
抗菌性の糖質及びそれを用いた方法
胃潰瘍を引き起こす細菌を含む細菌の増殖を阻害するのに有用な組成物を提供する。さらに、そのような組成物を製造できるトランスジェニック生体を提供する。また、この組成物を使用してヒトを含む個体の胃潰瘍を治療又は予防する方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
この出願は、米国特許法(35U.S.C.)119条に従い、2004年2月20日に出願された米国出願番号60/546,600に基づく優先権を主張するもので、その全内容が本明細書に含まれている。
【0002】
この発明は、国立癌研究所から授与されたCA71932及びCA33000の助成金に基づく政府支援により、なされたものである。米国政府は、本発明についてのある種の権利を有している。
【0003】
本発明は、概要すると抗菌性組成物に関するもので、特に、末端α1,4結合−N−アセチルグルコサミン残基を有するO−グリカン、末端α1,4結合−N−アセチルグルコサミン残基を有するこのような物質を含んでいる組成物、及びその組成物を用いて細菌の感染を予防及び/又は治療する方法に関するものである。
【背景技術】
【0004】
胃は、食物を消化し易くするために強い酸性環境となっている。これによって、栄養分が分解されて腸で吸収される。一般に、胃の内壁の細胞は、胃の細胞が産生する粘液層で保護されている。胃の内壁がむき出しとなったりびらんによる穴を生じたりして、胃壁が酸性環境に晒されると、胃潰瘍が惹き起こされる。
【0005】
胃潰瘍は、胃酸の分泌の増加や酸性物質の摂取によって惹き起こされる。アスピリンやイブプロフェンのような非ステロイド系抗炎症剤(NSAIDs)を服用したり、多量のアルコールやカフェインを飲用したりするヒトに特に惹き起こされ易い。胃潰瘍患者の約70%がヘリコバクター ピロリ(Helicobacter pylori: H.pylori)に感染している。これらのNSAIDs治療による胃潰瘍のうちの約50%は、ヘリコバクター ピロリによって胃炎が惹き起こされたものでないが、NSAIDsによる治療を行っていないほとんどの胃潰瘍は、胃内に穴が開いた胃炎やヘリコバクター ピロリ感染を伴う。NSAIDsによって惹き起こされた潰瘍患者を除けば、胃潰瘍患者のヘリコバクター ピロリ感染率は約96%にも達する。
【0006】
ヘリコバクター ピロリは、微好気性、グラム陰性の桿菌であり、全世界の人口のおよそ半分のヒトの胃の内壁上に棲息し、胃炎、慢性胃炎、胃潰瘍、十二指腸潰瘍などの主な原因であると言われている。また、胃癌等の疾患との関連性が報告されている。この為、ヘリコバクター ピロリによって惹き起こされる疾患に対する治療薬の開発は、ヒトが健康で快適な生活を維持していくために重要な課題の一つである。
【0007】
抗ヘリコバクター ピロリ剤として、例えば、ペニシリン系、テトラサイクリン系、セファロスポリン系、ニューロキノン系等の抗生物質が知られている。しかし、これらの抗生物質は、ヘリコバクター ピロリに対する抗菌作用を示すものの、単独での投与では十分な抗菌効果が得られない。このため、現在では、これらの抗生物質に加えて、ビスマス剤、プロトンポンプ阻害薬との3剤併用療法が主に行われている。また、特開平11−189529号公報に、ベンゾヒドロキサム酸を含有する抗ヘリコバクター ピロリ剤が提案されている。しかし、これらの場合、投与したときに下痢や吐き気などの副作用があり、安全性に劣るという問題がある。
【0008】
また、前記の治療薬とは別に、例えば、特開2003−73278号公報に、植物等から抽出したステリルグルコピラノシドを含有する抗ヘリコバクター ピロリ剤が提案されている。また、特開2002−370995号公報に、オウレン等の生薬から抽出された成分と、炭酸水素アルカリ金属塩とを含有する抗ヘリコバクター ピロリ薬が提案されている。しかし、これらの場合、ヒトの生体内で発現したものではないため副作用があり、さらに、ヘリコバクター ピロリの抗菌効果が十分ではないという問題がある。また、前記のいずれの提案にも、ヘリコバクター ピロリの運動性の低下、及び形態異常については何ら開示されていない。
【0009】
胃潰瘍治療の目的は、痛みの軽減、潰瘍の治癒、合併症の予防、及び潰瘍の再発の予防である。未治療の潰瘍による合併症には、出血、胃壁の穿孔、及び瘢痕による閉塞症などがある。さらに、胃潰瘍患者の約2〜3%が胃癌を発症する。ヘリコバクター ピロリに感染した胃潰瘍患者の治療に、抗生物質が使用される。抗生物質により治療すると、ヘリコバクター ピロリ感染を治癒した後に潰瘍を治癒することができる。また、治療が成功した場合、翌年に再発する患者は5%未満である(ヘリコバクター ピロリ感染が治癒していない場合の再発率は60〜80%である)。
【0010】
ヘリコバクター ピロリは胃の内壁を覆っている粘液層の深部に棲息するため、この細菌を胃内から根絶することは困難である。また、抗生物質が、細菌を殺すのに十分な濃度でこの粘液層を透過させるのも困難である。一般に、単剤療法は、感染してしまったヘリコバクター ピロリの除去に効果を奏しない。例えば、アモキシシリンを用いた場合、この細菌の根絶率は20%未満であり、メタロニダゾールを用いた場合、根絶率は10%未満であるが、クラリスロマイシンを併用すると、細菌の根絶率は約40%となる。そのため、2剤(例えば二種の抗生物質、又は一種の抗生物質と一種の酸低下薬)の併用療法や、3剤(例えば二種の抗生物質と一種の酸低下薬)の併用療法が行われる。二種の抗生物質の併用療法の根絶率は40〜60%である。また、最も優れた3剤併用療法で治療した患者のヘリコバクター ピロリ感染の根絶率は、現在のところ90%以上である。
【0011】
あいにく、ヘリコバクター ピロリも他の細菌のように、抗生物質に対して耐性を生じるようになるため、治療の成功率が低下してしまう。例えば、メタロニダゾールに対する耐性は部位によっては、40%の高率に達すると報告されている。また、クラリスロマイシンに対する耐性も僅か(2%未満)であるが報告されている。抗生物質に対する耐性は通常経時的に増加するので、それらがヘリコバクター ピロリの根絶と胃潰瘍の治療とに次第にそれほど有効でなくなるかもしれないという懸念がある。しかし、十分な抗菌効果を有し、副作用がなく、安定性に優れる物質やそれを用いた治療法は、いまだ提案されていない。そのため、胃潰瘍を予防し、また治療するための組成物と、そのための方法とが求められている。
【発明の開示】
【発明が解決しようとする課題】
【0012】
本発明は、胃に感染して胃潰瘍を引き起こすヘリコバクター ピロリの増殖を抑制する胃腺粘液細胞型ムチン、特に末端α1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有するムチンを見出したことに基づきなされたものである。この胃腺粘液細胞型ムチンにこのような活性があることが分かったので、α1,4結合−GlcNAc残基を有する典型的な糖タンパク質を調製したところ、それらがコレステリル−α−D−グルコピラノシド合成やヘリコバクター ピロリ増殖に対して抑制効果を有するということが、判明した。これによってなされた本発明は、胃潰瘍の治療、十二指腸潰瘍の治療、抗ヘリコバクター ピロリ剤、胃炎の治療、慢性胃炎の緩和に好適に使用される医薬物質;その物質の製造工程;その物質を用いた胃潰瘍の治療方法;及びその物質を製造できるトランスジェニック哺乳類を提供するというものである。
【課題を解決するための手段】
【0013】
本発明は、担体分子と相互作用して結合した少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する天然非存在の化合物に関するものである。担体分子は一つ以上のα1,4結合−GlcNAc残基に結合することができる分子であり、例えばO−グリカン(例えばコア2分岐型O−グリカン)、ポリペプチド、特に少なくとも一つのO−グリコシル化部位を有するポリペプチド、又は糖タンパク質(例えば、コア2分岐型O−グリカンを含むポリペプチドのような、O−グリカンを一つ以上含むポリペプチド)である。その一つの形態は、担体分子が、例えばCD43(ロイコシアリン)やCD34のようなムチン様ポリペプチドであるというものである。別な形態は、前記化合物が、例えば2、3、4、5又はそれ以上のα1,4結合−GlcNAc残基のような複数のα1,4結合−GlcNAc残基を有するというものである。さらに別な形態は、前記化合物が、合成されたオリゴ糖を有するというものである。
【0014】
また、本発明は、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質の製造方法に関するものである。その方法は例えば、ポリペプチドのグリコシル化に適した条件下で、α1,4−N−アセチルグルコサミン転移酵素(α4GnT)と、コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)又はコア1伸長酵素(例えばコア1伸長β1,3−N−アセチルグルコサミン転移酵素であるコア1−β3GlcNAcT又はC1−β3GnT)と、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドとを、接触させて、該ポリペプチドを、C2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質を製造するというものである。この担体ポリペプチドは、C2GnT−I(又はC1−β3GnT)とα4GnTとの活性によって一つ以上のα1,4結合−GlcNAc残基を結合し得るもので、例えば可溶性ポリペプチド(具体的には、可溶性CD43ポリペプチド又は可溶性CD34ポリペプチド)である。
【0015】
前記担体ポリペプチドの具体例の一つは、哺乳類の母乳中に通常存在するムチン型糖タンパク質を含むというものである。この具体例の一つの形態として、in vitroで、哺乳類の母乳と、少なくともα4GnTと、必要に応じてC2GnT−I及び/又はC1−β3GnTとを接触させ、母乳中のムチン型糖タンパク質を、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質に変換する方法が、挙げられる。この具体例の別な形態として、前記ムチン型糖タンパク質を哺乳類の母乳から単離し、哺乳類の母乳から単離されたムチン型糖タンパク質と、少なくともα4GnTと、必要に応じてC2GnT−I及び/又はC1−β3GnTとを接触させ、母乳中のムチン型糖タンパク質を、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質に変異させる方法が挙げられる。
【0016】
少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質の製造方法は、糖タンパク質を単離する工程を含んでいてもよい。それによって、本発明の方法で製造された少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質は、そのような方法で製造して単離された糖タンパク質として、得られる。
【0017】
さらに、本発明は、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質の製造方法に関するものである。この方法は例えば、少なくとも異型のα4GnTをエンコードするポリヌクレオチドを真核細胞内で発現させ、細胞により産生される内因性であってもよいポリペプチドを、細胞内に発現したC2GnT−I(又はC1−β3GnT)とα4GnTとによって、グリコシル化することができるというものである。例えば、哺乳類の細胞は、外因性(異型)発現α4GnTの存在下で、内因性C2GnT−Iと前記のようにグリコシル化され得るポリペプチドとを発現させて、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を産生して製造する。この方法で注意すべき点は、真核細胞内でα1,4結合−GlcNAc残基を有する天然非存在の糖タンパク質に変異されるため、糖タンパク質が“組換え型”糖タンパク質とみなされるが、この糖タンパク質は、細胞中の異型α4GnTの発現によってのみ産生されるということである。別な形態として、例えば真核細胞が天然に内因性C2GnT−Iや所望の担体ポリペプチドを発現してもしなくても、異型C2GnT−Iをエンコードするポリヌクレオチド及び/又は少なくとも一つのO−グリコシル化部位を有する異型担体ポリペプチドをエンコードするポリヌクレオチドが細胞内に発現して、担体ポリペプチドが内因性又は外因性(異型)であってもよいC2GnT−Iとα4GnTとによってグリコシル化されるというものである。
【0018】
前記担体ポリペプチドは、可溶性ポリペプチドであってもよく、例えば可溶性CD43又は可溶性CD34であってもよい。また、前記組換え型の糖タンパク質は、一つ又は複数のα1,4結合−GlcNAc残基を有していてもよい。前記真核細胞は、エンコードするポリヌクレオチドを発現できる如何なる真核細胞であってもよい。真核細胞は、内因性C2GnT−I、異型C2GnT−I、異型α4GnTのいずれかが、内因性担体ポリペプチドや異型ポリペプチドであってもよい担体ポリペプチドに一つ又はそれ以上のα1,4結合−GlcNAc残基を結合できるような酵素活性を有するものであってもよい。例えば前記真核細胞は、チャイニーズハムスター卵巣細胞のような哺乳類の細胞や、ヒトのような哺乳類の上皮細胞等の細胞であってもよい。さらに前記方法は、前記組換え型の糖タンパク質を単離する工程を含んでいてもよい。それによって、本発明の方法で製造されて、単離された組換え型の糖タンパク質を含む組換え型の糖タンパク質が、得られる。
【0019】
また、本発明は、少なくとも、α4GnTをエンコードする第一の外因性ポリヌクレオチドと、さらにC2GnT−Iをエンコードするポリヌクレオチドと、担体ポリペプチドをエンコードするポリヌクレオチドとを、ゲノム中に安定に組み込まれて有する非ヒトのトランスジェニック哺乳類であって、該第一の外因性ポリヌクレオチドが、細胞中又は関連する細胞中にエンコードされたC2GnTを発現させる5’制御配列に相互作用して結合されている非ヒトのトランスジェニック哺乳類に関するものである。5’制御配列は、活性な構成プロモータ、誘導プロモータ、組織特異プロモータ、又はそれと同様のものであってもよい。一つめの具体例として、前記5’制御配列は、乳腺特異的プロモータを含んでいるものである。そのC2GnTと担体ポリペプチドとをエンコードするポリヌクレオチドは、内因性ポリヌクレオチド(例えば非ヒトの動物のゲノムに含まれる遺伝子)であってもよいが、必ずしも必要ではない。二つめの具体例として、C2GnTをエンコードするポリヌクレオチドがC2GnT−Iをエンコードする第二の外因性ポリヌクレオチドであり;及び/又は、担体ポリペプチドをエンコードするポリヌクレオチドが少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチドであるものが挙げられる。この具体例の一つの形態として、前記第一の外因性ポリヌクレオチドと、共存するなら前記第二の外因性ポリヌクレオチドと、共存するなら前記第三の外因性ポリヌクレオチドとが夫々、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合しているものが挙げられる。この具体例の別な形態として、共存するなら前記第三の外因性ポリヌクレオチドは、担体ポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と相互作用して結合しているものが挙げられる。本発明のトランスジェニック哺乳類中のC2GnT−Iと、α4GnTと、担体ポリペプチドとの発現により、前記担体ポリペプチドは、C2GnT−Iとα4GnTとによってグリコシル化され、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質が産生される。
【0020】
本発明の目的に有用であって、乳腺特異的遺伝子の5’制御配列のすべて又は一部を構成するプロモータは、乳腺特異的な発現を示すプロモータであってもよい。そのようなプロモータとして、例えば乳清酸性タンパク質プロモータ、α−カゼインプロモータ、β−カゼインプロモータ、γ−カゼインプロモータ、α−ラクトアルブミンプロモータ、又はβ−ラクトグロブリンプロモータが挙げられる。前記第一の外因性ポリヌクレオチドと、前記第二の外因性ポリヌクレオチドと、及び前記第三の外因性ポリヌクレオチドとが、乳腺特異的遺伝子の3’制御配列、又は乳腺中の活性3’制御配列と、さらに相互作用して結合していてもよい。
【0021】
本発明の非ヒトのトランスジェニック哺乳類は、一つ、二つ、又は三つの導入遺伝子を用いて作製されて製造されるものであってもよい。これの一つの形態は、前記第一の外因性ポリヌクレオチドと、前記第二の外因性ポリヌクレオチドと、前記第三の外因性ポリヌクレオチドとが、夫々独立に、5’制御配列と、必要に応じ3’制御配列とに、前記のように相互作用して結合し得るというものである。別な形態は、前記第一の外因性ポリヌクレオチドと、前記第二の外因性ポリヌクレオチドと、前記第三の外因性ポリヌクレオチドとが、互いに相互作用して結合し、さらに5’制御配列と、必要に応じ3’制御配列とに、相互作用して結合し、単一の導入遺伝子を形成するというものである。さらに別な形態は、二つの導入遺伝子、例えば、第二の外因性ポリヌクレオチドに相互作用して結合した第一の外因性ポリヌクレオチドを含む第一の導入遺伝子と、第三の外因性ポリヌクレオチドを含む第二の導入遺伝子とであり、前記第一及び第二の導入遺伝子は夫々、5’制御配列と、必要に応じて3’制御配列とに、前記のように相互作用して結合し得るというものである。
【0022】
本発明の非ヒトのトランスジェニック哺乳類は、例えばマウス、ラット、ウサギ、ウマ、ブタ、ヒツジ、ヤギ、又はウシのような如何なる哺乳類であってもよい。具体例として、トランスジェニック哺乳類は、非ヒトの雌トランスジェニック哺乳類であって、抗菌活性型で、前記非ヒトの雌トランスジェニック哺乳類によって産生された組換え型糖タンパク質を、母乳中に分泌するというものである。それによって前記非ヒトの雌トランスジェニック哺乳類によって産生された母乳も、得られる。
【0023】
また、本発明は、非ヒトの雌トランスジェニック哺乳類の母乳中に、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を、産生させて製造する方法に関するものである。この方法は、例えば、非ヒトの哺乳類の胚にα4GnTをエンコードする第一の外因性ポリヌクレオチドと、必要に応じてC2GnT−I又はコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二の外因性ポリヌクレオチド;及び/又は必要に応じて少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチドを導入し、前記第一の外因性ポリヌクレオチドと、共存するなら前記第二の外因性ポリヌクレオチドと、共存するなら前記第三の外因性ポリヌクレオチドとが、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合され、共存するなら前記第三の外因性ポリヌクレオチドが、担体ポリペプチドを母乳中に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と相互作用して結合され;子孫を出産するような宿主の雌哺乳類に前記の胚を移植し;5’制御配列に相互作用して結合した第一の外因性ポリヌクレオチドと前記第二の外因性ポリヌクレオチドと前記第三の外因性ポリヌクレオチドとを含んで、ゲノム中に安定に組み込まれ、前記の雌の子孫に、母乳を産生させることによって行われるというものである。本発明の方法によって製造された非ヒトの雌トランスジェニック哺乳類中にC2GnT−Iとα4GnTと担体ポリペプチドとが発現し、前記担体ポリペプチドが、C2GnT−I(又はC1−β3GnT)とα4GnTとによってグリコシル化されて、少なくとも一つのα1,4結合−GlcNAc残基を有し、非ヒトの雌トランスジェニック哺乳類の母乳中に分泌される抗菌性の組換え型の糖タンパク質が、産生される。
【0024】
このような非ヒトの雌トランスジェニック哺乳類の製造方法は、さらに前記雌の子孫から搾乳して、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を含む母乳を得る工程を、含むものであってもよい。また前記の方法は、さらに前記母乳から少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を単離することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を得る工程を含んでいてもよい。それによって、本発明は、このような方法で製造される非ヒトの雌トランスジェニック哺乳類と、その雌トランスジェニック哺乳類から産生される母乳と、この方法で得られる少なくとも一つのα1,4結合−GlcNAc残基を有する単離された組換え型の糖タンパク質とを、提供することができるというものである。
【0025】
また、本発明は、コレステリル−α−D−グルコピラノシド(CGL)を発現する細菌の細胞壁の形成を、抑制又は阻害する方法に関するものである。この方法は、例えば、この細菌を、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物に接触させ、CGL合成を抑制又は阻害して、前記細菌の細胞壁の形成を抑制又は阻害するというものである。一つの形態として、前記化合物が、細菌のUDP−Glc:ステロール糖転移酵素活性を抑制又は阻害して、前記CGL合成を抑制又は阻害するというものが挙げられる。前記細菌は、GGLを発現する細菌であればよく、ヘリコバクター種の細菌、特にヘリコバクター ピロリも範疇に含んでいる。前記化合物は、本発明の化合物、例えば本発明の組換え型の糖タンパク質、又は胃腺粘膜細胞から単離された胃腺粘液細胞型ムチンも範疇に含んでいる。
【0026】
さらに本発明は、CGLを発現する細菌の増殖を抑制又は阻害する方法に関するものである。この方法は、例えば、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物に細菌を接触させ、CGL合成を抑制又は阻害して、前記細菌の増殖を抑制又は阻害するというものである。前記方法は例えば、培養中の細菌細胞に前記化合物を添加してin vitroで接触させたり、細菌に感染した個体、例えばヘリコバクター種の細菌に感染した哺乳類(例えばヘリコバクター ピロリに感染したヒト)の胃に前記化合物を直接投与し、in vivoで接触させたりして行なうというものである。前記化合物は、本明細書で示した方法を用いて得られるもので、例えばO−グリカンのような少なくとも一つのα1,4結合−GlcNAc残基を有する化合物であれば如何なるものであってもよく、胃腺粘膜細胞から単離された胃腺粘液細胞型ムチンであってもよい。
【0027】
さらに、本発明は、ヘリコバクター種の細菌の感染による胃潰瘍を治療又は予防する方法に関するものである。それによって本発明は、ヘリコバクター種の細菌が個体に感染することによる胃潰瘍の兆候又は症状を改善する方法を提供することができるというものである。この方法は、例えば、個体に少なくとも一つのα1,4結合−GlcNAc残基を有する化合物を投与し、前記化合物が前記ヘリコバクター種の細菌に接触し、前記ヘリコバクター種の細菌の増殖を抑制又は阻害して、胃潰瘍及び/又は胃癌の兆候又は症状を改善するというものである。
【0028】
本発明は、別な具体例として、胃潰瘍感受性の個体にヘリコバクター種の細菌が感染することによる胃潰瘍を、予防する方法を提供するというものである。この方法は、例えば、胃潰瘍感受性の個体に、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物を投与し、前記化合物が前記ヘリコバクター種の細菌の増殖を妨げて、前記ヘリコバクター種の細菌による胃潰瘍の形成を予防するというものである。
【0029】
本発明は、さらに他の具体例として、胃潰瘍又は胃癌を有する個体に少なくとも一つのα1,4結合−GlcNAc残基を有する化合物を投与して、前記ヘリコバクター種の細菌の感染による胃潰瘍又は胃癌を、改善する方法に関するというものである。前記化合物が、前記ヘリコバクター種の細菌の増殖を妨げて、前記ヘリコバクター種の細菌の感染による胃潰瘍又は胃癌が、改善される。例えば前記の方法は、本明細書中で示した非ヒトのトランスジェニック哺乳類から得られる前記化合物を含有する母乳を、前記個体が飲用することも範疇に含んでいる。
【0030】
本発明の方法により治療される個体は、胃粘膜がヘリコバクター感染しているかヘリコバクター感受性である如何なる個体であってもよい。それによる前記個体は例えばウマ、ブタ、ヤギ、イヌ、ネコ、フェレット、又はそれと同等のものであってもよい。具体例として、前記個体はヒトである。また、ヘリコバクター種の細菌は胃潰瘍形成に関係する如何なる細菌種であってもよく、例えばヒトの胃潰瘍形成に関係するヘリコバクター ピロリが挙げられる。
【0031】
前記化合物は、少なくとも一つの末端α1,4結合−GlcNAc残基を有するもので、本明細書で示したように、ヘリコバクター種の細菌の増殖を抑制又は阻害できるものである。前記化合物は、合成されたオリゴ糖、一つ又はそれ以上のα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質(例えば末端α1,4結合−GlcNAc残基を有する可溶性CD43ポリペプチド)、一つ又はそれ以上の末端α1,4結合−GlcNAc残基を有し、O−グリカンを含む化合物、又は胃腺粘膜細胞から単離することができる胃腺粘液細胞型ムチンを含む化合物であってもよい。少なくとも一つの末端α1,4結合−GlcNAc残基を有するそのような化合物を製造する方法は、慣用された公知の方法に準じる。例えば、担体分子のために有用な合成オリゴ糖を製造する方法として、コア2オリゴ糖を製造する方法(例えばSenguptaら、Tetrahedron Lett.44:6037-6042,2003;Ongら、J.Biol.Chem.277:18182-18190,2002参照;いずれも参考文献としてここに示す)、コア1伸長オリゴ糖を製造する方法(例えばBelotら、Tetrahedron Lett.74:2-6,2002参照;参考文献としてここに示す)、コア2分岐型O−グリカンにα1,4−GlcNAcを添加する方法(Nakayamaら、Proc.Natl.Acad.Sci.,USA 96:8991-8996,1999参照;参考文献としてここに示す)が挙げられる。さらに、前記化合物は、特に胃に直接投与できる経口投与のような便利な方法で投与されてもよい。一つの形態として、前記投与は、ヘリコバクター感染による胃潰瘍を有している個体又は前記胃潰瘍感受性の個体が、本明細書中で示した非ヒトのトランスジェニック哺乳類から得られる前記化合物を含有した母乳を、飲用することによって行われるというものである。
【0032】
また、本発明は、必須のα4GnTをエンコードする第一の外因性ポリヌクレオチドと;C2GnT−Iをエンコードし、又はC1−β3GnTをエンコードする第二のポリヌクレオチドと;少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとをゲノム中に安定に組み込んで有するトランスジェニック植物であって、該第一のポリヌクレオチド、該第二のポリヌクレオチド、及び該第三のポリヌクレオチドの少なくともいずれかが外因性ポリヌクレオチドであり;該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、該第三のポリヌクレオチドとが、プロモータを含む植物遺伝子の5’制御配列に相互作用して結合され;C2GnT−I又はC1−β3GnTと、α4GnTと、担体ポリペプチドとの発現により、該担体ポリペプチドを、C2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を産生して製造するトランスジェニック植物に関するものである。そのようなトランスジェニック植物中、前記第二のポリヌクレオチド及び/又は前記第三のポリヌクレオチドが、外因性のポリヌクレオチドであってもよく、一つの具体例として、前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、夫々外因性のポリヌクレオチドであるというものである。
【0033】
本発明のトランスジェニック植物中、外因性の核酸分子に相互作用して結合したプロモータは、構成プロモータ、誘導プロモータ、又は組織特異プロモータであってもよい。前記プロモータは、例えば種、花、葉及び/又は根のような植物の所望の細胞中でエンコードされたポリペプチド(例えばα4GnT)を転写する。例えば、前記プロモータは、構造的に活性な35Sカリフラワーモザイクウイルスプロモータ、ユビキチンプロモータ、コメアクチンプロモータ、誘導テトラサイクリン応答プロモータ、rbcS遺伝子光誘導プロモータ、又は種子特異的オレオシン遺伝子プロモータであってもよい。
【0034】
本発明のトランスジェニック植物は、如何なる植物であってもよく、単子葉植物、双子葉植物、裸子植物、被子植物も範疇に含むものである。さらに、前記トランスジェニック植物は、ヒトや動物の食材源として、用いられるものであってもよく、例えば農業経営(例えば遺伝子操作された作物の生産)のように商業上有用なものであってもよい。このような前記トランスジェニック植物は、例えば、ダイズ植物、コメ植物、トウモロコシ植物、又はコムギ植物であってもよい。それによって本発明は、例えばこのようなトランスジェニック植物によって産出された種;このようなトランスジェニック植物から産出された穀類のような産物;種や穀類に由来する産物、又はそれらの同等品例えば豆乳やライスミルクのような乳液を提供することができるというものである。
【0035】
また、本発明は、トランスジェニック植物中で、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を産生させて製造する方法に関するものである。この方法は例えば、植物細胞にα4GnTをエンコードする第一の外因性ポリヌクレオチドを少なくとも導入し;該第一の外因性ポリヌクレオチドが、プロモータを含んでいる植物遺伝子の5’制御配列に相互作用して結合され;α4GnTの発現により、該植物の細胞中の該担体ポリペプチドをα4GnTによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を産生させ;導入された外因性核酸分子を含む植物細胞から植物を生育させ、トランスジェニック植物を得て、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質をトランスジェニック植物中に産生させることができるというものである。この方法はさらに、外因性のC2GnT−Iをエンコードし、又は外因性のC1−β3GnTをエンコードする第二の外因性ポリヌクレオチド;及び/又は少なくとも一つのO−グリコシル化部位を有する外因性の担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチドをさらに植物細胞に導入する工程を含み、共存するなら該第二の外因性ポリヌクレオチド、及び共存するなら該第三の外因性ポリヌクレオチドが夫々、プロモータを含む植物遺伝子の5’制御配列に相互作用して結合され;共存するならC2GnT−I又はC1−β3GnTと、α4GnTと、共存するなら外因性の担体ポリペプチドとの発現により、該担体ポリペプチドを、共存するならC2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質をトランスジェニック植物中に産生させるものであってもよい。
【0036】
トランスジェニック植物中で、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を産生させて製造する方法は、さらに、例えば種子、葉、根、及び/又は花のようなトランスジェニック植物収穫物を、トランスジェニック植物から分取する工程を含んでいてもよい。前記の方法は、さらに、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を、前記トランスジェニック植物又は前記トランスジェニック植物収穫物から単離する工程を含んでいてもよい。一つの具体例として、この方法は、さらに前記トランスジェニック植物又は前記トランスジェニック植物収穫物を加工して、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を含む乳液製品を得る工程を含んでいてもよい。これにより、本発明の方法で産生されて単離された少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質、本発明の方法で産出されたトランスジェニック植物産出物、本発明の方法により得られた乳液も、供給することができる。
【発明を実施するための好ましい形態】
【0037】
本発明は、例えばヒトの胃潰瘍の原因となるヘリコバクター ピロリのようなヘリコバクター種の細菌に感染することによる胃潰瘍を、予防又は治療するのに有用な組成物と方法とを提供するというものである。本発明は、ヘリコバクター属に対する増殖性及び運動性を阻害し、また形態異常性を発現させるもので、抗菌活性を有し、ヘリコバクター属に対する抗菌活性が選択的及び特異的であるため副作用がなく、安全性に極めて優れる分子、機能性分子、組成物、医薬、飲食物、及び母乳、及びトランスジェニック哺乳類を提供することを目的とする。また、本発明は、前記機能性分子の効率的な製造方法、及び、機能性分子を投与することにより、副作用がなく、安全性に極めて優れる胃癌の治療方法、胃潰瘍の治療方法、及び胃潰瘍の予防方法を提供することを目的とする。
【0038】
図1に示すように、ヘリコバクター ピロリに感染した患者を検査したところ、細菌は、胃の内壁表面部を構成する表面粘液細胞から分泌される表面粘液細胞型ムチンにのみ関連しており、胃の内壁の深部に存在する腺粘液細胞部分にコロニーを形成しないということが観察される。腺粘液細胞から分泌される腺粘液細胞型ムチンは、末端でα−1,4−グルコシド結合(以下GlcNAcα(残基)と呼ぶことがある。)により結合されているN−アセチルグルコサミン残基を含んでいることも見出された。末端GlcNAcα残基は、ヘリコバクター ピロリの細胞壁に含まれるコレステリル−α−D−グルコピラノシド(CGL)の合成を阻害し、おそらくその結果、ヘリコバクター ピロリの増殖特性を阻害し、さらに、その運動性を阻害し、かつその形態異常性を発現させるということも見出された。図1中、「GlcNAcα(+)」は、GlcNAcα残基を含むムチンが存在することを意味し、「GlcNAcα(−)」は、GlcNAcα残基を含むムチンが存在しないことを意味する。このことは、α1,4−GlcNAc残基がPCSに染色されるものであり、またGOCTSがムチン表面を染色するという事実から、推察される(実施例1参照)。
【0039】
本発明は、複数のモノマーユニットからなり、その少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基である分子を提供するというものである。本明細書で開示したように、本発明の分子は、コレステリル−α−D−グルコピラノシドの合成を抑制及び阻害し、同様に前記コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び阻害する。さらに、前記分子は前記コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び阻害し、さらにはその細菌の形態異常を発現させる。一つの具体例として、前記分子は糖鎖を含み、その末端にN−アセチルグルコサミン残基を有していてもよい。一つの形態として、N−アセチルグルコサミン残基は、α1,4−N−アセチルグルコサミン残基である。このα1,4−N−アセチルグルコサミン残基は、コレステリル−α−D−グルコピラノシドの合成を抑制及び阻害することができる。同様に、前記コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び阻害することができる。さらにα1,4−N−アセチルグルコサミン残基は、前記コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び阻害し、また、その細菌の形態異常を発現させる。本発明の分子は、N−アセチルグルコサミン残基に隣接してガラクトース残基を有していてもよい。前記分子は直鎖又は分岐鎖であってもよい。
【0040】
本発明の分子は、N−アセチルグルコサミン(「GlcNAc」と称する)残基を一部に含み、必要に応じて適宜選択した分子構成ユニットを含む。前記GlcNAc残基は、化学合成した物質に由来する基であってもよく、天然物から抽出物に由来した基であってもよく、生体内に存在する基であってもよい。前記GlcNAcの製造方法は、特に制限はなく、例えば、キチンを塩酸で加水分解し、加水分解物を単離することにより製造することができる。
【0041】
前記分子構成ユニットは、本明細書に示したように目的に応じて適宜選択することができ、糖残基、樹脂ユニット等が挙げられる。これらを、1種単独で使用してもよく、2種以上併用してもよい。前記糖残基は、単糖残基であってもよいし、二糖以上の多糖残基であってもよい。前記単糖残基、又は多糖残基を構成する単糖残基として、例えば、ガラクトース残基、テトロース残基、エリトロース残基、トレオース残基、ペントース残基、リボース残基、アラビノース残基、キシロース残基、リキソース残基、ヘキソース残基、アロース残基、アルトロース残基、グルコース残基、マンノース残基、グロース残基、イドース残基、タロース残基、テトルロース残基、エリトルロース残基、2−ペンチロース残基、リブロース残基、キシルロース残基、プシコース残基、フルクトース残基、ソルボース残基、タガトース残基、2−ヘプツロース残基、セドヘプツロース残基、キノボース残基、ラムノース残基、フコース残基、ジギトキソース残基、アベクオース残基、チベロース残基、2−ベオキシリボース残基、2−デオキシグルコース残基、グルコサミン残基、マンノサミン残基、ガラクトサミン残基、フコサミン残基、キノボサミン残基、ラムノサミン残基、ノイラミン酸残基、ムラミン酸残基、ノジリマイシン残基などが挙げられる。これらの中でも、前記GlcNAc残基に隣接する糖残基として、ガラクトース残基が特に好ましい。前記糖残基は、1種単独でもよく、2種以上を併用してもよく、置換基が挿入されていてもよく、別な置換基でさらに置換されていてもよい。前記樹脂ユニットとして、例えば、L−ラクタイド、D−ラクタイド等が挙げられる。前記のユニットや別なユニットとして、生体適合性分子を構成する公知のユニットが用いられてもよい。
【0042】
前記GlcNAc残基は、本発明の前記分子のどこに位置していてもよく、例えば、分子の末端でもよく、末端以外の部分でもよい。しかし、前記GlcNAc残基が前記分子の末端に位置していると、優れた抗菌効果を示す。前記分子が糖鎖である場合、前記GlcNAc残基の結合はどのようなものでもよく、例えば、α−グルコシド結合やβ−グルコシド結合であってもよい。一つの形態として、前記結合はα1,4−グルコシド結合である。
【0043】
前記GlcNAc残基をα−1,4−グルコシド結合で前記分子に結合させるために酵素を使用する場合、その酵素を目的に応じて適宜選択することができる。前記酵素として、例えば、特開2001−46077号公報に記載のN−アセチルグルコサミン転移酵素(α4GnT)が挙げられる。前記α4GnTは、例えば、特開2001−46077号公報に記載の方法に準じて前記α4GnTをエンコードするDNAを形成させ、このDNAを組み込んだベクターを各種細胞にトランスフェクトさせることにより、前記α4GnTを発現させることによって、調製することができる(Nakayamaら、Proc.Natl.Acad.Sci.U.S.A.96,8991,1999参照)。
【0044】
前記分子は、前記GlcNAcα残基や付加ユニットを含む所望のモノマーユニットを多く含んでいてもよい。このモノマーユニットは、目的に応じて適宜選択される。例えば、前記分子は、2以上、特に2〜30(例えば5〜20)のモノマーユニットを含んでいてもよい。一般に、前記モノマーユニットの数が2未満であると、前記分子は抗菌効果を有しない。
【0045】
本発明の分子は、直鎖であっても分岐鎖であってもよい。前記分子が分岐である場合、分岐数は特に制限はなく、目的に応じて適宜選択することができる。一般に分岐数が多いほど、抗菌効果は高い。前記分子を分岐させるために酵素を使用してもよい。そのような酵素として、例えば、コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)を挙げることができ、このC2GnT−Iは例えば細胞への遺伝子の導入、遺伝子組み換えによるC2GnT−Iの調製、生物からの単離精製などにより調製することができる(例えばBierhuizen,Fukuda、Proc.Natl.Acad.Sci.U.S.A.89,9326,1992参照)。同様に、前記分子中で糖残基を結合させるために酵素を使用してもよい。そのような酵素として、例えば、コア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)を使用してもよい(Yehら、Cell,105:957-969,2001参照)。
【0046】
本発明の分子がGlcNAcα残基を含む糖鎖を有している場合、前記分子はヒトの胃壁深部に存在する腺粘液細胞から分泌される腺粘液細胞型ムチンが有する分子、又はその骨格構造を有する類似分子であってもよい。そのため、前記分子を、医薬や、食物や飲料のような飲食材として使用した場合、副作用がなく、極めて安全性が高い。
【0047】
また、本発明は、複数のモノマーユニットを含む分子であって、それの少なくとも一つのユニットがN−アセチルグルコサミン残基を有している分子を含んでいる組成物を提供するというものである。この組成物は、例えば薬物のような有用な医薬であってもよく、また食物や飲料のような飲食物用の組成物であってもよい。
【0048】
さらに、本発明は、機能性分子を提供するというものである。この機能性分子は、複数のモノマーユニットを含む分子であり、その少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基であるというものである。この機能性分子はコレステリル−α−D−グルコピラノシドの合成を抑制及び/又は阻害し、同様に前記コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び/又は阻害することができる。一つの形態として、前記機能性分子は、前記コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制又は阻害し、さらにはその細菌の形態異常を発現させる。そのような機能性分子は、主鎖と側鎖とを有する分子であってもよく、その側鎖が複数のモノマーユニットを含みその少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基である分子であってもよい。
【0049】
本発明の機能性分子は、本発明の前記の分子と、必要に応じて少なくとも一つの別な分子とで構成されている。一般に機能性分子は主鎖と側鎖とを有し、好ましくは本発明の分子は側鎖を有している。前記分子は主鎖に対してクラスター状に存在している。ここで用いられている前記「クラスター状」という用語は、ひとつの前記主鎖に対して、10以上の前記分子が末端で結合された状態をいう。前記主鎖の数と前記側鎖の数との比は特に制限はなく、目的に応じて適宜選択することができる。例えば、前記主鎖がタンパク質の場合、タンパク質を構成するアミノ酸残基の種類により比率は限定される。一般に、ひとつの主鎖に存する側鎖の数が多いほど、機能性分子の抗菌効果は高くなる。
【0050】
本発明の分子に結合される別な分子は、目的に応じて適宜選択することができる。前記その他の分子として、例えば、糖鎖、ポリペプチド、脂質、糖脂質、糖タンパク、ヌクレオチド、ポリ乳酸等の樹脂が挙げられ、これらは、1種単独で使用してもよく、2種以上を併用してもよい。一例として、前記その他の分子は、例えばCD43、CD34、及び/又はMuc−6を含むポリペプチドを有する分子である。前記分子と結合する前記ポリペプチド中のアミノ酸は、特に制限されない。前記アミノ酸として、例えば、フェニルアラニン(Phe)、ロイシン(Leu)、イソロイシン(Ile)、メチオニン(Met)、バリン(Val)、セリン(Ser)、プロリン(Pro)、トレオニン(Thr)、アラニン(Ala)、チロシン(Tyr)、ヒスチジン(His)、グルタミン(Gln)、アスパラギン(Asn)、リジン(Lys)、アスパラギン酸(Asp)、グルタミン酸(Glu)、システイン(Cys)、トリプトファン(Trp)、アルギニン(Arg)、グリシン(Gly)等が挙げられる。一般に、セリン及び/又はトレオニンが用いられる。
【0051】
本明細書に示したように、本発明の分子は、前記CGLの合成を抑制及び/又は阻害することができるものである。前記CGLの合成を抑制又は阻害することにより、前記CGLを必須とする細菌の増殖を抑制また阻害することができる。前記CGLを必須とする細菌として、例えば、ヘリコバクター種の細菌、特にヘリコバクター ピロリが挙げられる。前記ヘリコバクター種の細菌は、その細胞壁にα−コレステリルグルコシド(α−CGs)を含有する。このα−CGsの主要成分として、例えば、CGL、コレステリル−6−O−テトラデカノイル−α−D−グルコピラノシド(CAG)、及びコレステリル−6−O−ホスファチジル−α−D−グルコピラノシド(CPG)が挙げられる。このため、ヘリコバクター種の細菌は、CGLの合成が生存に必須となる細菌の一つである。
【0052】
本発明の機能性分子は、ヘリコバクター種の細菌に対して選択的及び特異的に抗菌活性を有し、ヒトの腸内の細菌層を損傷しない。さらに、本発明の機能性分子がGlcNAcα残基を含む場合、それは、ヒトの胃壁深部に存在する腺粘液細胞から分泌される腺粘液細胞型ムチン、又はムチンと類似の構造を有する他の機能性分子と同様なものである。したがって、本発明の機能性分子は、医薬、食物又は飲料として使用した場合でも副作用がなく、極めて安全性が高い点で有用である。
【0053】
機能性分子の主鎖が、例えばポリペプチドであってもよい。その一形態として、複数のモノマーユニットを含み、その少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基であって、主鎖に対してクラスター状に存在する分子が挙げられる。そのような機能性分子は、複数のモノマーユニットを含んでおり、その少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基であり主鎖に対してクラスター状に存在していると、前記コレステリル−α−D−グルコピラノシドの合成を抑制及び阻害することができ、同様に前記コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び阻害することができる。さらに前記機能性分子は、前記コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制又は阻害し、さらにはその細菌の形態異常を発現させる。
【0054】
このように、本発明の機能性分子は、コレステリル−α−D−グルコピラノシド(CGL)の合成を抑制及び/又は阻害することができ、及び/又はコレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び/又は阻害することができる。さらに本発明の機能性分子は、コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び/又は阻害し、また、その細菌の形態異常を発現させる。本発明の機能性分子は、例えばポリペプチドがCD43、CD34、又はMuc−6である分子が挙げられる。一つの形態として、主鎖に対する側鎖の数の比は10以上である。
【0055】
本発明の機能性分子は、胃潰瘍、十二指腸潰瘍、抗ヘリコバクター ピロリ剤、胃炎、及び/又は慢性胃炎の予防、治療、及び/又は改善のために、使用されるものであってもよい。また、機能性分子は、食物や飲料のような飲食物のために使用されるものであってもよい。これによって、このような機能性分子を含む組成物は、コレステリル−α−D−グルコピラノシドの合成を抑制及び/又は阻害することができ、コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び/又は阻害することができ、コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び/又は阻害することができ、及び/又はその細菌の形態異常を発現させることができる。また、本発明は、このような機能性分子を含むことにより前記のような抑制及び/又は阻害活性を奏する医薬を提供するというものである。この医薬組成物は、種々の形態で提供される。例えば、治療されるべき個体の胃及び/又は腸の粘膜層に前記医薬組成物を噴射又は塗布してもよい。また、別な形態として、前記医薬組成物をフィルム及び/又はシートに含浸又は含有させて、胃又は腸の粘膜層に付してもよい。本発明の医薬組成物は、コレステロールを分解するコレステロール分解酵素及びグルコースを分解するグルコース分解酵素の少なくともいずれかを有していてもよい。それにより、医薬組成物は前記したような抑制及び/又は阻害活性を有することができる。
【0056】
さらに、本発明は、機能性分子を含有する飲食物を提供するというものである。この機能性分子は、複数のモノマーユニットを含み、その少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基であるというものである。このような飲食物は、前記と同様に、コレステリル−α−D−グルコピラノシドの合成を抑制及び/又は阻害することができ、コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び/又は阻害することができ、生体内に存在しコレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び/又は阻害することができ、その細菌の形態異常を発現させることができる。
【0057】
また、本発明は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する機能性分子の製造方法に関するものである。この方法は例えば、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを接触させ、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する機能性分子を生成させて製造することができるというものである。本発明の方法の一つの具体例によれば、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとが接触すると、機能性分子が製造される。この具体例の一つの形態として、前記ポリペプチドは、可溶性ポリペプチド(例えばCD43、CD34、又はMuc−6)であるというものである。また、別な形態として、前記ポリペプチドは哺乳類の母乳中に分泌されるムチンであるというものである。
【0058】
本発明の機能性分子の製造方法は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する機能性分子を生成させた後に、単離する工程を含んでいてもよい。それによって本発明は、この方法により得られ、前記したような抑制及び/又は阻害活性を有する機能性分子を提供することができるというものである。例えば、単離された機能性分子を例えば医薬として投与することにより、生体内に存在しコレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び/又は阻害することができ、その細菌の形態異常を発現させることができる。
【0059】
また、本発明は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型の機能性分子の製造方法に関するものである。この方法は例えば、α4GnTをエンコードする第一のポリヌクレオチドと、C2GnT−IとC1−β3GnTとの少なくともいずれかをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとを真核細胞内で発現させるというものである。この方法によれば、第一のポリヌクレオチドと、第二のポリヌクレオチドと、第三のポリヌクレオチドとの少なくともいずれかは、真核細胞に導入される外因性ポリヌクレオチドである。例えばこの方法は、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとが発現することによって、行われるというものである。その後、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを互いに接触させて、機能性分子を生成させることができる。前記で示したように、前記ポリペプチドは可溶性ポリペプチド(例えばCD43、CD34、又はMuc−6)であってもよい。組換え型機能性分子がつくられる真核細胞は、例えば哺乳類の細胞であってもよい。
【0060】
本発明によれば、前記組換え型の機能性分子の製造方法は、さらに、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを接触させ、その発現後に、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型の機能性分子を生成させるというものであってもよい。この方法はさらに、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型機能性分子を生成後に単離させる工程を含んでいてもよい。それにより、本発明の方法で製造され、単離された組換え型機能性分子が、提供される。この単離された組換え型機能性分子は、前記したような抑制及び/又は阻害活性を有する。
【0061】
また、本発明は、母乳を分泌する非ヒトのトランスジェニック哺乳類を提供するというものである。本発明の非ヒトのトランスジェニック哺乳類は、α4GnTをエンコードする第一の外因性ポリヌクレオチドと、C2GnT−IとC1−β3GnTとの少なくともいずれかをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとをゲノム中に有することができるというものである。このようなトランスジェニック哺乳類は、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを発現することができる。その後、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを、互いに接触させ、機能性分子を生成することができる。
【0062】
本発明の非ヒトのトランスジェニック哺乳類は、外因性の第一のポリヌクレオチド、第二のポリヌクレオチド、及び/又は第三のポリヌクレオチドを有していてもよい。一つの形態として、前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとの少なくともいずれかが、ゲノム中に導入され、哺乳類の乳腺遺伝子の5’制御配列及び3’制御配列の少なくともいずれかに相互作用して結合する。別な形態として、第三のポリヌクレオチドが、ゲノム中に導入され、少なくとも一つのO−グリコシル化部位を有するポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と相互作用して結合する。ポリヌクレオチドが相互作用して結合する5’制御配列は、例えば乳清酸性タンパク質(WAP)プロモータ、α−カゼインプロモータ、β−カゼインプロモータ、γ−カゼインプロモータ、α−ラクトアルブミンプロモータ、又はβ−ラクトグロブリンプロモータのようなプロモータを含んでいてもよい。さらに別な形態として、前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、単一の核酸分子を形成し、この核酸分子が非ヒトのトランスジェニック哺乳類のゲノム中に導入されて、乳腺遺伝子の5’制御配列と3’制御配列とのいずれかに相互作用して結合する。トランスジェニック哺乳類は、母乳を分泌するものであれば如何なる哺乳類であってもよく、例えばマウス、ラット、ウサギ、ウマ、ブタ、ヒツジ、ヤギ、又はウシが挙げられる。
【0063】
前記トランスジェニック哺乳類が前記第二の外因性のポリヌクレオチドを必要としているか否かは、本発明の機能性分子を産生する細胞が、自然の状態で内因性の前記C2GnT−Iを発現するか否かによって決定され、それが発現する場合には導入しなくてもよい。また、前記トランスジェニック哺乳類が前記第三の外因性のポリヌクレオチドを導入する必要があるか否かは、本発明の機能性分子を産生する細胞が、自然の状態で前記少なくとも一つのO−グリコシル化部位を有する内因性のポリペプチドを発現するか否かによって決定され、それが発現する場合には導入しなくてもよい。例えば、前記細胞が乳腺上皮細胞である場合には、この乳腺上皮細胞は前記C2GnT−I及び前記少なくとも一つのO−グリコシル化部位を有するポリペプチドを発現するので、前記乳腺上皮細胞を有するトランスジェニック哺乳類は、前記第二及び第三のポリヌクレオチドを導入しなくてもよい。
【0064】
本発明のトランスジェニック哺乳類は、前記少なくとも一つのO−グリコシル化部位を有するポリペプチドが、前記α4GnTと、前記C2GnT−I及びC1−β3GnTの少なくともいずれかとによってグルコシル化されて、母乳を分泌する雌の場合には、その母乳中に本発明の機能性分子を分泌させることができる。組換え型のポリペプチドを母乳中に分泌できる非ヒトのトランスジェニック哺乳類の生産方法は、公知の方法に準じて製造することができるというものである(例えば、米国特許 第6,344,596号;第6,548,735号;第6,222,094号;第5,962,648号;第5,891,698号;第5,850,000号の各明細書参照)。
【0065】
また、本発明は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型の機能性分子を生成させて製造する方法に関するものである。その方法は例えば、α4GnTをエンコードする第一のポリヌクレオチドを非ヒトの哺乳類の胚に導入し;導入した胚を宿主の雌哺乳類に移植し;宿主の雌哺乳類に子孫を出産させ;出産した子孫の内、雌の子孫に母乳の産生を行わせるというものであってもよい。この方法によれば、この子孫の体内で、α4GnTが発現する。一つの形態として、α4GnTは子孫の体内に存在するポリペプチドと少なくとも接触し、それによって機能性分子を生成する。他の形態として、第一のポリヌクレオチドは、哺乳類の乳腺に関する遺伝子の5’制御配列及び3’制御配列の少なくともいずれかと互いに相互作用して結合するように胚に導入される。一つの具体例として、前記組換え型の機能性分子の製造方法は、C2GnT−I及びC1−β3GnTの少なくともいずれかをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有するポリペプチドをエンコードする第三のポリヌクレオチドとを胚に導入する工程を含む。別な具体例として、前記の方法は、雌の子孫に母乳を産生させた後、その母乳を搾乳する工程を含み、さらに前記母乳から組換え型の機能性分子を単離する工程を含んでもよい。これによって本発明は、前記の方法で得られ、特に前記組換え型の機能性分子を含み、前記したような抑制及び/又は阻害活性を有する母乳を提供することができるというものである。
【0066】
本発明の機能性分子は前記したいずれの方法によっても製造することができるが、本発明に基づいて公知で通常の別な方法で製造されてもよい。本発明の機能性分子の製造方法として、三つの形態を例示する。なお、本発明の母乳は、本発明の機能性分子の説明を通して明らかにする。前記第一の形態は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する機能性分子の製造方法であって、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを互いに接触させる工程を少なくとも含むものである。前記第二の形態は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型の機能性分子の製造方法であって、α4GnTをエンコードする第一のポリヌクレオチドと、C2GnT−I及びC1−β3GnTの少なくともいずれかをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有するポリペプチドをエンコードする第三のポリヌクレオチドとを真核細胞内で発現させる発現工程を少なくとも含むものである。前記第三の形態は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型の機能性分子の製造方法であって、α4GnTをエンコードする第一のポリヌクレオチドを非ヒトの哺乳類の胚に導入し、導入した胚を宿主の雌哺乳類に移植し、子孫を出産させ、出産した子孫の内、雌の子孫に母乳の産生を行わせる工程を含むものである。前記α4GnT、前記C2GnT−I、及び前記C1−β3GnTについては前記の通りである。前記ポリペプチドは、少なくとも一つのO−グリコシル化部位を有するポリペプチド(例えばCD43、CD34、Muc−6、ムチン)であって、哺乳類の母乳中に分泌される。
【0067】
前記第一の形態では、前記α4GnTと、前記C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとの他に、前記GlcNAcと糖とをさらに接触させてもよい。前記第二の形態では、前記発現工程の後、前記α4GnTと、前記C2GnT−I及びC1−β3GnTの少なくともいずれかと、前記少なくとも一つのO−グリコシル化部位を有するポリペプチドとを互いに接触させることにより前記機能性分子を製造することができる。前記GlcNAcと糖とをさらに接触させてもよい。また、前記第二の形態中、前記第一から第三のポリペプチドは、外因性であってもよく、内因性であってもよい。なお、本明細書中、「内因性」という用語は、ポリヌクレオチドに関して使用されている場合、ある特定の細胞内又は生物の細胞内で前記ポリヌクレオチドが自然状態で見出されることを意味し、「外因性」という用語は、ポリヌクレオチドに関して使用されている場合、ある特定の細胞内又は生物の細胞内で前記ポリヌクレオチドが自然状態では見出されないことを意味する。
【0068】
本発明の方法に、真核細胞を使用することができる。一般に、前記細胞は哺乳類の細胞であり、例えばチャイニーズハムスター卵巣細胞(CHO細胞)や乳腺細胞が挙げられる。前記真核細胞が乳腺細胞である場合には、前記C2GnT−Iと、前記少なくとも一つのO−グリコシル化部位を有するポリペプチドとを発現することができるため、前記α4GnTを外因性のものとして供給することにより前記機能性分子を産生することができる。また、前記真核細胞が前記C2GnT−Iを発現する場合には、前記α4GnTと、前記少なくとも一つのO−グリコシル化部位を有するポリペプチドとを外因性のものとして供給することにより前記機能性分子を産生することができる。
【0069】
前記第二の形態の製造方法の一例として、以下の方法がある。前記CD43の細胞外ドメイン全体を含むDNA断片(配列番号1におけるアミノ酸配列の20番目から254番目に相当する部分)をPCR法で増幅し、pSecTag2(インビトロジェン社製)のようなベクターにサブ・クローニングすることにより、Igκリーダー・ペプチドと、それに続くmycエピトープ、及び(His)6をエンコードするベクター(pSecTag2−sCD43)を作製する。次に、(1)C2GnT−Iをエンコードするポリヌクレオチド(cDNA)と、(2)α4GnTをエンコードするポリヌクレオチド(cDNA)と、(3)可溶性CD43をエンコードするポリヌクレオチド(cDNA)と、(4)ポリオーマ・ウイルス large T抗原をエンコードするポリヌクレオチド(cDNA)とを、夫々別々に前記pSecTag2−sCD43に組み込んだベクター4つを、LipofectAmine(登録商標、インビトロジェン社製)を用いて前記培養したCHO・Lec2細胞にトランスフェクトさせ、そのトランスフェクトさせたCHO・Lec2細胞内で、C2GnT−I、α4GnT、可溶性CD43、及びポリオーマ・ウイルス large T抗原を発現させ、接触させることにより、前記機能性分子として、末端GlcNAcα残基を含む可溶性CD43を発現させることができる。
【0070】
前記第三の形態の製造方法では、非ヒトの哺乳類の胚に、前記第一のポリヌクレオチドと、必要に応じて前記第二のポリヌクレオチド及び前記第三のポリヌクレオチドの少なくともいずれかを導入する。前記第一のポリヌクレオチドは、哺乳類の乳腺に関する遺伝子の5’制御配列及び3’制御配列の少なくともいずれかと互いに相互作用して胚に導入されてもよい。その場合、前記第二のポリヌクレオチドと前記第三のポリヌクレオチドとのいずれか又は両方が、哺乳類の乳腺に関連する遺伝子の5’制御配列及び3’制御配列の少なくともいずれかと互いに相互作用して胚に導入されてもよい。前記乳腺に関連する遺伝子は特に制限はないが、一般には乳腺特異的遺伝子である。前記5’制御配列は所望の制御要素であってもよく、前記したように一般にプロモータを有している。
【0071】
前記少なくとも一つのO−グリコシル化部位を有するポリペプチドをエンコードしている第三のポリヌクレオチドが、5’制御配列及び3’制御配列の少なくともいずれかの制御要素(例えば、乳腺特異的プロモータ等)に互いに相互作用して結合して、導入されると、前記制御要素は、前記ポリヌクレオチドに対して、通常の細胞内に関連付けられている別のポリヌクレオチド配列に対して行うように、その制御を行い、例えば、乳腺上皮細胞内で、前記エンコードされたポリペプチドが組織特異的に発現することができる。また、前記第一のポリヌクレオチド配列を前記第二のポリヌクレオチド配列に互いに相互作用させて結合させて導入し、二つ又はそれ以上のポリペプチドを前記相互作用させて導入したポリヌクレオチドから発現させることも可能である。また、必要に応じて、前記互いに相互作用させて導入したポリヌクレオチドからキメラの(組み合わされた)ポリペプチドを発現させることも可能である。前記キメラのポリペプチドは、融合ポリペプチドであってもよい。前記融合ポリペプチドとは、前記二つ又はそれ以上のエンコードされたポリペプチドが単一のポリペプチドに翻訳されたもの、つまり、それらがペプチド結合により共有結合したものである。
【0072】
前記第三のポリヌクレオチドは、少なくとも一つのO−グリコシル化部位を有するポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と互いに相互作用させて結合させて、胚に導入される。前記雌の子孫に母乳を産生させた後、母乳を搾乳してもよく、前記母乳から前記機能性分子を単離してもよい。前記第三の形態の製造方法では、前記雌の子孫の体内で前記第一のポリヌクレオチドから前記α4GnTが発現し、内因性又は外因性の前記少なくとも一つのO−グリコシル化部位を有するポリペプチドが前記C2GnT−I及び前記C1−β3GnTの少なくともいずれかによりグリコシル化され、前記機能性分子が産生される。前記機能性分子は、前記雌の子孫の母乳中に含まれ、該母乳を搾乳することにより効率的に産生することができる。
【0073】
前記したように、本発明の機能性分子は各種分野で使用可能である。前記機能性分子は、特に組成物、医薬及び/又は飲食物として供給される場合に、ヘリコバクター ピロリが原因である病気の治療薬、例えば、胃潰瘍治療薬、十二指腸潰瘍治療薬、胃炎治療薬、慢性胃炎緩和治療薬として好適に用いることができる。本発明の組成物、医薬及び飲食物は、本発明の分子及び機能性分子の少なくともいずれかを含有してなり、必要に応じて適宜選択したその他の成分を含有してなる。本発明の前記組成物、医薬、又は飲食物の全量中での前記分子及び機能性分子の少なくともいずれかの含有量は、特に制限はなく、目的に応じて適宜選択することができる。本発明の前記分子、前記機能性分子、前記医薬、又は前記組成物の全量に対する前記分子の含有量は、必要に応じて適宜選択することができるが、一般に、前記含有量が高ければ高いほど、抗菌効果が高くなる。
【0074】
種々の形状の前記組成物は、その他の成分を含んでいてもよい。前記その他の成分は特に制限はなく、目的に応じて適宜選択することができる。前記その他の成分として、例えば、賦形剤、滑沢剤、結合剤、水溶性高分子、塩基性無機塩、溶剤、溶解補助剤、懸濁化剤、等張化剤、緩衝剤、無痛化剤、防腐剤、抗酸化剤、着色剤、甘味剤、酸味剤、発泡剤、香料などが挙げられる。また、CGLの合成を効率的に抑制又は阻害する観点から、コレステロール分解酵素やグルコース分解酵素の添加も有用である。前記賦形剤として、例えば、乳糖、白糖、D−マンニトール、デンプン、コーンスターチ、結晶セルロース、軽質無水ケイ酸、酸化チタンなどが挙げられる。前記滑沢剤として、例えば、ステアリン酸マグネシウム、ショ糖脂肪酸エステル、ポリエチレングリコール、タルク、ステアリン酸などが挙げられる。前記結合剤として、例えば、ヒドロキシプロピルセルロース、ヒドロキシプロピルメチルセルロース、結晶セルロース、デンプン、ポリビニルピロリドン、アラビアゴム粉末、ゼラチン、プルラン、低置換度ヒドロキシプロピルセルロースなどが挙げられる。前記水溶性高分子として、例えば、ヒドロキシプロピルセルロース、ヒドロキシプロピルメチルセルロース、メチルセルロース、カルボキシメチルセルロースナトリウム等のセルロース誘導体、ポリビニルピロリドン、ポリアクリル酸ナトリウム、ポリビニルアルコール、アルギン酸ナトリウム、グアーガムなどが挙げられる。前記塩基性無機塩として、例えば、ナトリウム、カリウム、マグネシウム、カルシウムの塩基性無機塩が挙げられる。前記ナトリウムの塩基性無機塩として、例えば、炭酸ナトリウム、炭酸水素ナトリウム、リン酸水素二ナトリウムが挙げられる。前記カリウムの塩基性無機塩として、例えば、炭酸カリウム、炭酸水素カリウムが挙げられる。前記マグネシウムの塩基性無機塩として、例えば、重質炭酸マグネシウム、炭酸マグネシウム、酸化マグネシウム、水酸化マグネシウム、メタ珪酸アルミン酸マグネシウム、珪酸マグネシウム、アルミン酸マグネシウム、合成ヒドロタルサイト(Mg6Al2(OH)16・CO3・4H2O)、酸化アルミナ・マグネシウムが挙げられる。前記カルシウムの塩基性無機塩として、例えば、沈降炭酸カルシウム、水酸化カルシウムが挙げられる。前記溶剤として、例えば、注射用蒸留水、アルコール、プロピレングリコール、マクロゴール、ゴマ油、トウモロコシ油、オリーブ油などが挙げられる。前記溶解補助剤として、例えば、ポリエチレングリコール、プロピレングリコール、D−マンニトール、安息香酸ベンジル、エタノール、トリスアミノメタン、コレステロール、トリエタノールアミン、炭酸ナトリウム、クエン酸ナトリウムが挙げられる。前記懸濁化剤として、例えば、ステアリルトリエタノールアミン、ラウリル硫酸ナトリウム、ラウリルアミノプロピオン酸、レシチン、塩化ベンザルコニウム、塩化ベンゼトニウム、モノステアリン酸グリセリンのような界面活性剤;ポリビニルアルコール、ポリビニルピロリドン、カルボキシメチルセルロースナトリウム、メチルセルロース、ヒドロキシメチルセルロース、ヒドロキシエチルセルロース、ヒドロキシプロピルセルロースのような親水性高分子が挙げられる。前記等張化剤として、例えば、ブドウ糖、D−ソルビトール、塩化ナトリウム、グリセリン、D−マンニトールが挙げられる。前記緩衝剤として、例えば、リン酸塩、酢酸塩、炭酸塩、クエン酸塩が挙げられる。前記無痛化剤として、例えば、ベンジルアルコールが挙げられる。前記防腐剤として、例えば、パラオキシ安息香酸エステル類、クロロブタノール、ベンジルアルコール、フェネチルアルコール、デヒドロ酢酸、ソルビン酸が挙げられる。前記抗酸化剤として、例えば、亜硫酸塩、アスコルビン酸、α−トコフェロールが挙げられる。前記着色剤として、例えば、食用黄色5号、食用赤色2号、食用青色2号のような食用色素;食用レーキ色素;ベンガラが挙げられる。前記甘味料として、例えば、サッカリンナトリウム、グリチルリチン二カリウム、アスパルテーム、ステビア、ソーマチンが挙げられる。前記酸味料として、例えば、クエン酸(無水クエン酸)、酒石酸、リンゴ酸が挙げられる。前記発泡剤として、例えば、重曹が挙げられる。前記香料として、例えば、レモン、ライム、オレンジ、メントール、ストロベリーの抽出物が挙げられる。
【0075】
本発明の組成物及び/又は医薬は、投与の目的に合わせて、どのような形態、方法で投与されてもよい。その方法は、経口投与するものであってもよく、ヒトの胃及び腸の少なくともいずれかの粘膜層に前記組成物(又は機能性分子)を直接噴霧又は塗布するものであってもよい。また、本発明の組成物又は医薬を、フィルム又はシートに含浸させ、そのフィルム又はシートを胃及び/又は腸に付して使用してもよい。この投与方法は、胃又は腸の粘膜層表面で、噴霧又は塗布された前記組成物や医薬が流れ落ちることなく、一定時間保持されることにより、目的量の組成物又は医薬を投与でき、十分な抗菌効果を得ることができるため、有用である。また、前記使用方法は、内視鏡などの公知の医療器具を使用する場合に、好適に用いることができる。患部が胃である場合には、前記フィルム又はシートはキチン、糖、脂肪成分を含むことが好ましい。患部が腸である場合には、前記フィルム又はシートはキチンを含むことが好ましい。これにより、前記機能性分子は、胃や腸の中での胃液や腸液によって分解されることなく、長時間に亘って患部に接触することができる。
【0076】
本発明の組成物、医薬、及び飲食物は、十分な抗菌効果を有するが、抗菌効果をさらに向上させるために、抗生物質、生薬のような公知の胃潰瘍治療薬、十二指腸潰瘍治療薬、抗ヘリコバクター ピロリ薬、胃炎治療薬、及び慢性胃炎改善薬の少なくともいずれかと併用して使用してもよい。本発明の組成物、医薬及び飲食物は、本発明の機能性分子(又は分子)を含有するため副作用がなく、極めて安全性が高い点で有用であり、各種分野で使用可能である。本発明の組成物は例えば、ヘリコバクター ピロリに対する治療薬(例えば、胃潰瘍治療薬、十二指腸潰瘍治療薬、抗ヘリコバクター ピロリ薬、胃炎治療薬、慢性胃炎改善薬)に好適に用いることができ、本発明の医薬及び飲食物に特に好適に用いることができる。本発明の飲食物は、清涼飲料、炭酸飲料、栄養飲料、果実飲料、乳酸飲料、栄養補助食品、冷菓、麺類、菓子類、水産物、乳製品、加工食品、調味料、錠剤、カプセル剤などとして好適に使用することができる。
【0077】
このように、本発明は細菌の増殖抑制方法に関するものである。この方法は例えば、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する機能性分子に、CGLを発現する細菌を接触させ、細菌の増殖と、細菌の細胞壁の形成との少なくともいずれかを抑制又は阻害するものである。この方法によれば、前記機能性分子はコレステリル−α−D−グルコピラノシドの発現(即ち合成)を抑制又は阻害し、同様に、コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖と、細菌の細胞壁の形成との少なくともいずれかを抑制又は阻害する。さらに前記機能性分子は、コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制又は阻害し、さらにはその細菌の形態異常を発現させる。α1,4−N−アセチルグルコサミン残基を有する前記機能性分子は、UDP−Glc:ステロール糖転移酵素活性を抑制又は阻害することができる。増殖が抑制及び/又は阻害される細菌として、例えばヘリコバクター属の細菌(例えばヘリコバクター ピロリ)が挙げられる。
【0078】
さらに、本発明は胃潰瘍の治療方法を提供するというものである。この方法は例えば、ヘリコバクター種の細菌が個体に感染することによって起こる胃潰瘍の兆候又は症状を改善するために、α1,4−N−アセチルグルコサミン残基を有する機能性分子を投与するものである。この胃潰瘍治療方法では、前記機能性分子はコレステリル−α−D−グルコピラノシドの発現(合成)を抑制又は阻害し、同様に、コレステリル−α−D−グルコピラノシドを必須とするヘリコバクター種の細菌に感染することによって起こる胃潰瘍の兆候又は症状を改善する。前記ヘリコバクター種の細菌はヘリコバクター ピロリであってもよい。前記個体は例えばマウス、ラット、ウサギ、ウマ、ブタ、ヒツジ、ヤギ、ウシのような哺乳類であって、特にヒトであるとなおよい。
【0079】
胃潰瘍の治療方法では、胃腺粘液細胞型ムチンを含みα1,4−N−アセチルグルコサミン残基を有する機能性分子を利用することができる。前記機能性分子は種々の方法で投与される。投与方法として、例えば、経口投与、胃粘膜層への噴霧及び/又は塗布、前記機能性分子を含むフィルム及び/又はシートの貼付が挙げられる。一つの形態として、前記機能性分子は、前記のようにして生産された母乳を用いて投与される。
【0080】
さらに、本発明は胃潰瘍を予防する方法に関するものである。この方法は例えば、α1,4−N−アセチルグルコサミン残基を有する機能性分子の投与により、個体にヘリコバクター種の細菌が感染することによって起こる胃潰瘍の兆候又は症状を妨げ、前記機能性分子がコレステリル−α−D−グルコピラノシドの発現(合成)を抑制又は阻害し、コレステリル−α−D−グルコピラノシドを必須とするヘリコバクター種の細菌に感染することによって起こる胃潰瘍の兆候又は症状を予防するというものである。α1,4−N−アセチルグルコサミン残基を有する機能性分子は胃腺粘液細胞型ムチンを含んでいてもよい。また前記機能性分子は、前記した方法や公知の方法に準じて、投与される。
【0081】
さらに本発明は、胃癌を治療する方法に関するものである。この方法は例えば、α1,4−N−アセチルグルコサミン残基を有する機能性分子の投与により、個体にヘリコバクター種の細菌が感染することによって起こる胃癌の兆候又は症状を改善し、前記機能性分子がコレステリル−α−D−グルコピラノシドの発現(合成)を抑制又は阻害し、同様に、ヘリコバクター種の細菌に感染することによって起こる胃癌の兆候又は症状を改善するものである。
【0082】
本発明は、担体分子と相互作用して結合した少なくとも一つの末端α1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する非天然化合物を提供するというものである。「相互作用して結合」という用語は、本明細書中で、二つ以上の分子が一つの単位として働き、さらに一つ又は両方の分子、又はそれらの組み合わせによる機能効果を奏するような位置関係である二つ又はそれ以上の分子に関して広く使用される。例えば、担体分子と相互作用して結合したα1,4結合−GlcNAc残基とは、ヘリコバクター ピロリのようなヘリコバクター種の細菌の増殖を抑制又は阻害する効果を奏するように、α1,4結合−GlcNAc残基が位置しているということを意味している。そのように相互作用して結合したα1,4結合−GlcNAc残基は例えば、末端α1,4結合−GlcNAc残基を含むCD43分子やCD34分子で例示される。それらの化合物は他の担体分子へのα1,4結合−GlcNAc残基の相互作用した結合を確認する基準となる。
【0083】
二つ又はそれ以上のポリヌクレオチド配列もまた、相互作用して結合し得る。例えば、ポリペプチド(例えば担体ポリペプチド)をエンコードするポリヌクレオチドは、制御要素(例えば乳腺特異的プロモータ)に相互作用して結合することができ、前記制御要素は、通常細胞内に制御要素と関係するポリヌクレオチド配列をもたらすのと同様に、前記ポリヌクレオチドに対してその制御効果を与える(例えば、乳腺上皮細胞内で、前記エンコードされたポリペプチドが組織特異的に発現することができる)。また、前記第一のポリヌクレオチド配列を前記第二又はそれ以上のポリヌクレオチド配列に互いに相互作用させて導入し、二つ又はそれ以上のポリペプチドを前記相互作用させて導入したポリヌクレオチドから発現させることも可能である。また、必要に応じて、前記互いに相互作用させて導入したポリヌクレオチド配列からキメラのポリペプチドを発現させることも可能である。前記キメラのポリペプチドは、融合ポリペプチドであってもよい。前記融合ポリペプチドとは、前記二つ又はそれ以上のエンコードされたポリペプチドが単一のポリペプチドに翻訳されたもの、つまり、それらがペプチド結合により共有結合したものである。
【0084】
「担体分子」という用語は、ヘリコバクター増殖抑制活性を示す一つ又はそれ以上のα1,4結合−GlcNAc残基を相互作用させて導入できる分子に関して使われている。そのような担体分子として、ポリペプチド、ポリヌクレオチド、オリゴ糖、脂質、糖タンパク質、ヌクレオチドタンパク質、又は同様のものがある。担体分子は例えばO−グリカン(例えばコア2分岐型O−グリカン)で例示される。担体分子はさらにCD43やCD34のようなポリペプチドと相互作用して結合して、糖タンパク質を製造することができる。
【0085】
本明細書で示したように、一つ又はそれ以上のα1,4結合−GlcNAc残基を有する化合物は、コレステリル−α−D−グルコピラノシド(CGL)を発現する細菌の種々の機能、例えば細菌の増殖、細菌の運動性、細菌によるCGLの合成の一つ又はそれ以上の機能を抑制又は阻害することができる。「抑制又は阻害」という用語は、前記化合物の存在下で検査した機能が、前記化合物の非存在下での機能レベルと比較して、減少するか全く機能がなくなる状態となることを意味する。例えば一つ又はそれ以上のα1,4結合−GlcNAc残基を有する化合物は、その非存在下の場合と比べて、ヘリコバクター ピロリによるCGL合成の量を抑制又は阻害していることが本明細書に明示されている(実施例1参照)。「抑制又は阻害」という用語は、機能レベルが特別な分析によって検知できるレベル以下にまで減少すると認められるので、本明細書では通常一緒に使用している。例えば、前記化合物と接触したヘリコバクター ピロリによるCGL合成が完全に抑制されたかどうか、又は特別な分析による検知レベル以下のその合成レベルが維持されているかどうかという分析は確定できないかもしれない。しかし、制御レベルの減少は、前記化合物との接触によるものである。
【0086】
微好気性、グラム陰性の細菌であって、全世界の人口のおよそ半分のヒトの胃に感染しているヘリコバクター ピロリは、胃潰瘍の原因となり、また、胃の悪性腫瘍の原因となる(Marshall and Warren, Lancet 1311, 1984; Peek and Blaser,Nat.Rev.Cancer 2:28,2002;Du and Isaccson, Lancet Oncol.3:97, 2002参照)。感染しているほとんどのヒトは無症候群であるか、慢性胃炎のみを患い、患者の一部が消化性潰瘍、胃癌、悪性リンパ腫を患う(Cave,Semin.Gastrointest.Dis.12:196,2001参照、参考文献としてここに示す)。無症候群であるヒトは自己防御能を有しているのかもしれない。ヘリコバクター ピロリは、末端α1,4結合−GlcNAc残基を有するO−グリカンを発現する胃粘膜の深部でめったにみられないので、これらのO−グリカンがヘリコバクター ピロリの増殖に影響を及ぼすかどうか調べた。
【0087】
組織化学特性に基づき、胃粘素を二つに分類した(Otaら、Histochem.J.23:22,1991参照、参考文献としてここに示す)。一つは粘液細胞表面から分泌される表面粘液細胞型ムチンである。もう一つは粘液深部でみられ、粘液頚細胞、心腺細胞、幽門腺細胞のような腺粘液細胞から分泌される腺粘液細胞型ムチンである。ヘリコバクター ピロリに感染した胃粘液では、微生物が単独で表面粘液細胞型ムチンに接合しており、このムチンにヘリコバクター ピロリが付着するための糖質リガンドが存在する。ルイスbやシアリル−ルイスXとして知られる糖質構造はヘリコバクター付着特異リガンドBabAとSabAとを夫々供給する。両方の糖質リガンドは粘液細胞表面に専ら発現する(Ilverら、Science 279:373,1998; Mahdaviら、Science 297:573,2002)。対照的に、ヘリコバクター ピロリは、コア2分岐型O−グリカン(GlcNAca1→4Ga1β1→4GlcNAcβ1→6(GlcNAca1→4Galβ1→3)GalNAca→Ser/Thr)に結合した末端α1,4結合−GlcNAc残基を有する腺粘液細胞型ムチンを製造する胃粘液の深部に、コロニーを殆ど形成しない(Hidakaら、前出,2001)。腺粘液の損失に伴い洞萎縮すると、洞萎縮のない慢性胃炎に比べて、消化性潰瘍や胃癌の危険性が2〜3倍に高まってしまう(Sipponen and Hyvarinen, Scand.J.Gastroenterol.196(Suppl.3):3,1993参照、参考文献としてここに示す)。この観察と、ヘリコバクター ピロリ感染した大多数のヒトが無症候群である(Cave,前出,2001)という知見に基づき、深部ムチンに発現した末端α1,4結合−GlcNAc残基を有するO−グリカンがヘリコバクター ピロリ感染からヒトを保護できるかについて研究した。
【0088】
本明細書に示したように、末端α1,4結合−GlcNAc残基を有するO−グリカンはヘリコバクター ピロリに対する抗菌活性を有し、その細胞壁の主成分であるコレステリル−α−D−グルコピラノシドの生合成を抑制する。したがって、胃ムチン中のO−グリカン特有のサブセットは天然抗生物質としての機能を発揮し、ヘリコバクター ピロリ感染から宿主を保護する。これらのムチンがヘリコバクター ピロリ感染から宿主を保護することを確認できたことから、少なくとも一部がCGLの発現によって特徴づけられる他の細菌による感染だけでなく、ヘリコバクター種の細菌による感染を治療又は予防するのに有用な化合物を創り出すことができた(実施例1参照)。これによって、ヘリコバクター感染を治療するための組成物、その製造方法及びその使用方法が提供される。
【0089】
一つの具体例として、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質の製造方法を示す。それの一形態として、例えば、ポリペプチドのグリコシル化に適した条件下で、α4GnTと、C2GnT−Iと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドとを接触させ、このポリペプチドを、C2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化し、少なくとも一つのα1,4結合−N−アセチルグルコサミン残基を有する糖タンパク質を製造するものである。第二の形態として、前記C2GnT−Iとの組み合わせに代えて、C1−β3GnTのようなコア1伸長酵素と、C1−β3GnTに対する基質である担体ポリペプチドとを使用してもよい(Yahら、Cell 105:957-969,2001参照、参考文献としてここに示す)。この形態によれば、担体ポリペプチドはC1−β3GnTとα4GnTとによってグリコシル化され、少なくとも一つのα1,4結合−N−アセチルグルコサミン残基を有する糖タンパク質が製造される。前記担体ポリペプチドは、C2GnT−I(又はC1−β3GnT)とα4GnTとの活性により、一つ以上のα1,4結合−N−アセチルグルコサミン残基を結合できるものであればよく、例えば、可溶性ポリペプチド(例えば可溶性CD43ポリペプチド又は可溶性CD34ポリペプチド)が挙げられる。前記製造方法はさらに、前記糖タンパク質を単離する工程を含んでいてもよい。このようにして、本発明の方法で製造された糖タンパク質が単離された糖タンパク質として提供される。
【0090】
前記の通り、CGLを発現する細菌、特にヘリコバクター種の細菌の感染を治療又は予防するのに有用な組成成分は、担体分子と相互作用して結合した少なくとも一つの末端α1,4結合−GlcNAc残基を有する。このような組成成分はin vitro又はin vivoでつくられる。例えば、少なくとも一つのα1,4結合−GlcNAc残基を有するポリペプチド担体分子を含む糖タンパク質は、ポリペプチドのグリコシル化に適した条件下で、α1,4−N−アセチルグルコサミン転移酵素(α4GnT)と、コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)と、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドとを接触させて製造される。この方法によれば、前記ポリペプチドはC2GnT−Iとα4GnTとによってグリコシル化され、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質が製造される。
【0091】
このような糖タンパク質は、組み換えDNAの技術を用いて製造することもできる。例えば少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質は、α4GnTをエンコードする第一のポリヌクレオチドと、C2GnT−Iをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとを真核細胞内で発現させて製造できる。一般に、一つ、二つ、又は三つのポリヌクレオチドは真核細胞に対して内因性ではなく、一つ、二つ又は三つ全てのエンコードされたポリペプチド(α4GnT、C2GnT−I、担体ポリペプチド)は真核細胞に対して異型である。この方法で製造した組換え型の糖タンパク質は例えば、α1,4結合−GlcNAc残基を有するCD43(ロイコシアリン)分子、α1,4結合−GlcNAc残基を有するCD34分子で例示される(実施例1参照)。
【0092】
別な具体例として、本発明は、非ヒトの雌トランスジェニック哺乳類も範疇に含む非ヒトのトランスジェニック哺乳類に関するものである。この非ヒトのトランスジェニック哺乳類は、一つ又はそれ以上のα1,4結合−GlcNAc残基を有する組換え型糖タンパク質を製造する一つ又はそれ以上の導入遺伝子を含んでいる。前記糖タンパク質は非ヒトの雌トランスジェニック哺乳類の母乳中に分泌される。本発明の非ヒトのトランスジェニック哺乳類は、少なくともα4GnTをエンコードする第一の外因性ポリヌクレオチドと、必要に応じてC2GnT−Iをエンコードする第二の外因性ポリヌクレオチド及び/又は少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチドとをゲノム中に安定に組み込んで有している。本明細書に示したように、前記トランスジェニック哺乳類がさらに前記第二の外因性のポリヌクレオチド及び/又は前記第三の外因性のポリヌクレオチドを導入するか否かは、細胞、又は一つ又はそれ以上のα1,4結合−GlcNAc残基を有する組換え型糖タンパク質を産生する細胞が、自然の状態で内因性のC2GnT−I及び/又は所望の担体ポリペプチドを発現するか否かに依存する。例えば、乳腺上皮細胞は、C2GnT−Iとムチン型糖タンパク質とを発現する。よって、非ヒトのトランスジェニック哺乳類がその母乳中に一つ又はそれ以上のα1,4結合−GlcNAc残基を有する組換え型糖タンパク質を発現できる場合は、第二及び/又は第三のポリヌクレオチドを導入してもよいが、必ずしもその必要はない。したがって、種々の具体例では、本発明の非ヒトのトランスジェニック哺乳類は、一つの、又は二つの、又は三つの外因性ポリヌクレオチドを有している。一つの形態として、非ヒトのトランスジェニック哺乳類は、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合された第一の外因性ポリヌクレオチドを、ゲノム中に安定に組み込んで有している。第二の形態として、非ヒトのトランスジェニック哺乳類はさらに、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合された第二及び/又は第三のポリヌクレオチドを、夫々必要に応じてゲノム中に安定に組み込んで有している。さらに、第三のポリヌクレオチドは、共存するときには担体ポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と相互作用して結合している。
【0093】
さらに別な形態として、本発明は、少なくともα4GnTをエンコードする第一の外因性ポリヌクレオチドと、C2GnT−Iをエンコードし、又はC1−β3GnTをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとをゲノム中に安定に組み込んで有し、前記第一のポリヌクレオチドと前記第二のポリヌクレオチドと前記第三のポリヌクレオチドとの少なくともいずれかが外因性ポリヌクレオチドであり、前記第一のポリヌクレオチドと前記第二のポリヌクレオチドと前記第三のポリヌクレオチドとがプロモータを含む植物遺伝子の5’制御配列に相互作用して結合され、C2GnT−I又はC1−β3GnTとα4GnTと担体ポリペプチドとの発現により、前記担体ポリペプチドを、C2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化するトランスジェニック植物を提供するというものである。例えばエンハンサー及び/又は翻訳制御要素をさらに含む5’制御要素を構成する、相互作用して結合されたプロモータは、構成プロモータ、誘導プロモータ、又は組織特異プロモータであってもよく、例えば種、花、葉、及び/又は根のような植物の所望の細胞中のエンコードされたポリペプチド(例えばα4GnT)を転写するものである。さらに本発明は、前記トランスジェニック植物によって産出された種子、前記トランスジェニック植物から産生された穀類や小麦粉のような産物、前記トランスジェニック植物やその種、花、葉、又は根に由来する産物、豆乳やライスミルク等の乳液のような産物を提供するというものである。植物から前記のような産物を収穫して生産する方法は通常、公知の方法に準じて行われ、例えば豆乳を生産する方法(例えば、米国特許 第4,119,733号;第4,409,256号;第4,902,526号;第6,316,043号;第6,322,846号;第6,451,359号の各明細書参照、夫々参考文献としてここに示す)や、ライスミルクを生産する方法(例えば、米国特許 第4,744,992号;第4,894,242号;第5,609,895号;第6,599,552号の各明細書参照、夫々参考文献としてここに示す)がある。
【0094】
ポリヌクレオチドに関して使用されている「外因性」という用語は、ある特定の細胞内又は生物の細胞内で前記ポリヌクレオチドが自然状態では発見されないことを意味する。「異型」という用語も同様の意味で使用している。「異型」という用語はさらに、通常自然状態では発見されない二つ又はそれ以上のヌクレオチド配列を有するキメラのポリヌクレオチドに関して使用している。例えば、一つの遺伝子のコードされた配列が、第二の遺伝子のコードされた配列及び/又はプロモータ又は他の第二の制御配列(又は他の遺伝子)に結合(相互作用して結合)している場合、第一のコードされた配列は、第二のコードされた配列及び/又は制御配列に対して異型である。外因性のポリヌクレオチドは通常組換えDNA法を用いて細胞(例えば非ヒトの胚の細胞)中に導入される。本発明のトランスジェニック哺乳類中にC2GnT−Iと、α4GnTと、担体ポリペプチドとが発現することにより、担体ポリペプチドはC2GnT−Iとα4GnTとによってグリコシル化され、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質が製造される。製造された糖タンパク質は、雌の動物が生産する母乳中に分泌される。
【0095】
組換え型の糖タンパク質を含む組換え型のポリペプチドを母乳中に分泌可能な非ヒトのトランスジェニック哺乳類の生産方法は公知である(例えば、米国特許 第6,344,596号;第6,548,735号;第6,222,094号;第5,962,648号;第5,891,698号;第5,850,000号の各明細書参照、夫々参考文献としてここに示す)。そのような非ヒトのトランスジェニック哺乳類はどのような哺乳類でもよく、例えばマウス、ラット、ウサギ、ウマ、ブタ、ヒツジ、ヤギ、又はウシが挙げられる。一つの形態として、前記トランスジェニック哺乳類は非ヒトの雌トランスジェニック哺乳類であって、組換え型糖タンパク質が抗菌活性型で、前記非ヒトの雌トランスジェニック哺乳類によって生産された母乳中に分泌される。
【0096】
本発明は、非ヒトの雌トランスジェニック哺乳類の母乳中に、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造する方法を提供するというものである。「抗菌性」という用語は、ここでは少なくとも一つのα1,4結合−GlcNAc残基を有する化合物について使用しているが、前記化合物の非存在下での増殖レベル又は機能レベルと比較して、前記化合物がCGLを発現する細菌の増殖又は機能を抑制又は阻害することができることを意味している。ヘリコバクター ピロリのようなヘリコバクター種の細菌等の増殖又は機能が抑制又は阻害されていることは、本明細書に示した方法又は公知の方法を用いて確認できる。例えば化合物の抗菌活性は、その化合物の非存在下での運動性と比較して細菌の運動性が減少することを検出することにより、又は細菌増殖に適した培地で培養した細菌の数が減少することを検出することにより、確認できる。
【0097】
非ヒトの雌トランスジェニック哺乳類の母乳中に前記抗菌性の組換え型の糖タンパク質を製造する方法は次のようにして行われる。1)非ヒトの哺乳類の胚に、α4GnTをエンコードする第一のポリヌクレオチドと、C2GnT−Iをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとの少なくともいずれかを導入し、これら三つのポリヌクレオチドを全て前記胚に導入しないときには前記胚の細胞は対応する内因性のポリヌクレオチドを有し、前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合され、前記第三のポリヌクレオチドが、担体ポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と相互作用して結合し、2)前記胚を、子孫を生産するような宿主の雌哺乳類に移植し、3)前記雌の子孫に、5’制御配列に相互作用して結合しゲノム中に安定に組み込まれた第一の外因性ポリヌクレオチドと第二の外因性ポリヌクレオチドと第三の外因性ポリヌクレオチドとを含む母乳の生産を行わせる。母乳は、その動物をつがいにして妊娠させ、乳腺上皮で生産させるか、あるいは母乳を生産するように非ヒトの雌トランスジェニック哺乳類のホルモン処理を行うことで、非ヒトの雌トランスジェニック哺乳類により産生される。前記非ヒトの雌トランスジェニック哺乳類中にC2GnT−Iと、α4GnTと、担体ポリペプチドとが発現することにより、担体ポリペプチドはC2GnT−Iとα4GnTとによってグリコシル化され、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質が製造される。製造された糖タンパク質は、非ヒトの雌トランスジェニック哺乳類が産生する母乳中に分泌される。
【0098】
少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を母乳中に分泌する非ヒトの雌トランスジェニック哺乳類の生産方法はさらに、雌トランスジェニック哺乳類から搾乳することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を含む母乳を得る工程を含んでいてもよい。さらに前記方法は、前記母乳から少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を単離することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を得る工程を含んでいてもよい。本発明によれば、本発明の方法で生産される非ヒトの雌トランスジェニック哺乳類、その雌トランスジェニック哺乳類から生産される母乳、及びその母乳から得られる単離された少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を提供することができる。
【0099】
本発明は、ヘリコバクター種の細菌に感染することによる胃潰瘍の治療又は予防方法を提供するというものである。一つの具体例として、本発明はヘリコバクター種の細菌が個体に感染することによる胃潰瘍の兆候又は症状を改善する方法であって、個体に少なくとも一つのα1,4結合−GlcNAc残基を有する化合物が投与され、この化合物が前記ヘリコバクター種の細菌に接触して前記ヘリコバクター種の細菌の増殖を抑制又は阻害することにより、胃潰瘍の兆候又は症状を改善する方法を提供することができるというものである。第二の具体例として、本発明は、胃潰瘍感受性の個体にヘリコバクター種の細菌が感染することによる胃潰瘍の予防方法であって、前記胃潰瘍感受性の個体に少なくとも一つのα1,4結合−GlcNAc残基を有する化合物が投与され、この化合物が前記ヘリコバクター種の細菌の増殖を妨げることによって、前記ヘリコバクター種の細菌の感染による胃潰瘍を予防する方法を提供することができるというものである。
【0100】
ここで使用される「改善」という用語は、胃潰瘍の兆候や症状が緩和することを意味する。胃潰瘍の兆候や症状は、兆候や条件を監視する方法と同様に、熟練した臨床医によく知られている。胃潰瘍の症状として、例えば、腹部の灼熱痛、食べたり制酸剤を服用したりすると改善する痛み、食事の2〜3時間後にひどくなる痛みがある。潰瘍が出血している場合は、鮮血を含む嘔吐や、黒やタール色の排便のような症状がみられる。本発明の方法は、胃潰瘍の改善、例えば、患者の痛みを緩和させるものである。同様に、例えば、液体バリウムの摂取後に胃腸上部にX線を照射し、減少した潰瘍の大きさを検出したり、抗ヘリコバクター ピロリ抗体やヘリコバクター ピロリに対する血液検査を行い、抗体又は細菌の夫々の数の減少や欠乏を測定したり、検便中の潜血の減少又は欠乏を検出したり、出血性胃潰瘍による貧血の減少を検出したり、胃粘膜を内視鏡で検査したりすることによって、胃潰瘍の臨床学的兆候が緩和することを、熟練した臨床医は熟知している。
【0101】
ヘリコバクター感染は、本明細書で示した方法や、他の公知の方法で検知できる。例えば、らせん状細胞を直接観察するワルティン−スターリー銀染色のスタイネル変異による組織学的検査では、電子顕微鏡でヘリコバクター種の細菌を検知できる。糞粒又は盲端塗抹標本の微生物学的培養もまた、特異培地と微好気性培養条件とを用いたヘリコバクター種の細菌の確認のために使用される。ポリメラーゼ鎖反応(PCR)法もまたヘリコバクター種の細菌を検出するために使用され、制限エンドヌクレアーゼは、種々のヘリコバクター種の細菌を区別することができる。
【0102】
本発明の方法によって治療される個体は、胃粘膜がヘリコバクター種の細菌に感染しているか、又は感染しやすい感受性の個体である。したがって、前記個体は例えばウマ、ブタ、ヤギ、イヌ、ネコ、フェレット、又は同様のもののような哺乳類である。一つの具体例として、前記個体はヒトである。同様に、ヘリコバクター種の細菌は胃潰瘍形成に関与するもので、例えばヒトの胃潰瘍形成に関与するヘリコバクター ピロリである。さらに、胃潰瘍に関係するヘリコバクター ピロリ、ヘリコバクター種の細菌として、例えば胃炎のチーターから単離したH.acinonychis、シリアンハムスターの胃腸組織から培養したH.aurati、犬の胃にみられるH.bizzozeronii、ブタオザル(Macaca nemestrina)の胃にみられるH.nemestrinae、鳥やブタの排泄物から単離したH.pametensis、家禽や胃腸炎のヒト患者から単離したH.pullorum(Microbiology,140:3441-3449,1994参照)、Hericobacter felisやHericobacter bizzozeroniiに関係する犬胃ヘリコバクター種の細菌であるH.salomonis(JP Euzeby,List of Bacterial Names with Standing in Nomenclature Genus Helicobacter,URL:http/www/bacterio.cict.fr/h/helicobacter.html参照)、ペットのフェレットに広く存在するH.mustelae、ヘリコバクター ピロリに類似しブタに感染するH.heilmanniiなどが挙げられる。さらに、本発明の組成物と方法とは、細胞壁の段階でコレステリル−α−D−グルコピラノシド(CGL;Glcα1→コレステロール)を発現する細菌による病的状態を改善するために使用される。
【0103】
前記で示したように、本発明で使用される化合物は少なくとも一つの末端α1,4結合−GlcNAc残基を有する化合物であればよい。また前記化合物は、単独で使用してもよく、潰瘍患者の治療に有用であると臨床医が認めた一種又はそれ以上の治療剤や栄養剤と併用してもよい。前記化合物として、例えば、少なくとも一つの末端α1,4結合−GlcNAc残基を有する組換え型の糖タンパク質(例えば末端α1,4結合−GlcNAc残基を有する可溶性CD43ポリペプチド)、O−グリカンを含み、一つ又はそれ以上の末端α1,4結合−GlcNAc残基を有する化合物、胃腺粘液細胞型ムチンを含み、胃腺粘液細胞から単離され得る化合物が挙げられる。
【0104】
個体に投与される化合物は、その種類に応じた特有な如何なる方法により処方されてもよい。また、前記化合物は適切であれば如何なる方法で投与されてもよく、特に経口投与や胃へ直接挿管する方法であってもよい。治療方法のためだけではなく予防方法のためにも、例えば前記で示した非ヒトの雌トランスジェニック哺乳類から得られた母乳のような前記化合物を含む母乳を、ヘリコバクター感染による胃潰瘍を有していたり胃潰瘍になり易かったりする個体が、飲用することによって、前記化合物の投与が行われる。
【0105】
前記化合物が母乳に含まれる以外の形態の場合、前記化合物は個体への投与に適した組成に処方される。このように、本発明は少なくとも一つの末端α1,4結合−GlcNAc残基を有する化合物を含み、ヘリコバクター感染による胃潰瘍の予防又は改善に有効で、製剤学的に許容される担体をさらに含むことができる組成物を提供するというものである。そのような化合物やその化合物を含む組成物は、胃潰瘍を有しているか、又は胃潰瘍になりやすい個体の治療に用いる医薬として有用である。
【0106】
製剤学的に許容される担体は公知であり、例えば水、生理食塩水、他の溶媒、グリコールやグリセロールで例示されるビヒクルを含む水溶液;オリーブオイルや注射可能な有機エステルのような油状物が挙げられる。製剤学的に許容される担体は、例えば少なくとも一つの末端α1,4結合−GlcNAc残基を有する化合物(例えば前記組換え型の糖タンパク質)の吸収を安定化又は増大させる、生理的に許容される化合物を、含んでいてもよい。そのような生理的に許容される化合物として、例えば、グルコース、スクロース、デキストランのような糖質;アスコルビン酸、グルタチオンのような抗酸化剤;キレート化剤;低分子量タンパク質;他の安定化剤や賦形剤が挙げられる。例えば前記治療効果のある化合物や予防効果のある化合物の物理化学的特性や、一般的な経口投与のみならず静脈注射、注射、挿管、又は他の公知の方法のような非経口投与という組成物投与方法に応じて、生理的に許容される化合物を含む製剤学的に許容される担体を、選択すべきであることは、よく知られている。前記のように、前記組成物は、例えば、診断試薬、栄養物質、毒素、制酸剤及び/又は栄養補助剤等の治療有効剤のような別な薬剤を、さらに含んでいてもよい。
【0107】
本発明の方法で投与される、末端α1,4結合−GlcNAc残基を有する化合物が、治療目的で投与されるのか予防目的で投与されるのかに応じて、その総量を、一度に又は長期にわたって複数回に分けて、投与してもよい。個体の病状を治療するための前記化合物の量は、個体の歳、全身の健康状態、投与経路、投与回数、ヘリコバクター感染の重症度のような多くの要因によるべきであることが知られている。専門医は、これらの要因から、第1相及び第2相臨床試験結果に基づき、必要に応じて、投与量を調整し、さらに組成物の形状とその投与経路、投与量と投与頻度を、決定する。
【0108】
経口投与用として、前記化合物は錠剤、溶液又は懸濁液として処方され、通常無毒で製剤学的に許容される担体を含む錠剤、ペレット剤、カプセル、座薬、溶液、エマルジョン、懸濁液、又は使用に適した形状に調剤されてもよい。前記担体として前記のもの以外に、グルコース、ラクトース、マンノース、アカシアゴム、ゼラチン、マンニトール、デンプン糊、ケイ酸マグネシウム、タルク、コーンスターチ、ケラチン、コロイダルシリカ、ポテトスターチ、尿素、中鎖長トリグリセリド、デキストラン、又は使用に適した他の担体を固体、半固体、又は液体の形状で用いてもよい。さらに、補助剤、安定化剤、農厚化剤、着色剤、香料を使用することもできる。例えば、トリウロースのような安定乾燥剤(例えば米国特許 第5,314,695号明細書参照)を使用してもよい。
【0109】
また、本発明は、CGLを発現する細菌の細胞壁形成を抑制又は阻害する方法を提供するものである。その方法は例えば、前記細菌を、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物に接触させ、CGL合成を抑制又は阻害して、この細菌の細胞壁の形成を抑制又は阻害するというものである。前記化合物は、CGL合成の抑制又は阻害によって細菌のUDP−Glc:ステロール糖転移酵素活性を抑制又は阻害するものであってもよいが、必ずしも必要ではない。また、本発明はCGLを発現する細菌の増殖を抑制又は阻害する方法を提供するというものである。その方法は例えば、前記細菌を、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物に接触させ、CGL合成を抑制又は阻害して、前記細菌の増殖を抑制又は阻害するというものである。この方法では、前記細菌と前記化合物とをin vitroで接触させてもよく、例えば培養中の細菌細胞に前記化合物を添加して行われる。また、前記細菌と前記化合物とをin vivoで接触させてもよく、例えばヘリコバクター種の細菌に感染した哺乳類、特にヘリコバクター ピロリに感染したヒトのような、細菌感染した個体の胃に、前記化合物を直接投与して行われる。
【0110】
以下、本発明の実施例について説明するが、本発明はこの実施例に限定されるものではない。
【0111】
10%ウシ胎児血清(FCS)を添加したα−MEM培地中で、CMP−シアル酸トランスポーター欠損の突然変異体チャイニーズハムスター卵巣・Lec2細胞(CHO・Lec2細胞)を培養した。CD43の細胞外ドメイン全体を含むDNA断片(配列番号1におけるアミノ酸配列の20番目から254番目に相当する部分)をPCR法で増幅し、pSecTag2(インビトロジェン社製)にサブ・クローニングすることにより、Igκリーダー・ペプチドと、それに続くmycエピトープ、及び(His)6をエンコードするベクター(pSecTag2−sCD43)を作製した。次に、(1)コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)をエンコードするcDNAと、(2)α1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードするcDNAと、(3)ポリオーマ・ウイルス large T抗原をエンコードするcDNAとを、夫々別々に前記pSecTag2−sCD43に組み込んだベクター4つを、LipofectAmine(登録商標、インビトロジェン社製)を用いて前記培養したCHO・Lec2細胞にトランスフェクトさせた。トランスフェクトさせたCHO・Lec2細胞内で、末端GlcNAcα残基を含む可溶性CD43(以下、「GlcNAcα−CD43」と称する)を発現させることによりGlcNAcα−CD43を調製した。
【0112】
前記調製したGlcNAcα−CD43の活性は、前記GlcNAcα残基に特異的なHIK1083抗体(胃ムチン検知用HIK1083ラテックス、関東化学社製)(例えば悪性腺腫と病変の新診断方法:新モノクローナル抗体(HIK1083;Clinica Chimica Acta 312(2001)231-233)存在下でのラテックス凝集テスト)を用いて市販のELISAキット(胃腺粘液細胞型ムチン測定用:関東化学社製)により測定し、発色反応にはELISA ELAST増幅システム(パーキン エルマー社製)を用いた。この結果、発色を確認することができ、GlcNAcα残基が発現していることが確認できた。さらに、市販の定量システム(BCAタンパク質検定キット、ピエルスバイオテクノロジー社製、商品番号23225,23227)を用いてタンパク質の定量を行った。その結果、前記GlcNAcα−CD43中の側鎖(糖鎖)の数は、主鎖(CD43)夫々に対して80であり、該糖鎖は、該主鎖に対してクラスター状に存在していることが判った。さらに別な市販の定量キット(糖タンパク 糖質評価キット、ピエルスバイオテクノロジー社製、商品番号23260)を用いて糖鎖の分析を行った。前記糖鎖は、GlcNAcα1→4Galβ1→4GlcNAcβ1→6(GlcNAcα→4Galβ1→3)GalNAcα→Ser/Thrで構成され、該糖鎖中の分岐数は1であり、モノマーユニット数は6であることが判った。前記「Ser/Thr」は、主鎖としてのCD43中、セリン(Ser)及びトレオニン(Thr)のいずれかに結合していることを表す。
【0113】
なお、前記トランスフェクトしたCHO・Lec2細胞は、ポリオーマ・ウイルス large T抗原により、前記CD43を効率的に合成することができた。また、前記α4GnTをエンコードするcDNAは、特開平2001−46077号公報に記載の方法に準じて調製した。α型N−アセチルグルコサミン残基を含まないCD43の調製は以下のようにして行った。コントロール測定用として、前記α4GnTのcDNAのみを組み込んでいないpcDNAIベクター(インビトロジェン社製)を用いた以外は、GlcNAcα−CD43の調製と同様にして、O−グリカンのコア2分岐にGlcNAcα残基を含まない可溶性CD43を調製した。なお、前記調製した該GlcNAcαを含まないCD43をGlcNAcα残基に特異的なラテックス粒子抗体を用いて縮合反応を行ったところ、縮合は見られなかった。この結果、該GlcNAcαを含まないCD43はGlcNAcα残基を含まないことがわかった。さらに、前記GlcNAcα残基を含まないCD43中の側鎖(糖鎖)は、Galβ1→4GlcNAcβ1→6(Galβ1→3)GalNAcα→Ser/Thrで構成されることがわかった。前記「Ser/Thr」は、主鎖としてのCD43中のセリン(Ser)及びトレオニン(Thr)のいずれかに結合していることを表す。
【0114】
前記調製したGlcNAcα残基を含む可溶性CD43、及びGlcNAcα残基を含まない可溶性CD43を用いて、以下のようにして細菌増殖性、細菌の運動性、及び細菌の形態異常発現性、並びに、GlcNAcα残基によるCGL生合成の抑制又は阻害性についての評価を行った。細菌増殖性は以下のとおりである。微量液体希釈法(microbroth dilution method)により、10%ウマ血清を添加したブルセラ培地(ベクトン デッキンソン マイクロバイオテクノロジー システムズ社製)でヘリコバクター ピロリ(ATCC43504)を前培養した。続いて、1×107cells/mlに希釈した前記ヘリコバクター ピロリと、図2Aに示す濃度のGlcNAcα−CD43とを96穴のプレートに投入し、35℃、15%CO2の環境下で4日間培養した。図2A中、「αGlcNAc(+)」は、前記培地中に前記GlcNAcα−CD43が含まれていることを意味し、「αGlcNAc(−)」は、前記培地中に前記GlcNAcα残基を含まない可溶性CD43が含まれていることを意味し、前記培地中の前記GlcNAcα残基を含まない可溶性CD43の濃度は、125.0mU/mlである。培養したヘリコバクター ピロリの増殖は、OD600nmの条件下、マイクロプレート分光光度計SPECTRAmax PLUS384(モレキュラーデバイス社製)を用いて測定した。結果を図2Aに示した。なお、前記ブルセラ培地に代えて、ミューラーヒントン培地(栄研化学社製)を使用した以外は、前記の方法と同様にして最小発育阻止濃度を測定したところ、同様の結果を得ることができた。
【0115】
この結果、図2Aに示すように、GlcNacα残基の存在の有無に関わらず、最初の2.5日間での増殖はごくわずかであるが、これはヘリコバクター ピロリの増殖の誘導期の特徴である。培養が3日を超えると増殖期に入り、GlcNAcα残基を含まない可溶性CD43含有の培地では、ヘリコバクター ピロリの急速な増殖が観られた。しかし、GlcNAcα−CD43と共に培養した場合には、ヘリコバクター ピロリの増殖が抑制又は阻害されていることがわかった。なお、1mUは、1μgのp−ニトロフェニル−N−アセチルグルコサミン(GlcNAcα−PNP)と当量である。細菌運動性は以下のようにして測定した。31.2mU/mlのGlcNAcα−CD43を含有するブルセラ培地を用いてヘリコバクター ピロリを3日間培養した。培養したヘリコバクター ピロリの運動性をレーザー共焦点顕微鏡LSM510 META(カルル ツァィス社製)を用いて、1秒間隔で低速度写真を撮影し、代表的なヘリコバクター ピロリを矢印で示した。結果を図2Bに示した。また、GlcNAcα残基を含まない可溶性CD43についても同様に撮影をした。結果を図2Bに示した。
【0116】
なお、図2B中、「αGlcNAcα(+)」は、前記培地中に前記GlcNAcα−CD43が含まれていることを意味し、「αGlcNAcα(−)」は、前記培地中に前記GlcNAcα残基を含まない可溶性CD43が含まれていることを意味し、濃度は、夫々31.2mU/mlである。また、図2B右下に記載した下線は、50μmを表し、図はすべて等倍率で撮影した低速度写真である。
【0117】
この結果、GlcNAcα−CD43を含む培地(αGlcNAcα(+))で培養された7個のヘリコバクター ピロリの平均速度は3.1±3.5μm/secであり、GlcNAcα残基を含まないCD43を含む培地(αGlcNAcα(−))で培養された7個のヘリコバクター ピロリの平均速度は21.1±2.6μm/secであり、t検定による有意性は、P<0.001であった。従って、GlcNAcα−CD43を含む培地では、ヘリコバクター ピロリの運動性が著しく減少することがわかった。
【0118】
細菌形態異常は以下のようにして検出した。GlcNAcα−CD43、又はGlcNAcα残基を含まない可溶性CD43を31.2mU/ml濃度で含有するブルセラ培地を用いてヘリコバクター ピロリを3日間培養させた。培養したヘリコバクター ピロリは、走査電子顕微鏡(日本電子社製、JSM−6360LV)を用いて加速電圧15kVの条件でSEM写真を撮影した。結果を図2Cに示した。なお、図2C中、「αGlcNAc(+)」は、前記培地中に前記GlcNAcα−CD43が含まれていることを意味し、「αGlcNAc(−)」は、前記培地中に前記GlcNAcα残基を含まない可溶性CD43が含まれていることを意味し、濃度は、夫々31.2mU/mlである。また、図2C右下に記載した下線は、1μmを表し、図はすべて等倍率で撮影したSEM写真である。この結果、GlcNAcα−CD43を含む培地(αGlcNAcα(+))で培養されたヘリコバクター ピロリは、伸び、狭窄、屈曲などの形態異常が観られるのに対し、GlcNAcα残基を含まないCD43を含む培地(αGlcNAcα(−))で培養されたヘリコバクター ピロリは、形態異常が観られなかった。
【0119】
CGL生合成の阻害性を測定した。GlcNAcα残基によるCGL生合成の阻害性の分析は、以下のようにして、(1)ヘリコバクター ピロリのin vivoでのCGL生合成、及び、(2)in vitroでのCGL生合成の2つの場合について行った。ヘリコバクター ピロリ生体内でのCGL生合成の阻害性を測定した。GlcNAcα−CD43存在下でのヘリコバクター ピロリによるCGLの合成を行った。コレステロール及びUDP−グルコースを用いたヘリコバクター ピロリによるCGLの合成をGlcNAcα−CD43の存在下で行い、以下の様にしてヘリコバクター ピロリ生体内(in vivo)で生合成したCGLについてのMALDI−TOF質量分析を行った。10%ウマ血清を添加したブルセラ培地(ベクトン デッキンソン マイクロバイオテクノロジー システムズ社製)でヘリコバクター ピロリ(ATCC43504)を前培養した後、希釈した前記ヘリコバクター ピロリ(1×107cells/ml)と、4.0mU/ml濃度に調整したGlcNAcα−CD43とを96穴のプレートに投入して、35℃で2日間培養した。培養したヘリコバクター ピロリを回収し、Ca2+、Mg2+を含まないPBSで3回洗浄した後、1mlの蒸留水で懸濁し、懸濁した液を毎分6,000回転で10分間遠心することにより菌体を得た。得られた菌体を、2mlのクロロホルム・メタノール混合液(クロロホルム:メタノールの質量比=2:1)を用いて4℃で一晩かけて抽出し、抽出液をろ過して、窒素気流で乾燥させた。乾燥して得られた試料を4mlのクロロホルム・メタノール混合液(クロロホルム:メタノールの質量比=2:1)に溶解させた。
【0120】
Folchらの方法に準じ、混合液に1mlの水を添加した。得られた混合液の下相を窒素気流で乾燥させ、1mlの0.5N水酸化ナトリウム・メタノール溶液を用いて、50℃で1時間の処理を行った。さらに処理液を6N塩酸水溶液を用いて中和した後、反応管に1mlの石油エーテルを添加した。次に、上相を除去した後、さらに2mlの石油エーテルを下相に添加した。下相を乾燥させた後、1mlのクロロホルム・メタノール・水(クロロホルム:メタノール:水の質量比=86:14:1)の混合液(TLP)に溶解させ、同じ反応管に0.5mlの別のクロロホルム・メタノール・水(クロロホルム:メタノール:水の質量比=3:48:47)混合液(TUP)を添加した。得られた混合液の下相を回収し、窒素気流で乾燥させ、50μlのクロロホルムに溶解させた。次に、この試料を1μl量り取り、1μlの2,5−ジヒドロキシ安息香酸、又は1μlのtrans−3−インドールアクリル酸を添加し、これをマトリックスとして用いた。前記資料の質量スペクトルは、DE MALDI−TOF MSのVoyager−DE STR Biospectrometry Workstation(PE アプライド バイオシステムズ社製)を用い、リフレクターモード、レーザー強度2300の条件で、陽イオンモード又は負イオンモードで測定した。なお、ヘリコバクター ピロリに内生発現するホスファチジン酸を内部基準とし、2点外部較正を行った。結果を図3Bに示した。
【0121】
また、コントロールとして、前記GlcNAcα−CD43を、GlcNAcα残基を含まないCD43に代えた以外は前記の方法と同様にしてCGLの質量スペクトルを測定した。結果を図3Aに示した。図3A及び3Bに示したように、前記GlcNAcα−CD43が存在する場合、及びGlcNAcα残基を含まないCD43が存在する場合のいずれの場合についても、m/zが571.6の位置にCGLのピークが検出された。しかし、前記GlcNAcα−CD43の存在下でのCGLのピークは、コントロールで測定したCGLのピークと比較して29.5%と大きく減少していた。この結果、前記GlcNAcα−CD43が、ヘリコバクター ピロリ生体内でのCGLの生合成を阻害していることがわかった。
【0122】
in vitro(試験管内)でのCGL生合成の阻害性を測定した。GlcNAcα−CD43存在下でのヘリコバクター ピロリによるCGLの合成を行った。コレステロール及びUDP−グルコースを用いて、超音波処理したヘリコバクター ピロリによるCGLの合成を行い、以下の様にしてin vitroで合成したCGLについてのMALDI−TOF質量分析を行った。10%ウマ血清を添加したブルセラ培地(ベクトン デッキンソン マイクロバイオテクノロジー システムズ社製)でヘリコバクター ピロリ(ATCC43504)を前培養した後、希釈した前記ヘリコバクター ピロリ(5×107cells/ml)を5%ウマ血清添加のブルセラ培地中で、35℃で2日間培養した。培養した2mlのヘリコバクター ピロリ(5×107cells/ml)を回収し、Ca2+、Mg2+を含まないPBSで3回洗浄した後、1mlの蒸留水で懸濁し、懸濁した液を毎分6,000回転で10分間遠心することにより脂質成分を得た。
【0123】
次に、100mMのトリス緩衝液(pH7.5)と、15質量%グリセロールと、5mMのDTTと、200μMのPefabloc(メルク社製)と、0.5mg/mlのリゾチームとを含有する反応緩衝液1ml中にヘリコバクター ピロリを懸濁し、20℃で5分間培養した。これらのサンプルを氷浴上で、超音波ウォーターバスにて30秒間の超音波処理を30秒間隔で10回行った。次に、80μlの超音波処理済ヘリコバクター ピロリと、8mMコレステロールエタノール溶液5μlと、7.2μMのUDP−Glc(UDP−グルコース)5μlと、トリトンCF−54(シグマケミカル社製)1μlと、反応緩衝液9μlとを含有する100μlの反応混合液を、30℃で3時間定温放置(インキュベート)した。その後、0.45% NaCl溶液900μl、及びクロロホルム・メタノール混合液(2:1)4mlを添加して反応を停止させた。
【0124】
次に、添加後の混合液の下相をろ過し、窒素気流下で乾燥させた。乾燥して得られた脂質試料を4mlのクロロホルム・メタノール混合液(クロロホルム:メタノールの質量比=2:1)に溶解させ、Folchらの方法に準じ、混合液に1mlの水を添加した。得られた混合液の下相を窒素気流で乾燥させ、1mlの0.5N水酸化ナトリウム・メタノール溶液を用いて、50℃で1時間の処理を行った。さらに処理液を6N塩酸水溶液を用いて中和した後、反応管に1mlの石油エーテルを添加した。次に、上相を除去した後、さらに2mlの石油エーテルを下相に添加した。乾燥させた下相は、1mlのクロロホルム・メタノール・水(クロロホルム:メタノール:水の質量比=86:14:1)の混合液(TLP)に溶解させ、同じ反応管に0.5mlの別のクロロホルム・メタノール・水(クロロホルム:メタノール:水の質量比=3:48:47)混合液(TUP)を添加した。得られた混合液の下相を回収し、窒素気流で乾燥させ、50μlのクロロホルムに溶解させた。
【0125】
次に、この試料を1μl量り取り、1μlの2,5−ジヒドロキシ安息香酸、又は1μlのtrans−3−インドールアクリル酸に添加し、これをマトリックスとして用いた。DE MALDI−TOF MSのVoyager−DE STR Biospectrometry Workstation(PE アプライド バイオシステムズ社製)を用い、リフレクターモード、レーザー強度2500の条件で、陽イオンモードでのCGLの質量スペクトルを測定した。なお、ヘリコバクター ピロリに内生発現するホスファチジン酸を内部基準とし、2点外部較正を行った。結果を図3Cに示した。
【0126】
次に、前記反応緩衝液に5.0mUのGlcNAcα−CD43を含有させた以外は前記の方法と同様にしてCGLの質量スペクトルを測定した。結果を図3Dに示した。さらに、前記反応緩衝液に5.0mUのGlcNAcα残基を含まないCD43を含有させた以外は前記の方法と同様にしてCGLの質量スペクトルを測定した。結果を図3Eに示した。
【0127】
UDP−Glcとコレステロールとから超音波処理したヘリコバクター ピロリにより合成された試料についてMALDI−TOF質量分析を行ったところ、図3Cから明らかなように、m/zが571.6の位置にCGLのピークが検出された。これは、下記スキームに示すようにヘリコバクター ピロリが、UDP−GlcをコレステロールのC3の位置に転移するUDP−Glc:ステロール糖転移酵素活性を有するためであると考えられる。
【化1】

【0128】
しかし、前記GlcNAcα−CD43の存在下で同様にして培養した場合には、図3Dに示したように、m/zが571.6の位置にCGLのピークが検出されなかった。また、前記GlcNAcα−CD43に代えて、前記GlcNAcα残基を含まないCD43の存在下で同様にして培養した場合、m/zが571.6の位置にCGLのピークが検出され、前記反応緩衝液に前記GlcNAcα−CD43及び前記GlcNAcα残基を含まないCD43を添加しない場合、CGLのピークに変化が観られなかった。これらの結果から、GlcNAcα残基が、最終的にCGLの生合成を阻害していることが証明された。
【0129】
α−CGsの生合成に関わる糖転移酵素の同定を含むヘリコバクター ピロリの細胞壁の生成機構は解明されていないが、GlcNAcα残基と、CGLに含まれるα結合型Glcとの構造の類似性を考慮すると、前記分析結果から、GlcNAcα残基と、UDP−Glcとが互いに競合し、UDP−Glc:ステロール糖転移酵素の供与体基質を互いに奪い合っているか、又は、GlcNAcα残基が最終生成物阻害機構によって直接的にUDP−Glc:ステロール糖転移酵素活性を阻害し、CGLの生合成を阻害しているものと考えられる。前記細菌増殖性、細菌の運動性、及び細菌の形態異常発現性、並びに、GlcNAcα残基によるCGL生合成の抑制乃至阻害性の評価結果から、α1,4アセチルグルコサミン残基を有する機能性分子を投与することにより、個体がヘリコバクター属の細菌に感染して引き起こされる胃潰瘍及び胃癌の兆候及び症状を改善及び予防できることがわかった。
【0130】
コレステロールの有無に対するヘリコバクター ピロリの生存状況、及びヘリコバクター ピロリによるCGLの合成について実験を行った。コレステロールの有無に対するヘリコバクター ピロリの生存状況を調べた。コレステロールの生合成に関わる遺伝子は、ヘリコバクター ピロリのゲノムデータからは発見されていない。このため、ヘリコバクター ピロリは、外因性のコレステロールの存在がなければCGLを合成できない可能性があることから以下の実験を行った。即ち、コレステロールを含まないHam's F−12培地でヘリコバクター ピロリ(ATCC43504)を35℃の標準嫌気条件にて5日間培養すると、ヘリコバクター ピロリの増殖は同条件でコレステロールを含むHam’s F−12培地に対して50%のレベルにとどまった。結果を表1に示した。
【0131】
コレステロールを含まない培地では、ヘリコバクター ピロリは運動性を示さず、異常伸長(形態異常)として図2C左に見られるようなヘリコバクター ピロリの伸びがみられた。このヘリコバクター ピロリにコレステロールの非存在下でさらに21日間培養すると、ヘリコバクター ピロリは完全に死滅した。一方、コレステロールを加えた培地でヘリコバクター ピロリを培養すると、ヘリコバクター ピロリは順調に増殖し、形態異常は観られなかった(表1)。これらの結果から、ヘリコバクター ピロリの生存には、コレステロールを用いたCGLの合成が必要であることがわかった。
【0132】
ヘリコバクター ピロリによるCGLの合成を行った。コレステロールの有無に対するヘリコバクター ピロリでのCGLの発現レベルを調べるため、ヘリコバクター ピロリの糖脂質部のCGLを含むα−CGsの発現レベルの変化を以下の様にして薄膜クロマトグラフィー(TLC)により分析した。コレステロールを含むブルセラ培地と、コレステロールを含まない培地で培養されたヘリコバクター ピロリについて、夫々の培地から1×108個のヘリコバクター ピロリを回収し、クロロホルム・メタノール混合液(クロロホルム:メタノールの質量比=2:1)を用いて室温で1時間かけて糖脂質部を抽出した。抽出液を窒素気流下で乾燥させ、クロロホルム・メタノール・水混合液(クロロホルム:メタノール:水の質量比=16:6:1)を用いて薄膜クロマトグラフィー(TLC)を行った。単離した糖脂質部は、オルシノール硫酸緩衝液により110℃で染色し、可視化した。結果を図4に示した。なお、図4中、レーン1は、コレステロールを含まないブルセラ培地で培養されたヘリコバクター ピロリから抽出した糖脂質部の分析結果であり、レーン2は、コレステロールを含むブルセラ培地で培養されたヘリコバクター ピロリから抽出した糖脂質部の分析結果である。この結果、コレステロールを含む培地で培養したヘリコバクター ピロリの場合には、ヘリコバクター ピロリの特徴である、CGLを含むα−CGsの典型的な隣接三重線が検出された(レーン2)。しかし、コレステロールを含まない培地で培養したヘリコバクター ピロリでは、α−CGs(CGL)は検出されなかった(レーン1)。
【0133】
α型N−アセチルグルコサミン残基を含むCD34の調製を行った。実施例1中のpSecTag2に代えてpcDNA3(インビトロジェン社製)を用いたこと、及び、可溶性CD43をエンコードするcDNAに代えて、可溶性CD34−IgGキメラをエンコードするpcDNA3−CD34−IgGを用いた以外は以下の実施例1と同様にして、GlcNAcα残基を含む可溶性CD34を調製した。調製したGlcNAcα残基を含む可溶性CD34について、実施例1と同様にして細菌増殖性、細菌の運動性、及び細菌の形態異常発現性の分析、及びGlcNAcα残基によるCGL生合成の阻害性についての評価を前記のようにして行ったところ、実施例1と同様の結果が得られた。
【0134】
α型N−アセチルグルコサミン残基を含むCD43を発現するAGS細胞の調製を行った。実施例1中のCHO・Lec2細胞を胃腺癌細胞(AGS細胞)に代えた以外は以下の実施例1と同様にして、GlcNAcα−CD43を安定的に発現するAGS−α4GnT細胞を調製した。α型N−アセチルグルコサミン残基を含まないCD43を発現するAGS細胞の調製を行った。コントロール測定用として、前記α4GnTのcDNAのみを組み込んでいないpcDNAIベクター(インビトロジェン社製)を用いた以外は、AGS−α4GnT細胞の調製と同様にして、GlcNAcα残基を含まないCD43を安定的に発現するMock−トランスフェクトされたAGS細胞を調製した。前記調製したAGS−α4GnT細胞、又はMock−トランスフェクトされたAGS細胞は、1×107cells/mlのヘリコバクター ピロリと共に24時間共同培養を行った。培養後の状態をノマルスキー顕微鏡を用いて写真撮影した。結果を図5下段に示した。
【0135】
次に、20%緩衝ホルマリンで固定した後、ヘリコバクター ピロリに対するウサギ・ポリクローナル抗血清(ダコ社製)、及びマウス・モノクローナルHIK1083抗体(関東化学社製)の混合物と共に前記AGS−α4GnT細胞、又はMock−トランスフェクトされたAGS細胞をインキュベートし、前記マウス・モノクローナルHIK1083抗体により蛍光染色した。二次抗体として、ローダミン標識の抗ウサギ免疫グロブリン(抗ヘリコバクター ピロリ抗体用)と、イソチオシアン酸フルオレセイン標識抗マウスIgM(HIK1083抗体用)を添加してスライドを作製した。該スライドは、Vectashield(ベクター ラボラトリーズ社製)で封入し、レーザー共焦点顕微鏡LSM510 META(カルル ツァィス社製)を用いて写真撮影した。結果を図5上段に示した。
【0136】
図5右上段に示した図は、AGS−α4GnT細胞を蛍光染色した写真である。白色で表された部分は、蛍光発色した前記HIK1083抗体を表し、HIK1083抗体が、前記AGS−α4GnT細胞を取り囲むように位置していることが確認できた。前記HIK1083抗体は、GlcNAcα残基に特異的な抗体であることから、前記AGS−α4GnT細胞によりGlcNAcα残基が発現していることがわかる。また、図5右下段に示すように、GlcNAcα残基が発現している前記AGS−α4GnT細胞では、細胞の損傷がみられなかった。
【0137】
これに対し、図5左上段に示した図は、Mock−トランスフェクトされたAGS細胞をAGS−α4GnT細胞と同様に蛍光染色した写真である。白色で表された部分は、ヘリコバクター ピロリを表し、前記Mock−トランスフェクトされたAGS細胞を取り囲むようにヘリコバクター ピロリが位置していること、及び、蛍光発色した前記HIK1083抗体が存在しないことからGlcNAcα残基が発現していないことが確認できた。また、図5左下段に示すように、GlcNAcα残基が発現していない前記AGS−α4GnT細胞では、顕著に細胞の損傷がみられた。
【0138】
公知の酵素合成法(例えば、Murata, Usui、Trends Glycsci. Glycotech., 12,161-174,2000)に従って、6残基から成る糖鎖、GlcNAcα1→4Galβ1→4GlcNAcβ1→6(GlcNAcα→4Galβ1→3)GalNAcαを得た。前記の方法(以下の実施例1参照)と同様にして細菌増殖性、細菌の運動性、及び細菌の形態異常発現性の分析、並びに、GlcNAcα残基によるCGL生合成の阻害性についての評価を行ったところ、前記したものと同様の結果が得られた(以下の実施例1参照)。
【0139】
これらの結果から、ヘリコバクター ピロリの増殖にはCGLが必須であり、GlcNAcα残基は、CGLの生合成を阻害することにより、ヘリコバクター ピロリに対する抗菌性を示すことが明らかとなった。
【実施例】
【0140】
(実施例1)末端α1,4結合−N−アセチルグルコサミンを有するO−グリカンが持つ抗菌活性
この実施例では、胃腺粘液細胞型ムチン中で製造されたO−グリカンが、ヘリコバクター ピロリ増殖の天然由来阻害剤として作用すること、また末端α1,4結合−N−アセチルグルコサミンを有する合成組成物が抗菌活性を有することを実証する。
【0141】
材料及び方法
細菌株
ヘリコバクター ピロリの標準菌株(ATCC43504)は、アメリカンタイプカルチャーコレクション(ATCC;バージニア州ナマサス)から購入し、本明細書に示す実験に使用した。さらに細胞増殖評価に使用した細菌株は、次の通りである:ATCCから購入したヘリコバクター ピロリ(ATCC43526)及びヘリコバクター ピロリの臨床的単離物(#749、#750、#751)、大腸菌(Escherichia coli)、緑膿菌(Pseudomonas aeruginosa)、肺炎桿菌(Klebsiella pneumoniae)、黄色ブドウ球菌(Staphylococcus aureus)、I−レンサ球菌(I-Streptococcus)、肺炎レンサ球菌(Streptococcus pneumoniae)。全ての臨床的単離物は、信州大学医学部附属病院臨床検査部で確認されたものである。
【0142】
α1,4結合−GlcNAc残基を有する可溶性CD43、及びα1,4結合−GlcNAc残基を有しない可溶性CD43の製造
10%ウシ胎児血清(FCS)を添加したα−MEM培地中で、CMP−シアル酸トランスポーター欠損の突然変異体チャイニーズハムスター卵巣・Lec2細胞(Deutscherら、Cell 39;295,1984参照、参考文献としてここに示す)を、培養した。CD43の細胞外ドメイン全体を含むDNA断片(アミノ酸残基の20番目から254番目)をPCR法で増幅し、IgPリーダー・ペプチドと、それに続くmycエピトープ、及び(His)6をエンコードするpSecTag2ベクター(インビトロジェン社製;カリフォルニア州カールズバッド)にサブ・クローニングした(pSecTag2−sCD43)。次に、C2GnT−IをエンコードするcDNA(Bierhuizen、Fukuda、Proc.Natl.Acad.Sci.,USA89:9326,1992参照、参考文献としてここに示す)と、α4GnTをエンコードするcDNA(Nakayamaら、Proc.Natl.Acad.Sci.,USA96:8991,1999参照、参考文献としてここに示す)と、可溶性CD43をエンコードするcDNAと、ポリオーマ・ウイルス large T抗原をエンコードするcDNAとを、LipofectAmine(登録商標、インビトロジェン社製)を用いて前記Lec2細胞にトランスフェクトさせた。ポリオーマ・ウイルス large T抗原により、前記トランスフェクトされたLec2細胞は、効率よく組換え型タンパク質を産生した(Bierhuizen,Fukuda、前出,1992)。
【0143】
前記α4GnTのcDNAを組み込んでいないpcDNAIベクター(インビトロジェン社製)を用いて、O−グリカンのコア2分岐に末端α1,4結合−GlcNAc残基を有しない可溶性CD43を調製し、コントロール測定用とした。一週間後、CENTRIPREP YM30フィルター(ミリポア社製:マサチューセッツ州ベッドフォード)を用いて、10%ウシ胎児血清(FCS)を添加したα−MEM培地中に投入した可溶性CD43を濃縮し、測定用とした。前記調製した末端α−1,4−結合−GlcNAcを有する可溶性CD43の活性は、前記GlcNAcα残基に特異的なHIK1083抗体(関東化学社製、Ishiharaら、Biochem.J.318:409,1995参照、参考文献としてここに示す)を用いて市販のELISAキットにより測定し、発色反応にはELISA ELAST増幅システム(パーキン
エルマー社製;マサチューセッツ州ボストン)を用いた。この分析において、1ユニットは製造元の指示に従いp−ニトロフェニル−N−アセチル−α−グルコサミニド(GlcNAcα−PNP)の1mg(2.9Tmol)に相当する免疫反応力と定義した。これらの組換え体のタンパク質濃度はBCAタンパク質分析(ピエルス社製;イリノイ州ロックフォード)を用いて測定した。同様に、pSecTag2−sCD43に代えて可溶性CD34−IgGキメラをエンコードするpcDNA3−CD34−IgG(Yehら、Cell 104:957,2001参照、参考文献としてここに示す)を用いて、末端α1,4結合−GlcNAc残基を有する可溶性CD34タンパク質と、末端α1,4結合−GlcNAc残基を有しない可溶性CD34タンパク質とを調製した。
【0144】
細胞増殖試験
微量液体希釈法(Piccolominiら、J.Clin.Microbiol.35:1842,1997参照、参考文献としてここに示す)により、10%ウマ血清を添加したブルセラ培地(ベクトン ディッキンソン マイクロバイオロジー システムズ社製;メリーランド州スパークス)でヘリコバクター ピロリを前培養した。続いて96穴のプレート中、末端α1,4結合−GlcNAc残基を有する可溶性CD34又は末端α1,4結合−GlcNAc残基を有しない可溶性CD34の種々の量を含む、5%ウマ血清を添加したブルセラ培地で、1×107cells/mlに希釈した細菌を35℃、15%CO2の環境下で4日間培養した。細菌増殖は、OD600nmの条件下、マイクロプレート分光光度計SPECTRAmax PLUS384(モレキュラーデバイス社製;カリフォルニア州サニーベール)を用いて測定した。また、前記ブルセラ培地に代えて、微量液体希釈法のミューラーヒントン培地(栄研化学社製、東京都)を使用した以外は、前記の方法と同様にして最小発育阻止濃度を測定した。
【0145】
細胞運動性試験
31.2mU/mlの末端α1,4結合−GlcNAc残基を有する可溶性CD43又は同じタンパク質濃度の末端α1,4結合−GlcNAc残基を有しないCD43含有するブルセラ培地を用いてヘリコバクター ピロリを3日間培養した。培養したヘリコバクター ピロリの細胞運動性を、レーザー共焦点顕微鏡LSM510 META(カルル ツァィス社製;ドイツ国ジェナ)を用いて、1秒間隔で低速度写真を撮影し、評価した。
【0146】
走査電子顕微鏡
31.2mU/mlの末端α1,4結合−GlcNAc残基を有する可溶性CD43又は同じタンパク質濃度の末端α1,4結合−GlcNAc残基を有しないCD43を含有するブルセラ培地を用いてヘリコバクター ピロリを3日間培養した。培養したヘリコバクター ピロリを走査電子顕微鏡JSM−6360LV(日本電子社製;東京都)を用いて加速電圧15kVの条件で観察し、走査電子顕微鏡写真を撮影した。
【0147】
質量分析によるCGLの検出
10%ウマ血清を添加したブルセラ培地でヘリコバクター ピロリを前培養した後、5%ウマ血清と、4.0mU/mlの末端α1,4結合−GlcNAc残基を有する可溶性CD43又は同じタンパク質濃度の末端α1,4結合−GlcNAc残基を有しないCD43とを含有するブルセラ培地を用いて、1×107cells/mlの微生物を35℃で2日間培養した。培養したヘリコバクター ピロリを回収し、Ca2+、Mg2+を含まないリン酸緩衝生理食塩水(PBS)で3回洗浄した後、1mlの蒸留水で再度懸濁した。毎分6,000回転で10分間遠心した後、得られた脂質画分を、2mlのクロロホルム・メタノール混合液(2:1)を用いて4℃で一晩かけて抽出し、抽出液をろ紙でろ過して、窒素気流で乾燥させた。得られた脂質試料を4mlのクロロホルム・メタノール混合液(2:1)に溶解させ、Folchらの方法(J.Biol.Chem.226:497,1957参照、参考文献としてここに示す)に準じ、混合液に1mlの水を添加した。得られた混合液の下相を試験管中、窒素気流で乾燥させ、1mlの0.5N水酸化ナトリウム・メタノール溶液により50℃で1時間の処理し、処理液を6N塩酸水溶液で中和した後、1mlの石油エーテルを添加した。
【0148】
上相を除去した後、さらに2mlの石油エーテルを下相に添加した。下相を乾燥させた後、1mlのクロロホルム・メタノール・水(86:14:1)の混合液(TLP)に溶解させ、同じ反応管に0.5mlの別のクロロホルム・メタノール・水(3:48:47)混合液(TUP)を添加した。得られた混合液の下相を回収し、窒素気流で乾燥させ、50μlのクロロホルムに溶解させた。次に、この試料の1μlに、1μlの2,5−ジヒドロキシ安息香酸、又は1μlのtrans−3−インドールアクリル酸を添加し、これをマトリックスとして用いた。陽イオンモード又は負イオンモードでのCGLの質量スペクトルは、DE マトリックス支援イオン化−飛行時間型質量分析(DE MALDI−TOF MS)装置のVoyager−DE STR Biospectrometry Workstation(PE アプライド バイオシステムズ社製;カリフォルニア州フォスターシティ)を用い、前記のようにリフレクターモード、レーザー強度2300の条件で測定した(Taketomi,Sugiyama、Meth,Enzymol.312:80,2000参照、参考文献としてここに示す)。ヘリコバクター ピロリ中に内生発現するホスファチジン酸(Inamotoら、前出,1993)を内部基準とし、2点外部較正を行った。
【0149】
超音波処理したヘリコバクター ピロリによるin vitroでのCGL合成
in vitroでのCGL合成は、前記で示したように変異体を用いて分析した(Warneckeら、J.Biol.Chem.274:13048,1999参照、参考文献としてここに示す)。すなわち、10%ウマ血清を添加したブルセラ培地でヘリコバクター ピロリを前培養した。次に、5×107cells/mlに希釈した細菌を、5%ウマ血清を添加したブルセラ培地を用いて35℃で2日間培養した。培養した2mlのヘリコバクター ピロリ(5×108cells/mlを毎分6,000回転で10分間遠心し、Ca2+、Mg2+を含まないPBSで3回洗浄した。その後100mMのトリス緩衝液(pH7.5)と、15%グリセロールと、5mMのDTTと、200TMのPEFABLOCプロテアーゼ阻害剤(メルク社製;ドイツ国ダルムスタッド)と、0.5mg/mlのリゾチームとを含有する反応緩衝液1ml中にヘリコバクター ピロリを懸濁させ、20℃で5分間培養した。これらのサンプルを氷浴上で超音波ウォーターバスにて30秒間隔で30秒間の超音波処理を10回行った。
【0150】
次に、超音波処理したヘリコバクター ピロリの80μlと、8mMコレステロールのエタノール溶液5μlと、7.2μMのUDP−グルコース5μlと、1μlのCF−54と、9μlの反応緩衝液又は5.0mUの末端α1,4結合−GlcNAc残基を有する可溶性CD43又は同じタンパク質濃度の末端α1,4結合−GlcNAc残基を有しないCD43を含有する同様の緩衝液とを含有する100μlの反応混合液を、30℃で3時間培養した。その後、0.45%のNaCl溶液900μl、及びクロロホルム・メタノール混合液(2:1)の4mlを添加して反応を停止させた。その混合液の下相をろ過し、窒素気流下で乾燥させた。その後、乾燥して得られた試料を前記で示した方法と同様にして連続処理した。陽イオンモードでのCGLの質量スペクトルは、DE MALDI−TOF MS装置のVoyager−DE STR Biospectrometry Workstation(アプライド バイオシステムズ社製)を用い、リフレクターモード、レーザー強度2500の条件で、前記の方法(Taketomi, Sugiyama、前出,2000)で測定した。なお2点外部較正を行った。
【0151】
コレステロール存在下、又はコレステロール非存在下でのヘリコバクター ピロリの培養
10%ウマ血清を添加したブルセラ培地でヘリコバクター ピロリを前培養した。次に、5%ウマ血清を添加したブルセラ培地で、1×108cells/mlに希釈した細菌を40時間培養し、その後、標準培養条件下で、0.005%コレステロールを含み又はコレステロールを含まないウマ血清除去ブルセラ培地(TLC分析用)と、0.005%コレステロールを含んでいるか又はコレステロールを含まないF−12培地(細胞増殖分析用)とで、1×107cellのヘリコバクター ピロリを5〜21日間培養した。F−12培地での培養では、培地体積の半分量を注意深く除去し、除去した培地と同体積量の新鮮な培地で置き換えるという作業を2〜3日おきに21日間繰り返した。
【0152】
薄層クロマトグラフィー(TLC)
1×108cellのヘリコバクター ピロリからクロロホルム−メタノール混合液(2:1)を用いて室温で1時間かけて糖脂質の分画を抽出し、窒素気流で乾燥させ、クロロホルム−メタノール−水(16:6:1)混合液を用いて(8)で示したようにしてTLCを行った。単離した糖脂質は、オルシノール含有硫酸緩衝液を用い110℃で、目視できるように発色させた。
【0153】
ヘリコバクター ピロリと、末端α1,4結合−GlcNAcを安定に発現するAGS細胞との共生培養
末端α1,4結合−GlcNAcを安定に発現する胃腺癌AGS−α4GnT細胞を、前記の方法(Nakayamaら、前出,1999)で製造した。AGS−I4GnT細胞とMock−トランスフェクトしたAGS細胞とを、1×107cells/mlのヘリコバクター ピロリ(ATCC43504)と共に24時間、共培養した。20%の緩衝ホルマリンで固定した後、ヘリコバクター ピロリに対するウサギ・ポリクローナル抗血清(ダコ社製;デンマーク国グロストラップ)と、マウス・モノクローナルHIK1083抗体(関東化学社製;Ishiharaら、前出,1996)との混合物と共に前記細胞をインキュベートした。二次抗体として、ローダミン標識の抗ウサギ免疫グロブリン(抗ヘリコバクター ピロリ抗体用)と、イソチオシアン酸フルオレセイン標識抗マウスIgM(HIK1083抗体用)とを使用した。スライドは、VECTORSHIELDシールド溶液(ベクターラボラトリーズ社製;カリフォルニア州ブーリンガム)で封入し、レーザー共焦点顕微鏡LSM510 META(カルル ツァィス社製)を用いて観察した。
【0154】
結果
本明細書に示したように、培養時のヘリコバクター ピロリに対する効果を調べるために、α1,4結合−GlcNAc残基を有するムチン型糖タンパク質を調製した。前記のように、CD43(ロイコシアリン)は前記O−グリカンの優先的コアタンパク質としての役割を果たす。ヘリコバクター ピロリのO−グリカンのサブセットの活性を調べるために、α1,4−N−アセチルグルコサミン転移酵素(α4GnT;Nakayamaら、前出,1999)のcDNAと、コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I;Bierhuizen,
Fukuda、前出,1992)のcDNAと、可溶性CD43のcDNAとでトランスフェクトすることによって、末端α1,4結合−GlcNAc残基を有するO−グリカンを含む組換え型の可溶性CD43を、チャイニーズハムスター卵巣細胞(CHO)に発生させた。コントロールとして、α4GnTcDNAを使用しないこと以外は前記と同様の方法でCHOにトランスフェクトして、末端α1,4結合−GlcNAc残基を有しない可溶性CD43を製造した。
【0155】
異なる量の組換え型可溶性CD43を含む培地でヘリコバクター ピロリ(ATCC43504)をインキュベートした。最初の2.5日間での微生物の増殖は、末端α1,4結合−GlcNAc残基の有無に関係なくごくわずかであり(図2A)、これはヘリコバクター ピロリの増殖の遅滞期の特徴である。培養が3日を超えると増殖期に入り、微生物は急速に増殖した。しかし、末端α1,4結合−GlcNAc残基を有する可溶性CD43と共に細菌をインキュベートした場合は、ヘリコバクター ピロリの増殖速度が劇的に抑制された。この抑制効果は、可溶性CD43の濃度が62.5mU/mlを超えたときに顕著となった(1mUは、1μgのp−ニトロフェニル−N−アセチルグルコサミン(GlcNAcα−PNP)と当量である;Warneckeら、J.Biol.Chem.274:13048,1999参照、参考文献としてここに示す)。さらに、α1,4結合−GlcNAcを発現する可溶性CD43が31.2mU/mlで存在する培地で培養したヘリコバクター ピロリの運動性が、著しく抑制されることが、低速度写真からわかった(図2B)。また、電子顕微鏡写真は、ヘリコバクター ピロリの伸び、分節狭窄、屈曲のような顕著な形態異常を示した(図2C)。
【0156】
末端α1,4結合−GlcNAc残基を有する可溶性CD43の阻害効果について、15.6mU/mlから125.0mU/mlの最小阻害濃度で、別の標準菌株であるATCC43526や、3種の臨床的単離物(Warneckeら、前出,1999)を含む種々のヘリコバクター ピロリの菌株を用いて観察した。前記とは対照的に、末端α1,4結合−GlcNAc残基を有しない可溶性CD43と共にヘリコバクター ピロリをインキュベートした場合は、増殖阻害効果も形態異常もみられなかった(図2)。これらの結果は、末端α1,4結合−GlcNAc残基を有するO−グリカンが、抗菌剤と同様に、ヘリコバクター ピロリの増殖を抑制することを示している。また、ヘリコバクター ピロリに対する同様の阻害効果は末端α1,4結合−GlcNAc残基を結合した別のムチン様タンパク質であるCD34(Horiiら、Helicobacter 7:39,2002参照、参考文献としてここに示す)を使用した場合にも観察された。この結果は、骨格タンパク質ではなく末端α1,4結合−GlcNAc残基が、ヘリコバクター ピロリに対する増殖阻害活性のために必須であることを示している。
【0157】
末端α1,4結合−GlcNAc残基を有するO−グリカンによって引き起こされるヘリコバクター ピロリの形態異常は、β−ラクタマーゼ阻害剤のような抗生物質によって引き起こされる形態異常と類似しており、多くの細菌株の細胞壁の主成分であるペプチドグリカンの生合成を妨げる(Finalyら、J.Antimicrob.Chemother.52:18,2003参照、参考文献としてここに示す;Horiiら、前出,2002)。このように、これらのO−グリカンがヘリコバクター ピロリの細胞壁生合成を阻害する作用について調べた。ヘリコバクター種の細菌の細胞壁はα−コレステリルグルコシド(α−CGs)を特徴的に有しているが、その細胞壁の主成分はコレステリル−α−D−グルコピラノシド(CGL;Glcα1→コレステロール)、コレステリル−6−O−テトラデカノイル−α−D−グルコピラノシド(CAG)、コレステリル−6−O−ホスファチジル−α−D−グルコピラノシド(CPG;Hiraiら、J.Bacteriol.177:5327,1995;Haqueら、J.Bacteriol.178:2065,1996参照、夫々参考文献としてここに示す)である。
【0158】
末端α1,4結合−GlcNAc残基を有するO−グリカンがヘリコバクター ピロリのCGL生合成を妨げるかどうかを調べるために、O−グリカン存在下で培養したヘリコバクター ピロリ中のCGLの発現レベルを分析した。末端α1,4結合−GlcNAc残基を有しないO−グリカンを含む培地で培養したヘリコバクター ピロリの細胞壁成分を質量分析したところ、CGLに起因する明確なシグナルが現れた(図3A)。対照的に、末端α1,4結合−GlcNAc残基を有するO−グリカンを含む培地で培養したヘリコバクター ピロリサンプルの並行分析では、CGLは顕著な抑制を示し、末端α1,4結合−GlcNAc残基を有しないコントロール群の29.5%に減少した(図3B)。これらの結果は、末端α1,4結合−GlcNAc残基を有するO−グリカンが、ヘリコバクター ピロリのin vivoでのCGLの生合成を阻害していることを示している。
【0159】
α−CGsの生合成に関わる糖転移酵素の同定をはじめとしてヘリコバクター ピロリ細胞壁の根本的な生成機構はあまり解明されていないが、CGLはおそらく、グルコース(Glc)をUDP−GlcからコレステロールのC3位に転移させるUDP−Glc:ステロール糖転移酵素によって形成されていると考えられる。超音波処理されたヘリコバクター ピロリのUDP−Glc:ステロール糖転移酵素活性の発現を調べ、その発現が見られた場合は、それらの活性が末端α1,4結合−GlcNAc残基を有するO−グリカンによって阻害されるかどうかが分かる。コレステロールとUDP−Glcとを超音波処理したヘリコバクター ピロリと共にインキュベートし、生成物を質量分析した。多くの量のCGLが検出された(図3C)。これは、超音波処理したヘリコバクター ピロリが酵素学的にUDP−Glc:ステロール糖転移酵素活性を発現したことを示している。
【0160】
末端α1,4結合−GlcNAc残基を有する可溶性CD43を前記評価分析にかけたところ、CGL生成は抑制された(図3D)が、末端α1,4結合−GlcNAc残基を有しない可溶性CD43を前記評価分析にかけても何の効果もみられなかった(図3E)。腺粘液細胞型ムチン中に存在するα−結合型GlcNAcと、CGL中に存在するα−結合型Glcとの構造的類似性を考慮すると、これらの結果は、α1,4結合−GlcNAc残基がUDP−Glc:ステロール糖転移酵素のドナー基質のためにUDP−Glcと互いに競合し、又はα1,4結合−GlcNAc残基が最終生成物を阻害するメカニズムによって直接的にUDP−Glc:ステロール糖転移酵素活性を阻害する(Nakayamaら、J.Biol.Chem.271:3684,1996参照、参考文献としてここに示す)ことによって、CGLの生合成を減少させていることを示している。
【0161】
コレステロールはCGLの前駆体である。しかし、コレステロールの生合成に関係する遺伝子はヘリコバクター ピロリのゲノムデータベースにはみられない(Tombら、Nature 388:539,1997;Maraisら、Microbiol.Mol.Biol.Rev.63:642,1999)。よって、ヘリコバクター ピロリは外因性のコレステロールが存在しない状態ではCGLを合成できないかもしれない。コレステロールが存在しない培地でヘリコバクター ピロリを5日間培養した場合、微生物の細胞増殖はコレステロール存在下で培養した場合の約50%のレベルにとどまった(以下の表1参照)。コレステロール非存在下での培養において、運動性微生物は検出されず、形態異常、特にヘリコバクター ピロリの伸長がはっきりとみられた(図2Cで示したものと同様)。コレステロールが存在しない培地でヘリコバクター ピロリをさらに21日目まで培養すると、微生物は完全に死滅した。対照的に、コレステロールを補充した培地でヘリコバクター ピロリを培養した場合、細胞はよく増殖し形態異常の兆候はみられなかった(表1)。これらの結果は、ヘリコバクター ピロリの生存のために、外因的に補給されたコレステロールによるCGLの合成が必須であることを、示している。
【0162】
【表1】
【0163】
ヘリコバクター ピロリの標準菌株(ATCC43504)の1×107cells/mlを35℃の標準嫌気性条件下で、0.005%のコレステロールを補充したHam's F−12培地で5日間、及びコレステロールを含まない培地で21日間、夫々インキュベートした。CFUはコロニー形成ユニットである。
【0164】
コレステロールを含み、又は含まない培地で培養したヘリコバクター ピロリのCGL発現レベルを調べるために、ヘリコバクター ピロリの糖脂質分画のCGLを含むα−CGsの発現過程をTLCにより分析した。コレステロール存在下で培養したヘリコバクター ピロリでは、ヘリコバクター ピロリの特徴であるCGLを含むα−CGsの典型的な三本線が検出された(図4、レーン2)。しかしコレステロール非存在下で培養したヘリコバクター ピロリでは、α−CGsは検出されなかった(図4、レーン1)。これらの結果を合わせると、これらの結果は、CGLがヘリコバクター ピロリ細胞増殖に必須であること、また末端α1,4結合−GlcNAc残基を有するO−グリカンはCGLの生合成を阻害するためヘリコバクター ピロリに対して抗菌活性を有することを、示している。さらに、末端α1,4結合−GlcNAc残基を有するO−グリカンの抗菌活性は、CGLを発現する細菌の菌株に限定されるのかもしれない。実際に、ヘリコバクター ピロリ以外の菌株やCGL欠損菌株(例えば大腸菌、緑膿菌、肺炎桿菌、黄色ブドウ球菌、α−レンサ球菌、肺炎レンサ球菌)を末端α1,4結合−GlcNAc残基を有する可溶性CD43存在下でインキュベートしても、抗菌効果はみられなかった。
【0165】
前記の結果は、末端α1,4結合−GlcNAc残基を有するO−グリカンを発現する粘液細胞は、ヘリコバクター ピロリ感染から自分自身を保護していることを示している。この効果を確認するため、胃腺癌AGS−α4GnT細胞をα4GnT cDNAに安定にトランスフェクトし(Nakayamaら、前出,1999)、ヘリコバクター ピロリ感染による細胞の損傷を調べた。ヘリコバクター ピロリと共にインキュベートしてMock−トランスフェクトされた細胞は、細胞の扁平や萎縮のような顕著な荒廃を示した(図5矢印参照)。対照的に、ヘリコバクター ピロリと共に培養したAGS−α4GnT細胞ではごくわずかな損傷しか観察されなかった(図5)。これらの結果から、末端α1,4結合−GlcNAc残基を有するムチン型O−グリカンを発現する細胞はヘリコバクター ピロリ感染に対して抗力を示すことがわかった。
【0166】
グリカン鎖は、細胞間相互作用における細胞表面レセプターのリガンドとして(Lowe,Cell 104:809,2001;Akamaら、Science 295:124,2002;Perilloら、Nature 378:736,1995)、またレセプターや粘着タンパク質のモジュレーターとして(Moloneyら、Nature 406:369,2000;Demitriouら、Nature 409:733,2001;Nakayamaら、Proc.Natl.Acad.Sci.,USA 92:7031,1995)、種々の役割を果たしている。これらの結果は、哺乳類のグリカンが天然抗菌剤としても機能することを示している。末端α1,4結合−GlcNAc残基を有するO−グリカンはヒト胃腺粘液細胞により生成される。そのためこれらの結果は、副作用を引き起こすことなくヒトへのヘリコバクター ピロリ感染を予防及び治療する新規な医薬品を開発できるという、根拠となるものである。
【0167】
前記の実施例を参照しながら本発明を説明したが、本発明の本質や技術的範囲には、その改良や差異も包含される。したがって、本発明は、特許請求の範囲により定められるものである。
【図面の簡単な説明】
【0168】
【図1】図1は、通常のヒトの胃壁の断面概略図であって、GOCTSで表面のムチンを染色し、PCS染色剤により染色されたα1,4結合−GlcNAc残基を示すものである。
【0169】
【図2】図2Aから図2Cは、末端α1,4結合−GlcNAc残基を有するO−グリカンがヘリコバクター ピロリの増殖と運動性とを抑制することを示すものである。
【0170】
図2Aは末端α1,4結合−GlcNAcを有する(αGlcNAc(+))可溶性CD43、及び末端α1,4結合−GlcNAcを有しない(αGlcNAc(−))可溶性CD43を用いて培養したヘリコバクター ピロリの増殖曲線を示す。αGlcNAc(+)の1ミリユニット(mU)が、p−ニトロフェニル−N−アセチル−α−グルコサミン(GlcNAcα−PNP)の1Tgと等量の免疫反応性として定義される。αGlcNAc(−)のタンパク質濃度は、αGlcNAc(+)の125.0mU/mlでのタンパク質濃度と同等であった。
【0171】
図2Bは、末端α1,4結合−GlcNAcを有する(αGlcNAc(+))可溶性CD43の31.2mU/ml、又は同タンパク質濃度の末端α1,4結合−GlcNAcを有しない(αGlcNAc(−))可溶性CD43を用いて、3日間培養したヘリコバクター ピロリの運動性を1秒間隔で経時的に記録した図を示す。典型的なヘリコバクター ピロリを矢印で示している。αGlcNAc(+)の存在下で培養された7個のヘリコバクター ピロリの平均速度は3.1±3.5μm/sec(平均±標準偏差)であり、αGlcNAc(−)の存在下で培養された7個のヘリコバクター ピロリの平均速度は21.2±2.6μm/secであり、スチューデントt検定による有意性は、P<0.001であった。スケールバーは50μmである。
【0172】
図2Cは、末端α1,4結合−GlcNAcを有する(αGlcNAc(+))可溶性CD43の31.2mU/ml、又は同タンパク質濃度の末端α1,4結合−GlcNAcを有しない(αGlcNAc(−))可溶性CD43を用いて、3日間培養したヘリコバクター ピロリの走査電子顕微鏡写真である。αGlcNAc(+)で培養されたヘリコバクター ピロリは伸び、分節狭窄、屈曲などの形態異常がみられた。写真はすべて等倍率で撮影したものであり、スケールバーは1Tmを示す。
【0173】
【図3】図3Aから図3Eは、末端α1,4結合−GlcNAc残基を有する可溶性CD43が、ヘリコバクター ピロリのin vivoでのコレステリル−α−D−グルコピラノシド(CGL)の生合成(図3A、図3B)と、同様なヘリコバクター ピロリのinvitroでの生合成(図3C〜図3E)とを阻害することを、マトリックス支援レーザー脱離イオン化飛行時間型質量分析(MALDI−TOF−mass)で測定した結果を示す図である。
【0174】
図3Aは、末端α1,4結合−GlcNAc残基を有しない可溶性CD43を用いて培養したヘリコバクター ピロリの分画液中、m/z571.6の位置にナトリウム付加CGL{M+Na}+が検出されたことを示す(矢印)ものである。
【0175】
図3Bは、末端α1,4結合−GlcNAc残基を有する可溶性CD43の4.0mU/mlを用いて培養した試料のCGLを同様に測定した結果を示す。内部標準としてホスファチジン酸(Inamotoら、J.Clin.Gastroenterol. 17:S136,1993参照、参考文献としてここに示す)に対するCGLの相対量は対照実験(図2A矢印)の29.5%まで顕著に減少している。2回測定した平均値を示してある。
【0176】
図3Cは、超音波処理したヘリコバクター ピロリによってUDP−グルコース及びコレステロールから生合成された生成物のMALDI−TOF−massの測定結果を示す。CGL{M+Na}+がm/z571.6の位置が現れている。
【0177】
図3Dは、末端α1,4結合−GlcNAc残基を有する可溶性CD43の50.0mU/mlの存在下で、超音波処理したヘリコバクター ピロリによってUDP−グルコース及びコレステロールから生合成された生成物の質量分析の結果を示す。この条件下では、CGLのピークは検知できなかった。
【0178】
図3Eは、末端α1,4結合−GlcNAc残基を有しない可溶性CD43の存在下で、超音波処理したヘリコバクター ピロリによってUDP−グルコース及びコレステロールから生合成された生成物の質量分析の結果を示す。この条件下でCGLは合成されていた。
【0179】
【図4】図4は、コレステロール非存在下で培養したヘリコバクター ピロリにCGLが存在しないことを示す図である。コレステロールを含まないブルセラ培地(レーン1)又は0.005%のコレステロールを含むブルセラ培地(レーン2)で培養したヘリコバクター ピロリの1×108個から抽出した全糖脂質を薄層クロマトグラフィーで分析した。使用した溶媒系はクロロホルム・メタノール・水混合液(16:6:1)であり、糖脂質は、オルシノール硫酸緩衝液を用いて呈色させた。CAGはコレステリル−6−O−テトラデカノイル−α−D−グルコピラノシド;CGLはコレステリル−α−D−グルコピラノシド;CPGはコレステリル−6−O−ホスファチジル−α−D−グルコピラノシドである。
【0180】
【図5】図5は、末端α1,4結合−GlcNAc残基を有するO−グリカンが、胃腺癌細胞をヘリコバクター ピロリによる細胞損傷から保護していることを示すものである。AGS細胞に付着した多くのヘリコバクター ピロリはベクターのみにトランスフェクトされ、細胞の平面化や縮小のような細胞損傷がみられた(矢印)。一方、AGS−α4GnT細胞に不完全に付着したヘリコバクター ピロリはα4GnTに安定にトランスフェクトされ、細胞損傷はみられなかった。抗ヘリコバクター ピロリ抗体(赤)と、末端α1,4結合−GlcNAc残基特異的のHIK1083抗体(緑)(Ishiharaら、Biochem.J.318:409,1996参照、参考文献としてここに示す)とによりAGS細胞を二重の免疫組織化学的により示す図である。同じ領域のノマルスキー顕微鏡写真を図5下段に示す。スケールバーは50Tmである。
【技術分野】
【0001】
この出願は、米国特許法(35U.S.C.)119条に従い、2004年2月20日に出願された米国出願番号60/546,600に基づく優先権を主張するもので、その全内容が本明細書に含まれている。
【0002】
この発明は、国立癌研究所から授与されたCA71932及びCA33000の助成金に基づく政府支援により、なされたものである。米国政府は、本発明についてのある種の権利を有している。
【0003】
本発明は、概要すると抗菌性組成物に関するもので、特に、末端α1,4結合−N−アセチルグルコサミン残基を有するO−グリカン、末端α1,4結合−N−アセチルグルコサミン残基を有するこのような物質を含んでいる組成物、及びその組成物を用いて細菌の感染を予防及び/又は治療する方法に関するものである。
【背景技術】
【0004】
胃は、食物を消化し易くするために強い酸性環境となっている。これによって、栄養分が分解されて腸で吸収される。一般に、胃の内壁の細胞は、胃の細胞が産生する粘液層で保護されている。胃の内壁がむき出しとなったりびらんによる穴を生じたりして、胃壁が酸性環境に晒されると、胃潰瘍が惹き起こされる。
【0005】
胃潰瘍は、胃酸の分泌の増加や酸性物質の摂取によって惹き起こされる。アスピリンやイブプロフェンのような非ステロイド系抗炎症剤(NSAIDs)を服用したり、多量のアルコールやカフェインを飲用したりするヒトに特に惹き起こされ易い。胃潰瘍患者の約70%がヘリコバクター ピロリ(Helicobacter pylori: H.pylori)に感染している。これらのNSAIDs治療による胃潰瘍のうちの約50%は、ヘリコバクター ピロリによって胃炎が惹き起こされたものでないが、NSAIDsによる治療を行っていないほとんどの胃潰瘍は、胃内に穴が開いた胃炎やヘリコバクター ピロリ感染を伴う。NSAIDsによって惹き起こされた潰瘍患者を除けば、胃潰瘍患者のヘリコバクター ピロリ感染率は約96%にも達する。
【0006】
ヘリコバクター ピロリは、微好気性、グラム陰性の桿菌であり、全世界の人口のおよそ半分のヒトの胃の内壁上に棲息し、胃炎、慢性胃炎、胃潰瘍、十二指腸潰瘍などの主な原因であると言われている。また、胃癌等の疾患との関連性が報告されている。この為、ヘリコバクター ピロリによって惹き起こされる疾患に対する治療薬の開発は、ヒトが健康で快適な生活を維持していくために重要な課題の一つである。
【0007】
抗ヘリコバクター ピロリ剤として、例えば、ペニシリン系、テトラサイクリン系、セファロスポリン系、ニューロキノン系等の抗生物質が知られている。しかし、これらの抗生物質は、ヘリコバクター ピロリに対する抗菌作用を示すものの、単独での投与では十分な抗菌効果が得られない。このため、現在では、これらの抗生物質に加えて、ビスマス剤、プロトンポンプ阻害薬との3剤併用療法が主に行われている。また、特開平11−189529号公報に、ベンゾヒドロキサム酸を含有する抗ヘリコバクター ピロリ剤が提案されている。しかし、これらの場合、投与したときに下痢や吐き気などの副作用があり、安全性に劣るという問題がある。
【0008】
また、前記の治療薬とは別に、例えば、特開2003−73278号公報に、植物等から抽出したステリルグルコピラノシドを含有する抗ヘリコバクター ピロリ剤が提案されている。また、特開2002−370995号公報に、オウレン等の生薬から抽出された成分と、炭酸水素アルカリ金属塩とを含有する抗ヘリコバクター ピロリ薬が提案されている。しかし、これらの場合、ヒトの生体内で発現したものではないため副作用があり、さらに、ヘリコバクター ピロリの抗菌効果が十分ではないという問題がある。また、前記のいずれの提案にも、ヘリコバクター ピロリの運動性の低下、及び形態異常については何ら開示されていない。
【0009】
胃潰瘍治療の目的は、痛みの軽減、潰瘍の治癒、合併症の予防、及び潰瘍の再発の予防である。未治療の潰瘍による合併症には、出血、胃壁の穿孔、及び瘢痕による閉塞症などがある。さらに、胃潰瘍患者の約2〜3%が胃癌を発症する。ヘリコバクター ピロリに感染した胃潰瘍患者の治療に、抗生物質が使用される。抗生物質により治療すると、ヘリコバクター ピロリ感染を治癒した後に潰瘍を治癒することができる。また、治療が成功した場合、翌年に再発する患者は5%未満である(ヘリコバクター ピロリ感染が治癒していない場合の再発率は60〜80%である)。
【0010】
ヘリコバクター ピロリは胃の内壁を覆っている粘液層の深部に棲息するため、この細菌を胃内から根絶することは困難である。また、抗生物質が、細菌を殺すのに十分な濃度でこの粘液層を透過させるのも困難である。一般に、単剤療法は、感染してしまったヘリコバクター ピロリの除去に効果を奏しない。例えば、アモキシシリンを用いた場合、この細菌の根絶率は20%未満であり、メタロニダゾールを用いた場合、根絶率は10%未満であるが、クラリスロマイシンを併用すると、細菌の根絶率は約40%となる。そのため、2剤(例えば二種の抗生物質、又は一種の抗生物質と一種の酸低下薬)の併用療法や、3剤(例えば二種の抗生物質と一種の酸低下薬)の併用療法が行われる。二種の抗生物質の併用療法の根絶率は40〜60%である。また、最も優れた3剤併用療法で治療した患者のヘリコバクター ピロリ感染の根絶率は、現在のところ90%以上である。
【0011】
あいにく、ヘリコバクター ピロリも他の細菌のように、抗生物質に対して耐性を生じるようになるため、治療の成功率が低下してしまう。例えば、メタロニダゾールに対する耐性は部位によっては、40%の高率に達すると報告されている。また、クラリスロマイシンに対する耐性も僅か(2%未満)であるが報告されている。抗生物質に対する耐性は通常経時的に増加するので、それらがヘリコバクター ピロリの根絶と胃潰瘍の治療とに次第にそれほど有効でなくなるかもしれないという懸念がある。しかし、十分な抗菌効果を有し、副作用がなく、安定性に優れる物質やそれを用いた治療法は、いまだ提案されていない。そのため、胃潰瘍を予防し、また治療するための組成物と、そのための方法とが求められている。
【発明の開示】
【発明が解決しようとする課題】
【0012】
本発明は、胃に感染して胃潰瘍を引き起こすヘリコバクター ピロリの増殖を抑制する胃腺粘液細胞型ムチン、特に末端α1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有するムチンを見出したことに基づきなされたものである。この胃腺粘液細胞型ムチンにこのような活性があることが分かったので、α1,4結合−GlcNAc残基を有する典型的な糖タンパク質を調製したところ、それらがコレステリル−α−D−グルコピラノシド合成やヘリコバクター ピロリ増殖に対して抑制効果を有するということが、判明した。これによってなされた本発明は、胃潰瘍の治療、十二指腸潰瘍の治療、抗ヘリコバクター ピロリ剤、胃炎の治療、慢性胃炎の緩和に好適に使用される医薬物質;その物質の製造工程;その物質を用いた胃潰瘍の治療方法;及びその物質を製造できるトランスジェニック哺乳類を提供するというものである。
【課題を解決するための手段】
【0013】
本発明は、担体分子と相互作用して結合した少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する天然非存在の化合物に関するものである。担体分子は一つ以上のα1,4結合−GlcNAc残基に結合することができる分子であり、例えばO−グリカン(例えばコア2分岐型O−グリカン)、ポリペプチド、特に少なくとも一つのO−グリコシル化部位を有するポリペプチド、又は糖タンパク質(例えば、コア2分岐型O−グリカンを含むポリペプチドのような、O−グリカンを一つ以上含むポリペプチド)である。その一つの形態は、担体分子が、例えばCD43(ロイコシアリン)やCD34のようなムチン様ポリペプチドであるというものである。別な形態は、前記化合物が、例えば2、3、4、5又はそれ以上のα1,4結合−GlcNAc残基のような複数のα1,4結合−GlcNAc残基を有するというものである。さらに別な形態は、前記化合物が、合成されたオリゴ糖を有するというものである。
【0014】
また、本発明は、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質の製造方法に関するものである。その方法は例えば、ポリペプチドのグリコシル化に適した条件下で、α1,4−N−アセチルグルコサミン転移酵素(α4GnT)と、コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)又はコア1伸長酵素(例えばコア1伸長β1,3−N−アセチルグルコサミン転移酵素であるコア1−β3GlcNAcT又はC1−β3GnT)と、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドとを、接触させて、該ポリペプチドを、C2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質を製造するというものである。この担体ポリペプチドは、C2GnT−I(又はC1−β3GnT)とα4GnTとの活性によって一つ以上のα1,4結合−GlcNAc残基を結合し得るもので、例えば可溶性ポリペプチド(具体的には、可溶性CD43ポリペプチド又は可溶性CD34ポリペプチド)である。
【0015】
前記担体ポリペプチドの具体例の一つは、哺乳類の母乳中に通常存在するムチン型糖タンパク質を含むというものである。この具体例の一つの形態として、in vitroで、哺乳類の母乳と、少なくともα4GnTと、必要に応じてC2GnT−I及び/又はC1−β3GnTとを接触させ、母乳中のムチン型糖タンパク質を、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質に変換する方法が、挙げられる。この具体例の別な形態として、前記ムチン型糖タンパク質を哺乳類の母乳から単離し、哺乳類の母乳から単離されたムチン型糖タンパク質と、少なくともα4GnTと、必要に応じてC2GnT−I及び/又はC1−β3GnTとを接触させ、母乳中のムチン型糖タンパク質を、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質に変異させる方法が挙げられる。
【0016】
少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質の製造方法は、糖タンパク質を単離する工程を含んでいてもよい。それによって、本発明の方法で製造された少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質は、そのような方法で製造して単離された糖タンパク質として、得られる。
【0017】
さらに、本発明は、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質の製造方法に関するものである。この方法は例えば、少なくとも異型のα4GnTをエンコードするポリヌクレオチドを真核細胞内で発現させ、細胞により産生される内因性であってもよいポリペプチドを、細胞内に発現したC2GnT−I(又はC1−β3GnT)とα4GnTとによって、グリコシル化することができるというものである。例えば、哺乳類の細胞は、外因性(異型)発現α4GnTの存在下で、内因性C2GnT−Iと前記のようにグリコシル化され得るポリペプチドとを発現させて、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を産生して製造する。この方法で注意すべき点は、真核細胞内でα1,4結合−GlcNAc残基を有する天然非存在の糖タンパク質に変異されるため、糖タンパク質が“組換え型”糖タンパク質とみなされるが、この糖タンパク質は、細胞中の異型α4GnTの発現によってのみ産生されるということである。別な形態として、例えば真核細胞が天然に内因性C2GnT−Iや所望の担体ポリペプチドを発現してもしなくても、異型C2GnT−Iをエンコードするポリヌクレオチド及び/又は少なくとも一つのO−グリコシル化部位を有する異型担体ポリペプチドをエンコードするポリヌクレオチドが細胞内に発現して、担体ポリペプチドが内因性又は外因性(異型)であってもよいC2GnT−Iとα4GnTとによってグリコシル化されるというものである。
【0018】
前記担体ポリペプチドは、可溶性ポリペプチドであってもよく、例えば可溶性CD43又は可溶性CD34であってもよい。また、前記組換え型の糖タンパク質は、一つ又は複数のα1,4結合−GlcNAc残基を有していてもよい。前記真核細胞は、エンコードするポリヌクレオチドを発現できる如何なる真核細胞であってもよい。真核細胞は、内因性C2GnT−I、異型C2GnT−I、異型α4GnTのいずれかが、内因性担体ポリペプチドや異型ポリペプチドであってもよい担体ポリペプチドに一つ又はそれ以上のα1,4結合−GlcNAc残基を結合できるような酵素活性を有するものであってもよい。例えば前記真核細胞は、チャイニーズハムスター卵巣細胞のような哺乳類の細胞や、ヒトのような哺乳類の上皮細胞等の細胞であってもよい。さらに前記方法は、前記組換え型の糖タンパク質を単離する工程を含んでいてもよい。それによって、本発明の方法で製造されて、単離された組換え型の糖タンパク質を含む組換え型の糖タンパク質が、得られる。
【0019】
また、本発明は、少なくとも、α4GnTをエンコードする第一の外因性ポリヌクレオチドと、さらにC2GnT−Iをエンコードするポリヌクレオチドと、担体ポリペプチドをエンコードするポリヌクレオチドとを、ゲノム中に安定に組み込まれて有する非ヒトのトランスジェニック哺乳類であって、該第一の外因性ポリヌクレオチドが、細胞中又は関連する細胞中にエンコードされたC2GnTを発現させる5’制御配列に相互作用して結合されている非ヒトのトランスジェニック哺乳類に関するものである。5’制御配列は、活性な構成プロモータ、誘導プロモータ、組織特異プロモータ、又はそれと同様のものであってもよい。一つめの具体例として、前記5’制御配列は、乳腺特異的プロモータを含んでいるものである。そのC2GnTと担体ポリペプチドとをエンコードするポリヌクレオチドは、内因性ポリヌクレオチド(例えば非ヒトの動物のゲノムに含まれる遺伝子)であってもよいが、必ずしも必要ではない。二つめの具体例として、C2GnTをエンコードするポリヌクレオチドがC2GnT−Iをエンコードする第二の外因性ポリヌクレオチドであり;及び/又は、担体ポリペプチドをエンコードするポリヌクレオチドが少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチドであるものが挙げられる。この具体例の一つの形態として、前記第一の外因性ポリヌクレオチドと、共存するなら前記第二の外因性ポリヌクレオチドと、共存するなら前記第三の外因性ポリヌクレオチドとが夫々、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合しているものが挙げられる。この具体例の別な形態として、共存するなら前記第三の外因性ポリヌクレオチドは、担体ポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と相互作用して結合しているものが挙げられる。本発明のトランスジェニック哺乳類中のC2GnT−Iと、α4GnTと、担体ポリペプチドとの発現により、前記担体ポリペプチドは、C2GnT−Iとα4GnTとによってグリコシル化され、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質が産生される。
【0020】
本発明の目的に有用であって、乳腺特異的遺伝子の5’制御配列のすべて又は一部を構成するプロモータは、乳腺特異的な発現を示すプロモータであってもよい。そのようなプロモータとして、例えば乳清酸性タンパク質プロモータ、α−カゼインプロモータ、β−カゼインプロモータ、γ−カゼインプロモータ、α−ラクトアルブミンプロモータ、又はβ−ラクトグロブリンプロモータが挙げられる。前記第一の外因性ポリヌクレオチドと、前記第二の外因性ポリヌクレオチドと、及び前記第三の外因性ポリヌクレオチドとが、乳腺特異的遺伝子の3’制御配列、又は乳腺中の活性3’制御配列と、さらに相互作用して結合していてもよい。
【0021】
本発明の非ヒトのトランスジェニック哺乳類は、一つ、二つ、又は三つの導入遺伝子を用いて作製されて製造されるものであってもよい。これの一つの形態は、前記第一の外因性ポリヌクレオチドと、前記第二の外因性ポリヌクレオチドと、前記第三の外因性ポリヌクレオチドとが、夫々独立に、5’制御配列と、必要に応じ3’制御配列とに、前記のように相互作用して結合し得るというものである。別な形態は、前記第一の外因性ポリヌクレオチドと、前記第二の外因性ポリヌクレオチドと、前記第三の外因性ポリヌクレオチドとが、互いに相互作用して結合し、さらに5’制御配列と、必要に応じ3’制御配列とに、相互作用して結合し、単一の導入遺伝子を形成するというものである。さらに別な形態は、二つの導入遺伝子、例えば、第二の外因性ポリヌクレオチドに相互作用して結合した第一の外因性ポリヌクレオチドを含む第一の導入遺伝子と、第三の外因性ポリヌクレオチドを含む第二の導入遺伝子とであり、前記第一及び第二の導入遺伝子は夫々、5’制御配列と、必要に応じて3’制御配列とに、前記のように相互作用して結合し得るというものである。
【0022】
本発明の非ヒトのトランスジェニック哺乳類は、例えばマウス、ラット、ウサギ、ウマ、ブタ、ヒツジ、ヤギ、又はウシのような如何なる哺乳類であってもよい。具体例として、トランスジェニック哺乳類は、非ヒトの雌トランスジェニック哺乳類であって、抗菌活性型で、前記非ヒトの雌トランスジェニック哺乳類によって産生された組換え型糖タンパク質を、母乳中に分泌するというものである。それによって前記非ヒトの雌トランスジェニック哺乳類によって産生された母乳も、得られる。
【0023】
また、本発明は、非ヒトの雌トランスジェニック哺乳類の母乳中に、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を、産生させて製造する方法に関するものである。この方法は、例えば、非ヒトの哺乳類の胚にα4GnTをエンコードする第一の外因性ポリヌクレオチドと、必要に応じてC2GnT−I又はコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二の外因性ポリヌクレオチド;及び/又は必要に応じて少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチドを導入し、前記第一の外因性ポリヌクレオチドと、共存するなら前記第二の外因性ポリヌクレオチドと、共存するなら前記第三の外因性ポリヌクレオチドとが、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合され、共存するなら前記第三の外因性ポリヌクレオチドが、担体ポリペプチドを母乳中に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と相互作用して結合され;子孫を出産するような宿主の雌哺乳類に前記の胚を移植し;5’制御配列に相互作用して結合した第一の外因性ポリヌクレオチドと前記第二の外因性ポリヌクレオチドと前記第三の外因性ポリヌクレオチドとを含んで、ゲノム中に安定に組み込まれ、前記の雌の子孫に、母乳を産生させることによって行われるというものである。本発明の方法によって製造された非ヒトの雌トランスジェニック哺乳類中にC2GnT−Iとα4GnTと担体ポリペプチドとが発現し、前記担体ポリペプチドが、C2GnT−I(又はC1−β3GnT)とα4GnTとによってグリコシル化されて、少なくとも一つのα1,4結合−GlcNAc残基を有し、非ヒトの雌トランスジェニック哺乳類の母乳中に分泌される抗菌性の組換え型の糖タンパク質が、産生される。
【0024】
このような非ヒトの雌トランスジェニック哺乳類の製造方法は、さらに前記雌の子孫から搾乳して、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を含む母乳を得る工程を、含むものであってもよい。また前記の方法は、さらに前記母乳から少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を単離することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を得る工程を含んでいてもよい。それによって、本発明は、このような方法で製造される非ヒトの雌トランスジェニック哺乳類と、その雌トランスジェニック哺乳類から産生される母乳と、この方法で得られる少なくとも一つのα1,4結合−GlcNAc残基を有する単離された組換え型の糖タンパク質とを、提供することができるというものである。
【0025】
また、本発明は、コレステリル−α−D−グルコピラノシド(CGL)を発現する細菌の細胞壁の形成を、抑制又は阻害する方法に関するものである。この方法は、例えば、この細菌を、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物に接触させ、CGL合成を抑制又は阻害して、前記細菌の細胞壁の形成を抑制又は阻害するというものである。一つの形態として、前記化合物が、細菌のUDP−Glc:ステロール糖転移酵素活性を抑制又は阻害して、前記CGL合成を抑制又は阻害するというものが挙げられる。前記細菌は、GGLを発現する細菌であればよく、ヘリコバクター種の細菌、特にヘリコバクター ピロリも範疇に含んでいる。前記化合物は、本発明の化合物、例えば本発明の組換え型の糖タンパク質、又は胃腺粘膜細胞から単離された胃腺粘液細胞型ムチンも範疇に含んでいる。
【0026】
さらに本発明は、CGLを発現する細菌の増殖を抑制又は阻害する方法に関するものである。この方法は、例えば、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物に細菌を接触させ、CGL合成を抑制又は阻害して、前記細菌の増殖を抑制又は阻害するというものである。前記方法は例えば、培養中の細菌細胞に前記化合物を添加してin vitroで接触させたり、細菌に感染した個体、例えばヘリコバクター種の細菌に感染した哺乳類(例えばヘリコバクター ピロリに感染したヒト)の胃に前記化合物を直接投与し、in vivoで接触させたりして行なうというものである。前記化合物は、本明細書で示した方法を用いて得られるもので、例えばO−グリカンのような少なくとも一つのα1,4結合−GlcNAc残基を有する化合物であれば如何なるものであってもよく、胃腺粘膜細胞から単離された胃腺粘液細胞型ムチンであってもよい。
【0027】
さらに、本発明は、ヘリコバクター種の細菌の感染による胃潰瘍を治療又は予防する方法に関するものである。それによって本発明は、ヘリコバクター種の細菌が個体に感染することによる胃潰瘍の兆候又は症状を改善する方法を提供することができるというものである。この方法は、例えば、個体に少なくとも一つのα1,4結合−GlcNAc残基を有する化合物を投与し、前記化合物が前記ヘリコバクター種の細菌に接触し、前記ヘリコバクター種の細菌の増殖を抑制又は阻害して、胃潰瘍及び/又は胃癌の兆候又は症状を改善するというものである。
【0028】
本発明は、別な具体例として、胃潰瘍感受性の個体にヘリコバクター種の細菌が感染することによる胃潰瘍を、予防する方法を提供するというものである。この方法は、例えば、胃潰瘍感受性の個体に、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物を投与し、前記化合物が前記ヘリコバクター種の細菌の増殖を妨げて、前記ヘリコバクター種の細菌による胃潰瘍の形成を予防するというものである。
【0029】
本発明は、さらに他の具体例として、胃潰瘍又は胃癌を有する個体に少なくとも一つのα1,4結合−GlcNAc残基を有する化合物を投与して、前記ヘリコバクター種の細菌の感染による胃潰瘍又は胃癌を、改善する方法に関するというものである。前記化合物が、前記ヘリコバクター種の細菌の増殖を妨げて、前記ヘリコバクター種の細菌の感染による胃潰瘍又は胃癌が、改善される。例えば前記の方法は、本明細書中で示した非ヒトのトランスジェニック哺乳類から得られる前記化合物を含有する母乳を、前記個体が飲用することも範疇に含んでいる。
【0030】
本発明の方法により治療される個体は、胃粘膜がヘリコバクター感染しているかヘリコバクター感受性である如何なる個体であってもよい。それによる前記個体は例えばウマ、ブタ、ヤギ、イヌ、ネコ、フェレット、又はそれと同等のものであってもよい。具体例として、前記個体はヒトである。また、ヘリコバクター種の細菌は胃潰瘍形成に関係する如何なる細菌種であってもよく、例えばヒトの胃潰瘍形成に関係するヘリコバクター ピロリが挙げられる。
【0031】
前記化合物は、少なくとも一つの末端α1,4結合−GlcNAc残基を有するもので、本明細書で示したように、ヘリコバクター種の細菌の増殖を抑制又は阻害できるものである。前記化合物は、合成されたオリゴ糖、一つ又はそれ以上のα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質(例えば末端α1,4結合−GlcNAc残基を有する可溶性CD43ポリペプチド)、一つ又はそれ以上の末端α1,4結合−GlcNAc残基を有し、O−グリカンを含む化合物、又は胃腺粘膜細胞から単離することができる胃腺粘液細胞型ムチンを含む化合物であってもよい。少なくとも一つの末端α1,4結合−GlcNAc残基を有するそのような化合物を製造する方法は、慣用された公知の方法に準じる。例えば、担体分子のために有用な合成オリゴ糖を製造する方法として、コア2オリゴ糖を製造する方法(例えばSenguptaら、Tetrahedron Lett.44:6037-6042,2003;Ongら、J.Biol.Chem.277:18182-18190,2002参照;いずれも参考文献としてここに示す)、コア1伸長オリゴ糖を製造する方法(例えばBelotら、Tetrahedron Lett.74:2-6,2002参照;参考文献としてここに示す)、コア2分岐型O−グリカンにα1,4−GlcNAcを添加する方法(Nakayamaら、Proc.Natl.Acad.Sci.,USA 96:8991-8996,1999参照;参考文献としてここに示す)が挙げられる。さらに、前記化合物は、特に胃に直接投与できる経口投与のような便利な方法で投与されてもよい。一つの形態として、前記投与は、ヘリコバクター感染による胃潰瘍を有している個体又は前記胃潰瘍感受性の個体が、本明細書中で示した非ヒトのトランスジェニック哺乳類から得られる前記化合物を含有した母乳を、飲用することによって行われるというものである。
【0032】
また、本発明は、必須のα4GnTをエンコードする第一の外因性ポリヌクレオチドと;C2GnT−Iをエンコードし、又はC1−β3GnTをエンコードする第二のポリヌクレオチドと;少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとをゲノム中に安定に組み込んで有するトランスジェニック植物であって、該第一のポリヌクレオチド、該第二のポリヌクレオチド、及び該第三のポリヌクレオチドの少なくともいずれかが外因性ポリヌクレオチドであり;該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、該第三のポリヌクレオチドとが、プロモータを含む植物遺伝子の5’制御配列に相互作用して結合され;C2GnT−I又はC1−β3GnTと、α4GnTと、担体ポリペプチドとの発現により、該担体ポリペプチドを、C2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を産生して製造するトランスジェニック植物に関するものである。そのようなトランスジェニック植物中、前記第二のポリヌクレオチド及び/又は前記第三のポリヌクレオチドが、外因性のポリヌクレオチドであってもよく、一つの具体例として、前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、夫々外因性のポリヌクレオチドであるというものである。
【0033】
本発明のトランスジェニック植物中、外因性の核酸分子に相互作用して結合したプロモータは、構成プロモータ、誘導プロモータ、又は組織特異プロモータであってもよい。前記プロモータは、例えば種、花、葉及び/又は根のような植物の所望の細胞中でエンコードされたポリペプチド(例えばα4GnT)を転写する。例えば、前記プロモータは、構造的に活性な35Sカリフラワーモザイクウイルスプロモータ、ユビキチンプロモータ、コメアクチンプロモータ、誘導テトラサイクリン応答プロモータ、rbcS遺伝子光誘導プロモータ、又は種子特異的オレオシン遺伝子プロモータであってもよい。
【0034】
本発明のトランスジェニック植物は、如何なる植物であってもよく、単子葉植物、双子葉植物、裸子植物、被子植物も範疇に含むものである。さらに、前記トランスジェニック植物は、ヒトや動物の食材源として、用いられるものであってもよく、例えば農業経営(例えば遺伝子操作された作物の生産)のように商業上有用なものであってもよい。このような前記トランスジェニック植物は、例えば、ダイズ植物、コメ植物、トウモロコシ植物、又はコムギ植物であってもよい。それによって本発明は、例えばこのようなトランスジェニック植物によって産出された種;このようなトランスジェニック植物から産出された穀類のような産物;種や穀類に由来する産物、又はそれらの同等品例えば豆乳やライスミルクのような乳液を提供することができるというものである。
【0035】
また、本発明は、トランスジェニック植物中で、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を産生させて製造する方法に関するものである。この方法は例えば、植物細胞にα4GnTをエンコードする第一の外因性ポリヌクレオチドを少なくとも導入し;該第一の外因性ポリヌクレオチドが、プロモータを含んでいる植物遺伝子の5’制御配列に相互作用して結合され;α4GnTの発現により、該植物の細胞中の該担体ポリペプチドをα4GnTによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を産生させ;導入された外因性核酸分子を含む植物細胞から植物を生育させ、トランスジェニック植物を得て、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質をトランスジェニック植物中に産生させることができるというものである。この方法はさらに、外因性のC2GnT−Iをエンコードし、又は外因性のC1−β3GnTをエンコードする第二の外因性ポリヌクレオチド;及び/又は少なくとも一つのO−グリコシル化部位を有する外因性の担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチドをさらに植物細胞に導入する工程を含み、共存するなら該第二の外因性ポリヌクレオチド、及び共存するなら該第三の外因性ポリヌクレオチドが夫々、プロモータを含む植物遺伝子の5’制御配列に相互作用して結合され;共存するならC2GnT−I又はC1−β3GnTと、α4GnTと、共存するなら外因性の担体ポリペプチドとの発現により、該担体ポリペプチドを、共存するならC2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質をトランスジェニック植物中に産生させるものであってもよい。
【0036】
トランスジェニック植物中で、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を産生させて製造する方法は、さらに、例えば種子、葉、根、及び/又は花のようなトランスジェニック植物収穫物を、トランスジェニック植物から分取する工程を含んでいてもよい。前記の方法は、さらに、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を、前記トランスジェニック植物又は前記トランスジェニック植物収穫物から単離する工程を含んでいてもよい。一つの具体例として、この方法は、さらに前記トランスジェニック植物又は前記トランスジェニック植物収穫物を加工して、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を含む乳液製品を得る工程を含んでいてもよい。これにより、本発明の方法で産生されて単離された少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質、本発明の方法で産出されたトランスジェニック植物産出物、本発明の方法により得られた乳液も、供給することができる。
【発明を実施するための好ましい形態】
【0037】
本発明は、例えばヒトの胃潰瘍の原因となるヘリコバクター ピロリのようなヘリコバクター種の細菌に感染することによる胃潰瘍を、予防又は治療するのに有用な組成物と方法とを提供するというものである。本発明は、ヘリコバクター属に対する増殖性及び運動性を阻害し、また形態異常性を発現させるもので、抗菌活性を有し、ヘリコバクター属に対する抗菌活性が選択的及び特異的であるため副作用がなく、安全性に極めて優れる分子、機能性分子、組成物、医薬、飲食物、及び母乳、及びトランスジェニック哺乳類を提供することを目的とする。また、本発明は、前記機能性分子の効率的な製造方法、及び、機能性分子を投与することにより、副作用がなく、安全性に極めて優れる胃癌の治療方法、胃潰瘍の治療方法、及び胃潰瘍の予防方法を提供することを目的とする。
【0038】
図1に示すように、ヘリコバクター ピロリに感染した患者を検査したところ、細菌は、胃の内壁表面部を構成する表面粘液細胞から分泌される表面粘液細胞型ムチンにのみ関連しており、胃の内壁の深部に存在する腺粘液細胞部分にコロニーを形成しないということが観察される。腺粘液細胞から分泌される腺粘液細胞型ムチンは、末端でα−1,4−グルコシド結合(以下GlcNAcα(残基)と呼ぶことがある。)により結合されているN−アセチルグルコサミン残基を含んでいることも見出された。末端GlcNAcα残基は、ヘリコバクター ピロリの細胞壁に含まれるコレステリル−α−D−グルコピラノシド(CGL)の合成を阻害し、おそらくその結果、ヘリコバクター ピロリの増殖特性を阻害し、さらに、その運動性を阻害し、かつその形態異常性を発現させるということも見出された。図1中、「GlcNAcα(+)」は、GlcNAcα残基を含むムチンが存在することを意味し、「GlcNAcα(−)」は、GlcNAcα残基を含むムチンが存在しないことを意味する。このことは、α1,4−GlcNAc残基がPCSに染色されるものであり、またGOCTSがムチン表面を染色するという事実から、推察される(実施例1参照)。
【0039】
本発明は、複数のモノマーユニットからなり、その少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基である分子を提供するというものである。本明細書で開示したように、本発明の分子は、コレステリル−α−D−グルコピラノシドの合成を抑制及び阻害し、同様に前記コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び阻害する。さらに、前記分子は前記コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び阻害し、さらにはその細菌の形態異常を発現させる。一つの具体例として、前記分子は糖鎖を含み、その末端にN−アセチルグルコサミン残基を有していてもよい。一つの形態として、N−アセチルグルコサミン残基は、α1,4−N−アセチルグルコサミン残基である。このα1,4−N−アセチルグルコサミン残基は、コレステリル−α−D−グルコピラノシドの合成を抑制及び阻害することができる。同様に、前記コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び阻害することができる。さらにα1,4−N−アセチルグルコサミン残基は、前記コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び阻害し、また、その細菌の形態異常を発現させる。本発明の分子は、N−アセチルグルコサミン残基に隣接してガラクトース残基を有していてもよい。前記分子は直鎖又は分岐鎖であってもよい。
【0040】
本発明の分子は、N−アセチルグルコサミン(「GlcNAc」と称する)残基を一部に含み、必要に応じて適宜選択した分子構成ユニットを含む。前記GlcNAc残基は、化学合成した物質に由来する基であってもよく、天然物から抽出物に由来した基であってもよく、生体内に存在する基であってもよい。前記GlcNAcの製造方法は、特に制限はなく、例えば、キチンを塩酸で加水分解し、加水分解物を単離することにより製造することができる。
【0041】
前記分子構成ユニットは、本明細書に示したように目的に応じて適宜選択することができ、糖残基、樹脂ユニット等が挙げられる。これらを、1種単独で使用してもよく、2種以上併用してもよい。前記糖残基は、単糖残基であってもよいし、二糖以上の多糖残基であってもよい。前記単糖残基、又は多糖残基を構成する単糖残基として、例えば、ガラクトース残基、テトロース残基、エリトロース残基、トレオース残基、ペントース残基、リボース残基、アラビノース残基、キシロース残基、リキソース残基、ヘキソース残基、アロース残基、アルトロース残基、グルコース残基、マンノース残基、グロース残基、イドース残基、タロース残基、テトルロース残基、エリトルロース残基、2−ペンチロース残基、リブロース残基、キシルロース残基、プシコース残基、フルクトース残基、ソルボース残基、タガトース残基、2−ヘプツロース残基、セドヘプツロース残基、キノボース残基、ラムノース残基、フコース残基、ジギトキソース残基、アベクオース残基、チベロース残基、2−ベオキシリボース残基、2−デオキシグルコース残基、グルコサミン残基、マンノサミン残基、ガラクトサミン残基、フコサミン残基、キノボサミン残基、ラムノサミン残基、ノイラミン酸残基、ムラミン酸残基、ノジリマイシン残基などが挙げられる。これらの中でも、前記GlcNAc残基に隣接する糖残基として、ガラクトース残基が特に好ましい。前記糖残基は、1種単独でもよく、2種以上を併用してもよく、置換基が挿入されていてもよく、別な置換基でさらに置換されていてもよい。前記樹脂ユニットとして、例えば、L−ラクタイド、D−ラクタイド等が挙げられる。前記のユニットや別なユニットとして、生体適合性分子を構成する公知のユニットが用いられてもよい。
【0042】
前記GlcNAc残基は、本発明の前記分子のどこに位置していてもよく、例えば、分子の末端でもよく、末端以外の部分でもよい。しかし、前記GlcNAc残基が前記分子の末端に位置していると、優れた抗菌効果を示す。前記分子が糖鎖である場合、前記GlcNAc残基の結合はどのようなものでもよく、例えば、α−グルコシド結合やβ−グルコシド結合であってもよい。一つの形態として、前記結合はα1,4−グルコシド結合である。
【0043】
前記GlcNAc残基をα−1,4−グルコシド結合で前記分子に結合させるために酵素を使用する場合、その酵素を目的に応じて適宜選択することができる。前記酵素として、例えば、特開2001−46077号公報に記載のN−アセチルグルコサミン転移酵素(α4GnT)が挙げられる。前記α4GnTは、例えば、特開2001−46077号公報に記載の方法に準じて前記α4GnTをエンコードするDNAを形成させ、このDNAを組み込んだベクターを各種細胞にトランスフェクトさせることにより、前記α4GnTを発現させることによって、調製することができる(Nakayamaら、Proc.Natl.Acad.Sci.U.S.A.96,8991,1999参照)。
【0044】
前記分子は、前記GlcNAcα残基や付加ユニットを含む所望のモノマーユニットを多く含んでいてもよい。このモノマーユニットは、目的に応じて適宜選択される。例えば、前記分子は、2以上、特に2〜30(例えば5〜20)のモノマーユニットを含んでいてもよい。一般に、前記モノマーユニットの数が2未満であると、前記分子は抗菌効果を有しない。
【0045】
本発明の分子は、直鎖であっても分岐鎖であってもよい。前記分子が分岐である場合、分岐数は特に制限はなく、目的に応じて適宜選択することができる。一般に分岐数が多いほど、抗菌効果は高い。前記分子を分岐させるために酵素を使用してもよい。そのような酵素として、例えば、コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)を挙げることができ、このC2GnT−Iは例えば細胞への遺伝子の導入、遺伝子組み換えによるC2GnT−Iの調製、生物からの単離精製などにより調製することができる(例えばBierhuizen,Fukuda、Proc.Natl.Acad.Sci.U.S.A.89,9326,1992参照)。同様に、前記分子中で糖残基を結合させるために酵素を使用してもよい。そのような酵素として、例えば、コア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)を使用してもよい(Yehら、Cell,105:957-969,2001参照)。
【0046】
本発明の分子がGlcNAcα残基を含む糖鎖を有している場合、前記分子はヒトの胃壁深部に存在する腺粘液細胞から分泌される腺粘液細胞型ムチンが有する分子、又はその骨格構造を有する類似分子であってもよい。そのため、前記分子を、医薬や、食物や飲料のような飲食材として使用した場合、副作用がなく、極めて安全性が高い。
【0047】
また、本発明は、複数のモノマーユニットを含む分子であって、それの少なくとも一つのユニットがN−アセチルグルコサミン残基を有している分子を含んでいる組成物を提供するというものである。この組成物は、例えば薬物のような有用な医薬であってもよく、また食物や飲料のような飲食物用の組成物であってもよい。
【0048】
さらに、本発明は、機能性分子を提供するというものである。この機能性分子は、複数のモノマーユニットを含む分子であり、その少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基であるというものである。この機能性分子はコレステリル−α−D−グルコピラノシドの合成を抑制及び/又は阻害し、同様に前記コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び/又は阻害することができる。一つの形態として、前記機能性分子は、前記コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制又は阻害し、さらにはその細菌の形態異常を発現させる。そのような機能性分子は、主鎖と側鎖とを有する分子であってもよく、その側鎖が複数のモノマーユニットを含みその少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基である分子であってもよい。
【0049】
本発明の機能性分子は、本発明の前記の分子と、必要に応じて少なくとも一つの別な分子とで構成されている。一般に機能性分子は主鎖と側鎖とを有し、好ましくは本発明の分子は側鎖を有している。前記分子は主鎖に対してクラスター状に存在している。ここで用いられている前記「クラスター状」という用語は、ひとつの前記主鎖に対して、10以上の前記分子が末端で結合された状態をいう。前記主鎖の数と前記側鎖の数との比は特に制限はなく、目的に応じて適宜選択することができる。例えば、前記主鎖がタンパク質の場合、タンパク質を構成するアミノ酸残基の種類により比率は限定される。一般に、ひとつの主鎖に存する側鎖の数が多いほど、機能性分子の抗菌効果は高くなる。
【0050】
本発明の分子に結合される別な分子は、目的に応じて適宜選択することができる。前記その他の分子として、例えば、糖鎖、ポリペプチド、脂質、糖脂質、糖タンパク、ヌクレオチド、ポリ乳酸等の樹脂が挙げられ、これらは、1種単独で使用してもよく、2種以上を併用してもよい。一例として、前記その他の分子は、例えばCD43、CD34、及び/又はMuc−6を含むポリペプチドを有する分子である。前記分子と結合する前記ポリペプチド中のアミノ酸は、特に制限されない。前記アミノ酸として、例えば、フェニルアラニン(Phe)、ロイシン(Leu)、イソロイシン(Ile)、メチオニン(Met)、バリン(Val)、セリン(Ser)、プロリン(Pro)、トレオニン(Thr)、アラニン(Ala)、チロシン(Tyr)、ヒスチジン(His)、グルタミン(Gln)、アスパラギン(Asn)、リジン(Lys)、アスパラギン酸(Asp)、グルタミン酸(Glu)、システイン(Cys)、トリプトファン(Trp)、アルギニン(Arg)、グリシン(Gly)等が挙げられる。一般に、セリン及び/又はトレオニンが用いられる。
【0051】
本明細書に示したように、本発明の分子は、前記CGLの合成を抑制及び/又は阻害することができるものである。前記CGLの合成を抑制又は阻害することにより、前記CGLを必須とする細菌の増殖を抑制また阻害することができる。前記CGLを必須とする細菌として、例えば、ヘリコバクター種の細菌、特にヘリコバクター ピロリが挙げられる。前記ヘリコバクター種の細菌は、その細胞壁にα−コレステリルグルコシド(α−CGs)を含有する。このα−CGsの主要成分として、例えば、CGL、コレステリル−6−O−テトラデカノイル−α−D−グルコピラノシド(CAG)、及びコレステリル−6−O−ホスファチジル−α−D−グルコピラノシド(CPG)が挙げられる。このため、ヘリコバクター種の細菌は、CGLの合成が生存に必須となる細菌の一つである。
【0052】
本発明の機能性分子は、ヘリコバクター種の細菌に対して選択的及び特異的に抗菌活性を有し、ヒトの腸内の細菌層を損傷しない。さらに、本発明の機能性分子がGlcNAcα残基を含む場合、それは、ヒトの胃壁深部に存在する腺粘液細胞から分泌される腺粘液細胞型ムチン、又はムチンと類似の構造を有する他の機能性分子と同様なものである。したがって、本発明の機能性分子は、医薬、食物又は飲料として使用した場合でも副作用がなく、極めて安全性が高い点で有用である。
【0053】
機能性分子の主鎖が、例えばポリペプチドであってもよい。その一形態として、複数のモノマーユニットを含み、その少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基であって、主鎖に対してクラスター状に存在する分子が挙げられる。そのような機能性分子は、複数のモノマーユニットを含んでおり、その少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基であり主鎖に対してクラスター状に存在していると、前記コレステリル−α−D−グルコピラノシドの合成を抑制及び阻害することができ、同様に前記コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び阻害することができる。さらに前記機能性分子は、前記コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制又は阻害し、さらにはその細菌の形態異常を発現させる。
【0054】
このように、本発明の機能性分子は、コレステリル−α−D−グルコピラノシド(CGL)の合成を抑制及び/又は阻害することができ、及び/又はコレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び/又は阻害することができる。さらに本発明の機能性分子は、コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び/又は阻害し、また、その細菌の形態異常を発現させる。本発明の機能性分子は、例えばポリペプチドがCD43、CD34、又はMuc−6である分子が挙げられる。一つの形態として、主鎖に対する側鎖の数の比は10以上である。
【0055】
本発明の機能性分子は、胃潰瘍、十二指腸潰瘍、抗ヘリコバクター ピロリ剤、胃炎、及び/又は慢性胃炎の予防、治療、及び/又は改善のために、使用されるものであってもよい。また、機能性分子は、食物や飲料のような飲食物のために使用されるものであってもよい。これによって、このような機能性分子を含む組成物は、コレステリル−α−D−グルコピラノシドの合成を抑制及び/又は阻害することができ、コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び/又は阻害することができ、コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び/又は阻害することができ、及び/又はその細菌の形態異常を発現させることができる。また、本発明は、このような機能性分子を含むことにより前記のような抑制及び/又は阻害活性を奏する医薬を提供するというものである。この医薬組成物は、種々の形態で提供される。例えば、治療されるべき個体の胃及び/又は腸の粘膜層に前記医薬組成物を噴射又は塗布してもよい。また、別な形態として、前記医薬組成物をフィルム及び/又はシートに含浸又は含有させて、胃又は腸の粘膜層に付してもよい。本発明の医薬組成物は、コレステロールを分解するコレステロール分解酵素及びグルコースを分解するグルコース分解酵素の少なくともいずれかを有していてもよい。それにより、医薬組成物は前記したような抑制及び/又は阻害活性を有することができる。
【0056】
さらに、本発明は、機能性分子を含有する飲食物を提供するというものである。この機能性分子は、複数のモノマーユニットを含み、その少なくとも一つのモノマーユニットがN−アセチルグルコサミン残基であるというものである。このような飲食物は、前記と同様に、コレステリル−α−D−グルコピラノシドの合成を抑制及び/又は阻害することができ、コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖を抑制及び/又は阻害することができ、生体内に存在しコレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び/又は阻害することができ、その細菌の形態異常を発現させることができる。
【0057】
また、本発明は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する機能性分子の製造方法に関するものである。この方法は例えば、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを接触させ、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する機能性分子を生成させて製造することができるというものである。本発明の方法の一つの具体例によれば、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとが接触すると、機能性分子が製造される。この具体例の一つの形態として、前記ポリペプチドは、可溶性ポリペプチド(例えばCD43、CD34、又はMuc−6)であるというものである。また、別な形態として、前記ポリペプチドは哺乳類の母乳中に分泌されるムチンであるというものである。
【0058】
本発明の機能性分子の製造方法は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する機能性分子を生成させた後に、単離する工程を含んでいてもよい。それによって本発明は、この方法により得られ、前記したような抑制及び/又は阻害活性を有する機能性分子を提供することができるというものである。例えば、単離された機能性分子を例えば医薬として投与することにより、生体内に存在しコレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制及び/又は阻害することができ、その細菌の形態異常を発現させることができる。
【0059】
また、本発明は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型の機能性分子の製造方法に関するものである。この方法は例えば、α4GnTをエンコードする第一のポリヌクレオチドと、C2GnT−IとC1−β3GnTとの少なくともいずれかをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとを真核細胞内で発現させるというものである。この方法によれば、第一のポリヌクレオチドと、第二のポリヌクレオチドと、第三のポリヌクレオチドとの少なくともいずれかは、真核細胞に導入される外因性ポリヌクレオチドである。例えばこの方法は、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとが発現することによって、行われるというものである。その後、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを互いに接触させて、機能性分子を生成させることができる。前記で示したように、前記ポリペプチドは可溶性ポリペプチド(例えばCD43、CD34、又はMuc−6)であってもよい。組換え型機能性分子がつくられる真核細胞は、例えば哺乳類の細胞であってもよい。
【0060】
本発明によれば、前記組換え型の機能性分子の製造方法は、さらに、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを接触させ、その発現後に、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型の機能性分子を生成させるというものであってもよい。この方法はさらに、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型機能性分子を生成後に単離させる工程を含んでいてもよい。それにより、本発明の方法で製造され、単離された組換え型機能性分子が、提供される。この単離された組換え型機能性分子は、前記したような抑制及び/又は阻害活性を有する。
【0061】
また、本発明は、母乳を分泌する非ヒトのトランスジェニック哺乳類を提供するというものである。本発明の非ヒトのトランスジェニック哺乳類は、α4GnTをエンコードする第一の外因性ポリヌクレオチドと、C2GnT−IとC1−β3GnTとの少なくともいずれかをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとをゲノム中に有することができるというものである。このようなトランスジェニック哺乳類は、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを発現することができる。その後、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを、互いに接触させ、機能性分子を生成することができる。
【0062】
本発明の非ヒトのトランスジェニック哺乳類は、外因性の第一のポリヌクレオチド、第二のポリヌクレオチド、及び/又は第三のポリヌクレオチドを有していてもよい。一つの形態として、前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとの少なくともいずれかが、ゲノム中に導入され、哺乳類の乳腺遺伝子の5’制御配列及び3’制御配列の少なくともいずれかに相互作用して結合する。別な形態として、第三のポリヌクレオチドが、ゲノム中に導入され、少なくとも一つのO−グリコシル化部位を有するポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と相互作用して結合する。ポリヌクレオチドが相互作用して結合する5’制御配列は、例えば乳清酸性タンパク質(WAP)プロモータ、α−カゼインプロモータ、β−カゼインプロモータ、γ−カゼインプロモータ、α−ラクトアルブミンプロモータ、又はβ−ラクトグロブリンプロモータのようなプロモータを含んでいてもよい。さらに別な形態として、前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、単一の核酸分子を形成し、この核酸分子が非ヒトのトランスジェニック哺乳類のゲノム中に導入されて、乳腺遺伝子の5’制御配列と3’制御配列とのいずれかに相互作用して結合する。トランスジェニック哺乳類は、母乳を分泌するものであれば如何なる哺乳類であってもよく、例えばマウス、ラット、ウサギ、ウマ、ブタ、ヒツジ、ヤギ、又はウシが挙げられる。
【0063】
前記トランスジェニック哺乳類が前記第二の外因性のポリヌクレオチドを必要としているか否かは、本発明の機能性分子を産生する細胞が、自然の状態で内因性の前記C2GnT−Iを発現するか否かによって決定され、それが発現する場合には導入しなくてもよい。また、前記トランスジェニック哺乳類が前記第三の外因性のポリヌクレオチドを導入する必要があるか否かは、本発明の機能性分子を産生する細胞が、自然の状態で前記少なくとも一つのO−グリコシル化部位を有する内因性のポリペプチドを発現するか否かによって決定され、それが発現する場合には導入しなくてもよい。例えば、前記細胞が乳腺上皮細胞である場合には、この乳腺上皮細胞は前記C2GnT−I及び前記少なくとも一つのO−グリコシル化部位を有するポリペプチドを発現するので、前記乳腺上皮細胞を有するトランスジェニック哺乳類は、前記第二及び第三のポリヌクレオチドを導入しなくてもよい。
【0064】
本発明のトランスジェニック哺乳類は、前記少なくとも一つのO−グリコシル化部位を有するポリペプチドが、前記α4GnTと、前記C2GnT−I及びC1−β3GnTの少なくともいずれかとによってグルコシル化されて、母乳を分泌する雌の場合には、その母乳中に本発明の機能性分子を分泌させることができる。組換え型のポリペプチドを母乳中に分泌できる非ヒトのトランスジェニック哺乳類の生産方法は、公知の方法に準じて製造することができるというものである(例えば、米国特許 第6,344,596号;第6,548,735号;第6,222,094号;第5,962,648号;第5,891,698号;第5,850,000号の各明細書参照)。
【0065】
また、本発明は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型の機能性分子を生成させて製造する方法に関するものである。その方法は例えば、α4GnTをエンコードする第一のポリヌクレオチドを非ヒトの哺乳類の胚に導入し;導入した胚を宿主の雌哺乳類に移植し;宿主の雌哺乳類に子孫を出産させ;出産した子孫の内、雌の子孫に母乳の産生を行わせるというものであってもよい。この方法によれば、この子孫の体内で、α4GnTが発現する。一つの形態として、α4GnTは子孫の体内に存在するポリペプチドと少なくとも接触し、それによって機能性分子を生成する。他の形態として、第一のポリヌクレオチドは、哺乳類の乳腺に関する遺伝子の5’制御配列及び3’制御配列の少なくともいずれかと互いに相互作用して結合するように胚に導入される。一つの具体例として、前記組換え型の機能性分子の製造方法は、C2GnT−I及びC1−β3GnTの少なくともいずれかをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有するポリペプチドをエンコードする第三のポリヌクレオチドとを胚に導入する工程を含む。別な具体例として、前記の方法は、雌の子孫に母乳を産生させた後、その母乳を搾乳する工程を含み、さらに前記母乳から組換え型の機能性分子を単離する工程を含んでもよい。これによって本発明は、前記の方法で得られ、特に前記組換え型の機能性分子を含み、前記したような抑制及び/又は阻害活性を有する母乳を提供することができるというものである。
【0066】
本発明の機能性分子は前記したいずれの方法によっても製造することができるが、本発明に基づいて公知で通常の別な方法で製造されてもよい。本発明の機能性分子の製造方法として、三つの形態を例示する。なお、本発明の母乳は、本発明の機能性分子の説明を通して明らかにする。前記第一の形態は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する機能性分子の製造方法であって、α4GnTと、C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとを互いに接触させる工程を少なくとも含むものである。前記第二の形態は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型の機能性分子の製造方法であって、α4GnTをエンコードする第一のポリヌクレオチドと、C2GnT−I及びC1−β3GnTの少なくともいずれかをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有するポリペプチドをエンコードする第三のポリヌクレオチドとを真核細胞内で発現させる発現工程を少なくとも含むものである。前記第三の形態は、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する組換え型の機能性分子の製造方法であって、α4GnTをエンコードする第一のポリヌクレオチドを非ヒトの哺乳類の胚に導入し、導入した胚を宿主の雌哺乳類に移植し、子孫を出産させ、出産した子孫の内、雌の子孫に母乳の産生を行わせる工程を含むものである。前記α4GnT、前記C2GnT−I、及び前記C1−β3GnTについては前記の通りである。前記ポリペプチドは、少なくとも一つのO−グリコシル化部位を有するポリペプチド(例えばCD43、CD34、Muc−6、ムチン)であって、哺乳類の母乳中に分泌される。
【0067】
前記第一の形態では、前記α4GnTと、前記C2GnT−I及びC1−β3GnTの少なくともいずれかと、少なくとも一つのO−グリコシル化部位を有するポリペプチドとの他に、前記GlcNAcと糖とをさらに接触させてもよい。前記第二の形態では、前記発現工程の後、前記α4GnTと、前記C2GnT−I及びC1−β3GnTの少なくともいずれかと、前記少なくとも一つのO−グリコシル化部位を有するポリペプチドとを互いに接触させることにより前記機能性分子を製造することができる。前記GlcNAcと糖とをさらに接触させてもよい。また、前記第二の形態中、前記第一から第三のポリペプチドは、外因性であってもよく、内因性であってもよい。なお、本明細書中、「内因性」という用語は、ポリヌクレオチドに関して使用されている場合、ある特定の細胞内又は生物の細胞内で前記ポリヌクレオチドが自然状態で見出されることを意味し、「外因性」という用語は、ポリヌクレオチドに関して使用されている場合、ある特定の細胞内又は生物の細胞内で前記ポリヌクレオチドが自然状態では見出されないことを意味する。
【0068】
本発明の方法に、真核細胞を使用することができる。一般に、前記細胞は哺乳類の細胞であり、例えばチャイニーズハムスター卵巣細胞(CHO細胞)や乳腺細胞が挙げられる。前記真核細胞が乳腺細胞である場合には、前記C2GnT−Iと、前記少なくとも一つのO−グリコシル化部位を有するポリペプチドとを発現することができるため、前記α4GnTを外因性のものとして供給することにより前記機能性分子を産生することができる。また、前記真核細胞が前記C2GnT−Iを発現する場合には、前記α4GnTと、前記少なくとも一つのO−グリコシル化部位を有するポリペプチドとを外因性のものとして供給することにより前記機能性分子を産生することができる。
【0069】
前記第二の形態の製造方法の一例として、以下の方法がある。前記CD43の細胞外ドメイン全体を含むDNA断片(配列番号1におけるアミノ酸配列の20番目から254番目に相当する部分)をPCR法で増幅し、pSecTag2(インビトロジェン社製)のようなベクターにサブ・クローニングすることにより、Igκリーダー・ペプチドと、それに続くmycエピトープ、及び(His)6をエンコードするベクター(pSecTag2−sCD43)を作製する。次に、(1)C2GnT−Iをエンコードするポリヌクレオチド(cDNA)と、(2)α4GnTをエンコードするポリヌクレオチド(cDNA)と、(3)可溶性CD43をエンコードするポリヌクレオチド(cDNA)と、(4)ポリオーマ・ウイルス large T抗原をエンコードするポリヌクレオチド(cDNA)とを、夫々別々に前記pSecTag2−sCD43に組み込んだベクター4つを、LipofectAmine(登録商標、インビトロジェン社製)を用いて前記培養したCHO・Lec2細胞にトランスフェクトさせ、そのトランスフェクトさせたCHO・Lec2細胞内で、C2GnT−I、α4GnT、可溶性CD43、及びポリオーマ・ウイルス large T抗原を発現させ、接触させることにより、前記機能性分子として、末端GlcNAcα残基を含む可溶性CD43を発現させることができる。
【0070】
前記第三の形態の製造方法では、非ヒトの哺乳類の胚に、前記第一のポリヌクレオチドと、必要に応じて前記第二のポリヌクレオチド及び前記第三のポリヌクレオチドの少なくともいずれかを導入する。前記第一のポリヌクレオチドは、哺乳類の乳腺に関する遺伝子の5’制御配列及び3’制御配列の少なくともいずれかと互いに相互作用して胚に導入されてもよい。その場合、前記第二のポリヌクレオチドと前記第三のポリヌクレオチドとのいずれか又は両方が、哺乳類の乳腺に関連する遺伝子の5’制御配列及び3’制御配列の少なくともいずれかと互いに相互作用して胚に導入されてもよい。前記乳腺に関連する遺伝子は特に制限はないが、一般には乳腺特異的遺伝子である。前記5’制御配列は所望の制御要素であってもよく、前記したように一般にプロモータを有している。
【0071】
前記少なくとも一つのO−グリコシル化部位を有するポリペプチドをエンコードしている第三のポリヌクレオチドが、5’制御配列及び3’制御配列の少なくともいずれかの制御要素(例えば、乳腺特異的プロモータ等)に互いに相互作用して結合して、導入されると、前記制御要素は、前記ポリヌクレオチドに対して、通常の細胞内に関連付けられている別のポリヌクレオチド配列に対して行うように、その制御を行い、例えば、乳腺上皮細胞内で、前記エンコードされたポリペプチドが組織特異的に発現することができる。また、前記第一のポリヌクレオチド配列を前記第二のポリヌクレオチド配列に互いに相互作用させて結合させて導入し、二つ又はそれ以上のポリペプチドを前記相互作用させて導入したポリヌクレオチドから発現させることも可能である。また、必要に応じて、前記互いに相互作用させて導入したポリヌクレオチドからキメラの(組み合わされた)ポリペプチドを発現させることも可能である。前記キメラのポリペプチドは、融合ポリペプチドであってもよい。前記融合ポリペプチドとは、前記二つ又はそれ以上のエンコードされたポリペプチドが単一のポリペプチドに翻訳されたもの、つまり、それらがペプチド結合により共有結合したものである。
【0072】
前記第三のポリヌクレオチドは、少なくとも一つのO−グリコシル化部位を有するポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と互いに相互作用させて結合させて、胚に導入される。前記雌の子孫に母乳を産生させた後、母乳を搾乳してもよく、前記母乳から前記機能性分子を単離してもよい。前記第三の形態の製造方法では、前記雌の子孫の体内で前記第一のポリヌクレオチドから前記α4GnTが発現し、内因性又は外因性の前記少なくとも一つのO−グリコシル化部位を有するポリペプチドが前記C2GnT−I及び前記C1−β3GnTの少なくともいずれかによりグリコシル化され、前記機能性分子が産生される。前記機能性分子は、前記雌の子孫の母乳中に含まれ、該母乳を搾乳することにより効率的に産生することができる。
【0073】
前記したように、本発明の機能性分子は各種分野で使用可能である。前記機能性分子は、特に組成物、医薬及び/又は飲食物として供給される場合に、ヘリコバクター ピロリが原因である病気の治療薬、例えば、胃潰瘍治療薬、十二指腸潰瘍治療薬、胃炎治療薬、慢性胃炎緩和治療薬として好適に用いることができる。本発明の組成物、医薬及び飲食物は、本発明の分子及び機能性分子の少なくともいずれかを含有してなり、必要に応じて適宜選択したその他の成分を含有してなる。本発明の前記組成物、医薬、又は飲食物の全量中での前記分子及び機能性分子の少なくともいずれかの含有量は、特に制限はなく、目的に応じて適宜選択することができる。本発明の前記分子、前記機能性分子、前記医薬、又は前記組成物の全量に対する前記分子の含有量は、必要に応じて適宜選択することができるが、一般に、前記含有量が高ければ高いほど、抗菌効果が高くなる。
【0074】
種々の形状の前記組成物は、その他の成分を含んでいてもよい。前記その他の成分は特に制限はなく、目的に応じて適宜選択することができる。前記その他の成分として、例えば、賦形剤、滑沢剤、結合剤、水溶性高分子、塩基性無機塩、溶剤、溶解補助剤、懸濁化剤、等張化剤、緩衝剤、無痛化剤、防腐剤、抗酸化剤、着色剤、甘味剤、酸味剤、発泡剤、香料などが挙げられる。また、CGLの合成を効率的に抑制又は阻害する観点から、コレステロール分解酵素やグルコース分解酵素の添加も有用である。前記賦形剤として、例えば、乳糖、白糖、D−マンニトール、デンプン、コーンスターチ、結晶セルロース、軽質無水ケイ酸、酸化チタンなどが挙げられる。前記滑沢剤として、例えば、ステアリン酸マグネシウム、ショ糖脂肪酸エステル、ポリエチレングリコール、タルク、ステアリン酸などが挙げられる。前記結合剤として、例えば、ヒドロキシプロピルセルロース、ヒドロキシプロピルメチルセルロース、結晶セルロース、デンプン、ポリビニルピロリドン、アラビアゴム粉末、ゼラチン、プルラン、低置換度ヒドロキシプロピルセルロースなどが挙げられる。前記水溶性高分子として、例えば、ヒドロキシプロピルセルロース、ヒドロキシプロピルメチルセルロース、メチルセルロース、カルボキシメチルセルロースナトリウム等のセルロース誘導体、ポリビニルピロリドン、ポリアクリル酸ナトリウム、ポリビニルアルコール、アルギン酸ナトリウム、グアーガムなどが挙げられる。前記塩基性無機塩として、例えば、ナトリウム、カリウム、マグネシウム、カルシウムの塩基性無機塩が挙げられる。前記ナトリウムの塩基性無機塩として、例えば、炭酸ナトリウム、炭酸水素ナトリウム、リン酸水素二ナトリウムが挙げられる。前記カリウムの塩基性無機塩として、例えば、炭酸カリウム、炭酸水素カリウムが挙げられる。前記マグネシウムの塩基性無機塩として、例えば、重質炭酸マグネシウム、炭酸マグネシウム、酸化マグネシウム、水酸化マグネシウム、メタ珪酸アルミン酸マグネシウム、珪酸マグネシウム、アルミン酸マグネシウム、合成ヒドロタルサイト(Mg6Al2(OH)16・CO3・4H2O)、酸化アルミナ・マグネシウムが挙げられる。前記カルシウムの塩基性無機塩として、例えば、沈降炭酸カルシウム、水酸化カルシウムが挙げられる。前記溶剤として、例えば、注射用蒸留水、アルコール、プロピレングリコール、マクロゴール、ゴマ油、トウモロコシ油、オリーブ油などが挙げられる。前記溶解補助剤として、例えば、ポリエチレングリコール、プロピレングリコール、D−マンニトール、安息香酸ベンジル、エタノール、トリスアミノメタン、コレステロール、トリエタノールアミン、炭酸ナトリウム、クエン酸ナトリウムが挙げられる。前記懸濁化剤として、例えば、ステアリルトリエタノールアミン、ラウリル硫酸ナトリウム、ラウリルアミノプロピオン酸、レシチン、塩化ベンザルコニウム、塩化ベンゼトニウム、モノステアリン酸グリセリンのような界面活性剤;ポリビニルアルコール、ポリビニルピロリドン、カルボキシメチルセルロースナトリウム、メチルセルロース、ヒドロキシメチルセルロース、ヒドロキシエチルセルロース、ヒドロキシプロピルセルロースのような親水性高分子が挙げられる。前記等張化剤として、例えば、ブドウ糖、D−ソルビトール、塩化ナトリウム、グリセリン、D−マンニトールが挙げられる。前記緩衝剤として、例えば、リン酸塩、酢酸塩、炭酸塩、クエン酸塩が挙げられる。前記無痛化剤として、例えば、ベンジルアルコールが挙げられる。前記防腐剤として、例えば、パラオキシ安息香酸エステル類、クロロブタノール、ベンジルアルコール、フェネチルアルコール、デヒドロ酢酸、ソルビン酸が挙げられる。前記抗酸化剤として、例えば、亜硫酸塩、アスコルビン酸、α−トコフェロールが挙げられる。前記着色剤として、例えば、食用黄色5号、食用赤色2号、食用青色2号のような食用色素;食用レーキ色素;ベンガラが挙げられる。前記甘味料として、例えば、サッカリンナトリウム、グリチルリチン二カリウム、アスパルテーム、ステビア、ソーマチンが挙げられる。前記酸味料として、例えば、クエン酸(無水クエン酸)、酒石酸、リンゴ酸が挙げられる。前記発泡剤として、例えば、重曹が挙げられる。前記香料として、例えば、レモン、ライム、オレンジ、メントール、ストロベリーの抽出物が挙げられる。
【0075】
本発明の組成物及び/又は医薬は、投与の目的に合わせて、どのような形態、方法で投与されてもよい。その方法は、経口投与するものであってもよく、ヒトの胃及び腸の少なくともいずれかの粘膜層に前記組成物(又は機能性分子)を直接噴霧又は塗布するものであってもよい。また、本発明の組成物又は医薬を、フィルム又はシートに含浸させ、そのフィルム又はシートを胃及び/又は腸に付して使用してもよい。この投与方法は、胃又は腸の粘膜層表面で、噴霧又は塗布された前記組成物や医薬が流れ落ちることなく、一定時間保持されることにより、目的量の組成物又は医薬を投与でき、十分な抗菌効果を得ることができるため、有用である。また、前記使用方法は、内視鏡などの公知の医療器具を使用する場合に、好適に用いることができる。患部が胃である場合には、前記フィルム又はシートはキチン、糖、脂肪成分を含むことが好ましい。患部が腸である場合には、前記フィルム又はシートはキチンを含むことが好ましい。これにより、前記機能性分子は、胃や腸の中での胃液や腸液によって分解されることなく、長時間に亘って患部に接触することができる。
【0076】
本発明の組成物、医薬、及び飲食物は、十分な抗菌効果を有するが、抗菌効果をさらに向上させるために、抗生物質、生薬のような公知の胃潰瘍治療薬、十二指腸潰瘍治療薬、抗ヘリコバクター ピロリ薬、胃炎治療薬、及び慢性胃炎改善薬の少なくともいずれかと併用して使用してもよい。本発明の組成物、医薬及び飲食物は、本発明の機能性分子(又は分子)を含有するため副作用がなく、極めて安全性が高い点で有用であり、各種分野で使用可能である。本発明の組成物は例えば、ヘリコバクター ピロリに対する治療薬(例えば、胃潰瘍治療薬、十二指腸潰瘍治療薬、抗ヘリコバクター ピロリ薬、胃炎治療薬、慢性胃炎改善薬)に好適に用いることができ、本発明の医薬及び飲食物に特に好適に用いることができる。本発明の飲食物は、清涼飲料、炭酸飲料、栄養飲料、果実飲料、乳酸飲料、栄養補助食品、冷菓、麺類、菓子類、水産物、乳製品、加工食品、調味料、錠剤、カプセル剤などとして好適に使用することができる。
【0077】
このように、本発明は細菌の増殖抑制方法に関するものである。この方法は例えば、少なくとも一つのα1,4−N−アセチルグルコサミン残基を有する機能性分子に、CGLを発現する細菌を接触させ、細菌の増殖と、細菌の細胞壁の形成との少なくともいずれかを抑制又は阻害するものである。この方法によれば、前記機能性分子はコレステリル−α−D−グルコピラノシドの発現(即ち合成)を抑制又は阻害し、同様に、コレステリル−α−D−グルコピラノシドを必須とする細菌の増殖と、細菌の細胞壁の形成との少なくともいずれかを抑制又は阻害する。さらに前記機能性分子は、コレステリル−α−D−グルコピラノシドを必須とする細菌の運動性を抑制又は阻害し、さらにはその細菌の形態異常を発現させる。α1,4−N−アセチルグルコサミン残基を有する前記機能性分子は、UDP−Glc:ステロール糖転移酵素活性を抑制又は阻害することができる。増殖が抑制及び/又は阻害される細菌として、例えばヘリコバクター属の細菌(例えばヘリコバクター ピロリ)が挙げられる。
【0078】
さらに、本発明は胃潰瘍の治療方法を提供するというものである。この方法は例えば、ヘリコバクター種の細菌が個体に感染することによって起こる胃潰瘍の兆候又は症状を改善するために、α1,4−N−アセチルグルコサミン残基を有する機能性分子を投与するものである。この胃潰瘍治療方法では、前記機能性分子はコレステリル−α−D−グルコピラノシドの発現(合成)を抑制又は阻害し、同様に、コレステリル−α−D−グルコピラノシドを必須とするヘリコバクター種の細菌に感染することによって起こる胃潰瘍の兆候又は症状を改善する。前記ヘリコバクター種の細菌はヘリコバクター ピロリであってもよい。前記個体は例えばマウス、ラット、ウサギ、ウマ、ブタ、ヒツジ、ヤギ、ウシのような哺乳類であって、特にヒトであるとなおよい。
【0079】
胃潰瘍の治療方法では、胃腺粘液細胞型ムチンを含みα1,4−N−アセチルグルコサミン残基を有する機能性分子を利用することができる。前記機能性分子は種々の方法で投与される。投与方法として、例えば、経口投与、胃粘膜層への噴霧及び/又は塗布、前記機能性分子を含むフィルム及び/又はシートの貼付が挙げられる。一つの形態として、前記機能性分子は、前記のようにして生産された母乳を用いて投与される。
【0080】
さらに、本発明は胃潰瘍を予防する方法に関するものである。この方法は例えば、α1,4−N−アセチルグルコサミン残基を有する機能性分子の投与により、個体にヘリコバクター種の細菌が感染することによって起こる胃潰瘍の兆候又は症状を妨げ、前記機能性分子がコレステリル−α−D−グルコピラノシドの発現(合成)を抑制又は阻害し、コレステリル−α−D−グルコピラノシドを必須とするヘリコバクター種の細菌に感染することによって起こる胃潰瘍の兆候又は症状を予防するというものである。α1,4−N−アセチルグルコサミン残基を有する機能性分子は胃腺粘液細胞型ムチンを含んでいてもよい。また前記機能性分子は、前記した方法や公知の方法に準じて、投与される。
【0081】
さらに本発明は、胃癌を治療する方法に関するものである。この方法は例えば、α1,4−N−アセチルグルコサミン残基を有する機能性分子の投与により、個体にヘリコバクター種の細菌が感染することによって起こる胃癌の兆候又は症状を改善し、前記機能性分子がコレステリル−α−D−グルコピラノシドの発現(合成)を抑制又は阻害し、同様に、ヘリコバクター種の細菌に感染することによって起こる胃癌の兆候又は症状を改善するものである。
【0082】
本発明は、担体分子と相互作用して結合した少なくとも一つの末端α1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する非天然化合物を提供するというものである。「相互作用して結合」という用語は、本明細書中で、二つ以上の分子が一つの単位として働き、さらに一つ又は両方の分子、又はそれらの組み合わせによる機能効果を奏するような位置関係である二つ又はそれ以上の分子に関して広く使用される。例えば、担体分子と相互作用して結合したα1,4結合−GlcNAc残基とは、ヘリコバクター ピロリのようなヘリコバクター種の細菌の増殖を抑制又は阻害する効果を奏するように、α1,4結合−GlcNAc残基が位置しているということを意味している。そのように相互作用して結合したα1,4結合−GlcNAc残基は例えば、末端α1,4結合−GlcNAc残基を含むCD43分子やCD34分子で例示される。それらの化合物は他の担体分子へのα1,4結合−GlcNAc残基の相互作用した結合を確認する基準となる。
【0083】
二つ又はそれ以上のポリヌクレオチド配列もまた、相互作用して結合し得る。例えば、ポリペプチド(例えば担体ポリペプチド)をエンコードするポリヌクレオチドは、制御要素(例えば乳腺特異的プロモータ)に相互作用して結合することができ、前記制御要素は、通常細胞内に制御要素と関係するポリヌクレオチド配列をもたらすのと同様に、前記ポリヌクレオチドに対してその制御効果を与える(例えば、乳腺上皮細胞内で、前記エンコードされたポリペプチドが組織特異的に発現することができる)。また、前記第一のポリヌクレオチド配列を前記第二又はそれ以上のポリヌクレオチド配列に互いに相互作用させて導入し、二つ又はそれ以上のポリペプチドを前記相互作用させて導入したポリヌクレオチドから発現させることも可能である。また、必要に応じて、前記互いに相互作用させて導入したポリヌクレオチド配列からキメラのポリペプチドを発現させることも可能である。前記キメラのポリペプチドは、融合ポリペプチドであってもよい。前記融合ポリペプチドとは、前記二つ又はそれ以上のエンコードされたポリペプチドが単一のポリペプチドに翻訳されたもの、つまり、それらがペプチド結合により共有結合したものである。
【0084】
「担体分子」という用語は、ヘリコバクター増殖抑制活性を示す一つ又はそれ以上のα1,4結合−GlcNAc残基を相互作用させて導入できる分子に関して使われている。そのような担体分子として、ポリペプチド、ポリヌクレオチド、オリゴ糖、脂質、糖タンパク質、ヌクレオチドタンパク質、又は同様のものがある。担体分子は例えばO−グリカン(例えばコア2分岐型O−グリカン)で例示される。担体分子はさらにCD43やCD34のようなポリペプチドと相互作用して結合して、糖タンパク質を製造することができる。
【0085】
本明細書で示したように、一つ又はそれ以上のα1,4結合−GlcNAc残基を有する化合物は、コレステリル−α−D−グルコピラノシド(CGL)を発現する細菌の種々の機能、例えば細菌の増殖、細菌の運動性、細菌によるCGLの合成の一つ又はそれ以上の機能を抑制又は阻害することができる。「抑制又は阻害」という用語は、前記化合物の存在下で検査した機能が、前記化合物の非存在下での機能レベルと比較して、減少するか全く機能がなくなる状態となることを意味する。例えば一つ又はそれ以上のα1,4結合−GlcNAc残基を有する化合物は、その非存在下の場合と比べて、ヘリコバクター ピロリによるCGL合成の量を抑制又は阻害していることが本明細書に明示されている(実施例1参照)。「抑制又は阻害」という用語は、機能レベルが特別な分析によって検知できるレベル以下にまで減少すると認められるので、本明細書では通常一緒に使用している。例えば、前記化合物と接触したヘリコバクター ピロリによるCGL合成が完全に抑制されたかどうか、又は特別な分析による検知レベル以下のその合成レベルが維持されているかどうかという分析は確定できないかもしれない。しかし、制御レベルの減少は、前記化合物との接触によるものである。
【0086】
微好気性、グラム陰性の細菌であって、全世界の人口のおよそ半分のヒトの胃に感染しているヘリコバクター ピロリは、胃潰瘍の原因となり、また、胃の悪性腫瘍の原因となる(Marshall and Warren, Lancet 1311, 1984; Peek and Blaser,Nat.Rev.Cancer 2:28,2002;Du and Isaccson, Lancet Oncol.3:97, 2002参照)。感染しているほとんどのヒトは無症候群であるか、慢性胃炎のみを患い、患者の一部が消化性潰瘍、胃癌、悪性リンパ腫を患う(Cave,Semin.Gastrointest.Dis.12:196,2001参照、参考文献としてここに示す)。無症候群であるヒトは自己防御能を有しているのかもしれない。ヘリコバクター ピロリは、末端α1,4結合−GlcNAc残基を有するO−グリカンを発現する胃粘膜の深部でめったにみられないので、これらのO−グリカンがヘリコバクター ピロリの増殖に影響を及ぼすかどうか調べた。
【0087】
組織化学特性に基づき、胃粘素を二つに分類した(Otaら、Histochem.J.23:22,1991参照、参考文献としてここに示す)。一つは粘液細胞表面から分泌される表面粘液細胞型ムチンである。もう一つは粘液深部でみられ、粘液頚細胞、心腺細胞、幽門腺細胞のような腺粘液細胞から分泌される腺粘液細胞型ムチンである。ヘリコバクター ピロリに感染した胃粘液では、微生物が単独で表面粘液細胞型ムチンに接合しており、このムチンにヘリコバクター ピロリが付着するための糖質リガンドが存在する。ルイスbやシアリル−ルイスXとして知られる糖質構造はヘリコバクター付着特異リガンドBabAとSabAとを夫々供給する。両方の糖質リガンドは粘液細胞表面に専ら発現する(Ilverら、Science 279:373,1998; Mahdaviら、Science 297:573,2002)。対照的に、ヘリコバクター ピロリは、コア2分岐型O−グリカン(GlcNAca1→4Ga1β1→4GlcNAcβ1→6(GlcNAca1→4Galβ1→3)GalNAca→Ser/Thr)に結合した末端α1,4結合−GlcNAc残基を有する腺粘液細胞型ムチンを製造する胃粘液の深部に、コロニーを殆ど形成しない(Hidakaら、前出,2001)。腺粘液の損失に伴い洞萎縮すると、洞萎縮のない慢性胃炎に比べて、消化性潰瘍や胃癌の危険性が2〜3倍に高まってしまう(Sipponen and Hyvarinen, Scand.J.Gastroenterol.196(Suppl.3):3,1993参照、参考文献としてここに示す)。この観察と、ヘリコバクター ピロリ感染した大多数のヒトが無症候群である(Cave,前出,2001)という知見に基づき、深部ムチンに発現した末端α1,4結合−GlcNAc残基を有するO−グリカンがヘリコバクター ピロリ感染からヒトを保護できるかについて研究した。
【0088】
本明細書に示したように、末端α1,4結合−GlcNAc残基を有するO−グリカンはヘリコバクター ピロリに対する抗菌活性を有し、その細胞壁の主成分であるコレステリル−α−D−グルコピラノシドの生合成を抑制する。したがって、胃ムチン中のO−グリカン特有のサブセットは天然抗生物質としての機能を発揮し、ヘリコバクター ピロリ感染から宿主を保護する。これらのムチンがヘリコバクター ピロリ感染から宿主を保護することを確認できたことから、少なくとも一部がCGLの発現によって特徴づけられる他の細菌による感染だけでなく、ヘリコバクター種の細菌による感染を治療又は予防するのに有用な化合物を創り出すことができた(実施例1参照)。これによって、ヘリコバクター感染を治療するための組成物、その製造方法及びその使用方法が提供される。
【0089】
一つの具体例として、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質の製造方法を示す。それの一形態として、例えば、ポリペプチドのグリコシル化に適した条件下で、α4GnTと、C2GnT−Iと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドとを接触させ、このポリペプチドを、C2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化し、少なくとも一つのα1,4結合−N−アセチルグルコサミン残基を有する糖タンパク質を製造するものである。第二の形態として、前記C2GnT−Iとの組み合わせに代えて、C1−β3GnTのようなコア1伸長酵素と、C1−β3GnTに対する基質である担体ポリペプチドとを使用してもよい(Yahら、Cell 105:957-969,2001参照、参考文献としてここに示す)。この形態によれば、担体ポリペプチドはC1−β3GnTとα4GnTとによってグリコシル化され、少なくとも一つのα1,4結合−N−アセチルグルコサミン残基を有する糖タンパク質が製造される。前記担体ポリペプチドは、C2GnT−I(又はC1−β3GnT)とα4GnTとの活性により、一つ以上のα1,4結合−N−アセチルグルコサミン残基を結合できるものであればよく、例えば、可溶性ポリペプチド(例えば可溶性CD43ポリペプチド又は可溶性CD34ポリペプチド)が挙げられる。前記製造方法はさらに、前記糖タンパク質を単離する工程を含んでいてもよい。このようにして、本発明の方法で製造された糖タンパク質が単離された糖タンパク質として提供される。
【0090】
前記の通り、CGLを発現する細菌、特にヘリコバクター種の細菌の感染を治療又は予防するのに有用な組成成分は、担体分子と相互作用して結合した少なくとも一つの末端α1,4結合−GlcNAc残基を有する。このような組成成分はin vitro又はin vivoでつくられる。例えば、少なくとも一つのα1,4結合−GlcNAc残基を有するポリペプチド担体分子を含む糖タンパク質は、ポリペプチドのグリコシル化に適した条件下で、α1,4−N−アセチルグルコサミン転移酵素(α4GnT)と、コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)と、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドとを接触させて製造される。この方法によれば、前記ポリペプチドはC2GnT−Iとα4GnTとによってグリコシル化され、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質が製造される。
【0091】
このような糖タンパク質は、組み換えDNAの技術を用いて製造することもできる。例えば少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質は、α4GnTをエンコードする第一のポリヌクレオチドと、C2GnT−Iをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとを真核細胞内で発現させて製造できる。一般に、一つ、二つ、又は三つのポリヌクレオチドは真核細胞に対して内因性ではなく、一つ、二つ又は三つ全てのエンコードされたポリペプチド(α4GnT、C2GnT−I、担体ポリペプチド)は真核細胞に対して異型である。この方法で製造した組換え型の糖タンパク質は例えば、α1,4結合−GlcNAc残基を有するCD43(ロイコシアリン)分子、α1,4結合−GlcNAc残基を有するCD34分子で例示される(実施例1参照)。
【0092】
別な具体例として、本発明は、非ヒトの雌トランスジェニック哺乳類も範疇に含む非ヒトのトランスジェニック哺乳類に関するものである。この非ヒトのトランスジェニック哺乳類は、一つ又はそれ以上のα1,4結合−GlcNAc残基を有する組換え型糖タンパク質を製造する一つ又はそれ以上の導入遺伝子を含んでいる。前記糖タンパク質は非ヒトの雌トランスジェニック哺乳類の母乳中に分泌される。本発明の非ヒトのトランスジェニック哺乳類は、少なくともα4GnTをエンコードする第一の外因性ポリヌクレオチドと、必要に応じてC2GnT−Iをエンコードする第二の外因性ポリヌクレオチド及び/又は少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチドとをゲノム中に安定に組み込んで有している。本明細書に示したように、前記トランスジェニック哺乳類がさらに前記第二の外因性のポリヌクレオチド及び/又は前記第三の外因性のポリヌクレオチドを導入するか否かは、細胞、又は一つ又はそれ以上のα1,4結合−GlcNAc残基を有する組換え型糖タンパク質を産生する細胞が、自然の状態で内因性のC2GnT−I及び/又は所望の担体ポリペプチドを発現するか否かに依存する。例えば、乳腺上皮細胞は、C2GnT−Iとムチン型糖タンパク質とを発現する。よって、非ヒトのトランスジェニック哺乳類がその母乳中に一つ又はそれ以上のα1,4結合−GlcNAc残基を有する組換え型糖タンパク質を発現できる場合は、第二及び/又は第三のポリヌクレオチドを導入してもよいが、必ずしもその必要はない。したがって、種々の具体例では、本発明の非ヒトのトランスジェニック哺乳類は、一つの、又は二つの、又は三つの外因性ポリヌクレオチドを有している。一つの形態として、非ヒトのトランスジェニック哺乳類は、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合された第一の外因性ポリヌクレオチドを、ゲノム中に安定に組み込んで有している。第二の形態として、非ヒトのトランスジェニック哺乳類はさらに、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合された第二及び/又は第三のポリヌクレオチドを、夫々必要に応じてゲノム中に安定に組み込んで有している。さらに、第三のポリヌクレオチドは、共存するときには担体ポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と相互作用して結合している。
【0093】
さらに別な形態として、本発明は、少なくともα4GnTをエンコードする第一の外因性ポリヌクレオチドと、C2GnT−Iをエンコードし、又はC1−β3GnTをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとをゲノム中に安定に組み込んで有し、前記第一のポリヌクレオチドと前記第二のポリヌクレオチドと前記第三のポリヌクレオチドとの少なくともいずれかが外因性ポリヌクレオチドであり、前記第一のポリヌクレオチドと前記第二のポリヌクレオチドと前記第三のポリヌクレオチドとがプロモータを含む植物遺伝子の5’制御配列に相互作用して結合され、C2GnT−I又はC1−β3GnTとα4GnTと担体ポリペプチドとの発現により、前記担体ポリペプチドを、C2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化するトランスジェニック植物を提供するというものである。例えばエンハンサー及び/又は翻訳制御要素をさらに含む5’制御要素を構成する、相互作用して結合されたプロモータは、構成プロモータ、誘導プロモータ、又は組織特異プロモータであってもよく、例えば種、花、葉、及び/又は根のような植物の所望の細胞中のエンコードされたポリペプチド(例えばα4GnT)を転写するものである。さらに本発明は、前記トランスジェニック植物によって産出された種子、前記トランスジェニック植物から産生された穀類や小麦粉のような産物、前記トランスジェニック植物やその種、花、葉、又は根に由来する産物、豆乳やライスミルク等の乳液のような産物を提供するというものである。植物から前記のような産物を収穫して生産する方法は通常、公知の方法に準じて行われ、例えば豆乳を生産する方法(例えば、米国特許 第4,119,733号;第4,409,256号;第4,902,526号;第6,316,043号;第6,322,846号;第6,451,359号の各明細書参照、夫々参考文献としてここに示す)や、ライスミルクを生産する方法(例えば、米国特許 第4,744,992号;第4,894,242号;第5,609,895号;第6,599,552号の各明細書参照、夫々参考文献としてここに示す)がある。
【0094】
ポリヌクレオチドに関して使用されている「外因性」という用語は、ある特定の細胞内又は生物の細胞内で前記ポリヌクレオチドが自然状態では発見されないことを意味する。「異型」という用語も同様の意味で使用している。「異型」という用語はさらに、通常自然状態では発見されない二つ又はそれ以上のヌクレオチド配列を有するキメラのポリヌクレオチドに関して使用している。例えば、一つの遺伝子のコードされた配列が、第二の遺伝子のコードされた配列及び/又はプロモータ又は他の第二の制御配列(又は他の遺伝子)に結合(相互作用して結合)している場合、第一のコードされた配列は、第二のコードされた配列及び/又は制御配列に対して異型である。外因性のポリヌクレオチドは通常組換えDNA法を用いて細胞(例えば非ヒトの胚の細胞)中に導入される。本発明のトランスジェニック哺乳類中にC2GnT−Iと、α4GnTと、担体ポリペプチドとが発現することにより、担体ポリペプチドはC2GnT−Iとα4GnTとによってグリコシル化され、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質が製造される。製造された糖タンパク質は、雌の動物が生産する母乳中に分泌される。
【0095】
組換え型の糖タンパク質を含む組換え型のポリペプチドを母乳中に分泌可能な非ヒトのトランスジェニック哺乳類の生産方法は公知である(例えば、米国特許 第6,344,596号;第6,548,735号;第6,222,094号;第5,962,648号;第5,891,698号;第5,850,000号の各明細書参照、夫々参考文献としてここに示す)。そのような非ヒトのトランスジェニック哺乳類はどのような哺乳類でもよく、例えばマウス、ラット、ウサギ、ウマ、ブタ、ヒツジ、ヤギ、又はウシが挙げられる。一つの形態として、前記トランスジェニック哺乳類は非ヒトの雌トランスジェニック哺乳類であって、組換え型糖タンパク質が抗菌活性型で、前記非ヒトの雌トランスジェニック哺乳類によって生産された母乳中に分泌される。
【0096】
本発明は、非ヒトの雌トランスジェニック哺乳類の母乳中に、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造する方法を提供するというものである。「抗菌性」という用語は、ここでは少なくとも一つのα1,4結合−GlcNAc残基を有する化合物について使用しているが、前記化合物の非存在下での増殖レベル又は機能レベルと比較して、前記化合物がCGLを発現する細菌の増殖又は機能を抑制又は阻害することができることを意味している。ヘリコバクター ピロリのようなヘリコバクター種の細菌等の増殖又は機能が抑制又は阻害されていることは、本明細書に示した方法又は公知の方法を用いて確認できる。例えば化合物の抗菌活性は、その化合物の非存在下での運動性と比較して細菌の運動性が減少することを検出することにより、又は細菌増殖に適した培地で培養した細菌の数が減少することを検出することにより、確認できる。
【0097】
非ヒトの雌トランスジェニック哺乳類の母乳中に前記抗菌性の組換え型の糖タンパク質を製造する方法は次のようにして行われる。1)非ヒトの哺乳類の胚に、α4GnTをエンコードする第一のポリヌクレオチドと、C2GnT−Iをエンコードする第二のポリヌクレオチドと、少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとの少なくともいずれかを導入し、これら三つのポリヌクレオチドを全て前記胚に導入しないときには前記胚の細胞は対応する内因性のポリヌクレオチドを有し、前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合され、前記第三のポリヌクレオチドが、担体ポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列と相互作用して結合し、2)前記胚を、子孫を生産するような宿主の雌哺乳類に移植し、3)前記雌の子孫に、5’制御配列に相互作用して結合しゲノム中に安定に組み込まれた第一の外因性ポリヌクレオチドと第二の外因性ポリヌクレオチドと第三の外因性ポリヌクレオチドとを含む母乳の生産を行わせる。母乳は、その動物をつがいにして妊娠させ、乳腺上皮で生産させるか、あるいは母乳を生産するように非ヒトの雌トランスジェニック哺乳類のホルモン処理を行うことで、非ヒトの雌トランスジェニック哺乳類により産生される。前記非ヒトの雌トランスジェニック哺乳類中にC2GnT−Iと、α4GnTと、担体ポリペプチドとが発現することにより、担体ポリペプチドはC2GnT−Iとα4GnTとによってグリコシル化され、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質が製造される。製造された糖タンパク質は、非ヒトの雌トランスジェニック哺乳類が産生する母乳中に分泌される。
【0098】
少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を母乳中に分泌する非ヒトの雌トランスジェニック哺乳類の生産方法はさらに、雌トランスジェニック哺乳類から搾乳することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を含む母乳を得る工程を含んでいてもよい。さらに前記方法は、前記母乳から少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を単離することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を得る工程を含んでいてもよい。本発明によれば、本発明の方法で生産される非ヒトの雌トランスジェニック哺乳類、その雌トランスジェニック哺乳類から生産される母乳、及びその母乳から得られる単離された少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を提供することができる。
【0099】
本発明は、ヘリコバクター種の細菌に感染することによる胃潰瘍の治療又は予防方法を提供するというものである。一つの具体例として、本発明はヘリコバクター種の細菌が個体に感染することによる胃潰瘍の兆候又は症状を改善する方法であって、個体に少なくとも一つのα1,4結合−GlcNAc残基を有する化合物が投与され、この化合物が前記ヘリコバクター種の細菌に接触して前記ヘリコバクター種の細菌の増殖を抑制又は阻害することにより、胃潰瘍の兆候又は症状を改善する方法を提供することができるというものである。第二の具体例として、本発明は、胃潰瘍感受性の個体にヘリコバクター種の細菌が感染することによる胃潰瘍の予防方法であって、前記胃潰瘍感受性の個体に少なくとも一つのα1,4結合−GlcNAc残基を有する化合物が投与され、この化合物が前記ヘリコバクター種の細菌の増殖を妨げることによって、前記ヘリコバクター種の細菌の感染による胃潰瘍を予防する方法を提供することができるというものである。
【0100】
ここで使用される「改善」という用語は、胃潰瘍の兆候や症状が緩和することを意味する。胃潰瘍の兆候や症状は、兆候や条件を監視する方法と同様に、熟練した臨床医によく知られている。胃潰瘍の症状として、例えば、腹部の灼熱痛、食べたり制酸剤を服用したりすると改善する痛み、食事の2〜3時間後にひどくなる痛みがある。潰瘍が出血している場合は、鮮血を含む嘔吐や、黒やタール色の排便のような症状がみられる。本発明の方法は、胃潰瘍の改善、例えば、患者の痛みを緩和させるものである。同様に、例えば、液体バリウムの摂取後に胃腸上部にX線を照射し、減少した潰瘍の大きさを検出したり、抗ヘリコバクター ピロリ抗体やヘリコバクター ピロリに対する血液検査を行い、抗体又は細菌の夫々の数の減少や欠乏を測定したり、検便中の潜血の減少又は欠乏を検出したり、出血性胃潰瘍による貧血の減少を検出したり、胃粘膜を内視鏡で検査したりすることによって、胃潰瘍の臨床学的兆候が緩和することを、熟練した臨床医は熟知している。
【0101】
ヘリコバクター感染は、本明細書で示した方法や、他の公知の方法で検知できる。例えば、らせん状細胞を直接観察するワルティン−スターリー銀染色のスタイネル変異による組織学的検査では、電子顕微鏡でヘリコバクター種の細菌を検知できる。糞粒又は盲端塗抹標本の微生物学的培養もまた、特異培地と微好気性培養条件とを用いたヘリコバクター種の細菌の確認のために使用される。ポリメラーゼ鎖反応(PCR)法もまたヘリコバクター種の細菌を検出するために使用され、制限エンドヌクレアーゼは、種々のヘリコバクター種の細菌を区別することができる。
【0102】
本発明の方法によって治療される個体は、胃粘膜がヘリコバクター種の細菌に感染しているか、又は感染しやすい感受性の個体である。したがって、前記個体は例えばウマ、ブタ、ヤギ、イヌ、ネコ、フェレット、又は同様のもののような哺乳類である。一つの具体例として、前記個体はヒトである。同様に、ヘリコバクター種の細菌は胃潰瘍形成に関与するもので、例えばヒトの胃潰瘍形成に関与するヘリコバクター ピロリである。さらに、胃潰瘍に関係するヘリコバクター ピロリ、ヘリコバクター種の細菌として、例えば胃炎のチーターから単離したH.acinonychis、シリアンハムスターの胃腸組織から培養したH.aurati、犬の胃にみられるH.bizzozeronii、ブタオザル(Macaca nemestrina)の胃にみられるH.nemestrinae、鳥やブタの排泄物から単離したH.pametensis、家禽や胃腸炎のヒト患者から単離したH.pullorum(Microbiology,140:3441-3449,1994参照)、Hericobacter felisやHericobacter bizzozeroniiに関係する犬胃ヘリコバクター種の細菌であるH.salomonis(JP Euzeby,List of Bacterial Names with Standing in Nomenclature Genus Helicobacter,URL:http/www/bacterio.cict.fr/h/helicobacter.html参照)、ペットのフェレットに広く存在するH.mustelae、ヘリコバクター ピロリに類似しブタに感染するH.heilmanniiなどが挙げられる。さらに、本発明の組成物と方法とは、細胞壁の段階でコレステリル−α−D−グルコピラノシド(CGL;Glcα1→コレステロール)を発現する細菌による病的状態を改善するために使用される。
【0103】
前記で示したように、本発明で使用される化合物は少なくとも一つの末端α1,4結合−GlcNAc残基を有する化合物であればよい。また前記化合物は、単独で使用してもよく、潰瘍患者の治療に有用であると臨床医が認めた一種又はそれ以上の治療剤や栄養剤と併用してもよい。前記化合物として、例えば、少なくとも一つの末端α1,4結合−GlcNAc残基を有する組換え型の糖タンパク質(例えば末端α1,4結合−GlcNAc残基を有する可溶性CD43ポリペプチド)、O−グリカンを含み、一つ又はそれ以上の末端α1,4結合−GlcNAc残基を有する化合物、胃腺粘液細胞型ムチンを含み、胃腺粘液細胞から単離され得る化合物が挙げられる。
【0104】
個体に投与される化合物は、その種類に応じた特有な如何なる方法により処方されてもよい。また、前記化合物は適切であれば如何なる方法で投与されてもよく、特に経口投与や胃へ直接挿管する方法であってもよい。治療方法のためだけではなく予防方法のためにも、例えば前記で示した非ヒトの雌トランスジェニック哺乳類から得られた母乳のような前記化合物を含む母乳を、ヘリコバクター感染による胃潰瘍を有していたり胃潰瘍になり易かったりする個体が、飲用することによって、前記化合物の投与が行われる。
【0105】
前記化合物が母乳に含まれる以外の形態の場合、前記化合物は個体への投与に適した組成に処方される。このように、本発明は少なくとも一つの末端α1,4結合−GlcNAc残基を有する化合物を含み、ヘリコバクター感染による胃潰瘍の予防又は改善に有効で、製剤学的に許容される担体をさらに含むことができる組成物を提供するというものである。そのような化合物やその化合物を含む組成物は、胃潰瘍を有しているか、又は胃潰瘍になりやすい個体の治療に用いる医薬として有用である。
【0106】
製剤学的に許容される担体は公知であり、例えば水、生理食塩水、他の溶媒、グリコールやグリセロールで例示されるビヒクルを含む水溶液;オリーブオイルや注射可能な有機エステルのような油状物が挙げられる。製剤学的に許容される担体は、例えば少なくとも一つの末端α1,4結合−GlcNAc残基を有する化合物(例えば前記組換え型の糖タンパク質)の吸収を安定化又は増大させる、生理的に許容される化合物を、含んでいてもよい。そのような生理的に許容される化合物として、例えば、グルコース、スクロース、デキストランのような糖質;アスコルビン酸、グルタチオンのような抗酸化剤;キレート化剤;低分子量タンパク質;他の安定化剤や賦形剤が挙げられる。例えば前記治療効果のある化合物や予防効果のある化合物の物理化学的特性や、一般的な経口投与のみならず静脈注射、注射、挿管、又は他の公知の方法のような非経口投与という組成物投与方法に応じて、生理的に許容される化合物を含む製剤学的に許容される担体を、選択すべきであることは、よく知られている。前記のように、前記組成物は、例えば、診断試薬、栄養物質、毒素、制酸剤及び/又は栄養補助剤等の治療有効剤のような別な薬剤を、さらに含んでいてもよい。
【0107】
本発明の方法で投与される、末端α1,4結合−GlcNAc残基を有する化合物が、治療目的で投与されるのか予防目的で投与されるのかに応じて、その総量を、一度に又は長期にわたって複数回に分けて、投与してもよい。個体の病状を治療するための前記化合物の量は、個体の歳、全身の健康状態、投与経路、投与回数、ヘリコバクター感染の重症度のような多くの要因によるべきであることが知られている。専門医は、これらの要因から、第1相及び第2相臨床試験結果に基づき、必要に応じて、投与量を調整し、さらに組成物の形状とその投与経路、投与量と投与頻度を、決定する。
【0108】
経口投与用として、前記化合物は錠剤、溶液又は懸濁液として処方され、通常無毒で製剤学的に許容される担体を含む錠剤、ペレット剤、カプセル、座薬、溶液、エマルジョン、懸濁液、又は使用に適した形状に調剤されてもよい。前記担体として前記のもの以外に、グルコース、ラクトース、マンノース、アカシアゴム、ゼラチン、マンニトール、デンプン糊、ケイ酸マグネシウム、タルク、コーンスターチ、ケラチン、コロイダルシリカ、ポテトスターチ、尿素、中鎖長トリグリセリド、デキストラン、又は使用に適した他の担体を固体、半固体、又は液体の形状で用いてもよい。さらに、補助剤、安定化剤、農厚化剤、着色剤、香料を使用することもできる。例えば、トリウロースのような安定乾燥剤(例えば米国特許 第5,314,695号明細書参照)を使用してもよい。
【0109】
また、本発明は、CGLを発現する細菌の細胞壁形成を抑制又は阻害する方法を提供するものである。その方法は例えば、前記細菌を、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物に接触させ、CGL合成を抑制又は阻害して、この細菌の細胞壁の形成を抑制又は阻害するというものである。前記化合物は、CGL合成の抑制又は阻害によって細菌のUDP−Glc:ステロール糖転移酵素活性を抑制又は阻害するものであってもよいが、必ずしも必要ではない。また、本発明はCGLを発現する細菌の増殖を抑制又は阻害する方法を提供するというものである。その方法は例えば、前記細菌を、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物に接触させ、CGL合成を抑制又は阻害して、前記細菌の増殖を抑制又は阻害するというものである。この方法では、前記細菌と前記化合物とをin vitroで接触させてもよく、例えば培養中の細菌細胞に前記化合物を添加して行われる。また、前記細菌と前記化合物とをin vivoで接触させてもよく、例えばヘリコバクター種の細菌に感染した哺乳類、特にヘリコバクター ピロリに感染したヒトのような、細菌感染した個体の胃に、前記化合物を直接投与して行われる。
【0110】
以下、本発明の実施例について説明するが、本発明はこの実施例に限定されるものではない。
【0111】
10%ウシ胎児血清(FCS)を添加したα−MEM培地中で、CMP−シアル酸トランスポーター欠損の突然変異体チャイニーズハムスター卵巣・Lec2細胞(CHO・Lec2細胞)を培養した。CD43の細胞外ドメイン全体を含むDNA断片(配列番号1におけるアミノ酸配列の20番目から254番目に相当する部分)をPCR法で増幅し、pSecTag2(インビトロジェン社製)にサブ・クローニングすることにより、Igκリーダー・ペプチドと、それに続くmycエピトープ、及び(His)6をエンコードするベクター(pSecTag2−sCD43)を作製した。次に、(1)コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)をエンコードするcDNAと、(2)α1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードするcDNAと、(3)ポリオーマ・ウイルス large T抗原をエンコードするcDNAとを、夫々別々に前記pSecTag2−sCD43に組み込んだベクター4つを、LipofectAmine(登録商標、インビトロジェン社製)を用いて前記培養したCHO・Lec2細胞にトランスフェクトさせた。トランスフェクトさせたCHO・Lec2細胞内で、末端GlcNAcα残基を含む可溶性CD43(以下、「GlcNAcα−CD43」と称する)を発現させることによりGlcNAcα−CD43を調製した。
【0112】
前記調製したGlcNAcα−CD43の活性は、前記GlcNAcα残基に特異的なHIK1083抗体(胃ムチン検知用HIK1083ラテックス、関東化学社製)(例えば悪性腺腫と病変の新診断方法:新モノクローナル抗体(HIK1083;Clinica Chimica Acta 312(2001)231-233)存在下でのラテックス凝集テスト)を用いて市販のELISAキット(胃腺粘液細胞型ムチン測定用:関東化学社製)により測定し、発色反応にはELISA ELAST増幅システム(パーキン エルマー社製)を用いた。この結果、発色を確認することができ、GlcNAcα残基が発現していることが確認できた。さらに、市販の定量システム(BCAタンパク質検定キット、ピエルスバイオテクノロジー社製、商品番号23225,23227)を用いてタンパク質の定量を行った。その結果、前記GlcNAcα−CD43中の側鎖(糖鎖)の数は、主鎖(CD43)夫々に対して80であり、該糖鎖は、該主鎖に対してクラスター状に存在していることが判った。さらに別な市販の定量キット(糖タンパク 糖質評価キット、ピエルスバイオテクノロジー社製、商品番号23260)を用いて糖鎖の分析を行った。前記糖鎖は、GlcNAcα1→4Galβ1→4GlcNAcβ1→6(GlcNAcα→4Galβ1→3)GalNAcα→Ser/Thrで構成され、該糖鎖中の分岐数は1であり、モノマーユニット数は6であることが判った。前記「Ser/Thr」は、主鎖としてのCD43中、セリン(Ser)及びトレオニン(Thr)のいずれかに結合していることを表す。
【0113】
なお、前記トランスフェクトしたCHO・Lec2細胞は、ポリオーマ・ウイルス large T抗原により、前記CD43を効率的に合成することができた。また、前記α4GnTをエンコードするcDNAは、特開平2001−46077号公報に記載の方法に準じて調製した。α型N−アセチルグルコサミン残基を含まないCD43の調製は以下のようにして行った。コントロール測定用として、前記α4GnTのcDNAのみを組み込んでいないpcDNAIベクター(インビトロジェン社製)を用いた以外は、GlcNAcα−CD43の調製と同様にして、O−グリカンのコア2分岐にGlcNAcα残基を含まない可溶性CD43を調製した。なお、前記調製した該GlcNAcαを含まないCD43をGlcNAcα残基に特異的なラテックス粒子抗体を用いて縮合反応を行ったところ、縮合は見られなかった。この結果、該GlcNAcαを含まないCD43はGlcNAcα残基を含まないことがわかった。さらに、前記GlcNAcα残基を含まないCD43中の側鎖(糖鎖)は、Galβ1→4GlcNAcβ1→6(Galβ1→3)GalNAcα→Ser/Thrで構成されることがわかった。前記「Ser/Thr」は、主鎖としてのCD43中のセリン(Ser)及びトレオニン(Thr)のいずれかに結合していることを表す。
【0114】
前記調製したGlcNAcα残基を含む可溶性CD43、及びGlcNAcα残基を含まない可溶性CD43を用いて、以下のようにして細菌増殖性、細菌の運動性、及び細菌の形態異常発現性、並びに、GlcNAcα残基によるCGL生合成の抑制又は阻害性についての評価を行った。細菌増殖性は以下のとおりである。微量液体希釈法(microbroth dilution method)により、10%ウマ血清を添加したブルセラ培地(ベクトン デッキンソン マイクロバイオテクノロジー システムズ社製)でヘリコバクター ピロリ(ATCC43504)を前培養した。続いて、1×107cells/mlに希釈した前記ヘリコバクター ピロリと、図2Aに示す濃度のGlcNAcα−CD43とを96穴のプレートに投入し、35℃、15%CO2の環境下で4日間培養した。図2A中、「αGlcNAc(+)」は、前記培地中に前記GlcNAcα−CD43が含まれていることを意味し、「αGlcNAc(−)」は、前記培地中に前記GlcNAcα残基を含まない可溶性CD43が含まれていることを意味し、前記培地中の前記GlcNAcα残基を含まない可溶性CD43の濃度は、125.0mU/mlである。培養したヘリコバクター ピロリの増殖は、OD600nmの条件下、マイクロプレート分光光度計SPECTRAmax PLUS384(モレキュラーデバイス社製)を用いて測定した。結果を図2Aに示した。なお、前記ブルセラ培地に代えて、ミューラーヒントン培地(栄研化学社製)を使用した以外は、前記の方法と同様にして最小発育阻止濃度を測定したところ、同様の結果を得ることができた。
【0115】
この結果、図2Aに示すように、GlcNacα残基の存在の有無に関わらず、最初の2.5日間での増殖はごくわずかであるが、これはヘリコバクター ピロリの増殖の誘導期の特徴である。培養が3日を超えると増殖期に入り、GlcNAcα残基を含まない可溶性CD43含有の培地では、ヘリコバクター ピロリの急速な増殖が観られた。しかし、GlcNAcα−CD43と共に培養した場合には、ヘリコバクター ピロリの増殖が抑制又は阻害されていることがわかった。なお、1mUは、1μgのp−ニトロフェニル−N−アセチルグルコサミン(GlcNAcα−PNP)と当量である。細菌運動性は以下のようにして測定した。31.2mU/mlのGlcNAcα−CD43を含有するブルセラ培地を用いてヘリコバクター ピロリを3日間培養した。培養したヘリコバクター ピロリの運動性をレーザー共焦点顕微鏡LSM510 META(カルル ツァィス社製)を用いて、1秒間隔で低速度写真を撮影し、代表的なヘリコバクター ピロリを矢印で示した。結果を図2Bに示した。また、GlcNAcα残基を含まない可溶性CD43についても同様に撮影をした。結果を図2Bに示した。
【0116】
なお、図2B中、「αGlcNAcα(+)」は、前記培地中に前記GlcNAcα−CD43が含まれていることを意味し、「αGlcNAcα(−)」は、前記培地中に前記GlcNAcα残基を含まない可溶性CD43が含まれていることを意味し、濃度は、夫々31.2mU/mlである。また、図2B右下に記載した下線は、50μmを表し、図はすべて等倍率で撮影した低速度写真である。
【0117】
この結果、GlcNAcα−CD43を含む培地(αGlcNAcα(+))で培養された7個のヘリコバクター ピロリの平均速度は3.1±3.5μm/secであり、GlcNAcα残基を含まないCD43を含む培地(αGlcNAcα(−))で培養された7個のヘリコバクター ピロリの平均速度は21.1±2.6μm/secであり、t検定による有意性は、P<0.001であった。従って、GlcNAcα−CD43を含む培地では、ヘリコバクター ピロリの運動性が著しく減少することがわかった。
【0118】
細菌形態異常は以下のようにして検出した。GlcNAcα−CD43、又はGlcNAcα残基を含まない可溶性CD43を31.2mU/ml濃度で含有するブルセラ培地を用いてヘリコバクター ピロリを3日間培養させた。培養したヘリコバクター ピロリは、走査電子顕微鏡(日本電子社製、JSM−6360LV)を用いて加速電圧15kVの条件でSEM写真を撮影した。結果を図2Cに示した。なお、図2C中、「αGlcNAc(+)」は、前記培地中に前記GlcNAcα−CD43が含まれていることを意味し、「αGlcNAc(−)」は、前記培地中に前記GlcNAcα残基を含まない可溶性CD43が含まれていることを意味し、濃度は、夫々31.2mU/mlである。また、図2C右下に記載した下線は、1μmを表し、図はすべて等倍率で撮影したSEM写真である。この結果、GlcNAcα−CD43を含む培地(αGlcNAcα(+))で培養されたヘリコバクター ピロリは、伸び、狭窄、屈曲などの形態異常が観られるのに対し、GlcNAcα残基を含まないCD43を含む培地(αGlcNAcα(−))で培養されたヘリコバクター ピロリは、形態異常が観られなかった。
【0119】
CGL生合成の阻害性を測定した。GlcNAcα残基によるCGL生合成の阻害性の分析は、以下のようにして、(1)ヘリコバクター ピロリのin vivoでのCGL生合成、及び、(2)in vitroでのCGL生合成の2つの場合について行った。ヘリコバクター ピロリ生体内でのCGL生合成の阻害性を測定した。GlcNAcα−CD43存在下でのヘリコバクター ピロリによるCGLの合成を行った。コレステロール及びUDP−グルコースを用いたヘリコバクター ピロリによるCGLの合成をGlcNAcα−CD43の存在下で行い、以下の様にしてヘリコバクター ピロリ生体内(in vivo)で生合成したCGLについてのMALDI−TOF質量分析を行った。10%ウマ血清を添加したブルセラ培地(ベクトン デッキンソン マイクロバイオテクノロジー システムズ社製)でヘリコバクター ピロリ(ATCC43504)を前培養した後、希釈した前記ヘリコバクター ピロリ(1×107cells/ml)と、4.0mU/ml濃度に調整したGlcNAcα−CD43とを96穴のプレートに投入して、35℃で2日間培養した。培養したヘリコバクター ピロリを回収し、Ca2+、Mg2+を含まないPBSで3回洗浄した後、1mlの蒸留水で懸濁し、懸濁した液を毎分6,000回転で10分間遠心することにより菌体を得た。得られた菌体を、2mlのクロロホルム・メタノール混合液(クロロホルム:メタノールの質量比=2:1)を用いて4℃で一晩かけて抽出し、抽出液をろ過して、窒素気流で乾燥させた。乾燥して得られた試料を4mlのクロロホルム・メタノール混合液(クロロホルム:メタノールの質量比=2:1)に溶解させた。
【0120】
Folchらの方法に準じ、混合液に1mlの水を添加した。得られた混合液の下相を窒素気流で乾燥させ、1mlの0.5N水酸化ナトリウム・メタノール溶液を用いて、50℃で1時間の処理を行った。さらに処理液を6N塩酸水溶液を用いて中和した後、反応管に1mlの石油エーテルを添加した。次に、上相を除去した後、さらに2mlの石油エーテルを下相に添加した。下相を乾燥させた後、1mlのクロロホルム・メタノール・水(クロロホルム:メタノール:水の質量比=86:14:1)の混合液(TLP)に溶解させ、同じ反応管に0.5mlの別のクロロホルム・メタノール・水(クロロホルム:メタノール:水の質量比=3:48:47)混合液(TUP)を添加した。得られた混合液の下相を回収し、窒素気流で乾燥させ、50μlのクロロホルムに溶解させた。次に、この試料を1μl量り取り、1μlの2,5−ジヒドロキシ安息香酸、又は1μlのtrans−3−インドールアクリル酸を添加し、これをマトリックスとして用いた。前記資料の質量スペクトルは、DE MALDI−TOF MSのVoyager−DE STR Biospectrometry Workstation(PE アプライド バイオシステムズ社製)を用い、リフレクターモード、レーザー強度2300の条件で、陽イオンモード又は負イオンモードで測定した。なお、ヘリコバクター ピロリに内生発現するホスファチジン酸を内部基準とし、2点外部較正を行った。結果を図3Bに示した。
【0121】
また、コントロールとして、前記GlcNAcα−CD43を、GlcNAcα残基を含まないCD43に代えた以外は前記の方法と同様にしてCGLの質量スペクトルを測定した。結果を図3Aに示した。図3A及び3Bに示したように、前記GlcNAcα−CD43が存在する場合、及びGlcNAcα残基を含まないCD43が存在する場合のいずれの場合についても、m/zが571.6の位置にCGLのピークが検出された。しかし、前記GlcNAcα−CD43の存在下でのCGLのピークは、コントロールで測定したCGLのピークと比較して29.5%と大きく減少していた。この結果、前記GlcNAcα−CD43が、ヘリコバクター ピロリ生体内でのCGLの生合成を阻害していることがわかった。
【0122】
in vitro(試験管内)でのCGL生合成の阻害性を測定した。GlcNAcα−CD43存在下でのヘリコバクター ピロリによるCGLの合成を行った。コレステロール及びUDP−グルコースを用いて、超音波処理したヘリコバクター ピロリによるCGLの合成を行い、以下の様にしてin vitroで合成したCGLについてのMALDI−TOF質量分析を行った。10%ウマ血清を添加したブルセラ培地(ベクトン デッキンソン マイクロバイオテクノロジー システムズ社製)でヘリコバクター ピロリ(ATCC43504)を前培養した後、希釈した前記ヘリコバクター ピロリ(5×107cells/ml)を5%ウマ血清添加のブルセラ培地中で、35℃で2日間培養した。培養した2mlのヘリコバクター ピロリ(5×107cells/ml)を回収し、Ca2+、Mg2+を含まないPBSで3回洗浄した後、1mlの蒸留水で懸濁し、懸濁した液を毎分6,000回転で10分間遠心することにより脂質成分を得た。
【0123】
次に、100mMのトリス緩衝液(pH7.5)と、15質量%グリセロールと、5mMのDTTと、200μMのPefabloc(メルク社製)と、0.5mg/mlのリゾチームとを含有する反応緩衝液1ml中にヘリコバクター ピロリを懸濁し、20℃で5分間培養した。これらのサンプルを氷浴上で、超音波ウォーターバスにて30秒間の超音波処理を30秒間隔で10回行った。次に、80μlの超音波処理済ヘリコバクター ピロリと、8mMコレステロールエタノール溶液5μlと、7.2μMのUDP−Glc(UDP−グルコース)5μlと、トリトンCF−54(シグマケミカル社製)1μlと、反応緩衝液9μlとを含有する100μlの反応混合液を、30℃で3時間定温放置(インキュベート)した。その後、0.45% NaCl溶液900μl、及びクロロホルム・メタノール混合液(2:1)4mlを添加して反応を停止させた。
【0124】
次に、添加後の混合液の下相をろ過し、窒素気流下で乾燥させた。乾燥して得られた脂質試料を4mlのクロロホルム・メタノール混合液(クロロホルム:メタノールの質量比=2:1)に溶解させ、Folchらの方法に準じ、混合液に1mlの水を添加した。得られた混合液の下相を窒素気流で乾燥させ、1mlの0.5N水酸化ナトリウム・メタノール溶液を用いて、50℃で1時間の処理を行った。さらに処理液を6N塩酸水溶液を用いて中和した後、反応管に1mlの石油エーテルを添加した。次に、上相を除去した後、さらに2mlの石油エーテルを下相に添加した。乾燥させた下相は、1mlのクロロホルム・メタノール・水(クロロホルム:メタノール:水の質量比=86:14:1)の混合液(TLP)に溶解させ、同じ反応管に0.5mlの別のクロロホルム・メタノール・水(クロロホルム:メタノール:水の質量比=3:48:47)混合液(TUP)を添加した。得られた混合液の下相を回収し、窒素気流で乾燥させ、50μlのクロロホルムに溶解させた。
【0125】
次に、この試料を1μl量り取り、1μlの2,5−ジヒドロキシ安息香酸、又は1μlのtrans−3−インドールアクリル酸に添加し、これをマトリックスとして用いた。DE MALDI−TOF MSのVoyager−DE STR Biospectrometry Workstation(PE アプライド バイオシステムズ社製)を用い、リフレクターモード、レーザー強度2500の条件で、陽イオンモードでのCGLの質量スペクトルを測定した。なお、ヘリコバクター ピロリに内生発現するホスファチジン酸を内部基準とし、2点外部較正を行った。結果を図3Cに示した。
【0126】
次に、前記反応緩衝液に5.0mUのGlcNAcα−CD43を含有させた以外は前記の方法と同様にしてCGLの質量スペクトルを測定した。結果を図3Dに示した。さらに、前記反応緩衝液に5.0mUのGlcNAcα残基を含まないCD43を含有させた以外は前記の方法と同様にしてCGLの質量スペクトルを測定した。結果を図3Eに示した。
【0127】
UDP−Glcとコレステロールとから超音波処理したヘリコバクター ピロリにより合成された試料についてMALDI−TOF質量分析を行ったところ、図3Cから明らかなように、m/zが571.6の位置にCGLのピークが検出された。これは、下記スキームに示すようにヘリコバクター ピロリが、UDP−GlcをコレステロールのC3の位置に転移するUDP−Glc:ステロール糖転移酵素活性を有するためであると考えられる。
【化1】
【0128】
しかし、前記GlcNAcα−CD43の存在下で同様にして培養した場合には、図3Dに示したように、m/zが571.6の位置にCGLのピークが検出されなかった。また、前記GlcNAcα−CD43に代えて、前記GlcNAcα残基を含まないCD43の存在下で同様にして培養した場合、m/zが571.6の位置にCGLのピークが検出され、前記反応緩衝液に前記GlcNAcα−CD43及び前記GlcNAcα残基を含まないCD43を添加しない場合、CGLのピークに変化が観られなかった。これらの結果から、GlcNAcα残基が、最終的にCGLの生合成を阻害していることが証明された。
【0129】
α−CGsの生合成に関わる糖転移酵素の同定を含むヘリコバクター ピロリの細胞壁の生成機構は解明されていないが、GlcNAcα残基と、CGLに含まれるα結合型Glcとの構造の類似性を考慮すると、前記分析結果から、GlcNAcα残基と、UDP−Glcとが互いに競合し、UDP−Glc:ステロール糖転移酵素の供与体基質を互いに奪い合っているか、又は、GlcNAcα残基が最終生成物阻害機構によって直接的にUDP−Glc:ステロール糖転移酵素活性を阻害し、CGLの生合成を阻害しているものと考えられる。前記細菌増殖性、細菌の運動性、及び細菌の形態異常発現性、並びに、GlcNAcα残基によるCGL生合成の抑制乃至阻害性の評価結果から、α1,4アセチルグルコサミン残基を有する機能性分子を投与することにより、個体がヘリコバクター属の細菌に感染して引き起こされる胃潰瘍及び胃癌の兆候及び症状を改善及び予防できることがわかった。
【0130】
コレステロールの有無に対するヘリコバクター ピロリの生存状況、及びヘリコバクター ピロリによるCGLの合成について実験を行った。コレステロールの有無に対するヘリコバクター ピロリの生存状況を調べた。コレステロールの生合成に関わる遺伝子は、ヘリコバクター ピロリのゲノムデータからは発見されていない。このため、ヘリコバクター ピロリは、外因性のコレステロールの存在がなければCGLを合成できない可能性があることから以下の実験を行った。即ち、コレステロールを含まないHam's F−12培地でヘリコバクター ピロリ(ATCC43504)を35℃の標準嫌気条件にて5日間培養すると、ヘリコバクター ピロリの増殖は同条件でコレステロールを含むHam’s F−12培地に対して50%のレベルにとどまった。結果を表1に示した。
【0131】
コレステロールを含まない培地では、ヘリコバクター ピロリは運動性を示さず、異常伸長(形態異常)として図2C左に見られるようなヘリコバクター ピロリの伸びがみられた。このヘリコバクター ピロリにコレステロールの非存在下でさらに21日間培養すると、ヘリコバクター ピロリは完全に死滅した。一方、コレステロールを加えた培地でヘリコバクター ピロリを培養すると、ヘリコバクター ピロリは順調に増殖し、形態異常は観られなかった(表1)。これらの結果から、ヘリコバクター ピロリの生存には、コレステロールを用いたCGLの合成が必要であることがわかった。
【0132】
ヘリコバクター ピロリによるCGLの合成を行った。コレステロールの有無に対するヘリコバクター ピロリでのCGLの発現レベルを調べるため、ヘリコバクター ピロリの糖脂質部のCGLを含むα−CGsの発現レベルの変化を以下の様にして薄膜クロマトグラフィー(TLC)により分析した。コレステロールを含むブルセラ培地と、コレステロールを含まない培地で培養されたヘリコバクター ピロリについて、夫々の培地から1×108個のヘリコバクター ピロリを回収し、クロロホルム・メタノール混合液(クロロホルム:メタノールの質量比=2:1)を用いて室温で1時間かけて糖脂質部を抽出した。抽出液を窒素気流下で乾燥させ、クロロホルム・メタノール・水混合液(クロロホルム:メタノール:水の質量比=16:6:1)を用いて薄膜クロマトグラフィー(TLC)を行った。単離した糖脂質部は、オルシノール硫酸緩衝液により110℃で染色し、可視化した。結果を図4に示した。なお、図4中、レーン1は、コレステロールを含まないブルセラ培地で培養されたヘリコバクター ピロリから抽出した糖脂質部の分析結果であり、レーン2は、コレステロールを含むブルセラ培地で培養されたヘリコバクター ピロリから抽出した糖脂質部の分析結果である。この結果、コレステロールを含む培地で培養したヘリコバクター ピロリの場合には、ヘリコバクター ピロリの特徴である、CGLを含むα−CGsの典型的な隣接三重線が検出された(レーン2)。しかし、コレステロールを含まない培地で培養したヘリコバクター ピロリでは、α−CGs(CGL)は検出されなかった(レーン1)。
【0133】
α型N−アセチルグルコサミン残基を含むCD34の調製を行った。実施例1中のpSecTag2に代えてpcDNA3(インビトロジェン社製)を用いたこと、及び、可溶性CD43をエンコードするcDNAに代えて、可溶性CD34−IgGキメラをエンコードするpcDNA3−CD34−IgGを用いた以外は以下の実施例1と同様にして、GlcNAcα残基を含む可溶性CD34を調製した。調製したGlcNAcα残基を含む可溶性CD34について、実施例1と同様にして細菌増殖性、細菌の運動性、及び細菌の形態異常発現性の分析、及びGlcNAcα残基によるCGL生合成の阻害性についての評価を前記のようにして行ったところ、実施例1と同様の結果が得られた。
【0134】
α型N−アセチルグルコサミン残基を含むCD43を発現するAGS細胞の調製を行った。実施例1中のCHO・Lec2細胞を胃腺癌細胞(AGS細胞)に代えた以外は以下の実施例1と同様にして、GlcNAcα−CD43を安定的に発現するAGS−α4GnT細胞を調製した。α型N−アセチルグルコサミン残基を含まないCD43を発現するAGS細胞の調製を行った。コントロール測定用として、前記α4GnTのcDNAのみを組み込んでいないpcDNAIベクター(インビトロジェン社製)を用いた以外は、AGS−α4GnT細胞の調製と同様にして、GlcNAcα残基を含まないCD43を安定的に発現するMock−トランスフェクトされたAGS細胞を調製した。前記調製したAGS−α4GnT細胞、又はMock−トランスフェクトされたAGS細胞は、1×107cells/mlのヘリコバクター ピロリと共に24時間共同培養を行った。培養後の状態をノマルスキー顕微鏡を用いて写真撮影した。結果を図5下段に示した。
【0135】
次に、20%緩衝ホルマリンで固定した後、ヘリコバクター ピロリに対するウサギ・ポリクローナル抗血清(ダコ社製)、及びマウス・モノクローナルHIK1083抗体(関東化学社製)の混合物と共に前記AGS−α4GnT細胞、又はMock−トランスフェクトされたAGS細胞をインキュベートし、前記マウス・モノクローナルHIK1083抗体により蛍光染色した。二次抗体として、ローダミン標識の抗ウサギ免疫グロブリン(抗ヘリコバクター ピロリ抗体用)と、イソチオシアン酸フルオレセイン標識抗マウスIgM(HIK1083抗体用)を添加してスライドを作製した。該スライドは、Vectashield(ベクター ラボラトリーズ社製)で封入し、レーザー共焦点顕微鏡LSM510 META(カルル ツァィス社製)を用いて写真撮影した。結果を図5上段に示した。
【0136】
図5右上段に示した図は、AGS−α4GnT細胞を蛍光染色した写真である。白色で表された部分は、蛍光発色した前記HIK1083抗体を表し、HIK1083抗体が、前記AGS−α4GnT細胞を取り囲むように位置していることが確認できた。前記HIK1083抗体は、GlcNAcα残基に特異的な抗体であることから、前記AGS−α4GnT細胞によりGlcNAcα残基が発現していることがわかる。また、図5右下段に示すように、GlcNAcα残基が発現している前記AGS−α4GnT細胞では、細胞の損傷がみられなかった。
【0137】
これに対し、図5左上段に示した図は、Mock−トランスフェクトされたAGS細胞をAGS−α4GnT細胞と同様に蛍光染色した写真である。白色で表された部分は、ヘリコバクター ピロリを表し、前記Mock−トランスフェクトされたAGS細胞を取り囲むようにヘリコバクター ピロリが位置していること、及び、蛍光発色した前記HIK1083抗体が存在しないことからGlcNAcα残基が発現していないことが確認できた。また、図5左下段に示すように、GlcNAcα残基が発現していない前記AGS−α4GnT細胞では、顕著に細胞の損傷がみられた。
【0138】
公知の酵素合成法(例えば、Murata, Usui、Trends Glycsci. Glycotech., 12,161-174,2000)に従って、6残基から成る糖鎖、GlcNAcα1→4Galβ1→4GlcNAcβ1→6(GlcNAcα→4Galβ1→3)GalNAcαを得た。前記の方法(以下の実施例1参照)と同様にして細菌増殖性、細菌の運動性、及び細菌の形態異常発現性の分析、並びに、GlcNAcα残基によるCGL生合成の阻害性についての評価を行ったところ、前記したものと同様の結果が得られた(以下の実施例1参照)。
【0139】
これらの結果から、ヘリコバクター ピロリの増殖にはCGLが必須であり、GlcNAcα残基は、CGLの生合成を阻害することにより、ヘリコバクター ピロリに対する抗菌性を示すことが明らかとなった。
【実施例】
【0140】
(実施例1)末端α1,4結合−N−アセチルグルコサミンを有するO−グリカンが持つ抗菌活性
この実施例では、胃腺粘液細胞型ムチン中で製造されたO−グリカンが、ヘリコバクター ピロリ増殖の天然由来阻害剤として作用すること、また末端α1,4結合−N−アセチルグルコサミンを有する合成組成物が抗菌活性を有することを実証する。
【0141】
材料及び方法
細菌株
ヘリコバクター ピロリの標準菌株(ATCC43504)は、アメリカンタイプカルチャーコレクション(ATCC;バージニア州ナマサス)から購入し、本明細書に示す実験に使用した。さらに細胞増殖評価に使用した細菌株は、次の通りである:ATCCから購入したヘリコバクター ピロリ(ATCC43526)及びヘリコバクター ピロリの臨床的単離物(#749、#750、#751)、大腸菌(Escherichia coli)、緑膿菌(Pseudomonas aeruginosa)、肺炎桿菌(Klebsiella pneumoniae)、黄色ブドウ球菌(Staphylococcus aureus)、I−レンサ球菌(I-Streptococcus)、肺炎レンサ球菌(Streptococcus pneumoniae)。全ての臨床的単離物は、信州大学医学部附属病院臨床検査部で確認されたものである。
【0142】
α1,4結合−GlcNAc残基を有する可溶性CD43、及びα1,4結合−GlcNAc残基を有しない可溶性CD43の製造
10%ウシ胎児血清(FCS)を添加したα−MEM培地中で、CMP−シアル酸トランスポーター欠損の突然変異体チャイニーズハムスター卵巣・Lec2細胞(Deutscherら、Cell 39;295,1984参照、参考文献としてここに示す)を、培養した。CD43の細胞外ドメイン全体を含むDNA断片(アミノ酸残基の20番目から254番目)をPCR法で増幅し、IgPリーダー・ペプチドと、それに続くmycエピトープ、及び(His)6をエンコードするpSecTag2ベクター(インビトロジェン社製;カリフォルニア州カールズバッド)にサブ・クローニングした(pSecTag2−sCD43)。次に、C2GnT−IをエンコードするcDNA(Bierhuizen、Fukuda、Proc.Natl.Acad.Sci.,USA89:9326,1992参照、参考文献としてここに示す)と、α4GnTをエンコードするcDNA(Nakayamaら、Proc.Natl.Acad.Sci.,USA96:8991,1999参照、参考文献としてここに示す)と、可溶性CD43をエンコードするcDNAと、ポリオーマ・ウイルス large T抗原をエンコードするcDNAとを、LipofectAmine(登録商標、インビトロジェン社製)を用いて前記Lec2細胞にトランスフェクトさせた。ポリオーマ・ウイルス large T抗原により、前記トランスフェクトされたLec2細胞は、効率よく組換え型タンパク質を産生した(Bierhuizen,Fukuda、前出,1992)。
【0143】
前記α4GnTのcDNAを組み込んでいないpcDNAIベクター(インビトロジェン社製)を用いて、O−グリカンのコア2分岐に末端α1,4結合−GlcNAc残基を有しない可溶性CD43を調製し、コントロール測定用とした。一週間後、CENTRIPREP YM30フィルター(ミリポア社製:マサチューセッツ州ベッドフォード)を用いて、10%ウシ胎児血清(FCS)を添加したα−MEM培地中に投入した可溶性CD43を濃縮し、測定用とした。前記調製した末端α−1,4−結合−GlcNAcを有する可溶性CD43の活性は、前記GlcNAcα残基に特異的なHIK1083抗体(関東化学社製、Ishiharaら、Biochem.J.318:409,1995参照、参考文献としてここに示す)を用いて市販のELISAキットにより測定し、発色反応にはELISA ELAST増幅システム(パーキン
エルマー社製;マサチューセッツ州ボストン)を用いた。この分析において、1ユニットは製造元の指示に従いp−ニトロフェニル−N−アセチル−α−グルコサミニド(GlcNAcα−PNP)の1mg(2.9Tmol)に相当する免疫反応力と定義した。これらの組換え体のタンパク質濃度はBCAタンパク質分析(ピエルス社製;イリノイ州ロックフォード)を用いて測定した。同様に、pSecTag2−sCD43に代えて可溶性CD34−IgGキメラをエンコードするpcDNA3−CD34−IgG(Yehら、Cell 104:957,2001参照、参考文献としてここに示す)を用いて、末端α1,4結合−GlcNAc残基を有する可溶性CD34タンパク質と、末端α1,4結合−GlcNAc残基を有しない可溶性CD34タンパク質とを調製した。
【0144】
細胞増殖試験
微量液体希釈法(Piccolominiら、J.Clin.Microbiol.35:1842,1997参照、参考文献としてここに示す)により、10%ウマ血清を添加したブルセラ培地(ベクトン ディッキンソン マイクロバイオロジー システムズ社製;メリーランド州スパークス)でヘリコバクター ピロリを前培養した。続いて96穴のプレート中、末端α1,4結合−GlcNAc残基を有する可溶性CD34又は末端α1,4結合−GlcNAc残基を有しない可溶性CD34の種々の量を含む、5%ウマ血清を添加したブルセラ培地で、1×107cells/mlに希釈した細菌を35℃、15%CO2の環境下で4日間培養した。細菌増殖は、OD600nmの条件下、マイクロプレート分光光度計SPECTRAmax PLUS384(モレキュラーデバイス社製;カリフォルニア州サニーベール)を用いて測定した。また、前記ブルセラ培地に代えて、微量液体希釈法のミューラーヒントン培地(栄研化学社製、東京都)を使用した以外は、前記の方法と同様にして最小発育阻止濃度を測定した。
【0145】
細胞運動性試験
31.2mU/mlの末端α1,4結合−GlcNAc残基を有する可溶性CD43又は同じタンパク質濃度の末端α1,4結合−GlcNAc残基を有しないCD43含有するブルセラ培地を用いてヘリコバクター ピロリを3日間培養した。培養したヘリコバクター ピロリの細胞運動性を、レーザー共焦点顕微鏡LSM510 META(カルル ツァィス社製;ドイツ国ジェナ)を用いて、1秒間隔で低速度写真を撮影し、評価した。
【0146】
走査電子顕微鏡
31.2mU/mlの末端α1,4結合−GlcNAc残基を有する可溶性CD43又は同じタンパク質濃度の末端α1,4結合−GlcNAc残基を有しないCD43を含有するブルセラ培地を用いてヘリコバクター ピロリを3日間培養した。培養したヘリコバクター ピロリを走査電子顕微鏡JSM−6360LV(日本電子社製;東京都)を用いて加速電圧15kVの条件で観察し、走査電子顕微鏡写真を撮影した。
【0147】
質量分析によるCGLの検出
10%ウマ血清を添加したブルセラ培地でヘリコバクター ピロリを前培養した後、5%ウマ血清と、4.0mU/mlの末端α1,4結合−GlcNAc残基を有する可溶性CD43又は同じタンパク質濃度の末端α1,4結合−GlcNAc残基を有しないCD43とを含有するブルセラ培地を用いて、1×107cells/mlの微生物を35℃で2日間培養した。培養したヘリコバクター ピロリを回収し、Ca2+、Mg2+を含まないリン酸緩衝生理食塩水(PBS)で3回洗浄した後、1mlの蒸留水で再度懸濁した。毎分6,000回転で10分間遠心した後、得られた脂質画分を、2mlのクロロホルム・メタノール混合液(2:1)を用いて4℃で一晩かけて抽出し、抽出液をろ紙でろ過して、窒素気流で乾燥させた。得られた脂質試料を4mlのクロロホルム・メタノール混合液(2:1)に溶解させ、Folchらの方法(J.Biol.Chem.226:497,1957参照、参考文献としてここに示す)に準じ、混合液に1mlの水を添加した。得られた混合液の下相を試験管中、窒素気流で乾燥させ、1mlの0.5N水酸化ナトリウム・メタノール溶液により50℃で1時間の処理し、処理液を6N塩酸水溶液で中和した後、1mlの石油エーテルを添加した。
【0148】
上相を除去した後、さらに2mlの石油エーテルを下相に添加した。下相を乾燥させた後、1mlのクロロホルム・メタノール・水(86:14:1)の混合液(TLP)に溶解させ、同じ反応管に0.5mlの別のクロロホルム・メタノール・水(3:48:47)混合液(TUP)を添加した。得られた混合液の下相を回収し、窒素気流で乾燥させ、50μlのクロロホルムに溶解させた。次に、この試料の1μlに、1μlの2,5−ジヒドロキシ安息香酸、又は1μlのtrans−3−インドールアクリル酸を添加し、これをマトリックスとして用いた。陽イオンモード又は負イオンモードでのCGLの質量スペクトルは、DE マトリックス支援イオン化−飛行時間型質量分析(DE MALDI−TOF MS)装置のVoyager−DE STR Biospectrometry Workstation(PE アプライド バイオシステムズ社製;カリフォルニア州フォスターシティ)を用い、前記のようにリフレクターモード、レーザー強度2300の条件で測定した(Taketomi,Sugiyama、Meth,Enzymol.312:80,2000参照、参考文献としてここに示す)。ヘリコバクター ピロリ中に内生発現するホスファチジン酸(Inamotoら、前出,1993)を内部基準とし、2点外部較正を行った。
【0149】
超音波処理したヘリコバクター ピロリによるin vitroでのCGL合成
in vitroでのCGL合成は、前記で示したように変異体を用いて分析した(Warneckeら、J.Biol.Chem.274:13048,1999参照、参考文献としてここに示す)。すなわち、10%ウマ血清を添加したブルセラ培地でヘリコバクター ピロリを前培養した。次に、5×107cells/mlに希釈した細菌を、5%ウマ血清を添加したブルセラ培地を用いて35℃で2日間培養した。培養した2mlのヘリコバクター ピロリ(5×108cells/mlを毎分6,000回転で10分間遠心し、Ca2+、Mg2+を含まないPBSで3回洗浄した。その後100mMのトリス緩衝液(pH7.5)と、15%グリセロールと、5mMのDTTと、200TMのPEFABLOCプロテアーゼ阻害剤(メルク社製;ドイツ国ダルムスタッド)と、0.5mg/mlのリゾチームとを含有する反応緩衝液1ml中にヘリコバクター ピロリを懸濁させ、20℃で5分間培養した。これらのサンプルを氷浴上で超音波ウォーターバスにて30秒間隔で30秒間の超音波処理を10回行った。
【0150】
次に、超音波処理したヘリコバクター ピロリの80μlと、8mMコレステロールのエタノール溶液5μlと、7.2μMのUDP−グルコース5μlと、1μlのCF−54と、9μlの反応緩衝液又は5.0mUの末端α1,4結合−GlcNAc残基を有する可溶性CD43又は同じタンパク質濃度の末端α1,4結合−GlcNAc残基を有しないCD43を含有する同様の緩衝液とを含有する100μlの反応混合液を、30℃で3時間培養した。その後、0.45%のNaCl溶液900μl、及びクロロホルム・メタノール混合液(2:1)の4mlを添加して反応を停止させた。その混合液の下相をろ過し、窒素気流下で乾燥させた。その後、乾燥して得られた試料を前記で示した方法と同様にして連続処理した。陽イオンモードでのCGLの質量スペクトルは、DE MALDI−TOF MS装置のVoyager−DE STR Biospectrometry Workstation(アプライド バイオシステムズ社製)を用い、リフレクターモード、レーザー強度2500の条件で、前記の方法(Taketomi, Sugiyama、前出,2000)で測定した。なお2点外部較正を行った。
【0151】
コレステロール存在下、又はコレステロール非存在下でのヘリコバクター ピロリの培養
10%ウマ血清を添加したブルセラ培地でヘリコバクター ピロリを前培養した。次に、5%ウマ血清を添加したブルセラ培地で、1×108cells/mlに希釈した細菌を40時間培養し、その後、標準培養条件下で、0.005%コレステロールを含み又はコレステロールを含まないウマ血清除去ブルセラ培地(TLC分析用)と、0.005%コレステロールを含んでいるか又はコレステロールを含まないF−12培地(細胞増殖分析用)とで、1×107cellのヘリコバクター ピロリを5〜21日間培養した。F−12培地での培養では、培地体積の半分量を注意深く除去し、除去した培地と同体積量の新鮮な培地で置き換えるという作業を2〜3日おきに21日間繰り返した。
【0152】
薄層クロマトグラフィー(TLC)
1×108cellのヘリコバクター ピロリからクロロホルム−メタノール混合液(2:1)を用いて室温で1時間かけて糖脂質の分画を抽出し、窒素気流で乾燥させ、クロロホルム−メタノール−水(16:6:1)混合液を用いて(8)で示したようにしてTLCを行った。単離した糖脂質は、オルシノール含有硫酸緩衝液を用い110℃で、目視できるように発色させた。
【0153】
ヘリコバクター ピロリと、末端α1,4結合−GlcNAcを安定に発現するAGS細胞との共生培養
末端α1,4結合−GlcNAcを安定に発現する胃腺癌AGS−α4GnT細胞を、前記の方法(Nakayamaら、前出,1999)で製造した。AGS−I4GnT細胞とMock−トランスフェクトしたAGS細胞とを、1×107cells/mlのヘリコバクター ピロリ(ATCC43504)と共に24時間、共培養した。20%の緩衝ホルマリンで固定した後、ヘリコバクター ピロリに対するウサギ・ポリクローナル抗血清(ダコ社製;デンマーク国グロストラップ)と、マウス・モノクローナルHIK1083抗体(関東化学社製;Ishiharaら、前出,1996)との混合物と共に前記細胞をインキュベートした。二次抗体として、ローダミン標識の抗ウサギ免疫グロブリン(抗ヘリコバクター ピロリ抗体用)と、イソチオシアン酸フルオレセイン標識抗マウスIgM(HIK1083抗体用)とを使用した。スライドは、VECTORSHIELDシールド溶液(ベクターラボラトリーズ社製;カリフォルニア州ブーリンガム)で封入し、レーザー共焦点顕微鏡LSM510 META(カルル ツァィス社製)を用いて観察した。
【0154】
結果
本明細書に示したように、培養時のヘリコバクター ピロリに対する効果を調べるために、α1,4結合−GlcNAc残基を有するムチン型糖タンパク質を調製した。前記のように、CD43(ロイコシアリン)は前記O−グリカンの優先的コアタンパク質としての役割を果たす。ヘリコバクター ピロリのO−グリカンのサブセットの活性を調べるために、α1,4−N−アセチルグルコサミン転移酵素(α4GnT;Nakayamaら、前出,1999)のcDNAと、コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I;Bierhuizen,
Fukuda、前出,1992)のcDNAと、可溶性CD43のcDNAとでトランスフェクトすることによって、末端α1,4結合−GlcNAc残基を有するO−グリカンを含む組換え型の可溶性CD43を、チャイニーズハムスター卵巣細胞(CHO)に発生させた。コントロールとして、α4GnTcDNAを使用しないこと以外は前記と同様の方法でCHOにトランスフェクトして、末端α1,4結合−GlcNAc残基を有しない可溶性CD43を製造した。
【0155】
異なる量の組換え型可溶性CD43を含む培地でヘリコバクター ピロリ(ATCC43504)をインキュベートした。最初の2.5日間での微生物の増殖は、末端α1,4結合−GlcNAc残基の有無に関係なくごくわずかであり(図2A)、これはヘリコバクター ピロリの増殖の遅滞期の特徴である。培養が3日を超えると増殖期に入り、微生物は急速に増殖した。しかし、末端α1,4結合−GlcNAc残基を有する可溶性CD43と共に細菌をインキュベートした場合は、ヘリコバクター ピロリの増殖速度が劇的に抑制された。この抑制効果は、可溶性CD43の濃度が62.5mU/mlを超えたときに顕著となった(1mUは、1μgのp−ニトロフェニル−N−アセチルグルコサミン(GlcNAcα−PNP)と当量である;Warneckeら、J.Biol.Chem.274:13048,1999参照、参考文献としてここに示す)。さらに、α1,4結合−GlcNAcを発現する可溶性CD43が31.2mU/mlで存在する培地で培養したヘリコバクター ピロリの運動性が、著しく抑制されることが、低速度写真からわかった(図2B)。また、電子顕微鏡写真は、ヘリコバクター ピロリの伸び、分節狭窄、屈曲のような顕著な形態異常を示した(図2C)。
【0156】
末端α1,4結合−GlcNAc残基を有する可溶性CD43の阻害効果について、15.6mU/mlから125.0mU/mlの最小阻害濃度で、別の標準菌株であるATCC43526や、3種の臨床的単離物(Warneckeら、前出,1999)を含む種々のヘリコバクター ピロリの菌株を用いて観察した。前記とは対照的に、末端α1,4結合−GlcNAc残基を有しない可溶性CD43と共にヘリコバクター ピロリをインキュベートした場合は、増殖阻害効果も形態異常もみられなかった(図2)。これらの結果は、末端α1,4結合−GlcNAc残基を有するO−グリカンが、抗菌剤と同様に、ヘリコバクター ピロリの増殖を抑制することを示している。また、ヘリコバクター ピロリに対する同様の阻害効果は末端α1,4結合−GlcNAc残基を結合した別のムチン様タンパク質であるCD34(Horiiら、Helicobacter 7:39,2002参照、参考文献としてここに示す)を使用した場合にも観察された。この結果は、骨格タンパク質ではなく末端α1,4結合−GlcNAc残基が、ヘリコバクター ピロリに対する増殖阻害活性のために必須であることを示している。
【0157】
末端α1,4結合−GlcNAc残基を有するO−グリカンによって引き起こされるヘリコバクター ピロリの形態異常は、β−ラクタマーゼ阻害剤のような抗生物質によって引き起こされる形態異常と類似しており、多くの細菌株の細胞壁の主成分であるペプチドグリカンの生合成を妨げる(Finalyら、J.Antimicrob.Chemother.52:18,2003参照、参考文献としてここに示す;Horiiら、前出,2002)。このように、これらのO−グリカンがヘリコバクター ピロリの細胞壁生合成を阻害する作用について調べた。ヘリコバクター種の細菌の細胞壁はα−コレステリルグルコシド(α−CGs)を特徴的に有しているが、その細胞壁の主成分はコレステリル−α−D−グルコピラノシド(CGL;Glcα1→コレステロール)、コレステリル−6−O−テトラデカノイル−α−D−グルコピラノシド(CAG)、コレステリル−6−O−ホスファチジル−α−D−グルコピラノシド(CPG;Hiraiら、J.Bacteriol.177:5327,1995;Haqueら、J.Bacteriol.178:2065,1996参照、夫々参考文献としてここに示す)である。
【0158】
末端α1,4結合−GlcNAc残基を有するO−グリカンがヘリコバクター ピロリのCGL生合成を妨げるかどうかを調べるために、O−グリカン存在下で培養したヘリコバクター ピロリ中のCGLの発現レベルを分析した。末端α1,4結合−GlcNAc残基を有しないO−グリカンを含む培地で培養したヘリコバクター ピロリの細胞壁成分を質量分析したところ、CGLに起因する明確なシグナルが現れた(図3A)。対照的に、末端α1,4結合−GlcNAc残基を有するO−グリカンを含む培地で培養したヘリコバクター ピロリサンプルの並行分析では、CGLは顕著な抑制を示し、末端α1,4結合−GlcNAc残基を有しないコントロール群の29.5%に減少した(図3B)。これらの結果は、末端α1,4結合−GlcNAc残基を有するO−グリカンが、ヘリコバクター ピロリのin vivoでのCGLの生合成を阻害していることを示している。
【0159】
α−CGsの生合成に関わる糖転移酵素の同定をはじめとしてヘリコバクター ピロリ細胞壁の根本的な生成機構はあまり解明されていないが、CGLはおそらく、グルコース(Glc)をUDP−GlcからコレステロールのC3位に転移させるUDP−Glc:ステロール糖転移酵素によって形成されていると考えられる。超音波処理されたヘリコバクター ピロリのUDP−Glc:ステロール糖転移酵素活性の発現を調べ、その発現が見られた場合は、それらの活性が末端α1,4結合−GlcNAc残基を有するO−グリカンによって阻害されるかどうかが分かる。コレステロールとUDP−Glcとを超音波処理したヘリコバクター ピロリと共にインキュベートし、生成物を質量分析した。多くの量のCGLが検出された(図3C)。これは、超音波処理したヘリコバクター ピロリが酵素学的にUDP−Glc:ステロール糖転移酵素活性を発現したことを示している。
【0160】
末端α1,4結合−GlcNAc残基を有する可溶性CD43を前記評価分析にかけたところ、CGL生成は抑制された(図3D)が、末端α1,4結合−GlcNAc残基を有しない可溶性CD43を前記評価分析にかけても何の効果もみられなかった(図3E)。腺粘液細胞型ムチン中に存在するα−結合型GlcNAcと、CGL中に存在するα−結合型Glcとの構造的類似性を考慮すると、これらの結果は、α1,4結合−GlcNAc残基がUDP−Glc:ステロール糖転移酵素のドナー基質のためにUDP−Glcと互いに競合し、又はα1,4結合−GlcNAc残基が最終生成物を阻害するメカニズムによって直接的にUDP−Glc:ステロール糖転移酵素活性を阻害する(Nakayamaら、J.Biol.Chem.271:3684,1996参照、参考文献としてここに示す)ことによって、CGLの生合成を減少させていることを示している。
【0161】
コレステロールはCGLの前駆体である。しかし、コレステロールの生合成に関係する遺伝子はヘリコバクター ピロリのゲノムデータベースにはみられない(Tombら、Nature 388:539,1997;Maraisら、Microbiol.Mol.Biol.Rev.63:642,1999)。よって、ヘリコバクター ピロリは外因性のコレステロールが存在しない状態ではCGLを合成できないかもしれない。コレステロールが存在しない培地でヘリコバクター ピロリを5日間培養した場合、微生物の細胞増殖はコレステロール存在下で培養した場合の約50%のレベルにとどまった(以下の表1参照)。コレステロール非存在下での培養において、運動性微生物は検出されず、形態異常、特にヘリコバクター ピロリの伸長がはっきりとみられた(図2Cで示したものと同様)。コレステロールが存在しない培地でヘリコバクター ピロリをさらに21日目まで培養すると、微生物は完全に死滅した。対照的に、コレステロールを補充した培地でヘリコバクター ピロリを培養した場合、細胞はよく増殖し形態異常の兆候はみられなかった(表1)。これらの結果は、ヘリコバクター ピロリの生存のために、外因的に補給されたコレステロールによるCGLの合成が必須であることを、示している。
【0162】
【表1】
【0163】
ヘリコバクター ピロリの標準菌株(ATCC43504)の1×107cells/mlを35℃の標準嫌気性条件下で、0.005%のコレステロールを補充したHam's F−12培地で5日間、及びコレステロールを含まない培地で21日間、夫々インキュベートした。CFUはコロニー形成ユニットである。
【0164】
コレステロールを含み、又は含まない培地で培養したヘリコバクター ピロリのCGL発現レベルを調べるために、ヘリコバクター ピロリの糖脂質分画のCGLを含むα−CGsの発現過程をTLCにより分析した。コレステロール存在下で培養したヘリコバクター ピロリでは、ヘリコバクター ピロリの特徴であるCGLを含むα−CGsの典型的な三本線が検出された(図4、レーン2)。しかしコレステロール非存在下で培養したヘリコバクター ピロリでは、α−CGsは検出されなかった(図4、レーン1)。これらの結果を合わせると、これらの結果は、CGLがヘリコバクター ピロリ細胞増殖に必須であること、また末端α1,4結合−GlcNAc残基を有するO−グリカンはCGLの生合成を阻害するためヘリコバクター ピロリに対して抗菌活性を有することを、示している。さらに、末端α1,4結合−GlcNAc残基を有するO−グリカンの抗菌活性は、CGLを発現する細菌の菌株に限定されるのかもしれない。実際に、ヘリコバクター ピロリ以外の菌株やCGL欠損菌株(例えば大腸菌、緑膿菌、肺炎桿菌、黄色ブドウ球菌、α−レンサ球菌、肺炎レンサ球菌)を末端α1,4結合−GlcNAc残基を有する可溶性CD43存在下でインキュベートしても、抗菌効果はみられなかった。
【0165】
前記の結果は、末端α1,4結合−GlcNAc残基を有するO−グリカンを発現する粘液細胞は、ヘリコバクター ピロリ感染から自分自身を保護していることを示している。この効果を確認するため、胃腺癌AGS−α4GnT細胞をα4GnT cDNAに安定にトランスフェクトし(Nakayamaら、前出,1999)、ヘリコバクター ピロリ感染による細胞の損傷を調べた。ヘリコバクター ピロリと共にインキュベートしてMock−トランスフェクトされた細胞は、細胞の扁平や萎縮のような顕著な荒廃を示した(図5矢印参照)。対照的に、ヘリコバクター ピロリと共に培養したAGS−α4GnT細胞ではごくわずかな損傷しか観察されなかった(図5)。これらの結果から、末端α1,4結合−GlcNAc残基を有するムチン型O−グリカンを発現する細胞はヘリコバクター ピロリ感染に対して抗力を示すことがわかった。
【0166】
グリカン鎖は、細胞間相互作用における細胞表面レセプターのリガンドとして(Lowe,Cell 104:809,2001;Akamaら、Science 295:124,2002;Perilloら、Nature 378:736,1995)、またレセプターや粘着タンパク質のモジュレーターとして(Moloneyら、Nature 406:369,2000;Demitriouら、Nature 409:733,2001;Nakayamaら、Proc.Natl.Acad.Sci.,USA 92:7031,1995)、種々の役割を果たしている。これらの結果は、哺乳類のグリカンが天然抗菌剤としても機能することを示している。末端α1,4結合−GlcNAc残基を有するO−グリカンはヒト胃腺粘液細胞により生成される。そのためこれらの結果は、副作用を引き起こすことなくヒトへのヘリコバクター ピロリ感染を予防及び治療する新規な医薬品を開発できるという、根拠となるものである。
【0167】
前記の実施例を参照しながら本発明を説明したが、本発明の本質や技術的範囲には、その改良や差異も包含される。したがって、本発明は、特許請求の範囲により定められるものである。
【図面の簡単な説明】
【0168】
【図1】図1は、通常のヒトの胃壁の断面概略図であって、GOCTSで表面のムチンを染色し、PCS染色剤により染色されたα1,4結合−GlcNAc残基を示すものである。
【0169】
【図2】図2Aから図2Cは、末端α1,4結合−GlcNAc残基を有するO−グリカンがヘリコバクター ピロリの増殖と運動性とを抑制することを示すものである。
【0170】
図2Aは末端α1,4結合−GlcNAcを有する(αGlcNAc(+))可溶性CD43、及び末端α1,4結合−GlcNAcを有しない(αGlcNAc(−))可溶性CD43を用いて培養したヘリコバクター ピロリの増殖曲線を示す。αGlcNAc(+)の1ミリユニット(mU)が、p−ニトロフェニル−N−アセチル−α−グルコサミン(GlcNAcα−PNP)の1Tgと等量の免疫反応性として定義される。αGlcNAc(−)のタンパク質濃度は、αGlcNAc(+)の125.0mU/mlでのタンパク質濃度と同等であった。
【0171】
図2Bは、末端α1,4結合−GlcNAcを有する(αGlcNAc(+))可溶性CD43の31.2mU/ml、又は同タンパク質濃度の末端α1,4結合−GlcNAcを有しない(αGlcNAc(−))可溶性CD43を用いて、3日間培養したヘリコバクター ピロリの運動性を1秒間隔で経時的に記録した図を示す。典型的なヘリコバクター ピロリを矢印で示している。αGlcNAc(+)の存在下で培養された7個のヘリコバクター ピロリの平均速度は3.1±3.5μm/sec(平均±標準偏差)であり、αGlcNAc(−)の存在下で培養された7個のヘリコバクター ピロリの平均速度は21.2±2.6μm/secであり、スチューデントt検定による有意性は、P<0.001であった。スケールバーは50μmである。
【0172】
図2Cは、末端α1,4結合−GlcNAcを有する(αGlcNAc(+))可溶性CD43の31.2mU/ml、又は同タンパク質濃度の末端α1,4結合−GlcNAcを有しない(αGlcNAc(−))可溶性CD43を用いて、3日間培養したヘリコバクター ピロリの走査電子顕微鏡写真である。αGlcNAc(+)で培養されたヘリコバクター ピロリは伸び、分節狭窄、屈曲などの形態異常がみられた。写真はすべて等倍率で撮影したものであり、スケールバーは1Tmを示す。
【0173】
【図3】図3Aから図3Eは、末端α1,4結合−GlcNAc残基を有する可溶性CD43が、ヘリコバクター ピロリのin vivoでのコレステリル−α−D−グルコピラノシド(CGL)の生合成(図3A、図3B)と、同様なヘリコバクター ピロリのinvitroでの生合成(図3C〜図3E)とを阻害することを、マトリックス支援レーザー脱離イオン化飛行時間型質量分析(MALDI−TOF−mass)で測定した結果を示す図である。
【0174】
図3Aは、末端α1,4結合−GlcNAc残基を有しない可溶性CD43を用いて培養したヘリコバクター ピロリの分画液中、m/z571.6の位置にナトリウム付加CGL{M+Na}+が検出されたことを示す(矢印)ものである。
【0175】
図3Bは、末端α1,4結合−GlcNAc残基を有する可溶性CD43の4.0mU/mlを用いて培養した試料のCGLを同様に測定した結果を示す。内部標準としてホスファチジン酸(Inamotoら、J.Clin.Gastroenterol. 17:S136,1993参照、参考文献としてここに示す)に対するCGLの相対量は対照実験(図2A矢印)の29.5%まで顕著に減少している。2回測定した平均値を示してある。
【0176】
図3Cは、超音波処理したヘリコバクター ピロリによってUDP−グルコース及びコレステロールから生合成された生成物のMALDI−TOF−massの測定結果を示す。CGL{M+Na}+がm/z571.6の位置が現れている。
【0177】
図3Dは、末端α1,4結合−GlcNAc残基を有する可溶性CD43の50.0mU/mlの存在下で、超音波処理したヘリコバクター ピロリによってUDP−グルコース及びコレステロールから生合成された生成物の質量分析の結果を示す。この条件下では、CGLのピークは検知できなかった。
【0178】
図3Eは、末端α1,4結合−GlcNAc残基を有しない可溶性CD43の存在下で、超音波処理したヘリコバクター ピロリによってUDP−グルコース及びコレステロールから生合成された生成物の質量分析の結果を示す。この条件下でCGLは合成されていた。
【0179】
【図4】図4は、コレステロール非存在下で培養したヘリコバクター ピロリにCGLが存在しないことを示す図である。コレステロールを含まないブルセラ培地(レーン1)又は0.005%のコレステロールを含むブルセラ培地(レーン2)で培養したヘリコバクター ピロリの1×108個から抽出した全糖脂質を薄層クロマトグラフィーで分析した。使用した溶媒系はクロロホルム・メタノール・水混合液(16:6:1)であり、糖脂質は、オルシノール硫酸緩衝液を用いて呈色させた。CAGはコレステリル−6−O−テトラデカノイル−α−D−グルコピラノシド;CGLはコレステリル−α−D−グルコピラノシド;CPGはコレステリル−6−O−ホスファチジル−α−D−グルコピラノシドである。
【0180】
【図5】図5は、末端α1,4結合−GlcNAc残基を有するO−グリカンが、胃腺癌細胞をヘリコバクター ピロリによる細胞損傷から保護していることを示すものである。AGS細胞に付着した多くのヘリコバクター ピロリはベクターのみにトランスフェクトされ、細胞の平面化や縮小のような細胞損傷がみられた(矢印)。一方、AGS−α4GnT細胞に不完全に付着したヘリコバクター ピロリはα4GnTに安定にトランスフェクトされ、細胞損傷はみられなかった。抗ヘリコバクター ピロリ抗体(赤)と、末端α1,4結合−GlcNAc残基特異的のHIK1083抗体(緑)(Ishiharaら、Biochem.J.318:409,1996参照、参考文献としてここに示す)とによりAGS細胞を二重の免疫組織化学的により示す図である。同じ領域のノマルスキー顕微鏡写真を図5下段に示す。スケールバーは50Tmである。
【特許請求の範囲】
【請求項1】
担体分子に相互作用して結合した少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有することを特徴とする天然非存在の化合物。
【請求項2】
前記担体分子が、O−グリカンを有していることを特徴とする請求項1に記載の化合物。
【請求項3】
前記O−グリカンが、コア2分岐型のO−グリカンであることを特徴とする請求項2に記載の化合物。
【請求項4】
前記担体分子が、ポリペプチドを有していることを特徴とする請求項1に記載の化合物。
【請求項5】
前記ポリペプチドが、少なくとも一つのO−グリコシル化部位を有することを特徴とする請求項4に記載の化合物。
【請求項6】
前記ポリペプチドが、O−グリカンを有していることを特徴とする請求項4に記載の化合物。
【請求項7】
前記O−グリカンが、コア2分岐型のO−グリカンであることを特徴とする請求項5に記載の化合物。
【請求項8】
前記担体分子が、ムチン様ポリペプチドを有していることを特徴とする請求項1に記載の化合物。
【請求項9】
前記ムチン様ポリペプチドが、CD43(ロイコシアリン)、CD34、又はMuc6を有していることを特徴とする請求項8に記載の化合物。
【請求項10】
前記O−グリカンが、コア1型のO−グリカンであることを特徴とする請求項5に記載の化合物。
【請求項11】
複数のα1,4結合−GlcNAc残基を有することを特徴とする請求項1に記載の化合物。
【請求項12】
合成したオリゴ糖を有することを特徴とする請求項1に記載の化合物。
【請求項13】
少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する糖タンパク質を製造する方法であって、ポリペプチドのグリコシル化条件下で、
α1,4−N−アセチルグルコサミン転移酵素(α4GnT)と、
コア2型のβ1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)又はコア1型の伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)と、
少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドとを接触させ、
該ポリペプチドを、C2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化して、少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する糖タンパク質を製造する方法。
【請求項14】
前記担体ポリペプチドが、可溶性ポリペプチドであることを特徴とする請求項13に記載の方法。
【請求項15】
前記担体ポリペプチドが、CD43(ロイコシアリン)、CD34、又はMuc6であることを特徴とする請求項13に記載の方法。
【請求項16】
前記担体ポリペプチドが、哺乳類の母乳中に分泌されるムチン型糖タンパク質であることを特徴とする請求項13に記載の方法。
【請求項17】
前記ムチン型糖タンパク質を含む母乳に、α4GnTと、必要に応じてC2GnT−I又はC1−β3GnTとを接触させることを特徴とする請求項16に記載の方法。
【請求項18】
前記糖タンパク質が、複数のα1,4結合−GlcNAc残基を有することを特徴とする請求項13に記載の方法。
【請求項19】
前記糖タンパク質を単離することを特徴とする請求項13に記載の方法。
【請求項20】
請求項13に記載の方法により製造されることを特徴とする、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質。
【請求項21】
請求項17に記載の方法により製造される母乳であって、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質を含むことを特徴とする母乳。
【請求項22】
少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する組換え型の糖タンパク質を製造する方法であって、
α1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードする第一のポリヌクレオチドと、
コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)又はコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二のポリヌクレオチドと、
少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとを
真核細胞内で発現させる工程を含み、
該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、及び該第三のポリヌクレオチドとの少なくともいずれかが、真核細胞に導入された外因性ポリヌクレオチドであり、
該担体ポリペプチドを、該C2GnT−Iと該α4GnTとによって、又は該C1−β3GnTと該α4GnTとによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を製造する方法。
【請求項23】
前記担体ポリペプチドが、可溶性ポリペプチドであることを特徴とする請求項22に記載の方法。
【請求項24】
前記担体ポリペプチドが、CD43(ロイコシアリン)、CD34、又はMuc6であることを特徴とする請求項22に記載の方法。
【請求項25】
前記組換え型の糖タンパク質が、複数のα1,4結合−GlcNAc残基を有することを特徴とする請求項22に記載の方法。
【請求項26】
前記真核細胞が、哺乳類の細胞であることを特徴とする請求項22に記載の方法。
【請求項27】
前記組換え型の糖タンパク質を単離することを特徴とする請求項22に記載の方法。
【請求項28】
請求項22に記載の方法により製造され、少なくとも一つのα1,4結合−GlcNAc残基を有することを特徴とする組換え型の糖タンパク質。
【請求項29】
請求項27に記載の方法により製造され、少なくとも一つのα1,4結合−GlcNAc残基を有することを特徴とする単離された組換え型の糖タンパク質。
【請求項30】
少なくともα1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードする第一の外因性ポリヌクレオチドと;
コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)をエンコードし、又はコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二のポリヌクレオチドと;
少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとを、
少なくともゲノム中に、安定に組み込まれて有する非ヒトのトランスジェニック哺乳類であって、
該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、該第三のポリヌクレオチドとの少なくともいずれかが外因性ポリヌクレオチドであり;
該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、該第三のポリヌクレオチドとが、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合されており;
該第三のポリヌクレオチドが、該担体ポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列に相互作用して結合しており;
該C2GnT−I又は該C1−β3GnTと、該α4GnTと、該担体ポリペプチドとの発現により、該担体ポリペプチドが、該C2GnT−I又は該C1−β3GnTと該α4GnTとでグリコシル化されて、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を製造することを特徴とする非ヒトのトランスジェニック哺乳類。
【請求項31】
前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとの少なくともいずれかが、外因性のポリヌクレオチドを含んでいることを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項32】
前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、夫々外因性のポリヌクレオチドを含んでいることを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項33】
前記プロモータが、乳清酸性タンパク質(WAP)プロモータ、α−カゼインプロモータ、β−カゼインプロモータ、γ−カゼインプロモータ、α−ラクトアルブミンプロモータ、又はβ−ラクトグロブリンプロモータを含んでいることを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項34】
前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、及び前記第三のポリヌクレオチドとが、乳腺特異的遺伝子の3’制御配列、又は乳腺中で活性化した3’制御配列にも相互作用して結合していることを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項35】
前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが夫々、前記5’制御配列と前記3’制御配列とに相互作用して結合していることを特徴とする請求項34に記載の非ヒトのトランスジェニック哺乳類。
【請求項36】
前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、単一の核酸分子を構成しており、該核酸分子が前記5’制御配列と前記3’制御配列とに相互作用して結合していることを特徴とする請求項34に記載の非ヒトのトランスジェニック哺乳類。
【請求項37】
前記哺乳類が、マウス、ラット、ウサギ、ウマ、ブタ、ヒツジ、ヤギ、又はウシのいずれかであることを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項38】
前記トランスジェニック哺乳類が、非ヒトの雌トランスジェニック哺乳類であり、抗菌活性型の組換え型糖タンパク質を、非ヒトの雌トランスジェニック哺乳類により製造された母乳中に、分泌することを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項39】
請求項38に記載の非ヒトの雌トランスジェニック哺乳類によって製造された母乳。
【請求項40】
非ヒトの雌トランスジェニック哺乳類の母乳中に、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造させる方法であって、
a)α1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードする少なくとも第一の外因性ポリヌクレオチドを、非ヒトの哺乳類の胚に導入し;
該第一の外因性ポリヌクレオチドが、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合し、α4GnTの発現により、乳腺上皮細胞中の担体ポリペプチドが、α4GnTによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造するもので;
b)子孫を出産する宿主の雌哺乳類に、該a)の胚を移植し;
c)前記のb)の雌の子孫によって、5’制御配列に相互作用して結合しゲノム中で安定に組み換えられた第一の外因性ポリヌクレオチドを含む母乳を、製造させる工程を有することにより、
非ヒトの雌トランスジェニック哺乳類の母乳中に、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造させる方法。
【請求項41】
該非ヒトの哺乳類の胚に、
a)外因性のコア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)をエンコードし、又は外因性のコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二の外因性ポリヌクレオチド;又は
b)少なくとも一つのO−グリコシル化部位を有する外因性の担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチド;又は
c)該第二の外因性ポリヌクレオチド及び該第三の外因性ポリヌクレオチド
をさらに導入する工程を有しており、
該第二の外因性ポリヌクレオチドが共存する場合、該第三の外因性ポリヌクレオチドが共存する場合、又は該第二の外因性ポリヌクレオチド及び該第三の外因性ポリヌクレオチドが共存する場合、それが、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合しており;
該第三の外因性ポリヌクレオチドが共存する場合、それが、外因性の担体ポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列に相互作用して結合しており;
該C2GnT−I又は該C1−β3GnTが共存する場合、α4GnTと外因性の担体ポリペプチドとが共存する場合、C2GnT−Iとα4GnTがあるときはそれと、α4GnTとによって、該担体ポリペプチドが、グリコシル化されて、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造させることを特徴とする請求項40に記載の方法。
【請求項42】
前記c)の雌の子孫から搾乳する工程を有することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を含む母乳を得ることを特徴とする請求項40に記載の方法。
【請求項43】
前記母乳から少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を単離する工程をさらに有することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を得ることを特徴とする請求項42に記載の方法。
【請求項44】
請求項42に記載の方法により得られる母乳。
【請求項45】
請求項43に記載の方法により単離されて得られ、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質。
【請求項46】
コレステリル−α−D−グルコピラノシド(CGL)を発現する細菌の細胞壁の形成を抑制乃至阻害する方法であって、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物に接触させて、該CGL合成を抑制乃至阻害する工程を有することによって、該細菌の細胞壁の形成を抑制乃至阻害することを特徴とする方法。
【請求項47】
前記化合物が、グラム陰性細菌中のUDP−Glc:ステロール糖転移酵素活性を抑制乃至阻害することを特徴とする請求項46に記載の方法。
【請求項48】
前記細菌が、ヘリコバクター種の細菌であることを特徴とする請求項46に記載の方法。
【請求項49】
前記ヘリコバクター種の細菌が、ヘリコバクター ピロリであることを特徴とする請求項48に記載の方法。
【請求項50】
前記化合物が、単離された胃腺粘液細胞型ムチンであることを特徴とする請求項46に記載の方法。
【請求項51】
コレステリル−α−D−グルコピラノシド(CGL)を発現する細菌の増殖を抑制乃至阻害する方法であって、少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する化合物に、該細胞を接触させて、該CGL合成を抑制乃至阻害する工程を有することによって、該細菌の増殖を抑制乃至阻害することを特徴とする方法。
【請求項52】
前記化合物が、細菌のUDP−Glc:ステロール糖転移酵素活性を抑制乃至阻害することを特徴とする請求項51に記載の方法。
【請求項53】
前記細菌が、ヘリコバクター種の細菌であることを特徴とする請求項51に記載の方法。
【請求項54】
前記ヘリコバクター種の細菌が、ヘリコバクター ピロリであることを特徴とする請求項53に記載の方法。
【請求項55】
前記化合物が、単離された胃腺粘液細胞型ムチンであることを特徴とする請求項51に記載の方法。
【請求項56】
前記化合物が、O−グリカンを含んでいることを特徴とする請求項51に記載の方法。
【請求項57】
前記化合物が合成されたオリゴ糖を含んでいることを特徴とする請求項51に記載の方法。
【請求項58】
ヘリコバクター種の細菌が個体に感染することによる胃潰瘍の兆候又は症状を改善する方法であって、該個体に、少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する化合物が投与されて、該化合物が該ヘリコバクター種の細菌に接触し該ヘリコバクター種の細菌の増殖を抑制乃至阻害する工程を含むことによって、胃潰瘍の兆候又は症状を改善することを特徴とする方法。
【請求項59】
前記ヘリコバクター種の細菌が、ヘリコバクター ピロリであることを特徴とする請求項58に記載の方法。
【請求項60】
前記個体が哺乳類であることを特徴とする請求項58に記載の方法。
【請求項61】
前記哺乳類が、ウシ、ウマ、ブタ、ヤギ、イヌ、ネコ、又はフェレットであることを特徴とする請求項58に記載の方法。
【請求項62】
前記個体がヒトであることを特徴とする請求項58に記載の方法。
【請求項63】
前記化合物が、単離された胃腺粘液細胞型ムチンを含んでいることを特徴とする請求項58に記載の方法。
【請求項64】
前記化合物が、末端α1,4結合−GlcNAc残基を有するO−グリカンを含んでいることを特徴とする請求項58に記載の方法。
【請求項65】
前記化合物が、末端α1,4結合−GlcNAc残基を有する可溶性CD43(ロイコシアリン)を含んでいることを特徴とする請求項58に記載の方法。
【請求項66】
前記化合物が、合成されたオリゴ糖を含んでいることを特徴とする請求項58に記載の方法。
【請求項67】
前記化合物が経口投与されることを特徴とする請求項58に記載の方法。
【請求項68】
請求項21に記載の母乳を飲用することを特徴とする請求項67に記載の方法。
【請求項69】
胃潰瘍感受性の個体にヘリコバクター種の細菌が感染することによる胃潰瘍を予防する方法であって、該個体に少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する化合物が投与されて、該化合物が該ヘリコバクター種の細菌の増殖を妨げる工程を有することによって、該ヘリコバクター種の細菌の感染による胃潰瘍を予防することを特徴とする方法。
【請求項70】
前記個体への前記化合物の投与が、請求項21に記載の母乳を飲用する工程を有していることを特徴とする請求項69に記載の方法。
【請求項71】
ヘリコバクター種の細菌が個体に感染して胃潰瘍又は胃癌を発症した個体の胃潰瘍又は胃癌を改善する方法であって、該個体に少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する化合物が投与されて、該化合物が該ヘリコバクター種の細菌の増殖を妨げる工程を有することによって、該ヘリコバクター種の細菌の感染による胃潰瘍又は胃癌を改善することを特徴とする方法。
【請求項72】
前記個体への前記化合物の投与が、請求項21に記載の母乳を飲用する工程を有していることを特徴とする請求項71に記載の方法。
【請求項73】
少なくともα1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードする第一の外因性ポリヌクレオチドと;
コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)をエンコードし、又はコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二のポリヌクレオチドと;
少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとが、
ゲノム中に安定に組み込まれているトランスジェニック植物であって、
該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、該第三のポリヌクレオチドとの少なくともいずれかが外因性ポリヌクレオチドであり;
該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、該第三のポリヌクレオチドとが、プロモータを含む植物遺伝子の5’制御配列に相互作用して結合されており;
C2GnT−I又はC1−β3GnTと、α4GnTと、担体ポリペプチドとの発現により、該担体ポリペプチドが、該C2GnT−I又は該C1−β3GnTとα4GnTとによってグリコシル化されて、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を製造することを特徴とするトランスジェニック植物。
【請求項74】
前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとの少なくともいずれかが、外因性のポリヌクレオチドを含んでいることを特徴とする請求項73に記載のトランスジェニック植物。
【請求項75】
前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、夫々外因性のポリヌクレオチドであることを特徴とする請求項73に記載のトランスジェニック植物。
【請求項76】
前記プロモータが、構成プロモータ、誘導プロモータ、又は組織特異プロモータを含んでいることを特徴とする請求項73に記載のトランスジェニック植物。
【請求項77】
前記プロモータが、35Sカリフラワーモザイクウイルスプロモータ、ユビキチンプロモータ、テトラサイクリン応答プロモータ、rbcS遺伝子光誘導プロモータ、種子特異的オレオシン遺伝子プロモータ、又はコメアクチンプロモータを含んでいることを特徴とする請求項73に記載のトランスジェニック植物。
【請求項78】
前記植物がダイズ又はコメであることを特徴とする請求項73に記載のトランスジェニック植物。
【請求項79】
請求項73に記載のトランスジェニック植物から収穫された種子。
【請求項80】
請求項73に記載のトランスジェニック植物から収穫された穀類。
【請求項81】
請求項73に記載のトランスジェニック植物から収穫された乳液。
【請求項82】
トランスジェニック植物中に、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性組換え型の糖タンパク質を製造させる方法であって、
a)植物細胞に少なくとも、α1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードする第一の外因性ポリヌクレオチドを導入し;
該第一の外因性ポリヌクレオチドが、プロモータを含む植物遺伝子の5’制御配列に相互作用して結合されており;
該α4GnTの発現により、該植物の細胞中の該担体ポリペプチドが、該α4GnTによってグリコシル化され、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造するもので;
b)該a)の植物細胞から植物を生育させ、トランスジェニック植物を得ることによって、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質をトランスジェニック植物中に製造させる工程を有することを特徴とする、抗菌性の組換え型の糖タンパク質の製造方法。
【請求項83】
a)外因性のコア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)をエンコードし、又は外因性のコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二の外因性ポリヌクレオチド;又は
b)少なくとも一つのO−グリコシル化部位を有する外因性の担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチド;又は
c)該第二の外因性ポリヌクレオチド及び該第三の外因性ポリヌクレオチド
をさらに植物細胞に導入する工程を有しており、
該第二の外因性ポリヌクレオチドが共存する場合、該第三の外因性ポリヌクレオチドが共存する場合、又は該第二の外因性ポリヌクレオチド及び該第三の外因性ポリヌクレオチドが共存する場合、それが、プロモータを含む植物遺伝子の5’制御配列に相互作用して結合しており;
該C2GnT−I又は該C1−β3GnTが共存する場合、α4GnTと外因性の担体ポリペプチドとが共存する場合、C2GnT−Iとα4GnTがあるときはそれと、α4GnTとによって、該担体ポリペプチドが、グリコシル化されて、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質をトランスジェニック植物中に製造させることを特徴とする請求項82に記載の方法。
【請求項84】
前記トランスジェニック植物から、種子、葉、根、又は花の少なくともいずれかを含むトランスジェニック植物収穫物を分取することを特徴とする請求項82に記載の方法。
【請求項85】
前記トランスジェニック植物収穫物から、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を単離することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を得る工程を有することを特徴とする請求項84に記載の方法。
【請求項86】
前記トランスジェニック植物収穫物を加工して、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を含む乳液を得る工程を有することを特徴とする請求項84に記載の方法。
【請求項87】
請求項85に記載の方法により製造されることを特徴とする、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質。
【請求項88】
請求項86に記載の方法により得られることを特徴とする乳液。
【請求項1】
担体分子に相互作用して結合した少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有することを特徴とする天然非存在の化合物。
【請求項2】
前記担体分子が、O−グリカンを有していることを特徴とする請求項1に記載の化合物。
【請求項3】
前記O−グリカンが、コア2分岐型のO−グリカンであることを特徴とする請求項2に記載の化合物。
【請求項4】
前記担体分子が、ポリペプチドを有していることを特徴とする請求項1に記載の化合物。
【請求項5】
前記ポリペプチドが、少なくとも一つのO−グリコシル化部位を有することを特徴とする請求項4に記載の化合物。
【請求項6】
前記ポリペプチドが、O−グリカンを有していることを特徴とする請求項4に記載の化合物。
【請求項7】
前記O−グリカンが、コア2分岐型のO−グリカンであることを特徴とする請求項5に記載の化合物。
【請求項8】
前記担体分子が、ムチン様ポリペプチドを有していることを特徴とする請求項1に記載の化合物。
【請求項9】
前記ムチン様ポリペプチドが、CD43(ロイコシアリン)、CD34、又はMuc6を有していることを特徴とする請求項8に記載の化合物。
【請求項10】
前記O−グリカンが、コア1型のO−グリカンであることを特徴とする請求項5に記載の化合物。
【請求項11】
複数のα1,4結合−GlcNAc残基を有することを特徴とする請求項1に記載の化合物。
【請求項12】
合成したオリゴ糖を有することを特徴とする請求項1に記載の化合物。
【請求項13】
少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する糖タンパク質を製造する方法であって、ポリペプチドのグリコシル化条件下で、
α1,4−N−アセチルグルコサミン転移酵素(α4GnT)と、
コア2型のβ1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)又はコア1型の伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)と、
少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドとを接触させ、
該ポリペプチドを、C2GnT−I又はC1−β3GnTとα4GnTとによってグリコシル化して、少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する糖タンパク質を製造する方法。
【請求項14】
前記担体ポリペプチドが、可溶性ポリペプチドであることを特徴とする請求項13に記載の方法。
【請求項15】
前記担体ポリペプチドが、CD43(ロイコシアリン)、CD34、又はMuc6であることを特徴とする請求項13に記載の方法。
【請求項16】
前記担体ポリペプチドが、哺乳類の母乳中に分泌されるムチン型糖タンパク質であることを特徴とする請求項13に記載の方法。
【請求項17】
前記ムチン型糖タンパク質を含む母乳に、α4GnTと、必要に応じてC2GnT−I又はC1−β3GnTとを接触させることを特徴とする請求項16に記載の方法。
【請求項18】
前記糖タンパク質が、複数のα1,4結合−GlcNAc残基を有することを特徴とする請求項13に記載の方法。
【請求項19】
前記糖タンパク質を単離することを特徴とする請求項13に記載の方法。
【請求項20】
請求項13に記載の方法により製造されることを特徴とする、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質。
【請求項21】
請求項17に記載の方法により製造される母乳であって、少なくとも一つのα1,4結合−GlcNAc残基を有する糖タンパク質を含むことを特徴とする母乳。
【請求項22】
少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する組換え型の糖タンパク質を製造する方法であって、
α1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードする第一のポリヌクレオチドと、
コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)又はコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二のポリヌクレオチドと、
少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとを
真核細胞内で発現させる工程を含み、
該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、及び該第三のポリヌクレオチドとの少なくともいずれかが、真核細胞に導入された外因性ポリヌクレオチドであり、
該担体ポリペプチドを、該C2GnT−Iと該α4GnTとによって、又は該C1−β3GnTと該α4GnTとによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を製造する方法。
【請求項23】
前記担体ポリペプチドが、可溶性ポリペプチドであることを特徴とする請求項22に記載の方法。
【請求項24】
前記担体ポリペプチドが、CD43(ロイコシアリン)、CD34、又はMuc6であることを特徴とする請求項22に記載の方法。
【請求項25】
前記組換え型の糖タンパク質が、複数のα1,4結合−GlcNAc残基を有することを特徴とする請求項22に記載の方法。
【請求項26】
前記真核細胞が、哺乳類の細胞であることを特徴とする請求項22に記載の方法。
【請求項27】
前記組換え型の糖タンパク質を単離することを特徴とする請求項22に記載の方法。
【請求項28】
請求項22に記載の方法により製造され、少なくとも一つのα1,4結合−GlcNAc残基を有することを特徴とする組換え型の糖タンパク質。
【請求項29】
請求項27に記載の方法により製造され、少なくとも一つのα1,4結合−GlcNAc残基を有することを特徴とする単離された組換え型の糖タンパク質。
【請求項30】
少なくともα1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードする第一の外因性ポリヌクレオチドと;
コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)をエンコードし、又はコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二のポリヌクレオチドと;
少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとを、
少なくともゲノム中に、安定に組み込まれて有する非ヒトのトランスジェニック哺乳類であって、
該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、該第三のポリヌクレオチドとの少なくともいずれかが外因性ポリヌクレオチドであり;
該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、該第三のポリヌクレオチドとが、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合されており;
該第三のポリヌクレオチドが、該担体ポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列に相互作用して結合しており;
該C2GnT−I又は該C1−β3GnTと、該α4GnTと、該担体ポリペプチドとの発現により、該担体ポリペプチドが、該C2GnT−I又は該C1−β3GnTと該α4GnTとでグリコシル化されて、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を製造することを特徴とする非ヒトのトランスジェニック哺乳類。
【請求項31】
前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとの少なくともいずれかが、外因性のポリヌクレオチドを含んでいることを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項32】
前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、夫々外因性のポリヌクレオチドを含んでいることを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項33】
前記プロモータが、乳清酸性タンパク質(WAP)プロモータ、α−カゼインプロモータ、β−カゼインプロモータ、γ−カゼインプロモータ、α−ラクトアルブミンプロモータ、又はβ−ラクトグロブリンプロモータを含んでいることを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項34】
前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、及び前記第三のポリヌクレオチドとが、乳腺特異的遺伝子の3’制御配列、又は乳腺中で活性化した3’制御配列にも相互作用して結合していることを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項35】
前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが夫々、前記5’制御配列と前記3’制御配列とに相互作用して結合していることを特徴とする請求項34に記載の非ヒトのトランスジェニック哺乳類。
【請求項36】
前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、単一の核酸分子を構成しており、該核酸分子が前記5’制御配列と前記3’制御配列とに相互作用して結合していることを特徴とする請求項34に記載の非ヒトのトランスジェニック哺乳類。
【請求項37】
前記哺乳類が、マウス、ラット、ウサギ、ウマ、ブタ、ヒツジ、ヤギ、又はウシのいずれかであることを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項38】
前記トランスジェニック哺乳類が、非ヒトの雌トランスジェニック哺乳類であり、抗菌活性型の組換え型糖タンパク質を、非ヒトの雌トランスジェニック哺乳類により製造された母乳中に、分泌することを特徴とする請求項30に記載の非ヒトのトランスジェニック哺乳類。
【請求項39】
請求項38に記載の非ヒトの雌トランスジェニック哺乳類によって製造された母乳。
【請求項40】
非ヒトの雌トランスジェニック哺乳類の母乳中に、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造させる方法であって、
a)α1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードする少なくとも第一の外因性ポリヌクレオチドを、非ヒトの哺乳類の胚に導入し;
該第一の外因性ポリヌクレオチドが、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合し、α4GnTの発現により、乳腺上皮細胞中の担体ポリペプチドが、α4GnTによってグリコシル化して、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造するもので;
b)子孫を出産する宿主の雌哺乳類に、該a)の胚を移植し;
c)前記のb)の雌の子孫によって、5’制御配列に相互作用して結合しゲノム中で安定に組み換えられた第一の外因性ポリヌクレオチドを含む母乳を、製造させる工程を有することにより、
非ヒトの雌トランスジェニック哺乳類の母乳中に、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造させる方法。
【請求項41】
該非ヒトの哺乳類の胚に、
a)外因性のコア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)をエンコードし、又は外因性のコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二の外因性ポリヌクレオチド;又は
b)少なくとも一つのO−グリコシル化部位を有する外因性の担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチド;又は
c)該第二の外因性ポリヌクレオチド及び該第三の外因性ポリヌクレオチド
をさらに導入する工程を有しており、
該第二の外因性ポリヌクレオチドが共存する場合、該第三の外因性ポリヌクレオチドが共存する場合、又は該第二の外因性ポリヌクレオチド及び該第三の外因性ポリヌクレオチドが共存する場合、それが、プロモータを含む乳腺特異的遺伝子の5’制御配列に相互作用して結合しており;
該第三の外因性ポリヌクレオチドが共存する場合、それが、外因性の担体ポリペプチドを母乳内に分泌させる効果を有する信号配列をエンコードするヌクレオチド配列に相互作用して結合しており;
該C2GnT−I又は該C1−β3GnTが共存する場合、α4GnTと外因性の担体ポリペプチドとが共存する場合、C2GnT−Iとα4GnTがあるときはそれと、α4GnTとによって、該担体ポリペプチドが、グリコシル化されて、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造させることを特徴とする請求項40に記載の方法。
【請求項42】
前記c)の雌の子孫から搾乳する工程を有することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を含む母乳を得ることを特徴とする請求項40に記載の方法。
【請求項43】
前記母乳から少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を単離する工程をさらに有することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を得ることを特徴とする請求項42に記載の方法。
【請求項44】
請求項42に記載の方法により得られる母乳。
【請求項45】
請求項43に記載の方法により単離されて得られ、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質。
【請求項46】
コレステリル−α−D−グルコピラノシド(CGL)を発現する細菌の細胞壁の形成を抑制乃至阻害する方法であって、少なくとも一つのα1,4結合−GlcNAc残基を有する化合物に接触させて、該CGL合成を抑制乃至阻害する工程を有することによって、該細菌の細胞壁の形成を抑制乃至阻害することを特徴とする方法。
【請求項47】
前記化合物が、グラム陰性細菌中のUDP−Glc:ステロール糖転移酵素活性を抑制乃至阻害することを特徴とする請求項46に記載の方法。
【請求項48】
前記細菌が、ヘリコバクター種の細菌であることを特徴とする請求項46に記載の方法。
【請求項49】
前記ヘリコバクター種の細菌が、ヘリコバクター ピロリであることを特徴とする請求項48に記載の方法。
【請求項50】
前記化合物が、単離された胃腺粘液細胞型ムチンであることを特徴とする請求項46に記載の方法。
【請求項51】
コレステリル−α−D−グルコピラノシド(CGL)を発現する細菌の増殖を抑制乃至阻害する方法であって、少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する化合物に、該細胞を接触させて、該CGL合成を抑制乃至阻害する工程を有することによって、該細菌の増殖を抑制乃至阻害することを特徴とする方法。
【請求項52】
前記化合物が、細菌のUDP−Glc:ステロール糖転移酵素活性を抑制乃至阻害することを特徴とする請求項51に記載の方法。
【請求項53】
前記細菌が、ヘリコバクター種の細菌であることを特徴とする請求項51に記載の方法。
【請求項54】
前記ヘリコバクター種の細菌が、ヘリコバクター ピロリであることを特徴とする請求項53に記載の方法。
【請求項55】
前記化合物が、単離された胃腺粘液細胞型ムチンであることを特徴とする請求項51に記載の方法。
【請求項56】
前記化合物が、O−グリカンを含んでいることを特徴とする請求項51に記載の方法。
【請求項57】
前記化合物が合成されたオリゴ糖を含んでいることを特徴とする請求項51に記載の方法。
【請求項58】
ヘリコバクター種の細菌が個体に感染することによる胃潰瘍の兆候又は症状を改善する方法であって、該個体に、少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する化合物が投与されて、該化合物が該ヘリコバクター種の細菌に接触し該ヘリコバクター種の細菌の増殖を抑制乃至阻害する工程を含むことによって、胃潰瘍の兆候又は症状を改善することを特徴とする方法。
【請求項59】
前記ヘリコバクター種の細菌が、ヘリコバクター ピロリであることを特徴とする請求項58に記載の方法。
【請求項60】
前記個体が哺乳類であることを特徴とする請求項58に記載の方法。
【請求項61】
前記哺乳類が、ウシ、ウマ、ブタ、ヤギ、イヌ、ネコ、又はフェレットであることを特徴とする請求項58に記載の方法。
【請求項62】
前記個体がヒトであることを特徴とする請求項58に記載の方法。
【請求項63】
前記化合物が、単離された胃腺粘液細胞型ムチンを含んでいることを特徴とする請求項58に記載の方法。
【請求項64】
前記化合物が、末端α1,4結合−GlcNAc残基を有するO−グリカンを含んでいることを特徴とする請求項58に記載の方法。
【請求項65】
前記化合物が、末端α1,4結合−GlcNAc残基を有する可溶性CD43(ロイコシアリン)を含んでいることを特徴とする請求項58に記載の方法。
【請求項66】
前記化合物が、合成されたオリゴ糖を含んでいることを特徴とする請求項58に記載の方法。
【請求項67】
前記化合物が経口投与されることを特徴とする請求項58に記載の方法。
【請求項68】
請求項21に記載の母乳を飲用することを特徴とする請求項67に記載の方法。
【請求項69】
胃潰瘍感受性の個体にヘリコバクター種の細菌が感染することによる胃潰瘍を予防する方法であって、該個体に少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する化合物が投与されて、該化合物が該ヘリコバクター種の細菌の増殖を妨げる工程を有することによって、該ヘリコバクター種の細菌の感染による胃潰瘍を予防することを特徴とする方法。
【請求項70】
前記個体への前記化合物の投与が、請求項21に記載の母乳を飲用する工程を有していることを特徴とする請求項69に記載の方法。
【請求項71】
ヘリコバクター種の細菌が個体に感染して胃潰瘍又は胃癌を発症した個体の胃潰瘍又は胃癌を改善する方法であって、該個体に少なくとも一つのα1,4結合−N−アセチルグルコサミン(α1,4結合−GlcNAc)残基を有する化合物が投与されて、該化合物が該ヘリコバクター種の細菌の増殖を妨げる工程を有することによって、該ヘリコバクター種の細菌の感染による胃潰瘍又は胃癌を改善することを特徴とする方法。
【請求項72】
前記個体への前記化合物の投与が、請求項21に記載の母乳を飲用する工程を有していることを特徴とする請求項71に記載の方法。
【請求項73】
少なくともα1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードする第一の外因性ポリヌクレオチドと;
コア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)をエンコードし、又はコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二のポリヌクレオチドと;
少なくとも一つのO−グリコシル化部位を有する担体ポリペプチドをエンコードする第三のポリヌクレオチドとが、
ゲノム中に安定に組み込まれているトランスジェニック植物であって、
該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、該第三のポリヌクレオチドとの少なくともいずれかが外因性ポリヌクレオチドであり;
該第一のポリヌクレオチドと、該第二のポリヌクレオチドと、該第三のポリヌクレオチドとが、プロモータを含む植物遺伝子の5’制御配列に相互作用して結合されており;
C2GnT−I又はC1−β3GnTと、α4GnTと、担体ポリペプチドとの発現により、該担体ポリペプチドが、該C2GnT−I又は該C1−β3GnTとα4GnTとによってグリコシル化されて、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を製造することを特徴とするトランスジェニック植物。
【請求項74】
前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとの少なくともいずれかが、外因性のポリヌクレオチドを含んでいることを特徴とする請求項73に記載のトランスジェニック植物。
【請求項75】
前記第一のポリヌクレオチドと、前記第二のポリヌクレオチドと、前記第三のポリヌクレオチドとが、夫々外因性のポリヌクレオチドであることを特徴とする請求項73に記載のトランスジェニック植物。
【請求項76】
前記プロモータが、構成プロモータ、誘導プロモータ、又は組織特異プロモータを含んでいることを特徴とする請求項73に記載のトランスジェニック植物。
【請求項77】
前記プロモータが、35Sカリフラワーモザイクウイルスプロモータ、ユビキチンプロモータ、テトラサイクリン応答プロモータ、rbcS遺伝子光誘導プロモータ、種子特異的オレオシン遺伝子プロモータ、又はコメアクチンプロモータを含んでいることを特徴とする請求項73に記載のトランスジェニック植物。
【請求項78】
前記植物がダイズ又はコメであることを特徴とする請求項73に記載のトランスジェニック植物。
【請求項79】
請求項73に記載のトランスジェニック植物から収穫された種子。
【請求項80】
請求項73に記載のトランスジェニック植物から収穫された穀類。
【請求項81】
請求項73に記載のトランスジェニック植物から収穫された乳液。
【請求項82】
トランスジェニック植物中に、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性組換え型の糖タンパク質を製造させる方法であって、
a)植物細胞に少なくとも、α1,4−N−アセチルグルコサミン転移酵素(α4GnT)をエンコードする第一の外因性ポリヌクレオチドを導入し;
該第一の外因性ポリヌクレオチドが、プロモータを含む植物遺伝子の5’制御配列に相互作用して結合されており;
該α4GnTの発現により、該植物の細胞中の該担体ポリペプチドが、該α4GnTによってグリコシル化され、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質を製造するもので;
b)該a)の植物細胞から植物を生育させ、トランスジェニック植物を得ることによって、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質をトランスジェニック植物中に製造させる工程を有することを特徴とする、抗菌性の組換え型の糖タンパク質の製造方法。
【請求項83】
a)外因性のコア2β1,6−N−アセチルグルコサミン転移酵素−I(C2GnT−I)をエンコードし、又は外因性のコア1伸長β1,3−N−アセチルグルコサミン転移酵素(C1−β3GnT)をエンコードする第二の外因性ポリヌクレオチド;又は
b)少なくとも一つのO−グリコシル化部位を有する外因性の担体ポリペプチドをエンコードする第三の外因性ポリヌクレオチド;又は
c)該第二の外因性ポリヌクレオチド及び該第三の外因性ポリヌクレオチド
をさらに植物細胞に導入する工程を有しており、
該第二の外因性ポリヌクレオチドが共存する場合、該第三の外因性ポリヌクレオチドが共存する場合、又は該第二の外因性ポリヌクレオチド及び該第三の外因性ポリヌクレオチドが共存する場合、それが、プロモータを含む植物遺伝子の5’制御配列に相互作用して結合しており;
該C2GnT−I又は該C1−β3GnTが共存する場合、α4GnTと外因性の担体ポリペプチドとが共存する場合、C2GnT−Iとα4GnTがあるときはそれと、α4GnTとによって、該担体ポリペプチドが、グリコシル化されて、少なくとも一つのα1,4結合−GlcNAc残基を有する抗菌性の組換え型の糖タンパク質をトランスジェニック植物中に製造させることを特徴とする請求項82に記載の方法。
【請求項84】
前記トランスジェニック植物から、種子、葉、根、又は花の少なくともいずれかを含むトランスジェニック植物収穫物を分取することを特徴とする請求項82に記載の方法。
【請求項85】
前記トランスジェニック植物収穫物から、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を単離することにより、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を得る工程を有することを特徴とする請求項84に記載の方法。
【請求項86】
前記トランスジェニック植物収穫物を加工して、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質を含む乳液を得る工程を有することを特徴とする請求項84に記載の方法。
【請求項87】
請求項85に記載の方法により製造されることを特徴とする、少なくとも一つのα1,4結合−GlcNAc残基を有する組換え型の糖タンパク質。
【請求項88】
請求項86に記載の方法により得られることを特徴とする乳液。
【図1】

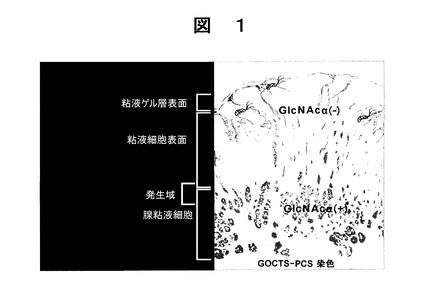
【図2】
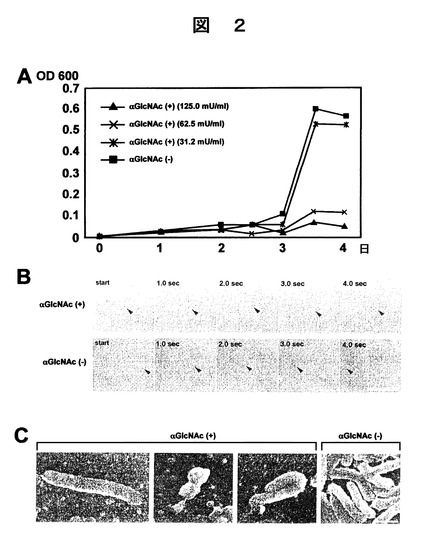
【図3】
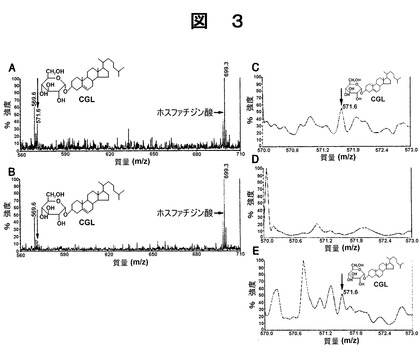
【図4】
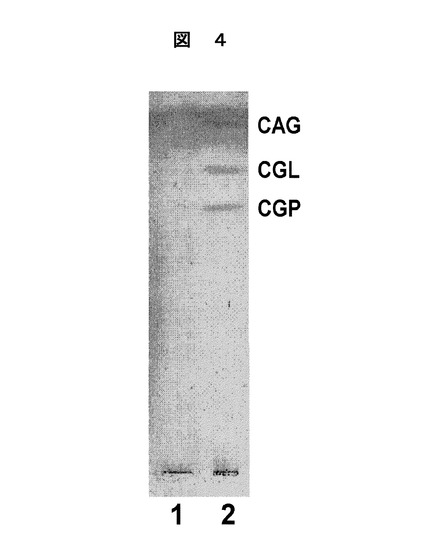
【図5】
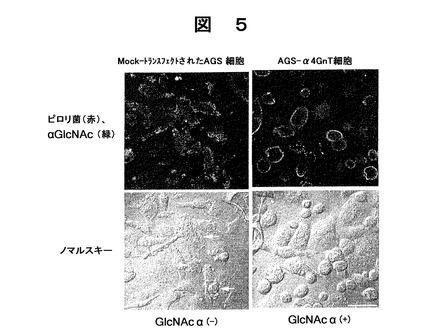
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2007−524693(P2007−524693A)
【公表日】平成19年8月30日(2007.8.30)
【国際特許分類】
【出願番号】特願2006−554266(P2006−554266)
【出願日】平成17年2月18日(2005.2.18)
【国際出願番号】PCT/US2005/005407
【国際公開番号】WO2005/081904
【国際公開日】平成17年9月9日(2005.9.9)
【出願人】(500114586)ザ バーナム インスティチュート (8)
【出願人】(504180239)国立大学法人信州大学 (759)
【Fターム(参考)】
【公表日】平成19年8月30日(2007.8.30)
【国際特許分類】
【出願日】平成17年2月18日(2005.2.18)
【国際出願番号】PCT/US2005/005407
【国際公開番号】WO2005/081904
【国際公開日】平成17年9月9日(2005.9.9)
【出願人】(500114586)ザ バーナム インスティチュート (8)
【出願人】(504180239)国立大学法人信州大学 (759)
【Fターム(参考)】
[ Back to top ]