
抗RHESUS−Dおよび抗HLA−DR抗体のフコース含量/ガラクトース含量の比とADCC活性の間の関連
本発明は、高いADCC活性を有し、Fc領域のグリコシル化部位にフコース含量/ガラクトース含量の比が0.6 以下であるグリカン構造を有するモノクローナル抗体に関する。本発明はまた、高いエフェクター活性をもつ前記モノクローナル抗体を含有する薬剤組成物にも関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、高いADCC活性を有し、Fc領域のグリコシル化部位に存在するグリカン構造のフコース含量/ガラクトース含量の比が0.6 以下であるモノクローナル抗体の組成物に関する。本発明はまた、高いエフェクター活性をもつ前記モノクローナル抗体を含有する薬剤組成物にも関する。
【背景技術】
【0002】
広く普及した受動免疫療法は、細胞または所定の物質に対する抗体、特にIgG型の免疫グロブリンの投与に基づく。しかし、モノクローナル抗体の使用が、感染性汚染がないことに関する生成物の安全性の保証などのいくつかの利点を有する場合、他方では有効なモノクローナル抗体を得ることが困難であると判明するかもしれない。
【0003】
G型の免疫グロブリン(IgG)は、ジスルフィド架橋で結合した2本の重鎖と2本の軽鎖からなるヘテロ二量体である。N末端位置の各鎖は、抗体が向けられる抗原に特異的な可変部分からなり、C末端位置の各鎖は、抗体のエフェクター特性を誘導する定常部分からなる。
【0004】
可変部分と、重鎖および軽鎖のCH1 およびCLドメインとの結合によりFab部分が形成され、これは格別の柔軟性をもった領域(転移領域)を介してFc領域(重鎖の定常部分)に結合しており、それによって各Fabがその抗原標的に固定されることを可能にし、一方Fc領域はFcγR受容体などのエフェクター分子に接近可能な状態のままである。
【0005】
Fc領域はCH2 およびCH3 と称される2つの球状ドメインからなる。両方の重鎖はCH3 ドメインにおいて接近して相互作用し、一方CH2 ドメインでは、Asn 297 に結合したラクトサミン型のバイアンテナリー(biantennary)N−グリカンが両鎖のそれぞれに存在することが、両ドメインの分離に寄与する。
【0006】
Fc領域のグリコシル化がIgGの生物学的活性、特に補体に媒介される細胞溶解 (CDC)および抗体依存性の細胞傷害(ADCC)にとって必須であることは多くの研究により示されてきた。このように、指定された突然変異誘発により、またはツニカマイシンの存在下で抗体産生細胞を培養することにより得られるを非グリコシル化IgGは、補体を活性化する能力およびFcγR受容体を固定する能力を失うことが実証された(NoseおよびWigzell, 1983; TaoおよびMorrison, 1989) 。
【0007】
各単糖の役割についてのより具体的研究により、二つに分かれる位置にN-アセチルグルコサミン (GlcNac) 残基が結合するとIgGのADCC活性を高める結果となることが示された (Umana 等, 1999;Davies, 2001) 。他方、ガラクトース残基がAsn 297 に結合するオリゴ糖中に存在するか否かの影響がより論議の的となっている。ガラクトース残基の存在がIgGのエフェクター機能に必須であるとして報告されると (Tsuchiya等, 1989;FurukawaおよびKobata, 1991;Kumpel等, 1994) 、別の著者等はガラクトース残基の欠如によりIgGの機能的活性は変化しなかったことを示した (Boyd等,1995 ;WrightおよびMorrison,1998) 。
【0008】
我々は特許出願WO 01/77181 において、Fc領域のグリコシル化がIgGの生物学的活性、特にCDCおよびADCC活性に必須であることを実証した。我々は、短鎖、低シアリル化、低フコシル化、末端マンノース残基および/または非挿入末端GlcNac残基を特徴とするラクトサミン型のバイアンテナリーN−グリカンがモノクローナル抗体に高いADCC活性を与えるグリカン構造の共通の特徴であることを示している。その後、我々の発見はShields 等 (2002) およびShinkawa等 (2003) の研究によって確認された。
【0009】
本発明において我々は、治療用抗Dポリクローナル抗体 (NATEAD, WinRho) が、そのフコース含量を考慮すると、非常に高いADCC活性を有することを認めた。
この観察は、低フコース含量それ自身が、FcγR受容体、特にFcγRIII を活性化する抗体の能力に影響する唯一の因子ではないことを示唆する。
【0010】
ポリクローナル抗体の全グリコシドのプロフィールを検討することによって、我々は (フコース含量/ガラクトース含量) の比と抗体のエフェクター活性との間に反比例の関係を見出した。
【0011】
事実、抗体が高度にフコシル化されていると、最適のエフェクター活性を得るには高度にガラクトシル化することが必要である。逆に、抗体が僅かしかフコシル化されていないと、最適のエフェクター活性を得るには、このガラクトース含量を、フコース含量/ガラクトース含量の比が0.6 より低く、好ましくは0.5 または0.4 よりも低くなるようにするべきである。
【0012】
従って、我々は実験結果に照らして、高いエフェクター活性が得られる、最適化されたフコース含量/ガラクトース含量の比を有する抗体を製造する方法を完成した。換言すれば、特異的オリゴ糖構造を有し (特にフコースおよびガラクトース残基に関して) 、高いエフェクター活性を付与する新規なモノクローナル抗体を提案する。他方、我々はまた、グリカン構造が細胞傷害活性の活性化を与えない抗体、およびそれを得る方法を提案する。
【発明の開示】
【0013】
従って、第1の面において、本発明は、下記工程を含むことを特徴とする、高いエフェクター活性を有するヒト化またはヒトキメラモノクローナル抗体を製造する方法に関する。
【0014】
a)各種供給源、特に、場合により遺伝的に改変されているか形質転換されている細胞、植物またはヒト以外の動物から得られるモノクローナル抗体を製造し精製すること、
b)該抗体のFc領域のグリコシル化部位が有するグリカン (多糖) 構造のフコース含量およびガラクトース含量を測定すること、および
c)フコース含量/ガラクトース含量の比が0.6 以下、好ましくは0.5 または0.4 以下である抗体を選択すること。
【0015】
「モノクローナル抗体」とは、自然に生じた突然変異を有する少割合の抗体を除き同じ主構造、同じ特異性、および翻訳後修飾、特に分子毎に変化しうるグリコシル化修飾を有するモノクローナル抗体を含む組成物を意味する。本発明の目的にとって、「モノクローナル抗体」または「モノクローナル抗体の組成物」なる表現は同義語である。
【0016】
本発明のモノクローナル抗体は、KohlerおよびMilstein (1975) により報告されたハイブリドーマの製造、エプスタイン・バーウィルス (EBV)によるヒトBリンパ球の不死化などの慣用の方法、またはファージ展示技術、ヒトもしくはトランスジェニック動物抗体 (特にマウス、Xenomouse TM由来) の組み合わせライブラリーの使用などのより最近の方法により製造できる;モノクローナル抗体は分子工学、特に抗体のキメラ化またはヒト化のための分子工学によっても製造できる。本発明の目的にとって、グリカンの分析は、例えばレーザー励起蛍光による高性能キャピラリー電気泳動 (High-Performance Capillary Electrophoresis with Laser-Induced Fluorescence, HPCE-LIF) 、または当業者に既知の任意のその他のグリカン分析方法により行うことができる。
【0017】
本発明の方法により、高いエフェクター活性、より詳しくはADCC型の高い機能活性を有するモノクローナル抗体を得ることができる。このため、エフェクター活性は抗体のFc領域に帰することができる生物学的活性を意味する。これらのエフェクター機能の例には、抗体依存性細胞傷害 (Antibody-Dependent Cell-mediated Cytotoxicity, ADCC) 活性、補体依存性細胞傷害 (Complement-Dependent Cytotoxicity , CDC) 活性、食細胞活性、エンドサイトーシス活性またはサイトカイン分泌の誘導が挙げられるが、これらに限定されない。
【0018】
「高い」エフェクター活性とは、同じ特異性を有するがフコース含量/ガラクトース含量の比が0. 6より大きい抗体のエフェクター活性に比べ、少なくとも20倍、50倍、60倍、70倍、80倍もしくは90倍、好ましくは100 倍まで、または優先的には500 倍のエフェクター活性を意味する。
【0019】
優先的には、フコース含量/ガラクトース含量の比は0.6 〜0.3 、好ましくは0.5 〜0.35の値である。事実、本発明の範囲内で行われた実験を考慮すると、制限的な比が存在する、即ち、比の減少に伴いそれ以下では機能的 (特にADCC) 活性がもはや直線的に増加しないフコース含量/ガラクトース含量の比が存在するようである。従って、これらの限定値の間となるように本発明方法を実施することが特に有利である。
【0020】
例えば、フコース含量が35%〜45%である場合、ガラクトース含量は70〜99%でありうる。フコース含量が20%〜35%である場合、ガラクトース含量は55%〜70%、または60%〜99%である。
【0021】
本発明の目的にとって、0.6 以下の比は、数百分の1、例えば400 〜500 分の1の単位で0.6 より大きい値も意味する。
本発明のある特定の面において、本発明方法により得られる抗体は、抗体の発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変した細胞において製造され、これらの細胞は真核または原核細胞、特に哺乳類、昆虫、植物、細菌または酵母由来の細胞である。
【0022】
有利には、得られる抗体はIgG型のヒト免疫グロブリンである。
より有利には、これらの細胞は、グリコシルトランスフェラーゼ活性を有する少なくとも1種のポリペプチドの発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変した細胞でありうる。
【0023】
優先的には、このグリコシルトランスフェラーゼ活性は、ガラクトシルトランスフェラーゼ活性、特にベータ(1,4)-ガラクトシルトランスフェラーゼまたはベータ(1,3)-ガラクトシルトランスフェラーゼ活性である。
【0024】
本発明の目的にとって、「ガラクトシルトランスフェラーゼ活性を有するポリペプチド」とは、N-グリカンの非還元位置においてガラクトース残基をUDP-ガラクトースからGlcNAc残基へ付加することを触媒しうる任意のポリペプチドを意味する。
【0025】
本発明の目的にとって、「ベータ(1,4)-ガラクトシルトランスフェラーゼ活性を有するポリペプチドの発現を可能にするベクター」とは、二糖型のGal ベータ(1,4)-GlcNacを合成しうるポリペプチドの発現を可能にするポリヌクレオチドを含む任意のベクターを意味し、このポリヌクレオチドはヒト、マウス、ハムスター、ウシ、ヒツジ、ヤギ、ブタ、ウマ、ラット、サル、ウサギ、トリなどの種に由来しうる。例えば、NM 001497 、AB 024434 、NM 003780 、BC 053006 、 XM 242992、 NM 177512などの配列 (これは網羅しているわけではない) が、Genbank などのヌクレオチドおよび/またはタンパク質配列のバンクにおいて利用できる。
【0026】
本発明の目的にとって、「ベータ(1,3)-ガラクトシルトランスフェラーゼ活性を有するポリペプチドの発現を可能にするベクター」とは、二糖型のGal ベータ(1,3)-GlcNacを合成しうるポリペプチドの発現を可能にするポリヌクレオチドを含む任意のベクターを意味し、このポリヌクレオチドはヒト、マウス、ハムスター、ウシ、ヒツジ、ヤギ、ブタ、ウマ、ラット、サル、ウサギ、トリなどの種に由来しうる。特に、ヒト、マウス、ハムスター、ウシ、ヒツジ、ヤギ、ブタ、ウマ、ラット、サル、ウサギ、トリなどの種に由来するベータ(1,3)-ガラクトシルトランスフェラーゼをコードする配列が特に適している。例えば、NM 020981 、AB 084170 、AY 043479(これらに制限されない) などの配列が、Genbank で利用できる。
【0027】
「抗体のFc領域のグリコシル化部位」とは一般的に、Kabat (Kabatデータベース、http://immuno.bme.nwu.edu)の番号付与による両方のAsn297残基を意味するが、本発明はアミノ酸配列が変化した抗体にも関する。
【0028】
本発明のある特定の態様において、細胞はさらに、GDP-フコースの合成および/または輸送に関与する活性、並びに/または抗体のグリコシル化部位のオリゴ糖へのフコースの付加に関与する酵素の活性が低減または欠失している。有利には、GDP-フコースの合成に関与する酵素は、GMD (GDP-D- マンノース 4,6- デヒドラターゼ) 、Fx (GDP-ケト-6- デオキシマンノース 3,5- エピメラーゼ、4-レダクターゼ) またはGFPP (GDP-ベータ-L- フコースピロホスホリラーゼ) であるが、これらは網羅するものではない。有利には、フコースの付加に関与する酵素はフコシルトランスフェラーゼである。GDP-フコースの輸送に関与するタンパク質は、有利にはヒトGDP-フコーストランスポーター1でありうる。
【0029】
本発明のある特定の態様において、工程b)で測定したフコースおよびガラクトースの含量が0.6 より大きい比を与える場合、抗体の機能活性を増加させるためには前記比が0.6 より小さく、好ましくは0.5 より小さく、0.4 より小さくさえなるように、工程c)の前に抗体を脱フコシル化する、および/またはガラクトース残基を抗体に付加することが可能である。この脱フコシル化は、保存培地であってよい抗体含有培地中にフコシダーゼを添加することにより行いうる。ガラクトース残基の付加は、抗体含有培地、または抗体およびUDP-ガラクトースなどの供与基質を含有する溶液にガラクトシルトランスフェラーゼを添加するなどの任意の適宜手段により行いうる。
【0030】
有利には、本発明方法を適用するために用いる細胞は、動物またはヒト細胞株由来であり、これらの株は特に、ラットのミエローマ株 (特にYB2/0 およびIR983F) 、ヒトミエローマ株 (例、Namalwa)、またはヒト起源のその他の任意の細胞 (PERC6, CHO株, 特にCHO-K, CHO-Lec10, CHO-Lec1, CHO Pro-5, CHO dhfr-, CHO Lec13)、または Wil-2, Jurkat, Vero, Molt-4, COS-7, 293-HEK, BHK, K6H6, NSO, SP2/0-Ag 14 およびP3X63Ag8.653から選択されたその他の株から選択される。
【0031】
有利には、抗体は抗-Rhesus D(抗-D) 、抗-CD 、抗−腫瘍、抗−ウイルス、抗-CD20 、または抗-HLA-DR であり、より詳細には以下の表0の抗体由来である。
【0032】
(表0)
抗体の名称 会社 標的 適用
および商品名 Edrecolomab Centocor 抗-Ep-CAM 結腸直腸がん
PANOREX
Rituximab Idec 抗CD20 B細胞リンパ腫
RITUXAN Genentech/ 血小板減少症
Hoffman La Roche 紫斑病
にライセンス
Trastuzumab Genentech 抗HER2 卵巣がん
HERCEPTIN Hoffmfan La Roche/
Immunogen にライセンス
Palivizumab Medimmune RSV
SYNAGIS Abott にライセンス
Alemtuzumab BTG 抗CD52 白血病
CAMPATH Scheringにライセンス
Ibritumomab IDEC 抗CD20 NHL
Tiuxetan Scheringにライセンス
ZEVALIN
Cetuximab Merck/BMS/ 抗EGFR がん
IMC-C225 Imclone
Bevacizumab Genentech/ 抗VEGFR がん
AVASTIN Hoffnan La Roche
Epratuzumab Immunmedics/ 抗CD22 がん:
Amgen 非ホジキンリンパ腫
Hu M195Mab Protein Design Labs 抗CD33 がん
MDX-210 Immuno-Designed ND がん
Molecules
BEC2 Imclone 抗GD3 がん
Mitunomab
Oregovomab Altarex 抗CA125 卵巣がん
OVAREX
Ecromeximab Kyowa-Hakko 抗GD3 悪性黒色腫
KW-2971
ABX-EGF Abgenix EGF がん
MDX010 Medarex 抗CD4R がん
XTL 002 XTL ND 抗ウイルス:HCV Bio-pharmaceuticals
H11SCFV viventia biotech ND がん
4B5 vivantia biotech 抗GD2 がん
XTL 001 XTL ND 抗ウィルス:HBV
Bio-pharmaceutical
MDX-070 MEDAREX 抗-PSMA 前立腺がん TNX-901 TANOX 抗IgE アレルギー
IDEC-114 IDEC プロテインC 非ホジキンリンパ腫
阻害
この一覧は制限するためのものではない。
【0033】
本発明の第2の目的は、免疫学的機能性分子の組成物のエフェクター活性、特にADCC活性を向上させる方法であり、分子組成物中のガラクトース含量を増加させ、および/またはフコース含量を減少させることを含む方法を提供することである。
【0034】
「免疫学的に機能性の分子」とは、免疫学的能力を実証することにより何らかの免疫原との何らかの接触に反応しうる分子を意味する。本来の状態のこれらの分子は良好なエフェクター活性、例えばADCC活性、または低いエフェクター活性を有するかもしれない。そられはグリコシル化部位を含むFc領域を有する。この目的にとって、これらの機能的免疫学的分子は優先的には抗体であり、有利にはモノクローナル抗体またはポリクローナル抗体である。
【0035】
本来の状態の分子は高いフコース含量を有するかもしれない。特に、この場合は、これらの分子または抗体のガラクトース含量を向上させることが有利である。
本発明のある態様において、フコース含量の減少はフコシダーゼの作用による組成物の分子の脱フコシル化によりなされる。この脱フコシル化はα1,6-フコシダーゼにより行うことができる。ウシの腎臓またはCharonia lampas から抽出したフコシダーゼはこの特異性を有する。
【0036】
本発明の別の態様では、組成物の分子のガラクトース含量における増加は、ガラクトシルトランスフェラーゼの作用による組成物のガラクトシル化による。
本発明のある特定の態様では、脱フコシル化のための酵素およびガラクトシル化のための酵素を両方とも作用させる。
【0037】
酵素処理に代わる方法として、免疫学的機能性分子の組成物を、低フコシル化抗体および/または高ガラクトシル化抗体を多くするレクチン上での一連のクロマトグラフィーにより精製しうる。
【0038】
一例として、有利には抗体である免疫学的機能性分子の組成物を含む溶液を、HPLCシステムに連結したレクチンカラム (例、LA-LCAまたはLA-AALカラム、Shimazu Corporation)上を通過させる。溶液を非吸着画分と吸着画分に分ける。非吸着画分および吸着画分のグリカン分析を行う:酵素作用によりタンパク質部分から開裂したオリゴ糖をAPTSで標識し、HPCE-LIFで分離して定量する。ピークの面積を計算する:フコースを含まないグリカンを有する抗体はそれにより分離され選択される。選択された画分を次いで、フェニル-5PW型 (Toso Corporation製) の疎水性カラムまたは第2レクチンカラム (LA-RCA 120またはLA-WGA、Seikagaku America)上を通過させる (非吸着画分または吸着画分から流出しうる) 。フコース含量/ガラクトース含量の比が0.6 以下の画分をそれによって正確に選択できる。
【0039】
本発明の第3の目的は、抗体分子をコードする少なくとも1つのベクターが導入された細胞、優先的にはYB2/0 細胞株由来の細胞であり、この細胞は、Fc領域のグリコシル化部位由来のオリゴ糖のフコース含量/ガラクトース含量の比が0.6 以下であるモノクローナル抗体を産生する。優先的にはこの比は0.5 より小さく、0.4 よりさえも小さい。本発明の好ましい面において、この比は0.6 〜0.3 である。
【0040】
本発明の好ましい面において、この細胞は、ガラクトシルトランスフェラーゼ、特にベータ(1,4)-ガラクトシルトランスフェラーゼまたはベータ(1,3)-ガラクトシルトランスフェラーゼをコードする発現ベクターでトランスフェクされる。有利には、この細胞は組換えガラクトシルトランスフェラーゼを発現または過発現する。
【0041】
YB2/0 株は本来、ベータ(1,4) およびベータ(1,3) のガラクトシルトランスフェラーゼファミリーを発現する。さらに、この細胞株は、低フコース含量の抗体を産生することが知られている (WO 01/77181, LFB) 。しかし、本発明による細胞は、非改変株により産生される抗体に比べて、改変された細胞により産生される抗体のフコース含量/ガラクトース含量の比を変化させる効果を有する、ガラクトシルトランスフェラーゼを過発現するという利点を有する。従って、抗体はもともとフコシル化が少ないので、そのガラクトース含量の向上はさらにそのフコース含量/ガラクトース含量の比を低くし、これはそのADCC活性をさらに最適化する効果がある。
【0042】
有利には、ガラクトシルトランスフェラーゼは、ヒト、マウス、ハムスター、ウシ、ヒツジ、ヤギ、ブタ、ウマ、ラット、サル、ウサギ、またはトリ (これに制限されない) 由来の配列によりコードされる。より詳しくは、コード配列はNM 001497, AB 024434, NM 003780, BC 053006, XM 242992 またはNM 177512 配列である。
【0043】
従って、本発明は、前記ベクターの発現を可能にする培地および条件下で前述の細胞を培養することを含む、Fc領域のグリコシル化部位に存在するグリカン構造が0.6 以下、優先的には0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有するモノクローナル抗体を製造する方法に関する。
【0044】
あるいは、上に規定したような抗体組成物は、フコース、ガラクトースまたはそれらを含むオリゴ糖を特異的に捕捉しうる任意の分子を用いた1または2以上のクロマトグラフィー工程により製造することもできる。前述のようにレクチン上での分離なども使用できる。
【0045】
また、本発明は、先に記載した方法により得られるか、または既述の方法で得られた、高いエフェクター活性をもった治療用抗体に関し、これらの抗体は、Fc領域のグリコシル化部位に、0.6 より小さい、優先的には0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有するグリカン構造をもつことを特徴とする。
【0046】
より有利には、これらは前述の方法で得ることができる治療用モノクローナル抗体であり、この抗体は増強されたADCC活性を有し、一例としてはポリクローナル抗体と等しいかそれより大きいADCC活性を有するモノクローナル抗-Dである。この増強されたADCC活性は、CHO DG44またはDxB11 株において発現されたポリクローナルまたはモノクローナル (同じ特異性を有する) 治療用抗体のものと少なくとも等しいか、好ましくはより大きい。
【0047】
有利には、これらはIgGであり、例えば、キメラ、ヒト化もしくはヒトのIgG1もしくはIgG3、またはヒトFc領域を有するIgGである。好ましくは、これらの抗体はヒトIgG、またはヒトFc領域を有する任意のキメラ分子である。
【0048】
同じ観点において、本発明は上述の抗体を含む薬剤組成物に関する。
また、本発明は、Fc領域のグリコシル化部位に存在するグリカン構造が0.6 より小さい、優先的には0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有するモノクローナルまたはポリクローナル抗体を少なくとも50%、好ましくは60%、70%、80%、または90%もしくは99%含む薬剤組成物に関する。好ましくは、この比は0.6 〜0.3 、特に0.5 〜0.35である。
【0049】
本発明の組成物は好ましくは、非遍在の通常抗原、特にRh因子 (例、ヒト赤血球細胞のRh因子(D))、またはヒトに対する病原性細胞もしくは病原性生物の抗原、特にがん細胞の抗原、に対する抗体を含む。抗体はさらに好ましくはIgGである。
【0050】
本発明の別の目的は、同種免疫、特に新生児溶血性疾患の治療用薬剤を製造するための本発明抗体の使用に関する。
本発明の別の目的は、自己免疫疾患、がん、および病原体による感染症の治療、特にセザリー症候群、固形がんから選択された免疫応答をのがれる疾患の治療、特に抗原性標的が弱く発現したもの、特に、乳がん、ポリ塩化ビフェニルに暴露されたヒトを対象とする環境に関連する病状、感染症 (特に結核) 、慢性疲労症候群(CFS) 、寄生虫感染症 (例、住血吸虫) およびウイルス感染症、の治療用薬剤を製造するための本発明抗体の使用に関する。
【0051】
さらに、本発明の抗体は、黒色腫などの陽性クラスII HLA細胞のがん、BおよびT細胞の急性リンパ性白血病、急性および慢性骨髄性白血病、バーキットリンパ腫、ホジキンリンパ腫、T細胞リンパ腫および非ホジキンリンパ腫の治療用薬剤を製造するのに使用できる。
【0052】
本発明の抗体は表0に示された抗体より選択することができる。
有利には、抗体は抗-HLA-DR または抗-CD20 である。
本発明の別の面において、本発明の抗体は、免疫システムの天然のエフェクター細胞によりIL-1α、IL-1β、IL-2、IL-3、IL-4、IL-5、IL-6、IL-12 、IL-18 、IL-21 、TGF β1 、TGF β2 、TNF α、TNF β、INF γ、およびIP10から選ばれた少なくとも1種のサイトカインの発現を誘導するための薬剤を製造するのに使用され、この薬剤はがんおよびウイルス、細菌もしくは寄生虫感染症を治療するのに特に有用である。
【0053】
本発明の別の特定の面では、本発明の抗体は、CD16多型の1つ、特にV/F158またはF/F158を有する患者、特に現在利用可能な抗体では治療できない症状の患者、または望ましくない二次的作用を受けている患者、の治療のための薬剤の製造に使用される。
【0054】
さらに別の面では、本発明はまた、下記工程を含むことを特徴とする、低いエフェクター活性、特に低いADCC型の機能活性を有する、キメラ、ヒト化またはヒトモノクローナル抗体を製造する方法に関する。
【0055】
a)各種供給源、特に、場合により遺伝的に改変されているか形質転換されている細胞、植物またはヒト以外の動物から得られるモノクローナル抗体を製造し精製すること、
b)該抗体のFc領域のグリコシル化部位に存在するグリカン (多糖) 構造のフコース含量およびガラクトース含量を測定すること、および
c)フコース含量/ガラクトース含量の比が0.6 より大きい、好ましくは1.2 より大きい抗体を選択すること。
【0056】
モノクローナル抗体のエフェクター活性の定義は前記した通りである。
さらに、「低いエフェクター活性」とは、同じ特異性を有するがフコース含量/ガラクトース含量の比が0. 6より小さい抗体のエフェクター活性、特にADCC型の機能活性に比べ、少なくとも20倍、50倍、60倍、70倍、80倍もしくは90倍、好ましくは100 倍まで、または優先的には500 倍低いエフェクター活性を意味する。
【0057】
従って、相補的な面において、本発明は、Fc領域のグリコシル化部位(Asn 297) が1.2 より大きいフコース含量/ガラクトース含量を有することを特徴とする、低いADCC活性を有する抗体、およびこれを含む組成物に関する。
【0058】
これらの抗体は、自己免疫疾患、特に免疫性血小板減少性紫斑病 (PTI)、同種免疫、移植片拒絶、アレルギー、喘息、皮膚炎、蕁麻疹、紅班および炎症性疾患の治療および/または予防のための薬剤の製造に有用である。
【0059】
本発明のある特定の面において、抗体は、この抗体の発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変した細胞において産生され、この細胞は真核細胞でも原核細胞でもよく、特に哺乳動物、昆虫、植物、細菌または酵母由来の細胞である。
【0060】
本発明のある態様において、細胞は、グリコシルトランスフェラーゼ活性、好ましくはフコシルトランスフェラーゼ活性、特にα1.6-フコシルトランスフェラーゼ活性を有する少なくとも1つのポリペプチドの発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変される。
【0061】
本発明の別の態様では、細胞は、UDP-ガラクトースの合成および/または輸送に関する活性、並びに/または抗体のグリコシル化部位のオリゴ糖へガラクトースを付加することに関与する酵素の活性が減少または欠失している。有利にはガラクトースの付加に関与するこの酵素は、β1,4-ガラクトシルトランスフェラーゼである。
【0062】
有利には、細胞はグリコシルトランスフェラーゼ活性、特にグリコシルトランスフェラーゼ活性と、UDP-ガラクトースの合成および/または輸送に関する活性、並びに/または抗体のグリコシル化部位のオリゴ糖へガラクトースを付加することに関与する酵素の活性が減少または欠失している。
【0063】
本発明のある態様において、工程b)で測定した比が0.6 より小さい場合、フコース含量/ガラクトース含量の比が0.6 より大きくなるように、工程c)の前にフコシル化を行う、および/またはガラクトース残基を抗体より除去することが可能である。有利には、脱ガラクトシル化は、抗体含有培地中にガラクトシダーゼを添加することにより行う。有利には、フコース残基の付加は、抗体含有培地にフコシルトランスフェラーゼを添加することにより行う。
【0064】
より有利には、抗体はIgG型のヒト免疫グロブリンである。有利には、抗体は、ヒト血球の分化のマーカーであるCDに対するか、またはバイオテロの場合には特に危険であるとして挙げられている病原体もしくはその毒素、特にバチルス・アントラシス (Bacillus anthracis) 、クロストリジウム・ボツリウム (Clostridium botulium) 、エルシニア・ペスティス (Yersinia pestis)、バリオラ・マジョル (Variola major)、フランシセラ・ツラレンシス(Francisella tularensis)、フィロウイルス(filovirus) 、アレナウイルス (arenavirus) 、ブルセラ・スピーシーズ (Brucella species) 、クロストリジウム・ペルフリンゲンス (Clostridium perfringens)、サルモネラ (Salmonellla)、大腸菌 (E.coli) 、赤痢菌 (Shigella) 、コクシエラ・バーネッティイ (Coxiella burnetii)、リシン毒素、リケッチア (Rickettsia) 、ウイルス性脳炎ウイルス、ビブリオコレラ (Vibrio cholerae)またはハンタウイルス、に対するものである。
【0065】
本発明の別の目的は、組成物のフコース含量の増加および/またはガラクトース含量の低下を含む、免疫機能性分子の組成物の活性を低下させる方法にも関する。
有利には、免疫機能性分子はモノクローナルまたはポリクローナル抗体である。
【0066】
ある特定の面において、フコース含量の増加は、フコシルトランスフェラーゼ、好ましくはα1,6-フコシルトランスフェラーゼの作用による前記組成物のフコシル化による。
別の特定の面において、前記組成物のガラクトース含量の低下は、ガラクトシダーゼ、好ましくは1または2以上のβ- ガラクトシダーゼの作用による組成物の脱ガラクトシル化による。
【0067】
より有利には、この組成物のフコシル化および脱ガラクトシル化の両方を行う。
従って、本発明の目的は、上記の本発明方法により得ることができる抗体の組成物、またはこれらの方法のいずれかにより得られた抗体組成物に関する。
【0068】
本発明の別の目的は、自己免疫疾患、特にPTI 、同種免疫、移植片拒絶、アレルギー、喘息、皮膚炎、蕁麻疹、紅班または炎症性疾患 (これらは網羅するものではない) を治療および/または予防する薬剤を製造するための、この抗体組成物の使用である。
【0069】
最後に、本発明は、抗体のFc領域のグリコシル化部位由来のオリゴ糖におけるフコース含量/ガラクトース含量の比を調節することを含む、免疫機能性分子の組成物の活性を制御する方法に関する。
【0070】
本発明のその他の面および利点は、ガラクトースによる「フコース効果」調節を示す実施例に記載されるであろう。これは非制限的と考えられるべきであり、本発明の範囲を制限するものではない。
【実施例1】
【0071】
一連の抗-Rh(D)抗体のフコース含量/ガラクトース含量の比とADCC活性との間の相関関係
Rhesus (Rh)(D)抗原に対する各種モノクローナルおよびポリクローナル抗体のフコース含量、次いでガラクトース含量を測定した。これより両者の間の比を算出し、各抗体についてADCC活性を測定した。
1.抗-Rh(D)モノクローナル抗体の産生
モノクローナル抗体は、Rh(D) 抗原を有する赤血球で免疫した、陰性のRh(D) ヒトドナーからのBリンパ球のEBV によるトランスフォーメーションに由来する。2つのクローンをこのトランスフォーメーションから選択した:
1)クローンの1つをK6H6-B5 ヒト/マウスヘテロミエローマと融合させ;この融合物よりクローンHH01を選択した。
【0072】
2)抗体の重鎖および軽鎖を発現させるためのベクターを作製するために、その他のクローンから抗-Rh(D)抗体をコードする RNAを抽出した。
この発現ベクターを、一方でEMAB1 、EMAB2 、EMAB3 およびEMAB4 抗体を産生させるYB2/0 細胞株、他方で次のCHQ 株をトランスフェクトするのに使用した:抗-D1 、抗-D2 および抗-D3 抗体をそれぞれ合成するDG44、K1およびLec13 。
2.ポリクローナル抗体の精製
抗-Rh(D)ポリクローナル抗体を、治療用製剤のWinRho (Cangene)から、Rh(D+)赤血球上での陽性選択により、次いでRhD(-)赤血球上での陰性選択により免疫精製し、最後に、セファロース−プロティンAゲルを用いたアフィニティークロマトグラフィー工程により、一方で赤血球上での免疫精製中に回収された汚染物質を除去し、他方でIgG1をIgG3から分離した。というのはIgG1のみが以下の試験において使用されるためである。
3.HPCE-LIFによるグリカン分析
抗-Rh(D)モノクローナルおよびポリクローナル抗体をセファデックスG-25 (HiTrap Desalting, Amersham Biosciences) カラム上で脱塩し、蒸発により乾燥させ、50mMのβ- メルカプトエタノールの存在下で加水分解 PNGアーゼF (Glyko) 用の緩衝液中に再懸濁する。37℃で16時間の温置後、無水エタノールを添加することによりタンパク質部分を沈殿させ、そしてN-グリカンを含有する上清を蒸発により乾燥させる。そうして得られたオリゴ糖は蛍光色素APTS (1-アミノピレン-3,6,8- トリスルホネート) で直接標識するか、またはAPTSで標識する前に特異的エキソグルコシダーゼの作用を受けさせる。次に、標識オリゴ糖をN-CHO キャピラリー上に注入し、レーザー励起蛍光の検出によるキャピラリー電気泳動 (HPCE-LIF) により分離、定量する。
【0073】
電気泳動図上にフコシル化されているかまたはされていない五糖 [GlcNac2-Man3] に対応する2つのピークが得られるように、分離されたフコシル化形態を、より詳しくはノイラミニダーゼ、β−ガラクトシダーゼおよびN-アセチルエキソサミニダーゼを同時に作用させた後添加することによりフコース含量の評価を行う。
【0074】
フコース含量 (%で表す) は以下の式を用いて算出される:
フコシル化[GlcNac2-Man3]×100
フコース含量=───────────────
[GlcNac2-Man3+フコシル化GlcNac2-Man3]
ガラクトース含量 (%で表す) は、末端位置にガラクトースを含有するオリゴ糖形態の割合を加えることにより算出される。使用する式は以下の通りである:
ガラクトース含量=[(G1+G1B +G1F +G1FB) +2×(G2 +G2F +G2B +G2FB)]
フコース含量/ガラクトース含量の比は、フコース含量をガラクトース含量で割ることにより得られ、これらの含量は上記のようにして算出される。
4.抗体の機能活性:ADCC
ADCC (抗体依存性細胞傷害) 法により、エフェクター細胞 (単核細胞またはリンパ球) の存在下でRh(D+)赤血球の溶解を引き起こす抗体の能力を評価することができる。
【0075】
簡単に述べると、赤血球細胞RhD(+)濃縮物の赤血球をパパインで処理し (1mg/ml 、37℃で10分) 、0.9 %NaClで洗浄する。エフェクター細胞を少なくとも3つの軟膜のプールからFicoll (Amersham) での遠心分離、次いで25%FCS の存在下での接着工程によりにより分離し9のオーダーのリンパ球/単球の比とする。マイクロタイトレーションプレート (96ウェル) において1ウェル当たり、精製抗-Rh(D)抗体の希釈物 (9.3 から150ng/ml) 100 μl、パパイン処理したRh(D+)赤血球25μl (即ち、1.106 ) 、エフェクター細胞25μl (即ち、2.106 ) および通常濃度2および10mg/ml の多価IgG (Tegeline, LFB)50μlを入れる。希釈物はウシ胎児血清 (FCS)の0.25% IMDM 中で作製する。37℃で1時間温置した後、プレートを遠心分離にかけ、次いで上清中の放出ヘモグロビンを発色性基質、2,7-ジアミノフルオレン (DAF)の存在下でそのペルオキシダーゼ活性を介して測定する。結果は溶解率 (%) として表され、100 %はNH4Cl 中の赤血球の全溶解に相当し (100 %対照) 、0%は抗体の存在しない反応混合物に相当する (0%対照) 。特異的溶解は下記式により%で算出する:
((試料のOD−0%対照のOD) ×100)
────────────────=ADCC%
(100%対照のOD−0%対照のOD)
抗-Rh(D)IgG1のFc領域のグリコシル化部位が有するオリゴ糖のHPCE-LIF分析を行った。
【0076】
【表1】

【0077】
* 免疫精製したポリクローナル抗-D
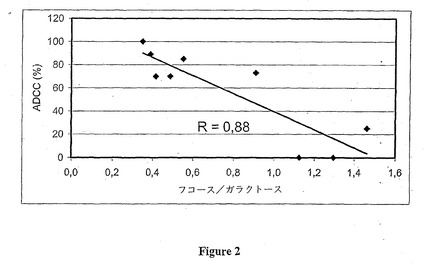
表1に含まれる、 [フコース含量/ガラクトース含量] の比およびADCC率の値は図2においてそれぞれ横座標および縦座標に記入されている。プロットされた一次回帰線の相関係数は0.92に等しい。
【0078】
このように、モノクローナルおよびポリクローナル抗-Rh(D)抗体の [フコース含量/ガラクトース含量] の比とADCC活性の間には相関関係がある。顕著なADCC活性を有する抗体はフコース含量/ガラクトース含量の比が0.6 より小さい。
【実施例2】
【0079】
脱ガラクトシル化前および後の抗-Rh(D)ポリクローナル抗体のADCC活性の比較
1.抗-Rh(D)ポリクローナル抗体の脱ガラクトシル化
免疫精製したポリクローナル抗体を加水分解緩衝液 (50mM 酢酸ナトリウム、pH5.5 、塩化カルシウム4mM含有) で透析する。抗体を、コレラ菌 (Vibrio cholerae)由来のノイラミニダーゼ (EC 3.2.1.18) (Calbiochem) 5mUおよびE.coli産生のβ−ガラクトシダーゼ (EC 3.2.1.23) (Roche)9mMの存在下で温置することにより脱シアリル化および脱ガラクトシル化する。「対照」と記載された対照は、上記のように、しかしノイラミニダーゼおよびβ−ガラクトシダーゼの不在下で処理した同じ抗体調製物からなる。37℃で25時間温置後、抗体を4℃で保存する。
【0080】
この実施例で製造された抗体を2つの部分に分ける:1つはグリカン分析に使用し、他はADCC活性の測定に取っておく。
2.HPCE-LIFによるグリカン分析
操作は、塩、また生成物に存在するかもしれない遊離の単糖類を除去するための、セファデックス-G25カラム上での、脱ガラクトシル化抗-Rh(D)ポリクローナル抗体の脱塩からなる。抗体の変性および還元の後、グリカンをエンドグリコシダーゼ、PNG アーゼF (Glyko)の作用を介して放出させる。37℃で16時間の温置後、タンパク質部分を無水エタノールの添加により沈殿させ、N-グリカンを含む上清を蒸発により乾燥させる。こうして得られたオリゴ糖中に含まれるガラクトースおよびフコースの量を評価するために、APTSでの標識前に、試料をそれぞれシアリダーゼおよびフコシダーゼ、またはシアリダーゼ、β−ガラクトシダーゼおよびN-アセチルヘキソサミニダーゼの同時の作用を受けさせる。次いで、標識オリゴ糖をレーザー励起蛍光の検出によるキャピラリー電気泳動 (HPCE-LIF) により分離、定量する。
3.ADCC活性の測定
実施例1に記載の方法に従い、β−ガラクトシダーゼ処理の前および後にポリクローナル抗体のADCC活性を測定する。
【0081】
こうして、β−ガラクトシダーゼ処理の後、抗-Rh(D)ポリクローナル抗体抗のFc領域のグリカンは残存ガラクトース含量が17.7%で、フコース含量が68.5%となる。従って、脱ガラクトシル化ポリクローナル抗体のフコース含量/ガラクトース含量の比は3.8 である。
【0082】
この試料中のテゲリン (Tegeline) などの多価IgGのADCC試験における存在は、高親和性受容体 (即ち、FcγRIまたはCD64) を阻害し、それによりRh(D+)赤血球溶解がエフェクター細胞上に存在するFcγRIII受容体と抗-Rh(D)抗体の相互作用により特異的となるようにする。
【0083】
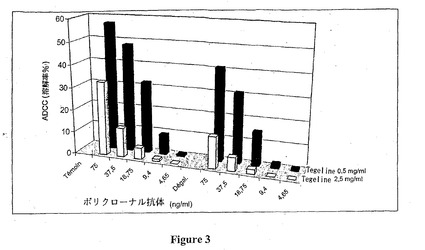
図3に示す結果は、抗-Rh(D)ポリクローナル抗体のADCC活性は一方で用量依存性であることを、そして反応混合物中の多価IgGの量が増加するとポリクローナル抗体の溶解性を減少させることを示している。さらに、脱ガラクトシル化ポリクローナル抗体は対照抗体に比べて低いADCC活性を有する。
【0084】
【表2】
【0085】
対照抗体、即ちノイラミニダーゼおよびβ−ガラクトシダーゼの不在下で同じように温置した抗体と比べた、脱ガラクトシル化抗-Rh(D)抗体のADCC活性の割合 (%) を表2に示す。
【0086】
このように、対照抗体に比べた、脱ガラクトシル化ポリクローナル抗体のADCC活性の減少は、抗体の量が少ない場合より有意である。さらに、脱ガラクトシル化ポリクローナル抗体の活性の減少は2.5mg/ml濃度の多価IgGの存在下でより有意である。
【実施例3】
【0087】
脱ガラクトシル化抗-Rh(D)モノクローナル抗体により生じるCD16受容体の活性化の測定
1.抗-Rh(D)モノクローナル抗体の脱ガラクトシル化
抗体を加水分解緩衝液 (50mM 酢酸ナトリウム、pH5.5 、塩化カルシウム4mM含有) で透析する。抗体を、コレラ菌 (Vibrio cholerae)由来のノイラミニダーゼ (EC 3.2.1.18) (Calbiochem) 5mUおよびE.coli産生のβ−ガラクトシダーゼ (EC 3.2.1.23) (Roche)9mUの存在下で温置することにより脱シアリル化および脱ガラクトシル化する。「対照」と記載された対照は、上記のように、しかしノイラミニダーゼおよびβ−ガラクトシダーゼの不在下で処理した同じ抗体調製物からなる。37℃で24時間温置後、抗体を4℃で保存する。
【0088】
この実施例で製造された抗体を2つの部分に分ける:1つはグリカン分析に使用し、他は機能活性の測定に取っておく。
2.CD16受容体活性化の測定
Jurkat CD16 細胞の活性化試験は、Fab を標的細胞上に存在する抗原に結合させた後、CD16 (FcγRIIIA)上に抗体のFcを固定することにより生じるインタロイキン-2 (IL-2) の分泌を測定する。Jurkat CD16 細胞により分泌されるIL-2のレベルはCD受容体の活性化に比例する。
【0089】
抗体希釈物50μl、6.105/mlの赤血球懸濁液50μl、1.106 /ml のJurkat CD16 細胞懸濁液50μlおよび40ng/ml のPMA 溶液50μlを順次96−ウェルマイクロタイトレーションプレートに入れる。希釈はすべて、5%FCS を含有するIMDE培地で行う。
【0090】
7%CO2 、37℃において16時間温置した後、マイクロタイトレーションプレートを遠心分離し、上清に含まれるIL-2の量を市販のキット (Duoset, R&D)で測定する。分泌されたIL-2の濃度をpg/ml で表す。
【0091】
結果はCD16活性化率 (%) として表し、対照モノクローナル抗体の存在下の分泌IL-2濃度は100 %に等しいと考えられる。
実施例2に記載したHPCE-LIFにより行ったグリカン分析の結果を表3にまとめる。
【0092】
【表3】
【0093】
このように、EMAB2 モノクローナル抗体はすべて脱ガラクトシル化され、HH01抗体は依然として17.3%のモノガラクトシル化形態を含有しているようである。従って、β−ガラクトシダーゼの作用の後、EMAB2 とHH01抗体のフコース含量/ガラクトース含量の比は0.6 よりずっと大きくなる。
【0094】
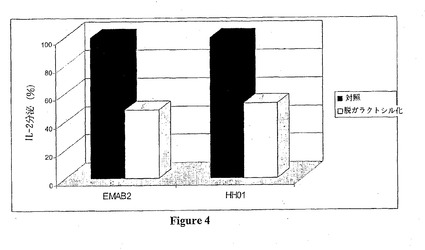
脱ガラクトシル化抗-Rh(D)モノクローナル抗体は、対照抗体に比べてCD16活性化が非常に低い (図4) 。このように、EMAB2 およびHH01モノクローナル抗体は、それぞれ52および47%のCD16活性化誘導能の低下を示す。
【実施例4】
【0095】
ガラクトシル化抗-Rh(D)モノクローナル抗体により生じるCD16活性化の測定
1.抗体のガラクトシル化
抗体を50mMヘペス緩衝液 (pH7.20) で透析する。反応混合物は、MnCl2 10mM、UDP-ガラクトース20mM、およびウシベータ1.4-ガラクトシルトランスフェラーゼク(Calbiochem)40mUを添加したモノクローナル抗体溶液からなる。37℃で24時間温置した後、試験管を使用まで4℃で保存する。
【0096】
対照は、反応培地中でUDP-Gal の不在を除き同じ条件下で温置した同種の抗体からなる。
この実施例で製造された抗体を2つの部分に分ける:1つはグリカン分析に使用し、他はADCC活性の測定に取っておく。
2.レクチンELISA によるガラクトース定量
レクチンはその認識特異性により生物学および医学の多くの用途、特にELISA 法によるグリカン分析に使用されてきた。β1,4 に結合したガラクトースを認識するレクチンRCA1を、抗体のN-グリカン中に存在するガラクトースを定量するのに使用した。
【0097】
モノクローナル抗体をマイクロタイトレーションプレートのウェルに固定する。Fc領域のN-グリカンを利用できるように、IgG分子を100 ℃で20分加熱して変性させた後、ウェルを室温、ビオチニル化RCA1溶液 (Vector) の存在下穏やかに攪拌しながら2時間温置する。未反応レクチンを除去するための洗浄の後、ストレプトアビジンペルオキシダーゼを各ウェルに添加し、1時間温置し、そして固定化レクチンをO-フェニレンジアミンの添加後 492nmで測定する。並行して、マイクロタイトレーションプレートのウェル中の固定化抗体の量を、ペルオキシダーゼで標識したヒト抗−IgG抗体により測定する。
【0098】
次いで、固定化レクチンの量をマイクロタイトレーションウェル中の固定化抗体の量により補正する。
3.CD16受容体活性化の測定
ガラクトシル化モノクローナル抗体のCD16受容体の活性化の測定に使用する操作条件は上に記載したものと同じである。
【0099】
この実施例に記載されるモノクローナル抗体は、同じ主要配列を有し、YB2/0 細胞により産生された抗-Rh(D)抗体である。それらは、EMAB2 については25%、EMAB3 については53%のα1,6-フコシル化率に関して、その機能活性において異なる。
【0100】
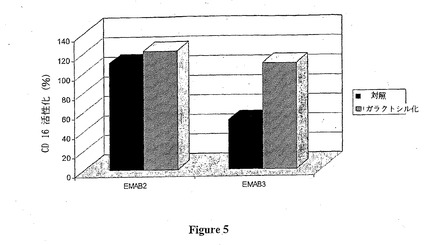
ベータ1,4-ガラクトシルトランスフェラーゼのin vitro作用の後、EMAB2 およびEMAB3 モノクローナル抗体により誘導されるCD16活性化はそれぞれ10および54%向上する (図5) 。このように、本来非常に良好なエフェクター活性を有するEMAB2 抗体のガラクトシル化の増加は、CD16活性化をわずかに向上させるだけであるが、高度にフコシル化されているEMAB3 抗体のガラクトシル化における増加は、CD16活性の非常に顕著な向上を示す。
【実施例5】
【0101】
EMAB2 抗-Rh(D)モノクローナル抗体により感作された赤血球のクリアランスの検討
EMAB2 抗-Rh(D)モノクローナル抗体により感作された赤血球のクリアランスを、診療所で使用されている抗-Rh(D)ポリクローナル抗体の治療用製剤であるRhophylac TMにより感作された赤血球のものと比較するために、EMAB2 抗-Rh(D)モノクローナル抗体を臨床試験第I相にて評価した。
【0102】
健康なボランティアの赤血球をクロム51 (51Cr) でex vivo にて標識し、感作する、即ち、ボランティアに再注入する前に抗原性部位の25%飽和レベルを得るために、抗-Rh(D)抗体であるEMAB2 またはRhophylac TMの存在下で温置する。
【0103】
51Crで標識した赤血球の血流中での消失を、標識した感作赤血球を注入して3、15、30分および1、2、4、6、8、10、24、48、72、96時間後に採取した血液サンプルにおいてガンマカウンターで放射能を測定することにより追跡した。赤血球の注入3分後に採取した血液サンプルは100 %の赤血球の生存を示す。
【0104】
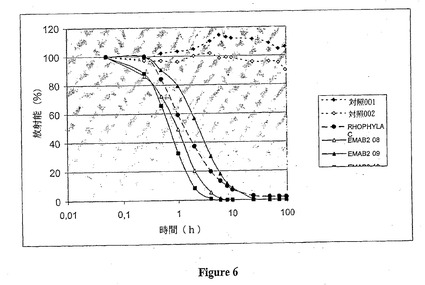
図6に示す結果は、抗体による放射標識赤血球の感作を行わない場合、100 時間より長い時間にわたり測定した放射能の減少は20%より少ないことを示している。しかし、赤血球がポリクローナル抗体治療用製剤またはEMAB2 モノクローナル抗体で感作される場合、血液の放射能は急激に減少し;注入後10時間は注入された放射能の10%より少ない状態である。このように、EMAB2 モノクローナル抗体で感作された赤血球の消失曲線は、Rhophylac TMポリクローナル抗体の治療用製剤により感作された赤血球のものと似たプロフィールを有する。
【0105】
フコース含量/ガラクトース含量が0.4 であるEMAB2 モノクローナル抗体は、予備感作Rh(D+)赤血球のクリアランスに関して、少なくともポリクローナル抗体の治療用製剤のものに匹敵するin vivo 活性を有する。同じ条件であるが、MonoD と称される別のモノクローナル抗体を用いて行った臨床検討では、非常に異なる結果を与えた;膜抗原性部位の25%飽和では、MonoD により生じるクリアランスは部分的にすぎない。MonoD 抗体のグリカン分析により、フコース含量が80%でガラクトース含量が86%である、即ち、比が0.93であることが明らかになった。
【0106】
従って、これらの臨床結果を比較すると、フコース含量/ガラクトース含量の比が0.6 以下である抗-Dモノクローナル抗体は、その比が1に近い抗体よりも赤血球のクリアラン
スに対してより高い効果を有することが分かる。
【実施例6】
【0107】
CHO およびYB2/0 細胞株により発現された抗-HLA DR モノクローナル抗体のガラクトース含量の改変
1.抗-HLA DR モノクローナル抗体の製造
1.1 発現ベクターの作製
これらの検討で用いる抗-HLA DR 抗体は、Lym-1 ハイブリドーマ (ATCC Hb-8621)により発現されるIgG2a アイソタイプマウス抗体のキメラ化に由来する。
【0108】
マウス抗体を産生するハイブリドーマから抽出した RNAをcDNAに変換した。マウスVK領域をK-Lym-Not1およびK-Lym-Dra3プライマーにより増幅し、次いで、予めNot1およびDra3で消化し、ヒト抗-D抗体のCK配列およびDHFR選択遺伝子を含有するキメラ化ベクターCK-Hu にクローン化した。
【0109】
マウスVH領域をH-Lym-Not1およびH-Lym-Apa1プライマーにより増幅し、次いで、予めNot1およびApa1で消化し、ヒト抗-D抗体のG1配列およびNEO 選択遺伝子を含有するキメラ化ベクターG1-Hu にクローン化した。
【0110】
hEF-1aプロモーターおよび非コーディングエクソン1および第1イントロンを含有するhEF-1a遺伝子の5'UTR 領域を市販のプラスミドpEF/Bsd (Invitrogen)からNhe 1 およびAcc 65 Iの二重消化により分離した。並行して、上記発現ベクターに存在するRSV プロモーターをBgl IIおよびSpe I の二重消化により除去し、次いでNhe I-Acc65 I 断片で置換した。
【0111】
1.2 安定な産生株の獲得
抗-HLA DR キメラ抗体の軽鎖および重鎖をコードする発現ベクターpEF-Lym-dhfr-K-10 およびpEF-Lym-neo-H-12を、エレクトロポレーションによりCHO-DXB11 (ATCC No. CRL-11397)およびYB2/0 (ATCC No. CRL-1662) 株をそれぞれコトランスフェクトするために使用した。
【0112】
トランスフェクション後、培養細胞を、一方では培地のヌクレオシドへの欠失および他方ではG418k 添加からなる二重選択圧にかける。この二重選択圧に耐性の形質転換体を次いで、希釈を制限することよりクローン化した。
【0113】
2つの選択されたクローンは、YB2/0 発現細胞株についてはYB2/0-DR-4B7であり、CHO-DXB11 発現細胞株についてはDXB11-DR-22A10である。
1.3 抗-HLA DR キメラ抗体の作製および精製
クローンYB2/0-DR-4B7を、EM-SF1.1培地、インスリン (1μg/ml)、クエン酸鉄 (50μg/ml)、HEPES (4mg/ml)およびPluronic F68 (0.5mg/ml) を添加したEMS 基本培地で10リットルの細胞培養バイオリアクター (Biolafitte) 中で増殖させた。
【0114】
クローンDXB11-DR-22A10を、2%ヒポキサンチンを添加したCHO SFM4ユーティリティ培地 (Perbio) で10リットルの細胞培養バイオリアクター (Biolafitte) 中で増殖させた。
細胞の生存率が50%より低くなったら、細胞を除去するために培地を集め、遠心分離し、上清に含まれるキメラ抗体をセファロース−プロティインAでのアフィニティークロマトグラフィーにより精製する。
2.脱ガラクトシル化
抗-HLA DR キメラ抗体を50mM 酢酸ナトリウム緩衝液 (pH5.5 、CaCl2 4mM含有) で透析した。抗体を、コレラ菌 (Vibrio cholerae)由来のノイラミニダーゼ (EC 3.2.1.18) (Calbiochem) 5mUおよびE.coli産生のβ−ガラクトシダーゼ (EC 3.2.1.23) (Roche)9mUの存在下で温置することにより脱ガラクトシル化した。対照は、上記のように、しかしノイラミニダーゼおよびβ−ガラクトシダーゼの不在下で処理した同じ抗体調製物からなる。抗体を37℃で25時間温置後、4℃で保存する。
【0115】
この実施例で製造された抗体を2つの部分に分ける:1つはグリカン分析に使用し、他は機能活性の測定に取っておく。
2.CD16活性化の測定
Raji細胞株を、その表面にHLA-DR主要組織適合遺伝子複合体の抗原決定基を有する標的として用いる。
【0116】
抗体希釈物50μl、6.105/mlのRaji細胞懸濁液50μl、1.106/mlのJurkat CD16 細胞懸濁液50μlおよび40ng/ml のPMA 溶液50μlを順次96−ウェルマイクロタイトレーションプレートに入れた。希釈はすべて、5%FCS を含有するEMS 培地で行った。
【0117】
7%CO2 、37℃において16時間温置した後、マイクロタイトレーションプレートを遠心分離し、上清に含まれるIL-2の量を市販のキット (Duoset, R&D)で測定する。分泌されたIL-2の濃度をpg/ml で表す。
【0118】
結果はCD16活性化の割合 (%) として表し、対照モノクローナル抗体の存在下の分泌IL-2濃度は100 %に等しいと考えられる。
抗-HLA DR キメラ抗体は、YB2/0 株で発現されるかCHO DXB11 株で発現されるかによっって非常に異なるグリカン構造を有する。このように、YB2/0 で発現された抗体のフコース含量/ガラクトース含量の比は0.37であり、CHO で発現された抗体については比は1.3 でありずっと高い。
【0119】
天然の抗体のCD16活性化は、フコース含量/ガラクトース含量比の値に一致する;このように、YB2/0 で合成され、比が0.37である抗-HLA DR 抗体により誘導されるIL-2分泌は、CHO DXB11 により合成され、比が1.3 である同じ種類の抗体により誘導される分泌の2倍である。
【0120】
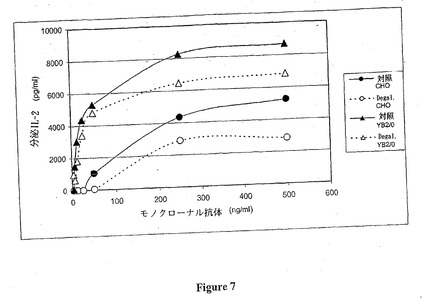
β−ガラクトシダーゼの作用の後、Fc領域のN-グリカン上に残るガラクトース含量をHPCE-LIFにより決定した。脱ガラクトシル化はほぼ完全であり、CHO により産生される抗体に対するG1形態のレベル、およびYB2/0 により産生される抗体についてのG1B 形態のレベルは、それぞれ7%および4.4 %である。このガラクトース含量の低下は、図7に示すように、対照抗体と比較してCD16活性化の著しい減少を示す。
参考文献
Boyd PN, Lines AC, Patel AK.「Campath-1Hの機能活性に及ぼすシアル酸、ガラクトースおよび全炭水化物の除去の影響」(1995) Mol. Immunol, 32, 1311-1318.
Davies J, Jiang L, Pan LY, Labarre MJ, Anderson D, Reff M.「組換え抗-CD20 CHO 産生細胞系におけるGnTIIIの発現:変化した糖形態を有する抗体の発現はFCガンマRIIIに対するより高い親和性を介したADCCの増加をもたらす」(2001) Biotechnol.Bioeng. 74, 288-294.
Furukawa K, Kodbata A.「IgGガラクトシル化−その生物学的意味および病理学」 (1991) Mol. Immunol.28, 1333-1340.
Kohler G, Milstein C. 「規定の特異性を有する抗体を分泌する融合細胞の連続培養」 (1995) Nature 256, 495-7.
Kumpel BM, Rademacher TW, Rook GA, Williams PJ, Wilson IB. 「EBV でトランスフォームしたB-リンパ芽球様細胞系により産生されたヒトIgGモノクローナル抗-Dのガラクトシル化は培養方法に依存し、Fc受容体媒介機能活性に影響する」 (1994) Hum. Antibodies Hybridomas 5, 143-451.
Nose M, Wigzell H.「モノクローナル抗体上の炭水化物鎖の生物学的意味」(1983) Proc.Natl.Acad.Sci.USA 80, 6632-6636.
Schields RL, Lai m Keck R, O'Connell LY, Hong K, Meng YG, Weiler SHA, Presta LG.「ヒトIgG1 N-結合オリゴ糖上のフコースの欠如はヒトFcγRIII への結合および抗体依存性細胞傷害を改善する」(2002) J.Biol.Chem. 277, 26733-26740.
Shinkawa T, Nakamura K, Yamane N, Shoji-Hosaka E, Kanda Y, Sakurada M, Uchida K, Anazawa H, Satoh M, Yamasaki M, Hanai N, Shitara K 「ヒトIgG1複合型オリゴ糖においてフコースが欠如しているが、ガラクトースまたは二分岐N-アセチルグルコサミンも存在しないことは、抗体依存性細胞傷害を増強する重要な役割を示す」 (2003) J. Biol.Chem. 278, 3466-3473.
Tao MH, Morrison SL 「非グリコシル化キメラマウス−ヒトIgGの検討:ヒトIgG定常領域を介した構造およびエフェクター機能における炭化水素の役割」(1989) J. Immunol. 143, 2595-2601.
Tsuchyia N, Endo T, Matsuta K, Yoshinoya S, Aikawa T, Kosuge E, Takeuchi F, Miyamoto T, Kobata A「ヒトIgGの免疫活性に及ぼすオリゴ糖からのガラクトース除去の影響」 (1989) J.Rheumatol. 16, 285-290.
Umana P, Jean-Mairet J, Moudry r, Amstutz H, Bailey JE「最適化抗体依存性細胞傷害活性を有する抗神経芽腫IgG1 の遺伝子工学処理したグリコ形態」(1999) Nat.Biotechnol. 17, 176-180.
Wright A, Morrioson SL「Igエフェクター機能に及ぼすC2関連炭水化物構造の影響:チャイニーズハムスター卵巣細胞1のグリコシル化変異体におけるキメラマウス−ヒトIgG1抗体を用いた検討」(1998) J.Immunol. 160, 3393-3402.
【図面の簡単な説明】
【0121】
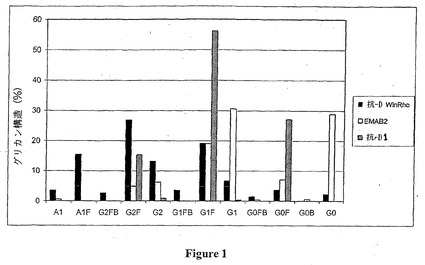
【図1】各種抗-Rh(D)抗体のFc領域のグリコシル化部位上に存在するグリカン構造を示す。この図は、次の3種類の抗-Rh(D)抗体のAsn297残基が有する各種グリカン形態の割合 (%) を図示する:WinRhoの抗-D IgG1(黒色) 、モノクローナルEMAB2 抗体 (白色) および抗-D1(斜線) 。
【図2】フコース含量/ガラクトース含量の比と抗-Rh(D)抗体のADCC活性の間の相関線を示す。
【図3】抗-Rh(D)抗体のADCC活性に及ぼすガラクトースの影響を示す。この図は、0.5 および2.5mg/ml濃度の多価IgG (Tegeline, LFB)の存在下での、脱ガラクトシル化 (Degal)または脱ガラクトシル化されない (対照) 抗-Rh(D)ポリクローナル抗体により生じるRh(D+)赤血球の溶解率 (%) を示す。
【図4】脱ガラクトシル化抗-Rh(D)モノクローナル抗体のCD16活性化を示す。この図は、脱ガラクトシル化 (白色) または脱ガラクトシル化されない (対照、黒色)EMAB2およびHH01抗-Rh(D)モノクローナル抗体の存在により生じるCD16活性化の割合 (%) を図示する。
【図5】ガラクトシル化抗-Rh(D)モノクローナル抗体のCD16活性化を示す。この図は、ウシβ1,4-ガラクトシルトランスフェラーゼによるin vitroガラクトシル化の前 (対照、黒色) および後 (白色) のEMAB2 およびEMAB3 抗-Rh(D)モノクローナル抗体により生じるCD16活性化を図示する。
【図6】抗-RH(D)抗体により感作された、またはされていない放射標識赤血球のクリアランス曲線を示す。この図は、Rhophylac TMポリクローナル抗体の治療用製剤 (●) またはEMAB2 モノクローナル抗体 (■、▲、△) により感作されたか、感作されない (◆、◇) 、Cr51放射標識赤血球の一定量を再注入したボランティアの血液中に含まれる放射能の追跡 (%で表示) を図示する。EMAB2 抗体は三人のボランティアで試験した (008 、009 および010)。
【図7】YB2/0 およびCHO-DG44細胞株で発現した抗-HLA DR モノクローナル抗体の脱ガラクトシル化のCD16活性化に及ぼす影響を示す。この図は、Jurkat CD16 細胞により分泌されるIl-2の量 (pg/mlで表示) を図示し、CD16受容体はその膜上にHLA DR分子を有するRaji細胞の存在下で天然の (実線) または脱ガラクトシル化 (点線) 抗-HLA DR キメラ抗体により活性化されている。
【技術分野】
【0001】
本発明は、高いADCC活性を有し、Fc領域のグリコシル化部位に存在するグリカン構造のフコース含量/ガラクトース含量の比が0.6 以下であるモノクローナル抗体の組成物に関する。本発明はまた、高いエフェクター活性をもつ前記モノクローナル抗体を含有する薬剤組成物にも関する。
【背景技術】
【0002】
広く普及した受動免疫療法は、細胞または所定の物質に対する抗体、特にIgG型の免疫グロブリンの投与に基づく。しかし、モノクローナル抗体の使用が、感染性汚染がないことに関する生成物の安全性の保証などのいくつかの利点を有する場合、他方では有効なモノクローナル抗体を得ることが困難であると判明するかもしれない。
【0003】
G型の免疫グロブリン(IgG)は、ジスルフィド架橋で結合した2本の重鎖と2本の軽鎖からなるヘテロ二量体である。N末端位置の各鎖は、抗体が向けられる抗原に特異的な可変部分からなり、C末端位置の各鎖は、抗体のエフェクター特性を誘導する定常部分からなる。
【0004】
可変部分と、重鎖および軽鎖のCH1 およびCLドメインとの結合によりFab部分が形成され、これは格別の柔軟性をもった領域(転移領域)を介してFc領域(重鎖の定常部分)に結合しており、それによって各Fabがその抗原標的に固定されることを可能にし、一方Fc領域はFcγR受容体などのエフェクター分子に接近可能な状態のままである。
【0005】
Fc領域はCH2 およびCH3 と称される2つの球状ドメインからなる。両方の重鎖はCH3 ドメインにおいて接近して相互作用し、一方CH2 ドメインでは、Asn 297 に結合したラクトサミン型のバイアンテナリー(biantennary)N−グリカンが両鎖のそれぞれに存在することが、両ドメインの分離に寄与する。
【0006】
Fc領域のグリコシル化がIgGの生物学的活性、特に補体に媒介される細胞溶解 (CDC)および抗体依存性の細胞傷害(ADCC)にとって必須であることは多くの研究により示されてきた。このように、指定された突然変異誘発により、またはツニカマイシンの存在下で抗体産生細胞を培養することにより得られるを非グリコシル化IgGは、補体を活性化する能力およびFcγR受容体を固定する能力を失うことが実証された(NoseおよびWigzell, 1983; TaoおよびMorrison, 1989) 。
【0007】
各単糖の役割についてのより具体的研究により、二つに分かれる位置にN-アセチルグルコサミン (GlcNac) 残基が結合するとIgGのADCC活性を高める結果となることが示された (Umana 等, 1999;Davies, 2001) 。他方、ガラクトース残基がAsn 297 に結合するオリゴ糖中に存在するか否かの影響がより論議の的となっている。ガラクトース残基の存在がIgGのエフェクター機能に必須であるとして報告されると (Tsuchiya等, 1989;FurukawaおよびKobata, 1991;Kumpel等, 1994) 、別の著者等はガラクトース残基の欠如によりIgGの機能的活性は変化しなかったことを示した (Boyd等,1995 ;WrightおよびMorrison,1998) 。
【0008】
我々は特許出願WO 01/77181 において、Fc領域のグリコシル化がIgGの生物学的活性、特にCDCおよびADCC活性に必須であることを実証した。我々は、短鎖、低シアリル化、低フコシル化、末端マンノース残基および/または非挿入末端GlcNac残基を特徴とするラクトサミン型のバイアンテナリーN−グリカンがモノクローナル抗体に高いADCC活性を与えるグリカン構造の共通の特徴であることを示している。その後、我々の発見はShields 等 (2002) およびShinkawa等 (2003) の研究によって確認された。
【0009】
本発明において我々は、治療用抗Dポリクローナル抗体 (NATEAD, WinRho) が、そのフコース含量を考慮すると、非常に高いADCC活性を有することを認めた。
この観察は、低フコース含量それ自身が、FcγR受容体、特にFcγRIII を活性化する抗体の能力に影響する唯一の因子ではないことを示唆する。
【0010】
ポリクローナル抗体の全グリコシドのプロフィールを検討することによって、我々は (フコース含量/ガラクトース含量) の比と抗体のエフェクター活性との間に反比例の関係を見出した。
【0011】
事実、抗体が高度にフコシル化されていると、最適のエフェクター活性を得るには高度にガラクトシル化することが必要である。逆に、抗体が僅かしかフコシル化されていないと、最適のエフェクター活性を得るには、このガラクトース含量を、フコース含量/ガラクトース含量の比が0.6 より低く、好ましくは0.5 または0.4 よりも低くなるようにするべきである。
【0012】
従って、我々は実験結果に照らして、高いエフェクター活性が得られる、最適化されたフコース含量/ガラクトース含量の比を有する抗体を製造する方法を完成した。換言すれば、特異的オリゴ糖構造を有し (特にフコースおよびガラクトース残基に関して) 、高いエフェクター活性を付与する新規なモノクローナル抗体を提案する。他方、我々はまた、グリカン構造が細胞傷害活性の活性化を与えない抗体、およびそれを得る方法を提案する。
【発明の開示】
【0013】
従って、第1の面において、本発明は、下記工程を含むことを特徴とする、高いエフェクター活性を有するヒト化またはヒトキメラモノクローナル抗体を製造する方法に関する。
【0014】
a)各種供給源、特に、場合により遺伝的に改変されているか形質転換されている細胞、植物またはヒト以外の動物から得られるモノクローナル抗体を製造し精製すること、
b)該抗体のFc領域のグリコシル化部位が有するグリカン (多糖) 構造のフコース含量およびガラクトース含量を測定すること、および
c)フコース含量/ガラクトース含量の比が0.6 以下、好ましくは0.5 または0.4 以下である抗体を選択すること。
【0015】
「モノクローナル抗体」とは、自然に生じた突然変異を有する少割合の抗体を除き同じ主構造、同じ特異性、および翻訳後修飾、特に分子毎に変化しうるグリコシル化修飾を有するモノクローナル抗体を含む組成物を意味する。本発明の目的にとって、「モノクローナル抗体」または「モノクローナル抗体の組成物」なる表現は同義語である。
【0016】
本発明のモノクローナル抗体は、KohlerおよびMilstein (1975) により報告されたハイブリドーマの製造、エプスタイン・バーウィルス (EBV)によるヒトBリンパ球の不死化などの慣用の方法、またはファージ展示技術、ヒトもしくはトランスジェニック動物抗体 (特にマウス、Xenomouse TM由来) の組み合わせライブラリーの使用などのより最近の方法により製造できる;モノクローナル抗体は分子工学、特に抗体のキメラ化またはヒト化のための分子工学によっても製造できる。本発明の目的にとって、グリカンの分析は、例えばレーザー励起蛍光による高性能キャピラリー電気泳動 (High-Performance Capillary Electrophoresis with Laser-Induced Fluorescence, HPCE-LIF) 、または当業者に既知の任意のその他のグリカン分析方法により行うことができる。
【0017】
本発明の方法により、高いエフェクター活性、より詳しくはADCC型の高い機能活性を有するモノクローナル抗体を得ることができる。このため、エフェクター活性は抗体のFc領域に帰することができる生物学的活性を意味する。これらのエフェクター機能の例には、抗体依存性細胞傷害 (Antibody-Dependent Cell-mediated Cytotoxicity, ADCC) 活性、補体依存性細胞傷害 (Complement-Dependent Cytotoxicity , CDC) 活性、食細胞活性、エンドサイトーシス活性またはサイトカイン分泌の誘導が挙げられるが、これらに限定されない。
【0018】
「高い」エフェクター活性とは、同じ特異性を有するがフコース含量/ガラクトース含量の比が0. 6より大きい抗体のエフェクター活性に比べ、少なくとも20倍、50倍、60倍、70倍、80倍もしくは90倍、好ましくは100 倍まで、または優先的には500 倍のエフェクター活性を意味する。
【0019】
優先的には、フコース含量/ガラクトース含量の比は0.6 〜0.3 、好ましくは0.5 〜0.35の値である。事実、本発明の範囲内で行われた実験を考慮すると、制限的な比が存在する、即ち、比の減少に伴いそれ以下では機能的 (特にADCC) 活性がもはや直線的に増加しないフコース含量/ガラクトース含量の比が存在するようである。従って、これらの限定値の間となるように本発明方法を実施することが特に有利である。
【0020】
例えば、フコース含量が35%〜45%である場合、ガラクトース含量は70〜99%でありうる。フコース含量が20%〜35%である場合、ガラクトース含量は55%〜70%、または60%〜99%である。
【0021】
本発明の目的にとって、0.6 以下の比は、数百分の1、例えば400 〜500 分の1の単位で0.6 より大きい値も意味する。
本発明のある特定の面において、本発明方法により得られる抗体は、抗体の発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変した細胞において製造され、これらの細胞は真核または原核細胞、特に哺乳類、昆虫、植物、細菌または酵母由来の細胞である。
【0022】
有利には、得られる抗体はIgG型のヒト免疫グロブリンである。
より有利には、これらの細胞は、グリコシルトランスフェラーゼ活性を有する少なくとも1種のポリペプチドの発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変した細胞でありうる。
【0023】
優先的には、このグリコシルトランスフェラーゼ活性は、ガラクトシルトランスフェラーゼ活性、特にベータ(1,4)-ガラクトシルトランスフェラーゼまたはベータ(1,3)-ガラクトシルトランスフェラーゼ活性である。
【0024】
本発明の目的にとって、「ガラクトシルトランスフェラーゼ活性を有するポリペプチド」とは、N-グリカンの非還元位置においてガラクトース残基をUDP-ガラクトースからGlcNAc残基へ付加することを触媒しうる任意のポリペプチドを意味する。
【0025】
本発明の目的にとって、「ベータ(1,4)-ガラクトシルトランスフェラーゼ活性を有するポリペプチドの発現を可能にするベクター」とは、二糖型のGal ベータ(1,4)-GlcNacを合成しうるポリペプチドの発現を可能にするポリヌクレオチドを含む任意のベクターを意味し、このポリヌクレオチドはヒト、マウス、ハムスター、ウシ、ヒツジ、ヤギ、ブタ、ウマ、ラット、サル、ウサギ、トリなどの種に由来しうる。例えば、NM 001497 、AB 024434 、NM 003780 、BC 053006 、 XM 242992、 NM 177512などの配列 (これは網羅しているわけではない) が、Genbank などのヌクレオチドおよび/またはタンパク質配列のバンクにおいて利用できる。
【0026】
本発明の目的にとって、「ベータ(1,3)-ガラクトシルトランスフェラーゼ活性を有するポリペプチドの発現を可能にするベクター」とは、二糖型のGal ベータ(1,3)-GlcNacを合成しうるポリペプチドの発現を可能にするポリヌクレオチドを含む任意のベクターを意味し、このポリヌクレオチドはヒト、マウス、ハムスター、ウシ、ヒツジ、ヤギ、ブタ、ウマ、ラット、サル、ウサギ、トリなどの種に由来しうる。特に、ヒト、マウス、ハムスター、ウシ、ヒツジ、ヤギ、ブタ、ウマ、ラット、サル、ウサギ、トリなどの種に由来するベータ(1,3)-ガラクトシルトランスフェラーゼをコードする配列が特に適している。例えば、NM 020981 、AB 084170 、AY 043479(これらに制限されない) などの配列が、Genbank で利用できる。
【0027】
「抗体のFc領域のグリコシル化部位」とは一般的に、Kabat (Kabatデータベース、http://immuno.bme.nwu.edu)の番号付与による両方のAsn297残基を意味するが、本発明はアミノ酸配列が変化した抗体にも関する。
【0028】
本発明のある特定の態様において、細胞はさらに、GDP-フコースの合成および/または輸送に関与する活性、並びに/または抗体のグリコシル化部位のオリゴ糖へのフコースの付加に関与する酵素の活性が低減または欠失している。有利には、GDP-フコースの合成に関与する酵素は、GMD (GDP-D- マンノース 4,6- デヒドラターゼ) 、Fx (GDP-ケト-6- デオキシマンノース 3,5- エピメラーゼ、4-レダクターゼ) またはGFPP (GDP-ベータ-L- フコースピロホスホリラーゼ) であるが、これらは網羅するものではない。有利には、フコースの付加に関与する酵素はフコシルトランスフェラーゼである。GDP-フコースの輸送に関与するタンパク質は、有利にはヒトGDP-フコーストランスポーター1でありうる。
【0029】
本発明のある特定の態様において、工程b)で測定したフコースおよびガラクトースの含量が0.6 より大きい比を与える場合、抗体の機能活性を増加させるためには前記比が0.6 より小さく、好ましくは0.5 より小さく、0.4 より小さくさえなるように、工程c)の前に抗体を脱フコシル化する、および/またはガラクトース残基を抗体に付加することが可能である。この脱フコシル化は、保存培地であってよい抗体含有培地中にフコシダーゼを添加することにより行いうる。ガラクトース残基の付加は、抗体含有培地、または抗体およびUDP-ガラクトースなどの供与基質を含有する溶液にガラクトシルトランスフェラーゼを添加するなどの任意の適宜手段により行いうる。
【0030】
有利には、本発明方法を適用するために用いる細胞は、動物またはヒト細胞株由来であり、これらの株は特に、ラットのミエローマ株 (特にYB2/0 およびIR983F) 、ヒトミエローマ株 (例、Namalwa)、またはヒト起源のその他の任意の細胞 (PERC6, CHO株, 特にCHO-K, CHO-Lec10, CHO-Lec1, CHO Pro-5, CHO dhfr-, CHO Lec13)、または Wil-2, Jurkat, Vero, Molt-4, COS-7, 293-HEK, BHK, K6H6, NSO, SP2/0-Ag 14 およびP3X63Ag8.653から選択されたその他の株から選択される。
【0031】
有利には、抗体は抗-Rhesus D(抗-D) 、抗-CD 、抗−腫瘍、抗−ウイルス、抗-CD20 、または抗-HLA-DR であり、より詳細には以下の表0の抗体由来である。
【0032】
(表0)
抗体の名称 会社 標的 適用
および商品名 Edrecolomab Centocor 抗-Ep-CAM 結腸直腸がん
PANOREX
Rituximab Idec 抗CD20 B細胞リンパ腫
RITUXAN Genentech/ 血小板減少症
Hoffman La Roche 紫斑病
にライセンス
Trastuzumab Genentech 抗HER2 卵巣がん
HERCEPTIN Hoffmfan La Roche/
Immunogen にライセンス
Palivizumab Medimmune RSV
SYNAGIS Abott にライセンス
Alemtuzumab BTG 抗CD52 白血病
CAMPATH Scheringにライセンス
Ibritumomab IDEC 抗CD20 NHL
Tiuxetan Scheringにライセンス
ZEVALIN
Cetuximab Merck/BMS/ 抗EGFR がん
IMC-C225 Imclone
Bevacizumab Genentech/ 抗VEGFR がん
AVASTIN Hoffnan La Roche
Epratuzumab Immunmedics/ 抗CD22 がん:
Amgen 非ホジキンリンパ腫
Hu M195Mab Protein Design Labs 抗CD33 がん
MDX-210 Immuno-Designed ND がん
Molecules
BEC2 Imclone 抗GD3 がん
Mitunomab
Oregovomab Altarex 抗CA125 卵巣がん
OVAREX
Ecromeximab Kyowa-Hakko 抗GD3 悪性黒色腫
KW-2971
ABX-EGF Abgenix EGF がん
MDX010 Medarex 抗CD4R がん
XTL 002 XTL ND 抗ウイルス:HCV Bio-pharmaceuticals
H11SCFV viventia biotech ND がん
4B5 vivantia biotech 抗GD2 がん
XTL 001 XTL ND 抗ウィルス:HBV
Bio-pharmaceutical
MDX-070 MEDAREX 抗-PSMA 前立腺がん TNX-901 TANOX 抗IgE アレルギー
IDEC-114 IDEC プロテインC 非ホジキンリンパ腫
阻害
この一覧は制限するためのものではない。
【0033】
本発明の第2の目的は、免疫学的機能性分子の組成物のエフェクター活性、特にADCC活性を向上させる方法であり、分子組成物中のガラクトース含量を増加させ、および/またはフコース含量を減少させることを含む方法を提供することである。
【0034】
「免疫学的に機能性の分子」とは、免疫学的能力を実証することにより何らかの免疫原との何らかの接触に反応しうる分子を意味する。本来の状態のこれらの分子は良好なエフェクター活性、例えばADCC活性、または低いエフェクター活性を有するかもしれない。そられはグリコシル化部位を含むFc領域を有する。この目的にとって、これらの機能的免疫学的分子は優先的には抗体であり、有利にはモノクローナル抗体またはポリクローナル抗体である。
【0035】
本来の状態の分子は高いフコース含量を有するかもしれない。特に、この場合は、これらの分子または抗体のガラクトース含量を向上させることが有利である。
本発明のある態様において、フコース含量の減少はフコシダーゼの作用による組成物の分子の脱フコシル化によりなされる。この脱フコシル化はα1,6-フコシダーゼにより行うことができる。ウシの腎臓またはCharonia lampas から抽出したフコシダーゼはこの特異性を有する。
【0036】
本発明の別の態様では、組成物の分子のガラクトース含量における増加は、ガラクトシルトランスフェラーゼの作用による組成物のガラクトシル化による。
本発明のある特定の態様では、脱フコシル化のための酵素およびガラクトシル化のための酵素を両方とも作用させる。
【0037】
酵素処理に代わる方法として、免疫学的機能性分子の組成物を、低フコシル化抗体および/または高ガラクトシル化抗体を多くするレクチン上での一連のクロマトグラフィーにより精製しうる。
【0038】
一例として、有利には抗体である免疫学的機能性分子の組成物を含む溶液を、HPLCシステムに連結したレクチンカラム (例、LA-LCAまたはLA-AALカラム、Shimazu Corporation)上を通過させる。溶液を非吸着画分と吸着画分に分ける。非吸着画分および吸着画分のグリカン分析を行う:酵素作用によりタンパク質部分から開裂したオリゴ糖をAPTSで標識し、HPCE-LIFで分離して定量する。ピークの面積を計算する:フコースを含まないグリカンを有する抗体はそれにより分離され選択される。選択された画分を次いで、フェニル-5PW型 (Toso Corporation製) の疎水性カラムまたは第2レクチンカラム (LA-RCA 120またはLA-WGA、Seikagaku America)上を通過させる (非吸着画分または吸着画分から流出しうる) 。フコース含量/ガラクトース含量の比が0.6 以下の画分をそれによって正確に選択できる。
【0039】
本発明の第3の目的は、抗体分子をコードする少なくとも1つのベクターが導入された細胞、優先的にはYB2/0 細胞株由来の細胞であり、この細胞は、Fc領域のグリコシル化部位由来のオリゴ糖のフコース含量/ガラクトース含量の比が0.6 以下であるモノクローナル抗体を産生する。優先的にはこの比は0.5 より小さく、0.4 よりさえも小さい。本発明の好ましい面において、この比は0.6 〜0.3 である。
【0040】
本発明の好ましい面において、この細胞は、ガラクトシルトランスフェラーゼ、特にベータ(1,4)-ガラクトシルトランスフェラーゼまたはベータ(1,3)-ガラクトシルトランスフェラーゼをコードする発現ベクターでトランスフェクされる。有利には、この細胞は組換えガラクトシルトランスフェラーゼを発現または過発現する。
【0041】
YB2/0 株は本来、ベータ(1,4) およびベータ(1,3) のガラクトシルトランスフェラーゼファミリーを発現する。さらに、この細胞株は、低フコース含量の抗体を産生することが知られている (WO 01/77181, LFB) 。しかし、本発明による細胞は、非改変株により産生される抗体に比べて、改変された細胞により産生される抗体のフコース含量/ガラクトース含量の比を変化させる効果を有する、ガラクトシルトランスフェラーゼを過発現するという利点を有する。従って、抗体はもともとフコシル化が少ないので、そのガラクトース含量の向上はさらにそのフコース含量/ガラクトース含量の比を低くし、これはそのADCC活性をさらに最適化する効果がある。
【0042】
有利には、ガラクトシルトランスフェラーゼは、ヒト、マウス、ハムスター、ウシ、ヒツジ、ヤギ、ブタ、ウマ、ラット、サル、ウサギ、またはトリ (これに制限されない) 由来の配列によりコードされる。より詳しくは、コード配列はNM 001497, AB 024434, NM 003780, BC 053006, XM 242992 またはNM 177512 配列である。
【0043】
従って、本発明は、前記ベクターの発現を可能にする培地および条件下で前述の細胞を培養することを含む、Fc領域のグリコシル化部位に存在するグリカン構造が0.6 以下、優先的には0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有するモノクローナル抗体を製造する方法に関する。
【0044】
あるいは、上に規定したような抗体組成物は、フコース、ガラクトースまたはそれらを含むオリゴ糖を特異的に捕捉しうる任意の分子を用いた1または2以上のクロマトグラフィー工程により製造することもできる。前述のようにレクチン上での分離なども使用できる。
【0045】
また、本発明は、先に記載した方法により得られるか、または既述の方法で得られた、高いエフェクター活性をもった治療用抗体に関し、これらの抗体は、Fc領域のグリコシル化部位に、0.6 より小さい、優先的には0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有するグリカン構造をもつことを特徴とする。
【0046】
より有利には、これらは前述の方法で得ることができる治療用モノクローナル抗体であり、この抗体は増強されたADCC活性を有し、一例としてはポリクローナル抗体と等しいかそれより大きいADCC活性を有するモノクローナル抗-Dである。この増強されたADCC活性は、CHO DG44またはDxB11 株において発現されたポリクローナルまたはモノクローナル (同じ特異性を有する) 治療用抗体のものと少なくとも等しいか、好ましくはより大きい。
【0047】
有利には、これらはIgGであり、例えば、キメラ、ヒト化もしくはヒトのIgG1もしくはIgG3、またはヒトFc領域を有するIgGである。好ましくは、これらの抗体はヒトIgG、またはヒトFc領域を有する任意のキメラ分子である。
【0048】
同じ観点において、本発明は上述の抗体を含む薬剤組成物に関する。
また、本発明は、Fc領域のグリコシル化部位に存在するグリカン構造が0.6 より小さい、優先的には0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有するモノクローナルまたはポリクローナル抗体を少なくとも50%、好ましくは60%、70%、80%、または90%もしくは99%含む薬剤組成物に関する。好ましくは、この比は0.6 〜0.3 、特に0.5 〜0.35である。
【0049】
本発明の組成物は好ましくは、非遍在の通常抗原、特にRh因子 (例、ヒト赤血球細胞のRh因子(D))、またはヒトに対する病原性細胞もしくは病原性生物の抗原、特にがん細胞の抗原、に対する抗体を含む。抗体はさらに好ましくはIgGである。
【0050】
本発明の別の目的は、同種免疫、特に新生児溶血性疾患の治療用薬剤を製造するための本発明抗体の使用に関する。
本発明の別の目的は、自己免疫疾患、がん、および病原体による感染症の治療、特にセザリー症候群、固形がんから選択された免疫応答をのがれる疾患の治療、特に抗原性標的が弱く発現したもの、特に、乳がん、ポリ塩化ビフェニルに暴露されたヒトを対象とする環境に関連する病状、感染症 (特に結核) 、慢性疲労症候群(CFS) 、寄生虫感染症 (例、住血吸虫) およびウイルス感染症、の治療用薬剤を製造するための本発明抗体の使用に関する。
【0051】
さらに、本発明の抗体は、黒色腫などの陽性クラスII HLA細胞のがん、BおよびT細胞の急性リンパ性白血病、急性および慢性骨髄性白血病、バーキットリンパ腫、ホジキンリンパ腫、T細胞リンパ腫および非ホジキンリンパ腫の治療用薬剤を製造するのに使用できる。
【0052】
本発明の抗体は表0に示された抗体より選択することができる。
有利には、抗体は抗-HLA-DR または抗-CD20 である。
本発明の別の面において、本発明の抗体は、免疫システムの天然のエフェクター細胞によりIL-1α、IL-1β、IL-2、IL-3、IL-4、IL-5、IL-6、IL-12 、IL-18 、IL-21 、TGF β1 、TGF β2 、TNF α、TNF β、INF γ、およびIP10から選ばれた少なくとも1種のサイトカインの発現を誘導するための薬剤を製造するのに使用され、この薬剤はがんおよびウイルス、細菌もしくは寄生虫感染症を治療するのに特に有用である。
【0053】
本発明の別の特定の面では、本発明の抗体は、CD16多型の1つ、特にV/F158またはF/F158を有する患者、特に現在利用可能な抗体では治療できない症状の患者、または望ましくない二次的作用を受けている患者、の治療のための薬剤の製造に使用される。
【0054】
さらに別の面では、本発明はまた、下記工程を含むことを特徴とする、低いエフェクター活性、特に低いADCC型の機能活性を有する、キメラ、ヒト化またはヒトモノクローナル抗体を製造する方法に関する。
【0055】
a)各種供給源、特に、場合により遺伝的に改変されているか形質転換されている細胞、植物またはヒト以外の動物から得られるモノクローナル抗体を製造し精製すること、
b)該抗体のFc領域のグリコシル化部位に存在するグリカン (多糖) 構造のフコース含量およびガラクトース含量を測定すること、および
c)フコース含量/ガラクトース含量の比が0.6 より大きい、好ましくは1.2 より大きい抗体を選択すること。
【0056】
モノクローナル抗体のエフェクター活性の定義は前記した通りである。
さらに、「低いエフェクター活性」とは、同じ特異性を有するがフコース含量/ガラクトース含量の比が0. 6より小さい抗体のエフェクター活性、特にADCC型の機能活性に比べ、少なくとも20倍、50倍、60倍、70倍、80倍もしくは90倍、好ましくは100 倍まで、または優先的には500 倍低いエフェクター活性を意味する。
【0057】
従って、相補的な面において、本発明は、Fc領域のグリコシル化部位(Asn 297) が1.2 より大きいフコース含量/ガラクトース含量を有することを特徴とする、低いADCC活性を有する抗体、およびこれを含む組成物に関する。
【0058】
これらの抗体は、自己免疫疾患、特に免疫性血小板減少性紫斑病 (PTI)、同種免疫、移植片拒絶、アレルギー、喘息、皮膚炎、蕁麻疹、紅班および炎症性疾患の治療および/または予防のための薬剤の製造に有用である。
【0059】
本発明のある特定の面において、抗体は、この抗体の発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変した細胞において産生され、この細胞は真核細胞でも原核細胞でもよく、特に哺乳動物、昆虫、植物、細菌または酵母由来の細胞である。
【0060】
本発明のある態様において、細胞は、グリコシルトランスフェラーゼ活性、好ましくはフコシルトランスフェラーゼ活性、特にα1.6-フコシルトランスフェラーゼ活性を有する少なくとも1つのポリペプチドの発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変される。
【0061】
本発明の別の態様では、細胞は、UDP-ガラクトースの合成および/または輸送に関する活性、並びに/または抗体のグリコシル化部位のオリゴ糖へガラクトースを付加することに関与する酵素の活性が減少または欠失している。有利にはガラクトースの付加に関与するこの酵素は、β1,4-ガラクトシルトランスフェラーゼである。
【0062】
有利には、細胞はグリコシルトランスフェラーゼ活性、特にグリコシルトランスフェラーゼ活性と、UDP-ガラクトースの合成および/または輸送に関する活性、並びに/または抗体のグリコシル化部位のオリゴ糖へガラクトースを付加することに関与する酵素の活性が減少または欠失している。
【0063】
本発明のある態様において、工程b)で測定した比が0.6 より小さい場合、フコース含量/ガラクトース含量の比が0.6 より大きくなるように、工程c)の前にフコシル化を行う、および/またはガラクトース残基を抗体より除去することが可能である。有利には、脱ガラクトシル化は、抗体含有培地中にガラクトシダーゼを添加することにより行う。有利には、フコース残基の付加は、抗体含有培地にフコシルトランスフェラーゼを添加することにより行う。
【0064】
より有利には、抗体はIgG型のヒト免疫グロブリンである。有利には、抗体は、ヒト血球の分化のマーカーであるCDに対するか、またはバイオテロの場合には特に危険であるとして挙げられている病原体もしくはその毒素、特にバチルス・アントラシス (Bacillus anthracis) 、クロストリジウム・ボツリウム (Clostridium botulium) 、エルシニア・ペスティス (Yersinia pestis)、バリオラ・マジョル (Variola major)、フランシセラ・ツラレンシス(Francisella tularensis)、フィロウイルス(filovirus) 、アレナウイルス (arenavirus) 、ブルセラ・スピーシーズ (Brucella species) 、クロストリジウム・ペルフリンゲンス (Clostridium perfringens)、サルモネラ (Salmonellla)、大腸菌 (E.coli) 、赤痢菌 (Shigella) 、コクシエラ・バーネッティイ (Coxiella burnetii)、リシン毒素、リケッチア (Rickettsia) 、ウイルス性脳炎ウイルス、ビブリオコレラ (Vibrio cholerae)またはハンタウイルス、に対するものである。
【0065】
本発明の別の目的は、組成物のフコース含量の増加および/またはガラクトース含量の低下を含む、免疫機能性分子の組成物の活性を低下させる方法にも関する。
有利には、免疫機能性分子はモノクローナルまたはポリクローナル抗体である。
【0066】
ある特定の面において、フコース含量の増加は、フコシルトランスフェラーゼ、好ましくはα1,6-フコシルトランスフェラーゼの作用による前記組成物のフコシル化による。
別の特定の面において、前記組成物のガラクトース含量の低下は、ガラクトシダーゼ、好ましくは1または2以上のβ- ガラクトシダーゼの作用による組成物の脱ガラクトシル化による。
【0067】
より有利には、この組成物のフコシル化および脱ガラクトシル化の両方を行う。
従って、本発明の目的は、上記の本発明方法により得ることができる抗体の組成物、またはこれらの方法のいずれかにより得られた抗体組成物に関する。
【0068】
本発明の別の目的は、自己免疫疾患、特にPTI 、同種免疫、移植片拒絶、アレルギー、喘息、皮膚炎、蕁麻疹、紅班または炎症性疾患 (これらは網羅するものではない) を治療および/または予防する薬剤を製造するための、この抗体組成物の使用である。
【0069】
最後に、本発明は、抗体のFc領域のグリコシル化部位由来のオリゴ糖におけるフコース含量/ガラクトース含量の比を調節することを含む、免疫機能性分子の組成物の活性を制御する方法に関する。
【0070】
本発明のその他の面および利点は、ガラクトースによる「フコース効果」調節を示す実施例に記載されるであろう。これは非制限的と考えられるべきであり、本発明の範囲を制限するものではない。
【実施例1】
【0071】
一連の抗-Rh(D)抗体のフコース含量/ガラクトース含量の比とADCC活性との間の相関関係
Rhesus (Rh)(D)抗原に対する各種モノクローナルおよびポリクローナル抗体のフコース含量、次いでガラクトース含量を測定した。これより両者の間の比を算出し、各抗体についてADCC活性を測定した。
1.抗-Rh(D)モノクローナル抗体の産生
モノクローナル抗体は、Rh(D) 抗原を有する赤血球で免疫した、陰性のRh(D) ヒトドナーからのBリンパ球のEBV によるトランスフォーメーションに由来する。2つのクローンをこのトランスフォーメーションから選択した:
1)クローンの1つをK6H6-B5 ヒト/マウスヘテロミエローマと融合させ;この融合物よりクローンHH01を選択した。
【0072】
2)抗体の重鎖および軽鎖を発現させるためのベクターを作製するために、その他のクローンから抗-Rh(D)抗体をコードする RNAを抽出した。
この発現ベクターを、一方でEMAB1 、EMAB2 、EMAB3 およびEMAB4 抗体を産生させるYB2/0 細胞株、他方で次のCHQ 株をトランスフェクトするのに使用した:抗-D1 、抗-D2 および抗-D3 抗体をそれぞれ合成するDG44、K1およびLec13 。
2.ポリクローナル抗体の精製
抗-Rh(D)ポリクローナル抗体を、治療用製剤のWinRho (Cangene)から、Rh(D+)赤血球上での陽性選択により、次いでRhD(-)赤血球上での陰性選択により免疫精製し、最後に、セファロース−プロティンAゲルを用いたアフィニティークロマトグラフィー工程により、一方で赤血球上での免疫精製中に回収された汚染物質を除去し、他方でIgG1をIgG3から分離した。というのはIgG1のみが以下の試験において使用されるためである。
3.HPCE-LIFによるグリカン分析
抗-Rh(D)モノクローナルおよびポリクローナル抗体をセファデックスG-25 (HiTrap Desalting, Amersham Biosciences) カラム上で脱塩し、蒸発により乾燥させ、50mMのβ- メルカプトエタノールの存在下で加水分解 PNGアーゼF (Glyko) 用の緩衝液中に再懸濁する。37℃で16時間の温置後、無水エタノールを添加することによりタンパク質部分を沈殿させ、そしてN-グリカンを含有する上清を蒸発により乾燥させる。そうして得られたオリゴ糖は蛍光色素APTS (1-アミノピレン-3,6,8- トリスルホネート) で直接標識するか、またはAPTSで標識する前に特異的エキソグルコシダーゼの作用を受けさせる。次に、標識オリゴ糖をN-CHO キャピラリー上に注入し、レーザー励起蛍光の検出によるキャピラリー電気泳動 (HPCE-LIF) により分離、定量する。
【0073】
電気泳動図上にフコシル化されているかまたはされていない五糖 [GlcNac2-Man3] に対応する2つのピークが得られるように、分離されたフコシル化形態を、より詳しくはノイラミニダーゼ、β−ガラクトシダーゼおよびN-アセチルエキソサミニダーゼを同時に作用させた後添加することによりフコース含量の評価を行う。
【0074】
フコース含量 (%で表す) は以下の式を用いて算出される:
フコシル化[GlcNac2-Man3]×100
フコース含量=───────────────
[GlcNac2-Man3+フコシル化GlcNac2-Man3]
ガラクトース含量 (%で表す) は、末端位置にガラクトースを含有するオリゴ糖形態の割合を加えることにより算出される。使用する式は以下の通りである:
ガラクトース含量=[(G1+G1B +G1F +G1FB) +2×(G2 +G2F +G2B +G2FB)]
フコース含量/ガラクトース含量の比は、フコース含量をガラクトース含量で割ることにより得られ、これらの含量は上記のようにして算出される。
4.抗体の機能活性:ADCC
ADCC (抗体依存性細胞傷害) 法により、エフェクター細胞 (単核細胞またはリンパ球) の存在下でRh(D+)赤血球の溶解を引き起こす抗体の能力を評価することができる。
【0075】
簡単に述べると、赤血球細胞RhD(+)濃縮物の赤血球をパパインで処理し (1mg/ml 、37℃で10分) 、0.9 %NaClで洗浄する。エフェクター細胞を少なくとも3つの軟膜のプールからFicoll (Amersham) での遠心分離、次いで25%FCS の存在下での接着工程によりにより分離し9のオーダーのリンパ球/単球の比とする。マイクロタイトレーションプレート (96ウェル) において1ウェル当たり、精製抗-Rh(D)抗体の希釈物 (9.3 から150ng/ml) 100 μl、パパイン処理したRh(D+)赤血球25μl (即ち、1.106 ) 、エフェクター細胞25μl (即ち、2.106 ) および通常濃度2および10mg/ml の多価IgG (Tegeline, LFB)50μlを入れる。希釈物はウシ胎児血清 (FCS)の0.25% IMDM 中で作製する。37℃で1時間温置した後、プレートを遠心分離にかけ、次いで上清中の放出ヘモグロビンを発色性基質、2,7-ジアミノフルオレン (DAF)の存在下でそのペルオキシダーゼ活性を介して測定する。結果は溶解率 (%) として表され、100 %はNH4Cl 中の赤血球の全溶解に相当し (100 %対照) 、0%は抗体の存在しない反応混合物に相当する (0%対照) 。特異的溶解は下記式により%で算出する:
((試料のOD−0%対照のOD) ×100)
────────────────=ADCC%
(100%対照のOD−0%対照のOD)
抗-Rh(D)IgG1のFc領域のグリコシル化部位が有するオリゴ糖のHPCE-LIF分析を行った。
【0076】
【表1】
【0077】
* 免疫精製したポリクローナル抗-D
表1に含まれる、 [フコース含量/ガラクトース含量] の比およびADCC率の値は図2においてそれぞれ横座標および縦座標に記入されている。プロットされた一次回帰線の相関係数は0.92に等しい。
【0078】
このように、モノクローナルおよびポリクローナル抗-Rh(D)抗体の [フコース含量/ガラクトース含量] の比とADCC活性の間には相関関係がある。顕著なADCC活性を有する抗体はフコース含量/ガラクトース含量の比が0.6 より小さい。
【実施例2】
【0079】
脱ガラクトシル化前および後の抗-Rh(D)ポリクローナル抗体のADCC活性の比較
1.抗-Rh(D)ポリクローナル抗体の脱ガラクトシル化
免疫精製したポリクローナル抗体を加水分解緩衝液 (50mM 酢酸ナトリウム、pH5.5 、塩化カルシウム4mM含有) で透析する。抗体を、コレラ菌 (Vibrio cholerae)由来のノイラミニダーゼ (EC 3.2.1.18) (Calbiochem) 5mUおよびE.coli産生のβ−ガラクトシダーゼ (EC 3.2.1.23) (Roche)9mMの存在下で温置することにより脱シアリル化および脱ガラクトシル化する。「対照」と記載された対照は、上記のように、しかしノイラミニダーゼおよびβ−ガラクトシダーゼの不在下で処理した同じ抗体調製物からなる。37℃で25時間温置後、抗体を4℃で保存する。
【0080】
この実施例で製造された抗体を2つの部分に分ける:1つはグリカン分析に使用し、他はADCC活性の測定に取っておく。
2.HPCE-LIFによるグリカン分析
操作は、塩、また生成物に存在するかもしれない遊離の単糖類を除去するための、セファデックス-G25カラム上での、脱ガラクトシル化抗-Rh(D)ポリクローナル抗体の脱塩からなる。抗体の変性および還元の後、グリカンをエンドグリコシダーゼ、PNG アーゼF (Glyko)の作用を介して放出させる。37℃で16時間の温置後、タンパク質部分を無水エタノールの添加により沈殿させ、N-グリカンを含む上清を蒸発により乾燥させる。こうして得られたオリゴ糖中に含まれるガラクトースおよびフコースの量を評価するために、APTSでの標識前に、試料をそれぞれシアリダーゼおよびフコシダーゼ、またはシアリダーゼ、β−ガラクトシダーゼおよびN-アセチルヘキソサミニダーゼの同時の作用を受けさせる。次いで、標識オリゴ糖をレーザー励起蛍光の検出によるキャピラリー電気泳動 (HPCE-LIF) により分離、定量する。
3.ADCC活性の測定
実施例1に記載の方法に従い、β−ガラクトシダーゼ処理の前および後にポリクローナル抗体のADCC活性を測定する。
【0081】
こうして、β−ガラクトシダーゼ処理の後、抗-Rh(D)ポリクローナル抗体抗のFc領域のグリカンは残存ガラクトース含量が17.7%で、フコース含量が68.5%となる。従って、脱ガラクトシル化ポリクローナル抗体のフコース含量/ガラクトース含量の比は3.8 である。
【0082】
この試料中のテゲリン (Tegeline) などの多価IgGのADCC試験における存在は、高親和性受容体 (即ち、FcγRIまたはCD64) を阻害し、それによりRh(D+)赤血球溶解がエフェクター細胞上に存在するFcγRIII受容体と抗-Rh(D)抗体の相互作用により特異的となるようにする。
【0083】
図3に示す結果は、抗-Rh(D)ポリクローナル抗体のADCC活性は一方で用量依存性であることを、そして反応混合物中の多価IgGの量が増加するとポリクローナル抗体の溶解性を減少させることを示している。さらに、脱ガラクトシル化ポリクローナル抗体は対照抗体に比べて低いADCC活性を有する。
【0084】
【表2】
【0085】
対照抗体、即ちノイラミニダーゼおよびβ−ガラクトシダーゼの不在下で同じように温置した抗体と比べた、脱ガラクトシル化抗-Rh(D)抗体のADCC活性の割合 (%) を表2に示す。
【0086】
このように、対照抗体に比べた、脱ガラクトシル化ポリクローナル抗体のADCC活性の減少は、抗体の量が少ない場合より有意である。さらに、脱ガラクトシル化ポリクローナル抗体の活性の減少は2.5mg/ml濃度の多価IgGの存在下でより有意である。
【実施例3】
【0087】
脱ガラクトシル化抗-Rh(D)モノクローナル抗体により生じるCD16受容体の活性化の測定
1.抗-Rh(D)モノクローナル抗体の脱ガラクトシル化
抗体を加水分解緩衝液 (50mM 酢酸ナトリウム、pH5.5 、塩化カルシウム4mM含有) で透析する。抗体を、コレラ菌 (Vibrio cholerae)由来のノイラミニダーゼ (EC 3.2.1.18) (Calbiochem) 5mUおよびE.coli産生のβ−ガラクトシダーゼ (EC 3.2.1.23) (Roche)9mUの存在下で温置することにより脱シアリル化および脱ガラクトシル化する。「対照」と記載された対照は、上記のように、しかしノイラミニダーゼおよびβ−ガラクトシダーゼの不在下で処理した同じ抗体調製物からなる。37℃で24時間温置後、抗体を4℃で保存する。
【0088】
この実施例で製造された抗体を2つの部分に分ける:1つはグリカン分析に使用し、他は機能活性の測定に取っておく。
2.CD16受容体活性化の測定
Jurkat CD16 細胞の活性化試験は、Fab を標的細胞上に存在する抗原に結合させた後、CD16 (FcγRIIIA)上に抗体のFcを固定することにより生じるインタロイキン-2 (IL-2) の分泌を測定する。Jurkat CD16 細胞により分泌されるIL-2のレベルはCD受容体の活性化に比例する。
【0089】
抗体希釈物50μl、6.105/mlの赤血球懸濁液50μl、1.106 /ml のJurkat CD16 細胞懸濁液50μlおよび40ng/ml のPMA 溶液50μlを順次96−ウェルマイクロタイトレーションプレートに入れる。希釈はすべて、5%FCS を含有するIMDE培地で行う。
【0090】
7%CO2 、37℃において16時間温置した後、マイクロタイトレーションプレートを遠心分離し、上清に含まれるIL-2の量を市販のキット (Duoset, R&D)で測定する。分泌されたIL-2の濃度をpg/ml で表す。
【0091】
結果はCD16活性化率 (%) として表し、対照モノクローナル抗体の存在下の分泌IL-2濃度は100 %に等しいと考えられる。
実施例2に記載したHPCE-LIFにより行ったグリカン分析の結果を表3にまとめる。
【0092】
【表3】
【0093】
このように、EMAB2 モノクローナル抗体はすべて脱ガラクトシル化され、HH01抗体は依然として17.3%のモノガラクトシル化形態を含有しているようである。従って、β−ガラクトシダーゼの作用の後、EMAB2 とHH01抗体のフコース含量/ガラクトース含量の比は0.6 よりずっと大きくなる。
【0094】
脱ガラクトシル化抗-Rh(D)モノクローナル抗体は、対照抗体に比べてCD16活性化が非常に低い (図4) 。このように、EMAB2 およびHH01モノクローナル抗体は、それぞれ52および47%のCD16活性化誘導能の低下を示す。
【実施例4】
【0095】
ガラクトシル化抗-Rh(D)モノクローナル抗体により生じるCD16活性化の測定
1.抗体のガラクトシル化
抗体を50mMヘペス緩衝液 (pH7.20) で透析する。反応混合物は、MnCl2 10mM、UDP-ガラクトース20mM、およびウシベータ1.4-ガラクトシルトランスフェラーゼク(Calbiochem)40mUを添加したモノクローナル抗体溶液からなる。37℃で24時間温置した後、試験管を使用まで4℃で保存する。
【0096】
対照は、反応培地中でUDP-Gal の不在を除き同じ条件下で温置した同種の抗体からなる。
この実施例で製造された抗体を2つの部分に分ける:1つはグリカン分析に使用し、他はADCC活性の測定に取っておく。
2.レクチンELISA によるガラクトース定量
レクチンはその認識特異性により生物学および医学の多くの用途、特にELISA 法によるグリカン分析に使用されてきた。β1,4 に結合したガラクトースを認識するレクチンRCA1を、抗体のN-グリカン中に存在するガラクトースを定量するのに使用した。
【0097】
モノクローナル抗体をマイクロタイトレーションプレートのウェルに固定する。Fc領域のN-グリカンを利用できるように、IgG分子を100 ℃で20分加熱して変性させた後、ウェルを室温、ビオチニル化RCA1溶液 (Vector) の存在下穏やかに攪拌しながら2時間温置する。未反応レクチンを除去するための洗浄の後、ストレプトアビジンペルオキシダーゼを各ウェルに添加し、1時間温置し、そして固定化レクチンをO-フェニレンジアミンの添加後 492nmで測定する。並行して、マイクロタイトレーションプレートのウェル中の固定化抗体の量を、ペルオキシダーゼで標識したヒト抗−IgG抗体により測定する。
【0098】
次いで、固定化レクチンの量をマイクロタイトレーションウェル中の固定化抗体の量により補正する。
3.CD16受容体活性化の測定
ガラクトシル化モノクローナル抗体のCD16受容体の活性化の測定に使用する操作条件は上に記載したものと同じである。
【0099】
この実施例に記載されるモノクローナル抗体は、同じ主要配列を有し、YB2/0 細胞により産生された抗-Rh(D)抗体である。それらは、EMAB2 については25%、EMAB3 については53%のα1,6-フコシル化率に関して、その機能活性において異なる。
【0100】
ベータ1,4-ガラクトシルトランスフェラーゼのin vitro作用の後、EMAB2 およびEMAB3 モノクローナル抗体により誘導されるCD16活性化はそれぞれ10および54%向上する (図5) 。このように、本来非常に良好なエフェクター活性を有するEMAB2 抗体のガラクトシル化の増加は、CD16活性化をわずかに向上させるだけであるが、高度にフコシル化されているEMAB3 抗体のガラクトシル化における増加は、CD16活性の非常に顕著な向上を示す。
【実施例5】
【0101】
EMAB2 抗-Rh(D)モノクローナル抗体により感作された赤血球のクリアランスの検討
EMAB2 抗-Rh(D)モノクローナル抗体により感作された赤血球のクリアランスを、診療所で使用されている抗-Rh(D)ポリクローナル抗体の治療用製剤であるRhophylac TMにより感作された赤血球のものと比較するために、EMAB2 抗-Rh(D)モノクローナル抗体を臨床試験第I相にて評価した。
【0102】
健康なボランティアの赤血球をクロム51 (51Cr) でex vivo にて標識し、感作する、即ち、ボランティアに再注入する前に抗原性部位の25%飽和レベルを得るために、抗-Rh(D)抗体であるEMAB2 またはRhophylac TMの存在下で温置する。
【0103】
51Crで標識した赤血球の血流中での消失を、標識した感作赤血球を注入して3、15、30分および1、2、4、6、8、10、24、48、72、96時間後に採取した血液サンプルにおいてガンマカウンターで放射能を測定することにより追跡した。赤血球の注入3分後に採取した血液サンプルは100 %の赤血球の生存を示す。
【0104】
図6に示す結果は、抗体による放射標識赤血球の感作を行わない場合、100 時間より長い時間にわたり測定した放射能の減少は20%より少ないことを示している。しかし、赤血球がポリクローナル抗体治療用製剤またはEMAB2 モノクローナル抗体で感作される場合、血液の放射能は急激に減少し;注入後10時間は注入された放射能の10%より少ない状態である。このように、EMAB2 モノクローナル抗体で感作された赤血球の消失曲線は、Rhophylac TMポリクローナル抗体の治療用製剤により感作された赤血球のものと似たプロフィールを有する。
【0105】
フコース含量/ガラクトース含量が0.4 であるEMAB2 モノクローナル抗体は、予備感作Rh(D+)赤血球のクリアランスに関して、少なくともポリクローナル抗体の治療用製剤のものに匹敵するin vivo 活性を有する。同じ条件であるが、MonoD と称される別のモノクローナル抗体を用いて行った臨床検討では、非常に異なる結果を与えた;膜抗原性部位の25%飽和では、MonoD により生じるクリアランスは部分的にすぎない。MonoD 抗体のグリカン分析により、フコース含量が80%でガラクトース含量が86%である、即ち、比が0.93であることが明らかになった。
【0106】
従って、これらの臨床結果を比較すると、フコース含量/ガラクトース含量の比が0.6 以下である抗-Dモノクローナル抗体は、その比が1に近い抗体よりも赤血球のクリアラン
スに対してより高い効果を有することが分かる。
【実施例6】
【0107】
CHO およびYB2/0 細胞株により発現された抗-HLA DR モノクローナル抗体のガラクトース含量の改変
1.抗-HLA DR モノクローナル抗体の製造
1.1 発現ベクターの作製
これらの検討で用いる抗-HLA DR 抗体は、Lym-1 ハイブリドーマ (ATCC Hb-8621)により発現されるIgG2a アイソタイプマウス抗体のキメラ化に由来する。
【0108】
マウス抗体を産生するハイブリドーマから抽出した RNAをcDNAに変換した。マウスVK領域をK-Lym-Not1およびK-Lym-Dra3プライマーにより増幅し、次いで、予めNot1およびDra3で消化し、ヒト抗-D抗体のCK配列およびDHFR選択遺伝子を含有するキメラ化ベクターCK-Hu にクローン化した。
【0109】
マウスVH領域をH-Lym-Not1およびH-Lym-Apa1プライマーにより増幅し、次いで、予めNot1およびApa1で消化し、ヒト抗-D抗体のG1配列およびNEO 選択遺伝子を含有するキメラ化ベクターG1-Hu にクローン化した。
【0110】
hEF-1aプロモーターおよび非コーディングエクソン1および第1イントロンを含有するhEF-1a遺伝子の5'UTR 領域を市販のプラスミドpEF/Bsd (Invitrogen)からNhe 1 およびAcc 65 Iの二重消化により分離した。並行して、上記発現ベクターに存在するRSV プロモーターをBgl IIおよびSpe I の二重消化により除去し、次いでNhe I-Acc65 I 断片で置換した。
【0111】
1.2 安定な産生株の獲得
抗-HLA DR キメラ抗体の軽鎖および重鎖をコードする発現ベクターpEF-Lym-dhfr-K-10 およびpEF-Lym-neo-H-12を、エレクトロポレーションによりCHO-DXB11 (ATCC No. CRL-11397)およびYB2/0 (ATCC No. CRL-1662) 株をそれぞれコトランスフェクトするために使用した。
【0112】
トランスフェクション後、培養細胞を、一方では培地のヌクレオシドへの欠失および他方ではG418k 添加からなる二重選択圧にかける。この二重選択圧に耐性の形質転換体を次いで、希釈を制限することよりクローン化した。
【0113】
2つの選択されたクローンは、YB2/0 発現細胞株についてはYB2/0-DR-4B7であり、CHO-DXB11 発現細胞株についてはDXB11-DR-22A10である。
1.3 抗-HLA DR キメラ抗体の作製および精製
クローンYB2/0-DR-4B7を、EM-SF1.1培地、インスリン (1μg/ml)、クエン酸鉄 (50μg/ml)、HEPES (4mg/ml)およびPluronic F68 (0.5mg/ml) を添加したEMS 基本培地で10リットルの細胞培養バイオリアクター (Biolafitte) 中で増殖させた。
【0114】
クローンDXB11-DR-22A10を、2%ヒポキサンチンを添加したCHO SFM4ユーティリティ培地 (Perbio) で10リットルの細胞培養バイオリアクター (Biolafitte) 中で増殖させた。
細胞の生存率が50%より低くなったら、細胞を除去するために培地を集め、遠心分離し、上清に含まれるキメラ抗体をセファロース−プロティインAでのアフィニティークロマトグラフィーにより精製する。
2.脱ガラクトシル化
抗-HLA DR キメラ抗体を50mM 酢酸ナトリウム緩衝液 (pH5.5 、CaCl2 4mM含有) で透析した。抗体を、コレラ菌 (Vibrio cholerae)由来のノイラミニダーゼ (EC 3.2.1.18) (Calbiochem) 5mUおよびE.coli産生のβ−ガラクトシダーゼ (EC 3.2.1.23) (Roche)9mUの存在下で温置することにより脱ガラクトシル化した。対照は、上記のように、しかしノイラミニダーゼおよびβ−ガラクトシダーゼの不在下で処理した同じ抗体調製物からなる。抗体を37℃で25時間温置後、4℃で保存する。
【0115】
この実施例で製造された抗体を2つの部分に分ける:1つはグリカン分析に使用し、他は機能活性の測定に取っておく。
2.CD16活性化の測定
Raji細胞株を、その表面にHLA-DR主要組織適合遺伝子複合体の抗原決定基を有する標的として用いる。
【0116】
抗体希釈物50μl、6.105/mlのRaji細胞懸濁液50μl、1.106/mlのJurkat CD16 細胞懸濁液50μlおよび40ng/ml のPMA 溶液50μlを順次96−ウェルマイクロタイトレーションプレートに入れた。希釈はすべて、5%FCS を含有するEMS 培地で行った。
【0117】
7%CO2 、37℃において16時間温置した後、マイクロタイトレーションプレートを遠心分離し、上清に含まれるIL-2の量を市販のキット (Duoset, R&D)で測定する。分泌されたIL-2の濃度をpg/ml で表す。
【0118】
結果はCD16活性化の割合 (%) として表し、対照モノクローナル抗体の存在下の分泌IL-2濃度は100 %に等しいと考えられる。
抗-HLA DR キメラ抗体は、YB2/0 株で発現されるかCHO DXB11 株で発現されるかによっって非常に異なるグリカン構造を有する。このように、YB2/0 で発現された抗体のフコース含量/ガラクトース含量の比は0.37であり、CHO で発現された抗体については比は1.3 でありずっと高い。
【0119】
天然の抗体のCD16活性化は、フコース含量/ガラクトース含量比の値に一致する;このように、YB2/0 で合成され、比が0.37である抗-HLA DR 抗体により誘導されるIL-2分泌は、CHO DXB11 により合成され、比が1.3 である同じ種類の抗体により誘導される分泌の2倍である。
【0120】
β−ガラクトシダーゼの作用の後、Fc領域のN-グリカン上に残るガラクトース含量をHPCE-LIFにより決定した。脱ガラクトシル化はほぼ完全であり、CHO により産生される抗体に対するG1形態のレベル、およびYB2/0 により産生される抗体についてのG1B 形態のレベルは、それぞれ7%および4.4 %である。このガラクトース含量の低下は、図7に示すように、対照抗体と比較してCD16活性化の著しい減少を示す。
参考文献
Boyd PN, Lines AC, Patel AK.「Campath-1Hの機能活性に及ぼすシアル酸、ガラクトースおよび全炭水化物の除去の影響」(1995) Mol. Immunol, 32, 1311-1318.
Davies J, Jiang L, Pan LY, Labarre MJ, Anderson D, Reff M.「組換え抗-CD20 CHO 産生細胞系におけるGnTIIIの発現:変化した糖形態を有する抗体の発現はFCガンマRIIIに対するより高い親和性を介したADCCの増加をもたらす」(2001) Biotechnol.Bioeng. 74, 288-294.
Furukawa K, Kodbata A.「IgGガラクトシル化−その生物学的意味および病理学」 (1991) Mol. Immunol.28, 1333-1340.
Kohler G, Milstein C. 「規定の特異性を有する抗体を分泌する融合細胞の連続培養」 (1995) Nature 256, 495-7.
Kumpel BM, Rademacher TW, Rook GA, Williams PJ, Wilson IB. 「EBV でトランスフォームしたB-リンパ芽球様細胞系により産生されたヒトIgGモノクローナル抗-Dのガラクトシル化は培養方法に依存し、Fc受容体媒介機能活性に影響する」 (1994) Hum. Antibodies Hybridomas 5, 143-451.
Nose M, Wigzell H.「モノクローナル抗体上の炭水化物鎖の生物学的意味」(1983) Proc.Natl.Acad.Sci.USA 80, 6632-6636.
Schields RL, Lai m Keck R, O'Connell LY, Hong K, Meng YG, Weiler SHA, Presta LG.「ヒトIgG1 N-結合オリゴ糖上のフコースの欠如はヒトFcγRIII への結合および抗体依存性細胞傷害を改善する」(2002) J.Biol.Chem. 277, 26733-26740.
Shinkawa T, Nakamura K, Yamane N, Shoji-Hosaka E, Kanda Y, Sakurada M, Uchida K, Anazawa H, Satoh M, Yamasaki M, Hanai N, Shitara K 「ヒトIgG1複合型オリゴ糖においてフコースが欠如しているが、ガラクトースまたは二分岐N-アセチルグルコサミンも存在しないことは、抗体依存性細胞傷害を増強する重要な役割を示す」 (2003) J. Biol.Chem. 278, 3466-3473.
Tao MH, Morrison SL 「非グリコシル化キメラマウス−ヒトIgGの検討:ヒトIgG定常領域を介した構造およびエフェクター機能における炭化水素の役割」(1989) J. Immunol. 143, 2595-2601.
Tsuchyia N, Endo T, Matsuta K, Yoshinoya S, Aikawa T, Kosuge E, Takeuchi F, Miyamoto T, Kobata A「ヒトIgGの免疫活性に及ぼすオリゴ糖からのガラクトース除去の影響」 (1989) J.Rheumatol. 16, 285-290.
Umana P, Jean-Mairet J, Moudry r, Amstutz H, Bailey JE「最適化抗体依存性細胞傷害活性を有する抗神経芽腫IgG1 の遺伝子工学処理したグリコ形態」(1999) Nat.Biotechnol. 17, 176-180.
Wright A, Morrioson SL「Igエフェクター機能に及ぼすC2関連炭水化物構造の影響:チャイニーズハムスター卵巣細胞1のグリコシル化変異体におけるキメラマウス−ヒトIgG1抗体を用いた検討」(1998) J.Immunol. 160, 3393-3402.
【図面の簡単な説明】
【0121】
【図1】各種抗-Rh(D)抗体のFc領域のグリコシル化部位上に存在するグリカン構造を示す。この図は、次の3種類の抗-Rh(D)抗体のAsn297残基が有する各種グリカン形態の割合 (%) を図示する:WinRhoの抗-D IgG1(黒色) 、モノクローナルEMAB2 抗体 (白色) および抗-D1(斜線) 。
【図2】フコース含量/ガラクトース含量の比と抗-Rh(D)抗体のADCC活性の間の相関線を示す。
【図3】抗-Rh(D)抗体のADCC活性に及ぼすガラクトースの影響を示す。この図は、0.5 および2.5mg/ml濃度の多価IgG (Tegeline, LFB)の存在下での、脱ガラクトシル化 (Degal)または脱ガラクトシル化されない (対照) 抗-Rh(D)ポリクローナル抗体により生じるRh(D+)赤血球の溶解率 (%) を示す。
【図4】脱ガラクトシル化抗-Rh(D)モノクローナル抗体のCD16活性化を示す。この図は、脱ガラクトシル化 (白色) または脱ガラクトシル化されない (対照、黒色)EMAB2およびHH01抗-Rh(D)モノクローナル抗体の存在により生じるCD16活性化の割合 (%) を図示する。
【図5】ガラクトシル化抗-Rh(D)モノクローナル抗体のCD16活性化を示す。この図は、ウシβ1,4-ガラクトシルトランスフェラーゼによるin vitroガラクトシル化の前 (対照、黒色) および後 (白色) のEMAB2 およびEMAB3 抗-Rh(D)モノクローナル抗体により生じるCD16活性化を図示する。
【図6】抗-RH(D)抗体により感作された、またはされていない放射標識赤血球のクリアランス曲線を示す。この図は、Rhophylac TMポリクローナル抗体の治療用製剤 (●) またはEMAB2 モノクローナル抗体 (■、▲、△) により感作されたか、感作されない (◆、◇) 、Cr51放射標識赤血球の一定量を再注入したボランティアの血液中に含まれる放射能の追跡 (%で表示) を図示する。EMAB2 抗体は三人のボランティアで試験した (008 、009 および010)。
【図7】YB2/0 およびCHO-DG44細胞株で発現した抗-HLA DR モノクローナル抗体の脱ガラクトシル化のCD16活性化に及ぼす影響を示す。この図は、Jurkat CD16 細胞により分泌されるIl-2の量 (pg/mlで表示) を図示し、CD16受容体はその膜上にHLA DR分子を有するRaji細胞の存在下で天然の (実線) または脱ガラクトシル化 (点線) 抗-HLA DR キメラ抗体により活性化されている。
【特許請求の範囲】
【請求項1】
下記工程を含むことを特徴とする、高いエフェクター活性を有するヒト化またはヒトキメラモノクローナル抗体を製造する方法:
a)各種供給源、特に、場合により遺伝的に改変されているか形質転換されている細胞、植物またはヒト以外の動物から得られるモノクローナル抗体を製造し精製すること、
b)該抗体のFc領域のグリコシル化部位が有するグリカン (多糖) 構造のフコース含量およびガラクトース含量を測定すること、および
c)フコース含量/ガラクトース含量の比が0.6 以下、好ましくは0.5 または0.4 以下である抗体を選択すること。
【請求項2】
前記抗体が、該抗体の発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変した細胞において製造され、該細胞は真核または原核細胞、特に哺乳類、昆虫、植物、細菌または酵母由来の細胞であることを特徴とする、請求項1記載の方法。
【請求項3】
前記細胞が、グリコシルトランスフェラーゼ活性を有する少なくとも1つのポリペプチドの発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変される、請求項1または2記載の方法。
【請求項4】
前記グリコシルトランスフェラーゼ活性がガラクトシルトランスフェラーゼ活性である請求項3記載の方法。
【請求項5】
前記ガラクトシルトランスフェラーゼ活性がベータ(1,4)-ガラクトシルトランスフェラーゼ活性またはベータ(1,3)-ガラクトシルトランスフェラーゼ活性であることを特徴とする、請求項4記載の方法。
【請求項6】
前記細胞が、GDP-フコースの合成および/または輸送に関与する活性、並びに/または抗体のグリコシル化部位のオリゴ糖へのフコースの付加に関与する酵素の活性が低減または欠失していることを特徴とする請求項1〜5のいずれかの項記載の方法。
【請求項7】
GDP-フコースの合成に関与する酵素が、GMD (GDP-D- マンノース 4,6- デヒドラターゼ) 、Fx (GDP-ケト-6- デオキシマンノース 3,5- エピメラーゼ、4-レダクターゼ) またはGFPP (GDP-ベータ-L- フコースピロホスホリラーゼ) であることを特徴とする請求項6記載の方法。
【請求項8】
フコースの付加に関与する酵素がフコシルトランスフェラーゼであることを特徴とする請求項6記載の方法。
【請求項9】
工程b)において測定した比が0.6 より大きい場合、工程c)の前に脱フコシル化を行う、および/またはガラクトース残基を前記抗体に付加することを特徴とする、請求項1〜8のいずれかの項記載の方法。
【請求項10】
前記脱フコシル化を抗体含有培地中にフコシダーゼを添加することにより行うことを特徴とする、請求項9記載の方法。
【請求項11】
ガラクトース残基の付加を、抗体含有培地にガラクトシルトランスフェラーゼを添加することにより行うことを特徴とする、請求項8または9記載の方法。
【請求項12】
前記細胞が、動物またはヒト細胞株由来であり、該株が特に、ラットのミエローマ株 (特にYB2/0 およびIR983F) 、ヒトミエローマ株 (例、Namalwa)、またはヒト起源のその他の任意の細胞 (例、PERC6)、 CHO株, 特にCHO-K, CHO-Lec10, CHO-Lec1, CHO Pro-5, CHO dhfr-, CHO Lec13 株、または Wil-2, Jurkat, Vero, Molt-4, COS-7, 293-HEK, BHK, K6H6, NSO, SP2/0-Ag 14 およびP3X63Ag8.653から選択されたその他の株から選択されることを特徴とする、請求項1〜11のいずれかの項記載の方法。
【請求項13】
前記細胞がIgG 型のヒト免疫グロブリンであることを特徴とする、請求項1〜12のいずれかの項記載の方法。
【請求項14】
抗体が抗-Rhesus 因子 (抗-D) 、抗-CD 、抗−腫瘍、抗−ウイルス、抗-CD20 、または抗-HLA-DR であることを特徴とする、請求項1〜13のいずれかの項記載の方法。
【請求項15】
前記エフェクター活性がADCC型機能活性であることを特徴とする、請求項1〜14のいずれかの項記載の方法。
【請求項16】
免疫学的機能性分子の組成物のエフェクター活性を向上させる方法であり、該分子組成物中のガラクトース含量の増加、および/またはフコース含量の減少を含む、前記方法。
【請求項17】
前記免疫学的機能性分子がモノクローナル抗体またはポリクローナル抗体であることを特徴とする請求項16記載の方法。
【請求項18】
前記分子が本来の状態では高いフコース含量を有することを特徴とする、請求項16または17記載の方法。
【請求項19】
フコース含量の減少が、フコシダーゼ、特にα1,6-フコシダーゼの作用による前記組成物の脱フコシル化によることを特徴とする、請求項16〜18のいずれかの項記載の方法。
【請求項20】
前記組成物のガラクトース含量の増加が、ガラクトシルトランスフェラーゼの作用による組成物のガラクトシル化によることを特徴とする、請求項16〜19のいずれかの項記載の方法。。
【請求項21】
抗体分子をコードする少なくとも1つのベクターが導入され、抗体Fc領域のグリコシル化部位のオリゴ糖のフコース含量/ガラクトース含量の比が0.6 以下である抗体を産生する、YB2/0 細胞株由来の細胞。
【請求項22】
ガラクトシルトランスフェラーゼをコードする発現ベクターでトランスフェクされることを特徴とする請求項21記載の細胞。
【請求項23】
前記ガラクトシルトランスフェラーゼがベータ(1,4)-ガラクトシルトランスフェラーゼまたはベータ(1,3)-ガラクトシルトランスフェラーゼであることを特徴とする、請求項21または22記載の細胞。
【請求項24】
前記細胞がガラクトシルトランスフェラーゼを過発現することを特徴とする、請求項21〜23のいずれかの項記載の細胞。
【請求項25】
前記ガラクトシルトランスフェラーゼが、ヒト、マウス、ハムスター、ウシ、ヒツジ、ヤギ、ブタ、ウマ、ラット、サル、ウサギ、またはトリ由来の配列によりコードされることを特徴とする請求項21〜24のいずれかの項記載の細胞。
【請求項26】
前記配列がNM 001497, AB 024434, NM 003780, BC 053006, XM 242992 またはNM 177512 配列であることを特徴とする請求項25記載の細胞。
【請求項27】
前記ベクターの発現を可能にする培地および条件下で請求項21〜26のいずれかの項記載の細胞を培養することを含む、Fc領域のグリコシル化部位に存在するグリカン構造が0.6 以下、好ましくは0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有する抗体を製造する方法。
【請求項28】
0.6 より小さい、好ましくは0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有するグリカン構造をFc領域のグリコシル化部位に含有することを特徴とする、請求項1〜20および27のいずれかの項記載の方法で得ることができる高いエフェクター活性を有する治療用抗体。
【請求項29】
請求項28記載の抗体および少なくとも1つの賦形剤を含む薬剤組成物。
【請求項30】
Fc領域のグリコシル化部位に存在するグリカン構造が0.6 より小さい、優先的には0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有するモノクローナル抗体を少なくとも50%、好ましくは60%、70%、80%、90%または99%含む薬剤組成物。
【請求項31】
抗体が、非遍在の通常抗原、特にRhesus因子 (例、ヒト赤血球細胞のRh因子(D))、またはヒトに対する病原性細胞もしくは病原性生物の抗原、特にがん細胞の抗原、に対する、請求項29または30記載の薬剤組成物。
【請求項32】
前記抗体がIgGである、請求項29〜31のいずれの項記載の薬剤組成物。
【請求項33】
同種免疫、特に新生児溶血性疾患の治療用薬剤を製造するための、請求項28記載の抗体の使用。
【請求項34】
自己免疫疾患、がん、および病原体による感染症の治療、特にセザリー症候群、固形がん、特に抗原性標的が弱く発現したもの、特に、乳がん、ポリ塩化ビフェニルに暴露されたヒトが罹患する環境に関連する病状、感染症 (特に結核) 、慢性疲労症候群(CFS) 、寄生虫感染症 (例、住血吸虫) およびウイルス感染症、の治療用薬剤を製造するための請求項28記載の抗体の使用。
【請求項35】
陽性クラスII HLA細胞のがん、BおよびT細胞の急性リンパ性白血病、急性および慢性骨髄性白血病、バーキットリンパ腫、ホジキンリンパ腫、骨髄性白血病、T細胞リンパ腫および非ホジキンリンパ腫の治療用薬剤を製造するための請求項28記載の抗体の使用。
【請求項36】
抗体が抗-HLA-DR または抗-CD20 であることを特徴とする、請求項33〜35のいずれかの項記載の使用。
【請求項37】
免疫システムの天然のエフェクター細胞によりIL-1α、IL-1β、IL-2、IL-3、IL-4、IL-5、IL-6、IL-12 、IL-18 、IL-21 、TGF β1 、TGF β2 、TNF α、TNF β、INF γ、およびIP10の発現を誘導するための薬剤を製造するための請求項28記載の抗体の使用。
【請求項38】
CD16多型の1つ、特にV/F158またはF/F158を有する患者、特に現在利用可能な抗体では治療できない症状の患者、または望ましくない二次的作用を受けている患者、の治療用の薬剤を製造するための請求項28記載の抗体の使用。
【請求項39】
下記工程を含むことを特徴とする、低いエフェクター活性を有する、ヒトまたはヒト化キメラモノクローナル抗体を製造する方法:
a)各種供給源、特に、場合により遺伝的に改変されているか形質転換されている細胞、植物またはヒト以外の動物から得られるモノクローナル抗体を製造し精製すること、
b)該抗体のFc領域のグリコシル化部位に存在するグリカン (多糖) 構造のフコース含量およびガラクトース含量を測定すること、および
c)フコース含量/ガラクトース含量の比が0.6 より大きい抗体を選択すること。
【請求項40】
前記抗体が、該抗体の発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変した細胞において産生され、この細胞は真核細胞でも原核細胞でもよく、特に哺乳動物、昆虫、植物、細菌または酵母由来の細胞であることを特徴とする請求項39記載の方法。
【請求項41】
前記細胞が、グリコシルトランスフェラーゼ活性を有する少なくとも1つのポリペプチドの発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変されることを特徴とする、請求項39または40記載の方法。
【請求項42】
前記グリコシルトランスフェラーゼ活性がフコシルトランスフェラーゼ活性、特にα1.6-フコシルトランスフェラーゼ活性であることを特徴とする請求項41記載の方法。
【請求項43】
前記細胞が、UDP-ガラクトースの合成および/または輸送に関する活性、並びに/または抗体のグリコシル化部位のオリゴ糖へガラクトースを付加することに関与する酵素の活性が低下または欠失している、請求項39〜42のいずれの項記載の方法。
【請求項44】
ガラクトースの付加に関与する前記酵素が、ガラクトシルトランスフェラーゼ、特にβ1,4-ガラクトシルトランスフェラーゼであることを特徴とする請求項43記載の方法。
【請求項45】
工程b)において測定した比が0.6 より小さい場合、工程c)の前にフコシル化を行う、および/またはガラクトース残基を抗体より除去することを特徴とする、請求項39〜44のいずれかの項記載の方法。
【請求項46】
前記脱ガラクトシル化を、抗体含有培地にガラクトシダーゼを添加することにより行うこと特徴とする請求項45記載の方法。
【請求項47】
フコース残基の付加を、抗体含有培地へのフコシルトランスフェラーゼの添加により行うことを特徴とする、請求項45または46記載の方法。
【請求項48】
前記抗体がIgG型のヒト免疫グロブリンであることを特徴とする請求項39〜47のいずれかの項記載の方法。
【請求項49】
前記抗体が、ヒト血球の分化マーカーであるCDに対するか、またはバイオテロの場合には特に危険であるとして挙げられている病原体もしくはその毒素、特にバチルス・アントラシス (Bacillus anthracis) 、クロストリジウム・ボツリウム (Clostridium botulium) 、エルシニア・ペスティス (Yersinia pestis)、バリオラ・マジョル (Variola major)、フランシセラ・ツラレンシス(Francisella tularensis)、フィロウイルス(filovirus) 、アレナウイルス (arenavirus) 、ブルセラ・スピーシーズ (Brucella species) 、クロストリジウム・ペルフリンゲンス (Clostridium perfringens)、サルモネラ (Salmonellla)、大腸菌 (E.coli) 、赤痢菌 (Shigella) 、コクシエラ・バーネッティイ (Coxiella burnetii)、リシン毒素、リケッチア (Rickettsia) 、ウイルス性脳炎ウイルス、ビブリオコレラ (Vibrio cholerae)またはハンタウイルス、に対するものであることを特徴とする、請求項39〜48のいずれかの項記載の方法。
【請求項50】
エフェクター活性がADCC型機能活性であることを特徴とする、請求項39〜49のいずれかの項記載の方法。
【請求項51】
免疫機能性分子の組成物の活性を低下させる方法であり、組成物のフコース含量の増加および/またはガラクトース含量の低下を含む、前記方法。
【請求項52】
免疫機能性分子がモノクローナルまたはポリクローナル抗体であることを特徴とする請求項51記載の方法。
【請求項53】
フコース含量の増加が、フコシルトランスフェラーゼの作用による前記組成物のフコシル化によることを特徴とする請求項51または52記載の方法。
【請求項54】
前記組成物のガラクトース含量の低下が、ガラクトシダーゼの作用による組成物の脱ガラクトシル化によることを特徴とする請求項51〜53のいずれかの項記載の方法。
【請求項55】
請求項39〜54のいずれかの項記載の方法により得ることができる抗体組成物。
【請求項56】
自己免疫疾患、同種免疫、特にPTI 、移植片拒絶、アレルギー、喘息、皮膚炎、蕁麻疹、紅班または炎症性疾患を治療および/または予防する薬剤を製造するための、請求項55記載の組成物の使用。
【請求項57】
抗体のFc領域のグリコシル化部位のオリゴ糖におけるフコース含量/ガラクトース含量の比を調節することを含む、免疫機能性分子の組成物の活性を制御する方法。
【請求項1】
下記工程を含むことを特徴とする、高いエフェクター活性を有するヒト化またはヒトキメラモノクローナル抗体を製造する方法:
a)各種供給源、特に、場合により遺伝的に改変されているか形質転換されている細胞、植物またはヒト以外の動物から得られるモノクローナル抗体を製造し精製すること、
b)該抗体のFc領域のグリコシル化部位が有するグリカン (多糖) 構造のフコース含量およびガラクトース含量を測定すること、および
c)フコース含量/ガラクトース含量の比が0.6 以下、好ましくは0.5 または0.4 以下である抗体を選択すること。
【請求項2】
前記抗体が、該抗体の発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変した細胞において製造され、該細胞は真核または原核細胞、特に哺乳類、昆虫、植物、細菌または酵母由来の細胞であることを特徴とする、請求項1記載の方法。
【請求項3】
前記細胞が、グリコシルトランスフェラーゼ活性を有する少なくとも1つのポリペプチドの発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変される、請求項1または2記載の方法。
【請求項4】
前記グリコシルトランスフェラーゼ活性がガラクトシルトランスフェラーゼ活性である請求項3記載の方法。
【請求項5】
前記ガラクトシルトランスフェラーゼ活性がベータ(1,4)-ガラクトシルトランスフェラーゼ活性またはベータ(1,3)-ガラクトシルトランスフェラーゼ活性であることを特徴とする、請求項4記載の方法。
【請求項6】
前記細胞が、GDP-フコースの合成および/または輸送に関与する活性、並びに/または抗体のグリコシル化部位のオリゴ糖へのフコースの付加に関与する酵素の活性が低減または欠失していることを特徴とする請求項1〜5のいずれかの項記載の方法。
【請求項7】
GDP-フコースの合成に関与する酵素が、GMD (GDP-D- マンノース 4,6- デヒドラターゼ) 、Fx (GDP-ケト-6- デオキシマンノース 3,5- エピメラーゼ、4-レダクターゼ) またはGFPP (GDP-ベータ-L- フコースピロホスホリラーゼ) であることを特徴とする請求項6記載の方法。
【請求項8】
フコースの付加に関与する酵素がフコシルトランスフェラーゼであることを特徴とする請求項6記載の方法。
【請求項9】
工程b)において測定した比が0.6 より大きい場合、工程c)の前に脱フコシル化を行う、および/またはガラクトース残基を前記抗体に付加することを特徴とする、請求項1〜8のいずれかの項記載の方法。
【請求項10】
前記脱フコシル化を抗体含有培地中にフコシダーゼを添加することにより行うことを特徴とする、請求項9記載の方法。
【請求項11】
ガラクトース残基の付加を、抗体含有培地にガラクトシルトランスフェラーゼを添加することにより行うことを特徴とする、請求項8または9記載の方法。
【請求項12】
前記細胞が、動物またはヒト細胞株由来であり、該株が特に、ラットのミエローマ株 (特にYB2/0 およびIR983F) 、ヒトミエローマ株 (例、Namalwa)、またはヒト起源のその他の任意の細胞 (例、PERC6)、 CHO株, 特にCHO-K, CHO-Lec10, CHO-Lec1, CHO Pro-5, CHO dhfr-, CHO Lec13 株、または Wil-2, Jurkat, Vero, Molt-4, COS-7, 293-HEK, BHK, K6H6, NSO, SP2/0-Ag 14 およびP3X63Ag8.653から選択されたその他の株から選択されることを特徴とする、請求項1〜11のいずれかの項記載の方法。
【請求項13】
前記細胞がIgG 型のヒト免疫グロブリンであることを特徴とする、請求項1〜12のいずれかの項記載の方法。
【請求項14】
抗体が抗-Rhesus 因子 (抗-D) 、抗-CD 、抗−腫瘍、抗−ウイルス、抗-CD20 、または抗-HLA-DR であることを特徴とする、請求項1〜13のいずれかの項記載の方法。
【請求項15】
前記エフェクター活性がADCC型機能活性であることを特徴とする、請求項1〜14のいずれかの項記載の方法。
【請求項16】
免疫学的機能性分子の組成物のエフェクター活性を向上させる方法であり、該分子組成物中のガラクトース含量の増加、および/またはフコース含量の減少を含む、前記方法。
【請求項17】
前記免疫学的機能性分子がモノクローナル抗体またはポリクローナル抗体であることを特徴とする請求項16記載の方法。
【請求項18】
前記分子が本来の状態では高いフコース含量を有することを特徴とする、請求項16または17記載の方法。
【請求項19】
フコース含量の減少が、フコシダーゼ、特にα1,6-フコシダーゼの作用による前記組成物の脱フコシル化によることを特徴とする、請求項16〜18のいずれかの項記載の方法。
【請求項20】
前記組成物のガラクトース含量の増加が、ガラクトシルトランスフェラーゼの作用による組成物のガラクトシル化によることを特徴とする、請求項16〜19のいずれかの項記載の方法。。
【請求項21】
抗体分子をコードする少なくとも1つのベクターが導入され、抗体Fc領域のグリコシル化部位のオリゴ糖のフコース含量/ガラクトース含量の比が0.6 以下である抗体を産生する、YB2/0 細胞株由来の細胞。
【請求項22】
ガラクトシルトランスフェラーゼをコードする発現ベクターでトランスフェクされることを特徴とする請求項21記載の細胞。
【請求項23】
前記ガラクトシルトランスフェラーゼがベータ(1,4)-ガラクトシルトランスフェラーゼまたはベータ(1,3)-ガラクトシルトランスフェラーゼであることを特徴とする、請求項21または22記載の細胞。
【請求項24】
前記細胞がガラクトシルトランスフェラーゼを過発現することを特徴とする、請求項21〜23のいずれかの項記載の細胞。
【請求項25】
前記ガラクトシルトランスフェラーゼが、ヒト、マウス、ハムスター、ウシ、ヒツジ、ヤギ、ブタ、ウマ、ラット、サル、ウサギ、またはトリ由来の配列によりコードされることを特徴とする請求項21〜24のいずれかの項記載の細胞。
【請求項26】
前記配列がNM 001497, AB 024434, NM 003780, BC 053006, XM 242992 またはNM 177512 配列であることを特徴とする請求項25記載の細胞。
【請求項27】
前記ベクターの発現を可能にする培地および条件下で請求項21〜26のいずれかの項記載の細胞を培養することを含む、Fc領域のグリコシル化部位に存在するグリカン構造が0.6 以下、好ましくは0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有する抗体を製造する方法。
【請求項28】
0.6 より小さい、好ましくは0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有するグリカン構造をFc領域のグリコシル化部位に含有することを特徴とする、請求項1〜20および27のいずれかの項記載の方法で得ることができる高いエフェクター活性を有する治療用抗体。
【請求項29】
請求項28記載の抗体および少なくとも1つの賦形剤を含む薬剤組成物。
【請求項30】
Fc領域のグリコシル化部位に存在するグリカン構造が0.6 より小さい、優先的には0.5 または0.4 より小さいフコース含量/ガラクトース含量の比を有するモノクローナル抗体を少なくとも50%、好ましくは60%、70%、80%、90%または99%含む薬剤組成物。
【請求項31】
抗体が、非遍在の通常抗原、特にRhesus因子 (例、ヒト赤血球細胞のRh因子(D))、またはヒトに対する病原性細胞もしくは病原性生物の抗原、特にがん細胞の抗原、に対する、請求項29または30記載の薬剤組成物。
【請求項32】
前記抗体がIgGである、請求項29〜31のいずれの項記載の薬剤組成物。
【請求項33】
同種免疫、特に新生児溶血性疾患の治療用薬剤を製造するための、請求項28記載の抗体の使用。
【請求項34】
自己免疫疾患、がん、および病原体による感染症の治療、特にセザリー症候群、固形がん、特に抗原性標的が弱く発現したもの、特に、乳がん、ポリ塩化ビフェニルに暴露されたヒトが罹患する環境に関連する病状、感染症 (特に結核) 、慢性疲労症候群(CFS) 、寄生虫感染症 (例、住血吸虫) およびウイルス感染症、の治療用薬剤を製造するための請求項28記載の抗体の使用。
【請求項35】
陽性クラスII HLA細胞のがん、BおよびT細胞の急性リンパ性白血病、急性および慢性骨髄性白血病、バーキットリンパ腫、ホジキンリンパ腫、骨髄性白血病、T細胞リンパ腫および非ホジキンリンパ腫の治療用薬剤を製造するための請求項28記載の抗体の使用。
【請求項36】
抗体が抗-HLA-DR または抗-CD20 であることを特徴とする、請求項33〜35のいずれかの項記載の使用。
【請求項37】
免疫システムの天然のエフェクター細胞によりIL-1α、IL-1β、IL-2、IL-3、IL-4、IL-5、IL-6、IL-12 、IL-18 、IL-21 、TGF β1 、TGF β2 、TNF α、TNF β、INF γ、およびIP10の発現を誘導するための薬剤を製造するための請求項28記載の抗体の使用。
【請求項38】
CD16多型の1つ、特にV/F158またはF/F158を有する患者、特に現在利用可能な抗体では治療できない症状の患者、または望ましくない二次的作用を受けている患者、の治療用の薬剤を製造するための請求項28記載の抗体の使用。
【請求項39】
下記工程を含むことを特徴とする、低いエフェクター活性を有する、ヒトまたはヒト化キメラモノクローナル抗体を製造する方法:
a)各種供給源、特に、場合により遺伝的に改変されているか形質転換されている細胞、植物またはヒト以外の動物から得られるモノクローナル抗体を製造し精製すること、
b)該抗体のFc領域のグリコシル化部位に存在するグリカン (多糖) 構造のフコース含量およびガラクトース含量を測定すること、および
c)フコース含量/ガラクトース含量の比が0.6 より大きい抗体を選択すること。
【請求項40】
前記抗体が、該抗体の発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変した細胞において産生され、この細胞は真核細胞でも原核細胞でもよく、特に哺乳動物、昆虫、植物、細菌または酵母由来の細胞であることを特徴とする請求項39記載の方法。
【請求項41】
前記細胞が、グリコシルトランスフェラーゼ活性を有する少なくとも1つのポリペプチドの発現を可能にする少なくとも1つのベクターを導入することにより遺伝的に改変されることを特徴とする、請求項39または40記載の方法。
【請求項42】
前記グリコシルトランスフェラーゼ活性がフコシルトランスフェラーゼ活性、特にα1.6-フコシルトランスフェラーゼ活性であることを特徴とする請求項41記載の方法。
【請求項43】
前記細胞が、UDP-ガラクトースの合成および/または輸送に関する活性、並びに/または抗体のグリコシル化部位のオリゴ糖へガラクトースを付加することに関与する酵素の活性が低下または欠失している、請求項39〜42のいずれの項記載の方法。
【請求項44】
ガラクトースの付加に関与する前記酵素が、ガラクトシルトランスフェラーゼ、特にβ1,4-ガラクトシルトランスフェラーゼであることを特徴とする請求項43記載の方法。
【請求項45】
工程b)において測定した比が0.6 より小さい場合、工程c)の前にフコシル化を行う、および/またはガラクトース残基を抗体より除去することを特徴とする、請求項39〜44のいずれかの項記載の方法。
【請求項46】
前記脱ガラクトシル化を、抗体含有培地にガラクトシダーゼを添加することにより行うこと特徴とする請求項45記載の方法。
【請求項47】
フコース残基の付加を、抗体含有培地へのフコシルトランスフェラーゼの添加により行うことを特徴とする、請求項45または46記載の方法。
【請求項48】
前記抗体がIgG型のヒト免疫グロブリンであることを特徴とする請求項39〜47のいずれかの項記載の方法。
【請求項49】
前記抗体が、ヒト血球の分化マーカーであるCDに対するか、またはバイオテロの場合には特に危険であるとして挙げられている病原体もしくはその毒素、特にバチルス・アントラシス (Bacillus anthracis) 、クロストリジウム・ボツリウム (Clostridium botulium) 、エルシニア・ペスティス (Yersinia pestis)、バリオラ・マジョル (Variola major)、フランシセラ・ツラレンシス(Francisella tularensis)、フィロウイルス(filovirus) 、アレナウイルス (arenavirus) 、ブルセラ・スピーシーズ (Brucella species) 、クロストリジウム・ペルフリンゲンス (Clostridium perfringens)、サルモネラ (Salmonellla)、大腸菌 (E.coli) 、赤痢菌 (Shigella) 、コクシエラ・バーネッティイ (Coxiella burnetii)、リシン毒素、リケッチア (Rickettsia) 、ウイルス性脳炎ウイルス、ビブリオコレラ (Vibrio cholerae)またはハンタウイルス、に対するものであることを特徴とする、請求項39〜48のいずれかの項記載の方法。
【請求項50】
エフェクター活性がADCC型機能活性であることを特徴とする、請求項39〜49のいずれかの項記載の方法。
【請求項51】
免疫機能性分子の組成物の活性を低下させる方法であり、組成物のフコース含量の増加および/またはガラクトース含量の低下を含む、前記方法。
【請求項52】
免疫機能性分子がモノクローナルまたはポリクローナル抗体であることを特徴とする請求項51記載の方法。
【請求項53】
フコース含量の増加が、フコシルトランスフェラーゼの作用による前記組成物のフコシル化によることを特徴とする請求項51または52記載の方法。
【請求項54】
前記組成物のガラクトース含量の低下が、ガラクトシダーゼの作用による組成物の脱ガラクトシル化によることを特徴とする請求項51〜53のいずれかの項記載の方法。
【請求項55】
請求項39〜54のいずれかの項記載の方法により得ることができる抗体組成物。
【請求項56】
自己免疫疾患、同種免疫、特にPTI 、移植片拒絶、アレルギー、喘息、皮膚炎、蕁麻疹、紅班または炎症性疾患を治療および/または予防する薬剤を製造するための、請求項55記載の組成物の使用。
【請求項57】
抗体のFc領域のグリコシル化部位のオリゴ糖におけるフコース含量/ガラクトース含量の比を調節することを含む、免疫機能性分子の組成物の活性を制御する方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2007−533299(P2007−533299A)
【公表日】平成19年11月22日(2007.11.22)
【国際特許分類】
【出願番号】特願2006−534807(P2006−534807)
【出願日】平成16年10月20日(2004.10.20)
【国際出願番号】PCT/FR2004/002686
【国際公開番号】WO2005/040221
【国際公開日】平成17年5月6日(2005.5.6)
【出願人】(506129544)ラボラトワール・フランセ・デュ・フラクシオンマン・エ・デ・ビョテクノロジー (5)
【氏名又は名称原語表記】LABORATOIRE FRANCAIS DU FRACTIONNEMENT ET DES BIOTECHNOLOGIES
【Fターム(参考)】
【公表日】平成19年11月22日(2007.11.22)
【国際特許分類】
【出願日】平成16年10月20日(2004.10.20)
【国際出願番号】PCT/FR2004/002686
【国際公開番号】WO2005/040221
【国際公開日】平成17年5月6日(2005.5.6)
【出願人】(506129544)ラボラトワール・フランセ・デュ・フラクシオンマン・エ・デ・ビョテクノロジー (5)
【氏名又は名称原語表記】LABORATOIRE FRANCAIS DU FRACTIONNEMENT ET DES BIOTECHNOLOGIES
【Fターム(参考)】
[ Back to top ]