
改善されたシトシンデアミナーゼ活性を有するポリペプチド
本発明は、アミノ酸配列を付加することによって天然のCDアーゼから誘導されることを特徴とする、CDアーゼ活性を有するポリペプチドであって、UPRTアーゼまたはチミジンキナーゼ活性を持たない、ポリペプチドに関する。
【発明の詳細な説明】
【発明の背景】
【0001】
発明の分野
本発明は、アミノ酸配列を付加することによって天然のCDアーゼから誘導されることを特徴とする、CDアーゼ活性を有するポリペプチドであって、UPRTアーゼまたはチミジンキナーゼ活性を持たない、ポリペプチドに関する。
【0002】
本発明は、CDアーゼ活性を有するこのポリペプチドをコードするヌクレオチド配列、CDアーゼ活性を有する上記ポリペプチドを発現するベクター、ウイルス粒子および宿主細胞、およびそれらを含んでなる組成物にも関する。最後に、本発明は、それらの治療使用、およびそれらを実行する治療の方法にも関する。本発明は、自殺遺伝子療法に関しては、特に増殖性および感染性疾患に関する応用に特に有用である。
【0003】
背景技術
遺伝子療法は、宿主細胞または生物へ遺伝情報を導入することであると定義される。ヒトに応用される第一のプロトコールは、アデニンデアミナーゼ(ADA)をコードする遺伝子に影響を及ぼす突然変異により遺伝的に免疫不全である患者について1990年9月に米国で開始された。この最初の実験が割合成功したことにより、遺伝病(欠陥遺伝子の機能不全を癒す目的の)および後天的疾患(癌、AIDSなどの感染症)の両方を含む様々な疾患に対するこの方法の開発が盛んになった。この手法では、そのとき以来発現産物が不活性物質(プロドラッグ)を細胞傷害性物質に形質転換することができる遺伝子を用いることによって、細胞死をもたらす「自殺遺伝子」療法など数多くの開発が行われてきた。1992年には、幾つかの研究グループが、この新規な方法の、腫瘍の治療およびAIDSに関与するHIVウイルスの伝播の阻害についての関連性を明らかにした。
【0004】
この点で、単純ヘルペス1型ウイルスチミジンキナーゼ(HSV−1 TK)は、自殺遺伝子のプロトタイプを構成している(Caruso et al., 1993, Proc. Natl. Acad. Sci. USA 90, 7024-7028; Culver et al., 1992, Science 256, 1550-1552; Ram et al., 1997, Nat. Med. 3, 1354-1361)。TKポリペプチドはそれ自体では毒性がないが、それはアシクロビールまたはガンシクロビール(GCV)のようなヌクレオシド類似体の形質転換を触媒する。改質ヌクレオシドは伸長過程にあるDNA鎖に組込まれ、結果として細胞分裂を阻害する。多数の自殺遺伝子/プロドラッグの対が、現在利用可能である。更に具体的に述べることができるものは、ラットチトクロームp450およびシクロホスファミド(Human Gene Therapy 5, 969-978)、Escherichia coli (E. Coli)プリンヌクレオシドホスホリラーゼおよび6−メチルプリンデオキシリボヌクレオシド(Sorscher et al., 1994, Gene Therapy 1, 223-238)、E. coliグアニンホスホリボシルトランスフェラーゼおよび6−チオキサンチン(Mzoz and Moolten, 1993, Human Gene Therapy 4,589-595)、およびシトシンデアミナーゼ(CDアーゼ)および5−フルオロシトシン(5FC)である。
【0005】
CDアーゼは、ピリミジン代謝経路に関与しており、これにより外来シトシンが加水分解的脱アミノ化によってウラシルに変換される。CDアーゼ活性は原核生物および下級真核生物では明らかにされているが(Jund and Lacroute, 1970, J. Bacteriol. 102, 607-615; Beck et al., 1972, J. Bacteriol. 110, 219-228; De Haan et al., 1972, Antonie van Leeuwenhoek 38, 257-263; Hoeprich et al., 1974, J. Inf. Dis. 130, 112-118; Esders and Lynn, 1985, J. Biol. Chem. 260, 3915-3922)、それらは哺乳類では見られない(Koechlin et al., 1966, Biochem Pharmacol. 15, 435-446; Polak et al., 1976, Chemotherapy 22, 137-153)。Saccharomyces cerevisiae (S. cerevisiae) FCY1およびE. coli codA遺伝子であって、これら2種類の生物のCDアーゼをそれぞれコードする遺伝子が知られており、それらの配列が公表されている(欧州特許第402 108号明細書;Erbs et al., 1997, Curr. Genet. 31, 1-6;WO93/01281号明細書)。
【0006】
CDアーゼはまた、シトシンの類似体、すなわち5−フルオロシトシン(5−FC)を脱アミノすることによって、5−フルオロウラシル(5−FU)を形成し、これは5−フルオロ−UMP(5−FUMP)に転換されると、細胞傷害性の高い化合物である。酵素をコードする遺伝子を不活性化する突然変異のためまたは哺乳類細胞のようにこの酵素を天然で欠損しているためにCDアーゼ活性を欠いている細胞は5−FC耐性である(Jund and Lacroute, 1970, J. Bacteriol, 102, 607-615; Kilstrup et al., 1989, J. Bacteriol. 1989 171, 2124-2127)。対照的に、CDアーゼ活性をコードする配列が導入された哺乳類細胞は、5−FCに感受性になった(Huber et al., 1993, Cancer Res. 53, 4619-4626; Mullen et al., 1992, Proc. Nati. Acad. Sci USA 89, 33-37;WO93/01281号明細書)。更に、隣接する形質転換されていない細胞も5−FCに感受性になる(Huber et al., 1994, Proc. Natl. Acad. Sci. USA 91, 8302-8306)。傍観者効果と呼ばれるこの現象は、細胞が5−FUを分泌するCDアーゼ活性を発現しており、これが次に血漿膜を通って直接的に拡散することによって近隣細胞を中毒させることによる。拡散における5−FUのこの特性は、tk/GCVリファレンス系と比較して消極的な意味で利点があり、傍観者効果はそこでtkを発現している細胞と接触する必要がある(Mesnil et al., 1996, Proc. Natl. Acad. Sci. USA 93, 1831-1835)。従って、遺伝子療法、特に抗癌遺伝子療法に関してCDアーゼが提供する総ての利点は、容易に理解することができる。
【0007】
CDアーゼ活性を用いる方法の効率を改良するため、従来技術書類であるWO−A−96/16183号明細書では、CDアーゼとUPRTアーゼ活性を有する2ドメイン酵素をコードする融合タンパク質の使用を推奨し、発現プラスミドによって運ばれるハイブリッドcodA::uppまたはFCY1::FUR1遺伝子の導入が、トランスフェクションしたB16細胞の5−FCに対する感作を増すことをイン・ビトロで明らかにしている。WO99/54481号明細書は、N末端部分に欠失があるUPRTアーゼをコードする突然変異したFUR1遺伝子を用いることによるこの発明を改良している。
【発明の概要】
【0008】
本発明は、アミノ酸配列の付加により天然CDアーゼから誘導されることを特徴とする、CDアーゼ活性を有するポリペプチドであって、UPRTアーゼまたはチミジンキナーゼ活性を持たない、ポリペプチドを用いた点において、初期の手法の改良がある。
【0009】
本発明は、一層効率的なポリペプチドであって、これによって5−FCに対する細胞の感受性を増加し且つ自殺遺伝子を用いる遺伝子療法の展望を改良することができるものを提供する。この突然変異体は、多数の用途、特に抗癌および抗ウイルス用途、および細胞死を必要とする総ての用途に用いることができる。
【発明の具体的説明】
【0010】
「シトシンデアミナーゼ活性」(CDアーゼ活性)は、シトシンまたはその類似体の一つの脱アミノ化を包含するものと理解される。
【0011】
「天然のCDアーゼに由来する」とは、広義には、CDアーゼ活性を有する上記ポリペプチドが、上記の天然のCDアーゼと70%を上回り、有利には80%を上回り、好ましくは90%を上回り、極めて好ましくは95%を上回る同一性の程度を有するアミノ酸配列を含んでなることを意味する。
【0012】
本発明の意味においては、天然のCDアーゼは、原核生物または真核生物起源のCDアーゼを表す。好ましくは、CDアーゼは酵母CDアーゼであり、特にSaccharomyces cerevisiae FCY1遺伝子によってコードされたものである。異なる起源のCDアーゼをコードする遺伝子のクローニングおよび配列は、文献や専門のデーターベースで入手することができる。参考として、FCY1遺伝子の配列は、Erbs et al.の文献に開示されている(1997, Curr. Genet. 31, 1-6)。
【0013】
好ましい例は、実質的に配列番号1の配列アイデンティファイアーに表されている通りであり、1位のMet残基から出発して158位のGlu残基で終了するアミノ酸配列を含んでなる天然のCDアーゼである。「実質的に」という用語は同一性の程度を表し、上記配列番号1の配列では、これは70%を上回り、有利には80%を上回り、好ましくは90%を上回り、極めて好ましくは95%を上回る。更に一層好ましくは、このポリペプチドは、1位のMet残基から出発して158位のGlu残基で終了する配列番号1の配列アイデンティファイアーに表されているアミノ酸配列を含んでなる。1位のMet残基から出発して158位のGlu残基で終了する配列番号1の配列アイデンティファイアーに表されているアミノ酸配列を有するポリペプチドは、本発明の実施に極めて適している。
【0014】
有利な態様によれば、本発明によるポリペプチドは上記の天然CDアーゼよりかなり高いCDアーゼ活性を示す。従って、下記の実施例は、UPRTアーゼ活性を持たないアミノ酸配列を付加することによって、5−FCに対する標的細胞の感作および/または治療を受けた動物に誘導される傍観者効果を増加することができる。感作を増加する因子は、有利には少なくとも2であり、好ましくは少なくとも5であり、極めて好ましくは10以上である。
【0015】
「UPRTアーゼ活性を持たないポリペプチド」は、5−FUを5−FUMPに転換することができないポリペプチドを包含する。5−FUを5−FUMPに転換するアミノ酸配列の能力は、本願明細書の実施例に開示されている方法を用いることによって評価することができる。
【0016】
「チミジンキナーゼ活性を持たないポリペプチド」は、ガンシクロビール(9−{[2−ヒドロキシ−1−(ヒドロキシメチル)エトキシルメチル}グアノシン)、ファムシクロビール、ブシクロビール、ペンシクロビール、バルシクロビール、アシクロビール(9−[2−ヒドロキシエトキシ)メチル]グアノシン)、トリフルオロチミジン、1−[2−デオキシ,2−フルオロ,β−D−アラビノフラノシル]−5−ヨードウラシル、ara−A (アデノシンアラビノシド,ビバラビン)、1−β−D−アラビノフラノキシルチミン、5−エチル−2’−デオキシウリジン、5−ヨード−5’−アミノ−2,5’−ジデオキシウリジン、イドクスウリジン(5−ヨード−2’−デオキシウリジン)、AZT(3’−アジド−3’チミジン)、ddC(ジデオキシシチジン)、AIU(5−ヨード−5’−アミノ−2’,5’−ジデオキシウリジン)、およびAraC(シチジンアラビノシド)へすることができないポリペプチドを包含する。
【0017】
好ましい態様によれば、天然CDアーゼに付加したアミノ酸配列は、10〜1000アミノ酸長であり、更に好ましくは100〜400、非常に好ましくは200〜300アミノ酸長である。付加は天然CDアーゼの任意の部位に起こることができるが、NまたはC末端が好ましく、特にC末端が好ましい。
【0018】
有利には、天然CDアーゼに付加するアミノ酸配列は、UPRTアーゼ活性を有するポリペプチドから誘導される。
【0019】
「UPRTアーゼ活性を有するポリペプチドから誘導される」とは、広義には、上記アミノ酸配列のUPRTアーゼ活性ポリペプチドのアミノ酸配列との同一性の程度が70%を上回り、有利には80%を上回り、好ましくは90%を上回り、極めて好ましくは95%を上回ることを意味する。
【0020】
本発明の意味においては、UPRTアーゼ活性を有するポリペプチドは、ウラシルまたはその誘導体の一つを一リン酸類似体に、特に5−FUを5−FUMPに転換することができるポリペプチドを表す。「突然変異」とは、上記ポリペプチドの任意の部位における1以上の残基の付加、欠失および/または置換であると理解すべきである。
【0021】
本発明によるアミノ酸配列が由来することがあるUPRTアーゼ活性を有するポリペプチドは、任意の起源、特に原核生物、真菌または酵母起源のものであることができる。例えば、E. coli (Anderson et al., 1992, Eur. J. Biochem 204, 51-56)、Lactococcus lactis (Martinussen and Hammer, 1994, J. Bacteriol. 176, 6457-6463)、Mycobacterium bovis (Kim et al., 1997, Biochem Mol. Biol. mt 41, 1117-1124)、およびBacillus subtilis (Martinussen et al., 1995, J. Bacteriol. 177, 271-274)由来のUPRTアーゼは、本発明に関して用いることができる。しかしながら、酵母UPRTアーゼ、特にS.cerevisiae FUR1遺伝子によってコードされるものを用いるのが特に好ましく、その配列はKern et al.の文献に開示されている(1990, Gene 88, 149-157)。更に詳細には、例えば、遺伝子の配列、および相当するUPRTアーゼの配列は、文献や専門のデーターベースに見出すことができる(SWISSPROT、EMBL、Genbank、Medlineなど)。更に詳細には、fur 1−7、fur 1−8およびfur 1−9によってコードされた突然変異体を、本発明で用いることができる。好ましい態様では、UPRTアーゼ活性を有するポリペプチドに由来するアミノ酸配列は、fur 1−8対立遺伝子によってコードされたアミノ酸配列である(Kern et al., 1990, Gene 88, 149-157)。
【0022】
特に有利な態様によれば、本発明の天然CDアーゼに付加したアミノ酸配列は、天然UPRTアーゼの欠失突然変異体に由来する。この欠失は、好ましくは元のUPRTアーゼのN末端領域に位置している。欠失は全体的(上記N末端領域の総ての残基に影響を与える)または部分的(一次構造において連続的であることもまたはないこともある1以上の残基に影響を与える)である可能性がある。一般的には、ポリペプチドはN末端部分、中心部分、およびC末端部分からなり、それぞれの部分は分子の約三分の一を占める。例えば、251アミノ酸を含むS. cerevisiae UPRTアーゼの場合には、N末端部分は天然形態の最初の位置にあるいわゆる開始メチオニンから始まる最初の83残基からなっている。E. coli UPRTアーゼの場合には、N末端部分は1−69位を包含する。
【0023】
FUR1Δ105と呼ばれる突然変異体遺伝子の発現産物はS. cerevisiae furl突然変異体を補足することができることが示されており、これにより機能性であることが立証されている。
【0024】
極めて好ましくは、本発明によるUPRTアーゼ活性を有するポリペプチドは、少なくとも天然のUPRTアーゼの第二のATGコドンの上流のN末端領域を全部または一部欠失することによって上記天然のUPRTアーゼから誘導される。上記領域の完全な欠失が好ましい。例えば、FURl遺伝子によってコードされるUPRTアーゼは、+1位に第一のATGコドン(開始ATGコドン)に続いて+36位に第二のコドンを含んでなる。従って、本発明に関しては残基+1−35を考えることによって、天然形態の+36位に通常に見られるメチオニンから始まるポリペプチドを得ることできる。FUR1Δ105と呼ばれる突然変異体遺伝子の発現産物はS. cerevisiae furl突然変異体を補足することができることが示されており、これにより機能性であることが立証されている。
【0025】
本発明による天然CDアーゼに付加した好ましいアミノ酸配列は、実質的に配列番号2の配列アイデンティファイアー(identifier)に表されているものであり、2位のSer残基から出発して216位のVal残基で終了するアミノ酸配列を含んでなる。「実質的に」という用語は同一性の程度を表し、上記配列では、これは70%を上回り、有利には80%を上回り、好ましくは90%を上回り、極めて好ましくは95%を上回る。更に一層好ましくは、天然のCDアーゼに付加したアミノ酸配列は、2位のSer残基から出発して216位のVal残基で終了する配列番号2の配列アイデンティファイアーに表されているアミノ酸配列を含んでなる。2位のSer残基から出発して216位のVal残基で終了する配列番号2の配列アイデンティファイアーに表されているアミノ酸配列を有する天然のCDアーゼに付加したアミノ酸配列は、本発明の実施に極めて適している。天然のCDアーゼに付加したアミノ酸配列は、追加の突然変異を含むことができる。配列番号2の配列アイデンティファイアーの2位におけるセリン残基をアラニン残基で置換することを、特に挙げることができる。
【0026】
本発明によるCDアーゼ活性を有する好ましいポリペプチドは、実質的に配列番号1の配列アイデンティファイアーに表されている通りであり、1位のMet残基から出発して373位のVal残基で終了するアミノ酸配列を含んでなる。「実質的に」という用語は同一性の程度を表し、上記配列では、上記配列番号1の配列では70%を上回り、有利には80%を上回り、好ましくは90%を上回り、極めて好ましくは95%を上回る。更に一層好ましくは、ポリペプチドは、1位のMet残基から出発して373位のVal残基で終了する配列番号1の配列アイデンティファイアーに表されているアミノ酸配列を含んでなる。1位のMet残基から出発して373位のVal残基で終了する配列番号2の配列アイデンティファイアーに表されているアミノ酸配列を有するポリペプチドが、本発明の実施に極めて適している。
【0027】
一般的には、本発明によるポリペプチドは、通常の化学合成の方法によって、または組換えDNA法によって産生することができる(例えば、Maniatis et al., 1989,「実験室便覧」,Cold Spring Harbor, Laboratory Press,コールド・スプリング・ハーバー,ニューヨークを参照されたい)この理由により、本発明は、上記ポリペプチドをコードするヌクレオチド配列を細胞に導入して形質転換細胞を生成させ、上記形質転換細胞を上記ポリペプチドを産生させるのに適当な条件下で培養し、上記ポリペプチドを細胞培養物から回収する、調製方法も包含する。プロデューサー細胞は、任意の起源のものであり、考慮中のヌクレオチド配列がそのゲノムに組込まれまたは複製することができる適当な発現ベクターに組込まれる限り制限がなく、細菌、酵母または哺乳類細胞であることができる。天然では、ヌクレオチド配列は、転写および翻訳シグナルの制御下に置かれることにより、プロデューサー細胞で発現させることができる。発現ベクターおよび制御シグナルは、熟練者に知られている。ポリペプチドは、培地または(リーシスした後の)細胞から回収し、通常の精製段階(クロマトグラフィー、電気泳動、濾過、免疫精製などによる)を施すことができる。
【0028】
本発明は、本発明によるポリペプチドをコードするヌクレオチド配列にも関する。ヌクレオチド配列は、cDNAまたはゲノム配列、または混合型であることができる。これは、場合によっては天然、異種(例えば、ウサギβ−グロブリン遺伝子のイントロンなど)、または合成起源の1種類以上のイントロンを含み、宿主細胞での発現を増加することができる。本発明に関して用いられる配列は、分子生物学の通常の手法によって、例えば、特定のプローブを有するライブラリーをスクリーニングすることによって、発現ライブラリーの免疫スクリーニングによって、または適当なプライマーを用いるPCRによって、あるいは化学合成によって得ることができる。突然変異体は、位置指定突然変異誘発、PCR、制限および連結酵素による消化の手法を用いて1以上のヌクレオチドを置換、欠失および/または付加することによって、あるいは化学合成によって、天然配列から生成させることができる。突然変異体および構築体が機能する能力は、酵素活性を評価することによって、または標的細胞の5−FCおよび/または5−FUに対する感受性を測定することによって明らかにすることができる。
【0029】
本発明は、宿主細胞中での発現に必要な要素の制御下に置かれた本発明によるヌクレオチド配列を有する組換えベクターにも関する。組換えベクターは、プラスミドまたはウイルス起源のものであることができ、場合によってはベクターのトランスフェクション効率および/または安定性を向上させる1以上の物質と組み合わせることができる。これらの物質は、熟練者が利用可能な文献に広汎に報告されている(例えば、Feigner et al., 1987, Proc. West. Pharmacol. Soc. 32,115-121; Hodgson and Solaiman, 1996, Nature Biotechnology 14, 339-342; Remy et al., 1994, Bioconjugate Chemistry, 5,647-654)。非制限的例としては、これらの物質は、ポリマー、脂質、特にカチオン性脂質、リポソーム、核タンパク質、または中性脂質であることができる。これらの物質は、単独でまたは組み合わせて用いることができる。考えられる組合せは、カチオン性脂質(DOGS、DC−CHOL、スペルミン−chol、スペルミジン−cholなど)、リゾリン脂質(例えば、ヘキサデシルホスホコリン)、および中性脂質(DOPE)と組み合わせる組換えプラスミドベクターの組合せである。
【0030】
好ましい態様によれば、本発明で用いることができるカチオン性脂質は、欧州特許第901463B1号明細書に記載のカチオン性脂質であり、更に好ましくはpcTG90である。
【0031】
本発明に関して用いることができるプラスミドの選択は、広大なものである。それらは、クローニングベクターおよび/または発現ベクターであることができる。一般的には、それらは熟練者に知られており、それらの多くは市販されているが、遺伝子操作の手法を用いてそれらを構築しまたはそれらを改質することもできる。挙げることができる例は、pBR322 (Gibco BRL)、pUC (Gibco BRL)、pBluescript (Stratagene)、pREP4、pCEP4 (Invitrogene)、またはpPoly (Lathe et al., 1987, Gene 57, 193-201)から誘導されるプラスミドである。好ましくは、本発明に関して用いられるプラスミドは、複製がプロデューサー細胞および/または宿主細胞で確実に開始されるようにする複製起源を含む(例えば、ColE1起源はE. coliで産生しようとするプラスミドについて選択され、oriP/EBNA1系は哺乳類宿主細胞で自己複製することが所望な場合に選択される。Lupton and Levine, 1985, Mol. Cell. Biol. 5, 2533-2542; Yates et al., Nature 313, 812-815)。プラスミドは、トランスフェクション細胞を選択または同定することができるようにする選択遺伝子をも含んでなる(栄養要求性変異、抗生物質耐性をコードする遺伝子などの補足)。天然では、プラスミドは、所定の細胞でその維持および/またはその安定性を改良する追加要素を含むことができる(cer配列であって、モノマー形態でのプラスミドの維持を促進する。(Summers and Sherrat, 1984, Cell 36, 1097-1103)細胞ゲノムへ組込む配列)。
【0032】
ウイルスベクターに関しては、ポックスウイルス(ワクシニアウイルス、特にMVA、カナリアポックスウイルスなど)、アデノウイルス、レトロウイルス、ヘルペスウイルス、アルファウイルス、フォーミーウイルス、またはアデノ随伴ウイルスから誘導されるベクターを考えることができる。複製コンピテントまたは複製欠失ベクターを用いることができる。組込みを行わないベクターを用いるのが好ましい。この点において、アデノウイルスベクターおよびMVAが、本発明の実施に極めて適している。
【0033】
好ましい態様によれば、本発明のウイルスベクターは、改質ワクシニアウイルスアンカラス(MVA)に由来する。MVAベクターおよびそれらのベクターを産生する方法は、欧州特許第83286号および欧州特許第206920号明細書、並びにMayr et al. (1975, Infection 3, 6-14)およびSutter et Moss (1992, Proc. Natl. Acad. Sci. USA 89,10847-10851)に詳細に記載されている。更に好ましい態様によれば、本発明のヌクレオチド配列をMVAベクターの欠失I、II、III、IV、VおよびVIへ、更に一層好ましくは欠失IIIに挿入することができる(Meyer et al., 1991, J. Gen. Virol. 72, 1031-1038 ; Sutter et al., 1994, Vaccine 12, 1032-1040)。
【0034】
レトロウイルスは、細胞に感染し、ほとんどの場合には細胞と結合して、分割させる特性を有し、この点において、癌に関連した使用に特に適している。本発明による組換えレトロウイルスは、一般にLTR配列、キャプシド化領域、および本発明によるヌクレオチド配列であって、レトロウイルスLTRまたは以下に記載されるような内部プロモーターの制御下に置かれているものを含む。組換えレトロウイルスは、任意の起源(ネズミ、霊長類、ネコ、ヒトなど)のレトロウイルスから、特にMOMuLV(Moloneyネズミ白血病ウイルス)、MVS(ネズミ肉腫ウイルス)、またはFriendネズミレトロウイルス(Fb29)から誘導することができる。これは、ウイルス粒子を構成するのに必要なウイルスポリペプチドgag、polおよび/またはenvを途中で供給することができるキャプシド化細胞系で増殖する。このような細胞系は、文献に記載されている(PA317、Psi CRIP GP +Am−12など)。本発明によるレトロウイルスベクターは、特にLTR(プロモーター領域の真核生物プロモーターによる置換)またはキャプシド化領域(異種キャプシド化領域、例えば、VL30型による置換)に修飾を含むことができる(仏国特許出願第94 08300号明細書および第97 05203号明細書を参照されたい)。
【0035】
複製に本質的であり且つE1、E2、E4およびL1−L5領域から選択される少なくとも1つの領域の全部または一部を欠くアデノウイルスベクターを用いて、国ベクターが宿主生物または環境中で増殖しないようにすることも好ましい。E1領域の欠失が好ましい。しかしながら、それを、特に欠陥のある本質的機能が補足細胞系および/またはヘルパーウイルスによって途中で補足される程度までE2、E4および/またはL1−L5領域の全部または一部に影響を与える他の(複数の)修飾/欠失と組み合わせることができる。この点では、当該技術分野の状況の第二世代ベクターを用いることができる(例えば、国際特許出願WO−A−94/28152号明細書およびWO−A−97/04119号明細書を参照されたい)。例えば、E1領域の主要部分およびE4転写単位の欠失が、特に有利である。クローニング容量を増加させるために、アデノウイルスベクターは更に非本質的なE3領域の全部または一部を欠くことができる。もう一つの代替例によれば、キャプシド化に本質的な配列、すなわち5’および3’ITR(逆方向末端反復配列)とキャプシド化領域を保持する最小アデノウイルスベクターを用いることができる。様々なアデノウイルスベクターおよびそれらを調製するための手法が知られている(例えば、Graham and Prevect, 1991, 「分子生物学の方法」,第7巻,p109-128; E. J. Murey監修,The Human Press mcを参照されたい)。
【0036】
更に、本発明によるアデノウイルスベクターの起源は、種の観点および血清型の観点の両方から変動することができる。ベクターは、ヒトおよび動物(イヌ、トリ、ウシ、ネズミ、ヒツジ、ブタ、サルなど)起源のアデノウイルスのゲノムから、または少なくとも2種類の異なる起源のアデノウイルスゲノム断片を含んでなるハイブリッドから誘導することができる。更に詳細には、イヌ起源のCAV−1またはCAV−2アデノウイルス、トリ起源のDAVアデノウイルス、またはウシ起源のBad 3型アデノウイルスを挙げることができる(Zakharchuk et al., Arch. Virol. , 1993, 128: 171-176; Spibey and Cavanagh, J. Gen. Virol. 1989, 70:165-172; Jouvenne et al., Gene, 1987, 60: 21-28; Mittal et al., J. Gen. Virol., 1995, 76: 93-102)。しかしながら、ヒト起源のアデノウイルスベクターであって、好ましくは血清型C−アデノウイルス、特に2または5型の血清型Cアデノウイルスから誘導されるものが好ましい。
【0037】
本明細書で用いられる「複製コンピテント」という用語は、任意のトランス相補性(transcomplementation)の非存在下における宿主細胞で複製することができるウイルスベクターを表す。本発明に関しては、この用語は、癌または過剰増殖性宿主細胞で一層良好にまたは選択的に複製するように遺伝子工学処理された複製選択的または条件複製アデノウイルスも包含する。
【0038】
本発明の好ましい態様によれば、複製コンピテントベクターは複製コンピテントアデノウイルスベクターである。これらの複製コンピテントアデノウイルスベクターは、当業者には周知である。これらの中で、ONYX−015ウイルスのように55kD P53インヒビターをコードするE1b領域で欠失したアデノウイルスベクターが特に好ましい(Bischoff et al, 1996; Heise et al., 2000;WO94/18992号明細書)。従って、このウイルスは、p53欠損腫瘍細胞に選択的に感染して殺すのに用いることができる。当業者であれば、公表された手法に従ってアデノウイルス5または他のウイルスにおけるp53インヒビター遺伝子を突然変異させ、崩壊させることもできる。E1A Rb結合領域において欠失したアデノウイルスを、本発明に用いることもできる。例えば、E1A領域に24塩基対の欠失を有する突然変異アデノウイルスであるデルタ24ウイルス(Fueyo et al., 2000)。デルタ24はRb結合領域に欠失を有し、Rbに結合しない。従って、突然変異ウイルスの複製は、正常な細胞ではRbによって阻害される。しかしながら、Rbが不活性化しており、細胞が腫瘍性になる場合には、デルタ24は最早阻害されない。代わりに、突然変異体ウイルスは効率的に複製し、Rb欠損細胞をリーシスする。
【0039】
本発明によるアデノウイルスベクターは、連結反応または相同組換えによって、または補足細胞系での組換えによってEscherichia coli(E. coli)でイン・ビトロで生成することができる(例えば、国際特許出願WO−A−96/17070号明細書を参照されたい)。
【0040】
発現に必要な要素は、ヌクレオチド配列をRNAおよびmRNAに転写して、ポリペプチドに翻訳することができる総ての要素からなっている。これらの要素は、特に調節可能であるかまたは構成的であることがあるプロモーターを含んでなる。天然では、このプロモーターは、選択されたベクターおよび宿主細胞に適している。挙げることができる例は、PGK(ホスホグリセレートキナーゼ)、MT(メタロチオネイン;Mclvor et al., 1987, Mol. Cell Biol. 7,838-848)、α−1アンチトリプシン、CFTR、界面活性剤、免疫グロブリン、β−アクチン(Tabin et al., 1982, Mol. Cell Biol. 2,426-436)およびSRa(Takebe et al., 1988, Mol. Cell Biol. 8, 466-472)遺伝子の真核生物プロモーター、SV40ウイルス(シミアンウイルス)の初期プロモーター、RSV(ラウス肉腫ウイルス)のLTR、HSV−1 TKプロモーター、CMVウイルス(サイトメガロウイルス)の初期プロモーター、ワクシニアウイルスのp7.5K pH5R、pKlL、p28およびp11プロモーター、およびE1AおよびMLPアデノウイルスプロモーターである。プロモーターは、腫瘍または癌細胞での発現を刺激するプロモーターであることもできる。特に、胸部および前立腺癌で過剰発現するMUC−1遺伝子(Chen et al., 1995, J. Clin. Invest. 96, 2775-2782)、結腸癌で過剰発現するCEA(癌胎児性抗原を表す)遺伝子(Schrewe et al., 1990, Mol. Cell. Biol. 10, 2738-2748)、黒色腫で過剰発現するチロシナーゼ遺伝子(Vile et al., 1993, Cancer Res. 53, 3860-3864)、胸部および膵臓癌で過剰発現するERBB−2遺伝子(Harris et al., 1994, Gene Therapy 1, 170-175)、肝臓癌で過剰発現するα−フェトプロテイン遺伝子(Kanai et al., 1997, Cancer Res. 57, 461-465)を挙げることができる。サイトメガロウイルス(CMV)初期プロモーターが、極めて好ましい。
【0041】
しかしながら、ワクシニアウイルスに由来するベクター(例えば、MVAベクターとして)を用いるときには、チミジンキナーゼ7.5K遺伝子のプロモーターが特に好ましい。
【0042】
必要な要素としては、更に、本発明によるヌクレオチド配列の発現または宿主細胞中でのその維持を向上させる追加要素が挙げられる。イントロン配列、分泌シグナル配列、核局在化配列、IRES型の翻訳の再開始のための内部部位、転写終結ポリA配列、トリパルタイト(tripartite)リーダーおよび複製の起源が、特に挙げられる。これらの要素は、熟練者に知られている。
【0043】
本発明による組換えベクターは、1個以上の目的とする追加遺伝子を含んでなることもでき、これらの遺伝子を同じ調節要素(ポリシストン性カセット)または独立した要素の制御下に置くことができる。特に挙げることができる遺伝子は、インターロイキンIL−2、IL−4、IL−7、IL−10およびIL−12、インターフェロン、腫瘍壊死因子(TNF)、コロニー刺激因子(CSF)、特にGM−CSF、および脈管形成に作用する因子(例えば、PAl−1、プラスミノーゲンアクチベーターインヒビターを表す)をコードする遺伝子である。一つの具体的態様では、本発明による組換えベクターは、IL−2をコードするまたはインターフェロンγ(INFγ)をコードする目的の遺伝子を含んでなる。本発明によるヌクレオチド配列を、HSV−1 TK遺伝子、リシン遺伝子、コレラ毒素遺伝子などの他の自殺遺伝子と組み合わせることも考えることができる。
【0044】
本発明は、本発明による組換えベクターを含んでなるウイルス粒子にも関する。このようなウイルス粒子は、当該技術分野で通常の任意の手法を用いてウイルスベクターから生成することができる。ウイルス粒子を、ベクターの欠損に適当な補足細胞で増殖させる。アデノウイルスベクターに関しては、例えば、ヒト胎児腎臓細胞を用いて確立され且つA549−E1細胞系(Imler et al., 1996, Gene Therapy 3, 75-84)または二重相補性が可能な細胞系(Yeh et al., 1996, J. Virol. 70, 559-565; Krougliak and Graham, 1995, Human Gene Therapy 6, 1575-1586; Wang et al., 1995 Gene Therapy 2, 775-783;国際特許出願第WO 97/04119号明細書)のE1機能(Graham et al., 1977, J. Gen. Virol. 36, 59-72)を効率的に補足する293細胞系が用いられる。ヘルパーウイルスを用いて、欠損機能を少なくとも部分的に補足することもできる。補足細胞は、ウイルスゲノムをウイルスキャプシドにキャプシド化して組換えベクターを含むウイルス粒子を生成するのに必要な初期および/または後期因子を途中で供給することができる細胞と理解される。上記細胞は、それ自身のベクターの欠損機能を総て補足することができないことがあり、この場合には、追加機能を供給するベクター/ヘルパーウイルスでトランスフェクション/形質導入を行うことができる。
【0045】
本発明は、ウイルス粒子の製造方法であって、
(i) 本発明による組換えベクターを、途中で上記ベクターを補足してトランスフェクションした補足細胞を得るようにすることができる補足細胞に導入し、
(ii) 上記トランスフェクションした補足細胞を、ウイルス粒子を産生することができるようにするのに適当な条件下で培養し、
(iii) ウイルス粒子を細胞培養物から回収する
ことを特徴とする、方法にも関する。
【0046】
ウイルス粒子は培養上清から回収することができるのは勿論であるが、細胞から回収することもできる。一般に用いられるポリの一つは、連続的凍結/融解サイクルによって細胞をリーシスして、リーシス上清のビリオンを回収することからなっている。次に、ビリオンを、当該技術分野の手法(クロマトグラフィー法、超遠心法、特に塩化セシウムグラディエントによる方法など)を用いて増幅し、精製することができる。
【0047】
本発明は、本発明によるヌクレオチド配列または組換えベクターを含んでなる、または本発明によるウイルス粒子に感染している宿主細胞にも関する。本発明の本発明の趣旨では、宿主細胞は、上記に定義されているように、組換えベクターでトランスフェクションすることができる、またはウイルス粒子に感染させることができる任意の細胞からなっている。哺乳類細胞、特にヒト細胞が極めて好ましい。この細胞は、上記ベクターをゲノムに組込まれているまたは組込まれていない形態(エピソーム)で含んでなることができる。この細胞は、原発性または任意の起源の腫瘍細胞、特に造血細胞(全能性幹細胞、白血球、リンパ球、単球またはマクロファージなど)、筋細胞(衛星細胞、筋細胞、筋芽細胞、平滑筋細胞など)、心臓細胞、肺細胞、気管細胞、肝細胞、上皮細胞または繊維芽細胞であることができる。
【0048】
本発明は、本発明によるポリペプチド、ヌクレオチド配列、組換えベクター、ウイルス粒子、または宿主細胞を、薬学上許容可能な賦形剤と組み合わせて含んでなる、組成物にも関する。
【0049】
本発明は、更に、本発明によるポリペプチドと目的とする上記遺伝子の一つによってコードされる目的とするポリペプチドを含んでなる組成物に関する。これらの目的とするポリペプチドの中で、特にインターロイキンIL−2、IL−4、IL−7、IL−10およびIL−12、インターフェロン、腫瘍壊死因子(TNF)、コロニー刺激因子(CSF)、特にGM−CSF、および脈管新生に作用する因子(例えば、PAl−1、プラスミノーゲンアクチベーターインヒビターを表す)を挙げることができる。IL−2またはINFγが特に考えられる。
【0050】
組成物は、宿主細胞中で上記ポリペプチドを発現させることができるヌクレオチド配列に基づくこともできる。ヌクレオチド配列は、1個の同じ発現ベクターによってまたは2個の独立したベクターによって運ぶことができる。上記組成物は、上記の(複数の)ヌクレオチド配列を発現する(複数の)ウイルスベクターから生成されるウイルス粒子を含んでなることができるのは勿論である。
【0051】
本発明は、更に、本発明によるヌクレオチド配列と、IL−2およびINFγから選択されるポリペプチドをコードする目的とする第二のヌクレオチド配列を含んでなる、組成物にも関する。
【0052】
本発明による組成物は、更に具体的には遺伝子療法による疾患の予防的または治癒的治療をしようとするものであり、更に具体的には増殖性疾患(癌、腫瘍、再狭窄など)および感染性起源、特にウイルス起源(BまたはC型肝炎ウイルス、HIV、ヘルペス、レトロウイルスなどによって誘発される)の疾患を目指すものである。
【0053】
本発明による組成物は、それを局所的に、非経口的にまたは消化器経路によって投与する目的で常法によって製造することができる。特に、この治療薬または予防薬の治療上有効量を、薬学上許容可能な賦形剤と組み合わせる。多数の投与経路を考えることができる。例えば、胃内、皮下、心臓内、筋肉内、静脈内、腹腔内、腫瘍内、肺内、および気管内経路を挙げることができる。これらの3つの後者の態様の場合には、エアゾールによってまたは点滴によって投与を行うのが有利である。投与は、単回用量として、または特定の時間間隔の後に1回以上反復される用量として行うことができる。適当な投与および投薬経路は、様々なパラメーター、例えば、個体、治療を行う疾患、または目的とする(複数の)遺伝子によって変化する。本発明によるウイルス粒子を基剤とする製剤は、104〜1014pfu(プラーク形成単位)、有利には105〜1013pfu、好ましくは106〜1012pfu、更に好ましくは、アデノウイルス粒子を用いるときには109〜1010、およびMVA粒子を用いるときには106〜107の用量の形態で処方することができる。本発明による組換えベクターに関する限り、DNA 0.01〜100mg、好ましくは0.05〜10mg、特に好ましくは0.5〜5mgを含んでなる用量が考えられる。ポリペプチドを基剤とする組成物は、好ましくは上記ポリペプチド0.05〜10g、特に好ましくは0.05〜5gを含んでなる。用量は、医師が調整することができるのは当然である。
【0054】
処方物は、薬学上の観点から許容可能な希釈剤、アジュバントまたは賦形剤、並びに可溶化剤、安定剤および防腐剤を含むこともできる。注射可能な投与の場合には、水性、非水性または等張溶液の処方物が好ましい。これは、適当な希釈剤を用いて使用時点に再構成することができる液状または乾燥(粉末、凍結乾燥物などの)形態で、単回用量としてまたは多数回用量として提示することができる。この処方物は、適当量のプロドラッグを含んでなることもできる。
【0055】
本発明は、遺伝子療法によってまたは組換え経路によって産生されたタンパク質を投与することによってヒトまたは動物体の治療を目的とする薬剤を調製するための、本発明によるポリペプチド、組換えベクター、ウイルス粒子、または宿主細胞の治療または予防使用にも関する。第一の可能性によれば、この薬剤は、イン・ビボで(例えば、静脈内注射により接近可能な腫瘍中に、エアゾールによって肺に、適当なカテーテルを用いて脈管系に、など)直接投与することができる。患者から細胞(骨髄幹細胞、末梢血リンパ球、筋細胞など)を採取し、当該技術分野の手法によりそれらをイン・ビトロでトランスフェクションまたは乾癬させた後、それらを患者に再投与することからなる、エクス・ビボ法を採用することもできる。好ましい使用は、癌、腫瘍、および望ましくない細胞増殖から生じる疾患の治療または予防にある。考えられる応用は、胸部、子宮(特に、パピローマウイルスによって誘発されるもの)、前立腺、肺、膀胱、肝臓、結腸、膵臓、胃、食道、喉頭、中枢神経系(例えば、神経膠芽細胞腫)、および血液(リンパ腫、白血病など)の癌を挙げることができる。これは、循環器疾患に関して用いて、例えば、血管壁の平滑筋の増殖(再狭窄)を阻害しまたは遅らせることもできる。最後に、感染性疾患の場合には、AIDSに応用される薬剤を考えることができる。
【0056】
本発明は、本発明によるヌクレオチド配列、組換えベクター、ウイルス粒子、または宿主細胞を、そのような治療を必要とする生体または宿主細胞に投与することを特徴とする、遺伝子療法による疾患の治療方法にも敷衍される。
【0057】
有利な態様によれば、治療使用または治療方法は、薬学上許容可能な量のプロドラッグ、有利にはシトシンの類似体、特に5−FCを宿主生物または細胞に投与する追加段階も含んでなる。例えば、50〜500mg/kg/日の用量を用いることができ、200mg/kg/日の用量が好ましい。本発明に関しては、プロドラッグを標準的実施(例えば、経口、体系的)に従って投与し、投与は本発明による治療薬の投与の前に、と同時に、あるいはの後に行う。経口投与が好ましい。プロドラッグの単回用量、または宿主生物または細胞中に毒性代謝物を産生できるほど十分長い時間反復する用量を投与することができる。
【0058】
更に、本発明による組成物または方法を、5−FUの細胞傷害性効果を増強する1種類以上の物質と組み合わせることができる。特に、ピリミジンのデ・ノボ生合成の経路を阻害する薬剤(例えば、下記のもの)、5−FUの代謝生成物(5−FdUMP)の存在下でチミジレートシンターゼの阻害を増加し、複製に必要なdTMPの貯留を減少させるロイコボリンのような薬剤(Waxman et al., 1982, Eur. J. Cancer Clin. Oncol. 18, 685-692)、および最後に、ジヒドロホレートレダクターゼを阻害し、PRPP(ホスホリボシルピロホスフェート)の貯留を増加することによって5−FUの細胞RNAへの組込みを増加させるメトトレキセートのような薬剤(Cadman et al., 1979, Science 250, 1135-1137)を挙げることができる。
【0059】
本発明による組成物または方法は、抗癌療法に有効な1種類以上の物質と組み合わせることができる。本発明による組成物と共にまたはと組み合わせて用いることができる抗癌療法に有効な医薬物質の中には、例えば、マイトマイシンC、シクロホスファミド、ブスルファン、イホスファミド、イソスファミド、メルファラン、ヘキサメチルメラミン、チオテパ、クロラムブシルまたはダカルバジンのようなアルキル化剤、例えば、ゲンシタビン、カペシタビン、5−フルオロウラシル、シタラビン、2−フルオロデオキシシチジン、メトトレキセート、イダトレキセート、トムデックスまたはトリムトレキセートのような代謝拮抗薬、例えば、ドキソルビシン、エピルビシン、エトポシド、テニポシドまたはミトキサントロンのようなトポイソメラーゼII阻害薬、例えば、イリノテカン(CPT−11)、7−エチル−10−ヒドロキシ−カンプトテシン(SN−38)またはトポテカンのようなトポイソメラーゼI阻害薬、例えば、パクリタキセル、ドセタキセル、ビンブラスチン、ビンクリスチンまたはビノレルビンのような抗有糸分裂薬、および例えば、シスプラチン、オキサリプラチン、スピロプラチナムまたはカルボプラチナムのような白金誘導体を挙げることができる。本発明による組成物または方法は、放射線療法と組み合わせて用いることもできる。
【0060】
本発明は、哺乳類細胞における正の選択マーカーとしての本発明による配列または組換えベクターの使用にも関する。有利には、細胞をトランスフェクションし、次に、細胞混合物を、PALA (N(ホスホノアセチル)−L−アスパルテート;Moore et al., 1982, Biochem. Pharmacol. 31, 3317-3321)、A77 1726 (レフルノミドの活性代謝物;Davis et al., 1996, Biochem. 35, 1270-1273)およびブレキナール(Chen et al., 1992, Cancer Res. 52, 3251-3257)のようなピリミジンのデ・ノボ生合成の経路の阻害薬の存在下で培養する。このような阻害薬の存在により、RNAおよびDNAの合成に必要なUMPのデ・ノボ合成がブロックされることによって、細胞死を生じる。この細胞傷害効果は、ウラシルの存在下にてUPRTアーゼ活性をコードする本発明によるヌクレオチド配列を発現させることによって、またはシトシンの存在下にてこの後者の配列をCDアーゼ活性をコードする配列と(場合によっては、融合形態で)同時発現させることによって回避することができる。結果として、トランスフェクションした細胞(UPRTアーゼ/CDアーゼ配列を組込んだ細胞)は、ピリミジン合成経路の阻害薬の存在下で増殖することができるようになる。従って本発明による使用によって、トランスフェクションした細胞を、細胞混合物中で効率的に同定しおよび/または細胞混合物から単離することができる。
【0061】
本発明は、本発明による配列または組換えベクターの、負の選択マーカーとしての使用にも関する。例えば、、本発明による配列または組換えベクターを用いて、胎児性幹細胞または遺伝子が中断されまたは改質される核導入の後に得られる細胞を選択することができる(例えば、トランスジェニック動物の製造方法)(例えば、Capecchi, 1989, Science 244, 1288-1292; Reid et al., 1990, Proc. Natl. Acad. Sci. USA 87, 4299-4303を参照されたい)。例えば、ネオマイシン耐性についての遺伝子と組み合わせたこのような使用により、相同組換え事象を行っており且つゲネティシンおよび相当するフッ素化ピリミジン(CDアーゼ活性をコードするヌクレオチド配列を用いるときには、5−FC)の存在下にて単独で増殖することができる細胞を選択することができる。非標的組換え事象を行った細胞は、ゲネティシンの存在下で増殖することができるが、フッ素化ピリミジンの存在下では増殖することができない。負の選択マーカーとしてのもう一つの有望な使用は、哺乳類細胞と同様に、植物は内因性のCDアーゼ活性を持たないので、植物の分野において見出されるものである。それらを、本発明によるヌクレオチド配列をトランスフェクションすることによって5−FCに増感して、外来CDアーゼを発現することができる(例えば、Perera et al. 1993, Plant. Mol. Biol. 23, 797-799を参照されたい)。
【0062】
本発明による配列または組換えベクターは、細菌または酵母の負の選択マーカーとして用いることもできる。この特定の態様では、天然のCDアーゼ遺伝子を欠いている酵母または細菌が好ましく用いられる。これらの細菌の中では、欧州特許第0792369BB1号明細書に開示されているものが、特に好ましい。例えば、本発明による配列を、本発明による配列を含んでなるプラスミド中で、相同組換えによってクローニング配列に用いることができる。これらの実験では、本発明による配列は、クローニングを行う配列を挿入しなければならない部位に配置される。従って、相同組換えの後には、所望なプラスミドを含んでなる細胞は、5−FCの存在下で増殖することができる。
【実施例】
【0063】
実施例1: Fcu1−8の構築
FCU1−8は、pCI−neoFCU1(pTG13046)から出発してFCU1の直接突然変異誘発によって構築した。183位のArgは、オリゴ5’−gtattcttattactgatgatgg−3’(A549en Tを改質するため)を有するSerによって置換して、プラスミドpCI−neoFCU1−8(pTG15546)を得た。
【0064】
実施例2: 哺乳類細胞でのFCY1、FCU1およびFcu1−8の発現、およびCDアーゼおよびUPRTアーゼ活性の測定
プラスミドpCIneo(Promega)、pCI−neoFCY1(pTG15916)、pCI−neoFCU1(pTG13046)およびpCI−neoFCU1−8(pTG15546)によって一時的にトランスフェクションした哺乳類COS7の株を、CDアーゼおよびUPRTアーゼ活性について試験した。
【0065】
プロトコールは下記の通りである: 5 105 COS7 細胞を、DMEM/10% FCS培地5mlを含む37℃の60mmフラスコに播種した。24時間後、細胞をプラスミド5μgの非存在下または存在下でリポフェクチン(Gibco BRL)20μlで処理した。DMEM培地2ml中にて37℃で20時間後、細胞をDMEM/10% FCS 5mlの存在下でインキュベーションした。37℃で48時間後、細胞をPBSで洗浄した後、リーシス緩衝液(Tris−HCl pH7.5 50mM/NaCl 150mM/EDTA 5mM/DTT 1mM/トリトン1%)25μlに懸濁した。4℃で30分間リーシスし、遠心分離した後、CDアーゼおよびUPRTアーゼ活性を細胞溶解物を含む上清中で測定した。
【0066】
CDアーゼ活性については、反応性CDアーゼ緩衝液(Tris−HCl pH7.5 100mM/[H3] 5−FC 3mM,0.25μCi/μl)2μlの存在下にて37℃で20分間インキュベーションする。
【0067】
UPRTアーゼ活性については、細胞溶解物4μlを、反応性UPRTアーゼ緩衝液(Tris−HCl pH7.5 100mM/MgCl2 10mM/5−PRPP 10mM/[C14] 5−FU 3mM,0.02μCi/μl)2μlの存在下にて37℃で20分間インキュベーションする。
【0068】
酵素反応を、100℃で1分間停止する。1μlの分量を、TLCポリエチレンイミン−セルロースプレート(Merck)上に置く。水/ブタノール−1混合物(14%/86%)を溶媒として用いて、5−FUを5−FCから分離し、5−FUMPと5−FUの分離は、水を溶媒として用いて行う。移動の後、TLCプレートをPosphorImager(445SI;Molecular Dynamics)で走査した。タンパク質濃度は、Bradfordの方法によって測定した。
【0069】
3回の独立した測定から計算される酵素活性の結果を、下表に示す。
【0070】
【表1】

【0071】
FCU1遺伝子と比較して、Fcu1−8突然変異体はUPRTアーゼ活性を失うが、CDアーゼ活性は保存され、天然のFCY1遺伝子のCDアーゼ活性より10倍高い。遺伝子FCU1でのArgl83のSerへの変化によりUPRTアーゼ活性が失われるが、CDアーゼ活性に変化はない。
【0072】
実施例3: FCU1を発現するアデノウイルス(AdTG14800)の構築
FCU1遺伝子を含むpCI−neoFCU1(pTG13046)のXhoI−MluI断片を単離し、これらの同じ酵素によって開裂したトランスファーベクターpTG13387に導入して、トランスファーベクターpTG14799を得る。アデノウイルスベクターAD−FCU1(pTG14800)は、pTG14799のPacI−BstEII断片とClaIによって線形化したベクターpTG6624との間のBJ 5183 E. coli中での相同組換えによって再構築する。最終構築物AD−FCU1は、E1およびE3を欠失したAd5ゲノム(ヌクレオチド459−3510および28249−30758)を含み、E1の代わりに、FCU1遺伝子を含んでなる発現カセットをCMVプロモーターとβ−グロリン/IgGに続いてbGHのポリA配列の制御下に置く。FCU1を発現するアデノウイルス粒子(AdTG14800)は、相補細胞系でのトランスフェクションによって生成する(例えば、PERC6系)。
【0073】
実施例4: Fcu1−8を発現するアデノウイルスの構築(AdTG15606)
Fcu1−8遺伝子を含むpCI−neoFCU1−8(pTG15546)のXhoI−MluI断片を単離し、これらの同じ酵素によって開裂したトランスファーベクターpTG13387に導入して、トランスファーベクターpTG15547を得る。アデノウイルスベクターAD−FCU1−8(pTG15606)は、pTG15547のPacI−BstEII断片とClaIによって線形化したベクターpTG6624との間のBJ 5183 E. coli中での相同組換えによって再構築する。最終構築物AD−FCU1−8は、E1(NT 459−3510)およびE3(NT 28249−30758)領域を欠失したAd5ゲノムを含み、E1の代わりに、FCU1−8遺伝子を含んでなる発現カセットをCMV初期プロモーターハイブリッドβ−グロリン/IgGおよびMVCに続いてbGHのポリA配列の制御下に置く。FCU1−8を発現するアデノウイルス粒子(AdTG15606)は、E1機能を補足する細胞系でのトランスフェクションによって生成する(例えば、PERC6系)。
【0074】
実施例5: FCU1(AdTG14800)およびFcul−8(AdTG15606)を発現するアデノウイルスによる感染:イン・ビトロでの結果、CDアーゼとUPRTアーゼの活性の測定
ヒト腫瘍細胞系A549(ATCC Ccl−185)を、中空アデノウイルス(AdTG15149)、FCU1を発現するアデノウイルス(AdTG14800)、およびFcu1−8を発現するアデノウイルス(AdTG15606)によって感染させた。感染プロトコールは下記の通りである:PBS−2% FCS−l%カチオン50μlの懸濁液で5x106個の細胞をMOI 5でのアデノウイルスの存在下にて37℃で30分間インキュベーションする。次に、総量が5x106個の細胞を、DMEM−10% FCS 5mlの存在下にて60mmフラスコ中でインキュベーションし、細胞をPBSで洗浄した後、リーシス緩衝液(Tris−HCl pH7.5 50mM/NaCl 150mM/EDTA 5mM/DTT 1mM/トリトン1%)25μlに再懸濁した。4℃で30分間リーシスし、遠心分離の後、CDアーゼおよびUPRTアーゼ活性を細胞溶解物を含む上清中で測定した(上記プロトコールによる)。
【0075】
3回の独立した測定から計算される酵素活性の結果を、下表に示す。
【0076】
【表2】
【0077】
Fcu1−8を発現するアデノウイルスによって感染した細胞について、結果はUPRTアーゼ活性の喪失を示唆しているが、CDアーゼ活性は保存される。
【0078】
実施例6: FCU1を発現するMVAの構築(MVATG15637)
トランスファーベクターMva−fcu1(pTG15637)を、pCI−neoFCU1(pTG13046)のHindIII−KpnI断片とXhoIによって線形化したベクターpTG14269の間のBJ 5183 E.coli株中での相同組換えによって再構築する。このトランスファーベクター MVA−fcu1(pTG15637)は、p11K7.5プロモーターの制御下に置かれ且つMVAの欠失IIIに接する配列によって枠どられたFCU1遺伝子の発現カセットを含む。FCU1を発現するMVA粒子(MVATG15637、FCU1は欠失IIIに挿入され且つp11K7.5プロモーターの制御下にある)は、胎児性のニワトリ細胞でのMVAN33とプラスミドpTG15637の間の相同組換えによって生成する。
【0079】
実施例7: Fcu1−8を発現するMvaの構築(MVATG15638)
トランスファーベクターMVA−FCU1−8(pTG15638)は、pCI−neoFCU1−8(pTG15546)のHindIII−KpnI断片とXhoIによって線形化されたベクターpTG14269との間のBJ 5183 E.coli株での相同組換えによって再構築される。このトランスファーベクターMVA−FCU1−8(pTG15638)は、p11K7.5プロモーターの制御下に置かれ且つMVAの欠失IIIに接する配列によって枠どられたFcu1−8遺伝子の発現カセットを含む。Fcu1−8を発現するMVA粒子(MVATG15638、Fcu1−8は欠失IIIに挿入され且つp11K7.5プロモーターの制御下にある)は、胎児性のニワトリ細胞でのMVAN33とプラスミドpTG15638の間の相同組換えによって生成する。
【0080】
実施例8: FCU1(MVATG15637)とFCU1−8(MVATG15638)を発現するMVAによ
ルスによる感染:イン・ビトロでの結果、CDアーゼとUPRTアーゼの活性の測定
ヒト腫瘍細胞系A549(ATCC CCCL−185)を、中空MVA(MVAN33)、FCU1を発現するMVA(MVA15637)、およびFCU1−8を発現するMVA(MVA15638)によって感染させた。感染プロトコールは下記の通りである:PBS−2% FCS−l%カチオン50μlの懸濁液で5x106個の細胞をMOI 5でのアデノウイルスの存在下にて37℃で30分間インキュベーションする。次に、総量が5x106個の細胞を、DMEM−10% FCS 5mlの存在下にて60mmフラスコ中でインキュベーションする。37℃で24時間後、細胞をPBSで洗浄した後、リーシス緩衝液(Tris−HCl pH7.5 50mM/NaCl 150mM/EDTA 5mM/DTT 1mM/トリトン1%)25μlに再懸濁した。4℃で30分間リーシスし、遠心分離の後、CDアーゼおよびUPRTアーゼ活性を細胞溶解物を含む上清中で測定した(上記プロトコールによる)。
【0081】
3回の独立した測定から計算される酵素活性の結果を、下表に示す。
【0082】
【表3】
【0083】
FCU1−8を発現するMVAによって感染した細胞について、結果はUPRTアーゼ活性の喪失を示唆しているが、CDアーゼ活性は保存される。
【0084】
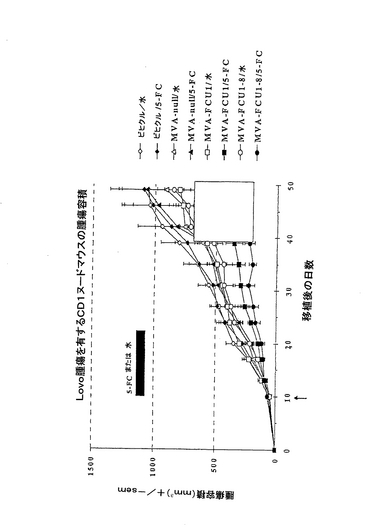
実施例9: イン・ビボでの実験
5x106個のヒト細胞LoVo(結腸腺癌)を、8群の15 CD1ヌードマウスにJ0に皮内投与する。腫瘍の容積が30−50mm3となったら直ぐに(J+10)、MVAを5x106 PFUの量を腫瘍内投与する。J+10から、0.5ml水または0.5ml 0.5% 5−FC溶液を経口で1日2回14日間投与する。腫瘍の容積は、群MVA−FCU1/5−FCと比較して群MVA−fCU1−8/5−FCでの腫瘍の大きさが一層良好に制御されることを強調している。
【図面の簡単な説明】
【0085】
【図1】本発明によるベクターを投与したCD1ヌードマウスでのLOVO腫瘍細胞の増殖。
【発明の背景】
【0001】
発明の分野
本発明は、アミノ酸配列を付加することによって天然のCDアーゼから誘導されることを特徴とする、CDアーゼ活性を有するポリペプチドであって、UPRTアーゼまたはチミジンキナーゼ活性を持たない、ポリペプチドに関する。
【0002】
本発明は、CDアーゼ活性を有するこのポリペプチドをコードするヌクレオチド配列、CDアーゼ活性を有する上記ポリペプチドを発現するベクター、ウイルス粒子および宿主細胞、およびそれらを含んでなる組成物にも関する。最後に、本発明は、それらの治療使用、およびそれらを実行する治療の方法にも関する。本発明は、自殺遺伝子療法に関しては、特に増殖性および感染性疾患に関する応用に特に有用である。
【0003】
背景技術
遺伝子療法は、宿主細胞または生物へ遺伝情報を導入することであると定義される。ヒトに応用される第一のプロトコールは、アデニンデアミナーゼ(ADA)をコードする遺伝子に影響を及ぼす突然変異により遺伝的に免疫不全である患者について1990年9月に米国で開始された。この最初の実験が割合成功したことにより、遺伝病(欠陥遺伝子の機能不全を癒す目的の)および後天的疾患(癌、AIDSなどの感染症)の両方を含む様々な疾患に対するこの方法の開発が盛んになった。この手法では、そのとき以来発現産物が不活性物質(プロドラッグ)を細胞傷害性物質に形質転換することができる遺伝子を用いることによって、細胞死をもたらす「自殺遺伝子」療法など数多くの開発が行われてきた。1992年には、幾つかの研究グループが、この新規な方法の、腫瘍の治療およびAIDSに関与するHIVウイルスの伝播の阻害についての関連性を明らかにした。
【0004】
この点で、単純ヘルペス1型ウイルスチミジンキナーゼ(HSV−1 TK)は、自殺遺伝子のプロトタイプを構成している(Caruso et al., 1993, Proc. Natl. Acad. Sci. USA 90, 7024-7028; Culver et al., 1992, Science 256, 1550-1552; Ram et al., 1997, Nat. Med. 3, 1354-1361)。TKポリペプチドはそれ自体では毒性がないが、それはアシクロビールまたはガンシクロビール(GCV)のようなヌクレオシド類似体の形質転換を触媒する。改質ヌクレオシドは伸長過程にあるDNA鎖に組込まれ、結果として細胞分裂を阻害する。多数の自殺遺伝子/プロドラッグの対が、現在利用可能である。更に具体的に述べることができるものは、ラットチトクロームp450およびシクロホスファミド(Human Gene Therapy 5, 969-978)、Escherichia coli (E. Coli)プリンヌクレオシドホスホリラーゼおよび6−メチルプリンデオキシリボヌクレオシド(Sorscher et al., 1994, Gene Therapy 1, 223-238)、E. coliグアニンホスホリボシルトランスフェラーゼおよび6−チオキサンチン(Mzoz and Moolten, 1993, Human Gene Therapy 4,589-595)、およびシトシンデアミナーゼ(CDアーゼ)および5−フルオロシトシン(5FC)である。
【0005】
CDアーゼは、ピリミジン代謝経路に関与しており、これにより外来シトシンが加水分解的脱アミノ化によってウラシルに変換される。CDアーゼ活性は原核生物および下級真核生物では明らかにされているが(Jund and Lacroute, 1970, J. Bacteriol. 102, 607-615; Beck et al., 1972, J. Bacteriol. 110, 219-228; De Haan et al., 1972, Antonie van Leeuwenhoek 38, 257-263; Hoeprich et al., 1974, J. Inf. Dis. 130, 112-118; Esders and Lynn, 1985, J. Biol. Chem. 260, 3915-3922)、それらは哺乳類では見られない(Koechlin et al., 1966, Biochem Pharmacol. 15, 435-446; Polak et al., 1976, Chemotherapy 22, 137-153)。Saccharomyces cerevisiae (S. cerevisiae) FCY1およびE. coli codA遺伝子であって、これら2種類の生物のCDアーゼをそれぞれコードする遺伝子が知られており、それらの配列が公表されている(欧州特許第402 108号明細書;Erbs et al., 1997, Curr. Genet. 31, 1-6;WO93/01281号明細書)。
【0006】
CDアーゼはまた、シトシンの類似体、すなわち5−フルオロシトシン(5−FC)を脱アミノすることによって、5−フルオロウラシル(5−FU)を形成し、これは5−フルオロ−UMP(5−FUMP)に転換されると、細胞傷害性の高い化合物である。酵素をコードする遺伝子を不活性化する突然変異のためまたは哺乳類細胞のようにこの酵素を天然で欠損しているためにCDアーゼ活性を欠いている細胞は5−FC耐性である(Jund and Lacroute, 1970, J. Bacteriol, 102, 607-615; Kilstrup et al., 1989, J. Bacteriol. 1989 171, 2124-2127)。対照的に、CDアーゼ活性をコードする配列が導入された哺乳類細胞は、5−FCに感受性になった(Huber et al., 1993, Cancer Res. 53, 4619-4626; Mullen et al., 1992, Proc. Nati. Acad. Sci USA 89, 33-37;WO93/01281号明細書)。更に、隣接する形質転換されていない細胞も5−FCに感受性になる(Huber et al., 1994, Proc. Natl. Acad. Sci. USA 91, 8302-8306)。傍観者効果と呼ばれるこの現象は、細胞が5−FUを分泌するCDアーゼ活性を発現しており、これが次に血漿膜を通って直接的に拡散することによって近隣細胞を中毒させることによる。拡散における5−FUのこの特性は、tk/GCVリファレンス系と比較して消極的な意味で利点があり、傍観者効果はそこでtkを発現している細胞と接触する必要がある(Mesnil et al., 1996, Proc. Natl. Acad. Sci. USA 93, 1831-1835)。従って、遺伝子療法、特に抗癌遺伝子療法に関してCDアーゼが提供する総ての利点は、容易に理解することができる。
【0007】
CDアーゼ活性を用いる方法の効率を改良するため、従来技術書類であるWO−A−96/16183号明細書では、CDアーゼとUPRTアーゼ活性を有する2ドメイン酵素をコードする融合タンパク質の使用を推奨し、発現プラスミドによって運ばれるハイブリッドcodA::uppまたはFCY1::FUR1遺伝子の導入が、トランスフェクションしたB16細胞の5−FCに対する感作を増すことをイン・ビトロで明らかにしている。WO99/54481号明細書は、N末端部分に欠失があるUPRTアーゼをコードする突然変異したFUR1遺伝子を用いることによるこの発明を改良している。
【発明の概要】
【0008】
本発明は、アミノ酸配列の付加により天然CDアーゼから誘導されることを特徴とする、CDアーゼ活性を有するポリペプチドであって、UPRTアーゼまたはチミジンキナーゼ活性を持たない、ポリペプチドを用いた点において、初期の手法の改良がある。
【0009】
本発明は、一層効率的なポリペプチドであって、これによって5−FCに対する細胞の感受性を増加し且つ自殺遺伝子を用いる遺伝子療法の展望を改良することができるものを提供する。この突然変異体は、多数の用途、特に抗癌および抗ウイルス用途、および細胞死を必要とする総ての用途に用いることができる。
【発明の具体的説明】
【0010】
「シトシンデアミナーゼ活性」(CDアーゼ活性)は、シトシンまたはその類似体の一つの脱アミノ化を包含するものと理解される。
【0011】
「天然のCDアーゼに由来する」とは、広義には、CDアーゼ活性を有する上記ポリペプチドが、上記の天然のCDアーゼと70%を上回り、有利には80%を上回り、好ましくは90%を上回り、極めて好ましくは95%を上回る同一性の程度を有するアミノ酸配列を含んでなることを意味する。
【0012】
本発明の意味においては、天然のCDアーゼは、原核生物または真核生物起源のCDアーゼを表す。好ましくは、CDアーゼは酵母CDアーゼであり、特にSaccharomyces cerevisiae FCY1遺伝子によってコードされたものである。異なる起源のCDアーゼをコードする遺伝子のクローニングおよび配列は、文献や専門のデーターベースで入手することができる。参考として、FCY1遺伝子の配列は、Erbs et al.の文献に開示されている(1997, Curr. Genet. 31, 1-6)。
【0013】
好ましい例は、実質的に配列番号1の配列アイデンティファイアーに表されている通りであり、1位のMet残基から出発して158位のGlu残基で終了するアミノ酸配列を含んでなる天然のCDアーゼである。「実質的に」という用語は同一性の程度を表し、上記配列番号1の配列では、これは70%を上回り、有利には80%を上回り、好ましくは90%を上回り、極めて好ましくは95%を上回る。更に一層好ましくは、このポリペプチドは、1位のMet残基から出発して158位のGlu残基で終了する配列番号1の配列アイデンティファイアーに表されているアミノ酸配列を含んでなる。1位のMet残基から出発して158位のGlu残基で終了する配列番号1の配列アイデンティファイアーに表されているアミノ酸配列を有するポリペプチドは、本発明の実施に極めて適している。
【0014】
有利な態様によれば、本発明によるポリペプチドは上記の天然CDアーゼよりかなり高いCDアーゼ活性を示す。従って、下記の実施例は、UPRTアーゼ活性を持たないアミノ酸配列を付加することによって、5−FCに対する標的細胞の感作および/または治療を受けた動物に誘導される傍観者効果を増加することができる。感作を増加する因子は、有利には少なくとも2であり、好ましくは少なくとも5であり、極めて好ましくは10以上である。
【0015】
「UPRTアーゼ活性を持たないポリペプチド」は、5−FUを5−FUMPに転換することができないポリペプチドを包含する。5−FUを5−FUMPに転換するアミノ酸配列の能力は、本願明細書の実施例に開示されている方法を用いることによって評価することができる。
【0016】
「チミジンキナーゼ活性を持たないポリペプチド」は、ガンシクロビール(9−{[2−ヒドロキシ−1−(ヒドロキシメチル)エトキシルメチル}グアノシン)、ファムシクロビール、ブシクロビール、ペンシクロビール、バルシクロビール、アシクロビール(9−[2−ヒドロキシエトキシ)メチル]グアノシン)、トリフルオロチミジン、1−[2−デオキシ,2−フルオロ,β−D−アラビノフラノシル]−5−ヨードウラシル、ara−A (アデノシンアラビノシド,ビバラビン)、1−β−D−アラビノフラノキシルチミン、5−エチル−2’−デオキシウリジン、5−ヨード−5’−アミノ−2,5’−ジデオキシウリジン、イドクスウリジン(5−ヨード−2’−デオキシウリジン)、AZT(3’−アジド−3’チミジン)、ddC(ジデオキシシチジン)、AIU(5−ヨード−5’−アミノ−2’,5’−ジデオキシウリジン)、およびAraC(シチジンアラビノシド)へすることができないポリペプチドを包含する。
【0017】
好ましい態様によれば、天然CDアーゼに付加したアミノ酸配列は、10〜1000アミノ酸長であり、更に好ましくは100〜400、非常に好ましくは200〜300アミノ酸長である。付加は天然CDアーゼの任意の部位に起こることができるが、NまたはC末端が好ましく、特にC末端が好ましい。
【0018】
有利には、天然CDアーゼに付加するアミノ酸配列は、UPRTアーゼ活性を有するポリペプチドから誘導される。
【0019】
「UPRTアーゼ活性を有するポリペプチドから誘導される」とは、広義には、上記アミノ酸配列のUPRTアーゼ活性ポリペプチドのアミノ酸配列との同一性の程度が70%を上回り、有利には80%を上回り、好ましくは90%を上回り、極めて好ましくは95%を上回ることを意味する。
【0020】
本発明の意味においては、UPRTアーゼ活性を有するポリペプチドは、ウラシルまたはその誘導体の一つを一リン酸類似体に、特に5−FUを5−FUMPに転換することができるポリペプチドを表す。「突然変異」とは、上記ポリペプチドの任意の部位における1以上の残基の付加、欠失および/または置換であると理解すべきである。
【0021】
本発明によるアミノ酸配列が由来することがあるUPRTアーゼ活性を有するポリペプチドは、任意の起源、特に原核生物、真菌または酵母起源のものであることができる。例えば、E. coli (Anderson et al., 1992, Eur. J. Biochem 204, 51-56)、Lactococcus lactis (Martinussen and Hammer, 1994, J. Bacteriol. 176, 6457-6463)、Mycobacterium bovis (Kim et al., 1997, Biochem Mol. Biol. mt 41, 1117-1124)、およびBacillus subtilis (Martinussen et al., 1995, J. Bacteriol. 177, 271-274)由来のUPRTアーゼは、本発明に関して用いることができる。しかしながら、酵母UPRTアーゼ、特にS.cerevisiae FUR1遺伝子によってコードされるものを用いるのが特に好ましく、その配列はKern et al.の文献に開示されている(1990, Gene 88, 149-157)。更に詳細には、例えば、遺伝子の配列、および相当するUPRTアーゼの配列は、文献や専門のデーターベースに見出すことができる(SWISSPROT、EMBL、Genbank、Medlineなど)。更に詳細には、fur 1−7、fur 1−8およびfur 1−9によってコードされた突然変異体を、本発明で用いることができる。好ましい態様では、UPRTアーゼ活性を有するポリペプチドに由来するアミノ酸配列は、fur 1−8対立遺伝子によってコードされたアミノ酸配列である(Kern et al., 1990, Gene 88, 149-157)。
【0022】
特に有利な態様によれば、本発明の天然CDアーゼに付加したアミノ酸配列は、天然UPRTアーゼの欠失突然変異体に由来する。この欠失は、好ましくは元のUPRTアーゼのN末端領域に位置している。欠失は全体的(上記N末端領域の総ての残基に影響を与える)または部分的(一次構造において連続的であることもまたはないこともある1以上の残基に影響を与える)である可能性がある。一般的には、ポリペプチドはN末端部分、中心部分、およびC末端部分からなり、それぞれの部分は分子の約三分の一を占める。例えば、251アミノ酸を含むS. cerevisiae UPRTアーゼの場合には、N末端部分は天然形態の最初の位置にあるいわゆる開始メチオニンから始まる最初の83残基からなっている。E. coli UPRTアーゼの場合には、N末端部分は1−69位を包含する。
【0023】
FUR1Δ105と呼ばれる突然変異体遺伝子の発現産物はS. cerevisiae furl突然変異体を補足することができることが示されており、これにより機能性であることが立証されている。
【0024】
極めて好ましくは、本発明によるUPRTアーゼ活性を有するポリペプチドは、少なくとも天然のUPRTアーゼの第二のATGコドンの上流のN末端領域を全部または一部欠失することによって上記天然のUPRTアーゼから誘導される。上記領域の完全な欠失が好ましい。例えば、FURl遺伝子によってコードされるUPRTアーゼは、+1位に第一のATGコドン(開始ATGコドン)に続いて+36位に第二のコドンを含んでなる。従って、本発明に関しては残基+1−35を考えることによって、天然形態の+36位に通常に見られるメチオニンから始まるポリペプチドを得ることできる。FUR1Δ105と呼ばれる突然変異体遺伝子の発現産物はS. cerevisiae furl突然変異体を補足することができることが示されており、これにより機能性であることが立証されている。
【0025】
本発明による天然CDアーゼに付加した好ましいアミノ酸配列は、実質的に配列番号2の配列アイデンティファイアー(identifier)に表されているものであり、2位のSer残基から出発して216位のVal残基で終了するアミノ酸配列を含んでなる。「実質的に」という用語は同一性の程度を表し、上記配列では、これは70%を上回り、有利には80%を上回り、好ましくは90%を上回り、極めて好ましくは95%を上回る。更に一層好ましくは、天然のCDアーゼに付加したアミノ酸配列は、2位のSer残基から出発して216位のVal残基で終了する配列番号2の配列アイデンティファイアーに表されているアミノ酸配列を含んでなる。2位のSer残基から出発して216位のVal残基で終了する配列番号2の配列アイデンティファイアーに表されているアミノ酸配列を有する天然のCDアーゼに付加したアミノ酸配列は、本発明の実施に極めて適している。天然のCDアーゼに付加したアミノ酸配列は、追加の突然変異を含むことができる。配列番号2の配列アイデンティファイアーの2位におけるセリン残基をアラニン残基で置換することを、特に挙げることができる。
【0026】
本発明によるCDアーゼ活性を有する好ましいポリペプチドは、実質的に配列番号1の配列アイデンティファイアーに表されている通りであり、1位のMet残基から出発して373位のVal残基で終了するアミノ酸配列を含んでなる。「実質的に」という用語は同一性の程度を表し、上記配列では、上記配列番号1の配列では70%を上回り、有利には80%を上回り、好ましくは90%を上回り、極めて好ましくは95%を上回る。更に一層好ましくは、ポリペプチドは、1位のMet残基から出発して373位のVal残基で終了する配列番号1の配列アイデンティファイアーに表されているアミノ酸配列を含んでなる。1位のMet残基から出発して373位のVal残基で終了する配列番号2の配列アイデンティファイアーに表されているアミノ酸配列を有するポリペプチドが、本発明の実施に極めて適している。
【0027】
一般的には、本発明によるポリペプチドは、通常の化学合成の方法によって、または組換えDNA法によって産生することができる(例えば、Maniatis et al., 1989,「実験室便覧」,Cold Spring Harbor, Laboratory Press,コールド・スプリング・ハーバー,ニューヨークを参照されたい)この理由により、本発明は、上記ポリペプチドをコードするヌクレオチド配列を細胞に導入して形質転換細胞を生成させ、上記形質転換細胞を上記ポリペプチドを産生させるのに適当な条件下で培養し、上記ポリペプチドを細胞培養物から回収する、調製方法も包含する。プロデューサー細胞は、任意の起源のものであり、考慮中のヌクレオチド配列がそのゲノムに組込まれまたは複製することができる適当な発現ベクターに組込まれる限り制限がなく、細菌、酵母または哺乳類細胞であることができる。天然では、ヌクレオチド配列は、転写および翻訳シグナルの制御下に置かれることにより、プロデューサー細胞で発現させることができる。発現ベクターおよび制御シグナルは、熟練者に知られている。ポリペプチドは、培地または(リーシスした後の)細胞から回収し、通常の精製段階(クロマトグラフィー、電気泳動、濾過、免疫精製などによる)を施すことができる。
【0028】
本発明は、本発明によるポリペプチドをコードするヌクレオチド配列にも関する。ヌクレオチド配列は、cDNAまたはゲノム配列、または混合型であることができる。これは、場合によっては天然、異種(例えば、ウサギβ−グロブリン遺伝子のイントロンなど)、または合成起源の1種類以上のイントロンを含み、宿主細胞での発現を増加することができる。本発明に関して用いられる配列は、分子生物学の通常の手法によって、例えば、特定のプローブを有するライブラリーをスクリーニングすることによって、発現ライブラリーの免疫スクリーニングによって、または適当なプライマーを用いるPCRによって、あるいは化学合成によって得ることができる。突然変異体は、位置指定突然変異誘発、PCR、制限および連結酵素による消化の手法を用いて1以上のヌクレオチドを置換、欠失および/または付加することによって、あるいは化学合成によって、天然配列から生成させることができる。突然変異体および構築体が機能する能力は、酵素活性を評価することによって、または標的細胞の5−FCおよび/または5−FUに対する感受性を測定することによって明らかにすることができる。
【0029】
本発明は、宿主細胞中での発現に必要な要素の制御下に置かれた本発明によるヌクレオチド配列を有する組換えベクターにも関する。組換えベクターは、プラスミドまたはウイルス起源のものであることができ、場合によってはベクターのトランスフェクション効率および/または安定性を向上させる1以上の物質と組み合わせることができる。これらの物質は、熟練者が利用可能な文献に広汎に報告されている(例えば、Feigner et al., 1987, Proc. West. Pharmacol. Soc. 32,115-121; Hodgson and Solaiman, 1996, Nature Biotechnology 14, 339-342; Remy et al., 1994, Bioconjugate Chemistry, 5,647-654)。非制限的例としては、これらの物質は、ポリマー、脂質、特にカチオン性脂質、リポソーム、核タンパク質、または中性脂質であることができる。これらの物質は、単独でまたは組み合わせて用いることができる。考えられる組合せは、カチオン性脂質(DOGS、DC−CHOL、スペルミン−chol、スペルミジン−cholなど)、リゾリン脂質(例えば、ヘキサデシルホスホコリン)、および中性脂質(DOPE)と組み合わせる組換えプラスミドベクターの組合せである。
【0030】
好ましい態様によれば、本発明で用いることができるカチオン性脂質は、欧州特許第901463B1号明細書に記載のカチオン性脂質であり、更に好ましくはpcTG90である。
【0031】
本発明に関して用いることができるプラスミドの選択は、広大なものである。それらは、クローニングベクターおよび/または発現ベクターであることができる。一般的には、それらは熟練者に知られており、それらの多くは市販されているが、遺伝子操作の手法を用いてそれらを構築しまたはそれらを改質することもできる。挙げることができる例は、pBR322 (Gibco BRL)、pUC (Gibco BRL)、pBluescript (Stratagene)、pREP4、pCEP4 (Invitrogene)、またはpPoly (Lathe et al., 1987, Gene 57, 193-201)から誘導されるプラスミドである。好ましくは、本発明に関して用いられるプラスミドは、複製がプロデューサー細胞および/または宿主細胞で確実に開始されるようにする複製起源を含む(例えば、ColE1起源はE. coliで産生しようとするプラスミドについて選択され、oriP/EBNA1系は哺乳類宿主細胞で自己複製することが所望な場合に選択される。Lupton and Levine, 1985, Mol. Cell. Biol. 5, 2533-2542; Yates et al., Nature 313, 812-815)。プラスミドは、トランスフェクション細胞を選択または同定することができるようにする選択遺伝子をも含んでなる(栄養要求性変異、抗生物質耐性をコードする遺伝子などの補足)。天然では、プラスミドは、所定の細胞でその維持および/またはその安定性を改良する追加要素を含むことができる(cer配列であって、モノマー形態でのプラスミドの維持を促進する。(Summers and Sherrat, 1984, Cell 36, 1097-1103)細胞ゲノムへ組込む配列)。
【0032】
ウイルスベクターに関しては、ポックスウイルス(ワクシニアウイルス、特にMVA、カナリアポックスウイルスなど)、アデノウイルス、レトロウイルス、ヘルペスウイルス、アルファウイルス、フォーミーウイルス、またはアデノ随伴ウイルスから誘導されるベクターを考えることができる。複製コンピテントまたは複製欠失ベクターを用いることができる。組込みを行わないベクターを用いるのが好ましい。この点において、アデノウイルスベクターおよびMVAが、本発明の実施に極めて適している。
【0033】
好ましい態様によれば、本発明のウイルスベクターは、改質ワクシニアウイルスアンカラス(MVA)に由来する。MVAベクターおよびそれらのベクターを産生する方法は、欧州特許第83286号および欧州特許第206920号明細書、並びにMayr et al. (1975, Infection 3, 6-14)およびSutter et Moss (1992, Proc. Natl. Acad. Sci. USA 89,10847-10851)に詳細に記載されている。更に好ましい態様によれば、本発明のヌクレオチド配列をMVAベクターの欠失I、II、III、IV、VおよびVIへ、更に一層好ましくは欠失IIIに挿入することができる(Meyer et al., 1991, J. Gen. Virol. 72, 1031-1038 ; Sutter et al., 1994, Vaccine 12, 1032-1040)。
【0034】
レトロウイルスは、細胞に感染し、ほとんどの場合には細胞と結合して、分割させる特性を有し、この点において、癌に関連した使用に特に適している。本発明による組換えレトロウイルスは、一般にLTR配列、キャプシド化領域、および本発明によるヌクレオチド配列であって、レトロウイルスLTRまたは以下に記載されるような内部プロモーターの制御下に置かれているものを含む。組換えレトロウイルスは、任意の起源(ネズミ、霊長類、ネコ、ヒトなど)のレトロウイルスから、特にMOMuLV(Moloneyネズミ白血病ウイルス)、MVS(ネズミ肉腫ウイルス)、またはFriendネズミレトロウイルス(Fb29)から誘導することができる。これは、ウイルス粒子を構成するのに必要なウイルスポリペプチドgag、polおよび/またはenvを途中で供給することができるキャプシド化細胞系で増殖する。このような細胞系は、文献に記載されている(PA317、Psi CRIP GP +Am−12など)。本発明によるレトロウイルスベクターは、特にLTR(プロモーター領域の真核生物プロモーターによる置換)またはキャプシド化領域(異種キャプシド化領域、例えば、VL30型による置換)に修飾を含むことができる(仏国特許出願第94 08300号明細書および第97 05203号明細書を参照されたい)。
【0035】
複製に本質的であり且つE1、E2、E4およびL1−L5領域から選択される少なくとも1つの領域の全部または一部を欠くアデノウイルスベクターを用いて、国ベクターが宿主生物または環境中で増殖しないようにすることも好ましい。E1領域の欠失が好ましい。しかしながら、それを、特に欠陥のある本質的機能が補足細胞系および/またはヘルパーウイルスによって途中で補足される程度までE2、E4および/またはL1−L5領域の全部または一部に影響を与える他の(複数の)修飾/欠失と組み合わせることができる。この点では、当該技術分野の状況の第二世代ベクターを用いることができる(例えば、国際特許出願WO−A−94/28152号明細書およびWO−A−97/04119号明細書を参照されたい)。例えば、E1領域の主要部分およびE4転写単位の欠失が、特に有利である。クローニング容量を増加させるために、アデノウイルスベクターは更に非本質的なE3領域の全部または一部を欠くことができる。もう一つの代替例によれば、キャプシド化に本質的な配列、すなわち5’および3’ITR(逆方向末端反復配列)とキャプシド化領域を保持する最小アデノウイルスベクターを用いることができる。様々なアデノウイルスベクターおよびそれらを調製するための手法が知られている(例えば、Graham and Prevect, 1991, 「分子生物学の方法」,第7巻,p109-128; E. J. Murey監修,The Human Press mcを参照されたい)。
【0036】
更に、本発明によるアデノウイルスベクターの起源は、種の観点および血清型の観点の両方から変動することができる。ベクターは、ヒトおよび動物(イヌ、トリ、ウシ、ネズミ、ヒツジ、ブタ、サルなど)起源のアデノウイルスのゲノムから、または少なくとも2種類の異なる起源のアデノウイルスゲノム断片を含んでなるハイブリッドから誘導することができる。更に詳細には、イヌ起源のCAV−1またはCAV−2アデノウイルス、トリ起源のDAVアデノウイルス、またはウシ起源のBad 3型アデノウイルスを挙げることができる(Zakharchuk et al., Arch. Virol. , 1993, 128: 171-176; Spibey and Cavanagh, J. Gen. Virol. 1989, 70:165-172; Jouvenne et al., Gene, 1987, 60: 21-28; Mittal et al., J. Gen. Virol., 1995, 76: 93-102)。しかしながら、ヒト起源のアデノウイルスベクターであって、好ましくは血清型C−アデノウイルス、特に2または5型の血清型Cアデノウイルスから誘導されるものが好ましい。
【0037】
本明細書で用いられる「複製コンピテント」という用語は、任意のトランス相補性(transcomplementation)の非存在下における宿主細胞で複製することができるウイルスベクターを表す。本発明に関しては、この用語は、癌または過剰増殖性宿主細胞で一層良好にまたは選択的に複製するように遺伝子工学処理された複製選択的または条件複製アデノウイルスも包含する。
【0038】
本発明の好ましい態様によれば、複製コンピテントベクターは複製コンピテントアデノウイルスベクターである。これらの複製コンピテントアデノウイルスベクターは、当業者には周知である。これらの中で、ONYX−015ウイルスのように55kD P53インヒビターをコードするE1b領域で欠失したアデノウイルスベクターが特に好ましい(Bischoff et al, 1996; Heise et al., 2000;WO94/18992号明細書)。従って、このウイルスは、p53欠損腫瘍細胞に選択的に感染して殺すのに用いることができる。当業者であれば、公表された手法に従ってアデノウイルス5または他のウイルスにおけるp53インヒビター遺伝子を突然変異させ、崩壊させることもできる。E1A Rb結合領域において欠失したアデノウイルスを、本発明に用いることもできる。例えば、E1A領域に24塩基対の欠失を有する突然変異アデノウイルスであるデルタ24ウイルス(Fueyo et al., 2000)。デルタ24はRb結合領域に欠失を有し、Rbに結合しない。従って、突然変異ウイルスの複製は、正常な細胞ではRbによって阻害される。しかしながら、Rbが不活性化しており、細胞が腫瘍性になる場合には、デルタ24は最早阻害されない。代わりに、突然変異体ウイルスは効率的に複製し、Rb欠損細胞をリーシスする。
【0039】
本発明によるアデノウイルスベクターは、連結反応または相同組換えによって、または補足細胞系での組換えによってEscherichia coli(E. coli)でイン・ビトロで生成することができる(例えば、国際特許出願WO−A−96/17070号明細書を参照されたい)。
【0040】
発現に必要な要素は、ヌクレオチド配列をRNAおよびmRNAに転写して、ポリペプチドに翻訳することができる総ての要素からなっている。これらの要素は、特に調節可能であるかまたは構成的であることがあるプロモーターを含んでなる。天然では、このプロモーターは、選択されたベクターおよび宿主細胞に適している。挙げることができる例は、PGK(ホスホグリセレートキナーゼ)、MT(メタロチオネイン;Mclvor et al., 1987, Mol. Cell Biol. 7,838-848)、α−1アンチトリプシン、CFTR、界面活性剤、免疫グロブリン、β−アクチン(Tabin et al., 1982, Mol. Cell Biol. 2,426-436)およびSRa(Takebe et al., 1988, Mol. Cell Biol. 8, 466-472)遺伝子の真核生物プロモーター、SV40ウイルス(シミアンウイルス)の初期プロモーター、RSV(ラウス肉腫ウイルス)のLTR、HSV−1 TKプロモーター、CMVウイルス(サイトメガロウイルス)の初期プロモーター、ワクシニアウイルスのp7.5K pH5R、pKlL、p28およびp11プロモーター、およびE1AおよびMLPアデノウイルスプロモーターである。プロモーターは、腫瘍または癌細胞での発現を刺激するプロモーターであることもできる。特に、胸部および前立腺癌で過剰発現するMUC−1遺伝子(Chen et al., 1995, J. Clin. Invest. 96, 2775-2782)、結腸癌で過剰発現するCEA(癌胎児性抗原を表す)遺伝子(Schrewe et al., 1990, Mol. Cell. Biol. 10, 2738-2748)、黒色腫で過剰発現するチロシナーゼ遺伝子(Vile et al., 1993, Cancer Res. 53, 3860-3864)、胸部および膵臓癌で過剰発現するERBB−2遺伝子(Harris et al., 1994, Gene Therapy 1, 170-175)、肝臓癌で過剰発現するα−フェトプロテイン遺伝子(Kanai et al., 1997, Cancer Res. 57, 461-465)を挙げることができる。サイトメガロウイルス(CMV)初期プロモーターが、極めて好ましい。
【0041】
しかしながら、ワクシニアウイルスに由来するベクター(例えば、MVAベクターとして)を用いるときには、チミジンキナーゼ7.5K遺伝子のプロモーターが特に好ましい。
【0042】
必要な要素としては、更に、本発明によるヌクレオチド配列の発現または宿主細胞中でのその維持を向上させる追加要素が挙げられる。イントロン配列、分泌シグナル配列、核局在化配列、IRES型の翻訳の再開始のための内部部位、転写終結ポリA配列、トリパルタイト(tripartite)リーダーおよび複製の起源が、特に挙げられる。これらの要素は、熟練者に知られている。
【0043】
本発明による組換えベクターは、1個以上の目的とする追加遺伝子を含んでなることもでき、これらの遺伝子を同じ調節要素(ポリシストン性カセット)または独立した要素の制御下に置くことができる。特に挙げることができる遺伝子は、インターロイキンIL−2、IL−4、IL−7、IL−10およびIL−12、インターフェロン、腫瘍壊死因子(TNF)、コロニー刺激因子(CSF)、特にGM−CSF、および脈管形成に作用する因子(例えば、PAl−1、プラスミノーゲンアクチベーターインヒビターを表す)をコードする遺伝子である。一つの具体的態様では、本発明による組換えベクターは、IL−2をコードするまたはインターフェロンγ(INFγ)をコードする目的の遺伝子を含んでなる。本発明によるヌクレオチド配列を、HSV−1 TK遺伝子、リシン遺伝子、コレラ毒素遺伝子などの他の自殺遺伝子と組み合わせることも考えることができる。
【0044】
本発明は、本発明による組換えベクターを含んでなるウイルス粒子にも関する。このようなウイルス粒子は、当該技術分野で通常の任意の手法を用いてウイルスベクターから生成することができる。ウイルス粒子を、ベクターの欠損に適当な補足細胞で増殖させる。アデノウイルスベクターに関しては、例えば、ヒト胎児腎臓細胞を用いて確立され且つA549−E1細胞系(Imler et al., 1996, Gene Therapy 3, 75-84)または二重相補性が可能な細胞系(Yeh et al., 1996, J. Virol. 70, 559-565; Krougliak and Graham, 1995, Human Gene Therapy 6, 1575-1586; Wang et al., 1995 Gene Therapy 2, 775-783;国際特許出願第WO 97/04119号明細書)のE1機能(Graham et al., 1977, J. Gen. Virol. 36, 59-72)を効率的に補足する293細胞系が用いられる。ヘルパーウイルスを用いて、欠損機能を少なくとも部分的に補足することもできる。補足細胞は、ウイルスゲノムをウイルスキャプシドにキャプシド化して組換えベクターを含むウイルス粒子を生成するのに必要な初期および/または後期因子を途中で供給することができる細胞と理解される。上記細胞は、それ自身のベクターの欠損機能を総て補足することができないことがあり、この場合には、追加機能を供給するベクター/ヘルパーウイルスでトランスフェクション/形質導入を行うことができる。
【0045】
本発明は、ウイルス粒子の製造方法であって、
(i) 本発明による組換えベクターを、途中で上記ベクターを補足してトランスフェクションした補足細胞を得るようにすることができる補足細胞に導入し、
(ii) 上記トランスフェクションした補足細胞を、ウイルス粒子を産生することができるようにするのに適当な条件下で培養し、
(iii) ウイルス粒子を細胞培養物から回収する
ことを特徴とする、方法にも関する。
【0046】
ウイルス粒子は培養上清から回収することができるのは勿論であるが、細胞から回収することもできる。一般に用いられるポリの一つは、連続的凍結/融解サイクルによって細胞をリーシスして、リーシス上清のビリオンを回収することからなっている。次に、ビリオンを、当該技術分野の手法(クロマトグラフィー法、超遠心法、特に塩化セシウムグラディエントによる方法など)を用いて増幅し、精製することができる。
【0047】
本発明は、本発明によるヌクレオチド配列または組換えベクターを含んでなる、または本発明によるウイルス粒子に感染している宿主細胞にも関する。本発明の本発明の趣旨では、宿主細胞は、上記に定義されているように、組換えベクターでトランスフェクションすることができる、またはウイルス粒子に感染させることができる任意の細胞からなっている。哺乳類細胞、特にヒト細胞が極めて好ましい。この細胞は、上記ベクターをゲノムに組込まれているまたは組込まれていない形態(エピソーム)で含んでなることができる。この細胞は、原発性または任意の起源の腫瘍細胞、特に造血細胞(全能性幹細胞、白血球、リンパ球、単球またはマクロファージなど)、筋細胞(衛星細胞、筋細胞、筋芽細胞、平滑筋細胞など)、心臓細胞、肺細胞、気管細胞、肝細胞、上皮細胞または繊維芽細胞であることができる。
【0048】
本発明は、本発明によるポリペプチド、ヌクレオチド配列、組換えベクター、ウイルス粒子、または宿主細胞を、薬学上許容可能な賦形剤と組み合わせて含んでなる、組成物にも関する。
【0049】
本発明は、更に、本発明によるポリペプチドと目的とする上記遺伝子の一つによってコードされる目的とするポリペプチドを含んでなる組成物に関する。これらの目的とするポリペプチドの中で、特にインターロイキンIL−2、IL−4、IL−7、IL−10およびIL−12、インターフェロン、腫瘍壊死因子(TNF)、コロニー刺激因子(CSF)、特にGM−CSF、および脈管新生に作用する因子(例えば、PAl−1、プラスミノーゲンアクチベーターインヒビターを表す)を挙げることができる。IL−2またはINFγが特に考えられる。
【0050】
組成物は、宿主細胞中で上記ポリペプチドを発現させることができるヌクレオチド配列に基づくこともできる。ヌクレオチド配列は、1個の同じ発現ベクターによってまたは2個の独立したベクターによって運ぶことができる。上記組成物は、上記の(複数の)ヌクレオチド配列を発現する(複数の)ウイルスベクターから生成されるウイルス粒子を含んでなることができるのは勿論である。
【0051】
本発明は、更に、本発明によるヌクレオチド配列と、IL−2およびINFγから選択されるポリペプチドをコードする目的とする第二のヌクレオチド配列を含んでなる、組成物にも関する。
【0052】
本発明による組成物は、更に具体的には遺伝子療法による疾患の予防的または治癒的治療をしようとするものであり、更に具体的には増殖性疾患(癌、腫瘍、再狭窄など)および感染性起源、特にウイルス起源(BまたはC型肝炎ウイルス、HIV、ヘルペス、レトロウイルスなどによって誘発される)の疾患を目指すものである。
【0053】
本発明による組成物は、それを局所的に、非経口的にまたは消化器経路によって投与する目的で常法によって製造することができる。特に、この治療薬または予防薬の治療上有効量を、薬学上許容可能な賦形剤と組み合わせる。多数の投与経路を考えることができる。例えば、胃内、皮下、心臓内、筋肉内、静脈内、腹腔内、腫瘍内、肺内、および気管内経路を挙げることができる。これらの3つの後者の態様の場合には、エアゾールによってまたは点滴によって投与を行うのが有利である。投与は、単回用量として、または特定の時間間隔の後に1回以上反復される用量として行うことができる。適当な投与および投薬経路は、様々なパラメーター、例えば、個体、治療を行う疾患、または目的とする(複数の)遺伝子によって変化する。本発明によるウイルス粒子を基剤とする製剤は、104〜1014pfu(プラーク形成単位)、有利には105〜1013pfu、好ましくは106〜1012pfu、更に好ましくは、アデノウイルス粒子を用いるときには109〜1010、およびMVA粒子を用いるときには106〜107の用量の形態で処方することができる。本発明による組換えベクターに関する限り、DNA 0.01〜100mg、好ましくは0.05〜10mg、特に好ましくは0.5〜5mgを含んでなる用量が考えられる。ポリペプチドを基剤とする組成物は、好ましくは上記ポリペプチド0.05〜10g、特に好ましくは0.05〜5gを含んでなる。用量は、医師が調整することができるのは当然である。
【0054】
処方物は、薬学上の観点から許容可能な希釈剤、アジュバントまたは賦形剤、並びに可溶化剤、安定剤および防腐剤を含むこともできる。注射可能な投与の場合には、水性、非水性または等張溶液の処方物が好ましい。これは、適当な希釈剤を用いて使用時点に再構成することができる液状または乾燥(粉末、凍結乾燥物などの)形態で、単回用量としてまたは多数回用量として提示することができる。この処方物は、適当量のプロドラッグを含んでなることもできる。
【0055】
本発明は、遺伝子療法によってまたは組換え経路によって産生されたタンパク質を投与することによってヒトまたは動物体の治療を目的とする薬剤を調製するための、本発明によるポリペプチド、組換えベクター、ウイルス粒子、または宿主細胞の治療または予防使用にも関する。第一の可能性によれば、この薬剤は、イン・ビボで(例えば、静脈内注射により接近可能な腫瘍中に、エアゾールによって肺に、適当なカテーテルを用いて脈管系に、など)直接投与することができる。患者から細胞(骨髄幹細胞、末梢血リンパ球、筋細胞など)を採取し、当該技術分野の手法によりそれらをイン・ビトロでトランスフェクションまたは乾癬させた後、それらを患者に再投与することからなる、エクス・ビボ法を採用することもできる。好ましい使用は、癌、腫瘍、および望ましくない細胞増殖から生じる疾患の治療または予防にある。考えられる応用は、胸部、子宮(特に、パピローマウイルスによって誘発されるもの)、前立腺、肺、膀胱、肝臓、結腸、膵臓、胃、食道、喉頭、中枢神経系(例えば、神経膠芽細胞腫)、および血液(リンパ腫、白血病など)の癌を挙げることができる。これは、循環器疾患に関して用いて、例えば、血管壁の平滑筋の増殖(再狭窄)を阻害しまたは遅らせることもできる。最後に、感染性疾患の場合には、AIDSに応用される薬剤を考えることができる。
【0056】
本発明は、本発明によるヌクレオチド配列、組換えベクター、ウイルス粒子、または宿主細胞を、そのような治療を必要とする生体または宿主細胞に投与することを特徴とする、遺伝子療法による疾患の治療方法にも敷衍される。
【0057】
有利な態様によれば、治療使用または治療方法は、薬学上許容可能な量のプロドラッグ、有利にはシトシンの類似体、特に5−FCを宿主生物または細胞に投与する追加段階も含んでなる。例えば、50〜500mg/kg/日の用量を用いることができ、200mg/kg/日の用量が好ましい。本発明に関しては、プロドラッグを標準的実施(例えば、経口、体系的)に従って投与し、投与は本発明による治療薬の投与の前に、と同時に、あるいはの後に行う。経口投与が好ましい。プロドラッグの単回用量、または宿主生物または細胞中に毒性代謝物を産生できるほど十分長い時間反復する用量を投与することができる。
【0058】
更に、本発明による組成物または方法を、5−FUの細胞傷害性効果を増強する1種類以上の物質と組み合わせることができる。特に、ピリミジンのデ・ノボ生合成の経路を阻害する薬剤(例えば、下記のもの)、5−FUの代謝生成物(5−FdUMP)の存在下でチミジレートシンターゼの阻害を増加し、複製に必要なdTMPの貯留を減少させるロイコボリンのような薬剤(Waxman et al., 1982, Eur. J. Cancer Clin. Oncol. 18, 685-692)、および最後に、ジヒドロホレートレダクターゼを阻害し、PRPP(ホスホリボシルピロホスフェート)の貯留を増加することによって5−FUの細胞RNAへの組込みを増加させるメトトレキセートのような薬剤(Cadman et al., 1979, Science 250, 1135-1137)を挙げることができる。
【0059】
本発明による組成物または方法は、抗癌療法に有効な1種類以上の物質と組み合わせることができる。本発明による組成物と共にまたはと組み合わせて用いることができる抗癌療法に有効な医薬物質の中には、例えば、マイトマイシンC、シクロホスファミド、ブスルファン、イホスファミド、イソスファミド、メルファラン、ヘキサメチルメラミン、チオテパ、クロラムブシルまたはダカルバジンのようなアルキル化剤、例えば、ゲンシタビン、カペシタビン、5−フルオロウラシル、シタラビン、2−フルオロデオキシシチジン、メトトレキセート、イダトレキセート、トムデックスまたはトリムトレキセートのような代謝拮抗薬、例えば、ドキソルビシン、エピルビシン、エトポシド、テニポシドまたはミトキサントロンのようなトポイソメラーゼII阻害薬、例えば、イリノテカン(CPT−11)、7−エチル−10−ヒドロキシ−カンプトテシン(SN−38)またはトポテカンのようなトポイソメラーゼI阻害薬、例えば、パクリタキセル、ドセタキセル、ビンブラスチン、ビンクリスチンまたはビノレルビンのような抗有糸分裂薬、および例えば、シスプラチン、オキサリプラチン、スピロプラチナムまたはカルボプラチナムのような白金誘導体を挙げることができる。本発明による組成物または方法は、放射線療法と組み合わせて用いることもできる。
【0060】
本発明は、哺乳類細胞における正の選択マーカーとしての本発明による配列または組換えベクターの使用にも関する。有利には、細胞をトランスフェクションし、次に、細胞混合物を、PALA (N(ホスホノアセチル)−L−アスパルテート;Moore et al., 1982, Biochem. Pharmacol. 31, 3317-3321)、A77 1726 (レフルノミドの活性代謝物;Davis et al., 1996, Biochem. 35, 1270-1273)およびブレキナール(Chen et al., 1992, Cancer Res. 52, 3251-3257)のようなピリミジンのデ・ノボ生合成の経路の阻害薬の存在下で培養する。このような阻害薬の存在により、RNAおよびDNAの合成に必要なUMPのデ・ノボ合成がブロックされることによって、細胞死を生じる。この細胞傷害効果は、ウラシルの存在下にてUPRTアーゼ活性をコードする本発明によるヌクレオチド配列を発現させることによって、またはシトシンの存在下にてこの後者の配列をCDアーゼ活性をコードする配列と(場合によっては、融合形態で)同時発現させることによって回避することができる。結果として、トランスフェクションした細胞(UPRTアーゼ/CDアーゼ配列を組込んだ細胞)は、ピリミジン合成経路の阻害薬の存在下で増殖することができるようになる。従って本発明による使用によって、トランスフェクションした細胞を、細胞混合物中で効率的に同定しおよび/または細胞混合物から単離することができる。
【0061】
本発明は、本発明による配列または組換えベクターの、負の選択マーカーとしての使用にも関する。例えば、、本発明による配列または組換えベクターを用いて、胎児性幹細胞または遺伝子が中断されまたは改質される核導入の後に得られる細胞を選択することができる(例えば、トランスジェニック動物の製造方法)(例えば、Capecchi, 1989, Science 244, 1288-1292; Reid et al., 1990, Proc. Natl. Acad. Sci. USA 87, 4299-4303を参照されたい)。例えば、ネオマイシン耐性についての遺伝子と組み合わせたこのような使用により、相同組換え事象を行っており且つゲネティシンおよび相当するフッ素化ピリミジン(CDアーゼ活性をコードするヌクレオチド配列を用いるときには、5−FC)の存在下にて単独で増殖することができる細胞を選択することができる。非標的組換え事象を行った細胞は、ゲネティシンの存在下で増殖することができるが、フッ素化ピリミジンの存在下では増殖することができない。負の選択マーカーとしてのもう一つの有望な使用は、哺乳類細胞と同様に、植物は内因性のCDアーゼ活性を持たないので、植物の分野において見出されるものである。それらを、本発明によるヌクレオチド配列をトランスフェクションすることによって5−FCに増感して、外来CDアーゼを発現することができる(例えば、Perera et al. 1993, Plant. Mol. Biol. 23, 797-799を参照されたい)。
【0062】
本発明による配列または組換えベクターは、細菌または酵母の負の選択マーカーとして用いることもできる。この特定の態様では、天然のCDアーゼ遺伝子を欠いている酵母または細菌が好ましく用いられる。これらの細菌の中では、欧州特許第0792369BB1号明細書に開示されているものが、特に好ましい。例えば、本発明による配列を、本発明による配列を含んでなるプラスミド中で、相同組換えによってクローニング配列に用いることができる。これらの実験では、本発明による配列は、クローニングを行う配列を挿入しなければならない部位に配置される。従って、相同組換えの後には、所望なプラスミドを含んでなる細胞は、5−FCの存在下で増殖することができる。
【実施例】
【0063】
実施例1: Fcu1−8の構築
FCU1−8は、pCI−neoFCU1(pTG13046)から出発してFCU1の直接突然変異誘発によって構築した。183位のArgは、オリゴ5’−gtattcttattactgatgatgg−3’(A549en Tを改質するため)を有するSerによって置換して、プラスミドpCI−neoFCU1−8(pTG15546)を得た。
【0064】
実施例2: 哺乳類細胞でのFCY1、FCU1およびFcu1−8の発現、およびCDアーゼおよびUPRTアーゼ活性の測定
プラスミドpCIneo(Promega)、pCI−neoFCY1(pTG15916)、pCI−neoFCU1(pTG13046)およびpCI−neoFCU1−8(pTG15546)によって一時的にトランスフェクションした哺乳類COS7の株を、CDアーゼおよびUPRTアーゼ活性について試験した。
【0065】
プロトコールは下記の通りである: 5 105 COS7 細胞を、DMEM/10% FCS培地5mlを含む37℃の60mmフラスコに播種した。24時間後、細胞をプラスミド5μgの非存在下または存在下でリポフェクチン(Gibco BRL)20μlで処理した。DMEM培地2ml中にて37℃で20時間後、細胞をDMEM/10% FCS 5mlの存在下でインキュベーションした。37℃で48時間後、細胞をPBSで洗浄した後、リーシス緩衝液(Tris−HCl pH7.5 50mM/NaCl 150mM/EDTA 5mM/DTT 1mM/トリトン1%)25μlに懸濁した。4℃で30分間リーシスし、遠心分離した後、CDアーゼおよびUPRTアーゼ活性を細胞溶解物を含む上清中で測定した。
【0066】
CDアーゼ活性については、反応性CDアーゼ緩衝液(Tris−HCl pH7.5 100mM/[H3] 5−FC 3mM,0.25μCi/μl)2μlの存在下にて37℃で20分間インキュベーションする。
【0067】
UPRTアーゼ活性については、細胞溶解物4μlを、反応性UPRTアーゼ緩衝液(Tris−HCl pH7.5 100mM/MgCl2 10mM/5−PRPP 10mM/[C14] 5−FU 3mM,0.02μCi/μl)2μlの存在下にて37℃で20分間インキュベーションする。
【0068】
酵素反応を、100℃で1分間停止する。1μlの分量を、TLCポリエチレンイミン−セルロースプレート(Merck)上に置く。水/ブタノール−1混合物(14%/86%)を溶媒として用いて、5−FUを5−FCから分離し、5−FUMPと5−FUの分離は、水を溶媒として用いて行う。移動の後、TLCプレートをPosphorImager(445SI;Molecular Dynamics)で走査した。タンパク質濃度は、Bradfordの方法によって測定した。
【0069】
3回の独立した測定から計算される酵素活性の結果を、下表に示す。
【0070】
【表1】
【0071】
FCU1遺伝子と比較して、Fcu1−8突然変異体はUPRTアーゼ活性を失うが、CDアーゼ活性は保存され、天然のFCY1遺伝子のCDアーゼ活性より10倍高い。遺伝子FCU1でのArgl83のSerへの変化によりUPRTアーゼ活性が失われるが、CDアーゼ活性に変化はない。
【0072】
実施例3: FCU1を発現するアデノウイルス(AdTG14800)の構築
FCU1遺伝子を含むpCI−neoFCU1(pTG13046)のXhoI−MluI断片を単離し、これらの同じ酵素によって開裂したトランスファーベクターpTG13387に導入して、トランスファーベクターpTG14799を得る。アデノウイルスベクターAD−FCU1(pTG14800)は、pTG14799のPacI−BstEII断片とClaIによって線形化したベクターpTG6624との間のBJ 5183 E. coli中での相同組換えによって再構築する。最終構築物AD−FCU1は、E1およびE3を欠失したAd5ゲノム(ヌクレオチド459−3510および28249−30758)を含み、E1の代わりに、FCU1遺伝子を含んでなる発現カセットをCMVプロモーターとβ−グロリン/IgGに続いてbGHのポリA配列の制御下に置く。FCU1を発現するアデノウイルス粒子(AdTG14800)は、相補細胞系でのトランスフェクションによって生成する(例えば、PERC6系)。
【0073】
実施例4: Fcu1−8を発現するアデノウイルスの構築(AdTG15606)
Fcu1−8遺伝子を含むpCI−neoFCU1−8(pTG15546)のXhoI−MluI断片を単離し、これらの同じ酵素によって開裂したトランスファーベクターpTG13387に導入して、トランスファーベクターpTG15547を得る。アデノウイルスベクターAD−FCU1−8(pTG15606)は、pTG15547のPacI−BstEII断片とClaIによって線形化したベクターpTG6624との間のBJ 5183 E. coli中での相同組換えによって再構築する。最終構築物AD−FCU1−8は、E1(NT 459−3510)およびE3(NT 28249−30758)領域を欠失したAd5ゲノムを含み、E1の代わりに、FCU1−8遺伝子を含んでなる発現カセットをCMV初期プロモーターハイブリッドβ−グロリン/IgGおよびMVCに続いてbGHのポリA配列の制御下に置く。FCU1−8を発現するアデノウイルス粒子(AdTG15606)は、E1機能を補足する細胞系でのトランスフェクションによって生成する(例えば、PERC6系)。
【0074】
実施例5: FCU1(AdTG14800)およびFcul−8(AdTG15606)を発現するアデノウイルスによる感染:イン・ビトロでの結果、CDアーゼとUPRTアーゼの活性の測定
ヒト腫瘍細胞系A549(ATCC Ccl−185)を、中空アデノウイルス(AdTG15149)、FCU1を発現するアデノウイルス(AdTG14800)、およびFcu1−8を発現するアデノウイルス(AdTG15606)によって感染させた。感染プロトコールは下記の通りである:PBS−2% FCS−l%カチオン50μlの懸濁液で5x106個の細胞をMOI 5でのアデノウイルスの存在下にて37℃で30分間インキュベーションする。次に、総量が5x106個の細胞を、DMEM−10% FCS 5mlの存在下にて60mmフラスコ中でインキュベーションし、細胞をPBSで洗浄した後、リーシス緩衝液(Tris−HCl pH7.5 50mM/NaCl 150mM/EDTA 5mM/DTT 1mM/トリトン1%)25μlに再懸濁した。4℃で30分間リーシスし、遠心分離の後、CDアーゼおよびUPRTアーゼ活性を細胞溶解物を含む上清中で測定した(上記プロトコールによる)。
【0075】
3回の独立した測定から計算される酵素活性の結果を、下表に示す。
【0076】
【表2】
【0077】
Fcu1−8を発現するアデノウイルスによって感染した細胞について、結果はUPRTアーゼ活性の喪失を示唆しているが、CDアーゼ活性は保存される。
【0078】
実施例6: FCU1を発現するMVAの構築(MVATG15637)
トランスファーベクターMva−fcu1(pTG15637)を、pCI−neoFCU1(pTG13046)のHindIII−KpnI断片とXhoIによって線形化したベクターpTG14269の間のBJ 5183 E.coli株中での相同組換えによって再構築する。このトランスファーベクター MVA−fcu1(pTG15637)は、p11K7.5プロモーターの制御下に置かれ且つMVAの欠失IIIに接する配列によって枠どられたFCU1遺伝子の発現カセットを含む。FCU1を発現するMVA粒子(MVATG15637、FCU1は欠失IIIに挿入され且つp11K7.5プロモーターの制御下にある)は、胎児性のニワトリ細胞でのMVAN33とプラスミドpTG15637の間の相同組換えによって生成する。
【0079】
実施例7: Fcu1−8を発現するMvaの構築(MVATG15638)
トランスファーベクターMVA−FCU1−8(pTG15638)は、pCI−neoFCU1−8(pTG15546)のHindIII−KpnI断片とXhoIによって線形化されたベクターpTG14269との間のBJ 5183 E.coli株での相同組換えによって再構築される。このトランスファーベクターMVA−FCU1−8(pTG15638)は、p11K7.5プロモーターの制御下に置かれ且つMVAの欠失IIIに接する配列によって枠どられたFcu1−8遺伝子の発現カセットを含む。Fcu1−8を発現するMVA粒子(MVATG15638、Fcu1−8は欠失IIIに挿入され且つp11K7.5プロモーターの制御下にある)は、胎児性のニワトリ細胞でのMVAN33とプラスミドpTG15638の間の相同組換えによって生成する。
【0080】
実施例8: FCU1(MVATG15637)とFCU1−8(MVATG15638)を発現するMVAによ
ルスによる感染:イン・ビトロでの結果、CDアーゼとUPRTアーゼの活性の測定
ヒト腫瘍細胞系A549(ATCC CCCL−185)を、中空MVA(MVAN33)、FCU1を発現するMVA(MVA15637)、およびFCU1−8を発現するMVA(MVA15638)によって感染させた。感染プロトコールは下記の通りである:PBS−2% FCS−l%カチオン50μlの懸濁液で5x106個の細胞をMOI 5でのアデノウイルスの存在下にて37℃で30分間インキュベーションする。次に、総量が5x106個の細胞を、DMEM−10% FCS 5mlの存在下にて60mmフラスコ中でインキュベーションする。37℃で24時間後、細胞をPBSで洗浄した後、リーシス緩衝液(Tris−HCl pH7.5 50mM/NaCl 150mM/EDTA 5mM/DTT 1mM/トリトン1%)25μlに再懸濁した。4℃で30分間リーシスし、遠心分離の後、CDアーゼおよびUPRTアーゼ活性を細胞溶解物を含む上清中で測定した(上記プロトコールによる)。
【0081】
3回の独立した測定から計算される酵素活性の結果を、下表に示す。
【0082】
【表3】
【0083】
FCU1−8を発現するMVAによって感染した細胞について、結果はUPRTアーゼ活性の喪失を示唆しているが、CDアーゼ活性は保存される。
【0084】
実施例9: イン・ビボでの実験
5x106個のヒト細胞LoVo(結腸腺癌)を、8群の15 CD1ヌードマウスにJ0に皮内投与する。腫瘍の容積が30−50mm3となったら直ぐに(J+10)、MVAを5x106 PFUの量を腫瘍内投与する。J+10から、0.5ml水または0.5ml 0.5% 5−FC溶液を経口で1日2回14日間投与する。腫瘍の容積は、群MVA−FCU1/5−FCと比較して群MVA−fCU1−8/5−FCでの腫瘍の大きさが一層良好に制御されることを強調している。
【図面の簡単な説明】
【0085】
【図1】本発明によるベクターを投与したCD1ヌードマウスでのLOVO腫瘍細胞の増殖。
【特許請求の範囲】
【請求項1】
アミノ酸配列を付加することによって天然のCDアーゼから誘導されることを特徴とする、CDアーゼ活性を有するポリペプチドであって、
UPRTアーゼまたはチミジンキナーゼ活性を持たない、ポリペプチド。
【請求項2】
天然CDアーゼに付加したアミノ酸配列が、天然CDアーゼのC末端に連結している、請求項1に記載のポリペプチド。
【請求項3】
天然CDアーゼに付加したアミノ酸配列が、10〜1000アミノ酸長である、請求項1または2に記載のポリペプチド。
【請求項4】
天然CDアーゼに付加したアミノ酸配列が、100〜400アミノ酸長である、請求項3に記載のポリペプチド。
【請求項5】
天然CDアーゼに付加したアミノ酸配列が、200〜300アミノ酸長である、請求項4に記載のポリペプチド。
【請求項6】
天然CDアーゼに付加したアミノ酸配列が、UPRTアーゼ活性を有するポリペプチドに由来する、請求項1〜5のいずれか一項に記載のポリペプチド。
【請求項7】
UPRTアーゼ活性を有するポリペプチドが、酵母UPRTアーゼ、特にSaccharomyces cerevisiae FUR1遺伝子によってコードされたものに由来する、請求項6に記載のポリペプチド。
【請求項8】
天然CDアーゼに付加したアミノ酸配列が、実質的に配列番号2の配列アイデンティファイアーに表されているものであって、2位のSer残基から出発して216位のVal残基で終了するアミノ酸配列に由来する、請求項7に記載のポリペプチド。
【請求項9】
天然CDアーゼに付加したアミノ酸配列が、配列番号2の2位のSer残基から出発して216位のVal残基で終了する配列アイデンティファイアーに表されている通りである、請求項8に記載のポリペプチド。
【請求項10】
上記天然CDアーゼが酵母CDアーゼ、特にSaccharomyces cerevisiae FCY1遺伝子によってコードされたものである、請求項1〜9のいずれか一項に記載のポリペプチド。
【請求項11】
天然CDアーゼが、実質的に配列番号1の配列アイデンティファイアーに表されている通りであって、1位のMet残基から出発して158位のGlu残基で終了するアミノ酸配列を含んでなる、請求項10に記載のポリペプチド。
【請求項12】
天然CDアーゼが配列番号1の配列アイデンティファイアーに表されている通りの、1位のMet残基から出発して158位のGlu残基で終了するアミノ酸配列を含んでなる、請求項11に記載のポリペプチド。
【請求項13】
実質的に配列番号1の配列アイデンティファイアーに表されている通りであって、1位のMet残基から出発して373位のVal残基で終了するアミノ酸配列を含んでなる、請求項10に記載のポリペプチド。
【請求項14】
配列番号1配列アイデンティファイアーに表されている通りの、1位のMet残基から出発して374位のVal残基で終了するアミノ酸配列を含んでなる、請求項13に記載のポリペプチド。
【請求項15】
天然CDアーゼの活性よりかなり高いCDアーゼ活性を示す、請求項1〜14のいずれか一項に記載のポリペプチド。
【請求項16】
請求項1〜15のいずれか一項に記載のポリペプチドをコードするヌクレオチド配列。
【請求項17】
請求項16に記載のヌクレオチド配列を有し、宿主細胞中での発現に必要な要素の制御下に置かれた、組換えベクター。
【請求項18】
ベクターがプラスミドとウイルスベクターとからなる群から選択されるものであって、必要に応じてはベクターのトランスフェクション効力および/または安定性を改良する1個以上の物質と組み合わされている、請求項17に記載の組換えベクター。
【請求項19】
ベクターのトランスフェクション効力および/または安定性を改良する前記物質が、カチオン性脂質、カチオン性ポリマー、リゾリン脂質およびポリペプチドを含んでなる群から選択される、請求項18に記載の組換えベクター。
【請求項20】
ポックスウイルス、アデノウイルス、レトロウイルス、ヘルペスウイルス、アルファウイルス、フォーミーウイルスまたはアデノ随伴ウイルスに由来するウイルスベクターである、請求項18に記載の組換えベクター。
【請求項21】
改質ワクシニアアンカラ(MVA)ウイルスに由来する、請求項20に記載の組換えベクター。
【請求項22】
請求項16に記載のヌクレオチド配列を、欠失I、II、III、IV、VおよびVIからなる群から選択されるMVAゲノム内の天然に存在する欠失の部位に挿入する、請求項21に記載の組換えベクター。
【請求項23】
天然に存在する欠失の部位が欠失IIIである、請求項22に記載の組換えベクター。
【請求項24】
発現に必要な要素がプロモーターを含んでなる、請求項17〜23のいずれか一項に記載の組換えベクター。
【請求項25】
プロモーターがチミジンキナーゼ7.5K遺伝子のプロモーターである、請求項24に記載の組換えベクター。
【請求項26】
複製に本質的な少なくとも1つの領域の全部または一部を欠き、El、E2、E4およびL1−L5領域から選択されるアデノウイルスベクターである、請求項20に記載の組換えベクター。
【請求項27】
非本質的E3領域の全部または一部を更に欠いているアデノウイルスベクターである、請求項26に記載の組換えベクター。
【請求項28】
プロモーターがサイトメガロウイルス(CMV)初期プロモーターである、請求項24に記載の組換えベクター。
【請求項29】
インターロイキンIL−2、IL−4、IL−7、IL−10およびIL−12、インターフェロン、腫瘍壊死因子(TNF)、コロニー刺激因子(CSF)、および脈管形成に作用する因子をコードする遺伝子から選択される1個以上の遺伝子を更に、含んでなる、請求項17〜28のいずれか一項に記載の組換えベクター。
【請求項30】
目的とする遺伝子が、IL−2およびINFγから選択されるポリペプチドをコードする、請求項29に記載の組換えベクター。
【請求項31】
ウイルス粒子の調製方法であって、
(i) 請求項17〜29のいずれか一項に記載の組換えベクターを、途中で前記ベクターを補足してトランスフェクションした補足細胞を得るようにすることができる補足細胞に導入し、
(ii) 上記トランスフェクションした補足細胞を、ウイルス粒子を産生することができるようにするのに適当な条件下で培養し、
(iii) ウイルス粒子を細胞培養物から回収する
ことを特徴とする、方法。
【請求項32】
請求項17〜30のいずれか一項に記載の組換えベクターを含んでなる、または請求項31に記載の方法によって得られる、ウイルス粒子。
【請求項33】
請求項16に記載のヌクレオチド配列または請求項17〜30のいずれか一項に記載の組換えベクターを含んでなる、または請求項32に記載のウイルス粒子に感染している、宿主細胞。
【請求項34】
請求項1〜15のいずれか一項に記載のポリペプチド、請求項16に記載のヌクレオチド配列、請求項17〜30のいずれか一項に記載の組換えベクター、請求項32に記載のウイルス粒子、または請求項33に記載の宿主細胞を、薬学上許容可能な賦形剤と組み合わせて含んでなる、組成物。
【請求項35】
請求項1〜15のいずれか一項に記載のポリペプチドと、目的とする第二のポリペプチド、特にIL−2およびINFγから選択されるポリペプチドとを含んでなる、請求項34に記載の組成物。
【請求項36】
請求項16に記載のヌクレオチド配列と、IL−2およびINFγから選択されるポリペプチドをコードする目的とする第二のヌクレオチド配列とを含んでなる、請求項34に記載の組成物。
【請求項37】
遺伝子療法によってまたは組換え経路によって産生されたタンパク質を投与することによってヒトまたは動物体の治療を目的とする薬剤を調製するための、請求項1〜15のいずれか一項に記載のポリペプチド、請求項16に記載のヌクレオチド配列、請求項17〜30のいずれか一項に記載の組換えベクター、請求項32に記載のウイルス粒子、または請求項32に記載の宿主細胞の、治療または予防的使用。
【請求項38】
癌、腫瘍、および望ましくない細胞増殖から生じる疾患の治療を目的とする薬剤を調製するための、請求項37に記載の治療的使用。
【請求項39】
請求項16に記載のヌクレオチド配列、請求項17〜30のいずれか一項に記載の組換えベクター、請求項32に記載のウイルス粒子、または請求項33に記載の宿主細胞を、そのような治療を必要とする生物または宿主細胞に投与することを特徴とする、遺伝子療法による疾患の治療方法。
【請求項40】
薬学上許容可能な量のプロドラッグ、有利にはシトシンの類似体、特に5−FCを宿主生物または細胞に投与する、請求項38に記載の方法、または請求項37もしくは38に記載の治療的使用。
【請求項1】
アミノ酸配列を付加することによって天然のCDアーゼから誘導されることを特徴とする、CDアーゼ活性を有するポリペプチドであって、
UPRTアーゼまたはチミジンキナーゼ活性を持たない、ポリペプチド。
【請求項2】
天然CDアーゼに付加したアミノ酸配列が、天然CDアーゼのC末端に連結している、請求項1に記載のポリペプチド。
【請求項3】
天然CDアーゼに付加したアミノ酸配列が、10〜1000アミノ酸長である、請求項1または2に記載のポリペプチド。
【請求項4】
天然CDアーゼに付加したアミノ酸配列が、100〜400アミノ酸長である、請求項3に記載のポリペプチド。
【請求項5】
天然CDアーゼに付加したアミノ酸配列が、200〜300アミノ酸長である、請求項4に記載のポリペプチド。
【請求項6】
天然CDアーゼに付加したアミノ酸配列が、UPRTアーゼ活性を有するポリペプチドに由来する、請求項1〜5のいずれか一項に記載のポリペプチド。
【請求項7】
UPRTアーゼ活性を有するポリペプチドが、酵母UPRTアーゼ、特にSaccharomyces cerevisiae FUR1遺伝子によってコードされたものに由来する、請求項6に記載のポリペプチド。
【請求項8】
天然CDアーゼに付加したアミノ酸配列が、実質的に配列番号2の配列アイデンティファイアーに表されているものであって、2位のSer残基から出発して216位のVal残基で終了するアミノ酸配列に由来する、請求項7に記載のポリペプチド。
【請求項9】
天然CDアーゼに付加したアミノ酸配列が、配列番号2の2位のSer残基から出発して216位のVal残基で終了する配列アイデンティファイアーに表されている通りである、請求項8に記載のポリペプチド。
【請求項10】
上記天然CDアーゼが酵母CDアーゼ、特にSaccharomyces cerevisiae FCY1遺伝子によってコードされたものである、請求項1〜9のいずれか一項に記載のポリペプチド。
【請求項11】
天然CDアーゼが、実質的に配列番号1の配列アイデンティファイアーに表されている通りであって、1位のMet残基から出発して158位のGlu残基で終了するアミノ酸配列を含んでなる、請求項10に記載のポリペプチド。
【請求項12】
天然CDアーゼが配列番号1の配列アイデンティファイアーに表されている通りの、1位のMet残基から出発して158位のGlu残基で終了するアミノ酸配列を含んでなる、請求項11に記載のポリペプチド。
【請求項13】
実質的に配列番号1の配列アイデンティファイアーに表されている通りであって、1位のMet残基から出発して373位のVal残基で終了するアミノ酸配列を含んでなる、請求項10に記載のポリペプチド。
【請求項14】
配列番号1配列アイデンティファイアーに表されている通りの、1位のMet残基から出発して374位のVal残基で終了するアミノ酸配列を含んでなる、請求項13に記載のポリペプチド。
【請求項15】
天然CDアーゼの活性よりかなり高いCDアーゼ活性を示す、請求項1〜14のいずれか一項に記載のポリペプチド。
【請求項16】
請求項1〜15のいずれか一項に記載のポリペプチドをコードするヌクレオチド配列。
【請求項17】
請求項16に記載のヌクレオチド配列を有し、宿主細胞中での発現に必要な要素の制御下に置かれた、組換えベクター。
【請求項18】
ベクターがプラスミドとウイルスベクターとからなる群から選択されるものであって、必要に応じてはベクターのトランスフェクション効力および/または安定性を改良する1個以上の物質と組み合わされている、請求項17に記載の組換えベクター。
【請求項19】
ベクターのトランスフェクション効力および/または安定性を改良する前記物質が、カチオン性脂質、カチオン性ポリマー、リゾリン脂質およびポリペプチドを含んでなる群から選択される、請求項18に記載の組換えベクター。
【請求項20】
ポックスウイルス、アデノウイルス、レトロウイルス、ヘルペスウイルス、アルファウイルス、フォーミーウイルスまたはアデノ随伴ウイルスに由来するウイルスベクターである、請求項18に記載の組換えベクター。
【請求項21】
改質ワクシニアアンカラ(MVA)ウイルスに由来する、請求項20に記載の組換えベクター。
【請求項22】
請求項16に記載のヌクレオチド配列を、欠失I、II、III、IV、VおよびVIからなる群から選択されるMVAゲノム内の天然に存在する欠失の部位に挿入する、請求項21に記載の組換えベクター。
【請求項23】
天然に存在する欠失の部位が欠失IIIである、請求項22に記載の組換えベクター。
【請求項24】
発現に必要な要素がプロモーターを含んでなる、請求項17〜23のいずれか一項に記載の組換えベクター。
【請求項25】
プロモーターがチミジンキナーゼ7.5K遺伝子のプロモーターである、請求項24に記載の組換えベクター。
【請求項26】
複製に本質的な少なくとも1つの領域の全部または一部を欠き、El、E2、E4およびL1−L5領域から選択されるアデノウイルスベクターである、請求項20に記載の組換えベクター。
【請求項27】
非本質的E3領域の全部または一部を更に欠いているアデノウイルスベクターである、請求項26に記載の組換えベクター。
【請求項28】
プロモーターがサイトメガロウイルス(CMV)初期プロモーターである、請求項24に記載の組換えベクター。
【請求項29】
インターロイキンIL−2、IL−4、IL−7、IL−10およびIL−12、インターフェロン、腫瘍壊死因子(TNF)、コロニー刺激因子(CSF)、および脈管形成に作用する因子をコードする遺伝子から選択される1個以上の遺伝子を更に、含んでなる、請求項17〜28のいずれか一項に記載の組換えベクター。
【請求項30】
目的とする遺伝子が、IL−2およびINFγから選択されるポリペプチドをコードする、請求項29に記載の組換えベクター。
【請求項31】
ウイルス粒子の調製方法であって、
(i) 請求項17〜29のいずれか一項に記載の組換えベクターを、途中で前記ベクターを補足してトランスフェクションした補足細胞を得るようにすることができる補足細胞に導入し、
(ii) 上記トランスフェクションした補足細胞を、ウイルス粒子を産生することができるようにするのに適当な条件下で培養し、
(iii) ウイルス粒子を細胞培養物から回収する
ことを特徴とする、方法。
【請求項32】
請求項17〜30のいずれか一項に記載の組換えベクターを含んでなる、または請求項31に記載の方法によって得られる、ウイルス粒子。
【請求項33】
請求項16に記載のヌクレオチド配列または請求項17〜30のいずれか一項に記載の組換えベクターを含んでなる、または請求項32に記載のウイルス粒子に感染している、宿主細胞。
【請求項34】
請求項1〜15のいずれか一項に記載のポリペプチド、請求項16に記載のヌクレオチド配列、請求項17〜30のいずれか一項に記載の組換えベクター、請求項32に記載のウイルス粒子、または請求項33に記載の宿主細胞を、薬学上許容可能な賦形剤と組み合わせて含んでなる、組成物。
【請求項35】
請求項1〜15のいずれか一項に記載のポリペプチドと、目的とする第二のポリペプチド、特にIL−2およびINFγから選択されるポリペプチドとを含んでなる、請求項34に記載の組成物。
【請求項36】
請求項16に記載のヌクレオチド配列と、IL−2およびINFγから選択されるポリペプチドをコードする目的とする第二のヌクレオチド配列とを含んでなる、請求項34に記載の組成物。
【請求項37】
遺伝子療法によってまたは組換え経路によって産生されたタンパク質を投与することによってヒトまたは動物体の治療を目的とする薬剤を調製するための、請求項1〜15のいずれか一項に記載のポリペプチド、請求項16に記載のヌクレオチド配列、請求項17〜30のいずれか一項に記載の組換えベクター、請求項32に記載のウイルス粒子、または請求項32に記載の宿主細胞の、治療または予防的使用。
【請求項38】
癌、腫瘍、および望ましくない細胞増殖から生じる疾患の治療を目的とする薬剤を調製するための、請求項37に記載の治療的使用。
【請求項39】
請求項16に記載のヌクレオチド配列、請求項17〜30のいずれか一項に記載の組換えベクター、請求項32に記載のウイルス粒子、または請求項33に記載の宿主細胞を、そのような治療を必要とする生物または宿主細胞に投与することを特徴とする、遺伝子療法による疾患の治療方法。
【請求項40】
薬学上許容可能な量のプロドラッグ、有利にはシトシンの類似体、特に5−FCを宿主生物または細胞に投与する、請求項38に記載の方法、または請求項37もしくは38に記載の治療的使用。
【図1】

【公表番号】特表2006−527994(P2006−527994A)
【公表日】平成18年12月14日(2006.12.14)
【国際特許分類】
【出願番号】特願2006−520936(P2006−520936)
【出願日】平成16年6月29日(2004.6.29)
【国際出願番号】PCT/IB2004/002505
【国際公開番号】WO2005/007857
【国際公開日】平成17年1月27日(2005.1.27)
【出願人】(599082883)トランジェーヌ、ソシエテ、アノニム (32)
【氏名又は名称原語表記】TRANSGENE S.A.
【Fターム(参考)】
【公表日】平成18年12月14日(2006.12.14)
【国際特許分類】
【出願日】平成16年6月29日(2004.6.29)
【国際出願番号】PCT/IB2004/002505
【国際公開番号】WO2005/007857
【国際公開日】平成17年1月27日(2005.1.27)
【出願人】(599082883)トランジェーヌ、ソシエテ、アノニム (32)
【氏名又は名称原語表記】TRANSGENE S.A.
【Fターム(参考)】
[ Back to top ]