
抗OX40L抗体
本発明は、抗OX40L抗体、特にヒト起源に由来するFc部分を含有しそして補体因子C1qに結合しない抗OX40L抗体およびその変異体に関する。これらの抗体は、炎症性疾患に罹っている患者に有益な新規で発明的な性質を有する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、抗OX40L抗体、特に、補体因子Clqに結合しない抗OX40L抗体、その医薬組成物および使用に関する。好ましくは、これらの抗体はヒト抗体又はヒト化抗体である。
【0002】
ヒトOX40L(gp34、SwissProt P23510)は、CD40/CD40Lライゲーションすると活性化されたB細胞および樹状細胞に発現されそして炎症性組織において内皮細胞に発現される(Review: Weinberg, A.D., Trends Immunol. 23(2002)102-109)。それは、HTLV−1感染したヒト白血病細胞(OX40を有するオートクリンループの発生によりこれらのT細胞の不死化)から単離された。OX40Lおよび対する抗体は、例えば、WO 95/12673; WO 95/21915; WO 99/15200; Baum, P.R., et al., EMBO J. 13(1994)3992-4001; Imura, A., et al., Blood 89(1997)2951-2958; Imura, A., et al., J. Exp.Med. 183(1996)2185-2195; Kjaergaard, J., et al., I. Immunol. 167(2001)6669-6677; Lane, P., J.Exp. Med. 191(2000)201-206; Mallett, S., and Barclay, A. N., Immunol. Today 12(1991)220-223; Mallett, S., et al., EMBO J. 9(1990)1063-1068; Ndhlovu, L.C., et al., J.Immunol. 167(2001)2991-2999; Ohshima, Y., et al., J.Immunol. 159(1997)3838-3848; Rogers, P.R., et al., Immunity 15(2001)445-455; Stueber, E., and Strober, W., J. Exp.Med. 183(1996)979-989; Stueber, E., et al., Gastroenterology 115(1998)1205-1215; Takahashi, Y., et al., J.Virol. 75(2001)6748-6757; Takasawa, N., et al., Jpn. J.Cancer Res. 92(2001)377-382; Taylor, L., and Schwarz, H., J.Immunol. Meth. 255(2001)67-72; Weinberg, A.D., et al., Nature Medicine 2(1996) 183-189; Weinberg, A.D., et al., Semin. Immunol. 10(1998) 471-480; Weinberg, A.D., Trends Immunol. 23(2002) 102-109; Wu, T., et al., Transplant. Proc. 33(2001) 217-218; Higgins, L.M., et al., J. Immunol. 162(1999) 486-493; and Yoshioka, T., et al., Eur. J. Immunol. 30(2000) 2815-2823)に記載されている。ヒトOX40Lは、活性化されたCD4+T細胞上に一過性に発現されるヒトOX40(CD134)に対するリガンドである。そのリガンドによるOX40のエンゲージメントは、T細胞活性化のための共刺激性シグナルをもたらす。OX40/OX40L相互作用は、両方向性シグナルを創り出すことが記載されている(Matsumura, Y., et al., J. Immunol. 163(1999) 3007-3011; Kotani, A., et al., Immunol. Lett. 84(2002) 1-7)。さらにOX40/OX40L相互作用は、炎症性組織における内皮細胞への活性化されたT細胞の付着を媒介する。OX40Lは、活性化されたB細胞、DCおよび内皮細胞上に一過性にのみ発現されるので、OX40Lに対する抗体は炎症性応答期間中T細胞活性化および内皮細胞付着を選択的にブロッキングするであろうが、不活性化されていない末梢T細胞を変化させない。Yoshioka, A等(Eur. J. Immunol. 30(2000)2815-2823)は、慢性関節リウマチのマウスモデルにおける中和性抗mOX40L mAbの治療的潜在力を証明した。その投与は、疾患の重度を劇的に軽減した。この抗体は、他の関連した疾患モデル、例えば、炎症性皮膚疾患、実験的自己免疫疾患(EAE)、GVHD、尿炎症性腸疾患における同様な活性を示した(Yoshioka, A., et al., Eur. J. Immunol 30(1999) 2815-2823; Salek-Ardakani, S., et al., J.Exp.Med. 198(2003) 315-324; Burgess, J.K., et al., J. Allergy Clin. Immunol. 113(2004) 683-689; Hoshino, A., et al., Eur. J. Immunol. 33(2003) 861-869; Arestides, R.S., et al., Eur. J. Immunol. 32(2002) 2874-2880; Nohara, C., et al., J. Immunol. 166(2001) 2108-2115; Weinberg, A. D., et al., J. Immunol. 162(1999)1818-1826; Higgins, L.M., et al., J. Immunol. 162(1999) 486-493 ; Humphreys, I.R., et al., J.Exp.Med. 198(2003) 1237-1242; Akiba, H. et al., J.Exp. Med. 191(2003) 375-380; Ishii, N., et al., Eur. J. Immunol. 33(2003) 2372-2381; Blazar, B. R., et al., Blood 101(2003) 3741-3748; Tsukada, N., et al., Blood 95(2000) 2434-2439; Akiba, H., et al., Biochem. Biophys. Res. Commun. 251(1998) 131-136。
【0003】
OX40Lに対する抗体は種々の疾患モデルにおけるそれらの抗炎症性効果について調べられた(Sugamura, K., et al., Nat. Rev. Immunol;. 4(2004)420-431)。
【0004】
Tanaka, Y., et al, Int. J. Cancer 36, (1985)549-555; Tozawa, H., et al., Int. J. Cancer 41(1998)231-238; and Miura, S., et al., Mol. Cell. Biol. 11(1991) 1313-1325は、ヒトT細胞白血病ウイルスI型(HTLV−I)を有するヒトリンパ球の系統の表面抗原と反応するTARM−34およびTAG−34と名づけられたマウスモノクローナル抗体を記載している。TAG−34抗体は、MBL International Corporationから市販されている。TAG−34はOX40Lにも結合する。
【0005】
本発明の要約
本発明は、抗体、好ましくはモノクローナル抗体であって、該抗体は、OX40Lに結合し、ヒト起源のFc部分を含有しそしてヒト補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合しないことを特徴とする抗体、好ましくはモノクローナル抗体に関する。
【0006】
本発明は、更に、抗体、好ましくはモノクローナル抗体であって、該抗体は、ヒト起源のFc部分を含有し、100ngの抗体濃度でOX40Lに結合しそして変性されたOX40Lに結合する(ウエスタンブロットにおいて)ことを特徴とする抗体、好ましくはモノクローナル抗体に関する。この抗体は、モノクローナル抗体LC.001が結合するエピトープと同じOX40Lポリペプチドエピトープに結合する。このような抗体は、例えば、LC.001、LC.033およびLC.060である。これらの抗体は、好ましくはヒトIgG1型(野生型)であるか、またはヒト補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合しない。
【0007】
本発明は、さらに可変軽鎖および可変重鎖を含むことを特徴とし、該可変重鎖はCDR1、CDR2およびCDR3を含むことを特徴とし、該CDR3は配列番号33〜38から選ばれることを特徴とするOX40Lに結合する抗体に関する。CDR1が配列番号21〜25から選ばれ、CDR2が配列番号26〜32から選ばれそしてCDR3が配列番号33〜38から選ばれることは特に好ましい。
【0008】
本発明に従う抗体は、好ましくは、可変軽鎖および可変重鎖を含むことを特徴とし、該可変軽鎖はCDR1、CDR2およびCDR3を含むことを特徴とし、該CDR3は配列番号51〜57から選ばれることを特徴とする。CDR1が配列番号39〜44から選ばれ、CDR2が配列番号45〜50から選ばれそしてCDR3が配列番号51〜57から選ばれることが特に好ましい。
【0009】
本発明に従う抗体は、好ましくは、可変重鎖および可変軽鎖を含むことを特徴とし、該可変重鎖はCDR1、CDR2およびCDR3を含むことを特徴とし、重鎖のCDR3は配列番号33〜38から選ばれそして軽鎖のCDR3は配列番号51〜57から選ばれることを特徴とする。可変重鎖は、配列番号21〜25から選ばれるCDR1、配列番号26〜32から選ばれるCDR2および配列番号33〜38から選ばれるCDR3を含み、そして可変軽鎖は、配列番号39〜44から選ばれるCDR1、配列番号45〜50から選ばれるCDR2および配列番号51〜57から選ばれるCDR3を含むことが特に好ましい。
【0010】
すべてのCDRsは互いに独立に、しかし当然のこととして、抗体がOX40Lに結合するような方法で選ばれる。したがって、同じLC抗体の軽鎖及び重鎖のCDRsを組み合わせることができ、またはLC.001の軽鎖CDRsはLC.001、LC.059またはLC.063の重鎖CDRsと組み合わせることができる。各鎖上のCDRsはフレームワークアミノ酸により分離されている。
【0011】
本発明に従う抗体は、好ましくは、抗体が、
a)配列番号1のアミノ酸配列の軽鎖(VL)可変CDRsおよび配列番号2の重鎖(VH)可変CDRs、
b)配列番号3のアミノ酸配列の軽鎖可変CDRsおよび配列番号4の重鎖可変CDRs、
c)配列番号5のアミノ酸配列の軽鎖可変CDRsおよび配列番号6の重鎖可変CDRs、
d)配列番号7のアミノ酸配列の軽鎖可変CDRsおよび配列番号8の重鎖可変CDRs、
e)配列番号9のアミノ酸配列の軽鎖可変CDRsおよび配列番号10の重鎖可変CDRs、
f)配列番号11または16のアミノ酸配列の軽鎖可変CDRsおよび配列番号12の重鎖可変CDRs、
g)配列番号1のアミノ酸配列により規定された軽鎖(VL)可変ドメインおよび配列番号17により規定された重鎖(VH)可変ドメイン、
h)配列番号18のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号19により規定された重鎖可変ドメイン、
i)配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号20により規定された重鎖可変ドメイン、または
そのOX40Lに結合性のフラグメント
からなる群より独立に選ばれるCDRsを含むことを特徴とする。
【0012】
本発明に従う抗体は、好ましくは、該抗体が、
a)配列番号1のアミノ酸配列により規定された軽鎖(VL)可変ドメインおよび配列番号2により規定された重鎖(VH)可変ドメイン、
b)配列番号3のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号4により規定された重鎖可変ドメイン、
c)配列番号5のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号6により規定された重鎖可変ドメイン、
d)配列番号7のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号8により規定された重鎖可変ドメイン、
e)配列番号9のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号10により規定された重鎖可変ドメイン、
f)配列番号11または16のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号12により規定された重鎖可変ドメイン、
g)配列番号1のアミノ酸配列により規定された軽鎖(VL)可変ドメインおよび配列番号17により規定された重鎖(VH)可変ドメイン、
h)配列番号18のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号19により規定された重鎖可変ドメイン、
i)配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号20により規定された重鎖可変ドメイン、
またはそのOX40Lに結合性のフラグメント、
からなる群より独立に選ばれる可変領域を含むことを特徴とする。
【0013】
本発明に従う抗体は、好ましくは、ヒト軽鎖可変領域が、配列番号1、3、5、7、9、11,16および18からなる群より独立に選ばれるアミノ酸配列を含むことを特徴とする。
【0014】
本発明に従う抗体は、好ましくは、ヒト重鎖可変領域が、配列番号2、4、6、8、10、12,17、19および20からなる群より独立に選ばれるアミノ酸配列を含むことを特徴とする。
【0015】
重鎖および軽鎖のCDR領域は配列番号21〜38および39〜57に示される。
【0016】
本発明に従う抗体は、好ましくは、該抗体が配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号2、17または20により規定された重鎖可変ドメインを含むことを特徴とする。
【0017】
本発明に従う抗体は、好ましくは、ヒト重鎖定常領域が配列番号14および15からなる群より独立に選ばれるアミノ酸配列または配列番号58の重鎖定常領域を含むことを特徴とする。
【0018】
本発明に従う抗体は、好ましくは、該抗体が配列番号13のκ軽鎖定常領域または配列番号61、65もしくは69の軽鎖定常領域を含むことを特徴とする。
【0019】
好ましくは、本発明に従う抗体は、OX40Lに結合することおよびヒトIgG1クラス(野生型)であることにより特徴付けられそしてγ重鎖として配列番号58、62または66を含む。特に好ましいのは、
a)γ重鎖として配列番号58およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号62およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号66およびκ軽鎖として配列番号69、
を含む抗体である。
【0020】
本発明の更なる態様は、OX40Lに結合する抗体であって、細胞系hu−Mab<hOX40L>LC.001、hu−Mab<hOX40L>LC.005、hu−Mab<hOX40L>LC.010、hu−Mab<hOX40L>LC.019、hu−Mab<hOX40L>LC.029、またはhu−Mab<hOX40L>LC.033により産生されることを特徴とする抗体である。
【0021】
本発明な従う抗体は、好ましくはキメラ抗体、ヒト抗体またはヒト化抗体である。
【0022】
本発明に従う抗体は、BIAcoreアッセイにおいて10-8M未満(10-12〜10-8M)のKD値でOX40Lに結合することにより特徴付けられ、更に好ましくは、BIAcoreアッセイにおいて10-12〜10-9MのKD範囲により特徴付けられる。
【0023】
本発明に従う抗体は、好ましくは、4nM以下のIC50値で0.5μg/mlのコーティング濃度で固定化されたOX40L(好ましくはストレプトアビジン表面に固定化されたビオチニル化OX40L)を使用するELISAにおいてOX40LのOX40との相互作用を阻害する。さらに好ましくは、IC50値は1〜4nMの範囲にある。
【0024】
本発明に従う抗体は、好ましくは、補体因子C1qへの抗体の非結合(non-binding)は、C1qに対する10μg/mlの濃度での抗体の最大結合(Bmax)が抗体LC.001のBmaxに比較して30%以下、好ましくは20%以下であるELISAアッセイ測定値を指すことを特徴とする。
【0025】
好ましくは、抗体は、ヒトFcγRI、FcγRIIAおよび/またはFcγRIIIAに結合しない。特に好ましくは、抗体はNKエフェクター細胞上のヒトFcγレセプターに結合しない。
【0026】
本発明に従う抗体は、好ましくは、NK細胞上のFcγレセプターへの抗体の非結合は、NK細胞に対する20μg/mlの濃度での抗体の最大結合(Bmax)が抗体LC.001のBmaxに比較して20%以下、好ましくは10%以下である、アッセイを指すことを特徴とする。
【0027】
本発明に従う抗体は、好ましくは、それがFcγRIに結合しないことを特徴とする。これは、FcγRIIAおよびFcγIIBを欠いているがリコンビナントFcγRIを発現しているB細胞リンパ腫細胞に対する0.078〜10μg/mlの範囲の濃度の抗体の結合を試験するアッセイで測定されるとき、LC.001のEC50値に比べて5倍以上、好ましくは7倍以上、例えば8倍以上であるEC50により抗体が特徴付けられることを意味する。
【0028】
本発明に従う抗体は、好ましくは、補体因子C1qへの非結合および/またはNK細胞上のヒトFcγレセプターへの非結合を引き起こす、好ましくはヒトFc部分における少なくとも1つのアミノ酸突然変異を含むIgG4抗体またはIgG1抗体であるとして特徴付けられる。
【0029】
本発明に従う抗体は、好ましくは、それが補体因子C3を活性化しないことを特徴とする。
【0030】
本発明に従う抗体は、好ましくは、ヒトサブクラスIgG4であることにより特徴付けられる。本発明のさらなる好ましい態様では、抗体は、E233、L234、L235、G236、D270、N297、E318、K320、K322、A327、A330、P331および/またはP329(EUインデックスに従う番号付け)における少なくとも1つの突然変異を含む、任意のIgGクラス、好ましくはIgG1またはIgG4であることにより特徴付けられる。特に好ましいのは、IgG1突然変異PVA236、L234A/L235Aおよび/またはGLPSS331ならびにIgG4突然変異L235Eである。IgG4サブクラスの抗体が、突然変異S228Pまたは突然変異S228PおよびL235Eを含有することが更に好ましい(Angal, S., et al., Mol. Immunol. 30(1993)105−108)。
【0031】
したがって、本発明に従う抗体は、好ましくは、PVA236、GLPSS331および/またはL234A/L235A(EUインデックスに従う番号付け)からの1つ以上の突然変異を含む、ヒトサブクラスIgG1の抗体である。
【0032】
好ましくは、本発明に従う抗体は、OX40Lに結合すること、突然変異L234A/L235Aを含有するIgG1クラスであることにより特徴付けられ、そしてγ重鎖として配列番号59、63または67を含む。
【0033】
特に好ましいのは、
a)γ重鎖として配列番号59およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号63およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号67およびκ軽鎖として配列番号69、
を含む抗体である。
【0034】
好ましくは、突然変異S228Pを含有するIgG4クラスであることにより特徴付けられる本発明に従う抗体は、γ重鎖として配列番号60、64または68を含む。
【0035】
特に好ましいものは、
a)γ重鎖として配列番号60およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号64およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号68およびκ軽鎖として配列番号69、
を含む抗体である。
【0036】
本発明に従う抗体は、好ましくは、それが補体依存性細胞傷害(CDC)を誘発しないことを特徴とする。
【0037】
本発明に従う抗体は、好ましくは、それが抗体依存性細胞性細胞傷害(ADCC)を誘発しないことを特徴とする。
【0038】
したがって、本発明は、抗OX40L抗体または単一重鎖もしくは単一軽鎖であって、それらのCDRs、可変領域、完全アミノ酸配列またはハイブリドーマにより特徴付けられ、そしてFc部分を含まないか、または任意の型のFc部分を含み、好ましくは、ヒト起源の改変されていないまたは上記した突然変異により改変されている、ヒトIgG1 FcもしくはヒトIgG4 Fcを含む、抗OX40L抗体または単一重鎖もしくは単一軽鎖を含む。
【0039】
したがって、本発明は、抗体、好ましくはモノクローナル抗体であって、該抗体は、OX40Lに結合し、ヒト由来のFc部分を含有しそしてヒト補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合せず、ヒトIgG4型であるかまたは上記した突然変異により改変されたヒトIgG1もしくはヒトIgG4であることを特徴とする抗体、好ましくはモノクローナル抗体も含む。
【0040】
従って、本発明は、抗体、好ましくはモノクローナル抗体であって、該抗体は、100ngの抗体濃度で、OX40Lにおよび変性されたOX40Lに結合する(ウエスタンブロットにおいて)ことを特徴とする抗体、好ましくはモノクローナル抗体も含む。この抗体は、モノクローナル抗体LC.001が結合するエピトープと同じOX40Lポリペプチドエピトープに結合する。抗体は、Fc部分含まないか、または任意の型のFc部分を含み、好ましくは、野生型もしくは上記した突然変異により修飾されたヒトIgG1もしくはヒトIgG4を含む。
【0041】
本発明に従う抗体は、OX40Lに対する抗体による治療を必要としている患者、特に炎症性疾患、特に慢性関節リウマチ、アレルギー性喘息および移植におけるGvHD(Sugamura, K., et al., Nat. Rev. Immunol. 4(2004)420-431も参照)に罹患している患者に有益な新規で発明的な性質を有する。
【0042】
本発明のさらなる態様は、本発明に従う抗体分子、その可変鎖またはそのCDRドメインをコードする核酸分子である。
【0043】
本発明の好ましい態様では、抗体は、Fab、F(ab’)2、または一本鎖フラグメントである。
【0044】
本発明のさらなる態様は、本発明に従う核酸分子を含むベクターである。
【0045】
本発明のさらなる態様は本発明に従うベクターを含む宿主細胞である。
【0046】
本発明のさらなる態様は、本発明に従う抗体の製造方法であって、該抗体分子の合成を可能とする条件下に本発明に従う宿主細胞を培養し、そしてこの培養物から該抗体分子を回収することを含む方法である。
【0047】
本発明のさらなる態様は、本発明に従う抗体の組成物、好ましくは医薬組成物または診断組成物である。
【0048】
本発明のさらなる態様は、本発明に従う抗体および少なくとも1種の薬学的に許容されうる賦形剤を含む医薬組成物である。
【0049】
本発明のさらなる態様は、本発明に従う治療的に有効量の抗体を患者に投与することを特徴とする治療を必要としている患者の処置のための方法である。
【0050】
本発明のさらなる態様は、治療、好ましくは炎症性疾患の処置のための、特に慢性関節リウマチ、喘息およびGvHD(宿主に対する移植片疾患)の処置および/または予防のための本発明に従う抗体の使用である。
【0051】
本発明のさらなる態様は、炎症性疾患の予防および/または処置のための、好ましくは慢性関節リウマチ、喘息およびGvHDの処置用の医薬の製造のための本発明に従う抗体の使用である。
【0052】
本発明のさらなる態様は、本発明に従う抗体、本発明に従う核酸分子、本発明に従うベクターまたは本発明に従う宿主細胞を含む診断キットである。
【0053】
発明の詳細な説明
用語「OX40L」は、TNFリガンドファミリーに属するII型膜タンパク質を指す。更なる名称は、ACT−4レセプター、CD134L、g34またはTNF4_Humanである。それは34kDaの分子量を有しそしてアクセッションナンバーP23510でSwissProtに保存されている。
【0054】
用語「OX40」は、OX40Lに結合するレセプターを指す。それはTNFレセプターファミリーに属するI型膜タンパク質である。さらなる名称は、ACT−4、OX40Lレセプター、CD134抗原、ACT35抗原、TNR4_Humanである。それは50kDaの分子量を有しそしてアクセッションナンバーP43489でSwissProtに保存されている。
【0055】
用語「抗体」は、種々の形態の抗体、好ましくは、完全抗体、抗体フラグメント、ヒト抗体、キメラ抗体、ヒト化抗体(humanized antibodies)および本発明に従う特性が保持されている限り、遺伝子工学的に作成された抗体(変異体または突然変異体抗体)を含むが、それらに限定されない種々の形態の抗体、好ましくはモノクローナル抗体を包含する。特に好ましいのは、ヒトモノクローナル抗体またはヒト化モノクローナル抗体、特にリコンビナントヒト抗体である。
【0056】
本明細書で使用された用語「モノクローナル抗体」または「モノクローナル抗体組成物」は、単一のアミノ酸組成の抗体分子の調製物を指す。
【0057】
用語「キメラ抗体」は、通常リコンビナントDNA技術により作成された、1種のソースもしくは種からの可変領域、即ち結合領域および異なるソースもしくは種に由来する定常領域の少なくとも一部を含むモノクローナル抗体を指す。マウス可変領域およびヒト定常領域を含むキメラ抗体が好ましい。本発明により包含される他の好ましい形態の「キメラ抗体」は、定常領域を元の抗体の定常領域から改変しまたは変化させて、特にC1q結合および/またはFcレセプター(FcR)結合に関して、本発明に従う性質を発生させたキメラ抗体である。このようなキメラ抗体は「クラススイッチ抗体(class-switched antibodies)」とも呼ばれる。キメラ抗体は、免疫グロブリン可変領域をコードするDNAセグメントおよび免疫グロブリン定常領域をコードするDNAセグメントを含む発現された免疫グロブリン遺伝子の産物である。キメラ抗体を産生するための方法は、慣用のリコンビナントDNAを含みそして遺伝子トランスフェクション技術は当技術分野で周知されている。例えば、Morrison, S.L., et al., Proc. Natl. Acad. Sci. USA 81(1984)6851-6855; US Patent Nos. 5,202,238 and 5,204,244参照。
【0058】
用語「ヒト化抗体」は、フレームワークまたは「相補性決定領域」(CDR)が改変されて親免疫グロブリンの特異性と比較して異なる特異性の免疫グロブリンのCDRを含む抗体を指す。好ましい態様では、マウスCDRを、ヒト抗体のフレームワーク領域に移植させて「ヒト化抗体」を調製する。例えば、Riechmann, L., et al., Nature 332(1988)323-327; and Neuberger, M.S., et al., Nature 314(1985)268-270参照。特に好ましいCDRsは、キメラおよび二官能性抗体について上記した抗原を認識する配列を示すCDRsに相当する。本発明により包含された他の形態の「ヒト化抗体」は、定常領域を元の抗体の定常領域からさらに改変もしくは変化させて、特にC1q結合および/またはFcレセプター(FcR)結合に関して、本発明に従う性質を発生させたヒト化抗体である。
【0059】
本明細書で使用された用語「ヒト抗体」は、ヒト生殖系列免疫グロブリン配列に由来する可変領域及び定常領域を有する抗体を含むことを意図する。ヒト抗体は、当技術水準で周知されている(van Dijk, M. A.., and van de Winkel, J. G., Curr. Opin. Chem. Biol. 5(2001)368-374)。ヒト抗体は、免疫感作されると、内因性免疫グロブリン産生の不存在下にヒト抗体のフルレパートリーまたは選択を生じさせることができるトランスジェニック動物(例えば、マウス)において産生させることもできる。このような生殖系列突然変異体マウスにおけるヒト生殖系列免疫グロブリン遺伝子アレーのトランスファーは、抗原チャレンジによるヒト抗体の産生をもたらすであろう(例えば、Jakobovits, A., et al., Proc. Natl. Acad. Sci. USA 90(1993)2551-2555; Jakobovits, A., et al., Nature 362(1993)255-258; Bruggemann, M., et al., Year Immunol. 7(1993)33-40参照)。ヒト抗体は、ファージディスプレーライブラリーにおいて産生させることもできる(Hoogenboom, H. R.., and Winter, G., J. Mil. Biol. 227(1992)381-388; Marks, J.D., et al., J. Mol. Biol. 222(1991)581-597)。Cole et al. and Boerner et al.,の技術も、ヒトモノクローナル抗体の産生のために利用可能である(Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77(1985); and Boerner, P., et al., J. Immunol. 147(1991)86-95)。本発明に従うキメラ抗体およびヒト化抗体について既に述べたとおり、本明細書で使用された用語「ヒト抗体」は、定常領域において改変されて特にC14q結合および/またはFcR結合に関して、例えば「クラススイッチ」、即ちFc部分の変化または突然変異(IgG1からIgG4へのおよび/またはIgG1/IgG4突然変異)により、本発明に従う性質を発生させたこのような抗体も含む。さらに、本発明は、C1qおよび/またはFcRに結合するOX40Lに対するモノクローナルヒト抗体を含む。このようなヒト抗体は、マウスOX40Lと比較してヒトOX40Lについて高い選択性により特徴付けられ(ヒトOX40Lへの結合よりもマウスOX40Lへの結合が>30倍低い)そして500nMの濃度のまでTNFαまたはCD40Lへの非特異的結合を示さない。このような抗体は、C1qおよび/またはFcRに結合しない抗体の発生のために有用である。
【0060】
本明細書で使用された用語「リコンビナントヒト抗体」は、リコンビナント手段により調製され、発現され、創生されまたは単離されるすべてのヒト抗体、例えばNSOもしくはCHO細胞などの宿主細胞からまたはヒト免疫グロブリン遺伝子用のトランスジェニック動物(例えばマウス)から単離された抗体、または宿主細胞にトランスフェクションされたリコンビナント発現ベクターを使用して発現された抗体を含むことを意図する。このようなリコンビナントヒト抗体は、再配列された形態において可変領域と定常領域を有する。本発明に従うリコンビナントヒト抗体は、in vivo体細胞超突然変異を受けている。したがって、リコンビナント抗体のVHおよびVL領域のアミノ酸配列は、ヒト生殖系列VHおよびVL配列に由来するおよび関連するが、in vivoでヒト抗体生殖系列レパートリーの範囲内に天然に存在しないことがありうる配列である。
【0061】
本明細書で使用された「可変領域」(軽鎖の可変領域(VL)、重鎖の可変領域(VH))は、抗体を抗原に結合させることに直接関与する軽鎖および重鎖のペアーの各々を示す。可変ヒト軽鎖および重鎖のドメインは、同じ一般的構造を有しそして各ドメインは、3つの「超可変領域」(または相補性決定領域、CDRs)により接続された、その配列が広く保存されている4つのフレームワーク(FR)領域を含む。フレームワーク領域は、βシートコンフォメーションを採用しておりそしてCDRsはβシート構造を接続するループを形成することができる。各鎖におけるCDRsは、フレームワーク領域によりそれらの三次元構造に保持されそして他の鎖からのCDRsと共に抗原結合部位を形成する。抗体重鎖および軽鎖CDR3領域は、本発明に従う抗体の結合特異性/アフィニティーにおいて特に重要な役割を演じ、したがって本発明のさらなる目的を与える。
【0062】
本明細書で使用されるとき用語「超可変領域」または「抗体の抗原結合部分」は、抗原結合の責任を担う抗体のアミノ酸残基を指す。超可変領域は、「相補性決定領域」または「CDRs」からのアミノ酸残基を含む。「フレームワーク」または「FR」領域は、本明細書で定義されたとおりの超可変領域残基以外のこれらの可変ドメイン領域である。したがって、抗体の軽鎖および重鎖は、N末端からC末端までドメインFR1、CDR1、FR2、CDR2、FR3、CDR3およびFR4を含む。各鎖上のCDRsは、このようなフレームワークアミノ酸により分離されている。特に、重鎖のCDR3は、抗原結合に最も寄与する領域である。CDRおよびFR領域は、Kabat等、Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD(1991)の標準定義に従って決定される。
【0063】
本明細書で使用された用語「核酸または核酸分子」は、DNA分子およびRNA分子を含むことを意図する。核酸分子は一本鎖または二本鎖であることができるが、好ましくは二本鎖DNAである。
【0064】
核酸は、それが他の核酸配列との機能的関係に置かれているとき「作用可能に連結されている(operably linked)」。例えば、プレ配列又は分泌リーダーのためのDNAは、もしもそれがポリペプチドの分泌に関与するプレタンパク質として発現されるならば、ポリペプチドのためのDNAに作用可能に連結されており;プロモーターまたはエンハンサーはもしもそれが配列の転写に影響を与えるならば、コード化配列に作用可能に連結されており;またはリボソーム結合部位は、もしそれが翻訳を促進するように位置しているならば、コード化配列に作用可能に連結されている。一般に、「作用可能に連結された」とは、連結されるDNA配列が連続的であり(contiguous)そして分泌リーダーの場合には、連続的でありそしてリーディングフレームにあることを意味する。しかしながら、エンハンサーは、連続的である必要はない。連結は都合の良い制限部位においてライゲーションにより達成される。このような部位が存在しないならば、合成オリゴヌクレオチドアダプターまたはリンカーが慣用の実施に従って使用される。
【0065】
本明細書に使用された、「細胞」、「細胞系」および「細胞培養物」という表現は、交換可能に使用されそしてすべてのこのような命名は子孫を含む。従って、語「トランスフォーマント」および「トランスフォーメーションされた細胞」は、トランスファーの数に関係なく初代対象細胞(primary subject cell)およびそれに由来する培養物を含む。すべての子孫は、意図的な突然変異または故意でない突然変異により、DNA内容において正確に同一でなくてもよいことも理解される。最初にトランスフォーメーションされた細胞においてスクリーニングされたのと同じ機能または生物学的活性を有する変異体子孫は含まれる。別の命名が意図される場合には、それは状況から明らかであろう。
【0066】
「定常ドメイン」は、抗体を抗原に結合することに直接関与していないが、種々のエフェクター機能を示す。抗体または免疫グロブリンは、それらの重鎖の定常領域のアミノ酸配列に依存して、クラス:IgA、IgD、IgE、IgGおよびIgMに分けられ、そしてこれらのいくつかはサブクラス(アイソタイプ)、例えばIgG1、IgG2、IgG3およびIgG4、IgA1およびIgA2に更に分けられうる。免疫グロブリンの異なるクラスに対応する重鎖定常領域は、それぞれα、ε、γおよびμと呼ばれる。本発明に従う抗体は好ましくはIgG型である。
【0067】
本発明に従う抗体は、Fc部分として、好ましくはヒト起源に由来するFc部分および好ましくはヒト定常領域のすべての他の部分を含有する。抗体のFc部分は、補体活性化、C1q結合、C3活性化およびFcレセプター結合に直接関与している。補体系に対する抗体の影響はある条件に依存するが、C1qへの結合はFc部分における規定された結合部位により引き起こされる。このような結合部位は、当技術分野の技術水準で知られておりそして例えば、Lukas, T.J., et al., J. Immunol. 127(1981)2555-2560; Brunhouse, R.., and Cebra, J. J., Mol. Immunol. 16(1979)907-917; Burton, D.R., et al., Nature 288(1980)338-344; Thommesen, J. E., et al., Mol. Immunol. 37(2000)995-1004; Idusogie, E.E., et al., J. Immunol. 264(2000)4178-4184; Hezareh, M.., et al., J. Virol. 75(2001)12161-12168; Morgan, A., et al., Immunology 86(1995)319-324; およびEP 0 307 434により記載されている。このような結合部位は、例えばL234、L235、D270、N297、E318、K320、K322、P331およびP329(KabatのEUインデックスに従う番号付け、下記参照)である。サブクラスIgG1、IgG2およびIgG3の抗体は、通常補体活性化、C1q結合およびC3活性化を示すが、これに対してIgG4は補体系を活性化せず、C1qに結合せずそしてC3を活性化しない。本明細書で使用された用語「ヒト起源由来のそしてヒト補体因子C1qおよび/またはNK細胞上のヒトFc□レセプターに結合しないFc部分」は、サブクラスIgG4のヒト抗体のFc部分、または下記に定義されたC1q結合、C3活性化および/またはFcR結合が検出できないような方法で改変されているサブクラスIgG1、IgG2およびIgG3のヒト抗体のFc部分である、Fc部分を示す。「抗体のFc部分」は、当業者に周知のそして抗体のパパイン開裂に基づいて定義された用語である。好ましくは、Fc部分はヒトFc部分でありそしてヒトIgG4サブクラスからの、好ましくはヒンジ領域において突然変異した(例えば、S228Pおよび/またはL235E)Fc部分またはヒトIgG1サブクラスからの突然変異したFc部分である。最も好ましいのは、配列番号14および15に示された領域あるいは配列番号58,59、60、突然変異L234AおよびL235Aを有する配列番号14または突然変異S228Pもしくは突然変異S228PおよびL235Eを有する配列番号15に含まれる領域から選ばれる重鎖定常領域を含むFc部分である。
【0068】
本発明は、OX40Lに結合しそして補体因子C1qおよび/またはFcレセプターに結合しない抗体を指す。本発明の好ましい態様では、これらの抗体は、補体依存性細胞傷害(CDC)および/または抗体依存性細胞性細胞傷害(ADCC)を誘発しない。好ましくは、この抗体は、それがOX40Lに結合し、ヒト起源に由来するFc部分を含有しそして補体因子C1qに結合しないことを特徴とする。さらに好ましくは、この抗体はヒトまたはヒト化モノクローナル抗体である。
【0069】
抗体Fc領域のFc部分により媒介されるエフェクター機能は、抗体の抗原への結合の後に作動するエフェクター機能を指す(これらの機能は、Fcレセプターによる補体カスケードの活性化および/または細胞活性化を含む)。
【0070】
補体カスケードの機能は、CH50アッセイにより評価されうる。抗赤血球抗体(EA)で感作されたヒツジ赤血球を試験血清に加えて溶血をもたらす古典的経路を活性化する。赤血球の50%を溶解するのに必要な血清の容積は、CH50単位を決定する。AP−CH50は、代替経路(alternative pathway)およびターミナル経路(terminal pathway)を測定する。手順はウサギ赤血球を使用することを除いては同様である。代替経路は試験血清を添加すると活性化される。
【0071】
C1qおよび2つのセリンプロテアーゼ、C1rおよびClsは、複合体C1、補体依存性細胞傷害(CDC)経路の第1成分を形成する。補体カスケードを活性化するために、C1qは、抗原性標的に結合されたIgG1の少なくとも2つの分子またはIgMの1つの分子に結合する(Ward, E. S.., and Ghetie, V., Ther. Immunol. 2(1995)77-94)。Burton, D. R.,は、アミノ酸残基318〜337を含む重鎖領域が、補体結合反応(complement fixation)に関与していると述べている(Mol. Immunol. 22(1985)161-206)。Duncan, A. R., およびWinter, G. (Nature 332(1988)738-740)は、部位特異的突然変異誘発を使用して、Glu318、Lys320およびLys322がC1qへの結合部位を形成することを報告した。C1qの結合におけるGlu318、Lys320およびLys322残基の役割は、これらの残基を含有する短い合成ペプチドの補体媒介溶解を阻害する能力により確かめられた。
【0072】
用語「補体依存性細胞傷害(CDC)」は、補体の存在下に本発明に従う抗体による、OX40L発現ヒト内皮細胞の溶解を指す。CDCは、好ましくは、補体の存在下に本発明に従う抗体によるOX40L発現ヒト内皮細胞の処理により測定される。細胞は、好ましくはカルセインで標識される。もしも抗体が30μg/mlの濃度で標的細胞の20%以上の溶解を誘導するならば、CDCが見出される。本発明者は、本発明に従う抗体の性質について、ELISAアッセイにおける補体因子C1qへの減少した結合が必須であることを見出した。このようなアッセイにおいて、一般に、ELISAプレートをある濃度範囲の抗体でコーティングし、それに精製されたヒトC1qまたはヒト血清を加える。C1q結合は、C1qに対して指向された抗体により、続いてペルオキシダーゼで標識されたコンジュケートにより検出される。結合の検出(最大結合Bmax)は、ペルオキシダーゼ基質、ABTS(登録商標)(2,2’−アジノ−ジ−[3−エチルベンゾチアゾリン−6−スルホネート(6)]について405nmにおける光学濃度(OD405)として測定される。したがって、本発明は、補体因子C1qへの抗体の非結合は、10μg/mlの抗体の濃度での本発明に従う抗体へのC1qの最大結合(Bmax)が抗体LC.001で観察されたBmaxの20%以下、好ましくは10%以下である、このようなELISAアッセイ測定値を指す。
【0073】
本発明に従う抗体は、ELISAアッセイにおける補体因子C3の減少した活性化を示すことが更に好ましい。このアッセイはC1qアッセイと同じ方式で行われる。このようなアッセイにおいて、一般にELISAプレートを所定の濃度範囲の抗体でコーティングし、それにヒト血清を加える。C3結合は、C3に対して指向された抗体により、次いでペルオキシダーゼで標識されたコンジュゲートにより検出される。結合(最大結合Bmax)の検出は、ペルオキシダーゼ基質ABTS(登録商標)について405nmにおける光学濃度(OD405)として測定される。したがって、本発明は、補体因子C3への抗体の非結合は、10μg/mlの抗体の濃度で抗体へのC3の最大結合(Bmax)が抗体LC.001のBmaxの10%以下、好ましくは5%以下である、このようなELISAアッセイ測定値を指す。
【0074】
用語「抗体依存性細胞性細胞傷害(ADCC)」は、Fcレセプター結合により媒介される機能であり、そしてエフェクター細胞の存在下に本発明に従う抗体によるOX40L発現標的細胞の溶解を指す。ADCCは、好ましくは、エフェクター細胞、例えば単離されたばかりのPBMC(末梢血単核細胞)またはバフィーコートから精製されたエフェクター細胞、例えば単球またはNK(ナチュラルキラー)細胞の存在下に、本発明に従う抗体によるOX40L発現赤血球系細胞(例えばリコンビナントヒトOX40Lを発現するK562細胞)の調製物の処理により測定される。標的細胞を、51Crで標識し、次いで抗体と共にインキュベーションする。標識された細胞をエフェクター細胞と共にインキュベーションし、そして上清を放出された51Crについて分析する。コントロールは、抗体なしで標的内皮細胞のエフェクター細胞とのインキュベーションを含む。ADCCを媒介する最初の段階を誘導する抗体の能力を、Fcγレセプター発現細胞、例えばFcγRIおよび/またはFcγRIIAをリコンビナント的に発現する細胞またはNK細胞(本質的にFcγRIIIAを発現する)へのそれらの結合を測定することにより調べた。好ましくはNK細胞上のFcγRへの結合を測定する。
【0075】
Fcレセプター結合性エフェクター機能は、造血細胞上の特殊化された細胞表面レセプターであるFcレセプター(FcRs)と抗体のFc領域の相互作用により媒介されうる。Fcレセプターは免疫グロブリンスーパーファミリーに属し、そして抗体依存性細胞性細胞傷害(ADCC)を介して、免疫複合体のファゴサイトーシスによる抗体でコーティングされた病原体の除去ならびに、対応する抗体でコーティングされた赤血球および種々の他の細胞標的(例えば腫瘍細胞)の溶解の両方を媒介することが示された。Van de Winkel, J. C., and Anderson, C. L., J. Leukoc. Biol. 49(1991)511-524)。FcRsは、免疫グロブリンアイソタイプに対するそれらの特異性により定義され;IgG抗体に対するFcレセプターは、FcγRと呼ばれ、IgEに対してはFcεRと呼ばれ、IgAに対してはFcαRと呼ばれる等である。Fcレセプター結合は、例えば、Ravetch, J. V., and Kinet, J. P., Annu. Rev. Immunol. 9(1991)457-492; Capel, P. J., et al., Immunomethods 4(1994)25-34; de Haas, M., et al., J. Lab. Clin. Med. 126(995)330-341; and Gessner, J. E., et al., Ann. Hematol. 76(1998)231-248に記載されている。
【0076】
IgG抗体のFcドメインに対するレセプター(FcγR)の架橋は、ファゴサイトーシス、抗体依存性細胞性細胞傷害および炎症性メディエーターの放出、ならびに免疫複合体クリアランスおよび抗体産生の調節を含む広範な種類のエフェクター機能をトリガーする。ヒトでは、3つのクラスのFcγRが特徴付けられており、それらは、FcγRI(CD64)、FcγRII(CD32)およびFcγRIII(CD16)であり、
−FcγRI(CD64)は、高いアフィニティーで一量体IgGに結合し、そしてマクロファージ、単球、好中球および好酸球上に発現される。E233〜G236、P238、D265、N297、A327およびP329の少なくとも1つのIgGにおける改変は、FcγRIへの結合を減少させる。IgG1およびIgG4中に置換された位置233〜236におけるIgG2残基はFcγRIへの結合を103倍減少させそして抗体感作された赤血球に対するヒト単球応答を排除した(Armour, K.L., et al., Eur. J. Immunol. 29(1999)2613-2624)。
−FcγRII(CD32)は、複合体化されたIgGに中〜低アフィニティーで結合しそして広く発現される。これらのレセプターは2つの重要なタイプ、FcγRIIAおよびFcγRIIBに分けることができる。FcγRIIAは、キリング(killing)に関与する多くの細胞(例えばマクロファージ、単球、好中球)上に見出されそしてキリングプロセスを活性化することができると思われる。FcγRIIBは、阻害プロセスにおいて役割を演じると思われ、そしてB細胞、マクロファージおよびマスト細胞および好酸球上に見出される。B細胞上では、それはさらなる免疫グロブリン産生および例えばIgEクラスへのアイソタイプスイッチを抑制するように機能すると思われる。マクロファージ上では、FcγRIIBは、FcγRIIAにより媒介されるファゴサイトーシスを抑制するように作用する。好酸球およびマスト細胞上では、該b形態は、その別のレセプターへのIgE結合を通じてこれらの細胞の活性化を阻害するのを助けることができる。FcγRIIAに対する減少した結合は、例えば、E233〜G236、P238、D265、N297、A327、P329、D270、Q295、A327、R292およびK414の少なくとも1つのIgG突然変異で見出される。
−FcγRIII(CD16)は、IgGに中〜低アフィニティーで結合しそして2つのタイプとして存在する。FcγRIIIAは、NK細胞、マクロファージ、好酸球およびいくらかの単球およびT細胞上に見出され、そしてADCCを媒介する。FcγRIIIBは、好中球上に高度に発現される。FcγRIIIAへの減少した結合は、例えば、E233〜G236、P238、D265、N297、A327、P329、D270、Q295、A327、S239、E269、E293、Y296、V303、A327、K338およびD376の少なくとも1つの突然変異で、見出される。
【0077】
Fcレセプターに対するヒトIgG1上の結合部位のマッピング、上記した突然変異部位およびFcγRIおよびFcγRIIAへの結合を測定するための方法は、Shields, R.L., et al. JBC276(2001)6591-6604に記載されている。
【0078】
本明細書で使用される用語「Fcレセプター」は、該レセプターと会合した(associated)細胞質ITAM配列の存在により特徴付けられた活性化レセプターを指す(例えば、Ravetch, J. V.., and Bolland, S., Annu. Rev. Immunol. 19(2001)275-290)。このようなレセプターは、FcγRI、FcγRIIAおよびFcγRIIIAである。本発明に従う抗体は、好ましくは、Fcγレセプター、好ましくはFcγIIIAへの減少した結合を示す。好ましくは、用語「FcγRの非結合」は、10μg/mlの抗体濃度においてNK細胞への本発明に従う抗体の結合が抗体LC.001について見出された結合の10%以下であることを意味する。
【0079】
IgG4は減少したFcR結合を示すが、他のIgGサブクラスの抗体は強い結合を示す。しかしながら、Pro238、Asp265、Asp270、Asn297(Fc炭水化物の損失)、Pro329および234、235、236および237 Ile253、Ser254、Lys288、Thr307、Gln311、Asn434およびHis435も、もし変えられるならば減少したFcR結合を与える残基である(Shields, R. L., et al. J. Biol. Chem. 276(2001)6591-6604; Lund, J., et al. FASEB J. 9(1995)115-119; Morgan, A., et al., Immunology 86(1995)319-324; およびEP 0 307 434)。好ましくは、IgG1またはIgG2サブクラスの本発明に従う抗体は、突然変異PVA236、GLPSS331および/またはL234A/L235Aを含む。IgG4サブクラスの本発明に従う抗体は、好ましくは突然変異L235Eを含む。さらなる好ましいIgG4突然変異はS228PまたはL235EおよびS228Pである(表1参照)。
【0080】
本明細書で使用される用語「OX40Lへの結合」は、BIAcoreアッセイにおけるヒトOX40Lへの抗体の結合を意味する(Pharmacia Biosensor AB, Uppsala, Sweden)。更なる確認のために、OX40Lへの結合は、精製されたOX40LがマイクロタイタープレートにコーティングされているELISAまたは直接もしくは間接に標識された抗体を、OX40Lを発現するK562細胞に結合させるFACSアッセイにおいて決定することもできる。
【0081】
BIAcoreアッセイにおいては、抗体を表面に結合させそしてOX40Lの結合は、表面プラズモン共鳴(SPR)により測定される。結合のアフィニティーは、用語ka(抗原への抗体の会合にたいする速度定数)、kd(解離に対する速度定数)およびKD(kd/ka)により定義される。本発明に従う抗体は、10-8以下、好ましくは約10-12〜10-9MのKDを示す(実施例参照)。したがって、本発明は、抗体がBIAcoreアッセイにおいて10-8M未満のKD値でOX40Lに結合する、好ましくは、KD範囲は10-12〜10-9Mである、上記した抗体を指す。
【0082】
OX40L特異的結合ELISAにおいて、OX40Lをマイクロタイタープレート上にコーティングし、そしてOX40Lへの抗体の結合は、HRPをコンジュゲーションした抗ヒトIgGおよびELISAの通常の段階により検出される。このアッセイにおけるEC50値は、好ましくは3nM〜8nMの範囲にある。
【0083】
本明細書で使用された「OX40LへのOX40の結合を阻害する」という用語は、ヒトOX40Lに本発明に記載の抗体を結合させ、それによりOX40/OX40L相互作用を阻害し、それによりOX40Lで誘導されるシグナル伝達を阻害することを指す。
【0084】
本発明の抗体は、hOX40L/OX40相互作用を、好ましくは、
i)1nM〜4nMの範囲のIC50値で0.5μg/mlのビオチニル化OX40Lの(固相)コーティング濃度で抗体により可溶性OX40とビオチニル化され固定化されたOX40Lとの相互作用をブロッキングすることによりELISAアッセイにより示されたin vitroレベルにおいて、
ii)1nM〜10nMの範囲のIC50値で0.78〜100nMの抗体濃度で抗体により可溶性OX40L(10nM、好ましくはhOX40L−Hisとして)と固定化されたOX40との相互作用をブロッキングすることによりBiacoreアッセイにより示されたin vitroレベルにおいて、
iii)抗体が、4〜20nMの範囲のIC50値でOX40と2×105細胞/サンプルの濃度のOX40Lを発現するK562細胞(K562_OX40L)との相互作用をブロッキングする、FACSアッセイにより示された細胞レベルにおいて、
iv)抗体が、1〜5nMの範囲のIC50でNFκB活性化のブロッキングをもたらす、サンプル当たり3×104個のOX40を発現するHeLa細胞への、K562_OX40Lにより誘導されるOX40シグナル伝達をブロッキングする、OX40シグナル伝達アッセイにより、
v)抗体が、1nM〜10nMの範囲のIC50値で1.5×105細胞/サンプルの濃度および0.75μg/mlのPHA濃度でK562_OX40LによるOX40Lで誘導されるT細胞活性化をブロッキングする、T細胞活性化アッセイにより、および/または
vi)抗体が、10μg/mlの抗体濃度で活性化されたB細胞または樹状細胞によるOX40Lで誘導されるT細胞活性化をブロッキングする、T細胞活性化アッセイ(Tetanus assay)により、阻害し、40%〜60%の阻害が得られた。
【0085】
1nM〜4nMの範囲のIC50値で0.5μg/mlOX40Lのコーティング濃度で可溶性OX40と固定化されたOX40Lとの相互作用をブロッキングすることによりELISAアッセイにおいて阻害を示す抗体が好ましい。
【0086】
したがって、本発明の更なる好ましい態様は、OX40/OX40L相互作用を阻害し、それによりOX40Lで誘導されるシグナル伝達を阻害することを特徴とする抗体を指す。
【0087】
本発明に従う抗体は、TNFαまたはCD40L 500nMの濃度までTNFαおよびCD40Lへの非特異的結合を示さないことが更に好ましい。
【0088】
本発明に従う抗体はヒトOX40Lに比較してマウスOX40Lへの少なくとも30倍低い結合を示すことが更に好ましい。
【0089】
10μg/mlの濃度の本発明に従う抗体は、HUVEC細胞上のOX40L発現のダウンレギュレーションを誘導しないことが更に好ましい。
【0090】
さらなる好ましい態様では、本発明の抗体は、それらが組合せ、
a)配列番号1のアミノ酸配列により規定された抗体LC.001の軽鎖可変ドメインおよび配列番号2により規定された抗体LC.001の重鎖可変ドメイン;
b)配列番号3のアミノ酸配列により規定された抗体LC.005の軽鎖可変ドメインおよび配列番号4により規定された抗体LC.005の重鎖可変ドメイン;
c)配列番号5のアミノ酸配列により規定された抗体LC.010の軽鎖可変ドメインおよび配列番号6により規定された抗体LC.010の重鎖可変ドメイン;
d)配列番号7のアミノ酸配列により規定された抗体LC.029の軽鎖可変ドメインおよび配列番号8により規定された抗体LC.029の重鎖可変ドメイン;
e)配列番号9のアミノ酸配列により規定された抗体LC.019の軽鎖可変ドメインおよび配列番号10により規定された抗体LC.019の重鎖可変ドメイン;
f)配列番号11または16のアミノ酸配列により規定された抗体LC.033の軽鎖可変ドメインおよび配列番号12により規定された抗体LC.033の重鎖可変ドメイン;
g)配列番号1のアミノ酸配列により規定された軽鎖(VL)可変ドメインおよび配列番号17により規定された重鎖(VH)可変ドメイン;
h)配列番号18のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号19により規定された重鎖可変ドメイン;
i)配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号20により規定された重鎖可変ドメイン;
からなる群より独立に選ばれる可変ドメインの組合せを含むことを特徴とする。
【0091】
さらなる好ましい態様では、本発明の抗体は、それらが、
k)配列番号13の配列により規定された軽/κ鎖;
l)L234AおよびL235A、PVA236またはGLPSS331から選ばれる1つ以上の突然変異を有するIgG1アイソタイプ配列番号14の重/γ鎖;
m)IgG4アイソタイプ配列番号15の重/γ鎖;
n)突然変異S228Pまたは突然変異S228PおよびL235Eを有するIgG4アイソタイプ配列番号15の重/γ鎖;
o)配列番号61、65または69に含まれる軽定常鎖(light constant chain)、
p)配列番号58、59、60、62、63、64、66、67または68に含まれる重定常鎖(heavy constant chain)、
からなる群より独立に選ばれる定常領域を含むことを特徴とする。
【0092】
更に好ましいのは、各可変抗体ドメイン組合せa)〜i)とγ鎖l)、m)、n)またはp)および好ましくはκ鎖k)またはo)とのすべての組合せである。特に好ましいのは、抗体LC.001、LC.005、LC.010、LC.019、LC.029、LC.033、LC.059、LC.060またはLC.063の可変鎖を、各々、配列番号13の配列により規定されたκ鎖または配列番号61、65もしくは69に含まれる軽定常鎖および突然変異L234AおよびL235Aを有するIgG1アイソタイプ配列番号14の重/γ鎖または配列番号59、63もしくは67に含まれる重定常鎖と共に含む抗体;抗体LC.001、LC.005、LC.010、LC.019、LC.029、LC.033、LC.059、LC.060またはLC.063の可変鎖を、各々、配列番号13の配列により規定されたκ鎖または配列番号59、63もしくは67に含まれる重定常鎖およびIgG4アイソタイプ配列番号15の重/γ鎖または配列番号60、64もしくは68(3つのすべてがS228P突然変異を有していない)に含まれる重定常鎖と共に含む抗体;抗体LC.001、LC.005、LC.010、LC.019、LC.029、LC.033、LC.059、LC.060またはLC.063の可変鎖を、各々、配列番号13の配列により規定されたκ鎖または配列番号61、65もしくは69に含まれる軽定常鎖および突然変異S228Pを有するIgG4アイソタイプ配列番号15の重/γ鎖または配列番号60、64もしくは68に含まれる重定常鎖と共に含む抗体である。
【0093】
好ましくは、抗体は、配列番号1のアミノ酸配列の軽鎖可変CDRおよび配列番号2、17もしくは20の重鎖可変CDR、または配列番号18のアミノ酸配列の軽鎖可変CDRおよび配列番号19の重鎖可変CDRを含む。
【0094】
好ましい抗体は、該抗体が、補体因子C1qへの非結合および/またはFCR結合の損失を引き起こす少なくとも1つのアミノ酸突然変異を含むヒトIgG4サブクラスまたは他のヒトサブクラス(好ましくはIgG1)であることを特徴とする。このような好ましい変異体抗体は、例えば、突然変異L234AおよびL235Aを有する配列番号14のアミノ酸配列または突然変異S228Pを有するもしくは有さない配列番号15を含む。
【0095】
本発明に従う好ましい抗体は、IgG1v1(E233P;L234V;L235A;ΔG236;A327G;A330S;P331Sにより特定されたPVA−236;GLPSS331)、IgG1v2(L234A;L235A)およびIgG4v1(S228P;L235E)およびIgG4x(S228P)として定義された抗体である。
【0096】
本発明に従うハイブリドーマ細胞系hu−Mab<hOX40L>LC.001は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の下に、寄託番号DSM ACC2672の下に2004年7月27日にDeutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSMZ)、Germanyに寄託された。
【0097】
本発明に従うハイブリドーマ細胞系hu−Mab<hOX40L>LC.005(DSM ACC 2685)、hu−Mab<hOX40L>LC.010(DSM ACC 2686)、hu−Mab<hOX40L>LC.019、hu−Mab<hOX40L>LC.029(DSM ACC 2688)およびhu−Mab<hOX40L>LC.033(DSM ACC 2689)は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の下に、2004年9月2日にDeutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSMZ)、Germanyに寄託された。
【0098】
前記細胞系から得られうる抗体は、本発明の好ましい態様でありそして補体因子C1qに結合しないおよび/またはヒトFcγレセプターに結合しない本発明に従う抗体の発生のための中間物質として特に有用である。
【0099】
本発明の更なる好ましい態様は、OX40Lに結合し、そして寄託されたハイブリドーマ細胞系により産生されたモノクローナル抗体LC.005、LC.010またはLC.029も結合する同じOX40Lエピトープに結合する、単離された抗OX40L抗体である。
【0100】
本発明の更なる態様は、補体因子C1qおよび/またはヒトFcγレセプターに結合しない、OX40Lに対する抗体の産生方法であって、10-8M未満のKD値でOX40Lに結合する抗体の重鎖をコードする核酸の配列を、改変された抗体が補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合しないような方式で改変し、該改変された核酸および該抗体の軽鎖をコードする核酸を発現ベクターに挿入し、該ベクターを原核生物もしくは真核生物宿主細胞に挿入し、コードされたタンパク質を発現しそして宿主細胞または上清から回収することを特徴とする方法である。
【0101】
本発明のさらなる態様は、補体因子C1qに結合しないおよび/またはヒトFcγレセプターに結合しない本発明に従う抗体の産生方法であって、前記細胞系の1つから得られうる抗体を、「クラススイッチ」、即ち、好ましくは、IgG1v1(E233P;L234V;L235A;ΔG236;A327G;A330S;P331Sにより特定されたPVA−236;GLPSS331)、IgG1v2(L234A;L235A)およびIgG4v1(S228P;L235E)およびIgG4x(S228P)として定義されたFc部分の変化または突然変異(例えばIgG1からIgG4へのおよび/またはIgG1/IgG4突然変異)により改変することを特徴とする方法である。
【0102】
更なる好ましい態様では、これらの抗体は、Fab、(Fab’)2および一本鎖フラグメントからなる群より選ばれる抗体フラグメントも含む。
【0103】
したがって、「変異体」抗OX40L抗体は、本明細書では、親抗体配列における1つ以上のアミノ酸残基の付加、欠失および/または置換により「親」抗OX40L抗体アミノ酸配列とはアミノ酸配列において異なる分子を指す。好ましい態様では、変異体は、親抗体の1つ以上の定常領域もしくは可変領域における、好ましくは定常領域における1つ以上のアミノ酸置換を含む。例えば、変異体は、親抗体の1つ以上の可変領域における少なくとも1つ、例えば約1〜約10、好ましくは約2〜約5の置換を含むことができる。通常、変異体は、親抗体定常および/または可変ドメイン配列との少なくとも90%、更に好ましくは少なくとも95%、最も好ましくは少なくとも99%のアミノ酸配列同一性を有するアミノ酸配列を有するであろう。
【0104】
本発明は、配列番号21〜38からなる群より選ばれる親抗体重鎖CDRおよび/または配列番号39〜57からなる群より選ばれる抗体軽鎖CDRの最初のアミノ酸配列を改変する方法であって、該最初のアミノ酸配列をコードする核酸を提供し、該核酸を、1つのアミノ酸が重鎖CDR1において改変され、1〜2つのアミノ酸が重鎖CDR2において改変され、1〜2つのアミノ酸が重鎖CDR3において改変され、1〜3つのアミノ酸が軽鎖CDR1において改変され、1〜3つのアミノ酸が軽鎖CDR2において改変されおよび/または1〜3つのアミノ酸が軽鎖CDR3において改変されるように改変し、該改変されたCDRアミノ酸配列を抗体構造において発現し、該抗体が10-8M未満のKDでOX40Lに結合するかどうかを測定し、そしてもしも該抗体が10-8M未満のKDでOX40Lに結合するならば、該改変されたCDRを選ぶことを特徴とする方法を含む。好ましくは、このような改変は保存的配列改変である。
【0105】
配列に関する同一性又は相同性は、本明細書では、配列をアラインメントしそして必要ならばギャップを導入して、最大百分率配列同一性を達成した後に、親抗体残基と同一である候補配列におけるアミノ酸残基の百分率として定義される。抗体配列におけるN末端、C末端もしくは内部の延長、欠失または挿入のどれも配列同一性または相同性に影響を与えるものとみなすべきではない。変異体は、ヒトOX40Lに結合する能力を保持し、そして好ましくは親抗体の性質より優れた性質を有する。例えば、変異体は、慢性関節リウマチおよび喘息の処置期間中減少した副作用を有することができる。何故ならば、OX40Lは、B細胞、樹状細胞およびマクロファージ上に一過性に発現されるのみならず内皮細胞(Kotani, A., et al., Immunol. Lett. 84(2002)1-7)、気道平滑筋細胞(=ASM)(Burgess, J. K., J. Allergy Clin. Immunol 113(2004)683-689)およびミクログリア細胞(Weinberg, A.D., et al., J.Immunol. 162(1999)1818-1826)上にも一過性に発現されるからである。内皮細胞、ASMおよびミクログリア細胞へのOX40Lに対する抗体の結合は、血管漏洩をもたらす内皮細胞の細胞損傷、肺破壊をもたらすASM細胞の細胞損傷、ミクログリアの損傷をもたらすミクログリア細胞の細胞損傷をもたらすことがありうる。
【0106】
本明細書における「親」抗体は、変異体の調製のために使用されるアミノ酸配列によりコードされている親抗体である。好ましくは、親抗体は、ヒトフレームワーク領域を有し、そしてもし存在するならば、ヒト抗体定常領域(1つまたは複数)を有する。例えば親抗体は、好ましくはIgG1型のヒト化抗体またはヒト抗体であることができる。
【0107】
本発明に従う抗体は、更に、本発明に従う抗体の上記した特徴に影響を与えないまたは該特徴を変えない、「保存性配列改変」、ヌクレオチドおよびアミノ酸配列改変を有するこのような抗体を含む。改変は、部位特異的突然変異誘発およびPCR媒介突然変異誘発などの当技術分野で知られている標準技術により導入することができる。保存性アミノ酸置換は、アミノ酸残基が同様な側鎖を有するアミノ酸残基で置き換えられているアミノ酸置換を含む。同様な側鎖を有するアミノ酸残基のファミリーは、当技術分野で定義されている。これらのファミリーは、塩基性側鎖(例えばリシン、アルギニン、ヒスチジン)、酸性側鎖(例えば、アスパラギン酸、グルタミン酸)、電荷のない極性側鎖(例えば、グリシン、アスパラギン、グルタミン、セリン、トレオニン、チロシン、システイン、トリプトファン)、非極性側鎖(例えば、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン)、β−分岐状側鎖(例えば、トレオニン、バリン、イソロイシン)および芳香族側鎖(例えば、チロシン、フェニルアラニン、トリプトファン、ヒスチジン)を有するアミノ酸を含む。したがって、ヒト抗OX40L抗体における予想される非必須アミノ酸残基は、好ましくは、同じ側鎖ファミリーからの他のアミノ酸残基で置き換えることができる。
【0108】
アミノ酸置換は、Riechmann, L., et al., Nature 332(1988)323-327 and Queen, C., et al., Proc. Natl. Acad. Aci. USA 86(1989)10029-10033により記載された分子モデル化に基づく突然変異誘発により達成されうる。
【0109】
本発明は、抗体の産生方法であって、10-8M未満のKD値でOX40Lに結合する抗体の重鎖をコードする第1核酸の配列を、改変された抗体が補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合しないような方法で改変し、該改変された第1核酸および該抗体の軽鎖をコードする第2核酸を発現ベクターに挿入し、該ベクターを原核生物または真核生物宿主細胞に挿入し、該抗体の合成を可能とする条件下に該宿主細胞を培養し、そしてこの培養物から該抗体を回収することを特徴とする方法を更に含む。
【0110】
本発明は、本発明に従うそしてヒト起源に由来するFc部分を含む抗体の産生方法であって、a)本発明に従う親ヒト抗体の軽鎖をコードする第1核酸配列およびFc部分が補体因子C1qおよび/またはFcレセプターに結合しないようにFc部分が改変されている該親ヒト抗体の重鎖をコードする第2DNA配列で宿主細胞をトランスフォーメーションし;b)該第1DNA配列及び第2DNA配列を、該抗体重鎖および軽鎖が産生されるように発現しそしてc)該抗体を宿主細胞または宿主細胞培養物から回収する段階を含む方法を更に含む。
【0111】
本発明は、上記した抗体をコードする核酸分子、これらの核酸を含む対応するベクターおよびこれらのベクターのための対応する宿主細胞も含む。本発明は、抗体の製造方法であって、該抗体分子の合成を可能とする条件下に対応する宿主細胞を培養しそしてこの培養物から該抗体を回収することを含む、例えば、原核生物または真核生物宿主細胞において重鎖をコードする核酸および軽鎖をコードする核酸を発現しそしてこのポリペプチドを該細胞から回収することによる、抗体の製造方法を包含する。
【0112】
抗体の診断的使用及び治療的使用が包含される。1つの診断用途では、本発明は、OX40Lを含有する疑いのあるサンプルを抗OX40L抗体に曝露しそしてサンプルへの抗体の結合を決定することを含む、OX40Lタンパク質の存在を決定する方法を提供する。OX40Lタンパク質は、OX40L発現細胞の細胞膜中にその膜貫通ドメインにより挿入されることができ、またはシェディング(shedding)もしくはタンパク質分解放出のような機構により放出された体液中の可溶性細胞外ドメインとして存在することができる。この使用のために、本発明は、抗体を使用してOX40Lタンパク質を検出するための抗体および指示を含むキットを提供する。
【0113】
本発明の抗体は、哺乳動物、好ましくは炎症性疾患を有するまたは炎症性疾患に罹っている疑いがある患者における炎症性疾患の予防および/または処置のために有用である。このような疾患は、アレルギー反応、例えば喘息を含む。他の適応は慢性関節リウマチを含む自己免疫疾患の処置である。
【0114】
本発明は、上記した炎症性疾患、特に喘息および慢性関節リウマチに罹っている哺乳動物を処置する方法を提供する。
【0115】
好ましくは、本発明の抗体は、その症状が吸入されたコルチコステロイドで十分にコントロールされない患者における重い持続性喘息の処置のために使用することができる。患者集団は、不十分にコントロールされた重い持続性喘息を有する成人および青年(12歳以上)を含む。抗体は、1月に1回または2回好ましくは皮下に送達されるであろう。主要な最終目標は、好ましくは急性増悪の減少であろう。他の最終目標は、ピークフロー、日中の喘息症状、夜間の目覚め、クオリティーオブライフ、救急室診察、喘息のない日、β−2アゴニスト使用、ステロイド減少または漸減および過敏性(hyper-responsiveness)に対する効果を含む。
【0116】
中程度〜重度の活性慢性関節リウマチを有する成人の処置のために単独療法のためまたはメトレキセートもしくは他のDMARDs(Disease Modifying Anti-Rheumatic Drugs)との併用において本発明に従う抗体を使用することは更に好ましい。それは2または4週間毎に皮下注射として投与されるであろう。それは、1種以上のDMARDsが効かなかった患者における慢性の治療であろう。最終目標は、活性な慢性関節リウマチを有する成人患者における兆候及び症状の減少および構造的損傷の進行の抑制を含むであろう。ACR基準により測定された障害の防止、兆候及び症状の改善(ACR20>60%、ACR50>35%、ACR70>15%;米国リウマチ学会からのインデックス;www.rheumatology.com)。
【0117】
本発明の更なる態様は、これらの疾患の処置用の医薬の製造のための本発明に従う抗体の使用である。
【0118】
本発明は、医薬組成物の製造のために上記に定義した抗体の使用にも関しそして薬学的有効量で本発明に従う抗体を、場合により緩衝液および/または薬学的目的での抗体の処方のために有用な佐剤と共に含有する医薬組成物を含む。
【0119】
本発明は、薬学的に許容されうる担体中にこのような抗体を含む医薬組成物を更に提供する。1つの態様では、医薬組成物は、製造物又はキットに含ませることもできる。
【0120】
本発明に従う抗体は好ましくはリコンビナント手段により産生される。このような方法は、当技術分野の技術水準で広く知られておりそして抗体ポリペプチドのその後の単離および通常薬学的に許容されうる純度への精製を伴う原核細胞及び真核細胞におけるタンパク質発現を含む。タンパク質発現のために、軽鎖および重鎖またはそのフラグメントをコードする核酸を標準方法により発現ベクターに挿入する。発現は、CHO細胞、NSO細胞、SP2/0細胞、HEK293細胞、COS細胞、酵母またはE.coli細胞などの適切な原核生物宿主細胞または真核生物宿主細胞において行われ、そして抗体は細胞(溶解後の上清または細胞)から回収される。
【0121】
抗体のリコンビナント産生は、当技術分野の技術水準でよく知られておりそして例えばMakrides, S. C., Protein Expr. Purif. 17(1999)183-202; Geisse, S., et al., Protein Expr. Purif. 8(1996) 271-282; Kaufman, R.J., Mol. Biotechnol. 16(2000) 151-161; Werner, R.C., et al., Arzneimittelforschung 48(1998)870-880の概説論文に記載されている。
【0122】
抗体は、完全細胞中に、細胞溶解物中にまたは部分的に精製された形態もしくは実質的に純粋な形態で存在することができる。精製は、他の細胞成分または他の汚染物、例えば他の細胞核酸もしくはタンパク質を除去するために、アルカリ/SDS処理、カラムクロマトグラフィー、および当技術分野で周知の他の技術を含む標準技術により行われる。Ausubel, F., et al., ed., Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York (1987)参照。
【0123】
NS0細胞における発現は、例えば、Barnes, L. M., et al., Cytotechnology 32(2000) 109-123; and Barnes, L. M., et al.,Biotech. Bioeng. 73(2001) 261-270により記載されている。一過性発現は、例えば、Durocher, Y., et al., Nucl. Acids. Res. 30(2002)E9により記載されている。可変ドメインのクローニングは、Orlandi, R., et al., Proc. Natl. Acad. Sci. USA 86(1989)3833-3837; Carter, P., et al., Proc. Natl. Acad. Sci. USA 89(1992) 4285-4289; およびNorderhaug, L., et al., J. Immunol. Methods 204(1997)77-87により記載されている。好ましい一過性発現系(HEK293)は、Schlaeger, E-J., and Christensen, K., in Cytotechnology 30(1999) 71-83 および Schlaeger, E-J., in J. Immunol. Methods 194(1996) 191-199により記載されている。
【0124】
原核生物に適当なコントロール配列は、例えばプロモーター、場合によりオペレーター配列およびリボソーム結合部位を含む。真核細胞は、プロモーター、エンハンサーおよびポリアデニル化シグナルを利用することが知られている。
【0125】
核酸は、それが他の核酸配列と機能的に関連させられているとき、「作用可能に連結されている」。例えば、プレ配列または分泌リーダーのためのDNAは、もしもそれがポリペプチドの分泌に関与するプレタンパク質として発現されるならば、ポリペプチドのためのDNAに作用可能に連結されており;プロモーターまたはエンハンサーは、それが配列の転写に影響を与えるならばコード化配列に作用可能に連結されており;またはリボソーム結合部位は、それが翻訳を促進するように配置されているならばコード化配列に作用可能に連結されている。一般に、「作用可能に連結されている」とは、連結されているDNA配列が連続的であり(contiguous)そして分泌リーダーの場合には、連続的であり且つリーディングフレーム内にある。しかしながら、エンハンサーは、連続的である必要はない。連結は、都合の良い制限部位におけるライゲーションにより達成される。もしもこのような部位が存在しないならば、合成オリゴヌクレオチドアダプターまたはリンカーが慣用の実施に従って使用される。
【0126】
モノクローナル抗体は、慣用の免疫グロブリン精製手順、例えば、プロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析またはアフィニティークロマトグラフィーにより培養培地から適当に分離される。モノクローナル抗体をコードするDNA及びRNAは、容易に単離されそして慣用の手順を使用して配列決定される。ハイブリドーマ細胞は、このようなDNAおよびRNAのソースとして役立つことができる。一旦単離されると、DNAを発現ベクターに挿入することができ、次いでこれを、宿主細胞、例えば他の場合には免疫グロブリンタンパク質を産生しないHEK293細胞、CHO細胞またはミエローマ細胞にトランスフェクションして、宿主細胞においてリコンビナントモノクローナル抗体の合成を達成する。
【0127】
ヒトOX40L抗体のアミノ酸配列変異体(または突然変異体)は、抗体DNAに適当なヌクレオチド変化を導入することまたはヌクレオチド合成により調製される。しかしながら、このような改変は、例えば上記した非常に限られた範囲においてのみ行うことができる。例えば、改変は、上記した抗体の特性、例えばIgGアイソタイプ及びエピトープ結合を変えないが、リコンビナント産生の収率、タンパク質安定性を改良することができまたは精製を容易にすることができる。
【0128】
抗OX40L抗体の特有のコンフォメーションを維持するのに関与しないいかなるシステイン残基も、一般にセリンにより置換することができ、それにより分子の酸化安定性を改良しそして異常な架橋を防止することができる。反対に、システイン結合(1つまたは複数)を抗体に加えて、その安定性(特に、抗体がFvフラグメントの如き抗体フラグメントである場合)を改良することができる。
【0129】
抗OX40L抗体のアミノ酸配列変異体をコードする核酸分子は、当技術分野で知られている種々の方法により調製される。これらの方法は、天然のソース(天然に存在するアミノ酸配列変異体の場合に)からの単離、またはオリゴヌクレオチド媒介(又は部位特異的)突然変異誘発、PCR突然変異誘発およびヒト化抗OX40L抗体の以前に調製された変異体または非変異体バージョンのカセット突然変異誘発による調製を含むが、それらに限定されない。
【0130】
本発明は、化学療法剤、毒素(例えばバクテリア、真菌、植物もしくは動物起源の酵素的に活性な毒素またはそのフラグメント)、放射性同位元素(即ち、放射性コンジュゲート)、などの細胞傷害剤にコンジュゲーションされた本発明に従う抗体を含む免疫コンジュゲートにも関する。抗体と細胞傷害剤とのコンジュゲートは、種々の二官能性タンパク質カップリング剤、例えばN−スクシンイミジル−3−(2−ピリジルジチオール)プロピオネート(SPDP)、イミノチオラン(IT)、イミドエステルの二官能性誘導体(例えばジメチルアジピミデートHCL)、活性エステル(例えばジスクシンイミジルスベレート)、アルデヒド(例えばグルタルアルデヒド)、ビス−アジド化合物(例えばビス(p−アジドベンゾイル)ヘキサンジアミン)、ビス−ジアゾニウム誘導体(例えばビス−(p−ジアゾニウムベンゾイル)−エチレンジアミン)、ジイソシアナート(例えばトリエン2,6−ジイソシアナート)およびビス−活性フッ素化合物(例えば1,5−ジフルオロ−2,4−ジニトロベンゼン)を使用して製造される。例えばリシンイムノトキシンは、Vitetta, E.S., et al., Science 238(1987) 1098-1104)に記載のとおりに製造することができる。炭素14で標識された1−イソチオシアナートベンジル−3−メチルジエチレントリアミンペンタ酢酸(MX−DTPA)は、抗体への放射性ヌクレオチドのコンジュゲートのための例示的キレート剤である。WO 94/11026参照。
【0131】
別のタイプの抗体の共有結合改変は、US Patent Nos. 4,640,835; 4,496,689; 4,301,144; 4,670,417; 4,791,192または4,179,337に記載された方法において、種々の非タンパク質性ポリマー、例えばポリエチレングリコール、ポリプロピレングリコールまたはポリオキシアルキレンの1つに抗体を連結することを含む。
【0132】
なお別の局面では、本発明は、ヒト抗OX40L抗体(本発明に従う抗体を産生するハイブリドーマ細胞からなる群より選ばれる細胞系により産生された親抗体)を発現するトランスジェニック非ヒト動物、例えばトランスジェニックマウスからの単離されたB細胞を提供する。好ましくは、単離されたB細胞は、精製されたもしくはリコンビナント形態のOX40L抗原および/またはOX40Lを発現する細胞で免疫感作されたトランスジェニック非ヒト動物、例えばトランスジェニックマウスから得られる。好ましくは、トランスジェニック非ヒト動物、例えばトランスジェニックマウスは、本発明の抗体のすべてもしくは一部をコードするヒト重鎖導入遺伝子およびヒト軽鎖導入遺伝子を含むゲノムを有する。単離されたB細胞を次いで不死化してヒト抗OX40L抗体のソース(例えばハイブリドーマ)を提供する。したがって、本発明は、本発明に従うヒトモノクローナル抗体を産生することができるハイブリドーマも提供する。1つの態様では、ハイブリドーマは、不死化された細胞に融合された、本発明の抗体のすべてもしくは一部をコードするヒト重鎖導入遺伝子およびヒト軽鎖導入遺伝子を含むゲノムを有するトランスジェニック非ヒト動物、例えばトランスジェニックマウスから得られるB細胞を含む。
【0133】
特定の態様では、トランスジェニック非ヒト動物は、本発明の抗体のすべてもしくは一部をコードするヒト重鎖導入遺伝子およびヒト軽鎖導入遺伝子を含むゲノムを有するトランスジェニックマウスである。トランスジェニック非ヒト動物は、OX40L抗原の精製されたもしくは濃縮された調製物および/またはOX40Lを発現する細胞で免疫感作されうる。好ましくは、トランスジェニック非ヒト動物、例えばトランスジェニックマウスは、OX40Lに対するヒトモノクローナル抗体のアイソタイプを産生することができる。
【0134】
本発明に従うヒトモノクローナル抗体は、本発明の抗体のすべてもしくは一部をコードするヒト重鎖導入遺伝子およびヒト軽鎖導入遺伝子を含むゲノムを有するトランスジェニック非ヒト動物、例えばトランスジェニックマウスを、OX40L抗原の精製されたもしくは濃縮された調製物および/またはOX40Lを発現する細胞で免疫感作させることにより産生することができる。次いで動物のB細胞(例えば、脾臓B細胞)を得そしてミエローマ細胞と融合して、OX40Lに対するヒトモノクローナル抗体を分泌する不死のハイブリドーマ細胞を形成する。
【0135】
好ましい態様では、OX40Lに対して指向されたヒトモノクローナル抗体は、マウス系よりはむしろヒト免疫系の一部を有するトランスジェニックマウスを使用して発生させることができる。本明細書で「HuMab」マウスと呼ばれるこれらのトランスジェニックマウスは、内在性μおよびκ鎖ローカスを不活性化するターゲティングされた突然変異と共に、重(μおよびγ)鎖およびκ軽鎖(定常領域遺伝子)を含む再配列されていないヒト免疫グロブリン遺伝子をコードするヒト免疫グロブリン遺伝子ミニローカスを含有する(Lonberg, N., et al., Nature 368(1994) 856-859)。したがって、このマウスは、マウスIgMまたはKの減少した発現を示し、そして免疫感作に応答して、導入されたヒト重鎖および軽鎖導入遺伝子は、クラススイッチおよび体細胞突然変異を受けて、高アフィニティーヒトIgGモノクローナル抗体を発生する(Lonberg, N., et al., Nature 368(1994)856-859; Lonberg, N., Handbook of Experimental Pharmacology 113(1994)49-101に概説されている; Lonberg, N., and Huszar, D., Intern. Rev. Immunol. 25(1995)65-93; およびHarding, F., and Lonberg, N., Ann. N. Acad. Sci. 764(1995) 536-546))。HuMAbマウスの調製は、Taylor, L., et al., Nucleic Acids Res. 20 (1992) 6287-6295; Chen, J., et al., Int. Immunol. 5(1993) 647-656; Tuaillon, N., et al., Proc. Natl. Acad. Sci. USA 90(1993) 3720-3724; Choi, T. K., et al., Nat. Genet. 4(1993)117-123; Chen, J., et al., EMBO J. 12(1993) 821-830; Tuaillon, N., et al.,J. Immunol. 152(1994) 2912-2920; Lonberg, N., et al., Nature 368(1994) 856-859; Lonberg, N., Handbook of Experimental Pharmacology 113(1994)49-101; Taylor, L., et al., Int. Immunol. 6(1994) 579-591; Lonberg, N., and Huszar, D., Intern. Rev. Immunol. 25(1995)65-93; およびHarding, F., and Lonberg, N., Ann. N. Acad. Sci. 764(1995) 536-546; Fishwild, D. M., et al., Nat. Biotechnol. 14(1996) 845-851;に記載されており、それらのすべての内容はそのまま参照により本明細書に組み込まれる。更に、US Patent Nos. 5,545,806; 5,569,825; 5,625,126; 5,633,425; 5,789,650; 5,877,397; 5,661,016; 5,814,318; 5,874,299; 5,545,807; 5,770,429; WO 98/24884; WO 94/25585; WO 93/1227; WO 92/22645; and WO 92/03918を参照のこと。
【0136】
OX40Lに対するヒトモノクローナル抗体を完全に発生させるために、HuMAbマウスを、Lonberg, N., et al., Nature 368(1994)856-859; Fishwild, D. M., et al., Nat. Biotechnol. 14(1996) 845-851およびWO 98/24884により記載のとおり、一般的方法に従ってOX40L抗原の精製されたもしくは濃縮された調製物および/またはOX40Lを発現する細胞で免疫感作させることができる。好ましくは、マウスは、最初の免疫感作時に6〜16週齢であろう。例えば、KLHにカップリングされたもしくはPBS中の可溶性OX40L抗原の精製されたもしくは濃縮された調製物(例えば、OX40L発現細胞から精製された)を使用して、HuMAbマウスを腹腔内に免疫感作させることができる。これは、単離されたOX40Lタンパク質による免疫感作とOX40Lを発現する細胞、例えば腫瘍細胞系による免疫感作を交互に行うことにより組み合わせて、免疫応答を促進させることができる。種々の抗原による累積した経験は、完全フロイントアジュバント中の抗原で腹腔内に(ip)最初に免疫感作され、次いで不完全フロイントアジュバント中の抗原で隔週毎にip免疫感作(例えば、総計6回まで)されると、HuMAbトランスジェニックマウスは最善に応答することを示した。免疫応答は、眼窩後方の出血(retroorbital bleeds)により得られる血漿サンプルにより、免疫感作プロトコールの経過にわたって監視することができる。血漿は、ELISAによりスクリーニングすることができ、そして十分な力価の抗OX40Lヒト免疫グロブリンを有するマウスを、対応するB細胞の不死化のために使用することができる。マウスを抗原で3〜4日静脈内に追加抗原刺激した後殺し、そして脾臓およびリンパ節を除去することができる。いくつかのマウスは、各抗原について免疫感作されるであろう。例えば、HCo7およびHCo12系統の総計12匹のHuMAbマウスを免疫感作させることができる。
【0137】
HCo7マウスは、それらの内在性軽鎖(κ)遺伝子におけるJKD破壊(JKD disruption)(Chen, J., et al., EMBO J. 12(1993)821-830)、それらの内在性重鎖遺伝子におけるCMD破壊(WO 01/14424の実施例1に記載のとおり)、KCo5ヒトκ軽鎖導入遺伝子(Fishwild, D. M., et al., Nat. Biotechnol. 14(1996) 845-851に記載のとおり)およびHCo7ヒト重鎖導入遺伝子(US Patent No.5,770,429に記載のとおり)を有する。
【0138】
HCo12マウスは、それらの内在性軽鎖(κ)遺伝子におけるJKD破壊(Chen, J., et al., EMBO J. 12(1993)821-830)、それらの内在性重鎖遺伝子におけるCMD破壊(WO 01/14424の実施例1に記載のとおり)、KCo5ヒトκ軽鎖導入遺伝子(Fishwild, D. M., et al., Nat. Biotechnol. 14(1996) 845-851に記載のとおり)およびHCo12ヒト重鎖導入遺伝子(WO 01/14424の実施例2に記載のとおり)を有する。マウスリンパ球を単離しそして標準プロトコールに基づいてPEGを使用してマウスミエローマ細胞系と融合させてハイブリドーマを発生させることができる。得られるハイブリドーマを、次いで抗原特異的抗体の産生についてスクリーニングする。例えば免疫感作されたマウスからの脾臓およびリンパ節由来のリンパ球の単一細胞懸濁液を、50%PEGによりSP2/0非分泌マウスミエローマ細胞(ATCC、CRL1581)の数の6分の1に融合させる。細胞を平底マイクロタイタープレート中に約2×105でプレートし、続いて選択培地中で約2週間インキュベーションする。
【0139】
次いで個々のウエルをヒト抗OX40LモノクローナルIgMおよびIgG抗体についてELISAによりスクリーニングする。一旦多量のハイブリドーマ増殖が起こると、培地を通常10〜14日後に分析する。抗体分泌ハイブリドーマを再びプレートし、再びスクリーニングし、そしてもしもヒトIgGについて依然としてポジティブであるならば、抗OX40Lモノクローナル抗体を限界希釈により少なくとも2回サブクローニングすることができる。次いで安定なサブクローンをin vitroで培養して特徴付けのために組織培養培地中で抗体を産生させる。
【0140】
アッセイツリー(assay tree)は、主として、IgGに関する非特異的アッセイ(「IgG−ELISA」)、次いで精製されたOX40Lタンパク質またはOX40L発現細胞への抗原結合の決定のための特異的ELISAおよび見かけのFACSアッセイからなる。次の段階は、抗OX40L抗体の、精製されたOX40Lまたは細胞上に発現されたOX40Lに対するその天然の相互作用パートナー、例えば可溶性の精製されたOX40との競合を決定する機能的アッセイ、例えば競合ELISAまたはFACSを含む。次の段階は、OX40シグナル伝達の抗OX40L抗体の阻止能力を決定する機能的アッセイ、例えばNFκB活性化(=「NFκBアッセイ」)を含む。次の段階は、T細胞活性化に関する抗OX40L抗体の阻止能力を決定する機能的アッセイ(「T細胞活性化アッセイ」および「TTアッセイ」)を含む。
【0141】
CDR配列は抗体−抗原相互作用について責任を担っているので、異なるヒト抗体からのフレームワーク配列上に本発明に従うCDR配列を含む発現ベクターを構築することにより本発明に従うリコンビナント抗体を発現することが可能である(例えば、Riechmann, L., et al., Nature 332(1998) 323-327; Jones, P., et al., Nature 321(1986) 522-525; and Queen, C., et al., Proc. Natl. Acad. Sci. USA 86(1989)10029-10033参照)。このようなフレームワーク配列は、生殖系列ヒト抗体遺伝子配列を含む公のDNAデータベースから得ることができる。これらの生殖系列配列は、成熟抗体遺伝子配列とは異なるであろう。何故ならば、それらはB細胞成熟期間中にV(D)J結合により形成される完全にアセンブリーされた可変遺伝子を含まないからである。生殖系列遺伝子配列は、各々可変領域にまんべんなく、高アフィニティー二次レパートリー抗体の配列とも異なるであろう。
【0142】
本発明は、好ましくはサンプルのOX40L(可溶性または膜に結合した)と本発明に従う抗体との結合を決定する免疫学的アッセイにより、in vitroでのOX40Lの診断のための本発明に従う抗体の使用をさらに含む。
【0143】
他の局面では、本発明は、薬学的に許容されうる担体と共に処方された、本発明のヒトモノクローナル抗体またはその抗原結合性部分の1つまたは組合せを含有する組成物、例えば医薬組成物を提供する。更に詳しくは、組成物は、医薬組成物または診断組成物であり、なおさらに詳しくは、医薬組成物は、上記に定義した抗体と少なくとも1種の薬学的に許容されうる賦形剤を含む。組成物は無菌でなければならず、そして組成物が注射器により送達可能である程度に流動性でなければならない。
【0144】
本明細書で使用された、「薬学的に許容される担体」は、生理学的に適合性の、いかなるおよびすべての溶媒、分散媒体、コーティング、抗バクテリア剤および抗真菌剤、等張剤および吸収遅延剤等を含む。好ましくは、担体は、静脈内、筋肉内、皮下、非経口、脊髄または表皮投与(例えば、注射または注入(infusion)により)のために適当である。好ましくは、このような担体は、好ましくは、等張性であり、好ましくは更に無機塩、糖、ポリオールおよび/または界面活性剤を含有する、水性のpH緩衝化された溶液(例えば酢酸塩、クエン酸塩、リン酸塩またはヒスチジン)である。薬学的に許容されうる担体は、Remington's Pharmaceutical Science, 16th edition, Osol, A. Ed.(1980)に記載されたようなものでもある。
【0145】
抗体濃度は、好ましくは0.1mg/ml〜50mg/mlである。好ましくは、緩衝化された溶液のpH値は、1mM〜200mMの緩衝剤濃度で4.0〜8.0の範囲にある。好ましい塩は、1mM〜200mMの範囲にある塩化ナトリウムおよび/またはリン酸ナトリウムである。好ましい糖は、1%〜15%(重量/容積)の範囲にあるスクロースおよび/またはトレハロースである。好ましいポリオールは、1%〜15%(重量/容積)の範囲にあるグリセロール、プロピレングリコール、液体ポリエチレングリコールおよび/または同様なものである。界面活性剤は、好ましくは、0.001%〜0.5%(重量/容積)の範囲のポリソルベート(例えばポリソルベート20または80)および/またはポロキサメール(poloxamere)である。このましい医薬組成物は、0.1mg/ml〜50mg/mlの抗体と1mM〜200mMのリン酸緩衝化生理食塩水pH4.0〜8.0を含有する。
【0146】
本発明の組成物は、それを必要としている患者に当技術分野で知られている多様の方法により投与することができる。当業者により認識されるとおり、投与の経路および/または方式は所望の結果に依存して変わるであろう。
【0147】
薬学的に許容されうる賦形剤または担体は、無菌の水性溶液剤または無菌の注射可能な溶液剤もしくは分散液剤の処方に応じて調合される製剤用の無菌の散剤を含む。このような媒体および薬学的有効物質のための作用物質の使用は当技術分野で知られている。
【0148】
本明細書で使用された語句「非経口投与」及び「非経口的に投与される」は、通常注射による腸内および局所的投与以外の投与方式を意味し、そして静脈内、筋肉内、動脈内、鞘内、カプセル内、眼窩内、心臓内、皮膚内、腹腔内、経気管、皮下、表皮下、関節内、被膜下、くも膜下、髄腔内、硬膜外、および胸骨内注射および注入を含むが、それらに限定されない。
【0149】
本発明の医薬組成物中の有効成分の実際の投与レベルは、患者に毒性であることなく、特定の患者、組成および投与方式について所望の治療応答を達成するのに有効な有効成分の量を得るように変えることができる。選ばれる投与レベルは、使用される本発明の特定の組成物、または又はそのエステル、塩もしくはアミドの活性、投与経路、投与の時間、使用される特定の化合物の排出の速度、処置の期間、使用される特定の組成物と組み合わせて使用される他の薬物、化合物および/または物質、処置される患者の年齢、性、体重、状態、一般に健康及び以前の医学的経歴および医学分野で周知の同様な因子を含む種々の薬物動態学的因子に依存するであろう。典型的な毎週の投与量(weekly dosage)は、上記した因子に依存して約0.1mg/kg〜約20mg/kg以上の範囲にあることができる。
【0150】
下記の実施例、参考文献、配列リストおよび図は本発明の理解を助けるために提供され、本発明の真の範囲は特許請求の範囲に記載されている。本発明の精神から逸脱することなく前記した手法において改変がなされうることは理解される。
【0151】
配列リストの説明
配列番号1 LC.001のκ軽鎖、可変領域
配列番号2 LC.001のγ重鎖可変領域
配列番号3 LC.005のκ軽鎖、可変領域
配列番号4 LC.005のγ重鎖可変領域
配列番号5 LC.010のκ軽鎖、可変領域
配列番号6 LC.010のγ重鎖可変領域
配列番号7 LC.029のκ軽鎖、可変領域
配列番号8 LC.029のγ重鎖可変領域
配列番号9 LC.019のκ軽鎖、可変領域
配列番号10 LC.019のγ重鎖可変領域
配列番号11 LC.033のκ軽鎖、可変領域
配列番号12 LC.033のγ重鎖可変領域
配列番号13 κ軽鎖定常領域
配列番号14 γ1重鎖定常領域
配列番号15 γ4重鎖定常領域
配列番号16 LC.033のκ軽鎖、突然変異体可変領域
配列番号1 LC.059のκ軽鎖、可変領域
配列番号17 LC.059のγ重鎖可変領域
配列番号18 LC.060のκ軽鎖、可変領域
配列番号19 LC.060のγ重鎖可変領域
配列番号1 LC.063のκ軽鎖、可変領域
配列番号20 LC.063のγ重鎖可変領域
配列番号21〜57 CDR配列
配列番号58 LC.001のγ重鎖(ヒトIgG1型)
配列番号59 LC.001のγ重鎖(L234A、L235AヒトIgG1突然変異体)
配列番号60 LC.001のγ重鎖(S228PヒトIgG4突然変異体)
配列番号61 LC.001のκ軽鎖
配列番号62 LC.005のγ重鎖(ヒトIgG1型)
配列番号63 LC.005のγ重鎖(L234A、L235AヒトIgG1突然変異体)
配列番号64 LC.005のγ重鎖(S228PヒトIgG4突然変異体)
配列番号65 LC.005のκ軽鎖
配列番号66 LC.060のγ重鎖(ヒトIgG1型)
配列番号67 LC.060のγ重鎖(L234A、L235AヒトIgG1突然変異体)
配列番号68 LC.060のγ重鎖(S228PヒトIgG4突然変異体)
配列番号69 LC.060のκ軽鎖
【0152】
略号
アミノ酸は、3文字コード(Leu)または1文字コード(L)において略記される。
S228Pは、IgG4重鎖の位置228におけるセリンのプロリンへの交換を意味する。
L234は、EU番号付け(Kabat)に従う位置234におけるアミノ酸ロイシンを意味する。
L234Aは、位置234におけるアミノ酸ロイシンがアラニンに変えられていることを意味する。
L235Aは、位置235におけるアミノ酸ロイシンがアラニンに変えられていることを意味する。
PVA236は、236領域において、IgG1のELLGまたはIgG4のEFLGがPVAに修正されていることを意味する。
GLPSS331は、331領域において、IgG1のALPAPまたはIgG2のGLPAPがGLPSSに変えられていることを意味する。
ΔG236は、位置236におけるアミノ酸が欠失していることを意味する。
IgG4xは、IgG4における突然変異S228Pを意味する。
LC2010−001は、LC.001と同義である。
Fcgは、Fcガンマ(Fcγ)の同義語である。
【0153】
抗体の他の配列修正は、同様に命名される。
ヒスチジンタグに融合されたリコンビナント可溶性ヒトOX40L
hOX40L−His
ヒスチジンタグに融合されたリコンビナント可溶性マウスOX40L
mOX40L−His
フラッグタグに融合されたリコンビナント可溶性ヒトOX40L
hOX40L−Flag
フラッグタグに融合されたリコンビナント可溶性マウスOX40L
mOX40L−Flag
ヒトFcγに融合されたリコンビナント可溶性ヒトOX40
hOX40−hFc
ウサギ抗マウスFcγモノクローナル抗体 Anti−mFc
ヤギ抗ヒトFcγモノクローナル抗体 Anti−hFc
マウス抗ヒスチジンモノクローナル抗体 Anti−His
マウスFcγに融合されたリコンビナント可溶性ヒトOX40
hOX40−mFc
マウス抗TNFαモノクローナル抗体 Anti−TNFα
マウス抗CD40Lモノクローナル抗体 Anti−CD40L
腫瘍壊死因子アルファ TNFα
CD40リガンド CD40L
ラット抗ヒトOX40Lモノクローナル抗体 TAG34
ヒト抗ヒトOX40Lモノクローナル抗体 LC.001、LC.005
LC.010、LC.019
LC.029、LC.033
LC.059、LC.060
LC.063
フィトヘマグルチニン PHA
【0154】
実施例
実施例1
抗OX40L抗体を産生するハイブリドーマ細胞系の発生
ハイブリドーマの培養
HuMAbハイブリドーマを、IMDM(Cambrex)、胎仔クローン1ウシ血清(Perbio Science)、起源ハイブリドーマクローニング因子(Igen)、ピルビン酸ナトリウム、ペニシリン/ストレプトマイシン、2−メルカプトエタノール、HAT(Sigma-Aldrich)およびカナマイシン(Invitrogen)中で37℃で、5%CO2において培養した。
【0155】
トランスジェニックマウスの免疫感作手順
LC.2010−001:6匹のHCo7(2匹のオスと4匹の雌)、系統GG2201(Medarex, San Jose, CA, USA)および4匹のHCo12(4匹の雄)、系統GG2198(Mesarex, San Jose, CA, USA)を、ヒトOX40L(hOX40L)のための発現ベクターで一過性にトランスフェクションされた1×106HEK293細胞、およびhOX40Lの可溶性細胞外ドメイン20μgで交互に免疫感作させた。8回の免疫感作を、hOX40L発現細胞による総計4回の腹腔内(i.p)免疫感作およびリコンビナントタンパク質による尾基部(tail base)での4回の皮下(s.c)免疫感作により行った。最初の免疫感作では、1×106HEK293−hOX40L細胞100μlを完全フロイントアジュバント(CFA; Difco Laboratories, Detroit, USA)100μlと混合した。すべての他の免疫感作では、PBS中の細胞100μlを使用するかまたはリコンビナントタンパク質を不完全フロイントアジュバント(ICFA;Difco)100μlと混合した。
【0156】
抗hOX40Lの血清力価が十分であることが見出されると、マウスを、融合の4日および3日前にPBS200μl中のhOX40L細胞外ドメイン15μgで静脈内に(i.v)2回さらに追加抗原刺激した。
【0157】
LC.2010−001、LC.059、LC.060およびLC.063はHCo12マウスに由来していた。
【0158】
LC.2010−005、−010、−019、−029および−033:5匹のHCo7(4匹の雄と1匹の雌)、系統GG2201(Medarex, San Jose, CA,USA)を、hOX40Lの可溶性細胞外ドメイン20μgで免疫感作させた。7回の免疫感作を、総計4回の腹腔内(i.p)免疫感作および尾基部での3回の皮下(s.c)免疫感作により行った。最初の免疫感作では、リコンビナントタンパク質100μlを、完全フロイントアジュバント(CFA; Difco Laboratories, Detroit, USA)100μlと混合した。すべての他の免疫感作では、リコンビナントタンパク質100μlを不完全フロイントアジュバント(ICFA; Difco)100μlと混合した。
【0159】
抗hOX40Lの血清力価が十分であることが見出されると、マウスを、融合の4日および3日前にPBS200μl中のhOX40L細胞外ドメイン15μgで静脈内に(i.v)2回さらに追加抗原刺激した。
【0160】
ハイブリドーマ発生
マウスを殺しそして脾臓および腹大動脈と大静脈をフランキングする(flanking)リンパ節を採集した。脾臓細胞およびリンパ節細胞の融合パートナーSP2.0細胞との融合は、標準操作手順に従って行った。
【0161】
抗原特異的ELISA
免疫感作されたマウスの血清における抗OX40L力価を、抗原特異的ELISAにより決定した。プレート(96平底ELISAプレート、Greiner)を、PBS中に溶解した精製されたOX40L0.1μg/mlでコーティングしそして室温で一夜コーティングした。次いで、ウエルを、PBSTC(Tween 20、0.05%(Sigma-Aldrich Chemie BV)および2%ニワトリ血清(Gibco)を含有するPBS)で室温で1時間ブロッキングした。
【0162】
試験した血清タップ(serum taps)を、PBSTC中に1:50に希釈しそしてウエルに加えた。免疫感作の前にマウスから得られた血清をPBSTC中に1:100で溶解しそしてネガティブコントロールとして使用した。ヒトOX40Lに対して指向されたマウス抗体をPBSTC中に1:50で溶解しそしてポジティブコントロールとして使用した。プレートを室温で1時間インキュベーションした。次いで、プレートをPBST(Tween 20、005%を含有するPBS)を使用して2回洗浄した。Gt−α−hulgG−HRP(Jackson)をPBSTC中に1:5000に希釈しそして試験されたタップおよびネガティブコントロールを含有するウエルに加えた。Rb−α−mIgG(Jackson)をPBSTC中に1:3000に希釈しそしてポジティブコントロールを含有するウエルに加えた。プレートを室温で1時間インキュベーションした。最後に、プレートをPBSTを使用して3回洗浄し、そして調製されたばかりのABTS(登録商標)溶液(1mg/ml)(ABTS:2,2’−アジノ ビス(3−エチルベンゾチアゾリン−6−スルホン酸)で暗所で室温(RT)で30分間発色させた。吸光度を405nmで測定した。
【0163】
κ−ELISA
融合から生じたハイブリドーマがヒト抗体を発生するかどうかを決定するために、κ−ELISAを行った。ELISAプレートを、PBS中に1/10000に希釈したラット抗ヒトIgGκ軽鎖抗体(DAKO)で、4℃での一夜のインキュベーションによりコーティングした。ウエルを捨てた後に、プレートを、PBSTC(Tween−20、0.05%を補充したPBSC(PBSTC))との室温で1時間のインキュベーションによりブロッキングした。次いでウエルをPBSTC中に1/2に希釈したハイブリドーマ培養上清とインキュベーションした。PBSTC中に1/2に希釈した培養培地をネガティブコントロールとして使用し、PBSTC中に1/100に希釈したκ鎖ポジティブマウス血清をポジティブコントロールとして使用した。次いで、ウエルを3回洗浄しそしてPBSTC中に1/2000に希釈したHRPをコンジュゲーションしたラット抗ヒトIgGF(ab’)2(DAKO)と37℃で1時間インキュベーションした。ウエルを3回洗浄しそしてアッセイを、調製されたばかりのABTS(登録商標)溶液(1mg/ml)で暗所で室温(RT)で30分間発色させた。吸光度をELISAプレートリーダーにおいて405nmで測定した。
【0164】
実施例2
抗OX40LHuMab可変ドメインのクローニングおよび配列分析
(κ軽鎖およびγ1重鎖)
OX40L HuMabsの軽鎖可変領域VLおよび重鎖可変領域VHをコードするヌクレオチド配列を、標準cDNA合成/PCR法により単離した。
【0165】
トータルRNAを、GeneRacer(商標)Kit(Invitrogen)を使用して1×106〜1×107ハイブリドーマ細胞から調製した。ハイブリドーマ由来のRNAを、第1鎖cDNA合成およびGeneRacer(商標)Oligo−dTプライマーのライゲーシヨンのためのテンプレートとして使用した。第2鎖cDNA合成およびVLおよびVHコード化cDNAフラグメントのさらなるPCR増幅を、それぞれ、カッパ軽鎖およびγ1重鎖定常領域のヌクレオチド配列に相補性のリバース軽鎖及び重鎖プライマーならびに5’−特異的GeneRacer(商標)プライマーにより行った。PCR産物を、Invitrogen(商標)Life TechnologiesからのTOPO(商標)TAクローニングキットおよびクローニングベクターとしてpCR4−TOPO(商標)を使用してクローニングした。クローニングされたPCR産物を、消化のためのEcoRIを使用して適切なプラスミドの制限マッピングにより同定し、そして、それぞれ、VLおよびVHのための約740および790bpのDNAフラグメントサイズを予測/計算した。
【0166】
クローニングされたPCRフラグメントのDNA配列を二本鎖配列決定により決定した。
【0167】
GCG(Genetics Computer Group, Madison, Wisconsin)ソフトウエアパッケージバージョン10.2およびVector−NTI8(InforMax,Inc)を一般的データプロセッシングのために使用した。DNAおよびタンパク質配列を、GCGモジュールCLUSTALWを使用してアライメントさせた。配列アライメントは、プログラムGENEDOC(バージョン2.1)を使用して作成された。
【0168】
実施例3
抗OX40L IgG1 HuMabのための発現プラスミドの構築
抗OX40L HuMab軽鎖及び重鎖コード化遺伝子を哺乳動物細胞発現ベクター中に別々にアセンブリーした。
【0169】
それにより、抗OX40L HuMab軽鎖可変領域(VL)およびヒトκ軽鎖定常領域(CL、配列番号13、または配列番号61、65もしくは69からの)をコードする遺伝子セグメントを、抗OX40L HuMab重鎖可変領域(VH)およびヒトγ1重鎖定常領域(CH1−Hinge−CH2−CH3、配列番号14または配列番号58、62もしくは66からの)のための遺伝子セグメントが接合されたと同様にして、接合させた。
【0170】
コドン使用頻度を推定することができるヒト軽鎖及び重鎖のヌクレオチド配列に関する一般的情報は、Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, fifth ed., NIH Publication No.91-3242(1991)に与えられる。
【0171】
抗OX40L HuMabκ軽鎖の転写単位は、下記のエレメント:
−ヒトサイトメガロウイルス(HCMV)からの前初期(immediate early)エンハンサーおよびプロモーター
−コザック配列を含む合成5’−UT、
−シグナル配列イントロンを含むマウス免疫グロブリン重鎖シグナル配列、
−5’端部における独特のBsmI制限部位およびスプライスドナー部位および3’端部における独特のNotI制限部位を持って配列された(arranged)クローニングされた抗OX40L HuMab可変軽鎖cDNA、
−イントロン2マウスIg−κエンハンサーを含むゲノムヒトκ遺伝子定常領域[Picard, D., and Schaffner, W., Nature 307(1984)80-82]および
−ヒト免疫グロブリンκポリアデニル化(「ポリA」シグナル配列、
からなる。
【0172】
抗OX40L HuMabγ1重鎖の転写単位は、下記のエレメント:
−ヒトサイトメガロウイルス(HCMV)からの前初期エンハンサーおよびプロモーター
−コザック配列を含む合成5’−UT、
−シグナル配列イントロンを含む改変されたマウス免疫グロブリン重鎖シグナル配列、
−5’における独特のBsmI制限部位およびスプライスドナー部位および3’端部における独特のNotI制限部位を持って配列されたクローニングされた抗OX40L HuMab可変重鎖cDNA、
−マウスIgμエンハンサーを含むゲノムヒトγ1重遺伝子定常領域(Neuberger, M.S., EMBO J.2(1983)1373-1378)、
−ヒトγ1免疫グロブリンポリアデニル化(「ポリA」シグナル配列、
からなる。
【0173】
抗OX40L HuMabκ軽鎖およびγ1重鎖発現プラスミドの機能的エレメント:抗OX40L HuMabκ軽鎖またはγ1重鎖発現カセットの外に、これらのプラスミドは、
−ヒグロマイシン耐性遺伝子
−エプスタイン−バールウイルス(EBV)の複製の起点、oriP
−E.coliにおいてこのプラスミドの複製を可能とするベクターpUC18からの複製の起点および
E.coliにおけるアンピシリン耐性を与えるβ−ラクタマーゼ遺伝子
を含有する。
【0174】
実施例4
抗OX40L IgG4 HuMabのための発現プラスミドの構築
抗OX40Lγ4重鎖プロトタイプ発現プラスミドは、ヒトゲノムγ1定常領域およびγ1免疫グロブリンポリアデニル化(「ポリA」)シグナル配列をヒトゲノムγ4定常領域(配列番号15、または配列番号60,64もしくは68からの突然変異していないγ4定常領域からの)およびγ4免疫グロブリンポリアデニル化シグナル配列により置き換えることにより、抗OX40Lγ1重鎖発現プラスミドから誘導された。
【0175】
抗OX40L HuMabκ軽鎖の発現のために、同じ発現プラスミドを、IgG1について説明したとおりに使用した(上記参照)。
【0176】
実施例5
LC.001に基づく突然変異体(変異体)抗OX40L IgG1およびIgG4のための発現プラスミドの構築
突然変異体抗OX40Lγ1−およびγ4−重鎖をコードする発現プラスミドは、QuickChange(商標)部位特異的突然変異誘発キット(Stratagene)を使用する野生型発現プラスミドの部位特異的突然変異誘発により創生された。アミノ酸は、EU番号付けに従って番号を付けられた(Edelman, G. M., et al., Proc. Natl. Acad. Sci. USA 63(1969) 78-85; Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, NIH Publication No. 91-3242, Bethesda, MD(1991))。
【0177】
【表1】

【0178】
実施例6
リコンビナント抗OX40L HuMabsの産生
10%超低IgG FCS(Gibco)、2mMグルタミン(Gibco)、1%容積/容積非必須アミノ酸(Gibco)および250μg/mlG418(Roche)を補充されたDMEM(Gibco)中で培養された付着HEK293−EBNA細胞(ATTC CRL−10852)の一過性トランスフェクションによりリコンビナントHuMabsを発生させた。トランスフェクションのために、Fugene(商標)6(Roche)トランスフェクション試薬を3:1〜6:1の範囲の試薬(μl)対DNA(μg)の割合で使用した。免疫グロブリン軽鎖および重鎖を、1:2〜2:1の軽鎖対重鎖コード化プラスミドのモル比を使用して2つの異なるプラスミドから発現させた。HuMab含有細胞培養上清を、トランスフェクションの4〜11日後に回収した。上清を精製まで−20℃に保存した。
【0179】
例えば、HEK293におけるヒト抗体のリコンビナント発現に関する一般的情報は、Meissner, P., et al., Biotechnol. Bioeng. 75(2001)197-203に与えられる。
【0180】
実施例7
抗体TAG−34、LC.001、LC.005、LC.010、LC.019、LC.029、LC.033のアフィニティー分析
機器:Biacore 3000、ランニングバッファーおよび反応バッファー:HBS−P(10mM HEPES、150mM NaCl、0.005%Tween20、ph7.4)、25℃。アナライトの注射は、0.78nM〜100nMの7つの濃度で3分間行いそしてHBS−Pで5分間洗浄した。表面(カルボキシメチル化されたデキストラン表面、CM)の再生は、10mMグリシンpH2.0の各々1分間の2回の注射により行った。チップ、アッセイフォーマットおよび注射のシーケンスおよび速度論的データは、下表における説明に対応する。速度論的データは、1:1ラングミュア結合モデルに速度論的データを適合させることにより計算された。
【0181】
【表2】
TAG34とmOX40Lとの相互作用は測定できなかった。
【0182】
すべてのBiacoreアッセイについてのデータ評価およびデータ寄託
ネガティブコントロールデータ(例えば、緩衝曲線)は、システム固有のベースラインドリフトの補正およびノイズシグナル減少のためにサンプル曲線から差し引いた。BiaEvaluationバージョン4.01をセンサーグラムの解析およびアフィニティーデータの計算のために使用した。
【0183】
実施例8
hOX40Lの固定化されたhOX40との相互作用を阻害する抗hOX40L抗体の阻害競合アッセイ
機器:Biacore 3000、ランニングバッファーおよび反応バッファー:HBS−P(10mM HEPES、150mM NaCl、0.005%Tween20、ph7.4)、25℃。注射の前に、アナライト(10mM)および競合剤(0.78nM〜100nMの8つの濃度)を22℃で少なくとも20分間プレインキュベーションした。アナライト+/−競合剤の注射を3分間行いそしてHBS−Pで3分間洗浄した。表面の再生は、10mMグリシンpH2.0の各1分の2回の注射により行った。チップ、アッセイフォーマットおよび注射のシーケンスおよび速度論的データは、下表3における説明に対応する。
【0184】
【表3】
【0185】
すべての抗体は、溶液中のOX40へのOX40Lの結合を阻害する(溶液アフィニティー)。LC.001およびLC.005は、TAG34よりも低いIC50値を示す。
【0186】
実施例9
抗OX40L抗体TAG34、LC.001、LC.005、LC.010、LC.019、LC.029、LC.033、LC.060のエピトープ特徴付け
機器:Biacore 3000、ランニングバッファーおよび反応バッファー:HBS−P(10mM HEPES、150mM NaCl、0.005%Tween20、ph7.4)、25℃。エピトープグループを、列挙された抗体間の交差競合(cross competition)により決定した。注射の前に、アナライト(50mM)および競合剤(100nM)を22℃で少なくとも20分間プレインキュベーションした。アナライト+/−競合剤の注射を2分間行いそしてHBS−Pで3分間洗浄した。表面の再生は、10mMグリシンpH2.0の各1分の2回の注射により行った。チップ、アッセイフォーマットおよび注射のシーケンスおよび速度論的データは、下表4における説明に対応する。
【0187】
【表4】
【0188】
TAG34により認識されたOX40Lエピトープは、エピトープAとして定義された。しかしながら、TAG34は、100ngの抗体濃度において変性されたOX40Lに結合しない(ウエスタンブロットにおいて)。1つのエピトープグループ(AまたはB)内の抗体は、交差阻害活性(cross inhibitory activity)を示すが、異なるグループからの抗体は、相加結合シグナル(additive binding signal)を示す。LC.019は、グループAからおよびグループBからの他の抗体を中和する。
【0189】
実施例10
TAG34、LC.001およびLC.005のCD40LおよびTNFαへの結合特異性
機器:Biacore 3000、ランニングバッファーおよび反応バッファー:HBS−P(10mM HEPES、150mM NaCl、0.005%Tween20、ph7.4)、25℃。アナライトの注射は、100nMおよび500nMで3分間行いそしてHBS−Pで2分間洗浄した。表面の再生は、100mMHClの各々1分間の2回の注射により行った。チップ、アッセイフォーマットおよび注射のシーケンスおよび速度論的データは、下表5における説明に対応する。
【0190】
【表5】
【0191】
このアッセイにおいて、CD40Lは、すべての抗体に対するまたはチップ表面に対するいくらかの非特異的結合を示したが、バックグラウンドシグナルを差し引いた後にこのアッセイは、固定化された抗体TAG34、LC.001およびLC.005に対するTNFαおよびCD40L(500nMまで)の非特異的結合がないことを示した。
【0192】
実施例11
抗体LC.001−IgG1およびLC.001−IgG4xのアフィニティー分析
機器:Biacore 3000、ランニングバッファーおよび反応バッファー:HBS−P(10mM HEPES、150mM NaCl、0.005%Tween20、ph7.4)、25℃。アナライトの注射は、0.78nM〜100nMの8つの濃度で3分間行いそしてHBS−Pで5分間洗浄した。表面の再生は、100mM HClの各々1分間の2回の注射により行った。チップ、アッセイフォーマットおよび注射のシーケンスおよび速度論的データは、下表における説明に対応する。速度論的データは、1:1ラングミュア結合モデルに速度論的データを適合させることにより計算された。
【0193】
【表6】
【0194】
LC.001およびLC.001−IgG4xは、hOX40L−Hisに対する同じアフィニティーを示す。
【0195】
実施例12
OX40Lに結合する抗体の検出のためのELISAアッセイ
SAコーティングしたプレート(96平底ELISAブレート、Microcoat)をインキュベーションバッファー(IB=0.1%Tween20(Serva)および1%遮断タンパク質を含有するPBS)中に溶解された0.5μg/mlビオチニル化OX40Lで室温で1時間コーティングした。次いでプレートを洗浄バッファー(WB=0.1%Tween20を含有する生理食塩水)を使用して2回洗浄した。
【0196】
サンプル(細胞培養上清または精製した抗体)をIB中に段階希釈しそしてウエルに加えた。プレートを室温で1時間インキュベーションした。次いで、プレートをWBを使用して2回洗浄した。次いで、ヒトIgGに対するヤギ抗体とPOD(Dianova)のコンジュゲートをIB中に50ng/mlに希釈しそしてウエルに加えた。プレートを室温で1時間インキュベーションした。最後に、プレートをWBを使用して2回洗浄しそしてすぐ使用できるABTS(登録商標)溶液(Roche)で暗所で室温(RT)で発色させた。最も高い濃度の吸光度が十分なODに達した後に、405nmで吸光度を測定した(図1a)。3nM〜8nMの範囲のEC50値が得られた。
【0197】
実施例13
ヒトOX40/ヒトOX40Lの相互作用を阻害する抗体の検出のためのELISAアッセイ
SAコーティングしたプレート(96平底ELISAプレート、Microcoat. Germany)をIB中に溶解された0.5μg/mlビオチニル化OX40Lで室温で1時間コーティングした。次いでプレートをWB(PBS緩衝液、0.1%(重量/容量)Tween(商標)20)を使用して2回洗浄した。
【0198】
サンプルをIB中に1μg/mlの濃度に希釈しそして段階希釈物においてウエルに加えた。OX40LへのOX40の最大結合を達成するために、いくらかのウエルにおいてIBのみを加えた。次いで各ウエルに、0.2μg/mlの濃度の、Digoxigenin(Roche Diagnostics GmbH, DE)とコンジュゲーションされたヒトOX40の溶液を加えた。プレートを室温で1時間インキュベーションした。次いで、プレートをWBを使用して2回洗浄した。ヒツジ<Digoxigenin>−POD(Roche)をIB中に50mU/mlに希釈しそしてウエルに加えた。プレートを室温で1時間インキュベーションした。最後に、プレートをWBを使用して2回洗浄しそしてすぐ使用できるABTS(登録商標)溶液(Roche)により暗所で室温(RT)で発色させた。吸光度を、10〜20分後に405nmで測定した(図1b)。1nM〜4nMの範囲のIC50を得た。
【0199】
実施例14
K562細胞(K562_OX40L細胞)上に発現されたヒトOX40LとヒトOX40の相互作用を阻害するHuMabsの検出のためのFACSアッセイ
目的:Dig標識されたhOX40:hFc融合タンパク質のhOX40L発現細胞系K562_hOX40Lとの相互作用をブロッキングするためのHuMab hOX40Lの性質の決定のためのアッセイ
【0200】
手順:このアッセイは、「アッセイ試薬」としてDig標識されたhOX40:hFcおよび「競合剤」としてHuMab hOX40Lを使用して行われた。
【0201】
アッセイ試薬:ストック0.5μg/μl−PBS中に1:10に希釈された)、100μl抗ジオキシゲニンFLUOS、PBS/0.5%BSA/1%ブロッキング試薬中に1:25に希釈された(Roche Diagnostics GmbH, DE)。
【0202】
2×105K562_OX40L細胞(ISF−0中で成長させた)を、PBS2ml中で洗浄しそしてPBS100μl中に再懸濁させた。この後PBS中の競合剤(0:1/1:1/1.5:1/2:1/2.5:2/5:1競合剤/試薬関係)を添加した。次いで室温で日光において30分間インキュベーションした。次いで試薬(PBS中の)を加えた;インキュベーション時間:30分、室温および日光。細胞をPBS2mlで洗浄しそして遠心によりペレット化した。染色用の二次抗体(抗ジゴキシゲニン−フルオレセイン)、Fabフラグメント(Roche, 1207741)を加えそして暗所で4℃で30分間インキュベーションした。細胞をPBS2mlで洗浄しそして遠心によりペレット化した。その後細胞をPBS0.5ml中に再懸濁させた。サンプルの測定をFACS−Scanにおいて行った(図2)。
【0203】
実施例15
hOX40/hOX40Lシグナリングについての抗体の阻害能力の決定のための機能的アッセイ(「NFκBアッセイ」)
Hela野生型(wt)およびヒトOX40を発現するHeLa細胞(HeLa_OX40)を、最小必須培地(MEM)、1×ピルビン酸Na、1×非必須アミノ酸(Gibco)、10%FCSにおいてそしてリコンビナント細胞の場合には+600μg/mlG418において成長させた。K562およびOX40Lを発現するK562をISF−O培地中で成長させそしてリコンビナント細胞の場合には、200μg/mlG418を加えた。
【0204】
HeLa_wtまたはHeLa_OX40細胞を、96ウエルプレートw/oG418中に3×104細胞/100μlの細胞密度で播種しそしてCO2インキュベータ中で一夜インキュベーションした。K562_wtまたはH562_OX40L細胞を1:1の細胞対細胞関係において加えた。OX40を発現するホルマリン固定された又はホルマリン固定されていないK562(−70℃で凍結された)を解凍しそしてMEM/10%FCS中で1:1に希釈し;K562_OX40L細胞を、OX40Lに対する抗体と共に室温で30分間プレインキュベーションした。K562_OX40L細胞による刺激時間は30分〜150分であった。細胞核からのタンパク質抽出をActive MotifからのNE−Kitにより供給者の指示に従って行った。Active MotifからのTransAM NFκB−ELISA(供給者の指示に従ってアッセイを行った)を使用して、NFκB活性化をもたらす、OX40シグナリングを決定した。Tecan MTP−Readerにおいて波長450/620nmで吸光度の測定を行った(図3)。すべてのLC抗体について0.6〜5nMの範囲のIC50値を得た。
【0205】
実施例16
T細胞活性化アッセイ
アッセイ原理
ヒト末梢血単核細胞(PBL)を、最適より低い濃度のT細胞マイトジェンフィトヘマグルチニン(PHA)で活性化し、そしてOX40Lを過発現するK562細胞で共刺激する(co-stimulated)。アッセイ条件下に、活性化されたT細胞を37℃で24時間インキュベーションして、IL−2を産生させる。ELISAアッセイを使用して上清中のサイトカインを測定する。Mabのブロッキグ効果を決定するために、K562_OX40L細胞をPBLとの共培養の前に抗体の適切な希釈物と1時間プレインキュベーションした。
【0206】
手順
Histopaque(登録商標)-1077(Sigma)における密度勾配遠心によりヘパリンを添加された全血から、ヒト末梢血単核細胞(PBL)を分離する。Hanksで洗浄した後に、細胞をTurkの溶液を使用して係数し、そして細胞を、ペニシリン、ストレプトマイシンおよびグルタミン(Gibco 10378-016)および10%FBSを補充されたRPMI 1640(Gibco)中に106/mlの濃度で再懸濁させる。K562コントロール細胞(野生型)を上記したとおりに補充された同じRPMI培地中に維持する。OX40LによりトランスフェクションされたK562細胞をGeneticin(G418、Gibco)を補充された同じ培地中に50mg/mlの最終濃度に維持する。K562細胞(WTまたはOX40L+)を、1.5×105細胞/mlに同じ培地で希釈しそして50μl/ウエル(0.75×104/ウエル)において96ウエル組織培養プレートの各ウエルに分配する。Mabの適切な希釈物を20μl/ウエルの容積において細胞に加えそして37℃で1時間インキュベーションする。各希釈物を二重測定ウエル(duplicate wells)において試験する。PBLを、100μl/ウエル(105細胞/ウエル)の容積で加える。PBL対K562細胞の最終比は〜13:1である。PHA(10X)(Sigma L-9132)を20μl/ウエルで加える(最終濃度0.75μg/ml)。ウエル当たりの総容積はRPMI/10%FCSで200μlにする。プレートを5%CO2−給湿されたインキュベータ中で37℃で24時間インキュベーションする。プレートの遠心の後、上清を集めそして製造者の指定に従ってELISA(BD,San Diego, CA, Cat No2627K1)によりIL−2を試験する(図4)。IC50(OX40Lで刺激されたPBLによるIL−2放出の50%を阻止するMab濃度)を計算するために、コントロール培養物(PBL+PHA+K562WT)において産生されたバックグラウンドIL−2濃度を、K562xOX40L+細胞により刺激されたPBLにより産生された総IL−2から差し引いた(図5)。IC50 TAG34:0.07μM;LC.001:2nM;LC.005:10nM。IC50値は2〜10nMの範囲にある。
【0207】
実施例17
破傷風トキソイドにより刺激された末梢血リンパ球に対する抗体の阻害効果を試験する破傷風アッセイ「TT−アッセイ」)
末梢血単核細胞(PBMC)をFicoll Hypaqueにより、ヘパリンを添加された血液から単離した。大部分の場合に、単離されたばかりのPBMCをこのアッセイのために使用した。ある場合には冷凍保存されたPBMCも使用した。このアッセイのための培地は、10%ヒト男性AB血清(Sigma-Aldrich);2mMグルタミンおよびPen/Strep(抗生物質ペニシリンおよびストレプトマイシンのすぐ使用できる混合物(Roche Diagnostics GmbH DE);20mlに再構成された凍結乾燥物;1000ml培地当たり2mlの使用)を含有するRPMIであった。
【0208】
プラスチックに付着させるために、ウエル当たり300.000PBMCを、96ウエル平底プレートにおいて一夜プレインキュベーションした。
【0209】
次の日に破傷風トキソイド(TT)(Chiron Behring)を2〜5μg/mlの最終濃度でウエルに加えた。ポジティブコントロールウエル(最大増殖/刺激)はTTのみを含有しており、すべての他のウエルに、抗体(精製されたIgG)を10μg/mlの最終濃度になるように加えた。マウスMabTAG34は、このアッセイにおいて含まれた(最終濃度10μg/ml)。非刺激性バックグラウンドとして、コントロール培地のみを使用した。すべてのアッセイは三重測定するように(in triplicates)セットアップされた。
【0210】
6日のさらなるインキュベーション(37℃、5%CO2、95%湿度)の後、3Hチミジンを1μキューリー/mlの最終濃度となるように加え、16時間の追加のインキュベーション期間の後に、プレートを回収しそして取り込まれた3Hチミジンをβ計数器において決定した(図6)。
【0211】
実施例18
OX40L抗体のマウスOX40Lとの交差反応性
本発明の抗体のマウスOX40Lと交差反応する能力を決定するために、段階的に希釈した抗体およびコントロール抗体を、mOX40Lを安定に発現するK562−mOX40L細胞と共にインキュベーションした。K562 WT細胞およびhOX40Lを安定に発現するK562−hOX40L細胞への結合も評価した。ネガティブコントロールとして、スカシ貝ヘモシアニン(Keyhole Limpet Hemocyanin)(α−KLH)に対して指向されたHuMab抗体を使用した。抗体RM134L、ラット抗mOX40L(eBioscience, San Diego, CA)が、mOX40L発現のためのポジティブコントロールとして含まれた。抗体TAG−34、マウス抗hOX40L(MBL, Nagoya, Japan)がhOX40L発現のためのポジティブコントロールとして含まれた。結合したヒト抗体の検出のために、フルオレセイン(FITC)コンジュゲーションされたヤギ抗ヒトIgG抗体を使用した。結合したRMI34Lの検出のために、ビオチニル化ウサギ抗ラットIgG抗体(DAKO、Glostrup, Denmark)を、フィコエリトリン(PE)(DAKO)とコンジュゲーションされたストレプトアビジンと組み合わせて使用した。結合したTAG−34の検出のために、FITCコンジュゲーションされたウサギ抗マウスIgG抗体を使用した。EC50値または試験したHuMabの20μg/mlでの最大結合(Bmax)に関する計算を、Graphpad Prismソフトウエアを使用する非線形回帰(可変スロープを有するS字状用量−応答)を使用して決定した。
【0212】
結果:
本発明に従うLC.001は、5.16±2.93μg/mlのEC50値および385.22のBmax(MFI)値により示されたとおりhOX40Lに結合することができたが、それぞれ11.41および9.67のBmax(MFI)値により示されたとおりmOX40LまたはWT細胞に結合することができなかった。さらに、本発明に従うLC.001(IgG4)は、8.19±1.05μg/mlのEC50値および311.30のBmax(MFI)値により示されたとおりhOX40Lにも有効に結合することができたが、それぞれ13.47および9.58のBmax(MFI)により示されたとおりmOX40LまたはWT細胞に結合することができなかった。予想されるとおり、ネガティブコントロールα−KLHはいかなる細胞にも結合しなかった(図7)。したがって、本発明に従うOX40L抗体は、ヒトOX40Lと比較してマウスOX40Lへの少なくとも30倍低い結合を示す。
【0213】
実施例19
OX40L HuMabの補体系を活性化する潜在力
C1qおよびC3c結合ELISA
本発明の抗体のC1q結合およびC3活性化を誘導する能力を決定するために、ELISAプレートを段階的に希釈した抗体およびコントロール抗体でコーティングした。ネガティブコントロールとして、C1qに非常に弱く結合するヒトIgG4(Tke Binding Site, Birmingham, England)を使用した。ヒトIgG1(The Binding Site)およびα−KLH(IgG1)は、ポジティブコントロールとして含まれた。次いで、コーティングされた抗体を、リコンビナントC1qおよびC3のソースとしてヒトのプールされた血清と共にインキュベーションした。結合したC1qの検出のために、セイヨウワサビペルオキシダーゼ(HRP)とコンジュゲーションされたブタ抗ウサギIgG抗体(DAKO)と組み合わせたC1qに対して指向されたウサギ抗体(DAKO)を使用した。活性化されたC3c(C3の活性化により発生した)の検出のために、HRPとコンジュゲーションしたウサギ抗マウスIgG抗体(Jackson ImmunoResearch Laboratories, West Grove, PA)と組み合わせてマウス抗ヒトC3c抗体(DAKO)を使用した。コーティング有効性の差を評価するために、コーティングされた抗体を、HRPとコンジュゲーションされたヤギ抗ヒトIgG抗体で可視化した。EC50値または試験されたHuMab10μg/mlにおける最大結合(Bmax)に関する計算を、Graphpad Prismソフトウエアを使用する非線形回帰(可変スロープを有するS字状用量−応答)を使用して決定した。
【0214】
結果:
本発明に従うLC.001は、2.19±0.42μg/mlのEC50値および3.089のBmax(OD405)値により示されたとおり、C1qに有効に結合することができた。さらに、両ポジティブコントロールヒトIgG1および抗KLHは、それぞれ4.17±1.08μg/mlおよび2.57±1.51μg/mlのEC50値、ならびにそれぞれ2.685および3.306のBmax(OD405)値により示されたとおり、C1qに有効に結合することができた。予想されるとおり、ネガティブコントロールヒトIgG4は、0.353のOD405Bmax値により示されたとおり、C1qに結合しなかった。さらに、本発明に従うLC.001IgG4xは、0.357のOD405Bmax値により示されたとおり、C1qに結合する能力を失っていた。
【0215】
C1q結合能力と一致して、LC.001によるC3c沈着(deposition)が、2.67±0.16μg/mlのEC50値および2.614のBmax(OD405)値で、抗体濃度依存性方式で起こった。さらに、両ポジティブコントロールヒトIgG1および抗KLHは、それぞれ5.45±0.36μg/mlおよび2.16±0.26μg/mlのEC50値、ならびにそれぞれ2.543および2.633のBmax(OD405)値により示されたとおり、C3cを有効に沈着することができた。予想されるとおり、ネガティブコントロールヒトIgG4は、0.095のOD405Bmax値により示されたとおり、C3cを沈着しなかった。さらに、本発明に従うLC.001IgG4xは、0.090のOD405Bmax値により示されたとおり、C3cを沈着する能力を失っていた(図8および図9)。
【0216】
実施例20
OX40L HuMabsのFcγレセプターI、IIaおよびIIbに結合する潜在力
IgGで誘導される抗体依存性細胞性細胞傷害(ADCC)は、エフェクター細胞に対するFcγレセプター(FcγR)により媒介される。本発明の抗体のFcγRsに結合する能力を決定するために、ヒトFcγRI、FcγRIIa、FcγRIIbで安定にトランスフェクションされたIIA1.6細胞(IIA1細胞から限界希釈により誘導された;Jones, B., et al., J. Immunol. 136(1986) 348-356)および野生型細胞を、段階希釈された抗体およびコントロール抗体と共にインキュベーションした。ネガティブコントロールとして、FcγRIに結合しないヒトIgG2(The Binding Site Ltd)およびFcγRIIに結合しないヒトIgG4(The Binding Site)を使用した。ヒトIgG1(The Binding Site)は、FcγRI結合のためのポジティブコントロールとして含まれ、そしてヒトIgG3(The Binding Site)はFcγRII結合のためのポジティブコントロールとして含まれた。結合した抗体を、フィコエリトリン(PE)とコンジュゲーションされたヒトIgGに対して指向された抗体を使用するFACS分析により検出した。EC50値または試験されたHuMab10μg/mlにおける最大結合(Bmax)に関する計算を、Graphpad Prismソフトウエアを使用する非線形回帰曲線適合(可変スロープ)を使用して決定した。
【0217】
LC.001は、0.11±0.03μg/mlのEC50値および8041.54のBmax(MFI)値により示されたとおりFcγRIに有効に(コントロールIgG1抗体に匹敵しうる)結合することができたが、それぞれ25.06および21.18のBmax(MFI)により示されたとおりFcγRIIaおよびFcγRIIbに結合することができなかった。
【0218】
LC.001IgG4xは、0.86±0.12μg/mlのEC50値および6030.07のBmax(MFI)で、LC.001に比較してFcγRIへの結合においてより少なく有効でありそしてコントロールIgG4抗体に匹敵していた。LC.001IgG4xのFcγRIIaおよびFcγRIIbへの結合は観察されなかったが(それぞれ21.40および19.27のBmax(MFI)値)、これに対してコントロールIgG3抗体は、結合することができた(それぞれ536.65および418.59のBmax(MFI)値)(図10)。したがって、FcγRIへの結合のEC50値は、抗体LC.001のEC50値に比較してLC.001IgG4xでは8倍である。
【0219】
実施例21
OX40L HuMabsのNK細胞上のFcγRIIIaに結合する潜在力
本発明の抗体のナチュラルキラー(NK)細胞上のFcγRIIIa(CD16)に結合する能力を決定するために、末梢血単核細胞(PBMCs)を単離しそして、FcγRIIIaに対するブロッキングマウス抗体(抗CD−16、クローン3G8、RDI、Flanders,NJ)20μg/mlの存在下または不存在下にHuMab抗体およびコントロール抗体20μg/mlとインキュベーションして、FcγRIIIaを介する結合を証明した。ネガティブコントロールとして、FcγRIIIaに結合しないヒトIgG2およびIgG4(The Binding Site)を使用した。ヒトIgG1およびIgG3(The Binding Site)は、FcγRIIIa結合のためのポジティブコントロールとして含まれた。NK細胞上の結合した抗体を、PE標識されたマウス抗ヒトCD56(NK細胞表面マーカー)抗体(BD Bioscience Pharmingen, San Diego,CA)を、FITC標識されたヤギF(ab)2抗ヒトIgG(Fc)抗体(Protos immunoresearch, Burlingame,CA)と組み合わせて使用するFACSにより検出した。試験されたHuMab20μg/mlにおける最大結合(Bmax)を決定した。
【0220】
LC.001は、641.37のBmax(MFI)値により示されたとおり、FcγRIIIaに有効に(コントロールIgG1抗体に匹敵しうる)結合することができた。FcγRIIIaに対するブロッキング抗体の添加は、NK細胞へのLC.001の結合を無効にした(145.38のバックグラウンド染色に比較して194.61のBmax(MFI)値)。LC.001 IgG4xは、FcγRIIIaに結合せずそして170.52のBmax(MFI)値で、コントロールIgG4抗体に匹敵しうる挙動をして、LC.001のBmaxの約10%にすぎないLC.001 IgG4xのBmaxをもたらした。FcγRIIIaに対するブロッキング抗体の添加は、LC.001 IgG4x結合に対する効果を示さなかった(174.26のBmax(MFI)値)(図11)。
【0221】
実施例22
HUVEC(初代ヒト臍帯静脈内皮細胞/Promo Cell)へのhMab_hOX40LおよびMab TAG34結合の効果
内皮細胞は、hOX40Lを発現することが記載されている(Kotani, A., et al., Immunol. Lett. 84(2002)1-7)。ヒト臍帯静脈内皮細胞(HUVEC)は、当然hOX40Lを発現し、したがって「内皮細胞モデル」として使用することができる。このアッセイの目的は、抗体TAG−34およびLC.001への結合後のHUVEC細胞上のhOX40Lの運命を決定することであった。
【0222】
HUVECを解凍しそしてT175フラスコ(Sarstedt)中でECG−M培地+2%FCS中で4日間増大させた。細胞を24ウエルプレート中にプレートした(10.000細胞/ウエル)。
【0223】
3日後、培地をECG−M培地+0.5%FCSに変えた。10μg/mlの抗体(<KLH>(スカシ貝ヘモシアニンに対する抗体)、ダウンモデュレーション(down-modulation)の誘導のためのTAG−34またはLC.001)の添加および2,5時間または24時間のインキュベーション。TAG−34またはLC.001によるHUVEC細胞の再染色。Alexa488(=<m>)で標識されたマウスIgGに対する二次抗体またはAlexa488(=<h>)で標識されたヒトIgGに対する二次抗体、各々10μg/mlによるFACS染色。FACS測定は、FACS−scan(Becton Dickinson)において行いそして平均蛍光強度(MFI)を計算した。
【0224】
前記<KLH>抗体を非特異的ネガティブコントロールとして使用した。
【0225】
表7は、LC.001の添加は、2.5時間後も24時間後もHUVEC細胞上のOX40Lのダウンモデュレーションをもたらさない(ライン4をライン5および6と比較する)ことを示す。しかしながら、TAG34の添加は、2.5時間後、そして24時間後にHUVEC細胞上のhOX40Lの強い(約3倍)ダウンモデュレーションを示す(ライン10をライン11および12と比較する)。
【0226】
10μg/mlの濃度の本発明に従う抗体は、HUVEC細胞上のOX40L発現のダウンレギュレーションを誘導しない。
【0227】
【表7】
【0228】
実施例23
TAG34、LC.001およびLC.005のウエスタンブロット分析
40および100nghOx40L−His(R&D Systems, 28〜34kDaの理論的サイズ)および分子量マーカーMagik Mark XP(Invitrogen; 20, 30, 40, 50, 60, 80 , 100, 120, 220 kDa)をゲル電気泳動のために調製した。したがって、xμlタンパク質、2.5μlNuPage LDS(ドデシルサルフェートのリチウム塩)サンプルバッファー(4x)、1μlNuPage還元剤(10x)および10μlとする量のH2Oを一緒にしそして10分間70℃で変性した。その後サンプルをNuPageゲル(Novex; 10%Bis-Tris)上にローディングしそして1×MOPSランニングバッファー(Novex)中で150Vで1時間ランニングした。
【0229】
ゲルを半乾燥チャンバ中で1時間/50mA、1×NuPageトランスファーバッファー(1×バッファー、0.1%Antioxidance、10%メタノール)を使用してPVDF膜(Millipore; メタノール中の5分間のインキュベーションおよび1×トランスファーバッファー中の10分のインキュベーションによる膜の活性化)上に半乾燥ブロットによりブロッティングした。膜を、室温で1時間振とう下に1×PBS/5%ミルク/0.5%Tween中でブロッキングした。一次抗体(pAB)を1×PBS/1%ミルク/0.5%Tween中に希釈し、加え、4℃で一夜インキュベーションした。
LC.001: 全容積4ml中の1.9μl(1.6μg)
LC.005: 1.1μl(1.6μg)/4ml
TAG34: 1.6μl(1.6μg)/4ml
【0230】
膜を1×PBS/0.5%Tween中で10分間3回洗浄した。二次抗体を(sAB)を1×PBS/1%ミルク/0.5%Tween中に希釈し、加えそして室温で1.5時間インキュベーションした。LC.001およびLC.005については、1:10000希釈におけるヒトIgGに対するポリクローナル抗体(Pierce)をsABとして使用し;TAG34では、1:400希釈におけるLumi Light Western Blotting Kit(Roche)からのマウスIgGに対するポリクローナル抗体をsABとして使用した。膜を1×PBS/0.5%Tweenで30分間2回洗浄した。検出のために、Lumi Light Western Blotting Kit(Roche)を、製造者の指示に従って使用した。ウエスタンブロットからの結果を図12に示す。LC.001は(ドデシルサルフェート)変性されたOX40Lを検出することができるが、これに対してLC.005およびTAG34は変性されたOX40Lに結合しない。
【0231】
【表8】
【図面の簡単な説明】
【0232】
【図1a】TAG−34、LC.001、LC.005、LC.010、LC.019、LC.029、LC.033についての「結合ELISA」を示す。
【図1b】TAG−34、LC.001、LC.005、LC.010、LC.019、LC.029、LC.033についての「阻止ELISA」+IC50データを示す。
【図2】TAG−34、LC.001、LC.005についての「阻止FACS」を示す。
【図3】TAG−34、LC.001、LC.019およびLC.024についての「NFκB−アッセイ」を示す(非結合抗体)。
【図4】TAG−34、LC.001およびLC.005についての「T細胞活性化アッセイ」およびIC50値(図4:IL−2放出、図5:阻害)を示す。
【図5】TAG−34、LC.001およびLC.005についての「T細胞活性化アッセイ」およびIC50値(図4:IL−2放出、図5:阻害)を示す。
【図6】TAG−34、LC.001およびLC.033についての「TT−アッセイ」データを示す。
【図7】本発明の抗体のマウスOX40Lとのクロス反応性を示す。A)トランスフェクションされた細胞上の及びWT細胞上のhOX40L発現のためのコントロール、B)hOX40L発現K562細胞への抗体の結合、C)トランスフェクションされた細胞およびWT細胞上のmOX40L発現のためのコントロール、D)mOX40L発現K562細胞への抗体の結合、およびE)WT K562細胞への抗体の結合(n=3)。
【図8】本発明の抗体のC1qに結合する能力を示す(n=3)。
【図9】本発明の抗体のC3cを活性化する能力を示す(n=3)。
【図10】本発明の抗体のFcγRI(n=4)、FcγRIIa(n=4)およびFcγRIIB(n=4)に結合する能力を示す。
【図11】本発明の抗体のNK細胞上のFcγRIIIA(CD16)に結合する能力を示す(6つのドナーの平均±SEM)。
【図12】ウエスタンブロット;レーン1、4、7:マーカー;レーン2,5,8:100ngOX40L;レーン3,6,9:40ngOX40L。
【技術分野】
【0001】
本発明は、抗OX40L抗体、特に、補体因子Clqに結合しない抗OX40L抗体、その医薬組成物および使用に関する。好ましくは、これらの抗体はヒト抗体又はヒト化抗体である。
【0002】
ヒトOX40L(gp34、SwissProt P23510)は、CD40/CD40Lライゲーションすると活性化されたB細胞および樹状細胞に発現されそして炎症性組織において内皮細胞に発現される(Review: Weinberg, A.D., Trends Immunol. 23(2002)102-109)。それは、HTLV−1感染したヒト白血病細胞(OX40を有するオートクリンループの発生によりこれらのT細胞の不死化)から単離された。OX40Lおよび対する抗体は、例えば、WO 95/12673; WO 95/21915; WO 99/15200; Baum, P.R., et al., EMBO J. 13(1994)3992-4001; Imura, A., et al., Blood 89(1997)2951-2958; Imura, A., et al., J. Exp.Med. 183(1996)2185-2195; Kjaergaard, J., et al., I. Immunol. 167(2001)6669-6677; Lane, P., J.Exp. Med. 191(2000)201-206; Mallett, S., and Barclay, A. N., Immunol. Today 12(1991)220-223; Mallett, S., et al., EMBO J. 9(1990)1063-1068; Ndhlovu, L.C., et al., J.Immunol. 167(2001)2991-2999; Ohshima, Y., et al., J.Immunol. 159(1997)3838-3848; Rogers, P.R., et al., Immunity 15(2001)445-455; Stueber, E., and Strober, W., J. Exp.Med. 183(1996)979-989; Stueber, E., et al., Gastroenterology 115(1998)1205-1215; Takahashi, Y., et al., J.Virol. 75(2001)6748-6757; Takasawa, N., et al., Jpn. J.Cancer Res. 92(2001)377-382; Taylor, L., and Schwarz, H., J.Immunol. Meth. 255(2001)67-72; Weinberg, A.D., et al., Nature Medicine 2(1996) 183-189; Weinberg, A.D., et al., Semin. Immunol. 10(1998) 471-480; Weinberg, A.D., Trends Immunol. 23(2002) 102-109; Wu, T., et al., Transplant. Proc. 33(2001) 217-218; Higgins, L.M., et al., J. Immunol. 162(1999) 486-493; and Yoshioka, T., et al., Eur. J. Immunol. 30(2000) 2815-2823)に記載されている。ヒトOX40Lは、活性化されたCD4+T細胞上に一過性に発現されるヒトOX40(CD134)に対するリガンドである。そのリガンドによるOX40のエンゲージメントは、T細胞活性化のための共刺激性シグナルをもたらす。OX40/OX40L相互作用は、両方向性シグナルを創り出すことが記載されている(Matsumura, Y., et al., J. Immunol. 163(1999) 3007-3011; Kotani, A., et al., Immunol. Lett. 84(2002) 1-7)。さらにOX40/OX40L相互作用は、炎症性組織における内皮細胞への活性化されたT細胞の付着を媒介する。OX40Lは、活性化されたB細胞、DCおよび内皮細胞上に一過性にのみ発現されるので、OX40Lに対する抗体は炎症性応答期間中T細胞活性化および内皮細胞付着を選択的にブロッキングするであろうが、不活性化されていない末梢T細胞を変化させない。Yoshioka, A等(Eur. J. Immunol. 30(2000)2815-2823)は、慢性関節リウマチのマウスモデルにおける中和性抗mOX40L mAbの治療的潜在力を証明した。その投与は、疾患の重度を劇的に軽減した。この抗体は、他の関連した疾患モデル、例えば、炎症性皮膚疾患、実験的自己免疫疾患(EAE)、GVHD、尿炎症性腸疾患における同様な活性を示した(Yoshioka, A., et al., Eur. J. Immunol 30(1999) 2815-2823; Salek-Ardakani, S., et al., J.Exp.Med. 198(2003) 315-324; Burgess, J.K., et al., J. Allergy Clin. Immunol. 113(2004) 683-689; Hoshino, A., et al., Eur. J. Immunol. 33(2003) 861-869; Arestides, R.S., et al., Eur. J. Immunol. 32(2002) 2874-2880; Nohara, C., et al., J. Immunol. 166(2001) 2108-2115; Weinberg, A. D., et al., J. Immunol. 162(1999)1818-1826; Higgins, L.M., et al., J. Immunol. 162(1999) 486-493 ; Humphreys, I.R., et al., J.Exp.Med. 198(2003) 1237-1242; Akiba, H. et al., J.Exp. Med. 191(2003) 375-380; Ishii, N., et al., Eur. J. Immunol. 33(2003) 2372-2381; Blazar, B. R., et al., Blood 101(2003) 3741-3748; Tsukada, N., et al., Blood 95(2000) 2434-2439; Akiba, H., et al., Biochem. Biophys. Res. Commun. 251(1998) 131-136。
【0003】
OX40Lに対する抗体は種々の疾患モデルにおけるそれらの抗炎症性効果について調べられた(Sugamura, K., et al., Nat. Rev. Immunol;. 4(2004)420-431)。
【0004】
Tanaka, Y., et al, Int. J. Cancer 36, (1985)549-555; Tozawa, H., et al., Int. J. Cancer 41(1998)231-238; and Miura, S., et al., Mol. Cell. Biol. 11(1991) 1313-1325は、ヒトT細胞白血病ウイルスI型(HTLV−I)を有するヒトリンパ球の系統の表面抗原と反応するTARM−34およびTAG−34と名づけられたマウスモノクローナル抗体を記載している。TAG−34抗体は、MBL International Corporationから市販されている。TAG−34はOX40Lにも結合する。
【0005】
本発明の要約
本発明は、抗体、好ましくはモノクローナル抗体であって、該抗体は、OX40Lに結合し、ヒト起源のFc部分を含有しそしてヒト補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合しないことを特徴とする抗体、好ましくはモノクローナル抗体に関する。
【0006】
本発明は、更に、抗体、好ましくはモノクローナル抗体であって、該抗体は、ヒト起源のFc部分を含有し、100ngの抗体濃度でOX40Lに結合しそして変性されたOX40Lに結合する(ウエスタンブロットにおいて)ことを特徴とする抗体、好ましくはモノクローナル抗体に関する。この抗体は、モノクローナル抗体LC.001が結合するエピトープと同じOX40Lポリペプチドエピトープに結合する。このような抗体は、例えば、LC.001、LC.033およびLC.060である。これらの抗体は、好ましくはヒトIgG1型(野生型)であるか、またはヒト補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合しない。
【0007】
本発明は、さらに可変軽鎖および可変重鎖を含むことを特徴とし、該可変重鎖はCDR1、CDR2およびCDR3を含むことを特徴とし、該CDR3は配列番号33〜38から選ばれることを特徴とするOX40Lに結合する抗体に関する。CDR1が配列番号21〜25から選ばれ、CDR2が配列番号26〜32から選ばれそしてCDR3が配列番号33〜38から選ばれることは特に好ましい。
【0008】
本発明に従う抗体は、好ましくは、可変軽鎖および可変重鎖を含むことを特徴とし、該可変軽鎖はCDR1、CDR2およびCDR3を含むことを特徴とし、該CDR3は配列番号51〜57から選ばれることを特徴とする。CDR1が配列番号39〜44から選ばれ、CDR2が配列番号45〜50から選ばれそしてCDR3が配列番号51〜57から選ばれることが特に好ましい。
【0009】
本発明に従う抗体は、好ましくは、可変重鎖および可変軽鎖を含むことを特徴とし、該可変重鎖はCDR1、CDR2およびCDR3を含むことを特徴とし、重鎖のCDR3は配列番号33〜38から選ばれそして軽鎖のCDR3は配列番号51〜57から選ばれることを特徴とする。可変重鎖は、配列番号21〜25から選ばれるCDR1、配列番号26〜32から選ばれるCDR2および配列番号33〜38から選ばれるCDR3を含み、そして可変軽鎖は、配列番号39〜44から選ばれるCDR1、配列番号45〜50から選ばれるCDR2および配列番号51〜57から選ばれるCDR3を含むことが特に好ましい。
【0010】
すべてのCDRsは互いに独立に、しかし当然のこととして、抗体がOX40Lに結合するような方法で選ばれる。したがって、同じLC抗体の軽鎖及び重鎖のCDRsを組み合わせることができ、またはLC.001の軽鎖CDRsはLC.001、LC.059またはLC.063の重鎖CDRsと組み合わせることができる。各鎖上のCDRsはフレームワークアミノ酸により分離されている。
【0011】
本発明に従う抗体は、好ましくは、抗体が、
a)配列番号1のアミノ酸配列の軽鎖(VL)可変CDRsおよび配列番号2の重鎖(VH)可変CDRs、
b)配列番号3のアミノ酸配列の軽鎖可変CDRsおよび配列番号4の重鎖可変CDRs、
c)配列番号5のアミノ酸配列の軽鎖可変CDRsおよび配列番号6の重鎖可変CDRs、
d)配列番号7のアミノ酸配列の軽鎖可変CDRsおよび配列番号8の重鎖可変CDRs、
e)配列番号9のアミノ酸配列の軽鎖可変CDRsおよび配列番号10の重鎖可変CDRs、
f)配列番号11または16のアミノ酸配列の軽鎖可変CDRsおよび配列番号12の重鎖可変CDRs、
g)配列番号1のアミノ酸配列により規定された軽鎖(VL)可変ドメインおよび配列番号17により規定された重鎖(VH)可変ドメイン、
h)配列番号18のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号19により規定された重鎖可変ドメイン、
i)配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号20により規定された重鎖可変ドメイン、または
そのOX40Lに結合性のフラグメント
からなる群より独立に選ばれるCDRsを含むことを特徴とする。
【0012】
本発明に従う抗体は、好ましくは、該抗体が、
a)配列番号1のアミノ酸配列により規定された軽鎖(VL)可変ドメインおよび配列番号2により規定された重鎖(VH)可変ドメイン、
b)配列番号3のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号4により規定された重鎖可変ドメイン、
c)配列番号5のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号6により規定された重鎖可変ドメイン、
d)配列番号7のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号8により規定された重鎖可変ドメイン、
e)配列番号9のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号10により規定された重鎖可変ドメイン、
f)配列番号11または16のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号12により規定された重鎖可変ドメイン、
g)配列番号1のアミノ酸配列により規定された軽鎖(VL)可変ドメインおよび配列番号17により規定された重鎖(VH)可変ドメイン、
h)配列番号18のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号19により規定された重鎖可変ドメイン、
i)配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号20により規定された重鎖可変ドメイン、
またはそのOX40Lに結合性のフラグメント、
からなる群より独立に選ばれる可変領域を含むことを特徴とする。
【0013】
本発明に従う抗体は、好ましくは、ヒト軽鎖可変領域が、配列番号1、3、5、7、9、11,16および18からなる群より独立に選ばれるアミノ酸配列を含むことを特徴とする。
【0014】
本発明に従う抗体は、好ましくは、ヒト重鎖可変領域が、配列番号2、4、6、8、10、12,17、19および20からなる群より独立に選ばれるアミノ酸配列を含むことを特徴とする。
【0015】
重鎖および軽鎖のCDR領域は配列番号21〜38および39〜57に示される。
【0016】
本発明に従う抗体は、好ましくは、該抗体が配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号2、17または20により規定された重鎖可変ドメインを含むことを特徴とする。
【0017】
本発明に従う抗体は、好ましくは、ヒト重鎖定常領域が配列番号14および15からなる群より独立に選ばれるアミノ酸配列または配列番号58の重鎖定常領域を含むことを特徴とする。
【0018】
本発明に従う抗体は、好ましくは、該抗体が配列番号13のκ軽鎖定常領域または配列番号61、65もしくは69の軽鎖定常領域を含むことを特徴とする。
【0019】
好ましくは、本発明に従う抗体は、OX40Lに結合することおよびヒトIgG1クラス(野生型)であることにより特徴付けられそしてγ重鎖として配列番号58、62または66を含む。特に好ましいのは、
a)γ重鎖として配列番号58およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号62およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号66およびκ軽鎖として配列番号69、
を含む抗体である。
【0020】
本発明の更なる態様は、OX40Lに結合する抗体であって、細胞系hu−Mab<hOX40L>LC.001、hu−Mab<hOX40L>LC.005、hu−Mab<hOX40L>LC.010、hu−Mab<hOX40L>LC.019、hu−Mab<hOX40L>LC.029、またはhu−Mab<hOX40L>LC.033により産生されることを特徴とする抗体である。
【0021】
本発明な従う抗体は、好ましくはキメラ抗体、ヒト抗体またはヒト化抗体である。
【0022】
本発明に従う抗体は、BIAcoreアッセイにおいて10-8M未満(10-12〜10-8M)のKD値でOX40Lに結合することにより特徴付けられ、更に好ましくは、BIAcoreアッセイにおいて10-12〜10-9MのKD範囲により特徴付けられる。
【0023】
本発明に従う抗体は、好ましくは、4nM以下のIC50値で0.5μg/mlのコーティング濃度で固定化されたOX40L(好ましくはストレプトアビジン表面に固定化されたビオチニル化OX40L)を使用するELISAにおいてOX40LのOX40との相互作用を阻害する。さらに好ましくは、IC50値は1〜4nMの範囲にある。
【0024】
本発明に従う抗体は、好ましくは、補体因子C1qへの抗体の非結合(non-binding)は、C1qに対する10μg/mlの濃度での抗体の最大結合(Bmax)が抗体LC.001のBmaxに比較して30%以下、好ましくは20%以下であるELISAアッセイ測定値を指すことを特徴とする。
【0025】
好ましくは、抗体は、ヒトFcγRI、FcγRIIAおよび/またはFcγRIIIAに結合しない。特に好ましくは、抗体はNKエフェクター細胞上のヒトFcγレセプターに結合しない。
【0026】
本発明に従う抗体は、好ましくは、NK細胞上のFcγレセプターへの抗体の非結合は、NK細胞に対する20μg/mlの濃度での抗体の最大結合(Bmax)が抗体LC.001のBmaxに比較して20%以下、好ましくは10%以下である、アッセイを指すことを特徴とする。
【0027】
本発明に従う抗体は、好ましくは、それがFcγRIに結合しないことを特徴とする。これは、FcγRIIAおよびFcγIIBを欠いているがリコンビナントFcγRIを発現しているB細胞リンパ腫細胞に対する0.078〜10μg/mlの範囲の濃度の抗体の結合を試験するアッセイで測定されるとき、LC.001のEC50値に比べて5倍以上、好ましくは7倍以上、例えば8倍以上であるEC50により抗体が特徴付けられることを意味する。
【0028】
本発明に従う抗体は、好ましくは、補体因子C1qへの非結合および/またはNK細胞上のヒトFcγレセプターへの非結合を引き起こす、好ましくはヒトFc部分における少なくとも1つのアミノ酸突然変異を含むIgG4抗体またはIgG1抗体であるとして特徴付けられる。
【0029】
本発明に従う抗体は、好ましくは、それが補体因子C3を活性化しないことを特徴とする。
【0030】
本発明に従う抗体は、好ましくは、ヒトサブクラスIgG4であることにより特徴付けられる。本発明のさらなる好ましい態様では、抗体は、E233、L234、L235、G236、D270、N297、E318、K320、K322、A327、A330、P331および/またはP329(EUインデックスに従う番号付け)における少なくとも1つの突然変異を含む、任意のIgGクラス、好ましくはIgG1またはIgG4であることにより特徴付けられる。特に好ましいのは、IgG1突然変異PVA236、L234A/L235Aおよび/またはGLPSS331ならびにIgG4突然変異L235Eである。IgG4サブクラスの抗体が、突然変異S228Pまたは突然変異S228PおよびL235Eを含有することが更に好ましい(Angal, S., et al., Mol. Immunol. 30(1993)105−108)。
【0031】
したがって、本発明に従う抗体は、好ましくは、PVA236、GLPSS331および/またはL234A/L235A(EUインデックスに従う番号付け)からの1つ以上の突然変異を含む、ヒトサブクラスIgG1の抗体である。
【0032】
好ましくは、本発明に従う抗体は、OX40Lに結合すること、突然変異L234A/L235Aを含有するIgG1クラスであることにより特徴付けられ、そしてγ重鎖として配列番号59、63または67を含む。
【0033】
特に好ましいのは、
a)γ重鎖として配列番号59およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号63およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号67およびκ軽鎖として配列番号69、
を含む抗体である。
【0034】
好ましくは、突然変異S228Pを含有するIgG4クラスであることにより特徴付けられる本発明に従う抗体は、γ重鎖として配列番号60、64または68を含む。
【0035】
特に好ましいものは、
a)γ重鎖として配列番号60およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号64およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号68およびκ軽鎖として配列番号69、
を含む抗体である。
【0036】
本発明に従う抗体は、好ましくは、それが補体依存性細胞傷害(CDC)を誘発しないことを特徴とする。
【0037】
本発明に従う抗体は、好ましくは、それが抗体依存性細胞性細胞傷害(ADCC)を誘発しないことを特徴とする。
【0038】
したがって、本発明は、抗OX40L抗体または単一重鎖もしくは単一軽鎖であって、それらのCDRs、可変領域、完全アミノ酸配列またはハイブリドーマにより特徴付けられ、そしてFc部分を含まないか、または任意の型のFc部分を含み、好ましくは、ヒト起源の改変されていないまたは上記した突然変異により改変されている、ヒトIgG1 FcもしくはヒトIgG4 Fcを含む、抗OX40L抗体または単一重鎖もしくは単一軽鎖を含む。
【0039】
したがって、本発明は、抗体、好ましくはモノクローナル抗体であって、該抗体は、OX40Lに結合し、ヒト由来のFc部分を含有しそしてヒト補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合せず、ヒトIgG4型であるかまたは上記した突然変異により改変されたヒトIgG1もしくはヒトIgG4であることを特徴とする抗体、好ましくはモノクローナル抗体も含む。
【0040】
従って、本発明は、抗体、好ましくはモノクローナル抗体であって、該抗体は、100ngの抗体濃度で、OX40Lにおよび変性されたOX40Lに結合する(ウエスタンブロットにおいて)ことを特徴とする抗体、好ましくはモノクローナル抗体も含む。この抗体は、モノクローナル抗体LC.001が結合するエピトープと同じOX40Lポリペプチドエピトープに結合する。抗体は、Fc部分含まないか、または任意の型のFc部分を含み、好ましくは、野生型もしくは上記した突然変異により修飾されたヒトIgG1もしくはヒトIgG4を含む。
【0041】
本発明に従う抗体は、OX40Lに対する抗体による治療を必要としている患者、特に炎症性疾患、特に慢性関節リウマチ、アレルギー性喘息および移植におけるGvHD(Sugamura, K., et al., Nat. Rev. Immunol. 4(2004)420-431も参照)に罹患している患者に有益な新規で発明的な性質を有する。
【0042】
本発明のさらなる態様は、本発明に従う抗体分子、その可変鎖またはそのCDRドメインをコードする核酸分子である。
【0043】
本発明の好ましい態様では、抗体は、Fab、F(ab’)2、または一本鎖フラグメントである。
【0044】
本発明のさらなる態様は、本発明に従う核酸分子を含むベクターである。
【0045】
本発明のさらなる態様は本発明に従うベクターを含む宿主細胞である。
【0046】
本発明のさらなる態様は、本発明に従う抗体の製造方法であって、該抗体分子の合成を可能とする条件下に本発明に従う宿主細胞を培養し、そしてこの培養物から該抗体分子を回収することを含む方法である。
【0047】
本発明のさらなる態様は、本発明に従う抗体の組成物、好ましくは医薬組成物または診断組成物である。
【0048】
本発明のさらなる態様は、本発明に従う抗体および少なくとも1種の薬学的に許容されうる賦形剤を含む医薬組成物である。
【0049】
本発明のさらなる態様は、本発明に従う治療的に有効量の抗体を患者に投与することを特徴とする治療を必要としている患者の処置のための方法である。
【0050】
本発明のさらなる態様は、治療、好ましくは炎症性疾患の処置のための、特に慢性関節リウマチ、喘息およびGvHD(宿主に対する移植片疾患)の処置および/または予防のための本発明に従う抗体の使用である。
【0051】
本発明のさらなる態様は、炎症性疾患の予防および/または処置のための、好ましくは慢性関節リウマチ、喘息およびGvHDの処置用の医薬の製造のための本発明に従う抗体の使用である。
【0052】
本発明のさらなる態様は、本発明に従う抗体、本発明に従う核酸分子、本発明に従うベクターまたは本発明に従う宿主細胞を含む診断キットである。
【0053】
発明の詳細な説明
用語「OX40L」は、TNFリガンドファミリーに属するII型膜タンパク質を指す。更なる名称は、ACT−4レセプター、CD134L、g34またはTNF4_Humanである。それは34kDaの分子量を有しそしてアクセッションナンバーP23510でSwissProtに保存されている。
【0054】
用語「OX40」は、OX40Lに結合するレセプターを指す。それはTNFレセプターファミリーに属するI型膜タンパク質である。さらなる名称は、ACT−4、OX40Lレセプター、CD134抗原、ACT35抗原、TNR4_Humanである。それは50kDaの分子量を有しそしてアクセッションナンバーP43489でSwissProtに保存されている。
【0055】
用語「抗体」は、種々の形態の抗体、好ましくは、完全抗体、抗体フラグメント、ヒト抗体、キメラ抗体、ヒト化抗体(humanized antibodies)および本発明に従う特性が保持されている限り、遺伝子工学的に作成された抗体(変異体または突然変異体抗体)を含むが、それらに限定されない種々の形態の抗体、好ましくはモノクローナル抗体を包含する。特に好ましいのは、ヒトモノクローナル抗体またはヒト化モノクローナル抗体、特にリコンビナントヒト抗体である。
【0056】
本明細書で使用された用語「モノクローナル抗体」または「モノクローナル抗体組成物」は、単一のアミノ酸組成の抗体分子の調製物を指す。
【0057】
用語「キメラ抗体」は、通常リコンビナントDNA技術により作成された、1種のソースもしくは種からの可変領域、即ち結合領域および異なるソースもしくは種に由来する定常領域の少なくとも一部を含むモノクローナル抗体を指す。マウス可変領域およびヒト定常領域を含むキメラ抗体が好ましい。本発明により包含される他の好ましい形態の「キメラ抗体」は、定常領域を元の抗体の定常領域から改変しまたは変化させて、特にC1q結合および/またはFcレセプター(FcR)結合に関して、本発明に従う性質を発生させたキメラ抗体である。このようなキメラ抗体は「クラススイッチ抗体(class-switched antibodies)」とも呼ばれる。キメラ抗体は、免疫グロブリン可変領域をコードするDNAセグメントおよび免疫グロブリン定常領域をコードするDNAセグメントを含む発現された免疫グロブリン遺伝子の産物である。キメラ抗体を産生するための方法は、慣用のリコンビナントDNAを含みそして遺伝子トランスフェクション技術は当技術分野で周知されている。例えば、Morrison, S.L., et al., Proc. Natl. Acad. Sci. USA 81(1984)6851-6855; US Patent Nos. 5,202,238 and 5,204,244参照。
【0058】
用語「ヒト化抗体」は、フレームワークまたは「相補性決定領域」(CDR)が改変されて親免疫グロブリンの特異性と比較して異なる特異性の免疫グロブリンのCDRを含む抗体を指す。好ましい態様では、マウスCDRを、ヒト抗体のフレームワーク領域に移植させて「ヒト化抗体」を調製する。例えば、Riechmann, L., et al., Nature 332(1988)323-327; and Neuberger, M.S., et al., Nature 314(1985)268-270参照。特に好ましいCDRsは、キメラおよび二官能性抗体について上記した抗原を認識する配列を示すCDRsに相当する。本発明により包含された他の形態の「ヒト化抗体」は、定常領域を元の抗体の定常領域からさらに改変もしくは変化させて、特にC1q結合および/またはFcレセプター(FcR)結合に関して、本発明に従う性質を発生させたヒト化抗体である。
【0059】
本明細書で使用された用語「ヒト抗体」は、ヒト生殖系列免疫グロブリン配列に由来する可変領域及び定常領域を有する抗体を含むことを意図する。ヒト抗体は、当技術水準で周知されている(van Dijk, M. A.., and van de Winkel, J. G., Curr. Opin. Chem. Biol. 5(2001)368-374)。ヒト抗体は、免疫感作されると、内因性免疫グロブリン産生の不存在下にヒト抗体のフルレパートリーまたは選択を生じさせることができるトランスジェニック動物(例えば、マウス)において産生させることもできる。このような生殖系列突然変異体マウスにおけるヒト生殖系列免疫グロブリン遺伝子アレーのトランスファーは、抗原チャレンジによるヒト抗体の産生をもたらすであろう(例えば、Jakobovits, A., et al., Proc. Natl. Acad. Sci. USA 90(1993)2551-2555; Jakobovits, A., et al., Nature 362(1993)255-258; Bruggemann, M., et al., Year Immunol. 7(1993)33-40参照)。ヒト抗体は、ファージディスプレーライブラリーにおいて産生させることもできる(Hoogenboom, H. R.., and Winter, G., J. Mil. Biol. 227(1992)381-388; Marks, J.D., et al., J. Mol. Biol. 222(1991)581-597)。Cole et al. and Boerner et al.,の技術も、ヒトモノクローナル抗体の産生のために利用可能である(Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p. 77(1985); and Boerner, P., et al., J. Immunol. 147(1991)86-95)。本発明に従うキメラ抗体およびヒト化抗体について既に述べたとおり、本明細書で使用された用語「ヒト抗体」は、定常領域において改変されて特にC14q結合および/またはFcR結合に関して、例えば「クラススイッチ」、即ちFc部分の変化または突然変異(IgG1からIgG4へのおよび/またはIgG1/IgG4突然変異)により、本発明に従う性質を発生させたこのような抗体も含む。さらに、本発明は、C1qおよび/またはFcRに結合するOX40Lに対するモノクローナルヒト抗体を含む。このようなヒト抗体は、マウスOX40Lと比較してヒトOX40Lについて高い選択性により特徴付けられ(ヒトOX40Lへの結合よりもマウスOX40Lへの結合が>30倍低い)そして500nMの濃度のまでTNFαまたはCD40Lへの非特異的結合を示さない。このような抗体は、C1qおよび/またはFcRに結合しない抗体の発生のために有用である。
【0060】
本明細書で使用された用語「リコンビナントヒト抗体」は、リコンビナント手段により調製され、発現され、創生されまたは単離されるすべてのヒト抗体、例えばNSOもしくはCHO細胞などの宿主細胞からまたはヒト免疫グロブリン遺伝子用のトランスジェニック動物(例えばマウス)から単離された抗体、または宿主細胞にトランスフェクションされたリコンビナント発現ベクターを使用して発現された抗体を含むことを意図する。このようなリコンビナントヒト抗体は、再配列された形態において可変領域と定常領域を有する。本発明に従うリコンビナントヒト抗体は、in vivo体細胞超突然変異を受けている。したがって、リコンビナント抗体のVHおよびVL領域のアミノ酸配列は、ヒト生殖系列VHおよびVL配列に由来するおよび関連するが、in vivoでヒト抗体生殖系列レパートリーの範囲内に天然に存在しないことがありうる配列である。
【0061】
本明細書で使用された「可変領域」(軽鎖の可変領域(VL)、重鎖の可変領域(VH))は、抗体を抗原に結合させることに直接関与する軽鎖および重鎖のペアーの各々を示す。可変ヒト軽鎖および重鎖のドメインは、同じ一般的構造を有しそして各ドメインは、3つの「超可変領域」(または相補性決定領域、CDRs)により接続された、その配列が広く保存されている4つのフレームワーク(FR)領域を含む。フレームワーク領域は、βシートコンフォメーションを採用しておりそしてCDRsはβシート構造を接続するループを形成することができる。各鎖におけるCDRsは、フレームワーク領域によりそれらの三次元構造に保持されそして他の鎖からのCDRsと共に抗原結合部位を形成する。抗体重鎖および軽鎖CDR3領域は、本発明に従う抗体の結合特異性/アフィニティーにおいて特に重要な役割を演じ、したがって本発明のさらなる目的を与える。
【0062】
本明細書で使用されるとき用語「超可変領域」または「抗体の抗原結合部分」は、抗原結合の責任を担う抗体のアミノ酸残基を指す。超可変領域は、「相補性決定領域」または「CDRs」からのアミノ酸残基を含む。「フレームワーク」または「FR」領域は、本明細書で定義されたとおりの超可変領域残基以外のこれらの可変ドメイン領域である。したがって、抗体の軽鎖および重鎖は、N末端からC末端までドメインFR1、CDR1、FR2、CDR2、FR3、CDR3およびFR4を含む。各鎖上のCDRsは、このようなフレームワークアミノ酸により分離されている。特に、重鎖のCDR3は、抗原結合に最も寄与する領域である。CDRおよびFR領域は、Kabat等、Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD(1991)の標準定義に従って決定される。
【0063】
本明細書で使用された用語「核酸または核酸分子」は、DNA分子およびRNA分子を含むことを意図する。核酸分子は一本鎖または二本鎖であることができるが、好ましくは二本鎖DNAである。
【0064】
核酸は、それが他の核酸配列との機能的関係に置かれているとき「作用可能に連結されている(operably linked)」。例えば、プレ配列又は分泌リーダーのためのDNAは、もしもそれがポリペプチドの分泌に関与するプレタンパク質として発現されるならば、ポリペプチドのためのDNAに作用可能に連結されており;プロモーターまたはエンハンサーはもしもそれが配列の転写に影響を与えるならば、コード化配列に作用可能に連結されており;またはリボソーム結合部位は、もしそれが翻訳を促進するように位置しているならば、コード化配列に作用可能に連結されている。一般に、「作用可能に連結された」とは、連結されるDNA配列が連続的であり(contiguous)そして分泌リーダーの場合には、連続的でありそしてリーディングフレームにあることを意味する。しかしながら、エンハンサーは、連続的である必要はない。連結は都合の良い制限部位においてライゲーションにより達成される。このような部位が存在しないならば、合成オリゴヌクレオチドアダプターまたはリンカーが慣用の実施に従って使用される。
【0065】
本明細書に使用された、「細胞」、「細胞系」および「細胞培養物」という表現は、交換可能に使用されそしてすべてのこのような命名は子孫を含む。従って、語「トランスフォーマント」および「トランスフォーメーションされた細胞」は、トランスファーの数に関係なく初代対象細胞(primary subject cell)およびそれに由来する培養物を含む。すべての子孫は、意図的な突然変異または故意でない突然変異により、DNA内容において正確に同一でなくてもよいことも理解される。最初にトランスフォーメーションされた細胞においてスクリーニングされたのと同じ機能または生物学的活性を有する変異体子孫は含まれる。別の命名が意図される場合には、それは状況から明らかであろう。
【0066】
「定常ドメイン」は、抗体を抗原に結合することに直接関与していないが、種々のエフェクター機能を示す。抗体または免疫グロブリンは、それらの重鎖の定常領域のアミノ酸配列に依存して、クラス:IgA、IgD、IgE、IgGおよびIgMに分けられ、そしてこれらのいくつかはサブクラス(アイソタイプ)、例えばIgG1、IgG2、IgG3およびIgG4、IgA1およびIgA2に更に分けられうる。免疫グロブリンの異なるクラスに対応する重鎖定常領域は、それぞれα、ε、γおよびμと呼ばれる。本発明に従う抗体は好ましくはIgG型である。
【0067】
本発明に従う抗体は、Fc部分として、好ましくはヒト起源に由来するFc部分および好ましくはヒト定常領域のすべての他の部分を含有する。抗体のFc部分は、補体活性化、C1q結合、C3活性化およびFcレセプター結合に直接関与している。補体系に対する抗体の影響はある条件に依存するが、C1qへの結合はFc部分における規定された結合部位により引き起こされる。このような結合部位は、当技術分野の技術水準で知られておりそして例えば、Lukas, T.J., et al., J. Immunol. 127(1981)2555-2560; Brunhouse, R.., and Cebra, J. J., Mol. Immunol. 16(1979)907-917; Burton, D.R., et al., Nature 288(1980)338-344; Thommesen, J. E., et al., Mol. Immunol. 37(2000)995-1004; Idusogie, E.E., et al., J. Immunol. 264(2000)4178-4184; Hezareh, M.., et al., J. Virol. 75(2001)12161-12168; Morgan, A., et al., Immunology 86(1995)319-324; およびEP 0 307 434により記載されている。このような結合部位は、例えばL234、L235、D270、N297、E318、K320、K322、P331およびP329(KabatのEUインデックスに従う番号付け、下記参照)である。サブクラスIgG1、IgG2およびIgG3の抗体は、通常補体活性化、C1q結合およびC3活性化を示すが、これに対してIgG4は補体系を活性化せず、C1qに結合せずそしてC3を活性化しない。本明細書で使用された用語「ヒト起源由来のそしてヒト補体因子C1qおよび/またはNK細胞上のヒトFc□レセプターに結合しないFc部分」は、サブクラスIgG4のヒト抗体のFc部分、または下記に定義されたC1q結合、C3活性化および/またはFcR結合が検出できないような方法で改変されているサブクラスIgG1、IgG2およびIgG3のヒト抗体のFc部分である、Fc部分を示す。「抗体のFc部分」は、当業者に周知のそして抗体のパパイン開裂に基づいて定義された用語である。好ましくは、Fc部分はヒトFc部分でありそしてヒトIgG4サブクラスからの、好ましくはヒンジ領域において突然変異した(例えば、S228Pおよび/またはL235E)Fc部分またはヒトIgG1サブクラスからの突然変異したFc部分である。最も好ましいのは、配列番号14および15に示された領域あるいは配列番号58,59、60、突然変異L234AおよびL235Aを有する配列番号14または突然変異S228Pもしくは突然変異S228PおよびL235Eを有する配列番号15に含まれる領域から選ばれる重鎖定常領域を含むFc部分である。
【0068】
本発明は、OX40Lに結合しそして補体因子C1qおよび/またはFcレセプターに結合しない抗体を指す。本発明の好ましい態様では、これらの抗体は、補体依存性細胞傷害(CDC)および/または抗体依存性細胞性細胞傷害(ADCC)を誘発しない。好ましくは、この抗体は、それがOX40Lに結合し、ヒト起源に由来するFc部分を含有しそして補体因子C1qに結合しないことを特徴とする。さらに好ましくは、この抗体はヒトまたはヒト化モノクローナル抗体である。
【0069】
抗体Fc領域のFc部分により媒介されるエフェクター機能は、抗体の抗原への結合の後に作動するエフェクター機能を指す(これらの機能は、Fcレセプターによる補体カスケードの活性化および/または細胞活性化を含む)。
【0070】
補体カスケードの機能は、CH50アッセイにより評価されうる。抗赤血球抗体(EA)で感作されたヒツジ赤血球を試験血清に加えて溶血をもたらす古典的経路を活性化する。赤血球の50%を溶解するのに必要な血清の容積は、CH50単位を決定する。AP−CH50は、代替経路(alternative pathway)およびターミナル経路(terminal pathway)を測定する。手順はウサギ赤血球を使用することを除いては同様である。代替経路は試験血清を添加すると活性化される。
【0071】
C1qおよび2つのセリンプロテアーゼ、C1rおよびClsは、複合体C1、補体依存性細胞傷害(CDC)経路の第1成分を形成する。補体カスケードを活性化するために、C1qは、抗原性標的に結合されたIgG1の少なくとも2つの分子またはIgMの1つの分子に結合する(Ward, E. S.., and Ghetie, V., Ther. Immunol. 2(1995)77-94)。Burton, D. R.,は、アミノ酸残基318〜337を含む重鎖領域が、補体結合反応(complement fixation)に関与していると述べている(Mol. Immunol. 22(1985)161-206)。Duncan, A. R., およびWinter, G. (Nature 332(1988)738-740)は、部位特異的突然変異誘発を使用して、Glu318、Lys320およびLys322がC1qへの結合部位を形成することを報告した。C1qの結合におけるGlu318、Lys320およびLys322残基の役割は、これらの残基を含有する短い合成ペプチドの補体媒介溶解を阻害する能力により確かめられた。
【0072】
用語「補体依存性細胞傷害(CDC)」は、補体の存在下に本発明に従う抗体による、OX40L発現ヒト内皮細胞の溶解を指す。CDCは、好ましくは、補体の存在下に本発明に従う抗体によるOX40L発現ヒト内皮細胞の処理により測定される。細胞は、好ましくはカルセインで標識される。もしも抗体が30μg/mlの濃度で標的細胞の20%以上の溶解を誘導するならば、CDCが見出される。本発明者は、本発明に従う抗体の性質について、ELISAアッセイにおける補体因子C1qへの減少した結合が必須であることを見出した。このようなアッセイにおいて、一般に、ELISAプレートをある濃度範囲の抗体でコーティングし、それに精製されたヒトC1qまたはヒト血清を加える。C1q結合は、C1qに対して指向された抗体により、続いてペルオキシダーゼで標識されたコンジュケートにより検出される。結合の検出(最大結合Bmax)は、ペルオキシダーゼ基質、ABTS(登録商標)(2,2’−アジノ−ジ−[3−エチルベンゾチアゾリン−6−スルホネート(6)]について405nmにおける光学濃度(OD405)として測定される。したがって、本発明は、補体因子C1qへの抗体の非結合は、10μg/mlの抗体の濃度での本発明に従う抗体へのC1qの最大結合(Bmax)が抗体LC.001で観察されたBmaxの20%以下、好ましくは10%以下である、このようなELISAアッセイ測定値を指す。
【0073】
本発明に従う抗体は、ELISAアッセイにおける補体因子C3の減少した活性化を示すことが更に好ましい。このアッセイはC1qアッセイと同じ方式で行われる。このようなアッセイにおいて、一般にELISAプレートを所定の濃度範囲の抗体でコーティングし、それにヒト血清を加える。C3結合は、C3に対して指向された抗体により、次いでペルオキシダーゼで標識されたコンジュゲートにより検出される。結合(最大結合Bmax)の検出は、ペルオキシダーゼ基質ABTS(登録商標)について405nmにおける光学濃度(OD405)として測定される。したがって、本発明は、補体因子C3への抗体の非結合は、10μg/mlの抗体の濃度で抗体へのC3の最大結合(Bmax)が抗体LC.001のBmaxの10%以下、好ましくは5%以下である、このようなELISAアッセイ測定値を指す。
【0074】
用語「抗体依存性細胞性細胞傷害(ADCC)」は、Fcレセプター結合により媒介される機能であり、そしてエフェクター細胞の存在下に本発明に従う抗体によるOX40L発現標的細胞の溶解を指す。ADCCは、好ましくは、エフェクター細胞、例えば単離されたばかりのPBMC(末梢血単核細胞)またはバフィーコートから精製されたエフェクター細胞、例えば単球またはNK(ナチュラルキラー)細胞の存在下に、本発明に従う抗体によるOX40L発現赤血球系細胞(例えばリコンビナントヒトOX40Lを発現するK562細胞)の調製物の処理により測定される。標的細胞を、51Crで標識し、次いで抗体と共にインキュベーションする。標識された細胞をエフェクター細胞と共にインキュベーションし、そして上清を放出された51Crについて分析する。コントロールは、抗体なしで標的内皮細胞のエフェクター細胞とのインキュベーションを含む。ADCCを媒介する最初の段階を誘導する抗体の能力を、Fcγレセプター発現細胞、例えばFcγRIおよび/またはFcγRIIAをリコンビナント的に発現する細胞またはNK細胞(本質的にFcγRIIIAを発現する)へのそれらの結合を測定することにより調べた。好ましくはNK細胞上のFcγRへの結合を測定する。
【0075】
Fcレセプター結合性エフェクター機能は、造血細胞上の特殊化された細胞表面レセプターであるFcレセプター(FcRs)と抗体のFc領域の相互作用により媒介されうる。Fcレセプターは免疫グロブリンスーパーファミリーに属し、そして抗体依存性細胞性細胞傷害(ADCC)を介して、免疫複合体のファゴサイトーシスによる抗体でコーティングされた病原体の除去ならびに、対応する抗体でコーティングされた赤血球および種々の他の細胞標的(例えば腫瘍細胞)の溶解の両方を媒介することが示された。Van de Winkel, J. C., and Anderson, C. L., J. Leukoc. Biol. 49(1991)511-524)。FcRsは、免疫グロブリンアイソタイプに対するそれらの特異性により定義され;IgG抗体に対するFcレセプターは、FcγRと呼ばれ、IgEに対してはFcεRと呼ばれ、IgAに対してはFcαRと呼ばれる等である。Fcレセプター結合は、例えば、Ravetch, J. V., and Kinet, J. P., Annu. Rev. Immunol. 9(1991)457-492; Capel, P. J., et al., Immunomethods 4(1994)25-34; de Haas, M., et al., J. Lab. Clin. Med. 126(995)330-341; and Gessner, J. E., et al., Ann. Hematol. 76(1998)231-248に記載されている。
【0076】
IgG抗体のFcドメインに対するレセプター(FcγR)の架橋は、ファゴサイトーシス、抗体依存性細胞性細胞傷害および炎症性メディエーターの放出、ならびに免疫複合体クリアランスおよび抗体産生の調節を含む広範な種類のエフェクター機能をトリガーする。ヒトでは、3つのクラスのFcγRが特徴付けられており、それらは、FcγRI(CD64)、FcγRII(CD32)およびFcγRIII(CD16)であり、
−FcγRI(CD64)は、高いアフィニティーで一量体IgGに結合し、そしてマクロファージ、単球、好中球および好酸球上に発現される。E233〜G236、P238、D265、N297、A327およびP329の少なくとも1つのIgGにおける改変は、FcγRIへの結合を減少させる。IgG1およびIgG4中に置換された位置233〜236におけるIgG2残基はFcγRIへの結合を103倍減少させそして抗体感作された赤血球に対するヒト単球応答を排除した(Armour, K.L., et al., Eur. J. Immunol. 29(1999)2613-2624)。
−FcγRII(CD32)は、複合体化されたIgGに中〜低アフィニティーで結合しそして広く発現される。これらのレセプターは2つの重要なタイプ、FcγRIIAおよびFcγRIIBに分けることができる。FcγRIIAは、キリング(killing)に関与する多くの細胞(例えばマクロファージ、単球、好中球)上に見出されそしてキリングプロセスを活性化することができると思われる。FcγRIIBは、阻害プロセスにおいて役割を演じると思われ、そしてB細胞、マクロファージおよびマスト細胞および好酸球上に見出される。B細胞上では、それはさらなる免疫グロブリン産生および例えばIgEクラスへのアイソタイプスイッチを抑制するように機能すると思われる。マクロファージ上では、FcγRIIBは、FcγRIIAにより媒介されるファゴサイトーシスを抑制するように作用する。好酸球およびマスト細胞上では、該b形態は、その別のレセプターへのIgE結合を通じてこれらの細胞の活性化を阻害するのを助けることができる。FcγRIIAに対する減少した結合は、例えば、E233〜G236、P238、D265、N297、A327、P329、D270、Q295、A327、R292およびK414の少なくとも1つのIgG突然変異で見出される。
−FcγRIII(CD16)は、IgGに中〜低アフィニティーで結合しそして2つのタイプとして存在する。FcγRIIIAは、NK細胞、マクロファージ、好酸球およびいくらかの単球およびT細胞上に見出され、そしてADCCを媒介する。FcγRIIIBは、好中球上に高度に発現される。FcγRIIIAへの減少した結合は、例えば、E233〜G236、P238、D265、N297、A327、P329、D270、Q295、A327、S239、E269、E293、Y296、V303、A327、K338およびD376の少なくとも1つの突然変異で、見出される。
【0077】
Fcレセプターに対するヒトIgG1上の結合部位のマッピング、上記した突然変異部位およびFcγRIおよびFcγRIIAへの結合を測定するための方法は、Shields, R.L., et al. JBC276(2001)6591-6604に記載されている。
【0078】
本明細書で使用される用語「Fcレセプター」は、該レセプターと会合した(associated)細胞質ITAM配列の存在により特徴付けられた活性化レセプターを指す(例えば、Ravetch, J. V.., and Bolland, S., Annu. Rev. Immunol. 19(2001)275-290)。このようなレセプターは、FcγRI、FcγRIIAおよびFcγRIIIAである。本発明に従う抗体は、好ましくは、Fcγレセプター、好ましくはFcγIIIAへの減少した結合を示す。好ましくは、用語「FcγRの非結合」は、10μg/mlの抗体濃度においてNK細胞への本発明に従う抗体の結合が抗体LC.001について見出された結合の10%以下であることを意味する。
【0079】
IgG4は減少したFcR結合を示すが、他のIgGサブクラスの抗体は強い結合を示す。しかしながら、Pro238、Asp265、Asp270、Asn297(Fc炭水化物の損失)、Pro329および234、235、236および237 Ile253、Ser254、Lys288、Thr307、Gln311、Asn434およびHis435も、もし変えられるならば減少したFcR結合を与える残基である(Shields, R. L., et al. J. Biol. Chem. 276(2001)6591-6604; Lund, J., et al. FASEB J. 9(1995)115-119; Morgan, A., et al., Immunology 86(1995)319-324; およびEP 0 307 434)。好ましくは、IgG1またはIgG2サブクラスの本発明に従う抗体は、突然変異PVA236、GLPSS331および/またはL234A/L235Aを含む。IgG4サブクラスの本発明に従う抗体は、好ましくは突然変異L235Eを含む。さらなる好ましいIgG4突然変異はS228PまたはL235EおよびS228Pである(表1参照)。
【0080】
本明細書で使用される用語「OX40Lへの結合」は、BIAcoreアッセイにおけるヒトOX40Lへの抗体の結合を意味する(Pharmacia Biosensor AB, Uppsala, Sweden)。更なる確認のために、OX40Lへの結合は、精製されたOX40LがマイクロタイタープレートにコーティングされているELISAまたは直接もしくは間接に標識された抗体を、OX40Lを発現するK562細胞に結合させるFACSアッセイにおいて決定することもできる。
【0081】
BIAcoreアッセイにおいては、抗体を表面に結合させそしてOX40Lの結合は、表面プラズモン共鳴(SPR)により測定される。結合のアフィニティーは、用語ka(抗原への抗体の会合にたいする速度定数)、kd(解離に対する速度定数)およびKD(kd/ka)により定義される。本発明に従う抗体は、10-8以下、好ましくは約10-12〜10-9MのKDを示す(実施例参照)。したがって、本発明は、抗体がBIAcoreアッセイにおいて10-8M未満のKD値でOX40Lに結合する、好ましくは、KD範囲は10-12〜10-9Mである、上記した抗体を指す。
【0082】
OX40L特異的結合ELISAにおいて、OX40Lをマイクロタイタープレート上にコーティングし、そしてOX40Lへの抗体の結合は、HRPをコンジュゲーションした抗ヒトIgGおよびELISAの通常の段階により検出される。このアッセイにおけるEC50値は、好ましくは3nM〜8nMの範囲にある。
【0083】
本明細書で使用された「OX40LへのOX40の結合を阻害する」という用語は、ヒトOX40Lに本発明に記載の抗体を結合させ、それによりOX40/OX40L相互作用を阻害し、それによりOX40Lで誘導されるシグナル伝達を阻害することを指す。
【0084】
本発明の抗体は、hOX40L/OX40相互作用を、好ましくは、
i)1nM〜4nMの範囲のIC50値で0.5μg/mlのビオチニル化OX40Lの(固相)コーティング濃度で抗体により可溶性OX40とビオチニル化され固定化されたOX40Lとの相互作用をブロッキングすることによりELISAアッセイにより示されたin vitroレベルにおいて、
ii)1nM〜10nMの範囲のIC50値で0.78〜100nMの抗体濃度で抗体により可溶性OX40L(10nM、好ましくはhOX40L−Hisとして)と固定化されたOX40との相互作用をブロッキングすることによりBiacoreアッセイにより示されたin vitroレベルにおいて、
iii)抗体が、4〜20nMの範囲のIC50値でOX40と2×105細胞/サンプルの濃度のOX40Lを発現するK562細胞(K562_OX40L)との相互作用をブロッキングする、FACSアッセイにより示された細胞レベルにおいて、
iv)抗体が、1〜5nMの範囲のIC50でNFκB活性化のブロッキングをもたらす、サンプル当たり3×104個のOX40を発現するHeLa細胞への、K562_OX40Lにより誘導されるOX40シグナル伝達をブロッキングする、OX40シグナル伝達アッセイにより、
v)抗体が、1nM〜10nMの範囲のIC50値で1.5×105細胞/サンプルの濃度および0.75μg/mlのPHA濃度でK562_OX40LによるOX40Lで誘導されるT細胞活性化をブロッキングする、T細胞活性化アッセイにより、および/または
vi)抗体が、10μg/mlの抗体濃度で活性化されたB細胞または樹状細胞によるOX40Lで誘導されるT細胞活性化をブロッキングする、T細胞活性化アッセイ(Tetanus assay)により、阻害し、40%〜60%の阻害が得られた。
【0085】
1nM〜4nMの範囲のIC50値で0.5μg/mlOX40Lのコーティング濃度で可溶性OX40と固定化されたOX40Lとの相互作用をブロッキングすることによりELISAアッセイにおいて阻害を示す抗体が好ましい。
【0086】
したがって、本発明の更なる好ましい態様は、OX40/OX40L相互作用を阻害し、それによりOX40Lで誘導されるシグナル伝達を阻害することを特徴とする抗体を指す。
【0087】
本発明に従う抗体は、TNFαまたはCD40L 500nMの濃度までTNFαおよびCD40Lへの非特異的結合を示さないことが更に好ましい。
【0088】
本発明に従う抗体はヒトOX40Lに比較してマウスOX40Lへの少なくとも30倍低い結合を示すことが更に好ましい。
【0089】
10μg/mlの濃度の本発明に従う抗体は、HUVEC細胞上のOX40L発現のダウンレギュレーションを誘導しないことが更に好ましい。
【0090】
さらなる好ましい態様では、本発明の抗体は、それらが組合せ、
a)配列番号1のアミノ酸配列により規定された抗体LC.001の軽鎖可変ドメインおよび配列番号2により規定された抗体LC.001の重鎖可変ドメイン;
b)配列番号3のアミノ酸配列により規定された抗体LC.005の軽鎖可変ドメインおよび配列番号4により規定された抗体LC.005の重鎖可変ドメイン;
c)配列番号5のアミノ酸配列により規定された抗体LC.010の軽鎖可変ドメインおよび配列番号6により規定された抗体LC.010の重鎖可変ドメイン;
d)配列番号7のアミノ酸配列により規定された抗体LC.029の軽鎖可変ドメインおよび配列番号8により規定された抗体LC.029の重鎖可変ドメイン;
e)配列番号9のアミノ酸配列により規定された抗体LC.019の軽鎖可変ドメインおよび配列番号10により規定された抗体LC.019の重鎖可変ドメイン;
f)配列番号11または16のアミノ酸配列により規定された抗体LC.033の軽鎖可変ドメインおよび配列番号12により規定された抗体LC.033の重鎖可変ドメイン;
g)配列番号1のアミノ酸配列により規定された軽鎖(VL)可変ドメインおよび配列番号17により規定された重鎖(VH)可変ドメイン;
h)配列番号18のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号19により規定された重鎖可変ドメイン;
i)配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号20により規定された重鎖可変ドメイン;
からなる群より独立に選ばれる可変ドメインの組合せを含むことを特徴とする。
【0091】
さらなる好ましい態様では、本発明の抗体は、それらが、
k)配列番号13の配列により規定された軽/κ鎖;
l)L234AおよびL235A、PVA236またはGLPSS331から選ばれる1つ以上の突然変異を有するIgG1アイソタイプ配列番号14の重/γ鎖;
m)IgG4アイソタイプ配列番号15の重/γ鎖;
n)突然変異S228Pまたは突然変異S228PおよびL235Eを有するIgG4アイソタイプ配列番号15の重/γ鎖;
o)配列番号61、65または69に含まれる軽定常鎖(light constant chain)、
p)配列番号58、59、60、62、63、64、66、67または68に含まれる重定常鎖(heavy constant chain)、
からなる群より独立に選ばれる定常領域を含むことを特徴とする。
【0092】
更に好ましいのは、各可変抗体ドメイン組合せa)〜i)とγ鎖l)、m)、n)またはp)および好ましくはκ鎖k)またはo)とのすべての組合せである。特に好ましいのは、抗体LC.001、LC.005、LC.010、LC.019、LC.029、LC.033、LC.059、LC.060またはLC.063の可変鎖を、各々、配列番号13の配列により規定されたκ鎖または配列番号61、65もしくは69に含まれる軽定常鎖および突然変異L234AおよびL235Aを有するIgG1アイソタイプ配列番号14の重/γ鎖または配列番号59、63もしくは67に含まれる重定常鎖と共に含む抗体;抗体LC.001、LC.005、LC.010、LC.019、LC.029、LC.033、LC.059、LC.060またはLC.063の可変鎖を、各々、配列番号13の配列により規定されたκ鎖または配列番号59、63もしくは67に含まれる重定常鎖およびIgG4アイソタイプ配列番号15の重/γ鎖または配列番号60、64もしくは68(3つのすべてがS228P突然変異を有していない)に含まれる重定常鎖と共に含む抗体;抗体LC.001、LC.005、LC.010、LC.019、LC.029、LC.033、LC.059、LC.060またはLC.063の可変鎖を、各々、配列番号13の配列により規定されたκ鎖または配列番号61、65もしくは69に含まれる軽定常鎖および突然変異S228Pを有するIgG4アイソタイプ配列番号15の重/γ鎖または配列番号60、64もしくは68に含まれる重定常鎖と共に含む抗体である。
【0093】
好ましくは、抗体は、配列番号1のアミノ酸配列の軽鎖可変CDRおよび配列番号2、17もしくは20の重鎖可変CDR、または配列番号18のアミノ酸配列の軽鎖可変CDRおよび配列番号19の重鎖可変CDRを含む。
【0094】
好ましい抗体は、該抗体が、補体因子C1qへの非結合および/またはFCR結合の損失を引き起こす少なくとも1つのアミノ酸突然変異を含むヒトIgG4サブクラスまたは他のヒトサブクラス(好ましくはIgG1)であることを特徴とする。このような好ましい変異体抗体は、例えば、突然変異L234AおよびL235Aを有する配列番号14のアミノ酸配列または突然変異S228Pを有するもしくは有さない配列番号15を含む。
【0095】
本発明に従う好ましい抗体は、IgG1v1(E233P;L234V;L235A;ΔG236;A327G;A330S;P331Sにより特定されたPVA−236;GLPSS331)、IgG1v2(L234A;L235A)およびIgG4v1(S228P;L235E)およびIgG4x(S228P)として定義された抗体である。
【0096】
本発明に従うハイブリドーマ細胞系hu−Mab<hOX40L>LC.001は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の下に、寄託番号DSM ACC2672の下に2004年7月27日にDeutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSMZ)、Germanyに寄託された。
【0097】
本発明に従うハイブリドーマ細胞系hu−Mab<hOX40L>LC.005(DSM ACC 2685)、hu−Mab<hOX40L>LC.010(DSM ACC 2686)、hu−Mab<hOX40L>LC.019、hu−Mab<hOX40L>LC.029(DSM ACC 2688)およびhu−Mab<hOX40L>LC.033(DSM ACC 2689)は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約の下に、2004年9月2日にDeutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSMZ)、Germanyに寄託された。
【0098】
前記細胞系から得られうる抗体は、本発明の好ましい態様でありそして補体因子C1qに結合しないおよび/またはヒトFcγレセプターに結合しない本発明に従う抗体の発生のための中間物質として特に有用である。
【0099】
本発明の更なる好ましい態様は、OX40Lに結合し、そして寄託されたハイブリドーマ細胞系により産生されたモノクローナル抗体LC.005、LC.010またはLC.029も結合する同じOX40Lエピトープに結合する、単離された抗OX40L抗体である。
【0100】
本発明の更なる態様は、補体因子C1qおよび/またはヒトFcγレセプターに結合しない、OX40Lに対する抗体の産生方法であって、10-8M未満のKD値でOX40Lに結合する抗体の重鎖をコードする核酸の配列を、改変された抗体が補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合しないような方式で改変し、該改変された核酸および該抗体の軽鎖をコードする核酸を発現ベクターに挿入し、該ベクターを原核生物もしくは真核生物宿主細胞に挿入し、コードされたタンパク質を発現しそして宿主細胞または上清から回収することを特徴とする方法である。
【0101】
本発明のさらなる態様は、補体因子C1qに結合しないおよび/またはヒトFcγレセプターに結合しない本発明に従う抗体の産生方法であって、前記細胞系の1つから得られうる抗体を、「クラススイッチ」、即ち、好ましくは、IgG1v1(E233P;L234V;L235A;ΔG236;A327G;A330S;P331Sにより特定されたPVA−236;GLPSS331)、IgG1v2(L234A;L235A)およびIgG4v1(S228P;L235E)およびIgG4x(S228P)として定義されたFc部分の変化または突然変異(例えばIgG1からIgG4へのおよび/またはIgG1/IgG4突然変異)により改変することを特徴とする方法である。
【0102】
更なる好ましい態様では、これらの抗体は、Fab、(Fab’)2および一本鎖フラグメントからなる群より選ばれる抗体フラグメントも含む。
【0103】
したがって、「変異体」抗OX40L抗体は、本明細書では、親抗体配列における1つ以上のアミノ酸残基の付加、欠失および/または置換により「親」抗OX40L抗体アミノ酸配列とはアミノ酸配列において異なる分子を指す。好ましい態様では、変異体は、親抗体の1つ以上の定常領域もしくは可変領域における、好ましくは定常領域における1つ以上のアミノ酸置換を含む。例えば、変異体は、親抗体の1つ以上の可変領域における少なくとも1つ、例えば約1〜約10、好ましくは約2〜約5の置換を含むことができる。通常、変異体は、親抗体定常および/または可変ドメイン配列との少なくとも90%、更に好ましくは少なくとも95%、最も好ましくは少なくとも99%のアミノ酸配列同一性を有するアミノ酸配列を有するであろう。
【0104】
本発明は、配列番号21〜38からなる群より選ばれる親抗体重鎖CDRおよび/または配列番号39〜57からなる群より選ばれる抗体軽鎖CDRの最初のアミノ酸配列を改変する方法であって、該最初のアミノ酸配列をコードする核酸を提供し、該核酸を、1つのアミノ酸が重鎖CDR1において改変され、1〜2つのアミノ酸が重鎖CDR2において改変され、1〜2つのアミノ酸が重鎖CDR3において改変され、1〜3つのアミノ酸が軽鎖CDR1において改変され、1〜3つのアミノ酸が軽鎖CDR2において改変されおよび/または1〜3つのアミノ酸が軽鎖CDR3において改変されるように改変し、該改変されたCDRアミノ酸配列を抗体構造において発現し、該抗体が10-8M未満のKDでOX40Lに結合するかどうかを測定し、そしてもしも該抗体が10-8M未満のKDでOX40Lに結合するならば、該改変されたCDRを選ぶことを特徴とする方法を含む。好ましくは、このような改変は保存的配列改変である。
【0105】
配列に関する同一性又は相同性は、本明細書では、配列をアラインメントしそして必要ならばギャップを導入して、最大百分率配列同一性を達成した後に、親抗体残基と同一である候補配列におけるアミノ酸残基の百分率として定義される。抗体配列におけるN末端、C末端もしくは内部の延長、欠失または挿入のどれも配列同一性または相同性に影響を与えるものとみなすべきではない。変異体は、ヒトOX40Lに結合する能力を保持し、そして好ましくは親抗体の性質より優れた性質を有する。例えば、変異体は、慢性関節リウマチおよび喘息の処置期間中減少した副作用を有することができる。何故ならば、OX40Lは、B細胞、樹状細胞およびマクロファージ上に一過性に発現されるのみならず内皮細胞(Kotani, A., et al., Immunol. Lett. 84(2002)1-7)、気道平滑筋細胞(=ASM)(Burgess, J. K., J. Allergy Clin. Immunol 113(2004)683-689)およびミクログリア細胞(Weinberg, A.D., et al., J.Immunol. 162(1999)1818-1826)上にも一過性に発現されるからである。内皮細胞、ASMおよびミクログリア細胞へのOX40Lに対する抗体の結合は、血管漏洩をもたらす内皮細胞の細胞損傷、肺破壊をもたらすASM細胞の細胞損傷、ミクログリアの損傷をもたらすミクログリア細胞の細胞損傷をもたらすことがありうる。
【0106】
本明細書における「親」抗体は、変異体の調製のために使用されるアミノ酸配列によりコードされている親抗体である。好ましくは、親抗体は、ヒトフレームワーク領域を有し、そしてもし存在するならば、ヒト抗体定常領域(1つまたは複数)を有する。例えば親抗体は、好ましくはIgG1型のヒト化抗体またはヒト抗体であることができる。
【0107】
本発明に従う抗体は、更に、本発明に従う抗体の上記した特徴に影響を与えないまたは該特徴を変えない、「保存性配列改変」、ヌクレオチドおよびアミノ酸配列改変を有するこのような抗体を含む。改変は、部位特異的突然変異誘発およびPCR媒介突然変異誘発などの当技術分野で知られている標準技術により導入することができる。保存性アミノ酸置換は、アミノ酸残基が同様な側鎖を有するアミノ酸残基で置き換えられているアミノ酸置換を含む。同様な側鎖を有するアミノ酸残基のファミリーは、当技術分野で定義されている。これらのファミリーは、塩基性側鎖(例えばリシン、アルギニン、ヒスチジン)、酸性側鎖(例えば、アスパラギン酸、グルタミン酸)、電荷のない極性側鎖(例えば、グリシン、アスパラギン、グルタミン、セリン、トレオニン、チロシン、システイン、トリプトファン)、非極性側鎖(例えば、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン)、β−分岐状側鎖(例えば、トレオニン、バリン、イソロイシン)および芳香族側鎖(例えば、チロシン、フェニルアラニン、トリプトファン、ヒスチジン)を有するアミノ酸を含む。したがって、ヒト抗OX40L抗体における予想される非必須アミノ酸残基は、好ましくは、同じ側鎖ファミリーからの他のアミノ酸残基で置き換えることができる。
【0108】
アミノ酸置換は、Riechmann, L., et al., Nature 332(1988)323-327 and Queen, C., et al., Proc. Natl. Acad. Aci. USA 86(1989)10029-10033により記載された分子モデル化に基づく突然変異誘発により達成されうる。
【0109】
本発明は、抗体の産生方法であって、10-8M未満のKD値でOX40Lに結合する抗体の重鎖をコードする第1核酸の配列を、改変された抗体が補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合しないような方法で改変し、該改変された第1核酸および該抗体の軽鎖をコードする第2核酸を発現ベクターに挿入し、該ベクターを原核生物または真核生物宿主細胞に挿入し、該抗体の合成を可能とする条件下に該宿主細胞を培養し、そしてこの培養物から該抗体を回収することを特徴とする方法を更に含む。
【0110】
本発明は、本発明に従うそしてヒト起源に由来するFc部分を含む抗体の産生方法であって、a)本発明に従う親ヒト抗体の軽鎖をコードする第1核酸配列およびFc部分が補体因子C1qおよび/またはFcレセプターに結合しないようにFc部分が改変されている該親ヒト抗体の重鎖をコードする第2DNA配列で宿主細胞をトランスフォーメーションし;b)該第1DNA配列及び第2DNA配列を、該抗体重鎖および軽鎖が産生されるように発現しそしてc)該抗体を宿主細胞または宿主細胞培養物から回収する段階を含む方法を更に含む。
【0111】
本発明は、上記した抗体をコードする核酸分子、これらの核酸を含む対応するベクターおよびこれらのベクターのための対応する宿主細胞も含む。本発明は、抗体の製造方法であって、該抗体分子の合成を可能とする条件下に対応する宿主細胞を培養しそしてこの培養物から該抗体を回収することを含む、例えば、原核生物または真核生物宿主細胞において重鎖をコードする核酸および軽鎖をコードする核酸を発現しそしてこのポリペプチドを該細胞から回収することによる、抗体の製造方法を包含する。
【0112】
抗体の診断的使用及び治療的使用が包含される。1つの診断用途では、本発明は、OX40Lを含有する疑いのあるサンプルを抗OX40L抗体に曝露しそしてサンプルへの抗体の結合を決定することを含む、OX40Lタンパク質の存在を決定する方法を提供する。OX40Lタンパク質は、OX40L発現細胞の細胞膜中にその膜貫通ドメインにより挿入されることができ、またはシェディング(shedding)もしくはタンパク質分解放出のような機構により放出された体液中の可溶性細胞外ドメインとして存在することができる。この使用のために、本発明は、抗体を使用してOX40Lタンパク質を検出するための抗体および指示を含むキットを提供する。
【0113】
本発明の抗体は、哺乳動物、好ましくは炎症性疾患を有するまたは炎症性疾患に罹っている疑いがある患者における炎症性疾患の予防および/または処置のために有用である。このような疾患は、アレルギー反応、例えば喘息を含む。他の適応は慢性関節リウマチを含む自己免疫疾患の処置である。
【0114】
本発明は、上記した炎症性疾患、特に喘息および慢性関節リウマチに罹っている哺乳動物を処置する方法を提供する。
【0115】
好ましくは、本発明の抗体は、その症状が吸入されたコルチコステロイドで十分にコントロールされない患者における重い持続性喘息の処置のために使用することができる。患者集団は、不十分にコントロールされた重い持続性喘息を有する成人および青年(12歳以上)を含む。抗体は、1月に1回または2回好ましくは皮下に送達されるであろう。主要な最終目標は、好ましくは急性増悪の減少であろう。他の最終目標は、ピークフロー、日中の喘息症状、夜間の目覚め、クオリティーオブライフ、救急室診察、喘息のない日、β−2アゴニスト使用、ステロイド減少または漸減および過敏性(hyper-responsiveness)に対する効果を含む。
【0116】
中程度〜重度の活性慢性関節リウマチを有する成人の処置のために単独療法のためまたはメトレキセートもしくは他のDMARDs(Disease Modifying Anti-Rheumatic Drugs)との併用において本発明に従う抗体を使用することは更に好ましい。それは2または4週間毎に皮下注射として投与されるであろう。それは、1種以上のDMARDsが効かなかった患者における慢性の治療であろう。最終目標は、活性な慢性関節リウマチを有する成人患者における兆候及び症状の減少および構造的損傷の進行の抑制を含むであろう。ACR基準により測定された障害の防止、兆候及び症状の改善(ACR20>60%、ACR50>35%、ACR70>15%;米国リウマチ学会からのインデックス;www.rheumatology.com)。
【0117】
本発明の更なる態様は、これらの疾患の処置用の医薬の製造のための本発明に従う抗体の使用である。
【0118】
本発明は、医薬組成物の製造のために上記に定義した抗体の使用にも関しそして薬学的有効量で本発明に従う抗体を、場合により緩衝液および/または薬学的目的での抗体の処方のために有用な佐剤と共に含有する医薬組成物を含む。
【0119】
本発明は、薬学的に許容されうる担体中にこのような抗体を含む医薬組成物を更に提供する。1つの態様では、医薬組成物は、製造物又はキットに含ませることもできる。
【0120】
本発明に従う抗体は好ましくはリコンビナント手段により産生される。このような方法は、当技術分野の技術水準で広く知られておりそして抗体ポリペプチドのその後の単離および通常薬学的に許容されうる純度への精製を伴う原核細胞及び真核細胞におけるタンパク質発現を含む。タンパク質発現のために、軽鎖および重鎖またはそのフラグメントをコードする核酸を標準方法により発現ベクターに挿入する。発現は、CHO細胞、NSO細胞、SP2/0細胞、HEK293細胞、COS細胞、酵母またはE.coli細胞などの適切な原核生物宿主細胞または真核生物宿主細胞において行われ、そして抗体は細胞(溶解後の上清または細胞)から回収される。
【0121】
抗体のリコンビナント産生は、当技術分野の技術水準でよく知られておりそして例えばMakrides, S. C., Protein Expr. Purif. 17(1999)183-202; Geisse, S., et al., Protein Expr. Purif. 8(1996) 271-282; Kaufman, R.J., Mol. Biotechnol. 16(2000) 151-161; Werner, R.C., et al., Arzneimittelforschung 48(1998)870-880の概説論文に記載されている。
【0122】
抗体は、完全細胞中に、細胞溶解物中にまたは部分的に精製された形態もしくは実質的に純粋な形態で存在することができる。精製は、他の細胞成分または他の汚染物、例えば他の細胞核酸もしくはタンパク質を除去するために、アルカリ/SDS処理、カラムクロマトグラフィー、および当技術分野で周知の他の技術を含む標準技術により行われる。Ausubel, F., et al., ed., Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York (1987)参照。
【0123】
NS0細胞における発現は、例えば、Barnes, L. M., et al., Cytotechnology 32(2000) 109-123; and Barnes, L. M., et al.,Biotech. Bioeng. 73(2001) 261-270により記載されている。一過性発現は、例えば、Durocher, Y., et al., Nucl. Acids. Res. 30(2002)E9により記載されている。可変ドメインのクローニングは、Orlandi, R., et al., Proc. Natl. Acad. Sci. USA 86(1989)3833-3837; Carter, P., et al., Proc. Natl. Acad. Sci. USA 89(1992) 4285-4289; およびNorderhaug, L., et al., J. Immunol. Methods 204(1997)77-87により記載されている。好ましい一過性発現系(HEK293)は、Schlaeger, E-J., and Christensen, K., in Cytotechnology 30(1999) 71-83 および Schlaeger, E-J., in J. Immunol. Methods 194(1996) 191-199により記載されている。
【0124】
原核生物に適当なコントロール配列は、例えばプロモーター、場合によりオペレーター配列およびリボソーム結合部位を含む。真核細胞は、プロモーター、エンハンサーおよびポリアデニル化シグナルを利用することが知られている。
【0125】
核酸は、それが他の核酸配列と機能的に関連させられているとき、「作用可能に連結されている」。例えば、プレ配列または分泌リーダーのためのDNAは、もしもそれがポリペプチドの分泌に関与するプレタンパク質として発現されるならば、ポリペプチドのためのDNAに作用可能に連結されており;プロモーターまたはエンハンサーは、それが配列の転写に影響を与えるならばコード化配列に作用可能に連結されており;またはリボソーム結合部位は、それが翻訳を促進するように配置されているならばコード化配列に作用可能に連結されている。一般に、「作用可能に連結されている」とは、連結されているDNA配列が連続的であり(contiguous)そして分泌リーダーの場合には、連続的であり且つリーディングフレーム内にある。しかしながら、エンハンサーは、連続的である必要はない。連結は、都合の良い制限部位におけるライゲーションにより達成される。もしもこのような部位が存在しないならば、合成オリゴヌクレオチドアダプターまたはリンカーが慣用の実施に従って使用される。
【0126】
モノクローナル抗体は、慣用の免疫グロブリン精製手順、例えば、プロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析またはアフィニティークロマトグラフィーにより培養培地から適当に分離される。モノクローナル抗体をコードするDNA及びRNAは、容易に単離されそして慣用の手順を使用して配列決定される。ハイブリドーマ細胞は、このようなDNAおよびRNAのソースとして役立つことができる。一旦単離されると、DNAを発現ベクターに挿入することができ、次いでこれを、宿主細胞、例えば他の場合には免疫グロブリンタンパク質を産生しないHEK293細胞、CHO細胞またはミエローマ細胞にトランスフェクションして、宿主細胞においてリコンビナントモノクローナル抗体の合成を達成する。
【0127】
ヒトOX40L抗体のアミノ酸配列変異体(または突然変異体)は、抗体DNAに適当なヌクレオチド変化を導入することまたはヌクレオチド合成により調製される。しかしながら、このような改変は、例えば上記した非常に限られた範囲においてのみ行うことができる。例えば、改変は、上記した抗体の特性、例えばIgGアイソタイプ及びエピトープ結合を変えないが、リコンビナント産生の収率、タンパク質安定性を改良することができまたは精製を容易にすることができる。
【0128】
抗OX40L抗体の特有のコンフォメーションを維持するのに関与しないいかなるシステイン残基も、一般にセリンにより置換することができ、それにより分子の酸化安定性を改良しそして異常な架橋を防止することができる。反対に、システイン結合(1つまたは複数)を抗体に加えて、その安定性(特に、抗体がFvフラグメントの如き抗体フラグメントである場合)を改良することができる。
【0129】
抗OX40L抗体のアミノ酸配列変異体をコードする核酸分子は、当技術分野で知られている種々の方法により調製される。これらの方法は、天然のソース(天然に存在するアミノ酸配列変異体の場合に)からの単離、またはオリゴヌクレオチド媒介(又は部位特異的)突然変異誘発、PCR突然変異誘発およびヒト化抗OX40L抗体の以前に調製された変異体または非変異体バージョンのカセット突然変異誘発による調製を含むが、それらに限定されない。
【0130】
本発明は、化学療法剤、毒素(例えばバクテリア、真菌、植物もしくは動物起源の酵素的に活性な毒素またはそのフラグメント)、放射性同位元素(即ち、放射性コンジュゲート)、などの細胞傷害剤にコンジュゲーションされた本発明に従う抗体を含む免疫コンジュゲートにも関する。抗体と細胞傷害剤とのコンジュゲートは、種々の二官能性タンパク質カップリング剤、例えばN−スクシンイミジル−3−(2−ピリジルジチオール)プロピオネート(SPDP)、イミノチオラン(IT)、イミドエステルの二官能性誘導体(例えばジメチルアジピミデートHCL)、活性エステル(例えばジスクシンイミジルスベレート)、アルデヒド(例えばグルタルアルデヒド)、ビス−アジド化合物(例えばビス(p−アジドベンゾイル)ヘキサンジアミン)、ビス−ジアゾニウム誘導体(例えばビス−(p−ジアゾニウムベンゾイル)−エチレンジアミン)、ジイソシアナート(例えばトリエン2,6−ジイソシアナート)およびビス−活性フッ素化合物(例えば1,5−ジフルオロ−2,4−ジニトロベンゼン)を使用して製造される。例えばリシンイムノトキシンは、Vitetta, E.S., et al., Science 238(1987) 1098-1104)に記載のとおりに製造することができる。炭素14で標識された1−イソチオシアナートベンジル−3−メチルジエチレントリアミンペンタ酢酸(MX−DTPA)は、抗体への放射性ヌクレオチドのコンジュゲートのための例示的キレート剤である。WO 94/11026参照。
【0131】
別のタイプの抗体の共有結合改変は、US Patent Nos. 4,640,835; 4,496,689; 4,301,144; 4,670,417; 4,791,192または4,179,337に記載された方法において、種々の非タンパク質性ポリマー、例えばポリエチレングリコール、ポリプロピレングリコールまたはポリオキシアルキレンの1つに抗体を連結することを含む。
【0132】
なお別の局面では、本発明は、ヒト抗OX40L抗体(本発明に従う抗体を産生するハイブリドーマ細胞からなる群より選ばれる細胞系により産生された親抗体)を発現するトランスジェニック非ヒト動物、例えばトランスジェニックマウスからの単離されたB細胞を提供する。好ましくは、単離されたB細胞は、精製されたもしくはリコンビナント形態のOX40L抗原および/またはOX40Lを発現する細胞で免疫感作されたトランスジェニック非ヒト動物、例えばトランスジェニックマウスから得られる。好ましくは、トランスジェニック非ヒト動物、例えばトランスジェニックマウスは、本発明の抗体のすべてもしくは一部をコードするヒト重鎖導入遺伝子およびヒト軽鎖導入遺伝子を含むゲノムを有する。単離されたB細胞を次いで不死化してヒト抗OX40L抗体のソース(例えばハイブリドーマ)を提供する。したがって、本発明は、本発明に従うヒトモノクローナル抗体を産生することができるハイブリドーマも提供する。1つの態様では、ハイブリドーマは、不死化された細胞に融合された、本発明の抗体のすべてもしくは一部をコードするヒト重鎖導入遺伝子およびヒト軽鎖導入遺伝子を含むゲノムを有するトランスジェニック非ヒト動物、例えばトランスジェニックマウスから得られるB細胞を含む。
【0133】
特定の態様では、トランスジェニック非ヒト動物は、本発明の抗体のすべてもしくは一部をコードするヒト重鎖導入遺伝子およびヒト軽鎖導入遺伝子を含むゲノムを有するトランスジェニックマウスである。トランスジェニック非ヒト動物は、OX40L抗原の精製されたもしくは濃縮された調製物および/またはOX40Lを発現する細胞で免疫感作されうる。好ましくは、トランスジェニック非ヒト動物、例えばトランスジェニックマウスは、OX40Lに対するヒトモノクローナル抗体のアイソタイプを産生することができる。
【0134】
本発明に従うヒトモノクローナル抗体は、本発明の抗体のすべてもしくは一部をコードするヒト重鎖導入遺伝子およびヒト軽鎖導入遺伝子を含むゲノムを有するトランスジェニック非ヒト動物、例えばトランスジェニックマウスを、OX40L抗原の精製されたもしくは濃縮された調製物および/またはOX40Lを発現する細胞で免疫感作させることにより産生することができる。次いで動物のB細胞(例えば、脾臓B細胞)を得そしてミエローマ細胞と融合して、OX40Lに対するヒトモノクローナル抗体を分泌する不死のハイブリドーマ細胞を形成する。
【0135】
好ましい態様では、OX40Lに対して指向されたヒトモノクローナル抗体は、マウス系よりはむしろヒト免疫系の一部を有するトランスジェニックマウスを使用して発生させることができる。本明細書で「HuMab」マウスと呼ばれるこれらのトランスジェニックマウスは、内在性μおよびκ鎖ローカスを不活性化するターゲティングされた突然変異と共に、重(μおよびγ)鎖およびκ軽鎖(定常領域遺伝子)を含む再配列されていないヒト免疫グロブリン遺伝子をコードするヒト免疫グロブリン遺伝子ミニローカスを含有する(Lonberg, N., et al., Nature 368(1994) 856-859)。したがって、このマウスは、マウスIgMまたはKの減少した発現を示し、そして免疫感作に応答して、導入されたヒト重鎖および軽鎖導入遺伝子は、クラススイッチおよび体細胞突然変異を受けて、高アフィニティーヒトIgGモノクローナル抗体を発生する(Lonberg, N., et al., Nature 368(1994)856-859; Lonberg, N., Handbook of Experimental Pharmacology 113(1994)49-101に概説されている; Lonberg, N., and Huszar, D., Intern. Rev. Immunol. 25(1995)65-93; およびHarding, F., and Lonberg, N., Ann. N. Acad. Sci. 764(1995) 536-546))。HuMAbマウスの調製は、Taylor, L., et al., Nucleic Acids Res. 20 (1992) 6287-6295; Chen, J., et al., Int. Immunol. 5(1993) 647-656; Tuaillon, N., et al., Proc. Natl. Acad. Sci. USA 90(1993) 3720-3724; Choi, T. K., et al., Nat. Genet. 4(1993)117-123; Chen, J., et al., EMBO J. 12(1993) 821-830; Tuaillon, N., et al.,J. Immunol. 152(1994) 2912-2920; Lonberg, N., et al., Nature 368(1994) 856-859; Lonberg, N., Handbook of Experimental Pharmacology 113(1994)49-101; Taylor, L., et al., Int. Immunol. 6(1994) 579-591; Lonberg, N., and Huszar, D., Intern. Rev. Immunol. 25(1995)65-93; およびHarding, F., and Lonberg, N., Ann. N. Acad. Sci. 764(1995) 536-546; Fishwild, D. M., et al., Nat. Biotechnol. 14(1996) 845-851;に記載されており、それらのすべての内容はそのまま参照により本明細書に組み込まれる。更に、US Patent Nos. 5,545,806; 5,569,825; 5,625,126; 5,633,425; 5,789,650; 5,877,397; 5,661,016; 5,814,318; 5,874,299; 5,545,807; 5,770,429; WO 98/24884; WO 94/25585; WO 93/1227; WO 92/22645; and WO 92/03918を参照のこと。
【0136】
OX40Lに対するヒトモノクローナル抗体を完全に発生させるために、HuMAbマウスを、Lonberg, N., et al., Nature 368(1994)856-859; Fishwild, D. M., et al., Nat. Biotechnol. 14(1996) 845-851およびWO 98/24884により記載のとおり、一般的方法に従ってOX40L抗原の精製されたもしくは濃縮された調製物および/またはOX40Lを発現する細胞で免疫感作させることができる。好ましくは、マウスは、最初の免疫感作時に6〜16週齢であろう。例えば、KLHにカップリングされたもしくはPBS中の可溶性OX40L抗原の精製されたもしくは濃縮された調製物(例えば、OX40L発現細胞から精製された)を使用して、HuMAbマウスを腹腔内に免疫感作させることができる。これは、単離されたOX40Lタンパク質による免疫感作とOX40Lを発現する細胞、例えば腫瘍細胞系による免疫感作を交互に行うことにより組み合わせて、免疫応答を促進させることができる。種々の抗原による累積した経験は、完全フロイントアジュバント中の抗原で腹腔内に(ip)最初に免疫感作され、次いで不完全フロイントアジュバント中の抗原で隔週毎にip免疫感作(例えば、総計6回まで)されると、HuMAbトランスジェニックマウスは最善に応答することを示した。免疫応答は、眼窩後方の出血(retroorbital bleeds)により得られる血漿サンプルにより、免疫感作プロトコールの経過にわたって監視することができる。血漿は、ELISAによりスクリーニングすることができ、そして十分な力価の抗OX40Lヒト免疫グロブリンを有するマウスを、対応するB細胞の不死化のために使用することができる。マウスを抗原で3〜4日静脈内に追加抗原刺激した後殺し、そして脾臓およびリンパ節を除去することができる。いくつかのマウスは、各抗原について免疫感作されるであろう。例えば、HCo7およびHCo12系統の総計12匹のHuMAbマウスを免疫感作させることができる。
【0137】
HCo7マウスは、それらの内在性軽鎖(κ)遺伝子におけるJKD破壊(JKD disruption)(Chen, J., et al., EMBO J. 12(1993)821-830)、それらの内在性重鎖遺伝子におけるCMD破壊(WO 01/14424の実施例1に記載のとおり)、KCo5ヒトκ軽鎖導入遺伝子(Fishwild, D. M., et al., Nat. Biotechnol. 14(1996) 845-851に記載のとおり)およびHCo7ヒト重鎖導入遺伝子(US Patent No.5,770,429に記載のとおり)を有する。
【0138】
HCo12マウスは、それらの内在性軽鎖(κ)遺伝子におけるJKD破壊(Chen, J., et al., EMBO J. 12(1993)821-830)、それらの内在性重鎖遺伝子におけるCMD破壊(WO 01/14424の実施例1に記載のとおり)、KCo5ヒトκ軽鎖導入遺伝子(Fishwild, D. M., et al., Nat. Biotechnol. 14(1996) 845-851に記載のとおり)およびHCo12ヒト重鎖導入遺伝子(WO 01/14424の実施例2に記載のとおり)を有する。マウスリンパ球を単離しそして標準プロトコールに基づいてPEGを使用してマウスミエローマ細胞系と融合させてハイブリドーマを発生させることができる。得られるハイブリドーマを、次いで抗原特異的抗体の産生についてスクリーニングする。例えば免疫感作されたマウスからの脾臓およびリンパ節由来のリンパ球の単一細胞懸濁液を、50%PEGによりSP2/0非分泌マウスミエローマ細胞(ATCC、CRL1581)の数の6分の1に融合させる。細胞を平底マイクロタイタープレート中に約2×105でプレートし、続いて選択培地中で約2週間インキュベーションする。
【0139】
次いで個々のウエルをヒト抗OX40LモノクローナルIgMおよびIgG抗体についてELISAによりスクリーニングする。一旦多量のハイブリドーマ増殖が起こると、培地を通常10〜14日後に分析する。抗体分泌ハイブリドーマを再びプレートし、再びスクリーニングし、そしてもしもヒトIgGについて依然としてポジティブであるならば、抗OX40Lモノクローナル抗体を限界希釈により少なくとも2回サブクローニングすることができる。次いで安定なサブクローンをin vitroで培養して特徴付けのために組織培養培地中で抗体を産生させる。
【0140】
アッセイツリー(assay tree)は、主として、IgGに関する非特異的アッセイ(「IgG−ELISA」)、次いで精製されたOX40Lタンパク質またはOX40L発現細胞への抗原結合の決定のための特異的ELISAおよび見かけのFACSアッセイからなる。次の段階は、抗OX40L抗体の、精製されたOX40Lまたは細胞上に発現されたOX40Lに対するその天然の相互作用パートナー、例えば可溶性の精製されたOX40との競合を決定する機能的アッセイ、例えば競合ELISAまたはFACSを含む。次の段階は、OX40シグナル伝達の抗OX40L抗体の阻止能力を決定する機能的アッセイ、例えばNFκB活性化(=「NFκBアッセイ」)を含む。次の段階は、T細胞活性化に関する抗OX40L抗体の阻止能力を決定する機能的アッセイ(「T細胞活性化アッセイ」および「TTアッセイ」)を含む。
【0141】
CDR配列は抗体−抗原相互作用について責任を担っているので、異なるヒト抗体からのフレームワーク配列上に本発明に従うCDR配列を含む発現ベクターを構築することにより本発明に従うリコンビナント抗体を発現することが可能である(例えば、Riechmann, L., et al., Nature 332(1998) 323-327; Jones, P., et al., Nature 321(1986) 522-525; and Queen, C., et al., Proc. Natl. Acad. Sci. USA 86(1989)10029-10033参照)。このようなフレームワーク配列は、生殖系列ヒト抗体遺伝子配列を含む公のDNAデータベースから得ることができる。これらの生殖系列配列は、成熟抗体遺伝子配列とは異なるであろう。何故ならば、それらはB細胞成熟期間中にV(D)J結合により形成される完全にアセンブリーされた可変遺伝子を含まないからである。生殖系列遺伝子配列は、各々可変領域にまんべんなく、高アフィニティー二次レパートリー抗体の配列とも異なるであろう。
【0142】
本発明は、好ましくはサンプルのOX40L(可溶性または膜に結合した)と本発明に従う抗体との結合を決定する免疫学的アッセイにより、in vitroでのOX40Lの診断のための本発明に従う抗体の使用をさらに含む。
【0143】
他の局面では、本発明は、薬学的に許容されうる担体と共に処方された、本発明のヒトモノクローナル抗体またはその抗原結合性部分の1つまたは組合せを含有する組成物、例えば医薬組成物を提供する。更に詳しくは、組成物は、医薬組成物または診断組成物であり、なおさらに詳しくは、医薬組成物は、上記に定義した抗体と少なくとも1種の薬学的に許容されうる賦形剤を含む。組成物は無菌でなければならず、そして組成物が注射器により送達可能である程度に流動性でなければならない。
【0144】
本明細書で使用された、「薬学的に許容される担体」は、生理学的に適合性の、いかなるおよびすべての溶媒、分散媒体、コーティング、抗バクテリア剤および抗真菌剤、等張剤および吸収遅延剤等を含む。好ましくは、担体は、静脈内、筋肉内、皮下、非経口、脊髄または表皮投与(例えば、注射または注入(infusion)により)のために適当である。好ましくは、このような担体は、好ましくは、等張性であり、好ましくは更に無機塩、糖、ポリオールおよび/または界面活性剤を含有する、水性のpH緩衝化された溶液(例えば酢酸塩、クエン酸塩、リン酸塩またはヒスチジン)である。薬学的に許容されうる担体は、Remington's Pharmaceutical Science, 16th edition, Osol, A. Ed.(1980)に記載されたようなものでもある。
【0145】
抗体濃度は、好ましくは0.1mg/ml〜50mg/mlである。好ましくは、緩衝化された溶液のpH値は、1mM〜200mMの緩衝剤濃度で4.0〜8.0の範囲にある。好ましい塩は、1mM〜200mMの範囲にある塩化ナトリウムおよび/またはリン酸ナトリウムである。好ましい糖は、1%〜15%(重量/容積)の範囲にあるスクロースおよび/またはトレハロースである。好ましいポリオールは、1%〜15%(重量/容積)の範囲にあるグリセロール、プロピレングリコール、液体ポリエチレングリコールおよび/または同様なものである。界面活性剤は、好ましくは、0.001%〜0.5%(重量/容積)の範囲のポリソルベート(例えばポリソルベート20または80)および/またはポロキサメール(poloxamere)である。このましい医薬組成物は、0.1mg/ml〜50mg/mlの抗体と1mM〜200mMのリン酸緩衝化生理食塩水pH4.0〜8.0を含有する。
【0146】
本発明の組成物は、それを必要としている患者に当技術分野で知られている多様の方法により投与することができる。当業者により認識されるとおり、投与の経路および/または方式は所望の結果に依存して変わるであろう。
【0147】
薬学的に許容されうる賦形剤または担体は、無菌の水性溶液剤または無菌の注射可能な溶液剤もしくは分散液剤の処方に応じて調合される製剤用の無菌の散剤を含む。このような媒体および薬学的有効物質のための作用物質の使用は当技術分野で知られている。
【0148】
本明細書で使用された語句「非経口投与」及び「非経口的に投与される」は、通常注射による腸内および局所的投与以外の投与方式を意味し、そして静脈内、筋肉内、動脈内、鞘内、カプセル内、眼窩内、心臓内、皮膚内、腹腔内、経気管、皮下、表皮下、関節内、被膜下、くも膜下、髄腔内、硬膜外、および胸骨内注射および注入を含むが、それらに限定されない。
【0149】
本発明の医薬組成物中の有効成分の実際の投与レベルは、患者に毒性であることなく、特定の患者、組成および投与方式について所望の治療応答を達成するのに有効な有効成分の量を得るように変えることができる。選ばれる投与レベルは、使用される本発明の特定の組成物、または又はそのエステル、塩もしくはアミドの活性、投与経路、投与の時間、使用される特定の化合物の排出の速度、処置の期間、使用される特定の組成物と組み合わせて使用される他の薬物、化合物および/または物質、処置される患者の年齢、性、体重、状態、一般に健康及び以前の医学的経歴および医学分野で周知の同様な因子を含む種々の薬物動態学的因子に依存するであろう。典型的な毎週の投与量(weekly dosage)は、上記した因子に依存して約0.1mg/kg〜約20mg/kg以上の範囲にあることができる。
【0150】
下記の実施例、参考文献、配列リストおよび図は本発明の理解を助けるために提供され、本発明の真の範囲は特許請求の範囲に記載されている。本発明の精神から逸脱することなく前記した手法において改変がなされうることは理解される。
【0151】
配列リストの説明
配列番号1 LC.001のκ軽鎖、可変領域
配列番号2 LC.001のγ重鎖可変領域
配列番号3 LC.005のκ軽鎖、可変領域
配列番号4 LC.005のγ重鎖可変領域
配列番号5 LC.010のκ軽鎖、可変領域
配列番号6 LC.010のγ重鎖可変領域
配列番号7 LC.029のκ軽鎖、可変領域
配列番号8 LC.029のγ重鎖可変領域
配列番号9 LC.019のκ軽鎖、可変領域
配列番号10 LC.019のγ重鎖可変領域
配列番号11 LC.033のκ軽鎖、可変領域
配列番号12 LC.033のγ重鎖可変領域
配列番号13 κ軽鎖定常領域
配列番号14 γ1重鎖定常領域
配列番号15 γ4重鎖定常領域
配列番号16 LC.033のκ軽鎖、突然変異体可変領域
配列番号1 LC.059のκ軽鎖、可変領域
配列番号17 LC.059のγ重鎖可変領域
配列番号18 LC.060のκ軽鎖、可変領域
配列番号19 LC.060のγ重鎖可変領域
配列番号1 LC.063のκ軽鎖、可変領域
配列番号20 LC.063のγ重鎖可変領域
配列番号21〜57 CDR配列
配列番号58 LC.001のγ重鎖(ヒトIgG1型)
配列番号59 LC.001のγ重鎖(L234A、L235AヒトIgG1突然変異体)
配列番号60 LC.001のγ重鎖(S228PヒトIgG4突然変異体)
配列番号61 LC.001のκ軽鎖
配列番号62 LC.005のγ重鎖(ヒトIgG1型)
配列番号63 LC.005のγ重鎖(L234A、L235AヒトIgG1突然変異体)
配列番号64 LC.005のγ重鎖(S228PヒトIgG4突然変異体)
配列番号65 LC.005のκ軽鎖
配列番号66 LC.060のγ重鎖(ヒトIgG1型)
配列番号67 LC.060のγ重鎖(L234A、L235AヒトIgG1突然変異体)
配列番号68 LC.060のγ重鎖(S228PヒトIgG4突然変異体)
配列番号69 LC.060のκ軽鎖
【0152】
略号
アミノ酸は、3文字コード(Leu)または1文字コード(L)において略記される。
S228Pは、IgG4重鎖の位置228におけるセリンのプロリンへの交換を意味する。
L234は、EU番号付け(Kabat)に従う位置234におけるアミノ酸ロイシンを意味する。
L234Aは、位置234におけるアミノ酸ロイシンがアラニンに変えられていることを意味する。
L235Aは、位置235におけるアミノ酸ロイシンがアラニンに変えられていることを意味する。
PVA236は、236領域において、IgG1のELLGまたはIgG4のEFLGがPVAに修正されていることを意味する。
GLPSS331は、331領域において、IgG1のALPAPまたはIgG2のGLPAPがGLPSSに変えられていることを意味する。
ΔG236は、位置236におけるアミノ酸が欠失していることを意味する。
IgG4xは、IgG4における突然変異S228Pを意味する。
LC2010−001は、LC.001と同義である。
Fcgは、Fcガンマ(Fcγ)の同義語である。
【0153】
抗体の他の配列修正は、同様に命名される。
ヒスチジンタグに融合されたリコンビナント可溶性ヒトOX40L
hOX40L−His
ヒスチジンタグに融合されたリコンビナント可溶性マウスOX40L
mOX40L−His
フラッグタグに融合されたリコンビナント可溶性ヒトOX40L
hOX40L−Flag
フラッグタグに融合されたリコンビナント可溶性マウスOX40L
mOX40L−Flag
ヒトFcγに融合されたリコンビナント可溶性ヒトOX40
hOX40−hFc
ウサギ抗マウスFcγモノクローナル抗体 Anti−mFc
ヤギ抗ヒトFcγモノクローナル抗体 Anti−hFc
マウス抗ヒスチジンモノクローナル抗体 Anti−His
マウスFcγに融合されたリコンビナント可溶性ヒトOX40
hOX40−mFc
マウス抗TNFαモノクローナル抗体 Anti−TNFα
マウス抗CD40Lモノクローナル抗体 Anti−CD40L
腫瘍壊死因子アルファ TNFα
CD40リガンド CD40L
ラット抗ヒトOX40Lモノクローナル抗体 TAG34
ヒト抗ヒトOX40Lモノクローナル抗体 LC.001、LC.005
LC.010、LC.019
LC.029、LC.033
LC.059、LC.060
LC.063
フィトヘマグルチニン PHA
【0154】
実施例
実施例1
抗OX40L抗体を産生するハイブリドーマ細胞系の発生
ハイブリドーマの培養
HuMAbハイブリドーマを、IMDM(Cambrex)、胎仔クローン1ウシ血清(Perbio Science)、起源ハイブリドーマクローニング因子(Igen)、ピルビン酸ナトリウム、ペニシリン/ストレプトマイシン、2−メルカプトエタノール、HAT(Sigma-Aldrich)およびカナマイシン(Invitrogen)中で37℃で、5%CO2において培養した。
【0155】
トランスジェニックマウスの免疫感作手順
LC.2010−001:6匹のHCo7(2匹のオスと4匹の雌)、系統GG2201(Medarex, San Jose, CA, USA)および4匹のHCo12(4匹の雄)、系統GG2198(Mesarex, San Jose, CA, USA)を、ヒトOX40L(hOX40L)のための発現ベクターで一過性にトランスフェクションされた1×106HEK293細胞、およびhOX40Lの可溶性細胞外ドメイン20μgで交互に免疫感作させた。8回の免疫感作を、hOX40L発現細胞による総計4回の腹腔内(i.p)免疫感作およびリコンビナントタンパク質による尾基部(tail base)での4回の皮下(s.c)免疫感作により行った。最初の免疫感作では、1×106HEK293−hOX40L細胞100μlを完全フロイントアジュバント(CFA; Difco Laboratories, Detroit, USA)100μlと混合した。すべての他の免疫感作では、PBS中の細胞100μlを使用するかまたはリコンビナントタンパク質を不完全フロイントアジュバント(ICFA;Difco)100μlと混合した。
【0156】
抗hOX40Lの血清力価が十分であることが見出されると、マウスを、融合の4日および3日前にPBS200μl中のhOX40L細胞外ドメイン15μgで静脈内に(i.v)2回さらに追加抗原刺激した。
【0157】
LC.2010−001、LC.059、LC.060およびLC.063はHCo12マウスに由来していた。
【0158】
LC.2010−005、−010、−019、−029および−033:5匹のHCo7(4匹の雄と1匹の雌)、系統GG2201(Medarex, San Jose, CA,USA)を、hOX40Lの可溶性細胞外ドメイン20μgで免疫感作させた。7回の免疫感作を、総計4回の腹腔内(i.p)免疫感作および尾基部での3回の皮下(s.c)免疫感作により行った。最初の免疫感作では、リコンビナントタンパク質100μlを、完全フロイントアジュバント(CFA; Difco Laboratories, Detroit, USA)100μlと混合した。すべての他の免疫感作では、リコンビナントタンパク質100μlを不完全フロイントアジュバント(ICFA; Difco)100μlと混合した。
【0159】
抗hOX40Lの血清力価が十分であることが見出されると、マウスを、融合の4日および3日前にPBS200μl中のhOX40L細胞外ドメイン15μgで静脈内に(i.v)2回さらに追加抗原刺激した。
【0160】
ハイブリドーマ発生
マウスを殺しそして脾臓および腹大動脈と大静脈をフランキングする(flanking)リンパ節を採集した。脾臓細胞およびリンパ節細胞の融合パートナーSP2.0細胞との融合は、標準操作手順に従って行った。
【0161】
抗原特異的ELISA
免疫感作されたマウスの血清における抗OX40L力価を、抗原特異的ELISAにより決定した。プレート(96平底ELISAプレート、Greiner)を、PBS中に溶解した精製されたOX40L0.1μg/mlでコーティングしそして室温で一夜コーティングした。次いで、ウエルを、PBSTC(Tween 20、0.05%(Sigma-Aldrich Chemie BV)および2%ニワトリ血清(Gibco)を含有するPBS)で室温で1時間ブロッキングした。
【0162】
試験した血清タップ(serum taps)を、PBSTC中に1:50に希釈しそしてウエルに加えた。免疫感作の前にマウスから得られた血清をPBSTC中に1:100で溶解しそしてネガティブコントロールとして使用した。ヒトOX40Lに対して指向されたマウス抗体をPBSTC中に1:50で溶解しそしてポジティブコントロールとして使用した。プレートを室温で1時間インキュベーションした。次いで、プレートをPBST(Tween 20、005%を含有するPBS)を使用して2回洗浄した。Gt−α−hulgG−HRP(Jackson)をPBSTC中に1:5000に希釈しそして試験されたタップおよびネガティブコントロールを含有するウエルに加えた。Rb−α−mIgG(Jackson)をPBSTC中に1:3000に希釈しそしてポジティブコントロールを含有するウエルに加えた。プレートを室温で1時間インキュベーションした。最後に、プレートをPBSTを使用して3回洗浄し、そして調製されたばかりのABTS(登録商標)溶液(1mg/ml)(ABTS:2,2’−アジノ ビス(3−エチルベンゾチアゾリン−6−スルホン酸)で暗所で室温(RT)で30分間発色させた。吸光度を405nmで測定した。
【0163】
κ−ELISA
融合から生じたハイブリドーマがヒト抗体を発生するかどうかを決定するために、κ−ELISAを行った。ELISAプレートを、PBS中に1/10000に希釈したラット抗ヒトIgGκ軽鎖抗体(DAKO)で、4℃での一夜のインキュベーションによりコーティングした。ウエルを捨てた後に、プレートを、PBSTC(Tween−20、0.05%を補充したPBSC(PBSTC))との室温で1時間のインキュベーションによりブロッキングした。次いでウエルをPBSTC中に1/2に希釈したハイブリドーマ培養上清とインキュベーションした。PBSTC中に1/2に希釈した培養培地をネガティブコントロールとして使用し、PBSTC中に1/100に希釈したκ鎖ポジティブマウス血清をポジティブコントロールとして使用した。次いで、ウエルを3回洗浄しそしてPBSTC中に1/2000に希釈したHRPをコンジュゲーションしたラット抗ヒトIgGF(ab’)2(DAKO)と37℃で1時間インキュベーションした。ウエルを3回洗浄しそしてアッセイを、調製されたばかりのABTS(登録商標)溶液(1mg/ml)で暗所で室温(RT)で30分間発色させた。吸光度をELISAプレートリーダーにおいて405nmで測定した。
【0164】
実施例2
抗OX40LHuMab可変ドメインのクローニングおよび配列分析
(κ軽鎖およびγ1重鎖)
OX40L HuMabsの軽鎖可変領域VLおよび重鎖可変領域VHをコードするヌクレオチド配列を、標準cDNA合成/PCR法により単離した。
【0165】
トータルRNAを、GeneRacer(商標)Kit(Invitrogen)を使用して1×106〜1×107ハイブリドーマ細胞から調製した。ハイブリドーマ由来のRNAを、第1鎖cDNA合成およびGeneRacer(商標)Oligo−dTプライマーのライゲーシヨンのためのテンプレートとして使用した。第2鎖cDNA合成およびVLおよびVHコード化cDNAフラグメントのさらなるPCR増幅を、それぞれ、カッパ軽鎖およびγ1重鎖定常領域のヌクレオチド配列に相補性のリバース軽鎖及び重鎖プライマーならびに5’−特異的GeneRacer(商標)プライマーにより行った。PCR産物を、Invitrogen(商標)Life TechnologiesからのTOPO(商標)TAクローニングキットおよびクローニングベクターとしてpCR4−TOPO(商標)を使用してクローニングした。クローニングされたPCR産物を、消化のためのEcoRIを使用して適切なプラスミドの制限マッピングにより同定し、そして、それぞれ、VLおよびVHのための約740および790bpのDNAフラグメントサイズを予測/計算した。
【0166】
クローニングされたPCRフラグメントのDNA配列を二本鎖配列決定により決定した。
【0167】
GCG(Genetics Computer Group, Madison, Wisconsin)ソフトウエアパッケージバージョン10.2およびVector−NTI8(InforMax,Inc)を一般的データプロセッシングのために使用した。DNAおよびタンパク質配列を、GCGモジュールCLUSTALWを使用してアライメントさせた。配列アライメントは、プログラムGENEDOC(バージョン2.1)を使用して作成された。
【0168】
実施例3
抗OX40L IgG1 HuMabのための発現プラスミドの構築
抗OX40L HuMab軽鎖及び重鎖コード化遺伝子を哺乳動物細胞発現ベクター中に別々にアセンブリーした。
【0169】
それにより、抗OX40L HuMab軽鎖可変領域(VL)およびヒトκ軽鎖定常領域(CL、配列番号13、または配列番号61、65もしくは69からの)をコードする遺伝子セグメントを、抗OX40L HuMab重鎖可変領域(VH)およびヒトγ1重鎖定常領域(CH1−Hinge−CH2−CH3、配列番号14または配列番号58、62もしくは66からの)のための遺伝子セグメントが接合されたと同様にして、接合させた。
【0170】
コドン使用頻度を推定することができるヒト軽鎖及び重鎖のヌクレオチド配列に関する一般的情報は、Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, fifth ed., NIH Publication No.91-3242(1991)に与えられる。
【0171】
抗OX40L HuMabκ軽鎖の転写単位は、下記のエレメント:
−ヒトサイトメガロウイルス(HCMV)からの前初期(immediate early)エンハンサーおよびプロモーター
−コザック配列を含む合成5’−UT、
−シグナル配列イントロンを含むマウス免疫グロブリン重鎖シグナル配列、
−5’端部における独特のBsmI制限部位およびスプライスドナー部位および3’端部における独特のNotI制限部位を持って配列された(arranged)クローニングされた抗OX40L HuMab可変軽鎖cDNA、
−イントロン2マウスIg−κエンハンサーを含むゲノムヒトκ遺伝子定常領域[Picard, D., and Schaffner, W., Nature 307(1984)80-82]および
−ヒト免疫グロブリンκポリアデニル化(「ポリA」シグナル配列、
からなる。
【0172】
抗OX40L HuMabγ1重鎖の転写単位は、下記のエレメント:
−ヒトサイトメガロウイルス(HCMV)からの前初期エンハンサーおよびプロモーター
−コザック配列を含む合成5’−UT、
−シグナル配列イントロンを含む改変されたマウス免疫グロブリン重鎖シグナル配列、
−5’における独特のBsmI制限部位およびスプライスドナー部位および3’端部における独特のNotI制限部位を持って配列されたクローニングされた抗OX40L HuMab可変重鎖cDNA、
−マウスIgμエンハンサーを含むゲノムヒトγ1重遺伝子定常領域(Neuberger, M.S., EMBO J.2(1983)1373-1378)、
−ヒトγ1免疫グロブリンポリアデニル化(「ポリA」シグナル配列、
からなる。
【0173】
抗OX40L HuMabκ軽鎖およびγ1重鎖発現プラスミドの機能的エレメント:抗OX40L HuMabκ軽鎖またはγ1重鎖発現カセットの外に、これらのプラスミドは、
−ヒグロマイシン耐性遺伝子
−エプスタイン−バールウイルス(EBV)の複製の起点、oriP
−E.coliにおいてこのプラスミドの複製を可能とするベクターpUC18からの複製の起点および
E.coliにおけるアンピシリン耐性を与えるβ−ラクタマーゼ遺伝子
を含有する。
【0174】
実施例4
抗OX40L IgG4 HuMabのための発現プラスミドの構築
抗OX40Lγ4重鎖プロトタイプ発現プラスミドは、ヒトゲノムγ1定常領域およびγ1免疫グロブリンポリアデニル化(「ポリA」)シグナル配列をヒトゲノムγ4定常領域(配列番号15、または配列番号60,64もしくは68からの突然変異していないγ4定常領域からの)およびγ4免疫グロブリンポリアデニル化シグナル配列により置き換えることにより、抗OX40Lγ1重鎖発現プラスミドから誘導された。
【0175】
抗OX40L HuMabκ軽鎖の発現のために、同じ発現プラスミドを、IgG1について説明したとおりに使用した(上記参照)。
【0176】
実施例5
LC.001に基づく突然変異体(変異体)抗OX40L IgG1およびIgG4のための発現プラスミドの構築
突然変異体抗OX40Lγ1−およびγ4−重鎖をコードする発現プラスミドは、QuickChange(商標)部位特異的突然変異誘発キット(Stratagene)を使用する野生型発現プラスミドの部位特異的突然変異誘発により創生された。アミノ酸は、EU番号付けに従って番号を付けられた(Edelman, G. M., et al., Proc. Natl. Acad. Sci. USA 63(1969) 78-85; Kabat, E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, NIH Publication No. 91-3242, Bethesda, MD(1991))。
【0177】
【表1】
【0178】
実施例6
リコンビナント抗OX40L HuMabsの産生
10%超低IgG FCS(Gibco)、2mMグルタミン(Gibco)、1%容積/容積非必須アミノ酸(Gibco)および250μg/mlG418(Roche)を補充されたDMEM(Gibco)中で培養された付着HEK293−EBNA細胞(ATTC CRL−10852)の一過性トランスフェクションによりリコンビナントHuMabsを発生させた。トランスフェクションのために、Fugene(商標)6(Roche)トランスフェクション試薬を3:1〜6:1の範囲の試薬(μl)対DNA(μg)の割合で使用した。免疫グロブリン軽鎖および重鎖を、1:2〜2:1の軽鎖対重鎖コード化プラスミドのモル比を使用して2つの異なるプラスミドから発現させた。HuMab含有細胞培養上清を、トランスフェクションの4〜11日後に回収した。上清を精製まで−20℃に保存した。
【0179】
例えば、HEK293におけるヒト抗体のリコンビナント発現に関する一般的情報は、Meissner, P., et al., Biotechnol. Bioeng. 75(2001)197-203に与えられる。
【0180】
実施例7
抗体TAG−34、LC.001、LC.005、LC.010、LC.019、LC.029、LC.033のアフィニティー分析
機器:Biacore 3000、ランニングバッファーおよび反応バッファー:HBS−P(10mM HEPES、150mM NaCl、0.005%Tween20、ph7.4)、25℃。アナライトの注射は、0.78nM〜100nMの7つの濃度で3分間行いそしてHBS−Pで5分間洗浄した。表面(カルボキシメチル化されたデキストラン表面、CM)の再生は、10mMグリシンpH2.0の各々1分間の2回の注射により行った。チップ、アッセイフォーマットおよび注射のシーケンスおよび速度論的データは、下表における説明に対応する。速度論的データは、1:1ラングミュア結合モデルに速度論的データを適合させることにより計算された。
【0181】
【表2】
TAG34とmOX40Lとの相互作用は測定できなかった。
【0182】
すべてのBiacoreアッセイについてのデータ評価およびデータ寄託
ネガティブコントロールデータ(例えば、緩衝曲線)は、システム固有のベースラインドリフトの補正およびノイズシグナル減少のためにサンプル曲線から差し引いた。BiaEvaluationバージョン4.01をセンサーグラムの解析およびアフィニティーデータの計算のために使用した。
【0183】
実施例8
hOX40Lの固定化されたhOX40との相互作用を阻害する抗hOX40L抗体の阻害競合アッセイ
機器:Biacore 3000、ランニングバッファーおよび反応バッファー:HBS−P(10mM HEPES、150mM NaCl、0.005%Tween20、ph7.4)、25℃。注射の前に、アナライト(10mM)および競合剤(0.78nM〜100nMの8つの濃度)を22℃で少なくとも20分間プレインキュベーションした。アナライト+/−競合剤の注射を3分間行いそしてHBS−Pで3分間洗浄した。表面の再生は、10mMグリシンpH2.0の各1分の2回の注射により行った。チップ、アッセイフォーマットおよび注射のシーケンスおよび速度論的データは、下表3における説明に対応する。
【0184】
【表3】
【0185】
すべての抗体は、溶液中のOX40へのOX40Lの結合を阻害する(溶液アフィニティー)。LC.001およびLC.005は、TAG34よりも低いIC50値を示す。
【0186】
実施例9
抗OX40L抗体TAG34、LC.001、LC.005、LC.010、LC.019、LC.029、LC.033、LC.060のエピトープ特徴付け
機器:Biacore 3000、ランニングバッファーおよび反応バッファー:HBS−P(10mM HEPES、150mM NaCl、0.005%Tween20、ph7.4)、25℃。エピトープグループを、列挙された抗体間の交差競合(cross competition)により決定した。注射の前に、アナライト(50mM)および競合剤(100nM)を22℃で少なくとも20分間プレインキュベーションした。アナライト+/−競合剤の注射を2分間行いそしてHBS−Pで3分間洗浄した。表面の再生は、10mMグリシンpH2.0の各1分の2回の注射により行った。チップ、アッセイフォーマットおよび注射のシーケンスおよび速度論的データは、下表4における説明に対応する。
【0187】
【表4】
【0188】
TAG34により認識されたOX40Lエピトープは、エピトープAとして定義された。しかしながら、TAG34は、100ngの抗体濃度において変性されたOX40Lに結合しない(ウエスタンブロットにおいて)。1つのエピトープグループ(AまたはB)内の抗体は、交差阻害活性(cross inhibitory activity)を示すが、異なるグループからの抗体は、相加結合シグナル(additive binding signal)を示す。LC.019は、グループAからおよびグループBからの他の抗体を中和する。
【0189】
実施例10
TAG34、LC.001およびLC.005のCD40LおよびTNFαへの結合特異性
機器:Biacore 3000、ランニングバッファーおよび反応バッファー:HBS−P(10mM HEPES、150mM NaCl、0.005%Tween20、ph7.4)、25℃。アナライトの注射は、100nMおよび500nMで3分間行いそしてHBS−Pで2分間洗浄した。表面の再生は、100mMHClの各々1分間の2回の注射により行った。チップ、アッセイフォーマットおよび注射のシーケンスおよび速度論的データは、下表5における説明に対応する。
【0190】
【表5】
【0191】
このアッセイにおいて、CD40Lは、すべての抗体に対するまたはチップ表面に対するいくらかの非特異的結合を示したが、バックグラウンドシグナルを差し引いた後にこのアッセイは、固定化された抗体TAG34、LC.001およびLC.005に対するTNFαおよびCD40L(500nMまで)の非特異的結合がないことを示した。
【0192】
実施例11
抗体LC.001−IgG1およびLC.001−IgG4xのアフィニティー分析
機器:Biacore 3000、ランニングバッファーおよび反応バッファー:HBS−P(10mM HEPES、150mM NaCl、0.005%Tween20、ph7.4)、25℃。アナライトの注射は、0.78nM〜100nMの8つの濃度で3分間行いそしてHBS−Pで5分間洗浄した。表面の再生は、100mM HClの各々1分間の2回の注射により行った。チップ、アッセイフォーマットおよび注射のシーケンスおよび速度論的データは、下表における説明に対応する。速度論的データは、1:1ラングミュア結合モデルに速度論的データを適合させることにより計算された。
【0193】
【表6】
【0194】
LC.001およびLC.001−IgG4xは、hOX40L−Hisに対する同じアフィニティーを示す。
【0195】
実施例12
OX40Lに結合する抗体の検出のためのELISAアッセイ
SAコーティングしたプレート(96平底ELISAブレート、Microcoat)をインキュベーションバッファー(IB=0.1%Tween20(Serva)および1%遮断タンパク質を含有するPBS)中に溶解された0.5μg/mlビオチニル化OX40Lで室温で1時間コーティングした。次いでプレートを洗浄バッファー(WB=0.1%Tween20を含有する生理食塩水)を使用して2回洗浄した。
【0196】
サンプル(細胞培養上清または精製した抗体)をIB中に段階希釈しそしてウエルに加えた。プレートを室温で1時間インキュベーションした。次いで、プレートをWBを使用して2回洗浄した。次いで、ヒトIgGに対するヤギ抗体とPOD(Dianova)のコンジュゲートをIB中に50ng/mlに希釈しそしてウエルに加えた。プレートを室温で1時間インキュベーションした。最後に、プレートをWBを使用して2回洗浄しそしてすぐ使用できるABTS(登録商標)溶液(Roche)で暗所で室温(RT)で発色させた。最も高い濃度の吸光度が十分なODに達した後に、405nmで吸光度を測定した(図1a)。3nM〜8nMの範囲のEC50値が得られた。
【0197】
実施例13
ヒトOX40/ヒトOX40Lの相互作用を阻害する抗体の検出のためのELISAアッセイ
SAコーティングしたプレート(96平底ELISAプレート、Microcoat. Germany)をIB中に溶解された0.5μg/mlビオチニル化OX40Lで室温で1時間コーティングした。次いでプレートをWB(PBS緩衝液、0.1%(重量/容量)Tween(商標)20)を使用して2回洗浄した。
【0198】
サンプルをIB中に1μg/mlの濃度に希釈しそして段階希釈物においてウエルに加えた。OX40LへのOX40の最大結合を達成するために、いくらかのウエルにおいてIBのみを加えた。次いで各ウエルに、0.2μg/mlの濃度の、Digoxigenin(Roche Diagnostics GmbH, DE)とコンジュゲーションされたヒトOX40の溶液を加えた。プレートを室温で1時間インキュベーションした。次いで、プレートをWBを使用して2回洗浄した。ヒツジ<Digoxigenin>−POD(Roche)をIB中に50mU/mlに希釈しそしてウエルに加えた。プレートを室温で1時間インキュベーションした。最後に、プレートをWBを使用して2回洗浄しそしてすぐ使用できるABTS(登録商標)溶液(Roche)により暗所で室温(RT)で発色させた。吸光度を、10〜20分後に405nmで測定した(図1b)。1nM〜4nMの範囲のIC50を得た。
【0199】
実施例14
K562細胞(K562_OX40L細胞)上に発現されたヒトOX40LとヒトOX40の相互作用を阻害するHuMabsの検出のためのFACSアッセイ
目的:Dig標識されたhOX40:hFc融合タンパク質のhOX40L発現細胞系K562_hOX40Lとの相互作用をブロッキングするためのHuMab hOX40Lの性質の決定のためのアッセイ
【0200】
手順:このアッセイは、「アッセイ試薬」としてDig標識されたhOX40:hFcおよび「競合剤」としてHuMab hOX40Lを使用して行われた。
【0201】
アッセイ試薬:ストック0.5μg/μl−PBS中に1:10に希釈された)、100μl抗ジオキシゲニンFLUOS、PBS/0.5%BSA/1%ブロッキング試薬中に1:25に希釈された(Roche Diagnostics GmbH, DE)。
【0202】
2×105K562_OX40L細胞(ISF−0中で成長させた)を、PBS2ml中で洗浄しそしてPBS100μl中に再懸濁させた。この後PBS中の競合剤(0:1/1:1/1.5:1/2:1/2.5:2/5:1競合剤/試薬関係)を添加した。次いで室温で日光において30分間インキュベーションした。次いで試薬(PBS中の)を加えた;インキュベーション時間:30分、室温および日光。細胞をPBS2mlで洗浄しそして遠心によりペレット化した。染色用の二次抗体(抗ジゴキシゲニン−フルオレセイン)、Fabフラグメント(Roche, 1207741)を加えそして暗所で4℃で30分間インキュベーションした。細胞をPBS2mlで洗浄しそして遠心によりペレット化した。その後細胞をPBS0.5ml中に再懸濁させた。サンプルの測定をFACS−Scanにおいて行った(図2)。
【0203】
実施例15
hOX40/hOX40Lシグナリングについての抗体の阻害能力の決定のための機能的アッセイ(「NFκBアッセイ」)
Hela野生型(wt)およびヒトOX40を発現するHeLa細胞(HeLa_OX40)を、最小必須培地(MEM)、1×ピルビン酸Na、1×非必須アミノ酸(Gibco)、10%FCSにおいてそしてリコンビナント細胞の場合には+600μg/mlG418において成長させた。K562およびOX40Lを発現するK562をISF−O培地中で成長させそしてリコンビナント細胞の場合には、200μg/mlG418を加えた。
【0204】
HeLa_wtまたはHeLa_OX40細胞を、96ウエルプレートw/oG418中に3×104細胞/100μlの細胞密度で播種しそしてCO2インキュベータ中で一夜インキュベーションした。K562_wtまたはH562_OX40L細胞を1:1の細胞対細胞関係において加えた。OX40を発現するホルマリン固定された又はホルマリン固定されていないK562(−70℃で凍結された)を解凍しそしてMEM/10%FCS中で1:1に希釈し;K562_OX40L細胞を、OX40Lに対する抗体と共に室温で30分間プレインキュベーションした。K562_OX40L細胞による刺激時間は30分〜150分であった。細胞核からのタンパク質抽出をActive MotifからのNE−Kitにより供給者の指示に従って行った。Active MotifからのTransAM NFκB−ELISA(供給者の指示に従ってアッセイを行った)を使用して、NFκB活性化をもたらす、OX40シグナリングを決定した。Tecan MTP−Readerにおいて波長450/620nmで吸光度の測定を行った(図3)。すべてのLC抗体について0.6〜5nMの範囲のIC50値を得た。
【0205】
実施例16
T細胞活性化アッセイ
アッセイ原理
ヒト末梢血単核細胞(PBL)を、最適より低い濃度のT細胞マイトジェンフィトヘマグルチニン(PHA)で活性化し、そしてOX40Lを過発現するK562細胞で共刺激する(co-stimulated)。アッセイ条件下に、活性化されたT細胞を37℃で24時間インキュベーションして、IL−2を産生させる。ELISAアッセイを使用して上清中のサイトカインを測定する。Mabのブロッキグ効果を決定するために、K562_OX40L細胞をPBLとの共培養の前に抗体の適切な希釈物と1時間プレインキュベーションした。
【0206】
手順
Histopaque(登録商標)-1077(Sigma)における密度勾配遠心によりヘパリンを添加された全血から、ヒト末梢血単核細胞(PBL)を分離する。Hanksで洗浄した後に、細胞をTurkの溶液を使用して係数し、そして細胞を、ペニシリン、ストレプトマイシンおよびグルタミン(Gibco 10378-016)および10%FBSを補充されたRPMI 1640(Gibco)中に106/mlの濃度で再懸濁させる。K562コントロール細胞(野生型)を上記したとおりに補充された同じRPMI培地中に維持する。OX40LによりトランスフェクションされたK562細胞をGeneticin(G418、Gibco)を補充された同じ培地中に50mg/mlの最終濃度に維持する。K562細胞(WTまたはOX40L+)を、1.5×105細胞/mlに同じ培地で希釈しそして50μl/ウエル(0.75×104/ウエル)において96ウエル組織培養プレートの各ウエルに分配する。Mabの適切な希釈物を20μl/ウエルの容積において細胞に加えそして37℃で1時間インキュベーションする。各希釈物を二重測定ウエル(duplicate wells)において試験する。PBLを、100μl/ウエル(105細胞/ウエル)の容積で加える。PBL対K562細胞の最終比は〜13:1である。PHA(10X)(Sigma L-9132)を20μl/ウエルで加える(最終濃度0.75μg/ml)。ウエル当たりの総容積はRPMI/10%FCSで200μlにする。プレートを5%CO2−給湿されたインキュベータ中で37℃で24時間インキュベーションする。プレートの遠心の後、上清を集めそして製造者の指定に従ってELISA(BD,San Diego, CA, Cat No2627K1)によりIL−2を試験する(図4)。IC50(OX40Lで刺激されたPBLによるIL−2放出の50%を阻止するMab濃度)を計算するために、コントロール培養物(PBL+PHA+K562WT)において産生されたバックグラウンドIL−2濃度を、K562xOX40L+細胞により刺激されたPBLにより産生された総IL−2から差し引いた(図5)。IC50 TAG34:0.07μM;LC.001:2nM;LC.005:10nM。IC50値は2〜10nMの範囲にある。
【0207】
実施例17
破傷風トキソイドにより刺激された末梢血リンパ球に対する抗体の阻害効果を試験する破傷風アッセイ「TT−アッセイ」)
末梢血単核細胞(PBMC)をFicoll Hypaqueにより、ヘパリンを添加された血液から単離した。大部分の場合に、単離されたばかりのPBMCをこのアッセイのために使用した。ある場合には冷凍保存されたPBMCも使用した。このアッセイのための培地は、10%ヒト男性AB血清(Sigma-Aldrich);2mMグルタミンおよびPen/Strep(抗生物質ペニシリンおよびストレプトマイシンのすぐ使用できる混合物(Roche Diagnostics GmbH DE);20mlに再構成された凍結乾燥物;1000ml培地当たり2mlの使用)を含有するRPMIであった。
【0208】
プラスチックに付着させるために、ウエル当たり300.000PBMCを、96ウエル平底プレートにおいて一夜プレインキュベーションした。
【0209】
次の日に破傷風トキソイド(TT)(Chiron Behring)を2〜5μg/mlの最終濃度でウエルに加えた。ポジティブコントロールウエル(最大増殖/刺激)はTTのみを含有しており、すべての他のウエルに、抗体(精製されたIgG)を10μg/mlの最終濃度になるように加えた。マウスMabTAG34は、このアッセイにおいて含まれた(最終濃度10μg/ml)。非刺激性バックグラウンドとして、コントロール培地のみを使用した。すべてのアッセイは三重測定するように(in triplicates)セットアップされた。
【0210】
6日のさらなるインキュベーション(37℃、5%CO2、95%湿度)の後、3Hチミジンを1μキューリー/mlの最終濃度となるように加え、16時間の追加のインキュベーション期間の後に、プレートを回収しそして取り込まれた3Hチミジンをβ計数器において決定した(図6)。
【0211】
実施例18
OX40L抗体のマウスOX40Lとの交差反応性
本発明の抗体のマウスOX40Lと交差反応する能力を決定するために、段階的に希釈した抗体およびコントロール抗体を、mOX40Lを安定に発現するK562−mOX40L細胞と共にインキュベーションした。K562 WT細胞およびhOX40Lを安定に発現するK562−hOX40L細胞への結合も評価した。ネガティブコントロールとして、スカシ貝ヘモシアニン(Keyhole Limpet Hemocyanin)(α−KLH)に対して指向されたHuMab抗体を使用した。抗体RM134L、ラット抗mOX40L(eBioscience, San Diego, CA)が、mOX40L発現のためのポジティブコントロールとして含まれた。抗体TAG−34、マウス抗hOX40L(MBL, Nagoya, Japan)がhOX40L発現のためのポジティブコントロールとして含まれた。結合したヒト抗体の検出のために、フルオレセイン(FITC)コンジュゲーションされたヤギ抗ヒトIgG抗体を使用した。結合したRMI34Lの検出のために、ビオチニル化ウサギ抗ラットIgG抗体(DAKO、Glostrup, Denmark)を、フィコエリトリン(PE)(DAKO)とコンジュゲーションされたストレプトアビジンと組み合わせて使用した。結合したTAG−34の検出のために、FITCコンジュゲーションされたウサギ抗マウスIgG抗体を使用した。EC50値または試験したHuMabの20μg/mlでの最大結合(Bmax)に関する計算を、Graphpad Prismソフトウエアを使用する非線形回帰(可変スロープを有するS字状用量−応答)を使用して決定した。
【0212】
結果:
本発明に従うLC.001は、5.16±2.93μg/mlのEC50値および385.22のBmax(MFI)値により示されたとおりhOX40Lに結合することができたが、それぞれ11.41および9.67のBmax(MFI)値により示されたとおりmOX40LまたはWT細胞に結合することができなかった。さらに、本発明に従うLC.001(IgG4)は、8.19±1.05μg/mlのEC50値および311.30のBmax(MFI)値により示されたとおりhOX40Lにも有効に結合することができたが、それぞれ13.47および9.58のBmax(MFI)により示されたとおりmOX40LまたはWT細胞に結合することができなかった。予想されるとおり、ネガティブコントロールα−KLHはいかなる細胞にも結合しなかった(図7)。したがって、本発明に従うOX40L抗体は、ヒトOX40Lと比較してマウスOX40Lへの少なくとも30倍低い結合を示す。
【0213】
実施例19
OX40L HuMabの補体系を活性化する潜在力
C1qおよびC3c結合ELISA
本発明の抗体のC1q結合およびC3活性化を誘導する能力を決定するために、ELISAプレートを段階的に希釈した抗体およびコントロール抗体でコーティングした。ネガティブコントロールとして、C1qに非常に弱く結合するヒトIgG4(Tke Binding Site, Birmingham, England)を使用した。ヒトIgG1(The Binding Site)およびα−KLH(IgG1)は、ポジティブコントロールとして含まれた。次いで、コーティングされた抗体を、リコンビナントC1qおよびC3のソースとしてヒトのプールされた血清と共にインキュベーションした。結合したC1qの検出のために、セイヨウワサビペルオキシダーゼ(HRP)とコンジュゲーションされたブタ抗ウサギIgG抗体(DAKO)と組み合わせたC1qに対して指向されたウサギ抗体(DAKO)を使用した。活性化されたC3c(C3の活性化により発生した)の検出のために、HRPとコンジュゲーションしたウサギ抗マウスIgG抗体(Jackson ImmunoResearch Laboratories, West Grove, PA)と組み合わせてマウス抗ヒトC3c抗体(DAKO)を使用した。コーティング有効性の差を評価するために、コーティングされた抗体を、HRPとコンジュゲーションされたヤギ抗ヒトIgG抗体で可視化した。EC50値または試験されたHuMab10μg/mlにおける最大結合(Bmax)に関する計算を、Graphpad Prismソフトウエアを使用する非線形回帰(可変スロープを有するS字状用量−応答)を使用して決定した。
【0214】
結果:
本発明に従うLC.001は、2.19±0.42μg/mlのEC50値および3.089のBmax(OD405)値により示されたとおり、C1qに有効に結合することができた。さらに、両ポジティブコントロールヒトIgG1および抗KLHは、それぞれ4.17±1.08μg/mlおよび2.57±1.51μg/mlのEC50値、ならびにそれぞれ2.685および3.306のBmax(OD405)値により示されたとおり、C1qに有効に結合することができた。予想されるとおり、ネガティブコントロールヒトIgG4は、0.353のOD405Bmax値により示されたとおり、C1qに結合しなかった。さらに、本発明に従うLC.001IgG4xは、0.357のOD405Bmax値により示されたとおり、C1qに結合する能力を失っていた。
【0215】
C1q結合能力と一致して、LC.001によるC3c沈着(deposition)が、2.67±0.16μg/mlのEC50値および2.614のBmax(OD405)値で、抗体濃度依存性方式で起こった。さらに、両ポジティブコントロールヒトIgG1および抗KLHは、それぞれ5.45±0.36μg/mlおよび2.16±0.26μg/mlのEC50値、ならびにそれぞれ2.543および2.633のBmax(OD405)値により示されたとおり、C3cを有効に沈着することができた。予想されるとおり、ネガティブコントロールヒトIgG4は、0.095のOD405Bmax値により示されたとおり、C3cを沈着しなかった。さらに、本発明に従うLC.001IgG4xは、0.090のOD405Bmax値により示されたとおり、C3cを沈着する能力を失っていた(図8および図9)。
【0216】
実施例20
OX40L HuMabsのFcγレセプターI、IIaおよびIIbに結合する潜在力
IgGで誘導される抗体依存性細胞性細胞傷害(ADCC)は、エフェクター細胞に対するFcγレセプター(FcγR)により媒介される。本発明の抗体のFcγRsに結合する能力を決定するために、ヒトFcγRI、FcγRIIa、FcγRIIbで安定にトランスフェクションされたIIA1.6細胞(IIA1細胞から限界希釈により誘導された;Jones, B., et al., J. Immunol. 136(1986) 348-356)および野生型細胞を、段階希釈された抗体およびコントロール抗体と共にインキュベーションした。ネガティブコントロールとして、FcγRIに結合しないヒトIgG2(The Binding Site Ltd)およびFcγRIIに結合しないヒトIgG4(The Binding Site)を使用した。ヒトIgG1(The Binding Site)は、FcγRI結合のためのポジティブコントロールとして含まれ、そしてヒトIgG3(The Binding Site)はFcγRII結合のためのポジティブコントロールとして含まれた。結合した抗体を、フィコエリトリン(PE)とコンジュゲーションされたヒトIgGに対して指向された抗体を使用するFACS分析により検出した。EC50値または試験されたHuMab10μg/mlにおける最大結合(Bmax)に関する計算を、Graphpad Prismソフトウエアを使用する非線形回帰曲線適合(可変スロープ)を使用して決定した。
【0217】
LC.001は、0.11±0.03μg/mlのEC50値および8041.54のBmax(MFI)値により示されたとおりFcγRIに有効に(コントロールIgG1抗体に匹敵しうる)結合することができたが、それぞれ25.06および21.18のBmax(MFI)により示されたとおりFcγRIIaおよびFcγRIIbに結合することができなかった。
【0218】
LC.001IgG4xは、0.86±0.12μg/mlのEC50値および6030.07のBmax(MFI)で、LC.001に比較してFcγRIへの結合においてより少なく有効でありそしてコントロールIgG4抗体に匹敵していた。LC.001IgG4xのFcγRIIaおよびFcγRIIbへの結合は観察されなかったが(それぞれ21.40および19.27のBmax(MFI)値)、これに対してコントロールIgG3抗体は、結合することができた(それぞれ536.65および418.59のBmax(MFI)値)(図10)。したがって、FcγRIへの結合のEC50値は、抗体LC.001のEC50値に比較してLC.001IgG4xでは8倍である。
【0219】
実施例21
OX40L HuMabsのNK細胞上のFcγRIIIaに結合する潜在力
本発明の抗体のナチュラルキラー(NK)細胞上のFcγRIIIa(CD16)に結合する能力を決定するために、末梢血単核細胞(PBMCs)を単離しそして、FcγRIIIaに対するブロッキングマウス抗体(抗CD−16、クローン3G8、RDI、Flanders,NJ)20μg/mlの存在下または不存在下にHuMab抗体およびコントロール抗体20μg/mlとインキュベーションして、FcγRIIIaを介する結合を証明した。ネガティブコントロールとして、FcγRIIIaに結合しないヒトIgG2およびIgG4(The Binding Site)を使用した。ヒトIgG1およびIgG3(The Binding Site)は、FcγRIIIa結合のためのポジティブコントロールとして含まれた。NK細胞上の結合した抗体を、PE標識されたマウス抗ヒトCD56(NK細胞表面マーカー)抗体(BD Bioscience Pharmingen, San Diego,CA)を、FITC標識されたヤギF(ab)2抗ヒトIgG(Fc)抗体(Protos immunoresearch, Burlingame,CA)と組み合わせて使用するFACSにより検出した。試験されたHuMab20μg/mlにおける最大結合(Bmax)を決定した。
【0220】
LC.001は、641.37のBmax(MFI)値により示されたとおり、FcγRIIIaに有効に(コントロールIgG1抗体に匹敵しうる)結合することができた。FcγRIIIaに対するブロッキング抗体の添加は、NK細胞へのLC.001の結合を無効にした(145.38のバックグラウンド染色に比較して194.61のBmax(MFI)値)。LC.001 IgG4xは、FcγRIIIaに結合せずそして170.52のBmax(MFI)値で、コントロールIgG4抗体に匹敵しうる挙動をして、LC.001のBmaxの約10%にすぎないLC.001 IgG4xのBmaxをもたらした。FcγRIIIaに対するブロッキング抗体の添加は、LC.001 IgG4x結合に対する効果を示さなかった(174.26のBmax(MFI)値)(図11)。
【0221】
実施例22
HUVEC(初代ヒト臍帯静脈内皮細胞/Promo Cell)へのhMab_hOX40LおよびMab TAG34結合の効果
内皮細胞は、hOX40Lを発現することが記載されている(Kotani, A., et al., Immunol. Lett. 84(2002)1-7)。ヒト臍帯静脈内皮細胞(HUVEC)は、当然hOX40Lを発現し、したがって「内皮細胞モデル」として使用することができる。このアッセイの目的は、抗体TAG−34およびLC.001への結合後のHUVEC細胞上のhOX40Lの運命を決定することであった。
【0222】
HUVECを解凍しそしてT175フラスコ(Sarstedt)中でECG−M培地+2%FCS中で4日間増大させた。細胞を24ウエルプレート中にプレートした(10.000細胞/ウエル)。
【0223】
3日後、培地をECG−M培地+0.5%FCSに変えた。10μg/mlの抗体(<KLH>(スカシ貝ヘモシアニンに対する抗体)、ダウンモデュレーション(down-modulation)の誘導のためのTAG−34またはLC.001)の添加および2,5時間または24時間のインキュベーション。TAG−34またはLC.001によるHUVEC細胞の再染色。Alexa488(=<m>)で標識されたマウスIgGに対する二次抗体またはAlexa488(=<h>)で標識されたヒトIgGに対する二次抗体、各々10μg/mlによるFACS染色。FACS測定は、FACS−scan(Becton Dickinson)において行いそして平均蛍光強度(MFI)を計算した。
【0224】
前記<KLH>抗体を非特異的ネガティブコントロールとして使用した。
【0225】
表7は、LC.001の添加は、2.5時間後も24時間後もHUVEC細胞上のOX40Lのダウンモデュレーションをもたらさない(ライン4をライン5および6と比較する)ことを示す。しかしながら、TAG34の添加は、2.5時間後、そして24時間後にHUVEC細胞上のhOX40Lの強い(約3倍)ダウンモデュレーションを示す(ライン10をライン11および12と比較する)。
【0226】
10μg/mlの濃度の本発明に従う抗体は、HUVEC細胞上のOX40L発現のダウンレギュレーションを誘導しない。
【0227】
【表7】
【0228】
実施例23
TAG34、LC.001およびLC.005のウエスタンブロット分析
40および100nghOx40L−His(R&D Systems, 28〜34kDaの理論的サイズ)および分子量マーカーMagik Mark XP(Invitrogen; 20, 30, 40, 50, 60, 80 , 100, 120, 220 kDa)をゲル電気泳動のために調製した。したがって、xμlタンパク質、2.5μlNuPage LDS(ドデシルサルフェートのリチウム塩)サンプルバッファー(4x)、1μlNuPage還元剤(10x)および10μlとする量のH2Oを一緒にしそして10分間70℃で変性した。その後サンプルをNuPageゲル(Novex; 10%Bis-Tris)上にローディングしそして1×MOPSランニングバッファー(Novex)中で150Vで1時間ランニングした。
【0229】
ゲルを半乾燥チャンバ中で1時間/50mA、1×NuPageトランスファーバッファー(1×バッファー、0.1%Antioxidance、10%メタノール)を使用してPVDF膜(Millipore; メタノール中の5分間のインキュベーションおよび1×トランスファーバッファー中の10分のインキュベーションによる膜の活性化)上に半乾燥ブロットによりブロッティングした。膜を、室温で1時間振とう下に1×PBS/5%ミルク/0.5%Tween中でブロッキングした。一次抗体(pAB)を1×PBS/1%ミルク/0.5%Tween中に希釈し、加え、4℃で一夜インキュベーションした。
LC.001: 全容積4ml中の1.9μl(1.6μg)
LC.005: 1.1μl(1.6μg)/4ml
TAG34: 1.6μl(1.6μg)/4ml
【0230】
膜を1×PBS/0.5%Tween中で10分間3回洗浄した。二次抗体を(sAB)を1×PBS/1%ミルク/0.5%Tween中に希釈し、加えそして室温で1.5時間インキュベーションした。LC.001およびLC.005については、1:10000希釈におけるヒトIgGに対するポリクローナル抗体(Pierce)をsABとして使用し;TAG34では、1:400希釈におけるLumi Light Western Blotting Kit(Roche)からのマウスIgGに対するポリクローナル抗体をsABとして使用した。膜を1×PBS/0.5%Tweenで30分間2回洗浄した。検出のために、Lumi Light Western Blotting Kit(Roche)を、製造者の指示に従って使用した。ウエスタンブロットからの結果を図12に示す。LC.001は(ドデシルサルフェート)変性されたOX40Lを検出することができるが、これに対してLC.005およびTAG34は変性されたOX40Lに結合しない。
【0231】
【表8】
【図面の簡単な説明】
【0232】
【図1a】TAG−34、LC.001、LC.005、LC.010、LC.019、LC.029、LC.033についての「結合ELISA」を示す。
【図1b】TAG−34、LC.001、LC.005、LC.010、LC.019、LC.029、LC.033についての「阻止ELISA」+IC50データを示す。
【図2】TAG−34、LC.001、LC.005についての「阻止FACS」を示す。
【図3】TAG−34、LC.001、LC.019およびLC.024についての「NFκB−アッセイ」を示す(非結合抗体)。
【図4】TAG−34、LC.001およびLC.005についての「T細胞活性化アッセイ」およびIC50値(図4:IL−2放出、図5:阻害)を示す。
【図5】TAG−34、LC.001およびLC.005についての「T細胞活性化アッセイ」およびIC50値(図4:IL−2放出、図5:阻害)を示す。
【図6】TAG−34、LC.001およびLC.033についての「TT−アッセイ」データを示す。
【図7】本発明の抗体のマウスOX40Lとのクロス反応性を示す。A)トランスフェクションされた細胞上の及びWT細胞上のhOX40L発現のためのコントロール、B)hOX40L発現K562細胞への抗体の結合、C)トランスフェクションされた細胞およびWT細胞上のmOX40L発現のためのコントロール、D)mOX40L発現K562細胞への抗体の結合、およびE)WT K562細胞への抗体の結合(n=3)。
【図8】本発明の抗体のC1qに結合する能力を示す(n=3)。
【図9】本発明の抗体のC3cを活性化する能力を示す(n=3)。
【図10】本発明の抗体のFcγRI(n=4)、FcγRIIa(n=4)およびFcγRIIB(n=4)に結合する能力を示す。
【図11】本発明の抗体のNK細胞上のFcγRIIIA(CD16)に結合する能力を示す(6つのドナーの平均±SEM)。
【図12】ウエスタンブロット;レーン1、4、7:マーカー;レーン2,5,8:100ngOX40L;レーン3,6,9:40ngOX40L。
【特許請求の範囲】
【請求項1】
OX40Lに結合し、ヒト起源に由来するFc部分を含有しそして補体因子C1qに結合しないことを特徴とする抗体。
【請求項2】
前記抗体は、NK細胞上のヒトFcγレセプターに結合しないことを特徴とする、請求項1に記載の抗体。
【請求項3】
前記抗体は、ヒト抗体であることを特徴とする、請求項1または2に記載の抗体。
【請求項4】
前記抗体は、キメラ抗体又はヒト化抗体であることを特徴とする、請求項1または2に記載の抗体。
【請求項5】
前記抗体は、10−8M未満のKD値でOX40Lに結合する、請求項1〜4のいずれかに記載の抗体。
【請求項6】
KDの範囲が10−12〜10−9Mである、請求項5に記載の抗体。
【請求項7】
前記抗体が、PVA236、GLPSS331および/またはL234A/L235A(EUインデックスに従う番号付け)からの1つ以上の突然変異を含むヒトサブクラスIgG1の抗体であることを特徴とする、請求項1〜6のいずれかに記載の抗体。
【請求項8】
前記抗体が、ヒトサブクラスIgG4の抗体であることを特徴とする、請求項1〜6のいずれかに記載の抗体。
【請求項9】
突然変異S228Pを含有することを特徴とする、請求項8に記載の抗体。
【請求項10】
突然変異L235Eを含有することを特徴とする、請求項8または9に記載の抗体。
【請求項11】
補体因子C3を活性化しないことを特徴とする、請求項1〜10のいずれかに記載の抗体。
【請求項12】
前記抗体が補体依存性細胞傷害(CDC)を誘発しないことを特徴とする、請求項1〜11のいずれかに記載の抗体。
【請求項13】
前記抗体が抗体依存性細胞性細胞傷害(ADCC)を誘発しないことを特徴とする、請求項1〜12のいずれかに記載の抗体。
【請求項14】
前記抗体が、1nM〜4nMの範囲のIC50値で0.5μg/mlOX40Lのコーティング濃度で可溶性OX40と固定化されたOX40Lとの相互作用をブロッキングすることにより、ELISAアッセイにおいて阻害を示すことを特徴とする、請求項1〜13のいずれかに記載の抗体。
【請求項15】
抗体であって、該抗体がOX40Lに結合することを特徴とし、そして該抗体が、下記の組合せ、
a)配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号2により規定された重鎖可変ドメイン、
b)配列番号3のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号4により規定された重鎖可変ドメイン、
c)配列番号5のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号6により規定された重鎖可変ドメイン、
d)配列番号7のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号8により規定された重鎖可変ドメイン、
e)配列番号9のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号10により規定された重鎖可変ドメイン、
f)配列番号11または16のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号12により規定された重鎖可変ドメイン、
g)配列番号1のアミノ酸配列により規定された軽鎖(VL)可変ドメインおよび配列番号17により規定された重鎖(VH)可変ドメイン、
h)配列番号18のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号19により規定された重鎖可変ドメイン、
i)配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号20により規定された重鎖可変ドメイン、
からなる群より独立に選ばれる可変領域組合せを含むことを特徴とする抗体。
【請求項16】
ヒト軽鎖可変領域が、配列番号1、3、5、7、9、11、16および18からなる群より独立に選ばれるアミノ酸配列を含むことを特徴とする、請求項1〜15のいずれかに記載の抗体。
【請求項17】
ヒト重鎖可変領域が、配列番号2、4、6、8、10、12、17、19および20からなる群より独立に選ばれるアミノ酸配列を含むことを特徴とする、請求項1〜16のいずれかに記載の抗体。
【請求項18】
前記抗体が、配列番号1のアミノ酸配列により規定される軽鎖可変ドメインおよび配列番号2、17または20により規定された重鎖可変ドメインを含むことを特徴とする、請求項1〜16のいずれかに記載の抗体。
【請求項19】
前記抗体が、補体因子C1qへの非結合を引き起こすFc部分における少なくとも1つのアミノ酸突然変異を含むことを特徴とする、請求項1〜18のいずれかに記載の抗体。
【請求項20】
前記抗体が、配列番号13により規定されたκ軽鎖定常領域または配列番号61、65もしくは69の軽鎖定常領域を含むことを特徴とする、請求項1〜19のいずれかに記載の抗体。
【請求項21】
前記抗体が、配列番号14または配列番号15により規定された重定常領域または配列番号58の重鎖定常領域を含むことを特徴とする、請求項1〜20のいずれかに記載の抗体。
【請求項22】
可変軽鎖および可変重鎖を含む、OX40Lに結合する抗体であって、該可変重鎖が配列番号33〜38から選ばれるCDR3を含むこと、および/または該可変軽鎖が、配列番号51〜57から選ばれるCDR3を含むことを特徴とする抗体。
【請求項23】
可変重鎖が、配列番号21〜25から選ばれるCDR1および配列番号26〜32から選ばれるCDR2を含むこと、および/または可変軽鎖が、配列番号39〜44から選ばれるCDR1および配列番号45〜50から選ばれるCDR2を含むことを特徴とする、請求項22に記載の抗体。
【請求項24】
細胞系、
hu−Mab<hOX40L>LC.001、
hu−Mab<hOX40L>LC.005、hu−Mab<hOX40L>LC.010、hu−Mab<hOX40L>LC.019、hu−Mab<hOX40L>LC.029、およびhu−Mab<hOX40L>LC.033
からなる群より選ばれる細胞系により産生されることを特徴とする抗体。
【請求項25】
前記抗体が、Fab、F(ab’)2、または一本鎖フラグメントであることを特徴とする、請求項1〜24のいずれかに記載の抗体。
【請求項26】
請求項1〜25に記載の抗体の産生方法であって、10−8M未満のKD値でOX40Lに結合する抗体の重鎖をコードする第1核酸の配列を、改変された抗体が補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合しないような方式で改変し、該改変された第1核酸および該抗体の軽鎖をコードする第2核酸を発現ベクターに挿入し、該ベクターを原核生物宿主細胞もしくは真核生物宿主細胞に挿入し、該宿主細胞を該抗体の合成を可能とする条件下に培養し、そして該抗体をこの培養物から回収することを特徴とする方法。
【請求項27】
前記抗体の重鎖をコードする前記第1核酸の配列が、請求項22に記載の抗体の重鎖をコードすることを特徴とする、請求項26に記載の抗体の産生方法。
【請求項28】
請求項1〜25のいずれかの抗体分子をコードする核酸分子。
【請求項29】
請求項28の核酸分子を含むベクター。
【請求項30】
請求項29のベクターを含む宿主細胞。
【請求項31】
請求項12〜25のいずれかに記載の抗体分子の製造方法であって、該抗体分子の合成を可能とする条件下に請求項30の宿主細胞を培養し、そして該抗体分子をこの培養物から回収することを含む方法。
【請求項32】
請求項1〜25のいずれかに記載の抗体分子又は請求項26もしくは27の方法により産生された抗体分子を含む組成物。
【請求項33】
医薬組成物又は診断組成物である、請求項32に記載の組成物。
【請求項34】
請求項1〜25のいずれかの抗体と少なくとも1種の薬学的に許容されうる賦形剤を含む医薬組成物。
【請求項35】
治療を必要としている患者に、治療的に有効量の請求項1〜23のいずれかに記載の抗体を投与することを特徴とする、治療を必要としている患者の処置方法。
【請求項36】
治療のための請求項1〜23に記載の抗体の使用。
【請求項37】
炎症性疾患の予防及び処置のための請求項1〜23のいずれかに記載の抗体の使用。
【請求項38】
炎症性疾患の予防及び処置用の医薬の製造のための請求項1〜23のいずれかに記載の抗体の使用。
【請求項39】
請求項1〜25のいずれかの抗体分子、請求項28の核酸分子、請求項29のベクターまたは請求項30の宿主細胞を含むキット。
【請求項40】
配列番号21〜38からなる群より選ばれる親抗体重鎖CDRおよび/または配列番号39〜57からなる群より選ばれる親抗体軽鎖CDRの最初のアミノ酸配列を改変する方法であって、該最初のアミノ酸配列をコードする核酸を提供し、1つのアミノ酸が重鎖CDR1において改変され、1〜2つのアミノ酸が重鎖CDR2において改変され、1〜2つのアミノ酸が重鎖CDR3において改変され、1〜3つのアミノ酸が軽鎖CDR1において改変され、1〜3つのアミノ酸が軽鎖CDR2において改変されおよび/または1〜3つのアミノ酸が軽鎖CDR3において改変されるように、該核酸を改変し、該改変されたCDRアミノ酸配列を抗体構造において発現し、該抗体が10−8M未満のKDでOX40Lに結合するかどうかを測定し、そしてもしも該抗体が10−8M未満のKDでOX40Lに結合するならば、該改変されたCDRを選ぶことを特徴とする方法。
【請求項41】
抗体であって、該抗体がOX40Lに結合し、ヒトIgG1クラスでありそしてγ重鎖として配列番号58、62又は66を含むことを特徴とする抗体。
【請求項42】
a)γ重鎖として配列番号58およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号62およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号66およびκ軽鎖として配列番号69、
を含む請求項41に記載の抗体。
【請求項43】
抗体であって、該抗体が、OX40Lに結合し、突然変異L234A/L235Aを含有するIgG1クラスであり、そしてγ重鎖として配列番号59、63または67を含むことを特徴とする抗体。
【請求項44】
a)γ重鎖として配列番号59およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号63およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号67およびκ軽鎖として配列番号69、
を含む、請求項43に記載の抗体。
【請求項45】
抗体であって、該抗体が、OX40Lに結合し、突然変異S228Pを含有するIgG4クラスであり、そしてγ重鎖として配列番号60、64または68を含むことを特徴とする抗体。
【請求項46】
a)γ重鎖として配列番号60およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号64およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号68およびκ軽鎖として配列番号69、
を含む、請求項45に記載の抗体。
【請求項1】
OX40Lに結合し、ヒト起源に由来するFc部分を含有しそして補体因子C1qに結合しないことを特徴とする抗体。
【請求項2】
前記抗体は、NK細胞上のヒトFcγレセプターに結合しないことを特徴とする、請求項1に記載の抗体。
【請求項3】
前記抗体は、ヒト抗体であることを特徴とする、請求項1または2に記載の抗体。
【請求項4】
前記抗体は、キメラ抗体又はヒト化抗体であることを特徴とする、請求項1または2に記載の抗体。
【請求項5】
前記抗体は、10−8M未満のKD値でOX40Lに結合する、請求項1〜4のいずれかに記載の抗体。
【請求項6】
KDの範囲が10−12〜10−9Mである、請求項5に記載の抗体。
【請求項7】
前記抗体が、PVA236、GLPSS331および/またはL234A/L235A(EUインデックスに従う番号付け)からの1つ以上の突然変異を含むヒトサブクラスIgG1の抗体であることを特徴とする、請求項1〜6のいずれかに記載の抗体。
【請求項8】
前記抗体が、ヒトサブクラスIgG4の抗体であることを特徴とする、請求項1〜6のいずれかに記載の抗体。
【請求項9】
突然変異S228Pを含有することを特徴とする、請求項8に記載の抗体。
【請求項10】
突然変異L235Eを含有することを特徴とする、請求項8または9に記載の抗体。
【請求項11】
補体因子C3を活性化しないことを特徴とする、請求項1〜10のいずれかに記載の抗体。
【請求項12】
前記抗体が補体依存性細胞傷害(CDC)を誘発しないことを特徴とする、請求項1〜11のいずれかに記載の抗体。
【請求項13】
前記抗体が抗体依存性細胞性細胞傷害(ADCC)を誘発しないことを特徴とする、請求項1〜12のいずれかに記載の抗体。
【請求項14】
前記抗体が、1nM〜4nMの範囲のIC50値で0.5μg/mlOX40Lのコーティング濃度で可溶性OX40と固定化されたOX40Lとの相互作用をブロッキングすることにより、ELISAアッセイにおいて阻害を示すことを特徴とする、請求項1〜13のいずれかに記載の抗体。
【請求項15】
抗体であって、該抗体がOX40Lに結合することを特徴とし、そして該抗体が、下記の組合せ、
a)配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号2により規定された重鎖可変ドメイン、
b)配列番号3のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号4により規定された重鎖可変ドメイン、
c)配列番号5のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号6により規定された重鎖可変ドメイン、
d)配列番号7のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号8により規定された重鎖可変ドメイン、
e)配列番号9のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号10により規定された重鎖可変ドメイン、
f)配列番号11または16のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号12により規定された重鎖可変ドメイン、
g)配列番号1のアミノ酸配列により規定された軽鎖(VL)可変ドメインおよび配列番号17により規定された重鎖(VH)可変ドメイン、
h)配列番号18のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号19により規定された重鎖可変ドメイン、
i)配列番号1のアミノ酸配列により規定された軽鎖可変ドメインおよび配列番号20により規定された重鎖可変ドメイン、
からなる群より独立に選ばれる可変領域組合せを含むことを特徴とする抗体。
【請求項16】
ヒト軽鎖可変領域が、配列番号1、3、5、7、9、11、16および18からなる群より独立に選ばれるアミノ酸配列を含むことを特徴とする、請求項1〜15のいずれかに記載の抗体。
【請求項17】
ヒト重鎖可変領域が、配列番号2、4、6、8、10、12、17、19および20からなる群より独立に選ばれるアミノ酸配列を含むことを特徴とする、請求項1〜16のいずれかに記載の抗体。
【請求項18】
前記抗体が、配列番号1のアミノ酸配列により規定される軽鎖可変ドメインおよび配列番号2、17または20により規定された重鎖可変ドメインを含むことを特徴とする、請求項1〜16のいずれかに記載の抗体。
【請求項19】
前記抗体が、補体因子C1qへの非結合を引き起こすFc部分における少なくとも1つのアミノ酸突然変異を含むことを特徴とする、請求項1〜18のいずれかに記載の抗体。
【請求項20】
前記抗体が、配列番号13により規定されたκ軽鎖定常領域または配列番号61、65もしくは69の軽鎖定常領域を含むことを特徴とする、請求項1〜19のいずれかに記載の抗体。
【請求項21】
前記抗体が、配列番号14または配列番号15により規定された重定常領域または配列番号58の重鎖定常領域を含むことを特徴とする、請求項1〜20のいずれかに記載の抗体。
【請求項22】
可変軽鎖および可変重鎖を含む、OX40Lに結合する抗体であって、該可変重鎖が配列番号33〜38から選ばれるCDR3を含むこと、および/または該可変軽鎖が、配列番号51〜57から選ばれるCDR3を含むことを特徴とする抗体。
【請求項23】
可変重鎖が、配列番号21〜25から選ばれるCDR1および配列番号26〜32から選ばれるCDR2を含むこと、および/または可変軽鎖が、配列番号39〜44から選ばれるCDR1および配列番号45〜50から選ばれるCDR2を含むことを特徴とする、請求項22に記載の抗体。
【請求項24】
細胞系、
hu−Mab<hOX40L>LC.001、
hu−Mab<hOX40L>LC.005、hu−Mab<hOX40L>LC.010、hu−Mab<hOX40L>LC.019、hu−Mab<hOX40L>LC.029、およびhu−Mab<hOX40L>LC.033
からなる群より選ばれる細胞系により産生されることを特徴とする抗体。
【請求項25】
前記抗体が、Fab、F(ab’)2、または一本鎖フラグメントであることを特徴とする、請求項1〜24のいずれかに記載の抗体。
【請求項26】
請求項1〜25に記載の抗体の産生方法であって、10−8M未満のKD値でOX40Lに結合する抗体の重鎖をコードする第1核酸の配列を、改変された抗体が補体因子C1qおよび/またはNK細胞上のヒトFcγレセプターに結合しないような方式で改変し、該改変された第1核酸および該抗体の軽鎖をコードする第2核酸を発現ベクターに挿入し、該ベクターを原核生物宿主細胞もしくは真核生物宿主細胞に挿入し、該宿主細胞を該抗体の合成を可能とする条件下に培養し、そして該抗体をこの培養物から回収することを特徴とする方法。
【請求項27】
前記抗体の重鎖をコードする前記第1核酸の配列が、請求項22に記載の抗体の重鎖をコードすることを特徴とする、請求項26に記載の抗体の産生方法。
【請求項28】
請求項1〜25のいずれかの抗体分子をコードする核酸分子。
【請求項29】
請求項28の核酸分子を含むベクター。
【請求項30】
請求項29のベクターを含む宿主細胞。
【請求項31】
請求項12〜25のいずれかに記載の抗体分子の製造方法であって、該抗体分子の合成を可能とする条件下に請求項30の宿主細胞を培養し、そして該抗体分子をこの培養物から回収することを含む方法。
【請求項32】
請求項1〜25のいずれかに記載の抗体分子又は請求項26もしくは27の方法により産生された抗体分子を含む組成物。
【請求項33】
医薬組成物又は診断組成物である、請求項32に記載の組成物。
【請求項34】
請求項1〜25のいずれかの抗体と少なくとも1種の薬学的に許容されうる賦形剤を含む医薬組成物。
【請求項35】
治療を必要としている患者に、治療的に有効量の請求項1〜23のいずれかに記載の抗体を投与することを特徴とする、治療を必要としている患者の処置方法。
【請求項36】
治療のための請求項1〜23に記載の抗体の使用。
【請求項37】
炎症性疾患の予防及び処置のための請求項1〜23のいずれかに記載の抗体の使用。
【請求項38】
炎症性疾患の予防及び処置用の医薬の製造のための請求項1〜23のいずれかに記載の抗体の使用。
【請求項39】
請求項1〜25のいずれかの抗体分子、請求項28の核酸分子、請求項29のベクターまたは請求項30の宿主細胞を含むキット。
【請求項40】
配列番号21〜38からなる群より選ばれる親抗体重鎖CDRおよび/または配列番号39〜57からなる群より選ばれる親抗体軽鎖CDRの最初のアミノ酸配列を改変する方法であって、該最初のアミノ酸配列をコードする核酸を提供し、1つのアミノ酸が重鎖CDR1において改変され、1〜2つのアミノ酸が重鎖CDR2において改変され、1〜2つのアミノ酸が重鎖CDR3において改変され、1〜3つのアミノ酸が軽鎖CDR1において改変され、1〜3つのアミノ酸が軽鎖CDR2において改変されおよび/または1〜3つのアミノ酸が軽鎖CDR3において改変されるように、該核酸を改変し、該改変されたCDRアミノ酸配列を抗体構造において発現し、該抗体が10−8M未満のKDでOX40Lに結合するかどうかを測定し、そしてもしも該抗体が10−8M未満のKDでOX40Lに結合するならば、該改変されたCDRを選ぶことを特徴とする方法。
【請求項41】
抗体であって、該抗体がOX40Lに結合し、ヒトIgG1クラスでありそしてγ重鎖として配列番号58、62又は66を含むことを特徴とする抗体。
【請求項42】
a)γ重鎖として配列番号58およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号62およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号66およびκ軽鎖として配列番号69、
を含む請求項41に記載の抗体。
【請求項43】
抗体であって、該抗体が、OX40Lに結合し、突然変異L234A/L235Aを含有するIgG1クラスであり、そしてγ重鎖として配列番号59、63または67を含むことを特徴とする抗体。
【請求項44】
a)γ重鎖として配列番号59およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号63およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号67およびκ軽鎖として配列番号69、
を含む、請求項43に記載の抗体。
【請求項45】
抗体であって、該抗体が、OX40Lに結合し、突然変異S228Pを含有するIgG4クラスであり、そしてγ重鎖として配列番号60、64または68を含むことを特徴とする抗体。
【請求項46】
a)γ重鎖として配列番号60およびκ軽鎖として配列番号61、
b)γ重鎖として配列番号64およびκ軽鎖として配列番号65、または
c)γ重鎖として配列番号68およびκ軽鎖として配列番号69、
を含む、請求項45に記載の抗体。
【図1a】

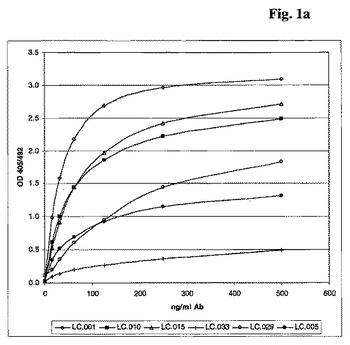
【図1b】
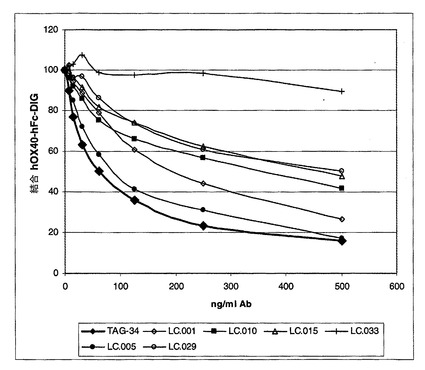
【図2】
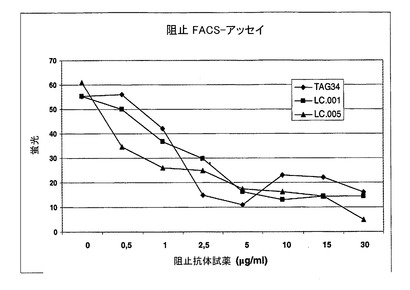
【図3】
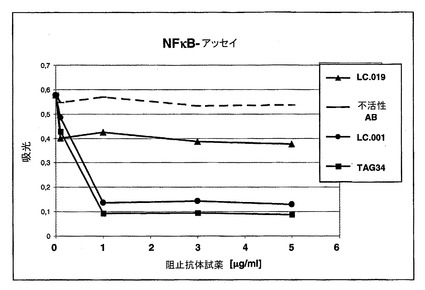
【図4】
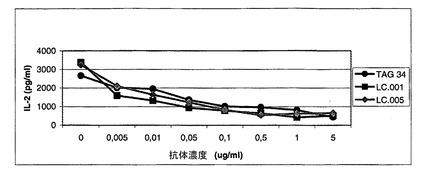
【図5】
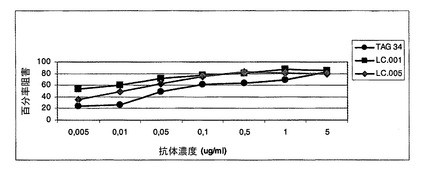
【図6】
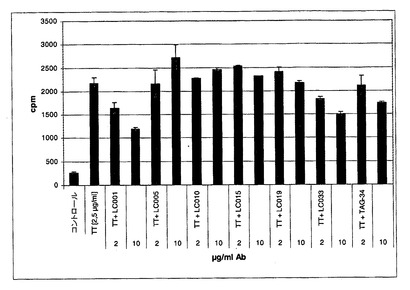
【図7】
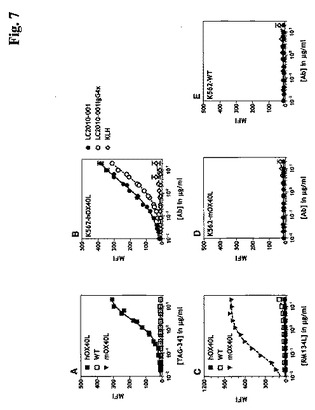
【図8】
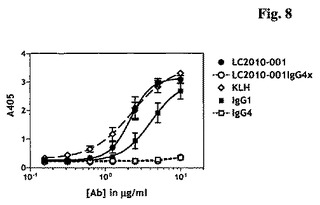
【図9】
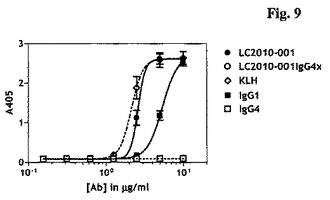
【図10】
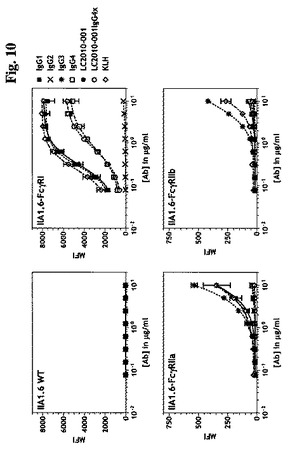
【図11】
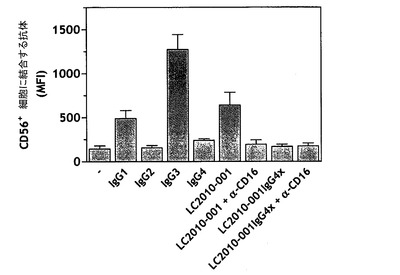
【図12】
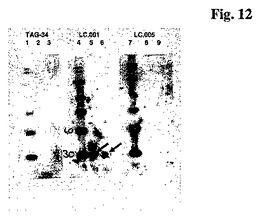
【図1b】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2008−512995(P2008−512995A)
【公表日】平成20年5月1日(2008.5.1)
【国際特許分類】
【出願番号】特願2007−531688(P2007−531688)
【出願日】平成17年9月16日(2005.9.16)
【国際出願番号】PCT/EP2005/009968
【国際公開番号】WO2006/029879
【国際公開日】平成18年3月23日(2006.3.23)
【出願人】(591003013)エフ.ホフマン−ラ ロシュ アーゲー (1,754)
【氏名又は名称原語表記】F. HOFFMANN−LA ROCHE AKTIENGESELLSCHAFT
【Fターム(参考)】
【公表日】平成20年5月1日(2008.5.1)
【国際特許分類】
【出願日】平成17年9月16日(2005.9.16)
【国際出願番号】PCT/EP2005/009968
【国際公開番号】WO2006/029879
【国際公開日】平成18年3月23日(2006.3.23)
【出願人】(591003013)エフ.ホフマン−ラ ロシュ アーゲー (1,754)
【氏名又は名称原語表記】F. HOFFMANN−LA ROCHE AKTIENGESELLSCHAFT
【Fターム(参考)】
[ Back to top ]