
抗P−セレクチン抗体
【課題】抗P−セレクチン抗体、特に補体因子C1qと結合しない抗P−セレクチン抗体の提供。
【解決手段】抗P−セレクチン抗体に、特にヒト起源由来のFc部分を含み補体因子C1qと結合しない抗P−セレクチン抗体およびそれらの変異体に関する。P−セレクチンとE−および/またはL−セレクチンとをマイクロタイタープレートに被覆する細胞性ELISAでのEC50値の測定では、E−またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍高く特異的に結合する。これらの抗体は、重症肢虚血または末梢動脈閉塞性疾患(CLI/PAOD)を患う患者に利益をもたらす。
【解決手段】抗P−セレクチン抗体に、特にヒト起源由来のFc部分を含み補体因子C1qと結合しない抗P−セレクチン抗体およびそれらの変異体に関する。P−セレクチンとE−および/またはL−セレクチンとをマイクロタイタープレートに被覆する細胞性ELISAでのEC50値の測定では、E−またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍高く特異的に結合する。これらの抗体は、重症肢虚血または末梢動脈閉塞性疾患(CLI/PAOD)を患う患者に利益をもたらす。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般的に抗P−セレクチン抗体、特に補体因子C1qと結合しない抗P−セレクチン抗体に関するものである。 好ましくは、これらの抗体はヒト抗体またはヒト化抗体である。
【0002】
P−セレクチン(CD62P、GMP−140、PADGEM、LECAM−3)は、トロンビンおよび他のアゴニストに応答して活性化血小板および内皮の表面に発現する140kDaのカルシウム依存性糖質結合タンパク質である(McEver et al, J Biol Chem 270:11025 (1995); Varki, Proc Natl Acad Sci USA 91:7390 (1994); Springer TA, Annu Rev Physiol 57:827 (1995))。両方の種類の細胞では、P−セレクチンは分泌顆粒、すなわち血小板のα顆粒および内皮細胞のWeibel−Palade小体に貯蔵される(McEver et al., J Clin Invest 84:92 (1984))。P−セレクチンは、NH2末端レクチンドメインの後にEGF様ドメイン、補体調節タンパク質に相同性を有する9個のショートコンセンサスリピート(short consensus repeat)、膜貫通ドメイン、および短鎖の細胞質側末端が続いたものから構成されるI型膜貫通糖タンパク質である(Johnston et al., Cell 56:1033 (1989))。P−セレクチンの構造は、セレクチンファミリーの他の二つのメンバーであるE−セレクチンおよびL−セレクチンと類似しており、これらのセレクチンは、サイトカインによって活性化した内皮細胞に発現している(E−セレクチン)か、または大部分のクラスの白血球上に構成的に発現している(L−セレクチン)かのいずれかである。
【0003】
全てのセレクチンは、シアリルルイスx(sLex; Foxall et al., J Cell Biol 117:895 (1992); Varki, Curr Opin Cell Biol 257:257 (1992))のような低分子シアリル化フコシル化オリゴ糖に低親和性で結合することが公知である。E−セレクチンではなくP−セレクチンおよびL−セレクチンは、ヘパリン硫酸のような特定の硫酸化糖質にも結合する(総説については、McEver and Cummings, J Clin Invest 100:S97 (1997)を参照されたい)。P−セレクチンに対する高親和性リガンドは、ムチン様糖タンパク質(McEver et al., J Biol Chem 270:11025 (1995))であり、その糖タンパク質は、シアリル化O−グリカンのクラスターを有するポリペプチド主鎖からなる。P−セレクチンが優先的に結合するあるシアロムチンリガンドは、循環している白血球によって、約120kDaの相対分子量を有する二つのジスルフィド結合サブユニットを有するホモ二量体として通常は発現されるP−セレクチン糖タンパク質リガンド1(PSGL−1、CD162)である。P−セレクチンの結合部位は、PSGL−1のNH2末端の最末端部分に局在する。そのリガンドに結合することによって、P−セレクチンは活性化血小板および内皮細胞上での白血球のローリングを仲介する。このローリング過程は、白血球の移動速度を効果的に低下させ、強固な接着および内皮下への白血球のその後の移行に、また血栓への白血球の蓄積にも欠かせない。
【0004】
P−セレクチン欠損マウスおよびP−セレクチン特異的遮断抗体を用いた研究から、虚血/再潅流傷害を含めた多数の急性および慢性炎症疾患の病態生理にP−セレクチンが関係することが示された(Winn et al., J Clin Invest 92:2042 (1993); Massberg et al., Blood 92:507 (1998))。さらに、アテローム性動脈硬化症(Collins et al., J Exp Med 191: 189 (2000); Johnson et al., J Clin Invest 99:1037 (1997))、再狭窄(Manka et al., Circulation 103:1000 (2001); Bienvenu et al., Circulation 103:1128 (2001))および血栓症(Kumar et al., Circulation 99:1363 (1999); Andre et al., Proc Natl Acad Sci USA 97:13835 (2000); Blann et al., Br. J. Haematol 108:191 (2000); Myers et al., Thromb Haemostasis 85: 423 (2001)のような炎症性構成要素を有する心血管疾患にP−セレクチンは明らかに寄与している。どうやら、P−セレクチン機能の阻害は、血管内皮または血小板に対する白血球の接着を伴う種々の疾患での治療として有効であろう(例えばWO93/06863を参照されたい)。
【0005】
P−セレクチンに対する抗体は、当技術分野の現状に記載され、それらの抗体の抗炎症効果および抗血栓効果に関して研究されてきた。米国特許第4,783,399号およびWO93/06863は、活性化血小板と反応性のP−セレクチンに対するマウスモノクローナル抗体を記載している。Geng J. G.ら(J. Biol. Chem., 266 (1991) 22313-22318)は、P−セレクチンのアミノ酸(aa)フラグメントaa60〜75(シグナル配列を含むスイスプロット配列P16109により計数してCysからGlu。WO93/21956はP−セレクチンに対するマウスモノクローナル抗体と、規定の抗体と競合してP−セレクチンフラグメントaa60〜75の存在下で結合するIgG1サブクラスのヒト化抗体とを参照している)にカルシウムイオンの不在下で結合するマウスモノクローナル抗体を記載している。挙げられた、ヒトP−セレクチンに対するマウスモノクローナル抗体のどれも、ヒト患者の処置に有用ではない。WO93/21956に挙げられたP−セレクチンに対するヒトIgG1サブクラスのヒト化抗体は、開発の前臨床段階にある(www.mrctechnology.org)。
【0006】
発明の開示
本発明は、P−セレクチンと結合して、ヒト補体因子C1qと結合しないことを特徴とする抗体に関するものである。好ましくは、この抗体はNK細胞上のヒトFcγ受容体にも結合しない。本発明による抗体は、ヒト起源由来のFc部分を含む。好ましくは、これらの抗体はヒト化抗体またはヒト抗体である。これらの抗体は、特に末梢動脈閉塞性疾患(PAOD)および重症肢虚血(CLI)による炎症障害および血栓障害を患う患者に利益をもたらす新しい発明的な性質を有する。
【図面の簡単な説明】
【0007】
【図1】本発明の抗体がマイクロタイタープレートに被覆した精製P−セレクチンに対する白血球様HL60細胞の接着を阻害することを示す図である。突然変異抗体は非突然変異親抗体よりも強力である。
【図2】HL60細胞に対するトロンビン活性化血小板の接着を測定するロゼット形成アッセイでの本発明の抗体の阻害活性を示す図である。
【図3a】本発明の抗体とラットおよびカニクイザルP−セレクチンとの交差反応性を示す図である。抗P−セレクチン抗体はHL60細胞に対するトロンビン活性化ラット血小板の接着に影響しないが、一方で商業的に入手できるポリクローナル抗P−セレクチン抗体(Pharmingen 09361A)はこの相互作用を阻害する。
【図3b】本発明の抗体とラットおよびカニクイザルP−セレクチンとの交差反応性を示す図である。本発明の抗体はHL60細胞に対するカニクイザル活性化血小板の接着を阻害する。
【図4a】P−、E−、およびL−セレクチントランスフェクタントに関する代表的な結合曲線によりE−およびL−セレクチンに比べてP−セレクチンに対する抗体の選択性を実証する図である。本発明による抗体は、0.01から0.07μg/mlの範囲のEC50値でP−セレクチンCHO細胞に結合する。E−セレクチンCHO細胞およびL−セレクチン300.19細胞に関するEC50値は好ましくは100μg/mlよりも大きい。
【図4b】P−、E−、およびL−セレクチントランスフェクタントに関する代表的な結合曲線によりE−およびL−セレクチンに比べてP−セレクチンに対する抗体の選択性を実証する図である。本発明による抗体は、0.01から0.07μg/mlの範囲のEC50値でP−セレクチンCHO細胞に結合する。E−セレクチンCHO細胞およびL−セレクチン300.19細胞に関するEC50値は好ましくは100μg/mlよりも大きい。
【図4c】P−、E−、およびL−セレクチントランスフェクタントに関する代表的な結合曲線によりE−およびL−セレクチンに比べてP−セレクチンに対する抗体の選択性を実証する図である。本発明による抗体は、0.01から0.07μg/mlの範囲のEC50値でP−セレクチンCHO細胞に結合する。E−セレクチンCHO細胞およびL−セレクチン300.19細胞に関するEC50値は好ましくは100μg/mlよりも大きい。
【図5】完全ヒト流動系での本発明の抗体の阻害活性を示す図である。これらの抗体は剪断速度65/sで濃度依存的に血小板単層へのヒト白血球の接着を阻害する。
【図6a】P−セレクチンを発現しているヒト内皮細胞に対する白血球の接着に及ぼす本発明の抗体の阻害効果を示す図である。対照に対する%で表した白血球の接着の合計阻害を明らかにする。
【図6b】P−セレクチンを発現しているヒト内皮細胞に対する白血球の接着に及ぼす本発明の抗体の阻害効果を示す図である。種々の白血球サブセットの絶対数に及ぼす抗体の一つの阻害効果を代表的に示す。
【0008】
発明の詳細な説明
I.定義
用語「P−セレクチン」はHsu-Lin et al., J Biol Chem 259: 9121 (1984)およびMc Ever et al., J Clin Invest 84:92 (1989)に記載されたような、ヒト血小板および内皮細胞によって発現される140kDaのタンパク質を指す。このI型膜貫通糖タンパク質は、NH2末端レクチンドメインの後に上皮増殖因子(EGF)様ドメインおよび9個のコンセンサスリピートドメインが続いたものから構成される。P−セレクチンは、単一の膜貫通ドメインによって膜に固定され、短鎖の細胞質側末端を含む。本発明は、P−セレクチンによって仲介される一つまたは複数の生物学的活性、例えば、炎症活性または血栓活性を阻害できる抗体を提供する。この抗体はP−セレクチンに結合し、P−セレクチンとそのリガンドとの結合を妨害することによって作用する。
【0009】
用語「P−セレクチンリガンド」は、Moore et al., J Cell Biol 118:2445 (1992)、Sako et al., Cell 75:1179 (1993)によって記載されたようなムチン様糖タンパク質であるP−セレクチンリガンド糖タンパク質1(PSGL−1)のような高親和性生物関連P−セレクチンリガンドに好ましくは関するものである。PSGL−1は、シアリル化O−グリカンのクラスターと連結した一連の10個のリピートを含む、セリン、トレオニン、およびプロリンに富む細胞外ドメインを有するI型膜タンパク質である。PSGL−1は、循環している白血球によって約120kDaの相対分子量を有する二つのジスルフィド結合サブユニットを有するホモ二量体として通常は発現される。P−セレクチンの結合部位は、PSGL−1のNH2末端の最末端部に局在する。血小板によって発現され、PSGL−1と構造類似性を有するシアロムチンGPIbαは、P−セレクチンに対する血小板のリガンドであると最近実証された(Romo et al, J Exp Med 190:803 (1999)。P−セレクチンに結合するGPIbαの生理学的重要性はまだ研究中であるが、その相互作用がローリングおよび活性化内皮細胞に対する血小板の接着に貢献しているようである(Berndt et al., Thromb Haemost 86:178 (2001)。P−セレクチンは、シアリルルイスxのような低分子シアル酸化フコシル化オリゴ糖(Foxall et al., J Cell Biol 117:895 (1992), Varki, Curr Opin Cell Biol 257 (1992)にも、特にヘパリン硫酸のような硫酸化糖質(McEver et al., J Biol Chem 270:11025 (1995)にも低親和性で結合する。
【0010】
用語「抗体」は、本発明による性質が保たれる限りは全抗体、抗体フラグメント、ヒト抗体、ヒト化抗体、キメラ抗体、および遺伝子操作抗体(変異抗体または突然変異抗体)を含むがそれに限定されるわけではない多様な形態の抗体、好ましくはモノクローナル抗体を包含する。特に組換えヒト抗体としてのヒトまたはヒト化モノクローナル抗体が特に好ましい。
【0011】
本明細書に使用するような用語「モノクローナル抗体」または「モノクローナル抗体組成物」は、単一のアミノ酸組成の抗体分子の調製物を指す。
【0012】
用語「キメラ抗体」は、ある起源または種からの可変部、すなわち結合部と、異なる起源または種由来の定常部の少なくとも部分とを含む、組換えDNA法によって通常は調製されたモノクローナル抗体を指す。マウス可変部とヒト定常部とを含むキメラ抗体が特に好ましい。そのようなマウス/ヒトキメラ抗体は、マウス免疫グロブリン可変部をコードするDNAセグメントと、ヒト免疫グロブリン定常部をコードするDNAセグメントとを含む発現された免疫グロブリン遺伝子の生成物である。本発明に包含される他の形態の「キメラ抗体」は、定常部が本来の抗体の定常部から修飾または変更されて、特にC1qの結合および/またはFc受容体(FcR)の結合に関して本発明による性質を発生する抗体である。そのような「キメラ」抗体は、「クラススイッチ抗体」とも称される。キメラ抗体を生成させる方法は、従来の組換えDNA法および当技術分野で現在周知である遺伝子トランスフェクション法を含む。例えばMorrison, S.L., et al., Proc. Natl. Acad. Sci. USA 81 (1984) 6851-6855;米国特許第5,202,238号および第5,204,244号を参照されたい。
【0013】
用語「ヒト化抗体」は、フレームワーク部または「相補性決定部」(CDR)が修飾されて親免疫グロブリンとは特異性が異なる免疫グロブリンCDRを含む抗体を指す。好ましい態様では、マウスCDRをヒト抗体のフレームワーク部にグラフトして、「ヒト化抗体」を調製する。例えばRiechmann, L., et al., Nature 332 (1988) 323-327;およびNeuberger, M.S., et al., Nature 314 (1985) 268-270を参照されたい。特に好ましいCDRは、キメラ抗体および二機能抗体に関して上に述べたように、抗原を認識する配列を表すCDRに対応する。本発明に包含される他の形態の「ヒト化抗体」は、定常部が本来の抗体から修飾または変更されて、特にC1qの結合および/またはFc受容体(FcR)の結合に関して本発明による性質を発生する抗体である。
【0014】
本明細書に使用するような用語「ヒト抗体」は、ヒト生殖細胞系免疫グロブリン配列由来の可変部および定常部を有する抗体を含むことを意図する。ヒト抗体は、当技術分野の現状で周知である (van Dijk and van de Winkel, Curr Opin Pharmacol 5:368 (2001)。内因性免疫グロブリンの生成の不在下で免疫処置に応じてヒト抗体の全レパートリーを生成できるトランスジェニック動物(例えばマウス)でヒト抗体を生成させることもできる。そのような生殖細胞系突然変異マウスへのヒト生殖細胞系免疫グロブリン遺伝子アレイの導入は、抗原による攻撃誘発に応答してヒト抗体の生成を招くであろう(例えばJakobovits et al., Proc. Natl. Acad. Sci. USA, 90: 2551-2555 (1993); Jakobovits et al., Nature, 362:255-258 (1993); Bruggemann et al., Year in Immuno., 7:33 (1993)を参照されたい)。ファージディスプレイライブラリーでもヒト抗体を生成させることができる(Hoogenboom and Winter, J. Mol. Biol., 227:381 (1992); Marks et al., J. Mol. Biol, 222:581 (19991))。ColeらおよびBoernerらの技法もヒトモノクローナル抗体の調製に利用できる(Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985)およびBoerner et al., J. Immunol., 147(l):86-95 (1991))。本発明によるキメラ抗体およびヒト化抗体に関してすでに触れたように、本明細書に使用するような用語「ヒト抗体」は、定常部で修飾されて、特にC1qの結合および/またはFcRの結合に関して本発明による性質を発生する抗体も含む。さらに、本発明はC1qおよび/またはFcRに結合するヒト抗体を含む。そのようなヒト抗体は、E−およびL−セレクチンよりもP−セレクチンに対する高い選択性を特徴とする。本発明によるそのような抗体は、0.01から0.07μg/mlの範囲のEC50値で、P−セレクチン発現細胞に結合する。E−セレクチンおよびL−セレクチン発現細胞に関するEC50値は、好ましくは100μg/mlよりも大きい。そのような抗体は、本発明による性質を有するヒト抗体を製造するための中間体として好ましくは有用である。
【0015】
本明細書に使用するような用語「組換えヒト抗体」は、NS0もしくはCHO細胞のような宿主細胞から、またはヒト免疫グロブリン遺伝子に関してトランスジェニックな動物(例えばマウス)から単離された抗体、あるいは宿主細胞にトランスフェクトされた組換え発現ベクターを使用して発現された抗体のような、組換え手段によって調製、発現、作出、または単離された全てのヒト抗体を含むことを意図する。本発明による組換えヒト抗体は、再編成された形態の可変部および定常部を有する。本発明による組換えヒト抗体は、インビボ体細胞高頻度突然変異に供されている。よって、これらの組換え抗体のVHおよびVL部のアミノ酸配列は、ヒト生殖細胞系VHおよびVL配列に由来して、それらの配列に関係する一方で、自然にはインビボでヒト抗体生殖細胞系レパートリーに存在しないおそれがある配列である。
【0016】
「可変部」(本明細書に使用するようなL鎖可変部(VL)、H鎖可変部(VH))は、抗原に対する抗体の結合に直接関与するL鎖およびH鎖のペアのそれぞれを示す。ヒト可変部L鎖およびH鎖のドメインは同じ一般構造を有し、各ドメインは四つのフレームワーク(FR)部を含み、その配列は広く保存され、三つの「超可変部」(または相補性決定部CDR)によって連結している。フレームワーク部はβシートコンホメーションを採り、CDRはβシート構造を連結しているループを形成する場合がある。各鎖のCDRは、フレームワーク部によって三次元構造に保持され、もう一方の鎖からのCDRと共に抗原結合部位を形成する。抗体H鎖およびL鎖のCDR3部は、本発明による抗体の結合特異性/親和性に特に重要な役割を演じ、よって本発明のさらなる目的を提供する。
【0017】
用語「超可変部」または「抗体の抗原結合部分」は、本明細書に使用する場合に、抗原結合を担う抗体のアミノ酸残基を指す。超可変部は、「相補性決定部」または「CDR」からのアミノ酸残基を含む。「フレームワーク」部または「FR」部は、本明細書に定義するような超可変部の残基以外の可変ドメイン部である。よって、抗体のL鎖およびH鎖は、N末端からC末端方向にFR1、CDR1、FR2、CDR2、FR3、CDR3およびFR4ドメインを含む。特に、H鎖CDR3は抗原結合に最も寄与する領域である。CDRおよびFR部は、Kabatら、Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991))の標準的な定義および/または「超可変ループ」の残基によって決定される。
【0018】
本明細書に使用するような用語「核酸または核酸分子」は、DNA分子およびRNA分子を含むことを意図する。核酸分子は、一本鎖または二本鎖の場合があるが、好ましくは二本鎖DNAである。
【0019】
核酸は、別の核酸配列と機能的関係に置かれた場合に「作動可能に連結」される。例えば、プレ配列または分泌リーダーがポリペプチドの分泌に関与するプレタンパク質としてポリペプチドが発現するならば、そのプレ配列または分泌リーダー配列に関するDNAはそのポリペプチドに関するDNAと作動可能に連結され、プロモーターまたはエンハンサーがコード配列の転写に影響するならば、そのプロモーターまたはエンハンサーはその配列に作動可能に連結され、また、リボソーム結合部位が翻訳を促進するように位置するならば、そのリボソーム結合部位はコード配列に作動可能に連結される。一般に「作動可能に連結した」は、連結されたDNA配列が隣接し、分泌リーダーの場合は隣接して読出し期であることを意味する。しかし、エンハンサーは隣接している必要がない。連結は、都合のよい制限部位での連結反応によって実現される。そのような制限部位が存在しないならば、従来の常法により合成オリゴヌクレオチドアダプターまたはリンカーを使用する。
【0020】
本明細書に使用するような表現「細胞」、「細胞系」、および「細胞培養物」は相互交換可能に使用され、そのような呼称の全ては子孫を含む。このように、語「形質転換体」および「形質転換細胞」は、初代対象細胞および継代数とは無関係にそれ由来の培養物を含む。全ての子孫は、故意または偶然の突然変異が原因でDNA含量が厳密には同一ではない場合があることも了解されている。本来形質転換された細胞でスクリーニングされたものと同じ機能または生物学的活性を有する変異子孫も含まれる。別個の呼称が意図される場合は、情況から明らかであろう。
【0021】
「定常ドメイン」は、抗原に対する抗体の結合に直接関与せず、様々なエフェクター機能を示す。H鎖定常部のアミノ酸配列に応じて、抗体または免疫グロブリンは、IgA、IgD、IgE、IgGおよびIgMのクラスに分かれ、これらのいくつかをサブクラス(アイソタイプ)、例えばIgG1、IgG2、IgG3、およびIgG4と、IgA1およびIgA2とにさらに分けることができる。種々のクラスの免疫グロブリンに対応するH鎖定常部は、それぞれα、δ、ε、γ、およびμと呼ばれる。本発明による抗体は、好ましくはIgG型である。
【0022】
抗体のFc部分は補体活性化、すなわちC1qの結合およびFc受容体の結合に直接関与する。補体系に及ぼす抗体の影響はある状態に依存するが、C1qに対する結合は、Fc部分での所定の結合部位によって起こる。そのような結合部位は、当技術分野の現状で公知であり、例えばBoakle et al., Nature 282 (1975) 742-743; Lukas et al., J. Immunol. 127 (1981) 2555-2560, Brunhouse and Cebra, Mol. Immunol. 16 (1979) 907-917; Burton et al., Nature 288 (1980) 338-344; Thommesen et al., Mol. Immunol. 37 (2000) 995-1004; Idusogie et al., J. Immunol.164 (2000) 4178-4184; Hezareh et al., J. Virology 75 (2001) 12161-12168; Morgan et al., Immunology 86 (1995) 319-324、EP0307434に記載されている。そのような結合部位は、例えばL234、L235、D270、N297、E318、K320、K322、P331、およびP329(Kabat、下記参照のEUインデックスにより付番)である。サブクラスIgG1、IgG2、およびIgG3の抗体は、通常は補体活性化とC1qおよびC3の結合を示すが、一方IgG4は補体系を活性化せず、C1qおよびC3を結合しない。本明細書に使用するような、用語「ヒト起源由来のFc部分」は、サブクラスIgG4のヒト抗体のFc部分、または下記に定義するようなC1qの結合および/もしくはFcRの結合を検出できないように修飾されたサブクラスIgG1、IgG2、もしくはIgG3のヒト抗体のFc部分のいずれかであるFc部分を示す。「抗体のFc部分」は、当業者に周知の用語であり、パパインによる抗体の切断に基づき定義される。本発明による抗体は、Fc部分としてヒト起源由来のFc部分と、好ましくはヒト定常部の他の全ての部分とを含む。好ましくは、Fc部分はヒトFc部分であり、特に好ましくはヒトIgG4サブクラス、またはヒトIgG1サブクラスからの突然変異Fc部分のいずれか由来である。Fc部分と、配列番号25〜28に示すか、またはPVA236突然変異を有さない配列番号25のH鎖定常部とが最も好ましい。
【0023】
II.本発明の好ましい態様
本発明は、P−セレクチンに結合する抗体であって、該抗体の可変H鎖アミノ酸配列CDR3がH鎖CDR3配列である配列番号38、39、40、41または42からなる群より選択されることを特徴とする、抗体を含む。
【0024】
本発明は、可変H鎖および可変L鎖を含む、P−セレクチンに結合する抗体を好ましくは提供し、その抗体は、可変H鎖がCDR配列であるCDR1、CDR2およびCDR3を含み、CDR1が、配列番号29、30、31、および32からなる群より選択され、CDR2が配列番号33、34、35、36、および37からなる群より選択され、CDR3が配列番号38、39、40、41、および42からなる群より選択されることを特徴とし、ここで、該CDRは相互に独立して選択される。
【0025】
本発明による抗体は、可変部L鎖がCDR配列であるCDR1、CDR2、およびCDR3を含み、CDR1が配列番号43および44より選択され、CDR2が配列番号45および46より選択され、CDR3が配列番号47、48、49、50、51、および52より選択されることを好ましくは特徴とし、ここで、該CDRは相互に独立して選択される。
【0026】
その抗体は、H鎖CDRとして配列番号2のCDRおよびL鎖CDRとして配列番号1のCDR、H鎖CDRとして配列番号4のCDRおよびL鎖CDRとして配列番号3のCDR、H鎖CDRとして配列番号6のCDRおよびL鎖CDRとして配列番号5のCDR、H鎖CDRとして配列番号8のCDRおよびL鎖CDRとして配列番号7のCDR、H鎖CDRとして配列番号10のCDRおよびL鎖CDRとして配列番号9のCDR、H鎖CDRとして配列番号12のCDRおよびL鎖CDRとして配列番号11のCDR、H鎖CDRとして配列番号14のCDRおよびL鎖CDRとして配列番号13のCDR、H鎖CDRとして配列番号16のCDRおよびL鎖CDRとして配列番号15のCDR、H鎖CDRとして配列番号18のCDRおよびL鎖CDRとして配列番号17のCDR、H鎖CDRとして配列番号20のCDRおよびL鎖CDRとして配列番号19のCDR、またはH鎖CDRとして配列番号22のCDRおよびL鎖CDRとして配列番号21のCDRを含むことを好ましくは特徴とする。
【0027】
Kabatら、Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)の標準的な定義によりCDR配列を決定できる。各鎖のCDRは、フレームワークアミノ酸によって引き離されている。配列番号1〜22のCDRを配列番号29〜52に示す。
【0028】
本発明による抗体は、該抗体がP−セレクチンと結合し、以下からなる群より独立して選択されるH鎖可変部およびL鎖可変部を含むことを好ましくは特徴とする:
a)配列番号2のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号1によって定義されるL鎖可変ドメイン;
b)配列番号4のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号3によって定義されるL鎖可変ドメイン;
c)配列番号6のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号5によって定義されるL鎖可変ドメイン;
d)配列番号8のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号7によって定義されるL鎖可変ドメイン;
e)配列番号10のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号9によって定義されるL鎖可変ドメイン;
f)配列番号12のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号11によって定義されるL鎖可変ドメイン;
g)配列番号14のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号13によって定義されるL鎖可変ドメイン;
h)配列番号16のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号15によって定義されるL鎖可変ドメイン;
i)配列番号18のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号17によって定義されるL鎖可変ドメイン;
j)配列番号20のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号19によって定義されるL鎖可変ドメイン;
k)配列番号22のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号21によって定義されるL鎖可変ドメイン。
【0029】
本発明による抗体は、H鎖可変部が配列番号2、4、6、8、10、12、14、16、18、20、および22からなる群より独立して選択されるアミノ酸配列を含むことを好ましくは特徴とする。
【0030】
本発明による抗体は、L鎖可変部が配列番号1、3、5、7、9、11、13、15、17、19、および21からなる群より独立して選択されるアミノ酸配列を含むことを好ましくは特徴とする。
【0031】
本発明は、P−セレクチンと結合し、補体因子C1qおよび/またはFc受容体と結合しない抗体を述べる。これらの抗体は、補体依存性細胞傷害作用(CDC)および/または抗体依存性細胞性細胞傷害作用(ADCC)を誘発しない。好ましくは、この抗体は、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しないことを特徴とする。さらに好ましくは、この抗体はヒト抗体またはヒト化抗体である。
【0032】
本発明による抗体は、定常鎖がヒト起源であることを好ましくは特徴とする。そのような定常鎖は当技術分野の現状において周知であり、例えばKabatに記載されている(例えばJohnson, G., and Wu, T.T., Nucleic Acids Res. 28 (2000) 214-218を参照されたい)。例えば有用なヒトH鎖定常部は、配列番号24、25、26、27、および28からなる群より独立して選択されるアミノ酸配列を含む。例えば有用なヒトL鎖定常部は、配列番号23のカッパL鎖定常部のアミノ酸配列を含む。
【0033】
抗体Fc部のFc部分によって仲介されるエフェクター機能は、抗原に対する抗体の結合後に作動するエフェクター機能を指す(これらの機能は、Fc受容体(FcR)による補体カスケードの活性化および/または細胞活性化を含む)。
【0034】
補体カスケードの機能をCH50アッセイによって評価できる。抗赤血球抗体(EA)で感作されたヒツジ赤血球を被験血清に加えて古典的経路を活性化する結果として、溶血が生じる。50%の赤血球を溶解させるのに必要な血清の容積がCH50ユニットを決定する。AP−CH50は代替経路および最終経路の尺度となる。その手順は、ウサギ赤血球を使用する以外は同様である。代替経路は被験血清の添加に応じて活性化する。
【0035】
C1qと、二つのセリンプロテアーゼであるC1rおよびC1sとは、補体依存性細胞傷害(CDC)経路の第一成分である複合体C1を形成する。補体カスケードを活性化するために、抗原性標的に付着したC1qは少なくとも2分子のIgG1または1分子のIgMと結合する(Ward and Ghetie, Therapeutic Immunology 2:77-94 (1995))。Burton(Molec. Immunol., 22(3): 161-206 (1985))は、アミノ酸残基318から337を含むH鎖部が補体の固定に関与していると記載した。DuncanおよびWinter(Nature 332:738-40 (1988))は部位特異的突然変異誘発を使用して、Glu318、Lys320、およびLys322がC1qに対する結合部位を形成すると報告した。Glu318、Lys320、およびLys322残基がC1qの結合に果たす役割は、これらの残基を含む短鎖合成ペプチドが補体介在性溶解を阻害できることによって確認された。
【0036】
用語「補体依存性細胞傷害作用(CDC)」は、本発明による抗体による、補体の存在下でのP−セレクチンを発現しているヒト内皮細胞および血小板の溶解を指す。CDCは、P−セレクチンを発現しているヒト内皮細胞および血小板を補体の存在下で本発明による抗体で処理することによって好ましくは測定される。細胞をカルセインで好ましくは標識する。抗体が30μg/mlの濃度で標的細胞の20%以上に溶解を誘導するならば、CDCが認められる。しかし、本発明者らは、本発明による抗体の性質にはELISAアッセイで補体因子C1qに対する減少した結合が不可欠であると見出した。そのようなアッセイでは、原則としてELISAプレートに様々な濃度範囲の抗体を被覆し、そのプレートに精製ヒトC1qまたはヒト血清を加える。C1qの結合をC1qに対する抗体に続いてペルオキシダーゼ標識コンジュゲートによって検出する。結合(最大結合Bmax)の検出をペルオキシダーゼの基質であるABTS(2,2’−アジノ−ジ−[3−エチルベンズチアゾリン−6−スルホナート(6)])に関する405nmでの吸光度(OD405)として測定する。したがって、本発明は抗体を説明し、その抗体は、補体因子C1qに対するその抗体の非結合がそのようなELISAアッセイ測定を参照することを特徴とし、ここで、抗体濃度10μg/mlでその抗体に対するC1qの最大結合(Bmax)が、細胞系hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)の抗体LC1004−002のBmaxの30%以下、好ましくは20%以下である。
【0037】
本発明による抗体がELISAアッセイにおいて補体因子C3に対して減少した結合を示すこともさらに好ましい。C1qアッセイと同様にしてアッセイを行う。そのようなアッセイでは原則としてELISAプレートを様々な濃度範囲の抗体で被覆し、そのプレートに精製ヒトC3またはヒト血清を加える。C3に対する抗体に続いてペルオキシダーゼ標識コンジュゲートによってC3の結合を検出する。結合(最大結合Bmax)の検出をペルオキシダーゼの基質であるABTS(2,2’−アジノ−ジ−[3−エチルベンズチアゾリンスルホナート(6)])に関する405nmでの吸光度(OD405)として測定する。したがって、本発明は抗体を説明し、その抗体は、補体因子C3に対する抗体の非結合がそのようなELISAアッセイ測定を参照することを特徴とし、ここで、抗体濃度10μg/mlでその抗体に対するC3の最大結合(Bmax)が、細胞系hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)の抗体LC1004−002のBmaxの10%、好ましくは5%以下である。
【0038】
用語「抗体依存性細胞性細胞傷害作用(ADCC)」は、Fc受容体の結合によって仲介される機能であり、本発明による抗体による、エフェクター細胞の存在下でのP−セレクチンを発現している標的細胞の溶解を指す。ADCCは、新鮮単離されたPBMC(末梢血単核細胞)のようなエフェクター細胞、または単球もしくはNK(ナチュラルキラー)細胞のようなバフィコート由来精製エフェクター細胞の存在下で、P−セレクチンを発現している内皮細胞調製物を本発明による抗体で処理することによって好ましくは測定される。標的細胞を51Crで標識してから、抗体と共にインキュベートする。標識された細胞をエフェクター細胞と共にインキュベートして、放出された51Crに関して上清を分析する。対照はエフェクター細胞存在下であるが抗体不在下で標的内皮細胞をインキュベートすることを含む。(FcγRIIおよびFcγRIIIを発現している)顆粒球、(FcγRIIIを発現している)NK細胞、および(FcγRIおよびFcγRIIを発現している)単球のようなFcγ受容体発現細胞に対するこれらの抗体の結合を測定することによって、これらの抗体がADCCを仲介する初段階を誘導する能力が決定された。
【0039】
抗体のFc部とFc受容体(FcR)との相互作用によって、Fc受容体と結合するエフェクター機能を仲介できる。FcRは造血細胞上の専門化した細胞表面受容体である。Fc受容体は免疫グロブリンスーパーファミリーに属し、抗体で被覆された病原体を免疫複合体の食作用によって除去すること、ならびに対応する抗体で被覆された赤血球および他の様々な細胞標的(例えば腫瘍細胞)の抗体依存性細胞介在性細胞傷害作用(ADCC)を経由した溶解の両方を仲介することが示された。Van de Winkel and Anderson, J. Leuk. Biol. 49:511-24 (1991)。FcRは、免疫グロブリンアイソタイプに対する特異性によって定義され、IgG抗体に対するFc受容体はFcγR、IgEに対してはFcεR、IgAに対してはFcαRなどと呼ばれる。Fc受容体の結合は、例えばRavetch and Kinet, Ann. Rev. Immunol. 9 (1991) 457-492; Capel et al., Immunomethods 4 (1994) 32-34; de Haas et al., J. Lab. Clin. Med. 126 (1995) 330-341およびGessner et al., Ann. Hematol. 76 1998) 231-248に記載されている。本発明による抗体は、Fcγ受容体、好ましくはFγcRI、FγcRIIA、FγcRIIB、および/またはFγcRIIIAに対して減少した結合を示す。
【0040】
本発明の抗体による抗体は、好ましくはエフェクター機能を全く誘発せず、NK細胞上に提示されたFcγRに結合しない。よって、用語「FcγRと結合しない」は、抗体濃度10μg/mlでのNK細胞に対する本発明による抗体の結合が、細胞系hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)の抗体LC1004−002に対して見いだされた結合の1%以下であることを意味する。
【0041】
IgG4はFcRとの減少した結合を示すが、他のIgGサブクラスの抗体は強い結合を示す。しかし、Pro238、Asp265、Asp270、Asn297(Fc糖質を欠如)、Pro329と、234、235、236および237と、Ile253、Ser254、Lys288、Thr307、Gln311、Asn434、およびHis435とは、改変されたならばFcRの減少した結合を提供する残基である(Shields et al. J. Biol. Chem. 276 (2001), 6591-6604; Lund et al. FASEB J. 9 (1995), 115-119; Morgan et al. Immunology 86 (1995) 319-324、EP0307434)。好ましくは本発明による抗体は、FcRと、S228、L234、L235および/もしくはD265に突然変異を有し、かつ/またはPVA236もしくはGLPSS331突然変異を含むIgG4サブクラスの、あるいはIgG1またはIgG2サブクラスの結合に関するものである。突然変異S228P(IgG4)、L234A(IgG1)、L235A(IgG1)、L235E(IgG4)、GLPSS331(IgG1)および/またはPVA236(IgG1)が特に好ましい。突然変異の好ましい組み合わせを表1にも示す。追加的な好ましい組み合わせはD265A/N297Aである。
【0042】
本明細書に使用するような用語「P−セレクチンに対する結合」は、BIAcoreアッセイ(Pharmacia Biosensor AB, Uppsala, Sweden)、またはマイクロタイタープレートに精製P−セレクチンもしくはP−セレクチンCHOトランスフェクタントのいずれかを被覆したELISAのいずれかにおけるP−セレクチンに対する抗体の結合を意味する。
【0043】
BIAcoreアッセイでは、抗体を表面に結合させ、P−セレクチンの結合を表面プラズモン共鳴法(SPR)によって測定する。結合の親和性を、用語ka(抗体/抗原複合体からの抗体の会合速度定数)、kd(解離定数)、およびKD(kd/ka)によって定義する。本発明による抗体は、10−8以下、好ましくは約10−11から10−9MのKDを示す(実施例を参照されたい)。したがって、本発明は、上記のような抗体を指し、ここで、その抗体はBIAcoreアッセイで10−8M未満のKD値でP−セレクチンに結合し、好ましくはKDの範囲は10−11から10−9Mである。
【0044】
好ましくは、その抗体はヒトIgG1またはIgG4サブタイプである。
【0045】
さらに好ましくは、その抗体はL234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体、またはL235およびS228(EUインデックスにより付番)に少なくとも一つの突然変異を含むヒトサブクラスIgG4の抗体であることを特徴とする。
【0046】
P−セレクチン特異的ELISAでは、精製P−セレクチンをマイクロタイタープレートに被覆し、ビオチン化抗ヒトIgGおよびELISAの通常の段階を用いて、P−セレクチンに対する抗体の結合を検出する。このアッセイにおけるEC50値は、P−セレクチンCHO細胞では0.002から0.03μg/mlの間の範囲に好ましくはあり、すなわち、本発明はELISAアッセイでP−セレクチンの結合に関するEC50値がP−セレクチンを提示しているCHO細胞に対して0.002から0.03μg/mlの間の範囲にある抗体を指す。P−セレクチン発現CHOトランスフェクタントをマイクロタイタープレートに被覆するアッセイでは、EC50値は0.01から0.08μg/mlの間、好ましくは0.01から0.04μg/mlの間の範囲にある。
【0047】
E−およびL−セレクチントランスフェクタントに対するEC50値は、好ましくは100μg/mlよりも大きい。本発明の抗体は、P−セレクチンと、E−および/またはL−セレクチンとをマイクロタイタープレートに被覆するELISAアッセイでのEC50値によって測定されるように、E−および/またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍大きく特異的に結合することを特徴とする。
【0048】
本明細書に使用するような用語「P−セレクチンに対するP−セレクチンリガンド結合の阻害」は、HL60細胞上に提示されたP−セレクチンリガンドに対する精製P−セレクチンまたは細胞発現P−セレクチンの結合を指す。リガンドに対するP−セレクチンの結合は本発明による抗体によって阻害される。抗体がリガンドに対するP−セレクチンの結合を阻害する能力を分析するインビトロアッセイでのIC50としてその阻害を測定する。そのようなアッセイについて実施例に記載する。これらのアッセイはP−セレクチンの適切な供給源として親和性精製されたP−セレクチンおよび活性化血小板を、リガンドの適切な供給源としてHL60細胞のような白血球様細胞を使用する。そのようなアッセイでは、P−セレクチンまたは活性化血小板に対するP−セレクチンの生理関連リガンドとしてのPSGL−1を発現しているHL60細胞の接着を、漸増する濃度の抗体の不在下および存在下で測定する。IC50値を少なくとも3回の独立した測定の平均値として測定する。阻害は、1μg/mlよりも大きくない、好ましくは0.5から0.08μg/mlのIC50値を意味する。
【0049】
本発明の抗体は、0.08から0.5μg/mlの範囲、好ましくは0.08から0.11μg/mlのIC50値で精製P−セレクチンに対する白血球様HL60細胞の接着を阻害する。活性化血小板に対する白血球様HL60細胞の接着は、0.05から0.3μg/mlの範囲のIC50値で阻害される。
【0050】
したがって、本発明のさらなる態様は、P−セレクチンの結合に関するEC50値がELISAアッセイでは0.01から0.08μg/mlの範囲であることを特徴とする抗体を述べ、ここで、P−セレクチンを発現するCHOトランスフェクタントをマイクロタイタープレートに被覆する。好ましい範囲は0.01から0.04μg/mlである。E−およびL−セレクチントランスフェクタントに関するEC50値は100μg/mlよりも大きい。さらなる態様では、本発明の抗体は、精製P−セレクチンに対する白血球様HL60細胞の接着を0.08から0.5μg/mlの間のIC50値で阻害する。好ましい範囲は、0.08から0.11μg/mlである。
【0051】
本発明の抗体は、完全にヒトの流動系で好ましくは70%よりも大きく白血球と血小板の単層との相互作用を(10μg/mlの濃度で)阻害する。さらに、これらの抗体は、3μg/mlの濃度で60〜90%の範囲でヒト流動系での活性化内皮細胞に対する白血球の接着を(白血球サブタイプに対する異なる効果を有して)阻害する。
【0052】
本発明の抗体は、P−セレクチンフラグメントaa60〜75(Swiss-Prot配列P16109)の存在下でP−セレクチンと好ましくは結合でき、かつ/またはATCCアクセッション番号HB11041と称する細胞系によって分泌される、抗体のP−セレクチンに対する結合を競合的に阻害しない。
【0053】
本発明の抗体は、ELISAアッセイ形式でP−セレクチンと血小板膜糖タンパク質GPIbαとの相互作用を好ましくは阻害しない。ELISAではGPIbαの可溶性細胞外部分であるグリコカリシンを、記載されたようにマイクロタイタープレートのウェルに固定化し(Romo et al., J Exp Med 190:803 (1999)、ポリクローナル抗P−セレクチン抗体を用いて、P−セレクチンHuMabと共に予備インキュベート後の精製P−セレクチンの結合を検出した。
【0054】
本発明のさらなる好ましい態様では、その抗体はC3タンパク質と結合しないことを特徴とし、さらに好ましくは補体依存性細胞傷害作用(CDC)を誘発しないことを特徴とする。さらに、その抗体はNKエフェクター細胞上のFcγ受容体に結合しないことを特徴とする場合がある。好ましくは、その抗体はL234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体、またはL235およびS228(EUインデックスにより付番)に少なくとも一つの突然変異を含むヒトサブクラスIgG4の抗体であることを特徴とする。さらなる好ましい態様では、その抗体は抗体依存性細胞性細胞傷害作用(ADCC)を誘発しないことを特徴とする。
【0055】
なおさらに好ましい態様では、本発明の抗体は、P−セレクチンと結合し、以下からなる群より独立して選択される可変部を含むことを特徴とする:
a)配列番号1のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号2によって定義されるH鎖可変ドメイン、
b)配列番号3のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号4によって定義されるH鎖可変ドメイン、
c)配列番号5のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号6によって定義されるH鎖可変ドメイン、
d)配列番号7のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号8によって定義されるH鎖可変ドメイン、
e)配列番号9のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号10によって定義されるH鎖可変ドメイン、
f)配列番号11のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号12によって定義されるH鎖可変ドメイン、
g)配列番号13のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号14によって定義されるH鎖可変ドメイン、
h)配列番号15のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号16によって定義されるH鎖可変ドメイン、
i)配列番号17のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号18によって定義されるH鎖可変ドメイン、
j)配列番号19のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号20によって定義されるH鎖可変ドメイン、
k)配列番号21のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号22によって定義されるH鎖可変ドメイン。
【0056】
好ましくは、その抗体はアミノ酸配列である配列番号3によって定義されるL鎖可変ドメインおよび配列番号4によって定義されるH鎖可変ドメインを含む。
【0057】
好ましい抗体は、ヒトIgG4サブクラスであるか、補体因子C1qに対する非結合を引き起こす少なくとも一つのアミノ酸突然変異を含むことを特徴とする。これらの変異抗体は、例えば配列番号25または配列番号26と、配列番号28とからなる群より独立して選択されるアミノ酸配列を含む。
【0058】
「変異」抗P−セレクチン抗体は、本明細書において親抗体配列での一つまたは複数のアミノ酸残基における付加、欠失および/または置換によって「親」抗P−セレクチン抗体のアミノ酸配列とアミノ酸配列が異なる分子を指す。好ましい態様では、その変異体は、親抗体の一つまたは複数の定常部または可変部に、好ましくは定常部に、一つまたは複数のアミノ酸置換を含む。例えば、その変異体は親抗体の一つまたは複数の可変部に少なくとも1個、例えば約1から約10個、好ましくは約2から約5個の置換を含む場合がある。通常は、その変異体は親抗体の定常ドメインおよび/または可変ドメイン配列と少なくとも90%、さらに好ましくは少なくとも95%、最も好ましくは少なくとも99%のアミノ酸配列同一性を有するアミノ酸配列を有するであろう。
【0059】
この配列に関係して同一性または相同性は、配列を整列させて必要ならばギャップを導入して最大率の配列同一性を実現した後で、候補配列において親抗体残基と同一であるアミノ酸残基の率として本明細書に定義される。N末端、C末端、または抗体配列への内部伸長、欠失、もしくは挿入のどれも配列同一性または相同性に影響するとして解釈してはならない。その変異体はヒトP−セレクチンと結合する能力を保有し、好ましくは親抗体よりも優れた性質を有する。例えば、その変異体は、さらに強い結合親和性、重症肢虚血または末梢動脈閉塞性疾患(CLI/PAOD)に関連する疾患を処置する能力の増強を有する場合がある。
【0060】
本明細書において特に関心対象である変異抗体は、Fcγ受容体との結合の排除が原因で、親抗体に比較したときに接着アッセイで阻害活性の少なくとも約4倍の増強を示す抗体である。
【0061】
本明細書における「親」抗体は、変異体の調製に使用されたアミノ酸配列にコードされる抗体である。好ましくは、親抗体はヒトフレームワーク部を有し、もしも存在するならばヒト抗体定常部を有する。例えば、親抗体はヒト化抗体またはヒト抗体でありうる。
【0062】
本発明による抗体は、本発明による抗体の上記性質に影響せず、それを改変もしない「保存的配列修飾」であるヌクレオチドおよびアミノ酸配列修飾を有するような抗体をさらに含む。部位特異的突然変異誘発およびPCR介在性突然変異誘発のような当技術分野で公知の標準的な技法によって修飾を導入できる。保存的アミノ酸置換には、アミノ酸残基が類似の側鎖を有するアミノ酸残基に交換された置換がある。類似した側鎖を有するアミノ酸残基のファミリーは、当技術分野で定義されている。これらのファミリーには、塩基性側鎖(例えばリジン、アルギニン、ヒスチジン)、酸性側鎖(例えばアスパラギン酸、グルタミン酸)、非荷電極性側鎖(例えばグリシン、アスパラギン、グルタミン、セリン、トレオニン、チロシン、システイン、トリプトファン)、非極性側鎖(例えばアラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン)、ベータ分岐側鎖(例えばトレオニン、バリン、イソロイシン)、および芳香族側鎖(例えばチロシン、フェニルアラニン、トリプトファン、ヒスチジン)を有するアミノ酸がある。このように、ヒト抗P−セレクチン抗体における予測される非必須アミノ酸残基を、同じ側鎖ファミリーからの別のアミノ酸残基に好ましくは交換できる。
【0063】
Riechmann, L., et al., Nature 332 (1988) 323-327およびQueen, C., et al., Proc. Natl. Acad. Sci. USA 86 (1989) 10029-10033によって記載されたような分子モデリングに基づいた突然変異誘発によってアミノ酸置換を行うことができる。
【0064】
さらなる好ましい態様では、抗体は配列番号23に定義されるようなκL鎖定常部を含む。
【0065】
本発明による好ましい抗体は、IgG1v1(E233P、L234V、L235A、デルタG236、A327G、A330S、P331Sによって規定されるPVA−236;GLPSS331)、IgG1v2(L234A、L235A)およびIgG4v1(S228P、L235E)として定義される抗体である。
【0066】
さらに好ましい態様では、これらの抗体はFab、F(ab’)2および一本鎖フラグメントからなる群より選択される抗体フラグメントも含む。
【0067】
本発明は、本発明による抗体の製造方法をさらに含み、その方法は、a)本発明による親ヒト抗体のL鎖をコードする第一核酸配列と、Fc部分が補体因子C1qおよび/またはFc受容体と結合しないために修飾されている該親ヒト抗体のH鎖をコードする第二DNA配列とを用いて宿主細胞を形質転換する段階、b)該抗体H鎖およびL鎖が生成するように該第一および第二DNA配列を発現させる段階、ならびにc)該抗体を宿主細胞または宿主細胞培養物から回収する段階とを含む。
【0068】
本発明は、中間体である抗体、すなわち抗P−セレクチン抗体についても述べ、それらの抗体は、ヒト抗体またはヒト化抗体であって、P−セレクチンとE−および/またはL−セレクチンとをマイクロタイタープレートに被覆するELISAアッセイで測定したときに、E−またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍大きく特異的に結合することを特徴とする。好ましくはこれらの抗体はIgG1またはIgG4抗体である。これらの抗体は、配列番号24のγ1H鎖定常部または配列番号27のγ4H鎖定常部によって定義されるようなアミノ酸配列も含む場合がある。特に、これらの抗体は、hu−Mab<P−セレクチン>LC1004−001(DSM ACC2640)、hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)、およびhu−Mab<P−セレクチン>LC1004−017(DSM ACC2642)からなる群より選択される細胞系によって生成される抗体を指す。
【0069】
本発明による抗体は、本発明による抗体の上記性質に影響せず、それを改変もしない「保存的配列修飾」であるヌクレオチドおよびアミノ酸配列修飾を有するような抗体をさらに含む。部位特異的突然変異誘発およびPCR介在性突然変異誘発のような当技術分野で公知の標準的な技法によって修飾を導入できる。保存的アミノ酸置換には、アミノ酸残基が類似の側鎖を有するアミノ酸残基に交換された置換がある。類似した側鎖を有するアミノ酸残基のファミリーは、当技術分野で定義されている。これらのファミリーには、塩基性側鎖(例えばリジン、アルギニン、ヒスチジン)、酸性側鎖(例えばアスパラギン酸、グルタミン酸)、非荷電極性側鎖(例えばグリシン、アスパラギン、グルタミン、セリン、トレオニン、チロシン、システイン、トリプトファン)、非極性側鎖(例えばアラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン)、ベータ−分岐側鎖(例えばトレオニン、バリン、イソロイシン)、および芳香族側鎖(例えばチロシン、フェニルアラニン、トリプトファン、ヒスチジン)を有するアミノ酸がある。このように、ヒト抗P−セレクチン抗体における予測される非必須アミノ酸残基を、同じ側鎖ファミリーからの別のアミノ酸残基に好ましくは交換できる。
【0070】
Riechmann, L.ら、Nature 332 (1988) 323-327およびQueen, C.ら、Proc. Natl. Acad. Sci. USA 86 (1989) 10029-10033によって記載された分子モデリングに基づいた突然変異誘発によってアミノ酸置換を行うことができる。
【0071】
本発明は、上に挙げた抗体をコードする核酸分子、これらの核酸を含む対応するベクター、およびこれらのベクターに関する対応する宿主細胞も含む。本発明は、該抗体分子の合成を可能にする条件で対応する宿主細胞を培養する段階と、該培養物から該抗体を回収する段階とを含む、例えばH鎖をコードする核酸およびL鎖をコードする核酸を原核または真核宿主細胞に発現させる段階と、該細胞から該ポリペプチドを回収する段階とによる、抗体の調製方法を包含する。
【0072】
診断および治療へのこの抗体の使用が考えられている。一つの診断応用では、本発明はP−セレクチンを含むと疑われる試料を抗P−セレクチン抗体に曝露する段階と、その試料に対する抗体の結合を決定する段階とを含む、P−セレクチンタンパク質の存在を決定する方法を提供する。この使用に関して、本発明は抗体と、その抗体を使用するための指示とを含む、P−セレクチンタンパク質を検出するためのキットを提供する。
【0073】
本発明の抗体は、炎症疾患および血栓疾患の処置に有用である。そのような疾患には,アテローム性動脈硬化症、動脈および深部静脈血栓症、血管形成術またはステント留置術後の再狭窄のような血管障害がある。好ましい適用は、末梢動脈閉塞性疾患(PAOD)および重症肢虚血(CLI)である。他の適用は、心筋梗塞、脳虚血イベント(例えば卒中)、腎梗塞などによって起こる虚血後白血球介在性組織損傷の処置である。これらの抗体は、敗血症、急性白血球介在性肺傷害、および喘息のようなアレルギー反応の処置にも適している。他の適用は、臓器移植の拒絶反応および慢性関節リウマチを含む自己免疫疾患の予防である。さらに、循環している癌細胞の接着を阻害することによって腫瘍転移を予防できる。
【0074】
本発明は、上記の炎症障害および血栓障害、特にPAODおよびCLI(末梢動脈閉塞性疾患または重症肢虚血)を患う哺乳動物を処置するための方法をさらに提供する。
【0075】
本発明は、治療、例えばこれらの疾患の処置のための薬を製造するための上記抗体の使用をさらに提供する。
【0076】
本発明は、薬学的組成物を製造するための上に定義したような抗体の使用にも関し、薬学的有効量で本発明による抗体を、場合によっては薬学的目的の抗体製剤に有用な緩衝剤および/または補助剤と共に含む薬学的組成物を含む。
【0077】
本発明は、薬学的に許容される担体中にそのような抗体を含む薬学的組成物をさらに提供する。一態様では、その薬学的組成物は製品またはキットに含まれる場合がある。
【0078】
本発明は、本発明によるそのようなアンタゴニストモノクローナル抗体、例えば親抗体を生成するハイブリドーマ細胞系をさらに提供する。
【0079】
本発明による好ましいハイブリドーマ細胞系であるhu−Mab<P−セレクチン>LC1004−001(抗体HuMab001)、hu−Mab<P−セレクチン>LC1004−002(抗体HuMab002)、およびhu−Mab<P−セレクチン>LC1004−017(抗体HuMab017)は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約に基づき、Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSMZ)、ドイツに寄託された。
【0080】
【表1】

【0081】
該細胞系から得ることができる抗体は、本発明の好ましい態様である。
【0082】
本発明による抗体は、組換え手段によって好ましくは生成される。そのような方法は当技術分野の現状で広く公知であり、原核および真核細胞でのタンパク質の発現と、それに続く抗体ポリペプチドの単離と、通常は薬学的に許容される純度までの精製とを含む。タンパク質の発現のために、L鎖およびH鎖、またはそれらのフラグメントをコードする核酸を標準的な方法によって発現ベクターに挿入する。CHO細胞、NS0細胞、SP2/0細胞、HEK293細胞、COS細胞、酵母、または大腸菌(E.coli)細胞のような適切な原核または真核宿主細胞で発現を行い、それらの細胞(上清または溶解後の細胞)から抗体を回収する。
【0083】
抗体の組換え生成は、当技術分野の現状で周知であり、例えばMakrides, S. C., Protein Expr. Purif. 17 (1999) 183-202; Geisse, S., et al., Protein Expr. Purif. 8 (1996) 271-282; Kaufman, R. J., Mol. Biotechnol. 16 (2000) 151-161; Werner, R. G., Drug Res. 48 (1998) 870-880の総説に記載されている。
【0084】
抗体は、全細胞中、細胞溶解液中、または部分精製された形もしくは実質的に純粋な形で存在しうる。他の細胞構成要素または他の混入物、例えば他の細胞性核酸またはタンパク質を除去するために、アルカリ/SDS処理、カラムクロマトグラフィー、および当技術分野で周知の他の技法を含めた標準的な技法によって精製を行う。Ausubel, F., et al., ed. Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York (1987)を参照されたい。
【0085】
NS0細胞での発現は、例えばBarnes, L.M., et al., Cytotechnology 32 (2000) 109-123およびBarnes, L.M., et al., Biotech. Bioeng. 73 (2001) 261-270に記載されている。一過性発現は、例えばDurocher, Y., et al., Nucl. Acids. Res. 30 (2002) E9に記載されている。可変ドメインのクローニングは、Orlandi, R., et al., Proc. Natl. Acad. Sci. USA 86 (1989) 3833-3837; Carter, P., et al., Proc. Natl. Acad. Sci. USA 89 (1992) 4285-4289;およびNorderhaug, L., et al., J. Immunol. Methods 204 (1997) 77-87に記載されている。好ましい一過性発現系(HEK293)は、Schlaeger, E.-J.およびChristensen, K.(Cytotechnology 30 (1999) 71-83)ならびにSchlaeger, E.-J.(J. Immunol. Methods 194 (1996) 191-199)によって記載されている。
【0086】
原核生物に適した制御配列には、例えばプロモーターと、場合によってはオペレーター配列と、リボソーム結合部位とがある。真核細胞はプロモーターと、エンハンサーと、ポリアデニル化シグナルとを利用することが公知である。
【0087】
別の核酸配列と機能的関係に置かれた場合に、核酸は「作動可能に連結」している。例えば、プレ配列もしくは分泌リーダーがポリペプチドの分泌に関与するプレタンパク質として発現するならば、そのプレ配列もしくは分泌リーダーに関するDNAはそのポリペプチドに関するDNAと作動可能に連結しており;プロモーターもしくはエンハンサーが配列の転写に影響するならば、そのプロモーターもしくはエンハンサーはコード配列に作動可能に連結し;またはリボソーム結合部位が翻訳を可能にするように位置するならば、そのリボソーム結合部位はコード配列に作動可能に連結する。一般的に「作動可能に連結」は、連結しているDNA配列が隣接し、分泌リーダーの場合は隣接してリーディングフレームに存在することを意味する。しかし、エンハンサーは隣接している必要はない。連結は、好都合な制限部位での連結反応によって実現される。そのような部位が存在しないならば、常法により合成オリゴヌクレオチドアダプターまたはリンカーを使用する。
【0088】
モノクローナル抗体は、例えばプロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、またはアフィニティクロマトグラフィーのような従来の免疫グロブリン精製法によって培地から適切に分離される。モノクローナル抗体をコードするDNAおよびRNAは、従来法を使用して容易に単離および配列決定される。ハイブリドーマ細胞はそのようなDNAおよびRNAの供給源として作用できる。いったん単離されたならば、そのDNAを発現ベクターに挿入でき、それを次にHEK293細胞、CHO細胞、または骨髄腫細胞のような宿主細胞にトランスフェクトして、宿主細胞での組換えモノクローナル抗体の合成を得る。それらの宿主細胞は、さもなければ免疫グロブリンタンパク質を生成しない。
【0089】
抗体DNAに適切なヌクレオチド変化を導入することによって、またはヌクレオチド合成によって、ヒトP−セレクチン抗体のアミノ酸配列変異体(または突然変異体)を調製する。しかし、そのような修飾を例えば上記のように非常に限定した範囲内でのみ行うことができる。例えば、これらの修飾はIgGアイソタイプおよびエピトープの結合のような先に述べた抗体特性を改変しないが、組換え生成の収率もしくはタンパク質の安定性を改善するか、または精製を容易にする場合がある。
【0090】
抗P−セレクチン抗体の固有のコンホメーションの維持に関与しない任意のシステイン残基を一般的にセリンに置換して、その分子の酸化安定性を高めて異所性の架橋を防止することもできる。逆に、システイン結合を抗体に加えて(特にその抗体がFvフラグメントのような抗体フラグメントである場合に)その安定性を高めることができる。
【0091】
その抗体の別の種類のアミノ酸変異体は、その抗体の本来のグリコシル化パターンを改変する。改変によって、その抗体にみられる一つもしくは複数の糖質部分を欠失させること、および/またはその抗体に存在しない一つもしくは複数のグリコシル化部位を付加することを意味する。抗体のグリコシル化は、概してN−結合である。N−結合は、アスパラギン残基の側鎖に対する糖質部分の付着を指す。トリペプチド配列であるアスパラギン−X−セリンおよびアスパラギン−X−トレオニン(Xはプロリン以外の任意のアミノ酸)は、アスパラギン側鎖に糖質部分が酵素的に付着するための認識配列である。このように、ポリペプチドにこれらのトリペプチド配列のいずれかが存在することによって、潜在的グリコシル化部位が生み出される。(N−結合グリコシル化部位に関して)先に述べた一つまたは複数のトリペプチド配列を含むようにアミノ酸配列を改変することによって、抗体へのグリコシル化部位の付加を好都合に実現する。
【0092】
抗P−セレクチン抗体のアミノ酸配列変異体をコードする核酸分子は、当技術分野で公知の様々な方法によって調製される。これらの方法には、(天然アミノ酸配列変異体の場合)天然供給源からの単離、または初期に調製した変異体もしくは非変異体版のヒト化抗P−セレクチン抗体のオリゴヌクレオチド介在性(または部位特異的)突然変異誘発と、PCR突然変異誘発と、カセット突然変異誘発とによる調製があるが、それに限定されるわけではない。
【0093】
本発明は、化学療法剤、毒素(例えば細菌、真菌、植物、もしくは動物起源の酵素的に活性な毒素、またはそれらのフラグメント)、放射性同位体(すなわち放射性コンジュゲート)、または炎症障害および血栓障害、特にPAODおよびCLIによる障害の予防もしくは処置のための薬剤のプロドラッグのような細胞傷害剤に結合した、本発明による抗体を含む免疫コンジュゲートにも関する。抗体と細胞傷害剤とのコンジュゲートは、N−スクシンイミジル−3−(2−ピリジルジチオール)プロピオネート(SPDP)、イミノチオラン(IT)、(ジメチルアジピミダートHCLのような)イミドエステルの二官能性誘導体、(スベリン酸ジスクシンイミジルのような)活性エステル、(グルタルアルデヒドのような)アルデヒド、(ビス(p−アジドベンゾイル)ヘキサンジアミンのような)ビス−アジド化合物、(ビス−(p−ジアゾニウムベンゾイル)−エチレンジアミン(bis-(p-diazoniumbenzoyl)-ethylenediatnine)のような)ビス−ジアゾニウム誘導体、(トリエン(tolyene)2,6−ジイソシアナートのような)ジイソシアナート、および(1,5−ジフルオロ−2,4−ジニトロベンゼンのような)ビス−活性フッ素化合物のような様々な二官能性タンパク質カップリング剤を使用して作製される。例えば、リシン免疫毒素をVitetta, E. S., et al., Science 238 (1987) 1098-1104)に記載されているように調製できる。炭素14標識1−イソチオシアナートベンジル−3−メチルジエチレントリアミン五酢酸(MX−DTPA)は抗体に放射性ヌクレオチドを結合させるための例示的なキレート剤である。WO94/11026を参照されたい。
【0094】
別の種類の共有結合性修飾は、その抗体にグリコシドを化学的または酵素的にカップリングすることを含む。これらの手順は、N−またはO−結合グリコシル化に関するグリコシル化能力を有する宿主細胞で抗体を生成させる必要がない点で有利である。使用したカップリング様式に応じて、(a)アルギニンおよびヒスチジン、(b)遊離カルボキシル基、(c)システインのもののような遊離スルフヒドリル基、(d)セリン、トレオニン、もしくはヒドロキシプロリンのもののような遊離ヒドロキシル基、(e)フェニルアラニン、チロシン、もしくはトリプトファンのもののような芳香族残基、または(f)グルタミンのアミド基に糖(類)を付着させることができる。これらの方法は、WO87/05330、およびAplin, J.D., and Wriston, J.C. Jr., CRC Crit. Rev. Biochem. (1981) 259-306に記載されている。
【0095】
抗体上に存在する任意の糖質部分の除去を化学的または酵素的に実現できる。化学的脱グリコシル化は、化合物であるトリフルオロメタンスルホン酸または同等の化合物にその抗体を曝露することを必要とする。この処理の結果として、結合糖(N−アセチルグルコサミンまたはN−アセチルガラクトサミン)以外の大部分または全ての糖の開裂が生じる一方で、抗体はインタクトのまま残る。化学的脱グリコシル化は、Sojahr, H. T., and Bahl, O. P., Arch. Biochem. Biophys. 259 (1987) 52-57およびEdge, A. S., et al. Anal. Biochem. 118 (1981) 131-137に記載されている。Thotakura, N.R., and Bahl, O. P., Meth. Enzymol. 138 (1987) 350-359に記載されたように様々なエンドグリコシダーゼおよびエキソグリコシダーゼの使用によって、抗体上の糖質部分の酵素的開裂を実現できる。
【0096】
別の種類の抗体の共有結合性修飾は、米国特許第4,640,835号、第4,496,689号、第4,301,144号、第4,670,417号、第4,791,192号、または第4,179,337号に記述されているように、様々な非タンパク質ポリマーの一つ、例えばポリエチレングリコール、ポリプロピレングリコール、またはポリオキシアルキレンに抗体を連結する段階を含む。
【0097】
なお別の局面では、本発明は、ヒト抗P−セレクチン抗体(例えば本発明によるhu−Mab<P−セレクチン>LC1004−001(DSM ACC2640)、hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)、およびhu−Mab<P−セレクチン>LC1004−017(DSM ACC2642)からなる群より選択される細胞系によって生成される親抗体)を発現する非ヒトトランスジェニック動物、例えばトランスジェニックマウスから単離されたB細胞を提供する。KMマウスは、適切な染色体導入マウスである。KMマウスは、ヒトH鎖導入染色体およびヒトカッパL鎖導入遺伝子を含む。KMマウスでは、内因性マウスH鎖およびL鎖遺伝子も破壊されている結果、マウスの免疫処置によってマウス免疫グロブリンよりもヒト免疫グロブリンの生成がもたらされる。KMマウスの構築およびヒト免疫グロブリンを発生させるためのそれらのマウスの使用はWO02/43478に詳細に記載されている。
【0098】
好ましくは、 単離されたB細胞は、精製型、もしくは組換え型のP−セレクチン抗原および/またはP−セレクチンを発現している細胞を用いて免疫処置された非ヒトトランスジェニック動物、例えばトランスジェニックマウスから得られる。好ましくは、非ヒトトランスジェニック動物、例えばトランスジェニックマウスは、本発明の抗体の全てまたは部分をコードする、ヒトH鎖導入遺伝子とヒトL鎖導入遺伝子とを含むゲノムを有する。次に、単離されたB細胞は不死化されてヒト抗P−セレクチン抗体の供給源(例えばハイブリドーマ)を提供する。したがって、本発明は本発明によるヒトモノクローナル抗体を生成できるハイブリドーマも提供する。一態様では、ハイブリドーマは、本発明の抗体の全てまたは部分をコードするヒトH鎖導入遺伝子とヒトL鎖導入遺伝子とを含むゲノムを有する非ヒトトランスジェニック動物、例えばトランスジェニックマウスから得られるB細胞が不死化細胞と融合したものを含む。
【0099】
特定の態様では、非ヒトトランスジェニック動物は、本発明の抗体の全てまたは部分をコードする、ヒトH鎖導入遺伝子とヒトL鎖導入遺伝子とを含むゲノムを有するトランスジェニックマウスである。精製もしくは濃縮されたP−セレクチン抗原調製物および/またはP−セレクチンを発現している細胞を用いて非ヒトトランスジェニック動物を免疫処置できる。好ましくは、非ヒトトランスジェニック動物、例えばトランスジェニックマウスは、P−セレクチンに対するヒトモノクローナル抗体のP−セレクチンアイソタイプを生成できる。
【0100】
精製もしくは濃縮されたP−セレクチン抗原調製物および/またはP−セレクチンを発現している細胞を用いて、本発明の抗体の全てまたは部分をコードする、ヒトH鎖導入遺伝子とヒトL鎖導入遺伝子とを含むゲノムを有する非ヒトトランスジェニック動物、例えばトランスジェニックマウスを免疫処置することによって、本発明によるヒトモノクローナル抗体を生成させることができる。次に、その動物のB細胞(例えば脾臓B細胞)を得て、骨髄腫細胞と融合させて、P−セレクチンに対するヒトモノクローナル抗体を分泌する不死化ハイブリドーマ細胞を形成させる。
【0101】
好ましい態様では、P−セレクチンに対するヒトモノクローナル抗体を、マウス免疫系よりもヒト免疫系の部分を保有するトランスジェニックマウスを使用して発生させることができる。本明細書において「HuMab」マウスと称されるこれらのトランスジェニックマウスは、H鎖(μおよびγ)およびκL鎖(定常部遺伝子)と、内因性μおよびκ鎖遺伝子座を不活性化する標的突然変異とを含む再編成されていないヒト免疫グロブリン遺伝子をコードするヒト免疫グロブリン遺伝子ミニ遺伝子座を含む(Lonberg, N., et al., Nature 368 (1994) 856-859)。したがって、そのマウスはマウスIgMまたはIgKの減少した発現を示し、導入されたヒトH鎖およびL鎖導入遺伝子は、免疫処置に応答してクラススイッチおよび体細胞突然変異を受けて、高親和性のヒトIgGモノクローナル抗体を発生する(Lonberg, N., et al., Nature 368 (1994) 856-859。Lonberg, N., Handbook of Experimental Pharmacology 113 (1994) 49-101; Lonberg, N., and Huszar, D., Intern. Rev. Immunol. 25 (1995) 65-93;およびHarding, F., and Lonberg, N., Ann. N. Acad. Sci 764 (1995) 536-546に総説されている)。HuMabマウスの調製は、Taylor, L., et al., Nucleic Acids Research 20 (1992) 6287-6295; Chen, J., et al., International Immunology 5 (1993) 647-656; Tuaillon, N., et al., Proc. Natl. Acad. Sci USA 90 (1993) 3720-3724; Choi, T.K., et al., Nature Genetics 4 (1993) 117-123; Chen, J., et al., EMBO J. 12 (1993) 821-830; Tuaillon, N., et al., Immunol. 152 (1994) 2912-2920; Lonberg, N., et al., Nature 368 (1994) 856-859; Lonberg, N., Handbook of Experimental Pharmacology 113 (1994) 49-101; Taylor, L., et al., Int. Immunol. 6 (1994) 579-591; Lonberg, N., and Huszar, D., Intern. Rev. Immunol. 25 (1995) 65-93; Harding, F., and Lonberg, N., Ann. N. Acad. Sci 764 (1995) 536-546; Fishwild, D.M., et al., Nat. Biotechnol. 14 (1996) 845-851に記載されており、これら全ての内容はその全体が参照により本明細書に組み入れられる。米国特許第5,545,806号、第5,569,825号、第5,625,126号、第5,633,425号、第5,789,650号、第5,877,397号、第5,661,016号、第5,814,318号、第5,874,299号、第5,545,807号、第5,770,429号、WO98/24884、WO94/25585、WO93/1227、WO92/22645、およびWO92/03918をさらに参照されたい。
【0102】
P−セレクチンに対する完全ヒトモノクローナル抗体を発生させるために、Lonberg, N. et al., Nature 368 (1994) 856-859; Fishwild, D. M. et al., Nat. Biotechnol. 14 (1996) 845-851、およびWO98/24884に記載されたような一般的な方法により、P−セレクチン抗原の精製もしくは濃縮された調製物および/またはP−セレクチンを発現している細胞を用いてHuMabマウスを免疫処置することができる。好ましくは、最初の免疫処置の際にマウスは6〜16週齢であろう。例えば、可溶性P−セレクチン抗原の精製または濃縮された(例えばP−セレクチン発現細胞から精製された)調製物を使用して、HuMabマウスを腹腔内免疫処置することができる。P−セレクチン抗原の精製または濃縮された調製物を使用した免疫処置の結果として抗体が生じない場合には、P−セレクチンを発現している細胞、例えば腫瘍細胞系を用いてマウスを免疫処置して免疫応答を促進することもできる。様々な抗原での累積的な経験は、最初フロイントの完全アジュバントに入れた抗原を用いて腹腔内(i.p.)免疫処置してから、フロイントの不完全アジュバントに入れた抗原を用いて2週間毎に(例えば合計6週間まで)i.p.免疫処置を行う場合に、HuMabトランスジェニックマウスが最もよく応答することを示した。眼窩採血によって得られた血漿試料を用いた、免疫処置プロトコールの過程にわたり免疫応答を監視できる。ELISAによって血漿をスクリーニングでき、十分な力価の抗P−セレクチンヒト免疫グロブリンを有するマウスを対応するB細胞の不死化に使用できる。屠殺して脾臓およびリンパ節を取り出す3から4日前に、マウスの静脈内に抗原を追加免疫できる。各抗原について2〜3回の融合を行う必要がありうると予想される。各抗原について数匹のマウスに免疫処置を行う。例えば、HCo7およびHCo12系統のHuMabマウス合計12匹に免疫処置を行うことができる。
【0103】
HCo7マウスは、(Chen, J. et al., EMBO J. 12 (1993) 821-830に記載されているように)内因性L鎖(カッパ)遺伝子にJKD破壊、(WO01/14424の実施例1に記載されているように)内因性H鎖遺伝子にCMD破壊、(Fishwild, D.M., et al., Nat. Biotechnol. 14 (1996) 845-851に記載されているように)KCo5ヒトカッパL鎖導入遺伝子、および(米国特許第5,770,429号に記載されているように)HCo7ヒトH鎖導入遺伝子を有する。
【0104】
HCo12マウスは、(Chen, J. et al., EMBO J. 12 (1993) 821-830に記載されているように)内因性L鎖(カッパ)遺伝子にJKD破壊、(WO01/14424の実施例1に記載されているように)内因性H鎖遺伝子にCMD破壊、(Fishwild, D.M., et al., Nat. Biotechnol. 14 (1996) 845-851に記載されているように)KCo5ヒトカッパL鎖導入遺伝子、および(WO01/14424の実施例2に記載されているように)HCo12ヒトH鎖導入遺伝子を有する。マウスリンパ球を単離して、標準的なプロトコールに基づいてPEGを使用してマウス骨髄腫細胞系と融合させてハイブリドーマを発生させることができる。結果として生じたハイブリドーマから、次に抗原特異的抗体の生成をスクリーニングする。例えば、免疫処置したマウスからの脾臓およびリンパ節由来リンパ球の単細胞懸濁物を、50%PEGを用いて6分の1の数のSP2/0非分泌型マウス骨髄腫細胞(ATCC、CRL1581)と融合させる。平底マイクロタイタープレートで約2×105個の細胞を平板培養してから、選択培地で約2週間インキュベートする。
【0105】
次に、個別のウェルからELISAによってヒト抗P−セレクチンモノクローナルIgMおよびIgG抗体をスクリーニングする。いったんハイブリドーマの大規模な成長が起こったならば、通常は10〜14日後に培地を分析する。抗体を分泌しているハイブリドーマをもう一度平板培養し、再度スクリーニングして、ヒトIgG抗P−セレクチンモノクローナル抗体がまだ陽性であれば、そのハイブリドーマを限界希釈によって少なくとも2回サブクローニングできる。次に、安定なサブクローンをインビトロで培養してキャラクタリゼーション用の組織培養培地で抗体を生成させる。
【0106】
CDR配列が抗体−抗原相互作用を担うことから、種々のヒト抗体からのフレームワーク配列上に本発明によるCDR配列を含む発現ベクターを構築することによって、本発明による組換え抗体を発現させることが可能である(例えばRiechmann, L., et al., Nature 332 (1998) 323-327; Jones, P., et al., Nature 321 (1986) 522-525;およびQueen, C, et al., Proc. Natl. Acad. See. U.S.A. 86 (1989)10029-10033を参照されたい)。そのようなフレームワーク配列を、生殖細胞系ヒト抗体遺伝子配列を含む公共のDNAデータベースから得ることができる。これらの生殖細胞系配列は、B細胞が成熟する間にV(D)J組換えによって形成する完全に集合した可変部遺伝子を含まないと思われることから、成熟抗体遺伝子配列とは異なるであろう。生殖細胞系遺伝子配列は、個別の高親和性二次レパートリー抗体の配列とも可変部に均等にわたって異なるであろう。
【0107】
本発明は、好ましくは試料のP−セレクチンと本発明による抗体との間の結合を決定する免疫アッセイによる、P−セレクチンのインビトロ診断のための本発明による抗体の使用をさらに含む。
【0108】
別の局面では、本発明は、本発明のヒトモノクローナル抗体の一つもしくは組み合わせ、またはその抗原結合部分が薬学的に許容される担体と共に製剤されたものを含む組成物、例えば薬学的組成物を提供する。さらに具体的には、その組成物は薬学的または診断用組成物であり、なおさらに具体的には、その薬学的組成物は上に定義されたような抗体と、少なくとも一つの薬学的に許容される賦形剤とを含む。
【0109】
本発明の薬学的組成物を、併用療法で、すなわち他の薬剤と組み合わせて投与することもできる。例えば、併用療法は、本発明の組成物を、重症肢虚血(CLI/PAOD)に関連した疾患の予防もしくは処置、または他の従来の治療法に有用な少なくとも一つの薬剤と共に含む場合がある。
【0110】
本明細書に使用するような「薬学的に許容される担体」には、生理学的に適合性の任意および全ての溶媒、分散媒、コーティング剤、抗細菌および抗真菌剤、等張化剤および吸収遅延剤などがある。好ましくは、担体は(例えば注射または注入による)静脈内、筋肉内、皮下、非経口、脊髄または上皮投与に適する。
【0111】
「薬学的に許容される塩」は、抗体の所望の生物学的活性を保持し、任意の望まれない毒性作用を与えない塩を指す(例えばBerge, S.M., et al., J. Pharm. Sci. 66 (1977) 1-19を参照されたい)。そのような塩は本発明に含まれる。そのような塩の例には、酸付加塩および塩基付加塩がある。酸付加塩には、塩酸塩のような無毒性無機酸由来の塩がある。
【0112】
本発明の組成物を、当技術分野で公知の様々な方法によって投与できる。当業者に認識されているであろうが、投与経路および/または投与様式は、所望の結果に応じて変動するであろう。
【0113】
ある投与経路で本発明の化合物を投与するために、その化合物をある物質で被覆するか、またはある物質と同時投与してその化合物の不活性化を防止することが必要な場合がある。例えば、適切な担体、例えばリポソーム、または希釈剤に入れてその化合物を被験体に投与できる。薬学的に許容される希釈剤には、生理食塩水および水性緩衝溶液がある。
【0114】
薬学的に許容される賦形剤または担体には、注射可能な滅菌溶液または分散物を即時調製するための滅菌水溶液または分散物と滅菌粉末とがある。薬学的に活性な物質のためのそのような媒質および薬剤の使用は、当技術分野で公知である。
【0115】
本明細書に使用する句「非経口投与」および「非経口的に投与する」は、経腸および局所投与以外の、通常は注射による投与様式を意味し、非限定的に静脈内、筋肉内、動脈内、髄腔内、関節包内、眼窩内、心臓内、皮内、腹腔内、経気管、皮下、上皮下、関節内、嚢下、クモ膜下、髄腔内、硬膜外、および胸骨内への注射および注入を含む。
【0116】
これらの組成物は、保存料、湿潤剤、乳化剤、および分散剤のような賦形剤または佐剤も含む場合もある。上記滅菌法と、例えばパラベン、クロロブタノール、フェノール、ソルビン酸などの様々な抗細菌剤および抗真菌剤の包含との両方によって、微生物の存在防止を確実にすることができる。組成物に糖類、塩化ナトリウムなどのような等張化剤を含ませることも理想的でありうる。さらに、モノステアリン酸アルミニウムおよびゼラチンのような、吸収を遅延させる薬剤の包含によって、注射可能な薬学的形態の持続性吸収をなし遂げることができる。
【0117】
選択した投与経路とは無関係に、適切な水和形態で使用できる本発明の化合物および/または本発明の薬学的組成物を、当業者に公知の通常の方法によって薬学的に許容される投薬剤形に製剤する。
【0118】
本発明の薬学的組成物中の有効成分の実際の投薬レベルは、特定の患者に有毒とならずにその患者、組成物、および投与様式に関して所望の治療応答を実現するために有効である有効成分の量を得るように変動しうる。選択された投薬レベルは、採用された本発明の特定の組成物、またはそのエステル、塩、もしくはアミドの活性と、投与経路と、投与時間と、採用される特定の化合物の排泄速度と、処置の持続時間と、採用された特定の組成物と併用される他の薬物、化合物、および/または物質と、処置される患者の年齢、性別、体重、状態、全身の健康状態、および以前の病歴と、医学の分野で周知の同様の因子を含めた様々な薬物動態因子とに依存するであろう。典型的な1週用量は、上に挙げた因子に応じて約0.1mg/kgから約20mg/kg以上まで変動しうるであろう。
【0119】
その組成物は無菌であって、シリンジによって送達できる程度に流動性でなければならない。水以外に、担体は等張緩衝生理食塩水、エタノール、ポリオール(例えばグリセロール、プロピレングリコール、および液体ポリエチレングリコールなど)、およびそれらの適切な混合物でありうる。
【0120】
例えばレシチンのようなコーティング剤の使用によって、分散物の場合は必要な粒子径を維持することによって、および界面活性剤の使用によって、妥当な流動性を維持できる。多くの場合で等張化剤、例えば糖類、マンニトールまたはソルビトールのようなポリアルコール、および塩化ナトリウムをその組成物に含ませることが好ましい。吸収を遅延させる薬剤、例えばモノステアリン酸アルミニウムまたはゼラチンをその組成物に含ませることによって、注射可能な組成物の長時間吸収をもたらすことができる。
【0121】
本発明は治療を必要とする患者を処置するための方法を含み、その方法は、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない、治療有効量の抗体をその患者に投与することを特徴とする。
【0122】
本発明は、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない抗体を治療に使用することを含む。
【0123】
本発明は、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない抗体を、炎症障害および血栓障害の予防および処置のための薬の調製に使用することを含む。
【0124】
本発明は、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない抗体を、PAODおよびCLIの処置のために使用することを含む。
【0125】
このように、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。一態様では、その抗体はヒト抗体である。別の態様では、その抗体はヒト化抗体である。
【0126】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、補体因子C1qにその抗体が結合しないことは、その抗体の濃度10μg/mlでその抗体に対するC1qの最大結合(Bmax)が、細胞系hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)の抗体LC1004−002のBmaxの30%以下であるELISAアッセイの測定値を指す。別の態様では、最大結合は細胞系hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)の抗体LC1004−002のBmaxの20%以下である。
【0127】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、その抗体はBIAcoreアッセイで10−8M未満のKD値でP−セレクチンに結合する。別の態様では、KDの範囲は10−11から10−9Mである。
【0128】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、その抗体は、P−セレクチンとE−および/またはL−セレクチンとがマクロタイタープレートに被覆されたELISAアッセイにおいてEC50値によって測定されたように、E−および/またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍大きく特異的に結合する。別の態様では、E−およびL−セレクチントランスフェクタントに対するEC50値は100μg/mlよりも大きい。
【0129】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、その抗体は1μg/mlよりも大きくないIC50値で精製P−セレクチンに対する白血球様HL60細胞の接着を阻害する。別の態様では、IC50値は0.08から0.5μg/mlの範囲である。なお別の態様では、IC50値は0.08から0.11μg/mlの範囲である。
【0130】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、
(a)活性化血小板に対する白血球様HL60細胞の接着は0.05から0.3μg/mlのIC50値で阻害されるか、
(b)その抗体は白血球と血小板単層との相互作用を70%よりも大きく阻害するか、
(c)その抗体は3μg/mlの濃度でヒト流動系中の活性化内皮細胞に対する白血球の接着を60から90%の範囲で阻害するか、
(d)その抗体はC3タンパク質と結合しないか、
(e)その抗体は補体依存性細胞傷害作用(CDC)を誘発しないか、
(f)その抗体はNKエフェクター細胞上のFcγ受容体に結合しないか、または
(g)その抗体は抗体依存性細胞性細胞傷害作用(ADCC)を誘発しない。
【0131】
一態様では、本発明はP−セレクチンに結合する抗体を提供し、該抗体は、可変部H鎖アミノ酸配列CDR3がH鎖CDR3配列である配列番号38、39、40、41、または42からなる群より選択されることを特徴とする。
【0132】
一態様では、本発明は、P−セレクチンに結合し、可変部H鎖および可変部L鎖を含む抗体を提供し、その抗体は、可変部H鎖がCDR配列であるCDR1、CDR2、およびCDR3を含み、CDR1が配列番号29、30、31、32からなる群より選択され、CDR2が配列番号33、34、35、36、37からなる群より独立して選択され、CDR3が配列番号38、39、40、41、42からなる群より選択されることを特徴とし、ここで、該CDRは相互に独立して選択される。
【0133】
一態様では、本発明は、可変部L鎖がCDR配列であるCDR1、CDR2、およびCDR3を含み、CDR1が配列番号43、44より選択され、CDR2が配列番号45、46より選択され、CDR3が配列番号47、48、49、50、51、52より選択されることを特徴とする抗体を提供し、ここで、該CDRは相互に独立して選択される。
【0134】
一態様では、本発明は、P−セレクチンと結合すること、および以下からなる群より独立して選択される可変部を含むことを特徴とする抗体を提供する:
a)配列番号1のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号2によって定義されるH鎖可変ドメイン、
b)配列番号3のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号4によって定義されるH鎖可変ドメイン、
c)配列番号5のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号6によって定義されるH鎖可変ドメイン、
d)配列番号7のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号8によって定義されるH鎖可変ドメイン、
e)配列番号9のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号10によって定義されるH鎖可変ドメイン、
f)配列番号11のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号12によって定義されるH鎖可変ドメイン、
g)配列番号13のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号14によって定義されるH鎖可変ドメイン、
h)配列番号15のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号16によって定義されるH鎖可変ドメイン、
i)配列番号17のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号18によって定義されるH鎖可変ドメイン、
j)配列番号19のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号20によって定義されるH鎖可変ドメイン、
k)配列番号21のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号22によって定義されるH鎖可変ドメイン。
【0135】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、その抗体は、アミノ酸配列である配列番号3によって定義されるL鎖可変ドメインのCDR1、CDR2、およびCDR3部と、配列番号4によって定義されるH鎖可変ドメインのCDR1、CDR2、およびCDR3部とを含む。別の態様では、その抗体は、アミノ酸配列である配列番号3によって定義されるL鎖可変ドメインと、配列番号4によって定義されるH鎖可変ドメインとを含む。
【0136】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、
(a)その抗体は補体因子C1qに結合しないようにさせる少なくとも一つのアミノ酸突然変異をFc部分に含むか、
(b)そのヒトH鎖定常部は配列番号25、配列番号26、および28からなる群より独立して選択されるアミノ酸配列を含むか、
(c)その抗体は配列番号23によって定義されるようなκL鎖定常部を含むか、
(d)その抗体は補体C1qに結合しないようにさせる少なくとも一つのアミノ酸突然変異を含むか、
(e)その抗体はIgG1v1、IgG1v2、およびIgG4v1からなる群より選択されるH鎖定常部を含むか、または
(f)その抗体はFab、F(ab’)2、もしくは一本鎖フラグメントである。
【0137】
一態様では、本発明は、a)ヒト抗体またはヒト化抗体であり、b)P−セレクチンとE−および/またはL−セレクチンとをマイクロタイタープレートに被覆するELISAアッセイにおいてEC50値によって測定されるようにE−またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍大きく特異的に結合することを特徴とする抗P−セレクチン抗体を提供する。別の態様では、その抗体は、配列番号24、25、もしくは26のγ1H鎖定常部、または配列番号27もしくは28のγ4H鎖定常部によって定義されるようなアミノ酸配列を含む。なお別の態様では、その抗体は、hu−Mab<P−セレクチン>LC1004−001(DSM ACC2640)、hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)、およびhu−+Mab<P−セレクチン>LC1004−017(DSM ACC2642)からなる群より選択される細胞系によって生成される。
【0138】
本発明は、段落[00129]から[00141]に記載されたような定義を有する態様およびそれらの組み合わせも提供するものとする。
【0139】
本発明の理解を助けるために以下の実施例、参照、配列リスト、および図面を提供し、その真の範囲を添付の特許請求の範囲に示す。本発明の精神から逸脱せずに、示された手順に変更を加えることができると了解されている。
【0140】
配列リストの説明
配列番号1 HuMab1004−001のLC1004−001L鎖可変ドメイン
配列番号2 HuMab1004−001のLC1004−001H鎖可変ドメイン
配列番号3 HuMab002のLC1004−002L鎖可変ドメイン
配列番号4 HuMab002のLC1004−002H鎖可変ドメイン
配列番号5 HuMab003のLC1004−003L鎖可変ドメイン
配列番号6 HuMab003のLC1004−003H鎖可変ドメイン
配列番号7 HuMab004(I)のLC1004−004L鎖(I)可変ドメイン
配列番号8 HuMab004(I)のLC1004−004H鎖(I)可変ドメイン
配列番号9 HuMab004(II)のLC1004−004L鎖(II)可変ドメイン
配列番号10 HuMab004(II)のLC1004−004H鎖(II)可変ドメイン
配列番号11 HuMab005のL鎖可変ドメイン
配列番号12 HuMab005のH鎖可変ドメイン
配列番号13 HuMab010(I)のL鎖可変ドメイン
配列番号14 HuMab010(I)のH鎖可変ドメイン
配列番号15 HuMab010(II)のL鎖可変ドメイン
配列番号16 HuMab010(II)のH鎖可変ドメイン
配列番号17 HuMab010(III)のL鎖可変ドメイン
配列番号18 HuMab010(III)のH鎖可変ドメイン
配列番号19 HuMab011のL鎖可変ドメイン
配列番号20 HuMab011のH鎖可変ドメイン
配列番号21 HuMab017のL鎖可変ドメイン
配列番号22 HuMab017のH鎖可変ドメイン
配列番号23 κL鎖定常部
配列番号24 γ1H鎖定常部
配列番号25 γ1H鎖定常部PVA236/GLPSS331(IgG1v1)
配列番号26 γ1H鎖定常部L234A/L235A(IgG1v2)
配列番号27 γ4H鎖定常部
配列番号28 γ4H鎖定常部S228/L235E(IgG4v1)
配列番号29〜32 H鎖CDR1
配列番号33〜37 H鎖CDR2
配列番号38〜42 H鎖CDR3
配列番号43〜44 L鎖CDR1
配列番号45〜46 L鎖CDR2
配列番号47〜52 L鎖CDR3
【0141】
省語:
アミノ酸を3文字(Leu)または1文字の略号(L)のいずれかに省略する
抗体HuMab00Xを抗体00Xとも呼ぶ
L234は、EU番号(Kabat)により位置234のアミノ酸ロイシンを意味する
L234Aは、位置234のアミノ酸ロイシンがアラニンに交換されたことを意味する
PVA236は、236領域においてIgG1のELLGまたはIgG4のEFLGがPVAに修正されたことを意味する
GLPSS331は、331領域におけるIgG1のALPAPまたはIgG2のGLPAPがGLPSSに交換されたことを意味する
他のIgGサブクラスも同様に修正する
【0142】
実施例
抗P−セレクチン抗体を生成するハイブリドーマ細胞系の発生
ハイブリドーマの培養
HuMabハイブリドーマをIMDM(Cambrex)、ウシ胎仔クローン1血清(Perbio Science)、ハイブリドーマクローニング因子原液(Igen)、ピルビン酸ナトリウム、ペニシリン/ストレプトマイシン、2−メルカプトエタノール、HAT(Sigma-Aldrich)、およびカナマイシン(Invitrogen)中で37℃および5%CO2で培養した。
【0143】
抗P−セレクチン抗体を生成するハイブリドーマ細胞系の発生
トランスジェニックマウスの免疫処置法
プロトコールA:
P−セレクチンの膜貫通ドメインおよび細胞質ドメインを欠如した組換え短縮型P−セレクチンをR&D Systemsから購入し、それを用いてHCo7トランスジェニックマウス10匹(雄5匹、雌5匹)GG2201系統(Medarex, San Jose, CA, USA)に免疫処置を行った。初回免疫処置には、PBS100μlに溶かした組換えP−セレクチン50μgをフロイントの完全アジュバント100μlと混合した。残りの免疫処置には、KLHとカップリングした組換えP−セレクチンを使用した。2回目の免疫処置には、KLHとカップリングした組換えP−セレクチン50μgをPBS100μlに溶かし、フロイントの不完全アジュバント100μlと混合した。残りの免疫処置には、KLHとカップリングした組換えP−セレクチン20μgをPBS100μlに溶かし、フロイントの不完全アジュバント100μlと混合した。腹腔内免疫処置から開始して、腹腔内投与と皮下投与とを交互にして免疫処置を行った。
【0144】
プロトコールB:
HCo7トランスジェニックマウス(全て雌)3匹およびKMトランスジェニックマウス(全て雄)3匹GG2489系統(Medarex, San Jose, CA, USA)に、イムノアフィニティークロマトグラフィー(下記参照)によって無効のヒト血小板から精製した完全長P−セレクチンを用いて免疫処置を行った。初回免疫処置には、PBS100μlに溶かした精製P−セレクチン50μgをフロイントの完全アジュバント(CFA; Difco Laboratories, Detroit, USA)100μlと混合した。2回目の免疫処置にはPBS100μlに溶かした精製P−セレクチン50μgをフロイントの不完全アジュバント100μl(ICFA; Difco)と混合した。
【0145】
他の全ての免疫処置については、精製P−セレクチン20μgを使用して、フロイントの不完全アジュバント100μlと混合した。
【0146】
抗原特異的ELISA
免疫処置されたマウスの血清中抗P−セレクチン力価を抗原特異的ELISAによって決定した。平板(96平底ELISAプレート、Greiner)に、PBSに溶かした0.1μg/ml精製P−セレクチンを被覆し、室温で一晩被覆した。その後、ウェルをPBSTC(0.05%Tween20(Sigma-Aldrich Chemie BV)および2%ニワトリ血清(Gibco)を含むPBS)を用いて室温で1時間ブロッキングした。
【0147】
被験血清タップをPBSTCで1:100に希釈してウェルに加えた。免疫処置前にマウスから得た血清をPBSTCで1:100の割合で溶かし、陰性対照として使用した。ヒトP−セレクチンに対するマウス抗体(1/7、Roche Baselが社内で製造)をPBSTCに1:100の割合で溶かし、陽性対照として使用した。プレートを室温で1時間インキュベートした。次に、PBST(0.05%Tween20を含むPBS)を使用してプレートを2回洗浄した。Gt−α−huIgG−HRP(Jackson)をPBSTCに1:5000に希釈して被験タップおよび陰性対照を含むウェルに加えた。Rb−α−mIgG(Jackson)をPBSTCに1:3000に希釈し、陽性対照を含むウェルに加えた。プレートを室温で1時間インキュベートした。最後にPBSTを使用してプレートを2回洗浄し、新鮮調製したABTS(登録商標)溶液(1mg/ml)(ABTS:2,2’−アジノビス(3−エチルベンズチアゾリン−6−スルホン酸)を用いて暗条件で室温(RT)で30分間発色させた。405nmの吸光度を測定した。
【0148】
マウスの追加免疫
血清中の抗P−セレクチン力価が十分な場合、PBS100μlに入れた組換えヒトP−セレクチン20μgを用いて、融合の4日および3日前にマウス静脈内に追加的に追加免疫処置を2回行った。
【0149】
ハイブリドーマの発生
マウスを屠殺し、腹大動脈および大静脈に隣接する脾臓およびリンパ節を集めた。脾臓細胞およびリンパ節細胞と、融合パートナーであるSP2.0細胞との融合を標準的な操作法により行った。
【0150】
配列番号1〜22の可変部H鎖およびL鎖配列を有するヒトモノクローナル抗体を免疫処置法によって得た。
【0151】
κ−ELISA
融合の結果生じたハイブリドーマがヒト抗体を発生するかどうかを決定するために、κ−ELISAを行った。4℃で一晩インキュベートすることによってPBSで1/10000に希釈したラット抗ヒトIgGκL鎖抗体(DAKO)をELISAプレートに被覆した。ウェルの中身を捨てた後で、室温で1時間PBSTCと共にインキュベートすることによってプレートをブロッキングした。その後、PBSTCで1/2に希釈したハイブリドーマ培養上清と共にウェルをインキュベートした。PBSTCで1/2に希釈した培地を陰性対照として使用し、PBSTCで1/100に希釈したκL鎖陽性マウス血清を陽性対照として役立てた。続いて、ウェルを3回洗浄して、PBSTCで1/2000に希釈したHRP結合ラット抗ヒトIgG F(ab’)2(DAKO)と共に37℃で1時間インキュベートした。ウェルを3回洗浄して、新鮮調製したABTS(登録商標)溶液(1mg/ml)を用いて暗条件で室温(RT)で30分間アッセイ液を発色させた。ELISAプレートリーダーを用いて405nmで吸光度を測定した。
【0152】
抗P−セレクチンHuMab可変ドメインのクローニングおよび配列分析(κL鎖およびγ1H鎖)
P−セレクチンHuMabのL鎖可変部VLおよびH鎖可変部VHをコードするヌクレオチド配列を標準的なcDNA合成/PCR法によって単離した。
【0153】
RNeasy(登録商標)ミニキット(Qiagen)を使用して1×106〜1×107個のハイブリドーマ細胞から総RNAを調製した。ハイブリドーマ由来RNAを第一鎖のcDNA合成のためのテンプレートとして使用し、その合成をオリゴdTプライマーを使用した従来法により行った。第二鎖cDNA合成と、VLおよびVHをコードするcDNAフラグメントのさらなるPCR増幅とを、κL鎖およびγ1H鎖定常部のヌクレオチド配列に相補的な逆方向L鎖およびH鎖プライマーと、5’特異的L鎖およびH鎖プライマーとをそれぞれ用いて行った。Invitrogen(商標)life technologiesからのTOPO TAクローニングキットと、クローニングベクターとしてpCR4−TOPOとを使用してPCR生成物をクローニングした。クローニングしたPCR生成物を、消化にEcoRIを使用して適切なプラスミドの制限地図を作成することによって同定し、VLおよびVHに対してそれぞれ約740および790bpのDNAフラグメントサイズであると予測/計算した。
【0154】
クローニングされたPCRフラグメントのDNA配列を二本鎖の配列決定により決定した。
【0155】
GCG(Genetics Computer Group, Madison, Wisconsin)ソフトウェアパッケージバージョン10.2をデータ処理全般に使用した。DNAおよびタンパク質の配列をGCGモジュールCLUSTALWを使用してアライメントを作成した。プログラムGENEDOC(バージョン2.1)を使用して配列アライメントを一覧にして、編集して、色分けした。
【0156】
抗P−セレクチンIgG1 HuMabに関する発現プラスミドの構築
抗P−セレクチンHuMabL鎖をコードしている遺伝子およびH鎖をコードしている遺伝子を哺乳動物細胞発現ベクターに別々に集めた。
【0157】
それによって、抗P−セレクチンHuMabL鎖可変部(VL)をコードする遺伝子セグメントおよびヒトκL鎖定常部(CL)をコードする遺伝子セグメントを、抗P−セレクチンHuMabH鎖可変部(VH)およびヒトγ1H鎖定常部(CH1−ヒンジ−CH2−CH3)に関する遺伝子セグメントと同様に連結した。
【0158】
コドン利用頻度を推定できるヒトL鎖およびH鎖のヌクレオチド配列に関する一般的な情報は、Kabat, E. A., Wu, T. T., Perry, H. M., Gottesman, K. S., and Foeller, C. (1991) Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242に示されている。
【0159】
抗P−セレクチンHuMab κL鎖の転写ユニットは、以下のエレメントから構成される:
・ ヒトサイトメガロウイルス(HCMV)由来前初期エンハンサーおよびプロモーター、
・ Kozak配列を含む合成5’−UT、
・ シグナル配列イントロンを含むマウス免疫グロブリンH鎖シグナル配列、
・ 5’末端に独特のBsmI制限部位と、スプライスドナー部位と、3’末端に独特のNotI制限部位とを有して配列した、クローニングされた抗P−セレクチンHuMab可変部L鎖cDNA、
・ イントロン2マウスIg−κエンハンサーを含むヒトゲノムκ遺伝子定常部(Picard, D., and Schaffner, W. (1984) Nature 307, 80-82)、ならびに
・ ヒト免疫グロブリンκ−ポリアデニル化(「ポリA」)シグナル配列。
【0160】
抗P−セレクチンHuMab γ1H鎖の転写ユニットは、以下のエレメントから構成される:
・ ヒトサイトメガロウイルス(HCMV)由来前初期エンハンサーおよびプロモーター、
・ Kozak配列を含む合成5’−UT、
・ シグナル配列イントロンを含む修飾されたマウス免疫グロブリンH鎖シグナル配列、
・ 5’末端に独特のBsmI制限部位と、スプライスドナー部位と、3’末端に独特のNotI制限部位とを有して配列した、クローニングされた抗P−セレクチンHuMab可変部H鎖cDNA、
・ マウスIgμ−エンハンサーを含むヒトゲノムγ1H鎖遺伝子定常部(Neuberger, M. S. (1983) Embo J2, 1373-1378)、
・ ヒトγ1−免疫グロブリンポリアデニル化(「ポリA」)シグナル配列。
【0161】
抗P−セレクチンHuMab κL鎖発現プラスミドおよびγ1H鎖発現プラスミドの機能的エレメント:抗P−セレクチンHuMab κL鎖またはγ1H鎖発現カセットの他に、これらのプラスミドは、以下を含む:
・ ヒグロマイシン耐性遺伝子
・ エプスタン・バールウイルス(EBV)の複製起点oriP
・ E.coliでのベクターpUC18の複製を可能にする、このプラスミド由来の複製起点、および
・ E.coliにアンピシリン耐性を付与するβ−ラクタマーゼ遺伝子。
【0162】
抗P−セレクチンIgG4 HuMabに関する発現プラスミドの構築
ヒトゲノムγ1定常部およびγ1免疫グロブリンポリアデニル化(「ポリA」)シグナル配列を、ヒトゲノムγ4定常部およびγ4−免疫グロブリンポリアデニル化シグナル配列に交換することによって、抗P−セレクチンγ1H鎖発現プラスミドから抗P−セレクチンγ4H鎖プロトタイプの発現プラスミドを得た。
【0163】
抗P−セレクチンHuMab κL鎖を発現させるために、IgG1について記載したのと同様の発現プラスミドを使用した(上記参照)。
【0164】
突然変異(変異)抗P−セレクチンIgG1およびIgG4に関する発現プラスミドの構築
突然変異抗P−セレクチンγ1およびγ4H鎖をコードする発現プラスミドを、QuickChange(商標)部位特異的突然変異誘発キット(Stratagene)を使用した野生型発現プラスミドの部位特異的突然変異誘発によって生み出した。
【0165】
LC1004−002に対して以下の突然変異体が発生した。EU付番によりアミノ酸に番号を付けた(Edelman, G. M., Cunningham, B. A., Gall, W. E., Gottlieb, P. D., Rutishauser, U., and Waxdal, M. J. (1969) Proc Natl Acad Sci USA 63, 78-85; Kabat, E. A., Wu, T. T., Perry, H. M., Gottesman, K. S., and Foeller, C. (1991) Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242)。
【0166】
【表2】
【0167】
組換え抗P−セレクチンHuMabの生成
10%超低濃度IgG FCS(Gibco)、2mMグルタミン(Gibco)、1%v/v 非必須アミノ酸(Gibco)、および250μg/mlG418(Roche)を補充したDMEM(Gibco)中で培養した接着HEK293−EBNA細胞(ATTC#CRL−10852)の一過性トランスフェクションによって組換えHuMabを発生させた。トランスフェクションにFugene(商標)6(Roche)トランスフェクション試薬を、試薬(μl)対DNA(μg)の比が3:1から6:1の範囲で使用した。L鎖をコードするプラスミド対H鎖をコードするプラスミドのモル比を1:2から2:1で用いて、二つの異なるプラスミドから免疫グロブリンL鎖およびH鎖を、発現させた。HuMabを含む細胞培養上清をトランスフェクションの4日後および11日後に採集した。精製まで上清を−20℃で保存した。
【0168】
例えばHEK293でのヒト抗体の組換え発現に関する一般的な情報は、Meissner, P., Pick, H., Kulangara, A., Chatellard, P., Friedrich, K., and Wurm, F. M. (2001) Biotechnol Bioeng 75, 197-203に示されている。
【0169】
抗P−セレクチンHuMabの親和性の決定
装置:
機器:BIACORE(登録商標)2000
チップ:CM5
カップリング法:アミンカップリング
緩衝液:HBS(HEPES、NaCl)、pH7.4、25℃
【0170】
親和性の測定のために、ウサギ抗ヒトFcγ抗体(Dianova)をアミンカップリング法によってチップ表面にカップリングし、P−セレクチンに対する抗体に提示させた。約400RUの抗P−セレクチン抗体が結合した。組換えP−セレクチン(R&D Systems)を0〜50nMの間の様々な濃度で加えた。P−セレクチンを120秒間インジェクトすることによって会合を測定し、チップ表面を緩衝液で180秒間洗浄することによって解離を測定した。種々のP−セレクチン抗体に関する親和性のデータを表2に示す。Biaevaluationソフトウェアを使用して、提示されたモノクローナル抗体に対するP−セレクチンの1:1ラングミュア型結合モデルに動態データをあてはめた。
【0171】
【表3】
【0172】
細胞性接着およびロゼット形成アッセイでのP−セレクチン抗体の阻害活性
材料および方法:
細胞接着アッセイ:接着アッセイではマイクロタイタープレートに被覆したP−セレクチンに対する白血球様HL60細胞(ATCC CCL240)の接着に及ぼすHuMabの効果を評価した。HL60細胞をBCECF−AM(2’,7’−ビス−(2−カルボキシエチル)−5−(および−6−)カルボキシフルオレセインアセトキシメチルエステル、カタログ番号216254、Calbiochem)で標識した。150mM NaCl、1mM CaCl2、1mM MgCl2、20mM Tris(pH7.4)、および0.0005%Tx100を含む緩衝液中の濃度1μg/mlの完全長精製P−セレクチン(精製法については上記参照)を96ウェルプレート(Nunc Immunoplate Maxisorp F96)に4℃で一晩被覆した。その後、3.5%ウシ血清アルブミン(BSA, Fluka)を含む上記緩衝液で室温(RT)で2時間ウェルをブロッキングした。1%BSAを含む上記緩衝液にP−セレクチンHuMabまたは参照マウスP−セレクチン抗体(WAPS12.2、各ハイブリドーマ細胞系はATCCが提供)を様々に希釈した液50μlと共に、これらのウェルをRTで20分間予備インキュベートした。標識HL60細胞(50μl、細胞70000個/ウェル)を加えて、RTで45分間結合させた。一部の実験では、Fc受容体をブロッキングするために20μg/mlのヒトIgG1と共にHL60細胞を30分間予備インキュベートしてから、それらの細胞をウェルに加えた。未結合のHL60細胞を静かに洗浄する(上記緩衝液で4回)ことによって除去してから、接着した細胞をNP−40(Fluka、H2Oに溶かした1%溶液)120μlで溶解させた。上清100μlをプレートに移して、発光分光分析装置LS50B(Perkin Elmer)を使用して励起波長485nmおよび発光538nmで各蛍光を測定した。
【0173】
ロゼット形成アッセイ:活性化血小板とHL60細胞との相互作用に及ぼす抗体の効果を評価するために、ロゼット形成アッセイ(Jungi et al, Blood 67:629 (1986))を二色細胞蛍光分析(Evangelista et al., Blood 88:4183 (1996))と組み合わせて適用した。記載されたように(Fox et al, Methods Enzymol 215:45 (1992))洗浄ヒト血小板を調製した。トロンビン(終濃度1U/ml)を用いて5分間それらの血小板を活性化させ、FITC結合抗ヒトGPIIb抗体pl−36で標識した(Kouns et al., J Biol Chem 267:18844 (1992))。次に、タイロード溶液70μl中の血小板1.4〜2×106個を、種々に希釈したHuMab(100μl)と共に暗条件でRTで30分間インキュベートした。20×106個/mlに調整した(タイロード溶液中の)HL60細胞懸濁液50μlを加えた。PE(フィコエリトリン)結合抗ヒトCD45Ab(コード番号555483、Pharmingen)20μlと共にインキュベートすることによってHL60細胞を標識した。標識したHL60細胞を血小板およびHuMabと共に室温で30分間FACS管(Becton Dickinson)中でインキュベートした後で、FACScan(Becton Dickinson)を使用して血小板とHL60細胞との両方のマーカー蛍光を測定することによって、混合凝集体またはロゼットの形成を分析した。前方および側方散乱、ならびに緑色(FITC)および赤色(PE)シグナルを、励起波長488nmならびにそれぞれ発光波長530nm(FITC)および570nm(PE)での対数増幅によって取得した。電子補正を使用して、スペクトルの重複を除いた。前方散乱および側方散乱に基づいてHL60細胞を同定した。HL60細胞と同定されたイベントに関してゲーティングを行って、単一の血小板を除外した。HL60細胞のゲートに入った5000イベントを各試料について測定した。非活性化またはトロンビン活性化血小板をEDTA(10mmol/l)の存在下でHL60細胞と混合した試料を使用して、緑色蛍光スケールでの閾値を設定した。閾値よりも大きいHL60細胞の率は、血小板と結合しているHL60細胞の率を表す。血小板マーカーの蛍光が低蛍光値に移動することは、多数の接着血小板を有する混合凝集体の数が減少して、少数の接着血小板を有する混合凝集体の数が増加するのに有利になっていることを反映する。
【0174】
結果:
HL60細胞接着アッセイでは、P−セレクチン抗体はHL60細胞が0.08〜0.5μg/mlの範囲のIC50値で精製P−セレクチンに接着するのを阻害した。抗体のFc部分に突然変異が導入されたが、HuMabのIgG4およびIgG1変異体の両方は、図1に示すように0.08〜0.11μg/mlのIC50値を有して、親抗体よりも強力であった。HL60細胞をヒトIgG1と予備インキュベートした場合、図1のHuMab002について実証されたようにIC50値の約3から4倍の低下となって非突然変異親抗体の効力も増加した。この結果は、接着アッセイでの突然変異体の有効性が増加したことの主な原因が、その抗体のFc部分がFcγ受容体に対することを介して、P−セレクチンに対するHL60細胞の接着が除去されることであると示唆している。
【0175】
HL60細胞に対する、P−セレクチンを発現しているヒト活性化血小板の接着を評価するロゼット形成アッセイでは、HuMabのIC50値は、このアッセイでのP−セレクチン受容体の数が少ないことが原因で接着アッセイでの値よりもなお低かった(IC50:0.05〜0.3μg/ml、好ましくは0.05〜0.2μg/ml)。各HuMabのFc変異体の有効性は、非突然変異親抗体に比べて増加する傾向にある(図2)。活性化血小板と共にインキュベートする前にHL60細胞をIgG1およびIgG4と予備インキュベーションしても、突然変異体と親抗体との両方の阻害活性に有意に影響しなかった。これは、接着アッセイに比べてロゼット形成アッセイではFcγ受容体介在性結合の役割があまり顕著でないことを示している。
【0176】
P−セレクチン抗体と動物種由来P−セレクチンとの交差反応性
材料および方法:P−セレクチンHuMabの交差反応性を、(i)FACS分析を用いてラットおよびカニクイザル由来活性化血小板に対するHuMabの結合、ならびに(ii)HL60細胞に対するラットおよびカニクイザル血小板の接着を評価するロゼット形成アッセイでのそれらの抗体の阻害活性を測定することによって評価した。
【0177】
ラットおよびカニクイザルの活性化血小板に対するHuMabの結合を測定するために、ラットおよびカニクイザルの洗浄した血小板を、ヒトの洗浄血小板(上記参照)の調製と同様に調製した。それらの血小板をトロンビンで5分間活性化させた(終濃度1U/ml)。活性化血小板を種々に希釈したHuMab(20μl)と共にRTで30分間インキュベートした。HuMabと結合させた後で、血小板を2%PFAを用いてRTで15分間固定した。試料をタイロード緩衝液で洗浄してタイロード300mlに再懸濁した。HuMabの結合を、ウサギ抗ヒトIgGのFITC結合F(ab’)2フラグメント(コード番号F0056、Dako)を用いて検出した。ラットP−セレクチンを阻害する対照抗体としてウサギ抗ヒトポリクローナル抗P−セレクチン抗体(コード番号09361A、Pharmingen)を使用した。
【0178】
ロゼット形成アッセイでのP−セレクチンHuMabの阻害効果を測定するために、ヒト血小板について上に記載したのと同様に、ラットおよびカニクイザルの洗浄血小板を調製した。本質的にヒト血小板について記載したようにロゼット形成アッセイを行った。カニクイザル血小板を標識するためにFITC結合抗ヒトGPIIb抗体pl−36を使用し、一方でラット血小板をFITC結合マウス抗ラットCD61抗体(コード番号554952、Pharmingen)で標識した。
【0179】
結果:図3aのいくつかの例に示すように、ヒトP−セレクチン介在性機能を阻害する本発明のP−セレクチン特異的抗体のどれも、ラットP−セレクチンに結合せず、ラット血小板およびHL60細胞からなる混合凝集体の形成も阻害しないことが示された。しかし、P−セレクチンHuMabはカニクイザルP−セレクチンに結合し、それを阻害する(図3b)。
【0180】
E−およびL−セレクチンと比べたP−セレクチン抗体の選択性
材料および方法:E−およびL−セレクチンと比べたP−セレクチンHuMabの選択性を、組換えE−およびL−セレクチンに対する抗体(ADP1およびADP2、R&D Systems)の結合を測定する無細胞ELISAと、E−セレクチンCHOトランスフェクタントおよびL−セレクチン300.19トランスフェクタントに対する抗体の結合を測定する細胞性ELISAとで決定した(Goetz et al., J Cell Biol 137:509 (1997); Ley et al., Blood 82:1632 (1993)に記載されているようにトランスフェクタントを発生させた)。
【0181】
無細胞ELISAでは、150mM NaCl、1mM CaCl2、1mM MgCl2、20mMトリス(pH7.4)、および0.0005%Tx100を含む緩衝液中濃度1μg/mlの組換えP−セレクチン、E−セレクチン、またはL−セレクチンを96ウェルプレート(Nunc Immunoplate Maxisorp F96)に4℃で一晩被覆した。その後、ウェルを3.5%ウシ血清アルブミン(BSA, Fluka)を含む上記緩衝液でRTで2時間ブロッキングした。1%BSAを含む上記緩衝液に種々に希釈したP−セレクチンHuMabまたは参照マウスP−、E−セレクチン抗体(BBA26;R&D Systems)およびヤギL−セレクチン抗体(AF728;R&D Systems)50μlと共にウェルをRTで一晩予備インキュベートした。ビオチン化抗ヒトIgG(Amersham、RPN1003、終濃度1:1000)または対照抗体については対応するビオチン化抗マウスもしくは抗ヤギIgGを使用することによって、HuMabの結合を検出した。1時間インキュベートした後で、ウェルを上記の緩衝液で(3回)洗浄し、0.1%BSAを含む上記緩衝液に1:750に希釈したストレプトアビジン−ビオチン化ペルオキシダーゼ複合体(Amersham、RPN1051)0.1mlを30分間加えた。次に、ウェルを洗浄して、ABTS(2,2’−アジノ−ジ−(3−エチルベンズチアゾリンスルホナート、Boehringer, Mannheim)を含むペルオキシダーゼ基質溶液0.2mlを加えた(ABTS原液;40mM ABTS 1ml、30%H2O25μl、および0.1M酢酸Na20ml、0.05 NaH2PO4)。約10分後に0.1Mクエン酸塩および0.01%NaN3の液50μlを使用して反応を停止させた。呈色反応を405nmで読み取った。
【0182】
細胞性ELISAでは、細胞解離溶液(Sigma C5914)で細胞をはがした後に、P−およびE−セレクチンCHO−トランスフェクタントを96ウェルプレート(TC Microwell F96 Nunc 167008)の各ウェルに細胞100000個/ウェルに調整して蒔き、各培地中で37℃で一晩培養した(P−CHO−トランスフェクタント用培地:DMEM+10%FCS+2mMグルタミン+ペニシリン100U/ml/ストレプトマイシン100μg/ml;E−セレクチントランスフェクタント用培地:HAM F12+10%FCS+2mMグルタミン+ペニシリン100U/ml/ストレプトマイシン100μg/ml+0.1%フンギゾン+100μg/mlネオマイシン)。培地を除き、3%TopBlock(コード番号TB232010; Juro)を含むA−T緩衝液(150mM NaCl、1mM CaCl2、1mM MgCl2、20mM トリス(pH7.4))で1時間ウェルをブロッキングした後で、1%TopBlockおよび0.1%アジドを含む上記緩衝液に種々に希釈したP−セレクチンHuMabまたは参照マウスP−およびE−セレクチン抗体(上記参照)50μlを加えて、RTで60分間インキュベートした。ウェルを(4回)洗浄後に、無細胞ELISAについて上に挙げたのと同じ段階を使用して、結合した抗体を検出した。
【0183】
L−セレクチン300.19細胞は懸濁細胞であることから、96ウェルポリスチレンフィルタープレート(Corning 3510)のウェルにL−セレクチン300.19トランスフェクタントを平板培養することによって細胞性ELISAの形式を変更しなければならなかった。フィルタープレートのブロッキングを使用して、インキュベーション溶液をプレートの底を通して濾過することによって除去したが、それ以外ではプロトコールはP−およびE−セレクチンCHO細胞を使用した場合と同様であった。対照としてトランスフェクトしていないCHOおよび300.19を使用した。
【0184】
結果:
本発明の抗体は、E−およびL−セレクチンに比べて高度に選択性であった。これらの抗体は0.01から0.08μg/ml、好ましくは0.01から0.04μg/mlの範囲のEC50値でP−セレクチンCHO細胞に結合したが、E−セレクチンCHO細胞およびL−セレクチン300.19に対するEC50値は50μg/mlよりも明らかに大きく、好ましくは100μg/mlよりも大きかった。HuMab002は細胞性ELISAでE−およびL−セレクチンに比べて4000倍よりも大きい選択係数を伴う最高の選択性を有した。さらに、HuMab002は、濃度100μg/mlまでのベースラインレベルを超えてE−およびL−セレクチントランスフェクタントに結合しない。HuMab002のFc変異体IgG4v1およびIgG1v1の選択性は、親HuMab002の選択性と類似している(図4a〜c)。
【0185】
完全ヒト血流系でのP−セレクチン抗体のex vivo阻害活性
血小板単層への白血球接着に及ぼすP−セレクチンHuMabの効果
材料および方法:
血管壁損傷部および血小板血栓の部位への白血球の動員に及ぼすP−セレクチン抗体の効果について取り組みために、異なる剪断速度でヒト白血球とヒト血小板との相互作用を測定できるヒト血流系を、本質的に記載されたように使用した(Kirchhofer et al., Blood 89:1270 (1997))。平行平板型潅流装置に健康ドナーの前肘静脈から採取したヒト全血で、損傷して露出した血管壁を模倣するコラーゲン表面に潅流した。以前に記載されたようにコラーゲンを被覆したカバースリップを調製した(Kirchhofer et al., Blood 89:1270 (1997))。それらのカバースリップを3個の平行平板型潅流チャンバーに配置した。異なる剪断速度(65/sおよび280/s)の測定を可能にするために、異なる寸法の潅流チャンバーを使用して、一定の血流1ml/minでコラーゲン被覆カバースリップ上に血液を潅流させた。潅流装置と離れて位置する個別のローラーポンプで血流を制御した。静脈から採血して3本の管にその血液を分注した直後に、GPIIb/IIIa阻害剤(0.5μmol/ラミフィバン)を加えて血小板の凝集を防止して血小板単層を発生させた。同時に、P−セレクチン抗体(HuMab、突然変異体、各参照抗体、または対照としてヒトIgG1およびIgG4)を種々の濃度で投与して、次に、コラーゲン被覆カバースリップを含む潅流チャンバーに血液−阻害剤混合物を導入した。5.5分間の潅流時間の後で、流動を中断せずにPBSで潅流チャンバーを3分間潅流した。流動を短時間中断した後で、チャンバーをPBSに溶かした3%パラホルムアルデヒドで1ml/minで2分間固定した。次に、カバースリップをチャンバーからはずし、PBSに溶かした3%パラホルムアルデヒドで4℃で1時間再び固定し、PBS−0.03%ナトリウムアジド中で保存した。血小板単層に接着している白血球数を評価するために、風乾した後でカバースリップをDiff-Quick溶液(Dade Behring AG)で染色してMerckoglas(Merck, Germany)に包埋した。画像解析システム(MCID, Imaging Research Inc.)を使用して、カバースリップの先端から1mm離れた、血流に垂直方向の標準領域に接着している白血球数を決定した。剪断速度65/sおよび280/sで白血球数を計数した面積は、それぞれ3.1mm2および2.1mm2を有した。
【0186】
結果:
P−セレクチンHuMabは血小板単層に対する白血球の接着を濃度依存的に阻害した。剪断速度65/sおよび濃度10μg/mlでHuMabは白血球の接着を60〜99%、好ましくは70〜99%阻害した。HuMabの阻害効果は、静脈の剪断速度65/sよりも高い剪断速度280/s(動脈の状況に近い)の方が顕著であった。全体的に、剪断速度280/sで接着している白血球数は、65/sの場合よりも小さかった。Fc変異体を各親抗体と比べると、HuMab002とその変異体IgG4v1およびIgG1v1とについて実証されたように、それらの抗体はex vivo潅流チャンバーで類似した阻害活性を有した(図5)。インビトロアッセイでみられた親抗体に比べて突然変異体で増加した阻害活性は、ex vivo潅流チャンバーでは観察されなかった。これは、ヒト全血で白血球のFcγ受容体が飽和していることが原因であろう。
【0187】
内皮細胞への白血球の接着に及ぼすP−セレクチンHuMabの効果
材料および方法:
剪断条件でのP−セレクチンHuMabの抗炎症能について取り組むために、内皮細胞をカバースリップ上に被覆した設定で、上に挙げたヒト血流系を使用した。Jaffeら、1993の方法(完全な引用を添えられたい)によりII型コラゲナーゼ(Roche Switzerland)で消化することによって臍帯からヒト臍静脈内皮細胞(HUVEC)を単離した。それらの細胞を20%ウシ胎仔血清(Gibco, Auckland)、100IU/mlペニシリン(Gibco, Auckland)、0.1mg/mlストレプトマイシン(Gibco, Auckland)、2mmol/l L−グルタミン(Gibco, Auckland)、10U/mlヘパリン(Sigma)、および50μg/ml EC成長補助剤(Sigma, Germany)を補充した199培地(M199, Sigma, Germany)中で1%ゼラチンを被覆した組織培養フラスコに入れて培養した。HUVECを集密まで(約4日)成長させ、トリプシン/エチレンジアミン四酢酸(Gibco, Auckland)と共に継代して、1%ゼラチン(Fluka, Germany)で予め被覆したThermanoxプラスチックカバースリップ上に蒔いた(EC約200000個/カバースリップ)。HUVECを沈降させ、HUVECは1〜2日で集密となった。潅流開始の24時間前に20ng/ml IL−4(R&D Systems)で、潅流の5〜10分前に10−4Mヒスタミン(Fluka, Germany)でこれらの細胞を刺激した。各実験を第1回継代のHUVECで行った。刺激されたHUVECの集密単層を有するカバースリップを上記のような平行平板型潅流チャンバーに配置した。上記の潅流実験と同様に、健康ドナーから全血を採取した。しかし、これらの実験ではトロンビン阻害剤Ro−46−6240(10μM)で血液の凝固を阻止して、種々の濃度のP−セレクチン抗体(HuMab、突然変異体、各参照抗体)または対照としてヒトIgG1およびIgG4と共に、活性化内皮細胞上を潅流する直前に5分間予備インキュベートした。血流を1ml/minに、剪断速度を65/sに、かつ潅流時間を5.5分間に調整した。PBSを用いた3分間の洗浄時間の後で、接着している白血球を有するHUVECを上記と同じ流動条件で3%パラホルムアルデヒドで2分間固定した。次に、チャンバーからカバースリップを取り出し、新鮮固定液に1時間浸し、PBS−0.02%アジ化ナトリウム中で保存した。形態分析のために、修飾ビオチン化抗マウス免疫グロブリン(Animal Research Kit, Dako, USA)を使用して予め標識した、白血球共通抗原CD45に対するマウス抗体で白血球を染色した。核をヘマトキシリン(J.T Baker, Holland)で対比染色した。
【0188】
結果:
IL−4およびヒスタミンの組み合わせを用いたHUVECの刺激は、P−セレクチンの発現と、種々の種類の白血球の接着とを招き、(PMNおよび好酸球を含めた)顆粒球が接着している白血球の主要な部分を構成した。本発明のHuMabは、3μg/mlで総白血球数の60〜90%の接着を阻害した。全体的に、Fc変異体の阻害活性は非突然変異HuMabの阻害活性と有意には異ならなかった。
【0189】
P−セレクチンHuMabは、種々の白血球サブタイプに異なる効果を及ぼすことを実証した。顆粒球に及ぼす効果は、単核性白血球に比べて顕著である。本発明による抗体は、(PMNおよび好酸球を含めた)顆粒球の接着を90〜99%、単球を50〜88%、リンパ球を5〜40%阻害した。種々の白血球サブタイプの絶対数の各減少を、図6にIgG4v1について代表的に示す。
【0190】
P−セレクチンHuMabが補体系を活性化する能力
C1qおよびC3c結合ELISA:
本発明の抗体がC1qの結合およびC3の活性化を誘導する能力を決定するために、ELISA法を使用した。C1qは適応免疫系の一部であり、免疫複合体と結合するといくつかの酵素前駆体の連続活性化を誘発する。それらの酵素は順番にC3分子の開裂を引き起こし、これは炎症反応の開始、外来または異常粒子のオプソニン化、および細胞膜の溶解を招く場合がある。
【0191】
原則として、ELISAプレートに種々の濃度範囲の抗体を被覆し、そこにヒトC1qか、またはC3の供給源としてヒトプール血清を加える。ヒトC1qまたはC3εに対する抗体によって、続いてペルオキシダーゼ標識コンジュゲートによってC1qまたはC3εの結合を検出する。
【0192】
HuMab002(ハイブリドーマおよび一過性トランスフェクトーマ由来材料、その突然変異体、ならびに対照抗体)を0.16〜20μg/mlの濃度で試験した。陰性対照として、C1qと非常に弱く結合するヒトIgG4(CLB, the Netherlands,0.5μg/ml原液)を使用した。ヒトIgG1(Sigma, 2ug/ml原液)を陽性対照として組み入れた。C1qを検出するために、C1qに対するウサギ抗体(Dako)と、セイヨウワサビペルオキシダーゼ(Sigma)と結合したブタ抗ウサギIgG抗体とを使用した。C3εを検出するために、マウス抗ヒトC3抗体と、セイヨウワサビペルオキシダーゼ(Sigma)と結合したウサギ抗マウスIgG抗体とを適用した。
【0193】
試験したHuMabのEC50値または10μg/mlでの最大結合(Bmax)に関する計算を、Graphpad Prismソフトウェアを使用した非線形回帰曲線あてはめ(1部位結合)を使用して決定した。
【0194】
結果:
本発明によるHuMab002は、ハイブリドーマおよびトランスフェクトーマ由来材料に関してそれぞれ0.946μg/mlおよび1.159μg/mlのEC50値と、0.987および0.711のBmax(OD405)値とから示されるように、効率的にC1qと結合できた。予想通り、OD405でのBmax値0.222により示されるように、陰性対照であるヒトIgG4はC1qと結合しなかった。しかし、試験した三つのFc変異体全て(IgG4v1、IgG1v1、IgG1v2)は、それぞれ0.132、0.119、および0.132のOD405 Bmax値によって示されるようにC1qと結合する能力を欠如した(表3)。C1qとの結合能と一致して、(ハイブリドーマおよびトランスフェクトーマ由来)HuMab002に対するC3の沈着は抗体濃度依存的に起こり、EC50値は2.7μg/mlから8.3μg/mlの範囲であった。しかし、三つのFc変異体全ては、それぞれ0.104、0.156、および0.133のOD405 Bmax値によって示されるように、C3の沈着を開始できなかった(表3)。
【0195】
HuMab002が補体成分と相互作用することから、この抗体はインビボでCDCを誘導する内因性能力を有する。したがって、この抗体のFc部分は本発明により修飾されている。
【0196】
【表4】
【0197】
P−セレクチンHuMabがFcγ受容体に結合する能力
IgG抗体依存性細胞傷害効果は、エフェクター細胞上のFcγ受容体によって仲介される。ヒト血液由来FcγR発現エフェクター細胞に対する、ハイブリドーマおよびトランスフェクトーマ由来HuMab002と、突然変異抗体と、対照抗体との結合をFACS分析によって検討した。
【0198】
材料および方法:
FcγRI IIA1.6トランスフェクタントまたは新鮮単離したエフェクター細胞を抗体と共にインキュベートして、抗体の結合をFITC標識ウサギ抗ヒトIgG F(ab)2(DAKO)、またはFITC標識ウサギ抗ヒトIgG F(ab)2(BD/Pharmingen)を用いて検出した。HuMab002(一過性トランスフェクトーマおよび/またはハイブリドーマ由来材料、ならびに突然変異体)を濃度1μg/ml(IIA1.6トランスフェクタント)または10μg/ml(エフェクター細胞)で試験した。一次抗体またはヒトIgG4(10μg/ml)の不在を陰性対照として使用した。IIA1.6細胞上のFcγRIの発現を検出するために、FITC標識マウス抗ヒトCD64(BD/Pharmingen)を使用した。NK細胞に富む末梢血単核細胞を使用した実験では、PE標識マウス抗ヒトCD56(BD/Pharmingen)を使用した二重染色によってNK細胞を同定した。FSC/SSCプロフィールに基づいて顆粒球および単球を同定した。
【0199】
IIA1.6細胞、IIA1.6−FcγRIトランスフェクタント、および新鮮単離したエフェクター細胞を抗体と共にインキュベートした。抗体の結合をFITC標識Rb−α−huIgG F(ab)2(DAKO)またはFITC標識Rb−α−huIgG F(ab)2(BD/Pharmingen)を用いて検出した。
【0200】
IIA1.6−FcγRIトランスフェクタント結合アッセイでは、HuMab002(一過性トランスフェクトーマ由来、ハイブリドーマ由来および突然変異体材料)を1μg/mlの濃度で試験した。IIA1.6野生型細胞を陰性対照として使用した。FcγRI発現の対照としてm−α−huCD64−FITC(BD/Pharmingen)を使用した。
【0201】
エフェクター細胞結合アッセイでは、HuMab002(一過性トランスフェクトーマ由来、ハイブリドーマ由来および突然変異体材料)を10μg/mlの濃度で試験した。顆粒球結合アッセイでは一過性トランスフェクトーマ材料を試験しなかった。顆粒球結合アッセイを除く全てのエフェクター細胞結合アッセイでIgG4(10μg/ml)を陰性対照として使用した。
【0202】
NK単離キット(Dynal Biotech ASA, Oslo, Norway)を使用して全血のNK細胞を濃縮した。m−α−huCD56−FITC染色によってNK細胞を同定した。
【0203】
NK単離キット(Dynal Biotech ASA, Oslo, Norway)に同封されているプロトコールに記載されているフィコール法を使用して、全血からPBMC(末梢血単核細胞)を得た。FSC/SSCプロフィールに基づいて単球を同定した。FACS溶解緩衝液を使用して全血から顆粒球を単離して、FSC/SSCプロフィールに基づいて同定した。
【0204】
新鮮単離したエフェクター細胞を抗体と共にインキュベートして、抗体の結合をFITC標識ウサギ抗ヒトIgG F(ab)2(DAKO)またはFITC標識ウサギ抗ヒトIgG F(ab)2(BD/Pharmingen)を用いて検出した。HuMab002(一過性トランスフェクトーマおよび/またはハイブリドーマ由来材料、ならびに突然変異体)を濃度10μg/mlで試験した。一次抗体またはヒトIgG4(10μg/ml)の不在を陰性対照として使用した。NK単離キット(Miltenyi Biotec, USA)によってMNC試料からNK細胞を単離した。NK細胞に富む末梢血単核細胞を使用した実験では、PE標識マウス抗ヒトCD56(BD/Pharmingen)を使用した二重染色によってNK細胞を同定した。当技術分野の現状により、PBMCから顆粒球および単球を単離した(例えば単球単離キット(Miltenyi、上記参照)。顆粒球および単球をFSC/SSCプロフィールに基づいて同定した。
【0205】
結果:
本発明によるHuMab002は、顆粒球、単球、およびNK細胞に対する結合によって示されるようにFcRに結合できた。試験した三つのFc変異体全て(IgG4v1、IgG1v1、およびIgG1v2)はNK細胞に結合する能力を完全に欠如していた(表4)。さらに、HuMab002は顆粒球および単球に効率的に結合したが、表5および6に抗体に結合している細胞の率によって示されるように、突然変異体はヒトIgG4に対する一次抗体の不在に匹敵する結合レベルを示した。これは、突然変異体がエフェクター細胞上のFcRと相互作用する能力を欠如したことを示している。
【0206】
HuMab002はFcRと効率的に相互作用できるので、この抗体はインビボで抗体依存性細胞性細胞傷害作用を誘導する内因性能力を有する。本発明によるFc変異体に関して行われたようなFcRとの相互作用の不活性化は、効果的にADCCを阻止する。
【0207】
【表5】
【0208】
【表6】
【0209】
【表7】
【技術分野】
【0001】
本発明は、一般的に抗P−セレクチン抗体、特に補体因子C1qと結合しない抗P−セレクチン抗体に関するものである。 好ましくは、これらの抗体はヒト抗体またはヒト化抗体である。
【0002】
P−セレクチン(CD62P、GMP−140、PADGEM、LECAM−3)は、トロンビンおよび他のアゴニストに応答して活性化血小板および内皮の表面に発現する140kDaのカルシウム依存性糖質結合タンパク質である(McEver et al, J Biol Chem 270:11025 (1995); Varki, Proc Natl Acad Sci USA 91:7390 (1994); Springer TA, Annu Rev Physiol 57:827 (1995))。両方の種類の細胞では、P−セレクチンは分泌顆粒、すなわち血小板のα顆粒および内皮細胞のWeibel−Palade小体に貯蔵される(McEver et al., J Clin Invest 84:92 (1984))。P−セレクチンは、NH2末端レクチンドメインの後にEGF様ドメイン、補体調節タンパク質に相同性を有する9個のショートコンセンサスリピート(short consensus repeat)、膜貫通ドメイン、および短鎖の細胞質側末端が続いたものから構成されるI型膜貫通糖タンパク質である(Johnston et al., Cell 56:1033 (1989))。P−セレクチンの構造は、セレクチンファミリーの他の二つのメンバーであるE−セレクチンおよびL−セレクチンと類似しており、これらのセレクチンは、サイトカインによって活性化した内皮細胞に発現している(E−セレクチン)か、または大部分のクラスの白血球上に構成的に発現している(L−セレクチン)かのいずれかである。
【0003】
全てのセレクチンは、シアリルルイスx(sLex; Foxall et al., J Cell Biol 117:895 (1992); Varki, Curr Opin Cell Biol 257:257 (1992))のような低分子シアリル化フコシル化オリゴ糖に低親和性で結合することが公知である。E−セレクチンではなくP−セレクチンおよびL−セレクチンは、ヘパリン硫酸のような特定の硫酸化糖質にも結合する(総説については、McEver and Cummings, J Clin Invest 100:S97 (1997)を参照されたい)。P−セレクチンに対する高親和性リガンドは、ムチン様糖タンパク質(McEver et al., J Biol Chem 270:11025 (1995))であり、その糖タンパク質は、シアリル化O−グリカンのクラスターを有するポリペプチド主鎖からなる。P−セレクチンが優先的に結合するあるシアロムチンリガンドは、循環している白血球によって、約120kDaの相対分子量を有する二つのジスルフィド結合サブユニットを有するホモ二量体として通常は発現されるP−セレクチン糖タンパク質リガンド1(PSGL−1、CD162)である。P−セレクチンの結合部位は、PSGL−1のNH2末端の最末端部分に局在する。そのリガンドに結合することによって、P−セレクチンは活性化血小板および内皮細胞上での白血球のローリングを仲介する。このローリング過程は、白血球の移動速度を効果的に低下させ、強固な接着および内皮下への白血球のその後の移行に、また血栓への白血球の蓄積にも欠かせない。
【0004】
P−セレクチン欠損マウスおよびP−セレクチン特異的遮断抗体を用いた研究から、虚血/再潅流傷害を含めた多数の急性および慢性炎症疾患の病態生理にP−セレクチンが関係することが示された(Winn et al., J Clin Invest 92:2042 (1993); Massberg et al., Blood 92:507 (1998))。さらに、アテローム性動脈硬化症(Collins et al., J Exp Med 191: 189 (2000); Johnson et al., J Clin Invest 99:1037 (1997))、再狭窄(Manka et al., Circulation 103:1000 (2001); Bienvenu et al., Circulation 103:1128 (2001))および血栓症(Kumar et al., Circulation 99:1363 (1999); Andre et al., Proc Natl Acad Sci USA 97:13835 (2000); Blann et al., Br. J. Haematol 108:191 (2000); Myers et al., Thromb Haemostasis 85: 423 (2001)のような炎症性構成要素を有する心血管疾患にP−セレクチンは明らかに寄与している。どうやら、P−セレクチン機能の阻害は、血管内皮または血小板に対する白血球の接着を伴う種々の疾患での治療として有効であろう(例えばWO93/06863を参照されたい)。
【0005】
P−セレクチンに対する抗体は、当技術分野の現状に記載され、それらの抗体の抗炎症効果および抗血栓効果に関して研究されてきた。米国特許第4,783,399号およびWO93/06863は、活性化血小板と反応性のP−セレクチンに対するマウスモノクローナル抗体を記載している。Geng J. G.ら(J. Biol. Chem., 266 (1991) 22313-22318)は、P−セレクチンのアミノ酸(aa)フラグメントaa60〜75(シグナル配列を含むスイスプロット配列P16109により計数してCysからGlu。WO93/21956はP−セレクチンに対するマウスモノクローナル抗体と、規定の抗体と競合してP−セレクチンフラグメントaa60〜75の存在下で結合するIgG1サブクラスのヒト化抗体とを参照している)にカルシウムイオンの不在下で結合するマウスモノクローナル抗体を記載している。挙げられた、ヒトP−セレクチンに対するマウスモノクローナル抗体のどれも、ヒト患者の処置に有用ではない。WO93/21956に挙げられたP−セレクチンに対するヒトIgG1サブクラスのヒト化抗体は、開発の前臨床段階にある(www.mrctechnology.org)。
【0006】
発明の開示
本発明は、P−セレクチンと結合して、ヒト補体因子C1qと結合しないことを特徴とする抗体に関するものである。好ましくは、この抗体はNK細胞上のヒトFcγ受容体にも結合しない。本発明による抗体は、ヒト起源由来のFc部分を含む。好ましくは、これらの抗体はヒト化抗体またはヒト抗体である。これらの抗体は、特に末梢動脈閉塞性疾患(PAOD)および重症肢虚血(CLI)による炎症障害および血栓障害を患う患者に利益をもたらす新しい発明的な性質を有する。
【図面の簡単な説明】
【0007】
【図1】本発明の抗体がマイクロタイタープレートに被覆した精製P−セレクチンに対する白血球様HL60細胞の接着を阻害することを示す図である。突然変異抗体は非突然変異親抗体よりも強力である。
【図2】HL60細胞に対するトロンビン活性化血小板の接着を測定するロゼット形成アッセイでの本発明の抗体の阻害活性を示す図である。
【図3a】本発明の抗体とラットおよびカニクイザルP−セレクチンとの交差反応性を示す図である。抗P−セレクチン抗体はHL60細胞に対するトロンビン活性化ラット血小板の接着に影響しないが、一方で商業的に入手できるポリクローナル抗P−セレクチン抗体(Pharmingen 09361A)はこの相互作用を阻害する。
【図3b】本発明の抗体とラットおよびカニクイザルP−セレクチンとの交差反応性を示す図である。本発明の抗体はHL60細胞に対するカニクイザル活性化血小板の接着を阻害する。
【図4a】P−、E−、およびL−セレクチントランスフェクタントに関する代表的な結合曲線によりE−およびL−セレクチンに比べてP−セレクチンに対する抗体の選択性を実証する図である。本発明による抗体は、0.01から0.07μg/mlの範囲のEC50値でP−セレクチンCHO細胞に結合する。E−セレクチンCHO細胞およびL−セレクチン300.19細胞に関するEC50値は好ましくは100μg/mlよりも大きい。
【図4b】P−、E−、およびL−セレクチントランスフェクタントに関する代表的な結合曲線によりE−およびL−セレクチンに比べてP−セレクチンに対する抗体の選択性を実証する図である。本発明による抗体は、0.01から0.07μg/mlの範囲のEC50値でP−セレクチンCHO細胞に結合する。E−セレクチンCHO細胞およびL−セレクチン300.19細胞に関するEC50値は好ましくは100μg/mlよりも大きい。
【図4c】P−、E−、およびL−セレクチントランスフェクタントに関する代表的な結合曲線によりE−およびL−セレクチンに比べてP−セレクチンに対する抗体の選択性を実証する図である。本発明による抗体は、0.01から0.07μg/mlの範囲のEC50値でP−セレクチンCHO細胞に結合する。E−セレクチンCHO細胞およびL−セレクチン300.19細胞に関するEC50値は好ましくは100μg/mlよりも大きい。
【図5】完全ヒト流動系での本発明の抗体の阻害活性を示す図である。これらの抗体は剪断速度65/sで濃度依存的に血小板単層へのヒト白血球の接着を阻害する。
【図6a】P−セレクチンを発現しているヒト内皮細胞に対する白血球の接着に及ぼす本発明の抗体の阻害効果を示す図である。対照に対する%で表した白血球の接着の合計阻害を明らかにする。
【図6b】P−セレクチンを発現しているヒト内皮細胞に対する白血球の接着に及ぼす本発明の抗体の阻害効果を示す図である。種々の白血球サブセットの絶対数に及ぼす抗体の一つの阻害効果を代表的に示す。
【0008】
発明の詳細な説明
I.定義
用語「P−セレクチン」はHsu-Lin et al., J Biol Chem 259: 9121 (1984)およびMc Ever et al., J Clin Invest 84:92 (1989)に記載されたような、ヒト血小板および内皮細胞によって発現される140kDaのタンパク質を指す。このI型膜貫通糖タンパク質は、NH2末端レクチンドメインの後に上皮増殖因子(EGF)様ドメインおよび9個のコンセンサスリピートドメインが続いたものから構成される。P−セレクチンは、単一の膜貫通ドメインによって膜に固定され、短鎖の細胞質側末端を含む。本発明は、P−セレクチンによって仲介される一つまたは複数の生物学的活性、例えば、炎症活性または血栓活性を阻害できる抗体を提供する。この抗体はP−セレクチンに結合し、P−セレクチンとそのリガンドとの結合を妨害することによって作用する。
【0009】
用語「P−セレクチンリガンド」は、Moore et al., J Cell Biol 118:2445 (1992)、Sako et al., Cell 75:1179 (1993)によって記載されたようなムチン様糖タンパク質であるP−セレクチンリガンド糖タンパク質1(PSGL−1)のような高親和性生物関連P−セレクチンリガンドに好ましくは関するものである。PSGL−1は、シアリル化O−グリカンのクラスターと連結した一連の10個のリピートを含む、セリン、トレオニン、およびプロリンに富む細胞外ドメインを有するI型膜タンパク質である。PSGL−1は、循環している白血球によって約120kDaの相対分子量を有する二つのジスルフィド結合サブユニットを有するホモ二量体として通常は発現される。P−セレクチンの結合部位は、PSGL−1のNH2末端の最末端部に局在する。血小板によって発現され、PSGL−1と構造類似性を有するシアロムチンGPIbαは、P−セレクチンに対する血小板のリガンドであると最近実証された(Romo et al, J Exp Med 190:803 (1999)。P−セレクチンに結合するGPIbαの生理学的重要性はまだ研究中であるが、その相互作用がローリングおよび活性化内皮細胞に対する血小板の接着に貢献しているようである(Berndt et al., Thromb Haemost 86:178 (2001)。P−セレクチンは、シアリルルイスxのような低分子シアル酸化フコシル化オリゴ糖(Foxall et al., J Cell Biol 117:895 (1992), Varki, Curr Opin Cell Biol 257 (1992)にも、特にヘパリン硫酸のような硫酸化糖質(McEver et al., J Biol Chem 270:11025 (1995)にも低親和性で結合する。
【0010】
用語「抗体」は、本発明による性質が保たれる限りは全抗体、抗体フラグメント、ヒト抗体、ヒト化抗体、キメラ抗体、および遺伝子操作抗体(変異抗体または突然変異抗体)を含むがそれに限定されるわけではない多様な形態の抗体、好ましくはモノクローナル抗体を包含する。特に組換えヒト抗体としてのヒトまたはヒト化モノクローナル抗体が特に好ましい。
【0011】
本明細書に使用するような用語「モノクローナル抗体」または「モノクローナル抗体組成物」は、単一のアミノ酸組成の抗体分子の調製物を指す。
【0012】
用語「キメラ抗体」は、ある起源または種からの可変部、すなわち結合部と、異なる起源または種由来の定常部の少なくとも部分とを含む、組換えDNA法によって通常は調製されたモノクローナル抗体を指す。マウス可変部とヒト定常部とを含むキメラ抗体が特に好ましい。そのようなマウス/ヒトキメラ抗体は、マウス免疫グロブリン可変部をコードするDNAセグメントと、ヒト免疫グロブリン定常部をコードするDNAセグメントとを含む発現された免疫グロブリン遺伝子の生成物である。本発明に包含される他の形態の「キメラ抗体」は、定常部が本来の抗体の定常部から修飾または変更されて、特にC1qの結合および/またはFc受容体(FcR)の結合に関して本発明による性質を発生する抗体である。そのような「キメラ」抗体は、「クラススイッチ抗体」とも称される。キメラ抗体を生成させる方法は、従来の組換えDNA法および当技術分野で現在周知である遺伝子トランスフェクション法を含む。例えばMorrison, S.L., et al., Proc. Natl. Acad. Sci. USA 81 (1984) 6851-6855;米国特許第5,202,238号および第5,204,244号を参照されたい。
【0013】
用語「ヒト化抗体」は、フレームワーク部または「相補性決定部」(CDR)が修飾されて親免疫グロブリンとは特異性が異なる免疫グロブリンCDRを含む抗体を指す。好ましい態様では、マウスCDRをヒト抗体のフレームワーク部にグラフトして、「ヒト化抗体」を調製する。例えばRiechmann, L., et al., Nature 332 (1988) 323-327;およびNeuberger, M.S., et al., Nature 314 (1985) 268-270を参照されたい。特に好ましいCDRは、キメラ抗体および二機能抗体に関して上に述べたように、抗原を認識する配列を表すCDRに対応する。本発明に包含される他の形態の「ヒト化抗体」は、定常部が本来の抗体から修飾または変更されて、特にC1qの結合および/またはFc受容体(FcR)の結合に関して本発明による性質を発生する抗体である。
【0014】
本明細書に使用するような用語「ヒト抗体」は、ヒト生殖細胞系免疫グロブリン配列由来の可変部および定常部を有する抗体を含むことを意図する。ヒト抗体は、当技術分野の現状で周知である (van Dijk and van de Winkel, Curr Opin Pharmacol 5:368 (2001)。内因性免疫グロブリンの生成の不在下で免疫処置に応じてヒト抗体の全レパートリーを生成できるトランスジェニック動物(例えばマウス)でヒト抗体を生成させることもできる。そのような生殖細胞系突然変異マウスへのヒト生殖細胞系免疫グロブリン遺伝子アレイの導入は、抗原による攻撃誘発に応答してヒト抗体の生成を招くであろう(例えばJakobovits et al., Proc. Natl. Acad. Sci. USA, 90: 2551-2555 (1993); Jakobovits et al., Nature, 362:255-258 (1993); Bruggemann et al., Year in Immuno., 7:33 (1993)を参照されたい)。ファージディスプレイライブラリーでもヒト抗体を生成させることができる(Hoogenboom and Winter, J. Mol. Biol., 227:381 (1992); Marks et al., J. Mol. Biol, 222:581 (19991))。ColeらおよびBoernerらの技法もヒトモノクローナル抗体の調製に利用できる(Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985)およびBoerner et al., J. Immunol., 147(l):86-95 (1991))。本発明によるキメラ抗体およびヒト化抗体に関してすでに触れたように、本明細書に使用するような用語「ヒト抗体」は、定常部で修飾されて、特にC1qの結合および/またはFcRの結合に関して本発明による性質を発生する抗体も含む。さらに、本発明はC1qおよび/またはFcRに結合するヒト抗体を含む。そのようなヒト抗体は、E−およびL−セレクチンよりもP−セレクチンに対する高い選択性を特徴とする。本発明によるそのような抗体は、0.01から0.07μg/mlの範囲のEC50値で、P−セレクチン発現細胞に結合する。E−セレクチンおよびL−セレクチン発現細胞に関するEC50値は、好ましくは100μg/mlよりも大きい。そのような抗体は、本発明による性質を有するヒト抗体を製造するための中間体として好ましくは有用である。
【0015】
本明細書に使用するような用語「組換えヒト抗体」は、NS0もしくはCHO細胞のような宿主細胞から、またはヒト免疫グロブリン遺伝子に関してトランスジェニックな動物(例えばマウス)から単離された抗体、あるいは宿主細胞にトランスフェクトされた組換え発現ベクターを使用して発現された抗体のような、組換え手段によって調製、発現、作出、または単離された全てのヒト抗体を含むことを意図する。本発明による組換えヒト抗体は、再編成された形態の可変部および定常部を有する。本発明による組換えヒト抗体は、インビボ体細胞高頻度突然変異に供されている。よって、これらの組換え抗体のVHおよびVL部のアミノ酸配列は、ヒト生殖細胞系VHおよびVL配列に由来して、それらの配列に関係する一方で、自然にはインビボでヒト抗体生殖細胞系レパートリーに存在しないおそれがある配列である。
【0016】
「可変部」(本明細書に使用するようなL鎖可変部(VL)、H鎖可変部(VH))は、抗原に対する抗体の結合に直接関与するL鎖およびH鎖のペアのそれぞれを示す。ヒト可変部L鎖およびH鎖のドメインは同じ一般構造を有し、各ドメインは四つのフレームワーク(FR)部を含み、その配列は広く保存され、三つの「超可変部」(または相補性決定部CDR)によって連結している。フレームワーク部はβシートコンホメーションを採り、CDRはβシート構造を連結しているループを形成する場合がある。各鎖のCDRは、フレームワーク部によって三次元構造に保持され、もう一方の鎖からのCDRと共に抗原結合部位を形成する。抗体H鎖およびL鎖のCDR3部は、本発明による抗体の結合特異性/親和性に特に重要な役割を演じ、よって本発明のさらなる目的を提供する。
【0017】
用語「超可変部」または「抗体の抗原結合部分」は、本明細書に使用する場合に、抗原結合を担う抗体のアミノ酸残基を指す。超可変部は、「相補性決定部」または「CDR」からのアミノ酸残基を含む。「フレームワーク」部または「FR」部は、本明細書に定義するような超可変部の残基以外の可変ドメイン部である。よって、抗体のL鎖およびH鎖は、N末端からC末端方向にFR1、CDR1、FR2、CDR2、FR3、CDR3およびFR4ドメインを含む。特に、H鎖CDR3は抗原結合に最も寄与する領域である。CDRおよびFR部は、Kabatら、Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991))の標準的な定義および/または「超可変ループ」の残基によって決定される。
【0018】
本明細書に使用するような用語「核酸または核酸分子」は、DNA分子およびRNA分子を含むことを意図する。核酸分子は、一本鎖または二本鎖の場合があるが、好ましくは二本鎖DNAである。
【0019】
核酸は、別の核酸配列と機能的関係に置かれた場合に「作動可能に連結」される。例えば、プレ配列または分泌リーダーがポリペプチドの分泌に関与するプレタンパク質としてポリペプチドが発現するならば、そのプレ配列または分泌リーダー配列に関するDNAはそのポリペプチドに関するDNAと作動可能に連結され、プロモーターまたはエンハンサーがコード配列の転写に影響するならば、そのプロモーターまたはエンハンサーはその配列に作動可能に連結され、また、リボソーム結合部位が翻訳を促進するように位置するならば、そのリボソーム結合部位はコード配列に作動可能に連結される。一般に「作動可能に連結した」は、連結されたDNA配列が隣接し、分泌リーダーの場合は隣接して読出し期であることを意味する。しかし、エンハンサーは隣接している必要がない。連結は、都合のよい制限部位での連結反応によって実現される。そのような制限部位が存在しないならば、従来の常法により合成オリゴヌクレオチドアダプターまたはリンカーを使用する。
【0020】
本明細書に使用するような表現「細胞」、「細胞系」、および「細胞培養物」は相互交換可能に使用され、そのような呼称の全ては子孫を含む。このように、語「形質転換体」および「形質転換細胞」は、初代対象細胞および継代数とは無関係にそれ由来の培養物を含む。全ての子孫は、故意または偶然の突然変異が原因でDNA含量が厳密には同一ではない場合があることも了解されている。本来形質転換された細胞でスクリーニングされたものと同じ機能または生物学的活性を有する変異子孫も含まれる。別個の呼称が意図される場合は、情況から明らかであろう。
【0021】
「定常ドメイン」は、抗原に対する抗体の結合に直接関与せず、様々なエフェクター機能を示す。H鎖定常部のアミノ酸配列に応じて、抗体または免疫グロブリンは、IgA、IgD、IgE、IgGおよびIgMのクラスに分かれ、これらのいくつかをサブクラス(アイソタイプ)、例えばIgG1、IgG2、IgG3、およびIgG4と、IgA1およびIgA2とにさらに分けることができる。種々のクラスの免疫グロブリンに対応するH鎖定常部は、それぞれα、δ、ε、γ、およびμと呼ばれる。本発明による抗体は、好ましくはIgG型である。
【0022】
抗体のFc部分は補体活性化、すなわちC1qの結合およびFc受容体の結合に直接関与する。補体系に及ぼす抗体の影響はある状態に依存するが、C1qに対する結合は、Fc部分での所定の結合部位によって起こる。そのような結合部位は、当技術分野の現状で公知であり、例えばBoakle et al., Nature 282 (1975) 742-743; Lukas et al., J. Immunol. 127 (1981) 2555-2560, Brunhouse and Cebra, Mol. Immunol. 16 (1979) 907-917; Burton et al., Nature 288 (1980) 338-344; Thommesen et al., Mol. Immunol. 37 (2000) 995-1004; Idusogie et al., J. Immunol.164 (2000) 4178-4184; Hezareh et al., J. Virology 75 (2001) 12161-12168; Morgan et al., Immunology 86 (1995) 319-324、EP0307434に記載されている。そのような結合部位は、例えばL234、L235、D270、N297、E318、K320、K322、P331、およびP329(Kabat、下記参照のEUインデックスにより付番)である。サブクラスIgG1、IgG2、およびIgG3の抗体は、通常は補体活性化とC1qおよびC3の結合を示すが、一方IgG4は補体系を活性化せず、C1qおよびC3を結合しない。本明細書に使用するような、用語「ヒト起源由来のFc部分」は、サブクラスIgG4のヒト抗体のFc部分、または下記に定義するようなC1qの結合および/もしくはFcRの結合を検出できないように修飾されたサブクラスIgG1、IgG2、もしくはIgG3のヒト抗体のFc部分のいずれかであるFc部分を示す。「抗体のFc部分」は、当業者に周知の用語であり、パパインによる抗体の切断に基づき定義される。本発明による抗体は、Fc部分としてヒト起源由来のFc部分と、好ましくはヒト定常部の他の全ての部分とを含む。好ましくは、Fc部分はヒトFc部分であり、特に好ましくはヒトIgG4サブクラス、またはヒトIgG1サブクラスからの突然変異Fc部分のいずれか由来である。Fc部分と、配列番号25〜28に示すか、またはPVA236突然変異を有さない配列番号25のH鎖定常部とが最も好ましい。
【0023】
II.本発明の好ましい態様
本発明は、P−セレクチンに結合する抗体であって、該抗体の可変H鎖アミノ酸配列CDR3がH鎖CDR3配列である配列番号38、39、40、41または42からなる群より選択されることを特徴とする、抗体を含む。
【0024】
本発明は、可変H鎖および可変L鎖を含む、P−セレクチンに結合する抗体を好ましくは提供し、その抗体は、可変H鎖がCDR配列であるCDR1、CDR2およびCDR3を含み、CDR1が、配列番号29、30、31、および32からなる群より選択され、CDR2が配列番号33、34、35、36、および37からなる群より選択され、CDR3が配列番号38、39、40、41、および42からなる群より選択されることを特徴とし、ここで、該CDRは相互に独立して選択される。
【0025】
本発明による抗体は、可変部L鎖がCDR配列であるCDR1、CDR2、およびCDR3を含み、CDR1が配列番号43および44より選択され、CDR2が配列番号45および46より選択され、CDR3が配列番号47、48、49、50、51、および52より選択されることを好ましくは特徴とし、ここで、該CDRは相互に独立して選択される。
【0026】
その抗体は、H鎖CDRとして配列番号2のCDRおよびL鎖CDRとして配列番号1のCDR、H鎖CDRとして配列番号4のCDRおよびL鎖CDRとして配列番号3のCDR、H鎖CDRとして配列番号6のCDRおよびL鎖CDRとして配列番号5のCDR、H鎖CDRとして配列番号8のCDRおよびL鎖CDRとして配列番号7のCDR、H鎖CDRとして配列番号10のCDRおよびL鎖CDRとして配列番号9のCDR、H鎖CDRとして配列番号12のCDRおよびL鎖CDRとして配列番号11のCDR、H鎖CDRとして配列番号14のCDRおよびL鎖CDRとして配列番号13のCDR、H鎖CDRとして配列番号16のCDRおよびL鎖CDRとして配列番号15のCDR、H鎖CDRとして配列番号18のCDRおよびL鎖CDRとして配列番号17のCDR、H鎖CDRとして配列番号20のCDRおよびL鎖CDRとして配列番号19のCDR、またはH鎖CDRとして配列番号22のCDRおよびL鎖CDRとして配列番号21のCDRを含むことを好ましくは特徴とする。
【0027】
Kabatら、Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)の標準的な定義によりCDR配列を決定できる。各鎖のCDRは、フレームワークアミノ酸によって引き離されている。配列番号1〜22のCDRを配列番号29〜52に示す。
【0028】
本発明による抗体は、該抗体がP−セレクチンと結合し、以下からなる群より独立して選択されるH鎖可変部およびL鎖可変部を含むことを好ましくは特徴とする:
a)配列番号2のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号1によって定義されるL鎖可変ドメイン;
b)配列番号4のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号3によって定義されるL鎖可変ドメイン;
c)配列番号6のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号5によって定義されるL鎖可変ドメイン;
d)配列番号8のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号7によって定義されるL鎖可変ドメイン;
e)配列番号10のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号9によって定義されるL鎖可変ドメイン;
f)配列番号12のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号11によって定義されるL鎖可変ドメイン;
g)配列番号14のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号13によって定義されるL鎖可変ドメイン;
h)配列番号16のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号15によって定義されるL鎖可変ドメイン;
i)配列番号18のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号17によって定義されるL鎖可変ドメイン;
j)配列番号20のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号19によって定義されるL鎖可変ドメイン;
k)配列番号22のアミノ酸配列によって定義されるH鎖可変ドメインおよび配列番号21によって定義されるL鎖可変ドメイン。
【0029】
本発明による抗体は、H鎖可変部が配列番号2、4、6、8、10、12、14、16、18、20、および22からなる群より独立して選択されるアミノ酸配列を含むことを好ましくは特徴とする。
【0030】
本発明による抗体は、L鎖可変部が配列番号1、3、5、7、9、11、13、15、17、19、および21からなる群より独立して選択されるアミノ酸配列を含むことを好ましくは特徴とする。
【0031】
本発明は、P−セレクチンと結合し、補体因子C1qおよび/またはFc受容体と結合しない抗体を述べる。これらの抗体は、補体依存性細胞傷害作用(CDC)および/または抗体依存性細胞性細胞傷害作用(ADCC)を誘発しない。好ましくは、この抗体は、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しないことを特徴とする。さらに好ましくは、この抗体はヒト抗体またはヒト化抗体である。
【0032】
本発明による抗体は、定常鎖がヒト起源であることを好ましくは特徴とする。そのような定常鎖は当技術分野の現状において周知であり、例えばKabatに記載されている(例えばJohnson, G., and Wu, T.T., Nucleic Acids Res. 28 (2000) 214-218を参照されたい)。例えば有用なヒトH鎖定常部は、配列番号24、25、26、27、および28からなる群より独立して選択されるアミノ酸配列を含む。例えば有用なヒトL鎖定常部は、配列番号23のカッパL鎖定常部のアミノ酸配列を含む。
【0033】
抗体Fc部のFc部分によって仲介されるエフェクター機能は、抗原に対する抗体の結合後に作動するエフェクター機能を指す(これらの機能は、Fc受容体(FcR)による補体カスケードの活性化および/または細胞活性化を含む)。
【0034】
補体カスケードの機能をCH50アッセイによって評価できる。抗赤血球抗体(EA)で感作されたヒツジ赤血球を被験血清に加えて古典的経路を活性化する結果として、溶血が生じる。50%の赤血球を溶解させるのに必要な血清の容積がCH50ユニットを決定する。AP−CH50は代替経路および最終経路の尺度となる。その手順は、ウサギ赤血球を使用する以外は同様である。代替経路は被験血清の添加に応じて活性化する。
【0035】
C1qと、二つのセリンプロテアーゼであるC1rおよびC1sとは、補体依存性細胞傷害(CDC)経路の第一成分である複合体C1を形成する。補体カスケードを活性化するために、抗原性標的に付着したC1qは少なくとも2分子のIgG1または1分子のIgMと結合する(Ward and Ghetie, Therapeutic Immunology 2:77-94 (1995))。Burton(Molec. Immunol., 22(3): 161-206 (1985))は、アミノ酸残基318から337を含むH鎖部が補体の固定に関与していると記載した。DuncanおよびWinter(Nature 332:738-40 (1988))は部位特異的突然変異誘発を使用して、Glu318、Lys320、およびLys322がC1qに対する結合部位を形成すると報告した。Glu318、Lys320、およびLys322残基がC1qの結合に果たす役割は、これらの残基を含む短鎖合成ペプチドが補体介在性溶解を阻害できることによって確認された。
【0036】
用語「補体依存性細胞傷害作用(CDC)」は、本発明による抗体による、補体の存在下でのP−セレクチンを発現しているヒト内皮細胞および血小板の溶解を指す。CDCは、P−セレクチンを発現しているヒト内皮細胞および血小板を補体の存在下で本発明による抗体で処理することによって好ましくは測定される。細胞をカルセインで好ましくは標識する。抗体が30μg/mlの濃度で標的細胞の20%以上に溶解を誘導するならば、CDCが認められる。しかし、本発明者らは、本発明による抗体の性質にはELISAアッセイで補体因子C1qに対する減少した結合が不可欠であると見出した。そのようなアッセイでは、原則としてELISAプレートに様々な濃度範囲の抗体を被覆し、そのプレートに精製ヒトC1qまたはヒト血清を加える。C1qの結合をC1qに対する抗体に続いてペルオキシダーゼ標識コンジュゲートによって検出する。結合(最大結合Bmax)の検出をペルオキシダーゼの基質であるABTS(2,2’−アジノ−ジ−[3−エチルベンズチアゾリン−6−スルホナート(6)])に関する405nmでの吸光度(OD405)として測定する。したがって、本発明は抗体を説明し、その抗体は、補体因子C1qに対するその抗体の非結合がそのようなELISAアッセイ測定を参照することを特徴とし、ここで、抗体濃度10μg/mlでその抗体に対するC1qの最大結合(Bmax)が、細胞系hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)の抗体LC1004−002のBmaxの30%以下、好ましくは20%以下である。
【0037】
本発明による抗体がELISAアッセイにおいて補体因子C3に対して減少した結合を示すこともさらに好ましい。C1qアッセイと同様にしてアッセイを行う。そのようなアッセイでは原則としてELISAプレートを様々な濃度範囲の抗体で被覆し、そのプレートに精製ヒトC3またはヒト血清を加える。C3に対する抗体に続いてペルオキシダーゼ標識コンジュゲートによってC3の結合を検出する。結合(最大結合Bmax)の検出をペルオキシダーゼの基質であるABTS(2,2’−アジノ−ジ−[3−エチルベンズチアゾリンスルホナート(6)])に関する405nmでの吸光度(OD405)として測定する。したがって、本発明は抗体を説明し、その抗体は、補体因子C3に対する抗体の非結合がそのようなELISAアッセイ測定を参照することを特徴とし、ここで、抗体濃度10μg/mlでその抗体に対するC3の最大結合(Bmax)が、細胞系hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)の抗体LC1004−002のBmaxの10%、好ましくは5%以下である。
【0038】
用語「抗体依存性細胞性細胞傷害作用(ADCC)」は、Fc受容体の結合によって仲介される機能であり、本発明による抗体による、エフェクター細胞の存在下でのP−セレクチンを発現している標的細胞の溶解を指す。ADCCは、新鮮単離されたPBMC(末梢血単核細胞)のようなエフェクター細胞、または単球もしくはNK(ナチュラルキラー)細胞のようなバフィコート由来精製エフェクター細胞の存在下で、P−セレクチンを発現している内皮細胞調製物を本発明による抗体で処理することによって好ましくは測定される。標的細胞を51Crで標識してから、抗体と共にインキュベートする。標識された細胞をエフェクター細胞と共にインキュベートして、放出された51Crに関して上清を分析する。対照はエフェクター細胞存在下であるが抗体不在下で標的内皮細胞をインキュベートすることを含む。(FcγRIIおよびFcγRIIIを発現している)顆粒球、(FcγRIIIを発現している)NK細胞、および(FcγRIおよびFcγRIIを発現している)単球のようなFcγ受容体発現細胞に対するこれらの抗体の結合を測定することによって、これらの抗体がADCCを仲介する初段階を誘導する能力が決定された。
【0039】
抗体のFc部とFc受容体(FcR)との相互作用によって、Fc受容体と結合するエフェクター機能を仲介できる。FcRは造血細胞上の専門化した細胞表面受容体である。Fc受容体は免疫グロブリンスーパーファミリーに属し、抗体で被覆された病原体を免疫複合体の食作用によって除去すること、ならびに対応する抗体で被覆された赤血球および他の様々な細胞標的(例えば腫瘍細胞)の抗体依存性細胞介在性細胞傷害作用(ADCC)を経由した溶解の両方を仲介することが示された。Van de Winkel and Anderson, J. Leuk. Biol. 49:511-24 (1991)。FcRは、免疫グロブリンアイソタイプに対する特異性によって定義され、IgG抗体に対するFc受容体はFcγR、IgEに対してはFcεR、IgAに対してはFcαRなどと呼ばれる。Fc受容体の結合は、例えばRavetch and Kinet, Ann. Rev. Immunol. 9 (1991) 457-492; Capel et al., Immunomethods 4 (1994) 32-34; de Haas et al., J. Lab. Clin. Med. 126 (1995) 330-341およびGessner et al., Ann. Hematol. 76 1998) 231-248に記載されている。本発明による抗体は、Fcγ受容体、好ましくはFγcRI、FγcRIIA、FγcRIIB、および/またはFγcRIIIAに対して減少した結合を示す。
【0040】
本発明の抗体による抗体は、好ましくはエフェクター機能を全く誘発せず、NK細胞上に提示されたFcγRに結合しない。よって、用語「FcγRと結合しない」は、抗体濃度10μg/mlでのNK細胞に対する本発明による抗体の結合が、細胞系hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)の抗体LC1004−002に対して見いだされた結合の1%以下であることを意味する。
【0041】
IgG4はFcRとの減少した結合を示すが、他のIgGサブクラスの抗体は強い結合を示す。しかし、Pro238、Asp265、Asp270、Asn297(Fc糖質を欠如)、Pro329と、234、235、236および237と、Ile253、Ser254、Lys288、Thr307、Gln311、Asn434、およびHis435とは、改変されたならばFcRの減少した結合を提供する残基である(Shields et al. J. Biol. Chem. 276 (2001), 6591-6604; Lund et al. FASEB J. 9 (1995), 115-119; Morgan et al. Immunology 86 (1995) 319-324、EP0307434)。好ましくは本発明による抗体は、FcRと、S228、L234、L235および/もしくはD265に突然変異を有し、かつ/またはPVA236もしくはGLPSS331突然変異を含むIgG4サブクラスの、あるいはIgG1またはIgG2サブクラスの結合に関するものである。突然変異S228P(IgG4)、L234A(IgG1)、L235A(IgG1)、L235E(IgG4)、GLPSS331(IgG1)および/またはPVA236(IgG1)が特に好ましい。突然変異の好ましい組み合わせを表1にも示す。追加的な好ましい組み合わせはD265A/N297Aである。
【0042】
本明細書に使用するような用語「P−セレクチンに対する結合」は、BIAcoreアッセイ(Pharmacia Biosensor AB, Uppsala, Sweden)、またはマイクロタイタープレートに精製P−セレクチンもしくはP−セレクチンCHOトランスフェクタントのいずれかを被覆したELISAのいずれかにおけるP−セレクチンに対する抗体の結合を意味する。
【0043】
BIAcoreアッセイでは、抗体を表面に結合させ、P−セレクチンの結合を表面プラズモン共鳴法(SPR)によって測定する。結合の親和性を、用語ka(抗体/抗原複合体からの抗体の会合速度定数)、kd(解離定数)、およびKD(kd/ka)によって定義する。本発明による抗体は、10−8以下、好ましくは約10−11から10−9MのKDを示す(実施例を参照されたい)。したがって、本発明は、上記のような抗体を指し、ここで、その抗体はBIAcoreアッセイで10−8M未満のKD値でP−セレクチンに結合し、好ましくはKDの範囲は10−11から10−9Mである。
【0044】
好ましくは、その抗体はヒトIgG1またはIgG4サブタイプである。
【0045】
さらに好ましくは、その抗体はL234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体、またはL235およびS228(EUインデックスにより付番)に少なくとも一つの突然変異を含むヒトサブクラスIgG4の抗体であることを特徴とする。
【0046】
P−セレクチン特異的ELISAでは、精製P−セレクチンをマイクロタイタープレートに被覆し、ビオチン化抗ヒトIgGおよびELISAの通常の段階を用いて、P−セレクチンに対する抗体の結合を検出する。このアッセイにおけるEC50値は、P−セレクチンCHO細胞では0.002から0.03μg/mlの間の範囲に好ましくはあり、すなわち、本発明はELISAアッセイでP−セレクチンの結合に関するEC50値がP−セレクチンを提示しているCHO細胞に対して0.002から0.03μg/mlの間の範囲にある抗体を指す。P−セレクチン発現CHOトランスフェクタントをマイクロタイタープレートに被覆するアッセイでは、EC50値は0.01から0.08μg/mlの間、好ましくは0.01から0.04μg/mlの間の範囲にある。
【0047】
E−およびL−セレクチントランスフェクタントに対するEC50値は、好ましくは100μg/mlよりも大きい。本発明の抗体は、P−セレクチンと、E−および/またはL−セレクチンとをマイクロタイタープレートに被覆するELISAアッセイでのEC50値によって測定されるように、E−および/またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍大きく特異的に結合することを特徴とする。
【0048】
本明細書に使用するような用語「P−セレクチンに対するP−セレクチンリガンド結合の阻害」は、HL60細胞上に提示されたP−セレクチンリガンドに対する精製P−セレクチンまたは細胞発現P−セレクチンの結合を指す。リガンドに対するP−セレクチンの結合は本発明による抗体によって阻害される。抗体がリガンドに対するP−セレクチンの結合を阻害する能力を分析するインビトロアッセイでのIC50としてその阻害を測定する。そのようなアッセイについて実施例に記載する。これらのアッセイはP−セレクチンの適切な供給源として親和性精製されたP−セレクチンおよび活性化血小板を、リガンドの適切な供給源としてHL60細胞のような白血球様細胞を使用する。そのようなアッセイでは、P−セレクチンまたは活性化血小板に対するP−セレクチンの生理関連リガンドとしてのPSGL−1を発現しているHL60細胞の接着を、漸増する濃度の抗体の不在下および存在下で測定する。IC50値を少なくとも3回の独立した測定の平均値として測定する。阻害は、1μg/mlよりも大きくない、好ましくは0.5から0.08μg/mlのIC50値を意味する。
【0049】
本発明の抗体は、0.08から0.5μg/mlの範囲、好ましくは0.08から0.11μg/mlのIC50値で精製P−セレクチンに対する白血球様HL60細胞の接着を阻害する。活性化血小板に対する白血球様HL60細胞の接着は、0.05から0.3μg/mlの範囲のIC50値で阻害される。
【0050】
したがって、本発明のさらなる態様は、P−セレクチンの結合に関するEC50値がELISAアッセイでは0.01から0.08μg/mlの範囲であることを特徴とする抗体を述べ、ここで、P−セレクチンを発現するCHOトランスフェクタントをマイクロタイタープレートに被覆する。好ましい範囲は0.01から0.04μg/mlである。E−およびL−セレクチントランスフェクタントに関するEC50値は100μg/mlよりも大きい。さらなる態様では、本発明の抗体は、精製P−セレクチンに対する白血球様HL60細胞の接着を0.08から0.5μg/mlの間のIC50値で阻害する。好ましい範囲は、0.08から0.11μg/mlである。
【0051】
本発明の抗体は、完全にヒトの流動系で好ましくは70%よりも大きく白血球と血小板の単層との相互作用を(10μg/mlの濃度で)阻害する。さらに、これらの抗体は、3μg/mlの濃度で60〜90%の範囲でヒト流動系での活性化内皮細胞に対する白血球の接着を(白血球サブタイプに対する異なる効果を有して)阻害する。
【0052】
本発明の抗体は、P−セレクチンフラグメントaa60〜75(Swiss-Prot配列P16109)の存在下でP−セレクチンと好ましくは結合でき、かつ/またはATCCアクセッション番号HB11041と称する細胞系によって分泌される、抗体のP−セレクチンに対する結合を競合的に阻害しない。
【0053】
本発明の抗体は、ELISAアッセイ形式でP−セレクチンと血小板膜糖タンパク質GPIbαとの相互作用を好ましくは阻害しない。ELISAではGPIbαの可溶性細胞外部分であるグリコカリシンを、記載されたようにマイクロタイタープレートのウェルに固定化し(Romo et al., J Exp Med 190:803 (1999)、ポリクローナル抗P−セレクチン抗体を用いて、P−セレクチンHuMabと共に予備インキュベート後の精製P−セレクチンの結合を検出した。
【0054】
本発明のさらなる好ましい態様では、その抗体はC3タンパク質と結合しないことを特徴とし、さらに好ましくは補体依存性細胞傷害作用(CDC)を誘発しないことを特徴とする。さらに、その抗体はNKエフェクター細胞上のFcγ受容体に結合しないことを特徴とする場合がある。好ましくは、その抗体はL234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体、またはL235およびS228(EUインデックスにより付番)に少なくとも一つの突然変異を含むヒトサブクラスIgG4の抗体であることを特徴とする。さらなる好ましい態様では、その抗体は抗体依存性細胞性細胞傷害作用(ADCC)を誘発しないことを特徴とする。
【0055】
なおさらに好ましい態様では、本発明の抗体は、P−セレクチンと結合し、以下からなる群より独立して選択される可変部を含むことを特徴とする:
a)配列番号1のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号2によって定義されるH鎖可変ドメイン、
b)配列番号3のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号4によって定義されるH鎖可変ドメイン、
c)配列番号5のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号6によって定義されるH鎖可変ドメイン、
d)配列番号7のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号8によって定義されるH鎖可変ドメイン、
e)配列番号9のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号10によって定義されるH鎖可変ドメイン、
f)配列番号11のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号12によって定義されるH鎖可変ドメイン、
g)配列番号13のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号14によって定義されるH鎖可変ドメイン、
h)配列番号15のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号16によって定義されるH鎖可変ドメイン、
i)配列番号17のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号18によって定義されるH鎖可変ドメイン、
j)配列番号19のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号20によって定義されるH鎖可変ドメイン、
k)配列番号21のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号22によって定義されるH鎖可変ドメイン。
【0056】
好ましくは、その抗体はアミノ酸配列である配列番号3によって定義されるL鎖可変ドメインおよび配列番号4によって定義されるH鎖可変ドメインを含む。
【0057】
好ましい抗体は、ヒトIgG4サブクラスであるか、補体因子C1qに対する非結合を引き起こす少なくとも一つのアミノ酸突然変異を含むことを特徴とする。これらの変異抗体は、例えば配列番号25または配列番号26と、配列番号28とからなる群より独立して選択されるアミノ酸配列を含む。
【0058】
「変異」抗P−セレクチン抗体は、本明細書において親抗体配列での一つまたは複数のアミノ酸残基における付加、欠失および/または置換によって「親」抗P−セレクチン抗体のアミノ酸配列とアミノ酸配列が異なる分子を指す。好ましい態様では、その変異体は、親抗体の一つまたは複数の定常部または可変部に、好ましくは定常部に、一つまたは複数のアミノ酸置換を含む。例えば、その変異体は親抗体の一つまたは複数の可変部に少なくとも1個、例えば約1から約10個、好ましくは約2から約5個の置換を含む場合がある。通常は、その変異体は親抗体の定常ドメインおよび/または可変ドメイン配列と少なくとも90%、さらに好ましくは少なくとも95%、最も好ましくは少なくとも99%のアミノ酸配列同一性を有するアミノ酸配列を有するであろう。
【0059】
この配列に関係して同一性または相同性は、配列を整列させて必要ならばギャップを導入して最大率の配列同一性を実現した後で、候補配列において親抗体残基と同一であるアミノ酸残基の率として本明細書に定義される。N末端、C末端、または抗体配列への内部伸長、欠失、もしくは挿入のどれも配列同一性または相同性に影響するとして解釈してはならない。その変異体はヒトP−セレクチンと結合する能力を保有し、好ましくは親抗体よりも優れた性質を有する。例えば、その変異体は、さらに強い結合親和性、重症肢虚血または末梢動脈閉塞性疾患(CLI/PAOD)に関連する疾患を処置する能力の増強を有する場合がある。
【0060】
本明細書において特に関心対象である変異抗体は、Fcγ受容体との結合の排除が原因で、親抗体に比較したときに接着アッセイで阻害活性の少なくとも約4倍の増強を示す抗体である。
【0061】
本明細書における「親」抗体は、変異体の調製に使用されたアミノ酸配列にコードされる抗体である。好ましくは、親抗体はヒトフレームワーク部を有し、もしも存在するならばヒト抗体定常部を有する。例えば、親抗体はヒト化抗体またはヒト抗体でありうる。
【0062】
本発明による抗体は、本発明による抗体の上記性質に影響せず、それを改変もしない「保存的配列修飾」であるヌクレオチドおよびアミノ酸配列修飾を有するような抗体をさらに含む。部位特異的突然変異誘発およびPCR介在性突然変異誘発のような当技術分野で公知の標準的な技法によって修飾を導入できる。保存的アミノ酸置換には、アミノ酸残基が類似の側鎖を有するアミノ酸残基に交換された置換がある。類似した側鎖を有するアミノ酸残基のファミリーは、当技術分野で定義されている。これらのファミリーには、塩基性側鎖(例えばリジン、アルギニン、ヒスチジン)、酸性側鎖(例えばアスパラギン酸、グルタミン酸)、非荷電極性側鎖(例えばグリシン、アスパラギン、グルタミン、セリン、トレオニン、チロシン、システイン、トリプトファン)、非極性側鎖(例えばアラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン)、ベータ分岐側鎖(例えばトレオニン、バリン、イソロイシン)、および芳香族側鎖(例えばチロシン、フェニルアラニン、トリプトファン、ヒスチジン)を有するアミノ酸がある。このように、ヒト抗P−セレクチン抗体における予測される非必須アミノ酸残基を、同じ側鎖ファミリーからの別のアミノ酸残基に好ましくは交換できる。
【0063】
Riechmann, L., et al., Nature 332 (1988) 323-327およびQueen, C., et al., Proc. Natl. Acad. Sci. USA 86 (1989) 10029-10033によって記載されたような分子モデリングに基づいた突然変異誘発によってアミノ酸置換を行うことができる。
【0064】
さらなる好ましい態様では、抗体は配列番号23に定義されるようなκL鎖定常部を含む。
【0065】
本発明による好ましい抗体は、IgG1v1(E233P、L234V、L235A、デルタG236、A327G、A330S、P331Sによって規定されるPVA−236;GLPSS331)、IgG1v2(L234A、L235A)およびIgG4v1(S228P、L235E)として定義される抗体である。
【0066】
さらに好ましい態様では、これらの抗体はFab、F(ab’)2および一本鎖フラグメントからなる群より選択される抗体フラグメントも含む。
【0067】
本発明は、本発明による抗体の製造方法をさらに含み、その方法は、a)本発明による親ヒト抗体のL鎖をコードする第一核酸配列と、Fc部分が補体因子C1qおよび/またはFc受容体と結合しないために修飾されている該親ヒト抗体のH鎖をコードする第二DNA配列とを用いて宿主細胞を形質転換する段階、b)該抗体H鎖およびL鎖が生成するように該第一および第二DNA配列を発現させる段階、ならびにc)該抗体を宿主細胞または宿主細胞培養物から回収する段階とを含む。
【0068】
本発明は、中間体である抗体、すなわち抗P−セレクチン抗体についても述べ、それらの抗体は、ヒト抗体またはヒト化抗体であって、P−セレクチンとE−および/またはL−セレクチンとをマイクロタイタープレートに被覆するELISAアッセイで測定したときに、E−またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍大きく特異的に結合することを特徴とする。好ましくはこれらの抗体はIgG1またはIgG4抗体である。これらの抗体は、配列番号24のγ1H鎖定常部または配列番号27のγ4H鎖定常部によって定義されるようなアミノ酸配列も含む場合がある。特に、これらの抗体は、hu−Mab<P−セレクチン>LC1004−001(DSM ACC2640)、hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)、およびhu−Mab<P−セレクチン>LC1004−017(DSM ACC2642)からなる群より選択される細胞系によって生成される抗体を指す。
【0069】
本発明による抗体は、本発明による抗体の上記性質に影響せず、それを改変もしない「保存的配列修飾」であるヌクレオチドおよびアミノ酸配列修飾を有するような抗体をさらに含む。部位特異的突然変異誘発およびPCR介在性突然変異誘発のような当技術分野で公知の標準的な技法によって修飾を導入できる。保存的アミノ酸置換には、アミノ酸残基が類似の側鎖を有するアミノ酸残基に交換された置換がある。類似した側鎖を有するアミノ酸残基のファミリーは、当技術分野で定義されている。これらのファミリーには、塩基性側鎖(例えばリジン、アルギニン、ヒスチジン)、酸性側鎖(例えばアスパラギン酸、グルタミン酸)、非荷電極性側鎖(例えばグリシン、アスパラギン、グルタミン、セリン、トレオニン、チロシン、システイン、トリプトファン)、非極性側鎖(例えばアラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン)、ベータ−分岐側鎖(例えばトレオニン、バリン、イソロイシン)、および芳香族側鎖(例えばチロシン、フェニルアラニン、トリプトファン、ヒスチジン)を有するアミノ酸がある。このように、ヒト抗P−セレクチン抗体における予測される非必須アミノ酸残基を、同じ側鎖ファミリーからの別のアミノ酸残基に好ましくは交換できる。
【0070】
Riechmann, L.ら、Nature 332 (1988) 323-327およびQueen, C.ら、Proc. Natl. Acad. Sci. USA 86 (1989) 10029-10033によって記載された分子モデリングに基づいた突然変異誘発によってアミノ酸置換を行うことができる。
【0071】
本発明は、上に挙げた抗体をコードする核酸分子、これらの核酸を含む対応するベクター、およびこれらのベクターに関する対応する宿主細胞も含む。本発明は、該抗体分子の合成を可能にする条件で対応する宿主細胞を培養する段階と、該培養物から該抗体を回収する段階とを含む、例えばH鎖をコードする核酸およびL鎖をコードする核酸を原核または真核宿主細胞に発現させる段階と、該細胞から該ポリペプチドを回収する段階とによる、抗体の調製方法を包含する。
【0072】
診断および治療へのこの抗体の使用が考えられている。一つの診断応用では、本発明はP−セレクチンを含むと疑われる試料を抗P−セレクチン抗体に曝露する段階と、その試料に対する抗体の結合を決定する段階とを含む、P−セレクチンタンパク質の存在を決定する方法を提供する。この使用に関して、本発明は抗体と、その抗体を使用するための指示とを含む、P−セレクチンタンパク質を検出するためのキットを提供する。
【0073】
本発明の抗体は、炎症疾患および血栓疾患の処置に有用である。そのような疾患には,アテローム性動脈硬化症、動脈および深部静脈血栓症、血管形成術またはステント留置術後の再狭窄のような血管障害がある。好ましい適用は、末梢動脈閉塞性疾患(PAOD)および重症肢虚血(CLI)である。他の適用は、心筋梗塞、脳虚血イベント(例えば卒中)、腎梗塞などによって起こる虚血後白血球介在性組織損傷の処置である。これらの抗体は、敗血症、急性白血球介在性肺傷害、および喘息のようなアレルギー反応の処置にも適している。他の適用は、臓器移植の拒絶反応および慢性関節リウマチを含む自己免疫疾患の予防である。さらに、循環している癌細胞の接着を阻害することによって腫瘍転移を予防できる。
【0074】
本発明は、上記の炎症障害および血栓障害、特にPAODおよびCLI(末梢動脈閉塞性疾患または重症肢虚血)を患う哺乳動物を処置するための方法をさらに提供する。
【0075】
本発明は、治療、例えばこれらの疾患の処置のための薬を製造するための上記抗体の使用をさらに提供する。
【0076】
本発明は、薬学的組成物を製造するための上に定義したような抗体の使用にも関し、薬学的有効量で本発明による抗体を、場合によっては薬学的目的の抗体製剤に有用な緩衝剤および/または補助剤と共に含む薬学的組成物を含む。
【0077】
本発明は、薬学的に許容される担体中にそのような抗体を含む薬学的組成物をさらに提供する。一態様では、その薬学的組成物は製品またはキットに含まれる場合がある。
【0078】
本発明は、本発明によるそのようなアンタゴニストモノクローナル抗体、例えば親抗体を生成するハイブリドーマ細胞系をさらに提供する。
【0079】
本発明による好ましいハイブリドーマ細胞系であるhu−Mab<P−セレクチン>LC1004−001(抗体HuMab001)、hu−Mab<P−セレクチン>LC1004−002(抗体HuMab002)、およびhu−Mab<P−セレクチン>LC1004−017(抗体HuMab017)は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約に基づき、Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSMZ)、ドイツに寄託された。
【0080】
【表1】
【0081】
該細胞系から得ることができる抗体は、本発明の好ましい態様である。
【0082】
本発明による抗体は、組換え手段によって好ましくは生成される。そのような方法は当技術分野の現状で広く公知であり、原核および真核細胞でのタンパク質の発現と、それに続く抗体ポリペプチドの単離と、通常は薬学的に許容される純度までの精製とを含む。タンパク質の発現のために、L鎖およびH鎖、またはそれらのフラグメントをコードする核酸を標準的な方法によって発現ベクターに挿入する。CHO細胞、NS0細胞、SP2/0細胞、HEK293細胞、COS細胞、酵母、または大腸菌(E.coli)細胞のような適切な原核または真核宿主細胞で発現を行い、それらの細胞(上清または溶解後の細胞)から抗体を回収する。
【0083】
抗体の組換え生成は、当技術分野の現状で周知であり、例えばMakrides, S. C., Protein Expr. Purif. 17 (1999) 183-202; Geisse, S., et al., Protein Expr. Purif. 8 (1996) 271-282; Kaufman, R. J., Mol. Biotechnol. 16 (2000) 151-161; Werner, R. G., Drug Res. 48 (1998) 870-880の総説に記載されている。
【0084】
抗体は、全細胞中、細胞溶解液中、または部分精製された形もしくは実質的に純粋な形で存在しうる。他の細胞構成要素または他の混入物、例えば他の細胞性核酸またはタンパク質を除去するために、アルカリ/SDS処理、カラムクロマトグラフィー、および当技術分野で周知の他の技法を含めた標準的な技法によって精製を行う。Ausubel, F., et al., ed. Current Protocols in Molecular Biology, Greene Publishing and Wiley Interscience, New York (1987)を参照されたい。
【0085】
NS0細胞での発現は、例えばBarnes, L.M., et al., Cytotechnology 32 (2000) 109-123およびBarnes, L.M., et al., Biotech. Bioeng. 73 (2001) 261-270に記載されている。一過性発現は、例えばDurocher, Y., et al., Nucl. Acids. Res. 30 (2002) E9に記載されている。可変ドメインのクローニングは、Orlandi, R., et al., Proc. Natl. Acad. Sci. USA 86 (1989) 3833-3837; Carter, P., et al., Proc. Natl. Acad. Sci. USA 89 (1992) 4285-4289;およびNorderhaug, L., et al., J. Immunol. Methods 204 (1997) 77-87に記載されている。好ましい一過性発現系(HEK293)は、Schlaeger, E.-J.およびChristensen, K.(Cytotechnology 30 (1999) 71-83)ならびにSchlaeger, E.-J.(J. Immunol. Methods 194 (1996) 191-199)によって記載されている。
【0086】
原核生物に適した制御配列には、例えばプロモーターと、場合によってはオペレーター配列と、リボソーム結合部位とがある。真核細胞はプロモーターと、エンハンサーと、ポリアデニル化シグナルとを利用することが公知である。
【0087】
別の核酸配列と機能的関係に置かれた場合に、核酸は「作動可能に連結」している。例えば、プレ配列もしくは分泌リーダーがポリペプチドの分泌に関与するプレタンパク質として発現するならば、そのプレ配列もしくは分泌リーダーに関するDNAはそのポリペプチドに関するDNAと作動可能に連結しており;プロモーターもしくはエンハンサーが配列の転写に影響するならば、そのプロモーターもしくはエンハンサーはコード配列に作動可能に連結し;またはリボソーム結合部位が翻訳を可能にするように位置するならば、そのリボソーム結合部位はコード配列に作動可能に連結する。一般的に「作動可能に連結」は、連結しているDNA配列が隣接し、分泌リーダーの場合は隣接してリーディングフレームに存在することを意味する。しかし、エンハンサーは隣接している必要はない。連結は、好都合な制限部位での連結反応によって実現される。そのような部位が存在しないならば、常法により合成オリゴヌクレオチドアダプターまたはリンカーを使用する。
【0088】
モノクローナル抗体は、例えばプロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析、またはアフィニティクロマトグラフィーのような従来の免疫グロブリン精製法によって培地から適切に分離される。モノクローナル抗体をコードするDNAおよびRNAは、従来法を使用して容易に単離および配列決定される。ハイブリドーマ細胞はそのようなDNAおよびRNAの供給源として作用できる。いったん単離されたならば、そのDNAを発現ベクターに挿入でき、それを次にHEK293細胞、CHO細胞、または骨髄腫細胞のような宿主細胞にトランスフェクトして、宿主細胞での組換えモノクローナル抗体の合成を得る。それらの宿主細胞は、さもなければ免疫グロブリンタンパク質を生成しない。
【0089】
抗体DNAに適切なヌクレオチド変化を導入することによって、またはヌクレオチド合成によって、ヒトP−セレクチン抗体のアミノ酸配列変異体(または突然変異体)を調製する。しかし、そのような修飾を例えば上記のように非常に限定した範囲内でのみ行うことができる。例えば、これらの修飾はIgGアイソタイプおよびエピトープの結合のような先に述べた抗体特性を改変しないが、組換え生成の収率もしくはタンパク質の安定性を改善するか、または精製を容易にする場合がある。
【0090】
抗P−セレクチン抗体の固有のコンホメーションの維持に関与しない任意のシステイン残基を一般的にセリンに置換して、その分子の酸化安定性を高めて異所性の架橋を防止することもできる。逆に、システイン結合を抗体に加えて(特にその抗体がFvフラグメントのような抗体フラグメントである場合に)その安定性を高めることができる。
【0091】
その抗体の別の種類のアミノ酸変異体は、その抗体の本来のグリコシル化パターンを改変する。改変によって、その抗体にみられる一つもしくは複数の糖質部分を欠失させること、および/またはその抗体に存在しない一つもしくは複数のグリコシル化部位を付加することを意味する。抗体のグリコシル化は、概してN−結合である。N−結合は、アスパラギン残基の側鎖に対する糖質部分の付着を指す。トリペプチド配列であるアスパラギン−X−セリンおよびアスパラギン−X−トレオニン(Xはプロリン以外の任意のアミノ酸)は、アスパラギン側鎖に糖質部分が酵素的に付着するための認識配列である。このように、ポリペプチドにこれらのトリペプチド配列のいずれかが存在することによって、潜在的グリコシル化部位が生み出される。(N−結合グリコシル化部位に関して)先に述べた一つまたは複数のトリペプチド配列を含むようにアミノ酸配列を改変することによって、抗体へのグリコシル化部位の付加を好都合に実現する。
【0092】
抗P−セレクチン抗体のアミノ酸配列変異体をコードする核酸分子は、当技術分野で公知の様々な方法によって調製される。これらの方法には、(天然アミノ酸配列変異体の場合)天然供給源からの単離、または初期に調製した変異体もしくは非変異体版のヒト化抗P−セレクチン抗体のオリゴヌクレオチド介在性(または部位特異的)突然変異誘発と、PCR突然変異誘発と、カセット突然変異誘発とによる調製があるが、それに限定されるわけではない。
【0093】
本発明は、化学療法剤、毒素(例えば細菌、真菌、植物、もしくは動物起源の酵素的に活性な毒素、またはそれらのフラグメント)、放射性同位体(すなわち放射性コンジュゲート)、または炎症障害および血栓障害、特にPAODおよびCLIによる障害の予防もしくは処置のための薬剤のプロドラッグのような細胞傷害剤に結合した、本発明による抗体を含む免疫コンジュゲートにも関する。抗体と細胞傷害剤とのコンジュゲートは、N−スクシンイミジル−3−(2−ピリジルジチオール)プロピオネート(SPDP)、イミノチオラン(IT)、(ジメチルアジピミダートHCLのような)イミドエステルの二官能性誘導体、(スベリン酸ジスクシンイミジルのような)活性エステル、(グルタルアルデヒドのような)アルデヒド、(ビス(p−アジドベンゾイル)ヘキサンジアミンのような)ビス−アジド化合物、(ビス−(p−ジアゾニウムベンゾイル)−エチレンジアミン(bis-(p-diazoniumbenzoyl)-ethylenediatnine)のような)ビス−ジアゾニウム誘導体、(トリエン(tolyene)2,6−ジイソシアナートのような)ジイソシアナート、および(1,5−ジフルオロ−2,4−ジニトロベンゼンのような)ビス−活性フッ素化合物のような様々な二官能性タンパク質カップリング剤を使用して作製される。例えば、リシン免疫毒素をVitetta, E. S., et al., Science 238 (1987) 1098-1104)に記載されているように調製できる。炭素14標識1−イソチオシアナートベンジル−3−メチルジエチレントリアミン五酢酸(MX−DTPA)は抗体に放射性ヌクレオチドを結合させるための例示的なキレート剤である。WO94/11026を参照されたい。
【0094】
別の種類の共有結合性修飾は、その抗体にグリコシドを化学的または酵素的にカップリングすることを含む。これらの手順は、N−またはO−結合グリコシル化に関するグリコシル化能力を有する宿主細胞で抗体を生成させる必要がない点で有利である。使用したカップリング様式に応じて、(a)アルギニンおよびヒスチジン、(b)遊離カルボキシル基、(c)システインのもののような遊離スルフヒドリル基、(d)セリン、トレオニン、もしくはヒドロキシプロリンのもののような遊離ヒドロキシル基、(e)フェニルアラニン、チロシン、もしくはトリプトファンのもののような芳香族残基、または(f)グルタミンのアミド基に糖(類)を付着させることができる。これらの方法は、WO87/05330、およびAplin, J.D., and Wriston, J.C. Jr., CRC Crit. Rev. Biochem. (1981) 259-306に記載されている。
【0095】
抗体上に存在する任意の糖質部分の除去を化学的または酵素的に実現できる。化学的脱グリコシル化は、化合物であるトリフルオロメタンスルホン酸または同等の化合物にその抗体を曝露することを必要とする。この処理の結果として、結合糖(N−アセチルグルコサミンまたはN−アセチルガラクトサミン)以外の大部分または全ての糖の開裂が生じる一方で、抗体はインタクトのまま残る。化学的脱グリコシル化は、Sojahr, H. T., and Bahl, O. P., Arch. Biochem. Biophys. 259 (1987) 52-57およびEdge, A. S., et al. Anal. Biochem. 118 (1981) 131-137に記載されている。Thotakura, N.R., and Bahl, O. P., Meth. Enzymol. 138 (1987) 350-359に記載されたように様々なエンドグリコシダーゼおよびエキソグリコシダーゼの使用によって、抗体上の糖質部分の酵素的開裂を実現できる。
【0096】
別の種類の抗体の共有結合性修飾は、米国特許第4,640,835号、第4,496,689号、第4,301,144号、第4,670,417号、第4,791,192号、または第4,179,337号に記述されているように、様々な非タンパク質ポリマーの一つ、例えばポリエチレングリコール、ポリプロピレングリコール、またはポリオキシアルキレンに抗体を連結する段階を含む。
【0097】
なお別の局面では、本発明は、ヒト抗P−セレクチン抗体(例えば本発明によるhu−Mab<P−セレクチン>LC1004−001(DSM ACC2640)、hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)、およびhu−Mab<P−セレクチン>LC1004−017(DSM ACC2642)からなる群より選択される細胞系によって生成される親抗体)を発現する非ヒトトランスジェニック動物、例えばトランスジェニックマウスから単離されたB細胞を提供する。KMマウスは、適切な染色体導入マウスである。KMマウスは、ヒトH鎖導入染色体およびヒトカッパL鎖導入遺伝子を含む。KMマウスでは、内因性マウスH鎖およびL鎖遺伝子も破壊されている結果、マウスの免疫処置によってマウス免疫グロブリンよりもヒト免疫グロブリンの生成がもたらされる。KMマウスの構築およびヒト免疫グロブリンを発生させるためのそれらのマウスの使用はWO02/43478に詳細に記載されている。
【0098】
好ましくは、 単離されたB細胞は、精製型、もしくは組換え型のP−セレクチン抗原および/またはP−セレクチンを発現している細胞を用いて免疫処置された非ヒトトランスジェニック動物、例えばトランスジェニックマウスから得られる。好ましくは、非ヒトトランスジェニック動物、例えばトランスジェニックマウスは、本発明の抗体の全てまたは部分をコードする、ヒトH鎖導入遺伝子とヒトL鎖導入遺伝子とを含むゲノムを有する。次に、単離されたB細胞は不死化されてヒト抗P−セレクチン抗体の供給源(例えばハイブリドーマ)を提供する。したがって、本発明は本発明によるヒトモノクローナル抗体を生成できるハイブリドーマも提供する。一態様では、ハイブリドーマは、本発明の抗体の全てまたは部分をコードするヒトH鎖導入遺伝子とヒトL鎖導入遺伝子とを含むゲノムを有する非ヒトトランスジェニック動物、例えばトランスジェニックマウスから得られるB細胞が不死化細胞と融合したものを含む。
【0099】
特定の態様では、非ヒトトランスジェニック動物は、本発明の抗体の全てまたは部分をコードする、ヒトH鎖導入遺伝子とヒトL鎖導入遺伝子とを含むゲノムを有するトランスジェニックマウスである。精製もしくは濃縮されたP−セレクチン抗原調製物および/またはP−セレクチンを発現している細胞を用いて非ヒトトランスジェニック動物を免疫処置できる。好ましくは、非ヒトトランスジェニック動物、例えばトランスジェニックマウスは、P−セレクチンに対するヒトモノクローナル抗体のP−セレクチンアイソタイプを生成できる。
【0100】
精製もしくは濃縮されたP−セレクチン抗原調製物および/またはP−セレクチンを発現している細胞を用いて、本発明の抗体の全てまたは部分をコードする、ヒトH鎖導入遺伝子とヒトL鎖導入遺伝子とを含むゲノムを有する非ヒトトランスジェニック動物、例えばトランスジェニックマウスを免疫処置することによって、本発明によるヒトモノクローナル抗体を生成させることができる。次に、その動物のB細胞(例えば脾臓B細胞)を得て、骨髄腫細胞と融合させて、P−セレクチンに対するヒトモノクローナル抗体を分泌する不死化ハイブリドーマ細胞を形成させる。
【0101】
好ましい態様では、P−セレクチンに対するヒトモノクローナル抗体を、マウス免疫系よりもヒト免疫系の部分を保有するトランスジェニックマウスを使用して発生させることができる。本明細書において「HuMab」マウスと称されるこれらのトランスジェニックマウスは、H鎖(μおよびγ)およびκL鎖(定常部遺伝子)と、内因性μおよびκ鎖遺伝子座を不活性化する標的突然変異とを含む再編成されていないヒト免疫グロブリン遺伝子をコードするヒト免疫グロブリン遺伝子ミニ遺伝子座を含む(Lonberg, N., et al., Nature 368 (1994) 856-859)。したがって、そのマウスはマウスIgMまたはIgKの減少した発現を示し、導入されたヒトH鎖およびL鎖導入遺伝子は、免疫処置に応答してクラススイッチおよび体細胞突然変異を受けて、高親和性のヒトIgGモノクローナル抗体を発生する(Lonberg, N., et al., Nature 368 (1994) 856-859。Lonberg, N., Handbook of Experimental Pharmacology 113 (1994) 49-101; Lonberg, N., and Huszar, D., Intern. Rev. Immunol. 25 (1995) 65-93;およびHarding, F., and Lonberg, N., Ann. N. Acad. Sci 764 (1995) 536-546に総説されている)。HuMabマウスの調製は、Taylor, L., et al., Nucleic Acids Research 20 (1992) 6287-6295; Chen, J., et al., International Immunology 5 (1993) 647-656; Tuaillon, N., et al., Proc. Natl. Acad. Sci USA 90 (1993) 3720-3724; Choi, T.K., et al., Nature Genetics 4 (1993) 117-123; Chen, J., et al., EMBO J. 12 (1993) 821-830; Tuaillon, N., et al., Immunol. 152 (1994) 2912-2920; Lonberg, N., et al., Nature 368 (1994) 856-859; Lonberg, N., Handbook of Experimental Pharmacology 113 (1994) 49-101; Taylor, L., et al., Int. Immunol. 6 (1994) 579-591; Lonberg, N., and Huszar, D., Intern. Rev. Immunol. 25 (1995) 65-93; Harding, F., and Lonberg, N., Ann. N. Acad. Sci 764 (1995) 536-546; Fishwild, D.M., et al., Nat. Biotechnol. 14 (1996) 845-851に記載されており、これら全ての内容はその全体が参照により本明細書に組み入れられる。米国特許第5,545,806号、第5,569,825号、第5,625,126号、第5,633,425号、第5,789,650号、第5,877,397号、第5,661,016号、第5,814,318号、第5,874,299号、第5,545,807号、第5,770,429号、WO98/24884、WO94/25585、WO93/1227、WO92/22645、およびWO92/03918をさらに参照されたい。
【0102】
P−セレクチンに対する完全ヒトモノクローナル抗体を発生させるために、Lonberg, N. et al., Nature 368 (1994) 856-859; Fishwild, D. M. et al., Nat. Biotechnol. 14 (1996) 845-851、およびWO98/24884に記載されたような一般的な方法により、P−セレクチン抗原の精製もしくは濃縮された調製物および/またはP−セレクチンを発現している細胞を用いてHuMabマウスを免疫処置することができる。好ましくは、最初の免疫処置の際にマウスは6〜16週齢であろう。例えば、可溶性P−セレクチン抗原の精製または濃縮された(例えばP−セレクチン発現細胞から精製された)調製物を使用して、HuMabマウスを腹腔内免疫処置することができる。P−セレクチン抗原の精製または濃縮された調製物を使用した免疫処置の結果として抗体が生じない場合には、P−セレクチンを発現している細胞、例えば腫瘍細胞系を用いてマウスを免疫処置して免疫応答を促進することもできる。様々な抗原での累積的な経験は、最初フロイントの完全アジュバントに入れた抗原を用いて腹腔内(i.p.)免疫処置してから、フロイントの不完全アジュバントに入れた抗原を用いて2週間毎に(例えば合計6週間まで)i.p.免疫処置を行う場合に、HuMabトランスジェニックマウスが最もよく応答することを示した。眼窩採血によって得られた血漿試料を用いた、免疫処置プロトコールの過程にわたり免疫応答を監視できる。ELISAによって血漿をスクリーニングでき、十分な力価の抗P−セレクチンヒト免疫グロブリンを有するマウスを対応するB細胞の不死化に使用できる。屠殺して脾臓およびリンパ節を取り出す3から4日前に、マウスの静脈内に抗原を追加免疫できる。各抗原について2〜3回の融合を行う必要がありうると予想される。各抗原について数匹のマウスに免疫処置を行う。例えば、HCo7およびHCo12系統のHuMabマウス合計12匹に免疫処置を行うことができる。
【0103】
HCo7マウスは、(Chen, J. et al., EMBO J. 12 (1993) 821-830に記載されているように)内因性L鎖(カッパ)遺伝子にJKD破壊、(WO01/14424の実施例1に記載されているように)内因性H鎖遺伝子にCMD破壊、(Fishwild, D.M., et al., Nat. Biotechnol. 14 (1996) 845-851に記載されているように)KCo5ヒトカッパL鎖導入遺伝子、および(米国特許第5,770,429号に記載されているように)HCo7ヒトH鎖導入遺伝子を有する。
【0104】
HCo12マウスは、(Chen, J. et al., EMBO J. 12 (1993) 821-830に記載されているように)内因性L鎖(カッパ)遺伝子にJKD破壊、(WO01/14424の実施例1に記載されているように)内因性H鎖遺伝子にCMD破壊、(Fishwild, D.M., et al., Nat. Biotechnol. 14 (1996) 845-851に記載されているように)KCo5ヒトカッパL鎖導入遺伝子、および(WO01/14424の実施例2に記載されているように)HCo12ヒトH鎖導入遺伝子を有する。マウスリンパ球を単離して、標準的なプロトコールに基づいてPEGを使用してマウス骨髄腫細胞系と融合させてハイブリドーマを発生させることができる。結果として生じたハイブリドーマから、次に抗原特異的抗体の生成をスクリーニングする。例えば、免疫処置したマウスからの脾臓およびリンパ節由来リンパ球の単細胞懸濁物を、50%PEGを用いて6分の1の数のSP2/0非分泌型マウス骨髄腫細胞(ATCC、CRL1581)と融合させる。平底マイクロタイタープレートで約2×105個の細胞を平板培養してから、選択培地で約2週間インキュベートする。
【0105】
次に、個別のウェルからELISAによってヒト抗P−セレクチンモノクローナルIgMおよびIgG抗体をスクリーニングする。いったんハイブリドーマの大規模な成長が起こったならば、通常は10〜14日後に培地を分析する。抗体を分泌しているハイブリドーマをもう一度平板培養し、再度スクリーニングして、ヒトIgG抗P−セレクチンモノクローナル抗体がまだ陽性であれば、そのハイブリドーマを限界希釈によって少なくとも2回サブクローニングできる。次に、安定なサブクローンをインビトロで培養してキャラクタリゼーション用の組織培養培地で抗体を生成させる。
【0106】
CDR配列が抗体−抗原相互作用を担うことから、種々のヒト抗体からのフレームワーク配列上に本発明によるCDR配列を含む発現ベクターを構築することによって、本発明による組換え抗体を発現させることが可能である(例えばRiechmann, L., et al., Nature 332 (1998) 323-327; Jones, P., et al., Nature 321 (1986) 522-525;およびQueen, C, et al., Proc. Natl. Acad. See. U.S.A. 86 (1989)10029-10033を参照されたい)。そのようなフレームワーク配列を、生殖細胞系ヒト抗体遺伝子配列を含む公共のDNAデータベースから得ることができる。これらの生殖細胞系配列は、B細胞が成熟する間にV(D)J組換えによって形成する完全に集合した可変部遺伝子を含まないと思われることから、成熟抗体遺伝子配列とは異なるであろう。生殖細胞系遺伝子配列は、個別の高親和性二次レパートリー抗体の配列とも可変部に均等にわたって異なるであろう。
【0107】
本発明は、好ましくは試料のP−セレクチンと本発明による抗体との間の結合を決定する免疫アッセイによる、P−セレクチンのインビトロ診断のための本発明による抗体の使用をさらに含む。
【0108】
別の局面では、本発明は、本発明のヒトモノクローナル抗体の一つもしくは組み合わせ、またはその抗原結合部分が薬学的に許容される担体と共に製剤されたものを含む組成物、例えば薬学的組成物を提供する。さらに具体的には、その組成物は薬学的または診断用組成物であり、なおさらに具体的には、その薬学的組成物は上に定義されたような抗体と、少なくとも一つの薬学的に許容される賦形剤とを含む。
【0109】
本発明の薬学的組成物を、併用療法で、すなわち他の薬剤と組み合わせて投与することもできる。例えば、併用療法は、本発明の組成物を、重症肢虚血(CLI/PAOD)に関連した疾患の予防もしくは処置、または他の従来の治療法に有用な少なくとも一つの薬剤と共に含む場合がある。
【0110】
本明細書に使用するような「薬学的に許容される担体」には、生理学的に適合性の任意および全ての溶媒、分散媒、コーティング剤、抗細菌および抗真菌剤、等張化剤および吸収遅延剤などがある。好ましくは、担体は(例えば注射または注入による)静脈内、筋肉内、皮下、非経口、脊髄または上皮投与に適する。
【0111】
「薬学的に許容される塩」は、抗体の所望の生物学的活性を保持し、任意の望まれない毒性作用を与えない塩を指す(例えばBerge, S.M., et al., J. Pharm. Sci. 66 (1977) 1-19を参照されたい)。そのような塩は本発明に含まれる。そのような塩の例には、酸付加塩および塩基付加塩がある。酸付加塩には、塩酸塩のような無毒性無機酸由来の塩がある。
【0112】
本発明の組成物を、当技術分野で公知の様々な方法によって投与できる。当業者に認識されているであろうが、投与経路および/または投与様式は、所望の結果に応じて変動するであろう。
【0113】
ある投与経路で本発明の化合物を投与するために、その化合物をある物質で被覆するか、またはある物質と同時投与してその化合物の不活性化を防止することが必要な場合がある。例えば、適切な担体、例えばリポソーム、または希釈剤に入れてその化合物を被験体に投与できる。薬学的に許容される希釈剤には、生理食塩水および水性緩衝溶液がある。
【0114】
薬学的に許容される賦形剤または担体には、注射可能な滅菌溶液または分散物を即時調製するための滅菌水溶液または分散物と滅菌粉末とがある。薬学的に活性な物質のためのそのような媒質および薬剤の使用は、当技術分野で公知である。
【0115】
本明細書に使用する句「非経口投与」および「非経口的に投与する」は、経腸および局所投与以外の、通常は注射による投与様式を意味し、非限定的に静脈内、筋肉内、動脈内、髄腔内、関節包内、眼窩内、心臓内、皮内、腹腔内、経気管、皮下、上皮下、関節内、嚢下、クモ膜下、髄腔内、硬膜外、および胸骨内への注射および注入を含む。
【0116】
これらの組成物は、保存料、湿潤剤、乳化剤、および分散剤のような賦形剤または佐剤も含む場合もある。上記滅菌法と、例えばパラベン、クロロブタノール、フェノール、ソルビン酸などの様々な抗細菌剤および抗真菌剤の包含との両方によって、微生物の存在防止を確実にすることができる。組成物に糖類、塩化ナトリウムなどのような等張化剤を含ませることも理想的でありうる。さらに、モノステアリン酸アルミニウムおよびゼラチンのような、吸収を遅延させる薬剤の包含によって、注射可能な薬学的形態の持続性吸収をなし遂げることができる。
【0117】
選択した投与経路とは無関係に、適切な水和形態で使用できる本発明の化合物および/または本発明の薬学的組成物を、当業者に公知の通常の方法によって薬学的に許容される投薬剤形に製剤する。
【0118】
本発明の薬学的組成物中の有効成分の実際の投薬レベルは、特定の患者に有毒とならずにその患者、組成物、および投与様式に関して所望の治療応答を実現するために有効である有効成分の量を得るように変動しうる。選択された投薬レベルは、採用された本発明の特定の組成物、またはそのエステル、塩、もしくはアミドの活性と、投与経路と、投与時間と、採用される特定の化合物の排泄速度と、処置の持続時間と、採用された特定の組成物と併用される他の薬物、化合物、および/または物質と、処置される患者の年齢、性別、体重、状態、全身の健康状態、および以前の病歴と、医学の分野で周知の同様の因子を含めた様々な薬物動態因子とに依存するであろう。典型的な1週用量は、上に挙げた因子に応じて約0.1mg/kgから約20mg/kg以上まで変動しうるであろう。
【0119】
その組成物は無菌であって、シリンジによって送達できる程度に流動性でなければならない。水以外に、担体は等張緩衝生理食塩水、エタノール、ポリオール(例えばグリセロール、プロピレングリコール、および液体ポリエチレングリコールなど)、およびそれらの適切な混合物でありうる。
【0120】
例えばレシチンのようなコーティング剤の使用によって、分散物の場合は必要な粒子径を維持することによって、および界面活性剤の使用によって、妥当な流動性を維持できる。多くの場合で等張化剤、例えば糖類、マンニトールまたはソルビトールのようなポリアルコール、および塩化ナトリウムをその組成物に含ませることが好ましい。吸収を遅延させる薬剤、例えばモノステアリン酸アルミニウムまたはゼラチンをその組成物に含ませることによって、注射可能な組成物の長時間吸収をもたらすことができる。
【0121】
本発明は治療を必要とする患者を処置するための方法を含み、その方法は、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない、治療有効量の抗体をその患者に投与することを特徴とする。
【0122】
本発明は、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない抗体を治療に使用することを含む。
【0123】
本発明は、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない抗体を、炎症障害および血栓障害の予防および処置のための薬の調製に使用することを含む。
【0124】
本発明は、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない抗体を、PAODおよびCLIの処置のために使用することを含む。
【0125】
このように、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。一態様では、その抗体はヒト抗体である。別の態様では、その抗体はヒト化抗体である。
【0126】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、補体因子C1qにその抗体が結合しないことは、その抗体の濃度10μg/mlでその抗体に対するC1qの最大結合(Bmax)が、細胞系hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)の抗体LC1004−002のBmaxの30%以下であるELISAアッセイの測定値を指す。別の態様では、最大結合は細胞系hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)の抗体LC1004−002のBmaxの20%以下である。
【0127】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、その抗体はBIAcoreアッセイで10−8M未満のKD値でP−セレクチンに結合する。別の態様では、KDの範囲は10−11から10−9Mである。
【0128】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、その抗体は、P−セレクチンとE−および/またはL−セレクチンとがマクロタイタープレートに被覆されたELISAアッセイにおいてEC50値によって測定されたように、E−および/またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍大きく特異的に結合する。別の態様では、E−およびL−セレクチントランスフェクタントに対するEC50値は100μg/mlよりも大きい。
【0129】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、その抗体は1μg/mlよりも大きくないIC50値で精製P−セレクチンに対する白血球様HL60細胞の接着を阻害する。別の態様では、IC50値は0.08から0.5μg/mlの範囲である。なお別の態様では、IC50値は0.08から0.11μg/mlの範囲である。
【0130】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、
(a)活性化血小板に対する白血球様HL60細胞の接着は0.05から0.3μg/mlのIC50値で阻害されるか、
(b)その抗体は白血球と血小板単層との相互作用を70%よりも大きく阻害するか、
(c)その抗体は3μg/mlの濃度でヒト流動系中の活性化内皮細胞に対する白血球の接着を60から90%の範囲で阻害するか、
(d)その抗体はC3タンパク質と結合しないか、
(e)その抗体は補体依存性細胞傷害作用(CDC)を誘発しないか、
(f)その抗体はNKエフェクター細胞上のFcγ受容体に結合しないか、または
(g)その抗体は抗体依存性細胞性細胞傷害作用(ADCC)を誘発しない。
【0131】
一態様では、本発明はP−セレクチンに結合する抗体を提供し、該抗体は、可変部H鎖アミノ酸配列CDR3がH鎖CDR3配列である配列番号38、39、40、41、または42からなる群より選択されることを特徴とする。
【0132】
一態様では、本発明は、P−セレクチンに結合し、可変部H鎖および可変部L鎖を含む抗体を提供し、その抗体は、可変部H鎖がCDR配列であるCDR1、CDR2、およびCDR3を含み、CDR1が配列番号29、30、31、32からなる群より選択され、CDR2が配列番号33、34、35、36、37からなる群より独立して選択され、CDR3が配列番号38、39、40、41、42からなる群より選択されることを特徴とし、ここで、該CDRは相互に独立して選択される。
【0133】
一態様では、本発明は、可変部L鎖がCDR配列であるCDR1、CDR2、およびCDR3を含み、CDR1が配列番号43、44より選択され、CDR2が配列番号45、46より選択され、CDR3が配列番号47、48、49、50、51、52より選択されることを特徴とする抗体を提供し、ここで、該CDRは相互に独立して選択される。
【0134】
一態様では、本発明は、P−セレクチンと結合すること、および以下からなる群より独立して選択される可変部を含むことを特徴とする抗体を提供する:
a)配列番号1のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号2によって定義されるH鎖可変ドメイン、
b)配列番号3のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号4によって定義されるH鎖可変ドメイン、
c)配列番号5のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号6によって定義されるH鎖可変ドメイン、
d)配列番号7のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号8によって定義されるH鎖可変ドメイン、
e)配列番号9のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号10によって定義されるH鎖可変ドメイン、
f)配列番号11のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号12によって定義されるH鎖可変ドメイン、
g)配列番号13のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号14によって定義されるH鎖可変ドメイン、
h)配列番号15のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号16によって定義されるH鎖可変ドメイン、
i)配列番号17のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号18によって定義されるH鎖可変ドメイン、
j)配列番号19のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号20によって定義されるH鎖可変ドメイン、
k)配列番号21のアミノ酸配列によって定義されるL鎖可変ドメインおよび配列番号22によって定義されるH鎖可変ドメイン。
【0135】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、その抗体は、アミノ酸配列である配列番号3によって定義されるL鎖可変ドメインのCDR1、CDR2、およびCDR3部と、配列番号4によって定義されるH鎖可変ドメインのCDR1、CDR2、およびCDR3部とを含む。別の態様では、その抗体は、アミノ酸配列である配列番号3によって定義されるL鎖可変ドメインと、配列番号4によって定義されるH鎖可変ドメインとを含む。
【0136】
一態様では、本発明は、P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体を提供し、該抗体は、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに交換され、L235がEに交換されたヒトサブクラスIgG4の抗体であることを特徴とする。ここで、
(a)その抗体は補体因子C1qに結合しないようにさせる少なくとも一つのアミノ酸突然変異をFc部分に含むか、
(b)そのヒトH鎖定常部は配列番号25、配列番号26、および28からなる群より独立して選択されるアミノ酸配列を含むか、
(c)その抗体は配列番号23によって定義されるようなκL鎖定常部を含むか、
(d)その抗体は補体C1qに結合しないようにさせる少なくとも一つのアミノ酸突然変異を含むか、
(e)その抗体はIgG1v1、IgG1v2、およびIgG4v1からなる群より選択されるH鎖定常部を含むか、または
(f)その抗体はFab、F(ab’)2、もしくは一本鎖フラグメントである。
【0137】
一態様では、本発明は、a)ヒト抗体またはヒト化抗体であり、b)P−セレクチンとE−および/またはL−セレクチンとをマイクロタイタープレートに被覆するELISAアッセイにおいてEC50値によって測定されるようにE−またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍大きく特異的に結合することを特徴とする抗P−セレクチン抗体を提供する。別の態様では、その抗体は、配列番号24、25、もしくは26のγ1H鎖定常部、または配列番号27もしくは28のγ4H鎖定常部によって定義されるようなアミノ酸配列を含む。なお別の態様では、その抗体は、hu−Mab<P−セレクチン>LC1004−001(DSM ACC2640)、hu−Mab<P−セレクチン>LC1004−002(DSM ACC2641)、およびhu−+Mab<P−セレクチン>LC1004−017(DSM ACC2642)からなる群より選択される細胞系によって生成される。
【0138】
本発明は、段落[00129]から[00141]に記載されたような定義を有する態様およびそれらの組み合わせも提供するものとする。
【0139】
本発明の理解を助けるために以下の実施例、参照、配列リスト、および図面を提供し、その真の範囲を添付の特許請求の範囲に示す。本発明の精神から逸脱せずに、示された手順に変更を加えることができると了解されている。
【0140】
配列リストの説明
配列番号1 HuMab1004−001のLC1004−001L鎖可変ドメイン
配列番号2 HuMab1004−001のLC1004−001H鎖可変ドメイン
配列番号3 HuMab002のLC1004−002L鎖可変ドメイン
配列番号4 HuMab002のLC1004−002H鎖可変ドメイン
配列番号5 HuMab003のLC1004−003L鎖可変ドメイン
配列番号6 HuMab003のLC1004−003H鎖可変ドメイン
配列番号7 HuMab004(I)のLC1004−004L鎖(I)可変ドメイン
配列番号8 HuMab004(I)のLC1004−004H鎖(I)可変ドメイン
配列番号9 HuMab004(II)のLC1004−004L鎖(II)可変ドメイン
配列番号10 HuMab004(II)のLC1004−004H鎖(II)可変ドメイン
配列番号11 HuMab005のL鎖可変ドメイン
配列番号12 HuMab005のH鎖可変ドメイン
配列番号13 HuMab010(I)のL鎖可変ドメイン
配列番号14 HuMab010(I)のH鎖可変ドメイン
配列番号15 HuMab010(II)のL鎖可変ドメイン
配列番号16 HuMab010(II)のH鎖可変ドメイン
配列番号17 HuMab010(III)のL鎖可変ドメイン
配列番号18 HuMab010(III)のH鎖可変ドメイン
配列番号19 HuMab011のL鎖可変ドメイン
配列番号20 HuMab011のH鎖可変ドメイン
配列番号21 HuMab017のL鎖可変ドメイン
配列番号22 HuMab017のH鎖可変ドメイン
配列番号23 κL鎖定常部
配列番号24 γ1H鎖定常部
配列番号25 γ1H鎖定常部PVA236/GLPSS331(IgG1v1)
配列番号26 γ1H鎖定常部L234A/L235A(IgG1v2)
配列番号27 γ4H鎖定常部
配列番号28 γ4H鎖定常部S228/L235E(IgG4v1)
配列番号29〜32 H鎖CDR1
配列番号33〜37 H鎖CDR2
配列番号38〜42 H鎖CDR3
配列番号43〜44 L鎖CDR1
配列番号45〜46 L鎖CDR2
配列番号47〜52 L鎖CDR3
【0141】
省語:
アミノ酸を3文字(Leu)または1文字の略号(L)のいずれかに省略する
抗体HuMab00Xを抗体00Xとも呼ぶ
L234は、EU番号(Kabat)により位置234のアミノ酸ロイシンを意味する
L234Aは、位置234のアミノ酸ロイシンがアラニンに交換されたことを意味する
PVA236は、236領域においてIgG1のELLGまたはIgG4のEFLGがPVAに修正されたことを意味する
GLPSS331は、331領域におけるIgG1のALPAPまたはIgG2のGLPAPがGLPSSに交換されたことを意味する
他のIgGサブクラスも同様に修正する
【0142】
実施例
抗P−セレクチン抗体を生成するハイブリドーマ細胞系の発生
ハイブリドーマの培養
HuMabハイブリドーマをIMDM(Cambrex)、ウシ胎仔クローン1血清(Perbio Science)、ハイブリドーマクローニング因子原液(Igen)、ピルビン酸ナトリウム、ペニシリン/ストレプトマイシン、2−メルカプトエタノール、HAT(Sigma-Aldrich)、およびカナマイシン(Invitrogen)中で37℃および5%CO2で培養した。
【0143】
抗P−セレクチン抗体を生成するハイブリドーマ細胞系の発生
トランスジェニックマウスの免疫処置法
プロトコールA:
P−セレクチンの膜貫通ドメインおよび細胞質ドメインを欠如した組換え短縮型P−セレクチンをR&D Systemsから購入し、それを用いてHCo7トランスジェニックマウス10匹(雄5匹、雌5匹)GG2201系統(Medarex, San Jose, CA, USA)に免疫処置を行った。初回免疫処置には、PBS100μlに溶かした組換えP−セレクチン50μgをフロイントの完全アジュバント100μlと混合した。残りの免疫処置には、KLHとカップリングした組換えP−セレクチンを使用した。2回目の免疫処置には、KLHとカップリングした組換えP−セレクチン50μgをPBS100μlに溶かし、フロイントの不完全アジュバント100μlと混合した。残りの免疫処置には、KLHとカップリングした組換えP−セレクチン20μgをPBS100μlに溶かし、フロイントの不完全アジュバント100μlと混合した。腹腔内免疫処置から開始して、腹腔内投与と皮下投与とを交互にして免疫処置を行った。
【0144】
プロトコールB:
HCo7トランスジェニックマウス(全て雌)3匹およびKMトランスジェニックマウス(全て雄)3匹GG2489系統(Medarex, San Jose, CA, USA)に、イムノアフィニティークロマトグラフィー(下記参照)によって無効のヒト血小板から精製した完全長P−セレクチンを用いて免疫処置を行った。初回免疫処置には、PBS100μlに溶かした精製P−セレクチン50μgをフロイントの完全アジュバント(CFA; Difco Laboratories, Detroit, USA)100μlと混合した。2回目の免疫処置にはPBS100μlに溶かした精製P−セレクチン50μgをフロイントの不完全アジュバント100μl(ICFA; Difco)と混合した。
【0145】
他の全ての免疫処置については、精製P−セレクチン20μgを使用して、フロイントの不完全アジュバント100μlと混合した。
【0146】
抗原特異的ELISA
免疫処置されたマウスの血清中抗P−セレクチン力価を抗原特異的ELISAによって決定した。平板(96平底ELISAプレート、Greiner)に、PBSに溶かした0.1μg/ml精製P−セレクチンを被覆し、室温で一晩被覆した。その後、ウェルをPBSTC(0.05%Tween20(Sigma-Aldrich Chemie BV)および2%ニワトリ血清(Gibco)を含むPBS)を用いて室温で1時間ブロッキングした。
【0147】
被験血清タップをPBSTCで1:100に希釈してウェルに加えた。免疫処置前にマウスから得た血清をPBSTCで1:100の割合で溶かし、陰性対照として使用した。ヒトP−セレクチンに対するマウス抗体(1/7、Roche Baselが社内で製造)をPBSTCに1:100の割合で溶かし、陽性対照として使用した。プレートを室温で1時間インキュベートした。次に、PBST(0.05%Tween20を含むPBS)を使用してプレートを2回洗浄した。Gt−α−huIgG−HRP(Jackson)をPBSTCに1:5000に希釈して被験タップおよび陰性対照を含むウェルに加えた。Rb−α−mIgG(Jackson)をPBSTCに1:3000に希釈し、陽性対照を含むウェルに加えた。プレートを室温で1時間インキュベートした。最後にPBSTを使用してプレートを2回洗浄し、新鮮調製したABTS(登録商標)溶液(1mg/ml)(ABTS:2,2’−アジノビス(3−エチルベンズチアゾリン−6−スルホン酸)を用いて暗条件で室温(RT)で30分間発色させた。405nmの吸光度を測定した。
【0148】
マウスの追加免疫
血清中の抗P−セレクチン力価が十分な場合、PBS100μlに入れた組換えヒトP−セレクチン20μgを用いて、融合の4日および3日前にマウス静脈内に追加的に追加免疫処置を2回行った。
【0149】
ハイブリドーマの発生
マウスを屠殺し、腹大動脈および大静脈に隣接する脾臓およびリンパ節を集めた。脾臓細胞およびリンパ節細胞と、融合パートナーであるSP2.0細胞との融合を標準的な操作法により行った。
【0150】
配列番号1〜22の可変部H鎖およびL鎖配列を有するヒトモノクローナル抗体を免疫処置法によって得た。
【0151】
κ−ELISA
融合の結果生じたハイブリドーマがヒト抗体を発生するかどうかを決定するために、κ−ELISAを行った。4℃で一晩インキュベートすることによってPBSで1/10000に希釈したラット抗ヒトIgGκL鎖抗体(DAKO)をELISAプレートに被覆した。ウェルの中身を捨てた後で、室温で1時間PBSTCと共にインキュベートすることによってプレートをブロッキングした。その後、PBSTCで1/2に希釈したハイブリドーマ培養上清と共にウェルをインキュベートした。PBSTCで1/2に希釈した培地を陰性対照として使用し、PBSTCで1/100に希釈したκL鎖陽性マウス血清を陽性対照として役立てた。続いて、ウェルを3回洗浄して、PBSTCで1/2000に希釈したHRP結合ラット抗ヒトIgG F(ab’)2(DAKO)と共に37℃で1時間インキュベートした。ウェルを3回洗浄して、新鮮調製したABTS(登録商標)溶液(1mg/ml)を用いて暗条件で室温(RT)で30分間アッセイ液を発色させた。ELISAプレートリーダーを用いて405nmで吸光度を測定した。
【0152】
抗P−セレクチンHuMab可変ドメインのクローニングおよび配列分析(κL鎖およびγ1H鎖)
P−セレクチンHuMabのL鎖可変部VLおよびH鎖可変部VHをコードするヌクレオチド配列を標準的なcDNA合成/PCR法によって単離した。
【0153】
RNeasy(登録商標)ミニキット(Qiagen)を使用して1×106〜1×107個のハイブリドーマ細胞から総RNAを調製した。ハイブリドーマ由来RNAを第一鎖のcDNA合成のためのテンプレートとして使用し、その合成をオリゴdTプライマーを使用した従来法により行った。第二鎖cDNA合成と、VLおよびVHをコードするcDNAフラグメントのさらなるPCR増幅とを、κL鎖およびγ1H鎖定常部のヌクレオチド配列に相補的な逆方向L鎖およびH鎖プライマーと、5’特異的L鎖およびH鎖プライマーとをそれぞれ用いて行った。Invitrogen(商標)life technologiesからのTOPO TAクローニングキットと、クローニングベクターとしてpCR4−TOPOとを使用してPCR生成物をクローニングした。クローニングしたPCR生成物を、消化にEcoRIを使用して適切なプラスミドの制限地図を作成することによって同定し、VLおよびVHに対してそれぞれ約740および790bpのDNAフラグメントサイズであると予測/計算した。
【0154】
クローニングされたPCRフラグメントのDNA配列を二本鎖の配列決定により決定した。
【0155】
GCG(Genetics Computer Group, Madison, Wisconsin)ソフトウェアパッケージバージョン10.2をデータ処理全般に使用した。DNAおよびタンパク質の配列をGCGモジュールCLUSTALWを使用してアライメントを作成した。プログラムGENEDOC(バージョン2.1)を使用して配列アライメントを一覧にして、編集して、色分けした。
【0156】
抗P−セレクチンIgG1 HuMabに関する発現プラスミドの構築
抗P−セレクチンHuMabL鎖をコードしている遺伝子およびH鎖をコードしている遺伝子を哺乳動物細胞発現ベクターに別々に集めた。
【0157】
それによって、抗P−セレクチンHuMabL鎖可変部(VL)をコードする遺伝子セグメントおよびヒトκL鎖定常部(CL)をコードする遺伝子セグメントを、抗P−セレクチンHuMabH鎖可変部(VH)およびヒトγ1H鎖定常部(CH1−ヒンジ−CH2−CH3)に関する遺伝子セグメントと同様に連結した。
【0158】
コドン利用頻度を推定できるヒトL鎖およびH鎖のヌクレオチド配列に関する一般的な情報は、Kabat, E. A., Wu, T. T., Perry, H. M., Gottesman, K. S., and Foeller, C. (1991) Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242に示されている。
【0159】
抗P−セレクチンHuMab κL鎖の転写ユニットは、以下のエレメントから構成される:
・ ヒトサイトメガロウイルス(HCMV)由来前初期エンハンサーおよびプロモーター、
・ Kozak配列を含む合成5’−UT、
・ シグナル配列イントロンを含むマウス免疫グロブリンH鎖シグナル配列、
・ 5’末端に独特のBsmI制限部位と、スプライスドナー部位と、3’末端に独特のNotI制限部位とを有して配列した、クローニングされた抗P−セレクチンHuMab可変部L鎖cDNA、
・ イントロン2マウスIg−κエンハンサーを含むヒトゲノムκ遺伝子定常部(Picard, D., and Schaffner, W. (1984) Nature 307, 80-82)、ならびに
・ ヒト免疫グロブリンκ−ポリアデニル化(「ポリA」)シグナル配列。
【0160】
抗P−セレクチンHuMab γ1H鎖の転写ユニットは、以下のエレメントから構成される:
・ ヒトサイトメガロウイルス(HCMV)由来前初期エンハンサーおよびプロモーター、
・ Kozak配列を含む合成5’−UT、
・ シグナル配列イントロンを含む修飾されたマウス免疫グロブリンH鎖シグナル配列、
・ 5’末端に独特のBsmI制限部位と、スプライスドナー部位と、3’末端に独特のNotI制限部位とを有して配列した、クローニングされた抗P−セレクチンHuMab可変部H鎖cDNA、
・ マウスIgμ−エンハンサーを含むヒトゲノムγ1H鎖遺伝子定常部(Neuberger, M. S. (1983) Embo J2, 1373-1378)、
・ ヒトγ1−免疫グロブリンポリアデニル化(「ポリA」)シグナル配列。
【0161】
抗P−セレクチンHuMab κL鎖発現プラスミドおよびγ1H鎖発現プラスミドの機能的エレメント:抗P−セレクチンHuMab κL鎖またはγ1H鎖発現カセットの他に、これらのプラスミドは、以下を含む:
・ ヒグロマイシン耐性遺伝子
・ エプスタン・バールウイルス(EBV)の複製起点oriP
・ E.coliでのベクターpUC18の複製を可能にする、このプラスミド由来の複製起点、および
・ E.coliにアンピシリン耐性を付与するβ−ラクタマーゼ遺伝子。
【0162】
抗P−セレクチンIgG4 HuMabに関する発現プラスミドの構築
ヒトゲノムγ1定常部およびγ1免疫グロブリンポリアデニル化(「ポリA」)シグナル配列を、ヒトゲノムγ4定常部およびγ4−免疫グロブリンポリアデニル化シグナル配列に交換することによって、抗P−セレクチンγ1H鎖発現プラスミドから抗P−セレクチンγ4H鎖プロトタイプの発現プラスミドを得た。
【0163】
抗P−セレクチンHuMab κL鎖を発現させるために、IgG1について記載したのと同様の発現プラスミドを使用した(上記参照)。
【0164】
突然変異(変異)抗P−セレクチンIgG1およびIgG4に関する発現プラスミドの構築
突然変異抗P−セレクチンγ1およびγ4H鎖をコードする発現プラスミドを、QuickChange(商標)部位特異的突然変異誘発キット(Stratagene)を使用した野生型発現プラスミドの部位特異的突然変異誘発によって生み出した。
【0165】
LC1004−002に対して以下の突然変異体が発生した。EU付番によりアミノ酸に番号を付けた(Edelman, G. M., Cunningham, B. A., Gall, W. E., Gottlieb, P. D., Rutishauser, U., and Waxdal, M. J. (1969) Proc Natl Acad Sci USA 63, 78-85; Kabat, E. A., Wu, T. T., Perry, H. M., Gottesman, K. S., and Foeller, C. (1991) Sequences of Proteins of Immunological Interest, Fifth Ed., NIH Publication No 91-3242)。
【0166】
【表2】
【0167】
組換え抗P−セレクチンHuMabの生成
10%超低濃度IgG FCS(Gibco)、2mMグルタミン(Gibco)、1%v/v 非必須アミノ酸(Gibco)、および250μg/mlG418(Roche)を補充したDMEM(Gibco)中で培養した接着HEK293−EBNA細胞(ATTC#CRL−10852)の一過性トランスフェクションによって組換えHuMabを発生させた。トランスフェクションにFugene(商標)6(Roche)トランスフェクション試薬を、試薬(μl)対DNA(μg)の比が3:1から6:1の範囲で使用した。L鎖をコードするプラスミド対H鎖をコードするプラスミドのモル比を1:2から2:1で用いて、二つの異なるプラスミドから免疫グロブリンL鎖およびH鎖を、発現させた。HuMabを含む細胞培養上清をトランスフェクションの4日後および11日後に採集した。精製まで上清を−20℃で保存した。
【0168】
例えばHEK293でのヒト抗体の組換え発現に関する一般的な情報は、Meissner, P., Pick, H., Kulangara, A., Chatellard, P., Friedrich, K., and Wurm, F. M. (2001) Biotechnol Bioeng 75, 197-203に示されている。
【0169】
抗P−セレクチンHuMabの親和性の決定
装置:
機器:BIACORE(登録商標)2000
チップ:CM5
カップリング法:アミンカップリング
緩衝液:HBS(HEPES、NaCl)、pH7.4、25℃
【0170】
親和性の測定のために、ウサギ抗ヒトFcγ抗体(Dianova)をアミンカップリング法によってチップ表面にカップリングし、P−セレクチンに対する抗体に提示させた。約400RUの抗P−セレクチン抗体が結合した。組換えP−セレクチン(R&D Systems)を0〜50nMの間の様々な濃度で加えた。P−セレクチンを120秒間インジェクトすることによって会合を測定し、チップ表面を緩衝液で180秒間洗浄することによって解離を測定した。種々のP−セレクチン抗体に関する親和性のデータを表2に示す。Biaevaluationソフトウェアを使用して、提示されたモノクローナル抗体に対するP−セレクチンの1:1ラングミュア型結合モデルに動態データをあてはめた。
【0171】
【表3】
【0172】
細胞性接着およびロゼット形成アッセイでのP−セレクチン抗体の阻害活性
材料および方法:
細胞接着アッセイ:接着アッセイではマイクロタイタープレートに被覆したP−セレクチンに対する白血球様HL60細胞(ATCC CCL240)の接着に及ぼすHuMabの効果を評価した。HL60細胞をBCECF−AM(2’,7’−ビス−(2−カルボキシエチル)−5−(および−6−)カルボキシフルオレセインアセトキシメチルエステル、カタログ番号216254、Calbiochem)で標識した。150mM NaCl、1mM CaCl2、1mM MgCl2、20mM Tris(pH7.4)、および0.0005%Tx100を含む緩衝液中の濃度1μg/mlの完全長精製P−セレクチン(精製法については上記参照)を96ウェルプレート(Nunc Immunoplate Maxisorp F96)に4℃で一晩被覆した。その後、3.5%ウシ血清アルブミン(BSA, Fluka)を含む上記緩衝液で室温(RT)で2時間ウェルをブロッキングした。1%BSAを含む上記緩衝液にP−セレクチンHuMabまたは参照マウスP−セレクチン抗体(WAPS12.2、各ハイブリドーマ細胞系はATCCが提供)を様々に希釈した液50μlと共に、これらのウェルをRTで20分間予備インキュベートした。標識HL60細胞(50μl、細胞70000個/ウェル)を加えて、RTで45分間結合させた。一部の実験では、Fc受容体をブロッキングするために20μg/mlのヒトIgG1と共にHL60細胞を30分間予備インキュベートしてから、それらの細胞をウェルに加えた。未結合のHL60細胞を静かに洗浄する(上記緩衝液で4回)ことによって除去してから、接着した細胞をNP−40(Fluka、H2Oに溶かした1%溶液)120μlで溶解させた。上清100μlをプレートに移して、発光分光分析装置LS50B(Perkin Elmer)を使用して励起波長485nmおよび発光538nmで各蛍光を測定した。
【0173】
ロゼット形成アッセイ:活性化血小板とHL60細胞との相互作用に及ぼす抗体の効果を評価するために、ロゼット形成アッセイ(Jungi et al, Blood 67:629 (1986))を二色細胞蛍光分析(Evangelista et al., Blood 88:4183 (1996))と組み合わせて適用した。記載されたように(Fox et al, Methods Enzymol 215:45 (1992))洗浄ヒト血小板を調製した。トロンビン(終濃度1U/ml)を用いて5分間それらの血小板を活性化させ、FITC結合抗ヒトGPIIb抗体pl−36で標識した(Kouns et al., J Biol Chem 267:18844 (1992))。次に、タイロード溶液70μl中の血小板1.4〜2×106個を、種々に希釈したHuMab(100μl)と共に暗条件でRTで30分間インキュベートした。20×106個/mlに調整した(タイロード溶液中の)HL60細胞懸濁液50μlを加えた。PE(フィコエリトリン)結合抗ヒトCD45Ab(コード番号555483、Pharmingen)20μlと共にインキュベートすることによってHL60細胞を標識した。標識したHL60細胞を血小板およびHuMabと共に室温で30分間FACS管(Becton Dickinson)中でインキュベートした後で、FACScan(Becton Dickinson)を使用して血小板とHL60細胞との両方のマーカー蛍光を測定することによって、混合凝集体またはロゼットの形成を分析した。前方および側方散乱、ならびに緑色(FITC)および赤色(PE)シグナルを、励起波長488nmならびにそれぞれ発光波長530nm(FITC)および570nm(PE)での対数増幅によって取得した。電子補正を使用して、スペクトルの重複を除いた。前方散乱および側方散乱に基づいてHL60細胞を同定した。HL60細胞と同定されたイベントに関してゲーティングを行って、単一の血小板を除外した。HL60細胞のゲートに入った5000イベントを各試料について測定した。非活性化またはトロンビン活性化血小板をEDTA(10mmol/l)の存在下でHL60細胞と混合した試料を使用して、緑色蛍光スケールでの閾値を設定した。閾値よりも大きいHL60細胞の率は、血小板と結合しているHL60細胞の率を表す。血小板マーカーの蛍光が低蛍光値に移動することは、多数の接着血小板を有する混合凝集体の数が減少して、少数の接着血小板を有する混合凝集体の数が増加するのに有利になっていることを反映する。
【0174】
結果:
HL60細胞接着アッセイでは、P−セレクチン抗体はHL60細胞が0.08〜0.5μg/mlの範囲のIC50値で精製P−セレクチンに接着するのを阻害した。抗体のFc部分に突然変異が導入されたが、HuMabのIgG4およびIgG1変異体の両方は、図1に示すように0.08〜0.11μg/mlのIC50値を有して、親抗体よりも強力であった。HL60細胞をヒトIgG1と予備インキュベートした場合、図1のHuMab002について実証されたようにIC50値の約3から4倍の低下となって非突然変異親抗体の効力も増加した。この結果は、接着アッセイでの突然変異体の有効性が増加したことの主な原因が、その抗体のFc部分がFcγ受容体に対することを介して、P−セレクチンに対するHL60細胞の接着が除去されることであると示唆している。
【0175】
HL60細胞に対する、P−セレクチンを発現しているヒト活性化血小板の接着を評価するロゼット形成アッセイでは、HuMabのIC50値は、このアッセイでのP−セレクチン受容体の数が少ないことが原因で接着アッセイでの値よりもなお低かった(IC50:0.05〜0.3μg/ml、好ましくは0.05〜0.2μg/ml)。各HuMabのFc変異体の有効性は、非突然変異親抗体に比べて増加する傾向にある(図2)。活性化血小板と共にインキュベートする前にHL60細胞をIgG1およびIgG4と予備インキュベーションしても、突然変異体と親抗体との両方の阻害活性に有意に影響しなかった。これは、接着アッセイに比べてロゼット形成アッセイではFcγ受容体介在性結合の役割があまり顕著でないことを示している。
【0176】
P−セレクチン抗体と動物種由来P−セレクチンとの交差反応性
材料および方法:P−セレクチンHuMabの交差反応性を、(i)FACS分析を用いてラットおよびカニクイザル由来活性化血小板に対するHuMabの結合、ならびに(ii)HL60細胞に対するラットおよびカニクイザル血小板の接着を評価するロゼット形成アッセイでのそれらの抗体の阻害活性を測定することによって評価した。
【0177】
ラットおよびカニクイザルの活性化血小板に対するHuMabの結合を測定するために、ラットおよびカニクイザルの洗浄した血小板を、ヒトの洗浄血小板(上記参照)の調製と同様に調製した。それらの血小板をトロンビンで5分間活性化させた(終濃度1U/ml)。活性化血小板を種々に希釈したHuMab(20μl)と共にRTで30分間インキュベートした。HuMabと結合させた後で、血小板を2%PFAを用いてRTで15分間固定した。試料をタイロード緩衝液で洗浄してタイロード300mlに再懸濁した。HuMabの結合を、ウサギ抗ヒトIgGのFITC結合F(ab’)2フラグメント(コード番号F0056、Dako)を用いて検出した。ラットP−セレクチンを阻害する対照抗体としてウサギ抗ヒトポリクローナル抗P−セレクチン抗体(コード番号09361A、Pharmingen)を使用した。
【0178】
ロゼット形成アッセイでのP−セレクチンHuMabの阻害効果を測定するために、ヒト血小板について上に記載したのと同様に、ラットおよびカニクイザルの洗浄血小板を調製した。本質的にヒト血小板について記載したようにロゼット形成アッセイを行った。カニクイザル血小板を標識するためにFITC結合抗ヒトGPIIb抗体pl−36を使用し、一方でラット血小板をFITC結合マウス抗ラットCD61抗体(コード番号554952、Pharmingen)で標識した。
【0179】
結果:図3aのいくつかの例に示すように、ヒトP−セレクチン介在性機能を阻害する本発明のP−セレクチン特異的抗体のどれも、ラットP−セレクチンに結合せず、ラット血小板およびHL60細胞からなる混合凝集体の形成も阻害しないことが示された。しかし、P−セレクチンHuMabはカニクイザルP−セレクチンに結合し、それを阻害する(図3b)。
【0180】
E−およびL−セレクチンと比べたP−セレクチン抗体の選択性
材料および方法:E−およびL−セレクチンと比べたP−セレクチンHuMabの選択性を、組換えE−およびL−セレクチンに対する抗体(ADP1およびADP2、R&D Systems)の結合を測定する無細胞ELISAと、E−セレクチンCHOトランスフェクタントおよびL−セレクチン300.19トランスフェクタントに対する抗体の結合を測定する細胞性ELISAとで決定した(Goetz et al., J Cell Biol 137:509 (1997); Ley et al., Blood 82:1632 (1993)に記載されているようにトランスフェクタントを発生させた)。
【0181】
無細胞ELISAでは、150mM NaCl、1mM CaCl2、1mM MgCl2、20mMトリス(pH7.4)、および0.0005%Tx100を含む緩衝液中濃度1μg/mlの組換えP−セレクチン、E−セレクチン、またはL−セレクチンを96ウェルプレート(Nunc Immunoplate Maxisorp F96)に4℃で一晩被覆した。その後、ウェルを3.5%ウシ血清アルブミン(BSA, Fluka)を含む上記緩衝液でRTで2時間ブロッキングした。1%BSAを含む上記緩衝液に種々に希釈したP−セレクチンHuMabまたは参照マウスP−、E−セレクチン抗体(BBA26;R&D Systems)およびヤギL−セレクチン抗体(AF728;R&D Systems)50μlと共にウェルをRTで一晩予備インキュベートした。ビオチン化抗ヒトIgG(Amersham、RPN1003、終濃度1:1000)または対照抗体については対応するビオチン化抗マウスもしくは抗ヤギIgGを使用することによって、HuMabの結合を検出した。1時間インキュベートした後で、ウェルを上記の緩衝液で(3回)洗浄し、0.1%BSAを含む上記緩衝液に1:750に希釈したストレプトアビジン−ビオチン化ペルオキシダーゼ複合体(Amersham、RPN1051)0.1mlを30分間加えた。次に、ウェルを洗浄して、ABTS(2,2’−アジノ−ジ−(3−エチルベンズチアゾリンスルホナート、Boehringer, Mannheim)を含むペルオキシダーゼ基質溶液0.2mlを加えた(ABTS原液;40mM ABTS 1ml、30%H2O25μl、および0.1M酢酸Na20ml、0.05 NaH2PO4)。約10分後に0.1Mクエン酸塩および0.01%NaN3の液50μlを使用して反応を停止させた。呈色反応を405nmで読み取った。
【0182】
細胞性ELISAでは、細胞解離溶液(Sigma C5914)で細胞をはがした後に、P−およびE−セレクチンCHO−トランスフェクタントを96ウェルプレート(TC Microwell F96 Nunc 167008)の各ウェルに細胞100000個/ウェルに調整して蒔き、各培地中で37℃で一晩培養した(P−CHO−トランスフェクタント用培地:DMEM+10%FCS+2mMグルタミン+ペニシリン100U/ml/ストレプトマイシン100μg/ml;E−セレクチントランスフェクタント用培地:HAM F12+10%FCS+2mMグルタミン+ペニシリン100U/ml/ストレプトマイシン100μg/ml+0.1%フンギゾン+100μg/mlネオマイシン)。培地を除き、3%TopBlock(コード番号TB232010; Juro)を含むA−T緩衝液(150mM NaCl、1mM CaCl2、1mM MgCl2、20mM トリス(pH7.4))で1時間ウェルをブロッキングした後で、1%TopBlockおよび0.1%アジドを含む上記緩衝液に種々に希釈したP−セレクチンHuMabまたは参照マウスP−およびE−セレクチン抗体(上記参照)50μlを加えて、RTで60分間インキュベートした。ウェルを(4回)洗浄後に、無細胞ELISAについて上に挙げたのと同じ段階を使用して、結合した抗体を検出した。
【0183】
L−セレクチン300.19細胞は懸濁細胞であることから、96ウェルポリスチレンフィルタープレート(Corning 3510)のウェルにL−セレクチン300.19トランスフェクタントを平板培養することによって細胞性ELISAの形式を変更しなければならなかった。フィルタープレートのブロッキングを使用して、インキュベーション溶液をプレートの底を通して濾過することによって除去したが、それ以外ではプロトコールはP−およびE−セレクチンCHO細胞を使用した場合と同様であった。対照としてトランスフェクトしていないCHOおよび300.19を使用した。
【0184】
結果:
本発明の抗体は、E−およびL−セレクチンに比べて高度に選択性であった。これらの抗体は0.01から0.08μg/ml、好ましくは0.01から0.04μg/mlの範囲のEC50値でP−セレクチンCHO細胞に結合したが、E−セレクチンCHO細胞およびL−セレクチン300.19に対するEC50値は50μg/mlよりも明らかに大きく、好ましくは100μg/mlよりも大きかった。HuMab002は細胞性ELISAでE−およびL−セレクチンに比べて4000倍よりも大きい選択係数を伴う最高の選択性を有した。さらに、HuMab002は、濃度100μg/mlまでのベースラインレベルを超えてE−およびL−セレクチントランスフェクタントに結合しない。HuMab002のFc変異体IgG4v1およびIgG1v1の選択性は、親HuMab002の選択性と類似している(図4a〜c)。
【0185】
完全ヒト血流系でのP−セレクチン抗体のex vivo阻害活性
血小板単層への白血球接着に及ぼすP−セレクチンHuMabの効果
材料および方法:
血管壁損傷部および血小板血栓の部位への白血球の動員に及ぼすP−セレクチン抗体の効果について取り組みために、異なる剪断速度でヒト白血球とヒト血小板との相互作用を測定できるヒト血流系を、本質的に記載されたように使用した(Kirchhofer et al., Blood 89:1270 (1997))。平行平板型潅流装置に健康ドナーの前肘静脈から採取したヒト全血で、損傷して露出した血管壁を模倣するコラーゲン表面に潅流した。以前に記載されたようにコラーゲンを被覆したカバースリップを調製した(Kirchhofer et al., Blood 89:1270 (1997))。それらのカバースリップを3個の平行平板型潅流チャンバーに配置した。異なる剪断速度(65/sおよび280/s)の測定を可能にするために、異なる寸法の潅流チャンバーを使用して、一定の血流1ml/minでコラーゲン被覆カバースリップ上に血液を潅流させた。潅流装置と離れて位置する個別のローラーポンプで血流を制御した。静脈から採血して3本の管にその血液を分注した直後に、GPIIb/IIIa阻害剤(0.5μmol/ラミフィバン)を加えて血小板の凝集を防止して血小板単層を発生させた。同時に、P−セレクチン抗体(HuMab、突然変異体、各参照抗体、または対照としてヒトIgG1およびIgG4)を種々の濃度で投与して、次に、コラーゲン被覆カバースリップを含む潅流チャンバーに血液−阻害剤混合物を導入した。5.5分間の潅流時間の後で、流動を中断せずにPBSで潅流チャンバーを3分間潅流した。流動を短時間中断した後で、チャンバーをPBSに溶かした3%パラホルムアルデヒドで1ml/minで2分間固定した。次に、カバースリップをチャンバーからはずし、PBSに溶かした3%パラホルムアルデヒドで4℃で1時間再び固定し、PBS−0.03%ナトリウムアジド中で保存した。血小板単層に接着している白血球数を評価するために、風乾した後でカバースリップをDiff-Quick溶液(Dade Behring AG)で染色してMerckoglas(Merck, Germany)に包埋した。画像解析システム(MCID, Imaging Research Inc.)を使用して、カバースリップの先端から1mm離れた、血流に垂直方向の標準領域に接着している白血球数を決定した。剪断速度65/sおよび280/sで白血球数を計数した面積は、それぞれ3.1mm2および2.1mm2を有した。
【0186】
結果:
P−セレクチンHuMabは血小板単層に対する白血球の接着を濃度依存的に阻害した。剪断速度65/sおよび濃度10μg/mlでHuMabは白血球の接着を60〜99%、好ましくは70〜99%阻害した。HuMabの阻害効果は、静脈の剪断速度65/sよりも高い剪断速度280/s(動脈の状況に近い)の方が顕著であった。全体的に、剪断速度280/sで接着している白血球数は、65/sの場合よりも小さかった。Fc変異体を各親抗体と比べると、HuMab002とその変異体IgG4v1およびIgG1v1とについて実証されたように、それらの抗体はex vivo潅流チャンバーで類似した阻害活性を有した(図5)。インビトロアッセイでみられた親抗体に比べて突然変異体で増加した阻害活性は、ex vivo潅流チャンバーでは観察されなかった。これは、ヒト全血で白血球のFcγ受容体が飽和していることが原因であろう。
【0187】
内皮細胞への白血球の接着に及ぼすP−セレクチンHuMabの効果
材料および方法:
剪断条件でのP−セレクチンHuMabの抗炎症能について取り組むために、内皮細胞をカバースリップ上に被覆した設定で、上に挙げたヒト血流系を使用した。Jaffeら、1993の方法(完全な引用を添えられたい)によりII型コラゲナーゼ(Roche Switzerland)で消化することによって臍帯からヒト臍静脈内皮細胞(HUVEC)を単離した。それらの細胞を20%ウシ胎仔血清(Gibco, Auckland)、100IU/mlペニシリン(Gibco, Auckland)、0.1mg/mlストレプトマイシン(Gibco, Auckland)、2mmol/l L−グルタミン(Gibco, Auckland)、10U/mlヘパリン(Sigma)、および50μg/ml EC成長補助剤(Sigma, Germany)を補充した199培地(M199, Sigma, Germany)中で1%ゼラチンを被覆した組織培養フラスコに入れて培養した。HUVECを集密まで(約4日)成長させ、トリプシン/エチレンジアミン四酢酸(Gibco, Auckland)と共に継代して、1%ゼラチン(Fluka, Germany)で予め被覆したThermanoxプラスチックカバースリップ上に蒔いた(EC約200000個/カバースリップ)。HUVECを沈降させ、HUVECは1〜2日で集密となった。潅流開始の24時間前に20ng/ml IL−4(R&D Systems)で、潅流の5〜10分前に10−4Mヒスタミン(Fluka, Germany)でこれらの細胞を刺激した。各実験を第1回継代のHUVECで行った。刺激されたHUVECの集密単層を有するカバースリップを上記のような平行平板型潅流チャンバーに配置した。上記の潅流実験と同様に、健康ドナーから全血を採取した。しかし、これらの実験ではトロンビン阻害剤Ro−46−6240(10μM)で血液の凝固を阻止して、種々の濃度のP−セレクチン抗体(HuMab、突然変異体、各参照抗体)または対照としてヒトIgG1およびIgG4と共に、活性化内皮細胞上を潅流する直前に5分間予備インキュベートした。血流を1ml/minに、剪断速度を65/sに、かつ潅流時間を5.5分間に調整した。PBSを用いた3分間の洗浄時間の後で、接着している白血球を有するHUVECを上記と同じ流動条件で3%パラホルムアルデヒドで2分間固定した。次に、チャンバーからカバースリップを取り出し、新鮮固定液に1時間浸し、PBS−0.02%アジ化ナトリウム中で保存した。形態分析のために、修飾ビオチン化抗マウス免疫グロブリン(Animal Research Kit, Dako, USA)を使用して予め標識した、白血球共通抗原CD45に対するマウス抗体で白血球を染色した。核をヘマトキシリン(J.T Baker, Holland)で対比染色した。
【0188】
結果:
IL−4およびヒスタミンの組み合わせを用いたHUVECの刺激は、P−セレクチンの発現と、種々の種類の白血球の接着とを招き、(PMNおよび好酸球を含めた)顆粒球が接着している白血球の主要な部分を構成した。本発明のHuMabは、3μg/mlで総白血球数の60〜90%の接着を阻害した。全体的に、Fc変異体の阻害活性は非突然変異HuMabの阻害活性と有意には異ならなかった。
【0189】
P−セレクチンHuMabは、種々の白血球サブタイプに異なる効果を及ぼすことを実証した。顆粒球に及ぼす効果は、単核性白血球に比べて顕著である。本発明による抗体は、(PMNおよび好酸球を含めた)顆粒球の接着を90〜99%、単球を50〜88%、リンパ球を5〜40%阻害した。種々の白血球サブタイプの絶対数の各減少を、図6にIgG4v1について代表的に示す。
【0190】
P−セレクチンHuMabが補体系を活性化する能力
C1qおよびC3c結合ELISA:
本発明の抗体がC1qの結合およびC3の活性化を誘導する能力を決定するために、ELISA法を使用した。C1qは適応免疫系の一部であり、免疫複合体と結合するといくつかの酵素前駆体の連続活性化を誘発する。それらの酵素は順番にC3分子の開裂を引き起こし、これは炎症反応の開始、外来または異常粒子のオプソニン化、および細胞膜の溶解を招く場合がある。
【0191】
原則として、ELISAプレートに種々の濃度範囲の抗体を被覆し、そこにヒトC1qか、またはC3の供給源としてヒトプール血清を加える。ヒトC1qまたはC3εに対する抗体によって、続いてペルオキシダーゼ標識コンジュゲートによってC1qまたはC3εの結合を検出する。
【0192】
HuMab002(ハイブリドーマおよび一過性トランスフェクトーマ由来材料、その突然変異体、ならびに対照抗体)を0.16〜20μg/mlの濃度で試験した。陰性対照として、C1qと非常に弱く結合するヒトIgG4(CLB, the Netherlands,0.5μg/ml原液)を使用した。ヒトIgG1(Sigma, 2ug/ml原液)を陽性対照として組み入れた。C1qを検出するために、C1qに対するウサギ抗体(Dako)と、セイヨウワサビペルオキシダーゼ(Sigma)と結合したブタ抗ウサギIgG抗体とを使用した。C3εを検出するために、マウス抗ヒトC3抗体と、セイヨウワサビペルオキシダーゼ(Sigma)と結合したウサギ抗マウスIgG抗体とを適用した。
【0193】
試験したHuMabのEC50値または10μg/mlでの最大結合(Bmax)に関する計算を、Graphpad Prismソフトウェアを使用した非線形回帰曲線あてはめ(1部位結合)を使用して決定した。
【0194】
結果:
本発明によるHuMab002は、ハイブリドーマおよびトランスフェクトーマ由来材料に関してそれぞれ0.946μg/mlおよび1.159μg/mlのEC50値と、0.987および0.711のBmax(OD405)値とから示されるように、効率的にC1qと結合できた。予想通り、OD405でのBmax値0.222により示されるように、陰性対照であるヒトIgG4はC1qと結合しなかった。しかし、試験した三つのFc変異体全て(IgG4v1、IgG1v1、IgG1v2)は、それぞれ0.132、0.119、および0.132のOD405 Bmax値によって示されるようにC1qと結合する能力を欠如した(表3)。C1qとの結合能と一致して、(ハイブリドーマおよびトランスフェクトーマ由来)HuMab002に対するC3の沈着は抗体濃度依存的に起こり、EC50値は2.7μg/mlから8.3μg/mlの範囲であった。しかし、三つのFc変異体全ては、それぞれ0.104、0.156、および0.133のOD405 Bmax値によって示されるように、C3の沈着を開始できなかった(表3)。
【0195】
HuMab002が補体成分と相互作用することから、この抗体はインビボでCDCを誘導する内因性能力を有する。したがって、この抗体のFc部分は本発明により修飾されている。
【0196】
【表4】
【0197】
P−セレクチンHuMabがFcγ受容体に結合する能力
IgG抗体依存性細胞傷害効果は、エフェクター細胞上のFcγ受容体によって仲介される。ヒト血液由来FcγR発現エフェクター細胞に対する、ハイブリドーマおよびトランスフェクトーマ由来HuMab002と、突然変異抗体と、対照抗体との結合をFACS分析によって検討した。
【0198】
材料および方法:
FcγRI IIA1.6トランスフェクタントまたは新鮮単離したエフェクター細胞を抗体と共にインキュベートして、抗体の結合をFITC標識ウサギ抗ヒトIgG F(ab)2(DAKO)、またはFITC標識ウサギ抗ヒトIgG F(ab)2(BD/Pharmingen)を用いて検出した。HuMab002(一過性トランスフェクトーマおよび/またはハイブリドーマ由来材料、ならびに突然変異体)を濃度1μg/ml(IIA1.6トランスフェクタント)または10μg/ml(エフェクター細胞)で試験した。一次抗体またはヒトIgG4(10μg/ml)の不在を陰性対照として使用した。IIA1.6細胞上のFcγRIの発現を検出するために、FITC標識マウス抗ヒトCD64(BD/Pharmingen)を使用した。NK細胞に富む末梢血単核細胞を使用した実験では、PE標識マウス抗ヒトCD56(BD/Pharmingen)を使用した二重染色によってNK細胞を同定した。FSC/SSCプロフィールに基づいて顆粒球および単球を同定した。
【0199】
IIA1.6細胞、IIA1.6−FcγRIトランスフェクタント、および新鮮単離したエフェクター細胞を抗体と共にインキュベートした。抗体の結合をFITC標識Rb−α−huIgG F(ab)2(DAKO)またはFITC標識Rb−α−huIgG F(ab)2(BD/Pharmingen)を用いて検出した。
【0200】
IIA1.6−FcγRIトランスフェクタント結合アッセイでは、HuMab002(一過性トランスフェクトーマ由来、ハイブリドーマ由来および突然変異体材料)を1μg/mlの濃度で試験した。IIA1.6野生型細胞を陰性対照として使用した。FcγRI発現の対照としてm−α−huCD64−FITC(BD/Pharmingen)を使用した。
【0201】
エフェクター細胞結合アッセイでは、HuMab002(一過性トランスフェクトーマ由来、ハイブリドーマ由来および突然変異体材料)を10μg/mlの濃度で試験した。顆粒球結合アッセイでは一過性トランスフェクトーマ材料を試験しなかった。顆粒球結合アッセイを除く全てのエフェクター細胞結合アッセイでIgG4(10μg/ml)を陰性対照として使用した。
【0202】
NK単離キット(Dynal Biotech ASA, Oslo, Norway)を使用して全血のNK細胞を濃縮した。m−α−huCD56−FITC染色によってNK細胞を同定した。
【0203】
NK単離キット(Dynal Biotech ASA, Oslo, Norway)に同封されているプロトコールに記載されているフィコール法を使用して、全血からPBMC(末梢血単核細胞)を得た。FSC/SSCプロフィールに基づいて単球を同定した。FACS溶解緩衝液を使用して全血から顆粒球を単離して、FSC/SSCプロフィールに基づいて同定した。
【0204】
新鮮単離したエフェクター細胞を抗体と共にインキュベートして、抗体の結合をFITC標識ウサギ抗ヒトIgG F(ab)2(DAKO)またはFITC標識ウサギ抗ヒトIgG F(ab)2(BD/Pharmingen)を用いて検出した。HuMab002(一過性トランスフェクトーマおよび/またはハイブリドーマ由来材料、ならびに突然変異体)を濃度10μg/mlで試験した。一次抗体またはヒトIgG4(10μg/ml)の不在を陰性対照として使用した。NK単離キット(Miltenyi Biotec, USA)によってMNC試料からNK細胞を単離した。NK細胞に富む末梢血単核細胞を使用した実験では、PE標識マウス抗ヒトCD56(BD/Pharmingen)を使用した二重染色によってNK細胞を同定した。当技術分野の現状により、PBMCから顆粒球および単球を単離した(例えば単球単離キット(Miltenyi、上記参照)。顆粒球および単球をFSC/SSCプロフィールに基づいて同定した。
【0205】
結果:
本発明によるHuMab002は、顆粒球、単球、およびNK細胞に対する結合によって示されるようにFcRに結合できた。試験した三つのFc変異体全て(IgG4v1、IgG1v1、およびIgG1v2)はNK細胞に結合する能力を完全に欠如していた(表4)。さらに、HuMab002は顆粒球および単球に効率的に結合したが、表5および6に抗体に結合している細胞の率によって示されるように、突然変異体はヒトIgG4に対する一次抗体の不在に匹敵する結合レベルを示した。これは、突然変異体がエフェクター細胞上のFcRと相互作用する能力を欠如したことを示している。
【0206】
HuMab002はFcRと効率的に相互作用できるので、この抗体はインビボで抗体依存性細胞性細胞傷害作用を誘導する内因性能力を有する。本発明によるFc変異体に関して行われたようなFcRとの相互作用の不活性化は、効果的にADCCを阻止する。
【0207】
【表5】
【0208】
【表6】
【0209】
【表7】
【特許請求の範囲】
【請求項1】
P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体であって、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに置換され、L235がEに置換されたヒトサブクラスIgG4の抗体であることを特徴とする抗体。
【請求項2】
ハイブリドーマ細胞系DSM ACC2640、DSM ACC2641、またはDSM ACC2642。
【請求項3】
ハイブリドーマ細胞系DSM ACC2640、DSM ACC2641、またはDSM ACC2642から入手できる抗P−セレクチン抗体。
【請求項4】
a)ヒト抗体またはヒト化抗体であり、かつ
b)P−セレクチンとE−および/またはL−セレクチンとをマイクロタイタープレートに被覆するELISAアッセイでのEC50値によって測定されるように、E−またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍高く特異的に結合することを特徴とする抗P−セレクチン抗体。
【請求項5】
請求項1、3、または4記載の抗体分子をコードする核酸分子。
【請求項6】
請求項5記載の核酸分子を含むベクター。
【請求項7】
請求項6記載のベクターを含む宿主細胞。
【請求項8】
請求項1、3、または4記載の抗体分子を調製するための方法であって、該抗体分子の合成を可能にする条件で請求項7記載の宿主細胞を培養する工程と、該培養物から該抗体分子を回収する工程とを含む方法。
【請求項9】
請求項1、3、もしくは4記載の抗体分子または請求項8記載の方法によって生成される抗体分子を含む組成物。
【請求項10】
薬学的組成物または診断用組成物である、請求項9記載の組成物。
【請求項11】
請求項1、3、または4記載の抗体と、少なくとも一つの薬学的に許容されうる賦形剤とを含む薬学的組成物。
【請求項12】
治療を必要とする患者を処置するための方法であって、請求項1、3、または4記載の治療有効量の抗体を該患者に投与することを特徴とする方法。
【請求項13】
治療のための請求項1、3、または4記載の抗体の使用。
【請求項14】
炎症障害および血栓障害を予防および処置するための医薬を調製するための、請求項1、3、または4記載の抗体の使用。
【請求項15】
PAODまたはCLIの処置のための、請求項14記載の使用。
【請求項16】
請求項1、3、または4記載の抗体分子、請求項5記載の核酸分子、請求項6記載のベクター、または請求項7記載の宿主細胞を含むキット。
【請求項17】
抗体を含む組成物であって、該抗体がP−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しないことを特徴とする組成物。
【請求項18】
薬学的組成物または診断用組成物である、請求項17記載の組成物。
【請求項19】
治療を必要とする患者を処置するための方法であって、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない治療有効量の抗体を該患者に投与することを特徴とする方法。
【請求項20】
治療のための、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない抗体の使用。
【請求項21】
炎症障害および血栓障害の予防および処置のための医薬を調製するための、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない抗体の使用。
【請求項22】
PAODおよびCLIの処置のための請求項21記載の使用。
【請求項23】
本明細書に記載した発明。
【請求項1】
P−セレクチンに結合し、補体因子C1qに結合せず、ヒト起源由来のFc部分を含む抗体であって、L234、L235、D270、N297、E318、K320、K322、P331、および/もしくはP329に少なくとも一つの突然変異を含むヒトサブクラスIgG1の抗体であるか、またはS228がPに置換され、L235がEに置換されたヒトサブクラスIgG4の抗体であることを特徴とする抗体。
【請求項2】
ハイブリドーマ細胞系DSM ACC2640、DSM ACC2641、またはDSM ACC2642。
【請求項3】
ハイブリドーマ細胞系DSM ACC2640、DSM ACC2641、またはDSM ACC2642から入手できる抗P−セレクチン抗体。
【請求項4】
a)ヒト抗体またはヒト化抗体であり、かつ
b)P−セレクチンとE−および/またはL−セレクチンとをマイクロタイタープレートに被覆するELISAアッセイでのEC50値によって測定されるように、E−またはL−セレクチンよりもP−セレクチンに対して少なくとも1000倍高く特異的に結合することを特徴とする抗P−セレクチン抗体。
【請求項5】
請求項1、3、または4記載の抗体分子をコードする核酸分子。
【請求項6】
請求項5記載の核酸分子を含むベクター。
【請求項7】
請求項6記載のベクターを含む宿主細胞。
【請求項8】
請求項1、3、または4記載の抗体分子を調製するための方法であって、該抗体分子の合成を可能にする条件で請求項7記載の宿主細胞を培養する工程と、該培養物から該抗体分子を回収する工程とを含む方法。
【請求項9】
請求項1、3、もしくは4記載の抗体分子または請求項8記載の方法によって生成される抗体分子を含む組成物。
【請求項10】
薬学的組成物または診断用組成物である、請求項9記載の組成物。
【請求項11】
請求項1、3、または4記載の抗体と、少なくとも一つの薬学的に許容されうる賦形剤とを含む薬学的組成物。
【請求項12】
治療を必要とする患者を処置するための方法であって、請求項1、3、または4記載の治療有効量の抗体を該患者に投与することを特徴とする方法。
【請求項13】
治療のための請求項1、3、または4記載の抗体の使用。
【請求項14】
炎症障害および血栓障害を予防および処置するための医薬を調製するための、請求項1、3、または4記載の抗体の使用。
【請求項15】
PAODまたはCLIの処置のための、請求項14記載の使用。
【請求項16】
請求項1、3、または4記載の抗体分子、請求項5記載の核酸分子、請求項6記載のベクター、または請求項7記載の宿主細胞を含むキット。
【請求項17】
抗体を含む組成物であって、該抗体がP−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しないことを特徴とする組成物。
【請求項18】
薬学的組成物または診断用組成物である、請求項17記載の組成物。
【請求項19】
治療を必要とする患者を処置するための方法であって、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない治療有効量の抗体を該患者に投与することを特徴とする方法。
【請求項20】
治療のための、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない抗体の使用。
【請求項21】
炎症障害および血栓障害の予防および処置のための医薬を調製するための、P−セレクチンと結合し、ヒト起源由来のFc部分を含み、かつ補体因子C1qと結合しない抗体の使用。
【請求項22】
PAODおよびCLIの処置のための請求項21記載の使用。
【請求項23】
本明細書に記載した発明。
【図1】

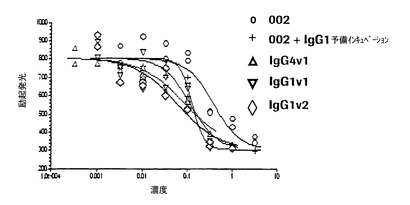
【図2】
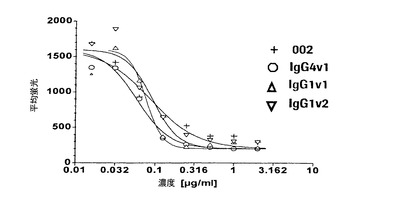
【図3a】
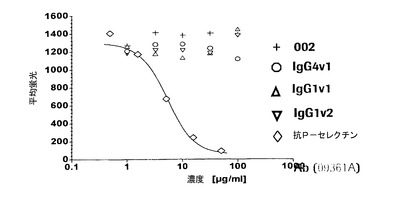
【図3b】
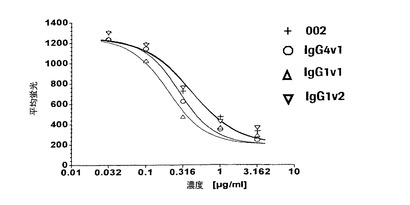
【図4a】
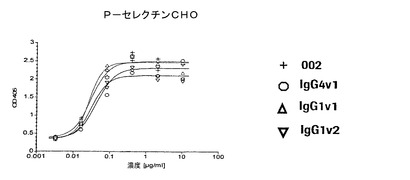
【図4b】
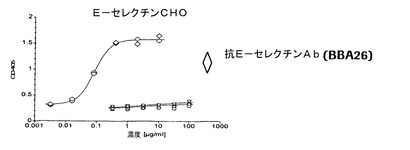
【図4c】
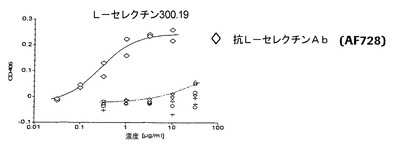
【図5】
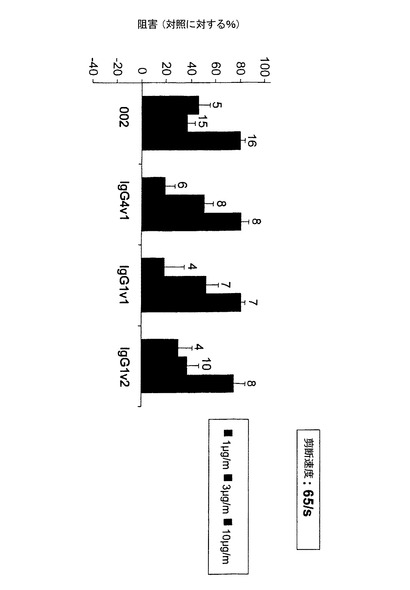
【図6a】
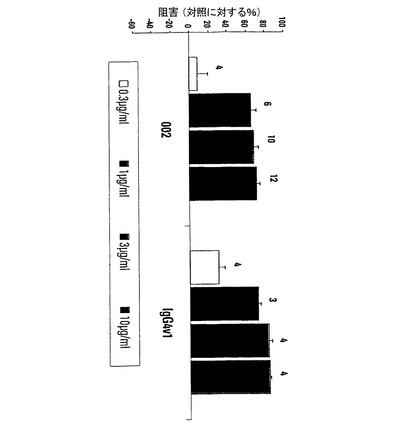
【図6b】
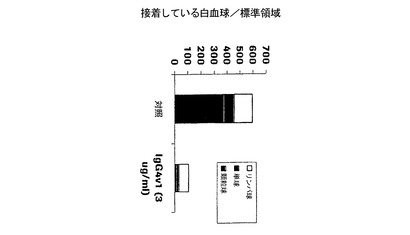
【図2】
【図3a】
【図3b】
【図4a】
【図4b】
【図4c】
【図5】
【図6a】
【図6b】
【公開番号】特開2013−63079(P2013−63079A)
【公開日】平成25年4月11日(2013.4.11)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−259616(P2012−259616)
【出願日】平成24年11月28日(2012.11.28)
【分割の表示】特願2010−67822(P2010−67822)の分割
【原出願日】平成17年4月5日(2005.4.5)
【出願人】(591003013)エフ.ホフマン−ラ ロシュ アーゲー (1,754)
【氏名又は名称原語表記】F. HOFFMANN−LA ROCHE AKTIENGESELLSCHAFT
【Fターム(参考)】
【公開日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願番号】特願2012−259616(P2012−259616)
【出願日】平成24年11月28日(2012.11.28)
【分割の表示】特願2010−67822(P2010−67822)の分割
【原出願日】平成17年4月5日(2005.4.5)
【出願人】(591003013)エフ.ホフマン−ラ ロシュ アーゲー (1,754)
【氏名又は名称原語表記】F. HOFFMANN−LA ROCHE AKTIENGESELLSCHAFT
【Fターム(参考)】
[ Back to top ]