
抗真菌性物質
【課題】安全であり、かつ、中性において抗真菌活性を有する抗真菌性物質、該抗真菌性物質を産生する菌、並びに、これらを用いるカビ及び/又は酵母の増殖阻害方法の提供。
【解決手段】キャンディダ属菌、サッカロマイセス属菌、ピキア属菌、ロドトルラ属菌、クラビスポラ属菌、及びトルラスポラ属菌からなる群より選ばれる1の酵母が産生する、胞子の発芽を阻害する抗真菌性物質、及び、下記の特徴を有する抗真菌性物質;1)pH7.0において、抗真菌活性を有すること、2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有すること、3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCO(Molecular Weight Cut Off、分画分子量)が10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持されること。
【解決手段】キャンディダ属菌、サッカロマイセス属菌、ピキア属菌、ロドトルラ属菌、クラビスポラ属菌、及びトルラスポラ属菌からなる群より選ばれる1の酵母が産生する、胞子の発芽を阻害する抗真菌性物質、及び、下記の特徴を有する抗真菌性物質;1)pH7.0において、抗真菌活性を有すること、2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有すること、3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCO(Molecular Weight Cut Off、分画分子量)が10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持されること。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、キャンディダ(Candida)属菌、サッカロマイセス(Saccharomyces)属菌、ピキア(Pichia)属菌、ロドトルラ(Rhodotorula)属菌、クラビスポラ(Clavispora)属菌、及びトルラスポラ(Torulaspora)属菌からなる群より選ばれる1の酵母が産生する、胞子の発芽を阻害する抗真菌性物質、該抗真菌性物質を含有する防カビ剤及び食品保存料、該抗真菌性物質又は該酵母を用いるカビ及び/又は酵母の増殖を阻害する方法、並びに、該抗真菌性物質を産生する新規キャンディダ属菌に関する。
【背景技術】
【0002】
微生物による腐敗や変敗を防止し、食品の保存性を高めるため、様々な抗菌性物質が、食品保存料として用いられている。食品の保存性を向上させることにより、流通過程における食品の腐敗等が効率的に防止されるため、製造コストの改善が期待でき、また、食品の安全性上も好ましい。このため、優れた抗菌性物質の開発が望まれている。ここで、抗菌性物質は、主に、プロピオン酸やソルビン酸等の化学合成物質と、微生物等から得られた天然由来の物質に分類できるが、食品保存料として用いる場合には、安全性の点から、天然由来の物質であることが好ましい。
【0003】
近年、天然由来の抗菌性物質として、ナイシン等の、乳酸菌由来の様々な種類のバクテリオシンが知られている(例えば、非特許文献1参照。)。バクテリオシンは、通常、ペプチド若しくはタンパク質であり、消化酵素等により容易に分解されるため、非常に安全性が高い。また、主にグラム陽性菌に対し優れた抗菌活性を有することが知られている。
【0004】
その他、グラム陽性菌とグラム陰性菌の両方に対し優れた抗菌活性を有する抗菌性物質を含む食品保存料の製造方法として、例えば、(1)酵母の増殖に必要な栄養素、無機塩類と炭素源としてグルコースを含む培地で酵母を定常期に達するまで増殖させた後、酵母を除去した培地に必要ならば炭素源を加え、再び定常期に達するまで増殖させる繰り返し培養を2回以上行って得られた水溶液であることを特徴とする酵母が産生する抗菌性物質を含む食品保存料の製造方法が開示されている(例えば、特許文献1参照。)。
【非特許文献1】園元ら、食品と開発、41巻3号、p73〜75、2006年
【特許文献1】特許第2545739号公報
【発明の開示】
【発明が解決しようとする課題】
【0005】
グラム陽性菌等のバクテリアのみならず、カビや酵母等の真菌も、食品の腐敗等を引き起こす。特にカビは、多くの食品の腐敗の主要な要因であることから、カビの増殖を抑制することができる抗真菌活性を有する物質を、食品保存料や抗菌剤として使用できることが好ましい。
しかしながら、ナイシン等のバクテリオシンは、一般にグラム陽性菌にのみ抗菌活性を示し、カビや酵母等の真菌の増殖を抑制することができないという問題がある。また、上記(1)の方法で得られる食品保存料は、酵母由来の抗菌性物質であるため、食品に対し安全に使用することができ、また、耐熱性も有しているが、特許文献1には、カビに対する抗菌活性については一切記載がない。
【0006】
一方、プロピオン酸等の有機酸は、真菌の増殖抑制に対し、一定の効果を有するものの、化学合成物質であるため、天然由来の物質に比べて安全性が劣ることに加え、独特のにおいを有するため、食品の風味上好ましくないという問題がある。
また、有機酸は、主に酸性下で抗菌活性を有し、中性の食品に対する抗菌性に乏しいという問題もある。有機酸の中性における抗菌活性の低下は、中性溶液中では、有機酸はほぼ完全に電離しているが、解離して負に帯電した有機酸は、微生物の細胞内に取り込まれ難いためと考えられている。
【0007】
本発明は、安全であり、かつ、中性においてカビや酵母等の真菌の増殖を抑制することのできる、抗真菌性物質を提供することを目的とする。
また、本発明は、該抗真菌性物質を含有する防カビ剤及び食品保存料、該抗真菌性物質を産生する菌、並びに、該抗真菌性物質又は該菌を用いるカビ及び/又は酵母の増殖を阻害する方法を提供することを目的とする。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決すべく鋭意研究した結果、食品由来の微生物が産生する抗真菌性物質であれば、食品保存料としても安全に用いることができると考え、様々な種類のチーズから分離した微生物の中から、pH7.0において、アスペルギルス・ニガー(Aspergillus niger)又はリゾパス・ストロニファー(Rhizopus stolonifer)の増殖を抑制することのできる抗真菌活性を有する微生物を見出し、さらに、該微生物が産生する抗真菌性物質を見出すことにより、本発明を完成させた。
【0009】
すなわち、本発明は、キャンディダ(Candida)属菌、サッカロマイセス(Saccharomyces)属菌、ピキア(Pichia)属菌、ロドトルラ(Rhodotorula)属菌、クラビスポラ(Clavispora)属菌、及びトルラスポラ(Torulaspora)属菌からなる群より選ばれる1の酵母が産生する、胞子の発芽を阻害する抗真菌性物質を提供するものである。
また、本発明は、下記の特徴を有する抗真菌性物質を提供するものである。
(1)pH7.0において、抗真菌活性を有すること、
(2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有すること、
(3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCO(Molecular Weight Cut Off、分画分子量)が10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持されること。
また、本発明は、下記の特徴を有する抗真菌性物質を提供するものである。
(1)pH7.0において、抗真菌活性を有すること、
(2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有すること、
(3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCO(Molecular Weight Cut Off、分画分子量)が10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持されること、
(4)55℃で15分間の加熱処理により、前記抗真菌活性が失活すること。
また、本発明は、前記キャンディダ属菌が、キャンディダ・マルトーサ(Candida maltosa)、キャンディダ・トロピカリス(Candida tropicalis)、又はキャンディダ・シュードインターメディア(Candida pseudintermedia)である抗真菌性物質を提供するものである。
また、本発明は、前記キャンディダ属菌が、キャンディダ・マルトーサO9−NP9(受託番号 NITE BP−297)、キャンディダ・マルトーサIAM12247、キャンディダ・マルトーサIAM12248、キャンディダ・トロピカリスIAM4965、又はキャンディダ・シュードインターメディアIAM12510である抗真菌性物質を提供するものである。
また、本発明は、前記サッカロマイセス属菌がサッカロマイセス・セレビシエ(Saccharomyces cerevisiae)であり、前記ピキア属菌がピキア・フェルメンタス(Pichia fermentans)であり、前記ロドトルラ属菌がロドトルラ・アクタ(Rhodotorula acuta)であり、前記クラビスポラ属菌がクラビスポラ・ルシタニエ(Clavispora lusitaniae)であり、前記トルラスポラ属菌がトルラスポラ・デルブレッキー(Torulaspora delbrueckii)である、抗真菌性物質を提供するものである。
また、本発明は、前記ピキア属菌がピキア・フェルメンタスJCM1824であり、前記ロドトルラ属菌がロドトルラ・アクタJCM9494であり、前記サッカロマイセス属菌がサッカロマイセス・セレビシエH−1(受領番号 NITE ABP−473)であり、前記クラビスポラ属菌がクラビスポラ・スピーシーズP−5(Clavispora sp. P−5、受領番号 NITE ABP−474)又はクラビスポラ・ルシタニエNBRC10059であり、前記トルラスポラ属菌がトルラスポラ・デルブルッキーNBRC0955である抗真菌性物質を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を含有する防カビ剤を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を含有する食品保存料を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を用いることを特徴とする、カビ及び/又は酵母の増殖を阻害する方法を提供するものである。
また、本発明は、前記いずれか記載の酵母を用いることを特徴とする、カビ及び/又は酵母の増殖を阻害する方法を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を産生することを特徴とするキャンディダ・マルトーサO9−NP9(受託番号 NITE BP−297)を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を産生することを特徴とするサッカロマイセス・セレビシエH−1(受領番号 NITE ABP−473)を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を産生することを特徴とするクラビスポラ・スピーシーズP−5(受領番号 NITE ABP−474)を提供するものである。
【発明の効果】
【0010】
本発明の抗真菌性物質により、効果的にカビや酵母等の真菌の増殖を抑制することができる。本発明の抗真菌性物質は、チーズから分離された酵母が産生する物質であるため、食品に対して安全に用いることができるのみならず、化粧品や医薬品等にも用いることが可能である。特に中性において、抗真菌活性を示すため、中性の食品等においても有効にカビ等の増殖を抑制することができる。本発明の抗真菌性物質により、食品等の保存期間を安全に延長することができるため、製造コストの改善も期待される。
また、本発明の抗真菌性物質を産生する酵母により、本発明の抗真菌性物質を効率よく産生することができる。
さらに、本発明のカビ及び/又は酵母の増殖を阻害する方法により、安全かつ簡便に、カビや酵母等の真菌の増殖を抑制することができる。
【発明を実施するための最良の形態】
【0011】
本発明の抗真菌性物質は、食品から得られた微生物が産生するものであって、pH7.0において、抗真菌活性を有するものである。このような抗真菌性物質であれば、食品等に安全に使用することができ、かつ、中性の食品等に対しても効果を発揮することができるためである。
【0012】
本発明の抗真菌性物質を産生する微生物は、例えば、各種の食品から分離した微生物の中から、pH7.0において、カビ等の増殖を抑制することができる抗真菌活性を有する菌株を選択することにより得ることができる。pH7.0において抗真菌活性を有する菌株は、本発明の抗真菌性物質を産生していると考えられるためである。以下、本発明の抗真菌性物質、及び該抗真菌性物質を産生する微生物を得る方法を、さらに詳細に説明する。
【0013】
1.微生物群の取得
本発明者らは、抗真菌性物質を産生する微生物を食品中から取得するにあたり、良好な発酵食品の一つであるチーズに着目し、常法により、微生物を分離した。具体的には、チーズを滅菌したチーズおろし金ですり下ろした後、0.05gを、スクリューキャップ付き試験管に分注した滅菌済10%(w/v)スキムミルク溶液5mLに加え、28℃で48時間静置培養した。該スクリューキャップ付き試験管を遠心分離して回収した沈殿物を、滅菌生理リン酸緩衝液で2回洗浄した後、生理リン酸緩衝液で希釈してサンプルを調製した。該サンプルを、MRS寒天培地に塗抹した後、28℃で培養することにより、微生物を分離した。チーズは、カマンベールチーズ(日本製)、カマンベールチーズ(フランス製)、フロマージュ(日本製)、モツァレラチーズ(ドイツ製)、3種類のゴーダチーズ(オランダ製)、クリームチーズ(イタリア製)、山羊乳チーズ(フランス製)、チーズフード(オーストラリア製)、北海道チェダーチーズ(日本製)、ローファットチェダーチーズ(オランダ製)、ホワイトチェダーチーズ(イギリス製)、カスピアンチーズ(イラン製)の、計14種類の市販されているチーズを用いた。一種類のチーズから分離された微生物は、1つの菌群にまとめた。
【0014】
2.抗真菌活性を有する菌が存在する菌群の選択
得られた14種類の菌群を、10%(w/w)スキムミルク培地に接種し、28℃で48時間培養して菌群培養液を調製した。10%(w/w)スキムミルク平板培地に、該菌群培養液を画線塗布した後、アスペルギルス・ニガーあるいはリゾパス・ストロニファーの胞子を懸濁した軟寒天培地を重層した。
該スキムミルク平板培地を、28℃で48時間培養した後に観察した結果、カスピアンチーズ(イラン製)由来のO−9菌群のスキムミルク平板培地において、一部のコロニー周辺で、アスペルギルス・ニガー等の増殖が抑制されていることが観察された。その他の13種類の菌群では、アスペルギルス・ニガー等の増殖抑制は観察されなかった。この結果から、O−9菌群中には、抗真菌性物質を産生する菌が存在していることが分かった。
【0015】
3.pH7.0において抗真菌活性を有する菌が存在する菌群の選択
(1)抗真菌活性を測定するサンプルの調整
O−9菌群を、10%(w/w)スキムミルク培地に接種し、28℃で48時間培養して菌群培養液を調製した。該菌群培養液を超音波処理することにより、菌体を均一に分散させた後、3,000×gで10分間遠心分離を行い、上清と沈殿物に分離した。得られた上清は約pH4であったため、5%(w/v)炭酸ナトリウム溶液を用いてpH7.0に調整したものを、上清サンプルとした。得られた沈殿物に、ほぼ等量の、pH7.0に調整した10%(w/w)スキムミルク溶液を添加して懸濁液としたものを、沈殿物サンプルとした。バクテリアの増殖を抑制して、抗真菌活性の測定を容易にするために、各サンプルには、10分の1相当量の後記実施例1に記載の抗菌剤を添加した。さらに、それぞれの抗菌剤含有サンプルに、9分の1相当量のサブロー培地(グルコース:40g/L、ペプトンS:10g/L)を添加して、抗真菌活性の測定用のサンプル溶液を調製した。
【0016】
(2)抗真菌活性の測定
96ウェルマイクロプレートの各ウェルに、上記3(1)で調整したサンプル溶液の2倍段階希釈系列(20〜29倍)を、それぞれ130μLずつ添加した。該サンプル溶液の希釈には、サブロー培地を用いた。コントロールとして、130μLのサブロー培地のみを添加したウェルを用いた。さらに、各ウェルに、後記参考例1に記載の方法で調整した1.0×104spores/mLのアスペルギルス・ニガーNBRC9455株胞子懸濁液を50μLずつそれぞれ添加した。該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルを顕微鏡で観察した。
【0017】
図1は、顕微鏡観察の結果を表したものである。図中のレーンAはO−9菌群由来の上清サンプル溶液の希釈系列を、レーンBはO−9菌群由来の沈殿物サンプル溶液の希釈系列を、レーンCはコントロールを、それぞれ示している。図中の○は、各ウェルを表したものであり、○の中の×は、アスペルギルス・ニガーの増殖が観察されたウェルを、○の中の○は、アスペルギルス・ニガーの増殖が観察されなかった、すなわち増殖抑制が観察されたウェルを、それぞれ表したものである。なお、アスペルギルス・ニガーの増殖は、アスペルギルス・ニガーの胞子の発芽や菌糸の伸長により判断した。つまり、アスペルギルス・ニガーの増殖抑制は、アスペルギルス・ニガーの胞子から、菌糸の発芽や伸長が阻止されている状態を意味する。
【0018】
図1に示されているように、コントロールのウェルにおいて、アスペルギルス・ニガーの増殖が観察された。一方、O−9菌群由来のサンプル溶液の場合には、沈殿物サンプル溶液を添加した全てのウェルと、上清サンプル溶液の8〜512倍希釈液を添加したウェルでは、増殖したアスペルギルス・ニガーが観察されたが、上清サンプル溶液の1〜4倍希釈液を添加したウェルでは、アスペルギルス・ニガーの増殖は観察されなかった。
【0019】
該観察の結果から、O−9菌群由来の上清サンプル溶液中には、本発明の抗真菌性物質が存在しており、該抗真菌性物質により、アスペルギルス・ニガーの増殖が抑制されたことが明らかである。したがって、O−9菌群には、pH7.0において抗真菌活性を有する菌、すなわち、本発明の抗真菌性物質を産生する菌が存在していることがわかった。
また、上清サンプル溶液を添加したウェルでアスペルギルス・ニガーの増殖抑制が観察されたことから、該菌が産生する本発明の抗真菌性物質は、10%(w/w)スキムミルク培地で培養した場合に、培養液中に移行し得る性質を有することが明らかである。
【0020】
4.抗真菌性物質の耐熱性
O−9菌群由来の上清サンプル溶液中に存在している本発明の抗真菌性物質の耐熱性を検討するために、上記3(1)で調整したO−9菌群由来の上清サンプルを、100℃で5分間の加熱処理後氷冷した後、上記3(1)と同様にpH7.0に調整後、抗菌剤とサブロー培地を添加して、加熱処理済上清サンプル溶液を調整した。上記3(2)と同様にして、該加熱処理済上清サンプル溶液の抗真菌活性を測定したところ、アスペルギルス・ニガーの増殖抑制は観察されなかった。該観察の結果から、本発明の抗真菌性物質は、100℃で5分間の加熱処理により、前記抗真菌活性が失活する性質を有することがわかった。なお、特許文献1記載の食品保存料に含有される抗菌性物質は、加熱処理によっては失活しないものであるため、本発明の抗真菌性物質とは異なる物質であることが明らかである。
【0021】
5.抗真菌性物質の大きさ
O−9菌群由来の上清サンプル溶液中に存在している本発明の抗真菌性物質の大きさを検討するために、上記3(1)で調整したO−9菌群由来の上清サンプルを、MWCOが10kDである限外濾過膜を用いて限外濾過を行い、限外濾過膜表面に保持された高分子画分と、限外濾過膜を通過した低分子画分に分画した。該高分子画分と該低分子画分をサンプルとして、上記3(1)と同様にpH7.0に調整後、抗菌剤とサブロー培地を添加して、上清高分子画分サンプル溶液と上清低分子画分サンプル溶液を調整した。上記3(2)と同様にして、該上清高分子画分サンプル溶液と該上清低分子画分サンプル溶液の抗真菌活性をそれぞれ測定したところ、該上清高分子画分サンプル溶液を添加したウェルではアスペルギルス・ニガーの増殖抑制が観察されたが、該上清低分子画分サンプル溶液を添加したウェルでは観察されなかった。該観察の結果から、本発明の抗真菌性物質は、該高分子画分に分画されており、MWCOが10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持される性質を有することが明らかである。
【0022】
なお、カスピアンチーズは他のチーズと比較して非常に塩分濃度の高いチーズである。そこで、本発明の抗真菌性物質を産生する菌が、好塩性である可能性を考慮し、O−9菌群を、2.5%NaCl含有の10%(w/w)スキムミルク培地に接種した以外は、全て上記3と同様にして抗真菌活性を測定した。その結果、上清サンプル溶液の1〜16倍希釈液を添加したウェルにおいて、アスペルギルス・ニガーの増殖抑制が観察された。つまり、2.5%NaCl含有培地で培養したことにより、抗真菌活性が約4倍上昇したことから、本発明の抗真菌性物質を産生する菌は、高い塩濃度下においても、十分な量の抗真菌性物質を産生し得ることがわかった。
【0023】
6.本発明の抗真菌性物質を産生する菌の単離
上記3(1)で調製したO−9菌群の菌群培養液を、BCP(ブロモクレゾールパープル)平板培地に塗布した後、37℃で48時間培養した。該BCP平板培地から、コロニーの周囲が黄変した6のコロニーと、黄変が観察されなかった42のコロニーの、計48コロニーを得た。BCP寒天培地入り48ウェルマイクロプレートの各ウェルに、得られた48コロニーから釣菌した菌を、それぞれ接種し、さらに37℃で48時間培養した。
【0024】
培養した48株から、培地が紫色のままであった42株の中から10株(P1〜P10株)を、培地が黄変した6株の中から3株(Y1〜Y3株)を、それぞれ得た。得られた13株を、それぞれ10%(w/w)スキムミルク培地に接種し、37℃で48時間培養して培養液を調製した。上記3(1)と同様にして、該培養液を遠心分離して得た上清を、pH7.0に調整後、抗菌剤とサブロー培地を添加して、それぞれの株由来の上清サンプル溶液を調整した。上記3(2)と同様にして、96ウェルマイクロプレートに該上清サンプル溶液の2倍段階希釈系列を添加して、抗真菌活性を測定した。
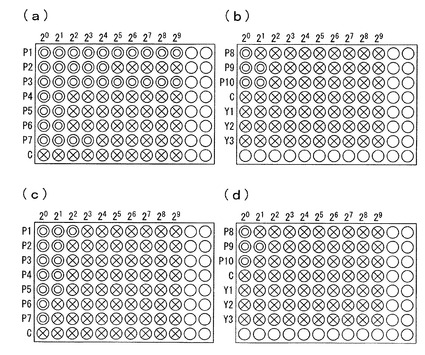
【0025】
図2の(a)と(b)は、該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルを顕微鏡で観察した結果を、(c)と(d)は同じく36時間培養した後の観察結果を、それぞれ表したものである。図中のCはコントロールを表している。また、図中の○は、各ウェルを表したものであり、○の中の×は、アスペルギルス・ニガーの増殖が観察されたウェルを、○の中の○は、アスペルギルス・ニガーの増殖抑制が観察されたウェルを、それぞれ表したものである。
該観察の結果から、Y1〜Y3株は、コントロールと同様に、抗真菌活性は全く観察されなかったが、P1〜P10株では、アスペルギルス・ニガーの増殖抑制が観察されたことから、それぞれ活性強度の差があるものの、P1〜P10株は全て、pH7.0において、抗真菌活性を有する抗真菌性物質を産生することができる菌株であることが明らかである。
【0026】
7.pH7.0において抗真菌性物質を産生する菌の同定
P1〜P10株のうち、P9株(以下、O9−NP9株)及びP5株(以下、O9−NP5株)の同定を試みた。
【0027】
7−1.O9−NP9株の同定
まず、遺伝学的性質を調べるため、O9−NP9株の26S rDNAの塩基配列を常法により同定した。具体的には、O9−NP9株の培養液から抽出したDNAと、NL1、NL2、NL3及びNL4プライマー(O’Donnell、Fusarium and its near relatives、CAB International、p225〜233、1993年)を用いて、26S rDNA−D1/D2領域の塩基配列を同定した。なお、DNAの抽出にはDNeasy Plant Mini Kit(QIAGEN社製)を、PCRにはpuReTaq Ready−To−Go PCR beads(GEヘルスケアバイオサイエンシズ社製)及びBigDye Terminator v3.1 Cycle Sequencing Kit(アプライドバイオシステムズ社製)を、塩基配列の同定にはABI PRISM3100Genetic Analyzer System(アプライドバイオシステムズ社製)を、それぞれ用いた。
国際塩基配列データベース(GenBank/DDBJ/EMBL)上で、該塩基配列について相同性検索を行ったところ、子嚢菌糸アナモルフ(無性時代)酵母の一種であるキャンディダ・マルトーサの基準株であるNRRL Y−17677T(アクセッション番号:U45745)と100%の相同性を示した。したがって、O9−NP9株は、キャンディダ・マルトーサに属する微生物であると推定された。
【0028】
以下にO9−NP9株の菌学的性質を示す。なお、菌学的性質の測定は、YM寒天培地(べクトンディッキンソン社製)で、25℃で2日〜1ヶ月間培養したO9−NP9株を用いた。また、糖類発酵性試験、炭素源資化性試験、窒素源資化性試験、ビタミン要求性試験、温度耐性試験、薬剤耐性試験は、Yeast: Characteristics and identification, 3rd edition(Barnett et al、Cambridge University Press、2000年)、及び、The Yeast, a taxonomic study, 4th edition(Kurtzman et al、Elsevier、1998年)に、ほぼ従って行った。
【0029】
菌学的性質
(1)コロニー観察(YM寒天培地平板で25℃、4日間培養した時の目視観察時)
周縁の形状:ほぼ全縁、隆起状態:扁平形、表面の形状:平滑、光沢及び性状:弱い光沢、湿性、色調:白色からクリーム色
(2)培養的・形態的性質(YM寒天培地平板で25℃、2日間培養した時の光学顕微鏡観察時)
栄養細胞の大きさと形状:直径3〜6μmの楕円形から円筒形
増殖の形式:多極出芽
菌糸等:偽菌糸の形成が有
(3)有性生殖器官の形成(YM寒天培地平板で25℃、1ヶ月間培養した時の光学顕微鏡観察時):なし
【0030】
【表1】

【0031】
(7)37℃下における生育性:陽性
(8)40℃下における生育性:陰性
(9)0.01%シクロヘキシミド含有培地における生育性:陽性
(10)10%塩化ナトリウム及び5%グルコース含有培地における生育性:弱陽性
(11)ビタミン欠乏培地での生育性:陰性
(12)デンプン様物質の形成:陰性
(13)尿素の分解:陰性
【0032】
図3は、YM寒天培地平板で25℃、2日間培養したO9−NP9株の電子顕微鏡写真である。
【0033】
O9−NP9株は、ガラクトース発酵性、イノシトール、エリスリトール、ラフィノース、セロビオース、メレジトース、ガラクトース、ラクトース、可溶性デンプン、L−ラムノース、DL−lactate、キシリトール、及び硝酸塩の資化性、ビタミン欠乏培地、0.01%シクロヘキシミド培地、37℃及び40℃下での生育性等において、キャンディダ・マルトーサと一致する菌学的性質を示した。一方、キャンディダ・マルトーサはトレハロースを発酵するとされているのに対し、O9−NP9株はトレハロース発酵性を示さなかった。
【0034】
これらの遺伝学的及び菌学的性質から、O9−NP9株はキャンディダ・マルトーサ菌種であることが確認された。26S rDNA−D1/D2領域の塩基配列は、キャンディダ・マルトーサNRRL Y−17677T株と100%の相同性があったが、トレハロース発酵能において異なるため、キャンディダ・マルトーサO9−NP9株はキャンディダ・マルトーサNRRL Y−17677T株とは異なる性質を有することが明らかである。
【0035】
そこで、出願人は、キャンディダ・マルトーサO9−NP9株を、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託した(寄託日:平成19年1月10日)。受託番号は、NITE BP−297である。
【0036】
7−2.O9−NP5株の同定
まず、遺伝学的性質を調べるため、O9−NP5株の26S rDNAの塩基配列を、O9−NP9株と同様にして同定した。国際塩基配列データベース(GenBank/DDBJ/EMBL)上で、該塩基配列について相同性検索を行ったところ、子嚢菌糸酵母の一種であるクラビスポラ・ルシタニエMTCC1001(アクセッション番号:AF538871)と100%の相同性を示したが、クラビスポラ・ルシタニエの基準株であるクラビスポラ・ルシタニエNRRL Y−11827T(アクセッション番号:U44817)とは93.8%の相同性を示した。また、該塩基配列と99%以上の相同性を示す既知種の基準株は存在しなかった。
以下にO9−NP5株の菌学的性質を示す。なお、菌学的性質の測定は、O9−NP9株と同様にして行った。
【0037】
菌学的性質
(1)コロニー観察(YM寒天培地平板で25℃、4日間培養した時の目視観察時)
周縁の形状:全縁、隆起状態:クッション形、表面の形状:平滑、光沢及び性状:バター様、湿性、色調:白色からクリーム色
(2)培養的・形態的性質(YM寒天培地平板で25℃、2日間培養した時の光学顕微鏡観察時)
栄養細胞の大きさと形状:直径2〜6μmの球形から広楕円形あるいは偏円形
増殖の形式:多極出芽
菌糸等:偽菌糸の形成が有
(3)有性生殖器官の形成(YM寒天培地平板で25℃、1ヶ月間培養した時の光学顕微鏡観察時):なし
【0038】
【表2】
【0039】
(7)37℃下における生育性:陽性
(8)40℃下における生育性:陽性
(9)10μg/mLシクロヘキシミド含有培地における生育性:陽性
(10)10%塩化ナトリウム及び5%グルコース含有培地における生育性:陽性
(11)50%グルコース含有培地における生育性:陽性
(12)ビタミン欠乏培地での生育性:陰性
(13)デンプン様物質の形成:陰性
(14)尿素の分解:陰性
(15)ゼラチンの液化性:陰性
【0040】
O9−NP5株は、グルコース発酵性を示し、硝酸塩の資化性を示さず、クラビスポラ属菌と一致する菌学的性質を示した。また、ほぼ全ての特徴において、クラビスポラ・ルシタニエと類似した菌学的性質を示したが、ラフィノース、可溶性デンプン、及びヘキサデカンの資化性において、クラビスポラ・ルシタニエとは異なる菌学的性質を示した。
【0041】
これらの遺伝学的及び菌学的性質から、O9−NP5株はクラビスポラ属菌であることが確認された。また、クラビスポラ・ルシタニエの基準株との相同性が低かったこと、及びラフィノース等の資化性が異なることから、クラビスポラ・ルシタニエO9−NP5株は、クラビスポラ・ルシタニエの新規菌株、若しくは、クラビスポラ・ルシタニエに近縁な新規な菌種であると推定された。
【0042】
そこで、出願人は、クラビスポラ・ルシタニエO9−NP5株を、『クラビスポラ・スピーシーズP−5株』として、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託した(受領日:2008年1月16日)。受領番号は、NITE ABP−474である。
なお、本願明細書中、「クラビスポラ・ルシタニエO9−NP5株」と「クラビスポラ・スピーシーズP−5株」は同一の菌株を表している。
【0043】
8.本発明の抗真菌性物質を産生する酵母の取得
MRS寒天培地に代えて、YM寒天培地を用いた以外は、全てキャンディダ・マルトーサO9−NP9株を単離した場合と同様にして、別のロットのカスピアンチーズから、抗真菌性物質を産生する酵母を1株得た。該1株を、O9−H1株と名付け、菌の同定を行った。
【0044】
まず、遺伝学的性質を調べるため、O9−H1株の26S rDNAの塩基配列を、O9−NP9株と同様にして同定した。国際塩基配列データベース(GenBank/DDBJ/EMBL)上で、該塩基配列について相同性検索を行ったところ、子嚢菌糸酵母の一種であるサッカロマイセス・セレビシエの基準株であるNRRL Y−12632T(アクセッション番号:AY048154)と100%の相同性を示した。したがって、O9−H1株は、サッカロマイセス・セレビシエに属する微生物であると推定された。
以下にO9−H1株の菌学的性質を示す。なお、菌学的性質の測定は、O9−NP9株と同様にして行った。
【0045】
菌学的性質
(1)コロニー観察(YM寒天培地平板で25℃、4日間培養した時の目視観察時)
周縁の形状:ほぼ全縁、隆起状態:中央突起形、表面の形状:平滑、光沢及び性状:バター様、湿性、色調:明るいクリーム色
(2)培養的・形態的性質(YM寒天培地平板で25℃、3日間培養した時の光学顕微鏡観察時)
栄養細胞の大きさと形状:直径3〜8μmの広楕円形から偏円形
増殖の形式:多極出芽
(3)有性生殖器官の形成(YM寒天培地平板で25℃、20日間培養した時の光学顕微鏡観察時):子嚢様の肥大した細胞は観察されたが、子嚢胞子の形成は認められなかった。
【0046】
【表3】
【0047】
(7)30℃下における生育性:陽性
(8)37℃下における生育性:陽性
(9)40℃下における生育性:陽性
(10)1μg/mLシクロヘキシミド含有培地における生育性:陽性
(11)10%塩化ナトリウム及び5%グルコース含有培地における生育性:陽性
(12)50%グルコース含有培地における生育性:陽性
(13)ビタミン欠乏培地での生育性:陰性
(14)デンプン様物質の形成:陰性
(15)尿素の分解:陰性
【0048】
O9−H1株は、グルコース、ガラクトース、スクロース等の急速な糖発酵性を示し、硝酸塩資化性を示さず、サッカロマイセス属菌と一致する菌学的性質を示した。また、ほぼ全ての特徴において、サッカロマイセス・セレビシエと類似した菌学的性質を示した。一方、ラフィノースを発酵せず、イヌリン資化性を示した点で、O9−H1株は、サッカロマイセス・セレビシエと異なる菌学的性質を示した。
【0049】
これらの遺伝学的及び菌学的性質から、O9−H1株はサッカロマイセス・セレビシエ菌種であることが確認された。26S rDNA−D1/D2領域の塩基配列は、サッカロマイセス・セレビシエNRRL Y−12632T株と100%の相同性があったが、ラフィノース発酵能及びイヌリン資化において異なるため、サッカロマイセス・セレビシエO9−H1株はサッカロマイセス・セレビシエNRRL Y−12632T株とは異なる性質を有することが明らかである。
【0050】
そこで、出願人は、サッカロマイセス・セレビシエO9−H1株を、『サッカロマイセス・セレビシエH−1株』として、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託した(受領日:2008年1月16日)。受領番号は、NITE ABP−473である。
なお、本願明細書中、「サッカロマイセス・セレビシエO9−H1株」と「サッカロマイセス・セレビシエH−1株」は同一の菌株を表している。
【0051】
本発明における抗真菌活性とは、カビや酵母等の真菌の増殖を抑制する活性のうち、胞子の発芽を阻害する活性を意味する。胞子の発芽を阻害することにより、効果的にカビ等の発生及び増殖を防止することができる。
【0052】
本発明の抗真菌性物質は、明確な物質名は未だ明らかではないが、キャンディダ属菌、サッカロマイセス属菌、ピキア属菌、ロドトルラ属菌、クラビスポラ属菌、及びトルラスポラ属菌からなる群より選ばれる1の酵母が産生する、胞子の発芽を阻害する抗真菌性物質である。
【0053】
また、本発明の抗真菌性物質は、(1)pH7.0において、抗真菌活性を有し、(2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有し、及び(3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCOが10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持される、という特徴を有するものである。さらに、(4)55℃で15分間の加熱処理により、前記抗真菌活性が失活する、という特徴を有するものであってもよい。
【0054】
本発明の抗真菌性物質を産生するキャンディダ属菌は、キャンディダ・マルトーサ、キャンディダ・トロピカリス、又はキャンディダ・シュードインターメディアであることが好ましく、キャンディダ・マルトーサであることが特に好ましい。該キャンディダ・マルトーサには、例えば、キャンディダ・マルトーサO9−NP9株、キャンディダ・マルトーサIAM12247、及びキャンディダ・マルトーサIAM12248等がある。該キャンディダ・トロピカリスには、例えば、キャンディダ・トロピカリスIAM4965等がある。該キャンディダ・シュードインターメディアには、例えば、キャンディダ・シュードインターメディアIAM12510等がある。
【0055】
本発明の抗真菌性物質を産生するピキア属菌は、ピキア・フェルメンタスであることが好ましく、ピキア・フェルメンタスJCM1824であることが特に好ましい。また、本発明の抗真菌性物質を産生するロドトルラ属菌は、ロドトルラ・アクタであることが好ましく、ロドトルラ・アクタJCM9494であることが特に好ましい。また、本発明の抗真菌性物質を産生するサッカロマイセス属菌は、サッカロマイセス・セレビシエであることが好ましく、サッカロマイセス・セレビシエH−1であることが特に好ましい。また、本発明の抗真菌性物質を産生するクラビスポラ属菌は、クラビスポラ・ルシタニエであることが好ましく、クラビスポラ・スピーシーズP−5又はクラビスポラ・ルシタニエNBRC10059であることが特に好ましい。また、本発明の抗真菌性物質を産生するトルラスポラ属菌は、トルラスポラ・デルブレッキーであることが好ましく、トルラスポラ・デルブルッキーNBRC0955であることが特に好ましい。
【0056】
本発明の酵母、すなわち、本発明の抗真菌性物質を産生するキャンディダ属菌、サッカロマイセス属菌、ピキア属菌、ロドトルラ属菌、クラビスポラ属菌、及びトルラスポラ属菌は、常法により培養することができ、培地や温度等の培養条件は、通常、キャンディダ属菌等の酵母を培養する条件であれば、特に限定されるものではない。該培地として、例えば、YM培地や10%(w/w)スキムミルク培地等がある。本発明の抗真菌性物質の活性が阻害されるおそれが少ないため、及び、該抗真菌性物質の精製が簡便になるため、10%(w/w)スキムミルク培地であることが好ましい。また、培養温度は、27〜30℃が好ましい。
【0057】
本発明の抗真菌性物質は、本発明の酵母を培養することにより得ることができる。例えば、本発明の酵母の培養液を、遠心分離処理をすることによって得た上清中に、本発明の抗真菌性物質は存在する。該遠心分離処理の条件は、該培養液中の菌体等の固体成分を除去できる条件であれば、特に限定されるものではないが、菌体由来の他の物質の混入を避けるため、2,500〜5,000×gの低速で遠心分離処理を行うことが好ましい。また、該上清を、MWCOが10kDである限外濾過膜を用いた限外濾過処理を行うことにより、該上清中に含まれる低分子の不純物を除去することもできる。このようにして得た該上清を、本発明の抗真菌性物質含有溶液として、防カビ剤や食品保存料として用いることもできる。
【0058】
本発明のカビ及び/又は酵母の増殖を阻害する方法は、本発明の抗真菌性物質又は本発明の酵母を用いる方法であって、本発明の抗真菌性物質の抗真菌活性が得られる方法であれば、特に限定されるものではない。本発明の抗真菌性物質を用いる方法として、例えば、該抗真菌性物質を含有する該酵母の培養液の上清等を、食品等に原料として添加する方法や、製造された食品に噴霧若しくは塗布する方法等がある。本発明の抗真菌性物質は、加熱処理により失活してしまうため、製造された食品に噴霧若しくは塗布する方法が好ましい。また、本発明の酵母を用いる方法として、例えば、発酵性食品の製造において、原料に本発明の酵母を添加した後、発酵させる方法等がある。該発酵性食品は、加熱処理を含まない発酵性食品であることが好ましい。
【実施例】
【0059】
次に実施例を示して本発明をさらに詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【0060】
[参考例1]
アスペルギルス・ニガーの胞子懸濁液の調整
アスペルギルス・ニガーNBRC9455株は、PDA(ポテトデキストロース寒天)平板培地に塗布した後、28℃で3日間培養して得られた胞子を、0.05%tween80添加生理食塩水に懸濁し、懸濁液を綿濾過後、−80℃で凍結保存したものを用いた。該凍結保存後のアスペルギルス・ニガーNBRC9455株を、生理食塩水を用いて希釈した後、サブロー培地を用いて、1.0×104spores/mLのアスペルギルス・ニガー胞子懸濁液を調製した。
【0061】
[実施例1]
キャンディダ・マルトーサO9−NP9株を、10%(w/w)スキムミルク培地に接種し、28℃で48時間培養して培養液を調製した。該培養液を超音波処理することにより、菌体を均一に分散させた後、3,000×gで10分間遠心分離を行って、上清を得た。
該上清を、5%(w/v)炭酸ナトリウム水溶液を用いてpHを7.0に調整した後、MWCOが10kDである限外濾過膜を用いて限外濾過を行った。限外濾過膜表面に保持された溶液を、抗真菌性物質溶液とした。
【0062】
1mLの該抗真菌性物質溶液を分注した1.5mLチューブを10本作製した。該チューブを、10、30、40、45、50、55、60、80、90℃の各温度で15分間加熱処理後、氷冷した。氷冷済チューブのそれぞれに対して、100μLの抗菌剤(ペニシリウムGK:0.7mg/mL、カナマイシン:60μg/mL、硫酸ストレプトマイシン:0.1mg/mL)を添加して混合した。その後、252μLの各チューブの抗真菌性物質溶液と、28μLのサブロー培地を混合して、活性測定用の抗真菌性物質溶液を調整した。
【0063】
96ウェルマイクロプレートの各ウェルに、調整した該抗真菌性物質溶液の2倍段階希釈系列(20〜29倍)を、それぞれ130μLずつ添加した。該抗真菌性物質溶液の希釈には、サブロー培地を用いた。具体的には、96ウェルマイクロプレートの1列目に該抗真菌性物質溶液を添加し、2列目に、該抗真菌性物質溶液の2倍希釈液を添加し、3列目に、該抗真菌性物質溶液の4(22)倍希釈溶液を添加した。各列に、順次、2倍段階希釈液をウェルに添加し、最後に10列目に、該加抗真菌性物質溶液の512(29)倍希釈溶液を添加した。コントロールとして、130μLのサブロー培地のみを添加した。さらに各ウェルに、前記参考例1に記載の方法で調整した1.0×104spores/mLのアスペルギルス・ニガー胞子懸濁液を50μLずつそれぞれ添加した。該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルを顕微鏡で観察した。
【0064】
【表4】
【0065】
表4は、顕微鏡観察の結果に基づき、各温度処理済抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「24」の記載は、抗真菌性物質溶液等の24倍希釈溶液以下の希釈倍率の溶液、すなわち、20〜24倍希釈溶液を添加したウェルにおいて、菌の増殖抑制が観察されたことを示している。また、表中「29」の記載は、抗真菌性物質溶液等の29倍希釈溶液以下の希釈倍率の溶液、すなわち、本実施例の全ての希釈溶液を添加したウェルにおいて、菌の増殖抑制が観察されたことを示している。さらに表中、「−」の記載は、20倍希釈溶液(原液)を添加したウェルにおいても活性が観察されなかったことを示している。
表4より明らかであるように、10℃と40℃の各温度処理済抗真菌性物質溶液では、本実施例の全ての希釈溶液を添加したウェルにおいて、菌の増殖抑制が観察された。したがって、10℃と40℃の各温度処理済抗真菌性物質溶液では、29倍率以上の希釈液においても、菌の増殖が抑制された可能性があり、高い抗真菌活性を有していることが明らかである。一方、加熱処理温度が40℃を超えると、急激に抗真菌活性は低下し、55℃15分間の加熱処理により、失活することが分かった。
【0066】
すなわち、実施例1の結果から、本発明の抗真菌性物質は、キャンディダ属菌等の酵母が産生するものであって、pH7.0において、抗真菌活性を有し、10%(w/w)スキムミルク培地で該酵母を培養した場合に、培養液中に分泌され、10%(w/w)スキムミルク培地で該酵母を培養した培養液の上清を、MWCOが10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持され、55℃で15分間の加熱処理により、該抗真菌活性が失活する、という特徴を有するものであることが明らかである。
【0067】
[実施例2]
本発明の抗真菌性物質の、様々な真菌に対する抗真菌活性を測定した。抗真菌活性のポジティブコントロールとして、真核細胞に対するタンパク質合成阻害剤であるシクロヘキシミドを用いた。
まず、加熱処理を行わないこと以外は、全て実施例1と同様にして、キャンディダ・マルトーサO9−NP9株の培養液から、抗真菌性物質溶液を調製した。また、表5記載の18菌株の1.0×104spores/mLの胞子懸濁液を、参考例1と同様にしてそれぞれ調製した。
【0068】
実施例1と同様にして、96ウェルマイクロプレートに、調製した抗真菌性物質溶液若しくは500ppmのシクロヘキシミド溶液の、2倍段階希釈系列(20〜29倍)を調製した。さらに各2倍段階希釈系列に、各菌株の胞子懸濁液を、1ウェル当たり50μLずつ、それぞれ添加した。該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルで菌の増殖が抑制されているかどうかを顕微鏡で観察した。
【0069】
表5は、実施例1と同様に、各抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。
【0070】
【表5】
【0071】
表5記載の18菌株のシクロヘキシミドに対する感受性は、それぞれ異なっていたが、全ての菌株において、500ppmのシクロヘキシミド溶液の添加により、増殖抑制が観察された。一方、本発明の抗真菌性物質溶液は、全ての菌株において、500ppmシクロヘキシミド溶液よりも大きい希釈倍率の希釈溶液で、菌の増殖抑制が観察された。
表5の結果から、本発明の抗真菌性物質は、多様な真菌に対して抗真菌活性を有していること、及び、実施例2記載の方法により調製した本発明の抗真菌性物質溶液は、500ppmシクロヘキシミド溶液よりも高い抗真菌活性を有していることが明らかである。
【0072】
[実施例3]
キャンディダ・マルトーサO9−NP9株以外のキャンディダ属菌や他の酵母が、キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を産生するかどうかを調べた。抗真菌活性のポジティブコントロールとして、実施例2と同様にシクロヘキシミドを用いた。
まず、実施例2と同様にして、表6記載の酵母の培養液から、抗真菌性物質溶液をそれぞれ調製した。また、参考例1と同様にして、1.0×104spores/mLのアスペルギルス・ニガー胞子懸濁液を調製した。
【0073】
実施例2と同様にして、96ウェルマイクロプレートに、調製した各酵母の抗真菌性物質溶液若しくは500ppmのシクロヘキシミド溶液の、2倍段階希釈系列(20〜29倍)を調製した。さらに各2倍段階希釈系列に、アスペルギルス・ニガー胞子懸濁液を、1ウェル当たり50μLずつ、それぞれ添加した。該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルで菌の増殖が抑制されているかどうかを顕微鏡で観察した。
【0074】
表6は、実施例2と同様に、各酵母の抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」の記載は、表4と同様に、20倍希釈溶液(原液)を添加したウェルにおいても活性が観察されなかったことを示している。
【0075】
【表6】
【0076】
表6の結果から、キャンディダ・マルトーサO9−NP9株以外の幾つかの酵母も、キャンディダ・マルトーサO9−NP9株と同様に、アスペルギルス・ニガーの増殖抑制が観察され、本発明の抗真菌性物質を産生することが明らかとなった。特に、キャンディダ・マルトーサIAM12247とキャンディダ・マルトーサIAM12248も、キャンディダ・マルトーサO9−NP9株と同様に、他の菌株に比べて、顕著に高い抗真菌活性を有していた。つまり、キャンディダ・マルトーサは、他の酵母に比べて、本発明の抗真菌性物質の産生能が非常に優れていることが明らかとなった。また、キャンディダ・トロピカリスIAM4965の抗真菌性物質溶液も、500ppmシクロヘキシミド溶液よりも高い抗真菌活性を有していた。その他、キャンディダ属菌のうち、キャンディダ・シュードインターメディアIAM12510も抗真菌活性を有していた。キャンディダ属菌以外にも、ピキア・フェルメンタスJCM1824やロドトルラ・アクタJCM9494も抗真菌活性を有していることが分かった。
【0077】
[実施例4]
実施例3で調べた酵母に加えて、さらに多くの酵母において、キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を産生するかどうかを調べた。具体的には、表7及び8記載の酵母を用いた以外は、全て実施例3と同様にして行った。
【0078】
【表7】
【0079】
【表8】
【0080】
表7及び8は、実施例3と同様に、各酵母の抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」の記載は、表6と同様である。
表7及び8の結果から、キャンディダ属菌のうち、さらにキャンディタ・サイトアナ(Candida saitoana)JCM1438等の幾つかの菌も抗真菌活性を有していることがわかった。
【0081】
[実施例5]
キャンディダ・マルトーサO9−NP9株、サッカロマイセス・セレビシエH−1株、及び、クラビスポラ・スピーシーズP−5株の抗真菌活性を比較した。
まず、実施例2と同様にして、該3菌株の培養液から、抗真菌性物質溶液をそれぞれ調製した。また、参考例1と同様にして、1.0×104spores/mLのアスペルギルス・ニガー胞子懸濁液を調製した。
【0082】
実施例2と同様にして、96ウェルマイクロプレートに、該3菌株の抗真菌性物質溶液の、2倍段階希釈系列(20〜29倍)を調製した。さらに各2倍段階希釈系列に、アスペルギルス・ニガー胞子懸濁液を、1ウェル当たり50μLずつ、それぞれ添加した。該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルで菌の増殖が抑制されているかどうかを顕微鏡で観察した。
【0083】
顕微鏡観察の結果、キャンディダ・マルトーサO9−NP9株では、抗真菌性物質溶液の29倍希釈液を添加したウェルであっても、アスペルギルス・ニガーの増殖抑制が観察された。一方、サッカロマイセス・セレビシエH−1株及びクラビスポラ・スピーシーズP−5株では、抗真菌性物質溶液の20〜24倍希釈液を添加したウェルにおいてのみ、アスペルギルス・ニガーの増殖抑制が観察された。
【0084】
実施例5の結果から、サッカロマイセス・セレビシエH−1株及びクラビスポラ・スピーシーズP−5株は、キャンディダ・マルトーサO9−NP9株と同様に本発明の抗真菌活性物質を産生すること、及び、該3株のなかでは、キャンディダ・マルトーサO9−NP9株が最も本発明の抗真菌性物質の産生能が高いことが明らかである。
【0085】
[実施例6]
サッカロマイセス属菌やその近縁菌が、キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を産生するかどうかを調べた。抗真菌活性のポジティブコントロールとして、実施例2と同様にシクロヘキシミドを用いた。
まず、実施例2と同様にして、表9記載の酵母の培養液から、抗真菌性物質溶液をそれぞれ調製した。また、参考例1と同様にして、1.0×104spores/mLのアスペルギルス・ニガー胞子懸濁液を調製した。
【0086】
実施例2と同様にして、96ウェルマイクロプレートに、調製した各酵母の抗真菌性物質溶液若しくは500ppmのシクロヘキシミド溶液の、2倍段階希釈系列(20〜29倍)を調製した。さらに各2倍段階希釈系列に、アスペルギルス・ニガー胞子懸濁液を、1ウェル当たり50μLずつ、それぞれ添加した。該96ウェルマイクロプレートを28℃で3日間又は7日間培養した後、各ウェルで菌の増殖が抑制されているかどうかを顕微鏡で観察した。
表9は、実施例2と同様に、各酵母の抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」の記載は、表4と同様に、20倍希釈溶液(原液)を添加したウェルにおいても活性が観察されなかったことを示しており、「ND」は測定しなかったことを意味している。また、表中「3日後」及び「7日後」は、それぞれ3日培養後又は7日培養後の結果を示している。
【0087】
【表9】
【0088】
表9の結果から、サッカロマイセス・セレビシエH−1株以外の幾つかの酵母においても、アスペルギルス・ニガーの増殖抑制が観察され、これらの酵母が本発明の抗真菌性物質を産生することが明らかとなった。特に、培養3日後においては、サッカロマイセス・セレビシエNBRC0216及びサッカロマイセス・セレビシエNBRC10181は、サッカロマイセス・セレビシエH−1と同様に、他の菌株に比べて、顕著に高い抗真菌活性を有していた。その他、トルラスポラ・デルブルッキーNBRC0955(受領番号 NITE ABP−475、受領日 2008年1月16日)、サッカロマイセス・パストリアナス(Saccharomyces pastprianus)NBRC11024、クリベロマイセス・ラクティス・バール・ドロソフィララム(Kluyveromyces lactis var.drosophilarum)NBRC1012、クリベロマイセス・ドブハンスキ(Kluyveromyces dobzhanskii)NBRC10603も、高い抗真菌活性を有していた。特に、トルロスポラ・デルブルッキーNBRC0955は、7日培養後であってもシクロヘキシミドよりも高い抗真菌活性を有しており、非常に優れていることが明らかとなった。
【0089】
[実施例7]
クラビスポラ属菌やその近縁菌が、キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を産生するかどうかを調べた。
具体的には、表9記載の酵母に代えて、表10記載の酵母を用いた以外は、全て実施例6と同様にして、各酵母の抗真菌性物質溶液を添加することにより、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察した。
表10は、各酵母の抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」、「ND」、「3日後」及び「7日後」の記載は、表9と同様である。
【0090】
【表10】
【0091】
表10の結果から、クラビスポラ・スピーシーズP−5株以外の幾つかの酵母も、キャンディダ・マルトーサO9−NP9株と同様に、アスペルギルス・ニガーの増殖抑制が観察され、本発明の抗真菌性物質を産生することが明らかとなった。培養3日後においては、クラビスポラ・スピーシーズP−5、クラビスポラ・ルシタニエNBRC10059は、キャンディダ・マルトーサO9−NP9と同様に、顕著に高い抗真菌活性を有していた。その他、クラビスポラ・ルシタニエNBRC10058もシクロヘキシミドよりも高い抗真菌活性を有していた。特に、クラビスポラ・ルシタニエNBRC10059(受領番号 NITE ABP−476、受領日 2008年1月16日)は、7日培養後であってもキャンディダ・マルトーサO9−NP9と同様に高い抗真菌活性を有しており、非常に優れていることが明らかとなった。
【0092】
[実施例8]
クラビスポラ・ルシタニエNBRC10059株及びトルロスポラ・デルブルッキーNBRC0955株が産生する本発明の抗真菌性物質の、様々な真菌に対する抗真菌活性を測定した。
具体的には、キャンディダ・マルトーサO9−NP9株に代えて、クラビスポラ・ルシタニエNBRC10059株又はトルロスポラ・デルブルッキーNBRC0955株を用いた以外は、全て実施例2と同様にして、各酵母の抗真菌性物質溶液を添加することにより、表11記載の菌の増殖が抑制されるかどうかを顕微鏡で観察した。
表11は、各酵母の抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」、「ND」、「3日後」及び「7日後」の記載は、表9と同様である。
【0093】
【表11】
【0094】
表11記載の13菌株の全てにおいて、クラビスポラ・ルシタニエNBRC10059株はシクロヘキシミドよりも高い抗真菌活性を示した。一方、トルロスポラ・デルブルッキーNBRC0955株は、幾つか増殖抑制効果を奏さない菌もあったが、真菌の種類によっては、シクロヘキシミドよりも高い抗真菌活性を示した。
【0095】
[実施例9]
各酵母が有する抗真菌活性物質の耐熱性を測定した。
まず、表12記載の各酵母をそれぞれ植菌した1LのPD(ポテトデキストロース)培地を、30℃48時間振とう培養したものを、3,000×gで5分間遠心分離処理し、上清を回収した。該上清を50℃、60℃、85℃、90℃の各温度で10分間処理後冷却したものを、抗真菌性物質溶液とした。なお、加熱処理をしなかった(非加熱処理)上清をコントロールとした。
実施例2と同様にして、これらの抗真菌性物質溶液を添加することにより、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察した。
表12は、各抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」の記載は、20倍希釈溶液(原液)を添加したウェルにおいても活性が観察されなかったことを示しており、「29」の記載は、29倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察されたことを示している。
この結果、表12記載の酵母が有する抗真菌活性物質は、いずれも60℃10分間以上の加熱処理で失活してしまうことが分かった。
【0096】
【表12】
【0097】
[実施例10]
キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を、原料としてパンに添加した場合の、抗真菌効果を観察した。
まず、キャンディダ・マルトーサO9−NP9株を植菌した1LのPD(ポテトデキストロース)培地を、30℃で3日間培養した。この培養液全体を、抗真菌性物質溶液として、実施例2と同様にして、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察したところ、該培養液の29倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察された。
この培養液を、水原料全量分に代えて食パンの原料とした。市販されている抗菌剤ネオカビンP(ジェイティーフーズ株式会社販売)を添加した食パンと、抗菌剤等を無添加の食パンを、それぞれ比較対象とした。具体的には、表12に示す配合量で、常法により食パンを製造した。表中、「%」は強力粉100%に対する重量部(%)を意味する。
【0098】
【表13】
【0099】
製造された食パンは、通常の6枚切りの厚さにスライスした。各食パン20枚ずつを、室内で10分間放置し、食パン表面上に落下菌を付着させた。その後、1枚ずつ袋に入れて、28℃で5日間保温した後、カビの有無や生え方等を観察した。カビの生え方は、コロニーの数が0個、1〜数10個、数10個以上、計数不能の4区分に区分けした。なお、「計数不能」とは、カビが表面の大部分を覆っていてコロニーの個数を計測できなかったものを意味する。
【0100】
【表14】
【0101】
表14は、観察により20枚の食パンを区分けした結果を、区分ごとの枚数で示したものである。この結果、全ての種類の食パンにおいて、20枚全てにカビが生えていた。無添加の食パンでは、1枚当たりのコロニー数が多く、計数不能なものが6枚もあったが、ネオカビンを添加した食パンは、1枚当たりのコロニー数が少なく、計数不能なものはなかった。これに対し、キャンディダ・マルトーサO9−NP9株の培養液を添加した食パンでは、ネオカビンを添加したものよりは多い傾向にあったが、無添加のものに比べて1枚当たりのコロニー数が非常に少なく、計数不能なものは1枚しかなかった。特に、培養液添加食パンでは、食パンの中心部にはほとんどカビが生えておらず、カビが生えているものには、食パンの耳のみにカビが生えているものが多かった。
このように、培養液を添加した食パンにおいて、カビが食パンの外側に多く、内側には少なかったのは、本発明の抗真菌性物質は加熱に弱いため、パンを焼成する工程で失活してしまったためと推察される。つまり、これらの結果から、本発明の抗真菌性物質は、加熱処理を施さない食品等への添加剤として用いることにより、従来の抗菌剤と同様に抗菌効果が期待できることが明らかである。
【0102】
[実施例11]
キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を、食パンに塗布した場合の、アスペルギルス・ニガーに対する抗真菌効果を観察した。
まず、キャンディダ・マルトーサO9−NP9株を接種した1LのPD培地を、30℃で3日間培養し、培養液全体を、抗真菌性物質溶液として、実施例2と同様にして、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察した。この結果、該培養液の29倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察された。この培養液を、市販の6枚切りの食パン(サンアロマ、山崎製パン社製)に、スプレーで全体に塗布し、コンラージ棒を用いて全体に塗り広げた。
培養液を塗布した食パンと、塗布していない食パン(コントロール)に、0.5mLのアスペルギルス・ニガー胞子懸濁液(2.0×104spores/mL)を、1枚当たり10点ずつ接種し、28℃で保温した。なお、アスペルギルス・ニガー胞子懸濁液は、参考例1と同様にして調製した。
この結果、接種から3日後には、コントロールの食パンにはカビが生えて緑色になっていたが、培養液を塗布した食パンにはカビは生えていなかった。さらに接種から5日後には、コントロールの食パンは全体的に黒いカビのコロニーが観察されたが、培養液を塗布した食パンでは、一箇所に白い毛状の菌糸が観察されただけであった。
これらの結果から、本発明の抗真菌性物質は、非加熱条件下で、非常に良好な抗真菌効果を奏することが明らかである。
【0103】
[実施例12]
キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を、食パンに塗布した場合の、落下菌に対する抗真菌効果を観察した。
実施例11と同様にして作製したキャンディダ・マルトーサO9−NP9株の培養液を塗布した食パンと、塗布していない食パン(コントロール)とを、各6枚ずつ、室内で10分間放置し、食パン表面上に落下菌を付着させた。その後、1枚ずつ袋に入れて、28℃で5日間保温した後、カビの有無を観察した。
この結果、コントロールの食パンは6枚ともカビが生えていたが、培養液を塗布した食パンにはいずれもカビは生えていなかった。これらの結果からも、本発明の抗真菌性物質は、非加熱条件下で、非常に良好な抗真菌効果を奏することが明らかである。
【0104】
[実施例13]
本発明の酵母を培養した培養液の抗真菌活性に対する培養条件の影響を観察した。具体的には、振とう培養と静置培養の、培養液の抗真菌活性に対する影響を調べた。
キャンディダ・マルトーサO9−NP9株を植菌した1LのPD培地を、25℃24時間又は48時間振とう培養して得た培養液と、同じくキャンディダ・マルトーサO9−NP9株を植菌した1LのPD培地を、25℃24時間又は48時間静置培養して得た培養液とを調製した。これらの培養液を、それぞれ3,000×gで5分間遠心分離処理し、上清と沈殿に分離して回収した。このようにして得られた上清は、そのまま抗真菌物質溶液として用いた。一方、菌体を含む沈殿は、上清と等量の蒸留水を加えて超音波処理することにより、菌体を均一に分散させた溶液を、抗真菌物質溶液として用いた。
各抗真菌物質溶液の抗真菌活性を、実施例2と同様にして、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察することにより測定した。
【0105】
【表15】
【0106】
表15は、各抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。この結果、培養時間や培養条件にかかわらず、沈殿由来の抗真菌性物質溶液では、この実験の最大希釈倍率である29倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察され、高い抗真菌活性を有していた。これに対して、上清由来の抗真菌性物質溶液では、培養時間による相違はあまりなく、いずれも、振とう培養由来の抗真菌性物質溶液は28倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察されたが、静置培養由来の抗真菌性物質溶液では22倍希釈溶液を添加したウェルまでしか菌の増殖抑制が観察されなかった。また、いずれの培養条件においても、上清由来の抗真菌性物質溶液よりも、沈殿由来の抗真菌性物質溶液のほうが高い抗真菌活性を有していた。
つまり、本発明の抗真菌性物質は、静置培養よりも振とう培養によって培養液に移行し易いこと、及び、沈殿(菌体)のほうが上清よりも抗真菌活性が高いことが分かった。したがって、本発明の抗真菌性物質は、静置培養よりも振とう培養時により積極的に酵母外へ分泌され易い可能性が示唆された。また、上清よりも沈殿のほうが高い抗真菌活性が検出されたことから、本発明の抗真菌性物質は、酵母表面に吸着し易い性質を有しているのではないかと推察される。
【0107】
[実施例14]
本発明の抗真菌性物質の性質を調べるため、粗精製した後、粗精製物中の物質の中性糖の定量分析を行った。
まず、キャンディダ・マルトーサO9−NP9株を接種した1LのPD培地を、30℃24時間振とう培養したものを、3,000×gで5分間遠心分離処理し、776.95gの上清を回収した。該上清の一部を抗真菌性物質溶液として、実施例2と同様にして、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察したところ、該培養液の210倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察された。
該上清の残り417.86gを、限外濾過モジュールSEP0031(MWCO:3kD、旭化成社製)を用いて限外濾過を行った。このとき、限外濾過膜を透過した溶液(S1)は314.87gであり、該限外濾過膜表面に保持された残渣(R1)は128.34gであった。このR1に脱イオン水を加えて500gとして攪拌した後、再度限外濾過モジュールSEP0031を用いて限外濾過を行った。このとき、限外濾過膜を透過した溶液(S2)は305.52gであり、該限外濾過膜表面に保持された残渣(R2)は138.87gであった。このR2に脱イオン水を加えて500gとして攪拌した後、再度限外濾過モジュールSEP0031を用いて限外濾過を行った。このとき、限外濾過膜を透過した溶液(S3)は402.36gであり、該限外濾過膜表面に保持された残渣(R3)は59.28gであった。
得られた残渣R3をそのまま均一に攪拌後、抗真菌性物質溶液として、実施例2と同様にして、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察したところ、該培養液の20〜25倍希釈溶液を添加したウェルにおいて菌の増殖抑制が観察された。
【0108】
さらに、残渣R3中の物質の中性糖の定量分析を行った。
具体的には、まず、抗真菌性物質溶液とした残渣R3を均一に攪拌した懸濁液を、ガラス試験管に100μL採取し、減圧乾固した。得られた残渣に200μLのトリフルオロ酢酸(2mol/L)を添加し、真空封管後、100℃で6時間加水分解した。加水分解物を減圧乾固して得られた残渣を、100μLの精製水に溶解させた後、孔径0.22μmのフィルターに通して濾過した濾液を測定試料原液とした。該測定試料原液を精製水で100倍又は1000倍希釈したものを測定試料とし、下記の条件で高速液体クロマトグラフ(HPLC)法で測定し、中性糖(ラムノース、リボース、マンノース、アラビノース、ガラクトース、キシロース、及びグルコース)の定量分析を行った。なお、ピークの同定は各中性糖の標準品との比較により行った。
HPLCシステム:LC−9Aシステム(島津製作所)
検出器:分光蛍光光度計RF−10AXL(島津製作所)
カラム:TSK−gel Sugar AXG 150×4.6mmI.D.(東ソー)
カラム温度:70℃
移動相:0.5Mホウ酸カリウム緩衝液pH8.7
移動相流速:0.4mL/min
ポストカラム標識:反応試薬:1%アルギニン/3%ホウ酸
反応試薬流速:0.5mL/min
反応温度:150℃
検出波長:Ex.320nm、Em.430nm
【0109】
この結果、残渣R3は、約1%のグルコース、及び少量のマンノースを含んでいることが分かった。つまり、残渣R3には多くの糖類が含まれていることから、本発明の抗真菌性物質は糖タンパク質や多糖類等の糖を多く含む化合物である可能性が示唆された。
また、この中性糖の分析結果と、静置培養よりも振とう培養によって培養液に移行し易い点、さらに後述の実施例16の結果からも、実施例13と同様に、本発明の抗真菌性物質は、酵母表面の糖タンパク質と、自身が有する中性糖を介して相互作用する等により、酵母表面に吸着され易い性質を有する可能性が示唆される。
【0110】
[実施例15]
500μLのキャンディダ・マルトーサO9−NP9株の培養液と、500μLの2.4×106spores/mLのアスペルギルス・ニガー胞子懸濁液を混合し、28℃で24時間インキュベートした。なお、キャンディダ・マルトーサO9−NP9株の培養液は、キャンディダ・マルトーサO9−NP9株を、10%(w/w)スキムミルク培地に接種し、28℃で48時間培養することにより調製した。また、アスペルギルス・ニガー胞子懸濁液は参考例1と同様にして調製した。
この培養液1mLに、200μLの蛍光色素0.5μM SYTOX−Green(タカラバイオ社製)を添加し、さらに0.5時間インキュベートした。その後、この培養液を3,000×gで5分間遠心分離処理し、上清を除去することにより蛍光色素を除去した。得られた沈殿に蒸留水を添加して懸濁させた後、再度遠心分離処理して上清を除去して沈殿を洗浄した。
この沈殿にさらに1mLの蒸留水を添加して、酵母と胞子の懸濁液を調製した。この懸濁液を通常の光学顕微鏡を用いて、蛍光及び透過光観察したところ、SYTOX−Greenで染色された胞子が観察された。また、この蛍光染色されている胞子は、吸水時と同等又はそれ以上の大きさに膨張していることが観察された。
ここで、SYTOX−Greenは、DNAと特異的に結合し得る色素であるが、細胞膜を通過することはできない。にもかかわらず、キャンディダ・マルトーサO9−NP9株とインキュベートしたアスペルギルス・ニガーの胞子が染色されたのは、キャンディダ・マルトーサO9−NP9株が有する抗真菌性物質の作用により、胞子の細胞壁が損傷を受けたためと推察される。また、蛍光染色された胞子が膨張していたことからも、胞子の細胞壁が損傷を受けたことが示唆される。
500μLの10%(w/w)スキムミルク培地に500μLの2.4×106spores/mLのアスペルギルス・ニガー胞子懸濁液を混合したものを同様にインキュベートし、SYTOX−Greenを添加したところ、蛍光染色された胞子は観察されなかった。また、キャンディダ・マルトーサO9−NP9株に代えて、本発明の抗真菌活性が観察されなかったキャンディダ・ユーティリスを用いたものを同様にインキュベートし、SYTOX−Greenを添加した場合にも、蛍光染色された胞子は観察されなかった。これらの結果からも、胞子が染色されたのは、キャンディダ・マルトーサO9−NP9株が有する本発明の抗真菌物質によるためであることが示唆される。
【0111】
[実施例16]
寒天培地中で培養した場合の本発明の抗真菌活性物質の活性を調べた。
まず、キャンディダ・マルトーサO9−NP9株を混釈したPDA培地を、28℃で48時間培養した後、滅菌済みの型を用いてくりぬき、キャンディダ・マルトーサO9−NP9株が生育している円柱形のPDA培地(以下、酵母混釈寒天培地という。)を作製した。
一方、同じ型を用いて、参考例1と同様にして調製したアスペルギルス・ニガー胞子懸濁液を混釈したPDA培地をくりぬき、アスペルギルス・ニガー胞子を含む円柱形のPDA培地(以下、胞子混釈寒天培地という。)を作製した。
これらの2種類の寒天培地を、(a)酵母混釈寒天培地の上に、胞子混釈寒天培地を直接重ね合わせた状態、(b)酵母混釈寒天培地の上に、ラップを挟んで胞子混釈寒天培地を重ね合わせた状態、(c)酵母混釈寒天培地の上に、孔径14kDの透析膜を挟んで胞子混釈寒天培地を重ね合わせた状態、の3つの状態で、それぞれ28℃で48〜72時間培養し、胞子の状態を観察した。
この結果、(b)の胞子混釈寒天培地では、胞子の増殖が観察された。一方、(a)と(c)の胞子混釈寒天培地では、胞子の増殖は観察されなかった。
また、培養後の胞子混釈寒天培地の一部を、液体のPD培地に添加し、28℃で24時間培養した後、培養液をPDA平板培地に塗抹して、さらに培養した。この結果、(a)の胞子混釈寒天培地由来のPDA平板培地には、酵母が増殖しており、カビは生えてこなかった。一方、(b)の胞子混釈寒天培地由来のPDA平板培地には、カビが生えてきたが、酵母は確認できなかった。さらに(c)の胞子混釈寒天培地由来のPDA平板培地には、カビが生えてきたが、酵母は確認できなかった。
これらの結果から、(b)では、2つの寒天培地がラップで挟まれていたため、胞子混釈寒天培地中のアスペルギルス・ニガー胞子は、酵母混釈寒天培地の影響を受けず、胞子が生育したが、(a)では、キャンディダ・マルトーサO9−NP9株が胞子混釈寒天培地へ移行して増殖した結果、アスペルギルス・ニガー胞子の増殖が阻害されたと考えられる。これに対して、(c)では、透析膜に阻まれて、(b)と同様に酵母は胞子混釈寒天培地へ移行できなかったが、酵母から産生された本発明の抗真菌物質が透析膜を透過して胞子混釈寒天培地へ移行したため、アスペルギルス・ニガー胞子の増殖が阻害された結果、胞子混釈寒天培地では胞子の増殖が観察されなかったと考えられる。一方、この胞子混釈寒天培地中に最初に混釈されていた胞子は死滅しなかったため、(c)の胞子混釈寒天培地由来のPDA平板培地にはカビが生育したと考えられる。
これらの結果からも、キャンディダ・マルトーサO9−NP9株が本発明の抗真菌性物質を産生していることが明らかである。
【産業上の利用可能性】
【0112】
本発明の抗真菌性物質は、チーズ由来の酵母が産生するため安全であり、中性において抗真菌活性を示すことから、特にカビの増殖が問題となる食品分野等で利用が可能である。
【図面の簡単な説明】
【0113】
【図1】O−9菌群培養液由来のサンプル溶液の2倍段階希釈系列と、アスペルギルス・ニガー胞子懸濁液を、96ウェルマイクロプレートの各ウェルにそれぞれ添加して、28℃で48時間培養した後に、各ウェルを顕微鏡で観察した結果を示したものである。図中のレーンAはO−9菌群由来の上清サンプル溶液の希釈系列を、レーンBはO−9菌群由来の沈殿物サンプル溶液の希釈系列を、レーンCはコントロールを、それぞれ示している。図中の○は、各ウェルを表したものであり、○の中の×は、アスペルギルス・ニガーの増殖が観察されたウェルを、○の中の○は、アスペルギルス・ニガーの増殖抑制が観察されたウェルを、それぞれ表したものである。
【図2】P1〜P10株及びY1〜Y3株をそれぞれ培養した培養液由来の上清サンプル溶液の2倍段階希釈系列と、アスペルギルス・ニガー胞子懸濁液を、96ウェルマイクロプレートの各ウェルにそれぞれ添加して培養した後に、各ウェルを顕微鏡で観察した結果を示したものである。(a)と(b)は、該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルを顕微鏡で観察した結果を、(c)と(d)は同じく36時間培養した後の観察結果を、それぞれ表したものである。図中の○は、各ウェルを表したものであり、○の中の×は、アスペルギルス・ニガーの増殖が観察されたウェルを、○の中の○は、アスペルギルス・ニガーの増殖抑制が観察されたウェルを、それぞれ表したものである。
【図3】YM寒天培地平板で25℃、2日間培養したO9−NP9株の電子顕微鏡写真である。
【技術分野】
【0001】
本発明は、キャンディダ(Candida)属菌、サッカロマイセス(Saccharomyces)属菌、ピキア(Pichia)属菌、ロドトルラ(Rhodotorula)属菌、クラビスポラ(Clavispora)属菌、及びトルラスポラ(Torulaspora)属菌からなる群より選ばれる1の酵母が産生する、胞子の発芽を阻害する抗真菌性物質、該抗真菌性物質を含有する防カビ剤及び食品保存料、該抗真菌性物質又は該酵母を用いるカビ及び/又は酵母の増殖を阻害する方法、並びに、該抗真菌性物質を産生する新規キャンディダ属菌に関する。
【背景技術】
【0002】
微生物による腐敗や変敗を防止し、食品の保存性を高めるため、様々な抗菌性物質が、食品保存料として用いられている。食品の保存性を向上させることにより、流通過程における食品の腐敗等が効率的に防止されるため、製造コストの改善が期待でき、また、食品の安全性上も好ましい。このため、優れた抗菌性物質の開発が望まれている。ここで、抗菌性物質は、主に、プロピオン酸やソルビン酸等の化学合成物質と、微生物等から得られた天然由来の物質に分類できるが、食品保存料として用いる場合には、安全性の点から、天然由来の物質であることが好ましい。
【0003】
近年、天然由来の抗菌性物質として、ナイシン等の、乳酸菌由来の様々な種類のバクテリオシンが知られている(例えば、非特許文献1参照。)。バクテリオシンは、通常、ペプチド若しくはタンパク質であり、消化酵素等により容易に分解されるため、非常に安全性が高い。また、主にグラム陽性菌に対し優れた抗菌活性を有することが知られている。
【0004】
その他、グラム陽性菌とグラム陰性菌の両方に対し優れた抗菌活性を有する抗菌性物質を含む食品保存料の製造方法として、例えば、(1)酵母の増殖に必要な栄養素、無機塩類と炭素源としてグルコースを含む培地で酵母を定常期に達するまで増殖させた後、酵母を除去した培地に必要ならば炭素源を加え、再び定常期に達するまで増殖させる繰り返し培養を2回以上行って得られた水溶液であることを特徴とする酵母が産生する抗菌性物質を含む食品保存料の製造方法が開示されている(例えば、特許文献1参照。)。
【非特許文献1】園元ら、食品と開発、41巻3号、p73〜75、2006年
【特許文献1】特許第2545739号公報
【発明の開示】
【発明が解決しようとする課題】
【0005】
グラム陽性菌等のバクテリアのみならず、カビや酵母等の真菌も、食品の腐敗等を引き起こす。特にカビは、多くの食品の腐敗の主要な要因であることから、カビの増殖を抑制することができる抗真菌活性を有する物質を、食品保存料や抗菌剤として使用できることが好ましい。
しかしながら、ナイシン等のバクテリオシンは、一般にグラム陽性菌にのみ抗菌活性を示し、カビや酵母等の真菌の増殖を抑制することができないという問題がある。また、上記(1)の方法で得られる食品保存料は、酵母由来の抗菌性物質であるため、食品に対し安全に使用することができ、また、耐熱性も有しているが、特許文献1には、カビに対する抗菌活性については一切記載がない。
【0006】
一方、プロピオン酸等の有機酸は、真菌の増殖抑制に対し、一定の効果を有するものの、化学合成物質であるため、天然由来の物質に比べて安全性が劣ることに加え、独特のにおいを有するため、食品の風味上好ましくないという問題がある。
また、有機酸は、主に酸性下で抗菌活性を有し、中性の食品に対する抗菌性に乏しいという問題もある。有機酸の中性における抗菌活性の低下は、中性溶液中では、有機酸はほぼ完全に電離しているが、解離して負に帯電した有機酸は、微生物の細胞内に取り込まれ難いためと考えられている。
【0007】
本発明は、安全であり、かつ、中性においてカビや酵母等の真菌の増殖を抑制することのできる、抗真菌性物質を提供することを目的とする。
また、本発明は、該抗真菌性物質を含有する防カビ剤及び食品保存料、該抗真菌性物質を産生する菌、並びに、該抗真菌性物質又は該菌を用いるカビ及び/又は酵母の増殖を阻害する方法を提供することを目的とする。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決すべく鋭意研究した結果、食品由来の微生物が産生する抗真菌性物質であれば、食品保存料としても安全に用いることができると考え、様々な種類のチーズから分離した微生物の中から、pH7.0において、アスペルギルス・ニガー(Aspergillus niger)又はリゾパス・ストロニファー(Rhizopus stolonifer)の増殖を抑制することのできる抗真菌活性を有する微生物を見出し、さらに、該微生物が産生する抗真菌性物質を見出すことにより、本発明を完成させた。
【0009】
すなわち、本発明は、キャンディダ(Candida)属菌、サッカロマイセス(Saccharomyces)属菌、ピキア(Pichia)属菌、ロドトルラ(Rhodotorula)属菌、クラビスポラ(Clavispora)属菌、及びトルラスポラ(Torulaspora)属菌からなる群より選ばれる1の酵母が産生する、胞子の発芽を阻害する抗真菌性物質を提供するものである。
また、本発明は、下記の特徴を有する抗真菌性物質を提供するものである。
(1)pH7.0において、抗真菌活性を有すること、
(2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有すること、
(3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCO(Molecular Weight Cut Off、分画分子量)が10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持されること。
また、本発明は、下記の特徴を有する抗真菌性物質を提供するものである。
(1)pH7.0において、抗真菌活性を有すること、
(2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有すること、
(3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCO(Molecular Weight Cut Off、分画分子量)が10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持されること、
(4)55℃で15分間の加熱処理により、前記抗真菌活性が失活すること。
また、本発明は、前記キャンディダ属菌が、キャンディダ・マルトーサ(Candida maltosa)、キャンディダ・トロピカリス(Candida tropicalis)、又はキャンディダ・シュードインターメディア(Candida pseudintermedia)である抗真菌性物質を提供するものである。
また、本発明は、前記キャンディダ属菌が、キャンディダ・マルトーサO9−NP9(受託番号 NITE BP−297)、キャンディダ・マルトーサIAM12247、キャンディダ・マルトーサIAM12248、キャンディダ・トロピカリスIAM4965、又はキャンディダ・シュードインターメディアIAM12510である抗真菌性物質を提供するものである。
また、本発明は、前記サッカロマイセス属菌がサッカロマイセス・セレビシエ(Saccharomyces cerevisiae)であり、前記ピキア属菌がピキア・フェルメンタス(Pichia fermentans)であり、前記ロドトルラ属菌がロドトルラ・アクタ(Rhodotorula acuta)であり、前記クラビスポラ属菌がクラビスポラ・ルシタニエ(Clavispora lusitaniae)であり、前記トルラスポラ属菌がトルラスポラ・デルブレッキー(Torulaspora delbrueckii)である、抗真菌性物質を提供するものである。
また、本発明は、前記ピキア属菌がピキア・フェルメンタスJCM1824であり、前記ロドトルラ属菌がロドトルラ・アクタJCM9494であり、前記サッカロマイセス属菌がサッカロマイセス・セレビシエH−1(受領番号 NITE ABP−473)であり、前記クラビスポラ属菌がクラビスポラ・スピーシーズP−5(Clavispora sp. P−5、受領番号 NITE ABP−474)又はクラビスポラ・ルシタニエNBRC10059であり、前記トルラスポラ属菌がトルラスポラ・デルブルッキーNBRC0955である抗真菌性物質を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を含有する防カビ剤を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を含有する食品保存料を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を用いることを特徴とする、カビ及び/又は酵母の増殖を阻害する方法を提供するものである。
また、本発明は、前記いずれか記載の酵母を用いることを特徴とする、カビ及び/又は酵母の増殖を阻害する方法を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を産生することを特徴とするキャンディダ・マルトーサO9−NP9(受託番号 NITE BP−297)を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を産生することを特徴とするサッカロマイセス・セレビシエH−1(受領番号 NITE ABP−473)を提供するものである。
また、本発明は、前記いずれか記載の抗真菌性物質を産生することを特徴とするクラビスポラ・スピーシーズP−5(受領番号 NITE ABP−474)を提供するものである。
【発明の効果】
【0010】
本発明の抗真菌性物質により、効果的にカビや酵母等の真菌の増殖を抑制することができる。本発明の抗真菌性物質は、チーズから分離された酵母が産生する物質であるため、食品に対して安全に用いることができるのみならず、化粧品や医薬品等にも用いることが可能である。特に中性において、抗真菌活性を示すため、中性の食品等においても有効にカビ等の増殖を抑制することができる。本発明の抗真菌性物質により、食品等の保存期間を安全に延長することができるため、製造コストの改善も期待される。
また、本発明の抗真菌性物質を産生する酵母により、本発明の抗真菌性物質を効率よく産生することができる。
さらに、本発明のカビ及び/又は酵母の増殖を阻害する方法により、安全かつ簡便に、カビや酵母等の真菌の増殖を抑制することができる。
【発明を実施するための最良の形態】
【0011】
本発明の抗真菌性物質は、食品から得られた微生物が産生するものであって、pH7.0において、抗真菌活性を有するものである。このような抗真菌性物質であれば、食品等に安全に使用することができ、かつ、中性の食品等に対しても効果を発揮することができるためである。
【0012】
本発明の抗真菌性物質を産生する微生物は、例えば、各種の食品から分離した微生物の中から、pH7.0において、カビ等の増殖を抑制することができる抗真菌活性を有する菌株を選択することにより得ることができる。pH7.0において抗真菌活性を有する菌株は、本発明の抗真菌性物質を産生していると考えられるためである。以下、本発明の抗真菌性物質、及び該抗真菌性物質を産生する微生物を得る方法を、さらに詳細に説明する。
【0013】
1.微生物群の取得
本発明者らは、抗真菌性物質を産生する微生物を食品中から取得するにあたり、良好な発酵食品の一つであるチーズに着目し、常法により、微生物を分離した。具体的には、チーズを滅菌したチーズおろし金ですり下ろした後、0.05gを、スクリューキャップ付き試験管に分注した滅菌済10%(w/v)スキムミルク溶液5mLに加え、28℃で48時間静置培養した。該スクリューキャップ付き試験管を遠心分離して回収した沈殿物を、滅菌生理リン酸緩衝液で2回洗浄した後、生理リン酸緩衝液で希釈してサンプルを調製した。該サンプルを、MRS寒天培地に塗抹した後、28℃で培養することにより、微生物を分離した。チーズは、カマンベールチーズ(日本製)、カマンベールチーズ(フランス製)、フロマージュ(日本製)、モツァレラチーズ(ドイツ製)、3種類のゴーダチーズ(オランダ製)、クリームチーズ(イタリア製)、山羊乳チーズ(フランス製)、チーズフード(オーストラリア製)、北海道チェダーチーズ(日本製)、ローファットチェダーチーズ(オランダ製)、ホワイトチェダーチーズ(イギリス製)、カスピアンチーズ(イラン製)の、計14種類の市販されているチーズを用いた。一種類のチーズから分離された微生物は、1つの菌群にまとめた。
【0014】
2.抗真菌活性を有する菌が存在する菌群の選択
得られた14種類の菌群を、10%(w/w)スキムミルク培地に接種し、28℃で48時間培養して菌群培養液を調製した。10%(w/w)スキムミルク平板培地に、該菌群培養液を画線塗布した後、アスペルギルス・ニガーあるいはリゾパス・ストロニファーの胞子を懸濁した軟寒天培地を重層した。
該スキムミルク平板培地を、28℃で48時間培養した後に観察した結果、カスピアンチーズ(イラン製)由来のO−9菌群のスキムミルク平板培地において、一部のコロニー周辺で、アスペルギルス・ニガー等の増殖が抑制されていることが観察された。その他の13種類の菌群では、アスペルギルス・ニガー等の増殖抑制は観察されなかった。この結果から、O−9菌群中には、抗真菌性物質を産生する菌が存在していることが分かった。
【0015】
3.pH7.0において抗真菌活性を有する菌が存在する菌群の選択
(1)抗真菌活性を測定するサンプルの調整
O−9菌群を、10%(w/w)スキムミルク培地に接種し、28℃で48時間培養して菌群培養液を調製した。該菌群培養液を超音波処理することにより、菌体を均一に分散させた後、3,000×gで10分間遠心分離を行い、上清と沈殿物に分離した。得られた上清は約pH4であったため、5%(w/v)炭酸ナトリウム溶液を用いてpH7.0に調整したものを、上清サンプルとした。得られた沈殿物に、ほぼ等量の、pH7.0に調整した10%(w/w)スキムミルク溶液を添加して懸濁液としたものを、沈殿物サンプルとした。バクテリアの増殖を抑制して、抗真菌活性の測定を容易にするために、各サンプルには、10分の1相当量の後記実施例1に記載の抗菌剤を添加した。さらに、それぞれの抗菌剤含有サンプルに、9分の1相当量のサブロー培地(グルコース:40g/L、ペプトンS:10g/L)を添加して、抗真菌活性の測定用のサンプル溶液を調製した。
【0016】
(2)抗真菌活性の測定
96ウェルマイクロプレートの各ウェルに、上記3(1)で調整したサンプル溶液の2倍段階希釈系列(20〜29倍)を、それぞれ130μLずつ添加した。該サンプル溶液の希釈には、サブロー培地を用いた。コントロールとして、130μLのサブロー培地のみを添加したウェルを用いた。さらに、各ウェルに、後記参考例1に記載の方法で調整した1.0×104spores/mLのアスペルギルス・ニガーNBRC9455株胞子懸濁液を50μLずつそれぞれ添加した。該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルを顕微鏡で観察した。
【0017】
図1は、顕微鏡観察の結果を表したものである。図中のレーンAはO−9菌群由来の上清サンプル溶液の希釈系列を、レーンBはO−9菌群由来の沈殿物サンプル溶液の希釈系列を、レーンCはコントロールを、それぞれ示している。図中の○は、各ウェルを表したものであり、○の中の×は、アスペルギルス・ニガーの増殖が観察されたウェルを、○の中の○は、アスペルギルス・ニガーの増殖が観察されなかった、すなわち増殖抑制が観察されたウェルを、それぞれ表したものである。なお、アスペルギルス・ニガーの増殖は、アスペルギルス・ニガーの胞子の発芽や菌糸の伸長により判断した。つまり、アスペルギルス・ニガーの増殖抑制は、アスペルギルス・ニガーの胞子から、菌糸の発芽や伸長が阻止されている状態を意味する。
【0018】
図1に示されているように、コントロールのウェルにおいて、アスペルギルス・ニガーの増殖が観察された。一方、O−9菌群由来のサンプル溶液の場合には、沈殿物サンプル溶液を添加した全てのウェルと、上清サンプル溶液の8〜512倍希釈液を添加したウェルでは、増殖したアスペルギルス・ニガーが観察されたが、上清サンプル溶液の1〜4倍希釈液を添加したウェルでは、アスペルギルス・ニガーの増殖は観察されなかった。
【0019】
該観察の結果から、O−9菌群由来の上清サンプル溶液中には、本発明の抗真菌性物質が存在しており、該抗真菌性物質により、アスペルギルス・ニガーの増殖が抑制されたことが明らかである。したがって、O−9菌群には、pH7.0において抗真菌活性を有する菌、すなわち、本発明の抗真菌性物質を産生する菌が存在していることがわかった。
また、上清サンプル溶液を添加したウェルでアスペルギルス・ニガーの増殖抑制が観察されたことから、該菌が産生する本発明の抗真菌性物質は、10%(w/w)スキムミルク培地で培養した場合に、培養液中に移行し得る性質を有することが明らかである。
【0020】
4.抗真菌性物質の耐熱性
O−9菌群由来の上清サンプル溶液中に存在している本発明の抗真菌性物質の耐熱性を検討するために、上記3(1)で調整したO−9菌群由来の上清サンプルを、100℃で5分間の加熱処理後氷冷した後、上記3(1)と同様にpH7.0に調整後、抗菌剤とサブロー培地を添加して、加熱処理済上清サンプル溶液を調整した。上記3(2)と同様にして、該加熱処理済上清サンプル溶液の抗真菌活性を測定したところ、アスペルギルス・ニガーの増殖抑制は観察されなかった。該観察の結果から、本発明の抗真菌性物質は、100℃で5分間の加熱処理により、前記抗真菌活性が失活する性質を有することがわかった。なお、特許文献1記載の食品保存料に含有される抗菌性物質は、加熱処理によっては失活しないものであるため、本発明の抗真菌性物質とは異なる物質であることが明らかである。
【0021】
5.抗真菌性物質の大きさ
O−9菌群由来の上清サンプル溶液中に存在している本発明の抗真菌性物質の大きさを検討するために、上記3(1)で調整したO−9菌群由来の上清サンプルを、MWCOが10kDである限外濾過膜を用いて限外濾過を行い、限外濾過膜表面に保持された高分子画分と、限外濾過膜を通過した低分子画分に分画した。該高分子画分と該低分子画分をサンプルとして、上記3(1)と同様にpH7.0に調整後、抗菌剤とサブロー培地を添加して、上清高分子画分サンプル溶液と上清低分子画分サンプル溶液を調整した。上記3(2)と同様にして、該上清高分子画分サンプル溶液と該上清低分子画分サンプル溶液の抗真菌活性をそれぞれ測定したところ、該上清高分子画分サンプル溶液を添加したウェルではアスペルギルス・ニガーの増殖抑制が観察されたが、該上清低分子画分サンプル溶液を添加したウェルでは観察されなかった。該観察の結果から、本発明の抗真菌性物質は、該高分子画分に分画されており、MWCOが10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持される性質を有することが明らかである。
【0022】
なお、カスピアンチーズは他のチーズと比較して非常に塩分濃度の高いチーズである。そこで、本発明の抗真菌性物質を産生する菌が、好塩性である可能性を考慮し、O−9菌群を、2.5%NaCl含有の10%(w/w)スキムミルク培地に接種した以外は、全て上記3と同様にして抗真菌活性を測定した。その結果、上清サンプル溶液の1〜16倍希釈液を添加したウェルにおいて、アスペルギルス・ニガーの増殖抑制が観察された。つまり、2.5%NaCl含有培地で培養したことにより、抗真菌活性が約4倍上昇したことから、本発明の抗真菌性物質を産生する菌は、高い塩濃度下においても、十分な量の抗真菌性物質を産生し得ることがわかった。
【0023】
6.本発明の抗真菌性物質を産生する菌の単離
上記3(1)で調製したO−9菌群の菌群培養液を、BCP(ブロモクレゾールパープル)平板培地に塗布した後、37℃で48時間培養した。該BCP平板培地から、コロニーの周囲が黄変した6のコロニーと、黄変が観察されなかった42のコロニーの、計48コロニーを得た。BCP寒天培地入り48ウェルマイクロプレートの各ウェルに、得られた48コロニーから釣菌した菌を、それぞれ接種し、さらに37℃で48時間培養した。
【0024】
培養した48株から、培地が紫色のままであった42株の中から10株(P1〜P10株)を、培地が黄変した6株の中から3株(Y1〜Y3株)を、それぞれ得た。得られた13株を、それぞれ10%(w/w)スキムミルク培地に接種し、37℃で48時間培養して培養液を調製した。上記3(1)と同様にして、該培養液を遠心分離して得た上清を、pH7.0に調整後、抗菌剤とサブロー培地を添加して、それぞれの株由来の上清サンプル溶液を調整した。上記3(2)と同様にして、96ウェルマイクロプレートに該上清サンプル溶液の2倍段階希釈系列を添加して、抗真菌活性を測定した。
【0025】
図2の(a)と(b)は、該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルを顕微鏡で観察した結果を、(c)と(d)は同じく36時間培養した後の観察結果を、それぞれ表したものである。図中のCはコントロールを表している。また、図中の○は、各ウェルを表したものであり、○の中の×は、アスペルギルス・ニガーの増殖が観察されたウェルを、○の中の○は、アスペルギルス・ニガーの増殖抑制が観察されたウェルを、それぞれ表したものである。
該観察の結果から、Y1〜Y3株は、コントロールと同様に、抗真菌活性は全く観察されなかったが、P1〜P10株では、アスペルギルス・ニガーの増殖抑制が観察されたことから、それぞれ活性強度の差があるものの、P1〜P10株は全て、pH7.0において、抗真菌活性を有する抗真菌性物質を産生することができる菌株であることが明らかである。
【0026】
7.pH7.0において抗真菌性物質を産生する菌の同定
P1〜P10株のうち、P9株(以下、O9−NP9株)及びP5株(以下、O9−NP5株)の同定を試みた。
【0027】
7−1.O9−NP9株の同定
まず、遺伝学的性質を調べるため、O9−NP9株の26S rDNAの塩基配列を常法により同定した。具体的には、O9−NP9株の培養液から抽出したDNAと、NL1、NL2、NL3及びNL4プライマー(O’Donnell、Fusarium and its near relatives、CAB International、p225〜233、1993年)を用いて、26S rDNA−D1/D2領域の塩基配列を同定した。なお、DNAの抽出にはDNeasy Plant Mini Kit(QIAGEN社製)を、PCRにはpuReTaq Ready−To−Go PCR beads(GEヘルスケアバイオサイエンシズ社製)及びBigDye Terminator v3.1 Cycle Sequencing Kit(アプライドバイオシステムズ社製)を、塩基配列の同定にはABI PRISM3100Genetic Analyzer System(アプライドバイオシステムズ社製)を、それぞれ用いた。
国際塩基配列データベース(GenBank/DDBJ/EMBL)上で、該塩基配列について相同性検索を行ったところ、子嚢菌糸アナモルフ(無性時代)酵母の一種であるキャンディダ・マルトーサの基準株であるNRRL Y−17677T(アクセッション番号:U45745)と100%の相同性を示した。したがって、O9−NP9株は、キャンディダ・マルトーサに属する微生物であると推定された。
【0028】
以下にO9−NP9株の菌学的性質を示す。なお、菌学的性質の測定は、YM寒天培地(べクトンディッキンソン社製)で、25℃で2日〜1ヶ月間培養したO9−NP9株を用いた。また、糖類発酵性試験、炭素源資化性試験、窒素源資化性試験、ビタミン要求性試験、温度耐性試験、薬剤耐性試験は、Yeast: Characteristics and identification, 3rd edition(Barnett et al、Cambridge University Press、2000年)、及び、The Yeast, a taxonomic study, 4th edition(Kurtzman et al、Elsevier、1998年)に、ほぼ従って行った。
【0029】
菌学的性質
(1)コロニー観察(YM寒天培地平板で25℃、4日間培養した時の目視観察時)
周縁の形状:ほぼ全縁、隆起状態:扁平形、表面の形状:平滑、光沢及び性状:弱い光沢、湿性、色調:白色からクリーム色
(2)培養的・形態的性質(YM寒天培地平板で25℃、2日間培養した時の光学顕微鏡観察時)
栄養細胞の大きさと形状:直径3〜6μmの楕円形から円筒形
増殖の形式:多極出芽
菌糸等:偽菌糸の形成が有
(3)有性生殖器官の形成(YM寒天培地平板で25℃、1ヶ月間培養した時の光学顕微鏡観察時):なし
【0030】
【表1】
【0031】
(7)37℃下における生育性:陽性
(8)40℃下における生育性:陰性
(9)0.01%シクロヘキシミド含有培地における生育性:陽性
(10)10%塩化ナトリウム及び5%グルコース含有培地における生育性:弱陽性
(11)ビタミン欠乏培地での生育性:陰性
(12)デンプン様物質の形成:陰性
(13)尿素の分解:陰性
【0032】
図3は、YM寒天培地平板で25℃、2日間培養したO9−NP9株の電子顕微鏡写真である。
【0033】
O9−NP9株は、ガラクトース発酵性、イノシトール、エリスリトール、ラフィノース、セロビオース、メレジトース、ガラクトース、ラクトース、可溶性デンプン、L−ラムノース、DL−lactate、キシリトール、及び硝酸塩の資化性、ビタミン欠乏培地、0.01%シクロヘキシミド培地、37℃及び40℃下での生育性等において、キャンディダ・マルトーサと一致する菌学的性質を示した。一方、キャンディダ・マルトーサはトレハロースを発酵するとされているのに対し、O9−NP9株はトレハロース発酵性を示さなかった。
【0034】
これらの遺伝学的及び菌学的性質から、O9−NP9株はキャンディダ・マルトーサ菌種であることが確認された。26S rDNA−D1/D2領域の塩基配列は、キャンディダ・マルトーサNRRL Y−17677T株と100%の相同性があったが、トレハロース発酵能において異なるため、キャンディダ・マルトーサO9−NP9株はキャンディダ・マルトーサNRRL Y−17677T株とは異なる性質を有することが明らかである。
【0035】
そこで、出願人は、キャンディダ・マルトーサO9−NP9株を、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託した(寄託日:平成19年1月10日)。受託番号は、NITE BP−297である。
【0036】
7−2.O9−NP5株の同定
まず、遺伝学的性質を調べるため、O9−NP5株の26S rDNAの塩基配列を、O9−NP9株と同様にして同定した。国際塩基配列データベース(GenBank/DDBJ/EMBL)上で、該塩基配列について相同性検索を行ったところ、子嚢菌糸酵母の一種であるクラビスポラ・ルシタニエMTCC1001(アクセッション番号:AF538871)と100%の相同性を示したが、クラビスポラ・ルシタニエの基準株であるクラビスポラ・ルシタニエNRRL Y−11827T(アクセッション番号:U44817)とは93.8%の相同性を示した。また、該塩基配列と99%以上の相同性を示す既知種の基準株は存在しなかった。
以下にO9−NP5株の菌学的性質を示す。なお、菌学的性質の測定は、O9−NP9株と同様にして行った。
【0037】
菌学的性質
(1)コロニー観察(YM寒天培地平板で25℃、4日間培養した時の目視観察時)
周縁の形状:全縁、隆起状態:クッション形、表面の形状:平滑、光沢及び性状:バター様、湿性、色調:白色からクリーム色
(2)培養的・形態的性質(YM寒天培地平板で25℃、2日間培養した時の光学顕微鏡観察時)
栄養細胞の大きさと形状:直径2〜6μmの球形から広楕円形あるいは偏円形
増殖の形式:多極出芽
菌糸等:偽菌糸の形成が有
(3)有性生殖器官の形成(YM寒天培地平板で25℃、1ヶ月間培養した時の光学顕微鏡観察時):なし
【0038】
【表2】
【0039】
(7)37℃下における生育性:陽性
(8)40℃下における生育性:陽性
(9)10μg/mLシクロヘキシミド含有培地における生育性:陽性
(10)10%塩化ナトリウム及び5%グルコース含有培地における生育性:陽性
(11)50%グルコース含有培地における生育性:陽性
(12)ビタミン欠乏培地での生育性:陰性
(13)デンプン様物質の形成:陰性
(14)尿素の分解:陰性
(15)ゼラチンの液化性:陰性
【0040】
O9−NP5株は、グルコース発酵性を示し、硝酸塩の資化性を示さず、クラビスポラ属菌と一致する菌学的性質を示した。また、ほぼ全ての特徴において、クラビスポラ・ルシタニエと類似した菌学的性質を示したが、ラフィノース、可溶性デンプン、及びヘキサデカンの資化性において、クラビスポラ・ルシタニエとは異なる菌学的性質を示した。
【0041】
これらの遺伝学的及び菌学的性質から、O9−NP5株はクラビスポラ属菌であることが確認された。また、クラビスポラ・ルシタニエの基準株との相同性が低かったこと、及びラフィノース等の資化性が異なることから、クラビスポラ・ルシタニエO9−NP5株は、クラビスポラ・ルシタニエの新規菌株、若しくは、クラビスポラ・ルシタニエに近縁な新規な菌種であると推定された。
【0042】
そこで、出願人は、クラビスポラ・ルシタニエO9−NP5株を、『クラビスポラ・スピーシーズP−5株』として、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託した(受領日:2008年1月16日)。受領番号は、NITE ABP−474である。
なお、本願明細書中、「クラビスポラ・ルシタニエO9−NP5株」と「クラビスポラ・スピーシーズP−5株」は同一の菌株を表している。
【0043】
8.本発明の抗真菌性物質を産生する酵母の取得
MRS寒天培地に代えて、YM寒天培地を用いた以外は、全てキャンディダ・マルトーサO9−NP9株を単離した場合と同様にして、別のロットのカスピアンチーズから、抗真菌性物質を産生する酵母を1株得た。該1株を、O9−H1株と名付け、菌の同定を行った。
【0044】
まず、遺伝学的性質を調べるため、O9−H1株の26S rDNAの塩基配列を、O9−NP9株と同様にして同定した。国際塩基配列データベース(GenBank/DDBJ/EMBL)上で、該塩基配列について相同性検索を行ったところ、子嚢菌糸酵母の一種であるサッカロマイセス・セレビシエの基準株であるNRRL Y−12632T(アクセッション番号:AY048154)と100%の相同性を示した。したがって、O9−H1株は、サッカロマイセス・セレビシエに属する微生物であると推定された。
以下にO9−H1株の菌学的性質を示す。なお、菌学的性質の測定は、O9−NP9株と同様にして行った。
【0045】
菌学的性質
(1)コロニー観察(YM寒天培地平板で25℃、4日間培養した時の目視観察時)
周縁の形状:ほぼ全縁、隆起状態:中央突起形、表面の形状:平滑、光沢及び性状:バター様、湿性、色調:明るいクリーム色
(2)培養的・形態的性質(YM寒天培地平板で25℃、3日間培養した時の光学顕微鏡観察時)
栄養細胞の大きさと形状:直径3〜8μmの広楕円形から偏円形
増殖の形式:多極出芽
(3)有性生殖器官の形成(YM寒天培地平板で25℃、20日間培養した時の光学顕微鏡観察時):子嚢様の肥大した細胞は観察されたが、子嚢胞子の形成は認められなかった。
【0046】
【表3】
【0047】
(7)30℃下における生育性:陽性
(8)37℃下における生育性:陽性
(9)40℃下における生育性:陽性
(10)1μg/mLシクロヘキシミド含有培地における生育性:陽性
(11)10%塩化ナトリウム及び5%グルコース含有培地における生育性:陽性
(12)50%グルコース含有培地における生育性:陽性
(13)ビタミン欠乏培地での生育性:陰性
(14)デンプン様物質の形成:陰性
(15)尿素の分解:陰性
【0048】
O9−H1株は、グルコース、ガラクトース、スクロース等の急速な糖発酵性を示し、硝酸塩資化性を示さず、サッカロマイセス属菌と一致する菌学的性質を示した。また、ほぼ全ての特徴において、サッカロマイセス・セレビシエと類似した菌学的性質を示した。一方、ラフィノースを発酵せず、イヌリン資化性を示した点で、O9−H1株は、サッカロマイセス・セレビシエと異なる菌学的性質を示した。
【0049】
これらの遺伝学的及び菌学的性質から、O9−H1株はサッカロマイセス・セレビシエ菌種であることが確認された。26S rDNA−D1/D2領域の塩基配列は、サッカロマイセス・セレビシエNRRL Y−12632T株と100%の相同性があったが、ラフィノース発酵能及びイヌリン資化において異なるため、サッカロマイセス・セレビシエO9−H1株はサッカロマイセス・セレビシエNRRL Y−12632T株とは異なる性質を有することが明らかである。
【0050】
そこで、出願人は、サッカロマイセス・セレビシエO9−H1株を、『サッカロマイセス・セレビシエH−1株』として、独立行政法人製品評価技術基盤機構特許微生物寄託センターに寄託した(受領日:2008年1月16日)。受領番号は、NITE ABP−473である。
なお、本願明細書中、「サッカロマイセス・セレビシエO9−H1株」と「サッカロマイセス・セレビシエH−1株」は同一の菌株を表している。
【0051】
本発明における抗真菌活性とは、カビや酵母等の真菌の増殖を抑制する活性のうち、胞子の発芽を阻害する活性を意味する。胞子の発芽を阻害することにより、効果的にカビ等の発生及び増殖を防止することができる。
【0052】
本発明の抗真菌性物質は、明確な物質名は未だ明らかではないが、キャンディダ属菌、サッカロマイセス属菌、ピキア属菌、ロドトルラ属菌、クラビスポラ属菌、及びトルラスポラ属菌からなる群より選ばれる1の酵母が産生する、胞子の発芽を阻害する抗真菌性物質である。
【0053】
また、本発明の抗真菌性物質は、(1)pH7.0において、抗真菌活性を有し、(2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有し、及び(3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCOが10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持される、という特徴を有するものである。さらに、(4)55℃で15分間の加熱処理により、前記抗真菌活性が失活する、という特徴を有するものであってもよい。
【0054】
本発明の抗真菌性物質を産生するキャンディダ属菌は、キャンディダ・マルトーサ、キャンディダ・トロピカリス、又はキャンディダ・シュードインターメディアであることが好ましく、キャンディダ・マルトーサであることが特に好ましい。該キャンディダ・マルトーサには、例えば、キャンディダ・マルトーサO9−NP9株、キャンディダ・マルトーサIAM12247、及びキャンディダ・マルトーサIAM12248等がある。該キャンディダ・トロピカリスには、例えば、キャンディダ・トロピカリスIAM4965等がある。該キャンディダ・シュードインターメディアには、例えば、キャンディダ・シュードインターメディアIAM12510等がある。
【0055】
本発明の抗真菌性物質を産生するピキア属菌は、ピキア・フェルメンタスであることが好ましく、ピキア・フェルメンタスJCM1824であることが特に好ましい。また、本発明の抗真菌性物質を産生するロドトルラ属菌は、ロドトルラ・アクタであることが好ましく、ロドトルラ・アクタJCM9494であることが特に好ましい。また、本発明の抗真菌性物質を産生するサッカロマイセス属菌は、サッカロマイセス・セレビシエであることが好ましく、サッカロマイセス・セレビシエH−1であることが特に好ましい。また、本発明の抗真菌性物質を産生するクラビスポラ属菌は、クラビスポラ・ルシタニエであることが好ましく、クラビスポラ・スピーシーズP−5又はクラビスポラ・ルシタニエNBRC10059であることが特に好ましい。また、本発明の抗真菌性物質を産生するトルラスポラ属菌は、トルラスポラ・デルブレッキーであることが好ましく、トルラスポラ・デルブルッキーNBRC0955であることが特に好ましい。
【0056】
本発明の酵母、すなわち、本発明の抗真菌性物質を産生するキャンディダ属菌、サッカロマイセス属菌、ピキア属菌、ロドトルラ属菌、クラビスポラ属菌、及びトルラスポラ属菌は、常法により培養することができ、培地や温度等の培養条件は、通常、キャンディダ属菌等の酵母を培養する条件であれば、特に限定されるものではない。該培地として、例えば、YM培地や10%(w/w)スキムミルク培地等がある。本発明の抗真菌性物質の活性が阻害されるおそれが少ないため、及び、該抗真菌性物質の精製が簡便になるため、10%(w/w)スキムミルク培地であることが好ましい。また、培養温度は、27〜30℃が好ましい。
【0057】
本発明の抗真菌性物質は、本発明の酵母を培養することにより得ることができる。例えば、本発明の酵母の培養液を、遠心分離処理をすることによって得た上清中に、本発明の抗真菌性物質は存在する。該遠心分離処理の条件は、該培養液中の菌体等の固体成分を除去できる条件であれば、特に限定されるものではないが、菌体由来の他の物質の混入を避けるため、2,500〜5,000×gの低速で遠心分離処理を行うことが好ましい。また、該上清を、MWCOが10kDである限外濾過膜を用いた限外濾過処理を行うことにより、該上清中に含まれる低分子の不純物を除去することもできる。このようにして得た該上清を、本発明の抗真菌性物質含有溶液として、防カビ剤や食品保存料として用いることもできる。
【0058】
本発明のカビ及び/又は酵母の増殖を阻害する方法は、本発明の抗真菌性物質又は本発明の酵母を用いる方法であって、本発明の抗真菌性物質の抗真菌活性が得られる方法であれば、特に限定されるものではない。本発明の抗真菌性物質を用いる方法として、例えば、該抗真菌性物質を含有する該酵母の培養液の上清等を、食品等に原料として添加する方法や、製造された食品に噴霧若しくは塗布する方法等がある。本発明の抗真菌性物質は、加熱処理により失活してしまうため、製造された食品に噴霧若しくは塗布する方法が好ましい。また、本発明の酵母を用いる方法として、例えば、発酵性食品の製造において、原料に本発明の酵母を添加した後、発酵させる方法等がある。該発酵性食品は、加熱処理を含まない発酵性食品であることが好ましい。
【実施例】
【0059】
次に実施例を示して本発明をさらに詳細に説明するが、本発明は以下の実施例に限定されるものではない。
【0060】
[参考例1]
アスペルギルス・ニガーの胞子懸濁液の調整
アスペルギルス・ニガーNBRC9455株は、PDA(ポテトデキストロース寒天)平板培地に塗布した後、28℃で3日間培養して得られた胞子を、0.05%tween80添加生理食塩水に懸濁し、懸濁液を綿濾過後、−80℃で凍結保存したものを用いた。該凍結保存後のアスペルギルス・ニガーNBRC9455株を、生理食塩水を用いて希釈した後、サブロー培地を用いて、1.0×104spores/mLのアスペルギルス・ニガー胞子懸濁液を調製した。
【0061】
[実施例1]
キャンディダ・マルトーサO9−NP9株を、10%(w/w)スキムミルク培地に接種し、28℃で48時間培養して培養液を調製した。該培養液を超音波処理することにより、菌体を均一に分散させた後、3,000×gで10分間遠心分離を行って、上清を得た。
該上清を、5%(w/v)炭酸ナトリウム水溶液を用いてpHを7.0に調整した後、MWCOが10kDである限外濾過膜を用いて限外濾過を行った。限外濾過膜表面に保持された溶液を、抗真菌性物質溶液とした。
【0062】
1mLの該抗真菌性物質溶液を分注した1.5mLチューブを10本作製した。該チューブを、10、30、40、45、50、55、60、80、90℃の各温度で15分間加熱処理後、氷冷した。氷冷済チューブのそれぞれに対して、100μLの抗菌剤(ペニシリウムGK:0.7mg/mL、カナマイシン:60μg/mL、硫酸ストレプトマイシン:0.1mg/mL)を添加して混合した。その後、252μLの各チューブの抗真菌性物質溶液と、28μLのサブロー培地を混合して、活性測定用の抗真菌性物質溶液を調整した。
【0063】
96ウェルマイクロプレートの各ウェルに、調整した該抗真菌性物質溶液の2倍段階希釈系列(20〜29倍)を、それぞれ130μLずつ添加した。該抗真菌性物質溶液の希釈には、サブロー培地を用いた。具体的には、96ウェルマイクロプレートの1列目に該抗真菌性物質溶液を添加し、2列目に、該抗真菌性物質溶液の2倍希釈液を添加し、3列目に、該抗真菌性物質溶液の4(22)倍希釈溶液を添加した。各列に、順次、2倍段階希釈液をウェルに添加し、最後に10列目に、該加抗真菌性物質溶液の512(29)倍希釈溶液を添加した。コントロールとして、130μLのサブロー培地のみを添加した。さらに各ウェルに、前記参考例1に記載の方法で調整した1.0×104spores/mLのアスペルギルス・ニガー胞子懸濁液を50μLずつそれぞれ添加した。該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルを顕微鏡で観察した。
【0064】
【表4】
【0065】
表4は、顕微鏡観察の結果に基づき、各温度処理済抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「24」の記載は、抗真菌性物質溶液等の24倍希釈溶液以下の希釈倍率の溶液、すなわち、20〜24倍希釈溶液を添加したウェルにおいて、菌の増殖抑制が観察されたことを示している。また、表中「29」の記載は、抗真菌性物質溶液等の29倍希釈溶液以下の希釈倍率の溶液、すなわち、本実施例の全ての希釈溶液を添加したウェルにおいて、菌の増殖抑制が観察されたことを示している。さらに表中、「−」の記載は、20倍希釈溶液(原液)を添加したウェルにおいても活性が観察されなかったことを示している。
表4より明らかであるように、10℃と40℃の各温度処理済抗真菌性物質溶液では、本実施例の全ての希釈溶液を添加したウェルにおいて、菌の増殖抑制が観察された。したがって、10℃と40℃の各温度処理済抗真菌性物質溶液では、29倍率以上の希釈液においても、菌の増殖が抑制された可能性があり、高い抗真菌活性を有していることが明らかである。一方、加熱処理温度が40℃を超えると、急激に抗真菌活性は低下し、55℃15分間の加熱処理により、失活することが分かった。
【0066】
すなわち、実施例1の結果から、本発明の抗真菌性物質は、キャンディダ属菌等の酵母が産生するものであって、pH7.0において、抗真菌活性を有し、10%(w/w)スキムミルク培地で該酵母を培養した場合に、培養液中に分泌され、10%(w/w)スキムミルク培地で該酵母を培養した培養液の上清を、MWCOが10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持され、55℃で15分間の加熱処理により、該抗真菌活性が失活する、という特徴を有するものであることが明らかである。
【0067】
[実施例2]
本発明の抗真菌性物質の、様々な真菌に対する抗真菌活性を測定した。抗真菌活性のポジティブコントロールとして、真核細胞に対するタンパク質合成阻害剤であるシクロヘキシミドを用いた。
まず、加熱処理を行わないこと以外は、全て実施例1と同様にして、キャンディダ・マルトーサO9−NP9株の培養液から、抗真菌性物質溶液を調製した。また、表5記載の18菌株の1.0×104spores/mLの胞子懸濁液を、参考例1と同様にしてそれぞれ調製した。
【0068】
実施例1と同様にして、96ウェルマイクロプレートに、調製した抗真菌性物質溶液若しくは500ppmのシクロヘキシミド溶液の、2倍段階希釈系列(20〜29倍)を調製した。さらに各2倍段階希釈系列に、各菌株の胞子懸濁液を、1ウェル当たり50μLずつ、それぞれ添加した。該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルで菌の増殖が抑制されているかどうかを顕微鏡で観察した。
【0069】
表5は、実施例1と同様に、各抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。
【0070】
【表5】
【0071】
表5記載の18菌株のシクロヘキシミドに対する感受性は、それぞれ異なっていたが、全ての菌株において、500ppmのシクロヘキシミド溶液の添加により、増殖抑制が観察された。一方、本発明の抗真菌性物質溶液は、全ての菌株において、500ppmシクロヘキシミド溶液よりも大きい希釈倍率の希釈溶液で、菌の増殖抑制が観察された。
表5の結果から、本発明の抗真菌性物質は、多様な真菌に対して抗真菌活性を有していること、及び、実施例2記載の方法により調製した本発明の抗真菌性物質溶液は、500ppmシクロヘキシミド溶液よりも高い抗真菌活性を有していることが明らかである。
【0072】
[実施例3]
キャンディダ・マルトーサO9−NP9株以外のキャンディダ属菌や他の酵母が、キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を産生するかどうかを調べた。抗真菌活性のポジティブコントロールとして、実施例2と同様にシクロヘキシミドを用いた。
まず、実施例2と同様にして、表6記載の酵母の培養液から、抗真菌性物質溶液をそれぞれ調製した。また、参考例1と同様にして、1.0×104spores/mLのアスペルギルス・ニガー胞子懸濁液を調製した。
【0073】
実施例2と同様にして、96ウェルマイクロプレートに、調製した各酵母の抗真菌性物質溶液若しくは500ppmのシクロヘキシミド溶液の、2倍段階希釈系列(20〜29倍)を調製した。さらに各2倍段階希釈系列に、アスペルギルス・ニガー胞子懸濁液を、1ウェル当たり50μLずつ、それぞれ添加した。該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルで菌の増殖が抑制されているかどうかを顕微鏡で観察した。
【0074】
表6は、実施例2と同様に、各酵母の抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」の記載は、表4と同様に、20倍希釈溶液(原液)を添加したウェルにおいても活性が観察されなかったことを示している。
【0075】
【表6】
【0076】
表6の結果から、キャンディダ・マルトーサO9−NP9株以外の幾つかの酵母も、キャンディダ・マルトーサO9−NP9株と同様に、アスペルギルス・ニガーの増殖抑制が観察され、本発明の抗真菌性物質を産生することが明らかとなった。特に、キャンディダ・マルトーサIAM12247とキャンディダ・マルトーサIAM12248も、キャンディダ・マルトーサO9−NP9株と同様に、他の菌株に比べて、顕著に高い抗真菌活性を有していた。つまり、キャンディダ・マルトーサは、他の酵母に比べて、本発明の抗真菌性物質の産生能が非常に優れていることが明らかとなった。また、キャンディダ・トロピカリスIAM4965の抗真菌性物質溶液も、500ppmシクロヘキシミド溶液よりも高い抗真菌活性を有していた。その他、キャンディダ属菌のうち、キャンディダ・シュードインターメディアIAM12510も抗真菌活性を有していた。キャンディダ属菌以外にも、ピキア・フェルメンタスJCM1824やロドトルラ・アクタJCM9494も抗真菌活性を有していることが分かった。
【0077】
[実施例4]
実施例3で調べた酵母に加えて、さらに多くの酵母において、キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を産生するかどうかを調べた。具体的には、表7及び8記載の酵母を用いた以外は、全て実施例3と同様にして行った。
【0078】
【表7】
【0079】
【表8】
【0080】
表7及び8は、実施例3と同様に、各酵母の抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」の記載は、表6と同様である。
表7及び8の結果から、キャンディダ属菌のうち、さらにキャンディタ・サイトアナ(Candida saitoana)JCM1438等の幾つかの菌も抗真菌活性を有していることがわかった。
【0081】
[実施例5]
キャンディダ・マルトーサO9−NP9株、サッカロマイセス・セレビシエH−1株、及び、クラビスポラ・スピーシーズP−5株の抗真菌活性を比較した。
まず、実施例2と同様にして、該3菌株の培養液から、抗真菌性物質溶液をそれぞれ調製した。また、参考例1と同様にして、1.0×104spores/mLのアスペルギルス・ニガー胞子懸濁液を調製した。
【0082】
実施例2と同様にして、96ウェルマイクロプレートに、該3菌株の抗真菌性物質溶液の、2倍段階希釈系列(20〜29倍)を調製した。さらに各2倍段階希釈系列に、アスペルギルス・ニガー胞子懸濁液を、1ウェル当たり50μLずつ、それぞれ添加した。該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルで菌の増殖が抑制されているかどうかを顕微鏡で観察した。
【0083】
顕微鏡観察の結果、キャンディダ・マルトーサO9−NP9株では、抗真菌性物質溶液の29倍希釈液を添加したウェルであっても、アスペルギルス・ニガーの増殖抑制が観察された。一方、サッカロマイセス・セレビシエH−1株及びクラビスポラ・スピーシーズP−5株では、抗真菌性物質溶液の20〜24倍希釈液を添加したウェルにおいてのみ、アスペルギルス・ニガーの増殖抑制が観察された。
【0084】
実施例5の結果から、サッカロマイセス・セレビシエH−1株及びクラビスポラ・スピーシーズP−5株は、キャンディダ・マルトーサO9−NP9株と同様に本発明の抗真菌活性物質を産生すること、及び、該3株のなかでは、キャンディダ・マルトーサO9−NP9株が最も本発明の抗真菌性物質の産生能が高いことが明らかである。
【0085】
[実施例6]
サッカロマイセス属菌やその近縁菌が、キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を産生するかどうかを調べた。抗真菌活性のポジティブコントロールとして、実施例2と同様にシクロヘキシミドを用いた。
まず、実施例2と同様にして、表9記載の酵母の培養液から、抗真菌性物質溶液をそれぞれ調製した。また、参考例1と同様にして、1.0×104spores/mLのアスペルギルス・ニガー胞子懸濁液を調製した。
【0086】
実施例2と同様にして、96ウェルマイクロプレートに、調製した各酵母の抗真菌性物質溶液若しくは500ppmのシクロヘキシミド溶液の、2倍段階希釈系列(20〜29倍)を調製した。さらに各2倍段階希釈系列に、アスペルギルス・ニガー胞子懸濁液を、1ウェル当たり50μLずつ、それぞれ添加した。該96ウェルマイクロプレートを28℃で3日間又は7日間培養した後、各ウェルで菌の増殖が抑制されているかどうかを顕微鏡で観察した。
表9は、実施例2と同様に、各酵母の抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」の記載は、表4と同様に、20倍希釈溶液(原液)を添加したウェルにおいても活性が観察されなかったことを示しており、「ND」は測定しなかったことを意味している。また、表中「3日後」及び「7日後」は、それぞれ3日培養後又は7日培養後の結果を示している。
【0087】
【表9】
【0088】
表9の結果から、サッカロマイセス・セレビシエH−1株以外の幾つかの酵母においても、アスペルギルス・ニガーの増殖抑制が観察され、これらの酵母が本発明の抗真菌性物質を産生することが明らかとなった。特に、培養3日後においては、サッカロマイセス・セレビシエNBRC0216及びサッカロマイセス・セレビシエNBRC10181は、サッカロマイセス・セレビシエH−1と同様に、他の菌株に比べて、顕著に高い抗真菌活性を有していた。その他、トルラスポラ・デルブルッキーNBRC0955(受領番号 NITE ABP−475、受領日 2008年1月16日)、サッカロマイセス・パストリアナス(Saccharomyces pastprianus)NBRC11024、クリベロマイセス・ラクティス・バール・ドロソフィララム(Kluyveromyces lactis var.drosophilarum)NBRC1012、クリベロマイセス・ドブハンスキ(Kluyveromyces dobzhanskii)NBRC10603も、高い抗真菌活性を有していた。特に、トルロスポラ・デルブルッキーNBRC0955は、7日培養後であってもシクロヘキシミドよりも高い抗真菌活性を有しており、非常に優れていることが明らかとなった。
【0089】
[実施例7]
クラビスポラ属菌やその近縁菌が、キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を産生するかどうかを調べた。
具体的には、表9記載の酵母に代えて、表10記載の酵母を用いた以外は、全て実施例6と同様にして、各酵母の抗真菌性物質溶液を添加することにより、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察した。
表10は、各酵母の抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」、「ND」、「3日後」及び「7日後」の記載は、表9と同様である。
【0090】
【表10】
【0091】
表10の結果から、クラビスポラ・スピーシーズP−5株以外の幾つかの酵母も、キャンディダ・マルトーサO9−NP9株と同様に、アスペルギルス・ニガーの増殖抑制が観察され、本発明の抗真菌性物質を産生することが明らかとなった。培養3日後においては、クラビスポラ・スピーシーズP−5、クラビスポラ・ルシタニエNBRC10059は、キャンディダ・マルトーサO9−NP9と同様に、顕著に高い抗真菌活性を有していた。その他、クラビスポラ・ルシタニエNBRC10058もシクロヘキシミドよりも高い抗真菌活性を有していた。特に、クラビスポラ・ルシタニエNBRC10059(受領番号 NITE ABP−476、受領日 2008年1月16日)は、7日培養後であってもキャンディダ・マルトーサO9−NP9と同様に高い抗真菌活性を有しており、非常に優れていることが明らかとなった。
【0092】
[実施例8]
クラビスポラ・ルシタニエNBRC10059株及びトルロスポラ・デルブルッキーNBRC0955株が産生する本発明の抗真菌性物質の、様々な真菌に対する抗真菌活性を測定した。
具体的には、キャンディダ・マルトーサO9−NP9株に代えて、クラビスポラ・ルシタニエNBRC10059株又はトルロスポラ・デルブルッキーNBRC0955株を用いた以外は、全て実施例2と同様にして、各酵母の抗真菌性物質溶液を添加することにより、表11記載の菌の増殖が抑制されるかどうかを顕微鏡で観察した。
表11は、各酵母の抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」、「ND」、「3日後」及び「7日後」の記載は、表9と同様である。
【0093】
【表11】
【0094】
表11記載の13菌株の全てにおいて、クラビスポラ・ルシタニエNBRC10059株はシクロヘキシミドよりも高い抗真菌活性を示した。一方、トルロスポラ・デルブルッキーNBRC0955株は、幾つか増殖抑制効果を奏さない菌もあったが、真菌の種類によっては、シクロヘキシミドよりも高い抗真菌活性を示した。
【0095】
[実施例9]
各酵母が有する抗真菌活性物質の耐熱性を測定した。
まず、表12記載の各酵母をそれぞれ植菌した1LのPD(ポテトデキストロース)培地を、30℃48時間振とう培養したものを、3,000×gで5分間遠心分離処理し、上清を回収した。該上清を50℃、60℃、85℃、90℃の各温度で10分間処理後冷却したものを、抗真菌性物質溶液とした。なお、加熱処理をしなかった(非加熱処理)上清をコントロールとした。
実施例2と同様にして、これらの抗真菌性物質溶液を添加することにより、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察した。
表12は、各抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。表中、「−」の記載は、20倍希釈溶液(原液)を添加したウェルにおいても活性が観察されなかったことを示しており、「29」の記載は、29倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察されたことを示している。
この結果、表12記載の酵母が有する抗真菌活性物質は、いずれも60℃10分間以上の加熱処理で失活してしまうことが分かった。
【0096】
【表12】
【0097】
[実施例10]
キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を、原料としてパンに添加した場合の、抗真菌効果を観察した。
まず、キャンディダ・マルトーサO9−NP9株を植菌した1LのPD(ポテトデキストロース)培地を、30℃で3日間培養した。この培養液全体を、抗真菌性物質溶液として、実施例2と同様にして、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察したところ、該培養液の29倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察された。
この培養液を、水原料全量分に代えて食パンの原料とした。市販されている抗菌剤ネオカビンP(ジェイティーフーズ株式会社販売)を添加した食パンと、抗菌剤等を無添加の食パンを、それぞれ比較対象とした。具体的には、表12に示す配合量で、常法により食パンを製造した。表中、「%」は強力粉100%に対する重量部(%)を意味する。
【0098】
【表13】
【0099】
製造された食パンは、通常の6枚切りの厚さにスライスした。各食パン20枚ずつを、室内で10分間放置し、食パン表面上に落下菌を付着させた。その後、1枚ずつ袋に入れて、28℃で5日間保温した後、カビの有無や生え方等を観察した。カビの生え方は、コロニーの数が0個、1〜数10個、数10個以上、計数不能の4区分に区分けした。なお、「計数不能」とは、カビが表面の大部分を覆っていてコロニーの個数を計測できなかったものを意味する。
【0100】
【表14】
【0101】
表14は、観察により20枚の食パンを区分けした結果を、区分ごとの枚数で示したものである。この結果、全ての種類の食パンにおいて、20枚全てにカビが生えていた。無添加の食パンでは、1枚当たりのコロニー数が多く、計数不能なものが6枚もあったが、ネオカビンを添加した食パンは、1枚当たりのコロニー数が少なく、計数不能なものはなかった。これに対し、キャンディダ・マルトーサO9−NP9株の培養液を添加した食パンでは、ネオカビンを添加したものよりは多い傾向にあったが、無添加のものに比べて1枚当たりのコロニー数が非常に少なく、計数不能なものは1枚しかなかった。特に、培養液添加食パンでは、食パンの中心部にはほとんどカビが生えておらず、カビが生えているものには、食パンの耳のみにカビが生えているものが多かった。
このように、培養液を添加した食パンにおいて、カビが食パンの外側に多く、内側には少なかったのは、本発明の抗真菌性物質は加熱に弱いため、パンを焼成する工程で失活してしまったためと推察される。つまり、これらの結果から、本発明の抗真菌性物質は、加熱処理を施さない食品等への添加剤として用いることにより、従来の抗菌剤と同様に抗菌効果が期待できることが明らかである。
【0102】
[実施例11]
キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を、食パンに塗布した場合の、アスペルギルス・ニガーに対する抗真菌効果を観察した。
まず、キャンディダ・マルトーサO9−NP9株を接種した1LのPD培地を、30℃で3日間培養し、培養液全体を、抗真菌性物質溶液として、実施例2と同様にして、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察した。この結果、該培養液の29倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察された。この培養液を、市販の6枚切りの食パン(サンアロマ、山崎製パン社製)に、スプレーで全体に塗布し、コンラージ棒を用いて全体に塗り広げた。
培養液を塗布した食パンと、塗布していない食パン(コントロール)に、0.5mLのアスペルギルス・ニガー胞子懸濁液(2.0×104spores/mL)を、1枚当たり10点ずつ接種し、28℃で保温した。なお、アスペルギルス・ニガー胞子懸濁液は、参考例1と同様にして調製した。
この結果、接種から3日後には、コントロールの食パンにはカビが生えて緑色になっていたが、培養液を塗布した食パンにはカビは生えていなかった。さらに接種から5日後には、コントロールの食パンは全体的に黒いカビのコロニーが観察されたが、培養液を塗布した食パンでは、一箇所に白い毛状の菌糸が観察されただけであった。
これらの結果から、本発明の抗真菌性物質は、非加熱条件下で、非常に良好な抗真菌効果を奏することが明らかである。
【0103】
[実施例12]
キャンディダ・マルトーサO9−NP9株が産生する本発明の抗真菌性物質を、食パンに塗布した場合の、落下菌に対する抗真菌効果を観察した。
実施例11と同様にして作製したキャンディダ・マルトーサO9−NP9株の培養液を塗布した食パンと、塗布していない食パン(コントロール)とを、各6枚ずつ、室内で10分間放置し、食パン表面上に落下菌を付着させた。その後、1枚ずつ袋に入れて、28℃で5日間保温した後、カビの有無を観察した。
この結果、コントロールの食パンは6枚ともカビが生えていたが、培養液を塗布した食パンにはいずれもカビは生えていなかった。これらの結果からも、本発明の抗真菌性物質は、非加熱条件下で、非常に良好な抗真菌効果を奏することが明らかである。
【0104】
[実施例13]
本発明の酵母を培養した培養液の抗真菌活性に対する培養条件の影響を観察した。具体的には、振とう培養と静置培養の、培養液の抗真菌活性に対する影響を調べた。
キャンディダ・マルトーサO9−NP9株を植菌した1LのPD培地を、25℃24時間又は48時間振とう培養して得た培養液と、同じくキャンディダ・マルトーサO9−NP9株を植菌した1LのPD培地を、25℃24時間又は48時間静置培養して得た培養液とを調製した。これらの培養液を、それぞれ3,000×gで5分間遠心分離処理し、上清と沈殿に分離して回収した。このようにして得られた上清は、そのまま抗真菌物質溶液として用いた。一方、菌体を含む沈殿は、上清と等量の蒸留水を加えて超音波処理することにより、菌体を均一に分散させた溶液を、抗真菌物質溶液として用いた。
各抗真菌物質溶液の抗真菌活性を、実施例2と同様にして、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察することにより測定した。
【0105】
【表15】
【0106】
表15は、各抗真菌性物質溶液の2倍段階希釈系列において、菌の増殖抑制が観察されたウェルのうち、最も抗真菌性物質濃度の低いウェルの希釈倍率、すなわち、最大希釈倍率を示したものである。この結果、培養時間や培養条件にかかわらず、沈殿由来の抗真菌性物質溶液では、この実験の最大希釈倍率である29倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察され、高い抗真菌活性を有していた。これに対して、上清由来の抗真菌性物質溶液では、培養時間による相違はあまりなく、いずれも、振とう培養由来の抗真菌性物質溶液は28倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察されたが、静置培養由来の抗真菌性物質溶液では22倍希釈溶液を添加したウェルまでしか菌の増殖抑制が観察されなかった。また、いずれの培養条件においても、上清由来の抗真菌性物質溶液よりも、沈殿由来の抗真菌性物質溶液のほうが高い抗真菌活性を有していた。
つまり、本発明の抗真菌性物質は、静置培養よりも振とう培養によって培養液に移行し易いこと、及び、沈殿(菌体)のほうが上清よりも抗真菌活性が高いことが分かった。したがって、本発明の抗真菌性物質は、静置培養よりも振とう培養時により積極的に酵母外へ分泌され易い可能性が示唆された。また、上清よりも沈殿のほうが高い抗真菌活性が検出されたことから、本発明の抗真菌性物質は、酵母表面に吸着し易い性質を有しているのではないかと推察される。
【0107】
[実施例14]
本発明の抗真菌性物質の性質を調べるため、粗精製した後、粗精製物中の物質の中性糖の定量分析を行った。
まず、キャンディダ・マルトーサO9−NP9株を接種した1LのPD培地を、30℃24時間振とう培養したものを、3,000×gで5分間遠心分離処理し、776.95gの上清を回収した。該上清の一部を抗真菌性物質溶液として、実施例2と同様にして、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察したところ、該培養液の210倍希釈溶液を添加したウェルにおいても菌の増殖抑制が観察された。
該上清の残り417.86gを、限外濾過モジュールSEP0031(MWCO:3kD、旭化成社製)を用いて限外濾過を行った。このとき、限外濾過膜を透過した溶液(S1)は314.87gであり、該限外濾過膜表面に保持された残渣(R1)は128.34gであった。このR1に脱イオン水を加えて500gとして攪拌した後、再度限外濾過モジュールSEP0031を用いて限外濾過を行った。このとき、限外濾過膜を透過した溶液(S2)は305.52gであり、該限外濾過膜表面に保持された残渣(R2)は138.87gであった。このR2に脱イオン水を加えて500gとして攪拌した後、再度限外濾過モジュールSEP0031を用いて限外濾過を行った。このとき、限外濾過膜を透過した溶液(S3)は402.36gであり、該限外濾過膜表面に保持された残渣(R3)は59.28gであった。
得られた残渣R3をそのまま均一に攪拌後、抗真菌性物質溶液として、実施例2と同様にして、アスペルギルス・ニガーの増殖が抑制されるかどうかを顕微鏡で観察したところ、該培養液の20〜25倍希釈溶液を添加したウェルにおいて菌の増殖抑制が観察された。
【0108】
さらに、残渣R3中の物質の中性糖の定量分析を行った。
具体的には、まず、抗真菌性物質溶液とした残渣R3を均一に攪拌した懸濁液を、ガラス試験管に100μL採取し、減圧乾固した。得られた残渣に200μLのトリフルオロ酢酸(2mol/L)を添加し、真空封管後、100℃で6時間加水分解した。加水分解物を減圧乾固して得られた残渣を、100μLの精製水に溶解させた後、孔径0.22μmのフィルターに通して濾過した濾液を測定試料原液とした。該測定試料原液を精製水で100倍又は1000倍希釈したものを測定試料とし、下記の条件で高速液体クロマトグラフ(HPLC)法で測定し、中性糖(ラムノース、リボース、マンノース、アラビノース、ガラクトース、キシロース、及びグルコース)の定量分析を行った。なお、ピークの同定は各中性糖の標準品との比較により行った。
HPLCシステム:LC−9Aシステム(島津製作所)
検出器:分光蛍光光度計RF−10AXL(島津製作所)
カラム:TSK−gel Sugar AXG 150×4.6mmI.D.(東ソー)
カラム温度:70℃
移動相:0.5Mホウ酸カリウム緩衝液pH8.7
移動相流速:0.4mL/min
ポストカラム標識:反応試薬:1%アルギニン/3%ホウ酸
反応試薬流速:0.5mL/min
反応温度:150℃
検出波長:Ex.320nm、Em.430nm
【0109】
この結果、残渣R3は、約1%のグルコース、及び少量のマンノースを含んでいることが分かった。つまり、残渣R3には多くの糖類が含まれていることから、本発明の抗真菌性物質は糖タンパク質や多糖類等の糖を多く含む化合物である可能性が示唆された。
また、この中性糖の分析結果と、静置培養よりも振とう培養によって培養液に移行し易い点、さらに後述の実施例16の結果からも、実施例13と同様に、本発明の抗真菌性物質は、酵母表面の糖タンパク質と、自身が有する中性糖を介して相互作用する等により、酵母表面に吸着され易い性質を有する可能性が示唆される。
【0110】
[実施例15]
500μLのキャンディダ・マルトーサO9−NP9株の培養液と、500μLの2.4×106spores/mLのアスペルギルス・ニガー胞子懸濁液を混合し、28℃で24時間インキュベートした。なお、キャンディダ・マルトーサO9−NP9株の培養液は、キャンディダ・マルトーサO9−NP9株を、10%(w/w)スキムミルク培地に接種し、28℃で48時間培養することにより調製した。また、アスペルギルス・ニガー胞子懸濁液は参考例1と同様にして調製した。
この培養液1mLに、200μLの蛍光色素0.5μM SYTOX−Green(タカラバイオ社製)を添加し、さらに0.5時間インキュベートした。その後、この培養液を3,000×gで5分間遠心分離処理し、上清を除去することにより蛍光色素を除去した。得られた沈殿に蒸留水を添加して懸濁させた後、再度遠心分離処理して上清を除去して沈殿を洗浄した。
この沈殿にさらに1mLの蒸留水を添加して、酵母と胞子の懸濁液を調製した。この懸濁液を通常の光学顕微鏡を用いて、蛍光及び透過光観察したところ、SYTOX−Greenで染色された胞子が観察された。また、この蛍光染色されている胞子は、吸水時と同等又はそれ以上の大きさに膨張していることが観察された。
ここで、SYTOX−Greenは、DNAと特異的に結合し得る色素であるが、細胞膜を通過することはできない。にもかかわらず、キャンディダ・マルトーサO9−NP9株とインキュベートしたアスペルギルス・ニガーの胞子が染色されたのは、キャンディダ・マルトーサO9−NP9株が有する抗真菌性物質の作用により、胞子の細胞壁が損傷を受けたためと推察される。また、蛍光染色された胞子が膨張していたことからも、胞子の細胞壁が損傷を受けたことが示唆される。
500μLの10%(w/w)スキムミルク培地に500μLの2.4×106spores/mLのアスペルギルス・ニガー胞子懸濁液を混合したものを同様にインキュベートし、SYTOX−Greenを添加したところ、蛍光染色された胞子は観察されなかった。また、キャンディダ・マルトーサO9−NP9株に代えて、本発明の抗真菌活性が観察されなかったキャンディダ・ユーティリスを用いたものを同様にインキュベートし、SYTOX−Greenを添加した場合にも、蛍光染色された胞子は観察されなかった。これらの結果からも、胞子が染色されたのは、キャンディダ・マルトーサO9−NP9株が有する本発明の抗真菌物質によるためであることが示唆される。
【0111】
[実施例16]
寒天培地中で培養した場合の本発明の抗真菌活性物質の活性を調べた。
まず、キャンディダ・マルトーサO9−NP9株を混釈したPDA培地を、28℃で48時間培養した後、滅菌済みの型を用いてくりぬき、キャンディダ・マルトーサO9−NP9株が生育している円柱形のPDA培地(以下、酵母混釈寒天培地という。)を作製した。
一方、同じ型を用いて、参考例1と同様にして調製したアスペルギルス・ニガー胞子懸濁液を混釈したPDA培地をくりぬき、アスペルギルス・ニガー胞子を含む円柱形のPDA培地(以下、胞子混釈寒天培地という。)を作製した。
これらの2種類の寒天培地を、(a)酵母混釈寒天培地の上に、胞子混釈寒天培地を直接重ね合わせた状態、(b)酵母混釈寒天培地の上に、ラップを挟んで胞子混釈寒天培地を重ね合わせた状態、(c)酵母混釈寒天培地の上に、孔径14kDの透析膜を挟んで胞子混釈寒天培地を重ね合わせた状態、の3つの状態で、それぞれ28℃で48〜72時間培養し、胞子の状態を観察した。
この結果、(b)の胞子混釈寒天培地では、胞子の増殖が観察された。一方、(a)と(c)の胞子混釈寒天培地では、胞子の増殖は観察されなかった。
また、培養後の胞子混釈寒天培地の一部を、液体のPD培地に添加し、28℃で24時間培養した後、培養液をPDA平板培地に塗抹して、さらに培養した。この結果、(a)の胞子混釈寒天培地由来のPDA平板培地には、酵母が増殖しており、カビは生えてこなかった。一方、(b)の胞子混釈寒天培地由来のPDA平板培地には、カビが生えてきたが、酵母は確認できなかった。さらに(c)の胞子混釈寒天培地由来のPDA平板培地には、カビが生えてきたが、酵母は確認できなかった。
これらの結果から、(b)では、2つの寒天培地がラップで挟まれていたため、胞子混釈寒天培地中のアスペルギルス・ニガー胞子は、酵母混釈寒天培地の影響を受けず、胞子が生育したが、(a)では、キャンディダ・マルトーサO9−NP9株が胞子混釈寒天培地へ移行して増殖した結果、アスペルギルス・ニガー胞子の増殖が阻害されたと考えられる。これに対して、(c)では、透析膜に阻まれて、(b)と同様に酵母は胞子混釈寒天培地へ移行できなかったが、酵母から産生された本発明の抗真菌物質が透析膜を透過して胞子混釈寒天培地へ移行したため、アスペルギルス・ニガー胞子の増殖が阻害された結果、胞子混釈寒天培地では胞子の増殖が観察されなかったと考えられる。一方、この胞子混釈寒天培地中に最初に混釈されていた胞子は死滅しなかったため、(c)の胞子混釈寒天培地由来のPDA平板培地にはカビが生育したと考えられる。
これらの結果からも、キャンディダ・マルトーサO9−NP9株が本発明の抗真菌性物質を産生していることが明らかである。
【産業上の利用可能性】
【0112】
本発明の抗真菌性物質は、チーズ由来の酵母が産生するため安全であり、中性において抗真菌活性を示すことから、特にカビの増殖が問題となる食品分野等で利用が可能である。
【図面の簡単な説明】
【0113】
【図1】O−9菌群培養液由来のサンプル溶液の2倍段階希釈系列と、アスペルギルス・ニガー胞子懸濁液を、96ウェルマイクロプレートの各ウェルにそれぞれ添加して、28℃で48時間培養した後に、各ウェルを顕微鏡で観察した結果を示したものである。図中のレーンAはO−9菌群由来の上清サンプル溶液の希釈系列を、レーンBはO−9菌群由来の沈殿物サンプル溶液の希釈系列を、レーンCはコントロールを、それぞれ示している。図中の○は、各ウェルを表したものであり、○の中の×は、アスペルギルス・ニガーの増殖が観察されたウェルを、○の中の○は、アスペルギルス・ニガーの増殖抑制が観察されたウェルを、それぞれ表したものである。
【図2】P1〜P10株及びY1〜Y3株をそれぞれ培養した培養液由来の上清サンプル溶液の2倍段階希釈系列と、アスペルギルス・ニガー胞子懸濁液を、96ウェルマイクロプレートの各ウェルにそれぞれ添加して培養した後に、各ウェルを顕微鏡で観察した結果を示したものである。(a)と(b)は、該96ウェルマイクロプレートを28℃で48時間培養した後、各ウェルを顕微鏡で観察した結果を、(c)と(d)は同じく36時間培養した後の観察結果を、それぞれ表したものである。図中の○は、各ウェルを表したものであり、○の中の×は、アスペルギルス・ニガーの増殖が観察されたウェルを、○の中の○は、アスペルギルス・ニガーの増殖抑制が観察されたウェルを、それぞれ表したものである。
【図3】YM寒天培地平板で25℃、2日間培養したO9−NP9株の電子顕微鏡写真である。
【特許請求の範囲】
【請求項1】
キャンディダ(Candida)属菌、サッカロマイセス(Saccharomyces)属菌、ピキア(Pichia)属菌、ロドトルラ(Rhodotorula)属菌、クラビスポラ(Clavispora)属菌、及びトルラスポラ(Torulaspora)属菌からなる群より選ばれる1の酵母が産生する、胞子の発芽を阻害する抗真菌性物質。
【請求項2】
下記の特徴を有する請求項1記載の抗真菌性物質。
(1)pH7.0において、抗真菌活性を有すること、
(2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有すること、
(3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCO(Molecular Weight Cut Off、分画分子量)が10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持されること。
【請求項3】
下記の特徴を有する請求項1記載の抗真菌性物質。
(1)pH7.0において、抗真菌活性を有すること、
(2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有すること、
(3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCO(Molecular Weight Cut Off、分画分子量)が10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持されること、
(4)55℃で15分間の加熱処理により、前記抗真菌活性が失活すること。
【請求項4】
前記キャンディダ属菌が、キャンディダ・マルトーサ(Candida maltosa)、キャンディダ・トロピカリス(Candida tropicalis)、又はキャンディダ・シュードインターメディア(Candida pseudintermedia)である、請求項1〜3のいずれか記載の抗真菌性物質。
【請求項5】
前記キャンディダ属菌が、キャンディダ・マルトーサO9−NP9(受託番号 NITE BP−297)、キャンディダ・マルトーサIAM12247、キャンディダ・マルトーサIAM12248、キャンディダ・トロピカリスIAM4965、又はキャンディダ・シュードインターメディアIAM12510である、請求項1〜3のいずれか記載の抗真菌性物質。
【請求項6】
前記サッカロマイセス属菌がサッカロマイセス・セレビシエ(Saccharomyces cerevisiae)であり、前記ピキア属菌がピキア・フェルメンタス(Pichia fermentans)であり、前記ロドトルラ属菌がロドトルラ・アクタ(Rhodotorula acuta)であり、前記クラビスポラ属菌がクラビスポラ・ルシタニエ(Clavispora lusitaniae)であり、前記トルラスポラ属菌がトルラスポラ・デルブレッキー(Torulaspora delbrueckii)である、請求項1〜3のいずれか記載の抗真菌性物質。
【請求項7】
前記ピキア属菌がピキア・フェルメンタスJCM1824であり、前記ロドトルラ属菌がロドトルラ・アクタJCM9494であり、前記サッカロマイセス属菌がサッカロマイセス・セレビシエH−1(受領番号 NITE ABP−473)であり、前記クラビスポラ属菌がクラビスポラ・スピーシーズP−5(Clavispora sp. P−5、受領番号 NITE ABP−474)又はクラビスポラ・ルシタニエNBRC10059であり、前記トルラスポラ属菌がトルラスポラ・デルブルッキーNBRC0955である、請求項1〜3のいずれか記載の抗真菌性物質。
【請求項8】
請求項1〜7のいずれか記載の抗真菌性物質を含有する防カビ剤。
【請求項9】
請求項1〜7のいずれか記載の抗真菌性物質を含有する食品保存料。
【請求項10】
請求項1〜7のいずれか記載の抗真菌性物質を用いることを特徴とする、カビ及び/又は酵母の増殖を阻害する方法。
【請求項11】
請求項1〜7のいずれか記載の抗真菌性物質を産生する酵母を用いることを特徴とする、カビ及び/又は酵母の増殖を阻害する方法。
【請求項12】
請求項1〜3のいずれか記載の抗真菌性物質を産生することを特徴とするキャンディダ・マルトーサO9−NP9(受託番号 NITE BP−297)。
【請求項13】
請求項1〜3のいずれか記載の抗真菌性物質を産生することを特徴とするサッカロマイセス・セレビシエH−1(受領番号 NITE ABP−473)。
【請求項14】
請求項1〜3のいずれか記載の抗真菌性物質を産生することを特徴とするクラビスポラ・スピーシーズP−5(受領番号 NITE ABP−474)。
【請求項1】
キャンディダ(Candida)属菌、サッカロマイセス(Saccharomyces)属菌、ピキア(Pichia)属菌、ロドトルラ(Rhodotorula)属菌、クラビスポラ(Clavispora)属菌、及びトルラスポラ(Torulaspora)属菌からなる群より選ばれる1の酵母が産生する、胞子の発芽を阻害する抗真菌性物質。
【請求項2】
下記の特徴を有する請求項1記載の抗真菌性物質。
(1)pH7.0において、抗真菌活性を有すること、
(2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有すること、
(3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCO(Molecular Weight Cut Off、分画分子量)が10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持されること。
【請求項3】
下記の特徴を有する請求項1記載の抗真菌性物質。
(1)pH7.0において、抗真菌活性を有すること、
(2)10%(w/w)スキムミルク培地で前記酵母を培養した培養液が、抗真菌活性を有すること、
(3)10%(w/w)スキムミルク培地で前記酵母を培養した培養液の上清を、MWCO(Molecular Weight Cut Off、分画分子量)が10kDである限外濾過膜を用いて限外濾過した場合に、限外濾過膜表面に保持されること、
(4)55℃で15分間の加熱処理により、前記抗真菌活性が失活すること。
【請求項4】
前記キャンディダ属菌が、キャンディダ・マルトーサ(Candida maltosa)、キャンディダ・トロピカリス(Candida tropicalis)、又はキャンディダ・シュードインターメディア(Candida pseudintermedia)である、請求項1〜3のいずれか記載の抗真菌性物質。
【請求項5】
前記キャンディダ属菌が、キャンディダ・マルトーサO9−NP9(受託番号 NITE BP−297)、キャンディダ・マルトーサIAM12247、キャンディダ・マルトーサIAM12248、キャンディダ・トロピカリスIAM4965、又はキャンディダ・シュードインターメディアIAM12510である、請求項1〜3のいずれか記載の抗真菌性物質。
【請求項6】
前記サッカロマイセス属菌がサッカロマイセス・セレビシエ(Saccharomyces cerevisiae)であり、前記ピキア属菌がピキア・フェルメンタス(Pichia fermentans)であり、前記ロドトルラ属菌がロドトルラ・アクタ(Rhodotorula acuta)であり、前記クラビスポラ属菌がクラビスポラ・ルシタニエ(Clavispora lusitaniae)であり、前記トルラスポラ属菌がトルラスポラ・デルブレッキー(Torulaspora delbrueckii)である、請求項1〜3のいずれか記載の抗真菌性物質。
【請求項7】
前記ピキア属菌がピキア・フェルメンタスJCM1824であり、前記ロドトルラ属菌がロドトルラ・アクタJCM9494であり、前記サッカロマイセス属菌がサッカロマイセス・セレビシエH−1(受領番号 NITE ABP−473)であり、前記クラビスポラ属菌がクラビスポラ・スピーシーズP−5(Clavispora sp. P−5、受領番号 NITE ABP−474)又はクラビスポラ・ルシタニエNBRC10059であり、前記トルラスポラ属菌がトルラスポラ・デルブルッキーNBRC0955である、請求項1〜3のいずれか記載の抗真菌性物質。
【請求項8】
請求項1〜7のいずれか記載の抗真菌性物質を含有する防カビ剤。
【請求項9】
請求項1〜7のいずれか記載の抗真菌性物質を含有する食品保存料。
【請求項10】
請求項1〜7のいずれか記載の抗真菌性物質を用いることを特徴とする、カビ及び/又は酵母の増殖を阻害する方法。
【請求項11】
請求項1〜7のいずれか記載の抗真菌性物質を産生する酵母を用いることを特徴とする、カビ及び/又は酵母の増殖を阻害する方法。
【請求項12】
請求項1〜3のいずれか記載の抗真菌性物質を産生することを特徴とするキャンディダ・マルトーサO9−NP9(受託番号 NITE BP−297)。
【請求項13】
請求項1〜3のいずれか記載の抗真菌性物質を産生することを特徴とするサッカロマイセス・セレビシエH−1(受領番号 NITE ABP−473)。
【請求項14】
請求項1〜3のいずれか記載の抗真菌性物質を産生することを特徴とするクラビスポラ・スピーシーズP−5(受領番号 NITE ABP−474)。
【図1】


【図2】
【図3】

【図2】
【図3】
【公開番号】特開2008−239604(P2008−239604A)
【公開日】平成20年10月9日(2008.10.9)
【国際特許分類】
【出願番号】特願2008−25692(P2008−25692)
【出願日】平成20年2月5日(2008.2.5)
【出願人】(000004569)日本たばこ産業株式会社 (406)
【Fターム(参考)】
【公開日】平成20年10月9日(2008.10.9)
【国際特許分類】
【出願日】平成20年2月5日(2008.2.5)
【出願人】(000004569)日本たばこ産業株式会社 (406)
【Fターム(参考)】
[ Back to top ]