラムフェニコールを含むLB培地で同様に増殖できることを示す。ショ糖存在下でプレートしたとき、30−37℃から42℃への温度の切換えは細胞死を引き起こす。
【図7】クロラムフェニコールアセチルトランスフェラーゼ(CatR)をコードする遺伝子で目印を付された、λcI遺伝子を含む両プラスミド(pPB829およびpPB838)を示す。pPB838はpMCS5の誘導体で、前記誘導体ではクロラムフェニコールアセチルトランスフェラーゼ(cat)がアンピシリン耐性遺伝子と入れ代わっている。このプラスミドは、カナマイシン遺伝子の弱いプロモータ(P1)の制御下に配置されたcI遺伝子を含む。pPB829はcat遺伝子を含むpVR1012プラスミド誘導体である。このプラスミドはcI遺伝子を含み、前記は弱いプロモーター(P1)の制御下に配置されている。ΔpurN(またはΔedA)親大腸菌株に導入されたとき、これらのプラスミドはショ糖の存在下での細胞の増殖を可能にする。クロラムフェニコールは概念の明白な立証を可能にする。
【図8】A−Eは宿主株の操作を示す。図8Aおよび8BはλPr::sacBΩkanカセットの作製を示す。前記カセットは、それぞれedAまたはpurN遺伝子の対立遺伝子置換によって大腸菌染色体に挿入される。図8Cは、ダブルsacBカセット大腸菌株を作製するために、ΔpurNΩλPr::sacB Km大腸菌宿主株にedA遺伝子を対立遺伝子置換するために用いられる完全なsacBカセット(ΔpurNΩλPr::sacBcat)の、PCRおよび結合PCRによる操作を示す。
【図9】A、BおよびCは、宿主株の操作、すなわち大腸菌のedAおよびpurN欠失のデモンストレーションのためにPCRの使用を示す。図9Aは、2つのPCR生成物(それぞれ151および729bp)を部分として示す。それら生成物を精製し、さらにPB1186およびPB1189プライマー並びにPhusion DNAポリメラーゼとともに第二のPCR工程で鋳型として用いる。図9BはPCRチェックを示し、λPr::sacB+Ωkanカセットの大腸菌染色体への導入を示す。図9Cは、λPr::sacB+ΩkanカセットがedAおよび/またはpurN遺伝子のいずれかにおける対立遺伝子置換によって遺伝子座に正確に挿入されることを示す。図9Dは、二重sacBカセット宿主株の操作、すなわち対立遺伝子置換による大腸菌のedAおよびpurNの欠失のデモンストレーションのためにPCRの使用を示す。PCRチェックは、λPr::sacB+ΩkanおよびλPr::sacB+ΩCatカセットの両方が、シークェンシングによって確認されるように、大腸菌の染色体の特定の遺伝子座に正確に導入されることを示す。図9Dに示すように、レーン1はBJ5183wt、レーン2はBJ5138ΔpurN、レーン3はBJ5138ΔedA、レーン4はBJ5183ΔpurNΔedAで希釈せずに1μL、レーン5はBJ5183ΔpurNΔedAで1/10希釈、レーン6は水で0bp(コントロール)、AはBJ5183wt、BはBJ5138ΔpurN、CはBJ5138ΔedA、DはBJ5183ΔpurNΔedAで希釈せずに3μL、EはBJ5183ΔpurNΔedAで希釈せずに1μL、FはBJ5183ΔpurNΔedAで1/10希釈、GはBJ5183ΔpurNΔedAで1/50希釈、Hは水で0bp(コントロール)である。
【図10】Aは、宿主株(ΔpurNλPr::sacB+Ωkan)はショ糖に対して高度に感受性であること(増殖しない)、pPB829またはpPB838プラスミドで形質転換した細胞はショ糖の存在下において30℃で良好に増殖したこと、各cIプラスミドから合成されたcI遺伝子生成物によってsacB遺伝子発現は完全に抑圧されたこと、およびクロラムフェニコールの存在下で増殖する同様な細胞を用いて平行して実施したコントロール実験によって明確に示されるようにプラスミドの維持は100%有効であったことを示す。42℃への温度上昇は、cI遺伝子生成物の不活化およびショ糖またはクロラムフェニコールのどちらかを含むLB寒天にプレートされた細胞の死をもたらす。この厳しいプラスミド維持実験は、ショ糖またはクロラムフェニコールの存在下で細胞を2回続けて継代して増殖させた後実施された。図10B−Cは、二重sacBカセット大腸菌株(ΔpurNλPr::sacB+Ωkan ΔedAλPr::sacB+ΩCat)は、30℃および42℃でインキュベートしたときショ糖の存在下で高度に感受性である(増殖しない)が、一方、ショ糖上にプレートされたとき1つのSacBカセットをもつ大腸菌株とともにいくつかの偶発的変異体が出現することを示している。図10Cはまた、1つのSacBカセットをもつ大腸菌株と比較した二重sacBカセット大腸菌株が要求する最低のショ糖濃度を示す(10%から最終の2%まで)。
【図11】増殖細胞中でのpPB829およびpPB838プラスミドの安定性を示す。
【図12】Aは、λPr::sacB 1つのsacBカセットでは30℃で偶発的変異が発生しないことを示す。図12B−Cは、2つのsacBカセット(1つではなくて)の存在は、30℃から37℃の範囲の温度ではショ糖感受性についてより強力な厳格性を付与することを示している(それでもなお2%ショ糖が最適である)。大腸菌染色体に1つのsacBカセットが存在する場合、同様な温度で2%から4%の範囲のショ糖濃度でより低い厳格性が存在する。この平板培養アッセイで確認されたように、2つのsacBカセットをもつ大腸菌株のショ糖に対する変異率は、37℃でインキュベートしたとき最低ショ糖濃度(2%)では検出不能であったが、1つのsacBカセット大腸菌株の変異率は3.8 x 10-6から5 x 10-5の範囲であった。別個の実験セットによって、2つのsacBカセットの大腸菌株における変異率は、2%ショ糖の存在下においては37℃でインキュベートしたときほぼ5 x 10-10であることが示された。
【図13】A−Bは、クロラムフェニコール(Cat)の目印を有するcIプラスミドで形質転換した細胞を選別圧力として抗生物質を使用することなく寒天プレート上でスクリーニングすることができる実験手順を示す。
【図14】A−Cは、cIプラスミドのみで(pPB829およびpPB838)形質転換した細胞の選別に対する、選別圧力として抗生物質を使用しない前記実験手順の有効性を示す。
【図15】A−Bは、1コピーのsacBカセットを含む増殖中の大腸菌細胞でのpPB829およびpPB838プラスミドの安定性および産生を示す。C−Eは、1つのsacBカセットおよび2つのsacBカセットをもつ大腸菌株(それぞれΔpurNλPr::sacB+ΩkanおよびΔpurNλPr::sacB+Ωkan ΔedAλPr::sacB+Ωcat)におけるcatフリーpPB885プラスミドの安定性および産生を示す。図15Fは、CHO細胞における抗生物質フリープラスミド(pPB896/選別圧としてショ糖)および抗生物質プラスミド(pCG105/選別圧としてCat)のトランスフェクション有効性(GFP発現)を示す。これら2つの選別圧間(ショ糖対抗生物質としてのCat)において発現GFPタンパク質に関する明白な相違は認められない。
【図16】pCRblunt+PB1184-1185プラスミドマップを示す。
【図17】pPB828の制限マップおよび特徴を示す。
【図18】pPB829の制限マップおよび特徴を示す。
【図19】pPB838のプラスミドマップおよび特徴を示す。
【図20】pPB844のプラスミドマップおよび特徴を示す。
【図21】pPB845のプラスミドマップおよび特徴を示す。
【図22】pPB846のプラスミドマップおよび特徴を示す。
【図23】pPB847のプラスミドマップおよび特徴を示す。
【図24】pPB885のプラスミドマップを示す。pPB885の特徴物マップは以下を示す:2つのCDS:cIリプレッサー:324−1037;AP(R):2321−2990(相補性)、本来の位置(補完物1626..2483);2つの複製起点:pUC起点:1500−2173(相補性)、本来の位置(補完物805..1478);f1起点:2991−3301(本来の位置2615−3053)。
【図25】pPB896のプラスミドマップを示す。pPB896の特徴物マップは以下を示す:3つのCDS:GFP:1675−2397(本来の位置1676…2398);cIリプレッサー:4089−4802;Cm(R):4828−5484(相補性)、補完物(950−1606);1つのイントロン:イントロンA:805−1625(本来の位置805−1625);1つの真核細胞プロモーター:CMVプロモーター:1−683;1つの複製起点:3133−3640(本来の位置2425..2932);1つのターミネーター:BGH転写ターミネーター/PolyA:2405−2951(本来の位置1697..2243);1つの5'UTR:CMV IE5'UTR:684−804(本来の位置684..804)。
【図26】cIリプレッサータンパク質の配列アラインメントおよび配列同一性のパーセンテージを示す。
【図27】sacBタンパク質の配列アラインメントおよび配列同一性のパーセンテージを示す。
【図28】ポリヌクレオチドおよびタンパク質に割り当てた配列番号を示す表である。
【図29】cIリプレッサータンパク質をコードするポリヌクレオチドの配列アラインメントおよびヌクレオチドレベルにおける配列同一性のパーセンテージを示す。
【図30】sacBタンパク質をコードするポリヌクレオチドの配列アラインメントおよびヌクレオチドレベルにおける配列同一性のパーセンテージを示す。
【図31】P1プロモーター、cIの天然のプロモーター、λPrプロモーター、λPrプロモーター+5'UTR+sacB遺伝子、P1プロモーター+cI遺伝子、およびcIの天然のプロモーター+cI遺伝子の配列を示す。
【発明を実施するための形態】
【0009】
詳細な説明
本開示および特に特許請求の範囲では、例えば“comprises”、“comprised”および“comprising”などのような用語は、米国特許法でそれらに与えられた意味を有し得る。例えば、それら用語は“includes”、“included”および“including”などを意味することができ、さらに、例えば“consisting essentially of”および“consists essentially of”のような用語は、米国特許法でそれらに与えられた意味を有し得る。例えば、それら用語は明示的に列挙されていない成分も許容するが、しかし従来技術で見出されている成分、または当該発明の基本的なまたは新規な特徴に影響を及ぼす成分は除かれる。
特段の説明がなければ、本明細書で用いられる全ての技術用語および学術用語は、本開示が属する分野の業者のある者が一般的に理解する意味と同じ意味を有する。単数用語の“a”、“an”および“the”は、文脈が明らかにそうでないことを示していないかぎり、対応する複数形を含む。同様に、“or”という語は、文脈が明らかにそうでないことを示していないかぎり、“and”を含むことが意図される。
【0010】
“動物”は哺乳動物、鳥類などが意図される。動物または宿主には、哺乳動物およびヒトが含まれる。動物は、ウマ科の動物(例えばウマ)、イヌ科の動物(例えばイヌ、オオカミ、キツネ、コヨーテ、ジャッカル)、ネコ科の動物(例えばライオン、トラ、イエネコ、野生のネコ、他の大型のネコおよび他のネコ科の動物(チーターおよびオオヤマネコを含む))、ヒツジ類(例えばヒツジ)、ウシ科の動物(例えばウシ)、ブタ類(例えばブタ)、鳥類(例えばニワトリ、アヒル、カモ、シチメンチョウ、ウズラ、キジ、オウム、ウソ、タカ、カラス、ダチョウ、エミュおよびヒクイドリ)、霊長類(例えばキツネザル、メガネザル、サル、テナガザル、ヒヒ)および魚類が含まれる。“動物”という用語はまた、全ての発育期(胚および胎児期を含む)の個々の動物を含む。
“タンパク質”、“ペプチド”、“ポリペプチド”および“ポリペプチドフラグメント”という用語は本明細書で互換的に用いられ、任意の長さのアミノ酸残基ポリマーを指す。ポリマーは線状でも分枝していてもよく、改変アミノ酸またはアミノ酸アナローグを含むことができ、さらにアミノ酸以外の化学的部分によって中断されていてもよい。前記用語はまた、天然に改変されているか、または介入(例えばジスルフィド結合形成、グリコシル化、リピド化、アセチル化、リン酸化)または他の任意の操作もしくは修飾(例えば標識または生物活性成分との結合)によって改変されているアミノ酸ポリマーを包含する。
【0011】
“核酸”または“ポリヌクレオチド”という用語は互換的に用いられ、線状もしくは分枝状、一本鎖状もしくは二本鎖状、または前記の混合であるRNAまたはDNAを指す。前記用語はまた、RNA/DNAハイブリッドを包含する。以下はポリヌクレオチドの非限定的な例である:遺伝子または遺伝子フラグメント、エキソン、イントロン、mRNA、tRNA、rRNA、リボザイム、cDNA、組換えポリヌクレオチド、分枝ポリヌクレオチド、プラスミド、ベクター、任意の配列の単離DNA、任意の配列の単離RNA、核酸プローブおよびプライマー。ポリヌクレオチドは、改変ヌクレオチド、例えばメチル化ヌクレオチドおよびヌクレオチドアナローグ、ウラシル、他の糖および連結基(例えばフルオロリボースおよびチオレート)、並びにヌクレオチド分枝を含むことができる。ヌクレオチドの配列は、重合の後で例えば標識成分との結合によってさらに改変することができる。本定義に含まれる他のタイプの改変は、キャップ、天然に存在する1つまたは2つ以上のヌクレオチドのアナローグによる置換、ポリヌクレオチドをタンパク質、金属イオン、標識成分、他のポリヌクレオチドまたは固相に結合させるための手段の導入である。ポリヌクレオチドは、化学的合成によって入手するか、または微生物から誘導することができる。
“ドラッグレスプラスミド”または“抗生物質フリープラスミド”という用語は互換的に用いられ、抗生物質選別遺伝子を含まないDNAプラスミドを指す。
“遺伝子”という用語は広範囲に用いられ、生物学的機能と結びついた任意のポリヌクレオチドセグメントを指す。したがって、遺伝子には、ゲノム配列の場合のイントロンおよびエキソン、cDNAの場合のコード配列および/または前記配列のための調節配列が含まれる。例えば、遺伝子はまた、mRNAまたは機能的RNAを発現するか、または具体的なタンパク質をコードする核酸フラグメントを指し、前記には調節配列が含まれる。
【0012】
本明細書に開示する主題は、抗生物質耐性遺伝子の使用を回避することによって細菌プラスミドDNAベクターを作製し、安全なワクチンおよび免疫原性組成物を製造する新規なアプローチに関する。
本明細書に開示する主題は、グラム陰性宿主細菌細胞内で多数のプラスミドコピーを維持するための新規な概念(3つの構成成分に基づく)を提示する。第一の構成成分はグラム陰性宿主細菌であり、前記細菌は規定の培養条件下で当該細菌にとって有毒な生成物を発現し、前記有毒遺伝子は当該細菌染色体の非必須領域に挿入されている。第二の構成成分は、当該グラム陰性細菌の染色体上の遺伝子の存在であり、前記遺伝子は、厳密に制御され得る構成的プロモーターの制御下で有毒生成物をコードする。第三の構成成分は、前記宿主染色体上の有毒遺伝子に作動できるように連結されたプロモーターを調節するプラスミド由来の特異的なリプレッサーの発現である。
本発明で意図されるグラム陰性細菌には、アビバクテリウム(Avibacterium)、ブルセラ(Brucella)、大腸菌(Escherichia coli)、ヘモフィルス(Haemophilus)(例えばヘモフィルス・スイス(Haemophilus suis))、サルモネラ(Salmonella)(例えばサルモネラ・エンテリディス(Salmonella enteridis)、ネズミチフス菌(Salmonella typhimurium)、サルモネラ・インファンチス(Salmonella infantis))、シゲラ(Shigella)、パスツレラ(Pasteurella)およびリメイレラ(Rimeirella)が含まれるが、ただしこれらに限定されない。
ある実施態様では、有毒遺伝子は、レバンスクラーゼをコードするsacB構造遺伝子である。別の実施態様では、枯草菌(Bacillus subtilis)のレバンスクラーゼをコードする枯草菌のsacB構造遺伝子である。その天然の環境でのsacBの発現はグラム陽性細菌にとっては無害であるが、グラム陰性細菌で発現される場合、ショ糖含有培地にグラム陰性細菌がプレートされたとき形質転換細菌の急速な死を引き起こす。
【0013】
ある特徴では、本発明はsacBタンパク質(レバンスクラーゼ)を提供する。別の特徴では、本発明は、配列番号:4、60、62、64、66、68、70、72または74に示す配列を有するsacBタンパク質およびその変種またはフラグメントを提供する。さらに別の特徴では、本発明は、配列番号:4、60、62、64、66、68、70、72または74に示す配列を有するポリペプチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するsacBタンパク質を提供する。さらに別の特徴では、本発明は、上記で認定したsacBタンパク質(配列番号:4、60、62、64、66、68、70、72または74)のフラグメントおよび変種を提供する(前記は周知の分子生物学的技術を用いて当業者により容易に調製され得る)。変種は、本発明のポリペプチド(特に配列番号:4、60、62、64、66、68、70、72または74に示すアミノ酸配列)と少なくとも約75%、80%、85%、90%、95%、96%、97%、98%または99%のアミノ酸配列同一性を有する相同なポリペプチドである。
【0014】
別の特徴では、本発明は、sacBタンパク質(レバンスクラーゼ)をコードするポリヌクレオチド(例えばsacB遺伝子)、例えば配列番号:4、60、62、64、66、68、70、72または74に示す配列を有するsacBタンパク質をコードするポリヌクレオチドを提供する。さらに別の特徴では、本発明は、配列番号:4、60、62、64、66、68、70、72または74に示す配列を有するポリペプチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するsacBタンパク質、または保存的変種、対立遺伝子変種、ホモローグ、またはこれらポリペプチドの1つの少なくとも8つまたは10の連続するアミノ酸を含むフラグメント、またはこれらポリペプチドの組み合わせをコードするポリヌクレオチドを提供する。別の特徴では、本発明は、配列番号:3、61、63、65、67、69、71または73に示すヌクレオチド配列を有するポリヌクレオチドまたはその変種を提供する。さらに別の特徴では、本発明は、配列番号:3、61、63、65、67、69、71または73に示す配列を有するポリヌクレオチドの1つと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するポリヌクレオチドまたはその変種を提供する。
【0015】
本発明で利用できる他の有毒な遺伝子にはリステリア(Listeria)またはスタフィロコッカス(Staphylococcus)遺伝子が含まれるが、ただしこれらに限定されない。有毒な遺伝子は、有毒な遺伝子生成物(有毒タンパク質またはRNA)として発現されるDNAであってもよいが、またそのもの自体が有毒であるものおよび単独で有毒であるものであってもよい。そのような有毒遺伝子生成物の例は当分野では周知であり、制限エンドヌクレアーゼ(例えばDpnI)、CcdA/CcdB(S. Maki et al. 1996, J Mol Biol 256:473-482)、および抑圧機能の非存在下で宿主を殺す遺伝子(例えばkicB)が含まれるが、ただしこれらに限定されない。また別には、有毒遺伝子はin vitroで選別可能であり得る(例えば制限部位)。
本明細書で用いられるように、“ホモローグ”という用語にはオルトローグ、アナローグおよびパラローグが含まれる。野生型ポリペプチドのアナローグ、オルトローグおよびパラローグは、翻訳後修飾により、アミノ酸配列の相違により、またはその両方により野生型ポリペプチドと相違し得る。特に、本発明のホモローグは、一般的には野生型ポリペプチドまたはポリヌクレオチド配列の全体または部分に関して少なくとも80−85%、85−90%、90−95%または95%、96%、97%、98%または99%の配列同一性を示し、かつ類似の機能を示すであろう。
【0016】
“変種”は実質的に類似する配列を意味することが意図される。ポリヌクレオチドについては、変種は、天然のポリヌクレオチド内の1つまたは2つ以上の部位の1つまたは2つ以上の欠失および/または付加、および/または天然のポリヌクレオチド内の1つまたは2つ以上の部位の1つまたは2つ以上のヌクレオチドの置換を含む。本明細書で用いられるように、“天然の”ポリヌクレオチドまたはポリペプチドは、それぞれ天然に存在するヌクレオチド配列またはアミノ酸配列を含む。本発明の特定のポリヌクレオチド(すなわち参照ポリヌクレオチド)の変種はまた、変種ポリヌクレオチドによってコードされるポリペプチドおよび参照ポリヌクレオチドによってコードされるポリペプチド間のパーセント配列同一性を比較することによって評価することができる。“変種”タンパク質は、天然のタンパク質内の1つまたは2つ以上の部位の1つまたは2つ以上のアミノ酸の欠失または付加、および/または天然のタンパク質内の1つまたは2つ以上の部位の1つまたは2つ以上のアミノ酸の置換によって天然のタンパク質から誘導されたタンパク質を意味する。本発明に包含される変種タンパク質は生物学的に活性を有する。すなわちそれらは免疫応答を誘引する能力を有する。
【0017】
変種には対立遺伝子変種が含まれる。“対立遺伝子変種”という用語は、タンパク質のアミノ酸配列に変化をもたらし、さらに天然の集団(例えばウイルス種またはウイルス変種)内に存在する多形性を含むポリヌクレオチドまたはポリペプチドを指す。そのような天然の対立遺伝子変種は、典型的にはポリヌクレオチドまたはポリペプチドで1−5%の変動を生じる。対立遺伝子変種は、多数の異なる種の中の問題の核酸配列をシークェンシングすることによって識別することができる。前記は、そのような種の中で同じ遺伝子座を識別するハイブリダイゼーションプローブを用いることによって容易に実施することができる。そのような核酸の多様性およびその結果生じたアミノ酸多形性または変種(それらは天然の対立遺伝子変種の結果であり、問題の遺伝子の機能的活性を変化させない)の任意のものおよび全てが本発明の範囲内に含まれる。
“保存的変種”という用語は、アミノ酸残基の生物学的に類似する別の残基による置換、またはコードされるアミノ酸残基が変化しないかまたは生物学的に類似する別の残基となるような核酸配列内のヌクレオチドの置換を指す。これに関して特に好ましい置換は、上記に記載したように一般的に性質が保存的であろう。
【0018】
開示のポリヌクレオチドには、遺伝暗号の結果として、例えば特定の宿主のための最適なコドン使用頻度の結果として縮退性である配列が含まれる。本明細書で用いられるように、“最適化”とは、ある種においてその発現を高めるために一般的に操作されるポリヌクレオチドを指す。本発明のポリペプチドをコードする最適化ポリヌクレオチドを提供するために、ポリペプチドのDNA配列は、1)個別の種において高度に発現される遺伝子の優先的コドンを含むように;2)前記種で実質的に見出されるヌクレオチド塩基組成のA+TまたはG+C含量と類似する含量を含むように;3)前記種の開始配列を形成するように;または4)脱安定化、不適切なポリアデニル化、RNAの分解および停止を引き起こす配列、または二次構造ヘアピンまたはRNAスプライス部位を形成する配列を除去するように改変できる。前記の種における本発明のポリペプチド(例えばsacBタンパク質またはcIリプレッサータンパク質)の増加は、真核生物および原核生物または個別の種のコドン使用頻度の頻度分布を利用することによって達成できる。“優先的コドン使用頻度”という用語は、あるアミノ酸を指定するためにヌクレオチドコドンの使用において個々の宿主細胞によって示される優先性を指す。20の天然のアミノ酸が存在し、そのほとんどが2つ以上のコドンによって指定される。したがって、ヌクレオチド配列によってコードされるポリペプチドのアミノ酸配列が機能的に変化しないかぎり、全ての縮退ヌクレオチド配列が本発明に含まれる。
【0019】
配列に関して“同一性”とは、2つの配列の短い方のヌクレオチドまたはアミノ酸の数で割った、同一ヌクレオチドまたはアミノ酸を有する位置の数を意味し、この場合、2つの配列のアラインメントはWilburとLipmannのアルゴリズム(Wilbur & Lipman)にしたがって決定できる。RNA配列がDNA配列と類似している、またはある程度の配列同一性もしくは配列相同性を有するというとき、DNA配列中のチミジン(T)はRNA配列中のウラシル(U)と等価であると考えられる。したがって、DNA配列中のチミジン(T)をRNA配列中のウラシル(U)と等価であると考えることによって、RNA配列は本発明の範囲内にあり、DNA配列から誘導することができる。2つのアミノ酸配列の配列同一性または配列類似性、または2つのヌクレオチド配列間の配列同一性は、Vector NTIソフトウェアパッケージ(Invitrogen, 1600 Faraday Ave., Carlsbad, CA)を用いて決定できる。
【0020】
別の実施態様では、本発明の有毒な遺伝子に作動できるように連結されるプロモーターは、λファージ由来のプロモーターである。λファージは、大腸菌内で生活する溶原性ファージである。ファージがその宿主細胞内に入ると、ファージは自身を宿主のDNAに組み入れる。この状態でλはプロファージと呼ばれ、宿主に大きな害を引き起こすことなく宿主ゲノム内に留まる。このようにして、プロファージは宿主の細胞分割のたびに二倍になる。この状態で発現されるプロファージのDNAは、宿主細胞のストレスの徴候に注意するタンパク質をコードする。ストレスは、飢餓、毒物(抗生物質のような)および宿主を損傷または破壊することができる他の因子の結果であり得る。ストレス状態が検出されたとき、プロファージは再び活性化し、宿主細胞のDNAからそれ自身を切り出し、その溶菌周期に入る。再活性化されたファージは宿主DNAから離れ、そのタンパク質工場を初期化して新ファージの多数のコピーを生産する。新ファージの構築によって宿主の全資源が枯渇したとき、細胞が溶解し(細胞膜が破壊され)、新ファージが放出される。
【0021】
ラムダリプレッサー遺伝子系は以下から成る(染色体の左から右へ):cI遺伝子、OR3、OR2、OR1、cro遺伝子。cI遺伝子はλリプレッサー(“cIリプレッサータンパク質”)をコードする。cIリプレッサータンパク質をコードするゲノム領域は免疫領域として知られている。cIリプレッサータンパク質は、遺伝子転写の正および負の両方の調節因子である。前記は、λファージがその宿主細菌の染色体上で“休眠状態”に留まることを可能にする。バクテリオファージλの溶原性状態は、調節タンパク質cIとOR(右オペレーター)およびOL(左オペレーター)(それぞれλPlおよびλPrプロモーターに由来し、溶菌状態に必要なタンパク質の転写を妨げる)が結合することによって維持される。溶原性λファージを保持する細菌細胞はλファージによる更なる感染に対して免疫である。cIリプレッサータンパク質はさらに別のファージ粒子の感染による溶菌の進行を阻害する。
この実施態様のある特徴では、プロモーターは配列番号:5に示す配列を有するポリヌクレオチドを含む。別の特徴では、プロモーターは、配列番号:5に示す配列を有するポリヌクレオチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するポリヌクレオチドを含む。
【0022】
ある実施態様では、本発明のリプレッサータンパク質は、cIリプレッサータンパク質、例えば温度感受性のcI857リプレッサーである。溶原としてcI857を保持するλファージは39℃より低い温度で増殖するが、続いて温度上昇によって溶菌性増殖を誘発するであろう。30℃では、cIリプレッサータンパク質は活性であり、感染ファージの右および左オペレーターと結合する。これによっていずれのファージタンパク質の転写も妨害され、したがって溶菌が妨げられる。しかしながら42℃では、cIリプレッサーは不活化され、プロモーターオペレーターと結合できない。
ある特徴では、本発明はcIリプレッサータンパク質を提供する。別の特徴では、本発明は、配列番号:2、44、46、48、50、52、54、56または58に示す配列を有するcIリプレッサータンパク質およびその変種またはフラグメントを提供する。さらに別の特徴では、本発明は、配列番号:2、44、46、48、50、52、54、56または58に示す配列を有するポリペプチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するcIリプレッサータンパク質を提供する。さらに別の特徴では、本発明は上記で認定したcIリプレッサータンパク質(配列番号:2、44、46、48、50、52、54、56または58)のフラグメントおよび変種を提供する(前記は周知の分子生物学的技術を用いて当業者によって容易に調製され得る)。変種は、本発明のポリペプチド、特に、配列番号:2、44、46、48、50、52、54、56または58に示すアミノ酸配列と少なくとも約75%、80%、85%、90%、95%、96%、97%、98%または99%の同一性を有する相同なポリペプチドである。
【0023】
別の特徴では、本発明は、cIリプレッサータンパク質をコードするポリヌクレオチド(例えばcI遺伝子)、例えば配列番号:2、44、46、48、50、52、54、56または58に示す配列を有するcIリプレッサータンパク質をコードするポリヌクレオチドを提供する。さらに別の特徴では、本発明は、配列番号:2、44、46、48、50、52、54、56または58に示す配列を有するポリペプチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するcIリプレッサータンパク質、または保存的変種、対立遺伝子変種、ホモローグ、またはこれらポリペプチドの1つの少なくとも8つまたは10の連続するアミノ酸を含むフラグメント、またはこれらポリペプチドの組み合わせをコードするポリヌクレオチドを提供する。別の特徴では、本発明は、配列番号:1、45、47、49、51、53、55、57または59に示すヌクレオチド配列を有するポリヌクレオチドまたはその変種を提供する。さらに別の特徴では、本発明は、配列番号:1、45、47、49、51、53、55、57または59に示す配列を有するポリヌクレオチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するポリヌクレオチド、またはその変種を提供する。
【0024】
ある実施態様では、cI遺伝子を駆動するプロモーターは天然のcI 遺伝子プロモーターである。別の実施態様では、cI遺伝子を駆動するプロモーターはカナマイシン遺伝子の弱いプロモーター(P1)である。別の実施態様では、プロモーターは配列番号:29、83または30に示す配列を有するポリヌクレオチドを含む。さらに別の実施態様では、プロモーターは、配列番号:29、83または30に示す配列を有するポリヌクレオチドと少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、少なくとも95%、96%、97%、98%または99%の配列同一性を有するポリヌクレオチドを含む。
本発明の方法で利用できる他のプロモーター/リプレッサー対には、CarA-CarSリプレッサー-アンチリプレッサー対により制御されるcarBプロモーター、成長ホルモン遺伝子プロモーターおよびリプレッサー、並びにLacリプレッサー(lacl)およびPtrc-2プロモーターが含まれるが、ただしこれらに限定されない。
【0025】
変異導入のための基質として機能する変異プロモーター/リプレッサー対は、現在入手可能であり当分野で報告されているようなものから選択することができる。変異リプレッサーとそれらのオペレーターとの結合に関する研究(以下を参照されたい:Nelson and Sauer, 1985, Cell 42:549;Nelson and Sauer, 1986, J Mol Biol 192:27;およびGussin et al. 1983, Lambda II, (Hendrix, Roberts, Stahl and Weisberg, eds) Cold Spring Harbor Press, Cold Spring Harbor, NY, p.93-123)は、減速されるが停止されることはない転写速度を提供する既知の変異プロモーターおよび/または変異リプレッサー配列の選別を可能にする。この選別プロモーター領域を異種発現プラスミドに取り込み、続いて通常の位置特異的変異導入によって前記領域を改変する(例えば以下を参照されたい:Morinaga et al. 1984, Biotechnology, 2:636)。また別には、リプレッサーコード配列を類似の態様で変更する。生じた変異リプレッサー/プロモーター対を、位置特異的変異導入に付されていない野生型プラスミドと発現を比較することによって、異種タンパク質の分泌を促進するその能力について解析する。
本明細書に開示する主題はドラッグレスまたは抗生物質フリーの概念を提供する。本概念では、プラスミドから発現されるリプレッサー活性(例えばcIリプレッサー)は、細胞宿主染色体上に存在するλファージプロモーターの下に配置された有毒遺伝子生成物(例えばsacB遺伝子生成物)の転写を阻害し、さらに基質(例えばショ糖)の存在下における宿主細胞の生存活性は、プラスミド由来リプレッサータンパク質(例えばλcIリプレッサータンパク質)がプラスミドから十分なレベルで発現されることによって担保される。抗生物質フリープラスミドを含む宿主株のショ糖存在下での増殖は、効率的な系のDNAプラスミドの維持および産生を担保する。
プラスミドという用語は、本発明のポリヌクレオチドおよび所望の宿主または標的の細胞における前記ポリヌクレオチドのin vivo発現に必要なエレメントを含む任意のDNA転写ユニットをカバーし、さらに前記ユニットに関して、スーパーコイルまたは非スーパーコイルプラスミド、環状プラスミドが、線状形と同様に本発明の範囲内に含まれることが特記される。
【0026】
本発明のある実施態様は、1つまたは2つ以上のプロモーターに作動できるように連結された1つまたは2つ以上のリプレッサー遺伝子を含むドラッグレスプラスミドに関する。ある特徴では、ドラッグレスプラスミドはcIリプレッサータンパク質をコードするポリヌクレオチドを含む。別の特徴では、ドラッグレスプラスミドはcIリプレッサータンパク質をコードするポリヌクレオチドに作動できるように連結されたプロモーターを含む。プロモーターは、天然のcI遺伝子プロモーターまたはカナマイシン遺伝子の弱いプロモーターであり得る。別の特徴では、ドラッグレスプラスミドはさらに、免疫原またはタンパク質をコードする異種ポリヌクレオチドを含む。別の特徴では、ドラッグレスプラスミドは、免疫原またはタンパク質をコードする異種ポリヌクレオチドに作動できるように連結されたプロモーターを含む。当分野で公知の任意のプロモーターを、本発明のドラッグレスプラスミドで用いることができる。前記には、細菌性、酵母、菌類、昆虫、哺乳動物および植物のプロモーターが含まれる。プロモーターは、ヒトまたはネズミ起源の、または場合によって別の起源(例えばラットまたはモルモット)を有する極初期サイトメガロウイルスプロモーター(CMV-IE)、スーパープロモーター(M. Ni et al. 1995, Plant J 7:661-676)であるが、ただし前記に限定されない。CMV-IEプロモーターは実際のプロモーター部分を含むことができるが、前記はエンハンサー部分と結合していてもいなくてもよい。EP-A-260148、EP-A-323597、米国特許5,168,062号、5,385,839号および4,968,615号をPCT出願WO87/03905と同様に参照することができる。グラム陰性細菌に由来する他のプロモーターもまた適切である。前記には、アビバクテリウム、ブルセラ、大腸菌、ヘモフィルス(例えばヘモフィルス・スイス)、サルモネラ(例えばサルモネラ・エンテリディス、ネズミチフス菌、サルモネラ・インファンチス)、シゲラ、パスツレラおよびリメイレラが含まれるが、ただしこれらに限定されない。
【0027】
プラスミドは他の発現制御エレメントを含むことができる。安定化配列、例えばイントロン配列を取り込むことは有利である。イントロン配列は、例えばhCMV-IEの最初のイントロン(PCT出願WO1,989/01036)、ウサギβグロビン遺伝子のイントロンII(van Ooyen et al. 1979)である。別の実施態様では、プラスミドは3'UTRを含むことができる。3'UTRは、アグロバクテリウムノパリンシンターゼ(Nos)3'UTRであるが、ただし前記に限定されない。
プラスミドおよびポックスウイルス以外のウイルスベクターのためのポリアデニル化(ポリA)シグナルに関しては、ウシ成長ホルモン(BGH)遺伝子のポリ(A)シグナル(US 5,122,458参照)、またはウサギβグロビン遺伝子ポリ(A)シグナル、またはSV40ウイルスのポリ(A)シグナルが有用であり得る。
“宿主細胞”は、外因性ポリヌクレオチド(例えば組換えプラスミドまたはベクター)の投与によって既に遺伝的に改変されているかまたは遺伝的に改変することができる原核細胞または真核細胞を意味する。遺伝的に改変された細胞というとき、前記用語は最初に改変された細胞およびその子孫の両方を指す。
【0028】
本発明の別の実施態様は、ドラッグレスプラスミドを含むグラム陰性細菌宿主株に関する。この実施態様のある特徴は、ドラッグレスプラスミドおよび宿主染色体の1つまたは2つ以上の非必須領域に挿入された異種ポリヌクレオチドを含むグラム陰性細菌宿主株に関し、この場合、前記異種ポリヌクレオチドは、リプレッサーにより厳密に調節され得るプロモーターに作動できるように連結されている。この実施態様の別の特徴では、宿主染色体に挿入された異種ポリヌクレオチドはsacB遺伝子である。この実施態様の別の特徴では、sacB遺伝子は、プロモーター(例えばλファージの右のプロモーター)に作動できるように連結される。sacB遺伝子およびプロモーターはマルチコピー(例えば2または3コピー)として宿主染色体に挿入できることは理解されよう。宿主株の非必須領域は、大腸菌染色体のdeAまたはpurN遺伝子であり得る。deAまたはpurN遺伝子は非必須であり、これら遺伝子の欠失は細菌の増殖速度に影響を与えない(J. Kim et al. Biochemistry. 46,44:12501-12511)。ある特徴では、sacB遺伝子およびプロモーターのカセットの1コピーが、deA遺伝子座またはpurN遺伝子座のどちらかに対立遺伝子置換によって挿入される。別の特徴では、sacB遺伝子およびプロモーターのカセットの2コピーが、deA遺伝子座またはpurN遺伝子座のどちらかに挿入される。さらに別の特徴では、sacB遺伝子およびプロモーターのカセットの2コピーが宿主染色体に挿入され、この場合sacB遺伝子およびプロモーターカセットの1つのコピーがdeA遺伝子座に挿入され、他方のコピーがpurN遺伝子座に挿入される。sacB遺伝子およびプロモーターのカセットは、プロモーターに作動できるように連結されたsacB遺伝子を含むポリヌクレオチドを指す。ある特徴では、sacB遺伝子およびプロモーターのカセットは、sacB遺伝子およびλファージの右のプロモーターを含む。別の特徴では、sacB遺伝子およびプロモーターのカセットは、配列番号:75に示す配列を有するポリヌクレオチドを含む。
【0029】
本明細書に開示する主題はさらに、免疫原またはタンパク質をコードする異種遺伝子を含むドラッグレスプラスミドを含むワクチンもしくは組成物、またはドラッグレスプラスミドを用いて発現させた免疫原またはタンパク質を含むワクチンもしくは組成物に関する。前記ワクチンまたは組成物はさらに医薬的に許容できる担体を含むことができる。
ある実施態様では、免疫原は、ネコの病原体、例えばネコヘルペスウイルス(FHV)、ネコカリシウイルス(FCV)、ネコ白血病ウイルス(FeLV)、ネコ免疫不全ウイルス(FIV)(ただしこれらに限定されない)、狂犬病ウイルスなどおよび前記の組み合わせから選択される。
別の実施態様では、免疫原は、イヌの病原体(狂犬病ウイルス、イヌヘルペスウイルス(CHV)、イヌパルボプイルス(CPV)、イヌコロナウイルス、レプトスピラ・カニコラ(Leptospira canicola)を含むが、ただしこれらに限定されない)、レプトスピラ・イクテロヘモラージアエ(L. ichterohaemorragiae)、レプトスピラ・グリッポチフォサ(L. grippotyphosa)、ボレリア・バーグドルフェリ(Borrelia burgdorferi)、ボルデテラ・ブロンキセプチカ(Bordetella bronchiseptica)など、および前記の組み合わせから選択される。
さらに別の実施態様では、免疫原は、ウマの病原体、例えばウマヘルペスウイルス(1型または4型)、ウマインフルエンザウイルス、破傷風、西ナイルウイルス、ストレプトコッカス・エクィ(Streptococcus equi)、ロドコッカス・エクィ(Rhodococcus equi)など、または前記の組合せから選択される。
【0030】
さらに別の実施態様では、免疫原は、ウシの病原体、例えば狂犬病ウイルス、ウシロタウイルス、ウシパラインフルエンザウイルス3型(bPIV-3)、ウシコロナウイルス、ウシウイルス性下痢ウイルス(BVDV)、口蹄疫ウイルス(FMDV)、ウシ呼吸器合胞体ウイルス(BRSV)、伝染性ウシ鼻気管炎ウイルス(IBR)、大腸菌、パスツレラ・マルトシダ(Pasteurella multocida)、パスツレラ・ヘモリチカ(P. haemolytica)、サルモネラ、クリプトスポリジウム(Cryptosporidium)など、および前記の組合せから選択される。
さらに別の実施態様では、免疫原は、ブタの病原体(例えばブタインフルエンザウイルス(SIV)、ブタシルコウイルス2型(PCV-2)、ブタ生殖器呼吸器症候群ウイルス(PRRS)、仮性狂犬病ウイルス(PRV)、ブタパルボウイルス(PPV)を含むが、ただしこれらに限定されない)、FMDV、マイコプラズマ・ヒオプニューモニアエ(Mycoplasma hyopneumoniae)、エリシペロトリクス・リューシオパシアエ(Erysipelothrix rhusiopathiae)、パスツレラ・マルトシダ、ボルデテラ・ブロンキセプチカ、大腸菌、ブルータングウイルス、アフリカウマ病ウイルス、リフトバレー熱、ニパ(Nipah)ウイルスなど、および前記の組合せから選択される。
【0031】
“抗原”または“免役原”という用語は互換的に用いられ、宿主動物で特異的な免疫応答をを誘発する物質を意味する。抗原は、免疫原性特性を有する挿入物を含む組換えベクター;宿主動物に提示されたとき免疫応答を誘発することができるDNAの断片またはフラグメント;タンパク質、ポリペプチド、ペプチド、エピトープ、ハプテンまたは前記の任意の組合せを含むことができる。また別には、免疫原または抗原は毒素または抗毒素を含むことができる。
本明細書で用いられるように、“医薬的に許容できる担体”および“医薬的に許容できるベヒクル”は相互に用いることができ、副作用を示さずに宿主に注射できる、ワクチン抗原収容のための液体ベヒクルを指す。当分野で公知の医薬的に許容できる適切な担体には、無菌的な水、食塩水、グルコース、デキストロースまたは緩衝溶液が含まれるが、ただしこれらに限定されない。担体は補助剤を含むことができ、前記には、希釈剤、安定化剤(すなわち糖類およびアミノ酸)、保存料、湿潤剤、乳化剤、pH緩衝剤、粘性強化添加剤、着色剤などが含まれるが、ただしこれらに限定されない。
【0032】
プラスミドに有益である(ただし絶対的に適切というわけではないが)第四アンモニウム塩を含む陽イオン性脂質は、有利には以下の式を有するものである:
【0033】
【化1】
【0034】
式中、R1は、12から18の炭素原子を有する飽和または不飽和の直鎖脂肪族ラジカルであり、R2は、2または3個の炭素原子を含む別の脂肪族ラジカルであり、Xは、アミンまたはヒドロキシル基(例えばDMRIE)である。別の実施態様では、陽イオン性脂質は、中性脂質(例えばDOPE)と結合することができる。
これらの陽イオン性脂質でとりわけ好ましいものはDNRIE(N-(2-ヒドロキシエチル)-N,N-ジメチル-2,3-ビス(テトラデシルオキシ)-1-プロパンアンモニウム(WO96/34109)であり、有利には中性脂質(有利にはDOPE(ジオレオイル-ホスファチジル-エタノールアミン))(Behr, 1994)と結合してDMRIE-DOPEを生成する。
有利には、アジュバントを含むプラスミド混合物は即席に、有利には調製物の投与と同時にまたは調製物の投与前近くに(例えば投与前近くまたは投与の前に)形成される。プラスミド-アジュバント混合物は、有利には投与前に十分な時間、例えば投与前に約10から約60分、例えば投与前にほぼ30分を与えて混合物が複合体を生じるように形成される。
DOPEが存在するときは、DMRIE:DOPE分子比は有利には約95:約5から約5:95、より有利には約1:約1、例えば1:1である。
DMRIEまたはDMRIE-DOPEアジュバント:プラスミド重量比は、約50:約1から約1:約10、例えば約10:約1から約1:約5、有利には約1:約1から約1:約2、例えば1:1から1:2である。
【0035】
別の実施態様では、医薬的にまたは獣医的に許容できる担体、賦形剤またはベヒクルは油中水エマルジョンであり得る。適切な油中水エマルジョンの例には、安定で4℃で液状である、油を基剤とする油中水ワクチンエマルジョンが含まれる。前記エマルジョンは、6から50v/v%、好ましくは12から25v/v%の抗原含有水相、50から94v/v%の油相(非代謝性油(例えばパラフィン油のような鉱物油)および/または代謝性油(例えば植物油)または脂肪酸、ポリオールまたはアルコールエステルを全部としてまたは部分として含む)、0.2から20p/v%、好ましくは3から8p/v%の界面活性剤(後者は全部としてまたは部分として含まれる)、または混合物としてポリグリセロールエステル(前記ポリグリセロールエステルは、好ましくはポリグリセロール(ポリ)リシンオレエート)またはポリオキシエチレンリシン油または他の水素添加ポリオキシエチレンリシン油を含む。油中水エマルジョンで用いることができる界面活性剤の例には、エトキシル化ソルビタンエステル(例えばポリオキシエチレン(20)ソルビタンモノオレエート(Tween80(登録商標))(AppliChem社(Cheshire, CT)から入手できる)およびソルビタンエステル(例えばソルビタンモノオレエート(Span 80(登録商標))(Sigma Aldrich(St. Louis, MO)から入手できる))が含まれる。さらに、油中水エマルジョンに関しては、米国特許6,919,084号(例えばその実施例8)を参照されたい(前記文献は参照により本明細書に含まれる)。いくつかの実施態様では、抗原含有水相は、1つまたは2つ以上の緩衝剤を含む食塩水溶液を含む。適切な緩衝溶液の例はリン酸緩衝食塩水である。有利な実施態様では、油中水エマルジョンは、水/油/水(W/O/W)三重エマルジョンである(例えば米国特許6,358,500号を参照されたい(前記文献は参照により本明細書に含まれる))。他の適切なエマルジョンの例は米国特許7,371,395号に記載されている(前記文献は参照により本明細書に含まれる)。
【0036】
ワクチンまたは組成物は、受容動物の齢、性別、体重、種および状態のような要件、および投与ルートを考慮しながら、医学または獣医学分野の業者によく知られた用量または技術によって投与することができる。投与ルートは、経皮、経粘膜投与(例えば経口、鼻内、肛門、膣投与)または非経口ルート(皮内、筋肉内、皮下、静脈内または腹腔内投与)であり得る。ワクチンまたは組成物は単独で投与できるが、また他の処置または治療と一緒にまたは連続して投与できる。投与形態には懸濁物、シロップまたはエリキシル、および非経口、皮下、皮内、筋肉内または静脈内投与用調製物(例えば注射可能投与物)、例えば無菌的懸濁物またはエマルジョンが含まれ得る。ワクチンまたは組成物は、スプレーとして投与するか、または食べ物および/または水に混合するか、または適切な担体、希釈剤または賦形剤(例えば無菌的な水、生理学的食塩水、グルコースなど)との混合物としてデリバ−することができる。組成物は、補助物質、例えば湿潤化もしくは乳化剤、pH緩衝剤、アジュバント、ゲル化もしくは粘性強化用添加剤、保存料、香料、着色剤などを、所望される投与経路および調製物に応じて含むことができる。標準的な薬学テキスト、例えば“Remington's Pharmaceutical Sciences”(1990)を参考にして、煩雑な実験を実施することなく適切な調製物を製造することができる。
【0037】
本明細書に開示する主題はさらに、以下の工程を含むドラッグプラスミドの製造方法に関する:1)宿主染色体の1つまたは2つ以上の非必須領域に対立遺伝子置換によって挿入された異種ポリヌクレオチドを含むグラム陰性細菌宿主株を操作する工程;2)cIリプレッサータンパク質をコードするポリヌクレオチドを含むDNAプラスミドを構築する工程;3)cIリプレッサータンパク質をコードする遺伝子を含むDNAプラスミドで細菌宿主株を形質転換する工程;4)形質転換細菌宿主株をショ糖の存在下において30℃から42℃の範囲の温度で増殖させる工程;および5)DNAプラスミドを回収する工程。
この実施態様のある特徴では、DNAプラスミドはさらに免疫原またはタンパク質をコードする異種ポリヌクレオチドを含み、このプラスミドでは異種ポリヌクレオチドは作動できるようにプロモーターに連結される。前記プロモーターは原核細胞または真核細胞で機能するプロモーター、例えばCMVプロモーターでもよい。
本明細書に開示する主題はさらに、以下の工程を含む、ドラッグレスプラスミドを用いてタンパク質または免疫原を製造する方法に関する:1)宿主染色体の1つまたは2つ以上の非必須領域に対立遺伝子置換によって挿入された異種ポリヌクレオチドを含むグラム陰性細菌宿主株を操作する工程;2)cIリプレッサータンパク質をコードするポリヌクレオチドおよび免疫原またはタンパク質をコードする遺伝子を含むDNAプラスミドを構築する工程;3)cIリプレッサータンパク質をコードするポリヌクレオチドおよび免疫原またはタンパク質をコードする遺伝子を含むDNAプラスミドで細菌宿主株を形質転換する工程;4)形質転換細菌宿主株をショ糖の存在下において30℃から42℃の範囲の温度で増殖させる工程;および5)免疫原またはタンパク質を回収する工程。
【0038】
この実施態様のある特徴では、免疫原またはタンパク質をコードする遺伝子は原核細胞で機能するプロモーターに作動できるように連結される。プロモーターはグラム陰性細菌由来のプロモーターでもよく、アビバクテリウム、ブルセラ、大腸菌、ヘモフィルス(例えばヘモフィルス・スイス)、サルモネラ(例えばサルモネラ・エンテリディス、ネズミチフス菌、サルモネラ・インファンチス)、シゲラ、パスツレラおよびリメイレラから単離されたプロモーターが含まれるが、ただしこれらに限定されない。
この実施態様のある特徴では、宿主染色体に挿入される異種ポリヌクレオチドはsacBタンパク質をコードする。別の特徴では、ショ糖濃度は、0%から約20%、約1%から約20%、約1%から約15%、約1%から約10%の範囲であり得る。さらに別の特徴では、ショ糖濃度は、約1%から約10%、約1%から約2%、約2%から約3%、約3%から約4%、約4%から約5%、約5%から約6%、約6%から約7%、約7%から約8%、約8%から約9%または約9%から約10%であり得る。さらに別の特徴では、ショ糖濃度は、約1%、約2%、約3%、約4%、約5%、約6%、約7%、約8%、約9%または約10%であり得る。
この実施態様のある特徴では、温度は約30℃から約42℃の範囲であり得る。別の特徴では、温度は、約30℃から約41℃、約30℃から約39℃、約30℃から約38℃、約30℃から約37℃の範囲であり得る。さらに別の特徴では、温度は、約30℃、約31℃、約33℃、約34℃、約35℃、約36℃、約37℃、約38℃、約39℃、約40℃または約41℃であり得る。
【0039】
“回収”という用語は、培養液から採集、抽出、採集または精製することを含むが、ただし前記に限定されない。
“単離された”生物学的成分(例えばポリヌクレオチド、DNAプラスミド、タンパク質または細胞小器官)とは、前記成分が通常存在する生物の細胞内の他の生物学的成分、例えば他の染色体性および染色体外DNAおよびRNA、タンパク質並びに細胞小器官から実質的に分離または精製されている成分を指す。“単離されている”ポリヌクレオチド(DNAプラスミドを含む)およびタンパク質には、標準的な精製方法によって精製されたポリヌクレオチドおよびタンパク質が含まれる(例えば以下を参照されたい:David W. Russell, Joseph Sambrook (2001), Molecular Cloning: a laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory)。前記用語はまた、化学的合成と同様に組み換え技術によって調製されたポリヌクレオチドおよびタンパク質を包含する。
本明細書で用いられる“精製された”という用語は完全に純粋であることを要求せず、むしろ相対的用語として意図される。したがって、例えば、精製されたポリペプチドまたはポリヌクレオチド調製物は、当該ポリペプチドまたはポリヌクレオチドがその天然の環境で存在するよりも濃縮されている調製物である。すなわち、当該ポリペプチドまたはポリヌクレオチドは細胞性成分から分離されてある。“実質的に精製された”とは、細胞性成分または物質の少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、または少なくとも98%、またはそれ以上が取り除かれているようなものが意図される。同様に、ポリペプチドまたはポリヌクレオチドは部分的に精製されてあってもよい。“部分的に精製”とは、細胞性成分または物質の60%未満が除去されることを意味する。
【0040】
本発明はさらに免疫または防御応答を動物で誘発する方法を提供する。前記方法は、本発明の免疫学的組成物、ワクチンまたは組成物を動物に投与する工程を含む。誘引される免疫応答は、特に抗体および/または細胞性免疫応答、および特にガンマインターフェロン応答である。
特に、本発明は、病原微生物による動物の感染(例えばウイルス、細菌、菌類または原生動物寄生による感染)に対して免疫するか、または前記を予防するか、または前記によって引き起こされる症状を緩和するための方法を提供する。本発明の方法は脊椎動物で有用であり、前記脊椎動物にはヒト、イヌ科の動物(例えばイヌ)、ネコ科の動物(例えばネコ)、ウマ科の動物(例えばウマ)、ウシ科の動物(例えばウシ)およびブタのような動物(例えばブタ)だけでなく鳥類(ニワトリ、シチメンチョウ、アヒル、カモ、ウズラ、キジ、オウム、ウソ、タカ、カラスおよび走鳥類(ダチョウ、エミュおよびヒクイドリ)を含むが、ただしこれらに限定されない)が含まれるが、ただしこれらに限定されない。本発明の方法はまた魚類にDNAワクチンを提供するために有用である。
本発明の具体的な特徴では、これらの方法は、本発明にしたがって製造したワクチン組成物を投与することによって分娩前に妊娠雌動物をワクチン接種することから成る。これらの方法はさらに、ワクチン接種プロトコルにより誘引される防御抗体の誘導およびワクチン接種妊娠雌動物からそれらの子孫への前記防御抗体の伝達を含む。そのような母性抗体の伝達は続いて当該子孫を病気から防御する。
【0041】
本発明にしたがって製造したワクチン組成物の投与量は、ワクチン接種される動物の種、品種、齢、サイズ、ワクチン接種歴および健康状態に左右されるであろう。抗原濃度、追加のワクチン成分および投与経路(すなわち皮下、皮内、経口、筋肉内または静脈内投与)のような他の要件もまた有効な投与量に影響を与えるであろう。ワクチン投与量は、ワクチンの抗原濃度、投与経路、並びにワクチン接種される動物の齢および状態に基づいて容易に決定することができる。各抗原バッチを個々に計測することができる。また別には、MPD(最少防御用量)試験および他のスクリーニング方法と同様に種々の投与量による秩序だった免疫原性試験を用いて、煩雑な実験を実施することなく本発明のワクチン組成物のための有効な投与量を決定することができる。以下に提供する実施例から、本明細書に記載したワクチン組成物の使用に適切な投与量および体積は極めて明白であろう。もっとも重要な要素は、当該投与量によって自然感染に対して少なくとも部分的な防御効果が提供されることである(前記は自然感染に付随する死亡率および罹患率の減少によって立証される)。適切な体積は、当業者によって同様に容易に確認される。例えば、鳥類の種では、1用量の体積は約0.1mLから約0.5mL、有利には約0.3mLから約0.5mLであり得る。ネコ科、イヌ科およびウマ科の種では、1用量の体積は約0.2mLから約3.0mL、有利には約0.3mLから約2.0mL、より有利には約0.5mLから約1.0mLであり得る。ウシ類およびブタ類の種では、前記用量体積は約0.2mLから約5.0mL、有利には約0.3mLから約3.0mL、より有利には0.5mLから約2.0mLであり得る。
【0042】
早期に免疫応答を強化するために、または最後の投与から長期間が経過したときには、好ましくは、周期的な期間間隔での反復ワクチン接種を実施することができる。本発明のある実施態様得は、ワクチン組成物は非経口注射として(すなわち皮下、皮内または筋肉内に)投与される。組成物は1回の投与として投与されるか、または別の実施態様では、約2から約6週間、好ましくは約2から約5週間の間隔で与えられる約2回から約5回の反復投与で与えることが。しかしながら、当業者は、投与回数およびワクチン接種の間の間隔は、多数の要件(ワクチン接種動物の齢、動物の状態、免疫経路、1回あたりに利用可能な抗原量などを含むが、ただしこれらに限定されない)に左右されることを承知しているであろう。最初のワクチン接種の場合、前記期間は一般的に1週間よりも長く、好ましくは約2から約5週間であろう。以前にワクチン接種された動物の場合、好ましくは、約1年間隔のブースターワクチン接種(妊娠前および妊娠中)が実施される。
本発明はまた、無針注入器(例えばPigjet(登録商標)、Avijet(登録商標)、Dermojet(登録商標)またはBiojector(登録商標)(Bioject, Oregon, USA))を用いてワクチン組成物を投与すること意図する。当業者は、煩雑な実験を実施することなく、複数の要件(例えばワクチン接種される動物の種、動物の齢および体重など)に関して要求されるように前記注入器の詳細を調節することができる。
本発明はさらに、活性成分(例えば免疫原または医薬組成物)を含む第一のバイアルおよび第二のバイアルに本発明にしたがって作製した希釈剤を含むキットに関する。前記免疫原は凍結乾燥形、乾燥形であっても、または本明細書に記載した水溶液中に存在してもよい。
本発明は、以下の非限定的な実施例の手段によってこれからさらに説明されるであろう。
【実施例1】
【0043】
抗生物質フリープラスミド維持の概念
以下の実施例は、グラム陰性宿主細胞内に多数のプラスミドコピーを維持するための、3つの成分に基づく新規な概念を示す:
1.所定の培養条件下でグラム陰性細菌にとって有毒な生成物を発現するグラム陰性細菌(前記有毒な遺伝子は、細菌宿主染色体の1つまたは2つ以上の非必須領域に挿入される);
2.厳密に調節することができる構成的プロモーターの制御下にある有毒生成物をコードする遺伝子が細菌染色体に存在すること;および
3.宿主染色体上で有毒遺伝子に作動できるように連結されたプロモーターを調節する特異的リプレッサーがプラスミドから発現されること。
この設計では、グラム陰性細菌宿主が所定の培養条件下(例えばショ糖の存在下)にあるときは、前記宿主の増殖を制御するのはプラスミドの存在である。
第一の成分では、グラム陰性細菌は形質転換され、レバンスクラーゼをコードするsacB遺伝子を発現する(図1参照)。第二および第三の成分は、cIリプレッサー遺伝子生成物および右λプロモーターに関する。λバクテリオファージのcI遺伝子はλリプレッサーをコードする。
cIリプレッサータンパク質をコードするゲノム領域は免疫領域として知られている。免疫領域は図2に示されている。cI遺伝子生成物を発現するプラスミド系は、λプロモーターの制御下に配置されたsacB遺伝子の転写を阻害するはずである。そのような抑圧はショ糖に対する耐性を付与し、プラスミドの維持および宿主細胞の増殖の両方を可能にするはずである(図1参照)。
図3は“ドラッグレス”の概念を示す。この概念では、cIリプレッサー活性(プラスミドから発現される)が、細胞宿主染色体に存在するλPrプロモーター下に配置されたsacB遺伝子生成物の転写を阻害し、さらにショ糖の存在下における宿主大腸菌の生存活性は、プラスミドから発現されるλcIリプレッサータンパク質の十分なレベルによって担保される。cIリプレッサーとその特異的プロモーターとの結合は、30℃から37℃の温度で最適である。
特定の事例では(例えば温度感受性cI857リプレッサーを用いるとき)、cIリプレッサーは40℃より高い温度ではプロモーターと結合しないであろう。したがって、42℃への切換えは、cIリプレッサーのλPrプロモーターとの結合活性を阻害し、結果として宿主細胞をショ糖(より具体的にはレバン副生成物)に感受性にするはずである。ショ糖の存在下における42℃でのインキュベーションは完全系における機能性の有効性を立証するためのad hoc条件であり、この系の完全な機能性を支持する“負の証明”を構成する(図4および図6C参照)。実際、Catまたはショ糖のどちらかの存在下で増殖するpPB829またはpPB838のどちらかで形質転換した親細胞λPr::sacBpurNΩkanは、LBショ糖寒天に播種し42℃でインキュベートしたとき生存しなかった。このことは、親細胞の生存活性はcI遺伝子を含むプラスミドの維持に依存することを示している。
【実施例2】
【0044】
λプロモーター/レバンスクラーゼ遺伝子を保持する大腸菌宿主株の作製
λPr::sacBpurNΩkanカセットを含む2つの操作宿主株を調製した。この場合、カセット(ラムダプロモーター(λPr)の制御下にsacB遺伝子を含む)にカナマイシンで目印を付し、続いてedAまたはpurN遺伝子の対立遺伝子置換によって大腸菌染色体に導入する。これらの遺伝子は必須ではなく、前記の欠失は増殖速度に影響を与えない。形質転換細胞はショ糖に対して高度に感受性になった。
図8A−Bは、大腸菌染色体にそれぞれedAまたはpurN遺伝子の対立遺伝子置換によって挿入されるべきλPr::sacBpurNΩkanカセットが、結合PCRによって融合される前にPCR増幅された6つの別個の成分を含むことを示す。
図8Dは、λPr::sacBpurNΩkanカセットが、edAまたはpurN遺伝子の対立遺伝子置換によって大腸菌の染色体に挿入されたことを示す。このカセット挿入は、線状DNA鋳型(染色体遺伝子座の上流および下流の相同領域によってフランキングされた欠失遺伝子を含む)で大腸菌を形質転換することによって実施し、さらに組換え熟練大腸菌株(例えばrecD, recB recC sbcA変異体)に伝達した。親株の操作のための大腸菌株はStratageneのBJ5183株(ref# 200154)で、遺伝子型はendA1 sbcBC recBC galK met thi-1 bioT hsdR(Strr)であった。提示したように、この大腸菌野生型の最初の表現型はKanS、CatSおよびSucroseRであった。この最新の表現型は、抗生物質フリープラスミドの機能性の提示に関する更なる実験に用いられた親株のものであった。
図8Cは、Prλ::sacBΩkan大腸菌染色体にedA遺伝子の対立遺伝子置換によって挿入されるべきλPr::sacBΩcatカセットが、結合PCRによって融合される前にPCR増幅された6つの別個の成分を含んでいたことを示す。対立遺伝子交換によりedA遺伝子の欠失を標的とする新規なλPr::sacBΩcatカセットの目的は、2つのsacBカセットを発現する二重欠失変異体ΔedAΔpurNを操作することであった。
図8Eは、親株をcIプラスミド(pPB829またはpPB838)で形質転換したとき、細胞がカナマイシン耐性(KanR)、クロラムフェニコール耐性(CatR)およびショ糖耐性(SucroseR)になったことを示す(親株+cIプラスミド[pPB829またはpPB838])。図は、概念を立証するために、sacBがラムダプロモーターの制御下に配置され、構築物がedAまたはpurN遺伝子の対立遺伝子置換によって大腸菌の染色体に導入されたときに、操作された宿主株(ΔpurNλPr::sacB+Ωkan)または(ΔedAλPr::sacB+Ωkan)およびドラッグプラスミドが同時に存在することを示している。形質転換細胞は高度にショ糖感受性になった。cIリプレッサーを含むクロラムフェニコールの目印をもつプラスミド(pPB838またはpPB829)がΔpurNλPr::sacB+ΩkanまたはΔedAλPr::sacB+Ωkan株に導入されたとき、形質転換細胞はカナマイシン、クロラムフェニコール、およびショ糖耐性になった。
【0045】
λプロモーター(λPr)が、purN欠失のためにPB1232およびPB1233を用いて増幅された。
PB1232プライマー(配列番号:6):
(CCGAACAACGCGTGGTTATCGACACCGCAAGGGATAAATATCTAACACCG) および
PB1232プライマー(配列番号:7)
(CAAACTTTTTGATGTTCATATCCATCTGATCCTCTTCAAAAGGCCACCTG)
λプロモーター(λPr)が、edA欠失のためにPB1234およびPB1233を用いて増幅された。
PB1234プライマー(配列番号:8):
(GACGACAAATTTGTAATCAGGCGAGAGCACCGCAAGGGATAAATATCTAACACCG)。
λプロモーター(λPr)(配列番号:5)の増幅はDNA鋳型としてpLDR8プラスミド(ATCC#77357)を用いて実施した。sacB遺伝子(配列番号:3)は、λPr::sacBΩkanカセットの操作のために結合PCRを用いたときは、PB1192(配列番号:9)(ATGGATATGAACATCAAAAAGTTTGC)およびPB1193(配列番号:10)(AAACAAATAGGGGTTCCGCGCACATTTATTTGTTAACTGTTAATTGTCCTTG)プライマーを用い、DNA鋳型としてはpNB350(Merialの特許物)を用いて増幅した(図8A−B参照)。λPr::sacBΩcatカセットの操作のためには、リバースプライマーPB1320(配列番号:80)(GCCGATCAACGTCTCATTTTCGCCGTTAACAGATCTTTATTTGTTAACTGTTAATTGTCCTTG)がPB1193の代わりに用いられた。blaプロモーターは、PB1194(配列番号:11)(ATGTGCGCGGAACCCCTATTTG)およびPB1195(配列番号:12)(GACGTTTCCCGTTGAATATGGCTCATACTCTTCCTTTTTCAATATTATTGAAGC)プライマーを用い、DNA鋳型としてはpCMVβを用いて増幅された。カナマイシン耐性遺伝子は、PB1196(配列番号:13)(ATGAGCCATATTCAACGGGAAACG)およびPB1197(配列番号:14)(GAAAAACGCCAGCGGCAGAGCTGGCGCTTAGAAAAACTCATCGAGCATCAAATG)プライマーを用い、DNA鋳型としてはプラスミドpLL14(カナマイシン遺伝子の弱いプロモーターを含む、Merialの特許物)を用いて増幅された。その天然のcatプロモーターの制御下に配置されたクロラムフェニコール(cat)耐性遺伝子は、PB1321(配列番号:81)((AGATCTGTTAACGGCGAAAATGAG)およびPB1322(配列番号:82)(AAAACGCTACAAAAATGCCCGATCCTTTACGCCCCGCCCTGCCACTCATCGC)プライマーを用い、DNA鋳型としてはpPB829を用いて増幅された。purN遺伝子の5'フランキング領域は、PB1237(配列番号:15)(TTTGCGGCCGCTGGTGGTGGTCGCCATGTGCGTTAATGACC)およびPB1199(配列番号:16)(TATTCGATAACCACGCGTTGTTCGG)を用い、DNA鋳型としては大腸菌SCS1株のゲノムDNAを用いて増幅された。purN遺伝子の3'フランキング領域は、PB1200(配列番号:17)(GCGCCAGCTCTGCCGCTGGCGTTTTTC)およびPB1238(配列番号:18)(TTTGGATCCGCTGGTGGATATCATCAAGGCAGTAACGCAGAATG)プライマーを用い、DNA鋳型としては大腸菌SCS1株のゲノムDNAを用いて増幅された。edA遺伝子の5'フランキング領域は、PB1235(配列番号:19)(TTTGCGGCCGCTGGTGGTTGAGAACCAGGTGATTGAAGCGCC)およびPB1209(配列番号:20)(CTCTCGCCTGATTACAAATTTGTCGTC)プライマーを用い、DNA鋳型としてはSCS1のゲノムDNAを用いて増幅された。edA遺伝子の3'フランキング領域は、以下のプライマーPB1210(配列番号:21)(AGGATCGGGCATTTTTGTAGCGT)およびPB1236(配列番号:22)(TTTCTAGAGCTGGTGGCGACTACCGTGAATCCTGGCAACC)を用い、DNA鋳型としてはプラスミドpLDR8を用いて増幅された。
【0046】
大腸菌染色体に伝達されるべきλPr::sacBΩkanカセットまたはλPr::sacBΩcatのこれら6つの個々のPCR生成物を結合PCRに続いて融合させた(図8A、B、C)これら6つの個々の増幅フラグメントをPCR clean-upキット(Geneclean turbo kit; MP Biomedicals, CA, USA)を用いて精製し、さらに6つ全てのフラグメントを用いプライマーは使用せずに第2ラウンドのPCRをセットした。PCRの条件は以下のとおりであった:PCR混合物(最終25μL)、1μLの各PCRフラグメント、2μLのdNTP(各々2.5mM、5μLの5x PCR緩衝液、11.5μLの滅菌蒸留水、0.5μLのPhusionDNAポリメラーゼ(Finenzymes, Finland);PCRサイクル:98℃で30秒、(98℃で10秒、60℃で30秒、72℃で5分)を15サイクル、および72℃で10分。第三ラウンドのPCR増幅は、1μLの精製した第2ラウンドの生成物で、purN(対立遺伝子置換)のためにはPB1235およびPB1236を用い、edA(対立遺伝子置換)のためにはPB1237およびPB1238を用いて実施した。PCR条件は以下のとおりであった:PCR混合物(最終50μL)、1μLの第二のPCRフラグメント、1μLのdNTP(各々2.5mM、10μLの5x PCR緩衝液、37μLの滅菌蒸留水、0.25μLのフォーワードプライマー、0.25μLのリバースプライマー、0.5μLのPhusionDNAポリメラーゼ(Finenzymes, Finland);PCRサイクル:98℃で30秒、(98℃で15秒、60℃で30秒、72℃で4分)を35サイクル、および72℃で10分。最終的なPCR結合アンプリコン(purN対立遺伝子置換については4007bp、およびedA対立遺伝子置換については3407bp)は、アガロースゲルでチェックし、大腸菌の形質転換前に精製した。各結合PCRアンプリコンは制限分析によってチェックした。
標的遺伝子座(edAまたはpurN)の境界のDNA配列に相同な2つの長い領域によってフランキングされた、λPrプロモーター(配列番号:75)の制御下に配置されたsacB遺伝子およびblaプロモーターの制御下に配置されたKan耐性遺伝子をコードする線状DNAフラグメント(λPr::sacBΩkan)が、エレクトロポレーションによって大腸菌の染色体に組み込まれた。ほぼ300の形質転換候補体(ΔpurNλPr::sacBΩkanまたはΔedAλPr::sacBΩkanシングル変異体)が、選別圧としてカナマイシンを含むLB寒天プレートで得られた。20個のコロニーをランダムに採取し、カナマイシン含有プレートでストリーキングすることにより精製し、PCRによってedAまたはpurN遺伝子座へのλPr::sacBΩkanカセットの挿入を立証した(図9B参照)。PB1196およびPB1197を用いたPCRを実施し、染色体へのKanamycinの挿入をチェックした。PB1192およびPB1193を用いてPCRを実施し、染色体にsacB遺伝子が存在することを立証した。PB1204(配列番号:23)(GTGGTGCTTATTTCCGGCAACGG)およびPB1205(配列番号:24)(CCAGCCACGCGGCGTTTTCGTGC)を用いてPCRを実施し、染色体にpurN遺伝子が存在しないことを立証した。PB1214(配列番号:25)(GACCACCGGCCCGGTTGTACCGG)およびPB1215(配列番号:26)(CGGACCCGCGATCGCCTGCAGG)を用いてPCRを実施し、edA遺伝子が存在しないことを立証した(図9B参照)。PB1213(配列番号:27)(GGTGGATGGCGTCCATTTCTGTGC)およびPB1196プライマーを用いてPCRを実施し、edA遺伝子座の右側に正しくカセットが挿入されていることを立証した。PB1212(配列番号:28)(CAAAAGTGTTAAGCGGTAACCTG)およびPB1233プライマーを用いてPCRを実施し、edA遺伝子座の左に正しくカセットが挿入されていることを立証した。PB1203およびPB1196プライマーを用いてPCRを実施し、purN遺伝子座の右に正しくカセットが挿入されていることを立証した。PB1202およびPB1233プライマーを用いてPCRを実施し、purN遺伝子座の左に正しくカセットが挿入されていることを立証した(図9C参照)。全てのPCRチェックによって正しい遺伝子座への組込みが確認された。さらにまた、sacB遺伝子の機能性もまた図10Bに示すように確認された。この操作した親株ΔpurNλPr::sacBΩkanはショ糖を含むLB寒天プレートでは増殖できなかった(図10)。
標的遺伝子座(edA)の境界のDNA配列に相同な2つの長い領域によってフランキングされた、sacB遺伝子(λPrプロモーターの制御下に配置されている)およびCat耐性遺伝子(その天然のcatプロモーターの制御下に配置されている)をコードする線状DNAフラグメント(λPr::sacBΩcat)が、エレクトロポレーションによってΔpurNλPr::sacBΩkan大腸菌株の染色体に組み込まれた。ほぼ200の形質転換候補体(ΔpurNλPr::sacBΩkanΔedAλPr::sacBΩcatダブル変異体)が、選別圧としてクロラムフェニコールを含むLB寒天プレートで得られた。20個のコロニーをランダムに採取し、クロラムフェニコール上でストリーキングすることにより精製し、プライマーセットPB1212およびPB1213を用いたPCRによって、edA遺伝子座へのλPr::sacBΩcatカセットの挿入を立証した(図9C参照)。PCR生成物はシークェンシングによってチェックし、λPr::sacBΩcatとの同一性を立証した。
宿主株の操作は図8Dに示されている。図8Eは、宿主株(1つのsacBカセット[λPr::sacBΩkan])およびドラッグレスプラスミドの同時存在を示す。最初の大腸菌の遺伝子型はKanS、CatS、SucroseRである。大腸菌ΔpurN Prλ::sacBΩkanの染色体へのカセットの組込みは、新規な遺伝子型KanR、CatS、SucroseSを生じた。cIリプレッサーを保持するプラスミドの導入によって、最終的な遺伝子型KanR、CatR、SucroseRが生成された。
【実施例3】
【0047】
cIリプレッサー発現カセットを保持するプラスミドの作製(pPB838-pPB844からpPB847-pPB885-pPB896)
プラスミド構築の要旨
pPB828から誘導したプラスミド、例えばpPB829およびpPB844-pPB847を作製した。前記は、それ自身のプロモーターまたはカナマイシン遺伝子の弱いプロモーター(P1)のどちらかの制御下にあるλファージのcI ORFを含む。カナマイシン遺伝子の弱いプロモーター(P1)の制御下に配置されたcI遺伝子を含むpPB383プラスミドを中間プラスミドの市販ベクターpMCS5を用いて作製した(pMCS5ではアンピシリン耐性遺伝子がクロラムフェニコールアセチルトランスフェラーゼ遺伝子(cat)に置換されていた)(図19)。
pPB844およびpPB845プラスミドはcI遺伝子(P1プロモーターの制御下に配置されている)を含む。前記cI遺伝子は、cat遺伝子を含むpVR1012誘導体(pVR1020または1012プラスミド、VICAL Inc.(Luke et al. 1997;Hartikka et al. 1996;例えば米国特許5,846,946号および6,451,769号参照))であるpPB828でクローニングされた。pPB844およびpPB845プラスミドは、cI遺伝子の当該プラスミド中での向きが異なっている(図20−21)。
pPB846およびpPB847プラスミドはcI遺伝子(それ自身の(天然の)プロモーター(配列番号:30)の制御下に配置されている)を含む(前記cI遺伝子は、cat遺伝子を含むpVR1012誘導体であるpPB828でクローニングされた)。これらのプラスミドは、cI遺伝子の当該プラスミド中での向きが異なっている(図22−23)。
pPB885はpPB838プラスミド(cI遺伝子はカナマイシン遺伝子の弱いプロモーター(P1)の制御下に配置されている)から調製され、前記では、クロラムフェニコールアセチルトランスフェラーゼ遺伝子(cat)はAvrII制限消化によって除去された(図24)。
pPB829から誘導されたプラスミド、例えばpPB896は、カナマイシン遺伝子の弱いプロモーター(P1)の制御下に配置されたcI遺伝子を含む。pPB896プラスミドは、真核細胞CMVプロモーターの制御下に配置されたGFPマーカー遺伝子を含む(図25)。
これらのプラスミドをドラッグレス系で異種ポリヌクレオチド、対象の遺伝子(トランスジーン)のクローニングのためにベクターとして用いた。各プラスミドは抗生物質耐性遺伝子(cat)を含み、前記は、固有の制限酵素によって切り出して除去し、適切な大腸菌sacB+株でショ糖の存在下において抗生物質選別圧(例えばクロラムフェニコール)によることなくプラスミドの増殖を可能にすることができる。
これらのプラスミドの抗生物質耐性遺伝子(cat)は概念の証明のためにのみ用いられた。最終的なドラッグレスプラスミドではこのcat遺伝子は最終的に除去される(図13−15および実施例4を参照)。
プラスミドpLDR8(ATCC No#77357)由来のcI遺伝子はリプレッサータンパク質cI857変異体を生成する(NCBI: AB248924, Cloning vector pND707, C.A. Love et al. Gene 1996, 17; 176(1-2):49-53)。
【0048】
A.弱いプロモーターP1の制御下に配置されたcIリプレッサーORFを含むプラスミドpPB829の構築
pPB828プラスミド:(pVR1012系プラスミド+Cat遺伝子)(図17)
pPB828誘導プラスミド、例えばpPB829を作製した。前記はカナマイシン遺伝子の弱いプロモーター(P1)の制御下にあるλファージのcI ORFおよび固有のEcoRI消化部位によって切り出すことができるCat遺伝子を含む。クロラムフェニコールアセチルトランスフェラーゼ(cat)遺伝子を含み、pPB828プラスミドを生じる、pVR1012系発現ベクターを作製した。pPB627(Merialの特許物質)由来のcat遺伝子はpNB335(Merialの特許物質)の誘導体であった(pNB335はそれ自体最初のプラスミドpSW23T(クローニングプラスミドGenBankアクセッション番号AY733066)の誘導体である)。cat遺伝子に対応するDNAフラグメント(798bp)(DNAについては配列番号:31、タンパク質については配列番号:32)は、プライマーPB1184(配列番号:33)(GAATTCCGGTCCGGGCGAAAATGAGACGTTGATCGGC3)(EcoRI部位(下線部)を含む)およびPB1185(配列番号:34)(CCTAGGCTGTGTTAATTAAGGCGCGCCGAATTCCGGTCCGTTACGCCCCGCCCTGCCACTCATCGC)(EcoRI部位(下線部)を含む)を用い、鋳型のpNB350およびPhusionTM High-Fidelity DNAポリメラーゼ(Finnzymes, 02150 Espoo, Finland)を用いてPCRによって入手した。得られたPCR生成物(832bp)をpCRblunt Topoベクター(Invitrogen, CA, USA)に連結し、プラスミドpCRblunt/PB1184-1185(4351bp)を得た(図16)。pCRblunt/PB1184-1185の実体は制限分析(PvuII消化)によって確認した。Cat遺伝子に対応するDNAフラグメントは、pCRblunt/PB1184-1185プラスミドのEcl136IIおよびEcoRVによる酵素消化によって入手した。897bpフラグメントをpVR1012系プラスミド(先にMscIおよびStuIで消化しておいた(3303bp))に連結してプラスミドpPB828を作製した(図17)。pPB828の実体は制限分析によって確認した(挿入物のサイズ決定のためにはPvuIIおよびNcoI消化、および遺伝子catが良好に除去されていることを立証するためにはEcoRI消化)。
pPB829プラスミド:(pPB828プラスミド+P1::cI遺伝子)(図18)
P1プロモーター(カナマイシン耐性遺伝子の弱いプロモーター)の制御下にあるcI遺伝子を含む構築物は融合PCRを用いて入手した。弱いプロモーターP1に対応するDNAフラグメントは、プライマーPB1186(配列番号:35)(TCATACCAGGCCTAGGTGATACGCCTATTTTTATAGGTTAATG)およびPB1187(配列番号:36)(AACACCCCTTGTATTACTGTTTATG)を用い、鋳型としてpLL14(Merialの特許出願US2005/0164946を参照されたい)、さらにPhusion DNAポリメラーゼを用いたPCRによって入手した。cI遺伝子(配列番号:1)に対応するDNAフラグメントは、プライマーPB1188(配列番号:37)(AATACAAGGGGTGTTATGAGCACAAAAAAGAAACCATTAACAC)(5'末端のP1配列の相補性領域(下線部)を含む)およびPB1189(配列番号:38)(CCGGAATTCGGCGCGTCAGCCAAACGTCTCTTCAGGCCACTG)を用い、鋳型としてpLDR8(ATCC#77357)、さらにPhusion DNAポリメラーゼを用いたPCRによって入手した。2つのPCR生成物(それぞれ151および729bp)を精製し、第二のPCR工程(PB1186およびPB1189プライマーおよびPhusion DNAポリメラーゼ(Finnzymes, Finland)を使用)で鋳型として用いた(図24参照)。2つのPCR生成物(それぞれ151および729bp)を精製し、AscIおよびAvrIIで消化したpPB828と連結して、プラスミドpPB829(4970bp)を作製した(図9A)。連結は、Clontech(Takara Bio Europe, St-Germain-en-Laye, France)の“In-Fusion PCR Cloning Kit”(Cat No. 740590.250)を用いて実施した。HindIII消化分析に続いてクローンpPB829を選別した。cI遺伝子およびP1プロモーターの配列の完全性は、プラスミドpPB829のシークェンシングによって確認した。
pPB896プラスミド:(pPB829プラスミド+P1::cI遺伝子+GFP)(図25)
GFPマーカー遺伝子は、真核細胞CMVプロモーター(配列番号:43)の制御下に配置した。GFP遺伝子は、pCG105プラスミド(真核細胞CMVプロモーターの制御下に配置されたGFP遺伝子を有するpPB828誘導プラスミド、Merialの特許物)のNotI/BglIIの両方による制限消化により調製した。この消化GFP遺伝子を、先にNotI/BglIIで切断しておいたpPB829プラスミドでクローニングしpPB896プラスミドを生成した。
【0049】
B.中間プラスミド(pPB838)およびP1プロモーター(カナマイシン遺伝子の弱いプロモーター)の制御下にあるcI遺伝子を含むプラスミドpPB844-pPB845の構築
pPB837.1プラスミド:(pMCS5プラスミド+Cat遺伝子)
Cat遺伝子に対応するDNAフラグメントは、プライマーPB1182(配列番号:39)(AGATCTGTTAACGGCGAAAATGAGACGTTGATCGGC)(5'末端にBglII部位を含む)およびPB1183(配列番号:40)(GTCGACGTTAACTTACGCCCCGCCCTGCCACTCATCGC)(5'末端にSalI部位を含む)、鋳型としてpPB791(プラスミドpNB350の誘導体)、およびPhusion DNAポリメラーゼを用いてPCRによって入手した。
PCRプライマーはpNB350のcat配列を土台にして設計した。3つの別個のPCR生成物(779bp)をプールし、pCRIIベクターに連結してプラスミドpCRII+PB1182-1183(4734bp)を入手した。pCRII+PB1182-1183のクローンをSalI/BglII消化に続いて選別した。DNAフラグメントA(2950bp)を得るために、pMCS5プラスミド(MoBiTec, Germany)をSalIおよびBglIIで消化した。pCRII+PB1182-1183をSalIおよびBglIIで消化し、フラグメントSalI-BglIIをアガロースゲルから単離した(797bp:フラグメントB)。フラグメントAおよびBを連結してプラスミドpPB837.1(3747bp)を生成した。
pPB837.2:blaプロモーターをもたない(pMCS5+Cat)
blaプロモーター領域を含まないプラスミドpPB837.2を作製した。プラスミドpPB837.1をNaeIおよびXmnIで消化し、3300bpのフラグメントNaeI-XmnIを単離し、アガロースゲルで精製した(フラグメントA)。このフラグメントAを連結してpPB837.2(3300bp)を得た。BglI消化に続いてクローンpPB837.2を選別した。
pPB838プラスミド(blaプロモーターのないpMCS5+Cat)+P1::cI(図22)
P1プロモーター(カナマイシン耐性遺伝子の弱いプロモーター)の制御下にあるcI遺伝子を含む構築物は融合PCRを用いて入手した。弱いプロモーターP1に対応するDNAフラグメントは、プライマーPB1186およびPB1187、鋳型としてPLL14、およびPhusion DNAポリメラーゼを用いてPCRにより入手した。cI遺伝子に対応するDNAフラグメントは、プライマーPB1188およびPB1189、鋳型としてpLDR8(ATCC#77357)、およびPhusion DNAポリメラーゼ(Finnzymes. Finland)を用いてPCRによって入手した。2つのPCR生成物(それぞれ151および729bp)を精製し、第二のPCR工程でPB1186およびPB1189プライマー並びにPhusion DNAポリメラーゼとともに鋳型として用いた。2つのPCR生成物(880bp)をプールし、pCRIIbluntベクターに連結してプラスミドpCRblunt+PB1186-1189(4400bp)を得た。クローンpCRblunt+PB1186-1189をEcoRV/SpeI消化に続いて選別した。
EcoRV-SpeIフラグメントA(936bp)を得るために、pCRblunt+PB1186-1189をEcoRVおよびSpeIで消化した。pPB837.2をEcoRVおよびSpeIで消化し、フラグメントEcoRV-SpeIをアガロースゲルから単離した(3464p:フラグメントB)。フラグメントAおよびBを連結してプラスミドpPB838(4216bp)を生成した。HindIII消化に続いてクローンpPB838を選別した。cI遺伝子およびP1プロモーターの配列の完全性はシークェンシングによって確認した。
pPB885プラスミド(blaプロモーターのないpMCS5+Cat)+P1::cI(図24)
このプラスミドはpPB838プラスミドから誘導した(前記では、クロラムフェニコールアセチルトランスフェラーゼ遺伝子(cat)はAvrII制限消化によって除去された)(図24)。pPB885プラスミドは、カナマイシン耐性遺伝子の弱いプロモーター(P1)の制御下にcI遺伝子を含む。pPB885プラスミドは、ΔpurNλPr::sacBΩkan大腸菌宿主株の形質転換に続いて、実施例4(下記に示す)に記載するように10%ショ糖補充LB寒天プレート上での直接選別によって単離された。
プラスミドpPB844-pPB845(図23−24)
P1プロモーターの制御下のcI遺伝子に対応するDNAフラグメントは、プライマーPB1186およびPB1263(配列番号:41)(TCAGCCAAACGTCTCTTCAGGCCAC)、鋳型としてpPB838、並びにPhusion DNAポリメラーゼを用いてPCRによって入手した。4つの別個のPCR生成物PB1186-PB1263(864bp)をプールし、pCRbluntベクターに連結してプラスミドpCRblunt+PB1186-1263(4383bp)を入手した。pCRblunt+PB1186-1263のクローンをHindII消化に続いて選別した。
pCRblunt+PB1186-1263プラスミドをEcoRIで消化し、末端をクレノーポリメラーゼで充填して平滑端フラグメント(929bp:フラグメントA)を生成した。pPB828プラスミドをXmnIで消化し、得られたXmnI-XmnIフラグメント(フラグメントB:4135bp)を精製した。フラグメントAおよびBを連結してプラスミドpPB844またはpPB845を生成した。候補物をBamHI/HindIII消化によってスクリーニングした。pPB844プラスミド(図20参照)では、cI遺伝子は、ベクターのMCS(マルチクローニング部位)でクローニングされるべき対象のトランスジーンとCMVプロモーター(配列番号:43)の制御下で同じ向きに存在する。pPB845プラスミド(図21参照)では、cIは反対方向に向いている。
【0050】
C.その天然のプロモーターをもつcI遺伝子プラスミドpPB846-pPB847の入手
それ自身のプロモーター(cIの天然のプロモーター:配列番号:30)の制御下のcI遺伝子に対応するDNAフラグメントは、プライマーPB1266(配列番号:42)(GCTGACTCATACCAGGCACGCACGGTGTTAGATATTTATCCC)およびPB1263、鋳型としてpLDR8、並びにPhusion DNAポリメラーゼを用いてPCRによって入手した。
pPB846-pPB847プラスミド
4つの別個のPCR生成物PB1266-PB1263(777bp)をプールし、pCRbluntベクターに連結してプラスミドpCRblunt+PB1266-1263(4296bp)を入手した。pCRblunt+PB1266-1263のクローンをHindII消化に続いて選別した。
pCRblunt+PB1266-1263プラスミドをEcoRIで消化し、末端をクレノーポリメラーゼで充填して平滑端フラグメント(842bp:フラグメントA)を生成した。pPB828プラスミドをXmnIで消化し、得られたXmnI-XmnIフラグメント(フラグメントB:4135bp)を精製した。フラグメントAおよびBを連結してプラスミドpPB846またはpPB847を生成した。候補物をBamHI/HindIII消化によってスクリーニングした。pPB846プラスミド(図22参照)では、cI遺伝子は、ベクターのMCS部位でクローニングされるべき対象のトランスジーンとCMVプロモーターの制御下で同じ向きに存在する。pPB847プラスミド(図23参照)では、cI遺伝子は反対方向に向いている。
D.構築の結果
図7は、ドラッグレスプラスミドの調製に用いられたcIリプレッサーを保持するpPB838およびpPB829の構築を示す。両プラスミドはクロラムフェニコール(CatR)遺伝子で目印を付されていた。これらのプラスミドのCat遺伝子は概念の証明のためにのみ要求されたが、最終的には除去することができる(図13−15および実施例4を参照されたい)。
【実施例4】
【0051】
大腸菌宿主細胞のcIプラスミドによる形質転換およびショ糖寒天プレートでの直接選別
クロラムフェニコール(Cat)の目印をもつcIプラスミドによって形質転換した細胞を選別圧として抗生物質を使用することなく寒天プレートでスクリーニングできる実験的方法は有効であることが示された(図13−15参照)。大腸菌λPr::sacB+ΔpurNΩkanのコンピテント細胞を1μgのpPB838またはpPB829プラスミドで形質転換し、LB液体ブロスにて30℃で3時間インキュベートした。希釈した(10-3から10-4)形質転換細胞(pPB838またはpPB829プラスミドで)の100μLアリコットを10%のショ糖を含むLB寒天プレートに播種した。10-3から10-4の範囲のこれら希釈物は本物の形質転換細胞(例えばcIプラスミドを有する)の選別に適切であるように思われる。なぜならば、ショ糖含有LB寒天プレートで増殖するショ糖耐性細胞の98−100%がCatRであったからである。このことは、これら形質転換細胞はcIプラスミド(pPB838またはpPB829)を有することを示唆している。
図13Aは、大腸菌λPr::sacB+ΔpurNΩkanのコンピテント細胞が、1μgのpPB838またはpPB829プラスミドで形質転換され、ドットスポット(各ドットに4μL)の前にLB液体ブロスにて30℃で3時間インキュベートされたことを示す。図は、クロラムフェニコール(15μg/mL)またはショ糖(10%)を含むLB寒天プレートでのLog希釈(未希釈から6Log希釈の範囲)を示す。大腸菌λPr::sacB+の非形質転換コンピテント親株細胞を同様な条件下での標準細胞増殖として用いた。期待したとおり、標準細胞はcatおよびショ糖のような選別プレートでは増殖しなかった。それにもかかわらず、長期間(3日間)のインキュベーションのために、いくつかの偶発的ショ糖耐性変異体がこの条件下で出現した。このレベルの偶発的変異体は、本物の形質転換細胞のスクリーニングについて考慮されるべきベースラインを規定する。この実験条件下では、2Log差がcatおよびショ糖で増殖する形質転換細胞で観察された。前記各規定のコンピテント細胞は、一時的に細胞壁の脆弱を示す。形質転換細胞をエレクトロポレーション後にLB液体培地でインキュベーションすることによって、エレクトロポレーションに続く前述の細胞壁の脆弱並びに細胞壁および細胞膜の誘発孔の復活が可能になる。コンピテント細胞は、λPr::sacB+ΔpurNΩkanカセットの挿入のために遺伝的にショ糖に感受性であるので、ショ糖の存在下での形質転換細胞の回復は、Catでの細胞の回復と比較して2Log低下した。これらの観察は、100μLの形質転換細胞を選別プレートに播種することによって確認された(図13B参照)。
図13Bは、catおよびショ糖上での形質転換有効率を決定し、さらにLBショ糖寒天プレートで増殖できる本物の形質転換細胞のスクリーニングの成功のために考慮すべき偶発的変異体出現率のベースラインをより厳密に規定するために、100μLの形質転換細胞および非形質転換標準細胞を適切なLog希釈を用いてcatまたはショ糖含有LB寒天プレートに播種したことを示す。以前のドットスポット細胞実験(図13A参照)と一致して、偶発的ショ糖耐性変異体は、形質転換細胞の10-3希釈をショ糖含有LB寒天プレートに播種したときは検出できなかった。以前に示したように(図13A参照)、この条件下ではcatおよびショ糖上での選別で形質転換効率に2Log差が存在した。10-3から10-4の範囲の希釈は、ショ糖含有LB寒天に播種したとき、本物の形質転換細胞(例えばcIプラスミドを有する)の選別に適切であるように思われた。
図14A−Cは、選別圧として抗生物質を使用することなく、プラスミド(pPB829およびpPB838)で形質転換した細胞を選別する実験的方法の有効性を示す。
図14Aは、cIプラスミドを有する本物の形質転換細胞のスクリーニングの有効性を立証するためにレプリカプレートアッセイを用いた実験の概要を示す。図13Bに示したものと同様に、コンピテント親細胞λPr::sacB+ΔpurNΩkanをcIプラスミド(pPB829およびpPB838)で形質転換し、30℃で3日間インキュベートする前に適切なLog希釈物を選別(Catまたはショ糖)LB寒天プレートに播種した。その後、20から50の範囲のCFU(コロニー形成単位)を示したプレートのレプリカを、30℃でさらに3日間インキュベートする前にCatまたはショ糖またはその組合せを含む別の新しいLB寒天プレートで作成した。理論的には、Cat耐性コロニーはcIプラスミドを増殖させる細胞と一致するはずである。
図14Bは、CatR CFUは、レプリカプレートアッセイ後にショ糖LB寒天プレートおよびCat/ショ糖LB寒天プレートで得られた数と同一であることを確認した実験結果を示す。図14B(パネルB)に示すように、CatR CFUは、i)全てがショ糖耐性、ii)全てがCatおよびショ糖の組合せに耐性であり、全ての親株は形質転換後にcIプラスミドを保持することを示している。パネルCは、ショ糖上で直接選別した10-3および10-4希釈の形質転換細胞はまた、ショ糖耐性であり、さらにCatおよびショ糖の組合せに耐性であることを示し、これらショ糖耐性CFUはまたcIプラスミドを含む形質転換細胞と一致することを示す。実際、非形質転換親細胞をショ糖含有LB寒天プレートに播種したとき、ショ糖耐性CFUは、ショ糖およびCatの組合せ上で複製させたとき期待したように増殖しなかった(クロラムフェニコール耐性はpPB829またはpPB838プラスミドのどちらかによって持ち込まれた表現型の属性であるからである)。
図14Cは、レプリカプレートアッセイ(図14B参照)から得られた統計学的データ、より具体的には、抗生物質を使用することなく選別されたcIプラスミド保有コロニーのパーセンテージを示す。この実験条件下では、cIプラスミド保有コロニーのパーセンテージは98%から100%の範囲である。
【実施例5】
【0052】
cIプラスミドを維持できることを示す、ショ糖が存在するまたは存在しない種々の培養条件
大腸菌の染色体に挿入されたλPr::sacB+カセットの機能性をチェックするために、 sacBを発現する大腸菌細胞、ΔpurNλPr::sacB+Ωkanをショ糖の存在下または非存在下で増殖させた。
図4はこの不活化系の“ショ糖感受性”を示す(前記不活化系では、42℃への切換えは、cIリプレッサーのλPrプロモーターとの結合活性を阻害し、宿主細胞をショ糖感受性にする)。cIリプレッサータンパク質(プラスミドから発現される)は、宿主染色体に位置するλPrプロモーターの制御下に置かれた有毒なsacB遺伝子生成物の転写を阻害する。ショ糖の存在下での宿主大腸菌細胞の生存活性は、プラスミドから発現されるλcIリプレッサータンパク質の十分なレベルによって担保される。図5は、ショ糖存在下での宿主細胞と抗生物質フリーpDL1プラスミドとの組合せによるドラッグレスプラスミド(pDL:DrugLess Plasmid)の生成を示す。概念の証明のための実験計画はさらに図6A、6Bおよび6Cに示される。
ショ糖の存在下または非存在下で増殖させたときのsacB発現宿主細胞、ΔpurNΩλPr::sacB+Ωkanの生理学的特徴を表1に示す。
表1:ショ糖の存在下および非存在下で増殖させたときのsacB+宿主細胞(ΔpurNΩλPr::sacB+)の生理学的特徴
【0053】
ショ糖の存在下または非存在下で増殖させたときの、pDL1プラスミド保持および非保持sacB発現宿主細胞(λPr::sacB+)の生理学的特徴を表2に示す。
表2:ショ糖の存在下および非存在下で増殖させたときのpDL1プラスミド保持および非保持(λPr::sacB+)宿主細胞の生理学的特徴
図10−11は、遺伝的に操作した宿主株は30−37℃でショ糖に対して高度に耐性であるが、42℃では増殖は認められないことが確認された結果を提供する。図10は、pPB829プラスミドまたはpPB838プラスミドを保持する大腸菌宿主株(ΔpurNλPr::sacB+Ωkan)の2回目の継代はショ糖の存在下において32℃で良好に増殖したことを示す。sacB遺伝子発現は、両方のcIプラスミドから合成されたcI遺伝子生成物によって完全に抑圧された。プラスミドの維持は100%有効で(CatRのコントロールに対して)、細胞はショ糖の存在下で生存できた。42℃では、cI遺伝子生成物は不活化された。プラスミドは維持されず、細胞はショ糖の存在下で死滅した。このことは、ショ糖の存在下における細胞の生存活性は機能的cIプラスミドの存在に帰せられることを示している。
【実施例6】
【0054】
抗生物質選別圧を使用せずに5継代にわたってcIプラスミドが安定であることを示す培養条件およびショ糖耐性の偶発的変異体の欠如
ドラッグレスプラスミドを生成したら、安定性およびショ糖またはCat含有新鮮ブロス培養液中での数代にわたるそれらの強固さについて試験した。
図11は、増殖細胞中でのpPB829およびpPB838プラスミドの安定性を示す。前記安定性には、クロラムフェニコールまたはショ糖による選別中に明白な遺伝的再編成が存在しないことが含まれる。ショ糖およびクロラムフェニコールの存在下で増殖する培養に匹敵し得るプラスミドの収量および維持が5連続継代の後でも認められた。抗生物質系および非抗生物質系は、プラスミドの維持および増殖細胞からの選別に関しては少なくとも等価であった。
図12は、細胞を30℃で増殖させたときは、親大腸菌に挿入されたλPr::sacBカセットで偶発的変異は発生しないことを示す。この観察は、細胞をショ糖の存在下で増殖させたとき、そのような大腸菌の遺伝的背景におけるcIプラスミドの安定性をはっきりと示している(図10参照)。この変異発生率実験は、親株(purNが欠失しsacB遺伝子を発現する)を用いて実施した。30℃では18時間インキュベートした後で変異は発生しないことが示された。染色体にsacB+カセットを保持する親細胞のクローン培養をショ糖含有LBで増殖させた。細胞を1.0のODで採集し、100μLアリコットをショ糖含有LB寒天プレートに播種した。sacB遺伝子の存在は細胞死を引き起こすので、偶発的変異発生率は、30℃での一晩インキュベーション後のショ糖耐性コロニーのスコアによって判定した。sacB+カセットを保持するがcIプラスミド(pPB829およびpPB838)で形質転換されていない標準細胞はショ糖耐性となり、スコア判定のために、ショ糖補充LB寒天プレートに播種(100μL)する前にOD1.0の培養を5Log希釈することが必要であった。一晩(18時間インキュベーション)培養の後で親細胞培養(ΔpurNλPr::sacBΩkan)に関してショ糖耐性コロニーは検出されず、一方、pPB829およびpPB838プラスミドで形質転換したΔpurNλPr::sacBΩkanの5Log希釈は増殖した。細胞を37℃でインキュベートしたとき、36時間がいくつかのショ糖耐性コロニー(ΔpurNλPr::sacBΩkan)の検出に必要であった。
37℃でインキュベートしたときには、偶発的変異発生は、36時間後に1.3 x 105細胞当たり1変異発生率で生じた。ショ糖の存在下で増殖している細胞でのcIプラスミドの維持は30℃での一晩インキュベーションで有効であるが、一方、親株の増殖は検出されなかった。
【実施例7】
【0055】
前述の1つのsacBカセット保有大腸菌宿主株(ΔpurNλPr::sacBΩkan)に第二のsacBカセット(ΔedAλPr::sacBΩcat)を付加することによる、ショ糖に対する偶発的変異発生率低下のための親大腸菌宿主株の改良
図8Cおよび図9Dの両図は、2つのsacBカセット(ΔpurNλPr::sacBΩkan ΔedAλPr::sacBΩcat)を保有する大腸菌の新規な親宿主株の操作態様を示す。
図10B−Cは、二重sacBカセット大腸菌株(ΔpurNλPr::sacB+ΩkanΔedAλPr::sacB+ΩCat)は、30℃および42℃でインキュベートしたとき、ショ糖の存在下で高度に感受性である(増殖しない)が、一方、1つのsacBカセットを有する大腸菌株に関しては10%ショ糖に播種したときでさえ、いくつかの偶発的変異体(ΔpurNλPr::sacBΩkan)が出現したことを示す。
図10Cはまた、1つのsacBカセットをもつ大腸菌(最終ショ糖濃度10%)と比較して二重sacBカセット大腸菌株が要求する最低ショ糖濃度(最終2%)を示す。さらにまた、低いショ糖%および最高温度(例えば42℃)で示されるように、2つのsacBカセットをもつ大腸菌株はショ糖に対してはるかに感受性のようである。このことは、約37℃の温度および約2%の(またはわずかに低い)ショ糖濃度が、増殖細胞でcI遺伝子リプレッサーを保持するプラスミドを維持するための最適条件と一致し得ることを示唆する。
図12B−Cは、2つの(1つではなく)sacBカセットの存在は、30℃から37℃の範囲の温度でショ糖感受性に対する強固さの向上を付与することを示す(しかもなお2%ショ糖が最適である)。2つのsacBカセットを含むプラスミドで形質転換した大腸菌細胞、BJ5183ΔpurNΩλPr::sacBKmΔedaΩλPr::sacBCatクローン#1およびBJ5183ΔpurNΩλPr::sacBKmΔedaΩλPr::sacBCatクローン#2、並びに1つのsacBカセットを含むプラスミドで形質転換した大腸菌細胞、BJ5183ΔpurNΩλPr::sacBKmクローン#16およびBJ5183ΔedaΩλPr::sacBKmクローン#11を約1.0のOD600まで増殖させた。各培養の(10-5希釈)約100μLを、0%、2%および4%のショ糖を含むLB/寒天プレートにCFUスコア判定のために播種した。このプレートを30℃から37℃でインキュベートした。37℃でのインキュベーションは、2%および4%のショ糖の存在下における二重sacBカセット発現親株の強固さを判定するために用いた。図12B−Cおよび下記の表3−7は、大腸菌染色体中の1つのsacBカセットの存在は、同様の温度(例えば30℃および37℃)での2%から4%の範囲のショ糖濃度では強固さは低下することを示す。このプレートアッセイで確認されたように、2つのsacBカセットをもつ大腸菌株に関するショ糖に対する変異発生率は、37℃でインキュベートしたとき最低ショ糖濃度(2%)では検出できなかったが、1つのsacBカセットをもつ大腸菌株に関する変異発生率は、3.8 10-6から5 10-5の範囲であった。別個の実験セットによって、2つのsacBカセットをもつ大腸菌株の変異発生率は、2%のショ糖の存在下において37℃でインキュベートしたときほぼ5.10-10であることが示された。
【0056】
表3
表4
表5
表6
表7
1*:BJ5183ΔpurNΩλPr::sacBKmΔedaΩλPr::sacBCatクローン#1
2*:BJ5183ΔpurNΩλPr::sacBKmΔedaΩλPr::sacBCatクローン#2
3*:BJ5183ΔpurNΩλPr::sacBKmクローン#16
4*:BJ5183ΔedaΩλPr::sacBKmクローン#11
【実施例8】
【0057】
抗生物質選別圧の非存在下で得られるプラスミドの維持および収量(cIプラスミドは宿主株内において高コピー数で維持される)
sacBを発現しcIプラスミド(pPB829またはpPB838)を保持する大腸菌宿主細胞を抗生物質選別圧の非存在下(例えばショ糖の存在下)で、またはコントロールとしてCatの存在下で増殖させた。
図11では、プラスミド収量は2および5継代で決定した。30℃で維持した培養からプラスミドを抽出し、ゲル電気泳動に付した。HindII制限消化は、増殖中およびCatまたはショ糖による選別中にpPB829またはpPB838で明白な遺伝的再編成は発生しなかったことを示している。さらにまた、Catまたはショ糖選別圧下でのcIプラスミド選別中に、同様なcIプラスミド維持が認められた。
図15A−Bは、培養におけるcIプラスミド(pPB829およびpPB838、sacBカセットの1コピーを含む)の安定性および維持の有効性を示す。前記安定性には、クロラムフェニコールまたはショ糖による選別後に遺伝的再編成が発生しないことが含まれる。ショ糖またはクロラムフェニコールの存在下で増殖させたcIプラスミドの効率的なプラスミド維持および安定性が観察された。さらにまた、プラスミド収量は、pPB829またはpPB838で形質転換した親宿主株を選別圧としてショ糖の存在下において30℃で増殖させたとき、選別圧としてCatの存在下で培養したときよりもはるかに高かった(約700/細胞)。プラスミドの維持および収量は極めて効率的であり、コピー数は、Catの存在下で培養した細胞と比較して2倍多かった。宿主細胞をショ糖の存在下で培養したときのこのcIプラスミド収量の向上は、cI発現を制御する弱いプロモーター(P1)によるものであった。実際、カナマイシン遺伝子の弱いプロモーター(P1)の制御下でのcIリプレッサーの発現は、λPrプロモーターの制御下に置かれたsacB遺伝子の毒性をより良好に無効にするためにプラスミド収量に対して正の作用を有していた。
以前に示したように(図11A参照)、抗生物質系および非抗生物質系は、プラスミドの維持およびcIプラスミド保持増殖細胞の選別に関しては等価である。図15Bは、プラスミド収量(コピー数)は、pPB829またはpPB838で形質転換した親宿主株を選別圧としてショ糖の存在下で増殖させたとき、選別圧としてCatの存在下で培養したときよりもはるかに高かった(約700/細胞)ことを示している。プラスミドの維持および収量は極めて効率的であり、コピー数は、抗生物質選別、例えばCatの存在下で培養した細胞と比較して2倍多かった
【実施例9】
【0058】
cIプラスミドに抗生物質遺伝子が存在しない場合のプラスミドの維持および収量:1つのsacBカセットおよび2つのsacBカセットをもつ大腸菌株間の比較解析
1つのsacBカセットおよび2つのsacBカセットをもつ大腸菌株(それぞれΔpurNλPr::sacB+ΩkanおよびΔpurNλPr::sacB+ΩkanΔedAλPr::sacB+Ωcat)の両方におけるcatフリーpPB885プラスミドの安定性および収量を示す。ΔpurNΩλPr::sacBKm(1つのsacBカセット)およびΔpurNΩλPr::sacBKmΔedaΩλPr::sacBcat(2つのsacBカセット)の両親株をcIcatフリーpPB885プラスミドで形質転換し、ショ糖プレートへの播種とその後の培養の前に3時間インキュベートした。プラスミド収量は、2つのsacBカセットの大腸菌株および1つのsacBカセットの大腸菌株に対してそれぞれ5%および10%ショ糖補充LBで3継代して決定した。37℃で維持した培養からpPB885を抽出し、ゲル電気泳動に付した。PvuII制限消化は、増殖およびショ糖選別の間にどちらのpPB885でも明白な遺伝的再編成は発生しないことを示した(図15D参照)。このゲルが示すように、10%および5%のどちらのショ糖選別後でもそれぞれ1つのsacBカセットおよび2つのsacBカセットをもつ両大腸菌で、明白な遺伝的再編成は生じなかった。
図15Eは、1つのsacBカセットおよび2つのsacBカセットをもつ両大腸菌株(それぞれΔpurNλPr::sacB+ΩkanおよびΔpurNλPr::sacB+ΩkanΔedAλPr::sacB+Ωcat)でのcatフリーpPB885プラスミドの収量を示す。興味深いことに、プラスミド収量(コピー数)は、1つのsacBカセットをもつ大腸菌株から得られたプラスミド収量(ほぼ142/細胞)と比較して2つのsacBカセットをもつ大腸菌株ではるかに高かった(ほぼ278/細胞).この実験で得られたプラスミド収量は以前の比較実験で得られたプラスミド収量より低いように思われる(図15B参照)。これは主として以下の事実によるものである:i)pPB885による形質転換後に、最良のクローンを同定するためにより多くのクローンを評価する必要があること、およびii)さらにcatフリーpPB885プラスミドで形質転換した2つのsacBカセットをもつ大腸菌株の増殖条件は最適ではなかったということ。実際、我々は、このもっとも新しい株の最適増殖条件は約37℃で約2%(またはわずかに低い)ショ糖であることを示した。しかしながら、この実験結果は、最適で強固な条件下では、プラスミド収量は、1つのsacBカセットをもつ大腸菌株と比較して2つのsacBカセットをもつ大腸菌株でより多量であり得ることを示唆している。
【実施例10】
【0059】
CHO細胞への抗生物質フリーcIプラスミドのトランスフェクション
図15Eおよび下記の表8は、抗生物質フリープラスミド(pPB896/選別圧としてショ糖)および抗生物質プラスミド(pCG105/選別圧としてCat)のCHO細胞へのプラスミドトランスフェクション効率(GFP発現)を示す。発現されるGFPに関してこれら2つの選別圧(ショ糖対抗生物質としてCat)間で明白な差異は認められない。pCG105プラスミド(CMVプロモーターの制御下に配置されたGFP遺伝子を有するpPB828誘導プラスミド(Merialの特許物))およびpPB896(pPB829プラスミド+P1::cI遺伝子+GFP)プラスミドをCHO細胞のトランスフェクションに用いて、従来の抗生物質プラスミドに対して抗生物質フリープラスミド概念の機能性を評価した。この実験は、1つのsacBカセットをもつ大腸菌宿主株の培養から単離したプラスミドを用いて実施した。pCG105は選別圧としてカナマイシンを用いて大腸菌で増殖させ、一方、pPB896は選別圧として10%ショ糖を用いて大腸菌で増殖させた。DNAプラスミドを抽出し、同じ濃度で投与した。
図15は、リポフェクタミン2000を用いたCHO-K1細胞の一過性トランスフェクション後の、pCG105およびpPB896によりコードされるGFPタンパク質のin vitro発現を示す。直径6cmのプレート中の90%コンフルエンシーのCHO-K1細胞に、各々5μgのプラスミドおよび10μLのリポフェクタミンを用い製造業者の指示にしたがいながらトランスフェクションを実施した。トランスフェクション後に、1%のSVFを含むMEM-glutamax培養液で細胞を培養した。培養を採集し、蛍光顕微鏡(図15)およびフローサイトメトリー(FACS Calibur(Becton Dickinson))(表8)を用いて解析した。
図15は、トランスフェクションを実施した両方のCHO-K1細胞で同様なGFPエピフルオレセンスが可視化されたことを示す。クロラムフェニコールおよびショ糖の選別圧下でpCG105およびpPB896プラスミドは比較可能な効率でCHO-K1細胞にトランスフェクトできた。
表8は、pCG105およびpPB896プラスミドのCHOトランスフェクション効率を示す。この表に示すように、CHO細胞にpCG105(59%のGFP発現)およびpPB896(48%のGFP発現)をトランスフェクトしたとき、比較可能なGFPタンパク質発現CHO細胞の%が測定された。これらの結果は、CHOのトランスフェクションおよび最終的にはDNAワクチンでの応用のためにショ糖系抗生物質フリープラスミドを使用することの有効性を立証した。
表8
これらの実験は、染色体上に存在する有毒遺伝子の制御はプラスミド上に存在するリプレッサーにより達成できることを示す。この系は、抗生物質の選別圧の非存在下で完全に機能を発揮する。この系は宿主細胞の5継代にわたって安定である。この系は、宿主細胞が細胞当たり高いプラスミドコピー数を達成することを可能にする。この系は、最少合成培養液と完ぺきに適合し得る。
これまで本発明の好ましい実施態様を詳細に説明してきたが、上記で規定した本発明は、その多くの変型が本発明の範囲を逸脱することなく可能であるので、上述の記載に示す具体的な内容に限定されないことは理解されよう。
本明細書に引用または参照した全ての文書(“本明細書引用文書”)、および本明細書引用文書中に引用または参照された全ての文書は、製造業者の指示、説明書、製品明細書および本明細書に記載したまたは参照により含まれる任意の文書中の任意の製品のプロダクトシートとともに、参照により本明細書に含まれ、本発明の実施で利用することができる。
【特許請求の範囲】
【請求項1】
cIリプレッサータンパク質をコードするポリヌクレオチドを含む、ドラッグレスプラスミド。
【請求項2】
ポリヌクレオチドが、配列番号:2、44、46、48、50、52、54、56または58に示される配列を有するポリペプチドと少なくとも90%の配列同一性を有するcIリプレッサータンパク質をコードする、請求項1に記載のドラッグレスプラスミド。
【請求項3】
ポリヌクレオチドが作動できるようにプロモーターに連結される、請求項1または2に記載のドラッグレスプラスミド。
【請求項4】
プロモーターがP1プロモーターまたは天然のcIプロモーターである、請求項3に記載のドラッグレスプラスミド。
【請求項5】
ポリヌクレオチドが、配列番号:1、45、47、49、51、53、55、57または59に示される配列を有するポリヌクレオチドと少なくとも90%の配列同一性を有する、請求項1−4のいずれか1項に記載のドラッグレスプラスミド。
【請求項6】
プラスミドがさらに、免疫原またはタンパク質をコードする異種ポリヌクレオチドを含む、請求項1−5のいずれか1項に記載のドラッグレスプラスミド。
【請求項7】
異種ポリヌクレオチドが、真核細胞または原核細胞で機能するプロモーターに作動できるように連結される、請求項6に記載のドラッグレスプラスミド。
【請求項8】
プロモーターがCMVプロモーターまたはグラム陰性細菌から単離されたプロモーターである、請求項7に記載のドラッグレスプラスミド。
【請求項9】
請求項1−8のいずれか1項に記載のドラッグレスプラスミドを含む、操作されたグラム陰性細菌宿主株。
【請求項10】
細菌宿主が、前記細菌の染色体の非必須領域に1つまたは2つ以上の異種ポリヌクレオチドを含む請求項9に記載の細菌宿主株であって、前記異種ポリヌクレオチドが宿主に有毒な生成物をコードする、前記細菌宿主株。
【請求項11】
ポリヌクレオチドがsacB遺伝子を含む、請求項10に記載の細菌株。
【請求項12】
sacB遺伝子が、配列番号:4、60、62、64、66、68、70、72または74に示される配列を有するポリペプチドと少なくとも90%の配列を有するSacBタンパク質をコードする、請求項11に記載の細菌株。
【請求項13】
異種ポリヌクレオチドがプロモーターに作動できるように連結される、請求項10−12のいずれか1項に記載の細菌株。
【請求項14】
プロモーターがλファージ由来プロモーターである、請求項13に記載の細菌株。
【請求項15】
非必須領域がdeA遺伝子またはpurN遺伝子を含む、請求項10−14のいずれか1項に記載の細菌株。
【請求項16】
sacB遺伝子が、配列番号:4に示される配列を有するポリペプチドと少なくとも90%の配列同一性を有するSacBタンパク質をコードし、さらにsacB遺伝子が、配列番号:5に示される配列を有するプロモーターに作動できるように連結される、請求項11−15のいずれか1項に記載の細菌株。
【請求項17】
細菌株が、配列番号:75に示される配列を有するポリヌクレオチドの1つまたは2つ以上のコピーを含む、請求項11−16のいずれか1項に記載の細菌株。
【請求項18】
細菌株が、宿主染色体のdeA遺伝子およびpurN遺伝子に挿入された配列番号:75のポリヌクレオチドの2つのコピーを含む、請求項17に記載の細菌株。
【請求項19】
細菌株が、アビバクテリウム、ブルセラ、大腸菌、ヘモフィルス(例えばヘモフィルス・スイス(Haemophilus suis))、サルモネラ(例えばサルモネラ・エンテリディス(Salmonella enteridis)、ネズミチフス菌(Salmonella typhimurium)、サルモネラ・インファンチス(Salmonella infantis))、シゲラ、パスツレラおよびリメイレラ(Rimeirella)から成る群から選択される、請求項10−18のいずれか1項に記載の細菌株。
【請求項20】
請求項1−8のいずれか1項に記載のドラッグレスプラスミドを含む組成物。
【請求項21】
さらに医薬的に許容できる担体を含む、請求項20に記載の組成物。
【請求項22】
以下の工程を含む、ドラッグレスプラスミドの製造方法:
1)宿主染色体の1つまたは2つ以上の非必須領域に対立遺伝子置換によって挿入された異種ポリヌクレオチドを含むグラム陰性細菌宿主株を操作する工程;
2)cIリプレッサータンパク質をコードするポリヌクレオチドを含むDNAプラスミドを構築する工程;
3)cIリプレッサータンパク質をコードする遺伝子を含むDNAプラスミドで細菌宿主株を形質転換する工程;
4)形質転換細菌宿主株をショ糖の存在下において30℃から42℃の範囲の温度で増殖させる工程;および
5)DNAプラスミドを回収する工程。
【請求項23】
以下の工程を含む、タンパク質または免疫原の製造方法:
1)宿主染色体の1つまたは2つ以上の非必須領域に対立遺伝子置換によって挿入された異種ポリヌクレオチドを含むグラム陰性細菌宿主株を操作する工程;
2)cIリプレッサータンパク質をコードするポリヌクレオチドおよび免疫原またはタンパク質をコードする遺伝子を含むDNAプラスミドを構築する工程;
3)cIリプレッサータンパク質をコードするポリヌクレオチドおよび免疫原またはタンパク質をコードする遺伝子を含むDNAプラスミドで細菌宿主株を形質転換する工程;
4)形質転換細菌宿主株をショ糖の存在下において30℃から42℃の範囲の温度で増殖させる工程;および
5)免疫原またはタンパク質を回収する工程。
【請求項24】
請求項23に記載の方法によって製造されたタンパク質または免疫原を含む組成物。
【請求項1】
cIリプレッサータンパク質をコードするポリヌクレオチドを含む、ドラッグレスプラスミド。
【請求項2】
ポリヌクレオチドが、配列番号:2、44、46、48、50、52、54、56または58に示される配列を有するポリペプチドと少なくとも90%の配列同一性を有するcIリプレッサータンパク質をコードする、請求項1に記載のドラッグレスプラスミド。
【請求項3】
ポリヌクレオチドが作動できるようにプロモーターに連結される、請求項1または2に記載のドラッグレスプラスミド。
【請求項4】
プロモーターがP1プロモーターまたは天然のcIプロモーターである、請求項3に記載のドラッグレスプラスミド。
【請求項5】
ポリヌクレオチドが、配列番号:1、45、47、49、51、53、55、57または59に示される配列を有するポリヌクレオチドと少なくとも90%の配列同一性を有する、請求項1−4のいずれか1項に記載のドラッグレスプラスミド。
【請求項6】
プラスミドがさらに、免疫原またはタンパク質をコードする異種ポリヌクレオチドを含む、請求項1−5のいずれか1項に記載のドラッグレスプラスミド。
【請求項7】
異種ポリヌクレオチドが、真核細胞または原核細胞で機能するプロモーターに作動できるように連結される、請求項6に記載のドラッグレスプラスミド。
【請求項8】
プロモーターがCMVプロモーターまたはグラム陰性細菌から単離されたプロモーターである、請求項7に記載のドラッグレスプラスミド。
【請求項9】
請求項1−8のいずれか1項に記載のドラッグレスプラスミドを含む、操作されたグラム陰性細菌宿主株。
【請求項10】
細菌宿主が、前記細菌の染色体の非必須領域に1つまたは2つ以上の異種ポリヌクレオチドを含む請求項9に記載の細菌宿主株であって、前記異種ポリヌクレオチドが宿主に有毒な生成物をコードする、前記細菌宿主株。
【請求項11】
ポリヌクレオチドがsacB遺伝子を含む、請求項10に記載の細菌株。
【請求項12】
sacB遺伝子が、配列番号:4、60、62、64、66、68、70、72または74に示される配列を有するポリペプチドと少なくとも90%の配列を有するSacBタンパク質をコードする、請求項11に記載の細菌株。
【請求項13】
異種ポリヌクレオチドがプロモーターに作動できるように連結される、請求項10−12のいずれか1項に記載の細菌株。
【請求項14】
プロモーターがλファージ由来プロモーターである、請求項13に記載の細菌株。
【請求項15】
非必須領域がdeA遺伝子またはpurN遺伝子を含む、請求項10−14のいずれか1項に記載の細菌株。
【請求項16】
sacB遺伝子が、配列番号:4に示される配列を有するポリペプチドと少なくとも90%の配列同一性を有するSacBタンパク質をコードし、さらにsacB遺伝子が、配列番号:5に示される配列を有するプロモーターに作動できるように連結される、請求項11−15のいずれか1項に記載の細菌株。
【請求項17】
細菌株が、配列番号:75に示される配列を有するポリヌクレオチドの1つまたは2つ以上のコピーを含む、請求項11−16のいずれか1項に記載の細菌株。
【請求項18】
細菌株が、宿主染色体のdeA遺伝子およびpurN遺伝子に挿入された配列番号:75のポリヌクレオチドの2つのコピーを含む、請求項17に記載の細菌株。
【請求項19】
細菌株が、アビバクテリウム、ブルセラ、大腸菌、ヘモフィルス(例えばヘモフィルス・スイス(Haemophilus suis))、サルモネラ(例えばサルモネラ・エンテリディス(Salmonella enteridis)、ネズミチフス菌(Salmonella typhimurium)、サルモネラ・インファンチス(Salmonella infantis))、シゲラ、パスツレラおよびリメイレラ(Rimeirella)から成る群から選択される、請求項10−18のいずれか1項に記載の細菌株。
【請求項20】
請求項1−8のいずれか1項に記載のドラッグレスプラスミドを含む組成物。
【請求項21】
さらに医薬的に許容できる担体を含む、請求項20に記載の組成物。
【請求項22】
以下の工程を含む、ドラッグレスプラスミドの製造方法:
1)宿主染色体の1つまたは2つ以上の非必須領域に対立遺伝子置換によって挿入された異種ポリヌクレオチドを含むグラム陰性細菌宿主株を操作する工程;
2)cIリプレッサータンパク質をコードするポリヌクレオチドを含むDNAプラスミドを構築する工程;
3)cIリプレッサータンパク質をコードする遺伝子を含むDNAプラスミドで細菌宿主株を形質転換する工程;
4)形質転換細菌宿主株をショ糖の存在下において30℃から42℃の範囲の温度で増殖させる工程;および
5)DNAプラスミドを回収する工程。
【請求項23】
以下の工程を含む、タンパク質または免疫原の製造方法:
1)宿主染色体の1つまたは2つ以上の非必須領域に対立遺伝子置換によって挿入された異種ポリヌクレオチドを含むグラム陰性細菌宿主株を操作する工程;
2)cIリプレッサータンパク質をコードするポリヌクレオチドおよび免疫原またはタンパク質をコードする遺伝子を含むDNAプラスミドを構築する工程;
3)cIリプレッサータンパク質をコードするポリヌクレオチドおよび免疫原またはタンパク質をコードする遺伝子を含むDNAプラスミドで細菌宿主株を形質転換する工程;
4)形質転換細菌宿主株をショ糖の存在下において30℃から42℃の範囲の温度で増殖させる工程;および
5)免疫原またはタンパク質を回収する工程。
【請求項24】
請求項23に記載の方法によって製造されたタンパク質または免疫原を含む組成物。
【図1】

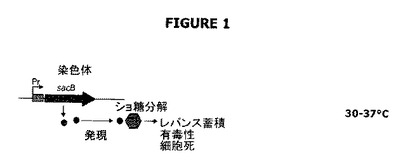
【図2】
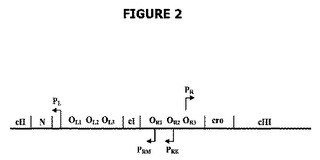
【図3】
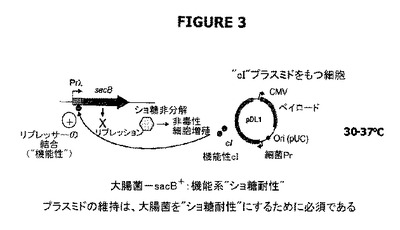
【図4】
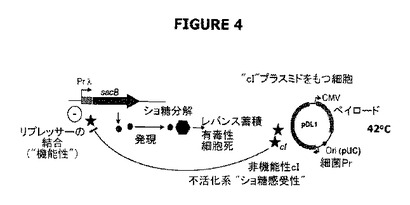
【図5】
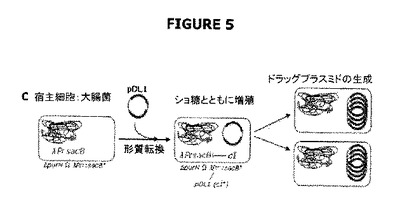
【図6A】
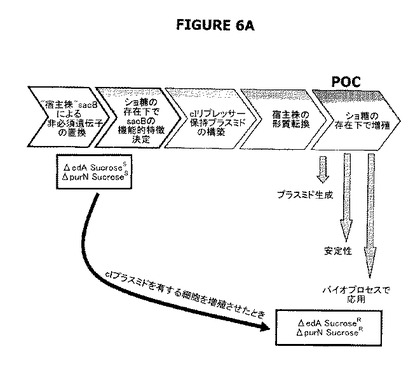
【図6B】
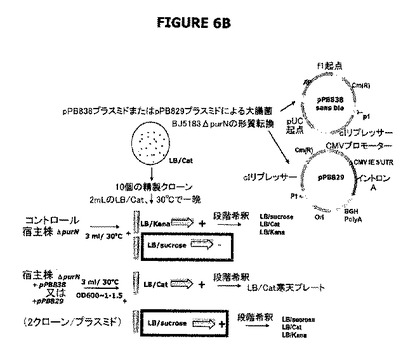
【図6C】
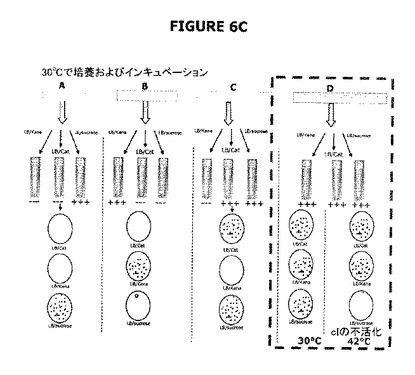
【図7】
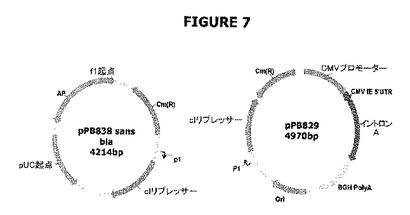
【図8A−B】
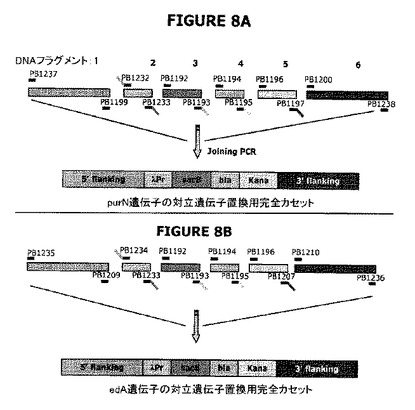
【図8C】
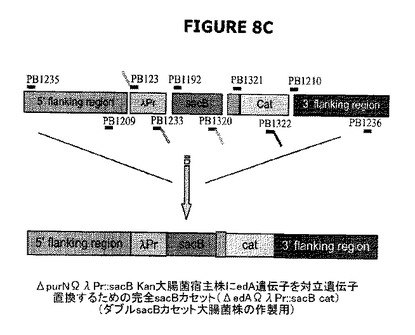
【図8D】
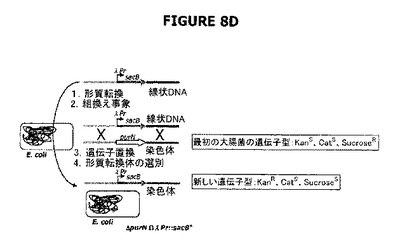
【図8E】
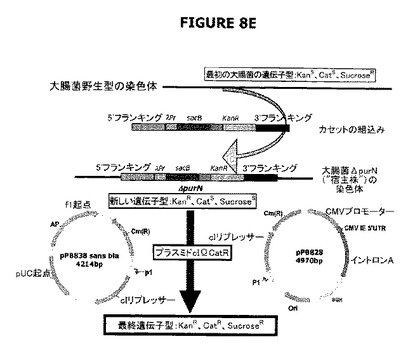
【図9A】
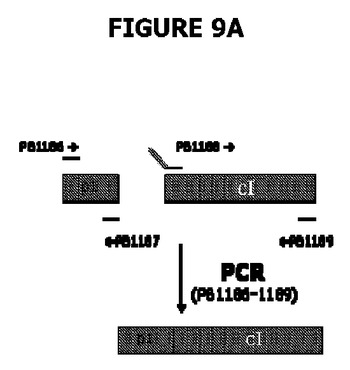
【図9B】
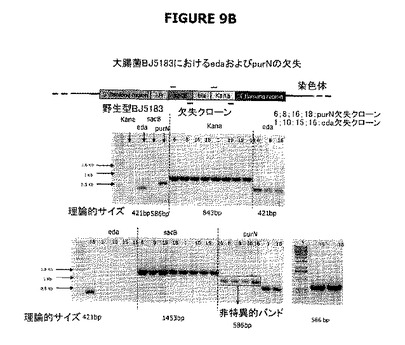
【図9C】
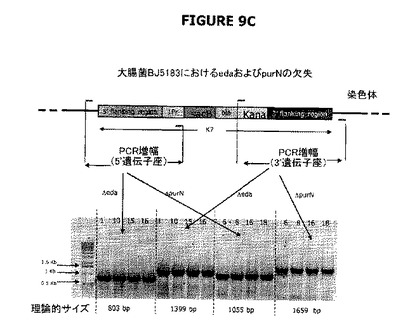
【図9D】
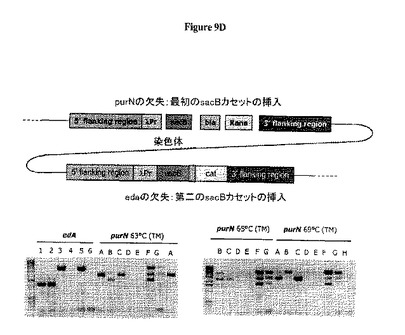
【図10A】
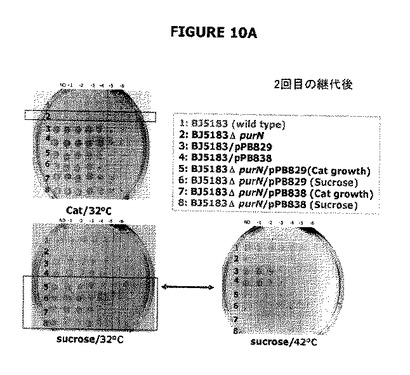
【図10B】
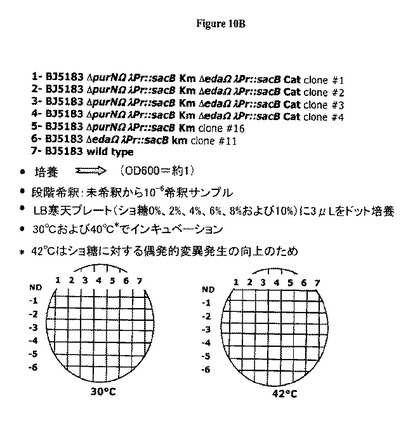
【図10C】
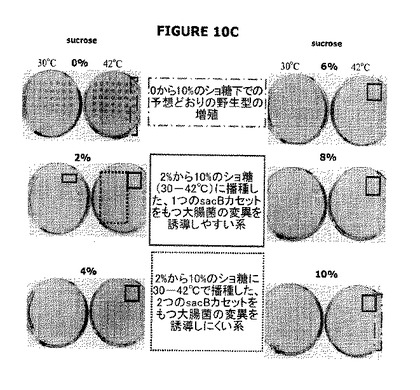
【図11】
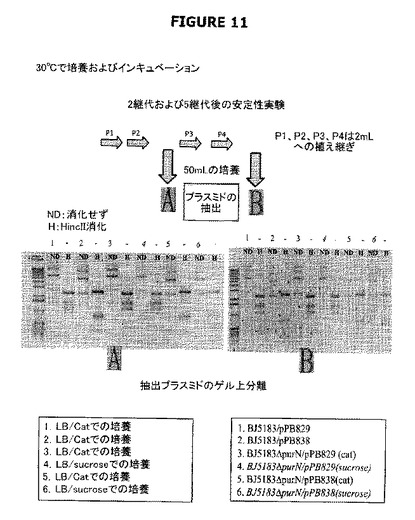
【図12A】
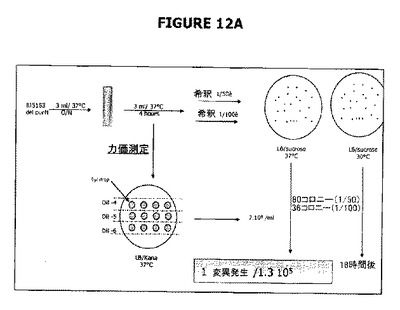
【図12B】
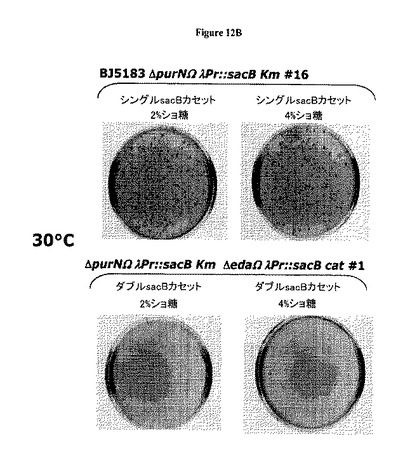
【図12C】
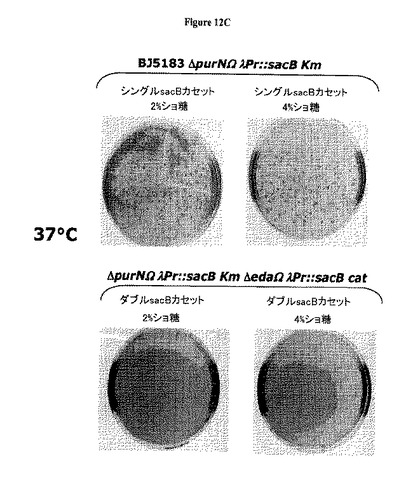
【図13A】

【図13B】
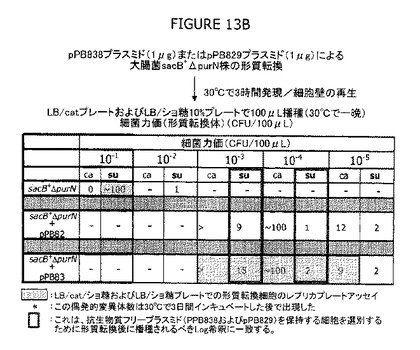
【図14A】
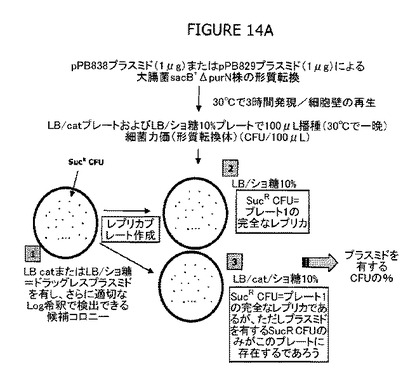
【図14B】
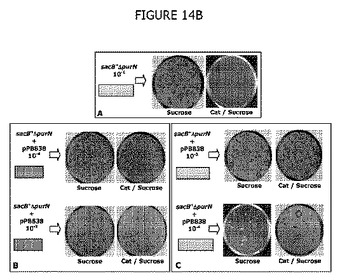
【図14C】
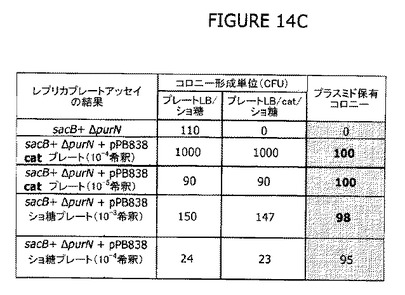
【図15A】
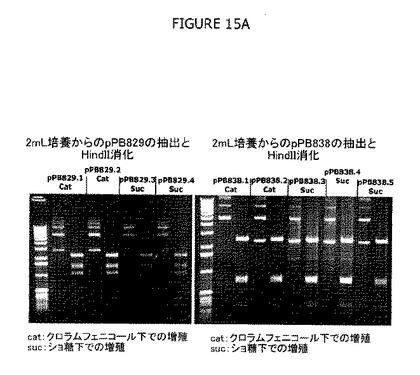
【図15B】
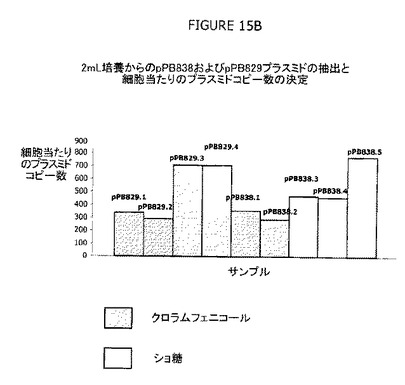
【図15C】
【図15D】
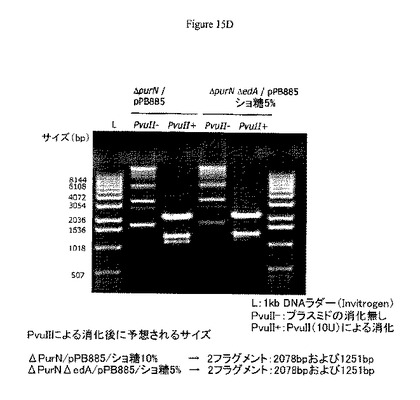
【図15E】
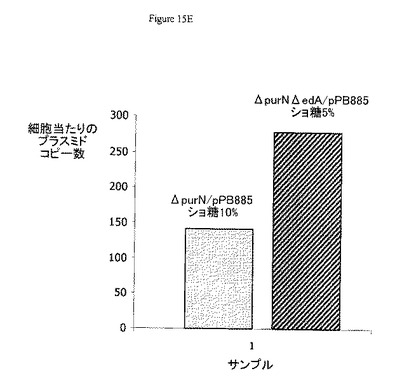
【図15F】

【図16】
【図17】
【図18】
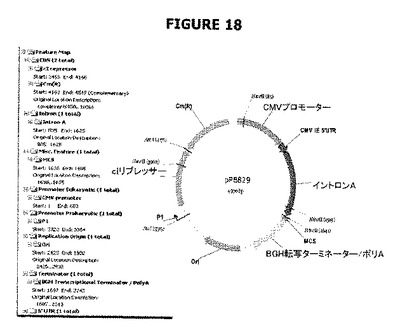
【図19】
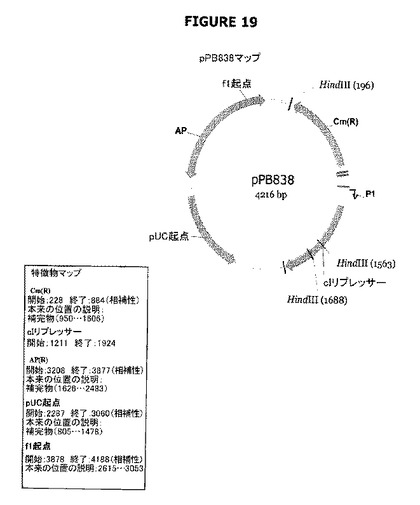
【図20】
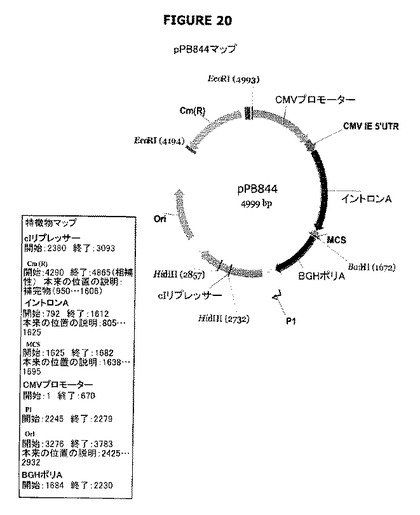
【図21】
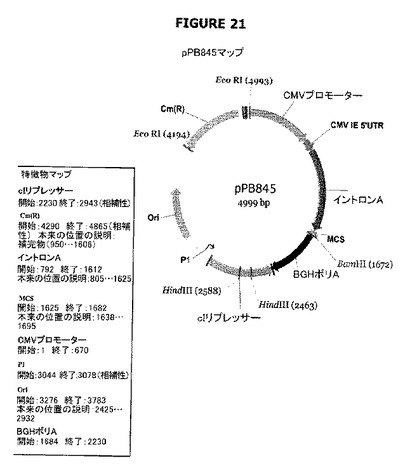
【図22】
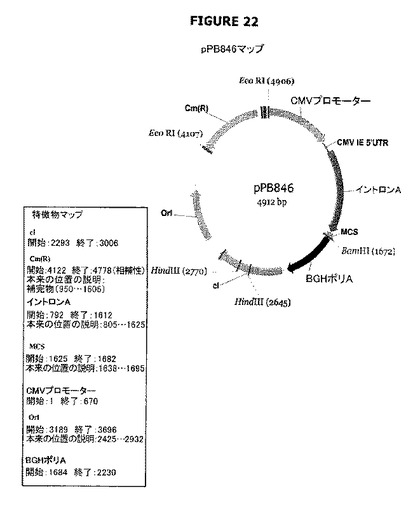
【図23】
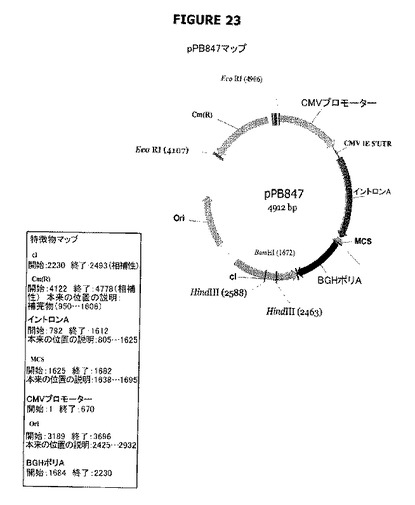
【図24】
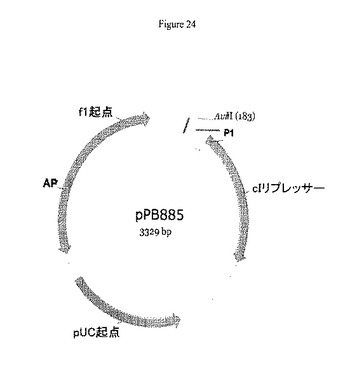
【図25】
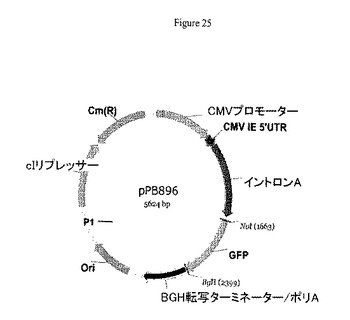
【図26−1】
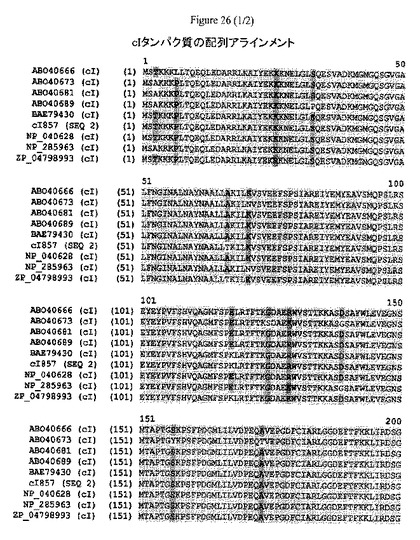
【図26−2】

【図27−1】
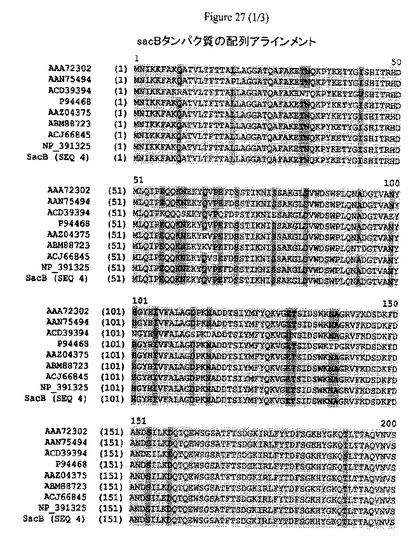
【図27−2】
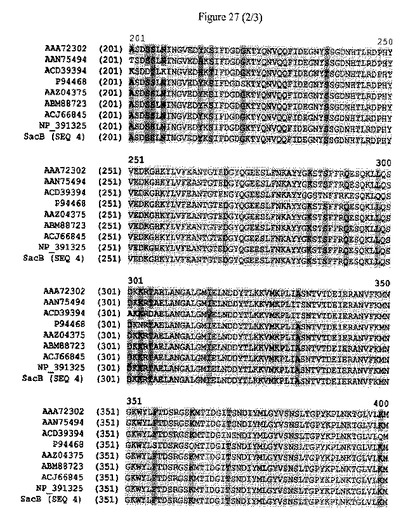
【図27−3】

【図28−1】

【図28−2】

【図29−1】
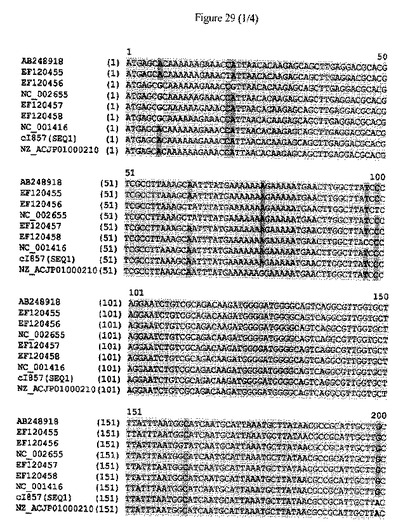
【図29−2】
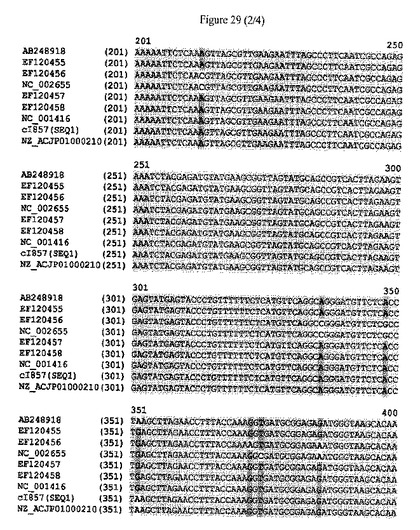
【図29−3】
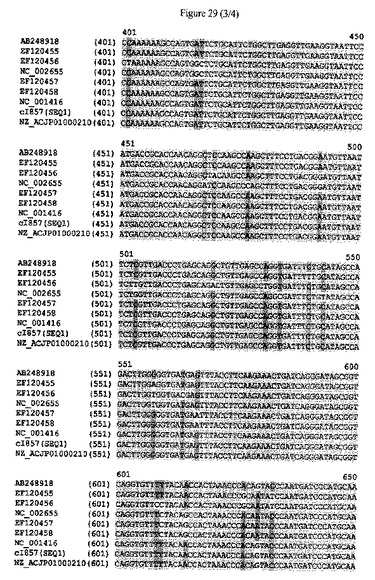
【図29−4】

【図30−1】
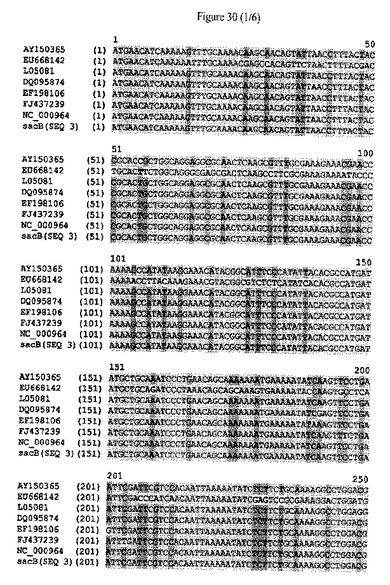
【図30−2】
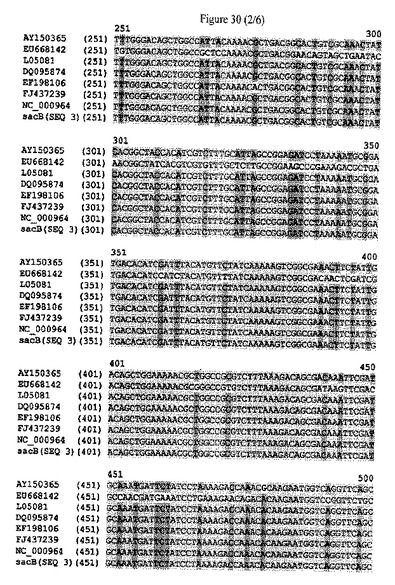
【図30−3】
【図30−4】

【図30−5】

【図30−6】

【図31−1】

【図31−2】

【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図6C】
【図7】
【図8A−B】
【図8C】
【図8D】
【図8E】
【図9A】
【図9B】
【図9C】
【図9D】
【図10A】
【図10B】
【図10C】
【図11】
【図12A】
【図12B】
【図12C】
【図13A】
【図13B】
【図14A】
【図14B】
【図14C】
【図15A】
【図15B】
【図15C】
【図15D】
【図15E】
【図15F】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26−1】
【図26−2】
【図27−1】
【図27−2】
【図27−3】
【図28−1】
【図28−2】
【図29−1】
【図29−2】
【図29−3】
【図29−4】
【図30−1】
【図30−2】
【図30−3】
【図30−4】
【図30−5】
【図30−6】
【図31−1】
【図31−2】
【公表番号】特表2012−527249(P2012−527249A)
【公表日】平成24年11月8日(2012.11.8)
【国際特許分類】
【出願番号】特願2012−512086(P2012−512086)
【出願日】平成22年5月24日(2010.5.24)
【国際出願番号】PCT/US2010/035979
【国際公開番号】WO2010/135742
【国際公開日】平成22年11月25日(2010.11.25)
【出願人】(304040692)メリアル リミテッド (73)
【Fターム(参考)】
【公表日】平成24年11月8日(2012.11.8)
【国際特許分類】
【出願日】平成22年5月24日(2010.5.24)
【国際出願番号】PCT/US2010/035979
【国際公開番号】WO2010/135742
【国際公開日】平成22年11月25日(2010.11.25)
【出願人】(304040692)メリアル リミテッド (73)
【Fターム(参考)】
[ Back to top ]