
改善されたヤロウイア・リポリティカ変異株によるジカルボン酸の製造
【課題】選択マーカーとして、LEU2およびURA3遺伝子による形質転換のために使用可能である原栄養性変異体MTLY37から栄養要求性ヤロウイア・リポリティカ変異株MTLY66,Leu−Ura−を得る方法を提供する。
【解決手段】生物変換条件下に、変異体MTLY66から、NADPH−シトクロムリダクターゼをコードするCPR遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY74 Leu+Ura−を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に選択マーカーLEU2およびCPR遺伝子を有する発現カセットを含有するJMP21−CPRベクターを変換することによって得る方法。
【解決手段】生物変換条件下に、変異体MTLY66から、NADPH−シトクロムリダクターゼをコードするCPR遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY74 Leu+Ura−を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に選択マーカーLEU2およびCPR遺伝子を有する発現カセットを含有するJMP21−CPRベクターを変換することによって得る方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、生物変換基質からの酵母ヤロウイア・リポリティカ(Yarrowia lipolytica)の変異株を用いた発酵によってジカルボン酸を製造する方法に関する。
【背景技術】
【0002】
ジカルボン酸(「二塩基酸」とも称される)は、ポリアミドおよびポリエステル、潤滑油、可塑剤または香料の合成のためのベース材料として用いられる。
【0003】
二塩基酸製造方法は、考慮される二塩基酸の炭素ネットワークの炭素原子の数に従って変動する(非特許文献1)。それ故に、アゼライン酸(C9二塩基酸)が、通常、オゾンによるオレイン酸の化学酸化によって得られる一方で、セバシン酸(C10二塩基酸)は、リシノール酸のアルカリ性酸化によって製造される。ドデカン二酸(C12二塩基酸)は、石油化学の製品である。細菌学は、トリデカンからブラッシル酸(C13二塩基酸)を製造するために用いられる。
【0004】
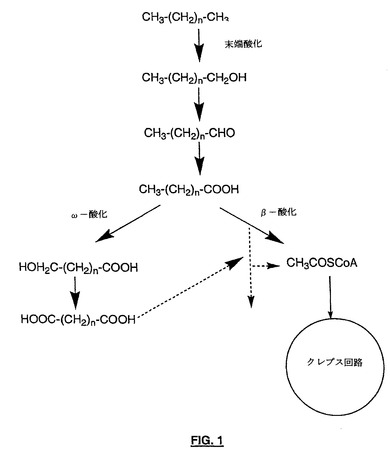
種々の適用において用いられる二塩基酸の多様性を考慮すると、最も広い範囲の考えられる二塩基酸に適用可能な製造経路の利点は疑う余地がない。化学製造より遅い反応速度によって特徴付けられるものの、生物学的製造は、非常に多様な基質に適用可能であるという利点を提供する(生物学的二塩基酸製造法は、概略的に図1に示される)。
【0005】
事実、多くの野生型の細菌種、例えば、クリプトコッカス・ネオフォルマンス(Cryptococcus neoformans)、緑膿菌(Pseudomonas aeruginosa)、キャンジダ・クロアカエ(Candida cloacae)等は、少量の二塩基酸を分泌することができる(非特許文献2〜3)。
【0006】
しかしながら、十分な量の二塩基酸分泌物を得るために、β酸化が阻止された変異体が用いられる必要がある。多くの種について、ランダムな突然変異生成およびその後の適切な選択によってこのような変異体が得られた(特許文献1(シイオ(Shiio)ら著)、非特許文献4)。
【0007】
しかしながら、特異的変異技術を用いるはるかによりエレガントではあるがより制限的な選択肢が、キャンジダ・トロピカリスについて開発された。この種に属する野生型株から、Picataggioらは、第1のβ酸化段階を触媒するアセチル−CoAオキシダーゼ(Aox)の2つのイソ酵素をコードする4つの遺伝子を連続的に破壊した(非特許文献5および特許文献2)。
【0008】
これらの著者は、その時、n−アルカンの二塩基酸への変換の反応速度論を制限する活性を構成していた、シトクロムP450モノオキシゲナーゼおよびNADPH−シトクロムリダクターゼをコードする遺伝子を過剰発現させた。(非特許文献6および特許文献3)。しかしながら、多コピー増幅システムに従って生じさせられたキャンジダ・トロピカリスの変異体は、全体的に安定でないようであり、それらは、場合によっては、復帰変異(reversions)を経る。これは、従来技術においてなぜ改良が行われなければならないかの理由である(特許文献4)。
【特許文献1】欧州特許第0229252号明細書
【特許文献2】米国特許第5254466号明細書
【特許文献3】米国特許第5648247号明細書
【特許文献4】米国特許出願公開第2004/0014198号明細書
【非特許文献1】ジョンソン・RW(Johnson RW)、ポロック・CM(Pollock CM)、キャントレル・RR(Cantrell RR)著、出版者「キルク・オスマー・エンサイクロペディア・オフ・ケミカル・テクノロジー(Kirk-Othmer Encyclopedia of Chemical Technology)」、第4版、1983年、p.118−136
【非特許文献2】チャン(Chan)ら著、「スティミュレーション・オフ・n−アルカン・コンバージョン・ツー・ジカルボキシリック・アシッド・バイ・オーガニック−ソルベント−アンド・デタージェント・トリーテッド・マイクローブス(Stimulation of n-alkane conversion to dicarboxylic acid by organic-solvent-and detergent-treated microbes)」、Appl.Microbiol.Biotechnol.、1991年、第34巻、p.772−777
【非特許文献3】シイオ(Shiio)ら著、「ミクロバイアル・プロダクション・オフ・ロング−チェイン・ジカルボキシリック・アシッド・フロム・n−アルカンズ(Microbial Production of Long-chain Dicarboxylic Acids from n-Alkanes) パートI(Part I) スクリーニング・アンド・プロパティーズ・オフ・ミクロオーガニスムズ・プロデューシング・ジカルボキシリック・アシッド(Screening and Properties of Microorganisms Producing Dicarboxylic Acids)」、Agr.Biol.Chem.、1971年、第35巻,第13号、p.2033−2042
【非特許文献4】ジアオ(Jiao)ら著、「アイソレーション・アンド・エンザイム・デターミネーション・オフ・キャンジダ・トロピカリス・ミュータンツ・フォー・DCA・プロダクション(Isolation and Enzyme Determination of Candida tropicalis Mutants for DCA Production)」、J.Gen.Appl.Microbiol.、2000年、第46巻,p.245−249
【非特許文献5】ピカタジオ(Picataggio)ら著、「デタミネーション・オフ・キャンジダ・トロピカリス・アシルコエンザイム・A・オキシダーゼ・イソエンザイム・ファンクション・バイ・シークエンシャル・ジーン・ディスラプション(Determination of Candida tropicalis Acylcoenzyme A Oxidase Isoenzyme Function by Sequential Gene Disruption)」、Mol.Cell.Biol.、1991年、第11巻、p.4333−4339
【非特許文献6】ピカタジオ(Picataggio)ら著、「メタボリック・エンジニアリング・オフ・キャンジダ・トロピカリス・フォー・ザ・プロダクション・オフ・ロング−チェイン・ジカルボキシリック・アシッド(Metabolic Engineering of Candida tropicalis for the Production of Long-chain Dicarboxylic Acids)」、Biotechnol.、1992年、第10巻、p.894−898
【発明の概要】
【発明が解決しようとする課題】
【0009】
(発明の目的)
本発明の目的は、従来技術の欠点を克服することである。実際に、アシル−CoAオキシダーゼをコードする遺伝子が破壊されたヤロウイア・リポリティカ変異体を用いて二塩基酸を生じさせることが有利には可能であることが発見された。
【課題を解決するための手段】
【0010】
本発明による方法が調製することを目的とする二塩基酸は、少なくとも10個の炭素原子を有する直線状炭化水素鎖を有し、鎖の各端部にカルボン酸基を含む有機化合物である。
【0011】
用いられる微生物は、ヤロウイア・リポリティカ変異体であって、β酸化において部分的に阻止するように少なくともPOX2、POX3、POX4およびPOX5遺伝子が破壊されたものである。
【0012】
POX2、POX3、POX4およびPOX5遺伝子によってコードされたアシル−CoAオキシダーゼとは別に、POX1およびPOX6遺伝子によってコードされる(未知機能の)2つの他のアシル−CoAオキシダーゼがヤロウイア・リポリティカのゲノムに存在する。
【0013】
これらの2つのアシル−CoAオキシダーゼは、2つの炭素原子の連続的脱離によって生合成された二塩基酸の再消費に必要とされる。POX1およびPOX6遺伝子のさらなる破壊は、それ故に、生物変換基質のプロファイルに相当する二塩基酸の産生をもたらす。例えば、18個の炭素原子を有する脂肪酸から主としてなるオレイン酸ヒマワリ油が生物変換基質として用いられる場合、POX遺伝子の全部が除かれた株は、18個の炭素原子を有する二塩基酸の大部分を生じさせるだろう。
【0014】
他方、生物変換基質は、脂肪体の形態の細胞内でトリグリセリドの形態で保存され得、それ故に、その二塩基酸への生物変換のために利用しにくくなる。遺伝子はプロテオーム解析によって、生物変換基質の蓄積において必要とされることが確認された。本発明者らは、下記遺伝子を確認した:DGA1(アシル−CoA:ジアシルグリセロール・アシルトランスフェラーゼ)、TGL1(トリアシルグリセロール・リパーゼ)、G3P(グリセロール−3−ホスファート・デヒドロゲナーゼ)、SCP2(推定のステロールキャリア)、LR01(推定のレシチン・コレステロール・アシルトランスフェラーゼ)並びにIFP 621 IPF(未知機能)、IPF 905(未知機能)およびIPF 2569(NADH−ユビキノン・リダクターゼ・サブユニット)。
【0015】
これらの遺伝子の活性の改変(破壊または過剰発現による)は、それ故に、ヤロウイア・リポリティカ細胞内の脂肪体の形態の生物変換基質の蓄積の減少をもたらす。
【0016】
ヤロウイア・リポリティカの遺伝系は、キャンジダ・トロピカリスのものとは全く異なることが留意され得る。二倍体酵母であるキャンジダ・トロピカリスとは異なり、ヤロウイア・リポリティカは、実際に、半数体種である。したがって、後者の微生物では、単一セットの染色体の存在のため、遺伝子削除操作は、より効果的かつ確実である。
【0017】
他方、アシル−CoAオキシダーゼをコードするPOX2遺伝子のプロモーターは、遺伝子、例えば、P450モノオキシゲナーゼ・シトクロムおよびNADPHシトクロムリダクターゼをコードする遺伝子を過剰発現させるために用いられる。プロモーターpPOX2は、生物変換基質によって高誘導性であるという特性を有する。それ故に、興味のある遺伝子の過剰発現は、プロモーターpPOX2の制御下に単一遺伝子コピーを付与することによって行われ、多コピー増幅システムによって得られるキャンジダ・トロピカリス変異体とは異なる、効率的、安定、かつ、非復帰性のヤロウイア・リポリティカ変異体を得ることが可能である。
【0018】
脂肪酸エステルまたは天然油(例えば、植物油)からの二塩基酸の産生についてのキャンジダ・トロピカリスを超えるヤロウイア・リポリティカの別の利点は、次の通りである:キャンジダ・トロピカリスによる天然油の二塩基酸への変換は、発酵に先行して基質の化学的な加水分解を少なくとも部分的に必要とする(米国特許5,962,285を参照)。この加水分解は、カルシウムまたはマグネシウムの水酸化物の存在中の鹸化によって行われる。それは、対応する脂肪酸塩(石鹸)を生じさせる。ここで、ヤロウイア・リポリティカは、トリグリセリドを炭素源として吸収する能力を有する。この異化の第1の段階は、1951年にPetersとNelsonにより確認された脂肪分解酵素(リパーゼ)によるトリグリセリドのフリーな脂肪酸とグリセロールへの加水分解を必要とする。細胞外のリパーゼ活性および39および44kDaの2つの膜リパーゼ(Barthら,ヤロウイア・リポリティカ、「Nonconventional Yeasts in Biotechnology A Handbook(Wolf,K.Ed.)Vol.1,1996,pp.313−388.Springer-Verlag」)がその後に記載された。ヤロウイア・リポリティカは、複数のリパーゼ(細胞外、膜および細胞内活性)を産生し得る。最近、上記のリパーゼに対応する遺伝子が特定された。LIP2遺伝子は、細胞外リパーゼLip2p(Pignedeら,2000)をコードする。それは好ましくはオレイン残基の長鎖トリグリセリドを加水分解することが示された(Barthら,1996)。
【0019】
ヤロウイア・リポリティカは、それ故に、発酵を行うのに適したpH条件下にエステルおよび天然油をフリーの脂肪酸とグリセロールに直接的に加水分解する。このような条件下に、エステルまたは油の加水分解および二塩基酸へのその変換は同時に起こり、これは、化学的加水分解段階が除かれるので、単純化された操作手順もたらすという利点を有する。
【発明を実施するための最良の形態】
【0020】
(詳細な説明)
より詳細な方法では、本発明は、少なくとも1種のジカルボン酸を製造する方法であって、
(a)生育段階であって、(アシル−CoAオキシダーゼをコードする)POX2、POX3、POX4およびPOX5遺伝子が少なくとも破壊されたヤロウイア・リポリティカの変異株が、炭素源および窒素源を少なくとも含むエネルギー性の基質から本質的になる培養培地において培養される、段階と、
(b)生物変換段階であって、前記株が、エネルギー性の基質の存在下に少なくとも10個の炭素原子を有するn−アルカン、少なくとも10個の炭素原子を有する脂肪酸、これらの脂肪酸の1〜4個の炭素原子を有するアルキルエステル(例えば、メチルまたはエチルエステルの混合物)および天然油(グリセロールの脂肪酸エステルの混合物)の中から選択される生物変換基質に供される、段階と、
(c)形成されたジカルボン酸を回収する段階と
を包含する方法を提供する。
【0021】
本発明による方法において用いられる株は、野生型ヤロウイア・リポリティカW29株(ATCC 20460,Collection de Levures d’Interet Biotechnologique(CLIB)においてCLIB89の下に記録された)に由来する。
【0022】
それ故に、G.Barthらの概説に記載されたPold株[ロイシン(leu−)およびウラシル(ura−)栄養要求性の株]によってヤロウイア・リポリティカ ATCC20460株に由来する新しい変異株を用いることが可能である。それは、CLIBにおいてCLIB139の下に記録されている。
【0023】
これらの新しい変異株(MTLY37、MTLY66、MTLY74、MTLY79、MTLY80、MTLY81、FT120およびFT130)が得られる方法は、下記に記載される。
【0024】
本発明による二塩基酸製造方法において、選択された変異株は、炭素源および窒素源を少なくとも含むエネルギー性の基質から本質的になる培地において生育が終わるまで培養される。次いで、生物変換基質(アルカンまたはアルカン混合物、脂肪酸または脂肪酸混合物、脂肪酸エステルまたは脂肪酸エステル混合物、天然油、またはこれらの種々の基質の混合物)が、二塩基酸への生物変換を開始するように加えられ、形成された二塩基酸は、当業者に知られる技術、例えば、カルシウム塩沈殿によって回収される。
【0025】
生物変換段階の間、培養培地は、一般的には、少なくとも1種のポリヒドロキシ化合物、例えば、グリセロール、糖等からなる二次的なエネルギー性の基質の供給を必要とし得る。
【0026】
(1.変異株MTLY37の取得)
この株を得るための手順は、次の通りであり得る:
1)興味のある遺伝子の配列を決定する(または、この遺伝子の配列は、データバンクにおいて入手可能である);
2)対抗選択マーカー(counter-selectable marker)、例えば、マーカーURA3(これにより、Ura+表現型またはUra−表現型について選択し得る)を用いるPCR(ポリメラーゼ連鎖反応)またはクローニングによって破壊用カセットを構築する;
3)削除された興味株を有する株を選択し(変換および軽質転換体の選択)(マーカーがURA3であれば有利にはUra+)、遺伝子崩壊をチェックする。
【0027】
変異株MTLY37を得る特定の方法は以降に記載される。
【0028】
野生型株のPOX遺伝子(その配列は、キャンジダ・トロピカリスの配列とは異なる)が、最初に、クローニングされかつ配列決定される。アシル−CoAオキシダーゼのイソ酵素をコードする遺伝子についての破壊用カセットが、次いで、構築される。アシル−CoAオキシダーゼの遺伝子は、選択マーカーURA3を用いて破壊される。プロモーターおよびターミネーター領域は、特定のオリゴヌクレオチド対を用いる第1のPCRによって増幅させられ、これは、オープンリーディングフレーム(ORF)から全配列を除去する。次いで、第2のPCRが外部のプライマーと、プロモーターおよびターミネーターのPCR産物とにより行われ、これらは、制限酵素I−Scelについての部位を含む20bpの共通の延長部を介して融合する。PCR産物がクローニングされて、プロモーター−ターミネーターモジュールを含有する一連のプラスミド(pPOX−PTによって設計される)が与えられる(破壊用カセット2)。
【0029】
URA3遺伝子が、POX−PTカセットのI−Scel部位に導入される。プロモーター−URA3−ターミネーターモジュールを含有する一連のpPOX−PUTプラスミドが構築される(破壊用カセット1)。これらの構築物は、一方で破壊用カセット1を含有するプラスミドについてpPOX1−PUT、pPOX2−PUT、pPOX3−PUT、pPOX4−PUTおよびpPOX5−PUTと称され、他方で、破壊用カセット2を含有するプラスミドについてpPOX1−PT、pPOX2−PT、pPOX3−PT、pPOX4−PTおよびpPOX5−PTと称される。
【0030】
破壊用カセットは、特定の外部プライマーによるPCRによって、例えば、Pfuポリメラーゼ(Stratagene,La Jolla,Californiaによって提供される)を用いて増幅させられる。タンパク質配列の最終分析は、ヤロウイア・リポリティカのアシル−CoAオキシダーゼが、他の酵母のものと45%の同一性度(50%類似性)を有することを示す。それらの間の同一性度は、55〜70%(65〜76%の類似性)である。
【0031】
アシル−CoAオキシダーゼをコードする遺伝子の破壊用カセットを構築した後、ヤロウイア・リポリティカの変換が、種々の方法によって行われ得る。エレクトロポレーションが行われ得、ここでは、DNAは、電気ショックによって導入される。より有利には、酢酸リチウムおよびポリエチレングリコール法が用いられ得る。それは、Gaillardinらによって記載されている:LEU2 Directed Expression of Beta-gallactosidase Activity and Phleomycin Resistance in Yarrowia lipolytica.Curr.Genet.11,1987,369−375。
【0032】
破壊の存在は、PCRによって、Gussowらの技術にしたがってチェックされ:Direct Clone Characterization from Plaques and Colonies by the Polymerase Chain Reaction.Nucleic Acids Res.17,1989,4000;次いで、サザンブロットハイブリッド形成によって確認される。
【0033】
Pold株の開始時に、破壊は2段階で行われる。
【0034】
Poldは、最初に、PCR PUT破壊用カセット1により変換され、選択される。次いで、Ura+クローンが、破壊用カセット2により変換され、URA3遺伝子が除かれ、それらが選択される。この手順は、四重崩壊MTLY37−pox2ΔPT−pox3ΔPT−pox4ΔPT−pox5ΔPUTを得ることを可能にする。この変異体の構築物の図表表示は、下記表1において要約される。
【0035】
【表1】

【0036】
遺伝子の破壊およびマーカーの切除はまた、組み換えを含む方法またはリコンビナーゼによってなされ得る。例えば、いずれかの側に、反復配列(選択された組み換えを可能にする)またはリコンビナーゼCreによって認識されるlox配列を有するマーカーを用いることが可能である。リコンビナーゼCreが発現させられた場合に切除は起こる(Fickersら,2003 New Disruption Cassettes for Rapid Gene Disruption and Marker Rescue in the Yeast Yarrowia lipolytica.J.Microbiol.Methods 55/3:727-737)。
【0037】
(2.ヤロウイア・リポリティカ変異株MTLY74の取得)
株MTLY74Leu+Ura−は変異株MTLY37から構築された。
【0038】
(2a)原栄養性変異体MTLY37(Leu+、Ura+)から、本発明者らは、ロイシンおよびウラシル栄養要求性ヤロウイア・リポリティカ変異株MTLY66(Leu−、Ura−)を構築する。第一段階は、マーカーura3−41によるマーカーURA3の変換、このマーカーを含有するPCRフラグメントを形質転換することおよび5FOAの存在下にUra−を選択することによるウラシル栄養要求性株MTLY40(Leu+、Ura−)の構築である。変異体MTLY40から、本発明者らは、ロイシン栄養要求性ヤロウイア・リポリティカ変異株MTLY64(Leu−、Ura−、Hyg+)を、マーカーLEU2を破壊すること、破壊用カセットPHTleu2を変換することおよび抵抗性ヒグロマイシン形質転換体(leu2::Hyg)を選択することによって構築する。MTLY64変異株から、本発明者らは、ロイシン栄養要求性ヤロウイア・リポリティカ変異株MTLY66(Leu−、Ura−)を、HYGマーカーを切除すること、リコンビナーゼCreおよびマーカーLEU2(Cre−LEU2)を含有する複製ベクターpRRQ2を軽質転換することおよび感受性ハイゴロマイシン形質転換体Leu+を選択することにより構築する。プラスミドpRRQ2の喪失は、富栄養培地YPD上での培養およびクローン(Leu−、Ura−、Hyg−)の単離によって達成される。
【0039】
(2b)変異体MTLY66から、本発明者らは、NADPH−シトクロムリダクターゼを過剰発現させるヤロウイア・リポリティカ変異株MTLY74 Leu+ Ura−を、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導される強いプロモーターpPOX2の制御下にそれを発現させることにより構築する。
【0040】
NADPH−シトクロムリダクターゼをコードする遺伝子(CPR)は、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に、選択遺伝子LEU2、JMP21(例)を含有するベクター中に導入される。マーカー−プロモータ−遺伝子のカセット(LEU2−pPOX2−CPR)は、変換によって導入される。
【0041】
この変異体の構築物の概略表示は、以下の表2に要約される。
【0042】
【表2】
【0043】
(3 共通の変異体MTLY74からの株MTLY79、MTLY80およびMTLY81の取得)
(3a)脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下にNADPH−シトクロムリダクターゼおよびシトクロムP450モノオキシゲナーゼALK1を発現させるMTLY79。
【0044】
変異体MTLY74から、本発明者らは、NADPH−シトクロムリダクターゼおよびシトクロムP450モノオキシゲナーゼALK1を過剰発現させるヤロウイア・リポリティカ変異株MTLY79を、生物変換条件下、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導される強いプロモーターpPOX2の制御下に構築する。
【0045】
本発明者らは、シトクロムP450モノオキシゲナーゼをコードするALK1遺伝子を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に、URA3選択遺伝子(例えば、JMP61)を含有するベクター中に導入する。マーカー−プロモーター−遺伝子のカセット(URA3−pPOX2−AKL1)は変換によって導入される。
【0046】
この変異体の構築物の概略表示は、以下の表3に要約される。
【0047】
【表3】
【0048】
(3b)脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下にNADPH−シトクロムリダクターゼおよびシトクロムP450モノオキシゲナーゼALK2を発現させるMTLY80。
【0049】
変異体MTLY74から、本発明者らは、NADPH−シトクロムリダクターゼ(CPR)およびシトクロムP450モノオキシゲナーゼ(ALK2)をコードする遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY80を、生物変換条件下、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導される強いプロモーターpPOX2の制御下に構築する。
【0050】
本発明者らは、シトクロムP450モノオキシゲナーゼをコードするALK2遺伝子を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に、URA3選択遺伝子(例えばJMP61)を含有するベクター中に導入する。マーカー−プロモーター−遺伝子のカセット(URA3−pPOX2−AKL2)は、変換によって導入される。
【0051】
この変異体の構築物の概略表示は、以下の表4に要約される。
【0052】
【表4】
【0053】
(3c)脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下にシトクロムP450モノオキシゲナーゼ(ALK1またはALK2)の遺伝子なしでNADPH−シトクロムリダクターゼを発現させるMTLY81。
【0054】
変異体MTLY74から、本発明者らは、NADPH−シトクロムリダクターゼ(CPR)をコードする遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY81を、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導される強いプロモーターpPOX2の制御下に構築する。
【0055】
変異体MTLY74は、マーカーURA3を有するプラスミドJMP61による形質転換によって無機物栄養性にされる。
【0056】
この変異体の構築物の概略表示は、以下の表5に要約される。
【0057】
【表5】
【0058】
(4 変異株FT120の取得)
この株は、MTLY37株またはMTLY66株の構築と同一の手順に従うことによって得られ得る。
【0059】
1)対抗選択マーカー、例えばマーカーURA3(これにより、Ura+表現型またはUra−表現型について選択し得る)を用いる、または、いずれかの側上に、反復配列(選択された組み換えを可能にする)またはリコンビナーゼCreによって認識されるlox配列を有するマーカー(選択された組み換えを可能にする)を用いる、PCR(ポリメラーゼ連鎖反応)またはクローニングにより破壊用カセットを構築する。
【0060】
2)削除された興味遺伝子を有する株を選択し(変換、および軽質転換体;マーカーがURA3ならば有利にはUra+の選択)および遺伝子破壊をチェックする。
【0061】
3)削除されたマーカーを有する株を選択し(変換および軽質転換体の選択);マーカーがURA3ならば有利には5FOARまたはマーカーがlox配列を示すならば有利にはリコンビナーゼを発現させるプラスミド;および遺伝子破壊をチェックする。
【0062】
変異株FT120を得るための特定の方法が以下に記載される。
【0063】
変異体MTLY66から、本発明者らは、株FT120Leu−Ura−,Δpox1−6を構築する。
【0064】
(4a)変異体MTLY66 Δpox2−5から、本発明者らは、ヤロウイア・リポリティカ変異株MTLY95 Δpox1−6を、POX1およびPOX6遺伝子の削除部およびマーカーの削除部の挿入により上記の方法に従って構築する。
【0065】
(4b)変異体MTLY95から、本発明者らは、生物変換条件下にNADPH−シトクロムリダクターゼ(CPR)をコードする遺伝子を過剰発現させるヤロウイア・リポリティカ変異株FT101 Leu+Ura−を、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導される強いプロモーターpPOX2の制御下にそれを発現させることによって構築する。
【0066】
本発明者らは、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に、NADPH−シトクロムリダクターゼをコードする遺伝子を、切除可能なLEU2選択遺伝子JMP21−LEU2ex(例)を含有するベクター内に導入した。マーカー−プロモーター−遺伝子のカセット(LEU2ex−pPOX2−CPR)は、形質転換によって導入される。FT120株は、マーカーLEU2exの切除後に、pUB4−CREプラスミド(Hyg+選択)による変換、YPD上のプラスミド喪失および最後のLeuクローンの単離によって得られる。
【0067】
この変異体の構築物の概略表示は、以下の表6に要約される。
【0068】
【表6】
【0069】
(5 変異株FT130の取得)
この株は、株FT120の構築物と同一の手順に従うことによって得られ得る。
【0070】
1)対抗選択マーカー、例えば、マーカーURA3(これにより、Ura+表現型またはUra−表現型を選択することができる)を用いる、または、いずれかの側上に、反復配列を有するマーカー(選択された組み換えを可能にする)またはリコンビナーゼCreによって認識されるlox配列を用いる、PCR(ポリメラーゼ連鎖反応)またはクローニングによって破壊用カセットを構築する。
【0071】
2)削除された興味遺伝子を有する株を選択し(変換および形質転換体の選択:マーカーがURA3であれば有利にはUra+)、遺伝子破壊をチェックする。
【0072】
3)削除されたマーカーを有する株を選択し(変換および形質転換体の選択;マーカーがURA3ならば有利には5FOAR、または、マーカーがlox配列を示すならば有利にはリコンビナーゼCreを発現させるプラスミド)、遺伝子破壊をチェックする。
【0073】
変異株FT130を得るための特定の方法が以下に記載される。
【0074】
変異体FT120から、本発明者らは、株FT130Leu−Ura−,Δpox1−6,Δdga1を構築する。
【0075】
(6)変異体FT120 Leu−Ura−,Δpox1−6,pPOX2−CPRから、本発明者らは、ヤロウイア・リポリティカ変異体FT130 Leu−Ura+Δpox1−6,pPOX2−CPR,Δdgalを、アシル−CoAジアシルグリセロール・アセチルトランスフェラーゼをコードするDGA1遺伝子の削除物の挿入によって構築する。
【0076】
【表7】
【0077】
株MTLY66、MTLY81、FT120およびFT130は、Collection Nationale de Cultures de Microorganismesに、それぞれ登録番号CNCM I−3319、CNCM I−3320、CNCM I−3527およびCNCM I−3528の下に登録されている。
【0078】
以下の実施例は本発明を説明するが、本発明の範囲を制限するものではない。
【0079】
(実施例)
これらの実施例において、本発明者らは、二塩基酸の産生についての培養条件および培地組成の影響を試験した。そして、本発明者らは、変異体MTLY37により、ペプトンの使用が、特にバクト−トリプトンと関連して、二塩基酸の産生に非常に有利であることを観察した(実施例1および2)。
【0080】
本発明者らはまた、同じ条件下に、変異体MTLY79、MTLY80およびMTLY81を試験した。そして、本発明者らは、NADPH−シトクロムリダクターゼが、二塩基酸の産生における制限段階を触媒することを観察した。実際に、この酵素の過剰発現のみが、二塩基酸の産生および産生能を大幅に増加させることを可能にする(実施例4〜6)。
【0081】
本発明者らは、同一条件下に、変異株FT120およびFT130を試験した(実施例7および8)。POX1およびPOX6遺伝子の削除は、FT120についての二塩基酸の分解を減少させることを可能にする。アシル−CoAジアシルグリセロール・アセチルトランスフェラーゼをコードするさらなるDGA1遺伝子の削除により、ヤロウイア・リポリティカの細胞内の脂肪体の形態にある生物変換基質の蓄積が減少する。それ故に、これらの実施例において得られた二塩基酸の大部分は、18個の炭素原子を有する脂肪酸を有する脂肪酸から本質的になる用いられる生物変換基質のような18個の炭素原子を有する二塩基酸からなる。
【0082】
(実施例1:変異体MTLY37によりオレインヒマワリ油からジカルボン酸を製造する方法)
組成物:酵母抽出物10g・L−1、ペプトン10g・L−1、グルコース10g・L−1、寒天(Agar)20g・L−1のグロース培地(gelosed medium)において維持される、変異体MTLY37の予備培養が、0.30近くの予備培養培地の初期吸光度を提供する播種によって行われた。予備培養は、軌道攪拌下(200rpm)に24時間にわたり30℃で、25mLの培地(10g・L−1の酵母抽出物、10g・L−1のペプトン、20g・L−1のグルコース)を含有する500mLのフランジ付きフラスコにおいて行われる。
【0083】
培養のために用いられた培地は、脱イオン水、10g・L−1の酵母抽出物、20g・L−1のトリプトン、40g・L−1のグルコースおよび30g・L−1のオレインヒマワリ油から構成される。
【0084】
発酵槽の播種は、予備培養フラスコの全てにより達成される。
【0085】
培養は、30℃で、2Lの培地を有し、複動半径流タービン(double-acting centripetal turbine)を備えた4Lの発酵槽において、0.5vvmの通気速度、800rpmの攪拌速度で行われる。
【0086】
17時間の培養後、培地のグルコースが除去されて直ぐに、60mLのオレインヒマワリ油(18個の炭素原子を有する脂肪酸から本質的になる)が、反応器に加えられ、1mL・h−1の速度での連続的なグリセロール供給に付される。その時、培養のpH値は、4Mのソーダの調整された添加によって8の一定値に維持される。発酵は130時間で終了する。
【0087】
培養手順の終了時に、遠心分離によって細胞バイオマスが取り除かれる。その後、6MのHClを添加することによって上澄は2.5のpH値まで酸性にされ、酸性ワート(wort)の遠心分離によって不溶性のジカルボン酸が集められ、次いで、乾燥させられる。
【0088】
混合物のジカルボン酸組成は、Uchioらの「Microbial Production of Long-chain Dicarboxylic Acids from n-Alkanes.Part II.Production by Candida cloacae Mutant Usable to Assimilate Dicarboxylic Acid」(Agr Biol.Chem.36,No.3,1972,426-433)によって記載された方法によるジカルボン酸のジエステルへの変換後に、カラムDB1におけるガスクロマトグラフィーによって測定される。クロマトグラフ・オーブンの温度は、150℃から280℃に8℃/分の速さでプログラムされる。
【0089】
結果は、130時間後に上澄における最大の二塩基酸産生度5.9g・L−1を示す。
【0090】
(実施例2)
培養培地において、同一濃度でトリプトンをペプトンに置き換えることによって実施例1が繰り返される。培養130時間後に、9.9g・L−1のジカルボン酸が得られた。すなわち、実施例1と関連して約68%産生度が増加した。
【0091】
(実施例3:連続的なオレインヒマワリ油の供給による変異体MTLY37によるジカルボン酸の産生)
オレインヒマワリ油が培養培地から除かれ、かつ、反応器内への1mLの二次制限(sublimiting)流量でのこの油の連続注入によって置き換えられて、実施例2が繰り返される。
【0092】
このような条件下に、130時間後に14.7g・L−1のジカルボン酸が培養培地において産生される。
【0093】
(実施例4:CPRおよびALK1を過剰発現させる変異体MTLY79によるオレインヒマワリ油からのジカルボン酸の産生)
変異体MTLY37を、CPRおよびALK1遺伝子を過剰発現させる変異体MTLY79によって置き換えることによって実施例3が繰り返される。130時間の培養後、16g・L−1のジカルボン酸が得られた。
【0094】
(実施例5:CPRおよびALK2を過剰発現させる変異体MTLY80によるオレインヒマワリ油からのジカルボン酸の産生)
変異体MTLY37を、CPRおよびALK2遺伝子を過剰発現させる変異体MTLY80によって置き換えることによって実施例3が繰り返される。130時間の培養後、16g・L−1のジカルボン酸が得られる。
【0095】
(実施例6)
変異体MTLY37を、CPR遺伝子のみを過剰発現させる変異体MTLY81によって置き換えることによって実施例3が繰り返される。130時間の培養後、16g・L−1のジカルボン酸が得られる。
【0096】
(実施例7:6のPOX(Δpox1−6)遺伝子が削除され、CPR遺伝子を過剰発現させる変異体FT120によるオレインヒマワリ油からのジカルボン酸の産生)
変異体MTLY37を、6のPOX遺伝子が削除され、CPR遺伝子のみを過剰発現させる変異体FT120によって置き換えることによって実施例3が繰り返される。130時間の培養後、18g・L−1のジカルボン酸が得られる。
【0097】
(実施例8:6のPOX(Δpox1−6)遺伝子が削除され、DGA1遺伝子(Δdga1)が削除され、かつ、CPR遺伝子を過剰発現させる変異体FT130によるオレインヒマワリ油からのジカルボン酸の産生)
変異体MTLY37を、6のPOX遺伝子およびDGA1遺伝子が削除され、CPR遺伝子を過剰発現させる変異体FT130によって置き換えることによって実施例3が繰り返される。130時間の培養後、23g・L−1のジカルボン酸が得られる。
【図面の簡単な説明】
【0098】
【図1】生物学的二塩基酸製造法を示す概略図である。
【技術分野】
【0001】
本発明は、生物変換基質からの酵母ヤロウイア・リポリティカ(Yarrowia lipolytica)の変異株を用いた発酵によってジカルボン酸を製造する方法に関する。
【背景技術】
【0002】
ジカルボン酸(「二塩基酸」とも称される)は、ポリアミドおよびポリエステル、潤滑油、可塑剤または香料の合成のためのベース材料として用いられる。
【0003】
二塩基酸製造方法は、考慮される二塩基酸の炭素ネットワークの炭素原子の数に従って変動する(非特許文献1)。それ故に、アゼライン酸(C9二塩基酸)が、通常、オゾンによるオレイン酸の化学酸化によって得られる一方で、セバシン酸(C10二塩基酸)は、リシノール酸のアルカリ性酸化によって製造される。ドデカン二酸(C12二塩基酸)は、石油化学の製品である。細菌学は、トリデカンからブラッシル酸(C13二塩基酸)を製造するために用いられる。
【0004】
種々の適用において用いられる二塩基酸の多様性を考慮すると、最も広い範囲の考えられる二塩基酸に適用可能な製造経路の利点は疑う余地がない。化学製造より遅い反応速度によって特徴付けられるものの、生物学的製造は、非常に多様な基質に適用可能であるという利点を提供する(生物学的二塩基酸製造法は、概略的に図1に示される)。
【0005】
事実、多くの野生型の細菌種、例えば、クリプトコッカス・ネオフォルマンス(Cryptococcus neoformans)、緑膿菌(Pseudomonas aeruginosa)、キャンジダ・クロアカエ(Candida cloacae)等は、少量の二塩基酸を分泌することができる(非特許文献2〜3)。
【0006】
しかしながら、十分な量の二塩基酸分泌物を得るために、β酸化が阻止された変異体が用いられる必要がある。多くの種について、ランダムな突然変異生成およびその後の適切な選択によってこのような変異体が得られた(特許文献1(シイオ(Shiio)ら著)、非特許文献4)。
【0007】
しかしながら、特異的変異技術を用いるはるかによりエレガントではあるがより制限的な選択肢が、キャンジダ・トロピカリスについて開発された。この種に属する野生型株から、Picataggioらは、第1のβ酸化段階を触媒するアセチル−CoAオキシダーゼ(Aox)の2つのイソ酵素をコードする4つの遺伝子を連続的に破壊した(非特許文献5および特許文献2)。
【0008】
これらの著者は、その時、n−アルカンの二塩基酸への変換の反応速度論を制限する活性を構成していた、シトクロムP450モノオキシゲナーゼおよびNADPH−シトクロムリダクターゼをコードする遺伝子を過剰発現させた。(非特許文献6および特許文献3)。しかしながら、多コピー増幅システムに従って生じさせられたキャンジダ・トロピカリスの変異体は、全体的に安定でないようであり、それらは、場合によっては、復帰変異(reversions)を経る。これは、従来技術においてなぜ改良が行われなければならないかの理由である(特許文献4)。
【特許文献1】欧州特許第0229252号明細書
【特許文献2】米国特許第5254466号明細書
【特許文献3】米国特許第5648247号明細書
【特許文献4】米国特許出願公開第2004/0014198号明細書
【非特許文献1】ジョンソン・RW(Johnson RW)、ポロック・CM(Pollock CM)、キャントレル・RR(Cantrell RR)著、出版者「キルク・オスマー・エンサイクロペディア・オフ・ケミカル・テクノロジー(Kirk-Othmer Encyclopedia of Chemical Technology)」、第4版、1983年、p.118−136
【非特許文献2】チャン(Chan)ら著、「スティミュレーション・オフ・n−アルカン・コンバージョン・ツー・ジカルボキシリック・アシッド・バイ・オーガニック−ソルベント−アンド・デタージェント・トリーテッド・マイクローブス(Stimulation of n-alkane conversion to dicarboxylic acid by organic-solvent-and detergent-treated microbes)」、Appl.Microbiol.Biotechnol.、1991年、第34巻、p.772−777
【非特許文献3】シイオ(Shiio)ら著、「ミクロバイアル・プロダクション・オフ・ロング−チェイン・ジカルボキシリック・アシッド・フロム・n−アルカンズ(Microbial Production of Long-chain Dicarboxylic Acids from n-Alkanes) パートI(Part I) スクリーニング・アンド・プロパティーズ・オフ・ミクロオーガニスムズ・プロデューシング・ジカルボキシリック・アシッド(Screening and Properties of Microorganisms Producing Dicarboxylic Acids)」、Agr.Biol.Chem.、1971年、第35巻,第13号、p.2033−2042
【非特許文献4】ジアオ(Jiao)ら著、「アイソレーション・アンド・エンザイム・デターミネーション・オフ・キャンジダ・トロピカリス・ミュータンツ・フォー・DCA・プロダクション(Isolation and Enzyme Determination of Candida tropicalis Mutants for DCA Production)」、J.Gen.Appl.Microbiol.、2000年、第46巻,p.245−249
【非特許文献5】ピカタジオ(Picataggio)ら著、「デタミネーション・オフ・キャンジダ・トロピカリス・アシルコエンザイム・A・オキシダーゼ・イソエンザイム・ファンクション・バイ・シークエンシャル・ジーン・ディスラプション(Determination of Candida tropicalis Acylcoenzyme A Oxidase Isoenzyme Function by Sequential Gene Disruption)」、Mol.Cell.Biol.、1991年、第11巻、p.4333−4339
【非特許文献6】ピカタジオ(Picataggio)ら著、「メタボリック・エンジニアリング・オフ・キャンジダ・トロピカリス・フォー・ザ・プロダクション・オフ・ロング−チェイン・ジカルボキシリック・アシッド(Metabolic Engineering of Candida tropicalis for the Production of Long-chain Dicarboxylic Acids)」、Biotechnol.、1992年、第10巻、p.894−898
【発明の概要】
【発明が解決しようとする課題】
【0009】
(発明の目的)
本発明の目的は、従来技術の欠点を克服することである。実際に、アシル−CoAオキシダーゼをコードする遺伝子が破壊されたヤロウイア・リポリティカ変異体を用いて二塩基酸を生じさせることが有利には可能であることが発見された。
【課題を解決するための手段】
【0010】
本発明による方法が調製することを目的とする二塩基酸は、少なくとも10個の炭素原子を有する直線状炭化水素鎖を有し、鎖の各端部にカルボン酸基を含む有機化合物である。
【0011】
用いられる微生物は、ヤロウイア・リポリティカ変異体であって、β酸化において部分的に阻止するように少なくともPOX2、POX3、POX4およびPOX5遺伝子が破壊されたものである。
【0012】
POX2、POX3、POX4およびPOX5遺伝子によってコードされたアシル−CoAオキシダーゼとは別に、POX1およびPOX6遺伝子によってコードされる(未知機能の)2つの他のアシル−CoAオキシダーゼがヤロウイア・リポリティカのゲノムに存在する。
【0013】
これらの2つのアシル−CoAオキシダーゼは、2つの炭素原子の連続的脱離によって生合成された二塩基酸の再消費に必要とされる。POX1およびPOX6遺伝子のさらなる破壊は、それ故に、生物変換基質のプロファイルに相当する二塩基酸の産生をもたらす。例えば、18個の炭素原子を有する脂肪酸から主としてなるオレイン酸ヒマワリ油が生物変換基質として用いられる場合、POX遺伝子の全部が除かれた株は、18個の炭素原子を有する二塩基酸の大部分を生じさせるだろう。
【0014】
他方、生物変換基質は、脂肪体の形態の細胞内でトリグリセリドの形態で保存され得、それ故に、その二塩基酸への生物変換のために利用しにくくなる。遺伝子はプロテオーム解析によって、生物変換基質の蓄積において必要とされることが確認された。本発明者らは、下記遺伝子を確認した:DGA1(アシル−CoA:ジアシルグリセロール・アシルトランスフェラーゼ)、TGL1(トリアシルグリセロール・リパーゼ)、G3P(グリセロール−3−ホスファート・デヒドロゲナーゼ)、SCP2(推定のステロールキャリア)、LR01(推定のレシチン・コレステロール・アシルトランスフェラーゼ)並びにIFP 621 IPF(未知機能)、IPF 905(未知機能)およびIPF 2569(NADH−ユビキノン・リダクターゼ・サブユニット)。
【0015】
これらの遺伝子の活性の改変(破壊または過剰発現による)は、それ故に、ヤロウイア・リポリティカ細胞内の脂肪体の形態の生物変換基質の蓄積の減少をもたらす。
【0016】
ヤロウイア・リポリティカの遺伝系は、キャンジダ・トロピカリスのものとは全く異なることが留意され得る。二倍体酵母であるキャンジダ・トロピカリスとは異なり、ヤロウイア・リポリティカは、実際に、半数体種である。したがって、後者の微生物では、単一セットの染色体の存在のため、遺伝子削除操作は、より効果的かつ確実である。
【0017】
他方、アシル−CoAオキシダーゼをコードするPOX2遺伝子のプロモーターは、遺伝子、例えば、P450モノオキシゲナーゼ・シトクロムおよびNADPHシトクロムリダクターゼをコードする遺伝子を過剰発現させるために用いられる。プロモーターpPOX2は、生物変換基質によって高誘導性であるという特性を有する。それ故に、興味のある遺伝子の過剰発現は、プロモーターpPOX2の制御下に単一遺伝子コピーを付与することによって行われ、多コピー増幅システムによって得られるキャンジダ・トロピカリス変異体とは異なる、効率的、安定、かつ、非復帰性のヤロウイア・リポリティカ変異体を得ることが可能である。
【0018】
脂肪酸エステルまたは天然油(例えば、植物油)からの二塩基酸の産生についてのキャンジダ・トロピカリスを超えるヤロウイア・リポリティカの別の利点は、次の通りである:キャンジダ・トロピカリスによる天然油の二塩基酸への変換は、発酵に先行して基質の化学的な加水分解を少なくとも部分的に必要とする(米国特許5,962,285を参照)。この加水分解は、カルシウムまたはマグネシウムの水酸化物の存在中の鹸化によって行われる。それは、対応する脂肪酸塩(石鹸)を生じさせる。ここで、ヤロウイア・リポリティカは、トリグリセリドを炭素源として吸収する能力を有する。この異化の第1の段階は、1951年にPetersとNelsonにより確認された脂肪分解酵素(リパーゼ)によるトリグリセリドのフリーな脂肪酸とグリセロールへの加水分解を必要とする。細胞外のリパーゼ活性および39および44kDaの2つの膜リパーゼ(Barthら,ヤロウイア・リポリティカ、「Nonconventional Yeasts in Biotechnology A Handbook(Wolf,K.Ed.)Vol.1,1996,pp.313−388.Springer-Verlag」)がその後に記載された。ヤロウイア・リポリティカは、複数のリパーゼ(細胞外、膜および細胞内活性)を産生し得る。最近、上記のリパーゼに対応する遺伝子が特定された。LIP2遺伝子は、細胞外リパーゼLip2p(Pignedeら,2000)をコードする。それは好ましくはオレイン残基の長鎖トリグリセリドを加水分解することが示された(Barthら,1996)。
【0019】
ヤロウイア・リポリティカは、それ故に、発酵を行うのに適したpH条件下にエステルおよび天然油をフリーの脂肪酸とグリセロールに直接的に加水分解する。このような条件下に、エステルまたは油の加水分解および二塩基酸へのその変換は同時に起こり、これは、化学的加水分解段階が除かれるので、単純化された操作手順もたらすという利点を有する。
【発明を実施するための最良の形態】
【0020】
(詳細な説明)
より詳細な方法では、本発明は、少なくとも1種のジカルボン酸を製造する方法であって、
(a)生育段階であって、(アシル−CoAオキシダーゼをコードする)POX2、POX3、POX4およびPOX5遺伝子が少なくとも破壊されたヤロウイア・リポリティカの変異株が、炭素源および窒素源を少なくとも含むエネルギー性の基質から本質的になる培養培地において培養される、段階と、
(b)生物変換段階であって、前記株が、エネルギー性の基質の存在下に少なくとも10個の炭素原子を有するn−アルカン、少なくとも10個の炭素原子を有する脂肪酸、これらの脂肪酸の1〜4個の炭素原子を有するアルキルエステル(例えば、メチルまたはエチルエステルの混合物)および天然油(グリセロールの脂肪酸エステルの混合物)の中から選択される生物変換基質に供される、段階と、
(c)形成されたジカルボン酸を回収する段階と
を包含する方法を提供する。
【0021】
本発明による方法において用いられる株は、野生型ヤロウイア・リポリティカW29株(ATCC 20460,Collection de Levures d’Interet Biotechnologique(CLIB)においてCLIB89の下に記録された)に由来する。
【0022】
それ故に、G.Barthらの概説に記載されたPold株[ロイシン(leu−)およびウラシル(ura−)栄養要求性の株]によってヤロウイア・リポリティカ ATCC20460株に由来する新しい変異株を用いることが可能である。それは、CLIBにおいてCLIB139の下に記録されている。
【0023】
これらの新しい変異株(MTLY37、MTLY66、MTLY74、MTLY79、MTLY80、MTLY81、FT120およびFT130)が得られる方法は、下記に記載される。
【0024】
本発明による二塩基酸製造方法において、選択された変異株は、炭素源および窒素源を少なくとも含むエネルギー性の基質から本質的になる培地において生育が終わるまで培養される。次いで、生物変換基質(アルカンまたはアルカン混合物、脂肪酸または脂肪酸混合物、脂肪酸エステルまたは脂肪酸エステル混合物、天然油、またはこれらの種々の基質の混合物)が、二塩基酸への生物変換を開始するように加えられ、形成された二塩基酸は、当業者に知られる技術、例えば、カルシウム塩沈殿によって回収される。
【0025】
生物変換段階の間、培養培地は、一般的には、少なくとも1種のポリヒドロキシ化合物、例えば、グリセロール、糖等からなる二次的なエネルギー性の基質の供給を必要とし得る。
【0026】
(1.変異株MTLY37の取得)
この株を得るための手順は、次の通りであり得る:
1)興味のある遺伝子の配列を決定する(または、この遺伝子の配列は、データバンクにおいて入手可能である);
2)対抗選択マーカー(counter-selectable marker)、例えば、マーカーURA3(これにより、Ura+表現型またはUra−表現型について選択し得る)を用いるPCR(ポリメラーゼ連鎖反応)またはクローニングによって破壊用カセットを構築する;
3)削除された興味株を有する株を選択し(変換および軽質転換体の選択)(マーカーがURA3であれば有利にはUra+)、遺伝子崩壊をチェックする。
【0027】
変異株MTLY37を得る特定の方法は以降に記載される。
【0028】
野生型株のPOX遺伝子(その配列は、キャンジダ・トロピカリスの配列とは異なる)が、最初に、クローニングされかつ配列決定される。アシル−CoAオキシダーゼのイソ酵素をコードする遺伝子についての破壊用カセットが、次いで、構築される。アシル−CoAオキシダーゼの遺伝子は、選択マーカーURA3を用いて破壊される。プロモーターおよびターミネーター領域は、特定のオリゴヌクレオチド対を用いる第1のPCRによって増幅させられ、これは、オープンリーディングフレーム(ORF)から全配列を除去する。次いで、第2のPCRが外部のプライマーと、プロモーターおよびターミネーターのPCR産物とにより行われ、これらは、制限酵素I−Scelについての部位を含む20bpの共通の延長部を介して融合する。PCR産物がクローニングされて、プロモーター−ターミネーターモジュールを含有する一連のプラスミド(pPOX−PTによって設計される)が与えられる(破壊用カセット2)。
【0029】
URA3遺伝子が、POX−PTカセットのI−Scel部位に導入される。プロモーター−URA3−ターミネーターモジュールを含有する一連のpPOX−PUTプラスミドが構築される(破壊用カセット1)。これらの構築物は、一方で破壊用カセット1を含有するプラスミドについてpPOX1−PUT、pPOX2−PUT、pPOX3−PUT、pPOX4−PUTおよびpPOX5−PUTと称され、他方で、破壊用カセット2を含有するプラスミドについてpPOX1−PT、pPOX2−PT、pPOX3−PT、pPOX4−PTおよびpPOX5−PTと称される。
【0030】
破壊用カセットは、特定の外部プライマーによるPCRによって、例えば、Pfuポリメラーゼ(Stratagene,La Jolla,Californiaによって提供される)を用いて増幅させられる。タンパク質配列の最終分析は、ヤロウイア・リポリティカのアシル−CoAオキシダーゼが、他の酵母のものと45%の同一性度(50%類似性)を有することを示す。それらの間の同一性度は、55〜70%(65〜76%の類似性)である。
【0031】
アシル−CoAオキシダーゼをコードする遺伝子の破壊用カセットを構築した後、ヤロウイア・リポリティカの変換が、種々の方法によって行われ得る。エレクトロポレーションが行われ得、ここでは、DNAは、電気ショックによって導入される。より有利には、酢酸リチウムおよびポリエチレングリコール法が用いられ得る。それは、Gaillardinらによって記載されている:LEU2 Directed Expression of Beta-gallactosidase Activity and Phleomycin Resistance in Yarrowia lipolytica.Curr.Genet.11,1987,369−375。
【0032】
破壊の存在は、PCRによって、Gussowらの技術にしたがってチェックされ:Direct Clone Characterization from Plaques and Colonies by the Polymerase Chain Reaction.Nucleic Acids Res.17,1989,4000;次いで、サザンブロットハイブリッド形成によって確認される。
【0033】
Pold株の開始時に、破壊は2段階で行われる。
【0034】
Poldは、最初に、PCR PUT破壊用カセット1により変換され、選択される。次いで、Ura+クローンが、破壊用カセット2により変換され、URA3遺伝子が除かれ、それらが選択される。この手順は、四重崩壊MTLY37−pox2ΔPT−pox3ΔPT−pox4ΔPT−pox5ΔPUTを得ることを可能にする。この変異体の構築物の図表表示は、下記表1において要約される。
【0035】
【表1】
【0036】
遺伝子の破壊およびマーカーの切除はまた、組み換えを含む方法またはリコンビナーゼによってなされ得る。例えば、いずれかの側に、反復配列(選択された組み換えを可能にする)またはリコンビナーゼCreによって認識されるlox配列を有するマーカーを用いることが可能である。リコンビナーゼCreが発現させられた場合に切除は起こる(Fickersら,2003 New Disruption Cassettes for Rapid Gene Disruption and Marker Rescue in the Yeast Yarrowia lipolytica.J.Microbiol.Methods 55/3:727-737)。
【0037】
(2.ヤロウイア・リポリティカ変異株MTLY74の取得)
株MTLY74Leu+Ura−は変異株MTLY37から構築された。
【0038】
(2a)原栄養性変異体MTLY37(Leu+、Ura+)から、本発明者らは、ロイシンおよびウラシル栄養要求性ヤロウイア・リポリティカ変異株MTLY66(Leu−、Ura−)を構築する。第一段階は、マーカーura3−41によるマーカーURA3の変換、このマーカーを含有するPCRフラグメントを形質転換することおよび5FOAの存在下にUra−を選択することによるウラシル栄養要求性株MTLY40(Leu+、Ura−)の構築である。変異体MTLY40から、本発明者らは、ロイシン栄養要求性ヤロウイア・リポリティカ変異株MTLY64(Leu−、Ura−、Hyg+)を、マーカーLEU2を破壊すること、破壊用カセットPHTleu2を変換することおよび抵抗性ヒグロマイシン形質転換体(leu2::Hyg)を選択することによって構築する。MTLY64変異株から、本発明者らは、ロイシン栄養要求性ヤロウイア・リポリティカ変異株MTLY66(Leu−、Ura−)を、HYGマーカーを切除すること、リコンビナーゼCreおよびマーカーLEU2(Cre−LEU2)を含有する複製ベクターpRRQ2を軽質転換することおよび感受性ハイゴロマイシン形質転換体Leu+を選択することにより構築する。プラスミドpRRQ2の喪失は、富栄養培地YPD上での培養およびクローン(Leu−、Ura−、Hyg−)の単離によって達成される。
【0039】
(2b)変異体MTLY66から、本発明者らは、NADPH−シトクロムリダクターゼを過剰発現させるヤロウイア・リポリティカ変異株MTLY74 Leu+ Ura−を、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導される強いプロモーターpPOX2の制御下にそれを発現させることにより構築する。
【0040】
NADPH−シトクロムリダクターゼをコードする遺伝子(CPR)は、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に、選択遺伝子LEU2、JMP21(例)を含有するベクター中に導入される。マーカー−プロモータ−遺伝子のカセット(LEU2−pPOX2−CPR)は、変換によって導入される。
【0041】
この変異体の構築物の概略表示は、以下の表2に要約される。
【0042】
【表2】
【0043】
(3 共通の変異体MTLY74からの株MTLY79、MTLY80およびMTLY81の取得)
(3a)脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下にNADPH−シトクロムリダクターゼおよびシトクロムP450モノオキシゲナーゼALK1を発現させるMTLY79。
【0044】
変異体MTLY74から、本発明者らは、NADPH−シトクロムリダクターゼおよびシトクロムP450モノオキシゲナーゼALK1を過剰発現させるヤロウイア・リポリティカ変異株MTLY79を、生物変換条件下、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導される強いプロモーターpPOX2の制御下に構築する。
【0045】
本発明者らは、シトクロムP450モノオキシゲナーゼをコードするALK1遺伝子を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に、URA3選択遺伝子(例えば、JMP61)を含有するベクター中に導入する。マーカー−プロモーター−遺伝子のカセット(URA3−pPOX2−AKL1)は変換によって導入される。
【0046】
この変異体の構築物の概略表示は、以下の表3に要約される。
【0047】
【表3】
【0048】
(3b)脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下にNADPH−シトクロムリダクターゼおよびシトクロムP450モノオキシゲナーゼALK2を発現させるMTLY80。
【0049】
変異体MTLY74から、本発明者らは、NADPH−シトクロムリダクターゼ(CPR)およびシトクロムP450モノオキシゲナーゼ(ALK2)をコードする遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY80を、生物変換条件下、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導される強いプロモーターpPOX2の制御下に構築する。
【0050】
本発明者らは、シトクロムP450モノオキシゲナーゼをコードするALK2遺伝子を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に、URA3選択遺伝子(例えばJMP61)を含有するベクター中に導入する。マーカー−プロモーター−遺伝子のカセット(URA3−pPOX2−AKL2)は、変換によって導入される。
【0051】
この変異体の構築物の概略表示は、以下の表4に要約される。
【0052】
【表4】
【0053】
(3c)脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下にシトクロムP450モノオキシゲナーゼ(ALK1またはALK2)の遺伝子なしでNADPH−シトクロムリダクターゼを発現させるMTLY81。
【0054】
変異体MTLY74から、本発明者らは、NADPH−シトクロムリダクターゼ(CPR)をコードする遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY81を、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導される強いプロモーターpPOX2の制御下に構築する。
【0055】
変異体MTLY74は、マーカーURA3を有するプラスミドJMP61による形質転換によって無機物栄養性にされる。
【0056】
この変異体の構築物の概略表示は、以下の表5に要約される。
【0057】
【表5】
【0058】
(4 変異株FT120の取得)
この株は、MTLY37株またはMTLY66株の構築と同一の手順に従うことによって得られ得る。
【0059】
1)対抗選択マーカー、例えばマーカーURA3(これにより、Ura+表現型またはUra−表現型について選択し得る)を用いる、または、いずれかの側上に、反復配列(選択された組み換えを可能にする)またはリコンビナーゼCreによって認識されるlox配列を有するマーカー(選択された組み換えを可能にする)を用いる、PCR(ポリメラーゼ連鎖反応)またはクローニングにより破壊用カセットを構築する。
【0060】
2)削除された興味遺伝子を有する株を選択し(変換、および軽質転換体;マーカーがURA3ならば有利にはUra+の選択)および遺伝子破壊をチェックする。
【0061】
3)削除されたマーカーを有する株を選択し(変換および軽質転換体の選択);マーカーがURA3ならば有利には5FOARまたはマーカーがlox配列を示すならば有利にはリコンビナーゼを発現させるプラスミド;および遺伝子破壊をチェックする。
【0062】
変異株FT120を得るための特定の方法が以下に記載される。
【0063】
変異体MTLY66から、本発明者らは、株FT120Leu−Ura−,Δpox1−6を構築する。
【0064】
(4a)変異体MTLY66 Δpox2−5から、本発明者らは、ヤロウイア・リポリティカ変異株MTLY95 Δpox1−6を、POX1およびPOX6遺伝子の削除部およびマーカーの削除部の挿入により上記の方法に従って構築する。
【0065】
(4b)変異体MTLY95から、本発明者らは、生物変換条件下にNADPH−シトクロムリダクターゼ(CPR)をコードする遺伝子を過剰発現させるヤロウイア・リポリティカ変異株FT101 Leu+Ura−を、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導される強いプロモーターpPOX2の制御下にそれを発現させることによって構築する。
【0066】
本発明者らは、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に、NADPH−シトクロムリダクターゼをコードする遺伝子を、切除可能なLEU2選択遺伝子JMP21−LEU2ex(例)を含有するベクター内に導入した。マーカー−プロモーター−遺伝子のカセット(LEU2ex−pPOX2−CPR)は、形質転換によって導入される。FT120株は、マーカーLEU2exの切除後に、pUB4−CREプラスミド(Hyg+選択)による変換、YPD上のプラスミド喪失および最後のLeuクローンの単離によって得られる。
【0067】
この変異体の構築物の概略表示は、以下の表6に要約される。
【0068】
【表6】
【0069】
(5 変異株FT130の取得)
この株は、株FT120の構築物と同一の手順に従うことによって得られ得る。
【0070】
1)対抗選択マーカー、例えば、マーカーURA3(これにより、Ura+表現型またはUra−表現型を選択することができる)を用いる、または、いずれかの側上に、反復配列を有するマーカー(選択された組み換えを可能にする)またはリコンビナーゼCreによって認識されるlox配列を用いる、PCR(ポリメラーゼ連鎖反応)またはクローニングによって破壊用カセットを構築する。
【0071】
2)削除された興味遺伝子を有する株を選択し(変換および形質転換体の選択:マーカーがURA3であれば有利にはUra+)、遺伝子破壊をチェックする。
【0072】
3)削除されたマーカーを有する株を選択し(変換および形質転換体の選択;マーカーがURA3ならば有利には5FOAR、または、マーカーがlox配列を示すならば有利にはリコンビナーゼCreを発現させるプラスミド)、遺伝子破壊をチェックする。
【0073】
変異株FT130を得るための特定の方法が以下に記載される。
【0074】
変異体FT120から、本発明者らは、株FT130Leu−Ura−,Δpox1−6,Δdga1を構築する。
【0075】
(6)変異体FT120 Leu−Ura−,Δpox1−6,pPOX2−CPRから、本発明者らは、ヤロウイア・リポリティカ変異体FT130 Leu−Ura+Δpox1−6,pPOX2−CPR,Δdgalを、アシル−CoAジアシルグリセロール・アセチルトランスフェラーゼをコードするDGA1遺伝子の削除物の挿入によって構築する。
【0076】
【表7】
【0077】
株MTLY66、MTLY81、FT120およびFT130は、Collection Nationale de Cultures de Microorganismesに、それぞれ登録番号CNCM I−3319、CNCM I−3320、CNCM I−3527およびCNCM I−3528の下に登録されている。
【0078】
以下の実施例は本発明を説明するが、本発明の範囲を制限するものではない。
【0079】
(実施例)
これらの実施例において、本発明者らは、二塩基酸の産生についての培養条件および培地組成の影響を試験した。そして、本発明者らは、変異体MTLY37により、ペプトンの使用が、特にバクト−トリプトンと関連して、二塩基酸の産生に非常に有利であることを観察した(実施例1および2)。
【0080】
本発明者らはまた、同じ条件下に、変異体MTLY79、MTLY80およびMTLY81を試験した。そして、本発明者らは、NADPH−シトクロムリダクターゼが、二塩基酸の産生における制限段階を触媒することを観察した。実際に、この酵素の過剰発現のみが、二塩基酸の産生および産生能を大幅に増加させることを可能にする(実施例4〜6)。
【0081】
本発明者らは、同一条件下に、変異株FT120およびFT130を試験した(実施例7および8)。POX1およびPOX6遺伝子の削除は、FT120についての二塩基酸の分解を減少させることを可能にする。アシル−CoAジアシルグリセロール・アセチルトランスフェラーゼをコードするさらなるDGA1遺伝子の削除により、ヤロウイア・リポリティカの細胞内の脂肪体の形態にある生物変換基質の蓄積が減少する。それ故に、これらの実施例において得られた二塩基酸の大部分は、18個の炭素原子を有する脂肪酸を有する脂肪酸から本質的になる用いられる生物変換基質のような18個の炭素原子を有する二塩基酸からなる。
【0082】
(実施例1:変異体MTLY37によりオレインヒマワリ油からジカルボン酸を製造する方法)
組成物:酵母抽出物10g・L−1、ペプトン10g・L−1、グルコース10g・L−1、寒天(Agar)20g・L−1のグロース培地(gelosed medium)において維持される、変異体MTLY37の予備培養が、0.30近くの予備培養培地の初期吸光度を提供する播種によって行われた。予備培養は、軌道攪拌下(200rpm)に24時間にわたり30℃で、25mLの培地(10g・L−1の酵母抽出物、10g・L−1のペプトン、20g・L−1のグルコース)を含有する500mLのフランジ付きフラスコにおいて行われる。
【0083】
培養のために用いられた培地は、脱イオン水、10g・L−1の酵母抽出物、20g・L−1のトリプトン、40g・L−1のグルコースおよび30g・L−1のオレインヒマワリ油から構成される。
【0084】
発酵槽の播種は、予備培養フラスコの全てにより達成される。
【0085】
培養は、30℃で、2Lの培地を有し、複動半径流タービン(double-acting centripetal turbine)を備えた4Lの発酵槽において、0.5vvmの通気速度、800rpmの攪拌速度で行われる。
【0086】
17時間の培養後、培地のグルコースが除去されて直ぐに、60mLのオレインヒマワリ油(18個の炭素原子を有する脂肪酸から本質的になる)が、反応器に加えられ、1mL・h−1の速度での連続的なグリセロール供給に付される。その時、培養のpH値は、4Mのソーダの調整された添加によって8の一定値に維持される。発酵は130時間で終了する。
【0087】
培養手順の終了時に、遠心分離によって細胞バイオマスが取り除かれる。その後、6MのHClを添加することによって上澄は2.5のpH値まで酸性にされ、酸性ワート(wort)の遠心分離によって不溶性のジカルボン酸が集められ、次いで、乾燥させられる。
【0088】
混合物のジカルボン酸組成は、Uchioらの「Microbial Production of Long-chain Dicarboxylic Acids from n-Alkanes.Part II.Production by Candida cloacae Mutant Usable to Assimilate Dicarboxylic Acid」(Agr Biol.Chem.36,No.3,1972,426-433)によって記載された方法によるジカルボン酸のジエステルへの変換後に、カラムDB1におけるガスクロマトグラフィーによって測定される。クロマトグラフ・オーブンの温度は、150℃から280℃に8℃/分の速さでプログラムされる。
【0089】
結果は、130時間後に上澄における最大の二塩基酸産生度5.9g・L−1を示す。
【0090】
(実施例2)
培養培地において、同一濃度でトリプトンをペプトンに置き換えることによって実施例1が繰り返される。培養130時間後に、9.9g・L−1のジカルボン酸が得られた。すなわち、実施例1と関連して約68%産生度が増加した。
【0091】
(実施例3:連続的なオレインヒマワリ油の供給による変異体MTLY37によるジカルボン酸の産生)
オレインヒマワリ油が培養培地から除かれ、かつ、反応器内への1mLの二次制限(sublimiting)流量でのこの油の連続注入によって置き換えられて、実施例2が繰り返される。
【0092】
このような条件下に、130時間後に14.7g・L−1のジカルボン酸が培養培地において産生される。
【0093】
(実施例4:CPRおよびALK1を過剰発現させる変異体MTLY79によるオレインヒマワリ油からのジカルボン酸の産生)
変異体MTLY37を、CPRおよびALK1遺伝子を過剰発現させる変異体MTLY79によって置き換えることによって実施例3が繰り返される。130時間の培養後、16g・L−1のジカルボン酸が得られた。
【0094】
(実施例5:CPRおよびALK2を過剰発現させる変異体MTLY80によるオレインヒマワリ油からのジカルボン酸の産生)
変異体MTLY37を、CPRおよびALK2遺伝子を過剰発現させる変異体MTLY80によって置き換えることによって実施例3が繰り返される。130時間の培養後、16g・L−1のジカルボン酸が得られる。
【0095】
(実施例6)
変異体MTLY37を、CPR遺伝子のみを過剰発現させる変異体MTLY81によって置き換えることによって実施例3が繰り返される。130時間の培養後、16g・L−1のジカルボン酸が得られる。
【0096】
(実施例7:6のPOX(Δpox1−6)遺伝子が削除され、CPR遺伝子を過剰発現させる変異体FT120によるオレインヒマワリ油からのジカルボン酸の産生)
変異体MTLY37を、6のPOX遺伝子が削除され、CPR遺伝子のみを過剰発現させる変異体FT120によって置き換えることによって実施例3が繰り返される。130時間の培養後、18g・L−1のジカルボン酸が得られる。
【0097】
(実施例8:6のPOX(Δpox1−6)遺伝子が削除され、DGA1遺伝子(Δdga1)が削除され、かつ、CPR遺伝子を過剰発現させる変異体FT130によるオレインヒマワリ油からのジカルボン酸の産生)
変異体MTLY37を、6のPOX遺伝子およびDGA1遺伝子が削除され、CPR遺伝子を過剰発現させる変異体FT130によって置き換えることによって実施例3が繰り返される。130時間の培養後、23g・L−1のジカルボン酸が得られる。
【図面の簡単な説明】
【0098】
【図1】生物学的二塩基酸製造法を示す概略図である。
【特許請求の範囲】
【請求項1】
選択マーカーとして、LEU2およびURA3遺伝子による形質転換のために使用可能である原栄養性変異体MTLY37から栄養要求性ヤロウイア・リポリティカ変異株MTLY66,Leu−Ura−を得る方法であって、
下記表の段階(1)〜(3):
【表1】
の変換操作が行われることを特徴とする方法。
【請求項2】
生物変換条件下に、変異体MTLY66から、NADPH−シトクロムリダクターゼをコードするCPR遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY74 Leu+Ura−を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に選択マーカーLEU2およびCPR遺伝子を有する発現カセットを含有するJMP21−CPRベクターを変換することによって得る方法。
【請求項3】
生物変換条件下に、ヤロウイア・リポリティカ変異株MTLY74から、NADPH−シトクロムリダクターゼをおよびシトクロムP450モノオキシゲナーゼをコードする遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY79を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に選択マーカーURA3と、ALK1遺伝子を有する発現カセットとを含有するJMP61−ALK1ベクターを変換することによって得る方法。
【請求項4】
生物変換条件下に、ヤロウイア・リポリティカ変異株MTLY74から、NADPH−シトクロムリダクターゼおよびシトクロムP450モノオキシゲナーゼALK2をコードする遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY80を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に選択マーカーURA3と、ALK2遺伝子を有する発現カセットとを含有するJMP61−ALK2ベクターを変換することによって得る方法。
【請求項5】
生物変換条件下に、変異株MTLY74から、NADPH−シトクロムリダクターゼをコードするCPR遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY81を、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導可能なプロモーターpPOX2の制御下にそれを発現させることによって得る方法であって、該ヤロウイア・リポリティカ変異株MTLY81は、マーカーURA3を有するJMP61プラスミドにより軽質転換させることによって変異体MTLY74を原栄養性にすることによって、原栄養性である、方法。
【請求項6】
ヤロウイア・リポリティカ変異株MTLY66から、ヤロウイア・リポリティカ変異株FT120 Leu−Ura−を得る方法であって、
下記表の段階(1)〜(4):
【表2】
の変換操作が行われ、生物変換条件下にNADPH−シトクロムリダクターゼをコードする遺伝子(CPR)を過剰発現させる変異株FT101 Leu+Ura−の構築は、ヤロウイア・リポリティカ株MTLY95から、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に切除可能な選択マーカーLEU2およびCPR遺伝子を含有するJMP21−LEU2ex−CPRベクターを変換することによって行われ、株FT120は、マーカーLEU2exの切除後に、pUB4−CREプラスミド(Hyg+選択)による変換、YPD上のプラスミドの喪失および最後のクローンLeu−の単離によって得られることを特徴とする方法。
【請求項7】
ヤロウイア・リポリティカ変異株FT120から、ヤロウイア・リポリティカ変異株FT130を得る方法であって、
下記変換段階:
【表3】
が行われることを特徴とする方法。
【請求項8】
新ヤロウイア・リポリティカ変異株MTLY66。
【請求項9】
新ヤロウイア・リポリティカ変異株MTLY81。
【請求項10】
新ヤロウイア・リポリティカ変異株FT120。
【請求項11】
新ヤロウイア・リポリティカ変異株FT130。
【請求項1】
選択マーカーとして、LEU2およびURA3遺伝子による形質転換のために使用可能である原栄養性変異体MTLY37から栄養要求性ヤロウイア・リポリティカ変異株MTLY66,Leu−Ura−を得る方法であって、
下記表の段階(1)〜(3):
【表1】
の変換操作が行われることを特徴とする方法。
【請求項2】
生物変換条件下に、変異体MTLY66から、NADPH−シトクロムリダクターゼをコードするCPR遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY74 Leu+Ura−を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に選択マーカーLEU2およびCPR遺伝子を有する発現カセットを含有するJMP21−CPRベクターを変換することによって得る方法。
【請求項3】
生物変換条件下に、ヤロウイア・リポリティカ変異株MTLY74から、NADPH−シトクロムリダクターゼをおよびシトクロムP450モノオキシゲナーゼをコードする遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY79を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に選択マーカーURA3と、ALK1遺伝子を有する発現カセットとを含有するJMP61−ALK1ベクターを変換することによって得る方法。
【請求項4】
生物変換条件下に、ヤロウイア・リポリティカ変異株MTLY74から、NADPH−シトクロムリダクターゼおよびシトクロムP450モノオキシゲナーゼALK2をコードする遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY80を、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に選択マーカーURA3と、ALK2遺伝子を有する発現カセットとを含有するJMP61−ALK2ベクターを変換することによって得る方法。
【請求項5】
生物変換条件下に、変異株MTLY74から、NADPH−シトクロムリダクターゼをコードするCPR遺伝子を過剰発現させるヤロウイア・リポリティカ変異株MTLY81を、脂肪酸、脂肪酸エステルまたは天然油タイプの生物変換基質によって誘導可能なプロモーターpPOX2の制御下にそれを発現させることによって得る方法であって、該ヤロウイア・リポリティカ変異株MTLY81は、マーカーURA3を有するJMP61プラスミドにより軽質転換させることによって変異体MTLY74を原栄養性にすることによって、原栄養性である、方法。
【請求項6】
ヤロウイア・リポリティカ変異株MTLY66から、ヤロウイア・リポリティカ変異株FT120 Leu−Ura−を得る方法であって、
下記表の段階(1)〜(4):
【表2】
の変換操作が行われ、生物変換条件下にNADPH−シトクロムリダクターゼをコードする遺伝子(CPR)を過剰発現させる変異株FT101 Leu+Ura−の構築は、ヤロウイア・リポリティカ株MTLY95から、脂肪酸、脂肪酸エステルまたは天然油によって誘導可能なプロモーターpPOX2の制御下に切除可能な選択マーカーLEU2およびCPR遺伝子を含有するJMP21−LEU2ex−CPRベクターを変換することによって行われ、株FT120は、マーカーLEU2exの切除後に、pUB4−CREプラスミド(Hyg+選択)による変換、YPD上のプラスミドの喪失および最後のクローンLeu−の単離によって得られることを特徴とする方法。
【請求項7】
ヤロウイア・リポリティカ変異株FT120から、ヤロウイア・リポリティカ変異株FT130を得る方法であって、
下記変換段階:
【表3】
が行われることを特徴とする方法。
【請求項8】
新ヤロウイア・リポリティカ変異株MTLY66。
【請求項9】
新ヤロウイア・リポリティカ変異株MTLY81。
【請求項10】
新ヤロウイア・リポリティカ変異株FT120。
【請求項11】
新ヤロウイア・リポリティカ変異株FT130。
【図1】

【公開番号】特開2012−130352(P2012−130352A)
【公開日】平成24年7月12日(2012.7.12)
【国際特許分類】
【出願番号】特願2012−49322(P2012−49322)
【出願日】平成24年3月6日(2012.3.6)
【分割の表示】特願2007−546115(P2007−546115)の分割
【原出願日】平成17年12月13日(2005.12.13)
【出願人】(500393413)アンスティテュ フランセ デュ ペトロール (32)
【出願人】(507199975)サーントゥル ナシオナル ドゥ ラ ルシェルシュ シャーンティフィク セエンエールエス (13)
【出願人】(507199997)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミク (4)
【Fターム(参考)】
【公開日】平成24年7月12日(2012.7.12)
【国際特許分類】
【出願日】平成24年3月6日(2012.3.6)
【分割の表示】特願2007−546115(P2007−546115)の分割
【原出願日】平成17年12月13日(2005.12.13)
【出願人】(500393413)アンスティテュ フランセ デュ ペトロール (32)
【出願人】(507199975)サーントゥル ナシオナル ドゥ ラ ルシェルシュ シャーンティフィク セエンエールエス (13)
【出願人】(507199997)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミク (4)
【Fターム(参考)】
[ Back to top ]