
改善された相同的組換え効率を有する糸状菌変異体

本発明は、NHR選好を有する糸状菌細胞のゲノムへのポリヌクレオチドの所定の部位に対する標的取込みの効率を増大させるための方法に関し、前記ポリヌクレオチドは、前記所定の部位との相同の領域を有し、取込み経路をHRへ導くステップを含んで成る。本発明は、親細胞に由来する変異体糸状菌にも関し、前記変異体は高い効率のHR経路および/または低い効率のNHR経路および/または同じ条件下の前記親細胞の前記HRおよび/またはNHR効率および/またはNHR/HR比と比べ減少した効率のNHR/HR比を有する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は分子生物学の分野に関する。詳しくは、糸状菌のゲノムへの核酸の方向づけられた取込みの効率を改善する方法、およびその使用に関する。
【背景技術】
【0002】
真核細胞は、ポリペプチドおよび二次代謝産物の(組換え)生産のための好ましい生物である。例えば、タンパク質生産菌株を構成する際には、生産されるタンパク質をコードする目的とする遺伝子の取込みの部位は、目的とする取込み遺伝子の転写および/または発現の調節に重要である。大部分の真核生物におけるDNAのゲノムへの取込みは無作為に高頻度で起こるため、組換えDNA技術によるタンパク質生産菌株の構成はしばしば、生産されるタンパク質をコードする遺伝子を含んで成る発現カセットの望ましくない無作為取込みをもたらす。発現カセットのこのコントロール不良の「無作為多重取込み」は潜在的に危険な過程であり、宿主のゲノムの望ましくな修飾をもたらしうる。したがって、高い効率で発現カセットの正確な標的を確実にすることによってタンパク質生産菌株を構成することが可能であることがきわめて望ましい。さらに、生物の増大する量の完全ゲノムの配列が入手可能になっている現在、これは過剰発現および削除ライブラリーの橋渡しをするゲノムを構成する機会を開く。かかるライブラリーの効率的な構成の重要な要件は、問題の生物が効率的に形質転換されうること、および核酸のゲノムへの直接の標的取込みに必要とされる必要な相同が比較的短いことである。
【0003】
真核細胞は少なくとも2つの別個の経路を有し(1つは相同的組換えを介し、また1つは非相同的組換えを介し)、それらを通じて核酸(特にDNAのコース)が宿主ゲノムへ取込まれうる。酵母サッカロミセス・セルビシエ(Saccharomyces cerevisiae)は、相同的組換え(HR)選好の生物である。この生物の非相同的組換えと相同的組換え(NHR/HR)の比は、約0.07〜0.007のばらつきがありうる。
【0004】
国際公開第02/052026号パンフレットは、DNA配列のそのゲノムへの改善された標的効率を有するサッカロミセス・セルビシエの変異体を開示している。かかる変異体菌株は、NHRに関与する遺伝子(KU70)を欠いている。
【0005】
サッカロミセス・セルビシエとは逆に、哺乳類細胞に至るまでの糸状菌細胞など大部分の真核生物はNHR選好を有する。糸状菌の中では、NHR/HR比は1〜100超の範囲である。かかる生物では、標的取込み頻度はやや低い。この頻度を改善するには、かかる生物のゲノムへ取込まれるポリヌクレオチド配列をフランキングする相同的領域の長さは、例えば単一遺伝子を分裂させるためには少なくとも2000bp、および推定的形質転換体をスクリーニングするためには少なくとも500bpと比較的長いことが必要である。かかるフランキング領域の必要性は、前記ポリヌクレオチドを含んで成るDNA構成物をクローン化する際、かつそれで生物を形質転換する際の大きな負担を表し、さらに、そのフランキング領域内にある隣接遺伝子は形質転換後の組換え過程中に容易に阻害され、それによって望ましくない予想外の副作用を引起こしうる。
【0006】
KU70を欠く哺乳類細胞はすでに単離されている(ピアス(Pierce)ら、Genes and Development、(2001年)、15:3237−3242頁)。これらの変異体は6倍高い相同指向修復頻度を示すが、相同指向標的取込みの効率の増加は示さない。これは、HR選好の生物(サッカロミセス・セルビシエ)において得られる結果が、NHR選好の生物には外挿されえないことを示す。
【0007】
意外にも、本発明者らは、糸状菌におけるHRへ核酸の取込み経路を導くステップが結果として糸状菌のゲノムへの核酸の標的取込みの改善された効率をもたらすことを見出した。
【発明の詳細な説明】
【0008】
本明細書で参照される、すべての特許および刊行物は、かかる特許および刊行物内で開示されたすべての配列および方法を含めて、参照により明示的に援用される。これらの特許および刊行物としては、欧州特許第357127B号明細書、同第635574B号明細書、国際公開第97/06261号パンフレット、同第98/46772号パンフレットが挙げられる。
【0009】
ポリヌクレオチドの糸状菌細胞のゲノムへの標的取込みの効率を増大させるための方法
本発明は、NHR選好を有する糸状菌細胞のゲノムへのポリヌクレオチドの所定の部位に対する標的取込みの効率を増大させるための方法に関し、前記ポリヌクレオチドは、前記所定の部位との相同の領域を有し、取込み経路をHRへ導くステップを含んで成る。本発明は、HR経路の効率を上昇させることによって、かつ/またはNHR経路の効率を低下させる(削減を意味する)ことによって、かつ/またはNHR/HR比を減少させることによって、かかる導くステップに達する。
【0010】
本発明に照らして、HR経路は、ポリヌクレオチドの宿主のゲノムへの標的取込みのコントロールに関与されるすべての遺伝子および構成要素と定義され、前記ポリヌクレオチドは、取込みが標的される宿主のゲノムの特定の所定部位と特定の相同を有する。NHR経路は、宿主のゲノム配列との前記ポリヌクレオチドの相同の程度と関係なく、ポリヌクレオチドの宿主のゲノムへの取込みのコントロールに関与されるすべての遺伝子および構成要素と定義される。
【0011】
好ましい実施形態によれば、導くステップは親糸状菌細胞の変異体を提供するステップを含んで成り、ここでNHR/HR比は、以下のアッセイによって測定された前記親生物における前記比と比べ少なくとも5%の変異体において減少する。より好ましくは、NHR/HR比は、前記親生物における前記比と比べ少なくとも10%、さらにより好ましくは少なくとも50%、かつ最も好ましくは少なくとも100%の変異体において減少する。
【0012】
別の好ましい実施形態によれば、本発明の糸状菌細胞は、以下のアッセイによって測定されうるように、少なくとも200、少なくとも50、少なくとも10であるNHR/HR比を有する。好ましくは、糸状菌細胞の比は少なくとも1、より好ましくは少なくとも0.5、さらにより好ましくは少なくとも0.1、さらにより好ましくは少なくとも0.05、さらにより好ましくは少なくとも0.01、さらにより好ましくは少なくとも0.005、さらにより好ましくは少なくとも0.001、さらにより好ましくは少なくとも0.0005、さらにより好ましくは少なくとも0.0001、かつ最も好ましくは少なくとも0.00001である。
【0013】
より好ましい実施形態によれば、本発明の糸状菌細胞は、以下のアッセイによって測定されるように200未満、さらにより好ましくは50未満、10未満であるNHR/HR比を有する。さらにより好ましくは、糸状菌細胞の比は1未満、さらにより好ましくは0.5未満、さらにより好ましくは0.1未満、さらにより好ましくは0.05未満、さらにより好ましくは0.01未満、さらにより好ましくは0.005未満、さらにより好ましくは0.001未満、さらにより好ましくは0.0005未満、さらにより好ましくは0.0001未満、かつ最も好ましくは0.00001未満である。
【0014】
NHR/HR比は、国際公開第02/052026号パンフレット(表2、23頁)に記載されたアッセイによって測定されることが好ましい。好ましい実施形態によれば、親生物は宿主細胞の部で定義された糸状菌細胞の1つである。別の好ましい実施形態によれば、本発明の糸状菌細胞は宿主細胞の部で定義された種由来である。
【0015】
あるいは、またより好ましくない実施形態によれば、糸状菌におけるNHR/HR比は、かかる経路に関与する以下の成分、すなわちKU70、KU80、MRE11、RAD50、RAD51、RAD52、XRS2、SIR4、LIG4の少なくとも1つの転写プロファイリングおよび/またはノーザンブロット法および/またはウェスタンブロット法など当業者に周知の方法を使用してモニタリングされる。
【0016】
本発明に照らして、「相同の領域」は「少なくとも1つ」の相同の領域を意味する。所定の部位が、本明細書では、宿主細胞によって含まれる遺伝物質内の部位と定義されるが、この同じ部位に対する相同を有するポリヌクレオチドが本発明による方法で取込まれる。
【0017】
好ましい実施形態において、本発明は、NHR選好を有する糸状菌細胞のゲノムへの所定部位に対するポリヌクレオチドの標的取込みの効率を増大させるための方法を提供するが、前記ポリヌクレオチドは、糸状菌を提供することによってHRへの取込み経路を導くステップを含んで成る前記所定の部位と相同の領域を有し、同じ条件下に由来する糸状菌のNHR経路の効率および/またはNHR/HR比と比べ、NHR経路の効率は低下し、かつ/またはNHR/HR比は減少している。好ましい実施形態によれば、親生物は宿主細胞の部で定義されている糸状菌の1つである。
【0018】
NHR経路の効率は、国際公開第02/052026号パンフレット(表2、23頁)に記載されたアッセイで測定されることが好ましい。
【0019】
あるいは、より好ましくない実施形態によれば、糸状菌におけるNHR経路の効率は、かかる経路に関与する成分の転写プロファイリングおよび/またはノーザンブロット法および/またはウェスタンブロット法など当業者に周知の方法を使用してモニタリングされる。より好ましくは、以下の成分、すなわちKU70、KU80、MRE11、RAD50、RAD51、RAD52、XRS2、SIR4、LIG4の少なくとも1つの発現レベルがモニタリングされる。さらにより好ましくは、KU複合体の相同的成分の発現レベルがモニタリングされる。最も好ましくは、相同的KU70および/またはKU80の発現レベルがモニタリングされる。
【0020】
NHR効率の低下は、少なくとも得られた細胞が由来する親細胞におけるよりも低いことを意味する。好ましくは、低下は2倍低く、より好ましくは10倍低く、さらにより好ましくは100倍低く、最も好ましくは1000倍超低く、かつさらに最も好ましくは、ノーザンまたはウェスタンブロット法、アレイ技術または表現型スクリーンを使用して検出可能ではないことを意味する。
【0021】
使用されうる典型的な表現型スクリーンは、以下のステップ、すなわち、所定のゲノム部位の相同的配列によってフランキングされる選択マーカー遺伝子を含んで成る発現カセットで推定的NHR変異体を形質転換するステップを含んで成る。この表現型スクリーンにおいて使用される選択マーカー遺伝子は、糸状菌の形質転換に有用である多くのマーカー遺伝子から選択されうる。一例として、これらのマーカーは、アセトアミダーゼ(amdS)遺伝子など優性および双方向選択マーカー遺伝子(欧州特許第635574B号明細書または国際公開第97/06261号パンフレット)、argB、trpC、またはpyrGなど栄養要求性マーカー遺伝子、および例えば、フレオマイシン(ble遺伝子によってコードされる産物はフレオマイシンに対する抵抗を与える)、ハイグロマイシンB、またはG418に対する抵抗を提供する抗生物質抵抗性遺伝子を含むがこれらに限定されない。好ましい選択マーカー遺伝子が、フレオマイシンに対する抵抗を与えるタンパク質をコードするble遺伝子である。推定的NHR変異体はすでにこの所定のゲノム部位で、amdS遺伝子、硝酸還元酵素遺伝子(niaD)、硫酸パーミアーゼ(Sut B)遺伝子、またはPyrG遺伝子など双方向選択マーカー遺伝子を含有する。niaD遺伝子はすでに他所で記載されている(ゴウカ(Gouka)RJ、ファン・ハルチングスフェルト(van Hartingsveldt)W、ボーベンベルグ(Bovenberg)RA、ファン・デン・ホンデル(van den Hondel)CA、ファン・ゴルコム(van Gorcom)RF、Cloning of the nitrate−nitrite reductase gene cluster of penicillium chrysogenum and use of the niaD gene as a homologous selection marker、J Biotechnol.、1991年9月、20(2):189−99頁)。niaD遺伝子は、細胞が塩素酸塩に耐性を示すようになると、塩素酸塩を含有するプレート上で形質転換体の直接選択を可能にする。sutB遺伝子はすでに他所で記載されている(ファン・デ・カンプ(van de Kamp)M、ピッツィニーニ(Pizzinini)E、フォス(Vos)A、ファン・デア・レンデ(van der Lende)TR、シュールス(Schuurs)TA、ニューベルト(Newbert)RW、ターナー(Turner)G、コーニングス(Konings)WN、ドリーゼン(Driessen)AJ、Sulfate transport in Penicillium chrysogenum:cloning and characterization of the sutA and sutB genes.、J. Bacteriol.1999年12月、181(23):7228−34頁)。好ましい選択マーカー遺伝子が、A.ニドゥランス(nidulans)gpdAプロモータに融合されるA.ニドゥランスamdSコード配列である(欧州特許第635574B号明細書)。他の糸状菌からのamdS遺伝子も使用されうる(国際公開第97/06261号パンフレット)。表現型スクリーンの好ましい形態において、amdS遺伝子は所定のゲノム部位に存在し、ble遺伝子は所定の部位に標的される遺伝子として使用される。非HR改善変異体において、bleカセットは無作為にゲノムに取込み、多くの形質転換体がアセトアミドとフレオマイシンの両方を有する二重選択培地で成長し、かつ比較的少ない形質転換体がフルオロアセトアミド−フレオマイシンプレートで成長することを可能にする。改善されたHRを有する変異体においては、amdSカセットがbleカセットで効率的に交換されると、アセトアミド−プレオマイシン二重選択プレートでの形質転換体の数は限られる。このカセットではより多くの変異体がフルオロアセトアミド−フレオマシン二重選択プレートで現れる。
【0022】
別の好ましい実施形態によれば、低下したNHR効率および/または減少したNHR/HR比を有する糸状菌は、NHRに関与する1つの成分が阻害されている糸状菌である。これに関連して、「1つの(a)」は「少なくとも1つ」を意味する。すなわち、NHRに関与する少なくとも1つの成分が一定の糸状菌で阻害されている。阻害は、NHRに関与する遺伝子の発現レベルをダウンレギュレートし、またはNHRに関与する成分をコードする遺伝子を不活性化することによって、かつ/またはNHRに関与する成分の発現レベルをダウンレギュレートし、かつ/またはNHRに関与する成分の(タンパク質)活性を(一時的に)減少させ、およびこれらの可能性の組合せによって達成されうる。
【0023】
好ましくは、得られた糸状菌は、同じ条件下に由来する親糸状菌細胞における前記遺伝子の発現との比較によってダウンレギュレートされたNHRに関与する遺伝子の発現を有する。好ましい実施形態によれば、親糸状菌は宿主細胞の部で定義される糸状菌の1つである。
【0024】
遺伝子の発現レベル、またはDNA配列は、得られた糸状菌におけるこの特定の遺伝子またはDNA配列の発現レベルが、それが由来する親糸状菌における同じ遺伝子またはDNA配列の発現レベルよりも低い、好ましくは3倍低く、より好ましくは4倍低く、最も好ましくは4倍超低く、さらに最も好ましくは、ノーザンまたはウェスタンブロット法、またはトランスクリプトミクスおよびプロテオミクスのような「オミクス」法を使用して検出可能ではない場合にダウンレギュレートされる。
【0025】
DNA配列の発現レベルのダウン/アップレギュレーションは、例えば、ノーザンブロット法によって細胞に存在する対応するmRNAの量を定量化することによって(Molecular Cloning:A Laboratory Manual、サンブルック(Sambrook)ら、ニューヨーク・コールド・スプリング・ハーバー・プレス(New York Cold Spring Harbour Press)、1989年)、かつ/または、例えば、ウェスタンブロット法によって細胞に存在する対応するタンパク質の量を定量化することによってモニタリングされうる。mRNAの量の差は、DNA配列分析(アイゼン(Eisen),M.B.およびブラウン(Brown),P.O.、DNA arrays for analysis of gene expression.Methods Enzymol.1993年:303:179−205頁)によっても定量化されうる。
【0026】
少なくとも1つの遺伝子またはDNA配列の発現レベルのダウンレギュレーションは、以下の方法の1つまたはその組合せによる遺伝子操作によって得られうる。すなわち、
a.組換え遺伝子操作法を使用するステップ、
b.糸状菌を変異誘発に提示するステップ。
【0027】
あるいは、または上記の方法と組合せて、かつ別の好ましい実施形態によれば、少なくとも1つの遺伝子またはDNA配列の少なくとも1つの発現レベルのダウンレギュレーションは、糸状菌を阻害化合物/組成物に提示することによって得られうる。
【0028】
得られた糸状菌はその後にDNA配列の前記遺伝子の発現レベルをモニタリングすることによって選択されうる。場合により、糸状菌はその後にそのNHR経路および/またはHR経路の効率および/またはそのNHR/HR比を測定することによって選択されうる。本発明に照らして、糸状菌のHR経路の効率は、一定の相同領域を使用することにより糸状菌のゲノムにおける一定のポリヌクレオチド配列の所定の部位への標的取込みの効率によって測定されうる。本発明に照らして、糸状菌のNHR経路の効率は、相同領域と関係なく糸状菌のゲノムにおける一定のポリヌクレオチド配列の非標的取込みの効率によって測定されうる。
【0029】
より好ましくは、少なくとも1つのDNA配列の発現のダウンレギュレーションは、例えばステップa.で定義された組換え遺伝子操作法で行われ、組換え糸状菌を得る。最も好ましくは、ステップa.は、DNA配列を削除するステップを含んで成り、さらに最も好ましくは、削除型DNA配列はその非機能的変異形によって置換され、かつさらに最も好ましくは、削除および置換は、好ましくは欧州特許第357127B号明細書に記載された遺伝子置換によって行われる。
【0030】
選ばれた糸状菌の少なくとも1つのDNA配列の削除または置換の場合、適切なDNA配列が標的遺伝子座で導入されなければならない。標的遺伝子座はこの場合、削除または置換されるNHR経路に関与するDNA配列である。適切なDNA配列は好ましくはクローニングベクター上に存在する。適切なクローニングベクターは、使用される糸状菌宿主細胞の染色体における所定の標的遺伝子座で取込むことが可能であるものである。
【0031】
好ましい統合的クローニングベクターは、この所定の遺伝子座に対するクローニングベクターの取込みを標的するために削除または置換されるDNA配列に対して相同的であるDNA断片を含んで成る。標的取込みを促進するために、クローニングベクターは好ましくは宿主細胞の形質転換前に線状化される。好ましくは、線状化は、クローニングベクターの少なくとも1つ、ただし好ましくはいずれかの末端が削除または置換されるDNA配列に対して相同的な配列によってフランキングされるように実行される。
【0032】
削除または置換されるDNA配列をフランキングする相同的配列の長さは好ましくは2kb未満、さらに好ましくは1kb未満、さらにより好ましくは0.5kb未満、さらにより好ましくは0.2kb未満、さらにより好ましくは0.1kb未満、さらにより好ましくは50bp未満、かつ最も好ましくは30bp未満である。
【0033】
クローニングベクターにおける選択マーカー遺伝子は、糸状菌の形質転換に有用である多くのマーカー遺伝子から選択されうる。一例として、これらのマーカーは、アセトアミダーゼ(amdS)遺伝子など優性および双方向選択マーカー遺伝子(欧州特許第635574B号明細書または国際公開第97/06261号パンフレット)、argB、trpC、またはpyrGなど栄養要求性マーカー遺伝子、および、例えば、フレオマイシン、ハイグロマイシンB、またはG418に対する抵抗を示す抗生物質抵抗性遺伝子を含むがこれらに限定されない。好ましい選択マーカー遺伝子が、A.ニドゥランスgpdAプロモータに融合したA.ニドゥランスamdSコード配列である(欧州特許第635574B号明細書)。他の糸状菌からのamdS遺伝子も使用されうる(国際公開第97/06261号パンフレット)。amdS選択マーカー遺伝子は、明確なDNA配列を置換および/または削除する同じ菌株における数倍で使用されうる利点を有する。
【0034】
欧州特許第635574B号明細書に記載されたフルオロアセトアミド培地の対抗選択を行うことで、結果として生じる菌株はマーカーを含まず、さらなる遺伝子修飾のために使用されうる。
【0035】
一定のDNA配列の発現をダウンレギュレートするための好ましい方法が、野生型DNA配列の削除および/または修飾DNA配列による置換を含んで成るが、その発現産物は機能的ではない。削除および置換は、好ましくは、欧州特許第0357127B1号明細書に記載された遺伝子置換法によって実行される。遺伝子の特定の削除は、好ましくは、欧州特許第635574B号明細書に記載されているように選択マーカー遺伝子としてamdS遺伝子を使用して実行される。
【0036】
あるいは、または他の言及された方法と組合せて、大腸菌(E.coli)におけるコスミドのインビボ組換えに基づく方法が以下に記載されているように使用されうる。すなわち、A rapid method for efficient gene replacement in the filamentous fungus Aspergillus nidulans(2000年)、シャーベロシュ(Chaveroche),MK、ジコ(Ghico),J.M.、およびダンフェルト(d’Enfert)C、Nucleic acids Research、28巻、22号。この方法は、例えばA.ニゲル(niger)のような他の糸状菌に適用可能である。
【0037】
DNA配列の発現のダウンレギュレーションは、アンチセンス核酸を使用することによって、もしくはUVまたは化学的変異誘発を使用することによっても達成されうる(マテム(Mattem),I.E.、ファン・ヌルト(van Noort)J.M.、ファン・デン・ベルグ(van den Berg),P.、アーチャー(Archer),D.B.、ロバーツ(Roberts),I.N.、およびファン・デン・ホンデル,C.A.、Isolation and characterization of mutans of Aspergillus niger deficient in extracellular proteases.Mol Gen Genet.1992年8月、234(2):332−6頁)。
【0038】
好ましくは、NHR経路にもたらされる欠損は誘導性のものである。これは、新しい調節領域によって、好ましくは抑制可能または調節可能なプロモータを使用することによって、より好ましくは、スイッチオン/オフされうるプロモータを使用することによって、すなわちグルコース抑制、もくしはアンモニア抑制、またはpH抑制によって、NHRに関与する成分をコードする遺伝子の内在性調節領域を置換することによって達成されうる。グルコース抑制されたプロモータの例は、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)pcbABプロモータ(マルチン(Martin)JF、カスケイロ(Casqueiro)J、コサルコバ(Kosalkova)K、マルコス(Marcos)AT、グチレツ(Gutierrez)S.、Penicillin and cephalosporin biosynthesis:mechanism of carbon catabolite regulation of penicillin production.アントニー・ファン・リューエンヘック(Antonie Van Leeuwenhoek)、1999年1−2月、75(1−2):21−31頁、総説)、またはアスペルギルス・ニゲル(Aspergillus niger)グルコアミラーゼプロモータである。オン/オフスイッチ可能なプロモータの例は以下の刊行物に記載されている。すなわち、
−An activator/repressor dual system allows tight tetracycline−regulated gene expression in budding yeast:ベリ(Belli)ら、(1998年)Nucl.Acid Research.26巻、4号:942−947頁、
−A light−switchable gene promoter system:シミズ−サトウ(Shimizu Sato)ら、(2002年)Nat.Biotech.20巻、10号、:1041−1044頁。
【0039】
好ましい実施形態によれば、糸状菌は、NHR経路に関与する以下の酵母遺伝子、KU70、KU80、RAD50、MRE11、XRS2、およびSIR4と相同的である、その内在性遺伝子の少なくとも1つを欠いている(ファン・デン・ボシュ(van den Bosch)ら(2002年):DNA double−strand break repair by homologous recombination.Biol.Chern.383巻:873−892頁、およびアレン(Allen)ら、(2003年):Interactive competition between homologous recombination and non−homologous end joining.Mol.Cancer Res.、1巻:913−920頁)。
【0040】
NHRに関与する少なくとも1つの成分を有する全種類の変異体は、NHRの過程においてその機能を実行することがもはや可能ではなく、または少なくとも相当に可能性が低く、本発明によって意図された変異体である。好ましくは、NHRに関与する成分は、得られた変異体におけるNHR経路の効率が前記定義のアッセイで測定されたように同じ条件下に由来する親細胞における活性の90%未満、さらに好ましくは85%未満、より好ましくは80%未満、さらにより好ましくは70%未満、最も好ましくは50%未満であるように阻害されている。
【0041】
好ましい実施形態によれば、親糸状菌は宿主細胞の部で定義される糸状菌の1つである。
【0042】
好ましくは、糸状菌細胞は以下の遺伝子の少なくとも1つを欠いている。すなわち、
−配列番号2または19で識別されるhdfAもしくはその相同体、または
−配列番号5または22で識別されるhdfBもしくはその相同体、またはその両方。
【0043】
別の好ましい実施形態によれば、糸状菌は、誘導と同時に減少するこれら遺伝子hdfAおよびhdfBによってコードされたタンパク質の少なくとも1つの量を有する。
【0044】
別の好ましい実施形態によれば、少なくとも1つの遺伝子またはDNA配列の発現レベルのダウンレギュレーションは、糸状菌を変異誘発にかけることによる遺伝子修飾によって得られうる。糸状菌細胞は、無作為変異誘発、およびその後に選択アッセイにかけられ、改善されたHRを有する変異体を変異体のすべての集団から単離することができる。
【0045】
本発明の好ましい実施形態によれば、宿主細胞の部で定義された糸状菌細胞の1つが変異誘発を実行する出発菌株として使用される。
【0046】
例えば、出発菌株はUV照射にかけられ、生存率が0.001%〜60%の範囲であるようにUV照射にかけられる。好ましくは、生存率は0.01%〜50%の範囲である。分生子が、物理的または化学的手段によって糸状菌を突然変異させる好ましい材料であることは当業者に公知である。しかし、変異体は菌糸細胞から得ることもできる。また、UV以外の他の変異誘発処置が化学的薬剤(例えば、NTG)として適用されうる。本明細書に記載された選択方法は、分生子または菌糸細胞のいずれかから得られる変異体を選択するためにも適用されうる。
【0047】
好ましくは、変異誘発は分生子に適用される。UV照射は、好ましくは、7.5、15、および30分など異なる時間にわたって適用され、細胞における軽度、中等度、および強度の突然変異率レベルを得る。突然変異試料は、直接、再胞子形成され、または胞子形成が誘発された前に(例えば、実施例9に記載)、YNBまたはYEPD(実施例9の定義)など富栄養培地において長い回復期間インキュベートされうる。
【0048】
次いで、胞子形成バッチは遺伝子標的におけるその効率について試験されうる。これは以下の方法によって試験されうる。プロトプラストは、機能的選択マーカーの発現カセットを有する少なくとも1つ、好ましくは2つもしくはそれ以上のDNA断片で形質転換されうる。発現カセットにおける選択マーカー遺伝子は、糸状菌の形質転換に有用である多くのマーカー遺伝子から選択されうる。一例として、これらのマーカーは、アセトアミダーゼ(amdS)遺伝子など優性および双方向選択マーカー遺伝子(欧州特許第635574B号明細書または国際公開第97/06261号パンフレット)、argB、trpC、またはpyrGなど栄養要求性マーカー遺伝子、および例えば、フレオマイシン、ハイグロマイシンB、またはG418に対する抵抗を提供する抗生物質抵抗性遺伝子含むがこれらに限定されない。好ましくは、使用される選択マーカーはbleおよびamdS遺伝子である。使用されるamdSカセットは、A.ニドゥランスgpdAプロモータに融合したA.ニドゥランスコード配列である(欧州特許第635574B号明細書)。他の糸状菌からのamdS遺伝子も使用されうる(国際公開第97/06261号パンフレット)。遺伝子bleは、フレオマイシンに対する抵抗を与えることが可能なタンパク質をコードする。遺伝子amdSは、細胞が唯一の窒素源としてアセトアミドで成長することを可能にするタンパク質をコードする(欧州特許第635574B号明細書に記載)。糸状菌のプロトプラストへのDNAの移動に適用される方法は当技術分野で周知であり、フィンケルシュタイン(Finkelstein)とボール(Ball)(編)、Biotechnology of filamentous fungi,technology and products,バターワース・ハイネマン(Butterworth−Heinemann)(1992年)、ベネット(Bennett)とラシュア(Lasure)(編)More Gene Manipulations in fungi、Academic Press(1991年)、ターナー、ピューラー(Puehler)(編)、Biotechnology、第2完全改訂版、VHC(1992年)を含む多くの文献に記載されている。Ca−PEG介在プロトプラスト形質転換は、欧州特許第635574B明細書に記載されているように使用される。
【0049】
これら2つの発現カセットの糸状菌ゲノムにおける2つの明確な特定の遺伝子座への標的取込みを選択するには、DNAの短い相同的ストレッチが、例えば、DNA断片の両側のPCRを介して添加されうる。いくつかの種類の構成物を作り、改善された標的効率を有する変異体を選択する機会を改善しうる。すなわち、DNAの相同的ストレッチは通常、30bp〜1000bp、好ましくは30b〜700bp、より好ましくは30bp〜500bp、さらにより好ましくは30bp〜300bp、より好ましく好ましくは30bp〜200bp、さらにより好ましくは30bp〜100bp、かつ最も好ましくは30bpでありうる。理論上、糸状菌ゲノムにおけるすべての遺伝子座が発現カセットの標的取込みのために選択されうる。好ましくは、標的が行われる遺伝子座は、この遺伝子座に存在する野生型遺伝子が発現カセットに含まれる遺伝子によって置換されている場合、得られた変異体が一定のアッセイによって検出可能である変化を示すようになっている。好ましくは、遺伝子座はniaD遺伝子座であり、それによって硝酸還元酵素遺伝子を破壊し(ゴウカ RJ、ファン・ハルチングスフェルト W、ボーフェンベルク(Bovenberg)RA、ファン・デン・ホンデル CA、ファン・ゴルコム RF、Cloning of the nitrate−nitrite reductase gene cluster of Penicillium chrysogenum and use of the niaD gene as a homologous selection marker.J Biotechnol.1991年9月、20(2):189−99頁)、細胞が塩素酸塩に対して抵抗性になると、塩素酸塩を含有するプレート上で形質転換体の直接選択を可能にする。別の好ましい遺伝子座はsutB遺伝子座であり、それによって硫酸パーミアーゼ遺伝子を破壊し(ファン・デ・カンプ M、ピッツィニーニ E、フォス A、ファン・デア・レンデ TR、シュールス TA、ニューベルト RW、ターナー G、コーニングス WN、ドリーゼン AJ、Sulfate transport in Penicillium chrysogenum:cloning and characterization of the sutA and sutB genes.、J. Bacteriol.1999年12月、181(23):7228−34頁)、セレン酸塩を含有するプレート上で形質転換体の直接選択を可能にする。存在する両方の選択マーカーを有し、かつ取込み遺伝子座に存在する遺伝子の不活性化に起因する2つの変化を有する変異体は、改善された標的取込みを有する菌株である。
【0050】
別の好ましい実施形態によれば、NHR経路における低下した効率、または減少したNHR/HR比および/またはHR経路の高い効率は、NHRに関与する成分を減少させることによって、より好ましくは部分的もしくは最も好ましくは完全に阻害することによって得られる。
【0051】
NHRに関与する成分の部分的または完全な阻害は異なる方法によって、例えば、かかる成分もしくは化学的阻害剤またはタンパク質阻害剤もしくは物理的阻害剤(ツール(Tour)O.ら(2003年)Nat.Biotech:Genetically targeted chromophore−assisted light inactivation.21巻、12号、1505−1508頁)またはペプチド阻害剤もしくはアンチセンス分子またはRNAi分子(R.S.カマス(Kamath)ら、(2003年)Nature:Systematic functional analysis of the Caenorhaboditis elegans genome using RNAi.421巻、231−237頁)に対する抗体によって得られうる。阻害の種類(部分的またはより好ましくは完全)に関係なく、NHRに関与する成分が上記定義されたNHRの過程におけるその機能を実行することがもはや不可能であり、少なくとも相当に可能性が低いことが重要である。
【0052】
NHRに関与する成分は、酵母KU70、RAD50、MREII、XRS2、LIG4、SIR4、KU80、LIFL、もしくはNEIL、または会合成分の糸状菌相同体を含んで成る。遺伝子の命名は生物間で異なるため、機能的等価物および/またはその機能的断片はすべて、酵母遺伝子KU70、RAD50、MREII、XRS2、LIG4、SIR4、KU80、LIFL、またはNEILの少なくとも1つの機能(機能であって、量ではなく)を実行するのが可能であると本明細書で定義されているが、これらは本発明にも含まれる。NHRに関与する成分を一時的に(部分的より好ましくは完全に)阻害することによって、核酸が所望の位置で取込まれるが、NHRに関与する成分を永続的に修飾することはなく、かつNHRに関与するかかる修飾成分の永続的存在によって引起こされる望ましくない副作用を予防する。
【0053】
上記の方法に加えて、または代替として、NHRに関与するタンパク質の活性を阻害することによって低下したNHR効率を得ること、または代わりのシグナル配列(ラモン・ド・ルーカス(Ramon de Lucas),J.、マルチネス(Martinez)O、ペレス(Perez)P.、イザベル・ロペス(Isabel Lopez),M.、バレンシアーノ(Valenciano),S.、およびラボルダ(Laborda),F.、The Aspergillus nidulans carnitine carrier encoded by the acuH gene is exclusively located in the mitochondria.FEMS Microbiol Lett.2001年7月24日、201(2):193−8頁)、または保持シグナル(デルクス(Derkx),P.M.とマドリッド(Madrid),S. M.、The foldase CYPB is a component of the secretory pathway of Aspergillus niger and contains the endoplasmic reticulum retention signal HEEL. Mol.Genet.Genomics.2001年12月、266(4):537−45頁)によってNHR関与タンパク質を再局在することも可能である。
【0054】
あるいは、または上記の方法と組合せて、タンパク質活性の阻害はUVまたは化学的変異誘発によって(マテム,I.E.、ファン・ヌルト J.M.、ファン・デン・ベルグ,P.、アーチャー,D.B.、ロバーツ,I.N.、およびファン・デン・ホンデル,C.A.、Isolation and characterization of mutans of Aspergillus niger deficient in extracellular proteases.Mol Gen Genet.1992年8月、234(2):332−6頁)、または親和性のプロテアソーム阻害剤のような阻害剤の使用によって(クラスト−ラクタクリスチン−β−ラクトン、アフィニティー・リサーチ・プロダクツ社(Affinity Research Products Ltd.)、CW8405−Z02185)も得られうる。
【0055】
別の好ましい実施形態によれば、HRへ導くステップは、取込まれるポリヌクレオチドは別にして、結合し、NHR成分の発現を制限することが可能な過剰な小さな二本鎖ポリヌクレオチドを添加するステップを含んで成る(アグラワル(Agrawal)N.ら:RNA interference:biology,mechanism and applications.Microbiol.Mol.Biol.Rev.、67巻、4号、657−685頁)。
【0056】
好ましい実施形態において、本発明は、所定の部位へのポリヌクレオチドの標的取込みの効率を増大させる方法を提供し、それによって前記ポリヌクレオチドは、糸状菌細胞を提供することによってHRへ取込み経路を導くステップを含んで成るNHR選好を有する糸状菌において、前記所定の部位で、またはその周りで相同性を有し、ここでHR経路の効率は同じ条件下に由来する親糸状菌の1つに比べ高められている。HR経路の効率は、好ましくは、NHR/HR比を測定するために使用されたものと同じアッセイによって測定される。好ましい実施形態によれば、親生物は宿主細胞の部で定義されている糸状菌の1つである。
【0057】
高められたというのは、得られる細胞が由来する親細胞におけるよりも少なくとも高いことを意味する。好ましくは、高められたというのは、2倍高い、より好ましくは3倍高い、さらにより好ましくは4倍高い、最も好ましくは、ノーザン、またはウェスタンブロット法、もしくはアレイ技術、または表現型スクリーンを使用して4倍超高いことを意味する。
【0058】
別の好ましい実施形態によれば、糸状菌は、それが由来する糸状菌細胞における同じ遺伝子の発現レベルと比較することによってアップレギュレートされている、HRに関与する少なくとも1つの遺伝子の発現レベルを有する。これは、HRに関与する成分をコードする遺伝子の発現レベルを増大させることによって、かつ/またはHRに関与する成分の発現レベルを増大させることによって、かつ/またはHRに関与する成分の活性を(一時的に)増大させることによって達成されうる。
【0059】
好ましくは、得られた糸状菌は、それが由来する糸状菌細胞における同じ遺伝子の発現レベルと比較することによってアップレギュレートされている、HRに関与する遺伝子の発現を有する。
【0060】
DNA配列の発現レベルは、得られた糸状菌におけるこの特定のDNA配列の発現レベルが、それが由来する親糸状菌における同じDNA配列の発現レベルよりも高い、好ましくは3倍高い、より好ましくは4倍高い、最も好ましくは、ノーザン、またはウェスタンブロット法、もしくはアレイ技術を使用して4倍超高い場合にアップレギュレートされる。好ましい実施形態によれば、親生物は宿主細胞で定義されている糸状菌の1つである。
【0061】
少なくとも1つのDNA配列の発現レベルのアップレギュレーションは、以下の方法の1つによって、またはその組合せによる遺伝子操作によって得られうる。すなわち、
c.組換え遺伝子操作法を使用するステップ、
d.糸状菌を変異誘発させるステップ、
あるいは、または上記の方法と組合せ、かつ別の好ましい実施形態によれば、少なくとも1つの遺伝子またはDNA配列の発現レベルのアップレギュレーションは、糸状菌を活性化化合物/組成物に提示することによって得られうる。
【0062】
糸状菌はその後に前記DNA配列の発現レベル、および場合により糸状菌のHR経路の効率をモニタリングすることによって選択されうる。糸状菌のHR効率は、一定の相同領域を使用することにより糸状菌のゲノムにおける所定の部位への一定のポリヌクレオチド配列の標的取込みの効率によって測定されうる。
【0063】
好ましくは、少なくとも1つのDNA配列の発現のアップレギュレーションは、組換え糸状菌を得るステップa.で定義されているような組換え遺伝子操作法で行われる。好ましくは、ステップa.は、DNA配列を含んで成るDNA構成物で糸状菌を形質転換するステップを含んで成り、好ましくは前記DNA配列は高発現遺伝子のプロモータに操作連結されている。選択プロモータは、過剰発現されるDNA配列の内在性プロモータよりも強い場合がある。DNA配列の発現のプロモータは、高発現真菌遺伝子由来であることが好ましい。
【0064】
多くの好ましい高発現真菌遺伝子が一例として挙げられている。すなわち、アスペルギルスまたはトリコルデルマからのアミラーゼ遺伝子、グルコアミラーゼ遺伝子、アルコールデヒドロゲナーゼ遺伝子、キシラナーゼ遺伝子、グリセルアルデヒドリン酸デヒドロゲナーゼ遺伝子、またはセロビオヒドロラーゼ遺伝子である。これらの目的のために最も好ましい高発現遺伝子は、アスペルギルス・ニゲル・グルコアミラーゼ遺伝子、アスペルギルス・オリゼ(Aspergillus oryzae)TAKAアミラーゼ遺伝子、アスペルギルス・ニドゥランス(Aspergillus nidulans)gpdA遺伝子、またはトリコデルマ・エセイ(Trichoderma reesei)セロビオヒドロラーゼ遺伝子である。グルコアミラーゼプロモータが、使用される最も好ましいプロモータである。これらの高発現遺伝子は、クローニングベクターの取込みのための標的遺伝子座として、および高発現真菌遺伝子の供給源として適切である。
【0065】
別の好ましい実施形態によれば、ステップa.は、好ましくは、DNA配列のそのゲノムコピー取込みによって、より好ましくは、高発現遺伝子座で、好ましくはグルコアミラーゼ遺伝子座でDNA配列の取込みを標的することによって、糸状菌細胞へのDNA配列のコピー数を増大させるステップを含んで成る。
【0066】
DNA配列の発現のアップレギュレーションは、DNA配列の少なくとも1つにコピーを糸状菌へ導入することによって、または強いプロモータのために変更し、または良好な動態および/または寿命を有するタンパク質をコードする遺伝子のために変更することによってDNA配列のコピー数を増大させることによって達成されうる。DNA配列はプラスミドに存在し、またはゲノムへ取込みうる。当業者は2つの代替の可能性のうちで選択しうる。すなわち、
−HR経路に関与する糸状菌の少なくとも1つの内在性DNA配列を過剰発現する。この場合、糸状菌はその内在性DNA配列のいくつかのコピーを含んで成る。
−HRに関与する少なくとも1つの非相同DNAを過剰発現する。この場合、糸状菌はHRに関与するその内在性DNA配列、およびさらにHRに関与する非相同DNA配列の少なくとも1つのコピーを有することになる。この非相同DNA配列はその対応する内在性DNA配列の相同体である。
【0067】
糸状菌は、DNA配列の1つもしくはそれ以上のコピーで形質転換されうる(とりわけ、チルブム(Tilbum)ら、1983年、Gene、26:205−221頁による)。DNA配列は、糸状菌のゲノムへ安定して取込まれ、または自発的複製が可能なDNA分子の一部として細胞へ導入されうる。DNA配列は、クローニングベクターに存在することが好ましい。糸状菌宿主細胞を形質転換することが可能なクローニングベクターが本発明における使用に適切である。したがって、本発明において使用するためのクローニングベクターは、糸状菌宿主細胞の染色体において無作為または所定の標的遺伝子座で取込む統合的クローニングベクター、およびAMA1配列を含んで成るベクターなど自発的に維持されるクローニングベクターを含んで成る。本発明の好ましい実施形態において、統合的クローニングベクターは、糸状菌宿主細胞のゲノムにおける所定の標的遺伝子座における、この所定の遺伝子座にクローニングベクターの取込みを標的するためのDNA断片を含んで成る。標的取込みを促進するために、クローニングベクターは宿主細胞の形質転換前に線状化されることが好ましい。線状化は、クローニングベクターの少なくとも1つであるが、好ましくはいずれかの末端が標的遺伝子座と相同的な配列によってフランキングされるように実行されることが好ましい。標的遺伝子座をフランキングする相同的配列の長さは好ましくは少なくとも30bp、好ましくは少なくとも50bp、好ましくは少なくとも0.1kb、さらに好ましくは少なくとも0.2kb、より好ましくは少なくとも0.5kb、さらにより好ましくは少なくとも1kb、最も好ましくは少なくとも2kbである。
【0068】
好ましくは、標的遺伝子座と相同的であるクローニングベクターにおけるDNA配列は、高発現遺伝子座由来であり、これは、糸状菌宿主細胞において高い発現レベルが可能である遺伝子由来であることを意味する。高い発現レベルが可能な遺伝子、すなわち高発現遺伝子が、本明細書では、そのmRNAが、例えば、誘導条件下に全細胞のmRNAの少なくとも0.5%(w/w)を構成しうる遺伝子、あるいは、その遺伝子産物が全細胞タンパク質の少なくとも1%(w/w)を構成し、または、分泌遺伝子産物の場合、少なくとも0.1g/lのレベルに分泌されうる遺伝子と定義されている(欧州特許第357127B1号明細書に記載)。
【0069】
過剰発現されるDNA配列のコピーの数を増加させるには、国際公開第98/46772号パンフレットに記載された遺伝子変換の方法が使用されうる。
【0070】
当業者は、標的のための相同的DNA配列およびプロモータ配列が1つのDNA断片で一致しうることを理解するであろう。上記の高発現遺伝子のリストも標的遺伝子座として適合されている。
【0071】
自発的に維持されるクローニングベクターの例が、AMA1配列を含んで成るクローニングベクターである。AMA1は、アスペルギルス・ニドゥランスから単離された6.0kbゲノムDNA断片であるが、これはアスペルギルスにおける自発的維持が可能である(例えば、アレスケンコ(Aleksenko)とクラッターバック(Clutterbuck)(1997年)、Fungal Genet.Biol.21:373−397頁を参照)。
【0072】
本発明の方法の別の好ましい実施形態によれば、ステップa.は、選択マーカー遺伝子を含んで成るDNA構成物で糸状菌を形質転換するステップを含んで成る。クローニングベクターにおける選択マーカー遺伝子は、糸状菌の形質転換に有用である多くのマーカー遺伝子から選択されうる。一例として、これらのマーカーは、amdS遺伝子など優性および双方向選択マーカー遺伝子(欧州特許第635574B号明細書または国際公開第97/06261号パンフレット)、argB、trpC、またはpyrGなど栄養要求性マーカー遺伝子、および例えば、フレオマイシン、ハイグロマイシンB、またはG418に対する抵抗を提供する抗生物質抵抗性遺伝子を含むがこれらに限定されない。優性および双方向選択マーカー遺伝子の使用が好ましい。好ましくは、amdS遺伝子が好ましく、より好ましくは、アスペルギルス・ニドゥランスまたはアスペルギルス・ニゲルからのamdS遺伝子である。最も好ましい選択マーカー遺伝子が、A.ニドゥランスgpdAプロモータに融合したA.ニドゥランスamdSコード配列である(EP635574号明細書を参照)。他の糸状菌からのamdS遺伝子も使用される(国際公開第97/06261号パンフレット)。amdS選択マーカー遺伝子は、それが同じ菌株において数倍で使用され、明確なDNA配列を導入し、過剰発現し、かつ/または削除しうる利点を有する。欧州特許第635574号明細書に記載されたフルオロアセトアミド培地での対抗選択を用いて、結果として生じる菌株はマーカーを含まず、さらなる遺伝子修飾のために使用されうる。
【0073】
あるいは、または上記の方法とともに、DNA配列の発現のアップレギュレーションがUVまたは化学的変異誘発を使用して達成されうる(マテム,I.E.、ファン・ヌルトJ.M.、ファン・デン・ベルグ,P.、アーチャー,D.B.、ロバーツ,I.N.、およびファン・デン・ホンデル,C.A.、Isolation and characterization of mutants of Aspergillus niger deficient in extracellular proteases.Mol Gen Genet.1992年8月、234(2):332−6頁)。
【0074】
さらに、かつ/またはHRに関与するDNA配列の発現のアップレギュレーションと組合せて、UVまたは化学的変異誘発によってHRに関与するタンパク質の活性を増大させることによってHR効率の増大を得ることも可能である(マテム,I.E.、ファン・ヌルト J.M.、ファン・デン・ベルグ,P.、アーチャー,D.B.、ロバーツ,I.N.、およびファン・デン・ホンデル,C.A.、Isolation and characterization of mutants of Aspergillus niger deficient in extracellular proteases.Mol Gen Genet.1992年8月、234(2):332−6頁)。
【0075】
当業者は、DNA配列の発現のアップレギュレーションを達成するには、記載された方法の各々を別々に、または組合せて使用しうることを理解するであろう。
【0076】
当業者は、増大したHR/NHR比、および/または低下したNHR効率、および/または高いHR効率を有する糸状菌を得るには、糸状菌における一定の遺伝子の発現をそれぞれダウンおよびアップレギュレートするために記載された各々の方法の少なくとも1つを使用することができる。好ましくは、低下したNHR効率および高いHR効率を有する組換え糸状菌を得るために糸状菌で実行される方法のすべては、優性および双方向選択マーカー、好ましくはamdS遺伝子、より好ましくは、アスペルギルス・ニドゥランスまたはアスペルギルス・ニゲルからのamdS遺伝子を使用して実行されている。
【0077】
得られた糸状菌はその後、前述されているように、例えばノーザンおよび/またはウェスタンブロット法および/またはアレイおよび/または表現型スクリーニングを使用することによって前記DNA配列の発現レベルをモニタリングすることによって選択されうる。場合により、細胞のNHRおよび/またはHR経路の効率がモニタリングされる。糸状菌のこれらの経路の効率は以前に規定されているようにモニタリングされうる。
【0078】
好ましくは、HR経路にもたらされる修飾は誘導性のものである。これは誘導性調節領域によってHRに関与する成分をコードする遺伝子の内在性調節領域を置換することによって、好ましくは誘導性プロモータを使用することによって達成されうる。誘導性プロモータの例は、アスペルギルス・ニゲルのグルコアミラーゼプロモータ、アスペルギルス・オリゼのTAKAアミラーゼプロモータ、pafプロモータ(マルクス(Marx),F.、ハース(Haas),H.、ラインドル(Reindl),M.、ストッフラー(Stoffler),G.、ロットスパイヒ(Lottspeich),F.、およびレドル(Redl),B.、Cloning, structural organization and regulation of expression of the Penicillium chrysogenum paf gene encoding an abundantly secreted protein with antifungal activity Gene 167(1−2頁)、167− 171頁 (1995年)、またはペニシリウム・クリソゲヌムのpcbCプロモータ(マルチン JF、カスケイロ J、コサルコワ K、マルコス AT、グチレス S.、Penicillin and cephalosporin biosynthesis:mechanism of carbon catabolite regulation of penicillin production.Antonie Van Leeuwenhoek. 1999年1−2月、75(1−2頁):21−31、総説)、またはNHRに関与する遺伝子の発現のダウンレギュレーションについて以前に引用されたスイッチオン/オフシステムである。
【0079】
好ましい実施形態によれば、修飾されるHR経路に関与する遺伝子は、以下の遺伝子またはその相同体、すなわち、RAD51、RAD52である。
【0080】
HRの過程においてその機能を実行することがより可能または少なくとも相当により可能である、HRに関与する少なくとも1つの成分を有する変異体のすべての種類が、本発明によって意図される変異体である。好ましくは、HRに関与する成分の活性は修飾されており、HR経路の効率は、以前に規定されたアッセイにおいて測定されるように、同じ条件下にそれが由来する親細胞における効率の110%超、より好ましくは200%超、最も好ましくは500%超である。好ましい実施形態によれば、親生物は宿主細胞の部で定義される糸状菌の1つである。本発明による方法は、広範囲であるが限定的に上述されているように、さまざまな用途において使用されうる。一部の特定の用途が以下に記載されている。
【0081】
宿主細胞
したがって、本発明はさらに糸状菌それ自体に関するが、これは好ましくは、前記糸状菌細胞のゲノムへの所定の部位に対するポリヌクレオチドの標的取込みの効率を増大させるための本発明の方法において使用され、前記糸状菌はNHR選好を有し、かつ前記ポリヌクレオチドは前記所定の部位と相同の領域を有するとともに、前記方法は取込み経路をHRへ導くステップを含んで成る。この方法において使用されうる糸状菌の特徴は以前に規定されている。
【0082】
本発明の方法において好ましく使用される糸状菌は親細胞に由来する変異体であり、NHR/HRの比が減少しており、かつ/またはNHR経路の効率が低下されており、かつ/またはHR経路の効率が同じ条件下に前記親生物における前記比および前記効率と比べ前記変異体細胞において高められている。NHR/HRの比および/またはNHR経路の効率および/またはHR経路の効率を測定するために使用されるアッセイは以前に記載されている。
【0083】
本発明の宿主細胞が糸状菌であるが、これはクローニングベクターで形質転換されることが可能である。したがって、試験される大部分の糸状菌では、それらはアスペルギルスのために開発された形質転換プロトコールを使用して形質転換されうることがわかった(とりわけ、チルブムら、1983年、Gene、26:205−221頁による)。当業者は、糸状菌宿主種の有効な形質転換が、本明細書で具体的に例示されたベクター、選択マーカー系、プロモータ、および形質転換プロトコールの使用に限定されていない。
【0084】
糸状菌は本明細書では、糸状形態における分類Eumycotinaの真核微生物と定義されており、すなわち、その栄養成長は菌糸伸長によって起こる。好ましい糸状菌宿主細胞は、アスペルギルス属、トリコデルマ属、フサリウム(Fusarium)属、ペニシリウム属、およびアクレモニウム(Acremonium)属からなる群から選択される。
【0085】
本発明のより好ましい実施形態において、糸状菌宿主細胞は、A.ニドゥランス、A.オリゼ、A.ソーヤ(sojae)、A.ニゲル群のアスペルギルス、トリコデルマ・エセイ、およびペニシリウム種からなる群から選択される。好ましくは、ペニシリウムはペニシリウム・クリソゲヌムまたはペニシリウム・シトリヌム(Penicillium citrinum)種である。
【0086】
A.ニゲル群は本明細書ではレイパー(Raper)とフェネル(Fennell)(1965年、The Genus Aspergillus,The Williams & Wilkins Company,Baltimore,293−344頁)に従って定義され、かつこれらの著者らによって含まれるすべての(黒色)アスペルギルスを含んで成る。最も好ましい糸状菌宿主細胞は、A.ニゲル群、A.オリゼのアスペルギルス、トリコデルマ・エセイ、およびペニシリウム・クリソゲヌムから成る群から選択される。
【0087】
好ましい実施形態によれば、親生物は寄託された糸状菌細胞アスペルギルス・ニゲルCBS513.88、アスペルギルス・オリゼATCC20423、IFO4177、ATCC1011、ATCC9576、ATCC14488−14491、ATCC11601、ATCC12892、ペニシリウム・クリソゲヌムCBS455.95、またはペニシリウム・シトリヌムATCC38065、ペニシリウム・クリソゲヌムP2、アクレモニウム・クリソゲヌム(Acremonium chrysogenum)ATCC36225またはATCC48272、トリコデルマ・エセイATCC26921またはATCC56765またはATCC26921、アスペルギルス・ソーヤATCC11906、クリソスポリウム・ルキノウェンゼ(Chrysosporium lucknowense)ATCC44006、クラビセプス・パスパリ(Claviceps paspali)CBS110.22、クラビセプス・プルプラ(Claviceps purpurea)CBS164.59、ペニシリウム・ブリビコンパクツム(Penicillium brevicompactum)ATCC 9056、アスペルギルス・テレウス(Aspergillus terreus)ATCC20542、アスペルギルス・ニドゥランスATCC28901およびまたはそれらの誘導体である。
【0088】
別の好ましい実施形態によれば、本発明の糸状菌細胞は、以下のアッセイによって測定されるように、少なくとも200、少なくとも50、少なくとも10であるNHR/HR比を有する。好ましくは、糸状菌細胞の比は少なくとも1、より好ましくは少なくとも0.5、さらにより好ましくは少なくとも0.1、さらにより好ましくは少なくとも0.05、さらにより好ましくは少なくとも0.01さらにより好ましくは少なくとも0.005さらにより好ましくは少なくとも0.001さらにより好ましくは少なくとも0.0005さらにより好ましくは少なくとも0.0001、かつ最も好ましくは少なくとも0.00001である。
【0089】
より好ましい実施形態によれば、本発明の糸状菌細胞は、以下のアッセイによって測定されるように200未満、さらにより好ましくは50未満、10未満であるNHR/HR比を有する。さらにより好ましくは、糸状菌細胞の比は1未満、さらにより好ましくは0.5未満、さらにより好ましくは0.1未満、さらにより好ましくは0.05未満、さらにより好ましくは0.01未満、さらにより好ましくは0.005未満、さらにより好ましくは0.001未満、さらにより好ましくは0.0005未満、さらにより好ましくは0.0001未満、かつ最も好ましくは0.00001未満である。
【0090】
NHR/HRの比は好ましくは国際公開第02/052026号パンフレット(表2、23頁)に記載されたアッセイによって測定される。
【0091】
好ましくは、糸状菌細胞は、NHRに関与する成分をコードする遺伝子を欠き、かつ/またはNHRに関与する成分の減少したレベルを有する。
【0092】
さらにより好ましくは、糸状菌細胞は以下の遺伝子の少なくとも1つを欠いている。すなわち、hdfAまたは配列番号2または19で識別されるその相同体、hdfBまたは配列番号5、もしくは22または両方で識別されるその相同体、かつ/または、好ましくは、これらの遺伝子によってコードされたタンパク質の少なくとも1つの減少した量を有する。
【0093】
最も好ましくは、糸状菌細胞は誘導性に以下の遺伝子の少なくとも1つを欠いている。すなわち、hdfAまたは配列番号2または19で識別されるその相同体、hdfBまたは配列番号5、もしくは22または両方で識別されるその相同体、かつ/または、好ましくは誘導性に、これらの遺伝子によってコードされたタンパク質の少なくとも1つの減少した量を有する。
【0094】
別の好ましい実施形態によれば、糸状菌細胞は、そのゲノムにおいて、NHRに関与する遺伝子が非機能的遺伝子によって、もしくは選択マーカーによって、または別の遺伝子によって置換されているようになっている。
【0095】
別の好ましい実施形態によれば、変異体はHRに関与する成分の増大したレベルを有する。
【0096】
本発明による糸状菌は、分子生物学法によって得られている。かかる遺伝子組換え法によって得られる糸状菌は組換え糸状菌と定義される。しかし本発明に即した組換え糸状菌は、早期に変異誘発法にかけられ、別の望ましい効果を達成しうる。最も好ましい実施形態によれば、得られる糸状菌は組換え糸状菌である。
【0097】
本発明の宿主細胞の使用
好ましい実施形態によれば、少なくとも、例えば、形質転換またはエレクトロポレーションの方法、および前記ポリヌクレオチドの前記細胞の遺伝物質への取込みによって、目的とするポリヌクレオチドを本発明の糸状菌へ導入するステップを含んで成る方法が提供される。取込みは複合方法であり、核酸配列が宿主細胞の遺伝物質の一部となる。核酸取込みの方法における1つのステップは組換えであり、組換えによって、核酸配列が交換または挿入され、導入された核酸は宿主細胞の遺伝物質の一部となる。原則として、2種類の組換え法が可能である。すなわち、相同的および非正統的すなわちNHRである。大部分の(高等)真核生物はHRを実行することはなく、または少なくとも顕著に実行することはないが、かかる方法を達成する必須タンパク質は利用可能である。この現象の1つの理由は、(高等)真核生物における相同的組換えの頻繁な使用が、繰返し核酸配列の存在により望ましくない染色体再配置をもたらしうることである。本発明による方法によってHRを達成するには、所定の部位との相同性を有するポリヌクレオチドを提供することが重要である。相同の割合および相同的領域の長さが相同的組換えにおいて重要な役割を果たすことは当業者に明らかである。相同の割合はほぼ100%であることが好ましい。当業者であれば、低い割合の相同も相同的組換えの分野で使用されるが、例えば、相同の領域、およびその全体的な分布に依存し、HRの低い効率をもたらしうるが、依然として有用であり、したがって本発明に含まれることを認識している。さらに、(ほぼ)相同的領域の長さは約3kbであり、これは相同的組換えを導くのに十分である。少なくとも1つの相同的領域が組換えには必要であるが、より好ましくは、目的とする核酸をフランキングする2つの相同的領域が標的取込みのために使用される。当業者は、相同の適切な割合、相同の長さ、および相同的領域の量を選択するやり方を知っている。かかる相同を提供することによって、核酸が宿主細胞の遺伝物質内のすべての所望の位置で取込まれる。本明細書で開示された本発明が、相同の長さおよび相同の割合がHRを提供/可能にするのに十分に高い限り、すべての核酸(好ましくはDNA)を所定の部位に導くために使用されることは当業者には明らかである。
【0098】
本発明がなされる前は、ポリヌクレオチドがつねに容易にすべての所望の位置で一定の糸状菌のゲノムへ取込まれたわけではない。本発明による方法は、例えば、さまざまな方法で遺伝子機能に影響を及ぼすために、完全な不活性化のためだけではなく、発現レベルまたは発現の調節における変化、タンパク質活性またはコードされたタンパク質の細胞内標的の変化を介在するためにも適用される。完全な不活性化は、通常、アンチセンス技術またはRNAi技術(ズレナー(Zrenner)R、ウィルミッツァー(Willmitzer)L、ソネワルド(Sonnewald)U、Analysis of the expression of potato Uridinediphosphate−glucose pyrophosphorylase and its inhibition by antisense RNA.Planta.(1993年)、190(2):247−52頁)など既存の方法によって達成されえないが、例えば、代謝経路の望ましくない側枝をコントロールし、例えば、(ベータ・ラクタム)抗生物質またはカルチノイドなど特定の二次代謝産物の産生を増大させる遺伝子の不活性化に有用である。完全な不活性化は、毒性または望ましくない化合物の産生を削減させるためにも有用である(chrysogenin in Penicillium;Aflatoxin in Aspergillus:マクドナルド(MacDonald)KDら、Heterokaryon studies and the genetic control of penicillin and chrysogenin production in Penicillium chrysogenum.J Gen Microbiol.(1963年)33:375−83頁)。完全な不活性化は、発酵過程および下流処理が改善されるような方法で生物の形態を変化させるのにも有用である。
【0099】
本発明は、代わりの調節配列によって既存の調節配列を置換し、内在性遺伝子の発現(例えば、特定の誘導剤に反応した発現)を変化させることを可能にする。
【0100】
本発明の一態様は、本発明の方法による活性遺伝子の不活性遺伝子による置換に関する。完全な不活性化は、通常、アンチセンス技術またはRNAi技術など既存の方法によって達成されえないが、例えば、代謝経路の望ましくない側枝をコントロールし、例えば、でんぷんなどバルク製品の品質を増大し、または特定の二次代謝産物の産生を増大させ、または望ましくない代謝産物の形成を阻害するために有用である。
【0101】
本発明の別の態様は、糸状菌細胞の広範囲の代謝再プログラミングまたはエンジニアリングに関する。完全に新しい経路の導入および/または望ましくない経路の修飾は、タンパク質または代謝産物など特定の化合物の生産に具体的に適合される細胞の取得をもたらす。
【0102】
本発明の別の態様は、不活性または変化した遺伝子の活性遺伝子による置換に関する。例えば、伝統的な変異誘発の有効な期間後、選択された糸状菌株が無作為変異誘発法の間に変化し、さらに不活性化した一部の内在性遺伝子を有する場合が多い。
【0103】
本発明のさらに別の態様では、抗生物質の糸状菌細胞に対する抵抗を与える物資を導入する方法が提供される。さらに本発明の別の態様では、所望の特性を糸状菌細胞に与える方法が提供される。好ましい実施形態では、遺伝子送達賦形剤が使用され、所望のポリヌクレオチドを所定の部位に送達する。遺伝子送達賦形剤は当技術分野で周知であり、説明で以前に記載されている。
【0104】
本発明の別の態様による別の好ましい方法も、例えば、所望の新規生成物のためのもので所望の発現プロフィールを与える遺伝子の既存のコード配列を置換することによって、新規生成物をコードする導入遺伝子の予測可能な発現を達成する。より好ましい実施形態によれば、本発明によって提供される糸状菌はさらに、産生される所望のタンパク質の所望の遺伝子コードを含んで成るDNA構成物を含んで成る。
【0105】
好ましくは、産生される所望のタンパク質をコードする所望の遺伝子は発現ベクターへ挿入され、これはその後に得られた宿主細胞を形質転換するために使用される。発現ベクターにおいて、DNA配列は、プロモータなどの適切な発現シグナル、場合によりシグナル配列および末端に操作連結されうるが、これらは宿主生物におけるタンパク質の発現および合成を導くことが可能である。
【0106】
より好ましくは、所望の遺伝子はプロモータおよび分泌シグナルに操作連結される。所望の遺伝子を発現させるために使用されうる方法は、DNA配列の発現のアップレギュレーションの部で記載されたものと同じであり、その発現産物はHR、すなわち、コピー数の増大、標的取込み、高発現遺伝子のプロモータの使用、選択マーカー遺伝子およびそれらの組合せの選択に関与する。
【0107】
所望のタンパク質は好ましくは酵素である。タンパク質が自然に分泌されない場合は、タンパク質をコードするポリヌクレオチドが当技術分野で周知の方法に従ってシグナル配列を有するように修飾されうる。分泌されるタンパク質は、自然に発現される内在性タンパク質でありうるが、非相同でもありうる。非相同は、タンパク質によってコードされる遺伝子が野生型糸状菌における自然条件下に産生されないことを意味する。本発明の糸状菌によって産生されうる酵素の例は、カルボヒドラーゼ、例えば、エンドグルカナーゼ、β−グルカナーゼ、セロビオヒドラーゼまたはβ−グルコシダーゼなどのセルラーゼ、ヘミセルラーゼ、またはキシラナーゼ、キシロシダーゼ、マナナース、ガラクタナース、ガラクトシダーゼ、ラムノガラクツロナーゼ、アラバナーゼ、ガラクツロナーゼ、リアーゼなどのペクチン分解酵素、またはアミノ分解酵素、フィターゼなどのホスファターゼ、リパーゼなどのエステラーゼ、タンパク質分解酵素、オキシダーゼ、トランスフェラーゼ、またはイソメラーゼなどの酸化還元酵素である。より好ましくは、所望の遺伝子はフィターゼをコードする。
【0108】
別の例として、既存のコード配列が修飾され、コードされたタンパク質は、例えば、改善された熱的特性および/または改善された動態特性(Km、Kcat)、および/または改善された酵素安定性、および/または基質範囲の延長、および/または寿命の増大等を有するタンパク質を作る最適特徴を有するようになる。
【0109】
本発明はさらに目的とするポリペプチドを製造するための本発明の糸状菌の使用に関する。あるいは、得られた糸状菌は二次代謝産物を製造するために使用されうる。好ましい二次代謝産物は、カルチノイド化合物、ベータ−ラクタム化合物、薬剤、抗腫瘍化合物等である。
【0110】
好ましくは、本発明で得られた糸状菌は、所望のタンパク質をコードするDNA配列の発現を促す条件下に形質転換された宿主細胞を培養し、かつ例えば以下の文献に記載された所望のタンパク質を回収することによって所望のタンパク質を製造するために使用される。すなわち、
−リー(Li),Z.J.、シュクラ(Shukla),V.、フォルダイス(Fordyce),A.P.、ペデルセン(ペデルセン),A.G.、ウェグナー(Wegner),K.S.、マルテン(Marten),M.R.、Fungal morphology and fragmentation behavior in a fed−batch Aspergillus oryzae fermentation at the production scale.Biotechnol Bioeng.2000年11月5日、70(3):300−12頁
−ウィザース(Withers),J.M.、スウィフト(Swift),R.J.、ウィーベ(Wiebe),M.G.、ロブソン(Robson),G.D.、プント(Punt),P.J.、ファン・デン・ホンデル,C.A.、Optimization and stability of glucoamylase production by recombinant strains of Aspergillus niger in chemostat culture.Biotechnol Bioeng.1998年8月20日、59(4):407−18頁。
−アマルラー(Amanullah),A.、クリステンセン(Christensen),L.H.、ハンセン(Hansen),K.、ニーナウ(Nienow),A.W.、トーマス(Thomas),R.C.、Dependence of morphology on agitation intensity in fed−batch cultures of Aspergillus oryzae and its implications for recombinant protein production.Biotechnol Bioeng.2002年3月30日、77(7):815−26頁。
【0111】
DNA配列およびこれらのDNA配列によってコードされたポリペプチド
本発明の別の態様によれば、以下の単離cDNA配列が提供される。すなわち、
配列番号2 A.ニゲルからのhdfA
配列番号19 ペニシリウム・クリソゲヌムからのhdfA
配列番号5 A.ニゲルからのhdfB
配列番号22 ペニシリウム・クリソゲヌムからのhdfB
およびそれらの相同体。
【0112】
配列番号1、18、4、および21の各々は、それぞれ、上記の各々のcDNA配列と関係があるゲノムDNA配列に対応する。
【0113】
配列番号3、20、6、および23の各々は、それぞれ、上記のそれぞれのcDNA配列によってコードされたタンパク質配列に対応する。
【0114】
本明細書で提供される配列情報は、誤って識別された塩基の包含を必要とするために狭義に考えるべきではない。本明細書で開示された特定の配列は、特にA.ニゲルまたはペニシリウム・クリソゲノムにおける糸状菌から完全な遺伝子を単離するために容易に使用されうるが、次いでさらに配列分析に容易にかけられ、それによって配列決定の誤りを識別することができる。
【0115】
別段の指示がない限り、本明細書でのDNA分子の配列決定によって決定される全ヌクレオチド配列は、自動化DNAシーケンサを使用して決定され、かつ本明細書で決定されるDNA分子によってコードされたポリペプチドの全アミノ酸配列は上記のように決定されたDNA配列の翻訳によって予測された。したがって、この自動化法によって決定されるDNA配列について当技術分野で周知であるように、本明細書で決定されたヌクレオチド配列はいつくかの誤りを含有する。自動化によって決定されたヌクレオチド配列は一般的に、配列決定されたDNA分子の実際のヌクレオチド配列と少なくとも約90%同一であり、より一般的には少なくとも約95%〜少なくとも約99.9%同一である。実際の配列は、当技術分野で周知の手動のDNA配列決定法を含む他の方法によってより正確に決定されうる。やはり当技術分野で周知であるように、実際の配列と比べ決定されたヌクレオチド配列における単一の挿入または削除が、ヌクレオチド配列の翻訳におけるフレームシフトをもたらし、決定されたヌクレオチド配列によってコードされた予測アミノ酸配列が、かかる挿入または削除の点で開始する、配列決定されたDNA分子によって実際にコードされたアミノ酸配列と完全に異なるようになる。
【0116】
当業者は、かかる誤って識別された塩基を識別することが可能であり、かかる誤りを正すやり方を知っている。
【0117】
「相同」は以下に定義されている。相同的はアスペルギルス・ニゲルまたはペニシリウム・クリソゲノム以外の他の糸状菌由来を意味すると理解されうる。
【0118】
他の生物からの全長DNAは、他の生物、例えば、糸状菌、具体的には、それらをスクリーニングすることによってアスペルギルス種またはペニシリウム種から構成されたcDNAまたはゲノムDNAライブラリーを使用して得られうる。
【0119】
本発明は、hdfAおよび/またはhdfBのパラログをも包含する。本発明に照らして、パラログは、配列番号1または配列番号4または配列番号18または配列番号21と相同的、かつそれぞれ、A.ニゲルまたはペニシリウム・クリソゲノム由来のDNA配列を意味する。
【0120】
例えば、アスペルギルスまたはペニシリウム菌株は、ノーザンブロット分析によって相同的hdfAおよび/またはhdfBポリヌクレオチドについてスクリーニングされうる。本発明によるポリヌクレオチドと相同的な転写産物の検出と同時に、cDNAライブラリーが、当業者に公知の標準の方法を利用することにより、適切な菌株から単離されるRNAで構成されうる。あるいは、全ゲノムDNAライブラリーが、本発明によるhdfAおよび/またはhdfBにハイブリダイズ可能なプローブを使用することによりスクリーニングされうる。
【0121】
相同的遺伝子配列は、例えば、本明細書で教示されたヌクレオチド配列に基づき設計された2つの退化オリゴヌクレオチドプライマープールを使用するPCRを実行することによって単離されうる。
【0122】
反応のためのテンプレートは、本発明によるポリヌクレオチドを発現することが知られ、またはそう思われる菌株から調製されるmRNAの逆転写によって得られるcDNAでありうる。PCR産物はサブクローン化されて配列決定され、増幅配列が新しいhdfAおよび/またはhdfB核酸配列、またはそれらの機能的等価物の配列を示すことを確実にしうる。
【0123】
次いで、PCR断片を使用し、さまざまな既知の方法によって行われる完全長cDNAを単離することができる。例えば、増幅断片は標識され、バクテリオファージまたはコスミドcDNAライブラリーをスクリーニングするために使用されうる。あるいは、標識断片を使用してゲノムライブラリーをスクリーニングすることができる。
【0124】
PCR技術をも使用し、他の生物から完全長cDNA配列を単離することができる。例えば、RNAは、標準の方法に従い、適切な細胞または組織源から単離されうる。逆転写反応は、最初のストランド合成のプライミングのために増幅断片の大部分の5’末端に特異的なオリゴヌクレオチドプライマーを使用してRNAで実行されうる。
【0125】
次いで、結果として生じるRNA/DNAハイブリッドは、標準の末端トランスフェラーゼ反応を使用して(例えば、グアニンで)「末端付加」されうるが、このハイブリッドはRNaseHで消化されうるとともに、次いで第2のストランド合成が(例えば、ポリCプライマーで)プライミングされうる。したがって、増幅断片の上流のcDNA配列は容易に単離されうる。有用なクローニング法の検討では、例えば、サンブルックら、上記参照、およびオーズベル(Ausubel)ら、下記参照を参照。
【0126】
「相同」は機能的等価物を意味するとも理解されうる。
【0127】
「機能的等価物」および「機能的変異形」という語は本明細書で同義で使用される。hdfAおよび/またはhdfB DNAの機能的等価物は、hdfAおよび/またはhdfBの特定の機能を示すポリペプチドをコードする単離DNA断片である。本発明によるhdfAおよび/またはhdfBポリペプチドの機能的等価物は、NHR複合体の一部として少なくとも1つの機能を示すポリペプチドである。したがって、機能的等価物も生体活性断片をも包含する。
【0128】
機能的タンパク質またはポリペプチド等価物は、配列番号3または6または20または23または非必須アミノ酸の置換、挿入、または削除を有する配列の1つもしくはそれ以上のアミノ酸の保存的置換のみを含有しうる。したがって、非必須アミノ酸は、生体機能を実質的に変化せずにこれらの配列の1つにおいて変化されうる残基である。例えば、本発明のhdfAおよび/またはhdfBタンパク質の中で保存されるアミノ酸残基は、特に変化の影響を受けにくいことが予測される。さらに、本発明のhdfAおよび/またはhdfBタンパク質の中で保存されるアミノ酸は、変化の影響を受ける可能性が低い。
【0129】
「保存的置換」という語は、アミノ酸残基が同様の側鎖を有するアミノ酸残基で置換される置換を意味することが意図されている。これらのファミリーは当技術分野で周知であり、塩基側鎖(例えば、リシン、アルギニン、およびヒスチジン)、酸側鎖(例えば、アスパラギン酸、グルタミン酸)、非荷電極性側鎖(例えば、グリシン、アスパラギン、グルタミン、セリン、トレオニン、チロシン、システイン)、非極性側鎖(例えば、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン)、ベータ−分岐側鎖(例えば、トレオニン、バリン、イソロイシン)、および芳香族側鎖(例えば、チロシン、フェニルアラニン、トリプトファン、ヒスチジン)を含むアミノ酸を含む。
【0130】
機能的核酸等価物は一般的に、サイレント突然変異、すなわちコードされたポリペプチドの生体機能を変化させることがない突然変異を含有しうる。したがって、本発明は、特定の生物活性に重要ではないアミノ酸残基における変化を含有するhdfAおよび/またはhdfBタンパク質をコードする核酸分子を提供する。かかるhdfAおよび/またはhdfBタンパク質はアミノ酸配列において配列番号3または6、または20または23と異なり、さらにその生物活性の少なくとも1つを保持する。一実施形態において、単離核酸分子はタンパク質をコードするヌクレオチド配列を含んで成るが、そのタンパク質は、配列番号3または6または20または23で示されたアミノ酸配列と少なくとも約60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%もしくはそれ以上相同の実質的に相同のアミノ酸配列を含んで成る。例えば、表現型的にサイレントアミノ酸置換するやり方に関する手引きは、ボウイ(Bowie),J.U.ら、Science 247:1306−1310頁(1990年)に示されており、ここで著者らは、変化に対するアミノ酸配列の耐性を研究する2つの主要な方法があることを示している。最初の方法は進化の方法に依拠し、ここで突然変異は自然な選択によって受入れられ、または拒絶される。第2の方法では、クローン化遺伝子の特定の位置でアミノ酸の変化を導入する遺伝子操作、および選択、すなわちスクリーニングが使用され、機能性を維持する配列を特定する。著者らが述べているように、これらの研究では、タンパク質が意外にもアミノ酸置換に耐えることが示された。著者らはさらにどの変化がタンパク質の特定の位置で許容的である可能性が高いかを示している。例えば、大部分の埋もれたアミノ酸残基は非極性側鎖を必要とするが、表面側鎖のほとんどの特徴は、ほとんどの場合、保存されていない。かかる表現型的にサイレントの他の置換がボウイ(Bowie)らおよびそれに引用された文献に記載されている。
【0131】
配列番号3または6または20または23によるタンパク質と相同的なhdfAおよび/またはhdfBタンパク質をコードする単離核酸分子が、1つもしくはそれ以上のアミノ酸置換、削除、または挿入がコードされたタンパク質へ導入されるように、1つもしくはそれ以上のヌクレオチド置換、付加、または削除を配列番号2または配列番号5、または配列番号19または配列番号22によるコードヌクレオチド配列へ導入しすることによって生成されうる。かかる突然変異は、部位特異的変異誘発およびPCR介在変異誘発など標準の方法によって導入されうる。
【0132】
「機能的等価物」という用語は、A.ニゲルのhdfAおよび/またはhdfBタンパク質のオルソログをも包含する。A.ニゲルのhdfAおよび/またはhdfBタンパク質のオルソログは、他の菌株または種から単離されうるとともに、同様または同一の生物活性を有するタンパク質である。かかるオルソログは、配列番号3または6または20または23と実質的に相同的であるアミノ酸配列を含んで成るとして容易に識別されうる。
【0133】
「相同」は「実質的に相同」の意味としても理解されうる。
【0134】
「実質的に相同」という用語は、第2のアミノ酸またはヌクレオチド配列と十分または最小の数の同一または等価の(例えば、同様の側鎖を有する)アミノ酸またはヌクレオチドを含有する第1のアミノ酸またはヌクレオチド配列を指し、第1および第2のアミノ酸またはヌクレオチド配列は共通のドメインを有するようになっている。例えば、約45%、好ましくは約50%、好ましくは約60%、好ましくは約65%、より好ましくは約70%、さらにより好ましくは約75%、80%、85%、90%、95%、96%、97%、98%、もしくは99%もしくはそれ以上の同一性を有する共通のドメイン含有するアミノ酸またはヌクレオチド配列が、十分に同一と本明細書で規定されている。
【0135】
また、配列番号2または5または19または22と異なるヌクレオチド配列を有する他のhdfAおよび/またはhdfBファミリーメンバーをコードする核酸も本発明の範囲内である。さらに、異なる種からのhdfAおよび/またはhdfBタンパク質をコードする核酸は、したがって配列番号2または5または19または22と異なるヌクレオチド配列を有する。
【0136】
本発明のhdfAおよび/またはhdfBDNAの変異形(例えば、天然対立遺伝子多型)および相同体に対応する核酸分子が、好ましくはきわめて厳格なハイブリダイゼーション条件下に標準のハイブリダイゼーション法によるハイブリダイゼーションプローブとして本明細書で開示されたcDNAまたはその適切な断片を使用する本明細書で開示されたhdfAおよび/またはhdfB核酸とのその相同に基づき単離されうる。
【0137】
ハイブリダイゼーション反応の「厳格性」は、当業者によって容易に決定可能であり、ほとんどの場合、プローブの長さ、洗浄温度、および塩濃度に依存した経験的計算である。一般に、長いプローブは適切なアニーリングのために高い温度を必要とするが、短いプローブは低い温度を必要とする。ハイブリダイゼーションはほとんどの場合、相補鎖がその融解温度未満の環境で存在する場合に再アニーリングする変性DNAの能力に依存する。プローブとハイブリダイズ可能な配列との間の所望の相同の程度が高いほど、使用されうる相対温度は高くなる。結果として、高い相対温度は、反応条件をより厳格にする傾向があるが、低い温度は厳格でなくなることになる。
【0138】
ハイブリダイゼーション反応の厳格性の追加の詳細および説明については、オーズベルら、Current Protocols in Molecular Biology、ワイリー・インターサイエンス・パブリッシャーズ(Wiley Interscience Publishers)(1995年)を参照されたい。
【0139】
本明細書で規定される「厳格な条件」または「高厳格性条件」は、以下によって識別されうる。すなわち、(1)洗浄のために低いイオン強度および高い温度、例えば、0.015M塩化ナトリウム/0.0015Mクエン酸ナトリウム/0.1%ドデシル硫酸ナトリウムを50℃下に使用し、(2)ハイブリダイゼーション中に、ホルムアミドなどの変性剤、例えば、50%(v/v)ホルムアミドを0.1%ウシ血清アルブミン/0.1%Ficdl/0.1%ポリビニルピロリドン/750mM塩化ナトリウムとともにpH6.5で50mMリン酸ナトリウム緩衝液、75mMクエン酸ナトリウムを42℃下に使用し、または(3)50%ホルムアミド、5×SSC(0.75M NaCl、0.075Mクエン酸ナトリウム)、50mMリン酸ナトリウム(pH6.8)、0.1%ピロリン酸ナトリウム、5×デンハルト液、超音波処理サケ精子DNA(50Rg/ml)、0.1%SDS、および10%硫酸デキストランを42℃下に使用するとともに、0.2×SSC(塩化ナトリウム/クエン酸ナトリウム)中42℃下および55℃下での50%ホルムアミド中での洗浄後、55℃下にEDTAを含有する0.1×SSCから成る高厳格性洗浄を行う。
【0140】
「中程度に厳格な条件」は、サンブルックら、Molecular Cloning:A Laboratory Manual、ニューヨーク:コールド・スプリング・ハーバー・プレス、1989年によって記載されているように識別されうるとともに、洗浄液および上記の厳格性の低いハイブリダイゼーション条件(例えば、温度、イオン強度、および%SDS)の使用を含む。中程度に厳格な条件の例は、20%ホルムアミド、5×SSC(150mM NaCl、5mMクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハルト液、10%硫酸デキストラン、および20mg/ml変性剪断サケ精子DNAを含んで成る溶液中37℃での一夜インキュベーション後、ろ過液の1×SSC中約37〜50℃下での洗浄である。当業者は、温度、イオン強度等を調節するやり方を、必要に応じて、プローブの長さなどの要因を適合させ、または配列類似性を決定するために適切なアルゴリズムを使用することによって認識するであろう。
【0141】
相同的(同様または同一)配列は、「配列比較アルゴリズム」を使用することによっても決定されうる。比較のための配列の最適なアラインメントは、例えば、スミス(Smith)とウォーターマン(Waterman)、Adv.Appl.Math.2:482頁(1981年)の局所相同アルゴリズムによって、ニードルマン(Needleman)とブンシュ(Wunsch)、J.Mol.Biol.48:443頁(1970年)の相同アラインメントアルゴリズムによって、ピアソン(Pearson)とリップマン(Lipman)、Proc.Nat’l Acad.Sci.米国(USA)85:2444頁(1988年)の類似性法の調査によって、これらのアルゴリズムのコンピュータによる実行(ウィスコンシン・ジェネティックス・ソフトウェア・パッケージ(Wisconsin Genetics Software Package)のGAP、BESTFIT、FASTA、およびTFASTA、ジェネティックス・コンピュータ・グループ(Genetics Computer Group)、575サイエンス(Science)Dr.、Madison、ウィスコンシン州(WI))によって、または目視検査によって実行されうる。配列類似性を決定するために適切であるアルゴリズムの例がBLASTアルゴリズムであり、これはアルトシュール(Altschul)ら、J.Mol.Biol.215:403−410頁(1990年)に記載されている。
【0142】
BLAST分析を実行するためのソフトウェアは、国立バイオテクノロジー情報センター(National Center for Biotechnology Information)によって公開されている。(http://www.ncbi.nlm.nih.gov/)。このアルゴリズムは最初に、データベース配列における同じ長さの単語と整列すると一部の正に評価された閾値スコアTに一致またはこれを満たすクエリー配列における長さWの短い単語を識別することによる高スコアリング配列ペア(HSP)の識別を含む。これら最初の近傍単語のヒットは、それらを含有する長いHSPを見出す出発点として作用する。単語のヒットは、累積的アラインメントスコアが増大しうる限り比較される2つの配列の各々に沿った両方の方向で拡大する。単語のヒットの拡張は、累積的アラインメントスコアが最大達成値から数量Xだけ減少すると停止し、累積的スコアはゼロもしくはそれ未満になり、またはどちらか一方の配列の末端に達する。
【0143】
BLASTアルゴリズムパラメータW、T、およびXは、アラインメントの感度および速度を決定する。BLASTプログラムでは初期設定として11の語長(W)、50のBLOSUM62スコアリングマトリクス(ヘニコフ(Henikoff)とヘニコフ、Proc.Natl.Acad.Sci.米国(USA)89:10915頁(1989年)を参照)50のアラインメント(B)、10の期待値(E)、M=5、N=4、および両方の鎖の比較が使用される。
【0144】
次いで、BLASTアルゴリズムでは2つの配列間の類似性の統計的分析を実行する(例えば、カーリン(Karlin)とアルトシュール、Proc.Nat’l.Acad.Sci.米国(USA)90:5873−5787頁(1993年)を参照)。BLASTアルゴリズムによって提供される類似性の1つの尺度は最小合計確率(P(N))であり、これはそれによって2つのヌクレオチドまたはアミノ酸配列の間の適合が偶然に起こりうる確率の指標を提供する。例えば、アミノ酸配列がプロテアーゼなどのタンパク質と同様とみなされるのは、プロテアーゼアミノ酸配列などのタンパク質との試験アミノ酸配列の比較における最小合計確率が約0.1未満、より好ましくは約0.01未満、かつ最も好ましくは約0.001未満である場合である。好ましくは、類似性は、配列番号2、5、19、および22を有するDNA配列の1つと少なくとも40%相同である。より好ましくは、類似性は少なくとも50%、より好ましくは少なくとも60%、より好ましくは少なくとも70%、より好ましくは少なくとも80%、より好ましくは少なくとも90%である。
【0145】
hdfAおよび/またはhdfB配列の自然発生的対立遺伝子多型に加えて、当業者は、突然変異によって配列番号2またh5または19または22のヌクレオチド配列へ変化が導入され、それによってhdfAおよび/またはhdfBタンパク質の機能を実質的に変化させることなく、hdfAおよび/またはhdfBタンパク質のアミノ酸配列の変化をもたらしうることを認識するであろう。
【0146】
本発明の別の態様において、劣化hdfAおよび/またはhdfBタンパク質が提供される。劣化hdfAおよび/またはhdfBタンパク質は、少なくとも1つの生物活性が減少しているタンパク質である。かかるタンパク質は、hdfAおよび/またはhdfBコード配列の全部または一部に沿って、飽和変異誘発によるなど、突然変異を無作為に導入することによって得られうるが、結果として生じる変異体は組換え発現され、生物活性についてスクリーニングされる。例えば、当業界は、その酵素活性を測定するための標準のアッセイを提供しており、したがって劣化タンパク質が容易に選択されうる。好ましくは、アッセイは以前に記載されているものである(例えば、国際公開第02/052026号パンフレット、23頁、または表現型スクリーニングアッセイを参照)。
【0147】
好ましい実施形態では、hdfAおよび/またはhdfBタンパク質は、配列番号3または6または20または23によるアミノ酸配列を有する。別の実施形態では、hdfAおよび/またはhdfBポリペプチドは、配列番号3または6または20または23によるアミノ酸配列と実質的に相同的であり、かつ配列番号3または6または20または23によるポリペプチドの少なくとも1つの生物活性を保持するが、自然の変動もしくは上述の変異誘発によりアミノ酸配列が異なる。
【0148】
別の好ましい実施形態によれば、hdfAおよび/またはhdfBタンパク質は、好ましくはきわめて厳格なハイブリダイゼーション条件下、配列番号2または5または19または22による核酸にハイブリダイズが可能な単離核酸断片によってコードされたアミノ酸配列を有する。
【0149】
したがって、hdfAおよび/またはhdfBタンパク質は、配列番号3または6または20または23で示されたアミノ酸配列と少なくとも約60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%もしくはそれ以上相同のアミノ酸配列を含んで成るタンパク質であり、配列番号3または6または20または23によるポリペプチドの少なくとも1つの機能的活性を保持する。
【0150】
本発明によるタンパク質の機能的等価物は、例えば、一定の活性の本発明のタンパク質の変異体、例えば、切断変異体のコンビナトリアルライブラリーをスクリーニングすることによっても識別されうる。一実施形態では、変異形の変化に富んだライブラリーが核酸レベルでコンビナトリアル変異誘発によって生成される。変異形の変化に富んだライブラリーが、例えば、退化セットの潜在的タンパク質配列が個別のポリペプチドとして、あるいは、一連の大きな融合タンパク質として(例えば、ファージディスプレイのために)発現可能であるように、合成オリゴヌクレオチドの混合物を遺伝子配列へ酵素的にライゲートすることによって生成されうる。退化オリゴヌクレオチド配列からの本発明のポリペプチドの潜在的な変異形のライブラリーを生成するために使用されうるさまざまな方法がある。退化オリゴヌクレオチドを合成するための方法が当技術分野で周知である(例えば、ナラング(Narang)(1983年)Tetrahedron 39:3頁、イタクラ(Itakura)ら(1984年)Annu.Rev.Biochem.53:323頁、イタクラら(1984年)Science 198:1056頁、イケ(Ike)ら(1983年)Nucleic Acid Res.11:477頁)を参照)。
【0151】
また、本発明のポリペプチドのコード配列の断片のライブラリーを使用し、変異形のその後の選択をスクリーニングするためのポリペプチドの変化に富んだ集団を生成することができる。例えば、コード配列断片のライブラリーが、目的とするコード配列の二本鎖PCR断片を切断が分子当たり約1回のみ生じる条件下にヌクレアーゼで処理し、二本鎖DNAを変性させ、DNAを再生させて異なる切断生成物からのセンス/アンチセンスペアを含みうる二本鎖DNAを形成し、S1ヌクレアーゼによる処理によって再形成二本鎖から一本鎖部分を除去し、結果として生じる断片ライブラリーを発現ベクターへライゲートすることによって生成されうる。この方法によって、目的とするタンパク質のさまざまなサイズのN末端および内部断片をコードする発現ライブラリーが得られうる。
【0152】
切断の点突然変異によって製造されるコンビナトリアルライブラリーの遺伝子産物をスクリーニングし、かつ選択された特性を有する遺伝子産物のDNAライブラリーをスクリーニングするためのいくつかの方法が当技術分野で周知である。大きな遺伝子ライブラリーをスクリーニングするための高スループット分析に適している最も広く使用される方法は一般的に遺伝子ライブラリーを確実な発現ベクターへクローニングし、適切な細胞を結果として生じるベクターのライブラリーで形質転換し、かつ所望の活性の検出がその生成物が検出された遺伝子をコードするベクターの単離を促進する条件下にコンビナトリアル遺伝子を発現させるステップを含む。ライブラリーにおける機能的変異体の頻度を増強する技術である再帰的アンサンブル変異誘発(REM)は、本発明のタンパク質の変異形を識別するスクリーニングアッセイと組合せて使用されうる(アーキン(Arkin)と(ユアバン(Yourvan)(1992年)Proc.Natl.Acad.Sci. 米国(USA)89:7811−7815頁、デルグレーブ(Delgrave)ら(1993年)Protein Engineering 6(3):327−331頁)。
【0153】
配列番号2および5および19および22で示されたhdfAおよび/またはhdfB遺伝子配列に加えて、hdfAおよび/またはhdfBタンパク質のアミノ酸の変化をもたらしうるDNA配列多型が、一定の集団内に存在しうることを当業者には明らかであろう。かかる遺伝子多型は、自然な対立遺伝子の変動により異なる集団からの、または集団内の細胞に存在しうる。対立遺伝子変異形も機能的等価物を含みうる。
【0154】
本発明によるポリヌクレオチドの断片は、機能的ポリペプチドをコードしないポリヌクレオチドをも含んで成る。かかるポリヌクレオチドはPCR反応のプローブまたはプライマーとして機能しうる。
【0155】
それらが機能的または非機能的ポリペプチドをコードするかどうかに関係なく本発明による核酸は、ハイブリダイゼーションプローブまたはポリメラーゼ連鎖反応(PCR)プライマーとして使用されうる。hdfAおよび/またはhdfB活性を有するポリペプチドをコードすることがない本発明の核酸分子の使用には、とりわけ、(1)hdfAおよび/またはhdfBタンパク質をコードする遺伝子、またその対立遺伝子変異形をcDNAライブラリーから、例えば、A.ニゲルまたはペニシリウム・クリソゲヌム以外の他の生物から単離、(2)フェルマ(Verma)ら、Human Chromosomes:a Manual of Basic Techniques、ペルガモン・プレス(Pergamon Press)、ニューヨーク(1988年)に記載されているhdfAおよび/またはhdfB遺伝子の正確な染色体位置を提供する中期染色体スプレッドへのインサイチュハイブリダイゼーション(例えば、FISH)、(3)特定の組織および/または細胞におけるhdfAおよび/またはhdfB mRNAの発現を検出するためのノーザンブロット分析、および4)一定の生体(例えば、組織)試料におけるhdfAおよび/またはhdfBプローブにハイブリダイズ可能な核酸の存在を分析する診断ツールとして使用されうるプローブおよびプライマーがある。
【0156】
hdfAおよび/またはhdfB遺伝子またはcDNAの機能的等価物を得る方法も本発明によって包含される。かかる方法は、配列番号2または5または19または22による配列またはその変異形の全部または一部をコードする単離核酸を含む標識プローブを得るステップ、ライブラリーにおける核酸断片に対するプローブのハイブリダイゼーションを可能にする条件下に標識プローブで核酸断片ライブラリーをスクリーニングするステップ、それによって核酸二本鎖を形成するステップ、および任意の標識二本鎖における核酸断片から完全長遺伝子配列を調製し、hdfAおよび/またはhdfB遺伝子に関係がある遺伝子を得るステップを必要とする。
【0157】
1つの実施形態において、本発明のhdfAおよび/またはhdfB核酸は、配列番号1、または2、または4、または5、または18、または19、または21、または22に示されている核酸配列と少なくとも45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、もしくはそれ以上相同である。
【0158】
別の好ましい実施形態において本発明のhdfAおよび/またはhdfBポリペプチドは、配列番号3または6または20または23に示されたアミノ酸配列と少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、もしくはそれ以上相同である。
【0159】
本発明は、配列番号1、または2、または4、または5、または18、または19、または21、または22それ自体を有するDNA配列、および上記のその相同体に関する。これらのDNA配列に関し、かつ遺伝子コードの退化によって得られるDNA配列も本発明の一部である。DNA配列番号2、5、19、および22に関し、かつハイブリダイゼーション(前段落参照)によって得られるDNA配列も発明の一部である。これらDNA配列によってコードされた単離ポリペプチドまたは上記のその相同体も本発明の一部である。ポリペプチドhdfAおよびhdfBは、NHRに関与する機能を有する。これらポリペプチドのすべてが本発明の方法において使用され、改善された標的効率を有しうる糸状菌を得ることができる。
【0160】
本発明は、以下の実施例でより詳細に例示される。かかる実施例は本発明の範囲を限定することが意図されていない。
【実施例】
【0161】
実施例1:hdfAおよび/またはhdfB遺伝子の識別および削除ベクターの構成。
アスペルギルス・ニゲル菌株CBS513.88のゲノムDNAを配列決定し、分析した。KU70およびKU80との相同体と注釈付きの翻訳タンパク質を有する2つの遺伝子を識別し、それぞれhdfAおよびhdfBと命名した。オープンリーディングフレーム(ORF)(イントロンを有する)および遺伝子の約1000bp5’および3’を含んで成るhdfAおよび/またはhdfB遺伝子座の配列が、配列表1および4に示されている。hdfAおよび/またはhdfBの遺伝子置換ベクターが、既知の原理に従って設計され、ルーチンのクローニング法に従って構成された(図1および2を参照)。本質的に、これらのベクターは、予定されたゲノム遺伝子座での相同的組換えのhdf ORFの約1000bpフランキング領域を含んで成る。また、それらは、A.ニドゥランス双方向amdS選択マーカー、中間直接繰返しを含有する。これら削除ベクターの一般設計は、欧州特許第635574B号明細書および国際公開第98/46772号パンフレットにおいて既述されている。
【0162】
実施例2:アスペルギルス・ニゲルにおけるhdfA遺伝子の不活性化。
削除ベクターpDEL−HDFAの直鎖DNA(図1)を単離して使用し、以前に記載された方法(Biotechnology of Filamentous fungi:Technology and Products.(1992年)リード・パブリッシング(Reed Publishing)(米国(USA))、第6章:Transformation 113〜156頁)を使用することによりアスペルギルス・ニゲルCBS513.88を形質転換した。この直鎖DNAはhdfA遺伝子座でゲノムへ取込むことができ、したがって、図3に示されているようにamdS遺伝子によってhdfA遺伝子を置換する。形質転換体をアセトアミド培地で選択し、欧州特許第635574B号明細書に記載された標準の方法に従ってコロニーを精製した。胞子をフルオロ・アセトアミド培地上にプレーティングし、amdSマーカーを失った菌株を選択した。発育コロニーをhdfA遺伝子座での取込みについてPCRで診断し、候補の菌株をhdfA遺伝子の削除についてサザン分析で試験した。hdfA遺伝子の削除は、遺伝子座全体をカバーするDNA断片の約2.2kbサイズの削減によって検出可能であり、適切なプローブにハイブリダイズされた。約8個の菌株が、約400の初期形質転換体のプールからゲノムhdfA遺伝子の除去を示した。
【0163】
菌株dHDFAを不活性化したhdfA遺伝子を有する代表的な菌株として選択した。
【0164】
実施例3:アスペルギルス・ニゲルにおけるhdfB遺伝子の不活性化。
削除ベクターpDEL−HDFBの直鎖DNA(図2)を単離して使用し、アスペルギルス・ニゲル菌株CBS513.88を形質転換した。この直鎖DNAはhdfB遺伝子座でゲノムへ取込むことができ、したがって、amdS遺伝子によってhdfB遺伝子を置換する(図4)。実施例2に記載されたものと同じ遺伝子置換法を使用した。形質転換体をアセトアミド培地で選択し、標準の方法に従ってコロニーを精製した。胞子をフルオロ・アセトアミド培地上にプレーティングし、amdSマーカーを失った菌株を選択した(欧州特許第635574B号明細書)。発育コロニーをhdfB遺伝子座での取込みについてPCRで診断し、候補の菌株をhdfB遺伝子の削除についてサザン分析で試験した。hdfB遺伝子の削除は、遺伝子座全体をカバーするDNA断片の約2.6kbサイズの削減によって検出可能であり、適切なプローブにハイブリダイズされた。約7個の菌株が、約370の初期形質転換体のプールからゲノムhdfB遺伝子の除去を示した。
【0165】
菌株dHDFBを不活性化したhdfB遺伝子を有する代表的な菌株として選択した。
【0166】
実施例4:アスペルギルス・ニゲルにおけるhdfAおよびhdfB遺伝子の不活性化。
削除ベクターpDEL−HDFBの直鎖DNA(図2)を単離して使用し、実施例2で得られた菌株dHDFAを形質転換した。この直鎖DNAはhdfB遺伝子座でゲノムへ取込むことができ、したがって、amdS遺伝子によってhdfB遺伝子を置換する(図4)。実施例2に記載されたものと同じ遺伝子置換法を使用した。形質転換体をアセトアミド培地で選択し、標準の方法に従ってコロニーを精製した。胞子をフルオロ・アセトアミド培地上にプレーティングし、amdSマーカーを失った菌株を選択した。発育コロニーをhdfB遺伝子座での取込みについてPCRで診断し、候補の菌株をhdfB遺伝子の削除についてサザン分析で試験した。hdfB遺伝子の削除は、遺伝子座全体をカバーするDNA断片の約2.6kbサイズの削減によって検出可能であり、適切なプローブにハイブリダイズされた。約15個の菌株が、約380の初期形質転換体のプールからゲノムhdfB遺伝子の除去を示した。
【0167】
菌株dHDFABを不活性化したhdfAおよびhdfB遺伝子を有する代表的な菌株として選択した。
【0168】
実施例5:単一相同的組換えイベントの改善された標的
DNAが予定された遺伝子座でアスペルギルス・ニゲルのゲノムへ取込みうる1つの機序は、単一相同的組換えによるものである。相同的DNAは、組換えによってゲノム位置で整列して取込む(図5を参照)。2つのベクターを使用し、実施例2、3、および4で得られたアスペルギルス・ニゲル菌株の単一相同的組換えによる標的効率を試験した。2つのベクターは、組換えおよび結果として生じる取込みを導くグルコアミラーゼ(glaA)遺伝子座と相同の領域を含んで成る(図5)。
【0169】
かかる相同的取込みのために設計された第1のベクターはすでに以前に国際公開第02/45524号パンフレットに記載されている(pGBFIN11−EPO)。このベクターは、プロリン特異的エンドプロテアーゼをコードする遺伝子を含有する。
【0170】
第2のベクター(pGBFIN11−PLA)は、A.オリゼからのホスホリパーゼA1(PLA1)をコードする遺伝子を含有する。この酵素をコードする遺伝子はすでに公表されている(ワタナベ(Watanabe)Iら、Biosci.Biotechnol. Biochem.(1999年)、63巻、5号、820−826頁)。この遺伝子は、pGBFIN11−EPOにおけるプロリン特異的エンドプロテアーゼ遺伝子のクローニングのために国際公開第02/45524号パンフレットに記載されたのと同じ方法を使用してpGBFIN11へクローン化された。
【0171】
菌株CBS513.88、dHDFA、dHDFB、およびdHDFABを実施例2で既述された形質転換法に従ってpGBFIN11−EPOまたはGBFIN11−PLAプラスミドのいずれかで形質転換した。得られた結果は、使用された両方のプラスミドで同じであった。発明者らはそれぞれ、標的遺伝子座で取込まれたプラスミドによる形質転換体の5%、10%、10%、および10%を見出した。したがって、発明者らは、アスペルギルス・ニゲルにおける少なくとも1つのhdf遺伝子の不活性化が単一の相同的組換えイベントによってこれらの菌株の標的効率の大幅な増大をもたらすと結論づけた。
【0172】
実施例6:数種類の遺伝子座での二重相同的組換えイベントの改善された標的。
さらに、ゲノムからの遺伝子をコードする多くのアミラーゼの不活性化のために設計された削除ベクターでのdHDFA菌株の形質転換によって標的効率を測定した。遺伝子フランキング領域を実質的に実施例1に記載されているようにクローン化し、結果として生じるベクターを線状化し、これを使用してCBS513−88およびdHDFA菌株のプロトプラストを形質転換した。標的頻度をPCR分析、および対応する遺伝子の不活性化を示す活性に基づくプレートアッセイによって測定した。後者は、0.4%寒天を補充したPDAプレート上で形質転換体を増殖させ、その後にヨード/カリウムヨード溶液(ルゴール(Lugol)、シグマ(Sigma)L6146)による染色によって行われた。以下の表1でわかるように、PCR分析および/または対応する遺伝子の不活性化を示す活性に基づくプレートアッセイによって判定された標的頻度は、CBS513.88菌株で観察されたものに比べ大幅に改善された。
【0173】
【表1】

【0174】
これらの所見により、アスペルギルス・ニゲルにおける少なくとも1つのhdf遺伝子の不活性化が結果として二重相同的組換えによる取込みに対するベクターの標的効率の大幅な増大をもたらすという本発明者らの結論がさらに裏付けられる。
【0175】
実施例7:標的頻度に対するamyBII遺伝子の相同的フランキング領域のサイズ削減の効果。
別個の一連の実験において、二重相同的組換えによる形質転換効率および標的頻度に対するフランキング領域長さの効果をさらに調査した。菌株CBS513.88およびdHDFAのプロトプラストを、可変長のamyBIIフランキング領域によってフランキングされたA.ニドゥランスamdSマーカーを含むPCR断片で形質転換した。表2に示されたデータは、増強された全体的な形質転換効率に加えて、統合的カセットの標的がdHDFA菌株においてはるかに改善されたことを明らかに示す。
【0176】
【表2】
【0177】
実施例8:ポリペプチドの表現型分析および製造。
dHDFA、dHDFB、またはdHDFAB菌株の固体培地または振盪フラスコでの発育中の表現型の差異は確認されなかった。プラスミドpGBFIN11−EPOまたはpBGFIN−PLAで形質転換されたdHDFA、dHDFB、およびdHDFAB菌株はすべて、以下の方法に従って判定されるように培地へ活性酵素を分泌した。
【0178】
固体培地はポテトデキストロース寒天(PDA)培地(ジフコ(Difco)、ポテトデキストロース寒天(POTATO DEXTROSE AGAR)、培養培地、カタログ番号213400、1996−1997年)であった。
【0179】
振盪フラスコ実験を欧州特許第635574B号明細書に記載されているように培地100ml中、34℃下、および500mlバッフル付き振盪フラスコを使用してインキュベーター振盪機中170rpmで行った。発酵の4日後、試料を取り、プロリン特異的エンドプロテアーゼ活性またはホスホリパーゼ活性のいずれかに判定した。
【0180】
プロリン特異的エンドプロテアーゼのタンパク質分解活性を分光光度法で時間内にpH5および約37℃で基質としてZ−グリ(Gly)(シン)−プロ(Pro)(リン)−pNAを使用して測定した。1Uプロリン特異的エンドプロテアーゼは、毎分Z−グリ(Gly)(シン)−プロ(Pro)(リン)−pNA1ミクロモルをpH5および37℃下に変換する酵素の量と定義されている。
【0181】
アスペルギルス・ニゲル(PLA1)からのホスホリパーゼPLA1活性を分光光度法で測定するために、人工的基質、すなわち、1,2−ジチオジオクタノイルホスファチジルコリン(diC8、基質)が使用される。PLA1はA1位で硫化物結合を加水分解し、チオオクタン酸を解離する。チオオクタン酸は、4,4ジチオピリジン(呈色試薬、4−DTDP)と反応し、4−チオピリドンを形成する。4−チオピリドンは、334nmの波長を有する放射線を吸収する4−メルカプトピリジンと互変異性平衡にある。同波長での吸光変化は測定される。1単位は、1nmolチオオクタン酸を1,2−ジチオジオクタノイルホスファチジルコリンから毎分37℃およびpH4.0で遊離する酵素の量である。
【0182】
基質溶液は、エタノール66ml当たりdiC8 1gを溶解し、酢酸バッファー264mlを添加することによって調製される。酢酸バッファーは、0.2%トリトン(Triton)−X100を含有する0.1M酢酸バッファーpH3.85を含んで成る。呈色試薬は、11mM 4,4−ジチオジピリジン溶液である。これは、2mlエッペンドルフ試料カップ中に4,4−ジチオジピリジン5.0mgを秤量し、エタノール1.00ml中に溶解することによって調製した。ミリQ水1.00mlを添加した。
【0183】
興味深いことに、色差またはコロニー外観など形態的変化は、CBS513.88から得られた形質転換体よりもdHDFA、dHDFB、およびdHDFAB菌株から得られた形質転換体で低頻度に生じた。これは無作為取込み(NHR)の削減によるものであり、したがって予想外の表現型の変化を阻止すると考えられる。
【0184】
実施例9:変異誘発による相同的組換えの改善された効率を有するペニシリウム変異体の単離
遺伝子標的の改善された効率を有する変異体を単離するために、典型的な変異誘発と分子生物学の組合せを適用した。ペニシリウム・クリソゲヌム(CBS 455.95)胞子をYEPD(ジフコ製の2%酵母抽出物、ジフコ製の1%ペプトン、2%グルコース)中で胞子形成するコロニーから得た。これらの胞子を滅菌水道水中で洗浄し、ml当たり108分生子を含有する懸濁液10mlを254nmでUV照射にかけた(シルバニア(Sylvania)、15ワット・ブラックライトブルー管、モデルFT15T8/BLB)。UV照射を7.5、15、または30分間当てると同時に、懸濁液をゆっくり振盪した。これらの異なる照射時間は、細胞における軽度、中等度、および強度の突然変異率レベルを得るために選択された。暗所での回収の1時間後、これら3時点からの細胞を2つの同等のアリコートに分けた。第1の試料を既述のように直接再胞子形成し(ヘルスバッハ(Hersbach),GJM、ファン・デン・ビーク(Van der Beek),CP、およびファン・ジユック(Van Dijck),PWM.The Penicillins:properties,biosynthesis and fermentation.In:Vandamme EJ(編) Biotechnology of Industrial Antibiotics(45−140頁)、マルセル・デッカー(Marcel Dekker)、ニューヨーク)、もう一方の試料をYNB培地(アミノ酸(ジフコ)、2.0%w/vグルコースとともに0.67%w/v酵母窒素塩基)中で長い回収期間、胞子形成を導入する前に4時間、25℃下にインキュベートした。
【0185】
第3の変異誘発試料をYNB中で一夜、野生型胞子を発生させた後、滅菌水道水での2つの洗浄ステップによって得て、滅菌水道水中に再懸濁した。さらにUV照射を7.5、15、および30分間当てると同時に、懸濁液をゆっくり振盪した。これらの試料を(上述したように)暗所での回収の1時間後に直接、再胞子形成した。
【0186】
これらの望ましい変異体をこれら変異誘発集団から選択するために、変異誘発集団をYEPD培地に接種した。発芽後、細胞の発達を標準の光学顕微鏡を使用して追跡した。培養の平均菌糸が申し分なく発達している場合、細胞を収穫し、溶解酵素でインキュベートし、プロトプラストを得た。プロトプラストを機能的選択マーカーの発現カセットを有する2つのDNA断片、bleおよびamdSで形質転換された。遺伝子bleはフレオマイシンへの耐性を与えることが可能なタンパク質をコードする(コラー(Kolar)M、プント(Punt)PJ、ファン・デン・ホンデル CA、シュワブ(Schwab)H.Transformation of Penicillium chrysogenum using dominant selection markers and expression of an Escherichia coli lacZ fusion gene.Gene.1988年、62(1):127−34頁)。遺伝子amdSは、細胞が唯一の窒素源としてアセトアミドで発育することを可能にするタンパク質をコードする(欧州特許第635574B号明細書に記載)。P.クリソゲヌムのプロトプラストへのDNAの移動に適用される方法は当技術分野で周知であり、フィンケルシュタインとボール(編)、Biotechnology of filamentous fungi,Technology and products,Butterworth−Heinemann(1992年)、ベネットとラシュア(編)More Gene Manipulations in fungi、Academic Press(1991年)、ターナー、ピューラー(編)、Biotechnology、第2完全改訂版、VHC(1992年)を含む多くの文献に記載されている。Ca−PEG介在プロトプラスト形質転換はEP635574号明細書に記載されているように使用される。
【0187】
ペニシリウムゲノムにおける特異的遺伝子座に対するこれらの発現カセットの標的取込みを選択するために、DNAの短い相同的伸長をDNA断片の両側でPCRによって添加した。3タイプの構成物を作った。すなわち、第1のタイプは30bpのDNAの相同的伸長を、第2のタイプは50bp、および第3は100bpを含有する。選択は、27の明確なバッチを規定する30、50、または100bpの相同的伸長を有する2つのDNA構成物(bleおよびamdS)で9つの胞子形成バッチから得られた変異体を形質転換することによって行った。ble遺伝子はniaD遺伝子座に標的され、それによって硝酸還元酵素遺伝子を破壊し(ゴウカ RJ、ファン・ハルチングスフェルト W、ボーベンベルグ RA、ファン・デン・ホンデル CA、ファン・ゴルコム RF、Cloning of the nitrate−nitrite reductase gene cluster of penicillium chrysogenum and use of the niaD gene as a homologous selection marker、J Biotechnol.、1991年9月、20(2):189−99頁)、細胞が塩素酸塩に耐性となると、塩素酸塩を含有するプレートへの形質転換体の選択を導くことを可能にした。amdS遺伝子はsutB遺伝子座に標的され、それによって硫酸パーミアーゼ遺伝子を破壊し(ファン・デ・カンプ M、ピッツィニーニ E、フォス A、ファン・デア・レンデ TR、シュールス TA、ニューベルト RW、ターナー G、コーニングス WN、ドリーゼン AJ、Sulfate transport in Penicillium chrysogenum:cloning and characterization of the sutA and sutB genes.、J. Bacteriol.1999年12月、181(23):7228−34頁)、セレン酸塩を含有するプレートへの形質転換体の選択を導くことを可能にした。形質転換体が最初に塩素酸塩で選択され、次いでセレン酸塩について試験された。さらに、選択マーカーの存在は、唯一の窒素源としてアセトアミドを含有するプレート(欧州特許第635574B号明細書)、次いでフレオマイシンを含有するプレートで証明された。対照野生型P.クリソゲヌムとしてCBS455.95も同じDNA断片で形質転換された。存在する両方の選択マーカーを有し、塩素酸塩およびセレン酸塩に対して耐性の変異体は改善された標的取込みを有する菌株である。
【0188】
【表3】
【0189】
【表4】
【図面の簡単な説明】
【0190】
【図1】アスペルギルス・ニゲルにおけるhdfA遺伝子を不活性化するために使用される置換ベクターpDEL−HDFAを示す図である。置換ベクターは、hdfAフランキング領域、amdSマーカー、および大腸菌DNAを含んで成る。大腸菌DNAは、A.ニゲル菌株の形質転換前に制限酵素AsclおよびNotlによる消化によって除去された。
【図2】A.ニゲルにおけるhdfB遺伝子を不活性化するために使用される置換ベクターpDEL−HDFBを示す図である。置換ベクターは、hdfBフランキング領域、amdSマーカー、および大腸菌DNAを含んで成る。大腸菌DNAは、A.ニゲル菌株の形質転換前に制限酵素AsclおよびNotlによる消化によって除去された。
【図3】A.ニゲルのhdfA遺伝子を削除するために使用される方法を示す図である。使用されるDNA構成物は、hdfA遺伝子の相同領域(5’および3’)によってフランキングされたamdS選択マーカーを含んで成る(1)。この構成物は、ゲノムhdfA遺伝子座で二重相同的組換え(X)を通じて取込み(2)、かつゲノムhdfA遺伝子を置換する(3)。その後、直接繰返し(U)にわたる組換えがamdSマーカーを除去し、結果としてhdfA遺伝子の正確な切除が生じる(4)。
【図4】A.ニゲルのhdfB遺伝子を削除するために使用される方法を示す図である。DNA構成物は、hdfB遺伝子の相同領域(5’および3’)によってフランキングされたamdS選択マーカーを含んで成る(1)。この構成物は、ゲノムhdfB遺伝子座で二重相同的組換え(X)を通じて取込み(2)、かつゲノムhdfB遺伝子コピーを置換する(3)。その後、直接繰返し(U)にわたる組換えがamdSマーカーを除去し、結果としてhdfB遺伝子の正確な切除が生じる(4)。
【図5】単一相同的組換えを通じてDNA構成物をA.ニゲルのゲノムへ取込むために使用される概略的な方法を示す図である。発現ベクターは、選択可能なamdSマーカーおよびglaA遺伝子座の相同的領域によってフランキングされた目的とする遺伝子(それぞれ、3’glaAおよび3’’glaA)を含んで成り、ゲノムglaA遺伝子座での取込みを導く。
【技術分野】
【0001】
本発明は分子生物学の分野に関する。詳しくは、糸状菌のゲノムへの核酸の方向づけられた取込みの効率を改善する方法、およびその使用に関する。
【背景技術】
【0002】
真核細胞は、ポリペプチドおよび二次代謝産物の(組換え)生産のための好ましい生物である。例えば、タンパク質生産菌株を構成する際には、生産されるタンパク質をコードする目的とする遺伝子の取込みの部位は、目的とする取込み遺伝子の転写および/または発現の調節に重要である。大部分の真核生物におけるDNAのゲノムへの取込みは無作為に高頻度で起こるため、組換えDNA技術によるタンパク質生産菌株の構成はしばしば、生産されるタンパク質をコードする遺伝子を含んで成る発現カセットの望ましくない無作為取込みをもたらす。発現カセットのこのコントロール不良の「無作為多重取込み」は潜在的に危険な過程であり、宿主のゲノムの望ましくな修飾をもたらしうる。したがって、高い効率で発現カセットの正確な標的を確実にすることによってタンパク質生産菌株を構成することが可能であることがきわめて望ましい。さらに、生物の増大する量の完全ゲノムの配列が入手可能になっている現在、これは過剰発現および削除ライブラリーの橋渡しをするゲノムを構成する機会を開く。かかるライブラリーの効率的な構成の重要な要件は、問題の生物が効率的に形質転換されうること、および核酸のゲノムへの直接の標的取込みに必要とされる必要な相同が比較的短いことである。
【0003】
真核細胞は少なくとも2つの別個の経路を有し(1つは相同的組換えを介し、また1つは非相同的組換えを介し)、それらを通じて核酸(特にDNAのコース)が宿主ゲノムへ取込まれうる。酵母サッカロミセス・セルビシエ(Saccharomyces cerevisiae)は、相同的組換え(HR)選好の生物である。この生物の非相同的組換えと相同的組換え(NHR/HR)の比は、約0.07〜0.007のばらつきがありうる。
【0004】
国際公開第02/052026号パンフレットは、DNA配列のそのゲノムへの改善された標的効率を有するサッカロミセス・セルビシエの変異体を開示している。かかる変異体菌株は、NHRに関与する遺伝子(KU70)を欠いている。
【0005】
サッカロミセス・セルビシエとは逆に、哺乳類細胞に至るまでの糸状菌細胞など大部分の真核生物はNHR選好を有する。糸状菌の中では、NHR/HR比は1〜100超の範囲である。かかる生物では、標的取込み頻度はやや低い。この頻度を改善するには、かかる生物のゲノムへ取込まれるポリヌクレオチド配列をフランキングする相同的領域の長さは、例えば単一遺伝子を分裂させるためには少なくとも2000bp、および推定的形質転換体をスクリーニングするためには少なくとも500bpと比較的長いことが必要である。かかるフランキング領域の必要性は、前記ポリヌクレオチドを含んで成るDNA構成物をクローン化する際、かつそれで生物を形質転換する際の大きな負担を表し、さらに、そのフランキング領域内にある隣接遺伝子は形質転換後の組換え過程中に容易に阻害され、それによって望ましくない予想外の副作用を引起こしうる。
【0006】
KU70を欠く哺乳類細胞はすでに単離されている(ピアス(Pierce)ら、Genes and Development、(2001年)、15:3237−3242頁)。これらの変異体は6倍高い相同指向修復頻度を示すが、相同指向標的取込みの効率の増加は示さない。これは、HR選好の生物(サッカロミセス・セルビシエ)において得られる結果が、NHR選好の生物には外挿されえないことを示す。
【0007】
意外にも、本発明者らは、糸状菌におけるHRへ核酸の取込み経路を導くステップが結果として糸状菌のゲノムへの核酸の標的取込みの改善された効率をもたらすことを見出した。
【発明の詳細な説明】
【0008】
本明細書で参照される、すべての特許および刊行物は、かかる特許および刊行物内で開示されたすべての配列および方法を含めて、参照により明示的に援用される。これらの特許および刊行物としては、欧州特許第357127B号明細書、同第635574B号明細書、国際公開第97/06261号パンフレット、同第98/46772号パンフレットが挙げられる。
【0009】
ポリヌクレオチドの糸状菌細胞のゲノムへの標的取込みの効率を増大させるための方法
本発明は、NHR選好を有する糸状菌細胞のゲノムへのポリヌクレオチドの所定の部位に対する標的取込みの効率を増大させるための方法に関し、前記ポリヌクレオチドは、前記所定の部位との相同の領域を有し、取込み経路をHRへ導くステップを含んで成る。本発明は、HR経路の効率を上昇させることによって、かつ/またはNHR経路の効率を低下させる(削減を意味する)ことによって、かつ/またはNHR/HR比を減少させることによって、かかる導くステップに達する。
【0010】
本発明に照らして、HR経路は、ポリヌクレオチドの宿主のゲノムへの標的取込みのコントロールに関与されるすべての遺伝子および構成要素と定義され、前記ポリヌクレオチドは、取込みが標的される宿主のゲノムの特定の所定部位と特定の相同を有する。NHR経路は、宿主のゲノム配列との前記ポリヌクレオチドの相同の程度と関係なく、ポリヌクレオチドの宿主のゲノムへの取込みのコントロールに関与されるすべての遺伝子および構成要素と定義される。
【0011】
好ましい実施形態によれば、導くステップは親糸状菌細胞の変異体を提供するステップを含んで成り、ここでNHR/HR比は、以下のアッセイによって測定された前記親生物における前記比と比べ少なくとも5%の変異体において減少する。より好ましくは、NHR/HR比は、前記親生物における前記比と比べ少なくとも10%、さらにより好ましくは少なくとも50%、かつ最も好ましくは少なくとも100%の変異体において減少する。
【0012】
別の好ましい実施形態によれば、本発明の糸状菌細胞は、以下のアッセイによって測定されうるように、少なくとも200、少なくとも50、少なくとも10であるNHR/HR比を有する。好ましくは、糸状菌細胞の比は少なくとも1、より好ましくは少なくとも0.5、さらにより好ましくは少なくとも0.1、さらにより好ましくは少なくとも0.05、さらにより好ましくは少なくとも0.01、さらにより好ましくは少なくとも0.005、さらにより好ましくは少なくとも0.001、さらにより好ましくは少なくとも0.0005、さらにより好ましくは少なくとも0.0001、かつ最も好ましくは少なくとも0.00001である。
【0013】
より好ましい実施形態によれば、本発明の糸状菌細胞は、以下のアッセイによって測定されるように200未満、さらにより好ましくは50未満、10未満であるNHR/HR比を有する。さらにより好ましくは、糸状菌細胞の比は1未満、さらにより好ましくは0.5未満、さらにより好ましくは0.1未満、さらにより好ましくは0.05未満、さらにより好ましくは0.01未満、さらにより好ましくは0.005未満、さらにより好ましくは0.001未満、さらにより好ましくは0.0005未満、さらにより好ましくは0.0001未満、かつ最も好ましくは0.00001未満である。
【0014】
NHR/HR比は、国際公開第02/052026号パンフレット(表2、23頁)に記載されたアッセイによって測定されることが好ましい。好ましい実施形態によれば、親生物は宿主細胞の部で定義された糸状菌細胞の1つである。別の好ましい実施形態によれば、本発明の糸状菌細胞は宿主細胞の部で定義された種由来である。
【0015】
あるいは、またより好ましくない実施形態によれば、糸状菌におけるNHR/HR比は、かかる経路に関与する以下の成分、すなわちKU70、KU80、MRE11、RAD50、RAD51、RAD52、XRS2、SIR4、LIG4の少なくとも1つの転写プロファイリングおよび/またはノーザンブロット法および/またはウェスタンブロット法など当業者に周知の方法を使用してモニタリングされる。
【0016】
本発明に照らして、「相同の領域」は「少なくとも1つ」の相同の領域を意味する。所定の部位が、本明細書では、宿主細胞によって含まれる遺伝物質内の部位と定義されるが、この同じ部位に対する相同を有するポリヌクレオチドが本発明による方法で取込まれる。
【0017】
好ましい実施形態において、本発明は、NHR選好を有する糸状菌細胞のゲノムへの所定部位に対するポリヌクレオチドの標的取込みの効率を増大させるための方法を提供するが、前記ポリヌクレオチドは、糸状菌を提供することによってHRへの取込み経路を導くステップを含んで成る前記所定の部位と相同の領域を有し、同じ条件下に由来する糸状菌のNHR経路の効率および/またはNHR/HR比と比べ、NHR経路の効率は低下し、かつ/またはNHR/HR比は減少している。好ましい実施形態によれば、親生物は宿主細胞の部で定義されている糸状菌の1つである。
【0018】
NHR経路の効率は、国際公開第02/052026号パンフレット(表2、23頁)に記載されたアッセイで測定されることが好ましい。
【0019】
あるいは、より好ましくない実施形態によれば、糸状菌におけるNHR経路の効率は、かかる経路に関与する成分の転写プロファイリングおよび/またはノーザンブロット法および/またはウェスタンブロット法など当業者に周知の方法を使用してモニタリングされる。より好ましくは、以下の成分、すなわちKU70、KU80、MRE11、RAD50、RAD51、RAD52、XRS2、SIR4、LIG4の少なくとも1つの発現レベルがモニタリングされる。さらにより好ましくは、KU複合体の相同的成分の発現レベルがモニタリングされる。最も好ましくは、相同的KU70および/またはKU80の発現レベルがモニタリングされる。
【0020】
NHR効率の低下は、少なくとも得られた細胞が由来する親細胞におけるよりも低いことを意味する。好ましくは、低下は2倍低く、より好ましくは10倍低く、さらにより好ましくは100倍低く、最も好ましくは1000倍超低く、かつさらに最も好ましくは、ノーザンまたはウェスタンブロット法、アレイ技術または表現型スクリーンを使用して検出可能ではないことを意味する。
【0021】
使用されうる典型的な表現型スクリーンは、以下のステップ、すなわち、所定のゲノム部位の相同的配列によってフランキングされる選択マーカー遺伝子を含んで成る発現カセットで推定的NHR変異体を形質転換するステップを含んで成る。この表現型スクリーンにおいて使用される選択マーカー遺伝子は、糸状菌の形質転換に有用である多くのマーカー遺伝子から選択されうる。一例として、これらのマーカーは、アセトアミダーゼ(amdS)遺伝子など優性および双方向選択マーカー遺伝子(欧州特許第635574B号明細書または国際公開第97/06261号パンフレット)、argB、trpC、またはpyrGなど栄養要求性マーカー遺伝子、および例えば、フレオマイシン(ble遺伝子によってコードされる産物はフレオマイシンに対する抵抗を与える)、ハイグロマイシンB、またはG418に対する抵抗を提供する抗生物質抵抗性遺伝子を含むがこれらに限定されない。好ましい選択マーカー遺伝子が、フレオマイシンに対する抵抗を与えるタンパク質をコードするble遺伝子である。推定的NHR変異体はすでにこの所定のゲノム部位で、amdS遺伝子、硝酸還元酵素遺伝子(niaD)、硫酸パーミアーゼ(Sut B)遺伝子、またはPyrG遺伝子など双方向選択マーカー遺伝子を含有する。niaD遺伝子はすでに他所で記載されている(ゴウカ(Gouka)RJ、ファン・ハルチングスフェルト(van Hartingsveldt)W、ボーベンベルグ(Bovenberg)RA、ファン・デン・ホンデル(van den Hondel)CA、ファン・ゴルコム(van Gorcom)RF、Cloning of the nitrate−nitrite reductase gene cluster of penicillium chrysogenum and use of the niaD gene as a homologous selection marker、J Biotechnol.、1991年9月、20(2):189−99頁)。niaD遺伝子は、細胞が塩素酸塩に耐性を示すようになると、塩素酸塩を含有するプレート上で形質転換体の直接選択を可能にする。sutB遺伝子はすでに他所で記載されている(ファン・デ・カンプ(van de Kamp)M、ピッツィニーニ(Pizzinini)E、フォス(Vos)A、ファン・デア・レンデ(van der Lende)TR、シュールス(Schuurs)TA、ニューベルト(Newbert)RW、ターナー(Turner)G、コーニングス(Konings)WN、ドリーゼン(Driessen)AJ、Sulfate transport in Penicillium chrysogenum:cloning and characterization of the sutA and sutB genes.、J. Bacteriol.1999年12月、181(23):7228−34頁)。好ましい選択マーカー遺伝子が、A.ニドゥランス(nidulans)gpdAプロモータに融合されるA.ニドゥランスamdSコード配列である(欧州特許第635574B号明細書)。他の糸状菌からのamdS遺伝子も使用されうる(国際公開第97/06261号パンフレット)。表現型スクリーンの好ましい形態において、amdS遺伝子は所定のゲノム部位に存在し、ble遺伝子は所定の部位に標的される遺伝子として使用される。非HR改善変異体において、bleカセットは無作為にゲノムに取込み、多くの形質転換体がアセトアミドとフレオマイシンの両方を有する二重選択培地で成長し、かつ比較的少ない形質転換体がフルオロアセトアミド−フレオマイシンプレートで成長することを可能にする。改善されたHRを有する変異体においては、amdSカセットがbleカセットで効率的に交換されると、アセトアミド−プレオマイシン二重選択プレートでの形質転換体の数は限られる。このカセットではより多くの変異体がフルオロアセトアミド−フレオマシン二重選択プレートで現れる。
【0022】
別の好ましい実施形態によれば、低下したNHR効率および/または減少したNHR/HR比を有する糸状菌は、NHRに関与する1つの成分が阻害されている糸状菌である。これに関連して、「1つの(a)」は「少なくとも1つ」を意味する。すなわち、NHRに関与する少なくとも1つの成分が一定の糸状菌で阻害されている。阻害は、NHRに関与する遺伝子の発現レベルをダウンレギュレートし、またはNHRに関与する成分をコードする遺伝子を不活性化することによって、かつ/またはNHRに関与する成分の発現レベルをダウンレギュレートし、かつ/またはNHRに関与する成分の(タンパク質)活性を(一時的に)減少させ、およびこれらの可能性の組合せによって達成されうる。
【0023】
好ましくは、得られた糸状菌は、同じ条件下に由来する親糸状菌細胞における前記遺伝子の発現との比較によってダウンレギュレートされたNHRに関与する遺伝子の発現を有する。好ましい実施形態によれば、親糸状菌は宿主細胞の部で定義される糸状菌の1つである。
【0024】
遺伝子の発現レベル、またはDNA配列は、得られた糸状菌におけるこの特定の遺伝子またはDNA配列の発現レベルが、それが由来する親糸状菌における同じ遺伝子またはDNA配列の発現レベルよりも低い、好ましくは3倍低く、より好ましくは4倍低く、最も好ましくは4倍超低く、さらに最も好ましくは、ノーザンまたはウェスタンブロット法、またはトランスクリプトミクスおよびプロテオミクスのような「オミクス」法を使用して検出可能ではない場合にダウンレギュレートされる。
【0025】
DNA配列の発現レベルのダウン/アップレギュレーションは、例えば、ノーザンブロット法によって細胞に存在する対応するmRNAの量を定量化することによって(Molecular Cloning:A Laboratory Manual、サンブルック(Sambrook)ら、ニューヨーク・コールド・スプリング・ハーバー・プレス(New York Cold Spring Harbour Press)、1989年)、かつ/または、例えば、ウェスタンブロット法によって細胞に存在する対応するタンパク質の量を定量化することによってモニタリングされうる。mRNAの量の差は、DNA配列分析(アイゼン(Eisen),M.B.およびブラウン(Brown),P.O.、DNA arrays for analysis of gene expression.Methods Enzymol.1993年:303:179−205頁)によっても定量化されうる。
【0026】
少なくとも1つの遺伝子またはDNA配列の発現レベルのダウンレギュレーションは、以下の方法の1つまたはその組合せによる遺伝子操作によって得られうる。すなわち、
a.組換え遺伝子操作法を使用するステップ、
b.糸状菌を変異誘発に提示するステップ。
【0027】
あるいは、または上記の方法と組合せて、かつ別の好ましい実施形態によれば、少なくとも1つの遺伝子またはDNA配列の少なくとも1つの発現レベルのダウンレギュレーションは、糸状菌を阻害化合物/組成物に提示することによって得られうる。
【0028】
得られた糸状菌はその後にDNA配列の前記遺伝子の発現レベルをモニタリングすることによって選択されうる。場合により、糸状菌はその後にそのNHR経路および/またはHR経路の効率および/またはそのNHR/HR比を測定することによって選択されうる。本発明に照らして、糸状菌のHR経路の効率は、一定の相同領域を使用することにより糸状菌のゲノムにおける一定のポリヌクレオチド配列の所定の部位への標的取込みの効率によって測定されうる。本発明に照らして、糸状菌のNHR経路の効率は、相同領域と関係なく糸状菌のゲノムにおける一定のポリヌクレオチド配列の非標的取込みの効率によって測定されうる。
【0029】
より好ましくは、少なくとも1つのDNA配列の発現のダウンレギュレーションは、例えばステップa.で定義された組換え遺伝子操作法で行われ、組換え糸状菌を得る。最も好ましくは、ステップa.は、DNA配列を削除するステップを含んで成り、さらに最も好ましくは、削除型DNA配列はその非機能的変異形によって置換され、かつさらに最も好ましくは、削除および置換は、好ましくは欧州特許第357127B号明細書に記載された遺伝子置換によって行われる。
【0030】
選ばれた糸状菌の少なくとも1つのDNA配列の削除または置換の場合、適切なDNA配列が標的遺伝子座で導入されなければならない。標的遺伝子座はこの場合、削除または置換されるNHR経路に関与するDNA配列である。適切なDNA配列は好ましくはクローニングベクター上に存在する。適切なクローニングベクターは、使用される糸状菌宿主細胞の染色体における所定の標的遺伝子座で取込むことが可能であるものである。
【0031】
好ましい統合的クローニングベクターは、この所定の遺伝子座に対するクローニングベクターの取込みを標的するために削除または置換されるDNA配列に対して相同的であるDNA断片を含んで成る。標的取込みを促進するために、クローニングベクターは好ましくは宿主細胞の形質転換前に線状化される。好ましくは、線状化は、クローニングベクターの少なくとも1つ、ただし好ましくはいずれかの末端が削除または置換されるDNA配列に対して相同的な配列によってフランキングされるように実行される。
【0032】
削除または置換されるDNA配列をフランキングする相同的配列の長さは好ましくは2kb未満、さらに好ましくは1kb未満、さらにより好ましくは0.5kb未満、さらにより好ましくは0.2kb未満、さらにより好ましくは0.1kb未満、さらにより好ましくは50bp未満、かつ最も好ましくは30bp未満である。
【0033】
クローニングベクターにおける選択マーカー遺伝子は、糸状菌の形質転換に有用である多くのマーカー遺伝子から選択されうる。一例として、これらのマーカーは、アセトアミダーゼ(amdS)遺伝子など優性および双方向選択マーカー遺伝子(欧州特許第635574B号明細書または国際公開第97/06261号パンフレット)、argB、trpC、またはpyrGなど栄養要求性マーカー遺伝子、および、例えば、フレオマイシン、ハイグロマイシンB、またはG418に対する抵抗を示す抗生物質抵抗性遺伝子を含むがこれらに限定されない。好ましい選択マーカー遺伝子が、A.ニドゥランスgpdAプロモータに融合したA.ニドゥランスamdSコード配列である(欧州特許第635574B号明細書)。他の糸状菌からのamdS遺伝子も使用されうる(国際公開第97/06261号パンフレット)。amdS選択マーカー遺伝子は、明確なDNA配列を置換および/または削除する同じ菌株における数倍で使用されうる利点を有する。
【0034】
欧州特許第635574B号明細書に記載されたフルオロアセトアミド培地の対抗選択を行うことで、結果として生じる菌株はマーカーを含まず、さらなる遺伝子修飾のために使用されうる。
【0035】
一定のDNA配列の発現をダウンレギュレートするための好ましい方法が、野生型DNA配列の削除および/または修飾DNA配列による置換を含んで成るが、その発現産物は機能的ではない。削除および置換は、好ましくは、欧州特許第0357127B1号明細書に記載された遺伝子置換法によって実行される。遺伝子の特定の削除は、好ましくは、欧州特許第635574B号明細書に記載されているように選択マーカー遺伝子としてamdS遺伝子を使用して実行される。
【0036】
あるいは、または他の言及された方法と組合せて、大腸菌(E.coli)におけるコスミドのインビボ組換えに基づく方法が以下に記載されているように使用されうる。すなわち、A rapid method for efficient gene replacement in the filamentous fungus Aspergillus nidulans(2000年)、シャーベロシュ(Chaveroche),MK、ジコ(Ghico),J.M.、およびダンフェルト(d’Enfert)C、Nucleic acids Research、28巻、22号。この方法は、例えばA.ニゲル(niger)のような他の糸状菌に適用可能である。
【0037】
DNA配列の発現のダウンレギュレーションは、アンチセンス核酸を使用することによって、もしくはUVまたは化学的変異誘発を使用することによっても達成されうる(マテム(Mattem),I.E.、ファン・ヌルト(van Noort)J.M.、ファン・デン・ベルグ(van den Berg),P.、アーチャー(Archer),D.B.、ロバーツ(Roberts),I.N.、およびファン・デン・ホンデル,C.A.、Isolation and characterization of mutans of Aspergillus niger deficient in extracellular proteases.Mol Gen Genet.1992年8月、234(2):332−6頁)。
【0038】
好ましくは、NHR経路にもたらされる欠損は誘導性のものである。これは、新しい調節領域によって、好ましくは抑制可能または調節可能なプロモータを使用することによって、より好ましくは、スイッチオン/オフされうるプロモータを使用することによって、すなわちグルコース抑制、もくしはアンモニア抑制、またはpH抑制によって、NHRに関与する成分をコードする遺伝子の内在性調節領域を置換することによって達成されうる。グルコース抑制されたプロモータの例は、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)pcbABプロモータ(マルチン(Martin)JF、カスケイロ(Casqueiro)J、コサルコバ(Kosalkova)K、マルコス(Marcos)AT、グチレツ(Gutierrez)S.、Penicillin and cephalosporin biosynthesis:mechanism of carbon catabolite regulation of penicillin production.アントニー・ファン・リューエンヘック(Antonie Van Leeuwenhoek)、1999年1−2月、75(1−2):21−31頁、総説)、またはアスペルギルス・ニゲル(Aspergillus niger)グルコアミラーゼプロモータである。オン/オフスイッチ可能なプロモータの例は以下の刊行物に記載されている。すなわち、
−An activator/repressor dual system allows tight tetracycline−regulated gene expression in budding yeast:ベリ(Belli)ら、(1998年)Nucl.Acid Research.26巻、4号:942−947頁、
−A light−switchable gene promoter system:シミズ−サトウ(Shimizu Sato)ら、(2002年)Nat.Biotech.20巻、10号、:1041−1044頁。
【0039】
好ましい実施形態によれば、糸状菌は、NHR経路に関与する以下の酵母遺伝子、KU70、KU80、RAD50、MRE11、XRS2、およびSIR4と相同的である、その内在性遺伝子の少なくとも1つを欠いている(ファン・デン・ボシュ(van den Bosch)ら(2002年):DNA double−strand break repair by homologous recombination.Biol.Chern.383巻:873−892頁、およびアレン(Allen)ら、(2003年):Interactive competition between homologous recombination and non−homologous end joining.Mol.Cancer Res.、1巻:913−920頁)。
【0040】
NHRに関与する少なくとも1つの成分を有する全種類の変異体は、NHRの過程においてその機能を実行することがもはや可能ではなく、または少なくとも相当に可能性が低く、本発明によって意図された変異体である。好ましくは、NHRに関与する成分は、得られた変異体におけるNHR経路の効率が前記定義のアッセイで測定されたように同じ条件下に由来する親細胞における活性の90%未満、さらに好ましくは85%未満、より好ましくは80%未満、さらにより好ましくは70%未満、最も好ましくは50%未満であるように阻害されている。
【0041】
好ましい実施形態によれば、親糸状菌は宿主細胞の部で定義される糸状菌の1つである。
【0042】
好ましくは、糸状菌細胞は以下の遺伝子の少なくとも1つを欠いている。すなわち、
−配列番号2または19で識別されるhdfAもしくはその相同体、または
−配列番号5または22で識別されるhdfBもしくはその相同体、またはその両方。
【0043】
別の好ましい実施形態によれば、糸状菌は、誘導と同時に減少するこれら遺伝子hdfAおよびhdfBによってコードされたタンパク質の少なくとも1つの量を有する。
【0044】
別の好ましい実施形態によれば、少なくとも1つの遺伝子またはDNA配列の発現レベルのダウンレギュレーションは、糸状菌を変異誘発にかけることによる遺伝子修飾によって得られうる。糸状菌細胞は、無作為変異誘発、およびその後に選択アッセイにかけられ、改善されたHRを有する変異体を変異体のすべての集団から単離することができる。
【0045】
本発明の好ましい実施形態によれば、宿主細胞の部で定義された糸状菌細胞の1つが変異誘発を実行する出発菌株として使用される。
【0046】
例えば、出発菌株はUV照射にかけられ、生存率が0.001%〜60%の範囲であるようにUV照射にかけられる。好ましくは、生存率は0.01%〜50%の範囲である。分生子が、物理的または化学的手段によって糸状菌を突然変異させる好ましい材料であることは当業者に公知である。しかし、変異体は菌糸細胞から得ることもできる。また、UV以外の他の変異誘発処置が化学的薬剤(例えば、NTG)として適用されうる。本明細書に記載された選択方法は、分生子または菌糸細胞のいずれかから得られる変異体を選択するためにも適用されうる。
【0047】
好ましくは、変異誘発は分生子に適用される。UV照射は、好ましくは、7.5、15、および30分など異なる時間にわたって適用され、細胞における軽度、中等度、および強度の突然変異率レベルを得る。突然変異試料は、直接、再胞子形成され、または胞子形成が誘発された前に(例えば、実施例9に記載)、YNBまたはYEPD(実施例9の定義)など富栄養培地において長い回復期間インキュベートされうる。
【0048】
次いで、胞子形成バッチは遺伝子標的におけるその効率について試験されうる。これは以下の方法によって試験されうる。プロトプラストは、機能的選択マーカーの発現カセットを有する少なくとも1つ、好ましくは2つもしくはそれ以上のDNA断片で形質転換されうる。発現カセットにおける選択マーカー遺伝子は、糸状菌の形質転換に有用である多くのマーカー遺伝子から選択されうる。一例として、これらのマーカーは、アセトアミダーゼ(amdS)遺伝子など優性および双方向選択マーカー遺伝子(欧州特許第635574B号明細書または国際公開第97/06261号パンフレット)、argB、trpC、またはpyrGなど栄養要求性マーカー遺伝子、および例えば、フレオマイシン、ハイグロマイシンB、またはG418に対する抵抗を提供する抗生物質抵抗性遺伝子含むがこれらに限定されない。好ましくは、使用される選択マーカーはbleおよびamdS遺伝子である。使用されるamdSカセットは、A.ニドゥランスgpdAプロモータに融合したA.ニドゥランスコード配列である(欧州特許第635574B号明細書)。他の糸状菌からのamdS遺伝子も使用されうる(国際公開第97/06261号パンフレット)。遺伝子bleは、フレオマイシンに対する抵抗を与えることが可能なタンパク質をコードする。遺伝子amdSは、細胞が唯一の窒素源としてアセトアミドで成長することを可能にするタンパク質をコードする(欧州特許第635574B号明細書に記載)。糸状菌のプロトプラストへのDNAの移動に適用される方法は当技術分野で周知であり、フィンケルシュタイン(Finkelstein)とボール(Ball)(編)、Biotechnology of filamentous fungi,technology and products,バターワース・ハイネマン(Butterworth−Heinemann)(1992年)、ベネット(Bennett)とラシュア(Lasure)(編)More Gene Manipulations in fungi、Academic Press(1991年)、ターナー、ピューラー(Puehler)(編)、Biotechnology、第2完全改訂版、VHC(1992年)を含む多くの文献に記載されている。Ca−PEG介在プロトプラスト形質転換は、欧州特許第635574B明細書に記載されているように使用される。
【0049】
これら2つの発現カセットの糸状菌ゲノムにおける2つの明確な特定の遺伝子座への標的取込みを選択するには、DNAの短い相同的ストレッチが、例えば、DNA断片の両側のPCRを介して添加されうる。いくつかの種類の構成物を作り、改善された標的効率を有する変異体を選択する機会を改善しうる。すなわち、DNAの相同的ストレッチは通常、30bp〜1000bp、好ましくは30b〜700bp、より好ましくは30bp〜500bp、さらにより好ましくは30bp〜300bp、より好ましく好ましくは30bp〜200bp、さらにより好ましくは30bp〜100bp、かつ最も好ましくは30bpでありうる。理論上、糸状菌ゲノムにおけるすべての遺伝子座が発現カセットの標的取込みのために選択されうる。好ましくは、標的が行われる遺伝子座は、この遺伝子座に存在する野生型遺伝子が発現カセットに含まれる遺伝子によって置換されている場合、得られた変異体が一定のアッセイによって検出可能である変化を示すようになっている。好ましくは、遺伝子座はniaD遺伝子座であり、それによって硝酸還元酵素遺伝子を破壊し(ゴウカ RJ、ファン・ハルチングスフェルト W、ボーフェンベルク(Bovenberg)RA、ファン・デン・ホンデル CA、ファン・ゴルコム RF、Cloning of the nitrate−nitrite reductase gene cluster of Penicillium chrysogenum and use of the niaD gene as a homologous selection marker.J Biotechnol.1991年9月、20(2):189−99頁)、細胞が塩素酸塩に対して抵抗性になると、塩素酸塩を含有するプレート上で形質転換体の直接選択を可能にする。別の好ましい遺伝子座はsutB遺伝子座であり、それによって硫酸パーミアーゼ遺伝子を破壊し(ファン・デ・カンプ M、ピッツィニーニ E、フォス A、ファン・デア・レンデ TR、シュールス TA、ニューベルト RW、ターナー G、コーニングス WN、ドリーゼン AJ、Sulfate transport in Penicillium chrysogenum:cloning and characterization of the sutA and sutB genes.、J. Bacteriol.1999年12月、181(23):7228−34頁)、セレン酸塩を含有するプレート上で形質転換体の直接選択を可能にする。存在する両方の選択マーカーを有し、かつ取込み遺伝子座に存在する遺伝子の不活性化に起因する2つの変化を有する変異体は、改善された標的取込みを有する菌株である。
【0050】
別の好ましい実施形態によれば、NHR経路における低下した効率、または減少したNHR/HR比および/またはHR経路の高い効率は、NHRに関与する成分を減少させることによって、より好ましくは部分的もしくは最も好ましくは完全に阻害することによって得られる。
【0051】
NHRに関与する成分の部分的または完全な阻害は異なる方法によって、例えば、かかる成分もしくは化学的阻害剤またはタンパク質阻害剤もしくは物理的阻害剤(ツール(Tour)O.ら(2003年)Nat.Biotech:Genetically targeted chromophore−assisted light inactivation.21巻、12号、1505−1508頁)またはペプチド阻害剤もしくはアンチセンス分子またはRNAi分子(R.S.カマス(Kamath)ら、(2003年)Nature:Systematic functional analysis of the Caenorhaboditis elegans genome using RNAi.421巻、231−237頁)に対する抗体によって得られうる。阻害の種類(部分的またはより好ましくは完全)に関係なく、NHRに関与する成分が上記定義されたNHRの過程におけるその機能を実行することがもはや不可能であり、少なくとも相当に可能性が低いことが重要である。
【0052】
NHRに関与する成分は、酵母KU70、RAD50、MREII、XRS2、LIG4、SIR4、KU80、LIFL、もしくはNEIL、または会合成分の糸状菌相同体を含んで成る。遺伝子の命名は生物間で異なるため、機能的等価物および/またはその機能的断片はすべて、酵母遺伝子KU70、RAD50、MREII、XRS2、LIG4、SIR4、KU80、LIFL、またはNEILの少なくとも1つの機能(機能であって、量ではなく)を実行するのが可能であると本明細書で定義されているが、これらは本発明にも含まれる。NHRに関与する成分を一時的に(部分的より好ましくは完全に)阻害することによって、核酸が所望の位置で取込まれるが、NHRに関与する成分を永続的に修飾することはなく、かつNHRに関与するかかる修飾成分の永続的存在によって引起こされる望ましくない副作用を予防する。
【0053】
上記の方法に加えて、または代替として、NHRに関与するタンパク質の活性を阻害することによって低下したNHR効率を得ること、または代わりのシグナル配列(ラモン・ド・ルーカス(Ramon de Lucas),J.、マルチネス(Martinez)O、ペレス(Perez)P.、イザベル・ロペス(Isabel Lopez),M.、バレンシアーノ(Valenciano),S.、およびラボルダ(Laborda),F.、The Aspergillus nidulans carnitine carrier encoded by the acuH gene is exclusively located in the mitochondria.FEMS Microbiol Lett.2001年7月24日、201(2):193−8頁)、または保持シグナル(デルクス(Derkx),P.M.とマドリッド(Madrid),S. M.、The foldase CYPB is a component of the secretory pathway of Aspergillus niger and contains the endoplasmic reticulum retention signal HEEL. Mol.Genet.Genomics.2001年12月、266(4):537−45頁)によってNHR関与タンパク質を再局在することも可能である。
【0054】
あるいは、または上記の方法と組合せて、タンパク質活性の阻害はUVまたは化学的変異誘発によって(マテム,I.E.、ファン・ヌルト J.M.、ファン・デン・ベルグ,P.、アーチャー,D.B.、ロバーツ,I.N.、およびファン・デン・ホンデル,C.A.、Isolation and characterization of mutans of Aspergillus niger deficient in extracellular proteases.Mol Gen Genet.1992年8月、234(2):332−6頁)、または親和性のプロテアソーム阻害剤のような阻害剤の使用によって(クラスト−ラクタクリスチン−β−ラクトン、アフィニティー・リサーチ・プロダクツ社(Affinity Research Products Ltd.)、CW8405−Z02185)も得られうる。
【0055】
別の好ましい実施形態によれば、HRへ導くステップは、取込まれるポリヌクレオチドは別にして、結合し、NHR成分の発現を制限することが可能な過剰な小さな二本鎖ポリヌクレオチドを添加するステップを含んで成る(アグラワル(Agrawal)N.ら:RNA interference:biology,mechanism and applications.Microbiol.Mol.Biol.Rev.、67巻、4号、657−685頁)。
【0056】
好ましい実施形態において、本発明は、所定の部位へのポリヌクレオチドの標的取込みの効率を増大させる方法を提供し、それによって前記ポリヌクレオチドは、糸状菌細胞を提供することによってHRへ取込み経路を導くステップを含んで成るNHR選好を有する糸状菌において、前記所定の部位で、またはその周りで相同性を有し、ここでHR経路の効率は同じ条件下に由来する親糸状菌の1つに比べ高められている。HR経路の効率は、好ましくは、NHR/HR比を測定するために使用されたものと同じアッセイによって測定される。好ましい実施形態によれば、親生物は宿主細胞の部で定義されている糸状菌の1つである。
【0057】
高められたというのは、得られる細胞が由来する親細胞におけるよりも少なくとも高いことを意味する。好ましくは、高められたというのは、2倍高い、より好ましくは3倍高い、さらにより好ましくは4倍高い、最も好ましくは、ノーザン、またはウェスタンブロット法、もしくはアレイ技術、または表現型スクリーンを使用して4倍超高いことを意味する。
【0058】
別の好ましい実施形態によれば、糸状菌は、それが由来する糸状菌細胞における同じ遺伝子の発現レベルと比較することによってアップレギュレートされている、HRに関与する少なくとも1つの遺伝子の発現レベルを有する。これは、HRに関与する成分をコードする遺伝子の発現レベルを増大させることによって、かつ/またはHRに関与する成分の発現レベルを増大させることによって、かつ/またはHRに関与する成分の活性を(一時的に)増大させることによって達成されうる。
【0059】
好ましくは、得られた糸状菌は、それが由来する糸状菌細胞における同じ遺伝子の発現レベルと比較することによってアップレギュレートされている、HRに関与する遺伝子の発現を有する。
【0060】
DNA配列の発現レベルは、得られた糸状菌におけるこの特定のDNA配列の発現レベルが、それが由来する親糸状菌における同じDNA配列の発現レベルよりも高い、好ましくは3倍高い、より好ましくは4倍高い、最も好ましくは、ノーザン、またはウェスタンブロット法、もしくはアレイ技術を使用して4倍超高い場合にアップレギュレートされる。好ましい実施形態によれば、親生物は宿主細胞で定義されている糸状菌の1つである。
【0061】
少なくとも1つのDNA配列の発現レベルのアップレギュレーションは、以下の方法の1つによって、またはその組合せによる遺伝子操作によって得られうる。すなわち、
c.組換え遺伝子操作法を使用するステップ、
d.糸状菌を変異誘発させるステップ、
あるいは、または上記の方法と組合せ、かつ別の好ましい実施形態によれば、少なくとも1つの遺伝子またはDNA配列の発現レベルのアップレギュレーションは、糸状菌を活性化化合物/組成物に提示することによって得られうる。
【0062】
糸状菌はその後に前記DNA配列の発現レベル、および場合により糸状菌のHR経路の効率をモニタリングすることによって選択されうる。糸状菌のHR効率は、一定の相同領域を使用することにより糸状菌のゲノムにおける所定の部位への一定のポリヌクレオチド配列の標的取込みの効率によって測定されうる。
【0063】
好ましくは、少なくとも1つのDNA配列の発現のアップレギュレーションは、組換え糸状菌を得るステップa.で定義されているような組換え遺伝子操作法で行われる。好ましくは、ステップa.は、DNA配列を含んで成るDNA構成物で糸状菌を形質転換するステップを含んで成り、好ましくは前記DNA配列は高発現遺伝子のプロモータに操作連結されている。選択プロモータは、過剰発現されるDNA配列の内在性プロモータよりも強い場合がある。DNA配列の発現のプロモータは、高発現真菌遺伝子由来であることが好ましい。
【0064】
多くの好ましい高発現真菌遺伝子が一例として挙げられている。すなわち、アスペルギルスまたはトリコルデルマからのアミラーゼ遺伝子、グルコアミラーゼ遺伝子、アルコールデヒドロゲナーゼ遺伝子、キシラナーゼ遺伝子、グリセルアルデヒドリン酸デヒドロゲナーゼ遺伝子、またはセロビオヒドロラーゼ遺伝子である。これらの目的のために最も好ましい高発現遺伝子は、アスペルギルス・ニゲル・グルコアミラーゼ遺伝子、アスペルギルス・オリゼ(Aspergillus oryzae)TAKAアミラーゼ遺伝子、アスペルギルス・ニドゥランス(Aspergillus nidulans)gpdA遺伝子、またはトリコデルマ・エセイ(Trichoderma reesei)セロビオヒドロラーゼ遺伝子である。グルコアミラーゼプロモータが、使用される最も好ましいプロモータである。これらの高発現遺伝子は、クローニングベクターの取込みのための標的遺伝子座として、および高発現真菌遺伝子の供給源として適切である。
【0065】
別の好ましい実施形態によれば、ステップa.は、好ましくは、DNA配列のそのゲノムコピー取込みによって、より好ましくは、高発現遺伝子座で、好ましくはグルコアミラーゼ遺伝子座でDNA配列の取込みを標的することによって、糸状菌細胞へのDNA配列のコピー数を増大させるステップを含んで成る。
【0066】
DNA配列の発現のアップレギュレーションは、DNA配列の少なくとも1つにコピーを糸状菌へ導入することによって、または強いプロモータのために変更し、または良好な動態および/または寿命を有するタンパク質をコードする遺伝子のために変更することによってDNA配列のコピー数を増大させることによって達成されうる。DNA配列はプラスミドに存在し、またはゲノムへ取込みうる。当業者は2つの代替の可能性のうちで選択しうる。すなわち、
−HR経路に関与する糸状菌の少なくとも1つの内在性DNA配列を過剰発現する。この場合、糸状菌はその内在性DNA配列のいくつかのコピーを含んで成る。
−HRに関与する少なくとも1つの非相同DNAを過剰発現する。この場合、糸状菌はHRに関与するその内在性DNA配列、およびさらにHRに関与する非相同DNA配列の少なくとも1つのコピーを有することになる。この非相同DNA配列はその対応する内在性DNA配列の相同体である。
【0067】
糸状菌は、DNA配列の1つもしくはそれ以上のコピーで形質転換されうる(とりわけ、チルブム(Tilbum)ら、1983年、Gene、26:205−221頁による)。DNA配列は、糸状菌のゲノムへ安定して取込まれ、または自発的複製が可能なDNA分子の一部として細胞へ導入されうる。DNA配列は、クローニングベクターに存在することが好ましい。糸状菌宿主細胞を形質転換することが可能なクローニングベクターが本発明における使用に適切である。したがって、本発明において使用するためのクローニングベクターは、糸状菌宿主細胞の染色体において無作為または所定の標的遺伝子座で取込む統合的クローニングベクター、およびAMA1配列を含んで成るベクターなど自発的に維持されるクローニングベクターを含んで成る。本発明の好ましい実施形態において、統合的クローニングベクターは、糸状菌宿主細胞のゲノムにおける所定の標的遺伝子座における、この所定の遺伝子座にクローニングベクターの取込みを標的するためのDNA断片を含んで成る。標的取込みを促進するために、クローニングベクターは宿主細胞の形質転換前に線状化されることが好ましい。線状化は、クローニングベクターの少なくとも1つであるが、好ましくはいずれかの末端が標的遺伝子座と相同的な配列によってフランキングされるように実行されることが好ましい。標的遺伝子座をフランキングする相同的配列の長さは好ましくは少なくとも30bp、好ましくは少なくとも50bp、好ましくは少なくとも0.1kb、さらに好ましくは少なくとも0.2kb、より好ましくは少なくとも0.5kb、さらにより好ましくは少なくとも1kb、最も好ましくは少なくとも2kbである。
【0068】
好ましくは、標的遺伝子座と相同的であるクローニングベクターにおけるDNA配列は、高発現遺伝子座由来であり、これは、糸状菌宿主細胞において高い発現レベルが可能である遺伝子由来であることを意味する。高い発現レベルが可能な遺伝子、すなわち高発現遺伝子が、本明細書では、そのmRNAが、例えば、誘導条件下に全細胞のmRNAの少なくとも0.5%(w/w)を構成しうる遺伝子、あるいは、その遺伝子産物が全細胞タンパク質の少なくとも1%(w/w)を構成し、または、分泌遺伝子産物の場合、少なくとも0.1g/lのレベルに分泌されうる遺伝子と定義されている(欧州特許第357127B1号明細書に記載)。
【0069】
過剰発現されるDNA配列のコピーの数を増加させるには、国際公開第98/46772号パンフレットに記載された遺伝子変換の方法が使用されうる。
【0070】
当業者は、標的のための相同的DNA配列およびプロモータ配列が1つのDNA断片で一致しうることを理解するであろう。上記の高発現遺伝子のリストも標的遺伝子座として適合されている。
【0071】
自発的に維持されるクローニングベクターの例が、AMA1配列を含んで成るクローニングベクターである。AMA1は、アスペルギルス・ニドゥランスから単離された6.0kbゲノムDNA断片であるが、これはアスペルギルスにおける自発的維持が可能である(例えば、アレスケンコ(Aleksenko)とクラッターバック(Clutterbuck)(1997年)、Fungal Genet.Biol.21:373−397頁を参照)。
【0072】
本発明の方法の別の好ましい実施形態によれば、ステップa.は、選択マーカー遺伝子を含んで成るDNA構成物で糸状菌を形質転換するステップを含んで成る。クローニングベクターにおける選択マーカー遺伝子は、糸状菌の形質転換に有用である多くのマーカー遺伝子から選択されうる。一例として、これらのマーカーは、amdS遺伝子など優性および双方向選択マーカー遺伝子(欧州特許第635574B号明細書または国際公開第97/06261号パンフレット)、argB、trpC、またはpyrGなど栄養要求性マーカー遺伝子、および例えば、フレオマイシン、ハイグロマイシンB、またはG418に対する抵抗を提供する抗生物質抵抗性遺伝子を含むがこれらに限定されない。優性および双方向選択マーカー遺伝子の使用が好ましい。好ましくは、amdS遺伝子が好ましく、より好ましくは、アスペルギルス・ニドゥランスまたはアスペルギルス・ニゲルからのamdS遺伝子である。最も好ましい選択マーカー遺伝子が、A.ニドゥランスgpdAプロモータに融合したA.ニドゥランスamdSコード配列である(EP635574号明細書を参照)。他の糸状菌からのamdS遺伝子も使用される(国際公開第97/06261号パンフレット)。amdS選択マーカー遺伝子は、それが同じ菌株において数倍で使用され、明確なDNA配列を導入し、過剰発現し、かつ/または削除しうる利点を有する。欧州特許第635574号明細書に記載されたフルオロアセトアミド培地での対抗選択を用いて、結果として生じる菌株はマーカーを含まず、さらなる遺伝子修飾のために使用されうる。
【0073】
あるいは、または上記の方法とともに、DNA配列の発現のアップレギュレーションがUVまたは化学的変異誘発を使用して達成されうる(マテム,I.E.、ファン・ヌルトJ.M.、ファン・デン・ベルグ,P.、アーチャー,D.B.、ロバーツ,I.N.、およびファン・デン・ホンデル,C.A.、Isolation and characterization of mutants of Aspergillus niger deficient in extracellular proteases.Mol Gen Genet.1992年8月、234(2):332−6頁)。
【0074】
さらに、かつ/またはHRに関与するDNA配列の発現のアップレギュレーションと組合せて、UVまたは化学的変異誘発によってHRに関与するタンパク質の活性を増大させることによってHR効率の増大を得ることも可能である(マテム,I.E.、ファン・ヌルト J.M.、ファン・デン・ベルグ,P.、アーチャー,D.B.、ロバーツ,I.N.、およびファン・デン・ホンデル,C.A.、Isolation and characterization of mutants of Aspergillus niger deficient in extracellular proteases.Mol Gen Genet.1992年8月、234(2):332−6頁)。
【0075】
当業者は、DNA配列の発現のアップレギュレーションを達成するには、記載された方法の各々を別々に、または組合せて使用しうることを理解するであろう。
【0076】
当業者は、増大したHR/NHR比、および/または低下したNHR効率、および/または高いHR効率を有する糸状菌を得るには、糸状菌における一定の遺伝子の発現をそれぞれダウンおよびアップレギュレートするために記載された各々の方法の少なくとも1つを使用することができる。好ましくは、低下したNHR効率および高いHR効率を有する組換え糸状菌を得るために糸状菌で実行される方法のすべては、優性および双方向選択マーカー、好ましくはamdS遺伝子、より好ましくは、アスペルギルス・ニドゥランスまたはアスペルギルス・ニゲルからのamdS遺伝子を使用して実行されている。
【0077】
得られた糸状菌はその後、前述されているように、例えばノーザンおよび/またはウェスタンブロット法および/またはアレイおよび/または表現型スクリーニングを使用することによって前記DNA配列の発現レベルをモニタリングすることによって選択されうる。場合により、細胞のNHRおよび/またはHR経路の効率がモニタリングされる。糸状菌のこれらの経路の効率は以前に規定されているようにモニタリングされうる。
【0078】
好ましくは、HR経路にもたらされる修飾は誘導性のものである。これは誘導性調節領域によってHRに関与する成分をコードする遺伝子の内在性調節領域を置換することによって、好ましくは誘導性プロモータを使用することによって達成されうる。誘導性プロモータの例は、アスペルギルス・ニゲルのグルコアミラーゼプロモータ、アスペルギルス・オリゼのTAKAアミラーゼプロモータ、pafプロモータ(マルクス(Marx),F.、ハース(Haas),H.、ラインドル(Reindl),M.、ストッフラー(Stoffler),G.、ロットスパイヒ(Lottspeich),F.、およびレドル(Redl),B.、Cloning, structural organization and regulation of expression of the Penicillium chrysogenum paf gene encoding an abundantly secreted protein with antifungal activity Gene 167(1−2頁)、167− 171頁 (1995年)、またはペニシリウム・クリソゲヌムのpcbCプロモータ(マルチン JF、カスケイロ J、コサルコワ K、マルコス AT、グチレス S.、Penicillin and cephalosporin biosynthesis:mechanism of carbon catabolite regulation of penicillin production.Antonie Van Leeuwenhoek. 1999年1−2月、75(1−2頁):21−31、総説)、またはNHRに関与する遺伝子の発現のダウンレギュレーションについて以前に引用されたスイッチオン/オフシステムである。
【0079】
好ましい実施形態によれば、修飾されるHR経路に関与する遺伝子は、以下の遺伝子またはその相同体、すなわち、RAD51、RAD52である。
【0080】
HRの過程においてその機能を実行することがより可能または少なくとも相当により可能である、HRに関与する少なくとも1つの成分を有する変異体のすべての種類が、本発明によって意図される変異体である。好ましくは、HRに関与する成分の活性は修飾されており、HR経路の効率は、以前に規定されたアッセイにおいて測定されるように、同じ条件下にそれが由来する親細胞における効率の110%超、より好ましくは200%超、最も好ましくは500%超である。好ましい実施形態によれば、親生物は宿主細胞の部で定義される糸状菌の1つである。本発明による方法は、広範囲であるが限定的に上述されているように、さまざまな用途において使用されうる。一部の特定の用途が以下に記載されている。
【0081】
宿主細胞
したがって、本発明はさらに糸状菌それ自体に関するが、これは好ましくは、前記糸状菌細胞のゲノムへの所定の部位に対するポリヌクレオチドの標的取込みの効率を増大させるための本発明の方法において使用され、前記糸状菌はNHR選好を有し、かつ前記ポリヌクレオチドは前記所定の部位と相同の領域を有するとともに、前記方法は取込み経路をHRへ導くステップを含んで成る。この方法において使用されうる糸状菌の特徴は以前に規定されている。
【0082】
本発明の方法において好ましく使用される糸状菌は親細胞に由来する変異体であり、NHR/HRの比が減少しており、かつ/またはNHR経路の効率が低下されており、かつ/またはHR経路の効率が同じ条件下に前記親生物における前記比および前記効率と比べ前記変異体細胞において高められている。NHR/HRの比および/またはNHR経路の効率および/またはHR経路の効率を測定するために使用されるアッセイは以前に記載されている。
【0083】
本発明の宿主細胞が糸状菌であるが、これはクローニングベクターで形質転換されることが可能である。したがって、試験される大部分の糸状菌では、それらはアスペルギルスのために開発された形質転換プロトコールを使用して形質転換されうることがわかった(とりわけ、チルブムら、1983年、Gene、26:205−221頁による)。当業者は、糸状菌宿主種の有効な形質転換が、本明細書で具体的に例示されたベクター、選択マーカー系、プロモータ、および形質転換プロトコールの使用に限定されていない。
【0084】
糸状菌は本明細書では、糸状形態における分類Eumycotinaの真核微生物と定義されており、すなわち、その栄養成長は菌糸伸長によって起こる。好ましい糸状菌宿主細胞は、アスペルギルス属、トリコデルマ属、フサリウム(Fusarium)属、ペニシリウム属、およびアクレモニウム(Acremonium)属からなる群から選択される。
【0085】
本発明のより好ましい実施形態において、糸状菌宿主細胞は、A.ニドゥランス、A.オリゼ、A.ソーヤ(sojae)、A.ニゲル群のアスペルギルス、トリコデルマ・エセイ、およびペニシリウム種からなる群から選択される。好ましくは、ペニシリウムはペニシリウム・クリソゲヌムまたはペニシリウム・シトリヌム(Penicillium citrinum)種である。
【0086】
A.ニゲル群は本明細書ではレイパー(Raper)とフェネル(Fennell)(1965年、The Genus Aspergillus,The Williams & Wilkins Company,Baltimore,293−344頁)に従って定義され、かつこれらの著者らによって含まれるすべての(黒色)アスペルギルスを含んで成る。最も好ましい糸状菌宿主細胞は、A.ニゲル群、A.オリゼのアスペルギルス、トリコデルマ・エセイ、およびペニシリウム・クリソゲヌムから成る群から選択される。
【0087】
好ましい実施形態によれば、親生物は寄託された糸状菌細胞アスペルギルス・ニゲルCBS513.88、アスペルギルス・オリゼATCC20423、IFO4177、ATCC1011、ATCC9576、ATCC14488−14491、ATCC11601、ATCC12892、ペニシリウム・クリソゲヌムCBS455.95、またはペニシリウム・シトリヌムATCC38065、ペニシリウム・クリソゲヌムP2、アクレモニウム・クリソゲヌム(Acremonium chrysogenum)ATCC36225またはATCC48272、トリコデルマ・エセイATCC26921またはATCC56765またはATCC26921、アスペルギルス・ソーヤATCC11906、クリソスポリウム・ルキノウェンゼ(Chrysosporium lucknowense)ATCC44006、クラビセプス・パスパリ(Claviceps paspali)CBS110.22、クラビセプス・プルプラ(Claviceps purpurea)CBS164.59、ペニシリウム・ブリビコンパクツム(Penicillium brevicompactum)ATCC 9056、アスペルギルス・テレウス(Aspergillus terreus)ATCC20542、アスペルギルス・ニドゥランスATCC28901およびまたはそれらの誘導体である。
【0088】
別の好ましい実施形態によれば、本発明の糸状菌細胞は、以下のアッセイによって測定されるように、少なくとも200、少なくとも50、少なくとも10であるNHR/HR比を有する。好ましくは、糸状菌細胞の比は少なくとも1、より好ましくは少なくとも0.5、さらにより好ましくは少なくとも0.1、さらにより好ましくは少なくとも0.05、さらにより好ましくは少なくとも0.01さらにより好ましくは少なくとも0.005さらにより好ましくは少なくとも0.001さらにより好ましくは少なくとも0.0005さらにより好ましくは少なくとも0.0001、かつ最も好ましくは少なくとも0.00001である。
【0089】
より好ましい実施形態によれば、本発明の糸状菌細胞は、以下のアッセイによって測定されるように200未満、さらにより好ましくは50未満、10未満であるNHR/HR比を有する。さらにより好ましくは、糸状菌細胞の比は1未満、さらにより好ましくは0.5未満、さらにより好ましくは0.1未満、さらにより好ましくは0.05未満、さらにより好ましくは0.01未満、さらにより好ましくは0.005未満、さらにより好ましくは0.001未満、さらにより好ましくは0.0005未満、さらにより好ましくは0.0001未満、かつ最も好ましくは0.00001未満である。
【0090】
NHR/HRの比は好ましくは国際公開第02/052026号パンフレット(表2、23頁)に記載されたアッセイによって測定される。
【0091】
好ましくは、糸状菌細胞は、NHRに関与する成分をコードする遺伝子を欠き、かつ/またはNHRに関与する成分の減少したレベルを有する。
【0092】
さらにより好ましくは、糸状菌細胞は以下の遺伝子の少なくとも1つを欠いている。すなわち、hdfAまたは配列番号2または19で識別されるその相同体、hdfBまたは配列番号5、もしくは22または両方で識別されるその相同体、かつ/または、好ましくは、これらの遺伝子によってコードされたタンパク質の少なくとも1つの減少した量を有する。
【0093】
最も好ましくは、糸状菌細胞は誘導性に以下の遺伝子の少なくとも1つを欠いている。すなわち、hdfAまたは配列番号2または19で識別されるその相同体、hdfBまたは配列番号5、もしくは22または両方で識別されるその相同体、かつ/または、好ましくは誘導性に、これらの遺伝子によってコードされたタンパク質の少なくとも1つの減少した量を有する。
【0094】
別の好ましい実施形態によれば、糸状菌細胞は、そのゲノムにおいて、NHRに関与する遺伝子が非機能的遺伝子によって、もしくは選択マーカーによって、または別の遺伝子によって置換されているようになっている。
【0095】
別の好ましい実施形態によれば、変異体はHRに関与する成分の増大したレベルを有する。
【0096】
本発明による糸状菌は、分子生物学法によって得られている。かかる遺伝子組換え法によって得られる糸状菌は組換え糸状菌と定義される。しかし本発明に即した組換え糸状菌は、早期に変異誘発法にかけられ、別の望ましい効果を達成しうる。最も好ましい実施形態によれば、得られる糸状菌は組換え糸状菌である。
【0097】
本発明の宿主細胞の使用
好ましい実施形態によれば、少なくとも、例えば、形質転換またはエレクトロポレーションの方法、および前記ポリヌクレオチドの前記細胞の遺伝物質への取込みによって、目的とするポリヌクレオチドを本発明の糸状菌へ導入するステップを含んで成る方法が提供される。取込みは複合方法であり、核酸配列が宿主細胞の遺伝物質の一部となる。核酸取込みの方法における1つのステップは組換えであり、組換えによって、核酸配列が交換または挿入され、導入された核酸は宿主細胞の遺伝物質の一部となる。原則として、2種類の組換え法が可能である。すなわち、相同的および非正統的すなわちNHRである。大部分の(高等)真核生物はHRを実行することはなく、または少なくとも顕著に実行することはないが、かかる方法を達成する必須タンパク質は利用可能である。この現象の1つの理由は、(高等)真核生物における相同的組換えの頻繁な使用が、繰返し核酸配列の存在により望ましくない染色体再配置をもたらしうることである。本発明による方法によってHRを達成するには、所定の部位との相同性を有するポリヌクレオチドを提供することが重要である。相同の割合および相同的領域の長さが相同的組換えにおいて重要な役割を果たすことは当業者に明らかである。相同の割合はほぼ100%であることが好ましい。当業者であれば、低い割合の相同も相同的組換えの分野で使用されるが、例えば、相同の領域、およびその全体的な分布に依存し、HRの低い効率をもたらしうるが、依然として有用であり、したがって本発明に含まれることを認識している。さらに、(ほぼ)相同的領域の長さは約3kbであり、これは相同的組換えを導くのに十分である。少なくとも1つの相同的領域が組換えには必要であるが、より好ましくは、目的とする核酸をフランキングする2つの相同的領域が標的取込みのために使用される。当業者は、相同の適切な割合、相同の長さ、および相同的領域の量を選択するやり方を知っている。かかる相同を提供することによって、核酸が宿主細胞の遺伝物質内のすべての所望の位置で取込まれる。本明細書で開示された本発明が、相同の長さおよび相同の割合がHRを提供/可能にするのに十分に高い限り、すべての核酸(好ましくはDNA)を所定の部位に導くために使用されることは当業者には明らかである。
【0098】
本発明がなされる前は、ポリヌクレオチドがつねに容易にすべての所望の位置で一定の糸状菌のゲノムへ取込まれたわけではない。本発明による方法は、例えば、さまざまな方法で遺伝子機能に影響を及ぼすために、完全な不活性化のためだけではなく、発現レベルまたは発現の調節における変化、タンパク質活性またはコードされたタンパク質の細胞内標的の変化を介在するためにも適用される。完全な不活性化は、通常、アンチセンス技術またはRNAi技術(ズレナー(Zrenner)R、ウィルミッツァー(Willmitzer)L、ソネワルド(Sonnewald)U、Analysis of the expression of potato Uridinediphosphate−glucose pyrophosphorylase and its inhibition by antisense RNA.Planta.(1993年)、190(2):247−52頁)など既存の方法によって達成されえないが、例えば、代謝経路の望ましくない側枝をコントロールし、例えば、(ベータ・ラクタム)抗生物質またはカルチノイドなど特定の二次代謝産物の産生を増大させる遺伝子の不活性化に有用である。完全な不活性化は、毒性または望ましくない化合物の産生を削減させるためにも有用である(chrysogenin in Penicillium;Aflatoxin in Aspergillus:マクドナルド(MacDonald)KDら、Heterokaryon studies and the genetic control of penicillin and chrysogenin production in Penicillium chrysogenum.J Gen Microbiol.(1963年)33:375−83頁)。完全な不活性化は、発酵過程および下流処理が改善されるような方法で生物の形態を変化させるのにも有用である。
【0099】
本発明は、代わりの調節配列によって既存の調節配列を置換し、内在性遺伝子の発現(例えば、特定の誘導剤に反応した発現)を変化させることを可能にする。
【0100】
本発明の一態様は、本発明の方法による活性遺伝子の不活性遺伝子による置換に関する。完全な不活性化は、通常、アンチセンス技術またはRNAi技術など既存の方法によって達成されえないが、例えば、代謝経路の望ましくない側枝をコントロールし、例えば、でんぷんなどバルク製品の品質を増大し、または特定の二次代謝産物の産生を増大させ、または望ましくない代謝産物の形成を阻害するために有用である。
【0101】
本発明の別の態様は、糸状菌細胞の広範囲の代謝再プログラミングまたはエンジニアリングに関する。完全に新しい経路の導入および/または望ましくない経路の修飾は、タンパク質または代謝産物など特定の化合物の生産に具体的に適合される細胞の取得をもたらす。
【0102】
本発明の別の態様は、不活性または変化した遺伝子の活性遺伝子による置換に関する。例えば、伝統的な変異誘発の有効な期間後、選択された糸状菌株が無作為変異誘発法の間に変化し、さらに不活性化した一部の内在性遺伝子を有する場合が多い。
【0103】
本発明のさらに別の態様では、抗生物質の糸状菌細胞に対する抵抗を与える物資を導入する方法が提供される。さらに本発明の別の態様では、所望の特性を糸状菌細胞に与える方法が提供される。好ましい実施形態では、遺伝子送達賦形剤が使用され、所望のポリヌクレオチドを所定の部位に送達する。遺伝子送達賦形剤は当技術分野で周知であり、説明で以前に記載されている。
【0104】
本発明の別の態様による別の好ましい方法も、例えば、所望の新規生成物のためのもので所望の発現プロフィールを与える遺伝子の既存のコード配列を置換することによって、新規生成物をコードする導入遺伝子の予測可能な発現を達成する。より好ましい実施形態によれば、本発明によって提供される糸状菌はさらに、産生される所望のタンパク質の所望の遺伝子コードを含んで成るDNA構成物を含んで成る。
【0105】
好ましくは、産生される所望のタンパク質をコードする所望の遺伝子は発現ベクターへ挿入され、これはその後に得られた宿主細胞を形質転換するために使用される。発現ベクターにおいて、DNA配列は、プロモータなどの適切な発現シグナル、場合によりシグナル配列および末端に操作連結されうるが、これらは宿主生物におけるタンパク質の発現および合成を導くことが可能である。
【0106】
より好ましくは、所望の遺伝子はプロモータおよび分泌シグナルに操作連結される。所望の遺伝子を発現させるために使用されうる方法は、DNA配列の発現のアップレギュレーションの部で記載されたものと同じであり、その発現産物はHR、すなわち、コピー数の増大、標的取込み、高発現遺伝子のプロモータの使用、選択マーカー遺伝子およびそれらの組合せの選択に関与する。
【0107】
所望のタンパク質は好ましくは酵素である。タンパク質が自然に分泌されない場合は、タンパク質をコードするポリヌクレオチドが当技術分野で周知の方法に従ってシグナル配列を有するように修飾されうる。分泌されるタンパク質は、自然に発現される内在性タンパク質でありうるが、非相同でもありうる。非相同は、タンパク質によってコードされる遺伝子が野生型糸状菌における自然条件下に産生されないことを意味する。本発明の糸状菌によって産生されうる酵素の例は、カルボヒドラーゼ、例えば、エンドグルカナーゼ、β−グルカナーゼ、セロビオヒドラーゼまたはβ−グルコシダーゼなどのセルラーゼ、ヘミセルラーゼ、またはキシラナーゼ、キシロシダーゼ、マナナース、ガラクタナース、ガラクトシダーゼ、ラムノガラクツロナーゼ、アラバナーゼ、ガラクツロナーゼ、リアーゼなどのペクチン分解酵素、またはアミノ分解酵素、フィターゼなどのホスファターゼ、リパーゼなどのエステラーゼ、タンパク質分解酵素、オキシダーゼ、トランスフェラーゼ、またはイソメラーゼなどの酸化還元酵素である。より好ましくは、所望の遺伝子はフィターゼをコードする。
【0108】
別の例として、既存のコード配列が修飾され、コードされたタンパク質は、例えば、改善された熱的特性および/または改善された動態特性(Km、Kcat)、および/または改善された酵素安定性、および/または基質範囲の延長、および/または寿命の増大等を有するタンパク質を作る最適特徴を有するようになる。
【0109】
本発明はさらに目的とするポリペプチドを製造するための本発明の糸状菌の使用に関する。あるいは、得られた糸状菌は二次代謝産物を製造するために使用されうる。好ましい二次代謝産物は、カルチノイド化合物、ベータ−ラクタム化合物、薬剤、抗腫瘍化合物等である。
【0110】
好ましくは、本発明で得られた糸状菌は、所望のタンパク質をコードするDNA配列の発現を促す条件下に形質転換された宿主細胞を培養し、かつ例えば以下の文献に記載された所望のタンパク質を回収することによって所望のタンパク質を製造するために使用される。すなわち、
−リー(Li),Z.J.、シュクラ(Shukla),V.、フォルダイス(Fordyce),A.P.、ペデルセン(ペデルセン),A.G.、ウェグナー(Wegner),K.S.、マルテン(Marten),M.R.、Fungal morphology and fragmentation behavior in a fed−batch Aspergillus oryzae fermentation at the production scale.Biotechnol Bioeng.2000年11月5日、70(3):300−12頁
−ウィザース(Withers),J.M.、スウィフト(Swift),R.J.、ウィーベ(Wiebe),M.G.、ロブソン(Robson),G.D.、プント(Punt),P.J.、ファン・デン・ホンデル,C.A.、Optimization and stability of glucoamylase production by recombinant strains of Aspergillus niger in chemostat culture.Biotechnol Bioeng.1998年8月20日、59(4):407−18頁。
−アマルラー(Amanullah),A.、クリステンセン(Christensen),L.H.、ハンセン(Hansen),K.、ニーナウ(Nienow),A.W.、トーマス(Thomas),R.C.、Dependence of morphology on agitation intensity in fed−batch cultures of Aspergillus oryzae and its implications for recombinant protein production.Biotechnol Bioeng.2002年3月30日、77(7):815−26頁。
【0111】
DNA配列およびこれらのDNA配列によってコードされたポリペプチド
本発明の別の態様によれば、以下の単離cDNA配列が提供される。すなわち、
配列番号2 A.ニゲルからのhdfA
配列番号19 ペニシリウム・クリソゲヌムからのhdfA
配列番号5 A.ニゲルからのhdfB
配列番号22 ペニシリウム・クリソゲヌムからのhdfB
およびそれらの相同体。
【0112】
配列番号1、18、4、および21の各々は、それぞれ、上記の各々のcDNA配列と関係があるゲノムDNA配列に対応する。
【0113】
配列番号3、20、6、および23の各々は、それぞれ、上記のそれぞれのcDNA配列によってコードされたタンパク質配列に対応する。
【0114】
本明細書で提供される配列情報は、誤って識別された塩基の包含を必要とするために狭義に考えるべきではない。本明細書で開示された特定の配列は、特にA.ニゲルまたはペニシリウム・クリソゲノムにおける糸状菌から完全な遺伝子を単離するために容易に使用されうるが、次いでさらに配列分析に容易にかけられ、それによって配列決定の誤りを識別することができる。
【0115】
別段の指示がない限り、本明細書でのDNA分子の配列決定によって決定される全ヌクレオチド配列は、自動化DNAシーケンサを使用して決定され、かつ本明細書で決定されるDNA分子によってコードされたポリペプチドの全アミノ酸配列は上記のように決定されたDNA配列の翻訳によって予測された。したがって、この自動化法によって決定されるDNA配列について当技術分野で周知であるように、本明細書で決定されたヌクレオチド配列はいつくかの誤りを含有する。自動化によって決定されたヌクレオチド配列は一般的に、配列決定されたDNA分子の実際のヌクレオチド配列と少なくとも約90%同一であり、より一般的には少なくとも約95%〜少なくとも約99.9%同一である。実際の配列は、当技術分野で周知の手動のDNA配列決定法を含む他の方法によってより正確に決定されうる。やはり当技術分野で周知であるように、実際の配列と比べ決定されたヌクレオチド配列における単一の挿入または削除が、ヌクレオチド配列の翻訳におけるフレームシフトをもたらし、決定されたヌクレオチド配列によってコードされた予測アミノ酸配列が、かかる挿入または削除の点で開始する、配列決定されたDNA分子によって実際にコードされたアミノ酸配列と完全に異なるようになる。
【0116】
当業者は、かかる誤って識別された塩基を識別することが可能であり、かかる誤りを正すやり方を知っている。
【0117】
「相同」は以下に定義されている。相同的はアスペルギルス・ニゲルまたはペニシリウム・クリソゲノム以外の他の糸状菌由来を意味すると理解されうる。
【0118】
他の生物からの全長DNAは、他の生物、例えば、糸状菌、具体的には、それらをスクリーニングすることによってアスペルギルス種またはペニシリウム種から構成されたcDNAまたはゲノムDNAライブラリーを使用して得られうる。
【0119】
本発明は、hdfAおよび/またはhdfBのパラログをも包含する。本発明に照らして、パラログは、配列番号1または配列番号4または配列番号18または配列番号21と相同的、かつそれぞれ、A.ニゲルまたはペニシリウム・クリソゲノム由来のDNA配列を意味する。
【0120】
例えば、アスペルギルスまたはペニシリウム菌株は、ノーザンブロット分析によって相同的hdfAおよび/またはhdfBポリヌクレオチドについてスクリーニングされうる。本発明によるポリヌクレオチドと相同的な転写産物の検出と同時に、cDNAライブラリーが、当業者に公知の標準の方法を利用することにより、適切な菌株から単離されるRNAで構成されうる。あるいは、全ゲノムDNAライブラリーが、本発明によるhdfAおよび/またはhdfBにハイブリダイズ可能なプローブを使用することによりスクリーニングされうる。
【0121】
相同的遺伝子配列は、例えば、本明細書で教示されたヌクレオチド配列に基づき設計された2つの退化オリゴヌクレオチドプライマープールを使用するPCRを実行することによって単離されうる。
【0122】
反応のためのテンプレートは、本発明によるポリヌクレオチドを発現することが知られ、またはそう思われる菌株から調製されるmRNAの逆転写によって得られるcDNAでありうる。PCR産物はサブクローン化されて配列決定され、増幅配列が新しいhdfAおよび/またはhdfB核酸配列、またはそれらの機能的等価物の配列を示すことを確実にしうる。
【0123】
次いで、PCR断片を使用し、さまざまな既知の方法によって行われる完全長cDNAを単離することができる。例えば、増幅断片は標識され、バクテリオファージまたはコスミドcDNAライブラリーをスクリーニングするために使用されうる。あるいは、標識断片を使用してゲノムライブラリーをスクリーニングすることができる。
【0124】
PCR技術をも使用し、他の生物から完全長cDNA配列を単離することができる。例えば、RNAは、標準の方法に従い、適切な細胞または組織源から単離されうる。逆転写反応は、最初のストランド合成のプライミングのために増幅断片の大部分の5’末端に特異的なオリゴヌクレオチドプライマーを使用してRNAで実行されうる。
【0125】
次いで、結果として生じるRNA/DNAハイブリッドは、標準の末端トランスフェラーゼ反応を使用して(例えば、グアニンで)「末端付加」されうるが、このハイブリッドはRNaseHで消化されうるとともに、次いで第2のストランド合成が(例えば、ポリCプライマーで)プライミングされうる。したがって、増幅断片の上流のcDNA配列は容易に単離されうる。有用なクローニング法の検討では、例えば、サンブルックら、上記参照、およびオーズベル(Ausubel)ら、下記参照を参照。
【0126】
「相同」は機能的等価物を意味するとも理解されうる。
【0127】
「機能的等価物」および「機能的変異形」という語は本明細書で同義で使用される。hdfAおよび/またはhdfB DNAの機能的等価物は、hdfAおよび/またはhdfBの特定の機能を示すポリペプチドをコードする単離DNA断片である。本発明によるhdfAおよび/またはhdfBポリペプチドの機能的等価物は、NHR複合体の一部として少なくとも1つの機能を示すポリペプチドである。したがって、機能的等価物も生体活性断片をも包含する。
【0128】
機能的タンパク質またはポリペプチド等価物は、配列番号3または6または20または23または非必須アミノ酸の置換、挿入、または削除を有する配列の1つもしくはそれ以上のアミノ酸の保存的置換のみを含有しうる。したがって、非必須アミノ酸は、生体機能を実質的に変化せずにこれらの配列の1つにおいて変化されうる残基である。例えば、本発明のhdfAおよび/またはhdfBタンパク質の中で保存されるアミノ酸残基は、特に変化の影響を受けにくいことが予測される。さらに、本発明のhdfAおよび/またはhdfBタンパク質の中で保存されるアミノ酸は、変化の影響を受ける可能性が低い。
【0129】
「保存的置換」という語は、アミノ酸残基が同様の側鎖を有するアミノ酸残基で置換される置換を意味することが意図されている。これらのファミリーは当技術分野で周知であり、塩基側鎖(例えば、リシン、アルギニン、およびヒスチジン)、酸側鎖(例えば、アスパラギン酸、グルタミン酸)、非荷電極性側鎖(例えば、グリシン、アスパラギン、グルタミン、セリン、トレオニン、チロシン、システイン)、非極性側鎖(例えば、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン)、ベータ−分岐側鎖(例えば、トレオニン、バリン、イソロイシン)、および芳香族側鎖(例えば、チロシン、フェニルアラニン、トリプトファン、ヒスチジン)を含むアミノ酸を含む。
【0130】
機能的核酸等価物は一般的に、サイレント突然変異、すなわちコードされたポリペプチドの生体機能を変化させることがない突然変異を含有しうる。したがって、本発明は、特定の生物活性に重要ではないアミノ酸残基における変化を含有するhdfAおよび/またはhdfBタンパク質をコードする核酸分子を提供する。かかるhdfAおよび/またはhdfBタンパク質はアミノ酸配列において配列番号3または6、または20または23と異なり、さらにその生物活性の少なくとも1つを保持する。一実施形態において、単離核酸分子はタンパク質をコードするヌクレオチド配列を含んで成るが、そのタンパク質は、配列番号3または6または20または23で示されたアミノ酸配列と少なくとも約60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%もしくはそれ以上相同の実質的に相同のアミノ酸配列を含んで成る。例えば、表現型的にサイレントアミノ酸置換するやり方に関する手引きは、ボウイ(Bowie),J.U.ら、Science 247:1306−1310頁(1990年)に示されており、ここで著者らは、変化に対するアミノ酸配列の耐性を研究する2つの主要な方法があることを示している。最初の方法は進化の方法に依拠し、ここで突然変異は自然な選択によって受入れられ、または拒絶される。第2の方法では、クローン化遺伝子の特定の位置でアミノ酸の変化を導入する遺伝子操作、および選択、すなわちスクリーニングが使用され、機能性を維持する配列を特定する。著者らが述べているように、これらの研究では、タンパク質が意外にもアミノ酸置換に耐えることが示された。著者らはさらにどの変化がタンパク質の特定の位置で許容的である可能性が高いかを示している。例えば、大部分の埋もれたアミノ酸残基は非極性側鎖を必要とするが、表面側鎖のほとんどの特徴は、ほとんどの場合、保存されていない。かかる表現型的にサイレントの他の置換がボウイ(Bowie)らおよびそれに引用された文献に記載されている。
【0131】
配列番号3または6または20または23によるタンパク質と相同的なhdfAおよび/またはhdfBタンパク質をコードする単離核酸分子が、1つもしくはそれ以上のアミノ酸置換、削除、または挿入がコードされたタンパク質へ導入されるように、1つもしくはそれ以上のヌクレオチド置換、付加、または削除を配列番号2または配列番号5、または配列番号19または配列番号22によるコードヌクレオチド配列へ導入しすることによって生成されうる。かかる突然変異は、部位特異的変異誘発およびPCR介在変異誘発など標準の方法によって導入されうる。
【0132】
「機能的等価物」という用語は、A.ニゲルのhdfAおよび/またはhdfBタンパク質のオルソログをも包含する。A.ニゲルのhdfAおよび/またはhdfBタンパク質のオルソログは、他の菌株または種から単離されうるとともに、同様または同一の生物活性を有するタンパク質である。かかるオルソログは、配列番号3または6または20または23と実質的に相同的であるアミノ酸配列を含んで成るとして容易に識別されうる。
【0133】
「相同」は「実質的に相同」の意味としても理解されうる。
【0134】
「実質的に相同」という用語は、第2のアミノ酸またはヌクレオチド配列と十分または最小の数の同一または等価の(例えば、同様の側鎖を有する)アミノ酸またはヌクレオチドを含有する第1のアミノ酸またはヌクレオチド配列を指し、第1および第2のアミノ酸またはヌクレオチド配列は共通のドメインを有するようになっている。例えば、約45%、好ましくは約50%、好ましくは約60%、好ましくは約65%、より好ましくは約70%、さらにより好ましくは約75%、80%、85%、90%、95%、96%、97%、98%、もしくは99%もしくはそれ以上の同一性を有する共通のドメイン含有するアミノ酸またはヌクレオチド配列が、十分に同一と本明細書で規定されている。
【0135】
また、配列番号2または5または19または22と異なるヌクレオチド配列を有する他のhdfAおよび/またはhdfBファミリーメンバーをコードする核酸も本発明の範囲内である。さらに、異なる種からのhdfAおよび/またはhdfBタンパク質をコードする核酸は、したがって配列番号2または5または19または22と異なるヌクレオチド配列を有する。
【0136】
本発明のhdfAおよび/またはhdfBDNAの変異形(例えば、天然対立遺伝子多型)および相同体に対応する核酸分子が、好ましくはきわめて厳格なハイブリダイゼーション条件下に標準のハイブリダイゼーション法によるハイブリダイゼーションプローブとして本明細書で開示されたcDNAまたはその適切な断片を使用する本明細書で開示されたhdfAおよび/またはhdfB核酸とのその相同に基づき単離されうる。
【0137】
ハイブリダイゼーション反応の「厳格性」は、当業者によって容易に決定可能であり、ほとんどの場合、プローブの長さ、洗浄温度、および塩濃度に依存した経験的計算である。一般に、長いプローブは適切なアニーリングのために高い温度を必要とするが、短いプローブは低い温度を必要とする。ハイブリダイゼーションはほとんどの場合、相補鎖がその融解温度未満の環境で存在する場合に再アニーリングする変性DNAの能力に依存する。プローブとハイブリダイズ可能な配列との間の所望の相同の程度が高いほど、使用されうる相対温度は高くなる。結果として、高い相対温度は、反応条件をより厳格にする傾向があるが、低い温度は厳格でなくなることになる。
【0138】
ハイブリダイゼーション反応の厳格性の追加の詳細および説明については、オーズベルら、Current Protocols in Molecular Biology、ワイリー・インターサイエンス・パブリッシャーズ(Wiley Interscience Publishers)(1995年)を参照されたい。
【0139】
本明細書で規定される「厳格な条件」または「高厳格性条件」は、以下によって識別されうる。すなわち、(1)洗浄のために低いイオン強度および高い温度、例えば、0.015M塩化ナトリウム/0.0015Mクエン酸ナトリウム/0.1%ドデシル硫酸ナトリウムを50℃下に使用し、(2)ハイブリダイゼーション中に、ホルムアミドなどの変性剤、例えば、50%(v/v)ホルムアミドを0.1%ウシ血清アルブミン/0.1%Ficdl/0.1%ポリビニルピロリドン/750mM塩化ナトリウムとともにpH6.5で50mMリン酸ナトリウム緩衝液、75mMクエン酸ナトリウムを42℃下に使用し、または(3)50%ホルムアミド、5×SSC(0.75M NaCl、0.075Mクエン酸ナトリウム)、50mMリン酸ナトリウム(pH6.8)、0.1%ピロリン酸ナトリウム、5×デンハルト液、超音波処理サケ精子DNA(50Rg/ml)、0.1%SDS、および10%硫酸デキストランを42℃下に使用するとともに、0.2×SSC(塩化ナトリウム/クエン酸ナトリウム)中42℃下および55℃下での50%ホルムアミド中での洗浄後、55℃下にEDTAを含有する0.1×SSCから成る高厳格性洗浄を行う。
【0140】
「中程度に厳格な条件」は、サンブルックら、Molecular Cloning:A Laboratory Manual、ニューヨーク:コールド・スプリング・ハーバー・プレス、1989年によって記載されているように識別されうるとともに、洗浄液および上記の厳格性の低いハイブリダイゼーション条件(例えば、温度、イオン強度、および%SDS)の使用を含む。中程度に厳格な条件の例は、20%ホルムアミド、5×SSC(150mM NaCl、5mMクエン酸三ナトリウム)、50mMリン酸ナトリウム(pH7.6)、5×デンハルト液、10%硫酸デキストラン、および20mg/ml変性剪断サケ精子DNAを含んで成る溶液中37℃での一夜インキュベーション後、ろ過液の1×SSC中約37〜50℃下での洗浄である。当業者は、温度、イオン強度等を調節するやり方を、必要に応じて、プローブの長さなどの要因を適合させ、または配列類似性を決定するために適切なアルゴリズムを使用することによって認識するであろう。
【0141】
相同的(同様または同一)配列は、「配列比較アルゴリズム」を使用することによっても決定されうる。比較のための配列の最適なアラインメントは、例えば、スミス(Smith)とウォーターマン(Waterman)、Adv.Appl.Math.2:482頁(1981年)の局所相同アルゴリズムによって、ニードルマン(Needleman)とブンシュ(Wunsch)、J.Mol.Biol.48:443頁(1970年)の相同アラインメントアルゴリズムによって、ピアソン(Pearson)とリップマン(Lipman)、Proc.Nat’l Acad.Sci.米国(USA)85:2444頁(1988年)の類似性法の調査によって、これらのアルゴリズムのコンピュータによる実行(ウィスコンシン・ジェネティックス・ソフトウェア・パッケージ(Wisconsin Genetics Software Package)のGAP、BESTFIT、FASTA、およびTFASTA、ジェネティックス・コンピュータ・グループ(Genetics Computer Group)、575サイエンス(Science)Dr.、Madison、ウィスコンシン州(WI))によって、または目視検査によって実行されうる。配列類似性を決定するために適切であるアルゴリズムの例がBLASTアルゴリズムであり、これはアルトシュール(Altschul)ら、J.Mol.Biol.215:403−410頁(1990年)に記載されている。
【0142】
BLAST分析を実行するためのソフトウェアは、国立バイオテクノロジー情報センター(National Center for Biotechnology Information)によって公開されている。(http://www.ncbi.nlm.nih.gov/)。このアルゴリズムは最初に、データベース配列における同じ長さの単語と整列すると一部の正に評価された閾値スコアTに一致またはこれを満たすクエリー配列における長さWの短い単語を識別することによる高スコアリング配列ペア(HSP)の識別を含む。これら最初の近傍単語のヒットは、それらを含有する長いHSPを見出す出発点として作用する。単語のヒットは、累積的アラインメントスコアが増大しうる限り比較される2つの配列の各々に沿った両方の方向で拡大する。単語のヒットの拡張は、累積的アラインメントスコアが最大達成値から数量Xだけ減少すると停止し、累積的スコアはゼロもしくはそれ未満になり、またはどちらか一方の配列の末端に達する。
【0143】
BLASTアルゴリズムパラメータW、T、およびXは、アラインメントの感度および速度を決定する。BLASTプログラムでは初期設定として11の語長(W)、50のBLOSUM62スコアリングマトリクス(ヘニコフ(Henikoff)とヘニコフ、Proc.Natl.Acad.Sci.米国(USA)89:10915頁(1989年)を参照)50のアラインメント(B)、10の期待値(E)、M=5、N=4、および両方の鎖の比較が使用される。
【0144】
次いで、BLASTアルゴリズムでは2つの配列間の類似性の統計的分析を実行する(例えば、カーリン(Karlin)とアルトシュール、Proc.Nat’l.Acad.Sci.米国(USA)90:5873−5787頁(1993年)を参照)。BLASTアルゴリズムによって提供される類似性の1つの尺度は最小合計確率(P(N))であり、これはそれによって2つのヌクレオチドまたはアミノ酸配列の間の適合が偶然に起こりうる確率の指標を提供する。例えば、アミノ酸配列がプロテアーゼなどのタンパク質と同様とみなされるのは、プロテアーゼアミノ酸配列などのタンパク質との試験アミノ酸配列の比較における最小合計確率が約0.1未満、より好ましくは約0.01未満、かつ最も好ましくは約0.001未満である場合である。好ましくは、類似性は、配列番号2、5、19、および22を有するDNA配列の1つと少なくとも40%相同である。より好ましくは、類似性は少なくとも50%、より好ましくは少なくとも60%、より好ましくは少なくとも70%、より好ましくは少なくとも80%、より好ましくは少なくとも90%である。
【0145】
hdfAおよび/またはhdfB配列の自然発生的対立遺伝子多型に加えて、当業者は、突然変異によって配列番号2またh5または19または22のヌクレオチド配列へ変化が導入され、それによってhdfAおよび/またはhdfBタンパク質の機能を実質的に変化させることなく、hdfAおよび/またはhdfBタンパク質のアミノ酸配列の変化をもたらしうることを認識するであろう。
【0146】
本発明の別の態様において、劣化hdfAおよび/またはhdfBタンパク質が提供される。劣化hdfAおよび/またはhdfBタンパク質は、少なくとも1つの生物活性が減少しているタンパク質である。かかるタンパク質は、hdfAおよび/またはhdfBコード配列の全部または一部に沿って、飽和変異誘発によるなど、突然変異を無作為に導入することによって得られうるが、結果として生じる変異体は組換え発現され、生物活性についてスクリーニングされる。例えば、当業界は、その酵素活性を測定するための標準のアッセイを提供しており、したがって劣化タンパク質が容易に選択されうる。好ましくは、アッセイは以前に記載されているものである(例えば、国際公開第02/052026号パンフレット、23頁、または表現型スクリーニングアッセイを参照)。
【0147】
好ましい実施形態では、hdfAおよび/またはhdfBタンパク質は、配列番号3または6または20または23によるアミノ酸配列を有する。別の実施形態では、hdfAおよび/またはhdfBポリペプチドは、配列番号3または6または20または23によるアミノ酸配列と実質的に相同的であり、かつ配列番号3または6または20または23によるポリペプチドの少なくとも1つの生物活性を保持するが、自然の変動もしくは上述の変異誘発によりアミノ酸配列が異なる。
【0148】
別の好ましい実施形態によれば、hdfAおよび/またはhdfBタンパク質は、好ましくはきわめて厳格なハイブリダイゼーション条件下、配列番号2または5または19または22による核酸にハイブリダイズが可能な単離核酸断片によってコードされたアミノ酸配列を有する。
【0149】
したがって、hdfAおよび/またはhdfBタンパク質は、配列番号3または6または20または23で示されたアミノ酸配列と少なくとも約60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%もしくはそれ以上相同のアミノ酸配列を含んで成るタンパク質であり、配列番号3または6または20または23によるポリペプチドの少なくとも1つの機能的活性を保持する。
【0150】
本発明によるタンパク質の機能的等価物は、例えば、一定の活性の本発明のタンパク質の変異体、例えば、切断変異体のコンビナトリアルライブラリーをスクリーニングすることによっても識別されうる。一実施形態では、変異形の変化に富んだライブラリーが核酸レベルでコンビナトリアル変異誘発によって生成される。変異形の変化に富んだライブラリーが、例えば、退化セットの潜在的タンパク質配列が個別のポリペプチドとして、あるいは、一連の大きな融合タンパク質として(例えば、ファージディスプレイのために)発現可能であるように、合成オリゴヌクレオチドの混合物を遺伝子配列へ酵素的にライゲートすることによって生成されうる。退化オリゴヌクレオチド配列からの本発明のポリペプチドの潜在的な変異形のライブラリーを生成するために使用されうるさまざまな方法がある。退化オリゴヌクレオチドを合成するための方法が当技術分野で周知である(例えば、ナラング(Narang)(1983年)Tetrahedron 39:3頁、イタクラ(Itakura)ら(1984年)Annu.Rev.Biochem.53:323頁、イタクラら(1984年)Science 198:1056頁、イケ(Ike)ら(1983年)Nucleic Acid Res.11:477頁)を参照)。
【0151】
また、本発明のポリペプチドのコード配列の断片のライブラリーを使用し、変異形のその後の選択をスクリーニングするためのポリペプチドの変化に富んだ集団を生成することができる。例えば、コード配列断片のライブラリーが、目的とするコード配列の二本鎖PCR断片を切断が分子当たり約1回のみ生じる条件下にヌクレアーゼで処理し、二本鎖DNAを変性させ、DNAを再生させて異なる切断生成物からのセンス/アンチセンスペアを含みうる二本鎖DNAを形成し、S1ヌクレアーゼによる処理によって再形成二本鎖から一本鎖部分を除去し、結果として生じる断片ライブラリーを発現ベクターへライゲートすることによって生成されうる。この方法によって、目的とするタンパク質のさまざまなサイズのN末端および内部断片をコードする発現ライブラリーが得られうる。
【0152】
切断の点突然変異によって製造されるコンビナトリアルライブラリーの遺伝子産物をスクリーニングし、かつ選択された特性を有する遺伝子産物のDNAライブラリーをスクリーニングするためのいくつかの方法が当技術分野で周知である。大きな遺伝子ライブラリーをスクリーニングするための高スループット分析に適している最も広く使用される方法は一般的に遺伝子ライブラリーを確実な発現ベクターへクローニングし、適切な細胞を結果として生じるベクターのライブラリーで形質転換し、かつ所望の活性の検出がその生成物が検出された遺伝子をコードするベクターの単離を促進する条件下にコンビナトリアル遺伝子を発現させるステップを含む。ライブラリーにおける機能的変異体の頻度を増強する技術である再帰的アンサンブル変異誘発(REM)は、本発明のタンパク質の変異形を識別するスクリーニングアッセイと組合せて使用されうる(アーキン(Arkin)と(ユアバン(Yourvan)(1992年)Proc.Natl.Acad.Sci. 米国(USA)89:7811−7815頁、デルグレーブ(Delgrave)ら(1993年)Protein Engineering 6(3):327−331頁)。
【0153】
配列番号2および5および19および22で示されたhdfAおよび/またはhdfB遺伝子配列に加えて、hdfAおよび/またはhdfBタンパク質のアミノ酸の変化をもたらしうるDNA配列多型が、一定の集団内に存在しうることを当業者には明らかであろう。かかる遺伝子多型は、自然な対立遺伝子の変動により異なる集団からの、または集団内の細胞に存在しうる。対立遺伝子変異形も機能的等価物を含みうる。
【0154】
本発明によるポリヌクレオチドの断片は、機能的ポリペプチドをコードしないポリヌクレオチドをも含んで成る。かかるポリヌクレオチドはPCR反応のプローブまたはプライマーとして機能しうる。
【0155】
それらが機能的または非機能的ポリペプチドをコードするかどうかに関係なく本発明による核酸は、ハイブリダイゼーションプローブまたはポリメラーゼ連鎖反応(PCR)プライマーとして使用されうる。hdfAおよび/またはhdfB活性を有するポリペプチドをコードすることがない本発明の核酸分子の使用には、とりわけ、(1)hdfAおよび/またはhdfBタンパク質をコードする遺伝子、またその対立遺伝子変異形をcDNAライブラリーから、例えば、A.ニゲルまたはペニシリウム・クリソゲヌム以外の他の生物から単離、(2)フェルマ(Verma)ら、Human Chromosomes:a Manual of Basic Techniques、ペルガモン・プレス(Pergamon Press)、ニューヨーク(1988年)に記載されているhdfAおよび/またはhdfB遺伝子の正確な染色体位置を提供する中期染色体スプレッドへのインサイチュハイブリダイゼーション(例えば、FISH)、(3)特定の組織および/または細胞におけるhdfAおよび/またはhdfB mRNAの発現を検出するためのノーザンブロット分析、および4)一定の生体(例えば、組織)試料におけるhdfAおよび/またはhdfBプローブにハイブリダイズ可能な核酸の存在を分析する診断ツールとして使用されうるプローブおよびプライマーがある。
【0156】
hdfAおよび/またはhdfB遺伝子またはcDNAの機能的等価物を得る方法も本発明によって包含される。かかる方法は、配列番号2または5または19または22による配列またはその変異形の全部または一部をコードする単離核酸を含む標識プローブを得るステップ、ライブラリーにおける核酸断片に対するプローブのハイブリダイゼーションを可能にする条件下に標識プローブで核酸断片ライブラリーをスクリーニングするステップ、それによって核酸二本鎖を形成するステップ、および任意の標識二本鎖における核酸断片から完全長遺伝子配列を調製し、hdfAおよび/またはhdfB遺伝子に関係がある遺伝子を得るステップを必要とする。
【0157】
1つの実施形態において、本発明のhdfAおよび/またはhdfB核酸は、配列番号1、または2、または4、または5、または18、または19、または21、または22に示されている核酸配列と少なくとも45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、もしくはそれ以上相同である。
【0158】
別の好ましい実施形態において本発明のhdfAおよび/またはhdfBポリペプチドは、配列番号3または6または20または23に示されたアミノ酸配列と少なくとも60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、もしくはそれ以上相同である。
【0159】
本発明は、配列番号1、または2、または4、または5、または18、または19、または21、または22それ自体を有するDNA配列、および上記のその相同体に関する。これらのDNA配列に関し、かつ遺伝子コードの退化によって得られるDNA配列も本発明の一部である。DNA配列番号2、5、19、および22に関し、かつハイブリダイゼーション(前段落参照)によって得られるDNA配列も発明の一部である。これらDNA配列によってコードされた単離ポリペプチドまたは上記のその相同体も本発明の一部である。ポリペプチドhdfAおよびhdfBは、NHRに関与する機能を有する。これらポリペプチドのすべてが本発明の方法において使用され、改善された標的効率を有しうる糸状菌を得ることができる。
【0160】
本発明は、以下の実施例でより詳細に例示される。かかる実施例は本発明の範囲を限定することが意図されていない。
【実施例】
【0161】
実施例1:hdfAおよび/またはhdfB遺伝子の識別および削除ベクターの構成。
アスペルギルス・ニゲル菌株CBS513.88のゲノムDNAを配列決定し、分析した。KU70およびKU80との相同体と注釈付きの翻訳タンパク質を有する2つの遺伝子を識別し、それぞれhdfAおよびhdfBと命名した。オープンリーディングフレーム(ORF)(イントロンを有する)および遺伝子の約1000bp5’および3’を含んで成るhdfAおよび/またはhdfB遺伝子座の配列が、配列表1および4に示されている。hdfAおよび/またはhdfBの遺伝子置換ベクターが、既知の原理に従って設計され、ルーチンのクローニング法に従って構成された(図1および2を参照)。本質的に、これらのベクターは、予定されたゲノム遺伝子座での相同的組換えのhdf ORFの約1000bpフランキング領域を含んで成る。また、それらは、A.ニドゥランス双方向amdS選択マーカー、中間直接繰返しを含有する。これら削除ベクターの一般設計は、欧州特許第635574B号明細書および国際公開第98/46772号パンフレットにおいて既述されている。
【0162】
実施例2:アスペルギルス・ニゲルにおけるhdfA遺伝子の不活性化。
削除ベクターpDEL−HDFAの直鎖DNA(図1)を単離して使用し、以前に記載された方法(Biotechnology of Filamentous fungi:Technology and Products.(1992年)リード・パブリッシング(Reed Publishing)(米国(USA))、第6章:Transformation 113〜156頁)を使用することによりアスペルギルス・ニゲルCBS513.88を形質転換した。この直鎖DNAはhdfA遺伝子座でゲノムへ取込むことができ、したがって、図3に示されているようにamdS遺伝子によってhdfA遺伝子を置換する。形質転換体をアセトアミド培地で選択し、欧州特許第635574B号明細書に記載された標準の方法に従ってコロニーを精製した。胞子をフルオロ・アセトアミド培地上にプレーティングし、amdSマーカーを失った菌株を選択した。発育コロニーをhdfA遺伝子座での取込みについてPCRで診断し、候補の菌株をhdfA遺伝子の削除についてサザン分析で試験した。hdfA遺伝子の削除は、遺伝子座全体をカバーするDNA断片の約2.2kbサイズの削減によって検出可能であり、適切なプローブにハイブリダイズされた。約8個の菌株が、約400の初期形質転換体のプールからゲノムhdfA遺伝子の除去を示した。
【0163】
菌株dHDFAを不活性化したhdfA遺伝子を有する代表的な菌株として選択した。
【0164】
実施例3:アスペルギルス・ニゲルにおけるhdfB遺伝子の不活性化。
削除ベクターpDEL−HDFBの直鎖DNA(図2)を単離して使用し、アスペルギルス・ニゲル菌株CBS513.88を形質転換した。この直鎖DNAはhdfB遺伝子座でゲノムへ取込むことができ、したがって、amdS遺伝子によってhdfB遺伝子を置換する(図4)。実施例2に記載されたものと同じ遺伝子置換法を使用した。形質転換体をアセトアミド培地で選択し、標準の方法に従ってコロニーを精製した。胞子をフルオロ・アセトアミド培地上にプレーティングし、amdSマーカーを失った菌株を選択した(欧州特許第635574B号明細書)。発育コロニーをhdfB遺伝子座での取込みについてPCRで診断し、候補の菌株をhdfB遺伝子の削除についてサザン分析で試験した。hdfB遺伝子の削除は、遺伝子座全体をカバーするDNA断片の約2.6kbサイズの削減によって検出可能であり、適切なプローブにハイブリダイズされた。約7個の菌株が、約370の初期形質転換体のプールからゲノムhdfB遺伝子の除去を示した。
【0165】
菌株dHDFBを不活性化したhdfB遺伝子を有する代表的な菌株として選択した。
【0166】
実施例4:アスペルギルス・ニゲルにおけるhdfAおよびhdfB遺伝子の不活性化。
削除ベクターpDEL−HDFBの直鎖DNA(図2)を単離して使用し、実施例2で得られた菌株dHDFAを形質転換した。この直鎖DNAはhdfB遺伝子座でゲノムへ取込むことができ、したがって、amdS遺伝子によってhdfB遺伝子を置換する(図4)。実施例2に記載されたものと同じ遺伝子置換法を使用した。形質転換体をアセトアミド培地で選択し、標準の方法に従ってコロニーを精製した。胞子をフルオロ・アセトアミド培地上にプレーティングし、amdSマーカーを失った菌株を選択した。発育コロニーをhdfB遺伝子座での取込みについてPCRで診断し、候補の菌株をhdfB遺伝子の削除についてサザン分析で試験した。hdfB遺伝子の削除は、遺伝子座全体をカバーするDNA断片の約2.6kbサイズの削減によって検出可能であり、適切なプローブにハイブリダイズされた。約15個の菌株が、約380の初期形質転換体のプールからゲノムhdfB遺伝子の除去を示した。
【0167】
菌株dHDFABを不活性化したhdfAおよびhdfB遺伝子を有する代表的な菌株として選択した。
【0168】
実施例5:単一相同的組換えイベントの改善された標的
DNAが予定された遺伝子座でアスペルギルス・ニゲルのゲノムへ取込みうる1つの機序は、単一相同的組換えによるものである。相同的DNAは、組換えによってゲノム位置で整列して取込む(図5を参照)。2つのベクターを使用し、実施例2、3、および4で得られたアスペルギルス・ニゲル菌株の単一相同的組換えによる標的効率を試験した。2つのベクターは、組換えおよび結果として生じる取込みを導くグルコアミラーゼ(glaA)遺伝子座と相同の領域を含んで成る(図5)。
【0169】
かかる相同的取込みのために設計された第1のベクターはすでに以前に国際公開第02/45524号パンフレットに記載されている(pGBFIN11−EPO)。このベクターは、プロリン特異的エンドプロテアーゼをコードする遺伝子を含有する。
【0170】
第2のベクター(pGBFIN11−PLA)は、A.オリゼからのホスホリパーゼA1(PLA1)をコードする遺伝子を含有する。この酵素をコードする遺伝子はすでに公表されている(ワタナベ(Watanabe)Iら、Biosci.Biotechnol. Biochem.(1999年)、63巻、5号、820−826頁)。この遺伝子は、pGBFIN11−EPOにおけるプロリン特異的エンドプロテアーゼ遺伝子のクローニングのために国際公開第02/45524号パンフレットに記載されたのと同じ方法を使用してpGBFIN11へクローン化された。
【0171】
菌株CBS513.88、dHDFA、dHDFB、およびdHDFABを実施例2で既述された形質転換法に従ってpGBFIN11−EPOまたはGBFIN11−PLAプラスミドのいずれかで形質転換した。得られた結果は、使用された両方のプラスミドで同じであった。発明者らはそれぞれ、標的遺伝子座で取込まれたプラスミドによる形質転換体の5%、10%、10%、および10%を見出した。したがって、発明者らは、アスペルギルス・ニゲルにおける少なくとも1つのhdf遺伝子の不活性化が単一の相同的組換えイベントによってこれらの菌株の標的効率の大幅な増大をもたらすと結論づけた。
【0172】
実施例6:数種類の遺伝子座での二重相同的組換えイベントの改善された標的。
さらに、ゲノムからの遺伝子をコードする多くのアミラーゼの不活性化のために設計された削除ベクターでのdHDFA菌株の形質転換によって標的効率を測定した。遺伝子フランキング領域を実質的に実施例1に記載されているようにクローン化し、結果として生じるベクターを線状化し、これを使用してCBS513−88およびdHDFA菌株のプロトプラストを形質転換した。標的頻度をPCR分析、および対応する遺伝子の不活性化を示す活性に基づくプレートアッセイによって測定した。後者は、0.4%寒天を補充したPDAプレート上で形質転換体を増殖させ、その後にヨード/カリウムヨード溶液(ルゴール(Lugol)、シグマ(Sigma)L6146)による染色によって行われた。以下の表1でわかるように、PCR分析および/または対応する遺伝子の不活性化を示す活性に基づくプレートアッセイによって判定された標的頻度は、CBS513.88菌株で観察されたものに比べ大幅に改善された。
【0173】
【表1】
【0174】
これらの所見により、アスペルギルス・ニゲルにおける少なくとも1つのhdf遺伝子の不活性化が結果として二重相同的組換えによる取込みに対するベクターの標的効率の大幅な増大をもたらすという本発明者らの結論がさらに裏付けられる。
【0175】
実施例7:標的頻度に対するamyBII遺伝子の相同的フランキング領域のサイズ削減の効果。
別個の一連の実験において、二重相同的組換えによる形質転換効率および標的頻度に対するフランキング領域長さの効果をさらに調査した。菌株CBS513.88およびdHDFAのプロトプラストを、可変長のamyBIIフランキング領域によってフランキングされたA.ニドゥランスamdSマーカーを含むPCR断片で形質転換した。表2に示されたデータは、増強された全体的な形質転換効率に加えて、統合的カセットの標的がdHDFA菌株においてはるかに改善されたことを明らかに示す。
【0176】
【表2】
【0177】
実施例8:ポリペプチドの表現型分析および製造。
dHDFA、dHDFB、またはdHDFAB菌株の固体培地または振盪フラスコでの発育中の表現型の差異は確認されなかった。プラスミドpGBFIN11−EPOまたはpBGFIN−PLAで形質転換されたdHDFA、dHDFB、およびdHDFAB菌株はすべて、以下の方法に従って判定されるように培地へ活性酵素を分泌した。
【0178】
固体培地はポテトデキストロース寒天(PDA)培地(ジフコ(Difco)、ポテトデキストロース寒天(POTATO DEXTROSE AGAR)、培養培地、カタログ番号213400、1996−1997年)であった。
【0179】
振盪フラスコ実験を欧州特許第635574B号明細書に記載されているように培地100ml中、34℃下、および500mlバッフル付き振盪フラスコを使用してインキュベーター振盪機中170rpmで行った。発酵の4日後、試料を取り、プロリン特異的エンドプロテアーゼ活性またはホスホリパーゼ活性のいずれかに判定した。
【0180】
プロリン特異的エンドプロテアーゼのタンパク質分解活性を分光光度法で時間内にpH5および約37℃で基質としてZ−グリ(Gly)(シン)−プロ(Pro)(リン)−pNAを使用して測定した。1Uプロリン特異的エンドプロテアーゼは、毎分Z−グリ(Gly)(シン)−プロ(Pro)(リン)−pNA1ミクロモルをpH5および37℃下に変換する酵素の量と定義されている。
【0181】
アスペルギルス・ニゲル(PLA1)からのホスホリパーゼPLA1活性を分光光度法で測定するために、人工的基質、すなわち、1,2−ジチオジオクタノイルホスファチジルコリン(diC8、基質)が使用される。PLA1はA1位で硫化物結合を加水分解し、チオオクタン酸を解離する。チオオクタン酸は、4,4ジチオピリジン(呈色試薬、4−DTDP)と反応し、4−チオピリドンを形成する。4−チオピリドンは、334nmの波長を有する放射線を吸収する4−メルカプトピリジンと互変異性平衡にある。同波長での吸光変化は測定される。1単位は、1nmolチオオクタン酸を1,2−ジチオジオクタノイルホスファチジルコリンから毎分37℃およびpH4.0で遊離する酵素の量である。
【0182】
基質溶液は、エタノール66ml当たりdiC8 1gを溶解し、酢酸バッファー264mlを添加することによって調製される。酢酸バッファーは、0.2%トリトン(Triton)−X100を含有する0.1M酢酸バッファーpH3.85を含んで成る。呈色試薬は、11mM 4,4−ジチオジピリジン溶液である。これは、2mlエッペンドルフ試料カップ中に4,4−ジチオジピリジン5.0mgを秤量し、エタノール1.00ml中に溶解することによって調製した。ミリQ水1.00mlを添加した。
【0183】
興味深いことに、色差またはコロニー外観など形態的変化は、CBS513.88から得られた形質転換体よりもdHDFA、dHDFB、およびdHDFAB菌株から得られた形質転換体で低頻度に生じた。これは無作為取込み(NHR)の削減によるものであり、したがって予想外の表現型の変化を阻止すると考えられる。
【0184】
実施例9:変異誘発による相同的組換えの改善された効率を有するペニシリウム変異体の単離
遺伝子標的の改善された効率を有する変異体を単離するために、典型的な変異誘発と分子生物学の組合せを適用した。ペニシリウム・クリソゲヌム(CBS 455.95)胞子をYEPD(ジフコ製の2%酵母抽出物、ジフコ製の1%ペプトン、2%グルコース)中で胞子形成するコロニーから得た。これらの胞子を滅菌水道水中で洗浄し、ml当たり108分生子を含有する懸濁液10mlを254nmでUV照射にかけた(シルバニア(Sylvania)、15ワット・ブラックライトブルー管、モデルFT15T8/BLB)。UV照射を7.5、15、または30分間当てると同時に、懸濁液をゆっくり振盪した。これらの異なる照射時間は、細胞における軽度、中等度、および強度の突然変異率レベルを得るために選択された。暗所での回収の1時間後、これら3時点からの細胞を2つの同等のアリコートに分けた。第1の試料を既述のように直接再胞子形成し(ヘルスバッハ(Hersbach),GJM、ファン・デン・ビーク(Van der Beek),CP、およびファン・ジユック(Van Dijck),PWM.The Penicillins:properties,biosynthesis and fermentation.In:Vandamme EJ(編) Biotechnology of Industrial Antibiotics(45−140頁)、マルセル・デッカー(Marcel Dekker)、ニューヨーク)、もう一方の試料をYNB培地(アミノ酸(ジフコ)、2.0%w/vグルコースとともに0.67%w/v酵母窒素塩基)中で長い回収期間、胞子形成を導入する前に4時間、25℃下にインキュベートした。
【0185】
第3の変異誘発試料をYNB中で一夜、野生型胞子を発生させた後、滅菌水道水での2つの洗浄ステップによって得て、滅菌水道水中に再懸濁した。さらにUV照射を7.5、15、および30分間当てると同時に、懸濁液をゆっくり振盪した。これらの試料を(上述したように)暗所での回収の1時間後に直接、再胞子形成した。
【0186】
これらの望ましい変異体をこれら変異誘発集団から選択するために、変異誘発集団をYEPD培地に接種した。発芽後、細胞の発達を標準の光学顕微鏡を使用して追跡した。培養の平均菌糸が申し分なく発達している場合、細胞を収穫し、溶解酵素でインキュベートし、プロトプラストを得た。プロトプラストを機能的選択マーカーの発現カセットを有する2つのDNA断片、bleおよびamdSで形質転換された。遺伝子bleはフレオマイシンへの耐性を与えることが可能なタンパク質をコードする(コラー(Kolar)M、プント(Punt)PJ、ファン・デン・ホンデル CA、シュワブ(Schwab)H.Transformation of Penicillium chrysogenum using dominant selection markers and expression of an Escherichia coli lacZ fusion gene.Gene.1988年、62(1):127−34頁)。遺伝子amdSは、細胞が唯一の窒素源としてアセトアミドで発育することを可能にするタンパク質をコードする(欧州特許第635574B号明細書に記載)。P.クリソゲヌムのプロトプラストへのDNAの移動に適用される方法は当技術分野で周知であり、フィンケルシュタインとボール(編)、Biotechnology of filamentous fungi,Technology and products,Butterworth−Heinemann(1992年)、ベネットとラシュア(編)More Gene Manipulations in fungi、Academic Press(1991年)、ターナー、ピューラー(編)、Biotechnology、第2完全改訂版、VHC(1992年)を含む多くの文献に記載されている。Ca−PEG介在プロトプラスト形質転換はEP635574号明細書に記載されているように使用される。
【0187】
ペニシリウムゲノムにおける特異的遺伝子座に対するこれらの発現カセットの標的取込みを選択するために、DNAの短い相同的伸長をDNA断片の両側でPCRによって添加した。3タイプの構成物を作った。すなわち、第1のタイプは30bpのDNAの相同的伸長を、第2のタイプは50bp、および第3は100bpを含有する。選択は、27の明確なバッチを規定する30、50、または100bpの相同的伸長を有する2つのDNA構成物(bleおよびamdS)で9つの胞子形成バッチから得られた変異体を形質転換することによって行った。ble遺伝子はniaD遺伝子座に標的され、それによって硝酸還元酵素遺伝子を破壊し(ゴウカ RJ、ファン・ハルチングスフェルト W、ボーベンベルグ RA、ファン・デン・ホンデル CA、ファン・ゴルコム RF、Cloning of the nitrate−nitrite reductase gene cluster of penicillium chrysogenum and use of the niaD gene as a homologous selection marker、J Biotechnol.、1991年9月、20(2):189−99頁)、細胞が塩素酸塩に耐性となると、塩素酸塩を含有するプレートへの形質転換体の選択を導くことを可能にした。amdS遺伝子はsutB遺伝子座に標的され、それによって硫酸パーミアーゼ遺伝子を破壊し(ファン・デ・カンプ M、ピッツィニーニ E、フォス A、ファン・デア・レンデ TR、シュールス TA、ニューベルト RW、ターナー G、コーニングス WN、ドリーゼン AJ、Sulfate transport in Penicillium chrysogenum:cloning and characterization of the sutA and sutB genes.、J. Bacteriol.1999年12月、181(23):7228−34頁)、セレン酸塩を含有するプレートへの形質転換体の選択を導くことを可能にした。形質転換体が最初に塩素酸塩で選択され、次いでセレン酸塩について試験された。さらに、選択マーカーの存在は、唯一の窒素源としてアセトアミドを含有するプレート(欧州特許第635574B号明細書)、次いでフレオマイシンを含有するプレートで証明された。対照野生型P.クリソゲヌムとしてCBS455.95も同じDNA断片で形質転換された。存在する両方の選択マーカーを有し、塩素酸塩およびセレン酸塩に対して耐性の変異体は改善された標的取込みを有する菌株である。
【0188】
【表3】
【0189】
【表4】
【図面の簡単な説明】
【0190】
【図1】アスペルギルス・ニゲルにおけるhdfA遺伝子を不活性化するために使用される置換ベクターpDEL−HDFAを示す図である。置換ベクターは、hdfAフランキング領域、amdSマーカー、および大腸菌DNAを含んで成る。大腸菌DNAは、A.ニゲル菌株の形質転換前に制限酵素AsclおよびNotlによる消化によって除去された。
【図2】A.ニゲルにおけるhdfB遺伝子を不活性化するために使用される置換ベクターpDEL−HDFBを示す図である。置換ベクターは、hdfBフランキング領域、amdSマーカー、および大腸菌DNAを含んで成る。大腸菌DNAは、A.ニゲル菌株の形質転換前に制限酵素AsclおよびNotlによる消化によって除去された。
【図3】A.ニゲルのhdfA遺伝子を削除するために使用される方法を示す図である。使用されるDNA構成物は、hdfA遺伝子の相同領域(5’および3’)によってフランキングされたamdS選択マーカーを含んで成る(1)。この構成物は、ゲノムhdfA遺伝子座で二重相同的組換え(X)を通じて取込み(2)、かつゲノムhdfA遺伝子を置換する(3)。その後、直接繰返し(U)にわたる組換えがamdSマーカーを除去し、結果としてhdfA遺伝子の正確な切除が生じる(4)。
【図4】A.ニゲルのhdfB遺伝子を削除するために使用される方法を示す図である。DNA構成物は、hdfB遺伝子の相同領域(5’および3’)によってフランキングされたamdS選択マーカーを含んで成る(1)。この構成物は、ゲノムhdfB遺伝子座で二重相同的組換え(X)を通じて取込み(2)、かつゲノムhdfB遺伝子コピーを置換する(3)。その後、直接繰返し(U)にわたる組換えがamdSマーカーを除去し、結果としてhdfB遺伝子の正確な切除が生じる(4)。
【図5】単一相同的組換えを通じてDNA構成物をA.ニゲルのゲノムへ取込むために使用される概略的な方法を示す図である。発現ベクターは、選択可能なamdSマーカーおよびglaA遺伝子座の相同的領域によってフランキングされた目的とする遺伝子(それぞれ、3’glaAおよび3’’glaA)を含んで成り、ゲノムglaA遺伝子座での取込みを導く。
【特許請求の範囲】
【請求項1】
NHR選好の糸状菌細胞のゲノムへの所定の部位に対するポリヌクレオチドの標的取込みの効率を増大させるための方法であって、前記ポリヌクレオチドが、前記所定の部位との相同の領域を有し、HRの方へ取込み経路を導くステップを含んで成る、方法。
【請求項2】
前記導くステップが、親糸状菌細胞の変異体を提供するステップを含んで成り、NHR/HR比が、同じ条件下で測定された前記親生物における前記比と比べ変異体において減少している、請求項1に記載の方法。
【請求項3】
前記導くステップが、NHRに関与する成分をコードする遺伝子に欠ける、かつ/またはNHRに関与する成分の減少したレベルを有する変異体を提供するステップを含んで成る請求項1または2に記載の方法。
【請求項4】
前記変異体が、好ましくは誘導性に、以下の遺伝子、すなわちhdfAまたはその相同体、hdfBまたはその相同体、あるいはその両方の少なくとも1つに欠け、かつ/または、好ましくは誘導性に、これらの遺伝子によってコードされたタンパク質の少なくとも1つの減少した量を有する、請求項3に記載の方法。
【請求項5】
NHRに関与する遺伝子が非機能的変異形によって置換されている、請求項3または4に記載の方法。
【請求項6】
前記導くステップが、取込まれる前記ポリヌクレオチドに過剰な小さな二本鎖ポリヌクレオチドを添加するステップを含んで成る、請求項1〜5のいずれか一項に記載の方法。
【請求項7】
前記導くステップが、前記タンパク質の阻害剤を添加することによってNHRにおいて活性の少なくとも1つのタンパク質の活性を減少させるステップを含んで成る、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
前記変異体がHRに関与する成分の増加したレベルを有する、請求項1〜7のいずれか一項に記載の方法。
【請求項9】
NHR/HR比が50未満、好ましくは10未満、さらにより好ましくは1未満、かつ最も好ましくは0.001未満である糸状菌が使用される、請求項1〜8のいずれか一項に記載の方法。
【請求項10】
親糸状菌細胞の変異体であって、前記親生物がNHR選好を有し、NHR/HR比が、同じ条件下で測定された前記親生物における前記比と比べ変異体が減少している変異体。
【請求項11】
前記変異体が、NHRに関与する成分をコードする遺伝子を欠き、かつ/またはNHRに関与する成分の減少したレベルを有する、請求項10に記載の変異体。
【請求項12】
前記変異体が、好ましくは誘導性に、以下の遺伝子、すなわちhdfAまたはその相同体、hdfBまたはその相同体、あるいはその両方の少なくとも1つに欠け、かつ/または、好ましくは誘導性に、これらの遺伝子によってコードされたタンパク質の少なくとも1つの減少した量を有する、請求項10または11に記載の変異体。
【請求項13】
前記生物のゲノムにおいて、NHRに関与する遺伝子が非機能的変異形によって置換されている請求項10〜12のいずれか一項に記載の変異体。
【請求項14】
前記変異体がHRに関与する成分の増加したレベルを有する、請求項10〜13のいずれか一項に記載の変異体。
【請求項15】
前記変異体が組換え変異体である、請求項10〜14のいずれか一項に記載の変異体。
【請求項16】
NHR/HR比が50未満、好ましくは10未満、さらにより好ましくは1未満、かつ最も好ましくは0.001未満である糸状菌。
【請求項17】
目的とするポリペプチドをコードする目的とする遺伝子を含んで成るDNA配列を含んで成るDNA構成物で形質転換された請求項10〜16のいずれか一項に記載の糸状菌。
【請求項18】
前記糸状菌が、アスペルギルス種、ペニシリウム種、またはトリコデルマ種である、請求項10〜17のいずれか一項に記載の糸状菌。
【請求項19】
前記アスペルギルスが、アスペルギルス・ニゲル種またはアスペルギルス・オリゼ種である、請求項18に記載の糸状菌。
【請求項20】
前記ペニシリウムが、ペニシリウム・クリソゲヌム種またはペニシリウム・シトリナム種である、請求項18に記載の糸状菌。
【請求項21】
請求項10〜20のいずれか一項に記載の糸状菌が使用される、目的とするポリペプチドを産生するための方法。
【請求項22】
請求項10〜21のいずれか一項に記載の糸状菌が使用される、代謝産物を産生するための方法。
【請求項23】
前記代謝産物がカルチノイド化合物またはベータ・ラクタム化合物である、請求項22に記載の方法。
【請求項24】
配列番号2または5または19または22を有する単離DNA配列またはその相同体。
【請求項25】
請求項24に記載のDNA配列によってコードされた単離ポリペプチドまたはその相同体。
【請求項1】
NHR選好の糸状菌細胞のゲノムへの所定の部位に対するポリヌクレオチドの標的取込みの効率を増大させるための方法であって、前記ポリヌクレオチドが、前記所定の部位との相同の領域を有し、HRの方へ取込み経路を導くステップを含んで成る、方法。
【請求項2】
前記導くステップが、親糸状菌細胞の変異体を提供するステップを含んで成り、NHR/HR比が、同じ条件下で測定された前記親生物における前記比と比べ変異体において減少している、請求項1に記載の方法。
【請求項3】
前記導くステップが、NHRに関与する成分をコードする遺伝子に欠ける、かつ/またはNHRに関与する成分の減少したレベルを有する変異体を提供するステップを含んで成る請求項1または2に記載の方法。
【請求項4】
前記変異体が、好ましくは誘導性に、以下の遺伝子、すなわちhdfAまたはその相同体、hdfBまたはその相同体、あるいはその両方の少なくとも1つに欠け、かつ/または、好ましくは誘導性に、これらの遺伝子によってコードされたタンパク質の少なくとも1つの減少した量を有する、請求項3に記載の方法。
【請求項5】
NHRに関与する遺伝子が非機能的変異形によって置換されている、請求項3または4に記載の方法。
【請求項6】
前記導くステップが、取込まれる前記ポリヌクレオチドに過剰な小さな二本鎖ポリヌクレオチドを添加するステップを含んで成る、請求項1〜5のいずれか一項に記載の方法。
【請求項7】
前記導くステップが、前記タンパク質の阻害剤を添加することによってNHRにおいて活性の少なくとも1つのタンパク質の活性を減少させるステップを含んで成る、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
前記変異体がHRに関与する成分の増加したレベルを有する、請求項1〜7のいずれか一項に記載の方法。
【請求項9】
NHR/HR比が50未満、好ましくは10未満、さらにより好ましくは1未満、かつ最も好ましくは0.001未満である糸状菌が使用される、請求項1〜8のいずれか一項に記載の方法。
【請求項10】
親糸状菌細胞の変異体であって、前記親生物がNHR選好を有し、NHR/HR比が、同じ条件下で測定された前記親生物における前記比と比べ変異体が減少している変異体。
【請求項11】
前記変異体が、NHRに関与する成分をコードする遺伝子を欠き、かつ/またはNHRに関与する成分の減少したレベルを有する、請求項10に記載の変異体。
【請求項12】
前記変異体が、好ましくは誘導性に、以下の遺伝子、すなわちhdfAまたはその相同体、hdfBまたはその相同体、あるいはその両方の少なくとも1つに欠け、かつ/または、好ましくは誘導性に、これらの遺伝子によってコードされたタンパク質の少なくとも1つの減少した量を有する、請求項10または11に記載の変異体。
【請求項13】
前記生物のゲノムにおいて、NHRに関与する遺伝子が非機能的変異形によって置換されている請求項10〜12のいずれか一項に記載の変異体。
【請求項14】
前記変異体がHRに関与する成分の増加したレベルを有する、請求項10〜13のいずれか一項に記載の変異体。
【請求項15】
前記変異体が組換え変異体である、請求項10〜14のいずれか一項に記載の変異体。
【請求項16】
NHR/HR比が50未満、好ましくは10未満、さらにより好ましくは1未満、かつ最も好ましくは0.001未満である糸状菌。
【請求項17】
目的とするポリペプチドをコードする目的とする遺伝子を含んで成るDNA配列を含んで成るDNA構成物で形質転換された請求項10〜16のいずれか一項に記載の糸状菌。
【請求項18】
前記糸状菌が、アスペルギルス種、ペニシリウム種、またはトリコデルマ種である、請求項10〜17のいずれか一項に記載の糸状菌。
【請求項19】
前記アスペルギルスが、アスペルギルス・ニゲル種またはアスペルギルス・オリゼ種である、請求項18に記載の糸状菌。
【請求項20】
前記ペニシリウムが、ペニシリウム・クリソゲヌム種またはペニシリウム・シトリナム種である、請求項18に記載の糸状菌。
【請求項21】
請求項10〜20のいずれか一項に記載の糸状菌が使用される、目的とするポリペプチドを産生するための方法。
【請求項22】
請求項10〜21のいずれか一項に記載の糸状菌が使用される、代謝産物を産生するための方法。
【請求項23】
前記代謝産物がカルチノイド化合物またはベータ・ラクタム化合物である、請求項22に記載の方法。
【請求項24】
配列番号2または5または19または22を有する単離DNA配列またはその相同体。
【請求項25】
請求項24に記載のDNA配列によってコードされた単離ポリペプチドまたはその相同体。
【図1】

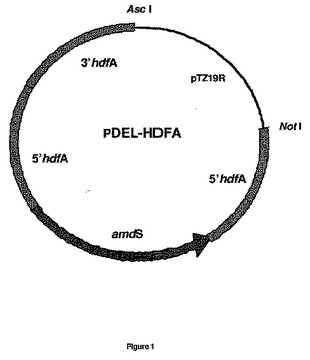
【図2】

【図3】
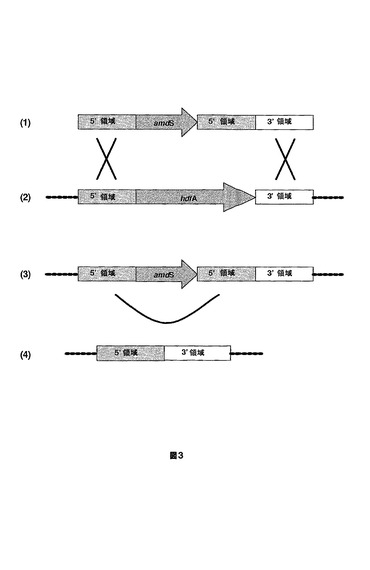
【図4】
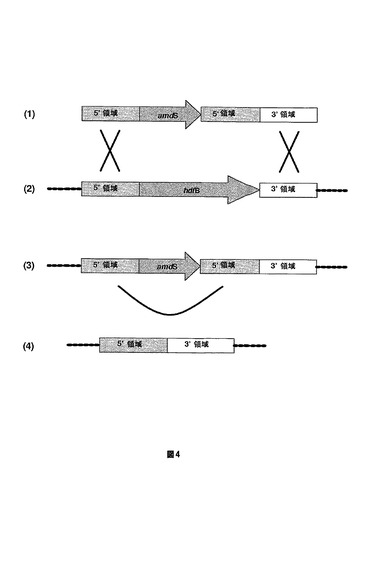
【図5】
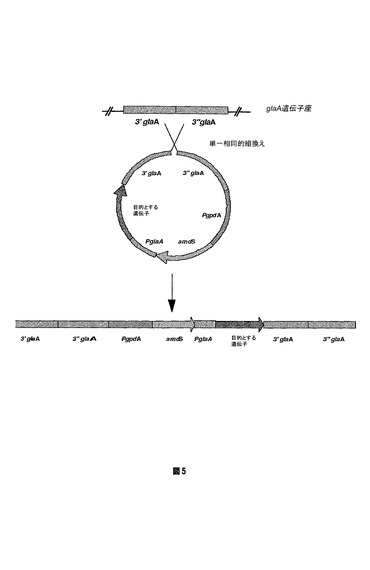
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2007−530065(P2007−530065A)
【公表日】平成19年11月1日(2007.11.1)
【国際特許分類】
【出願番号】特願2007−505561(P2007−505561)
【出願日】平成17年3月31日(2005.3.31)
【国際出願番号】PCT/EP2005/051464
【国際公開番号】WO2005/095624
【国際公開日】平成17年10月13日(2005.10.13)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公表日】平成19年11月1日(2007.11.1)
【国際特許分類】
【出願日】平成17年3月31日(2005.3.31)
【国際出願番号】PCT/EP2005/051464
【国際公開番号】WO2005/095624
【国際公開日】平成17年10月13日(2005.10.13)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]