
抗PDGF−DD抗体を用いた腎炎の処置方法
本明細書に記載の本発明の実施態様は、血小板由来成長因子DD(PDGF−DD)およびかかる抗体の使用に関する。本発明の抗体は、PDGF−DDの過剰産生と関連する疾患の診断および治療としての使用が見出されている。特に、本発明の実施態様と関連して、抗PDGF−DD抗体の、メサンギウム増殖により引き起こされる疾患を含む腎炎および関連疾患の処置のための使用が提供される。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
技術分野
本明細書に記載の本発明の実施態様は、血小板由来成長因子DD(PDGF−DD)に対する抗体、およびかかる抗体の使用に関する。本発明の抗体は、PDGF−DDの過剰産生に関連する疾患の診断および処置としての使用が見出されている。具体的には、本発明の実施態様をふまえて、メサンギウム増殖により引き起こされる疾患を含む腎炎および関連疾患の処置のための抗PDGF−DD抗体の使用が提供される。
【0002】
発明の背景
腎炎は、アメリカ合衆国および世界中で関心の増している問題である、腎臓疾患の1つの群である。腎炎は、透析処置または腎移植を受けなければ最終的には致命的となる、腎不全に徐々に進行し得る。異なる型の腎炎は、異なる遺伝パターン、および異なる進行速度を有する。遺伝性腎炎は、尿中の血液細胞およびタンパク質の顕微鏡トレースにより明らかにされ、出生時に存在するが、概して穏やかである。別の型の腎炎である糸球体腎炎は、腎臓の濾過単位である糸球体の炎症である。他の型の腎炎は、単球増加症および連鎖球菌(Streptococcus)(感染後)のような感染性疾患の後遺症であり得る。
【0003】
メサンギウム細胞の増殖に関連する腎炎および他の関連疾患の症状は、腎炎の特定の型に依存して変動するが、典型的には尿中の血液またはタンパク質の存在が挙げられる。該疾患の初期段階では、徴候または症状がないこともある。疾患が進行するにつれて、以下の症状のいくつかまたは全てが生じ得る:高血圧、尿の過剰な泡立ち、尿の色の(赤または黒褐色への)変化、眼、手および足の腫脹、悪心および嘔吐、呼吸困難、および頭痛。これらの症状は、疾患を同定し、処置コースに従い、そしてどのタイプの処置が必要かを同定するために用いられ得る。
【0004】
糸球体の損傷は、タンパク質尿、血尿、高窒素血症、乏尿、無尿、浮腫、および高血圧を含むがこれらに限定されない、種々の疾患の徴候を引き起こす。該疾患はまた、この疾患はまた、腎炎症候群、急性腎炎、および急性進行性糸球体腎炎を引き起こし得る。
【0005】
糖尿病性腎症を含む多くの進行性腎疾患、ならびにIgA腎症のような最も頻繁な型の糸球体腎炎は、糸球体メサンギウム細胞増殖および/または基質蓄積により特徴付けられる。Striker et al., Lab Invest 64: 446-456(1991)参照。現時点で、血小板由来成長因子(PDGF)および関連するPDGF系が、メサンギウム細胞増殖および基質蓄積に関与し得ることを示唆する証拠がいくつかある。Floege et al., supra(2001)およびFloege et al., Am.J. Pathol. 154: 169-79(1999); Gilbert et al., Kidney Int. 59: 1324-32(2001); Nakamura et al., Kidney Int., 59: 2134-45(2001)参照。さらに、PDGFβ受容体サブユニットおよびPDGF B鎖の両方が腎間質性線維症において過剰発現する。Kliem et al., Kidney Int. 49: 666-78(1996)参照。2量体PDGF−BB単独の多用量の注射は、正常ラット腎臓において間質性線維性変化を誘発し得る。Tang et al., Am. J. Pathol. 148: 1169-80(1996)参照。
【0006】
20年間、血小板由来成長因子系は、たった2つのPDGF鎖(PDGF−AおよびPDGF−B)からなるとされ、これは、ホモまたはヘテロ2量体として分泌され、α鎖および/またはβ鎖を含む2量体PDGF受容体に結合する。PDGF−Aはα鎖のみに結合するが、PDGF−βは全ての受容体タイプのリガンドである。Floege et al., "Growth factors and cytokines," in Immunologic Renal Diseases(Neilson E. G., and Couser W. G., eds., 2d ed. 2001)参照。最近、PDGF−CおよびPDGF−Dと命名された2つの他のPDGFアイソフォームが、ホモ2量体としてのみ放出されることが述べられた。現行の専門用語によると、PDGF−Cのホモ2量体形は、「PDGF−CC」として知られ、PDGF−Dのホモ2量体形は、「PDGF−DD」として知られている。LaRochelle et al.,Nat. Cell Biol. 3: 517-21(2001); Li et al.,Nat. Cell Biol. 2: 302-09(2000);およびBergsten et al.,Nat. Cell Biol. 3: 512-16(2001)を参照。PDGF−CCの中心鎖は、主にαα−PDGF受容体のリガンドであるようだが、PDGF−DDは、主にββ−PDGF受容体と結合する(同上)。両方の場合において、αβ受容体との結合もいくつか述べられている。LaRochelle et al., supra(2001); Bergsten et al., supra(2001); Gilbertson et al., J. Biol. Chem. 276: 27406-14(2001)参照。4つのPDGFアイソフォーム全て、ならびに両受容体鎖は、異なる空間配置ではあるが腎臓で発現する。Floege et al., supra(2001);Changsirikulchai et al., Kidney Int. 62(6): 2043-54(2002); Eitner et al., J. Am. Soc. Nephrol. 13(4): 910-17(2002)を参照。
【0007】
PDGF−Dは、ジスルフィド結合したホモ2量体PDGF−DDとして分泌され、そしてPDGF−ββ−およびαβ−受容体の特異的作用リガンドとなるために、そのCUBドメインの解離による制限的タンパク質分解で活性化される。発生において、および成人正常腎臓において、PDGF−DDは、内臓の糸球体上皮細胞およびいくつかの血管平滑筋細胞で発現する。Changsirikulchai et al., supra(2002)参照。マウス腎臓の発生において、分枝尿管(branching ureter)の細胞のみがPDGF−DD免疫反応性を示した。Bergsten et al., supra(2001)参照。
【0008】
腎炎の診断は、典型的には、家族歴、および/または赤血球およびタンパク質の存在、特に血尿またはアルブミン尿についての尿沈査検査による。残念なことに、根元的な病理過程に作用する、または臨床コースを変化させる特定の処置は知られていない。抗生物質、抗凝血剤、ステロイド剤、および免疫抑制剤は何ら利益をもたらさなかった。高血圧の制御が示唆されており、タンパク質の制限がいくぶん役に立つかもしれない。末期の尿毒症が起こった場合、透析およびさらには腎移植が必要とされる。従って、腎炎の処置のための新規アプローチが必要である。
【0009】
発明の要約
本発明の実施態様は、抗PDGF−DD抗体の投与が、糸球体細胞の増殖を低減し、そしてその増殖と関連する疾患を処置するのに非常に効果的であったという発見に関する。
【0010】
従って、本発明の1つの実施態様は、腎炎および関連疾患の進行を阻害するための完全ヒト抗PDGF−DD抗体、および治療展望から所望した特性を有する抗PDGF−DD抗体製剤の使用である。好ましくは、当該抗体は、配列番号2、6、10、14、18、22、26、30、34、38、42、46、50、54、58、62、66、70および74からなる群から選択される配列を有する重鎖アミノ酸を有する。より好ましくは、当該抗体はさらに、配列番号4、8、12、16、20、24、28、32、36、40、44、48、52、56、60、64、68および72からなる群から選択される配列を有する軽鎖アミノ酸を有する。
【0011】
本発明の実施態様は、任意の特定の抗PDGF−DD抗体、または任意の特定型の抗体に限定されないことは理解されるだろう。例えば、抗PDGF−DD抗体は、全長抗体(例えば、完全ヒトFc領域を有する)、または抗体フラグメント(例えば、Fab、Fab’またはF(ab’)2)であってもよい。さらに、抗体は、抗体を分泌するハイブリドーマから、または遺伝子または抗体をコードする遺伝子で形質転換またはトランスフェクションされた組換え産生細胞から製造されてもよい。
【0012】
好ましい実施態様において、本発明は、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎不全、および糖尿病性腎症を含む(これらに限定されない)、ヒトの腎炎および関連疾患の処置を含む。
【0013】
1つの実施態様において、抗PDGF−DD抗体は、有効量の抗体またはそのフラグメントを、医薬的に許容される担体または希釈剤と共に含む医薬組成物を成す。代わりの実施態様において、抗PDGF−DD抗体は、放射性同位元素、または毒素と結合している。別の実施態様において、抗PDGF−DD抗体またはそのフラグメントは、治療法と結合していてもよい。該治療法は、毒素または放射性同位元素であり得る。好ましくは、かかる抗体は、例えば、腎炎、進行性腎疾患、および関連疾患(例えば、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎間質性線維症、腎不全および糖尿病性腎症)のような疾患の処置のために用いられ得る。
【0014】
別の実施態様において、本発明は、患者に有効量の抗PDGF−DD抗体を投与することにより、患者におけるPDGF−DDの発現と関連する疾患または状態を処置する方法を含む。該患者は、哺乳動物患者、好ましくはヒト患者である。該疾患または状態は、例えば、腎炎、進行性腎疾患、および関連疾患(例えば、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎間質性線維症、腎不全または糖尿病性腎症)であり得る。さらなる実施態様は、腎炎の処置が必要な哺乳動物を同定すること、次に該哺乳動物に治療上有効用量の抗PDGF−DD抗体を投与することにより、哺乳動物のPDGF−DDの発現と関連する疾患または状態を処置する方法を含む。
【0015】
あるいは、抗PDGF−DD抗体は、哺乳動物が腎炎または関連疾患を含む(これに制限されない)PDGF−DDの発現に関係する疾患または状態、およびメサンギウム増殖により引き起こされる疾患にかかるのを予防するために投与されてもよい。好ましくは、抗PDGF−DD抗体は完全ヒト抗体である。疾患または状態は、腎炎、進行性腎疾患、および関連疾患(例えば、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎間質性線維症、腎不全、および糖尿病性腎症)含むが、これらに限らない、腎炎および関連疾患であり得る。
【0016】
さらに別の実施態様において、本発明は、細胞PDGF−DD発現細胞を有効量の抗PDGF−DD抗体またはそのフラグメントと接触させること、次に該細胞および抗体をインキュベートすることによる(インキュベーションが細胞増殖を阻害する)、PDGF−DDの発現に関連するかまたは当該発現により引き起こされる細胞増殖を阻害するための方法を含む。1つの実施態様において、細胞増殖は、メサンギウム細胞増殖である。さらに、メサンギウム細胞は、ヒトメサンギウム細胞であり得る。さらに、方法は、インビボで行われ得る。
【0017】
別の実施態様において、本発明は、抗PDGF−DD抗体を含有する組成物を有する容器、および該組成物がPDGF−Dの過剰発現により特徴付けられる状態を処置するために用いられ得ることを示すパッケージの記載またはラベルを含む製品である。好ましくは哺乳動物、より好ましくはヒトが、抗PDGF−DD抗体を受ける。好ましい実施態様において、ヒトの腎炎および関連疾患が処置され、これは、腎炎、進行性腎疾患、および関連疾患、例えば、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎間質性線維症、腎不全、および糖尿病性腎症を含むが、これらに限らない。
【0018】
別の実施態様は、抗PDGF−DD抗体を用いて、糸球体中のメサンギウム細胞の過剰増殖を同定することを含む、疾患の危険因子を同定するための方法、疾患の診断方法、および疾患の病期診断方法である。
【0019】
1つの実施態様において、本発明は、細胞を抗PDGF−DD抗体と接触させ、次にPDGF−DDの存在を検出することによる、細胞でのPDGF−DDの発現の発現と関連する状態の診断方法を含む。好ましい状態は、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎不全、および糖尿病性腎症を含むが、これに限定されない。
【0020】
さらに別の実施態様において、本発明は、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎不全、および糖尿病性腎症を含む(これらに限定されない)、ヒトにおいて腎炎および関連疾患をスクリーニングするための、哺乳動物組織または細胞におけるPDGF−DDの検出のためのアッセイキットを含む。該キットは、PDGF−DDと結合する抗体、および存在するなら、該抗体のPDGF−DDとの反応を示す手段を含む。好ましくは、当該抗体はモノクローナル抗体である。1つの実施態様において、PDGF−DDと結合する抗体が標識される。別の実施態様において、抗体は非標識の1次抗体であり、反応を示すための手段が、標識抗免疫グロブリン抗体である。好ましくは、抗体は、蛍光色素、酵素、放射性核種、および放射線不透過材料からなる群から選択されるマーカーで標識される。
【0021】
なお別の実施態様は、腎炎および関連疾患の処置のための医薬の製造における抗PDGF−DD抗体の使用である。1つの実施態様において、該疾患は、腎炎、進行性腎疾患、およびメサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎間質性線維症、腎不全、および糖尿病性腎症のような関連疾患からなる群から選択される。
【0022】
図面の簡単な説明
図1は、ELISAによる抗PDGF−DD mAb 6.4特異性の特徴決定を示す。
図2は、ELISAによる抗PDGF−DD mAb 6.4特異性のさらなる特徴決定を示す。
図3は、ウエスタンブロット分析による抗PDGF−DD mAb特異性の特徴決定を示す。
【0023】
図4は、抗PDGF−DD mAb 6.4が、NIH3T3細胞でのPDGF−DD誘導性BrdU取込みを、約75ng/mlのIC50で中和できることを示す線グラフである。
図5は、PDGF−DDがインビトロにおいてメサンギウム細胞の成長因子となることを示す棒グラフである。データは、4つの独立した実験の平均±SDである。*は、p<0.05対非刺激対照を示す。
【0024】
図6は、ヒトメサンギウム細胞におけるPDGF−DD誘導性BrdU取込みの結果を示す棒グラフである。
図7は、種々の型の腎炎を有する患者のヒト血清中でのPDGF−DD発現を示すグラフである。黒丸は、個々の臨床血清試料のPDGF−DD濃度を示す。PDGF−DD血清濃度は、患者の疾患の徴候に従いグループ分けした。所定の臨床徴候の患者数(n)が、平均PDGF−DD濃度(ng/ml)とともに提供される。
【0025】
図8は、正常ラットメサンギウム細胞および抗Thy−1誘導性腎炎を有するラットのメサンギウム細胞の免疫組織化学分析を示す。抗PDGF−DD染色性の上昇が、抗Thy−1誘導性腎炎のラットで見出された。メサンギウム、尿細管、および周辺脈管構造が示されている。メサンギウム細胞は、周辺細胞および腎尿細管を含んでいた。白色および灰色の矢印は、それぞれ毛細管および尿細管染色を示す。
【0026】
図9は、ラットにおいて成された完全ヒトmAbカイネティクスのシュミレーションを示す棒グラフである。示されるように、1回用量の後であっても4日にわたり、予測された小さなピークからトラフへの変動のみが生じている。
図10は、無処置のラットでの発現に対する、抗Thy1.1腎炎の経過におけるPDGF−A、−B、−Cおよび−Dの転写発現を示す線グラフである。
【0027】
図11は、PDGF−DDタンパク質が、糸球体での抗Thy1.1腎炎において過剰発現されることを示す。正常糸球体ではPDGF−DD発現は見られない(図11(A))が、疾患を誘導した7日後にメサンギウム増殖性腎炎において容易に検出され得る(図11(B))。抗PDGF−DD抗体を等濃度の対照IgGで置き換えた場合、糸球体染色性はない(図11(C))。倍率は600倍である。
【0028】
図12A〜Hは、抗PDGF−DD抗体、無関係の対照IgGまたはPBS単独のいずれかで処置した、メサンギウム増殖性抗Thy1.1腎炎を有するラットにおいて、糸球体が疾患の誘導5日後および8日後に変化していることを示す棒グラフである。
【0029】
図13は、ラットでのBrdU取込により測定された、糸球体増殖の結果を示す棒グラフである。腎炎のラットを、抗PDGF−DD mAb 6.4、対照抗体またはPBSで処置した。健常ラットを、抗PDGF−DD mAb 6.4または対照抗体で処置した。
【0030】
図14は、ラットにおけるPAS染色により測定された糸球体増殖および有糸分裂の結果を示す棒グラフである。腎炎のラットを、抗PDGF−DD mAb 6.4、対照抗体またはPBSで処置した。健常ラットを、抗PDGF−DD mAb 6.4または対照抗体で処置した。
【0031】
図15は、急性ラット抗Thy−1モデルにおけるメサンギウム細胞有糸分裂に対する抗PDGF−DD mAb 6.4の作用を示す棒グラフである。抗Thy−1ラットを、抗PDGF−DD mAb 6.4、または対照抗体またはPBSで処置した。健常ラットを、抗PDGF−DD mAb 6.4または対照抗体で処置した。
【0032】
図16は、急性ラット抗Thy−1モデルにおける糸球体細胞の有糸分裂に対する抗PDGF−DD mAb 6.4の用量応答性作用を示す棒グラフである。
図17は、急性ラットThy−1モデルにおけるBrdU取込みに対する抗PDGF−DD mAb 6.4の用量応答性作用を示す棒グラフである。
【0033】
図18は、正常および疾患のヒト腎臓組織上の免疫組織化学分析を示す。メサンギウム、尿細管および周辺脈管構造が示されている。白の矢印および灰色の矢印は、それぞれ毛細管および尿細管の染色を示す。小さな黒の矢印は、メサンギウムにおける点状の炎症性細胞の蓄積を示す。
【0034】
詳細な説明
本明細書に記載の発明は、腎炎および関連状態を効果的に処置、診断および/または病期診断する方法に関する。かかる状態は、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎不全、および糖尿病性腎症を含む。1つの特定の実施態様において、本発明は、腎炎および関連状態の処置として、治療上有効量の抗PDGF−DD抗体を投与することを含む。好ましい実施態様において、抗体は、2量体PDGF−DDに対する完全ヒト抗体である。
【0035】
本発明の他の実施態様は、インビボでメサンギウム増殖性変化の低減をもたらす他の化合物に関する。従って、PDGF−DDのレベルを低減する化合物は、腎炎の処置に有用であろう。PDGF−D核酸、ポリペプチド、抗体、アゴニスト、アンタゴニストおよび他の関連化合物の使用を、以下により詳細に開示する。
【0036】
上述のように、PDGF−DはPDGF−B受容体を介してシグナル伝達し、ラットのメサンギウム細胞(MC)の分裂を促進する。低レベルのPDGF−D mRNAが、正常ラット糸球体で検出された。しかしながら、培養したラットMCを100ng/ml PDGF−DDと共にインキュベーションすることにより、MC増殖が7倍に増加し、これは24時間後に最大となった。リアルタイムPCRにより、PDGF−D mRNAは、培養メサンギウム細胞および正常ラット腎臓から単離された糸球体の両方で検出された。ラットにおけるメサンギウム増殖性抗Thy1.1腎炎の誘導後、糸球体のPDGF−D mRNAおよびタンパク質発現は、リアルタイムPCRで測定された非腎炎ラットと比較して、4から9日間有意に増加した。PDGF−D mRNA発現のピークは、PDGF−B mRNA発現ピークの2日後に生じた。さらに、PDGF−DD血清レベルは、7日目の腎炎動物において有意に増加した。
【0037】
腎炎におけるPDGF−DDの機能的役割を調べるため、Xenomouse[登録商標](Abgenix, Inc., Fremont, CA)において中和完全ヒトモノクローナル抗PDGF−DD抗体を産生させた。抗Thy1.1腎炎の誘導後、ラットは、疾患誘導の3日後および5日後に、10および4mg/kg 完全ヒト抗PDGF−DD抗体 mAb 6.4(n=15)、または無関係なヒトモノクローナル抗体(n=15)またはPBS(n=15)の通常の腹腔内注射により処置された。疾患誘導の8日後に、PDGF−DDの拮抗作用により、100個の糸球体当たりの有糸分裂像の有意な低減(抗PDGF−DD:9.9±0.9;無関係なIgG:13.9±0.9;PBS:14.7±1.0;p<0.0014)、ならびにチミジン類似体BrdUを取り込んだ糸球体細胞の有意な低減(抗PDGF−DD mAb 6.4:1.62±0.23;無関係なIgG:2.88±0.28;PBS:2.91±0.18;p<0.00169)を生じた。抗PDGF−DDを投与されたラットにおける糸球体細胞増殖の低減は、リアルタイムPCRにより決定された、低減したPDGF−B mRNAの糸球体発現とは関連しなかった。
【0038】
抗PDGF−DD抗体の正常ラットへの注射は、無関係なIgGを投与された正常ラットと比較して、生理的な糸球体細胞のターンオーバーに影響を及ぼさなかった。従って、糸球体メサンギウム細胞により産生されるPDGF−DDは、インビトロおよびインビボの両方で糸球体細胞の分裂促進因子となる。
【0039】
配列表
代表的なヒト抗PDGF−DD抗体の重鎖および軽鎖の可変領域ヌクレオチドおよびアミノ酸配列が、配列表において提供され、その内容が以下の表1に要約されている。
【0040】
表1
【表1】

【0041】
【表2】
【0042】
【表3】
【0043】
定義
他に定義しない限り、本明細書に記載の本発明と関連して用いられる科学用語および技術用語は、当業者に通常理解される意味を有するものとする。さらに、文脈により他に必要とされない限り、単数形の用語は複数形も含み、複数形の用語は単数形も含むものとする。一般に、本明細書に記載の細胞および組織培養、分子生物学、ならびにタンパク質およびオリゴヌクレオチドまたはポリヌクレオチド化学およびハイブリダイゼーションと関連して利用される学名および技術は、よく知られたものであり、当該技術分野において通常用いられるものである。標準的技術は、組み換えDNA、オリゴヌクレオチド、ならびに組織培養および形質転換(例えば、エレクトロポレーション、リポフェクション)に用いられる。酵素反応および精製技術は、製造元の仕様書に従って、あるいは当該技術分野において通常成されるか、または本明細書に記載のように行われる。上記技術および手順は、一般に、当該技術分野でよく知られた通常の方法に従い、かつ本明細書を通じて引用されて考察された種々の一般的かつより具体的な引例に記載のように、行われる。例えば、Sambrook et al.Molecular Cloning : A Laboratory Manual(3rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y.(2001))を参照されたい。本明細書に記載の分析化学、合成有機化学、および医療化学および医薬化学と関連して利用される学名、および実験室手段および技術は、当該技術分野でよく知られ、かつ通常用いられるものである。標準的な技術は、化学合成、化学分析、医薬製造、製剤、および送達、および患者の処置のために用いられる。
【0044】
本明細書で提供される実施態様と関連して用いられるように、以下の用語は、他に示されない限り、以下の意味を有すると理解されるものとする。
メサンギウム細胞は、哺乳動物の腎臓の糸球体小葉(lubule)内で見出される細胞であり、構造支持体となり、血流を制御し得、食細胞であり、かつ免疫応答において抗原を提示する補助細胞となる。
【0045】
メサンギウム増殖性腎炎は、糸球体メサンギウム細胞または基質の増加、またはメサンギウムの蓄積を伴う糸球体腎炎である。
【0046】
メサンギウム増殖性糸球体腎炎は、糸球体毛細管のメサンギウム層でのIgM抗体の異常蓄積による腎糸球体(腎臓の血液濾過部分)の炎症である。
【0047】
メサンギウム毛細管性糸球体腎炎は、腎不全となる腎障害である。糸球体の炎症は、異常な免疫応答および腎臓(糸球体)内での抗体の蓄積から生じる。症状は、混濁尿(膿尿)、排尿減少、腫脹および高血圧が挙げられる。この障害は、たいてい末期の腎疾患となる。
【0048】
メサンギウムは、毛細管の間の糸球体の中心部である。メサンギウム細胞は、食細胞であり、大部分が内皮細胞により毛細管の管腔と分けられている。糸球体外メサンギウムは、密集斑と傍糸球体装置の輸入細動脈および輸出細動脈との間の三角形の空間を満たすメサンギウム細胞である。
【0049】
糸球体腎炎は、腎臓の糸球体内の毛細管ループの炎症により特徴付けられる、種々の腎炎である。これは、急性、亜急性および慢性の形で生じ、感染症または自己免疫疾患が続発し得る。
【0050】
用語「PDGF−DD」は、変異体と共に全長かつ成熟形のPDGF−DD、およびそれらのフラグメントを含む。従って、PDGF−DDは、変異体CG52053−01、CG52053−02、CG52053−03、CG52053−04、CG52053−05、CG52053−06、およびCG52053−07(CuraGen, New Haven, CT)を含むが、これらに限らない。より多くの情報が、PCT出願の公開番号第WO01/25433号(1999年10月7日出願)において見出され得る。
【0051】
本明細書で用いられる用語「単離されたポリヌクレオチド」は、ゲノム、cDNAまたは合成起源のポリヌクレオチド、あるいはこれらのいくつかの組合せを意味し、その起源に基づき、「単離されたポリヌクレオチド」は、(1)天然で見出される「単離されたポリヌクレオチド」の全てまたは部分と関連しないか、(2)天然で結合しないポリヌクレオチドに作動可能に結合するか、あるいは(3)より長い配列の一部分としては天然に存在しない。
【0052】
本明細書で言及される用語「単離されたタンパク質」は、cDNA、組換えRNAまたは合成起源のタンパク質、あるいはこれらのいくつかの組合せを意味し、その起源または変異体の供給源に基づき、「単離されたタンパク質」は、(1)天然で見出されるタンパク質とは関連しないか、(2)同じ供給源由来の他のタンパク質を含まない(例えば、マウスタンパク質を含まない)、(3)異なる種由来の細胞により発現されるか、あるいは(4)天然に存在しない。
【0053】
用語「ポリペプチド」は、本明細書で一般名称として用いられ、天然タンパク質、フラグメント、またはポリペプチド配列の類似体を意味する。従って、天然タンパク質、フラグメント、および類似体は、ポリペプチド属の種である。本発明をふまえて、好ましいポリペプチドは、ヒト免疫グロブリンの重鎖分子、およびヒト免疫グロブリンのκ軽鎖分子、ならびに免疫グロブリンの重鎖分子を免疫グロブリンの軽鎖(例えば、免疫グロブリンのκ軽鎖)と共に含む組合せ、またはその逆により形成される抗体分子、ならびそのフラグメントおよび類似体を含む。
【0054】
対象に適用して本明細書で用いられる用語「天然に存在する」は、対象が天然に見出され得るという事実を意味する。例えば、天然供給源から単離されることができ、実験室またはその他の方法で意図的に修飾されていない、生物(ウイルスを含む)に存在するポリペプチドまたはポリヌクレオチド配列は、天然に存在する。
【0055】
本明細書で用いられる用語「作動可能に結合した」は、成分の位置を意味し、意図された方法でそれらが機能することを許可する関係にあることを記載したものである。コーディング配列に「作動可能に結合した」対照配列は、コーディング配列の発現が対照配列と適合可能な条件下で達成されるような方法でライゲーションされている。
【0056】
本明細書で用いられる用語「対照配列」は、これらがライゲーションしているコーディング配列の発現およびプロセッシングに作用する必要のあるポリヌクレオチド配列を意味する。かかる対照配列の性質は、宿主生物に依存して異なる;原核生物では、かかる対照配列は、一般に、プロモーター、リボソーム結合部位、および転写終結配列を含み;真核生物では、一般に、かかる対照配列は、プロモーター、および転写終結配列を含む。用語「対照配列」は、最低でも、その存在が発現およびプロセッシングに必須である要素の全てを含むことが意図され、その存在が有利であるさらなる要素、例えば、リーダー配列および融合パートナー配列も含み得る。
【0057】
本明細書で言及される用語「ポリヌクレオチド」は、リボヌクレオチドまたはデオキシヌクレオチドのいずれかである、少なくとも10塩基長の重合体形ヌクレオチド、またはいずれかのタイプのヌクレオチドの修飾形を意味する。この用語は、1本鎖および2本鎖形のDNAを含む。
【0058】
本明細書で言及される用語「オリゴヌクレオチド」は、天然に存在するもの、および天然に存在するものと一体となって結合した修飾されたヌクレオチド、および天然に存在しないオリゴヌクレオチド結合体を含む。オリゴヌクレオチドは、一般に、200塩基長またはそれより短いものを含むポリヌクレオチドサブセットである。好ましくは、オリゴヌクレオチドは、10から60塩基長であり、最も好ましくは、12、13、14、15、16、17、18、19、または20から40塩基長である。オリゴヌクレオチドは通常1本鎖(例えば、プローブ用)であるが、オリゴヌクレオチドは、2本鎖(例えば、遺伝子変異の構築用)であってもよい。本発明のオリゴヌクレオチドは、センスオリゴヌクレオチドまたはアンチセンスオリゴヌクレオチドのいずれであってもよい。
【0059】
本明細書で言及される用語「天然に存在するヌクレオチド」は、デオキシリボヌクレオチドおよびリボヌクレオチドを含む。本明細書で言及される用語「修飾されたヌクレオチド」は、修飾または置換された糖基等を有するヌクレオチドを含む。本明細書で言及される用語「オリゴヌクレオチド結合」は、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホロアニラデート、ホスホロアミデート等のようなオリゴヌクレオチド結合を含む。例えば、LaPlanche et al.Nucl. Acids Res. 14: 9081(1986); Stec et al. J. Am. Chenu. Soc. 106: 6077(1984); Stein et al. Nucl. Acids Res. 16: 3209(1988); Zon et al. Anti-Cancer Drug Design 6: 539(1991); Zon et al. Oligonucreotides and analogues : A Practical Approach, pp. 87-108(F. Eckstein, Ed., Oxford University Press, Oxford England(1991)); Stec et al. 米国特許第5,151,510号; Uhlmann and Peyman Chemical Reviews 90: 543(1990)を参照されたい。オリゴヌクレオチドは、所望に応じて、検出用の標識を含むことができる。
【0060】
本明細書で言及される用語「選択的にハイブリダイズする」は、検出可能かつ特異的な結合を意味する。本発明をふまえて、ポリヌクレオチド、オリゴヌクレオチド、およびそれらのフラグメントは、非特異的な核酸との検出可能な結合の感知可能な量を最小化するハイブリダイゼーションおよび洗浄条件下で、核酸に選択的にハイブリダイズする。高ストリンジェントな条件を用いて、当該技術分野で知られ、かつ本明細書において考察される選択的ハイブリダイゼーション条件が達成され得る。一般に、本発明のポリヌクレオチド、オリゴヌクレオチド、およびフラグメントと対象の核酸配列間の核酸配列相同性は、少なくとも80%であり、より典型的には、好ましくは、少なくとも85%、90%、95%、99%および100%の増加した相同性を有するであろう。2つのアミノ酸配列は、それらの配列間に部分的または完全な同一性が存在するなら、相同である。例えば、85%の相同は、2つの配列がマッチングを最大とするようアラインメントされたとき、85%のアミノ酸が同一であることを意味する。(マッチングさせられている2配列のいずれかにおける)ギャップは、マッチングを最大化する際に許容される;5以下のギャップ長が好ましく、2以下のギャップ長がより好ましい。あるいは、そして好ましくは、2つのタンパク質配列(または少なくとも30アミノ酸長のもの由来のポリペプチド配列)は、変異データマトリックスおよび6以上のギャップペナルティーによるプログラムALIGNを用いて、5より多くのアラインメントスコア(標準偏差単位)を有する場合、相同である(この用語は本明細書で用いられる)。M. O. Dayhoff, in Atlas of Protein Sequence and Structure, Vol. 5,101-110、およびSupplement 2 to Vol. 5,1-10(National Biomedical Research Foundation 1972)を参照されたい。2つの配列またはその一部分の相同性は、そのアミノ酸がALIGNプログラムを用いて最適にアラインメントされた場合に50%以上同一であるなら、より好ましい。本明細書で用いられる用語「対応する」は、ポリヌクレオチド配列が、参照ポリヌクレオチド配列の全てまたは一部分に対して相同である(すなわち、厳密には進化上関連しないが、同一である)か、あるいはポリペプチド配列が、参照ポリペプチド配列と同一であることを意味する。対照的に、用語「相補である」は、相補的配列が、参照ポリヌクレオチド配列の全てまたは一部分に対して相同であることを意味するために本明細書で用いられる。例えば、ヌクレオチド配列「TATAC」は、参照配列「TATAC」に対応し、かつ「GTATA」に相補性である。
【0061】
以下の用語:「参照配列」、「比較ウインドウ」、「配列同一性」、「配列同一性の割合」、および「実質的な同一性」は、2以上のポリヌクレオチドまたはアミノ酸配列間の配列関係を記載するために用いられる。「参照配列」は、配列比較の基礎として用いられる所定の配列である;参照配列は、例えば、配列表で示される全長cDNAまたは遺伝子配列のセグメントとしてのより長い配列のサブセットであってもよく、あるいは完全なcDNAまたは遺伝子配列を含んでいてもよい。一般に、参照配列は、少なくとも18ヌクレオチド長または6アミノ酸長であり、しばしば少なくとも24ヌクレオチド長または8アミノ酸長であり、そしてたいてい少なくとも48ヌクレオチド長または16アミノ酸長である。2つのポリヌクレオチドまたはアミノ酸配列は、それぞれ(1)2分子間で類似する配列(すなわち、完全なポリヌクレオチドまたはアミノ酸配列の一部)を含み得、かつ(2)2つのポリヌクレオチドまたはアミノ酸配列間で異なる配列をさらに含み得るため、2(またはそれ以上)分子間の配列比較は、典型的には、配列類似性の局所領域を同定し、そして比較するための「比較ウインドウ」について2分子の配列を比較することで行われる。本明細書で用いられる「比較ウインドウ」は、ポリヌクレオチド配列またはアミノ酸配列が、少なくとも18の連続するヌクレオチドまたは6アミノ酸配列の参照配列と比較され得、かつ比較ウインドウのポリヌクレオチド配列の部分が、2配列の最適なアラインメントのための参照配列(付加または欠損を含まない)と比較して、20%以下の付加、欠損、置換などを含み得る、少なくとも18の連続するヌクレオチド位置または6アミノ酸である論理的セグメントを意味する。比較ウインドウをアラインメントするための配列の最適なアラインメントは、Smith and Watermanの局所相同性アルゴリズム(Adv. Appl. Math. 2: 482(1981))、Needleman and Wunschの相同性アラインメントアルゴリズム(J. Mol.Biol. 48: 443(1970))、Pearson and Lipmanの同様の検索方法(Proc. Natl. Acad. Sci.(U. S. A.)85: 2444(1988))、これらのアルゴリズムのコンピュータ上での実行(Wisconsin Genetics Software Package Release 7.0のGAP、BESTFIT、FASTA、およびTFASTA(Genetics Computer Group, 575 Science Dr., Madison, Wis.))、Genework、またはMacVector Software Package)、または調査により実行されてもよく、そして種々の方法で得られた最良のアラインメント(すなわち、比較ウインドウについて最も高い割合の相同性となる)が選択される。
【0062】
用語「配列同一性」は、比較ウインドウについて2つのポリヌクレオチドまたはアミノ酸配列が同一である(すなわち、ヌクレオチド対ヌクレオチドまたは残基対残基ベース)ことを意味する。用語「配列同一性の割合」は、比較ウインドウについて最適にアラインメントされた2配列を比較し、同一の核酸塩基(例えば、A、T、C、G、UまたはI)または残基が、両配列で生じる位置数を決定し、一致位置数を得、一致位置数を比較ウインドウの位置の総数(すなわち、ウインドウサイズ)で割り、次にこの結果に100を掛け、配列同一性の割合を得ることにより、算出される。本明細書で用いられる用語「実質的な同一性」は、ポリヌクレオチドまたはアミノ酸が、少なくとも18ヌクレオチド(6アミノ酸)位置の比較ウインドウ、しばしば少なくとも24〜48ヌクレオチド(8〜16アミノ酸)位置のウインドウについて参照配列と比較して、少なくとも85%の配列同一性、好ましくは、少なくとも90から95%の配列同一性、より一般的には、少なくとも99%の配列同一性を有するポリヌクレオチドまたはアミノ酸配列の特徴を示し、ここで、配列同一性の割合は、参照配列を、比較ウインドウについて参照配列の全20%以下が欠失または付加を含み得る配列と比較することにより算出される。参照配列は、より長い配列のサブセットであってもよい。
【0063】
本明細書で用いられる20の従来アミノ酸およびそれらの略語は、従来の使用に従う。Immunology-A Synthesis(2nd ed., Golub, E. S. and Gren, D. R. eds. , Sinauer Associates, Sunderland, Mass. 1991)を参照されたい。20の従来アミノ酸の立体異性体(例えば、D−アミノ酸)、α,α−2置換アミノ酸のような非天然アミノ酸、N−アルキルアミノ酸、乳酸、および他の従来にないアミノ酸もまた、本明細書に記載の本発明のポリペプチドの適切な要素であり得る。従来にないアミノ酸の例としては、4−ヒドロキシプロリン、γ−カルボキシグルタメート、ε−N,N,N−トリメチルリジン、ε−N−アセチルリジン、O−ホスホセリン、N−アセチルセリン、N−ホルミルメチオニン、3−メチルヒスチジン、5−ヒドロキシリジン、α−N−メチルアルギニン、および他の類似のアミノ酸およびイミノ酸(例えば、4−ヒドロキシプロリン)が挙げられる。本明細書で用いられるポリペプチド表記では、標準的使用および慣習に従い、左手方向がアミノ末端方向、そして右手方向がカルボキシ末端方向である。
【0064】
同様に、他に特定しない限り、1本鎖ポリヌクレオチド配列の左手末端は、5’末端である;2本鎖ポリヌクレオチド配列の左手方向は、5’方向として言及される。新生RNA転写物の5’から3’方向への付加は、転写方向として言及される;RNAと同じ配列を有し、かつRNA転写物の5’から5’末端であるDNA鎖の配列領域は、「上流配列」として言及される;RNAと同じ配列を有し、RNA転写物の3’から3’末端であるDNA鎖の配列領域は、「下流配列」として言及される。
【0065】
ポリペプチドに適用される用語「実質的な同一性」は、例えば、デフォルトギャップウエイトを用いたプログラムGAPまたはBESTFITにより、最適にアラインメントされた場合、2ペプチド配列は、少なくとも80%の配列同一性、好ましくは、少なくとも90%の配列同一性、より好ましくは、少なくとも95%の配列同一性、そして最も好ましくは、少なくとも99%の配列同一性を有することを意味する。好ましくは、同一でない残基の位置は、保存的アミノ酸置換により異なる。保存的アミノ酸置換は、互換性のある類似の側鎖を有する残基に言及する。例えば、脂肪族側鎖を有するアミノ酸基は、グリシン、アラニン、バリン、ロイシン、およびイソロイシンであり;脂肪族ヒドロキシル側鎖を有するアミノ酸基は、セリン、およびスレオニンであり;アミド含有側鎖を有するアミノ酸基は、アスパラギン、およびグルタミンであり;芳香族側鎖アミノ酸基を有するアミノ酸基は、フェニルアラニン、チロシン、およびトリプトファンであり;塩基性側鎖を有するアミノ酸基は、リジン、アルギニン、およびヒスチジンであり;そして硫黄含有側鎖を有するアミノ酸基は、システイン、およびメチオニンである。好ましい保存的アミノ酸の置換基は、バリン−ロイシン−イソロイシン、フェニルアラニン−チロシン、リジン−アルギニン、アラニン−バリン、グルタミン酸−アスパラギン酸、およびアスパラギン−グルタミンである。
【0066】
本明細書で考察される、抗体または免疫グロブリン分子のアミノ酸配列のマイナーバリエーションは、アミノ酸配列のバリエーションが、少なくとも75%、より好ましくは、少なくとも80%、90%、95%、そして最も好ましくは99%のオリジナル配列を保持している場合、本明細書に記載の発明に包含されることが意図される。特に、保存的アミノ酸置換が意図される。保存的置換は、それらの側鎖に関連性がある、アミノ酸ファミリー内で生じるものである。遺伝子的にコードされたアミノ酸は、一般に、以下のファミリー:(1)酸性=アスパラギン酸、グルタミン酸;(2)塩基性=リジン、アルギニン、ヒスチジン;(3)非極性=アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン;および(4)非荷電極性=グリシン、アスパラギン、グルタミン、システイン、セリン、スレオニン、チロシンに分けられる。より好ましいファミリーは以下である:セリンおよびスレオニンが脂肪族ヒドロキシファミリーであり;アスパラギンおよびグルタミンがアミド含有ファミリーであり;アラニン、バリン、ロイシン、およびイソロイシンが脂肪族ファミリーであり;そしてフェニルアラニン、トリプトファン、およびチロシンが芳香族ファミリーである。例えば、ロイシンのイソロイシンまたはバリンでの単離された置換、アスパラギン酸のグルタミン酸での単離された置換、スレオニンのセリンでの単離された置換、またはアミノ酸の構造的に関連するアミノ酸での同様の置換は、特に該置換がフレームワーク内にアミノ酸を含まない場合、得られた分子の結合または特性に大きく影響することはないと予測することはもっともである。アミノ酸変更が機能的ペプチドとなるか否かは、ポリペプチド誘導体の特異的活性をアッセイすることで、容易に決定され得る。アッセイは本明細書で詳細に説明されている。抗体または免疫グロブリン分子のフラグメントまたは類似体は、当業者により容易に製造され得る。フラグメントまたは類似体の好ましいアミノ末端およびカルボキシ末端は、機能ドメインの境界付近に存在する。構造ドメインおよび機能的ドメインは、ヌクレオチドおよび/またはアミノ酸配列データを公開または独自の配列データベースと比較することにより、同定され得る。好ましくは、コンピューター上での比較方法が、既知の構造および/または機能の他のタンパク質に存在する配列モチーフまたは予測タンパク質構造ドメインを同定するために用いられる。既知の3次元構造に折りたたまれたタンパク質配列を同定するための方法が知られている。Bowie et al., Science 253: 164(1991)。従って、上記の例は、当業者が、本発明をふまえて構造ドメインおよび機能ドメインを定義するために用いられ得る配列モチーフおよび構造を認識することができることを示している。
【0067】
好ましいアミノ酸置換は、(1)タンパク質分解に対する感受性を低減し、(2)酸化に対する感受性を低減し、(3)タンパク質複合体形成のための結合親和性を変化させ、(4)結合親和性を変化させ、かつ(4)かかる類似体の他の物理化学的または機能的特性を与えるか、または修飾するものである。類似体は、天然に存在するペプチド配列以外の配列の種々の変異体を含み得る。例えば、1つまたは複数のアミノ酸置換(好ましくは、保存的アミノ酸置換)は、天然に存在する配列内(好ましくは、分子間接触を形成するドメイン外のポリペプチドの部分において)で生じてもよい。保存的アミノ酸置換は、実質的には、親配列の構造特徴を変更してはいけない(例えば、置換アミノ酸は、親配列中に存在するらせん構造を破壊、あるいは親配列を特徴付ける他のタイプの二次構造を破壊する傾向があってはならない)。当該技術分野で認識されているポリペプチドの二次元および三次元構造の例は、Proteins, Structures and Molecular Principles(Creighton, ed., W. H. Freeman and Company, New York 1984); Introduction to Protein Structure(Branden, C. and Tooze, J. eds., Garland Publishing, New York, N. Y. 1991);およびThornton et al., Nature 354: 105(1991)に記載されている。
【0068】
本明細書で用いられる用語「ポリペプチドフラグメント」は、アミノ末端および/またはカルボキシ末端の欠失したポリペプチドを意味するが、残存アミノ酸配列は、例えば、全長cDNA配列から推定される天然に存在する配列の対応する位置と同一である。フラグメントは、典型的には、少なくとも5、6、8または10アミノ酸長であり、好ましくは、少なくとも14アミノ酸長であり、より好ましくは、少なくとも20アミノ酸長であり、通常は、少なくとも50アミノ酸長であり、そしてさらにより好ましくは、少なくとも70アミノ酸長である。本明細書で用いられる用語「類似体」は、推定されたアミノ酸配列の一部分と実質的に同一であり、かつ以下の特性:(1)適切な結合条件下でPDGF−DD2量体と特異的に結合すること、(2)適切なPDGF−DD結合を崩壊させる能力、または(3)インビトロまたはインビボでPDGF−DDを発現する細胞の増殖を阻害する能力のうち少なくとも1つを有する、少なくとも25アミノ酸のセグメントを含むポリペプチドに言及する。典型的には、ポリペプチド類似体は、天然に存在する配列に対して、保存的アミノ酸置換(または、付加または欠失)を含む。類似体は、典型的には、少なくとも20アミノ酸長、好ましくは、少なくとも50アミノ酸長、またはそれより長く、しばしば天然に存在するポリペプチドの全長と同じ長さであり得る。
【0069】
ペプチド類似体は、一般に、テンプレートペプチドのものと類似する特性を有する非ペプチド薬として医薬産業において用いられる。この種の非ペプチド化合物は、「ペプチドミメティック(peptide mimetic)」または「ペプチドミメティック(peptidomimetic)」と呼ばれる。Fauchere, J. Adv. Drug Res. 15: 29(1986); Veber and Freidinger, TINS p. 392(1985);およびEvans et al.,J. Med. Chem. 30: 1229(1987)を参照されたい。かかる化合物は、たいていコンピュータ分子モデリングの助けにより開発される。治療上有用なペプチドと構造が類似したペプチドミメティックは、均等な治療効果または予防効果を生み出すために用いられ得る。一般に、ペプチドミメティックは、ヒト抗体の様なパラダイムポリペプチド(すなわち、生化学特性または薬理学活性を有するポリペプチド)と構造が類似しているが、当該技術分野でよく知られた方法により、−CH2NH−、−CH2S−、−CH2−CH2−、−CH=CH−(シスおよびトランス)、−COCH2−、−CH(OH)CH2−、および−CH2SO−からなる群から選択される結合で必要に応じて置換された1以上のペプチド結合を有している。一致配列の1以上のアミノ酸の同種のD−アミノ酸での系統的置換が、より安定なペプチドを製造するために用いられてもよい。さらに、一致配列または実質的に同一の一致配列バリエーションを含む制約されたペプチドは、当該技術分野で既知の方法(Rizo and Gierasch Ann. Rev. Biochem. 61: 387(1992))、例えば、ペプチドを環化させる分子内ジスルフィド架橋を形成し得る内部システイン残基を付加することにより、製造されてもよい。
【0070】
「抗体」または「抗体ペプチド」は、未変性の抗体、または未変性の抗体と特異的結合に対して競合するその結合フラグメントを意味する。結合フラグメントは、組換えDNA技術、または未変性の抗体の酵素的分解または化学的分解により作製される。結合フラグメントは、Fab、Fab’、F(ab’)2、Fvおよび1本鎖抗体が挙げられる。「二重特異性」または「二機能性」抗体以外の他の抗体は、その結合部位のそれぞれが同一であることが理解される。過剰の抗体が、対受容体と結合する受容体の量を少なくとも約20%、40%、60%または80%、より一般的には約85%(インビトロ競合結合アッセイにて測定)より多く低減させるとき、抗体は、受容体の対受容体との接着を実質的に阻害する。
【0071】
用語「エピトープ」は、免疫グロブリンまたはT細胞受容体と特異的に結合し得る任意のタンパク質決定基を含む。エピトープ決定基は、一般的には、アミノ酸または糖側鎖のような分子の化学的に活性な表面群からなり、一般的には、特異的な三次元構造特徴、ならびに特異的荷電特徴を有する。抗体は、解離定数が1μM以下、好ましくは100nM以下、そして最も好ましくは、10nM以下であるときに、抗原と特異的に結合するといわれている。
【0072】
用語「剤」は、化学化合物、化学化合物の混合物、生物学的マクロ分子、または生物学的材料から作られた抽出物を示すために、本明細書で用いられる。
【0073】
本明細書での目的上、「活性な」または「活性」は、天然のPDGF−DDポリペプチドまたは天然に存在するPDGF−DDポリペプチドの生物学的活性および/または免疫学的活性を保持しているPDGF−DDポリペプチドの形態を意味し、ここで「生物学的」活性とは、天然のPDGF−DDまたは天然に存在するPDGF−DDポリペプチドが有する抗原性エピトープに対する抗体産生を誘導する能力以外の、天然のPDGF−DDまたは天然に存在するPDGF−DDポリペプチドにより引き起こされる生物学的機能(阻害性または刺激性のいずれか)を意味し、そして「免疫学的」活性とは、天然のPDGF−DDまたは天然に存在するPDGF−DDポリペプチドが有する抗原性エピトープに対する抗体産生を誘導する能力を意味する。
【0074】
「処置」は、治療上の処置、および予防的(prophylactic)または予防上(preventive)の測定の両方を意味し、目的は、標的の病状または疾患を予防するか、あるいは遅延(緩和)することである。処置の必要なものは、既に疾患を有する者、ならびに該疾患に罹りやすいものまたは当該疾患の予防が必要とされるものを含む。
【0075】
「哺乳動物」は、哺乳動物に分類される任意の動物を意味し、ヒト、サル、チンパンジーおよびゴリラの様な他の霊長類、イヌ、ネコ、ウシ、ウマ、ヒツジ、ブタ、ヤギ、ウサギ、げっ歯類等の様な家庭用および農場用動物(家畜)、ならびに動物園、スポーツ、実験、または愛玩動物が挙げられる。処置の目的上、哺乳動物は、好ましくはヒトである。
【0076】
本明細書で用いられる「担体」は、利用される投薬量および濃度でそれらに曝露された細胞または哺乳動物にとって毒性のない、医薬的に許容される担体、賦形剤、または安定化剤を含む。生理的に許容される担体は、たいてい水性のpH緩衝溶液である。生理的に許容される担体の例は、リン酸、クエン酸、および他の有機酸のような緩衝液;アスコルビン酸を含む抗酸化剤;低分子量(約10残基未満)のポリペプチド;血清アルブミン、ゼラチン、または免疫グロブリンのようなタンパク質;ポリビニルピロリドンのような親水性ポリマー;グリシン、グルタミン、アスパラギン、アルギニン、またはリジンのようなアミノ酸;グルコース、マンノース、またはデキストリンを含む単糖、二糖および他の多糖類;EDTAのようなキレート剤;マンニトールまたはソルビトールのような糖アルコール;ナトリウムのような塩形成対イオン;および/またはTWEEN(登録商標)、ポリエチレングリコール(PEG)およびPLURONICS(登録商標)のような非イオン性界面活性剤を含む。
【0077】
抗体のパパイン消化により、それぞれ単一の抗原結合部位を有する「Fab」フラグメントと呼ばれる2つの同一の抗原結合フラグメントが得られ、そして残りの「Fc」フラグメントは、容易に結晶化される能力を反映した表示である。ペプシン処理により、「F(ab’)2」フラグメントが得られる、これは2つの抗原結合部位を有し、かつ抗原と架橋することができる。
【0078】
「Fv」は、抗体の完全な抗原認識部位および結合部位を含有する最小の抗体フラグメントである。この領域は、非共有結合で強く会合した、1つの重鎖と1つの軽鎖の可変ドメインの2量体からなる。この構成で、各可変ドメインの3つのCDRが相互作用してVH−VL2量体の表面の抗原結合部位を規定する。総合すると、6つのCDRが、抗原結合特異性を抗体に与える。しかしながら、例えば、1つの可変ドメイン(例えば、Fv2量体のVHまたはVL部分、または抗原特異的な3つのCDRのみを含むFvの半分)でさえも、おそらく完全な結合部位よりも低い親和性ではあるが、抗原を認識し結合する能力を有し得る。
【0079】
Fabフラグメントはまた、軽鎖の定常ドメイン、および重鎖の第一の定常ドメイン(CH1)を含有する。Fabフラグメントは、抗体のヒンジ領域由来の1つ以上のシステインを含む重鎖CH1ドメインのカルボキシ末端での2、3の残基の付加により、Fab’フラグメントとは異なっている。F(ab’)2抗体フラグメントは、本来、それらの間にヒンジシステインを有するFab’フラグメント対として製造された。抗体フラグメントの他の化学結合もまた知られている。
【0080】
「固相」は、本明細書に記載の抗体と接着できる、非水性マトリックスを意味する。本明細書において包含される固相の例は、部分的または全体的に、ガラス(例えば、孔制御ガラス)、多糖(例えば、アガロース)、ポリアクリルアミド、ポリスチレン、ポリビニルアルコール、およびシリコーンから形成されたものが挙げられる。ある種の実施態様において、状況に応じて、固相は、アッセイプレート、いいかえれると、精製カラム(例、アフィニティークロマトグラフィーカラム)のウェルを含むこともできる。この用語はまた、米国特許第4,275,149号に記載されるものの様な個別の粒子の非連続固相も含む。
【0081】
用語「リポソーム」は、哺乳動物への薬(例えば、PDGF−DDポリペプチド、またはこれに対する抗体)の輸送に有用な、種々のタイプの脂質、リン脂質、および/または界面活性剤を含む、小胞を示すために本明細書において用いられる。リポソームの成分は、一般的に、生物学的な膜の脂質配列と類似した二層の構造で配列される。
【0082】
用語「小分子」は、約500ダルトン未満の分子量を有する分子を記載するために、本明細書で用いられる。
【0083】
本明細書で用いられる用語「標識」または「標識された」は、例えば、放射標識されたアミノ酸の取込による、検出可能なマーカーの取込、または印の付いたアビジンにより検出され得るビオチン化部分のポリペプチドとの接着(例えば、光学的方法または比色方法により検出され得る蛍光マーカーまたは酵素活性を含有するストレプトアビジン)。ある種の状況において、標識またはマーカーは治療用であり得る。ポリペプチドおよび糖タンパク質を標識する種々の方法が当該技術分野で知られており、用いられ得る。ポリペプチドの標識の例は、以下のものが挙げられるが、これに限らない:放射性同位元素または放射性核種(例えば、3H、14C、15N、35S、90Y、99Tc、111In、125I、131I)、蛍光標識(例えば、FITC、ローダミン、ランタニド蛍光体)、酵素標識(例えば、西洋ワサビペルオキシダーゼ、β−ガラクトシダーゼ、ルシフェラーゼ、アルカリホスファターゼ)、化学発光、ビオチニル基、二次レポーターにより認識される、事前に決定されたポリペプチドエピトープ(例えば、ロイシンジッパー対配列、二次抗体の結合部位、金属結合ドメイン、エピトープタグ)。ある実施態様において、標識は、種々の長さのスペーサーアームに結合して、可能性のある立体障害が低減される。
【0084】
本明細書で用いられる用語「医薬剤または薬」は、患者に適切に投与した場合に所望の効果を誘導し得る化学化合物または組成物を意味する。本明細書の他の化学用語は、The McGraw-Hill Dictionary of Chemical Terms(Parker, S. , Ed., McGraw-Hill, San Francisco(1985))に例示される、当該技術分野での従来の使用に従い、用いられる。
【0085】
本明細書で用いられる「実質的に純粋」は、目的の種が、存在する優勢種であること(すなわち、モル基準で、組成物において他のいかなる個々の種よりも豊富に存在すること)を意味し、好ましくは、実質的に精製された画分は、目的の種が、存在する全てのマクロ分子種の少なくとも約50%(モル基準)を含む、組成物である。一般的には、実質的に純粋な組成物は、存在する全てのマクロ分子種の約80%より多く、より好ましくは、約85%、90%、95%および99%より多くを含むであろう。最も好ましくは、目的の種は、精製されて本質的に均一になり(コンタミ種は、従来の検出方法によって組成物中で検出されない)、ここで、組成物は本質的には単一のマクロ分子種からなる。
用語「患者」は、ヒト患者および獣医対象を含む。
【0086】
抗PDGF−DD抗体
抗体、または部分、フラグメント、ミメティック、またはその誘導体は、PDGF−DD2量体を認識する、任意のタイプの抗体または部分であってもよい。ある種の実施態様において、好ましくは、抗体、またはその部分は、PDGF−DDを中和できる。さらなる実施態様において、好ましくは、抗体、またはその部分は、PDGF−DDおよび腎炎と関連する症状(炎症、体液鬱滞、組織腫脹、疼痛、腫脹、高血圧、脳腫脹、視覚障害、低尿量、および血尿が挙げられるが、これに限らない)を低減し得る。1つの実施態様によれば、抗体は、例えば、抗PDGF−DD mAb 6.4であり得る。かかる抗体のさらなる例は、関連米国特許出願第10/041,860号(出願日:2,002年1月7日)において、見出すことができる。
【0087】
抗体構造
基本的な抗体構造単位は、4量体を含むことが知られている。それぞれの4量体は、2つの同一対ポリペプチド鎖から構成され、それぞれの対は、1つの「軽」鎖(約25kDa)および1つの「重」鎖(約50〜70kDa)を有している。それぞれの鎖のアミノ末端部分は、主に抗原認識に関与する約100〜110またはそれ以上のアミノ酸からなる可変領域を含む。それぞれの鎖のカルボキシ末端部分は、主にエフェクター機能に関与する定常領域を規定する。ヒト軽鎖は、κおよびλ軽鎖に分類される。重鎖は、μ、δ、γ、α、またはεに分類され、アイソトープをそれぞれIgM、IgD、IgAおよびIgEと規定する。軽鎖および重鎖内の可変領域および定常領域は、約12以上のアミノ酸の「J」領域により結合されており、重鎖はまた、約10より多くのアミノ酸からなる「D」領域を含む。概論として、Fundamental Immunology Ch. 7(Paul, W., ed., 2d ed. Raven Press, N. Y.(1989))を参照されたい。それぞれの軽鎖/重鎖対の可変領域は、抗体結合部位を形成する。従って、未変性の抗体は、2つの結合部位を有する。例外的に、二機能性または二重特異性抗体では、2つの結合部位は同一である。
【0088】
鎖は全て、相補性決定領域またはCDRとも呼ばれる、3つの超可変領域により結合された比較的保存されたフレームワーク領域(FR)からなる同じ一般構造を有する。それぞれの対の2つの鎖由来のCDRは、フレームワーク領域によりアライメントされ、特異的エピトープと結合することが可能になる。N末端からC末端まで、軽鎖および重鎖は両方ともドメインFR1、CDR1、FR2、CDR2、FR3、CDR3およびFR4を含む。それぞれのドメインに対するアミノ酸指定は、Kabat Sequences of Proteins of Immunological Interest(National Institutes of Health, Bethesda, Md.(1987 and 1991))、またはChothia & Lesk J. Mol. Biol. 196: 901-917(1987);Chothia et al. Nature 342:878-883(1989)の定義に従う。
【0089】
二重特異性抗体または二機能性抗体は、2つの異なる重鎖/軽鎖対および2つの異なる結合部位を有する人工ハイブリッド抗体である。二重特異性抗体は、ハイブリドーマの融合またはFab’フラグメントの結合を含む、種々の方法により製造され得る。例えば、Songsivilai & Lachmann,Clin. Exp. Immunol. 79: 315-321(1990)、Kostelny et al., J. Immunol. 148: 1547-1553(1992)を参照されたい。二重特異性抗体の製造は、従来の抗体製造と比較してかなり苦労が多い方法であり得るが、二重特異性抗体の収率および純度は一般に低い。二重特異性抗体は、1つの結合部位を有するフラグメント(例えば、Fab、Fab’およびFv)の形では存在しない。
かかる二機能性抗体または二重特異性抗体が本発明で意図され、そして包含されることが理解されるであろう。
【0090】
ヒト抗体および抗体のヒト化
本明細書に記載の発明の実施態様はまた、ヒト抗体も意図し包含する。ヒトの処置のため、ヒト抗体は、マウスまたはラットの可変領域および/または定常領域を有する抗体に関連するある種の問題を回避する。かかるマウスまたはラット由来タンパク質の存在は、抗体の迅速なクリアランスをもたらし得るか、あるいは患者の抗体に対する免疫応答を発症し得る。マウスまたはラット由来抗体の利用を回避するため、ヒト化抗体を開発するか、あるいはげっ歯類が完全ヒト抗体を産生するように、ヒト抗体機能をげっ歯類に導入することで完全ヒト抗体を製造できると仮定されてきた。
【0091】
ヒト抗体
完全ヒト抗体を製造する1つの方法は、ゲノム内にヒト重鎖および軽鎖遺伝子を含有するように操作された、XenoMouse(登録商標)系マウスを用いることによる。例えば、ヒト重鎖遺伝子座およびκ軽鎖遺伝子座の245kbおよび190kbサイズの生殖系列立体配置フラグメントを含有するXenoMouse(登録商標)マウスは、Green et al., Nature Genetics 7: 13-21(1994)に記載されている。Green等の研究は、それぞれメガ塩基サイズのヒト重鎖遺伝子座およびκ軽鎖遺伝子座の生殖系列立体配置YACフラグメントの利用により、約80%より多くのヒト抗体レパートリーの導入にまで拡大した。Mendez et al., Nature Genetics 15: 146-56(1997)および米国特許出願第08/759,620号(出願日:1,996年12月3日)を参照のこと。さらに、全λ軽鎖遺伝子座を含有するXenoMouse(登録商標)マウスが作製された(米国特許出願第60/334,508号(出願日:2,001年11月30日))。さらに、複数のアイソタイプを産生するXenoMouse(登録商標)マウスが作製された(例えば、WO00/76310参照)。XenoMouse(登録商標)系は、Abgenix, Inc.(Fremont, CA)から入手可能である。
【0092】
XenoMouse(登録商標)の作製は、さらに、以下で考察され記載されている:米国特許出願第07/466,008号(出願日:1990年1月12日)、同第07/610,515号(出願日:1990年11月8日)、同第07/919,297号(出願日:1992年7月24日)、同第07/922,649号(出願日:1992年7月30日)同第08/031,801号(出願日:1993年3月15日)、同第08/112,848号(出願日:1993年8月27日)、同第08/234,145号(出願日:1994年4月28日)、同第08/376,279号(出願日:1995年1月20日)、同第08/430,938号(出願日:1995年4月27日)、同第08/464,584号(出願日:1995年6月5日)、同第08/464,582号(出願日:1995年6月5日)、同第08/463,191号(出願日:1995年6月5日)、同第08/462,837号(出願日:1995年6月5日)、同第08/486,853号(出願日:1995年6月5日)、同第08/486,857号(出願日:1995年6月5日)、同第08/486,859号(出願日:1995年6月5日)、同第08/462,513号(出願日:1995年6月5日)、同第08/724,752号(出願日:1996年10月2日)および同第08/759,620号(出願日:1996年12月3日)、ならびに米国特許第6,162,963号、同第6,150,584号、同第6,114,598号、同第6,075,181号および同第5,939,598号、ならびに日本国特許第3 068 180 B2号、同第3 068 506 B2号および同第3 068 507 B2号。Mendez et al. Nature Genetics 15: 146-156(1997)およびGreen and Jakobovits J.Exp. Med., 188: 483-495(1998)もまた参照されたい。欧州特許第463,151 B1号(特許発行日:1996年6月12日)、国際特許出願第WO94/02602号(公開日:1994年2月3日)、国際特許出願第WO 96/34096号(公開日:1996年10月31日)同第WO98/24893号(公開日:1998年6月11日)、同第WO00/76310号(公開日:2000年12月21日)もまた参照されたい。
【0093】
代わりのアプローチで、GenPharm International, Inc.を含む他社は、「ミニローカス(minilocus)」アプローチを利用した。ミニローカスアプローチにおいて、外因性のIg遺伝子座は、Ig遺伝子座由来の種(個々の遺伝子)を含むことにより模倣される。従って、1つ以上のVH遺伝子、1つ以上のDH遺伝子、1つ以上のJH遺伝子、μ定常領域および第二の定常領域(好ましくは、γ定常領域)が、動物に挿入するための構築物となる。このアプローチは、米国特許第5,545,807号(Surani et al.)および米国特許第5,545,806号、同第5,625,825号、同第5,625,126号、同第5,633,425号、同第5,661,016号、同第5,770,429号、同第5,789,650号、同第5,814,318号、同第5,877,397号、同第5,874,299号および同第6,255,458号(それぞれLonberg and Kay)、米国特許第5,591,669号および同第6,023,010号(Krimpenfort and Berns)、米国特許第5,612,205号、同第5,721,367号および同第5,789,215(Berns et al.)および米国特許第5,643,763号(Choi and Dunn)、ならびにGen Pharm Internationalの米国特許出願第07/574,748号(出願日:1990年8月29)、同第07/575,962号(出願日:1990年8月31日)、同第07/810,279号(出願日:1991年12月17日)、同第07/853,408号(出願日:1992年3月18日)、同第07/904,068号(出願日:1992年6月23日)、同第07/990,860号(出願日:1992年12月16日)、同第08/053,131号(出願日:1993年4月26日)、同第08/096,762号(出願日:1993年7月22日)、同第08/155,301号(出願日:1993年11月18日)、同第08/161,739号(出願日:1993年12月3日)、同第08/165,699号(出願日:1993年12月10日)、同第08/209,741号(出願日:1994年3月9日)を参照されたい。欧州特許第0 546 073 B1号、国際公開パンフレット第WO92/03918号、同第WO92/22645号、同第WO92/22647号、同第WO92/22670号、同第WO93/12227号、同第WO94/00569号、同第WO94/25585号、同第WO96/14436号、同第WO97/13852号および同第WO98/24884、ならびに米国特許第5,981,175号も参照されたい。さらに、Taylor et al., 1992, Chen et al., 1993, Tuaillon et al., 1993, Choi et al., 1993, Lonberg et al.,(1994), Taylor et al.,(1994),およびTuaillon et al.,(1995), Fishwild et al.,(1996)も参照されたい。
【0094】
上記で引用し、Medical Research Counsel(「MRC」)に帰属しているSurani等発明者は、ミニローカスアプローチの使用により、Ig遺伝子座を有するトランスジェニックマウスを作製した。上記で引用したGenPharm International workの発明者であるLonbergおよびKayは、本発明者らの指示に従って、Surani等の研究の実質的に重複する内因性マウスIg遺伝子座の不活性化を提案した。
【0095】
ミニローカスアプローチの利点は、Ig遺伝子座の一部分を含む構築物が製造され、動物に導入されるという迅速性である。しかしながら、同様にミニローカスアプローチの重大な欠点は、理論上、少数のV、DおよびJ遺伝子を含むことにより不十分な多様性が導入されることである。実際、公開された研究は、この懸念を支持しているようである。ミニローカスアプローチを用いて作製した動物のB細胞発生および抗体産生は、成長が止まっているように思われる。従って、本明細書に記載の発明の周辺研究は、より多くの多様性を達成するためにIg遺伝子座の大部分の導入を目差し続け、かつ動物の免疫レパートリーを再構築するために努力し続けている。
【0096】
Kirinはまた、マイクロ細胞融合により、染色体の巨大片または全染色体が導入されたマウス由来のヒト抗体産生を示している。欧州特許出願第773 288号および同第843 961号を参照されたい。
【0097】
Lidak Pharmaceuticals(現在はXenorex)はまた、ヒトドナー由来の非悪性成熟末梢白血球の注射によって修飾されたSCIDマウスにおけるヒト抗体の産生を示している。修飾されたマウスは、ヒト抗体の産生からなる、免疫原での刺激の際にヒトドナーの免疫応答特徴を示す。米国特許第5,476,996号および同第5,698,767号を参照されたい。
【0098】
ヒト抗マウス抗体(HAMA)応答は、キメラまたはそうでなければヒト化抗体を製造するための産業となる。キメラ抗体はヒト定常領域およびマウス可変領域を有するが、ある種のヒト抗キメラ抗体(HACA)応答は、特に抗体の慢性的または複数回投与の利用において観察されることが予測される。従って、HAMAまたはHACA応答の懸念および/または影響を低減するために、PDGF−DDに対する完全ヒト抗体を提供することが望まれるであろう。
【0099】
ヒト化およびディスプレイ技術
ヒト抗体の製造と関連して上述したように、免疫原性の低減した抗体を製造することには利点がある。これは、ある程度、適切なライブラリを用いたヒト化技術およびディスプレイ技術と関連して成され得る。マウス抗体または他の種由来の抗体は、当該技術分野でよく知られた技術を用いてヒト化または霊長類化し得ることは理解されるであろう。例えば、Winter and Harris,immunol Today 14: 43-46(1993)およびWright et al., Crit, Reviews in Immunol. 12: 125-168(1992)を参照されたい。目的の抗体は、組換えDNA技術により操作され、CH1、CH2、CH3、ヒンジドメインおよび/またはフレームワークドメインを対応するヒト配列と置き換えるられてもよい(例、WO92/02190および米国特許第5,530,101号、同第5,585,089号、同第5,693,761号、同第5,693,792号、同第5,714,350号および同第5,777,085号参照)。また、キメラ免疫グロブリン遺伝子の構築のためのIg cDNAの使用が当該技術分野で知られている(Liu et al., P.N. A. S. 84: 3439(1987)およびJ. Immunol. 139: 3521(1987))。mRNAは、抗体を産生するハイブリドーマまたは他の細胞から単離され、cDNAを製造するために用いられる。目的のcDNAは、特異的なプライマーを用いたポリメラーゼ連鎖反応により増幅されてもよい(米国特許第4,683,195号および同第4,683,202号)。あるいは、ライブラリを作成し、スクリーニングして、目的とする配列を単離する。次いで、抗体の可変領域をコードするDNA配列を、ヒト定常領域配列と融合させる。ヒト定常領域遺伝子の配列は、Kabat et al.,”Sequences of Protein of Immunological Interest”, N. I. H. publication no. 91-3242(1991)において見出され得る。ヒトC領域遺伝子は、既知のクローンから容易に得られる。アイソタイプの選択は、相補的固定の様な所望のエフェクター機能、または抗体依存性細胞毒性の活性により、導かれるであろう。好ましいアイソタイプは、IgG1、IgG3およびIgG4である。ヒト軽鎖定常領域、κまたはλのいずれを用いてもよい。次いで、キメラヒト化抗体は従来の方法により発現される。
【0100】
Fv、F(ab’).sub.2、およびFabのような抗体フラグメントは、未変性のタンパク質の分解、例えば、プロテアーゼまたは化学分解により、製造されてもよい。あるいは、切断された遺伝子が設計される。例えば、F(ab’)2フラグメントの一部分をコードするキメラ遺伝子は、CH1ドメインおよびH鎖のヒンジ領域をコードするDNA配列、次いで、翻訳停止コドンを含む、切断された分子が得られる。
【0101】
重鎖および軽鎖J領域の一致配列は、V領域セグメントのヒトC領域セグメントとの連続結合のため、有用な制限部位をJ領域に導入するためのプライマーとしての使用のためのオリゴヌクレオチドを設計するために用いられてもよい。C領域cDNAを、部位指定の変異誘発により修飾し、制限部位をヒト配列の類似位置に配置することができる。
【0102】
発現ベクターには、プラスミド、レトロウイルス、YAC、EBV由来エピソーム等が挙げられる。好都合なベクターは、任意のVHまたはVL配列が容易に挿入されて発現され得るように、操作された適切な制限部位を有する、機能的に完全ヒトCHまたはCL免疫グロブリン配列をコードするものである。かかるベクターでは、挿入されたJ領域のスプライスドナー部位とヒトC領域に先立つスプライスアクセプター部位との間、およびヒトCHエキソン内で生じるスプライス領域でもスプライシングが起こる。ポリアデニル化および転写の終結は、コーディング領域の下流の天然染色体部位で起こる。得られたキメラ抗体は、任意の強力なプロモーター(例えば、SV−40初期プロモーター(Okayama et al., Mol. Cell. Bio. 3: 280(1983))、Rous肉腫ウイルスLTR(Gorman et al., P. N. A. S. 79: 6777(1982))、およびモロニーマウス白血病ウイルスLTR(Grosschedl et al., Cell 41 : 885(1985))等のレトロウイルスLTRが挙げられる)と結合し得る。また、天然のIgプロモーター等も用いられ得ることは、理解されるだろう。
【0103】
さらに、ヒト抗体または他の種由来の抗体は、ディスプレイタイプ技術(ファージディスプレイ、レトロウイルスディスプレイ、リボソームディスプレイが挙げられるが、これに限らない)、および当該技術分野でよく知られた技術を用いる他の技術により製造され、得られた分子は、親和性成熟のようなさらなる成熟に供することができ、かかる技術は当該技術分野においてよく知られている。Wright and Harris, supra., Hanes and Plucthau, PNAS USA 94: 4937-4942(1997)(リボソームディスプレイ)、Parmley and Smith, Gene 73: 305-318(1988)(ファージディスプレイ)、Scott,TIBS 17: 241-245(1992)、Cwirla et al., PNAS USA 87: 6378-6382(1990)、Russel et al., Nucl. Acid Res. 21: 1081-1085(1993)、Hoganboom et al., Immunol. Reviews 130: 43-68(1992), Chiswell and McCafferty, TIBTECH 10: 80-84(1992)、および米国特許第5,733,743号を参照。ディスプレイ技術がヒトでない抗体を製造するために用いられる場合、かかる抗体は上述のヒト化抗体であり得る。
【0104】
これらの技術を用いて、PDGF−DD発現細胞、PDGF−DD自体、PDGF−DDの形態、それらのエピトープまたはペプチドに対する抗体が製造され、その後、それらの発現ライブラリー(例えば、米国特許第5,703,057号参照)は、上述の活性について上述のようにスクリーニングされ得る。
【0105】
抗体の調製
XenoMouse(登録商標)テクノロジーを用いることにより、PDGF−Dの2量体形に特異的な完全ヒトモノクローナル抗体が製造された。原則的に、マウスのXenoMouse(登録商標)系は、PDGF−DDまたはそれらのフラグメントで免疫され、リンパ球(例えば、B細胞)が、抗体を発現するマウスから回収され、回収された細胞がミエロイド型細胞株と融合されて不死化ハイブリドーマ細胞株が製造され、かかるハイブリドーマ細胞株がスクリーニングされて選択され、PDGF−DDに特異的な抗体を産生するハイブリドーマ細胞株が同定された。さらに、かかる細胞株により産生された抗体の特徴決定が、本明細書に記載され、これは、かかる抗体の重鎖および軽鎖のヌクレオチドおよびアミノ酸配列解析を含む。
【0106】
好ましい実施態様において、抗体は、本明細書に記載の抗PDGF−DD mAb 1.6、1.9、1.18、1.19、1.22、1.29、1.33、1.40.1、1.45、1.46、1.51、1.59、および6.4を中和するものから選択される。国際公開公報WO03/057,857号(2003年7月17日公開)を参照されたい。もちろん、開示された方法は任意の特定の抗PDGF−DDモノクローナル抗体の使用に限られず、むしろ任意のかかる抗体の使用を包含する。
【0107】
あるいは、ハイブリドーマを作成するためにミエローマ細胞と融合される代わりに、免疫されたXenoMouse(登録商標)系マウスから回収され単離された細胞は、最初の抗原、好ましくは、PDGF−DDタンパク質に対する反応性についてさらにスクリーニングされ得る。かかるスクリーニングには、PDGF−DD−Hisタンパク質を用いるELISA、目的の抗原と結合する既知の抗体を用いる競合アッセイ、および一過性にトランスフェクションされた全長PDGF−DD発現CHO細胞とのインビトロでの結合が挙げられる。次いで、目的の抗体を分泌する1つのB細胞が、PDGF−DD特異的溶血性プラークアッセイ(Babcook et al., Proc.Natl. Acad. Sci. USA, 93: 7843-7848(1996))を用いて単離される。溶解の標的細胞は、好ましくは、PDGF−DD抗原でコーティングされたヒツジ赤血球(SRBC)である。目的の免疫グロブリンおよび相補体を分泌するB細胞培養液の存在下で、プラークの形成は、標的細胞の特異的なPDGF−DD仲介性溶解を示す。プラークの中央にある単一抗原特異的血漿細胞が単離され、次いで、抗体をコードするDNAが、1つの血漿細胞から単離され得る。逆転写PCRを用いて、抗体の可変領域をコードするDNAが特異的にクローン化され得る。次いで、かかるクローン化されたDNAが、適当な発現ベクター、好ましくは、pcDNAのようなベクターカセット、より好ましくは、免疫グロブリン重鎖および軽鎖の定常ドメインを含有するpcDNAベクターにさらに挿入され得る。次いで、作成されたベクターが、宿主細胞、好ましくは、CHO細胞にトランスフェクションされ、プロモーターの誘導、形質転換体の選択、または所望の配列をコードする遺伝子の増幅のため必要に応じて改変された、通常は栄養培地中で培養され得る。PDGF−DDに特異的な抗体を産生する複数の単一血漿細胞の単離は、本明細書に記載されている。さらに、抗PDGF−DD抗体の特異性をコードする遺伝子材料が単離され、適切な発現ベクターに導入され、次いで宿主細胞にトランスフェクションされる。
【0108】
一般に、上述の細胞株により産生される抗体は、完全ヒトIgG2重鎖をヒトκ軽鎖と共に有することが見出された。該抗体は高い親和性、典型的には、固相および液相のいずれかで測定される場合、約10−6ないし約10−11MのKdを有していた。これらのmAbは、抗原結合競合研究に基づき、グループまたは「ビン(bin)」に分類され得る。国際公開公報第WO03/048,731号(2003年6月12日公開)参照。
【0109】
抗PDGF−DD抗体の治療上の有用性に対する親和性の重要性については、例えば、コンビナトリアルに抗PDGF−DD抗体を製造し、かかる抗体を結合親和性について評価し得ることは理解されるだろう。利用され得る1つのアプローチは、上述のように製造され、PDGF−DDに対する良好な親和性を有していることが見出された抗体由来の重鎖cDNAを取り出し、上述のように製造され、これを、PDGF−DDに対して良好な親和性を有していることが見出された2次抗体由来の軽鎖cDNAと組合せ、3次抗体が製造され得るものである。得られた3次抗体の親和性は、本明細書に記載のように測定され得、望ましい解離定数を有するものが単離され特徴決定される。あるいは、上述の任意の抗体の軽鎖が、当該軽鎖と対にしたときPDGF−DDに対して高親和性を示す重鎖、あるいはその逆のものの産生を補助するツールとして用いられ得る。このライブラリのこれらの重鎖可変領域は、未感作の動物から単離され、過免疫動物から単離され、CDR領域の異なる重鎖可変配列を含有するライブラリーから人工的に製造され、あるいは任意の重鎖可変領域遺伝子(例えば、ランダムまたは指定変異誘発)のCDR領域内で多様性を生み出す任意の他の方法により製造され得る。これらのCDR領域、および特定のCDR3は、元の抗体と最初に対になった重鎖とは有意に異なる長さまたは配列同一性であってもよい。次いで、得られたライブラリーが、PDGF−DDとの高親和性結合についてスクリーニングされ、元の抗体(高親和性および中和)と類似する特性を有する、治療関連抗体分子が製造され得る。重鎖または重鎖可変領域を用いた同様の方法を用いて、固有の軽鎖可変領域を有する治療関連抗体分子が製造され得る。さらに、次いで、新規重鎖可変領域または軽鎖可変領域が、上述と類似の様式で用いられ、新規な抗体分子の製造を可能にする新規軽鎖可変領域または重鎖可変領域が同定され得る。
【0110】
利用され得る別のコンビナトリアルアプローチは、本明細書に記載の発明の抗体、特に相補決定領域(CDR)において利用されるために示されている生殖系の重鎖および/または軽鎖上で変異誘発を実行することである。得られた抗体の親和性は、本明細書に記載のように測定され、望ましい解離定数を有するものが単離されて特徴決定される。好ましい結合体の選択の際には、配列またはそれをコードする配列を用いて、上述の組換え抗体が製造され得る。オリゴヌクレオチド上での変異誘発を実行するための適切な方法は、当業者に知られており、化学的変異誘発(例えば、亜硫酸ナトリウムの使用)、酵素誤取込み、および放射線への曝露が挙げられる。本明細書に記載の発明は、本明細書で定義された抗体を、変異誘発または他の任意の方法により製造された本明細書で明確に説明された抗体に包含することは理解される。さらに、本明細書で定義された保存的または非保存的アミノ酸置換を有する抗体は、本明細書で明確に説明された抗体において製造され、本明細書に記載の発明の実施態様に含まれる。
【0111】
用いられ得る別のコンビナトリアルアプローチは、他の可変領域の遺伝子由来のフレームワーク領域と関連して、上記の抗体のCDR領域、特にCDR3を発現させることである。例えば、1つの抗PDGF−DD抗体の重鎖のCDR1、CDR2およびCDR3は、他の重鎖可変遺伝子のフレームワーク領域と関連して、発現され得る。同様に、抗PDGF−DD抗体の軽鎖のCDR1、CDR2およびCDR3は、他の軽鎖可変遺伝子のフレームワーク領域と関連して、発現され得る。さらに、これらのCDR領域の生殖系配列は、他の重鎖または軽鎖可変領域遺伝子と関連して発現され得る。得られた抗体は、特異性および親和性についてアッセイされ、新規抗体分子の作製が可能になる。
【0112】
本明細書に記載の発明をふまえて製造される抗体が、ハイブリドーマ細胞株以外の細胞株でも発現され得ることは理解されるであろう。特定の抗体をコードする配列は、適当な哺乳動物宿主細胞の形質転換に用いられ得る。形質転換は、ポリヌクレオチドを宿主細胞に導入するための任意の既知の方法によるものであり、例えば、ポリヌクレオチドをウイルス(またはウイルスベクター)パッケージングすること、次に宿主細胞をウイルス(またはベクター)に形質導入すること、あるいは米国特許第4,399,216号、同第4,912,040号、同第4,740,461号および同第4,959,455号に例示される当該技術分野で既知のトランスフェクション方法によってもよい。用いられる形質転換方法は、形質転換されるべき宿主に依存する。異種性のポリヌクレオチドを哺乳動物細胞に導入する方法は当該技術分野で知られており、デキストラン仲介性トランスフェクション、リン酸カルシウム沈殿、ポリブレン仲介性トランスフェクション、原形質融合、エレクトロポレーション、ポリヌクレオチドのリポソームへのカプセル化、およびDNAの核への直接マイクロインジェクションが挙げられる。
【0113】
発現のための宿主哺乳動物細胞株として利用可能な哺乳動物細胞株は、当該技術分野でよく知られており、American Type Culture Collection(ATCC)から入手可能な不死化細胞株を含み、これは、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、ベビーハムスター腎臓(BHK)細胞、サル腎細胞(COS)、ヒト肝細胞癌腫細胞(例えば、Hep G2)、および多数の他の細胞株を含むが、これに限らない。特に好ましい細胞株は、どの細胞株が高い発現レベルを有し、保存的PDGF−DD結合特性を有する抗体を産生するかを決定することにより、選択される。
【0114】
抗体治療のさらなる基準
PDGF−DD抗体の機能は、その操作方法の少なくとも一部分に対して重要であるように思われることが、本明細書において考察される。機能とは、例えば、PDGF−DDに対する応答における抗PDGF−DD抗体の活性を意味する。従って、ある種の場合において、抗体が、補体依存細胞毒性(CDC)および抗体依存性細胞の細胞毒性(ADCC)を含むエフェクター機能を発揮し得ることが、PDGF−DDに対する治療候補としての抗体の製造に関連して望まれてもよい。同様のことが可能な抗体のアイソタイプは多数存在し、これは、マウスIgM、マウスIgG2a、マウスIgG2b、マウスIgG3、ヒトIgM、ヒトIgG1およびヒトIgG3を含むが、これに限らない。製造された抗体が、当初からかかるアイソタイプを有する必要はないが、製造された抗体が任意のアイソタイプを有し得、そして抗体が当該技術分野でよく知られた従来技術を用いてその後アイソタイプに切り替えられ得ることは、理解されるだろう。かかる技術は、直接組換え技術(例えば、米国特許第4,816,397号および同第6,331,415号を参照されたい)、細胞・細胞融合技術(例えば、米国特許第5,916,771号および同第6,207,418号を参照されたい)の使用を含む。
【0115】
細胞・細胞融合技術では、任意の所望のアイソタイプを有する重鎖を有する、ミエローマ細胞株または他の細胞株が製造され、そして軽鎖を有する別のミエローマ細胞株または他の細胞株が製造される。その後、かかる細胞は融合され、未変性の抗体を発現する細胞株が単離され得る。
【0116】
例えば、本明細書で考察される抗PDGF−DD抗体は、ヒト抗PDGF−DD IgG2およびIgG4抗体である。かかる抗体が、PDGF−DD分子との所望の結合性を有する場合、これは、容易には、ヒトIgM、ヒトIgG1またはヒトIgG3、IgGA1またはIgGA2アイソタイプを製造するたに切り換えられたアイソタイプであり得るが、これは、依然として同じ可変領域を有している(抗体の特異性およびその親和性のいくらかを有する)。次いで、かかる分子は、CDCにおいて補体を固定し、関与することができるであろう。
【0117】
従って、抗体候補は、上記の所望の「構造」属性に合致するように製造されるので、これらは一般的には、アイソタイプスイッチングにより、少なくともある種の所望の「機能的」属性を付与され得る。
【0118】
エピトープマッピング
免疫ブロット分析
本明細書に記載の抗体のPDGF−DDとの結合は、多数の方法で試験され得る。例えば、PDGF−DDは、SDS−PAGEに供され、免疫ブロッティングにより分析され得る。SDS−PAGEは、還元剤の不存下または存在下のいずれかで行われてもよい。かかる化学修飾は、システイン残基のメチル化をもたらし得る。従って、本明細書に記載のPDGF−DD抗体がPDGF−DDの線状エピトープと結合しているかどうかを決定することが可能である。
【0119】
表面増強レーザー脱離/イオン化
本明細書に記載のPDGF−DD抗体に対するエピトープのエピトープマッピングはまた、SELDIを用いて行われ得る。SELDI ProteinChip(登録商標)アレイは、タンパク質・タンパク質相互作用部位を定義するために用いられる。抗原は、最初のインキュベーションおよび洗浄によって、Protein Chipアレイ表面に共有結合で固定された抗体に特異的に捕獲される。結合された抗原は、レーザー誘導性脱離方法により検出され、そしてそれらの質量が直接決定され得る。結合した抗原のかかるフラグメントは、タンパク質の「エピトープ」と命名されている。
【0120】
SELDI方法は、複合体分子組成物内の個々の成分を直接検出し、迅速、高感度、かつ測定可能な方法で他の成分と比較して定量的にマッピングすることを可能にする。SELDIは、表面化学の多様なアレイを用いて、レーザー誘導性脱離方法による検出する多数の個々のタンパク質分子を捕獲し提示する。SELDI方法の成功は、それぞれが異なる技術に依存する、複数の機能の表面(「チップ」)上での複数機能の小型化および一体化により部分的に定義される。SELDI BioChipおよび他の型のSELDIプローブは、これらが個々の標的分子(例えば、タンパク質)または評価されるべき分子集団の捕獲、精製(分離)、提示、検出および特徴決定に積極的に関与するように、表面が「増強」されている。
【0121】
オリジナルのサンプルのみを充填された単一のSELDIタンパク質BioChipは、何千回も読み取られ得る。LumiCyteのSELDIタンパク質BioChipは、1平方センチメートル当たり10,000個もの指定可能なタンパク質結合位置を有している。それぞれの位置が、十数個の個々のタンパク質の存在を明らかにできる。それぞれの位置からのタンパク質組成物情報が比較され、独自の情報セットが組み合わされると、得られた組成物マップは、特異的パターンまたは分子「フィンガープリント」を定義するために総括的に用いられる、特徴セットを有するイメージを明らかにする。異なるフィンガープリントは、健常な種々の段階、疾患の発症、または適当な治療法の投与と関連する疾患の緩和と関係し得る。
【0122】
SELDI方法は、4つの部分に分けてより詳細に説明され得る。最初に、目的の1つ以上のタンパク質が、サンプルを調製および標識することなく、オリジナルの供給源材料から直接的にProteinChipアレイ上に捕獲または「ドッキング(docked)」される。第2の工程では、化学的および生体分子的「ノイズ」を低減することにより、「シグナル対ノイズ」比が増強される。かかる「ノイズ」は、非所望の材料を洗浄して除くことによるチップ上の標的の選択的保持により低減される。さらに、捕獲された1つ以上の標的タンパク質が、迅速かつ高感度のレーザー誘導性方法(SELDI)により読み取られ、これは、標的に関する直接的情報(分子量)を提供する。最後に、アレイ内の任意にの1以上の位置の標的タンパク質が、1つ以上のチップ上の結合またはタンパク質構造および機能を特徴決定するための修飾を行うことにより、インサイツで特徴決定され得る。
【0123】
ファージディスプレイ
本明細書に記載のPDGF−DD抗体のエピトープは、Filamentousファージ(New England Biolabs)上にディスプレイされたランダムペプチドである12merのコンビナトリアルライブラリにProteinChipアレイを曝露することにより決定され得る。
【0124】
ファージディスプレイは、選択技術を記載し、ここで、ペプチドは、バクテリオファージのコートタンパク質との融合として発現され、ウイルス粒子の表面に融合タンパク質のディスプレイとなる。パニングは、ペプチドを現したファージのライブラリーを標的でコートされたプレートおよびチューブとインキュベートし、非結合ファージを洗浄し、次に、特異的結合ファージを溶出することにより、行われる。次いで、溶出されたファージが増幅され、さらなる結合および増幅サイクルにより取り出され、所望の結合配列プールが富化される。3回または4回の後、個々の結合クローンが、抗体コートウェル上で行われるファージELISAアッセイにより結合についてさらに試験され、そしてポジティブクローンの特異的DNA配列決定により特徴決定される。
【0125】
本明細書に記載のPDGF−DD抗体に対するかかるパニングを複数回おこなった後、結合ファージは溶出され、結合ペプチドの同定および特徴決定のためのさらなる研究に供され得る。
【0126】
PDGF−DDアゴニストおよびアンタゴニスト
本明細書に記載の発明の実施態様はまた、PDGF−DDアゴニスト(ミメティック)またはPDGF−DDアンタゴニストのいずれかとして機能する、PDGF−DDタンパク質の変異体に関与する。好ましくは、PDGF−DDタンパク質の変異体は、腎炎の処置に有用である。PDGF−DDタンパク質の変異体は、変異誘発、例えば、PDGF−DDタンパク質の異なる点変異または切断により製造され得る。PDGF−DDタンパク質のアゴニストは、天然に存在する形のPDGF−DDの実質的な同一性、サブセット、生物学的活性を保持し得る。PDGF−DDタンパク質のアンタゴニストは、例えば、PDGF−DDタンパク質を含む細胞シグナル伝達カスケードの下流または上流メンバーと競合的に結合することにより、天然に存在する形態のPDGF−DDタンパク質の1つ以上の活性を阻害することができる。従って、特異的生物学的作用は、機能制限された変異体による処置により誘発され得る。1つの実施態様において、天然に存在する形態のタンパク質の生物学的活性のサブセットを有する変異体による対象の処置は、天然に存在する形態のPDGF−DDタンパク質での処置と比較して、副作用が少ない。
【0127】
PDGF−DDアゴニスト(ミメティック)またはPDGF−DDアンタゴニストのいずれかとして機能するPDGF−DDタンパク質の変異体は、PDGF−DDタンパク質の変異(例えば、切断変異)のコンビナトリアルライブラリーをタンパク質アゴニストまたはアンタゴニスト活性についてスクリーニングすることにより同定され得る。1つの実施態様において、PDGF−D変異体の変化に富むライブラリーは、核酸レベルでのコンビナトリアル変異誘発により製造され、これは、変化に富む遺伝子ライブラリーによりコードされる。PDGF−D変異体の変化に富むライブラリーは、可能性のあるPDGF−D配列の変性セットが個々のポリペプチド、またはそこにPDGF−D配列セットを含有する巨大融合タンパク質のセット(例えば、ファージディスプレイ)として発現可能なように、例えば、合成オリゴヌクレオチド混合物の遺伝子配列への酵素的ライゲーションにより生み出され得る。変性オリゴヌクレオチド配列由来の可能性のあるPDGF−D変異体ライブラリーを作製するために用いられ得る種々の方法が存在する。変性遺伝子配列の化学合成は、自動DNA合成装置で行われることができ、次いで、合成遺伝子は、適当な発現ベクタとライゲーションされる。遺伝子の変性セットの使用により、可能性のあるPDGF−D変異体配列の所望のセットをコードする配列の全てを1つの混合物で提供することが可能となる。変性オリゴヌクレオチドの合成方法は、当該技術分野で知られている(例えば、Narang, Tetrahedron 39: 3(1983); Itakura et al.,Annu. Rev.Biochem. 53: 323(1984); Itakura et al., Science 198: 1056(1984); Ike et al.,Nucl. Acid Res. 11: 477(1983)を参照されたい)。
【0128】
他の治療法の設計および創出
さらに、PDGF−DDと関連して本明細書で製造され、特徴決定される抗体の活性に基づき、抗体部分を超える他の治療法様式の設計が容易になる。かかる様式は、二重特異性抗体、免疫毒素および放射標識治療法、ペプチド治療法の創出、遺伝子治療剤(特に体内)、アンチセンス治療法、および小分子のような進歩した抗体治療法を含むが、これに限らない。
【0129】
進歩した抗体治療法の創出と関連して、補体固定が所望の属性である場合、例えば、二重特異性、免疫毒素または放射標識の使用により細胞殺傷のための補体依存性を回避することが可能であり得る。
【0130】
例えば、二重特異性抗体に関して、(i)一方はPDGF−DDに対する特異性を有し、他方は、一体となって結合した第2の分子に対する特異性を有する2つの抗体、(ii)PDGF−DDに特異的な1つの鎖および第2の分子に特異的な第2の鎖を有する単一抗体、または(iii)PDGF−DDおよび他の分子に対する特異性を有する1本鎖抗体を含む、二重特異性抗体が製造され得る。かかる二重特異性抗体は、例えば、よく知られた技術を用いて製造され得る((i)および(ii)に関しては、例えば、Fanger et al., Immunol Methods 4: 72-81(1994)およびWright and Harris, supraを、また、(iii)に関しては、例えば、Traunecker et al., Itit. J. Cancer(Suppl.)7: 51-52(1992)を参照されたい)。それぞれの場合で、第2の特異性は、重鎖活性化受容体に対して作りだされ、これは、CD16またはCD64(例えば、Deo et al., 18: 127(1997)参照)、またはCD89(例えば、Valerius et al., Blood 90: 4485-4492(1997)参照)を含むが、これに限らない。上記のように製造された二重特異性抗体は、PDGF−DD発現細胞、特に、本明細書に記載のPDGF−DD抗体が有効な細胞を殺傷するであろう。
【0131】
免疫毒素に関し、抗体は、当該技術分野でよく知られた技術を利用して免疫毒素として作用するように修飾され得る。例えば、Vitetta, Immunol Today 14: 252(1993)を参照されたい。また、米国特許第5,194,594を参照されたい。放射標識抗体の製造と関連し、かかる修飾抗体もまた、当該技術分野でよく知られた技術を用いて容易に製造され得る。例えば、Junghans et al., in Cancer Chemotherapy and Biotherapy 655-686(2d ed., Chafner and Longo, eds., Lippincott Raven(1996))を参照。また、米国特許第4,681,581号、同第4,735,210号、同第5,101,827号、同第5,102,990(RE35,500)号、同第5,648,471号および同第,697,902号も参照されたい。免疫毒素および放射標識分子はそれぞれ、PDGF−DD発現細胞、特に、本明細書に記載のPDGF−DD抗体が有効な細胞を殺傷するであろう。
【0132】
治療ペプチドの製造と関連し、PDGF−DD、および本明細書に記載の抗体(小分子に関しては後述)の様なこれに対する抗体と関連する構造情報を利用、またはペプチドライブラリーのスクリーニングにより、PDGF−DDに対して指定された治療用ペプチドが製造され得る。ペプチド治療法の設計およびスクリーニングは、Houghten et al., Biotechniques 13: 412-421(1992)、Houghten, PNAS USA82: 5131-5135(1985), Pinalla et al., Biotechniques 13: 901-905(1992)、Blake and Litzi-Davis, BioConjugate Chem. 3: 510-513(1992)と関連付けて考察される。免疫毒素および放射標識分子もまた、抗体と関連して上記で考察されたペプチド部分に関連して、類似の方法で製造され得る。
【0133】
PDGF−DD分子(またはスプライス変異体のような形態または代わりの形態)が、疾患過程において機能的に活性であると仮定すると、従来の技術により、遺伝子およびそのアンチセンス治療法を設計することが可能であろう。かかる様式は、PDGF−DDの機能を調節するのに利用され得る。これと関連して、本明細書に記載の抗体は、これと関連する機能的アッセイの設計および使用を可能にする。アンチセンス治療法の設計およびストラテジーは、国際特許出願第WO94/29444号に詳細に論じられている。遺伝子療法の設計およびストラテジーはよく知られいてる。しかしながら、特に、体内に関与する遺伝子治療技術の使用が特に有利であることを明らかにすることができた。例えば、Chen et al., Human Gene Therapy 5: 595-601(1994)およびMarasco, Gene Therapy 4: 11-15(1997)を参照されたい。遺伝子治療法に関する一般的設計および考慮はまた、国際特許出願第WO97/38137号でも考察されている。
【0134】
小分子治療法もまた想定され得る。薬剤は、本明細書に記載のPDGF−DDの活性を調節するように設計され得る。本明細書に記載のPDGF−DD分子の構造、本明細書に記載の抗体のような他の分子とのその相互作用から収集した知識、およびその他を利用して、さらなる治療の方法が合理的に設計され得る。これと関連し、X線結晶学、コンピュータ補助(またはアシスト)分子モデリング(CAMM)、量的または質的構造活性相関(QSAR)のような合理的薬剤設計技術および類似の技術を用いて、薬剤発見努力に焦点が当てられ得る。合理的設計により、PDGF−DD活性を修飾または調節するために用いられ得る、分子またはその特異的形態と相互作用し得るタンパク質または合成構造の予測が可能になる。かかる構造は化学的に合成されるか、または生物学的システムで発現させることができる。このアプローチは、Capsey et al, Genetically Engineered Human Therapeutic Drugs(Stockton Press, NY(1988))で概説されている。さらに、コンビナトリアルライブラリを設計し、合成して、ハイスループットスクリーニング作用の様なスクリーニングプログラムにおいて用いられ得る。
【0135】
治療投与および処方
抗体およびそのフラグメントを含むが、これに限らない抗PDGF−DD化合物は、PDGF−DDを調節する化合物を必要とする生物を処置する医薬への取込に適している。これらの薬理学的に活性な化合物は、生物、例えば、動物およびヒトを含む哺乳動物に投与するための医薬剤を製造するための製剤学の通常の方法と関連して、製造され得る。ある種の実施態様において、有効成分は、修飾してまたは修飾することなく医薬製品に取り込まれ得る。さらなる実施態様は、本明細書に記載の医薬的に活性な化合物をいくつかの経路で送達する医薬または治療剤の製造を含む。例えば、これに限らないが、抗体またはそのフラグメントをコードする配列を有するDNA、RNA、およびウイルスベクターが、ある種の実施態様において用いられ得る。さらに、抗体またはそのフラグメントをコードする核酸は、単独または他の有効成分と組み合わせて投与され得る。
【0136】
本明細書に記載の治療剤の投与は、移動、輸送、耐容性などの改善をもたらすために製剤に取込まれる適当な担体、賦形剤、安定化剤、および他の剤との混合物で投与され得ることは、理解されるだろう。医薬的に許容される担体は、本発明の医薬的有効成分と有害に反応しない非経腸、経腸(例えば、経口)、または局所投与に適した有機または無機担体物質を含む。適当な医薬的に許容される担体は、水、食塩水、アルコール、アラビアゴム、植物油、ベンジルアルコール、ポリエチレングリコール、ゼラチン、ラクトース、アミロースまたはデンプンのような炭水化物、ステアリン酸マグネシウム、タルク、ケイ酸、粘性パラフィン、香料油、脂肪酸モノグリセリドおよびジグリセリド、ペンタエリスリトール脂肪酸エステル、ヒドロキシメチルセルロース、ポリビニルピロリドン等が挙げられるが、これらに限定されない。さらなる担体、賦形剤および安定剤は、Tris−HClのような緩衝液、リン酸塩、クエン酸塩、酢酸塩および他の有機酸塩;アスコルビン酸のような抗酸化剤;ポリアルギニンのような低分子量(約10残基未満)ペプチド、血清アルブミンのようなタンパク質、ゼラチンまたは免疫グロブリン;ポリビニルピロリジノンのような親水性ポリマー;グリシン、グルタミン酸、アスパラギン酸またはアルギニンのようなアミノ酸;セルロースまたはその誘導体、グルコース、マンノースまたはデキストリンを含む単糖、二糖および他の炭水化物を含む他の炭水化物;EDTAのようなキレート剤;マンニトールまたはソルビトールのような糖アルコール;ナトリウムのような対イオンおよび/またはTWEEN、PLURONICSまたはポリエチレングリコールのような非イオン性界面活性剤を含む。多くのより適当なビークルは、Remmington's Pharmaceutical Sciences,15th Edition, Easton: Mack Publishing Company, pages 1405-1412 and 1461-1487(1975)およびThe National Formulary XIV, 14th Edition, Washington, American Pharmaceutical Association(1975)に記載されている。
【0137】
抗体の投与経路は、例えば、これに限定されないが、局所的、経皮的、非経腸、経消化管、経気管および経肺胞を含む既知の方法に従うことができる。非経腸経路投与は、中心静脈への注射、静脈内、脳内、筋肉内、腹腔内、皮内、動脈内、くも膜下内または病変内経路のような電気的または直接的注射または点滴を含むが、これに限らない。抗体は、好ましくは、点滴、ボーラス注射、あるいは下記の徐放システムにより、連続的に投与される。好ましい実施態様において、投与経路は皮下注射であり得る。代わりの実施態様において、抗体は腎動脈を介して投与される。投与の消化管経路には、摂取および直腸経路が挙げられるが、これらに限定されない。投与の気管および肺胞経路には、経口または経鼻経路が挙げられるが、これらに限定されない。
【0138】
インビボでの投与に用いる場合、抗体製剤は無菌であり得る。これは、凍結乾燥および再構成の前および後に滅菌フィルトレーションメンブレンを通してフィルトレーションすることにより、容易に行われ得る。抗体は、通常は、凍結乾燥形または溶液中に貯蔵されるであろう。さらに、治療用組成物は、発熱物質を含まず、pH、等張性および安定性に関して非経腸で許容される溶液であり得る。治療用抗体組成物は、一般に、無菌アクセスポートを有する容器、例えば、皮下注射用針を刺すことが可能なストッパーを有する静脈注射用バッグまたはバイアルに入れられる。
【0139】
注射用無菌組成物は、Remington's Pharmaceutical Sciences(18th ed., Mack Publishing Company, Easton, PA(1990))に記載の従来の医薬実施に従い、製剤され得る。医薬製剤は、所望に応じて、活性化合物と有害な反応をしない滑沢剤、保存剤、安定剤、湿潤剤、乳化剤、浸透圧に影響する塩、緩衝液、抗酸化剤、着色剤、香味剤、または芳香族物質等と混合して、無菌化され得る。例えば、水、またはゴマ油、ピーナッツ油または綿実油のような天然に存在する植物油、またはオレイン酸エチルのような合成脂質ビークル等の様なビークル中の活性化合物の溶解または懸濁が所望されてもよい。
【0140】
非経腸投与に適した本発明の薬理学的に活性な化合物を有する適当な組成物は、医薬的に許容される無菌等張溶液を含むが、これに限らない。かかる溶液は、中心静脈ラインへの注射、静脈内、筋肉内、腹腔内、皮内または皮下注射用の生理食塩水およびリン酸緩衝生理食塩水を含むが、これに限らない。
【0141】
消化管投与に適した、本発明の薬理的に活性な化合物を有する組成物は、医薬的に許容される消化用粉剤、丸薬、または液剤、または直腸投与用座剤を含むが、これに限らない。
【0142】
徐放性製剤の適当な例は、ポリペプチドを含有する固体疎水性ポリマーの半透性マトリックスが挙げられ、ここでマトリックスは、成形された物品、フィルムまたはマイクロカプセルの形態である。徐放性マトリックスの例は、ポリエステル、Langer et al., J. Biomed Mater. Res., 15: 167-277(1981)およびLanger, Chem. Tech. , 12: 98-105(1982)に記載のハイドロゲル、例えば、ポリ(2−ヒドロキシエチルメタクリレート)、またはポリ(ビニルアルコール))、ポリラクチド(米国特許第3,773,919号、欧州特許第58,481号)、L−グルタミン酸とγ−エチル−L−グルタメートとのコポリマー(Sidman et al.,Biopolymer, 22: 547-556(1983))、非分解性エチレン−酢酸ビニル(Langer et al., supra)、LUPRON Depot(登録商標)のような分解性乳酸−グリコール酸コポリマー(乳酸−グリコール酸コポリマーおよび酢酸リュープロリンからなる注射可能な微粒子)、およびポリ−D−(−)−3−ヒドロキシ酪酸(欧州特許第133,988号)が挙げられる。
【0143】
エチレン−酢酸ビニルおよび乳酸−グリコール酸のようなポリマーは、100日間にわたり分子を放出することができるが、特定のハイドロゲルは、タンパク質をより短い期間放出する。カプセル化されたタンパク質が体内に長時間残存すると、37℃で水分に曝露された結果、変性または凝集し得、結果として生物学的活性が損なわれ、免疫原性が変化する可能性がある。関与するメカニズムに応じて、タンパク質安定化のための合理的ストラテジーが考えられ得る。例えば、凝集メカニズムがジスルフィド結合間の変化による分子内S−S結合形成であることが見出された場合、安定化は、スルフヒドリル残基の修飾、酸性溶液からの凍結乾燥、水分含有量の制御、適当な添加剤の使用、および特別なポリマーマトリックス組成物の開発により達成され得る。
【0144】
徐放性組成物はまた、リポソームで包まれた本発明の抗体を含む。かかる抗体を含有するリポソームは、それ自体既知の方法により製造される:米国特許第DE3,218,121号;Epstein et al.,Proc.Natl.Acad. Sci. USA, 82: 3688-3692(1985);Hwang et al.,Proc. Natl. Acad. Sci. USA, 77: 4030-4034(1980);欧州特許第52,322号;欧州特許第36,676号;欧州特許第88,046号;欧州特許第143,949号;142,641号;日本国特許出願第83−118008号(特願昭59−118008号);米国特許第4,485,045号および同第4,544,545号;ならびに欧州特許第102,324号参照。
【0145】
治療に用いられる抗体の有効量は、例えば、治療の対象、投与経路、患者の状態に依存する。抗体用量は、疾患の重症度、種類、体重、性別、食事、投与時間および投与経路、他の医薬、および他の関連臨床因子を含む、薬剤の作用を改変することが知られている種々の因子を考慮しながら、担当医によって決定されるであろう。従って、治療者は、最適な治療効果を得るのに必要とされる用量を滴定し、投与経路を改変する必要があるであろう。典型的には、臨床医は、所望の効果を達成する投薬量に至るまで、抗体を投与するであろう。治療上有効用量は、インビトロまたはインビボでの方法のいずれかにより決定され得る。この療法の進行は、従来のアッセイまたは本明細書に記載のアッセイにより容易にモニターすることができる。
【0146】
かかる化合物の治療有効性および毒性は、細胞培養または実験動物における標準的な医薬手順、例えば、ED50(集団の50%が治療上効果的な用量)により決定され得る。腎炎ラットモデルまたは代わりのモデルの処置から得られたデータは、ヒトを含む他の生物に用いるための用量範囲の創出の際に用いられ得る。かかる化合物の投薬量は、好ましくは、ED50を毒性なく含む、循環濃度の範囲内である。投薬量は、出差(evectin)、ハイブリッド、結合パートナーまたはそのフラグメントの種類、利用される投薬形態、生物種の感度、および投与経路に依存して、この範囲を変動する。
【0147】
種々の抗体またはそのフラグメントの正常な投薬濃度は、約0.1〜100mg/kgで変動し得る。望ましい投薬量濃度は、例えば、0.lmg/kg、0.2mg/kg、0.3mg/kg、0.4mg/kg、0.5mg/kg、0.6mg/kg、0.7mg/kg、0.8mg/kg、0.9mg/kg、l.0mg/kg、1.5mg/kg、2.0mg/kg、2.5mg/kg、3.0mg/kg、3.5mg/kg、4.0mg/kg、4.5mg/kg、5.0mg/kg、5.5mg/kg、6.0mg/kg、6.5mg/kg、7.0mg/kg、7.5mg/kg、8.0mg/kg、8.5mg/kg、9.0mg/kg、10mg/kg、15mg/kg、20mg/kg、25mg/kg、30mg/kg、35mg/kg、40mg/kg、45mg/kg、50mg/kg、55mg/kg、60mg/kg、65mg/kg、70mg/kg、75mg/kg、80mg/kg、85mg/kg、90mg/kg、95mg/kgおよび100mg/kg、またはそれ以上を含む。1つの好ましい投薬量は、1〜10mg/kgである。
【0148】
ある実施態様において、抗体またはそのフラグメントの用量は、約0.1μM〜500mM、好ましくは約1〜800μM、そしてより好ましくは、約10μM〜約500μMの組織または血液濃度あるいはその両方を生じる。好ましい用量は、例えば、10μM、15μM、20μM、25μM、30μM、35μM、40μM、45μM、50μM、55μM、60μM、65μM、70μM、75μM、80μM、85μM、90μM、95μM、100μM、110μM、120μM、130μM、140μM、145μM、150μM、160μM、170μM、180μM、190μM、200μM、220μM、240μM、250μM、260μM、280μM、300μM、320μM、340μM、360μM、380μM、400μM、420μM、440μM、460μM、480μM、および500μMの組織または血液濃度あるいはその両方を達成するのに必要とされる量である。代わりの実施態様において、800μMより高い組織濃度を生じる用量が用いられ得る。抗体、ハイブリッド、結合対、またはそのフラグメントの一定の点滴もまた、血液レベルにより測定される組織での安定な濃度を維持するために提供され得る。
【0149】
投薬量および投与は、活性部分の十分なレベルをもたらすため、あるいは所望の効果を維持するために調整され得る。本明細書の実施態様は、短期作用する医薬組成物、および長期作用する医薬組成物の両方を含む。従って、実施態様は、医薬組成物が、約1、2、3、4、5または6日毎、1週間毎、2週間に1回、3週間に1回、4週間に1回、5週間に1回、6週間に1回、7週間に1回、または8週間に1回投与されるスケジュールを含む。特定の製剤の半減期およびクリアランス率に依存して、本明細書に記載の医薬組成物は、1日あたり約1、2、3、4、5、6、7、8、9および10回、またはそれ以上投与され得る。
【0150】
さらなる治療法が、抗PDGF−DD抗体の投与と組み合わせて、当該投与前または後に行われてもよい。これらの治療法は、疾患の症状を処置するために用いられてもよいし、あるいは抗PDGF−DD抗体の副作用を低減するために用いられてもよい。これらはまた、抗PDGF−DD抗体の活性を増強するために用いられてもよい。これらに限らないが、例えば、抗生物質、利尿剤、麻酔剤、鎮痛剤、抗炎症剤、およびインスリンを含む、任意の種類の治療法が用いられる。糸球体腎炎を処置するために典型的に用いられ、かつ抗体と組合せて用いられ得る剤の例は、プレドニゾン、シクロホスファミド、クロラムブシル、および例えば、ワーファリン、ジピリダモールおよびアスピリンのような血液希釈剤を含む。
【0151】
診断上の使用
PDGF−DDは、正常腎臓では低レベルで発現されるが、発現は虚血後の腎臓で劇的に増加されることが見出された(Ichimura T, Bonventre JV, Bailly V, Wei H, Hession CA, Cate RL, Sanicola M., J.Biol. Chem. 273(7): 4135-42(1998))。抗PDGF−DD抗体による免疫組織化学染色は、腎臓(renal)癌腫、腎臓(kidney)癌腫、前立腺癌腫、および卵巣癌腫での陽性染色を示し(以下参照)、これは、正常組織と比較してPDGF−DDの過剰発現が、かかる疾患の診断マーカーとなり得ることを示す。
【0152】
従って、本発明の実施態様はまた、アッセイ、特にインビトロ診断アッセイ(例えば、患者サンプルにおいてPDGF−DDレベルを決定するための使用)にも有用である。かかるアッセイは、PDGF−DDの過剰発現に関連する疾患の診断に有用であり得る。いくつかの実施態様において、疾患は腎炎である。患者サンプルは、例えば、体液、好ましくは血液、より好ましくは血液血清、滑液、組織溶解物、および疾患組織から調製した抽出物であってもよい。本発明の他の実施態様は、腎炎およびメサンギウム増殖性関連疾患の診断および段階決定において有用である。PDGF−DDのレベルのモニタリングは、疾患に対する患者応答の代わりの測定手段および患者の疾患の重症度のモニタリング方法として用いられ得る。他の溶解性マーカーレベルと比較して、高レベルのPDGF−DDが、虚血後腎臓の存在を示すであろう。患者サンプルに存在するPDGF−DD抗原の濃度は、存在する抗原の量を特異的に決定する方法を用いて、決定され得る。かかる方法には、ELISA法が挙げられ、当該方法では、例えば、本発明の抗体がポリマーマトリックスのような不溶性マトリックス上に簡便に固定され得る。あるいは、抗PDGF−DD抗体を用いる免疫組織学的化学染色アッセイは、サンプル中でPDGF−DDのレベルを決定するために用いられ得る。進行または治療のそれぞれの段階についての統計的に有意な結果をもたらすサンプル集団を用いて、疾患の各段階の特性であると考えられ得る抗原の濃度範囲が設計され得る。
【0153】
1つの実施態様において、血液サンプルが被検者から採取され、サンプル中に存在するPDGF−DD抗原の濃度が決定され、研究下の被検者の疾患の段階が評価されるか、あるいは一連の治療に対する応答が特徴決定される。このようにして得られた濃度を用いて、その値がどの濃度範囲に含まれるかが同定される。このようにして同定された範囲は、診断された対象からなる種々の集団で同定された疾患の進行段階または治療段階と相関し、これにより、研究下の対象の疾患段階を提供する。
【0154】
遺伝子増幅および/または発現は、適当に標識されたプローブを用い、本明細書で提供される配列に基づき、例えば、従来のサザンブロッティング、ノーザンブロッティング(Thomas,Proc. Natl. Acad. Sci. USA, 77: 5201-5205(1980))、ドットブロッティング(DNA分析)またはインサイツハイブリダイゼーションによりサンプル中で直接測定され、mRNAの転写が定量され得る。あるいは、特異的2本鎖(DNA2本鎖、RNA2本鎖、およびDNA・RNAハイブリッド2本鎖またはDNA・タンパク質2本鎖)を認識し得る抗体が用いられ得る。次いで、抗体は標識され得、アッセイが行われ得るが、この際、2量体が表面に結合し、その結果、表面での2本鎖の形成により、2本鎖に結合した抗体の存在が検出され得る。
【0155】
例えば、抗体フラグメントを含む抗体は、PDGF−DDタンパク質の発現を定性的または定量的に検出するために用いられ得る。上述のように、抗体には、好ましくは、検出可能な蛍光標識等が付与され、結合は、光学顕微鏡、フローサイトメトリー、蛍光分析または他の当該技術分野で既知の技術を用いてモニタリングされ得る。これらの技術は、増幅遺伝子が細胞表面タンパク質(例えば、成長因子)をコードするときに特に適している。かかる結合アッセイは、当該技術分野で知られた方法で行われる。
【0156】
PDGF−DDタンパク質と結合した抗体のインサイツ検出は、例えば、免疫蛍光顕微鏡または免疫電子顕微鏡により行われ得る。このため、組織標本が患者から採取され、標識された抗体が標本に塗布される(好ましくは、抗体を生物学的サンプルに重ねることによる)。この手順はまた、試験組織中のマーカー遺伝子産物の分布の決定も可能にする。インサイツでの検出に広範囲の種々の組織学的方法が容易に利用可能であることは、当業者には明らかであろう。
【0157】
異なる遺伝子発現を定量する最も高感度かつフレキシブルな定量方法の1つは、RT−PCRであり、これを用いて、異なるサンプル集団、正常組織および腫瘍組織、薬剤処置ありまたはなしでのmRNAレベルが比較され、遺伝子発現のパターンが特徴決定され、密接に関連するmRNA間が見出され、そしてRNA構造が分析され得る。
【0158】
第一の工程は、標的サンプルからmRNAを単離することである。出発材料は、典型的には、疾患組織および対応する正常組織からそれぞれ単離されたトータルRNAである。従って、mRNAは、例えば、凍結し保存された、またはパラフィン包埋して固定された(例えば、ホルマリン固定)疾患組織のサンプルから、同種の正常組織との比較のために抽出され得る。RNA抽出物の調製方法は、当該技術分野でよく知られており、Ausubel et al., Current Protocols of Molecular Biology, John Wiley and Sons(1997)を含む、分子生物学の標準的教科書に記載されている。パラフィン包埋からのRNA抽出方法は、例えば、Rupp and Locker, Lab Invest., 56: A67(1987)およびDe Andres et al., BioTechniques, 18: 42044(1995)に開示されている。特に、RNAの単離は、Qiagenのような市販の精製キット、緩衝液セットおよびプロテアーゼを用いて、製造者の指示に従い行われ得る。例えば、培養液中の細胞由来のトータルRNAは、QiagenのRNeasyミニカラムを用いて単離され得る。組織サンプル由来のトータルRNAは、RNA Stat-60(Tel-Test)を用いて単離され得る。
【0159】
RNAはPCRのテンプレートとはならないので、RT−PCRによる異なる遺伝子発現分析の第1の工程は、RNAテンプレートのcDNAへの逆転写であり、次いで、PCR反応においてそれが指数関数的に増幅される。通常用いられる2つの主な逆転写酵素は、Avilo骨髄芽球症ウイルス逆転写酵素(AMV−RT)およびMoloneyマウス白血病ウイルス逆転写酵素(MMLV−RT)である。逆転写工程は、典型的には、状況および発現プロファイリング上の目的に依存して、特異的プライマー、ランダムな6量体またはオリゴ-dTプライマーを用いて調製される。例えば、抽出されたRNAは、GeneAmp RNA PCRキット(Perkin Elmer, CA, USA)を用いて、製造元の指示に従い逆転写され得る。次いで、誘導されたcDNAは、引き続いてPCR反応のテンプレートとして用いられ得る。
【0160】
PCR工程は、種々の熱安定性DNA依存性DNAポリメラーゼを用い得る、典型的には、5’−3’ヌクレアーゼ活性を有するが3’−5’末端ヌクレアーゼ活性を有しないTaq DNAポリメラーゼが用いられる。従って、TaqMan PCRは、典型的には、TaqまたはTthポリメラーゼの5’−ヌクレアーゼ活性を用いて、その標的増幅物と結合したハイブリダイゼーションプローブが加水分解されるが、均等な5’ヌクレアーゼ活性を有する任意の酵素を用いることもできる。2つのオリゴヌクレオチドプライマーを用いて、PCR反応の代表的増幅物(amplicontypical)が産生される。第3のオリゴヌクレオチドまたはプローブは、2つのPCRプライマーの間に位置するヌクレオチド配列を検出するよう設計される。プローブは、Taq DNAポリメラーゼ酵素によって伸長されないが、レポーター蛍光色素およびクエンチャー蛍光色素で標識される。レポーター色素からの任意のレーザー励起放射は、2つの色素がプローブ上で密接に位置したときにクエンチング色素により消光される。増幅反応の間、Taq DNAポリメラーゼ酵素は、テンプレート依存性の方法でプローブを切断する。得られたプローブフラグメントは、溶液中で解離し、解離したレポーター色素からのシグナルは、第2の蛍光発光団の消光作用を有しない。レポーター色素の1つの分子は、合成したそれぞれの新しい分子に対して放出され、消光されていないレポーター色素の検出により、データの定量的解釈の基礎が提供される。
【0161】
TaqMan RT−PCRは、例えば、ABI PRIZM 7700(登録商標)Sequence Detection System(登録商標)(Perkin-Elmer-Applied Biosystems, Foster City, CA, USA)またはLight Cycler(Roche Molecular Biochemicals, Mannheim, Germany)のような市販の装置を用いて行われ得る。好ましい実施態様において、5’ヌクレアーゼ方法は、ABI PRIZM 7700(登録商標)Sequence Detection System(登録商標)のようなリアルタイム定量的PCR装置を用いて実行される。当該システムは、サーモサイクラー、レーザー、電荷結合素子(CCD)、カメラおよびコンピュータからなる。当該システムは、サンプルを、サーモサイクラー上の96ウェル形態で増幅する。増幅の間、レーザー誘導蛍光シグナルが、光ファイバーケーブルにより96ウェル全てに集積され、CCDで検出される。当該システムは、装置を実行させ、データを解析するためのソフトウェアを含む。
【0162】
5’−ヌクレアーゼアッセイデータは、最初はCtまたは閾値サイクルとして現される。上記で考察された蛍光値は、全サイクルで記録され、増幅反応のある点まで増幅された産物の量を現す。統計的に有意であるとして蛍光シグナルが最初に記録された点が、閾値サイクル(Ct)である。ΔCt値は、疾患組織中のRNAの発現を正常細胞のそれと比較した場合に、核酸サンプル中の特定の標的配列の出発コピーの相対数を定量的に測定するために用いられる。
【0163】
サンプルとサンプルの変動の影響を最小化するために、RT−PCRは、通常、内部標準を用いて実行される。理想的な内部標準は、異なる組織間で一定のレベルで発現され、実験処理による影響を受けないものである。遺伝子発現のパターンを標準化するために最もよく用いられるRNAは、ハウスキーピング遺伝子グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(GAPDH)およびβ−アクチンのmRNAである。
【0164】
異なる遺伝子発現もまた、マイクロアレイ技術を用いて同定または確認され得る。この方法では、目的のヌクレオチド配列は、マイクロチップ基板上にプレート化またはアレイ化される。次いで、アレイ化された配列を、目的の細胞または組織由来の特異的DNAプローブによりハイブリダイズする。
【0165】
マイクロアレイ技術の特定の実施態様において、cDNAクローンのPCR増幅挿入物は、高密度アレイの基板上に付与される。好ましくは、少なくとも10,000ヌクレオチド配列が基板上に付与される。10,000素子でそれぞれマイクロチップ上に固定されたマイクロアレイ遺伝子は、ストリンジェントな条件下でのハイブリダイゼーションに適している。蛍光標識されたcDNAプローブは、目的の組織から抽出されたRNAの逆転写による蛍光ヌクレオチドの取込みにより産生され得る。チップに付与された標識cDNAプローブは、アレイ上のDNAのそれぞれのスポットに選択的にハイブリダイズされる。ストリンジェントな洗浄により非特異的結合プローブを除去した後、チップを共焦点レーザー顕微鏡でスキャンする。それぞれのアレイ化エレメントのハイブリダイゼーションの定量により、対応するmRNA量の評価が可能になる。2色の蛍光により、RNAの2つの供給源から製造された特異的に標識されたcDNAプローブがアレイに対でハイブリダイズする。このように、2つの供給源からの転写において、それぞれ特定された遺伝子に対応する相対量が同時に決定される。ハイブリダイゼーションのスケールの最小化により、多数の遺伝子についての発現パターンの簡便かつ迅速な評価が得られる。かかる方法は、細胞1個あたり数コピーで発現される稀な転写物を検出するため、および発現レベルの少なくとも約2倍の差異を再現可能に検出するために必要とされる感度を有することが示されている(Schena et al., Proc. Natl. Acad. Sci. USA, 93(20)L106-49)。核酸およびマイクロアレイ技術のハイブリダイゼーションの方法論は、当該技術分野でよく知られている。
抗体および方法の選択した実施態様を、以下の実施例で説明する。
【0166】
実施例
行った実験および得られた結果を含む以下の実施例は、例示のみの目的で提供されており、本明細書に記載の発明の実施態様を限定するものとして解釈されるべきではない。
【0167】
実施例1
PDGF−DD抗原の調製
生物学的に活性なPDGF−DD p35である、CUBドメインを欠く、組換えヒトおよびマウスPDGF−DDを、LaRochelle et al., Nat Cell Biol 3: 517-521(2001)に記載のように製造した。ヒトPDGF−CCを、同様のプロトコールで製造した。PDGF−AAおよびPDGF−BBは、R&D Systems(Minneapolis,MN)から購入した。
【0168】
実施例2
PDGFに対するアプタマーベースのアンタゴニスト
PDGF−Bアプタマー(NX1975)の合成および特徴決定は、Green et al., Biochemistry 35: 14413-14424(1996)に詳細に記載されている。オリジナルのDNAアプタマーの修飾は、特定のヌクレオチドを2−フルオロピリミジンおよび2’−O−メチルプリンで置換し、ヌクレアーゼ抵抗性を改善すること、ならびにアプタマーを40kDaのポリエチレングリコール(PEG)と結合させてインビボでの血漿残存時間を延長させることを含む。Floege et al., Am J Pathol 154: 169-179(1999)。
【0169】
実施例3
抗PDGF−DD抗体
完全ヒト抗PDGF−DDモノクローナル抗体を、Yang et al., J. Leukoc. Biol. 66: 401-410(1999)に記載の方法を以下のように変更して産生した。簡潔にいうと、ヒトIgG2を有するXenoMouse(登録商標)系(8〜10週齢)に、週に2回、完全フロイントアジュバント中のV5−標識溶解性PDGF−DD(LaRochelle et al.,Nat Cell Biol 3: 517-521(2001)) 10μgを足蹠に注射して免疫した。Yang et al. supraを参照。電気的細胞融合を用いて、ハイブリドーマを製造した。完全ヒトアイソタイプと一致したPK16.3を、負の対照として用いた。
【0170】
実施例4
完全ヒト抗PDGF−DD mAB 6.4の特徴決定
PDGFのなかでも、PDGF−DDに対する完全ヒト抗PDGF−DD mAb 6.4の特異性を、固相ELISA、ウエスタンブロット分析、およびNIH 3T3 BrdU取込分析により特徴決定した。
【0171】
固相ELISA
完全ヒト抗PDGF−DDの特異性を、固相ELISAにより特徴決定した。簡潔に言うと、Corning96ウェル平底高タンパク質結合ポリスチレンマイクロタイタープレートを、500ng/mlのPDGF−AA、PDGF−BB、PDGF−CCまたはPDGF−DDで一晩コーティングした。プレートを、アッセイ希釈剤(Pharmingen, San Diego, CA)で1時間ブロッキングした。次いで、抗PDGF−DD mAb 6.4または対照mAb PK16.3を添加し、示した濃度で2時間インキュベートした。第1のmAb結合を、TMB試薬(Pharmingen, San Diego, CA)を用いて、2次抗体に結合した抗ヒト西洋ワサビペルオキシダーゼを用いて検出した。マイクロタイタープレートを、450nmにてKinetic Microplate Reader(Molecular Devices, Sunnyvale, CA)を用いて読み取った。
【0172】
図1に示すように、抗PDGF−DD mAb 6.4は、PDGF−DDを認識したが、PDGF−AA、PDGF−BB、またはPDGF−CCは認識しなかった。対照mAb PK16.3は、PDGF−DDの認識しなかった。ELISA結果を確認するため、ウエスタンブロット分析も行った。
【0173】
さらに、Corning96ウェル平底高タンパク質結合ポリスチレンマイクロタイタープレートを、500ng/ml ヒトまたはマウスPDGF−DDを用いて一晩コーティングすることにより、PDGF固相ELISAを行った。プレートを、アッセイ希釈剤(Pharmingen, San Diego, CA)で1時間ブロッキングした。次いで、抗PDGF−DD mAb 6.4を添加し、示した濃度で2時間インキュベートした。第1のmAb結合を、TMB試薬(Pharmingen)を用いて、二次抗体に結合した抗ヒト西洋ワサビペルオキシダーゼを用いて検出した。マイクロタイタープレートを、450nmでKinetic Microplate Reader(Molecular Devices, Menlo Park, CA)を用いて読み取った。
【0174】
図2に示すように、抗PDGF−DD mAb 6.4抗体はヒトおよびマウスPDGF−DDの両方を認識する。
【0175】
ウエスタンブロット分析
ウエスタンブロット分析用に、PDGF−AA、PDGF−BB、PDGF−CC、およびPDGF−DD(250μg)をSDS−PAGEサンプル緩衝液中で希釈し、沸騰させ、16% SDSポリアクリルアミドゲルを用いるSDS−PAGEゲル電気泳動に供した。タンパク質をHybond-Pメンブレン(Amersham, Piscataway, NJ)に移し、フィルターをPDGF−DD mAb 6.4または対照mAb PK16.3(0.85μg/ml)で12時間プローブ結合させた。洗浄後、フィルターを、抗ヒト西洋ワサビペルオキシダーゼ結合二次抗体と共にインキュベーションした。バンドを、増強した化学発光により視覚化した(Amersham, Piscataway, NJ)。
【0176】
図3は、抗PDGF−DD mAb 6.4がPDGF−DD、p35を免疫ブロットしたが、PDGF−AA、PDGF−BB、またはPDGF−CCは免疫ブロットしなかったことを示している。対照mAb PK16.3は、PDGFを認識しなかった。BIACorの動力学的測定を用いて、抗PDGF−DD抗体6.4のヒトPDGF−DDとの親和性が170PMであり、抗PDGF−DD mAb 6.4がマウスPDGF−DDに対して少なくとも20倍少ない親和性を示す(データは示していない)ことを決定した。
【0177】
NIH 3T3 BrdU取込みアッセイ
抗PDGF−DD mAb 6.4のPDGF−DD誘導性細胞分裂活性を中和する能力を試験するため、NIH 3T3 BrdU取込みアッセイを用いた。NIH 3T3中和アッセイは、LaRochelle et al.,Nat Cell Biol 3: 517-521(2001)に記載の方法を以下のように変更して行った。簡潔にいうと、NIH 3T3細胞を、24時間血清枯渇状態におき、モノクローナル抗体を示した濃度で添加した。次いで、PDGF−DDを18時間後に100ng/mlで添加した。18時間後、BrdUを5時間にわたって添加し、BrdUアッセイを、製造元(Roche)の指示に従い行った。
【0178】
図4に示すように、抗PDGF−DD mAb 6.4は、PDGF−DD誘導性BrdU取込みを、約75ng/mlのIC50で中和した。PDGF−DD誘導性BrdU取込みは、試験した最高濃度(5μg/ml、データは示していない)で影響を受けなかった。対照mAb PK16.3は、PDGF−DD誘導性BrdU取り込みに影響しなかった。これらの結果を総合して考えると、抗PDGF−DD mAb 6.4が、PDGF−DDに対して高度に特異的であるが、他のPDGFファミリーメンバーは認識せず、PDGF−DD誘導性BrdU取り込みを強く中和することを示した。
【0179】
実施例5
インビトロでのPDGF−DDのメサンギウム細胞増殖に対する作用
メサンギウム細胞培養実験
インビトロでのPDGF−DDのメサンギウム細胞増殖に対する作用を研究するため、ラットメサンギウム細胞を培地中で樹立し、特徴決定し、上述のように維持した。Radeke et al.,J Inzmunol 153 : 1281-1292,(1994)を参照。簡潔に言うと、ラットメサンギウム細胞を96ウェルプレート(Nunc, Wiesbaden, Germany)に播種し、密集させて、サブコンフルエントまで増殖させ、1%ウシ血清アルブミンを含むRPMI1640にて48時間増殖を停止させた。48時間後、PDGF−DD(10〜200ng/ml)およびPDGF−BB(l0ng/mlおよび50ng/ml)と共に、PDGF−B鎖アプタマー(100ng/ml)または配列スクランブルしたアプタマー(100ng/ml)を添加し、細胞を24時間インキュベートした。DNA合成を、BrdU取込みにより決定し、製造元の指示に従い、熱量測定細胞増殖ELISA(Roche, Mannheim, Germany)により測定した。
【0180】
増殖を停止させた培養ラットメサンギウム細胞のPDGF−DDとのインキュベーションは、増殖における用量依存性をもたらした(図5)。データは、4つの独立実験の平均±標準偏差である。統計上の有意性(p<0.05と定義した)を、ANOVAおよびBonferroni t検定を用いて評価した。アスタリスクは、非刺激対照に対するp<0.05を示す。
【0181】
細胞分裂誘導性PDGF−DD活性のPDGF−Bへの非依存性を、PDGF−DDと同時に、細胞をアンタゴニストPDGF−Bアプタマーまたは配列スクランブル対照アプタマーとインキュベーションすることにより示した。アプタマーは、PDGF−BB誘導性増殖をブロックしたが、PDGF−DDの細胞分裂誘導性の可能性には影響を及ぼさなかった(図5)。同様のデータ(示していない)が、ヒトメサンギウム細胞を用いて得られた。
【0182】
実施例6
PDGF−DDおよび抗PDGF−DD抗体のヒトメサンギウム細胞(HMC)に対する作用
ヒトメサンギウム細胞を血清枯渇状態におき、以下の100ng/mL、250ng/mL、1μg/mLの濃度でPDGF−DDまたはPDGF−BBと共にBrdUで一晩処理した。比較のため、他の間葉系細胞、例えば、NIH 3T3線維芽細胞、CCD 1070包皮線維芽細胞、および初代平滑筋細胞を、BrdUおよび完全な血清で処理した。BrdU取込みを、抗BrdU抗体ELISAを用いるアッセイにより検出した。図6が示すように、PDGF−DDは、l00ng/mLより高い濃度で初期ヒトメサンギウム細胞の増殖を誘発する。図6はさらに、ヒトメサンギウム細胞におけるBrdU取込みの同様の誘導に必要とされるPDGF−DDおよびPDGF−BBの濃度に、10倍の差を見出したことを示す。
【0183】
実施例7
腎炎の血清におけるPDGF−DDレベル
腎炎血清におけるPDGF−DDレベルを定量するために、サンドウィッチELISAを開発した。サンドウィッチELISAに用いた2つの完全ヒトmAb(抗PDGF−DD mAb 1.6および1.17)は、PDGF−DD分子に対する異なるエピトープを認識した(データは示していない)。抗PDGF−DD mAb 1.6を捕獲抗体として用い、抗PDGF−DD mAb 1.17を検出抗体として用いた。
【0184】
ELISAを以下のように行った。2μg/mlの濃度のコーティング緩衝液(0.1M NaHCO3、pH9.6)中の捕獲抗体(抗PDGF−DD mAb 1.6) 50μlを、ELISAプレート(Fisher)にコートした。4℃で一晩インキュベーションした後、プレートを、ブロッキング緩衝液(PBS中、0.5% BSA、0.1% Tween20、0.01% Thimerosal)200μlで1時間、25℃にて処理した。PBS中の0.05% Tween20(洗浄緩衝液、WB)を用いて、プレートを洗浄した(3回)。正常血清または患者血清(Clinomics, Bioreclamation, Cooperative Human Tissue Network)を、ヒト血清を50%含むブロッキング緩衝液で希釈した。プレートを血清サンプルと共に4℃にて一晩インキュベーションし、WBで洗浄し、次いで100μ1/ウェルのビオチンル化検出抗PDGF−DD mAb 1.17で1時間、25℃にてインキュベートした。洗浄後、プレートをHRP−ストレプトアビジンと共に15分間、インキュベートし、上述のように洗浄し、次いで、発色のため100μ1/ウェルのH2O2中のO−フェニレンジアミン(Sigma開発中溶液)で処理した。反応を2M H2SO4で停止させ、ELISAプレートリーダーを用いて492nmで分析した。血清サンプル中のPDGF−DDの濃度を、4つのパラメータ曲線フィッティングプログラムを用いてPDGF−DD標準曲線と比較することにより計算した。
【0185】
腎炎を有するII型糖尿病患者におけるPDGF−DD血清レベル
PDGF−DDが腎炎に関与するかどうかを決定するため、II型糖尿病を有する種々の型の腎炎患者由来の血清レベルを調べた。血清PDGF−DD濃度を、上述の定量的PDGF−DDサンドウィッチELISAを用いて評価した。ELISAはPDGF−DDに特異的であり、4ng/mlの感度を有していた。図7は、研究結果を概要する。黒丸は、個々の臨床血清サンプルのPDGF−DD濃度を示す。PDGF−DD血清濃度を、患者の疾患指標に従いグループ化した。所定の臨床指標を有する患者の数(n)を、平均PDGF−DD濃度(ng/ml)とともに提供する。
【0186】
図7に示すように、PDGF−DDは、II型糖尿病患者由来の10個の血清サンプルのうち8個で、正常血清(n=50)のと比較して6%上昇した(平均=11.4ng/ml、p<.001)。II型糖尿病患者におけるPDGF−DDの平均血清レベルは、正常個体の濃度が4ng/ml未満であるのに対し、約4〜24ng/mlの範囲にあった。これらのデータは、PDGF−DDがII型糖尿病の患者の血清中で増加し、そしてこれがPDGF−DDがII型糖尿病に関連する腎臓疾患/腎不全の発症を遅らせるための標的であり得ることを示唆している。これらの結果は、腎炎患者の血清では、正常個体の血清と比較して、PDGF−DDレベルが4〜7倍増加することを示している。
【0187】
実施例8
ラットメサンギウムの免疫組織化学分析
正常ラットメサンギウム細胞を、抗Thy−1誘導性腎炎を有するラットのメサンギウム細胞と比較した。Wistarラットは、Charles River社から入手した。免疫組織化学染色を、抗PDGF−DD血清を用いて行い、次いで、西洋ワサビペルオキシダーゼを結合したヤギ抗ウサギ抗体を用いて検出した。簡潔に言うと、組織を従来の技術によりパラフィン除去し、トリプシン(0.15%)で37℃にて10分間処理した。1次抗体および抗ウサギHRP結合体と、それぞれ10分間インキュベーションした後、ジアミノベンジジン(DAB)溶液を切片に添加し、免疫活性を視覚化した。図8に示すように、免疫組織化学分析により、抗Thy−1誘導性腎炎ラットでは、抗PDGF−DDレベルが上昇することが明らかとなった。メサンギウム、尿細管および周辺脈管構造を示す。メサンギウム細胞は、周皮細胞および腎尿細管を含んでいる。白色および灰色の矢印は、それぞれ毛細管および尿細管染色を示す。
【0188】
実施例9
完全ヒト抗PDGF−DD mAb 6.4のシミュレートした薬物動態学
ラットにおいて、完全ヒトmAb動態学のシミュレーションを行った。雄性Wistarラットに、抗PDGF−DD mAb 6.4を、3日目および5日目にそれぞれl0mg/kgおよび5mg/kgで投与した。血清を回収し、ヒト抗PDGF−DD mAbレベルを、ヒト特異的IgG ELISAで定量した。図9に示すように、単回投与後でさえも、4日間予想された大きなピークからトラフへの変動は生じ無かった。これらのデータは、ラットにおけるヒト抗体のクリアランスのpKシミュレーションモデルと好適に関連し、多くの抗PDGF−DD mAb 6.4は、一旦投与されると循環し続けることを示している。
【0189】
抗体クリアランス速度を分析するためのさらなる実験において、49匹のラットを以下の種々のレベルの抗PDGF−DD抗体、対照抗体またはPBSで処置した。
【表4】
【0190】
以下の「循環抗体」につての表において、左の列は49匹の動物についての5日目の結果を、右の列は、対応する動物の8日目の結果を示す。
抗PDGF−DD抗体クリアランス
【表5】
【0191】
【表6】
【0192】
上述のように、抗PDGF−DD mAb 6.4は、薬物動態学モデルにおいて計算したように、予測循環半減期を示した。
【0193】
実施例10
メサンギウム増殖性腎炎における糸球体中でのPDGF−DD発現
抗Thy1.1腎炎における糸球体でのPDGF−DD発現の動態学を研究するため、体重180gの雄性Wistarラット(Charles River, Sulzfeld, Germany)に1mg/kgのモノクローナル抗Thy1.1抗体(Clone OX-7; European Collection of Animal Cell Cultures, Salisbury, England)を注射することにより、抗Thy1.1メサンギウム増殖性糸球体腎炎を誘導した。45匹のラットに抗Thy1.1抗体を投与し、抗体注射の4時間後、1、2、4、7、9、14、21および28日の時点で屠殺した(それぞれ5匹ずつ)。屠殺後、腎組織および単離した糸球体を得た。糸球体の単離は、異なるふるいわけにより行った。Johnson et al., J Clin Invest 87: 847-858(1991)参照。全ての糸球体単離物を顕微鏡でチェックしたところ、98%より高い純度を示した。さらに、副腎組織を得た。
【0194】
糸球体のRNAの抽出および分析
RNAを糸球体から単離し、発現をリアルタイム定量的PCRで測定した。簡潔に言うと、単離したラット糸球体および副腎から、標準的手法を用いて、グアニジウムイソチオシアネート/フェノール/クロロホルム法により、トータルRNAを抽出した。Chomczynski et al., Anal Biochem 162: 156-159(1987)参照。得られたサンプルのRNA含有量および純度を、UV分光測定法により、260および280nmで決定した。
【0195】
cDNA合成は、トータルRNA 1μg、ランダムプライマー(6nt、250ng/μl、Roche)lμl、M−MLV逆転写酵素緩衝液(Invitrogen, Groningen, The Netherlands)6μ1、dNTP混合物(それぞれl0mM、Amersham Pharmacia Biotech, Freiburg, Germany)1.5μl、RNase阻害剤(40U/μl、Promega, Mannheim, Germany)0.7μl、M−MLV逆転写酵素(200U/μ1、Invitrogen)1μl、およびDEPC処理H20を含む、反応混合物 30μlにて行った。この混合物を、25℃で10分間、次いで42℃で1時間インキュベートした。
【0196】
リアルタイム定量的PCRを、ABI prism 7700配列検出装置(Applied Biosystems, Weiterstadt, Germany)を用いて行った。それぞれの反応において、cDNA 0.75μ1、およびPCR Master Mix(ROX Reference Dyeを用いるPlatinum Quantitative PCR SuperMix-UDG;Invitrogen)12.5μ1を、計25μlの体積で用いた。PCR条件は、50℃で2分、次いで95℃で15秒を40サイクル、そして60℃で1分であった。Taqmanプライマーおよびプローブを、Primer Express ソフトウェア(Applied Biosystems)を用い、Genbankデータベースの配列から設計した。本研究で用いたプライマーおよびプローブを、以下の表3に列挙する。
【0197】
表3
プライマーおよびプローブ
【表7】
【0198】
PGDF−DDはメサンギウム増殖性腎炎の糸球体において過剰発現する
ラットにメサンギウム増殖性抗Thy1.1腎炎を誘導した後、糸球体のPDGF−D mRNA発現は、当初は、疾患誘導4時間後に36%減少したが、次いで、4日〜9日目に、非腎炎ラットと比較して2.4〜2.9倍増加した(図10)。この後者のピークは、糸球体のPDGF−A mRNA発現のそれと平行し、PDGF−B mRNA最大発現後に若干遅れて生じた(図10)。これらの3つのPDGFアイソフォームとは対照的に、PDGF−C mRNAは、抗Thy1.1腎炎の最初の28日間にはアップレギュレートされなかった。
【0199】
抗Thy1.1腎炎におけるPDGF−D mRNAアップレギュレーションが腎臓に特異的であるかどうかを評価するために、副腎でのmRNAレベルもまた調査した。なぜなら、副腎はPDGF−Dの重要な供給源であることが見出されているからである。LaRochelle et al., Nat Cell Biol 3:517-521(2001)参照。糸球体とは対照的に、抗Thy1.1腎炎の最初の28日間において、副腎では、PDGF−D mRNA発現レベルの有意な変化は観察されなかった(データは示していない)。この後者の知見にもかかわらず、正常動物のPDGF−DDタンパク質レベルが常に検出限界未満(<0.02ng/ml、n=5)であったのに対し、疾患誘導8日後の腎炎ラットの血清中では、当該タンパク質レベルの劇的なアップレギュレーション(27.7±14.5ng PDGF−D/ml、n=9)を検出した。
【0200】
PDGF−DD発現の免疫組織化学
正常ラット腎臓における免疫組織化学PDGF−DD発現は、動脈および細動脈の血管平滑筋細胞に限られるが、糸球体では免疫反応は見られなかった(図11(A))。拡張したメサンギウムにおけるPDGF−DDの顕著な糸球体過剰発現は、疾患誘導の7日後には存在した(図11(B))が、残りの腎臓の染色パターンには影響を及ぼさなかった。抗PDGF−DD抗体を等濃度の対照IgGと置き換えた場合、糸球体染色は存在しなかった(図11(C))。
【0201】
実施例11
PDGF−DDとPDGF−BBの相互作用
PDGF−BBおよびPDGF−DDの両方が抗Thy1.1腎炎において過剰産生され(図10および11)、かついずれかの拮抗作用がメサンギウム増殖性変化の減少をもたらすと仮定した場合の、2つのPDGFアイソフォームの可能性のある相互作用を評価した。
【0202】
PDGF−DDの抗PDGF−DD mAb 6.4との拮抗作用は、疾患の8日目において、糸球体PDGF−BおよびPDGF−D mRNAレベルに対する有意な影響をもたらさなかった(表11)。また、このモデルにおける、特異的アプタマーによるPDGF−Bの拮抗作用は、8日目におけるPDGF−D mRNAの糸球体発現に何ら違いをもたらさなかった(PEG40対照群で3.10±1.30増加したのに対し、アプタマー群における非腎炎ラットについては3.18±0.58増加した(それぞれn=5))。しかしながら、後者の実験における糸球体PDGF−B mRNA発現は、PDGF−B拮抗作用によりわずかに誘導された(正常ラットにおける発現と比較して、PEG40対照群では発現が2.52±0.64であるのに対し、アプタマー群では発現が3.31±1.1(それぞれn=5)であった)。測定をそれぞれのサンプルについて2回行った。
【0203】
実施例12
インビボでのPDGF−DD拮抗作用
インビボでのPDGF−DD拮抗作用の効果を研究するため、ラットを、抗Thy1.1腎炎の誘導の3日および5日後に、抗PDGF−DD抗体 6.4、対照IgG PK16.3またはPBSで処置した。処置は、20mM Tris−HCl/100mM NaCl(pH7.4) 800μlに溶解した、抗体の腹腔内注射からなるものであった。処置のタイミングを、メサンギウム細胞増殖の開始後約1日目からピーク(OX−7−誘導性抗Thy1.1腎炎モデルでは、疾患誘導の5日と8日との間で生じる)までラットを処置するように選択した。抗PDGF−DD抗体の3つの異なる用量のインビボ効果を研究した。
【0204】
50μg/mlより高い血清レベルまたはインビトロでのPDGF−DDの最大阻害の2分の1を生じるという計算に基づき、体重1kgあたりl0mg(3日目)および4mg(5日目)の平均用量の抗PDGF−DD mAb 6.4を選択した。抗PDGF−DD mAb 6.4または無関係な対照IgG2 PK16.3と関係するレベルの達成を確認するため、5日目および第8日目において、疾患群1〜4におけるヒトIgG2血清レベルを測定した。5日目に30μg/ml未満のレベルを示す動物を分析から除いた。
【0205】
上記と共に、抗体で処置した群における十分なヒト血清IgG2を有する7つの群のラットを研究した:(1)3日目に5mg/kg体重、そして5日目に2mg/kg体重にて抗PDGF−DD mAb 6.4を投与した7匹の腎炎ラット;(2)3日目にl0mg/kg体重、そして5日目に4mg/kg体重にて抗PDGF−DD mAb 6.4投与した7匹の腎炎ラット;(3)3日目に20mg/kg体重、そして5日目に8mg/kg体重にて抗PDGF−DD mAb 6.4を投与した8匹の腎炎ラット;(4)3日目に20mg/kg体重、そして5日目に8mg/kg体重にて無関係な対照IgGを投与した8匹の腎炎ラット;(5)等量のPBSのみを注射した9匹の腎炎ラット;(6)3日目にl0mg/kg体重、そして5日目に4mg/kg体重にて抗PDGF−DD mAb 6.4を投与した5匹の非腎炎正常ラット;ならびに(7)等量の無関係な対照IgGを投与した5匹の非腎炎正常ラット。
【0206】
群1〜5のそれぞれから4匹のランダムに選択したラットにおいて、5日目に、Floege et al., Am J Pathol 154 : 169-179(1999)に記載の生体内バイオプシーにより、組織学的評価のための腎バイオプシーを得た。全てのラットにおいて、疾患誘導8日後に、死後バイオプシーを得た。次いで、全ての群の2または3匹のラットの残りの皮質組織をプールし、糸球体の単離に用いた(上記を参照されたい)。疾患誘導7日後に、尿を回収した。8日目、屠殺の4時間前にチミジン類似体5−ブロモ−2’−デオキシウリジン(BrdU; Sigma, Deisenhofen, Germany;l00 mg/kg体重)を腹腔内注射した。
【0207】
PDGF−DD阻害はインビボでの病理学的メサンギウム細胞増殖を低減する
抗Thy1.1抗体の注射後、PBS処置した動物は、腎炎の代表的な過程を発症し、これは初期のメサンギウム分解、および引き続き5日および8日目において、メサンギウム細胞増殖相およびマトリックスの蓄積により特徴付けられた。抗PDGF−DD mAb 6.4を反復注射した後に、目に見える悪影響は見られず、全てのラットが生存し研究の終了まで正常であるように見えた。腎炎群で至った抗体血清レベルを、表4に示す。腎炎群におけるアルブミン/クレアチニン比および全ての処置群における収縮期血圧は、有意な差はなかった。
【0208】
尿中アルブミンレベルは、ラットアルブミンに特異的なELISAキット(Nephrat, Exocell, Philadelphia, PA)を用いて決定した。尿中クレアチニンは、Vitros 250分析装置(Orthoclinical Diagnostics,Neckargmund, Germany)を用いて、2点間速度法で決定した。全ての測定を2回行った。血圧測定は、プログラム血圧計BP-98A(Softron, Tokyo, Japan)を用いて、tail cuff法により行った。Kitahara et al.,J Am Soc Nephrol 13 : 1261-1270(2002)参照。
【0209】
腎炎ラットでは、7日目に、非腎炎ラットと比較してアルブミン尿のかなりの増加が生じた(対照IgGを投与した非腎炎ラットではアルブミン/クレアチニン比:0.3±0.lmg/μmolであるのに対して、PBSを投与した腎炎ラットでは15.5±5.6mg/μmol;(p<0.01))。PBS、対照IgG、または3種類の用量の抗PDGF−DD mAb 6.4を投与した腎炎群の間には有意な差異は見られなかった(表4)。抗PDGF−DD mAb 6.4は、非腎炎ラットにおいてタンパク質尿を誘発しなかった。
【0210】
種々の抗PDGF−DD mAb 6.4用量でも、無関係な対照IgGでも、全身血圧レベルに有意な作用は観察されず、7日目において、全ての動物が正常血圧を維持した(表4)。
表4
インビボで達成されたヒトIgG2抗体(抗PDGF−DD mAb 6.4または無関係な対照IgG2)レベル、尿中アルブミン/クレアチニンおよび収縮期血圧
【表8】
**データは平均値±標準偏差である。n.d.は、測定しなかったことを示す。
【0211】
糸球体有糸分裂数を数えることにより評価する糸球体細胞増殖は、無関係なIgGまたはPBSを投与したラットと比較して、抗PDGF−DD mAb 6.4を投与したラットにおける8日目の用量依存性を有意に低減した(図12(A))。処置を3日目および5日目に行った。正常または非腎炎ラットを、抗PDGF−DD抗体または無関係な対照IgGで処置した。アスタリスクはp<0.05を示す。BrdU陽性の核のカウントにより、20+8mgの抗PDGF−DD抗体/kgでの処置群では、8日目に増殖が最も顕著に抑制されているという知見を確認した(図12(B))。抗PDGF−DD処置を受けた全3群のデータを集めたところ、インビボで至った抗体レベルとBrdU取込み核とは、5日目(r=−0.53;p=0.018)および8日目(r=−0.40;p=0.081)においてネガティブに相関した。
【0212】
メサンギウム細胞に対する処置の効果を評価するため、活性化メサンギウム細胞のみによって発現するα平滑筋アクチンを、腎断面で免疫染色した。Johnson et al., J Clin Invest 87: 847-858(1991)参照。α平滑筋アクチンの糸球体発現は、10+4mg/kgおよび20+8mg/kgの抗PDGF−DD mAb 6.4を投与したラットにおいて、8日目に、無関係なIgGまたはPBSを投与したラットと比較して有意に減少した(図12(C))。メサンギウム細胞増殖が減少しているかどうかを特異的に決定するため、抗PDGF−DD mAb 6.4で処置したラットおよび対照IgGまたはPBSで処置したラットのBrdUおよびα平滑筋アクチンを2重免疫染色した(図12(D))。このデータは、全3つの抗PDGF−DD抗体処置群で、疾患誘導の8日後にメサンギウム細胞の増殖の顕著な低減が観察され、メサンギウム細胞増殖が最大で57%低減されたことを確認した。メサンギウム分解スコアは、抗PDGF−DD mAb 6.4処置ラットおよび対照IgG処置ラットにおいて同じであった(図12(E))。
【0213】
抗PDGF−DD mAb 6.4の正常ラットへの注射は、無関係なIgGを投与した正常ラットと比較して、生理的な糸球体細胞ターンオーバーに影響しなかった。
【0214】
インビボでのPDGF−DDの阻害は、糸球体の単球/マクロファージ流動を低減する
5日目において、抗PDGF−DD mAb 6.4の3種類の用量全てが糸球体単球/マクロファージ流動の顕著な低減を生じたが、8日後には生じなかった(図12(G))。正常ラットの特異的抗PDGF−DD抗体または無関係なIgGのいずれかでの処置は、糸球体単球/マクロファージ流動に影響しなかった。
【0215】
インビボでのPDGF−DD阻害は、糸球体マトリックスの蓄積を低減する
ラットを10+4mg/kgの抗PDGF−DD mAb 6.4または20+8mg/kgの抗PDGF−DD mAb 6.4のいずれかで処置した結果、腎炎対照と比較して、糸球体フィブロネクチン蓄積が低減された(図12(H))。一方、I型コラーゲンの糸球体蓄積は、3つの腎炎群のいずれにおいても、対照IgGまたはPBSで処置したラットと比較して、抗PDGF−DD抗体処置により影響されなかった(図12(F))。
【0216】
正常ラットでは、糸球体マトリックス発現は、抗PDGF−DD抗体または無関係なIgGでの処置によって影響されなかった(図12(H))。
【0217】
実施例13
インビボでの抗PDGF−DD抗体の効力
ラットにおける抗Thy−1.1抗体誘導性メサンギウム増殖性糸球体腎炎モデルにおけるPDGF−DDと結合する、抗PDGF−DD mAb 6.4の効力を、インビボで以下のように評価した。
【0218】
正常な生理的状態を有し、体重が約150〜200gの雄性Wistarラット(10週齢)(Charles River, Sulzfeld, Germany)を得た。まず、ラットを、正常および抗Thy1.1メサンギウム増殖性糸球体腎炎を誘導する、2群に分けた。
【0219】
動物を、地方の動物施設で以下のように飼育した:ラットを7日間順応させ、食物および水道水を自由に与えた。研究を開始する前に動物を検査し、適度な健康および適合性を確認した。疾患を有すること、または不適切であることが判明した動物は本研究に供しなかった。この一連の研究において、12時間毎の明/暗サイクルを維持した。名目上の温度範囲は、20〜23℃であり、30%〜70%の相対湿度もまた維持した。
【0220】
完全ヒト抗PDGF−DD mAb 6.4を、上述のXenomouse(登録商標)技術を用いて産生した。1mg/kgのモノクローナル抗Thy1.1抗体を注射することにより、抗Thy1.1メサンギウム増殖性糸球体腎炎を雄性Wistarラットにて誘導した。抗Thy1.1腎炎を誘導した後、疾患誘導の3日後および5日後に、10mg/kgおよび4mg/kgのmAb 6.4(n=15)または無関係なヒトモノクローナル抗体(n=15)またはPBS(n=15)を毎日腹腔内注射することにより、ラットを処置した。残りのラットは処置しなかった。全部で5群のラットを研究した。処置後、ラットを、腎バイオプシー、尿アルブミンおよび屠殺後の組織回収により分析した。
【0221】
1)合計14mg/kg(疾患誘導の3日後にl0mg/kg、そして5日後に4mg/kg)の抗PDGF−DD mAb 6.4を投与した15匹の腎炎ラット;
2)合計14mg/kg(疾患誘導の3日後にl0mg/kg、そして5日後に4mg/kg)の無関係なアイソタイプ一致対照抗体(抗PDGF抗体ではない)を投与した15匹の腎炎ラット;
3)トリス緩衝生理食塩水のみ800μLのボーラス注射を受けた15匹の腎炎ラット;
4)合計14mg/kg(疾患誘導の3日後にl0mg/kg、そして5日後に4mg/kg)の抗PDGF−DD mAb 6.4を投与して5匹の腎炎ラット;および
5)5匹の正常無処置のラット。
【0222】
表5により、試験した5群についての研究設計のリストを提供する。
表5
研究設計
【表9】
【0223】
抗PDGF−DD mAb 6.4の純度は、90%より高いか、または同等であった。全てのバイアルを、使用するまで冷蔵庫中で4℃にて保管した。予備のサンプルを、−80℃で保管した。トリス緩衝生理食塩水中の10Mg/Kg体重、および4mg/kg体重を腹腔内注射した。トリス緩衝生理食塩水での希釈は、l0mg/kgおよび4mg/kg用量が、800μLの体積で投与できるように行った。これらの用量を、3日後および5日後にのみ、1日1回投与した。
【0224】
処置の持続を、メサンギウム細胞増殖の開始の約1日後から増殖のピーク(OX−7誘導性抗Thy1.1腎炎モデルでは、疾患誘導の6日目と9日目との間で生じる)までラットを処置するように選択した。
【0225】
ラットに投薬した約60分後、ラットの有意な臨床徴候、罹患率、および死亡率について、ラットを毎日観察した。研究の開始後は、体重測定を行わなかった。動物が検視前に死亡した場合、検死および組織データには含めず、組織も回収しなかった。
【0226】
動物が検視中に死亡した場合、死亡していたものと記録し、検視データを用いなかった。しかしながら、評価の可能性のため、組織をホルマリン中に回収した。瀕死の動物は、屠殺し、同様に処置した。
【0227】
8日目まで生存した動物全てを、頚椎脱臼により殺し、総合的に観察し、予定した全ての組織を組織形態学的評価のため、10%中性緩衝ホルマリン、Methacarn溶液、および液体窒素中に回収した。
【0228】
染色方法および組織の調製:光学顕微鏡および免疫ペルオキシダーゼ染色のための組織をメチルCarnoy溶液で固定し、パラフィンに包埋した。4μmの断面を過ヨウ素酸Schiff(PAS)試薬で染色し、ヘマトキシリンで対比染色した。PAS染色された断面において、有糸分裂数を、20より多い毛細管切片を含有する連続的皮質糸球体である30を超える断面(30〜100の範囲)を、公平な観察者により評価した。メサンギウム溶解を、Burg et al., Lab Invest 76: 505-516(1997)に記載の半定量スケールで階級付けした:0=メサンギウム溶解なし、1=部分的メサンギウム溶解、2=全体的メサンギウム溶解、3=微細動脈瘤。
【0229】
免疫ペルオキシダーゼ染色:メチルCarnoyで固定したバイオプシー組織の4μm断面を、Johnson et al, 1990に記載の間接免疫ペルオキシダーゼ技術により処理した。PDGF−DDは、ヒトPDGF−Dに対するポリクローナルウサギ抗体により検出した。一次抗体は、既に記載のもの(Burg et al, 1997; Yoshimura et al, 1991)と同一であり、α平滑筋アクチンに対するマウスモノクローナル抗体(クローン1A4);PDGF−B鎖に対するマウスモノクローナル抗体(クローンPGF−007);単球、マクロファージ、および樹状細胞に存在する細胞質抗原に対するマウスモノクローナルIgG抗体(クローンED1);ラット赤血球であらかじめ吸着した、アフィニティー精製ポリクローナルヤギ抗ヒト/ウシIV型コラーゲンIgG;ヒトI型コラーゲンに対するポリクローナルヤギ抗体(Southern Biotech Associates, Birmingham, AL, USA);アフィニティー精製ポリクローナルウサギ抗ラットフィブロネクチン抗体IgG分画(Chemicon, Temecula, CA, USA);および上述の適当な負の対照(Burg et al, 1997; Yoshimura et al, 1991)を含む。PDGF−DDは、ヒトPDGF−Dに対するポリクローナルウサギ抗体により検出した。血清を、Protein A Sepharoseクロマトグラフィーにより精製した。PDGF−Cアフィニティーカラムに吸着させることにより、PDGF−C交差活性を取り除いた。得られた免疫グロブリンフローを濃縮し、これはELISAまたはウエスタンブロット分析によりPDGF−A、BまたはCとは反応しないことがわかった。全てのスライドを、スライドの出処を知らされていない観察者により評価した。
【0230】
糸球体中の浸潤白血球の平均数を得るため、糸球体の50より多くの連続的断面を評価し、腎臓あたりの平均値を計算した。I型コラーゲン、フィブロネクチンおよびα平滑筋アクチンの免疫ペルオキシダーゼ染色を評価するため、それぞれの糸球体領域を、半定量的に段階付けし、1バイオプシー当たりの平均スコアを計算した。それぞれのスコアは、主に、染色強度ではなく染色程度の変化を反映し、これは、陽性染色が集中的に増強された糸球体房領域の割合に依存する:
I=0〜25%
II=25〜50%
III=50〜75%
IV=>75%
【0231】
免疫組織化学的二重染色:増殖細胞の型を同定するための二重免疫染色は、以前に報告(Kliem et al, 1996; Hugo et al, 1996)されているように、まず、増殖細胞の断面を、トリス緩衝生理食塩水中のヌクレアーゼ含有ブロモデオキシウリジン(Amersham, Braunschweig, Germany)に対するマウスモノクローナル抗体(クローンBU−1)による間接的免疫ペルオキシダーゼ方法を用いて、染色した。断面を、α平滑筋アクチンに対するIgG1 mAb 1A4および単球/マクロファージに対するED1と共にインキュベーションした。細胞がBrdUの陽性核染色を示し、当該核がα平滑筋アクチンまたはED1抗原陽性細胞質により完全に包囲されている場合、増殖メサンギウム細胞または単球/マクロファージであると同定した。負の対照では、いずれかの1次抗体を省き、この場合、二重染色は観察されなかった。
【0232】
尿測定:尿中タンパク質(アルブミン尿)は、Bio-Radタンパク質アッセイ(Bio-Rad Laboratories GmbH, Munchen, Germany)を用い、ウシ血清アルブミン(Sigma)を標準として用いることにより測定した。血圧は、テール血圧計により測定した。
【0233】
得られた多数のデータから統計的分析を行い、平均値および標準偏差を数学的に計算した。統計的分析は、予定屠殺時まで生存した動物由来のデータに基づき行った。全ての値を平均±標準偏差として表す。統計上の有意性(p<0.05と定義)を、Student t検定またはANOVAおよびBonferroni t検定を用いて評価した。データの解釈を助けるため、研究責任者の裁量で補助的分析も行った。
【0234】
結果:図13は、糸球体腎炎のラットモデルにおけるBrdU取込みにより測定した糸球体増殖の結果を示す。ラットを屠殺前にBrdUで6時間処置した。核のBrdU染色は、抗BrdU抗体を用いて測定した。腎炎ラットモデルで観察された糸球体100個あたりの有糸分裂数を計測した。表6は、試験した5群に基づき、糸球体100個あたりのBrdU陽性核数をまとめている。表6は、図13の対応するグラフと共に、抗PDGF−DD mAb 6.4を腎炎動物に投与することにより、チミジン類似体BrdUを取込んだ糸球体細胞が、無関係のIgGおよびPBSを投与した場合と比べて少なくなることを示している。1列は、疾患糸球体の有糸分裂指数を示す。2列は、対照抗体で処置した腎炎のラットモデルのものである。3列は、PBSで処置した腎炎ラットモデルのものである。4列は、抗PDGF−DD mAb 6.4で処置した健常ラット対照のものである。5列は、対照抗体で処置した健常ラット対照のものである。
【0235】
表6
糸球体によるBrdU取込み
【表10】
【0236】
同様に、図14は、糸球体腎炎のラットモデルを、抗PDGF−DD mAb 6.4で処置した場合の有糸分裂をPAS染色および定量測定した糸球体増殖の結果を示す。4μMの断面を過ヨウ素酸Schiff試薬で染色し、ヘマトキシリンで対比染色した。有糸分裂を、糸球体細胞100個当たりについて測定した。30〜50個の糸球体房の有糸分裂数を決定した。表7は、抗PDGF−DD mAb 6.4投与が、無関係なIgG抗体およびPBSと比較して、100個当たりの有糸分裂を顕著に減少させることを示している。図14の対応グラフは、抗PDGF−DD mAb 6.4を腎炎動物に投与することにより、無関係なIgG抗体およびPBSを腎炎動物に投与したときと比べて、有糸分裂する糸球体細胞がより少なくなることを示す。1列は、抗PDGF−DD mAb 6.4で処置した腎炎ラットモデルで観察された結果を示す。2列は、対照抗体で処置した腎炎ラットモデルを示す。3列は、PBSで処置した腎炎ラットモデルを示す。4列は、抗PDGF−DD mAb 6.4で処置した健常ラット対照を示す。5列は、対照抗体で処置した健常ラット対照を示す。
【0237】
表7
糸球体での有糸分裂の測定(PAS使用)
【表11】
【0238】
表6および7において示すように、抗PDGF−DD mAb 6.4での腎炎ラットの処置は、有糸分裂の手動計数(表7)およびBrdU取込みによる測定(表6)のいずれにおいても糸球体増殖を低減した。
【0239】
また、メサンギウム細胞を、抗平滑筋アクチンで対比染色した。所定セットのメサンギウム細胞(30〜50個の糸球体タフト)における有糸分裂数を決定した。二重染色アッセイの結果を、以下の表8に示す。表8は、対応する図15と共に、抗PDGF−DD mAb 6.4は、無関係な(非PDGF−D)抗体(PK16.3 IgG)およびPBSと比較して、糸球体有糸分裂の低減に有効あることを示している。
【0240】
表8
BrdU+抗sm−アクチン
【表12】
【0241】
実施例14
急性ラットThy−1モデルに対するmAb 6.4の用量応答性作用
抗PDGF−DD mAb 6.4のThy−1誘導腎炎に対する作用もまた、用量応答性方法で研究した。抗Thy1.1腎炎誘導後、ラットを疾患誘導の3日〜8日後に、それぞれ、5、10および20mg/kg、次いで2、4および8mg/kgのヒト抗PDGF−DD mAb 6.4(n=15)または無関係なヒトPK16.3モノクローナル抗体(n=15)またはPBS(n=15)のいずれかで腹腔内注射により毎日処置した。図16に示しかつ表9にまとめるように、8日後、抗PDGF−DD mAb 6.4を用いて処置した疾患誘導拮抗作用は、100個の糸球体当たりの有糸分裂数を有意に低減した。図17に示しかつ表10にまとめるように、抗PDGF−DD mAb 6.4での処置はまた、チミジン類似体BrdUを取込む糸球体細胞を有意に低減した。従って、ラット抗Thy−1モデルの10mg/kgの抗PDGF−DD mAb 6.4での処置は、若干用量応答性方法で増殖を40〜70%阻害する。
【0242】
表9
【表13】
【0243】
表10
【表14】
【0244】
表11
抗PDGF−D mAb処置ラットにおける糸球体PDGF−BおよびPDGF−D mRNA発現
【表15】
**データは、それぞれの処置群におけいてプールした分画の平均(および範囲)である。
【0245】
実施例15
ヒト腎臓疾患組織の免疫組織化学的分析
免疫組織化学的分析により、ヒト腎臓疾患組織におけるPDGF−DDの存在を試験した。
【0246】
PDGF−AA、PDGF−BBまたはPDGF−CCを認識しないウサギ抗PDGF−DD IgGを用いて、免疫組織化学染色を行った。染色に続いて、西洋ワサビペルオキシダーゼと結合したヤギ抗ウサギ複合体(抗ウサギHRP複合体)を用いて検出した。抗ウサギHRP複合体とのインキュベーション後、ジアミノベンジジン(DAB)溶液を断面に添加し、免疫反応を視覚化した。
【0247】
活発な糸球体腎炎において、尿細管(ほとんどはおそらく近位尿細管である)の染色が明確に認められた。いくつかの糸球体も、陽性に染色されていた(約10〜20%のメサンギウム/視野)。慢性的な同種移植拒絶を有する患者由来組織もまた陽性に染色されており、これは尿細管および血管染色を示していた。細胞蓄積物もまたメサンギウム中で検出され、これは炎症誘導性肥満細胞および細胞沈着を示唆した。対照ウサギIgGでは染色されなかった。
【0248】
PDGF−DD染色はまた、薬剤誘導性間質性腎炎でも明確に認められた。ここで、尿細管の染色増加、メサンギウム細胞の顕著な染色、およびいくつかの浸潤性炎症誘導性細胞染色が観察された。存在する可能性のある非常に弱い尿細管の染色を除いては、正常ヒト腎臓の顕著な染色は観察されなかった。腎硬化症においては、尿細管のPDGF−DD染色が見られた(データは示していない)。虚血性尿細管損傷においては、染色は観察されなかった(データは示していない)。これらの結果は、多くのヒト腎臓の病理においてPDGF−DDが増加することを示唆しており、腎臓疾患におけるPDGF−DDの役割を示唆する。PDGF−DDは、尿細管間質、メサンギウム増殖および活発な炎症過程における変化に関与し得る(図18を参照)。白色および灰色の矢印は、それぞれ毛細管および尿細管染色を示している。小さな黒色の矢印は、メサンギウム中の点状炎症性細胞蓄積物を示す。
【0249】
実施例16
ELISAを用いた腎炎の発症リスクの分析、診断および段階決定
腎炎に罹患した患者由来のPDGF−DD血清レベルを分析した。PDGF−DD濃度は、定量的サンドウィッチELISA法を用い、PDGF−DDに対して生じた全2つのヒトmAbを用いて評価した。腎炎患者の血清中では、正常患者と比べて、PDGF−DDレベルが4〜7倍に上昇していることがわかる。従って、PDGF−DDレベルのこれらの差異は、診断を成すための助けとなり得、かつ医師が腎炎および関連疾患の段階決定するための助けとなり得る。
【0250】
実施例17
抗PDGF−DD抗体によるヒトの腎炎の処置
医師は、有効量の抗PDGF−DD抗体をそれを必要とする患者に投与し、それによって当該必要とする患者の症状が緩和されるかまたは腎炎が効果的に治療される。投与および用量は患者に特有である。抗PDGF−DD抗体の投与は、皮下注射による。
【0251】
上述の種々の方法および技術は、多くの実施態様を行うための多数の方法を提供する。当然のことながら、本明細書に記載の任意の特定の実施態様に従って必ずしも記載された全ての目的または利点が達成され得るわけではないことは理解されるべきである。従って、例えば、当業者は、当該方法は、本明細書で教示される1つの利点または利点群を達成または最適化する方法で、本明細書で教示または示唆され得る他の目的または利点を必ずしも達成することなく実行され得ることを理解するであろう。
【0252】
さらに、当業者は、異なる実施態様の種々の特性は互換可能であることを理解するであろう。同様に、上述の種々の特性および工程、ならびにそのような特性または工程の他の公知の均等物は、本明細書に記載の原理に従った方法を実行するために、当業者によって結び付けられ適合され得る。
【0253】
本明細書に記載の方法は、ある種の実施態様および実施例の文脈によって開示されてきたが、これらの方法が、特定の開示された実施態様を超えて、他の代わりの実施態様および/または使用および明らかな改変ならびにそれらの均等物まで拡張されることは当業者に理解されるであろう。従って、本明細書に記載の方法は、本明細書で好ましい実施態様の特定の開示によって限定されないが、添付の特許請求の範囲の参照となることが意図されている。
【図面の簡単な説明】
【0254】
【図1】図1は、ELISAによる抗PDGF−DD mAb 6.4特異性の特徴決定を示す。
【図2】図2は、ELISAによる抗PDGF−DD mAb 6.4特異性のさらなる特徴決定を示す。
【図3】図3は、ウエスタンブロット分析による抗PDGF−DD mAb特異性の特徴決定を示す。
【図4】図4は、抗PDGF−DD mAb 6.4が、NIH3T3細胞でのPDGF−DD誘導性BrdU取込みを、約75ng/mlのIC50で中和できることを示す線グラフである。
【図5】図5は、PDGF−DDがインビトロにおいてメサンギウム細胞の成長因子となることを示す棒グラフである。
【図6】図6は、ヒトメサンギウム細胞におけるPDGF−DD誘導性BrdU取込みの結果を示す棒グラフである。
【図7】図7は、種々の型の腎炎を有する患者のヒト血清中でのPDGF−DD発現を示すグラフである。
【図8】図8は、正常ラットメサンギウム細胞および抗Thy−1誘導性腎炎を有するラットのメサンギウム細胞の免疫組織化学分析を示す。
【図9】図9は、ラットにおいて成された完全ヒトmAbカイネティクスのシュミレーションを示す棒グラフである。
【図10】図10は、無処置のラットでの発現に対する、抗Thy1.1腎炎の経過におけるPDGF−A、−B、−Cおよび−Dの転写発現を示す線グラフである。
【図11】図11は、PDGF−DDタンパク質が、糸球体での抗Thy1.1腎炎において過剰発現されることを示す。
【図12−1】図12A〜Hは、抗PDGF−DD抗体、無関係の対照IgGまたはPBS単独のいずれかで処置した、メサンギウム増殖性抗Thy1.1腎炎を有するラットにおいて、糸球体が疾患の誘導5日後および8日後に変化していることを示す棒グラフである。
【図12−2】図12A〜Hは、抗PDGF−DD抗体、無関係の対照IgGまたはPBS単独のいずれかで処置した、メサンギウム増殖性抗Thy1.1腎炎を有するラットにおいて、糸球体が疾患の誘導5日後および8日後に変化していることを示す棒グラフである。
【図12−3】図12A〜Hは、抗PDGF−DD抗体、無関係の対照IgGまたはPBS単独のいずれかで処置した、メサンギウム増殖性抗Thy1.1腎炎を有するラットにおいて、糸球体が疾患の誘導5日後および8日後に変化していることを示す棒グラフである。
【図12−4】図12A〜Hは、抗PDGF−DD抗体、無関係の対照IgGまたはPBS単独のいずれかで処置した、メサンギウム増殖性抗Thy1.1腎炎を有するラットにおいて、糸球体が疾患の誘導5日後および8日後に変化していることを示す棒グラフである。
【図13】図13は、ラットでのBrdU取込により測定された、糸球体増殖の結果を示す棒グラフである。
【図14】図14は、ラットにおけるPAS染色により測定された糸球体増殖および有糸分裂の結果を示す棒グラフである。
【図15】図15は、急性ラット抗Thy−1モデルにおけるメサンギウム細胞有糸分裂に対する抗PDGF−DD mAb 6.4の作用を示す棒グラフである。
【図16】図16は、急性ラット抗Thy−1モデルにおける糸球体細胞の有糸分裂に対する抗PDGF−DD mAb 6.4の用量応答性作用を示す棒グラフである。
【図17】図17は、急性ラットThy−1モデルにおけるBrdU取込みに対する抗PDGF−DD mAb 6.4の用量応答性作用を示す棒グラフである。
【図18】図18は、正常および疾患のヒト腎臓組織上の免疫組織化学分析を示す。
【配列表】
【発明の詳細な説明】
【0001】
技術分野
本明細書に記載の本発明の実施態様は、血小板由来成長因子DD(PDGF−DD)に対する抗体、およびかかる抗体の使用に関する。本発明の抗体は、PDGF−DDの過剰産生に関連する疾患の診断および処置としての使用が見出されている。具体的には、本発明の実施態様をふまえて、メサンギウム増殖により引き起こされる疾患を含む腎炎および関連疾患の処置のための抗PDGF−DD抗体の使用が提供される。
【0002】
発明の背景
腎炎は、アメリカ合衆国および世界中で関心の増している問題である、腎臓疾患の1つの群である。腎炎は、透析処置または腎移植を受けなければ最終的には致命的となる、腎不全に徐々に進行し得る。異なる型の腎炎は、異なる遺伝パターン、および異なる進行速度を有する。遺伝性腎炎は、尿中の血液細胞およびタンパク質の顕微鏡トレースにより明らかにされ、出生時に存在するが、概して穏やかである。別の型の腎炎である糸球体腎炎は、腎臓の濾過単位である糸球体の炎症である。他の型の腎炎は、単球増加症および連鎖球菌(Streptococcus)(感染後)のような感染性疾患の後遺症であり得る。
【0003】
メサンギウム細胞の増殖に関連する腎炎および他の関連疾患の症状は、腎炎の特定の型に依存して変動するが、典型的には尿中の血液またはタンパク質の存在が挙げられる。該疾患の初期段階では、徴候または症状がないこともある。疾患が進行するにつれて、以下の症状のいくつかまたは全てが生じ得る:高血圧、尿の過剰な泡立ち、尿の色の(赤または黒褐色への)変化、眼、手および足の腫脹、悪心および嘔吐、呼吸困難、および頭痛。これらの症状は、疾患を同定し、処置コースに従い、そしてどのタイプの処置が必要かを同定するために用いられ得る。
【0004】
糸球体の損傷は、タンパク質尿、血尿、高窒素血症、乏尿、無尿、浮腫、および高血圧を含むがこれらに限定されない、種々の疾患の徴候を引き起こす。該疾患はまた、この疾患はまた、腎炎症候群、急性腎炎、および急性進行性糸球体腎炎を引き起こし得る。
【0005】
糖尿病性腎症を含む多くの進行性腎疾患、ならびにIgA腎症のような最も頻繁な型の糸球体腎炎は、糸球体メサンギウム細胞増殖および/または基質蓄積により特徴付けられる。Striker et al., Lab Invest 64: 446-456(1991)参照。現時点で、血小板由来成長因子(PDGF)および関連するPDGF系が、メサンギウム細胞増殖および基質蓄積に関与し得ることを示唆する証拠がいくつかある。Floege et al., supra(2001)およびFloege et al., Am.J. Pathol. 154: 169-79(1999); Gilbert et al., Kidney Int. 59: 1324-32(2001); Nakamura et al., Kidney Int., 59: 2134-45(2001)参照。さらに、PDGFβ受容体サブユニットおよびPDGF B鎖の両方が腎間質性線維症において過剰発現する。Kliem et al., Kidney Int. 49: 666-78(1996)参照。2量体PDGF−BB単独の多用量の注射は、正常ラット腎臓において間質性線維性変化を誘発し得る。Tang et al., Am. J. Pathol. 148: 1169-80(1996)参照。
【0006】
20年間、血小板由来成長因子系は、たった2つのPDGF鎖(PDGF−AおよびPDGF−B)からなるとされ、これは、ホモまたはヘテロ2量体として分泌され、α鎖および/またはβ鎖を含む2量体PDGF受容体に結合する。PDGF−Aはα鎖のみに結合するが、PDGF−βは全ての受容体タイプのリガンドである。Floege et al., "Growth factors and cytokines," in Immunologic Renal Diseases(Neilson E. G., and Couser W. G., eds., 2d ed. 2001)参照。最近、PDGF−CおよびPDGF−Dと命名された2つの他のPDGFアイソフォームが、ホモ2量体としてのみ放出されることが述べられた。現行の専門用語によると、PDGF−Cのホモ2量体形は、「PDGF−CC」として知られ、PDGF−Dのホモ2量体形は、「PDGF−DD」として知られている。LaRochelle et al.,Nat. Cell Biol. 3: 517-21(2001); Li et al.,Nat. Cell Biol. 2: 302-09(2000);およびBergsten et al.,Nat. Cell Biol. 3: 512-16(2001)を参照。PDGF−CCの中心鎖は、主にαα−PDGF受容体のリガンドであるようだが、PDGF−DDは、主にββ−PDGF受容体と結合する(同上)。両方の場合において、αβ受容体との結合もいくつか述べられている。LaRochelle et al., supra(2001); Bergsten et al., supra(2001); Gilbertson et al., J. Biol. Chem. 276: 27406-14(2001)参照。4つのPDGFアイソフォーム全て、ならびに両受容体鎖は、異なる空間配置ではあるが腎臓で発現する。Floege et al., supra(2001);Changsirikulchai et al., Kidney Int. 62(6): 2043-54(2002); Eitner et al., J. Am. Soc. Nephrol. 13(4): 910-17(2002)を参照。
【0007】
PDGF−Dは、ジスルフィド結合したホモ2量体PDGF−DDとして分泌され、そしてPDGF−ββ−およびαβ−受容体の特異的作用リガンドとなるために、そのCUBドメインの解離による制限的タンパク質分解で活性化される。発生において、および成人正常腎臓において、PDGF−DDは、内臓の糸球体上皮細胞およびいくつかの血管平滑筋細胞で発現する。Changsirikulchai et al., supra(2002)参照。マウス腎臓の発生において、分枝尿管(branching ureter)の細胞のみがPDGF−DD免疫反応性を示した。Bergsten et al., supra(2001)参照。
【0008】
腎炎の診断は、典型的には、家族歴、および/または赤血球およびタンパク質の存在、特に血尿またはアルブミン尿についての尿沈査検査による。残念なことに、根元的な病理過程に作用する、または臨床コースを変化させる特定の処置は知られていない。抗生物質、抗凝血剤、ステロイド剤、および免疫抑制剤は何ら利益をもたらさなかった。高血圧の制御が示唆されており、タンパク質の制限がいくぶん役に立つかもしれない。末期の尿毒症が起こった場合、透析およびさらには腎移植が必要とされる。従って、腎炎の処置のための新規アプローチが必要である。
【0009】
発明の要約
本発明の実施態様は、抗PDGF−DD抗体の投与が、糸球体細胞の増殖を低減し、そしてその増殖と関連する疾患を処置するのに非常に効果的であったという発見に関する。
【0010】
従って、本発明の1つの実施態様は、腎炎および関連疾患の進行を阻害するための完全ヒト抗PDGF−DD抗体、および治療展望から所望した特性を有する抗PDGF−DD抗体製剤の使用である。好ましくは、当該抗体は、配列番号2、6、10、14、18、22、26、30、34、38、42、46、50、54、58、62、66、70および74からなる群から選択される配列を有する重鎖アミノ酸を有する。より好ましくは、当該抗体はさらに、配列番号4、8、12、16、20、24、28、32、36、40、44、48、52、56、60、64、68および72からなる群から選択される配列を有する軽鎖アミノ酸を有する。
【0011】
本発明の実施態様は、任意の特定の抗PDGF−DD抗体、または任意の特定型の抗体に限定されないことは理解されるだろう。例えば、抗PDGF−DD抗体は、全長抗体(例えば、完全ヒトFc領域を有する)、または抗体フラグメント(例えば、Fab、Fab’またはF(ab’)2)であってもよい。さらに、抗体は、抗体を分泌するハイブリドーマから、または遺伝子または抗体をコードする遺伝子で形質転換またはトランスフェクションされた組換え産生細胞から製造されてもよい。
【0012】
好ましい実施態様において、本発明は、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎不全、および糖尿病性腎症を含む(これらに限定されない)、ヒトの腎炎および関連疾患の処置を含む。
【0013】
1つの実施態様において、抗PDGF−DD抗体は、有効量の抗体またはそのフラグメントを、医薬的に許容される担体または希釈剤と共に含む医薬組成物を成す。代わりの実施態様において、抗PDGF−DD抗体は、放射性同位元素、または毒素と結合している。別の実施態様において、抗PDGF−DD抗体またはそのフラグメントは、治療法と結合していてもよい。該治療法は、毒素または放射性同位元素であり得る。好ましくは、かかる抗体は、例えば、腎炎、進行性腎疾患、および関連疾患(例えば、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎間質性線維症、腎不全および糖尿病性腎症)のような疾患の処置のために用いられ得る。
【0014】
別の実施態様において、本発明は、患者に有効量の抗PDGF−DD抗体を投与することにより、患者におけるPDGF−DDの発現と関連する疾患または状態を処置する方法を含む。該患者は、哺乳動物患者、好ましくはヒト患者である。該疾患または状態は、例えば、腎炎、進行性腎疾患、および関連疾患(例えば、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎間質性線維症、腎不全または糖尿病性腎症)であり得る。さらなる実施態様は、腎炎の処置が必要な哺乳動物を同定すること、次に該哺乳動物に治療上有効用量の抗PDGF−DD抗体を投与することにより、哺乳動物のPDGF−DDの発現と関連する疾患または状態を処置する方法を含む。
【0015】
あるいは、抗PDGF−DD抗体は、哺乳動物が腎炎または関連疾患を含む(これに制限されない)PDGF−DDの発現に関係する疾患または状態、およびメサンギウム増殖により引き起こされる疾患にかかるのを予防するために投与されてもよい。好ましくは、抗PDGF−DD抗体は完全ヒト抗体である。疾患または状態は、腎炎、進行性腎疾患、および関連疾患(例えば、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎間質性線維症、腎不全、および糖尿病性腎症)含むが、これらに限らない、腎炎および関連疾患であり得る。
【0016】
さらに別の実施態様において、本発明は、細胞PDGF−DD発現細胞を有効量の抗PDGF−DD抗体またはそのフラグメントと接触させること、次に該細胞および抗体をインキュベートすることによる(インキュベーションが細胞増殖を阻害する)、PDGF−DDの発現に関連するかまたは当該発現により引き起こされる細胞増殖を阻害するための方法を含む。1つの実施態様において、細胞増殖は、メサンギウム細胞増殖である。さらに、メサンギウム細胞は、ヒトメサンギウム細胞であり得る。さらに、方法は、インビボで行われ得る。
【0017】
別の実施態様において、本発明は、抗PDGF−DD抗体を含有する組成物を有する容器、および該組成物がPDGF−Dの過剰発現により特徴付けられる状態を処置するために用いられ得ることを示すパッケージの記載またはラベルを含む製品である。好ましくは哺乳動物、より好ましくはヒトが、抗PDGF−DD抗体を受ける。好ましい実施態様において、ヒトの腎炎および関連疾患が処置され、これは、腎炎、進行性腎疾患、および関連疾患、例えば、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎間質性線維症、腎不全、および糖尿病性腎症を含むが、これらに限らない。
【0018】
別の実施態様は、抗PDGF−DD抗体を用いて、糸球体中のメサンギウム細胞の過剰増殖を同定することを含む、疾患の危険因子を同定するための方法、疾患の診断方法、および疾患の病期診断方法である。
【0019】
1つの実施態様において、本発明は、細胞を抗PDGF−DD抗体と接触させ、次にPDGF−DDの存在を検出することによる、細胞でのPDGF−DDの発現の発現と関連する状態の診断方法を含む。好ましい状態は、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎不全、および糖尿病性腎症を含むが、これに限定されない。
【0020】
さらに別の実施態様において、本発明は、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎不全、および糖尿病性腎症を含む(これらに限定されない)、ヒトにおいて腎炎および関連疾患をスクリーニングするための、哺乳動物組織または細胞におけるPDGF−DDの検出のためのアッセイキットを含む。該キットは、PDGF−DDと結合する抗体、および存在するなら、該抗体のPDGF−DDとの反応を示す手段を含む。好ましくは、当該抗体はモノクローナル抗体である。1つの実施態様において、PDGF−DDと結合する抗体が標識される。別の実施態様において、抗体は非標識の1次抗体であり、反応を示すための手段が、標識抗免疫グロブリン抗体である。好ましくは、抗体は、蛍光色素、酵素、放射性核種、および放射線不透過材料からなる群から選択されるマーカーで標識される。
【0021】
なお別の実施態様は、腎炎および関連疾患の処置のための医薬の製造における抗PDGF−DD抗体の使用である。1つの実施態様において、該疾患は、腎炎、進行性腎疾患、およびメサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎間質性線維症、腎不全、および糖尿病性腎症のような関連疾患からなる群から選択される。
【0022】
図面の簡単な説明
図1は、ELISAによる抗PDGF−DD mAb 6.4特異性の特徴決定を示す。
図2は、ELISAによる抗PDGF−DD mAb 6.4特異性のさらなる特徴決定を示す。
図3は、ウエスタンブロット分析による抗PDGF−DD mAb特異性の特徴決定を示す。
【0023】
図4は、抗PDGF−DD mAb 6.4が、NIH3T3細胞でのPDGF−DD誘導性BrdU取込みを、約75ng/mlのIC50で中和できることを示す線グラフである。
図5は、PDGF−DDがインビトロにおいてメサンギウム細胞の成長因子となることを示す棒グラフである。データは、4つの独立した実験の平均±SDである。*は、p<0.05対非刺激対照を示す。
【0024】
図6は、ヒトメサンギウム細胞におけるPDGF−DD誘導性BrdU取込みの結果を示す棒グラフである。
図7は、種々の型の腎炎を有する患者のヒト血清中でのPDGF−DD発現を示すグラフである。黒丸は、個々の臨床血清試料のPDGF−DD濃度を示す。PDGF−DD血清濃度は、患者の疾患の徴候に従いグループ分けした。所定の臨床徴候の患者数(n)が、平均PDGF−DD濃度(ng/ml)とともに提供される。
【0025】
図8は、正常ラットメサンギウム細胞および抗Thy−1誘導性腎炎を有するラットのメサンギウム細胞の免疫組織化学分析を示す。抗PDGF−DD染色性の上昇が、抗Thy−1誘導性腎炎のラットで見出された。メサンギウム、尿細管、および周辺脈管構造が示されている。メサンギウム細胞は、周辺細胞および腎尿細管を含んでいた。白色および灰色の矢印は、それぞれ毛細管および尿細管染色を示す。
【0026】
図9は、ラットにおいて成された完全ヒトmAbカイネティクスのシュミレーションを示す棒グラフである。示されるように、1回用量の後であっても4日にわたり、予測された小さなピークからトラフへの変動のみが生じている。
図10は、無処置のラットでの発現に対する、抗Thy1.1腎炎の経過におけるPDGF−A、−B、−Cおよび−Dの転写発現を示す線グラフである。
【0027】
図11は、PDGF−DDタンパク質が、糸球体での抗Thy1.1腎炎において過剰発現されることを示す。正常糸球体ではPDGF−DD発現は見られない(図11(A))が、疾患を誘導した7日後にメサンギウム増殖性腎炎において容易に検出され得る(図11(B))。抗PDGF−DD抗体を等濃度の対照IgGで置き換えた場合、糸球体染色性はない(図11(C))。倍率は600倍である。
【0028】
図12A〜Hは、抗PDGF−DD抗体、無関係の対照IgGまたはPBS単独のいずれかで処置した、メサンギウム増殖性抗Thy1.1腎炎を有するラットにおいて、糸球体が疾患の誘導5日後および8日後に変化していることを示す棒グラフである。
【0029】
図13は、ラットでのBrdU取込により測定された、糸球体増殖の結果を示す棒グラフである。腎炎のラットを、抗PDGF−DD mAb 6.4、対照抗体またはPBSで処置した。健常ラットを、抗PDGF−DD mAb 6.4または対照抗体で処置した。
【0030】
図14は、ラットにおけるPAS染色により測定された糸球体増殖および有糸分裂の結果を示す棒グラフである。腎炎のラットを、抗PDGF−DD mAb 6.4、対照抗体またはPBSで処置した。健常ラットを、抗PDGF−DD mAb 6.4または対照抗体で処置した。
【0031】
図15は、急性ラット抗Thy−1モデルにおけるメサンギウム細胞有糸分裂に対する抗PDGF−DD mAb 6.4の作用を示す棒グラフである。抗Thy−1ラットを、抗PDGF−DD mAb 6.4、または対照抗体またはPBSで処置した。健常ラットを、抗PDGF−DD mAb 6.4または対照抗体で処置した。
【0032】
図16は、急性ラット抗Thy−1モデルにおける糸球体細胞の有糸分裂に対する抗PDGF−DD mAb 6.4の用量応答性作用を示す棒グラフである。
図17は、急性ラットThy−1モデルにおけるBrdU取込みに対する抗PDGF−DD mAb 6.4の用量応答性作用を示す棒グラフである。
【0033】
図18は、正常および疾患のヒト腎臓組織上の免疫組織化学分析を示す。メサンギウム、尿細管および周辺脈管構造が示されている。白の矢印および灰色の矢印は、それぞれ毛細管および尿細管の染色を示す。小さな黒の矢印は、メサンギウムにおける点状の炎症性細胞の蓄積を示す。
【0034】
詳細な説明
本明細書に記載の発明は、腎炎および関連状態を効果的に処置、診断および/または病期診断する方法に関する。かかる状態は、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、腎不全、および糖尿病性腎症を含む。1つの特定の実施態様において、本発明は、腎炎および関連状態の処置として、治療上有効量の抗PDGF−DD抗体を投与することを含む。好ましい実施態様において、抗体は、2量体PDGF−DDに対する完全ヒト抗体である。
【0035】
本発明の他の実施態様は、インビボでメサンギウム増殖性変化の低減をもたらす他の化合物に関する。従って、PDGF−DDのレベルを低減する化合物は、腎炎の処置に有用であろう。PDGF−D核酸、ポリペプチド、抗体、アゴニスト、アンタゴニストおよび他の関連化合物の使用を、以下により詳細に開示する。
【0036】
上述のように、PDGF−DはPDGF−B受容体を介してシグナル伝達し、ラットのメサンギウム細胞(MC)の分裂を促進する。低レベルのPDGF−D mRNAが、正常ラット糸球体で検出された。しかしながら、培養したラットMCを100ng/ml PDGF−DDと共にインキュベーションすることにより、MC増殖が7倍に増加し、これは24時間後に最大となった。リアルタイムPCRにより、PDGF−D mRNAは、培養メサンギウム細胞および正常ラット腎臓から単離された糸球体の両方で検出された。ラットにおけるメサンギウム増殖性抗Thy1.1腎炎の誘導後、糸球体のPDGF−D mRNAおよびタンパク質発現は、リアルタイムPCRで測定された非腎炎ラットと比較して、4から9日間有意に増加した。PDGF−D mRNA発現のピークは、PDGF−B mRNA発現ピークの2日後に生じた。さらに、PDGF−DD血清レベルは、7日目の腎炎動物において有意に増加した。
【0037】
腎炎におけるPDGF−DDの機能的役割を調べるため、Xenomouse[登録商標](Abgenix, Inc., Fremont, CA)において中和完全ヒトモノクローナル抗PDGF−DD抗体を産生させた。抗Thy1.1腎炎の誘導後、ラットは、疾患誘導の3日後および5日後に、10および4mg/kg 完全ヒト抗PDGF−DD抗体 mAb 6.4(n=15)、または無関係なヒトモノクローナル抗体(n=15)またはPBS(n=15)の通常の腹腔内注射により処置された。疾患誘導の8日後に、PDGF−DDの拮抗作用により、100個の糸球体当たりの有糸分裂像の有意な低減(抗PDGF−DD:9.9±0.9;無関係なIgG:13.9±0.9;PBS:14.7±1.0;p<0.0014)、ならびにチミジン類似体BrdUを取り込んだ糸球体細胞の有意な低減(抗PDGF−DD mAb 6.4:1.62±0.23;無関係なIgG:2.88±0.28;PBS:2.91±0.18;p<0.00169)を生じた。抗PDGF−DDを投与されたラットにおける糸球体細胞増殖の低減は、リアルタイムPCRにより決定された、低減したPDGF−B mRNAの糸球体発現とは関連しなかった。
【0038】
抗PDGF−DD抗体の正常ラットへの注射は、無関係なIgGを投与された正常ラットと比較して、生理的な糸球体細胞のターンオーバーに影響を及ぼさなかった。従って、糸球体メサンギウム細胞により産生されるPDGF−DDは、インビトロおよびインビボの両方で糸球体細胞の分裂促進因子となる。
【0039】
配列表
代表的なヒト抗PDGF−DD抗体の重鎖および軽鎖の可変領域ヌクレオチドおよびアミノ酸配列が、配列表において提供され、その内容が以下の表1に要約されている。
【0040】
表1
【表1】
【0041】
【表2】
【0042】
【表3】
【0043】
定義
他に定義しない限り、本明細書に記載の本発明と関連して用いられる科学用語および技術用語は、当業者に通常理解される意味を有するものとする。さらに、文脈により他に必要とされない限り、単数形の用語は複数形も含み、複数形の用語は単数形も含むものとする。一般に、本明細書に記載の細胞および組織培養、分子生物学、ならびにタンパク質およびオリゴヌクレオチドまたはポリヌクレオチド化学およびハイブリダイゼーションと関連して利用される学名および技術は、よく知られたものであり、当該技術分野において通常用いられるものである。標準的技術は、組み換えDNA、オリゴヌクレオチド、ならびに組織培養および形質転換(例えば、エレクトロポレーション、リポフェクション)に用いられる。酵素反応および精製技術は、製造元の仕様書に従って、あるいは当該技術分野において通常成されるか、または本明細書に記載のように行われる。上記技術および手順は、一般に、当該技術分野でよく知られた通常の方法に従い、かつ本明細書を通じて引用されて考察された種々の一般的かつより具体的な引例に記載のように、行われる。例えば、Sambrook et al.Molecular Cloning : A Laboratory Manual(3rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y.(2001))を参照されたい。本明細書に記載の分析化学、合成有機化学、および医療化学および医薬化学と関連して利用される学名、および実験室手段および技術は、当該技術分野でよく知られ、かつ通常用いられるものである。標準的な技術は、化学合成、化学分析、医薬製造、製剤、および送達、および患者の処置のために用いられる。
【0044】
本明細書で提供される実施態様と関連して用いられるように、以下の用語は、他に示されない限り、以下の意味を有すると理解されるものとする。
メサンギウム細胞は、哺乳動物の腎臓の糸球体小葉(lubule)内で見出される細胞であり、構造支持体となり、血流を制御し得、食細胞であり、かつ免疫応答において抗原を提示する補助細胞となる。
【0045】
メサンギウム増殖性腎炎は、糸球体メサンギウム細胞または基質の増加、またはメサンギウムの蓄積を伴う糸球体腎炎である。
【0046】
メサンギウム増殖性糸球体腎炎は、糸球体毛細管のメサンギウム層でのIgM抗体の異常蓄積による腎糸球体(腎臓の血液濾過部分)の炎症である。
【0047】
メサンギウム毛細管性糸球体腎炎は、腎不全となる腎障害である。糸球体の炎症は、異常な免疫応答および腎臓(糸球体)内での抗体の蓄積から生じる。症状は、混濁尿(膿尿)、排尿減少、腫脹および高血圧が挙げられる。この障害は、たいてい末期の腎疾患となる。
【0048】
メサンギウムは、毛細管の間の糸球体の中心部である。メサンギウム細胞は、食細胞であり、大部分が内皮細胞により毛細管の管腔と分けられている。糸球体外メサンギウムは、密集斑と傍糸球体装置の輸入細動脈および輸出細動脈との間の三角形の空間を満たすメサンギウム細胞である。
【0049】
糸球体腎炎は、腎臓の糸球体内の毛細管ループの炎症により特徴付けられる、種々の腎炎である。これは、急性、亜急性および慢性の形で生じ、感染症または自己免疫疾患が続発し得る。
【0050】
用語「PDGF−DD」は、変異体と共に全長かつ成熟形のPDGF−DD、およびそれらのフラグメントを含む。従って、PDGF−DDは、変異体CG52053−01、CG52053−02、CG52053−03、CG52053−04、CG52053−05、CG52053−06、およびCG52053−07(CuraGen, New Haven, CT)を含むが、これらに限らない。より多くの情報が、PCT出願の公開番号第WO01/25433号(1999年10月7日出願)において見出され得る。
【0051】
本明細書で用いられる用語「単離されたポリヌクレオチド」は、ゲノム、cDNAまたは合成起源のポリヌクレオチド、あるいはこれらのいくつかの組合せを意味し、その起源に基づき、「単離されたポリヌクレオチド」は、(1)天然で見出される「単離されたポリヌクレオチド」の全てまたは部分と関連しないか、(2)天然で結合しないポリヌクレオチドに作動可能に結合するか、あるいは(3)より長い配列の一部分としては天然に存在しない。
【0052】
本明細書で言及される用語「単離されたタンパク質」は、cDNA、組換えRNAまたは合成起源のタンパク質、あるいはこれらのいくつかの組合せを意味し、その起源または変異体の供給源に基づき、「単離されたタンパク質」は、(1)天然で見出されるタンパク質とは関連しないか、(2)同じ供給源由来の他のタンパク質を含まない(例えば、マウスタンパク質を含まない)、(3)異なる種由来の細胞により発現されるか、あるいは(4)天然に存在しない。
【0053】
用語「ポリペプチド」は、本明細書で一般名称として用いられ、天然タンパク質、フラグメント、またはポリペプチド配列の類似体を意味する。従って、天然タンパク質、フラグメント、および類似体は、ポリペプチド属の種である。本発明をふまえて、好ましいポリペプチドは、ヒト免疫グロブリンの重鎖分子、およびヒト免疫グロブリンのκ軽鎖分子、ならびに免疫グロブリンの重鎖分子を免疫グロブリンの軽鎖(例えば、免疫グロブリンのκ軽鎖)と共に含む組合せ、またはその逆により形成される抗体分子、ならびそのフラグメントおよび類似体を含む。
【0054】
対象に適用して本明細書で用いられる用語「天然に存在する」は、対象が天然に見出され得るという事実を意味する。例えば、天然供給源から単離されることができ、実験室またはその他の方法で意図的に修飾されていない、生物(ウイルスを含む)に存在するポリペプチドまたはポリヌクレオチド配列は、天然に存在する。
【0055】
本明細書で用いられる用語「作動可能に結合した」は、成分の位置を意味し、意図された方法でそれらが機能することを許可する関係にあることを記載したものである。コーディング配列に「作動可能に結合した」対照配列は、コーディング配列の発現が対照配列と適合可能な条件下で達成されるような方法でライゲーションされている。
【0056】
本明細書で用いられる用語「対照配列」は、これらがライゲーションしているコーディング配列の発現およびプロセッシングに作用する必要のあるポリヌクレオチド配列を意味する。かかる対照配列の性質は、宿主生物に依存して異なる;原核生物では、かかる対照配列は、一般に、プロモーター、リボソーム結合部位、および転写終結配列を含み;真核生物では、一般に、かかる対照配列は、プロモーター、および転写終結配列を含む。用語「対照配列」は、最低でも、その存在が発現およびプロセッシングに必須である要素の全てを含むことが意図され、その存在が有利であるさらなる要素、例えば、リーダー配列および融合パートナー配列も含み得る。
【0057】
本明細書で言及される用語「ポリヌクレオチド」は、リボヌクレオチドまたはデオキシヌクレオチドのいずれかである、少なくとも10塩基長の重合体形ヌクレオチド、またはいずれかのタイプのヌクレオチドの修飾形を意味する。この用語は、1本鎖および2本鎖形のDNAを含む。
【0058】
本明細書で言及される用語「オリゴヌクレオチド」は、天然に存在するもの、および天然に存在するものと一体となって結合した修飾されたヌクレオチド、および天然に存在しないオリゴヌクレオチド結合体を含む。オリゴヌクレオチドは、一般に、200塩基長またはそれより短いものを含むポリヌクレオチドサブセットである。好ましくは、オリゴヌクレオチドは、10から60塩基長であり、最も好ましくは、12、13、14、15、16、17、18、19、または20から40塩基長である。オリゴヌクレオチドは通常1本鎖(例えば、プローブ用)であるが、オリゴヌクレオチドは、2本鎖(例えば、遺伝子変異の構築用)であってもよい。本発明のオリゴヌクレオチドは、センスオリゴヌクレオチドまたはアンチセンスオリゴヌクレオチドのいずれであってもよい。
【0059】
本明細書で言及される用語「天然に存在するヌクレオチド」は、デオキシリボヌクレオチドおよびリボヌクレオチドを含む。本明細書で言及される用語「修飾されたヌクレオチド」は、修飾または置換された糖基等を有するヌクレオチドを含む。本明細書で言及される用語「オリゴヌクレオチド結合」は、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホロアニラデート、ホスホロアミデート等のようなオリゴヌクレオチド結合を含む。例えば、LaPlanche et al.Nucl. Acids Res. 14: 9081(1986); Stec et al. J. Am. Chenu. Soc. 106: 6077(1984); Stein et al. Nucl. Acids Res. 16: 3209(1988); Zon et al. Anti-Cancer Drug Design 6: 539(1991); Zon et al. Oligonucreotides and analogues : A Practical Approach, pp. 87-108(F. Eckstein, Ed., Oxford University Press, Oxford England(1991)); Stec et al. 米国特許第5,151,510号; Uhlmann and Peyman Chemical Reviews 90: 543(1990)を参照されたい。オリゴヌクレオチドは、所望に応じて、検出用の標識を含むことができる。
【0060】
本明細書で言及される用語「選択的にハイブリダイズする」は、検出可能かつ特異的な結合を意味する。本発明をふまえて、ポリヌクレオチド、オリゴヌクレオチド、およびそれらのフラグメントは、非特異的な核酸との検出可能な結合の感知可能な量を最小化するハイブリダイゼーションおよび洗浄条件下で、核酸に選択的にハイブリダイズする。高ストリンジェントな条件を用いて、当該技術分野で知られ、かつ本明細書において考察される選択的ハイブリダイゼーション条件が達成され得る。一般に、本発明のポリヌクレオチド、オリゴヌクレオチド、およびフラグメントと対象の核酸配列間の核酸配列相同性は、少なくとも80%であり、より典型的には、好ましくは、少なくとも85%、90%、95%、99%および100%の増加した相同性を有するであろう。2つのアミノ酸配列は、それらの配列間に部分的または完全な同一性が存在するなら、相同である。例えば、85%の相同は、2つの配列がマッチングを最大とするようアラインメントされたとき、85%のアミノ酸が同一であることを意味する。(マッチングさせられている2配列のいずれかにおける)ギャップは、マッチングを最大化する際に許容される;5以下のギャップ長が好ましく、2以下のギャップ長がより好ましい。あるいは、そして好ましくは、2つのタンパク質配列(または少なくとも30アミノ酸長のもの由来のポリペプチド配列)は、変異データマトリックスおよび6以上のギャップペナルティーによるプログラムALIGNを用いて、5より多くのアラインメントスコア(標準偏差単位)を有する場合、相同である(この用語は本明細書で用いられる)。M. O. Dayhoff, in Atlas of Protein Sequence and Structure, Vol. 5,101-110、およびSupplement 2 to Vol. 5,1-10(National Biomedical Research Foundation 1972)を参照されたい。2つの配列またはその一部分の相同性は、そのアミノ酸がALIGNプログラムを用いて最適にアラインメントされた場合に50%以上同一であるなら、より好ましい。本明細書で用いられる用語「対応する」は、ポリヌクレオチド配列が、参照ポリヌクレオチド配列の全てまたは一部分に対して相同である(すなわち、厳密には進化上関連しないが、同一である)か、あるいはポリペプチド配列が、参照ポリペプチド配列と同一であることを意味する。対照的に、用語「相補である」は、相補的配列が、参照ポリヌクレオチド配列の全てまたは一部分に対して相同であることを意味するために本明細書で用いられる。例えば、ヌクレオチド配列「TATAC」は、参照配列「TATAC」に対応し、かつ「GTATA」に相補性である。
【0061】
以下の用語:「参照配列」、「比較ウインドウ」、「配列同一性」、「配列同一性の割合」、および「実質的な同一性」は、2以上のポリヌクレオチドまたはアミノ酸配列間の配列関係を記載するために用いられる。「参照配列」は、配列比較の基礎として用いられる所定の配列である;参照配列は、例えば、配列表で示される全長cDNAまたは遺伝子配列のセグメントとしてのより長い配列のサブセットであってもよく、あるいは完全なcDNAまたは遺伝子配列を含んでいてもよい。一般に、参照配列は、少なくとも18ヌクレオチド長または6アミノ酸長であり、しばしば少なくとも24ヌクレオチド長または8アミノ酸長であり、そしてたいてい少なくとも48ヌクレオチド長または16アミノ酸長である。2つのポリヌクレオチドまたはアミノ酸配列は、それぞれ(1)2分子間で類似する配列(すなわち、完全なポリヌクレオチドまたはアミノ酸配列の一部)を含み得、かつ(2)2つのポリヌクレオチドまたはアミノ酸配列間で異なる配列をさらに含み得るため、2(またはそれ以上)分子間の配列比較は、典型的には、配列類似性の局所領域を同定し、そして比較するための「比較ウインドウ」について2分子の配列を比較することで行われる。本明細書で用いられる「比較ウインドウ」は、ポリヌクレオチド配列またはアミノ酸配列が、少なくとも18の連続するヌクレオチドまたは6アミノ酸配列の参照配列と比較され得、かつ比較ウインドウのポリヌクレオチド配列の部分が、2配列の最適なアラインメントのための参照配列(付加または欠損を含まない)と比較して、20%以下の付加、欠損、置換などを含み得る、少なくとも18の連続するヌクレオチド位置または6アミノ酸である論理的セグメントを意味する。比較ウインドウをアラインメントするための配列の最適なアラインメントは、Smith and Watermanの局所相同性アルゴリズム(Adv. Appl. Math. 2: 482(1981))、Needleman and Wunschの相同性アラインメントアルゴリズム(J. Mol.Biol. 48: 443(1970))、Pearson and Lipmanの同様の検索方法(Proc. Natl. Acad. Sci.(U. S. A.)85: 2444(1988))、これらのアルゴリズムのコンピュータ上での実行(Wisconsin Genetics Software Package Release 7.0のGAP、BESTFIT、FASTA、およびTFASTA(Genetics Computer Group, 575 Science Dr., Madison, Wis.))、Genework、またはMacVector Software Package)、または調査により実行されてもよく、そして種々の方法で得られた最良のアラインメント(すなわち、比較ウインドウについて最も高い割合の相同性となる)が選択される。
【0062】
用語「配列同一性」は、比較ウインドウについて2つのポリヌクレオチドまたはアミノ酸配列が同一である(すなわち、ヌクレオチド対ヌクレオチドまたは残基対残基ベース)ことを意味する。用語「配列同一性の割合」は、比較ウインドウについて最適にアラインメントされた2配列を比較し、同一の核酸塩基(例えば、A、T、C、G、UまたはI)または残基が、両配列で生じる位置数を決定し、一致位置数を得、一致位置数を比較ウインドウの位置の総数(すなわち、ウインドウサイズ)で割り、次にこの結果に100を掛け、配列同一性の割合を得ることにより、算出される。本明細書で用いられる用語「実質的な同一性」は、ポリヌクレオチドまたはアミノ酸が、少なくとも18ヌクレオチド(6アミノ酸)位置の比較ウインドウ、しばしば少なくとも24〜48ヌクレオチド(8〜16アミノ酸)位置のウインドウについて参照配列と比較して、少なくとも85%の配列同一性、好ましくは、少なくとも90から95%の配列同一性、より一般的には、少なくとも99%の配列同一性を有するポリヌクレオチドまたはアミノ酸配列の特徴を示し、ここで、配列同一性の割合は、参照配列を、比較ウインドウについて参照配列の全20%以下が欠失または付加を含み得る配列と比較することにより算出される。参照配列は、より長い配列のサブセットであってもよい。
【0063】
本明細書で用いられる20の従来アミノ酸およびそれらの略語は、従来の使用に従う。Immunology-A Synthesis(2nd ed., Golub, E. S. and Gren, D. R. eds. , Sinauer Associates, Sunderland, Mass. 1991)を参照されたい。20の従来アミノ酸の立体異性体(例えば、D−アミノ酸)、α,α−2置換アミノ酸のような非天然アミノ酸、N−アルキルアミノ酸、乳酸、および他の従来にないアミノ酸もまた、本明細書に記載の本発明のポリペプチドの適切な要素であり得る。従来にないアミノ酸の例としては、4−ヒドロキシプロリン、γ−カルボキシグルタメート、ε−N,N,N−トリメチルリジン、ε−N−アセチルリジン、O−ホスホセリン、N−アセチルセリン、N−ホルミルメチオニン、3−メチルヒスチジン、5−ヒドロキシリジン、α−N−メチルアルギニン、および他の類似のアミノ酸およびイミノ酸(例えば、4−ヒドロキシプロリン)が挙げられる。本明細書で用いられるポリペプチド表記では、標準的使用および慣習に従い、左手方向がアミノ末端方向、そして右手方向がカルボキシ末端方向である。
【0064】
同様に、他に特定しない限り、1本鎖ポリヌクレオチド配列の左手末端は、5’末端である;2本鎖ポリヌクレオチド配列の左手方向は、5’方向として言及される。新生RNA転写物の5’から3’方向への付加は、転写方向として言及される;RNAと同じ配列を有し、かつRNA転写物の5’から5’末端であるDNA鎖の配列領域は、「上流配列」として言及される;RNAと同じ配列を有し、RNA転写物の3’から3’末端であるDNA鎖の配列領域は、「下流配列」として言及される。
【0065】
ポリペプチドに適用される用語「実質的な同一性」は、例えば、デフォルトギャップウエイトを用いたプログラムGAPまたはBESTFITにより、最適にアラインメントされた場合、2ペプチド配列は、少なくとも80%の配列同一性、好ましくは、少なくとも90%の配列同一性、より好ましくは、少なくとも95%の配列同一性、そして最も好ましくは、少なくとも99%の配列同一性を有することを意味する。好ましくは、同一でない残基の位置は、保存的アミノ酸置換により異なる。保存的アミノ酸置換は、互換性のある類似の側鎖を有する残基に言及する。例えば、脂肪族側鎖を有するアミノ酸基は、グリシン、アラニン、バリン、ロイシン、およびイソロイシンであり;脂肪族ヒドロキシル側鎖を有するアミノ酸基は、セリン、およびスレオニンであり;アミド含有側鎖を有するアミノ酸基は、アスパラギン、およびグルタミンであり;芳香族側鎖アミノ酸基を有するアミノ酸基は、フェニルアラニン、チロシン、およびトリプトファンであり;塩基性側鎖を有するアミノ酸基は、リジン、アルギニン、およびヒスチジンであり;そして硫黄含有側鎖を有するアミノ酸基は、システイン、およびメチオニンである。好ましい保存的アミノ酸の置換基は、バリン−ロイシン−イソロイシン、フェニルアラニン−チロシン、リジン−アルギニン、アラニン−バリン、グルタミン酸−アスパラギン酸、およびアスパラギン−グルタミンである。
【0066】
本明細書で考察される、抗体または免疫グロブリン分子のアミノ酸配列のマイナーバリエーションは、アミノ酸配列のバリエーションが、少なくとも75%、より好ましくは、少なくとも80%、90%、95%、そして最も好ましくは99%のオリジナル配列を保持している場合、本明細書に記載の発明に包含されることが意図される。特に、保存的アミノ酸置換が意図される。保存的置換は、それらの側鎖に関連性がある、アミノ酸ファミリー内で生じるものである。遺伝子的にコードされたアミノ酸は、一般に、以下のファミリー:(1)酸性=アスパラギン酸、グルタミン酸;(2)塩基性=リジン、アルギニン、ヒスチジン;(3)非極性=アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン;および(4)非荷電極性=グリシン、アスパラギン、グルタミン、システイン、セリン、スレオニン、チロシンに分けられる。より好ましいファミリーは以下である:セリンおよびスレオニンが脂肪族ヒドロキシファミリーであり;アスパラギンおよびグルタミンがアミド含有ファミリーであり;アラニン、バリン、ロイシン、およびイソロイシンが脂肪族ファミリーであり;そしてフェニルアラニン、トリプトファン、およびチロシンが芳香族ファミリーである。例えば、ロイシンのイソロイシンまたはバリンでの単離された置換、アスパラギン酸のグルタミン酸での単離された置換、スレオニンのセリンでの単離された置換、またはアミノ酸の構造的に関連するアミノ酸での同様の置換は、特に該置換がフレームワーク内にアミノ酸を含まない場合、得られた分子の結合または特性に大きく影響することはないと予測することはもっともである。アミノ酸変更が機能的ペプチドとなるか否かは、ポリペプチド誘導体の特異的活性をアッセイすることで、容易に決定され得る。アッセイは本明細書で詳細に説明されている。抗体または免疫グロブリン分子のフラグメントまたは類似体は、当業者により容易に製造され得る。フラグメントまたは類似体の好ましいアミノ末端およびカルボキシ末端は、機能ドメインの境界付近に存在する。構造ドメインおよび機能的ドメインは、ヌクレオチドおよび/またはアミノ酸配列データを公開または独自の配列データベースと比較することにより、同定され得る。好ましくは、コンピューター上での比較方法が、既知の構造および/または機能の他のタンパク質に存在する配列モチーフまたは予測タンパク質構造ドメインを同定するために用いられる。既知の3次元構造に折りたたまれたタンパク質配列を同定するための方法が知られている。Bowie et al., Science 253: 164(1991)。従って、上記の例は、当業者が、本発明をふまえて構造ドメインおよび機能ドメインを定義するために用いられ得る配列モチーフおよび構造を認識することができることを示している。
【0067】
好ましいアミノ酸置換は、(1)タンパク質分解に対する感受性を低減し、(2)酸化に対する感受性を低減し、(3)タンパク質複合体形成のための結合親和性を変化させ、(4)結合親和性を変化させ、かつ(4)かかる類似体の他の物理化学的または機能的特性を与えるか、または修飾するものである。類似体は、天然に存在するペプチド配列以外の配列の種々の変異体を含み得る。例えば、1つまたは複数のアミノ酸置換(好ましくは、保存的アミノ酸置換)は、天然に存在する配列内(好ましくは、分子間接触を形成するドメイン外のポリペプチドの部分において)で生じてもよい。保存的アミノ酸置換は、実質的には、親配列の構造特徴を変更してはいけない(例えば、置換アミノ酸は、親配列中に存在するらせん構造を破壊、あるいは親配列を特徴付ける他のタイプの二次構造を破壊する傾向があってはならない)。当該技術分野で認識されているポリペプチドの二次元および三次元構造の例は、Proteins, Structures and Molecular Principles(Creighton, ed., W. H. Freeman and Company, New York 1984); Introduction to Protein Structure(Branden, C. and Tooze, J. eds., Garland Publishing, New York, N. Y. 1991);およびThornton et al., Nature 354: 105(1991)に記載されている。
【0068】
本明細書で用いられる用語「ポリペプチドフラグメント」は、アミノ末端および/またはカルボキシ末端の欠失したポリペプチドを意味するが、残存アミノ酸配列は、例えば、全長cDNA配列から推定される天然に存在する配列の対応する位置と同一である。フラグメントは、典型的には、少なくとも5、6、8または10アミノ酸長であり、好ましくは、少なくとも14アミノ酸長であり、より好ましくは、少なくとも20アミノ酸長であり、通常は、少なくとも50アミノ酸長であり、そしてさらにより好ましくは、少なくとも70アミノ酸長である。本明細書で用いられる用語「類似体」は、推定されたアミノ酸配列の一部分と実質的に同一であり、かつ以下の特性:(1)適切な結合条件下でPDGF−DD2量体と特異的に結合すること、(2)適切なPDGF−DD結合を崩壊させる能力、または(3)インビトロまたはインビボでPDGF−DDを発現する細胞の増殖を阻害する能力のうち少なくとも1つを有する、少なくとも25アミノ酸のセグメントを含むポリペプチドに言及する。典型的には、ポリペプチド類似体は、天然に存在する配列に対して、保存的アミノ酸置換(または、付加または欠失)を含む。類似体は、典型的には、少なくとも20アミノ酸長、好ましくは、少なくとも50アミノ酸長、またはそれより長く、しばしば天然に存在するポリペプチドの全長と同じ長さであり得る。
【0069】
ペプチド類似体は、一般に、テンプレートペプチドのものと類似する特性を有する非ペプチド薬として医薬産業において用いられる。この種の非ペプチド化合物は、「ペプチドミメティック(peptide mimetic)」または「ペプチドミメティック(peptidomimetic)」と呼ばれる。Fauchere, J. Adv. Drug Res. 15: 29(1986); Veber and Freidinger, TINS p. 392(1985);およびEvans et al.,J. Med. Chem. 30: 1229(1987)を参照されたい。かかる化合物は、たいていコンピュータ分子モデリングの助けにより開発される。治療上有用なペプチドと構造が類似したペプチドミメティックは、均等な治療効果または予防効果を生み出すために用いられ得る。一般に、ペプチドミメティックは、ヒト抗体の様なパラダイムポリペプチド(すなわち、生化学特性または薬理学活性を有するポリペプチド)と構造が類似しているが、当該技術分野でよく知られた方法により、−CH2NH−、−CH2S−、−CH2−CH2−、−CH=CH−(シスおよびトランス)、−COCH2−、−CH(OH)CH2−、および−CH2SO−からなる群から選択される結合で必要に応じて置換された1以上のペプチド結合を有している。一致配列の1以上のアミノ酸の同種のD−アミノ酸での系統的置換が、より安定なペプチドを製造するために用いられてもよい。さらに、一致配列または実質的に同一の一致配列バリエーションを含む制約されたペプチドは、当該技術分野で既知の方法(Rizo and Gierasch Ann. Rev. Biochem. 61: 387(1992))、例えば、ペプチドを環化させる分子内ジスルフィド架橋を形成し得る内部システイン残基を付加することにより、製造されてもよい。
【0070】
「抗体」または「抗体ペプチド」は、未変性の抗体、または未変性の抗体と特異的結合に対して競合するその結合フラグメントを意味する。結合フラグメントは、組換えDNA技術、または未変性の抗体の酵素的分解または化学的分解により作製される。結合フラグメントは、Fab、Fab’、F(ab’)2、Fvおよび1本鎖抗体が挙げられる。「二重特異性」または「二機能性」抗体以外の他の抗体は、その結合部位のそれぞれが同一であることが理解される。過剰の抗体が、対受容体と結合する受容体の量を少なくとも約20%、40%、60%または80%、より一般的には約85%(インビトロ競合結合アッセイにて測定)より多く低減させるとき、抗体は、受容体の対受容体との接着を実質的に阻害する。
【0071】
用語「エピトープ」は、免疫グロブリンまたはT細胞受容体と特異的に結合し得る任意のタンパク質決定基を含む。エピトープ決定基は、一般的には、アミノ酸または糖側鎖のような分子の化学的に活性な表面群からなり、一般的には、特異的な三次元構造特徴、ならびに特異的荷電特徴を有する。抗体は、解離定数が1μM以下、好ましくは100nM以下、そして最も好ましくは、10nM以下であるときに、抗原と特異的に結合するといわれている。
【0072】
用語「剤」は、化学化合物、化学化合物の混合物、生物学的マクロ分子、または生物学的材料から作られた抽出物を示すために、本明細書で用いられる。
【0073】
本明細書での目的上、「活性な」または「活性」は、天然のPDGF−DDポリペプチドまたは天然に存在するPDGF−DDポリペプチドの生物学的活性および/または免疫学的活性を保持しているPDGF−DDポリペプチドの形態を意味し、ここで「生物学的」活性とは、天然のPDGF−DDまたは天然に存在するPDGF−DDポリペプチドが有する抗原性エピトープに対する抗体産生を誘導する能力以外の、天然のPDGF−DDまたは天然に存在するPDGF−DDポリペプチドにより引き起こされる生物学的機能(阻害性または刺激性のいずれか)を意味し、そして「免疫学的」活性とは、天然のPDGF−DDまたは天然に存在するPDGF−DDポリペプチドが有する抗原性エピトープに対する抗体産生を誘導する能力を意味する。
【0074】
「処置」は、治療上の処置、および予防的(prophylactic)または予防上(preventive)の測定の両方を意味し、目的は、標的の病状または疾患を予防するか、あるいは遅延(緩和)することである。処置の必要なものは、既に疾患を有する者、ならびに該疾患に罹りやすいものまたは当該疾患の予防が必要とされるものを含む。
【0075】
「哺乳動物」は、哺乳動物に分類される任意の動物を意味し、ヒト、サル、チンパンジーおよびゴリラの様な他の霊長類、イヌ、ネコ、ウシ、ウマ、ヒツジ、ブタ、ヤギ、ウサギ、げっ歯類等の様な家庭用および農場用動物(家畜)、ならびに動物園、スポーツ、実験、または愛玩動物が挙げられる。処置の目的上、哺乳動物は、好ましくはヒトである。
【0076】
本明細書で用いられる「担体」は、利用される投薬量および濃度でそれらに曝露された細胞または哺乳動物にとって毒性のない、医薬的に許容される担体、賦形剤、または安定化剤を含む。生理的に許容される担体は、たいてい水性のpH緩衝溶液である。生理的に許容される担体の例は、リン酸、クエン酸、および他の有機酸のような緩衝液;アスコルビン酸を含む抗酸化剤;低分子量(約10残基未満)のポリペプチド;血清アルブミン、ゼラチン、または免疫グロブリンのようなタンパク質;ポリビニルピロリドンのような親水性ポリマー;グリシン、グルタミン、アスパラギン、アルギニン、またはリジンのようなアミノ酸;グルコース、マンノース、またはデキストリンを含む単糖、二糖および他の多糖類;EDTAのようなキレート剤;マンニトールまたはソルビトールのような糖アルコール;ナトリウムのような塩形成対イオン;および/またはTWEEN(登録商標)、ポリエチレングリコール(PEG)およびPLURONICS(登録商標)のような非イオン性界面活性剤を含む。
【0077】
抗体のパパイン消化により、それぞれ単一の抗原結合部位を有する「Fab」フラグメントと呼ばれる2つの同一の抗原結合フラグメントが得られ、そして残りの「Fc」フラグメントは、容易に結晶化される能力を反映した表示である。ペプシン処理により、「F(ab’)2」フラグメントが得られる、これは2つの抗原結合部位を有し、かつ抗原と架橋することができる。
【0078】
「Fv」は、抗体の完全な抗原認識部位および結合部位を含有する最小の抗体フラグメントである。この領域は、非共有結合で強く会合した、1つの重鎖と1つの軽鎖の可変ドメインの2量体からなる。この構成で、各可変ドメインの3つのCDRが相互作用してVH−VL2量体の表面の抗原結合部位を規定する。総合すると、6つのCDRが、抗原結合特異性を抗体に与える。しかしながら、例えば、1つの可変ドメイン(例えば、Fv2量体のVHまたはVL部分、または抗原特異的な3つのCDRのみを含むFvの半分)でさえも、おそらく完全な結合部位よりも低い親和性ではあるが、抗原を認識し結合する能力を有し得る。
【0079】
Fabフラグメントはまた、軽鎖の定常ドメイン、および重鎖の第一の定常ドメイン(CH1)を含有する。Fabフラグメントは、抗体のヒンジ領域由来の1つ以上のシステインを含む重鎖CH1ドメインのカルボキシ末端での2、3の残基の付加により、Fab’フラグメントとは異なっている。F(ab’)2抗体フラグメントは、本来、それらの間にヒンジシステインを有するFab’フラグメント対として製造された。抗体フラグメントの他の化学結合もまた知られている。
【0080】
「固相」は、本明細書に記載の抗体と接着できる、非水性マトリックスを意味する。本明細書において包含される固相の例は、部分的または全体的に、ガラス(例えば、孔制御ガラス)、多糖(例えば、アガロース)、ポリアクリルアミド、ポリスチレン、ポリビニルアルコール、およびシリコーンから形成されたものが挙げられる。ある種の実施態様において、状況に応じて、固相は、アッセイプレート、いいかえれると、精製カラム(例、アフィニティークロマトグラフィーカラム)のウェルを含むこともできる。この用語はまた、米国特許第4,275,149号に記載されるものの様な個別の粒子の非連続固相も含む。
【0081】
用語「リポソーム」は、哺乳動物への薬(例えば、PDGF−DDポリペプチド、またはこれに対する抗体)の輸送に有用な、種々のタイプの脂質、リン脂質、および/または界面活性剤を含む、小胞を示すために本明細書において用いられる。リポソームの成分は、一般的に、生物学的な膜の脂質配列と類似した二層の構造で配列される。
【0082】
用語「小分子」は、約500ダルトン未満の分子量を有する分子を記載するために、本明細書で用いられる。
【0083】
本明細書で用いられる用語「標識」または「標識された」は、例えば、放射標識されたアミノ酸の取込による、検出可能なマーカーの取込、または印の付いたアビジンにより検出され得るビオチン化部分のポリペプチドとの接着(例えば、光学的方法または比色方法により検出され得る蛍光マーカーまたは酵素活性を含有するストレプトアビジン)。ある種の状況において、標識またはマーカーは治療用であり得る。ポリペプチドおよび糖タンパク質を標識する種々の方法が当該技術分野で知られており、用いられ得る。ポリペプチドの標識の例は、以下のものが挙げられるが、これに限らない:放射性同位元素または放射性核種(例えば、3H、14C、15N、35S、90Y、99Tc、111In、125I、131I)、蛍光標識(例えば、FITC、ローダミン、ランタニド蛍光体)、酵素標識(例えば、西洋ワサビペルオキシダーゼ、β−ガラクトシダーゼ、ルシフェラーゼ、アルカリホスファターゼ)、化学発光、ビオチニル基、二次レポーターにより認識される、事前に決定されたポリペプチドエピトープ(例えば、ロイシンジッパー対配列、二次抗体の結合部位、金属結合ドメイン、エピトープタグ)。ある実施態様において、標識は、種々の長さのスペーサーアームに結合して、可能性のある立体障害が低減される。
【0084】
本明細書で用いられる用語「医薬剤または薬」は、患者に適切に投与した場合に所望の効果を誘導し得る化学化合物または組成物を意味する。本明細書の他の化学用語は、The McGraw-Hill Dictionary of Chemical Terms(Parker, S. , Ed., McGraw-Hill, San Francisco(1985))に例示される、当該技術分野での従来の使用に従い、用いられる。
【0085】
本明細書で用いられる「実質的に純粋」は、目的の種が、存在する優勢種であること(すなわち、モル基準で、組成物において他のいかなる個々の種よりも豊富に存在すること)を意味し、好ましくは、実質的に精製された画分は、目的の種が、存在する全てのマクロ分子種の少なくとも約50%(モル基準)を含む、組成物である。一般的には、実質的に純粋な組成物は、存在する全てのマクロ分子種の約80%より多く、より好ましくは、約85%、90%、95%および99%より多くを含むであろう。最も好ましくは、目的の種は、精製されて本質的に均一になり(コンタミ種は、従来の検出方法によって組成物中で検出されない)、ここで、組成物は本質的には単一のマクロ分子種からなる。
用語「患者」は、ヒト患者および獣医対象を含む。
【0086】
抗PDGF−DD抗体
抗体、または部分、フラグメント、ミメティック、またはその誘導体は、PDGF−DD2量体を認識する、任意のタイプの抗体または部分であってもよい。ある種の実施態様において、好ましくは、抗体、またはその部分は、PDGF−DDを中和できる。さらなる実施態様において、好ましくは、抗体、またはその部分は、PDGF−DDおよび腎炎と関連する症状(炎症、体液鬱滞、組織腫脹、疼痛、腫脹、高血圧、脳腫脹、視覚障害、低尿量、および血尿が挙げられるが、これに限らない)を低減し得る。1つの実施態様によれば、抗体は、例えば、抗PDGF−DD mAb 6.4であり得る。かかる抗体のさらなる例は、関連米国特許出願第10/041,860号(出願日:2,002年1月7日)において、見出すことができる。
【0087】
抗体構造
基本的な抗体構造単位は、4量体を含むことが知られている。それぞれの4量体は、2つの同一対ポリペプチド鎖から構成され、それぞれの対は、1つの「軽」鎖(約25kDa)および1つの「重」鎖(約50〜70kDa)を有している。それぞれの鎖のアミノ末端部分は、主に抗原認識に関与する約100〜110またはそれ以上のアミノ酸からなる可変領域を含む。それぞれの鎖のカルボキシ末端部分は、主にエフェクター機能に関与する定常領域を規定する。ヒト軽鎖は、κおよびλ軽鎖に分類される。重鎖は、μ、δ、γ、α、またはεに分類され、アイソトープをそれぞれIgM、IgD、IgAおよびIgEと規定する。軽鎖および重鎖内の可変領域および定常領域は、約12以上のアミノ酸の「J」領域により結合されており、重鎖はまた、約10より多くのアミノ酸からなる「D」領域を含む。概論として、Fundamental Immunology Ch. 7(Paul, W., ed., 2d ed. Raven Press, N. Y.(1989))を参照されたい。それぞれの軽鎖/重鎖対の可変領域は、抗体結合部位を形成する。従って、未変性の抗体は、2つの結合部位を有する。例外的に、二機能性または二重特異性抗体では、2つの結合部位は同一である。
【0088】
鎖は全て、相補性決定領域またはCDRとも呼ばれる、3つの超可変領域により結合された比較的保存されたフレームワーク領域(FR)からなる同じ一般構造を有する。それぞれの対の2つの鎖由来のCDRは、フレームワーク領域によりアライメントされ、特異的エピトープと結合することが可能になる。N末端からC末端まで、軽鎖および重鎖は両方ともドメインFR1、CDR1、FR2、CDR2、FR3、CDR3およびFR4を含む。それぞれのドメインに対するアミノ酸指定は、Kabat Sequences of Proteins of Immunological Interest(National Institutes of Health, Bethesda, Md.(1987 and 1991))、またはChothia & Lesk J. Mol. Biol. 196: 901-917(1987);Chothia et al. Nature 342:878-883(1989)の定義に従う。
【0089】
二重特異性抗体または二機能性抗体は、2つの異なる重鎖/軽鎖対および2つの異なる結合部位を有する人工ハイブリッド抗体である。二重特異性抗体は、ハイブリドーマの融合またはFab’フラグメントの結合を含む、種々の方法により製造され得る。例えば、Songsivilai & Lachmann,Clin. Exp. Immunol. 79: 315-321(1990)、Kostelny et al., J. Immunol. 148: 1547-1553(1992)を参照されたい。二重特異性抗体の製造は、従来の抗体製造と比較してかなり苦労が多い方法であり得るが、二重特異性抗体の収率および純度は一般に低い。二重特異性抗体は、1つの結合部位を有するフラグメント(例えば、Fab、Fab’およびFv)の形では存在しない。
かかる二機能性抗体または二重特異性抗体が本発明で意図され、そして包含されることが理解されるであろう。
【0090】
ヒト抗体および抗体のヒト化
本明細書に記載の発明の実施態様はまた、ヒト抗体も意図し包含する。ヒトの処置のため、ヒト抗体は、マウスまたはラットの可変領域および/または定常領域を有する抗体に関連するある種の問題を回避する。かかるマウスまたはラット由来タンパク質の存在は、抗体の迅速なクリアランスをもたらし得るか、あるいは患者の抗体に対する免疫応答を発症し得る。マウスまたはラット由来抗体の利用を回避するため、ヒト化抗体を開発するか、あるいはげっ歯類が完全ヒト抗体を産生するように、ヒト抗体機能をげっ歯類に導入することで完全ヒト抗体を製造できると仮定されてきた。
【0091】
ヒト抗体
完全ヒト抗体を製造する1つの方法は、ゲノム内にヒト重鎖および軽鎖遺伝子を含有するように操作された、XenoMouse(登録商標)系マウスを用いることによる。例えば、ヒト重鎖遺伝子座およびκ軽鎖遺伝子座の245kbおよび190kbサイズの生殖系列立体配置フラグメントを含有するXenoMouse(登録商標)マウスは、Green et al., Nature Genetics 7: 13-21(1994)に記載されている。Green等の研究は、それぞれメガ塩基サイズのヒト重鎖遺伝子座およびκ軽鎖遺伝子座の生殖系列立体配置YACフラグメントの利用により、約80%より多くのヒト抗体レパートリーの導入にまで拡大した。Mendez et al., Nature Genetics 15: 146-56(1997)および米国特許出願第08/759,620号(出願日:1,996年12月3日)を参照のこと。さらに、全λ軽鎖遺伝子座を含有するXenoMouse(登録商標)マウスが作製された(米国特許出願第60/334,508号(出願日:2,001年11月30日))。さらに、複数のアイソタイプを産生するXenoMouse(登録商標)マウスが作製された(例えば、WO00/76310参照)。XenoMouse(登録商標)系は、Abgenix, Inc.(Fremont, CA)から入手可能である。
【0092】
XenoMouse(登録商標)の作製は、さらに、以下で考察され記載されている:米国特許出願第07/466,008号(出願日:1990年1月12日)、同第07/610,515号(出願日:1990年11月8日)、同第07/919,297号(出願日:1992年7月24日)、同第07/922,649号(出願日:1992年7月30日)同第08/031,801号(出願日:1993年3月15日)、同第08/112,848号(出願日:1993年8月27日)、同第08/234,145号(出願日:1994年4月28日)、同第08/376,279号(出願日:1995年1月20日)、同第08/430,938号(出願日:1995年4月27日)、同第08/464,584号(出願日:1995年6月5日)、同第08/464,582号(出願日:1995年6月5日)、同第08/463,191号(出願日:1995年6月5日)、同第08/462,837号(出願日:1995年6月5日)、同第08/486,853号(出願日:1995年6月5日)、同第08/486,857号(出願日:1995年6月5日)、同第08/486,859号(出願日:1995年6月5日)、同第08/462,513号(出願日:1995年6月5日)、同第08/724,752号(出願日:1996年10月2日)および同第08/759,620号(出願日:1996年12月3日)、ならびに米国特許第6,162,963号、同第6,150,584号、同第6,114,598号、同第6,075,181号および同第5,939,598号、ならびに日本国特許第3 068 180 B2号、同第3 068 506 B2号および同第3 068 507 B2号。Mendez et al. Nature Genetics 15: 146-156(1997)およびGreen and Jakobovits J.Exp. Med., 188: 483-495(1998)もまた参照されたい。欧州特許第463,151 B1号(特許発行日:1996年6月12日)、国際特許出願第WO94/02602号(公開日:1994年2月3日)、国際特許出願第WO 96/34096号(公開日:1996年10月31日)同第WO98/24893号(公開日:1998年6月11日)、同第WO00/76310号(公開日:2000年12月21日)もまた参照されたい。
【0093】
代わりのアプローチで、GenPharm International, Inc.を含む他社は、「ミニローカス(minilocus)」アプローチを利用した。ミニローカスアプローチにおいて、外因性のIg遺伝子座は、Ig遺伝子座由来の種(個々の遺伝子)を含むことにより模倣される。従って、1つ以上のVH遺伝子、1つ以上のDH遺伝子、1つ以上のJH遺伝子、μ定常領域および第二の定常領域(好ましくは、γ定常領域)が、動物に挿入するための構築物となる。このアプローチは、米国特許第5,545,807号(Surani et al.)および米国特許第5,545,806号、同第5,625,825号、同第5,625,126号、同第5,633,425号、同第5,661,016号、同第5,770,429号、同第5,789,650号、同第5,814,318号、同第5,877,397号、同第5,874,299号および同第6,255,458号(それぞれLonberg and Kay)、米国特許第5,591,669号および同第6,023,010号(Krimpenfort and Berns)、米国特許第5,612,205号、同第5,721,367号および同第5,789,215(Berns et al.)および米国特許第5,643,763号(Choi and Dunn)、ならびにGen Pharm Internationalの米国特許出願第07/574,748号(出願日:1990年8月29)、同第07/575,962号(出願日:1990年8月31日)、同第07/810,279号(出願日:1991年12月17日)、同第07/853,408号(出願日:1992年3月18日)、同第07/904,068号(出願日:1992年6月23日)、同第07/990,860号(出願日:1992年12月16日)、同第08/053,131号(出願日:1993年4月26日)、同第08/096,762号(出願日:1993年7月22日)、同第08/155,301号(出願日:1993年11月18日)、同第08/161,739号(出願日:1993年12月3日)、同第08/165,699号(出願日:1993年12月10日)、同第08/209,741号(出願日:1994年3月9日)を参照されたい。欧州特許第0 546 073 B1号、国際公開パンフレット第WO92/03918号、同第WO92/22645号、同第WO92/22647号、同第WO92/22670号、同第WO93/12227号、同第WO94/00569号、同第WO94/25585号、同第WO96/14436号、同第WO97/13852号および同第WO98/24884、ならびに米国特許第5,981,175号も参照されたい。さらに、Taylor et al., 1992, Chen et al., 1993, Tuaillon et al., 1993, Choi et al., 1993, Lonberg et al.,(1994), Taylor et al.,(1994),およびTuaillon et al.,(1995), Fishwild et al.,(1996)も参照されたい。
【0094】
上記で引用し、Medical Research Counsel(「MRC」)に帰属しているSurani等発明者は、ミニローカスアプローチの使用により、Ig遺伝子座を有するトランスジェニックマウスを作製した。上記で引用したGenPharm International workの発明者であるLonbergおよびKayは、本発明者らの指示に従って、Surani等の研究の実質的に重複する内因性マウスIg遺伝子座の不活性化を提案した。
【0095】
ミニローカスアプローチの利点は、Ig遺伝子座の一部分を含む構築物が製造され、動物に導入されるという迅速性である。しかしながら、同様にミニローカスアプローチの重大な欠点は、理論上、少数のV、DおよびJ遺伝子を含むことにより不十分な多様性が導入されることである。実際、公開された研究は、この懸念を支持しているようである。ミニローカスアプローチを用いて作製した動物のB細胞発生および抗体産生は、成長が止まっているように思われる。従って、本明細書に記載の発明の周辺研究は、より多くの多様性を達成するためにIg遺伝子座の大部分の導入を目差し続け、かつ動物の免疫レパートリーを再構築するために努力し続けている。
【0096】
Kirinはまた、マイクロ細胞融合により、染色体の巨大片または全染色体が導入されたマウス由来のヒト抗体産生を示している。欧州特許出願第773 288号および同第843 961号を参照されたい。
【0097】
Lidak Pharmaceuticals(現在はXenorex)はまた、ヒトドナー由来の非悪性成熟末梢白血球の注射によって修飾されたSCIDマウスにおけるヒト抗体の産生を示している。修飾されたマウスは、ヒト抗体の産生からなる、免疫原での刺激の際にヒトドナーの免疫応答特徴を示す。米国特許第5,476,996号および同第5,698,767号を参照されたい。
【0098】
ヒト抗マウス抗体(HAMA)応答は、キメラまたはそうでなければヒト化抗体を製造するための産業となる。キメラ抗体はヒト定常領域およびマウス可変領域を有するが、ある種のヒト抗キメラ抗体(HACA)応答は、特に抗体の慢性的または複数回投与の利用において観察されることが予測される。従って、HAMAまたはHACA応答の懸念および/または影響を低減するために、PDGF−DDに対する完全ヒト抗体を提供することが望まれるであろう。
【0099】
ヒト化およびディスプレイ技術
ヒト抗体の製造と関連して上述したように、免疫原性の低減した抗体を製造することには利点がある。これは、ある程度、適切なライブラリを用いたヒト化技術およびディスプレイ技術と関連して成され得る。マウス抗体または他の種由来の抗体は、当該技術分野でよく知られた技術を用いてヒト化または霊長類化し得ることは理解されるであろう。例えば、Winter and Harris,immunol Today 14: 43-46(1993)およびWright et al., Crit, Reviews in Immunol. 12: 125-168(1992)を参照されたい。目的の抗体は、組換えDNA技術により操作され、CH1、CH2、CH3、ヒンジドメインおよび/またはフレームワークドメインを対応するヒト配列と置き換えるられてもよい(例、WO92/02190および米国特許第5,530,101号、同第5,585,089号、同第5,693,761号、同第5,693,792号、同第5,714,350号および同第5,777,085号参照)。また、キメラ免疫グロブリン遺伝子の構築のためのIg cDNAの使用が当該技術分野で知られている(Liu et al., P.N. A. S. 84: 3439(1987)およびJ. Immunol. 139: 3521(1987))。mRNAは、抗体を産生するハイブリドーマまたは他の細胞から単離され、cDNAを製造するために用いられる。目的のcDNAは、特異的なプライマーを用いたポリメラーゼ連鎖反応により増幅されてもよい(米国特許第4,683,195号および同第4,683,202号)。あるいは、ライブラリを作成し、スクリーニングして、目的とする配列を単離する。次いで、抗体の可変領域をコードするDNA配列を、ヒト定常領域配列と融合させる。ヒト定常領域遺伝子の配列は、Kabat et al.,”Sequences of Protein of Immunological Interest”, N. I. H. publication no. 91-3242(1991)において見出され得る。ヒトC領域遺伝子は、既知のクローンから容易に得られる。アイソタイプの選択は、相補的固定の様な所望のエフェクター機能、または抗体依存性細胞毒性の活性により、導かれるであろう。好ましいアイソタイプは、IgG1、IgG3およびIgG4である。ヒト軽鎖定常領域、κまたはλのいずれを用いてもよい。次いで、キメラヒト化抗体は従来の方法により発現される。
【0100】
Fv、F(ab’).sub.2、およびFabのような抗体フラグメントは、未変性のタンパク質の分解、例えば、プロテアーゼまたは化学分解により、製造されてもよい。あるいは、切断された遺伝子が設計される。例えば、F(ab’)2フラグメントの一部分をコードするキメラ遺伝子は、CH1ドメインおよびH鎖のヒンジ領域をコードするDNA配列、次いで、翻訳停止コドンを含む、切断された分子が得られる。
【0101】
重鎖および軽鎖J領域の一致配列は、V領域セグメントのヒトC領域セグメントとの連続結合のため、有用な制限部位をJ領域に導入するためのプライマーとしての使用のためのオリゴヌクレオチドを設計するために用いられてもよい。C領域cDNAを、部位指定の変異誘発により修飾し、制限部位をヒト配列の類似位置に配置することができる。
【0102】
発現ベクターには、プラスミド、レトロウイルス、YAC、EBV由来エピソーム等が挙げられる。好都合なベクターは、任意のVHまたはVL配列が容易に挿入されて発現され得るように、操作された適切な制限部位を有する、機能的に完全ヒトCHまたはCL免疫グロブリン配列をコードするものである。かかるベクターでは、挿入されたJ領域のスプライスドナー部位とヒトC領域に先立つスプライスアクセプター部位との間、およびヒトCHエキソン内で生じるスプライス領域でもスプライシングが起こる。ポリアデニル化および転写の終結は、コーディング領域の下流の天然染色体部位で起こる。得られたキメラ抗体は、任意の強力なプロモーター(例えば、SV−40初期プロモーター(Okayama et al., Mol. Cell. Bio. 3: 280(1983))、Rous肉腫ウイルスLTR(Gorman et al., P. N. A. S. 79: 6777(1982))、およびモロニーマウス白血病ウイルスLTR(Grosschedl et al., Cell 41 : 885(1985))等のレトロウイルスLTRが挙げられる)と結合し得る。また、天然のIgプロモーター等も用いられ得ることは、理解されるだろう。
【0103】
さらに、ヒト抗体または他の種由来の抗体は、ディスプレイタイプ技術(ファージディスプレイ、レトロウイルスディスプレイ、リボソームディスプレイが挙げられるが、これに限らない)、および当該技術分野でよく知られた技術を用いる他の技術により製造され、得られた分子は、親和性成熟のようなさらなる成熟に供することができ、かかる技術は当該技術分野においてよく知られている。Wright and Harris, supra., Hanes and Plucthau, PNAS USA 94: 4937-4942(1997)(リボソームディスプレイ)、Parmley and Smith, Gene 73: 305-318(1988)(ファージディスプレイ)、Scott,TIBS 17: 241-245(1992)、Cwirla et al., PNAS USA 87: 6378-6382(1990)、Russel et al., Nucl. Acid Res. 21: 1081-1085(1993)、Hoganboom et al., Immunol. Reviews 130: 43-68(1992), Chiswell and McCafferty, TIBTECH 10: 80-84(1992)、および米国特許第5,733,743号を参照。ディスプレイ技術がヒトでない抗体を製造するために用いられる場合、かかる抗体は上述のヒト化抗体であり得る。
【0104】
これらの技術を用いて、PDGF−DD発現細胞、PDGF−DD自体、PDGF−DDの形態、それらのエピトープまたはペプチドに対する抗体が製造され、その後、それらの発現ライブラリー(例えば、米国特許第5,703,057号参照)は、上述の活性について上述のようにスクリーニングされ得る。
【0105】
抗体の調製
XenoMouse(登録商標)テクノロジーを用いることにより、PDGF−Dの2量体形に特異的な完全ヒトモノクローナル抗体が製造された。原則的に、マウスのXenoMouse(登録商標)系は、PDGF−DDまたはそれらのフラグメントで免疫され、リンパ球(例えば、B細胞)が、抗体を発現するマウスから回収され、回収された細胞がミエロイド型細胞株と融合されて不死化ハイブリドーマ細胞株が製造され、かかるハイブリドーマ細胞株がスクリーニングされて選択され、PDGF−DDに特異的な抗体を産生するハイブリドーマ細胞株が同定された。さらに、かかる細胞株により産生された抗体の特徴決定が、本明細書に記載され、これは、かかる抗体の重鎖および軽鎖のヌクレオチドおよびアミノ酸配列解析を含む。
【0106】
好ましい実施態様において、抗体は、本明細書に記載の抗PDGF−DD mAb 1.6、1.9、1.18、1.19、1.22、1.29、1.33、1.40.1、1.45、1.46、1.51、1.59、および6.4を中和するものから選択される。国際公開公報WO03/057,857号(2003年7月17日公開)を参照されたい。もちろん、開示された方法は任意の特定の抗PDGF−DDモノクローナル抗体の使用に限られず、むしろ任意のかかる抗体の使用を包含する。
【0107】
あるいは、ハイブリドーマを作成するためにミエローマ細胞と融合される代わりに、免疫されたXenoMouse(登録商標)系マウスから回収され単離された細胞は、最初の抗原、好ましくは、PDGF−DDタンパク質に対する反応性についてさらにスクリーニングされ得る。かかるスクリーニングには、PDGF−DD−Hisタンパク質を用いるELISA、目的の抗原と結合する既知の抗体を用いる競合アッセイ、および一過性にトランスフェクションされた全長PDGF−DD発現CHO細胞とのインビトロでの結合が挙げられる。次いで、目的の抗体を分泌する1つのB細胞が、PDGF−DD特異的溶血性プラークアッセイ(Babcook et al., Proc.Natl. Acad. Sci. USA, 93: 7843-7848(1996))を用いて単離される。溶解の標的細胞は、好ましくは、PDGF−DD抗原でコーティングされたヒツジ赤血球(SRBC)である。目的の免疫グロブリンおよび相補体を分泌するB細胞培養液の存在下で、プラークの形成は、標的細胞の特異的なPDGF−DD仲介性溶解を示す。プラークの中央にある単一抗原特異的血漿細胞が単離され、次いで、抗体をコードするDNAが、1つの血漿細胞から単離され得る。逆転写PCRを用いて、抗体の可変領域をコードするDNAが特異的にクローン化され得る。次いで、かかるクローン化されたDNAが、適当な発現ベクター、好ましくは、pcDNAのようなベクターカセット、より好ましくは、免疫グロブリン重鎖および軽鎖の定常ドメインを含有するpcDNAベクターにさらに挿入され得る。次いで、作成されたベクターが、宿主細胞、好ましくは、CHO細胞にトランスフェクションされ、プロモーターの誘導、形質転換体の選択、または所望の配列をコードする遺伝子の増幅のため必要に応じて改変された、通常は栄養培地中で培養され得る。PDGF−DDに特異的な抗体を産生する複数の単一血漿細胞の単離は、本明細書に記載されている。さらに、抗PDGF−DD抗体の特異性をコードする遺伝子材料が単離され、適切な発現ベクターに導入され、次いで宿主細胞にトランスフェクションされる。
【0108】
一般に、上述の細胞株により産生される抗体は、完全ヒトIgG2重鎖をヒトκ軽鎖と共に有することが見出された。該抗体は高い親和性、典型的には、固相および液相のいずれかで測定される場合、約10−6ないし約10−11MのKdを有していた。これらのmAbは、抗原結合競合研究に基づき、グループまたは「ビン(bin)」に分類され得る。国際公開公報第WO03/048,731号(2003年6月12日公開)参照。
【0109】
抗PDGF−DD抗体の治療上の有用性に対する親和性の重要性については、例えば、コンビナトリアルに抗PDGF−DD抗体を製造し、かかる抗体を結合親和性について評価し得ることは理解されるだろう。利用され得る1つのアプローチは、上述のように製造され、PDGF−DDに対する良好な親和性を有していることが見出された抗体由来の重鎖cDNAを取り出し、上述のように製造され、これを、PDGF−DDに対して良好な親和性を有していることが見出された2次抗体由来の軽鎖cDNAと組合せ、3次抗体が製造され得るものである。得られた3次抗体の親和性は、本明細書に記載のように測定され得、望ましい解離定数を有するものが単離され特徴決定される。あるいは、上述の任意の抗体の軽鎖が、当該軽鎖と対にしたときPDGF−DDに対して高親和性を示す重鎖、あるいはその逆のものの産生を補助するツールとして用いられ得る。このライブラリのこれらの重鎖可変領域は、未感作の動物から単離され、過免疫動物から単離され、CDR領域の異なる重鎖可変配列を含有するライブラリーから人工的に製造され、あるいは任意の重鎖可変領域遺伝子(例えば、ランダムまたは指定変異誘発)のCDR領域内で多様性を生み出す任意の他の方法により製造され得る。これらのCDR領域、および特定のCDR3は、元の抗体と最初に対になった重鎖とは有意に異なる長さまたは配列同一性であってもよい。次いで、得られたライブラリーが、PDGF−DDとの高親和性結合についてスクリーニングされ、元の抗体(高親和性および中和)と類似する特性を有する、治療関連抗体分子が製造され得る。重鎖または重鎖可変領域を用いた同様の方法を用いて、固有の軽鎖可変領域を有する治療関連抗体分子が製造され得る。さらに、次いで、新規重鎖可変領域または軽鎖可変領域が、上述と類似の様式で用いられ、新規な抗体分子の製造を可能にする新規軽鎖可変領域または重鎖可変領域が同定され得る。
【0110】
利用され得る別のコンビナトリアルアプローチは、本明細書に記載の発明の抗体、特に相補決定領域(CDR)において利用されるために示されている生殖系の重鎖および/または軽鎖上で変異誘発を実行することである。得られた抗体の親和性は、本明細書に記載のように測定され、望ましい解離定数を有するものが単離されて特徴決定される。好ましい結合体の選択の際には、配列またはそれをコードする配列を用いて、上述の組換え抗体が製造され得る。オリゴヌクレオチド上での変異誘発を実行するための適切な方法は、当業者に知られており、化学的変異誘発(例えば、亜硫酸ナトリウムの使用)、酵素誤取込み、および放射線への曝露が挙げられる。本明細書に記載の発明は、本明細書で定義された抗体を、変異誘発または他の任意の方法により製造された本明細書で明確に説明された抗体に包含することは理解される。さらに、本明細書で定義された保存的または非保存的アミノ酸置換を有する抗体は、本明細書で明確に説明された抗体において製造され、本明細書に記載の発明の実施態様に含まれる。
【0111】
用いられ得る別のコンビナトリアルアプローチは、他の可変領域の遺伝子由来のフレームワーク領域と関連して、上記の抗体のCDR領域、特にCDR3を発現させることである。例えば、1つの抗PDGF−DD抗体の重鎖のCDR1、CDR2およびCDR3は、他の重鎖可変遺伝子のフレームワーク領域と関連して、発現され得る。同様に、抗PDGF−DD抗体の軽鎖のCDR1、CDR2およびCDR3は、他の軽鎖可変遺伝子のフレームワーク領域と関連して、発現され得る。さらに、これらのCDR領域の生殖系配列は、他の重鎖または軽鎖可変領域遺伝子と関連して発現され得る。得られた抗体は、特異性および親和性についてアッセイされ、新規抗体分子の作製が可能になる。
【0112】
本明細書に記載の発明をふまえて製造される抗体が、ハイブリドーマ細胞株以外の細胞株でも発現され得ることは理解されるであろう。特定の抗体をコードする配列は、適当な哺乳動物宿主細胞の形質転換に用いられ得る。形質転換は、ポリヌクレオチドを宿主細胞に導入するための任意の既知の方法によるものであり、例えば、ポリヌクレオチドをウイルス(またはウイルスベクター)パッケージングすること、次に宿主細胞をウイルス(またはベクター)に形質導入すること、あるいは米国特許第4,399,216号、同第4,912,040号、同第4,740,461号および同第4,959,455号に例示される当該技術分野で既知のトランスフェクション方法によってもよい。用いられる形質転換方法は、形質転換されるべき宿主に依存する。異種性のポリヌクレオチドを哺乳動物細胞に導入する方法は当該技術分野で知られており、デキストラン仲介性トランスフェクション、リン酸カルシウム沈殿、ポリブレン仲介性トランスフェクション、原形質融合、エレクトロポレーション、ポリヌクレオチドのリポソームへのカプセル化、およびDNAの核への直接マイクロインジェクションが挙げられる。
【0113】
発現のための宿主哺乳動物細胞株として利用可能な哺乳動物細胞株は、当該技術分野でよく知られており、American Type Culture Collection(ATCC)から入手可能な不死化細胞株を含み、これは、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、ベビーハムスター腎臓(BHK)細胞、サル腎細胞(COS)、ヒト肝細胞癌腫細胞(例えば、Hep G2)、および多数の他の細胞株を含むが、これに限らない。特に好ましい細胞株は、どの細胞株が高い発現レベルを有し、保存的PDGF−DD結合特性を有する抗体を産生するかを決定することにより、選択される。
【0114】
抗体治療のさらなる基準
PDGF−DD抗体の機能は、その操作方法の少なくとも一部分に対して重要であるように思われることが、本明細書において考察される。機能とは、例えば、PDGF−DDに対する応答における抗PDGF−DD抗体の活性を意味する。従って、ある種の場合において、抗体が、補体依存細胞毒性(CDC)および抗体依存性細胞の細胞毒性(ADCC)を含むエフェクター機能を発揮し得ることが、PDGF−DDに対する治療候補としての抗体の製造に関連して望まれてもよい。同様のことが可能な抗体のアイソタイプは多数存在し、これは、マウスIgM、マウスIgG2a、マウスIgG2b、マウスIgG3、ヒトIgM、ヒトIgG1およびヒトIgG3を含むが、これに限らない。製造された抗体が、当初からかかるアイソタイプを有する必要はないが、製造された抗体が任意のアイソタイプを有し得、そして抗体が当該技術分野でよく知られた従来技術を用いてその後アイソタイプに切り替えられ得ることは、理解されるだろう。かかる技術は、直接組換え技術(例えば、米国特許第4,816,397号および同第6,331,415号を参照されたい)、細胞・細胞融合技術(例えば、米国特許第5,916,771号および同第6,207,418号を参照されたい)の使用を含む。
【0115】
細胞・細胞融合技術では、任意の所望のアイソタイプを有する重鎖を有する、ミエローマ細胞株または他の細胞株が製造され、そして軽鎖を有する別のミエローマ細胞株または他の細胞株が製造される。その後、かかる細胞は融合され、未変性の抗体を発現する細胞株が単離され得る。
【0116】
例えば、本明細書で考察される抗PDGF−DD抗体は、ヒト抗PDGF−DD IgG2およびIgG4抗体である。かかる抗体が、PDGF−DD分子との所望の結合性を有する場合、これは、容易には、ヒトIgM、ヒトIgG1またはヒトIgG3、IgGA1またはIgGA2アイソタイプを製造するたに切り換えられたアイソタイプであり得るが、これは、依然として同じ可変領域を有している(抗体の特異性およびその親和性のいくらかを有する)。次いで、かかる分子は、CDCにおいて補体を固定し、関与することができるであろう。
【0117】
従って、抗体候補は、上記の所望の「構造」属性に合致するように製造されるので、これらは一般的には、アイソタイプスイッチングにより、少なくともある種の所望の「機能的」属性を付与され得る。
【0118】
エピトープマッピング
免疫ブロット分析
本明細書に記載の抗体のPDGF−DDとの結合は、多数の方法で試験され得る。例えば、PDGF−DDは、SDS−PAGEに供され、免疫ブロッティングにより分析され得る。SDS−PAGEは、還元剤の不存下または存在下のいずれかで行われてもよい。かかる化学修飾は、システイン残基のメチル化をもたらし得る。従って、本明細書に記載のPDGF−DD抗体がPDGF−DDの線状エピトープと結合しているかどうかを決定することが可能である。
【0119】
表面増強レーザー脱離/イオン化
本明細書に記載のPDGF−DD抗体に対するエピトープのエピトープマッピングはまた、SELDIを用いて行われ得る。SELDI ProteinChip(登録商標)アレイは、タンパク質・タンパク質相互作用部位を定義するために用いられる。抗原は、最初のインキュベーションおよび洗浄によって、Protein Chipアレイ表面に共有結合で固定された抗体に特異的に捕獲される。結合された抗原は、レーザー誘導性脱離方法により検出され、そしてそれらの質量が直接決定され得る。結合した抗原のかかるフラグメントは、タンパク質の「エピトープ」と命名されている。
【0120】
SELDI方法は、複合体分子組成物内の個々の成分を直接検出し、迅速、高感度、かつ測定可能な方法で他の成分と比較して定量的にマッピングすることを可能にする。SELDIは、表面化学の多様なアレイを用いて、レーザー誘導性脱離方法による検出する多数の個々のタンパク質分子を捕獲し提示する。SELDI方法の成功は、それぞれが異なる技術に依存する、複数の機能の表面(「チップ」)上での複数機能の小型化および一体化により部分的に定義される。SELDI BioChipおよび他の型のSELDIプローブは、これらが個々の標的分子(例えば、タンパク質)または評価されるべき分子集団の捕獲、精製(分離)、提示、検出および特徴決定に積極的に関与するように、表面が「増強」されている。
【0121】
オリジナルのサンプルのみを充填された単一のSELDIタンパク質BioChipは、何千回も読み取られ得る。LumiCyteのSELDIタンパク質BioChipは、1平方センチメートル当たり10,000個もの指定可能なタンパク質結合位置を有している。それぞれの位置が、十数個の個々のタンパク質の存在を明らかにできる。それぞれの位置からのタンパク質組成物情報が比較され、独自の情報セットが組み合わされると、得られた組成物マップは、特異的パターンまたは分子「フィンガープリント」を定義するために総括的に用いられる、特徴セットを有するイメージを明らかにする。異なるフィンガープリントは、健常な種々の段階、疾患の発症、または適当な治療法の投与と関連する疾患の緩和と関係し得る。
【0122】
SELDI方法は、4つの部分に分けてより詳細に説明され得る。最初に、目的の1つ以上のタンパク質が、サンプルを調製および標識することなく、オリジナルの供給源材料から直接的にProteinChipアレイ上に捕獲または「ドッキング(docked)」される。第2の工程では、化学的および生体分子的「ノイズ」を低減することにより、「シグナル対ノイズ」比が増強される。かかる「ノイズ」は、非所望の材料を洗浄して除くことによるチップ上の標的の選択的保持により低減される。さらに、捕獲された1つ以上の標的タンパク質が、迅速かつ高感度のレーザー誘導性方法(SELDI)により読み取られ、これは、標的に関する直接的情報(分子量)を提供する。最後に、アレイ内の任意にの1以上の位置の標的タンパク質が、1つ以上のチップ上の結合またはタンパク質構造および機能を特徴決定するための修飾を行うことにより、インサイツで特徴決定され得る。
【0123】
ファージディスプレイ
本明細書に記載のPDGF−DD抗体のエピトープは、Filamentousファージ(New England Biolabs)上にディスプレイされたランダムペプチドである12merのコンビナトリアルライブラリにProteinChipアレイを曝露することにより決定され得る。
【0124】
ファージディスプレイは、選択技術を記載し、ここで、ペプチドは、バクテリオファージのコートタンパク質との融合として発現され、ウイルス粒子の表面に融合タンパク質のディスプレイとなる。パニングは、ペプチドを現したファージのライブラリーを標的でコートされたプレートおよびチューブとインキュベートし、非結合ファージを洗浄し、次に、特異的結合ファージを溶出することにより、行われる。次いで、溶出されたファージが増幅され、さらなる結合および増幅サイクルにより取り出され、所望の結合配列プールが富化される。3回または4回の後、個々の結合クローンが、抗体コートウェル上で行われるファージELISAアッセイにより結合についてさらに試験され、そしてポジティブクローンの特異的DNA配列決定により特徴決定される。
【0125】
本明細書に記載のPDGF−DD抗体に対するかかるパニングを複数回おこなった後、結合ファージは溶出され、結合ペプチドの同定および特徴決定のためのさらなる研究に供され得る。
【0126】
PDGF−DDアゴニストおよびアンタゴニスト
本明細書に記載の発明の実施態様はまた、PDGF−DDアゴニスト(ミメティック)またはPDGF−DDアンタゴニストのいずれかとして機能する、PDGF−DDタンパク質の変異体に関与する。好ましくは、PDGF−DDタンパク質の変異体は、腎炎の処置に有用である。PDGF−DDタンパク質の変異体は、変異誘発、例えば、PDGF−DDタンパク質の異なる点変異または切断により製造され得る。PDGF−DDタンパク質のアゴニストは、天然に存在する形のPDGF−DDの実質的な同一性、サブセット、生物学的活性を保持し得る。PDGF−DDタンパク質のアンタゴニストは、例えば、PDGF−DDタンパク質を含む細胞シグナル伝達カスケードの下流または上流メンバーと競合的に結合することにより、天然に存在する形態のPDGF−DDタンパク質の1つ以上の活性を阻害することができる。従って、特異的生物学的作用は、機能制限された変異体による処置により誘発され得る。1つの実施態様において、天然に存在する形態のタンパク質の生物学的活性のサブセットを有する変異体による対象の処置は、天然に存在する形態のPDGF−DDタンパク質での処置と比較して、副作用が少ない。
【0127】
PDGF−DDアゴニスト(ミメティック)またはPDGF−DDアンタゴニストのいずれかとして機能するPDGF−DDタンパク質の変異体は、PDGF−DDタンパク質の変異(例えば、切断変異)のコンビナトリアルライブラリーをタンパク質アゴニストまたはアンタゴニスト活性についてスクリーニングすることにより同定され得る。1つの実施態様において、PDGF−D変異体の変化に富むライブラリーは、核酸レベルでのコンビナトリアル変異誘発により製造され、これは、変化に富む遺伝子ライブラリーによりコードされる。PDGF−D変異体の変化に富むライブラリーは、可能性のあるPDGF−D配列の変性セットが個々のポリペプチド、またはそこにPDGF−D配列セットを含有する巨大融合タンパク質のセット(例えば、ファージディスプレイ)として発現可能なように、例えば、合成オリゴヌクレオチド混合物の遺伝子配列への酵素的ライゲーションにより生み出され得る。変性オリゴヌクレオチド配列由来の可能性のあるPDGF−D変異体ライブラリーを作製するために用いられ得る種々の方法が存在する。変性遺伝子配列の化学合成は、自動DNA合成装置で行われることができ、次いで、合成遺伝子は、適当な発現ベクタとライゲーションされる。遺伝子の変性セットの使用により、可能性のあるPDGF−D変異体配列の所望のセットをコードする配列の全てを1つの混合物で提供することが可能となる。変性オリゴヌクレオチドの合成方法は、当該技術分野で知られている(例えば、Narang, Tetrahedron 39: 3(1983); Itakura et al.,Annu. Rev.Biochem. 53: 323(1984); Itakura et al., Science 198: 1056(1984); Ike et al.,Nucl. Acid Res. 11: 477(1983)を参照されたい)。
【0128】
他の治療法の設計および創出
さらに、PDGF−DDと関連して本明細書で製造され、特徴決定される抗体の活性に基づき、抗体部分を超える他の治療法様式の設計が容易になる。かかる様式は、二重特異性抗体、免疫毒素および放射標識治療法、ペプチド治療法の創出、遺伝子治療剤(特に体内)、アンチセンス治療法、および小分子のような進歩した抗体治療法を含むが、これに限らない。
【0129】
進歩した抗体治療法の創出と関連して、補体固定が所望の属性である場合、例えば、二重特異性、免疫毒素または放射標識の使用により細胞殺傷のための補体依存性を回避することが可能であり得る。
【0130】
例えば、二重特異性抗体に関して、(i)一方はPDGF−DDに対する特異性を有し、他方は、一体となって結合した第2の分子に対する特異性を有する2つの抗体、(ii)PDGF−DDに特異的な1つの鎖および第2の分子に特異的な第2の鎖を有する単一抗体、または(iii)PDGF−DDおよび他の分子に対する特異性を有する1本鎖抗体を含む、二重特異性抗体が製造され得る。かかる二重特異性抗体は、例えば、よく知られた技術を用いて製造され得る((i)および(ii)に関しては、例えば、Fanger et al., Immunol Methods 4: 72-81(1994)およびWright and Harris, supraを、また、(iii)に関しては、例えば、Traunecker et al., Itit. J. Cancer(Suppl.)7: 51-52(1992)を参照されたい)。それぞれの場合で、第2の特異性は、重鎖活性化受容体に対して作りだされ、これは、CD16またはCD64(例えば、Deo et al., 18: 127(1997)参照)、またはCD89(例えば、Valerius et al., Blood 90: 4485-4492(1997)参照)を含むが、これに限らない。上記のように製造された二重特異性抗体は、PDGF−DD発現細胞、特に、本明細書に記載のPDGF−DD抗体が有効な細胞を殺傷するであろう。
【0131】
免疫毒素に関し、抗体は、当該技術分野でよく知られた技術を利用して免疫毒素として作用するように修飾され得る。例えば、Vitetta, Immunol Today 14: 252(1993)を参照されたい。また、米国特許第5,194,594を参照されたい。放射標識抗体の製造と関連し、かかる修飾抗体もまた、当該技術分野でよく知られた技術を用いて容易に製造され得る。例えば、Junghans et al., in Cancer Chemotherapy and Biotherapy 655-686(2d ed., Chafner and Longo, eds., Lippincott Raven(1996))を参照。また、米国特許第4,681,581号、同第4,735,210号、同第5,101,827号、同第5,102,990(RE35,500)号、同第5,648,471号および同第,697,902号も参照されたい。免疫毒素および放射標識分子はそれぞれ、PDGF−DD発現細胞、特に、本明細書に記載のPDGF−DD抗体が有効な細胞を殺傷するであろう。
【0132】
治療ペプチドの製造と関連し、PDGF−DD、および本明細書に記載の抗体(小分子に関しては後述)の様なこれに対する抗体と関連する構造情報を利用、またはペプチドライブラリーのスクリーニングにより、PDGF−DDに対して指定された治療用ペプチドが製造され得る。ペプチド治療法の設計およびスクリーニングは、Houghten et al., Biotechniques 13: 412-421(1992)、Houghten, PNAS USA82: 5131-5135(1985), Pinalla et al., Biotechniques 13: 901-905(1992)、Blake and Litzi-Davis, BioConjugate Chem. 3: 510-513(1992)と関連付けて考察される。免疫毒素および放射標識分子もまた、抗体と関連して上記で考察されたペプチド部分に関連して、類似の方法で製造され得る。
【0133】
PDGF−DD分子(またはスプライス変異体のような形態または代わりの形態)が、疾患過程において機能的に活性であると仮定すると、従来の技術により、遺伝子およびそのアンチセンス治療法を設計することが可能であろう。かかる様式は、PDGF−DDの機能を調節するのに利用され得る。これと関連して、本明細書に記載の抗体は、これと関連する機能的アッセイの設計および使用を可能にする。アンチセンス治療法の設計およびストラテジーは、国際特許出願第WO94/29444号に詳細に論じられている。遺伝子療法の設計およびストラテジーはよく知られいてる。しかしながら、特に、体内に関与する遺伝子治療技術の使用が特に有利であることを明らかにすることができた。例えば、Chen et al., Human Gene Therapy 5: 595-601(1994)およびMarasco, Gene Therapy 4: 11-15(1997)を参照されたい。遺伝子治療法に関する一般的設計および考慮はまた、国際特許出願第WO97/38137号でも考察されている。
【0134】
小分子治療法もまた想定され得る。薬剤は、本明細書に記載のPDGF−DDの活性を調節するように設計され得る。本明細書に記載のPDGF−DD分子の構造、本明細書に記載の抗体のような他の分子とのその相互作用から収集した知識、およびその他を利用して、さらなる治療の方法が合理的に設計され得る。これと関連し、X線結晶学、コンピュータ補助(またはアシスト)分子モデリング(CAMM)、量的または質的構造活性相関(QSAR)のような合理的薬剤設計技術および類似の技術を用いて、薬剤発見努力に焦点が当てられ得る。合理的設計により、PDGF−DD活性を修飾または調節するために用いられ得る、分子またはその特異的形態と相互作用し得るタンパク質または合成構造の予測が可能になる。かかる構造は化学的に合成されるか、または生物学的システムで発現させることができる。このアプローチは、Capsey et al, Genetically Engineered Human Therapeutic Drugs(Stockton Press, NY(1988))で概説されている。さらに、コンビナトリアルライブラリを設計し、合成して、ハイスループットスクリーニング作用の様なスクリーニングプログラムにおいて用いられ得る。
【0135】
治療投与および処方
抗体およびそのフラグメントを含むが、これに限らない抗PDGF−DD化合物は、PDGF−DDを調節する化合物を必要とする生物を処置する医薬への取込に適している。これらの薬理学的に活性な化合物は、生物、例えば、動物およびヒトを含む哺乳動物に投与するための医薬剤を製造するための製剤学の通常の方法と関連して、製造され得る。ある種の実施態様において、有効成分は、修飾してまたは修飾することなく医薬製品に取り込まれ得る。さらなる実施態様は、本明細書に記載の医薬的に活性な化合物をいくつかの経路で送達する医薬または治療剤の製造を含む。例えば、これに限らないが、抗体またはそのフラグメントをコードする配列を有するDNA、RNA、およびウイルスベクターが、ある種の実施態様において用いられ得る。さらに、抗体またはそのフラグメントをコードする核酸は、単独または他の有効成分と組み合わせて投与され得る。
【0136】
本明細書に記載の治療剤の投与は、移動、輸送、耐容性などの改善をもたらすために製剤に取込まれる適当な担体、賦形剤、安定化剤、および他の剤との混合物で投与され得ることは、理解されるだろう。医薬的に許容される担体は、本発明の医薬的有効成分と有害に反応しない非経腸、経腸(例えば、経口)、または局所投与に適した有機または無機担体物質を含む。適当な医薬的に許容される担体は、水、食塩水、アルコール、アラビアゴム、植物油、ベンジルアルコール、ポリエチレングリコール、ゼラチン、ラクトース、アミロースまたはデンプンのような炭水化物、ステアリン酸マグネシウム、タルク、ケイ酸、粘性パラフィン、香料油、脂肪酸モノグリセリドおよびジグリセリド、ペンタエリスリトール脂肪酸エステル、ヒドロキシメチルセルロース、ポリビニルピロリドン等が挙げられるが、これらに限定されない。さらなる担体、賦形剤および安定剤は、Tris−HClのような緩衝液、リン酸塩、クエン酸塩、酢酸塩および他の有機酸塩;アスコルビン酸のような抗酸化剤;ポリアルギニンのような低分子量(約10残基未満)ペプチド、血清アルブミンのようなタンパク質、ゼラチンまたは免疫グロブリン;ポリビニルピロリジノンのような親水性ポリマー;グリシン、グルタミン酸、アスパラギン酸またはアルギニンのようなアミノ酸;セルロースまたはその誘導体、グルコース、マンノースまたはデキストリンを含む単糖、二糖および他の炭水化物を含む他の炭水化物;EDTAのようなキレート剤;マンニトールまたはソルビトールのような糖アルコール;ナトリウムのような対イオンおよび/またはTWEEN、PLURONICSまたはポリエチレングリコールのような非イオン性界面活性剤を含む。多くのより適当なビークルは、Remmington's Pharmaceutical Sciences,15th Edition, Easton: Mack Publishing Company, pages 1405-1412 and 1461-1487(1975)およびThe National Formulary XIV, 14th Edition, Washington, American Pharmaceutical Association(1975)に記載されている。
【0137】
抗体の投与経路は、例えば、これに限定されないが、局所的、経皮的、非経腸、経消化管、経気管および経肺胞を含む既知の方法に従うことができる。非経腸経路投与は、中心静脈への注射、静脈内、脳内、筋肉内、腹腔内、皮内、動脈内、くも膜下内または病変内経路のような電気的または直接的注射または点滴を含むが、これに限らない。抗体は、好ましくは、点滴、ボーラス注射、あるいは下記の徐放システムにより、連続的に投与される。好ましい実施態様において、投与経路は皮下注射であり得る。代わりの実施態様において、抗体は腎動脈を介して投与される。投与の消化管経路には、摂取および直腸経路が挙げられるが、これらに限定されない。投与の気管および肺胞経路には、経口または経鼻経路が挙げられるが、これらに限定されない。
【0138】
インビボでの投与に用いる場合、抗体製剤は無菌であり得る。これは、凍結乾燥および再構成の前および後に滅菌フィルトレーションメンブレンを通してフィルトレーションすることにより、容易に行われ得る。抗体は、通常は、凍結乾燥形または溶液中に貯蔵されるであろう。さらに、治療用組成物は、発熱物質を含まず、pH、等張性および安定性に関して非経腸で許容される溶液であり得る。治療用抗体組成物は、一般に、無菌アクセスポートを有する容器、例えば、皮下注射用針を刺すことが可能なストッパーを有する静脈注射用バッグまたはバイアルに入れられる。
【0139】
注射用無菌組成物は、Remington's Pharmaceutical Sciences(18th ed., Mack Publishing Company, Easton, PA(1990))に記載の従来の医薬実施に従い、製剤され得る。医薬製剤は、所望に応じて、活性化合物と有害な反応をしない滑沢剤、保存剤、安定剤、湿潤剤、乳化剤、浸透圧に影響する塩、緩衝液、抗酸化剤、着色剤、香味剤、または芳香族物質等と混合して、無菌化され得る。例えば、水、またはゴマ油、ピーナッツ油または綿実油のような天然に存在する植物油、またはオレイン酸エチルのような合成脂質ビークル等の様なビークル中の活性化合物の溶解または懸濁が所望されてもよい。
【0140】
非経腸投与に適した本発明の薬理学的に活性な化合物を有する適当な組成物は、医薬的に許容される無菌等張溶液を含むが、これに限らない。かかる溶液は、中心静脈ラインへの注射、静脈内、筋肉内、腹腔内、皮内または皮下注射用の生理食塩水およびリン酸緩衝生理食塩水を含むが、これに限らない。
【0141】
消化管投与に適した、本発明の薬理的に活性な化合物を有する組成物は、医薬的に許容される消化用粉剤、丸薬、または液剤、または直腸投与用座剤を含むが、これに限らない。
【0142】
徐放性製剤の適当な例は、ポリペプチドを含有する固体疎水性ポリマーの半透性マトリックスが挙げられ、ここでマトリックスは、成形された物品、フィルムまたはマイクロカプセルの形態である。徐放性マトリックスの例は、ポリエステル、Langer et al., J. Biomed Mater. Res., 15: 167-277(1981)およびLanger, Chem. Tech. , 12: 98-105(1982)に記載のハイドロゲル、例えば、ポリ(2−ヒドロキシエチルメタクリレート)、またはポリ(ビニルアルコール))、ポリラクチド(米国特許第3,773,919号、欧州特許第58,481号)、L−グルタミン酸とγ−エチル−L−グルタメートとのコポリマー(Sidman et al.,Biopolymer, 22: 547-556(1983))、非分解性エチレン−酢酸ビニル(Langer et al., supra)、LUPRON Depot(登録商標)のような分解性乳酸−グリコール酸コポリマー(乳酸−グリコール酸コポリマーおよび酢酸リュープロリンからなる注射可能な微粒子)、およびポリ−D−(−)−3−ヒドロキシ酪酸(欧州特許第133,988号)が挙げられる。
【0143】
エチレン−酢酸ビニルおよび乳酸−グリコール酸のようなポリマーは、100日間にわたり分子を放出することができるが、特定のハイドロゲルは、タンパク質をより短い期間放出する。カプセル化されたタンパク質が体内に長時間残存すると、37℃で水分に曝露された結果、変性または凝集し得、結果として生物学的活性が損なわれ、免疫原性が変化する可能性がある。関与するメカニズムに応じて、タンパク質安定化のための合理的ストラテジーが考えられ得る。例えば、凝集メカニズムがジスルフィド結合間の変化による分子内S−S結合形成であることが見出された場合、安定化は、スルフヒドリル残基の修飾、酸性溶液からの凍結乾燥、水分含有量の制御、適当な添加剤の使用、および特別なポリマーマトリックス組成物の開発により達成され得る。
【0144】
徐放性組成物はまた、リポソームで包まれた本発明の抗体を含む。かかる抗体を含有するリポソームは、それ自体既知の方法により製造される:米国特許第DE3,218,121号;Epstein et al.,Proc.Natl.Acad. Sci. USA, 82: 3688-3692(1985);Hwang et al.,Proc. Natl. Acad. Sci. USA, 77: 4030-4034(1980);欧州特許第52,322号;欧州特許第36,676号;欧州特許第88,046号;欧州特許第143,949号;142,641号;日本国特許出願第83−118008号(特願昭59−118008号);米国特許第4,485,045号および同第4,544,545号;ならびに欧州特許第102,324号参照。
【0145】
治療に用いられる抗体の有効量は、例えば、治療の対象、投与経路、患者の状態に依存する。抗体用量は、疾患の重症度、種類、体重、性別、食事、投与時間および投与経路、他の医薬、および他の関連臨床因子を含む、薬剤の作用を改変することが知られている種々の因子を考慮しながら、担当医によって決定されるであろう。従って、治療者は、最適な治療効果を得るのに必要とされる用量を滴定し、投与経路を改変する必要があるであろう。典型的には、臨床医は、所望の効果を達成する投薬量に至るまで、抗体を投与するであろう。治療上有効用量は、インビトロまたはインビボでの方法のいずれかにより決定され得る。この療法の進行は、従来のアッセイまたは本明細書に記載のアッセイにより容易にモニターすることができる。
【0146】
かかる化合物の治療有効性および毒性は、細胞培養または実験動物における標準的な医薬手順、例えば、ED50(集団の50%が治療上効果的な用量)により決定され得る。腎炎ラットモデルまたは代わりのモデルの処置から得られたデータは、ヒトを含む他の生物に用いるための用量範囲の創出の際に用いられ得る。かかる化合物の投薬量は、好ましくは、ED50を毒性なく含む、循環濃度の範囲内である。投薬量は、出差(evectin)、ハイブリッド、結合パートナーまたはそのフラグメントの種類、利用される投薬形態、生物種の感度、および投与経路に依存して、この範囲を変動する。
【0147】
種々の抗体またはそのフラグメントの正常な投薬濃度は、約0.1〜100mg/kgで変動し得る。望ましい投薬量濃度は、例えば、0.lmg/kg、0.2mg/kg、0.3mg/kg、0.4mg/kg、0.5mg/kg、0.6mg/kg、0.7mg/kg、0.8mg/kg、0.9mg/kg、l.0mg/kg、1.5mg/kg、2.0mg/kg、2.5mg/kg、3.0mg/kg、3.5mg/kg、4.0mg/kg、4.5mg/kg、5.0mg/kg、5.5mg/kg、6.0mg/kg、6.5mg/kg、7.0mg/kg、7.5mg/kg、8.0mg/kg、8.5mg/kg、9.0mg/kg、10mg/kg、15mg/kg、20mg/kg、25mg/kg、30mg/kg、35mg/kg、40mg/kg、45mg/kg、50mg/kg、55mg/kg、60mg/kg、65mg/kg、70mg/kg、75mg/kg、80mg/kg、85mg/kg、90mg/kg、95mg/kgおよび100mg/kg、またはそれ以上を含む。1つの好ましい投薬量は、1〜10mg/kgである。
【0148】
ある実施態様において、抗体またはそのフラグメントの用量は、約0.1μM〜500mM、好ましくは約1〜800μM、そしてより好ましくは、約10μM〜約500μMの組織または血液濃度あるいはその両方を生じる。好ましい用量は、例えば、10μM、15μM、20μM、25μM、30μM、35μM、40μM、45μM、50μM、55μM、60μM、65μM、70μM、75μM、80μM、85μM、90μM、95μM、100μM、110μM、120μM、130μM、140μM、145μM、150μM、160μM、170μM、180μM、190μM、200μM、220μM、240μM、250μM、260μM、280μM、300μM、320μM、340μM、360μM、380μM、400μM、420μM、440μM、460μM、480μM、および500μMの組織または血液濃度あるいはその両方を達成するのに必要とされる量である。代わりの実施態様において、800μMより高い組織濃度を生じる用量が用いられ得る。抗体、ハイブリッド、結合対、またはそのフラグメントの一定の点滴もまた、血液レベルにより測定される組織での安定な濃度を維持するために提供され得る。
【0149】
投薬量および投与は、活性部分の十分なレベルをもたらすため、あるいは所望の効果を維持するために調整され得る。本明細書の実施態様は、短期作用する医薬組成物、および長期作用する医薬組成物の両方を含む。従って、実施態様は、医薬組成物が、約1、2、3、4、5または6日毎、1週間毎、2週間に1回、3週間に1回、4週間に1回、5週間に1回、6週間に1回、7週間に1回、または8週間に1回投与されるスケジュールを含む。特定の製剤の半減期およびクリアランス率に依存して、本明細書に記載の医薬組成物は、1日あたり約1、2、3、4、5、6、7、8、9および10回、またはそれ以上投与され得る。
【0150】
さらなる治療法が、抗PDGF−DD抗体の投与と組み合わせて、当該投与前または後に行われてもよい。これらの治療法は、疾患の症状を処置するために用いられてもよいし、あるいは抗PDGF−DD抗体の副作用を低減するために用いられてもよい。これらはまた、抗PDGF−DD抗体の活性を増強するために用いられてもよい。これらに限らないが、例えば、抗生物質、利尿剤、麻酔剤、鎮痛剤、抗炎症剤、およびインスリンを含む、任意の種類の治療法が用いられる。糸球体腎炎を処置するために典型的に用いられ、かつ抗体と組合せて用いられ得る剤の例は、プレドニゾン、シクロホスファミド、クロラムブシル、および例えば、ワーファリン、ジピリダモールおよびアスピリンのような血液希釈剤を含む。
【0151】
診断上の使用
PDGF−DDは、正常腎臓では低レベルで発現されるが、発現は虚血後の腎臓で劇的に増加されることが見出された(Ichimura T, Bonventre JV, Bailly V, Wei H, Hession CA, Cate RL, Sanicola M., J.Biol. Chem. 273(7): 4135-42(1998))。抗PDGF−DD抗体による免疫組織化学染色は、腎臓(renal)癌腫、腎臓(kidney)癌腫、前立腺癌腫、および卵巣癌腫での陽性染色を示し(以下参照)、これは、正常組織と比較してPDGF−DDの過剰発現が、かかる疾患の診断マーカーとなり得ることを示す。
【0152】
従って、本発明の実施態様はまた、アッセイ、特にインビトロ診断アッセイ(例えば、患者サンプルにおいてPDGF−DDレベルを決定するための使用)にも有用である。かかるアッセイは、PDGF−DDの過剰発現に関連する疾患の診断に有用であり得る。いくつかの実施態様において、疾患は腎炎である。患者サンプルは、例えば、体液、好ましくは血液、より好ましくは血液血清、滑液、組織溶解物、および疾患組織から調製した抽出物であってもよい。本発明の他の実施態様は、腎炎およびメサンギウム増殖性関連疾患の診断および段階決定において有用である。PDGF−DDのレベルのモニタリングは、疾患に対する患者応答の代わりの測定手段および患者の疾患の重症度のモニタリング方法として用いられ得る。他の溶解性マーカーレベルと比較して、高レベルのPDGF−DDが、虚血後腎臓の存在を示すであろう。患者サンプルに存在するPDGF−DD抗原の濃度は、存在する抗原の量を特異的に決定する方法を用いて、決定され得る。かかる方法には、ELISA法が挙げられ、当該方法では、例えば、本発明の抗体がポリマーマトリックスのような不溶性マトリックス上に簡便に固定され得る。あるいは、抗PDGF−DD抗体を用いる免疫組織学的化学染色アッセイは、サンプル中でPDGF−DDのレベルを決定するために用いられ得る。進行または治療のそれぞれの段階についての統計的に有意な結果をもたらすサンプル集団を用いて、疾患の各段階の特性であると考えられ得る抗原の濃度範囲が設計され得る。
【0153】
1つの実施態様において、血液サンプルが被検者から採取され、サンプル中に存在するPDGF−DD抗原の濃度が決定され、研究下の被検者の疾患の段階が評価されるか、あるいは一連の治療に対する応答が特徴決定される。このようにして得られた濃度を用いて、その値がどの濃度範囲に含まれるかが同定される。このようにして同定された範囲は、診断された対象からなる種々の集団で同定された疾患の進行段階または治療段階と相関し、これにより、研究下の対象の疾患段階を提供する。
【0154】
遺伝子増幅および/または発現は、適当に標識されたプローブを用い、本明細書で提供される配列に基づき、例えば、従来のサザンブロッティング、ノーザンブロッティング(Thomas,Proc. Natl. Acad. Sci. USA, 77: 5201-5205(1980))、ドットブロッティング(DNA分析)またはインサイツハイブリダイゼーションによりサンプル中で直接測定され、mRNAの転写が定量され得る。あるいは、特異的2本鎖(DNA2本鎖、RNA2本鎖、およびDNA・RNAハイブリッド2本鎖またはDNA・タンパク質2本鎖)を認識し得る抗体が用いられ得る。次いで、抗体は標識され得、アッセイが行われ得るが、この際、2量体が表面に結合し、その結果、表面での2本鎖の形成により、2本鎖に結合した抗体の存在が検出され得る。
【0155】
例えば、抗体フラグメントを含む抗体は、PDGF−DDタンパク質の発現を定性的または定量的に検出するために用いられ得る。上述のように、抗体には、好ましくは、検出可能な蛍光標識等が付与され、結合は、光学顕微鏡、フローサイトメトリー、蛍光分析または他の当該技術分野で既知の技術を用いてモニタリングされ得る。これらの技術は、増幅遺伝子が細胞表面タンパク質(例えば、成長因子)をコードするときに特に適している。かかる結合アッセイは、当該技術分野で知られた方法で行われる。
【0156】
PDGF−DDタンパク質と結合した抗体のインサイツ検出は、例えば、免疫蛍光顕微鏡または免疫電子顕微鏡により行われ得る。このため、組織標本が患者から採取され、標識された抗体が標本に塗布される(好ましくは、抗体を生物学的サンプルに重ねることによる)。この手順はまた、試験組織中のマーカー遺伝子産物の分布の決定も可能にする。インサイツでの検出に広範囲の種々の組織学的方法が容易に利用可能であることは、当業者には明らかであろう。
【0157】
異なる遺伝子発現を定量する最も高感度かつフレキシブルな定量方法の1つは、RT−PCRであり、これを用いて、異なるサンプル集団、正常組織および腫瘍組織、薬剤処置ありまたはなしでのmRNAレベルが比較され、遺伝子発現のパターンが特徴決定され、密接に関連するmRNA間が見出され、そしてRNA構造が分析され得る。
【0158】
第一の工程は、標的サンプルからmRNAを単離することである。出発材料は、典型的には、疾患組織および対応する正常組織からそれぞれ単離されたトータルRNAである。従って、mRNAは、例えば、凍結し保存された、またはパラフィン包埋して固定された(例えば、ホルマリン固定)疾患組織のサンプルから、同種の正常組織との比較のために抽出され得る。RNA抽出物の調製方法は、当該技術分野でよく知られており、Ausubel et al., Current Protocols of Molecular Biology, John Wiley and Sons(1997)を含む、分子生物学の標準的教科書に記載されている。パラフィン包埋からのRNA抽出方法は、例えば、Rupp and Locker, Lab Invest., 56: A67(1987)およびDe Andres et al., BioTechniques, 18: 42044(1995)に開示されている。特に、RNAの単離は、Qiagenのような市販の精製キット、緩衝液セットおよびプロテアーゼを用いて、製造者の指示に従い行われ得る。例えば、培養液中の細胞由来のトータルRNAは、QiagenのRNeasyミニカラムを用いて単離され得る。組織サンプル由来のトータルRNAは、RNA Stat-60(Tel-Test)を用いて単離され得る。
【0159】
RNAはPCRのテンプレートとはならないので、RT−PCRによる異なる遺伝子発現分析の第1の工程は、RNAテンプレートのcDNAへの逆転写であり、次いで、PCR反応においてそれが指数関数的に増幅される。通常用いられる2つの主な逆転写酵素は、Avilo骨髄芽球症ウイルス逆転写酵素(AMV−RT)およびMoloneyマウス白血病ウイルス逆転写酵素(MMLV−RT)である。逆転写工程は、典型的には、状況および発現プロファイリング上の目的に依存して、特異的プライマー、ランダムな6量体またはオリゴ-dTプライマーを用いて調製される。例えば、抽出されたRNAは、GeneAmp RNA PCRキット(Perkin Elmer, CA, USA)を用いて、製造元の指示に従い逆転写され得る。次いで、誘導されたcDNAは、引き続いてPCR反応のテンプレートとして用いられ得る。
【0160】
PCR工程は、種々の熱安定性DNA依存性DNAポリメラーゼを用い得る、典型的には、5’−3’ヌクレアーゼ活性を有するが3’−5’末端ヌクレアーゼ活性を有しないTaq DNAポリメラーゼが用いられる。従って、TaqMan PCRは、典型的には、TaqまたはTthポリメラーゼの5’−ヌクレアーゼ活性を用いて、その標的増幅物と結合したハイブリダイゼーションプローブが加水分解されるが、均等な5’ヌクレアーゼ活性を有する任意の酵素を用いることもできる。2つのオリゴヌクレオチドプライマーを用いて、PCR反応の代表的増幅物(amplicontypical)が産生される。第3のオリゴヌクレオチドまたはプローブは、2つのPCRプライマーの間に位置するヌクレオチド配列を検出するよう設計される。プローブは、Taq DNAポリメラーゼ酵素によって伸長されないが、レポーター蛍光色素およびクエンチャー蛍光色素で標識される。レポーター色素からの任意のレーザー励起放射は、2つの色素がプローブ上で密接に位置したときにクエンチング色素により消光される。増幅反応の間、Taq DNAポリメラーゼ酵素は、テンプレート依存性の方法でプローブを切断する。得られたプローブフラグメントは、溶液中で解離し、解離したレポーター色素からのシグナルは、第2の蛍光発光団の消光作用を有しない。レポーター色素の1つの分子は、合成したそれぞれの新しい分子に対して放出され、消光されていないレポーター色素の検出により、データの定量的解釈の基礎が提供される。
【0161】
TaqMan RT−PCRは、例えば、ABI PRIZM 7700(登録商標)Sequence Detection System(登録商標)(Perkin-Elmer-Applied Biosystems, Foster City, CA, USA)またはLight Cycler(Roche Molecular Biochemicals, Mannheim, Germany)のような市販の装置を用いて行われ得る。好ましい実施態様において、5’ヌクレアーゼ方法は、ABI PRIZM 7700(登録商標)Sequence Detection System(登録商標)のようなリアルタイム定量的PCR装置を用いて実行される。当該システムは、サーモサイクラー、レーザー、電荷結合素子(CCD)、カメラおよびコンピュータからなる。当該システムは、サンプルを、サーモサイクラー上の96ウェル形態で増幅する。増幅の間、レーザー誘導蛍光シグナルが、光ファイバーケーブルにより96ウェル全てに集積され、CCDで検出される。当該システムは、装置を実行させ、データを解析するためのソフトウェアを含む。
【0162】
5’−ヌクレアーゼアッセイデータは、最初はCtまたは閾値サイクルとして現される。上記で考察された蛍光値は、全サイクルで記録され、増幅反応のある点まで増幅された産物の量を現す。統計的に有意であるとして蛍光シグナルが最初に記録された点が、閾値サイクル(Ct)である。ΔCt値は、疾患組織中のRNAの発現を正常細胞のそれと比較した場合に、核酸サンプル中の特定の標的配列の出発コピーの相対数を定量的に測定するために用いられる。
【0163】
サンプルとサンプルの変動の影響を最小化するために、RT−PCRは、通常、内部標準を用いて実行される。理想的な内部標準は、異なる組織間で一定のレベルで発現され、実験処理による影響を受けないものである。遺伝子発現のパターンを標準化するために最もよく用いられるRNAは、ハウスキーピング遺伝子グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(GAPDH)およびβ−アクチンのmRNAである。
【0164】
異なる遺伝子発現もまた、マイクロアレイ技術を用いて同定または確認され得る。この方法では、目的のヌクレオチド配列は、マイクロチップ基板上にプレート化またはアレイ化される。次いで、アレイ化された配列を、目的の細胞または組織由来の特異的DNAプローブによりハイブリダイズする。
【0165】
マイクロアレイ技術の特定の実施態様において、cDNAクローンのPCR増幅挿入物は、高密度アレイの基板上に付与される。好ましくは、少なくとも10,000ヌクレオチド配列が基板上に付与される。10,000素子でそれぞれマイクロチップ上に固定されたマイクロアレイ遺伝子は、ストリンジェントな条件下でのハイブリダイゼーションに適している。蛍光標識されたcDNAプローブは、目的の組織から抽出されたRNAの逆転写による蛍光ヌクレオチドの取込みにより産生され得る。チップに付与された標識cDNAプローブは、アレイ上のDNAのそれぞれのスポットに選択的にハイブリダイズされる。ストリンジェントな洗浄により非特異的結合プローブを除去した後、チップを共焦点レーザー顕微鏡でスキャンする。それぞれのアレイ化エレメントのハイブリダイゼーションの定量により、対応するmRNA量の評価が可能になる。2色の蛍光により、RNAの2つの供給源から製造された特異的に標識されたcDNAプローブがアレイに対でハイブリダイズする。このように、2つの供給源からの転写において、それぞれ特定された遺伝子に対応する相対量が同時に決定される。ハイブリダイゼーションのスケールの最小化により、多数の遺伝子についての発現パターンの簡便かつ迅速な評価が得られる。かかる方法は、細胞1個あたり数コピーで発現される稀な転写物を検出するため、および発現レベルの少なくとも約2倍の差異を再現可能に検出するために必要とされる感度を有することが示されている(Schena et al., Proc. Natl. Acad. Sci. USA, 93(20)L106-49)。核酸およびマイクロアレイ技術のハイブリダイゼーションの方法論は、当該技術分野でよく知られている。
抗体および方法の選択した実施態様を、以下の実施例で説明する。
【0166】
実施例
行った実験および得られた結果を含む以下の実施例は、例示のみの目的で提供されており、本明細書に記載の発明の実施態様を限定するものとして解釈されるべきではない。
【0167】
実施例1
PDGF−DD抗原の調製
生物学的に活性なPDGF−DD p35である、CUBドメインを欠く、組換えヒトおよびマウスPDGF−DDを、LaRochelle et al., Nat Cell Biol 3: 517-521(2001)に記載のように製造した。ヒトPDGF−CCを、同様のプロトコールで製造した。PDGF−AAおよびPDGF−BBは、R&D Systems(Minneapolis,MN)から購入した。
【0168】
実施例2
PDGFに対するアプタマーベースのアンタゴニスト
PDGF−Bアプタマー(NX1975)の合成および特徴決定は、Green et al., Biochemistry 35: 14413-14424(1996)に詳細に記載されている。オリジナルのDNAアプタマーの修飾は、特定のヌクレオチドを2−フルオロピリミジンおよび2’−O−メチルプリンで置換し、ヌクレアーゼ抵抗性を改善すること、ならびにアプタマーを40kDaのポリエチレングリコール(PEG)と結合させてインビボでの血漿残存時間を延長させることを含む。Floege et al., Am J Pathol 154: 169-179(1999)。
【0169】
実施例3
抗PDGF−DD抗体
完全ヒト抗PDGF−DDモノクローナル抗体を、Yang et al., J. Leukoc. Biol. 66: 401-410(1999)に記載の方法を以下のように変更して産生した。簡潔にいうと、ヒトIgG2を有するXenoMouse(登録商標)系(8〜10週齢)に、週に2回、完全フロイントアジュバント中のV5−標識溶解性PDGF−DD(LaRochelle et al.,Nat Cell Biol 3: 517-521(2001)) 10μgを足蹠に注射して免疫した。Yang et al. supraを参照。電気的細胞融合を用いて、ハイブリドーマを製造した。完全ヒトアイソタイプと一致したPK16.3を、負の対照として用いた。
【0170】
実施例4
完全ヒト抗PDGF−DD mAB 6.4の特徴決定
PDGFのなかでも、PDGF−DDに対する完全ヒト抗PDGF−DD mAb 6.4の特異性を、固相ELISA、ウエスタンブロット分析、およびNIH 3T3 BrdU取込分析により特徴決定した。
【0171】
固相ELISA
完全ヒト抗PDGF−DDの特異性を、固相ELISAにより特徴決定した。簡潔に言うと、Corning96ウェル平底高タンパク質結合ポリスチレンマイクロタイタープレートを、500ng/mlのPDGF−AA、PDGF−BB、PDGF−CCまたはPDGF−DDで一晩コーティングした。プレートを、アッセイ希釈剤(Pharmingen, San Diego, CA)で1時間ブロッキングした。次いで、抗PDGF−DD mAb 6.4または対照mAb PK16.3を添加し、示した濃度で2時間インキュベートした。第1のmAb結合を、TMB試薬(Pharmingen, San Diego, CA)を用いて、2次抗体に結合した抗ヒト西洋ワサビペルオキシダーゼを用いて検出した。マイクロタイタープレートを、450nmにてKinetic Microplate Reader(Molecular Devices, Sunnyvale, CA)を用いて読み取った。
【0172】
図1に示すように、抗PDGF−DD mAb 6.4は、PDGF−DDを認識したが、PDGF−AA、PDGF−BB、またはPDGF−CCは認識しなかった。対照mAb PK16.3は、PDGF−DDの認識しなかった。ELISA結果を確認するため、ウエスタンブロット分析も行った。
【0173】
さらに、Corning96ウェル平底高タンパク質結合ポリスチレンマイクロタイタープレートを、500ng/ml ヒトまたはマウスPDGF−DDを用いて一晩コーティングすることにより、PDGF固相ELISAを行った。プレートを、アッセイ希釈剤(Pharmingen, San Diego, CA)で1時間ブロッキングした。次いで、抗PDGF−DD mAb 6.4を添加し、示した濃度で2時間インキュベートした。第1のmAb結合を、TMB試薬(Pharmingen)を用いて、二次抗体に結合した抗ヒト西洋ワサビペルオキシダーゼを用いて検出した。マイクロタイタープレートを、450nmでKinetic Microplate Reader(Molecular Devices, Menlo Park, CA)を用いて読み取った。
【0174】
図2に示すように、抗PDGF−DD mAb 6.4抗体はヒトおよびマウスPDGF−DDの両方を認識する。
【0175】
ウエスタンブロット分析
ウエスタンブロット分析用に、PDGF−AA、PDGF−BB、PDGF−CC、およびPDGF−DD(250μg)をSDS−PAGEサンプル緩衝液中で希釈し、沸騰させ、16% SDSポリアクリルアミドゲルを用いるSDS−PAGEゲル電気泳動に供した。タンパク質をHybond-Pメンブレン(Amersham, Piscataway, NJ)に移し、フィルターをPDGF−DD mAb 6.4または対照mAb PK16.3(0.85μg/ml)で12時間プローブ結合させた。洗浄後、フィルターを、抗ヒト西洋ワサビペルオキシダーゼ結合二次抗体と共にインキュベーションした。バンドを、増強した化学発光により視覚化した(Amersham, Piscataway, NJ)。
【0176】
図3は、抗PDGF−DD mAb 6.4がPDGF−DD、p35を免疫ブロットしたが、PDGF−AA、PDGF−BB、またはPDGF−CCは免疫ブロットしなかったことを示している。対照mAb PK16.3は、PDGFを認識しなかった。BIACorの動力学的測定を用いて、抗PDGF−DD抗体6.4のヒトPDGF−DDとの親和性が170PMであり、抗PDGF−DD mAb 6.4がマウスPDGF−DDに対して少なくとも20倍少ない親和性を示す(データは示していない)ことを決定した。
【0177】
NIH 3T3 BrdU取込みアッセイ
抗PDGF−DD mAb 6.4のPDGF−DD誘導性細胞分裂活性を中和する能力を試験するため、NIH 3T3 BrdU取込みアッセイを用いた。NIH 3T3中和アッセイは、LaRochelle et al.,Nat Cell Biol 3: 517-521(2001)に記載の方法を以下のように変更して行った。簡潔にいうと、NIH 3T3細胞を、24時間血清枯渇状態におき、モノクローナル抗体を示した濃度で添加した。次いで、PDGF−DDを18時間後に100ng/mlで添加した。18時間後、BrdUを5時間にわたって添加し、BrdUアッセイを、製造元(Roche)の指示に従い行った。
【0178】
図4に示すように、抗PDGF−DD mAb 6.4は、PDGF−DD誘導性BrdU取込みを、約75ng/mlのIC50で中和した。PDGF−DD誘導性BrdU取込みは、試験した最高濃度(5μg/ml、データは示していない)で影響を受けなかった。対照mAb PK16.3は、PDGF−DD誘導性BrdU取り込みに影響しなかった。これらの結果を総合して考えると、抗PDGF−DD mAb 6.4が、PDGF−DDに対して高度に特異的であるが、他のPDGFファミリーメンバーは認識せず、PDGF−DD誘導性BrdU取り込みを強く中和することを示した。
【0179】
実施例5
インビトロでのPDGF−DDのメサンギウム細胞増殖に対する作用
メサンギウム細胞培養実験
インビトロでのPDGF−DDのメサンギウム細胞増殖に対する作用を研究するため、ラットメサンギウム細胞を培地中で樹立し、特徴決定し、上述のように維持した。Radeke et al.,J Inzmunol 153 : 1281-1292,(1994)を参照。簡潔に言うと、ラットメサンギウム細胞を96ウェルプレート(Nunc, Wiesbaden, Germany)に播種し、密集させて、サブコンフルエントまで増殖させ、1%ウシ血清アルブミンを含むRPMI1640にて48時間増殖を停止させた。48時間後、PDGF−DD(10〜200ng/ml)およびPDGF−BB(l0ng/mlおよび50ng/ml)と共に、PDGF−B鎖アプタマー(100ng/ml)または配列スクランブルしたアプタマー(100ng/ml)を添加し、細胞を24時間インキュベートした。DNA合成を、BrdU取込みにより決定し、製造元の指示に従い、熱量測定細胞増殖ELISA(Roche, Mannheim, Germany)により測定した。
【0180】
増殖を停止させた培養ラットメサンギウム細胞のPDGF−DDとのインキュベーションは、増殖における用量依存性をもたらした(図5)。データは、4つの独立実験の平均±標準偏差である。統計上の有意性(p<0.05と定義した)を、ANOVAおよびBonferroni t検定を用いて評価した。アスタリスクは、非刺激対照に対するp<0.05を示す。
【0181】
細胞分裂誘導性PDGF−DD活性のPDGF−Bへの非依存性を、PDGF−DDと同時に、細胞をアンタゴニストPDGF−Bアプタマーまたは配列スクランブル対照アプタマーとインキュベーションすることにより示した。アプタマーは、PDGF−BB誘導性増殖をブロックしたが、PDGF−DDの細胞分裂誘導性の可能性には影響を及ぼさなかった(図5)。同様のデータ(示していない)が、ヒトメサンギウム細胞を用いて得られた。
【0182】
実施例6
PDGF−DDおよび抗PDGF−DD抗体のヒトメサンギウム細胞(HMC)に対する作用
ヒトメサンギウム細胞を血清枯渇状態におき、以下の100ng/mL、250ng/mL、1μg/mLの濃度でPDGF−DDまたはPDGF−BBと共にBrdUで一晩処理した。比較のため、他の間葉系細胞、例えば、NIH 3T3線維芽細胞、CCD 1070包皮線維芽細胞、および初代平滑筋細胞を、BrdUおよび完全な血清で処理した。BrdU取込みを、抗BrdU抗体ELISAを用いるアッセイにより検出した。図6が示すように、PDGF−DDは、l00ng/mLより高い濃度で初期ヒトメサンギウム細胞の増殖を誘発する。図6はさらに、ヒトメサンギウム細胞におけるBrdU取込みの同様の誘導に必要とされるPDGF−DDおよびPDGF−BBの濃度に、10倍の差を見出したことを示す。
【0183】
実施例7
腎炎の血清におけるPDGF−DDレベル
腎炎血清におけるPDGF−DDレベルを定量するために、サンドウィッチELISAを開発した。サンドウィッチELISAに用いた2つの完全ヒトmAb(抗PDGF−DD mAb 1.6および1.17)は、PDGF−DD分子に対する異なるエピトープを認識した(データは示していない)。抗PDGF−DD mAb 1.6を捕獲抗体として用い、抗PDGF−DD mAb 1.17を検出抗体として用いた。
【0184】
ELISAを以下のように行った。2μg/mlの濃度のコーティング緩衝液(0.1M NaHCO3、pH9.6)中の捕獲抗体(抗PDGF−DD mAb 1.6) 50μlを、ELISAプレート(Fisher)にコートした。4℃で一晩インキュベーションした後、プレートを、ブロッキング緩衝液(PBS中、0.5% BSA、0.1% Tween20、0.01% Thimerosal)200μlで1時間、25℃にて処理した。PBS中の0.05% Tween20(洗浄緩衝液、WB)を用いて、プレートを洗浄した(3回)。正常血清または患者血清(Clinomics, Bioreclamation, Cooperative Human Tissue Network)を、ヒト血清を50%含むブロッキング緩衝液で希釈した。プレートを血清サンプルと共に4℃にて一晩インキュベーションし、WBで洗浄し、次いで100μ1/ウェルのビオチンル化検出抗PDGF−DD mAb 1.17で1時間、25℃にてインキュベートした。洗浄後、プレートをHRP−ストレプトアビジンと共に15分間、インキュベートし、上述のように洗浄し、次いで、発色のため100μ1/ウェルのH2O2中のO−フェニレンジアミン(Sigma開発中溶液)で処理した。反応を2M H2SO4で停止させ、ELISAプレートリーダーを用いて492nmで分析した。血清サンプル中のPDGF−DDの濃度を、4つのパラメータ曲線フィッティングプログラムを用いてPDGF−DD標準曲線と比較することにより計算した。
【0185】
腎炎を有するII型糖尿病患者におけるPDGF−DD血清レベル
PDGF−DDが腎炎に関与するかどうかを決定するため、II型糖尿病を有する種々の型の腎炎患者由来の血清レベルを調べた。血清PDGF−DD濃度を、上述の定量的PDGF−DDサンドウィッチELISAを用いて評価した。ELISAはPDGF−DDに特異的であり、4ng/mlの感度を有していた。図7は、研究結果を概要する。黒丸は、個々の臨床血清サンプルのPDGF−DD濃度を示す。PDGF−DD血清濃度を、患者の疾患指標に従いグループ化した。所定の臨床指標を有する患者の数(n)を、平均PDGF−DD濃度(ng/ml)とともに提供する。
【0186】
図7に示すように、PDGF−DDは、II型糖尿病患者由来の10個の血清サンプルのうち8個で、正常血清(n=50)のと比較して6%上昇した(平均=11.4ng/ml、p<.001)。II型糖尿病患者におけるPDGF−DDの平均血清レベルは、正常個体の濃度が4ng/ml未満であるのに対し、約4〜24ng/mlの範囲にあった。これらのデータは、PDGF−DDがII型糖尿病の患者の血清中で増加し、そしてこれがPDGF−DDがII型糖尿病に関連する腎臓疾患/腎不全の発症を遅らせるための標的であり得ることを示唆している。これらの結果は、腎炎患者の血清では、正常個体の血清と比較して、PDGF−DDレベルが4〜7倍増加することを示している。
【0187】
実施例8
ラットメサンギウムの免疫組織化学分析
正常ラットメサンギウム細胞を、抗Thy−1誘導性腎炎を有するラットのメサンギウム細胞と比較した。Wistarラットは、Charles River社から入手した。免疫組織化学染色を、抗PDGF−DD血清を用いて行い、次いで、西洋ワサビペルオキシダーゼを結合したヤギ抗ウサギ抗体を用いて検出した。簡潔に言うと、組織を従来の技術によりパラフィン除去し、トリプシン(0.15%)で37℃にて10分間処理した。1次抗体および抗ウサギHRP結合体と、それぞれ10分間インキュベーションした後、ジアミノベンジジン(DAB)溶液を切片に添加し、免疫活性を視覚化した。図8に示すように、免疫組織化学分析により、抗Thy−1誘導性腎炎ラットでは、抗PDGF−DDレベルが上昇することが明らかとなった。メサンギウム、尿細管および周辺脈管構造を示す。メサンギウム細胞は、周皮細胞および腎尿細管を含んでいる。白色および灰色の矢印は、それぞれ毛細管および尿細管染色を示す。
【0188】
実施例9
完全ヒト抗PDGF−DD mAb 6.4のシミュレートした薬物動態学
ラットにおいて、完全ヒトmAb動態学のシミュレーションを行った。雄性Wistarラットに、抗PDGF−DD mAb 6.4を、3日目および5日目にそれぞれl0mg/kgおよび5mg/kgで投与した。血清を回収し、ヒト抗PDGF−DD mAbレベルを、ヒト特異的IgG ELISAで定量した。図9に示すように、単回投与後でさえも、4日間予想された大きなピークからトラフへの変動は生じ無かった。これらのデータは、ラットにおけるヒト抗体のクリアランスのpKシミュレーションモデルと好適に関連し、多くの抗PDGF−DD mAb 6.4は、一旦投与されると循環し続けることを示している。
【0189】
抗体クリアランス速度を分析するためのさらなる実験において、49匹のラットを以下の種々のレベルの抗PDGF−DD抗体、対照抗体またはPBSで処置した。
【表4】
【0190】
以下の「循環抗体」につての表において、左の列は49匹の動物についての5日目の結果を、右の列は、対応する動物の8日目の結果を示す。
抗PDGF−DD抗体クリアランス
【表5】
【0191】
【表6】
【0192】
上述のように、抗PDGF−DD mAb 6.4は、薬物動態学モデルにおいて計算したように、予測循環半減期を示した。
【0193】
実施例10
メサンギウム増殖性腎炎における糸球体中でのPDGF−DD発現
抗Thy1.1腎炎における糸球体でのPDGF−DD発現の動態学を研究するため、体重180gの雄性Wistarラット(Charles River, Sulzfeld, Germany)に1mg/kgのモノクローナル抗Thy1.1抗体(Clone OX-7; European Collection of Animal Cell Cultures, Salisbury, England)を注射することにより、抗Thy1.1メサンギウム増殖性糸球体腎炎を誘導した。45匹のラットに抗Thy1.1抗体を投与し、抗体注射の4時間後、1、2、4、7、9、14、21および28日の時点で屠殺した(それぞれ5匹ずつ)。屠殺後、腎組織および単離した糸球体を得た。糸球体の単離は、異なるふるいわけにより行った。Johnson et al., J Clin Invest 87: 847-858(1991)参照。全ての糸球体単離物を顕微鏡でチェックしたところ、98%より高い純度を示した。さらに、副腎組織を得た。
【0194】
糸球体のRNAの抽出および分析
RNAを糸球体から単離し、発現をリアルタイム定量的PCRで測定した。簡潔に言うと、単離したラット糸球体および副腎から、標準的手法を用いて、グアニジウムイソチオシアネート/フェノール/クロロホルム法により、トータルRNAを抽出した。Chomczynski et al., Anal Biochem 162: 156-159(1987)参照。得られたサンプルのRNA含有量および純度を、UV分光測定法により、260および280nmで決定した。
【0195】
cDNA合成は、トータルRNA 1μg、ランダムプライマー(6nt、250ng/μl、Roche)lμl、M−MLV逆転写酵素緩衝液(Invitrogen, Groningen, The Netherlands)6μ1、dNTP混合物(それぞれl0mM、Amersham Pharmacia Biotech, Freiburg, Germany)1.5μl、RNase阻害剤(40U/μl、Promega, Mannheim, Germany)0.7μl、M−MLV逆転写酵素(200U/μ1、Invitrogen)1μl、およびDEPC処理H20を含む、反応混合物 30μlにて行った。この混合物を、25℃で10分間、次いで42℃で1時間インキュベートした。
【0196】
リアルタイム定量的PCRを、ABI prism 7700配列検出装置(Applied Biosystems, Weiterstadt, Germany)を用いて行った。それぞれの反応において、cDNA 0.75μ1、およびPCR Master Mix(ROX Reference Dyeを用いるPlatinum Quantitative PCR SuperMix-UDG;Invitrogen)12.5μ1を、計25μlの体積で用いた。PCR条件は、50℃で2分、次いで95℃で15秒を40サイクル、そして60℃で1分であった。Taqmanプライマーおよびプローブを、Primer Express ソフトウェア(Applied Biosystems)を用い、Genbankデータベースの配列から設計した。本研究で用いたプライマーおよびプローブを、以下の表3に列挙する。
【0197】
表3
プライマーおよびプローブ
【表7】
【0198】
PGDF−DDはメサンギウム増殖性腎炎の糸球体において過剰発現する
ラットにメサンギウム増殖性抗Thy1.1腎炎を誘導した後、糸球体のPDGF−D mRNA発現は、当初は、疾患誘導4時間後に36%減少したが、次いで、4日〜9日目に、非腎炎ラットと比較して2.4〜2.9倍増加した(図10)。この後者のピークは、糸球体のPDGF−A mRNA発現のそれと平行し、PDGF−B mRNA最大発現後に若干遅れて生じた(図10)。これらの3つのPDGFアイソフォームとは対照的に、PDGF−C mRNAは、抗Thy1.1腎炎の最初の28日間にはアップレギュレートされなかった。
【0199】
抗Thy1.1腎炎におけるPDGF−D mRNAアップレギュレーションが腎臓に特異的であるかどうかを評価するために、副腎でのmRNAレベルもまた調査した。なぜなら、副腎はPDGF−Dの重要な供給源であることが見出されているからである。LaRochelle et al., Nat Cell Biol 3:517-521(2001)参照。糸球体とは対照的に、抗Thy1.1腎炎の最初の28日間において、副腎では、PDGF−D mRNA発現レベルの有意な変化は観察されなかった(データは示していない)。この後者の知見にもかかわらず、正常動物のPDGF−DDタンパク質レベルが常に検出限界未満(<0.02ng/ml、n=5)であったのに対し、疾患誘導8日後の腎炎ラットの血清中では、当該タンパク質レベルの劇的なアップレギュレーション(27.7±14.5ng PDGF−D/ml、n=9)を検出した。
【0200】
PDGF−DD発現の免疫組織化学
正常ラット腎臓における免疫組織化学PDGF−DD発現は、動脈および細動脈の血管平滑筋細胞に限られるが、糸球体では免疫反応は見られなかった(図11(A))。拡張したメサンギウムにおけるPDGF−DDの顕著な糸球体過剰発現は、疾患誘導の7日後には存在した(図11(B))が、残りの腎臓の染色パターンには影響を及ぼさなかった。抗PDGF−DD抗体を等濃度の対照IgGと置き換えた場合、糸球体染色は存在しなかった(図11(C))。
【0201】
実施例11
PDGF−DDとPDGF−BBの相互作用
PDGF−BBおよびPDGF−DDの両方が抗Thy1.1腎炎において過剰産生され(図10および11)、かついずれかの拮抗作用がメサンギウム増殖性変化の減少をもたらすと仮定した場合の、2つのPDGFアイソフォームの可能性のある相互作用を評価した。
【0202】
PDGF−DDの抗PDGF−DD mAb 6.4との拮抗作用は、疾患の8日目において、糸球体PDGF−BおよびPDGF−D mRNAレベルに対する有意な影響をもたらさなかった(表11)。また、このモデルにおける、特異的アプタマーによるPDGF−Bの拮抗作用は、8日目におけるPDGF−D mRNAの糸球体発現に何ら違いをもたらさなかった(PEG40対照群で3.10±1.30増加したのに対し、アプタマー群における非腎炎ラットについては3.18±0.58増加した(それぞれn=5))。しかしながら、後者の実験における糸球体PDGF−B mRNA発現は、PDGF−B拮抗作用によりわずかに誘導された(正常ラットにおける発現と比較して、PEG40対照群では発現が2.52±0.64であるのに対し、アプタマー群では発現が3.31±1.1(それぞれn=5)であった)。測定をそれぞれのサンプルについて2回行った。
【0203】
実施例12
インビボでのPDGF−DD拮抗作用
インビボでのPDGF−DD拮抗作用の効果を研究するため、ラットを、抗Thy1.1腎炎の誘導の3日および5日後に、抗PDGF−DD抗体 6.4、対照IgG PK16.3またはPBSで処置した。処置は、20mM Tris−HCl/100mM NaCl(pH7.4) 800μlに溶解した、抗体の腹腔内注射からなるものであった。処置のタイミングを、メサンギウム細胞増殖の開始後約1日目からピーク(OX−7−誘導性抗Thy1.1腎炎モデルでは、疾患誘導の5日と8日との間で生じる)までラットを処置するように選択した。抗PDGF−DD抗体の3つの異なる用量のインビボ効果を研究した。
【0204】
50μg/mlより高い血清レベルまたはインビトロでのPDGF−DDの最大阻害の2分の1を生じるという計算に基づき、体重1kgあたりl0mg(3日目)および4mg(5日目)の平均用量の抗PDGF−DD mAb 6.4を選択した。抗PDGF−DD mAb 6.4または無関係な対照IgG2 PK16.3と関係するレベルの達成を確認するため、5日目および第8日目において、疾患群1〜4におけるヒトIgG2血清レベルを測定した。5日目に30μg/ml未満のレベルを示す動物を分析から除いた。
【0205】
上記と共に、抗体で処置した群における十分なヒト血清IgG2を有する7つの群のラットを研究した:(1)3日目に5mg/kg体重、そして5日目に2mg/kg体重にて抗PDGF−DD mAb 6.4を投与した7匹の腎炎ラット;(2)3日目にl0mg/kg体重、そして5日目に4mg/kg体重にて抗PDGF−DD mAb 6.4投与した7匹の腎炎ラット;(3)3日目に20mg/kg体重、そして5日目に8mg/kg体重にて抗PDGF−DD mAb 6.4を投与した8匹の腎炎ラット;(4)3日目に20mg/kg体重、そして5日目に8mg/kg体重にて無関係な対照IgGを投与した8匹の腎炎ラット;(5)等量のPBSのみを注射した9匹の腎炎ラット;(6)3日目にl0mg/kg体重、そして5日目に4mg/kg体重にて抗PDGF−DD mAb 6.4を投与した5匹の非腎炎正常ラット;ならびに(7)等量の無関係な対照IgGを投与した5匹の非腎炎正常ラット。
【0206】
群1〜5のそれぞれから4匹のランダムに選択したラットにおいて、5日目に、Floege et al., Am J Pathol 154 : 169-179(1999)に記載の生体内バイオプシーにより、組織学的評価のための腎バイオプシーを得た。全てのラットにおいて、疾患誘導8日後に、死後バイオプシーを得た。次いで、全ての群の2または3匹のラットの残りの皮質組織をプールし、糸球体の単離に用いた(上記を参照されたい)。疾患誘導7日後に、尿を回収した。8日目、屠殺の4時間前にチミジン類似体5−ブロモ−2’−デオキシウリジン(BrdU; Sigma, Deisenhofen, Germany;l00 mg/kg体重)を腹腔内注射した。
【0207】
PDGF−DD阻害はインビボでの病理学的メサンギウム細胞増殖を低減する
抗Thy1.1抗体の注射後、PBS処置した動物は、腎炎の代表的な過程を発症し、これは初期のメサンギウム分解、および引き続き5日および8日目において、メサンギウム細胞増殖相およびマトリックスの蓄積により特徴付けられた。抗PDGF−DD mAb 6.4を反復注射した後に、目に見える悪影響は見られず、全てのラットが生存し研究の終了まで正常であるように見えた。腎炎群で至った抗体血清レベルを、表4に示す。腎炎群におけるアルブミン/クレアチニン比および全ての処置群における収縮期血圧は、有意な差はなかった。
【0208】
尿中アルブミンレベルは、ラットアルブミンに特異的なELISAキット(Nephrat, Exocell, Philadelphia, PA)を用いて決定した。尿中クレアチニンは、Vitros 250分析装置(Orthoclinical Diagnostics,Neckargmund, Germany)を用いて、2点間速度法で決定した。全ての測定を2回行った。血圧測定は、プログラム血圧計BP-98A(Softron, Tokyo, Japan)を用いて、tail cuff法により行った。Kitahara et al.,J Am Soc Nephrol 13 : 1261-1270(2002)参照。
【0209】
腎炎ラットでは、7日目に、非腎炎ラットと比較してアルブミン尿のかなりの増加が生じた(対照IgGを投与した非腎炎ラットではアルブミン/クレアチニン比:0.3±0.lmg/μmolであるのに対して、PBSを投与した腎炎ラットでは15.5±5.6mg/μmol;(p<0.01))。PBS、対照IgG、または3種類の用量の抗PDGF−DD mAb 6.4を投与した腎炎群の間には有意な差異は見られなかった(表4)。抗PDGF−DD mAb 6.4は、非腎炎ラットにおいてタンパク質尿を誘発しなかった。
【0210】
種々の抗PDGF−DD mAb 6.4用量でも、無関係な対照IgGでも、全身血圧レベルに有意な作用は観察されず、7日目において、全ての動物が正常血圧を維持した(表4)。
表4
インビボで達成されたヒトIgG2抗体(抗PDGF−DD mAb 6.4または無関係な対照IgG2)レベル、尿中アルブミン/クレアチニンおよび収縮期血圧
【表8】
**データは平均値±標準偏差である。n.d.は、測定しなかったことを示す。
【0211】
糸球体有糸分裂数を数えることにより評価する糸球体細胞増殖は、無関係なIgGまたはPBSを投与したラットと比較して、抗PDGF−DD mAb 6.4を投与したラットにおける8日目の用量依存性を有意に低減した(図12(A))。処置を3日目および5日目に行った。正常または非腎炎ラットを、抗PDGF−DD抗体または無関係な対照IgGで処置した。アスタリスクはp<0.05を示す。BrdU陽性の核のカウントにより、20+8mgの抗PDGF−DD抗体/kgでの処置群では、8日目に増殖が最も顕著に抑制されているという知見を確認した(図12(B))。抗PDGF−DD処置を受けた全3群のデータを集めたところ、インビボで至った抗体レベルとBrdU取込み核とは、5日目(r=−0.53;p=0.018)および8日目(r=−0.40;p=0.081)においてネガティブに相関した。
【0212】
メサンギウム細胞に対する処置の効果を評価するため、活性化メサンギウム細胞のみによって発現するα平滑筋アクチンを、腎断面で免疫染色した。Johnson et al., J Clin Invest 87: 847-858(1991)参照。α平滑筋アクチンの糸球体発現は、10+4mg/kgおよび20+8mg/kgの抗PDGF−DD mAb 6.4を投与したラットにおいて、8日目に、無関係なIgGまたはPBSを投与したラットと比較して有意に減少した(図12(C))。メサンギウム細胞増殖が減少しているかどうかを特異的に決定するため、抗PDGF−DD mAb 6.4で処置したラットおよび対照IgGまたはPBSで処置したラットのBrdUおよびα平滑筋アクチンを2重免疫染色した(図12(D))。このデータは、全3つの抗PDGF−DD抗体処置群で、疾患誘導の8日後にメサンギウム細胞の増殖の顕著な低減が観察され、メサンギウム細胞増殖が最大で57%低減されたことを確認した。メサンギウム分解スコアは、抗PDGF−DD mAb 6.4処置ラットおよび対照IgG処置ラットにおいて同じであった(図12(E))。
【0213】
抗PDGF−DD mAb 6.4の正常ラットへの注射は、無関係なIgGを投与した正常ラットと比較して、生理的な糸球体細胞ターンオーバーに影響しなかった。
【0214】
インビボでのPDGF−DDの阻害は、糸球体の単球/マクロファージ流動を低減する
5日目において、抗PDGF−DD mAb 6.4の3種類の用量全てが糸球体単球/マクロファージ流動の顕著な低減を生じたが、8日後には生じなかった(図12(G))。正常ラットの特異的抗PDGF−DD抗体または無関係なIgGのいずれかでの処置は、糸球体単球/マクロファージ流動に影響しなかった。
【0215】
インビボでのPDGF−DD阻害は、糸球体マトリックスの蓄積を低減する
ラットを10+4mg/kgの抗PDGF−DD mAb 6.4または20+8mg/kgの抗PDGF−DD mAb 6.4のいずれかで処置した結果、腎炎対照と比較して、糸球体フィブロネクチン蓄積が低減された(図12(H))。一方、I型コラーゲンの糸球体蓄積は、3つの腎炎群のいずれにおいても、対照IgGまたはPBSで処置したラットと比較して、抗PDGF−DD抗体処置により影響されなかった(図12(F))。
【0216】
正常ラットでは、糸球体マトリックス発現は、抗PDGF−DD抗体または無関係なIgGでの処置によって影響されなかった(図12(H))。
【0217】
実施例13
インビボでの抗PDGF−DD抗体の効力
ラットにおける抗Thy−1.1抗体誘導性メサンギウム増殖性糸球体腎炎モデルにおけるPDGF−DDと結合する、抗PDGF−DD mAb 6.4の効力を、インビボで以下のように評価した。
【0218】
正常な生理的状態を有し、体重が約150〜200gの雄性Wistarラット(10週齢)(Charles River, Sulzfeld, Germany)を得た。まず、ラットを、正常および抗Thy1.1メサンギウム増殖性糸球体腎炎を誘導する、2群に分けた。
【0219】
動物を、地方の動物施設で以下のように飼育した:ラットを7日間順応させ、食物および水道水を自由に与えた。研究を開始する前に動物を検査し、適度な健康および適合性を確認した。疾患を有すること、または不適切であることが判明した動物は本研究に供しなかった。この一連の研究において、12時間毎の明/暗サイクルを維持した。名目上の温度範囲は、20〜23℃であり、30%〜70%の相対湿度もまた維持した。
【0220】
完全ヒト抗PDGF−DD mAb 6.4を、上述のXenomouse(登録商標)技術を用いて産生した。1mg/kgのモノクローナル抗Thy1.1抗体を注射することにより、抗Thy1.1メサンギウム増殖性糸球体腎炎を雄性Wistarラットにて誘導した。抗Thy1.1腎炎を誘導した後、疾患誘導の3日後および5日後に、10mg/kgおよび4mg/kgのmAb 6.4(n=15)または無関係なヒトモノクローナル抗体(n=15)またはPBS(n=15)を毎日腹腔内注射することにより、ラットを処置した。残りのラットは処置しなかった。全部で5群のラットを研究した。処置後、ラットを、腎バイオプシー、尿アルブミンおよび屠殺後の組織回収により分析した。
【0221】
1)合計14mg/kg(疾患誘導の3日後にl0mg/kg、そして5日後に4mg/kg)の抗PDGF−DD mAb 6.4を投与した15匹の腎炎ラット;
2)合計14mg/kg(疾患誘導の3日後にl0mg/kg、そして5日後に4mg/kg)の無関係なアイソタイプ一致対照抗体(抗PDGF抗体ではない)を投与した15匹の腎炎ラット;
3)トリス緩衝生理食塩水のみ800μLのボーラス注射を受けた15匹の腎炎ラット;
4)合計14mg/kg(疾患誘導の3日後にl0mg/kg、そして5日後に4mg/kg)の抗PDGF−DD mAb 6.4を投与して5匹の腎炎ラット;および
5)5匹の正常無処置のラット。
【0222】
表5により、試験した5群についての研究設計のリストを提供する。
表5
研究設計
【表9】
【0223】
抗PDGF−DD mAb 6.4の純度は、90%より高いか、または同等であった。全てのバイアルを、使用するまで冷蔵庫中で4℃にて保管した。予備のサンプルを、−80℃で保管した。トリス緩衝生理食塩水中の10Mg/Kg体重、および4mg/kg体重を腹腔内注射した。トリス緩衝生理食塩水での希釈は、l0mg/kgおよび4mg/kg用量が、800μLの体積で投与できるように行った。これらの用量を、3日後および5日後にのみ、1日1回投与した。
【0224】
処置の持続を、メサンギウム細胞増殖の開始の約1日後から増殖のピーク(OX−7誘導性抗Thy1.1腎炎モデルでは、疾患誘導の6日目と9日目との間で生じる)までラットを処置するように選択した。
【0225】
ラットに投薬した約60分後、ラットの有意な臨床徴候、罹患率、および死亡率について、ラットを毎日観察した。研究の開始後は、体重測定を行わなかった。動物が検視前に死亡した場合、検死および組織データには含めず、組織も回収しなかった。
【0226】
動物が検視中に死亡した場合、死亡していたものと記録し、検視データを用いなかった。しかしながら、評価の可能性のため、組織をホルマリン中に回収した。瀕死の動物は、屠殺し、同様に処置した。
【0227】
8日目まで生存した動物全てを、頚椎脱臼により殺し、総合的に観察し、予定した全ての組織を組織形態学的評価のため、10%中性緩衝ホルマリン、Methacarn溶液、および液体窒素中に回収した。
【0228】
染色方法および組織の調製:光学顕微鏡および免疫ペルオキシダーゼ染色のための組織をメチルCarnoy溶液で固定し、パラフィンに包埋した。4μmの断面を過ヨウ素酸Schiff(PAS)試薬で染色し、ヘマトキシリンで対比染色した。PAS染色された断面において、有糸分裂数を、20より多い毛細管切片を含有する連続的皮質糸球体である30を超える断面(30〜100の範囲)を、公平な観察者により評価した。メサンギウム溶解を、Burg et al., Lab Invest 76: 505-516(1997)に記載の半定量スケールで階級付けした:0=メサンギウム溶解なし、1=部分的メサンギウム溶解、2=全体的メサンギウム溶解、3=微細動脈瘤。
【0229】
免疫ペルオキシダーゼ染色:メチルCarnoyで固定したバイオプシー組織の4μm断面を、Johnson et al, 1990に記載の間接免疫ペルオキシダーゼ技術により処理した。PDGF−DDは、ヒトPDGF−Dに対するポリクローナルウサギ抗体により検出した。一次抗体は、既に記載のもの(Burg et al, 1997; Yoshimura et al, 1991)と同一であり、α平滑筋アクチンに対するマウスモノクローナル抗体(クローン1A4);PDGF−B鎖に対するマウスモノクローナル抗体(クローンPGF−007);単球、マクロファージ、および樹状細胞に存在する細胞質抗原に対するマウスモノクローナルIgG抗体(クローンED1);ラット赤血球であらかじめ吸着した、アフィニティー精製ポリクローナルヤギ抗ヒト/ウシIV型コラーゲンIgG;ヒトI型コラーゲンに対するポリクローナルヤギ抗体(Southern Biotech Associates, Birmingham, AL, USA);アフィニティー精製ポリクローナルウサギ抗ラットフィブロネクチン抗体IgG分画(Chemicon, Temecula, CA, USA);および上述の適当な負の対照(Burg et al, 1997; Yoshimura et al, 1991)を含む。PDGF−DDは、ヒトPDGF−Dに対するポリクローナルウサギ抗体により検出した。血清を、Protein A Sepharoseクロマトグラフィーにより精製した。PDGF−Cアフィニティーカラムに吸着させることにより、PDGF−C交差活性を取り除いた。得られた免疫グロブリンフローを濃縮し、これはELISAまたはウエスタンブロット分析によりPDGF−A、BまたはCとは反応しないことがわかった。全てのスライドを、スライドの出処を知らされていない観察者により評価した。
【0230】
糸球体中の浸潤白血球の平均数を得るため、糸球体の50より多くの連続的断面を評価し、腎臓あたりの平均値を計算した。I型コラーゲン、フィブロネクチンおよびα平滑筋アクチンの免疫ペルオキシダーゼ染色を評価するため、それぞれの糸球体領域を、半定量的に段階付けし、1バイオプシー当たりの平均スコアを計算した。それぞれのスコアは、主に、染色強度ではなく染色程度の変化を反映し、これは、陽性染色が集中的に増強された糸球体房領域の割合に依存する:
I=0〜25%
II=25〜50%
III=50〜75%
IV=>75%
【0231】
免疫組織化学的二重染色:増殖細胞の型を同定するための二重免疫染色は、以前に報告(Kliem et al, 1996; Hugo et al, 1996)されているように、まず、増殖細胞の断面を、トリス緩衝生理食塩水中のヌクレアーゼ含有ブロモデオキシウリジン(Amersham, Braunschweig, Germany)に対するマウスモノクローナル抗体(クローンBU−1)による間接的免疫ペルオキシダーゼ方法を用いて、染色した。断面を、α平滑筋アクチンに対するIgG1 mAb 1A4および単球/マクロファージに対するED1と共にインキュベーションした。細胞がBrdUの陽性核染色を示し、当該核がα平滑筋アクチンまたはED1抗原陽性細胞質により完全に包囲されている場合、増殖メサンギウム細胞または単球/マクロファージであると同定した。負の対照では、いずれかの1次抗体を省き、この場合、二重染色は観察されなかった。
【0232】
尿測定:尿中タンパク質(アルブミン尿)は、Bio-Radタンパク質アッセイ(Bio-Rad Laboratories GmbH, Munchen, Germany)を用い、ウシ血清アルブミン(Sigma)を標準として用いることにより測定した。血圧は、テール血圧計により測定した。
【0233】
得られた多数のデータから統計的分析を行い、平均値および標準偏差を数学的に計算した。統計的分析は、予定屠殺時まで生存した動物由来のデータに基づき行った。全ての値を平均±標準偏差として表す。統計上の有意性(p<0.05と定義)を、Student t検定またはANOVAおよびBonferroni t検定を用いて評価した。データの解釈を助けるため、研究責任者の裁量で補助的分析も行った。
【0234】
結果:図13は、糸球体腎炎のラットモデルにおけるBrdU取込みにより測定した糸球体増殖の結果を示す。ラットを屠殺前にBrdUで6時間処置した。核のBrdU染色は、抗BrdU抗体を用いて測定した。腎炎ラットモデルで観察された糸球体100個あたりの有糸分裂数を計測した。表6は、試験した5群に基づき、糸球体100個あたりのBrdU陽性核数をまとめている。表6は、図13の対応するグラフと共に、抗PDGF−DD mAb 6.4を腎炎動物に投与することにより、チミジン類似体BrdUを取込んだ糸球体細胞が、無関係のIgGおよびPBSを投与した場合と比べて少なくなることを示している。1列は、疾患糸球体の有糸分裂指数を示す。2列は、対照抗体で処置した腎炎のラットモデルのものである。3列は、PBSで処置した腎炎ラットモデルのものである。4列は、抗PDGF−DD mAb 6.4で処置した健常ラット対照のものである。5列は、対照抗体で処置した健常ラット対照のものである。
【0235】
表6
糸球体によるBrdU取込み
【表10】
【0236】
同様に、図14は、糸球体腎炎のラットモデルを、抗PDGF−DD mAb 6.4で処置した場合の有糸分裂をPAS染色および定量測定した糸球体増殖の結果を示す。4μMの断面を過ヨウ素酸Schiff試薬で染色し、ヘマトキシリンで対比染色した。有糸分裂を、糸球体細胞100個当たりについて測定した。30〜50個の糸球体房の有糸分裂数を決定した。表7は、抗PDGF−DD mAb 6.4投与が、無関係なIgG抗体およびPBSと比較して、100個当たりの有糸分裂を顕著に減少させることを示している。図14の対応グラフは、抗PDGF−DD mAb 6.4を腎炎動物に投与することにより、無関係なIgG抗体およびPBSを腎炎動物に投与したときと比べて、有糸分裂する糸球体細胞がより少なくなることを示す。1列は、抗PDGF−DD mAb 6.4で処置した腎炎ラットモデルで観察された結果を示す。2列は、対照抗体で処置した腎炎ラットモデルを示す。3列は、PBSで処置した腎炎ラットモデルを示す。4列は、抗PDGF−DD mAb 6.4で処置した健常ラット対照を示す。5列は、対照抗体で処置した健常ラット対照を示す。
【0237】
表7
糸球体での有糸分裂の測定(PAS使用)
【表11】
【0238】
表6および7において示すように、抗PDGF−DD mAb 6.4での腎炎ラットの処置は、有糸分裂の手動計数(表7)およびBrdU取込みによる測定(表6)のいずれにおいても糸球体増殖を低減した。
【0239】
また、メサンギウム細胞を、抗平滑筋アクチンで対比染色した。所定セットのメサンギウム細胞(30〜50個の糸球体タフト)における有糸分裂数を決定した。二重染色アッセイの結果を、以下の表8に示す。表8は、対応する図15と共に、抗PDGF−DD mAb 6.4は、無関係な(非PDGF−D)抗体(PK16.3 IgG)およびPBSと比較して、糸球体有糸分裂の低減に有効あることを示している。
【0240】
表8
BrdU+抗sm−アクチン
【表12】
【0241】
実施例14
急性ラットThy−1モデルに対するmAb 6.4の用量応答性作用
抗PDGF−DD mAb 6.4のThy−1誘導腎炎に対する作用もまた、用量応答性方法で研究した。抗Thy1.1腎炎誘導後、ラットを疾患誘導の3日〜8日後に、それぞれ、5、10および20mg/kg、次いで2、4および8mg/kgのヒト抗PDGF−DD mAb 6.4(n=15)または無関係なヒトPK16.3モノクローナル抗体(n=15)またはPBS(n=15)のいずれかで腹腔内注射により毎日処置した。図16に示しかつ表9にまとめるように、8日後、抗PDGF−DD mAb 6.4を用いて処置した疾患誘導拮抗作用は、100個の糸球体当たりの有糸分裂数を有意に低減した。図17に示しかつ表10にまとめるように、抗PDGF−DD mAb 6.4での処置はまた、チミジン類似体BrdUを取込む糸球体細胞を有意に低減した。従って、ラット抗Thy−1モデルの10mg/kgの抗PDGF−DD mAb 6.4での処置は、若干用量応答性方法で増殖を40〜70%阻害する。
【0242】
表9
【表13】
【0243】
表10
【表14】
【0244】
表11
抗PDGF−D mAb処置ラットにおける糸球体PDGF−BおよびPDGF−D mRNA発現
【表15】
**データは、それぞれの処置群におけいてプールした分画の平均(および範囲)である。
【0245】
実施例15
ヒト腎臓疾患組織の免疫組織化学的分析
免疫組織化学的分析により、ヒト腎臓疾患組織におけるPDGF−DDの存在を試験した。
【0246】
PDGF−AA、PDGF−BBまたはPDGF−CCを認識しないウサギ抗PDGF−DD IgGを用いて、免疫組織化学染色を行った。染色に続いて、西洋ワサビペルオキシダーゼと結合したヤギ抗ウサギ複合体(抗ウサギHRP複合体)を用いて検出した。抗ウサギHRP複合体とのインキュベーション後、ジアミノベンジジン(DAB)溶液を断面に添加し、免疫反応を視覚化した。
【0247】
活発な糸球体腎炎において、尿細管(ほとんどはおそらく近位尿細管である)の染色が明確に認められた。いくつかの糸球体も、陽性に染色されていた(約10〜20%のメサンギウム/視野)。慢性的な同種移植拒絶を有する患者由来組織もまた陽性に染色されており、これは尿細管および血管染色を示していた。細胞蓄積物もまたメサンギウム中で検出され、これは炎症誘導性肥満細胞および細胞沈着を示唆した。対照ウサギIgGでは染色されなかった。
【0248】
PDGF−DD染色はまた、薬剤誘導性間質性腎炎でも明確に認められた。ここで、尿細管の染色増加、メサンギウム細胞の顕著な染色、およびいくつかの浸潤性炎症誘導性細胞染色が観察された。存在する可能性のある非常に弱い尿細管の染色を除いては、正常ヒト腎臓の顕著な染色は観察されなかった。腎硬化症においては、尿細管のPDGF−DD染色が見られた(データは示していない)。虚血性尿細管損傷においては、染色は観察されなかった(データは示していない)。これらの結果は、多くのヒト腎臓の病理においてPDGF−DDが増加することを示唆しており、腎臓疾患におけるPDGF−DDの役割を示唆する。PDGF−DDは、尿細管間質、メサンギウム増殖および活発な炎症過程における変化に関与し得る(図18を参照)。白色および灰色の矢印は、それぞれ毛細管および尿細管染色を示している。小さな黒色の矢印は、メサンギウム中の点状炎症性細胞蓄積物を示す。
【0249】
実施例16
ELISAを用いた腎炎の発症リスクの分析、診断および段階決定
腎炎に罹患した患者由来のPDGF−DD血清レベルを分析した。PDGF−DD濃度は、定量的サンドウィッチELISA法を用い、PDGF−DDに対して生じた全2つのヒトmAbを用いて評価した。腎炎患者の血清中では、正常患者と比べて、PDGF−DDレベルが4〜7倍に上昇していることがわかる。従って、PDGF−DDレベルのこれらの差異は、診断を成すための助けとなり得、かつ医師が腎炎および関連疾患の段階決定するための助けとなり得る。
【0250】
実施例17
抗PDGF−DD抗体によるヒトの腎炎の処置
医師は、有効量の抗PDGF−DD抗体をそれを必要とする患者に投与し、それによって当該必要とする患者の症状が緩和されるかまたは腎炎が効果的に治療される。投与および用量は患者に特有である。抗PDGF−DD抗体の投与は、皮下注射による。
【0251】
上述の種々の方法および技術は、多くの実施態様を行うための多数の方法を提供する。当然のことながら、本明細書に記載の任意の特定の実施態様に従って必ずしも記載された全ての目的または利点が達成され得るわけではないことは理解されるべきである。従って、例えば、当業者は、当該方法は、本明細書で教示される1つの利点または利点群を達成または最適化する方法で、本明細書で教示または示唆され得る他の目的または利点を必ずしも達成することなく実行され得ることを理解するであろう。
【0252】
さらに、当業者は、異なる実施態様の種々の特性は互換可能であることを理解するであろう。同様に、上述の種々の特性および工程、ならびにそのような特性または工程の他の公知の均等物は、本明細書に記載の原理に従った方法を実行するために、当業者によって結び付けられ適合され得る。
【0253】
本明細書に記載の方法は、ある種の実施態様および実施例の文脈によって開示されてきたが、これらの方法が、特定の開示された実施態様を超えて、他の代わりの実施態様および/または使用および明らかな改変ならびにそれらの均等物まで拡張されることは当業者に理解されるであろう。従って、本明細書に記載の方法は、本明細書で好ましい実施態様の特定の開示によって限定されないが、添付の特許請求の範囲の参照となることが意図されている。
【図面の簡単な説明】
【0254】
【図1】図1は、ELISAによる抗PDGF−DD mAb 6.4特異性の特徴決定を示す。
【図2】図2は、ELISAによる抗PDGF−DD mAb 6.4特異性のさらなる特徴決定を示す。
【図3】図3は、ウエスタンブロット分析による抗PDGF−DD mAb特異性の特徴決定を示す。
【図4】図4は、抗PDGF−DD mAb 6.4が、NIH3T3細胞でのPDGF−DD誘導性BrdU取込みを、約75ng/mlのIC50で中和できることを示す線グラフである。
【図5】図5は、PDGF−DDがインビトロにおいてメサンギウム細胞の成長因子となることを示す棒グラフである。
【図6】図6は、ヒトメサンギウム細胞におけるPDGF−DD誘導性BrdU取込みの結果を示す棒グラフである。
【図7】図7は、種々の型の腎炎を有する患者のヒト血清中でのPDGF−DD発現を示すグラフである。
【図8】図8は、正常ラットメサンギウム細胞および抗Thy−1誘導性腎炎を有するラットのメサンギウム細胞の免疫組織化学分析を示す。
【図9】図9は、ラットにおいて成された完全ヒトmAbカイネティクスのシュミレーションを示す棒グラフである。
【図10】図10は、無処置のラットでの発現に対する、抗Thy1.1腎炎の経過におけるPDGF−A、−B、−Cおよび−Dの転写発現を示す線グラフである。
【図11】図11は、PDGF−DDタンパク質が、糸球体での抗Thy1.1腎炎において過剰発現されることを示す。
【図12−1】図12A〜Hは、抗PDGF−DD抗体、無関係の対照IgGまたはPBS単独のいずれかで処置した、メサンギウム増殖性抗Thy1.1腎炎を有するラットにおいて、糸球体が疾患の誘導5日後および8日後に変化していることを示す棒グラフである。
【図12−2】図12A〜Hは、抗PDGF−DD抗体、無関係の対照IgGまたはPBS単独のいずれかで処置した、メサンギウム増殖性抗Thy1.1腎炎を有するラットにおいて、糸球体が疾患の誘導5日後および8日後に変化していることを示す棒グラフである。
【図12−3】図12A〜Hは、抗PDGF−DD抗体、無関係の対照IgGまたはPBS単独のいずれかで処置した、メサンギウム増殖性抗Thy1.1腎炎を有するラットにおいて、糸球体が疾患の誘導5日後および8日後に変化していることを示す棒グラフである。
【図12−4】図12A〜Hは、抗PDGF−DD抗体、無関係の対照IgGまたはPBS単独のいずれかで処置した、メサンギウム増殖性抗Thy1.1腎炎を有するラットにおいて、糸球体が疾患の誘導5日後および8日後に変化していることを示す棒グラフである。
【図13】図13は、ラットでのBrdU取込により測定された、糸球体増殖の結果を示す棒グラフである。
【図14】図14は、ラットにおけるPAS染色により測定された糸球体増殖および有糸分裂の結果を示す棒グラフである。
【図15】図15は、急性ラット抗Thy−1モデルにおけるメサンギウム細胞有糸分裂に対する抗PDGF−DD mAb 6.4の作用を示す棒グラフである。
【図16】図16は、急性ラット抗Thy−1モデルにおける糸球体細胞の有糸分裂に対する抗PDGF−DD mAb 6.4の用量応答性作用を示す棒グラフである。
【図17】図17は、急性ラットThy−1モデルにおけるBrdU取込みに対する抗PDGF−DD mAb 6.4の用量応答性作用を示す棒グラフである。
【図18】図18は、正常および疾患のヒト腎臓組織上の免疫組織化学分析を示す。
【配列表】
【特許請求の範囲】
【請求項1】
腎炎を効果的に処置するための医薬の製造における抗体またはその結合フラグメントの使用であって、該抗体またはその結合フラグメントが血小板由来成長因子DD(PDGF−DD)と結合する、使用。
【請求項2】
該抗体が完全ヒトモノクローナル抗体である、請求項1記載の使用。
【請求項3】
該腎炎が、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、進行性腎疾患、腎間質性線維症、腎不全、および糖尿病性腎症からなる群から選択される、請求項1記載の使用。
【請求項4】
該腎炎が糸球体またはメサンギウム細胞の増殖と関連する、請求項1記載の使用。
【請求項5】
該医薬が生理的に許容される緩衝液をさらに含む、請求項1記載の使用。
【請求項6】
メサンギウム細胞の増殖を阻害するための医薬の製造における抗体またはその結合フラグメントの使用であって、該抗体またはその結合フラグメントが血小板由来成長因子DD(PDGF−DD)と結合する、使用。
【請求項7】
該抗体が完全ヒトモノクローナル抗体である、請求項6記載の使用。
【請求項8】
該メサンギウム細胞がヒトメサンギウム細胞である、請求項6記載の使用。
【請求項9】
該医薬が生理的に許容される緩衝液をさらに含む、請求項6記載の使用。
【請求項10】
腎炎の検出方法であって、
腎炎に対するリスクのある患者を選択すること;
該患者由来の腎臓細胞をPDGF−DDと結合する抗体またはその結合フラグメントと接触させること;次に
該細胞と該抗体との結合を検出し、検出可能な結合が腎炎の指標となる、方法。
【請求項11】
該抗体がモノクローナル抗体である、請求項10記載の使用。
【請求項12】
該抗体が完全ヒトモノクローナル抗体である、請求項10記載の使用。
【請求項13】
該抗体が、蛍光色素、酵素、放射性核種、および放射線不透過物質からなる群から選択されるマーカーで標識されている、請求項10記載の使用。
【請求項14】
該結合フラグメントがFab’フラグメントを含む、請求項10記載の使用。
【請求項15】
該腎炎が、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、進行性腎疾患、腎間質性線維症、腎不全、および糖尿病性腎症からなる群から選択される、請求項10記載の使用。
【請求項16】
腎炎を効果的に処置する方法であって、
腎炎の処置を必要とする動物を選択すること、次に
該動物に血小板由来成長因子DD(PDGF−DD)と結合する抗体またはその結合フラグメントを治療上有効用量投与すること
を含む、方法。
【請求項17】
該動物がヒトである、請求項16記載の方法。
【請求項18】
該抗体が完全ヒトモノクローナル抗体である、請求項16記載の方法。
【請求項19】
該腎炎が、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、進行性腎疾患、腎間質性線維症、腎不全、および糖尿病性腎症からなる群から選択される、請求項16記載の方法。
【請求項20】
該腎炎が、糸球体またはメサンギウム細胞の増殖と関連する、請求項16記載の方法。
【請求項21】
該投与が皮下注射によるものである、請求項16記載の方法。
【請求項22】
該投与が筋肉内注射によるものである、請求項16記載の方法。
【請求項23】
メサンギウム細胞の増殖を阻害する方法であって、
血小板由来成長因子DD(PDGF−DD)と結合するモノクローナル抗体またはその結合フラグメントを用意すること;次に
増殖メサンギウム細胞を、該細胞の阻害された増殖となる条件下で該モノクローナル抗体と接触させること
を含む、方法。
【請求項24】
該抗体が完全ヒトモノクローナル抗体である、請求項23記載の方法。
【請求項25】
該メサンギウム細胞がヒトメサンギウム細胞である、請求項23記載の方法。
【請求項26】
メサンギウム増殖性糸球体腎炎を効果的に処置する方法であって、
メサンギウム増殖性糸球体腎炎の処置を必要とする動物を選択すること、次に
該動物に、血小板由来成長因子DD(PDGF−DD)と結合する抗体またはその結合フラグメントを治療上有効量投与すること
を含む、方法。
【請求項27】
該動物がヒトである、請求項26記載の方法。
【請求項28】
該抗体が完全ヒトモノクローナル抗体である、請求項26記載の方法。
【請求項29】
該投与が皮下注射によるものである、請求項26記載の方法。
【請求項30】
該投与が筋肉内注射によるものである、請求項26記載の方法。
【請求項1】
腎炎を効果的に処置するための医薬の製造における抗体またはその結合フラグメントの使用であって、該抗体またはその結合フラグメントが血小板由来成長因子DD(PDGF−DD)と結合する、使用。
【請求項2】
該抗体が完全ヒトモノクローナル抗体である、請求項1記載の使用。
【請求項3】
該腎炎が、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、進行性腎疾患、腎間質性線維症、腎不全、および糖尿病性腎症からなる群から選択される、請求項1記載の使用。
【請求項4】
該腎炎が糸球体またはメサンギウム細胞の増殖と関連する、請求項1記載の使用。
【請求項5】
該医薬が生理的に許容される緩衝液をさらに含む、請求項1記載の使用。
【請求項6】
メサンギウム細胞の増殖を阻害するための医薬の製造における抗体またはその結合フラグメントの使用であって、該抗体またはその結合フラグメントが血小板由来成長因子DD(PDGF−DD)と結合する、使用。
【請求項7】
該抗体が完全ヒトモノクローナル抗体である、請求項6記載の使用。
【請求項8】
該メサンギウム細胞がヒトメサンギウム細胞である、請求項6記載の使用。
【請求項9】
該医薬が生理的に許容される緩衝液をさらに含む、請求項6記載の使用。
【請求項10】
腎炎の検出方法であって、
腎炎に対するリスクのある患者を選択すること;
該患者由来の腎臓細胞をPDGF−DDと結合する抗体またはその結合フラグメントと接触させること;次に
該細胞と該抗体との結合を検出し、検出可能な結合が腎炎の指標となる、方法。
【請求項11】
該抗体がモノクローナル抗体である、請求項10記載の使用。
【請求項12】
該抗体が完全ヒトモノクローナル抗体である、請求項10記載の使用。
【請求項13】
該抗体が、蛍光色素、酵素、放射性核種、および放射線不透過物質からなる群から選択されるマーカーで標識されている、請求項10記載の使用。
【請求項14】
該結合フラグメントがFab’フラグメントを含む、請求項10記載の使用。
【請求項15】
該腎炎が、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、進行性腎疾患、腎間質性線維症、腎不全、および糖尿病性腎症からなる群から選択される、請求項10記載の使用。
【請求項16】
腎炎を効果的に処置する方法であって、
腎炎の処置を必要とする動物を選択すること、次に
該動物に血小板由来成長因子DD(PDGF−DD)と結合する抗体またはその結合フラグメントを治療上有効用量投与すること
を含む、方法。
【請求項17】
該動物がヒトである、請求項16記載の方法。
【請求項18】
該抗体が完全ヒトモノクローナル抗体である、請求項16記載の方法。
【請求項19】
該腎炎が、メサンギウム増殖性腎炎、メサンギウム増殖性糸球体腎炎、メサンギウム毛細管性糸球体腎炎、全身性エリテマトーデス、糸球体腎炎、進行性腎疾患、腎間質性線維症、腎不全、および糖尿病性腎症からなる群から選択される、請求項16記載の方法。
【請求項20】
該腎炎が、糸球体またはメサンギウム細胞の増殖と関連する、請求項16記載の方法。
【請求項21】
該投与が皮下注射によるものである、請求項16記載の方法。
【請求項22】
該投与が筋肉内注射によるものである、請求項16記載の方法。
【請求項23】
メサンギウム細胞の増殖を阻害する方法であって、
血小板由来成長因子DD(PDGF−DD)と結合するモノクローナル抗体またはその結合フラグメントを用意すること;次に
増殖メサンギウム細胞を、該細胞の阻害された増殖となる条件下で該モノクローナル抗体と接触させること
を含む、方法。
【請求項24】
該抗体が完全ヒトモノクローナル抗体である、請求項23記載の方法。
【請求項25】
該メサンギウム細胞がヒトメサンギウム細胞である、請求項23記載の方法。
【請求項26】
メサンギウム増殖性糸球体腎炎を効果的に処置する方法であって、
メサンギウム増殖性糸球体腎炎の処置を必要とする動物を選択すること、次に
該動物に、血小板由来成長因子DD(PDGF−DD)と結合する抗体またはその結合フラグメントを治療上有効量投与すること
を含む、方法。
【請求項27】
該動物がヒトである、請求項26記載の方法。
【請求項28】
該抗体が完全ヒトモノクローナル抗体である、請求項26記載の方法。
【請求項29】
該投与が皮下注射によるものである、請求項26記載の方法。
【請求項30】
該投与が筋肉内注射によるものである、請求項26記載の方法。
【図1】

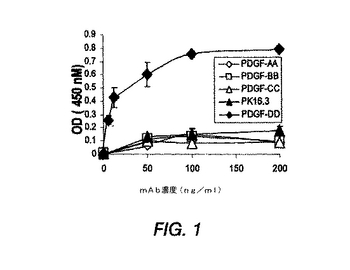
【図2】
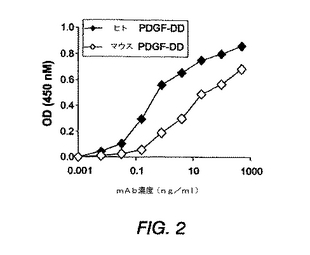
【図3】
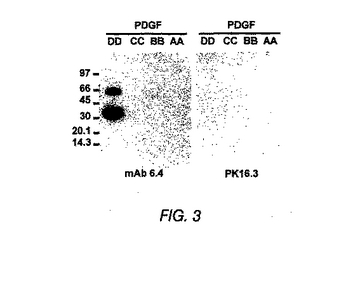
【図4】
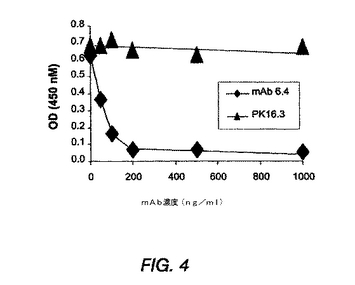
【図5】
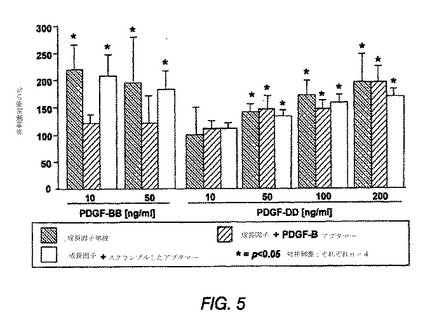
【図6】
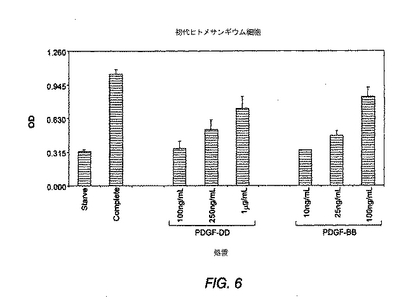
【図7】
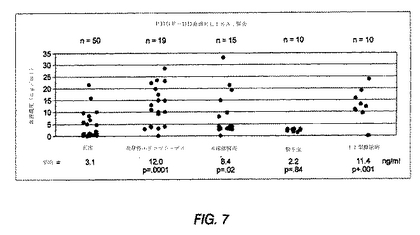
【図8】
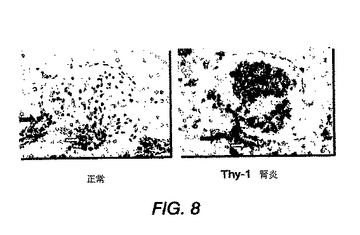
【図9】
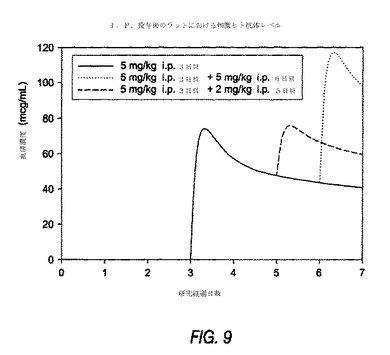
【図10】
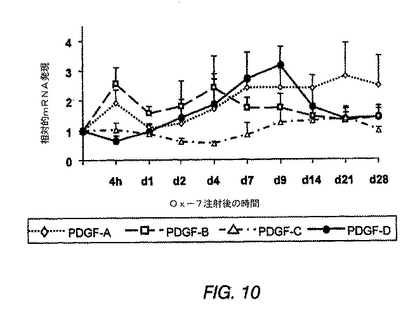
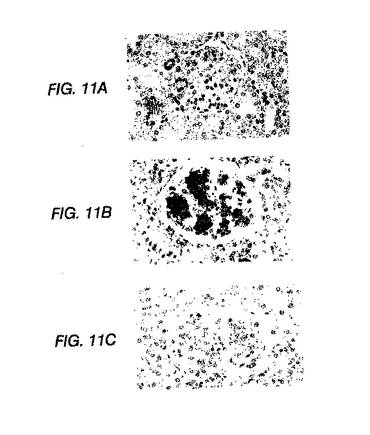
【図12−1】
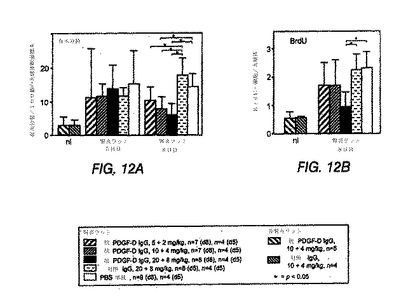
【図12−2】
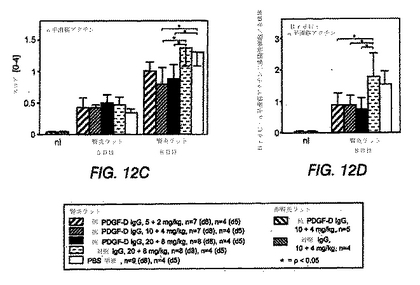
【図12−3】
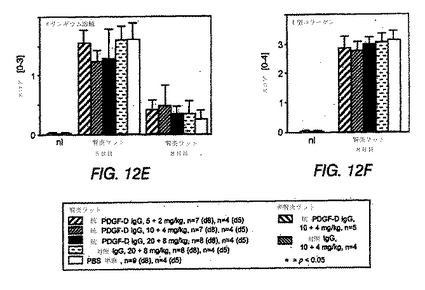
【図12−4】
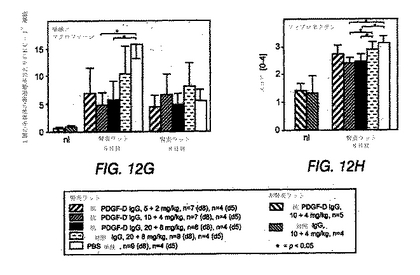
【図13】
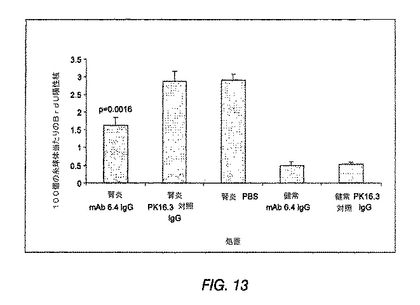
【図14】
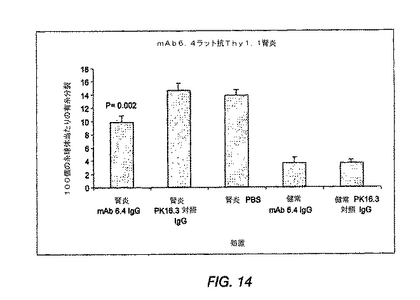
【図15】
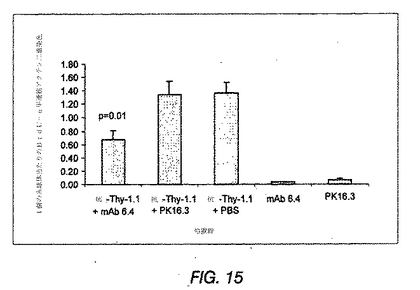
【図16】
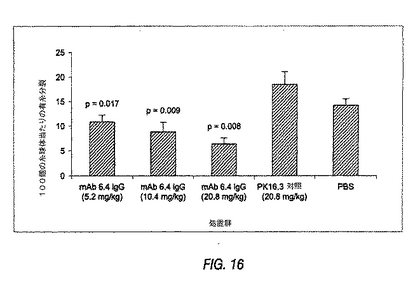
【図17】
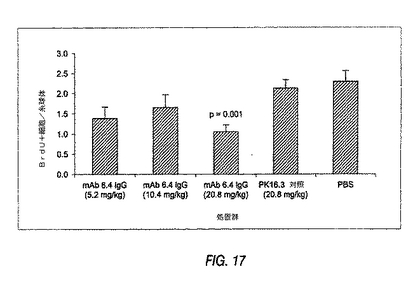
【図18】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図12−1】
【図12−2】
【図12−3】
【図12−4】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公表番号】特表2006−511471(P2006−511471A)
【公表日】平成18年4月6日(2006.4.6)
【国際特許分類】
【出願番号】特願2004−536624(P2004−536624)
【出願日】平成15年9月16日(2003.9.16)
【国際出願番号】PCT/US2003/029414
【国際公開番号】WO2004/024098
【国際公開日】平成16年3月25日(2004.3.25)
【出願人】(501343271)アブジェニックス インコーポレイテッド (5)
【出願人】(301062363)キュラジェン コーポレイション (18)
【Fターム(参考)】
【公表日】平成18年4月6日(2006.4.6)
【国際特許分類】
【出願日】平成15年9月16日(2003.9.16)
【国際出願番号】PCT/US2003/029414
【国際公開番号】WO2004/024098
【国際公開日】平成16年3月25日(2004.3.25)
【出願人】(501343271)アブジェニックス インコーポレイテッド (5)
【出願人】(301062363)キュラジェン コーポレイション (18)
【Fターム(参考)】
[ Back to top ]