
播種性カンジダ症および他の感染性因子に対するワクチン接種をするための医薬組成物および方法
本発明は、医薬上許容され得る媒体中のアジュバントと共に、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンを提供する。本発明は、血行的に播種性カンジダ症または粘膜皮膚カンジダ症を治療または予防する方法も提供する。該方法は、医薬上許容され得る媒体中の、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンの免疫原性量を投与することを含む。

【発明の詳細な説明】
【技術分野】
【0001】
本発明はCandida albicans表面アドヘシンタンパク質、C.albicans表面接着タンパク質でのワクチン接種に対する免疫応答から得られた抗体、およびC.albicans表面接着タンパク質でカンジダ症および他の細菌感染を予防および/または治療する方法に関する。
【背景技術】
【0002】
(発明の背景)
近年、Candida種によって引き起こされた院内感染の発生率の劇的な増大があった。血行的に散在したカンジダ感染の発生率は1980年から1989年にかけて11倍増加した。この増大しつつある発生率は1990年代まで続いた。Candida種による感染は、今日、院内敗血症の第四番目の最も一般的な原因であり、Escherichia coliのそれと同等であり、Klebsiella種によって引き起こされた発生率を凌ぐ。さらに、Candida種は、ひどい怪我を有する患者において執拗な真菌感染の最も一般的な原因である。骨髄移植を受けている個人の11%まで、および同所性肝臓移植体を有する固体の13%までが侵入性カンジダ感染を発生するであろう。
【0003】
Candida albicans、この属における主な病原体は、2つの形態:出芽胞子で(出芽酵母)およびフィラメント状(菌糸および偽菌糸)相の間をスイッチすることができる。フィラメント化を調節する遺伝子に欠陥があるCandida突然変異体は、動物モデルにおいて低下したビルレンスを有すると報告されている。この低下したビルレンスは、出芽胞子からフィラメントへ変化する能力がC.albicansの鍵となるビルレンス因子であることを示唆する。今日まで、これらのフィラメント化経路の必須のエフェクタはC.albicansにおいて同定されていない。非特許文献1を参照。
【0004】
黄色ブドウ球菌(Staphylococcus aureus)感染もまた一般的であり、益々、抗生物質に対する薬物耐性をもたらす。例えば、黄色ブドウ球菌は合衆国および世界中で皮膚および皮膚構造感染、心内膜炎および菌血症の通常の原因である。以前、地域感染型の黄色ブドウ球菌(CA−S.aureus)感染は、セファゾリン、オキサシリン、メチシリン、ペニシリンおよびアモキシシリンのようなペニシリナーゼ−耐性ベータラクタムに対してほとんど均一に感受性であった。しかしながら、過去10年間に渡って、ベータ−ラクタム耐性黄色ブドウ球菌(MRSA)感染の伝染病は、世界中で多数の地域で見られている。特に、地域感染型MRSA(CA−MRSA)。多くの場所において、MRSAは、CA感染を引き起こす支配的な黄色ブドウ球菌株となった。最近の合衆国における3つの州での予測される集団−ベースの概観は、CA−MRSA感染の発生率が100,000集団当たり500症例であると見積もっており、これは合衆国単独において1年あたりほぼ150万症例に換算される。薬物−耐性黄色ブドウ球菌感染の増大する頻度は、これらの感染を予防し、治療するための新しい方法に対する要望に焦点を当てている。
【0005】
ビルレンスに寄与する生物の調節経路におけるエフェクタの同定は、現存の抗真菌剤よりもすぐれた方法および組成物での治療的介入に対する機会を提供する。ビルレンスに関与する調節経路に影響する細胞表面タンパク質の同定は特に有望である。なぜならば、タンパク質の特徴付けは、カンジダ感染と戦う場合に、現存の抗真菌剤よりも優れた免疫療法技術を可能とするからである。
【0006】
Candida albicansのビルレンスは、宿主構成要素へのその接着、および酵母から菌糸へ変換する能力が病原性を決定するにおいて最も臨界的なうちにあるいくつかの推定ビルレンス因子によって調節される。Candidaに対して殺菌性である優れた抗真菌剤が存在するが、カンジダ血症に帰される死亡率は、アンフォテリシンBのような優れた抗真菌剤での治療をもってさえほぼ38%である。また、アンフォテリシンBのような現存の剤は望ましくない毒性を呈する傾向がある。アンフォテリシンBよりも毒性が低いさらなる抗真菌剤を開発することができるが、より優れた剤は開発されないであろう。従って、播種性カンジダ症を治療または予防するための受動的または能動的免疫療法は、標準的な抗真菌療法に代えて有望である。
【0007】
かくして、Candida、黄色ブドウ球菌および他の免疫学的に関連する病原体に対する宿主免疫保護および受動的免疫保護を提供する効果的な免疫原に対する要望が存在する。本発明はこの要望を満足し、同様に、関連する利点を提供する。
【非特許文献1】Caesar−TonThat,T.C.and J.E.Cutler,“A monoclonal antibody to Candida albicans enhances mouse neutrophil candidacidal activity,”Infect.Immun.65:5354−5357,1997
【発明の開示】
【課題を解決するための手段】
【0008】
(発明の要旨)
本発明は、医薬上許容され得る媒体中のアジュバントと共に、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバーまたはその免疫原性断片を含むワクチンを提供する。また、本発明は、播種性カンジダ症を治療または予防する方法も提供する。該方法は、医薬上許容され得る媒体中の、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンの免疫原性量を投与することを含む。宿主細胞または組織へのCandidaの結合または侵入を阻害するのに有効な量の細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその機能的断片を投与することを含む播種性カンジダ症を治療または予防する方法も提供される。該Alsタンパク質ファミリーメンバーはCandida albicans、Candida krusei、Candida tropicalis、Candida glabrataおよびCandida parapsilosisよりなる群から選択されるCandida株に由来することができ、該Alsタンパク質ファミリーメンバーはAls1p、Als3p、Als5p、Als6p、Als7pまたはAls9pを含む。また、黄色ブドウ球菌感染を治療また予防する方法も提供される。該方法は、医薬上許容され得る媒体中の、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンの免疫原性量を投与することを含む。
【発明を実施するための最良の形態】
【0009】
(発明の詳細な説明)
Candida albicansおよび黄色ブドウ球菌(Staphylococcus aureus)はヒトにおける一般的な病原体である。例えば、C.albicansは通常は無害な片利共生生物であるが、この生物は膣および/または口腔咽頭カンジダ症のような表層粘膜皮膚感染から、播種性カンジダ症における深器官関与の範囲の種々の疾患を引き起こし得る。病気を引き起こすに先立って、真菌は胃腸管および、ある場合には皮膚および粘膜でコロニー化する。宿主粘膜表面への接着はこの初期工程の鍵となる要件である。コロニー化後に、C.albicansは感染された血管内デバイスを介して、または化学療法、またはストレス潰瘍によって危うくされる胃腸粘膜を通じる移動によって血流に入る。次いで、生物が血流を介して散在し、血管内皮に結合し、それに貫入して、血管ツリーから脱出し、肝臓、脾臓および腎臓のような深い器官に侵入する。
【0010】
本明細書中に記載された種々の例示的なAlsタンパク質ファミリーメンバーの同定および機能的特徴付けは、タンパク質のこのメンバーがカンジダ症の治療で効果的に利用されるのを可能とする。多用な基材および他の選択的細胞接着機能に対する特異的結合活性は、能動的または受動免疫のためのワクチンの生産において、宿主細胞の結合、吸着または侵入を阻害することによって初期感染を低下または予防するための細胞接着のペプチド、ミメティック阻害剤のアナログにおいて開発することができる。さらに、異なる結合および侵入プロフィールは、Alsタンパク質ファミリーメンバーの活性の広いスペクトルまたは標的化阻害の設計および使用を可能とする。加えて、結合および/または侵入活性を付与する機能的断片は、望まない外来性タンパク質配列の排除を可能とし、かくして、Alsファミリータンパク質メンバーワクチンまたは治療阻害剤の効力を増大させる。
【0011】
内皮細胞への接着によるC.albicansの病原性の性質は、ここに具体的に引用してその全体を援用する米国特許第5,578,309において議論されている。アドヘシンとしての遺伝子産物の特徴付けを含めた、Als1遺伝子およびその特徴の記載については、Fu,Y.,G.Rieg,W.A.Forizi,P.H.Belanger,J.E.J.Edwards,and S.G.Filler.1998.Expression of the Candida albicans gene ALS1 in Saccharomyces cerevisiae induces adherence to endothelial and epithelial cells.Infect.Immun.66:1783−1786;Hoyer,L.L.1997.Fu Y,Ibrahim AS,Sheppard DC,Chen Y−C,French SW,Cutler JE,Filler SG,Edwards,JE,Jr.2002.Candida albicans Als1p:an adhesin that is a downstream effector of the EFG1 filamentation pathway.Molecular Microbiology 44:61−72,Sheppard DC,Yeaman MR,Welch WH,Phan QT,Fu Y,Ibrahim AS,Filler SG,Zhang M,Waring AJ,Edwards,Jr.,JE 2004.Functional and Structural Diversity in the Als Protein Family of Candida albicans.Journal Biological Chemistry.279:30480−30489.The ALS gene family of Candida albicans.International Society for Human and Animal Mycology Salsimorge,Italy:(Abstract);Hoyer,L.L.,S.Scherer,A.R.Shatzman,and G.P.Livi.1995.Candida albicans ALSI:domains related to a Saccharonzyces cerevisiae sexual agglutinin separated by a repeating motif.Mol.Microbiol.15:39−54参照。
【0012】
この点に関しては、ヒト真菌病原体Candida albicansはコロニー化し、広く種々の宿主の組織に侵入する。宿主構成要素への接着はこのプロセスにおいて重要な役割を演じる。C.albicans Alsタンパク質ファミリーの2つのメンバー(Als1pおよびAls5p)は、接着を媒介することが判明しており、Alsタンパク質ファミリーメンバーの結合、接着および細胞侵入活性が例示される。本明細書中に記載されるように、Als遺伝子ファミリーの膜をクローン化し、S.cerevisiaeにおいて発現させて、それらの個々の機能を特徴付けた。区別されるAlsタンパク質は区別される接着プロフィールを多様な宿主基材に付与した。キメラAls5p−Als6p構築物を用い、基材特異的接着を媒介する領域はAlsタンパク質におけるN末端ドメインと突き止められた。具体的には、Alsタンパク質のサブセットもまた、細胞内皮侵入を媒介したが、これはこのファミリーのこれまでは未知であった機能である。これらの結果と合致して、相同モデリングは、Alsメンバーが、免疫グロブリンスーパーファミリーの接着または侵入と類似の拡大された領域が間に存在する反−平行β−シートモチーフを含有することを明らかにした。この知見は、Als1pのN末端ドメインの円二色およびフーリエ変換赤外分光分析を用いて確認された。アミノ酸過敏の特異的領域がAlsタンパク質のN末端ドメイン内に見出され、エネルギー−ベースのモデルは、恐らくはAlsファミリーメンバーの多様な機能を支配するN末端ドメインにおける同様性および差を予測した。まとめると、これらの結果は、Alsファミリー内の構造的および機能的多様性は、C.albicansに、感染の間に広い範囲の宿主構成要素を認識し、それと相互作用できる細胞壁タンパク質のアレイを供することを示す。
【0013】
本発明は、医薬上許容され得る媒体中の、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片、およびアジュバントを有するワクチンを提供する。該ワクチンは、Candida albicans、Candida krusei、Candida tropicalis、Candida glabrataまたはCandida、parapsilosisのようなCandida種に由来するAlsタンパク質ファミリーメンバーであり得る。Alsタンパク質ファミリーメンバーは、例えば、Als1p、Als3p、Als5p、Als6p、Als7pおよびAls9p、またはその免疫原性断片であり得る。Candida種内の全ての他のAlsタンパク質ファミリーメンバーは、同様に、本発明のワクチンとして使用することができる。
【0014】
本発明は、播種性カンジダ症を治療し、予防し、または緩和するためのワクチンとしてのC.albicansアグルチニン様配列タンパク質ファミリーメンバーの遺伝子産物を利用する。該ワクチンはC.albicansの異なる株に対して、ならびに異なるCandida種に対して効果的である。Alsタンパク質ファミリーメンバーは、例えば、Als1p、Als3p、Als5p、Als6p、Als7pおよびAls9pであり得る。本発明は、C.albicansによる内皮細胞および/または上皮細胞への接着および侵入、および生物の病因を遅らせるためのワクチンとして使用されるAlsタンパク質ファミリーメンバー−発現表面タンパク質の罹患性におけるALS遺伝子の役割を探求する。
【0015】
本発明に従うと、ALSファミリーメンバー遺伝子は、C.albicansに対する免疫治療戦略の標的として選択される表面アドヘシンをコードする。ALS1遺伝子の発現産物であるAls1pタンパク質は表面タンパク質に典型的な構造的特徴を有し、事実、C.albicansの細胞表面で発現されることを示すのは、宿主組織へのアドヘシンとして作用するタンパク質についての1つの基準である。Alsタンパク質ファミリーメンバーは、N末端におけるシグナルペプチド、C−末端におけるグリコシルホスファチジルイノシン(CPI)繋留配列、およびスレオニンおよびセリンがリッチな反復を含む中央領域を有するものとして構造的に特徴付けることができる。また、Alsタンパク質ファミリーメンバーは、細胞表面に発現されるタンパク質に典型的なN−およびO−グリコシル化部位を有する。ALs1pのN末端に対して向けられたモノクローナル抗体を用いる間接的免疫蛍光は、例えば、ALs1pが出芽胞子の対数期の間に発現されることを明らかにした。ALs1pのこの発現は菌糸形成の間に増加し、菌糸エレメントが拡散した表面染色によって示されるように出芽胞子から伸びる接合点に位置する。さらに、このモノクローナル抗体は、C.albicans過剰発現突然変異体の内皮細胞への増強された接着をブロックし、それにより、ALs1pを用いる免疫療法適用についての原理を確立する。機能的特徴は、それが他のAlsファミリーメンバーの細胞接着および侵入に関するので、実施例VIにおいて後にさらに記載する。
【0016】
かくして、1つの態様によると、本発明は、医薬組成物に処方し、アジュバントを含むまたは含まないワクチンとして投与した場合に有用な特性を有する、例えば、Als1p、Als3p、Als5p、Als6p、Als7pおよびAls9pと命名されたAlsファミリーメンバー表面接着タンパク質またはその機能的断片、コンジュゲートまたはアナログを提供する。Alsタンパク質ファミリーメンバー、2以上のAlsタンパク質ファミリーメンバーまたは1以上のその機能的断片、アナログ、コンジュゲートまたは誘導体の組合せは、たとえば、Candida albicansから得ることができる。同様なアドヘシンまたはインバシン分子、またはそのアナログまたは誘導体はカンジダ起源のものとすることができ、たとえば、属Candida、たとえば、Candida parapsilosis、Candida krusei、Candida glabrataおよびCandida tropicalisに属する種から得ることができる。本発明による表面アドヘシンまたはインバシンタンパク質は単離されたまたは純粋な形態で得ることができ、かくして、本発明の1つの実施形態によると、実質的に純粋なAlsタンパク質ファミリーメンバーCandida表面アドヘシンタンパク質、またはその機能的断片、免疫原性断片、アナログ、コンジュゲートまたは誘導体は、ワクチンとして処方されて、患者において免疫応答を引き起こして、Candidaに対して免疫応答を誘導し、および/または該生物の内皮細胞への接着をブロックする。無傷Alsタンパク質ファミリーメンバーと同様な結合、接着または侵入活性を呈するAlsタンパク質ファミリーメンバーの断片を、本明細書中においては、機能的断片という。Candida種に対する抗体または細胞性免疫応答を誘導することができるAlsタンパク質ファミリーメンバーの断片を、本明細書中においては、免疫原性断片という。例示的な機能的断片は実施例VIにおいて後にさらに記載するAlsタンパク質ファミリーメンバーのN末端ポリペプチド領域を含む。例示的な免疫原性断片はN末端Alsポリペプチド領域、C−末端Alsポリペプチド領域、ならびに抗体免疫応答、細胞性免疫応答、あるいは抗体および細胞性両免疫応答を生じさせるのに十分ないずれかの他のAls断片を含む。そのような免疫原性断片は、約4アミノ酸と小さく、かつ無傷ポリペプチド程度に大きくすることができ、ならびにその間の全てのポリペプチド長さを含む。
【0017】
本発明による表面接着タンパク質のアナログまたは誘導体は、ALSファミリーメンバー遺伝子および/または遺伝子産物について本明細書中に記載された基準によって同定し、さらに特徴付けることができる。例えば、アナログまたは誘導体のヌル突然変異体は、対照と比較して、内皮細胞に対して顕著に低下した接着を有するであろう。同様に、適切なモデルにおけるアナログまたは誘導体の過剰発現は、対照と比較して内皮細胞への増大した接着を示し、前記した基準に従うと細胞表面アドヘシンとして確認される。また、アナログまたは誘導体に対する抗血清は抗Alsタンパク質ファミリーメンバー抗体と交差反応することができ、本明細書中に開示されたような播種性カンジダ症のマウスモデルに投与した場合に、増大した生存時間を呈することができる。
【0018】
また、本発明は、播種性カンジダ症を治療または予防する方法を提供する。該方法は、医薬上許容され得る媒体中の、細胞接着または侵入活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンの免疫原性量を投与することを含む。該ワクチンはアジュバントと共に、またはそれ無くして投与することができる。Alsタンパク質ファミリーメンバーは異なるCandida株から、ならびにCandida albicans、Candida krusei、Candida tropicalis、Candida glabrataおよびCandida、parapsilosisのような異なるCandida種から由来することができる。播種性カンジダ症を治療または予防する方法で用いるAlsタンパク質ファミリーメンバーはAls1p、Als3p、Als5p、Als6p、Als7pおよびAls9pを含む。
【0019】
異なるCandida株、異なるCandida種、他の細菌および感染剤に対する本発明のワクチンの有効性、およびその広い範囲の免疫活性は後にさらに記載し、実施例において例示する。例えば、実施例Vは、抗ALS抗体が粘膜および血行的に播種性のカンジダ感染に対して有効的であることを示す。実施例VIIは、細胞−媒介免疫を高めることによって、マウス播種性カンジダ症の間における生存を、rAls1p−Nでのワクチン接種が改良することを示す。実施例VIIIは、本発明のワクチンは真菌負荷を低下させ、免疫応答性および免疫無防備マウス双方において生存を改良することを示す。実施例IXは、黄色ブドウ球菌感染に対する本発明のALSワクチンの有効性を示す。実施例Xは、本発明のワクチンがC.albicansの異なる株に対して、およびC.glabrata、C.krusei、C.parapsilosisおよびC.tropicalisのような異なる種に対して効果的であることを例示し、異なる動物モデルにおける有効性を例示する。また、実施例XIは、異なる動物モデルにおける本発明の異なるワクチンの有効性を例示し、ならびに誘導された異なる応答点および2つの代表的なALSワクチンの能力の比較を提供する。
【0020】
さらに提供される発明は、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその機能的断片の有効量を投与して、Candidaの宿主細胞または組織への結合または侵入を阻害することを含む、播種性カンジダ症を治療または予防する方法である。Alsタンパク質ファミリーメンバーはCandida albicans、Candida krusei、Candida tropicalis、Candida glabrataおよびCandida parapsilosisに由来することができる。播種性カンジダ症を治療または予防する方法で用いるAlsタンパク質ファミリーメンバーは、Als1p、Als3p、Als5p、Als6p、Als7pおよびAls9pを含む。細胞接着活性は、ゼラチン、フィブロネクチン、ラミニン、上皮細胞または内皮細胞への結合、および/または細胞侵入への促進を含む。
【0021】
加えて、本発明は、本明細書中に記載されたAlsタンパク質ファミリーメンバーを用いて黄色ブドウ球菌感染を治療または予防する方法も提供する。特に、黄色ブドウ球菌感染を治療または予防する方法は、医薬上許容され得る媒体中の、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンの免疫原性量を投与することを含む。
【0022】
Als1pおよびAls3pは、黄色ブドウ球菌細胞表面タンパク質に対する有意な相同性のため特に効果的である。例えば、Als1pおよびAls3pの配列および構造的相同性は、実施例IXにおいて後にさらに記載する。本明細書中で提供される教示および指針があれば、当業者は、本発明のワクチンおよび方法を同様なCandidaおよびブドウ球菌(Staphylococcus)感染の治療に適用することができるのを理解するであろう。同様に、本明細書中に記載された教示および方法があれば、当業者であれば、本発明のワクチンおよび方法は、真菌、細菌などを含めた、本明細書中に記載されたAlsタンパク質ファミリーメンバーに対して同様な免疫原性、配列および/または構造的相同性を持つ細胞表面ポリペプチドを有する他の病原体にやはり適用することができるのを理解するであろう。
【0023】
Candidaまたはブドウ球菌感染に対する細胞接着または侵入戦略の免疫療法および/またはAlsポリペプチド阻害は、血管内皮細胞への結合のレベルにおいて、ならびにフィラメント化調節経路の下流エフェクタを介して働くことができる。可溶性Alsタンパク質ファミリーメンバーまたは機能的断片を用いる結合の免疫療法戦略または阻害はこの関係で有用である。なぜならば:(i)血行的に播種性のカンジダ症および他の感染病因に関連する罹患率および死亡率は、現在利用可能な抗真菌療法でもってしても許容できないくらい高いままである;(ii)抗真菌および抗生物質耐性の上昇する発生率は、抗真菌および抗菌剤の増大する使用に関連し、(iii)ひどいCandidaおよびブドウ球菌感染の危険がある患者の集団はよく定義されており、非常に大きく、かつ手術後患者、移植体患者、癌患者、および低体重で誕生した幼児を含み;および(iv)ひどいCandida感染を発生する患者の高いパーセンテージは好中球減少症ではなく、かくして、ワクチンまたは競合ポリペプチドまたは化合物阻害剤に応答できるからである。これらの理由で、Candidaおよびブドウ球菌は、受動的免疫療法、能動的免疫療法、または受動的または能動的免疫療法の組合せについて魅力的な真菌および細菌標的である。加えて、Candidaは、Alsタンパク質ファミリーメンバーポリペプチド、その機能的誘導体、および/または1以上のAlsファミリーメンバーに結合し、Candidaの宿主細胞受容体への結合を妨げるその化合物またはミメティックを用いる競合阻害で魅力的である。
【0024】
本明細書中で提供された教示および指針があれば、当業者であれば、当該分野でよく知られた免疫療法を、本発明のAlsタンパク質ファミリーメンバー、その免疫原性断片、アナログ、コンジュゲート、および/または誘導体と共に使用して、アジュバントと共に、またはそれなくしてワクチンとして投与された医薬上許容され得る組成物中の免疫原として該分子を1以上に用いることができるのを理解するであろう。本発明の目的では、用語「医薬の」または「医薬上許容され得る」とは、非毒性であり、望まれた場合、安全にヒトに投与できる担体または添加剤と共に用いられる公知の技術によって処方された組成物をいう。投与は、例えば、静脈内、筋肉内、腹腔内、または皮下注射を含めた良く知られた経路を用いて行うことができる。本発明のそのようなワクチンは当業者に知られた緩衝液、塩または他の溶媒を含んで、溶液中でのワクチンの活性を維持することができる。同様に、当該分野でよく知られた広い範囲のアジュバントのいずれかを本発明のワクチンと共に使用して、Candidaまたはブドウ球菌の罹患性宿主細胞への結合、侵入および/または感染を低下し、またはブロックすることができる治療上有効な免疫応答を誘導し、促進し、または高めることができる。
【0025】
同様に、本明細書中に提供された教示および指針があれば、当業者であれば、投与し、細胞表面分子のその同族受容体への結合を選択的にブロックするための当該分野でよく知られた治療方法を、本発明のAlsタンパク質ファミリーメンバー、その機能的断片、アナログ、コンジュゲート、および/または誘導体と共に使用して、Alsタンパク質ファミリーメンバーの1以上を医薬上許容され得る組成物中で阻害剤として用いることができるのを理解するであろう。ワクチン処方に関しては、阻害性処方は、例えば、静脈内、筋肉内、腹腔内、または皮下注射を含めた当該分野でよく知られた方法を用いて同様に投与することができる。Alsファミリーメンバー受容体に結合し、Alsタンパク質ファミリーメンバー結合をブロックするそのような阻害性組成物は当業者に知られた緩衝液、塩または他の溶媒を含んで、溶液中でのワクチンの活性を維持することができる。さらに、当該分野でよく知られた広い範囲の処方のいずれかを本発明の阻害性組成物で使用して、Candidaまたはブドウ球菌の罹患性宿主細胞への結合、侵入および/または感染を低下させ、または阻害するように、送達または摂取を標的化し、および/または高めることができる。
【0026】
本発明に従う治療免疫原または受容体結合阻害剤として用いる分子に関しては、当業者であれば、Alsタンパク質ファミリーメンバー分子を、免疫原性ワクチンまたは細胞接着もしくは侵入阻害剤としての必須の性質を失うことなく切形または断片化することができるのを認識するであろう。例えば、Alsタンパク質ファミリーメンバーを切形して、前記した、かつ実施例において後にさらに記載する機能的特性を維持したC−末端からの切形によってN末端断片を生じさせることができる。同様に、C−末端断片は、その機能的特性を維持したN末端からの切形によって生じさせることができる。本明細書中で提供された教示および指針に従った他の修飾を本発明に従って行って、他のAlsタンパク質ファミリーメンバーの機能的断片、その免疫原性断片、アナログまたは誘導体を作成して、天然タンパク質に関して本明細書中に記載された治療上有用な特性を達成することができる。
【0027】
本発明のAlsタンパク質ファミリーメンバーおよび方法の治療的有効性の1つの態様はフィラメント化の調節への干渉を達成して、生物の宿主構成要素への接着をブロックし、およびイムノエフェクタ細胞および他の生理学的メカニズムによって生物のクリアランスを高める。内皮細胞は血管系の大部分を被覆するので、抗体、Alsファミリーメンバータンパク質、ポリペプチド、またはペプチドまたはそのいずれかの組合せを用いて内皮細胞への生物の接着、侵入および/または双方をブロックする戦略は、本発明の有用な実施形態を含む。既に記載したように、そのような接着および/または侵入をブロックする戦略は、カンジダアドヘシン、インバシン、または本明細書中に開示された同族受容体に対して向けられた能動的または受動的免疫療法、あるいは阻害性結合を含む。かくして、例えば、いずれの適当な宿主もタンパク質を注射し、血清を集めて、適当な精製および/または濃縮後に所望の抗アドヘシン抗体を得ることができる。注射に先立って、アドヘシンまたはインバシンタンパク質またはその組合せを適当なビヒクル、好ましくは、多糖のような公知の免疫刺激剤、またはリポソームのような送達処方、または時間−放出組成物中に処方することができる。かくして、さらなる態様によると、本発明は、ワクチンまたはAls受容体阻害剤として用いるため、処方中に、1以上の医薬上許容され得る賦形剤と共にカンジダアドヘシンまたはインバシンタンパク質を含む医薬組成物を提供する。
【0028】
本発明の方法は、C.albicansの宿主構成要素の内皮または上皮細胞への接着をブロックすることによって、あるいは例えば抗体をブドウ球菌に結合させ、免疫メカニズムに病原体を除去させることによって、カンジダまたはブドウ球菌感染を緩和し、および/または阻害するものである。かくして、本発明の1つの態様によると、Alsタンパク質ファミリーメンバーのアドヘシンまたはインバシンタンパク質、その機能的または免疫原性断片、誘導体、アナログ、またはコンジュゲートを含む医薬組成物を、注射または注入用の生体適合性担体を含有する医薬組成物中のワクチンまたはAls受容体阻害剤として処方し、患者に投与される。また、Alsファミリーメンバータンパク質または単離されたまたは組換えAlsファミリーメンバータンパク質に対して生起された抗血清の直接的投与を用いて、C.albicansの哺乳動物宿主の構成要素への接着をブロックし、またはブドウ球菌病原体の除去を行うことができる。アドヘシンタンパク質に対する抗血清は公知の技術、Kohler and Milstein, Nature 256:495−499(1975)によって得ることができ、それをヒト化して、抗原生を低下させる(米国特許第5,693,762参照)。またはトランスジェニックマウスにおいて生産し、編集されていないヒト免疫グロブリン遺伝子を得ることができる(米国特許第5,877,397参照)。同様に、単離されたまたは組換えAlsタンパク質ファミリーメンバーもまた、例えば、後に実施例に記載される組換え生産を含めた、当業者によく知られた方法を用いて生産することもできる。
【0029】
本発明のなおさらなる使用は、Alsタンパク質ファミリーメンバーのアドヘシンまたはインバシンタンパク質を用いて、カンジダまたはブドウ球菌感染の予防および/または緩和のためのワクチン戦略を開発する。かくして、本発明の1つの態様によると、例えば、標準免疫学技術を使用してマルチ−成分ワクチン戦略を構築することができ、これは、C.albicansの接着をブロックし、またはブドウ球菌病原体の排除を行うために宿主構成要素からの免疫応答を高め、および/または誘導することができる。
【0030】
本発明のなおさらなる使用は、例えば、DNAワクチン戦略を開発することである。かくして、本発明の1つの態様によると、例えば、Alsタンパク質ファミリーメンバーのアドヘシンまたはインバシン、またはその機能的断片をコードするALSファミリーメンバーポリヌクレオチドを、遺伝子産物に対する免疫応答を得るように設計されたプロトコルに従って投与される。例えば、Felgner 米国特許第5,703,055参照。
【0031】
本発明のなおさらなる使用は、例えば、組合せワクチン戦略を開発することである。かくして、本発明の1つの態様によると、例えば、抗ALSタンパク質ファミリーメンバー抗体は、カンジダまたはブドウ球菌感染を治療し、および/または予防する抗体と共に用いることができる。米国特許第5,578,309参照。
【0032】
以下の実施例は、播種性カンジダ症の予防的尺度または治療のための基礎としてのALS1アドヘシンの免疫治療的利用性を説明する。実施例1は、ALS1の過剰発現によって特徴付けられるALS1ヌル突然変異体およびC.albicansの株を調製して、内皮細胞への接着の媒介を確認することを記載する。実施例2は、Als1pの局所化、およびefgフィラメント化調節経路の関与を記載する。実施例3は、ALS1アドヘシンタンパク質の精製を記載する。実施例4は、表面アドヘシンタンパク質のブロッキングを示すために用いられる、ALS1表面アドヘシンタンパク質に対して生起されたウサギポリクローナル抗体の調製を記載する。実施例5は、本発明に従って本明細書中に記載されたようにALS1表面接着タンパク質に対して生起されたポリクローナル抗体を用いてインビボにて接着をブロックして、マウスモデルにおいて播種性カンジダ症に対して保護することを記載する。実施例VIは、Alsタンパク質ファミリーメンバーの構造的および機能的特徴を記載する。
【0033】
本発明の種々の実施形態の活性に実質的に影響しない修飾もまた本明細書中で提供される発明の定義内に含まれることは理解される。従って、以下の実施例は本発明を説明するものであり、限定することを意図しない。
【0034】
(実施例I)
(Als1はC.albicansの内皮細胞への接着を媒介する)
URAブラスター技術を用いて、Als1pの発現を欠如するC.albicansのヌル突然変異体を構築した。als1/als1突然変異体は、Ura−ブラスター技術(Fonzi and Irwin,Genetics 134, 717(1993))の修飾を用いてC.albicans株CAI4において以下のように構築した。2つの別々のals1/hisG−IRA3−hisG−als1構築物を利用して、遺伝子の2つの異なる対立遺伝子を破壊した。4.9kbのAsLS1コーディング配列は、以下のプライマーを用いて高い忠実度PCR(Boehringer Mannheim,Indianapolis, IN)でもって生じさせた:
【0035】
【化1】
次に、PCR断片をpGEM−Tベクター(Promega,Madison,WI)にクローン化し、かくして、pGEM−T−ALS1を得た。hisG−URA3−hisG構築物をKpn1およびHind3での消化によってpMG−7から放出させ、これを用いて、pGEM−T−ALS1のKpn1およびHind3消化によって放出されたALS1の一部を置き換えた。最終のals1−hisG−URA3−hisG−als1構築物を、Xholでの消化によってプラスミドから放出させ、これを用いて株CAI−4の形質転換によってALS1の最初の対立遺伝子を破壊した。
【0036】
第2のals1−hisG−URA3−hisG−als1構築物を2つの工程で生じさせた。まず、pMB7のBg12−Hind3 hisG−URA3−hisG断片をpUC19のBamH1−Hind3部位にクローン化し、それにより、pYC2を得た。次いで、pYC2をHind3で消化し、T4 DNAポリメラーゼを用いてdATPおよびdGTPで部分的に満たし、次いで、Sma1で消化して新しいhisGURA3−hisGで断片を得た。第二に、ALS1相補的フランキング領域を生じさせるために、pGEM−T−ALS1をXba1で消化し、次いで、dCTPおよびdTTPで部分的に満たした。この断片をHpa1で消化して、ALS1の中央部分を欠失させ、次いで、hisG−URA3−hisG断片に連結させ、pYC3を得た。次いで、このプラスミドをXholによって消化して、構築物を放出させ、これを用いて、ALS1の第二の対立遺伝子を破壊した。実験を通じて増殖曲線を作成して、生じた突然変異が増殖速度に対して影響を有しないことを確認した。すべての組込みは、XbaIおよびHindIIIでのpYF5の消化によって生じた0.9kbのALS1特異的プローブを用いてサザーンブロット分析によって確認した。
【0037】
ヌル突然変異体を、ヒト臍静脈内皮細胞にインビトロで接着するその能力についてC.albicans CAI−12(URA+リバータント株)と比較した。接着実験では、YPD(2%グルコース、2%ペプトン、および1%酵母エキス)での一晩の培養からの酵母細胞を、グルタミンを含むRPM1中で25℃にて1時間増殖させて、Als1p発現を誘導した。ハンクス平衡塩溶液(HBSS)(Irvine Scientific, Irvine, CA)中の3×102生物を内皮細胞の各ウェルに加え、その後、プレートを37℃にて30分間インキュベートした。接種物のサイズは、YPD寒天中での定量的培養によって確認した。インキュベーション時間の最後に、非接着性生物を吸引し、内皮細胞単層を標準的な方法にてHBSSで2回すすいだ。ウェルをYPD寒天と重ね、接着性生物の数をコロニーカウンティングによって決定した。統計学的処理はWilcoxin順位和検定によって得られ、Bonferroni修正にて多数の比較のために修正した。P<0.001。
【0038】
図1を参照し、ALS1/ALS1およびals1/als1株の比較は、ALS1ヌル突然変異体はC.albicans CAI−12よりも内皮細胞に35%未満接着した。バックグラウンド接着を低下させるために、非ALS1発現条件下で増殖させた野生型株の接着を、Als1pを自律的に発現する突然変異体と比較した。この突然変異体は、構成的ADH1プロモーターの制御下のALS1の第三のコピーを野生型C.albicansに組込むことによって構築した。C.albicansにおけるALS1の構成的発現を達成するために、平滑末端PCR生成URA3遺伝子を、pOCUS−2ベクター(Novagen,Madison,WI)の平滑末端Bg12部位へ連結して、pOU−2を得た。(pLH−ADHptから単離され、親切にも、A.Brown,Aberdeen,UKによって提供された)C.albicansアルコールデヒドロゲナーゼ遺伝子(ADH1)プロモーターおよびターミネーターを含有した2.4kbのNot1−Stu1断片を、Not1およびStu1での消化後にpOU−2にクローン化した。pOAU−3と命名された新しいプラスミドは、ADH1プロモーターおよびターミネーターの間に唯一のBg12部位を有した。BamH1制限酵素部位が近接するALS1コーディング配列は、鋳型としてのpYF−5、および以下のプライマーを用いて高忠実度PCRによって生じさせた:
【0039】
【化2】
このPCR断片をBamH1で消化し、次いで、pOAU−3の適合するBg12部位にクローン化して、pAU−1を得た。最後に、pAU−1を、C.albicans CAI−4を形質転換するに先立ってXbalによって線状化した。部位−特異的組込みは、サザーンブロット分析によって確認した。図1Bを参照し、このPADH1−ALS1株におけるALS1の過剰発現の結果、野生型C.albicansと比較して内皮細胞への接着の76%増加がもたらされた。野生型の内皮細胞接着を過剰発現突然変異体と比較するにおいて、酵母細胞を25℃においてYPD中で一晩増殖させた。(Als1p非誘導性条件)。Als1p発現は、野生型のバックグラウンド接着を低下させるように誘導されず、かくして、PADH1−ALS1ハイブリッド遺伝子を通じての接着においてAls1pの役割を拡大した。接着性アッセイは前記したように行った。統計学的処理はWilcoxin順位和検定によって得られ、Bonferroni修正を施して多数の比較のために修正した。P<0.001。
【0040】
モノクローナル抗Als1pマウスIgG抗体を、Clontech YEXpress(TM)酵母発現系(Palo Alto,CA)を用いて発現させたAls1pの精製され切形されたN末端(アミノ酸#17から#432)に対して生起させた。これらのモノクローナル抗Als1p抗体の接着性ブロッキング能力は、1:50希釈にて、C.albicansを抗Als1抗体またはマウスIgG(Sigma,St.Louis,MO)いずれかと共にC.albicans細胞をインキュベートすることによって評価した。その後、酵母細胞を前記した接着アッセイで用いた。統計学的処理は、Wilcoxin順位和検定によって得られ、Bonferroni修正を施して多数比較のために修正した。P<0.001。結果は、PADH1−ALS1株の接着が26.8%±3.5%から14.7%±5.3%まで低下したことを明らかにした。かくして、ALS1欠失および過剰発現の効果は、Als1pが、C.albicansの内皮細胞への接着を媒介することを示す。
【0041】
(実施例II)
(Als1pの局所化)
アドヘシンとして機能するAls1pについては、それは細胞表面に位置しなければならない。Als1pの細胞表面位置決定は、抗Als1pモノクローナル抗体と共に間接的に免疫蛍光を用いて確認した。拡散染色は、指数関数的増殖の間に出芽胞子の表面で検出された。この染色は静止相において出芽胞子では検出不可能であった。図2Aを参照し、出芽胞子を、フィラメントを生産するように誘導すると、強い染色が観察され、これは出現するフィラメントの基部に専ら位置付けられた。免疫蛍光はals1−als1突然変異体で観察されず、これは、Als1pについてのこの抗体の特異性を確認する。図2B参照。これらの結果は、Als1pが細胞表面タンパク質であることを確立する。
【0042】
出芽胞子−フィラメント接合へのAls1pの特異的位置決定は、フィラメント化プロセスにおけるAls1pを示唆する。メカニズムを決定するために、C.albicans ALS1突然変異体のフィラメント化表現型を分析した。図3Aを参照し、als1/als1突然変異体はLee固体培地上での4日間のインキュベーション後にフィラメントを形成できず、他方ALS1/ALS1およびALS1/als1株ならびにALS1−捕捉突然変異体は共にこの時点において豊富なフィラメントを生産した。als1/als1突然変異体は、より長いインキュベーション時間の後にフィラメントを形成することができた。さらに、ALS1の過剰発現はフィラメント化を増大させ:PADH1−ALS1株は3日間のインキュベーションの後に豊富なフィラメントを形成し、他方、野生型株はこの時点において乏しいフィラメントを生産した。図3B参照。フィラメント化におけるAls1pの役割をさらに確認するために、ALS1のコーディング配列を反対向きに挿入した以外は、ALS1過剰発現突然変異体と同様な突然変異体を用いて陰性対照を提供した。得られた株のフィラメント化表現型は、野生型株のそれと同様であることが示された。Als1pのフィラメント−誘導特性は固体媒体上で増殖させた細胞に特異的である。なぜならば、前記した株の全ては液体培地中で匹敵するほどフィラメント化したからである。データは、Als1pがフィラメント化を促進し、フィラメント化制御経路の調節におけるALS1発現を示唆することを示す。efg1/efg1、cph1/cph1、efg1/efg cph1/cph1、tup1/tup1、およびcla4/cla4突然変異体を含めた、これらの経路の各々における欠点を持つ突然変異体でのALS1発現のノーザンブロット分析を行った。図4Aを参照し、EFG1の双方の対立遺伝子が破壊された突然変異体はALS1を発現できなかった。野生型EFG1のコピーのefg1/efg1突然変異体への導入はALS1発現を回復したが、低下したレベルのものであった。図4B参照。また、図4Aで見られるように、他のフィラメント化調節突然変異のいずれも、ALS1発現を有意に改変しなかった(図4A)。かくして、Efg1pはALS1発現で要求される。
【0043】
もしEfg1pがALS1の発現を刺激するならば、これは今度はフィラメント化を誘導するのであるが、efg1/efg1株におけるALS1の発現はフィラメント化を回復するはずである。ADH1プロモーターの制御下にあるALS1の機能的対立遺伝子はefg1/efg1株に組込まれた。ALS1遺伝子産物がefg1ヌル突然変異体におけるフィラメント化における欠陥を補足するという確率を調べるために、Ura efg1ヌル突然変異体を、線状化pAU−1で形質転換した。Ura+クローンを選択し、ALS1の第三のコピーの組込みは、プライマーを用いるPCRで確認した。得られた株はALS1を自律的に発現し、Lee寒天上でフィラメント化する能力を回復した:
【0044】
【化3】
図4BおよびC参照。従って、Efg1pはALS1発現の活性化を通じてのフィラメント化を誘導する。
【0045】
フィラメント化はC.albicinsにおける臨界的なビルレンス因子であるゆえに、フィラメント化を調節する経路の表示は病因についての重要な示唆を有する。ALS1に先立って、これらの調節経路の下流エフェクタをコードする遺伝子は同定されていない。細胞表面タンパク質をコードする2つの他の遺伝子、HWP1およびINTIの破壊の結果、フィラメント化欠陥を持つ突然変異体がもたらされる。HWP1発現もまたEfg1pによってやはり調節されるが、efg1/efg1突然変異体におけるHWP1の自律発現はフィラメント化を回復できなかった。従って、Hwp1p単独はEFG1の下流のフィラメント化のエフェクタとして機能しない。また、INT1発現を制御する調節エレメントは知られていない。かくして、Als1pは、フィラメント化の下流エフェクタとして機能する同定された最初の細胞−表面タンパク質であり、それにより、C.albicansのビルレンスにおけるこのタンパク質についての中枢的な役割を示唆する。
【0046】
C.albicansビルレンスに対するAls1pの寄与は、血行的に播種性のカンジダ症のモデルでテストした。A.S.Ibrahimら,Infect.Immun.63,1993(1995)。図5Aを参照し、als1/als1ヌル突然変異体で感染させたマウスは、ALS1/ALS1株、ALS1/als1突然変異体またはALS1−捕捉突然変異体で感染させたマウスよりも有意に長く生存した。感染から28時間後に、als1/als1突然変異体で感染させたマウスの腎臓は有意に少数の生物を含有した(5.70±0.46 log10CFU/g)(双方の比較について(P<0.0006)。テストした時点のいずれかにおいて株のいずれかで感染させたマウスの脾臓、肺、または肝臓から回収された生物のコロニーカウントにおいて、差は検出されなかった。これらの結果は、Als1pが、感染の最初の28時間の間のC.albicans増殖および腎臓における執拗性で重要であることを示す。図5Bを参照し、感染から28時間後のマウスの腎臓の調査は、als1/als1突然変異体が、ALS1−捕捉株の野生型いずれかよりも、有意により短いフィラメントを生産し、より弱い免疫応答を誘導したことを明らかとした。しかしながら、感染から40時間までは、フィラメントの長さ、およびそれらを囲う白血球の数は全ての3つの株について同様であった。
【0047】
組織病理学で見られたals1/als1突然変異体のフィラメント化欠陥は、固体培地でのインビトロフィラメント化アッセイと並行した。この突然変異体は、インビボおよびインビトロ双方において初期の時点で欠陥があるフィラメント化を示した。この欠陥は、結局は、延長された感染/インキュベーションで解決された。これらの結果が、ALS1から独立したフィラメント化調節経路がより遅い時点で操作可能となり得ることを示唆する。感染から40時間までのこの代替フィラメント化経路の活性化は、なぜals1/als1突然変異体で感染させたマウスが引き続いて次の2から3日において死滅するかの理由のようである。
【0048】
まとめると、これらのデータは、C.albicans ALS1が、内皮細胞への接着およびフィラメント化双方を媒介する細胞表面タンパク質をコードすることを示す。Als1pは、C.albicansにおけるいずれかの既知のフィラメント化調節経路の唯一の同定された下流エフェクタである。加えて、Als1pは血行性カンジダ感染におけるビルレンスに寄与する。Als1pの細胞表面位置およびデュアル機能性は、それを、薬物および免疫−ベースの療法双方にとって魅力的な標的とする。
【0049】
(実施例III)
(ALS1アドヘシンタンパク質精製)
E.coliによって合成されたALS1タンパク質は免疫原として適切である。しかしながら、E.coliによって合成された真核生物タンパク質は、不適切な折畳またはグリコシル化の欠如のため機能的ではないであろう。従って、ALS1タンパク質が、C.albicansの内皮細胞への接着をブロックできるかを決定するために、該タンパク質を好ましくは遺伝子工学により作成されたC.albicansから精製する。
【0050】
PCRを用いて、ヌクレオチド52から1296のALS1の断片を増幅した。この1246bp断片は、シグナルペプチドの端部からタンデムリピートの始まりまでの予測されたALS1タンパク質のN末端を含む。ALS1の領域を増幅した。というのは、それは、S.cerevisiae Aga1遺伝子産物の結合領域に対するその相同性に基づいて、アドヘシンの結合部位をコードするようだからである。加えて、予測されたALS1タンパク質のこの部分はほとんどグリコシル化部位を有さず、そのサイズはE.coliにおける効果的な発現で適切である。
【0051】
ALS1の断片をpQE32へ連結してpINS5を得た。このプラスミドにおいて、タンパク質をlacプロモーターの制御下で発現させ、それは、それがアフィニティー精製できるように、N末端に融合した6−hisタグを有する。我々は、pINS5でE.coliを形質転換し、それを(IPTGの存在下で)誘導条件下でそれを成長させ、次いで、細胞を溶解させた。細胞溶解物をNi2+−アガロースカラムを通して、ALS1−6His融合タンパク質をアフィニティー精製した。この手法により、実質的な量のALS1−6Hisを得た。融合タンパク質をさらにSDS−PAGEによって精製した。ポリクローナルウサギ抗血清がそれに対して生起できるように該タンパク質を含有するバンドをゲルから切り出した。本発明による表面アドヘシンタンパク質は、本発明の精神逸脱することなく種々の公知のプロセスによって調製し、精製できることは当業者によって認識されるであろう。Als1pの配列を図7にリストする。
【0052】
(実施例IV)
(ALS1タンパク質に対するポリクローナル抗血清の生起)
ALS1タンパク質に対する抗体がCandida albicansの内皮および上皮細胞、選択された宿主構成要素への接着をインビトロにてブロックするか否かを決定するために、ALS1タンパク質で形質転換したS.cerevisiaeでウサギを接種した。用いた免疫化プロトコルは、Candidaの種々の種内で抗原性関係を同定した抗血清の生産用のHasencleverおよびMitchellによって用いられた用量およびスケジュールであった。Hasenclever,H.F および W.O.Mitchell.1960。Antigenic relationships of Torulopsis glabrata and seven speceis of the genus Candida.J.Bacteriol.79:677−681。対照抗血清もまた、空のプラスミドで形質転換されたS.cerevisiaeに対して生起させた。全ての酵母細胞をガラクトース中で増殖させて、ALS遺伝子の発現を誘導した。接着実験でテストする前に、血清を56Cにおいて加熱−不活化させて、全ての補体活性を除去した。
【0053】
免疫化されたウサギからの血清を、空のプラスミドで形質転換されたS.cerevisiaeの全細胞で吸収させて、ALS1タンパク質以外の酵母の成分と反応性である抗体を除去した。抗血清の力価は、ALS1遺伝子を発現するS.cerevisiaeを用いる免疫蛍光によって決定した。FITC−標識抗ウサギ抗体は商業的源(Southern Biotechnology,Inc)から購入した。アフィニティー−精製二次抗体は必須であった。なぜならば、多くの市販の血清は、酵母グルカンおよびマンナンと反応性である抗体を含有するからである。二次抗体はプラスミドで形質転換されたCandida albicansならびにS.cerevisiaeを用いて予めテストし、必要に応じて吸収して、いずれの抗S.cerevisiaeまたは抗Candida抗体も除去した。陰性対照は1)免疫前血清、2)空のプラスミドで形質転換させたS.cerevisiae、および3)ALS(グルコース)遺伝子の発現を抑制する条件下で増殖させた以外はALS遺伝子で形質転換されたS.cerevisiaeであった。
【0054】
前記実験に加えて、ウエスタンブロッティングを用いて、抗血清が、それがそれに対して生起したALSタンパク質に特異的に結合するというさらなる確認を供した。ALS1で形質転換されたS.cerevisiaeを誘導条件下で増殖し、その原形質膜は標準的な方法によって単離した。Panaretou R and P.Piper.1996.Isolation of Yeast Plasma Membranes.p.117−121.InI.H.Evans.(ed.),Yeast Protocols.Methods in Cell and Molecular Biology.Humana Press,Totowa,New Jersey。原形質膜は空のプラスミドを形質転換し、かつ同一条件下で増殖させたS.cerevisiaeからも調製した。膜タンパク質をSDS−PAGEによって分離し、次いで、エレクトロブロッティングによってPVDF膜に移した。Harlow,EおよびD.Lane.1988.Antibodies:a laboratory manual.Cold Spring Harbor Laboratory Press。無脂肪乳でブロックした後、ブロットをALS抗血清と共にインキュベートした。予め吸収された抗血清は、空のプラスミドを含有するS.cerevisiaeから抽出されたタンパク質と反応しなかった。この抗血清はS.cerevisiae pYF5(Candida albicans ALS1を発現するクローン)の内皮細胞への接着をブロックした。
【0055】
(実施例V)
(特異的ALSタンパク質に対するポリクローナル抗体は、粘膜および血行的に播種性のカンジダ感染からマウスを予防的に保護する)
前記条件下で、ALS遺伝子で形質転換されたS.cerevisiaeのクローンの接着をブロックする抗血清を同定した後、これらの抗血清は、Candida albicans での静脈内チャレンジからマウスを保護することが示された。
【0056】
ALSタンパク質に対する抗血清を、まず、血行的に播種性のカンジダ症のネズミモデルでテストした。アフィニティー−精製抗ALS抗体は、酵母細胞の種々の基材への接着を妨げる点において効果的である(実施例3参照)。アフィニティー−精製はこの系で有用である。なぜならば、抗体は正確に決定できるからである。さらに、分画されていない抗血清は、疑いもなく、S.cerevisiaeキャリアー細胞上の抗原に向けられた多量の抗体を含有するであろう。これらの抗Saccharomyces抗体の多くはC.albicansに結合するようであり、結果の解釈を不可能とするようである。加えて、ALSタンパク質を発現するS.cerevisiaeから抗体を溶出させるのに用いる手法もまた、アジュバント−様活性を有し得る少量の酵母マンナンまたはグルカンを溶出することもできる可能性が高い。免疫アフィニティー−精製抗体は使用前にさらに精製される。それはマウス脾臓細胞で予め吸着することもできる。
【0057】
抗体用量を投与して、最も活性な免疫化プロトコルにおいて予測することができる血清抗体のレベルを画定する範囲をカバーし、およびカンジダ症のネズミモデルにおいて受動免疫で典型的に用いる抗体用量の範囲をカバーすることができる。Dromer,F.,J.Charreire,A.Contrepois,C.Carbon,およびP.Yeni.1987,Protection,of mice against experimental cryptococcosis by anti−Cryptococcus neofornwns monoclonal antibody,Infect.Immun.55:749−752;Han,Y.and J.E.Cutler.1995,Antibody reponse that protects against disseminated candidiasis,Infect.Immun.63:2714−2719;Mukherjee,J.,M.D.Scharff,and A.Casadevall.1992,Protective murine monoclonal antibodies to Cryptococcus neofornwns,Infect.Immun.60:4534−4541;Sanford,J.E.,D.M.Lupan,A.M.Schlageter,and T.R.Kozel.1990,Passive immunication against Cryptococcus neoformans with an isotype−switch family ofmonoclonal antibodies reactive with cryptococcal polysaccharide,Infect.Immun.58:1919−1923参照。BALB/cマウス(雌、7週齢、NCI)に、腹腔内(i.p.)注射によって、マウス脾臓細胞で吸収されている抗ALSを与えた。対照マウスは、各々、マウス脾臓細胞で吸収されている予め採血した血清、無傷抗ALS血清、またはDPBSを受けた。予備的吸収のために、2mlの抗ALSまたは予め採血した血清を、100μlのマウス(BALB/c、7週齢の雌、NCI)脾臓細胞(ml当たりほぼ9×106細胞)と室温にて20分間混合した。混合物を遠心(@300xg)によって温かい滅菌DPBSで3分間洗浄した。この手法を3回反復した。i.p.注射の用量はマウス当たり0.4mlであった。4時間後、マウスをC.albicans(株CA−1;i.v.注射によりマウス当たり5×105親水性酵母細胞)でチャレンジした。次いで、その生存時間を測定した。図6参照。
【0058】
従前の実験は、腹腔内経路を介して投与された抗体が血清へ迅速(数分内)かつほとんど完全に移ることを示している(Kozel and Casadevall、未公表観察)。抗体製剤を投与する効力についての対照として、マウスの平行群を、ALS遺伝子で形質転換されたS.cerevisiaで吸収されたプレ−免疫前血清から単離された抗体で処理した。生存時間および腎臓のグラム当たりの酵母の数を測定した。再度、図6を参照し、ALS1ヌル突然変異体の106出芽胞子を静脈内注射によって感染したマウスは、ALS1の1つの対立遺伝子が欠失されているCandida albicans CAI−12またはCandid albicansで感染させたマウスと比較した場合、より長いメジアン生存時間を有した(p=0.003)。
【0059】
これらの結果は、ワクチンとしてALS1タンパク質を用いる免疫療法戦略が、播種性カンジダ症に対して保護的予防効果を有することを示す。
【0060】
(実施例VI)
(Candida albicans のAlsタンパク質ファミリーにおける機能および構造の多様性)
非接着性S.cerevisiaeの異種補足によるC.albicansALS1遺伝子の単離および特徴付けは従前に記載されている(Fuら,Infect.Immun.66:1783−1786(1998))。ALS1は、内皮細胞および上皮細胞への接着を媒介する細胞表面タンパク質をコードする。C.albicansにおけるこの遺伝子の双方のコピーの破壊は、内皮細胞への接着の35%低下に関連しており、ALS1の過剰発現は接着を125%増大させる(Fuら,Mol.Microbiol.44:61−72(2002))。
【0061】
ALSIは、Hoyerらによって元来記載された少なくとも8つのメンバーよりなる多数の大きなC.albicans遺伝子ファミリーである(Hoyerら,Trends Microbiol.9:176−180(2001),Zhaoら,Microbiology 149:2947−2960(2003))。これらの遺伝子は、3つのドメインによって特徴付けられる細胞表面タンパク質をコードする。N末端領域は推定シグナルペプチドを含有し、Alsタンパク質の間で比較的保存されている。この領域は、貧弱にしかグリコシル化されていないと予測される(Zhaoら,Microbiology 149:2947−2960(2003),Hoyerら,Genetics 157:1555−1567(2001))。これらのタンパク質の中央部分は可変数のタンデムリピート(長さは〜36アミノ酸)よりなり、グリコシルホスファチジルイノシトールアンカー配列を含有するセリン−スレオニン−リッチなC−末端領域がある(上記)が続く。この遺伝子ファミリーによってコードされるタンパク質は感染の間に発現することが知られているが(Hoyerら,Infect.Immun.67:4251−4255(1999),Zhangら,Genome Res.13:2005−2017(2003))、異なるAlsタンパク質の機能は詳細に調べられていない。
【0062】
非接着性S.cerevisiaeにおけるAlsタンパク質の異種発現を行って、Alsタンパク質の機能を評価し、C.albicansによって発現された多数の他のアドヘシンによって媒介された高いバックグラウンド接着を回避した。この異種発現系は、アドヘシンALS1,ALS5およびEAP1の単離および特徴付けを含めた、C.albicans遺伝子の実験で広範に用いられてきた(Liら,Eukaryot Cell 2:1266−1273(2003),Fuら,Infect.Immun.66:1783−1786(1998),Gaurら,Infect.Immun.65:5289−5294(1997))。後にさらに記載するように、このモデル系を用い、Alsタンパク質は、多様な接着性および侵入性機能を有することが示された。これらの結果と合致して、相同性モデリングは、Alsタンパク質が、タンパク質の免疫グロブリンスーパーファミリーのアドヘシンおよびインバシンメンバーに対して構造が密接に関連していることを示した。CDおよびフーリエ変換赤外(FTIR)1分光分析を用いる構造解析により、Als1pのN末端ドメインが反−平行βシート、ターン、α−ラセン、および免疫グロブリンスーパーファミリーの他のメンバーの構造と合致する構造決定されていないドメインから構成されることが確認された。最後に、比較エネルギー−ベースのモデルは、それらの区別される接着および侵入性生物学的機能を支配し得る異なるAlsタンパク質の間でN末端ドメインの鍵となる物理化学的特性の差を示唆する。
【0063】
ALSファミリーメンバーをクローン化し、およびそれらをS.cerevisiaeにおいて発現させるために、ALSI、−3、−5、−6、−7、および−9を後に記載するように首尾よく増幅し、発現させた。簡単に述べれば、クローニングおよび他の培養工程について、S.cerevisiae株S150−2B(leu2 his3 trp1 ura3)を、従前に記載されているように異種発現で用いた(Fuら,Infect.Immun.66:2078−2084(1998))。C.albicans株SC5314をゲノムクローニングで用いた。全ての株を、必要に応じて1.5%バクト−寒天(Difco)で固化させた最小規定培地(100μg/mlのL−ロイシン、−Lトリプトファン、L−ヒスチジン、および硫酸アデニンを補足した1×酵母窒素ベースブロス(Difco)、2%グルコース、および0.5%硫酸アンモニウム)中で増殖させた。ura−株の増殖は、80μg/mlウリジン(Sigma)の添加によって支持された。ASL5を含有するプラスミドpGK103、ALS1を含有するpYF5、およびALS9を含有するpALSnは従前に記載されている(Fuら,Infect.Immune.66:1783−1786(1998),Gaurら,Infect.Immune.65:5289−5297(1997),Lucinodら,Proceedings of the 102nd Annual Meeting of the American Society for Microbiology,pp.204,American Society for Microbiology,Salt Lake City,Ut.(2002))。A.Brown(Aberdeen,UK)から得られたプラスミドpADH1はC.albicansアルコールデヒドロゲナーゼ遺伝子(ADHI)プロモーターおよびターミネーターを含有し、それはS.cerevisiaeで(機能的である)Baileyら,J.Bacteriol.178:5353−5360(1996))。このプラスミドを、S.cerevisiaeにおけるALS遺伝子の構成的発現で用いた。
【0064】
ヒト口腔上皮および血管内皮細胞を入手し、以下のように培養した。咽頭癌腫から単離されたFaDu口腔上皮細胞系はAmerican Type Culture Collection(ATCC)から購入し、その推奨されるプロトコルに従って維持した。内皮細胞は臍帯静脈から単離し、Jaffeらの方法の我々の従前に記載した修飾によって維持した(Fuら,Mol.Microbiol.44:61−72(2002),Jaffeら,J.Clin.Invest.52:2745−2756(1973))。全ての細胞培養を、5%CO2を含有する湿潤環境中で37℃に維持した。
【0065】
ALS遺伝子をクローニングするために、ALSファミリーのメンバーのゲノム配列を、(the World Wide Web at URL:sequence.stanford.edu/group/candida/search.htmlで入手可能な)StanfordデータベースのBLASTサーチングによって同定した。PCRプライマーは、5’BgIIIおよび3’Xhol制限酵素部位を取り組み、表Iにて以下に示されたオープンリーディングフレームの各々を特異的に増幅するように作成した(配列番号14から19(各々、ALS1、3、5、6、7および9センスプライマー);配列番号20から25(各々、ALS1、3、5、6、7および9アンチセンスプライマー))。各遺伝子は、Expand(登録商標)High Fidelity(高忠実度)PCR系(Roche Applied Science)を用いるPCRによってクローン化した。ALS3、ALS6、およびALS7をC.albicans SC5314ゲノムDNAから増幅し、他方、ALS1、ALS5、およびALS9は、C.albicansゲノムライブラリーから従前に検索されているプラスミドから増幅した(Fuら,Infect.Immune.66:1783−1786(1998),Gaurら,Infect.Immune.65:5289−5297(1997),Lucinodら,Proceedings of the 102nd Annual Meeting of the American Society for Microbiology,pp.204,American Society for Microbiology,Salt Lake City,Ut.(2002))。配列決定のために、PCR産物をpGEM−T−Easy(Promega)に連結した。次いで、配列が確認されたALSオープンリーディングフレームを、BglII−Xhol共−消化によってpGEM−T−Easyから放出させ、目的のALS遺伝子がADH1プロモーターの制御下となるように、pADH1に連結した。S.cerevisiae株S150−2Bを、酢酸リチウム方法を用い、ALS過剰発現構築物ならびに空のpADH1構築物の各々で形質転換した。S.cerevisiaeにおける各ALS遺伝子の発現をノーザンブロット分析によって確認し、しかる後、表現型分析を行った。
【0066】
(表I)
(S.cerevisiaにおける異種発現のためのALS遺伝子のコーディング領域を増幅するのに用いるPCRプライマー)
【0067】
【表1】
ALS mRNA発現は各構築物につきノーザンブロット分析によって検出した。プライマーの3つの組の使用に拘わらず、C.albicans SC5314のゲノムDNAからのALS2およびALS4の増幅は不成功であった。ALS遺伝子のタンデムリピートを横切った配列決定および組立の困難性を仮定すれば、この結果は、公表されたゲノムデーターベースで現在入手可能な配列アセンブリーにおける誤差を反映する可能性がある。
【0068】
フローサイトメトリーにより、Alsタンパク質の各々がその各S.cerevisiae宿主の表面で発現されたことが確認された。簡単に述べれば、Als構築物の各々についての細胞表面発現の確認は、2つの異なるポリクローナル抗Als抗血清を使用する間接的免疫蛍光を用いて決定した。抗血清Aは、Als1pの417−アミノ酸N末端断片でのウサギの免疫化によって生じた、抗Als1p抗体よりなるものであった。抗血清Bは、C.albicans細胞壁成分を認識するが、S.cerevisiae株(latron Laboratories)と交差反応しないウサギ抗C.albicansマンナン因子5であった。
【0069】
各株については、107出芽胞子を一晩培養から単離し、100μlのヤギ血清でブロックし、次いで、1:25希釈のポリクローナル抗血清AまたはBいずれかで染色し、続いて、1:100のフルオレセインイソチオシアネート−標識ヤギ抗ウサギIgGで染色した。488 nmにおいて発光するアルゴンレーザーを備えたFACSCaliber(Becton Dickinson)機器をフローサイトメトリー分析で用いた。蛍光発光は515/40−nmバンドバスフィルターで検出した。10,000事象についての蛍光を集め、ベースライン(すなわち、空のプラスミドで形質転換されたS.cerevisiae)を超える蛍光を持つ細胞の分布を、CELLQUESTソフトウェア(Becton Dickinson)を用いて各株について分析した。
【0070】
表IIに示すように、2つの区別される抗血清は、Alsp−発現株の全ては、空のプラスミドで形質転換されたS.cerevisiae と比較した場合に、蛍光の少なくとも4倍増加を呈したことを示した。Alsファミリーのメンバーの中の予測された構造多様性と合致して、抗血清は、個々のAls発現株の認識において差を呈した。
【0071】
(表II)
(フローサイトメトリー分析によるS.cerevisiaeの表面上のAlsタンパク質の検出)
ポリクローナル抗Als1p抗血清(A)またはポリクローナル抗C.albicans細胞壁抗血清(B)いずれかでの間接的免疫蛍光を用い、各株の出芽胞子を染色し、次いで、フローサイトメトリーを用いて分析した。結果は、括弧に入れた−倍増加と共に、バックグラウンド(空のプラスミドで形質転換されたS.cerevisiae)を超える陽性細胞のパーセンテージとして表す。
【0072】
【表2】
種々のAlsタンパク質を発現したS.cerevisiaeクローンを、種々の宿主基材に接着するそれらの能力について調べた。後に記載するように、結果は、Alsタンパク質が基材特異的接着の異なるプロフィールを呈することを示す。
【0073】
真菌接着アッセイを行って、形質転換されたS.cerevisiae株の接着特性を決定した。簡単に述べると、従前に記載された接着アッセイ(8)の修飾を以下のように使用した。ゼラチン(Sigma)、ラミニン(Sigma)、またはフィブロネクチン(Becton Dickinson)の0.01 mg/ml溶液の1mlを、6−ウェル組織培養プレート(Costar)の各ウェルに加え、37℃で一晩インキュベートすることによって、接着プレートを被覆した。内皮細胞については、第二継代細胞を、0.2%ゼラチンマトリックスを被覆した6−ウェル組織培養プレート中で密集するまで増幅させ、上皮細胞については、FaDU細胞を、0.1%フィブロネクチンマトリックスで被覆した6−ウェル組織培養プレート中で密集するまで増殖させた(3日)。接着テストの前に、ウェルを1mlの温かいハンクス平衡塩溶液(HBSS)で2回洗浄した。テストすべきS.cerevisiae株を30℃の最小規定培地中で一晩増殖させ、次いで、遠心によって収穫し、HBSS(Irvine Scientific)で洗浄し、ヘマサイトメーターを用いて数えた。300の生物を、目的の基材で被覆した6−ウェル組織培養プレートの各ウェルに加え、CO2中で37℃にて30分間インキュベートした。非接着性生物を、10mlのHBSSで標準化された方法にて2回洗浄することによって除去した。ウェルにYPD寒天(1%酵母エキス(Difco)、2%バクト−ペプトン(Difco)、2%D−グルコース、1.5%寒天)を重ね、接種物を定量的培養によって確認した。プレートを30℃にて48時間インキュベートし、コロニーをカウントした。接着は初期接種物のパーセンテージとして表した。接着の差は、単一因子偏差分析検定を用いて比較し、p<0.01で有意と考えられた。
【0074】
異なる基材へのS.cerevisiae形質転換体の接着プロフィールの顕著な差があった(図8)。Als1p−、Als3p−、およびAls5p−発現株はテストした全ての基材に結合したが、Als6p−発現S.cerevisiaeはゼラチンのみに接着し、およびAls9p−発現S.cerevisiaeはバックグラウンドレベルを超えてラミニンのみに接着した。さらに、種々の基材への接着に定量的差があった。例えば、Als3pと比較した場合、Als1pはより大きな接着をゼラチンに付与したが、上皮細胞にはより小さな接着を付与した(p<0.01、単一因子偏差分析)。Als7pを発現するS.cerevisiaeだけがテストした基材のいずれにも接着しなかった。Alsタンパク質発現のレベルの小さな差は表IIに示された免疫蛍光実験によって排除できないが、そのような差は本実験で見出された基材特異的結合パターンが原因であるようではない。細胞表面に発現されたAlsタンパク質の量のそのような全体的増加または減少は、基材を横切っての接着の釣り合った増加または減少を生じ、観察された基材特異的差をもたらすものではないと予測されるであろう。
【0075】
後に記載されるように、Alsタンパク質に対する基材結合特異性は、Alsタンパク質のN末端配列に存在する。簡単に述べれば、S.cerevisiaeにおけるAls5p発現は、ゼラチンおよび内皮細胞を含めた多数の基材へ接着を付与し、他方、Als6p発現はゼラチン単独への接着をもたらした。機能のこの顕著な差にかかわらず、Als5pおよびAls6pはアミノ酸レベルにおいて80%を超えて同一である。これらのタンパク質のタンデムリピートおよびC−末端部分は実質的に同一であり、配列の差の大部分はこれらの2つのタンパク質のN末端に集中している。これらのデータはN末端配列の変動性が基材に特異性を付与することを示す。
【0076】
前記した結果は、キメラALS5/ALS6構築物の接着表現型を決定する実験の結果によって裏付けられる。簡単に述べれば、キメラAls5/Als6タンパク質は、各タンパク質のN末端を交換することによって構築された。キメラALS5/6遺伝子は以下のように構築された。5’2117bpの遺伝子を含むALS5のBglII−HpaI断片を単離した。次いで、pGEM−T−ALS6をBglIIおよびHpaIで消化して、ALS6の対応する5’2126bpを放出させ、pGEM−T−Easy+ALS6の3’配列よりなる断片を単離し、5’ALS5断片に連結して、プラスミドpGEM−T−5N6Cを得た。対応するALS6の5’断片およびALS5の3’断片を用いる同一アプローチを用いて、プラスミドp−GEM−T−6N5Cを得た。配列確認の後、各キメラALS遺伝子をBglII−XhoI消化によって放出し、前記したようにpATH1にサブクローンした。次いで、S.cerevisiae S150−2Bをこれらの構築物で形質転換し、それらの接着特性の特徴付けの前に、発現をノーザンブロット分析によって確認した。
【0077】
Als6pのC末端へのAls5pのN末端のキメラ融合を発現するS.cerevisiaeは、Als5pと同様にゼラチンおよび内皮細胞双方に接着した(図9)。同様に、Als5pのC末端へのAls6のN末端のキメラ融合を発現する株は、Als6pを発現するS.cerevisiaeがそうであったように、ゼラチンのみに接着した(図9)。さらに、Als5pおよびキメラAls5N6Cタンパク質を発現する株は、フィブロネクチン−被覆ビーズを凝集させ、他方、Als6pおよびキメラAls6N5Cタンパク質を発現するものは、これらのビーズに対して親和性はほとんどまたは全く有しなかった。まとめると、これらのデータは、これらの形質転換されたS.cerevisiae株の接着プロフィールがAlsタンパク質のN末端部分によって支配されていたことを示す。
【0078】
Alsタンパク質ファミリーメンバーの間で示された基材特異性の差に加えて、他の生物学的機能の差も観察された。例えば、Alsタンパク質のサブセットは、S.cerevisiaeによる内皮細胞侵入を媒介することが示された。C.albicansはそれ自身のエンドサイトーシスを誘導することによって内皮細胞に侵入する(Fillerら,Infect Immune.63976−983(1995),Belangerら,Cell Microbiol.,in press(2002))。このエンドサイトーシスは、生物が内皮細胞に接着する後に起こる;しかしながら、このプロセスで必要とされるC.albicansリガンドは未知である。さらに、区別されるカンジダリガンドは接着およびエンドサイトーシス双方で必要であるかは明らかでない。非接着性であることに加え、S.cerevisiaeは内皮細胞による有意なエンドサイトーシスを受けない。従って、Alsタンパク質がインバシンならびにアドヘシンとして働くことができるか否かをテストするために、内皮細胞に侵入するAlsタンパク質を発現するS.cerevisiae株の能力を決定した。
【0079】
内皮細胞侵入を媒介するAlsタンパク質の能力は、従前に記載された差分蛍光アッセイの修飾を用いて決定された(Phanら,Infect.Immun.68:3485−3490(2000))。簡単に述べれば、内皮細胞を、フィブロネクチンで被覆された12−mm直径のカバーグラス上で密集するまで増殖させ、24−ウェル組織培養プレート(Corning)に入れた。次いで、細胞を、RPMI 1640培地(Irvine Scientific)中の各S.cerevisiae株の105出芽胞子で感染させた。陽性対照として、細胞を同様な数のC.albicans出芽胞子で感染させた。90分間のインキュベーションの後、細胞を0.5mlのHBSSで標準的な方法にて2回濯ぎ、3%パラホルムアルデヒドで固定した。内皮細胞の表面に接着したままである生物を、Alexa 568(Molecular Probes,Inc.,Eugene,OR)とコンジュゲートされている、ウサギ抗C.albicans抗血清(バイオデザイン)で1時間染色し、これは赤色の蛍光を発した。この抗血清は、2倍より高い希釈にてS.cerevisiaeと交差反応する。次いで、内皮細胞を、リン酸−緩衝化生理食塩水中の0.2%Triton X−100中で10分間浸透させ、その後、細胞−会合生物(内部化された+接着性の生物)を、Alexa488でコンジュゲートされた抗C.albicans抗血清で再度染色し、これは緑色の蛍光を発した。次いで、カバーグラスをエピ蛍光で観察した。内皮細胞によって内部化された生物の数を、(緑色の蛍光を発する)細胞−会合生物の数から(赤色の蛍光を発する)接着性生物の数を差し引くことによって決定した。少なくとも100の生物が各カバーグラス上でカウントされ、全ての実験は少なくとも3つの別々の状況で三連にて行った。
【0080】
フィブロネクチンビーズ接着アッセイを行って、特定のAlsタンパク質の結合特徴をさらに特徴付けた。この点に関し、Als5pは、S.cerevisiaeの表面で発現された場合に、フィブロネクチン−被覆ビーズの凝集を誘導するタンパク質の能力によって元来同定された(Gaurら,Infect.Immune.65:5289−5297(1997))。従って、フィブロネクチンのためにALS5,ALS6,5N6Cおよび6N5Cで形質転換されたS.cerevisiae株を、この方法を用いてビーズ接着についてテストした(Gaurら,Infect.Immune.65:5289−5297(1997),(Gaurら,Infect.Immun.67:6040−6047(1999))。簡単に述べれば、トシル化磁性ビーズ(Dynal Biotech)を製造業者の指示に従ってフィブロネクチンで被覆した。次に、10μlの被覆ビーズ(106ビーズ)を、1mlのS.cerevisiae 1 × Tris−EDTA(TE)緩衝液、pH7.0中の1×108形質転換S.cerevisiaeと混合し、温和に混合しつつ45分間インキュベートした。チューブを磁石中において、ビーズおよび接着性S.cerevisiaeを非接着性生物から分離した。非接着性生物を含有する上清を吸引によって取り出し、残存するビーズを1mlのTE緩衝液中に再懸濁させることによって3回洗浄し、続いて、磁気分離および上清の吸引を行った。最後に、洗浄されたビーズおよび接着性生物を100μlのTE緩衝液に再懸濁させ、共−凝集について顕微鏡で調べた。
【0081】
結果は、Als1p Als3pおよびAls5pを発現するS.cerevisiaeが、細胞−会合生物のパーセンテージの有意な増加を呈することを示し、これは、内皮細胞に接着するそれらの能力を反映する。加えて、Als3p、および程度は低いが、Als1pおよびAls5pを発現する生物は有意な内皮細胞侵入を示した(図10)。
【0082】
前記した機能試験に加えて、Alsタンパク質は、免疫グロブリンスーパーファミリーのアドヘシンおよびインバシンに相同であることも判明した。Alsタンパク質の分子モデリングにおける初期工程として、知識に基づくサーチアルゴリズムを用いて、Alsファミリーメンバーと有意な構造的同様性を共有する分子を同定した。簡単に述べれば、相同性およびエネルギー−ベースのモデリングを行って、Alsタンパク質の総じての物理化学的特徴を比較した。まず、知識に基づく方法(SWISS−MODEL)(Guexら,Electrophoresis 18:2714−2723(1997),Schwedeら,Nucleic Acids Res.31:3381−3385(2003))を用いて、相同な立体配座を持つタンパク質についてのSwissおよびBrookhavenタンパク質データベースにおいて構造のコンビナトーリアル拡張構造配列を分析し、比較した(Shindyalovら,Protein Eng.11:739−747(1998))。このアプローチは、ExNRL−3Dデータベースにおいて一次配列同様性についてサーチするためのBLASTP2アルゴリズム(Altschulら,Mol.Biol.215:403−410(1990))を含んだ。平行して、動的配列整列アルゴリズムSIM(Huangら,Adv.Appl.Math.12:337−367(1991))を用いて、最大の配列同一性を持つ候補鋳型を選択した。引き続いて、ProModllを用いて、一次および洗練されたマッチ分析を行った。得られたタンパク質を、Alsタンパク質骨格軌跡の相同性モデリングのための鋳型として用いた。
【0083】
Alsタンパク質のN末端ドメイン(例えば、アミノ酸1から480;初期タンデムリピートに先行)の頑強なモデルを、相補性アプローチを介して作り出した。Alsタンパク質のN末端ドメインを、Silicon Graphicsワークステーション(SGI,Inc.)でオペレートするSYBYL 6.9.1ソフトウェア(Tripos Associates)を用いて配列相同性(Composer(Tophamら,Biochem.Soc.Symp.57:1−9(1990))およびネジ処理方法(Matchmaker (Godzikら,J.Mol.Biol.227:227−238(1992))および遺伝子−折畳(Jaroszewskiら,Protein Sci.7:1431−1440(1998),Godzikら、Protein Eng.8:409−416(1995),Godzikら,Proc.Natl.Acad.Sci.U.S.A.89:12098−12102(1992),Godzikら,J.Comput.Aided Mol.Des.7:397−438(1993))によって推定溶液立体配座に変換した。得られたコンフォーマーおよび標的Alsドメインのアミノ酸側鎖を分子ダイナミックスによって洗練し、AMBER95強制場方法(Duanら,J.Comput.Chem.24:1999−2012(2003))およびPowellミニマイザー(Powellら,Math.Program 12:241−254(1977))を用いて株エネルギーを最小化した。
【0084】
これらのアプローチは、ペプチド骨格原子の位置が固定された側鎖相互作用を最適化する。好ましい立体配座は、水性溶媒中の拡大された分子ダイナミックスから決定された。次に、全てのペプチド結合のねじれ角を最小の拘束にて180±15°に調整した。いくつかの場合において、拘束なしで、またはα−ラセン領域を0.4−kJペナルティーをカノニカルラマチャンドラン(Ramachandran)φおよびΨ角度に適用することによって拘束して、分子ダイナミックスを実行した。最終全エネルギー最小化を、全ての拘束および凝集体の除去の後に各モデルについて行った。得られたAlsのN末端ドメインモデルは3つの基準:(i)最も好都合な歪みエネルギー(分子メカニックス);(ii)経験的位置エネルギー関数;および(iii)潜在的ジスルフィドブリッチングの空間的配置の保存に基づいて優先させた(Godzikら,J.Mol.Biol.227:227−238(1992),Bowieら,Science 253:164−170(1991),Eisenbergら,Methods Enzymol.227:396−404(1997),Fischerら,FASEB J.10:126−136(1996),Luthyら,Nature 356:83−85(1992))。標準的な尺度を用いる相同性鋳型に対する関係においてAlsモデルを確証のために評価した(e−値(Welchら,Biochemistry 35:7165−7173(1996),Welchら,Biochemistry 33:6074−6085(1994))。最後に、Alsモデルの物理化学的特性を、物理的特性がAlsのN末端ドメインの水接近可能表面へ投影されるように、SYBYLおよびHINTプラットフォーム(Kellogら,J.Comput.Aided Mol.Des.5:545−552(1991))において実行して、MOLCAD(Heidenら,J.Comput.Chem.14:246−250(1993))によって可視化した。
【0085】
これらのモデルは、全てのAlsタンパク質のN末端ドメインが、免疫グロブリンスーパーファミリーのメンバーと合致して、多数の反−平行β−シートドメインを含有することを示す。結果は表IIIにて以下にまとめる。これらのタンパク質は、典型的には、それからループ/コイル構造が投影される、複雑な7−ストランド反−平行β−シートドメインよりなる。β−シートドメインは間に存在する領域によって相互から分離される。この構造は、しばしば、ビーズ−オン−ア−ストリングモチーフという。特に注意すべきは、Alsタンパク質の実質的に全てが公知のアドヘシンまたはインバシンホモログに対してモデル化されたことである(表III)。同様性の異なるパターンが分析したAlsタンパク質内で観察された。例えば、Als7p以外の調べた全てのAlsタンパク質は、黄色ブドウ球菌のコラーゲン−結合タンパク質との有意な相同性を共に有した。しかしながら、特異的な一次、二次および三次ホモログはほとんどのファミリーメンバーについて変化した(表III)。例えば、Als2pおよびAls9pは同一の一次、二次、および三次ホモログを共有した。
【0086】
(表III)
(Alsタンパク質間のホモログの比較)
各Alsタンパク質のホモログは記載された知識に基づくアルゴリズムによって同定し、1から3の構造相関の下降順でランク付けした。NS、相同性モデリングについて有意なモデルは同定されなかった(相関係数((r2)≦70%。PDB、バイオテクノロジー情報様式でのNational Centerでのタンパク質データーバンクコード(Protein Data Bank)。
【0087】
【表3−1】
【0088】
【表3−2】
Alsタンパク質は、予測されたループ/コイル構造にマップされるN末端超可変領域を含有するとも決定された。この点に関し、個々のAlsタンパク質によって媒介される基材特異的接着の観察された差にも拘わらず、N末端ドメインにおける大きな配列の領域はこのファミリーを横切って保存されている。しかしながら、超可変領域(HVR)1から7と命名されたAlsタンパク質内の有意な多様性の7つの領域が見出された。(8以上のアミノ酸から構成される)これらの領域は、Alsタンパク質を横切っての明らかなコンセンサス同一性は含まず、かつ50%未満のコンセンサス保存を含んだ。対照的に、介入する保存領域(CR)1から7は、Alsタンパク質を横切って30%を超えるコンセンサス同一性、および50%を超えるコンセンサス保存を呈した。既知の機能を持つAlsタンパク質のN末端ドメイン(残基1から420)を含むこれらのアミノ酸配列の同一性プロットおよび模式的整列を図11、AおよびBに掲げる。特に、相同性モデリングは、異なるAlsタンパク質のHVRが、配列は識別可能でありつつ、CRのβ−シート成分から投影される同様なループ/コイル構造に適合すると予測されることを明らかとした。かくして、これらの保存されたHVRの存在は、それらが宿主構成要素と相互作用するのに入手可能であることを示す。
【0089】
前記した相同性モデリングおよび関連決定に加えて、経験的決定は、加えて、Als1pのN末端ドメインの予測される構造を確認する。我々の相同性モデリングによって作り出される仮説を検定するために、CDおよびFTIR分光分析の補充的なアプローチを用い、Als1pのN末端ドメインの構造的特徴を決定した。Als1pのアミノ酸17から432を含むこのタンパク質はS.cerevisiaeで生産され、Fuら,Molecular Microbiology,44:61−72(2002)によって従前に記載されている。
【0090】
簡単に述べれば、円二色スペクトルを、熱電気温度コントローラーを備えたAVIV62DSスペクトロポラリメーター(Aviv Biomedical Inc.)で記録した。0.1−mm光路脱着可能水晶セルを用い、Als1pの水溶液(リン酸−緩衝化生理食塩水中10μM)を260から185nmまで10nm/分の速度および0.2nmの試料間隔でスキャンした。ペプチドを欠く緩衝液からのスペクトルを試料溶液から差し引いて、光散乱人工物を最小化し、最終スペクトルは25℃で記録した8スキャンの平均であった。該機器は(+)−10−ショウノウスルホン酸(1−mm路長セル中1mg/ml)(Johnsonら,Proteins 7:205−214(1990))でルーチン的にキャリブレートされており、楕円度は平均残基楕円度(1)MRE(度−cm2 dmol−1)として表した。タンパク質濃度は、発現されたAls1pドメインの芳香族アミノ酸組成に基づく280nmにおける吸収によって決定された(Paceら,Protein Sci 4:2411−2423(1995))。CDスペクトルは、インターネットに基づくDichroweb(Lobleyら,Bioinformatics 18:211−212(2002))インターフェース(cryst.bbk.ac.uk/cdweb/html/home.html)を介してSelcon(Sreeramaら,Protein Sci.8:370−380(1999))を用い、ラセン、β−シート、ターン、および乱れた構造に分割した。
【0091】
Als1p自己−フィルムの赤外スペクトルは、4の利得の、256スキャンにわたって平均され、かつ2cm−1の分解能にて、ジューテリウムトリグリシン硫酸ディテクターを備えたBruker Vector 22 FTIRスペクトロメーター(Bruker Optics)で25℃にて記録した。50μlのリン酸−緩衝化生理食塩水中の50マイクログラムのタンパク質を、50×20×2−mmゲルマニウム減衰全反射試料結晶(Pike Technologies)の表面に広げ、乾燥させた。次いで、赤外スペクトルを記録するに先立って、乾燥タンパク質自己−フィルムをD2Oで1時間水和させた。赤外スペクトルのアミドIバンドを、曲線−フィッティングソフトウェア(GRAMS/32,Version 5;Galactic)を備えた成分ピークの面積計算によって二次立体配座について分析した。種々の立体配座についての周波数限界は以下の通りであった:α−ラセン(1662−1645 cm−1)、β−シート(1637−1613および1710−1682 cm−1)、β−ターンループ(1682−1662 cm−1)、および乱れた構造(1645−1637 cm−1)(50−52)。
【0092】
Als1pのN末端ドメインの円二色結果を図12Aに示し、これは、217nmにおける二色最小、および200nm近くの強い正の二色最大を明らかとする。これらの特徴は、支配的な反−平行βシート成分を有するタンパク質の特徴である。CDスペクトルの分解は、タンパク質は50.1%βシートの立体配座を採り、他方、他の構造クラス寄与は乱れた構造(26.9%)、ターン構造(19.3%)、およびα−ラセン(3.7%)を含む。
【0093】
図12Bに示すように、水和したAls1pの自己−フィルムのFTIR測定により、試料が支配的なβ−シート立体配座を有することが強く確認された。これらのスペクトルにより、1634および1628cm1に中心があるピークを持つ強い低周波数アミドIバンド、および1685cm−1に中心がある弱い高周波数バンドが明らかにされた。タンパク質アミドI赤外スペクトルの高および低周波数成分へのこの周波数分裂は、分子間反−平行β−シートの間の転移二極結合の効果に典型的であることが示されている(Halversonら,J.Am.Chem.Soc.113:6701−6703(1991))。スペクトルの曲線フィッティングは、タンパク質構築物が〜57.2%反平行β−シートであることを示した。IRスペクトルの曲線フィッティングからの他の二次構造立体配座は、乱れた構造(20.5%)、ターン成分(13.3%)、およびα−ラセン(9.0%)を含む。
【0094】
一緒にすると、FTIRおよびCDデータは、Als1pのN末端が、構造度が低い領域が中間に存在する、従たるα−ラセンおよびターン成分を含有する反−平行β−シート構造の支配的なドメインを含有することをさらに裏付ける。
【0095】
三次元モデルは、さらに、AlsのN末端ドメインの間の物理化学的区別を示す。この点に関し、分子モデルは、宿主細胞およびいくつかの基材とのそれらの相互作用に影響するようであるAlsタンパク質のN末端ドメインの予測された物理化学的属性の差を示した。図13に示すように、Alsタンパク質は、疎水性、電荷、および水素結合ポテンシャルの表面分布に基づいて3つの区別される群に分離可能である。Als1p、Als3pおよびAls5pは、各々、これらの特性の同様なパターンを共に有し、かくして、Als群Aと考えられる。対照的に、Als6pおよびAls7pのN末端ドメイン(Als群B)の予測された物理化学的特性は、Als群Aのそれからの顕著な差を有する(図13)。Als群Aのメンバーにおけるカチオンポテンシャルは、典型的には、それらの中性またはアニオン面から分離されるが、正の電荷はAls群BのメンバーAls6pおよびAls7pの全表面を横切って広く分布している。最後に、Als2p、Als4p、およびAls9pのN末端は、Als群AまたはBタンパク質いずれかとは構造的に異なるAlsタンパク質の第三の群(Als群C)を構成するようである。Als群Cタンパク質は、疎水性または静電分布の点でAls群Bタンパク質よりもAls群Aにより同様なように見えるであろう。
【0096】
接着機能を持ついくつかのタンパク質はC.albicansにおいて同定されている。Hwp1pは、哺乳動物トランスグルタミナーゼに対する基質として作用することによって、頬上皮細胞への接着を媒介することが示されている(5)。EAP1は、最近、S.cerevisiaeにおける異種発現によって同定されており、ポリスチレンおよび腎臓上皮細胞への接着をインビトロで媒介する(7)。Alsタンパク質ファミリーの8つのメンバーのうち、Als1pおよびAls5pのみが機能的見込みから研究されている。Als1pの異種発現は、ヒト血管内皮細胞および上皮細胞への結合を媒介することが示されており、遺伝子分布実験を介してC.albicansにおいて確認されているという知見である(Fuら,Mol.Microbiol.44:61−72(2002),Fuら,Infect.Immune.66:1783−1786(1998))。S.cerevisiaeにおけるALS5の異種発現は接着性をコラーゲン、フィブロネクチン、ウシ血清アルブミン、およびラミニンに付与する(Gaurら,Infect.Immune.65:5289−5297(1997),Gaurら,Infect.Immun.67:6040−6047(1999),Gaurら,Cell Commun.Adhes.9:45−57(2002))。C.albicansのアドヘシンの基材特異性の大規模な比較は行われたことがない。本実験において、我々は、Alsタンパク質ファミリーメンバーの構造的に多様な群の接着特性を比較した。我々のデータは、Alsタンパク質が、種々のヒト基材への接着に対する特異性の重複するスペクトルを持つ表面タンパク質の多様なファミリーを含むことを示す(図8)。さらに、本ドメイン交換実験からの結果は、Alsタンパク質のN末端ドメインが、それらの基材接着性プロフィールの特性を付与することを示す。
【0097】
接着性を媒介するに加え、我々のデータは、Alsタンパク質はインバシンとしても機能できることを示唆する。興味あることに、Als1pおよびAls3p双方を発現するS.cerevisiaeは同様な内皮細胞接着を示したが、Als3p−発現S.cerevisiaeはかなり高い速度での内部化を受けた。これらの結果は、エンドサイトーシスが接着の延長であるばかりか、むしろ、リガンド−受容体相互作用によって影響され得る区別されるプロセスであることを示す。Alsタンパク質中のN末端配列の差は、接着で当てはまるように、これらの区別される機能を媒介するようである。
【0098】
三次元空間に分布したタンパク質ドメインの物理化学的特性は、受容体−リガンド相互作用を支配する非常に重要な構造的特徴である(Eisenbergら,J.Mol.Biol.179:125−142(1984),Waringら,Protein Peptidew Lett.3:177−184(1996),Hancockら,Lancet349:418−422(1997))。Alsタンパク質は、免疫グロブリンスーパーファミリーの他のアドヘシンおよびインバシンに特徴的な立体配座特徴を共有する。しかしながら、ここのAlsタンパク質はそれらの一次ホモログが異なり、これは、Alsファミリーのメンバーが異なる基材結合プロフィールを呈することを示す実験データと合致する知見である。まとめると、Als相同性のこれらのパターンは、Alsタンパク質メンバーは構造および予測された折畳において全体的同様性を共有するが、機能のそれらの差の原因である区別されるAlsタンパク質の間で構造的差が存在することを示す。
【0099】
Alsファミリーメンバー構造決定に関する前記した結果は、Alsp1pのN末端領域が、少量の比較的構造化されていない領域と共に、ループ/コイル構造を含有する反−平行β−シートドメインから圧倒的に構成されることを示す相同性モデリングを裏付ける。これらの特徴は、免疫グロブリンスーパーファミリーのメンバーのモチーフを示す。これらの結果は、Als5pの円二色実験との有意な予測的裏付け(Hoyerら,Yeast 18:49−60(2001))を示し、これは、Als5pのN末端が反−平行β−シートおよびループ/コイル領域の相対的圧倒性によって特徴付けられることを示す。かくして、Alsタンパク質ファミリーの全てのメンバーはこの総じての構造をかなり呈するようである。特に、前記した構造の結果もまた、HVRの多くが区別されるAlsタンパク質のN末端中のβ−シートドメインから投影するフレキシブルなループ/コイル構造に対応することを示す相同性モデルとやはり合致する。まとめると、これらの結果は、これらの構造がAlsタンパク質による基材特異的結合に一体化されることを示す(図14)。前記した結果と合致して、マンノース−結合レクチンの類似領域、α−アグルチニン、および免疫グロブリンスーパーファミリーの他のメンバーが、基材結合特異性を付与するようである(Zhaoら,Hybrid Hybridomics21:25−36(2002),Wojciechowiczら,Mol.Cell.Biol.13:2554−2563(1993))。さらに、これらの可変ループ領域の突然変異が、これらの相同タンパク質において基材結合を有意に改変する(Renzら,J.Cell Biol.125:1395−1406(1994),Vineyら,J.Immunol.157:2488−2497(1996))。
【0100】
三次元モデリングの結果は、さらに、個々のAlsタンパク質のN末端ドメインが、それらの接着プロフィールに関連する区別される分子シグニチュアを保有することを示す。これらのシグニチュアは、Alsタンパク質の中で構造的関係を区別する立体配置を生じる、表面領域、疎水性、および静電気電荷のようなパラメーターを取り込む。例えば、Als群Aメンバー(Als1p,Als3pおよびAls5p)のような多数の基材に結合するAlsタンパク質が、立体バルク、疎水性分布、および静電ポテンシャルの点で同様な予測されるL−末端プロフィールを有する。なお、この群内においてさえ、該群内の機能的差を支配することができる特異的物理化学的区別が存在する(図13)。対照的に、低下した接着能力を持つAlsタンパク質は、疎水性および静電ポテンシャルを含めた、多数の物理化学的特性においてAls群Aタンパク質から区別されると予測される表面特徴を有する。これらの構造的特徴の差の集合効果は、区別されるAlsタンパク質の特異的機能的特性をかなり付与するようである。
【0101】
広範な遺伝的変動性がALS遺伝子ファミリー内で示されているC.albicansの異なる単離体の特異的ALS遺伝子における配列変動が観察されており(Zhangら,Genome Res.13:2005−2017(2003),Hoyerら,Yeast18:49−60(2001))、ALSファミリーの全てのメンバーが全ての単離体に存在するのではない。単一単離体における2つの異なる対立遺伝子の間の有意な配列多様性さえ見出されている(Zhaoら,Microbiology149:2947−2960(2003),Zhangら,Genome Res.13:2005−2017(2003))。この程度の遺伝的変動性は、これらのタンパク質が比較的高い周波数において再編成または突然変異を受け得ることを示唆する。そのようなメカニズムは、本実験において示された高度な構造的および機能的多様性を生じさせる能力を生物に供するであろう。この仮説に対する間接的裏付けは、この遺伝子が共に高突然変異性であり、これらの突然変異は選択圧に従うという双方を示唆したALS7の対立遺伝子変動の最近の研究によって供される(Zhangら,Genome Res.13:2005−2017(2003))。
【0102】
まとめると、前記結果は、構造および機能的レベルの双方において抗体およびAlsタンパク質の間の類似性を示す。例えば、相同性モデリングは、これらのファミリーの構造的立体配置における同様性を過小評価しており、過剰変動はそうでなければ安定なフレームワーク内の局所化ドメインに標的化されている(例えば、Alsタンパク質のHVRおよび免疫グロブリンにおけるFab領域)。さらに、抗体に関しては、ALS遺伝子ファミリーの遺伝的変動性は、Candidaが、接着および侵入における特異性のスペクトルを持つタンパク質の多様なアレイを呈する機会を供するであろう。関連タンパク質のそのような群の入手可能性は、感染の間に異なる解剖学的および生理学的ニッチをコロニー化し、およびそれに侵入する生物の能力を改良するようである。
【0103】
本出願を通じて、種々の刊行物を括弧内に入れて引用した。これらの刊行物の開示の全体をここに引用して本明細書中に組込んで、本発明が属する分野の最先端技術を十分に記載する。
【0104】
本発明を開示された実施形態を参照して記載してきたが、当業者であれば、具体的実施例および前記した詳細な実験は本発明を説明するだけであることを容易に認識するであろう。本発明の精神を逸脱することなく種々の修飾を成すことができることは理解されるべきである。従って、本発明は以下の請求の範囲によってのみ限定される。
【0105】
(実施例VII)
(細胞−媒介の液性でない免疫を高めることによって、rAls1p−Nでのワクチン接種はマウス播種性カンジダ症の間の生存を改良する)
本実施例は、Als1pの組換えN末端(rAls1p−N)でのBALB/cマウスの免疫化が、C.albicansの致死接種物での引き続いてのチャレンジの間に生存を改良したことを示す。保護的20μg用量のrAls1p−NはTh1脾臓細胞のCandida−刺激を有意に増加させ、インビボ遅延型過敏を増加させた。対照的に、抗体力価は保護に相関しなかった。最後に、ワクチンはT細胞−欠乏マウスにおいて保護的ではなかったが、B細胞−欠乏マウスにおいては保護的であった。これらのデータは、rAls1p−Nワクチンの作用メカニズムが、C.albicansに対する液性よりはむしろ細胞媒介免疫の刺激であることを示す。
【0106】
該実験で用いたC.albicansはSC5314、すなわち、W.Fonzi(Georgetown University)によって供給された動物モデルにおいて高度にビルレントなよく特徴付けられた臨床単離体であった(Spellbergら,Infect Immun.71:5756−5764(2003))。該生物は、感染に先立って酵母ペプトンデキストロースブロス(Difco)において3回系列的に継代された。
【0107】
該実験で用いたマウス株はNational Cancer Institute(Bethesda,MD)から入手した雌BALB/cマウスであった。ワクチン効力に対する年齢のインパクトを探求するために、若年性マウス(8から10週)およびリタイアしたブリーダー(≧6月)の双方を利用した。igh遺伝子座のホモ接合性欠失を担う雌B細胞−欠乏マウス(C.129B6−IgH−Jhdtm1Dhu)、T細胞−欠乏ヌードマウス(C.Cg/AnBomTac−FoxnlnuN20)、およびコンジェニック野生型BALB/c同腹子はTaconic農場(Germantown,NY)から入手した。マウスを、照射した食物およびオートクレーブ処理した水を自由に備えた濾過ケイジに収容した。生存実験では、マウスを種々の用量の抗原(後記参照)で免疫化し、引き続いて、C.albicans SC5314出芽胞子の適当な接種物、またはPBS(Irvine Scientific,Irvine,CA)対照で尾静脈を介して感染させた。もし個々のデータベースが統計学的不均一を示さなければ、複連生存実験の結果を合わせた(後記参照)。マウスに関連する全ての手法は、動物収容および看護のためのNational Institutes of Health指針に従い、制度的動物使用および看護委員会によって認可された。
【0108】
以下に記載されたrAls1p−N免疫化手法は以下のように行った。簡単に述べれば、rAls1p−N(Als1pのアミノ酸17から432)はS.cerevisiaeにおいて生産され、ゲル濾過およびNi−NTAマトリックスアフィニティー精製によって精製した(Fuら,Molec.Microbiol.44:61−72(2002))。タンパク質の量は修飾されたLowryアッセイによって定量した。高度な純度(約90%)が、前記したような、SDS−ポリアクリルアミドゲル電気泳動ならびに円二色性およびFTIRによって確認された。マウスは、0日目にフロイントの完全アジュバント(CFA−Sigma−Aldrich)と混合されたrAls1p−Nの腹腔内(ip)注射によって免疫化し、21日目にフロイントの不完全アジュバント(IFA,Sigma−Aldrich)を含むもう1つの用量の抗原でブースター注射し、次いで、ブーストから2週間後に感染した。
【0109】
得られた抗体力価は96ウェルプレートにてELISAによって決定した。簡単に述べれば、ウェルをPBS中の5μg/ml rAls1p−Nのウェル当たり100μlで被覆した。3%ウシ血清アルブミンを含有するTris緩衝化生理食塩水(TBS)(0.01 MTrisHCI,pH7.4,0.15 M NaCl)でのブロッキング工程に続いて、マウス血清を室温にて1時間インキュベートした。ウェルを、0.05%Tween20を含有するTBSで3回洗浄し、続いて、TBSでさらに3回洗浄した。ホースラディッシュペルオキシダーゼ(Sigma)とコンジュゲートさせたヤギ抗マウス2次抗体を1:5000の最終希釈にて加え、プレートを室温にて1時間さらにインキュベートした。ウェルをTBSで洗浄し、0.1Mクエン酸緩衝液(pH5.0)、50mg/mlのo−フェニレンジアミン(Sigma)、および10μlの30%H2O2を含有する基質と共にインキュベートした。色を30分間で発色させ、その後、10%H2SO4を加えることによって反応を停止させ、光学密度(OD)をマイクロタイタープレートリーダーにて490nmで決定した。陰性対照ウェルは希釈剤のみを受け、バックグラウンド吸収をテストウェルから差し引いて、最終OD読みを得た。ELISA力価は、正のOD読みを与えた最後の血清希釈の逆数を取った(すなわち、≧陰性対照試料の平均OD+2標準偏差)。
【0110】
以下に記載した他の方法は以下のように行った。簡単に述べると、C.albicans−誘導サイトカインプロフィールを行って、細胞−媒介免疫性およびインビボサイトカインプロフィールに対するrAls1p−Nワクチンの効果を決定した。マウスを前記したように免疫化した。最終ブーストから2週間後に、脾臓細胞を収穫し、従前に記載されているように4×106細胞/mlの密度にて完全培地中で培養した(Spellbergら,Infect.Immun.71:5756−5764(2003))。サイトカイン生産を刺激するために、脾臓細胞を熱で殺したC.albicans SC5314芽菅と共に共培養した。我々は、rAls1p−Nの代わりに加熱して殺したC.albicansを用いて、脾臓細胞を刺激して、感染の間にインビボ状況を模倣した。C.albicans細胞を、グルタミンを含むRPMI−1640(Gibco BRL)中で予め90分間発芽させて、Als1pの発現を誘導した(Fuら,Molec.Microbiol.44:61−72(2002))。得られたC.albicans芽管を60℃における90分間のインキュベーションによって加熱により殺した(Ibrahimら,Infect.Immun.63:4368−74(1995))。加熱して殺した真菌を2×107偽菌糸/mlの密度にて脾臓培養に加えた(1白血球に対する5真菌の比率)。48時間後に、従前に記載されているように、脾臓細胞を、細胞内サイトカイン検出およびフローサイトメトリーによってTh1(CD4+IFN−” +IL−4−)、Th2(CD4+IFN− −IL−4+)、またはCD4+IL−10+周波数についてプロファイリングした(Spellbergら, Infect. Immun. 71:5756−5764(2003))。3−色フローサイトメトリーは、製造業者の推奨に従ってFACSCompソフトウェアを用い、CaliBRITEビーズ(Becton Dickinson,San Jose,CA)でキャリブレートしたBecton−Dickson FACScan機器で行った。データ獲得の間、CD4+リンパ球は前方および側方散乱の連結、およびFITC−抗CD4抗体蛍光特性によって開閉した。各試料についてのデータは、10,000のCD4+リンパ球を分析するまで獲得した。結果は、Th1またはTh2細胞であった全ての開閉したリンパ球のパーセンテージのメジアン±25および75四分位として示す。
【0111】
足蹠膨潤はOomuraらの方法によって決定した(41)。簡単に述べれば、マウスを前記したように適当な用量のrAls1p−NまたはCFA単独で免疫化した。ブーストから二週間後に、免疫化マウスのベースライン足蹠サイズを、電子デジタルキャリパーを用いて測定した。25μlのPBS中の50μgのrAls1p−Nを右側足蹠に注射し、PBS単独は免疫化マウスの左側足蹠に注射した。24時間後に、足蹠を再度測定した。抗原−特異的足蹠膨潤を:(24時間における右側足蹠厚み−ベースラインにおける右側足蹠厚み)−(24時間後における左側足蹠厚み−ベースラインにおける左側足蹠厚み)として計算した。
【0112】
非パラメーターLog順位テストを利用して、マウスの生存時間の差を決定した。抗体の力価、Th1またはTh2リンパ球の頻度、および足蹠膨潤を、適切には、非パラメーター多数比較のためにSteel検定(Rhyneら, Biometrics 23:539−49(1967))、および不対比較のためにMann Whitney U検定によって比較した。相関はSpearman順位和検定で計算した。不均一性が複連生存試験で存在するかを決定するために、Kolmogorov−Smirnov検定を利用した。P値<0.05は有意と考えられた。
【0113】
rAls1p−N免疫原の最も有効な用量を決定するために、107−倍用量範囲を評価した(マウス当たり20pgから200μg)。雌のリタイアしたブリーダーBALB/cマウスをrAls1p−N+アジュバント(CFA/IFA)またはアジュバント単独で免疫化した。免疫化したマウスをブースティングから二週間後に採血して、抗rAls1p−N抗体力価を決定した(後記参照)。マウスを、引き続いて、C.albicans(2×105出芽胞子)の致死接種物で感染させた。反復実験からの生存データを合わせた。というのは、個々の実験は統計学的不均一性を示さなかったからである(Kolmogorov−Smirnov検定によるとp.>0.05)。20μg用量のrAls1p−Nは、感染マウスの25%の長期生存、およびアジュバント単独と比較した全生存の有意な増加(Log順位検定によるとp=0.044、図1)をもたらした。10倍高い(図15)または低い(データは示さず)用量いずれも、アジュバント単独と比較して生存を有意に増加させなかった。この結果は、中程度の用量のrAls1p−Nワクチンがネズミ播種性カンジダ症に対して保護を誘導することを示す。
【0114】
前記知見はrAls1p−Nワクチンについての保護的用量を確立した。次に、ワクチンの有効性を、106出芽胞子で感染させたより迅速に致死的なマウスのモデルにおいて評価した(各々、メジアン生存3 vs.11日、ワクチン接種されていないマウスにおいて106 vs.2×105接種物)。再度、反復実験からのデータを組み合わせた。というのは、個々の実験の結果は統計学的不均一性を示さなかったからである(Kolmogorov−Smirnov検定によるとp>0.05)。106C.albicans出芽胞子で感染させたBalb/cマウスに20μg用量+CFAとして投与した場合、rAls1p−Nワクチンはメジアン生存の2倍を超え、その結果、全生存の有意な増大−vs−ワクチン接種していない対照をもたらした(Log順位テストによるとp=0.001、図16A)。マウスの年齢がrAls1p−Nワクチンに対するそれらの応答に影響するかを決定するために、我々は若年マウスにおいてそれをテストした。若年マウスをワクチン接種し、同一の高い接種物で感染させた場合、同様な生存利益が見出された(Log順位検定によるとp=0.02、図16B)。
【0115】
200μg用量のrAls1p−Nの結果、20μg用量と比較して劣った保護がもたらされたが、200μg用量の抗原のみが血清抗Als1p抗体力価の有意な増大を誘導した(200μg用量 vs.全ての他の群についてp≦0.005、図17)。抗Als1p抗体力価の有意な増加は、中程度の保護的抗原用量で検出されなかった(20μg vs.アジュバントについてp=0.1)。個々のマウスの血清抗Als1p抗体力価を各マウスの生存時間に対してプロットすると、抗体力価および生存の間で相関は見出されなかった(R2=0.03、Spearman順位和検定によるとp>0.05)。事実、最高用量の抗原(200μg)で免疫化したマウスは、1:100,000過剰の抗rAls1p−N抗体力価を有したが、その力価が検出の下限である(〜1:100)抗原のより低い用量で免疫化したマウスから異ならない生存持続を有した。これらの結果は、rAls1p−Nワクチンによって誘導された保護が抗体力価に相関するように見えないことを示す。
【0116】
液性免疫はrAls1p−N−誘導保護と相関しなかったので、我々は、保護および非保護用量のrAls1p−Nによって誘導された細胞−媒介免疫応答を調べた。マウスを、前記したように、0.2、20、または200μgのrAls1p−N、またはアジュバント単独で免疫化した。ブーストから2週間後に、脾臓細胞を収穫し、Als1pを発現することが知られている、熱で殺した予め発芽させたC.albicansの存在下で培養した(Fuら,Molec.Microbiol.44:61−72(2002))。培養から48時間後に、脾臓細胞をフローサイトメトリーによって細胞内サイトカイン検出のために収穫した。保護的20μg用量の抗原で免疫化したマウスからのリンパ球のみが、アジュバント単独を与えられたマウスと比較してTh1細胞の有意に増加した頻度を生じた(P=0.03、図18)。Th2の頻度(図18)およびIL−10+Tリンパ球の頻度(データは示さず)の有意な差は、アジュバントまたは該用量のいずれかの抗原で免疫化されたマウスの間で検出されなかった。
【0117】
タイプ1免疫がインビボでrAls1p−Nによって刺激されたことを確認するために、足蹠膨潤によって遅延タイプ過敏をテストした。保護的20μg用量のrAls1p−Nでワクチン接種したマウスのみが、対照と比較して有意に増大した遅延型過敏反応を発生し、この応答は、非保護的0.2および200μg用量によって誘導されたものよりもやはり有意に高かった(図19、非パラメーターSteel検定によると、全ての比較−vs−20μg用量についてp<0.05)。まとめると、これらの結果は、保護的用量のrAls1p−N抗原が有意なTh1偏光および遅延型過敏反応を誘導したことを示す。
【0118】
ワクチン−媒介保護における抗体およびt−細胞の役割を規定するために、B細胞欠乏、T細胞欠乏ヌード、またはコンジェニックBALB/c野生型対照マウスを20μgのrAls1p−Nの+アジュバントまたはアジュバント単独で免疫化し、C.albicansの致死接種物(8×105出芽胞子)で感染させた。B細胞−欠乏マウスは感染に対してより耐性である傾向があり、他方、T細胞−欠乏マウスはアジュバントのみが与えられた野生型対照マウスよりも感受性であった(各々、B細胞欠乏およびT細胞欠乏マウス−vs−野生型アジュバント−処理についてp=0.065および0.01、図20)。最後に、rAls1p−NワクチンはB細胞−欠乏マウスにおいてその効果を維持した(rAls1p−Nワクチン接種−vs−アジュバント単独についてp=0.04、図6)が、T細胞−欠乏マウスにおいて効果的でなかった(rAls1p−Nワクチン接種−vs−アジュバント単独についてp=0.4、図20)。これらの結果は、Als1pワクチンがB細胞欠乏マウスにおいて効果的であったがT細胞欠乏ヌードマウスにおいてはそうではなかったことを示す。
【0119】
前記したのは、このタンパク質のN末端での免疫化が、引き続いての血行的に播種性のカンジダ症の間に若年性および成熟BALB/cマウス双方の生存を改良したことを示す結果である。特に、中間用量のrAls1p−N(20 μg)は、より低い用量およびより高い用量(200μg)双方と比較して優れた保護を供した。それにも拘わらず、非保護的200μg用量のrAls1p−Nは免疫原性であった。というのは、それは保護的20μg用量よりも100倍高い抗体の力価を誘導したからである。
【0120】
最高用量のrAls1p−Nにおけるより低い保護での、逆U−形状用量−応答効力曲線は、最初に、与えられた用量の抗原による液性および細胞−媒介免疫の誘導の間の逆関係を記載したParishら,の古典的研究を思い出させる。Parishの精液データとの関係で、逆U−形状用量−応答効力曲線は、もし:1)ワクチン効力が細胞−媒介免疫に依存し、および2)中程度の用量のrAls1p−Nが高い抗体−刺激用量と比較して優れた細胞−媒介免疫を刺激したならば説明することができよう。我々は、従って、rAls1p−Nワクチンで見られた逆U−形状用量応答効力曲線は、保護的な中程度の用量の抗原による細胞−媒介免疫の優れた誘導によるものであったと仮定した。
【0121】
この仮説を検定するために、Th1細胞および遅延型過敏を刺激する高い、中程度、および低い用量の抗原の能力を決定した。脾臓細胞からのサイトカイン−生産を刺激するために、我々は、rAls1p−Nの代わりに熱で殺したC.albicansへの暴露によって細胞を特異的に活性化して、感染の間のインビボ状況を模倣した。保護的20μgの用量だけがC.albicans−刺激脾臓Th1リンパ球の頻度を有意に増加させた。エクスビボC.albicans−刺激脾臓細胞で見られたTh1の頻度は、マウスにおける播種性カンジダ症の間にインビボで検出されたのと同様であり(59)、このアプローチの関連性を過小評価している。
【0122】
検出されたエクスビボTh1細胞がインビボにて機能的重要性のものであるかを決定するために、我々は、異なる用量のrAls1p−N免疫化によって誘導された遅延型過敏を比較した。Th1細胞の頻度と一致して、保護的20μg用量のrAls1p−Nだけが有意なインビボ遅延型過敏反応を刺激した。これらの結果は、ワクチン−誘導保護がタイプ1の誘導、細胞媒介免疫によるものであるという仮説と合致する。驚くべきことに、200μg用量のrAls1p−Nによる顕著に上昇した抗体力価の誘導にも拘わらず、我々は、この用量でワクチン接種したマウスにおいて脾臓Th2リンパ球の増加は見出さなかった。1つの可能な説明は、Th2細胞が脾臓細胞よりはむしろ末梢リンパ節において活性化されたというものである。あるいは、他のT細胞集団(例えば、NKT細胞)が、200μg用量のrAls1p−Nに応答して見られた高い抗体力価を誘導する原因であっただろう。
【0123】
抗体力価および保護の間の相関の欠如は、ワクチン−誘導保護を媒介するにおける抗体の役割を完全には排除しなかった。例えば、ELISA力価は、種々の特異性および親和性を持つ抗体の列挙の結果である。従って、ワクチン−媒介保護に参画した抗体の小さなサブセットが生じたという可能性は、抗体力価を測定することによって排除することができなかった。rAls1p−Nワクチン−媒介保護における細胞−媒介の液性でない免疫の役割を確認するために、我々は、B細胞−およびT細胞−欠乏マウスにおけるワクチンの効果をテストした。B細胞−欠乏マウスは、野生型対照よりも播種性カンジダ症に対してより耐性である傾向があり、ワクチンの効果はB細胞−欠乏マウスにおいて無効にならなかった。対照的に、T細胞−欠乏マウスは野生型対照よりも播種性カンジダ症に対してより感受性であり、ワクチンの効果はT細胞−欠乏マウスにおいて失われた。我々の知見は、従って、rAls1p−Nワクチンの効果はT細胞−媒介の主として液性ではない免疫の誘導に依存していることを確認する。同様に、B細胞―欠乏マウスはコンジェニック野生型同腹子よりも播種性カンジダ症に対してより感受性ではなかったゆえに、抗体はこのモデルにおいて播種性カンジダ症に対して支配的なエフェクタではない。
【0124】
まとめると、我々は、新規なrAls1p−Nワクチンは、液性免疫よりはむしろ細胞−媒介免疫を誘導することによって実験的散在カンジダ症に対する保護を媒介すると報告する。rAls1p−Nワクチンの中程度の保護的効果の増強は、従って、最適化アジュバントおよび/またはサイトカインを用いる細胞−媒介免疫のさらなるプライミング、または代替免疫化経路で達成することができる。事実、我々の進行している研究においては、我々は、腹腔内と比較してrAls1p−Nを皮下投与することによって効力の顕著な増大を既に見出している。
【0125】
(実施例VIII)
(抗Candida albicans rAls1p−Nワクチンは真菌負荷を低下させ、免疫応答性および免疫無防備マウスの双方において生存を改善する)
本実施例は、免疫応答性および免疫無防備マウスの双方における皮下(SQ)経路によって投与した場合の、実施例VIIに記載されたrAls1p−Nワクチンの効力の増強を記載する。まず、免疫応答性マウスにおけるrAls1p−Nワクチンの効力。アミノ酸19から433の全長タンパク質を含むrAls1p−Nを前記したようにS.cerevisiaeにおいて生産し、精製した。対象調製物を、同様に、空のプラスミドで形質転換したS.cerevisiaeから精製した。BALB/cリタイアしたブリーダーマウス(25から30g)を、0日目に、フロイントの完全アジュバント(CFA)と混合したrAls1p−N(20μg)または対照調製物のSQ注射によって、続いて、21日目にフロイントの不完全アジュバント(IFA)中のブースター用量によって免疫化した。ブーストから2週間後に、ワクチンの免疫原性は、従前に記載されているように、遅延型過敏(DTH)のマーカーとしての足蹠膨潤反応の強度を評価することによって確認した。ワクチン接種したマウスは、rAls1p−N特異的DTHの顕著な増加を有した(図21)。
【0126】
rAls1p−Nワクチンの効果は、感染したBALB/cマウスにおける生存に対するrAls1p−Nワクチン接種のインパクトを決定することによって評価した(図22A)。ワクチン接種したまたは対照マウスを、C.albicansの迅速に致死的な接種物(2.5から5×105出芽胞子)で尾静脈を介して感染させた。我々は、そのような接種物で感染させたマウスがひどい敗血症ショックで死亡したことを従前に示した(Spellbergら,J.Infect.Dis.In press(2005))。ワクチン接種は死亡までの時間を顕著に延長させ(Log順位検定によると双方の接種物についてp<0.05)、30日生存を改善した(50から57% vs.0%、Fisher’s Exact検定によると双方の接種物についてp<0.05)。
【0127】
次いで、血行的に播種性のカンジダ症の間における組織真菌負荷に対するワクチン接種のインパクトを決定した。ブーストから14日後に、ワクチン接種したおよび対照BALB/cマウスをC.albicans SC5314の5×105出芽胞子で尾−静脈を介して感染させた。感染から6日後に、対照実験群における最初の死亡の開始の前に、腎臓を収穫し、ホモゲナイズし、Sabouraudデキストロース寒天(Difco)中で定量的に培養した(18)。rAls1p−NでのSQワクチン接種の結果、対照と比較して腎臓真菌負荷のメジアン1.5 log CFU/g減少がもたらされた(Wilcoxon順位和検定によるとp=0.01、図22B)。
【0128】
rAls1p−Nワクチンの効力もまた、免疫無防備マウスにおいて評価した。免疫応答性マウスにおいて効力を証明したら、好中球減少症マウスにおいて免疫を誘導し、好中球減少症マウスを播種性カンジダ症から保護するrAls1p−Nワクチンの能力も評価した。ワクチン接種したBALB/cマウスを、シクロホスファミドの投与によって好中球減少症とした(記載されたように(Sheppardら,Antimicrob.Agents.Chemother.48:1908−11(2004)、ほぼ12日の好中球減少症をもたらす、感染に対して−2日に200mg/kg ip、および+9日に100mg/kg ip)。足蹠膨潤反応をシクロホスファミドの最初の量から2日後に行った。ワクチン接種した好中球減少症マウスは免疫応答性マウスと同様な大きさのDTH反応を発生した(図23A vs.1、平行して行った実験)。C.albicansの2.5×104出芽胞子で尾−静脈を介して感染させた好中球減少症マウスにおいて、ワクチン接種の結果、死亡までの時間(Log順位検定 vs.対照によるとp=0.007)、メジアン生存時間(Wilcoxon順位和検定によると>21 vs 12 d,p=0.008)、および全生存(88% vs.38%,Fisher’s Exact検定によるとp=0.005)の有意な改善がもたらされた(図23B)。
【0129】
粘膜感染におけるrAls1p−Nワクチン接種の効力を決定するために、ワクチンをマウス口腔咽頭カンジダ症(OPC)モデルでテストした(Kamaiら,Infect.Immun.70:5256−8(2002) and Kamaiら,Antimicrob.Agents Chemother.45:3195−97(2001))。ワクチン接種したマウスをコルチゾン酢酸(感染に対して−1、1および3日に225mg/kg SQ)で処理し、記載したように舌下感染させた。舌を感染から5日後に切り出した。ホモゲナイズした舌のコロニー形成単位は侵入性感染および表面−接着性コロニー化の間を区別できなかったゆえに、我々は組織病理学によって侵入の程度を評価した。盲検観察者(BJS)は、舌の全長に沿ってスキャンし、40×高−出力場当たりの真菌病巣のひどさを定量することによって各セクションをスコア取りした(0=病巣なし、1+=中程度の粘膜炎症、2+=上皮に制限される有意な炎症、3+=全上皮層を通じて伸びる炎症、4+=上皮下まで伸びる炎症)。サンプリングの偏りを回避するために、少なくとも生きた介入組織セクションによって隔てられた各舌の2つのセクションをスコア取りした。全ての対照マウスは多数の場所においてその舌の顕著な真菌侵入を発生し、他方、唯2匹のワクチン接種したマウスがいずれかの舌病巣を発生した。全体として、対照マウスにおける舌当たりの病巣のメジアン数(75、25四分位)はワクチン接種したマウスについての1(2.5、0)と比較して6.5(8、5.75)であった(Wilcoxon順位和検定によるとp=0.03)。感染のひどさの半定量的評価は、対照と比較してワクチン接種マウスにおいて有意な低下を示した(図24、Wilcoxon順位和検定によるとp=0.03)。
【0130】
粘膜感染におけるrAls1p−NまたはrAls3p−Nワクチン接種の効力を決定するために、膣コロニー化のマウスモデルにおいてこれら2つのワクチンを評価した(Clemonsら,Infect.Immun.72:4878−80(2004);Fidel.Int Rev Immunol.21:515−48(2002) and Wozniakら,InfectImmun.70:5790−9(2002))。ワクチン接種したマウスを感染に対して−3日目にエストロゲン(30μg,与えられたSQ)で処理し、次いで、C.albicansの106出芽胞子を含有する10μlのリン酸緩衝化生理食塩水で膣においてチャレンジさせた。膣を接種から3日後に切り出し、ホモゲナイズし、系列希釈をYPDプレート上で平板培養した。コロニー形成単位(CFU)を30から35℃においてプレートのインキュベーションから20から48時間後に数えた。rAls3p−Nでワクチン接種したマウスから集めた膣は、対照マウス(すなわち、CFA単独でワクチン接種したマウス)から集めた膣よりも低いCFUを有したが、rAls1p−Nでワクチン接種したマウスから集めたものはそうでなかった(図25、Wilcoxon順位和検定によるとp=0.01)。
【0131】
カンジダ症の発生率およびその継続する高い死亡率に徴すると、Candida spp.に対するワクチンの開発は非常に重要なものである。前記した結果は、rAls1p−NでのSQワクチン接種は、免疫応答性および免疫無防備マウス双方におけるそうでなければ迅速に致死的な血行的に播種性のカンジダ症の間において、生存の顕著な改善、および真菌負荷の有意な低下がもたらされた。興味深いのは個々のワクチン接種したマウスから得られた腎臓真菌負荷の結果であり、これは、マウスのほぼ半分が5 log CFU/g下で腎臓真菌負荷を有したことを示す。我々は、致死的感染を示す腎臓真菌負荷の閾値は5 log CFU/gであることを従前に見出しており;このレベルを超える腎臓真菌負荷を持つマウスは典型的には感染で死亡し、他方、この負荷未満の腎臓真菌負荷を持つマウスは感染から生存した(Spellbergら,J.Infect.Dis.In press (2005)) and (Spellbergら,Infect.Immun.71:5756−5764(2003))。従って、ワクチン接種した群における突発的死亡はワクチン接種に関わらない高い真菌負荷を反映するようである。組織真菌負荷におけるマウス間変動は、宿主−病原相互作用および/または可変ワクチン応答性の複雑性を反映するであろう。
【0132】
まとめると、rAls1p−Nワクチンは増大する通常のおよびかなり致死的な播種性カンジダ症の治療、ひどさの低下および/または予防で用いることができる。該ワクチンは免疫応答性マウスにおいて効果的であり、かつ効果は好中球減少症およびコルチコステロイド−処理宿主においてさえ保持される。最後に、ワクチンは膣および口腔咽頭カンジダ症を含めた粘膜皮膚カンジダ症を保護することができる。
【0133】
(実施例IX)
(黄色ブドウ球菌感染に対するAlsワクチンの効果)
本実施例は、C.albicansからのAlsタンパク質が黄色ブドウ球菌で感染させた動物モデルの生存を改善することを示す。
【0134】
C.albicansのAlsアドヘシンは、黄色ブドウ球菌でのアドヘシンに対して有意に相同あると同定された。この特徴を用いてAlsアドヘシンを用い黄色ブドウ球菌に対する効果的なワクチンを設計し実施した。簡単に述べると、C.albicans ALSファミリーは少なくとも9つの遺伝子よりなる(Hoyerら,Genetics 157:1555−67(2001); Hoyer LL.,Trends Microbiol.9:176−80(2001))。前記したように、Alsタンパク質は生物学的に関連する基材へのアドヘシンとして機能する(Fuら,Molec.Microbiol.44:61−72(2002);Gaur and Klotz,Infect.Immun 65:5289−94(1997);Zhao et al.,Microbiology 150:2415−28(2004);Ohら,Microbiology 151:673−81(2005);Zhaoら,Microbiology 151:1619−30(2005)); Hoyerら,Mol.Microbiol.15:39−54(1995))。特に、rAls1p−NおよびrAls3p−NのN末端は、コラーゲン結合タンパク質およびクランピング因子を含めた、病原性黄色ブドウ球菌によって発現された表面タンパク質に対して有意に相同である(表IV;Sheppardら J.Biol.Chem.279:30480−89(2004))。
【0135】
【表4】
表IVにおいて上記に掲げた相同性の計算は、配列整列および三次元表面構造の双方の特徴を考慮するものである。Als1pの相同性は、黄色ブドウ球菌のコラーゲン結合タンパク質またはクランピング因子と比較して95%または90%よりも大きいと計算された(r2≧90%;Sheppardら,上記)。同様に、Als3pの相同性は、黄色ブドウ球菌のコラーゲン結合タンパク質またはクランピング因子と比較して95%または80%よりも大と計算された(r2≧90%)。
【0136】
前記知見を裏付けるために、相同性およびねじ切り法を使用して、Als1pおよび黄色ブドウ球菌クランピング因子Aの間の構造−機能調和をモデル化した(ClfA − PDB code:c1n67A)。これらの方法は一次構造における特異的相同性を評価し、3−D立体配座およびパターン分析を行って、類似の機能モチーフを求めた。例えば、BLASTP、PROSITEおよびJALVIEW方法を使用して、ALS vs ClfA一次配列の同様性および差を分析した(Yountら Antimicrob.Agents Chemother.48:4395−4404(2004)and Yount and Yeaman.Proc.Natl.Acad.Sci.USA 101:7363−7368(2004))。次いで、3−D PSSMを含めたインターネットベースの適用を用いて、さらなる分析のための潜在的ALSホモログを優先付けた(Sheppardら J.Biol.Chem.279:30480−30489(2004))。得られたデータに沿って、PHYRE適用(Kelley,L.,R.Bennett−Lovsey,A.Herbert,and K.Fleming;ウェブサイトは以下の通りである:http://www.sbg.bio.ic.ac.uk/〜phyre/)を用いて、トポロジーマッピングを行い、および推定共有機能モチーフを同定する目的で、選択されたALSタンパク質に対する最大の構造的または機能的相同性を持つタンパク質によって共有された三次元モチーフを同定した。前記方法は公のドメインで広く入手可能であり、種々のプロテオミックおよび構造的生物学適用で用いられる。前記相同性およびねじ切り法結果に基づき、Als1pおよびClfAの間の機能的部位相同性のコンセンサスを生じさせ、ClfAに構築されたAls1pモデルの特異的残基にマッピングした。いくつかの特別な知見は、以下に記載するこれらのモデリング分析から生じた。
【0137】
まず、Als1pおよびClfAのN末端領域の間で、二次構造およびアミノ酸保存において、特に、アミノ酸30から300によって含まれる領域(双方のタンパク質のN末端)において、有意な相同性が同定された。
【0138】
第二に、確立されたClfAアドヘシン決定基に基づく相同機能部位のコンセンサスマッピングは、Als1pにおける特異的トポロジー的モチーフに集中した。このトポロジー的モチーフを、2つのβ−シートドメインの隣接面の屈曲によって形成された溝として図26に示される。
【0139】
第三に、一次構造相同性と合致して、Als1pにおける予測された機能的溝モチーフは、アミノ酸残基30から300を含むN末端領域における超可変領域に由来する特異的残基にマッピングされる。
【0140】
これらの結果は、調和した生物学的機能に対する構造的基礎、並びにAls1pおよびClfAに対する免疫学的応答を供した。これらの結果はAls1p構造−活性の我々の総じてのモデルをさらに裏付け、突然変異分析およびエピトープマッピングに対する標的化アプローチをさらに容易とする。最後に、これらの結果は、Als1pおよびClfAが、多様な微生物病原体に存在する類似の構造および機能のアドヘシンであることを示す。
【0141】
黄色ブドウ球菌に対するモノクローナル抗体もまた同定され、これはC.albicansによって引き起こされた感染を低下させることができる。前記構造的知見に関して、この特徴も用いて、Alsアドヘシンを用い、黄色ブドウ球菌に対する効果的なワクチンを設計し実施した。
【0142】
簡単に述べると、黄色ブドウ球菌上の表面アドヘシンを認識することが知られているヒト化抗ブドウ球菌モノクローナル抗体(Aurexis(登録商標))は現在臨床試験中である。このモノクローナル抗体もやはりAlsファミリーメンバーと交差反応する。ブドウ球菌血流感染の治療用のAurexis(登録商標)の相II臨床試験の好都合な結果が報告されている(Inhibitex Inc.,2005;http://phx.corporate−ir.net/phoenix.zhtml?c=176944&p=irol−newsArticle&ID=707322&highlight=において、2005年9月19日にアクセスされた)。簡単に述べると、この報告においては、血液中の公知の黄色ブドウ球菌を持つ患者に活性な感染に対する治療としてAurexis(登録商標)抗体を投与した(すなわち、これは能動的ワクチン戦略または予防的実験ではない)。プラセボを受ける9人の患者はCandida によって引き起こされた突発的な血流感染を経験し、他方、Aurexis(登録商標)実験群においては、3人の患者のみがCandida血流感染を経験した。前記相同性および構造的知見と組合せて、黄色ブドウ球菌に対する抗体で治療した患者についてのCandida血液感染の減少を認識すると、免疫原性エピトープはCandidaおよび黄色ブドウ球菌の間で共有されており、これらの免疫原性エピトープはおよび他の種の治療用の1つの種に対して生起された免疫応答、抗体またはエフェクタメカニズムを用いて治療的利益について標的化できることを示す。従って、前記データを一緒にすると、Candida spp.と交差反応する黄色ブドウ球菌上の表面アドヘシンに対する免疫応答を提供する。
【0143】
前記戦略に従い、例示的なAlsアドヘシンワクチンを設計し、それは、黄色ブドウ球菌に感染したマウスの生存を改善することが示された。ワクチン接種するのに用いた例示的なAlsアドヘシンはrAls1p−NまたはrAls3p−Nであり、これは前記したように生産し、用いた。簡単に述べれば、Candida、rAls1p−NおよびrAls3p−Nに対するこれらのAlsワクチンが黄色ブドウ球菌に対して交差−種保護を媒介できるかを決定するために、雌Balb/cマウスを従前に記載された養生法でワクチン接種した(0日目におけるフロイントの完全アジュバント+20μgのrAls1p−NまたはrAls3p−N、続いての、3週間におけるフロイントの不完全アジュバント中のブースター用量;共に皮下投与した)。ワクチン接種から2週間後に、マウスを、メチリシリン−耐性であって、動物モデルにおいてビルレントであることが知られている、致死用量の黄色ブドウ球菌株67−0で尾−静脈を介して感染させた。マウスの生存を示す結果を図26に示す。示したように、rAls1p−NおよびrAls3p−Nワクチンは、これらの感染したマウスにおいて改善された長期生存を媒介した(図27)。加えて、保護のメカニズムはTh2よりはむしろTh1の増強であるようである。というのは、Ab力価、およびrAls1p−NまたはrAls3p−Nいずれかでワクチン接種したマウスの生存の間に相関は観察されなかったからである(図28)。
【0144】
(実施例X)
(抗Candida rAls1p−Nワクチンは播種性カンジダ症に対する広い範囲の保護を媒介する)
本実施例は、rAls1p−Nワクチンが非近交系マウスを播種性カンジダ症から保護し、Balb/cマウスをC.albicansおよび非albicans Candidaの他のビルレント株に対して保護する。
【0145】
本実験を行って、C.albicansの他の株に対する、およびCandidaの非albicans種に対する、フロイントのアジュバント以外の第二のアジュバントと組み合わせた、非近交系マウスにおけるその効力を特異的に評価することによりrAls1p−Nによって誘導される保護の広さを説明する。
【0146】
rAls1p−Nでのワクチン接種は、播種性カンジダ症から非近交系マウスを保護した。簡単に述べると、非近交系マウスCD1マウスはNational Cancer Institute(Bethesda, MD)から入手した。マウスに関連する全ての手法は、動物収容および介護についてのNational Institutes of Health指針に従い、動物使用および看護制度委員会によって認可された。前記にて、および例えば、Ibrahimら,Infect.Immun.73:999−1005(2005);Spellbergら,Infect.Immun.73:6191−93(2005)に記載されているように、マウスをrAls1p−N+フロイントアジュバントでワクチン接種した。rAls1p−N(Als1pのアミノ酸17から432)はS.cerevisiaeにおいて生産され、ゲル濾過およびNi−NTAマトリックスアフィニティー精製によって精製した。高度な純度(約90%)は、前記にて、および例えば、Sheppardら,J Biol Chem 279:30480−89(2004)に記載されているように、SDS−ポリアクリルアミドゲル電気詠動並びに円二色およびFTIRによって確認した。マウスは、0日目においてフロイントの完全アジュバント(CFA;Sigma−Aldrich,St.Louis,MO)と混合されたrAls1p−N(20μg)のSQ注射、続いての21日目におけるフロイントの不完全アジュバント(IFA;Sigma−Aldrich)中のブースター用量によって免疫化した。対照マウスはCFA/IFA単独で免疫化した。ブーストから14日後に、我々は既に記載したように、免疫化されたマウスをC.albicans SC5314で尾−静脈を介して感染させた。Ibrahimら,(2005)上記;およびSpellbergら(2005)上記。Balb/cマウスにおける我々の従前の知見と同様に、rAls1p−Nワクチンは感染されたCD1マウスの生存を顕著に改良した(図29A)。
【0147】
フロイントのアジュバントはヒトで用いるのにはあまりにも毒性であるゆえに、我々はミョウバン(2% Alhydrogel,Brenntag Biosector,Frederikssund,Denmark)中のrAls1p−Nワクチン、ヒトで用いる合衆国食品薬品局(FDA)によって現在認可された唯一のワクチンの用量応答を行った。ミョウバンでのワクチン接種は、1日目における免疫化、21日目におけるブースト、および2週間後における感染にて、フロイントのアジュバントと同一のスケジュールで行った。我々は、ミョウバンと組合せたより高い用量のrAls1p−Nの結果、播種性カンジダ症を持つマウスの生存の有意な改善をもたらしたことを見出した(図29B)。また、用量応答の関係があるようであり、ミョウバンと組み合わせた場合に、より高い用量のrAls1p−Nにおいて改良された生存の傾向がある。
【0148】
また、rAls1p−Nワクチンは、C.albicansのいくつかの株で感染させたBalb/cマウスの生存を改善することが示された。特に有用なワクチンは、免疫系が標的病原体の多数の株を認識するように開始することができる免疫原を利用する。DNA配列分析によって、我々は、Als1pのN末端領域の予測されたアミノ酸配列が、血流(5株)、尿(5株)および口腔咽頭(10株)感染(データは示さず)からの臨床C.albicans単離体の多様な群の中で99.9%保存されていたことを見出した。これらの結果は、rals1p−NワクチンがC.albicans株の広いアレイに対して効果的であり得ることを示す。C.albicansの他の株に対するrAls1p−Nワクチンの保護の広さを確認するために、マウスをrAls1p−N+フロイントのアジュバントで前記したようにワクチン接種し、C.albicansのいくつかの臨床単離体の1つで感染させた(Ibrahimら, Infect Immun 63:1993−98(1995))。図30に示したように、rAls1p−Nワクチンは、これらの株の各々で感染させたマウスの生存を有意に改善した。
【0149】
また、rAls1p−Nワクチンは、Candidaのいくつかの非albicans種で感染させたマウスにおいて組織真菌負荷を低下させることが示された。簡単に述べると、ALS遺伝子ファミリーはC.dubliniensisおよびC.tropicalisを含めた他のCandida種に存在する(Hoyerら,Genetics 157:1555−67(2001))。同様に、Alsファミリーメンバーに類似するアドヘシンはC.glabrataに記載されている(Cormackら,Science 285:578−82(1999);Friemanら,Mol Microbiol 46:479−92(2002))。非albicans種に対するrAls1p−Nの効力を確認するために、Balb/cマウスを前記したようにrAls1p−N+フロイントのアジュバントでワクチン接種し、C.glabrata 31028(Harbor−UCLA Medical Centerにおける微生物学研究所からの臨床的血流単離体)、C.krusei 91−1159(親切にもMichael Rinaldi,San Antonio,Texasによって提供された)、C.parapsilosis 22019(Harbor−UCLA Medical Centerからの臨床的血流単離体)、またはC.tropicalis 4243(Harbor−UCLA Medical Centerからの臨床血流単離体)で尾−静脈を介して感染させた。図31に示すように、rAls1p−Nワクチンはこれらの種の各々で感染させたマウスの腎臓真菌負荷を低下させた。
【0150】
まとめると、rAls1p−Nワクチンは、増大している通常のおよびかなり致死的な播種性カンジダ症のひどさを予防し、および/または低下させることができる。該ワクチンは、アジュバントとしてのミョウバンと混合した場合、C.albicansの多数の株に対して、およびCandidaのいくつかの非albicans種に対して、同系交配および非近交系マウスの双方において有効である。これらの結果は、本発明のALDSワクチンが広く種々のカンジダおよび他の感染に対して有効であることをさらに裏付ける。
【0151】
(実施例XI)
(抗Candida rAls3p−Nワクチンは播種性カンジダ症に対してrAls1p−Nと同等に効果的であり、粘膜カンジダ症に対してより効果的である)
本実施例は、血行的に散在した口腔咽頭および膣カンジダ症のネズミモデルにおけるrAls3p−Nワクチンの効力をrAls1p−Nと比較する。
【0152】
ALSファミリーメンバーのうち、ALS1およびALS3遺伝子は基材親和性の最も広いアレイにてアドヘシンをコードする。相互に比較した場合、Als1pは内皮細胞およびゼラチンに対するより大きな接着を媒介したが、上皮細胞に対しては劣った接着を媒介した(Sheppardら,J Biol Chem 279:30480−89(2004))。接着の質の個の差は、rAls3p−NがrAls1p−Nと比較してワクチン免疫原として異なる有効性を有することができることを示唆した。
【0153】
ワクチンおよびワクチン接種は前記したように行った。簡単に述べれば、rAls1p−NおよびrAls3p−N(Als1pまたはAls3pのアミノ酸17から432)はS.cerevisiaeで生産され、前記にて、およびIbrahimら,(2005),上記;Spellbergら,(2005),上記;に記載されたように、ゲル濾過およびNi−NTAマトリックスアフィニティー精製によって精製した。タンパク質の量は修飾されたLowryアッセイによって定量した。高度な純度(約90%)は、前記にて、およびIbrahimら.,(2005),上記;Spellbergら.,(2005),上記;に記載されているように、SDS−ポリアクリルアミドゲル電気詠動ならびに円二色性およびFTIRによって確認した。マウスを、0日目においてフロイントの完全アジュバント(CFA,Sigma−Aldrich,St.Louis,MO)と混合された20μgのrAls1p−NまたはrAls3p−Nの皮下(SQ注射によって免疫化し、21日目においてもう1つの用量の抗原とフロイントの不完全なアジュバント(IFA,Sigma−Aldrich)でブースト注射し、次いで、ブーストから2週間後に感染させた。
【0154】
統計学的解析は以下のように行った。非パラメーターLog順位検定を利用して、マウスの生存時間の差を決定した。抗体力価および足蹠膨潤は、適切であれば、非パラメーター多数比較のためのSteel検定(Rhyneら.,Biometrics 23:539−49(1967))、または負対比較のためのMann Whitney U検定によって比較した。相関はSpearman順位検定で計算した。不均一が複連生存試験で存在するかを決定するために、Kolmogorov−Smimov検定を利用した。P値<0.05は有意と考えられた。
【0155】
rAls3p−Nでのワクチン接種は、rAls1p−Nとの比較において抗体応答のより広いアレイを刺激することが示された。この点に関して、図32に示される結果は、CFA + rAls1p−NまたはrAls3p−Nでワクチン接種したマウスは、CFA単独を受けるマウスよりも有意に大きな抗体力価を発生させたことを示す。注目すべきことには、rAls3p−Nワクチン接種したマウスは、rAls1p−Nワクチン接種したマウスに対して同等な力価にて抗rAls1p−N抗体を生じた(図32,上)。対照的には、rAls1p−Nワクチン接種したマウスは、rAls3p−Nでワクチン接種したマウスよりもrAls3p−Nに対してより小さな力価を生じた(図32,下)。しかしながら、rAls1p−NおよびrAls3p−Nは、図33に示したように、同様な遅延型過敏応答をインビボにてもたらした。
【0156】
rAls1p−NおよびrAls3p−Nワクチンは、播種性カンジダ症に対して同様な効果を媒介することも示されていた。簡単に述べれば、rAls3p−Nワクチンが、血行的に播種性のカンジダ症に対してrAls1p−Nと同程度に効果的であることをさらに裏付けるために、マウスをCFA、CFA+rAls1p−N、またはCFA+rAls3p−Nをワクチン接種し、続いて、C.albicansで尾−静脈を介して感染させた。図34に示す結果は、rAls1p−NおよびrAls3p−Nワクチン双方の結果、生存の有意な改良がもたらされたことを示す。
【0157】
抗Alsp抗体力価および遅延型過敏反応と、引き続いてC.albicansで感染させたワクチン接種したマウスにおける生存との相関も決定した。簡単に述べれば、抗体力価を、先に、およびIbrahimら,(2005),上記;Spellbergら,(2005),上記に我々が記載したように、96ウェルプレートにおいてELISAによって決定した。ウェルを、PBS中の5μg/mlのrAls1p−NまたはrAls3p−Nのウェル当たり100μlで被覆した。マウス血清を、3%ウシ血清アルブミンを含有するtris緩衝生理食塩水(TBS)(0.01 M TrisHCl,pH 7.4,0.15M NaCl)でのブロッキング工程に続いて室温にて1時間インキュベートした。ウェルを0.05%Tween20を含有するTBSで3回洗浄し、続いて、Tweenを含まないTBSでさらに3回洗浄した。ホースラディッシュペルオキシダーゼ(Sigma−Aldrich)でコンジュゲートしたヤギ抗マウスIgG二次抗体を1:5000の最終希釈で加え、プレートを室温にて1時間さらにインキュベートした。ウェルをTBSで洗浄し、0.1Mクエン酸緩衝液(pH 5.0);50mg/mlのo−フェニレンジアニン(Sigma)、および10μlの30%H2O2を含有する基質と共にインキュベートした。30分間発色させ、その後、10%H2SO4を加えることによって反応を停止させ、490nmにてマイクロタイタープレートにおいて光学密度(OD)を決定した。陰性対照ウェルは無関係な抗体を受け、バックグラウンド吸光度をテストウェルから差し引いて、最終OD読みを得た。ELISA力価は、陽性OD読みを与えた最後の血清希釈の逆数として取られた(すなわち、>は陰性対照試料の平均OD+(標準偏差*2)を意味する)。
【0158】
遅延型過敏反応は、足蹠膨潤テストを測定することによって評価した。簡単に述べれば、マウスをrAls1p−N、rAls3p−N、またはCFA単独で免疫化した。ブースとから2週間後に、免疫化マウスのベースライン足蹠サイズを電子デジタルキャリパーを用いて測定した。25μlのPBS中の50μgのrAls1p−NまたはrAls3p−Nを右側足蹠に注射し、PBS単独を免疫化マウスの左側足蹠に注射した。24時間後に、足蹠を再度測定した。抗原−特異的足蹠膨潤は:(24時間における右側足蹠厚み−ベースラインにおける右側足蹠)−(24時間における左側足蹠厚み−ベースラインにおける左側足蹠厚み)として計算された。
【0159】
ワクチン接種したマウスから力価決定のために採血し、感染に2日先立って足蹠膨潤テストを受けさせた。感染で生き残らなかったワクチン接種マウスは、それにもかかわらず、図35に示すように広い範囲の抗体力価を有した。多くのそのようなマウスは≧1:50,000(≧4.5log10)の抗rAls1p−Nおよび抗rAls3p−N抗体力価を有した。その結果、抗体力価は生存に有意には相関しなかった。対照的に、足蹠膨潤反応の強さは生存と相関した(図35、Spearman順位相関テストによるとp=0.6&p=0.009)。
【0160】
rAls3p−Nワクチンは、粘膜カンジダ症の2つのモデルにおいてrAls1p−Nよりも大きな効果を示した。Als3pはAls1pと比較して上皮細胞に対して優れた接着を媒介したので、この観察は、rAls3p−Nが感染の粘膜モデルにおいてユニークな効果を呈することができることを示す。rAls3p−Nと比較したrAls1p−Nの効力を、感染のステロイド−処理口腔咽頭モデルにおいて、およびカンジダ膣炎のモデルにおいて評価した。
【0161】
簡単に述べれば、前記マウス口腔咽頭でカンジダ症(OPC)モデルにおけるワクチン実験を従前に記載されているように、およびSpellbergら,(2005),上記;Kamaiら,Antimicrob Agents Chemother 45:3195−57(2001),and Kamaiら,Infect Immun 70:5256−58(2002)に記載されているように行った。ワクチン接種マウスをコルチゾン酢酸(感染に対して−1、1および3日目にて225mg/kg SQ)での処理によって免疫無防備とした。感染の日に、kg当たり8mgのキシラジンおよび110mgケタミンでの腹腔内注射によってマウスを麻酔した。アルギン酸カルシウム尿道スワブを、それを30℃のHBSS中のml当たり106生物の懸濁液に入れることによって、C.albicansで飽和させた。飽和したスワブを75分間でマウスの口腔中で舌下に入れた。感染の5日後に、舌および舌下神経組織を切り出し、秤量し、ホモゲナイズし、次いで、定量的に培養して、経口真菌負荷を決定した。
【0162】
マウス膣カンジダ症に対するワクチンの有効性は、感染に対して−3日目に(共にSigma−Aldrichからの)落花生油に溶解させた30μgの皮下エストラジオールバレレートで雌Balb/cマウスをワクチン接種して、プソイドエストラス(pseudoestrus)を誘導することによって行った。感染の日に、100mg/kgのケタミンのip投与によってマウスを鎮静させた。鎮静させたマウスを10μlのHBSS中のC.albicansの106出芽胞子で膣内感染させた。感染から3日後に、膣およびほぼ1センチメートルの各子宮角をブロック上に切開し、ホモゲナイズし、定量的に培養した。
【0163】
図36に示すように、口腔咽頭カンジダ症を持つコルチゾン−処理マウスにおいて、rAls1p−Nワクチンは低下した舌真菌負荷に対する強い傾向を媒介した(p=0.054)。総じての利点の大きさは<0.3logCFU/グラムであった(図36)。比較において、rAls3p−Nワクチンは統計学的に有意な舌真菌負荷の>0.6logCFU/グラム減少を媒介した(p=0.005,図36)。同様に、カンジダ膣炎の非免疫無防備モデルにおいて、rAls3p−Nワクチンは、図37に示すようにCFA単独と比較して膣真菌負荷の0.7logCFU/グラム減少を媒介した(p=0.02)。比較において、rAls1p−Nは膣炎モデルにおいて利点を全く媒介せず、およびrAls3p−NはrAls1p−Nよりも有意に効果的であった(p=0.01)。
【0164】
前記した結果は、アミノ酸レベルにおいてrAls1p−Nに対して85%相同であるrAls3p−Nに基づくワクチンは播種性カンジダ症に対して同等に効果的であるが、粘膜感染に対してrAls1p−Nよりも効果的であった。rAls3p−Nの増大した有効性は口腔咽頭カンジダ症のステロイド−処理モデルおよびカンジダ膣炎の免疫応答性モデル双方で見られた。前期結果は、補助的抗真菌療法を伴わないカンジダ敗血症ショックのマウスモデルにおいて≧50%長期生存の達成が刺激的であったことを示し、さらに、本発明の治療的利点の全てのALSワクチンをさらに裏付ける。
【0165】
抗体力価は、播種性カンジダ症の間におけるいずれかのワクチンの保護的効果と相関しなかったが、遅延型過敏の誘導はインビボにて保護と相関した。これらのデータは、さらに、ワクチン−感染保護のメカニズムがタイプ1の誘導、真菌に対する細胞−媒介免疫を裏付ける。rAls1p−NおよびrAls3p−Nの双方はrAls1p−Nに対する同等な力価の抗体を誘導したが、rAls3p−NがrAls1p−Nよりも有意により高い力価の抗rAls3p−N抗体を誘導した。これらのデータは、その高度なアミノ酸配列相同性(85%)にも拘わらず、液性免疫系はrAls1p−NおよびrAls3p−Nの間を識別できることを示す。前記結果は、さらに、Als1pおよびAls3p上皮細胞接着特徴の差に拘わらず、rAls1p−NおよびrAls3p−Nワクチンが血行的に播種性の(すなわち、血管内)カンジダ症に対して保護するのに同等に効果的であることを裏付ける。
【0166】
まとめると、抗カンジダrAls3p−Nワクチンは、rAls1p−Nワクチンよりも同等な細胞−媒介のものであるがより広い抗体−ベースの応答を誘導した。免疫原は、血行的に播種性のカンジダ症に対して同等な程度の保護をもたらしたが、rAls3p−Nは口腔咽頭および膣カンジダ症双方に対してより大きな保護を媒介した。
【0167】
本出願を通じて、種々の刊行物を括弧に入れて引用した。これらの刊行物の全ての開示をここに引用して本出願に取り込んで、本発明が属する分野の最先端技術をより十分に記載する。
【0168】
本発明を開示された実施形態を参照して記載してきたが、当業者であれば、前記にて詳細に記載した特別な実施例および実験は本発明を説明するものに過ぎないことを容易に認識するであろう。本発明の精神を逸脱することなく種々の修飾をなすことができるのは理解されるべきであろう。従って、本発明は特許請求の範囲によってのみ限定される。
【図面の簡単な説明】
【0169】
【図1A】図1Aは、ヒト臍静脈内皮細胞へのC.albicansのAlslp接着の媒介を示す。値は、各々三連で行われた少なくとも3つの独立した実験の平均±SDを表す。(A)ALS11/als2 als1/als1およびALS1−補足突然変異体および野生型CAIl2(30)の内皮細胞接着。統計学的処理はWilcoxon順位和検定によって得られ、Bonferroni修正での多数の比較について修正した。全ての比較について*P<0.001。
【図1B】図1Bは、ヒト臍静脈内皮細胞へのC.albicansのAlslp接着の媒介を示す。値は、各々三連で行われた少なくとも3つの独立した実験の平均±SDを表す。(B)野生型C.albicansと比較した、ALS1を過剰発現するPADHl−ALS1突然変異体の内皮細胞接着。統計学的処理はWilcoxon順位和検定によって得られ、Bonferroni修正での多数の比較について修正した。全ての比較について*P<0.001。
【図2A】図2Aは、C.albicans間接的免疫蛍光のフィラメントへのAls1pの細胞表面局所化を示す。C.albicansのフィラメント化は、グルタミンと共にRPMI l640培地中で酵母細胞を37℃で1.5時間インキュベートすることによって誘導された。Als1pは、まず、抗Als1pマウスmAb、続いて、FITC−標識ヤギ抗マウスIgGと共に生物をインキュベートすることによって検出された。また、C.albicans細胞表面を、Alexa 594(Molecular Probes,Eugene,OR)とコンジュゲートした抗C.albicansポリクローナルAbで染色した。黄色に染色される領域はAls1p局所化を表す(A)C.albicans野生型。
【図2B】図2Bは、C.albicans間接的免疫蛍光のフィラメントへのAls1pの細胞表面局所化を示す。C.albicansのフィラメント化は、グルタミンと共にRPMI l640培地中で酵母細胞を37℃で1.5時間インキュベートすることによって誘導された。Als1pは、まず、抗Als1pマウスmAb、続いて、FITC−標識ヤギ抗マウスIgGと共に生物をインキュベートすることによって検出された。また、C.albicans細胞表面を、Alexa 594(Molecular Probes,Eugene,OR)とコンジュゲートした抗C.albicansポリクローナルAbで染色した。黄色に染色される領域はAls1p局所化を表す(B)als1/als1突然変異体株。
【図2C】図2Cは、C.albicans間接的免疫蛍光のフィラメントへのAls1pの細胞表面局所化を示す。C.albicansのフィラメント化は、グルタミンと共にRPMI l640培地中で酵母細胞を37℃で1.5時間インキュベートすることによって誘導された。Als1pは、まず、抗Als1pマウスmAb、続いて、FITC−標識ヤギ抗マウスIgGと共に生物をインキュベートすることによって検出された。また、C.albicans細胞表面を、Alexa 594(Molecular Probes,Eugene,OR)とコンジュゲートした抗C.albicansポリクローナルAbで染色した。黄色に染色される領域はAls1p局所化を表す(C)野生型ALS1を補足したals1/als1。
【図2D】図2Dは、C.albicans間接的免疫蛍光のフィラメントへのAls1pの細胞表面局所化を示す。C.albicansのフィラメント化は、グルタミンと共にRPMI l640培地中で酵母細胞を37℃で1.5時間インキュベートすることによって誘導された。Als1pは、まず、抗Als1pマウスmAb、続いて、FITC−標識ヤギ抗マウスIgGと共に生物をインキュベートすることによって検出された。また、C.albicans細胞表面を、Alexa 594(Molecular Probes,Eugene,OR)とコンジュゲートした抗C.albicansポリクローナルAbで染色した。黄色に染色される領域はAls1p局所化を表す(D)PADHl−ALS1過剰発現突然変異体。
【図3A】図3Aは、固体培地上でのC.albicansフィラメント化に対するAls1pの媒介を示す。C.albicans出芽胞子をLee寒天プレート上にスポットし、37℃にて4日間(A)インキュベートした。
【図3B】図3Bは、固体培地上でのC.albicansフィラメント化に対するAls1pの媒介を示す。C.albicans出芽胞子をLee寒天プレート上にスポットし、37℃にて3日間(B)インキュベートした。
【図4A】図4Aは、EFG1フィラメント化調節経路によるALS1発現の制御を示す。異なるフィラメント化調節経路に欠陥がある突然変異体におけるALS1の発現を示すノーザンブロット分析。全RNAは、37℃にて90分間、RPM1 1640+グルタイン培地中で増殖させてフィラメント化を誘導した細胞から抽出した。ブロットはALS1およびTEF1でプローブした。
【図4B】図4Bは、EFG1フィラメント化調節経路によるALS1発現の制御を示す。EFG1またはPADHl−ALS1いずれかを補足したefgl/efgl突然変異体におけるALS1の発現を示すノーザンブロット分析。全RNAは、37℃にて90分間、RPM1 1640+グルタイン培地中で増殖させてフィラメント化を誘導した細胞から抽出した。ブロットはALS1およびTEF1でプローブした。
【図4C】図4Cは、C.albicansフィラメント化の媒介を示す。Lee寒天プレート上で37℃にて4日間増殖させたefgl/efgl突然変異体、およびPADHl−ALS1を補足したefgl/efgl突然変異体の顕微鏡写真。
【図5A】図5Aは、(A)雄Balb/Cマウス(各酵母株についてn=30)に静止相出芽胞子(0.5mlのPBS中マウスあたり106)で注射することによる、血行的に播種性のカンジダ症のマウスモデルにおけるビルレンスの低下を示す。曲線は三連実験の編集した結果である(各株についてn=30マウス)。30℃にてYPD中で増殖させた全ての株の倍加時間は1.29から1.52時間の間の範囲であり、相互に統計学的に異ならなかった。全染色体DNAのサザーンブロット分析を用いて、感染した生物から検索したC.albicans株の遺伝子型の同一性を、マウスを感染させるのに用いたC.albicans株のそれにマッチさせた。統計学的解析は、Wilcoxon順位和検定によって得られ、Bonferroni修正にて多数の比較のために修正した。als1/als1突然変異体−vs−他の株の各々について*P<0.002。
【図5B】図5B:C.albicans野生型、ホモ接合性als1 null突然変異体またはヘテロ接合性ALS1補足突然変異体で感染させた腎臓の組織学的顕微鏡写真。腎臓試料を感染から28時間後に検索し、パラホルムアルデヒド中で固定し、断面を銀で染色した(倍率×400)。矢印はC.albicans細胞を示す。
【図5C】図5C:C.albicans野生型、ホモ接合性als1 null突然変異体またはヘテロ接合性ALS1補足突然変異体で感染させた腎臓の組織学的顕微鏡写真。腎臓試料を感染から40時間後に検索し、パラホルムアルデヒド中で固定し、断面を銀で染色した(倍率×400)。矢印はC.albicans細胞を示す。
【図6】図6は、抗Als1pポリ血清を注入した動物についての30日間に渡って生存する動物の関数としての播種性カンジダ症に対する抗ALS抗体の予防効果を示す。
【図7】図7は、接着表現型によって配置された選択されたALSポリペプチドのN末端部分のポリペプチド配列整列である。頂部の3つの線はALS1、3および5ポリペプチド(各々、配列番号1から3)からの配列であり、これは内皮細胞に結合する。底部の3つはALS6、7および9ポリペプチドからの配列であり(各々、配列番号4から6)、これは内皮細胞に結合しない。最後の線はALSポリペプチドファミリーコンセンサス配列(配列番号7)を表す。
【図8】図8は、Saccharonzyces cerevisiaeにおいて不均一に発現させた場合に、Alsタンパク質が基材特異的接着特性を付与することを示す。各パネルは、C.albicansが接着することが知られている種々の基材への1つのAlsp発現株のパーセント接着を示す(塗りつぶした棒線)。空のベクターで形質転換されたS.cerevisiaeの接着(塗りつぶしていない棒線)は陰性対照として各パネルに含ませる。Gel、ゼラチン;FN、フィブロネクチン;LN、ラミニン;FaDu、FaDU上皮細胞;EC、内皮細胞。単一因子偏差分析によって空のプラスミド対照と比較した場合、*p<0.01。結果は三連で行った少なくとも3つの実験の平均±S.D.である。
【図9】図9は、ドメインスワッピングが、基材特異的接着が、Alsタンパク質のN末端ドメインの組成によって決定されることを示すことを示す。テストすべきAls遺伝子または構築物の表示は、ALS5(黒色)またはALS6(白色)からの配列よりなる棒線として描かれる。各突然変異体の接着特性は、フィブロネクチン−被覆ビーズへの形質転換されたS.cerevisiaeの接着を示す顕微鏡写真、および6−ウェルプレートアッセイで測定されたゼラチン(黒色棒線)および内皮細胞(灰色棒線)への接着を示すグラフとして表示される。結果は、各々が三連で行った少なくとも3つの実験の平均±S.D.である。
【図10】図10は、S.cerevisiaeにおいて発現された場合に、Alsタンパク質のサブセットが内皮細胞侵入を媒介することを示す。A、Alsタンパク質を発現する、または空のプラスミド(対照)で形質転換されたS.cerevisiae株の内皮細胞接着。データは、内皮細胞−関連生物の合計数を表し、高電力場当たりの細胞として表す。B、高電力場当たりの細胞内生物の数として表されたAlsp発現S.cerevisiae株の内皮細胞侵入の程度。単一因子偏差分析によって空のプラスミド対照と比較した場合、*p<0.01。結果は、三連で行った少なくとも3つの実験の平均±S.D.である。
【図11】図11は、公知の機能のAlsタンパク質のN末端アミノ酸配列の整列が、CRおよびHVRの交互パターンを示すことを示す。A、公知の機能のAlsタンパク質のN末端領域内のコンセンサス同一性のパーセンテージ。シグナルペプチド領域(アミノ酸1から20)は示されないことに注意されたい。塗りつぶしていないボックスは、HVR1から7として命名された領域を示す。B、個々のHVRの組成を示すAlsタンパク質(各々、配列番号1から6)の模式的整列。該配列を配置して、少数の同定された基材に結合する、または全く同定された基材に結合しないタンパク質と、多数の基材に対して親和性を持つタンパク質とを比較する。各保存された領域におけるアミノ酸配列の数を括弧に入れて示す。
【図12】図12は、Als1タンパク質N末端ドメインのCDおよびFTIRスペクトルを示す。A、リン酸緩衝化生理食塩水中の10μM Als1pの円二色スペクトル。B、D2O蒸気で水和されたAls1p自己−フィルムのFTIRスペクトル。
【図13】図13は、Alsタンパク質ファミリー内のN末端ドメインの予測された物理化学的特性の比較を示す。疎水性、静電性、または水素−結合特徴は各ドメインの水−接近可能表面に投影される。疎水性は以下のように示される:茶色、最も疎水性;青色、最も親水性。静電性(スペクトル連続)は以下のように示される:赤色、最も正の電荷(+10 kcal/モル);青色、最も負の電荷(−10 kcal/モル)。水素−結合ポテンシャル(H−結合)は以下のように示される:赤色、ドナー;青色、アクセプター。Alsタンパク質は、これらの特性の複合に基づいて3つの群に区別可能である。例えば、Als群Aタンパク質内の同様な疎水性、静電性および水素−結合プロフィール、Als1p、Als3p、およびAls5pに注意されたい。対照的に、Als群Bメンバー、Als6pおよびAls7pは、Als群Aのそれからの疎水性および静電性特徴の顕著な差を提示する。生化学的プロフィールに加えて、これらのドメイン内の予測される構造の差に注意されたい。
【図14】Alsファミリータンパク質における構造−機能関係の概念的モデル。Alsタンパク質は3つの一般的成分:N末端ドメイン、タンデムリピート、およびC.albicans細胞壁と界面を形成するグリコシルホスファチジルイノシトールアンカーを含有するセリン/スレオニン−リッチなC−末端ドメインから構成される。説明したように、Alsタンパク質は、免疫グロブリンスーパーファミリーに特徴的な、拡大されたスパンが間に存在する多数の保存された反―平行β−シート領域(CR1−n)を含有する。β−シートドメインから突出するのは、HVRを含有するループ/コイル構造である。特異的Alsタンパク質HVRの三次元物理化学的特性は、恐らくは、接着および侵入機能をCandidaに付与する宿主基材との相互作用を支配する。説明目的で、3つのN末端β−シート/コイルドメインおよびそれらの各CR/HVR成分のみを示す。この突出は、図13に示した構造イメージに対して直角で見られることに注意されたい。
【図15】rAls1p−Nでのマウス(リタイアしたブリーダー)の免疫化は、引き続いての播種性カンジダ症の間の生存を改良する。Als1p+アジュバントで免疫化されたマウスの生存。N=異なる日での二連実験における群当たり16匹のマウス;Adj.=アジュバント。*p<0.05 vsアジュバント。
【図16】rAls1p−Nでの免疫化は、リタイアしたブリーダーおよび若年マウス双方の生存を改良する。C.albicansの迅速に致死的な106接種物で感染させたリタイアしたブリーダー(A)および若年性(B)マウスの生存。N=異なる日の二連実験における群当たり16匹のマウス;Adj.=アジュバント。*p<0.05 vsアジュバント対照。
【図17】抗rAls1p−N力価は生存と相関しない。アジュバントと共に、またはそれなくして、変化する用量のrAls1p−Nで免疫化されたBalb/cマウスにおいて生起した抗rAls1p−Nポリクローナル抗体の力価。Adj.=アジュバント。200μg vs.全ての他のものについて*p≦0.005。
【図18】rAls1p−Nの保護用量のみがC.albicans−刺激Th1脾臓細胞の増加を誘導する。異なる用量のrAls1p−NワクチンによるTh1(CD4+IFN−γ+IL−4−)およびTh2(CD4+IFN−γ−IL−4+)脾臓細胞の誘導。免疫化されたマウス(群当たりn=9)からの脾臓細胞を熱で殺した予め発芽したC.albicansで48時間刺激し、次いで、3−色フローサイトメトリーによって分析した。*p=0.03 vs.アジュバント。
【図19】rAls1p−Nの保護用量のみがrAls1p−N刺激遅延タイプの過敏の増加を誘導する。rAls1p−NまたはCFA単独でワクチン接種したマウス(群当たりn=9から12)における、足蹠膨潤によって評価した遅延タイプの過敏。マウスを示した量のrAls1p−Nで免疫化し、次いで、50μgのrAls1p−Nを足蹠に注射した。足蹠膨潤は24時間後に評価した。*p<0.05−vs−アジュバント、0.2μg、および200μg。
【図20】rAls1p−Nワクチンは、保護的免疫性を誘導するのにB細胞ではなくT細胞を必要とする。B細胞−欠乏、T細胞−欠乏(ヌード)およびコンジェニック野生型Balb/c対照マウス(群当たりn=7または8)の生存を、rAls1p−N+アジュバントまたはアジュバント単独でのワクチン接種後に同時に評価した。*p<0.04−vs−アジュバント単独、¶p=0.003−vs−野生型アジュバント−処理。
【図21】rAls1p−NでのSQワクチン接種は免疫コンピテントマウスにおいてインビボDTH応答を誘導する。足蹠膨潤は、50μgのrAls1p−NのBALB/cマウス(群あたりn=10)における足蹠への注射から24時間後に評価した。メジアン値は黒色棒線として表示する。Wilcoxon順位和検定によって*p=0.002 vs.対照。
【図22】rAls1p−Nワクチンは、血行的に散在したカンジダ症を持つ免疫コンピテントマウスの生存を改良し、組織真菌負荷を低下させる。A)C.albicansで尾−静脈を介して引き続いて感染させた、ワクチン接種したまたは対照BALB/cマウス(各々、2.5または5 × 105接種物について群当たりn=7または10)の生存。各実験は感染から30日後に停止し、全ての残りのマウスは良好に見えた。Log順位検定により*p<0.05 vs.対照。B)C.albicansの5×105出芽胞子で尾静脈を介して感染させたBALB/cマウス(群当たりn=7)における腎臓真菌負荷。y軸はアッセイの検出の下限を反映する。メジアン値は黒色棒線として表示する。Wilxocon順位和検定によって*p=0.01 vs対照。
【図23】rAls1p−Nワクチンは好中球減少症マウスにおいてDTH反応を誘導し、引き続いての血行的に播種性のカンジダ症の間のそれらの生存を改良する。A)足蹠膨潤は、50μgのrAls1p−NのBALB/cマウス(対照についてはn=10、rAls1p−Nについてはn=8)の足蹠への注射から24時間後に評価した。Wilcoxon順位和検定により*p=0.006 vs対照。B)C.albicansの2.5 × 104出芽胞子で感染させた好中球減少症BALB/cマウス(2つの実験からの群当たりn=16)の生存。Log順位検定により*p=0.007 vsアジュバント対照。
【図24】rAls1p−Nワクチンは、口腔咽頭カンジダ症を持つマウスの舌上での組織病理学的真菌病相のひどさを低下させる。群当たりN=4匹マウス。テキスト中に記載されたような盲検観察者によって生じた炎症スコア。Wilcoxon順位和検定により*p=0.03。
【図25】図25は、rAls1p−NワクチンではなくrAls3p−Nワクチンが、C.albicansを接種したマウスの膣の真菌集落形成を減少させることを示す(Wlicoxon順位和検定により*p=0.01 vs CFA単独でワクチン接種したマウス)群当たりN=11匹マウス。
【図26】図26は、Als1p相同性マップ−vs−黄色ブドウ球菌クランピング因子A(c1n67A)を示す。C.albicansのAls1pおよび黄色ブドウ球菌のClfAからのコンセンサス機能的部位はAls1p相同性モデルにマッピングされた。Als1pおよびClfAのN末端からの多数の残基はコンセンサス溝モチーフにマッピングされ、これは、基材への結合が双方のアドヘシンについて起こると予測されるものである。
【図27】図27は、rAls1p−NおよびrAls3p−Nワクチンは、スタフィロコッカス血症マウスの生存を改良することを示す(Log順位検定により*p<0.003 vs CFA単独でワクチン接種したマウス)。群当たりN=22匹マウス。
【図28】図28は、抗体力価が個々のワクチン接種したマウスにおける保護の程度と相関しないが、それらはワクチン接種していないマウスをワクチン接種したマウスから区別することを示す。各々、CFA単独、またはrAls1p−NまたはrAls3p−NのCFA+20μgで免疫化されたBalb/cマウスにおいて生起した抗rAls1p−Nまたは抗rAls3p−Nポリクローナル抗体の力価。総じて、抗体力価および生存の間に有意な相関があり(rho=0.474,p=0.0057)、これは、抗体力価をワクチン保護のための同族マーカーとして用いることができることを示す。しかしながら、CFA単独を受けるマウスからのデータを排除すると、抗体−力価、およびrAls1p−NまたはrAls3p−Nをワクチン接種したマウスの生存の間に相関はなく(rho=0.041143,p=0.847)、これは、抗体がワクチンの保護の支配的メカニズムではないようであることを示す。
【図29】図29は、rAls1p−Nワクチンが非近交系CD1マウスを血行的に播種性のカンジダ症から保護することを示す。A)CD1マウス(群当たりn=8)を、rAls1p−N(20μg)+CFA、またはCFA単独でSQワクチン接種し、ブーストから14日後にC.albicans SC5314で尾−静脈を介して感染させた。B)CD1マウス(群当たりn=8)を、ミョウバンと共に種々の用量のrAls1p−Nまたはミョウバン単独でSQワクチン接種し、ブーストから14日後に、C.albicans SC5314で尾−静脈を介して感染させた。Log順位検定により*p<0.05 vs.アジュバント対照。
【図30】図30は、rAls1p−Nワクチンが、C.albicansのいくつかの株の1つで感染させたBalb/cマウスの生存を改良することを示す。rAls1p−N+CFA−vs−CFA単独で免疫化し、C.albicans 15563(7×105出芽胞子)、16240(4×105出芽胞子)、または36082(4×105出芽胞子)で尾−静脈を介して感染させたBalb/cマウスの生存(群当たりN=8匹マウス)。Log順位検定により*p<0.05 vs.アジュバント対照。
【図31】図31は、rAls1p−Nワクチンが、Candidaのいくつかの非albicans種で感染させたBalb/cマウスにおける組織真菌負荷を低下させることを示す。Balb/cマウス(群当たりn=5)をCFAまたはCFA+rAls1p−N(20μg)でワクチン接種し、C.glabrata、C.krusei、C.parapsilosisまたはC.tropicalisで尾−静脈を介して感染させた。感染接種物を種の名称の下に括弧に入れて示す。腎臓真菌負荷は感染から5日後に決定した。y軸はアッセイの検出の下限を反映する。多数の比較のための非パラメーターSteel検定により*p<0.05 vs.アジュバント対照。
【図32】図32は、rAls1p−Nに対して交差反応した抗体を、rAls3p−N−免疫化マウスが生じたことを示す。CFA単独、CFA+rAls1p−N、またはCFA+rAls3p−Nで免疫化した個々のマウスの力価。CFAおよびCFA+rAls3p−Nについて群当たりN=7匹マウス;CFA+rAls1p−Nについてn=6匹マウス。Mann Whitney U検定により*p<0.05 vs.CFA単独;**p<0.002 vs.CFA単独& p<0.011vs.CFA+rAls1p−N。棒線はメジアンを示す。
【図33】図33は、rAls1p−NおよびrAls3p−N双方がインビボ遅延型過敏応答につきマウスをプライムしたことを示す。マウス(CFAおよびCFA+rAls3p−Nについて群当たりn=7;CFA+rAls1p−Nについてn=6)をCFA単独、CFA+rAls1p−N、またはCFA+rAls3p−Nワクチン接種した。インビボでの遅延型過敏を足蹠膨潤によって測定した。Mann Whitney U検定によって*p<0.05 vs.CFA単独。棒線はメジアンを示す。
【図34】図34は、rAls1p−NおよびrAls3p−Nワクチンは、ネズミの血行的に播種性のカンジダ症に対する同様な有効性を媒介したことを示す。C.albicansの5×105出芽胞子で尾−静脈を介して感染させたBalb/cマウス(CFAおよびCFA+rAls3p−Nについての2つの実験からの群当たりn=15、およびCFA+rAls1p−Nについて2つの実験からn=14)の生存。実験は感染から28日後に停止させ、全ての残りのマウスは良好に見えた。Log順位検定により*p≦0.0001 vs.CFA対照。
【図35】図35は、インビボ遅延型過敏が播種性カンジダ症の間の生存と相関したことを示す。抗rAls1p−Nまたは抗rAls3p−N抗体力価および足蹠膨潤反応を、C.albicansでの尾−静脈を介する感染の2日前にマウス(CFAまたはCFA+rAls3p−Nについて群当たりn=7、CFA+rAls1p−Nについてn=6)において測定した。Spearman順位和検定で決定した相関。
【図36】図36は、rAls3p−Nワクチンがネズミ口腔咽頭カンジダ症の間において組織真菌負荷を有意に低下させたことを示す。口腔咽頭カンジダ症を持つマウス(CFAについてn=7、およびrAls1p−NまたはrAls3p−Nワクチン接種群について8)における舌真菌負荷。y軸はアッセイの検出の下限を反映する。Mann Whitney U検定により*p=0.005 vs.CFA。
【図37】図37は、ネズミカンジダ膣炎におけるCFA単独およびCFA+rAls1p−N双方と比較して、rAls3p−Nが膣真菌負荷を低下させたことを示す。CFA、CFA+rAls1p−N、またはCFA+rAls3p−Nワクチン接種したマウス(2つの実験からの群当たりn=11)における膣真菌負荷。y軸はアッセイの検出の下限を反映する。多数の比較のためのSteel検定により*p≦0.02 vs CFAおよびCFA+rAls1p−N。
【技術分野】
【0001】
本発明はCandida albicans表面アドヘシンタンパク質、C.albicans表面接着タンパク質でのワクチン接種に対する免疫応答から得られた抗体、およびC.albicans表面接着タンパク質でカンジダ症および他の細菌感染を予防および/または治療する方法に関する。
【背景技術】
【0002】
(発明の背景)
近年、Candida種によって引き起こされた院内感染の発生率の劇的な増大があった。血行的に散在したカンジダ感染の発生率は1980年から1989年にかけて11倍増加した。この増大しつつある発生率は1990年代まで続いた。Candida種による感染は、今日、院内敗血症の第四番目の最も一般的な原因であり、Escherichia coliのそれと同等であり、Klebsiella種によって引き起こされた発生率を凌ぐ。さらに、Candida種は、ひどい怪我を有する患者において執拗な真菌感染の最も一般的な原因である。骨髄移植を受けている個人の11%まで、および同所性肝臓移植体を有する固体の13%までが侵入性カンジダ感染を発生するであろう。
【0003】
Candida albicans、この属における主な病原体は、2つの形態:出芽胞子で(出芽酵母)およびフィラメント状(菌糸および偽菌糸)相の間をスイッチすることができる。フィラメント化を調節する遺伝子に欠陥があるCandida突然変異体は、動物モデルにおいて低下したビルレンスを有すると報告されている。この低下したビルレンスは、出芽胞子からフィラメントへ変化する能力がC.albicansの鍵となるビルレンス因子であることを示唆する。今日まで、これらのフィラメント化経路の必須のエフェクタはC.albicansにおいて同定されていない。非特許文献1を参照。
【0004】
黄色ブドウ球菌(Staphylococcus aureus)感染もまた一般的であり、益々、抗生物質に対する薬物耐性をもたらす。例えば、黄色ブドウ球菌は合衆国および世界中で皮膚および皮膚構造感染、心内膜炎および菌血症の通常の原因である。以前、地域感染型の黄色ブドウ球菌(CA−S.aureus)感染は、セファゾリン、オキサシリン、メチシリン、ペニシリンおよびアモキシシリンのようなペニシリナーゼ−耐性ベータラクタムに対してほとんど均一に感受性であった。しかしながら、過去10年間に渡って、ベータ−ラクタム耐性黄色ブドウ球菌(MRSA)感染の伝染病は、世界中で多数の地域で見られている。特に、地域感染型MRSA(CA−MRSA)。多くの場所において、MRSAは、CA感染を引き起こす支配的な黄色ブドウ球菌株となった。最近の合衆国における3つの州での予測される集団−ベースの概観は、CA−MRSA感染の発生率が100,000集団当たり500症例であると見積もっており、これは合衆国単独において1年あたりほぼ150万症例に換算される。薬物−耐性黄色ブドウ球菌感染の増大する頻度は、これらの感染を予防し、治療するための新しい方法に対する要望に焦点を当てている。
【0005】
ビルレンスに寄与する生物の調節経路におけるエフェクタの同定は、現存の抗真菌剤よりもすぐれた方法および組成物での治療的介入に対する機会を提供する。ビルレンスに関与する調節経路に影響する細胞表面タンパク質の同定は特に有望である。なぜならば、タンパク質の特徴付けは、カンジダ感染と戦う場合に、現存の抗真菌剤よりも優れた免疫療法技術を可能とするからである。
【0006】
Candida albicansのビルレンスは、宿主構成要素へのその接着、および酵母から菌糸へ変換する能力が病原性を決定するにおいて最も臨界的なうちにあるいくつかの推定ビルレンス因子によって調節される。Candidaに対して殺菌性である優れた抗真菌剤が存在するが、カンジダ血症に帰される死亡率は、アンフォテリシンBのような優れた抗真菌剤での治療をもってさえほぼ38%である。また、アンフォテリシンBのような現存の剤は望ましくない毒性を呈する傾向がある。アンフォテリシンBよりも毒性が低いさらなる抗真菌剤を開発することができるが、より優れた剤は開発されないであろう。従って、播種性カンジダ症を治療または予防するための受動的または能動的免疫療法は、標準的な抗真菌療法に代えて有望である。
【0007】
かくして、Candida、黄色ブドウ球菌および他の免疫学的に関連する病原体に対する宿主免疫保護および受動的免疫保護を提供する効果的な免疫原に対する要望が存在する。本発明はこの要望を満足し、同様に、関連する利点を提供する。
【非特許文献1】Caesar−TonThat,T.C.and J.E.Cutler,“A monoclonal antibody to Candida albicans enhances mouse neutrophil candidacidal activity,”Infect.Immun.65:5354−5357,1997
【発明の開示】
【課題を解決するための手段】
【0008】
(発明の要旨)
本発明は、医薬上許容され得る媒体中のアジュバントと共に、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバーまたはその免疫原性断片を含むワクチンを提供する。また、本発明は、播種性カンジダ症を治療または予防する方法も提供する。該方法は、医薬上許容され得る媒体中の、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンの免疫原性量を投与することを含む。宿主細胞または組織へのCandidaの結合または侵入を阻害するのに有効な量の細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその機能的断片を投与することを含む播種性カンジダ症を治療または予防する方法も提供される。該Alsタンパク質ファミリーメンバーはCandida albicans、Candida krusei、Candida tropicalis、Candida glabrataおよびCandida parapsilosisよりなる群から選択されるCandida株に由来することができ、該Alsタンパク質ファミリーメンバーはAls1p、Als3p、Als5p、Als6p、Als7pまたはAls9pを含む。また、黄色ブドウ球菌感染を治療また予防する方法も提供される。該方法は、医薬上許容され得る媒体中の、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンの免疫原性量を投与することを含む。
【発明を実施するための最良の形態】
【0009】
(発明の詳細な説明)
Candida albicansおよび黄色ブドウ球菌(Staphylococcus aureus)はヒトにおける一般的な病原体である。例えば、C.albicansは通常は無害な片利共生生物であるが、この生物は膣および/または口腔咽頭カンジダ症のような表層粘膜皮膚感染から、播種性カンジダ症における深器官関与の範囲の種々の疾患を引き起こし得る。病気を引き起こすに先立って、真菌は胃腸管および、ある場合には皮膚および粘膜でコロニー化する。宿主粘膜表面への接着はこの初期工程の鍵となる要件である。コロニー化後に、C.albicansは感染された血管内デバイスを介して、または化学療法、またはストレス潰瘍によって危うくされる胃腸粘膜を通じる移動によって血流に入る。次いで、生物が血流を介して散在し、血管内皮に結合し、それに貫入して、血管ツリーから脱出し、肝臓、脾臓および腎臓のような深い器官に侵入する。
【0010】
本明細書中に記載された種々の例示的なAlsタンパク質ファミリーメンバーの同定および機能的特徴付けは、タンパク質のこのメンバーがカンジダ症の治療で効果的に利用されるのを可能とする。多用な基材および他の選択的細胞接着機能に対する特異的結合活性は、能動的または受動免疫のためのワクチンの生産において、宿主細胞の結合、吸着または侵入を阻害することによって初期感染を低下または予防するための細胞接着のペプチド、ミメティック阻害剤のアナログにおいて開発することができる。さらに、異なる結合および侵入プロフィールは、Alsタンパク質ファミリーメンバーの活性の広いスペクトルまたは標的化阻害の設計および使用を可能とする。加えて、結合および/または侵入活性を付与する機能的断片は、望まない外来性タンパク質配列の排除を可能とし、かくして、Alsファミリータンパク質メンバーワクチンまたは治療阻害剤の効力を増大させる。
【0011】
内皮細胞への接着によるC.albicansの病原性の性質は、ここに具体的に引用してその全体を援用する米国特許第5,578,309において議論されている。アドヘシンとしての遺伝子産物の特徴付けを含めた、Als1遺伝子およびその特徴の記載については、Fu,Y.,G.Rieg,W.A.Forizi,P.H.Belanger,J.E.J.Edwards,and S.G.Filler.1998.Expression of the Candida albicans gene ALS1 in Saccharomyces cerevisiae induces adherence to endothelial and epithelial cells.Infect.Immun.66:1783−1786;Hoyer,L.L.1997.Fu Y,Ibrahim AS,Sheppard DC,Chen Y−C,French SW,Cutler JE,Filler SG,Edwards,JE,Jr.2002.Candida albicans Als1p:an adhesin that is a downstream effector of the EFG1 filamentation pathway.Molecular Microbiology 44:61−72,Sheppard DC,Yeaman MR,Welch WH,Phan QT,Fu Y,Ibrahim AS,Filler SG,Zhang M,Waring AJ,Edwards,Jr.,JE 2004.Functional and Structural Diversity in the Als Protein Family of Candida albicans.Journal Biological Chemistry.279:30480−30489.The ALS gene family of Candida albicans.International Society for Human and Animal Mycology Salsimorge,Italy:(Abstract);Hoyer,L.L.,S.Scherer,A.R.Shatzman,and G.P.Livi.1995.Candida albicans ALSI:domains related to a Saccharonzyces cerevisiae sexual agglutinin separated by a repeating motif.Mol.Microbiol.15:39−54参照。
【0012】
この点に関しては、ヒト真菌病原体Candida albicansはコロニー化し、広く種々の宿主の組織に侵入する。宿主構成要素への接着はこのプロセスにおいて重要な役割を演じる。C.albicans Alsタンパク質ファミリーの2つのメンバー(Als1pおよびAls5p)は、接着を媒介することが判明しており、Alsタンパク質ファミリーメンバーの結合、接着および細胞侵入活性が例示される。本明細書中に記載されるように、Als遺伝子ファミリーの膜をクローン化し、S.cerevisiaeにおいて発現させて、それらの個々の機能を特徴付けた。区別されるAlsタンパク質は区別される接着プロフィールを多様な宿主基材に付与した。キメラAls5p−Als6p構築物を用い、基材特異的接着を媒介する領域はAlsタンパク質におけるN末端ドメインと突き止められた。具体的には、Alsタンパク質のサブセットもまた、細胞内皮侵入を媒介したが、これはこのファミリーのこれまでは未知であった機能である。これらの結果と合致して、相同モデリングは、Alsメンバーが、免疫グロブリンスーパーファミリーの接着または侵入と類似の拡大された領域が間に存在する反−平行β−シートモチーフを含有することを明らかにした。この知見は、Als1pのN末端ドメインの円二色およびフーリエ変換赤外分光分析を用いて確認された。アミノ酸過敏の特異的領域がAlsタンパク質のN末端ドメイン内に見出され、エネルギー−ベースのモデルは、恐らくはAlsファミリーメンバーの多様な機能を支配するN末端ドメインにおける同様性および差を予測した。まとめると、これらの結果は、Alsファミリー内の構造的および機能的多様性は、C.albicansに、感染の間に広い範囲の宿主構成要素を認識し、それと相互作用できる細胞壁タンパク質のアレイを供することを示す。
【0013】
本発明は、医薬上許容され得る媒体中の、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片、およびアジュバントを有するワクチンを提供する。該ワクチンは、Candida albicans、Candida krusei、Candida tropicalis、Candida glabrataまたはCandida、parapsilosisのようなCandida種に由来するAlsタンパク質ファミリーメンバーであり得る。Alsタンパク質ファミリーメンバーは、例えば、Als1p、Als3p、Als5p、Als6p、Als7pおよびAls9p、またはその免疫原性断片であり得る。Candida種内の全ての他のAlsタンパク質ファミリーメンバーは、同様に、本発明のワクチンとして使用することができる。
【0014】
本発明は、播種性カンジダ症を治療し、予防し、または緩和するためのワクチンとしてのC.albicansアグルチニン様配列タンパク質ファミリーメンバーの遺伝子産物を利用する。該ワクチンはC.albicansの異なる株に対して、ならびに異なるCandida種に対して効果的である。Alsタンパク質ファミリーメンバーは、例えば、Als1p、Als3p、Als5p、Als6p、Als7pおよびAls9pであり得る。本発明は、C.albicansによる内皮細胞および/または上皮細胞への接着および侵入、および生物の病因を遅らせるためのワクチンとして使用されるAlsタンパク質ファミリーメンバー−発現表面タンパク質の罹患性におけるALS遺伝子の役割を探求する。
【0015】
本発明に従うと、ALSファミリーメンバー遺伝子は、C.albicansに対する免疫治療戦略の標的として選択される表面アドヘシンをコードする。ALS1遺伝子の発現産物であるAls1pタンパク質は表面タンパク質に典型的な構造的特徴を有し、事実、C.albicansの細胞表面で発現されることを示すのは、宿主組織へのアドヘシンとして作用するタンパク質についての1つの基準である。Alsタンパク質ファミリーメンバーは、N末端におけるシグナルペプチド、C−末端におけるグリコシルホスファチジルイノシン(CPI)繋留配列、およびスレオニンおよびセリンがリッチな反復を含む中央領域を有するものとして構造的に特徴付けることができる。また、Alsタンパク質ファミリーメンバーは、細胞表面に発現されるタンパク質に典型的なN−およびO−グリコシル化部位を有する。ALs1pのN末端に対して向けられたモノクローナル抗体を用いる間接的免疫蛍光は、例えば、ALs1pが出芽胞子の対数期の間に発現されることを明らかにした。ALs1pのこの発現は菌糸形成の間に増加し、菌糸エレメントが拡散した表面染色によって示されるように出芽胞子から伸びる接合点に位置する。さらに、このモノクローナル抗体は、C.albicans過剰発現突然変異体の内皮細胞への増強された接着をブロックし、それにより、ALs1pを用いる免疫療法適用についての原理を確立する。機能的特徴は、それが他のAlsファミリーメンバーの細胞接着および侵入に関するので、実施例VIにおいて後にさらに記載する。
【0016】
かくして、1つの態様によると、本発明は、医薬組成物に処方し、アジュバントを含むまたは含まないワクチンとして投与した場合に有用な特性を有する、例えば、Als1p、Als3p、Als5p、Als6p、Als7pおよびAls9pと命名されたAlsファミリーメンバー表面接着タンパク質またはその機能的断片、コンジュゲートまたはアナログを提供する。Alsタンパク質ファミリーメンバー、2以上のAlsタンパク質ファミリーメンバーまたは1以上のその機能的断片、アナログ、コンジュゲートまたは誘導体の組合せは、たとえば、Candida albicansから得ることができる。同様なアドヘシンまたはインバシン分子、またはそのアナログまたは誘導体はカンジダ起源のものとすることができ、たとえば、属Candida、たとえば、Candida parapsilosis、Candida krusei、Candida glabrataおよびCandida tropicalisに属する種から得ることができる。本発明による表面アドヘシンまたはインバシンタンパク質は単離されたまたは純粋な形態で得ることができ、かくして、本発明の1つの実施形態によると、実質的に純粋なAlsタンパク質ファミリーメンバーCandida表面アドヘシンタンパク質、またはその機能的断片、免疫原性断片、アナログ、コンジュゲートまたは誘導体は、ワクチンとして処方されて、患者において免疫応答を引き起こして、Candidaに対して免疫応答を誘導し、および/または該生物の内皮細胞への接着をブロックする。無傷Alsタンパク質ファミリーメンバーと同様な結合、接着または侵入活性を呈するAlsタンパク質ファミリーメンバーの断片を、本明細書中においては、機能的断片という。Candida種に対する抗体または細胞性免疫応答を誘導することができるAlsタンパク質ファミリーメンバーの断片を、本明細書中においては、免疫原性断片という。例示的な機能的断片は実施例VIにおいて後にさらに記載するAlsタンパク質ファミリーメンバーのN末端ポリペプチド領域を含む。例示的な免疫原性断片はN末端Alsポリペプチド領域、C−末端Alsポリペプチド領域、ならびに抗体免疫応答、細胞性免疫応答、あるいは抗体および細胞性両免疫応答を生じさせるのに十分ないずれかの他のAls断片を含む。そのような免疫原性断片は、約4アミノ酸と小さく、かつ無傷ポリペプチド程度に大きくすることができ、ならびにその間の全てのポリペプチド長さを含む。
【0017】
本発明による表面接着タンパク質のアナログまたは誘導体は、ALSファミリーメンバー遺伝子および/または遺伝子産物について本明細書中に記載された基準によって同定し、さらに特徴付けることができる。例えば、アナログまたは誘導体のヌル突然変異体は、対照と比較して、内皮細胞に対して顕著に低下した接着を有するであろう。同様に、適切なモデルにおけるアナログまたは誘導体の過剰発現は、対照と比較して内皮細胞への増大した接着を示し、前記した基準に従うと細胞表面アドヘシンとして確認される。また、アナログまたは誘導体に対する抗血清は抗Alsタンパク質ファミリーメンバー抗体と交差反応することができ、本明細書中に開示されたような播種性カンジダ症のマウスモデルに投与した場合に、増大した生存時間を呈することができる。
【0018】
また、本発明は、播種性カンジダ症を治療または予防する方法を提供する。該方法は、医薬上許容され得る媒体中の、細胞接着または侵入活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンの免疫原性量を投与することを含む。該ワクチンはアジュバントと共に、またはそれ無くして投与することができる。Alsタンパク質ファミリーメンバーは異なるCandida株から、ならびにCandida albicans、Candida krusei、Candida tropicalis、Candida glabrataおよびCandida、parapsilosisのような異なるCandida種から由来することができる。播種性カンジダ症を治療または予防する方法で用いるAlsタンパク質ファミリーメンバーはAls1p、Als3p、Als5p、Als6p、Als7pおよびAls9pを含む。
【0019】
異なるCandida株、異なるCandida種、他の細菌および感染剤に対する本発明のワクチンの有効性、およびその広い範囲の免疫活性は後にさらに記載し、実施例において例示する。例えば、実施例Vは、抗ALS抗体が粘膜および血行的に播種性のカンジダ感染に対して有効的であることを示す。実施例VIIは、細胞−媒介免疫を高めることによって、マウス播種性カンジダ症の間における生存を、rAls1p−Nでのワクチン接種が改良することを示す。実施例VIIIは、本発明のワクチンは真菌負荷を低下させ、免疫応答性および免疫無防備マウス双方において生存を改良することを示す。実施例IXは、黄色ブドウ球菌感染に対する本発明のALSワクチンの有効性を示す。実施例Xは、本発明のワクチンがC.albicansの異なる株に対して、およびC.glabrata、C.krusei、C.parapsilosisおよびC.tropicalisのような異なる種に対して効果的であることを例示し、異なる動物モデルにおける有効性を例示する。また、実施例XIは、異なる動物モデルにおける本発明の異なるワクチンの有効性を例示し、ならびに誘導された異なる応答点および2つの代表的なALSワクチンの能力の比較を提供する。
【0020】
さらに提供される発明は、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその機能的断片の有効量を投与して、Candidaの宿主細胞または組織への結合または侵入を阻害することを含む、播種性カンジダ症を治療または予防する方法である。Alsタンパク質ファミリーメンバーはCandida albicans、Candida krusei、Candida tropicalis、Candida glabrataおよびCandida parapsilosisに由来することができる。播種性カンジダ症を治療または予防する方法で用いるAlsタンパク質ファミリーメンバーは、Als1p、Als3p、Als5p、Als6p、Als7pおよびAls9pを含む。細胞接着活性は、ゼラチン、フィブロネクチン、ラミニン、上皮細胞または内皮細胞への結合、および/または細胞侵入への促進を含む。
【0021】
加えて、本発明は、本明細書中に記載されたAlsタンパク質ファミリーメンバーを用いて黄色ブドウ球菌感染を治療または予防する方法も提供する。特に、黄色ブドウ球菌感染を治療または予防する方法は、医薬上許容され得る媒体中の、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンの免疫原性量を投与することを含む。
【0022】
Als1pおよびAls3pは、黄色ブドウ球菌細胞表面タンパク質に対する有意な相同性のため特に効果的である。例えば、Als1pおよびAls3pの配列および構造的相同性は、実施例IXにおいて後にさらに記載する。本明細書中で提供される教示および指針があれば、当業者は、本発明のワクチンおよび方法を同様なCandidaおよびブドウ球菌(Staphylococcus)感染の治療に適用することができるのを理解するであろう。同様に、本明細書中に記載された教示および方法があれば、当業者であれば、本発明のワクチンおよび方法は、真菌、細菌などを含めた、本明細書中に記載されたAlsタンパク質ファミリーメンバーに対して同様な免疫原性、配列および/または構造的相同性を持つ細胞表面ポリペプチドを有する他の病原体にやはり適用することができるのを理解するであろう。
【0023】
Candidaまたはブドウ球菌感染に対する細胞接着または侵入戦略の免疫療法および/またはAlsポリペプチド阻害は、血管内皮細胞への結合のレベルにおいて、ならびにフィラメント化調節経路の下流エフェクタを介して働くことができる。可溶性Alsタンパク質ファミリーメンバーまたは機能的断片を用いる結合の免疫療法戦略または阻害はこの関係で有用である。なぜならば:(i)血行的に播種性のカンジダ症および他の感染病因に関連する罹患率および死亡率は、現在利用可能な抗真菌療法でもってしても許容できないくらい高いままである;(ii)抗真菌および抗生物質耐性の上昇する発生率は、抗真菌および抗菌剤の増大する使用に関連し、(iii)ひどいCandidaおよびブドウ球菌感染の危険がある患者の集団はよく定義されており、非常に大きく、かつ手術後患者、移植体患者、癌患者、および低体重で誕生した幼児を含み;および(iv)ひどいCandida感染を発生する患者の高いパーセンテージは好中球減少症ではなく、かくして、ワクチンまたは競合ポリペプチドまたは化合物阻害剤に応答できるからである。これらの理由で、Candidaおよびブドウ球菌は、受動的免疫療法、能動的免疫療法、または受動的または能動的免疫療法の組合せについて魅力的な真菌および細菌標的である。加えて、Candidaは、Alsタンパク質ファミリーメンバーポリペプチド、その機能的誘導体、および/または1以上のAlsファミリーメンバーに結合し、Candidaの宿主細胞受容体への結合を妨げるその化合物またはミメティックを用いる競合阻害で魅力的である。
【0024】
本明細書中で提供された教示および指針があれば、当業者であれば、当該分野でよく知られた免疫療法を、本発明のAlsタンパク質ファミリーメンバー、その免疫原性断片、アナログ、コンジュゲート、および/または誘導体と共に使用して、アジュバントと共に、またはそれなくしてワクチンとして投与された医薬上許容され得る組成物中の免疫原として該分子を1以上に用いることができるのを理解するであろう。本発明の目的では、用語「医薬の」または「医薬上許容され得る」とは、非毒性であり、望まれた場合、安全にヒトに投与できる担体または添加剤と共に用いられる公知の技術によって処方された組成物をいう。投与は、例えば、静脈内、筋肉内、腹腔内、または皮下注射を含めた良く知られた経路を用いて行うことができる。本発明のそのようなワクチンは当業者に知られた緩衝液、塩または他の溶媒を含んで、溶液中でのワクチンの活性を維持することができる。同様に、当該分野でよく知られた広い範囲のアジュバントのいずれかを本発明のワクチンと共に使用して、Candidaまたはブドウ球菌の罹患性宿主細胞への結合、侵入および/または感染を低下し、またはブロックすることができる治療上有効な免疫応答を誘導し、促進し、または高めることができる。
【0025】
同様に、本明細書中に提供された教示および指針があれば、当業者であれば、投与し、細胞表面分子のその同族受容体への結合を選択的にブロックするための当該分野でよく知られた治療方法を、本発明のAlsタンパク質ファミリーメンバー、その機能的断片、アナログ、コンジュゲート、および/または誘導体と共に使用して、Alsタンパク質ファミリーメンバーの1以上を医薬上許容され得る組成物中で阻害剤として用いることができるのを理解するであろう。ワクチン処方に関しては、阻害性処方は、例えば、静脈内、筋肉内、腹腔内、または皮下注射を含めた当該分野でよく知られた方法を用いて同様に投与することができる。Alsファミリーメンバー受容体に結合し、Alsタンパク質ファミリーメンバー結合をブロックするそのような阻害性組成物は当業者に知られた緩衝液、塩または他の溶媒を含んで、溶液中でのワクチンの活性を維持することができる。さらに、当該分野でよく知られた広い範囲の処方のいずれかを本発明の阻害性組成物で使用して、Candidaまたはブドウ球菌の罹患性宿主細胞への結合、侵入および/または感染を低下させ、または阻害するように、送達または摂取を標的化し、および/または高めることができる。
【0026】
本発明に従う治療免疫原または受容体結合阻害剤として用いる分子に関しては、当業者であれば、Alsタンパク質ファミリーメンバー分子を、免疫原性ワクチンまたは細胞接着もしくは侵入阻害剤としての必須の性質を失うことなく切形または断片化することができるのを認識するであろう。例えば、Alsタンパク質ファミリーメンバーを切形して、前記した、かつ実施例において後にさらに記載する機能的特性を維持したC−末端からの切形によってN末端断片を生じさせることができる。同様に、C−末端断片は、その機能的特性を維持したN末端からの切形によって生じさせることができる。本明細書中で提供された教示および指針に従った他の修飾を本発明に従って行って、他のAlsタンパク質ファミリーメンバーの機能的断片、その免疫原性断片、アナログまたは誘導体を作成して、天然タンパク質に関して本明細書中に記載された治療上有用な特性を達成することができる。
【0027】
本発明のAlsタンパク質ファミリーメンバーおよび方法の治療的有効性の1つの態様はフィラメント化の調節への干渉を達成して、生物の宿主構成要素への接着をブロックし、およびイムノエフェクタ細胞および他の生理学的メカニズムによって生物のクリアランスを高める。内皮細胞は血管系の大部分を被覆するので、抗体、Alsファミリーメンバータンパク質、ポリペプチド、またはペプチドまたはそのいずれかの組合せを用いて内皮細胞への生物の接着、侵入および/または双方をブロックする戦略は、本発明の有用な実施形態を含む。既に記載したように、そのような接着および/または侵入をブロックする戦略は、カンジダアドヘシン、インバシン、または本明細書中に開示された同族受容体に対して向けられた能動的または受動的免疫療法、あるいは阻害性結合を含む。かくして、例えば、いずれの適当な宿主もタンパク質を注射し、血清を集めて、適当な精製および/または濃縮後に所望の抗アドヘシン抗体を得ることができる。注射に先立って、アドヘシンまたはインバシンタンパク質またはその組合せを適当なビヒクル、好ましくは、多糖のような公知の免疫刺激剤、またはリポソームのような送達処方、または時間−放出組成物中に処方することができる。かくして、さらなる態様によると、本発明は、ワクチンまたはAls受容体阻害剤として用いるため、処方中に、1以上の医薬上許容され得る賦形剤と共にカンジダアドヘシンまたはインバシンタンパク質を含む医薬組成物を提供する。
【0028】
本発明の方法は、C.albicansの宿主構成要素の内皮または上皮細胞への接着をブロックすることによって、あるいは例えば抗体をブドウ球菌に結合させ、免疫メカニズムに病原体を除去させることによって、カンジダまたはブドウ球菌感染を緩和し、および/または阻害するものである。かくして、本発明の1つの態様によると、Alsタンパク質ファミリーメンバーのアドヘシンまたはインバシンタンパク質、その機能的または免疫原性断片、誘導体、アナログ、またはコンジュゲートを含む医薬組成物を、注射または注入用の生体適合性担体を含有する医薬組成物中のワクチンまたはAls受容体阻害剤として処方し、患者に投与される。また、Alsファミリーメンバータンパク質または単離されたまたは組換えAlsファミリーメンバータンパク質に対して生起された抗血清の直接的投与を用いて、C.albicansの哺乳動物宿主の構成要素への接着をブロックし、またはブドウ球菌病原体の除去を行うことができる。アドヘシンタンパク質に対する抗血清は公知の技術、Kohler and Milstein, Nature 256:495−499(1975)によって得ることができ、それをヒト化して、抗原生を低下させる(米国特許第5,693,762参照)。またはトランスジェニックマウスにおいて生産し、編集されていないヒト免疫グロブリン遺伝子を得ることができる(米国特許第5,877,397参照)。同様に、単離されたまたは組換えAlsタンパク質ファミリーメンバーもまた、例えば、後に実施例に記載される組換え生産を含めた、当業者によく知られた方法を用いて生産することもできる。
【0029】
本発明のなおさらなる使用は、Alsタンパク質ファミリーメンバーのアドヘシンまたはインバシンタンパク質を用いて、カンジダまたはブドウ球菌感染の予防および/または緩和のためのワクチン戦略を開発する。かくして、本発明の1つの態様によると、例えば、標準免疫学技術を使用してマルチ−成分ワクチン戦略を構築することができ、これは、C.albicansの接着をブロックし、またはブドウ球菌病原体の排除を行うために宿主構成要素からの免疫応答を高め、および/または誘導することができる。
【0030】
本発明のなおさらなる使用は、例えば、DNAワクチン戦略を開発することである。かくして、本発明の1つの態様によると、例えば、Alsタンパク質ファミリーメンバーのアドヘシンまたはインバシン、またはその機能的断片をコードするALSファミリーメンバーポリヌクレオチドを、遺伝子産物に対する免疫応答を得るように設計されたプロトコルに従って投与される。例えば、Felgner 米国特許第5,703,055参照。
【0031】
本発明のなおさらなる使用は、例えば、組合せワクチン戦略を開発することである。かくして、本発明の1つの態様によると、例えば、抗ALSタンパク質ファミリーメンバー抗体は、カンジダまたはブドウ球菌感染を治療し、および/または予防する抗体と共に用いることができる。米国特許第5,578,309参照。
【0032】
以下の実施例は、播種性カンジダ症の予防的尺度または治療のための基礎としてのALS1アドヘシンの免疫治療的利用性を説明する。実施例1は、ALS1の過剰発現によって特徴付けられるALS1ヌル突然変異体およびC.albicansの株を調製して、内皮細胞への接着の媒介を確認することを記載する。実施例2は、Als1pの局所化、およびefgフィラメント化調節経路の関与を記載する。実施例3は、ALS1アドヘシンタンパク質の精製を記載する。実施例4は、表面アドヘシンタンパク質のブロッキングを示すために用いられる、ALS1表面アドヘシンタンパク質に対して生起されたウサギポリクローナル抗体の調製を記載する。実施例5は、本発明に従って本明細書中に記載されたようにALS1表面接着タンパク質に対して生起されたポリクローナル抗体を用いてインビボにて接着をブロックして、マウスモデルにおいて播種性カンジダ症に対して保護することを記載する。実施例VIは、Alsタンパク質ファミリーメンバーの構造的および機能的特徴を記載する。
【0033】
本発明の種々の実施形態の活性に実質的に影響しない修飾もまた本明細書中で提供される発明の定義内に含まれることは理解される。従って、以下の実施例は本発明を説明するものであり、限定することを意図しない。
【0034】
(実施例I)
(Als1はC.albicansの内皮細胞への接着を媒介する)
URAブラスター技術を用いて、Als1pの発現を欠如するC.albicansのヌル突然変異体を構築した。als1/als1突然変異体は、Ura−ブラスター技術(Fonzi and Irwin,Genetics 134, 717(1993))の修飾を用いてC.albicans株CAI4において以下のように構築した。2つの別々のals1/hisG−IRA3−hisG−als1構築物を利用して、遺伝子の2つの異なる対立遺伝子を破壊した。4.9kbのAsLS1コーディング配列は、以下のプライマーを用いて高い忠実度PCR(Boehringer Mannheim,Indianapolis, IN)でもって生じさせた:
【0035】
【化1】
次に、PCR断片をpGEM−Tベクター(Promega,Madison,WI)にクローン化し、かくして、pGEM−T−ALS1を得た。hisG−URA3−hisG構築物をKpn1およびHind3での消化によってpMG−7から放出させ、これを用いて、pGEM−T−ALS1のKpn1およびHind3消化によって放出されたALS1の一部を置き換えた。最終のals1−hisG−URA3−hisG−als1構築物を、Xholでの消化によってプラスミドから放出させ、これを用いて株CAI−4の形質転換によってALS1の最初の対立遺伝子を破壊した。
【0036】
第2のals1−hisG−URA3−hisG−als1構築物を2つの工程で生じさせた。まず、pMB7のBg12−Hind3 hisG−URA3−hisG断片をpUC19のBamH1−Hind3部位にクローン化し、それにより、pYC2を得た。次いで、pYC2をHind3で消化し、T4 DNAポリメラーゼを用いてdATPおよびdGTPで部分的に満たし、次いで、Sma1で消化して新しいhisGURA3−hisGで断片を得た。第二に、ALS1相補的フランキング領域を生じさせるために、pGEM−T−ALS1をXba1で消化し、次いで、dCTPおよびdTTPで部分的に満たした。この断片をHpa1で消化して、ALS1の中央部分を欠失させ、次いで、hisG−URA3−hisG断片に連結させ、pYC3を得た。次いで、このプラスミドをXholによって消化して、構築物を放出させ、これを用いて、ALS1の第二の対立遺伝子を破壊した。実験を通じて増殖曲線を作成して、生じた突然変異が増殖速度に対して影響を有しないことを確認した。すべての組込みは、XbaIおよびHindIIIでのpYF5の消化によって生じた0.9kbのALS1特異的プローブを用いてサザーンブロット分析によって確認した。
【0037】
ヌル突然変異体を、ヒト臍静脈内皮細胞にインビトロで接着するその能力についてC.albicans CAI−12(URA+リバータント株)と比較した。接着実験では、YPD(2%グルコース、2%ペプトン、および1%酵母エキス)での一晩の培養からの酵母細胞を、グルタミンを含むRPM1中で25℃にて1時間増殖させて、Als1p発現を誘導した。ハンクス平衡塩溶液(HBSS)(Irvine Scientific, Irvine, CA)中の3×102生物を内皮細胞の各ウェルに加え、その後、プレートを37℃にて30分間インキュベートした。接種物のサイズは、YPD寒天中での定量的培養によって確認した。インキュベーション時間の最後に、非接着性生物を吸引し、内皮細胞単層を標準的な方法にてHBSSで2回すすいだ。ウェルをYPD寒天と重ね、接着性生物の数をコロニーカウンティングによって決定した。統計学的処理はWilcoxin順位和検定によって得られ、Bonferroni修正にて多数の比較のために修正した。P<0.001。
【0038】
図1を参照し、ALS1/ALS1およびals1/als1株の比較は、ALS1ヌル突然変異体はC.albicans CAI−12よりも内皮細胞に35%未満接着した。バックグラウンド接着を低下させるために、非ALS1発現条件下で増殖させた野生型株の接着を、Als1pを自律的に発現する突然変異体と比較した。この突然変異体は、構成的ADH1プロモーターの制御下のALS1の第三のコピーを野生型C.albicansに組込むことによって構築した。C.albicansにおけるALS1の構成的発現を達成するために、平滑末端PCR生成URA3遺伝子を、pOCUS−2ベクター(Novagen,Madison,WI)の平滑末端Bg12部位へ連結して、pOU−2を得た。(pLH−ADHptから単離され、親切にも、A.Brown,Aberdeen,UKによって提供された)C.albicansアルコールデヒドロゲナーゼ遺伝子(ADH1)プロモーターおよびターミネーターを含有した2.4kbのNot1−Stu1断片を、Not1およびStu1での消化後にpOU−2にクローン化した。pOAU−3と命名された新しいプラスミドは、ADH1プロモーターおよびターミネーターの間に唯一のBg12部位を有した。BamH1制限酵素部位が近接するALS1コーディング配列は、鋳型としてのpYF−5、および以下のプライマーを用いて高忠実度PCRによって生じさせた:
【0039】
【化2】
このPCR断片をBamH1で消化し、次いで、pOAU−3の適合するBg12部位にクローン化して、pAU−1を得た。最後に、pAU−1を、C.albicans CAI−4を形質転換するに先立ってXbalによって線状化した。部位−特異的組込みは、サザーンブロット分析によって確認した。図1Bを参照し、このPADH1−ALS1株におけるALS1の過剰発現の結果、野生型C.albicansと比較して内皮細胞への接着の76%増加がもたらされた。野生型の内皮細胞接着を過剰発現突然変異体と比較するにおいて、酵母細胞を25℃においてYPD中で一晩増殖させた。(Als1p非誘導性条件)。Als1p発現は、野生型のバックグラウンド接着を低下させるように誘導されず、かくして、PADH1−ALS1ハイブリッド遺伝子を通じての接着においてAls1pの役割を拡大した。接着性アッセイは前記したように行った。統計学的処理はWilcoxin順位和検定によって得られ、Bonferroni修正を施して多数の比較のために修正した。P<0.001。
【0040】
モノクローナル抗Als1pマウスIgG抗体を、Clontech YEXpress(TM)酵母発現系(Palo Alto,CA)を用いて発現させたAls1pの精製され切形されたN末端(アミノ酸#17から#432)に対して生起させた。これらのモノクローナル抗Als1p抗体の接着性ブロッキング能力は、1:50希釈にて、C.albicansを抗Als1抗体またはマウスIgG(Sigma,St.Louis,MO)いずれかと共にC.albicans細胞をインキュベートすることによって評価した。その後、酵母細胞を前記した接着アッセイで用いた。統計学的処理は、Wilcoxin順位和検定によって得られ、Bonferroni修正を施して多数比較のために修正した。P<0.001。結果は、PADH1−ALS1株の接着が26.8%±3.5%から14.7%±5.3%まで低下したことを明らかにした。かくして、ALS1欠失および過剰発現の効果は、Als1pが、C.albicansの内皮細胞への接着を媒介することを示す。
【0041】
(実施例II)
(Als1pの局所化)
アドヘシンとして機能するAls1pについては、それは細胞表面に位置しなければならない。Als1pの細胞表面位置決定は、抗Als1pモノクローナル抗体と共に間接的に免疫蛍光を用いて確認した。拡散染色は、指数関数的増殖の間に出芽胞子の表面で検出された。この染色は静止相において出芽胞子では検出不可能であった。図2Aを参照し、出芽胞子を、フィラメントを生産するように誘導すると、強い染色が観察され、これは出現するフィラメントの基部に専ら位置付けられた。免疫蛍光はals1−als1突然変異体で観察されず、これは、Als1pについてのこの抗体の特異性を確認する。図2B参照。これらの結果は、Als1pが細胞表面タンパク質であることを確立する。
【0042】
出芽胞子−フィラメント接合へのAls1pの特異的位置決定は、フィラメント化プロセスにおけるAls1pを示唆する。メカニズムを決定するために、C.albicans ALS1突然変異体のフィラメント化表現型を分析した。図3Aを参照し、als1/als1突然変異体はLee固体培地上での4日間のインキュベーション後にフィラメントを形成できず、他方ALS1/ALS1およびALS1/als1株ならびにALS1−捕捉突然変異体は共にこの時点において豊富なフィラメントを生産した。als1/als1突然変異体は、より長いインキュベーション時間の後にフィラメントを形成することができた。さらに、ALS1の過剰発現はフィラメント化を増大させ:PADH1−ALS1株は3日間のインキュベーションの後に豊富なフィラメントを形成し、他方、野生型株はこの時点において乏しいフィラメントを生産した。図3B参照。フィラメント化におけるAls1pの役割をさらに確認するために、ALS1のコーディング配列を反対向きに挿入した以外は、ALS1過剰発現突然変異体と同様な突然変異体を用いて陰性対照を提供した。得られた株のフィラメント化表現型は、野生型株のそれと同様であることが示された。Als1pのフィラメント−誘導特性は固体媒体上で増殖させた細胞に特異的である。なぜならば、前記した株の全ては液体培地中で匹敵するほどフィラメント化したからである。データは、Als1pがフィラメント化を促進し、フィラメント化制御経路の調節におけるALS1発現を示唆することを示す。efg1/efg1、cph1/cph1、efg1/efg cph1/cph1、tup1/tup1、およびcla4/cla4突然変異体を含めた、これらの経路の各々における欠点を持つ突然変異体でのALS1発現のノーザンブロット分析を行った。図4Aを参照し、EFG1の双方の対立遺伝子が破壊された突然変異体はALS1を発現できなかった。野生型EFG1のコピーのefg1/efg1突然変異体への導入はALS1発現を回復したが、低下したレベルのものであった。図4B参照。また、図4Aで見られるように、他のフィラメント化調節突然変異のいずれも、ALS1発現を有意に改変しなかった(図4A)。かくして、Efg1pはALS1発現で要求される。
【0043】
もしEfg1pがALS1の発現を刺激するならば、これは今度はフィラメント化を誘導するのであるが、efg1/efg1株におけるALS1の発現はフィラメント化を回復するはずである。ADH1プロモーターの制御下にあるALS1の機能的対立遺伝子はefg1/efg1株に組込まれた。ALS1遺伝子産物がefg1ヌル突然変異体におけるフィラメント化における欠陥を補足するという確率を調べるために、Ura efg1ヌル突然変異体を、線状化pAU−1で形質転換した。Ura+クローンを選択し、ALS1の第三のコピーの組込みは、プライマーを用いるPCRで確認した。得られた株はALS1を自律的に発現し、Lee寒天上でフィラメント化する能力を回復した:
【0044】
【化3】
図4BおよびC参照。従って、Efg1pはALS1発現の活性化を通じてのフィラメント化を誘導する。
【0045】
フィラメント化はC.albicinsにおける臨界的なビルレンス因子であるゆえに、フィラメント化を調節する経路の表示は病因についての重要な示唆を有する。ALS1に先立って、これらの調節経路の下流エフェクタをコードする遺伝子は同定されていない。細胞表面タンパク質をコードする2つの他の遺伝子、HWP1およびINTIの破壊の結果、フィラメント化欠陥を持つ突然変異体がもたらされる。HWP1発現もまたEfg1pによってやはり調節されるが、efg1/efg1突然変異体におけるHWP1の自律発現はフィラメント化を回復できなかった。従って、Hwp1p単独はEFG1の下流のフィラメント化のエフェクタとして機能しない。また、INT1発現を制御する調節エレメントは知られていない。かくして、Als1pは、フィラメント化の下流エフェクタとして機能する同定された最初の細胞−表面タンパク質であり、それにより、C.albicansのビルレンスにおけるこのタンパク質についての中枢的な役割を示唆する。
【0046】
C.albicansビルレンスに対するAls1pの寄与は、血行的に播種性のカンジダ症のモデルでテストした。A.S.Ibrahimら,Infect.Immun.63,1993(1995)。図5Aを参照し、als1/als1ヌル突然変異体で感染させたマウスは、ALS1/ALS1株、ALS1/als1突然変異体またはALS1−捕捉突然変異体で感染させたマウスよりも有意に長く生存した。感染から28時間後に、als1/als1突然変異体で感染させたマウスの腎臓は有意に少数の生物を含有した(5.70±0.46 log10CFU/g)(双方の比較について(P<0.0006)。テストした時点のいずれかにおいて株のいずれかで感染させたマウスの脾臓、肺、または肝臓から回収された生物のコロニーカウントにおいて、差は検出されなかった。これらの結果は、Als1pが、感染の最初の28時間の間のC.albicans増殖および腎臓における執拗性で重要であることを示す。図5Bを参照し、感染から28時間後のマウスの腎臓の調査は、als1/als1突然変異体が、ALS1−捕捉株の野生型いずれかよりも、有意により短いフィラメントを生産し、より弱い免疫応答を誘導したことを明らかとした。しかしながら、感染から40時間までは、フィラメントの長さ、およびそれらを囲う白血球の数は全ての3つの株について同様であった。
【0047】
組織病理学で見られたals1/als1突然変異体のフィラメント化欠陥は、固体培地でのインビトロフィラメント化アッセイと並行した。この突然変異体は、インビボおよびインビトロ双方において初期の時点で欠陥があるフィラメント化を示した。この欠陥は、結局は、延長された感染/インキュベーションで解決された。これらの結果が、ALS1から独立したフィラメント化調節経路がより遅い時点で操作可能となり得ることを示唆する。感染から40時間までのこの代替フィラメント化経路の活性化は、なぜals1/als1突然変異体で感染させたマウスが引き続いて次の2から3日において死滅するかの理由のようである。
【0048】
まとめると、これらのデータは、C.albicans ALS1が、内皮細胞への接着およびフィラメント化双方を媒介する細胞表面タンパク質をコードすることを示す。Als1pは、C.albicansにおけるいずれかの既知のフィラメント化調節経路の唯一の同定された下流エフェクタである。加えて、Als1pは血行性カンジダ感染におけるビルレンスに寄与する。Als1pの細胞表面位置およびデュアル機能性は、それを、薬物および免疫−ベースの療法双方にとって魅力的な標的とする。
【0049】
(実施例III)
(ALS1アドヘシンタンパク質精製)
E.coliによって合成されたALS1タンパク質は免疫原として適切である。しかしながら、E.coliによって合成された真核生物タンパク質は、不適切な折畳またはグリコシル化の欠如のため機能的ではないであろう。従って、ALS1タンパク質が、C.albicansの内皮細胞への接着をブロックできるかを決定するために、該タンパク質を好ましくは遺伝子工学により作成されたC.albicansから精製する。
【0050】
PCRを用いて、ヌクレオチド52から1296のALS1の断片を増幅した。この1246bp断片は、シグナルペプチドの端部からタンデムリピートの始まりまでの予測されたALS1タンパク質のN末端を含む。ALS1の領域を増幅した。というのは、それは、S.cerevisiae Aga1遺伝子産物の結合領域に対するその相同性に基づいて、アドヘシンの結合部位をコードするようだからである。加えて、予測されたALS1タンパク質のこの部分はほとんどグリコシル化部位を有さず、そのサイズはE.coliにおける効果的な発現で適切である。
【0051】
ALS1の断片をpQE32へ連結してpINS5を得た。このプラスミドにおいて、タンパク質をlacプロモーターの制御下で発現させ、それは、それがアフィニティー精製できるように、N末端に融合した6−hisタグを有する。我々は、pINS5でE.coliを形質転換し、それを(IPTGの存在下で)誘導条件下でそれを成長させ、次いで、細胞を溶解させた。細胞溶解物をNi2+−アガロースカラムを通して、ALS1−6His融合タンパク質をアフィニティー精製した。この手法により、実質的な量のALS1−6Hisを得た。融合タンパク質をさらにSDS−PAGEによって精製した。ポリクローナルウサギ抗血清がそれに対して生起できるように該タンパク質を含有するバンドをゲルから切り出した。本発明による表面アドヘシンタンパク質は、本発明の精神逸脱することなく種々の公知のプロセスによって調製し、精製できることは当業者によって認識されるであろう。Als1pの配列を図7にリストする。
【0052】
(実施例IV)
(ALS1タンパク質に対するポリクローナル抗血清の生起)
ALS1タンパク質に対する抗体がCandida albicansの内皮および上皮細胞、選択された宿主構成要素への接着をインビトロにてブロックするか否かを決定するために、ALS1タンパク質で形質転換したS.cerevisiaeでウサギを接種した。用いた免疫化プロトコルは、Candidaの種々の種内で抗原性関係を同定した抗血清の生産用のHasencleverおよびMitchellによって用いられた用量およびスケジュールであった。Hasenclever,H.F および W.O.Mitchell.1960。Antigenic relationships of Torulopsis glabrata and seven speceis of the genus Candida.J.Bacteriol.79:677−681。対照抗血清もまた、空のプラスミドで形質転換されたS.cerevisiaeに対して生起させた。全ての酵母細胞をガラクトース中で増殖させて、ALS遺伝子の発現を誘導した。接着実験でテストする前に、血清を56Cにおいて加熱−不活化させて、全ての補体活性を除去した。
【0053】
免疫化されたウサギからの血清を、空のプラスミドで形質転換されたS.cerevisiaeの全細胞で吸収させて、ALS1タンパク質以外の酵母の成分と反応性である抗体を除去した。抗血清の力価は、ALS1遺伝子を発現するS.cerevisiaeを用いる免疫蛍光によって決定した。FITC−標識抗ウサギ抗体は商業的源(Southern Biotechnology,Inc)から購入した。アフィニティー−精製二次抗体は必須であった。なぜならば、多くの市販の血清は、酵母グルカンおよびマンナンと反応性である抗体を含有するからである。二次抗体はプラスミドで形質転換されたCandida albicansならびにS.cerevisiaeを用いて予めテストし、必要に応じて吸収して、いずれの抗S.cerevisiaeまたは抗Candida抗体も除去した。陰性対照は1)免疫前血清、2)空のプラスミドで形質転換させたS.cerevisiae、および3)ALS(グルコース)遺伝子の発現を抑制する条件下で増殖させた以外はALS遺伝子で形質転換されたS.cerevisiaeであった。
【0054】
前記実験に加えて、ウエスタンブロッティングを用いて、抗血清が、それがそれに対して生起したALSタンパク質に特異的に結合するというさらなる確認を供した。ALS1で形質転換されたS.cerevisiaeを誘導条件下で増殖し、その原形質膜は標準的な方法によって単離した。Panaretou R and P.Piper.1996.Isolation of Yeast Plasma Membranes.p.117−121.InI.H.Evans.(ed.),Yeast Protocols.Methods in Cell and Molecular Biology.Humana Press,Totowa,New Jersey。原形質膜は空のプラスミドを形質転換し、かつ同一条件下で増殖させたS.cerevisiaeからも調製した。膜タンパク質をSDS−PAGEによって分離し、次いで、エレクトロブロッティングによってPVDF膜に移した。Harlow,EおよびD.Lane.1988.Antibodies:a laboratory manual.Cold Spring Harbor Laboratory Press。無脂肪乳でブロックした後、ブロットをALS抗血清と共にインキュベートした。予め吸収された抗血清は、空のプラスミドを含有するS.cerevisiaeから抽出されたタンパク質と反応しなかった。この抗血清はS.cerevisiae pYF5(Candida albicans ALS1を発現するクローン)の内皮細胞への接着をブロックした。
【0055】
(実施例V)
(特異的ALSタンパク質に対するポリクローナル抗体は、粘膜および血行的に播種性のカンジダ感染からマウスを予防的に保護する)
前記条件下で、ALS遺伝子で形質転換されたS.cerevisiaeのクローンの接着をブロックする抗血清を同定した後、これらの抗血清は、Candida albicans での静脈内チャレンジからマウスを保護することが示された。
【0056】
ALSタンパク質に対する抗血清を、まず、血行的に播種性のカンジダ症のネズミモデルでテストした。アフィニティー−精製抗ALS抗体は、酵母細胞の種々の基材への接着を妨げる点において効果的である(実施例3参照)。アフィニティー−精製はこの系で有用である。なぜならば、抗体は正確に決定できるからである。さらに、分画されていない抗血清は、疑いもなく、S.cerevisiaeキャリアー細胞上の抗原に向けられた多量の抗体を含有するであろう。これらの抗Saccharomyces抗体の多くはC.albicansに結合するようであり、結果の解釈を不可能とするようである。加えて、ALSタンパク質を発現するS.cerevisiaeから抗体を溶出させるのに用いる手法もまた、アジュバント−様活性を有し得る少量の酵母マンナンまたはグルカンを溶出することもできる可能性が高い。免疫アフィニティー−精製抗体は使用前にさらに精製される。それはマウス脾臓細胞で予め吸着することもできる。
【0057】
抗体用量を投与して、最も活性な免疫化プロトコルにおいて予測することができる血清抗体のレベルを画定する範囲をカバーし、およびカンジダ症のネズミモデルにおいて受動免疫で典型的に用いる抗体用量の範囲をカバーすることができる。Dromer,F.,J.Charreire,A.Contrepois,C.Carbon,およびP.Yeni.1987,Protection,of mice against experimental cryptococcosis by anti−Cryptococcus neofornwns monoclonal antibody,Infect.Immun.55:749−752;Han,Y.and J.E.Cutler.1995,Antibody reponse that protects against disseminated candidiasis,Infect.Immun.63:2714−2719;Mukherjee,J.,M.D.Scharff,and A.Casadevall.1992,Protective murine monoclonal antibodies to Cryptococcus neofornwns,Infect.Immun.60:4534−4541;Sanford,J.E.,D.M.Lupan,A.M.Schlageter,and T.R.Kozel.1990,Passive immunication against Cryptococcus neoformans with an isotype−switch family ofmonoclonal antibodies reactive with cryptococcal polysaccharide,Infect.Immun.58:1919−1923参照。BALB/cマウス(雌、7週齢、NCI)に、腹腔内(i.p.)注射によって、マウス脾臓細胞で吸収されている抗ALSを与えた。対照マウスは、各々、マウス脾臓細胞で吸収されている予め採血した血清、無傷抗ALS血清、またはDPBSを受けた。予備的吸収のために、2mlの抗ALSまたは予め採血した血清を、100μlのマウス(BALB/c、7週齢の雌、NCI)脾臓細胞(ml当たりほぼ9×106細胞)と室温にて20分間混合した。混合物を遠心(@300xg)によって温かい滅菌DPBSで3分間洗浄した。この手法を3回反復した。i.p.注射の用量はマウス当たり0.4mlであった。4時間後、マウスをC.albicans(株CA−1;i.v.注射によりマウス当たり5×105親水性酵母細胞)でチャレンジした。次いで、その生存時間を測定した。図6参照。
【0058】
従前の実験は、腹腔内経路を介して投与された抗体が血清へ迅速(数分内)かつほとんど完全に移ることを示している(Kozel and Casadevall、未公表観察)。抗体製剤を投与する効力についての対照として、マウスの平行群を、ALS遺伝子で形質転換されたS.cerevisiaで吸収されたプレ−免疫前血清から単離された抗体で処理した。生存時間および腎臓のグラム当たりの酵母の数を測定した。再度、図6を参照し、ALS1ヌル突然変異体の106出芽胞子を静脈内注射によって感染したマウスは、ALS1の1つの対立遺伝子が欠失されているCandida albicans CAI−12またはCandid albicansで感染させたマウスと比較した場合、より長いメジアン生存時間を有した(p=0.003)。
【0059】
これらの結果は、ワクチンとしてALS1タンパク質を用いる免疫療法戦略が、播種性カンジダ症に対して保護的予防効果を有することを示す。
【0060】
(実施例VI)
(Candida albicans のAlsタンパク質ファミリーにおける機能および構造の多様性)
非接着性S.cerevisiaeの異種補足によるC.albicansALS1遺伝子の単離および特徴付けは従前に記載されている(Fuら,Infect.Immun.66:1783−1786(1998))。ALS1は、内皮細胞および上皮細胞への接着を媒介する細胞表面タンパク質をコードする。C.albicansにおけるこの遺伝子の双方のコピーの破壊は、内皮細胞への接着の35%低下に関連しており、ALS1の過剰発現は接着を125%増大させる(Fuら,Mol.Microbiol.44:61−72(2002))。
【0061】
ALSIは、Hoyerらによって元来記載された少なくとも8つのメンバーよりなる多数の大きなC.albicans遺伝子ファミリーである(Hoyerら,Trends Microbiol.9:176−180(2001),Zhaoら,Microbiology 149:2947−2960(2003))。これらの遺伝子は、3つのドメインによって特徴付けられる細胞表面タンパク質をコードする。N末端領域は推定シグナルペプチドを含有し、Alsタンパク質の間で比較的保存されている。この領域は、貧弱にしかグリコシル化されていないと予測される(Zhaoら,Microbiology 149:2947−2960(2003),Hoyerら,Genetics 157:1555−1567(2001))。これらのタンパク質の中央部分は可変数のタンデムリピート(長さは〜36アミノ酸)よりなり、グリコシルホスファチジルイノシトールアンカー配列を含有するセリン−スレオニン−リッチなC−末端領域がある(上記)が続く。この遺伝子ファミリーによってコードされるタンパク質は感染の間に発現することが知られているが(Hoyerら,Infect.Immun.67:4251−4255(1999),Zhangら,Genome Res.13:2005−2017(2003))、異なるAlsタンパク質の機能は詳細に調べられていない。
【0062】
非接着性S.cerevisiaeにおけるAlsタンパク質の異種発現を行って、Alsタンパク質の機能を評価し、C.albicansによって発現された多数の他のアドヘシンによって媒介された高いバックグラウンド接着を回避した。この異種発現系は、アドヘシンALS1,ALS5およびEAP1の単離および特徴付けを含めた、C.albicans遺伝子の実験で広範に用いられてきた(Liら,Eukaryot Cell 2:1266−1273(2003),Fuら,Infect.Immun.66:1783−1786(1998),Gaurら,Infect.Immun.65:5289−5294(1997))。後にさらに記載するように、このモデル系を用い、Alsタンパク質は、多様な接着性および侵入性機能を有することが示された。これらの結果と合致して、相同性モデリングは、Alsタンパク質が、タンパク質の免疫グロブリンスーパーファミリーのアドヘシンおよびインバシンメンバーに対して構造が密接に関連していることを示した。CDおよびフーリエ変換赤外(FTIR)1分光分析を用いる構造解析により、Als1pのN末端ドメインが反−平行βシート、ターン、α−ラセン、および免疫グロブリンスーパーファミリーの他のメンバーの構造と合致する構造決定されていないドメインから構成されることが確認された。最後に、比較エネルギー−ベースのモデルは、それらの区別される接着および侵入性生物学的機能を支配し得る異なるAlsタンパク質の間でN末端ドメインの鍵となる物理化学的特性の差を示唆する。
【0063】
ALSファミリーメンバーをクローン化し、およびそれらをS.cerevisiaeにおいて発現させるために、ALSI、−3、−5、−6、−7、および−9を後に記載するように首尾よく増幅し、発現させた。簡単に述べれば、クローニングおよび他の培養工程について、S.cerevisiae株S150−2B(leu2 his3 trp1 ura3)を、従前に記載されているように異種発現で用いた(Fuら,Infect.Immun.66:2078−2084(1998))。C.albicans株SC5314をゲノムクローニングで用いた。全ての株を、必要に応じて1.5%バクト−寒天(Difco)で固化させた最小規定培地(100μg/mlのL−ロイシン、−Lトリプトファン、L−ヒスチジン、および硫酸アデニンを補足した1×酵母窒素ベースブロス(Difco)、2%グルコース、および0.5%硫酸アンモニウム)中で増殖させた。ura−株の増殖は、80μg/mlウリジン(Sigma)の添加によって支持された。ASL5を含有するプラスミドpGK103、ALS1を含有するpYF5、およびALS9を含有するpALSnは従前に記載されている(Fuら,Infect.Immune.66:1783−1786(1998),Gaurら,Infect.Immune.65:5289−5297(1997),Lucinodら,Proceedings of the 102nd Annual Meeting of the American Society for Microbiology,pp.204,American Society for Microbiology,Salt Lake City,Ut.(2002))。A.Brown(Aberdeen,UK)から得られたプラスミドpADH1はC.albicansアルコールデヒドロゲナーゼ遺伝子(ADHI)プロモーターおよびターミネーターを含有し、それはS.cerevisiaeで(機能的である)Baileyら,J.Bacteriol.178:5353−5360(1996))。このプラスミドを、S.cerevisiaeにおけるALS遺伝子の構成的発現で用いた。
【0064】
ヒト口腔上皮および血管内皮細胞を入手し、以下のように培養した。咽頭癌腫から単離されたFaDu口腔上皮細胞系はAmerican Type Culture Collection(ATCC)から購入し、その推奨されるプロトコルに従って維持した。内皮細胞は臍帯静脈から単離し、Jaffeらの方法の我々の従前に記載した修飾によって維持した(Fuら,Mol.Microbiol.44:61−72(2002),Jaffeら,J.Clin.Invest.52:2745−2756(1973))。全ての細胞培養を、5%CO2を含有する湿潤環境中で37℃に維持した。
【0065】
ALS遺伝子をクローニングするために、ALSファミリーのメンバーのゲノム配列を、(the World Wide Web at URL:sequence.stanford.edu/group/candida/search.htmlで入手可能な)StanfordデータベースのBLASTサーチングによって同定した。PCRプライマーは、5’BgIIIおよび3’Xhol制限酵素部位を取り組み、表Iにて以下に示されたオープンリーディングフレームの各々を特異的に増幅するように作成した(配列番号14から19(各々、ALS1、3、5、6、7および9センスプライマー);配列番号20から25(各々、ALS1、3、5、6、7および9アンチセンスプライマー))。各遺伝子は、Expand(登録商標)High Fidelity(高忠実度)PCR系(Roche Applied Science)を用いるPCRによってクローン化した。ALS3、ALS6、およびALS7をC.albicans SC5314ゲノムDNAから増幅し、他方、ALS1、ALS5、およびALS9は、C.albicansゲノムライブラリーから従前に検索されているプラスミドから増幅した(Fuら,Infect.Immune.66:1783−1786(1998),Gaurら,Infect.Immune.65:5289−5297(1997),Lucinodら,Proceedings of the 102nd Annual Meeting of the American Society for Microbiology,pp.204,American Society for Microbiology,Salt Lake City,Ut.(2002))。配列決定のために、PCR産物をpGEM−T−Easy(Promega)に連結した。次いで、配列が確認されたALSオープンリーディングフレームを、BglII−Xhol共−消化によってpGEM−T−Easyから放出させ、目的のALS遺伝子がADH1プロモーターの制御下となるように、pADH1に連結した。S.cerevisiae株S150−2Bを、酢酸リチウム方法を用い、ALS過剰発現構築物ならびに空のpADH1構築物の各々で形質転換した。S.cerevisiaeにおける各ALS遺伝子の発現をノーザンブロット分析によって確認し、しかる後、表現型分析を行った。
【0066】
(表I)
(S.cerevisiaにおける異種発現のためのALS遺伝子のコーディング領域を増幅するのに用いるPCRプライマー)
【0067】
【表1】
ALS mRNA発現は各構築物につきノーザンブロット分析によって検出した。プライマーの3つの組の使用に拘わらず、C.albicans SC5314のゲノムDNAからのALS2およびALS4の増幅は不成功であった。ALS遺伝子のタンデムリピートを横切った配列決定および組立の困難性を仮定すれば、この結果は、公表されたゲノムデーターベースで現在入手可能な配列アセンブリーにおける誤差を反映する可能性がある。
【0068】
フローサイトメトリーにより、Alsタンパク質の各々がその各S.cerevisiae宿主の表面で発現されたことが確認された。簡単に述べれば、Als構築物の各々についての細胞表面発現の確認は、2つの異なるポリクローナル抗Als抗血清を使用する間接的免疫蛍光を用いて決定した。抗血清Aは、Als1pの417−アミノ酸N末端断片でのウサギの免疫化によって生じた、抗Als1p抗体よりなるものであった。抗血清Bは、C.albicans細胞壁成分を認識するが、S.cerevisiae株(latron Laboratories)と交差反応しないウサギ抗C.albicansマンナン因子5であった。
【0069】
各株については、107出芽胞子を一晩培養から単離し、100μlのヤギ血清でブロックし、次いで、1:25希釈のポリクローナル抗血清AまたはBいずれかで染色し、続いて、1:100のフルオレセインイソチオシアネート−標識ヤギ抗ウサギIgGで染色した。488 nmにおいて発光するアルゴンレーザーを備えたFACSCaliber(Becton Dickinson)機器をフローサイトメトリー分析で用いた。蛍光発光は515/40−nmバンドバスフィルターで検出した。10,000事象についての蛍光を集め、ベースライン(すなわち、空のプラスミドで形質転換されたS.cerevisiae)を超える蛍光を持つ細胞の分布を、CELLQUESTソフトウェア(Becton Dickinson)を用いて各株について分析した。
【0070】
表IIに示すように、2つの区別される抗血清は、Alsp−発現株の全ては、空のプラスミドで形質転換されたS.cerevisiae と比較した場合に、蛍光の少なくとも4倍増加を呈したことを示した。Alsファミリーのメンバーの中の予測された構造多様性と合致して、抗血清は、個々のAls発現株の認識において差を呈した。
【0071】
(表II)
(フローサイトメトリー分析によるS.cerevisiaeの表面上のAlsタンパク質の検出)
ポリクローナル抗Als1p抗血清(A)またはポリクローナル抗C.albicans細胞壁抗血清(B)いずれかでの間接的免疫蛍光を用い、各株の出芽胞子を染色し、次いで、フローサイトメトリーを用いて分析した。結果は、括弧に入れた−倍増加と共に、バックグラウンド(空のプラスミドで形質転換されたS.cerevisiae)を超える陽性細胞のパーセンテージとして表す。
【0072】
【表2】
種々のAlsタンパク質を発現したS.cerevisiaeクローンを、種々の宿主基材に接着するそれらの能力について調べた。後に記載するように、結果は、Alsタンパク質が基材特異的接着の異なるプロフィールを呈することを示す。
【0073】
真菌接着アッセイを行って、形質転換されたS.cerevisiae株の接着特性を決定した。簡単に述べると、従前に記載された接着アッセイ(8)の修飾を以下のように使用した。ゼラチン(Sigma)、ラミニン(Sigma)、またはフィブロネクチン(Becton Dickinson)の0.01 mg/ml溶液の1mlを、6−ウェル組織培養プレート(Costar)の各ウェルに加え、37℃で一晩インキュベートすることによって、接着プレートを被覆した。内皮細胞については、第二継代細胞を、0.2%ゼラチンマトリックスを被覆した6−ウェル組織培養プレート中で密集するまで増幅させ、上皮細胞については、FaDU細胞を、0.1%フィブロネクチンマトリックスで被覆した6−ウェル組織培養プレート中で密集するまで増殖させた(3日)。接着テストの前に、ウェルを1mlの温かいハンクス平衡塩溶液(HBSS)で2回洗浄した。テストすべきS.cerevisiae株を30℃の最小規定培地中で一晩増殖させ、次いで、遠心によって収穫し、HBSS(Irvine Scientific)で洗浄し、ヘマサイトメーターを用いて数えた。300の生物を、目的の基材で被覆した6−ウェル組織培養プレートの各ウェルに加え、CO2中で37℃にて30分間インキュベートした。非接着性生物を、10mlのHBSSで標準化された方法にて2回洗浄することによって除去した。ウェルにYPD寒天(1%酵母エキス(Difco)、2%バクト−ペプトン(Difco)、2%D−グルコース、1.5%寒天)を重ね、接種物を定量的培養によって確認した。プレートを30℃にて48時間インキュベートし、コロニーをカウントした。接着は初期接種物のパーセンテージとして表した。接着の差は、単一因子偏差分析検定を用いて比較し、p<0.01で有意と考えられた。
【0074】
異なる基材へのS.cerevisiae形質転換体の接着プロフィールの顕著な差があった(図8)。Als1p−、Als3p−、およびAls5p−発現株はテストした全ての基材に結合したが、Als6p−発現S.cerevisiaeはゼラチンのみに接着し、およびAls9p−発現S.cerevisiaeはバックグラウンドレベルを超えてラミニンのみに接着した。さらに、種々の基材への接着に定量的差があった。例えば、Als3pと比較した場合、Als1pはより大きな接着をゼラチンに付与したが、上皮細胞にはより小さな接着を付与した(p<0.01、単一因子偏差分析)。Als7pを発現するS.cerevisiaeだけがテストした基材のいずれにも接着しなかった。Alsタンパク質発現のレベルの小さな差は表IIに示された免疫蛍光実験によって排除できないが、そのような差は本実験で見出された基材特異的結合パターンが原因であるようではない。細胞表面に発現されたAlsタンパク質の量のそのような全体的増加または減少は、基材を横切っての接着の釣り合った増加または減少を生じ、観察された基材特異的差をもたらすものではないと予測されるであろう。
【0075】
後に記載されるように、Alsタンパク質に対する基材結合特異性は、Alsタンパク質のN末端配列に存在する。簡単に述べれば、S.cerevisiaeにおけるAls5p発現は、ゼラチンおよび内皮細胞を含めた多数の基材へ接着を付与し、他方、Als6p発現はゼラチン単独への接着をもたらした。機能のこの顕著な差にかかわらず、Als5pおよびAls6pはアミノ酸レベルにおいて80%を超えて同一である。これらのタンパク質のタンデムリピートおよびC−末端部分は実質的に同一であり、配列の差の大部分はこれらの2つのタンパク質のN末端に集中している。これらのデータはN末端配列の変動性が基材に特異性を付与することを示す。
【0076】
前記した結果は、キメラALS5/ALS6構築物の接着表現型を決定する実験の結果によって裏付けられる。簡単に述べれば、キメラAls5/Als6タンパク質は、各タンパク質のN末端を交換することによって構築された。キメラALS5/6遺伝子は以下のように構築された。5’2117bpの遺伝子を含むALS5のBglII−HpaI断片を単離した。次いで、pGEM−T−ALS6をBglIIおよびHpaIで消化して、ALS6の対応する5’2126bpを放出させ、pGEM−T−Easy+ALS6の3’配列よりなる断片を単離し、5’ALS5断片に連結して、プラスミドpGEM−T−5N6Cを得た。対応するALS6の5’断片およびALS5の3’断片を用いる同一アプローチを用いて、プラスミドp−GEM−T−6N5Cを得た。配列確認の後、各キメラALS遺伝子をBglII−XhoI消化によって放出し、前記したようにpATH1にサブクローンした。次いで、S.cerevisiae S150−2Bをこれらの構築物で形質転換し、それらの接着特性の特徴付けの前に、発現をノーザンブロット分析によって確認した。
【0077】
Als6pのC末端へのAls5pのN末端のキメラ融合を発現するS.cerevisiaeは、Als5pと同様にゼラチンおよび内皮細胞双方に接着した(図9)。同様に、Als5pのC末端へのAls6のN末端のキメラ融合を発現する株は、Als6pを発現するS.cerevisiaeがそうであったように、ゼラチンのみに接着した(図9)。さらに、Als5pおよびキメラAls5N6Cタンパク質を発現する株は、フィブロネクチン−被覆ビーズを凝集させ、他方、Als6pおよびキメラAls6N5Cタンパク質を発現するものは、これらのビーズに対して親和性はほとんどまたは全く有しなかった。まとめると、これらのデータは、これらの形質転換されたS.cerevisiae株の接着プロフィールがAlsタンパク質のN末端部分によって支配されていたことを示す。
【0078】
Alsタンパク質ファミリーメンバーの間で示された基材特異性の差に加えて、他の生物学的機能の差も観察された。例えば、Alsタンパク質のサブセットは、S.cerevisiaeによる内皮細胞侵入を媒介することが示された。C.albicansはそれ自身のエンドサイトーシスを誘導することによって内皮細胞に侵入する(Fillerら,Infect Immune.63976−983(1995),Belangerら,Cell Microbiol.,in press(2002))。このエンドサイトーシスは、生物が内皮細胞に接着する後に起こる;しかしながら、このプロセスで必要とされるC.albicansリガンドは未知である。さらに、区別されるカンジダリガンドは接着およびエンドサイトーシス双方で必要であるかは明らかでない。非接着性であることに加え、S.cerevisiaeは内皮細胞による有意なエンドサイトーシスを受けない。従って、Alsタンパク質がインバシンならびにアドヘシンとして働くことができるか否かをテストするために、内皮細胞に侵入するAlsタンパク質を発現するS.cerevisiae株の能力を決定した。
【0079】
内皮細胞侵入を媒介するAlsタンパク質の能力は、従前に記載された差分蛍光アッセイの修飾を用いて決定された(Phanら,Infect.Immun.68:3485−3490(2000))。簡単に述べれば、内皮細胞を、フィブロネクチンで被覆された12−mm直径のカバーグラス上で密集するまで増殖させ、24−ウェル組織培養プレート(Corning)に入れた。次いで、細胞を、RPMI 1640培地(Irvine Scientific)中の各S.cerevisiae株の105出芽胞子で感染させた。陽性対照として、細胞を同様な数のC.albicans出芽胞子で感染させた。90分間のインキュベーションの後、細胞を0.5mlのHBSSで標準的な方法にて2回濯ぎ、3%パラホルムアルデヒドで固定した。内皮細胞の表面に接着したままである生物を、Alexa 568(Molecular Probes,Inc.,Eugene,OR)とコンジュゲートされている、ウサギ抗C.albicans抗血清(バイオデザイン)で1時間染色し、これは赤色の蛍光を発した。この抗血清は、2倍より高い希釈にてS.cerevisiaeと交差反応する。次いで、内皮細胞を、リン酸−緩衝化生理食塩水中の0.2%Triton X−100中で10分間浸透させ、その後、細胞−会合生物(内部化された+接着性の生物)を、Alexa488でコンジュゲートされた抗C.albicans抗血清で再度染色し、これは緑色の蛍光を発した。次いで、カバーグラスをエピ蛍光で観察した。内皮細胞によって内部化された生物の数を、(緑色の蛍光を発する)細胞−会合生物の数から(赤色の蛍光を発する)接着性生物の数を差し引くことによって決定した。少なくとも100の生物が各カバーグラス上でカウントされ、全ての実験は少なくとも3つの別々の状況で三連にて行った。
【0080】
フィブロネクチンビーズ接着アッセイを行って、特定のAlsタンパク質の結合特徴をさらに特徴付けた。この点に関し、Als5pは、S.cerevisiaeの表面で発現された場合に、フィブロネクチン−被覆ビーズの凝集を誘導するタンパク質の能力によって元来同定された(Gaurら,Infect.Immune.65:5289−5297(1997))。従って、フィブロネクチンのためにALS5,ALS6,5N6Cおよび6N5Cで形質転換されたS.cerevisiae株を、この方法を用いてビーズ接着についてテストした(Gaurら,Infect.Immune.65:5289−5297(1997),(Gaurら,Infect.Immun.67:6040−6047(1999))。簡単に述べれば、トシル化磁性ビーズ(Dynal Biotech)を製造業者の指示に従ってフィブロネクチンで被覆した。次に、10μlの被覆ビーズ(106ビーズ)を、1mlのS.cerevisiae 1 × Tris−EDTA(TE)緩衝液、pH7.0中の1×108形質転換S.cerevisiaeと混合し、温和に混合しつつ45分間インキュベートした。チューブを磁石中において、ビーズおよび接着性S.cerevisiaeを非接着性生物から分離した。非接着性生物を含有する上清を吸引によって取り出し、残存するビーズを1mlのTE緩衝液中に再懸濁させることによって3回洗浄し、続いて、磁気分離および上清の吸引を行った。最後に、洗浄されたビーズおよび接着性生物を100μlのTE緩衝液に再懸濁させ、共−凝集について顕微鏡で調べた。
【0081】
結果は、Als1p Als3pおよびAls5pを発現するS.cerevisiaeが、細胞−会合生物のパーセンテージの有意な増加を呈することを示し、これは、内皮細胞に接着するそれらの能力を反映する。加えて、Als3p、および程度は低いが、Als1pおよびAls5pを発現する生物は有意な内皮細胞侵入を示した(図10)。
【0082】
前記した機能試験に加えて、Alsタンパク質は、免疫グロブリンスーパーファミリーのアドヘシンおよびインバシンに相同であることも判明した。Alsタンパク質の分子モデリングにおける初期工程として、知識に基づくサーチアルゴリズムを用いて、Alsファミリーメンバーと有意な構造的同様性を共有する分子を同定した。簡単に述べれば、相同性およびエネルギー−ベースのモデリングを行って、Alsタンパク質の総じての物理化学的特徴を比較した。まず、知識に基づく方法(SWISS−MODEL)(Guexら,Electrophoresis 18:2714−2723(1997),Schwedeら,Nucleic Acids Res.31:3381−3385(2003))を用いて、相同な立体配座を持つタンパク質についてのSwissおよびBrookhavenタンパク質データベースにおいて構造のコンビナトーリアル拡張構造配列を分析し、比較した(Shindyalovら,Protein Eng.11:739−747(1998))。このアプローチは、ExNRL−3Dデータベースにおいて一次配列同様性についてサーチするためのBLASTP2アルゴリズム(Altschulら,Mol.Biol.215:403−410(1990))を含んだ。平行して、動的配列整列アルゴリズムSIM(Huangら,Adv.Appl.Math.12:337−367(1991))を用いて、最大の配列同一性を持つ候補鋳型を選択した。引き続いて、ProModllを用いて、一次および洗練されたマッチ分析を行った。得られたタンパク質を、Alsタンパク質骨格軌跡の相同性モデリングのための鋳型として用いた。
【0083】
Alsタンパク質のN末端ドメイン(例えば、アミノ酸1から480;初期タンデムリピートに先行)の頑強なモデルを、相補性アプローチを介して作り出した。Alsタンパク質のN末端ドメインを、Silicon Graphicsワークステーション(SGI,Inc.)でオペレートするSYBYL 6.9.1ソフトウェア(Tripos Associates)を用いて配列相同性(Composer(Tophamら,Biochem.Soc.Symp.57:1−9(1990))およびネジ処理方法(Matchmaker (Godzikら,J.Mol.Biol.227:227−238(1992))および遺伝子−折畳(Jaroszewskiら,Protein Sci.7:1431−1440(1998),Godzikら、Protein Eng.8:409−416(1995),Godzikら,Proc.Natl.Acad.Sci.U.S.A.89:12098−12102(1992),Godzikら,J.Comput.Aided Mol.Des.7:397−438(1993))によって推定溶液立体配座に変換した。得られたコンフォーマーおよび標的Alsドメインのアミノ酸側鎖を分子ダイナミックスによって洗練し、AMBER95強制場方法(Duanら,J.Comput.Chem.24:1999−2012(2003))およびPowellミニマイザー(Powellら,Math.Program 12:241−254(1977))を用いて株エネルギーを最小化した。
【0084】
これらのアプローチは、ペプチド骨格原子の位置が固定された側鎖相互作用を最適化する。好ましい立体配座は、水性溶媒中の拡大された分子ダイナミックスから決定された。次に、全てのペプチド結合のねじれ角を最小の拘束にて180±15°に調整した。いくつかの場合において、拘束なしで、またはα−ラセン領域を0.4−kJペナルティーをカノニカルラマチャンドラン(Ramachandran)φおよびΨ角度に適用することによって拘束して、分子ダイナミックスを実行した。最終全エネルギー最小化を、全ての拘束および凝集体の除去の後に各モデルについて行った。得られたAlsのN末端ドメインモデルは3つの基準:(i)最も好都合な歪みエネルギー(分子メカニックス);(ii)経験的位置エネルギー関数;および(iii)潜在的ジスルフィドブリッチングの空間的配置の保存に基づいて優先させた(Godzikら,J.Mol.Biol.227:227−238(1992),Bowieら,Science 253:164−170(1991),Eisenbergら,Methods Enzymol.227:396−404(1997),Fischerら,FASEB J.10:126−136(1996),Luthyら,Nature 356:83−85(1992))。標準的な尺度を用いる相同性鋳型に対する関係においてAlsモデルを確証のために評価した(e−値(Welchら,Biochemistry 35:7165−7173(1996),Welchら,Biochemistry 33:6074−6085(1994))。最後に、Alsモデルの物理化学的特性を、物理的特性がAlsのN末端ドメインの水接近可能表面へ投影されるように、SYBYLおよびHINTプラットフォーム(Kellogら,J.Comput.Aided Mol.Des.5:545−552(1991))において実行して、MOLCAD(Heidenら,J.Comput.Chem.14:246−250(1993))によって可視化した。
【0085】
これらのモデルは、全てのAlsタンパク質のN末端ドメインが、免疫グロブリンスーパーファミリーのメンバーと合致して、多数の反−平行β−シートドメインを含有することを示す。結果は表IIIにて以下にまとめる。これらのタンパク質は、典型的には、それからループ/コイル構造が投影される、複雑な7−ストランド反−平行β−シートドメインよりなる。β−シートドメインは間に存在する領域によって相互から分離される。この構造は、しばしば、ビーズ−オン−ア−ストリングモチーフという。特に注意すべきは、Alsタンパク質の実質的に全てが公知のアドヘシンまたはインバシンホモログに対してモデル化されたことである(表III)。同様性の異なるパターンが分析したAlsタンパク質内で観察された。例えば、Als7p以外の調べた全てのAlsタンパク質は、黄色ブドウ球菌のコラーゲン−結合タンパク質との有意な相同性を共に有した。しかしながら、特異的な一次、二次および三次ホモログはほとんどのファミリーメンバーについて変化した(表III)。例えば、Als2pおよびAls9pは同一の一次、二次、および三次ホモログを共有した。
【0086】
(表III)
(Alsタンパク質間のホモログの比較)
各Alsタンパク質のホモログは記載された知識に基づくアルゴリズムによって同定し、1から3の構造相関の下降順でランク付けした。NS、相同性モデリングについて有意なモデルは同定されなかった(相関係数((r2)≦70%。PDB、バイオテクノロジー情報様式でのNational Centerでのタンパク質データーバンクコード(Protein Data Bank)。
【0087】
【表3−1】
【0088】
【表3−2】
Alsタンパク質は、予測されたループ/コイル構造にマップされるN末端超可変領域を含有するとも決定された。この点に関し、個々のAlsタンパク質によって媒介される基材特異的接着の観察された差にも拘わらず、N末端ドメインにおける大きな配列の領域はこのファミリーを横切って保存されている。しかしながら、超可変領域(HVR)1から7と命名されたAlsタンパク質内の有意な多様性の7つの領域が見出された。(8以上のアミノ酸から構成される)これらの領域は、Alsタンパク質を横切っての明らかなコンセンサス同一性は含まず、かつ50%未満のコンセンサス保存を含んだ。対照的に、介入する保存領域(CR)1から7は、Alsタンパク質を横切って30%を超えるコンセンサス同一性、および50%を超えるコンセンサス保存を呈した。既知の機能を持つAlsタンパク質のN末端ドメイン(残基1から420)を含むこれらのアミノ酸配列の同一性プロットおよび模式的整列を図11、AおよびBに掲げる。特に、相同性モデリングは、異なるAlsタンパク質のHVRが、配列は識別可能でありつつ、CRのβ−シート成分から投影される同様なループ/コイル構造に適合すると予測されることを明らかとした。かくして、これらの保存されたHVRの存在は、それらが宿主構成要素と相互作用するのに入手可能であることを示す。
【0089】
前記した相同性モデリングおよび関連決定に加えて、経験的決定は、加えて、Als1pのN末端ドメインの予測される構造を確認する。我々の相同性モデリングによって作り出される仮説を検定するために、CDおよびFTIR分光分析の補充的なアプローチを用い、Als1pのN末端ドメインの構造的特徴を決定した。Als1pのアミノ酸17から432を含むこのタンパク質はS.cerevisiaeで生産され、Fuら,Molecular Microbiology,44:61−72(2002)によって従前に記載されている。
【0090】
簡単に述べれば、円二色スペクトルを、熱電気温度コントローラーを備えたAVIV62DSスペクトロポラリメーター(Aviv Biomedical Inc.)で記録した。0.1−mm光路脱着可能水晶セルを用い、Als1pの水溶液(リン酸−緩衝化生理食塩水中10μM)を260から185nmまで10nm/分の速度および0.2nmの試料間隔でスキャンした。ペプチドを欠く緩衝液からのスペクトルを試料溶液から差し引いて、光散乱人工物を最小化し、最終スペクトルは25℃で記録した8スキャンの平均であった。該機器は(+)−10−ショウノウスルホン酸(1−mm路長セル中1mg/ml)(Johnsonら,Proteins 7:205−214(1990))でルーチン的にキャリブレートされており、楕円度は平均残基楕円度(1)MRE(度−cm2 dmol−1)として表した。タンパク質濃度は、発現されたAls1pドメインの芳香族アミノ酸組成に基づく280nmにおける吸収によって決定された(Paceら,Protein Sci 4:2411−2423(1995))。CDスペクトルは、インターネットに基づくDichroweb(Lobleyら,Bioinformatics 18:211−212(2002))インターフェース(cryst.bbk.ac.uk/cdweb/html/home.html)を介してSelcon(Sreeramaら,Protein Sci.8:370−380(1999))を用い、ラセン、β−シート、ターン、および乱れた構造に分割した。
【0091】
Als1p自己−フィルムの赤外スペクトルは、4の利得の、256スキャンにわたって平均され、かつ2cm−1の分解能にて、ジューテリウムトリグリシン硫酸ディテクターを備えたBruker Vector 22 FTIRスペクトロメーター(Bruker Optics)で25℃にて記録した。50μlのリン酸−緩衝化生理食塩水中の50マイクログラムのタンパク質を、50×20×2−mmゲルマニウム減衰全反射試料結晶(Pike Technologies)の表面に広げ、乾燥させた。次いで、赤外スペクトルを記録するに先立って、乾燥タンパク質自己−フィルムをD2Oで1時間水和させた。赤外スペクトルのアミドIバンドを、曲線−フィッティングソフトウェア(GRAMS/32,Version 5;Galactic)を備えた成分ピークの面積計算によって二次立体配座について分析した。種々の立体配座についての周波数限界は以下の通りであった:α−ラセン(1662−1645 cm−1)、β−シート(1637−1613および1710−1682 cm−1)、β−ターンループ(1682−1662 cm−1)、および乱れた構造(1645−1637 cm−1)(50−52)。
【0092】
Als1pのN末端ドメインの円二色結果を図12Aに示し、これは、217nmにおける二色最小、および200nm近くの強い正の二色最大を明らかとする。これらの特徴は、支配的な反−平行βシート成分を有するタンパク質の特徴である。CDスペクトルの分解は、タンパク質は50.1%βシートの立体配座を採り、他方、他の構造クラス寄与は乱れた構造(26.9%)、ターン構造(19.3%)、およびα−ラセン(3.7%)を含む。
【0093】
図12Bに示すように、水和したAls1pの自己−フィルムのFTIR測定により、試料が支配的なβ−シート立体配座を有することが強く確認された。これらのスペクトルにより、1634および1628cm1に中心があるピークを持つ強い低周波数アミドIバンド、および1685cm−1に中心がある弱い高周波数バンドが明らかにされた。タンパク質アミドI赤外スペクトルの高および低周波数成分へのこの周波数分裂は、分子間反−平行β−シートの間の転移二極結合の効果に典型的であることが示されている(Halversonら,J.Am.Chem.Soc.113:6701−6703(1991))。スペクトルの曲線フィッティングは、タンパク質構築物が〜57.2%反平行β−シートであることを示した。IRスペクトルの曲線フィッティングからの他の二次構造立体配座は、乱れた構造(20.5%)、ターン成分(13.3%)、およびα−ラセン(9.0%)を含む。
【0094】
一緒にすると、FTIRおよびCDデータは、Als1pのN末端が、構造度が低い領域が中間に存在する、従たるα−ラセンおよびターン成分を含有する反−平行β−シート構造の支配的なドメインを含有することをさらに裏付ける。
【0095】
三次元モデルは、さらに、AlsのN末端ドメインの間の物理化学的区別を示す。この点に関し、分子モデルは、宿主細胞およびいくつかの基材とのそれらの相互作用に影響するようであるAlsタンパク質のN末端ドメインの予測された物理化学的属性の差を示した。図13に示すように、Alsタンパク質は、疎水性、電荷、および水素結合ポテンシャルの表面分布に基づいて3つの区別される群に分離可能である。Als1p、Als3pおよびAls5pは、各々、これらの特性の同様なパターンを共に有し、かくして、Als群Aと考えられる。対照的に、Als6pおよびAls7pのN末端ドメイン(Als群B)の予測された物理化学的特性は、Als群Aのそれからの顕著な差を有する(図13)。Als群Aのメンバーにおけるカチオンポテンシャルは、典型的には、それらの中性またはアニオン面から分離されるが、正の電荷はAls群BのメンバーAls6pおよびAls7pの全表面を横切って広く分布している。最後に、Als2p、Als4p、およびAls9pのN末端は、Als群AまたはBタンパク質いずれかとは構造的に異なるAlsタンパク質の第三の群(Als群C)を構成するようである。Als群Cタンパク質は、疎水性または静電分布の点でAls群Bタンパク質よりもAls群Aにより同様なように見えるであろう。
【0096】
接着機能を持ついくつかのタンパク質はC.albicansにおいて同定されている。Hwp1pは、哺乳動物トランスグルタミナーゼに対する基質として作用することによって、頬上皮細胞への接着を媒介することが示されている(5)。EAP1は、最近、S.cerevisiaeにおける異種発現によって同定されており、ポリスチレンおよび腎臓上皮細胞への接着をインビトロで媒介する(7)。Alsタンパク質ファミリーの8つのメンバーのうち、Als1pおよびAls5pのみが機能的見込みから研究されている。Als1pの異種発現は、ヒト血管内皮細胞および上皮細胞への結合を媒介することが示されており、遺伝子分布実験を介してC.albicansにおいて確認されているという知見である(Fuら,Mol.Microbiol.44:61−72(2002),Fuら,Infect.Immune.66:1783−1786(1998))。S.cerevisiaeにおけるALS5の異種発現は接着性をコラーゲン、フィブロネクチン、ウシ血清アルブミン、およびラミニンに付与する(Gaurら,Infect.Immune.65:5289−5297(1997),Gaurら,Infect.Immun.67:6040−6047(1999),Gaurら,Cell Commun.Adhes.9:45−57(2002))。C.albicansのアドヘシンの基材特異性の大規模な比較は行われたことがない。本実験において、我々は、Alsタンパク質ファミリーメンバーの構造的に多様な群の接着特性を比較した。我々のデータは、Alsタンパク質が、種々のヒト基材への接着に対する特異性の重複するスペクトルを持つ表面タンパク質の多様なファミリーを含むことを示す(図8)。さらに、本ドメイン交換実験からの結果は、Alsタンパク質のN末端ドメインが、それらの基材接着性プロフィールの特性を付与することを示す。
【0097】
接着性を媒介するに加え、我々のデータは、Alsタンパク質はインバシンとしても機能できることを示唆する。興味あることに、Als1pおよびAls3p双方を発現するS.cerevisiaeは同様な内皮細胞接着を示したが、Als3p−発現S.cerevisiaeはかなり高い速度での内部化を受けた。これらの結果は、エンドサイトーシスが接着の延長であるばかりか、むしろ、リガンド−受容体相互作用によって影響され得る区別されるプロセスであることを示す。Alsタンパク質中のN末端配列の差は、接着で当てはまるように、これらの区別される機能を媒介するようである。
【0098】
三次元空間に分布したタンパク質ドメインの物理化学的特性は、受容体−リガンド相互作用を支配する非常に重要な構造的特徴である(Eisenbergら,J.Mol.Biol.179:125−142(1984),Waringら,Protein Peptidew Lett.3:177−184(1996),Hancockら,Lancet349:418−422(1997))。Alsタンパク質は、免疫グロブリンスーパーファミリーの他のアドヘシンおよびインバシンに特徴的な立体配座特徴を共有する。しかしながら、ここのAlsタンパク質はそれらの一次ホモログが異なり、これは、Alsファミリーのメンバーが異なる基材結合プロフィールを呈することを示す実験データと合致する知見である。まとめると、Als相同性のこれらのパターンは、Alsタンパク質メンバーは構造および予測された折畳において全体的同様性を共有するが、機能のそれらの差の原因である区別されるAlsタンパク質の間で構造的差が存在することを示す。
【0099】
Alsファミリーメンバー構造決定に関する前記した結果は、Alsp1pのN末端領域が、少量の比較的構造化されていない領域と共に、ループ/コイル構造を含有する反−平行β−シートドメインから圧倒的に構成されることを示す相同性モデリングを裏付ける。これらの特徴は、免疫グロブリンスーパーファミリーのメンバーのモチーフを示す。これらの結果は、Als5pの円二色実験との有意な予測的裏付け(Hoyerら,Yeast 18:49−60(2001))を示し、これは、Als5pのN末端が反−平行β−シートおよびループ/コイル領域の相対的圧倒性によって特徴付けられることを示す。かくして、Alsタンパク質ファミリーの全てのメンバーはこの総じての構造をかなり呈するようである。特に、前記した構造の結果もまた、HVRの多くが区別されるAlsタンパク質のN末端中のβ−シートドメインから投影するフレキシブルなループ/コイル構造に対応することを示す相同性モデルとやはり合致する。まとめると、これらの結果は、これらの構造がAlsタンパク質による基材特異的結合に一体化されることを示す(図14)。前記した結果と合致して、マンノース−結合レクチンの類似領域、α−アグルチニン、および免疫グロブリンスーパーファミリーの他のメンバーが、基材結合特異性を付与するようである(Zhaoら,Hybrid Hybridomics21:25−36(2002),Wojciechowiczら,Mol.Cell.Biol.13:2554−2563(1993))。さらに、これらの可変ループ領域の突然変異が、これらの相同タンパク質において基材結合を有意に改変する(Renzら,J.Cell Biol.125:1395−1406(1994),Vineyら,J.Immunol.157:2488−2497(1996))。
【0100】
三次元モデリングの結果は、さらに、個々のAlsタンパク質のN末端ドメインが、それらの接着プロフィールに関連する区別される分子シグニチュアを保有することを示す。これらのシグニチュアは、Alsタンパク質の中で構造的関係を区別する立体配置を生じる、表面領域、疎水性、および静電気電荷のようなパラメーターを取り込む。例えば、Als群Aメンバー(Als1p,Als3pおよびAls5p)のような多数の基材に結合するAlsタンパク質が、立体バルク、疎水性分布、および静電ポテンシャルの点で同様な予測されるL−末端プロフィールを有する。なお、この群内においてさえ、該群内の機能的差を支配することができる特異的物理化学的区別が存在する(図13)。対照的に、低下した接着能力を持つAlsタンパク質は、疎水性および静電ポテンシャルを含めた、多数の物理化学的特性においてAls群Aタンパク質から区別されると予測される表面特徴を有する。これらの構造的特徴の差の集合効果は、区別されるAlsタンパク質の特異的機能的特性をかなり付与するようである。
【0101】
広範な遺伝的変動性がALS遺伝子ファミリー内で示されているC.albicansの異なる単離体の特異的ALS遺伝子における配列変動が観察されており(Zhangら,Genome Res.13:2005−2017(2003),Hoyerら,Yeast18:49−60(2001))、ALSファミリーの全てのメンバーが全ての単離体に存在するのではない。単一単離体における2つの異なる対立遺伝子の間の有意な配列多様性さえ見出されている(Zhaoら,Microbiology149:2947−2960(2003),Zhangら,Genome Res.13:2005−2017(2003))。この程度の遺伝的変動性は、これらのタンパク質が比較的高い周波数において再編成または突然変異を受け得ることを示唆する。そのようなメカニズムは、本実験において示された高度な構造的および機能的多様性を生じさせる能力を生物に供するであろう。この仮説に対する間接的裏付けは、この遺伝子が共に高突然変異性であり、これらの突然変異は選択圧に従うという双方を示唆したALS7の対立遺伝子変動の最近の研究によって供される(Zhangら,Genome Res.13:2005−2017(2003))。
【0102】
まとめると、前記結果は、構造および機能的レベルの双方において抗体およびAlsタンパク質の間の類似性を示す。例えば、相同性モデリングは、これらのファミリーの構造的立体配置における同様性を過小評価しており、過剰変動はそうでなければ安定なフレームワーク内の局所化ドメインに標的化されている(例えば、Alsタンパク質のHVRおよび免疫グロブリンにおけるFab領域)。さらに、抗体に関しては、ALS遺伝子ファミリーの遺伝的変動性は、Candidaが、接着および侵入における特異性のスペクトルを持つタンパク質の多様なアレイを呈する機会を供するであろう。関連タンパク質のそのような群の入手可能性は、感染の間に異なる解剖学的および生理学的ニッチをコロニー化し、およびそれに侵入する生物の能力を改良するようである。
【0103】
本出願を通じて、種々の刊行物を括弧内に入れて引用した。これらの刊行物の開示の全体をここに引用して本明細書中に組込んで、本発明が属する分野の最先端技術を十分に記載する。
【0104】
本発明を開示された実施形態を参照して記載してきたが、当業者であれば、具体的実施例および前記した詳細な実験は本発明を説明するだけであることを容易に認識するであろう。本発明の精神を逸脱することなく種々の修飾を成すことができることは理解されるべきである。従って、本発明は以下の請求の範囲によってのみ限定される。
【0105】
(実施例VII)
(細胞−媒介の液性でない免疫を高めることによって、rAls1p−Nでのワクチン接種はマウス播種性カンジダ症の間の生存を改良する)
本実施例は、Als1pの組換えN末端(rAls1p−N)でのBALB/cマウスの免疫化が、C.albicansの致死接種物での引き続いてのチャレンジの間に生存を改良したことを示す。保護的20μg用量のrAls1p−NはTh1脾臓細胞のCandida−刺激を有意に増加させ、インビボ遅延型過敏を増加させた。対照的に、抗体力価は保護に相関しなかった。最後に、ワクチンはT細胞−欠乏マウスにおいて保護的ではなかったが、B細胞−欠乏マウスにおいては保護的であった。これらのデータは、rAls1p−Nワクチンの作用メカニズムが、C.albicansに対する液性よりはむしろ細胞媒介免疫の刺激であることを示す。
【0106】
該実験で用いたC.albicansはSC5314、すなわち、W.Fonzi(Georgetown University)によって供給された動物モデルにおいて高度にビルレントなよく特徴付けられた臨床単離体であった(Spellbergら,Infect Immun.71:5756−5764(2003))。該生物は、感染に先立って酵母ペプトンデキストロースブロス(Difco)において3回系列的に継代された。
【0107】
該実験で用いたマウス株はNational Cancer Institute(Bethesda,MD)から入手した雌BALB/cマウスであった。ワクチン効力に対する年齢のインパクトを探求するために、若年性マウス(8から10週)およびリタイアしたブリーダー(≧6月)の双方を利用した。igh遺伝子座のホモ接合性欠失を担う雌B細胞−欠乏マウス(C.129B6−IgH−Jhdtm1Dhu)、T細胞−欠乏ヌードマウス(C.Cg/AnBomTac−FoxnlnuN20)、およびコンジェニック野生型BALB/c同腹子はTaconic農場(Germantown,NY)から入手した。マウスを、照射した食物およびオートクレーブ処理した水を自由に備えた濾過ケイジに収容した。生存実験では、マウスを種々の用量の抗原(後記参照)で免疫化し、引き続いて、C.albicans SC5314出芽胞子の適当な接種物、またはPBS(Irvine Scientific,Irvine,CA)対照で尾静脈を介して感染させた。もし個々のデータベースが統計学的不均一を示さなければ、複連生存実験の結果を合わせた(後記参照)。マウスに関連する全ての手法は、動物収容および看護のためのNational Institutes of Health指針に従い、制度的動物使用および看護委員会によって認可された。
【0108】
以下に記載されたrAls1p−N免疫化手法は以下のように行った。簡単に述べれば、rAls1p−N(Als1pのアミノ酸17から432)はS.cerevisiaeにおいて生産され、ゲル濾過およびNi−NTAマトリックスアフィニティー精製によって精製した(Fuら,Molec.Microbiol.44:61−72(2002))。タンパク質の量は修飾されたLowryアッセイによって定量した。高度な純度(約90%)が、前記したような、SDS−ポリアクリルアミドゲル電気泳動ならびに円二色性およびFTIRによって確認された。マウスは、0日目にフロイントの完全アジュバント(CFA−Sigma−Aldrich)と混合されたrAls1p−Nの腹腔内(ip)注射によって免疫化し、21日目にフロイントの不完全アジュバント(IFA,Sigma−Aldrich)を含むもう1つの用量の抗原でブースター注射し、次いで、ブーストから2週間後に感染した。
【0109】
得られた抗体力価は96ウェルプレートにてELISAによって決定した。簡単に述べれば、ウェルをPBS中の5μg/ml rAls1p−Nのウェル当たり100μlで被覆した。3%ウシ血清アルブミンを含有するTris緩衝化生理食塩水(TBS)(0.01 MTrisHCI,pH7.4,0.15 M NaCl)でのブロッキング工程に続いて、マウス血清を室温にて1時間インキュベートした。ウェルを、0.05%Tween20を含有するTBSで3回洗浄し、続いて、TBSでさらに3回洗浄した。ホースラディッシュペルオキシダーゼ(Sigma)とコンジュゲートさせたヤギ抗マウス2次抗体を1:5000の最終希釈にて加え、プレートを室温にて1時間さらにインキュベートした。ウェルをTBSで洗浄し、0.1Mクエン酸緩衝液(pH5.0)、50mg/mlのo−フェニレンジアミン(Sigma)、および10μlの30%H2O2を含有する基質と共にインキュベートした。色を30分間で発色させ、その後、10%H2SO4を加えることによって反応を停止させ、光学密度(OD)をマイクロタイタープレートリーダーにて490nmで決定した。陰性対照ウェルは希釈剤のみを受け、バックグラウンド吸収をテストウェルから差し引いて、最終OD読みを得た。ELISA力価は、正のOD読みを与えた最後の血清希釈の逆数を取った(すなわち、≧陰性対照試料の平均OD+2標準偏差)。
【0110】
以下に記載した他の方法は以下のように行った。簡単に述べると、C.albicans−誘導サイトカインプロフィールを行って、細胞−媒介免疫性およびインビボサイトカインプロフィールに対するrAls1p−Nワクチンの効果を決定した。マウスを前記したように免疫化した。最終ブーストから2週間後に、脾臓細胞を収穫し、従前に記載されているように4×106細胞/mlの密度にて完全培地中で培養した(Spellbergら,Infect.Immun.71:5756−5764(2003))。サイトカイン生産を刺激するために、脾臓細胞を熱で殺したC.albicans SC5314芽菅と共に共培養した。我々は、rAls1p−Nの代わりに加熱して殺したC.albicansを用いて、脾臓細胞を刺激して、感染の間にインビボ状況を模倣した。C.albicans細胞を、グルタミンを含むRPMI−1640(Gibco BRL)中で予め90分間発芽させて、Als1pの発現を誘導した(Fuら,Molec.Microbiol.44:61−72(2002))。得られたC.albicans芽管を60℃における90分間のインキュベーションによって加熱により殺した(Ibrahimら,Infect.Immun.63:4368−74(1995))。加熱して殺した真菌を2×107偽菌糸/mlの密度にて脾臓培養に加えた(1白血球に対する5真菌の比率)。48時間後に、従前に記載されているように、脾臓細胞を、細胞内サイトカイン検出およびフローサイトメトリーによってTh1(CD4+IFN−” +IL−4−)、Th2(CD4+IFN− −IL−4+)、またはCD4+IL−10+周波数についてプロファイリングした(Spellbergら, Infect. Immun. 71:5756−5764(2003))。3−色フローサイトメトリーは、製造業者の推奨に従ってFACSCompソフトウェアを用い、CaliBRITEビーズ(Becton Dickinson,San Jose,CA)でキャリブレートしたBecton−Dickson FACScan機器で行った。データ獲得の間、CD4+リンパ球は前方および側方散乱の連結、およびFITC−抗CD4抗体蛍光特性によって開閉した。各試料についてのデータは、10,000のCD4+リンパ球を分析するまで獲得した。結果は、Th1またはTh2細胞であった全ての開閉したリンパ球のパーセンテージのメジアン±25および75四分位として示す。
【0111】
足蹠膨潤はOomuraらの方法によって決定した(41)。簡単に述べれば、マウスを前記したように適当な用量のrAls1p−NまたはCFA単独で免疫化した。ブーストから二週間後に、免疫化マウスのベースライン足蹠サイズを、電子デジタルキャリパーを用いて測定した。25μlのPBS中の50μgのrAls1p−Nを右側足蹠に注射し、PBS単独は免疫化マウスの左側足蹠に注射した。24時間後に、足蹠を再度測定した。抗原−特異的足蹠膨潤を:(24時間における右側足蹠厚み−ベースラインにおける右側足蹠厚み)−(24時間後における左側足蹠厚み−ベースラインにおける左側足蹠厚み)として計算した。
【0112】
非パラメーターLog順位テストを利用して、マウスの生存時間の差を決定した。抗体の力価、Th1またはTh2リンパ球の頻度、および足蹠膨潤を、適切には、非パラメーター多数比較のためにSteel検定(Rhyneら, Biometrics 23:539−49(1967))、および不対比較のためにMann Whitney U検定によって比較した。相関はSpearman順位和検定で計算した。不均一性が複連生存試験で存在するかを決定するために、Kolmogorov−Smirnov検定を利用した。P値<0.05は有意と考えられた。
【0113】
rAls1p−N免疫原の最も有効な用量を決定するために、107−倍用量範囲を評価した(マウス当たり20pgから200μg)。雌のリタイアしたブリーダーBALB/cマウスをrAls1p−N+アジュバント(CFA/IFA)またはアジュバント単独で免疫化した。免疫化したマウスをブースティングから二週間後に採血して、抗rAls1p−N抗体力価を決定した(後記参照)。マウスを、引き続いて、C.albicans(2×105出芽胞子)の致死接種物で感染させた。反復実験からの生存データを合わせた。というのは、個々の実験は統計学的不均一性を示さなかったからである(Kolmogorov−Smirnov検定によるとp.>0.05)。20μg用量のrAls1p−Nは、感染マウスの25%の長期生存、およびアジュバント単独と比較した全生存の有意な増加(Log順位検定によるとp=0.044、図1)をもたらした。10倍高い(図15)または低い(データは示さず)用量いずれも、アジュバント単独と比較して生存を有意に増加させなかった。この結果は、中程度の用量のrAls1p−Nワクチンがネズミ播種性カンジダ症に対して保護を誘導することを示す。
【0114】
前記知見はrAls1p−Nワクチンについての保護的用量を確立した。次に、ワクチンの有効性を、106出芽胞子で感染させたより迅速に致死的なマウスのモデルにおいて評価した(各々、メジアン生存3 vs.11日、ワクチン接種されていないマウスにおいて106 vs.2×105接種物)。再度、反復実験からのデータを組み合わせた。というのは、個々の実験の結果は統計学的不均一性を示さなかったからである(Kolmogorov−Smirnov検定によるとp>0.05)。106C.albicans出芽胞子で感染させたBalb/cマウスに20μg用量+CFAとして投与した場合、rAls1p−Nワクチンはメジアン生存の2倍を超え、その結果、全生存の有意な増大−vs−ワクチン接種していない対照をもたらした(Log順位テストによるとp=0.001、図16A)。マウスの年齢がrAls1p−Nワクチンに対するそれらの応答に影響するかを決定するために、我々は若年マウスにおいてそれをテストした。若年マウスをワクチン接種し、同一の高い接種物で感染させた場合、同様な生存利益が見出された(Log順位検定によるとp=0.02、図16B)。
【0115】
200μg用量のrAls1p−Nの結果、20μg用量と比較して劣った保護がもたらされたが、200μg用量の抗原のみが血清抗Als1p抗体力価の有意な増大を誘導した(200μg用量 vs.全ての他の群についてp≦0.005、図17)。抗Als1p抗体力価の有意な増加は、中程度の保護的抗原用量で検出されなかった(20μg vs.アジュバントについてp=0.1)。個々のマウスの血清抗Als1p抗体力価を各マウスの生存時間に対してプロットすると、抗体力価および生存の間で相関は見出されなかった(R2=0.03、Spearman順位和検定によるとp>0.05)。事実、最高用量の抗原(200μg)で免疫化したマウスは、1:100,000過剰の抗rAls1p−N抗体力価を有したが、その力価が検出の下限である(〜1:100)抗原のより低い用量で免疫化したマウスから異ならない生存持続を有した。これらの結果は、rAls1p−Nワクチンによって誘導された保護が抗体力価に相関するように見えないことを示す。
【0116】
液性免疫はrAls1p−N−誘導保護と相関しなかったので、我々は、保護および非保護用量のrAls1p−Nによって誘導された細胞−媒介免疫応答を調べた。マウスを、前記したように、0.2、20、または200μgのrAls1p−N、またはアジュバント単独で免疫化した。ブーストから2週間後に、脾臓細胞を収穫し、Als1pを発現することが知られている、熱で殺した予め発芽させたC.albicansの存在下で培養した(Fuら,Molec.Microbiol.44:61−72(2002))。培養から48時間後に、脾臓細胞をフローサイトメトリーによって細胞内サイトカイン検出のために収穫した。保護的20μg用量の抗原で免疫化したマウスからのリンパ球のみが、アジュバント単独を与えられたマウスと比較してTh1細胞の有意に増加した頻度を生じた(P=0.03、図18)。Th2の頻度(図18)およびIL−10+Tリンパ球の頻度(データは示さず)の有意な差は、アジュバントまたは該用量のいずれかの抗原で免疫化されたマウスの間で検出されなかった。
【0117】
タイプ1免疫がインビボでrAls1p−Nによって刺激されたことを確認するために、足蹠膨潤によって遅延タイプ過敏をテストした。保護的20μg用量のrAls1p−Nでワクチン接種したマウスのみが、対照と比較して有意に増大した遅延型過敏反応を発生し、この応答は、非保護的0.2および200μg用量によって誘導されたものよりもやはり有意に高かった(図19、非パラメーターSteel検定によると、全ての比較−vs−20μg用量についてp<0.05)。まとめると、これらの結果は、保護的用量のrAls1p−N抗原が有意なTh1偏光および遅延型過敏反応を誘導したことを示す。
【0118】
ワクチン−媒介保護における抗体およびt−細胞の役割を規定するために、B細胞欠乏、T細胞欠乏ヌード、またはコンジェニックBALB/c野生型対照マウスを20μgのrAls1p−Nの+アジュバントまたはアジュバント単独で免疫化し、C.albicansの致死接種物(8×105出芽胞子)で感染させた。B細胞−欠乏マウスは感染に対してより耐性である傾向があり、他方、T細胞−欠乏マウスはアジュバントのみが与えられた野生型対照マウスよりも感受性であった(各々、B細胞欠乏およびT細胞欠乏マウス−vs−野生型アジュバント−処理についてp=0.065および0.01、図20)。最後に、rAls1p−NワクチンはB細胞−欠乏マウスにおいてその効果を維持した(rAls1p−Nワクチン接種−vs−アジュバント単独についてp=0.04、図6)が、T細胞−欠乏マウスにおいて効果的でなかった(rAls1p−Nワクチン接種−vs−アジュバント単独についてp=0.4、図20)。これらの結果は、Als1pワクチンがB細胞欠乏マウスにおいて効果的であったがT細胞欠乏ヌードマウスにおいてはそうではなかったことを示す。
【0119】
前記したのは、このタンパク質のN末端での免疫化が、引き続いての血行的に播種性のカンジダ症の間に若年性および成熟BALB/cマウス双方の生存を改良したことを示す結果である。特に、中間用量のrAls1p−N(20 μg)は、より低い用量およびより高い用量(200μg)双方と比較して優れた保護を供した。それにも拘わらず、非保護的200μg用量のrAls1p−Nは免疫原性であった。というのは、それは保護的20μg用量よりも100倍高い抗体の力価を誘導したからである。
【0120】
最高用量のrAls1p−Nにおけるより低い保護での、逆U−形状用量−応答効力曲線は、最初に、与えられた用量の抗原による液性および細胞−媒介免疫の誘導の間の逆関係を記載したParishら,の古典的研究を思い出させる。Parishの精液データとの関係で、逆U−形状用量−応答効力曲線は、もし:1)ワクチン効力が細胞−媒介免疫に依存し、および2)中程度の用量のrAls1p−Nが高い抗体−刺激用量と比較して優れた細胞−媒介免疫を刺激したならば説明することができよう。我々は、従って、rAls1p−Nワクチンで見られた逆U−形状用量応答効力曲線は、保護的な中程度の用量の抗原による細胞−媒介免疫の優れた誘導によるものであったと仮定した。
【0121】
この仮説を検定するために、Th1細胞および遅延型過敏を刺激する高い、中程度、および低い用量の抗原の能力を決定した。脾臓細胞からのサイトカイン−生産を刺激するために、我々は、rAls1p−Nの代わりに熱で殺したC.albicansへの暴露によって細胞を特異的に活性化して、感染の間のインビボ状況を模倣した。保護的20μgの用量だけがC.albicans−刺激脾臓Th1リンパ球の頻度を有意に増加させた。エクスビボC.albicans−刺激脾臓細胞で見られたTh1の頻度は、マウスにおける播種性カンジダ症の間にインビボで検出されたのと同様であり(59)、このアプローチの関連性を過小評価している。
【0122】
検出されたエクスビボTh1細胞がインビボにて機能的重要性のものであるかを決定するために、我々は、異なる用量のrAls1p−N免疫化によって誘導された遅延型過敏を比較した。Th1細胞の頻度と一致して、保護的20μg用量のrAls1p−Nだけが有意なインビボ遅延型過敏反応を刺激した。これらの結果は、ワクチン−誘導保護がタイプ1の誘導、細胞媒介免疫によるものであるという仮説と合致する。驚くべきことに、200μg用量のrAls1p−Nによる顕著に上昇した抗体力価の誘導にも拘わらず、我々は、この用量でワクチン接種したマウスにおいて脾臓Th2リンパ球の増加は見出さなかった。1つの可能な説明は、Th2細胞が脾臓細胞よりはむしろ末梢リンパ節において活性化されたというものである。あるいは、他のT細胞集団(例えば、NKT細胞)が、200μg用量のrAls1p−Nに応答して見られた高い抗体力価を誘導する原因であっただろう。
【0123】
抗体力価および保護の間の相関の欠如は、ワクチン−誘導保護を媒介するにおける抗体の役割を完全には排除しなかった。例えば、ELISA力価は、種々の特異性および親和性を持つ抗体の列挙の結果である。従って、ワクチン−媒介保護に参画した抗体の小さなサブセットが生じたという可能性は、抗体力価を測定することによって排除することができなかった。rAls1p−Nワクチン−媒介保護における細胞−媒介の液性でない免疫の役割を確認するために、我々は、B細胞−およびT細胞−欠乏マウスにおけるワクチンの効果をテストした。B細胞−欠乏マウスは、野生型対照よりも播種性カンジダ症に対してより耐性である傾向があり、ワクチンの効果はB細胞−欠乏マウスにおいて無効にならなかった。対照的に、T細胞−欠乏マウスは野生型対照よりも播種性カンジダ症に対してより感受性であり、ワクチンの効果はT細胞−欠乏マウスにおいて失われた。我々の知見は、従って、rAls1p−Nワクチンの効果はT細胞−媒介の主として液性ではない免疫の誘導に依存していることを確認する。同様に、B細胞―欠乏マウスはコンジェニック野生型同腹子よりも播種性カンジダ症に対してより感受性ではなかったゆえに、抗体はこのモデルにおいて播種性カンジダ症に対して支配的なエフェクタではない。
【0124】
まとめると、我々は、新規なrAls1p−Nワクチンは、液性免疫よりはむしろ細胞−媒介免疫を誘導することによって実験的散在カンジダ症に対する保護を媒介すると報告する。rAls1p−Nワクチンの中程度の保護的効果の増強は、従って、最適化アジュバントおよび/またはサイトカインを用いる細胞−媒介免疫のさらなるプライミング、または代替免疫化経路で達成することができる。事実、我々の進行している研究においては、我々は、腹腔内と比較してrAls1p−Nを皮下投与することによって効力の顕著な増大を既に見出している。
【0125】
(実施例VIII)
(抗Candida albicans rAls1p−Nワクチンは真菌負荷を低下させ、免疫応答性および免疫無防備マウスの双方において生存を改善する)
本実施例は、免疫応答性および免疫無防備マウスの双方における皮下(SQ)経路によって投与した場合の、実施例VIIに記載されたrAls1p−Nワクチンの効力の増強を記載する。まず、免疫応答性マウスにおけるrAls1p−Nワクチンの効力。アミノ酸19から433の全長タンパク質を含むrAls1p−Nを前記したようにS.cerevisiaeにおいて生産し、精製した。対象調製物を、同様に、空のプラスミドで形質転換したS.cerevisiaeから精製した。BALB/cリタイアしたブリーダーマウス(25から30g)を、0日目に、フロイントの完全アジュバント(CFA)と混合したrAls1p−N(20μg)または対照調製物のSQ注射によって、続いて、21日目にフロイントの不完全アジュバント(IFA)中のブースター用量によって免疫化した。ブーストから2週間後に、ワクチンの免疫原性は、従前に記載されているように、遅延型過敏(DTH)のマーカーとしての足蹠膨潤反応の強度を評価することによって確認した。ワクチン接種したマウスは、rAls1p−N特異的DTHの顕著な増加を有した(図21)。
【0126】
rAls1p−Nワクチンの効果は、感染したBALB/cマウスにおける生存に対するrAls1p−Nワクチン接種のインパクトを決定することによって評価した(図22A)。ワクチン接種したまたは対照マウスを、C.albicansの迅速に致死的な接種物(2.5から5×105出芽胞子)で尾静脈を介して感染させた。我々は、そのような接種物で感染させたマウスがひどい敗血症ショックで死亡したことを従前に示した(Spellbergら,J.Infect.Dis.In press(2005))。ワクチン接種は死亡までの時間を顕著に延長させ(Log順位検定によると双方の接種物についてp<0.05)、30日生存を改善した(50から57% vs.0%、Fisher’s Exact検定によると双方の接種物についてp<0.05)。
【0127】
次いで、血行的に播種性のカンジダ症の間における組織真菌負荷に対するワクチン接種のインパクトを決定した。ブーストから14日後に、ワクチン接種したおよび対照BALB/cマウスをC.albicans SC5314の5×105出芽胞子で尾−静脈を介して感染させた。感染から6日後に、対照実験群における最初の死亡の開始の前に、腎臓を収穫し、ホモゲナイズし、Sabouraudデキストロース寒天(Difco)中で定量的に培養した(18)。rAls1p−NでのSQワクチン接種の結果、対照と比較して腎臓真菌負荷のメジアン1.5 log CFU/g減少がもたらされた(Wilcoxon順位和検定によるとp=0.01、図22B)。
【0128】
rAls1p−Nワクチンの効力もまた、免疫無防備マウスにおいて評価した。免疫応答性マウスにおいて効力を証明したら、好中球減少症マウスにおいて免疫を誘導し、好中球減少症マウスを播種性カンジダ症から保護するrAls1p−Nワクチンの能力も評価した。ワクチン接種したBALB/cマウスを、シクロホスファミドの投与によって好中球減少症とした(記載されたように(Sheppardら,Antimicrob.Agents.Chemother.48:1908−11(2004)、ほぼ12日の好中球減少症をもたらす、感染に対して−2日に200mg/kg ip、および+9日に100mg/kg ip)。足蹠膨潤反応をシクロホスファミドの最初の量から2日後に行った。ワクチン接種した好中球減少症マウスは免疫応答性マウスと同様な大きさのDTH反応を発生した(図23A vs.1、平行して行った実験)。C.albicansの2.5×104出芽胞子で尾−静脈を介して感染させた好中球減少症マウスにおいて、ワクチン接種の結果、死亡までの時間(Log順位検定 vs.対照によるとp=0.007)、メジアン生存時間(Wilcoxon順位和検定によると>21 vs 12 d,p=0.008)、および全生存(88% vs.38%,Fisher’s Exact検定によるとp=0.005)の有意な改善がもたらされた(図23B)。
【0129】
粘膜感染におけるrAls1p−Nワクチン接種の効力を決定するために、ワクチンをマウス口腔咽頭カンジダ症(OPC)モデルでテストした(Kamaiら,Infect.Immun.70:5256−8(2002) and Kamaiら,Antimicrob.Agents Chemother.45:3195−97(2001))。ワクチン接種したマウスをコルチゾン酢酸(感染に対して−1、1および3日に225mg/kg SQ)で処理し、記載したように舌下感染させた。舌を感染から5日後に切り出した。ホモゲナイズした舌のコロニー形成単位は侵入性感染および表面−接着性コロニー化の間を区別できなかったゆえに、我々は組織病理学によって侵入の程度を評価した。盲検観察者(BJS)は、舌の全長に沿ってスキャンし、40×高−出力場当たりの真菌病巣のひどさを定量することによって各セクションをスコア取りした(0=病巣なし、1+=中程度の粘膜炎症、2+=上皮に制限される有意な炎症、3+=全上皮層を通じて伸びる炎症、4+=上皮下まで伸びる炎症)。サンプリングの偏りを回避するために、少なくとも生きた介入組織セクションによって隔てられた各舌の2つのセクションをスコア取りした。全ての対照マウスは多数の場所においてその舌の顕著な真菌侵入を発生し、他方、唯2匹のワクチン接種したマウスがいずれかの舌病巣を発生した。全体として、対照マウスにおける舌当たりの病巣のメジアン数(75、25四分位)はワクチン接種したマウスについての1(2.5、0)と比較して6.5(8、5.75)であった(Wilcoxon順位和検定によるとp=0.03)。感染のひどさの半定量的評価は、対照と比較してワクチン接種マウスにおいて有意な低下を示した(図24、Wilcoxon順位和検定によるとp=0.03)。
【0130】
粘膜感染におけるrAls1p−NまたはrAls3p−Nワクチン接種の効力を決定するために、膣コロニー化のマウスモデルにおいてこれら2つのワクチンを評価した(Clemonsら,Infect.Immun.72:4878−80(2004);Fidel.Int Rev Immunol.21:515−48(2002) and Wozniakら,InfectImmun.70:5790−9(2002))。ワクチン接種したマウスを感染に対して−3日目にエストロゲン(30μg,与えられたSQ)で処理し、次いで、C.albicansの106出芽胞子を含有する10μlのリン酸緩衝化生理食塩水で膣においてチャレンジさせた。膣を接種から3日後に切り出し、ホモゲナイズし、系列希釈をYPDプレート上で平板培養した。コロニー形成単位(CFU)を30から35℃においてプレートのインキュベーションから20から48時間後に数えた。rAls3p−Nでワクチン接種したマウスから集めた膣は、対照マウス(すなわち、CFA単独でワクチン接種したマウス)から集めた膣よりも低いCFUを有したが、rAls1p−Nでワクチン接種したマウスから集めたものはそうでなかった(図25、Wilcoxon順位和検定によるとp=0.01)。
【0131】
カンジダ症の発生率およびその継続する高い死亡率に徴すると、Candida spp.に対するワクチンの開発は非常に重要なものである。前記した結果は、rAls1p−NでのSQワクチン接種は、免疫応答性および免疫無防備マウス双方におけるそうでなければ迅速に致死的な血行的に播種性のカンジダ症の間において、生存の顕著な改善、および真菌負荷の有意な低下がもたらされた。興味深いのは個々のワクチン接種したマウスから得られた腎臓真菌負荷の結果であり、これは、マウスのほぼ半分が5 log CFU/g下で腎臓真菌負荷を有したことを示す。我々は、致死的感染を示す腎臓真菌負荷の閾値は5 log CFU/gであることを従前に見出しており;このレベルを超える腎臓真菌負荷を持つマウスは典型的には感染で死亡し、他方、この負荷未満の腎臓真菌負荷を持つマウスは感染から生存した(Spellbergら,J.Infect.Dis.In press (2005)) and (Spellbergら,Infect.Immun.71:5756−5764(2003))。従って、ワクチン接種した群における突発的死亡はワクチン接種に関わらない高い真菌負荷を反映するようである。組織真菌負荷におけるマウス間変動は、宿主−病原相互作用および/または可変ワクチン応答性の複雑性を反映するであろう。
【0132】
まとめると、rAls1p−Nワクチンは増大する通常のおよびかなり致死的な播種性カンジダ症の治療、ひどさの低下および/または予防で用いることができる。該ワクチンは免疫応答性マウスにおいて効果的であり、かつ効果は好中球減少症およびコルチコステロイド−処理宿主においてさえ保持される。最後に、ワクチンは膣および口腔咽頭カンジダ症を含めた粘膜皮膚カンジダ症を保護することができる。
【0133】
(実施例IX)
(黄色ブドウ球菌感染に対するAlsワクチンの効果)
本実施例は、C.albicansからのAlsタンパク質が黄色ブドウ球菌で感染させた動物モデルの生存を改善することを示す。
【0134】
C.albicansのAlsアドヘシンは、黄色ブドウ球菌でのアドヘシンに対して有意に相同あると同定された。この特徴を用いてAlsアドヘシンを用い黄色ブドウ球菌に対する効果的なワクチンを設計し実施した。簡単に述べると、C.albicans ALSファミリーは少なくとも9つの遺伝子よりなる(Hoyerら,Genetics 157:1555−67(2001); Hoyer LL.,Trends Microbiol.9:176−80(2001))。前記したように、Alsタンパク質は生物学的に関連する基材へのアドヘシンとして機能する(Fuら,Molec.Microbiol.44:61−72(2002);Gaur and Klotz,Infect.Immun 65:5289−94(1997);Zhao et al.,Microbiology 150:2415−28(2004);Ohら,Microbiology 151:673−81(2005);Zhaoら,Microbiology 151:1619−30(2005)); Hoyerら,Mol.Microbiol.15:39−54(1995))。特に、rAls1p−NおよびrAls3p−NのN末端は、コラーゲン結合タンパク質およびクランピング因子を含めた、病原性黄色ブドウ球菌によって発現された表面タンパク質に対して有意に相同である(表IV;Sheppardら J.Biol.Chem.279:30480−89(2004))。
【0135】
【表4】
表IVにおいて上記に掲げた相同性の計算は、配列整列および三次元表面構造の双方の特徴を考慮するものである。Als1pの相同性は、黄色ブドウ球菌のコラーゲン結合タンパク質またはクランピング因子と比較して95%または90%よりも大きいと計算された(r2≧90%;Sheppardら,上記)。同様に、Als3pの相同性は、黄色ブドウ球菌のコラーゲン結合タンパク質またはクランピング因子と比較して95%または80%よりも大と計算された(r2≧90%)。
【0136】
前記知見を裏付けるために、相同性およびねじ切り法を使用して、Als1pおよび黄色ブドウ球菌クランピング因子Aの間の構造−機能調和をモデル化した(ClfA − PDB code:c1n67A)。これらの方法は一次構造における特異的相同性を評価し、3−D立体配座およびパターン分析を行って、類似の機能モチーフを求めた。例えば、BLASTP、PROSITEおよびJALVIEW方法を使用して、ALS vs ClfA一次配列の同様性および差を分析した(Yountら Antimicrob.Agents Chemother.48:4395−4404(2004)and Yount and Yeaman.Proc.Natl.Acad.Sci.USA 101:7363−7368(2004))。次いで、3−D PSSMを含めたインターネットベースの適用を用いて、さらなる分析のための潜在的ALSホモログを優先付けた(Sheppardら J.Biol.Chem.279:30480−30489(2004))。得られたデータに沿って、PHYRE適用(Kelley,L.,R.Bennett−Lovsey,A.Herbert,and K.Fleming;ウェブサイトは以下の通りである:http://www.sbg.bio.ic.ac.uk/〜phyre/)を用いて、トポロジーマッピングを行い、および推定共有機能モチーフを同定する目的で、選択されたALSタンパク質に対する最大の構造的または機能的相同性を持つタンパク質によって共有された三次元モチーフを同定した。前記方法は公のドメインで広く入手可能であり、種々のプロテオミックおよび構造的生物学適用で用いられる。前記相同性およびねじ切り法結果に基づき、Als1pおよびClfAの間の機能的部位相同性のコンセンサスを生じさせ、ClfAに構築されたAls1pモデルの特異的残基にマッピングした。いくつかの特別な知見は、以下に記載するこれらのモデリング分析から生じた。
【0137】
まず、Als1pおよびClfAのN末端領域の間で、二次構造およびアミノ酸保存において、特に、アミノ酸30から300によって含まれる領域(双方のタンパク質のN末端)において、有意な相同性が同定された。
【0138】
第二に、確立されたClfAアドヘシン決定基に基づく相同機能部位のコンセンサスマッピングは、Als1pにおける特異的トポロジー的モチーフに集中した。このトポロジー的モチーフを、2つのβ−シートドメインの隣接面の屈曲によって形成された溝として図26に示される。
【0139】
第三に、一次構造相同性と合致して、Als1pにおける予測された機能的溝モチーフは、アミノ酸残基30から300を含むN末端領域における超可変領域に由来する特異的残基にマッピングされる。
【0140】
これらの結果は、調和した生物学的機能に対する構造的基礎、並びにAls1pおよびClfAに対する免疫学的応答を供した。これらの結果はAls1p構造−活性の我々の総じてのモデルをさらに裏付け、突然変異分析およびエピトープマッピングに対する標的化アプローチをさらに容易とする。最後に、これらの結果は、Als1pおよびClfAが、多様な微生物病原体に存在する類似の構造および機能のアドヘシンであることを示す。
【0141】
黄色ブドウ球菌に対するモノクローナル抗体もまた同定され、これはC.albicansによって引き起こされた感染を低下させることができる。前記構造的知見に関して、この特徴も用いて、Alsアドヘシンを用い、黄色ブドウ球菌に対する効果的なワクチンを設計し実施した。
【0142】
簡単に述べると、黄色ブドウ球菌上の表面アドヘシンを認識することが知られているヒト化抗ブドウ球菌モノクローナル抗体(Aurexis(登録商標))は現在臨床試験中である。このモノクローナル抗体もやはりAlsファミリーメンバーと交差反応する。ブドウ球菌血流感染の治療用のAurexis(登録商標)の相II臨床試験の好都合な結果が報告されている(Inhibitex Inc.,2005;http://phx.corporate−ir.net/phoenix.zhtml?c=176944&p=irol−newsArticle&ID=707322&highlight=において、2005年9月19日にアクセスされた)。簡単に述べると、この報告においては、血液中の公知の黄色ブドウ球菌を持つ患者に活性な感染に対する治療としてAurexis(登録商標)抗体を投与した(すなわち、これは能動的ワクチン戦略または予防的実験ではない)。プラセボを受ける9人の患者はCandida によって引き起こされた突発的な血流感染を経験し、他方、Aurexis(登録商標)実験群においては、3人の患者のみがCandida血流感染を経験した。前記相同性および構造的知見と組合せて、黄色ブドウ球菌に対する抗体で治療した患者についてのCandida血液感染の減少を認識すると、免疫原性エピトープはCandidaおよび黄色ブドウ球菌の間で共有されており、これらの免疫原性エピトープはおよび他の種の治療用の1つの種に対して生起された免疫応答、抗体またはエフェクタメカニズムを用いて治療的利益について標的化できることを示す。従って、前記データを一緒にすると、Candida spp.と交差反応する黄色ブドウ球菌上の表面アドヘシンに対する免疫応答を提供する。
【0143】
前記戦略に従い、例示的なAlsアドヘシンワクチンを設計し、それは、黄色ブドウ球菌に感染したマウスの生存を改善することが示された。ワクチン接種するのに用いた例示的なAlsアドヘシンはrAls1p−NまたはrAls3p−Nであり、これは前記したように生産し、用いた。簡単に述べれば、Candida、rAls1p−NおよびrAls3p−Nに対するこれらのAlsワクチンが黄色ブドウ球菌に対して交差−種保護を媒介できるかを決定するために、雌Balb/cマウスを従前に記載された養生法でワクチン接種した(0日目におけるフロイントの完全アジュバント+20μgのrAls1p−NまたはrAls3p−N、続いての、3週間におけるフロイントの不完全アジュバント中のブースター用量;共に皮下投与した)。ワクチン接種から2週間後に、マウスを、メチリシリン−耐性であって、動物モデルにおいてビルレントであることが知られている、致死用量の黄色ブドウ球菌株67−0で尾−静脈を介して感染させた。マウスの生存を示す結果を図26に示す。示したように、rAls1p−NおよびrAls3p−Nワクチンは、これらの感染したマウスにおいて改善された長期生存を媒介した(図27)。加えて、保護のメカニズムはTh2よりはむしろTh1の増強であるようである。というのは、Ab力価、およびrAls1p−NまたはrAls3p−Nいずれかでワクチン接種したマウスの生存の間に相関は観察されなかったからである(図28)。
【0144】
(実施例X)
(抗Candida rAls1p−Nワクチンは播種性カンジダ症に対する広い範囲の保護を媒介する)
本実施例は、rAls1p−Nワクチンが非近交系マウスを播種性カンジダ症から保護し、Balb/cマウスをC.albicansおよび非albicans Candidaの他のビルレント株に対して保護する。
【0145】
本実験を行って、C.albicansの他の株に対する、およびCandidaの非albicans種に対する、フロイントのアジュバント以外の第二のアジュバントと組み合わせた、非近交系マウスにおけるその効力を特異的に評価することによりrAls1p−Nによって誘導される保護の広さを説明する。
【0146】
rAls1p−Nでのワクチン接種は、播種性カンジダ症から非近交系マウスを保護した。簡単に述べると、非近交系マウスCD1マウスはNational Cancer Institute(Bethesda, MD)から入手した。マウスに関連する全ての手法は、動物収容および介護についてのNational Institutes of Health指針に従い、動物使用および看護制度委員会によって認可された。前記にて、および例えば、Ibrahimら,Infect.Immun.73:999−1005(2005);Spellbergら,Infect.Immun.73:6191−93(2005)に記載されているように、マウスをrAls1p−N+フロイントアジュバントでワクチン接種した。rAls1p−N(Als1pのアミノ酸17から432)はS.cerevisiaeにおいて生産され、ゲル濾過およびNi−NTAマトリックスアフィニティー精製によって精製した。高度な純度(約90%)は、前記にて、および例えば、Sheppardら,J Biol Chem 279:30480−89(2004)に記載されているように、SDS−ポリアクリルアミドゲル電気詠動並びに円二色およびFTIRによって確認した。マウスは、0日目においてフロイントの完全アジュバント(CFA;Sigma−Aldrich,St.Louis,MO)と混合されたrAls1p−N(20μg)のSQ注射、続いての21日目におけるフロイントの不完全アジュバント(IFA;Sigma−Aldrich)中のブースター用量によって免疫化した。対照マウスはCFA/IFA単独で免疫化した。ブーストから14日後に、我々は既に記載したように、免疫化されたマウスをC.albicans SC5314で尾−静脈を介して感染させた。Ibrahimら,(2005)上記;およびSpellbergら(2005)上記。Balb/cマウスにおける我々の従前の知見と同様に、rAls1p−Nワクチンは感染されたCD1マウスの生存を顕著に改良した(図29A)。
【0147】
フロイントのアジュバントはヒトで用いるのにはあまりにも毒性であるゆえに、我々はミョウバン(2% Alhydrogel,Brenntag Biosector,Frederikssund,Denmark)中のrAls1p−Nワクチン、ヒトで用いる合衆国食品薬品局(FDA)によって現在認可された唯一のワクチンの用量応答を行った。ミョウバンでのワクチン接種は、1日目における免疫化、21日目におけるブースト、および2週間後における感染にて、フロイントのアジュバントと同一のスケジュールで行った。我々は、ミョウバンと組合せたより高い用量のrAls1p−Nの結果、播種性カンジダ症を持つマウスの生存の有意な改善をもたらしたことを見出した(図29B)。また、用量応答の関係があるようであり、ミョウバンと組み合わせた場合に、より高い用量のrAls1p−Nにおいて改良された生存の傾向がある。
【0148】
また、rAls1p−Nワクチンは、C.albicansのいくつかの株で感染させたBalb/cマウスの生存を改善することが示された。特に有用なワクチンは、免疫系が標的病原体の多数の株を認識するように開始することができる免疫原を利用する。DNA配列分析によって、我々は、Als1pのN末端領域の予測されたアミノ酸配列が、血流(5株)、尿(5株)および口腔咽頭(10株)感染(データは示さず)からの臨床C.albicans単離体の多様な群の中で99.9%保存されていたことを見出した。これらの結果は、rals1p−NワクチンがC.albicans株の広いアレイに対して効果的であり得ることを示す。C.albicansの他の株に対するrAls1p−Nワクチンの保護の広さを確認するために、マウスをrAls1p−N+フロイントのアジュバントで前記したようにワクチン接種し、C.albicansのいくつかの臨床単離体の1つで感染させた(Ibrahimら, Infect Immun 63:1993−98(1995))。図30に示したように、rAls1p−Nワクチンは、これらの株の各々で感染させたマウスの生存を有意に改善した。
【0149】
また、rAls1p−Nワクチンは、Candidaのいくつかの非albicans種で感染させたマウスにおいて組織真菌負荷を低下させることが示された。簡単に述べると、ALS遺伝子ファミリーはC.dubliniensisおよびC.tropicalisを含めた他のCandida種に存在する(Hoyerら,Genetics 157:1555−67(2001))。同様に、Alsファミリーメンバーに類似するアドヘシンはC.glabrataに記載されている(Cormackら,Science 285:578−82(1999);Friemanら,Mol Microbiol 46:479−92(2002))。非albicans種に対するrAls1p−Nの効力を確認するために、Balb/cマウスを前記したようにrAls1p−N+フロイントのアジュバントでワクチン接種し、C.glabrata 31028(Harbor−UCLA Medical Centerにおける微生物学研究所からの臨床的血流単離体)、C.krusei 91−1159(親切にもMichael Rinaldi,San Antonio,Texasによって提供された)、C.parapsilosis 22019(Harbor−UCLA Medical Centerからの臨床的血流単離体)、またはC.tropicalis 4243(Harbor−UCLA Medical Centerからの臨床血流単離体)で尾−静脈を介して感染させた。図31に示すように、rAls1p−Nワクチンはこれらの種の各々で感染させたマウスの腎臓真菌負荷を低下させた。
【0150】
まとめると、rAls1p−Nワクチンは、増大している通常のおよびかなり致死的な播種性カンジダ症のひどさを予防し、および/または低下させることができる。該ワクチンは、アジュバントとしてのミョウバンと混合した場合、C.albicansの多数の株に対して、およびCandidaのいくつかの非albicans種に対して、同系交配および非近交系マウスの双方において有効である。これらの結果は、本発明のALDSワクチンが広く種々のカンジダおよび他の感染に対して有効であることをさらに裏付ける。
【0151】
(実施例XI)
(抗Candida rAls3p−Nワクチンは播種性カンジダ症に対してrAls1p−Nと同等に効果的であり、粘膜カンジダ症に対してより効果的である)
本実施例は、血行的に散在した口腔咽頭および膣カンジダ症のネズミモデルにおけるrAls3p−Nワクチンの効力をrAls1p−Nと比較する。
【0152】
ALSファミリーメンバーのうち、ALS1およびALS3遺伝子は基材親和性の最も広いアレイにてアドヘシンをコードする。相互に比較した場合、Als1pは内皮細胞およびゼラチンに対するより大きな接着を媒介したが、上皮細胞に対しては劣った接着を媒介した(Sheppardら,J Biol Chem 279:30480−89(2004))。接着の質の個の差は、rAls3p−NがrAls1p−Nと比較してワクチン免疫原として異なる有効性を有することができることを示唆した。
【0153】
ワクチンおよびワクチン接種は前記したように行った。簡単に述べれば、rAls1p−NおよびrAls3p−N(Als1pまたはAls3pのアミノ酸17から432)はS.cerevisiaeで生産され、前記にて、およびIbrahimら,(2005),上記;Spellbergら,(2005),上記;に記載されたように、ゲル濾過およびNi−NTAマトリックスアフィニティー精製によって精製した。タンパク質の量は修飾されたLowryアッセイによって定量した。高度な純度(約90%)は、前記にて、およびIbrahimら.,(2005),上記;Spellbergら.,(2005),上記;に記載されているように、SDS−ポリアクリルアミドゲル電気詠動ならびに円二色性およびFTIRによって確認した。マウスを、0日目においてフロイントの完全アジュバント(CFA,Sigma−Aldrich,St.Louis,MO)と混合された20μgのrAls1p−NまたはrAls3p−Nの皮下(SQ注射によって免疫化し、21日目においてもう1つの用量の抗原とフロイントの不完全なアジュバント(IFA,Sigma−Aldrich)でブースト注射し、次いで、ブーストから2週間後に感染させた。
【0154】
統計学的解析は以下のように行った。非パラメーターLog順位検定を利用して、マウスの生存時間の差を決定した。抗体力価および足蹠膨潤は、適切であれば、非パラメーター多数比較のためのSteel検定(Rhyneら.,Biometrics 23:539−49(1967))、または負対比較のためのMann Whitney U検定によって比較した。相関はSpearman順位検定で計算した。不均一が複連生存試験で存在するかを決定するために、Kolmogorov−Smimov検定を利用した。P値<0.05は有意と考えられた。
【0155】
rAls3p−Nでのワクチン接種は、rAls1p−Nとの比較において抗体応答のより広いアレイを刺激することが示された。この点に関して、図32に示される結果は、CFA + rAls1p−NまたはrAls3p−Nでワクチン接種したマウスは、CFA単独を受けるマウスよりも有意に大きな抗体力価を発生させたことを示す。注目すべきことには、rAls3p−Nワクチン接種したマウスは、rAls1p−Nワクチン接種したマウスに対して同等な力価にて抗rAls1p−N抗体を生じた(図32,上)。対照的には、rAls1p−Nワクチン接種したマウスは、rAls3p−Nでワクチン接種したマウスよりもrAls3p−Nに対してより小さな力価を生じた(図32,下)。しかしながら、rAls1p−NおよびrAls3p−Nは、図33に示したように、同様な遅延型過敏応答をインビボにてもたらした。
【0156】
rAls1p−NおよびrAls3p−Nワクチンは、播種性カンジダ症に対して同様な効果を媒介することも示されていた。簡単に述べれば、rAls3p−Nワクチンが、血行的に播種性のカンジダ症に対してrAls1p−Nと同程度に効果的であることをさらに裏付けるために、マウスをCFA、CFA+rAls1p−N、またはCFA+rAls3p−Nをワクチン接種し、続いて、C.albicansで尾−静脈を介して感染させた。図34に示す結果は、rAls1p−NおよびrAls3p−Nワクチン双方の結果、生存の有意な改良がもたらされたことを示す。
【0157】
抗Alsp抗体力価および遅延型過敏反応と、引き続いてC.albicansで感染させたワクチン接種したマウスにおける生存との相関も決定した。簡単に述べれば、抗体力価を、先に、およびIbrahimら,(2005),上記;Spellbergら,(2005),上記に我々が記載したように、96ウェルプレートにおいてELISAによって決定した。ウェルを、PBS中の5μg/mlのrAls1p−NまたはrAls3p−Nのウェル当たり100μlで被覆した。マウス血清を、3%ウシ血清アルブミンを含有するtris緩衝生理食塩水(TBS)(0.01 M TrisHCl,pH 7.4,0.15M NaCl)でのブロッキング工程に続いて室温にて1時間インキュベートした。ウェルを0.05%Tween20を含有するTBSで3回洗浄し、続いて、Tweenを含まないTBSでさらに3回洗浄した。ホースラディッシュペルオキシダーゼ(Sigma−Aldrich)でコンジュゲートしたヤギ抗マウスIgG二次抗体を1:5000の最終希釈で加え、プレートを室温にて1時間さらにインキュベートした。ウェルをTBSで洗浄し、0.1Mクエン酸緩衝液(pH 5.0);50mg/mlのo−フェニレンジアニン(Sigma)、および10μlの30%H2O2を含有する基質と共にインキュベートした。30分間発色させ、その後、10%H2SO4を加えることによって反応を停止させ、490nmにてマイクロタイタープレートにおいて光学密度(OD)を決定した。陰性対照ウェルは無関係な抗体を受け、バックグラウンド吸光度をテストウェルから差し引いて、最終OD読みを得た。ELISA力価は、陽性OD読みを与えた最後の血清希釈の逆数として取られた(すなわち、>は陰性対照試料の平均OD+(標準偏差*2)を意味する)。
【0158】
遅延型過敏反応は、足蹠膨潤テストを測定することによって評価した。簡単に述べれば、マウスをrAls1p−N、rAls3p−N、またはCFA単独で免疫化した。ブースとから2週間後に、免疫化マウスのベースライン足蹠サイズを電子デジタルキャリパーを用いて測定した。25μlのPBS中の50μgのrAls1p−NまたはrAls3p−Nを右側足蹠に注射し、PBS単独を免疫化マウスの左側足蹠に注射した。24時間後に、足蹠を再度測定した。抗原−特異的足蹠膨潤は:(24時間における右側足蹠厚み−ベースラインにおける右側足蹠)−(24時間における左側足蹠厚み−ベースラインにおける左側足蹠厚み)として計算された。
【0159】
ワクチン接種したマウスから力価決定のために採血し、感染に2日先立って足蹠膨潤テストを受けさせた。感染で生き残らなかったワクチン接種マウスは、それにもかかわらず、図35に示すように広い範囲の抗体力価を有した。多くのそのようなマウスは≧1:50,000(≧4.5log10)の抗rAls1p−Nおよび抗rAls3p−N抗体力価を有した。その結果、抗体力価は生存に有意には相関しなかった。対照的に、足蹠膨潤反応の強さは生存と相関した(図35、Spearman順位相関テストによるとp=0.6&p=0.009)。
【0160】
rAls3p−Nワクチンは、粘膜カンジダ症の2つのモデルにおいてrAls1p−Nよりも大きな効果を示した。Als3pはAls1pと比較して上皮細胞に対して優れた接着を媒介したので、この観察は、rAls3p−Nが感染の粘膜モデルにおいてユニークな効果を呈することができることを示す。rAls3p−Nと比較したrAls1p−Nの効力を、感染のステロイド−処理口腔咽頭モデルにおいて、およびカンジダ膣炎のモデルにおいて評価した。
【0161】
簡単に述べれば、前記マウス口腔咽頭でカンジダ症(OPC)モデルにおけるワクチン実験を従前に記載されているように、およびSpellbergら,(2005),上記;Kamaiら,Antimicrob Agents Chemother 45:3195−57(2001),and Kamaiら,Infect Immun 70:5256−58(2002)に記載されているように行った。ワクチン接種マウスをコルチゾン酢酸(感染に対して−1、1および3日目にて225mg/kg SQ)での処理によって免疫無防備とした。感染の日に、kg当たり8mgのキシラジンおよび110mgケタミンでの腹腔内注射によってマウスを麻酔した。アルギン酸カルシウム尿道スワブを、それを30℃のHBSS中のml当たり106生物の懸濁液に入れることによって、C.albicansで飽和させた。飽和したスワブを75分間でマウスの口腔中で舌下に入れた。感染の5日後に、舌および舌下神経組織を切り出し、秤量し、ホモゲナイズし、次いで、定量的に培養して、経口真菌負荷を決定した。
【0162】
マウス膣カンジダ症に対するワクチンの有効性は、感染に対して−3日目に(共にSigma−Aldrichからの)落花生油に溶解させた30μgの皮下エストラジオールバレレートで雌Balb/cマウスをワクチン接種して、プソイドエストラス(pseudoestrus)を誘導することによって行った。感染の日に、100mg/kgのケタミンのip投与によってマウスを鎮静させた。鎮静させたマウスを10μlのHBSS中のC.albicansの106出芽胞子で膣内感染させた。感染から3日後に、膣およびほぼ1センチメートルの各子宮角をブロック上に切開し、ホモゲナイズし、定量的に培養した。
【0163】
図36に示すように、口腔咽頭カンジダ症を持つコルチゾン−処理マウスにおいて、rAls1p−Nワクチンは低下した舌真菌負荷に対する強い傾向を媒介した(p=0.054)。総じての利点の大きさは<0.3logCFU/グラムであった(図36)。比較において、rAls3p−Nワクチンは統計学的に有意な舌真菌負荷の>0.6logCFU/グラム減少を媒介した(p=0.005,図36)。同様に、カンジダ膣炎の非免疫無防備モデルにおいて、rAls3p−Nワクチンは、図37に示すようにCFA単独と比較して膣真菌負荷の0.7logCFU/グラム減少を媒介した(p=0.02)。比較において、rAls1p−Nは膣炎モデルにおいて利点を全く媒介せず、およびrAls3p−NはrAls1p−Nよりも有意に効果的であった(p=0.01)。
【0164】
前記した結果は、アミノ酸レベルにおいてrAls1p−Nに対して85%相同であるrAls3p−Nに基づくワクチンは播種性カンジダ症に対して同等に効果的であるが、粘膜感染に対してrAls1p−Nよりも効果的であった。rAls3p−Nの増大した有効性は口腔咽頭カンジダ症のステロイド−処理モデルおよびカンジダ膣炎の免疫応答性モデル双方で見られた。前期結果は、補助的抗真菌療法を伴わないカンジダ敗血症ショックのマウスモデルにおいて≧50%長期生存の達成が刺激的であったことを示し、さらに、本発明の治療的利点の全てのALSワクチンをさらに裏付ける。
【0165】
抗体力価は、播種性カンジダ症の間におけるいずれかのワクチンの保護的効果と相関しなかったが、遅延型過敏の誘導はインビボにて保護と相関した。これらのデータは、さらに、ワクチン−感染保護のメカニズムがタイプ1の誘導、真菌に対する細胞−媒介免疫を裏付ける。rAls1p−NおよびrAls3p−Nの双方はrAls1p−Nに対する同等な力価の抗体を誘導したが、rAls3p−NがrAls1p−Nよりも有意により高い力価の抗rAls3p−N抗体を誘導した。これらのデータは、その高度なアミノ酸配列相同性(85%)にも拘わらず、液性免疫系はrAls1p−NおよびrAls3p−Nの間を識別できることを示す。前記結果は、さらに、Als1pおよびAls3p上皮細胞接着特徴の差に拘わらず、rAls1p−NおよびrAls3p−Nワクチンが血行的に播種性の(すなわち、血管内)カンジダ症に対して保護するのに同等に効果的であることを裏付ける。
【0166】
まとめると、抗カンジダrAls3p−Nワクチンは、rAls1p−Nワクチンよりも同等な細胞−媒介のものであるがより広い抗体−ベースの応答を誘導した。免疫原は、血行的に播種性のカンジダ症に対して同等な程度の保護をもたらしたが、rAls3p−Nは口腔咽頭および膣カンジダ症双方に対してより大きな保護を媒介した。
【0167】
本出願を通じて、種々の刊行物を括弧に入れて引用した。これらの刊行物の全ての開示をここに引用して本出願に取り込んで、本発明が属する分野の最先端技術をより十分に記載する。
【0168】
本発明を開示された実施形態を参照して記載してきたが、当業者であれば、前記にて詳細に記載した特別な実施例および実験は本発明を説明するものに過ぎないことを容易に認識するであろう。本発明の精神を逸脱することなく種々の修飾をなすことができるのは理解されるべきであろう。従って、本発明は特許請求の範囲によってのみ限定される。
【図面の簡単な説明】
【0169】
【図1A】図1Aは、ヒト臍静脈内皮細胞へのC.albicansのAlslp接着の媒介を示す。値は、各々三連で行われた少なくとも3つの独立した実験の平均±SDを表す。(A)ALS11/als2 als1/als1およびALS1−補足突然変異体および野生型CAIl2(30)の内皮細胞接着。統計学的処理はWilcoxon順位和検定によって得られ、Bonferroni修正での多数の比較について修正した。全ての比較について*P<0.001。
【図1B】図1Bは、ヒト臍静脈内皮細胞へのC.albicansのAlslp接着の媒介を示す。値は、各々三連で行われた少なくとも3つの独立した実験の平均±SDを表す。(B)野生型C.albicansと比較した、ALS1を過剰発現するPADHl−ALS1突然変異体の内皮細胞接着。統計学的処理はWilcoxon順位和検定によって得られ、Bonferroni修正での多数の比較について修正した。全ての比較について*P<0.001。
【図2A】図2Aは、C.albicans間接的免疫蛍光のフィラメントへのAls1pの細胞表面局所化を示す。C.albicansのフィラメント化は、グルタミンと共にRPMI l640培地中で酵母細胞を37℃で1.5時間インキュベートすることによって誘導された。Als1pは、まず、抗Als1pマウスmAb、続いて、FITC−標識ヤギ抗マウスIgGと共に生物をインキュベートすることによって検出された。また、C.albicans細胞表面を、Alexa 594(Molecular Probes,Eugene,OR)とコンジュゲートした抗C.albicansポリクローナルAbで染色した。黄色に染色される領域はAls1p局所化を表す(A)C.albicans野生型。
【図2B】図2Bは、C.albicans間接的免疫蛍光のフィラメントへのAls1pの細胞表面局所化を示す。C.albicansのフィラメント化は、グルタミンと共にRPMI l640培地中で酵母細胞を37℃で1.5時間インキュベートすることによって誘導された。Als1pは、まず、抗Als1pマウスmAb、続いて、FITC−標識ヤギ抗マウスIgGと共に生物をインキュベートすることによって検出された。また、C.albicans細胞表面を、Alexa 594(Molecular Probes,Eugene,OR)とコンジュゲートした抗C.albicansポリクローナルAbで染色した。黄色に染色される領域はAls1p局所化を表す(B)als1/als1突然変異体株。
【図2C】図2Cは、C.albicans間接的免疫蛍光のフィラメントへのAls1pの細胞表面局所化を示す。C.albicansのフィラメント化は、グルタミンと共にRPMI l640培地中で酵母細胞を37℃で1.5時間インキュベートすることによって誘導された。Als1pは、まず、抗Als1pマウスmAb、続いて、FITC−標識ヤギ抗マウスIgGと共に生物をインキュベートすることによって検出された。また、C.albicans細胞表面を、Alexa 594(Molecular Probes,Eugene,OR)とコンジュゲートした抗C.albicansポリクローナルAbで染色した。黄色に染色される領域はAls1p局所化を表す(C)野生型ALS1を補足したals1/als1。
【図2D】図2Dは、C.albicans間接的免疫蛍光のフィラメントへのAls1pの細胞表面局所化を示す。C.albicansのフィラメント化は、グルタミンと共にRPMI l640培地中で酵母細胞を37℃で1.5時間インキュベートすることによって誘導された。Als1pは、まず、抗Als1pマウスmAb、続いて、FITC−標識ヤギ抗マウスIgGと共に生物をインキュベートすることによって検出された。また、C.albicans細胞表面を、Alexa 594(Molecular Probes,Eugene,OR)とコンジュゲートした抗C.albicansポリクローナルAbで染色した。黄色に染色される領域はAls1p局所化を表す(D)PADHl−ALS1過剰発現突然変異体。
【図3A】図3Aは、固体培地上でのC.albicansフィラメント化に対するAls1pの媒介を示す。C.albicans出芽胞子をLee寒天プレート上にスポットし、37℃にて4日間(A)インキュベートした。
【図3B】図3Bは、固体培地上でのC.albicansフィラメント化に対するAls1pの媒介を示す。C.albicans出芽胞子をLee寒天プレート上にスポットし、37℃にて3日間(B)インキュベートした。
【図4A】図4Aは、EFG1フィラメント化調節経路によるALS1発現の制御を示す。異なるフィラメント化調節経路に欠陥がある突然変異体におけるALS1の発現を示すノーザンブロット分析。全RNAは、37℃にて90分間、RPM1 1640+グルタイン培地中で増殖させてフィラメント化を誘導した細胞から抽出した。ブロットはALS1およびTEF1でプローブした。
【図4B】図4Bは、EFG1フィラメント化調節経路によるALS1発現の制御を示す。EFG1またはPADHl−ALS1いずれかを補足したefgl/efgl突然変異体におけるALS1の発現を示すノーザンブロット分析。全RNAは、37℃にて90分間、RPM1 1640+グルタイン培地中で増殖させてフィラメント化を誘導した細胞から抽出した。ブロットはALS1およびTEF1でプローブした。
【図4C】図4Cは、C.albicansフィラメント化の媒介を示す。Lee寒天プレート上で37℃にて4日間増殖させたefgl/efgl突然変異体、およびPADHl−ALS1を補足したefgl/efgl突然変異体の顕微鏡写真。
【図5A】図5Aは、(A)雄Balb/Cマウス(各酵母株についてn=30)に静止相出芽胞子(0.5mlのPBS中マウスあたり106)で注射することによる、血行的に播種性のカンジダ症のマウスモデルにおけるビルレンスの低下を示す。曲線は三連実験の編集した結果である(各株についてn=30マウス)。30℃にてYPD中で増殖させた全ての株の倍加時間は1.29から1.52時間の間の範囲であり、相互に統計学的に異ならなかった。全染色体DNAのサザーンブロット分析を用いて、感染した生物から検索したC.albicans株の遺伝子型の同一性を、マウスを感染させるのに用いたC.albicans株のそれにマッチさせた。統計学的解析は、Wilcoxon順位和検定によって得られ、Bonferroni修正にて多数の比較のために修正した。als1/als1突然変異体−vs−他の株の各々について*P<0.002。
【図5B】図5B:C.albicans野生型、ホモ接合性als1 null突然変異体またはヘテロ接合性ALS1補足突然変異体で感染させた腎臓の組織学的顕微鏡写真。腎臓試料を感染から28時間後に検索し、パラホルムアルデヒド中で固定し、断面を銀で染色した(倍率×400)。矢印はC.albicans細胞を示す。
【図5C】図5C:C.albicans野生型、ホモ接合性als1 null突然変異体またはヘテロ接合性ALS1補足突然変異体で感染させた腎臓の組織学的顕微鏡写真。腎臓試料を感染から40時間後に検索し、パラホルムアルデヒド中で固定し、断面を銀で染色した(倍率×400)。矢印はC.albicans細胞を示す。
【図6】図6は、抗Als1pポリ血清を注入した動物についての30日間に渡って生存する動物の関数としての播種性カンジダ症に対する抗ALS抗体の予防効果を示す。
【図7】図7は、接着表現型によって配置された選択されたALSポリペプチドのN末端部分のポリペプチド配列整列である。頂部の3つの線はALS1、3および5ポリペプチド(各々、配列番号1から3)からの配列であり、これは内皮細胞に結合する。底部の3つはALS6、7および9ポリペプチドからの配列であり(各々、配列番号4から6)、これは内皮細胞に結合しない。最後の線はALSポリペプチドファミリーコンセンサス配列(配列番号7)を表す。
【図8】図8は、Saccharonzyces cerevisiaeにおいて不均一に発現させた場合に、Alsタンパク質が基材特異的接着特性を付与することを示す。各パネルは、C.albicansが接着することが知られている種々の基材への1つのAlsp発現株のパーセント接着を示す(塗りつぶした棒線)。空のベクターで形質転換されたS.cerevisiaeの接着(塗りつぶしていない棒線)は陰性対照として各パネルに含ませる。Gel、ゼラチン;FN、フィブロネクチン;LN、ラミニン;FaDu、FaDU上皮細胞;EC、内皮細胞。単一因子偏差分析によって空のプラスミド対照と比較した場合、*p<0.01。結果は三連で行った少なくとも3つの実験の平均±S.D.である。
【図9】図9は、ドメインスワッピングが、基材特異的接着が、Alsタンパク質のN末端ドメインの組成によって決定されることを示すことを示す。テストすべきAls遺伝子または構築物の表示は、ALS5(黒色)またはALS6(白色)からの配列よりなる棒線として描かれる。各突然変異体の接着特性は、フィブロネクチン−被覆ビーズへの形質転換されたS.cerevisiaeの接着を示す顕微鏡写真、および6−ウェルプレートアッセイで測定されたゼラチン(黒色棒線)および内皮細胞(灰色棒線)への接着を示すグラフとして表示される。結果は、各々が三連で行った少なくとも3つの実験の平均±S.D.である。
【図10】図10は、S.cerevisiaeにおいて発現された場合に、Alsタンパク質のサブセットが内皮細胞侵入を媒介することを示す。A、Alsタンパク質を発現する、または空のプラスミド(対照)で形質転換されたS.cerevisiae株の内皮細胞接着。データは、内皮細胞−関連生物の合計数を表し、高電力場当たりの細胞として表す。B、高電力場当たりの細胞内生物の数として表されたAlsp発現S.cerevisiae株の内皮細胞侵入の程度。単一因子偏差分析によって空のプラスミド対照と比較した場合、*p<0.01。結果は、三連で行った少なくとも3つの実験の平均±S.D.である。
【図11】図11は、公知の機能のAlsタンパク質のN末端アミノ酸配列の整列が、CRおよびHVRの交互パターンを示すことを示す。A、公知の機能のAlsタンパク質のN末端領域内のコンセンサス同一性のパーセンテージ。シグナルペプチド領域(アミノ酸1から20)は示されないことに注意されたい。塗りつぶしていないボックスは、HVR1から7として命名された領域を示す。B、個々のHVRの組成を示すAlsタンパク質(各々、配列番号1から6)の模式的整列。該配列を配置して、少数の同定された基材に結合する、または全く同定された基材に結合しないタンパク質と、多数の基材に対して親和性を持つタンパク質とを比較する。各保存された領域におけるアミノ酸配列の数を括弧に入れて示す。
【図12】図12は、Als1タンパク質N末端ドメインのCDおよびFTIRスペクトルを示す。A、リン酸緩衝化生理食塩水中の10μM Als1pの円二色スペクトル。B、D2O蒸気で水和されたAls1p自己−フィルムのFTIRスペクトル。
【図13】図13は、Alsタンパク質ファミリー内のN末端ドメインの予測された物理化学的特性の比較を示す。疎水性、静電性、または水素−結合特徴は各ドメインの水−接近可能表面に投影される。疎水性は以下のように示される:茶色、最も疎水性;青色、最も親水性。静電性(スペクトル連続)は以下のように示される:赤色、最も正の電荷(+10 kcal/モル);青色、最も負の電荷(−10 kcal/モル)。水素−結合ポテンシャル(H−結合)は以下のように示される:赤色、ドナー;青色、アクセプター。Alsタンパク質は、これらの特性の複合に基づいて3つの群に区別可能である。例えば、Als群Aタンパク質内の同様な疎水性、静電性および水素−結合プロフィール、Als1p、Als3p、およびAls5pに注意されたい。対照的に、Als群Bメンバー、Als6pおよびAls7pは、Als群Aのそれからの疎水性および静電性特徴の顕著な差を提示する。生化学的プロフィールに加えて、これらのドメイン内の予測される構造の差に注意されたい。
【図14】Alsファミリータンパク質における構造−機能関係の概念的モデル。Alsタンパク質は3つの一般的成分:N末端ドメイン、タンデムリピート、およびC.albicans細胞壁と界面を形成するグリコシルホスファチジルイノシトールアンカーを含有するセリン/スレオニン−リッチなC−末端ドメインから構成される。説明したように、Alsタンパク質は、免疫グロブリンスーパーファミリーに特徴的な、拡大されたスパンが間に存在する多数の保存された反―平行β−シート領域(CR1−n)を含有する。β−シートドメインから突出するのは、HVRを含有するループ/コイル構造である。特異的Alsタンパク質HVRの三次元物理化学的特性は、恐らくは、接着および侵入機能をCandidaに付与する宿主基材との相互作用を支配する。説明目的で、3つのN末端β−シート/コイルドメインおよびそれらの各CR/HVR成分のみを示す。この突出は、図13に示した構造イメージに対して直角で見られることに注意されたい。
【図15】rAls1p−Nでのマウス(リタイアしたブリーダー)の免疫化は、引き続いての播種性カンジダ症の間の生存を改良する。Als1p+アジュバントで免疫化されたマウスの生存。N=異なる日での二連実験における群当たり16匹のマウス;Adj.=アジュバント。*p<0.05 vsアジュバント。
【図16】rAls1p−Nでの免疫化は、リタイアしたブリーダーおよび若年マウス双方の生存を改良する。C.albicansの迅速に致死的な106接種物で感染させたリタイアしたブリーダー(A)および若年性(B)マウスの生存。N=異なる日の二連実験における群当たり16匹のマウス;Adj.=アジュバント。*p<0.05 vsアジュバント対照。
【図17】抗rAls1p−N力価は生存と相関しない。アジュバントと共に、またはそれなくして、変化する用量のrAls1p−Nで免疫化されたBalb/cマウスにおいて生起した抗rAls1p−Nポリクローナル抗体の力価。Adj.=アジュバント。200μg vs.全ての他のものについて*p≦0.005。
【図18】rAls1p−Nの保護用量のみがC.albicans−刺激Th1脾臓細胞の増加を誘導する。異なる用量のrAls1p−NワクチンによるTh1(CD4+IFN−γ+IL−4−)およびTh2(CD4+IFN−γ−IL−4+)脾臓細胞の誘導。免疫化されたマウス(群当たりn=9)からの脾臓細胞を熱で殺した予め発芽したC.albicansで48時間刺激し、次いで、3−色フローサイトメトリーによって分析した。*p=0.03 vs.アジュバント。
【図19】rAls1p−Nの保護用量のみがrAls1p−N刺激遅延タイプの過敏の増加を誘導する。rAls1p−NまたはCFA単独でワクチン接種したマウス(群当たりn=9から12)における、足蹠膨潤によって評価した遅延タイプの過敏。マウスを示した量のrAls1p−Nで免疫化し、次いで、50μgのrAls1p−Nを足蹠に注射した。足蹠膨潤は24時間後に評価した。*p<0.05−vs−アジュバント、0.2μg、および200μg。
【図20】rAls1p−Nワクチンは、保護的免疫性を誘導するのにB細胞ではなくT細胞を必要とする。B細胞−欠乏、T細胞−欠乏(ヌード)およびコンジェニック野生型Balb/c対照マウス(群当たりn=7または8)の生存を、rAls1p−N+アジュバントまたはアジュバント単独でのワクチン接種後に同時に評価した。*p<0.04−vs−アジュバント単独、¶p=0.003−vs−野生型アジュバント−処理。
【図21】rAls1p−NでのSQワクチン接種は免疫コンピテントマウスにおいてインビボDTH応答を誘導する。足蹠膨潤は、50μgのrAls1p−NのBALB/cマウス(群あたりn=10)における足蹠への注射から24時間後に評価した。メジアン値は黒色棒線として表示する。Wilcoxon順位和検定によって*p=0.002 vs.対照。
【図22】rAls1p−Nワクチンは、血行的に散在したカンジダ症を持つ免疫コンピテントマウスの生存を改良し、組織真菌負荷を低下させる。A)C.albicansで尾−静脈を介して引き続いて感染させた、ワクチン接種したまたは対照BALB/cマウス(各々、2.5または5 × 105接種物について群当たりn=7または10)の生存。各実験は感染から30日後に停止し、全ての残りのマウスは良好に見えた。Log順位検定により*p<0.05 vs.対照。B)C.albicansの5×105出芽胞子で尾静脈を介して感染させたBALB/cマウス(群当たりn=7)における腎臓真菌負荷。y軸はアッセイの検出の下限を反映する。メジアン値は黒色棒線として表示する。Wilxocon順位和検定によって*p=0.01 vs対照。
【図23】rAls1p−Nワクチンは好中球減少症マウスにおいてDTH反応を誘導し、引き続いての血行的に播種性のカンジダ症の間のそれらの生存を改良する。A)足蹠膨潤は、50μgのrAls1p−NのBALB/cマウス(対照についてはn=10、rAls1p−Nについてはn=8)の足蹠への注射から24時間後に評価した。Wilcoxon順位和検定により*p=0.006 vs対照。B)C.albicansの2.5 × 104出芽胞子で感染させた好中球減少症BALB/cマウス(2つの実験からの群当たりn=16)の生存。Log順位検定により*p=0.007 vsアジュバント対照。
【図24】rAls1p−Nワクチンは、口腔咽頭カンジダ症を持つマウスの舌上での組織病理学的真菌病相のひどさを低下させる。群当たりN=4匹マウス。テキスト中に記載されたような盲検観察者によって生じた炎症スコア。Wilcoxon順位和検定により*p=0.03。
【図25】図25は、rAls1p−NワクチンではなくrAls3p−Nワクチンが、C.albicansを接種したマウスの膣の真菌集落形成を減少させることを示す(Wlicoxon順位和検定により*p=0.01 vs CFA単独でワクチン接種したマウス)群当たりN=11匹マウス。
【図26】図26は、Als1p相同性マップ−vs−黄色ブドウ球菌クランピング因子A(c1n67A)を示す。C.albicansのAls1pおよび黄色ブドウ球菌のClfAからのコンセンサス機能的部位はAls1p相同性モデルにマッピングされた。Als1pおよびClfAのN末端からの多数の残基はコンセンサス溝モチーフにマッピングされ、これは、基材への結合が双方のアドヘシンについて起こると予測されるものである。
【図27】図27は、rAls1p−NおよびrAls3p−Nワクチンは、スタフィロコッカス血症マウスの生存を改良することを示す(Log順位検定により*p<0.003 vs CFA単独でワクチン接種したマウス)。群当たりN=22匹マウス。
【図28】図28は、抗体力価が個々のワクチン接種したマウスにおける保護の程度と相関しないが、それらはワクチン接種していないマウスをワクチン接種したマウスから区別することを示す。各々、CFA単独、またはrAls1p−NまたはrAls3p−NのCFA+20μgで免疫化されたBalb/cマウスにおいて生起した抗rAls1p−Nまたは抗rAls3p−Nポリクローナル抗体の力価。総じて、抗体力価および生存の間に有意な相関があり(rho=0.474,p=0.0057)、これは、抗体力価をワクチン保護のための同族マーカーとして用いることができることを示す。しかしながら、CFA単独を受けるマウスからのデータを排除すると、抗体−力価、およびrAls1p−NまたはrAls3p−Nをワクチン接種したマウスの生存の間に相関はなく(rho=0.041143,p=0.847)、これは、抗体がワクチンの保護の支配的メカニズムではないようであることを示す。
【図29】図29は、rAls1p−Nワクチンが非近交系CD1マウスを血行的に播種性のカンジダ症から保護することを示す。A)CD1マウス(群当たりn=8)を、rAls1p−N(20μg)+CFA、またはCFA単独でSQワクチン接種し、ブーストから14日後にC.albicans SC5314で尾−静脈を介して感染させた。B)CD1マウス(群当たりn=8)を、ミョウバンと共に種々の用量のrAls1p−Nまたはミョウバン単独でSQワクチン接種し、ブーストから14日後に、C.albicans SC5314で尾−静脈を介して感染させた。Log順位検定により*p<0.05 vs.アジュバント対照。
【図30】図30は、rAls1p−Nワクチンが、C.albicansのいくつかの株の1つで感染させたBalb/cマウスの生存を改良することを示す。rAls1p−N+CFA−vs−CFA単独で免疫化し、C.albicans 15563(7×105出芽胞子)、16240(4×105出芽胞子)、または36082(4×105出芽胞子)で尾−静脈を介して感染させたBalb/cマウスの生存(群当たりN=8匹マウス)。Log順位検定により*p<0.05 vs.アジュバント対照。
【図31】図31は、rAls1p−Nワクチンが、Candidaのいくつかの非albicans種で感染させたBalb/cマウスにおける組織真菌負荷を低下させることを示す。Balb/cマウス(群当たりn=5)をCFAまたはCFA+rAls1p−N(20μg)でワクチン接種し、C.glabrata、C.krusei、C.parapsilosisまたはC.tropicalisで尾−静脈を介して感染させた。感染接種物を種の名称の下に括弧に入れて示す。腎臓真菌負荷は感染から5日後に決定した。y軸はアッセイの検出の下限を反映する。多数の比較のための非パラメーターSteel検定により*p<0.05 vs.アジュバント対照。
【図32】図32は、rAls1p−Nに対して交差反応した抗体を、rAls3p−N−免疫化マウスが生じたことを示す。CFA単独、CFA+rAls1p−N、またはCFA+rAls3p−Nで免疫化した個々のマウスの力価。CFAおよびCFA+rAls3p−Nについて群当たりN=7匹マウス;CFA+rAls1p−Nについてn=6匹マウス。Mann Whitney U検定により*p<0.05 vs.CFA単独;**p<0.002 vs.CFA単独& p<0.011vs.CFA+rAls1p−N。棒線はメジアンを示す。
【図33】図33は、rAls1p−NおよびrAls3p−N双方がインビボ遅延型過敏応答につきマウスをプライムしたことを示す。マウス(CFAおよびCFA+rAls3p−Nについて群当たりn=7;CFA+rAls1p−Nについてn=6)をCFA単独、CFA+rAls1p−N、またはCFA+rAls3p−Nワクチン接種した。インビボでの遅延型過敏を足蹠膨潤によって測定した。Mann Whitney U検定によって*p<0.05 vs.CFA単独。棒線はメジアンを示す。
【図34】図34は、rAls1p−NおよびrAls3p−Nワクチンは、ネズミの血行的に播種性のカンジダ症に対する同様な有効性を媒介したことを示す。C.albicansの5×105出芽胞子で尾−静脈を介して感染させたBalb/cマウス(CFAおよびCFA+rAls3p−Nについての2つの実験からの群当たりn=15、およびCFA+rAls1p−Nについて2つの実験からn=14)の生存。実験は感染から28日後に停止させ、全ての残りのマウスは良好に見えた。Log順位検定により*p≦0.0001 vs.CFA対照。
【図35】図35は、インビボ遅延型過敏が播種性カンジダ症の間の生存と相関したことを示す。抗rAls1p−Nまたは抗rAls3p−N抗体力価および足蹠膨潤反応を、C.albicansでの尾−静脈を介する感染の2日前にマウス(CFAまたはCFA+rAls3p−Nについて群当たりn=7、CFA+rAls1p−Nについてn=6)において測定した。Spearman順位和検定で決定した相関。
【図36】図36は、rAls3p−Nワクチンがネズミ口腔咽頭カンジダ症の間において組織真菌負荷を有意に低下させたことを示す。口腔咽頭カンジダ症を持つマウス(CFAについてn=7、およびrAls1p−NまたはrAls3p−Nワクチン接種群について8)における舌真菌負荷。y軸はアッセイの検出の下限を反映する。Mann Whitney U検定により*p=0.005 vs.CFA。
【図37】図37は、ネズミカンジダ膣炎におけるCFA単独およびCFA+rAls1p−N双方と比較して、rAls3p−Nが膣真菌負荷を低下させたことを示す。CFA、CFA+rAls1p−N、またはCFA+rAls3p−Nワクチン接種したマウス(2つの実験からの群当たりn=11)における膣真菌負荷。y軸はアッセイの検出の下限を反映する。多数の比較のためのSteel検定により*p≦0.02 vs CFAおよびCFA+rAls1p−N。
【特許請求の範囲】
【請求項1】
医薬上許容され得る媒体中の、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンの免疫原性量を投与することを含む、黄色ブドウ球菌感染を治療または予防する方法。
【請求項2】
前記Alsタンパク質ファミリーメンバーが、Candida albicans、Candida krusei、Candida tropicalis、Candida glabrata、およびCandida parapsilosisよりなる群から選択されるCandida株に由来するAlsタンパク質を含む、請求項1に記載の方法。
【請求項3】
前記Alsタンパク質ファミリーメンバーがAls1pまたはAls3pから選択される、請求項1に記載の方法。
【請求項4】
前記免疫原性断片がAlsタンパク質ファミリーメンバーのN末端領域断片を含む、請求項1に記載の方法。
【請求項5】
前記投与が能動免疫、受動免疫またはその組合せを含む、請求項1に記載の方法。
【請求項6】
細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその機能的断片の有効量を投与して、Candidaが宿主の細胞または組織へと結合または侵入するのを阻害することを含む、黄色ブドウ球菌感染を治療または予防する方法。
【請求項7】
前記Alsタンパク質ファミリーメンバーが、Candida albicans、Candida krusei、Candida tropicalis、Candida glabrata、およびCandida parapsilosisよりなる群から選択されるCandida株に由来するAlsタンパク質を含む、請求項6に記載の方法。
【請求項8】
前記Alsタンパク質ファミリーメンバーがAls1pまたはAls3pから選択される、請求項6に記載の方法。
【請求項9】
前記細胞接着活性がゼラチン、フィブロネクチン、ラミニン、上皮細胞または内皮細胞への結合を含む、請求項8に記載の方法。
【請求項10】
前記機能的断片がAlsタンパク質ファミリーメンバーのN末端領域断片を含む、請求項6に記載の方法。
【請求項11】
前記宿主細胞が内皮細胞または上皮細胞起源の細胞を含む、請求項6に記載の方法。
【請求項1】
医薬上許容され得る媒体中の、細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその免疫原性断片を含むワクチンの免疫原性量を投与することを含む、黄色ブドウ球菌感染を治療または予防する方法。
【請求項2】
前記Alsタンパク質ファミリーメンバーが、Candida albicans、Candida krusei、Candida tropicalis、Candida glabrata、およびCandida parapsilosisよりなる群から選択されるCandida株に由来するAlsタンパク質を含む、請求項1に記載の方法。
【請求項3】
前記Alsタンパク質ファミリーメンバーがAls1pまたはAls3pから選択される、請求項1に記載の方法。
【請求項4】
前記免疫原性断片がAlsタンパク質ファミリーメンバーのN末端領域断片を含む、請求項1に記載の方法。
【請求項5】
前記投与が能動免疫、受動免疫またはその組合せを含む、請求項1に記載の方法。
【請求項6】
細胞接着活性を有する単離されたAlsタンパク質ファミリーメンバー、またはその機能的断片の有効量を投与して、Candidaが宿主の細胞または組織へと結合または侵入するのを阻害することを含む、黄色ブドウ球菌感染を治療または予防する方法。
【請求項7】
前記Alsタンパク質ファミリーメンバーが、Candida albicans、Candida krusei、Candida tropicalis、Candida glabrata、およびCandida parapsilosisよりなる群から選択されるCandida株に由来するAlsタンパク質を含む、請求項6に記載の方法。
【請求項8】
前記Alsタンパク質ファミリーメンバーがAls1pまたはAls3pから選択される、請求項6に記載の方法。
【請求項9】
前記細胞接着活性がゼラチン、フィブロネクチン、ラミニン、上皮細胞または内皮細胞への結合を含む、請求項8に記載の方法。
【請求項10】
前記機能的断片がAlsタンパク質ファミリーメンバーのN末端領域断片を含む、請求項6に記載の方法。
【請求項11】
前記宿主細胞が内皮細胞または上皮細胞起源の細胞を含む、請求項6に記載の方法。
【図1A】

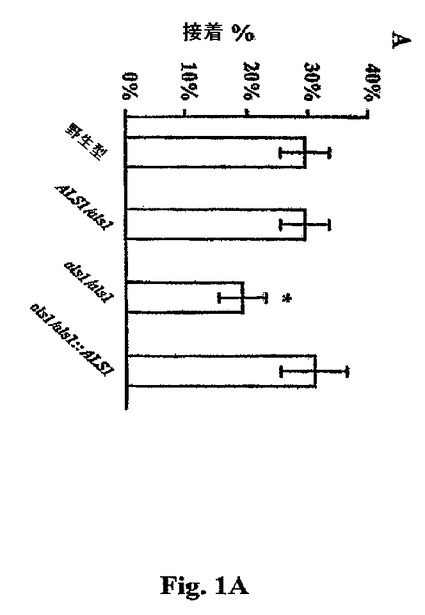
【図1B】
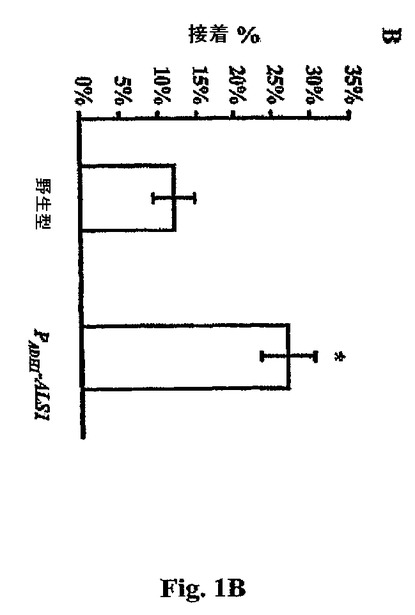
【図2A】

【図2B】

【図2C】
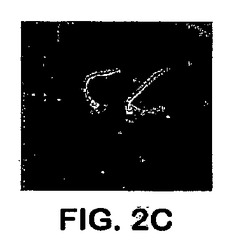
【図2D】
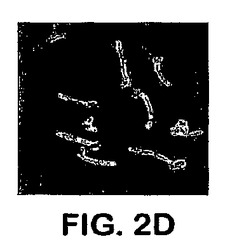
【図3A】
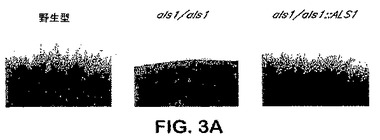
【図3B】
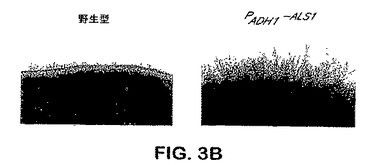
【図4A】
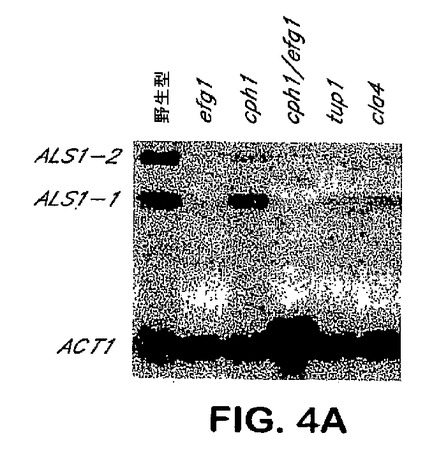
【図4B】
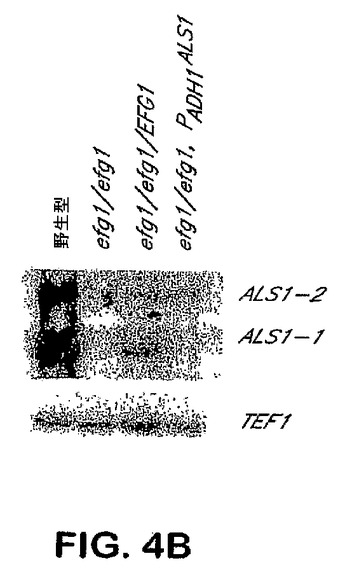
【図4C】

【図5A】
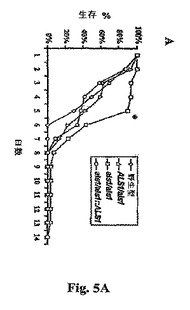
【図5B】

【図5C】
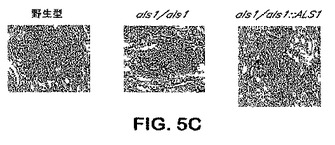
【図6】
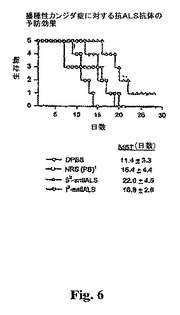
【図7】

【図8】
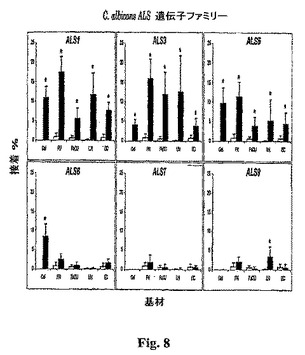
【図9】
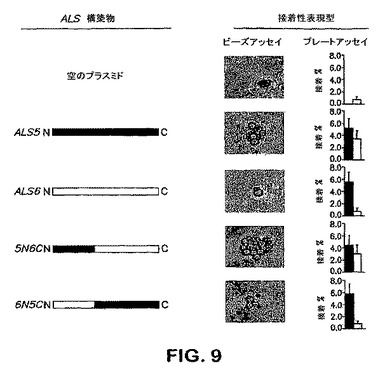
【図10】
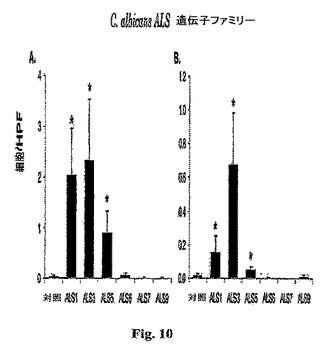
【図11】
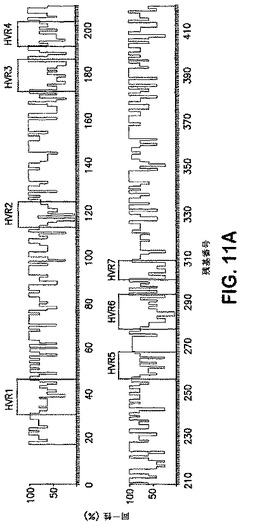
【図11B】

【図12】
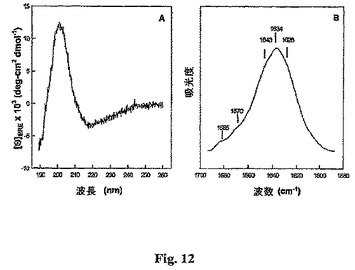
【図13】

【図14】
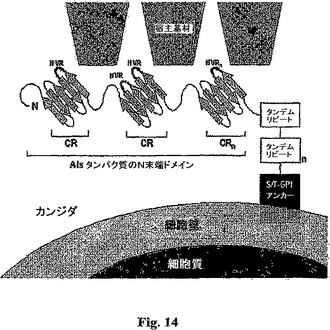
【図15】
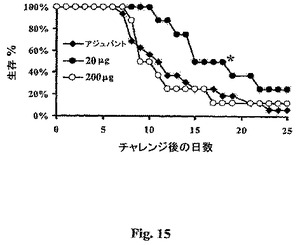
【図16】
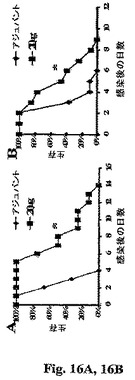
【図17】
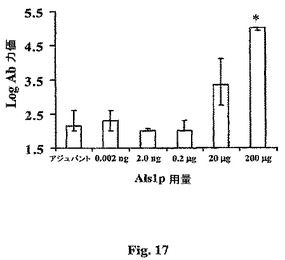
【図18】
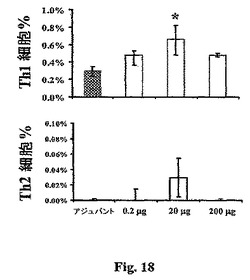
【図19】
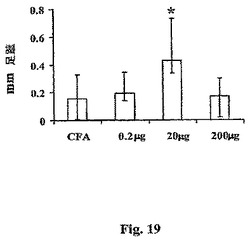
【図20】
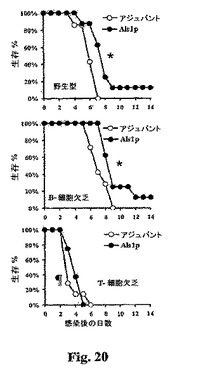
【図21】
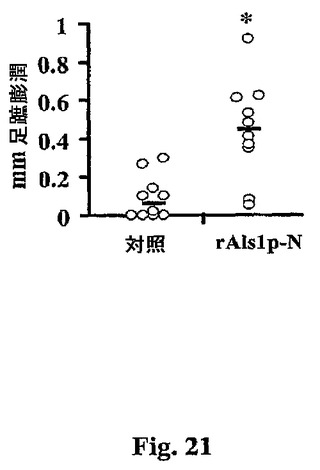
【図22】
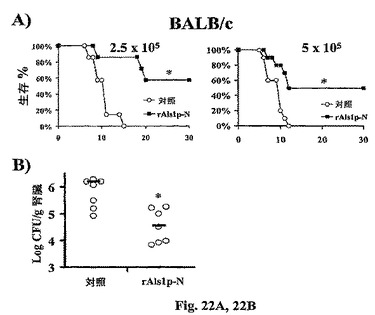
【図23】
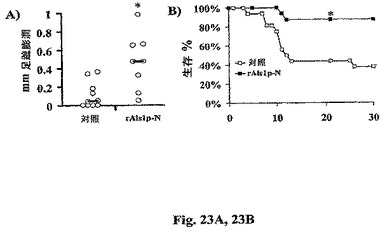
【図24】
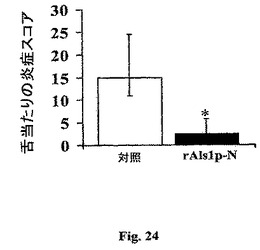
【図25】
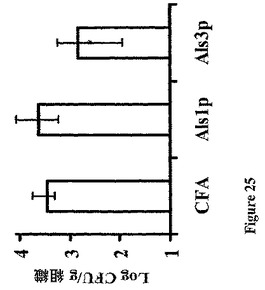
【図26】
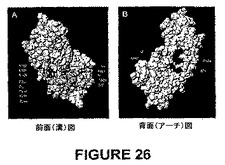
【図27】
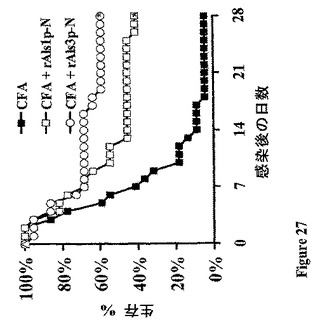
【図28】
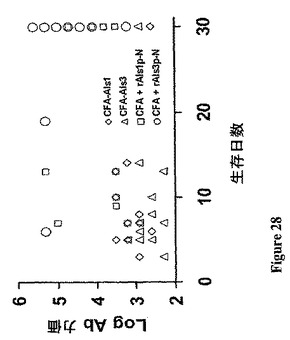
【図29】
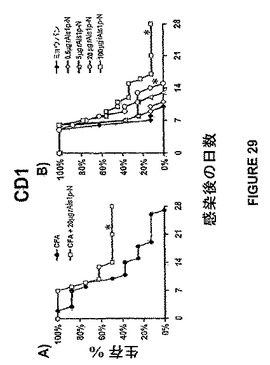
【図30】
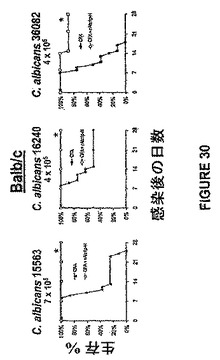
【図31】
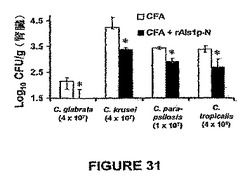
【図32】
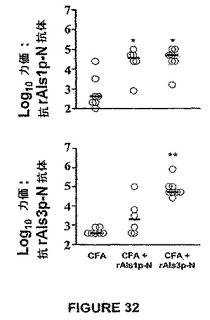
【図33】
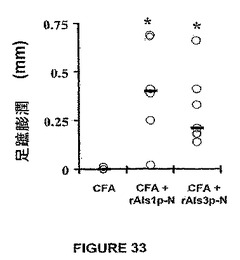
【図34】
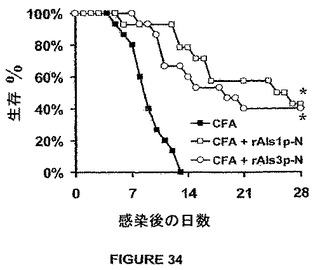
【図35】
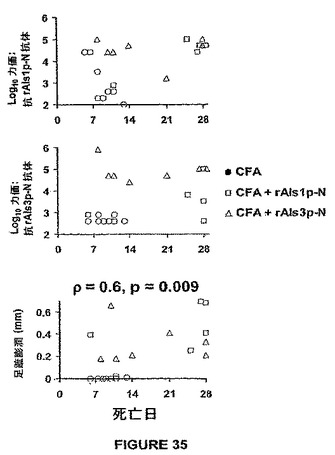
【図36】
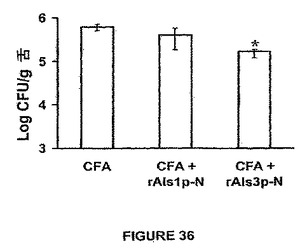
【図37】
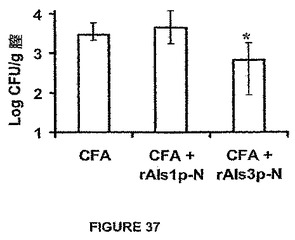
【図1B】
【図2A】
【図2B】
【図2C】
【図2D】
【図3A】
【図3B】
【図4A】
【図4B】
【図4C】
【図5A】
【図5B】
【図5C】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図11B】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【公表番号】特表2009−524601(P2009−524601A)
【公表日】平成21年7月2日(2009.7.2)
【国際特許分類】
【出願番号】特願2008−549598(P2008−549598)
【出願日】平成19年1月5日(2007.1.5)
【国際出願番号】PCT/US2007/000433
【国際公開番号】WO2007/081896
【国際公開日】平成19年7月19日(2007.7.19)
【出願人】(502170175)ロサンゼルス バイオメディカル リサーチ インスティテュート アット ハーバー− ユーシーエルエー メディカル センター (11)
【Fターム(参考)】
【公表日】平成21年7月2日(2009.7.2)
【国際特許分類】
【出願日】平成19年1月5日(2007.1.5)
【国際出願番号】PCT/US2007/000433
【国際公開番号】WO2007/081896
【国際公開日】平成19年7月19日(2007.7.19)
【出願人】(502170175)ロサンゼルス バイオメディカル リサーチ インスティテュート アット ハーバー− ユーシーエルエー メディカル センター (11)
【Fターム(参考)】
[ Back to top ]