
抗HSV抗体
特許請求の範囲に記載の通り、本発明は、特許請求の範囲に記載の抗HSV抗体、該抗体の有効量を含む薬学的組成物、該抗体をコードするヌクレオチド配列を含む発現ベクター、該ヌクレオチド配列を含む宿主細胞、該抗体を産生することができるハイブリドーマ細胞、および薬物としての、特に、対象におけるHSV関連疾患の予防的または治療的処置のための薬物の製造のための、該抗体の使用に関する。

【発明の詳細な説明】
【技術分野】
【0001】
特許請求の範囲に記載の通り、本発明は、特許請求の範囲に記載の抗HSV抗体、該抗体の有効量を含む薬学的組成物、該抗体をコードするヌクレオチド配列を含む発現ベクター、該ヌクレオチド配列を含む宿主細胞、該抗体を産生することができるハイブリドーマ細胞、および薬物としての、特に、対象におけるHSV関連疾患の予防的または治療的処置のための薬物の製造のための、該抗体の使用に関する。
【背景技術】
【0002】
ヒト病原性単純ヘルペスウイルス(HSV)は、皮膚親和性および神経親和性DNAウイルスであり、その臨床症状は皮膚およびその近傍の粘膜から主に生じ、とりわけ神経炎、髄膜炎、脳炎、脊髄炎、多発性神経根炎などの神経学的合併症を二次的に引き起こす。ある程度、高い致死性を伴う重度進行の、先天性、後天性およびまた、医原性の免疫不全が報告されている。1型HSV (HSV-1、95%; 口唇ヘルペス、角膜ヘルペス、ヘルペス性湿疹)および2型HSV (HSV-2、10〜30%; 陰部ヘルペス、新生児ヘルペス)による集団の高い感染率のため、ならびにウイルスの頻繁な再活性化、知覚神経節および自律神経節中で潜伏的に持続する存続期間のため、HSVは特に臨床的関連性がある。ウイルスのタイプとは無関係に、一次または再発HSV感染の対症療法の目標は、ウイルス複製の阻害、苦痛の時間の短縮、および再発の頻度に影響を与える全身合併症の予防である。
【0003】
初期認識および的確な投与量で、ウイルス抑制剤は抗ウイルス療法に成功裏に利用されている。最も一般的なウイルス抑制剤(例えばアシクロビル(acyclovir)、ペンシクロビル(penciclovir)、ホスカネット(foscarnet)、イドクスウリジン(idoxuridin))は、ヌクレオシド類似体またはピロリン酸類似体であり、その共通の活性原理はウイルス感染細胞におけるDNA合成の阻害に基づく。
【0004】
HSV感染の処置のための最も重要な治療剤の一つがプリンヌクレオシド類似体のアシクロビルである。これはウイルスチミジンキナーゼによりリン酸化され、その後、ウイルスDNAポリメラーゼを妨害する。対照的に、ヒトDNAポリメラーゼは30〜50倍アシクロビルに対する感受性が低く、このため、微々たる副作用しか観察されない。
【0005】
しかし、選択的に作用するウイルス抑制剤の開発にもかかわらず、ウイルス性疾患の化学療法処置は依然として深刻な問題である。具体的には、免疫抑制患者の長期にわたる予防的および治療的処置の間に観察される一般的な化学療法剤に対する耐性株の出現は解決が難しい。結果として、症例の10%超で、有効なウイルス抑制剤の欠如のため、致死的な進行を伴う急速進行性の全身感染が認められる。
【0006】
現在、ピロリン酸類似体のホスカネットがアシクロビル耐性ヘルペスウイルスに対して免疫抑制患者で特に利用されている。この薬剤はウイルスDNAポリメラーゼの直接的な阻害を引き起こし、チミジンキナーゼに影響を与えない。しかしながら、ホスカネットの使用は腎不全、心臓の問題などの重篤な望ましくない副作用をもたらし、骨髄に対する毒性を有し、皮膚潰瘍を引き起こすこともある。ホスカネットは、その催奇形効果のため、妊娠中に投与することもできない。さらに、交差耐性株の形成が認められ、これにより代替的な治療剤の開発が大いに必要となる。受動免疫学的予防は現在、利用できない。HSV1およびHSV2に対する能動免疫用の2〜3種の実験的ワクチンは、検証可能な成功を示さなかった。
【0007】
抗体はがん、自己免疫障害およびウイルス感染の処置にかなり有望である。JP 6135854には、HSVに対するヒトモノクローナル抗体およびアシクロビル(ACV) などの抗ウイルス核酸類似体が点滴静注により注射の形態で同時投与または連続投与される単純ヘルペスウイルス感染用の治療剤が記述されている。DK 187286には、HSV-1およびHSV-2由来の糖タンパク質D (gD) (HSV gD-1およびgD-2)に関して多特異的な免疫反応性を示す抗体が開示されている。WO 1997/26329には、HSV-1およびHSV-2の診断および処置に有用なヒトモノクローナル抗体が記述されている。後者の抗体は、HSV-1およびHSV-2の糖タンパク質D抗原との結合でHSV 863モノクローナル抗体と競合する。US 4,950,595には、抗ウイルス-ヒト抗体を産生するマウス-ヒトハイブリドーマ、その調製の過程、および抗ウイルス-ヒトモノクローナル抗体について開示されている。
【0008】
さらに、HSV 1/2に特異的な別のマウスモノクローナル抗体(Fd79) (Kogae et al., 1986)のヒト化が記述されている(Co, M.S. et al., 1991; Zeitlin L. et al., 1998)。この抗体は、HSV1およびHSV2の糖タンパク質B (gB)の共有エピトープを認識する。さらに、ヒト化Fd79がトランスジェニック植物においても真核生物細胞株SP2/0においても産生され、その後、特徴付けられており、53 nMの親和性を示している。モノクローナルマウス抗体H1815は、糖タンパク質B (gB)のアミノ酸263〜283の領域における類似の、しかし同一ではないエピトープを認識する(Navarro et al., 1992)。しかしながら、H1815はウイルス中和または「細胞間伝播」の阻害ができない。
【0009】
最後に、US 6,180,370には、ヒト化免疫グロブリンおよびそれを作出する方法について記述されている。さらに、WO 2003/105782は、ヒトフレームワーク上にマウス抗体をグラフトすることの特異性に関連する。
【0010】
このように、化学療法剤は望ましくない副作用を有し、ますます多くの耐性株が認められている。
【0011】
それゆえ、HSV感染を中和でき、かつ細胞間伝播を阻害できる(ヒト化)抗HSV抗体を提供することが本発明の目的である。さらに、従来適用される化学療法剤の上記の不利点を克服するHSV関連疾患の処置のための予防剤および/または治療剤を提供することが本発明の目的である。
【0012】
驚いたことに、本発明による抗体はこの目的を解決することが分かった。したがって、本発明は、ヒト化されてもよい、組換え産生抗体に基づくHSV感染の処置に向けた当技術分野において公知の治療剤に代わる有望な選択肢を提供する。これらの抗体は、宿主内で伝播するためのウイルス機構の両方を遮断することができる。それらは効果的に、無細胞ウイルス粒子を中和し、ウイルスの直接的な細胞間伝播を阻害する。これらの抗体は、ウイルス複製サイクルに不可欠な、HSV1およびHSV2外被の表面糖タンパク質B (gB)の高度に保存されたエピトープに特異的に結合するので、薬物耐性が出現する可能性はほとんどない。
【0013】
本発明のマウス抗体の効果は、Eis-Hubinger et al., Intervirology (1991); 32:351-360およびEis-Hubinger et al., Journal of General Virology (1993); 74: 379-385を参照されたく、部分的には、既に記述されているとはいえ、抗体それ自体または本発明の抗体の相補性決定領域(CDR)の配列およびそれが結合するエピトープは、決して公開されておらず、または公衆が利用可能なものではない。
【0014】
要約すれば、(ヒト化)抗体は以下の利点の1つまたは複数を提供する:
・本発明のマウスモノクローナル抗体の効力は既に証明されている(Eis-Hubinger et al., 1991; Eis-Hubinger et al., 1993を参照のこと)。さらに、本発明者らは、本発明のヒト化抗体がまた、インビトロでのHSV1およびHSV2ウイルス感染を中和でき、「細胞間伝播」機構を阻害することによりウイルス伝播を阻害できることを実施例の項において示す。ヒトでの感染進行との関連で、ヒト免疫系は、HSV 1/2に典型的な細胞間伝播を効率的に予防するための抗体特異性を生み出すことができない。
・アシクロビルおよびホスカネットなどの保守的な化学療法剤の頻繁かつ長期にわたる予防的および治療的適用は、耐性ウイルス株の形成の増加をもたらす。この耐性の問題は、単独でまたはアシクロビルおよび/もしくはホスカネットなどのウイルス抑制剤との組み合わせで投与される、本明細書に記述の(ヒト化)抗HSV抗体によって克服することができる。これは、この抗体が異なる作用機序に依るからである。
・本明細書において記述される抗体はHSV gBタンパク質のエピトープに特異的に結合する。本発明の抗体に対するHSV耐性の出現は、gB-タンパク質中の変異がウイルス感染価の喪失をもたらすので、予想されるものではない。
・従来のウイルス抑制剤の全身投与が禁忌とされる患者は、本明細書において記述される(ヒト化)抗体から特に利益を得る。
【発明の概要】
【0015】
したがって、SEQ ID NO: 1、SEQ ID NO: 2、SEQ ID NO: 3、SEQ ID NO: 4、SEQ ID NO: 5、およびSEQ ID NO: 6に示される相補性決定領域を含む抗体が提供される。さらに、SEQ ID NO: 1〜6に示される相補性決定領域を含む抗体と同じエピトープを認識する抗体が提供される。
【0016】
さらに、本明細書に記述の抗体の有効量と、少なくとも1つの薬学的に許容される賦形剤とを含む、薬学的組成物も提供される。
【0017】
さらに、特許請求の範囲に記載の抗体をコードするヌクレオチド配列を含む発現ベクター、該ヌクレオチド配列を含む細胞、および該抗体を産生することができるハイブリドーマ細胞が提供される。
【0018】
最後に、医薬における使用のための特許請求の範囲に記載の抗体、および対象におけるHSV関連疾患の予防的または治療的処置のための薬物の製造のための、特許請求の範囲に記載の抗体の使用が同様に本明細書において提供される。
【0019】
独立請求項に記載の本発明のさまざまな局面および従属請求項に含まれる好ましい態様は、参照により本明細書に組み入れられる。
【図面の簡単な説明】
【0020】
【図1】膣粘膜において確立された感染後のモノクローナル抗体(mAb) 2cの投与によるインビボでのHSV複製の低減を示す。免疫応答性(黒記号)マウスおよびCD4+枯渇(白記号)マウスを感染後3日目および11日目(矢印)に腹腔内(i.p.)注射によりポリクローナルHSV免疫血清(三角)で、mAb 2c (丸)、または対照培地(四角)で処置した。
【図2A】免疫抑制(CD4-/CD8-)マウスにおけるHSV感染の進行に及ぼすmAb 2cの効果を示す。感染後1日目および3日目の時点(矢印で示した)で、動物に対照培地(黒四角)、ポリクローナルHSV免疫血清(白四角)、またはmAb 2c (黒丸)のいずれかをi.p.投与した。(A)は生存率を示す。
【図2B】免疫抑制(CD4-/CD8-)マウスにおけるHSV感染の進行に及ぼすmAb 2cの効果を示す。感染後1日目および3日目の時点(矢印で示した)で、動物に対照培地(黒四角)、ポリクローナルHSV免疫血清(白四角)、またはmAb 2c (黒丸)のいずれかをi.p.投与した。(B)は膣粘膜におけるウイルス複製を示す。
【図3】mAb 2c由来抗体および抗体断片の略図である。(A) 完全な抗体: マウス可変領域(VL、VH; 左)とヒト定常ドメイン(CL、CH1、CH2、CH3; 中央)との遺伝的融合はキメラ抗体をもたらす。ヒト化IgG抗体(右)では、マウスモノクローナル抗体の超可変領域がヒト抗体のフレームワーク上にグラフトされる。(B) 抗体断片: 軽鎖(VL + CL)および重鎖の2つのN末端ドメイン(VH + CH1)からなる一価Fab断片(Fab)、または2つの不対C末端システイン残基によって共有結合的に連結されている二価F(ab')2断片を従来のプロテアーゼ消化によって産生することができる。マウスscFv抗体(「一本鎖可変断片」)の作出のため、可変ドメインVHおよびVLをコードする遺伝子を、2cハイブリドーマ細胞株から単離し、可動性の連結ペプチド(「リンカー」)をコードする遺伝子セグメントによって連結した。
【図4】可変重鎖および軽鎖ドメイン(VHおよびVL)の配列アライメントを示す。抗原結合部位はChothia (Chothia and Lesk, 1987; Chothia et al., 1989)(点線)およびKabat (Kabat and Wu, 1991)(破線)により規定される。ヒト生殖細胞系配列DP28およびDPK13はV-Baseデータベース(http://vbase.mrc-cpe.cam.ac.uk/)から入手し、マウスmAb 2cのCDRのためのアクセプター配列として役立てた。(A) 「不変の残基」(Kabat and Wu, 1991); (B) 「鍵となる残基」(Chothia et al., 1989)および(C) VH/VL界面の残基(Chothia et al., 1985)は、マウスおよびヒト配列の間で、それぞれ、適合残基の場合には(+)または非適合残基の場合には(-)で印されている。(D) それぞれ不変(i)残基部位; 類似(r)残基部位; 表面(s)残基R、K、E、D、Q、N; 中性(n)残基P、H、Y、G、A、S、T; および埋没(b)残基C、V、L、I、M、F、WとしてChothia (Chothia et al., 1998)により定義されている中心部位の残基; 埋没中性残基はxで印されている; 表面中性残基はyで印されている; マウスおよびヒト配列間の非適合残基部位は太文字で印されている; VHhum2cおよびVLhum2c、太文字で示されたマウスCDR残基を有する特異性グラフト配列。全ての残基が一文字表記で示され、Kabat (Kabat et al., 1991)にしたがって付番されている。
【図5】モノクローナル抗体2c、ch2cおよびヒト化バリアントhu2c-V1-V4の平衡結合曲線を示す。HSV-1 F感染Vero細胞の表面上の糖タンパク質Bとの特異的な結合をフローサイトメトリーによって判定した。表示濃度での結合活性を、バックグラウンドの蛍光を差し引いた平均蛍光強度(MFI)として示す。測定は三つ組で行った; 標準偏差をバーとして示す。抗原結合データをLevenberg-Marquard法による非線形回帰モデルに適合させることによって結合親和性定数KDを決定した。
【図6】マウス親mAb 2cと同じ、キメラ抗体およびヒト化抗体のエピトープ特異性を実証する競合研究を示す。キメラmAb ch2c (白丸)およびヒト化mAbバリアントhu2c-V1 (四角)、hu2c-V2 (三角)、hu2c-V3 (星印)、hu2c-V4 (菱形)は、HSV-1感染Vero細胞の表面上に存在するgBとの結合で親mAb 2cと競合する。感染Vero細胞を漸増濃度のmAb ch2cまたはヒト化mAb hu2c-V1-V4とまず初めにインキュベートし、その後、競合物質として100 nMのmAb 2cとのインキュベーションを行った。平均蛍光強度(MFI)は適用された競合物質の結合を示す。
【図7】親mAb 2cおよびヒト化mAb hu2c-V1のHSV中和活性が補体非依存的であることを示す。HSV-1を、Vero細胞に適用する前に10%ヒト補体の存在下または非存在下で培地(対照)、ポリクローナルIgG Cytotect(登録商標) (120μg/ml)、mAb 2c (2μg/ml)またはmAb hu2c-V1 (2μg/ml)とともにプレインキュベートする。2日後にプラークの出現をスコア化した。
【図8】非耐性実験室株(HSV-1 F、HSv-1 324hv、HSV-1 17 syn+、HSV-2 G)、ACV耐性実験室株HSV-1 TK-および耐性について詳しく調べられていない臨床分離株と比較した、アシクロビル(ACV)、ACVおよびホスカネット(PFA)、またはACV、PFAおよびシドビル(CDV)に対して臨床的に耐性の患者分離株に由来するHSV-1およびHSV-2を中和するヒト化mAb h2c-V1の効率を示す。完全なウイルス中和のためのmAb h2c-V1の力価を決定するために、いくつかの抗体濃度のものを100 TCID50のHSV-1またはHSV-2分離株とともに37℃で1時間インキュベートし、Vero細胞とともに3日間インキュベートした。mAb hu2c-V1は7.8〜15.6 nMの濃度で完全に、HSV-1実験室株HSV-1 F、HSV-1 324 hv、HSV-1 17 syn+、HSV-1 TK-を中和する。HSV-1臨床分離株はその耐性プロファイルに関係なく同様にmAb h2c-V1によって中和される。さらに、株HSV-2 GおよびACV耐性HSV-2分離株に対して31.3〜62.5 nMの濃度でmAb hu2c-V1の、同じ中和効率が示された。
【図9】本発明の抗HSV抗体によるウイルスの細胞間伝播の阻害を示す。4時間HSV-1 Fを感染させたVero細胞を2回洗浄し、過剰の、ヒトポリクローナル抗HSV対照血清(1:20)、マウスmAb 2c (500 nM)、酵素消化によって調製された2c由来の抗体断片F(ab')2 (500 nM)またはFab (3000 nM)、キメラmAb ch2c (500 nM)を含有する培地とともに、またはヒト化mAbバリアント1のhu2c-V1 (500 nM)とともに、それぞれ、インキュベートした。感染から2日後に、Leica DM IRE2共焦点顕微鏡を用い40倍の倍率で蛍光標識ポリクローナルヤギ抗HSV血清でウイルスの伝播を検出した。ヒトポリクローナル抗HSVの中和価は、100 TCID50を100μl容量中用いて1:160と以前に決定された。1:20の希釈で抗HSV血清は隣接細胞へのウイルスの伝播を抑止することができない。細胞間伝播を500 nMのマウスmAb 2c、2c-F(ab')2抗体断片、キメラmAbおよびヒト化mAbによって成功裏に阻害することができた。3.000 nMの最も高い試験濃度で一価2c-Fab断片は細胞間伝播を完全に阻害することができなかった。
【図10】抗HSV mAbによる受動免疫後にHSV-1を膣内感染させたNOD/SCIDマウスの生存を示す。マウスに感染の24時間前にPBS (十字)、2.5 mg/kg (四角)、5 mg/kg (三角)または15 mg/kg (丸)の親mAb 2c (白記号)またはヒト化mAb hu2c-V1 (黒記号)のいずれかを静脈内投与した。マウスに1×106 TCID50/20μlの神経病原性HSV-1株Fを膣内感染させた。体重減少、外陰炎/膣炎または神経疾患の症状を有する感染マウスを殺処理し、その臓器を、既述のようにVero細胞単層上での力価測定により感染性ウイルスについて調べた。非感染マウスを30日目の時点で殺処理した。1群あたりの動物数n=7。
【図11】HSV-1伝播に対しての全身適用抗体によるNOD/SCIDマウスの保護を示す。感染後24時間から始めて、マウスに、矢印で示した時点(24時間、40時間、56時間)で静脈内に3回15 mg/kgのmAb 2cまたはヒト化mAb hu2c-V1のいずれかを投与した。1群あたり感染動物数n=7。
【図12】アシクロビル、ホスカネットおよびシドボビルに対して耐性の患者由来HSV-1分離株を膣内感染させたNOD/SCIDマウスは、15 mg/kgのヒト化mAb hu2c-V1による感染後24時間、40時間および56時間の時点での静脈内処置によって致死性脳炎から有意に保護されたことを示す。1日2回アシクロビルによる標準的な処置を受けたマウスは、28日以内に全て死んだ。
【図13A−1】gBに対するmAb 2cのエピトープの局在性を示す。(A)はHSV1およびHSV2の糖タンパク質B (gB)のアミノ酸配列アライメントを示す。示されるのは、以下の株のgBタンパク質アミノ酸配列のアライメントである(括弧内は対応するNCBIアクセッション番号): HSV1株KOS (P06437)、F (P06436)、gC-39-R6 (ABM66850)、ならびにHSV2株333 (ABU45423)、HG52 (P08666)およびMMA (AAB60547)。gBのシグナル配列に下線を引いてある。成熟gBは31位からアミノ酸APで始まる。アミノ酸の付番は、シグナルペプチド配列を含むgBについて示されている。エピトープの付番はそれに従っている。mAb 2cは、ペプチドマイクロアレイによって示されるようにgB内の2つの別々の領域(囲まれた配列領域)に結合する。アミノ酸299PFYGYRE305はmAb 2cの結合に不可欠であることが示されている。
【図13A−2】gBに対するmAb 2cのエピトープの局在性を示す。(A)はHSV1およびHSV2の糖タンパク質B (gB)のアミノ酸配列アライメントを示す。示されるのは、以下の株のgBタンパク質アミノ酸配列のアライメントである(括弧内は対応するNCBIアクセッション番号): HSV1株KOS (P06437)、F (P06436)、gC-39-R6 (ABM66850)、ならびにHSV2株333 (ABU45423)、HG52 (P08666)およびMMA (AAB60547)。gBのシグナル配列に下線を引いてある。成熟gBは31位からアミノ酸APで始まる。アミノ酸の付番は、シグナルペプチド配列を含むgBについて示されている。エピトープの付番はそれに従っている。mAb 2cは、ペプチドマイクロアレイによって示されるようにgB内の2つの別々の領域(囲まれた配列領域)に結合する。アミノ酸299PFYGYRE305はmAb 2cの結合に不可欠であることが示されている。
【図13A−3】gBに対するmAb 2cのエピトープの局在性を示す。(A)はHSV1およびHSV2の糖タンパク質B (gB)のアミノ酸配列アライメントを示す。示されるのは、以下の株のgBタンパク質アミノ酸配列のアライメントである(括弧内は対応するNCBIアクセッション番号): HSV1株KOS (P06437)、F (P06436)、gC-39-R6 (ABM66850)、ならびにHSV2株333 (ABU45423)、HG52 (P08666)およびMMA (AAB60547)。gBのシグナル配列に下線を引いてある。成熟gBは31位からアミノ酸APで始まる。アミノ酸の付番は、シグナルペプチド配列を含むgBについて示されている。エピトープの付番はそれに従っている。mAb 2cは、ペプチドマイクロアレイによって示されるようにgB内の2つの別々の領域(囲まれた配列領域)に結合する。アミノ酸299PFYGYRE305はmAb 2cの結合に不可欠であることが示されている。
【図13B】gBに対するmAb 2cのエピトープの局在性を示す。(B) 異なるウエスタンブロット条件下での組換えgBとのmAb 2cの反応性によるmAb 2cの特徴付け。組換えgB (730t)を未変性(N)または変性(D)条件下での8% SDS-PAGEにて分離し、ニトロセルロース膜に転写し、2%ミルクを含有するTNTブロッキング用緩衝液中で1時間インキュベートした。この膜をgB特異的モノクローナル抗体mAb H1817、mAb H126またはmAb 2cでプローブし、gBとの結合をHRPコンジュゲートポリクローナルヤギ抗マウス血清および化学発光によって検出した。対照の場合には、連続的なエピトープ(Bender et al., 2007)および不連続的なエピトープ(Kousoulas et al., 1988)を認識する、それぞれ、mAb H1817およびmAb H126を用いた。線状エピトープに典型的な染色パターンがmAb H1817によるウエスタンブロット分析において得られ、非還元条件下でのgBの単量体型および三量体型の検出ならびに還元条件下でのgB単量体の唯一の主たる染色が示された。予想通り、mAb H126は未変性条件下でのみgBと反応した。170 kDa超(>170 kDa)の、唯一の、上方のgBタンパク質のバンドの認識から、mAb H126が三量体gBに特異的に結合することが示唆される。しかしながら、mAb 2cは未変性および変性gBと反応し、変性条件下での反応性はmAb H1817と比べてはるかに弱く、このことは、変性に対して耐性であるか、またはSDS-PAGE電気泳動の間に再び折り畳まれるように見え、それゆえ「偽連続」エピトープと呼ばれる、不連続なエピトープ(Bender et al., 2007)にmAb 2cが結合することを示唆している。分子量(kDa)が左側に示され、gB三量体および単量体の移動が右側に示されている。
【図14】gBに対するmAb 2cのペプチドマッピングを示す。(A) アミノ酸31〜505のgBの細胞外ドメインに及ぶペプチドライブラリに対して特定された結合領域AおよびBの図式的な局在。13 merのペプチドを3アミノ酸のずれで連続的なセルロース膜上で合成し、結合したmAb 2cをペルオキシダーゼコンジュゲート二次抗体で化学発光により検出した。機能ドメインI〜Vは、HeldweinらによるgBの結晶構造に対応し、結晶構造において解析されていない領域は灰色で示されている(24)。S, シグナル配列。(B) 可動性リンカーを介してスライドグラス上に固定化された13 merのペプチドによる高分解能レーザースキャンからの蛍光シグナル強度。
【図15】gB結晶構造(PDB-ID 2GUM)上の中和mAb 2cエピトープの局在を示す。gB三量体のリボンダイヤグラムが示されている。星印は2つのプロトマーの融合ループを示し、第3のプロトマーの融合ループは見えない。不連続的なmAb 2cエピトープF175〜A190およびF300〜E305のマッピング残基は、片方のプロトマーについては濃い灰色によりおよび他の両方のプロトマーについては薄い灰色により表面表現で示されている。
【図16】mAb 2cのデュオトープスキャニングを示す。mAb 2c結合領域AおよびB (ダッシュ記号付きのバー)のコンセンサス配列(下線を引いた)を、直接的にか、または1つもしくは2つのβアラニンスペーサー(B、B-B)によって離すかのいずれかで連結させたデュオトープ(白バーおよび黒バー)として合成した。mAb 2cとデュオトープとの反応性を高分解能レーザースキャンからの蛍光シグナル強度によって記録した。
【図17】フローサイトメトリーによって決定されたmAb 2c、2c-F(ab')2、2c-Fabおよび2c-scFvに対する平衡結合曲線を示す。表示濃度での(A) HSV-1 Fまたは(B) HSV-2 G感染Vero細胞に対する結合活性を最大の平均蛍光強度の百分率として示す。測定は三つ組で2回行った; バーは標準偏差を表す。
【図18】mAb 2cによる標的細胞へのHSV-1ウイルス粒子の付着の阻害を示す。(A) mAb 2c (0.98〜125 nM)または(B) 多価ヒトγグロブリン(Intratect(登録商標)) (0.33〜42μM)の連続希釈液を96ウェルマイクロタイタープレート中のVero細胞単層に添加し、その後、100 TCID50 HSV-1とのプレインキュベーション(付着前中和)処理または標的細胞への100 TCID50 HSV-1の吸着後(付着後中和)処理を行った。10個の個別の接種細胞単層におけるウイルス誘導性の細胞変性効果(CPE)を対照と比べて100%および50%抑止する、それぞれ、最も高い抗体および多価ヒトIgGの力価を37℃で72時間のインキュベーションの後に、終点と見なして決定した。独立した3回の実験の平均の標準誤差は0.1未満(< 0.1)であった。
【図19】HSVのインビトロでの中和に及ぼす抗gB抗体の結合価の影響を示す。(A) 二価抗体mAb 2c (IgG)および2c-F(ab')2、ならびに一価2c-Fabの希釈液をVero細胞への接種前に100 TCID50のHSV-1 FまたはHSV-2とともに1時間インキュベートした。図3に記述されるように72時間後にCPEをスコア化した。示されているのは、3回のうち1回の代表的な繰り返し実験からの、ウイルス接種材料の100%を中和するのに必要な抗体濃度である。(B) マウス抗Fab IgGで架橋された2c-Fab断片の抗ウイルス活性。
【図20】mAb 2c処置免疫不全マウスの用量依存的な生存を示す。NOD/SCIDマウスに1×106 TCID50 HSV-1での膣内攻撃の24時間前に異なる単回投与量のmAb 2cを静脈内投与した。1群あたりの動物数はPBSの場合n = 7、他の全群の場合n = 9。
【図21】HSV-1伝播に対して全身に適用されたmAb 2cまたはヒト化mAb hu2c-V1によるNOD/SCIDマウスの性器粘膜からの確立したHSV-1感染の除去を示す。感染後24時間から始めて、マウスに、矢印で示した時点(24時間、40時間、56時間)で静脈内に3回15 mg/kgのmAb 2c (白記号)またはヒト化mAb hu2c-V1 (黒記号)を投与した。抗体または対照処置マウスの膣ウイルス力価をVero細胞単層上で培養された膣洗浄液から決定した。誤差バーは標準偏差を示す。
【図22】アミノ酸31〜505のHSV-1 gB配列に対するペプチドスキャニングを示す。12アミノ酸の重複を有し、計155種の異なるペプチドスポットをもたらす、セルロース膜に結合させた15merのペプチド(15/12スキャン)をMAb 2cとともにインキュベートした。ペプチドに結合したMAb 2cを、ペルオキシダーゼ標識抗マウスIgG Fab断片およびルミノール様の化学発光基質を用いて検出した。MAb 2cは結合部位AおよびBと名付けた連続的なgBペプチドのトリプレット(ペプチド49〜51)およびダブレット(ペプチド90〜91)に結合することが分かった。各部位からのペプチドに共通するgB配列は太文字で強調されており(下部)、残基181〜189および残基301〜312のgB配列を表す。
【図23】アミノ酸296〜315のHSV-1 gB配列に対するペプチドスキャニングを示す。各ペプチドを1アミノ酸ずつ配列に沿ってシフトさせ(13/12スキャン)、二つ組で合成し、セルロース膜に結合させた13merのペプチドをMAb 2cとともにインキュベートし、その後、化学発光ウエスタンブロッティング検出を行った。一連の5つのペプチドに対してMAb 2cの結合を認めた。5つの反応性ペプチドに共通する配列は300FYGYREGSH308である。
【図24】残基295〜315のgB配列に対する鍵となるモチーフのスキャンを示す。アミノ酸295〜315のHSV-1 gB配列を、それぞれが1アミノ酸ずつ配列に沿ってシフトする六量体ペプチドに切断し、計16種のペプチドを得た。gB由来配列を各N末端およびC末端の位置で4つの無作為化残基により取り囲んだ。gB配列300FYGYRE305および301YGYREG306に相当する2つの連続ペプチドは、MAb 2cに結合することが特定された。これらのペプチドに共通する配列は太文字で強調されている(ペプチド6および7)。
【図25】ウイルス接種の24時間前に受動的に移入されたポリクローナル免疫血清(白四角、黒四角)、MAb 2c (白丸、黒丸)または対照として沈降培地(白三角、黒三角)を投与されたC57BL/6マウスの性器粘膜におけるHSV-1の除去動態を示す。野生型(wt)株Fおよびその変異派生体R126 (Y303N)、R1375 (R304Q)、R1435 (H308Y)およびR233 (R328H)を接種したマウスからのデータを白記号で示し、野生型株KOS 321およびその変異派生体B4.1 (E305K)を接種したマウスからのデータを黒記号で示す。S.E.の範囲(log10)および接種したマウスの数: wt株F、白四角、±1.5〜0、マウス12匹; 白丸、±0.4〜0、マウス7匹; 白三角、±1.3〜0、マウス9匹; wt株KOS、黒四角、±1.4〜0、マウス8匹; 黒丸、±0.6〜0、マウス8匹; 黒三角、±1.4〜0、マウス8匹; F変異体株R126 (Y303N)、白四角、±0.6〜0、マウス5匹; 白丸、±0.6〜0、マウス6匹; 白三角、±0.5〜0、マウス6匹; F変異体株R1375 (R304Q)、白四角、±1.2〜0、マウス11匹; 白丸、±1.3〜0、マウス10匹; 白三角、±1.2〜0、マウス11匹; KOS変異体株B4.1 (E305K)、黒四角、±0.9〜0、マウス12匹; 黒丸、±0.7〜0、マウス12匹; 黒三角、±0.9〜0、マウス10匹; F変異体株R1435 (H308Y)、白四角、±1.4〜0.6、マウス6匹; 白丸、0、マウス5匹; 白三角、±1.0〜0.6、マウス6匹; F変異体株R233 (R328H)、白四角、±1.0〜0、マウス5匹; 白丸、±0.1〜0、マウス5匹; 白三角、±1.1〜0、マウス6匹。
【図26】ペプチド90 (図22参照、gB配列298SPFYGYREGSHTEHT312; 左側)との、ならびに部位Bの重要な残基、グリシンリンカーおよび部位Aに由来するモチーフFEDFを含むようにデザインされたペプチド(PFYGYRE-G-FEDF; 右側)とのMAb 2cの反応性の比較を示す。
【発明を実施するための形態】
【0021】
発明の詳細な説明
したがって、第1の局面において、本発明は、
に示される相補性決定領域を含む抗体に関する。
【0022】
抗体または免疫グロブリンは、ジスルフィド結合によって連結された2本の大きな重鎖および2本の小さな軽鎖の、その天然形態にあるγグロブリンタンパク質である(図3参照)。5種類の哺乳類Ig重鎖: α、δ、ε、γおよびμが存在する。存在する重鎖の種類が抗体のクラス(アイソタイプ)を規定し; これらは、それぞれ、IgA、IgD、IgE、IgGおよびIgM抗体である。各重鎖は2つの領域、つまり定常領域および可変領域を有する。定常領域は同じ種の同じアイソタイプの全ての天然抗体においてほぼ同一である。軽鎖も1つの定常ドメインおよび1つの可変ドメインからなる。哺乳類では、2種類の免疫グロブリン軽鎖ラムダ(λ)およびカッパ(κ)が存在する。
【0023】
全ての抗体の一般構造は非常に似ているが、所与の抗体の特異的性質は可変(V)領域によって決定される。より具体的には、軽(VL)鎖上の各3つのおよび重(VH)鎖上の3つの可変ループが抗原との結合に、すなわち、その抗原特異性に関与する。これらのループは相補性決定領域(CDR)といわれる。VHおよびVLの両ドメイン由来のCDRが抗原結合部位に寄与するので、最終の抗原特異性を決定するのは重鎖および軽鎖の組み合わせであり、どちらか一方だけではない。
【0024】
本明細書において用いられる「抗体」という用語は、結合特異性がSEQ ID NO: 1〜6に示されるCDRによって決定される、抗原に結合できる任意のポリペプチドを意味する。ゆえに、「抗体」は、少なくとも1つの抗原結合断片を含む任意のポリペプチドに関するように意図される。抗原結合断片は、重鎖の可変ドメインおよび軽鎖の可変ドメインの両方がともに、特異的な抗原に結合できる形で配列された少なくともその両方のドメインからなる。「抗体」は完全な抗体、または抗体断片、例えばFab、F(ab)2もしくはscFv断片を含む(図3も参照のこと)。
【0025】
「完全な抗体」という用語に関して、各ドメインが、全体のドメイン構造を変化させない、変異、欠失または挿入などの、さらなる修飾をたとえ含んでいようとも、天然抗体(すなわち、3つまたは4つの定常ドメインの重鎖および1つの定常ドメインの軽鎖ならびに各可変ドメインを含む)に典型的な全体のドメイン構造を有する任意の抗体が意図される。
【0026】
「抗体断片」はまた、上記に定義される少なくとも1つの抗原結合断片を含み、断片が由来する完全な抗体と同じ機能および特異性を示す。Fab断片は、酵素パパインを用いて免疫グロブリンを切断することにより作出することができる。酵素パパインは、蝶番領域よりも下部、かくして、ジスルフィド結合よりも下部を切断し、その結果、F(ab)2断片が形成される。さらに、重鎖および軽鎖の可変領域をともに融合させて、一本鎖可変断片(scFv)を形成させることができる。
【0027】
さらに、「抗体」という用語は全ての上記の免疫グロブリンアイソタイプを含むよう意図され、すなわち、抗体はこれらのアイソタイプの任意のサブクラスを含めて、IgA、IgD、IgE、IgGまたはIgM抗体であってよい。好ましくは、抗体はIgG抗体、より好ましくはIgG1またはIgG2抗体である。抗体は組換えにより発現され産生されうるので、抗体は重鎖、例えば、1つのIgG1および1つのIgG2の重鎖、または異なる種由来の重鎖の、2つの異なる定常領域を含んでもよい。しかしながら、重鎖は同じ種に由来することが好ましい。さらに、抗体はラムダまたはカッパ軽鎖のどちらかを含むことができる。
【0028】
実施例2に示されるように、抗体の結合価は、ウイルス中和を媒介かつ細胞間伝播を阻害する効力に大きな影響を及ぼし、最良の結果は二価抗体で、すなわち、2つの抗原結合領域を有する抗体で示された。二価抗体の例は完全な抗体またはF(ab)2断片などの、二価抗体断片である。それゆえ、好ましい態様において、抗体は二価抗体であり、好ましくはここで抗体は完全な抗体または抗体断片であり、具体的にはここで抗体断片はF(ab)2断片である。その代わりに好ましい態様において、抗体は多価抗体、すなわち、組換え抗体もしくはその断片、好ましくは三重特異性抗体もしくは四重特異性抗体、またはIgM五量体もしくは連結された免疫グロブリンなどの全免疫グロブリンを含む、三つ以上の結合部位を有する抗体である。これらの抗体の形式は当技術分野において公知である。
【0029】
別の好ましい態様において、抗体はモノクローナル抗体であり、好ましくはここで抗体はマウス抗体、キメラ抗体またはヒト化抗体であり、より好ましくはここでヒト化抗体は以下で論じられるように、ヒト生殖細胞系配列に由来する。キメラ抗体は、その免疫原性を低減するために遺伝子操作によってある種の免疫グロブリンの少なくとも1つの領域が別の種の免疫グロブリンの別の領域に融合されている抗体である。キメラ抗体の例は図3Aに示されており、この図には、ヒト免疫グロブリンの残存部分に融合されたマウスVLおよびVH領域が描かれている。キメラ抗体の特定の種類はヒト化抗体である。ヒト化抗体は、非ヒト抗体のCDRをコードするDNAを、ヒト抗体フレームワークコーディングDNAにグラフトすることによって産生される。次に、得られたDNA構築物を用いて、単にCDRがヒト以外のものであるだけなので、非ヒト親抗体よりも、またはキメラ抗体よりも、通常、免疫原性ではない抗体を、発現かつ産生させることができる。
【0030】
1つの好ましい態様において、抗体は感染細胞から、隣接した第2の非感染細胞へのHSV感染の伝播(細胞間伝播)を阻害することができる。細胞間伝播は、無細胞粒子を放出させずに、ある感染細胞から、隣接した非感染細胞へと伝播するヘルペスウイルスの能力である。抗体が、感染細胞から、隣接した第2の非感染細胞へのHSVの伝播(細胞間伝播)を阻害できるかどうかを調べるために、以下のアッセイ法を用いることができる。
【0031】
24ウェル組織培養プレート中のカバーガラス上で集密まで増殖されたVero細胞を400 TCID50/ウェルの一定のウイルス量で37℃にて4時間感染させる。1メディアンの組織培養感染用量(1 TCID50)は、接種された細胞培養物の50%において細胞変性効果をもたらすと考えられる、ウイルスなどの細胞変性因子の量である。ウイルス接種材料を引き続いて除去し、細胞をPBSで2回洗浄し、上清によるウイルス伝播を防ぐために過剰の異なる抗HSV抗体またはポリクローナル抗HSV対照血清を含有するDMEM, 2% FCS, Pen/Strep 1 ml中37℃で2日間さらにインキュベートする。HSV感染細胞のウイルス抗原を蛍光標識ポリクローナルヤギ抗HSV血清(BETHYL Labolatories, Montgomery, TX USA, カタログ番号A190-136F, ロット番号A190-136F-2)で検出する。
【0032】
好ましくは、抗体は、隣接細胞の20%未満の感染であれば、細胞間伝播を阻害することができ、好ましくはここで上記のアッセイ法において隣接細胞の15%未満、10%未満、5%未満、より好ましくは3%未満および最も好ましくは1%未満が感染している。
【0033】
なお、さらに好ましい態様において、抗体は最大でも40 nM、好ましくは最大でも30 nM、より好ましくは最大でも20 nM、さらにより好ましくは最大でも15 nM、例えば最大でも13 nM、最大でも10 nM、および最も好ましくは最大でも7 nMの解離定数KDを有する。KDは、複合体がその構成要素へ可逆的に解離するその傾向(すなわち抗原に対する抗体の親和性)の尺度としての解離定数を表し、結合定数の逆のものである。KDはScatchard方程式から計算することができ、KDを決定するための方法は当技術分野において周知である。
【0034】
さらに好ましい態様において、最大でも20 nMの、好ましくは最大でも16 nMの、より好ましくは最大でも12 nMの、例えば最大でも10 nM、例えば最大でも8 nMまたは最大でも6 nMの、および最も好ましくは最大でも4 nMの濃度の抗体は、100 TCID50のHSVの規定量を80%超まで、好ましくは90%超、例えば95%超、より好ましくは96%超、例えば97%超、および最も好ましくは98%超、例えば99%超または場合により100%中和することができる。本明細書において「中和(する)」とは、ウイルスが任意のさらなる細胞に感染できないように抗体がウイルスをオプソニン化することを意味する。最大でも20 nMの濃度の抗体が100 TCID50のHSVの規定量を中和できるかどうか試験するためのアッセイ法は、Eis-Hubinger et al , 1991., およびEis-Hubinger et al., 1993に、ならびに以下の実施例1および2に示されている。
【0035】
さらに、1つの好ましい態様において、抗体は、図4に示されるように、SEQ ID NO: 9の1〜30位、38〜51位、68〜99位および112〜122位(またはKabatによる付番によれば、それぞれ、1〜30位、36〜49位、66〜94位および103〜113位)ならびにSEQ ID NO: 10の1〜23位、40〜54位、62〜93位および103〜113位(またはKabatによる付番によれば、それぞれ、1〜23位、35〜49位、57〜88位および98〜108位)に示されるアミノ酸残基と比べてフレームワーク領域中で少なくとも70%、好ましくは少なくとも75%、少なくとも80%、より好ましくは少なくとも85%、少なくとも90%、さらにより好ましくは少なくとも95%および最も好ましくは98%の(全体の)配列同一性を有するアミノ酸配列を含む。
【0036】
ポリペプチドは、SEQ ID NO: 9または10が関心対象のポリペプチドの最も適合している配列と整列され、整列されたそれらの2つの配列間のアミノ酸同一性が、SEQ ID NO: 9の1〜30位、38〜51位、68〜99位および112〜122位(またはKabatによる付番によれば、それぞれ、1〜30位、36〜49位、66〜94位および103〜113位)、ならびにSEQ ID NO: 10の1〜23位、40〜54位、62〜93位および103〜113位(またはKabatによる付番によれば、それぞれ、1〜23位、35〜49位、57〜88位および98〜108位)にわたって少なくともX %であるなら、SEQ ID NO: 9または10に対してフレームワーク領域中で「少なくともX %の配列同一性」を有する。かかるアミノ酸配列のアライメントは、例えば、国立バイオテクノロジー情報センター(NCBI)のホームページ上、http://www.ncbi.nlm.nih.gov/blast/blast.cgiで提供されている「BLAST」プログラムなどの公的に入手可能なコンピュータ相同性プログラムを使い、その中で提供されている初期設定を用いて行うことができる。アミノ酸配列または核酸配列のセットの配列同一性の割合を計算するさらなる方法は、当技術分野において公知である。
【0037】
あるいは、別の好ましい態様において、抗体は、図4に示されるように、SEQ ID NO: 7の1〜30位、38〜51位、68〜99位および112〜122位(またはKabatによる付番によれば、それぞれ、1〜30位、36〜49位、66〜94位および103〜113位)、ならびにSEQ ID NO: 8の1〜23位、41〜55位、63〜94位および104〜114位(またはKabatによる付番によれば、それぞれ、1〜23位、35〜49位、57〜88位および98〜108位)に示されるアミノ酸残基と比べてフレームワーク領域中で少なくとも80%、好ましくは少なくとも85%、より好ましくは少なくとも90%、さらにより好ましくは少なくとも95%、例えば98%および最も好ましくは100%の(全体の)配列同一性を有するアミノ酸配列を含む。ポリペプチドは、SEQ ID NO: 7または8が関心対象のポリペプチドの最も適合している配列と整列され、整列されたそれらの2つの配列間のアミノ酸同一性が、SEQ ID NO: 7の1〜30位、38〜51位、68〜99位および112〜122位(またはKabatによる付番によれば、それぞれ、1〜30位、36〜49位、66〜94位および103〜113位)、ならびにSEQ ID NO: 8の1〜23位、41〜55位、63〜94位および104〜114位(またはKabatによる付番によれば、それぞれ、1〜23位、35〜49位、57〜88位および98〜108位)にわたって少なくともX %であるなら、SEQ ID NO: 7または8に対してフレームワーク領域中で「少なくともX %の配列同一性」を有する。
【0038】
SEQ ID NO: 7および8はヒト生殖細胞系配列に由来する。たとえ非生殖細胞系列のヒト免疫グロブリンフレームワーク配列がヒト由来のものであっても、それらが免疫原性でないことを、通常、除外することはできない。それゆえ、本発明者らは、生殖細胞系配列を調べた。というのは、それらが高頻度変異しておらず、それゆえ、免疫原性ではないものと予想されるからである。したがって、ヒト化抗体は好ましくは、ヒト生殖細胞系配列に、例えばSEQ ID NO: 7および/または8に由来する。SEQ ID NO: 7および8は次の通りである。
【0039】
本発明との関連で、本発明のヒト化抗体の作出のためにSEQ ID NO:7および8を用いることで、親抗体に対するのと同じ親和性を達成するために帰突然変異は必要とされないことが分かったが、このことから、対応するヒト化抗体が非常に低い免疫原性を示すことが示唆されうる。その結果、本発明との関連で、SEQ ID NO: 9および10またはSEQ ID NO 7および8をそれぞれ含む抗体と同じ特異性を示す、抗体が含まれることが好ましい。
【0040】
さらに好ましい態様において、抗体はエフェクター部分、治療部分、または検出可能な部分とコンジュゲートしている。これに関連して、「コンジュゲート(している)」という用語は、非限定的に介在ドメインありまたはなしでの組換え融合、インテイン媒介融合、非共有結合性会合および共有結合、例えば、ジスルフィド結合、ペプチド結合、水素結合、静電結合、および高次構造的結合、例えば、ビオチン-アビジン会合を含む、タンパク質ドメインを機能的に結び付けるための当技術分野において公知の任意の方法をいう。エフェクター部分とのコンジュゲーションは化学的手段によるかまたは組換え手段によるかのどちらかであってよい。化学的手段は、1分子を形成するように2分子間で形成される共有結合が存在するような、抗体とエフェクター部分との間の反応をいう。
【0041】
「エフェクター部分」という用語は、抗体によって標的化される細胞に影響を及ぼすように意図された化合物を意味する。エフェクター部分は、例えば、治療部分または検出可能な部分でありうる。
【0042】
「治療部分」は、細胞毒性剤または細胞毒性薬などの、治療剤として作用するように意図された化合物である。化合物の例は以下、薬学的組成物に示される。
【0043】
「検出可能な標識」は、蛍光標識などの、分光学的、光化学的、生化学的、免疫化学的、電気的、光学的または化学的手段により検出可能な任意の化合物またはタンパク質タグを含む。
【0044】
抗体の特異性はCDRによりまたは抗体が結合するエピトープにより発現されうる。したがって、第2の局面において、本発明は、第1の局面の抗体と同じエピトープを認識する抗体に関する。実施例の項に示されるように、ならびに図13Aおよび13Bに例示されるように、このエピトープは、HSV1およびHSV2の糖タンパク質Bのアミノ酸172〜195および295〜313に位置する、変性に対して部分的に耐性の不連続な、というよりむしろ偽連続なエピトープである。
【0045】
本出願との関連で、mAb 2c抗体のエピトープはgBタンパク質の最初の487アミノ末端残基のなかに位置しうる。好ましくは、エピトープは、gBタンパク質の172位と307位との間のアミノ酸配列のなかに位置する少なくとも1つのアミノ酸配列を含みうる。
【0046】
エピトープは、gBタンパク質の連続アミノ酸配列301YGYRE305、好ましくは連続アミノ酸配列301YGYREG306または300FYGYRE305を含むことがあり、より好ましくは配列は、末端の位置でさらに伸長されうる(すなわち、299PFYGYRE305または300FYGYREGS307)。本発明の抗体のエピトープは、gBの連続アミノ酸配列298〜313 (298SPFYGYREGSHTEHTS313)を含みうる。
【0047】
あるいは、エピトープは連続アミノ酸配列172QVWFGHRYSQFMGIFED188に位置しうる。エピトープは連続アミノ酸配列172QVWFGHRYSQFMG184を含みうる。
【0048】
好ましくは、エピトープは2つ以上の連続アミノ酸配列からなりうる。エピトープは部分的に、不連続なエピトープであってもよい。より好ましくは、エピトープは2つの連続アミノ酸配列を含みうる。2つのアミノ酸配列からなるそのようなエピトープは、「デュオトープ」と指定することができる。抗体は両方のアミノ酸配列に結合することができる。
【0049】
より好ましくは、デュオトープのアミノ酸配列は、アミノ酸配列300FYGYRE305およびアミノ酸172位と188位との間に位置するアミノ酸配列を含みうる。さらにより好ましくは、エピトープは、gBタンパク質のアミノ酸配列300FYGYRE305およびアミノ酸配列179YSQFMG184を含みうる。あるいは、エピトープまたはデュオトープは、化学的に合成されてもよい。エピトープは、配列YSQFMG-βA-FYGYREを有する化学的に合成されたエピトープでありうる。本明細書において用いられる略語βAはβ-アラニンをいう。
【0050】
より好ましくは、エピトープは、gBタンパク質のアミノ酸配列FYGYREおよびアミノ酸配列FEDを含みうる。エピトープは、配列FED-βA-βA-FYGYREまたはPFYGYREGFEDFを有する化学的に合成されたエピトープでありうる。
【0051】
エピトープはgBタンパク質中に含まれうるが、その分解産物中に含まれてもよく、または化学的に合成されたペプチドであってよいことを当業者は理解することができる。アミノ酸の位置は、gBタンパク質の配列中の対応するアミノ酸配列の位置を明示するために示されるにすぎない。本発明は、該エピトープを含む全てのペプチドを包含する。ペプチドは、長さが100アミノ酸超のポリペプチドの一部であってよく、または100アミノ酸未満、好ましくは50アミノ酸未満、より好ましくは25アミノ酸未満、さらにより好ましくは16アミノ酸未満の小さなペプチドであってよい。そのようなペプチドのアミノ酸は天然アミノ酸もしくは非天然アミノ酸(例えば、β-アミノ酸、γ-アミノ酸、D-アミノ酸)またはその組み合わせであってよい。さらに、本発明はエピトープの各レトロインベルソペプチドを包含することができる。ペプチドは結合していてもよくまたは結合していなくてもよい。ペプチドは、例えば、小分子(例えば、薬物もしくはフルオロフォア)と結合していてもよく、高分子量重合体(例えば、ポリエチレングリコール(PEG)、ポリエチレンイミン(PEI)、ヒドロキシプロピルメタクリレート(HPMA)など)と結合していてもよく、またはタンパク質、脂肪酸、糖部分と結合していてもよく、あるいは膜中に挿入されていてもよい。
【0052】
当技術分野において公知の抗体H126とは対照的に、本発明のmAb 2c抗体によって認識されるエピトープは必ずしも不連続ではない。抗体H126とは対照的に、本発明の抗体は連続的なエピトープ、かくして、連続アミノ酸配列に結合することができ、または不連続なエピトープに結合しうる。それゆえ、本発明の抗体の特性は改善されている。例えば、mAb 2c抗体は、標的タンパク質が変性される方法(例えば、SDSページ電気泳動)に用いることができ、または小さな直鎖ペプチドの検出に用いることができる。
【0053】
問題の抗体および第1の局面の抗体が同じエピトープを認識するかどうかを調べるために、以下の競合試験を行うことができる: 3 moi (感染の多重度)で感染されたVero細胞を20時間後、競合物質としてさまざまな濃度の問題の抗体とともに1時間インキュベートする。第2のインキュベーション段階では、第1の局面の抗体を100 nMの定濃度で適用し、その結合を、第1の局面の抗体の定常ドメインに対して作製された蛍光標識抗体を用いてフローサイトメトリーにより検出する(実施例の項および図6も参照のこと)。問題の抗体の濃度に反比例する結合は、両方の抗体が同じエピトープを認識することを示す。しかしながら、使用できる多くの他のアッセイ法が当技術分野において公知である。
【0054】
第2の局面の好ましい態様は、上記の、第1の局面の場合と同じものである。
【0055】
第3の局面において、本発明は、第1または第2の局面による抗体の有効量と、少なくとも1つの薬学的に許容される賦形剤とを含む、薬学的組成物に関する。しかしながら、「薬学的組成物」という用語は「薬物」という用語と本明細書において互換的に用いることができる。
【0056】
薬学的組成物中の抗体の含量は、処置または予防に有用である限り、限定されないが、好ましくは全組成あたり0.0000001〜10重量%を含有する。さらに、本明細書において記述される抗体は、好ましくは担体中で利用される。担体の選択は投与の経路および活性薬剤の濃度に依ることができ、担体は凍結乾燥された組成物または水溶液の形態であることができる。一般的に、適切な量の薬学的に許容される塩を担体中に用いて、組成物を等張にする。担体の例としては、生理食塩水、リンゲル溶液およびデキストロース溶液が挙げられるが、これらに限定されることはない。好ましくは、クエン酸塩、リン酸塩および他の有機酸などの緩衝液; 塩形成対イオン、例えばナトリウムおよびカリウム; 低分子量(10アミノ酸残基未満)のポリペプチド; タンパク質、例えば血清アルブミンもしくはゼラチン; 親水性重合体、例えばポリビニルピロリドン; ヒスチジン、グルタミン、リジン、アスパラギン、アルギニンもしくはグリシンなどのアミノ酸; グルコース、マンノースもしくはデキストリンを含む糖質; 単糖類; 二糖類; 他の糖、例えばスクロース、マンニトール、トレハロースもしくはソルビトール; キレート剤、例えばEDTA; 非イオン性界面活性剤、例えばTween、プルロニック(Pluronic)もしくはポリエチレングリコール; メチオニン、アスコルビン酸およびトコフェロールを含む酸化防止剤; ならびに/または保存料、例えば塩化オクタデシルジメチルベンジルアンモニウム; 塩化ヘキサメトニウム; 塩化ベンザルコニウム、塩化ベンゼトニウム; フェノール、ブチルもしくはベンジルアルコール; アルキルパラベン、例えばメチルもしくはプロピルパラベン; カテコール; レゾルシノール; シクロヘキサノール; 3-ペンタノール; およびm-クレゾール)を含めて、許容される賦形剤、担体または安定剤は、利用される投与量および濃度で無毒性である。適当な担体およびその製剤はRemington's Pharmaceutical Sciences, 17th ed., 1985, Mack Publishing Coにさらに詳細に記述されている。
【0057】
組成物は、アシクロビル、ペンシクロビル、イドクスウリジンおよびホスカネットを含む、化学療法剤またはウイルス抑制剤などの、2つ以上の活性化合物を含むこともできる。
【0058】
アシクログアノシン(ACV)または2-アミノ-9-(2-ヒドロシキエトキシメチル)-3H-プリン-6-オンとしても公知のアシクロビルは、アセプレス(ACERPES)(登録商標)、アシック(Acic)(登録商標)、アシクロベータ(Aciclobeta)(登録商標)、アシクロCT(AcicloCT)(登録商標)、アシクロスタッド(Aciclostad)(登録商標)、アシクロビル、アシック(Acic)(登録商標)、オフタール(Ophtal)(登録商標)、アシビル(Acivir)(登録商標)、アシビジョン(AciVision)、アシクロビル(Acyclovir)(登録商標)、アウィラル(Aviral)(登録商標)、シクロビル、ヘルベビル(Helvevir)(登録商標)、ヘルペックス(Herpex)、スプラビラン(Supraviran)(登録商標)、ウィルカルム(Virucalm)(登録商標)、ウィルポス(Virupos)(登録商標)、ウィルジン(Virzin)、ゾリパリン(Zoliparin)(登録商標)、ゾビル(Zovir)およびゾビラックス(Zovirax)(登録商標)などの商品名で販売されているグアノシン類似体の抗ウイルス薬である。ペンシクロビル(2-アミノ-9-[4-ヒドロキシ-3-(ヒドロキシメチル)ブチル]-6,9-ジヒドロ-3H-プリン-6-オン)は、デナビル(Denavir)およびフェニスティル(Fenistil)などの商品名で販売されているグアニン類似体の抗ウイルス薬である。ファムシクロビル(Famciclovir)(酢酸2-[(アセチルオキシ)メチル]-4-(2-アミノ-9H-プリン-9-イル)ブチル)はペンシクロビルのプロドラッグであり、経口バイオアベイラビリティが改善されている。イドクスウリジン(2'-デオキシ-5-ヨードウリジン)は、ヌクレオシド・ウリジンの生化学的類似体であり、ウィルングェント(Virunguent)(登録商標)およびゾストラム(Zostrum)(登録商標)などの商品名で販売されている。ホスカネットは、式HO2CPO3H2を有する化合物の共役塩基であり、ホスカビル(Foscavir)(登録商標)およびトリアプテン(Triapten)(登録商標)などの商品名で販売されている。
【0059】
好ましくは、抗体および/または活性化合物は有効量で含まれる。
【0060】
「有効量」という用語は、薬学的組成物を投与しようとする対象において検出可能な治療反応を誘導するのに十分な量をいう。
【0061】
第4の局面において、本発明は、本発明の抗体をコードする核酸配列を含む発現ベクターを提供する。一般に、発現ベクターは、問題の遺伝子を標的細胞に導入し、その結果、該遺伝子によってコードされるタンパク質、すなわち抗体の転写および翻訳をもたらすために用いられるプラスミドである。したがって、発現ベクターは、発現ベクター上で運搬される遺伝子の効率的な転写を指令するためにポリアデニル化シグナル部位のほかに、プロモーターおよびエンハンサー領域などの調節配列を含む。発現ベクターはさらなる必要なまたは有用な領域、例えば真核生物または原核生物細胞における選択用の選択可能なマーカー、複製起点などを含むこともできる。
【0062】
そのため、第5の局面において、本発明は、本発明の抗体をコードするヌクレオチド配列を含む宿主細胞に関する。宿主細胞は、本発明の抗体を発現させるのに適した任意の細胞であることができ、哺乳類細胞、酵母細胞および昆虫細胞、好ましくは哺乳類細胞、より好ましくは骨髄腫細胞株などの不死細胞株を含む。適当な細胞株はアメリカンティッシュカルチャーコレクションATCCで入手可能である。
【0063】
さらに、第6の局面において、第1および/または第2の局面の抗体を産生することができるハイブリドーマ細胞が提供される。ハイブリドーマ細胞は、迅速かつ無制限に増殖し、所望の抗体を大量に産生しうる遺伝子操作細胞である。ハイブリドーマ細胞は、関連抗原で刺激された動物の脾臓から抗体産生B細胞を取り出し、これを次いで、不死骨髄腫腫瘍細胞と融合させることによって調製される。
【0064】
非常に重要な第7の局面において、本発明は、薬物としての使用のための本発明による抗体に関する。より具体的には、本発明は、対象におけるHSV関連疾患の予防的または治療的処置のための薬物の製造のための本発明の抗体の使用に関する。等しくは、本発明は、対象におけるHSV関連疾患の予防的または治療的処置における使用のための本発明の抗体に関する。さらに、本発明は、本発明の抗体が治療的有効量で対象に投与される、対象におけるHSV関連疾患の予防的または治療的処置の方法に関する。HSV感染はいくつかの異なる疾患を引き起こすことがある。よくある皮膚または粘膜の感染は、顔および口(口腔顔面ヘルペス)、生殖器(性器ヘルペス)または手(ヘルペス性ひょう疽)に影響を与えることがある。ウイルスが眼に感染し、損傷を与える場合(ヘルペス性角膜炎)、または中枢神経系に侵入し、脳に損傷を与える場合(ヘルペス脳炎)、より重篤な障害が発生する。新生児、移植レシピエント、またはAIDS患者などの、未成熟なまたは抑制された免疫系を有する患者は、HSV感染から重篤な合併症を起こす傾向がある。HSV関連疾患は剣状ヘルペス、モラレ髄膜炎、おそらくベル麻痺、躁鬱病、双極性感情障害または双極性気分障害としても公知の、双極性障害の認知障害と関連している障害、およびアルツハイマー病も含む。アルツハイマー病に関して、最近の科学的刊行物からβアミロイドプラーク内の1型単純ヘルペスウイルスDNAの著しい局在が実証され、このウイルスがプラークの原因である可能性が示唆されている。最後に、本発明による抗体の使用は、例えば免疫抑制患者の長期にわたる予防的および治療的処置の間に、よく用いられる化学療法のウイルス抑制剤に対する耐性株の出現が観察されるならば有用である。
【0065】
したがって、好ましい態様において、HSV関連疾患は以下の特徴の1つまたは複数を伴う: 経口再発の存在、生殖器再発の存在、ヘルペス性湿疹、新生児ヘルペス、免疫不全(免疫減弱状態の患者)、免疫抑制、脳炎、髄膜炎、髄膜脳炎、眼感染症、汎発性HSV感染症、および/またはウイルス抑制剤に対する耐性。
【0066】
別の好ましい態様において、HSV関連疾患は化学療法のウイルス抑制剤に対する不耐性を伴う。
【0067】
さらに好ましい態様において、薬物は少なくとも1つのさらなる活性剤を含み、好ましくはここでさらなる活性剤は化学療法剤またはウイルス抑制剤であり、より好ましくはここでさらなる活性剤は上記のように、アシクロビル、ペンシクロビル、イドクスウリジンおよびホスカネットからなる群より選択される。
【0068】
最後の好ましい態様において、対象はイヌ、ネコ、ブタ、ウシ、ヒツジ、ウマ、げっ歯類、例えばラット、マウスおよびモルモット、または霊長類、例えばゴリラ、チンパンジーおよびヒトなどの哺乳類であり、好ましくは対象はヒトである。
【実施例】
【0069】
本発明を以下の例によってさらに説明するが、これらは本発明の範囲を限定するものと解釈されるべきではない。
【0070】
実施例1
1型および2型単純ヘルペスウイルス(HSV1、HSV2)の糖タンパク質B (gB)に対する特異性を有するマウスモノクローナル抗体(mAb) 2cの調製
抗HSV特異的なmAbの作出のため、BALB/cマウスをUV不活化HSV1-株342 hvで免疫した。その後、マウス脾細胞を骨髄腫細胞株X63-Ag8.653との体細胞融合によって不死化し、抗HSV特異的なmAb 2c (IgG2a)を分泌するハイブリドーマ細胞株を、酵素免疫アッセイ法、免疫蛍光アッセイ法およびHSV中和アッセイ法を用いて単細胞クローンの上清をスクリーニングすることによって単離した。結合試験によって、mAb 2cがHSV1およびHSV2の糖タンパク質gBの共用の、不連続なエピトープを認識することが明らかになった。糖タンパク質Bは約904アミノ酸長であり、ウイルス膜中におよびHSV 1/2感染細胞の膜中に三量体として挿入される。mAb 2cが細胞外ウイルス粒子の伝播を中和するだけでなく、HSVに特徴的な、最初に感染した細胞から、隣接した非感染細胞への感染(細胞間伝播)の直接経路を効率的に阻害もすることは特に関係がある。後者の過程は、通常、ヒトにおいて天然のHSV特異的抗体のレパートリーによって阻害されることはない。
【0071】
mAb 2cのインビボでの効力を調べるために、マウスモデルにおける感染の経路を選択したが、これはヒトにおける自然感染のかかわり合いに酷似している。それゆえ、C57BL/6Jマウスに、その処女膣粘膜へのHSV1株342 hvの適用により感染させた。インビボでのHSV複製の阻害のため、mAb 2cを感染後の異なる時点でマウスに腹腔内(i.p.)投与した。免疫応答性動物においてもCD4+ T細胞枯渇動物においても、mAb 2cは、膣粘膜におけるウイルス増殖および短期間内での炎症性病変の形成を阻害することができる(図1)。
【0072】
ポリクローナルHSV血清とは対照的に、mAb 2cはウイルス複製を阻害することができただけでなく、T細胞(CD4+およびCD8+)が完全に枯渇されている免疫抑制動物において高い効率で全身の、致命的な進行性疾患を予防することができた(図2)。ウイルス接種24時間前のmAb 2cの投与は、感染から動物を効率的に保護した(Eis-Hubinger et al., 1993)。
【0073】
中和特性に及ぼす抗原結合部位の数(結合価)およびFc部分の影響を調べるために、mAb 2cのFab断片およびF(ab')2断片を従来のプロテアーゼ消化によって作出し、そのほかにも、当技術分野において周知の方法を用いてmAb 2cの組換え「一本鎖Fv」(scFv)をクローニングし、産生かつ精製した(図3)。
【0074】
当技術分野において周知の方法のように、自然のHSV複製サイクルの間に細胞表面に膜会合糖タンパク質としてgB-タンパク質を発現するHSV感染Vero細胞を用いてフローサイトメトリーにより親和性定数(KD)を決定した。この結果を下記表1に示す。
【0075】
(表1)マウスmAb 2cの親和性定数(KD)および作出された2c抗体断片の親和性定数(KD)
【0076】
親mAb 2cとは対照的に、F(ab')2断片はわずかに高い親和性を示す。Fab断片およびscFv断片はほぼ同一の親和性を有するが、しかし、これは親mAbの親和性よりも約1.7倍〜1.9倍弱いその一結合価によるものである。
【0077】
マイクロタイタープレート中で単層として増殖させたVero細胞を用いて標準的な終点希釈アッセイ法により抗体の中和活性を決定した。手短に言えば、100 TCID50のHSVをVero細胞の接種前に抗体の連続希釈液(2c-IgGおよびF(ab)2: 0.98 nM〜125 nM; Fab: 23 nM〜3000 nM)とともに37℃で1時間細胞培地100μl中でプレインキュベートした。37℃で72時間のインキュベーションの後、ウイルス誘導CPEの形成を100%まで抑止するのに必要な抗体濃度を完全中和価として決定した。さらに、一価2c-Fab断片のウイルス中和能を架橋抗体の存在下で決定した。
【0078】
(表2)100 TCID50の規定のウイルス量の完全中和
【0079】
親mAb 2cならびにそのF(ab')2断片およびFab断片はHSV1およびHSV2の完全中和の能力を有することを実証することができた。しかし、一価抗体断片はmAb 2cに比べてHSV 1/2に対して顕著に低減した中性化効率を示す。HSV1およびHSV2の100%の中和には、それぞれ、375倍および97倍高いFab断片濃度が必要である。scFvは、プラーク低減効果を示したが、3000 nMの最も高い試験濃度でCPEを完全に阻害することはできなかった(データ不掲載)。過剰の抗Fab特異的IgG (Jackson ImmunoResearch, Newmarket, England)をプレインキュベーション段階に加えることによって、一価2c Fab断片のウイルス中和能を2倍増強することができた(不掲載)。対照的に、二価F(ab')2断片は親mAb 2cと比べてHSV1に対してもHSV2に対しても実質的に2倍高い効率的な中和活性を示す。結論として、抗体価はその中和特性に重要な役割を果たす。HSV-2の完全な中和に必要な抗体濃度がいっそう高いことは、HSV-1およびHSV-2のDNAコピー数を決定するRT-PCRによって確認されるように、HSV-1に比べてHSV-2により産生される非感染性粒子の量がいっそう多いことで説明することができる(データ不掲載)。
【0080】
mAb 2cの特性およびその産生に関するさらなる詳細は、Eis-Hubinger et al.. 1991、およびEis-Hubinger et al.. 1993に示されている。
【0081】
実施例2
mAb 2cのキメラ化およびヒト化
マウスモノクローナル抗体2cを治療剤として利用するために、親抗体の特異性を完全に保持しながらも、ヒトへの投与中のその免疫原性を低減または排除することを目的として遺伝子操作の方法を用いて抗体を修飾した。したがって、mAb 2cと同じ特異性を有するキメラおよびヒト化モノクローナル抗体を作出した(図3)。
【0082】
最初に、mAb 2cの可変重鎖および軽鎖(VH、VL)の真正遺伝子を、5'-RACE (Rapid Amplification of cDNA Ends; cDNA末端の迅速増幅)-PCRによりハイブリドーマ細胞株から単離および増幅した。本発明者らの協力パートナーのGrosse-Hovest博士(Universitat Tubingen)が構築した発現ベクターであって、ヒトIgG1アイソタイプの軽鎖および重鎖の定常領域を含む該ベクターの中に、増幅されたVH-遺伝子およびVL-遺伝子をクローニングすることによってキメラIgG1抗体(ch2c)を作出した。この抗体は、安定的にトランスフェクトされたSp2/0マウス骨髄腫細胞の細胞培養上清へ最終的に分泌された。
【0083】
免疫原性をさらに低減するために、ヒト化抗体を構築した。それゆえ、mAb 2cの6つの相補性決定領域(CDR)をコードする遺伝子セグメント(2c VL-CDR1/2/3および2c VH-CDR1/2/3)をそれぞれ、ヒトVHおよびVL生殖細胞遺伝子の適当なヒトフレームワーク免疫グロブリンアクセプター骨格へクローニングした(CDRグラフト)。mAb 2cの軽鎖および重鎖のCDR領域をクローニングするのに適したヒト生殖細胞アクセプター骨格は、V-Baseデータベース(http://vbase.mrc-cpe.cam.ac.uk/)のうちの対応するヒトフレームワーク領域との配列のアライメントによって決定された。DP28は、対応するマウスmAb 2v重鎖VH配列に対して最も高いフレームワーク配列同一性(88.5%の配列同一性)を示した; DPK13は、対応するマウスmAb 2c軽鎖VL配列に対して最も高いフレームワーク配列同一性(88.9%の配列同一性)を示した。したがって、マウスドナー抗体2cのCDRコード遺伝子セグメント(すなわち2c VL-CDR1/2/3および2c VH-CDR1/2/3)をそれぞれ、DP28およびDPK13をコードするアクセプターフレームワークへクローニングした。
【0084】
モノクローナル抗体のヒト化との関連で、導入されたマウスCDRの構造的完全性にとって有害な可能性がある、したがって抗原結合特性にとって有害な可能性がある、ヒトフレームワーク領域中のアミノ酸を特定することが必要である。通常、そのようなアミノ酸はコンピュータで作った相同性モデルを用いて特定され、立体的に重要であるようにみえる位置を、マウスドナーmAbの抗原結合特性を保持するために、マウスの対応配列に変異させる(Queen el al., 1989参照)。しかしながら、潜在的に重要なアミノ酸は、抗体レパートリーのデータベースを用い、公知の三次元構造を有する参照抗体に基づきその臨界的意義を評価して特定することもできる(Krauss et al., 2003参照)。したがって、VH-およびVL-フレームワーク領域中のいくつかの潜在的に重要なアミノ酸を決定し(図4)、これらの潜在的に重要なアミノ酸を、マウスのその対応残基に連続的に復帰突然変異させた4種のヒト化mAb 2cバリアントを重複伸長PCRによって作出した(下記表3参照)。
【0085】
(表3)マウスドナー配列へのフレームワーク領域中の復帰突然変異を有するヒト化mAb 2cバリアント
【0086】
ヒト化抗体バリアントh2c-V1-4は、本発明者らの協力パートナーのGrosse-Hovest博士(Universitat Tubingen)が構築した上記の発現ベクターの中にヒト化VH-遺伝子およびVL-遺伝子をクローニングすることによって構築された。この抗体は、マウス骨髄腫細胞株Sp2/0の安定なトランスフェクションの後に最終的に発現された。高い特異的産生率を有するクローンの選択後、抗体が定量的に産生され、さらなる特徴付けのために細胞培養上清から精製された。
【0087】
キメラおよびヒト化抗HSV IgG1抗体の特徴付け
HSV感染Vero細胞を用いて、親mAb 2cおよび2c抗体断片の場合に記述したようにフローサイトメトリーによって同様に、親和性定数(KD)を決定した(実施例1参照)。この結果を図5に示す。
【0088】
キメラ抗体ch2cは親抗体mAb 2cの親和性を保持していた。ヒト化バリアントの場合、バリアントh2c-V1に関して唯一のCDRグラフトはマウスmAb 2cに匹敵する親和性を保存するのに十分であった。それゆえ、各マウス配列へのヒトフレームワーク残基のさらなる連続的な復帰突然変異は、抗原結合部位の構造的完全性を改善するのに必要ではなかった。バリアントh2c-V2はさらに、mAb 2cと比べて2倍低い親和性を示す。
【0089】
キメラmAb 2cおよびヒト化抗体バリアントmAb h2c1-4が親mAb 2cと同じエピトープを認識することを実証するために、競合試験を行った。HSV-1感染(3 moi、20時間) Vero細胞を最初に、漸増濃度の、それぞれ、キメラmAb ch2cまたはヒト化mAb hu2c V1-V4とともに1時間インキュベートした。第2のインキュベーション段階では、100 nMの親mAb 2cを添加し、その結合を、定常ドメインに対して作製された蛍光標識抗体を用いてフローサイトメトリーにより検出した(図6)。
【0090】
競合試験から、競合物質の結合を表す蛍光シグナルは第1のインキュベーション段階で適用された非標識抗体の濃度に反比例することが明らかである。このことは、キメラmAbおよびヒト化mAbバリアントが同じ特異的結合部位に向けてmAb 2cと競合し、ゆえに同じエピトープを認識することを証明している。
【0091】
mAb ch2cおよび4種のヒト化バリアントh2cのウイルス中和能を上記のように、精製された抗体調製物で調べた。この結果を下記表4に示す。100 TCID50の規定のウイルス量の50%のおよび完全なHSV中和に必要な各濃度を示す。
【0092】
(表4)100 TCID50の規定のウイルス量の50%のまたは完全な中和に必要な抗体濃度
【0093】
キメラmAb ch2c、およびmA h2c-V2以外の全てのヒト化mAb h2cは、親mAb 2cと同じ効率でHSVを中和する。mAb h2c-V2の2倍低い中和効率は、このバリアントのいっそう低い親和性と相関する。さらなる実験的特性評価および前臨床評価のため、mAb h2c-V1を選択したが、これは、このバリアントが親抗体mAb 2cと同じ親和性およびウイルス中和特性を保有するからである。さらに、mAb h2c-V1はフレームワーク領域中にマウスドナー配列への復帰突然変異を持たず、それゆえ、ヒトにおいて低い免疫原性能を保有するものと予想される。
【0094】
ヒト化mAb h2c-V1およびマウスmAb 2cの中和活性に及ぼす補体の影響を、プラーク低減アッセイ法を用いて詳しく調べた。ヒト高力価免疫グロブリン血清(Cytotect(登録商標), Biotest AG)とは対照的に、親mAb 2cおよびヒト化バリアントmAb h2c-V1はHSVを補体非依存的に中和する(図7)。HSV-1 gBに対する補体非依存的な中和抗体は、0.8〜160μg/mlの力価でウイルス投入量の50%を中和することが文献に記述されている(Navarro et al, 1992, Virology 186)。マウスmAb 2c、キメラmAb ch2cおよびヒト化mAbバリアントの終点希釈アッセイ法によりHSV-1 (F)を50%中和するのに必要な力価は3.3〜5.1 nMであり、これは0.49〜0.78μg/mlに相当する(表4参照)。
【0095】
HSV-1およびHSV-2の臨床分離株の中和アッセイ法から、HSV-1およびHSV-2の実験室株と比べてヒト化mAbバリアントh2c-V1による不活化に対する同じ感受性が実証される。さらに、mAb h2c-V1は非耐性実験室株または未知の耐性を有する臨床分離株と同じ効率で、アシクロビル(ACV)、ACVおよびホスカネット(PFA)、またはACV、PFAおよびシドビル(Cidovir) (CDV)に対して臨床的に耐性の患者分離株由来のHSVを中和する(図8)。ゆえに、ヒト化mAb h2c-V1は、耐性HSV株を誘導する従来の抗ヘルペス薬による限界を克服する新しい強力な抗ウイルス剤となる。
【0096】
宿主内での伝播のために、HSVは2つの機構: 無細胞粒子の放出および直接的な細胞間伝播を用いる。HSVの細胞間伝播は、より迅速なウイルス複製および播種、ならびに液性免疫反応の要素に対する耐性のような、無細胞伝播に対して優位性を保ちうる。抗HSV抗体による細胞間伝播を調べるために、Vero細胞を24ウェル組織培養プレート中のカバーガラス上に播き、集密まで増殖させ、これに37℃で4時間1ウェルあたり400 TCID50のHSV-1 Fを接種した。ウイルス接種材料を吸引し、Vero細胞をPBSで2回洗浄し、2% FBS、抗生物質を含有する、かつ過剰の中和抗体、つまりヒトポリクローナル抗HSV血清を含有するか、または対照の目的で中和抗体を含有しないかのどちらかのDMEM 1 ml中にて37℃で48時間さらに培養した。48時間後、培地を除去し、Vero細胞単層をHEPES緩衝生理食塩水で2回洗浄し、室温で15分間PBS中4%のパラホルムアルデヒド中で固定した。細胞単層をPBSで2回すすぎ、HEPES緩衝生理食塩水中0.05%のTween 20を含有するブロッキング緩衝液500μl中で15分間インキュベートした。ブロッキング緩衝液中で1:100に希釈されたFITCコンジュゲートポリクローナルヤギ抗HSV血清(BETHYL, Montgomery, TX, USA)でのHSV-1感染細胞単層の染色によりウイルス抗原を検出した。細胞単層をPBSで3回洗浄し、Mowiol (Calbiochem, San Diego, CA, USA)でマウントした。免疫蛍光陽性細胞をLeica DM IRE2共焦点顕微鏡により40倍の倍率で捕捉した(図9)。
【0097】
ヒトポリクローナル抗HSV血清(1:20)は、HSVの細胞間伝播に阻害効果を及ぼさない(図9A)。親mAb 2cおよびそのF(ab)2断片は500 nMの濃度で細胞間伝播を完全に阻害し、1つのHSV感染細胞しか検出することができない(図9BおよびC)。親mAb 2cの6倍高い濃度で適用されたFab断片は、ヒトポリクローナル抗HSV血清と比べて細胞間伝播をわずかに低減するが、しかし細胞間伝播を完全に阻害することができない(図9D)。中和アッセイ法において既に示されたように、これらの結果から、中和抗体の二価はHSV伝播を阻害するその能力に重要な役割を果たすことが確認される。キメラmAb ch2cおよびヒト化バリアントmAb h2c-V1は、親mAb 2cと同じくらい効率的に500 nMの濃度でHSVの細胞間伝播を阻害する(図9EおよびF)。さらに行われた従来のプラーク阻害アッセイ法(Highlander el al., 1988)から、共焦点顕微鏡での評価によって得た結果が確認された。
【0098】
最初のインビボHSV保護実験から、親mAb 2cと同様に5 mg/kg i.v.単回用量のヒト化mAb h2c-V1が膣内HSV-1 F感染した重症免疫不全マウス(NOD-SCID)の生存を顕著に引き延ばすことが明らかである(図10)。15 mg/kgのヒト化mAb h2c-V1または親mAb 2cのどちらかを投与されたマウスは、致死性脳炎から完全に保護される(図10)。さらに、ヒト化mAb 2c-V1はまた、樹立された末梢HSV感染の存在下でウイルス伝播および致死性脳炎からの保護をもたらす。ウイルス攻撃後24時間の時点で膣洗浄において高いHSV-1力価を有するNOD/SCIDマウスは、15 mg/kgのヒト化mAb 2c-V1またはmAb 2cにより24時間、40時間および56時間の時点で繰り返し静脈内処置された場合、感染の致命的結末から完全に保護された(図11)。さらに、ヒト化抗体mAb 2c-V1はまた、多耐性HSV株の樹立感染を有するNOD/SCIDにおいて致死性脳炎を抑止した。対照的に、アシクロビルを投与されたマウスは全て死んだ(図12)。
【0099】
エピトープマッピング
全長gB (31〜904)または720、630、503、487および470の位置でC末端の切断を伴うgB変異体のどちらかをコードする発現プラスミドでトランスフェクトされたCOS-1細胞を用いた結合試験により、マウス親mAb 2cによって認識されるエピトープがgBの最初の487アミノ酸のなかに位置付けられた。連続ペプチドの間に12 aaの重複を有する、固相に結合された15アミノ酸(aa)長の合成ペプチドを用いたさらなる研究によって、mAb 2cが立体構造エピトープに位置することが示された。mAb 2cはaa 175〜195の糖タンパク質B配列(領域A)に相当する3つの連続ペプチドに結合する。さらに、mAb 2cは、アミノ酸298〜312に相当するペプチド(298SPFYGYREGSHTEHT312) (SEQ ID NO: 17)に強力に結合し、アミノ酸301〜315に相当する後続のペプチドに中程度に結合する(図13Aおよび図22)。未変性または変性SDS-PAGE条件の下で分離された組換えgB (gB(730)t、Florent Bender, University of Pensylvania, Philadelphia, USAから親切にも提供していただいた)を用いたウエスタンブロットに関するその反応性によるmAb 2cの特徴付けからは、変性に対して部分的に耐性であるか、またはウエスタンブロット条件の間に再編成され、それゆえ、「偽連続」エピトープと呼ばれる(Bender, F et al. J. Virol. 2007, 81 p3872-3841を参照のこと)不連続なエピトープを、mAb 2cが認識することが確認される(図13B)。
【0100】
mAb2cのウイルス中和活性に関連するHSV-1 gBタンパク質中のエピトープを特定するために、その糖タンパク質中に単一のアミノ酸(aa)交換を有するモノクローナル抗体耐性(mar) HSV-1変異体について研究した(表5)。
【0101】
(表5)モノクローナル抗体耐性(mar) HSV-1変異体に対するmAb 2cの中和および結合活性
(a) シグナル配列を含む成熟糖タンパク質Bによる付番(図13A)
(1) Kousoulas et al., 1984
(2) Pellett et la., 1985
(3) Kousoulas et al., 1988
(4) Highlander et al., 1989
【0102】
mAb 2cはmar変異体R126、R1375、およびB4.1を中和しなかったが、それぞれ、変異体R1435およびR233の感染性を完全に中和した。さらに、免疫蛍光アッセイ法から、mAb 2cがmar変異体R126、R1375、およびB4.1に感染したVero細胞に結合しないことが確認された。これらの結果から、アミノ酸Y303、R304およびE305がmAb2cの中和活性に不可欠であることが示唆される。mar変異体R1435およびR233に感染したVero細胞を用いて強力な蛍光シグナルが得られた。
【0103】
特に、gBのアミノ酸303〜305に対するmAb 2cの認識および結合がその機能(ウイルス中和、細胞間伝播の阻害)に不可欠であることが分かった。gBのこの領域はHSV-1株とHSV-2株との間で高度に保存されているので、これらのアミノ酸はgBのコアの融合機構に属し、ウイルス侵入に不可欠であることが想定される。それゆえ、mAb 2cが結合しない天然のgB変異体の出現は、高い選択圧の下でさえ可能性が低い。
【0104】
実施例3
抗体親和性の判定
Vero細胞の単層に80〜90%の集密でMOI 3のHSV-1またはHSV-2を感染させ、これを翌日、トリプシン処理により、その後、PBS中での洗浄により収集した。2c抗体の細胞表面結合測定は既報(1)のように行った。手短に言えば、精製されたmAb 2cまたはmAb 2c由来抗体断片2c-F(ab')2、2c-Fabおよび2c-scFvを室温で1時間、FACS緩衝液(PBS、2% FBS、0.1%アジ化ナトリウム) 100μl中5×105個のVero細胞とともに、0.03 nM〜500 nMの濃度にて三つ組でインキュベートした。細胞をFACS緩衝液200μlで2回洗浄し、結合したmAb 2c、2c-F(ab')2、および2c-Fabの検出のためFITC標識Fab特異的ヤギ抗マウスIgG, (15μg/ml, Jackson ImmunoResearch, Newmarket, Suffolk, England)とともにインキュベートした。結合したscFvは飽和濃度の抗c-myc mAb 9E10 (10μg/ml; Roche, Indianapolis, IN, USA)と最初にインキュベートし、その後、2回の洗浄およびFcγ特異的FITC標識ヤギ抗マウスIgG (15μg/ml; Jackson ImmunoResearch)とのインキュベーションを行うことによって検出された。細胞を2回洗浄し、FACS緩衝液中に再懸濁した。蛍光をFACScalibur (BD Bioscience, San Jose, CA, USA)にて測定し、CellQuest(商標)ソフトウェア(BD Biosciences)を用いて平均蛍光強度(MFI)を計算した。バックグラウンドの蛍光を差し引き、GraphPad Prism 4.0版(GraphPad Software, La Jolla, CA)で非線形回帰のためのMarquardtおよびLevenbergの方法を用いることにより平衡結合定数を決定した。
【0105】
エピトープの特徴付け
Roselyn J. EisenbergおよびGary H. Cohen (University of Pennsylvania, Philadelphia, USA)から親切にも提供していただいた、未変性または変性切断型糖タンパク質B、つまりgB(730)t (4)とのmAb 2cの免疫反応性は、本質的には既報(4)のように行った: 精製gB(730)t (0,75μg)を非還元(0.2% SDSを含有するサンプル用緩衝液)条件または変性(2% SDSおよび155 mM β-メルカプトエタノールを含有するサンプル用緩衝液、95℃で2分)条件のどちらかの下で8% SDS-PAGEにて分離し、ニトロセルロース膜上に転写した。膜ストリップをTNT緩衝液(0.1 M Tris.HCl, pH 7.5, 0.15 M NaCl, 0.05% Tween-20)中2%のミルクで1時間ブロッキングし、その後、2%ミルク/TNT緩衝液中5μg/mlの糖タンパク質B特異的抗体mAb 2c、H126 (Novus Biologicals, Littleton, CO, USA)およびH1817 (Novus)との室温で2時間のインキュベーションを行った。結合した抗体を西洋ワサビペルオキシダーゼコンジュゲートポリクローナルヤギ抗マウス抗体(1:20,000 QED Bioscience Inc. San Diego, CA, USA)および化学発光(Thermo Scientific,)で、LAS 3000 Luminescent Image Analyzer (Fujifilm, Tokyo, Japan)を用いて検出した。
【0106】
COS-1細胞を、全長HSV-1 gB (31〜904, pRB9221)または720位(pTS690)、630位(pPS600)、503位(pRB9510)、487位(pRB9509)および470位(pRB9508)で切断されたC末端欠失変異体をコードするプラスミドでDEAE-デキストラン法により一過的にトランスフェクトした。これらのプラスミドはL. Pereira (52, 55)から親切にも提供していただいた。mAb2cまたは対照抗体を用いたトランスフェクト細胞での免疫蛍光アッセイ法は、他で既報(53)のように行った。
【0107】
ペプチドマッピング
セルロースに結合された重複する13 merのペプチドおよびデュオトープは既報(20, 34)のように標準的なSPOT合成プロトコルにしたがって自動的に調製された(JPT Peptide Technologies, Berlin, Germany)。さらに、反応性タグおよびリンカーとカップリングされたペプチドを3つの同一のサブアレイ中の修飾ガラス表面上に化学選択的に固定化し、切断されアセチル化された配列の除去、その後の洗浄段階によって精製した。ペプチドマイクロアレイを、TBS含有ブロッキング用緩衝液(Pierce International)で2時間ブロッキングし、ブロッキング用緩衝液中10μg/mlのmAb 2cとともに2時間インキュベートした。ペプチドマイクロアレイを、0.1% Tweenを含有するTBS緩衝液(T-TBS)で洗浄し、ペプチド膜上のペプチドに結合した抗体をPVDF膜上に転写した。ペルオキシダーゼ標識された抗マウスIgG (Sigma)または蛍光標識された抗マウスIgG (Pierce)をブロッキング用緩衝液中1μg/mlの終濃度で二次抗体として用いた。2時間のインキュベーションおよびT-TBSによる最終の洗浄の後、化学発光基質(Roche Diagnostics)を用いてPVDF膜を分析した。スライドガラスペプチドマイクロアレイをT-TBSおよび3 mM SSC緩衝液(JPT Peptide Technologies)で徹底的に洗浄し、窒素下で乾燥し、高分解能蛍光スキャナ(Axon GenePix 4200 AL)を用いてスキャンした。蛍光シグナル強度(発光量, LU)を、スポット認識ソフトウェア(GenePix 6.0)を用いて分析し、二次抗マウスIgGとの対照のインキュベーションからのバックグラウンドの強度に対して補正した。
【0108】
ウイルス中和アッセイ法
抗体の中和活性を既報(16)のように終点希釈アッセイ法によって決定した。手短に言えば、抗体の連続希釈液を細胞培地中37℃で1時間100 TCID50のHSV-1またはHSV-2とともにインキュベートした。抗体ウイルス接種材料を、マイクロタイタープレート中で増殖させたVero細胞単層に適用し、37℃で72時間のインキュベーション後に細胞変性効果(CPE)をスコア化した。ウイルス誘導性CPEを100%低減するのに必要な抗体濃度を完全中和価として決定した。さらに、一価2c-Fab断片のウイルス中和能を架橋抗体の存在下で、過剰の抗マウスFab IgG (2600 nM, Jackson ImmunoResearch, Newmarket, Suffolk, England)をプレインキュベーション段階に加えることによって調べた。対照の目的で、抗体なしのウイルスおよび抗体のみを用いて、それぞれ、最大CPEまたはCPEなしを誘導した。ウイルス中和アッセイ法を少なくとも2回繰り返し、同じ結果となった。
【0109】
付着後中和アッセイ法
予冷されたVero細胞単層(15分間4℃)に1時間4℃で100 TCID50 HSV-1 Fを感染させてウイルス吸収を可能とした後に、mAb 2cまたはヒト血漿由来の多価IgG調製物(Intratect(登録商標), Biotest AG, Dreieich, Germany)のどちらかの連続希釈液を添加した(付着後中和)。同一の実験条件下でのmAb 2cの付着前 vs 付着後中和効率を比較するため、予冷されたVero細胞単層に添加する前に、100 TCID50 HSV-1 Fを同じ抗体希釈液とともに4℃で1時間インキュベートした。両アッセイ由来の接種済みVero細胞を37℃に移す前に、4℃でさらに1時間インキュベートした。上記の標準的な中和アッセイ法に記述されているように、72時間後に中和価を決定した。
【0110】
細胞間伝播アッセイ法
集密までカバーガラス上で増殖されたVero細胞に37℃で4時間500μl中400 TCID50/ウェルでHSV-1 Fを接種した。ウイルス接種材料を吸引し、Vero細胞をPBSで2回洗浄し、2% FBSを含む増殖培地1 ml中、過剰の中和抗体の存在下または非存在下において37℃で48時間さらに培養した。高力価の抗HSV-1免疫グロブリンを有する免疫ドナーに由来するプール済みヒト血清を1:20の希釈で対照として用い、二価抗体mAb 2cおよび2c-F(ab')2の濃度は500 nMとし、一価2c-Fabの濃度は3000 nMとした。48時間後、培地を除去し、Vero細胞単層をHEPES緩衝生理食塩水で2回洗浄し、室温で15分間PBS中4%のパラホルムアルデヒドで固定した。ウイルス伝播を可視化するため、細胞をPBSで2回すすぎ、0.05% Tween 20を含むHEPES緩衝生理食塩水500μl中で15分間インキュベートし、FITCコンジュゲートポリクローナルヤギ抗HSV血清(1:100, BETHYL, Montgomery, TX, USA)で染色した。PBSによって3回洗浄した染色細胞を、0.2 g/ml Mowiol 4-88 (Calbiochem, San Diego, CA, USA)を含有するマウンティング培地中でマウントした。蛍光像をLeica DM IRE2共焦点顕微鏡により40倍の倍率で捕捉した。細胞間伝播の阻害を吸収後ウイルス中和アッセイ法によってさらに試験した。6ウェルプレート中で集密まで増殖されたVero細胞を、2% FBSおよび抗生物質を含有するDMEM 3 ml中、200 TCID50のHSV-1 Fとともに37℃で4時間インキュベートした。細胞単層をPBSで2回洗浄し、過剰の中和抗体またはポリクローナルヒトHSV-1中和血清を含有する温かいプラーク形成用培地(DMEM, 5% (w/v)アガロース, 10% FBS, 抗生物質)で重層した。37℃で48時間のインキュベーション後、光学顕微鏡検査によってプラーク形成を分析した。
【0111】
DNA定量
リアルタイム(RT) PCRを行ってHSV-1およびHSV-2ゲノムを定量化した。自動核酸抽出システムMagNA Pure LC System (Roche)を製造元の指示にしたがって用い、等量のHSV-1およびHSV-2感染粒子を含有するサンプルからDNAを精製した。ウイルスDNAを次いで、Real Art HSV-1 / HSV-2定量キット(Qiagen)を用いRT-PCR (Lightcycler, Roche)を行って定量した。
【0112】
マウス保護実験
麻酔した6〜8週齢の、雌性非肥満糖尿病/重症複合免疫不全(NOD-SCID)マウス(NOD.CB17-Prkdcscid/J, Charles River Laboratories, Research Models and Services, Sulzfeld, Germany)を1×106 TCID50 HSV-1 F/マウスの接種材料20μlで膣内から攻撃した。皮膚接着剤(Epiglu, Meyer-Haake Medical Innovations, Wehrheim, Germany)を外陰部に適当して、ウイルス接種材料の流出(dircharge)を抑止した。送達された接種材料は膣洗浄液の培養によって評価した場合に94%超(>94%)の感染率を誘導した。マウスをウイルス接種後毎日、体重減少、外陰炎/膣炎(発赤、粘液膿性排出物および炎症の兆候)ならびに神経疾患について調べた。これらの症状のいずれかを呈するマウスをすぐに殺処理した。免疫予防の場合にはウイルス接種の24時間前に、または治療的処置の場合にはウイルス感染から24時間、40時間および56時間後に、精製mAb 2cの静脈内(i.v.)注射によってマウスを受動的に免疫した。Vero細胞に対する終点希釈アッセイ法を用いて感染後1、2、4、6および8日目に、ならびに死亡時に得られた膣洗浄液からのウイルス力価の判定により感染についてマウスを評価した。殺処理したマウスの臓器(脾臓、副腎、肺、心臓、肝臓、腎臓、脊髄および脳)中のウイルス量を、他で既報(41)のようにVero細胞単層に対する力価測定によって臓器のホモジナイゼーション後に判定した。各試験および対照群には、検出可能なHSV-1感染を伴う動物9〜10匹を含めた。
【0113】
結果
mAb 2cによって認識されるgBエピトープのマッピングおよび分析
最近になって決定された1型HSV (HSV-1)由来gB細胞外ドメイン結晶構造から、5つの異なる構造ドメイン: ドメインI (基部)、ドメインII (中央部)、ドメインIII (コア部)、ドメインIV (冠部)およびドメインV (腕部)を有する多ドメイン三量体が明らかになった(24)。mAb 2cの中和エピトープを特徴付けるために、本発明者らは、還元条件下または非還元条件下のどちらかのウエスタンブロット分析において組換えgB(730t)(4)とのその反応性について試験した。対照として、本発明者らは、線状エピトープを認識するmAb H1817 (4)および不連続なエピトープを認識するmAb H126 (33)を用いた。線状エピトープに典型的な染色パターンがmAb H1817によるウエスタンブロット分析において得られ、非還元条件下でのgBの単量体型および三量体型の検出ならびに還元条件下でのgB単量体の唯一の主たる染色が示された(図13B)。予想通り、mAb H126は未変性条件下でのみgBと反応した。驚いたことに、170 kDa超(>170 kDa)の、唯一の、上方のgBタンパク質のバンドの認識から、mAb H126が三量体特異的であることが示唆される(図13B)。しかしながら、mAb 2cは未変性および変性gBと反応し、変性条件下での反応性はmAb H1817と比べてはるかに弱かった(図13B)。変性条件下でのgB単量体との弱い反応性は、変性に対して耐性であるか、またはSDS-PAGE電気泳動の間に再び折り畳まれるように見え、それゆえ「偽連続」エピトープと呼ばれる、不連続なエピトープに結合する一連の他の中和抗体についてこれまでに報告されている(4)。
【0114】
mAb 2cの結合に関与する特異的なエピトープを特定するため、本発明者らは、ペプチドマイクロアレイ上に提示されたgB由来ペプチドを用いた。最初に、重複する13 merのペプチドが3アミノ酸のずれで連続的なセルロース膜に非荷電性アセチル化アミノ末端で結合されるようにSPOT合成により、アミノ酸31〜505を提示するgB配列を調製した。非複合体化抗体に対する結合平衡の移動を回避するため、mAb 2cペプチドスキャンを化学発光による検出の前にPVDF膜上に固定化した。表示の機能ドメインを有する全長gBの略図(図14A)に示されるように、mAb 2cの反応性は、ドメインI内の2つの別個の領域にまたがるペプチド、つまり残基175〜193 (結合領域A)を含む3つの連続ペプチドおよび残基295〜310 (結合領域B)を含む2つの重複ペプチドに限定された。特定された両方の結合領域を検証するため、本発明者らは、可動性リンカーを介してスライドグラス上に固定化された13 merの精製ペプチドのさらなるセットを用いた。セルロースのスクリーニングと比べて、蛍光を介したこのマイクロアレイのスキャニングの読み出しから、同じエピトープ結合領域が確認された(図14B)。精製ペプチドおよび高分解能マイクロアレイスキャニングシステムの適用により、両結合部位のさらなる連続ペプチドがこのペプチドマイクロアレイにおいてmAb2cにより認識された(図14B)。
【0115】
本発明者らは、解析したgB構造に対して特定されたmAb 2cの結合部位をマッピングした(24)。興味深いことに、mAb 2cと最も強い反応性を示すペプチド172QVWFGHRYSQFMG184 (SEQ ID NO: 18)は、ドメインIの曲線状サブドメインに位置する2つの推定上の融合ループの1つ(融合ループ1 173VWFGHRY179) (SEQ ID NO: 19)と重なった(22) (図15)。しかしながら、gB三量体の基部に結合部位Aが局在化することで、融合後の立体構造を表す可能性が最も高い、利用可能なgB構造においてはmAb 2cにアクセスできなくなる(24)。結合部位Bの残基は曝露され、ドメインIの上部に位置付けられる(図15)。
【0116】
mAb 2cの立体構造依存的なエピトープをさらに評価するために、両方の結合領域のコンセンサス配列を直接的にまたは1つもしくは2つのβ-アラニンスペーサーによって分け隔てデュオトープとしてさまざまな組み合わせでつなぎ合わせた(図16)。残基E187の後の融合ループ1のすぐ近くでのリンカーの挿入は、gBが融合後の立体構造へ折り畳まれてさえも、融合欠損gB変異体(40,61)を生ずることが最近になって示された(61)。それゆえ、本発明者らは、別のデュオトープスキャンの中に、結合領域Aのコンセンサスモチーフ179YSQFMG184 (SEQ ID NO: 20)に加えて結合領域Aの186FED188モチーフを含めた。13 merのペプチドスキャンにおいてmAb 2cと最も強い結合反応性を示すペプチド172QVWFGHRYSQFMG184 (SEQ ID NO: 18)と比べて(図16)、結合部位Bのコンセンサスペプチド300FYGYRE305 (SEQ ID NO: 21)との両方の結合部位Aのモチーフの組み合わせが、シグナル強度の増強された2つのデュオトープを生じた(図16、デュオトープセットIおよびII)。デュオトープ179YSQFMG184-βA-300FYGYRE305 (SEQ ID NO: 22)に対するmAb 2cの結合強度はほんの少しだけ増したが、蛍光シグナル強度の大体の飽和は、デュオトープ186FED188-βA-βA-300FYGYRE305 (SEQ ID NO: 23)で得られた。
【0117】
したがって、ペプチドマイクロアレイからの結果は、ウエスタンブロッティングの結果に一致し、mAb 2cが立体構造依存的なエピトープを認識することを実証している。ウイルス粒子外被と細胞膜との融合を抑止するため、mAb 2cはgBの融合前の立体構造に結合するはずである。しかしながら、mAb 2cの中和エピトープは、利用可能なgB結晶構造(24)に存在するgB立体構造の表面に部分的にしかマッピングされず、gBが侵入の間に異なる立体構造を取りうることを示唆している。
【0118】
mAb 2c由来の二価および一価抗体の特徴付け
ウイルス侵入におけるgBの機能に不可欠なgB上の領域を特定するために幾人の研究者らによってモノクローナル抗体が用いられている(4, 25, 39, 52)。ドメインVのC末端の残基および近接プロモーターのドメインIの残基を含むgB三量体の基部の特異な機能領域にマッピングされている、中和抗体がgBの融合活性を妨害することが示唆されている(4)。本発明者らはそれゆえ、ドメインVのC末端に近いドメインI内のmAb 2cエピトープに結合する一価抗体がgBの活性化による協同的な立体構造変化を十分に遮断するはずであると仮定した。mAb 2cはインビトロにおいて補体なしでHSV-1を中和する(16)ので、本発明者らは、mAb 2cが媒介する仮説的機構を研究するのに有益なツールとして従来のF(ab')2およびFab断片ならびに組換え一本鎖可変断片(scFv)を作出した。作出した抗体調製物の均一性をサイズ排除クロマトグラフィーによってモニタリングした(データ不掲載)。それぞれ、HSV-1もしくはHSV-2で感染された、または感染されなかったVero細胞を用いたフローサイトメトリー分析から、mAb 2cおよびmAb 2c由来抗体断片の特異的な結合が実証された(データ不掲載)。本発明者らはさらに蛍光サイトメトリーを用いて、HSV-1およびHSV-2感染Vero細胞に対する抗体の平衡結合曲線を決定した(図17)。これらの研究の結果から、それぞれ、FabおよびscFvに対するよりも全IgGおよびF(ab')2断片に対するいっそう高い見掛け上の親和性が実証された(表6)。決定された一価抗体親和性に対する二価抗体の機能的親和性(親和力)の増加から、二価抗体が細胞表面上の2つのgBエピトープに同時に結合できたことが示唆される。二価mAb 2cおよび2c-F(ab')2は、その一価対応物と比べて1.7〜2.8倍高い見掛け上の親和性を示した。IgGに比べてF(ab')2断片の見掛け上のKDのわずかな増加は、F(ab')2構築物内の抗原結合部位のいっそう高い可動性によるものでありうる。両方のHSV-1およびHSV-2感染Vero細胞に対するmAb 2c、2c-F(ab')2および2c-Fabの同様の見掛け上の親和性から、認識されるgBエピトープが両方のウイルス間で構造的に違わないことが確認された(表6)。
【0119】
(表6)HSV-1 FまたはHSV-2 G感染Vero細胞との結合に対するmAb 2cおよびmAb 2c由来抗体断片の見掛け上の平衡定数
aHSV感染細胞上のgBとの結合に対するKD値は、フローサイトメトリーによって決定された平衡結合曲線からのデータ(図17)をMarquardt-Levenberg式に適合させることによって決定された。
【0120】
インビトロにおける一価および二価抗体の中和活性
HSV-1ウイルス粒子がVero細胞と相互作用する前(付着前)または後(付着後)に抗体が加えられるかどうかにかかわりなくmAb 2cの等しい中和効力(図18A)から、mAb 2cが標的細胞とのウイルスの結合を妨害しないことが示唆された。対照的に、ポリクローナルヒトγグロブリンIntratect(登録商標)は、標的細胞へのウイルス粒子の付着の阻害によって明らかに中和をしていた(図18B)。mAb 2c由来断片F(ab')2、FabおよびscFvの中和活性を、Vero細胞に対する標準的な中和アッセイ法においてその親IgG対応物と比較した。親mAb 2cは8 nMの濃度でHSV-1誘導性の細胞変性効果(CPE)を100%低減した。興味深いことに、HSV-2誘導性のCPEを完全に低減するには、4倍高いmAb 2c濃度が必要とされた(図19A)。二価2c-F(ab')2はHSV-1およびHSV-2誘導性のCPEの両方を親mAb 2cよりも2倍効率的に低減した。驚いたことに、本発明者らは、HSV-1およびHSV-2を中和するための一価2c抗体断片の能力の基本的相違を認めた。親mAb 2cと比べて、HSV-1およびHSV-2誘導性のCPEを100%低減するには、それぞれ、およそ375倍および94倍高い濃度の2c-Fabが必要であった(図19A)。組換え2c-scFvは光学顕微鏡の下でプラーク低減効果を示したが、3,000 nMの最も高い試験濃度でさえもHSV誘導性のCPEを100%低減することはできなかった(データ不掲載)。
【0121】
両方の二価抗体mAb 2cおよび2c-F(ab')2はHSV-1よりも約4倍低い効率でHSV-2を中和した(図19A)ので、本発明者らは、等量の感染粒子を含有するHSV-1およびHSV-2調製物のゲノムコピー数を定量的リアルタイムPCRによって分析した。HSV-1と比べて、HSV-2の場合には4倍多い数のゲノム相当物が認められ(データ不掲載)、HSV-2の中和に必要とされるmAb 2cおよび2c-F(ab')2の抗体力価がさらに高いこととよく相関していた。
【0122】
図19Aに示されるように中和アッセイ法から、抗体価と中和効率との間の強い相関が示唆された。その結果、本発明者らは、Fab断片の架橋によってウイルス感染を取り除くための2c-Fab断片の能力を回復できるかどうか調べた。ウイルス中和アッセイ法を、マウスFab断片と反応するIgGの非存在下または存在下で2c-Fabに対して繰り返した。図19Bに示されるように、2c-Fabの架橋結合は中和活性を劇的に増大したが、それを、親mAb 2cの場合と同じ効力まで回復することができなかった。抗マウスFab IgGのみではウイルス中和に対する効果が示されなかった(データ不掲載)。
【0123】
細胞間伝播の阻害
2c-Fab断片は遊離ウイルス粒子を効率的に中和しなかったが、それでも、小さなサイズの抗体断片はより好ましい拡散特性を有することが報告されており(66)、本発明者らは、HSV-1が細胞間結合を、感染細胞から非感染細胞へと横断することを抑止するその活性について調べた。両方の二価抗体mAb 2cおよび2c-F(ab')2は、Vero細胞単層中で伝播されたHSV-1を完全に抑制し、間接的な免疫蛍光によってただ1つの感染細胞を可視化できただけであった(図9)。遊離ウイルス粒子を中和するポリクローナルヒト血清の能力にもかかわらず、それはウイルスの細胞間伝播を阻害することが全くできなかった。これは、多数のHSVエピトープに対して作製された中和抗体の不均一集団の結果である可能性が最も高い。ポリクローナルヒト免疫血清と比べて、一価2c-Fab断片は細胞間伝播をある程度制御することができた。しかしながら、一価2c-Fab断片は、その二価対応物とは対照的に、6倍高い濃度で試験してさえもウイルス伝播を完全に排除することができなかった(図9)。ゆえに、抗体価は隣接細胞の間でのHSV-1の伝播の阻害においても鍵となる役割を果たした。
【0124】
播種性HSV感染に対する免疫不全マウスの免疫保護
本発明者らは、CD4+およびCD8+ T細胞の両方が枯渇したマウスが、膣内HSV-1感染の後にmAb 2cの受動移入によって致死性脳炎から完全に保護されることを以前に示した(17)。ヒトにおいてHSV-2感染の部位に蓄積するナチュラルキラー(NK)細胞(28)はインターフェロン-γの初期供給源であり(45)、インターフェロン-γはHSV感染の制御に不可欠な役割を果たす(2, 45, 62)。つい最近、初めて、ヒトNK細胞が先天性免疫応答としてヒト化マウスで初感染性器HSV感染に対する保護を媒介することが実証された(37)。mAb 2cが抗体媒介性の免疫反応とは独立して抗ウイルス活性を付与するかどうか調べるために、本発明者らは、SCID T細胞欠乏およびB細胞欠乏に加えて、NK細胞およびマクロファージの機能ならびに補体経路を刺激する能力を欠くNOD/SCIDマウスモデルを利用した。NOD/SCIDマウスの膣内HSV-1感染(1×106 TCID50)は、急速進行性の全身疾患を引き起こし、9日の生存期間中央値であった。臓器におけるHSV力価を終点希釈アッセイ法によって判定し、脊髄(2.3×106 TCID50)、脳(3.8×105 TCID50)および膣粘膜(1.4×106 TCID50)において高ウイルス力価、腎臓(1.7×104 TCID50)および副腎(1.1×104 TCID50)において中力価ならびに肺(1.1×103 TCID50)および心臓(1.9×102 TCID50)において低力価を示した(データ不掲載)。mAb 2cの治療効率を評価するため、NOD/SCIDマウスを膣内HSV-1攻撃の24時間前に2.5 mg/kg、5 mg/kgまたは15 mg/kgの抗体で静脈内処置した(図20)。低抗体用量を投与されたマウスは、HSV-1による致命的感染から十分には保護されなかった。しかしながら、5 mg/kgのmAb 2cで処置されたマウスの生存期間中央値は、PBSを投与された対照マウスと比べた場合に2.6倍引き延ばされた。致死性脳炎から保護されなかったマウスより調べた臓器におけるHSV-1の力価は未処置対照群に匹敵していた。対照的に、動物の完全保護は15 mg/kgのmAb 2cの用量で達成された。抗体によって保護されたマウスの臓器におけるウイルス力価は、1×102 TCID50の検出限界未満であった。
【0125】
本発明者らは次に、mAb 2cによる曝露後免疫がまた、樹立された末梢HSV感染の存在下でウイルス伝播および致死性脳炎からの保護を付与するかどうか評価した。ウイルス攻撃後24時間の時点で膣洗浄液中に高いHSV-1力価を有するNOD/SCIDマウスを24時間、40時間および56時間の時点で15 mg/kgのmAb 2cにより静脈内から繰り返し処置した(図11および図21)。PBS処置対照群は、神経症状を有するマウスを7日目〜9日目に殺処理しなければならなくなるまで一定の経膣的なウイルスの出芽を示した。対照的に、mAb 2cは8日目までに、樹立されたHSV-1感染を除去し、感染の致命的結末を完全に抑止した(3×300μg; PBSと比べてP = 0.0003)。さらに、感染から1ヶ月後にmAb 2c処置動物の感覚ニューロンおよび各臓器においてウイルス粒子は検出されなかった(データ不掲載)。
【0126】
考察
ウイルスが標的細胞に侵入するために取る段階に続いて、ウイルス中和mAbはいくつかの機構によって侵入を阻害することができる。ウイルス表面タンパク質と細胞のタンパク質、脂質または糖質との特異的な相互作用は、感染の初期段階に相当し、これを中和抗体によって遮断することができる。ウイルス付着を阻害する抗体は、HIV-1 gp120のCD4結合部位と反応するmAb F105およびインフルエンザ血球凝集素(HA)の受容体結合部位を網羅するFab HC19など、ウイルス粒子の受容体結合部位に直接結合するか(6, 19, 54)、またはHA受容体結合部位の近く17Åで結合するFab HC45など、受容体連結を立体的に妨害するか(18)のどちらかである。HSV gDとその細胞受容体の1つとの不可欠な結合に加えて、gBは標的細胞へのウイルス粒子の付着に関与している。最近、HSV gBに対する2つのへパラン硫酸プロテオグリカン非依存的な真の細胞表面受容体および/または付着因子の存在が報告された(5, 23, 60)。対になった免疫グロブリン様2型受容体(PILRα)は少なくともある種の細胞型において、gBの潜在的タンパク質受容体の1つと特徴付けられている(60)。mAb 2cに対し、比較による付着前 vs 付着後中和アッセイ法から、この抗体は細胞表面へのウイルスの結合を阻害できないが、ウイルス侵入を遮断することが示された。gBと、その融合ドメインの鍵となる疎水性および親水性残基を介した脂質膜との相互作用(22, 23)は、融合ループのすぐ近くのエピトープを認識するmAbによって遮断されうることがこれまでに示されている(4, 22)。mAb 2cの立体構造エピトープは融合ループ1と部分的に重複するので、本発明者らは、mAb 2cの結合が融合シグナルの伝達を妨害する可能性が最も高いと推論し、本発明者らはさらに、考えられる作用機序として結合後/融合前の段階で中和を評価した。
【0127】
構造的再配列の誘導はウイルス融合糖タンパク質の鍵となる特徴であり、異なる融合前および融合後立体構造を引き起こす。異なる中和mAbのエピトープがスパイクの側面ドメインに沿っておよびgB結晶構造の冠部の先端にマッピングされた(4, 24)。mAb 2cのエピトープは、ドメインVのC末端ヘリックスαF内の残基および近接プロモーターのドメインI内の残基からなるgB三量体の基部の特異な機能領域(FR1)に位置する(4)。本発明者らの相同性モデルは、mAb 2cによって認識される不連続なエピトープの一部分(F300〜E305)が、プレクストリン相同性(PH)ドメインの特徴を有する、gBのドメインIの上部に局在することを示す(7, 38)。しかしながら、ドメインIに同様に位置するエピトープの他の部分(F175〜A190)は埋もれており、gBが主要な立体構造変化を起こさなければ、mAb 2cの結合にアクセスできないと考えられる。本発明者らはそれゆえ、mAb 2cが融合前の立体構造において選択的にgBの転移を妨げると仮定した。mAb 2cエピトープの局在、および活性化による立体構造変化が協調的であるという仮定に基づき、本発明者らは、gBの融合ドメインおよび細胞膜の並置の遮断にはmAb 2cの一価相互作用で十分であろうと推論した。しかしながら、驚いたことに、作出された一価抗体断片(FabおよびscFv)のどれも遊離ウイルス粒子を効率的に中和すること、またはウイルスの細胞間伝播を阻害することができなかった。対照的に、両方の二価分子mAb 2cおよび2c-F(ab')2は、ウイルス中和および細胞間伝播の阻害について非常に効果的であった。本研究における全てのmAb 2c由来抗体の特異的かつ同等な結合活性の保持から、抗原認識の障害による一価抗体および二価抗体の機能的差異は除外される。免疫グロブリンの多価結合はその機能的親和性を増加させる(26)。しかしながら、機能的親和性の増加は抗体結合部位の固有の親和性と逆の相関がある(49)。二価2c抗体IgGおよびF(ab')2に対する1.7〜2.8の平衡定数の増分が、その一価対応物scFvおよびFabと比較した場合に、わずかでしかないのは、かくして、低いナノモル範囲において固有の親和性を有する抗体にとって珍しいことではない。このように、いっそう高い見掛け上の親和性は実のところ、gB抗原との多価(いっそう高い親和力)の結合が実際に起きていることを示し、mAb 2cおよび2c-F(ab')2の抗ウイルス活性がgB架橋結合の結果であることを示唆している。水痘帯状疱疹ウイルス(VZV)のgH抗原に対して特異性を有する二価抗体または三価抗体に比べて見劣りする一価抗体の中和効率は、これらの抗体のサイズの違いによる立体障害の問題として論じられている(15)。本発明者らは、mAb 2cバリアントに対する潜在的なさらなる中和機構としてのこの可能性を完全に除外することはできないが、これは、抗体のサイズ、中和効率および細胞間伝播の阻害の間に直接的な相関関係が認められなかったので、可能性が低いようである。さらに、本発明者らのデータは、いっそう小さい2c-F(ab')2がいっそう大きな2c-IgGよりもさらに良好なウイルス中和活性を有していたことを示している。ゆえに、本発明の所見は、gB架橋結合がmAb 2cの抗ウイルス活性に対する鍵となる機構であることを示し、gB三量体の固定化によるgB融合前立体構造の安定化が融合シグナルの活性化を阻害することを示唆している。Silvermanらによる最新の研究(61)から、融合ループ1に近い残基E187の後の5つのアミノ酸の挿入によるHSV-1 gB細胞外ドメインの融合欠損表現型がgBの立体構造変化の妨害から生じるのではなく、むしろ他の機構によるgB機能の妨害から生じうることが提唱された。本発明者らのデュオトープスキャンにおいて、mAb 2cは、gBの機能に極めて重要であるようにみえる、特定の挿入部位E187を網羅する結合部位A/Bデュオトープ186FED188-βA-βA-300FYGYRE305 (SEQ ID NO: 23)と最も強く反応した。それゆえ、mAb 2cの架橋結合がHSV融合機構の他の成分と相互作用するgBの能力を損なうと推測したくなる。しかしながら、本発明者らの結果は、架橋結合がgBそれ自体の立体構造変化を遮断するか、または細胞融合の間に起こり(3)、融合過程の完了に不可欠な(65)、gB、gDおよびgH/gLの間の相互作用を遮断するかどうかを区別することを可能にするものではないので、将来の研究が必要である。解析された結晶において観察されたHSV-1 gBの立体構造(24)は、融合後の形態に相当することを示唆しており、gBの融合前のモデルは未だ特徴付けられていない。それゆえ、gBと複合体化したmAb 2cまたはそのF(ab')2のX線結晶学的研究は、gBの未変性の立体構造に関する見識および融合シグナルの伝達に関するさらに良好な理解をもたらす可能性がある。
【0128】
マウスにおいてHSV-2感染の膣感染を防ぐために局所に適用された抗gD抗体および抗gB抗体の保護効力を評価する研究から、新しい膣消毒薬としての遺伝子操作組換え抗体の実現可能性が実証された(67〜69)。重篤な、そして致命的でさえあるHSV感染は、母親から感染した新生児において、再発性眼感染を有する患者において、または重度免疫減弱状態の患者において起こりうる。本発明者らの抗gB抗体の全身適用が高度に免疫不全なインビボの設定においても保護を付与するかどうか調べるために、本発明者らは、NOD/SCIDマウスモデルを利用した。本発明者らは、免疫応答性マウスにおいておよびT細胞枯渇マウスにおいて後肢麻痺および致命的疱疹脳炎を引き起こすウイルスの軸索伝播による確立された神経節感染経路として膣内HSV-1接種を用いた(16, 17)。ここで本発明者らは、mAb 2cが一次性HSV-1感染の急性期においてNOD/SCIDを完全に保護するだけでなく、末梢ウイルス伝播の開始後でさえも神経疾患および死を完全に抑止するのに有効でもあることを実証する。HSVの細胞間伝播は、ニューロンのシナプスおよび密着結合をまたいだウイルスの移動のために、ならびに適応免疫系の免疫学的障壁を回避するために非常に効率的な手段である。mAb 2cは、感染した膣組織のウイルス発現を低減もし、HSVの軸索伝播を阻害もする。他の報告から、ウイルス攻撃後の抗HSV IgGの投与が動物において急性神経節感染の量を低減できることが示された(16, 42)。一貫して、角膜HSV-1感染を有するマウスに腹腔内投与された組換えヒト抗gD IgGがHSVに感染した神経線維および感覚ニューロンに局在することが示された(59)。さらに、適切な時点で曝露後に投与されたHSV gD、gCまたはgBに特異的なmAbによる免疫応答性動物の受動免疫は、HSV誘導性の神経疾患からの保護を示した(13, 16)。しかし、いくつかの動物実験から、液性免疫だけではHSV感染の制御には効果がないとも結論付けられている。
【0129】
本発明者らの抗体のインビボでの保護効力は、免疫エフェクター機能から独立している。文献中のいくつかの動物試験から、液性免疫だけではHSV感染の制御には効果がないとも結論付けられている。この考えと一致して、抗HSV-1過免疫血清の投与は免疫抑制マウスまたは免疫不全マウスの保護に効果がないと報告されている(47, 48, 50, 51, 56)。HSV-1感染後24時間の無胸腺ヌードマウスをヒト抗gD mAbで全身処置することにより、未処置対照と比べて生存時間が引き延ばされたが、死が抑止されることはなかった(58)。別の研究では、HSV-1誘導性の間質性角膜炎マウスモデルにおいて、抗gD mAbがCD4+またはCD8+ T細胞のどちらかを枯渇させたマウスの死を抑止したものの、マウスの両方のT細胞サブセットを同時に枯渇させた場合には、死を抑止できないことが示された(64)。
【0130】
本発明者らの知る限りでは、本発明者らは、細胞エフェクター機構および補体から完全に独立してニューロンのHSV-1伝播を抑止する、全身に適用された抗gB架橋mAbの保護効力を初めて実証した。HSV複製に不可欠なgBの一種の共通エピトープに対するmAb 2cの特異性およびさらなる免疫エフェクター機能の動員の必要性なくして高いその保護効率は、新規の免疫治療薬としてのこの抗体の大いなる潜在性を示す。
【0131】
実施例3に関する参考文献一覧
【0132】
実施例4
ウイルス中和アッセイ法
ウイルスの中和感受性を決定するための過剰量の抗体によるプラーク低減アッセイ法としてまたは抗体溶液の中和価を決定するための終点希釈法として、中和アッセイ法をVero細胞に対してマイクロタイタープレート中で行った。プラーク低減アッセイ法は、20μgのMAb 2cとの250プラーク形成単位のインキュベーションによって行った。2時間後、50μL/ウェルのVero細胞懸濁液(細胞1.5×105個/mL)を添加した。3日後、細胞をクリスタルバイオレットで染色した。終点力価測定のため、希釈した抗体溶液(0.025 mL)を0.025 mL中100 TCID50のHSV-1、および1:10に希釈したモルモット補体0.025 mLとともにインキュベートした。培養物の50%においてウイルス誘導性の細胞変性効果を抑止する最大血清希釈の逆数として力価を表した。
【0133】
gB欠失変異体の構築およびCOS-1細胞における発現
全長HSV-1 gB (gB(1〜904) = pMT2gB)、gB(1〜720)、gB(1〜630)、gB(1〜505)、gB(1〜503)、gB(1〜487)およびgB(1〜470)をコードするプラスミドの構築は他で記述されている[30, 31]。プラスミドはLeonore Pereira, University of California, San Franciscoから親切にも提供していただいた。gB(1〜130)、gB(1〜223)、gB(183〜488)およびgB(436〜642)をコードするプラスミドは、制限酵素部位Bam HIおよびXho Iが隣接するPCR単位複製配列を、真核生物発現ベクターpSVL (Amersham Pharmacia, Freiburg, Germany)の中にクローニングすることによって構築した。gBヌクレオチド52588〜60362を含んだHSV-1株17+ [33; GenBank X14112]のサブゲノムプラスミドクローンをPCRにおける鋳型として用いた。N末端切断型のgB構築物の発現のため、gBシグナル配列をコードするDNAを、5'末端の位置にXhoI部位を含むプライマーを用いたPCRによって増幅し、小断片プラスミドgB(183〜488)およびgB(436〜642)のgBをコードするDNAの5'側に挿入した。挿入断片の正しい組み込みおよびその配列をヌクレオチド配列決定によって確認した。COS-1細胞を、24ウェルプレートの中に配置されたカバーガラス(直径10 mm)上で増殖させ、DEAE-デキストラン法によりプラスミドでトランスフェクトした[34]。gBおよびその切断型誘導体の発現を、よく特徴付けられた抗HSV-1 gBマウスモノクローナル抗体H1396およびH1781の混合物を用いた間接的免疫蛍光顕微鏡検査法により検証した。トランスフェクトされ固定されたCOS-1細胞をMAb 2cと反応させ、免疫蛍光顕微鏡検査法により分析した。
【0134】
gBの部位特異的突然変異誘発および組換えウイルスの構築
単一のアミノ酸変異を、Altered Sites(商標) in vitro Mutagenesis System (Promega, Mannheim, Germany)を用いてオリゴヌクレオチド特異的突然変異誘発によりHSV-1 gBに導入した。手短に言えば、pMT2gB [31]内のgBコード配列を大腸菌(E. coli)ファージミド突然変異誘発ベクターpAlter-1の中に移入した。pALTER-lgBで形質転換された大腸菌JM109細胞にファージR408を感染させることによって、一本鎖pALTER-lgB DNA分子を調製した。以下のようにミスマッチプライマー(変異させた位置に下線を引いた)を用い製造元のプロトコルにしたがって部位特異的突然変異誘発を行った。
【0135】
完全に二重鎖の、共有結合閉環状分子を形成させるためのプライマー伸長および核酸連結の後に、このヘテロ二重鎖DNAを、ヌクレオチドミスマッチを修復することができない大腸菌BMH 71-18 mutSに形質転換した。アンピシリン耐性クローンから得られたプラスミドを大腸菌JM109 (recA-)の形質転換に用いた。得られたpAlter-lgBプラスミドにおける変異の同一性をgB DNA配列決定によって確認した。突然変異誘発の後、変異させた挿入断片を発現ベクターpMT2gBにクローニングし戻し、各pMT2gBmut構築物を得た。単一のアミノ酸変異を有するgBに対するMAb 2cの反応性を分析するために、カバーガラス(10×30 mm)上で増殖させたCOS-1細胞をリポフェクトアミン(商標)/OPTI-MEM(商標)法(Invitrogen, Karlsruhe, Germany)により製造元の指示にしたがってpMT2gBmut構築物でトランスフェクトした。細胞をメタノール/アセトンにより培養44時間後に固定した。ポリクローナルウサギ抗HSV-1 IgG (Dako, Hamburg, Germany)を用いた間接的免疫蛍光分析により、gBmutの発現を検証した。
【0136】
単一のアミノ酸の交換を有する糖タンパク質Bを、HSV-1ゲノムとプラスミドpMT2gBmut DNAとの間の相同組換えによってHSV-1野生型株Fの中に挿入した。ゲノムHSV-1 DNAを56℃で1〜3時間SDS/プロテイナーゼK溶解によって、その後、フェノール/クロロホルム/イソアミルアルコール抽出および65時間の透析(緩衝液I, 10 mM NaCl, 10 mM EDTA, 50 mM Tris-HCl, pH 8; 緩衝液II, 10 mM NaCl, 1 mM EDTA, 10 mM Tris-HCl, pH 7.5)によってウイルスストック液から調製した。pMT2gBmutプラスミドをQiagen Plasmid Midiキット(Qiagen, Hilden, Germany)によって抽出した。組換えウイルスはVero細胞を、StowおよびWilkie [36]により改変されたGrahamおよびVan der Eb [35]の方法にしたがってリン酸カルシウム沈殿させたDNAで、または製造元の指示にしたがってリポフェクトアミン(商標)/OPTI-MEM(商標)法(Invitrogen, Karlsruhe, Germany)でトランスフェクトすることにより調製した。手短に言えば、リン酸カルシウム法によるトランスフェクションの場合、精製HSV-1 F DNAおよそ100 ngを5分間1 M CaCl2 64μlの添加によりA. bidest. 0.436 mL中500 ngのプラスミドDNAおよび5μgの仔ウシ胸腺DNAで沈殿させた。45分間DNA沈殿物とのおよび3時間10%ウシ胎仔血清を含むEMEMとのペトリ皿(直径25 cm)の中で増殖させた細胞のインキュベーションの後、2分間HEBS緩衝液中25%のDMSOによって細胞にショックを与えた。その後、培養を、10%ウシ胎仔血清を含むEMEMおよびMAb 2cで継続した。リポフェクトアミンによるトランスフェクションの場合、HSV-1 DNA 1μgおよびプラスミドDNA 1μgを用いた。十分に分離されたプラークを選び取り、PCR単位複製配列のサイクル配列決定により各変異についてスクリーニングした。全体としては、プラスミドコード化gBの存在下において全くプラークを産生することに成功しなかった実験では、得られるプラークの数は低かった(gBをコードするプラスミドおよびMAb 2cの存在なしでの対照実験では多数のプラークを生じた)。抗体反応性を試験するために、カバーガラス(8×16 mm)上で増殖させた3×105個のVero細胞にEMEM 1 mL中200〜300プラーク形成単位のウイルスを感染させた。1時間後、10%ウシ胎仔血清を含むEMEMによって細胞培地を交換した。2日間のインキュベーションの後、細胞を-20℃でメタノール/アセトンにより固定した。
【0137】
免疫蛍光アッセイ法
細胞とのHSV gB特異的マウスモノクローナル抗体の結合を、DTAFコンジュゲートヤギ抗マウスIgG (Jackson ImmunoResearch Laboratories, Dianova, Hamburg, Germany)を用いて検出した。ウサギ抗HSV-1 IgGの結合の検出のため、TexasRedまたはCy3コンジュゲートヤギ抗ウサギIgG (Jackson ImmunoResearch Laboratories, Dianova)を用いた。ヒト抗体の検出のため、DTAFコンジュゲートヤギ抗ヒトIgG (Jackson ImmunoResearch Laboratories, Dianova)を用いた。
【0138】
セルロース結合ペプチドの合成およびスキャニング(ペプチドスキャン、鍵となるモチーフのスキャン、置換分析)
全てのペプチドは既報[37, 38]のようにWhatman 50セルロース膜上での半自動SPOT(商標)合成により作出した。合成後、ブロッキング用試薬(1X, Cambridge Research Biochemicals, Northwich, UK)をTris緩衝生理食塩水(50 mM Tris, 125 mM NaCl, 4 mM KCl, pH 8.0)、0.05% (v/v) Tween 20および5% (w/v)スクロース(TBST)の中に含有するブロッキング用緩衝液中で終夜、この膜をブロッキングした。TBST中で1回洗浄した後に、このシートを室温で3時間ブロッキング用緩衝液中で同時にMAb 2c (0.5〜1.0μg/mL)およびPODコンジュゲート抗マウスIgG Fab断片(5倍過剰; Roche Applied Science, Mannheim, Germany)とともにインキュベートした。TBST中で2回洗浄した後に、Chemiluminescence Western Blotting Kit (Roche Applied Science, Mannheim, Germany)を製造元のプロトコルにしたがって用いることにより、抗体結合を検出した。
【0139】
重複するペプチドのスキャン(ペプチドスキャン) [39]は、アミノ酸31〜505のgB残基(HSV-1株17+ [33] GenBank X14112)に対応した、12アミノ酸の重複を有する(すなわち、3アミノ酸ずつgB配列に沿ってシフトする) 15merとして、およびgB残基296〜315に対応した12アミノ酸の重複を有する(すなわち、1アミノ酸ずつgB配列に沿ってシフトする) 13merとして合成された。
【0140】
MAb 2c結合部位B内の最小の結合モチーフを特定するために、N末端およびC末端のそれぞれの位置で4つの無作為化残基により囲まれた中央の、gB由来六量体から構成される14mer (x1x2x3x4B1B2B3B4B5B6x11x12x13x14; x, 無作為化される位置、B, 固定化されるgB由来の位置)を用いて鍵となるモチーフのスキャンを行った[40]。各ペプチドで、六量体配列は1アミノ酸ずつシフトさせて、gBのV295位から始まり、gBのA315位に至るまでの配列に及んだ。
【0141】
gB結合部位A内のMAb 2cの結合について単一アミノ酸の関連性を、既報[40〜42]のようにその他全ての19種の天然アミノ酸によって各アミノ酸を連続的に交換することにより行ったペプチド178RYSQFMGIFEDRAPV192 (SEQ ID NO: 43)の置換分析によって分析した。
【0142】
マウスおよびマウス保護実験
雌性C57BL/6J (H-2b)マウスを、Charles River Wiga (Charles River Laboratories, Sulzfeld, Germany)から入手し、33〜37日齢の時点で使用した。実験を既報[1, 2]のように行った。手短に言えば、マウスに、10%ウシ胎仔血清を含むEMEM 0.1 mL中2×106 TCID50のHSV-1を膣内から接種した。ウイルス接種の24時間前に、腹腔内注射によってMAb 2c、ポリクローナル免疫血清または沈殿させた培地のいずれか0.5 mLをマウスに投与した。使用したヒト標準的免疫血清調製物(Beriglobin S(商標), CSL Behring, Germany)は、0.025 mL中でHSV-1に対して1:1280の補体非依存性の中和価を有し、適用のためIscoveの培地中で4倍希釈した。MAb 2cのストック調製液は1:640の補体非依存性の中和価を有し、ポリクローナル免疫血清と同じ中和活性を含むように2倍希釈した。適用した抗体希釈液のELISA力価は、マウスおよびヒトIgGに対するペルオキシダーゼコンジュゲートウサギを用いKahlon & Whitleyの方法[43]にしたがって決定した場合に104.5〜105.5であった。対照の場合、等量のIscoveの培地を同じように処理した。培地を投与した対照マウスは、非HSV特異的なMAbを投与した対照と同等であった[1]。膣拭き取り検体をウイルス接種後1日おきに採取し、Vero細胞単層上のウイルスについてアッセイした。感染性ウイルスの力価をReed & Muenchの方法[44]にしたがって0.05 mLあたりのTCID50によりマイクロタイタープレート中で決定した。
【0143】
結果
gBの最初の487アミノ酸はMAb 2cの結合に必要である
MAb 2cエピトープの適切な折り畳みに必要な糖タンパク質B領域に関する最初の見識を得るために、全長HSV-1 gB構築物および一連のカルボキシ末端切断型gB構築物を方法の項に示したようにCOS-1細胞において発現させた。gBの発現を、マウスHSV gB特異的MAb H1396およびH1781の混合物を用いた間接的免疫蛍光顕微鏡検査法により検証した。間接的免疫蛍光アッセイ法によりMAb 2cの結合も可視化した。表7に示されるように、全長タンパク質および切断型誘導体gB(1〜720)、gB(1〜630)、gB(1〜505)、gB(1〜503)およびgB(1〜487)はMAb 2cによって認識された。対照的に、MAb 2cはgB(1〜470)、gB(1〜223)およびgB(1〜130)を結合することができなかった。さらに、N末端およびC末端の両方の切断のある2つの構築物(gB(183〜488)、gB(436〜642))で反応は認められなかった。これらの結果から、MAb 2cのエピトープが最初の487アミノ末端残基のなかに位置することが示唆された。
【0144】
(表7)COS-1細胞において発現された切断型HSV-1糖タンパク質B (gB)とのMAb 2cの結合
* 全てのgB構築物の発現をHSV gB特異的モノクローナル抗体H1396およびH1781を用いた間接的免疫蛍光によって確認した[30〜32]。
** MAb 2cの結合を間接的免疫蛍光によって検出した。
+ はMAb 2c結合を示し、- はMAb 2c結合の失敗を示す。
*** pSVL, 陰性対照として用いた発現ベクター。
【0145】
MAb 2cは2つの異なるgB領域の配列を認識する
MAb 2cによって認識されるエピトープの精密マッピングはCOS-1細胞において発現されたgB欠失構築物によって可能ではなかったので、gBに由来する重複ペプチドのスキャン(ペプチドスキャン)をSPOT(商標)合成により連続的なセルロース膜支持体上で合成した。アミノ酸31〜505のgB領域に及ぶペプチドを、12アミノ酸の重複を有する(すなわち、3アミノ酸ずつgB配列に沿ってシフトする) 15merとして合成し、計155種のペプチドを得た。MAb 2cの結合は一次(MAb 2c)および二次(PODコンジュゲート抗マウスIgG Fab)抗体との同時インキュベーション、ならびに化学発光による検出で示された。
【0146】
図22に示されるように、MAb 2cは、部位Aおよび部位Bと名付けられた、2つの異なるgB領域のなかの5つのペプチドに結合することが分かった。部位Aは、gB残基175FGHRYSQFMGIFEDRAPVPFE195 (SEQ ID NO: 44)に対応する3つの連続ペプチド(共通配列181QFMGIFEDR189 (SEQ ID NO: 45)を含み、そして部位Bは、残基298SPFYGYREGSHTEHTSYA315 (SEQ ID NO: 46)を包含する2つの連続ペプチド(共通配列301YGYREGSHTEHT312 (SEQ ID NO: 47)を含む。
【0147】
MAb 2cに対する結合部位Bの最小の長さの特定
ペプチド90 (図22; 298SPFYGYREGSHTEHT312 (SEQ ID NO: 48)は最も強いシグナル強度を示したので、本発明者らは、結合部位BがMAb 2c結合のための主要な決定因子であると仮定した。かくして、本発明者らは、さらに高分解能のセルロース結合ペプチドスキャンを用いてMAb 2c結合に必要な部位Bの最小の長さを特定した。12アミノ酸の重複を有する(すなわち、アミノ酸ただ1つずつgB配列に沿ってシフトする) gB由来の残基296〜315に及ぶ13merのペプチドを二つ組で合成した。5つの連続ペプチドとのMAb 2cの反応性を上記のようにインキュベーションおよび検出手順の後に認めた。5つの反応性ペプチドの配列アライメントを図23に示す。全5種のペプチドに対する共通配列は300FYGYREGSH308 (SEQ ID NO: 49)であった。
【0148】
第2の手法では、両端に無作為化される4つの位置が隣接した6個のgB由来アミノ酸からそれぞれがなる14merを用いた鍵となるモチーフのスキャニング法を適用した。このアッセイ法において、各ペプチド分子の4つの末端位置は、アミノ酸が統計的に組み入れられた無作為配列に相当していた。各スポットにはかくして、外側のペプチド位置で多数の配列を有するが、しかし全て5〜10番目のペプチド位置に同じgB由来配列を有するペプチドの膨大な混合物が含まれた。gBに由来する中央の六量体は残基V295〜A315のgB領域にまたがり、1アミノ酸ずつ沿うようにしてシフトした。六量体のgB配列を選択したが、これは、非線状エピトープの75%超が最大4〜7残基長の連続的なストレッチを含むことが知られているからである[45]。MAb 2cの反応性は2つの連続ペプチドxxxx300FYGYRE305xxxx (SEQ ID NO: 21)およびxxxx301YGYREG306xxxx (SEQ ID NO: 50)で見られた(図24)。したがって、配列300FYGYREG306 (SEQ ID NO: 51)はMAb 2cとの相互作用に必要な部位Bペプチドの最小の結合モチーフであるものと考えられた。
【0149】
部位BでのMAb 2cの結合に重要な個々の残基の、変異型gBによる特定
全体的なおよび自然に折り畳まれたタンパク質という状況でMAb 2cに対する結合部位Bを確認するために、本発明者らは、結合部位B内の単一アミノ酸の交換によって、pMT2gBにクローニングされた、全長gBのアミノ酸配列を改変した。部位特異的突然変異誘発用のファージミドに基づく系を用いることにより、単一アミノ酸の交換を有するいくつかのgB構築物を作出した。COS-1細胞における変異型gBの発現の後、免疫蛍光アッセイ法によってMAb 2cの結合を分析した。表8に示されるように、MAb 2cの結合に重要であることが証明された一連のgB残基が特定された。詳細には、SによるgB 299位の残基Pの置換、それぞれYおよびIによるF300の置換、NによるY301の置換、それぞれRおよびVによるG302の置換、NによるY303の置換、それぞれGおよびLによるR304の置換、ならびにKによるE305の置換は、MAb 2c結合の完全な喪失をもたらし、したがって、299〜305位の残基はそれぞれ、その側鎖を介して抗体と相互作用する主要な残基となることにより、または抗体によって認識されるエピトープの立体構造の形成に必要なgBタンパク質の適切な全体的または局部的折り畳みに影響を与えることにより、エピトープの形成に決定的に関与していることを示唆していた。COS-1細胞における変異型gBの発現は、マウスMAb 2cおよびポリクローナルウサギ抗HSV IgG免疫血清との細胞の同時インキュベーションにより、その後、DTAFコンジュゲート抗マウスIgG (MAb 2cが結合したなら緑色の蛍光)およびTexasRedまたはCy3コンジュゲート抗ウサギ抗体(同じ細胞の赤色の蛍光)との同時インキュベーションを介した特定により検証した。反対に、単一のgB残基Y296の、それぞれ、NおよびFへの交換、M297の、それぞれ、L、TおよびVへの交換、ならびにS298のAへの交換のほか、G306の、それぞれ、AおよびVへの交換、ならびにS307のAへの交換はMAb 2cの結合に影響を与えなかった。
【0150】
(表8)COS-1細胞において発現された、単一アミノ酸の交換を含むHSV-1全長糖タンパク質B (gB) バリアントとのMAb 2cの結合
* 野生型アミノ酸はgBの位置番号の前に示されており、導入された残基は位置番号の後に示されている。
全てのgBバリアントの発現をポリクローナルウサギ抗HSV-1 IgG血清との細胞の同時インキュベーションによって得た免疫蛍光で確認した。
** MAb 2cの結合を間接的免疫蛍光によって試験した。
+ はMAb 2c結合を示し、- はMAb 2c結合の失敗を示す。
*** この特定の結果の評価については、考察の項を参照されたい。
【0151】
部位BでのMAb 2cの結合に重要な個々の残基の、変異体ウイルスによる特定
インビボに最も近い状況に近づけるため、それぞれがgBの中にアミノ酸変異を含む、よく特徴付けられた5つのHSV-1バリアント(R126、R1375、B4.1、R1435、R233) [27〜29]を用いることにより、および方法の項に示されるように、本研究において作出された変異体ウイルス(vY301N [301位のYをNによって置換した]、vG302R、vG302V)により、部位Bの個々のアミノ酸の影響をさらに分析した。カバースライド上のVero細胞に200〜300プラーク形成単位のこれらの変異体または親の野生型ウイルスHSV-1 FおよびKOS 321のどちらかを感染させた。間接的免疫蛍光アッセイ法から、MAb 2cはウイルスvY301N、vG302R、vG302V、R126 (Nによって置換されたY303)、R1375 (R304Q)およびB4.1 (E305K)に感染した細胞に結合できないが、MAb 2cは変異体R1435 (H308Y)、R233 (R328H)を感染させた細胞、および野生型ウイルスを感染させた細胞に対して反応性を示すことが実証された(表9)。
【0152】
(表9)MAb 2cの結合およびMAb 2cによる中和に対するHSV-1野生型ウイルスおよびウイルスgB変異体の感受性
* gBの発現をポリクローナルウサギ抗HSV-1 IgG血清で得た免疫蛍光により全てのウイルスについて確認した。+ は間接的免疫蛍光によって検出された感染Vero細胞とのMAb 2cの結合を示す。- はMAb 2c結合の失敗を示す。
** + はMAb 2cによるウイルス中和を示し、- はウイルス中和の失敗を示す。
*** 野生型アミノ酸はgBの位置番号の前に示されており、導入された残基は位置番号の後に示されている。
【0153】
gBとのMAb 2cの結合に重要であると特定された残基が抗体の中和活性にも重要であったかどうかを判定するために、250プラーク形成単位のウイルス変異体または野生型ウイルスを用いて中和アッセイ法を行った。表9に示されるように、野生型株FおよびKOS 321ならびに変異体R1435 (H308Y)およびR233 (R328H)はMAb 2cによって完全に中和された。対照的に、MAb 2cは変異体ウイルスvY301N、vG302R、vG302V、R126 (Y303N)、R1375 (R304Q)およびB4.1 (E305K)を完全に中和することができず、これらの残基の各々がMAb 2cの中和能に必要なエピトープを形成させるのに不可欠な標的であることを示唆していた。総合すれば、ペプチド分析および変異型タンパク質によって得られた結果から、残基299〜305がエピトープ形成に、およびMAb 2cのインビトロでの生物活性に重要であることが示された。
【0154】
マウス保護実験によるエピトープマッピング
インビボでのMAb 2cの保護効果がまた、部位Bの特定のアミノ酸に依存するかどうか分析するために、計168匹のC57BL/6マウスにMAb 2cの腹腔内注射から24時間後に変異体ウイルスまたは親野生型株のどちらかを膣内から接種した。比較のため、同じ中和能力に調整されたポリクローナル免疫血清を投与した。この実験は既報[1, 2]のように行った。図25に示されるように、MAb 2cは、変異体R126 (Y303N)、R1375 (R304Q)およびB4.1 (E305K)を接種したマウスでは効果がなかったが、変異体R1435 (H308Y)もしくはR233 (R328H)または野生型ウイルスを接種したマウスでは有効であった。しかし、ウイルス変異体R126、B4.1およびR233を用いた実験は、マウスの粘膜におけるこれらの変異体のウイルス複製が非効率的であるという事実によってわずかに妨げられた。特に、変異体R126は非常に低い複製能を示した。ゆえに、感染の経過は、MAb 2c、ポリクローナル免疫血清または対照液で処置したR126感染マウスの間で違わなかった。総合すれば、マウス保護実験の結果から、MAb 2cがインビボでその保護効果を示すにはgB残基Y303、R304およびE305が不可欠であることが明らかに実証された。
【0155】
MAb 2c結合のための部位Aの特徴付け
ペプチドスキャン法によるMAb 2cエピトープマッピングの結果(図22)から、共通配列181QFMGIFEDR189を有する結合部位Aは、部位B、およびセグメント化されたペプチドによって検出できない可能性がある潜在的に他の領域とともに形成される不連続なエピトープの構成要素であるが、自然に折り畳まれたタンパク質において機能するエピトープの部分であることが最初に示唆された。しかしながら、部位Aは驚いたことに、三次元gB三量体構造の表面に位置していなかった[21]。さらに、部位AおよびBはgB表面上ですぐ近くになく、抗体の抗原結合部位の平均領域によって同時に網羅することができなかった。
【0156】
部位Aの関連性を調べるために、本発明者らは、gB構築物および単一アミノ酸の交換を有するウイルス変異体を作出することを開始した。しかし、gB構築物でのおよびウイルス変異体でのこれらの残基のいずれの置換も、ほぼ間違いなく分子中の部位Bモチーフの存在により、続けて抗体結合の喪失が起こらなかったので、生体系におけるこれらのアミノ酸の重要性を実証する努力は報いられなかった(データおよび突然変異誘発プライマーは示されていない)。
【0157】
ペプチドスキャン法によって特定されたペプチド50 178RYSQFMGIFEDRAPV192 (SEQ ID NO: 43)の各アミノ酸の相対的重要性を研究するために、本発明者らは、全置換分析を行った。それゆえ、可能な全ての単一部位置換類似体(すなわち、各位置がその他の全19種のタンパク新生アミノ酸によって置換されている)をSPOT(商標)合成によって合成し、MAb 2cの結合について試験した。結合の喪失なしにペプチドの大部分の位置をいくつかの物理化学的に異なるアミノ酸によって交換することができた。しかしながら、モチーフ186FED188は保存された。すなわち、これらのアミノ酸の改変は抗体結合の喪失と関連していた(データ不掲載)。本発明者らはそれゆえ、部位Aまたは主にモチーフ186FED188が不連続なMAb 2cエピトープの一部を模倣すると仮定した。
【0158】
この前提について調べるため、本発明者らは、生物学的に重要であると分かったMAb 2cの結合部位Bに位置する残基(299PFYGYRE305 (SEQ ID NO: 53)) (表8)、グリシンリンカーおよび部位Aの最も重要な配列に由来するC末端のFEDFモチーフから構成される、12merのペプチドPFYGYRE-G-FEDF (SEQ ID NO: 52)をコンピュータによりデザインした。このペプチドとのMAb 2cの結合を、15merのペプチドスキャンにおいて最も強い反応性を示したペプチド298SPFYGYREGSHTEHT312 (SEQ ID NO: 48) (図22)と比べて測定した。両方のペプチドを、方法に記述したようにセルロース膜上で合成し、MAb 2cでプローブした。図26に示されるように、組み合わせペプチドPFYGYRE-G-FEDF (SEQ ID NO: 52)のシグナルは、部位Bのペプチドのみと比べてはるかに強く、しかもX線フィルムの曝露時間は15merのペプチドスキャンの曝露時間の4分の1でしかなかった。この所見は、MAb 2cが、i) 残基299〜残基305 (PFYGYRE) (SEQ ID NO: 53)の部位B、およびii) 配列FEDF (SEQ ID NO: 54)によって模倣されうる1つまたは複数のさらなる不連続な領域から構成される不連続なエピトープを認識するという前提を強力に支持する。
【0159】
考察
現研究の目的は単純ヘルペスウイルス糖タンパク質B上のMAb 2cに対する結合部位をマッピングすること、および鍵となるエピトープ残基を特定することであった。一連のC末端切断型の組換え発現gBタンパク質を用いて、N末端の487残基がMAb2cの結合に必要であることが分かった。C末端からの17個またはそれ以上のアミノ酸のさらなる欠失は、全ての欠失型gBの一過性トランスフェクト細胞での合成を容易に確認できたという事実にもかかわらず、抗体結合の喪失をもたらした。エピトープの位置を絞り込むために、それぞれがシグナルペプチド配列(アミノ酸1〜30)に融合された2つのさらなる欠失変異体gB(183〜488)およびgB(436〜642)を構築した。MAb2cはこれらの後者の切断型gBタンパク質のいずれにも結合することができず、かくしてMAb 2cのエピトープが残基470〜487の間に局在している可能性があるという本発明者らの初期の前提に矛盾していたので、本発明者らは、合成ペプチドを利用する代替的なエピトープマッピング戦略に切り替えることに決めた。
【0160】
多くのB細胞エピトープがもともと不連続的である[46]。タンパク質の断片、すなわち、化学的にまたは生物学的に作出されたペプチドを用いたこれらの不連続的なエピトープのマッピングでは、単一の結合領域に由来するペプチドが結合パートナーに対して、ELISAまたは表面プラズモン共鳴アッセイ法では通常測定できない非常に低い親和性を一般的に有するという難点に悩まされる。ここ20年のうちに、連続表面上で合成されたペプチドを用いた不連続的なエピトープのマッピング研究にかかわるいくつかの調査例が記述されている[47〜50]。感受性の点で、SPOT(商標)法はセルロース膜上の高いペプチド密度(およそ50 nmol/cm2)によって特に適している[37]。これは親和力および再結合効果をもたらし、それゆえ、さらに低親和性のペプチド-抗体間の相互作用の特定を可能にする。2006年までの600超の引用文を網羅する包括的な見直しから、ペプチド合成のためにSPOT(商標)技術を用いて線状かつ不連続のエピトープをマッピングすることに関する研究が大々的に要約されている[51]。
【0161】
N末端の487個のgB残基内でのMAb 2cエピトープの位置決めの後、SPOT(商標)合成法と組み合わせてペプチドスキャン法を用い、抗体結合に重要なアミノ酸の精密マッピングを達成した。図20に示されるように、MAb 2cは、結合部位Bに局在する15merのペプチド298SPFYGYREGSHTEHT312 (SEQ ID NO: 48)に最も強く反応し、この配列がMAb 2c認識に主に関わるという前提につながった。その後、部位Bの認識配列をさらに高分解能のペプチドスキャニングによってgBアミノ酸300〜308にまで縮めることができた。エピトープの鍵となる残基の最終確認を次いで、単一の点突然変異を持つ全長gB構築物でトランスフェクトされた細胞とのMAb 2cの結合を研究する生物学的手法によって達成し、配列300FYGYRE305 (SEQ ID NO: 21)を得た。さらに、プロリン残基299は、三次元gB構造におけるその非常に限られた表面曝露にもかかわらずインビボにおいて重要であることが分かった[21]。しかしながら、プロリン側鎖による立体構造の制限のため、つまりプロリンは、その環状性の結果、固定化された構造としてタンパク質配列を安定化することが多いため、この残基はまた、抗体結合部位でのgBの局部的な折り畳みに不可欠であるものと考えられる。特定された鍵となる残基の影響は、鍵となる残基を変異させた場合にインビボでのMAb 2cの保護効果が無効化されることを実証するマウス保護実験によって裏付けられた。全体的に見て、これらのデータは、アミノ酸300〜305がエネルギー性MAb 2cエピトープの必須部分を形成することを証明する。
【0162】
予想通り、部位B内の鍵となる残基の単一の点突然変異を有するHSV変異体は、MAb 2c結合および中和に耐性であった。しかし、ウイルス変異体の大部分が細胞培養または粘膜において増殖不良を示し、重度に無能なことが分かった。とりわけ、300位の残基フェニルアラニンは、その位置にアミノ酸の交換がある生存ウイルス変異体を作出しようとする試みがこれまで失敗に終わっているので、gBの生物学的機能に重要であるようである。この事実はウイルスの溶解サイクルにおけるこのgBモチーフの重要な役割を示唆しうる。したがって、MAb2cのエピトープがいくぶんかgBの弱点になりうると推測することは興味深い。
【0163】
ペプチドスキャニングによる初期のエピトープマッピングから、MAb 2cが部位Aと名付けられた、さらなるgB領域を認識することが示唆された。その部位で鍵となる残基の判定をペプチド178RYSQFMGIFEDRAPV192に関する置換分析によって行い、残基F186、E187およびD188が非常に置き換え感受性が高いことが実証された。生体系におけるこれらのアミノ酸の関連性は、gB構築物でのおよびウイルス変異体でのこれらの残基の置換が、ほぼ間違いなく分子中の部位Bモチーフの存在により、抗体結合に影響を与えなかったので、実証することができなかった。
【0164】
最近になって決定された外部gBドメイン結晶構造[21]に基づいて、gB単量体を6つの異なる構造ドメインに分けた。ドメインIはアミノ酸154〜363を含む。本明細書で提示した結果によれば、MAb 2cの最も重要なエネルギー性エピトープの残基(部位B)は構造ドメインIに存在する。部位Bの鍵となる残基をgB結晶構造に重ね合わせることにより、これらの残基は構造ドメインIの上部3分の1にある2つのβ鎖(β13, β14)間の22アミノ酸のループ様ストレッチの範囲内でgBの表面に位置することが明らかである。部位Aの残基186FED188も構造ドメインIに、ただし、ほとんど曝露されない小腔中のgBドメインIの基部に局在していることから、この2つの部位が空間的に近接していないことが示唆された。ところがgBと他のウイルス糖タンパク質との構造的類似性によりおよびリンカー挿入突然変異誘発の結果により、その結晶構造はgBの融合後の形態に相当することが示唆される[52〜54]。しかし、ウイルス粒子はgBの融合前の形態を含んでおり、中和抗体はgBの融合前の立体構造を認識すべきであると提案されている[21]。とはいえ、最近の研究から、試験された全てのgB特異的MAbによる融合前および融合後gBの両方の認識が示唆されている[55]。
【0165】
三次元gB構造[21]の範囲内でのおよび部位Bに対しての部位Aの局在と組み合わせた実験データに基づく、代替の、さらに魅力的な説明は、部位Aが不連続的なMAb 2cエピトープの構成要素ではないというものである。ペプチドスキャンおよび置換分析の結果から、適用された方法論によっては明らかに検出できない不連続的なエピトープの1つまたは複数の領域の模倣が示唆される。抗体-抗原複合体のX線またはNMR技術によっては、部位Bの残基300〜305に主に存在するエネルギー性エピトープとは対照的に、機能的エピトープの全体、すなわちMAb 2cと接触する全てのアミノ酸が検出されうるにすぎない[56, 57]。
【0166】
部位AおよびとりわけFEDモチーフが不連続的なエピトープの別の部分を模倣するという仮説は、可動性スペーサー要素としてのグリシン残基を介して共有結合している1つの分子内で部位Bの重要な残基と部位Aに由来する配列FEDFとを組み合わせるという試みによって明らかに支持された(図26)。この遺伝子操作ペプチドは、親和性の増加と相関する部位B由来のペプチド90と比べて(図22参照)シグナル強度の極めて大きな増加をもたらした。例えば、不連続なエピトープを認識する抗体に対するインターロイキン-10模倣体など、一つの合成分子に集められた単一の結合領域を網羅するペプチドによる不連続な結合部位の模倣がいくつかの刊行物に記述されている[49]。
【0167】
幾人かの研究者らがHSV gBの機能ドメインを特定するために何年にもわたってモノクローナル抗体を用いてきた[30、32、58〜60]。最近の研究から、gBに対する中和MAbの結合パターンによって規定されるようにgB構造全体に分散している、少なくとも4つの機能領域の存在が示唆された[55]。これらの結果によれば、MAb 2cのエピトープは、構造ドメインI、および構造ドメインVの残基697〜725由来の配列によって形成される機能領域(FR) 1のなかに位置しており、構造ドメインVは残基670〜727にまで及ぶ。興味深いことに、Benderら[55]によって作出された最も強力な中和MAbの3つとも、残基98〜472を含むgBタンパク質分解的切断断片との反応性によって決定した場合にFR1内の構造ドメインIにマッピングされている。1型HSVによって誘発されたMAb 2cは、2型HSVと交差反応性である[1, 2]。本発明者らはそれゆえ、HSV-1の部位AおよびBのアミノ酸配列をHSV-2 gBのそれと比べた。NCBIタンパク質データベース(日付2010年7月30日)中で見出された全53種の全長HSV-2 gBの中に、HSV-1 gB配列178RYSQFMGIFEDRAPV192 (SEQ ID NO: 43)および298SPFYGYREGSHTEHT312 (SEQ ID NO: 48)が存在していた。
【0168】
抗HSV療法の主な目標は、ウイルス複製を素早く取り除くことであるので、MAb 2cは1型および2型HSVを処置するための潜在的な手段となりうる。原理上は、2つの戦略を追求できる。まず第一に、MAb 2cの特異性および生物活性の抗体を、部位B由来のペプチドまたは模倣エピトープPFYGYRE-G-FEDF (SEQ ID NO: 52)によって誘導できることを実証できたなら、能動免疫が考えられうる。MAb 2cの予防可能性および治療可能性を活用する代替的手法は、十分に確立された抗ウイルス化学療法に加えて、受動免疫用にマウス抗体をヒト化分子へ変換することである。
【0169】
実施例4に関する参考文献
【0170】
参考文献
【技術分野】
【0001】
特許請求の範囲に記載の通り、本発明は、特許請求の範囲に記載の抗HSV抗体、該抗体の有効量を含む薬学的組成物、該抗体をコードするヌクレオチド配列を含む発現ベクター、該ヌクレオチド配列を含む宿主細胞、該抗体を産生することができるハイブリドーマ細胞、および薬物としての、特に、対象におけるHSV関連疾患の予防的または治療的処置のための薬物の製造のための、該抗体の使用に関する。
【背景技術】
【0002】
ヒト病原性単純ヘルペスウイルス(HSV)は、皮膚親和性および神経親和性DNAウイルスであり、その臨床症状は皮膚およびその近傍の粘膜から主に生じ、とりわけ神経炎、髄膜炎、脳炎、脊髄炎、多発性神経根炎などの神経学的合併症を二次的に引き起こす。ある程度、高い致死性を伴う重度進行の、先天性、後天性およびまた、医原性の免疫不全が報告されている。1型HSV (HSV-1、95%; 口唇ヘルペス、角膜ヘルペス、ヘルペス性湿疹)および2型HSV (HSV-2、10〜30%; 陰部ヘルペス、新生児ヘルペス)による集団の高い感染率のため、ならびにウイルスの頻繁な再活性化、知覚神経節および自律神経節中で潜伏的に持続する存続期間のため、HSVは特に臨床的関連性がある。ウイルスのタイプとは無関係に、一次または再発HSV感染の対症療法の目標は、ウイルス複製の阻害、苦痛の時間の短縮、および再発の頻度に影響を与える全身合併症の予防である。
【0003】
初期認識および的確な投与量で、ウイルス抑制剤は抗ウイルス療法に成功裏に利用されている。最も一般的なウイルス抑制剤(例えばアシクロビル(acyclovir)、ペンシクロビル(penciclovir)、ホスカネット(foscarnet)、イドクスウリジン(idoxuridin))は、ヌクレオシド類似体またはピロリン酸類似体であり、その共通の活性原理はウイルス感染細胞におけるDNA合成の阻害に基づく。
【0004】
HSV感染の処置のための最も重要な治療剤の一つがプリンヌクレオシド類似体のアシクロビルである。これはウイルスチミジンキナーゼによりリン酸化され、その後、ウイルスDNAポリメラーゼを妨害する。対照的に、ヒトDNAポリメラーゼは30〜50倍アシクロビルに対する感受性が低く、このため、微々たる副作用しか観察されない。
【0005】
しかし、選択的に作用するウイルス抑制剤の開発にもかかわらず、ウイルス性疾患の化学療法処置は依然として深刻な問題である。具体的には、免疫抑制患者の長期にわたる予防的および治療的処置の間に観察される一般的な化学療法剤に対する耐性株の出現は解決が難しい。結果として、症例の10%超で、有効なウイルス抑制剤の欠如のため、致死的な進行を伴う急速進行性の全身感染が認められる。
【0006】
現在、ピロリン酸類似体のホスカネットがアシクロビル耐性ヘルペスウイルスに対して免疫抑制患者で特に利用されている。この薬剤はウイルスDNAポリメラーゼの直接的な阻害を引き起こし、チミジンキナーゼに影響を与えない。しかしながら、ホスカネットの使用は腎不全、心臓の問題などの重篤な望ましくない副作用をもたらし、骨髄に対する毒性を有し、皮膚潰瘍を引き起こすこともある。ホスカネットは、その催奇形効果のため、妊娠中に投与することもできない。さらに、交差耐性株の形成が認められ、これにより代替的な治療剤の開発が大いに必要となる。受動免疫学的予防は現在、利用できない。HSV1およびHSV2に対する能動免疫用の2〜3種の実験的ワクチンは、検証可能な成功を示さなかった。
【0007】
抗体はがん、自己免疫障害およびウイルス感染の処置にかなり有望である。JP 6135854には、HSVに対するヒトモノクローナル抗体およびアシクロビル(ACV) などの抗ウイルス核酸類似体が点滴静注により注射の形態で同時投与または連続投与される単純ヘルペスウイルス感染用の治療剤が記述されている。DK 187286には、HSV-1およびHSV-2由来の糖タンパク質D (gD) (HSV gD-1およびgD-2)に関して多特異的な免疫反応性を示す抗体が開示されている。WO 1997/26329には、HSV-1およびHSV-2の診断および処置に有用なヒトモノクローナル抗体が記述されている。後者の抗体は、HSV-1およびHSV-2の糖タンパク質D抗原との結合でHSV 863モノクローナル抗体と競合する。US 4,950,595には、抗ウイルス-ヒト抗体を産生するマウス-ヒトハイブリドーマ、その調製の過程、および抗ウイルス-ヒトモノクローナル抗体について開示されている。
【0008】
さらに、HSV 1/2に特異的な別のマウスモノクローナル抗体(Fd79) (Kogae et al., 1986)のヒト化が記述されている(Co, M.S. et al., 1991; Zeitlin L. et al., 1998)。この抗体は、HSV1およびHSV2の糖タンパク質B (gB)の共有エピトープを認識する。さらに、ヒト化Fd79がトランスジェニック植物においても真核生物細胞株SP2/0においても産生され、その後、特徴付けられており、53 nMの親和性を示している。モノクローナルマウス抗体H1815は、糖タンパク質B (gB)のアミノ酸263〜283の領域における類似の、しかし同一ではないエピトープを認識する(Navarro et al., 1992)。しかしながら、H1815はウイルス中和または「細胞間伝播」の阻害ができない。
【0009】
最後に、US 6,180,370には、ヒト化免疫グロブリンおよびそれを作出する方法について記述されている。さらに、WO 2003/105782は、ヒトフレームワーク上にマウス抗体をグラフトすることの特異性に関連する。
【0010】
このように、化学療法剤は望ましくない副作用を有し、ますます多くの耐性株が認められている。
【0011】
それゆえ、HSV感染を中和でき、かつ細胞間伝播を阻害できる(ヒト化)抗HSV抗体を提供することが本発明の目的である。さらに、従来適用される化学療法剤の上記の不利点を克服するHSV関連疾患の処置のための予防剤および/または治療剤を提供することが本発明の目的である。
【0012】
驚いたことに、本発明による抗体はこの目的を解決することが分かった。したがって、本発明は、ヒト化されてもよい、組換え産生抗体に基づくHSV感染の処置に向けた当技術分野において公知の治療剤に代わる有望な選択肢を提供する。これらの抗体は、宿主内で伝播するためのウイルス機構の両方を遮断することができる。それらは効果的に、無細胞ウイルス粒子を中和し、ウイルスの直接的な細胞間伝播を阻害する。これらの抗体は、ウイルス複製サイクルに不可欠な、HSV1およびHSV2外被の表面糖タンパク質B (gB)の高度に保存されたエピトープに特異的に結合するので、薬物耐性が出現する可能性はほとんどない。
【0013】
本発明のマウス抗体の効果は、Eis-Hubinger et al., Intervirology (1991); 32:351-360およびEis-Hubinger et al., Journal of General Virology (1993); 74: 379-385を参照されたく、部分的には、既に記述されているとはいえ、抗体それ自体または本発明の抗体の相補性決定領域(CDR)の配列およびそれが結合するエピトープは、決して公開されておらず、または公衆が利用可能なものではない。
【0014】
要約すれば、(ヒト化)抗体は以下の利点の1つまたは複数を提供する:
・本発明のマウスモノクローナル抗体の効力は既に証明されている(Eis-Hubinger et al., 1991; Eis-Hubinger et al., 1993を参照のこと)。さらに、本発明者らは、本発明のヒト化抗体がまた、インビトロでのHSV1およびHSV2ウイルス感染を中和でき、「細胞間伝播」機構を阻害することによりウイルス伝播を阻害できることを実施例の項において示す。ヒトでの感染進行との関連で、ヒト免疫系は、HSV 1/2に典型的な細胞間伝播を効率的に予防するための抗体特異性を生み出すことができない。
・アシクロビルおよびホスカネットなどの保守的な化学療法剤の頻繁かつ長期にわたる予防的および治療的適用は、耐性ウイルス株の形成の増加をもたらす。この耐性の問題は、単独でまたはアシクロビルおよび/もしくはホスカネットなどのウイルス抑制剤との組み合わせで投与される、本明細書に記述の(ヒト化)抗HSV抗体によって克服することができる。これは、この抗体が異なる作用機序に依るからである。
・本明細書において記述される抗体はHSV gBタンパク質のエピトープに特異的に結合する。本発明の抗体に対するHSV耐性の出現は、gB-タンパク質中の変異がウイルス感染価の喪失をもたらすので、予想されるものではない。
・従来のウイルス抑制剤の全身投与が禁忌とされる患者は、本明細書において記述される(ヒト化)抗体から特に利益を得る。
【発明の概要】
【0015】
したがって、SEQ ID NO: 1、SEQ ID NO: 2、SEQ ID NO: 3、SEQ ID NO: 4、SEQ ID NO: 5、およびSEQ ID NO: 6に示される相補性決定領域を含む抗体が提供される。さらに、SEQ ID NO: 1〜6に示される相補性決定領域を含む抗体と同じエピトープを認識する抗体が提供される。
【0016】
さらに、本明細書に記述の抗体の有効量と、少なくとも1つの薬学的に許容される賦形剤とを含む、薬学的組成物も提供される。
【0017】
さらに、特許請求の範囲に記載の抗体をコードするヌクレオチド配列を含む発現ベクター、該ヌクレオチド配列を含む細胞、および該抗体を産生することができるハイブリドーマ細胞が提供される。
【0018】
最後に、医薬における使用のための特許請求の範囲に記載の抗体、および対象におけるHSV関連疾患の予防的または治療的処置のための薬物の製造のための、特許請求の範囲に記載の抗体の使用が同様に本明細書において提供される。
【0019】
独立請求項に記載の本発明のさまざまな局面および従属請求項に含まれる好ましい態様は、参照により本明細書に組み入れられる。
【図面の簡単な説明】
【0020】
【図1】膣粘膜において確立された感染後のモノクローナル抗体(mAb) 2cの投与によるインビボでのHSV複製の低減を示す。免疫応答性(黒記号)マウスおよびCD4+枯渇(白記号)マウスを感染後3日目および11日目(矢印)に腹腔内(i.p.)注射によりポリクローナルHSV免疫血清(三角)で、mAb 2c (丸)、または対照培地(四角)で処置した。
【図2A】免疫抑制(CD4-/CD8-)マウスにおけるHSV感染の進行に及ぼすmAb 2cの効果を示す。感染後1日目および3日目の時点(矢印で示した)で、動物に対照培地(黒四角)、ポリクローナルHSV免疫血清(白四角)、またはmAb 2c (黒丸)のいずれかをi.p.投与した。(A)は生存率を示す。
【図2B】免疫抑制(CD4-/CD8-)マウスにおけるHSV感染の進行に及ぼすmAb 2cの効果を示す。感染後1日目および3日目の時点(矢印で示した)で、動物に対照培地(黒四角)、ポリクローナルHSV免疫血清(白四角)、またはmAb 2c (黒丸)のいずれかをi.p.投与した。(B)は膣粘膜におけるウイルス複製を示す。
【図3】mAb 2c由来抗体および抗体断片の略図である。(A) 完全な抗体: マウス可変領域(VL、VH; 左)とヒト定常ドメイン(CL、CH1、CH2、CH3; 中央)との遺伝的融合はキメラ抗体をもたらす。ヒト化IgG抗体(右)では、マウスモノクローナル抗体の超可変領域がヒト抗体のフレームワーク上にグラフトされる。(B) 抗体断片: 軽鎖(VL + CL)および重鎖の2つのN末端ドメイン(VH + CH1)からなる一価Fab断片(Fab)、または2つの不対C末端システイン残基によって共有結合的に連結されている二価F(ab')2断片を従来のプロテアーゼ消化によって産生することができる。マウスscFv抗体(「一本鎖可変断片」)の作出のため、可変ドメインVHおよびVLをコードする遺伝子を、2cハイブリドーマ細胞株から単離し、可動性の連結ペプチド(「リンカー」)をコードする遺伝子セグメントによって連結した。
【図4】可変重鎖および軽鎖ドメイン(VHおよびVL)の配列アライメントを示す。抗原結合部位はChothia (Chothia and Lesk, 1987; Chothia et al., 1989)(点線)およびKabat (Kabat and Wu, 1991)(破線)により規定される。ヒト生殖細胞系配列DP28およびDPK13はV-Baseデータベース(http://vbase.mrc-cpe.cam.ac.uk/)から入手し、マウスmAb 2cのCDRのためのアクセプター配列として役立てた。(A) 「不変の残基」(Kabat and Wu, 1991); (B) 「鍵となる残基」(Chothia et al., 1989)および(C) VH/VL界面の残基(Chothia et al., 1985)は、マウスおよびヒト配列の間で、それぞれ、適合残基の場合には(+)または非適合残基の場合には(-)で印されている。(D) それぞれ不変(i)残基部位; 類似(r)残基部位; 表面(s)残基R、K、E、D、Q、N; 中性(n)残基P、H、Y、G、A、S、T; および埋没(b)残基C、V、L、I、M、F、WとしてChothia (Chothia et al., 1998)により定義されている中心部位の残基; 埋没中性残基はxで印されている; 表面中性残基はyで印されている; マウスおよびヒト配列間の非適合残基部位は太文字で印されている; VHhum2cおよびVLhum2c、太文字で示されたマウスCDR残基を有する特異性グラフト配列。全ての残基が一文字表記で示され、Kabat (Kabat et al., 1991)にしたがって付番されている。
【図5】モノクローナル抗体2c、ch2cおよびヒト化バリアントhu2c-V1-V4の平衡結合曲線を示す。HSV-1 F感染Vero細胞の表面上の糖タンパク質Bとの特異的な結合をフローサイトメトリーによって判定した。表示濃度での結合活性を、バックグラウンドの蛍光を差し引いた平均蛍光強度(MFI)として示す。測定は三つ組で行った; 標準偏差をバーとして示す。抗原結合データをLevenberg-Marquard法による非線形回帰モデルに適合させることによって結合親和性定数KDを決定した。
【図6】マウス親mAb 2cと同じ、キメラ抗体およびヒト化抗体のエピトープ特異性を実証する競合研究を示す。キメラmAb ch2c (白丸)およびヒト化mAbバリアントhu2c-V1 (四角)、hu2c-V2 (三角)、hu2c-V3 (星印)、hu2c-V4 (菱形)は、HSV-1感染Vero細胞の表面上に存在するgBとの結合で親mAb 2cと競合する。感染Vero細胞を漸増濃度のmAb ch2cまたはヒト化mAb hu2c-V1-V4とまず初めにインキュベートし、その後、競合物質として100 nMのmAb 2cとのインキュベーションを行った。平均蛍光強度(MFI)は適用された競合物質の結合を示す。
【図7】親mAb 2cおよびヒト化mAb hu2c-V1のHSV中和活性が補体非依存的であることを示す。HSV-1を、Vero細胞に適用する前に10%ヒト補体の存在下または非存在下で培地(対照)、ポリクローナルIgG Cytotect(登録商標) (120μg/ml)、mAb 2c (2μg/ml)またはmAb hu2c-V1 (2μg/ml)とともにプレインキュベートする。2日後にプラークの出現をスコア化した。
【図8】非耐性実験室株(HSV-1 F、HSv-1 324hv、HSV-1 17 syn+、HSV-2 G)、ACV耐性実験室株HSV-1 TK-および耐性について詳しく調べられていない臨床分離株と比較した、アシクロビル(ACV)、ACVおよびホスカネット(PFA)、またはACV、PFAおよびシドビル(CDV)に対して臨床的に耐性の患者分離株に由来するHSV-1およびHSV-2を中和するヒト化mAb h2c-V1の効率を示す。完全なウイルス中和のためのmAb h2c-V1の力価を決定するために、いくつかの抗体濃度のものを100 TCID50のHSV-1またはHSV-2分離株とともに37℃で1時間インキュベートし、Vero細胞とともに3日間インキュベートした。mAb hu2c-V1は7.8〜15.6 nMの濃度で完全に、HSV-1実験室株HSV-1 F、HSV-1 324 hv、HSV-1 17 syn+、HSV-1 TK-を中和する。HSV-1臨床分離株はその耐性プロファイルに関係なく同様にmAb h2c-V1によって中和される。さらに、株HSV-2 GおよびACV耐性HSV-2分離株に対して31.3〜62.5 nMの濃度でmAb hu2c-V1の、同じ中和効率が示された。
【図9】本発明の抗HSV抗体によるウイルスの細胞間伝播の阻害を示す。4時間HSV-1 Fを感染させたVero細胞を2回洗浄し、過剰の、ヒトポリクローナル抗HSV対照血清(1:20)、マウスmAb 2c (500 nM)、酵素消化によって調製された2c由来の抗体断片F(ab')2 (500 nM)またはFab (3000 nM)、キメラmAb ch2c (500 nM)を含有する培地とともに、またはヒト化mAbバリアント1のhu2c-V1 (500 nM)とともに、それぞれ、インキュベートした。感染から2日後に、Leica DM IRE2共焦点顕微鏡を用い40倍の倍率で蛍光標識ポリクローナルヤギ抗HSV血清でウイルスの伝播を検出した。ヒトポリクローナル抗HSVの中和価は、100 TCID50を100μl容量中用いて1:160と以前に決定された。1:20の希釈で抗HSV血清は隣接細胞へのウイルスの伝播を抑止することができない。細胞間伝播を500 nMのマウスmAb 2c、2c-F(ab')2抗体断片、キメラmAbおよびヒト化mAbによって成功裏に阻害することができた。3.000 nMの最も高い試験濃度で一価2c-Fab断片は細胞間伝播を完全に阻害することができなかった。
【図10】抗HSV mAbによる受動免疫後にHSV-1を膣内感染させたNOD/SCIDマウスの生存を示す。マウスに感染の24時間前にPBS (十字)、2.5 mg/kg (四角)、5 mg/kg (三角)または15 mg/kg (丸)の親mAb 2c (白記号)またはヒト化mAb hu2c-V1 (黒記号)のいずれかを静脈内投与した。マウスに1×106 TCID50/20μlの神経病原性HSV-1株Fを膣内感染させた。体重減少、外陰炎/膣炎または神経疾患の症状を有する感染マウスを殺処理し、その臓器を、既述のようにVero細胞単層上での力価測定により感染性ウイルスについて調べた。非感染マウスを30日目の時点で殺処理した。1群あたりの動物数n=7。
【図11】HSV-1伝播に対しての全身適用抗体によるNOD/SCIDマウスの保護を示す。感染後24時間から始めて、マウスに、矢印で示した時点(24時間、40時間、56時間)で静脈内に3回15 mg/kgのmAb 2cまたはヒト化mAb hu2c-V1のいずれかを投与した。1群あたり感染動物数n=7。
【図12】アシクロビル、ホスカネットおよびシドボビルに対して耐性の患者由来HSV-1分離株を膣内感染させたNOD/SCIDマウスは、15 mg/kgのヒト化mAb hu2c-V1による感染後24時間、40時間および56時間の時点での静脈内処置によって致死性脳炎から有意に保護されたことを示す。1日2回アシクロビルによる標準的な処置を受けたマウスは、28日以内に全て死んだ。
【図13A−1】gBに対するmAb 2cのエピトープの局在性を示す。(A)はHSV1およびHSV2の糖タンパク質B (gB)のアミノ酸配列アライメントを示す。示されるのは、以下の株のgBタンパク質アミノ酸配列のアライメントである(括弧内は対応するNCBIアクセッション番号): HSV1株KOS (P06437)、F (P06436)、gC-39-R6 (ABM66850)、ならびにHSV2株333 (ABU45423)、HG52 (P08666)およびMMA (AAB60547)。gBのシグナル配列に下線を引いてある。成熟gBは31位からアミノ酸APで始まる。アミノ酸の付番は、シグナルペプチド配列を含むgBについて示されている。エピトープの付番はそれに従っている。mAb 2cは、ペプチドマイクロアレイによって示されるようにgB内の2つの別々の領域(囲まれた配列領域)に結合する。アミノ酸299PFYGYRE305はmAb 2cの結合に不可欠であることが示されている。
【図13A−2】gBに対するmAb 2cのエピトープの局在性を示す。(A)はHSV1およびHSV2の糖タンパク質B (gB)のアミノ酸配列アライメントを示す。示されるのは、以下の株のgBタンパク質アミノ酸配列のアライメントである(括弧内は対応するNCBIアクセッション番号): HSV1株KOS (P06437)、F (P06436)、gC-39-R6 (ABM66850)、ならびにHSV2株333 (ABU45423)、HG52 (P08666)およびMMA (AAB60547)。gBのシグナル配列に下線を引いてある。成熟gBは31位からアミノ酸APで始まる。アミノ酸の付番は、シグナルペプチド配列を含むgBについて示されている。エピトープの付番はそれに従っている。mAb 2cは、ペプチドマイクロアレイによって示されるようにgB内の2つの別々の領域(囲まれた配列領域)に結合する。アミノ酸299PFYGYRE305はmAb 2cの結合に不可欠であることが示されている。
【図13A−3】gBに対するmAb 2cのエピトープの局在性を示す。(A)はHSV1およびHSV2の糖タンパク質B (gB)のアミノ酸配列アライメントを示す。示されるのは、以下の株のgBタンパク質アミノ酸配列のアライメントである(括弧内は対応するNCBIアクセッション番号): HSV1株KOS (P06437)、F (P06436)、gC-39-R6 (ABM66850)、ならびにHSV2株333 (ABU45423)、HG52 (P08666)およびMMA (AAB60547)。gBのシグナル配列に下線を引いてある。成熟gBは31位からアミノ酸APで始まる。アミノ酸の付番は、シグナルペプチド配列を含むgBについて示されている。エピトープの付番はそれに従っている。mAb 2cは、ペプチドマイクロアレイによって示されるようにgB内の2つの別々の領域(囲まれた配列領域)に結合する。アミノ酸299PFYGYRE305はmAb 2cの結合に不可欠であることが示されている。
【図13B】gBに対するmAb 2cのエピトープの局在性を示す。(B) 異なるウエスタンブロット条件下での組換えgBとのmAb 2cの反応性によるmAb 2cの特徴付け。組換えgB (730t)を未変性(N)または変性(D)条件下での8% SDS-PAGEにて分離し、ニトロセルロース膜に転写し、2%ミルクを含有するTNTブロッキング用緩衝液中で1時間インキュベートした。この膜をgB特異的モノクローナル抗体mAb H1817、mAb H126またはmAb 2cでプローブし、gBとの結合をHRPコンジュゲートポリクローナルヤギ抗マウス血清および化学発光によって検出した。対照の場合には、連続的なエピトープ(Bender et al., 2007)および不連続的なエピトープ(Kousoulas et al., 1988)を認識する、それぞれ、mAb H1817およびmAb H126を用いた。線状エピトープに典型的な染色パターンがmAb H1817によるウエスタンブロット分析において得られ、非還元条件下でのgBの単量体型および三量体型の検出ならびに還元条件下でのgB単量体の唯一の主たる染色が示された。予想通り、mAb H126は未変性条件下でのみgBと反応した。170 kDa超(>170 kDa)の、唯一の、上方のgBタンパク質のバンドの認識から、mAb H126が三量体gBに特異的に結合することが示唆される。しかしながら、mAb 2cは未変性および変性gBと反応し、変性条件下での反応性はmAb H1817と比べてはるかに弱く、このことは、変性に対して耐性であるか、またはSDS-PAGE電気泳動の間に再び折り畳まれるように見え、それゆえ「偽連続」エピトープと呼ばれる、不連続なエピトープ(Bender et al., 2007)にmAb 2cが結合することを示唆している。分子量(kDa)が左側に示され、gB三量体および単量体の移動が右側に示されている。
【図14】gBに対するmAb 2cのペプチドマッピングを示す。(A) アミノ酸31〜505のgBの細胞外ドメインに及ぶペプチドライブラリに対して特定された結合領域AおよびBの図式的な局在。13 merのペプチドを3アミノ酸のずれで連続的なセルロース膜上で合成し、結合したmAb 2cをペルオキシダーゼコンジュゲート二次抗体で化学発光により検出した。機能ドメインI〜Vは、HeldweinらによるgBの結晶構造に対応し、結晶構造において解析されていない領域は灰色で示されている(24)。S, シグナル配列。(B) 可動性リンカーを介してスライドグラス上に固定化された13 merのペプチドによる高分解能レーザースキャンからの蛍光シグナル強度。
【図15】gB結晶構造(PDB-ID 2GUM)上の中和mAb 2cエピトープの局在を示す。gB三量体のリボンダイヤグラムが示されている。星印は2つのプロトマーの融合ループを示し、第3のプロトマーの融合ループは見えない。不連続的なmAb 2cエピトープF175〜A190およびF300〜E305のマッピング残基は、片方のプロトマーについては濃い灰色によりおよび他の両方のプロトマーについては薄い灰色により表面表現で示されている。
【図16】mAb 2cのデュオトープスキャニングを示す。mAb 2c結合領域AおよびB (ダッシュ記号付きのバー)のコンセンサス配列(下線を引いた)を、直接的にか、または1つもしくは2つのβアラニンスペーサー(B、B-B)によって離すかのいずれかで連結させたデュオトープ(白バーおよび黒バー)として合成した。mAb 2cとデュオトープとの反応性を高分解能レーザースキャンからの蛍光シグナル強度によって記録した。
【図17】フローサイトメトリーによって決定されたmAb 2c、2c-F(ab')2、2c-Fabおよび2c-scFvに対する平衡結合曲線を示す。表示濃度での(A) HSV-1 Fまたは(B) HSV-2 G感染Vero細胞に対する結合活性を最大の平均蛍光強度の百分率として示す。測定は三つ組で2回行った; バーは標準偏差を表す。
【図18】mAb 2cによる標的細胞へのHSV-1ウイルス粒子の付着の阻害を示す。(A) mAb 2c (0.98〜125 nM)または(B) 多価ヒトγグロブリン(Intratect(登録商標)) (0.33〜42μM)の連続希釈液を96ウェルマイクロタイタープレート中のVero細胞単層に添加し、その後、100 TCID50 HSV-1とのプレインキュベーション(付着前中和)処理または標的細胞への100 TCID50 HSV-1の吸着後(付着後中和)処理を行った。10個の個別の接種細胞単層におけるウイルス誘導性の細胞変性効果(CPE)を対照と比べて100%および50%抑止する、それぞれ、最も高い抗体および多価ヒトIgGの力価を37℃で72時間のインキュベーションの後に、終点と見なして決定した。独立した3回の実験の平均の標準誤差は0.1未満(< 0.1)であった。
【図19】HSVのインビトロでの中和に及ぼす抗gB抗体の結合価の影響を示す。(A) 二価抗体mAb 2c (IgG)および2c-F(ab')2、ならびに一価2c-Fabの希釈液をVero細胞への接種前に100 TCID50のHSV-1 FまたはHSV-2とともに1時間インキュベートした。図3に記述されるように72時間後にCPEをスコア化した。示されているのは、3回のうち1回の代表的な繰り返し実験からの、ウイルス接種材料の100%を中和するのに必要な抗体濃度である。(B) マウス抗Fab IgGで架橋された2c-Fab断片の抗ウイルス活性。
【図20】mAb 2c処置免疫不全マウスの用量依存的な生存を示す。NOD/SCIDマウスに1×106 TCID50 HSV-1での膣内攻撃の24時間前に異なる単回投与量のmAb 2cを静脈内投与した。1群あたりの動物数はPBSの場合n = 7、他の全群の場合n = 9。
【図21】HSV-1伝播に対して全身に適用されたmAb 2cまたはヒト化mAb hu2c-V1によるNOD/SCIDマウスの性器粘膜からの確立したHSV-1感染の除去を示す。感染後24時間から始めて、マウスに、矢印で示した時点(24時間、40時間、56時間)で静脈内に3回15 mg/kgのmAb 2c (白記号)またはヒト化mAb hu2c-V1 (黒記号)を投与した。抗体または対照処置マウスの膣ウイルス力価をVero細胞単層上で培養された膣洗浄液から決定した。誤差バーは標準偏差を示す。
【図22】アミノ酸31〜505のHSV-1 gB配列に対するペプチドスキャニングを示す。12アミノ酸の重複を有し、計155種の異なるペプチドスポットをもたらす、セルロース膜に結合させた15merのペプチド(15/12スキャン)をMAb 2cとともにインキュベートした。ペプチドに結合したMAb 2cを、ペルオキシダーゼ標識抗マウスIgG Fab断片およびルミノール様の化学発光基質を用いて検出した。MAb 2cは結合部位AおよびBと名付けた連続的なgBペプチドのトリプレット(ペプチド49〜51)およびダブレット(ペプチド90〜91)に結合することが分かった。各部位からのペプチドに共通するgB配列は太文字で強調されており(下部)、残基181〜189および残基301〜312のgB配列を表す。
【図23】アミノ酸296〜315のHSV-1 gB配列に対するペプチドスキャニングを示す。各ペプチドを1アミノ酸ずつ配列に沿ってシフトさせ(13/12スキャン)、二つ組で合成し、セルロース膜に結合させた13merのペプチドをMAb 2cとともにインキュベートし、その後、化学発光ウエスタンブロッティング検出を行った。一連の5つのペプチドに対してMAb 2cの結合を認めた。5つの反応性ペプチドに共通する配列は300FYGYREGSH308である。
【図24】残基295〜315のgB配列に対する鍵となるモチーフのスキャンを示す。アミノ酸295〜315のHSV-1 gB配列を、それぞれが1アミノ酸ずつ配列に沿ってシフトする六量体ペプチドに切断し、計16種のペプチドを得た。gB由来配列を各N末端およびC末端の位置で4つの無作為化残基により取り囲んだ。gB配列300FYGYRE305および301YGYREG306に相当する2つの連続ペプチドは、MAb 2cに結合することが特定された。これらのペプチドに共通する配列は太文字で強調されている(ペプチド6および7)。
【図25】ウイルス接種の24時間前に受動的に移入されたポリクローナル免疫血清(白四角、黒四角)、MAb 2c (白丸、黒丸)または対照として沈降培地(白三角、黒三角)を投与されたC57BL/6マウスの性器粘膜におけるHSV-1の除去動態を示す。野生型(wt)株Fおよびその変異派生体R126 (Y303N)、R1375 (R304Q)、R1435 (H308Y)およびR233 (R328H)を接種したマウスからのデータを白記号で示し、野生型株KOS 321およびその変異派生体B4.1 (E305K)を接種したマウスからのデータを黒記号で示す。S.E.の範囲(log10)および接種したマウスの数: wt株F、白四角、±1.5〜0、マウス12匹; 白丸、±0.4〜0、マウス7匹; 白三角、±1.3〜0、マウス9匹; wt株KOS、黒四角、±1.4〜0、マウス8匹; 黒丸、±0.6〜0、マウス8匹; 黒三角、±1.4〜0、マウス8匹; F変異体株R126 (Y303N)、白四角、±0.6〜0、マウス5匹; 白丸、±0.6〜0、マウス6匹; 白三角、±0.5〜0、マウス6匹; F変異体株R1375 (R304Q)、白四角、±1.2〜0、マウス11匹; 白丸、±1.3〜0、マウス10匹; 白三角、±1.2〜0、マウス11匹; KOS変異体株B4.1 (E305K)、黒四角、±0.9〜0、マウス12匹; 黒丸、±0.7〜0、マウス12匹; 黒三角、±0.9〜0、マウス10匹; F変異体株R1435 (H308Y)、白四角、±1.4〜0.6、マウス6匹; 白丸、0、マウス5匹; 白三角、±1.0〜0.6、マウス6匹; F変異体株R233 (R328H)、白四角、±1.0〜0、マウス5匹; 白丸、±0.1〜0、マウス5匹; 白三角、±1.1〜0、マウス6匹。
【図26】ペプチド90 (図22参照、gB配列298SPFYGYREGSHTEHT312; 左側)との、ならびに部位Bの重要な残基、グリシンリンカーおよび部位Aに由来するモチーフFEDFを含むようにデザインされたペプチド(PFYGYRE-G-FEDF; 右側)とのMAb 2cの反応性の比較を示す。
【発明を実施するための形態】
【0021】
発明の詳細な説明
したがって、第1の局面において、本発明は、
に示される相補性決定領域を含む抗体に関する。
【0022】
抗体または免疫グロブリンは、ジスルフィド結合によって連結された2本の大きな重鎖および2本の小さな軽鎖の、その天然形態にあるγグロブリンタンパク質である(図3参照)。5種類の哺乳類Ig重鎖: α、δ、ε、γおよびμが存在する。存在する重鎖の種類が抗体のクラス(アイソタイプ)を規定し; これらは、それぞれ、IgA、IgD、IgE、IgGおよびIgM抗体である。各重鎖は2つの領域、つまり定常領域および可変領域を有する。定常領域は同じ種の同じアイソタイプの全ての天然抗体においてほぼ同一である。軽鎖も1つの定常ドメインおよび1つの可変ドメインからなる。哺乳類では、2種類の免疫グロブリン軽鎖ラムダ(λ)およびカッパ(κ)が存在する。
【0023】
全ての抗体の一般構造は非常に似ているが、所与の抗体の特異的性質は可変(V)領域によって決定される。より具体的には、軽(VL)鎖上の各3つのおよび重(VH)鎖上の3つの可変ループが抗原との結合に、すなわち、その抗原特異性に関与する。これらのループは相補性決定領域(CDR)といわれる。VHおよびVLの両ドメイン由来のCDRが抗原結合部位に寄与するので、最終の抗原特異性を決定するのは重鎖および軽鎖の組み合わせであり、どちらか一方だけではない。
【0024】
本明細書において用いられる「抗体」という用語は、結合特異性がSEQ ID NO: 1〜6に示されるCDRによって決定される、抗原に結合できる任意のポリペプチドを意味する。ゆえに、「抗体」は、少なくとも1つの抗原結合断片を含む任意のポリペプチドに関するように意図される。抗原結合断片は、重鎖の可変ドメインおよび軽鎖の可変ドメインの両方がともに、特異的な抗原に結合できる形で配列された少なくともその両方のドメインからなる。「抗体」は完全な抗体、または抗体断片、例えばFab、F(ab)2もしくはscFv断片を含む(図3も参照のこと)。
【0025】
「完全な抗体」という用語に関して、各ドメインが、全体のドメイン構造を変化させない、変異、欠失または挿入などの、さらなる修飾をたとえ含んでいようとも、天然抗体(すなわち、3つまたは4つの定常ドメインの重鎖および1つの定常ドメインの軽鎖ならびに各可変ドメインを含む)に典型的な全体のドメイン構造を有する任意の抗体が意図される。
【0026】
「抗体断片」はまた、上記に定義される少なくとも1つの抗原結合断片を含み、断片が由来する完全な抗体と同じ機能および特異性を示す。Fab断片は、酵素パパインを用いて免疫グロブリンを切断することにより作出することができる。酵素パパインは、蝶番領域よりも下部、かくして、ジスルフィド結合よりも下部を切断し、その結果、F(ab)2断片が形成される。さらに、重鎖および軽鎖の可変領域をともに融合させて、一本鎖可変断片(scFv)を形成させることができる。
【0027】
さらに、「抗体」という用語は全ての上記の免疫グロブリンアイソタイプを含むよう意図され、すなわち、抗体はこれらのアイソタイプの任意のサブクラスを含めて、IgA、IgD、IgE、IgGまたはIgM抗体であってよい。好ましくは、抗体はIgG抗体、より好ましくはIgG1またはIgG2抗体である。抗体は組換えにより発現され産生されうるので、抗体は重鎖、例えば、1つのIgG1および1つのIgG2の重鎖、または異なる種由来の重鎖の、2つの異なる定常領域を含んでもよい。しかしながら、重鎖は同じ種に由来することが好ましい。さらに、抗体はラムダまたはカッパ軽鎖のどちらかを含むことができる。
【0028】
実施例2に示されるように、抗体の結合価は、ウイルス中和を媒介かつ細胞間伝播を阻害する効力に大きな影響を及ぼし、最良の結果は二価抗体で、すなわち、2つの抗原結合領域を有する抗体で示された。二価抗体の例は完全な抗体またはF(ab)2断片などの、二価抗体断片である。それゆえ、好ましい態様において、抗体は二価抗体であり、好ましくはここで抗体は完全な抗体または抗体断片であり、具体的にはここで抗体断片はF(ab)2断片である。その代わりに好ましい態様において、抗体は多価抗体、すなわち、組換え抗体もしくはその断片、好ましくは三重特異性抗体もしくは四重特異性抗体、またはIgM五量体もしくは連結された免疫グロブリンなどの全免疫グロブリンを含む、三つ以上の結合部位を有する抗体である。これらの抗体の形式は当技術分野において公知である。
【0029】
別の好ましい態様において、抗体はモノクローナル抗体であり、好ましくはここで抗体はマウス抗体、キメラ抗体またはヒト化抗体であり、より好ましくはここでヒト化抗体は以下で論じられるように、ヒト生殖細胞系配列に由来する。キメラ抗体は、その免疫原性を低減するために遺伝子操作によってある種の免疫グロブリンの少なくとも1つの領域が別の種の免疫グロブリンの別の領域に融合されている抗体である。キメラ抗体の例は図3Aに示されており、この図には、ヒト免疫グロブリンの残存部分に融合されたマウスVLおよびVH領域が描かれている。キメラ抗体の特定の種類はヒト化抗体である。ヒト化抗体は、非ヒト抗体のCDRをコードするDNAを、ヒト抗体フレームワークコーディングDNAにグラフトすることによって産生される。次に、得られたDNA構築物を用いて、単にCDRがヒト以外のものであるだけなので、非ヒト親抗体よりも、またはキメラ抗体よりも、通常、免疫原性ではない抗体を、発現かつ産生させることができる。
【0030】
1つの好ましい態様において、抗体は感染細胞から、隣接した第2の非感染細胞へのHSV感染の伝播(細胞間伝播)を阻害することができる。細胞間伝播は、無細胞粒子を放出させずに、ある感染細胞から、隣接した非感染細胞へと伝播するヘルペスウイルスの能力である。抗体が、感染細胞から、隣接した第2の非感染細胞へのHSVの伝播(細胞間伝播)を阻害できるかどうかを調べるために、以下のアッセイ法を用いることができる。
【0031】
24ウェル組織培養プレート中のカバーガラス上で集密まで増殖されたVero細胞を400 TCID50/ウェルの一定のウイルス量で37℃にて4時間感染させる。1メディアンの組織培養感染用量(1 TCID50)は、接種された細胞培養物の50%において細胞変性効果をもたらすと考えられる、ウイルスなどの細胞変性因子の量である。ウイルス接種材料を引き続いて除去し、細胞をPBSで2回洗浄し、上清によるウイルス伝播を防ぐために過剰の異なる抗HSV抗体またはポリクローナル抗HSV対照血清を含有するDMEM, 2% FCS, Pen/Strep 1 ml中37℃で2日間さらにインキュベートする。HSV感染細胞のウイルス抗原を蛍光標識ポリクローナルヤギ抗HSV血清(BETHYL Labolatories, Montgomery, TX USA, カタログ番号A190-136F, ロット番号A190-136F-2)で検出する。
【0032】
好ましくは、抗体は、隣接細胞の20%未満の感染であれば、細胞間伝播を阻害することができ、好ましくはここで上記のアッセイ法において隣接細胞の15%未満、10%未満、5%未満、より好ましくは3%未満および最も好ましくは1%未満が感染している。
【0033】
なお、さらに好ましい態様において、抗体は最大でも40 nM、好ましくは最大でも30 nM、より好ましくは最大でも20 nM、さらにより好ましくは最大でも15 nM、例えば最大でも13 nM、最大でも10 nM、および最も好ましくは最大でも7 nMの解離定数KDを有する。KDは、複合体がその構成要素へ可逆的に解離するその傾向(すなわち抗原に対する抗体の親和性)の尺度としての解離定数を表し、結合定数の逆のものである。KDはScatchard方程式から計算することができ、KDを決定するための方法は当技術分野において周知である。
【0034】
さらに好ましい態様において、最大でも20 nMの、好ましくは最大でも16 nMの、より好ましくは最大でも12 nMの、例えば最大でも10 nM、例えば最大でも8 nMまたは最大でも6 nMの、および最も好ましくは最大でも4 nMの濃度の抗体は、100 TCID50のHSVの規定量を80%超まで、好ましくは90%超、例えば95%超、より好ましくは96%超、例えば97%超、および最も好ましくは98%超、例えば99%超または場合により100%中和することができる。本明細書において「中和(する)」とは、ウイルスが任意のさらなる細胞に感染できないように抗体がウイルスをオプソニン化することを意味する。最大でも20 nMの濃度の抗体が100 TCID50のHSVの規定量を中和できるかどうか試験するためのアッセイ法は、Eis-Hubinger et al , 1991., およびEis-Hubinger et al., 1993に、ならびに以下の実施例1および2に示されている。
【0035】
さらに、1つの好ましい態様において、抗体は、図4に示されるように、SEQ ID NO: 9の1〜30位、38〜51位、68〜99位および112〜122位(またはKabatによる付番によれば、それぞれ、1〜30位、36〜49位、66〜94位および103〜113位)ならびにSEQ ID NO: 10の1〜23位、40〜54位、62〜93位および103〜113位(またはKabatによる付番によれば、それぞれ、1〜23位、35〜49位、57〜88位および98〜108位)に示されるアミノ酸残基と比べてフレームワーク領域中で少なくとも70%、好ましくは少なくとも75%、少なくとも80%、より好ましくは少なくとも85%、少なくとも90%、さらにより好ましくは少なくとも95%および最も好ましくは98%の(全体の)配列同一性を有するアミノ酸配列を含む。
【0036】
ポリペプチドは、SEQ ID NO: 9または10が関心対象のポリペプチドの最も適合している配列と整列され、整列されたそれらの2つの配列間のアミノ酸同一性が、SEQ ID NO: 9の1〜30位、38〜51位、68〜99位および112〜122位(またはKabatによる付番によれば、それぞれ、1〜30位、36〜49位、66〜94位および103〜113位)、ならびにSEQ ID NO: 10の1〜23位、40〜54位、62〜93位および103〜113位(またはKabatによる付番によれば、それぞれ、1〜23位、35〜49位、57〜88位および98〜108位)にわたって少なくともX %であるなら、SEQ ID NO: 9または10に対してフレームワーク領域中で「少なくともX %の配列同一性」を有する。かかるアミノ酸配列のアライメントは、例えば、国立バイオテクノロジー情報センター(NCBI)のホームページ上、http://www.ncbi.nlm.nih.gov/blast/blast.cgiで提供されている「BLAST」プログラムなどの公的に入手可能なコンピュータ相同性プログラムを使い、その中で提供されている初期設定を用いて行うことができる。アミノ酸配列または核酸配列のセットの配列同一性の割合を計算するさらなる方法は、当技術分野において公知である。
【0037】
あるいは、別の好ましい態様において、抗体は、図4に示されるように、SEQ ID NO: 7の1〜30位、38〜51位、68〜99位および112〜122位(またはKabatによる付番によれば、それぞれ、1〜30位、36〜49位、66〜94位および103〜113位)、ならびにSEQ ID NO: 8の1〜23位、41〜55位、63〜94位および104〜114位(またはKabatによる付番によれば、それぞれ、1〜23位、35〜49位、57〜88位および98〜108位)に示されるアミノ酸残基と比べてフレームワーク領域中で少なくとも80%、好ましくは少なくとも85%、より好ましくは少なくとも90%、さらにより好ましくは少なくとも95%、例えば98%および最も好ましくは100%の(全体の)配列同一性を有するアミノ酸配列を含む。ポリペプチドは、SEQ ID NO: 7または8が関心対象のポリペプチドの最も適合している配列と整列され、整列されたそれらの2つの配列間のアミノ酸同一性が、SEQ ID NO: 7の1〜30位、38〜51位、68〜99位および112〜122位(またはKabatによる付番によれば、それぞれ、1〜30位、36〜49位、66〜94位および103〜113位)、ならびにSEQ ID NO: 8の1〜23位、41〜55位、63〜94位および104〜114位(またはKabatによる付番によれば、それぞれ、1〜23位、35〜49位、57〜88位および98〜108位)にわたって少なくともX %であるなら、SEQ ID NO: 7または8に対してフレームワーク領域中で「少なくともX %の配列同一性」を有する。
【0038】
SEQ ID NO: 7および8はヒト生殖細胞系配列に由来する。たとえ非生殖細胞系列のヒト免疫グロブリンフレームワーク配列がヒト由来のものであっても、それらが免疫原性でないことを、通常、除外することはできない。それゆえ、本発明者らは、生殖細胞系配列を調べた。というのは、それらが高頻度変異しておらず、それゆえ、免疫原性ではないものと予想されるからである。したがって、ヒト化抗体は好ましくは、ヒト生殖細胞系配列に、例えばSEQ ID NO: 7および/または8に由来する。SEQ ID NO: 7および8は次の通りである。
【0039】
本発明との関連で、本発明のヒト化抗体の作出のためにSEQ ID NO:7および8を用いることで、親抗体に対するのと同じ親和性を達成するために帰突然変異は必要とされないことが分かったが、このことから、対応するヒト化抗体が非常に低い免疫原性を示すことが示唆されうる。その結果、本発明との関連で、SEQ ID NO: 9および10またはSEQ ID NO 7および8をそれぞれ含む抗体と同じ特異性を示す、抗体が含まれることが好ましい。
【0040】
さらに好ましい態様において、抗体はエフェクター部分、治療部分、または検出可能な部分とコンジュゲートしている。これに関連して、「コンジュゲート(している)」という用語は、非限定的に介在ドメインありまたはなしでの組換え融合、インテイン媒介融合、非共有結合性会合および共有結合、例えば、ジスルフィド結合、ペプチド結合、水素結合、静電結合、および高次構造的結合、例えば、ビオチン-アビジン会合を含む、タンパク質ドメインを機能的に結び付けるための当技術分野において公知の任意の方法をいう。エフェクター部分とのコンジュゲーションは化学的手段によるかまたは組換え手段によるかのどちらかであってよい。化学的手段は、1分子を形成するように2分子間で形成される共有結合が存在するような、抗体とエフェクター部分との間の反応をいう。
【0041】
「エフェクター部分」という用語は、抗体によって標的化される細胞に影響を及ぼすように意図された化合物を意味する。エフェクター部分は、例えば、治療部分または検出可能な部分でありうる。
【0042】
「治療部分」は、細胞毒性剤または細胞毒性薬などの、治療剤として作用するように意図された化合物である。化合物の例は以下、薬学的組成物に示される。
【0043】
「検出可能な標識」は、蛍光標識などの、分光学的、光化学的、生化学的、免疫化学的、電気的、光学的または化学的手段により検出可能な任意の化合物またはタンパク質タグを含む。
【0044】
抗体の特異性はCDRによりまたは抗体が結合するエピトープにより発現されうる。したがって、第2の局面において、本発明は、第1の局面の抗体と同じエピトープを認識する抗体に関する。実施例の項に示されるように、ならびに図13Aおよび13Bに例示されるように、このエピトープは、HSV1およびHSV2の糖タンパク質Bのアミノ酸172〜195および295〜313に位置する、変性に対して部分的に耐性の不連続な、というよりむしろ偽連続なエピトープである。
【0045】
本出願との関連で、mAb 2c抗体のエピトープはgBタンパク質の最初の487アミノ末端残基のなかに位置しうる。好ましくは、エピトープは、gBタンパク質の172位と307位との間のアミノ酸配列のなかに位置する少なくとも1つのアミノ酸配列を含みうる。
【0046】
エピトープは、gBタンパク質の連続アミノ酸配列301YGYRE305、好ましくは連続アミノ酸配列301YGYREG306または300FYGYRE305を含むことがあり、より好ましくは配列は、末端の位置でさらに伸長されうる(すなわち、299PFYGYRE305または300FYGYREGS307)。本発明の抗体のエピトープは、gBの連続アミノ酸配列298〜313 (298SPFYGYREGSHTEHTS313)を含みうる。
【0047】
あるいは、エピトープは連続アミノ酸配列172QVWFGHRYSQFMGIFED188に位置しうる。エピトープは連続アミノ酸配列172QVWFGHRYSQFMG184を含みうる。
【0048】
好ましくは、エピトープは2つ以上の連続アミノ酸配列からなりうる。エピトープは部分的に、不連続なエピトープであってもよい。より好ましくは、エピトープは2つの連続アミノ酸配列を含みうる。2つのアミノ酸配列からなるそのようなエピトープは、「デュオトープ」と指定することができる。抗体は両方のアミノ酸配列に結合することができる。
【0049】
より好ましくは、デュオトープのアミノ酸配列は、アミノ酸配列300FYGYRE305およびアミノ酸172位と188位との間に位置するアミノ酸配列を含みうる。さらにより好ましくは、エピトープは、gBタンパク質のアミノ酸配列300FYGYRE305およびアミノ酸配列179YSQFMG184を含みうる。あるいは、エピトープまたはデュオトープは、化学的に合成されてもよい。エピトープは、配列YSQFMG-βA-FYGYREを有する化学的に合成されたエピトープでありうる。本明細書において用いられる略語βAはβ-アラニンをいう。
【0050】
より好ましくは、エピトープは、gBタンパク質のアミノ酸配列FYGYREおよびアミノ酸配列FEDを含みうる。エピトープは、配列FED-βA-βA-FYGYREまたはPFYGYREGFEDFを有する化学的に合成されたエピトープでありうる。
【0051】
エピトープはgBタンパク質中に含まれうるが、その分解産物中に含まれてもよく、または化学的に合成されたペプチドであってよいことを当業者は理解することができる。アミノ酸の位置は、gBタンパク質の配列中の対応するアミノ酸配列の位置を明示するために示されるにすぎない。本発明は、該エピトープを含む全てのペプチドを包含する。ペプチドは、長さが100アミノ酸超のポリペプチドの一部であってよく、または100アミノ酸未満、好ましくは50アミノ酸未満、より好ましくは25アミノ酸未満、さらにより好ましくは16アミノ酸未満の小さなペプチドであってよい。そのようなペプチドのアミノ酸は天然アミノ酸もしくは非天然アミノ酸(例えば、β-アミノ酸、γ-アミノ酸、D-アミノ酸)またはその組み合わせであってよい。さらに、本発明はエピトープの各レトロインベルソペプチドを包含することができる。ペプチドは結合していてもよくまたは結合していなくてもよい。ペプチドは、例えば、小分子(例えば、薬物もしくはフルオロフォア)と結合していてもよく、高分子量重合体(例えば、ポリエチレングリコール(PEG)、ポリエチレンイミン(PEI)、ヒドロキシプロピルメタクリレート(HPMA)など)と結合していてもよく、またはタンパク質、脂肪酸、糖部分と結合していてもよく、あるいは膜中に挿入されていてもよい。
【0052】
当技術分野において公知の抗体H126とは対照的に、本発明のmAb 2c抗体によって認識されるエピトープは必ずしも不連続ではない。抗体H126とは対照的に、本発明の抗体は連続的なエピトープ、かくして、連続アミノ酸配列に結合することができ、または不連続なエピトープに結合しうる。それゆえ、本発明の抗体の特性は改善されている。例えば、mAb 2c抗体は、標的タンパク質が変性される方法(例えば、SDSページ電気泳動)に用いることができ、または小さな直鎖ペプチドの検出に用いることができる。
【0053】
問題の抗体および第1の局面の抗体が同じエピトープを認識するかどうかを調べるために、以下の競合試験を行うことができる: 3 moi (感染の多重度)で感染されたVero細胞を20時間後、競合物質としてさまざまな濃度の問題の抗体とともに1時間インキュベートする。第2のインキュベーション段階では、第1の局面の抗体を100 nMの定濃度で適用し、その結合を、第1の局面の抗体の定常ドメインに対して作製された蛍光標識抗体を用いてフローサイトメトリーにより検出する(実施例の項および図6も参照のこと)。問題の抗体の濃度に反比例する結合は、両方の抗体が同じエピトープを認識することを示す。しかしながら、使用できる多くの他のアッセイ法が当技術分野において公知である。
【0054】
第2の局面の好ましい態様は、上記の、第1の局面の場合と同じものである。
【0055】
第3の局面において、本発明は、第1または第2の局面による抗体の有効量と、少なくとも1つの薬学的に許容される賦形剤とを含む、薬学的組成物に関する。しかしながら、「薬学的組成物」という用語は「薬物」という用語と本明細書において互換的に用いることができる。
【0056】
薬学的組成物中の抗体の含量は、処置または予防に有用である限り、限定されないが、好ましくは全組成あたり0.0000001〜10重量%を含有する。さらに、本明細書において記述される抗体は、好ましくは担体中で利用される。担体の選択は投与の経路および活性薬剤の濃度に依ることができ、担体は凍結乾燥された組成物または水溶液の形態であることができる。一般的に、適切な量の薬学的に許容される塩を担体中に用いて、組成物を等張にする。担体の例としては、生理食塩水、リンゲル溶液およびデキストロース溶液が挙げられるが、これらに限定されることはない。好ましくは、クエン酸塩、リン酸塩および他の有機酸などの緩衝液; 塩形成対イオン、例えばナトリウムおよびカリウム; 低分子量(10アミノ酸残基未満)のポリペプチド; タンパク質、例えば血清アルブミンもしくはゼラチン; 親水性重合体、例えばポリビニルピロリドン; ヒスチジン、グルタミン、リジン、アスパラギン、アルギニンもしくはグリシンなどのアミノ酸; グルコース、マンノースもしくはデキストリンを含む糖質; 単糖類; 二糖類; 他の糖、例えばスクロース、マンニトール、トレハロースもしくはソルビトール; キレート剤、例えばEDTA; 非イオン性界面活性剤、例えばTween、プルロニック(Pluronic)もしくはポリエチレングリコール; メチオニン、アスコルビン酸およびトコフェロールを含む酸化防止剤; ならびに/または保存料、例えば塩化オクタデシルジメチルベンジルアンモニウム; 塩化ヘキサメトニウム; 塩化ベンザルコニウム、塩化ベンゼトニウム; フェノール、ブチルもしくはベンジルアルコール; アルキルパラベン、例えばメチルもしくはプロピルパラベン; カテコール; レゾルシノール; シクロヘキサノール; 3-ペンタノール; およびm-クレゾール)を含めて、許容される賦形剤、担体または安定剤は、利用される投与量および濃度で無毒性である。適当な担体およびその製剤はRemington's Pharmaceutical Sciences, 17th ed., 1985, Mack Publishing Coにさらに詳細に記述されている。
【0057】
組成物は、アシクロビル、ペンシクロビル、イドクスウリジンおよびホスカネットを含む、化学療法剤またはウイルス抑制剤などの、2つ以上の活性化合物を含むこともできる。
【0058】
アシクログアノシン(ACV)または2-アミノ-9-(2-ヒドロシキエトキシメチル)-3H-プリン-6-オンとしても公知のアシクロビルは、アセプレス(ACERPES)(登録商標)、アシック(Acic)(登録商標)、アシクロベータ(Aciclobeta)(登録商標)、アシクロCT(AcicloCT)(登録商標)、アシクロスタッド(Aciclostad)(登録商標)、アシクロビル、アシック(Acic)(登録商標)、オフタール(Ophtal)(登録商標)、アシビル(Acivir)(登録商標)、アシビジョン(AciVision)、アシクロビル(Acyclovir)(登録商標)、アウィラル(Aviral)(登録商標)、シクロビル、ヘルベビル(Helvevir)(登録商標)、ヘルペックス(Herpex)、スプラビラン(Supraviran)(登録商標)、ウィルカルム(Virucalm)(登録商標)、ウィルポス(Virupos)(登録商標)、ウィルジン(Virzin)、ゾリパリン(Zoliparin)(登録商標)、ゾビル(Zovir)およびゾビラックス(Zovirax)(登録商標)などの商品名で販売されているグアノシン類似体の抗ウイルス薬である。ペンシクロビル(2-アミノ-9-[4-ヒドロキシ-3-(ヒドロキシメチル)ブチル]-6,9-ジヒドロ-3H-プリン-6-オン)は、デナビル(Denavir)およびフェニスティル(Fenistil)などの商品名で販売されているグアニン類似体の抗ウイルス薬である。ファムシクロビル(Famciclovir)(酢酸2-[(アセチルオキシ)メチル]-4-(2-アミノ-9H-プリン-9-イル)ブチル)はペンシクロビルのプロドラッグであり、経口バイオアベイラビリティが改善されている。イドクスウリジン(2'-デオキシ-5-ヨードウリジン)は、ヌクレオシド・ウリジンの生化学的類似体であり、ウィルングェント(Virunguent)(登録商標)およびゾストラム(Zostrum)(登録商標)などの商品名で販売されている。ホスカネットは、式HO2CPO3H2を有する化合物の共役塩基であり、ホスカビル(Foscavir)(登録商標)およびトリアプテン(Triapten)(登録商標)などの商品名で販売されている。
【0059】
好ましくは、抗体および/または活性化合物は有効量で含まれる。
【0060】
「有効量」という用語は、薬学的組成物を投与しようとする対象において検出可能な治療反応を誘導するのに十分な量をいう。
【0061】
第4の局面において、本発明は、本発明の抗体をコードする核酸配列を含む発現ベクターを提供する。一般に、発現ベクターは、問題の遺伝子を標的細胞に導入し、その結果、該遺伝子によってコードされるタンパク質、すなわち抗体の転写および翻訳をもたらすために用いられるプラスミドである。したがって、発現ベクターは、発現ベクター上で運搬される遺伝子の効率的な転写を指令するためにポリアデニル化シグナル部位のほかに、プロモーターおよびエンハンサー領域などの調節配列を含む。発現ベクターはさらなる必要なまたは有用な領域、例えば真核生物または原核生物細胞における選択用の選択可能なマーカー、複製起点などを含むこともできる。
【0062】
そのため、第5の局面において、本発明は、本発明の抗体をコードするヌクレオチド配列を含む宿主細胞に関する。宿主細胞は、本発明の抗体を発現させるのに適した任意の細胞であることができ、哺乳類細胞、酵母細胞および昆虫細胞、好ましくは哺乳類細胞、より好ましくは骨髄腫細胞株などの不死細胞株を含む。適当な細胞株はアメリカンティッシュカルチャーコレクションATCCで入手可能である。
【0063】
さらに、第6の局面において、第1および/または第2の局面の抗体を産生することができるハイブリドーマ細胞が提供される。ハイブリドーマ細胞は、迅速かつ無制限に増殖し、所望の抗体を大量に産生しうる遺伝子操作細胞である。ハイブリドーマ細胞は、関連抗原で刺激された動物の脾臓から抗体産生B細胞を取り出し、これを次いで、不死骨髄腫腫瘍細胞と融合させることによって調製される。
【0064】
非常に重要な第7の局面において、本発明は、薬物としての使用のための本発明による抗体に関する。より具体的には、本発明は、対象におけるHSV関連疾患の予防的または治療的処置のための薬物の製造のための本発明の抗体の使用に関する。等しくは、本発明は、対象におけるHSV関連疾患の予防的または治療的処置における使用のための本発明の抗体に関する。さらに、本発明は、本発明の抗体が治療的有効量で対象に投与される、対象におけるHSV関連疾患の予防的または治療的処置の方法に関する。HSV感染はいくつかの異なる疾患を引き起こすことがある。よくある皮膚または粘膜の感染は、顔および口(口腔顔面ヘルペス)、生殖器(性器ヘルペス)または手(ヘルペス性ひょう疽)に影響を与えることがある。ウイルスが眼に感染し、損傷を与える場合(ヘルペス性角膜炎)、または中枢神経系に侵入し、脳に損傷を与える場合(ヘルペス脳炎)、より重篤な障害が発生する。新生児、移植レシピエント、またはAIDS患者などの、未成熟なまたは抑制された免疫系を有する患者は、HSV感染から重篤な合併症を起こす傾向がある。HSV関連疾患は剣状ヘルペス、モラレ髄膜炎、おそらくベル麻痺、躁鬱病、双極性感情障害または双極性気分障害としても公知の、双極性障害の認知障害と関連している障害、およびアルツハイマー病も含む。アルツハイマー病に関して、最近の科学的刊行物からβアミロイドプラーク内の1型単純ヘルペスウイルスDNAの著しい局在が実証され、このウイルスがプラークの原因である可能性が示唆されている。最後に、本発明による抗体の使用は、例えば免疫抑制患者の長期にわたる予防的および治療的処置の間に、よく用いられる化学療法のウイルス抑制剤に対する耐性株の出現が観察されるならば有用である。
【0065】
したがって、好ましい態様において、HSV関連疾患は以下の特徴の1つまたは複数を伴う: 経口再発の存在、生殖器再発の存在、ヘルペス性湿疹、新生児ヘルペス、免疫不全(免疫減弱状態の患者)、免疫抑制、脳炎、髄膜炎、髄膜脳炎、眼感染症、汎発性HSV感染症、および/またはウイルス抑制剤に対する耐性。
【0066】
別の好ましい態様において、HSV関連疾患は化学療法のウイルス抑制剤に対する不耐性を伴う。
【0067】
さらに好ましい態様において、薬物は少なくとも1つのさらなる活性剤を含み、好ましくはここでさらなる活性剤は化学療法剤またはウイルス抑制剤であり、より好ましくはここでさらなる活性剤は上記のように、アシクロビル、ペンシクロビル、イドクスウリジンおよびホスカネットからなる群より選択される。
【0068】
最後の好ましい態様において、対象はイヌ、ネコ、ブタ、ウシ、ヒツジ、ウマ、げっ歯類、例えばラット、マウスおよびモルモット、または霊長類、例えばゴリラ、チンパンジーおよびヒトなどの哺乳類であり、好ましくは対象はヒトである。
【実施例】
【0069】
本発明を以下の例によってさらに説明するが、これらは本発明の範囲を限定するものと解釈されるべきではない。
【0070】
実施例1
1型および2型単純ヘルペスウイルス(HSV1、HSV2)の糖タンパク質B (gB)に対する特異性を有するマウスモノクローナル抗体(mAb) 2cの調製
抗HSV特異的なmAbの作出のため、BALB/cマウスをUV不活化HSV1-株342 hvで免疫した。その後、マウス脾細胞を骨髄腫細胞株X63-Ag8.653との体細胞融合によって不死化し、抗HSV特異的なmAb 2c (IgG2a)を分泌するハイブリドーマ細胞株を、酵素免疫アッセイ法、免疫蛍光アッセイ法およびHSV中和アッセイ法を用いて単細胞クローンの上清をスクリーニングすることによって単離した。結合試験によって、mAb 2cがHSV1およびHSV2の糖タンパク質gBの共用の、不連続なエピトープを認識することが明らかになった。糖タンパク質Bは約904アミノ酸長であり、ウイルス膜中におよびHSV 1/2感染細胞の膜中に三量体として挿入される。mAb 2cが細胞外ウイルス粒子の伝播を中和するだけでなく、HSVに特徴的な、最初に感染した細胞から、隣接した非感染細胞への感染(細胞間伝播)の直接経路を効率的に阻害もすることは特に関係がある。後者の過程は、通常、ヒトにおいて天然のHSV特異的抗体のレパートリーによって阻害されることはない。
【0071】
mAb 2cのインビボでの効力を調べるために、マウスモデルにおける感染の経路を選択したが、これはヒトにおける自然感染のかかわり合いに酷似している。それゆえ、C57BL/6Jマウスに、その処女膣粘膜へのHSV1株342 hvの適用により感染させた。インビボでのHSV複製の阻害のため、mAb 2cを感染後の異なる時点でマウスに腹腔内(i.p.)投与した。免疫応答性動物においてもCD4+ T細胞枯渇動物においても、mAb 2cは、膣粘膜におけるウイルス増殖および短期間内での炎症性病変の形成を阻害することができる(図1)。
【0072】
ポリクローナルHSV血清とは対照的に、mAb 2cはウイルス複製を阻害することができただけでなく、T細胞(CD4+およびCD8+)が完全に枯渇されている免疫抑制動物において高い効率で全身の、致命的な進行性疾患を予防することができた(図2)。ウイルス接種24時間前のmAb 2cの投与は、感染から動物を効率的に保護した(Eis-Hubinger et al., 1993)。
【0073】
中和特性に及ぼす抗原結合部位の数(結合価)およびFc部分の影響を調べるために、mAb 2cのFab断片およびF(ab')2断片を従来のプロテアーゼ消化によって作出し、そのほかにも、当技術分野において周知の方法を用いてmAb 2cの組換え「一本鎖Fv」(scFv)をクローニングし、産生かつ精製した(図3)。
【0074】
当技術分野において周知の方法のように、自然のHSV複製サイクルの間に細胞表面に膜会合糖タンパク質としてgB-タンパク質を発現するHSV感染Vero細胞を用いてフローサイトメトリーにより親和性定数(KD)を決定した。この結果を下記表1に示す。
【0075】
(表1)マウスmAb 2cの親和性定数(KD)および作出された2c抗体断片の親和性定数(KD)
【0076】
親mAb 2cとは対照的に、F(ab')2断片はわずかに高い親和性を示す。Fab断片およびscFv断片はほぼ同一の親和性を有するが、しかし、これは親mAbの親和性よりも約1.7倍〜1.9倍弱いその一結合価によるものである。
【0077】
マイクロタイタープレート中で単層として増殖させたVero細胞を用いて標準的な終点希釈アッセイ法により抗体の中和活性を決定した。手短に言えば、100 TCID50のHSVをVero細胞の接種前に抗体の連続希釈液(2c-IgGおよびF(ab)2: 0.98 nM〜125 nM; Fab: 23 nM〜3000 nM)とともに37℃で1時間細胞培地100μl中でプレインキュベートした。37℃で72時間のインキュベーションの後、ウイルス誘導CPEの形成を100%まで抑止するのに必要な抗体濃度を完全中和価として決定した。さらに、一価2c-Fab断片のウイルス中和能を架橋抗体の存在下で決定した。
【0078】
(表2)100 TCID50の規定のウイルス量の完全中和
【0079】
親mAb 2cならびにそのF(ab')2断片およびFab断片はHSV1およびHSV2の完全中和の能力を有することを実証することができた。しかし、一価抗体断片はmAb 2cに比べてHSV 1/2に対して顕著に低減した中性化効率を示す。HSV1およびHSV2の100%の中和には、それぞれ、375倍および97倍高いFab断片濃度が必要である。scFvは、プラーク低減効果を示したが、3000 nMの最も高い試験濃度でCPEを完全に阻害することはできなかった(データ不掲載)。過剰の抗Fab特異的IgG (Jackson ImmunoResearch, Newmarket, England)をプレインキュベーション段階に加えることによって、一価2c Fab断片のウイルス中和能を2倍増強することができた(不掲載)。対照的に、二価F(ab')2断片は親mAb 2cと比べてHSV1に対してもHSV2に対しても実質的に2倍高い効率的な中和活性を示す。結論として、抗体価はその中和特性に重要な役割を果たす。HSV-2の完全な中和に必要な抗体濃度がいっそう高いことは、HSV-1およびHSV-2のDNAコピー数を決定するRT-PCRによって確認されるように、HSV-1に比べてHSV-2により産生される非感染性粒子の量がいっそう多いことで説明することができる(データ不掲載)。
【0080】
mAb 2cの特性およびその産生に関するさらなる詳細は、Eis-Hubinger et al.. 1991、およびEis-Hubinger et al.. 1993に示されている。
【0081】
実施例2
mAb 2cのキメラ化およびヒト化
マウスモノクローナル抗体2cを治療剤として利用するために、親抗体の特異性を完全に保持しながらも、ヒトへの投与中のその免疫原性を低減または排除することを目的として遺伝子操作の方法を用いて抗体を修飾した。したがって、mAb 2cと同じ特異性を有するキメラおよびヒト化モノクローナル抗体を作出した(図3)。
【0082】
最初に、mAb 2cの可変重鎖および軽鎖(VH、VL)の真正遺伝子を、5'-RACE (Rapid Amplification of cDNA Ends; cDNA末端の迅速増幅)-PCRによりハイブリドーマ細胞株から単離および増幅した。本発明者らの協力パートナーのGrosse-Hovest博士(Universitat Tubingen)が構築した発現ベクターであって、ヒトIgG1アイソタイプの軽鎖および重鎖の定常領域を含む該ベクターの中に、増幅されたVH-遺伝子およびVL-遺伝子をクローニングすることによってキメラIgG1抗体(ch2c)を作出した。この抗体は、安定的にトランスフェクトされたSp2/0マウス骨髄腫細胞の細胞培養上清へ最終的に分泌された。
【0083】
免疫原性をさらに低減するために、ヒト化抗体を構築した。それゆえ、mAb 2cの6つの相補性決定領域(CDR)をコードする遺伝子セグメント(2c VL-CDR1/2/3および2c VH-CDR1/2/3)をそれぞれ、ヒトVHおよびVL生殖細胞遺伝子の適当なヒトフレームワーク免疫グロブリンアクセプター骨格へクローニングした(CDRグラフト)。mAb 2cの軽鎖および重鎖のCDR領域をクローニングするのに適したヒト生殖細胞アクセプター骨格は、V-Baseデータベース(http://vbase.mrc-cpe.cam.ac.uk/)のうちの対応するヒトフレームワーク領域との配列のアライメントによって決定された。DP28は、対応するマウスmAb 2v重鎖VH配列に対して最も高いフレームワーク配列同一性(88.5%の配列同一性)を示した; DPK13は、対応するマウスmAb 2c軽鎖VL配列に対して最も高いフレームワーク配列同一性(88.9%の配列同一性)を示した。したがって、マウスドナー抗体2cのCDRコード遺伝子セグメント(すなわち2c VL-CDR1/2/3および2c VH-CDR1/2/3)をそれぞれ、DP28およびDPK13をコードするアクセプターフレームワークへクローニングした。
【0084】
モノクローナル抗体のヒト化との関連で、導入されたマウスCDRの構造的完全性にとって有害な可能性がある、したがって抗原結合特性にとって有害な可能性がある、ヒトフレームワーク領域中のアミノ酸を特定することが必要である。通常、そのようなアミノ酸はコンピュータで作った相同性モデルを用いて特定され、立体的に重要であるようにみえる位置を、マウスドナーmAbの抗原結合特性を保持するために、マウスの対応配列に変異させる(Queen el al., 1989参照)。しかしながら、潜在的に重要なアミノ酸は、抗体レパートリーのデータベースを用い、公知の三次元構造を有する参照抗体に基づきその臨界的意義を評価して特定することもできる(Krauss et al., 2003参照)。したがって、VH-およびVL-フレームワーク領域中のいくつかの潜在的に重要なアミノ酸を決定し(図4)、これらの潜在的に重要なアミノ酸を、マウスのその対応残基に連続的に復帰突然変異させた4種のヒト化mAb 2cバリアントを重複伸長PCRによって作出した(下記表3参照)。
【0085】
(表3)マウスドナー配列へのフレームワーク領域中の復帰突然変異を有するヒト化mAb 2cバリアント
【0086】
ヒト化抗体バリアントh2c-V1-4は、本発明者らの協力パートナーのGrosse-Hovest博士(Universitat Tubingen)が構築した上記の発現ベクターの中にヒト化VH-遺伝子およびVL-遺伝子をクローニングすることによって構築された。この抗体は、マウス骨髄腫細胞株Sp2/0の安定なトランスフェクションの後に最終的に発現された。高い特異的産生率を有するクローンの選択後、抗体が定量的に産生され、さらなる特徴付けのために細胞培養上清から精製された。
【0087】
キメラおよびヒト化抗HSV IgG1抗体の特徴付け
HSV感染Vero細胞を用いて、親mAb 2cおよび2c抗体断片の場合に記述したようにフローサイトメトリーによって同様に、親和性定数(KD)を決定した(実施例1参照)。この結果を図5に示す。
【0088】
キメラ抗体ch2cは親抗体mAb 2cの親和性を保持していた。ヒト化バリアントの場合、バリアントh2c-V1に関して唯一のCDRグラフトはマウスmAb 2cに匹敵する親和性を保存するのに十分であった。それゆえ、各マウス配列へのヒトフレームワーク残基のさらなる連続的な復帰突然変異は、抗原結合部位の構造的完全性を改善するのに必要ではなかった。バリアントh2c-V2はさらに、mAb 2cと比べて2倍低い親和性を示す。
【0089】
キメラmAb 2cおよびヒト化抗体バリアントmAb h2c1-4が親mAb 2cと同じエピトープを認識することを実証するために、競合試験を行った。HSV-1感染(3 moi、20時間) Vero細胞を最初に、漸増濃度の、それぞれ、キメラmAb ch2cまたはヒト化mAb hu2c V1-V4とともに1時間インキュベートした。第2のインキュベーション段階では、100 nMの親mAb 2cを添加し、その結合を、定常ドメインに対して作製された蛍光標識抗体を用いてフローサイトメトリーにより検出した(図6)。
【0090】
競合試験から、競合物質の結合を表す蛍光シグナルは第1のインキュベーション段階で適用された非標識抗体の濃度に反比例することが明らかである。このことは、キメラmAbおよびヒト化mAbバリアントが同じ特異的結合部位に向けてmAb 2cと競合し、ゆえに同じエピトープを認識することを証明している。
【0091】
mAb ch2cおよび4種のヒト化バリアントh2cのウイルス中和能を上記のように、精製された抗体調製物で調べた。この結果を下記表4に示す。100 TCID50の規定のウイルス量の50%のおよび完全なHSV中和に必要な各濃度を示す。
【0092】
(表4)100 TCID50の規定のウイルス量の50%のまたは完全な中和に必要な抗体濃度
【0093】
キメラmAb ch2c、およびmA h2c-V2以外の全てのヒト化mAb h2cは、親mAb 2cと同じ効率でHSVを中和する。mAb h2c-V2の2倍低い中和効率は、このバリアントのいっそう低い親和性と相関する。さらなる実験的特性評価および前臨床評価のため、mAb h2c-V1を選択したが、これは、このバリアントが親抗体mAb 2cと同じ親和性およびウイルス中和特性を保有するからである。さらに、mAb h2c-V1はフレームワーク領域中にマウスドナー配列への復帰突然変異を持たず、それゆえ、ヒトにおいて低い免疫原性能を保有するものと予想される。
【0094】
ヒト化mAb h2c-V1およびマウスmAb 2cの中和活性に及ぼす補体の影響を、プラーク低減アッセイ法を用いて詳しく調べた。ヒト高力価免疫グロブリン血清(Cytotect(登録商標), Biotest AG)とは対照的に、親mAb 2cおよびヒト化バリアントmAb h2c-V1はHSVを補体非依存的に中和する(図7)。HSV-1 gBに対する補体非依存的な中和抗体は、0.8〜160μg/mlの力価でウイルス投入量の50%を中和することが文献に記述されている(Navarro et al, 1992, Virology 186)。マウスmAb 2c、キメラmAb ch2cおよびヒト化mAbバリアントの終点希釈アッセイ法によりHSV-1 (F)を50%中和するのに必要な力価は3.3〜5.1 nMであり、これは0.49〜0.78μg/mlに相当する(表4参照)。
【0095】
HSV-1およびHSV-2の臨床分離株の中和アッセイ法から、HSV-1およびHSV-2の実験室株と比べてヒト化mAbバリアントh2c-V1による不活化に対する同じ感受性が実証される。さらに、mAb h2c-V1は非耐性実験室株または未知の耐性を有する臨床分離株と同じ効率で、アシクロビル(ACV)、ACVおよびホスカネット(PFA)、またはACV、PFAおよびシドビル(Cidovir) (CDV)に対して臨床的に耐性の患者分離株由来のHSVを中和する(図8)。ゆえに、ヒト化mAb h2c-V1は、耐性HSV株を誘導する従来の抗ヘルペス薬による限界を克服する新しい強力な抗ウイルス剤となる。
【0096】
宿主内での伝播のために、HSVは2つの機構: 無細胞粒子の放出および直接的な細胞間伝播を用いる。HSVの細胞間伝播は、より迅速なウイルス複製および播種、ならびに液性免疫反応の要素に対する耐性のような、無細胞伝播に対して優位性を保ちうる。抗HSV抗体による細胞間伝播を調べるために、Vero細胞を24ウェル組織培養プレート中のカバーガラス上に播き、集密まで増殖させ、これに37℃で4時間1ウェルあたり400 TCID50のHSV-1 Fを接種した。ウイルス接種材料を吸引し、Vero細胞をPBSで2回洗浄し、2% FBS、抗生物質を含有する、かつ過剰の中和抗体、つまりヒトポリクローナル抗HSV血清を含有するか、または対照の目的で中和抗体を含有しないかのどちらかのDMEM 1 ml中にて37℃で48時間さらに培養した。48時間後、培地を除去し、Vero細胞単層をHEPES緩衝生理食塩水で2回洗浄し、室温で15分間PBS中4%のパラホルムアルデヒド中で固定した。細胞単層をPBSで2回すすぎ、HEPES緩衝生理食塩水中0.05%のTween 20を含有するブロッキング緩衝液500μl中で15分間インキュベートした。ブロッキング緩衝液中で1:100に希釈されたFITCコンジュゲートポリクローナルヤギ抗HSV血清(BETHYL, Montgomery, TX, USA)でのHSV-1感染細胞単層の染色によりウイルス抗原を検出した。細胞単層をPBSで3回洗浄し、Mowiol (Calbiochem, San Diego, CA, USA)でマウントした。免疫蛍光陽性細胞をLeica DM IRE2共焦点顕微鏡により40倍の倍率で捕捉した(図9)。
【0097】
ヒトポリクローナル抗HSV血清(1:20)は、HSVの細胞間伝播に阻害効果を及ぼさない(図9A)。親mAb 2cおよびそのF(ab)2断片は500 nMの濃度で細胞間伝播を完全に阻害し、1つのHSV感染細胞しか検出することができない(図9BおよびC)。親mAb 2cの6倍高い濃度で適用されたFab断片は、ヒトポリクローナル抗HSV血清と比べて細胞間伝播をわずかに低減するが、しかし細胞間伝播を完全に阻害することができない(図9D)。中和アッセイ法において既に示されたように、これらの結果から、中和抗体の二価はHSV伝播を阻害するその能力に重要な役割を果たすことが確認される。キメラmAb ch2cおよびヒト化バリアントmAb h2c-V1は、親mAb 2cと同じくらい効率的に500 nMの濃度でHSVの細胞間伝播を阻害する(図9EおよびF)。さらに行われた従来のプラーク阻害アッセイ法(Highlander el al., 1988)から、共焦点顕微鏡での評価によって得た結果が確認された。
【0098】
最初のインビボHSV保護実験から、親mAb 2cと同様に5 mg/kg i.v.単回用量のヒト化mAb h2c-V1が膣内HSV-1 F感染した重症免疫不全マウス(NOD-SCID)の生存を顕著に引き延ばすことが明らかである(図10)。15 mg/kgのヒト化mAb h2c-V1または親mAb 2cのどちらかを投与されたマウスは、致死性脳炎から完全に保護される(図10)。さらに、ヒト化mAb 2c-V1はまた、樹立された末梢HSV感染の存在下でウイルス伝播および致死性脳炎からの保護をもたらす。ウイルス攻撃後24時間の時点で膣洗浄において高いHSV-1力価を有するNOD/SCIDマウスは、15 mg/kgのヒト化mAb 2c-V1またはmAb 2cにより24時間、40時間および56時間の時点で繰り返し静脈内処置された場合、感染の致命的結末から完全に保護された(図11)。さらに、ヒト化抗体mAb 2c-V1はまた、多耐性HSV株の樹立感染を有するNOD/SCIDにおいて致死性脳炎を抑止した。対照的に、アシクロビルを投与されたマウスは全て死んだ(図12)。
【0099】
エピトープマッピング
全長gB (31〜904)または720、630、503、487および470の位置でC末端の切断を伴うgB変異体のどちらかをコードする発現プラスミドでトランスフェクトされたCOS-1細胞を用いた結合試験により、マウス親mAb 2cによって認識されるエピトープがgBの最初の487アミノ酸のなかに位置付けられた。連続ペプチドの間に12 aaの重複を有する、固相に結合された15アミノ酸(aa)長の合成ペプチドを用いたさらなる研究によって、mAb 2cが立体構造エピトープに位置することが示された。mAb 2cはaa 175〜195の糖タンパク質B配列(領域A)に相当する3つの連続ペプチドに結合する。さらに、mAb 2cは、アミノ酸298〜312に相当するペプチド(298SPFYGYREGSHTEHT312) (SEQ ID NO: 17)に強力に結合し、アミノ酸301〜315に相当する後続のペプチドに中程度に結合する(図13Aおよび図22)。未変性または変性SDS-PAGE条件の下で分離された組換えgB (gB(730)t、Florent Bender, University of Pensylvania, Philadelphia, USAから親切にも提供していただいた)を用いたウエスタンブロットに関するその反応性によるmAb 2cの特徴付けからは、変性に対して部分的に耐性であるか、またはウエスタンブロット条件の間に再編成され、それゆえ、「偽連続」エピトープと呼ばれる(Bender, F et al. J. Virol. 2007, 81 p3872-3841を参照のこと)不連続なエピトープを、mAb 2cが認識することが確認される(図13B)。
【0100】
mAb2cのウイルス中和活性に関連するHSV-1 gBタンパク質中のエピトープを特定するために、その糖タンパク質中に単一のアミノ酸(aa)交換を有するモノクローナル抗体耐性(mar) HSV-1変異体について研究した(表5)。
【0101】
(表5)モノクローナル抗体耐性(mar) HSV-1変異体に対するmAb 2cの中和および結合活性
(a) シグナル配列を含む成熟糖タンパク質Bによる付番(図13A)
(1) Kousoulas et al., 1984
(2) Pellett et la., 1985
(3) Kousoulas et al., 1988
(4) Highlander et al., 1989
【0102】
mAb 2cはmar変異体R126、R1375、およびB4.1を中和しなかったが、それぞれ、変異体R1435およびR233の感染性を完全に中和した。さらに、免疫蛍光アッセイ法から、mAb 2cがmar変異体R126、R1375、およびB4.1に感染したVero細胞に結合しないことが確認された。これらの結果から、アミノ酸Y303、R304およびE305がmAb2cの中和活性に不可欠であることが示唆される。mar変異体R1435およびR233に感染したVero細胞を用いて強力な蛍光シグナルが得られた。
【0103】
特に、gBのアミノ酸303〜305に対するmAb 2cの認識および結合がその機能(ウイルス中和、細胞間伝播の阻害)に不可欠であることが分かった。gBのこの領域はHSV-1株とHSV-2株との間で高度に保存されているので、これらのアミノ酸はgBのコアの融合機構に属し、ウイルス侵入に不可欠であることが想定される。それゆえ、mAb 2cが結合しない天然のgB変異体の出現は、高い選択圧の下でさえ可能性が低い。
【0104】
実施例3
抗体親和性の判定
Vero細胞の単層に80〜90%の集密でMOI 3のHSV-1またはHSV-2を感染させ、これを翌日、トリプシン処理により、その後、PBS中での洗浄により収集した。2c抗体の細胞表面結合測定は既報(1)のように行った。手短に言えば、精製されたmAb 2cまたはmAb 2c由来抗体断片2c-F(ab')2、2c-Fabおよび2c-scFvを室温で1時間、FACS緩衝液(PBS、2% FBS、0.1%アジ化ナトリウム) 100μl中5×105個のVero細胞とともに、0.03 nM〜500 nMの濃度にて三つ組でインキュベートした。細胞をFACS緩衝液200μlで2回洗浄し、結合したmAb 2c、2c-F(ab')2、および2c-Fabの検出のためFITC標識Fab特異的ヤギ抗マウスIgG, (15μg/ml, Jackson ImmunoResearch, Newmarket, Suffolk, England)とともにインキュベートした。結合したscFvは飽和濃度の抗c-myc mAb 9E10 (10μg/ml; Roche, Indianapolis, IN, USA)と最初にインキュベートし、その後、2回の洗浄およびFcγ特異的FITC標識ヤギ抗マウスIgG (15μg/ml; Jackson ImmunoResearch)とのインキュベーションを行うことによって検出された。細胞を2回洗浄し、FACS緩衝液中に再懸濁した。蛍光をFACScalibur (BD Bioscience, San Jose, CA, USA)にて測定し、CellQuest(商標)ソフトウェア(BD Biosciences)を用いて平均蛍光強度(MFI)を計算した。バックグラウンドの蛍光を差し引き、GraphPad Prism 4.0版(GraphPad Software, La Jolla, CA)で非線形回帰のためのMarquardtおよびLevenbergの方法を用いることにより平衡結合定数を決定した。
【0105】
エピトープの特徴付け
Roselyn J. EisenbergおよびGary H. Cohen (University of Pennsylvania, Philadelphia, USA)から親切にも提供していただいた、未変性または変性切断型糖タンパク質B、つまりgB(730)t (4)とのmAb 2cの免疫反応性は、本質的には既報(4)のように行った: 精製gB(730)t (0,75μg)を非還元(0.2% SDSを含有するサンプル用緩衝液)条件または変性(2% SDSおよび155 mM β-メルカプトエタノールを含有するサンプル用緩衝液、95℃で2分)条件のどちらかの下で8% SDS-PAGEにて分離し、ニトロセルロース膜上に転写した。膜ストリップをTNT緩衝液(0.1 M Tris.HCl, pH 7.5, 0.15 M NaCl, 0.05% Tween-20)中2%のミルクで1時間ブロッキングし、その後、2%ミルク/TNT緩衝液中5μg/mlの糖タンパク質B特異的抗体mAb 2c、H126 (Novus Biologicals, Littleton, CO, USA)およびH1817 (Novus)との室温で2時間のインキュベーションを行った。結合した抗体を西洋ワサビペルオキシダーゼコンジュゲートポリクローナルヤギ抗マウス抗体(1:20,000 QED Bioscience Inc. San Diego, CA, USA)および化学発光(Thermo Scientific,)で、LAS 3000 Luminescent Image Analyzer (Fujifilm, Tokyo, Japan)を用いて検出した。
【0106】
COS-1細胞を、全長HSV-1 gB (31〜904, pRB9221)または720位(pTS690)、630位(pPS600)、503位(pRB9510)、487位(pRB9509)および470位(pRB9508)で切断されたC末端欠失変異体をコードするプラスミドでDEAE-デキストラン法により一過的にトランスフェクトした。これらのプラスミドはL. Pereira (52, 55)から親切にも提供していただいた。mAb2cまたは対照抗体を用いたトランスフェクト細胞での免疫蛍光アッセイ法は、他で既報(53)のように行った。
【0107】
ペプチドマッピング
セルロースに結合された重複する13 merのペプチドおよびデュオトープは既報(20, 34)のように標準的なSPOT合成プロトコルにしたがって自動的に調製された(JPT Peptide Technologies, Berlin, Germany)。さらに、反応性タグおよびリンカーとカップリングされたペプチドを3つの同一のサブアレイ中の修飾ガラス表面上に化学選択的に固定化し、切断されアセチル化された配列の除去、その後の洗浄段階によって精製した。ペプチドマイクロアレイを、TBS含有ブロッキング用緩衝液(Pierce International)で2時間ブロッキングし、ブロッキング用緩衝液中10μg/mlのmAb 2cとともに2時間インキュベートした。ペプチドマイクロアレイを、0.1% Tweenを含有するTBS緩衝液(T-TBS)で洗浄し、ペプチド膜上のペプチドに結合した抗体をPVDF膜上に転写した。ペルオキシダーゼ標識された抗マウスIgG (Sigma)または蛍光標識された抗マウスIgG (Pierce)をブロッキング用緩衝液中1μg/mlの終濃度で二次抗体として用いた。2時間のインキュベーションおよびT-TBSによる最終の洗浄の後、化学発光基質(Roche Diagnostics)を用いてPVDF膜を分析した。スライドガラスペプチドマイクロアレイをT-TBSおよび3 mM SSC緩衝液(JPT Peptide Technologies)で徹底的に洗浄し、窒素下で乾燥し、高分解能蛍光スキャナ(Axon GenePix 4200 AL)を用いてスキャンした。蛍光シグナル強度(発光量, LU)を、スポット認識ソフトウェア(GenePix 6.0)を用いて分析し、二次抗マウスIgGとの対照のインキュベーションからのバックグラウンドの強度に対して補正した。
【0108】
ウイルス中和アッセイ法
抗体の中和活性を既報(16)のように終点希釈アッセイ法によって決定した。手短に言えば、抗体の連続希釈液を細胞培地中37℃で1時間100 TCID50のHSV-1またはHSV-2とともにインキュベートした。抗体ウイルス接種材料を、マイクロタイタープレート中で増殖させたVero細胞単層に適用し、37℃で72時間のインキュベーション後に細胞変性効果(CPE)をスコア化した。ウイルス誘導性CPEを100%低減するのに必要な抗体濃度を完全中和価として決定した。さらに、一価2c-Fab断片のウイルス中和能を架橋抗体の存在下で、過剰の抗マウスFab IgG (2600 nM, Jackson ImmunoResearch, Newmarket, Suffolk, England)をプレインキュベーション段階に加えることによって調べた。対照の目的で、抗体なしのウイルスおよび抗体のみを用いて、それぞれ、最大CPEまたはCPEなしを誘導した。ウイルス中和アッセイ法を少なくとも2回繰り返し、同じ結果となった。
【0109】
付着後中和アッセイ法
予冷されたVero細胞単層(15分間4℃)に1時間4℃で100 TCID50 HSV-1 Fを感染させてウイルス吸収を可能とした後に、mAb 2cまたはヒト血漿由来の多価IgG調製物(Intratect(登録商標), Biotest AG, Dreieich, Germany)のどちらかの連続希釈液を添加した(付着後中和)。同一の実験条件下でのmAb 2cの付着前 vs 付着後中和効率を比較するため、予冷されたVero細胞単層に添加する前に、100 TCID50 HSV-1 Fを同じ抗体希釈液とともに4℃で1時間インキュベートした。両アッセイ由来の接種済みVero細胞を37℃に移す前に、4℃でさらに1時間インキュベートした。上記の標準的な中和アッセイ法に記述されているように、72時間後に中和価を決定した。
【0110】
細胞間伝播アッセイ法
集密までカバーガラス上で増殖されたVero細胞に37℃で4時間500μl中400 TCID50/ウェルでHSV-1 Fを接種した。ウイルス接種材料を吸引し、Vero細胞をPBSで2回洗浄し、2% FBSを含む増殖培地1 ml中、過剰の中和抗体の存在下または非存在下において37℃で48時間さらに培養した。高力価の抗HSV-1免疫グロブリンを有する免疫ドナーに由来するプール済みヒト血清を1:20の希釈で対照として用い、二価抗体mAb 2cおよび2c-F(ab')2の濃度は500 nMとし、一価2c-Fabの濃度は3000 nMとした。48時間後、培地を除去し、Vero細胞単層をHEPES緩衝生理食塩水で2回洗浄し、室温で15分間PBS中4%のパラホルムアルデヒドで固定した。ウイルス伝播を可視化するため、細胞をPBSで2回すすぎ、0.05% Tween 20を含むHEPES緩衝生理食塩水500μl中で15分間インキュベートし、FITCコンジュゲートポリクローナルヤギ抗HSV血清(1:100, BETHYL, Montgomery, TX, USA)で染色した。PBSによって3回洗浄した染色細胞を、0.2 g/ml Mowiol 4-88 (Calbiochem, San Diego, CA, USA)を含有するマウンティング培地中でマウントした。蛍光像をLeica DM IRE2共焦点顕微鏡により40倍の倍率で捕捉した。細胞間伝播の阻害を吸収後ウイルス中和アッセイ法によってさらに試験した。6ウェルプレート中で集密まで増殖されたVero細胞を、2% FBSおよび抗生物質を含有するDMEM 3 ml中、200 TCID50のHSV-1 Fとともに37℃で4時間インキュベートした。細胞単層をPBSで2回洗浄し、過剰の中和抗体またはポリクローナルヒトHSV-1中和血清を含有する温かいプラーク形成用培地(DMEM, 5% (w/v)アガロース, 10% FBS, 抗生物質)で重層した。37℃で48時間のインキュベーション後、光学顕微鏡検査によってプラーク形成を分析した。
【0111】
DNA定量
リアルタイム(RT) PCRを行ってHSV-1およびHSV-2ゲノムを定量化した。自動核酸抽出システムMagNA Pure LC System (Roche)を製造元の指示にしたがって用い、等量のHSV-1およびHSV-2感染粒子を含有するサンプルからDNAを精製した。ウイルスDNAを次いで、Real Art HSV-1 / HSV-2定量キット(Qiagen)を用いRT-PCR (Lightcycler, Roche)を行って定量した。
【0112】
マウス保護実験
麻酔した6〜8週齢の、雌性非肥満糖尿病/重症複合免疫不全(NOD-SCID)マウス(NOD.CB17-Prkdcscid/J, Charles River Laboratories, Research Models and Services, Sulzfeld, Germany)を1×106 TCID50 HSV-1 F/マウスの接種材料20μlで膣内から攻撃した。皮膚接着剤(Epiglu, Meyer-Haake Medical Innovations, Wehrheim, Germany)を外陰部に適当して、ウイルス接種材料の流出(dircharge)を抑止した。送達された接種材料は膣洗浄液の培養によって評価した場合に94%超(>94%)の感染率を誘導した。マウスをウイルス接種後毎日、体重減少、外陰炎/膣炎(発赤、粘液膿性排出物および炎症の兆候)ならびに神経疾患について調べた。これらの症状のいずれかを呈するマウスをすぐに殺処理した。免疫予防の場合にはウイルス接種の24時間前に、または治療的処置の場合にはウイルス感染から24時間、40時間および56時間後に、精製mAb 2cの静脈内(i.v.)注射によってマウスを受動的に免疫した。Vero細胞に対する終点希釈アッセイ法を用いて感染後1、2、4、6および8日目に、ならびに死亡時に得られた膣洗浄液からのウイルス力価の判定により感染についてマウスを評価した。殺処理したマウスの臓器(脾臓、副腎、肺、心臓、肝臓、腎臓、脊髄および脳)中のウイルス量を、他で既報(41)のようにVero細胞単層に対する力価測定によって臓器のホモジナイゼーション後に判定した。各試験および対照群には、検出可能なHSV-1感染を伴う動物9〜10匹を含めた。
【0113】
結果
mAb 2cによって認識されるgBエピトープのマッピングおよび分析
最近になって決定された1型HSV (HSV-1)由来gB細胞外ドメイン結晶構造から、5つの異なる構造ドメイン: ドメインI (基部)、ドメインII (中央部)、ドメインIII (コア部)、ドメインIV (冠部)およびドメインV (腕部)を有する多ドメイン三量体が明らかになった(24)。mAb 2cの中和エピトープを特徴付けるために、本発明者らは、還元条件下または非還元条件下のどちらかのウエスタンブロット分析において組換えgB(730t)(4)とのその反応性について試験した。対照として、本発明者らは、線状エピトープを認識するmAb H1817 (4)および不連続なエピトープを認識するmAb H126 (33)を用いた。線状エピトープに典型的な染色パターンがmAb H1817によるウエスタンブロット分析において得られ、非還元条件下でのgBの単量体型および三量体型の検出ならびに還元条件下でのgB単量体の唯一の主たる染色が示された(図13B)。予想通り、mAb H126は未変性条件下でのみgBと反応した。驚いたことに、170 kDa超(>170 kDa)の、唯一の、上方のgBタンパク質のバンドの認識から、mAb H126が三量体特異的であることが示唆される(図13B)。しかしながら、mAb 2cは未変性および変性gBと反応し、変性条件下での反応性はmAb H1817と比べてはるかに弱かった(図13B)。変性条件下でのgB単量体との弱い反応性は、変性に対して耐性であるか、またはSDS-PAGE電気泳動の間に再び折り畳まれるように見え、それゆえ「偽連続」エピトープと呼ばれる、不連続なエピトープに結合する一連の他の中和抗体についてこれまでに報告されている(4)。
【0114】
mAb 2cの結合に関与する特異的なエピトープを特定するため、本発明者らは、ペプチドマイクロアレイ上に提示されたgB由来ペプチドを用いた。最初に、重複する13 merのペプチドが3アミノ酸のずれで連続的なセルロース膜に非荷電性アセチル化アミノ末端で結合されるようにSPOT合成により、アミノ酸31〜505を提示するgB配列を調製した。非複合体化抗体に対する結合平衡の移動を回避するため、mAb 2cペプチドスキャンを化学発光による検出の前にPVDF膜上に固定化した。表示の機能ドメインを有する全長gBの略図(図14A)に示されるように、mAb 2cの反応性は、ドメインI内の2つの別個の領域にまたがるペプチド、つまり残基175〜193 (結合領域A)を含む3つの連続ペプチドおよび残基295〜310 (結合領域B)を含む2つの重複ペプチドに限定された。特定された両方の結合領域を検証するため、本発明者らは、可動性リンカーを介してスライドグラス上に固定化された13 merの精製ペプチドのさらなるセットを用いた。セルロースのスクリーニングと比べて、蛍光を介したこのマイクロアレイのスキャニングの読み出しから、同じエピトープ結合領域が確認された(図14B)。精製ペプチドおよび高分解能マイクロアレイスキャニングシステムの適用により、両結合部位のさらなる連続ペプチドがこのペプチドマイクロアレイにおいてmAb2cにより認識された(図14B)。
【0115】
本発明者らは、解析したgB構造に対して特定されたmAb 2cの結合部位をマッピングした(24)。興味深いことに、mAb 2cと最も強い反応性を示すペプチド172QVWFGHRYSQFMG184 (SEQ ID NO: 18)は、ドメインIの曲線状サブドメインに位置する2つの推定上の融合ループの1つ(融合ループ1 173VWFGHRY179) (SEQ ID NO: 19)と重なった(22) (図15)。しかしながら、gB三量体の基部に結合部位Aが局在化することで、融合後の立体構造を表す可能性が最も高い、利用可能なgB構造においてはmAb 2cにアクセスできなくなる(24)。結合部位Bの残基は曝露され、ドメインIの上部に位置付けられる(図15)。
【0116】
mAb 2cの立体構造依存的なエピトープをさらに評価するために、両方の結合領域のコンセンサス配列を直接的にまたは1つもしくは2つのβ-アラニンスペーサーによって分け隔てデュオトープとしてさまざまな組み合わせでつなぎ合わせた(図16)。残基E187の後の融合ループ1のすぐ近くでのリンカーの挿入は、gBが融合後の立体構造へ折り畳まれてさえも、融合欠損gB変異体(40,61)を生ずることが最近になって示された(61)。それゆえ、本発明者らは、別のデュオトープスキャンの中に、結合領域Aのコンセンサスモチーフ179YSQFMG184 (SEQ ID NO: 20)に加えて結合領域Aの186FED188モチーフを含めた。13 merのペプチドスキャンにおいてmAb 2cと最も強い結合反応性を示すペプチド172QVWFGHRYSQFMG184 (SEQ ID NO: 18)と比べて(図16)、結合部位Bのコンセンサスペプチド300FYGYRE305 (SEQ ID NO: 21)との両方の結合部位Aのモチーフの組み合わせが、シグナル強度の増強された2つのデュオトープを生じた(図16、デュオトープセットIおよびII)。デュオトープ179YSQFMG184-βA-300FYGYRE305 (SEQ ID NO: 22)に対するmAb 2cの結合強度はほんの少しだけ増したが、蛍光シグナル強度の大体の飽和は、デュオトープ186FED188-βA-βA-300FYGYRE305 (SEQ ID NO: 23)で得られた。
【0117】
したがって、ペプチドマイクロアレイからの結果は、ウエスタンブロッティングの結果に一致し、mAb 2cが立体構造依存的なエピトープを認識することを実証している。ウイルス粒子外被と細胞膜との融合を抑止するため、mAb 2cはgBの融合前の立体構造に結合するはずである。しかしながら、mAb 2cの中和エピトープは、利用可能なgB結晶構造(24)に存在するgB立体構造の表面に部分的にしかマッピングされず、gBが侵入の間に異なる立体構造を取りうることを示唆している。
【0118】
mAb 2c由来の二価および一価抗体の特徴付け
ウイルス侵入におけるgBの機能に不可欠なgB上の領域を特定するために幾人の研究者らによってモノクローナル抗体が用いられている(4, 25, 39, 52)。ドメインVのC末端の残基および近接プロモーターのドメインIの残基を含むgB三量体の基部の特異な機能領域にマッピングされている、中和抗体がgBの融合活性を妨害することが示唆されている(4)。本発明者らはそれゆえ、ドメインVのC末端に近いドメインI内のmAb 2cエピトープに結合する一価抗体がgBの活性化による協同的な立体構造変化を十分に遮断するはずであると仮定した。mAb 2cはインビトロにおいて補体なしでHSV-1を中和する(16)ので、本発明者らは、mAb 2cが媒介する仮説的機構を研究するのに有益なツールとして従来のF(ab')2およびFab断片ならびに組換え一本鎖可変断片(scFv)を作出した。作出した抗体調製物の均一性をサイズ排除クロマトグラフィーによってモニタリングした(データ不掲載)。それぞれ、HSV-1もしくはHSV-2で感染された、または感染されなかったVero細胞を用いたフローサイトメトリー分析から、mAb 2cおよびmAb 2c由来抗体断片の特異的な結合が実証された(データ不掲載)。本発明者らはさらに蛍光サイトメトリーを用いて、HSV-1およびHSV-2感染Vero細胞に対する抗体の平衡結合曲線を決定した(図17)。これらの研究の結果から、それぞれ、FabおよびscFvに対するよりも全IgGおよびF(ab')2断片に対するいっそう高い見掛け上の親和性が実証された(表6)。決定された一価抗体親和性に対する二価抗体の機能的親和性(親和力)の増加から、二価抗体が細胞表面上の2つのgBエピトープに同時に結合できたことが示唆される。二価mAb 2cおよび2c-F(ab')2は、その一価対応物と比べて1.7〜2.8倍高い見掛け上の親和性を示した。IgGに比べてF(ab')2断片の見掛け上のKDのわずかな増加は、F(ab')2構築物内の抗原結合部位のいっそう高い可動性によるものでありうる。両方のHSV-1およびHSV-2感染Vero細胞に対するmAb 2c、2c-F(ab')2および2c-Fabの同様の見掛け上の親和性から、認識されるgBエピトープが両方のウイルス間で構造的に違わないことが確認された(表6)。
【0119】
(表6)HSV-1 FまたはHSV-2 G感染Vero細胞との結合に対するmAb 2cおよびmAb 2c由来抗体断片の見掛け上の平衡定数
aHSV感染細胞上のgBとの結合に対するKD値は、フローサイトメトリーによって決定された平衡結合曲線からのデータ(図17)をMarquardt-Levenberg式に適合させることによって決定された。
【0120】
インビトロにおける一価および二価抗体の中和活性
HSV-1ウイルス粒子がVero細胞と相互作用する前(付着前)または後(付着後)に抗体が加えられるかどうかにかかわりなくmAb 2cの等しい中和効力(図18A)から、mAb 2cが標的細胞とのウイルスの結合を妨害しないことが示唆された。対照的に、ポリクローナルヒトγグロブリンIntratect(登録商標)は、標的細胞へのウイルス粒子の付着の阻害によって明らかに中和をしていた(図18B)。mAb 2c由来断片F(ab')2、FabおよびscFvの中和活性を、Vero細胞に対する標準的な中和アッセイ法においてその親IgG対応物と比較した。親mAb 2cは8 nMの濃度でHSV-1誘導性の細胞変性効果(CPE)を100%低減した。興味深いことに、HSV-2誘導性のCPEを完全に低減するには、4倍高いmAb 2c濃度が必要とされた(図19A)。二価2c-F(ab')2はHSV-1およびHSV-2誘導性のCPEの両方を親mAb 2cよりも2倍効率的に低減した。驚いたことに、本発明者らは、HSV-1およびHSV-2を中和するための一価2c抗体断片の能力の基本的相違を認めた。親mAb 2cと比べて、HSV-1およびHSV-2誘導性のCPEを100%低減するには、それぞれ、およそ375倍および94倍高い濃度の2c-Fabが必要であった(図19A)。組換え2c-scFvは光学顕微鏡の下でプラーク低減効果を示したが、3,000 nMの最も高い試験濃度でさえもHSV誘導性のCPEを100%低減することはできなかった(データ不掲載)。
【0121】
両方の二価抗体mAb 2cおよび2c-F(ab')2はHSV-1よりも約4倍低い効率でHSV-2を中和した(図19A)ので、本発明者らは、等量の感染粒子を含有するHSV-1およびHSV-2調製物のゲノムコピー数を定量的リアルタイムPCRによって分析した。HSV-1と比べて、HSV-2の場合には4倍多い数のゲノム相当物が認められ(データ不掲載)、HSV-2の中和に必要とされるmAb 2cおよび2c-F(ab')2の抗体力価がさらに高いこととよく相関していた。
【0122】
図19Aに示されるように中和アッセイ法から、抗体価と中和効率との間の強い相関が示唆された。その結果、本発明者らは、Fab断片の架橋によってウイルス感染を取り除くための2c-Fab断片の能力を回復できるかどうか調べた。ウイルス中和アッセイ法を、マウスFab断片と反応するIgGの非存在下または存在下で2c-Fabに対して繰り返した。図19Bに示されるように、2c-Fabの架橋結合は中和活性を劇的に増大したが、それを、親mAb 2cの場合と同じ効力まで回復することができなかった。抗マウスFab IgGのみではウイルス中和に対する効果が示されなかった(データ不掲載)。
【0123】
細胞間伝播の阻害
2c-Fab断片は遊離ウイルス粒子を効率的に中和しなかったが、それでも、小さなサイズの抗体断片はより好ましい拡散特性を有することが報告されており(66)、本発明者らは、HSV-1が細胞間結合を、感染細胞から非感染細胞へと横断することを抑止するその活性について調べた。両方の二価抗体mAb 2cおよび2c-F(ab')2は、Vero細胞単層中で伝播されたHSV-1を完全に抑制し、間接的な免疫蛍光によってただ1つの感染細胞を可視化できただけであった(図9)。遊離ウイルス粒子を中和するポリクローナルヒト血清の能力にもかかわらず、それはウイルスの細胞間伝播を阻害することが全くできなかった。これは、多数のHSVエピトープに対して作製された中和抗体の不均一集団の結果である可能性が最も高い。ポリクローナルヒト免疫血清と比べて、一価2c-Fab断片は細胞間伝播をある程度制御することができた。しかしながら、一価2c-Fab断片は、その二価対応物とは対照的に、6倍高い濃度で試験してさえもウイルス伝播を完全に排除することができなかった(図9)。ゆえに、抗体価は隣接細胞の間でのHSV-1の伝播の阻害においても鍵となる役割を果たした。
【0124】
播種性HSV感染に対する免疫不全マウスの免疫保護
本発明者らは、CD4+およびCD8+ T細胞の両方が枯渇したマウスが、膣内HSV-1感染の後にmAb 2cの受動移入によって致死性脳炎から完全に保護されることを以前に示した(17)。ヒトにおいてHSV-2感染の部位に蓄積するナチュラルキラー(NK)細胞(28)はインターフェロン-γの初期供給源であり(45)、インターフェロン-γはHSV感染の制御に不可欠な役割を果たす(2, 45, 62)。つい最近、初めて、ヒトNK細胞が先天性免疫応答としてヒト化マウスで初感染性器HSV感染に対する保護を媒介することが実証された(37)。mAb 2cが抗体媒介性の免疫反応とは独立して抗ウイルス活性を付与するかどうか調べるために、本発明者らは、SCID T細胞欠乏およびB細胞欠乏に加えて、NK細胞およびマクロファージの機能ならびに補体経路を刺激する能力を欠くNOD/SCIDマウスモデルを利用した。NOD/SCIDマウスの膣内HSV-1感染(1×106 TCID50)は、急速進行性の全身疾患を引き起こし、9日の生存期間中央値であった。臓器におけるHSV力価を終点希釈アッセイ法によって判定し、脊髄(2.3×106 TCID50)、脳(3.8×105 TCID50)および膣粘膜(1.4×106 TCID50)において高ウイルス力価、腎臓(1.7×104 TCID50)および副腎(1.1×104 TCID50)において中力価ならびに肺(1.1×103 TCID50)および心臓(1.9×102 TCID50)において低力価を示した(データ不掲載)。mAb 2cの治療効率を評価するため、NOD/SCIDマウスを膣内HSV-1攻撃の24時間前に2.5 mg/kg、5 mg/kgまたは15 mg/kgの抗体で静脈内処置した(図20)。低抗体用量を投与されたマウスは、HSV-1による致命的感染から十分には保護されなかった。しかしながら、5 mg/kgのmAb 2cで処置されたマウスの生存期間中央値は、PBSを投与された対照マウスと比べた場合に2.6倍引き延ばされた。致死性脳炎から保護されなかったマウスより調べた臓器におけるHSV-1の力価は未処置対照群に匹敵していた。対照的に、動物の完全保護は15 mg/kgのmAb 2cの用量で達成された。抗体によって保護されたマウスの臓器におけるウイルス力価は、1×102 TCID50の検出限界未満であった。
【0125】
本発明者らは次に、mAb 2cによる曝露後免疫がまた、樹立された末梢HSV感染の存在下でウイルス伝播および致死性脳炎からの保護を付与するかどうか評価した。ウイルス攻撃後24時間の時点で膣洗浄液中に高いHSV-1力価を有するNOD/SCIDマウスを24時間、40時間および56時間の時点で15 mg/kgのmAb 2cにより静脈内から繰り返し処置した(図11および図21)。PBS処置対照群は、神経症状を有するマウスを7日目〜9日目に殺処理しなければならなくなるまで一定の経膣的なウイルスの出芽を示した。対照的に、mAb 2cは8日目までに、樹立されたHSV-1感染を除去し、感染の致命的結末を完全に抑止した(3×300μg; PBSと比べてP = 0.0003)。さらに、感染から1ヶ月後にmAb 2c処置動物の感覚ニューロンおよび各臓器においてウイルス粒子は検出されなかった(データ不掲載)。
【0126】
考察
ウイルスが標的細胞に侵入するために取る段階に続いて、ウイルス中和mAbはいくつかの機構によって侵入を阻害することができる。ウイルス表面タンパク質と細胞のタンパク質、脂質または糖質との特異的な相互作用は、感染の初期段階に相当し、これを中和抗体によって遮断することができる。ウイルス付着を阻害する抗体は、HIV-1 gp120のCD4結合部位と反応するmAb F105およびインフルエンザ血球凝集素(HA)の受容体結合部位を網羅するFab HC19など、ウイルス粒子の受容体結合部位に直接結合するか(6, 19, 54)、またはHA受容体結合部位の近く17Åで結合するFab HC45など、受容体連結を立体的に妨害するか(18)のどちらかである。HSV gDとその細胞受容体の1つとの不可欠な結合に加えて、gBは標的細胞へのウイルス粒子の付着に関与している。最近、HSV gBに対する2つのへパラン硫酸プロテオグリカン非依存的な真の細胞表面受容体および/または付着因子の存在が報告された(5, 23, 60)。対になった免疫グロブリン様2型受容体(PILRα)は少なくともある種の細胞型において、gBの潜在的タンパク質受容体の1つと特徴付けられている(60)。mAb 2cに対し、比較による付着前 vs 付着後中和アッセイ法から、この抗体は細胞表面へのウイルスの結合を阻害できないが、ウイルス侵入を遮断することが示された。gBと、その融合ドメインの鍵となる疎水性および親水性残基を介した脂質膜との相互作用(22, 23)は、融合ループのすぐ近くのエピトープを認識するmAbによって遮断されうることがこれまでに示されている(4, 22)。mAb 2cの立体構造エピトープは融合ループ1と部分的に重複するので、本発明者らは、mAb 2cの結合が融合シグナルの伝達を妨害する可能性が最も高いと推論し、本発明者らはさらに、考えられる作用機序として結合後/融合前の段階で中和を評価した。
【0127】
構造的再配列の誘導はウイルス融合糖タンパク質の鍵となる特徴であり、異なる融合前および融合後立体構造を引き起こす。異なる中和mAbのエピトープがスパイクの側面ドメインに沿っておよびgB結晶構造の冠部の先端にマッピングされた(4, 24)。mAb 2cのエピトープは、ドメインVのC末端ヘリックスαF内の残基および近接プロモーターのドメインI内の残基からなるgB三量体の基部の特異な機能領域(FR1)に位置する(4)。本発明者らの相同性モデルは、mAb 2cによって認識される不連続なエピトープの一部分(F300〜E305)が、プレクストリン相同性(PH)ドメインの特徴を有する、gBのドメインIの上部に局在することを示す(7, 38)。しかしながら、ドメインIに同様に位置するエピトープの他の部分(F175〜A190)は埋もれており、gBが主要な立体構造変化を起こさなければ、mAb 2cの結合にアクセスできないと考えられる。本発明者らはそれゆえ、mAb 2cが融合前の立体構造において選択的にgBの転移を妨げると仮定した。mAb 2cエピトープの局在、および活性化による立体構造変化が協調的であるという仮定に基づき、本発明者らは、gBの融合ドメインおよび細胞膜の並置の遮断にはmAb 2cの一価相互作用で十分であろうと推論した。しかしながら、驚いたことに、作出された一価抗体断片(FabおよびscFv)のどれも遊離ウイルス粒子を効率的に中和すること、またはウイルスの細胞間伝播を阻害することができなかった。対照的に、両方の二価分子mAb 2cおよび2c-F(ab')2は、ウイルス中和および細胞間伝播の阻害について非常に効果的であった。本研究における全てのmAb 2c由来抗体の特異的かつ同等な結合活性の保持から、抗原認識の障害による一価抗体および二価抗体の機能的差異は除外される。免疫グロブリンの多価結合はその機能的親和性を増加させる(26)。しかしながら、機能的親和性の増加は抗体結合部位の固有の親和性と逆の相関がある(49)。二価2c抗体IgGおよびF(ab')2に対する1.7〜2.8の平衡定数の増分が、その一価対応物scFvおよびFabと比較した場合に、わずかでしかないのは、かくして、低いナノモル範囲において固有の親和性を有する抗体にとって珍しいことではない。このように、いっそう高い見掛け上の親和性は実のところ、gB抗原との多価(いっそう高い親和力)の結合が実際に起きていることを示し、mAb 2cおよび2c-F(ab')2の抗ウイルス活性がgB架橋結合の結果であることを示唆している。水痘帯状疱疹ウイルス(VZV)のgH抗原に対して特異性を有する二価抗体または三価抗体に比べて見劣りする一価抗体の中和効率は、これらの抗体のサイズの違いによる立体障害の問題として論じられている(15)。本発明者らは、mAb 2cバリアントに対する潜在的なさらなる中和機構としてのこの可能性を完全に除外することはできないが、これは、抗体のサイズ、中和効率および細胞間伝播の阻害の間に直接的な相関関係が認められなかったので、可能性が低いようである。さらに、本発明者らのデータは、いっそう小さい2c-F(ab')2がいっそう大きな2c-IgGよりもさらに良好なウイルス中和活性を有していたことを示している。ゆえに、本発明の所見は、gB架橋結合がmAb 2cの抗ウイルス活性に対する鍵となる機構であることを示し、gB三量体の固定化によるgB融合前立体構造の安定化が融合シグナルの活性化を阻害することを示唆している。Silvermanらによる最新の研究(61)から、融合ループ1に近い残基E187の後の5つのアミノ酸の挿入によるHSV-1 gB細胞外ドメインの融合欠損表現型がgBの立体構造変化の妨害から生じるのではなく、むしろ他の機構によるgB機能の妨害から生じうることが提唱された。本発明者らのデュオトープスキャンにおいて、mAb 2cは、gBの機能に極めて重要であるようにみえる、特定の挿入部位E187を網羅する結合部位A/Bデュオトープ186FED188-βA-βA-300FYGYRE305 (SEQ ID NO: 23)と最も強く反応した。それゆえ、mAb 2cの架橋結合がHSV融合機構の他の成分と相互作用するgBの能力を損なうと推測したくなる。しかしながら、本発明者らの結果は、架橋結合がgBそれ自体の立体構造変化を遮断するか、または細胞融合の間に起こり(3)、融合過程の完了に不可欠な(65)、gB、gDおよびgH/gLの間の相互作用を遮断するかどうかを区別することを可能にするものではないので、将来の研究が必要である。解析された結晶において観察されたHSV-1 gBの立体構造(24)は、融合後の形態に相当することを示唆しており、gBの融合前のモデルは未だ特徴付けられていない。それゆえ、gBと複合体化したmAb 2cまたはそのF(ab')2のX線結晶学的研究は、gBの未変性の立体構造に関する見識および融合シグナルの伝達に関するさらに良好な理解をもたらす可能性がある。
【0128】
マウスにおいてHSV-2感染の膣感染を防ぐために局所に適用された抗gD抗体および抗gB抗体の保護効力を評価する研究から、新しい膣消毒薬としての遺伝子操作組換え抗体の実現可能性が実証された(67〜69)。重篤な、そして致命的でさえあるHSV感染は、母親から感染した新生児において、再発性眼感染を有する患者において、または重度免疫減弱状態の患者において起こりうる。本発明者らの抗gB抗体の全身適用が高度に免疫不全なインビボの設定においても保護を付与するかどうか調べるために、本発明者らは、NOD/SCIDマウスモデルを利用した。本発明者らは、免疫応答性マウスにおいておよびT細胞枯渇マウスにおいて後肢麻痺および致命的疱疹脳炎を引き起こすウイルスの軸索伝播による確立された神経節感染経路として膣内HSV-1接種を用いた(16, 17)。ここで本発明者らは、mAb 2cが一次性HSV-1感染の急性期においてNOD/SCIDを完全に保護するだけでなく、末梢ウイルス伝播の開始後でさえも神経疾患および死を完全に抑止するのに有効でもあることを実証する。HSVの細胞間伝播は、ニューロンのシナプスおよび密着結合をまたいだウイルスの移動のために、ならびに適応免疫系の免疫学的障壁を回避するために非常に効率的な手段である。mAb 2cは、感染した膣組織のウイルス発現を低減もし、HSVの軸索伝播を阻害もする。他の報告から、ウイルス攻撃後の抗HSV IgGの投与が動物において急性神経節感染の量を低減できることが示された(16, 42)。一貫して、角膜HSV-1感染を有するマウスに腹腔内投与された組換えヒト抗gD IgGがHSVに感染した神経線維および感覚ニューロンに局在することが示された(59)。さらに、適切な時点で曝露後に投与されたHSV gD、gCまたはgBに特異的なmAbによる免疫応答性動物の受動免疫は、HSV誘導性の神経疾患からの保護を示した(13, 16)。しかし、いくつかの動物実験から、液性免疫だけではHSV感染の制御には効果がないとも結論付けられている。
【0129】
本発明者らの抗体のインビボでの保護効力は、免疫エフェクター機能から独立している。文献中のいくつかの動物試験から、液性免疫だけではHSV感染の制御には効果がないとも結論付けられている。この考えと一致して、抗HSV-1過免疫血清の投与は免疫抑制マウスまたは免疫不全マウスの保護に効果がないと報告されている(47, 48, 50, 51, 56)。HSV-1感染後24時間の無胸腺ヌードマウスをヒト抗gD mAbで全身処置することにより、未処置対照と比べて生存時間が引き延ばされたが、死が抑止されることはなかった(58)。別の研究では、HSV-1誘導性の間質性角膜炎マウスモデルにおいて、抗gD mAbがCD4+またはCD8+ T細胞のどちらかを枯渇させたマウスの死を抑止したものの、マウスの両方のT細胞サブセットを同時に枯渇させた場合には、死を抑止できないことが示された(64)。
【0130】
本発明者らの知る限りでは、本発明者らは、細胞エフェクター機構および補体から完全に独立してニューロンのHSV-1伝播を抑止する、全身に適用された抗gB架橋mAbの保護効力を初めて実証した。HSV複製に不可欠なgBの一種の共通エピトープに対するmAb 2cの特異性およびさらなる免疫エフェクター機能の動員の必要性なくして高いその保護効率は、新規の免疫治療薬としてのこの抗体の大いなる潜在性を示す。
【0131】
実施例3に関する参考文献一覧
【0132】
実施例4
ウイルス中和アッセイ法
ウイルスの中和感受性を決定するための過剰量の抗体によるプラーク低減アッセイ法としてまたは抗体溶液の中和価を決定するための終点希釈法として、中和アッセイ法をVero細胞に対してマイクロタイタープレート中で行った。プラーク低減アッセイ法は、20μgのMAb 2cとの250プラーク形成単位のインキュベーションによって行った。2時間後、50μL/ウェルのVero細胞懸濁液(細胞1.5×105個/mL)を添加した。3日後、細胞をクリスタルバイオレットで染色した。終点力価測定のため、希釈した抗体溶液(0.025 mL)を0.025 mL中100 TCID50のHSV-1、および1:10に希釈したモルモット補体0.025 mLとともにインキュベートした。培養物の50%においてウイルス誘導性の細胞変性効果を抑止する最大血清希釈の逆数として力価を表した。
【0133】
gB欠失変異体の構築およびCOS-1細胞における発現
全長HSV-1 gB (gB(1〜904) = pMT2gB)、gB(1〜720)、gB(1〜630)、gB(1〜505)、gB(1〜503)、gB(1〜487)およびgB(1〜470)をコードするプラスミドの構築は他で記述されている[30, 31]。プラスミドはLeonore Pereira, University of California, San Franciscoから親切にも提供していただいた。gB(1〜130)、gB(1〜223)、gB(183〜488)およびgB(436〜642)をコードするプラスミドは、制限酵素部位Bam HIおよびXho Iが隣接するPCR単位複製配列を、真核生物発現ベクターpSVL (Amersham Pharmacia, Freiburg, Germany)の中にクローニングすることによって構築した。gBヌクレオチド52588〜60362を含んだHSV-1株17+ [33; GenBank X14112]のサブゲノムプラスミドクローンをPCRにおける鋳型として用いた。N末端切断型のgB構築物の発現のため、gBシグナル配列をコードするDNAを、5'末端の位置にXhoI部位を含むプライマーを用いたPCRによって増幅し、小断片プラスミドgB(183〜488)およびgB(436〜642)のgBをコードするDNAの5'側に挿入した。挿入断片の正しい組み込みおよびその配列をヌクレオチド配列決定によって確認した。COS-1細胞を、24ウェルプレートの中に配置されたカバーガラス(直径10 mm)上で増殖させ、DEAE-デキストラン法によりプラスミドでトランスフェクトした[34]。gBおよびその切断型誘導体の発現を、よく特徴付けられた抗HSV-1 gBマウスモノクローナル抗体H1396およびH1781の混合物を用いた間接的免疫蛍光顕微鏡検査法により検証した。トランスフェクトされ固定されたCOS-1細胞をMAb 2cと反応させ、免疫蛍光顕微鏡検査法により分析した。
【0134】
gBの部位特異的突然変異誘発および組換えウイルスの構築
単一のアミノ酸変異を、Altered Sites(商標) in vitro Mutagenesis System (Promega, Mannheim, Germany)を用いてオリゴヌクレオチド特異的突然変異誘発によりHSV-1 gBに導入した。手短に言えば、pMT2gB [31]内のgBコード配列を大腸菌(E. coli)ファージミド突然変異誘発ベクターpAlter-1の中に移入した。pALTER-lgBで形質転換された大腸菌JM109細胞にファージR408を感染させることによって、一本鎖pALTER-lgB DNA分子を調製した。以下のようにミスマッチプライマー(変異させた位置に下線を引いた)を用い製造元のプロトコルにしたがって部位特異的突然変異誘発を行った。
【0135】
完全に二重鎖の、共有結合閉環状分子を形成させるためのプライマー伸長および核酸連結の後に、このヘテロ二重鎖DNAを、ヌクレオチドミスマッチを修復することができない大腸菌BMH 71-18 mutSに形質転換した。アンピシリン耐性クローンから得られたプラスミドを大腸菌JM109 (recA-)の形質転換に用いた。得られたpAlter-lgBプラスミドにおける変異の同一性をgB DNA配列決定によって確認した。突然変異誘発の後、変異させた挿入断片を発現ベクターpMT2gBにクローニングし戻し、各pMT2gBmut構築物を得た。単一のアミノ酸変異を有するgBに対するMAb 2cの反応性を分析するために、カバーガラス(10×30 mm)上で増殖させたCOS-1細胞をリポフェクトアミン(商標)/OPTI-MEM(商標)法(Invitrogen, Karlsruhe, Germany)により製造元の指示にしたがってpMT2gBmut構築物でトランスフェクトした。細胞をメタノール/アセトンにより培養44時間後に固定した。ポリクローナルウサギ抗HSV-1 IgG (Dako, Hamburg, Germany)を用いた間接的免疫蛍光分析により、gBmutの発現を検証した。
【0136】
単一のアミノ酸の交換を有する糖タンパク質Bを、HSV-1ゲノムとプラスミドpMT2gBmut DNAとの間の相同組換えによってHSV-1野生型株Fの中に挿入した。ゲノムHSV-1 DNAを56℃で1〜3時間SDS/プロテイナーゼK溶解によって、その後、フェノール/クロロホルム/イソアミルアルコール抽出および65時間の透析(緩衝液I, 10 mM NaCl, 10 mM EDTA, 50 mM Tris-HCl, pH 8; 緩衝液II, 10 mM NaCl, 1 mM EDTA, 10 mM Tris-HCl, pH 7.5)によってウイルスストック液から調製した。pMT2gBmutプラスミドをQiagen Plasmid Midiキット(Qiagen, Hilden, Germany)によって抽出した。組換えウイルスはVero細胞を、StowおよびWilkie [36]により改変されたGrahamおよびVan der Eb [35]の方法にしたがってリン酸カルシウム沈殿させたDNAで、または製造元の指示にしたがってリポフェクトアミン(商標)/OPTI-MEM(商標)法(Invitrogen, Karlsruhe, Germany)でトランスフェクトすることにより調製した。手短に言えば、リン酸カルシウム法によるトランスフェクションの場合、精製HSV-1 F DNAおよそ100 ngを5分間1 M CaCl2 64μlの添加によりA. bidest. 0.436 mL中500 ngのプラスミドDNAおよび5μgの仔ウシ胸腺DNAで沈殿させた。45分間DNA沈殿物とのおよび3時間10%ウシ胎仔血清を含むEMEMとのペトリ皿(直径25 cm)の中で増殖させた細胞のインキュベーションの後、2分間HEBS緩衝液中25%のDMSOによって細胞にショックを与えた。その後、培養を、10%ウシ胎仔血清を含むEMEMおよびMAb 2cで継続した。リポフェクトアミンによるトランスフェクションの場合、HSV-1 DNA 1μgおよびプラスミドDNA 1μgを用いた。十分に分離されたプラークを選び取り、PCR単位複製配列のサイクル配列決定により各変異についてスクリーニングした。全体としては、プラスミドコード化gBの存在下において全くプラークを産生することに成功しなかった実験では、得られるプラークの数は低かった(gBをコードするプラスミドおよびMAb 2cの存在なしでの対照実験では多数のプラークを生じた)。抗体反応性を試験するために、カバーガラス(8×16 mm)上で増殖させた3×105個のVero細胞にEMEM 1 mL中200〜300プラーク形成単位のウイルスを感染させた。1時間後、10%ウシ胎仔血清を含むEMEMによって細胞培地を交換した。2日間のインキュベーションの後、細胞を-20℃でメタノール/アセトンにより固定した。
【0137】
免疫蛍光アッセイ法
細胞とのHSV gB特異的マウスモノクローナル抗体の結合を、DTAFコンジュゲートヤギ抗マウスIgG (Jackson ImmunoResearch Laboratories, Dianova, Hamburg, Germany)を用いて検出した。ウサギ抗HSV-1 IgGの結合の検出のため、TexasRedまたはCy3コンジュゲートヤギ抗ウサギIgG (Jackson ImmunoResearch Laboratories, Dianova)を用いた。ヒト抗体の検出のため、DTAFコンジュゲートヤギ抗ヒトIgG (Jackson ImmunoResearch Laboratories, Dianova)を用いた。
【0138】
セルロース結合ペプチドの合成およびスキャニング(ペプチドスキャン、鍵となるモチーフのスキャン、置換分析)
全てのペプチドは既報[37, 38]のようにWhatman 50セルロース膜上での半自動SPOT(商標)合成により作出した。合成後、ブロッキング用試薬(1X, Cambridge Research Biochemicals, Northwich, UK)をTris緩衝生理食塩水(50 mM Tris, 125 mM NaCl, 4 mM KCl, pH 8.0)、0.05% (v/v) Tween 20および5% (w/v)スクロース(TBST)の中に含有するブロッキング用緩衝液中で終夜、この膜をブロッキングした。TBST中で1回洗浄した後に、このシートを室温で3時間ブロッキング用緩衝液中で同時にMAb 2c (0.5〜1.0μg/mL)およびPODコンジュゲート抗マウスIgG Fab断片(5倍過剰; Roche Applied Science, Mannheim, Germany)とともにインキュベートした。TBST中で2回洗浄した後に、Chemiluminescence Western Blotting Kit (Roche Applied Science, Mannheim, Germany)を製造元のプロトコルにしたがって用いることにより、抗体結合を検出した。
【0139】
重複するペプチドのスキャン(ペプチドスキャン) [39]は、アミノ酸31〜505のgB残基(HSV-1株17+ [33] GenBank X14112)に対応した、12アミノ酸の重複を有する(すなわち、3アミノ酸ずつgB配列に沿ってシフトする) 15merとして、およびgB残基296〜315に対応した12アミノ酸の重複を有する(すなわち、1アミノ酸ずつgB配列に沿ってシフトする) 13merとして合成された。
【0140】
MAb 2c結合部位B内の最小の結合モチーフを特定するために、N末端およびC末端のそれぞれの位置で4つの無作為化残基により囲まれた中央の、gB由来六量体から構成される14mer (x1x2x3x4B1B2B3B4B5B6x11x12x13x14; x, 無作為化される位置、B, 固定化されるgB由来の位置)を用いて鍵となるモチーフのスキャンを行った[40]。各ペプチドで、六量体配列は1アミノ酸ずつシフトさせて、gBのV295位から始まり、gBのA315位に至るまでの配列に及んだ。
【0141】
gB結合部位A内のMAb 2cの結合について単一アミノ酸の関連性を、既報[40〜42]のようにその他全ての19種の天然アミノ酸によって各アミノ酸を連続的に交換することにより行ったペプチド178RYSQFMGIFEDRAPV192 (SEQ ID NO: 43)の置換分析によって分析した。
【0142】
マウスおよびマウス保護実験
雌性C57BL/6J (H-2b)マウスを、Charles River Wiga (Charles River Laboratories, Sulzfeld, Germany)から入手し、33〜37日齢の時点で使用した。実験を既報[1, 2]のように行った。手短に言えば、マウスに、10%ウシ胎仔血清を含むEMEM 0.1 mL中2×106 TCID50のHSV-1を膣内から接種した。ウイルス接種の24時間前に、腹腔内注射によってMAb 2c、ポリクローナル免疫血清または沈殿させた培地のいずれか0.5 mLをマウスに投与した。使用したヒト標準的免疫血清調製物(Beriglobin S(商標), CSL Behring, Germany)は、0.025 mL中でHSV-1に対して1:1280の補体非依存性の中和価を有し、適用のためIscoveの培地中で4倍希釈した。MAb 2cのストック調製液は1:640の補体非依存性の中和価を有し、ポリクローナル免疫血清と同じ中和活性を含むように2倍希釈した。適用した抗体希釈液のELISA力価は、マウスおよびヒトIgGに対するペルオキシダーゼコンジュゲートウサギを用いKahlon & Whitleyの方法[43]にしたがって決定した場合に104.5〜105.5であった。対照の場合、等量のIscoveの培地を同じように処理した。培地を投与した対照マウスは、非HSV特異的なMAbを投与した対照と同等であった[1]。膣拭き取り検体をウイルス接種後1日おきに採取し、Vero細胞単層上のウイルスについてアッセイした。感染性ウイルスの力価をReed & Muenchの方法[44]にしたがって0.05 mLあたりのTCID50によりマイクロタイタープレート中で決定した。
【0143】
結果
gBの最初の487アミノ酸はMAb 2cの結合に必要である
MAb 2cエピトープの適切な折り畳みに必要な糖タンパク質B領域に関する最初の見識を得るために、全長HSV-1 gB構築物および一連のカルボキシ末端切断型gB構築物を方法の項に示したようにCOS-1細胞において発現させた。gBの発現を、マウスHSV gB特異的MAb H1396およびH1781の混合物を用いた間接的免疫蛍光顕微鏡検査法により検証した。間接的免疫蛍光アッセイ法によりMAb 2cの結合も可視化した。表7に示されるように、全長タンパク質および切断型誘導体gB(1〜720)、gB(1〜630)、gB(1〜505)、gB(1〜503)およびgB(1〜487)はMAb 2cによって認識された。対照的に、MAb 2cはgB(1〜470)、gB(1〜223)およびgB(1〜130)を結合することができなかった。さらに、N末端およびC末端の両方の切断のある2つの構築物(gB(183〜488)、gB(436〜642))で反応は認められなかった。これらの結果から、MAb 2cのエピトープが最初の487アミノ末端残基のなかに位置することが示唆された。
【0144】
(表7)COS-1細胞において発現された切断型HSV-1糖タンパク質B (gB)とのMAb 2cの結合
* 全てのgB構築物の発現をHSV gB特異的モノクローナル抗体H1396およびH1781を用いた間接的免疫蛍光によって確認した[30〜32]。
** MAb 2cの結合を間接的免疫蛍光によって検出した。
+ はMAb 2c結合を示し、- はMAb 2c結合の失敗を示す。
*** pSVL, 陰性対照として用いた発現ベクター。
【0145】
MAb 2cは2つの異なるgB領域の配列を認識する
MAb 2cによって認識されるエピトープの精密マッピングはCOS-1細胞において発現されたgB欠失構築物によって可能ではなかったので、gBに由来する重複ペプチドのスキャン(ペプチドスキャン)をSPOT(商標)合成により連続的なセルロース膜支持体上で合成した。アミノ酸31〜505のgB領域に及ぶペプチドを、12アミノ酸の重複を有する(すなわち、3アミノ酸ずつgB配列に沿ってシフトする) 15merとして合成し、計155種のペプチドを得た。MAb 2cの結合は一次(MAb 2c)および二次(PODコンジュゲート抗マウスIgG Fab)抗体との同時インキュベーション、ならびに化学発光による検出で示された。
【0146】
図22に示されるように、MAb 2cは、部位Aおよび部位Bと名付けられた、2つの異なるgB領域のなかの5つのペプチドに結合することが分かった。部位Aは、gB残基175FGHRYSQFMGIFEDRAPVPFE195 (SEQ ID NO: 44)に対応する3つの連続ペプチド(共通配列181QFMGIFEDR189 (SEQ ID NO: 45)を含み、そして部位Bは、残基298SPFYGYREGSHTEHTSYA315 (SEQ ID NO: 46)を包含する2つの連続ペプチド(共通配列301YGYREGSHTEHT312 (SEQ ID NO: 47)を含む。
【0147】
MAb 2cに対する結合部位Bの最小の長さの特定
ペプチド90 (図22; 298SPFYGYREGSHTEHT312 (SEQ ID NO: 48)は最も強いシグナル強度を示したので、本発明者らは、結合部位BがMAb 2c結合のための主要な決定因子であると仮定した。かくして、本発明者らは、さらに高分解能のセルロース結合ペプチドスキャンを用いてMAb 2c結合に必要な部位Bの最小の長さを特定した。12アミノ酸の重複を有する(すなわち、アミノ酸ただ1つずつgB配列に沿ってシフトする) gB由来の残基296〜315に及ぶ13merのペプチドを二つ組で合成した。5つの連続ペプチドとのMAb 2cの反応性を上記のようにインキュベーションおよび検出手順の後に認めた。5つの反応性ペプチドの配列アライメントを図23に示す。全5種のペプチドに対する共通配列は300FYGYREGSH308 (SEQ ID NO: 49)であった。
【0148】
第2の手法では、両端に無作為化される4つの位置が隣接した6個のgB由来アミノ酸からそれぞれがなる14merを用いた鍵となるモチーフのスキャニング法を適用した。このアッセイ法において、各ペプチド分子の4つの末端位置は、アミノ酸が統計的に組み入れられた無作為配列に相当していた。各スポットにはかくして、外側のペプチド位置で多数の配列を有するが、しかし全て5〜10番目のペプチド位置に同じgB由来配列を有するペプチドの膨大な混合物が含まれた。gBに由来する中央の六量体は残基V295〜A315のgB領域にまたがり、1アミノ酸ずつ沿うようにしてシフトした。六量体のgB配列を選択したが、これは、非線状エピトープの75%超が最大4〜7残基長の連続的なストレッチを含むことが知られているからである[45]。MAb 2cの反応性は2つの連続ペプチドxxxx300FYGYRE305xxxx (SEQ ID NO: 21)およびxxxx301YGYREG306xxxx (SEQ ID NO: 50)で見られた(図24)。したがって、配列300FYGYREG306 (SEQ ID NO: 51)はMAb 2cとの相互作用に必要な部位Bペプチドの最小の結合モチーフであるものと考えられた。
【0149】
部位BでのMAb 2cの結合に重要な個々の残基の、変異型gBによる特定
全体的なおよび自然に折り畳まれたタンパク質という状況でMAb 2cに対する結合部位Bを確認するために、本発明者らは、結合部位B内の単一アミノ酸の交換によって、pMT2gBにクローニングされた、全長gBのアミノ酸配列を改変した。部位特異的突然変異誘発用のファージミドに基づく系を用いることにより、単一アミノ酸の交換を有するいくつかのgB構築物を作出した。COS-1細胞における変異型gBの発現の後、免疫蛍光アッセイ法によってMAb 2cの結合を分析した。表8に示されるように、MAb 2cの結合に重要であることが証明された一連のgB残基が特定された。詳細には、SによるgB 299位の残基Pの置換、それぞれYおよびIによるF300の置換、NによるY301の置換、それぞれRおよびVによるG302の置換、NによるY303の置換、それぞれGおよびLによるR304の置換、ならびにKによるE305の置換は、MAb 2c結合の完全な喪失をもたらし、したがって、299〜305位の残基はそれぞれ、その側鎖を介して抗体と相互作用する主要な残基となることにより、または抗体によって認識されるエピトープの立体構造の形成に必要なgBタンパク質の適切な全体的または局部的折り畳みに影響を与えることにより、エピトープの形成に決定的に関与していることを示唆していた。COS-1細胞における変異型gBの発現は、マウスMAb 2cおよびポリクローナルウサギ抗HSV IgG免疫血清との細胞の同時インキュベーションにより、その後、DTAFコンジュゲート抗マウスIgG (MAb 2cが結合したなら緑色の蛍光)およびTexasRedまたはCy3コンジュゲート抗ウサギ抗体(同じ細胞の赤色の蛍光)との同時インキュベーションを介した特定により検証した。反対に、単一のgB残基Y296の、それぞれ、NおよびFへの交換、M297の、それぞれ、L、TおよびVへの交換、ならびにS298のAへの交換のほか、G306の、それぞれ、AおよびVへの交換、ならびにS307のAへの交換はMAb 2cの結合に影響を与えなかった。
【0150】
(表8)COS-1細胞において発現された、単一アミノ酸の交換を含むHSV-1全長糖タンパク質B (gB) バリアントとのMAb 2cの結合
* 野生型アミノ酸はgBの位置番号の前に示されており、導入された残基は位置番号の後に示されている。
全てのgBバリアントの発現をポリクローナルウサギ抗HSV-1 IgG血清との細胞の同時インキュベーションによって得た免疫蛍光で確認した。
** MAb 2cの結合を間接的免疫蛍光によって試験した。
+ はMAb 2c結合を示し、- はMAb 2c結合の失敗を示す。
*** この特定の結果の評価については、考察の項を参照されたい。
【0151】
部位BでのMAb 2cの結合に重要な個々の残基の、変異体ウイルスによる特定
インビボに最も近い状況に近づけるため、それぞれがgBの中にアミノ酸変異を含む、よく特徴付けられた5つのHSV-1バリアント(R126、R1375、B4.1、R1435、R233) [27〜29]を用いることにより、および方法の項に示されるように、本研究において作出された変異体ウイルス(vY301N [301位のYをNによって置換した]、vG302R、vG302V)により、部位Bの個々のアミノ酸の影響をさらに分析した。カバースライド上のVero細胞に200〜300プラーク形成単位のこれらの変異体または親の野生型ウイルスHSV-1 FおよびKOS 321のどちらかを感染させた。間接的免疫蛍光アッセイ法から、MAb 2cはウイルスvY301N、vG302R、vG302V、R126 (Nによって置換されたY303)、R1375 (R304Q)およびB4.1 (E305K)に感染した細胞に結合できないが、MAb 2cは変異体R1435 (H308Y)、R233 (R328H)を感染させた細胞、および野生型ウイルスを感染させた細胞に対して反応性を示すことが実証された(表9)。
【0152】
(表9)MAb 2cの結合およびMAb 2cによる中和に対するHSV-1野生型ウイルスおよびウイルスgB変異体の感受性
* gBの発現をポリクローナルウサギ抗HSV-1 IgG血清で得た免疫蛍光により全てのウイルスについて確認した。+ は間接的免疫蛍光によって検出された感染Vero細胞とのMAb 2cの結合を示す。- はMAb 2c結合の失敗を示す。
** + はMAb 2cによるウイルス中和を示し、- はウイルス中和の失敗を示す。
*** 野生型アミノ酸はgBの位置番号の前に示されており、導入された残基は位置番号の後に示されている。
【0153】
gBとのMAb 2cの結合に重要であると特定された残基が抗体の中和活性にも重要であったかどうかを判定するために、250プラーク形成単位のウイルス変異体または野生型ウイルスを用いて中和アッセイ法を行った。表9に示されるように、野生型株FおよびKOS 321ならびに変異体R1435 (H308Y)およびR233 (R328H)はMAb 2cによって完全に中和された。対照的に、MAb 2cは変異体ウイルスvY301N、vG302R、vG302V、R126 (Y303N)、R1375 (R304Q)およびB4.1 (E305K)を完全に中和することができず、これらの残基の各々がMAb 2cの中和能に必要なエピトープを形成させるのに不可欠な標的であることを示唆していた。総合すれば、ペプチド分析および変異型タンパク質によって得られた結果から、残基299〜305がエピトープ形成に、およびMAb 2cのインビトロでの生物活性に重要であることが示された。
【0154】
マウス保護実験によるエピトープマッピング
インビボでのMAb 2cの保護効果がまた、部位Bの特定のアミノ酸に依存するかどうか分析するために、計168匹のC57BL/6マウスにMAb 2cの腹腔内注射から24時間後に変異体ウイルスまたは親野生型株のどちらかを膣内から接種した。比較のため、同じ中和能力に調整されたポリクローナル免疫血清を投与した。この実験は既報[1, 2]のように行った。図25に示されるように、MAb 2cは、変異体R126 (Y303N)、R1375 (R304Q)およびB4.1 (E305K)を接種したマウスでは効果がなかったが、変異体R1435 (H308Y)もしくはR233 (R328H)または野生型ウイルスを接種したマウスでは有効であった。しかし、ウイルス変異体R126、B4.1およびR233を用いた実験は、マウスの粘膜におけるこれらの変異体のウイルス複製が非効率的であるという事実によってわずかに妨げられた。特に、変異体R126は非常に低い複製能を示した。ゆえに、感染の経過は、MAb 2c、ポリクローナル免疫血清または対照液で処置したR126感染マウスの間で違わなかった。総合すれば、マウス保護実験の結果から、MAb 2cがインビボでその保護効果を示すにはgB残基Y303、R304およびE305が不可欠であることが明らかに実証された。
【0155】
MAb 2c結合のための部位Aの特徴付け
ペプチドスキャン法によるMAb 2cエピトープマッピングの結果(図22)から、共通配列181QFMGIFEDR189を有する結合部位Aは、部位B、およびセグメント化されたペプチドによって検出できない可能性がある潜在的に他の領域とともに形成される不連続なエピトープの構成要素であるが、自然に折り畳まれたタンパク質において機能するエピトープの部分であることが最初に示唆された。しかしながら、部位Aは驚いたことに、三次元gB三量体構造の表面に位置していなかった[21]。さらに、部位AおよびBはgB表面上ですぐ近くになく、抗体の抗原結合部位の平均領域によって同時に網羅することができなかった。
【0156】
部位Aの関連性を調べるために、本発明者らは、gB構築物および単一アミノ酸の交換を有するウイルス変異体を作出することを開始した。しかし、gB構築物でのおよびウイルス変異体でのこれらの残基のいずれの置換も、ほぼ間違いなく分子中の部位Bモチーフの存在により、続けて抗体結合の喪失が起こらなかったので、生体系におけるこれらのアミノ酸の重要性を実証する努力は報いられなかった(データおよび突然変異誘発プライマーは示されていない)。
【0157】
ペプチドスキャン法によって特定されたペプチド50 178RYSQFMGIFEDRAPV192 (SEQ ID NO: 43)の各アミノ酸の相対的重要性を研究するために、本発明者らは、全置換分析を行った。それゆえ、可能な全ての単一部位置換類似体(すなわち、各位置がその他の全19種のタンパク新生アミノ酸によって置換されている)をSPOT(商標)合成によって合成し、MAb 2cの結合について試験した。結合の喪失なしにペプチドの大部分の位置をいくつかの物理化学的に異なるアミノ酸によって交換することができた。しかしながら、モチーフ186FED188は保存された。すなわち、これらのアミノ酸の改変は抗体結合の喪失と関連していた(データ不掲載)。本発明者らはそれゆえ、部位Aまたは主にモチーフ186FED188が不連続なMAb 2cエピトープの一部を模倣すると仮定した。
【0158】
この前提について調べるため、本発明者らは、生物学的に重要であると分かったMAb 2cの結合部位Bに位置する残基(299PFYGYRE305 (SEQ ID NO: 53)) (表8)、グリシンリンカーおよび部位Aの最も重要な配列に由来するC末端のFEDFモチーフから構成される、12merのペプチドPFYGYRE-G-FEDF (SEQ ID NO: 52)をコンピュータによりデザインした。このペプチドとのMAb 2cの結合を、15merのペプチドスキャンにおいて最も強い反応性を示したペプチド298SPFYGYREGSHTEHT312 (SEQ ID NO: 48) (図22)と比べて測定した。両方のペプチドを、方法に記述したようにセルロース膜上で合成し、MAb 2cでプローブした。図26に示されるように、組み合わせペプチドPFYGYRE-G-FEDF (SEQ ID NO: 52)のシグナルは、部位Bのペプチドのみと比べてはるかに強く、しかもX線フィルムの曝露時間は15merのペプチドスキャンの曝露時間の4分の1でしかなかった。この所見は、MAb 2cが、i) 残基299〜残基305 (PFYGYRE) (SEQ ID NO: 53)の部位B、およびii) 配列FEDF (SEQ ID NO: 54)によって模倣されうる1つまたは複数のさらなる不連続な領域から構成される不連続なエピトープを認識するという前提を強力に支持する。
【0159】
考察
現研究の目的は単純ヘルペスウイルス糖タンパク質B上のMAb 2cに対する結合部位をマッピングすること、および鍵となるエピトープ残基を特定することであった。一連のC末端切断型の組換え発現gBタンパク質を用いて、N末端の487残基がMAb2cの結合に必要であることが分かった。C末端からの17個またはそれ以上のアミノ酸のさらなる欠失は、全ての欠失型gBの一過性トランスフェクト細胞での合成を容易に確認できたという事実にもかかわらず、抗体結合の喪失をもたらした。エピトープの位置を絞り込むために、それぞれがシグナルペプチド配列(アミノ酸1〜30)に融合された2つのさらなる欠失変異体gB(183〜488)およびgB(436〜642)を構築した。MAb2cはこれらの後者の切断型gBタンパク質のいずれにも結合することができず、かくしてMAb 2cのエピトープが残基470〜487の間に局在している可能性があるという本発明者らの初期の前提に矛盾していたので、本発明者らは、合成ペプチドを利用する代替的なエピトープマッピング戦略に切り替えることに決めた。
【0160】
多くのB細胞エピトープがもともと不連続的である[46]。タンパク質の断片、すなわち、化学的にまたは生物学的に作出されたペプチドを用いたこれらの不連続的なエピトープのマッピングでは、単一の結合領域に由来するペプチドが結合パートナーに対して、ELISAまたは表面プラズモン共鳴アッセイ法では通常測定できない非常に低い親和性を一般的に有するという難点に悩まされる。ここ20年のうちに、連続表面上で合成されたペプチドを用いた不連続的なエピトープのマッピング研究にかかわるいくつかの調査例が記述されている[47〜50]。感受性の点で、SPOT(商標)法はセルロース膜上の高いペプチド密度(およそ50 nmol/cm2)によって特に適している[37]。これは親和力および再結合効果をもたらし、それゆえ、さらに低親和性のペプチド-抗体間の相互作用の特定を可能にする。2006年までの600超の引用文を網羅する包括的な見直しから、ペプチド合成のためにSPOT(商標)技術を用いて線状かつ不連続のエピトープをマッピングすることに関する研究が大々的に要約されている[51]。
【0161】
N末端の487個のgB残基内でのMAb 2cエピトープの位置決めの後、SPOT(商標)合成法と組み合わせてペプチドスキャン法を用い、抗体結合に重要なアミノ酸の精密マッピングを達成した。図20に示されるように、MAb 2cは、結合部位Bに局在する15merのペプチド298SPFYGYREGSHTEHT312 (SEQ ID NO: 48)に最も強く反応し、この配列がMAb 2c認識に主に関わるという前提につながった。その後、部位Bの認識配列をさらに高分解能のペプチドスキャニングによってgBアミノ酸300〜308にまで縮めることができた。エピトープの鍵となる残基の最終確認を次いで、単一の点突然変異を持つ全長gB構築物でトランスフェクトされた細胞とのMAb 2cの結合を研究する生物学的手法によって達成し、配列300FYGYRE305 (SEQ ID NO: 21)を得た。さらに、プロリン残基299は、三次元gB構造におけるその非常に限られた表面曝露にもかかわらずインビボにおいて重要であることが分かった[21]。しかしながら、プロリン側鎖による立体構造の制限のため、つまりプロリンは、その環状性の結果、固定化された構造としてタンパク質配列を安定化することが多いため、この残基はまた、抗体結合部位でのgBの局部的な折り畳みに不可欠であるものと考えられる。特定された鍵となる残基の影響は、鍵となる残基を変異させた場合にインビボでのMAb 2cの保護効果が無効化されることを実証するマウス保護実験によって裏付けられた。全体的に見て、これらのデータは、アミノ酸300〜305がエネルギー性MAb 2cエピトープの必須部分を形成することを証明する。
【0162】
予想通り、部位B内の鍵となる残基の単一の点突然変異を有するHSV変異体は、MAb 2c結合および中和に耐性であった。しかし、ウイルス変異体の大部分が細胞培養または粘膜において増殖不良を示し、重度に無能なことが分かった。とりわけ、300位の残基フェニルアラニンは、その位置にアミノ酸の交換がある生存ウイルス変異体を作出しようとする試みがこれまで失敗に終わっているので、gBの生物学的機能に重要であるようである。この事実はウイルスの溶解サイクルにおけるこのgBモチーフの重要な役割を示唆しうる。したがって、MAb2cのエピトープがいくぶんかgBの弱点になりうると推測することは興味深い。
【0163】
ペプチドスキャニングによる初期のエピトープマッピングから、MAb 2cが部位Aと名付けられた、さらなるgB領域を認識することが示唆された。その部位で鍵となる残基の判定をペプチド178RYSQFMGIFEDRAPV192に関する置換分析によって行い、残基F186、E187およびD188が非常に置き換え感受性が高いことが実証された。生体系におけるこれらのアミノ酸の関連性は、gB構築物でのおよびウイルス変異体でのこれらの残基の置換が、ほぼ間違いなく分子中の部位Bモチーフの存在により、抗体結合に影響を与えなかったので、実証することができなかった。
【0164】
最近になって決定された外部gBドメイン結晶構造[21]に基づいて、gB単量体を6つの異なる構造ドメインに分けた。ドメインIはアミノ酸154〜363を含む。本明細書で提示した結果によれば、MAb 2cの最も重要なエネルギー性エピトープの残基(部位B)は構造ドメインIに存在する。部位Bの鍵となる残基をgB結晶構造に重ね合わせることにより、これらの残基は構造ドメインIの上部3分の1にある2つのβ鎖(β13, β14)間の22アミノ酸のループ様ストレッチの範囲内でgBの表面に位置することが明らかである。部位Aの残基186FED188も構造ドメインIに、ただし、ほとんど曝露されない小腔中のgBドメインIの基部に局在していることから、この2つの部位が空間的に近接していないことが示唆された。ところがgBと他のウイルス糖タンパク質との構造的類似性によりおよびリンカー挿入突然変異誘発の結果により、その結晶構造はgBの融合後の形態に相当することが示唆される[52〜54]。しかし、ウイルス粒子はgBの融合前の形態を含んでおり、中和抗体はgBの融合前の立体構造を認識すべきであると提案されている[21]。とはいえ、最近の研究から、試験された全てのgB特異的MAbによる融合前および融合後gBの両方の認識が示唆されている[55]。
【0165】
三次元gB構造[21]の範囲内でのおよび部位Bに対しての部位Aの局在と組み合わせた実験データに基づく、代替の、さらに魅力的な説明は、部位Aが不連続的なMAb 2cエピトープの構成要素ではないというものである。ペプチドスキャンおよび置換分析の結果から、適用された方法論によっては明らかに検出できない不連続的なエピトープの1つまたは複数の領域の模倣が示唆される。抗体-抗原複合体のX線またはNMR技術によっては、部位Bの残基300〜305に主に存在するエネルギー性エピトープとは対照的に、機能的エピトープの全体、すなわちMAb 2cと接触する全てのアミノ酸が検出されうるにすぎない[56, 57]。
【0166】
部位AおよびとりわけFEDモチーフが不連続的なエピトープの別の部分を模倣するという仮説は、可動性スペーサー要素としてのグリシン残基を介して共有結合している1つの分子内で部位Bの重要な残基と部位Aに由来する配列FEDFとを組み合わせるという試みによって明らかに支持された(図26)。この遺伝子操作ペプチドは、親和性の増加と相関する部位B由来のペプチド90と比べて(図22参照)シグナル強度の極めて大きな増加をもたらした。例えば、不連続なエピトープを認識する抗体に対するインターロイキン-10模倣体など、一つの合成分子に集められた単一の結合領域を網羅するペプチドによる不連続な結合部位の模倣がいくつかの刊行物に記述されている[49]。
【0167】
幾人かの研究者らがHSV gBの機能ドメインを特定するために何年にもわたってモノクローナル抗体を用いてきた[30、32、58〜60]。最近の研究から、gBに対する中和MAbの結合パターンによって規定されるようにgB構造全体に分散している、少なくとも4つの機能領域の存在が示唆された[55]。これらの結果によれば、MAb 2cのエピトープは、構造ドメインI、および構造ドメインVの残基697〜725由来の配列によって形成される機能領域(FR) 1のなかに位置しており、構造ドメインVは残基670〜727にまで及ぶ。興味深いことに、Benderら[55]によって作出された最も強力な中和MAbの3つとも、残基98〜472を含むgBタンパク質分解的切断断片との反応性によって決定した場合にFR1内の構造ドメインIにマッピングされている。1型HSVによって誘発されたMAb 2cは、2型HSVと交差反応性である[1, 2]。本発明者らはそれゆえ、HSV-1の部位AおよびBのアミノ酸配列をHSV-2 gBのそれと比べた。NCBIタンパク質データベース(日付2010年7月30日)中で見出された全53種の全長HSV-2 gBの中に、HSV-1 gB配列178RYSQFMGIFEDRAPV192 (SEQ ID NO: 43)および298SPFYGYREGSHTEHT312 (SEQ ID NO: 48)が存在していた。
【0168】
抗HSV療法の主な目標は、ウイルス複製を素早く取り除くことであるので、MAb 2cは1型および2型HSVを処置するための潜在的な手段となりうる。原理上は、2つの戦略を追求できる。まず第一に、MAb 2cの特異性および生物活性の抗体を、部位B由来のペプチドまたは模倣エピトープPFYGYRE-G-FEDF (SEQ ID NO: 52)によって誘導できることを実証できたなら、能動免疫が考えられうる。MAb 2cの予防可能性および治療可能性を活用する代替的手法は、十分に確立された抗ウイルス化学療法に加えて、受動免疫用にマウス抗体をヒト化分子へ変換することである。
【0169】
実施例4に関する参考文献
【0170】
参考文献
【特許請求の範囲】
【請求項1】
SEQ ID NO: 1、SEQ ID NO: 2、SEQ ID NO: 3、SEQ ID NO: 4、SEQ ID NO: 5、およびSEQ ID NO: 6に示される相補性決定領域を含む、抗体。
【請求項2】
請求項1記載の抗体と同じエピトープを認識する抗体。
【請求項3】
感染細胞から、隣接する第2の非感染細胞へのHSVの伝播(細胞間伝播)を阻害することができる、請求項1または2のいずれか一項記載の抗体。
【請求項4】
最大でも40 nM、好ましくは最大でも30 nM、より好ましくは最大でも20 nM、さらにより好ましくは最大でも15 nM、最大でも13 nM、最大でも10 nM、および最も好ましくは最大でも7 nMの解離定数KDを有する、請求項1〜3のいずれか一項記載の抗体。
【請求項5】
最大でも20 nM、好ましくは最大でも16 nM、より好ましくは最大でも12 nM、最大でも10 nM、最大でも8 nM、最大でも6 nM、および最も好ましくは最大でも4 nMの濃度で100 TCID50のHSVの規定量を中和することができる、請求項1〜4のいずれか一項記載の抗体。
【請求項6】
SEQ ID NO: 9の1〜30位、38〜51位、68〜99位、および112〜122位、ならびにSEQ ID NO: 10の1〜23位、40〜54位、62〜93位、および103〜113位に示されるアミノ酸残基に対して少なくとも70%の配列同一性を有するアミノ酸配列を含む、請求項1〜5のいずれか一項記載の抗体。
【請求項7】
SEQ ID NO: 7の1〜30位、38〜51位、68〜99位、および112〜122位、ならびにSEQ ID NO: 8の1〜23位、41〜55位、63〜94位、および104〜114位に示されるアミノ酸残基対して少なくとも80%、好ましくは100%の配列同一性を有するアミノ酸配列を含む、請求項1〜6のいずれか一項記載の抗体。
【請求項8】
二価抗体または多価抗体である、請求項1〜7のいずれか一項記載の抗体。
【請求項9】
エフェクター部分、治療部分、または検出可能な標識とコンジュゲートしている、請求項1〜8のいずれか一項記載の抗体。
【請求項10】
請求項1〜9のいずれか一項記載の抗体の有効量と、少なくとも1つの薬学的に許容される賦形剤とを含む、薬学的組成物。
【請求項11】
請求項1〜8のいずれか一項記載の抗体をコードする核酸配列を含む、発現ベクター。
【請求項12】
請求項1〜8のいずれか一項記載の抗体をコードするヌクレオチド配列を含む、宿主細胞。
【請求項13】
請求項1〜8のいずれか一項記載の抗体を産生することができる、ハイブリドーマ細胞。
【請求項14】
薬物としての使用のための請求項1〜9のいずれか一項記載の抗体。
【請求項15】
対象におけるHSV関連疾患の予防的または治療的処置のための方法における使用のための請求項1〜9のいずれか一項記載の抗体。
【請求項16】
HSV関連疾患が以下の特徴の1つまたは複数を伴う、請求項15記載の使用のための抗体:
(a) 経口再発の存在、
(b) 生殖器再発の存在、
(c) ヘルペス性湿疹、
(d) 新生児ヘルペス、
(e) 免疫不全、免疫減弱状態の患者、
(f) ウイルス抑制剤に対する耐性、
(g) 脳炎、
(h) 髄膜炎、
(i) 髄膜脳炎、
(J) 眼感染症、および/または
(k) 汎発性HSV感染症。
【請求項1】
SEQ ID NO: 1、SEQ ID NO: 2、SEQ ID NO: 3、SEQ ID NO: 4、SEQ ID NO: 5、およびSEQ ID NO: 6に示される相補性決定領域を含む、抗体。
【請求項2】
請求項1記載の抗体と同じエピトープを認識する抗体。
【請求項3】
感染細胞から、隣接する第2の非感染細胞へのHSVの伝播(細胞間伝播)を阻害することができる、請求項1または2のいずれか一項記載の抗体。
【請求項4】
最大でも40 nM、好ましくは最大でも30 nM、より好ましくは最大でも20 nM、さらにより好ましくは最大でも15 nM、最大でも13 nM、最大でも10 nM、および最も好ましくは最大でも7 nMの解離定数KDを有する、請求項1〜3のいずれか一項記載の抗体。
【請求項5】
最大でも20 nM、好ましくは最大でも16 nM、より好ましくは最大でも12 nM、最大でも10 nM、最大でも8 nM、最大でも6 nM、および最も好ましくは最大でも4 nMの濃度で100 TCID50のHSVの規定量を中和することができる、請求項1〜4のいずれか一項記載の抗体。
【請求項6】
SEQ ID NO: 9の1〜30位、38〜51位、68〜99位、および112〜122位、ならびにSEQ ID NO: 10の1〜23位、40〜54位、62〜93位、および103〜113位に示されるアミノ酸残基に対して少なくとも70%の配列同一性を有するアミノ酸配列を含む、請求項1〜5のいずれか一項記載の抗体。
【請求項7】
SEQ ID NO: 7の1〜30位、38〜51位、68〜99位、および112〜122位、ならびにSEQ ID NO: 8の1〜23位、41〜55位、63〜94位、および104〜114位に示されるアミノ酸残基対して少なくとも80%、好ましくは100%の配列同一性を有するアミノ酸配列を含む、請求項1〜6のいずれか一項記載の抗体。
【請求項8】
二価抗体または多価抗体である、請求項1〜7のいずれか一項記載の抗体。
【請求項9】
エフェクター部分、治療部分、または検出可能な標識とコンジュゲートしている、請求項1〜8のいずれか一項記載の抗体。
【請求項10】
請求項1〜9のいずれか一項記載の抗体の有効量と、少なくとも1つの薬学的に許容される賦形剤とを含む、薬学的組成物。
【請求項11】
請求項1〜8のいずれか一項記載の抗体をコードする核酸配列を含む、発現ベクター。
【請求項12】
請求項1〜8のいずれか一項記載の抗体をコードするヌクレオチド配列を含む、宿主細胞。
【請求項13】
請求項1〜8のいずれか一項記載の抗体を産生することができる、ハイブリドーマ細胞。
【請求項14】
薬物としての使用のための請求項1〜9のいずれか一項記載の抗体。
【請求項15】
対象におけるHSV関連疾患の予防的または治療的処置のための方法における使用のための請求項1〜9のいずれか一項記載の抗体。
【請求項16】
HSV関連疾患が以下の特徴の1つまたは複数を伴う、請求項15記載の使用のための抗体:
(a) 経口再発の存在、
(b) 生殖器再発の存在、
(c) ヘルペス性湿疹、
(d) 新生児ヘルペス、
(e) 免疫不全、免疫減弱状態の患者、
(f) ウイルス抑制剤に対する耐性、
(g) 脳炎、
(h) 髄膜炎、
(i) 髄膜脳炎、
(J) 眼感染症、および/または
(k) 汎発性HSV感染症。
【図1】

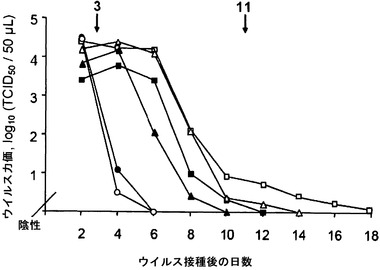
【図2A】
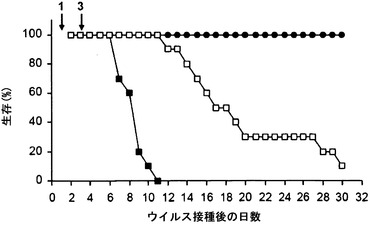
【図2B】
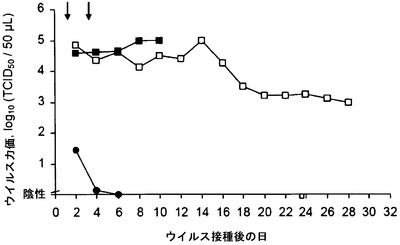
【図3】
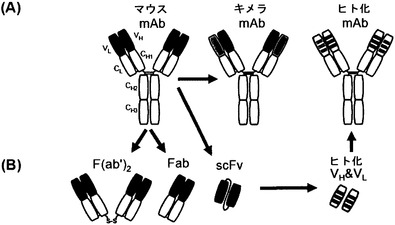
【図4】

【図5】
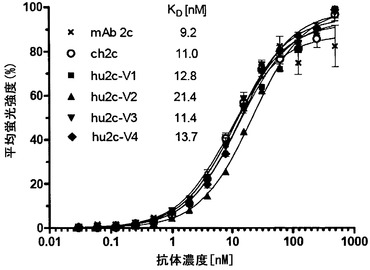
【図6】
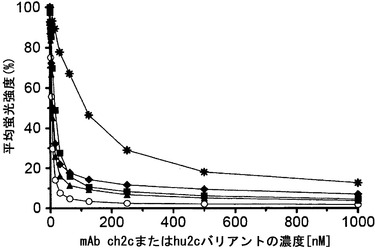
【図7】
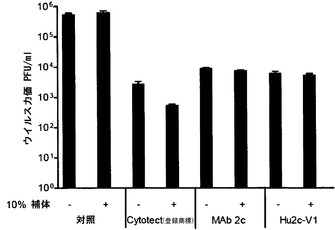
【図8】
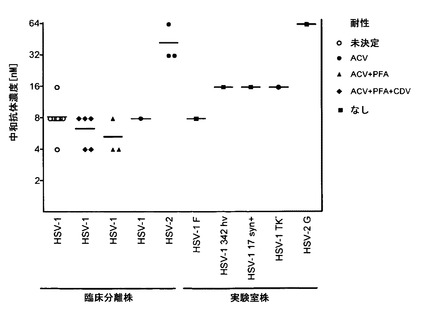
【図9】
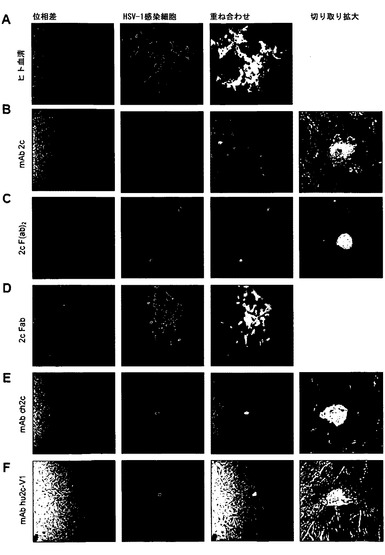
【図10】
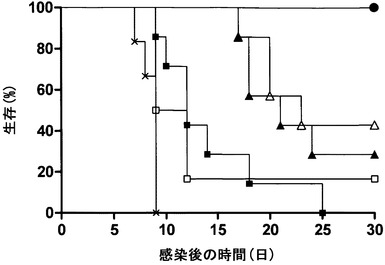
【図11】
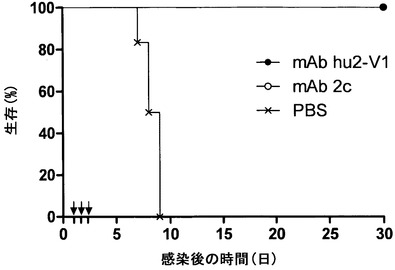
【図12】
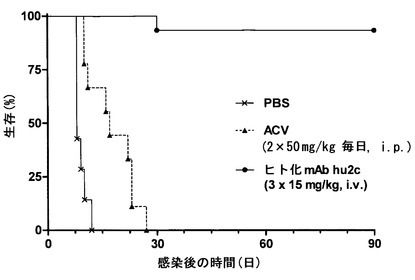
【図13A−1】

【図13A−2】
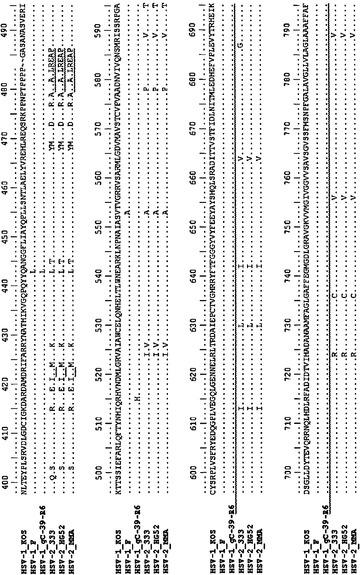
【図13A−3】

【図13B】

【図14】
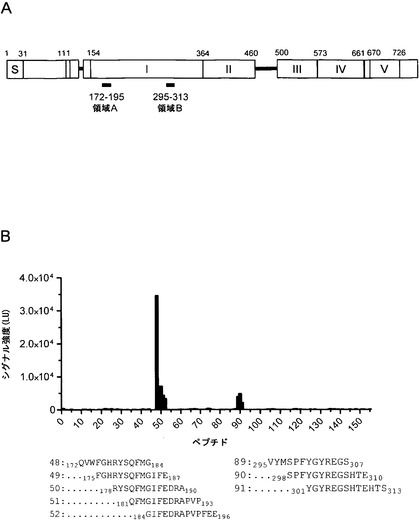
【図15】

【図16】
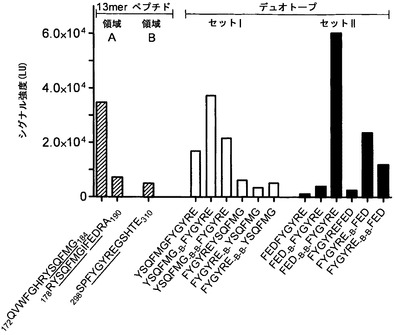
【図17】
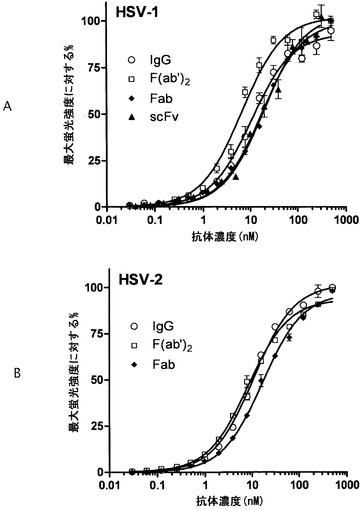
【図18】
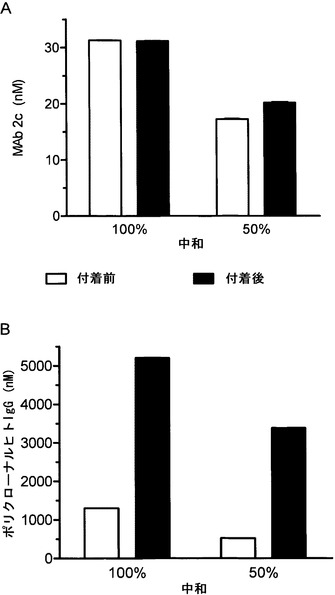
【図19】
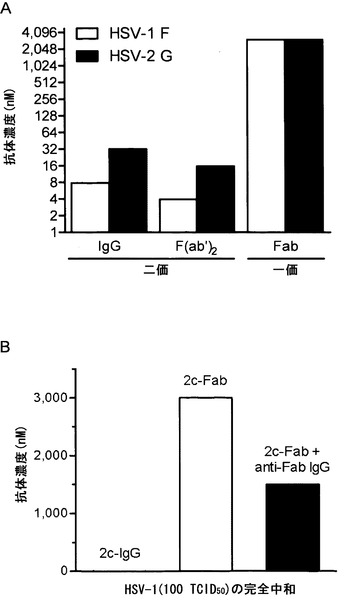
【図20】
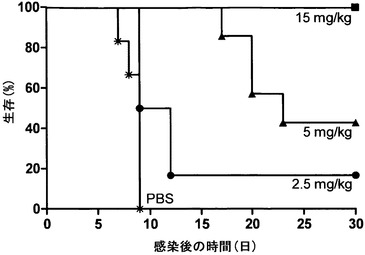
【図21】
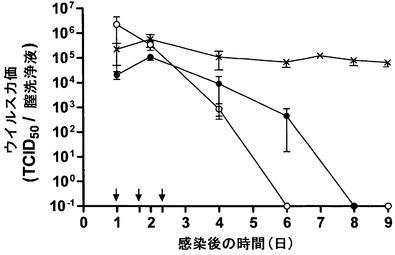
【図22】
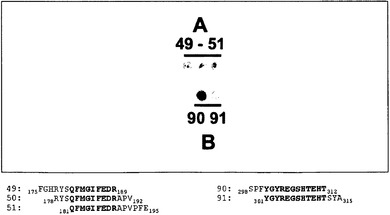
【図23】
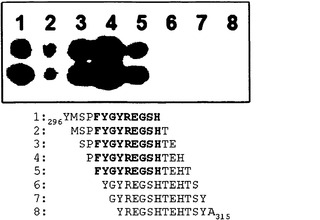
【図24】
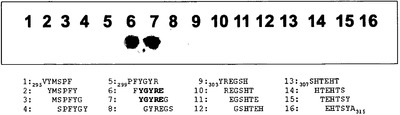
【図25】
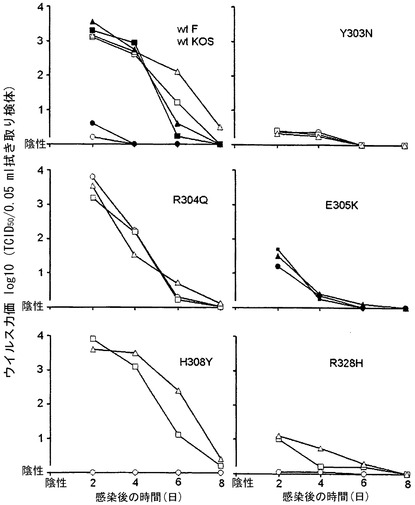
【図26】

【図2A】
【図2B】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13A−1】
【図13A−2】
【図13A−3】
【図13B】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【公表番号】特表2013−506403(P2013−506403A)
【公表日】平成25年2月28日(2013.2.28)
【国際特許分類】
【出願番号】特願2012−531286(P2012−531286)
【出願日】平成22年10月1日(2010.10.1)
【国際出願番号】PCT/EP2010/006020
【国際公開番号】WO2011/038933
【国際公開日】平成23年4月7日(2011.4.7)
【出願人】(512084422)ユニバーシタット ダイズバーグ−エッセン (1)
【出願人】(512084433)ライニッシュ フリードリッヒ−ウィルヘルムズ−ユニバーシタット ボン (1)
【Fターム(参考)】
【公表日】平成25年2月28日(2013.2.28)
【国際特許分類】
【出願日】平成22年10月1日(2010.10.1)
【国際出願番号】PCT/EP2010/006020
【国際公開番号】WO2011/038933
【国際公開日】平成23年4月7日(2011.4.7)
【出願人】(512084422)ユニバーシタット ダイズバーグ−エッセン (1)
【出願人】(512084433)ライニッシュ フリードリッヒ−ウィルヘルムズ−ユニバーシタット ボン (1)
【Fターム(参考)】
[ Back to top ]