
抗細菌感染、炎症および白血病組成物ならびにその使用
【課題】有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、グラム陽性菌の増殖を抑制する方法が提供される。
【解決手段】グラム陽性菌はメチシリン耐性黄色ブドウ球菌を含む。有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、敗血症を治療する方法も提供される。有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、白血病を治療する方法もさらに提供される。
【解決手段】グラム陽性菌はメチシリン耐性黄色ブドウ球菌を含む。有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、敗血症を治療する方法も提供される。有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、白血病を治療する方法もさらに提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、グラム陽性菌増殖の抑制、抗炎症および白血病の治療のためのテリマグランジン(Tellimagrandin)IIの使用に関する。
【背景技術】
【0002】
抗細菌感染
黄色ブドウ球菌(Staphylococcus aureus)(SA)はグラム陽性球菌に属し、臨床感染(たとえば、皮膚外傷感染、食中毒、敗血症性ショックなど)および院内感染の一般的な原因である。1950年のペニシリンの発見により、黄色ブドウ球菌との戦いに大きな進歩がもたらされた。しかし、1950年後半にはペニシリン耐性黄色ブドウ球菌が単離され同定された。メチシリンおよびオキサシリンなどの他の抗生物質がちょうど開発された時、抗生物質の乱用によりこれら2つの化合物に対する薬物耐性のある黄色ブドウ球菌が出現し、これはメチシリン耐性黄色ブドウ球菌(MRSA)としても知られている。抗生物質耐性の脅威は急速に増加した。2003年までに、米国において臨床試料から単離された黄色ブドウ球菌の50%がMRSAと見なされた。2009年、疾病管理予防センター(R.O.C.)による分析によれば、MRSAは台湾における院内感染の主因でもある(70〜80%)。
【0003】
抗生物質に抵抗するために、黄色ブドウ球菌はいくつかの根底にある機構を示す。βラクタマーゼを産生することが、βラクタム抗生物質(たとえば、ペニシリン、メチシリンおよびオキサシリン)耐性の主要な機構である。しかし、黄色ブドウ球菌のβラクタム抗生物質耐性には、βラクタマーゼを含有することよりもむしろmecA遺伝子発現が主に寄与している。mecA遺伝子は、βラクタムに対して親和性が低い、ペニシリン結合タンパク質2a(PBP2aまたはPBP2')と呼ばれる追加のタンパク質をコードしている。PBP2aは、メチシリンが黄色ブドウ球菌に結合し反応するのを困難にする。
【0004】
メチシリン耐性黄色ブドウ球菌の臨床指標は、4μg/mL以上のオキサシリンの最小抑制濃度(MIC)およびmecA遺伝子またはその産物を含有していることである。他の点では、欧州における多くのメチシリン耐性黄色ブドウ球菌は、テトラサイクリン抗生物質に対する耐性を示す。
【0005】
薬物耐性に加えて、黄色ブドウ球菌はバイオフィルムを形成する傾向があり、臨床コンダクター感染を引き起こす。したがって、バイオフィルム抑制化合物が開発されたら、コンダクターおよび他の臨床関連感染を著しく減少することになる。
【0006】
いくつかのタンニン天然抽出物の抗菌効果が報告されている。タンニンは2つのカテゴリー、加水分解性タンニンと濃縮タンニンに分類することができる。ポリフェノールはタンニンの一種であり、一般には様々な果実、野菜および赤ワイン中に存在する。ポリフェノールのもっとも一般的な機能は、酸化防止、抗真菌性、抗菌性などである。天然抽出物由来のポリフェノールは正常細胞には低細胞毒性を有するが、癌細胞には高細胞毒性を有する。いくつかの研究により、ポリフェノール天然抽出物はMRSAも死滅させることが可能であることが報告されている。これらの天然抽出物は、およそ3種類:低分子量のポリフェノール、酸化生成物産生ポリフェノールおよび高分子量のポリフェノールに分けることが可能である。ポリフェノール、特に(-)-エピカテキンガレート(Epicatechin gallate)(ECg)、コリラジン(Corilagin)およびテリマグランジンI(TG I)の抗菌効果はその分子量に比例していることが報告されている。様々なポリフェノール天然抽出物またはタンニン天然抽出物は、βラクタム抗生物質との相乗的殺菌効果を有し、たとえば、ECgは細胞壁の透過性とDアラニル化を減少させることできると示す複数の論文がある。さらに、遺伝的研究により、神経薬のチオリダジンもメチシリン耐性黄色ブドウ球菌の感受性を回復することが可能であることが指摘されている。上記効果は、mecA遺伝子発現およびPBP2aタンパク質産生の抑制を通じて作用する。しかし、チオリダジンはその毒性のために現在では使用されることは稀である。
【0007】
メチシリン耐性黄色ブドウ球菌感染の薬物耐性の深刻さは最近増大しており、新たな抗生物質の研究と開発には長い時間がかかる。天然抽出物には、正常細胞に対する低毒性、人体への害がほとんどないことおよびさらには他の治療的効果という利点がある。
【0008】
TG Iの誘導体であるテリマグランジンII(TG II)は、TG Iよりも1つ多く官能基、すなわち、没食子酸を有する。ポリフェノールでは、没食子酸は真菌およびウイルス感染に抵抗し酸化ストレスから細胞を保護する。さらに、没食子酸はより優れた癌細胞殺細胞能力も有する。テリマグランジンIIは、アセトンによってオニビシ(Trapa natans)Lまたはハマナス(Rosa rugosa Thunb)から均質に抽出することが可能である。
【0009】
抗炎症
敗血症、敗血症性ショックおよび多臓器不全症候群(MODS/MOF)は、ICUにおける患者の主要な死亡原因である。敗血症の病理発生および治療ならびにその合併症についての研究は大いに前進しているが、それでも敗血症の死亡率は改善が不十分であることを示している。米国での統計分析では、毎年75万人の敗血症患者が出ており、そのうちのわずか50%〜70%が生存することができる。敗血症とその合併症は13番目の死亡主要原因であった。敗血症とその合併症は毎年600以上の死亡例の一因になっている。
【0010】
この10年、グラム陰性桿菌は敗血症のもっとも一般的な病原体であった。大半のグラム陰性桿菌は消化管では正常な細菌叢であり、ある種の原因、たとえば、穿孔性虫垂炎に次ぐ腹膜炎および桿菌が会陰部を尿道または膀胱まで上がっていくことによる尿路感染の下で、隣接組織に広がる。敗血症の原病巣は通常、肺、尿生殖路、肝臓、胆嚢および消化管、皮膚または軟組織において発生する。原病巣は、約20〜30%の敗血症患者、特に、肝硬変、癌および他の慢性的な衰弱性疾患の患者では特定することができない。
【0011】
敗血症の病因および治療に関する最近の研究により、敗血症の病因は、グラム陰性桿菌の細胞壁から放出される内毒素としても知られるリポ多糖(LPS)と密接な関係があることが示されている。体細胞がリポ多糖類および内毒素などの微生物組成物により刺激されると、補体、白血球および血管内皮細胞はサイトカイン、TNFなどの放出により活性化される。免疫系が感染により活性化されると、血管緊張の調節因子である一酸化窒素が豊富に放出され、スーパーオキシドアニオンと相互作用してペルオキシナイトライトを形成するが、このペルオキシナイトライトは硝酸に再結合して、有毒なヒドロキシルイオンを生成するかまたは破壊的な酸化性ペルオキシ亜硝酸の形成を促進しうる。
【0012】
さらに、一酸化窒素は、潰瘍性大腸炎、乾癬および敗血症などの一部の炎症性疾患の調節において重要である。一酸化窒素は、一酸化窒素シンターゼ(NOS)がLアルギニンの酸化を調節する時に形成される。NOSは2つのカテゴリーに分けられる。1つは、神経型cNOS(NOS1)と内皮型cNOS(NOS3)などのカルシウム依存性構成的NOS(cNOS)である。もう1つはカルシウム非依存性誘導型NOS(NOS2)であり、大部分がマクロファージ、肝細胞および軟骨細胞により放出される。
【0013】
NOS2に加えて、シクロオキシゲナーゼII(COX-2)も炎症応答の初期段階において重要な役割を果たしている。COX-1と同様に、COX-2はアラキドン酸を、PGD2、PGE2、PGI2などの前駆体であるプロスタギランジンエンドペルオキシドH2(PGH2)に加水分解する。大半の動物細胞において構成的であるCOX-1と違って、COX-2発現は内毒素およびIL-1、IL-2またはTNF-αなどのサイトカインにより刺激されなければならない。COX-2は血流速度、浮腫および炎症応答における疼痛に対する身体感受性を調節する。
【0014】
研究によれば、ヘムオキシゲナーゼ-1(HO-1)は抗炎症性であり、敗血症における死亡率を減少させる。HO-1はヘムを一酸化炭素および二価イオンに分解する。一酸化炭素はマクロファージを調節して炎症応答の発生および悪化を抑制する。二価イオンは遊離ラジカルに結合し、したがって抗酸化性である。HO-1は血管拡張などの複数の機能を果たして、炎症応答における炎症機構から人体を保護する。
【0015】
テリマグランジンIIはポリフェノール化合物の加水分解性タンニンに属する。ポリフェノール化合物は大半の高等植物から抽出される。タンニンは動物皮膚に存在し、タンパク質沈殿を促進し、優れた金属キレート剤として機能する。タンニンは遊離ラジカルを除去することができるので、生物における炎症により誘導される過酸化を軽減するのに使用することができる。
【0016】
抗白血病
白血病は血液の癌としても知られている。プログラムされた細胞死は正常な細胞生理の一部であり、アポトーシスは不適切な生存条件下におけるプログラムされた細胞死である。実験におけるアポトーシスまたはネクローシスなどの他の細胞死の初期の識別は、顕微鏡を用いた細胞形態形成による。アポトーシスと他の細胞死の主な違いは、血清やオルガネラが漏出するのを妨げるための後期段階におけるより完全な細胞膜であり、独特なアポトーシス小体を生じる。
【0017】
このプロセスには、特定のプロテアーゼによる形状維持タンパク質細胞骨格の分解、核酸断片化、クロマチン凝縮、および細胞膜上のブレブも含まれる。ポリフェノール化合物の癌細胞抑制については多くの研究が発表されてきた。ヒト白血病細胞、HL-60の増殖の抑制および細胞周期停滞は立証されている。著者は、ある種の濃度のタンニン酸を用いて処置された細胞においてプログラムされたアポトーシスを観察した。最後に、関連するタンパク質の変動を使用して、大半がミトコンドリアによる内部シグナル伝達によって誘導されるアポトーシスを予想した。白血病細胞におけるそのアポトーシス誘導機構については、茶ポリフェノールに関するさらに多くの研究が行われた。
【0018】
アポトーシスの原因は様々であり、内部経路と外部経路に分けられる。細胞活性酸素種または細胞生存にとって不利な他の要因が誘導されて増加した結果として、ミトコンドリア膜電位が変化し、アポトーシスタンパク質Bcl-2が減少し、Baxが増加すると、内部経路が誘発される。ミトコンドリア中のチトクロムCがミトコンドリア膜の外側の細胞質に放出され、それに続いてアポトーシスタンパク質、大半がカスパーゼ9の活性化が誘導される。最後に、細胞DNAが断片化しアポトーシスが完了する。外部経路は、Fas-Lなどの死亡因子の存在により誘発される。Fas-Lは、アポトーシスタンパク質、大半がカスパーゼ8を活性化する前にFas受容体に結合し、次にプログラムされたアポトーシスが起こる。
【0019】
多くの研究が、タンニンが腫瘍細胞増殖の抑制と関係があることを確認している。タンニン化合物は、加水分解性タンニンと凝縮タンニンに分けられる。精製された植物ポリフェノールは加水分解性タンニンの一種に属する。加水分解性タンニンは、活性酸素種により媒介される腫瘍細胞におけるプログラムされた死に関係することが現在では知られている。
【先行技術文献】
【非特許文献】
【0020】
【非特許文献1】Srdjanら(APMIS 115: 891〜9頁、2007)
【発明の概要】
【発明が解決しようとする課題】
【0021】
黄色ブドウ球菌感染、敗血症および白血病を克服するために、本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、グラム陽性菌の増殖を抑制する方法を提供する。グラム陽性菌はメチシリン耐性黄色ブドウ球菌を含む。本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、敗血症などの抗炎症の方法も提供する。本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、白血病を治療する方法をさらに提供する。
【課題を解決するための手段】
【0022】
本明細書で使用されるように、用語「テリマグランジンII」とは、たとえば、トウビシ(Trapa bispinosa)の殻(しかし、これに限定されない)から抽出される天然のポリフェノール化合物のことである。
【0023】
本明細書で使用されるように、用語「対象」とは、哺乳動物、さらに好ましくはヒトのことである。哺乳動物には、ヒト、霊長類、家畜、スポーツ動物、げっ歯類および愛玩動物が挙げられるがこれらに限定されない。
【0024】
本明細書で使用されるように、用語「1つの(a)」または「1つの(an)」は、本発明の抗生物質または要素または成分のカテゴリーまたは種を説明するのに用いられる。これは便宜上のためだけに、本発明の一般的意味を与えるために行われる。この記述は、1つまたは少なくとも1つを含むと理解されるべきであり、単数形は、それが他の意味であることが明白でない限り、その複数形も含む。
【0025】
本明細書で使用されるように、用語「または(or)」は「および/または(and/or)」を記述するのに用いられる。
【図面の簡単な説明】
【0026】
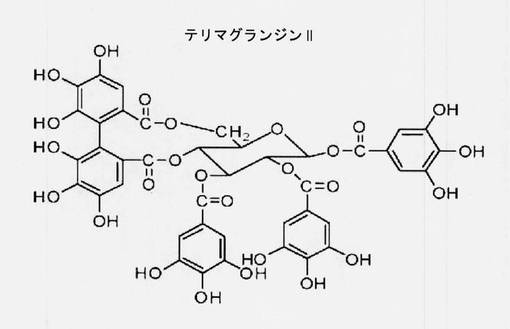
【図1】テリマグランジンII(TG II)の構造を示す図である。
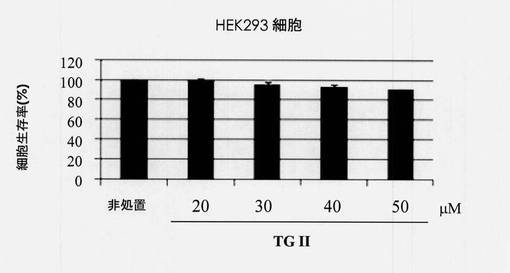
【図2】ヒト胚腎臓細胞系統HEK293における細胞生存率に対するテリマグランジンIIの効果を示す図である。
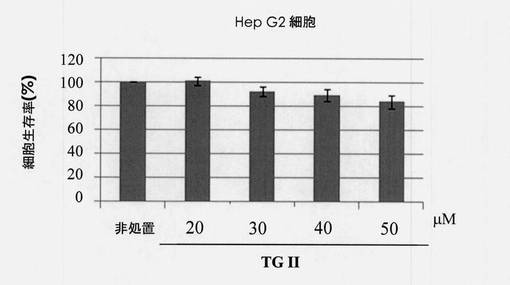
【図3】ヒト肝細胞腫細胞系統Hep G2における細胞生存率に対するテリマグランジンIIの効果を示す図である。
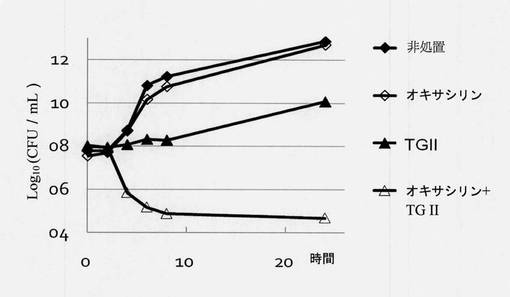
【図4】細菌に対する相乗的抗菌活性のためのオキサシリンとテリマグランジンIIの組合せを示す図である。Y軸:平均コロニー数(Log10cfu/mL)、X軸:培養時間。4つの群があり、◆対照群、薬物を何も添加しない;◇5μg/mLオキサシリンの試験群;▲40μg/mLテリマグランジンIIの試験群;Δ40μg/mLテリマグランジンIIと5μg/mLオキサシリンの試験群。
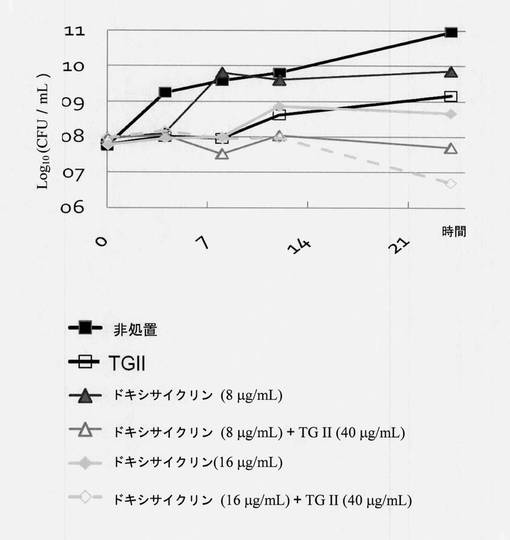
【図5】細菌に対する相乗的抗菌活性のためのドキシサイクリンとテリマグランジンIIの組合せを示す図である。Y軸:平均コロニー数(Log10cfu/mL)、X軸:培養時間。6つの群があり、■対照群、薬物を何も添加しない;□40μg/mLテリマグランジンIIの試験群;▲8μg/mLドキシサイクリンの試験群;Δ8μg/mLドキシサイクリンと40μg/mLテリマグランジンIIの試験群、◆16μg/mLドキシサイクリンの試験群、◇16μg/mLドキシサイクリンと40μg/mLテリマグランジンIIの試験群。
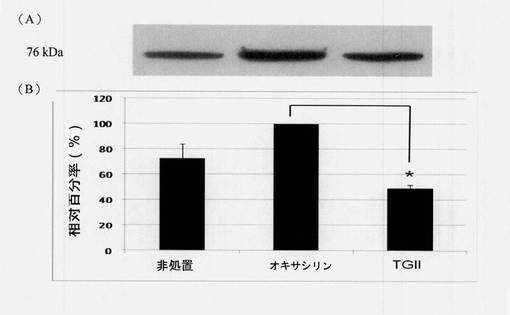
【図6】ペニシリン結合タンパク質2a(PBP2a)の発現に対するテリマグランジンIIの効果を示す図である(N=3)。PBP2a発現:(A)ウェスタンブロット分析結果(B)図6(A)からのPBP2aタンパク質のウェスタンブロットの定量的分析。各タンパク質バンドの強度はAlphaEase FCソフトウェアを使用して定量され、2μg/mLオキサシリンの試験群を100%として設定した。第1のバーは対照群を表し、第2のバーは2μg/mLオキサシリンの試験群を表し、第3のバーは40μg/mLテリマグランジンIIの試験群を表す。左から右へ標準偏差(SD)は、11.77、0および2.5;*p<0.01。
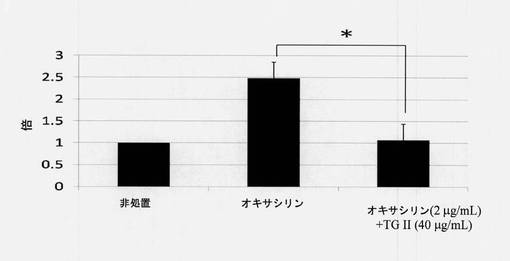
【図7】mecA遺伝子の発現に対するテリマグランジンIIの効果を示す図である(N=3、*p<0.05)。
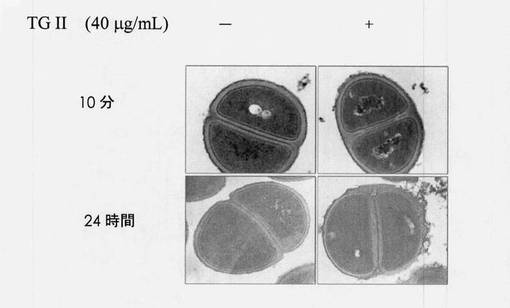
【図8】走査型電子顕微鏡(SEM)から観察される表現型変化を示す図である。
【図9】マウスマクロファージ細胞系統RAW264.7における細胞毒性に対するテリマグランジンIIの効果を示す図である(N=3、平均±SD)。
【図10】リポ多糖(LPS)により誘導される一酸化窒素の生成に対するテリマグランジンIIの効果を示す図である(N=3、平均±SD)。
【図11】リポ多糖(LPS)により誘導されるプロスタグランジンE2(PGE2)の発現に対するテリマグランジンIIの効果を示す図である(N=3、平均±SD、*p<0.05)。
【図12】リポ多糖(LPS)により誘導されるヘムオキシゲナーゼ-1(HO-1)の発現に対するテリマグランジンIIの効果を示す図である(N=3、平均±SD)。
【図13】WST-1アッセイを使用して測定された定量的細胞生存率結果を示す図である。テリマグランジンIIを用いてまたは用いずに処置されたヒト赤骨髄芽球様細胞白血病K-562細胞の細胞生存率は、WST-1アッセイを使用して決定された(N=3、平均±SD、#p<0.01)。
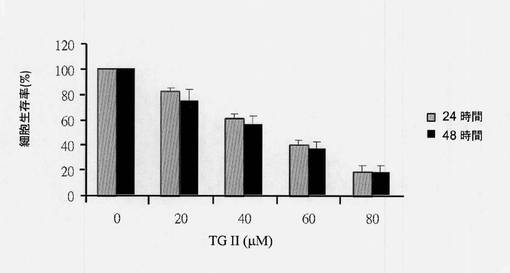
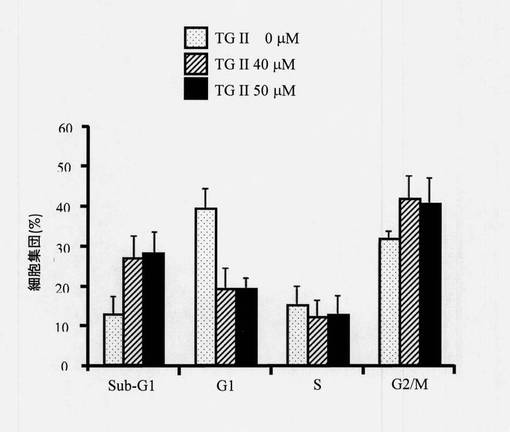
【図14】フローサイトメトリーを使用することによる細胞周期分布を示す図である。テリマグランジンIIを用いてまたは用いずに24時間処置されたヒト赤骨髄芽球様細胞白血病K-562細胞の細胞周期研究はフローサイトメトリーを使用して決定された(N=3、平均±SD、*p<0.05)。
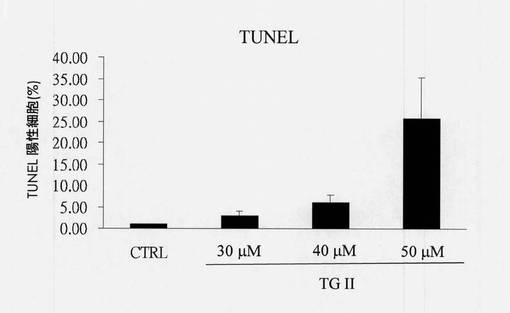
【図15】非処置細胞と比べた、テリマグランジンIIを用いて48時間処置されたヒト赤骨髄芽球様細胞白血病K-562細胞におけるDNA断片化の量を示す図である(N=3、平均±SD、#p<0.05)。
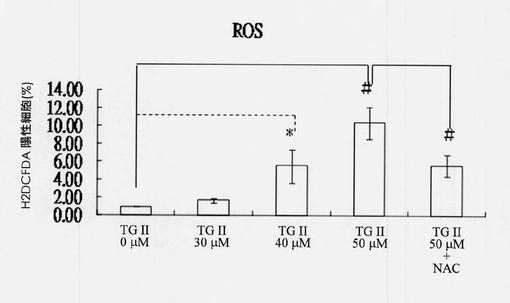
【図17】テリマグランジンIIを用いて2時間処置されたヒト赤骨髄芽球様細胞白血病K-562細胞のROS生成を示す図である(N=3、*p<0.05、#p<0.01)。
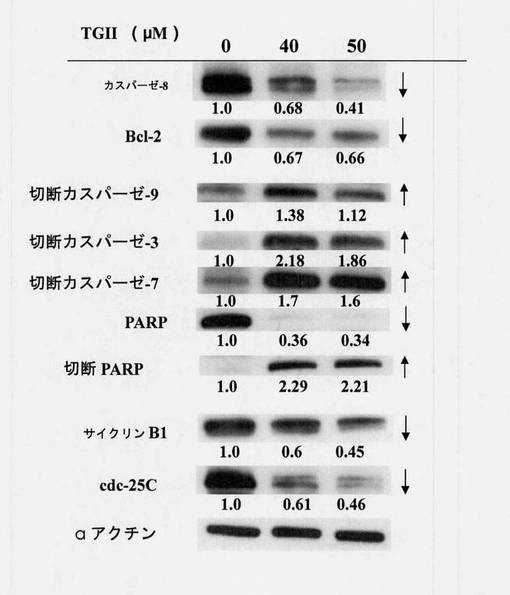
【図18】テリマグランジンIIを用いてまたは用いずに48時間処置されたヒト赤骨髄芽球様細胞白血病K-562細胞におけるウェスタン免疫ブロットによるカスパーゼ8、Bcl-2、カスパーゼ9、カスパーゼ3、カスパーゼ7、PARP、サイクリンB1およびCdc-25Cの発現を示す図である(N=3、タンパク質定量化はαアクチンに基づいていた)。
【発明を実施するための形態】
【0027】
したがって、本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、グラム陽性菌の増殖を抑制する方法を提供する。一実施形態では、方法は、有効量のβラクタム抗生物質またはポリケチド抗生物質を対象に投与するステップをさらに含む。別の実施形態では、細菌は黄色ブドウ球菌またはメチシリン耐性黄色ブドウ球菌である。
【0028】
一実施形態では、テリマグランジンIIはβラクタム抗生物質またはポリケチド抗生物質と相乗的に作用して、メチシリン耐性黄色ブドウ球菌の増殖を抑制する。別の実施形態では、βラクタム抗生物質には、ペニシリン、たとえば、ベンザチンペニシリン、ベンジルペニシリン、フェノキシメチルペニシリン、プロカインペニシリン、メチシリン、オキサシリン、ナフシリン、クロキサシリン、ジクロキサシリン、フルクロキサシリン、テモシリン、アモキシシリン、アンピシリン、コアモキシクラブ(co-amoxiclav)、アズロシリン、カルベニシリン、チカルシリン、メズロシリンおよびピペラシリン;セファロスポリン、たとえば、セファレキシン、セファロチン、セファゾリン、セファクロル、セフロキシム、セファマンドール、セフォテタン、セフォキシチン、セフトリアキソン、セフォタキシム、セフポドキシム、セフィキシム、セフタジジム、セフェピムおよびセフピロム;カルバペネムもしくはペネム、たとえば、イミペネム、メロペネム、パニペネム、ビアペネム、エルタペネム、ファロペネムおよびドリペネム;モノバクタム、たとえば、アズトレオナム、チゲモナム(tigemonam)、ノカルディシンAおよびタブトキシニン-βラクタム;またはクラブラン酸、タゾバクタムおよびスルバクタムなどのβラクタマーゼ阻害剤が挙げられるが、これらに限定されない。さらに別の実施形態では、ポリケチド抗生物質には、テトラサイクリン、ドキシサイクリン、ミノサイクリン、エリスロマイシン、モネンシンA(ルメンシン)、タイロシン、ナラシン、リファマイシン、セルビマイシンC、ジャドマイシンB、キロマイシンまたはムピロシンが挙げられるが、これらに限定されない。一実施形態では、βラクタム抗生物質は、メチシリン、オキサシリン、ナフシリン、クロキサシリン、ジクロキサシリン、フルクロキサシリン、テモシリンまたはアモキシシリンである。別の実施形態では、ポリケチド抗生物質は、テトラサイクリン、ドキシサイクリンまたはミノサイクリンである。さらに別の実施形態では、βラクタム抗生物質はオキサシリンであり、ポリケチド抗生物質はドキシサイクリンである。
【0029】
一実施形態では、テリマグランジンIIは、30μg/mLを超える濃度を有する。別の実施形態では、テリマグランジンIIは、40μg/mLを超える濃度を有する。一実施形態では、対象はヒトである。
【0030】
【表1】

【0031】
1つのMSSA株、3つのMRSA株(19615、18271および18631)ならびに1つの標準MRSA株(ATCC 33591)を使用した。試験された試薬は、オキサシリン、エリスロマイシン、ペニシリン、カナマイシン、レボフロキサシン、ドキシサイクリンおよびテリマグランジンIIである。
【0032】
【表2】
【0033】
テリマグランジンIIとの相乗効果が実証された抗生物質はオキサシリン、ドキシサイクリンおよびレボフロキサシンである。最小抑制濃度(MIC)と部分抑制濃度インデックス(FIC:fractional inhibitory concentration index)が計算された。相乗効果、相加効果、無差別効果およびアンタゴニスト効果のFIC値は、それぞれ、<0.5、0.5〜<1、1〜≦4、および4である。
【0034】
【表3】
【0035】
1つのMSSA株と3つのMRSA株(19615、18271および18631)が使用された。試験された抗生物質は、オキサシリン、アンピシリン、レボフロキサシン、エリスロマイシン、カナマイシン、ドキシサイクリンおよびバンコマイシンであった。最小抑制濃度(MIC)および部分抑制濃度インデックス(FIC)が計算された。相乗効果、相加効果、無差別効果およびアンタゴニスト効果のFIC値は、それぞれ、<0.5、0.5〜<1、1〜≦4、および4である。
【0036】
【表4】
【0037】
14の臨床的に単離されたMRSA株を、オキサシリンおよびテリマグランジンII(40μg/mL)を用いて試験し、FICインデックスを計算した。相乗効果、相加効果、無差別効果およびアンタゴニスト効果のFIC値は、それぞれ、<0.5、0.5〜<1、1〜≦4、および4である。
【0038】
一実施形態では、テリマグランジンIIとポリケチド抗生物質は抗酸化機構を通じて相乗的に作用する。
【0039】
一実施形態では、βラクタム抗生物質は、テリマグランジンIIと一緒に作用して、黄色ブドウ球菌においてペニシリン結合タンパク質2a発現を減少させる。別の実施形態では、βラクタム抗生物質は、テリマグランジンIIと一緒に作用して、黄色ブドウ球菌においてmecA遺伝子発現を抑制する。
【0040】
【表5】
【0041】
【表6】
【0042】
テリマグランジンIIとドキシサイクリンのMICは、FeCl2の存在下と比べた場合、FeCl2の非存在下のほうが高い(Table 6(表6))。これは、テリマグランジンIIとドキシサイクリンの抗MRSA活性がFeCl2の存在下では抑制されることを示唆している。
【0043】
一実施形態では、テリマグランジンIIは黄色ブドウ球菌のバイオフィルム形成を抑制する。
【0044】
本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、抗炎症の方法も提供する。一実施形態では、抗炎症の方法を使用して敗血症を治療する。
【0045】
一実施形態では、テリマグランジンIIは一酸化窒素およびプロスタグランジンE2産生を減少させる。別の実施形態では、テリマグランジンIIはヘムオキシゲナーゼ-1の産生を増加させる。
【0046】
一実施形態では、テリマグランジンIIは10〜50μMの範囲の濃度を有する。別の実施形態では、テリマグランジンIIは25〜50μMの範囲の濃度を有する。一実施形態では、対象はヒトである。
【0047】
本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、白血病を治療する方法をさらに提供する。
【0048】
一実施形態では、テリマグランジンIIは、白血病細胞の細胞アポトーシスおよび細胞周期停止を誘導する。別の実施形態では、テリマグランジンIIは、細胞内活性酸素種を増加させることにより細胞アポトーシスを誘導する。さらに別の実施形態では、白血病細胞の細胞アポトーシスは、カスパーゼ3およびカスパーゼ7を活性化することにより誘導される。別の実施形態では、白血病細胞の細胞周期停止はG2/M期に停止される。
【0049】
一実施形態では、テリマグランジンIIは、20〜80μMの範囲の濃度を有する。別の実施形態では、テリマグランジンIIは、40〜80μMの範囲の濃度を有する。別の実施形態では、テリマグランジンIIは、50〜80μMの範囲の濃度を有する。一実施形態では、対象はヒトである。
【0050】
次の実施例は、以下の通り、本発明のいくつかの例となる実施形態を提供する。
【0051】
(実施例)
以下の実施例は非限定的であり、本発明の様々な態様および特徴の典型にすぎない。
【0052】
(実施例1)
細胞生存率アッセイ
細胞生存率はアラマーブルーを使用して測定された。細胞生存率アッセイでは、ヒト胚腎臓細胞系統HEK293およびヒト肝細胞腫細胞系統Hep G2を、96ウェルプレート上の各ウェルに180μLプレーティングした(HEK293では、約10,000細胞/ウェルの密度で;Hep G2では約7500細胞/ウェルの密度で)。細胞を一晩(16〜18時間)増殖させ、付着させた。一晩のインキュベーション後、20μLのテリマグランジンIIを添加して、0、20、30、40および50μg/mLの最終濃度を得た。0μg/mLのテリマグランジンIIと追加の薬物を添加しない(培地のみ)培養物は対照であった。次に、細胞を、5%CO2、37℃インキュベーター中24時間インキュベートした。24時間後、100μLのアラマーブルーが各ウェルに添加され、5%CO2、37℃インキュベーター中4時間培養された。570nmおよび600nmで吸光度がモニターされ、細胞生存率が決定された。
【0053】
図2は、ヒト胚腎臓細胞系統HEK293における細胞生存率に対するテリマグランジンIIの効果を示している。Y軸は生存細胞の割合を表し、X軸は対照と20、30、40および50μg/mLのテリマグランジンIIの濃度に対応する。
【0054】
X軸は、対照と20、30、40および50μg/mLのテリマグランジンIIの濃度に対応している。標準偏差は、それぞれ0、1.59、2.01、1.64および0.87と決定された。図3は、ヒト肝細胞腫細胞系統Hep G2における細胞生存率に対するテリマグランジンIIの効果を示した。Y軸は、生存細胞の割合を表し、X軸は対照と20、30、40および50μg/mLのテリマグランジンIIの濃度に対応する。標準偏差は、それぞれ3.480、3.95、4.83および5.73と決定された。図2および図3に示されるように、テリマグランジンIIは、ヒト胚腎臓細胞系統とヒト肝細胞腫細胞系統において低い毒性を示した。細胞生存率は、40μg/mLのテリマグランジンIIで処置された細胞では少なくとも80%維持された。
【0055】
最小抑制濃度、MIC
試験される抗生物質(オキサシリン、エリスロマイシン、アンピシリン、カナマイシン、レボフロキサシン、バンコマイシンおよびドキシサイクリン)の連続2倍希釈は微量液体希釈法(Broth microdilution method)により、最終濃度2〜512μg/mLまで実施された。細菌の一晩培養物を収集し、Muller-Hilton(M-H)ブロスに再懸濁した。1アリコートの細胞懸濁液(OD600=0.1)を調製した。細菌懸濁液を96ウェルプレートにプレーティングし、試験抗生物質と一緒に37℃、5%CO2インキュベーター中でインキュベートした(Table 1(表1)〜Table 4(表4))。
【0056】
部分抑制濃度インデックス(FICインデックス)
2つの薬物間の相乗作用、相乗作用と拮抗作用は、以下の式により評価された。
【0057】
【数1】
【0058】
【表7】
【0059】
Table 1(表1)〜Table 4(表4)に示されるように、テリマグランジンIIとオキサシリンまたはドキシサイクリンは相乗作用として定義され、そのFICインデックスは0.5未満であった。
【0060】
1,1-ジフェニル-2-ピクリルヒドラジル(DPPH)捕捉アッセイ
試験試薬、天然抽出物およびビタミンC(アスコルビン酸)はメタノールに溶解させ、10X保存液を作製した。ビタミンCは陽性対照として使用された。120μMのDPPH溶液を調製した。96ウェルマイクロプレート上に保存試薬、天然抽出物およびビタミンCを20μL取って、総容量100μLまでになるようにメタノールを添加した。対照は100μLメタノールであった。各試料を100μL DPPH溶液と混合させ、暗所で40分間インキュベートした。各ウェルのDPPHレベルは、517nmでの各ウェルの光学密度を検出することにより評価し、結果はTable 5(表5)に示した。
【0061】
殺菌曲線(Time-Killing Curve)
MRSA 19615細菌を一晩培養し、収集し、指示最終濃度のTG IIおよび抗生物質を含有する25mLのMHB中に再懸濁した。試料を希釈するために、指示された時間間隔で試料を懸濁液から取り、濾過されたPBSと混合し、続いて、2プレート/各時点でプレーティングして希釈試料をMHA上に広げた。150〜500コロニー間の最初の時点のコロニー数を使用して希釈倍率を決定した。殺菌曲線はコロニー計数法から計算され、データ解析は一晩培養後に実施された。細菌殺菌曲線は、時間に対する平均コロニー数(log10 cfu/mL)をプロットすることにより作成された(図4と図5)。図4が示すように、オキサシリン(5μg/mL)処置単独では、細菌増殖を抑制できなかった。オキサシリン(5μg/mL)とテリマグランジンII(40μg/mL)を一緒に使用すると、24時間後に細菌の目に見える増殖を抑制した。図5が示すように、テリマグランジンII/ドキシサイクリン組合せ(40μg/mLテリマグランジンIIと8μg/mLドキシサイクリン; 40μg/mLテリマグランジンIIと16μg/mLドキシサイクリン)では、相乗効果が観察された。したがって、40μg/mLテリマグランジンIIと16μg/mLドキシサイクリン組合せの殺菌効率のほうがはるかに強力であった。
【0062】
ウェスタンブロッティング分析
タンパク質試料(15μg)は5×試料緩衝液と混合し、1×試料緩衝液中で5分間煮沸し、次にタンパク質がその成分ポリペプチド鎖に変性されるように、急速冷却した。等量のタンパク質(15μg)を17mAで2時間、10%SDS-PAGE上で分離し、ウェット式転写手順を使用してPVDF膜へ移した。PVDF膜は、非特異的結合を防ぐために5%(w/v)脱脂粉乳を含有するt-BST緩衝液中、室温で30分間ブロックし、次に5%脱脂粉乳中、4℃で16〜18時間、一次抗体(ウサギ抗ペニシリン結合タンパク質、希釈度1:5000)を用いてプローブした。抗グロイエール(Groel)は内部標準として使用された(希釈度1:80,000; Sigmaより購入)。それぞれ5分間で3回t-BSTを用いて洗浄後、膜を、5%(w/v)脱脂粉乳中、西洋ワサビペルオキシダーゼ(HRP)-コンジュゲート二次抗体と一緒に室温で30分間インキュベートした。膜をそれぞれ5分間で3回洗浄し、続いて増強された化学発光システムを使用して、フィルムに曝露する。ウェスタンブロットの定量的結果は図6に示した。図6に示されるように、40μg/mLのテリマグランジンII単独で、細菌中においてペニシリン結合タンパク質2a(PBP2a)の発現を抑制することができた。
【0063】
RNA単離および定量化
RNA単離手順は、Roche High Pure RNA Isolation Kitにより実施された。手短に言えば、各処置された細菌試料は、13,000rpmで2分間の遠心分離により収集し、37℃で15分間20μMリゾスタフィン処置により溶解した。試料を、400μL溶解結合緩衝液混合試料を添加した後に高純度フィルター管に充填し、次に9,600gで1分間遠心分離した。通過画分は除かれフィルター管は収集管に入れた。90μLの混合前デオキシリボヌクレアーゼインキュベーション緩衝液と10μLのデオキシリボヌクレアーゼIをフィルター管に添加し、続いて1時間反応させた。洗浄緩衝液I、IIおよびIIIは連続してフィルター管に添加し、通過画分は、各ステップ間に遠心分離(9,600gで1分間)により取り除いた。最終RNA産物は、30μL溶出緩衝液により溶出し、清潔なエッペンドルフ内に収集した。
【0064】
RNA調製物は、Backman DU800を使用して定量した。2マイクロリットル(2μL)のRNA試料はPCRチューブに98μLの溶出緩衝液と一緒に添加し、よく混合した。RNA試料の濃度と純度は、Backman DU800分光光度計を使用して評価した(希釈係数は50である)。
【0065】
逆転写PCR
SuperScript III逆転写酵素(Invitrogen製)が本研究で使用された。2マイクログラム(2μg)の精製RNAを、PCR反応管中で1μLのオリゴdT、1μLのランダムプライマー(250ng)および1μLの10mM dNTP混合物(dATP、dGTP、dCTPおよびdTTP、それぞれ10mM)と混合させた。混合液を5分間で65℃まで加熱し、氷上で急速冷却させた。4マイクロリットル(4μL)の5×Fist-stand緩衝液、1μLのDTT(0.1M)および0.5μLのSuperScript III逆転写酵素を添加し、よく混合させた。混合液は25℃で5分間、55℃で60分間、および70℃で5分間インキュベートした。
【0066】
リアルタイムPCR
標的遺伝子の発現を調べるために、Roche LightCyclerシステムを使用してRT-PCRを実施した。2マイクログラム(2μg)のcDNAを、反応プレートにおいて4μM MgCl2、5μMプライマーおよび2μLの10×LightCycler FastStart DNA Master SYBR Green I混合液(Roche製)と混合させ、よく混合させた。増幅条件は以下の通りであった。変性は1サイクル95℃で0.5分間、95℃で15秒間、60℃で30秒間および72℃で30秒間であった。PCR実行後、追加の融解分析が実施され(0.1℃/秒の温度転移速度で60℃から95℃まで)、これにより増幅反応の特異性を確認した。RNA遺伝子発現の倍率変化は、式2-ΔΔCt、Ctはサイクル閾値により計算された。サイクル閾値(Ct)は、RT-PCRにおいて蛍光シグナルが閾値を超えるのに必要とされるサイクル数と定義される。ΔCtは、標的遺伝子のCt値から内在性対照のCt値を引くことにより計算された。次に、ΔΔCtは、試料のΔCtから対照のΔCtを引くことにより計算された。図7に示されるように、第1のバーは対照群を表し、第2のバーは2μg/mLのオキサシリンを有する試験群を表し、第3のバーは40μg/mLテリマグランジンIIと2μg/mLのオキサシリンを有する試験群を表す。左から右へ標準偏差(SD)は、0、0.37および0.37であり、*p<0.05。図7に示される結果では、2μg/mLのオキサシリンと40μg/mLテリマグランジンIIの組合せはmecA遺伝子発現を抑制した。
【0067】
【表8】
【0068】
透過型電子顕微鏡(TEM)
TEMにより観察する前に、細菌をプレート上で一晩培養し、収集し、10mL MHBまたは40μg/mLテリマグランジンIIを含有する10mL MHBのどちらかに懸濁した。細菌溶液は、600nm(OD600)で0.1の光学密度まで希釈し、10分間または24時間培養した。次に、細胞を10,000rpm/分で遠心分離し、固定液に懸濁し、固定時間は24時間を超えることはできない。TEM用の試料は、高雄医学大学病院(Kaohsiung、Taiwan、ROC)の実験医学臨床研究部に送られた。図8に示されるように、40μg/mLテリマグランジンIIを用いて処置し10分間培養した細菌試料は、対照群と比べると滑らかではないざらざらした表面を示した。表現型は、40μg/mLテリマグランジンIIを用いて処置され24時間培養された細菌試料との著しい相関を示した。
【0069】
バイオフィルム形成
細菌を寒天プレート上で一晩(16〜18時間)増殖した。次に、収集し、0.45%無菌PBS中に再懸濁し、無菌PBSに600nm(OD600)で0.1の最終光学密度まで希釈した。ポリスチレン96ウェルマイクロタイタープレートを、ウェルあたり150μLの培養液と20μLの細菌溶液で満たした。培養物を37℃で18時間静置させた。インキュベーション期間後、培養物を取り除き、マイクロタイタープレートウェルは300μLの無菌PBSを用いて3回穏やかに洗浄し、ゆるく会合した細菌を取り除き、次に60℃オーブンで1時間乾燥させた。試料は、各ウェルに2%色素溶液(150μL)を最初の播種レベルより上まで添加することにより染色し、15分間インキュベートした。次にプレートを新鮮な水で洗浄した。染色強度は、各乾燥ウェルに95%アルコール(150μL)を添加後に測定された。試料を室温で30分間インキュベートし、その後OD517値をプレートリーダー上で測定した。
【0070】
【表9】
【0071】
光学密度カットオフ値(ODc)=陰性対照の平均OD+3×陰性対照の標準偏差(SD)(バイオフィルム生成の解釈は、Srdjanら(APMIS 115: 891〜9頁、2007)の基準に従って実行された)。MRSA17199は対照株として使用された。
バイオフィルム生成の解釈:
平均OD値 バイオフィルム生成
OD≦ODc なし
ODc<OD≦2×ODc 弱
2×ODc<OD≦4×ODc 中
4×ODc<OD 強
【0072】
(実施例2)
以下の研究ではマウスマクロファージ細胞系統RAW264.7が使用された。
【0073】
細胞生存率アッセイ
アラマーブルー試薬を使用して細胞生存率を評価した。RAW264.7細胞(200μLの培地中2×104)を96ウェルプレート上に播種した。一晩のインキュベーション後、細胞を2%ウシ胎児血清(FBS)を含有するDMEM培地中で3時間血清飢餓状態にした。いくつかの群があり、すなわちDMSO処置もテリマグランジンII処置もない細胞(対照); DMSOを用いて処置される細胞(対照);異なる濃度(10、25、50、75および100μM、DMSO中に希釈される)のテリマグランジンIIを用いて処置される細胞(試験群)であった。各群は2通りに決定された。
【0074】
24時間後、細胞培地を取り除き破棄し、細胞を180μLの新鮮な培養液で再懸濁した。各細胞懸濁液に20μLアラマーブルーを添加した。次に、プレートをさらに4時間インキュベーター内に置いた。インキュベーション後、培養液の吸光度を570と600nmで測定し、細胞生存率を、以下の式を使用して解析した。
【0075】
【数2】
【0076】
O1: 570nmでの酸化アラマーブルーのモル吸光係数(E)
O2: 600nmでの酸化アラマーブルーのモル吸光係数(E)
A1: 570nmでの試験ウェルの吸光度
A2: 600nmでの試験ウェルの吸光度
P1: 570nmでの陽性増殖対照ウェルの吸光度
P2: 600nmでの陽性増殖対照ウェルの吸光度
【0077】
図9では、x軸はテリマグランジンIIの濃度を表し、y軸は細胞生存率を表す。図9の結果により、細胞生存率は、濃度50μg/mLまたはそれよりも低い用量でテリマグランジンIIを用いて処置された場合、著しい影響を受けないことが示された。
【0078】
一酸化窒素生成解析
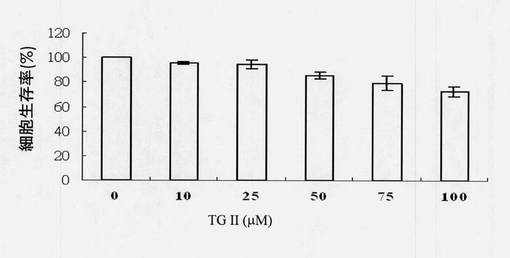
マウスマクロファージ細胞を、約4×105 RAW264.7細胞/mLの密度で12ウェルプレートの各ウェルにプレーティングした。一晩のインキュベーション後、細胞を2%ウシ胎児血清(FBS)を含有するDMEM培地中で3時間血清飢餓状態にした。リポ多糖(LPS)を用いて24時間刺激された細胞は、培養液中の亜硝酸レベルを著しく増加させた。内在性一酸化窒素生成はLPSにより誘導した。細胞を、LPSを用いて処置される前および後に様々な濃度(10、25および50μM)のテリマグランジンIIで処置した。一酸化窒素生成解析では、培養上清を96ウェルプレートに運び、グリース試薬と混合した。プレートをインキュベートし、次に吸光度を520nmで測定した。DMEMで調製された亜硝酸ナトリウム(NaNO2)溶液(0M〜0.1M、8ポイント)を使用して標準曲線を作成した。試料の亜硝酸濃度は標準曲線の内挿により決定した。図10に示されるように、テリマグランジンIIの導入(LPSを用いて処置される前または後のいずれでも)により、LPS誘導一酸化窒素の生成は減少した。試験群のテリマグランジンII濃度(25および50μM)は統計的有意を示した。図10は、相対的順序でLPSまたはテリマグランジンII処置および濃度を標識されたx軸ならびに一酸化窒素濃度を標識されたy軸により枠組みされている。対照(LPSのみ)および試験群はt検定を使用して比較した、*p<0.05(有意)および#p<0.01(高度に有意)。
【0079】
プロスタグランジンE2(PGE2)アッセイ
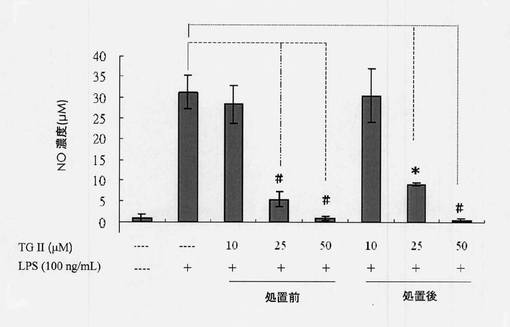
1ミリリットル(1mL)のマウスマクロファージ細胞系統RAW264.7(4×105細胞/ウェル)を12ウェルプレート上に一晩播種した。一晩のインキュベーション後、細胞を2%ウシ胎児血清(FBS)を含有するDMEM培地中で3時間血清飢餓状態にした。次に、テリマグランジンIIが、最終濃度の10、25および50μMが得られるように添加された。このようにして、30分のインキュベーション後、マウスマクロファージ細胞をLPSを用いて16時間処置し、細胞培養物上清を収集した。無細胞上清を希釈し、培養上清中のPGE2はPGE2アッセイキット(R&D system製)を使用することにより検出した。本研究の目的は、テリマグランジンIIを用いて共処置することによりLPS誘導PGE2発現を評価することであった。図11に示されるように、25または50μMのテリマグランジンIIを、LPSで処置する30分前に細胞に添加すると、PGE2の発現は著しく減少した(実験結果はLPS群と比較し、0.05未満のP値は有意と解釈した)。
【0080】
ヘムオキシゲナーゼ-1(HO-1)のウェスタンブロット分析
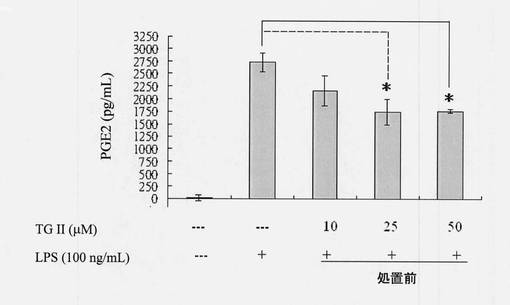
マウスマクロファージRAW264.7細胞を、播種密度5×106細胞/mLで6cm細胞皿にプレーティングし、一晩インキュベートした。一晩のインキュベーション後、細胞を異なった濃度のテリマグランジンII(10、25および50μM)およびLPSを用いて処置し、24時間インキュベートした。細胞はPBSを用いて2回洗浄し、こすり取り、氷上でRIPA緩衝液に溶解し、タンパク質を得た。ヘムオキシゲナーゼ-1発現は全細胞溶解物のウェスタンブロッティングにより分析した。ウェスタンブロットデータおよび定量的解析の結果は図12に示されている。図12では、第1のバーは対照群を表し、第2のバーはLPSのみを用いた試験群を表し、第3、第4および第5のバーは10、25および50μg/mLテリマグランジンIIを用いた試験群を表している(N=3、平均±SD)。図12に示されるように、ヘムオキシゲナーゼ-1の発現は、LPS刺激細胞上でわずかに増加した。テリマグランジンII処置(25または50μM)処置により、ヘムオキシゲナーゼ-1の発現に著しい増加が観察された。
【0081】
(実施例3)
細胞生存率アッセイ
定量的細胞生存率はWST-1アッセイを使用して評価された。WST-1は、代謝活性細胞上で高度に水溶性のホルマザンを生成し、細胞生存率の直接比色測定を可能にする試薬である。
【0082】
K-562細胞を、播種密度6,000細胞/ウェル(100μL)で96ウェル培養プレートにプレーティングし、37℃で5%CO2を用い16〜18時間インキュベートした。細胞を、最終濃度を0、20、40、60および80μMにするように20μLテリマグランジンIIを用いて処置して、37℃で5%CO2を用い24時間および48時間インキュベートした。テリマグランジンII処置のない細胞は対照として使用された。プレートの各ウェルに12μLのWST-1試薬(10:1)を添加し、よく混合し、次に37℃で5%CO2インキュベーターにおいて2時間インキュベートした。インキュベーション後、450nm(試験波長)および630nm(基準波長)で吸光度を測定した。データは450nm測定値から630nmバックグラウンド吸光度を引くことにより計算された。
【0083】
結果は図13に示されており、異なる濃度で24時間または48時間テリマグランジンIIを用いて処置されたK-562細胞の細胞生存率は、WST-1アッセイを使用して測定された。細胞生存は、対照培養物増殖(100%)に対して存在する生細胞の割合として表された。IC50値は、曲線から外挿された。24時間試験群のIC50の値は57.01μMであり、48時間試験群のIC50の値は48.11μMであった。濃度40μMまたは50μMのテリマグランジンIIを使用したフォローアップ実験は、この結果に基づいていた(N=3、平均±SD、*p<0.05)。
【0084】
図13は、WST-1アッセイを使用して測定された定量的細胞生存率の結果を示していた。テリマグランジンIIを用いて処置されたまたは処置されないヒト赤骨髄芽球様細胞白血病K-562細胞の細胞生存率は、WST-1アッセイを使用して決定された(N=3、平均±SD、#p<0.01)。
【0085】
細胞周期解析
K-562細胞を、密度5×105細胞/mL(15mL)で15cm皿にプレーティングし、37℃で5%CO2を用い16〜18時間インキュベートした。細胞を収集し、室温、700rpmで3分間遠心分離し、0.04%ウシ胎児血清および8mMのLグルタミンを含有する培養液に再懸濁した。次に、細胞を37℃で5%CO2を用い16〜18時間インキュベートした。
【0086】
上記細胞を収集し、開始密度5×105細胞/mL(3mL)で3cm皿にプレーティングした。細胞を、最終濃度0、10、20、30、40および50μMまでテリマグランジンIIを用いて処置し、37℃で5%CO2を用いて12時間および24時間インキュベートした。
【0087】
細胞を4℃、700rpmの速度で5分間遠心分離し、PBSを用いて1回洗浄した。細胞を、70%エタノールの滴下添加によって細胞懸濁液に固定した。16〜18時間後、エタノール懸濁細胞を4℃、3,500rpm/分の速度で5分間遠心分離し、エタノールを完全にデカントした。細胞を、リボヌクレアーゼA(10μg/mL)および1% Triton X-100を含有するPBS中に再懸濁し、37℃で5%CO2インキュベーターにおいて30分〜1時間インキュベートした。ヨウ化プロピジウム(PI)染色では、細胞は室温、3,500rpmで5分間遠心分離し、600μLのPBSに再懸濁した。25μLのPI(20μg/mL)溶液を混合物に添加し、室温で暗所に1時間インキュベートした。ヨウ化プロピジウム染色細胞のDNA含量分析はフローサイトメーターにおいて実施された。フローサイトメトリーによる細胞周期解析は、図14に示される通りであった。結果により、サブG1およびG2/M期細胞の割合はテリマグランジンII処置において用量依存的に増加することが示された(N=3、平均±SD、*p<0.05)。
【0088】
アポトーシス検出試験
TUNEL試験を使用して、細胞がアポトーシス過程に入っているかどうかを判定するためにDNA断片化を検出した。
【0089】
1mLあたり1×106 K-562細胞の濃度の合計で10mLの細胞溶液を、細胞が環境に順応できるように、5%CO2を供給されたインキュベーターにおいて37℃で30分間10cm細胞培養皿を用いて培養した。異なる濃度のテリマグランジンII溶液を、最終濃度がそれぞれ0、10、20、30、40および50μMになるように順に添加した。溶液は、37℃、5%CO2を用いたインキュベーター中に48時間置き、その後4℃、3,500rpmで5分間遠心分離した。上清を取り除いた後、細胞をPBSにより1回すすいだ。合計で1mLの1%Trioxane(PBSを用いて10%Trioxane溶液から希釈された)を添加し、溶液を室温で30分間〜1時間反応させておいた。次に、溶液を4℃、3,500rpmで5分間遠心分離した。上清を取り除いた後、70%エタノールを添加し、溶液を細胞固定のために16〜18時間-20℃に置いた。
【0090】
細胞固定後、溶液を4℃、400rpmで10分間遠心分離し、その後上清を取り除いた。TUNEL緩衝液80μLを添加し、溶液を室温で5分間反応させておいた。次に、EDTA 1mLを溶液に添加し、その後これを穏やかに混合した。溶液は4℃、3,500rpmで5分間遠心分離し、その後上清を取り除いた。次に、TUNEL試験のために50μLの蛍光検出試薬を溶液中に添加し、その後よく混合させ、37℃で1時間、暗所でインキュベートした。溶液は4℃、3,500rpmで5分間遠心分離し、PBSを使用して細胞を1回すすいだ。最後に、1mLのPBSを使用して細胞を再懸濁させ、その後BD FACScanを使用して蛍光シグナルを検出し、定量結果は図15である。TUNEL試験の定量図(図15)から、蛍光シグナルはK-562細胞が30μMのテリマグランジンIIと反応した48時間後に検出されたことが分かる。さらに、シグナル変化の変動は、濃度が50μMまで増加した場合のほうが顕著であった(N=3、平均±SD、*p<0.05)。
【0091】
DNAラダー試験
テリマグランジンIIを用いて処置されたK-562細胞を、DNA断片化を同定するため、電気泳動のために収集した。
【0092】
1mLあたり1×106 K-562細胞の濃度の合計で3mLの細胞溶液を、細胞が環境に順応できるように、37℃で30分間、5%CO2を供給されたインキュベーターにおいて6cm細胞培養プレートを用いて培養した。異なる濃度のテリマグランジンII溶液を、最終濃度がそれぞれ0、20、30、40、50および60μMになるように順に添加した。溶液を、37℃、5%CO2を用いたインキュベーター中に48時間置き、その後4℃、2,500rpmで5分間遠心分離した。上清を取り除いた後、細胞をPBSにより1回すすぐ。次に、細胞溶解緩衝液100μL(20μM EDTAおよび50μM Tris-HClに溶解した1% NP-40、pH8.0)を添加し、溶液を高速で10秒間浸盪させた。
【0093】
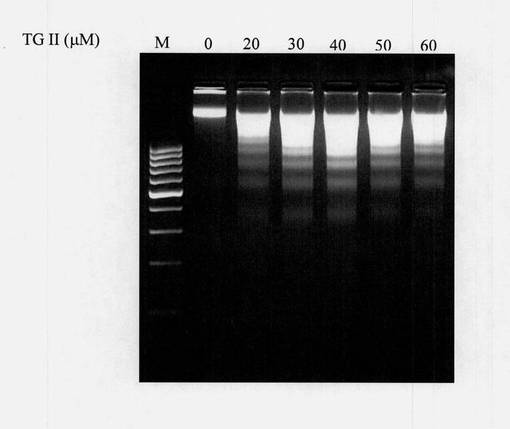
溶液は3,500rpmで5分間遠心分離し、その後90μLの上清を収集し、SDS溶液10μLとよく混合させた。リボヌクレアーゼA(4mg/mL)50μLを混合溶液に添加し、次に56℃の水浴中に1時間置いてRNAを取り除いた。次に、プロテイナーゼK(10mg/mL)30μLを添加して、その後さらに1時間56℃の水浴においてタンパク質を取り除いた。全溶液容積の半分の10M酢酸アンモニウム(90μL)を添加しよく混合させて、その後全容積の2倍の無水エタノール(540μL)を添加してよく混合させた。溶液を、DNA沈殿のために-80℃下に置いた。溶液は低温で30分間、12,000rpmで遠心分離し、その後DNA沈殿物の消失を避けるために上清を慎重に取り除いた。DNA沈殿物は70%エタノール200μLにより1回すすぎ、その後全上清を取り除いて、チューブキャップを開けて室温で乾燥させた。脱イオン水50μLまたは0.5×TBE緩衝液を添加して、DNAを完全に分解した。最後に、DNAゲル電気泳動を上述のDNA試料について実施した。図16のDNAラダー電気泳動図は、TUNEL試験の結果を裏付けた。異なる濃度のテリマグランジンIIを用いて処置した48時間後にK-562細胞においてDNA断片化が起きた。
【0094】
ROS試験
ラジカル特異的蛍光色素、2',7'-ジクロロフルオレセイン(DCF-DA)を使用して、それぞれ細胞中のH2O2/NOベースのラジカルまたはO2ベースのラジカルを検出した。
【0095】
1mLあたり1×106 K-562細胞の濃度の合計3mLの細胞溶液を、細胞が環境に順応できるように、37℃で30分間、5%CO2を供給されたインキュベーターにおいて、6cm細胞培養プレートを用いて培養した。異なる濃度のテリマグランジンII溶液を、最終濃度がそれぞれ0、30、40および50μMになるように順に添加した。溶液を、37℃、5%CO2を用いたインキュベーター中に2時間置き、その後4℃、3,500rpmで5分間遠心分離し、次にPBSにより1回すすいだ。2',7'-ジクロロフルオレセイン(DCF-DA)を添加して、その後溶液を暗所インキュベーターに45分間置き、次にPBSにより1回すすいだ。最後に、1mLのPBSを使用して細胞を再懸濁させ、その後BD FACScanを使用して蛍光シグナルを検出した。図17に示される結果から、K-562細胞中のROSは、ROS試験において40μMテリマグランジンIIとの相互作用の2時間後にBD FACScanにより増加しているのが観察された。その上、細胞がテリマグランジンII 50μLと2時間相互作用する前に抗酸化剤、N-アセチル-L-システイン10μMを用いて前処理された場合には、ROS生成の抑制が観察された(N=3、*p<0.05、#p<0.01)。
【0096】
ウェスタンブロッティング
1mLあたり1×106 K-562細胞の濃度の合計3mLの細胞溶液を、細胞が環境に順応できるように、37℃で30分間、5%CO2を供給されたインキュベーターにおいて、6cm細胞培養皿を用いて培養した。異なる濃度のテリマグランジンII溶液を、最終濃度がそれぞれ0、40および50μMになるように順に添加した。溶液を、37℃、5%CO2を用いたインキュベーター中に48時間置いた。インキュベーション後、溶液を4℃、1,000rpmで5分間遠心分離し、PBSにより1回すすいだ。プロテアーゼ阻害剤を含有するRIPA緩衝液を添加して細胞を分解した。
【0097】
溶液は、細胞が確実に物理的損傷により分解されているように、3回液体窒素を用いて急速凍結させ、解凍した。溶液を4℃、30分間15,000rpmで遠心分離し、全細胞タンパク質を含有する上清を収集した。タンパク質濃度はBCAタンパク質アッセイにより決定された。最後に、等量のタンパク質試料(40μg)は1×添加液中で3分間煮沸し、さらにタンパク質ゲル電気泳動にかけるためにタンパク質は一次構造まで分解した(または-80℃で保存した)。
【0098】
SDS-PAGE 10%または12%が、タンパク質ゲル電気泳動(150V、17mA、2時間5分)において目的のタンパク質分子量により選択された。タンパク質は、ウェット式転写手順(100V、300mA、30分間)を使用してゲルからPVDF膜に移した。膜は、非特異的結合を除去するために2%ウシ胎児血清(BSA)を含有するトリス緩衝生理食塩水-Tween(TBS-T)緩衝液中室温で1時間ブロックした。
【0099】
PVDF膜は、2%BSA-TBS-T緩衝液中4℃で16〜18時間、製造業者に従った推奨される量で希釈された一次抗体を用いてプローブした。相互作用の完了後、TBS-T緩衝液を使用して、非特異的に結合した一次抗体をすすぎ落した。TBS-T緩衝液は、約30分間のうちに10分ごとに3回交換した。次に、特異的二次抗体を添加して、室温で1時間相互作用させた。TBS-T緩衝液を使用して、非特異的に結合した二次抗体をすすぎ落した。TBS-T緩衝液は約60分間のうちに10分ごとに6回交換された。ECLシステム(増強された化学発光システム)を使用し、フィルムを光に曝露させて現像した。最後に定量的解析が実施された。図18は、ウェスタンブロッティングの結果から、先ず、おそらく切断されたカスパーゼ8に活性化されて、外部死滅経路におけるカスパーゼ8の不活性が減少することを示した。ROS増加という前の結果に示されるように、この結果も、抗アポトーシスのタンパク質、Bcl-2が減少して、カスパーゼ9、カスパーゼ3およびカスパーゼ7の活性化およびPARPの究極の活性化をさらに誘導することを示した。細胞周期調節に関連するタンパク質には、G2/Mチェックポイント調節因子、サイクリンB1およびCdc-25Cが挙げられる。上の結果により、(非リン酸化)活性はテリマグランジンIIとの相互作用後に減少し、細胞周期の停滞を裏付けていることも見出された(N=3、タンパク質定量化はαアクチンに基づいていた)。
【0100】
統計的解析
スチューデントt検定(不対の)を実施した。結果は、標準誤差(平均±S.D.)付きの少なくとも3つの個々の実験の平均として表されており、P値≦0.01または≦0.05を有意と見なした。
【0101】
当業者であれば、本発明が、目標を遂行するため、および言及された目的と利点に加えてその固有の目的と利点を得るために十分適合していることを容易に認識する。動物、プロセス、およびそのプロセスを生み出すための方法は、好ましい実施形態の典型であり、例となるものであり、本発明の範囲を限定するものとして意図されてはいない。その改変および他の使用を当業者であれば思い付くであろう。これらの改変は本発明の精神の範囲内に包含されており、特許請求の範囲により定義されている。
【技術分野】
【0001】
本発明は、グラム陽性菌増殖の抑制、抗炎症および白血病の治療のためのテリマグランジン(Tellimagrandin)IIの使用に関する。
【背景技術】
【0002】
抗細菌感染
黄色ブドウ球菌(Staphylococcus aureus)(SA)はグラム陽性球菌に属し、臨床感染(たとえば、皮膚外傷感染、食中毒、敗血症性ショックなど)および院内感染の一般的な原因である。1950年のペニシリンの発見により、黄色ブドウ球菌との戦いに大きな進歩がもたらされた。しかし、1950年後半にはペニシリン耐性黄色ブドウ球菌が単離され同定された。メチシリンおよびオキサシリンなどの他の抗生物質がちょうど開発された時、抗生物質の乱用によりこれら2つの化合物に対する薬物耐性のある黄色ブドウ球菌が出現し、これはメチシリン耐性黄色ブドウ球菌(MRSA)としても知られている。抗生物質耐性の脅威は急速に増加した。2003年までに、米国において臨床試料から単離された黄色ブドウ球菌の50%がMRSAと見なされた。2009年、疾病管理予防センター(R.O.C.)による分析によれば、MRSAは台湾における院内感染の主因でもある(70〜80%)。
【0003】
抗生物質に抵抗するために、黄色ブドウ球菌はいくつかの根底にある機構を示す。βラクタマーゼを産生することが、βラクタム抗生物質(たとえば、ペニシリン、メチシリンおよびオキサシリン)耐性の主要な機構である。しかし、黄色ブドウ球菌のβラクタム抗生物質耐性には、βラクタマーゼを含有することよりもむしろmecA遺伝子発現が主に寄与している。mecA遺伝子は、βラクタムに対して親和性が低い、ペニシリン結合タンパク質2a(PBP2aまたはPBP2')と呼ばれる追加のタンパク質をコードしている。PBP2aは、メチシリンが黄色ブドウ球菌に結合し反応するのを困難にする。
【0004】
メチシリン耐性黄色ブドウ球菌の臨床指標は、4μg/mL以上のオキサシリンの最小抑制濃度(MIC)およびmecA遺伝子またはその産物を含有していることである。他の点では、欧州における多くのメチシリン耐性黄色ブドウ球菌は、テトラサイクリン抗生物質に対する耐性を示す。
【0005】
薬物耐性に加えて、黄色ブドウ球菌はバイオフィルムを形成する傾向があり、臨床コンダクター感染を引き起こす。したがって、バイオフィルム抑制化合物が開発されたら、コンダクターおよび他の臨床関連感染を著しく減少することになる。
【0006】
いくつかのタンニン天然抽出物の抗菌効果が報告されている。タンニンは2つのカテゴリー、加水分解性タンニンと濃縮タンニンに分類することができる。ポリフェノールはタンニンの一種であり、一般には様々な果実、野菜および赤ワイン中に存在する。ポリフェノールのもっとも一般的な機能は、酸化防止、抗真菌性、抗菌性などである。天然抽出物由来のポリフェノールは正常細胞には低細胞毒性を有するが、癌細胞には高細胞毒性を有する。いくつかの研究により、ポリフェノール天然抽出物はMRSAも死滅させることが可能であることが報告されている。これらの天然抽出物は、およそ3種類:低分子量のポリフェノール、酸化生成物産生ポリフェノールおよび高分子量のポリフェノールに分けることが可能である。ポリフェノール、特に(-)-エピカテキンガレート(Epicatechin gallate)(ECg)、コリラジン(Corilagin)およびテリマグランジンI(TG I)の抗菌効果はその分子量に比例していることが報告されている。様々なポリフェノール天然抽出物またはタンニン天然抽出物は、βラクタム抗生物質との相乗的殺菌効果を有し、たとえば、ECgは細胞壁の透過性とDアラニル化を減少させることできると示す複数の論文がある。さらに、遺伝的研究により、神経薬のチオリダジンもメチシリン耐性黄色ブドウ球菌の感受性を回復することが可能であることが指摘されている。上記効果は、mecA遺伝子発現およびPBP2aタンパク質産生の抑制を通じて作用する。しかし、チオリダジンはその毒性のために現在では使用されることは稀である。
【0007】
メチシリン耐性黄色ブドウ球菌感染の薬物耐性の深刻さは最近増大しており、新たな抗生物質の研究と開発には長い時間がかかる。天然抽出物には、正常細胞に対する低毒性、人体への害がほとんどないことおよびさらには他の治療的効果という利点がある。
【0008】
TG Iの誘導体であるテリマグランジンII(TG II)は、TG Iよりも1つ多く官能基、すなわち、没食子酸を有する。ポリフェノールでは、没食子酸は真菌およびウイルス感染に抵抗し酸化ストレスから細胞を保護する。さらに、没食子酸はより優れた癌細胞殺細胞能力も有する。テリマグランジンIIは、アセトンによってオニビシ(Trapa natans)Lまたはハマナス(Rosa rugosa Thunb)から均質に抽出することが可能である。
【0009】
抗炎症
敗血症、敗血症性ショックおよび多臓器不全症候群(MODS/MOF)は、ICUにおける患者の主要な死亡原因である。敗血症の病理発生および治療ならびにその合併症についての研究は大いに前進しているが、それでも敗血症の死亡率は改善が不十分であることを示している。米国での統計分析では、毎年75万人の敗血症患者が出ており、そのうちのわずか50%〜70%が生存することができる。敗血症とその合併症は13番目の死亡主要原因であった。敗血症とその合併症は毎年600以上の死亡例の一因になっている。
【0010】
この10年、グラム陰性桿菌は敗血症のもっとも一般的な病原体であった。大半のグラム陰性桿菌は消化管では正常な細菌叢であり、ある種の原因、たとえば、穿孔性虫垂炎に次ぐ腹膜炎および桿菌が会陰部を尿道または膀胱まで上がっていくことによる尿路感染の下で、隣接組織に広がる。敗血症の原病巣は通常、肺、尿生殖路、肝臓、胆嚢および消化管、皮膚または軟組織において発生する。原病巣は、約20〜30%の敗血症患者、特に、肝硬変、癌および他の慢性的な衰弱性疾患の患者では特定することができない。
【0011】
敗血症の病因および治療に関する最近の研究により、敗血症の病因は、グラム陰性桿菌の細胞壁から放出される内毒素としても知られるリポ多糖(LPS)と密接な関係があることが示されている。体細胞がリポ多糖類および内毒素などの微生物組成物により刺激されると、補体、白血球および血管内皮細胞はサイトカイン、TNFなどの放出により活性化される。免疫系が感染により活性化されると、血管緊張の調節因子である一酸化窒素が豊富に放出され、スーパーオキシドアニオンと相互作用してペルオキシナイトライトを形成するが、このペルオキシナイトライトは硝酸に再結合して、有毒なヒドロキシルイオンを生成するかまたは破壊的な酸化性ペルオキシ亜硝酸の形成を促進しうる。
【0012】
さらに、一酸化窒素は、潰瘍性大腸炎、乾癬および敗血症などの一部の炎症性疾患の調節において重要である。一酸化窒素は、一酸化窒素シンターゼ(NOS)がLアルギニンの酸化を調節する時に形成される。NOSは2つのカテゴリーに分けられる。1つは、神経型cNOS(NOS1)と内皮型cNOS(NOS3)などのカルシウム依存性構成的NOS(cNOS)である。もう1つはカルシウム非依存性誘導型NOS(NOS2)であり、大部分がマクロファージ、肝細胞および軟骨細胞により放出される。
【0013】
NOS2に加えて、シクロオキシゲナーゼII(COX-2)も炎症応答の初期段階において重要な役割を果たしている。COX-1と同様に、COX-2はアラキドン酸を、PGD2、PGE2、PGI2などの前駆体であるプロスタギランジンエンドペルオキシドH2(PGH2)に加水分解する。大半の動物細胞において構成的であるCOX-1と違って、COX-2発現は内毒素およびIL-1、IL-2またはTNF-αなどのサイトカインにより刺激されなければならない。COX-2は血流速度、浮腫および炎症応答における疼痛に対する身体感受性を調節する。
【0014】
研究によれば、ヘムオキシゲナーゼ-1(HO-1)は抗炎症性であり、敗血症における死亡率を減少させる。HO-1はヘムを一酸化炭素および二価イオンに分解する。一酸化炭素はマクロファージを調節して炎症応答の発生および悪化を抑制する。二価イオンは遊離ラジカルに結合し、したがって抗酸化性である。HO-1は血管拡張などの複数の機能を果たして、炎症応答における炎症機構から人体を保護する。
【0015】
テリマグランジンIIはポリフェノール化合物の加水分解性タンニンに属する。ポリフェノール化合物は大半の高等植物から抽出される。タンニンは動物皮膚に存在し、タンパク質沈殿を促進し、優れた金属キレート剤として機能する。タンニンは遊離ラジカルを除去することができるので、生物における炎症により誘導される過酸化を軽減するのに使用することができる。
【0016】
抗白血病
白血病は血液の癌としても知られている。プログラムされた細胞死は正常な細胞生理の一部であり、アポトーシスは不適切な生存条件下におけるプログラムされた細胞死である。実験におけるアポトーシスまたはネクローシスなどの他の細胞死の初期の識別は、顕微鏡を用いた細胞形態形成による。アポトーシスと他の細胞死の主な違いは、血清やオルガネラが漏出するのを妨げるための後期段階におけるより完全な細胞膜であり、独特なアポトーシス小体を生じる。
【0017】
このプロセスには、特定のプロテアーゼによる形状維持タンパク質細胞骨格の分解、核酸断片化、クロマチン凝縮、および細胞膜上のブレブも含まれる。ポリフェノール化合物の癌細胞抑制については多くの研究が発表されてきた。ヒト白血病細胞、HL-60の増殖の抑制および細胞周期停滞は立証されている。著者は、ある種の濃度のタンニン酸を用いて処置された細胞においてプログラムされたアポトーシスを観察した。最後に、関連するタンパク質の変動を使用して、大半がミトコンドリアによる内部シグナル伝達によって誘導されるアポトーシスを予想した。白血病細胞におけるそのアポトーシス誘導機構については、茶ポリフェノールに関するさらに多くの研究が行われた。
【0018】
アポトーシスの原因は様々であり、内部経路と外部経路に分けられる。細胞活性酸素種または細胞生存にとって不利な他の要因が誘導されて増加した結果として、ミトコンドリア膜電位が変化し、アポトーシスタンパク質Bcl-2が減少し、Baxが増加すると、内部経路が誘発される。ミトコンドリア中のチトクロムCがミトコンドリア膜の外側の細胞質に放出され、それに続いてアポトーシスタンパク質、大半がカスパーゼ9の活性化が誘導される。最後に、細胞DNAが断片化しアポトーシスが完了する。外部経路は、Fas-Lなどの死亡因子の存在により誘発される。Fas-Lは、アポトーシスタンパク質、大半がカスパーゼ8を活性化する前にFas受容体に結合し、次にプログラムされたアポトーシスが起こる。
【0019】
多くの研究が、タンニンが腫瘍細胞増殖の抑制と関係があることを確認している。タンニン化合物は、加水分解性タンニンと凝縮タンニンに分けられる。精製された植物ポリフェノールは加水分解性タンニンの一種に属する。加水分解性タンニンは、活性酸素種により媒介される腫瘍細胞におけるプログラムされた死に関係することが現在では知られている。
【先行技術文献】
【非特許文献】
【0020】
【非特許文献1】Srdjanら(APMIS 115: 891〜9頁、2007)
【発明の概要】
【発明が解決しようとする課題】
【0021】
黄色ブドウ球菌感染、敗血症および白血病を克服するために、本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、グラム陽性菌の増殖を抑制する方法を提供する。グラム陽性菌はメチシリン耐性黄色ブドウ球菌を含む。本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、敗血症などの抗炎症の方法も提供する。本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、白血病を治療する方法をさらに提供する。
【課題を解決するための手段】
【0022】
本明細書で使用されるように、用語「テリマグランジンII」とは、たとえば、トウビシ(Trapa bispinosa)の殻(しかし、これに限定されない)から抽出される天然のポリフェノール化合物のことである。
【0023】
本明細書で使用されるように、用語「対象」とは、哺乳動物、さらに好ましくはヒトのことである。哺乳動物には、ヒト、霊長類、家畜、スポーツ動物、げっ歯類および愛玩動物が挙げられるがこれらに限定されない。
【0024】
本明細書で使用されるように、用語「1つの(a)」または「1つの(an)」は、本発明の抗生物質または要素または成分のカテゴリーまたは種を説明するのに用いられる。これは便宜上のためだけに、本発明の一般的意味を与えるために行われる。この記述は、1つまたは少なくとも1つを含むと理解されるべきであり、単数形は、それが他の意味であることが明白でない限り、その複数形も含む。
【0025】
本明細書で使用されるように、用語「または(or)」は「および/または(and/or)」を記述するのに用いられる。
【図面の簡単な説明】
【0026】
【図1】テリマグランジンII(TG II)の構造を示す図である。
【図2】ヒト胚腎臓細胞系統HEK293における細胞生存率に対するテリマグランジンIIの効果を示す図である。
【図3】ヒト肝細胞腫細胞系統Hep G2における細胞生存率に対するテリマグランジンIIの効果を示す図である。
【図4】細菌に対する相乗的抗菌活性のためのオキサシリンとテリマグランジンIIの組合せを示す図である。Y軸:平均コロニー数(Log10cfu/mL)、X軸:培養時間。4つの群があり、◆対照群、薬物を何も添加しない;◇5μg/mLオキサシリンの試験群;▲40μg/mLテリマグランジンIIの試験群;Δ40μg/mLテリマグランジンIIと5μg/mLオキサシリンの試験群。
【図5】細菌に対する相乗的抗菌活性のためのドキシサイクリンとテリマグランジンIIの組合せを示す図である。Y軸:平均コロニー数(Log10cfu/mL)、X軸:培養時間。6つの群があり、■対照群、薬物を何も添加しない;□40μg/mLテリマグランジンIIの試験群;▲8μg/mLドキシサイクリンの試験群;Δ8μg/mLドキシサイクリンと40μg/mLテリマグランジンIIの試験群、◆16μg/mLドキシサイクリンの試験群、◇16μg/mLドキシサイクリンと40μg/mLテリマグランジンIIの試験群。
【図6】ペニシリン結合タンパク質2a(PBP2a)の発現に対するテリマグランジンIIの効果を示す図である(N=3)。PBP2a発現:(A)ウェスタンブロット分析結果(B)図6(A)からのPBP2aタンパク質のウェスタンブロットの定量的分析。各タンパク質バンドの強度はAlphaEase FCソフトウェアを使用して定量され、2μg/mLオキサシリンの試験群を100%として設定した。第1のバーは対照群を表し、第2のバーは2μg/mLオキサシリンの試験群を表し、第3のバーは40μg/mLテリマグランジンIIの試験群を表す。左から右へ標準偏差(SD)は、11.77、0および2.5;*p<0.01。
【図7】mecA遺伝子の発現に対するテリマグランジンIIの効果を示す図である(N=3、*p<0.05)。
【図8】走査型電子顕微鏡(SEM)から観察される表現型変化を示す図である。
【図9】マウスマクロファージ細胞系統RAW264.7における細胞毒性に対するテリマグランジンIIの効果を示す図である(N=3、平均±SD)。
【図10】リポ多糖(LPS)により誘導される一酸化窒素の生成に対するテリマグランジンIIの効果を示す図である(N=3、平均±SD)。
【図11】リポ多糖(LPS)により誘導されるプロスタグランジンE2(PGE2)の発現に対するテリマグランジンIIの効果を示す図である(N=3、平均±SD、*p<0.05)。
【図12】リポ多糖(LPS)により誘導されるヘムオキシゲナーゼ-1(HO-1)の発現に対するテリマグランジンIIの効果を示す図である(N=3、平均±SD)。
【図13】WST-1アッセイを使用して測定された定量的細胞生存率結果を示す図である。テリマグランジンIIを用いてまたは用いずに処置されたヒト赤骨髄芽球様細胞白血病K-562細胞の細胞生存率は、WST-1アッセイを使用して決定された(N=3、平均±SD、#p<0.01)。
【図14】フローサイトメトリーを使用することによる細胞周期分布を示す図である。テリマグランジンIIを用いてまたは用いずに24時間処置されたヒト赤骨髄芽球様細胞白血病K-562細胞の細胞周期研究はフローサイトメトリーを使用して決定された(N=3、平均±SD、*p<0.05)。
【図15】非処置細胞と比べた、テリマグランジンIIを用いて48時間処置されたヒト赤骨髄芽球様細胞白血病K-562細胞におけるDNA断片化の量を示す図である(N=3、平均±SD、#p<0.05)。
【図17】テリマグランジンIIを用いて2時間処置されたヒト赤骨髄芽球様細胞白血病K-562細胞のROS生成を示す図である(N=3、*p<0.05、#p<0.01)。
【図18】テリマグランジンIIを用いてまたは用いずに48時間処置されたヒト赤骨髄芽球様細胞白血病K-562細胞におけるウェスタン免疫ブロットによるカスパーゼ8、Bcl-2、カスパーゼ9、カスパーゼ3、カスパーゼ7、PARP、サイクリンB1およびCdc-25Cの発現を示す図である(N=3、タンパク質定量化はαアクチンに基づいていた)。
【発明を実施するための形態】
【0027】
したがって、本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、グラム陽性菌の増殖を抑制する方法を提供する。一実施形態では、方法は、有効量のβラクタム抗生物質またはポリケチド抗生物質を対象に投与するステップをさらに含む。別の実施形態では、細菌は黄色ブドウ球菌またはメチシリン耐性黄色ブドウ球菌である。
【0028】
一実施形態では、テリマグランジンIIはβラクタム抗生物質またはポリケチド抗生物質と相乗的に作用して、メチシリン耐性黄色ブドウ球菌の増殖を抑制する。別の実施形態では、βラクタム抗生物質には、ペニシリン、たとえば、ベンザチンペニシリン、ベンジルペニシリン、フェノキシメチルペニシリン、プロカインペニシリン、メチシリン、オキサシリン、ナフシリン、クロキサシリン、ジクロキサシリン、フルクロキサシリン、テモシリン、アモキシシリン、アンピシリン、コアモキシクラブ(co-amoxiclav)、アズロシリン、カルベニシリン、チカルシリン、メズロシリンおよびピペラシリン;セファロスポリン、たとえば、セファレキシン、セファロチン、セファゾリン、セファクロル、セフロキシム、セファマンドール、セフォテタン、セフォキシチン、セフトリアキソン、セフォタキシム、セフポドキシム、セフィキシム、セフタジジム、セフェピムおよびセフピロム;カルバペネムもしくはペネム、たとえば、イミペネム、メロペネム、パニペネム、ビアペネム、エルタペネム、ファロペネムおよびドリペネム;モノバクタム、たとえば、アズトレオナム、チゲモナム(tigemonam)、ノカルディシンAおよびタブトキシニン-βラクタム;またはクラブラン酸、タゾバクタムおよびスルバクタムなどのβラクタマーゼ阻害剤が挙げられるが、これらに限定されない。さらに別の実施形態では、ポリケチド抗生物質には、テトラサイクリン、ドキシサイクリン、ミノサイクリン、エリスロマイシン、モネンシンA(ルメンシン)、タイロシン、ナラシン、リファマイシン、セルビマイシンC、ジャドマイシンB、キロマイシンまたはムピロシンが挙げられるが、これらに限定されない。一実施形態では、βラクタム抗生物質は、メチシリン、オキサシリン、ナフシリン、クロキサシリン、ジクロキサシリン、フルクロキサシリン、テモシリンまたはアモキシシリンである。別の実施形態では、ポリケチド抗生物質は、テトラサイクリン、ドキシサイクリンまたはミノサイクリンである。さらに別の実施形態では、βラクタム抗生物質はオキサシリンであり、ポリケチド抗生物質はドキシサイクリンである。
【0029】
一実施形態では、テリマグランジンIIは、30μg/mLを超える濃度を有する。別の実施形態では、テリマグランジンIIは、40μg/mLを超える濃度を有する。一実施形態では、対象はヒトである。
【0030】
【表1】
【0031】
1つのMSSA株、3つのMRSA株(19615、18271および18631)ならびに1つの標準MRSA株(ATCC 33591)を使用した。試験された試薬は、オキサシリン、エリスロマイシン、ペニシリン、カナマイシン、レボフロキサシン、ドキシサイクリンおよびテリマグランジンIIである。
【0032】
【表2】
【0033】
テリマグランジンIIとの相乗効果が実証された抗生物質はオキサシリン、ドキシサイクリンおよびレボフロキサシンである。最小抑制濃度(MIC)と部分抑制濃度インデックス(FIC:fractional inhibitory concentration index)が計算された。相乗効果、相加効果、無差別効果およびアンタゴニスト効果のFIC値は、それぞれ、<0.5、0.5〜<1、1〜≦4、および4である。
【0034】
【表3】
【0035】
1つのMSSA株と3つのMRSA株(19615、18271および18631)が使用された。試験された抗生物質は、オキサシリン、アンピシリン、レボフロキサシン、エリスロマイシン、カナマイシン、ドキシサイクリンおよびバンコマイシンであった。最小抑制濃度(MIC)および部分抑制濃度インデックス(FIC)が計算された。相乗効果、相加効果、無差別効果およびアンタゴニスト効果のFIC値は、それぞれ、<0.5、0.5〜<1、1〜≦4、および4である。
【0036】
【表4】
【0037】
14の臨床的に単離されたMRSA株を、オキサシリンおよびテリマグランジンII(40μg/mL)を用いて試験し、FICインデックスを計算した。相乗効果、相加効果、無差別効果およびアンタゴニスト効果のFIC値は、それぞれ、<0.5、0.5〜<1、1〜≦4、および4である。
【0038】
一実施形態では、テリマグランジンIIとポリケチド抗生物質は抗酸化機構を通じて相乗的に作用する。
【0039】
一実施形態では、βラクタム抗生物質は、テリマグランジンIIと一緒に作用して、黄色ブドウ球菌においてペニシリン結合タンパク質2a発現を減少させる。別の実施形態では、βラクタム抗生物質は、テリマグランジンIIと一緒に作用して、黄色ブドウ球菌においてmecA遺伝子発現を抑制する。
【0040】
【表5】
【0041】
【表6】
【0042】
テリマグランジンIIとドキシサイクリンのMICは、FeCl2の存在下と比べた場合、FeCl2の非存在下のほうが高い(Table 6(表6))。これは、テリマグランジンIIとドキシサイクリンの抗MRSA活性がFeCl2の存在下では抑制されることを示唆している。
【0043】
一実施形態では、テリマグランジンIIは黄色ブドウ球菌のバイオフィルム形成を抑制する。
【0044】
本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、抗炎症の方法も提供する。一実施形態では、抗炎症の方法を使用して敗血症を治療する。
【0045】
一実施形態では、テリマグランジンIIは一酸化窒素およびプロスタグランジンE2産生を減少させる。別の実施形態では、テリマグランジンIIはヘムオキシゲナーゼ-1の産生を増加させる。
【0046】
一実施形態では、テリマグランジンIIは10〜50μMの範囲の濃度を有する。別の実施形態では、テリマグランジンIIは25〜50μMの範囲の濃度を有する。一実施形態では、対象はヒトである。
【0047】
本発明は、有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、白血病を治療する方法をさらに提供する。
【0048】
一実施形態では、テリマグランジンIIは、白血病細胞の細胞アポトーシスおよび細胞周期停止を誘導する。別の実施形態では、テリマグランジンIIは、細胞内活性酸素種を増加させることにより細胞アポトーシスを誘導する。さらに別の実施形態では、白血病細胞の細胞アポトーシスは、カスパーゼ3およびカスパーゼ7を活性化することにより誘導される。別の実施形態では、白血病細胞の細胞周期停止はG2/M期に停止される。
【0049】
一実施形態では、テリマグランジンIIは、20〜80μMの範囲の濃度を有する。別の実施形態では、テリマグランジンIIは、40〜80μMの範囲の濃度を有する。別の実施形態では、テリマグランジンIIは、50〜80μMの範囲の濃度を有する。一実施形態では、対象はヒトである。
【0050】
次の実施例は、以下の通り、本発明のいくつかの例となる実施形態を提供する。
【0051】
(実施例)
以下の実施例は非限定的であり、本発明の様々な態様および特徴の典型にすぎない。
【0052】
(実施例1)
細胞生存率アッセイ
細胞生存率はアラマーブルーを使用して測定された。細胞生存率アッセイでは、ヒト胚腎臓細胞系統HEK293およびヒト肝細胞腫細胞系統Hep G2を、96ウェルプレート上の各ウェルに180μLプレーティングした(HEK293では、約10,000細胞/ウェルの密度で;Hep G2では約7500細胞/ウェルの密度で)。細胞を一晩(16〜18時間)増殖させ、付着させた。一晩のインキュベーション後、20μLのテリマグランジンIIを添加して、0、20、30、40および50μg/mLの最終濃度を得た。0μg/mLのテリマグランジンIIと追加の薬物を添加しない(培地のみ)培養物は対照であった。次に、細胞を、5%CO2、37℃インキュベーター中24時間インキュベートした。24時間後、100μLのアラマーブルーが各ウェルに添加され、5%CO2、37℃インキュベーター中4時間培養された。570nmおよび600nmで吸光度がモニターされ、細胞生存率が決定された。
【0053】
図2は、ヒト胚腎臓細胞系統HEK293における細胞生存率に対するテリマグランジンIIの効果を示している。Y軸は生存細胞の割合を表し、X軸は対照と20、30、40および50μg/mLのテリマグランジンIIの濃度に対応する。
【0054】
X軸は、対照と20、30、40および50μg/mLのテリマグランジンIIの濃度に対応している。標準偏差は、それぞれ0、1.59、2.01、1.64および0.87と決定された。図3は、ヒト肝細胞腫細胞系統Hep G2における細胞生存率に対するテリマグランジンIIの効果を示した。Y軸は、生存細胞の割合を表し、X軸は対照と20、30、40および50μg/mLのテリマグランジンIIの濃度に対応する。標準偏差は、それぞれ3.480、3.95、4.83および5.73と決定された。図2および図3に示されるように、テリマグランジンIIは、ヒト胚腎臓細胞系統とヒト肝細胞腫細胞系統において低い毒性を示した。細胞生存率は、40μg/mLのテリマグランジンIIで処置された細胞では少なくとも80%維持された。
【0055】
最小抑制濃度、MIC
試験される抗生物質(オキサシリン、エリスロマイシン、アンピシリン、カナマイシン、レボフロキサシン、バンコマイシンおよびドキシサイクリン)の連続2倍希釈は微量液体希釈法(Broth microdilution method)により、最終濃度2〜512μg/mLまで実施された。細菌の一晩培養物を収集し、Muller-Hilton(M-H)ブロスに再懸濁した。1アリコートの細胞懸濁液(OD600=0.1)を調製した。細菌懸濁液を96ウェルプレートにプレーティングし、試験抗生物質と一緒に37℃、5%CO2インキュベーター中でインキュベートした(Table 1(表1)〜Table 4(表4))。
【0056】
部分抑制濃度インデックス(FICインデックス)
2つの薬物間の相乗作用、相乗作用と拮抗作用は、以下の式により評価された。
【0057】
【数1】
【0058】
【表7】
【0059】
Table 1(表1)〜Table 4(表4)に示されるように、テリマグランジンIIとオキサシリンまたはドキシサイクリンは相乗作用として定義され、そのFICインデックスは0.5未満であった。
【0060】
1,1-ジフェニル-2-ピクリルヒドラジル(DPPH)捕捉アッセイ
試験試薬、天然抽出物およびビタミンC(アスコルビン酸)はメタノールに溶解させ、10X保存液を作製した。ビタミンCは陽性対照として使用された。120μMのDPPH溶液を調製した。96ウェルマイクロプレート上に保存試薬、天然抽出物およびビタミンCを20μL取って、総容量100μLまでになるようにメタノールを添加した。対照は100μLメタノールであった。各試料を100μL DPPH溶液と混合させ、暗所で40分間インキュベートした。各ウェルのDPPHレベルは、517nmでの各ウェルの光学密度を検出することにより評価し、結果はTable 5(表5)に示した。
【0061】
殺菌曲線(Time-Killing Curve)
MRSA 19615細菌を一晩培養し、収集し、指示最終濃度のTG IIおよび抗生物質を含有する25mLのMHB中に再懸濁した。試料を希釈するために、指示された時間間隔で試料を懸濁液から取り、濾過されたPBSと混合し、続いて、2プレート/各時点でプレーティングして希釈試料をMHA上に広げた。150〜500コロニー間の最初の時点のコロニー数を使用して希釈倍率を決定した。殺菌曲線はコロニー計数法から計算され、データ解析は一晩培養後に実施された。細菌殺菌曲線は、時間に対する平均コロニー数(log10 cfu/mL)をプロットすることにより作成された(図4と図5)。図4が示すように、オキサシリン(5μg/mL)処置単独では、細菌増殖を抑制できなかった。オキサシリン(5μg/mL)とテリマグランジンII(40μg/mL)を一緒に使用すると、24時間後に細菌の目に見える増殖を抑制した。図5が示すように、テリマグランジンII/ドキシサイクリン組合せ(40μg/mLテリマグランジンIIと8μg/mLドキシサイクリン; 40μg/mLテリマグランジンIIと16μg/mLドキシサイクリン)では、相乗効果が観察された。したがって、40μg/mLテリマグランジンIIと16μg/mLドキシサイクリン組合せの殺菌効率のほうがはるかに強力であった。
【0062】
ウェスタンブロッティング分析
タンパク質試料(15μg)は5×試料緩衝液と混合し、1×試料緩衝液中で5分間煮沸し、次にタンパク質がその成分ポリペプチド鎖に変性されるように、急速冷却した。等量のタンパク質(15μg)を17mAで2時間、10%SDS-PAGE上で分離し、ウェット式転写手順を使用してPVDF膜へ移した。PVDF膜は、非特異的結合を防ぐために5%(w/v)脱脂粉乳を含有するt-BST緩衝液中、室温で30分間ブロックし、次に5%脱脂粉乳中、4℃で16〜18時間、一次抗体(ウサギ抗ペニシリン結合タンパク質、希釈度1:5000)を用いてプローブした。抗グロイエール(Groel)は内部標準として使用された(希釈度1:80,000; Sigmaより購入)。それぞれ5分間で3回t-BSTを用いて洗浄後、膜を、5%(w/v)脱脂粉乳中、西洋ワサビペルオキシダーゼ(HRP)-コンジュゲート二次抗体と一緒に室温で30分間インキュベートした。膜をそれぞれ5分間で3回洗浄し、続いて増強された化学発光システムを使用して、フィルムに曝露する。ウェスタンブロットの定量的結果は図6に示した。図6に示されるように、40μg/mLのテリマグランジンII単独で、細菌中においてペニシリン結合タンパク質2a(PBP2a)の発現を抑制することができた。
【0063】
RNA単離および定量化
RNA単離手順は、Roche High Pure RNA Isolation Kitにより実施された。手短に言えば、各処置された細菌試料は、13,000rpmで2分間の遠心分離により収集し、37℃で15分間20μMリゾスタフィン処置により溶解した。試料を、400μL溶解結合緩衝液混合試料を添加した後に高純度フィルター管に充填し、次に9,600gで1分間遠心分離した。通過画分は除かれフィルター管は収集管に入れた。90μLの混合前デオキシリボヌクレアーゼインキュベーション緩衝液と10μLのデオキシリボヌクレアーゼIをフィルター管に添加し、続いて1時間反応させた。洗浄緩衝液I、IIおよびIIIは連続してフィルター管に添加し、通過画分は、各ステップ間に遠心分離(9,600gで1分間)により取り除いた。最終RNA産物は、30μL溶出緩衝液により溶出し、清潔なエッペンドルフ内に収集した。
【0064】
RNA調製物は、Backman DU800を使用して定量した。2マイクロリットル(2μL)のRNA試料はPCRチューブに98μLの溶出緩衝液と一緒に添加し、よく混合した。RNA試料の濃度と純度は、Backman DU800分光光度計を使用して評価した(希釈係数は50である)。
【0065】
逆転写PCR
SuperScript III逆転写酵素(Invitrogen製)が本研究で使用された。2マイクログラム(2μg)の精製RNAを、PCR反応管中で1μLのオリゴdT、1μLのランダムプライマー(250ng)および1μLの10mM dNTP混合物(dATP、dGTP、dCTPおよびdTTP、それぞれ10mM)と混合させた。混合液を5分間で65℃まで加熱し、氷上で急速冷却させた。4マイクロリットル(4μL)の5×Fist-stand緩衝液、1μLのDTT(0.1M)および0.5μLのSuperScript III逆転写酵素を添加し、よく混合させた。混合液は25℃で5分間、55℃で60分間、および70℃で5分間インキュベートした。
【0066】
リアルタイムPCR
標的遺伝子の発現を調べるために、Roche LightCyclerシステムを使用してRT-PCRを実施した。2マイクログラム(2μg)のcDNAを、反応プレートにおいて4μM MgCl2、5μMプライマーおよび2μLの10×LightCycler FastStart DNA Master SYBR Green I混合液(Roche製)と混合させ、よく混合させた。増幅条件は以下の通りであった。変性は1サイクル95℃で0.5分間、95℃で15秒間、60℃で30秒間および72℃で30秒間であった。PCR実行後、追加の融解分析が実施され(0.1℃/秒の温度転移速度で60℃から95℃まで)、これにより増幅反応の特異性を確認した。RNA遺伝子発現の倍率変化は、式2-ΔΔCt、Ctはサイクル閾値により計算された。サイクル閾値(Ct)は、RT-PCRにおいて蛍光シグナルが閾値を超えるのに必要とされるサイクル数と定義される。ΔCtは、標的遺伝子のCt値から内在性対照のCt値を引くことにより計算された。次に、ΔΔCtは、試料のΔCtから対照のΔCtを引くことにより計算された。図7に示されるように、第1のバーは対照群を表し、第2のバーは2μg/mLのオキサシリンを有する試験群を表し、第3のバーは40μg/mLテリマグランジンIIと2μg/mLのオキサシリンを有する試験群を表す。左から右へ標準偏差(SD)は、0、0.37および0.37であり、*p<0.05。図7に示される結果では、2μg/mLのオキサシリンと40μg/mLテリマグランジンIIの組合せはmecA遺伝子発現を抑制した。
【0067】
【表8】
【0068】
透過型電子顕微鏡(TEM)
TEMにより観察する前に、細菌をプレート上で一晩培養し、収集し、10mL MHBまたは40μg/mLテリマグランジンIIを含有する10mL MHBのどちらかに懸濁した。細菌溶液は、600nm(OD600)で0.1の光学密度まで希釈し、10分間または24時間培養した。次に、細胞を10,000rpm/分で遠心分離し、固定液に懸濁し、固定時間は24時間を超えることはできない。TEM用の試料は、高雄医学大学病院(Kaohsiung、Taiwan、ROC)の実験医学臨床研究部に送られた。図8に示されるように、40μg/mLテリマグランジンIIを用いて処置し10分間培養した細菌試料は、対照群と比べると滑らかではないざらざらした表面を示した。表現型は、40μg/mLテリマグランジンIIを用いて処置され24時間培養された細菌試料との著しい相関を示した。
【0069】
バイオフィルム形成
細菌を寒天プレート上で一晩(16〜18時間)増殖した。次に、収集し、0.45%無菌PBS中に再懸濁し、無菌PBSに600nm(OD600)で0.1の最終光学密度まで希釈した。ポリスチレン96ウェルマイクロタイタープレートを、ウェルあたり150μLの培養液と20μLの細菌溶液で満たした。培養物を37℃で18時間静置させた。インキュベーション期間後、培養物を取り除き、マイクロタイタープレートウェルは300μLの無菌PBSを用いて3回穏やかに洗浄し、ゆるく会合した細菌を取り除き、次に60℃オーブンで1時間乾燥させた。試料は、各ウェルに2%色素溶液(150μL)を最初の播種レベルより上まで添加することにより染色し、15分間インキュベートした。次にプレートを新鮮な水で洗浄した。染色強度は、各乾燥ウェルに95%アルコール(150μL)を添加後に測定された。試料を室温で30分間インキュベートし、その後OD517値をプレートリーダー上で測定した。
【0070】
【表9】
【0071】
光学密度カットオフ値(ODc)=陰性対照の平均OD+3×陰性対照の標準偏差(SD)(バイオフィルム生成の解釈は、Srdjanら(APMIS 115: 891〜9頁、2007)の基準に従って実行された)。MRSA17199は対照株として使用された。
バイオフィルム生成の解釈:
平均OD値 バイオフィルム生成
OD≦ODc なし
ODc<OD≦2×ODc 弱
2×ODc<OD≦4×ODc 中
4×ODc<OD 強
【0072】
(実施例2)
以下の研究ではマウスマクロファージ細胞系統RAW264.7が使用された。
【0073】
細胞生存率アッセイ
アラマーブルー試薬を使用して細胞生存率を評価した。RAW264.7細胞(200μLの培地中2×104)を96ウェルプレート上に播種した。一晩のインキュベーション後、細胞を2%ウシ胎児血清(FBS)を含有するDMEM培地中で3時間血清飢餓状態にした。いくつかの群があり、すなわちDMSO処置もテリマグランジンII処置もない細胞(対照); DMSOを用いて処置される細胞(対照);異なる濃度(10、25、50、75および100μM、DMSO中に希釈される)のテリマグランジンIIを用いて処置される細胞(試験群)であった。各群は2通りに決定された。
【0074】
24時間後、細胞培地を取り除き破棄し、細胞を180μLの新鮮な培養液で再懸濁した。各細胞懸濁液に20μLアラマーブルーを添加した。次に、プレートをさらに4時間インキュベーター内に置いた。インキュベーション後、培養液の吸光度を570と600nmで測定し、細胞生存率を、以下の式を使用して解析した。
【0075】
【数2】
【0076】
O1: 570nmでの酸化アラマーブルーのモル吸光係数(E)
O2: 600nmでの酸化アラマーブルーのモル吸光係数(E)
A1: 570nmでの試験ウェルの吸光度
A2: 600nmでの試験ウェルの吸光度
P1: 570nmでの陽性増殖対照ウェルの吸光度
P2: 600nmでの陽性増殖対照ウェルの吸光度
【0077】
図9では、x軸はテリマグランジンIIの濃度を表し、y軸は細胞生存率を表す。図9の結果により、細胞生存率は、濃度50μg/mLまたはそれよりも低い用量でテリマグランジンIIを用いて処置された場合、著しい影響を受けないことが示された。
【0078】
一酸化窒素生成解析
マウスマクロファージ細胞を、約4×105 RAW264.7細胞/mLの密度で12ウェルプレートの各ウェルにプレーティングした。一晩のインキュベーション後、細胞を2%ウシ胎児血清(FBS)を含有するDMEM培地中で3時間血清飢餓状態にした。リポ多糖(LPS)を用いて24時間刺激された細胞は、培養液中の亜硝酸レベルを著しく増加させた。内在性一酸化窒素生成はLPSにより誘導した。細胞を、LPSを用いて処置される前および後に様々な濃度(10、25および50μM)のテリマグランジンIIで処置した。一酸化窒素生成解析では、培養上清を96ウェルプレートに運び、グリース試薬と混合した。プレートをインキュベートし、次に吸光度を520nmで測定した。DMEMで調製された亜硝酸ナトリウム(NaNO2)溶液(0M〜0.1M、8ポイント)を使用して標準曲線を作成した。試料の亜硝酸濃度は標準曲線の内挿により決定した。図10に示されるように、テリマグランジンIIの導入(LPSを用いて処置される前または後のいずれでも)により、LPS誘導一酸化窒素の生成は減少した。試験群のテリマグランジンII濃度(25および50μM)は統計的有意を示した。図10は、相対的順序でLPSまたはテリマグランジンII処置および濃度を標識されたx軸ならびに一酸化窒素濃度を標識されたy軸により枠組みされている。対照(LPSのみ)および試験群はt検定を使用して比較した、*p<0.05(有意)および#p<0.01(高度に有意)。
【0079】
プロスタグランジンE2(PGE2)アッセイ
1ミリリットル(1mL)のマウスマクロファージ細胞系統RAW264.7(4×105細胞/ウェル)を12ウェルプレート上に一晩播種した。一晩のインキュベーション後、細胞を2%ウシ胎児血清(FBS)を含有するDMEM培地中で3時間血清飢餓状態にした。次に、テリマグランジンIIが、最終濃度の10、25および50μMが得られるように添加された。このようにして、30分のインキュベーション後、マウスマクロファージ細胞をLPSを用いて16時間処置し、細胞培養物上清を収集した。無細胞上清を希釈し、培養上清中のPGE2はPGE2アッセイキット(R&D system製)を使用することにより検出した。本研究の目的は、テリマグランジンIIを用いて共処置することによりLPS誘導PGE2発現を評価することであった。図11に示されるように、25または50μMのテリマグランジンIIを、LPSで処置する30分前に細胞に添加すると、PGE2の発現は著しく減少した(実験結果はLPS群と比較し、0.05未満のP値は有意と解釈した)。
【0080】
ヘムオキシゲナーゼ-1(HO-1)のウェスタンブロット分析
マウスマクロファージRAW264.7細胞を、播種密度5×106細胞/mLで6cm細胞皿にプレーティングし、一晩インキュベートした。一晩のインキュベーション後、細胞を異なった濃度のテリマグランジンII(10、25および50μM)およびLPSを用いて処置し、24時間インキュベートした。細胞はPBSを用いて2回洗浄し、こすり取り、氷上でRIPA緩衝液に溶解し、タンパク質を得た。ヘムオキシゲナーゼ-1発現は全細胞溶解物のウェスタンブロッティングにより分析した。ウェスタンブロットデータおよび定量的解析の結果は図12に示されている。図12では、第1のバーは対照群を表し、第2のバーはLPSのみを用いた試験群を表し、第3、第4および第5のバーは10、25および50μg/mLテリマグランジンIIを用いた試験群を表している(N=3、平均±SD)。図12に示されるように、ヘムオキシゲナーゼ-1の発現は、LPS刺激細胞上でわずかに増加した。テリマグランジンII処置(25または50μM)処置により、ヘムオキシゲナーゼ-1の発現に著しい増加が観察された。
【0081】
(実施例3)
細胞生存率アッセイ
定量的細胞生存率はWST-1アッセイを使用して評価された。WST-1は、代謝活性細胞上で高度に水溶性のホルマザンを生成し、細胞生存率の直接比色測定を可能にする試薬である。
【0082】
K-562細胞を、播種密度6,000細胞/ウェル(100μL)で96ウェル培養プレートにプレーティングし、37℃で5%CO2を用い16〜18時間インキュベートした。細胞を、最終濃度を0、20、40、60および80μMにするように20μLテリマグランジンIIを用いて処置して、37℃で5%CO2を用い24時間および48時間インキュベートした。テリマグランジンII処置のない細胞は対照として使用された。プレートの各ウェルに12μLのWST-1試薬(10:1)を添加し、よく混合し、次に37℃で5%CO2インキュベーターにおいて2時間インキュベートした。インキュベーション後、450nm(試験波長)および630nm(基準波長)で吸光度を測定した。データは450nm測定値から630nmバックグラウンド吸光度を引くことにより計算された。
【0083】
結果は図13に示されており、異なる濃度で24時間または48時間テリマグランジンIIを用いて処置されたK-562細胞の細胞生存率は、WST-1アッセイを使用して測定された。細胞生存は、対照培養物増殖(100%)に対して存在する生細胞の割合として表された。IC50値は、曲線から外挿された。24時間試験群のIC50の値は57.01μMであり、48時間試験群のIC50の値は48.11μMであった。濃度40μMまたは50μMのテリマグランジンIIを使用したフォローアップ実験は、この結果に基づいていた(N=3、平均±SD、*p<0.05)。
【0084】
図13は、WST-1アッセイを使用して測定された定量的細胞生存率の結果を示していた。テリマグランジンIIを用いて処置されたまたは処置されないヒト赤骨髄芽球様細胞白血病K-562細胞の細胞生存率は、WST-1アッセイを使用して決定された(N=3、平均±SD、#p<0.01)。
【0085】
細胞周期解析
K-562細胞を、密度5×105細胞/mL(15mL)で15cm皿にプレーティングし、37℃で5%CO2を用い16〜18時間インキュベートした。細胞を収集し、室温、700rpmで3分間遠心分離し、0.04%ウシ胎児血清および8mMのLグルタミンを含有する培養液に再懸濁した。次に、細胞を37℃で5%CO2を用い16〜18時間インキュベートした。
【0086】
上記細胞を収集し、開始密度5×105細胞/mL(3mL)で3cm皿にプレーティングした。細胞を、最終濃度0、10、20、30、40および50μMまでテリマグランジンIIを用いて処置し、37℃で5%CO2を用いて12時間および24時間インキュベートした。
【0087】
細胞を4℃、700rpmの速度で5分間遠心分離し、PBSを用いて1回洗浄した。細胞を、70%エタノールの滴下添加によって細胞懸濁液に固定した。16〜18時間後、エタノール懸濁細胞を4℃、3,500rpm/分の速度で5分間遠心分離し、エタノールを完全にデカントした。細胞を、リボヌクレアーゼA(10μg/mL)および1% Triton X-100を含有するPBS中に再懸濁し、37℃で5%CO2インキュベーターにおいて30分〜1時間インキュベートした。ヨウ化プロピジウム(PI)染色では、細胞は室温、3,500rpmで5分間遠心分離し、600μLのPBSに再懸濁した。25μLのPI(20μg/mL)溶液を混合物に添加し、室温で暗所に1時間インキュベートした。ヨウ化プロピジウム染色細胞のDNA含量分析はフローサイトメーターにおいて実施された。フローサイトメトリーによる細胞周期解析は、図14に示される通りであった。結果により、サブG1およびG2/M期細胞の割合はテリマグランジンII処置において用量依存的に増加することが示された(N=3、平均±SD、*p<0.05)。
【0088】
アポトーシス検出試験
TUNEL試験を使用して、細胞がアポトーシス過程に入っているかどうかを判定するためにDNA断片化を検出した。
【0089】
1mLあたり1×106 K-562細胞の濃度の合計で10mLの細胞溶液を、細胞が環境に順応できるように、5%CO2を供給されたインキュベーターにおいて37℃で30分間10cm細胞培養皿を用いて培養した。異なる濃度のテリマグランジンII溶液を、最終濃度がそれぞれ0、10、20、30、40および50μMになるように順に添加した。溶液は、37℃、5%CO2を用いたインキュベーター中に48時間置き、その後4℃、3,500rpmで5分間遠心分離した。上清を取り除いた後、細胞をPBSにより1回すすいだ。合計で1mLの1%Trioxane(PBSを用いて10%Trioxane溶液から希釈された)を添加し、溶液を室温で30分間〜1時間反応させておいた。次に、溶液を4℃、3,500rpmで5分間遠心分離した。上清を取り除いた後、70%エタノールを添加し、溶液を細胞固定のために16〜18時間-20℃に置いた。
【0090】
細胞固定後、溶液を4℃、400rpmで10分間遠心分離し、その後上清を取り除いた。TUNEL緩衝液80μLを添加し、溶液を室温で5分間反応させておいた。次に、EDTA 1mLを溶液に添加し、その後これを穏やかに混合した。溶液は4℃、3,500rpmで5分間遠心分離し、その後上清を取り除いた。次に、TUNEL試験のために50μLの蛍光検出試薬を溶液中に添加し、その後よく混合させ、37℃で1時間、暗所でインキュベートした。溶液は4℃、3,500rpmで5分間遠心分離し、PBSを使用して細胞を1回すすいだ。最後に、1mLのPBSを使用して細胞を再懸濁させ、その後BD FACScanを使用して蛍光シグナルを検出し、定量結果は図15である。TUNEL試験の定量図(図15)から、蛍光シグナルはK-562細胞が30μMのテリマグランジンIIと反応した48時間後に検出されたことが分かる。さらに、シグナル変化の変動は、濃度が50μMまで増加した場合のほうが顕著であった(N=3、平均±SD、*p<0.05)。
【0091】
DNAラダー試験
テリマグランジンIIを用いて処置されたK-562細胞を、DNA断片化を同定するため、電気泳動のために収集した。
【0092】
1mLあたり1×106 K-562細胞の濃度の合計で3mLの細胞溶液を、細胞が環境に順応できるように、37℃で30分間、5%CO2を供給されたインキュベーターにおいて6cm細胞培養プレートを用いて培養した。異なる濃度のテリマグランジンII溶液を、最終濃度がそれぞれ0、20、30、40、50および60μMになるように順に添加した。溶液を、37℃、5%CO2を用いたインキュベーター中に48時間置き、その後4℃、2,500rpmで5分間遠心分離した。上清を取り除いた後、細胞をPBSにより1回すすぐ。次に、細胞溶解緩衝液100μL(20μM EDTAおよび50μM Tris-HClに溶解した1% NP-40、pH8.0)を添加し、溶液を高速で10秒間浸盪させた。
【0093】
溶液は3,500rpmで5分間遠心分離し、その後90μLの上清を収集し、SDS溶液10μLとよく混合させた。リボヌクレアーゼA(4mg/mL)50μLを混合溶液に添加し、次に56℃の水浴中に1時間置いてRNAを取り除いた。次に、プロテイナーゼK(10mg/mL)30μLを添加して、その後さらに1時間56℃の水浴においてタンパク質を取り除いた。全溶液容積の半分の10M酢酸アンモニウム(90μL)を添加しよく混合させて、その後全容積の2倍の無水エタノール(540μL)を添加してよく混合させた。溶液を、DNA沈殿のために-80℃下に置いた。溶液は低温で30分間、12,000rpmで遠心分離し、その後DNA沈殿物の消失を避けるために上清を慎重に取り除いた。DNA沈殿物は70%エタノール200μLにより1回すすぎ、その後全上清を取り除いて、チューブキャップを開けて室温で乾燥させた。脱イオン水50μLまたは0.5×TBE緩衝液を添加して、DNAを完全に分解した。最後に、DNAゲル電気泳動を上述のDNA試料について実施した。図16のDNAラダー電気泳動図は、TUNEL試験の結果を裏付けた。異なる濃度のテリマグランジンIIを用いて処置した48時間後にK-562細胞においてDNA断片化が起きた。
【0094】
ROS試験
ラジカル特異的蛍光色素、2',7'-ジクロロフルオレセイン(DCF-DA)を使用して、それぞれ細胞中のH2O2/NOベースのラジカルまたはO2ベースのラジカルを検出した。
【0095】
1mLあたり1×106 K-562細胞の濃度の合計3mLの細胞溶液を、細胞が環境に順応できるように、37℃で30分間、5%CO2を供給されたインキュベーターにおいて、6cm細胞培養プレートを用いて培養した。異なる濃度のテリマグランジンII溶液を、最終濃度がそれぞれ0、30、40および50μMになるように順に添加した。溶液を、37℃、5%CO2を用いたインキュベーター中に2時間置き、その後4℃、3,500rpmで5分間遠心分離し、次にPBSにより1回すすいだ。2',7'-ジクロロフルオレセイン(DCF-DA)を添加して、その後溶液を暗所インキュベーターに45分間置き、次にPBSにより1回すすいだ。最後に、1mLのPBSを使用して細胞を再懸濁させ、その後BD FACScanを使用して蛍光シグナルを検出した。図17に示される結果から、K-562細胞中のROSは、ROS試験において40μMテリマグランジンIIとの相互作用の2時間後にBD FACScanにより増加しているのが観察された。その上、細胞がテリマグランジンII 50μLと2時間相互作用する前に抗酸化剤、N-アセチル-L-システイン10μMを用いて前処理された場合には、ROS生成の抑制が観察された(N=3、*p<0.05、#p<0.01)。
【0096】
ウェスタンブロッティング
1mLあたり1×106 K-562細胞の濃度の合計3mLの細胞溶液を、細胞が環境に順応できるように、37℃で30分間、5%CO2を供給されたインキュベーターにおいて、6cm細胞培養皿を用いて培養した。異なる濃度のテリマグランジンII溶液を、最終濃度がそれぞれ0、40および50μMになるように順に添加した。溶液を、37℃、5%CO2を用いたインキュベーター中に48時間置いた。インキュベーション後、溶液を4℃、1,000rpmで5分間遠心分離し、PBSにより1回すすいだ。プロテアーゼ阻害剤を含有するRIPA緩衝液を添加して細胞を分解した。
【0097】
溶液は、細胞が確実に物理的損傷により分解されているように、3回液体窒素を用いて急速凍結させ、解凍した。溶液を4℃、30分間15,000rpmで遠心分離し、全細胞タンパク質を含有する上清を収集した。タンパク質濃度はBCAタンパク質アッセイにより決定された。最後に、等量のタンパク質試料(40μg)は1×添加液中で3分間煮沸し、さらにタンパク質ゲル電気泳動にかけるためにタンパク質は一次構造まで分解した(または-80℃で保存した)。
【0098】
SDS-PAGE 10%または12%が、タンパク質ゲル電気泳動(150V、17mA、2時間5分)において目的のタンパク質分子量により選択された。タンパク質は、ウェット式転写手順(100V、300mA、30分間)を使用してゲルからPVDF膜に移した。膜は、非特異的結合を除去するために2%ウシ胎児血清(BSA)を含有するトリス緩衝生理食塩水-Tween(TBS-T)緩衝液中室温で1時間ブロックした。
【0099】
PVDF膜は、2%BSA-TBS-T緩衝液中4℃で16〜18時間、製造業者に従った推奨される量で希釈された一次抗体を用いてプローブした。相互作用の完了後、TBS-T緩衝液を使用して、非特異的に結合した一次抗体をすすぎ落した。TBS-T緩衝液は、約30分間のうちに10分ごとに3回交換した。次に、特異的二次抗体を添加して、室温で1時間相互作用させた。TBS-T緩衝液を使用して、非特異的に結合した二次抗体をすすぎ落した。TBS-T緩衝液は約60分間のうちに10分ごとに6回交換された。ECLシステム(増強された化学発光システム)を使用し、フィルムを光に曝露させて現像した。最後に定量的解析が実施された。図18は、ウェスタンブロッティングの結果から、先ず、おそらく切断されたカスパーゼ8に活性化されて、外部死滅経路におけるカスパーゼ8の不活性が減少することを示した。ROS増加という前の結果に示されるように、この結果も、抗アポトーシスのタンパク質、Bcl-2が減少して、カスパーゼ9、カスパーゼ3およびカスパーゼ7の活性化およびPARPの究極の活性化をさらに誘導することを示した。細胞周期調節に関連するタンパク質には、G2/Mチェックポイント調節因子、サイクリンB1およびCdc-25Cが挙げられる。上の結果により、(非リン酸化)活性はテリマグランジンIIとの相互作用後に減少し、細胞周期の停滞を裏付けていることも見出された(N=3、タンパク質定量化はαアクチンに基づいていた)。
【0100】
統計的解析
スチューデントt検定(不対の)を実施した。結果は、標準誤差(平均±S.D.)付きの少なくとも3つの個々の実験の平均として表されており、P値≦0.01または≦0.05を有意と見なした。
【0101】
当業者であれば、本発明が、目標を遂行するため、および言及された目的と利点に加えてその固有の目的と利点を得るために十分適合していることを容易に認識する。動物、プロセス、およびそのプロセスを生み出すための方法は、好ましい実施形態の典型であり、例となるものであり、本発明の範囲を限定するものとして意図されてはいない。その改変および他の使用を当業者であれば思い付くであろう。これらの改変は本発明の精神の範囲内に包含されており、特許請求の範囲により定義されている。
【特許請求の範囲】
【請求項1】
有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、グラム陽性菌の増殖を抑制する方法。
【請求項2】
有効量のβラクタム抗生物質またはポリケチド抗生物質を前記対象に投与するステップをさらに含む、請求項1に記載の方法。
【請求項3】
前記細菌が黄色ブドウ球菌またはメチシリン耐性黄色ブドウ球菌である、請求項1に記載の方法。
【請求項4】
テリマグランジンIIがβラクタム抗生物質またはポリケチド抗生物質と相乗的に作用して、メチシリン耐性黄色ブドウ球菌の増殖を抑制する、請求項1に記載の方法。
【請求項5】
βラクタム抗生物質が、メチシリン、オキサシリン、ナフシリン、クロキサシリン、ジクロキサシリン、フルクロキサシリン、テモシリンまたはアモキシシリンを含む、請求項2に記載の方法。
【請求項6】
ポリケチド抗生物質が、テトラサイクリン、ドキシサイクリンまたはミノサイクリンを含む、請求項2に記載の方法。
【請求項7】
テリマグランジンIIが30μg/mLを超える濃度を有する、請求項1に記載の方法。
【請求項8】
テリマグランジンIIとポリケチド抗生物質が抗酸化機構を通じて相乗的に作用する、請求項4に記載の方法。
【請求項9】
βラクタム抗生物質が、テリマグランジンIIと一緒に作用して、黄色ブドウ球菌においてペニシリン結合タンパク質2a発現を減少させる、請求項2に記載の方法。
【請求項10】
βラクタム抗生物質が、テリマグランジンIIと一緒に作用して、黄色ブドウ球菌においてmecA遺伝子発現を抑制する、請求項2に記載の方法。
【請求項11】
テリマグランジンIIが黄色ブドウ球菌のバイオフィルム形成を抑制する、請求項1に記載の方法。
【請求項12】
有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、抗炎症の方法。
【請求項13】
抗炎症が敗血症治療を含む、請求項12に記載の方法。
【請求項14】
テリマグランジンIIが一酸化窒素およびプロスタグランジンE2産生を減少させる、請求項13に記載の方法。
【請求項15】
テリマグランジンIIがヘムオキシゲナーゼ1の産生を増加させる、請求項13に記載の方法。
【請求項16】
テリマグランジンIIが10〜50μMの範囲の濃度を有する、請求項13に記載の方法。
【請求項17】
有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、白血病を治療する方法。
【請求項18】
テリマグランジンIIが、白血病細胞の細胞アポトーシスおよび細胞周期停止を誘導する、請求項17に記載の方法。
【請求項19】
テリマグランジンIIが20〜80μMの範囲の濃度を有する、請求項17に記載の方法。
【請求項20】
テリマグランジンIIが、細胞内活性酸素種を増加させることにより細胞アポトーシスを誘導する、請求項18に記載の方法。
【請求項21】
白血病細胞の細胞アポトーシスが、カスパーゼ3およびカスパーゼ7を活性化することにより誘導される、請求項18に記載の方法。
【請求項22】
白血病細胞の細胞周期停止がG2/M期に停止される、請求項18に記載の方法。
【請求項1】
有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、グラム陽性菌の増殖を抑制する方法。
【請求項2】
有効量のβラクタム抗生物質またはポリケチド抗生物質を前記対象に投与するステップをさらに含む、請求項1に記載の方法。
【請求項3】
前記細菌が黄色ブドウ球菌またはメチシリン耐性黄色ブドウ球菌である、請求項1に記載の方法。
【請求項4】
テリマグランジンIIがβラクタム抗生物質またはポリケチド抗生物質と相乗的に作用して、メチシリン耐性黄色ブドウ球菌の増殖を抑制する、請求項1に記載の方法。
【請求項5】
βラクタム抗生物質が、メチシリン、オキサシリン、ナフシリン、クロキサシリン、ジクロキサシリン、フルクロキサシリン、テモシリンまたはアモキシシリンを含む、請求項2に記載の方法。
【請求項6】
ポリケチド抗生物質が、テトラサイクリン、ドキシサイクリンまたはミノサイクリンを含む、請求項2に記載の方法。
【請求項7】
テリマグランジンIIが30μg/mLを超える濃度を有する、請求項1に記載の方法。
【請求項8】
テリマグランジンIIとポリケチド抗生物質が抗酸化機構を通じて相乗的に作用する、請求項4に記載の方法。
【請求項9】
βラクタム抗生物質が、テリマグランジンIIと一緒に作用して、黄色ブドウ球菌においてペニシリン結合タンパク質2a発現を減少させる、請求項2に記載の方法。
【請求項10】
βラクタム抗生物質が、テリマグランジンIIと一緒に作用して、黄色ブドウ球菌においてmecA遺伝子発現を抑制する、請求項2に記載の方法。
【請求項11】
テリマグランジンIIが黄色ブドウ球菌のバイオフィルム形成を抑制する、請求項1に記載の方法。
【請求項12】
有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、抗炎症の方法。
【請求項13】
抗炎症が敗血症治療を含む、請求項12に記載の方法。
【請求項14】
テリマグランジンIIが一酸化窒素およびプロスタグランジンE2産生を減少させる、請求項13に記載の方法。
【請求項15】
テリマグランジンIIがヘムオキシゲナーゼ1の産生を増加させる、請求項13に記載の方法。
【請求項16】
テリマグランジンIIが10〜50μMの範囲の濃度を有する、請求項13に記載の方法。
【請求項17】
有効量のテリマグランジンII、その薬学的に許容される塩、鏡像異性体、異性体または互換異性体を対象に投与するステップを含む、白血病を治療する方法。
【請求項18】
テリマグランジンIIが、白血病細胞の細胞アポトーシスおよび細胞周期停止を誘導する、請求項17に記載の方法。
【請求項19】
テリマグランジンIIが20〜80μMの範囲の濃度を有する、請求項17に記載の方法。
【請求項20】
テリマグランジンIIが、細胞内活性酸素種を増加させることにより細胞アポトーシスを誘導する、請求項18に記載の方法。
【請求項21】
白血病細胞の細胞アポトーシスが、カスパーゼ3およびカスパーゼ7を活性化することにより誘導される、請求項18に記載の方法。
【請求項22】
白血病細胞の細胞周期停止がG2/M期に停止される、請求項18に記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公開番号】特開2012−153687(P2012−153687A)
【公開日】平成24年8月16日(2012.8.16)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−12792(P2012−12792)
【出願日】平成24年1月25日(2012.1.25)
【出願人】(510137489)カオシュン メディカル ユニヴァーシティ (2)
【Fターム(参考)】
【公開日】平成24年8月16日(2012.8.16)
【国際特許分類】
【出願番号】特願2012−12792(P2012−12792)
【出願日】平成24年1月25日(2012.1.25)
【出願人】(510137489)カオシュン メディカル ユニヴァーシティ (2)
【Fターム(参考)】
[ Back to top ]