
抗肥満用免疫原性ハイブリッドポリペプチド及びこれを含む抗肥満ワクチン組成物
アポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドとヘルパーT細胞エピトープを含み、前記ミメティックペプチドのC末端がヘルパーT細胞エピトープのN末端と融合した免疫原ハイブリッドポリペプチド、及びこれを含む肥満予防または治療用ワクチン組成物を開示する。

【発明の詳細な説明】
【発明の詳細な説明】
【0001】
〔技術分野〕
本発明は、アポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドとヘルパーT細胞エピトープを含み、前記ミメティックペプチドのC末端がヘルパーT細胞エピトープ部分のN末端と融合した免疫原性ハイブリッドポリペプチド、及びこれを含む肥満予防または治療用ワクチン組成物に関する。
【0002】
〔背景技術〕
最近、韓国では、西欧的食生活の影響で動脈硬化症及び冠状動脈疾患(coronary atherosclerotic disease:CAD)が益々増加しつつあり、これによる死亡率も高くなってきている。この種の疾患の原因となる血清脂質としては、コレステロール(cholesterol)、トリグリセリド(triglyceride:TG)、遊離脂肪酸(free fatty acid)、リン脂質(phospholipid)などがあり、アポリポ蛋白質(apolipoprotein)と共にリポ蛋白質を形成して血液循環によって運搬されている。中でも、LDL(low density lipoprotein)は主にTGやコレステロールの運搬を担当しており、LDL−コレステロール数値の変化は前述したような疾病の予後を示す尺度となる。
【0003】
脂質代謝に関連した成人病の主要因子であるLDL−コレステロールは、各組織の細胞膜のLDL受容体に結合して組織内に蓄積、活用されるか、スカベンジャー細胞により捕獲されてから加水分解されて遊離コレステロールの形でアポEリポ蛋白質と共にHDLへ伝達されて肝で再活用されるか、胆汁酸の形に処理されて排泄される経路が明らかになっている。この過程において、アポリポ蛋白質は、リポ蛋白質の構造的恒常性を保ち、脂肪質分解酵素の補助因子機能、及び細胞膜上の特定の受容体と結合する非常に重要な機能を行っている。
【0004】
アポリポ蛋白質B−100(Apo B−100)は、このような低密度リポ蛋白質(LDL)の主な蛋白質成分であり、またIDL、VLDL、キロミクロン(chylomicron)に存在しているため、血中にある抗体がApo B−100を認識するように誘導すると、食細胞によるLDL−消去(clearance)が容易に起こり得るであろう。これを根拠とし、最近、ワクチンを用いてLDL−コレステロールの数値を低め且つ動脈硬化を減少させるための研究が行われている。このような抗−コレステロールワクチン療法により誘導された抗体はIgMタイプであり、VLDL、IDL及びLDLと結合するものと思われるが、このことから高コレステロール及びアテローム性動脈硬化症に対する予防及び治療用ワクチンの可能性が提示されている(Bailey, et al., Cholesterol vaccines. Science 264, 1067-1068, 1994; Palinski W et al., Proc Natl Acad Sci U S A. 92, 821-5, 1995; Wu R. de Faire U et al., Hypertension. 33, 53-9, 1999)。また、アポリポ蛋白質B−100は、4560個のアミノ酸残基(24個のシグナルペプチドを含む)からなる巨大な蛋白質分子であって、分子量は500kDaを超過する(Elovson J et al., Biochemistry, 24: 1569-1578, 1985)。主に肝から分泌される両親媒性の分子なので、リポ蛋白質のリピドとの相互作用が可能であり、水溶液の環境とも相互作用が可能である(Segrest J. P et al., Adv. Protein Chem., 45: 303-369, 1994)。アポリポ蛋白質B−100は、LDL粒子の大きさと構造を安定化させる役割と、収容基との結合によって血漿内LDLコレステロールの恒常性を調節するのに重要な役割をする(Brown MS et al., Science, 232 : 34-47, 1986)。
【0005】
本願の発明者により出願された韓国特許公開第2002−0018971号では、Apo B−100のエピトープに対するミメティックペプチド(mimetic peptide)が抗肥満効果を持つと記述している。ところが、前記出願では、単に、前述したB細胞のエピトープであるミメティックペプチドが抗肥満効果を示すことを開示している。
【0006】
本発明の前には、アポリポ蛋白質のB細胞エピトープとT細胞エピトープとを融合させて免疫原性を増強させようとしたことがなく、免疫反応を増強させるために蛋白質キャリアまたは補助剤として使用した試みのみがあった。
【0007】
米国特許第5,843,446号では、LHRHの免疫原性を増強させるために、LHRHにこれとは異なる蛋白質を結合させることができるが、この場合、主要免疫反応がLHRHに対して誘導されるよりは、結合したキャリア蛋白質に対して誘導され、却ってLHRHに対する免疫抑制が誘導されるおそれがあるという問題について記述している。これから分かるように、B細胞エピトープの免疫原性を増強させるための追加物質の選別、結合方式、結合位置などについての研究開発は、持続的な努力が必要である。
【0008】
ハプテンの免疫原性を高めるために媒体蛋白質と融合させようとする試みがたくさんあったが、一律的な相乗効果を得ることはできなかった。特に、本発明のようなB細胞エピトープとT細胞エピトープとの線形連結の際に、これらの方向性、それぞれのエピトープの種類などによって寧ろ免疫原性が消失してしまう場合もあり(Francis, M. J. et al., Nature 330: 168-170, 1987)、リンカーの存在により抗原性が減少する場合も発生した(Partidos, C. et al., Mol. Immunol. 29:651-658, 1992)。このようにペプチドワクチンのデザインに一貫的に適用することが可能な様態が存在せず、デザインされたワクチンの効能も予測することができない。同一の理由から、疎水性が非常に強いPB14ペプチドを、異種のペプチドであるT細胞エピトープと融合させる場合、抗原性を示す部位を分子の内部に取り入れて抗体誘導能の低下をもたらす可能性も排除することができない。
【0009】
このような背景の下で、本発明者らは、抗肥満効果を示すアポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドであるPB14を増強させるために様々な試みを重ねた結果、ミメティックエピトープのC末端にヘルパーT細胞のエピトープのN末端を融合させたハイブリッドポリペプチドが優れた免疫増強効果を示し、肥満の予防または治療に効果的に使用できることを確認した。このようなハイブリッドポリペプチドが、アポリポ蛋白質のB細胞エピトープの有利な活性または効果を中和させる免疫反応を誘導するか有害な副作用を起すことなく、優れた抗肥満活性を示すことができるということは、全く類推できない効果であった。
【0010】
〔発明の開示〕
一つの様態として、本発明は、アポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドのアミノ酸配列を含み、前記ミメティックペプチドのC末端がヘルパーT細胞エピトープのN末端と融合した免疫原性ハイブリッドポリペプチドを提供する。
【0011】
他の様態として、本発明は、アポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドのアミノ酸配列を含み、前記ミメティックペプチドのC末端がヘルパーT細胞エピトープのN末端と融合した免疫原性ハイブリッドポリペプチドを含む肥満予防または治療用ワクチンを提供する。
【0012】
別の様態として、本発明は、前述した免疫原性ハイブリッドポリペプチドをコードする遺伝子を含む組み換えベクター、前記組み換えベクターを含む形質転換体、及び前記組み換えベクターで形質転換された宿主細胞を培養して前記ハイブリッドポリペプチドを生産する方法を提供する。
【0013】
〔図面の簡単な説明〕
本発明の前記および他の目的、特徴および利点は、添付図面を参照する以降の詳細な説明からより明らかになるであろう。
【0014】
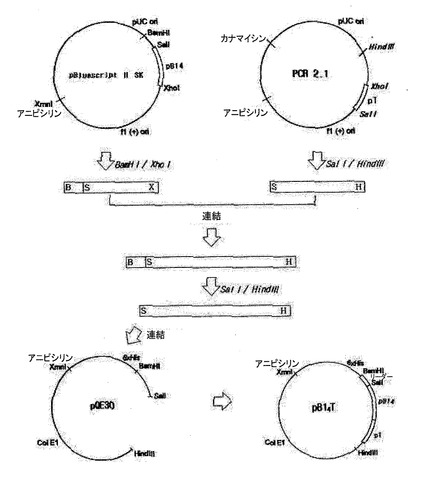
図1はpB14Tの製作図を示す。
【0015】
図2はpB14Tの制限酵素切断を示す。
【0016】
図3はDNA配列から類推したpB14Tの一次構造を示す。
【0017】
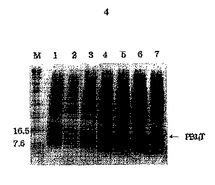
図4は形質転換された大腸菌(Escherichia coli)M15/pB14Tを、IPTGでpB14Tの発現を誘導した後、SDS−PAGE分析した結果である。発現された組み換えPB14Tは矢印で表示した。レインM:予め染色された蛋白質サイズマーカー;レイン1:IPTGで誘導されていない大腸菌M15;レイン3〜7:IPTG誘導1、2、3、4及び5時間後の大腸菌M15/pB14T。
【0018】
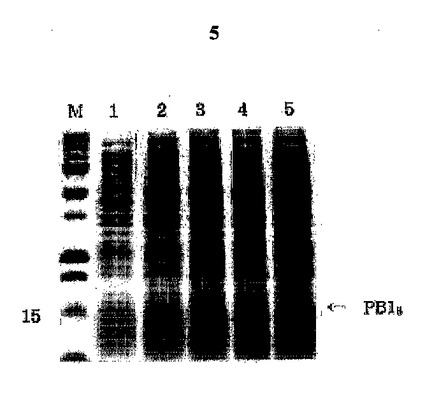
図5は形質転換された大腸菌M15/pB18を、IPTGでPB18の発現を誘導した後、SDS−PAGE分析した結果である。発現された組み換えPB18は矢印で表示した。レインM:予め染色された蛋白質サイズマーカー、レイン1:IPTGで誘導されていない大腸菌M15;レイン2〜5:IPTG誘導1、2、3及び4時間後の大腸菌M15/pB18。
【0019】
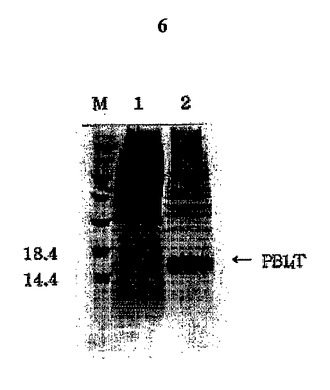
図6は大腸菌溶解物(lysate)のSDS−PAGE分析結果である。バクテリア溶解物を遠心分離して上澄液と沈殿物に分離した後、上澄液(レイン1)と沈殿物(レイン2)をSDS−PAGE分析した。PB14Tは、矢印で表示したように沈澱物に現われた。
【0020】
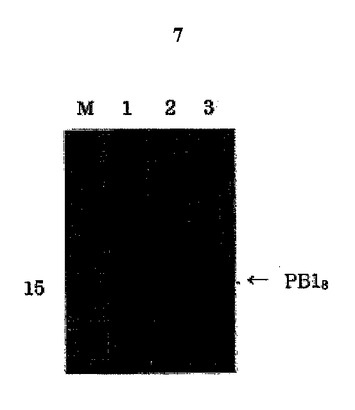
図7は大腸菌溶解物のSDS−PAGE分析結果である。バクテリア溶解物を遠心分離して上澄液と沈殿物に分離した後、全体溶解物(レイン1)、上澄液(レイン2)及び沈澱物(レイン3)をSDS−PAGE分析した。PB18は、矢印で表示したように沈殿物に現われた。
【0021】
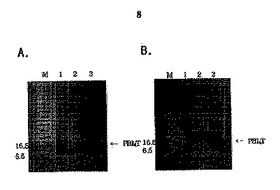
図8は精製されたPB14Tペプチドをウェスタンブロット分析した結果である。抗PB14ウサギ抗体(A)と抗preS2モノクローナル抗体(B)を用いてPB14Tを分析した。レイン1:大腸菌M5、レイン2:IPTGで誘導されていない大腸菌M15/pB14T、レイン3:IPTG誘導3時間後の大腸菌M15/pB14T。
【0022】
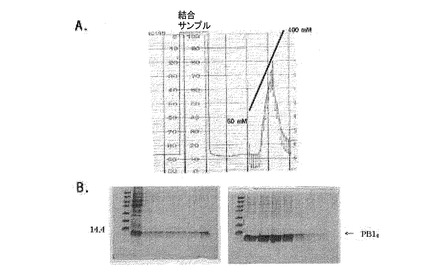
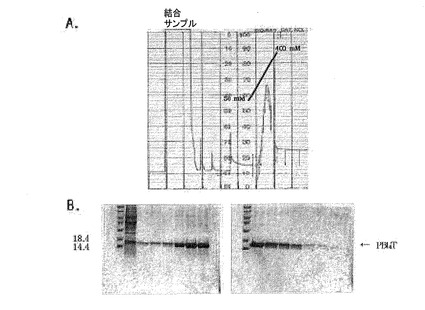
図9はイミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB14溶出プロファイルを示す。
【0023】
図10はイミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB14T溶出プロファイルを示す。
【0024】
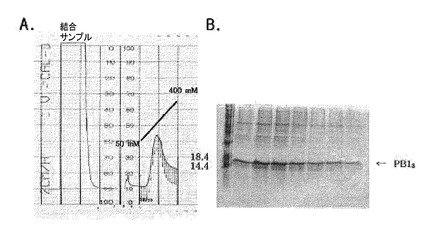
図11はイミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB18溶出プロファイルを示す。
【0025】
図12はpTB14の製作図を示す。
【0026】
図13は精製されたPB14、PB14T及びPTB14をウェスタンブロット分析した結果である。パネルAはマウス抗−preS2モノクローナル抗体と山羊抗−マウスIgG−HRP結合抗体と確認し、パネルBはウサギ抗−PB14抗血清と山羊抗−ウサギIgG−HRP結合抗体と確認した。
【0027】
図14はDNA配列から類推したTB14/pQE30の一次構造を示す。
【0028】
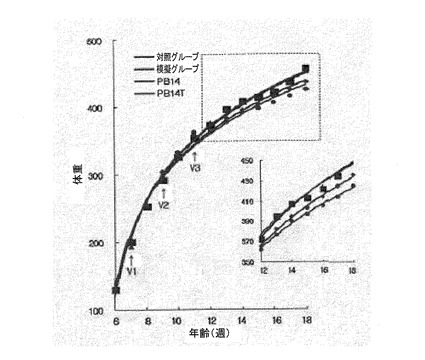
図15は対照グループ(normal)、模擬グループ(mock)、ワクチン処理グループにおけるSD系白ネズミの体重増加を比較したグラフを示す。対照グループ(■)はPBSで注射され、模擬グループ(▲)はオボアルブミンで注射された。ワクチン処理グループはPB14を媒体蛋白質OVAに抱合させたPB14+OVA(◆)ペプチドとPB14T(●)ペプチドで注射された。このようなペプチド注射は、2週間隔で3回ずつ行われた。矢印は、ワクチン処理の時点を示す。
【0029】
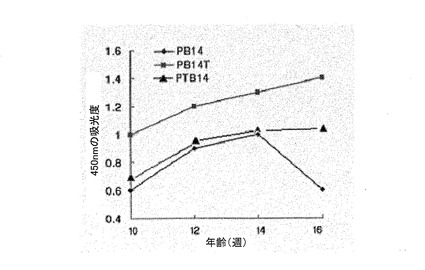
図16は抗−PB14、抗−PB14T及び抗−PTB14の抗体力価変化を測定した結果である。
【0030】
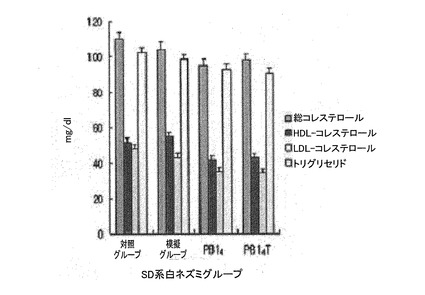
図17は血清内トリグリセリド、HDL、LDL、総コレステロール量を測定した結果である。
【0031】
〔発明を実施するための最良の様態〕
一つの様態として、本発明は、アポリポ蛋白質B−100のB細胞エピトープのミメティックペプチのアミノ酸配列を含み、前記ミメティックペプチドのC末端とヘルパーT細胞のエピトープのN末端とが融合した免疫原性ハイブリッドポリペプチドを提供する。
【0032】
本発明は、アポリポ蛋白質に対する免疫原性を強化させるための一つの方法として、アポリポ蛋白質、特にアポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドにT細胞エピトープが融合した形の免疫原性ハイブリッドペプチドを提供しようとする。このように、アポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドにT細胞エピトープを連結した結果、抗体誘導能が強化されたうえ、ワクチン効能期間が延長されて優れた抗肥満効果を示した。
【0033】
本発明で使用される用語「エピトープのミメティックペプチド」は、エピトープの最小部分を模倣するペプチドであって、抗体によって認識できるように天然エピトープと十分類似であるか、天然エピトープと交差反応する抗体を増加させることが可能なエピトープを意味し、ミモトープ(mimotope)ともいう。このようなミメティックペプチドは、生体内で非自己(non-self)と認知されるという利点があり、その場合、免疫反応において自己寛容(self tolerance)の問題を克服することができるという長所がある。前述したアポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドは、アポリボ蛋白質B−100と特異的に結合する抗体によって認識され、アポリポ蛋白質B−100と特異的に結合する抗体は、アポリポ蛋白質B−100を特異的に認識して結合するポリクローナル抗体とモノクローナル抗体を含み、これらの断片、例えばFc、Fab、F(ab’)2なども含む。特にモノクローナル抗体が好ましく、より好ましくはMab B9及びMab B23が使用される。
【0034】
本発明において、アポリポ蛋白質B−100のエピトープのミメティックペプチドは、配列番号1、2及び3の中から選択されたアミノ酸配列を含む。したがって、好ましい様態において、本発明は、配列番号1、2及び3の中から選択されたアミノ酸配列を含み、アポリポ蛋白質B−100と特異的に結合する抗体によって認識されるペプチドのC末端とヘルパーT細胞のエピトープのN末端とが融合した免疫原性ポリペプチドに関する。
【0035】
本発明者らは、ファージディスプレイペプチドライブラリ(phage displayed peptide library)からバイオパニング(biopanning)を用いてアポリポ蛋白質B−100に対するモノクローナル抗体Mab B9またはMab B23によって認識できるミメティックペプチド(配列番号1、2及び3)を究明した。
【0036】
前述した配列番号1、2及び3の中から選択されたアミノ酸配列を含むアポリポ蛋白質B−100のエピトープのミメティックペプチドは、前記配列番号のアミノ酸配列1個からなる単量体の形であってもよいが、免疫原性を一層増強させるために前記配列番号のアミノ酸配列が2個以上、好ましくは3〜8個、より好ましくは3〜6個が連結された多量体の形を取ってもよい。最も好ましくは、4個が連結された四量体(配列番号4)である。多量体の形を取る場合、単量体を形成するアミノ酸配列は、直接またはリンカーを介して共有結合で連結できる。リンカーを介して連結される場合、アミノ酸、例えばグリシン、アラニン、バリン、ロイシン、イソロイシン、プロリン、セリン、トレオニン、アスバラギン、アスパラギン酸、システイン、グルタミン、グルタミン酸、リジン、アルギニンを1個〜5個ずつ使用して連結させることができ、使用可能な好ましいアミノン酸としては、バリン、ロイシン、アスパラギン酸、グリシン、アラニン、プロリンを例示することができる。より好ましくは遺伝子操作の容易性を考慮してバリン、ロイシン、アスパラギン酸などの中から選択された2つのアミノ酸を連結して使用することができる。好ましくは、前記配列番号1、2及び3の中から選択されたアミノ酸配列が前記リンカーを介して2個以上連結されたミメティックペプチドが使用される。
【0037】
本発明において「T細胞エピトープ」という用語は、適当な効率でMHCクラスII分子に結合することができ、T細胞を刺激し或いはMHCクラスIIとの複合体でT細胞と結合することができるアミノ酸配列を意味する。その場合、T細胞エピトープは、T細胞上に存在する特定の受容体によって認識され、B細胞が抗体生産細胞に分化するのに要求される信号を提供する役割も果たし、細胞毒性T細胞(CTL)を誘導して標的細胞の溶菌を誘導する。T細胞エピトープは、T細胞を刺激させ且つ免疫反応を強化する限り特定なものと限定されず、目的に適した多様な蛋白質、ペプチドなどが使用可能である。本発明の目的上、T細胞エピトープは、ヘルパーT細胞エピトープが好ましく使用される。例えば、B型肝炎表面抗原ヘルパーT細胞エピトープ、クラミジア・トラコマティス(Chlamydia trachomatis)主要外膜蛋白質ヘルパーT細胞エピトープ、プラスモジウム・ファルシパルム・サーカムスポロゾイト(Plasmodium falciparum circumsporozoite)ヘルパーT細胞エピトープ、大腸菌TraTヘルパーT細胞エピトープ、破傷風トキソイド(Tetanus toxoid)ヘルパーT細胞エピトープ、ジフテリアトキソイド(diphtheria toxoid)ヘルパーT細胞エピトープ、マンソン住血吸虫(Schistosoma mansoni)トリオースリン酸イソメラーゼヘルパーT細胞エピトープ、麻疹(measles)ウィルスF蛋白質ヘルパーT細胞エピトープ、 百日咳ワクチン(pertussis vaccine)、BCG(Bacile Calmette-Guerin)、ポリオワクチン(polio vaccine)、流行性耳下腺炎ワクチン(mumps vaccine)、 風疹ワクチン(rubella vaccine)、狂犬病ワクチン(rabies vaccine)、ツベルクリンの精製蛋白質誘導体、キーホールリンペットヘモシアニン(keyhole limpet hemocyanin)、これらの断片または配合物などに由来したT細胞エピトープ配列を含むことができる。前述したT細胞エピトープは、特定の目的に応じて選択されたアミノ酸残基を添加、欠失、置換して使用することができ、また2つ以上の相異なるT細胞エピトープが結合している多量体の形でも提供できる。本発明の具体例では、B型肝炎ウィルスの表面抗原を使用した。B型肝炎ウィルス(HBV)遺伝子の大きさは3.2kbであって、大きく4種の重要な蛋白質に対する情報を持っており、S遺伝子(表現抗原蛋白質)、C遺伝子(コア蛋白質)、P遺伝子(DNA重合酵素)及びX遺伝子から構成されている。S遺伝子の場合、HBsAgをコードしているS領域とpreS領域に分けられる。preS領域は、HBVの菌株に応じて108または119個のアミノ酸をコードしているpreS1と、サブタイプに関係なく55個のアミノ酸残基からなるpreS2とに分けられる。HBV preS2蛋白質は、体内免疫反応過程でヘルパーT細胞を活性化させ、これはHBVに対する抗体の形成を促進させることができる。
【0038】
本発明で使用された用語「ハイブリッドポリペプチド」とは、一般に、起源の異なる異種ペプチドが連結された形のペプチドをいい、本発明では、B細胞エピトープとT細胞エピトープとを連結した形のペプチドを意味する。このようなハイブリッドポリペプチドは、それぞれのパトナーが決定された後、化学的に合成するか、遺伝子組み換え方法で発現及び精製して得ることができる。好ましくは、B細胞エピトープをコードする遺伝子配列とT細胞エピトープをコードする遺伝子配列とを連結したハイブリッド遺伝子を細胞発現システムで発現させて提供される。このようなハイブリッドポリペプチドは、B細胞エピトープとT細胞エピトープとが直接或いはリンカーなどの連結子を介して連結できる。リンカーを介して連結される場合、これによる免疫反応の誘導に不利な影響を及ぼさないように選定されるべきである。
【0039】
本発明で使用された用語「ポリペプチド」は、2つ以上のアミノ酸を含む残基が共有ペプチド結合によって結合した全長のアミノ酸鎖を含む用語であって、ジペプチド、トリペプチド、オリゴペプチド、ポリペプチドなどがこれに含まれるが、特に、本発明では、数個〜数十個のアミノ酸が共有結合したペプチド2種以上が互いに連結されたハイブリッドポリペプチドを意味する。本発明のハイブリッドポリペプチドは、2種以上のペプチド、例えばB細胞エピトープとT細胞エピトープとが連結されたポリペプチドである。ポリペプチドを構成するそれぞれのペプチド配列は、前述したようなエピトープに該当する配列を含み、それに隣接して配置された配列も共に含むことができる。これらのペプチドは、L−アミノ酸、D−アミノ酸、または2つの相異なる配置のアミノ酸の多様な組合せを取ることができる。本発明のハイブリッドポリペプチドは、全的に前述したようなB細胞エピトープ、T細胞エピトープ、及び任意にその隣接配列を含んだ抗原性部分のみからなってもよく、付加配列を含んでもよい。ところが、このような付加配列は、全体免疫原性(immunogenicity)を減少させないことが好ましい。このような付加配列としては、リンカー配列を例示することができる。
【0040】
本発明で使用された用語「免疫原性」とは、細胞性免疫及び体液性免疫反応を誘導して異物に対して対処する能力をいう。このような免疫反応を誘導する物質を免疫原と呼ぶ。本発明は、免疫原性物質として、B細胞エピトープとT細胞エピトープを両方とも備えるポリペプチドを使用する。
【0041】
本発明者らは、B細胞エピトープを持っているが、それ自体ではT細胞エピトープ部分が欠如している抗肥満機能的ペプチドであるApo B−100ミメティックペプチドの四量体PB14のC末端部分と、T細胞エピトープを持っているHBVのpreS2の一部(T断片)とを連結し、PB14Tの発現用遺伝子断片を製作した(図1)。PB14断片を得るためにBamHIとXhoIを、T断片を得るためにSalIとHindIIIを使用し、PB14Tの遺伝子断片とベクターpQE30とを結合して大腸菌JM109に形質転換した。制限酵素切断(図2)とDNA配列分析(図3)によってT細胞エピトープとB細胞エピトープ部分とが連結されている正しいクローンであることを確認し、このクローンをpB14Tと命名した。PB14T及びPB18の発現のためにpQE30ベクターを使用したが、ベクター自体内にある開始コドンから蛋白質の発現が始まり、精製を便利にするための6個のヒスチジンも共に発現し、エンテロキナーゼ(enterokinase)部位も発現する。このように発現する蛋白質の大きさは、PB14Tは16.2kDaであり、PB18は16.5kDaであった。時間帯別にサンプルを得て、SDS−PAGEによって発現を確認した(図4、図5)。
【0042】
したがって、本発明の具体的な実施様態によって四量体のアポリポ蛋白質B−100のミメティックペプチドとB型肝炎ウィルスの表面抗原preS2とが連結された配列番号9の免疫原性ハイブリッドポリペプチドが提供できる。
【0043】
本発明の免疫原性ハイブリッドポリペプチドは、化学的に合成するか、遺伝子組み換え方法によって生産することができ、好ましくは組み換えベクターを用いてこれを宿主細胞に形質転換させ、これから発現されたポリペプチドを分離、精製して生産することができる。
【0044】
したがって、他の様態として、本発明は、前記免疫原性ハイブリッドポリペプチドをコードする遺伝子を含む組み換えベクトル、及び前記組み換えベクターで形質転換された宿主細胞を提供する。
【0045】
別の様態として、本発明は、前記組み換えベクターで形質転換された宿主細胞を培養して前記免疫原性ハイブリッドポリペプチドを生産する方法を提供する。
【0046】
遺伝子組み換え方法によって本発明の免疫原性ハイブリッドポリペプチドを生産する過程は、次の段階を含む。
【0047】
第1段階は、ハイブリッドペプチドをコードする遺伝子をベクターに挿入して組み換えベクターを製造する段階である。外来遺伝子を挿入するためのベクターとしては、プラスミド、ウィルス、コスミドなど多様な形のベクターを使用することができる。組み換えベクターは、クローニングベクター及び発現ベクターを含む。クローニングベクターは、複製基点、例えばプラスミド、ファージまたはコスミドの複製基点を含み、他のDNA切片が付着し、付着している切片が複製できる「レプリコン」である。発現ベクターは、蛋白質の合成に使用されるように開発された。組み換えベクターは、通常、外来DNAの断片が挿入されたキャリアであって、一般に二本鎖のDNAの断片を意味する。本願において、「外来DNA」とは、外来種から起源するDNAを意味し、或いは同一の種から起源する場合には本来の形から実質的に変形された形を意味する。また、細胞で正常的に発現しない非変化DNA配列も含む。この場合、外来遺伝子は、転写される特定の目的の核酸でポリペプチドを暗号する。組み換えベクターは、宿主細胞における形質感染遺伝子の発現水準を高めるためには、当該遺伝子が選択された発現宿主内で機能を発揮する転写及び解読発現調節配列に作動可能に連結されなければならない。前記組み換えベクターは、個体の細胞内で遺伝子挿入物が発現されるように作動可能に連結された必須的な調節要素を含む遺伝子作製物であって、このような遺伝子作製物を製造するために、標準組み換えDNA技術を利用することができる。組み換えベクターの種類は、原核細胞または真核細胞の各種宿主細胞で目的の遺伝子を発現させ、目的の蛋白質を生産する機能を行うものであれば、特に限定されないが、強力な活性を示すプロモータと強い発現力を保有しながら自然状態と類似な形の外来蛋白質を大量生産することが可能なベクターが好ましい。組み換えベクターは、少なくともプロモータ、開始コドン、目的の蛋白質を暗号化する遺伝子、終結コドンターミネータを含んでいることが好ましい。その他に、シグナルペプチドをコードするDNA、エンハンサー配列、目的の遺伝子の5’側と3’側の非解読領域、選別マーカー領域、または複製可能単位などを適切に含むこともできる。
【0048】
第2段階は、前記組み換えベクターを用いて宿主細胞を形質転換させた後、培養する段階である。組み換えベクターを宿主細胞に導入して形質転換体を製造するための方法としては、文献(Sambrook, J. et al., Molecular Cloning, A Laboratory Manual(2版)、Cold Spring Harbor Laboratory, 1. 74, 1989)に記載のリン酸カルシウム法または塩化カルシウム/塩化ルビジウム法、エレクトロポレーション法、エレクトロインジェクション、PEGなどの化学的処理方法、遺伝子銃(gene gun)などを利用することができる。前記組み換えベクターが発現する形質転換体を栄養培地で培養すると、有用な蛋白質を大量に製造、分離可能である。培地と培養条件は、宿主細胞に応じて、寛容されるものを適切に選択して用いることができる。培養の際に細胞の生育と蛋白質の大量生産に適するように温度、培地のpH及び培養時間などの条件を適切に調節しなければならない。本発明に係る組み換えベクターで形質転換できる宿主細胞は、原核細胞と真核細胞を両方とも含み、DNAの導入効率が高く、導入されたDNAの発現効率が高い宿主が通常用いられる。細菌、例えばエシェリキア、シュードモナス、バシラス、ストレプトミセス、真菌、酵母などの周知の真核及び原核宿主、SF9(Spodoptera frugiperda)などの昆虫細胞、CHO、COS1、COS7、BSC1、BSC40、BMT10などの動物細胞などが使用できる宿主細胞の例である。好ましくは、大腸菌が使用できる。
【0049】
第3段階は、ハイブリッドペプチドの発現を誘導、蓄積する段階である。本発明では、誘導因子IPTGを用いてペプチドの発現を誘導し、誘導時間は蛋白質の量を最大化されるように調節した。
【0050】
第4段階は、ハイブリッドペプチドを分離、精製する段階である。一般に、組み換え的に生産されたペプチドは、培地または細胞分解物から回収できる。膜結合型の場合、適切な界面活性剤溶液(たとえばTriton−X 100)を使用し、或いは酵素的切断によって膜から遊離できる。ハイブリッドペプチドの発現に使用された細胞は、凍結−解凍反復、音波処理、機械的破壊または細胞分解剤などの多様な物質的または化学的手段によって破壊でき、通常の生化学分離技術によって分離、精製可能である(Sambrook et al., Molecular Cloning: A laborarory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, 1989; Deuscher, M., Guide to Protein Purification Methods Enzymology, Vol. 182. Academic Press. Inc., San Diego, CA, 1990)。電気泳動、遠心分離、ゲルろ過、沈澱、透析、クロマトグラフィー(イオン交換クロマトグラフィー、アフィニティクロマトグラフィー、免疫吸着アフィニティクロマトグラフィー、逆相HPLC、ゲル浸透HPLC)、等電点電気泳動法、並びにその多様な変化及び複合方法を含むが、これらに局限されない。
【0051】
本発明では、具体的にPB14Tの遺伝子断片をベクターpQE30と結合して大腸菌に形質転換した。pQE30は、ファージT5プロモータからなるプロモータとIPTGを誘導剤として使用するlacオペレーターシステムを持っている、大腸菌から蛋白質を大量生産するのに有用なベクターである。PB14Tを認識することが可能な2つの抗体、1次抗体はウサギ抗−PB14ポリクローナル抗体とマウス抗−preS2モノクローナル抗体を使用するウェスタンブロットによって正しく発現することを確認し、蛋白質の精製を始めた。PB14とPB14Tは不溶性なので、8M尿素に変性させた後、ヒスチジンタグ(tag)蛋白質用Ni−NTA樹脂を利用するアフィニティクロマトグラフィーを用いてPB14とPB14Tを精製した。
【0052】
上述したように発現及び精製したポリペプチドをマウスに接種した後、それによる体重の増加、血清内抗体力価、血清内脂質含量の変化を分析した。その結果、ハイブリッドポリペプチドでワクチン化されたグループが、対照グループ、またはハイブリッドされていないミメティックペプチドでワクチン化されたグループに比べて体重増加が抑制され、ミメティックペプチドに対する抗体力価が高く、残存期間が長く、TG及びLDL−コレステロールの数値も低くなることを確認した。
【0053】
ペプチドワクチンのデザインに一貫的に適用することが可能な様態が存在せず、デザインされたワクチンの効能も予測することができない。同一の理由から、疎水性が非常に強いPB14ペプチドを異種ペプチドであるT細胞エピトープと融合させる場合、抗原性を示す部位を分子の内部に取り入れて抗体誘導能の低下をもたらす可能性も排除することができない。このようにその結果を類推し難い状況で、本発明者らは、アポリポ蛋白質のB細胞エピトープのミメティックペプチドにT細胞エピトープを連結したハイブリッドポリペプチドを設計し、抗肥満に対する増加した免疫原性を立証した。
【0054】
人工合成ハイブリッドポリペプチド及びこれを含むワクチンの免疫原性は、B細胞エピトープとヘルパーT細胞エピトープが存在しなければならず、また、B細胞エピトープとヘルパーT細胞エピトープとの連結順序(orientation)によってワクチンの効能が異なる。すなわち、ヘルパーT細胞のエピトープがB細胞エピトープのN末端に位置するか、C末端に位置するかによって抗体誘導能が異なる(Partidos, C, Stanley, C, and Steward, M, The effect of orientation of epitope on the immunogenicity of chimeric synthetic peptides representing measles virus protein sequences, Molecular Immunology,29(5),651-658,1992)。
【0055】
本発明者らは、B細胞エピトープとヘルパーT細胞エピトープとの連結順序による免疫誘導効果の差異を確認するために、前記PB14のC末端部分とT断片とを連結して製作したB14T遺伝子断片とは異なり、PB14のN末端部分とT断片とを連結してTB14遺伝子断片を製作した(図12)。具体的に、実施例9による方法によってベクターpTB14を製作し、大腸菌M15に形質転換させて発現させ、発現したハイブリッドポリペプチドPTB14は、His−Tagが存在するので、Ni−NTA His−結合レジンを用いたアフィニティクロマトグラフィーを行って精製した。
【0056】
PTB14とPB14Tの抗体誘導反応性及び免疫原性を比較するために、白ネズミを用いて免疫化した後、採血してこれを比較した。PTB14の抗体誘導能はPB14よりは多少向上しており、残存期間の長期化現象を示したが、PB14Tに比べて約50〜60%程度に著しく弱いことを確認することができた(図16)。これは、体重増加抑制効能においても同一の結果を示した(表2)。前述したように、本発明のPB14のC末端部分とT断片とを連結して製作したPB14Tポリペプチドの免疫原性及び肥満抑制効能がより優れることを確認することができた。
【0057】
したがって、別の様態として、本発明は、アポリポ蛋白質B−100のエピトープのミメティックペプチドのアミノ酸配列を含み、前記ミメティックペプチドのC末端とヘルパーT細胞のエピトープのN末端とが融合した免疫原性ハイブリッドポリペプチドを含む肥満予防または治療用ワクチンを提供する。
【0058】
免疫原が示す反応の大きさ及び観察された個体の比率を比較すると、優れた効果を示すワクチンとして使用できるかどうかを判断することができる。本発明では、肥満予防及び治療用ワクチンという元々の発明の目的に合うように、(a)体重の増加推移観察、(b)血清内抗体力価の観察、及び(c)血清内脂質含量の変化によって免疫反応効果を検定し、抗原の優れた形態を立証した。
【0059】
具体的に、7週齢SD系白ネズミ(rat)の抗原注射による体重の変化を、精製PB14、PB14Tのペプチド100μgを腹腔に2週間隔3回注射した後、体重増加グラフで確認した(図15)。1次注射とブーストのときまでは各グループの身体質量は292〜297gと同様であったが、2回注射の1週後から、ワクチン化されたグループが対照グループ及び模擬グループとは差異を示し始めた。これは、最初注射による微弱な免疫反応が2次注射によってブーストされた後、増強した免疫応答反応によって体重増加が抑制されることを示す。対照グループ及び模擬グループよりはワクチン処理したときに体重の増加程度が減少し、また、PB14形態のペプチドよりPB14T形態のペプチドがより優れた体重増加抑制効果を示す(表2)。3次注射後にも、体重増加の偏差は維持された。また、キャリア蛋白質であるオボアルブミンを抱合させた形のPB14よりは均質なキメラ抗原PB14Tが免疫応答反応を効率よく誘導することを確認することができた。ワクチン処理されたグループにおけるSD系白ネズミの抗体力価は、10、12、14、16週齢でELISA分析した(図16)。PB14Tを注射したグループがPB14を注射したグループに比べて抗体力価が増加し、14週齢では、PB14Tでワクチン処理されたグループが、PB14ワクチン処理されたグループよりO.D数値が1.5倍高かった。16週齢では、PB14グループにおいて力価の減少が現われたが、PB14Tグループは力価の増加が維持された。血清内脂質水準は、ワクチン処理されたグループが、対照グループ及び模擬グループより全体的にTG及びコレステロールの数値が低かった。特に、LDL−コレステロールは正常値の60%水準と低くなった(図17)。
【0060】
前記結果は、B細胞エピトープであるPB14のみが在る形態より、T細胞エピトープが共に融合している形態が一層高い免疫原性を示し、効果的なワクチン組成物として使用できることを立証する。
【0061】
また、本発明者らは、PB14Tの効能を愛玩犬を対象として臨床実験した。PB14Tをアルミナと混合して10匹の愛玩犬に2週間隔で2回接種した後、体重変化を観察した結果、間食を始めとした高脂肪食を無制限提供しても、体重の増加は全く観察されなかった(表4)。抗体誘導の度合いにおいても、2次接種後、各愛玩犬の血清を採取して抗体力価をELISA実験法によって測定した結果、5000〜50000倍希釈した血清でも高い吸光度を示して優れた効果を発揮した。
【0062】
本発明の抗肥満ワクチンは、抗原、薬剤学的に許容可能な担体、適切な補助剤、その他の通常の物質からなり、免疫学的効果量で投与する。本発明において、「免疫学的効果量」とは、肥満治療及び予防効果を示すことができる程度の十分な量と、副作用や深刻なまたは過度な免疫反応を起さない程度の量を意味し、正確な投与濃度は投与される特定の免疫原によって異なり、免疫反応の発生を検査するために、当業者が公知の方法を用いてこれを決定することができる。また、投与形態及び経路、収容者の年齢、健康及び体重、症状の特性及び程度、現在治療法の種類、及び治療回数によって変化できる。担体は、当分野に公知になっているもので、安定化剤、希釈剤、緩衝液を含む。適切な安定化剤は、ソルビトール、ラクトース、マンニトール、澱粉、糖、デキストラン及びブドウ糖などの炭水化物;アルブミンまたはカゼインなどの蛋白質などを含む。適切な希釈剤には塩、ハンクス平衡塩、リンガー液などを含む。適切な緩衝液には、アルカリ金属リン酸塩、アルカリ金属炭酸塩、アルカリ土金属炭酸塩などを含む。また、ワクチンには、免疫反応を改善または強化させるために、少なくとも一つの補助剤などを含む。適切な補助剤には、ペプチド;水酸化アルミニウム;リン酸アルミニウム;酸化アルミニウム;Marcol 52などのミネラル油または植物性油及び少なくとも一つの乳化剤からなる組成物、またはリゾレシチン、多価陽イオン、多価陰イオンなどの表面活性物質などが含まれる。本発明のワクチン組成物は、個別治療剤として投与してもよく、他の治療剤と併用して投与してもよく、従来の治療剤とは順次または同時に投与してもよい。ワクチン組成物は、公知の投与経路によって投与される。このような方法には、経口、経皮、筋肉、腹膜、静脈、皮下、鼻腔経路を利用することができるが、これらに局限されない。また、製薬組成物は、活性物質が標的細胞へ移動することが可能な任意の装置によって投与できる。
【0063】
以下、実施例によって本発明をより具体的に説明する。これらの実施例は本発明を例示するためのもので、本発明の範囲を限定するものではない。
【0064】
〔実施例〕
<実験材料>
DNAミニプレップキット(miniprep kit)とゲルからDNAを抽出するために使用されたキットはNucelogen製を、細胞培養に必要なバクト−トリプトン(bacto-trypton)、バクト−酵素抽出物(bacto-yeast extract)、アガ(agar)などは、Difco社(Detroti、MI)製を、制限酵素はTakara製を、T4DNAリガーゼはNEB製をそれぞれ使用した。ベクターはpB1uescriptIISK(Stratagene社)、PCR2.1(Invitrogen、Carlsbad、CA)、pQE30(Qiagen社)を使用し、菌株は大腸菌JM109及びM15(Qiagen社)を使用した。
【0065】
蛋白質の生成を誘導するためのIPTGはSigma社製を、発現された蛋白質を精製するためのNi−NTAレジンはNovagen社製を使用した。SDS−PAGE及びウェスタンブロット、ECLなどに用いられた予備染色マーカー(prestained marker)はNEB社製であった。蛋白質を変性状態にするために使用された尿素はDuchefa社製であり、精製時に使用されたイミダゾールはUSB製であった。透析に使用された膜はSpectrum製のMWCO 3500であり、蛋白質凝集防止のために使用された試薬はAmresco製のCHAPSであった。ELISAに使用された抗体は、Sigma製のHRP結合抗−白ネズミIgGであった。ウェスタンブロットとECLに使用された基質溶液として、BCIP/NBTはSigma製を、ECL+plusウェスタンブロット検出試薬はAmersham製を使用した。補助剤は、フロイントアジュバント(Freund Adjuvant;Sigma社製)と水酸化アルミニウム(Reheis社製)を使用した。蛋白質の定量は、BCA蛋白質検査法(Pierce)とBradford法(Biorad)を使用した。
【0066】
血清内中性脂肪はトリグリザイム−Vを、総コレステロールはコレステザイム−Vを、HDLコレステロールはHDL−C555(シンヤン化学社製)を、LDLコレステロールはEZLDLコレステロール(Sigma社製)を用いて測定した。LDL測定器は、Randox製を使用した。
【0067】
SD(Sprague Dawley)系白ネズミは、(株)大韓バイオリンクから購入し、5週齢以上の雄を使用した。白ネズミの飼料としては韓国の(株)Samtako製(成分:天然蛋白質18%以上、粗脂肪5.3%、粗繊維4.5%、回分8.0%)を使用した。
【0068】
組み換えPB14T及びPB14ペプチドの精製のために、超音波粉砕緩衝液(5mMイミダゾール、0.5M NaCl、20mM tris−Cl、pH7.9)、結合緩衝液(5mMイミダゾール、0.5M NaCl、20mM tris−Cl、8M尿素、pH7.9)、洗浄緩衝液(50mMイミダゾール、0.5M NaCl、20mM tris−Cl、8M尿素、pH7.9)、溶出緩衝液(400mMイミダゾール、0.5M NaCl、20mM tris−Cl、8M尿素、pH7.9)を使用した。
【0069】
<実施例1:抗肥満作用ペプチドPB14T人工遺伝子の作成>
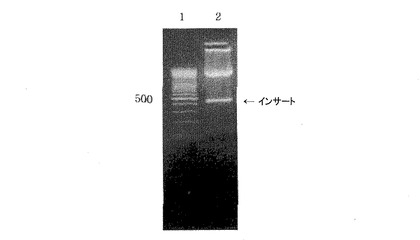
pB1uescritIISKベクターに入っているB14断片を得るために、BamHIとXhoI制限酵素を用いて切断し、PCR2.1ベクターに入っているT断片を得るために、SalIとHindIII制限酵素を用いて切断した。2つのベクターから得られた切片、B14とTに対して、B14のXhoI粘着末端とTのSalI粘着末端が相補的なので、T4 DNAリガーゼを用いて16℃で12時間結合させた。その結果、連結部位はSalIまたはXhoIによって切られなくなり、B14T断片を得るためにもう1回SalIとHindIIIで切った。蛋白質を発現するために選択されたベクターシステムとしてpQE30プラスミドを使用した。このベクターは、発現させようとする蛋白質が6Xヒスチジンと融合蛋白質の形で発現して精製が容易となるように考案された。B14T遺伝子断片をpQE30ベクターのSalI及びHindIII部位と結合した。こして作られたクローンをpB14Tと命名した(図1)。大腸菌JM109を形質転換させた後、プラスミドを分離してSalIとHindIIIで切って、450bp断片が挿入されたことを確認した(図2)。
【0070】
前述した組み換えベクターpB14Tは、大腸菌M15/pB14Tの形であって、2004年3月4日付けでKCCM(Korean Culture Center of Microorganisms、韓国ソウル市西大門区弘済1洞ユリムビル361−221)に寄託番号KCCM−10562で寄託された。
【0071】
<実施例2:抗肥満作用ペプチドPB18人工遺伝子の作成>
pB1uescriptIISKベクターに入っているB14断片をSalIとXhoIで切って得た。pBX4ベクター(B14断片が挿入されているpQX30ベクター、韓国特許公開第2002−0018971号)をSalIで切って前記B14断片とT4DNAリガーゼを用いて16℃で一晩中結合させた。
【0072】
<実施例3:遺伝子塩基配列の決定>
クローニングされたpB14Tの塩基配列を確認するために、プラスミド濃度を300〜500ng/μLとして韓国の(株)コアバイオシステムにDNA塩基配列分析を依頼し、正しいクローンであることを確認した(図3)。
【0073】
<実施例4:組み換えPB14Tペプチドの発現>
PB14T及びPB18は、pQE30ベクターで発現するので、ベクター自体内の開始コドンから蛋白質の発現が始まり、精製を便利にするために6個のヒスチジンも共に発現し、エンテロキナーゼ部位も含んでいる。ペプチドの発現に使用した宿主細胞は、大腸菌M15であって、アンピシリンとカナマイシンが含有されたLBプレートに塗抹してコロニーを得た後、Amp(100μg/mL)とKan(25μg/mL)が含有されたLB培地10mLで一晩中培養した。時間の変化に伴う蛋白質の誘導を調べるために、一晩中培養した培養液中の1mLを新鮮なLB培地50mLに接種した。本培養液を600nmで吸光度が0.4〜0.5となるまで37℃で1時間30分振とう培養した後、IPTGを最終1mM濃度となるように加え、5時間培養を行い続けて1時間ごとに1mLずつ分取した。IPTGを加える前に1mLを予め取って非誘導対照グループとして使用した。それぞれの培養液は、14,000rpmで1分間遠心分離してペレットを得た後、2×SDSサンプル緩衝液30μLに溶かしてSDS−PAGEの際にサンプルとして使用した。こうして計算された蛋白質の大きさは、それぞれPB14Tは16.2kDa、PB18は16.5kDaであった。時間帯別にサンプルを得てSDS−PAGEによって発現を確認した結果を、図4及び図5に示す。
【0074】
<実施例5:組み換えPB14Tペプチドのウェスタンブロット>
SDS−PAGEにおいて大きさの分析によってPB14Tを確認したが、正確な蛋白質が発現するかを確認するために、PB14Tを認識することが可能な2つの抗体を用いてウェスタンブロットを行った。PB14Tウェスタンブロットの対照グループには、M15にB14TがサブクローニングされていないpQE30ベクターのみを形質転換させて使用した。サンプルは、IPTGで誘導する前のものと、IPTG誘導3時間後のものにした。1次抗体は、ウサギ抗−PB14ポリクローナル抗体、マウス抗−preS2モノクローナル抗体をPBSに1:10000で希釈して使用した。1次抗体を認識することが可能な2次抗体は、ペルオキシダーゼが結合している山羊抗−ウサギIgG、山羊抗−マウスIgGをPBSに1:10000で希釈して使用した。発色反応は、ECL+Plusウェスタンブロットキットを使用し、膜をカセットに入れて富士メディカルのX線フィルムを入れた後、10秒間露出させて現像した。ウサギ抗−PB14ポリクローナル抗体はPB14TのPB14断片を認識する抗体であり、マウス抗−preS2モノクローナル抗体は、PB14TのT断片を認識する抗体なので、正確に蛋白質が発現すると、2つの抗体を使用したブロックの全てでバンドが現われるべきである。図8に示すように、それぞれの抗体がPB14TのPB14とTを認識したので、PB14Tが正確に発現したことを確認できた。
【0075】
<実施例6:組み換えPB14T及びPB18ペプチドのバクテリア細胞位置の確認>
PB14T及びPB18の細胞位置確認は、IPTGで誘導してから3時間が経過したサンプルを遠心分離し、それを超音波緩衝液で再懸濁させた後、超音波により得られたペレットと上澄み液それぞれSDS−PAGEで確認した。具体的に、IPTGで発現が誘導された細胞を4℃、9000rpmで30分間遠心分離した後、得られたペレットをしばらく−20℃で凍らせてから氷上で溶かし、その後超音波分解緩衝液をペレット1gに対して5mLの割合で入れて再懸濁した。細胞を溶解させるために、30秒ずつ15サイクル(サイクルの間に1分間休止)で超音波粉砕した。4℃、9000rpmで30分間遠心分離して上澄み液を分離し、これを可溶性蛋白質の入っている非加工抽出物A(粗抽出物A)として使用し、残っているペレットは、不溶性蛋白質の入っている非加工抽出物B(粗抽出物B)として使用した。それぞれのサンプルに2×SDSサンプル緩衝液を混ぜて95℃で5分間沸かした後、SDS−PAGEを行った。上澄み液よりはペレットに所望の蛋白質が多量現われることから、不溶性が強いことを確認した(図6及び図7)。
【0076】
<実施例7:組み換えPB14、PB14T及びPB18の精製>
ペプチドを精製するためにヒスチジンタグ蛋白質用Ni−NTAレジンを使用した。レジンに結合しているNi+と末端にヒスチジンヘキサマー融合蛋白質が相互作用する性質を利用することにより親和力を利用したアフィニティクロマトグラフィーの一つの方法である。遺伝子組み換えされた大腸菌を接種したLB培地10mLを一晩中前培養した後、500mLのLB培地に全量接種して600nmで吸光度0.4〜0.5となるまで37℃で培養した。IPTGを1mMで加え、4時間培養した後、9000rpmで30分間遠心分離して細胞ペレット得て−20℃に置いた。氷上で溶かしたペレットに超音波粉砕緩衝液を湿潤細胞と5mL/gの割合で入れて再懸濁させて超音波粉砕を行った。超音波粉砕をして細胞を溶解させ、4℃、9000rpmで30分間遠心分離してペレットを得た。結合緩衝液を、残っている上澄み液の体積と同一の体積となるようにペレットに入れた後、再懸濁させ、細胞残骸を除去するために超音波粉砕を3サイクル行った。これを再び4℃、9000rpmで30分間遠心分離して得た上澄み液を精製用サンプルとして使用した。
【0077】
Ni−NTAレジン充填カラムの大きさは直径1cm、高さ15cmであり、レジンは2mL、流速は2mL/minとした。レジンをカラムに充填した後、3〜5倍体積の蒸留水で洗浄し、5倍体積の1×荷電された緩衝液(50mM NiSO4)を流してNi2+を結合させた後、結合緩衝液で平衡化してNi−キレート化親和性樹脂を製造した。レジンにサンプルローディングを2回行った。こうしてサンプルローディングが完了すると、さらに結合緩衝液で1.0範囲の280nm吸光度で基底線に到達するまで流した。その後、洗浄緩衝液で10分間流し、平衡化が完了すると、洗浄緩衝液から溶出緩衝液にイミダゾールの濃度勾配をかけ、レジンに残留する蛋白質を完璧に溶出させるために溶出緩衝液のみを10分間さらに流した。分画は層20個であり、それぞれ2mLずつであった。精製されたペプチドは8M尿素状態であり、尿素を除去するために、PBSを用いて一晩中透析を行った。
【0078】
前述したように、それぞれの蛋白質が強い不溶性を持つため、精製は8M尿素入り緩衝液で蛋白質を変性させてから行い、樹脂に結合した蛋白質を溶出させるためにイミダゾールを使用し、濃度は50mM〜400mMの濃度勾配で精製した。その結果が図9、図10及び図11に示されている。約300mMのイミダゾール濃度のとき、大部分の蛋白質が溶出した。それぞれの蛋白質の収率は1L培養の際にPB18の場合には3〜3.5mg、PB14Tの場合には4〜4.5mgであった。
【0079】
<実施例8:組み換えPB14T、PB14及びPB18ペプチドの定量>
PB14T、PB14、PB18は、PBSで透析を行うと、尿素が除去されるので、蛋白質の凝集による沈殿が発生し、この状態では蛋白質の定量を正確に行うことができなかった。精製された蛋白質の凝集状態を除去するために、50mM CHAPSを使用した。定量は、BCA蛋白質定量法、ブラッドフォード法を用いて行った。2mg/mLのBSAを1000、500、250、125、62.5μg/mLと逐次希釈して標準として用いた。BCA分析は、Pierce社製のプロトコールによって行い、37℃で30分間反応させ、562nmで吸光度を測定し、ブラッドフォード試薬はサンプルと室温で10分間反応させた後、595nmで吸光度を測定し、BSAの逐次希釈液のBCA或いはブラッドフォード蛋白質定量発色反応の吸光度を標準定量線として用いてサンプルの蛋白質の濃度を定量した。
【0080】
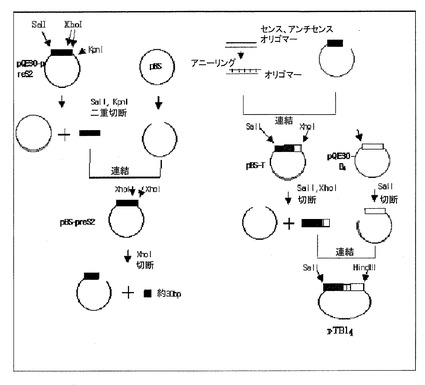
<実施例9:PTB14の発現のためのベクターpTB14の製作>
M15大腸菌株に形質転換されている発現用ベクターpQE30においてTカセット(preS2)をKpnI(Takara)、SalI(Takara)制限酵素で二重切断によって分離し、pB1uescriptも同一の酵素で処理した。分離したTカセットとpB1uescriptをそれぞれゲル上で確認して分離溶出させ、溶出させたベクターとTカセットはT4 DNAリガーゼで連結させた。pB1uescript4μL、Tカセット4μL、リガーゼ1μL(MBI Fermentas、1Weissμ/mL)、10×緩衝液(MBI Fermentas)1μLを1.5mLのチューブに入れて16℃で一晩中培養させた。組み換えされたベクターをJM109細胞と混ぜて42℃で90秒間熱衝撃を与えた後、LB培地を入れて37℃で1時間培養した。これをLB(Amp)培地に接種し、37℃で培養してコロニーを確認した。確認されたコロニー中の幾つかを任意選択して培養過程、プラスミド分離精製、制限酵素切断を経てアガロースゲル上でDNA断片のサイズによって確認した。Tカセット内にはXhoI部位が存在するが、これを無くさなければ、連結されたTB4カセットを得ることができなかった。すなわち、Tとは、Tカセット(HBVウィルスのpreS2遺伝子、183bp)の3’末端近く(Tカセットの5’を基準として約150bp)にXhoI部位が存在してクローニングに使用することができないので、内部のXhoI部位を点突然変異させたXhoIで切られない新しい配列のTカセットの形態である。pB1uescript−preS2は、Tカセット内部のXhoI部位により短いDNA断片(30bp)が切り出された。この部分に合成オリゴマーを挿入した。セルフライゲーション(Self-ligation)を防ぐために、ベクターは37℃、30分間アルカリ性ホスファターゼ(Boehringer Mannheim、GembH、Germany)で処理し、95℃で5分間脱リン酸させてゲル上で溶出させた。オリゴマーは、37℃で30分、65℃で20分間処理して5’−リン酸化させた。95℃で5分間静置反応させた後、熱遮断(heat block)をして徐々に冷やしながらアニーリングさせた。オリゴマーとpB1uescrpt−Tリガーゼを用いて16℃で一晩中培養した。組み換えされたpB1uescrtip−TをJM109細胞に形質転換導入してLB(Amp)培地でコロニーを確認し、コロニー選別過程を経て、所望のプラスミドを持った細胞を確保した。オリゴマーはpresS2の当該部分の28個のヌクレオチドからなっており、但し、XhoI部位を無くすために、センス−オリゴマー5’の5番目のヌクレオチドをGからAに変え、これによりアミノ酸配列がアルギニンからリジンに変わった。センスとアンチセンスでそれぞれ28merを製作してアニーリングさせ、これを、XhoIで処理したpB1uescript−preS2内に挿入した。pB1uescript−TをSalI、XhoIで二重切断し、pQE30−B4をSalIで処理してそれぞれをゲル上で分離溶出させた。5’側が切り出されたpQE30−B4に、分離されたTを挿入し、pQE30−pTB4を作成した。組み換えされたTB4をSalIとHindIIIで切ってバンドの大きさを確認した。こうして作成されたベクターをpTB14と命名した(図13)。
【0081】
<実施例10:PTB14の発現及び精製>
発現ベクターpTB14をM15に導入してLB(Amp、Kan)培地2Lに培養した。これを7000rpmで10分間遠心分離して湿潤細胞9gを回収した。遺伝子組み換えで発現されたハイブリッドポリペプチドPTB4は、His−Tagが存在するので、Ni−NTA His−結合レジンを用いたアフィニティクロマトグラフィーを行った。カラムは、レジン体積4mLで直径1.8cm、高さ8cmのものを使用した。Econo systemの吸光度範囲は0.5、記録計の用紙速度は2cm/hour、サンプルローディング速度は2mL/minに合わせて使用した。まず、湿潤細胞に超音波粉砕緩衝液(sonication buffer)を入れて超音波破砕によって細胞を壊し、4℃、10000rpmで30分間遠心分離した。ペレットのみを結合緩衝液に溶かした後、アフィニティクロマトグラフィーで精製した。まず、結合溶液をカラムに流しながらレジンを沈殿させた。検出器を見ながら、ベースラインを定め、一定の値を指すと、超音波粉砕したサンプルを入れた。サンプルがカラム内に入って再び一定の値を指すまで待ってから洗浄溶液を流し、その後さらに一定の値を指すと、溶出溶液も流してPTB14を分離した。発現及び精製されたハイブリッドポリペプチドは、SDS−PAGE後、ウェスタンブロットで確認した。SDS−PAGEしたPTB14を半乾燥トランスファ(semi-dry transfer)を用いて膜に移した。ブロッキング溶液(0.5% casein−phosphate buffered saline-tween、0.02%NaN3)を用いて37℃で2時間恒温処理した後、Tris-buffered saline-tween(TBS−T、pH7.6)で2分ずつ2回洗浄した。その後、1次抗体を37℃で1時間処理し、TBS−Tで5分ずつ4回洗浄した。2時抗体も1時間処理した後、同一の方法で洗浄した。Tカセットの確認には抗−preS2モノクローナル抗体(1:10000)、山羊抗−マウスIgG−HRP結合抗体(1:10000)を使用し、Bカセットの確認にはウサギ抗−PB14抗血清(1:10000)、山羊抗−ウサギIgG−HRP結合抗体(1:10000)を使用した。ECLキット溶液を乾燥膜上に散布して5分間反応させ、これをECLキットとしてバンドを確認した。その結果、Bカセットは、ウサギ抗−PB14抗血清と山羊抗−ウサギIgG−HRP結合抗体を用いてPB14TとPTB14でのみ約16KDaのバンドが現われたことを見ることができ、Tカセットは、抗−preS2モノクローナル抗体と山羊抗−マウスIgG−HRP結合抗体を用いて、PB14では約8kDaのバンドが観察され、PB14TとPTB14では16kDaのバンドが見えてそれぞれのハイブリッドポリペプチドが正確に発現、精製されたことを確認した(図14)。
【0082】
<実施例11:PB14とオボアルブミンとの連結>
PB14をキャリア蛋白質としてのオブアルブミンと連結した。キャリア蛋白質とPB14のモル比が約1:10となるようにした。約1時間4℃で反応バイアルで攪拌し、2%グルタルアルデヒド溶液を入れた後、3時間攪拌し、PBS緩衝液でMWCO3000透析膜を用いて一晩中透析して残余のグルタルアルデヒドを除去した。
【0083】
<実施例12:ワクチン処理(免疫化)>
7週齢のSD系白ネズミを6グループに分けてワクチン処理を行った(表1)。実施例7及び実施例10で精製及び定量したペプチド100μgを、表1に示すように、それぞれの補助剤と混合して100μLとなるようにして腹腔に注射した。注射は2週間隔で7、9及び11週齢に3回にわたって行った。補助剤はフロイントアジュバントと水酸化アルミニウムを使用した。フロイントアジュバントはペプチドと同量で混ぜ、水酸化アルミニウムは5.8mg/mLを最終濃度が0.2mg/mLとなるように調整した後、ペプチドと室温で攪拌しながら恒温処理した。最初ブースト5日目、最終ブースト5日目、2週目及び4週目に尻から採血した。
【0084】
【表1】
【0085】
前記ワクチン処理後の体重変化を図15にグラフで示した。最初注射してからブーストするまでは、各グループ別体重が292〜297gと同様であったが、2回注射後1週前からは、ワクチン処理グループ、対照グループおよび模擬グループの体重の差異が見られた。18週齢のとき、対照グループおよび模擬グループとPB14注射グループとの体重の差異は16gであり、PB14T注射グループとの体重の差異は27gであった(表2)。これは、最初注射による微弱な免疫反応が2次注射によってブーストされた後、増強した免疫応答反応によって体重の増加が抑制されることを示す。3次注射後にも、体重増加の偏差は維持された。
【0086】
【表2】
【0087】
表2において、標準偏差は、5匹のSD系白ネズミに対して計算されたものであり、それぞれの単位はgで表わす。
【0088】
<実施例13:抗体力価の測定>
血清サンプルを用いて間接的ELISA法によって抗体力価を測定した。微細力価平板(microtiter plate)にPB14を各ウェルに100ngの濃度で100μLずつ仕込んだ。4℃で一晩中処理した後、ブロッキング溶液(PBS、0.5%カゼイン、0.02%NaN3)を添加し、37℃で1時間恒温処理した。PBSTで3回洗い出し、SD系白ネズミワクチン処理によって得た血清をPBSに1/500〜1/8000倍で希釈して100μLずつ使用し、37℃で1時間恒温処理した。各ウェルをPBSTで3回洗い出し、2次抗体としての山羊抗−白ネズミIgGの1/1000倍希釈液で培養した。OPD発色反応を行い、吸光度を450nmで測定した。
【0089】
SD系白ネズミが10、12、14、16週齢のとき、ワクチン処理グループで白ネズミの抗体力価を確認した結果が図16に示されている。力価はそれぞれの血清希釈比を1/2000にしたとき、ELISA結果、吸光度が0.6を基準とした。1/500〜1/8000倍で希釈してPB14、PB14T及びPTB14を注射したグループは、14週齢までは抗体力価が増加し、PB14TグループはPB14よりO.D数値で1.5倍高かったが、PTB14グループはPB14グループに比べて多少向上した。16週齢のときは、PB14グループでは力価が減少し、PB14T及びPTB14のグループでは力価が維持されたが、PTB14はPB14Tに比べて約50〜60%と著しく弱いことを確認することができた。
【0090】
<実施例14:血清脂質含量の測定>
TGと総コレステロールの測定は、発色試薬200μLに血清4μLを入れて37℃で5分間高温処理した後、それぞれ505nm、500nmで吸光度を測定した。HDLの測定は、血清と沈澱試薬を1:1で混ぜた後、室温で10分間放置し、その後3000rpm以上で10分間遠心分離して得た上澄液を発色試薬200μLに4μL入れて37℃で5分間恒温処理した後、555mmで吸光度を測定した。LDLコレステロールの測定はEZ LDLコレステロールキット(Sigma社製)を使用し、LDLカリブレータはRandox社製を使用し、製造社から提供されたプロトコールによってキット内の試薬1150μLに血清4μLを入れて37℃で5分間反応させた後、試薬250μLを添加してさらに37℃で5分間反応し、その後600nmで吸光度を測定した。各測定シートから得た吸光度と標準試液を比較して濃度を計算した。
SD系白ネズミへの3次注射後5週目の血清で分析した結果が表3に示されている。
【0091】
【表3】
【0092】
表3において、標準偏差は5匹のSD系白ネズミに対して計算されたものであり、それぞれの単位はmg/dLで示した。
【0093】
対照グループおよび模擬グループでは、全体的にTG及びコレステロールの数値がワクチン処理グループより約10mg/dL(10mg/100mL)高かった。同じワクチン処理グループにおけるTGとLDLコレステロールの数値は、PB14で高かったが、その差異は大きくなかった(図17)。
【0094】
<実施例15:愛玩犬を対象とした臨床実験結果>
PB14Tをアジュバントとしてのアルミナと混合した混合物(2mg/mL)0.5mLを10匹の愛玩犬(韓国アンサン動物病院で肥満管理を受けている愛玩犬)に筋肉注射又は皮下注射の形で2週間隔で2回接種した後、1.5〜3ヶ月間体重の変化を観察した。その結果、抗体が徐々に減少する形に維持され(半減期:3ヶ月)、間食を始めとした高脂肪食を無制限提供しても、体重は全く増加していない。具体的に、接種した10匹の愛玩犬の全てが、間食及び高カロリー食餌を摂取する場合でも体重の増加が抑制されることを確認した。特に、ヨークシャーテリア(Yorkshire terrier)種の場合、性別及び年齢による体重増加が予想される状況でも、PB14Tの接種により体重が増加しないことを確認することができた。
【0095】
また、各愛玩犬の血清を採取して抗体誘導の度合いを確認するために、2次接種後1週目に、PB14T或いはPB14を認識する血清中の抗体力価をELISA実験法で測定した。その結果、5000〜50000倍で希釈した血清においても0.5の高い吸光度を示して抗体誘導能も非常に優れた。
【0096】
【表4】
【0097】
〔産業上の利用可能性〕
本発明の抗肥満効果を示すアポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドのC末端にヘルパーT細胞のエピトープのN末端を融合させたハイブリッドポリペプチドは、B細胞エピトープの有利な活性または効果を中和させる免疫反応を誘導するか有害な副作用を起すことなく、優れた免疫増強効果を示して肥満の予防または治療に効果的に使用できる。
特許手続き上微生物寄託の国際的承認に関するブダペスト条約
国際様式
このページの下端に確認された国際寄託当局から規則7.1に基づいて発行された原寄託申請の受託証
To:Hyo Joon Kim.
韓国(〒425−791)京畿道安山市常緑樹区四1洞漢陽大学校HBI604
┌──────────────────────────────────────┐
I.微生物表示 │
├─────────────────┬────────────────────┤
寄託者によって与えられた識別番号: 国際寄託当局によって与えられた受託番号 │
M15/pB14T : │
│ KCCM−10562 │
├─────────────────┴────────────────────┤
II. 科学的性質および/または提示された分類学的表示 │
├──────────────────────────────────────┤
前記Iで識別された微生物は、下記のとおりである。 │
[ ]科学的性質 │
[ ]提示された分類学的表示 │
(該当欄にX表示) │
├──────────────────────────────────────┤
III.受付および受託 │
├──────────────────────────────────────┤
本国際寄託当局は、前記Iに表示された微生物が2004年3月4日付で受付され │
たことを確認する。(原寄託日)1 │
├──────────────────────────────────────┤
IV.国際寄託当局 │
├───────────────────┬──────────────────┤
名称:韓国微生物保存センター(KCCM 本国際寄託当局を代表する委任状を有 │
) する者又は権限保有職員の署名 │
住所:韓国(〒120−091)ソウル市 │ │
西大門区弘済1洞361−221 日付:2004年3月12日 │
ユリムビル │ │
└───────────────────┴──────────────────┘
1 規則6.4(d)適用、この日付は国際寄託当局の資格を得た日付であるが、国際寄託当局の資格の獲得後、ブダペスト条約の外でなされた寄託がブダペスト条約下の寄託に転換される場合、この日付は国際寄託当局が微生物を受付けた日付である。
書式BP/4 単一ページ
【図面の簡単な説明】
【0098】
【図1】pB14Tの製作図を示す。
【図2】pB14Tの制限酵素切断を示す。
【図3】DNA配列から類推したpB14Tの一次構造を示す。
【図4】形質転換された大腸菌(Escherichia coli)M15/pB14Tを、IPTGでpB14Tの発現を誘導した後、SDS−PAGE分析した結果である。発現された組み換えPB14Tは矢印で表示した。レインM:予め染色された蛋白質サイズマーカー;レイン1:IPTGで誘導されていない大腸菌M15;レイン3〜7:IPTG誘導1、2、3、4及び5時間後の大腸菌M15/pB14T。
【図5】形質転換された大腸菌M15/pB18を、IPTGでPB18の発現を誘導した後、SDS−PAGE分析した結果である。発現された組み換えPB18は矢印で表示した。レインM:予め染色された蛋白質サイズマーカー、レイン1:IPTGで誘導されていない大腸菌M15;レイン2〜5:IPTG誘導1、2、3及び4時間後の大腸菌M15/pB18。
【図6】大腸菌溶解物(lysate)のSDS−PAGE分析結果である。バクテリア溶解物を遠心分離して上澄液と沈殿物に分離した後、上澄液(レイン1)と沈殿物(レイン2)をSDS−PAGE分析した。PB14Tは、矢印で表示したように沈澱物に現われた。
【図7】大腸菌溶解物のSDS−PAGE分析結果である。バクテリア溶解物を遠心分離して上澄液と沈殿物に分離した後、全体溶解物(レイン1)、上澄液(レイン2)及び沈澱物(レイン3)をSDS−PAGE分析した。PB18は、矢印で表示したように沈殿物に現われた。
【図8】精製されたPB14Tペプチドをウェスタンブロット分析した結果である。抗PB14ウサギ抗体(A)と抗preS2モノクローナル抗体(B)を用いてPB14Tを分析した。レイン1:大腸菌M5、レイン2:IPTGで誘導されていない大腸菌M15/pB14T、レイン3:IPTG誘導3時間後の大腸菌M15/pB14T。
【図9】イミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB14溶出プロファイルを示す。
【図10】イミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB14T溶出プロファイルを示す。
【図11】イミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB18溶出プロファイルを示す。
【図12】pTB14の製作図を示す。
【図13】精製されたPB14、PB14T及びPTB14をウェスタンブロット分析した結果である。パネルAはマウス抗−preS2モノクローナル抗体と山羊抗−マウスIgG−HRP結合抗体と確認し、パネルBはウサギ抗−PB14抗血清と山羊抗−ウサギIgG−HRP結合抗体と確認した。
【図14】DNA配列から類推したTB14/pQE30の一次構造を示す。
【図15】対照グループ(normal)、模擬グループ(mock)、ワクチン処理グループにおけるSD系白ネズミの体重増加を比較したグラフを示す。対照グループ(■)はPBSで注射され、模擬グループ(▲)はオボアルブミンで注射された。ワクチン処理グループはPB14を媒体蛋白質OVAに抱合させたPB14+OVA(◆)ペプチドとPB14T(●)ペプチドで注射された。このようなペプチド注射は、2週間隔で3回ずつ行われた。矢印は、ワクチン処理の時点を示す。
【図16】抗−PB14、抗−PB14T及び抗−PTB14の抗体力価変化を測定した結果である。
【図17】血清内トリグリセリド、HDL、LDL、総コレステロール量を測定した結果である。
【発明の詳細な説明】
【0001】
〔技術分野〕
本発明は、アポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドとヘルパーT細胞エピトープを含み、前記ミメティックペプチドのC末端がヘルパーT細胞エピトープ部分のN末端と融合した免疫原性ハイブリッドポリペプチド、及びこれを含む肥満予防または治療用ワクチン組成物に関する。
【0002】
〔背景技術〕
最近、韓国では、西欧的食生活の影響で動脈硬化症及び冠状動脈疾患(coronary atherosclerotic disease:CAD)が益々増加しつつあり、これによる死亡率も高くなってきている。この種の疾患の原因となる血清脂質としては、コレステロール(cholesterol)、トリグリセリド(triglyceride:TG)、遊離脂肪酸(free fatty acid)、リン脂質(phospholipid)などがあり、アポリポ蛋白質(apolipoprotein)と共にリポ蛋白質を形成して血液循環によって運搬されている。中でも、LDL(low density lipoprotein)は主にTGやコレステロールの運搬を担当しており、LDL−コレステロール数値の変化は前述したような疾病の予後を示す尺度となる。
【0003】
脂質代謝に関連した成人病の主要因子であるLDL−コレステロールは、各組織の細胞膜のLDL受容体に結合して組織内に蓄積、活用されるか、スカベンジャー細胞により捕獲されてから加水分解されて遊離コレステロールの形でアポEリポ蛋白質と共にHDLへ伝達されて肝で再活用されるか、胆汁酸の形に処理されて排泄される経路が明らかになっている。この過程において、アポリポ蛋白質は、リポ蛋白質の構造的恒常性を保ち、脂肪質分解酵素の補助因子機能、及び細胞膜上の特定の受容体と結合する非常に重要な機能を行っている。
【0004】
アポリポ蛋白質B−100(Apo B−100)は、このような低密度リポ蛋白質(LDL)の主な蛋白質成分であり、またIDL、VLDL、キロミクロン(chylomicron)に存在しているため、血中にある抗体がApo B−100を認識するように誘導すると、食細胞によるLDL−消去(clearance)が容易に起こり得るであろう。これを根拠とし、最近、ワクチンを用いてLDL−コレステロールの数値を低め且つ動脈硬化を減少させるための研究が行われている。このような抗−コレステロールワクチン療法により誘導された抗体はIgMタイプであり、VLDL、IDL及びLDLと結合するものと思われるが、このことから高コレステロール及びアテローム性動脈硬化症に対する予防及び治療用ワクチンの可能性が提示されている(Bailey, et al., Cholesterol vaccines. Science 264, 1067-1068, 1994; Palinski W et al., Proc Natl Acad Sci U S A. 92, 821-5, 1995; Wu R. de Faire U et al., Hypertension. 33, 53-9, 1999)。また、アポリポ蛋白質B−100は、4560個のアミノ酸残基(24個のシグナルペプチドを含む)からなる巨大な蛋白質分子であって、分子量は500kDaを超過する(Elovson J et al., Biochemistry, 24: 1569-1578, 1985)。主に肝から分泌される両親媒性の分子なので、リポ蛋白質のリピドとの相互作用が可能であり、水溶液の環境とも相互作用が可能である(Segrest J. P et al., Adv. Protein Chem., 45: 303-369, 1994)。アポリポ蛋白質B−100は、LDL粒子の大きさと構造を安定化させる役割と、収容基との結合によって血漿内LDLコレステロールの恒常性を調節するのに重要な役割をする(Brown MS et al., Science, 232 : 34-47, 1986)。
【0005】
本願の発明者により出願された韓国特許公開第2002−0018971号では、Apo B−100のエピトープに対するミメティックペプチド(mimetic peptide)が抗肥満効果を持つと記述している。ところが、前記出願では、単に、前述したB細胞のエピトープであるミメティックペプチドが抗肥満効果を示すことを開示している。
【0006】
本発明の前には、アポリポ蛋白質のB細胞エピトープとT細胞エピトープとを融合させて免疫原性を増強させようとしたことがなく、免疫反応を増強させるために蛋白質キャリアまたは補助剤として使用した試みのみがあった。
【0007】
米国特許第5,843,446号では、LHRHの免疫原性を増強させるために、LHRHにこれとは異なる蛋白質を結合させることができるが、この場合、主要免疫反応がLHRHに対して誘導されるよりは、結合したキャリア蛋白質に対して誘導され、却ってLHRHに対する免疫抑制が誘導されるおそれがあるという問題について記述している。これから分かるように、B細胞エピトープの免疫原性を増強させるための追加物質の選別、結合方式、結合位置などについての研究開発は、持続的な努力が必要である。
【0008】
ハプテンの免疫原性を高めるために媒体蛋白質と融合させようとする試みがたくさんあったが、一律的な相乗効果を得ることはできなかった。特に、本発明のようなB細胞エピトープとT細胞エピトープとの線形連結の際に、これらの方向性、それぞれのエピトープの種類などによって寧ろ免疫原性が消失してしまう場合もあり(Francis, M. J. et al., Nature 330: 168-170, 1987)、リンカーの存在により抗原性が減少する場合も発生した(Partidos, C. et al., Mol. Immunol. 29:651-658, 1992)。このようにペプチドワクチンのデザインに一貫的に適用することが可能な様態が存在せず、デザインされたワクチンの効能も予測することができない。同一の理由から、疎水性が非常に強いPB14ペプチドを、異種のペプチドであるT細胞エピトープと融合させる場合、抗原性を示す部位を分子の内部に取り入れて抗体誘導能の低下をもたらす可能性も排除することができない。
【0009】
このような背景の下で、本発明者らは、抗肥満効果を示すアポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドであるPB14を増強させるために様々な試みを重ねた結果、ミメティックエピトープのC末端にヘルパーT細胞のエピトープのN末端を融合させたハイブリッドポリペプチドが優れた免疫増強効果を示し、肥満の予防または治療に効果的に使用できることを確認した。このようなハイブリッドポリペプチドが、アポリポ蛋白質のB細胞エピトープの有利な活性または効果を中和させる免疫反応を誘導するか有害な副作用を起すことなく、優れた抗肥満活性を示すことができるということは、全く類推できない効果であった。
【0010】
〔発明の開示〕
一つの様態として、本発明は、アポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドのアミノ酸配列を含み、前記ミメティックペプチドのC末端がヘルパーT細胞エピトープのN末端と融合した免疫原性ハイブリッドポリペプチドを提供する。
【0011】
他の様態として、本発明は、アポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドのアミノ酸配列を含み、前記ミメティックペプチドのC末端がヘルパーT細胞エピトープのN末端と融合した免疫原性ハイブリッドポリペプチドを含む肥満予防または治療用ワクチンを提供する。
【0012】
別の様態として、本発明は、前述した免疫原性ハイブリッドポリペプチドをコードする遺伝子を含む組み換えベクター、前記組み換えベクターを含む形質転換体、及び前記組み換えベクターで形質転換された宿主細胞を培養して前記ハイブリッドポリペプチドを生産する方法を提供する。
【0013】
〔図面の簡単な説明〕
本発明の前記および他の目的、特徴および利点は、添付図面を参照する以降の詳細な説明からより明らかになるであろう。
【0014】
図1はpB14Tの製作図を示す。
【0015】
図2はpB14Tの制限酵素切断を示す。
【0016】
図3はDNA配列から類推したpB14Tの一次構造を示す。
【0017】
図4は形質転換された大腸菌(Escherichia coli)M15/pB14Tを、IPTGでpB14Tの発現を誘導した後、SDS−PAGE分析した結果である。発現された組み換えPB14Tは矢印で表示した。レインM:予め染色された蛋白質サイズマーカー;レイン1:IPTGで誘導されていない大腸菌M15;レイン3〜7:IPTG誘導1、2、3、4及び5時間後の大腸菌M15/pB14T。
【0018】
図5は形質転換された大腸菌M15/pB18を、IPTGでPB18の発現を誘導した後、SDS−PAGE分析した結果である。発現された組み換えPB18は矢印で表示した。レインM:予め染色された蛋白質サイズマーカー、レイン1:IPTGで誘導されていない大腸菌M15;レイン2〜5:IPTG誘導1、2、3及び4時間後の大腸菌M15/pB18。
【0019】
図6は大腸菌溶解物(lysate)のSDS−PAGE分析結果である。バクテリア溶解物を遠心分離して上澄液と沈殿物に分離した後、上澄液(レイン1)と沈殿物(レイン2)をSDS−PAGE分析した。PB14Tは、矢印で表示したように沈澱物に現われた。
【0020】
図7は大腸菌溶解物のSDS−PAGE分析結果である。バクテリア溶解物を遠心分離して上澄液と沈殿物に分離した後、全体溶解物(レイン1)、上澄液(レイン2)及び沈澱物(レイン3)をSDS−PAGE分析した。PB18は、矢印で表示したように沈殿物に現われた。
【0021】
図8は精製されたPB14Tペプチドをウェスタンブロット分析した結果である。抗PB14ウサギ抗体(A)と抗preS2モノクローナル抗体(B)を用いてPB14Tを分析した。レイン1:大腸菌M5、レイン2:IPTGで誘導されていない大腸菌M15/pB14T、レイン3:IPTG誘導3時間後の大腸菌M15/pB14T。
【0022】
図9はイミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB14溶出プロファイルを示す。
【0023】
図10はイミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB14T溶出プロファイルを示す。
【0024】
図11はイミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB18溶出プロファイルを示す。
【0025】
図12はpTB14の製作図を示す。
【0026】
図13は精製されたPB14、PB14T及びPTB14をウェスタンブロット分析した結果である。パネルAはマウス抗−preS2モノクローナル抗体と山羊抗−マウスIgG−HRP結合抗体と確認し、パネルBはウサギ抗−PB14抗血清と山羊抗−ウサギIgG−HRP結合抗体と確認した。
【0027】
図14はDNA配列から類推したTB14/pQE30の一次構造を示す。
【0028】
図15は対照グループ(normal)、模擬グループ(mock)、ワクチン処理グループにおけるSD系白ネズミの体重増加を比較したグラフを示す。対照グループ(■)はPBSで注射され、模擬グループ(▲)はオボアルブミンで注射された。ワクチン処理グループはPB14を媒体蛋白質OVAに抱合させたPB14+OVA(◆)ペプチドとPB14T(●)ペプチドで注射された。このようなペプチド注射は、2週間隔で3回ずつ行われた。矢印は、ワクチン処理の時点を示す。
【0029】
図16は抗−PB14、抗−PB14T及び抗−PTB14の抗体力価変化を測定した結果である。
【0030】
図17は血清内トリグリセリド、HDL、LDL、総コレステロール量を測定した結果である。
【0031】
〔発明を実施するための最良の様態〕
一つの様態として、本発明は、アポリポ蛋白質B−100のB細胞エピトープのミメティックペプチのアミノ酸配列を含み、前記ミメティックペプチドのC末端とヘルパーT細胞のエピトープのN末端とが融合した免疫原性ハイブリッドポリペプチドを提供する。
【0032】
本発明は、アポリポ蛋白質に対する免疫原性を強化させるための一つの方法として、アポリポ蛋白質、特にアポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドにT細胞エピトープが融合した形の免疫原性ハイブリッドペプチドを提供しようとする。このように、アポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドにT細胞エピトープを連結した結果、抗体誘導能が強化されたうえ、ワクチン効能期間が延長されて優れた抗肥満効果を示した。
【0033】
本発明で使用される用語「エピトープのミメティックペプチド」は、エピトープの最小部分を模倣するペプチドであって、抗体によって認識できるように天然エピトープと十分類似であるか、天然エピトープと交差反応する抗体を増加させることが可能なエピトープを意味し、ミモトープ(mimotope)ともいう。このようなミメティックペプチドは、生体内で非自己(non-self)と認知されるという利点があり、その場合、免疫反応において自己寛容(self tolerance)の問題を克服することができるという長所がある。前述したアポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドは、アポリボ蛋白質B−100と特異的に結合する抗体によって認識され、アポリポ蛋白質B−100と特異的に結合する抗体は、アポリポ蛋白質B−100を特異的に認識して結合するポリクローナル抗体とモノクローナル抗体を含み、これらの断片、例えばFc、Fab、F(ab’)2なども含む。特にモノクローナル抗体が好ましく、より好ましくはMab B9及びMab B23が使用される。
【0034】
本発明において、アポリポ蛋白質B−100のエピトープのミメティックペプチドは、配列番号1、2及び3の中から選択されたアミノ酸配列を含む。したがって、好ましい様態において、本発明は、配列番号1、2及び3の中から選択されたアミノ酸配列を含み、アポリポ蛋白質B−100と特異的に結合する抗体によって認識されるペプチドのC末端とヘルパーT細胞のエピトープのN末端とが融合した免疫原性ポリペプチドに関する。
【0035】
本発明者らは、ファージディスプレイペプチドライブラリ(phage displayed peptide library)からバイオパニング(biopanning)を用いてアポリポ蛋白質B−100に対するモノクローナル抗体Mab B9またはMab B23によって認識できるミメティックペプチド(配列番号1、2及び3)を究明した。
【0036】
前述した配列番号1、2及び3の中から選択されたアミノ酸配列を含むアポリポ蛋白質B−100のエピトープのミメティックペプチドは、前記配列番号のアミノ酸配列1個からなる単量体の形であってもよいが、免疫原性を一層増強させるために前記配列番号のアミノ酸配列が2個以上、好ましくは3〜8個、より好ましくは3〜6個が連結された多量体の形を取ってもよい。最も好ましくは、4個が連結された四量体(配列番号4)である。多量体の形を取る場合、単量体を形成するアミノ酸配列は、直接またはリンカーを介して共有結合で連結できる。リンカーを介して連結される場合、アミノ酸、例えばグリシン、アラニン、バリン、ロイシン、イソロイシン、プロリン、セリン、トレオニン、アスバラギン、アスパラギン酸、システイン、グルタミン、グルタミン酸、リジン、アルギニンを1個〜5個ずつ使用して連結させることができ、使用可能な好ましいアミノン酸としては、バリン、ロイシン、アスパラギン酸、グリシン、アラニン、プロリンを例示することができる。より好ましくは遺伝子操作の容易性を考慮してバリン、ロイシン、アスパラギン酸などの中から選択された2つのアミノ酸を連結して使用することができる。好ましくは、前記配列番号1、2及び3の中から選択されたアミノ酸配列が前記リンカーを介して2個以上連結されたミメティックペプチドが使用される。
【0037】
本発明において「T細胞エピトープ」という用語は、適当な効率でMHCクラスII分子に結合することができ、T細胞を刺激し或いはMHCクラスIIとの複合体でT細胞と結合することができるアミノ酸配列を意味する。その場合、T細胞エピトープは、T細胞上に存在する特定の受容体によって認識され、B細胞が抗体生産細胞に分化するのに要求される信号を提供する役割も果たし、細胞毒性T細胞(CTL)を誘導して標的細胞の溶菌を誘導する。T細胞エピトープは、T細胞を刺激させ且つ免疫反応を強化する限り特定なものと限定されず、目的に適した多様な蛋白質、ペプチドなどが使用可能である。本発明の目的上、T細胞エピトープは、ヘルパーT細胞エピトープが好ましく使用される。例えば、B型肝炎表面抗原ヘルパーT細胞エピトープ、クラミジア・トラコマティス(Chlamydia trachomatis)主要外膜蛋白質ヘルパーT細胞エピトープ、プラスモジウム・ファルシパルム・サーカムスポロゾイト(Plasmodium falciparum circumsporozoite)ヘルパーT細胞エピトープ、大腸菌TraTヘルパーT細胞エピトープ、破傷風トキソイド(Tetanus toxoid)ヘルパーT細胞エピトープ、ジフテリアトキソイド(diphtheria toxoid)ヘルパーT細胞エピトープ、マンソン住血吸虫(Schistosoma mansoni)トリオースリン酸イソメラーゼヘルパーT細胞エピトープ、麻疹(measles)ウィルスF蛋白質ヘルパーT細胞エピトープ、 百日咳ワクチン(pertussis vaccine)、BCG(Bacile Calmette-Guerin)、ポリオワクチン(polio vaccine)、流行性耳下腺炎ワクチン(mumps vaccine)、 風疹ワクチン(rubella vaccine)、狂犬病ワクチン(rabies vaccine)、ツベルクリンの精製蛋白質誘導体、キーホールリンペットヘモシアニン(keyhole limpet hemocyanin)、これらの断片または配合物などに由来したT細胞エピトープ配列を含むことができる。前述したT細胞エピトープは、特定の目的に応じて選択されたアミノ酸残基を添加、欠失、置換して使用することができ、また2つ以上の相異なるT細胞エピトープが結合している多量体の形でも提供できる。本発明の具体例では、B型肝炎ウィルスの表面抗原を使用した。B型肝炎ウィルス(HBV)遺伝子の大きさは3.2kbであって、大きく4種の重要な蛋白質に対する情報を持っており、S遺伝子(表現抗原蛋白質)、C遺伝子(コア蛋白質)、P遺伝子(DNA重合酵素)及びX遺伝子から構成されている。S遺伝子の場合、HBsAgをコードしているS領域とpreS領域に分けられる。preS領域は、HBVの菌株に応じて108または119個のアミノ酸をコードしているpreS1と、サブタイプに関係なく55個のアミノ酸残基からなるpreS2とに分けられる。HBV preS2蛋白質は、体内免疫反応過程でヘルパーT細胞を活性化させ、これはHBVに対する抗体の形成を促進させることができる。
【0038】
本発明で使用された用語「ハイブリッドポリペプチド」とは、一般に、起源の異なる異種ペプチドが連結された形のペプチドをいい、本発明では、B細胞エピトープとT細胞エピトープとを連結した形のペプチドを意味する。このようなハイブリッドポリペプチドは、それぞれのパトナーが決定された後、化学的に合成するか、遺伝子組み換え方法で発現及び精製して得ることができる。好ましくは、B細胞エピトープをコードする遺伝子配列とT細胞エピトープをコードする遺伝子配列とを連結したハイブリッド遺伝子を細胞発現システムで発現させて提供される。このようなハイブリッドポリペプチドは、B細胞エピトープとT細胞エピトープとが直接或いはリンカーなどの連結子を介して連結できる。リンカーを介して連結される場合、これによる免疫反応の誘導に不利な影響を及ぼさないように選定されるべきである。
【0039】
本発明で使用された用語「ポリペプチド」は、2つ以上のアミノ酸を含む残基が共有ペプチド結合によって結合した全長のアミノ酸鎖を含む用語であって、ジペプチド、トリペプチド、オリゴペプチド、ポリペプチドなどがこれに含まれるが、特に、本発明では、数個〜数十個のアミノ酸が共有結合したペプチド2種以上が互いに連結されたハイブリッドポリペプチドを意味する。本発明のハイブリッドポリペプチドは、2種以上のペプチド、例えばB細胞エピトープとT細胞エピトープとが連結されたポリペプチドである。ポリペプチドを構成するそれぞれのペプチド配列は、前述したようなエピトープに該当する配列を含み、それに隣接して配置された配列も共に含むことができる。これらのペプチドは、L−アミノ酸、D−アミノ酸、または2つの相異なる配置のアミノ酸の多様な組合せを取ることができる。本発明のハイブリッドポリペプチドは、全的に前述したようなB細胞エピトープ、T細胞エピトープ、及び任意にその隣接配列を含んだ抗原性部分のみからなってもよく、付加配列を含んでもよい。ところが、このような付加配列は、全体免疫原性(immunogenicity)を減少させないことが好ましい。このような付加配列としては、リンカー配列を例示することができる。
【0040】
本発明で使用された用語「免疫原性」とは、細胞性免疫及び体液性免疫反応を誘導して異物に対して対処する能力をいう。このような免疫反応を誘導する物質を免疫原と呼ぶ。本発明は、免疫原性物質として、B細胞エピトープとT細胞エピトープを両方とも備えるポリペプチドを使用する。
【0041】
本発明者らは、B細胞エピトープを持っているが、それ自体ではT細胞エピトープ部分が欠如している抗肥満機能的ペプチドであるApo B−100ミメティックペプチドの四量体PB14のC末端部分と、T細胞エピトープを持っているHBVのpreS2の一部(T断片)とを連結し、PB14Tの発現用遺伝子断片を製作した(図1)。PB14断片を得るためにBamHIとXhoIを、T断片を得るためにSalIとHindIIIを使用し、PB14Tの遺伝子断片とベクターpQE30とを結合して大腸菌JM109に形質転換した。制限酵素切断(図2)とDNA配列分析(図3)によってT細胞エピトープとB細胞エピトープ部分とが連結されている正しいクローンであることを確認し、このクローンをpB14Tと命名した。PB14T及びPB18の発現のためにpQE30ベクターを使用したが、ベクター自体内にある開始コドンから蛋白質の発現が始まり、精製を便利にするための6個のヒスチジンも共に発現し、エンテロキナーゼ(enterokinase)部位も発現する。このように発現する蛋白質の大きさは、PB14Tは16.2kDaであり、PB18は16.5kDaであった。時間帯別にサンプルを得て、SDS−PAGEによって発現を確認した(図4、図5)。
【0042】
したがって、本発明の具体的な実施様態によって四量体のアポリポ蛋白質B−100のミメティックペプチドとB型肝炎ウィルスの表面抗原preS2とが連結された配列番号9の免疫原性ハイブリッドポリペプチドが提供できる。
【0043】
本発明の免疫原性ハイブリッドポリペプチドは、化学的に合成するか、遺伝子組み換え方法によって生産することができ、好ましくは組み換えベクターを用いてこれを宿主細胞に形質転換させ、これから発現されたポリペプチドを分離、精製して生産することができる。
【0044】
したがって、他の様態として、本発明は、前記免疫原性ハイブリッドポリペプチドをコードする遺伝子を含む組み換えベクトル、及び前記組み換えベクターで形質転換された宿主細胞を提供する。
【0045】
別の様態として、本発明は、前記組み換えベクターで形質転換された宿主細胞を培養して前記免疫原性ハイブリッドポリペプチドを生産する方法を提供する。
【0046】
遺伝子組み換え方法によって本発明の免疫原性ハイブリッドポリペプチドを生産する過程は、次の段階を含む。
【0047】
第1段階は、ハイブリッドペプチドをコードする遺伝子をベクターに挿入して組み換えベクターを製造する段階である。外来遺伝子を挿入するためのベクターとしては、プラスミド、ウィルス、コスミドなど多様な形のベクターを使用することができる。組み換えベクターは、クローニングベクター及び発現ベクターを含む。クローニングベクターは、複製基点、例えばプラスミド、ファージまたはコスミドの複製基点を含み、他のDNA切片が付着し、付着している切片が複製できる「レプリコン」である。発現ベクターは、蛋白質の合成に使用されるように開発された。組み換えベクターは、通常、外来DNAの断片が挿入されたキャリアであって、一般に二本鎖のDNAの断片を意味する。本願において、「外来DNA」とは、外来種から起源するDNAを意味し、或いは同一の種から起源する場合には本来の形から実質的に変形された形を意味する。また、細胞で正常的に発現しない非変化DNA配列も含む。この場合、外来遺伝子は、転写される特定の目的の核酸でポリペプチドを暗号する。組み換えベクターは、宿主細胞における形質感染遺伝子の発現水準を高めるためには、当該遺伝子が選択された発現宿主内で機能を発揮する転写及び解読発現調節配列に作動可能に連結されなければならない。前記組み換えベクターは、個体の細胞内で遺伝子挿入物が発現されるように作動可能に連結された必須的な調節要素を含む遺伝子作製物であって、このような遺伝子作製物を製造するために、標準組み換えDNA技術を利用することができる。組み換えベクターの種類は、原核細胞または真核細胞の各種宿主細胞で目的の遺伝子を発現させ、目的の蛋白質を生産する機能を行うものであれば、特に限定されないが、強力な活性を示すプロモータと強い発現力を保有しながら自然状態と類似な形の外来蛋白質を大量生産することが可能なベクターが好ましい。組み換えベクターは、少なくともプロモータ、開始コドン、目的の蛋白質を暗号化する遺伝子、終結コドンターミネータを含んでいることが好ましい。その他に、シグナルペプチドをコードするDNA、エンハンサー配列、目的の遺伝子の5’側と3’側の非解読領域、選別マーカー領域、または複製可能単位などを適切に含むこともできる。
【0048】
第2段階は、前記組み換えベクターを用いて宿主細胞を形質転換させた後、培養する段階である。組み換えベクターを宿主細胞に導入して形質転換体を製造するための方法としては、文献(Sambrook, J. et al., Molecular Cloning, A Laboratory Manual(2版)、Cold Spring Harbor Laboratory, 1. 74, 1989)に記載のリン酸カルシウム法または塩化カルシウム/塩化ルビジウム法、エレクトロポレーション法、エレクトロインジェクション、PEGなどの化学的処理方法、遺伝子銃(gene gun)などを利用することができる。前記組み換えベクターが発現する形質転換体を栄養培地で培養すると、有用な蛋白質を大量に製造、分離可能である。培地と培養条件は、宿主細胞に応じて、寛容されるものを適切に選択して用いることができる。培養の際に細胞の生育と蛋白質の大量生産に適するように温度、培地のpH及び培養時間などの条件を適切に調節しなければならない。本発明に係る組み換えベクターで形質転換できる宿主細胞は、原核細胞と真核細胞を両方とも含み、DNAの導入効率が高く、導入されたDNAの発現効率が高い宿主が通常用いられる。細菌、例えばエシェリキア、シュードモナス、バシラス、ストレプトミセス、真菌、酵母などの周知の真核及び原核宿主、SF9(Spodoptera frugiperda)などの昆虫細胞、CHO、COS1、COS7、BSC1、BSC40、BMT10などの動物細胞などが使用できる宿主細胞の例である。好ましくは、大腸菌が使用できる。
【0049】
第3段階は、ハイブリッドペプチドの発現を誘導、蓄積する段階である。本発明では、誘導因子IPTGを用いてペプチドの発現を誘導し、誘導時間は蛋白質の量を最大化されるように調節した。
【0050】
第4段階は、ハイブリッドペプチドを分離、精製する段階である。一般に、組み換え的に生産されたペプチドは、培地または細胞分解物から回収できる。膜結合型の場合、適切な界面活性剤溶液(たとえばTriton−X 100)を使用し、或いは酵素的切断によって膜から遊離できる。ハイブリッドペプチドの発現に使用された細胞は、凍結−解凍反復、音波処理、機械的破壊または細胞分解剤などの多様な物質的または化学的手段によって破壊でき、通常の生化学分離技術によって分離、精製可能である(Sambrook et al., Molecular Cloning: A laborarory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, 1989; Deuscher, M., Guide to Protein Purification Methods Enzymology, Vol. 182. Academic Press. Inc., San Diego, CA, 1990)。電気泳動、遠心分離、ゲルろ過、沈澱、透析、クロマトグラフィー(イオン交換クロマトグラフィー、アフィニティクロマトグラフィー、免疫吸着アフィニティクロマトグラフィー、逆相HPLC、ゲル浸透HPLC)、等電点電気泳動法、並びにその多様な変化及び複合方法を含むが、これらに局限されない。
【0051】
本発明では、具体的にPB14Tの遺伝子断片をベクターpQE30と結合して大腸菌に形質転換した。pQE30は、ファージT5プロモータからなるプロモータとIPTGを誘導剤として使用するlacオペレーターシステムを持っている、大腸菌から蛋白質を大量生産するのに有用なベクターである。PB14Tを認識することが可能な2つの抗体、1次抗体はウサギ抗−PB14ポリクローナル抗体とマウス抗−preS2モノクローナル抗体を使用するウェスタンブロットによって正しく発現することを確認し、蛋白質の精製を始めた。PB14とPB14Tは不溶性なので、8M尿素に変性させた後、ヒスチジンタグ(tag)蛋白質用Ni−NTA樹脂を利用するアフィニティクロマトグラフィーを用いてPB14とPB14Tを精製した。
【0052】
上述したように発現及び精製したポリペプチドをマウスに接種した後、それによる体重の増加、血清内抗体力価、血清内脂質含量の変化を分析した。その結果、ハイブリッドポリペプチドでワクチン化されたグループが、対照グループ、またはハイブリッドされていないミメティックペプチドでワクチン化されたグループに比べて体重増加が抑制され、ミメティックペプチドに対する抗体力価が高く、残存期間が長く、TG及びLDL−コレステロールの数値も低くなることを確認した。
【0053】
ペプチドワクチンのデザインに一貫的に適用することが可能な様態が存在せず、デザインされたワクチンの効能も予測することができない。同一の理由から、疎水性が非常に強いPB14ペプチドを異種ペプチドであるT細胞エピトープと融合させる場合、抗原性を示す部位を分子の内部に取り入れて抗体誘導能の低下をもたらす可能性も排除することができない。このようにその結果を類推し難い状況で、本発明者らは、アポリポ蛋白質のB細胞エピトープのミメティックペプチドにT細胞エピトープを連結したハイブリッドポリペプチドを設計し、抗肥満に対する増加した免疫原性を立証した。
【0054】
人工合成ハイブリッドポリペプチド及びこれを含むワクチンの免疫原性は、B細胞エピトープとヘルパーT細胞エピトープが存在しなければならず、また、B細胞エピトープとヘルパーT細胞エピトープとの連結順序(orientation)によってワクチンの効能が異なる。すなわち、ヘルパーT細胞のエピトープがB細胞エピトープのN末端に位置するか、C末端に位置するかによって抗体誘導能が異なる(Partidos, C, Stanley, C, and Steward, M, The effect of orientation of epitope on the immunogenicity of chimeric synthetic peptides representing measles virus protein sequences, Molecular Immunology,29(5),651-658,1992)。
【0055】
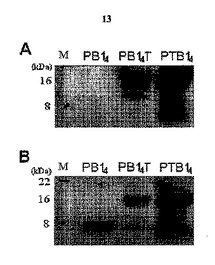
本発明者らは、B細胞エピトープとヘルパーT細胞エピトープとの連結順序による免疫誘導効果の差異を確認するために、前記PB14のC末端部分とT断片とを連結して製作したB14T遺伝子断片とは異なり、PB14のN末端部分とT断片とを連結してTB14遺伝子断片を製作した(図12)。具体的に、実施例9による方法によってベクターpTB14を製作し、大腸菌M15に形質転換させて発現させ、発現したハイブリッドポリペプチドPTB14は、His−Tagが存在するので、Ni−NTA His−結合レジンを用いたアフィニティクロマトグラフィーを行って精製した。
【0056】
PTB14とPB14Tの抗体誘導反応性及び免疫原性を比較するために、白ネズミを用いて免疫化した後、採血してこれを比較した。PTB14の抗体誘導能はPB14よりは多少向上しており、残存期間の長期化現象を示したが、PB14Tに比べて約50〜60%程度に著しく弱いことを確認することができた(図16)。これは、体重増加抑制効能においても同一の結果を示した(表2)。前述したように、本発明のPB14のC末端部分とT断片とを連結して製作したPB14Tポリペプチドの免疫原性及び肥満抑制効能がより優れることを確認することができた。
【0057】
したがって、別の様態として、本発明は、アポリポ蛋白質B−100のエピトープのミメティックペプチドのアミノ酸配列を含み、前記ミメティックペプチドのC末端とヘルパーT細胞のエピトープのN末端とが融合した免疫原性ハイブリッドポリペプチドを含む肥満予防または治療用ワクチンを提供する。
【0058】
免疫原が示す反応の大きさ及び観察された個体の比率を比較すると、優れた効果を示すワクチンとして使用できるかどうかを判断することができる。本発明では、肥満予防及び治療用ワクチンという元々の発明の目的に合うように、(a)体重の増加推移観察、(b)血清内抗体力価の観察、及び(c)血清内脂質含量の変化によって免疫反応効果を検定し、抗原の優れた形態を立証した。
【0059】
具体的に、7週齢SD系白ネズミ(rat)の抗原注射による体重の変化を、精製PB14、PB14Tのペプチド100μgを腹腔に2週間隔3回注射した後、体重増加グラフで確認した(図15)。1次注射とブーストのときまでは各グループの身体質量は292〜297gと同様であったが、2回注射の1週後から、ワクチン化されたグループが対照グループ及び模擬グループとは差異を示し始めた。これは、最初注射による微弱な免疫反応が2次注射によってブーストされた後、増強した免疫応答反応によって体重増加が抑制されることを示す。対照グループ及び模擬グループよりはワクチン処理したときに体重の増加程度が減少し、また、PB14形態のペプチドよりPB14T形態のペプチドがより優れた体重増加抑制効果を示す(表2)。3次注射後にも、体重増加の偏差は維持された。また、キャリア蛋白質であるオボアルブミンを抱合させた形のPB14よりは均質なキメラ抗原PB14Tが免疫応答反応を効率よく誘導することを確認することができた。ワクチン処理されたグループにおけるSD系白ネズミの抗体力価は、10、12、14、16週齢でELISA分析した(図16)。PB14Tを注射したグループがPB14を注射したグループに比べて抗体力価が増加し、14週齢では、PB14Tでワクチン処理されたグループが、PB14ワクチン処理されたグループよりO.D数値が1.5倍高かった。16週齢では、PB14グループにおいて力価の減少が現われたが、PB14Tグループは力価の増加が維持された。血清内脂質水準は、ワクチン処理されたグループが、対照グループ及び模擬グループより全体的にTG及びコレステロールの数値が低かった。特に、LDL−コレステロールは正常値の60%水準と低くなった(図17)。
【0060】
前記結果は、B細胞エピトープであるPB14のみが在る形態より、T細胞エピトープが共に融合している形態が一層高い免疫原性を示し、効果的なワクチン組成物として使用できることを立証する。
【0061】
また、本発明者らは、PB14Tの効能を愛玩犬を対象として臨床実験した。PB14Tをアルミナと混合して10匹の愛玩犬に2週間隔で2回接種した後、体重変化を観察した結果、間食を始めとした高脂肪食を無制限提供しても、体重の増加は全く観察されなかった(表4)。抗体誘導の度合いにおいても、2次接種後、各愛玩犬の血清を採取して抗体力価をELISA実験法によって測定した結果、5000〜50000倍希釈した血清でも高い吸光度を示して優れた効果を発揮した。
【0062】
本発明の抗肥満ワクチンは、抗原、薬剤学的に許容可能な担体、適切な補助剤、その他の通常の物質からなり、免疫学的効果量で投与する。本発明において、「免疫学的効果量」とは、肥満治療及び予防効果を示すことができる程度の十分な量と、副作用や深刻なまたは過度な免疫反応を起さない程度の量を意味し、正確な投与濃度は投与される特定の免疫原によって異なり、免疫反応の発生を検査するために、当業者が公知の方法を用いてこれを決定することができる。また、投与形態及び経路、収容者の年齢、健康及び体重、症状の特性及び程度、現在治療法の種類、及び治療回数によって変化できる。担体は、当分野に公知になっているもので、安定化剤、希釈剤、緩衝液を含む。適切な安定化剤は、ソルビトール、ラクトース、マンニトール、澱粉、糖、デキストラン及びブドウ糖などの炭水化物;アルブミンまたはカゼインなどの蛋白質などを含む。適切な希釈剤には塩、ハンクス平衡塩、リンガー液などを含む。適切な緩衝液には、アルカリ金属リン酸塩、アルカリ金属炭酸塩、アルカリ土金属炭酸塩などを含む。また、ワクチンには、免疫反応を改善または強化させるために、少なくとも一つの補助剤などを含む。適切な補助剤には、ペプチド;水酸化アルミニウム;リン酸アルミニウム;酸化アルミニウム;Marcol 52などのミネラル油または植物性油及び少なくとも一つの乳化剤からなる組成物、またはリゾレシチン、多価陽イオン、多価陰イオンなどの表面活性物質などが含まれる。本発明のワクチン組成物は、個別治療剤として投与してもよく、他の治療剤と併用して投与してもよく、従来の治療剤とは順次または同時に投与してもよい。ワクチン組成物は、公知の投与経路によって投与される。このような方法には、経口、経皮、筋肉、腹膜、静脈、皮下、鼻腔経路を利用することができるが、これらに局限されない。また、製薬組成物は、活性物質が標的細胞へ移動することが可能な任意の装置によって投与できる。
【0063】
以下、実施例によって本発明をより具体的に説明する。これらの実施例は本発明を例示するためのもので、本発明の範囲を限定するものではない。
【0064】
〔実施例〕
<実験材料>
DNAミニプレップキット(miniprep kit)とゲルからDNAを抽出するために使用されたキットはNucelogen製を、細胞培養に必要なバクト−トリプトン(bacto-trypton)、バクト−酵素抽出物(bacto-yeast extract)、アガ(agar)などは、Difco社(Detroti、MI)製を、制限酵素はTakara製を、T4DNAリガーゼはNEB製をそれぞれ使用した。ベクターはpB1uescriptIISK(Stratagene社)、PCR2.1(Invitrogen、Carlsbad、CA)、pQE30(Qiagen社)を使用し、菌株は大腸菌JM109及びM15(Qiagen社)を使用した。
【0065】
蛋白質の生成を誘導するためのIPTGはSigma社製を、発現された蛋白質を精製するためのNi−NTAレジンはNovagen社製を使用した。SDS−PAGE及びウェスタンブロット、ECLなどに用いられた予備染色マーカー(prestained marker)はNEB社製であった。蛋白質を変性状態にするために使用された尿素はDuchefa社製であり、精製時に使用されたイミダゾールはUSB製であった。透析に使用された膜はSpectrum製のMWCO 3500であり、蛋白質凝集防止のために使用された試薬はAmresco製のCHAPSであった。ELISAに使用された抗体は、Sigma製のHRP結合抗−白ネズミIgGであった。ウェスタンブロットとECLに使用された基質溶液として、BCIP/NBTはSigma製を、ECL+plusウェスタンブロット検出試薬はAmersham製を使用した。補助剤は、フロイントアジュバント(Freund Adjuvant;Sigma社製)と水酸化アルミニウム(Reheis社製)を使用した。蛋白質の定量は、BCA蛋白質検査法(Pierce)とBradford法(Biorad)を使用した。
【0066】
血清内中性脂肪はトリグリザイム−Vを、総コレステロールはコレステザイム−Vを、HDLコレステロールはHDL−C555(シンヤン化学社製)を、LDLコレステロールはEZLDLコレステロール(Sigma社製)を用いて測定した。LDL測定器は、Randox製を使用した。
【0067】
SD(Sprague Dawley)系白ネズミは、(株)大韓バイオリンクから購入し、5週齢以上の雄を使用した。白ネズミの飼料としては韓国の(株)Samtako製(成分:天然蛋白質18%以上、粗脂肪5.3%、粗繊維4.5%、回分8.0%)を使用した。
【0068】
組み換えPB14T及びPB14ペプチドの精製のために、超音波粉砕緩衝液(5mMイミダゾール、0.5M NaCl、20mM tris−Cl、pH7.9)、結合緩衝液(5mMイミダゾール、0.5M NaCl、20mM tris−Cl、8M尿素、pH7.9)、洗浄緩衝液(50mMイミダゾール、0.5M NaCl、20mM tris−Cl、8M尿素、pH7.9)、溶出緩衝液(400mMイミダゾール、0.5M NaCl、20mM tris−Cl、8M尿素、pH7.9)を使用した。
【0069】
<実施例1:抗肥満作用ペプチドPB14T人工遺伝子の作成>
pB1uescritIISKベクターに入っているB14断片を得るために、BamHIとXhoI制限酵素を用いて切断し、PCR2.1ベクターに入っているT断片を得るために、SalIとHindIII制限酵素を用いて切断した。2つのベクターから得られた切片、B14とTに対して、B14のXhoI粘着末端とTのSalI粘着末端が相補的なので、T4 DNAリガーゼを用いて16℃で12時間結合させた。その結果、連結部位はSalIまたはXhoIによって切られなくなり、B14T断片を得るためにもう1回SalIとHindIIIで切った。蛋白質を発現するために選択されたベクターシステムとしてpQE30プラスミドを使用した。このベクターは、発現させようとする蛋白質が6Xヒスチジンと融合蛋白質の形で発現して精製が容易となるように考案された。B14T遺伝子断片をpQE30ベクターのSalI及びHindIII部位と結合した。こして作られたクローンをpB14Tと命名した(図1)。大腸菌JM109を形質転換させた後、プラスミドを分離してSalIとHindIIIで切って、450bp断片が挿入されたことを確認した(図2)。
【0070】
前述した組み換えベクターpB14Tは、大腸菌M15/pB14Tの形であって、2004年3月4日付けでKCCM(Korean Culture Center of Microorganisms、韓国ソウル市西大門区弘済1洞ユリムビル361−221)に寄託番号KCCM−10562で寄託された。
【0071】
<実施例2:抗肥満作用ペプチドPB18人工遺伝子の作成>
pB1uescriptIISKベクターに入っているB14断片をSalIとXhoIで切って得た。pBX4ベクター(B14断片が挿入されているpQX30ベクター、韓国特許公開第2002−0018971号)をSalIで切って前記B14断片とT4DNAリガーゼを用いて16℃で一晩中結合させた。
【0072】
<実施例3:遺伝子塩基配列の決定>
クローニングされたpB14Tの塩基配列を確認するために、プラスミド濃度を300〜500ng/μLとして韓国の(株)コアバイオシステムにDNA塩基配列分析を依頼し、正しいクローンであることを確認した(図3)。
【0073】
<実施例4:組み換えPB14Tペプチドの発現>
PB14T及びPB18は、pQE30ベクターで発現するので、ベクター自体内の開始コドンから蛋白質の発現が始まり、精製を便利にするために6個のヒスチジンも共に発現し、エンテロキナーゼ部位も含んでいる。ペプチドの発現に使用した宿主細胞は、大腸菌M15であって、アンピシリンとカナマイシンが含有されたLBプレートに塗抹してコロニーを得た後、Amp(100μg/mL)とKan(25μg/mL)が含有されたLB培地10mLで一晩中培養した。時間の変化に伴う蛋白質の誘導を調べるために、一晩中培養した培養液中の1mLを新鮮なLB培地50mLに接種した。本培養液を600nmで吸光度が0.4〜0.5となるまで37℃で1時間30分振とう培養した後、IPTGを最終1mM濃度となるように加え、5時間培養を行い続けて1時間ごとに1mLずつ分取した。IPTGを加える前に1mLを予め取って非誘導対照グループとして使用した。それぞれの培養液は、14,000rpmで1分間遠心分離してペレットを得た後、2×SDSサンプル緩衝液30μLに溶かしてSDS−PAGEの際にサンプルとして使用した。こうして計算された蛋白質の大きさは、それぞれPB14Tは16.2kDa、PB18は16.5kDaであった。時間帯別にサンプルを得てSDS−PAGEによって発現を確認した結果を、図4及び図5に示す。
【0074】
<実施例5:組み換えPB14Tペプチドのウェスタンブロット>
SDS−PAGEにおいて大きさの分析によってPB14Tを確認したが、正確な蛋白質が発現するかを確認するために、PB14Tを認識することが可能な2つの抗体を用いてウェスタンブロットを行った。PB14Tウェスタンブロットの対照グループには、M15にB14TがサブクローニングされていないpQE30ベクターのみを形質転換させて使用した。サンプルは、IPTGで誘導する前のものと、IPTG誘導3時間後のものにした。1次抗体は、ウサギ抗−PB14ポリクローナル抗体、マウス抗−preS2モノクローナル抗体をPBSに1:10000で希釈して使用した。1次抗体を認識することが可能な2次抗体は、ペルオキシダーゼが結合している山羊抗−ウサギIgG、山羊抗−マウスIgGをPBSに1:10000で希釈して使用した。発色反応は、ECL+Plusウェスタンブロットキットを使用し、膜をカセットに入れて富士メディカルのX線フィルムを入れた後、10秒間露出させて現像した。ウサギ抗−PB14ポリクローナル抗体はPB14TのPB14断片を認識する抗体であり、マウス抗−preS2モノクローナル抗体は、PB14TのT断片を認識する抗体なので、正確に蛋白質が発現すると、2つの抗体を使用したブロックの全てでバンドが現われるべきである。図8に示すように、それぞれの抗体がPB14TのPB14とTを認識したので、PB14Tが正確に発現したことを確認できた。
【0075】
<実施例6:組み換えPB14T及びPB18ペプチドのバクテリア細胞位置の確認>
PB14T及びPB18の細胞位置確認は、IPTGで誘導してから3時間が経過したサンプルを遠心分離し、それを超音波緩衝液で再懸濁させた後、超音波により得られたペレットと上澄み液それぞれSDS−PAGEで確認した。具体的に、IPTGで発現が誘導された細胞を4℃、9000rpmで30分間遠心分離した後、得られたペレットをしばらく−20℃で凍らせてから氷上で溶かし、その後超音波分解緩衝液をペレット1gに対して5mLの割合で入れて再懸濁した。細胞を溶解させるために、30秒ずつ15サイクル(サイクルの間に1分間休止)で超音波粉砕した。4℃、9000rpmで30分間遠心分離して上澄み液を分離し、これを可溶性蛋白質の入っている非加工抽出物A(粗抽出物A)として使用し、残っているペレットは、不溶性蛋白質の入っている非加工抽出物B(粗抽出物B)として使用した。それぞれのサンプルに2×SDSサンプル緩衝液を混ぜて95℃で5分間沸かした後、SDS−PAGEを行った。上澄み液よりはペレットに所望の蛋白質が多量現われることから、不溶性が強いことを確認した(図6及び図7)。
【0076】
<実施例7:組み換えPB14、PB14T及びPB18の精製>
ペプチドを精製するためにヒスチジンタグ蛋白質用Ni−NTAレジンを使用した。レジンに結合しているNi+と末端にヒスチジンヘキサマー融合蛋白質が相互作用する性質を利用することにより親和力を利用したアフィニティクロマトグラフィーの一つの方法である。遺伝子組み換えされた大腸菌を接種したLB培地10mLを一晩中前培養した後、500mLのLB培地に全量接種して600nmで吸光度0.4〜0.5となるまで37℃で培養した。IPTGを1mMで加え、4時間培養した後、9000rpmで30分間遠心分離して細胞ペレット得て−20℃に置いた。氷上で溶かしたペレットに超音波粉砕緩衝液を湿潤細胞と5mL/gの割合で入れて再懸濁させて超音波粉砕を行った。超音波粉砕をして細胞を溶解させ、4℃、9000rpmで30分間遠心分離してペレットを得た。結合緩衝液を、残っている上澄み液の体積と同一の体積となるようにペレットに入れた後、再懸濁させ、細胞残骸を除去するために超音波粉砕を3サイクル行った。これを再び4℃、9000rpmで30分間遠心分離して得た上澄み液を精製用サンプルとして使用した。
【0077】
Ni−NTAレジン充填カラムの大きさは直径1cm、高さ15cmであり、レジンは2mL、流速は2mL/minとした。レジンをカラムに充填した後、3〜5倍体積の蒸留水で洗浄し、5倍体積の1×荷電された緩衝液(50mM NiSO4)を流してNi2+を結合させた後、結合緩衝液で平衡化してNi−キレート化親和性樹脂を製造した。レジンにサンプルローディングを2回行った。こうしてサンプルローディングが完了すると、さらに結合緩衝液で1.0範囲の280nm吸光度で基底線に到達するまで流した。その後、洗浄緩衝液で10分間流し、平衡化が完了すると、洗浄緩衝液から溶出緩衝液にイミダゾールの濃度勾配をかけ、レジンに残留する蛋白質を完璧に溶出させるために溶出緩衝液のみを10分間さらに流した。分画は層20個であり、それぞれ2mLずつであった。精製されたペプチドは8M尿素状態であり、尿素を除去するために、PBSを用いて一晩中透析を行った。
【0078】
前述したように、それぞれの蛋白質が強い不溶性を持つため、精製は8M尿素入り緩衝液で蛋白質を変性させてから行い、樹脂に結合した蛋白質を溶出させるためにイミダゾールを使用し、濃度は50mM〜400mMの濃度勾配で精製した。その結果が図9、図10及び図11に示されている。約300mMのイミダゾール濃度のとき、大部分の蛋白質が溶出した。それぞれの蛋白質の収率は1L培養の際にPB18の場合には3〜3.5mg、PB14Tの場合には4〜4.5mgであった。
【0079】
<実施例8:組み換えPB14T、PB14及びPB18ペプチドの定量>
PB14T、PB14、PB18は、PBSで透析を行うと、尿素が除去されるので、蛋白質の凝集による沈殿が発生し、この状態では蛋白質の定量を正確に行うことができなかった。精製された蛋白質の凝集状態を除去するために、50mM CHAPSを使用した。定量は、BCA蛋白質定量法、ブラッドフォード法を用いて行った。2mg/mLのBSAを1000、500、250、125、62.5μg/mLと逐次希釈して標準として用いた。BCA分析は、Pierce社製のプロトコールによって行い、37℃で30分間反応させ、562nmで吸光度を測定し、ブラッドフォード試薬はサンプルと室温で10分間反応させた後、595nmで吸光度を測定し、BSAの逐次希釈液のBCA或いはブラッドフォード蛋白質定量発色反応の吸光度を標準定量線として用いてサンプルの蛋白質の濃度を定量した。
【0080】
<実施例9:PTB14の発現のためのベクターpTB14の製作>
M15大腸菌株に形質転換されている発現用ベクターpQE30においてTカセット(preS2)をKpnI(Takara)、SalI(Takara)制限酵素で二重切断によって分離し、pB1uescriptも同一の酵素で処理した。分離したTカセットとpB1uescriptをそれぞれゲル上で確認して分離溶出させ、溶出させたベクターとTカセットはT4 DNAリガーゼで連結させた。pB1uescript4μL、Tカセット4μL、リガーゼ1μL(MBI Fermentas、1Weissμ/mL)、10×緩衝液(MBI Fermentas)1μLを1.5mLのチューブに入れて16℃で一晩中培養させた。組み換えされたベクターをJM109細胞と混ぜて42℃で90秒間熱衝撃を与えた後、LB培地を入れて37℃で1時間培養した。これをLB(Amp)培地に接種し、37℃で培養してコロニーを確認した。確認されたコロニー中の幾つかを任意選択して培養過程、プラスミド分離精製、制限酵素切断を経てアガロースゲル上でDNA断片のサイズによって確認した。Tカセット内にはXhoI部位が存在するが、これを無くさなければ、連結されたTB4カセットを得ることができなかった。すなわち、Tとは、Tカセット(HBVウィルスのpreS2遺伝子、183bp)の3’末端近く(Tカセットの5’を基準として約150bp)にXhoI部位が存在してクローニングに使用することができないので、内部のXhoI部位を点突然変異させたXhoIで切られない新しい配列のTカセットの形態である。pB1uescript−preS2は、Tカセット内部のXhoI部位により短いDNA断片(30bp)が切り出された。この部分に合成オリゴマーを挿入した。セルフライゲーション(Self-ligation)を防ぐために、ベクターは37℃、30分間アルカリ性ホスファターゼ(Boehringer Mannheim、GembH、Germany)で処理し、95℃で5分間脱リン酸させてゲル上で溶出させた。オリゴマーは、37℃で30分、65℃で20分間処理して5’−リン酸化させた。95℃で5分間静置反応させた後、熱遮断(heat block)をして徐々に冷やしながらアニーリングさせた。オリゴマーとpB1uescrpt−Tリガーゼを用いて16℃で一晩中培養した。組み換えされたpB1uescrtip−TをJM109細胞に形質転換導入してLB(Amp)培地でコロニーを確認し、コロニー選別過程を経て、所望のプラスミドを持った細胞を確保した。オリゴマーはpresS2の当該部分の28個のヌクレオチドからなっており、但し、XhoI部位を無くすために、センス−オリゴマー5’の5番目のヌクレオチドをGからAに変え、これによりアミノ酸配列がアルギニンからリジンに変わった。センスとアンチセンスでそれぞれ28merを製作してアニーリングさせ、これを、XhoIで処理したpB1uescript−preS2内に挿入した。pB1uescript−TをSalI、XhoIで二重切断し、pQE30−B4をSalIで処理してそれぞれをゲル上で分離溶出させた。5’側が切り出されたpQE30−B4に、分離されたTを挿入し、pQE30−pTB4を作成した。組み換えされたTB4をSalIとHindIIIで切ってバンドの大きさを確認した。こうして作成されたベクターをpTB14と命名した(図13)。
【0081】
<実施例10:PTB14の発現及び精製>
発現ベクターpTB14をM15に導入してLB(Amp、Kan)培地2Lに培養した。これを7000rpmで10分間遠心分離して湿潤細胞9gを回収した。遺伝子組み換えで発現されたハイブリッドポリペプチドPTB4は、His−Tagが存在するので、Ni−NTA His−結合レジンを用いたアフィニティクロマトグラフィーを行った。カラムは、レジン体積4mLで直径1.8cm、高さ8cmのものを使用した。Econo systemの吸光度範囲は0.5、記録計の用紙速度は2cm/hour、サンプルローディング速度は2mL/minに合わせて使用した。まず、湿潤細胞に超音波粉砕緩衝液(sonication buffer)を入れて超音波破砕によって細胞を壊し、4℃、10000rpmで30分間遠心分離した。ペレットのみを結合緩衝液に溶かした後、アフィニティクロマトグラフィーで精製した。まず、結合溶液をカラムに流しながらレジンを沈殿させた。検出器を見ながら、ベースラインを定め、一定の値を指すと、超音波粉砕したサンプルを入れた。サンプルがカラム内に入って再び一定の値を指すまで待ってから洗浄溶液を流し、その後さらに一定の値を指すと、溶出溶液も流してPTB14を分離した。発現及び精製されたハイブリッドポリペプチドは、SDS−PAGE後、ウェスタンブロットで確認した。SDS−PAGEしたPTB14を半乾燥トランスファ(semi-dry transfer)を用いて膜に移した。ブロッキング溶液(0.5% casein−phosphate buffered saline-tween、0.02%NaN3)を用いて37℃で2時間恒温処理した後、Tris-buffered saline-tween(TBS−T、pH7.6)で2分ずつ2回洗浄した。その後、1次抗体を37℃で1時間処理し、TBS−Tで5分ずつ4回洗浄した。2時抗体も1時間処理した後、同一の方法で洗浄した。Tカセットの確認には抗−preS2モノクローナル抗体(1:10000)、山羊抗−マウスIgG−HRP結合抗体(1:10000)を使用し、Bカセットの確認にはウサギ抗−PB14抗血清(1:10000)、山羊抗−ウサギIgG−HRP結合抗体(1:10000)を使用した。ECLキット溶液を乾燥膜上に散布して5分間反応させ、これをECLキットとしてバンドを確認した。その結果、Bカセットは、ウサギ抗−PB14抗血清と山羊抗−ウサギIgG−HRP結合抗体を用いてPB14TとPTB14でのみ約16KDaのバンドが現われたことを見ることができ、Tカセットは、抗−preS2モノクローナル抗体と山羊抗−マウスIgG−HRP結合抗体を用いて、PB14では約8kDaのバンドが観察され、PB14TとPTB14では16kDaのバンドが見えてそれぞれのハイブリッドポリペプチドが正確に発現、精製されたことを確認した(図14)。
【0082】
<実施例11:PB14とオボアルブミンとの連結>
PB14をキャリア蛋白質としてのオブアルブミンと連結した。キャリア蛋白質とPB14のモル比が約1:10となるようにした。約1時間4℃で反応バイアルで攪拌し、2%グルタルアルデヒド溶液を入れた後、3時間攪拌し、PBS緩衝液でMWCO3000透析膜を用いて一晩中透析して残余のグルタルアルデヒドを除去した。
【0083】
<実施例12:ワクチン処理(免疫化)>
7週齢のSD系白ネズミを6グループに分けてワクチン処理を行った(表1)。実施例7及び実施例10で精製及び定量したペプチド100μgを、表1に示すように、それぞれの補助剤と混合して100μLとなるようにして腹腔に注射した。注射は2週間隔で7、9及び11週齢に3回にわたって行った。補助剤はフロイントアジュバントと水酸化アルミニウムを使用した。フロイントアジュバントはペプチドと同量で混ぜ、水酸化アルミニウムは5.8mg/mLを最終濃度が0.2mg/mLとなるように調整した後、ペプチドと室温で攪拌しながら恒温処理した。最初ブースト5日目、最終ブースト5日目、2週目及び4週目に尻から採血した。
【0084】
【表1】
【0085】
前記ワクチン処理後の体重変化を図15にグラフで示した。最初注射してからブーストするまでは、各グループ別体重が292〜297gと同様であったが、2回注射後1週前からは、ワクチン処理グループ、対照グループおよび模擬グループの体重の差異が見られた。18週齢のとき、対照グループおよび模擬グループとPB14注射グループとの体重の差異は16gであり、PB14T注射グループとの体重の差異は27gであった(表2)。これは、最初注射による微弱な免疫反応が2次注射によってブーストされた後、増強した免疫応答反応によって体重の増加が抑制されることを示す。3次注射後にも、体重増加の偏差は維持された。
【0086】
【表2】
【0087】
表2において、標準偏差は、5匹のSD系白ネズミに対して計算されたものであり、それぞれの単位はgで表わす。
【0088】
<実施例13:抗体力価の測定>
血清サンプルを用いて間接的ELISA法によって抗体力価を測定した。微細力価平板(microtiter plate)にPB14を各ウェルに100ngの濃度で100μLずつ仕込んだ。4℃で一晩中処理した後、ブロッキング溶液(PBS、0.5%カゼイン、0.02%NaN3)を添加し、37℃で1時間恒温処理した。PBSTで3回洗い出し、SD系白ネズミワクチン処理によって得た血清をPBSに1/500〜1/8000倍で希釈して100μLずつ使用し、37℃で1時間恒温処理した。各ウェルをPBSTで3回洗い出し、2次抗体としての山羊抗−白ネズミIgGの1/1000倍希釈液で培養した。OPD発色反応を行い、吸光度を450nmで測定した。
【0089】
SD系白ネズミが10、12、14、16週齢のとき、ワクチン処理グループで白ネズミの抗体力価を確認した結果が図16に示されている。力価はそれぞれの血清希釈比を1/2000にしたとき、ELISA結果、吸光度が0.6を基準とした。1/500〜1/8000倍で希釈してPB14、PB14T及びPTB14を注射したグループは、14週齢までは抗体力価が増加し、PB14TグループはPB14よりO.D数値で1.5倍高かったが、PTB14グループはPB14グループに比べて多少向上した。16週齢のときは、PB14グループでは力価が減少し、PB14T及びPTB14のグループでは力価が維持されたが、PTB14はPB14Tに比べて約50〜60%と著しく弱いことを確認することができた。
【0090】
<実施例14:血清脂質含量の測定>
TGと総コレステロールの測定は、発色試薬200μLに血清4μLを入れて37℃で5分間高温処理した後、それぞれ505nm、500nmで吸光度を測定した。HDLの測定は、血清と沈澱試薬を1:1で混ぜた後、室温で10分間放置し、その後3000rpm以上で10分間遠心分離して得た上澄液を発色試薬200μLに4μL入れて37℃で5分間恒温処理した後、555mmで吸光度を測定した。LDLコレステロールの測定はEZ LDLコレステロールキット(Sigma社製)を使用し、LDLカリブレータはRandox社製を使用し、製造社から提供されたプロトコールによってキット内の試薬1150μLに血清4μLを入れて37℃で5分間反応させた後、試薬250μLを添加してさらに37℃で5分間反応し、その後600nmで吸光度を測定した。各測定シートから得た吸光度と標準試液を比較して濃度を計算した。
SD系白ネズミへの3次注射後5週目の血清で分析した結果が表3に示されている。
【0091】
【表3】
【0092】
表3において、標準偏差は5匹のSD系白ネズミに対して計算されたものであり、それぞれの単位はmg/dLで示した。
【0093】
対照グループおよび模擬グループでは、全体的にTG及びコレステロールの数値がワクチン処理グループより約10mg/dL(10mg/100mL)高かった。同じワクチン処理グループにおけるTGとLDLコレステロールの数値は、PB14で高かったが、その差異は大きくなかった(図17)。
【0094】
<実施例15:愛玩犬を対象とした臨床実験結果>
PB14Tをアジュバントとしてのアルミナと混合した混合物(2mg/mL)0.5mLを10匹の愛玩犬(韓国アンサン動物病院で肥満管理を受けている愛玩犬)に筋肉注射又は皮下注射の形で2週間隔で2回接種した後、1.5〜3ヶ月間体重の変化を観察した。その結果、抗体が徐々に減少する形に維持され(半減期:3ヶ月)、間食を始めとした高脂肪食を無制限提供しても、体重は全く増加していない。具体的に、接種した10匹の愛玩犬の全てが、間食及び高カロリー食餌を摂取する場合でも体重の増加が抑制されることを確認した。特に、ヨークシャーテリア(Yorkshire terrier)種の場合、性別及び年齢による体重増加が予想される状況でも、PB14Tの接種により体重が増加しないことを確認することができた。
【0095】
また、各愛玩犬の血清を採取して抗体誘導の度合いを確認するために、2次接種後1週目に、PB14T或いはPB14を認識する血清中の抗体力価をELISA実験法で測定した。その結果、5000〜50000倍で希釈した血清においても0.5の高い吸光度を示して抗体誘導能も非常に優れた。
【0096】
【表4】
【0097】
〔産業上の利用可能性〕
本発明の抗肥満効果を示すアポリポ蛋白質B−100のB細胞エピトープのミメティックペプチドのC末端にヘルパーT細胞のエピトープのN末端を融合させたハイブリッドポリペプチドは、B細胞エピトープの有利な活性または効果を中和させる免疫反応を誘導するか有害な副作用を起すことなく、優れた免疫増強効果を示して肥満の予防または治療に効果的に使用できる。
特許手続き上微生物寄託の国際的承認に関するブダペスト条約
国際様式
このページの下端に確認された国際寄託当局から規則7.1に基づいて発行された原寄託申請の受託証
To:Hyo Joon Kim.
韓国(〒425−791)京畿道安山市常緑樹区四1洞漢陽大学校HBI604
┌──────────────────────────────────────┐
I.微生物表示 │
├─────────────────┬────────────────────┤
寄託者によって与えられた識別番号: 国際寄託当局によって与えられた受託番号 │
M15/pB14T : │
│ KCCM−10562 │
├─────────────────┴────────────────────┤
II. 科学的性質および/または提示された分類学的表示 │
├──────────────────────────────────────┤
前記Iで識別された微生物は、下記のとおりである。 │
[ ]科学的性質 │
[ ]提示された分類学的表示 │
(該当欄にX表示) │
├──────────────────────────────────────┤
III.受付および受託 │
├──────────────────────────────────────┤
本国際寄託当局は、前記Iに表示された微生物が2004年3月4日付で受付され │
たことを確認する。(原寄託日)1 │
├──────────────────────────────────────┤
IV.国際寄託当局 │
├───────────────────┬──────────────────┤
名称:韓国微生物保存センター(KCCM 本国際寄託当局を代表する委任状を有 │
) する者又は権限保有職員の署名 │
住所:韓国(〒120−091)ソウル市 │ │
西大門区弘済1洞361−221 日付:2004年3月12日 │
ユリムビル │ │
└───────────────────┴──────────────────┘
1 規則6.4(d)適用、この日付は国際寄託当局の資格を得た日付であるが、国際寄託当局の資格の獲得後、ブダペスト条約の外でなされた寄託がブダペスト条約下の寄託に転換される場合、この日付は国際寄託当局が微生物を受付けた日付である。
書式BP/4 単一ページ
【図面の簡単な説明】
【0098】
【図1】pB14Tの製作図を示す。
【図2】pB14Tの制限酵素切断を示す。
【図3】DNA配列から類推したpB14Tの一次構造を示す。
【図4】形質転換された大腸菌(Escherichia coli)M15/pB14Tを、IPTGでpB14Tの発現を誘導した後、SDS−PAGE分析した結果である。発現された組み換えPB14Tは矢印で表示した。レインM:予め染色された蛋白質サイズマーカー;レイン1:IPTGで誘導されていない大腸菌M15;レイン3〜7:IPTG誘導1、2、3、4及び5時間後の大腸菌M15/pB14T。
【図5】形質転換された大腸菌M15/pB18を、IPTGでPB18の発現を誘導した後、SDS−PAGE分析した結果である。発現された組み換えPB18は矢印で表示した。レインM:予め染色された蛋白質サイズマーカー、レイン1:IPTGで誘導されていない大腸菌M15;レイン2〜5:IPTG誘導1、2、3及び4時間後の大腸菌M15/pB18。
【図6】大腸菌溶解物(lysate)のSDS−PAGE分析結果である。バクテリア溶解物を遠心分離して上澄液と沈殿物に分離した後、上澄液(レイン1)と沈殿物(レイン2)をSDS−PAGE分析した。PB14Tは、矢印で表示したように沈澱物に現われた。
【図7】大腸菌溶解物のSDS−PAGE分析結果である。バクテリア溶解物を遠心分離して上澄液と沈殿物に分離した後、全体溶解物(レイン1)、上澄液(レイン2)及び沈澱物(レイン3)をSDS−PAGE分析した。PB18は、矢印で表示したように沈殿物に現われた。
【図8】精製されたPB14Tペプチドをウェスタンブロット分析した結果である。抗PB14ウサギ抗体(A)と抗preS2モノクローナル抗体(B)を用いてPB14Tを分析した。レイン1:大腸菌M5、レイン2:IPTGで誘導されていない大腸菌M15/pB14T、レイン3:IPTG誘導3時間後の大腸菌M15/pB14T。
【図9】イミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB14溶出プロファイルを示す。
【図10】イミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB14T溶出プロファイルを示す。
【図11】イミダゾールの線形濃度勾配によるNi−NTAアフィニティクロマトグラフィーのPB18溶出プロファイルを示す。
【図12】pTB14の製作図を示す。
【図13】精製されたPB14、PB14T及びPTB14をウェスタンブロット分析した結果である。パネルAはマウス抗−preS2モノクローナル抗体と山羊抗−マウスIgG−HRP結合抗体と確認し、パネルBはウサギ抗−PB14抗血清と山羊抗−ウサギIgG−HRP結合抗体と確認した。
【図14】DNA配列から類推したTB14/pQE30の一次構造を示す。
【図15】対照グループ(normal)、模擬グループ(mock)、ワクチン処理グループにおけるSD系白ネズミの体重増加を比較したグラフを示す。対照グループ(■)はPBSで注射され、模擬グループ(▲)はオボアルブミンで注射された。ワクチン処理グループはPB14を媒体蛋白質OVAに抱合させたPB14+OVA(◆)ペプチドとPB14T(●)ペプチドで注射された。このようなペプチド注射は、2週間隔で3回ずつ行われた。矢印は、ワクチン処理の時点を示す。
【図16】抗−PB14、抗−PB14T及び抗−PTB14の抗体力価変化を測定した結果である。
【図17】血清内トリグリセリド、HDL、LDL、総コレステロール量を測定した結果である。
【特許請求の範囲】
【請求項1】
配列番号1、2及び3の中から選択されたアミノ酸配列を含むペプチドのC末端とヘルパーT細胞エピトープのN末端とが融合する、免疫原性ハイブリッドポリペプチド。
【請求項2】
ペプチドは、配列番号1、2及び3の中から選択されたアミノ酸配列が2〜8個連結されてなることを特徴とする、請求項1に記載のポリペプチド。
【請求項3】
ペプチドは、配列番号1、2及び3の中から選択されたアミノ酸配列が4個連結されてなることを特徴とする、請求項2に記載のポリペプチド。
【請求項4】
ペプチドは、配列番号1のアミノ酸配列が4個連結されてなることを特徴とする、請求項3に記載のポリペプチド。
【請求項5】
ペプチドは、配列番号5のアミノ酸配列を持つことを特徴とする、請求項4に記載のポリペプチド。
【請求項6】
ヘルパーT細胞エピトープは、B型肝炎ウィルス表面抗原ヘルパーT細胞エピトープ、クラミジア・トラコマティス(Chlamydia trachomatis)主要外膜蛋白質ヘルパーT細胞エピトープ、プラスモジウム・ファルシパルム・サーカムスポロゾイト(Plasmodium falciparum circumsporozoite)ヘルパーT細胞エピトープ、大腸菌TraTヘルパーT細胞エピトープ、破傷風トキソイド(Tetanus toxoid)ヘルパーT細胞エピトープ、ジフテリアトキソイド(diphtheria toxoid)ヘルパーT細胞エピトープ、マンソン住血吸虫(Schistosoma mansoni)トリオースリン酸イソメラーゼヘルパーT細胞エピトープ、麻疹(measles)ウィルスF蛋白質ヘルパーT細胞エピトープ及び狂犬病ウィルスヘルパーT細胞エピトープよりなるグループの中から選択されることを特徴とする、請求項1に記載のポリペプチド。
【請求項7】
T細胞エピトープがB型肝炎ウィルス表面抗原のヘルパーT細胞エピトープであることを特徴とする、請求項6に記載のポリペプチド。
【請求項8】
T細胞エピトープがB型肝炎ウィルス表面抗原のpreS2ヘルパーT細胞エピトープであることを特徴とする、請求項7に記載のポリペプチド。
【請求項9】
T細胞エピトープが配列番号7のアミノ酸配列を持つことを特徴とする、請求項8に記載のポリペプチド。
【請求項10】
配列番号9のアミノ酸配列を持つことを特徴とする、請求項1に記載のポリペプチド。
【請求項11】
請求項1〜10のいずれか1項に記載のポリペプチドを含む肥満予防または治療用ワクチン。
【請求項12】
請求項1〜10のいずれか1項に記載のポリペプチドをコードする遺伝子を含む組み換えベクター。
【請求項13】
pB14T(KCCM−10562)であることを特徴とする、請求項12に記載の組み換えベクター。
【請求項14】
請求項12に記載の組み換えベクターで形質転換された宿主細胞。
【請求項15】
請求項12に記載の組み換えベクターで形質転換された宿主細胞を培養して請求項1に記載のポリペプチドを製造する方法。
【請求項1】
配列番号1、2及び3の中から選択されたアミノ酸配列を含むペプチドのC末端とヘルパーT細胞エピトープのN末端とが融合する、免疫原性ハイブリッドポリペプチド。
【請求項2】
ペプチドは、配列番号1、2及び3の中から選択されたアミノ酸配列が2〜8個連結されてなることを特徴とする、請求項1に記載のポリペプチド。
【請求項3】
ペプチドは、配列番号1、2及び3の中から選択されたアミノ酸配列が4個連結されてなることを特徴とする、請求項2に記載のポリペプチド。
【請求項4】
ペプチドは、配列番号1のアミノ酸配列が4個連結されてなることを特徴とする、請求項3に記載のポリペプチド。
【請求項5】
ペプチドは、配列番号5のアミノ酸配列を持つことを特徴とする、請求項4に記載のポリペプチド。
【請求項6】
ヘルパーT細胞エピトープは、B型肝炎ウィルス表面抗原ヘルパーT細胞エピトープ、クラミジア・トラコマティス(Chlamydia trachomatis)主要外膜蛋白質ヘルパーT細胞エピトープ、プラスモジウム・ファルシパルム・サーカムスポロゾイト(Plasmodium falciparum circumsporozoite)ヘルパーT細胞エピトープ、大腸菌TraTヘルパーT細胞エピトープ、破傷風トキソイド(Tetanus toxoid)ヘルパーT細胞エピトープ、ジフテリアトキソイド(diphtheria toxoid)ヘルパーT細胞エピトープ、マンソン住血吸虫(Schistosoma mansoni)トリオースリン酸イソメラーゼヘルパーT細胞エピトープ、麻疹(measles)ウィルスF蛋白質ヘルパーT細胞エピトープ及び狂犬病ウィルスヘルパーT細胞エピトープよりなるグループの中から選択されることを特徴とする、請求項1に記載のポリペプチド。
【請求項7】
T細胞エピトープがB型肝炎ウィルス表面抗原のヘルパーT細胞エピトープであることを特徴とする、請求項6に記載のポリペプチド。
【請求項8】
T細胞エピトープがB型肝炎ウィルス表面抗原のpreS2ヘルパーT細胞エピトープであることを特徴とする、請求項7に記載のポリペプチド。
【請求項9】
T細胞エピトープが配列番号7のアミノ酸配列を持つことを特徴とする、請求項8に記載のポリペプチド。
【請求項10】
配列番号9のアミノ酸配列を持つことを特徴とする、請求項1に記載のポリペプチド。
【請求項11】
請求項1〜10のいずれか1項に記載のポリペプチドを含む肥満予防または治療用ワクチン。
【請求項12】
請求項1〜10のいずれか1項に記載のポリペプチドをコードする遺伝子を含む組み換えベクター。
【請求項13】
pB14T(KCCM−10562)であることを特徴とする、請求項12に記載の組み換えベクター。
【請求項14】
請求項12に記載の組み換えベクターで形質転換された宿主細胞。
【請求項15】
請求項12に記載の組み換えベクターで形質転換された宿主細胞を培養して請求項1に記載のポリペプチドを製造する方法。
【図1】

【図2】
【図3】

【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】

【図15】
【図16】
【図17】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公表番号】特表2008−500813(P2008−500813A)
【公表日】平成20年1月17日(2008.1.17)
【国際特許分類】
【出願番号】特願2007−503840(P2007−503840)
【出願日】平成17年3月18日(2005.3.18)
【国際出願番号】PCT/KR2005/000784
【国際公開番号】WO2005/087800
【国際公開日】平成17年9月22日(2005.9.22)
【出願人】(506315343)エスジェイ バイオメッド インコーポレイテッド (1)
【Fターム(参考)】
【公表日】平成20年1月17日(2008.1.17)
【国際特許分類】
【出願日】平成17年3月18日(2005.3.18)
【国際出願番号】PCT/KR2005/000784
【国際公開番号】WO2005/087800
【国際公開日】平成17年9月22日(2005.9.22)
【出願人】(506315343)エスジェイ バイオメッド インコーポレイテッド (1)
【Fターム(参考)】
[ Back to top ]