
改善された生体内半減期を有する生物学的に活性なタンパク質
【課題】生物学的に活性なタンパク質の半減期を変更する。
【解決手段】2〜約500単位の反復ペプチド・モチーフを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含み、非複合型の生物学的に活性なポリペプチド又はタンパク質の固有の半減期と比較して改善された血漿中半減期を示す。生物学的に活性なポリペプチドは、サイトカイン(例えば、顆粒球コロニー刺激因子(G-CSF))などである。また、主成分としてGly、Asn、及びGlnから選択される1種類のアミノ酸2残基以上、並びに副次成分としてAla、Ser、Thr、Asp、Gln、Glu、His、及びAsnから選択される1種類のアミノ酸1残基以上が、上記アミノ酸が主成分及び副次成分の中に存在しないという条件で含まれる、
【解決手段】2〜約500単位の反復ペプチド・モチーフを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含み、非複合型の生物学的に活性なポリペプチド又はタンパク質の固有の半減期と比較して改善された血漿中半減期を示す。生物学的に活性なポリペプチドは、サイトカイン(例えば、顆粒球コロニー刺激因子(G-CSF))などである。また、主成分としてGly、Asn、及びGlnから選択される1種類のアミノ酸2残基以上、並びに副次成分としてAla、Ser、Thr、Asp、Gln、Glu、His、及びAsnから選択される1種類のアミノ酸1残基以上が、上記アミノ酸が主成分及び副次成分の中に存在しないという条件で含まれる、
【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
当該出願は、合衆国法典第35巻第119(e)条の下、2005年8月30日に出願された米国出願第60/712,585号及び2005年1月25日に出願された米国出願第60/647,119号による優先権を主張し、上記出願の内容を本明細書中に援用する。
【0002】
技術分野
本願発明は、広く、生物学的に活性なタンパク質に関し、より詳しく述べると、生物学的に活性なタンパク質の半減期を変更することに関する。
【背景技術】
【0003】
ヒトの治療法として投与される時、生物学的に活性なタンパク質は、多くの場合、望ましくない半減期を有する。それらの固有の半減期は、多くの場合、最適に満たない治療効能、服薬遵守の問題、及び患者の不便を招く投薬スケジュール及び投与計画を強いる。
【0004】
ヒトの治療法向けの生物学的に活性なタンパク質の製造において、生物学的に活性なタンパク質の半減期の延長は、物理的手段(例えば、投与経路の変更、ナノ粒子カプセル化、及びリポソーム封入)、化学修飾(例えば、エマルジョン、ペグ化、及び高グリコシル化)、並びに遺伝的修飾(例えば、一次タンパク質構造の修飾、高分子タグ、ヒト血清アルブミン融合、翻訳後修飾の取り込み)を通じて試みられた。例えば、Lordら、Clin. Cancer Res. 7:2085-2090ページ(2001年)及びvan Der Auweraら、Am. J. Hematol. 55:245-251ページ(2001年)を参照のこと。しかしながら、そのようなアプローチは他の問題を招いた。物理的手段による生物学的に活性なタンパク質の半減期の延長は、多くの場合、高価で、且つ、時間のかかる製造中の追加の後続加工を伴う、さらなる製剤原料の複雑さを持ち込む。化学修飾は、生物学的に活性なタンパク質の生物活性又は安全性の特性を変えるかもしれない。生物学的に活性なタンパク質が組み換えDNA合成方法論により作製された場合には、特定の細胞発現系におけるタンパク質収量及び純度に対する遺伝的修飾の効果が、その意図される使用に関して評価される必要性が生じる。
【0005】
従って、生物学的に活性なタンパク質の固有の半減期を改善するための他のアプローチの必要性が存在する。
【発明の概要】
【0006】
発明の概要
本発明の1つの側面は、2〜約500反復単位のペプチド・モチーフを含むポリペプチド(アミノ酸伸長)にペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体に向けられる。前記モチーフは、Gly(G)、Asn(N)、及びGln(Q)から選択される1種類のアミノ酸2残基以上である主成分、並びにAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から選択される1種類のアミノ酸1残基以上である副次成分を、上記アミノ酸が主成分と副次成分の両方に存在していないという条件で含み、ここで、上記複合体の血漿中半減期は非複合型の生物学的に活性なポリペプチドの固有の半減期に比べて改善される。
本明細書中に使用される用語「改善された」は、非複合型の生物学的に活性なポリペプチド又はタンパク質自体の血漿中半減期(すなわち、固有の半減期)に比べて延びたか又は短くなった半減期を表す。語句「固有の半減期」は、天然の生物学的に活性なポリペプチドの半減期、又は非複合型のポリペプチド(よって、上記天然ポリペプチドの組み換え型を含む)の半減期を意味する。
【0007】
いくつかの態様において、ペプチド・モチーフは、3〜6個のアミノ酸残基(すなわち、3、4、5、又は6個)を含む。ペプチド・モチーフが5又は6個のアミノ酸残基を含むいくつかの態様において、副次成分は上記ペプチドの中の1個のアミノ酸残基を含む。いくつかの態様において、ペプチド・モチーフは、N及びTアミノ酸残基、N及びEアミノ酸残基、Q及びSアミノ酸残基、又はN及びQアミノ酸残基から成る配列を持つ。いくつかの態様において、アミノ酸伸長は、前記生物学的に活性なポリペプチドに対してN末端側であり;いくつかの実施例において、それは、前記生物学的に活性なポリペプチドに対してC末端側であり;そして、他の態様において、それは、前記生物学的に活性なポリペプチドに対してN及びC末端側の両方に位置する。いくつかの態様において、生物学的に活性なポリペプチドは、サイトカイン(例えば、顆粒球コロニー刺激因子(G-CSF)、ヒト成長ホルモン、又はインターフェロン、例えばβ-インターフェロン若しくはγ-インターフェロンなど)、抗体、抗体断片、タンパク質分解性抗体断片若しくはドメイン、一本鎖抗体、遺伝的若しくは化学的に最適化された抗体若しくはその断片、可溶性のgp120若しくはgp160糖タンパク質、血液凝固因子、可溶性受容体、例えば、腫瘍壊死因子(TNF)-αのII型受容体など、治療用酵素、又はエリスロポエチン(EPO)である。いくつかの態様において、タンパク質複合体は、非複合型の生物学的に活性なポリペプチドの固有の半減期に比べて短い改善された半減期を有する、例えば、ここで、上記生物学的に活性なポリペプチドは組み換え型の活性化プロテインC又は組み換え型の第VII因子を含む。
【0008】
本発明の他の側面は、タンパク質複合体及び担体を含む組成物に向けられる。いくつかの態様において、組成物は医薬組成物であり、そして、担体は医薬として許容される担体である。
【0009】
本発明の他の側面は、先に記載のタンパク質複合体をコードするキメラDNA分子、並びに上記キメラDNA分子を一緒に含むベクター、そして、上記キメラDNA分子又はそれを含むベクターで形質転換された細胞に向けられる。いくつかの態様において、ベクターは、プラスミド、例えば、pCE2である。いくつかの態様において、細胞は、哺乳動物細胞、例えば、チャイニーズハムスター卵巣(CHO)細胞、細菌、例えば、E.コリ(E. coli)、又は酵母である。
【0010】
本発明のさらに他の側面は、主成分としてGly(G)、Asn(N)、及びGln(Q)から選択される1種類のアミノ酸2残基以上を含み、且つ、副次成分としてAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から選択される1種類のアミノ酸1残基以上が、上記アミノ酸が上記主成分と上記副次成分に存在しないという条件で含まれる、2〜約500のペプチド単位から成るポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含む、上記生物学的に活性なタンパク質が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を有するような生物学的に活性なタンパク質複合体の作製方法であって、以下のステップ:上記タンパク質複合体をコードするキメラDNA分子で形質転換した細胞を上記DNAが発現される条件下で培養し、それによって、上記タンパク質複合体を産生し;そして、上記細胞から上記キメラDNA分子の発現産物を抽出する、を含む上記作製方法に向けられる。
【0011】
本発明の更なる側面は、所定のタンパク質複合体が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を示すかどうかを測定する方法であって、以下のステップ:a)Gly(G)、Asn(N)、及びGln(Q)から成る群から選択される1種類のアミノ酸2残基以上を含むか若しくはそれらから成る主成分、並びにAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から成る群から選択される1種類のアミノ酸1残基以上を含むか若しくはそれらから成るが、上記アミノ酸が上記主成分と上記副次成分の両方に存在することのない2〜約500反復単位のペプチド・モチーフを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体を調製し、そしてb)上記タンパク質複合体を試験して、そのタンパク質複合体が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を有するかどうか測定する、を含む上記方法に向けられる。
詳細な説明
【0012】
本明細書中に使用される用語「ポリペプチド」は、別段の指示のない限り、特定の長さを持たないアミノ酸の重合体を意味する。よって、ぺプチド及びタンパク質は、「ポリペプチド」の定義に含まれ、そして、これらの用語は、明細書を通して、並びに特許請求の範囲内で互換性を持って使用される。用語「ポリペプチド」は、翻訳後修飾、例えば、グリコシル基、アセチル基、リン酸基、脂質基の共有結合、プロリン又はリジンのヒドロキシル化などを持つポリペプチドを除外しない。「ポリペプチド」のこの定義によって、その相同体も同様に組み込まれる。
【0013】
用語「精製される」は、本明細書中では、生物学的に活性なタンパク質複合体がその意図される使用に適当なレベルまで精製されることを意味する。
【0014】
本発明は、全般的な側面において、Gly(G)、Asn(N)、及びGln(Q)から選択される1種類のアミノ酸2残基以上である主成分、並びにAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から選択される1種類のアミノ酸1残基以上である副次成分が、上記アミノ酸が上記主成分と上記副次成分の両方に存在しないという条件で含まれる、2〜約500単位のペプチド・モチーフを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体に向けられる。本発明のタンパク質複合体は、相当する非複合型の生物学的に活性なポリペプチド又はタンパク質より長い血漿中半減期を有する。
【0015】
モチーフの反復単位は、通常、3〜7個(3、4、5、6、又は7個)のアミノ酸残基を含む。主成分としてN(Asn)を持つ代表的なペプチド・モチーフが、表1に記載されている。
【0016】
主成分としてG(Gly)を持つ代表的なペプチド・モチーフが、表2に記載されている。
【0017】
主成分としてQ(Glu)を持つ代表的なペプチド・モチーフが、表3に記載されている。
【0018】
ペプチド・モチーフの数は、2〜約500におよぶ。よって、本明細書中に詳しく説明された特定のペプチド・モチーフを含めたモチーフは、ポリペプチド内に以下の単位数:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192、193、194、195、196、197、198、199、200、201、202、203、204、205、206、207、208、209、210、211、212、213、214、215、216、217、218、219、220、221、222、223、224、225、226、227、228、229、230、231、232、233、234、235、236、237、238、239、240、241、242、243、244、245、246、247、248、249、250、251、252、253、254、255、256、257、258、259、260、261、262、263、264、265、266、267、268、269、270、271、272、273、274、275、276、277、278、279、280、281、282、283、284、285、286、287、288、289、290、291、292、293、294、295、296、297、298、299、300、301、302、303、304、305、306、307、308、309、310、311、312、313、314、315、316、317、318、319、320、321、322、323、324、325、326、327、328、329、330、331、332、333、334、335、336、337、338、339、340、341、342、343、344、345、346、347、348、349、350、351、352、353、354、355、356、357、358、359、360、361、362、363、364、365、366、367、368、369、370、371、372、373、374、375、376、377、378、379、380、381、382、383、384、385、386、387、388、389、390、391、392、393、394、395、396、397、398、399、400、401、402、403、404、405、406、407、408、409、410、411、412、413、414、415、416、417、418、419、420、421、422、423、424、425、426、427、428、429、430、431、432、433、434、435、436、437、438、439、440、441、442、443、444、445、446、447、448、449、450、451、452、453、454、455、456、457、458、459、460、461、462、463、464、465、466、467、468、469、470、471、472、473、474、475、476、477、478、479、480、481、482、483、484、485、486、487、488、489、490、491、492、493、494、495、496、497、498、499、及び500で存在するかもしれない(よって、あらゆるその部分的範囲も同様に含む)。
【0019】
先に記載のとおり、生物学的に活性なポリペプチドに結合したポリペプチドは、生物学的に活性なポリペプチドのアミノ酸又はポリアミノ伸長(以降「アミノ酸伸長」)とも呼ばれるかもしれない。アミノ酸伸長は、生物学的に活性なポリペプチド配列に対して、N末端側に、C末端側に、又はN及びC末端側の両方に位置するかもしれない。
【0020】
理論に縛られるつもりはないが、出願人は、アミノ酸伸長が安定な立体構造をとらないので、それ自体はタンパク質の活性を妨げないか、そうでなければ、影響を及ぼさないと考える。また、アミノ酸伸長を2つの異なるアミノ酸に制限することは、免疫原性の可能性を最小限にするのに役立つアミノ酸伸長の化学的な複雑さを軽減し、並びにたった1つのタイプ又は種類のアミノ酸の使用を通じて可能になるよりもはるかに大規模な生理化学的性質の調節を許容すると考えられる。
【0021】
広く、生物学的に活性なポリペプチドは、天然のポリペプチド(すなわち、それらが生体内に存在する場合)、あるいは、例えば、改善された血漿中半減期がいくつかの観点、特に、脊椎動物体にデリバリーされた時に、その脊椎動物の所定の疾患の症状を治療すること、例えば、治癒させること、改善すること、又は軽減すること、あるいは、末期症状の経過を遅らせることによって脊椎動物の寿命を延ばす意味がある治療的観点から望ましいであろう組み換えヒトG-CSF(rh-G-CSF)などのように組み換えによって生み出されたポリペプチドを含めた、あらゆるタンパク質を含む。生物学的に活性なタンパク質の種類には、サイトカイン、ケモカイン、リンフォカイン、リガンド、受容体、ホルモン、アポトーシス誘導ポリペプチド、酵素、抗体及び抗体断片、並びに成長因子が含まれる。受容体の実例には、TNFのI型受容体、IL-1のII型受容体、IL-1受容体アンタゴニスト、IL-4受容体、及びいずれかの化学的に若しくは遺伝的に修飾された可溶性受容体も同様に含まれる。酵素の実例には、活性化プロテインC、第VII因子、(例えば、SANTYL(登録商標)という名称でAdvance Biofactures Corporationによって販売されている)コラゲナーゼ;(例えば、FABRAZYME(登録商標)という名称でGenzymeによって販売されている)アガルシダーゼ-β;(例えば、PULMOZYME(登録商標)という名称でGenentechによって販売されている)ドルナーゼ-α;(例えば、ACTIVASE(登録商標)という名称でGenentechによって販売されている)アルテプラーゼ;(例えば、ONCASPAR(登録商標)という名称でEnzonによって販売されている)ペグ化アスパラギナーゼ;(例えば、ELSPAR(登録商標)という名称でMerckによって販売されている)アスパラギナーゼ;及び(例えば、CEREDASE(登録商標)という名称でGenzymeによって販売されている)イミグルセラーゼが含まれる。特定のポリペプチド又はタンパク質の実例には、これだけに制限されることなく、顆粒球マクロファージ・コロニー刺激因子(GM-CSF)、顆粒球コロニー刺激因子(G-CSF)、マクロファージ・コロニー刺激因子(M-CSF)、コロニー刺激因子(CSF)、インターフェロンβ(IFN-β)、インターフェロンγ(IFN-γ)、I型インターフェロンγ誘導因子(IGIF)、トランスフォーミング成長因子β(TGF-β)、RANTES(活性化によって調節され、正常T細胞が発現し、そして、分泌したと思われる)、マクロファージ炎症タンパク質(例えば、MIP-1-α及びMIP-1-β)、リーシュマニア伸長開始因子(LEIF)、血小板誘導成長因子(PDGF)、腫瘍壊死因子(TNF)、成長因子、例えば、表皮増殖因子(EGF)、血管内皮成長因子(VEGF)、繊維芽細胞成長因子(FGF)、神経成長因子(NGF)、脳由来神経栄養因子(BDNF)、ニューロトロフィン-2(NT-2)、ニューロトロフィン-3(NT-3)、ニューロトロフィン-4(NT-4)、ニューロトロフィン-5(NT-5)、グリア細胞株由来神経栄養因子(GDNF)、繊毛様神経栄養因子(CNTF)、TNF-αのII型受容体、エリスロポエチン(EPO)、インスリン、並びに可溶性糖タンパク質、例えば、gp120及びgp160糖タンパク質が含まれる。gp120糖タンパク質はヒト免疫不全ウィルス(HIV)エンベロープ・タンパク質であり、そして、gp160糖タンパク質はgp120糖タンパク質の既知の前駆体である。
【0022】
いくつかの態様において、生物学的に活性なポリペプチドの半減期を、固有の半減期に対して短縮されるように改善することが望ましい。そのような態様には、(例えば、XIGRIS(登録商標)という名称でEli Lillyによって販売されている)組み換え型の活性化プロテインC、及び(NOVOSEVEN(登録商標)という名称でNovo Nordiskによって販売されている)組み換え型の第VII因子が含まれる。
【0023】
生物学的に活性なポリペプチドは、疾患、例えば、パーキンソン病、癌、及び心臓病などを治療するのに使用できる。加えて、治療用ポリペプチドは、自己免疫異常、例えば、多発硬化症;シェーグレン症候群;サルコイドーシス;インシュリン依存性糖尿病;自己免疫性甲状腺炎;関節炎(例えば、変形性関節症、関節リウマチ、反応性関節炎、及び乾癬性関節炎);強直性脊椎炎;並びに強皮症などを治療するのに使用できる。また、本発明の治療用ポリペプチドは、急性及び慢性炎症性障害を治療するのに、身長が伸びるのを促進するのに、創傷治癒を促進するのに、及び細胞、組織、又は臓器の移殖後の拒否反応を予防するのに使用できる。
【0024】
いくつかの好ましい態様において、ポリペプチドはG-CSFである。G-CSFは、好中性顆粒球の急速な増殖、そして、血流への放出を引き起こし、その結果、感染との闘いにおいて治療効果を与える。米国特許第6,831,158号で説明されているように、組み換えヒト(rh)-G-CSFは、一般に、様々な形態の白血球減少症(低レベルの白血球)及び好中球減少症(低レベルの好中球)を治療するのに使用される。白血球減少症及び好中球減少症は、様々な感染に対する高い感受性を引き起こす。
【0025】
rh-G-CSFの市販の調製品は、フィルグラスチム(GRAN(登録商標)及びNEUPOGEN(登録商標))、レノグラスチム(NEUTROGIN(登録商標)及びGRANOCYTE(登録商標))、及びナルトグラスチム(NEU-UP(登録商標))という名称で入手可能である。GRAN(登録商標)及びNEUPOGEN(登録商標)は、グリコシル化されておらず、組み換えE.コリ細胞により産生される。NEUTROGIN(登録商標)及びGRANOCYTE(登録商標)は、グリコシル化されており、組み換えCHO細胞により産生される。NEU-UP(登録商標)は、グリコシル化されておらず、組み換えE.コリ細胞で産生された完全なrh-G-CSFのN末端基領域にて置換された5つのアミノ酸を持つ。
【0026】
G-CSF自体に加えて、生物学的に機能的であるか、又は生物活性があるG-CSF類似体も有用である。rh-G-CSFの調製方法は、米国特許第4,810,643号で開示されている。様々なG-CSF類似体も米国特許第4,810,643号で報告されている。rh-G-CSFをコードするポリヌクレオチド、及びrh-G-CSFのアミノ酸構造物は、共に、米国特許第5,985,265号に提供されている。
【0027】
本発明のタンパク質複合体のアミノ酸配列(配列番号2450)(及び対応するポリヌクレオチド配列(配列番号2451))の代表例が、表4に示されている。PHOリーダー配列が含まれている。
【0028】
前記アミノ酸配列において、PHOリーダー配列は第1〜18アミノ酸であり、G-CSFは第19〜192アミノ酸であり、そして、NNT155アミノ酸伸長は第193〜347アミノ酸である。終止コドンは「*」記号によって示される。
【0029】
ポリヌクレオチド配列(配列番号2451)で表される場合、核酸1〜54がPHOリーダー配列をコードし、核酸55〜576がG-CSFをコードし、そして、核酸577〜1041がNNT155アミノ酸伸長をコードする。核酸1042〜1044(タグ)が終止コドンを構成する。
【0030】
いくつかの態様において、反復ペプチド単位の数は75〜225である。よって、配列NNTを持つペプチド・モチーフの反復単位を含むポリペプチドに連結されたG-CSFを含むタンパク質複合体の場合に、本発明の態様は、以下のタンパク質複合体:G-CSF-(NNT)75、G-CSF-(NNT)76、G-CSF-(NNT)77、G-CSF-(NNT)78、G-CSF-(NNT)79、G-CSF-(NNT)80、G-CSF-(NNT)81、G-CSF-(NNT)82、G-CSF-(NNT)83、G-CSF-(NNT)84、G-CSF-(NNT)85、G-CSF-(NNT)86、G-CSF-(NNT)87、G-CSF-(NNT)88、G-CSF-(NNT)89、G-CSF-(NNT)90、G-CSF-(NNT)91、G-CSF-(NNT)92、G-CSF-(NNT)93、G-CSF-(NNT)94、G-CSF-(NNT)95、G-CSF-(NNT)96、G-CSF-(NNT)97、G-CSF-(NNT)98、G-CSF-(NNT)99、G-CSF-(NNT)100、G-CSF-(NNT)101、G-CSF-(NNT)102、G-CSF-(NNT)103、G-CSF-(NNT)104、G-CSF-(NNT)105、G-CSF-(NNT)106、G-CSF-(NNT)107、G-CSF-(NNT)108、G-CSF-(NNT)109、G-CSF-(NNT)110、G-CSF-(NNT)111、G-CSF-(NNT)112、G-CSF-(NNT)113、G-CSF-(NNT)114、G-CSF-(NNT)115、G-CSF-(NNT)116、G-CSF-(NNT)117、G-CSF-(NNT)118、G-CSF-(NNT)119、G-CSF-(NNT)120、G-CSF-(NNT)121、G-CSF-(NNT)122、G-CSF-(NNT)123、G-CSF-(NNT)124、G-CSF-(NNT)125、G-CSF-(NNT)126、G-CSF-(NNT)127、G-CSF-(NNT)128、G-CSF-(NNT)129、G-CSF-(NNT)130、G-CSF(NNT)131、G-CSF-(NNT)132、G-CSF-(NNT)133、G-CSF-(NNT)134、G-CSF-(NNT)135、G-CSF-(NNT)136、G-CSF-(NNT)137、G-CSF-(NNT)138、G-CSF-(NNT)139、G-CSF-(NNT)140、G-CSF-(NNT)141、G-CSF-(NNT)142、G-CSF-(NNT)143、G-CSF-(NNT)144、G-CSF-(NNT)145、G-CSF-(NNT)146、G-CSF-(NNT)147、G-CSF-(NNT)148、G-CSF-(NNT)149、G-CSF-(NNT)150、G-CSF-(NNT)151、G-CSF-(NNT)152、G-CSF-(NNT)153、G-CSF-(NNT)154、G-CSF-(NNT)155、G-CSF-(NNT)156、G-CSF-(NNT)157、G-CSF-(NNT)158、G-CSF-(NNT)159、G-CSF-(NNT)160、G-CSF-(NNT)161、G-CSF-(NNT)162、G-CSF-(NNT)163、G-CSF-(NNT)164、G-CSF(NNT)165、G-CSF-(NNT)166、G-CSF-(NNT)167、G-CSF(NNT)168、G-CSF-(NNT)169、G-CSF-(NNT)170、G-CSF-(NNT)171、G-CSF-(NNT)172、G-CSF-(NNT)173、G-CSF-(NNT)174、G-CSF-(NNT)175、G-CSF-(NNT)176、G-CSF-(NNT)177、G-CSF-(NNT)178、G-CSF-(NNT)179、G-CSF-(NNT)180、G-CSF-(NNT)181、G-CSF-(NNT)182、G-CSF-(NNT)183、G-CSF-(NNT)184、G-CSF-(NNT)185、G-CSF-(NNT)186、G-CSF-(NNT)187、G-CSF-(NNT)188、G-CSF-(NNT)189、G-CSF-(NNT)190、G-CSF-(NNT)191、G-CSF-(NNT)192、G-CSF-(NNT)193、G-CSF-(NNT)194、G-CSF-(NNT)195、G-CSF-(NNT)196、G-CSF-(NNT)197、G-CSF-(NNT)198、G-CSF-(NNT)199、G-CSF-(NNT)200、G-CSF-(NNT)201、G-CSF-(NNT)202、G-CSF-(NNT)203、G-CSF-(NNT)204、G-CSF-(NNT)205、G-CSF-(NNT)206、G-CSF-(NNT)207、G-CSF-(NNT)208、G-CSF-(NNT)209、G-CSF-(NNT)210、G-CSF-(NNT)211、G-CSF-(NNT)212、G-CSF-(NNT)213、G-CSF-(NNT)214、G-CSF-(NNT)215、G-CSF-(NNT)216、G-CSF-(NNT)217、G-CSF-(NNT)218、G-CSF-(NNT)219、G-CSF-(NNT)220、G-CSF-(NNT)221、G-CSF-(NNT)222、G-CSF-(NNT)223、G-CSF-(NNT)224、及びG-CSF-(NNT)225、のいずれかを含むかもしれない。
【0031】
他の好ましい態様において、ポリペプチドはEPOである。Krantz、Blood 77:419ページ(1991年)で説明されているように、天然のEPOは、骨髄内で赤血球前駆細胞の分裂と分化を刺激し、そして、受容体及び赤血球前駆体に結合することによってその生物活性を発揮する。
EPOは、チャイニーズハムスター卵巣(CHO)細胞内に挿入され、そして、発現されたクローン化ヒトEPO(hEPO)遺伝子の産物として組み換え技術を使用して生合成的に製造された。Egrieら、Immunobiol. 72:213-224ページ(1986年)を参照のこと。hEPOの主要な、完全に処理された形態の一次構造(すなわち、アミノ酸配列)は、米国特許第6,583,272号で例証されている。EPOには、Cys7-Cys161及びCys29-Cys33の間に2個のジスルフィド架橋が存在する。糖部分を含まないEPOのポリペプチド鎖の分子量は、18,236DAである。完全なEPO分子において、分子量の約40%は、タンパク質上のグリコシル化部位でタンパク質をグリコシル化する炭水化物基で占められる。Sasakiら、J. Biol. Chem. 262:12059ページ(1987年)を参照のこと。
【0032】
hEPOは赤血球形成に不可欠なので、当該ホルモンは赤血球の産生が低いか若しくは欠陥があることを特徴とする血液疾患の治療に有用である。臨床的に、EPOは、慢性腎不全(CRF)患者の貧血の治療に使用される。Eschbachら、NEJM 316:73-78ページ(1987年);Eschbachら、Ann. Intern. Med. 111:992ページ(1989年);Egrieら、Kidney Intl. 33:262ページ(1988年);及びLimら、Ann. Intern. Med. 110:108-114ページ(1989年)を参照のこと。EPOは、後天性免疫不全症候群(AIDS)及び癌の化学療法中の患者の貧血の治療にも使用された。R. P. Dannaら、Erythropoietin In Clinical Applications - An International Perspective 301-324ページ(M. B. Garnick編、Marcel Dekker 1990年)を参照のこと。
【0033】
EPOのアミノ酸及び相当するヌクレオチド配列、並びに本発明において有用な他の生物学的に活性なポリペプチドを表5に示す。EPOの他の2種類のアミノ酸配列を表6に示す。
【0034】
タンパク質複合体は、1個以上のアフィニティー・タグをさらに含むことができる。一般に、アフィニティー・タグは、当該アフィニティー・タグを含む融合タンパク質の分離、精製、又は検出を容易にするポリペプチド断片である。原則として、抗体又は他の特異的結合作用物質が利用可能であるあらゆるペプチド又はタンパク質がアフィニティー・タグとして使用できる。代表的なアフィニティー・タグには、ポリヒスチジン・トラクト(poly-histidine tract)、プロテインA(Nilssonら、EMBO J. 4:1075ページ(1985年);Nilssonら、Methods Enzymol. 198:3ページ(1991年))、グルタチオンS転移酵素(Smithら、Gene 67:31ページ(1988年))、マルトース結合タンパク質(Kellermanら、Methods Enzymol. 90:459-463ページ(1982年);Guanら、Gene 67:21-30ページ(1987年))、Glu-Glu親和性タグ(Grussenmeyerら、Proc. Natl. Acad. Sci. USA 52:7952-4ページ(1985年);oNDEPHO-1Rを参照のこと)、サブスタンスP、Flag(商標)ペプチド(Hoppら、Biotechnology 6:1204-10(1988))、ストレプトアビジン結合ペプチド、チオレドキシン、ユビキチン、セルロース結合タンパク質、T7ポリメラーゼ、又は他の抗原エピトープ若しくは結合ドメインが含まれる。一般的に、Fordら、Protein Expression and Purification 2:95-107ページ(1991年)を参照のこと。アフィニティー・タグをコードするDNAは、商業的な供給者(例えば、Pharmacia Biotech、Piscataway, New Jersey;New England Biolabs、Beverly, Massachusetts;及びEastman Kodak、New Haven, Connecticut)から入手可能である。
【0035】
アミノ酸伸長の場合のように、アフィニティー・タグは、生物学的に活性なポリペプチド配列に対してN末端側に、C末端側に、又はN末端側とC末端側の両方に位置することができる。
【0036】
本発明は、タンパク質複合体の作製方法にも向けられる。その方法は、タンパク質複合体をコードするキメラDNA分子で形質転換された細胞を、上記DNAが発現される条件下で培養し、それによって、上記タンパク質複合体を産生し;そして、細胞又は培地(又は、細胞培養によるミレアウ(milieu))からキメラDNA分子の発現産物を抽出することを含む。化学的手段によって形成されたタンパク質複合体(例えば、組み換えメチオニル・ヒトG-CSFとモノメチオニル・ポリエチレングリコールの共有結合複合体である、市販品NEULASTA(PEG-G-CSF))と対照的に、本発明における結合は、連続したタンパク質から成る成分として生物学的に活性なタンパク質とポリペプチドの産生をもたらす、物理的又は化学的な手段とは対照的な組み換えにより実施される。アミノ酸伸長にタンパク質を繋ぎ合わせるのに、例えば、約10〜20アミノ酸の長さのリンカーが使用できる。
【0037】
キメラDNA分子は、タンパク質部分をコードする遺伝子又はポリヌクレオチド断片、及び1個以上の遺伝子断片、例えば、ポリペプチド又はアミノ酸伸長を一緒にコードするオリゴヌクレオチドを含む。アミノ酸伸長に含まれるペプチド・モチーフをコードする(そして、本明細書中に具体的に開示したペプチド・モチーフをコードする)オリゴヌクレオチドを表7に示す。(表7で使用される1字表記を表8で説明する。)DNA分子は、アフィニティー・タグ、リンカー、並びに5’及び3’調節エレメントをコードする断片をさらに含むことができる。タンパク質部分をコードする遺伝子又はポリヌクレオチドは、所望のタンパク質又はタンパク質部分のポリペプチドをコードすることが知られているあらゆる遺伝子又はポリヌクレオチドであってもよい。そのような遺伝子及びポリヌクレオチド、そしてそれらを作り出すのに使用されるプライマーは、タンパク質又はポリペプチドに特異的であり、当該技術分野で周知である。ポリペプチド部分をコードする連結されたオリゴヌクレオチドは、先に説明された、及び実施例1に記載される手順に従って産生できる。オリゴヌクレオチドは、どんな長さであってもよいが、好ましくは、転写を抑制することが知られている反復DNA配列の使用を避けるように設計される。例えば、2つのグルタミン酸コドンの組合せ物を含む連結されたオリゴヌクレオチドに、1個だけのグルタミン酸コドンから作られたポリヌクレオチドよりも遺伝子発現を妨げる構造的配置を採用する可能性は低い。本発明のタンパク質複合体をコードするキメラDNA分子は、発現を容易にするために、その5’末端にメチオニン(M)、及び/又はプロリン(P)アミノ酸をコードするコドンを含むように設計されるかもしれない。
【0038】
本発明の複合体は、分子生物学の標準的な組み換え技術によって作製される。いくつかの態様において、生物学的に活性なタンパク質をコードする遺伝子又はポリヌクレオチドは、最初に、構築物、例えば、プラスミド又は他のベクターにクローン化される。次に、ポリペプチド部分の反復単位をコードするオリゴヌクレオチドは、オリゴヌクレオチドがポリペプチド部分をコードするポリヌクレオチドを形成するように一緒に連結される連結又は多量体化スキームを通じて構築物内にクローン化される。このように、オリゴヌクレオチドは、タンパク質部分をコードする遺伝子又はポリヌクレオチドに加えられ、それによって、構築物内にキメラDNA分子を作り出す。選択肢として、より適切な発現ベクターである別の構築物に、キメラDNA分子を移すか、又はクローン化することができる。この時点で、キメラDNA分子を発現することができる宿主細胞がキメラDNA分子で形質転換される。形質転換は、場合によっては、例えば、発現ベクターなどの担体を利用して起こり得る。次に、形質転換された宿主細胞は、タンパク質複合体のコード化をもたらすキメラDNA分子の発現に好適な条件下で培養される。
【0039】
本発明において有効な連結又は多量体化の方法は周知である。Joseph Sambrookら、Molecular Cloning:A Laboratory Manual、第2版、1.53(Cold Spring Harbor Laboratory Press 1989年)を参照のこと。
【0040】
クローニング方法は、当該技術分野で周知の「定方向性クローニング(directional cloning)」を通して行われてもよい。定方向性クローニングは、特定の、且つ、事前に規定された方向での、プラスミド又はベクター内へのポリヌクレオチドの挿入を指す。いったんベクター内にクローン化されると、ポリヌクレオチド配列は、その3’末端側にて、あるいはその5’、及び/又は3’末端側に挿入される他のポリヌクレオチドで伸ばされるかもしれない。そのような設計は、連結の複数の過程を実施する必要なしに大きな重合体を作るのに効率的、且つ、比較的簡単な方法を提供する。第2のポリヌクレオチドの挿入を容易にするために、ベクターは、好ましくは、クローン化ポリヌクレオチドの上流に制限部位を含むが、発現のために必要とされる調節エレメントの下流には含まない。
【0041】
’定方向性クローニングを容易にするために、「アダプター・オリゴヌクレオチド」が、タンパク質複合体をコードするキメラDNA分子の5’及び3’末端に連結されるかもしれない。好ましくは、アダプターは、発現ベクター内に存在するものと両立し得る制限部位を含む。3’アダプター・オリゴヌクレオチドは、それが連結されているコード配列の末端を示すための終止コドンも含むかもしれない。長いポリヌクレオチドが連結後に作り出される可能性を高めるために、ポリペプチド部分をコードするオリゴヌクレオチドが、好ましくは、アダプター・オリゴヌクレオチドより多く加えられる。
【0042】
方法論は、いずれか特定のクローニング・ストラテジーに限定されない。当業者は、本発明のキメラDNA分子を含む構築物を産生するためのあらゆる種類のクローニング・ストラテジーを使用することができる。
【0043】
キメラDNA分子は、当業者に周知である既知の技術により宿主細胞内に挿入されることができる。これらの技術には、これだけに制限されることなく、リン酸カルシウム共沈キメラDNA分子を使用した形質転換、脂質試薬共トランスフェクション(すなわち、Lipofectamine)、エレクトロポレーション、細胞をウイルスと接触させることによる形質導入、又は細胞内へのキメラDNA分子のマイクロインジェクションが含まれる。宿主細胞は、高等真核細胞、例えば、哺乳動物細胞など、又は下等真核細胞、例えば、酵母細胞などであるか、あるいは宿主細胞は、原核細胞、例えば、細菌細胞などであるかもしれない。他のものが好みで利用されるかもしれないが、形質転換のために好適な原核生物宿主には、E.コリ(E. coli)、バチルス・ズブチリス(Bacillus subtillis)、サルモネラ・ティフィムリウム(Salmonella typhimurium)、並びにシュードモナス属(the genera of Pseudomonas)、ストレプトマイセス属(Streptomyces)、及びスタフィロコッカス属(Staphylococcus)の中の多様な種が含まれる。
【0044】
さまざまな宿主/発現ベクターの組合せが、本発明のタンパク質複合体を発現する際に利用される。一般に、組み換え発現ベクターには、複製開始点、及び宿主細胞の形質転換を可能にする選択マーカー(例えば、真核細胞培養のためのジヒドロ葉酸レダクターゼ若しくはネオマイシン耐性、又は、例えば、E.コリにおけるテトラサイクリン若しくはアンピシリン耐性、又はS.セレビシエ(S. cerevisiae)TRP1遺伝子)、及び下流の構造配列の転写を指示する高度に発現される遺伝子から得られるプロモーターが含まれる。前述のプロモーターは、解糖系酵素、例えば、とりわけ、3-ホスホグリセリン酸キナーゼ(PGK)、A因子、酸性ホスファターゼ、又は熱ショック・タンパク質などをコードするオペロンから得ることができる。異種構造配列は、適切な段階で、翻訳開始及び終止配列で構築され、そして、いくつかの態様において、リーダー配列が翻訳されたタンパク質複合体の分泌を指示できる。ベクターは複製開始点領域をさらに含み、ベクターの維持を確保し、そして、所望であれば、宿主内での増幅を提供する。
【0045】
使用できる有用な発現ベクターには、例えば、染色体、非染色体、及び合成DNA配列の断片が含まれる。好適なベクターには、これだけに制限されることなく、SV40及びpcDNAの誘導体、並びに既知の細菌プラスミド、例えば、col EI、pCR1、pBR322、pMal-C2、pET、Smithら、Gene 67:31-40ページ(1988年)によって説明されているpGEX、pMB9、及びその誘導体など、プラスミド、例えば、RP4など、ファージDNA、例えば、ファージIの多数の誘導体など、例えば、NM989など、並びに他のファージDNA、例えば、M13及び糸状単鎖ファージDNAなど;酵母プラスミド、例えば、2ミクロン・プラスミド及び2ミクロン・プラスミドの誘導体など、並びにセントメリック(centomeric)、且つ、集約的な酵母シャトルベクター;真核細胞において有用なベクター、例えば、昆虫又は哺乳動物細胞において有用なベクターなど;プラスミドとファージDNAの組合せから得られたベクター、例えば、ファージDNA又は発現制御配列を利用するように修飾されたプラスミドなど;等が含まれる。必要条件は、ベクターが最適な宿主細胞内で複製可能であり、且つ、存続可能であることである。低又は高コピー数のベクターを要望に応じて使用できる。
【0046】
例えば、バキュロウイルス発現系において、非融合トランスファー・ベクター、例えば、これだけに制限されることなく、pVL941(BamHIクローニング部位、Summersら、Virology 84:390-402ページ(1978年)から入手可能)、pVL1393(BamHI、SmaI、XbaI、EcoRI、NotI、XmaIII、BglII、及びPstIクローニング部位;Invitrogen)、pVL1392(BglII、PstI、NotI、XmaIII、EcoRI、XbaI、SmaI、及びBamHIクローニング部位;Summersら、Virology 54:390-402ページ(1978年)及びInvitrogen)、並びにpBlueBacIII(BamHI、BglII、PstI、NcoI、及びHindIIIクローニング部位、ブルー/ホワイト組み換えスクリーニングによる、Invitrogen)など、そして、融合トランスファー・ベクター、例えば、これだけに制限されることなく、pAc700(BamHI及びKpnIクローニング部位、BamHI認識部位が開始コドンで始まる;Summersら、Virology 54:390-402ページ(1978年))、pAc701及びpAc702(pAc700と同じ、異なるリーディング・フレームを持つ)、pAc360(ポリヘドリン開始コドンの36塩基対下流のBamHIクローニング部位;Invitrogen(1995年))、並びにpBlueBacHisA、B、C(BamHI、BglII、PstI、NcoI、及びHindIIIクローニング部位を有する3つの異なるリーディング・フレーム、ProBond精製及びブルー/ホワイト組み換えスクリーニングのためのN末端ペプチド;Invitrogen(220))の両方を使用できる。
【0047】
哺乳動物発現ベクターには、複製開始点、好適なプロモーター、及びエンハンサーを含み、そして、いずれかの必要なリボソーム結合部位、ポリアデニル化部位、スプライス・ドナー及びアクセプター部位、転写終止配列、並びに5’フランキング非転写配列が含まれる。SV40スプライスに由来するDNA配列、及びポリアデニル化部位は、必要な非転写遺伝要素を提供するのに使用できる。本発明における使用のために想定される哺乳動物発現ベクターには、誘導プロモーター、例えば、ジヒドロ葉酸レダクターゼ・プロモーターなどを有するベクター、DHFR発現カセットを有するあらゆる発現ベクター、又はDHFR/メトトレキサート共増幅ベクター、例えば、pED(PstI、SalI、SbaI、SmaI、及びEcoRIクローニング部位、クローン化遺伝子及びDHFRの両方を発現するベクターを有する;Randal J. Kaufman、1991年、Randal J. Kaufman, Current Protocols in Molecular Biology, 16.12(1991年))などが含まれる。あるいは、グルタミン・シンテターゼ/メチオニン・スルホキシミン共増幅ベクター、例えば、pEE14(HindIII、XbaI、SmaI、SbaI、EcoRI、及びBclIクローニング部位、ベクターがグルタミン・シンテターゼとクローン化遺伝子を発現する;Celltech)などである。エプスタイン・バー・ウイルス(EBV)又は核抗原(EBNA)の制御下でのエピソーム発現を指示するベクターには、例えば、pREP4(BamHI、SfiI、XhoI、NotI、NheI、HindIII、NheI、PvuII、及びKpnIクローニング部位、構成的RSV-LTRプロモーター、ハイグロマイシン選択マーカー;Invitrogen)、pCEP4(BamHI、SfiI、XhoI、NotI、NheI、HindIII、NheI、PvuII、及びKpnIクローニング部位、構成的hCMV前初期遺伝子プロモーター、ハイグロマイシン選択マーカー;Invitrogen)、pMEP4(KpnI、PvuI、NheI、HindIII、NotI、XhoI、SfiI、BamHIクローニング部位、誘導性メタロチオネインIIa遺伝子プロモーター、ハイグロマイシン選択マーカー、Invitrogen)、pREP8(BamHI、XhoI、NotI、HindIII、NheI、及びKpnIクローニング部位、RSV-LTRプロモーター、ヒスチジノール選択マーカー;Invitrogen)、pREP9(KpnI、NheI、HindIII、NotI、XhoI、SfiI、BamHIクローニング部位、RSV-LTRプロモーター、G418選択マーカー;Invitrogen)、並びにpEBVHis(RSV-LTRプロモーター、ハイグロマイシン選択マーカー、N末端ペプチドはProBond樹脂により精製されることができ、そしてエンテロキナーゼによって切断される;Invitrogen)などが使用できる。
【0048】
本発明における使用のために選択可能な哺乳動物発現ベクターには、これだけに制限されることなく、pRc/CMV(HindIII、BstXI、NotI、SbaI、及びApaIクローニング部位、G418選択、Invitrogen)、pRc/RSV(HindII、SpeI、BstXI、NotI、XbaIクローニング部位、G418選択、Invitrogen)等が含まれる。本発明に使用できるワクシニア・ウイルス哺乳動物発現ベクター(例えば、Randall J. Kaufman, Current Protocols in Molecular Biology 16.12(Frederick M. Ausubelら編、Wiley、1991年)を参照のこと)には、これだけに制限されることなく、pSC11(SmaIクローニング部位、TK-及びβ-gal選択)、pMJ601(SalI、SmaI、AflI、NarI、BspMII、BamHI、ApaI、NheI、SacII、KpnI、及びHindIIIクローニング部位;TK-及びβ-gal選択)、pTKgptF1S(EcoRI、PstI、SalII、AccI、HindII、SbaI、BamHI、及びHpaクローニング部位、TK又はXPRT選択)等が含まれる。
【0049】
本発明でも使用できる酵母発現系には、これだけに制限されることなく、非融合pYES2ベクター(XbaI、SphI、ShoI、NotI、GstXI、EcoRI、BstXI、BamHI、SacI、KpnI、及びHindIIIクローニング部位、Invitrogen)、融合pYESHisA、B、C(XbaI、SphI、ShoI、NotI、BstXI、EcoRI、BamHI、SacI、KpnI、及びHindIIIクローニング部位、N末端ペプチドはProBond樹脂で精製され、そして、エンテロキナーゼで切断される;Invitrogen)、pRSベクター等も含むことができる。
【0050】
本発明における使用のための1つの特に好ましいベクターは、プラスミドpCE2である。pCE2プラスミドは、当該技術分野で知られているいずれかの方法によっても得ることができる。実施例2.A.で利用された、そういった方法の1つは、Leungら、Proc. Natl. Acad. Sci. USA 92:4813-4817ページ(1995年)に記載されている。
【0051】
好ましい態様において、キメラDNA分子は、挿入されたキメラDNA分子の転写と翻訳に必要な要素を既に含む発現ベクター内に挿入される可能性がある。
【0052】
加えて、キメラDNA分子を含む発現ベクターは、薬物選択マーカーを含むかもしれない。そのようなマーカーは、クローニングにおいて、及びキメラDNA分子を含むベクターの選別又は検証において助けとなる。例えば、ネオマイシン、ピューロマイシン、ハイグロマイシン、ジヒドロ葉酸レダクターゼ(DHFR)、グアニン・ホスホリボシル転移酵素(GPT)、ゼオシン、及びヒスチジノールに対する耐性を与える遺伝子は有用な選択マーカーである。あるいは、酵素、例えば、単純ヘルペス・ウイルス・チミジンキナーゼ(tk)又はクロラムフェニコール・アセチルトランスフェラーゼ(CAT)などを利用することができる。免疫学的マーカーも利用することができる。遺伝子産物をコードする核酸を同時に発現することができる限り、あらゆる既知の選択マーカーも用いることができる。選択マーカーの更なる実例は、当業者にとって周知であり、そして、レポーター、例えば、高感度緑色蛍光タンパク質(EGFP)、β-ガラクトシダーゼ(β-gal)、又はクロラムフェニコール・アセチルトランスフェラーゼ(CAT)を含む。
【0053】
このため、哺乳動物、主として、ヒト細胞、並びに細菌、酵母、真菌、昆虫、線虫、及び植物細胞が宿主細胞として本発明に使用することができ、そして、本明細書中に規定されるように発現ベクターによって形質転換されることができる。いくつかの細胞宿主、例えば、哺乳動物細胞などにおいて、それが元の環境(例えば、それが自然発生的なものであれば、自然環境)から取り出されるので、キメラDNA分子を含む細胞は「分離」されるかもしれない。他の態様、例えば、植物などにおいて、植物細胞又は部分の培養物よりも植物全体を使用できるので、細胞が分離される必要はない。
【0054】
好適な細胞の実例は、これだけに制限されることなく、VERO細胞、HeLa細胞、例えば、ATCC番号CCL2など、CHO細胞株、例えば、ATCC番号CCL61など、COS細胞、例えば、COS-7細胞及びATCC番号CRL1650の細胞など、W138、BHK、HepG2、3T3、例えば、ATCC番号CRL6361など、A549、PC12、K562細胞、293細胞、Sf9細胞、例えば、ATCC番号CRL1711など、並びにCv1細胞、例えば、ATCC番号CCL70などが含まれる。
【0055】
本発明において使用できる他の好適な細胞には、これだけに制限されることなく、原核生物宿主細胞菌株、例えば、エシェリキア・コリ(Escherichia coli)、(例えば、菌株DH5-α)、バチルス・ズブチリス、サルモネラ・ティフィムリウム、又はシュードオモナス属、ストレプトコッカス属、及びスタフィロコッカス属の菌株などが含まれるかもしれない。本発明に使用できるさらに好適な細胞には、酵母細胞、例えば、サッカロマイセス属(Saccharomyces)のものなど、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)などが含まれる。
【0056】
着目のポリヌクレオチドを含む宿主細胞は、プロモーターを活性化するのに、形質転換体を選択するのに、又は遺伝子を増幅するのに適切なように改善された従来の栄養培地(例えば、Ham’s栄養混合物)中で培養されうる。培養条件、例えば、温度、pH等は、発現のために選択された宿主細胞でこれまで使用されたものであり、当業者には明白である。細胞は、通常、遠心分離によって採取され、物理的又は化学的手段によって破壊され、そして、更なる精製のために保持される粗抽出物となる。タンパク質発現に利用される微生物細胞は、その全てが当業者にとって周知である凍結融解サイクル、超音波処理、機械的破砕、又は細胞溶解剤の使用を含めたいずれか都合のよい方法で破壊されるかもしれない。細胞分解に関係する態様は、キメラDNA分子の発現後に分解を制限するプロテアーゼ・インヒビターを含むバッファーの使用を伴うかもしれない。好適なプロテアーゼ・インヒビターには、ロイペプチン、ペプスタチン、又はアプロチニンが含まれる。上清は、次に、飽和硫酸アンモニウムの濃度を連続的に高めて沈殿させる。
【0057】
タンパク質複合体生成物は、1種類以上の技術によって精製されることができる。通常、精製は、個別の手順、例えば、ゲル濾過、アフィニティー精製、塩分別、イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、ヒドロキシルアパタイト吸着クロマトグラフィー、疎水性相互作用クロマトグラフィー、及びゲル電気泳動などの組合せを伴う。タンパク質リホールディング・ステップが、タンパク質複合体の立体構造を完成するのに、適宜、使用されてもよい。高速液体クロマトグラフィー(HPLC)は、多くの場合、最終精製ステップに有用である。一般に、Robert K. Scopes, Protein Purification:Principles and Practice(Charles R. Castor編、Springer-Verlag 1994年)及びJoseph Sambrook, Molecular Cloning:A Laboratory Manual、第2版(Cold Spring Harbor Laboratory Press 1989年)を参照のこと。多段階精製分離の実例は、Baronら、Crit. Rev. Biotechnol. 10:179-90ページ(1990年)及びBelowら、J. Chromatogr. A. 679:67-83ページ(1994年)にも記載されている。
【0058】
複合体は、それらが非複合型タンパク質と比べて改善された血漿中半減期を示すかどうか測定するために使用前に試験される。例えば、G-CSFと様々なアミノ酸伸長、例えば、(NNT)などを用いて実施した実験において、出願人は、ある場合において半減期が延び、そして他の場合において短くなることがわかった。試験を、実施例3に示されているように、薬物動力学の標準的な技術に従って実施することができる。この手順は、予定された用量の複合体を動物、好ましくは、実験動物、例えば、齧歯動物など、例えば、マウスに投与し、予定された間隔でその動物から血漿を採取し、そして、濃度が測定できなくなるまで、例えば、酵素結合免疫吸着検定法(「ELISA」)によって血漿を分析して、上記複合体の濃度を測定することを伴う。半減期は、(例えば、WINNonLinソフトウェア・バージョン4.1を使用した)非コンパートメント薬物動力学的解析によって計算できる。複合体の濃度が計測できる最後の時間(tf)に加えて、分析は、以下の主要なパラメーター:λz、見かけの終末段階に関連した見かけの終末速度定数、血漿中の濃度の対数、対、曲線の単一指数型終末部分の時間に関する線形回帰分析によって概算される;t1/2,z、見かけの終末半減期、以下の方程式:t1/2,z=ln(2)/λzにより計算される;AUC、0時間〜無限の血漿中濃度−時間曲線下面積;AUC/D、用量単位あたりの血漿中濃度−時間曲線下面積;MRT、1次モーメント曲線、AUMC、とAUC下面積の間の比として計算される平均滞留時間;CL、全身クリアランス、CL=D/AUCとして計算される;及びVss、分布の定常状態容量、VSS=CL*MRTとして計算される、の観測又は計算を含む。
【0059】
本発明の更なる側面は、タンパク質複合体と担体を含む組成物に関する。広く、担体は、培地又はマトリツクス(例えば、精製マトリツクス)であるかもしれない。いくつかの態様において、担体は、医薬として許容される担体であり、その場合、その組成物はヒト又は動物、最も好ましくは、哺乳動物の障害、及び/又は疾患を予防又は治療するために、あるいは診断目的のために有用である。組成物の有効成分として、タンパク質複合体は、好ましくは、可溶性型の状態である。
【0060】
一般に、組成物は、例えば、治療又は診断に関する期待される効果を発揮する医薬として有効な量のタンパク質複合体を含む。医薬として有効な量は、細胞培養アッセイから推定することができる。例えば、生体外の系において所望の効果を挙げる濃度点若しくは範囲を含むか又は網羅する血中濃度範囲を達成するために、用量が動物モデルにおいて処方されるかもしれない。例えば、Molineuxら、Exp. Hematol. 27:1724-34ページ(1999年)を参照のこと。この情報は、よって、ヒトを含めた他の哺乳動物、及び動物において用量を正確に測定するために使用できる。一般に、投与量は、患者の体重に基づいて約1ng/kg〜約10mg/kgの範囲におよぶ。
【0061】
そのような化合物の有毒性と治療効能は、細胞培養又は実験動物における標準的な医薬の手順によって測定することができる。例えば、Molineuxら、Exp. Hematol. 27:1724-34ページ(1999年)を参照のこと。例えば、LD50(集団の50%に対して致死的な用量)、並びにED50(集団の50%において治療的に有効な用量)は、当該技術分野で知られている方法を使用して測定できる。毒性と治療効果の間の用量比は、高い治療指標を提供する化合物のLD50とED50の間の比として示すことができる治療係数である。
【0062】
細胞培養及び動物の研究から得られるデータは、好ましくは、わずかの若しくは全く毒性のないED50の中に含めた血中濃度の範囲内にある前述の化合物の一連の投与量を処方する際に使用できる。
【0063】
組成物は、いずれかの好適な経路、例えば、局所的に、経口的に、全身的に、静脈内に、筋肉内に、粘膜的に、(例えば、パッチによって)経皮的に投与できる。それらは、リポソーム、微小粒子、マイクロカプセル、ナノ粒子等の中にカプセル化することができる。生物学的に活性なポリペプチドを処方し、そして、投与する技術は、Remington:The Science and Practice of Pharmacy(Alfonso R. Gennaroら編、Philadelphia College of Pharmacy and Science 2000年)の中にも開示されている。
【0064】
本発明とその利点について完全に説明するために、以下の具体的な実施例が与えられるが、当該実施例は単なる一例としてのものであるので、決して制限するものではないことは理解されている。
【図面の簡単な説明】
【0065】
【図1】PHO×pBSKベクターの略図である。
【図2】PHO-G-CSF- NN×pBSKベクターの略図である。
【図3】PHO-G-CSF-NNT65×pBSKベクターの略図である。
【図4】PHO-G-CSF-NNT155×pBSKベクターの略図である。
【図5】PHO-G-CSF-NNT155×pCE2ベクターの略図である。
【図6】いくつかの異なるG-CSF-ポリペプチド複合体と非複合型G-CSFのウエスタンブロットである。
【図7】電気泳動前にPNGase Fで処理したいくつかの異なるG-CSF-ポリペプチド複合体と非複合型G-CSFのウエスタンブロットである。
【図8】PROCHO4-CDM培地で培養したCHOK1細胞において発現された(二重反復試験の)G-CSF-NNT155とG-CSF-NNT65構築物のウエスタンブロットである。
【実施例】
【0066】
実施例1
キメラDNA分子のクローニング
下流タンパク質精製の簡略化のために、G-CSF重合体タンパク質を細胞培地中に分泌されることにした。様々なサイトカイン−重合体構築物での細菌(ST2)分泌シグナルを使用したこれまでの経験では、原核生物及び真核細胞系の両方において低い分泌効能しか示さなかった。しかしながら、異種タンパク質(GFP及びHPV 16 E7)の培地中への分泌のためのpho1+酸ホスファターゼ(PHO)のシゾサッカロマイセス・ポンベ(Schizosaccharomycen pombe)分泌シグナル配列の使用が知られていた。このため、この分泌シグナルを、CHO細胞発現系において以下に記載のG-CSF重合体構築物を用いて試験した。また、以下に記載のものは、G-CSF重合体構築物の製造と、G-CSF重合体の発現のために合成したベクターである。
1.A. PHO×pBSK構築物の製造
PHO×pBSK構築物
【0067】
合成された最初の構築物は、単に、細菌クローニング・ベクターpBSK内にクローン化したPHOリーダー配列であった。PHO分泌シグナルのアミノ酸配列を以下に挙げた。
【0068】
【化1】

【0069】
PHO分泌シグナルを、2組の相補DNAオリゴヌクレオチドを一緒に融合し、そして、それらをpBSK内にクローン化することによって合成した。オリゴヌクレオチドの設計に通じる最も重要な考慮すべき事柄は、G-CSF重合体のN末端へのリーダー配列の融合があらゆる介在配列なしに直接的であるということであった。これは、分泌シグナル全体が加工中に分子から切り取られ、天然及び臨床的に入手可能なバージョンのG-CSFと比べた時に、N末端修飾を持たないG-CSFの分泌型を生じるのを確実にする。このような様式で構築物を調製することによって、G-CSFとPEG-G-CSFを直接比較することができるだけではなく、これは組み換えG-CSFタンパク質に追加アミノ酸を挿入することによってあらゆる潜在的な免疫原性をも制限した。PHOリーダーのpBSK内へのクローニング、並びにその後のPHO×pBSK内へのG-CSFのクローニングのための制限部位をオリゴヌクレオチド内に組み入れることによって、これらの必要条件は満たされた。使用したオリゴヌクレオチドを、以下に挙げる。
【0070】
【化2】
【0071】
PHO×pBSKベクターの図を図1に示す。PHOリーダー(PhoA)を、NotIとBamHIを使用してpBSK内にクローン化した。成熟G-CSFの配列を、BspMIとBamHI部位を利用してこの構築物内にクローン化した。このベクターは、細菌内で増えると、アンピシリン耐性(bla)を与える。
構築方法
【0072】
オリゴヌクレオチド(oNDEPHO-1FとoNDEPHO-1R)及び(oNDEPHO-2FとoNDEPHO-2R)の相補対を、T4ポリヌクレオチド・キナーゼ(PNK)を使用してリン酸化した。T4 PNKを熱不活化し、そして、オリゴヌクレオチド対を氷上でゆっくりとアニーリングさせた。反応物を、TE中に希釈し、そして、NotIとBamHIを使用して前もって消化したpBSKとの連結に使用した。連結産物を、Top10コンピテント細胞内にエレクトポレーション処理し、そして、LB-アンピシリン・プレートで培養した。ミニプレップを、アンピシリン抵抗性コロニーにて実施して、DNAを分離し、そして、特徴的な消化で推定上のクローンを識別した。配列分析で推定上のクローンが正しいと判定した。
1.B. PHO-G-CSF-NN×pBSK構築物の製造
PHO-G-CSF-NN×pBSK構築物
【0073】
この構築物は、BspMIとBamHIを使用したPHO×pBSKベクター内への、成熟G-CSFのコード配列に相当するPCR増幅物のクローニングから生じた。先に述べたとおり、この目的で使用されるオリゴヌクレオチドを、制限酵素BspMIを利用することによってPHOとG-CSFの間の接合点に介在配列を全く持たないことを確実にするように設計した。さらに、G-CSFのC末端は、2つのアスパラギン残基から成る直接的な融合(NN)、及び酵素BbsIとBamHIのための制限部位を持っていた。G-CSFのC末端へのNNT重合体の直接的な付加のためにBbsIを続いて利用すると同時に、ベクター内へのGCSF-NNのクローン化のためにBamHIを使用した。
【0074】
G-CSFを増幅するために合成したオリゴヌクレオチドは、以下のとおりである:
【0075】
【化3】
【0076】
PHO-G-CSF-NN×pBSKベクターの図を、図2に示す。成熟G-CSF-NNの配列を、BspMI(破壊される)とBamHIを使用してPHO×pBSK内にクローン化した。NNT重合体のその後の付加は、プラスミド地図上に示したBbsIとBamHI部位を利用した。このベクターは、細菌で増えると、アンピシリン耐性(bla)を与える。
構築方法
【0077】
G-CSFを、oBspMIGCSFとoGCSFBbsBamオリゴヌクレオチドを使用したPCR法によって増幅した。成熟G-CSFに相当する約520bpのバンドを、アガロース・ゲルから切り出し、そして、精製した。精製した断片を、BspMIとBamHIで消化し、精製し、そして、同じ酵素で切断したPRO×pBSK内に連結した。連結産物を、Top10コンピテント細胞内にエレクトポレーション処理し、LB-アンピシリン・プレート上で培養した。ミニプレップを、アンピシリン抵抗性コロニーに対して実施して、DNAを分離し、そして、特徴的な消化で推定上のクローンを識別した。配列分析で推定上のクローンが正しいと判定した。
1.C. PHO-G-CSF-NNT65×pBSK構築物の製造
PHO-G-CSF-NNT65×pBSK構築物
【0078】
この重合体のアミノ酸組成物は、コンセンサス哺乳動物N連結型グリコシル化部位、N-X-(S/T)をコードする。これにより、この構築物がCHO細胞内で発現される時、重合体はその重合体伸長のスレオニン残基でグリコシル化されることができる。期待されることは、翻訳産物サイズにおける重合増加と翻訳後修飾がG-CSFのpKパラメーターを調節し、その生物活性を低下させることなくタンパク質の延長された血清中半減期を与えるであろうことであった。
【0079】
この重合体の構築を、オリゴヌクレオチド連結/多量体化スキームを使用することで達成した。PHO-G-CSF-NN×pBSK構築物をBbsIで切断することによって、4塩基のアンダーハング(GTTG)をG-CSFのC末端に付加された2個のアスパラギン残基において作り出した。反復NNTトリプレットをコードし、並びにGTTGアンダーハングにアニールする相補的なオリゴヌクレオチドの組合せを設計することによって、より長い鎖に9つのアミノ酸をコードするオリゴヌクレオチドが多量体を形成することが可能であった。終止コドンを含む短いアダプター、重合体の将来的な伸長のためのBbsI部位、及びBamHI部位を、低い割合で付加して、多量体化を終了させ、そして、BbsI-BamHI重合体断片をPHO-G-CSF-NN×pBSK内にクローニングした。
【0080】
NNT65重合体を合成するのに使用したオリゴヌクレオチドを、以下に挙げる:
重合体骨格オリゴヌクレオチド
【0081】
【化4】
【0082】
アダプター・オリゴヌクレオチド
【0083】
【化5】
【0084】
以下の配列(NTNNTNNTN)を、重合体の多量体を形成するのに使用した、それぞれ、CAACオーバハングとGTTGアンダーハングを示すo3NNTFとo3NNTRの重合体骨格オリゴヌクレオチドを使用して作り出したNNT重合体の反復単位であった。
【0085】
【化6】
【0086】
以下に示すものは、重合体を完成させ、そして、終止コドン、重合体の将来的な伸長のためのBbsI部位、及びクローニングに必要なBamHI部位を含む終結アダプター分子である。
終結アダプター分子
【0087】
【化7】
【0088】
PHO-G-CSF-NNT65×pBSKベクターの図を、図3に示す。NNT65重合体の付加には、G-CSFのC末端に位置するBbsI部位、及びBamHI(破壊される)を利用した。図3に示されているように、将来的な長さの伸長のためにNNT65重合体の端にBbsI部位を作り直した。このベクターは、細菌内で増えると、アンピシリン耐性(bla)を与える。
構築方法
【0089】
重合体骨格オリゴヌクレオチド(o3NNTFとo3NNTR)及びアダプター・オリゴヌクレオチド(oDent3FとoDent3R)の相補的な対を、T4 PNKを使用してリン酸化した。T4 PNKを熱不活化し、そして、オリゴヌクレオチド対を氷上でゆっくりとアニーるさせた。重合体とアダプターの二体鎖をT4リガーゼと20:1及び40:1の比で混合することによって、重合体の多量体化を実施した。T4リガーゼを、熱不活性化し、そして、連結反応物全体をBamHIで一晩、消化した。両反応物を沈殿させ、そして、アクリルアミドゲル上に泳動した。250bp〜800bpの間の成分を切り出し、そして、ゲルを精製した。
【0090】
この成分を、BbsIとBamHIで消化したPHO-G-CSF-NN×pBSKとの連結に使用した。化学的コンピテントStbl2細胞を、連結産物で形質転換し、そして、LB-アンピシリン・プレート上で培養した。ミニプレップを実施して、DNAを分離し、そして、特徴的な消化で推定上のクローンを識別した。配列分析で推定上のクローンが正しいと判定した。
【0091】
このストラテジーから分離された中で最も長いクローンは、PHO-G-CSF-NNT65×pBSKであった。
1.D. PHO-G-CSF-NNT155×pBSK構築物の製造
PHO-G-CSF-NNT155×pBSK構築物
【0092】
このクローンの構築には、同じ多量体化スキームを使用したPHO-G-CSF-NNT65×pBSK構築物への追加のNNT残基の付加を必要とした。元々の重合体と伸長の間の接合点を識別するために、この伸長に使用されるオリゴヌクレオチドのヌクレオチド組成を変更した。これらの変更は、重合体の元々のNNT組成を維持し、且つ、同じGTTGアンダーハングとCAACオーバハング・ストラテジーを利用した。BbsIとBamHIでPHO-G-CSF-NNT65を消化することによって、同じオリゴヌクレオチド多量体化ストラテジーを使用して重合体の長さを伸長することが可能であった。
【0093】
NNT65重合体をNNT155に伸長するのに使用したオリゴヌクレオチドを、以下に挙げる:
重合体骨格オリゴヌクレオチド
【0094】
【化8】
【0095】
アダプター・オリゴヌクレオチド
【0096】
【化9】
【0097】
PHO-G-CSF-NNT65×pBSK構築物の製造と同様に、NTNNTNNTNが、o3NNTextFとo3NNTextRの重合体骨格オリゴヌクレオチドを使用して生み出したNNT重合体伸長の反復単位であり、CAACオーバハングとGTTGアンダーハングを重合体の多量体を形成するのにさらに使用し、そして、アミノ酸組成を元々のNNT65重合体から変えることはなかった。
【0098】
【化10】
【0099】
上記のo3NNTextFとo3NNTextRの中の下線を引いたヌクレオチドが、それぞれ、o3NNTFとo3NNTRの同じ位置のヌクレオチドと異なっていた。
【0100】
以下のものは、NNT155重合体を完成させ、且つ、クローニングに必要なBamHI部位を含む終結アダプター分子である。oDent3FとoDent3Rは、最初のNNT65重合体に使用したオリゴヌクレオチドと同じものである。
終結アダプター分子
【0101】
【化11】
【0102】
PHO-G-CSF-NNT155×pBSKベクターの図を、図4に示す。90AAによるWNT65重合体の伸長は、PHO-G-CSF-NNT65×pBSKのC末端に位置するBbsI部位と、BamHI(破壊される)を利用した。将来的な長さの伸長のためにNNT155重合体の端にBbsI部位を作り直したことに注目した。このベクターは、細菌内で増えると、アンピシリン耐性(bla)を与える。
構築方法
【0103】
重合体骨格オリゴヌクレオチドo3NNTextFとo3NNTextR、及びアダプター・オリゴヌクレオチドoDent3FとoDent3Rの相補的な対を、T4 PNKを使用してリン酸化した。T4 PNKを熱不活化し、そして、オリゴヌクレオチド対を氷上でゆっくりとアニーるさせた。重合体とアダプターの二体鎖をT4リガーゼと20:1及び40:1の比で混合することによって、重合体のオリゴマー化を実施した。T4リガーゼを、熱不活性化し、そして、連結反応物全体をBamHIで一晩、消化した。両反応物を沈殿させ、そして、アクリルアミドゲル上に泳動した。250bp〜800bpの間の成分を切り出し、そして、ゲルを精製した。この成分を、BbsIとBamHIで消化したPHO-G-CSF-NNT65×pBSK内に連結した。化学的コンピテントStbl2細胞を、連結産物で形質転換し、そして、LB-アンピシリン・プレート上で培養した。ミニプレップを実施して、DNAを分離し、そして、特徴的な消化で推定上のクローンを識別した。配列分析で推定上のクローンが正しいと判定した。
【0104】
このストラテジーから分離された最も長いクローンは、PHO-G-CSF-NNT155×pBSKであった。
実施例2
宿主細胞の形質転換とキメラDNA分子の発現
2.A. PHO-G-CSF-NNT155×pCE2構築物の製造
ベクターpCE2の調製
【0105】
プラスミドpCE2は、以下の操作でプラスミドpREP7bから得られた。まず最初に、2000bpのEBNAコード領域を取り除いた。次に、pBR322oriをpKS-oriによって置き換えた。そして、RSVプロモーター領域を、CMVエンハンサー、及び伸長因子-1a(EF-1a)プロモーターとイントロンによって置き換えた。
CMVエンハンサーを、以下のプライマー:
【0106】
【化12】
【0107】
を使用してpCEP4(Invitrogen、San Diego, CA)からポリメラーゼ連鎖反応(PCR)によって作り出した380bpのXbaI-SphI断片から得た。
EF-1aプロモーターとイントロン(Uetsukiら、J. Biol. Chem. 254:5791-5798ページ(1989年))を、以下のプライマー:
【0108】
【化13】
【0109】
を使用してヒト・ゲノムDNAからPCR法によって作り出した1200bpのSphI-Asp718I断片から得た。
【0110】
2つの断片を、pREP7bから得られたXbaI/Asp718I消化ベクター内に連結して、pCE2を作り出した。
PHO-G-CSF-NNT155×pCE2構築物
【0111】
哺乳動物細胞内でPHO-G-CSF-NNT155を発現するために、構築物をベクターpCE2内に移行することが必要であった。このベクターは、最低限のプロモーター、及び前初期CMVエンハンサーが隣接するヒト伸長因子-1-αの第1イントロンを含んだ。ベクターは、哺乳動物選択のためのヒグロマイシンB耐性マーカーをも含んだ。
【0112】
PHO-G-CSF-NNT155×pCE2ベクターの図を、図5に示す。CHO細胞内で発現を駆動するプロモーターは、Pcmv/ef-1aである。CHO細胞内へのエレクトロポレーションのために、SalIを使用してベクターを線状にした。このベクターは、哺乳動物細胞においてハイグロマイシンBに対する耐性を与え、そして、細菌内で増えると、アンピシリン耐性(bla)を与える。
構築方法
【0113】
この構築物は、PHO-G-CSF-NNT155×pBSKから1100bpのNotI-BamHI断片を分離し、そして、ゲル精製することによって製造した。この成分を、NotIとBamHIで消化したpCE2内に連結した。化学的コンピテントStbl2細胞を、連結産物で形質転換し、LB-アンピシリン・プレート上で培養した。ミニプレップを実施して、DNAを分離し、そして、特徴的な消化で推定上のクローンを識別した。大規模なマキシプレップを実施して、CHO細胞の形質転換のためのマイクログラム単位のプラスミドを分離した。
2.B. 形質移入細胞株の製造
【0114】
先に記載した重合体を製造するために、哺乳動物細胞を、発現のための宿主細胞として使用した。今回は、チャイニーズハムスター卵巣(CHO)細胞株を選択した。CHO細胞は、G-CSF及びEPOを含めた組み換えタンパク質治療薬を発現するために製薬業界で広く使用されている。以下で説明するように、CHO細胞におけるG-CSF重合体構築物の発現培養を確立した。
接着培養の確立
【0115】
PHO-G-CSF-NNT155×pCE2ベクターを、酵素SalIを使用して線状にした。消化物を、沈殿させ、そして、50μLのTE中に再懸濁した。このプラスミド1(1)μgを、5×106個のCHO細胞と共にエレクトロポレーションに使用した。接着CHO細胞を、10%のFBSを含むHam’s栄養混合物F-12(F-12 Ham’s培地)で培養した。細胞を、一晩、回復させ、そして、翌日、700μg/mLのハイグロマイシンBを含む培地を加えて、耐性菌の選択を開始した。2〜3週間の間に必要に応じて2〜3日おきに培地を交換した。耐性菌の大量のプールを、分離し、そして、ハイグロマイシンBの存在下でさらに2代継代した。
【0116】
培地中へのPHO-G-CSF-NNT155の分泌についてこれらの細胞を試験するために、細胞を、3〜5日間、低血清培地(0.5%)を含むHam’s培地中で培養した。培地を、分離し、そして、G-CSFに対するポリクローナル抗体を使用して調べるウエスタンブロットに使用した。ブロットを、図6及び7で再現する。ブロット間の相違点は、図7のサンプルがペプチド:N-グリコシダーゼF(PNGase F)処理をさらに受けた点である。図6に示されているように、ブロットは、免疫反応成分が見かけ上の分子量約95kDaの所に移動することを示した。図7に示されているように、N-結合型グリコシル化鎖を取り除くグリコシダーゼのPNGase Fを用いたこれらの同じサンプルの消化は、免疫反応成分を見かけ上の分子量、約49kDaまで低下させた。これは、改善されていないG-CSF-NNT155タンパク質の理論上のサイズにほぼ一致した。天然のG-CSFは、約18kDaの所に電気泳動により移動し;C末端へのNNT重合体の付加は、遺伝子組み換え型の、重合体分子にかなりの質量を加える。発現もかなり安定していると思われ;条件培地中に存在する未精製タンパク質について、それほどのタンパク分解は観察されなかった。さらに、図7に示されているように、PNGase F処理後の構築物のサイズの減少は、組み換えタンパク質構築物が相当量の糖付加を受けることを示唆した。
浮遊培養の確立
【0117】
確立したCHO培養の条件培地中に見られるG-CSF-NNT155の精製を容易にするために、細胞を、化学的に規定した、低タンパク質、無血清培地(PROCHO4-CDM, Cambrex)中で培養した。細胞をこの培地に適合させる過程において、細胞は接着から浮遊増殖に適合した。
【0118】
細胞を、T-185フラスコ内で、F-12 Ham’s+10%のFBS中で密な集密度まで培養し、トリプシン処理し、そして、PROCHO4-CDM培地中に懸濁させた。107個の細胞を、50mLのPROCHO4-CDM培地に加え、そして、T-185フラスコ内でインキューベートした。4〜5代の継代の後に、約90%の細胞が、もはや接着するものではなく、単独細胞の混合物及び凝集した細胞塊として増殖した。条件培地の回収のために、通常、60mLの新しいPROCHO4-CDM培地を、5〜6日間、7〜8mLの高細胞密度培養と一緒にインキューベートした。細胞を遠心分離によって培地から取り除き、そして、培地を、それに続いて、0.2ミクロン(μm)のフイルターを使用して不純物を除いた。培地は、ウェスタンブロットによってG-CSF-NNT155の収量を定量化し(通常、100〜200μg/L)、必要とされるまで保存した。
2.C. PROCHO4-CDM中でのG-CSF-NNT構築物の発現
【0119】
サンプルは、PROCHO4-CDM中で6日間、培養した前述の大量のCHO細胞株からの条件培地であった。ウエスタンブロットを、G-CSFに対するポリクローナル抗体を使用することで調べた。図8に示されているように、両構築物のサイズは、(先に説明した)Ham’s培地中に見られたものと同じであり、かつ、G-CSF-NNT155細胞株で観察された分解が無かった。
2.D. 発現の最適化
【0120】
所望の生成物の発現を最大にするのに必要な培養条件を決定するための試みにおいて、様々な化学的及び栄養添加物、並びに様々な環境パラメーターを試験した。以下のアプローチは、タンパク質蓄積の増大をもたらした。
化学添加物
【0121】
全ての化学添加物がタンパク質産生の増加をもたらすわけではないことが知られているが、刊行された文献は、2.5mMまでのアデノシン又はAMPの添加が細胞周期停止につながり;効果的に、タンパク質蓄積の付随した増大を伴って細胞培養の生存を長引かせることを示した。AMPの効果を、ProCHO4既知組成培地(CDM)中、G-CSF-NNT155産生のCHOを介した産生について試験した。AMPを、1mMの終濃度でProCHO4-CDM中に加えた。対照と比較して細胞生存率の減少は観察されなかった。AMPの添加はタンパク質産生のわずかな増加をもたらした。
環境パラメーター
温度変化
【0122】
CHO培養のための低い培養温度は、著しく高められたレベルのタンパク質産生つながる可能性がある。他のタンパク質産生向上の方法に見られるように、観察された効果は、細胞周期停止中の高められたタンパク質産生に起因するものである。37℃から28℃に組み換えCHO株の培養温度を下げることの効果を調べた。28℃で維持した培養物は37℃のものよりも顕著に長い生存のままであったが、低温は、高い組み換えタンパク質産生をもたらさなかった。
浮遊培養への適応
【0123】
大量のCHO細胞集団をスピナー・フラスコ培養条件に切り替えた。浮遊培養への適応は、ProCHO4-CDM中での培養によって仲介された。適応に続いて、細胞を、T型フラスコ内に広げ、そして、様々な密度でProCHO5 CDMを含むスピナー・フラスコ内に播種した。細胞が静置又は撹拌培養かどうかにかかわらず、タンパク質産生の著しい増加をProCHO5 CDM中での浮遊培養への適応中に観察した。
宿主としての異種細胞株の使用
【0124】
発現の最適化への1つのアプローチは、宿主細胞に新しい細胞株を使用することであるかもしれない。従って、ATCCからのCHO-K1の新しい発送品を調達した。新しい細胞集団を、増殖させ、そして今後の使用のために初期継代にて貯蔵した。この規定の培養物を、着目の組み換えタンパク質を発現する安定したクローン細胞株を作り出すために使用した。簡単に言えば、CHO-K1細胞を着目のベクターと一緒にエレクトポレーション処理し、そして、ハイグロマイシンの安定発現に関して選択した。92個のコロニーを、分離し、そして、着目の組み換えタンパク質の発現について評価した。これらのコロニーの多くが、先に記載したとおり、元々のトランスフェクト集団のものより顕著に高い発現レベルを示した。
結果
【0125】
培養物を1mMのAMPで処理した時、発現の非常にわずかな増強を観察した。低い培養温度は、培養物の寿命を延ばしたが、タンパク質発現の増加は無かった。ProCHO5 CDM中での懸濁培養条件への適応は、組み換えタンパク質の発現を劇的に高めた。また、ATCCによって規定されたCHO-K1細胞株から分離したクローンは、元々の分離集団よりも顕著に高いレベルの所望のタンパク質を発現した。
実施例3
G-CSF-NNT155複合体の薬物動態学的評価
【0126】
G-CSF-NNT155タンパク質複合体の薬物動力学的パラメーターを評価した。他の3つの化合物を正の対照として盛り込んだ。対照は、NEULASTA(PEG-G-CSF)、NEUPOGEN(rh-G-CSF)、及びG-CSF化合物であった。化合物を、マウスに単回静脈内(i.v.)投与し、そして、血液を、投薬の72時間後まで採取することによって試験した。血漿をELISA法によって分析した。
3.A. 分析方法
【0127】
アッセイには定量的なサンドイッチ酵素免疫分析技術を利用した。G-CSFに特異的なモノクローナル抗体を、マイクロプレート上にプレコートした。標準物質とサンプルをウェル内にピペットで計り取った。存在する全てのG-CSFを、固定化抗体によって結合した。全ての非結合物質を洗い流した後に、G-CSFに特異的な酵素結合ポリクローナル抗体をウェルに加えた。全ての非結合抗体−酵素試薬を取り除くための洗浄に続いて、基質溶液をウェルに加え、そして、初期段階で結合したG-CSFの量に比例して発色させた。発色を止め、そして、色の強さを計測した。
試薬
【0128】
以下の試薬と材料を使用した:
− G-CSFマイクロプレート:G-CSFに対するマウス・モノクローナル抗体でコートした96ウェル・ポリスチレン・マイクロプレート(8ウェルから成る12の細片)。
− G-CSF複合体:21mLの、保存料を含む西洋ワサビペルオキシダーゼに結合した、G-CSFに対するポリクローナル抗体。
− G-CSF標準物質:2バイアル(25ng/mL)の、凍結乾燥された、保存料を含む緩衝化タンパク質基材中の組み換えヒトG-CSF。
− アッセイ希釈剤RD1A:11mLの、保存料を含む緩衝化タンパク質基材。
− キャリブレーター希釈剤RD5:21mLの、保存料を含む緩衝化タンパク質基材。細胞培養上清サンプル用。
− キャリブレーター希釈剤RD6A:21mLの、保存料を含む動物血清。血清/血漿サンプル用。
− 洗浄バッファー濃縮物:21mLの、保存料を含む緩衝化界面活性剤の25倍濃縮溶液。
− 発色試薬A:12.5mLの安定化過酸化水素。
− 発色試薬B:12.5mLの安定化色素原(テトラメチルベンジジン)。
− 停止溶液:6mLの2N硫酸。
− プレート・カバー:4つの粘着性細片。
試薬の調製
【0129】
20mLの洗浄バッファー濃縮物を、脱イオン水又は蒸留水中に希釈して、500mLの洗浄バッファーを調製した。G-CSF標準物質を1mLの脱イオン水又は蒸留水によって再構成した。この再構成で25000pg/mLの原液を生じた。G-CSF標準物質を、希釈液を作製する前に、軽く撹拌しながら最低15分間、静置した。900(900)μLのキャリブレーター希釈剤RD6Aを、2500pg/mL用チューブ内にピペットで計り取った。600μLの前述の希釈剤を、残りの4個のチューブ内にピペットで計り取った。連続希釈:25000pg/mL(1:10)→2500pg/mL及び25000pg/mL(1:3)→833.3pg/mL(1:3)→277.8pg/mL(1:3)→92.6pg/mL(1:3)→30.9pg/mLを作り出すために、原液を使用した。次の移しかえの前に、各チューブを十分に混合した。2500pg/mLの標準物質は、高値の標準物質としての役割を果たした。キャリブレーター希釈剤RD6A(0pg/mL)は、ゼロ標準物質としての役割を果たした。検量線を、6点:2500-833.3-277.8-92.6-30.9-0pg/mL、各点について2つのウェルで実施した。基質溶液について、発色試薬A及びBを、使用前15分以内に等量で一緒に混合した。試薬を遮光した。
アッセイ手順
【0130】
全ての試薬とサンプルを、使用前に室温にした。全ての試薬と標準試薬を調製した。100(100)μLのアッセイ希釈剤RD1Aを各ウェルに加え、100μLの標準物質及び適量のサンプルも同様に各ウェルに加えた。ウェルを、粘着性細片で覆い、そして、室温で2時間インキューベートした。各ウェルを吸引し、そして、400μLの洗浄バッファーで洗浄した。合計3回の洗浄のためにそれを2回繰り返した。最後の洗浄後に、吸引又はデカントすることによって残っている洗浄バッファーを全て取り除いた。プレートを、裏返しにし、そして、きれいなペーパータオルに吸い取らせた。200μLのG-CSF複合体を各ウェルに加えた。ウェルを新しい粘着性細片で覆った。ウェルをに室温で2時間インキューベートした。先に説明した吸引/洗浄を繰り返した。200(200)μLの基質溶液を各ウェルに加えた。ウェルを室温で20分間、インキューベートした。ウェルを遮光した。そして、50μLの停止溶液を各ウェルに加えた。最後に、30分以内にそれぞれのウェルの吸光度を、570nmの補正を伴う450nmに設定したマイクロプレート・リーダー(VersaMax-Molecular Device)を使用して測定した(OD 450nm-OD 570nm)。
薬物動力学的解析
【0131】
非コンパートメント薬物動力学的解析(WINNonLinソフトウェア・バージョン4.1)を適用した。以下の主要なパラメーター:λz、見かけの終末段階に関連した見かけの終末速度定数、血漿中の濃度の対数、対、曲線の単一指数型終末部分の時間に関する線形回帰分析によって概算される;t1/2,z、見かけの終末半減期、以下の方程式:t1/2,z=ln(2)/λzにより計算される;AUC、0時間〜無限の血漿中濃度−時間曲線下面積;AUC/D、用量単位あたりの血漿中濃度−時間曲線下面積;MRT、1次モーメント曲線、AUMC、とAUC下面積の間の比として計算される平均滞留時間;CL、全身クリアランス、CL=D/AUCとして計算される;及びVss、分布の定常状態容量、VSS=CL*MRTとして計算される、を各G-CSF構築物について観察又は計算した。
結果
【0132】
表9に示されるように、マウスにおける薬物動力学的研究は、G-CSF-NNT155が、G-CSF対照(1.53時間)、NEUPOGEN化合物(1.13時間)、又はNEULASTA化合物(3.04時間)のいずれよりも長い半減期(6.2時間)を持つことを示した。G-CSF-NNT155のより長い半減期は、3種類の対照の全てと比べてより持続する効果の継続期間に対応する。9.5mL/時間/kgの全身クリアランスを占める全身クリアランスの対応する実質的な減少がG-CSF-NNT155に関して観察された。本明細書中に開示したアミノ酸伸長を含む他の複合体は、対照と比較して短かくなった半減期を示した。
実施例4
G-CSF-NNT155複合体の生物学的利用能と効能
【0133】
低い好中球絶対数である好中球減少症は、感染に対する闘いを妨げるかもしれない重篤な状態である。好中球数の末梢的な増加を刺激するために、好中球減少症の治療として、又は移植のための細胞採取における他の刺激因子と組み合わせて、顆粒球コロニー刺激因子(G-CSF)を使用できる。部分的に精製したG-CSF-NNT155、PEG-G-CSF(NEULASTA)、及びrh-G-CSF(NEUPOGEN)の生物学的利用能を、i.v.(静脈内)及びs.c.(皮下)投与を通じて比較した。NEUPOGEN化合物は、組み換えメチオニル・ヒトG-CSFである。NEULASTA化合物は、組み換えメチオニル・ヒトG-CSFとモノメトキシポリエチレングリコールの共有結合複合体である。先に記載のとおり、G-CSF-NNT155は、G-CSFのアミノ酸伸長、及び/又はグリコシル化形態である。
4.A. ELISAによる生物学的利用能の試験
【0134】
G-CSF-NNT155の生物学的利用能を評価するために、i.v.及びs.c.投与を通じてG-CSF-NNT155、PEG-G-CSF、及びrh-G-CSFの投与を受けたマウスから得られた血漿サンプルに対してELISA分析を実施した。
【0135】
試験物と対照物は、以下のとおりであった:部分的に精製したG-CSF-NNT115、PEG-G-CSF(NEULASTA)、rh-G-CSF(NEUPOGEN)、及び溶媒対照(150mMの塩化ナトリウム+20mMのNaOAc+0.004%のTween-20);それぞれを、指定された経路i.v.及びs.c投与を通じた動物における使用に許容される様式で製造した。
【0136】
マウスに、i.v.及びs.c.投与を通じて(それぞれ125μg/kgの量で)G-CSF-NNT155、PEG-G-CSF、rh-G-CSF、及び溶媒対照を投与した。マウス血漿サンプルを分離した。所望であれば、マウス血漿サンプルを希釈した。
【0137】
次に、マウス・サンプルをG-CSFのELISAで分析した。ELISAにより血漿サンプルを分析することの主な目的は、定量的な目的だけで、定性的な目的ではなかった(すなわち、過度に正確な血漿レベル濃度を得るためではない)。G-CSFのELISAの結果は、試験物が血漿中に存在したという定性的な肯定か、又は試験物が血漿中に無かったという定性的な否定のいずれかを提供した。血漿サンプルを、2種類の希釈剤を使用して三重反復試験で分析した。
【0138】
静脈内投与したG-CSF-NNT155は72時間まで検出できた一方で、PEG-G-CSFは24時間までしか検出できなかったという結果であった。加えて、皮下投与したG-CSF-NNT155とPEG-G-CSFの両方は、72時間まで検出できた。皮下投与したrh-G-CSFは、15分及び2時間のサンプリング時点において検出できた。
4.B. マウスにおける造血効果に関するG-CSF-NNT155とPEG-G-CSFの単回投与比較
【0139】
G-CSF-NNT155の生物学的利用能及び効能の他の評価において、白血球及び好中球数の計測を伴う血液学分析を、i.v.及びs.c.投与を通じてG-CSF-NNT155、PEG-G-CSF、及びrh-G-CSFの投与を受けたマウスから得られた血漿サンプルに対して実施した。
【0140】
試験物と対照物は、前記G-CSFのELISA評価と同じであった。血漿サンプルを得るための手順もG-CSFのELISA評価と同じであった。血液学分析の結果は、以下のとおりであった。
【0141】
溶媒対照に関する白血球数の平均データは、それぞれ、i.v.及びs.c.投与について、0.76×103/μL〜4.35×103/μL、及び1.27×103/μL〜4.25×103/μLにおよんだ。溶媒対照に関する平均好中球絶対数のデータは、それぞれ、i.v.及びs.c.投与について、0.11×103/μL〜0.55×103/μL、及び0.29×103/μL〜0.35×103/μLにおよんだ。
【0142】
G-CSF-NNT155に関する平均白血球数は、投与15分後の1.44×103/μLからi.v.投与後72時間の11.45×103/μLまで増加した。G-CSF-NNT155に関する平均白血球数は、投与後15分の3.15×103/μLからs.c投与後72時間の11.45×103/μLまで増加した。同じように、G-CSF-NNT155に関する平均好中球絶対数データは、投与後15分の0.04×103/μLからi.v.投与後72時間の4.24×103/μLまで増加した。G-CSF-NNT155に関する平均好中球絶対数は、投与後15分の0.25×103/μLから投与後72時間の2.04×103/μLまで増加し、そして、s.c.投与経路について、投与後120時間の0.14×103/μLまで減少した。
【0143】
PEG-G-CSF(NEULASTA)に関する平均白血球数は、投与後15分の1.36×103/μLから投与後24時間の6.03×103/μLまで増加し、その後、i.v.投与後72時間の3.01×103/μLまで減少した。PEG-G-CSF(NEULASTA)に関する平均白血球数は、投与後15分の1.47×103/μLから投与後24時間の4.58×103/μLまで増加し、その後、s.c.投与後120時間の2.47×103/μLまで減少した。PEG-G-CSF(NEULASTA)に関する平均好中球絶対数は、投与後15分の0.05×103/μLから投与後24時間の1.83×103/μLまで増加し、その後、i.v.投与後72時間の0.20×103/μLまで減少した。PEG-G-CSF(NEULASTA)に関する平均好中球絶対数は、投与後15分の0.18×103/μLから投与後24時間の1.04×103/μLまで増加し、その後、s.c.投与後120時間の0.22×103/μLまで減少した。
【0144】
rh-G-CSF(NEUPOGEN)に関する平均白血球数は、投与後15分の1.30×103/μLからs.c.投与後2時間の1.19×103/μLまで減少した。rh-G-CSF(NEUPOGEN)に関する平均好中球絶対数は、投与後15分の0.11×103/μLからs.c.経路での投与後2時間の0.49×103/μLまで増加した。
【0145】
結果は、G-CSF-NNT155の静脈内投与が好中球絶対数の増加に直接関係する投与後72時間の間中の白血球数の著しい増加を証明したことを示した。この平均増加は、溶媒対照の白血球数を約260%上回った。同じように、G-CSF-NNT155のs.c.投与に関する白血球数の平均増加は、投与後72時間で、溶媒対照及びPEG-G-CSFの両方を約295%上回った。しかしながら、平均白血球数と平均好中球絶対数の双方は、投与後120時間には溶媒対照及びPEG-G-CSFの両方の数値に戻った。
【0146】
いくつかの態様において、ペプチドは、配列NNT、NNNNT、又はNNNNNTを含み、且つ、nは約150〜約160の整数である。これらの実施例の一部において、生物学的に活性なタンパク質はG-CSFである。
【0147】
明細書中で引用した全ての刊行物が、本願発明が関係する当業者の技術レベルを示している。それぞれの個々の刊行物が、援用されたのと同じくらい明確に、且つ、個別に示されるのと同程度に、これらの刊行物の全てを本明細書中に援用する。
【0148】
特定の実施例を参照して本明細書中の本発明を説明してきたが、これらの実施例が本発明の原理と適用をただ例証するにすぎないことを理解すべきである。そのため、多数の修飾が例証のための実施例に対してもたらされること、並びに他の配列が添付の特許請求の範囲によって規定される本発明の本質及び範囲から逸脱することなく考え出される可能性があることを理解すべきである。
【表1】
【表2】
【表3】
【表4】
【表5】
【表6】
【表7】
【表8】
【表9】
【表10】
【表11】
【表12】
【表13】
【表14】
【表15】
【表16】
【表17】
【表18】
【表19】
【表20】
【表21】
【表22】
【表23】
【表24】
【表25】
【表26】
【表27】
【表28】
【表29】
【表30】
【表31】
【表32】
【表33】
【表34】
【表35】
【表36】
【表37】
【表38】
【表39】
【表40】
【表41】
【表42】
【表43】
【表44】
【表45】
【表46】
【表47】
【表48】
【表49】
【表50】
【表51】
【表52】
【表53】
【表54】
【表55】
【表56】
【表57】
【表58】
【表59】
【表60】
【表61】
【表62】
【表63】
【表64】
【表65】
【表66】
【表67】
【表68】
【表69】
【表70】
【表71】
【表72】
【表73】
【表74】
【表75】
【表76】
【表77】
【表78】
【表79】
【表80】
【表81】
【表82】
【表83】
【表84】
【表85】
【表86】
【表87】
【表88】
【表89】
【表90】
【表91】
【表92】
【表93】
【表94】
【表95】
【表96】
【表97】
【表98】
【表99】
【表100】
【技術分野】
【0001】
関連出願の相互参照
当該出願は、合衆国法典第35巻第119(e)条の下、2005年8月30日に出願された米国出願第60/712,585号及び2005年1月25日に出願された米国出願第60/647,119号による優先権を主張し、上記出願の内容を本明細書中に援用する。
【0002】
技術分野
本願発明は、広く、生物学的に活性なタンパク質に関し、より詳しく述べると、生物学的に活性なタンパク質の半減期を変更することに関する。
【背景技術】
【0003】
ヒトの治療法として投与される時、生物学的に活性なタンパク質は、多くの場合、望ましくない半減期を有する。それらの固有の半減期は、多くの場合、最適に満たない治療効能、服薬遵守の問題、及び患者の不便を招く投薬スケジュール及び投与計画を強いる。
【0004】
ヒトの治療法向けの生物学的に活性なタンパク質の製造において、生物学的に活性なタンパク質の半減期の延長は、物理的手段(例えば、投与経路の変更、ナノ粒子カプセル化、及びリポソーム封入)、化学修飾(例えば、エマルジョン、ペグ化、及び高グリコシル化)、並びに遺伝的修飾(例えば、一次タンパク質構造の修飾、高分子タグ、ヒト血清アルブミン融合、翻訳後修飾の取り込み)を通じて試みられた。例えば、Lordら、Clin. Cancer Res. 7:2085-2090ページ(2001年)及びvan Der Auweraら、Am. J. Hematol. 55:245-251ページ(2001年)を参照のこと。しかしながら、そのようなアプローチは他の問題を招いた。物理的手段による生物学的に活性なタンパク質の半減期の延長は、多くの場合、高価で、且つ、時間のかかる製造中の追加の後続加工を伴う、さらなる製剤原料の複雑さを持ち込む。化学修飾は、生物学的に活性なタンパク質の生物活性又は安全性の特性を変えるかもしれない。生物学的に活性なタンパク質が組み換えDNA合成方法論により作製された場合には、特定の細胞発現系におけるタンパク質収量及び純度に対する遺伝的修飾の効果が、その意図される使用に関して評価される必要性が生じる。
【0005】
従って、生物学的に活性なタンパク質の固有の半減期を改善するための他のアプローチの必要性が存在する。
【発明の概要】
【0006】
発明の概要
本発明の1つの側面は、2〜約500反復単位のペプチド・モチーフを含むポリペプチド(アミノ酸伸長)にペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体に向けられる。前記モチーフは、Gly(G)、Asn(N)、及びGln(Q)から選択される1種類のアミノ酸2残基以上である主成分、並びにAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から選択される1種類のアミノ酸1残基以上である副次成分を、上記アミノ酸が主成分と副次成分の両方に存在していないという条件で含み、ここで、上記複合体の血漿中半減期は非複合型の生物学的に活性なポリペプチドの固有の半減期に比べて改善される。
本明細書中に使用される用語「改善された」は、非複合型の生物学的に活性なポリペプチド又はタンパク質自体の血漿中半減期(すなわち、固有の半減期)に比べて延びたか又は短くなった半減期を表す。語句「固有の半減期」は、天然の生物学的に活性なポリペプチドの半減期、又は非複合型のポリペプチド(よって、上記天然ポリペプチドの組み換え型を含む)の半減期を意味する。
【0007】
いくつかの態様において、ペプチド・モチーフは、3〜6個のアミノ酸残基(すなわち、3、4、5、又は6個)を含む。ペプチド・モチーフが5又は6個のアミノ酸残基を含むいくつかの態様において、副次成分は上記ペプチドの中の1個のアミノ酸残基を含む。いくつかの態様において、ペプチド・モチーフは、N及びTアミノ酸残基、N及びEアミノ酸残基、Q及びSアミノ酸残基、又はN及びQアミノ酸残基から成る配列を持つ。いくつかの態様において、アミノ酸伸長は、前記生物学的に活性なポリペプチドに対してN末端側であり;いくつかの実施例において、それは、前記生物学的に活性なポリペプチドに対してC末端側であり;そして、他の態様において、それは、前記生物学的に活性なポリペプチドに対してN及びC末端側の両方に位置する。いくつかの態様において、生物学的に活性なポリペプチドは、サイトカイン(例えば、顆粒球コロニー刺激因子(G-CSF)、ヒト成長ホルモン、又はインターフェロン、例えばβ-インターフェロン若しくはγ-インターフェロンなど)、抗体、抗体断片、タンパク質分解性抗体断片若しくはドメイン、一本鎖抗体、遺伝的若しくは化学的に最適化された抗体若しくはその断片、可溶性のgp120若しくはgp160糖タンパク質、血液凝固因子、可溶性受容体、例えば、腫瘍壊死因子(TNF)-αのII型受容体など、治療用酵素、又はエリスロポエチン(EPO)である。いくつかの態様において、タンパク質複合体は、非複合型の生物学的に活性なポリペプチドの固有の半減期に比べて短い改善された半減期を有する、例えば、ここで、上記生物学的に活性なポリペプチドは組み換え型の活性化プロテインC又は組み換え型の第VII因子を含む。
【0008】
本発明の他の側面は、タンパク質複合体及び担体を含む組成物に向けられる。いくつかの態様において、組成物は医薬組成物であり、そして、担体は医薬として許容される担体である。
【0009】
本発明の他の側面は、先に記載のタンパク質複合体をコードするキメラDNA分子、並びに上記キメラDNA分子を一緒に含むベクター、そして、上記キメラDNA分子又はそれを含むベクターで形質転換された細胞に向けられる。いくつかの態様において、ベクターは、プラスミド、例えば、pCE2である。いくつかの態様において、細胞は、哺乳動物細胞、例えば、チャイニーズハムスター卵巣(CHO)細胞、細菌、例えば、E.コリ(E. coli)、又は酵母である。
【0010】
本発明のさらに他の側面は、主成分としてGly(G)、Asn(N)、及びGln(Q)から選択される1種類のアミノ酸2残基以上を含み、且つ、副次成分としてAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から選択される1種類のアミノ酸1残基以上が、上記アミノ酸が上記主成分と上記副次成分に存在しないという条件で含まれる、2〜約500のペプチド単位から成るポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含む、上記生物学的に活性なタンパク質が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を有するような生物学的に活性なタンパク質複合体の作製方法であって、以下のステップ:上記タンパク質複合体をコードするキメラDNA分子で形質転換した細胞を上記DNAが発現される条件下で培養し、それによって、上記タンパク質複合体を産生し;そして、上記細胞から上記キメラDNA分子の発現産物を抽出する、を含む上記作製方法に向けられる。
【0011】
本発明の更なる側面は、所定のタンパク質複合体が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を示すかどうかを測定する方法であって、以下のステップ:a)Gly(G)、Asn(N)、及びGln(Q)から成る群から選択される1種類のアミノ酸2残基以上を含むか若しくはそれらから成る主成分、並びにAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から成る群から選択される1種類のアミノ酸1残基以上を含むか若しくはそれらから成るが、上記アミノ酸が上記主成分と上記副次成分の両方に存在することのない2〜約500反復単位のペプチド・モチーフを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体を調製し、そしてb)上記タンパク質複合体を試験して、そのタンパク質複合体が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を有するかどうか測定する、を含む上記方法に向けられる。
詳細な説明
【0012】
本明細書中に使用される用語「ポリペプチド」は、別段の指示のない限り、特定の長さを持たないアミノ酸の重合体を意味する。よって、ぺプチド及びタンパク質は、「ポリペプチド」の定義に含まれ、そして、これらの用語は、明細書を通して、並びに特許請求の範囲内で互換性を持って使用される。用語「ポリペプチド」は、翻訳後修飾、例えば、グリコシル基、アセチル基、リン酸基、脂質基の共有結合、プロリン又はリジンのヒドロキシル化などを持つポリペプチドを除外しない。「ポリペプチド」のこの定義によって、その相同体も同様に組み込まれる。
【0013】
用語「精製される」は、本明細書中では、生物学的に活性なタンパク質複合体がその意図される使用に適当なレベルまで精製されることを意味する。
【0014】
本発明は、全般的な側面において、Gly(G)、Asn(N)、及びGln(Q)から選択される1種類のアミノ酸2残基以上である主成分、並びにAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から選択される1種類のアミノ酸1残基以上である副次成分が、上記アミノ酸が上記主成分と上記副次成分の両方に存在しないという条件で含まれる、2〜約500単位のペプチド・モチーフを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体に向けられる。本発明のタンパク質複合体は、相当する非複合型の生物学的に活性なポリペプチド又はタンパク質より長い血漿中半減期を有する。
【0015】
モチーフの反復単位は、通常、3〜7個(3、4、5、6、又は7個)のアミノ酸残基を含む。主成分としてN(Asn)を持つ代表的なペプチド・モチーフが、表1に記載されている。
【0016】
主成分としてG(Gly)を持つ代表的なペプチド・モチーフが、表2に記載されている。
【0017】
主成分としてQ(Glu)を持つ代表的なペプチド・モチーフが、表3に記載されている。
【0018】
ペプチド・モチーフの数は、2〜約500におよぶ。よって、本明細書中に詳しく説明された特定のペプチド・モチーフを含めたモチーフは、ポリペプチド内に以下の単位数:2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192、193、194、195、196、197、198、199、200、201、202、203、204、205、206、207、208、209、210、211、212、213、214、215、216、217、218、219、220、221、222、223、224、225、226、227、228、229、230、231、232、233、234、235、236、237、238、239、240、241、242、243、244、245、246、247、248、249、250、251、252、253、254、255、256、257、258、259、260、261、262、263、264、265、266、267、268、269、270、271、272、273、274、275、276、277、278、279、280、281、282、283、284、285、286、287、288、289、290、291、292、293、294、295、296、297、298、299、300、301、302、303、304、305、306、307、308、309、310、311、312、313、314、315、316、317、318、319、320、321、322、323、324、325、326、327、328、329、330、331、332、333、334、335、336、337、338、339、340、341、342、343、344、345、346、347、348、349、350、351、352、353、354、355、356、357、358、359、360、361、362、363、364、365、366、367、368、369、370、371、372、373、374、375、376、377、378、379、380、381、382、383、384、385、386、387、388、389、390、391、392、393、394、395、396、397、398、399、400、401、402、403、404、405、406、407、408、409、410、411、412、413、414、415、416、417、418、419、420、421、422、423、424、425、426、427、428、429、430、431、432、433、434、435、436、437、438、439、440、441、442、443、444、445、446、447、448、449、450、451、452、453、454、455、456、457、458、459、460、461、462、463、464、465、466、467、468、469、470、471、472、473、474、475、476、477、478、479、480、481、482、483、484、485、486、487、488、489、490、491、492、493、494、495、496、497、498、499、及び500で存在するかもしれない(よって、あらゆるその部分的範囲も同様に含む)。
【0019】
先に記載のとおり、生物学的に活性なポリペプチドに結合したポリペプチドは、生物学的に活性なポリペプチドのアミノ酸又はポリアミノ伸長(以降「アミノ酸伸長」)とも呼ばれるかもしれない。アミノ酸伸長は、生物学的に活性なポリペプチド配列に対して、N末端側に、C末端側に、又はN及びC末端側の両方に位置するかもしれない。
【0020】
理論に縛られるつもりはないが、出願人は、アミノ酸伸長が安定な立体構造をとらないので、それ自体はタンパク質の活性を妨げないか、そうでなければ、影響を及ぼさないと考える。また、アミノ酸伸長を2つの異なるアミノ酸に制限することは、免疫原性の可能性を最小限にするのに役立つアミノ酸伸長の化学的な複雑さを軽減し、並びにたった1つのタイプ又は種類のアミノ酸の使用を通じて可能になるよりもはるかに大規模な生理化学的性質の調節を許容すると考えられる。
【0021】
広く、生物学的に活性なポリペプチドは、天然のポリペプチド(すなわち、それらが生体内に存在する場合)、あるいは、例えば、改善された血漿中半減期がいくつかの観点、特に、脊椎動物体にデリバリーされた時に、その脊椎動物の所定の疾患の症状を治療すること、例えば、治癒させること、改善すること、又は軽減すること、あるいは、末期症状の経過を遅らせることによって脊椎動物の寿命を延ばす意味がある治療的観点から望ましいであろう組み換えヒトG-CSF(rh-G-CSF)などのように組み換えによって生み出されたポリペプチドを含めた、あらゆるタンパク質を含む。生物学的に活性なタンパク質の種類には、サイトカイン、ケモカイン、リンフォカイン、リガンド、受容体、ホルモン、アポトーシス誘導ポリペプチド、酵素、抗体及び抗体断片、並びに成長因子が含まれる。受容体の実例には、TNFのI型受容体、IL-1のII型受容体、IL-1受容体アンタゴニスト、IL-4受容体、及びいずれかの化学的に若しくは遺伝的に修飾された可溶性受容体も同様に含まれる。酵素の実例には、活性化プロテインC、第VII因子、(例えば、SANTYL(登録商標)という名称でAdvance Biofactures Corporationによって販売されている)コラゲナーゼ;(例えば、FABRAZYME(登録商標)という名称でGenzymeによって販売されている)アガルシダーゼ-β;(例えば、PULMOZYME(登録商標)という名称でGenentechによって販売されている)ドルナーゼ-α;(例えば、ACTIVASE(登録商標)という名称でGenentechによって販売されている)アルテプラーゼ;(例えば、ONCASPAR(登録商標)という名称でEnzonによって販売されている)ペグ化アスパラギナーゼ;(例えば、ELSPAR(登録商標)という名称でMerckによって販売されている)アスパラギナーゼ;及び(例えば、CEREDASE(登録商標)という名称でGenzymeによって販売されている)イミグルセラーゼが含まれる。特定のポリペプチド又はタンパク質の実例には、これだけに制限されることなく、顆粒球マクロファージ・コロニー刺激因子(GM-CSF)、顆粒球コロニー刺激因子(G-CSF)、マクロファージ・コロニー刺激因子(M-CSF)、コロニー刺激因子(CSF)、インターフェロンβ(IFN-β)、インターフェロンγ(IFN-γ)、I型インターフェロンγ誘導因子(IGIF)、トランスフォーミング成長因子β(TGF-β)、RANTES(活性化によって調節され、正常T細胞が発現し、そして、分泌したと思われる)、マクロファージ炎症タンパク質(例えば、MIP-1-α及びMIP-1-β)、リーシュマニア伸長開始因子(LEIF)、血小板誘導成長因子(PDGF)、腫瘍壊死因子(TNF)、成長因子、例えば、表皮増殖因子(EGF)、血管内皮成長因子(VEGF)、繊維芽細胞成長因子(FGF)、神経成長因子(NGF)、脳由来神経栄養因子(BDNF)、ニューロトロフィン-2(NT-2)、ニューロトロフィン-3(NT-3)、ニューロトロフィン-4(NT-4)、ニューロトロフィン-5(NT-5)、グリア細胞株由来神経栄養因子(GDNF)、繊毛様神経栄養因子(CNTF)、TNF-αのII型受容体、エリスロポエチン(EPO)、インスリン、並びに可溶性糖タンパク質、例えば、gp120及びgp160糖タンパク質が含まれる。gp120糖タンパク質はヒト免疫不全ウィルス(HIV)エンベロープ・タンパク質であり、そして、gp160糖タンパク質はgp120糖タンパク質の既知の前駆体である。
【0022】
いくつかの態様において、生物学的に活性なポリペプチドの半減期を、固有の半減期に対して短縮されるように改善することが望ましい。そのような態様には、(例えば、XIGRIS(登録商標)という名称でEli Lillyによって販売されている)組み換え型の活性化プロテインC、及び(NOVOSEVEN(登録商標)という名称でNovo Nordiskによって販売されている)組み換え型の第VII因子が含まれる。
【0023】
生物学的に活性なポリペプチドは、疾患、例えば、パーキンソン病、癌、及び心臓病などを治療するのに使用できる。加えて、治療用ポリペプチドは、自己免疫異常、例えば、多発硬化症;シェーグレン症候群;サルコイドーシス;インシュリン依存性糖尿病;自己免疫性甲状腺炎;関節炎(例えば、変形性関節症、関節リウマチ、反応性関節炎、及び乾癬性関節炎);強直性脊椎炎;並びに強皮症などを治療するのに使用できる。また、本発明の治療用ポリペプチドは、急性及び慢性炎症性障害を治療するのに、身長が伸びるのを促進するのに、創傷治癒を促進するのに、及び細胞、組織、又は臓器の移殖後の拒否反応を予防するのに使用できる。
【0024】
いくつかの好ましい態様において、ポリペプチドはG-CSFである。G-CSFは、好中性顆粒球の急速な増殖、そして、血流への放出を引き起こし、その結果、感染との闘いにおいて治療効果を与える。米国特許第6,831,158号で説明されているように、組み換えヒト(rh)-G-CSFは、一般に、様々な形態の白血球減少症(低レベルの白血球)及び好中球減少症(低レベルの好中球)を治療するのに使用される。白血球減少症及び好中球減少症は、様々な感染に対する高い感受性を引き起こす。
【0025】
rh-G-CSFの市販の調製品は、フィルグラスチム(GRAN(登録商標)及びNEUPOGEN(登録商標))、レノグラスチム(NEUTROGIN(登録商標)及びGRANOCYTE(登録商標))、及びナルトグラスチム(NEU-UP(登録商標))という名称で入手可能である。GRAN(登録商標)及びNEUPOGEN(登録商標)は、グリコシル化されておらず、組み換えE.コリ細胞により産生される。NEUTROGIN(登録商標)及びGRANOCYTE(登録商標)は、グリコシル化されており、組み換えCHO細胞により産生される。NEU-UP(登録商標)は、グリコシル化されておらず、組み換えE.コリ細胞で産生された完全なrh-G-CSFのN末端基領域にて置換された5つのアミノ酸を持つ。
【0026】
G-CSF自体に加えて、生物学的に機能的であるか、又は生物活性があるG-CSF類似体も有用である。rh-G-CSFの調製方法は、米国特許第4,810,643号で開示されている。様々なG-CSF類似体も米国特許第4,810,643号で報告されている。rh-G-CSFをコードするポリヌクレオチド、及びrh-G-CSFのアミノ酸構造物は、共に、米国特許第5,985,265号に提供されている。
【0027】
本発明のタンパク質複合体のアミノ酸配列(配列番号2450)(及び対応するポリヌクレオチド配列(配列番号2451))の代表例が、表4に示されている。PHOリーダー配列が含まれている。
【0028】
前記アミノ酸配列において、PHOリーダー配列は第1〜18アミノ酸であり、G-CSFは第19〜192アミノ酸であり、そして、NNT155アミノ酸伸長は第193〜347アミノ酸である。終止コドンは「*」記号によって示される。
【0029】
ポリヌクレオチド配列(配列番号2451)で表される場合、核酸1〜54がPHOリーダー配列をコードし、核酸55〜576がG-CSFをコードし、そして、核酸577〜1041がNNT155アミノ酸伸長をコードする。核酸1042〜1044(タグ)が終止コドンを構成する。
【0030】
いくつかの態様において、反復ペプチド単位の数は75〜225である。よって、配列NNTを持つペプチド・モチーフの反復単位を含むポリペプチドに連結されたG-CSFを含むタンパク質複合体の場合に、本発明の態様は、以下のタンパク質複合体:G-CSF-(NNT)75、G-CSF-(NNT)76、G-CSF-(NNT)77、G-CSF-(NNT)78、G-CSF-(NNT)79、G-CSF-(NNT)80、G-CSF-(NNT)81、G-CSF-(NNT)82、G-CSF-(NNT)83、G-CSF-(NNT)84、G-CSF-(NNT)85、G-CSF-(NNT)86、G-CSF-(NNT)87、G-CSF-(NNT)88、G-CSF-(NNT)89、G-CSF-(NNT)90、G-CSF-(NNT)91、G-CSF-(NNT)92、G-CSF-(NNT)93、G-CSF-(NNT)94、G-CSF-(NNT)95、G-CSF-(NNT)96、G-CSF-(NNT)97、G-CSF-(NNT)98、G-CSF-(NNT)99、G-CSF-(NNT)100、G-CSF-(NNT)101、G-CSF-(NNT)102、G-CSF-(NNT)103、G-CSF-(NNT)104、G-CSF-(NNT)105、G-CSF-(NNT)106、G-CSF-(NNT)107、G-CSF-(NNT)108、G-CSF-(NNT)109、G-CSF-(NNT)110、G-CSF-(NNT)111、G-CSF-(NNT)112、G-CSF-(NNT)113、G-CSF-(NNT)114、G-CSF-(NNT)115、G-CSF-(NNT)116、G-CSF-(NNT)117、G-CSF-(NNT)118、G-CSF-(NNT)119、G-CSF-(NNT)120、G-CSF-(NNT)121、G-CSF-(NNT)122、G-CSF-(NNT)123、G-CSF-(NNT)124、G-CSF-(NNT)125、G-CSF-(NNT)126、G-CSF-(NNT)127、G-CSF-(NNT)128、G-CSF-(NNT)129、G-CSF-(NNT)130、G-CSF(NNT)131、G-CSF-(NNT)132、G-CSF-(NNT)133、G-CSF-(NNT)134、G-CSF-(NNT)135、G-CSF-(NNT)136、G-CSF-(NNT)137、G-CSF-(NNT)138、G-CSF-(NNT)139、G-CSF-(NNT)140、G-CSF-(NNT)141、G-CSF-(NNT)142、G-CSF-(NNT)143、G-CSF-(NNT)144、G-CSF-(NNT)145、G-CSF-(NNT)146、G-CSF-(NNT)147、G-CSF-(NNT)148、G-CSF-(NNT)149、G-CSF-(NNT)150、G-CSF-(NNT)151、G-CSF-(NNT)152、G-CSF-(NNT)153、G-CSF-(NNT)154、G-CSF-(NNT)155、G-CSF-(NNT)156、G-CSF-(NNT)157、G-CSF-(NNT)158、G-CSF-(NNT)159、G-CSF-(NNT)160、G-CSF-(NNT)161、G-CSF-(NNT)162、G-CSF-(NNT)163、G-CSF-(NNT)164、G-CSF(NNT)165、G-CSF-(NNT)166、G-CSF-(NNT)167、G-CSF(NNT)168、G-CSF-(NNT)169、G-CSF-(NNT)170、G-CSF-(NNT)171、G-CSF-(NNT)172、G-CSF-(NNT)173、G-CSF-(NNT)174、G-CSF-(NNT)175、G-CSF-(NNT)176、G-CSF-(NNT)177、G-CSF-(NNT)178、G-CSF-(NNT)179、G-CSF-(NNT)180、G-CSF-(NNT)181、G-CSF-(NNT)182、G-CSF-(NNT)183、G-CSF-(NNT)184、G-CSF-(NNT)185、G-CSF-(NNT)186、G-CSF-(NNT)187、G-CSF-(NNT)188、G-CSF-(NNT)189、G-CSF-(NNT)190、G-CSF-(NNT)191、G-CSF-(NNT)192、G-CSF-(NNT)193、G-CSF-(NNT)194、G-CSF-(NNT)195、G-CSF-(NNT)196、G-CSF-(NNT)197、G-CSF-(NNT)198、G-CSF-(NNT)199、G-CSF-(NNT)200、G-CSF-(NNT)201、G-CSF-(NNT)202、G-CSF-(NNT)203、G-CSF-(NNT)204、G-CSF-(NNT)205、G-CSF-(NNT)206、G-CSF-(NNT)207、G-CSF-(NNT)208、G-CSF-(NNT)209、G-CSF-(NNT)210、G-CSF-(NNT)211、G-CSF-(NNT)212、G-CSF-(NNT)213、G-CSF-(NNT)214、G-CSF-(NNT)215、G-CSF-(NNT)216、G-CSF-(NNT)217、G-CSF-(NNT)218、G-CSF-(NNT)219、G-CSF-(NNT)220、G-CSF-(NNT)221、G-CSF-(NNT)222、G-CSF-(NNT)223、G-CSF-(NNT)224、及びG-CSF-(NNT)225、のいずれかを含むかもしれない。
【0031】
他の好ましい態様において、ポリペプチドはEPOである。Krantz、Blood 77:419ページ(1991年)で説明されているように、天然のEPOは、骨髄内で赤血球前駆細胞の分裂と分化を刺激し、そして、受容体及び赤血球前駆体に結合することによってその生物活性を発揮する。
EPOは、チャイニーズハムスター卵巣(CHO)細胞内に挿入され、そして、発現されたクローン化ヒトEPO(hEPO)遺伝子の産物として組み換え技術を使用して生合成的に製造された。Egrieら、Immunobiol. 72:213-224ページ(1986年)を参照のこと。hEPOの主要な、完全に処理された形態の一次構造(すなわち、アミノ酸配列)は、米国特許第6,583,272号で例証されている。EPOには、Cys7-Cys161及びCys29-Cys33の間に2個のジスルフィド架橋が存在する。糖部分を含まないEPOのポリペプチド鎖の分子量は、18,236DAである。完全なEPO分子において、分子量の約40%は、タンパク質上のグリコシル化部位でタンパク質をグリコシル化する炭水化物基で占められる。Sasakiら、J. Biol. Chem. 262:12059ページ(1987年)を参照のこと。
【0032】
hEPOは赤血球形成に不可欠なので、当該ホルモンは赤血球の産生が低いか若しくは欠陥があることを特徴とする血液疾患の治療に有用である。臨床的に、EPOは、慢性腎不全(CRF)患者の貧血の治療に使用される。Eschbachら、NEJM 316:73-78ページ(1987年);Eschbachら、Ann. Intern. Med. 111:992ページ(1989年);Egrieら、Kidney Intl. 33:262ページ(1988年);及びLimら、Ann. Intern. Med. 110:108-114ページ(1989年)を参照のこと。EPOは、後天性免疫不全症候群(AIDS)及び癌の化学療法中の患者の貧血の治療にも使用された。R. P. Dannaら、Erythropoietin In Clinical Applications - An International Perspective 301-324ページ(M. B. Garnick編、Marcel Dekker 1990年)を参照のこと。
【0033】
EPOのアミノ酸及び相当するヌクレオチド配列、並びに本発明において有用な他の生物学的に活性なポリペプチドを表5に示す。EPOの他の2種類のアミノ酸配列を表6に示す。
【0034】
タンパク質複合体は、1個以上のアフィニティー・タグをさらに含むことができる。一般に、アフィニティー・タグは、当該アフィニティー・タグを含む融合タンパク質の分離、精製、又は検出を容易にするポリペプチド断片である。原則として、抗体又は他の特異的結合作用物質が利用可能であるあらゆるペプチド又はタンパク質がアフィニティー・タグとして使用できる。代表的なアフィニティー・タグには、ポリヒスチジン・トラクト(poly-histidine tract)、プロテインA(Nilssonら、EMBO J. 4:1075ページ(1985年);Nilssonら、Methods Enzymol. 198:3ページ(1991年))、グルタチオンS転移酵素(Smithら、Gene 67:31ページ(1988年))、マルトース結合タンパク質(Kellermanら、Methods Enzymol. 90:459-463ページ(1982年);Guanら、Gene 67:21-30ページ(1987年))、Glu-Glu親和性タグ(Grussenmeyerら、Proc. Natl. Acad. Sci. USA 52:7952-4ページ(1985年);oNDEPHO-1Rを参照のこと)、サブスタンスP、Flag(商標)ペプチド(Hoppら、Biotechnology 6:1204-10(1988))、ストレプトアビジン結合ペプチド、チオレドキシン、ユビキチン、セルロース結合タンパク質、T7ポリメラーゼ、又は他の抗原エピトープ若しくは結合ドメインが含まれる。一般的に、Fordら、Protein Expression and Purification 2:95-107ページ(1991年)を参照のこと。アフィニティー・タグをコードするDNAは、商業的な供給者(例えば、Pharmacia Biotech、Piscataway, New Jersey;New England Biolabs、Beverly, Massachusetts;及びEastman Kodak、New Haven, Connecticut)から入手可能である。
【0035】
アミノ酸伸長の場合のように、アフィニティー・タグは、生物学的に活性なポリペプチド配列に対してN末端側に、C末端側に、又はN末端側とC末端側の両方に位置することができる。
【0036】
本発明は、タンパク質複合体の作製方法にも向けられる。その方法は、タンパク質複合体をコードするキメラDNA分子で形質転換された細胞を、上記DNAが発現される条件下で培養し、それによって、上記タンパク質複合体を産生し;そして、細胞又は培地(又は、細胞培養によるミレアウ(milieu))からキメラDNA分子の発現産物を抽出することを含む。化学的手段によって形成されたタンパク質複合体(例えば、組み換えメチオニル・ヒトG-CSFとモノメチオニル・ポリエチレングリコールの共有結合複合体である、市販品NEULASTA(PEG-G-CSF))と対照的に、本発明における結合は、連続したタンパク質から成る成分として生物学的に活性なタンパク質とポリペプチドの産生をもたらす、物理的又は化学的な手段とは対照的な組み換えにより実施される。アミノ酸伸長にタンパク質を繋ぎ合わせるのに、例えば、約10〜20アミノ酸の長さのリンカーが使用できる。
【0037】
キメラDNA分子は、タンパク質部分をコードする遺伝子又はポリヌクレオチド断片、及び1個以上の遺伝子断片、例えば、ポリペプチド又はアミノ酸伸長を一緒にコードするオリゴヌクレオチドを含む。アミノ酸伸長に含まれるペプチド・モチーフをコードする(そして、本明細書中に具体的に開示したペプチド・モチーフをコードする)オリゴヌクレオチドを表7に示す。(表7で使用される1字表記を表8で説明する。)DNA分子は、アフィニティー・タグ、リンカー、並びに5’及び3’調節エレメントをコードする断片をさらに含むことができる。タンパク質部分をコードする遺伝子又はポリヌクレオチドは、所望のタンパク質又はタンパク質部分のポリペプチドをコードすることが知られているあらゆる遺伝子又はポリヌクレオチドであってもよい。そのような遺伝子及びポリヌクレオチド、そしてそれらを作り出すのに使用されるプライマーは、タンパク質又はポリペプチドに特異的であり、当該技術分野で周知である。ポリペプチド部分をコードする連結されたオリゴヌクレオチドは、先に説明された、及び実施例1に記載される手順に従って産生できる。オリゴヌクレオチドは、どんな長さであってもよいが、好ましくは、転写を抑制することが知られている反復DNA配列の使用を避けるように設計される。例えば、2つのグルタミン酸コドンの組合せ物を含む連結されたオリゴヌクレオチドに、1個だけのグルタミン酸コドンから作られたポリヌクレオチドよりも遺伝子発現を妨げる構造的配置を採用する可能性は低い。本発明のタンパク質複合体をコードするキメラDNA分子は、発現を容易にするために、その5’末端にメチオニン(M)、及び/又はプロリン(P)アミノ酸をコードするコドンを含むように設計されるかもしれない。
【0038】
本発明の複合体は、分子生物学の標準的な組み換え技術によって作製される。いくつかの態様において、生物学的に活性なタンパク質をコードする遺伝子又はポリヌクレオチドは、最初に、構築物、例えば、プラスミド又は他のベクターにクローン化される。次に、ポリペプチド部分の反復単位をコードするオリゴヌクレオチドは、オリゴヌクレオチドがポリペプチド部分をコードするポリヌクレオチドを形成するように一緒に連結される連結又は多量体化スキームを通じて構築物内にクローン化される。このように、オリゴヌクレオチドは、タンパク質部分をコードする遺伝子又はポリヌクレオチドに加えられ、それによって、構築物内にキメラDNA分子を作り出す。選択肢として、より適切な発現ベクターである別の構築物に、キメラDNA分子を移すか、又はクローン化することができる。この時点で、キメラDNA分子を発現することができる宿主細胞がキメラDNA分子で形質転換される。形質転換は、場合によっては、例えば、発現ベクターなどの担体を利用して起こり得る。次に、形質転換された宿主細胞は、タンパク質複合体のコード化をもたらすキメラDNA分子の発現に好適な条件下で培養される。
【0039】
本発明において有効な連結又は多量体化の方法は周知である。Joseph Sambrookら、Molecular Cloning:A Laboratory Manual、第2版、1.53(Cold Spring Harbor Laboratory Press 1989年)を参照のこと。
【0040】
クローニング方法は、当該技術分野で周知の「定方向性クローニング(directional cloning)」を通して行われてもよい。定方向性クローニングは、特定の、且つ、事前に規定された方向での、プラスミド又はベクター内へのポリヌクレオチドの挿入を指す。いったんベクター内にクローン化されると、ポリヌクレオチド配列は、その3’末端側にて、あるいはその5’、及び/又は3’末端側に挿入される他のポリヌクレオチドで伸ばされるかもしれない。そのような設計は、連結の複数の過程を実施する必要なしに大きな重合体を作るのに効率的、且つ、比較的簡単な方法を提供する。第2のポリヌクレオチドの挿入を容易にするために、ベクターは、好ましくは、クローン化ポリヌクレオチドの上流に制限部位を含むが、発現のために必要とされる調節エレメントの下流には含まない。
【0041】
’定方向性クローニングを容易にするために、「アダプター・オリゴヌクレオチド」が、タンパク質複合体をコードするキメラDNA分子の5’及び3’末端に連結されるかもしれない。好ましくは、アダプターは、発現ベクター内に存在するものと両立し得る制限部位を含む。3’アダプター・オリゴヌクレオチドは、それが連結されているコード配列の末端を示すための終止コドンも含むかもしれない。長いポリヌクレオチドが連結後に作り出される可能性を高めるために、ポリペプチド部分をコードするオリゴヌクレオチドが、好ましくは、アダプター・オリゴヌクレオチドより多く加えられる。
【0042】
方法論は、いずれか特定のクローニング・ストラテジーに限定されない。当業者は、本発明のキメラDNA分子を含む構築物を産生するためのあらゆる種類のクローニング・ストラテジーを使用することができる。
【0043】
キメラDNA分子は、当業者に周知である既知の技術により宿主細胞内に挿入されることができる。これらの技術には、これだけに制限されることなく、リン酸カルシウム共沈キメラDNA分子を使用した形質転換、脂質試薬共トランスフェクション(すなわち、Lipofectamine)、エレクトロポレーション、細胞をウイルスと接触させることによる形質導入、又は細胞内へのキメラDNA分子のマイクロインジェクションが含まれる。宿主細胞は、高等真核細胞、例えば、哺乳動物細胞など、又は下等真核細胞、例えば、酵母細胞などであるか、あるいは宿主細胞は、原核細胞、例えば、細菌細胞などであるかもしれない。他のものが好みで利用されるかもしれないが、形質転換のために好適な原核生物宿主には、E.コリ(E. coli)、バチルス・ズブチリス(Bacillus subtillis)、サルモネラ・ティフィムリウム(Salmonella typhimurium)、並びにシュードモナス属(the genera of Pseudomonas)、ストレプトマイセス属(Streptomyces)、及びスタフィロコッカス属(Staphylococcus)の中の多様な種が含まれる。
【0044】
さまざまな宿主/発現ベクターの組合せが、本発明のタンパク質複合体を発現する際に利用される。一般に、組み換え発現ベクターには、複製開始点、及び宿主細胞の形質転換を可能にする選択マーカー(例えば、真核細胞培養のためのジヒドロ葉酸レダクターゼ若しくはネオマイシン耐性、又は、例えば、E.コリにおけるテトラサイクリン若しくはアンピシリン耐性、又はS.セレビシエ(S. cerevisiae)TRP1遺伝子)、及び下流の構造配列の転写を指示する高度に発現される遺伝子から得られるプロモーターが含まれる。前述のプロモーターは、解糖系酵素、例えば、とりわけ、3-ホスホグリセリン酸キナーゼ(PGK)、A因子、酸性ホスファターゼ、又は熱ショック・タンパク質などをコードするオペロンから得ることができる。異種構造配列は、適切な段階で、翻訳開始及び終止配列で構築され、そして、いくつかの態様において、リーダー配列が翻訳されたタンパク質複合体の分泌を指示できる。ベクターは複製開始点領域をさらに含み、ベクターの維持を確保し、そして、所望であれば、宿主内での増幅を提供する。
【0045】
使用できる有用な発現ベクターには、例えば、染色体、非染色体、及び合成DNA配列の断片が含まれる。好適なベクターには、これだけに制限されることなく、SV40及びpcDNAの誘導体、並びに既知の細菌プラスミド、例えば、col EI、pCR1、pBR322、pMal-C2、pET、Smithら、Gene 67:31-40ページ(1988年)によって説明されているpGEX、pMB9、及びその誘導体など、プラスミド、例えば、RP4など、ファージDNA、例えば、ファージIの多数の誘導体など、例えば、NM989など、並びに他のファージDNA、例えば、M13及び糸状単鎖ファージDNAなど;酵母プラスミド、例えば、2ミクロン・プラスミド及び2ミクロン・プラスミドの誘導体など、並びにセントメリック(centomeric)、且つ、集約的な酵母シャトルベクター;真核細胞において有用なベクター、例えば、昆虫又は哺乳動物細胞において有用なベクターなど;プラスミドとファージDNAの組合せから得られたベクター、例えば、ファージDNA又は発現制御配列を利用するように修飾されたプラスミドなど;等が含まれる。必要条件は、ベクターが最適な宿主細胞内で複製可能であり、且つ、存続可能であることである。低又は高コピー数のベクターを要望に応じて使用できる。
【0046】
例えば、バキュロウイルス発現系において、非融合トランスファー・ベクター、例えば、これだけに制限されることなく、pVL941(BamHIクローニング部位、Summersら、Virology 84:390-402ページ(1978年)から入手可能)、pVL1393(BamHI、SmaI、XbaI、EcoRI、NotI、XmaIII、BglII、及びPstIクローニング部位;Invitrogen)、pVL1392(BglII、PstI、NotI、XmaIII、EcoRI、XbaI、SmaI、及びBamHIクローニング部位;Summersら、Virology 54:390-402ページ(1978年)及びInvitrogen)、並びにpBlueBacIII(BamHI、BglII、PstI、NcoI、及びHindIIIクローニング部位、ブルー/ホワイト組み換えスクリーニングによる、Invitrogen)など、そして、融合トランスファー・ベクター、例えば、これだけに制限されることなく、pAc700(BamHI及びKpnIクローニング部位、BamHI認識部位が開始コドンで始まる;Summersら、Virology 54:390-402ページ(1978年))、pAc701及びpAc702(pAc700と同じ、異なるリーディング・フレームを持つ)、pAc360(ポリヘドリン開始コドンの36塩基対下流のBamHIクローニング部位;Invitrogen(1995年))、並びにpBlueBacHisA、B、C(BamHI、BglII、PstI、NcoI、及びHindIIIクローニング部位を有する3つの異なるリーディング・フレーム、ProBond精製及びブルー/ホワイト組み換えスクリーニングのためのN末端ペプチド;Invitrogen(220))の両方を使用できる。
【0047】
哺乳動物発現ベクターには、複製開始点、好適なプロモーター、及びエンハンサーを含み、そして、いずれかの必要なリボソーム結合部位、ポリアデニル化部位、スプライス・ドナー及びアクセプター部位、転写終止配列、並びに5’フランキング非転写配列が含まれる。SV40スプライスに由来するDNA配列、及びポリアデニル化部位は、必要な非転写遺伝要素を提供するのに使用できる。本発明における使用のために想定される哺乳動物発現ベクターには、誘導プロモーター、例えば、ジヒドロ葉酸レダクターゼ・プロモーターなどを有するベクター、DHFR発現カセットを有するあらゆる発現ベクター、又はDHFR/メトトレキサート共増幅ベクター、例えば、pED(PstI、SalI、SbaI、SmaI、及びEcoRIクローニング部位、クローン化遺伝子及びDHFRの両方を発現するベクターを有する;Randal J. Kaufman、1991年、Randal J. Kaufman, Current Protocols in Molecular Biology, 16.12(1991年))などが含まれる。あるいは、グルタミン・シンテターゼ/メチオニン・スルホキシミン共増幅ベクター、例えば、pEE14(HindIII、XbaI、SmaI、SbaI、EcoRI、及びBclIクローニング部位、ベクターがグルタミン・シンテターゼとクローン化遺伝子を発現する;Celltech)などである。エプスタイン・バー・ウイルス(EBV)又は核抗原(EBNA)の制御下でのエピソーム発現を指示するベクターには、例えば、pREP4(BamHI、SfiI、XhoI、NotI、NheI、HindIII、NheI、PvuII、及びKpnIクローニング部位、構成的RSV-LTRプロモーター、ハイグロマイシン選択マーカー;Invitrogen)、pCEP4(BamHI、SfiI、XhoI、NotI、NheI、HindIII、NheI、PvuII、及びKpnIクローニング部位、構成的hCMV前初期遺伝子プロモーター、ハイグロマイシン選択マーカー;Invitrogen)、pMEP4(KpnI、PvuI、NheI、HindIII、NotI、XhoI、SfiI、BamHIクローニング部位、誘導性メタロチオネインIIa遺伝子プロモーター、ハイグロマイシン選択マーカー、Invitrogen)、pREP8(BamHI、XhoI、NotI、HindIII、NheI、及びKpnIクローニング部位、RSV-LTRプロモーター、ヒスチジノール選択マーカー;Invitrogen)、pREP9(KpnI、NheI、HindIII、NotI、XhoI、SfiI、BamHIクローニング部位、RSV-LTRプロモーター、G418選択マーカー;Invitrogen)、並びにpEBVHis(RSV-LTRプロモーター、ハイグロマイシン選択マーカー、N末端ペプチドはProBond樹脂により精製されることができ、そしてエンテロキナーゼによって切断される;Invitrogen)などが使用できる。
【0048】
本発明における使用のために選択可能な哺乳動物発現ベクターには、これだけに制限されることなく、pRc/CMV(HindIII、BstXI、NotI、SbaI、及びApaIクローニング部位、G418選択、Invitrogen)、pRc/RSV(HindII、SpeI、BstXI、NotI、XbaIクローニング部位、G418選択、Invitrogen)等が含まれる。本発明に使用できるワクシニア・ウイルス哺乳動物発現ベクター(例えば、Randall J. Kaufman, Current Protocols in Molecular Biology 16.12(Frederick M. Ausubelら編、Wiley、1991年)を参照のこと)には、これだけに制限されることなく、pSC11(SmaIクローニング部位、TK-及びβ-gal選択)、pMJ601(SalI、SmaI、AflI、NarI、BspMII、BamHI、ApaI、NheI、SacII、KpnI、及びHindIIIクローニング部位;TK-及びβ-gal選択)、pTKgptF1S(EcoRI、PstI、SalII、AccI、HindII、SbaI、BamHI、及びHpaクローニング部位、TK又はXPRT選択)等が含まれる。
【0049】
本発明でも使用できる酵母発現系には、これだけに制限されることなく、非融合pYES2ベクター(XbaI、SphI、ShoI、NotI、GstXI、EcoRI、BstXI、BamHI、SacI、KpnI、及びHindIIIクローニング部位、Invitrogen)、融合pYESHisA、B、C(XbaI、SphI、ShoI、NotI、BstXI、EcoRI、BamHI、SacI、KpnI、及びHindIIIクローニング部位、N末端ペプチドはProBond樹脂で精製され、そして、エンテロキナーゼで切断される;Invitrogen)、pRSベクター等も含むことができる。
【0050】
本発明における使用のための1つの特に好ましいベクターは、プラスミドpCE2である。pCE2プラスミドは、当該技術分野で知られているいずれかの方法によっても得ることができる。実施例2.A.で利用された、そういった方法の1つは、Leungら、Proc. Natl. Acad. Sci. USA 92:4813-4817ページ(1995年)に記載されている。
【0051】
好ましい態様において、キメラDNA分子は、挿入されたキメラDNA分子の転写と翻訳に必要な要素を既に含む発現ベクター内に挿入される可能性がある。
【0052】
加えて、キメラDNA分子を含む発現ベクターは、薬物選択マーカーを含むかもしれない。そのようなマーカーは、クローニングにおいて、及びキメラDNA分子を含むベクターの選別又は検証において助けとなる。例えば、ネオマイシン、ピューロマイシン、ハイグロマイシン、ジヒドロ葉酸レダクターゼ(DHFR)、グアニン・ホスホリボシル転移酵素(GPT)、ゼオシン、及びヒスチジノールに対する耐性を与える遺伝子は有用な選択マーカーである。あるいは、酵素、例えば、単純ヘルペス・ウイルス・チミジンキナーゼ(tk)又はクロラムフェニコール・アセチルトランスフェラーゼ(CAT)などを利用することができる。免疫学的マーカーも利用することができる。遺伝子産物をコードする核酸を同時に発現することができる限り、あらゆる既知の選択マーカーも用いることができる。選択マーカーの更なる実例は、当業者にとって周知であり、そして、レポーター、例えば、高感度緑色蛍光タンパク質(EGFP)、β-ガラクトシダーゼ(β-gal)、又はクロラムフェニコール・アセチルトランスフェラーゼ(CAT)を含む。
【0053】
このため、哺乳動物、主として、ヒト細胞、並びに細菌、酵母、真菌、昆虫、線虫、及び植物細胞が宿主細胞として本発明に使用することができ、そして、本明細書中に規定されるように発現ベクターによって形質転換されることができる。いくつかの細胞宿主、例えば、哺乳動物細胞などにおいて、それが元の環境(例えば、それが自然発生的なものであれば、自然環境)から取り出されるので、キメラDNA分子を含む細胞は「分離」されるかもしれない。他の態様、例えば、植物などにおいて、植物細胞又は部分の培養物よりも植物全体を使用できるので、細胞が分離される必要はない。
【0054】
好適な細胞の実例は、これだけに制限されることなく、VERO細胞、HeLa細胞、例えば、ATCC番号CCL2など、CHO細胞株、例えば、ATCC番号CCL61など、COS細胞、例えば、COS-7細胞及びATCC番号CRL1650の細胞など、W138、BHK、HepG2、3T3、例えば、ATCC番号CRL6361など、A549、PC12、K562細胞、293細胞、Sf9細胞、例えば、ATCC番号CRL1711など、並びにCv1細胞、例えば、ATCC番号CCL70などが含まれる。
【0055】
本発明において使用できる他の好適な細胞には、これだけに制限されることなく、原核生物宿主細胞菌株、例えば、エシェリキア・コリ(Escherichia coli)、(例えば、菌株DH5-α)、バチルス・ズブチリス、サルモネラ・ティフィムリウム、又はシュードオモナス属、ストレプトコッカス属、及びスタフィロコッカス属の菌株などが含まれるかもしれない。本発明に使用できるさらに好適な細胞には、酵母細胞、例えば、サッカロマイセス属(Saccharomyces)のものなど、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)などが含まれる。
【0056】
着目のポリヌクレオチドを含む宿主細胞は、プロモーターを活性化するのに、形質転換体を選択するのに、又は遺伝子を増幅するのに適切なように改善された従来の栄養培地(例えば、Ham’s栄養混合物)中で培養されうる。培養条件、例えば、温度、pH等は、発現のために選択された宿主細胞でこれまで使用されたものであり、当業者には明白である。細胞は、通常、遠心分離によって採取され、物理的又は化学的手段によって破壊され、そして、更なる精製のために保持される粗抽出物となる。タンパク質発現に利用される微生物細胞は、その全てが当業者にとって周知である凍結融解サイクル、超音波処理、機械的破砕、又は細胞溶解剤の使用を含めたいずれか都合のよい方法で破壊されるかもしれない。細胞分解に関係する態様は、キメラDNA分子の発現後に分解を制限するプロテアーゼ・インヒビターを含むバッファーの使用を伴うかもしれない。好適なプロテアーゼ・インヒビターには、ロイペプチン、ペプスタチン、又はアプロチニンが含まれる。上清は、次に、飽和硫酸アンモニウムの濃度を連続的に高めて沈殿させる。
【0057】
タンパク質複合体生成物は、1種類以上の技術によって精製されることができる。通常、精製は、個別の手順、例えば、ゲル濾過、アフィニティー精製、塩分別、イオン交換クロマトグラフィー、サイズ排除クロマトグラフィー、ヒドロキシルアパタイト吸着クロマトグラフィー、疎水性相互作用クロマトグラフィー、及びゲル電気泳動などの組合せを伴う。タンパク質リホールディング・ステップが、タンパク質複合体の立体構造を完成するのに、適宜、使用されてもよい。高速液体クロマトグラフィー(HPLC)は、多くの場合、最終精製ステップに有用である。一般に、Robert K. Scopes, Protein Purification:Principles and Practice(Charles R. Castor編、Springer-Verlag 1994年)及びJoseph Sambrook, Molecular Cloning:A Laboratory Manual、第2版(Cold Spring Harbor Laboratory Press 1989年)を参照のこと。多段階精製分離の実例は、Baronら、Crit. Rev. Biotechnol. 10:179-90ページ(1990年)及びBelowら、J. Chromatogr. A. 679:67-83ページ(1994年)にも記載されている。
【0058】
複合体は、それらが非複合型タンパク質と比べて改善された血漿中半減期を示すかどうか測定するために使用前に試験される。例えば、G-CSFと様々なアミノ酸伸長、例えば、(NNT)などを用いて実施した実験において、出願人は、ある場合において半減期が延び、そして他の場合において短くなることがわかった。試験を、実施例3に示されているように、薬物動力学の標準的な技術に従って実施することができる。この手順は、予定された用量の複合体を動物、好ましくは、実験動物、例えば、齧歯動物など、例えば、マウスに投与し、予定された間隔でその動物から血漿を採取し、そして、濃度が測定できなくなるまで、例えば、酵素結合免疫吸着検定法(「ELISA」)によって血漿を分析して、上記複合体の濃度を測定することを伴う。半減期は、(例えば、WINNonLinソフトウェア・バージョン4.1を使用した)非コンパートメント薬物動力学的解析によって計算できる。複合体の濃度が計測できる最後の時間(tf)に加えて、分析は、以下の主要なパラメーター:λz、見かけの終末段階に関連した見かけの終末速度定数、血漿中の濃度の対数、対、曲線の単一指数型終末部分の時間に関する線形回帰分析によって概算される;t1/2,z、見かけの終末半減期、以下の方程式:t1/2,z=ln(2)/λzにより計算される;AUC、0時間〜無限の血漿中濃度−時間曲線下面積;AUC/D、用量単位あたりの血漿中濃度−時間曲線下面積;MRT、1次モーメント曲線、AUMC、とAUC下面積の間の比として計算される平均滞留時間;CL、全身クリアランス、CL=D/AUCとして計算される;及びVss、分布の定常状態容量、VSS=CL*MRTとして計算される、の観測又は計算を含む。
【0059】
本発明の更なる側面は、タンパク質複合体と担体を含む組成物に関する。広く、担体は、培地又はマトリツクス(例えば、精製マトリツクス)であるかもしれない。いくつかの態様において、担体は、医薬として許容される担体であり、その場合、その組成物はヒト又は動物、最も好ましくは、哺乳動物の障害、及び/又は疾患を予防又は治療するために、あるいは診断目的のために有用である。組成物の有効成分として、タンパク質複合体は、好ましくは、可溶性型の状態である。
【0060】
一般に、組成物は、例えば、治療又は診断に関する期待される効果を発揮する医薬として有効な量のタンパク質複合体を含む。医薬として有効な量は、細胞培養アッセイから推定することができる。例えば、生体外の系において所望の効果を挙げる濃度点若しくは範囲を含むか又は網羅する血中濃度範囲を達成するために、用量が動物モデルにおいて処方されるかもしれない。例えば、Molineuxら、Exp. Hematol. 27:1724-34ページ(1999年)を参照のこと。この情報は、よって、ヒトを含めた他の哺乳動物、及び動物において用量を正確に測定するために使用できる。一般に、投与量は、患者の体重に基づいて約1ng/kg〜約10mg/kgの範囲におよぶ。
【0061】
そのような化合物の有毒性と治療効能は、細胞培養又は実験動物における標準的な医薬の手順によって測定することができる。例えば、Molineuxら、Exp. Hematol. 27:1724-34ページ(1999年)を参照のこと。例えば、LD50(集団の50%に対して致死的な用量)、並びにED50(集団の50%において治療的に有効な用量)は、当該技術分野で知られている方法を使用して測定できる。毒性と治療効果の間の用量比は、高い治療指標を提供する化合物のLD50とED50の間の比として示すことができる治療係数である。
【0062】
細胞培養及び動物の研究から得られるデータは、好ましくは、わずかの若しくは全く毒性のないED50の中に含めた血中濃度の範囲内にある前述の化合物の一連の投与量を処方する際に使用できる。
【0063】
組成物は、いずれかの好適な経路、例えば、局所的に、経口的に、全身的に、静脈内に、筋肉内に、粘膜的に、(例えば、パッチによって)経皮的に投与できる。それらは、リポソーム、微小粒子、マイクロカプセル、ナノ粒子等の中にカプセル化することができる。生物学的に活性なポリペプチドを処方し、そして、投与する技術は、Remington:The Science and Practice of Pharmacy(Alfonso R. Gennaroら編、Philadelphia College of Pharmacy and Science 2000年)の中にも開示されている。
【0064】
本発明とその利点について完全に説明するために、以下の具体的な実施例が与えられるが、当該実施例は単なる一例としてのものであるので、決して制限するものではないことは理解されている。
【図面の簡単な説明】
【0065】
【図1】PHO×pBSKベクターの略図である。
【図2】PHO-G-CSF- NN×pBSKベクターの略図である。
【図3】PHO-G-CSF-NNT65×pBSKベクターの略図である。
【図4】PHO-G-CSF-NNT155×pBSKベクターの略図である。
【図5】PHO-G-CSF-NNT155×pCE2ベクターの略図である。
【図6】いくつかの異なるG-CSF-ポリペプチド複合体と非複合型G-CSFのウエスタンブロットである。
【図7】電気泳動前にPNGase Fで処理したいくつかの異なるG-CSF-ポリペプチド複合体と非複合型G-CSFのウエスタンブロットである。
【図8】PROCHO4-CDM培地で培養したCHOK1細胞において発現された(二重反復試験の)G-CSF-NNT155とG-CSF-NNT65構築物のウエスタンブロットである。
【実施例】
【0066】
実施例1
キメラDNA分子のクローニング
下流タンパク質精製の簡略化のために、G-CSF重合体タンパク質を細胞培地中に分泌されることにした。様々なサイトカイン−重合体構築物での細菌(ST2)分泌シグナルを使用したこれまでの経験では、原核生物及び真核細胞系の両方において低い分泌効能しか示さなかった。しかしながら、異種タンパク質(GFP及びHPV 16 E7)の培地中への分泌のためのpho1+酸ホスファターゼ(PHO)のシゾサッカロマイセス・ポンベ(Schizosaccharomycen pombe)分泌シグナル配列の使用が知られていた。このため、この分泌シグナルを、CHO細胞発現系において以下に記載のG-CSF重合体構築物を用いて試験した。また、以下に記載のものは、G-CSF重合体構築物の製造と、G-CSF重合体の発現のために合成したベクターである。
1.A. PHO×pBSK構築物の製造
PHO×pBSK構築物
【0067】
合成された最初の構築物は、単に、細菌クローニング・ベクターpBSK内にクローン化したPHOリーダー配列であった。PHO分泌シグナルのアミノ酸配列を以下に挙げた。
【0068】
【化1】
【0069】
PHO分泌シグナルを、2組の相補DNAオリゴヌクレオチドを一緒に融合し、そして、それらをpBSK内にクローン化することによって合成した。オリゴヌクレオチドの設計に通じる最も重要な考慮すべき事柄は、G-CSF重合体のN末端へのリーダー配列の融合があらゆる介在配列なしに直接的であるということであった。これは、分泌シグナル全体が加工中に分子から切り取られ、天然及び臨床的に入手可能なバージョンのG-CSFと比べた時に、N末端修飾を持たないG-CSFの分泌型を生じるのを確実にする。このような様式で構築物を調製することによって、G-CSFとPEG-G-CSFを直接比較することができるだけではなく、これは組み換えG-CSFタンパク質に追加アミノ酸を挿入することによってあらゆる潜在的な免疫原性をも制限した。PHOリーダーのpBSK内へのクローニング、並びにその後のPHO×pBSK内へのG-CSFのクローニングのための制限部位をオリゴヌクレオチド内に組み入れることによって、これらの必要条件は満たされた。使用したオリゴヌクレオチドを、以下に挙げる。
【0070】
【化2】
【0071】
PHO×pBSKベクターの図を図1に示す。PHOリーダー(PhoA)を、NotIとBamHIを使用してpBSK内にクローン化した。成熟G-CSFの配列を、BspMIとBamHI部位を利用してこの構築物内にクローン化した。このベクターは、細菌内で増えると、アンピシリン耐性(bla)を与える。
構築方法
【0072】
オリゴヌクレオチド(oNDEPHO-1FとoNDEPHO-1R)及び(oNDEPHO-2FとoNDEPHO-2R)の相補対を、T4ポリヌクレオチド・キナーゼ(PNK)を使用してリン酸化した。T4 PNKを熱不活化し、そして、オリゴヌクレオチド対を氷上でゆっくりとアニーリングさせた。反応物を、TE中に希釈し、そして、NotIとBamHIを使用して前もって消化したpBSKとの連結に使用した。連結産物を、Top10コンピテント細胞内にエレクトポレーション処理し、そして、LB-アンピシリン・プレートで培養した。ミニプレップを、アンピシリン抵抗性コロニーにて実施して、DNAを分離し、そして、特徴的な消化で推定上のクローンを識別した。配列分析で推定上のクローンが正しいと判定した。
1.B. PHO-G-CSF-NN×pBSK構築物の製造
PHO-G-CSF-NN×pBSK構築物
【0073】
この構築物は、BspMIとBamHIを使用したPHO×pBSKベクター内への、成熟G-CSFのコード配列に相当するPCR増幅物のクローニングから生じた。先に述べたとおり、この目的で使用されるオリゴヌクレオチドを、制限酵素BspMIを利用することによってPHOとG-CSFの間の接合点に介在配列を全く持たないことを確実にするように設計した。さらに、G-CSFのC末端は、2つのアスパラギン残基から成る直接的な融合(NN)、及び酵素BbsIとBamHIのための制限部位を持っていた。G-CSFのC末端へのNNT重合体の直接的な付加のためにBbsIを続いて利用すると同時に、ベクター内へのGCSF-NNのクローン化のためにBamHIを使用した。
【0074】
G-CSFを増幅するために合成したオリゴヌクレオチドは、以下のとおりである:
【0075】
【化3】
【0076】
PHO-G-CSF-NN×pBSKベクターの図を、図2に示す。成熟G-CSF-NNの配列を、BspMI(破壊される)とBamHIを使用してPHO×pBSK内にクローン化した。NNT重合体のその後の付加は、プラスミド地図上に示したBbsIとBamHI部位を利用した。このベクターは、細菌で増えると、アンピシリン耐性(bla)を与える。
構築方法
【0077】
G-CSFを、oBspMIGCSFとoGCSFBbsBamオリゴヌクレオチドを使用したPCR法によって増幅した。成熟G-CSFに相当する約520bpのバンドを、アガロース・ゲルから切り出し、そして、精製した。精製した断片を、BspMIとBamHIで消化し、精製し、そして、同じ酵素で切断したPRO×pBSK内に連結した。連結産物を、Top10コンピテント細胞内にエレクトポレーション処理し、LB-アンピシリン・プレート上で培養した。ミニプレップを、アンピシリン抵抗性コロニーに対して実施して、DNAを分離し、そして、特徴的な消化で推定上のクローンを識別した。配列分析で推定上のクローンが正しいと判定した。
1.C. PHO-G-CSF-NNT65×pBSK構築物の製造
PHO-G-CSF-NNT65×pBSK構築物
【0078】
この重合体のアミノ酸組成物は、コンセンサス哺乳動物N連結型グリコシル化部位、N-X-(S/T)をコードする。これにより、この構築物がCHO細胞内で発現される時、重合体はその重合体伸長のスレオニン残基でグリコシル化されることができる。期待されることは、翻訳産物サイズにおける重合増加と翻訳後修飾がG-CSFのpKパラメーターを調節し、その生物活性を低下させることなくタンパク質の延長された血清中半減期を与えるであろうことであった。
【0079】
この重合体の構築を、オリゴヌクレオチド連結/多量体化スキームを使用することで達成した。PHO-G-CSF-NN×pBSK構築物をBbsIで切断することによって、4塩基のアンダーハング(GTTG)をG-CSFのC末端に付加された2個のアスパラギン残基において作り出した。反復NNTトリプレットをコードし、並びにGTTGアンダーハングにアニールする相補的なオリゴヌクレオチドの組合せを設計することによって、より長い鎖に9つのアミノ酸をコードするオリゴヌクレオチドが多量体を形成することが可能であった。終止コドンを含む短いアダプター、重合体の将来的な伸長のためのBbsI部位、及びBamHI部位を、低い割合で付加して、多量体化を終了させ、そして、BbsI-BamHI重合体断片をPHO-G-CSF-NN×pBSK内にクローニングした。
【0080】
NNT65重合体を合成するのに使用したオリゴヌクレオチドを、以下に挙げる:
重合体骨格オリゴヌクレオチド
【0081】
【化4】
【0082】
アダプター・オリゴヌクレオチド
【0083】
【化5】
【0084】
以下の配列(NTNNTNNTN)を、重合体の多量体を形成するのに使用した、それぞれ、CAACオーバハングとGTTGアンダーハングを示すo3NNTFとo3NNTRの重合体骨格オリゴヌクレオチドを使用して作り出したNNT重合体の反復単位であった。
【0085】
【化6】
【0086】
以下に示すものは、重合体を完成させ、そして、終止コドン、重合体の将来的な伸長のためのBbsI部位、及びクローニングに必要なBamHI部位を含む終結アダプター分子である。
終結アダプター分子
【0087】
【化7】
【0088】
PHO-G-CSF-NNT65×pBSKベクターの図を、図3に示す。NNT65重合体の付加には、G-CSFのC末端に位置するBbsI部位、及びBamHI(破壊される)を利用した。図3に示されているように、将来的な長さの伸長のためにNNT65重合体の端にBbsI部位を作り直した。このベクターは、細菌内で増えると、アンピシリン耐性(bla)を与える。
構築方法
【0089】
重合体骨格オリゴヌクレオチド(o3NNTFとo3NNTR)及びアダプター・オリゴヌクレオチド(oDent3FとoDent3R)の相補的な対を、T4 PNKを使用してリン酸化した。T4 PNKを熱不活化し、そして、オリゴヌクレオチド対を氷上でゆっくりとアニーるさせた。重合体とアダプターの二体鎖をT4リガーゼと20:1及び40:1の比で混合することによって、重合体の多量体化を実施した。T4リガーゼを、熱不活性化し、そして、連結反応物全体をBamHIで一晩、消化した。両反応物を沈殿させ、そして、アクリルアミドゲル上に泳動した。250bp〜800bpの間の成分を切り出し、そして、ゲルを精製した。
【0090】
この成分を、BbsIとBamHIで消化したPHO-G-CSF-NN×pBSKとの連結に使用した。化学的コンピテントStbl2細胞を、連結産物で形質転換し、そして、LB-アンピシリン・プレート上で培養した。ミニプレップを実施して、DNAを分離し、そして、特徴的な消化で推定上のクローンを識別した。配列分析で推定上のクローンが正しいと判定した。
【0091】
このストラテジーから分離された中で最も長いクローンは、PHO-G-CSF-NNT65×pBSKであった。
1.D. PHO-G-CSF-NNT155×pBSK構築物の製造
PHO-G-CSF-NNT155×pBSK構築物
【0092】
このクローンの構築には、同じ多量体化スキームを使用したPHO-G-CSF-NNT65×pBSK構築物への追加のNNT残基の付加を必要とした。元々の重合体と伸長の間の接合点を識別するために、この伸長に使用されるオリゴヌクレオチドのヌクレオチド組成を変更した。これらの変更は、重合体の元々のNNT組成を維持し、且つ、同じGTTGアンダーハングとCAACオーバハング・ストラテジーを利用した。BbsIとBamHIでPHO-G-CSF-NNT65を消化することによって、同じオリゴヌクレオチド多量体化ストラテジーを使用して重合体の長さを伸長することが可能であった。
【0093】
NNT65重合体をNNT155に伸長するのに使用したオリゴヌクレオチドを、以下に挙げる:
重合体骨格オリゴヌクレオチド
【0094】
【化8】
【0095】
アダプター・オリゴヌクレオチド
【0096】
【化9】
【0097】
PHO-G-CSF-NNT65×pBSK構築物の製造と同様に、NTNNTNNTNが、o3NNTextFとo3NNTextRの重合体骨格オリゴヌクレオチドを使用して生み出したNNT重合体伸長の反復単位であり、CAACオーバハングとGTTGアンダーハングを重合体の多量体を形成するのにさらに使用し、そして、アミノ酸組成を元々のNNT65重合体から変えることはなかった。
【0098】
【化10】
【0099】
上記のo3NNTextFとo3NNTextRの中の下線を引いたヌクレオチドが、それぞれ、o3NNTFとo3NNTRの同じ位置のヌクレオチドと異なっていた。
【0100】
以下のものは、NNT155重合体を完成させ、且つ、クローニングに必要なBamHI部位を含む終結アダプター分子である。oDent3FとoDent3Rは、最初のNNT65重合体に使用したオリゴヌクレオチドと同じものである。
終結アダプター分子
【0101】
【化11】
【0102】
PHO-G-CSF-NNT155×pBSKベクターの図を、図4に示す。90AAによるWNT65重合体の伸長は、PHO-G-CSF-NNT65×pBSKのC末端に位置するBbsI部位と、BamHI(破壊される)を利用した。将来的な長さの伸長のためにNNT155重合体の端にBbsI部位を作り直したことに注目した。このベクターは、細菌内で増えると、アンピシリン耐性(bla)を与える。
構築方法
【0103】
重合体骨格オリゴヌクレオチドo3NNTextFとo3NNTextR、及びアダプター・オリゴヌクレオチドoDent3FとoDent3Rの相補的な対を、T4 PNKを使用してリン酸化した。T4 PNKを熱不活化し、そして、オリゴヌクレオチド対を氷上でゆっくりとアニーるさせた。重合体とアダプターの二体鎖をT4リガーゼと20:1及び40:1の比で混合することによって、重合体のオリゴマー化を実施した。T4リガーゼを、熱不活性化し、そして、連結反応物全体をBamHIで一晩、消化した。両反応物を沈殿させ、そして、アクリルアミドゲル上に泳動した。250bp〜800bpの間の成分を切り出し、そして、ゲルを精製した。この成分を、BbsIとBamHIで消化したPHO-G-CSF-NNT65×pBSK内に連結した。化学的コンピテントStbl2細胞を、連結産物で形質転換し、そして、LB-アンピシリン・プレート上で培養した。ミニプレップを実施して、DNAを分離し、そして、特徴的な消化で推定上のクローンを識別した。配列分析で推定上のクローンが正しいと判定した。
【0104】
このストラテジーから分離された最も長いクローンは、PHO-G-CSF-NNT155×pBSKであった。
実施例2
宿主細胞の形質転換とキメラDNA分子の発現
2.A. PHO-G-CSF-NNT155×pCE2構築物の製造
ベクターpCE2の調製
【0105】
プラスミドpCE2は、以下の操作でプラスミドpREP7bから得られた。まず最初に、2000bpのEBNAコード領域を取り除いた。次に、pBR322oriをpKS-oriによって置き換えた。そして、RSVプロモーター領域を、CMVエンハンサー、及び伸長因子-1a(EF-1a)プロモーターとイントロンによって置き換えた。
CMVエンハンサーを、以下のプライマー:
【0106】
【化12】
【0107】
を使用してpCEP4(Invitrogen、San Diego, CA)からポリメラーゼ連鎖反応(PCR)によって作り出した380bpのXbaI-SphI断片から得た。
EF-1aプロモーターとイントロン(Uetsukiら、J. Biol. Chem. 254:5791-5798ページ(1989年))を、以下のプライマー:
【0108】
【化13】
【0109】
を使用してヒト・ゲノムDNAからPCR法によって作り出した1200bpのSphI-Asp718I断片から得た。
【0110】
2つの断片を、pREP7bから得られたXbaI/Asp718I消化ベクター内に連結して、pCE2を作り出した。
PHO-G-CSF-NNT155×pCE2構築物
【0111】
哺乳動物細胞内でPHO-G-CSF-NNT155を発現するために、構築物をベクターpCE2内に移行することが必要であった。このベクターは、最低限のプロモーター、及び前初期CMVエンハンサーが隣接するヒト伸長因子-1-αの第1イントロンを含んだ。ベクターは、哺乳動物選択のためのヒグロマイシンB耐性マーカーをも含んだ。
【0112】
PHO-G-CSF-NNT155×pCE2ベクターの図を、図5に示す。CHO細胞内で発現を駆動するプロモーターは、Pcmv/ef-1aである。CHO細胞内へのエレクトロポレーションのために、SalIを使用してベクターを線状にした。このベクターは、哺乳動物細胞においてハイグロマイシンBに対する耐性を与え、そして、細菌内で増えると、アンピシリン耐性(bla)を与える。
構築方法
【0113】
この構築物は、PHO-G-CSF-NNT155×pBSKから1100bpのNotI-BamHI断片を分離し、そして、ゲル精製することによって製造した。この成分を、NotIとBamHIで消化したpCE2内に連結した。化学的コンピテントStbl2細胞を、連結産物で形質転換し、LB-アンピシリン・プレート上で培養した。ミニプレップを実施して、DNAを分離し、そして、特徴的な消化で推定上のクローンを識別した。大規模なマキシプレップを実施して、CHO細胞の形質転換のためのマイクログラム単位のプラスミドを分離した。
2.B. 形質移入細胞株の製造
【0114】
先に記載した重合体を製造するために、哺乳動物細胞を、発現のための宿主細胞として使用した。今回は、チャイニーズハムスター卵巣(CHO)細胞株を選択した。CHO細胞は、G-CSF及びEPOを含めた組み換えタンパク質治療薬を発現するために製薬業界で広く使用されている。以下で説明するように、CHO細胞におけるG-CSF重合体構築物の発現培養を確立した。
接着培養の確立
【0115】
PHO-G-CSF-NNT155×pCE2ベクターを、酵素SalIを使用して線状にした。消化物を、沈殿させ、そして、50μLのTE中に再懸濁した。このプラスミド1(1)μgを、5×106個のCHO細胞と共にエレクトロポレーションに使用した。接着CHO細胞を、10%のFBSを含むHam’s栄養混合物F-12(F-12 Ham’s培地)で培養した。細胞を、一晩、回復させ、そして、翌日、700μg/mLのハイグロマイシンBを含む培地を加えて、耐性菌の選択を開始した。2〜3週間の間に必要に応じて2〜3日おきに培地を交換した。耐性菌の大量のプールを、分離し、そして、ハイグロマイシンBの存在下でさらに2代継代した。
【0116】
培地中へのPHO-G-CSF-NNT155の分泌についてこれらの細胞を試験するために、細胞を、3〜5日間、低血清培地(0.5%)を含むHam’s培地中で培養した。培地を、分離し、そして、G-CSFに対するポリクローナル抗体を使用して調べるウエスタンブロットに使用した。ブロットを、図6及び7で再現する。ブロット間の相違点は、図7のサンプルがペプチド:N-グリコシダーゼF(PNGase F)処理をさらに受けた点である。図6に示されているように、ブロットは、免疫反応成分が見かけ上の分子量約95kDaの所に移動することを示した。図7に示されているように、N-結合型グリコシル化鎖を取り除くグリコシダーゼのPNGase Fを用いたこれらの同じサンプルの消化は、免疫反応成分を見かけ上の分子量、約49kDaまで低下させた。これは、改善されていないG-CSF-NNT155タンパク質の理論上のサイズにほぼ一致した。天然のG-CSFは、約18kDaの所に電気泳動により移動し;C末端へのNNT重合体の付加は、遺伝子組み換え型の、重合体分子にかなりの質量を加える。発現もかなり安定していると思われ;条件培地中に存在する未精製タンパク質について、それほどのタンパク分解は観察されなかった。さらに、図7に示されているように、PNGase F処理後の構築物のサイズの減少は、組み換えタンパク質構築物が相当量の糖付加を受けることを示唆した。
浮遊培養の確立
【0117】
確立したCHO培養の条件培地中に見られるG-CSF-NNT155の精製を容易にするために、細胞を、化学的に規定した、低タンパク質、無血清培地(PROCHO4-CDM, Cambrex)中で培養した。細胞をこの培地に適合させる過程において、細胞は接着から浮遊増殖に適合した。
【0118】
細胞を、T-185フラスコ内で、F-12 Ham’s+10%のFBS中で密な集密度まで培養し、トリプシン処理し、そして、PROCHO4-CDM培地中に懸濁させた。107個の細胞を、50mLのPROCHO4-CDM培地に加え、そして、T-185フラスコ内でインキューベートした。4〜5代の継代の後に、約90%の細胞が、もはや接着するものではなく、単独細胞の混合物及び凝集した細胞塊として増殖した。条件培地の回収のために、通常、60mLの新しいPROCHO4-CDM培地を、5〜6日間、7〜8mLの高細胞密度培養と一緒にインキューベートした。細胞を遠心分離によって培地から取り除き、そして、培地を、それに続いて、0.2ミクロン(μm)のフイルターを使用して不純物を除いた。培地は、ウェスタンブロットによってG-CSF-NNT155の収量を定量化し(通常、100〜200μg/L)、必要とされるまで保存した。
2.C. PROCHO4-CDM中でのG-CSF-NNT構築物の発現
【0119】
サンプルは、PROCHO4-CDM中で6日間、培養した前述の大量のCHO細胞株からの条件培地であった。ウエスタンブロットを、G-CSFに対するポリクローナル抗体を使用することで調べた。図8に示されているように、両構築物のサイズは、(先に説明した)Ham’s培地中に見られたものと同じであり、かつ、G-CSF-NNT155細胞株で観察された分解が無かった。
2.D. 発現の最適化
【0120】
所望の生成物の発現を最大にするのに必要な培養条件を決定するための試みにおいて、様々な化学的及び栄養添加物、並びに様々な環境パラメーターを試験した。以下のアプローチは、タンパク質蓄積の増大をもたらした。
化学添加物
【0121】
全ての化学添加物がタンパク質産生の増加をもたらすわけではないことが知られているが、刊行された文献は、2.5mMまでのアデノシン又はAMPの添加が細胞周期停止につながり;効果的に、タンパク質蓄積の付随した増大を伴って細胞培養の生存を長引かせることを示した。AMPの効果を、ProCHO4既知組成培地(CDM)中、G-CSF-NNT155産生のCHOを介した産生について試験した。AMPを、1mMの終濃度でProCHO4-CDM中に加えた。対照と比較して細胞生存率の減少は観察されなかった。AMPの添加はタンパク質産生のわずかな増加をもたらした。
環境パラメーター
温度変化
【0122】
CHO培養のための低い培養温度は、著しく高められたレベルのタンパク質産生つながる可能性がある。他のタンパク質産生向上の方法に見られるように、観察された効果は、細胞周期停止中の高められたタンパク質産生に起因するものである。37℃から28℃に組み換えCHO株の培養温度を下げることの効果を調べた。28℃で維持した培養物は37℃のものよりも顕著に長い生存のままであったが、低温は、高い組み換えタンパク質産生をもたらさなかった。
浮遊培養への適応
【0123】
大量のCHO細胞集団をスピナー・フラスコ培養条件に切り替えた。浮遊培養への適応は、ProCHO4-CDM中での培養によって仲介された。適応に続いて、細胞を、T型フラスコ内に広げ、そして、様々な密度でProCHO5 CDMを含むスピナー・フラスコ内に播種した。細胞が静置又は撹拌培養かどうかにかかわらず、タンパク質産生の著しい増加をProCHO5 CDM中での浮遊培養への適応中に観察した。
宿主としての異種細胞株の使用
【0124】
発現の最適化への1つのアプローチは、宿主細胞に新しい細胞株を使用することであるかもしれない。従って、ATCCからのCHO-K1の新しい発送品を調達した。新しい細胞集団を、増殖させ、そして今後の使用のために初期継代にて貯蔵した。この規定の培養物を、着目の組み換えタンパク質を発現する安定したクローン細胞株を作り出すために使用した。簡単に言えば、CHO-K1細胞を着目のベクターと一緒にエレクトポレーション処理し、そして、ハイグロマイシンの安定発現に関して選択した。92個のコロニーを、分離し、そして、着目の組み換えタンパク質の発現について評価した。これらのコロニーの多くが、先に記載したとおり、元々のトランスフェクト集団のものより顕著に高い発現レベルを示した。
結果
【0125】
培養物を1mMのAMPで処理した時、発現の非常にわずかな増強を観察した。低い培養温度は、培養物の寿命を延ばしたが、タンパク質発現の増加は無かった。ProCHO5 CDM中での懸濁培養条件への適応は、組み換えタンパク質の発現を劇的に高めた。また、ATCCによって規定されたCHO-K1細胞株から分離したクローンは、元々の分離集団よりも顕著に高いレベルの所望のタンパク質を発現した。
実施例3
G-CSF-NNT155複合体の薬物動態学的評価
【0126】
G-CSF-NNT155タンパク質複合体の薬物動力学的パラメーターを評価した。他の3つの化合物を正の対照として盛り込んだ。対照は、NEULASTA(PEG-G-CSF)、NEUPOGEN(rh-G-CSF)、及びG-CSF化合物であった。化合物を、マウスに単回静脈内(i.v.)投与し、そして、血液を、投薬の72時間後まで採取することによって試験した。血漿をELISA法によって分析した。
3.A. 分析方法
【0127】
アッセイには定量的なサンドイッチ酵素免疫分析技術を利用した。G-CSFに特異的なモノクローナル抗体を、マイクロプレート上にプレコートした。標準物質とサンプルをウェル内にピペットで計り取った。存在する全てのG-CSFを、固定化抗体によって結合した。全ての非結合物質を洗い流した後に、G-CSFに特異的な酵素結合ポリクローナル抗体をウェルに加えた。全ての非結合抗体−酵素試薬を取り除くための洗浄に続いて、基質溶液をウェルに加え、そして、初期段階で結合したG-CSFの量に比例して発色させた。発色を止め、そして、色の強さを計測した。
試薬
【0128】
以下の試薬と材料を使用した:
− G-CSFマイクロプレート:G-CSFに対するマウス・モノクローナル抗体でコートした96ウェル・ポリスチレン・マイクロプレート(8ウェルから成る12の細片)。
− G-CSF複合体:21mLの、保存料を含む西洋ワサビペルオキシダーゼに結合した、G-CSFに対するポリクローナル抗体。
− G-CSF標準物質:2バイアル(25ng/mL)の、凍結乾燥された、保存料を含む緩衝化タンパク質基材中の組み換えヒトG-CSF。
− アッセイ希釈剤RD1A:11mLの、保存料を含む緩衝化タンパク質基材。
− キャリブレーター希釈剤RD5:21mLの、保存料を含む緩衝化タンパク質基材。細胞培養上清サンプル用。
− キャリブレーター希釈剤RD6A:21mLの、保存料を含む動物血清。血清/血漿サンプル用。
− 洗浄バッファー濃縮物:21mLの、保存料を含む緩衝化界面活性剤の25倍濃縮溶液。
− 発色試薬A:12.5mLの安定化過酸化水素。
− 発色試薬B:12.5mLの安定化色素原(テトラメチルベンジジン)。
− 停止溶液:6mLの2N硫酸。
− プレート・カバー:4つの粘着性細片。
試薬の調製
【0129】
20mLの洗浄バッファー濃縮物を、脱イオン水又は蒸留水中に希釈して、500mLの洗浄バッファーを調製した。G-CSF標準物質を1mLの脱イオン水又は蒸留水によって再構成した。この再構成で25000pg/mLの原液を生じた。G-CSF標準物質を、希釈液を作製する前に、軽く撹拌しながら最低15分間、静置した。900(900)μLのキャリブレーター希釈剤RD6Aを、2500pg/mL用チューブ内にピペットで計り取った。600μLの前述の希釈剤を、残りの4個のチューブ内にピペットで計り取った。連続希釈:25000pg/mL(1:10)→2500pg/mL及び25000pg/mL(1:3)→833.3pg/mL(1:3)→277.8pg/mL(1:3)→92.6pg/mL(1:3)→30.9pg/mLを作り出すために、原液を使用した。次の移しかえの前に、各チューブを十分に混合した。2500pg/mLの標準物質は、高値の標準物質としての役割を果たした。キャリブレーター希釈剤RD6A(0pg/mL)は、ゼロ標準物質としての役割を果たした。検量線を、6点:2500-833.3-277.8-92.6-30.9-0pg/mL、各点について2つのウェルで実施した。基質溶液について、発色試薬A及びBを、使用前15分以内に等量で一緒に混合した。試薬を遮光した。
アッセイ手順
【0130】
全ての試薬とサンプルを、使用前に室温にした。全ての試薬と標準試薬を調製した。100(100)μLのアッセイ希釈剤RD1Aを各ウェルに加え、100μLの標準物質及び適量のサンプルも同様に各ウェルに加えた。ウェルを、粘着性細片で覆い、そして、室温で2時間インキューベートした。各ウェルを吸引し、そして、400μLの洗浄バッファーで洗浄した。合計3回の洗浄のためにそれを2回繰り返した。最後の洗浄後に、吸引又はデカントすることによって残っている洗浄バッファーを全て取り除いた。プレートを、裏返しにし、そして、きれいなペーパータオルに吸い取らせた。200μLのG-CSF複合体を各ウェルに加えた。ウェルを新しい粘着性細片で覆った。ウェルをに室温で2時間インキューベートした。先に説明した吸引/洗浄を繰り返した。200(200)μLの基質溶液を各ウェルに加えた。ウェルを室温で20分間、インキューベートした。ウェルを遮光した。そして、50μLの停止溶液を各ウェルに加えた。最後に、30分以内にそれぞれのウェルの吸光度を、570nmの補正を伴う450nmに設定したマイクロプレート・リーダー(VersaMax-Molecular Device)を使用して測定した(OD 450nm-OD 570nm)。
薬物動力学的解析
【0131】
非コンパートメント薬物動力学的解析(WINNonLinソフトウェア・バージョン4.1)を適用した。以下の主要なパラメーター:λz、見かけの終末段階に関連した見かけの終末速度定数、血漿中の濃度の対数、対、曲線の単一指数型終末部分の時間に関する線形回帰分析によって概算される;t1/2,z、見かけの終末半減期、以下の方程式:t1/2,z=ln(2)/λzにより計算される;AUC、0時間〜無限の血漿中濃度−時間曲線下面積;AUC/D、用量単位あたりの血漿中濃度−時間曲線下面積;MRT、1次モーメント曲線、AUMC、とAUC下面積の間の比として計算される平均滞留時間;CL、全身クリアランス、CL=D/AUCとして計算される;及びVss、分布の定常状態容量、VSS=CL*MRTとして計算される、を各G-CSF構築物について観察又は計算した。
結果
【0132】
表9に示されるように、マウスにおける薬物動力学的研究は、G-CSF-NNT155が、G-CSF対照(1.53時間)、NEUPOGEN化合物(1.13時間)、又はNEULASTA化合物(3.04時間)のいずれよりも長い半減期(6.2時間)を持つことを示した。G-CSF-NNT155のより長い半減期は、3種類の対照の全てと比べてより持続する効果の継続期間に対応する。9.5mL/時間/kgの全身クリアランスを占める全身クリアランスの対応する実質的な減少がG-CSF-NNT155に関して観察された。本明細書中に開示したアミノ酸伸長を含む他の複合体は、対照と比較して短かくなった半減期を示した。
実施例4
G-CSF-NNT155複合体の生物学的利用能と効能
【0133】
低い好中球絶対数である好中球減少症は、感染に対する闘いを妨げるかもしれない重篤な状態である。好中球数の末梢的な増加を刺激するために、好中球減少症の治療として、又は移植のための細胞採取における他の刺激因子と組み合わせて、顆粒球コロニー刺激因子(G-CSF)を使用できる。部分的に精製したG-CSF-NNT155、PEG-G-CSF(NEULASTA)、及びrh-G-CSF(NEUPOGEN)の生物学的利用能を、i.v.(静脈内)及びs.c.(皮下)投与を通じて比較した。NEUPOGEN化合物は、組み換えメチオニル・ヒトG-CSFである。NEULASTA化合物は、組み換えメチオニル・ヒトG-CSFとモノメトキシポリエチレングリコールの共有結合複合体である。先に記載のとおり、G-CSF-NNT155は、G-CSFのアミノ酸伸長、及び/又はグリコシル化形態である。
4.A. ELISAによる生物学的利用能の試験
【0134】
G-CSF-NNT155の生物学的利用能を評価するために、i.v.及びs.c.投与を通じてG-CSF-NNT155、PEG-G-CSF、及びrh-G-CSFの投与を受けたマウスから得られた血漿サンプルに対してELISA分析を実施した。
【0135】
試験物と対照物は、以下のとおりであった:部分的に精製したG-CSF-NNT115、PEG-G-CSF(NEULASTA)、rh-G-CSF(NEUPOGEN)、及び溶媒対照(150mMの塩化ナトリウム+20mMのNaOAc+0.004%のTween-20);それぞれを、指定された経路i.v.及びs.c投与を通じた動物における使用に許容される様式で製造した。
【0136】
マウスに、i.v.及びs.c.投与を通じて(それぞれ125μg/kgの量で)G-CSF-NNT155、PEG-G-CSF、rh-G-CSF、及び溶媒対照を投与した。マウス血漿サンプルを分離した。所望であれば、マウス血漿サンプルを希釈した。
【0137】
次に、マウス・サンプルをG-CSFのELISAで分析した。ELISAにより血漿サンプルを分析することの主な目的は、定量的な目的だけで、定性的な目的ではなかった(すなわち、過度に正確な血漿レベル濃度を得るためではない)。G-CSFのELISAの結果は、試験物が血漿中に存在したという定性的な肯定か、又は試験物が血漿中に無かったという定性的な否定のいずれかを提供した。血漿サンプルを、2種類の希釈剤を使用して三重反復試験で分析した。
【0138】
静脈内投与したG-CSF-NNT155は72時間まで検出できた一方で、PEG-G-CSFは24時間までしか検出できなかったという結果であった。加えて、皮下投与したG-CSF-NNT155とPEG-G-CSFの両方は、72時間まで検出できた。皮下投与したrh-G-CSFは、15分及び2時間のサンプリング時点において検出できた。
4.B. マウスにおける造血効果に関するG-CSF-NNT155とPEG-G-CSFの単回投与比較
【0139】
G-CSF-NNT155の生物学的利用能及び効能の他の評価において、白血球及び好中球数の計測を伴う血液学分析を、i.v.及びs.c.投与を通じてG-CSF-NNT155、PEG-G-CSF、及びrh-G-CSFの投与を受けたマウスから得られた血漿サンプルに対して実施した。
【0140】
試験物と対照物は、前記G-CSFのELISA評価と同じであった。血漿サンプルを得るための手順もG-CSFのELISA評価と同じであった。血液学分析の結果は、以下のとおりであった。
【0141】
溶媒対照に関する白血球数の平均データは、それぞれ、i.v.及びs.c.投与について、0.76×103/μL〜4.35×103/μL、及び1.27×103/μL〜4.25×103/μLにおよんだ。溶媒対照に関する平均好中球絶対数のデータは、それぞれ、i.v.及びs.c.投与について、0.11×103/μL〜0.55×103/μL、及び0.29×103/μL〜0.35×103/μLにおよんだ。
【0142】
G-CSF-NNT155に関する平均白血球数は、投与15分後の1.44×103/μLからi.v.投与後72時間の11.45×103/μLまで増加した。G-CSF-NNT155に関する平均白血球数は、投与後15分の3.15×103/μLからs.c投与後72時間の11.45×103/μLまで増加した。同じように、G-CSF-NNT155に関する平均好中球絶対数データは、投与後15分の0.04×103/μLからi.v.投与後72時間の4.24×103/μLまで増加した。G-CSF-NNT155に関する平均好中球絶対数は、投与後15分の0.25×103/μLから投与後72時間の2.04×103/μLまで増加し、そして、s.c.投与経路について、投与後120時間の0.14×103/μLまで減少した。
【0143】
PEG-G-CSF(NEULASTA)に関する平均白血球数は、投与後15分の1.36×103/μLから投与後24時間の6.03×103/μLまで増加し、その後、i.v.投与後72時間の3.01×103/μLまで減少した。PEG-G-CSF(NEULASTA)に関する平均白血球数は、投与後15分の1.47×103/μLから投与後24時間の4.58×103/μLまで増加し、その後、s.c.投与後120時間の2.47×103/μLまで減少した。PEG-G-CSF(NEULASTA)に関する平均好中球絶対数は、投与後15分の0.05×103/μLから投与後24時間の1.83×103/μLまで増加し、その後、i.v.投与後72時間の0.20×103/μLまで減少した。PEG-G-CSF(NEULASTA)に関する平均好中球絶対数は、投与後15分の0.18×103/μLから投与後24時間の1.04×103/μLまで増加し、その後、s.c.投与後120時間の0.22×103/μLまで減少した。
【0144】
rh-G-CSF(NEUPOGEN)に関する平均白血球数は、投与後15分の1.30×103/μLからs.c.投与後2時間の1.19×103/μLまで減少した。rh-G-CSF(NEUPOGEN)に関する平均好中球絶対数は、投与後15分の0.11×103/μLからs.c.経路での投与後2時間の0.49×103/μLまで増加した。
【0145】
結果は、G-CSF-NNT155の静脈内投与が好中球絶対数の増加に直接関係する投与後72時間の間中の白血球数の著しい増加を証明したことを示した。この平均増加は、溶媒対照の白血球数を約260%上回った。同じように、G-CSF-NNT155のs.c.投与に関する白血球数の平均増加は、投与後72時間で、溶媒対照及びPEG-G-CSFの両方を約295%上回った。しかしながら、平均白血球数と平均好中球絶対数の双方は、投与後120時間には溶媒対照及びPEG-G-CSFの両方の数値に戻った。
【0146】
いくつかの態様において、ペプチドは、配列NNT、NNNNT、又はNNNNNTを含み、且つ、nは約150〜約160の整数である。これらの実施例の一部において、生物学的に活性なタンパク質はG-CSFである。
【0147】
明細書中で引用した全ての刊行物が、本願発明が関係する当業者の技術レベルを示している。それぞれの個々の刊行物が、援用されたのと同じくらい明確に、且つ、個別に示されるのと同程度に、これらの刊行物の全てを本明細書中に援用する。
【0148】
特定の実施例を参照して本明細書中の本発明を説明してきたが、これらの実施例が本発明の原理と適用をただ例証するにすぎないことを理解すべきである。そのため、多数の修飾が例証のための実施例に対してもたらされること、並びに他の配列が添付の特許請求の範囲によって規定される本発明の本質及び範囲から逸脱することなく考え出される可能性があることを理解すべきである。
【表1】
【表2】
【表3】
【表4】
【表5】
【表6】
【表7】
【表8】
【表9】
【表10】
【表11】
【表12】
【表13】
【表14】
【表15】
【表16】
【表17】
【表18】
【表19】
【表20】
【表21】
【表22】
【表23】
【表24】
【表25】
【表26】
【表27】
【表28】
【表29】
【表30】
【表31】
【表32】
【表33】
【表34】
【表35】
【表36】
【表37】
【表38】
【表39】
【表40】
【表41】
【表42】
【表43】
【表44】
【表45】
【表46】
【表47】
【表48】
【表49】
【表50】
【表51】
【表52】
【表53】
【表54】
【表55】
【表56】
【表57】
【表58】
【表59】
【表60】
【表61】
【表62】
【表63】
【表64】
【表65】
【表66】
【表67】
【表68】
【表69】
【表70】
【表71】
【表72】
【表73】
【表74】
【表75】
【表76】
【表77】
【表78】
【表79】
【表80】
【表81】
【表82】
【表83】
【表84】
【表85】
【表86】
【表87】
【表88】
【表89】
【表90】
【表91】
【表92】
【表93】
【表94】
【表95】
【表96】
【表97】
【表98】
【表99】
【表100】
【特許請求の範囲】
【請求項1】
生物学的に活性なタンパク質複合体であって、上記生物学的に活性なタンパク質複合体が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を有するように、主成分としてGly(G)、Asn(N)、及びGln(Q)から選択される1種類のアミノ酸2残基以上、並びに副次成分としてAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から選択される1種類のアミノ酸1残基以上が、上記アミノ酸が主成分及び副次成分の中に存在しないという条件で含まれる、2〜約500単位のペプチドを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体。
【請求項2】
前記ペプチドが、3〜6個のアミノ酸残基を含む、請求項1に記載のタンパク質複合体。
【請求項3】
前記ペプチドが、3個のアミノ酸残基を含む、請求項1に記載のタンパク質複合体。
【請求項4】
前記ペプチドが、4個のアミノ酸残基を含む、請求項1に記載のタンパク質複合体。
【請求項5】
前記ペプチドが、5個のアミノ酸残基を含む、請求項1に記載のタンパク質複合体。
【請求項6】
前記副次成分が、前記ペプチドの中の1個のアミノ酸残基を含む、請求項5に記載のタンパク質複合体。
【請求項7】
前記ペプチドが、6個のアミノ酸残基を含む、請求項1に記載のタンパク質複合体。
【請求項8】
前記副次成分が、前記ペプチドの中の1個のアミノ酸残基を含む、請求項7に記載のタンパク質複合体。
【請求項9】
前記ペプチドが、N及びTアミノ酸残基から成る配列を持つ、請求項1に記載のタンパク質複合体。
【請求項10】
前記ペプチドが、N及びEアミノ酸残基から成る配列を持つ、請求項1に記載のタンパク質複合体。
【請求項11】
前記ペプチドが、Q及びSアミノ酸残基から成る配列を持つ、請求項1に記載のタンパク質複合体。
【請求項12】
前記ペプチドが、N及びQアミノ酸残基から成る配列を持つ、請求項1に記載のタンパク質複合体。
【請求項13】
前記ポリペプチドが、前記生物学的に活性なポリペプチドに対してN末端側にある、請求項1に記載のタンパク質複合体。
【請求項14】
前記ポリペプチドが、前記生物学的に活性なポリペプチドに対してC末端側にある、請求項1に記載のタンパク質複合体。
【請求項15】
前記ポリペプチドが、前記生物学的に活性なポリペプチドに対してN及びC末端側の両方に位置している、請求項1に記載のタンパク質複合体。
【請求項16】
前記生物学的に活性なポリペプチドが、サイトカインである、請求項1に記載のタンパク質複合体。
【請求項17】
前記サイトカインが、顆粒球コロニー刺激因子(G-CSF)である、請求項16に記載のタンパク質複合体。
【請求項18】
前記サイトカインが、ヒト成長ホルモンである、請求項16に記載のタンパク質複合体。
【請求項19】
前記サイトカインが、インターフェロンである、請求項16に記載のタンパク質複合体。
【請求項20】
前記インターフェロンが、β-インターフェロンである、請求項19に記載のタンパク質複合体。
【請求項21】
前記インターフェロンが、γ-インターフェロンである、請求項19に記載のタンパク質複合体。
【請求項22】
前記生物学的に活性なポリペプチドが、抗体、抗体断片、タンパク質分解性抗体断片若しくはドメイン、一本鎖抗体、遺伝的若しくは化学的に最適化した抗体、又はその断片を含む、請求項1に記載のタンパク質複合体。
【請求項23】
前記生物学的に活性なポリペプチドが、可溶性gp120又はgp160糖タンパク質を含む、請求項1に記載のタンパク質複合体。
【請求項24】
前記生物学的に活性なポリペプチドが、血液凝固因子を含む、請求項1に記載のタンパク質複合体。
【請求項25】
前記生物学的に活性なポリペプチドが、可溶性受容体を含む、請求項1に記載のタンパク質複合体。
【請求項26】
前記可溶性受容体が、腫瘍壊死因子(TNF)-αのII型受容体を含む、請求項25に記載のタンパク質複合体。
【請求項27】
前記生物学的に活性なポリペプチドが、エリスロポエチンを含む、請求項1に記載のタンパク質複合体。
【請求項28】
非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して相対的に短縮をされた改善された半減期を有する、請求項1に記載のタンパク質複合体。
【請求項29】
前記生物学的に活性なポリペプチドが、組み換え活性化プロテインCを含む、請求項28に記載のタンパク質複合体。
【請求項30】
前記生物学的に活性なポリペプチドが、組み換え第VII因子を含む、請求項28に記載のタンパク質複合体。
【請求項31】
前記生物学的に活性なポリペプチドが、治療用酵素である、請求項1に記載のタンパク質複合体。
【請求項32】
請求項1に記載のタンパク質複合体と担体を含む組成物。
【請求項33】
前記担体が、医薬として有効な担体である、請求項32に記載の組成物。
【請求項34】
請求項1に記載のタンパク質複合体をコードするキメラDNA分子。
【請求項35】
請求項34に記載のキメラDNA分子を含むベクター。
【請求項36】
プラスミドである、請求項35に記載のベクター。
【請求項37】
前記プラスミドが、pCE2である、請求項36に記載のベクター。
【請求項38】
請求項35に記載のベクターで形質転換した細胞。
【請求項39】
哺乳動物細胞である、請求項38に記載の細胞。
【請求項40】
前記哺乳動物細胞が、チャイニーズハムスター卵巣(CHO)細胞である、請求項39に記載の細胞。
【請求項41】
細菌である、請求項38に記載の細胞。
【請求項42】
前記細菌が、E.コリである、請求項41に記載の細胞。
【請求項43】
酵母である、請求項38に記載の細胞。
【請求項44】
生物学的に活性なタンパク質複合体であり、上記生物学的に活性なタンパク質が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を有するように、主成分としてGly(G)、Asn(N)、及びGln(Q)から選択される1種類のアミノ酸2残基以上、並びに副次成分としてAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から選択される1種類のアミノ酸1残基以上が、上記アミノ酸が主成分及び副次成分の中に存在しないという条件で含まれる、2〜約500単位のペプチドを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体の製造方法であって、以下のステップ:
上記タンパク質複合体をコードするキメラDNA分子で形質転換した細胞を、上記DNAが発現される条件下で培養し、それによって、上記タンパク質複合体を産生し;そして
上記細胞から上記キメラDNA分子の発現産物を抽出する、
を含む前記製造方法。
【請求項45】
前記キメラDNA分子を、その後で前記細胞内に形質転換されるベクター内への前記キメラDNA分子の挿入によって前記細胞内に挿入する、請求項44に記載の方法。
【請求項46】
所定のタンパク質複合体が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を示すかどうかを判定する方法であって、以下のステップ:
a)Gly(G)、Asn(N)、及びGln(Q)から成る群から選択される1種類のアミノ酸2残基以上を含むか、又はそれらから成る主成分、並びにAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から成る群から選択される1種類のアミノ酸1残基以上を含むか、又はそれらから成る副次成分を含むが、上記アミノ酸が上記主成分及び副次成分の両方に存在することのない、2〜約500反復単位のペプチド・モチーフを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体を調製し、そして
b)上記タンパク質複合体を試験して、そのタンパク質複合体が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を有するかどうかを判定する、
を含む前記方法。
【請求項1】
生物学的に活性なタンパク質複合体であって、上記生物学的に活性なタンパク質複合体が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を有するように、主成分としてGly(G)、Asn(N)、及びGln(Q)から選択される1種類のアミノ酸2残基以上、並びに副次成分としてAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から選択される1種類のアミノ酸1残基以上が、上記アミノ酸が主成分及び副次成分の中に存在しないという条件で含まれる、2〜約500単位のペプチドを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体。
【請求項2】
前記ペプチドが、3〜6個のアミノ酸残基を含む、請求項1に記載のタンパク質複合体。
【請求項3】
前記ペプチドが、3個のアミノ酸残基を含む、請求項1に記載のタンパク質複合体。
【請求項4】
前記ペプチドが、4個のアミノ酸残基を含む、請求項1に記載のタンパク質複合体。
【請求項5】
前記ペプチドが、5個のアミノ酸残基を含む、請求項1に記載のタンパク質複合体。
【請求項6】
前記副次成分が、前記ペプチドの中の1個のアミノ酸残基を含む、請求項5に記載のタンパク質複合体。
【請求項7】
前記ペプチドが、6個のアミノ酸残基を含む、請求項1に記載のタンパク質複合体。
【請求項8】
前記副次成分が、前記ペプチドの中の1個のアミノ酸残基を含む、請求項7に記載のタンパク質複合体。
【請求項9】
前記ペプチドが、N及びTアミノ酸残基から成る配列を持つ、請求項1に記載のタンパク質複合体。
【請求項10】
前記ペプチドが、N及びEアミノ酸残基から成る配列を持つ、請求項1に記載のタンパク質複合体。
【請求項11】
前記ペプチドが、Q及びSアミノ酸残基から成る配列を持つ、請求項1に記載のタンパク質複合体。
【請求項12】
前記ペプチドが、N及びQアミノ酸残基から成る配列を持つ、請求項1に記載のタンパク質複合体。
【請求項13】
前記ポリペプチドが、前記生物学的に活性なポリペプチドに対してN末端側にある、請求項1に記載のタンパク質複合体。
【請求項14】
前記ポリペプチドが、前記生物学的に活性なポリペプチドに対してC末端側にある、請求項1に記載のタンパク質複合体。
【請求項15】
前記ポリペプチドが、前記生物学的に活性なポリペプチドに対してN及びC末端側の両方に位置している、請求項1に記載のタンパク質複合体。
【請求項16】
前記生物学的に活性なポリペプチドが、サイトカインである、請求項1に記載のタンパク質複合体。
【請求項17】
前記サイトカインが、顆粒球コロニー刺激因子(G-CSF)である、請求項16に記載のタンパク質複合体。
【請求項18】
前記サイトカインが、ヒト成長ホルモンである、請求項16に記載のタンパク質複合体。
【請求項19】
前記サイトカインが、インターフェロンである、請求項16に記載のタンパク質複合体。
【請求項20】
前記インターフェロンが、β-インターフェロンである、請求項19に記載のタンパク質複合体。
【請求項21】
前記インターフェロンが、γ-インターフェロンである、請求項19に記載のタンパク質複合体。
【請求項22】
前記生物学的に活性なポリペプチドが、抗体、抗体断片、タンパク質分解性抗体断片若しくはドメイン、一本鎖抗体、遺伝的若しくは化学的に最適化した抗体、又はその断片を含む、請求項1に記載のタンパク質複合体。
【請求項23】
前記生物学的に活性なポリペプチドが、可溶性gp120又はgp160糖タンパク質を含む、請求項1に記載のタンパク質複合体。
【請求項24】
前記生物学的に活性なポリペプチドが、血液凝固因子を含む、請求項1に記載のタンパク質複合体。
【請求項25】
前記生物学的に活性なポリペプチドが、可溶性受容体を含む、請求項1に記載のタンパク質複合体。
【請求項26】
前記可溶性受容体が、腫瘍壊死因子(TNF)-αのII型受容体を含む、請求項25に記載のタンパク質複合体。
【請求項27】
前記生物学的に活性なポリペプチドが、エリスロポエチンを含む、請求項1に記載のタンパク質複合体。
【請求項28】
非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して相対的に短縮をされた改善された半減期を有する、請求項1に記載のタンパク質複合体。
【請求項29】
前記生物学的に活性なポリペプチドが、組み換え活性化プロテインCを含む、請求項28に記載のタンパク質複合体。
【請求項30】
前記生物学的に活性なポリペプチドが、組み換え第VII因子を含む、請求項28に記載のタンパク質複合体。
【請求項31】
前記生物学的に活性なポリペプチドが、治療用酵素である、請求項1に記載のタンパク質複合体。
【請求項32】
請求項1に記載のタンパク質複合体と担体を含む組成物。
【請求項33】
前記担体が、医薬として有効な担体である、請求項32に記載の組成物。
【請求項34】
請求項1に記載のタンパク質複合体をコードするキメラDNA分子。
【請求項35】
請求項34に記載のキメラDNA分子を含むベクター。
【請求項36】
プラスミドである、請求項35に記載のベクター。
【請求項37】
前記プラスミドが、pCE2である、請求項36に記載のベクター。
【請求項38】
請求項35に記載のベクターで形質転換した細胞。
【請求項39】
哺乳動物細胞である、請求項38に記載の細胞。
【請求項40】
前記哺乳動物細胞が、チャイニーズハムスター卵巣(CHO)細胞である、請求項39に記載の細胞。
【請求項41】
細菌である、請求項38に記載の細胞。
【請求項42】
前記細菌が、E.コリである、請求項41に記載の細胞。
【請求項43】
酵母である、請求項38に記載の細胞。
【請求項44】
生物学的に活性なタンパク質複合体であり、上記生物学的に活性なタンパク質が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を有するように、主成分としてGly(G)、Asn(N)、及びGln(Q)から選択される1種類のアミノ酸2残基以上、並びに副次成分としてAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から選択される1種類のアミノ酸1残基以上が、上記アミノ酸が主成分及び副次成分の中に存在しないという条件で含まれる、2〜約500単位のペプチドを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体の製造方法であって、以下のステップ:
上記タンパク質複合体をコードするキメラDNA分子で形質転換した細胞を、上記DNAが発現される条件下で培養し、それによって、上記タンパク質複合体を産生し;そして
上記細胞から上記キメラDNA分子の発現産物を抽出する、
を含む前記製造方法。
【請求項45】
前記キメラDNA分子を、その後で前記細胞内に形質転換されるベクター内への前記キメラDNA分子の挿入によって前記細胞内に挿入する、請求項44に記載の方法。
【請求項46】
所定のタンパク質複合体が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を示すかどうかを判定する方法であって、以下のステップ:
a)Gly(G)、Asn(N)、及びGln(Q)から成る群から選択される1種類のアミノ酸2残基以上を含むか、又はそれらから成る主成分、並びにAla(A)、Ser(S)、Thr(T)、Asp(D)、Gln(Q)、Glu(E)、His(H)、及びAsn(N)から成る群から選択される1種類のアミノ酸1残基以上を含むか、又はそれらから成る副次成分を含むが、上記アミノ酸が上記主成分及び副次成分の両方に存在することのない、2〜約500反復単位のペプチド・モチーフを含むポリペプチドにペプチド結合を介して結びついた生物学的に活性なポリペプチドを含むタンパク質複合体を調製し、そして
b)上記タンパク質複合体を試験して、そのタンパク質複合体が非複合型の生物学的に活性なポリペプチドの固有の半減期と比較して改善された血漿中半減期を有するかどうかを判定する、
を含む前記方法。
【図1】

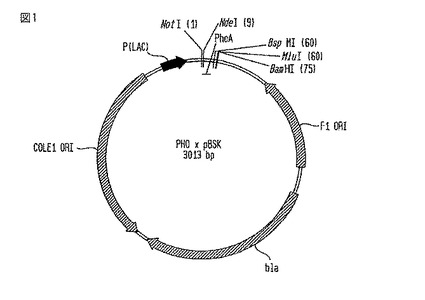
【図2】
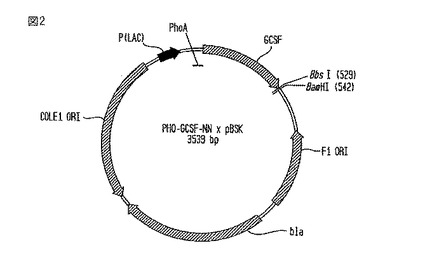
【図3】
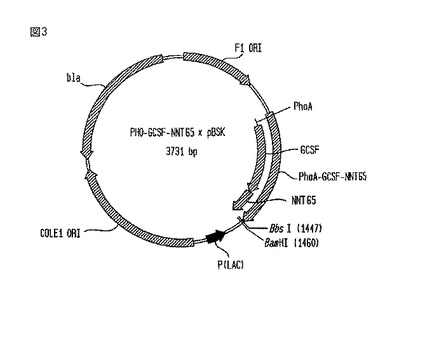
【図4】
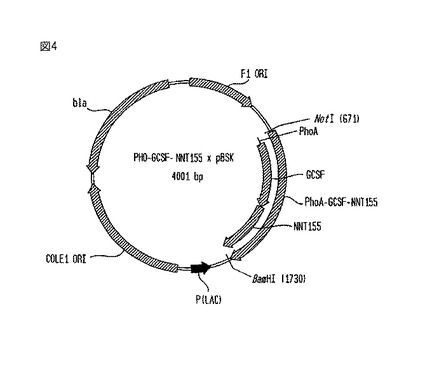
【図5】
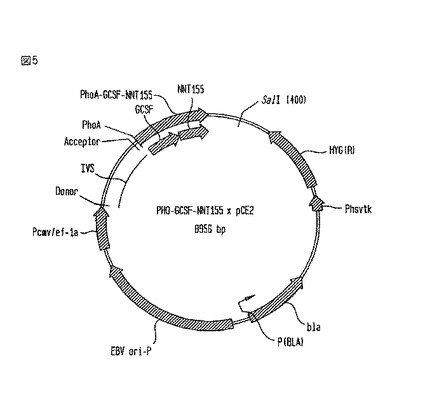
【図6】
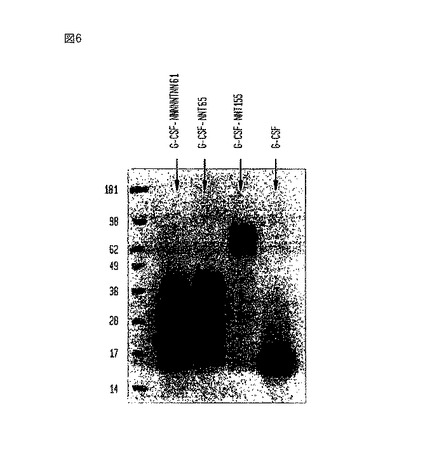
【図7】
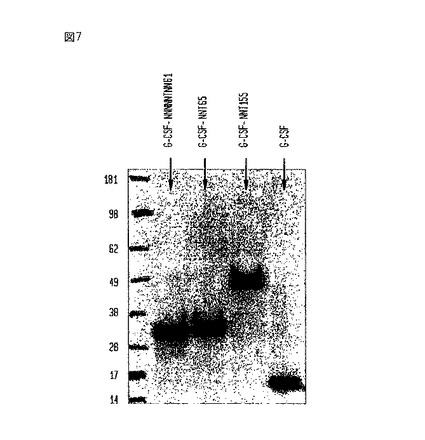
【図8】
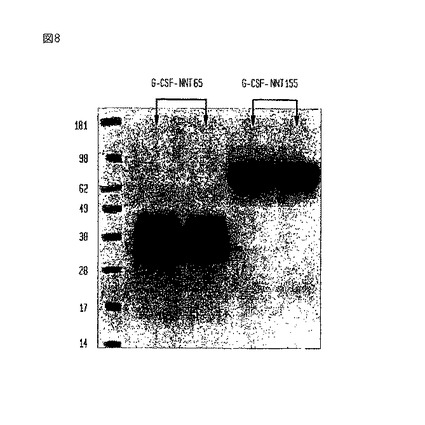
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2013−100292(P2013−100292A)
【公開日】平成25年5月23日(2013.5.23)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−263866(P2012−263866)
【出願日】平成24年11月30日(2012.11.30)
【分割の表示】特願2007−552379(P2007−552379)の分割
【原出願日】平成18年1月25日(2006.1.25)
【出願人】(501394804)セル セラピューティクス インコーポレーテッド (3)
【Fターム(参考)】
【公開日】平成25年5月23日(2013.5.23)
【国際特許分類】
【出願番号】特願2012−263866(P2012−263866)
【出願日】平成24年11月30日(2012.11.30)
【分割の表示】特願2007−552379(P2007−552379)の分割
【原出願日】平成18年1月25日(2006.1.25)
【出願人】(501394804)セル セラピューティクス インコーポレーテッド (3)
【Fターム(参考)】
[ Back to top ]