
改変プロモーターを使用する植物老化の操作
本発明は、植物の老化を操作する方法に関する。また、本発明は、かかる方法に有用なベクター、変更された老化特性を有する形質転換植物、ならびに植物細胞、種子およびかかる植物の他の部分に関する。一態様において、本発明は、植物に、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたは機能的に活性な断片またはその改変体を含む遺伝子構築物を導入することを含む、植物の老化を操作する方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
この出願は、2003年3月5日に出願された表題「Manipulation of plant senescence using an MYB gene promoter and cytokinin biosynthesis gene」の米国特許出願第10/363,723号の一部継続であり、米国特許出願第10/363,723号は、2001年8月30日に出願された表題「Manipulation of Plant Senescence Using An MYB Gene Promoter and Cytokinin Biosynthesis Genes」の国際特許出願PCT/AU01/01092からの優先権を主張し、国際特許出願PCT/AU01/01092は、2000年9月6日に出願された表題「Manipulation of Plant Senescence Using An MYB Gene Promoter and Cytokinin Biosynthesis Genes」の豪国特許出願PQ9946からの優先権を主張する。これらの内容は、それらの全体が参考として本明細書に援用される。
【0002】
本発明は、植物の老化を操作する方法に関する。また、本発明は、かかる方法に有用なベクター、変更された老化特性を有する形質転換植物、ならびに植物細胞、種子およびかかる植物の他の部分に関する。
【背景技術】
【0003】
葉の老化は、細胞死の前に細胞の代謝的および構造的変化を伴う。また、活発に生長している領域への栄養素の再循環も伴う。
【0004】
サイトカイニンによる植物および植物器官の老化の調節は、農業上重要な成果をもたらす。葉内のサイトカイニンレベルの上昇は、老化を遅延させる傾向がある。その産物(イソペンテニルトランスフェラーゼ)がサイトカイニン合成の重要な段階を触媒するipt遺伝子の発現を調節するために、いくつかのプロモーターが使用されている。しかしながら、一般に、ipt遺伝子を過剰発現するトランスジェニック植物は、地下部および苗条の生長の遅延、地下部の無形成、頂芽優性の低下、および葉面積の減少を有することが報告されている。
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明の目的は、先行技術に伴う問題点または欠点の1つ以上を解決または軽減することである。
【課題を解決するための手段】
【0006】
一態様において、本発明は、植物に、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたは機能的に活性な断片またはその改変体を含む遺伝子構築物を導入することを含む、植物の老化を操作する方法を提供する。
【0007】
老化の操作は、植物および/または特定の植物の器官に関連するものである。種々の植物の器官、例えば、葉、地下部、苗条、茎、塊茎、花、走根、および果実などの老化が操作され得る。植物および植物器官の老化の操作は、例えば、園芸製品および切り花における果実、花、葉および塊茎の貯蔵寿命の増大、園芸作物の腐り易さの低減、老化遅延葉内の炭素固定の増大による収穫率の増大、飼料植物におけるバイオマス生産の向上、採種の向上などの農業上重要な成果をもたらし得る。
【0008】
「老化の操作」は、一般的に、非形質転換対照植物と比べて形質転換植物における老化の遅滞に関する。しかしながら、一部の適用用途では、植物の老化を促進あるいは変更することが望ましい場合があり得る。老化は、例えば、アンチセンス遺伝子を利用することにより促進あるいは変更され得る。
【0009】
有効量の前記遺伝子構築物は、任意の適当な手法、例えば、形質導入、トランスフェクションまたは形質転換によって植物に導入され得る。「有効量」とは、前記植物、または該植物に由来する植物、植物種子もしくは他の植物部分において同定可能な表現型形質がもたらされるのに充分な量を意図する。かかる量は、適切な当業者が植物の種類、投与経路および他の関連要素を考慮することによって容易に決定され得る。かかる当業者は、適当な量および投与方法を容易に決定することができよう。例えば、Maniatisら,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory,Cold Spring Harbor(その全開示内容は、引用により本明細書に組み込まれる)を参照のこと。
【0010】
「修飾myb遺伝子プロモーター」とは、通常myb遺伝子と関連しているプロモーターであって、前記プロモーター内の1つ以上の地下部特異的モチーフおよび/または花粉特異的モチーフが欠失もしくは不活化されるように修飾されたプロモーターを意図する。
【0011】
本出願人は理論に拘束されることを望まないが、前記myb遺伝子プロモーター内の1つ以上の地下部特異的モチーフの欠失もしくは不活化により、一部の植物種で地下部の発育に影響し得る植物の成長点におけるサイトカイニン生合成酵素をコードする遺伝子の基底(leaky)発現の問題が軽減または解決され得ると仮定する。また、前記myb遺伝子プロモーター内の1つ以上の花粉特異的モチーフの欠失もしくは不活化により、一部の植物種で花粉の発育に影響し得る花粉におけるサイトカイニン生合成酵素をコードする遺伝子の基底発現の問題が軽減または解決され得ると仮定する。
【0012】
好ましくは、修飾myb遺伝子プロモーターは、修飾myb32遺伝子プロモーターである。好ましくは、修飾myb遺伝子プロモーターはArabidopsis由来のもの、より好ましくはシロイヌナズナ(Arabidopsis thaliana)由来のものである。
【0013】
本発明に従って修飾され得る好適なプロモーターは、Liら,Cloning of three MYB−like genes from Arabidopsis(PGR 99−138)Plant Physiology 121 :313(1999)(その全開示内容は、引用により本明細書に組み込まれる)に記載されたものである。
【0014】
「地下部特異的モチーフ」とは、植物の地下部内の関連遺伝子の発現を指令する3〜7ヌクレオチド、好ましくは4〜6ヌクレオチド、より好ましくは5ヌクレオチドの配列を意図する。
【0015】
好ましくは、地下部特異的モチーフは、コンセンサス配列ATATTまたはAATATを含む。
【0016】
好ましくは、前記myb遺伝子プロモーター内の1〜10個、より好ましくは3〜8個、さらにより好ましくは5〜7個の地下部特異的モチーフを欠失もしくは不活化させ、好ましくは欠失させる。
【0017】
地下部特異的モチーフは、個々のモチーフを取り出すこと、または1つ以上のモチーフを含む該プロモーターの断片を取り出すことにより欠失させ得る。例えば、シロイヌナズナmyb遺伝子プロモーターのヌクレオチド1〜530、好ましくはヌクレオチド110〜530の領域の全部または一部を欠失させ得る。
【0018】
該欠失は、例えば制限エンドヌクレアーゼで核酸を切断し、切断末端をリゲートし、取り出した断片を有するプロモーターを作製することにより行なわれ得る。
【0019】
例えば、修飾シロイヌナズナmyb遺伝子プロモーターは、162〜176位のXcmI部位と520〜525位のSspI部位の間の断片を取り出すことにより作製され得る。これにより、7つの地下部特異的モチーフのうち6つが欠失した修飾myb遺伝子プロモーターが作製される。あるいはまた、例えば、520〜525位のSspI部位の上流の領域を欠失させること、またはヌクレオチド1〜120の領域を、162〜176位のXcmI部位と520〜525位のSspI部位の間の領域とともに欠失させることにより、7つの地下部特異的モチーフすべてを欠失させ得る。
【0020】
地下部特異的モチーフは、該モチーフ内の1つ以上のヌクレオチドを、好ましいコンセンサス配列を有しなくなるように付加、欠失、置換または誘導体化することにより不活化させ得る。
【0021】
好ましくは、修飾myb遺伝子プロモーターは、添付の図2、3および4に示す配列(それぞれ、配列番号2、3および4)からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む。
【0022】
「花粉特異的モチーフ」とは、植物の花粉内の関連遺伝子の発現を指令する3〜7ヌクレオチド、好ましくは4〜6ヌクレオチド、より好ましくは4または5ヌクレオチドの配列を意図する。
【0023】
好ましくは、花粉特異的モチーフは、TTCTおよびAGAAからなる群より選択されるコンセンサス配列を含む。
【0024】
好ましくは、前記myb遺伝子プロモーター内の1〜30個、より好ましくは3〜15個、さらにより好ましくは4〜10個の花粉特異的モチーフを欠失もしくは不活化させ、好ましくは欠失させる。
【0025】
花粉特異的モチーフは、個々のモチーフを取り出すこと、または1つ以上のモチーフを含む該プロモーターの断片を取り出すことにより欠失させ得る。例えば、ヌクレオチド1〜540、好ましくはヌクレオチド390〜540のシロイヌナズナmyb遺伝子プロモーターの領域の全部または一部を欠失させ得る。
【0026】
該欠失は、例えば制限エンドヌクレアーゼで核酸を切断し、切断末端をリゲートし、取り出した断片を有するプロモーターを作製することにより行なわれ得る。
【0027】
例えば、修飾シロイヌナズナmyb遺伝子プロモーターは、162〜176位のXcmI部位と520〜525位のSspI部位の間の断片を取り出すことにより作製され得る。これにより、23個の花粉特異的モチーフのうち4個が欠失した修飾myb遺伝子プロモーターが作製される。あるいはまた、例えば、520〜525位のSspI部位の上流の領域を欠失させることにより10個の花粉特異的モチーフを欠失させ得る。
【0028】
花粉特異的モチーフは、該モチーフ内の1つ以上のヌクレオチドを、好ましいコンセンサス配列がなくなるように付加、欠失、置換または誘導体化することにより不活化させ得る。
【0029】
好ましくは、修飾myb遺伝子プロモーターは、添付の図2、3および4に示す配列(それぞれ、配列番号2、3および4)からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む。
【0030】
本発明のさらなる態様において、植物に、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結されたmyb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む遺伝子構築物を導入することを含む、植物のバイオマスを増大させる方法が提供される。
【0031】
myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体は、完全長のmyb遺伝子プロモーターまたは修飾myb遺伝子プロモーターであり得る。
【0032】
完全長のmyb遺伝子プロモーターは、myb32遺伝子プロモーターであり得る。好ましくは、myb遺伝子プロモーターはArabidopsis由来のもの、より好ましくはシロイヌナズナ由来のものである。最も好ましくはmyb遺伝子プロモーターは、添付の図1に示す配列(配列番号1)からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む。
【0033】
好適なプロモーターは、Liら,Cloning of three MYB−like genes from Arabidopsis(PGR 99−138)Plant Physiology 121 :313(1999)に記載されたものである。
【0034】
修飾myb遺伝子プロモーターは、本明細書において前述の修飾myb遺伝子プロモーターであり得る。
【0035】
「バイオマスの向上」とは、非形質転換対照植物と比べて形質転換植物における、総葉面積、累積葉面積、葉の生長力学(すなわち、経時的な葉の数)、走根長さ、および花1つまたは種蒔き面積あたりの種子収量からなる群より選択される生長特性の向上または増大を意図する。また、「バイオマスの向上」は、非形質転換対照植物と比べて形質転換植物における走根死の割合の低下または減少を包含する。
【0036】
特に、本発明によるトランスジェニック植物の種子重量(すなわち、千個の種子の重量)は非トランスジェニック対照植物と識別不可能であるが、同等の開花強度の非トランスジェニック対照植物と比較すると、花1つまたは種蒔き面積あたりの基準で示す総種子収量は、トランスジェニック植物より有意に高いことを、本出願人らは見い出した。
【0037】
myb遺伝子プロモーターまたは修飾myb遺伝子プロモーターに関連する「機能的に活性な」とは、その断片または改変体(類縁体、誘導体もしくは変異型など)が、本発明の方法による植物の老化の操作能を有することを意図する。かかる改変体には、天然に存在する対立遺伝子改変体および天然に存在しない改変体が包含される。1つ以上のヌクレオチドの付加、欠失、置換および誘導体化が想定されるが、修飾によって該断片または改変体の機能的活性の低下がもたらさないものとする。
【0038】
好ましくは、機能的に活性な断片または改変体は、該断片または改変体に相当する上記配列の関連部分と少なくともほぼ80%の同一性を有し、より好ましくは少なくともほぼ90%の同一性、最も好ましくは少なくともほぼ95%の同一性を有する。好ましくは、該断片は、少なくとも20ヌクレオチド、より好ましくは少なくとも50ヌクレオチド、より好ましくは少なくとも100ヌクレオチド、より好ましくは少なくとも200ヌクレオチド、最も好ましくは少なくとも300ヌクレオチドのサイズを有する。
【0039】
「サイトカイニンの生合成に関与している酵素をコードする遺伝子」とは、カイネチン、ゼアチンおよびベンジルアデニンなどのサイトカインの合成に関与している酵素をコードする遺伝子、例えば、イソペンチルトランスフェラーゼ(ipt)をコードする遺伝子、またはsho遺伝子(例えば、Petunia由来のもの)などのipt様遺伝子を意図する。好ましくは、該遺伝子は、イソペンテニルトランスフェラーゼ(ipt)遺伝子またはsho遺伝子である。好ましい実施形態において、該遺伝子は、Agrobacterium、より好ましくはAgrobacterium tumefaciens;Lotus、より好ましくはミヤコグサ(Lotus japonicus);およびPetunia、より好ましくはツクバネアサガオ(Petunia hybrida)からなる群より選択される種に由来するものである。
【0040】
最も好ましくは、該遺伝子は、添付の図6、8および10に示す配列(配列番号5、7および9)、添付の図7、9および11に示すポリペプチドをコードする配列(配列番号6、8および10)からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む。
【0041】
サイトカイニン生合成酵素をコードする遺伝子に関連する「機能的に活性な」とは、その断片または改変体(類縁体、誘導体または変異型など)が、本発明の方法による植物の老化の操作能を有することを意図する。かかる改変体には、天然に存在する対立遺伝子改変体および天然に存在しない改変体が包含される。ヌクレオチドの1つ以上の付加、欠失、置換および誘導体化が想定されるが、該修飾によって該断片または改変体の機能的活性の低下をもたらさないものとする。好ましくは、機能的に活性な断片または改変体は、該断片または改変体に相当する上記配列の関連部分と少なくともほぼ80%の同一性を有し、より好ましくは少なくともほぼ90%の同一性、最も好ましくは少なくともほぼ95%の同一性を有する。かかる機能的に活性な改変体および断片としては、例えば、同類核酸変化、または対応するアミノ酸配列内の1つ以上の残基の同類アミノ酸置換をもたらす核酸変化を有するものが挙げられる。例えば、機能的に活性な改変体は、図6、8または10に示す配列の1つ以上の同類核酸置換を含むものであり得、得られる機能的に活性な改変体は、それぞれ図7、9または11に示すアミノ酸配列をコードする。好ましくは、該断片は、少なくとも20ヌクレオチド、より好ましくは少なくとも50ヌクレオチド、より好ましくは少なくとも100ヌクレオチド、より好ましくは少なくとも500ヌクレオチドのサイズを有する。
【0042】
遺伝子構築物は植物に、任意の適当な手法によって導入され得る。本発明の遺伝子構築物を植物細胞内に組み込むための手法(例えば、形質導入、トランスフェクションまたは形質転換)は、当業者によく知られたものである。かかる手法としては、アグロバクテリウム媒介型誘導、組織、細胞およびプロトプラストへのエレクトロポレーション、プロトプラスト融合、生殖器官内への注入、未熟胚内への注入ならびに細胞、組織、カルス、未熟および成熟胚に対する高速プロジェクタイル誘導、微粒子銃形質転換ならびにその組合せが挙げられる。手法の選択は、形質転換対象の植物の種類に大きく依存し、適切な当業者によって容易に決定され得る。
【0043】
本発明の遺伝子構築物が組み込まれる細胞は、後述するようにして選択し、次いで、当該技術分野でよく知られた手法を用いて適切な培地中で培養すると、形質転換植物が再生し得る。例えば温度、pHなどの培養条件は当業者には自明であろう。得られた植物は、当該技術分野でよく知られた方法を用い、有性生殖または無性生殖のいずれかによって形質転換植物の後続世代を生成してもよい。
【0044】
本発明の方法は、さまざまな植物、例えば、単子葉[草(例えば、牧草、芝生およびバイオエネルギー草、例えば、ペレニアルライグラス、トールフェスク、イタリアンライグラス、レッドフェスキュー、リードカナリーグラス、ビッグブルーステム、コードグラス、ネピアグラス、野生ライムギ、野生サトウキビ、ススキ属(Miscanthus))、トウモロコシ、オートムギ、小麦および大麦)など]、双子葉[Arabidopsis、タバコ、大豆、クローバ(例えば、シロツメクサ、ムラサキツメクサ、サブタレニアンクローバ)、アルファルファ、アブラナ、アブラナ科野菜、レタス、ホウレンソウなど]および裸子植物に適用され得る。
【0045】
本発明のさらなる態様において、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む植物の老化を操作することができるベクターが提供される。
【0046】
本発明のまたさらなる態様において、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結されたmyb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む、植物のバイオマスを増大させることができるベクターが提供される。
【0047】
myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体は、本明細書において上記の完全長のmyb遺伝子プロモーターまたは修飾myb遺伝子プロモーターであり得る。
【0048】
本発明のこの態様の好ましい実施形態において、ベクターは、さらにターミネーターを含むものであり得、前記プロモーター、遺伝子およびターミネーターは、作動可能に連結されている。
【0049】
「作動可能に連結されている」とは、前記プロモーターが植物細胞において前記遺伝子の発現をもたらし得ること、および前記ターミネーターが植物細胞において前記遺伝子の発現を終了させ得ることを意図する。好ましくは、前記プロモーターは前記遺伝子の上流であり、前記ターミネーターは前記遺伝子の下流である。
【0050】
ベクターは、任意の適当な型のものであり得、ウイルス系または非ウイルス系であり得る。ベクターは、発現ベクターであり得る。かかるベクターとしては、染色体、非染色体および合成の核酸配列、例えば、植物ウイルスの誘導体;細菌系プラスミド;Agrobacterium tumefaciens由来のTiプラスミドの誘導体;Agrobacterium rhizogenes由来のRiプラスミドの誘導体;ファージDNA;酵母人工染色体;細菌系人工染色体;バイナリー細菌系人工染色体;プラスミドとファージDNAの組合せから誘導したベクターが挙げられる。しかしながら、植物細胞内で複製可能または組込み可能またはバイアブルである限り、任意の他のベクターが使用され得る。
【0051】
プロモーター、遺伝子およびターミネーターは、任意の適当な型のものであり得、標的植物細胞に対して内在性であってもよく、外来性であってもよいが、標的植物細胞において機能性であるものとする。
【0052】
また、本発明のベクターに使用され得るさまざまなターミネーターは、当業者に充分知られている。ターミネーターは、プロモーター配列と同じ遺伝子由来のものであってよく、異なる遺伝子由来のものであってもよい。特に好適なターミネーターは、ポリアデニル化シグナル(CaMV 35S ポリAなど)ならびにノパリンシンターゼ(nos)遺伝子およびオクトピンシンターゼ(ocs)遺伝子由来の他のターミネーターである。
【0053】
ベクターには、プロモーター、遺伝子およびターミネーターに加えて、さらに、遺伝子の発現に必要なエレメント、例えば、ベクターバックボーン、複製起点(ori)、マルチクローニング部位、スペーサー配列、エンハンサー、イントロン(トウモロコシユビキチンUbiイントロンなど)、抗生物質耐性遺伝子および他の選択可能マーカー遺伝子[ネオマイシンホスホトランスフェラーゼ(nptll)遺伝子、ハイグロマイシンホスホトランスフェラーゼ(hph)遺伝子、ホスフィノトリシンアセチルトランスフェラーゼ(barまたはpat)遺伝子など]、ならびにレポーター遺伝子(β−グルクロニダーゼ(GUS)遺伝子(gusA)など]が種々の組合せで含まれることがあり得る。また、ベクターには、翻訳開始のためのリボソーム結合部位を含めてもよい。また、ベクターには、発現の増幅のための適切な配列を含めてもよい。
【0054】
形質転換宿主細胞選択のための表現型形質をもたらすのに選択可能マーカー遺伝子を使用する択一法として、形質転換細胞内のベクターの存在を、当該技術分野でよく知られた他の手法、例えば、PCR(ポリメラーゼ連鎖反応)、サザンブロットハイブリダイゼーション解析、組織化学的アッセイ(例えば、GUSアッセイ)、薄層クロマトグラフィー(TLC)、ノーザンおよびウエスタンブロットハイブリダイゼーション解析などによって調べてもよい。
【0055】
当業者には、ベクターの種々の成分は、前記遺伝子の発現がもたらされるように作動可能に連結されていることが認識されよう。本発明のベクターの成分を作動可能に連結させるための手法は、当業者には充分知られている。かかる手法としては、リンカー(例えば、1つ以上の制限酵素部位を含む合成リンカーなど)の使用が挙げられる。
【0056】
本発明のさらなる態様において、変更された老化特性または向上したバイオマスを有するトランスジェニック植物細胞、植物、植物種子、または他の植物部分が提供される。好ましくは、前記植物細胞、植物、植物種子、または他の植物部分は、本発明によるベクターを含む。好ましくは、トランスジェニック植物細胞、植物、植物種子、または他の植物部分は、本発明による方法によって生成されるものである。
【0057】
また、本発明は、本発明の植物細胞由来のトランスジェニック植物、植物種子、または他の植物部分を提供する。
【0058】
また、本発明は、本発明の植物由来のトランスジェニック植物、植物種子、または他の植物部分を提供する。
【0059】
次に、本発明を、以下の実施例および添付の図面に関して、より充分に説明する。しかしながら、以下の説明は例示にすぎず、上記の本発明の一般説明をなんら制限するものと解釈されるべきでないことを理解されたい。
【図面の簡単な説明】
【0060】
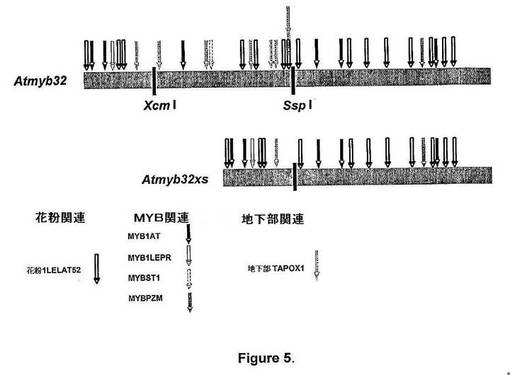
【図1】図1は、シロイヌナズナのmyb32遺伝子(atmyb32)由来のプロモーターのヌクレオチド配列であるAtmyb32プロモーター配列を示し(配列番号1)、MYB型、花粉特異的および地下部特異的モチーフを強調表示している。WAACCA(下線/イタリック体)MYB1AT;GTTAGTT(太字/線で囲んだ範囲)MYB1LEPR;CCWACC(線で囲んだ範囲)MYBPZM;GGATA(イタリック体)MYBST1;AGAAA(下線)花粉1LELAT52;ATATT(太字)地下部モチーフAPOX1。
【図2】図2は、XcmI−SspI植物配列欠失型Atmyb32プロモーター配列改変体(Atmyb32xs)を示す。MYB型、花粉特異的および地下部特異的モチーフを強調表示している。WAACCA(下線/イタリック体)MYB1AT;GTTAGTT(太字/線で囲んだ範囲)MYB1LEPR;CCWACC(線で囲んだ範囲)MYBPZM;AGAAA(下線)花粉1LELAT52;ATATT(太字)地下部モチーフAPOX1(配列番号2)。
【図3】図3は、すべての地下部モチーフを欠失させたAtmyb32プロモーター改変体配列を示す。MYB型、花粉特異的および地下部特異的モチーフを強調表示している。WAACCA(下線/イタリック体)MYB1AT;CCWACC(線で囲んだ範囲)MYBPZM;AGAAA(下線)花粉1LELAT52(配列番号3)。
【図4】図4は、SspI部位上流配列を欠失させたAtmyb32プロモーター改変体配列を示す。MYB型、花粉特異的および地下部特異的モチーフを強調表示している。WAACCA(下線/イタリック体)MYB1AT;CCWACC(線で囲んだ範囲)MYBPZM;AGAAA(下線)花粉1LELAT52(配列番号4)。
【図5】図5は、Atmyb32プロモーター配列およびAtmyb32xsプロモーター配列内のモチーフを示す。
【図6】図6は、Agrobacterium tumefaciens由来のイソペンテニルトランスフェラーゼ(ipt)遺伝子のヌクレオチド配列(配列番号5)を示す。
【図7】図7は、Agrobacterium tumefaciens由来のイソペンチルトランスフェラーゼ遺伝子の推定アミノ酸配列(配列番号6)を示す。
【図8】図8は、ミヤコグサ由来のイソペンチルトランスフェラーゼ遺伝子のヌクレオチド配列(配列番号7)を示す。
【図9】図9は、ミヤコグサ由来のイソペンチルトランスフェラーゼ遺伝子の推定アミノ酸配列(配列番号8)を示す。
【図10】図10は、ツクバネアサガオ由来のサイトカイニン生合成Sho遺伝子のヌクレオチド配列(配列番号9)を示す。
【図11】図11は、ツクバネアサガオ由来のサイトカイニン生合成Sho遺伝子の推定アミノ酸配列(配列番号10)を示す。
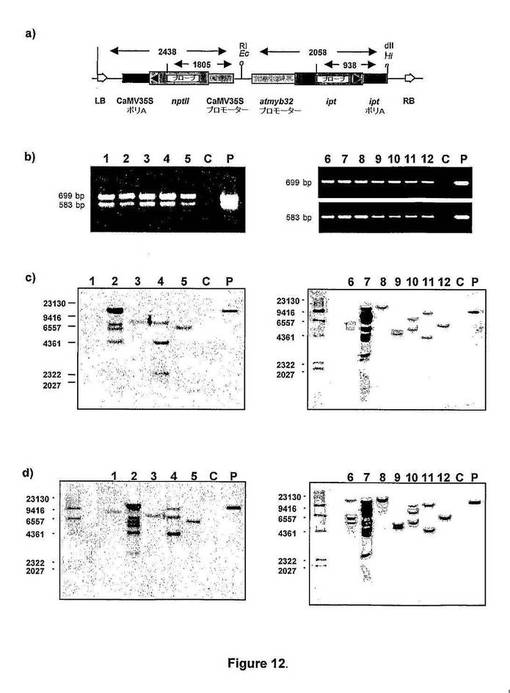
【図12】図12は、atmyb32::iptトランスジェニックシロツメクサ(Trifolium repens)植物のPCRおよびサザンDNA解析を示す。a)制限酵素部位およびサザンハイブリダイゼーション解析に使用されたプローブの位置を示すpatmyb32:iptのT−DNA領域、b)PCR増幅させた599bp nptll産物および583bp ipt産物の臭化エチジウム染色1%アガロースゲル、c)iptプローブとハイブリダイズさせたPCR陽性シロツメクサ植物から単離したHindIII消化全ゲノムDNAとのサザンブロットハイブリダイゼーション、d)nptllプローブとハイブリダイズさせたPCR陽性シロツメクサ植物から単離したHindIII消化全ゲノムDNAとのサザンブロットハイブリダイゼーション。レーン1〜2:2つの独立したカナマイシン耐性cv.Haifa 再生体(regenerant)、コード:それぞれ、Hmi01、Hmi08;レーン3〜12:12個の独立したカナマイシン耐性cv.Irrigation再生体、コード:それぞれ、Imi06、Imi07、Imi08、Imi09、Imi10、Imi11、Imi12、Imi14、Imi16、Imi18;レーンC:非形質転換シロツメクサ;レーンP:陽性対照プラスミドpatmyb32ipt。
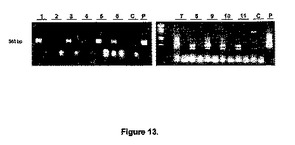
【図13】図13は、atmyb32::iptトランスジェニックシロツメクサ(T.repens)植物におけるipt mRNA発現のRT−PCR解析を示す。レーン1〜11は、図4.8の場合の対応植物コードを有する11種類の独立したトランスジェニック系統由来の試料である;レーンC、対照非形質転換植物;レーンP、陽性対照としてのプラスミド。全RNAを葉組織から単離した。全RNA(13μg)を各逆転写反応に使用し、RT産物の1/5をPCRによって増幅させた。右側のゲル上のDNA産物は、2×30サイクルの集中的(intensive)PCRによって増幅させた。交互のレーンに負荷した対応RT−PCR反応液には、逆転写酵素は添加しなかった。
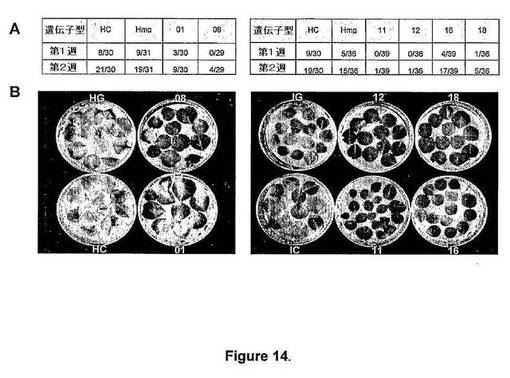
【図14】図14は、atmyb32::iptトランスジェニックシロツメクサ(T.repens)植物から切除した葉の老化バイオアッセイを示す。各植物系統から少なくとも30枚の葉を、各系統の走根上の類似した位置から収集した。A.切除した葉の総数に対する分数としての黄変葉の数。B.光の下で水上に2週間維持した葉の典型的な外観。植物系統に対するキー:それぞれ、HC、ICおよびHmg、Img、非形質転換およびatmyb32::gusAトランスジェニック植物(cv.HaifaおよびIrrigation);01および08、atmyb32::iptトランスジェニックHaifa系統は、それぞれ、Hmi01およびHmi08;11、12、16および18atmyb32::iptトランスジェニックIrrigation系統は、それぞれ、Imi11、Imi12、Imi16およびImi18。
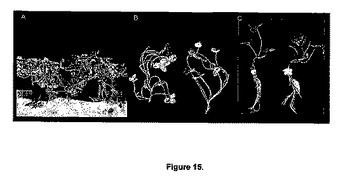
【図15】図15は、対照植物(左)と比較したatmyb32::iptトランスジェニックシロツメクサ(T.repens)(右)植物における、A)一般的な植物の形態、B)正常な苗条の発育、およびC)正常な地下部の発育を示す。
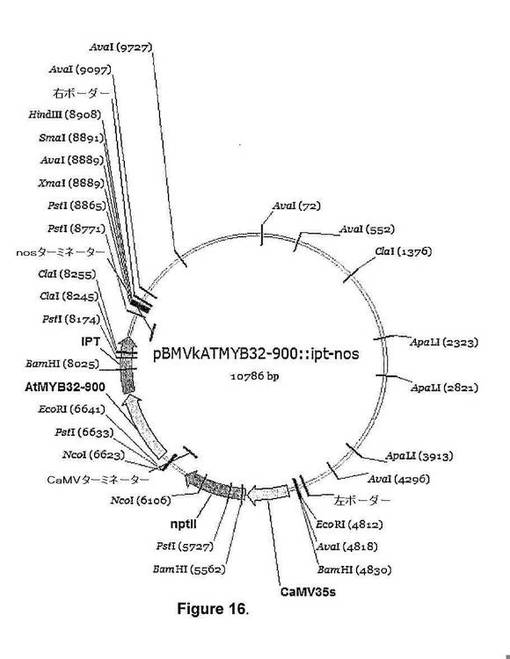
【図16】図16は、pBMVkAtMYB32−900::ipt[遺伝子:イソペンチルトランスフェラーゼ(IPT);ベクター:pBMVkAtMYB32−900::ipt−nos(バックボーンpPZPRCS2);選択可能マーカー:spec;植物選択可能マーカーカセット:35S::kan::35ST;遺伝子プロモーター:AtMYB32−900;遺伝子ターミネーター:nos.]のベクターの詳細を示す。
【図17】図17は、ベクターpBMVkAtMYB32−900::ipt−nosのヌクレオチド配列(配列番号11)を示す。
【図18】図18は、pBMVkAtMYB32xs::ipt−nos[遺伝子:イソペンチルトランスフェラーゼ(IPT);ベクター:pBMVkAtMYB32XS::ipt−nos(バックボーンpPZPRCS2);選択可能マーカー:spec;植物選択可能マーカーカセット:35S::kan::35ST;遺伝子プロモーター:AtMYB32−xs;遺伝子ターミネーター:nos.]のベクターの詳細を示す。
【図19】図19は、ベクターpBMVkAtMYB32xs::ipt−nosのヌクレオチド配列(配列番号12)を示す。
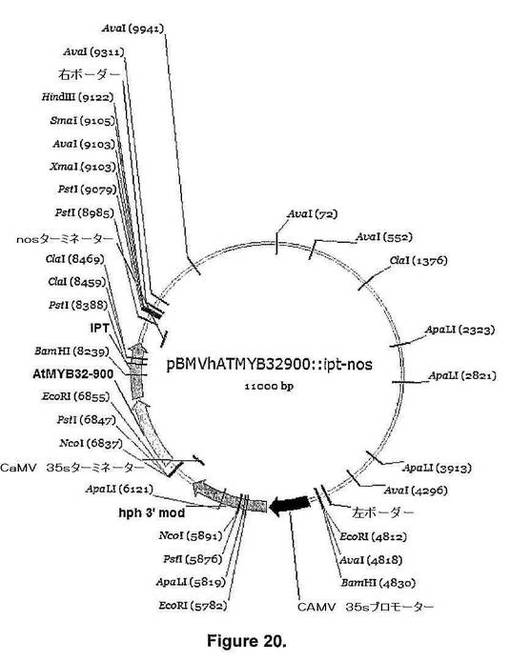
【図20】図20は、pBMVhAtMYB32−900::ipt−nos[遺伝子:イソペンチルトランスフェラーゼ(IPT);ベクター:pBMVhAtMYB32−900::ipt−nos(バックボーンpPZPRCS2);選択可能マーカー:spec;植物選択可能マーカーカセット:35S::hph::35ST;遺伝子プロモーター:AtMYB32−900;遺伝子ターミネーター:nos.]のベクターの詳細を示す。
【図21】図21は、ベクターpBMVhAtMYB32−900::ipt−nosのヌクレオチド配列(配列番号13)を示す。
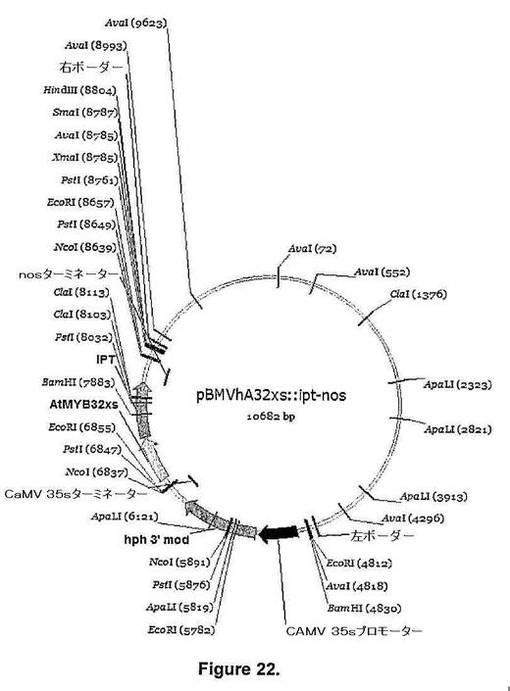
【図22】図22は、pBMVhAtMYB32xs::ipt−nos[遺伝子:イソペンチルトランスフェラーゼ(IPT);ベクター:pBMVhAtMYB32XS::ipt−nos(バックボーンpPZPRCS2);選択可能マーカー:spec;植物選択可能マーカーカセット:35S::hph::35ST;遺伝子プロモーター:AtMYB32−xs;遺伝子ターミネーター:nos.]のベクターの詳細を示す。
【図23】図23は、ベクターpBMVhAtMYB32xs::ipt−nosのヌクレオチド配列(配列番号14)を示す。
【図24】図24は、pBSubn−AtMYB32−900::ipt−nos[遺伝子:イソペンチルトランスフェラーゼ(IPT);ベクター:pBSubn−AtMYB32−900::ipt−nos(バックボーンpPZPRCS2);選択可能マーカー:spec;植物選択可能マーカーカセット:Ubi::bar::nos;遺伝子プロモーター:AtMYB32−900;遺伝子ターミネーター:nos.]のベクターの詳細を示す。
【図25】図25は、ベクターpBSAtMYB32900::ipt−nosのヌクレオチド配列(配列番号15)を示す。
【図26】図26は、pBMVhATMYB3−900::ipt−nosおよびpBMVhATMYB32xs::ipt−nosを含むトランスジェニックアブラナの生成を示す。A.アブラナ種子をインビトロで発芽させる;B.胚軸部を7日齢の苗木から切除し、アグロバクテリウム懸濁液を接種する;CおよびD.ハイグロマイシン選択下、接種胚軸部からの再生 ;E〜J.pBMVhATMYB3−900::ipt−nosおよびpBMVhATMYB32xs::ipt−nosベクターを有するトランスジェニックT0アブラナ植物。
【図27】図27は、小麦の微粒子銃形質転換のためのプロセスを示す。
【図28】図28は、小麦(Triticum aestivum L.MPB Bobwhite 26)の微粒子銃形質転換を示す。ドナー植物生産(AおよびB);接合子胚単離(CおよびD);グルホシネート選択下での再生(E〜G);選択下での地下部形成(H);トランスジェニック子孫の回収のための閉鎖温室条件下でのT0植物栽培(I)。
【図29】図29は、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物の隔離圃場試験を示す。
【図30】図30は、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物と、非トランスジェニック対照シロツメクサ植物での生長速度と生長力学の比較評価を示す。A)生長速度 B)生長力学 C)生長特性(45日後) 白抜きの四角 トランスジェニックシロツメクサ 黒塗りの四角 非トランスジェニック対照シロツメクサ (明) (暗)
【図31】図31は、隔離圃場条件下での、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12、LXR 18およびLXR 11)と、非トランスジェニック対照シロツメクサ植物(すなわち、WT)の開花強度(すなわち、1m2あたりの成熟花の数)を示す。
【図32】図32は、隔離圃場条件下での、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12、LXR 18およびLXR 11)と、非トランスジェニック対照シロツメクサ植物(すなわち、WT)の種子重量(すなわち、千個の種子の重量、単位:グラム)を示す。
【図33】図33は、隔離圃場条件下での、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12、LXR 18およびLXR 11)と、非トランスジェニック対照シロツメクサ植物(すなわち、WT)の花1つあたりの種子収量(単位:ミリグラム)を示す。
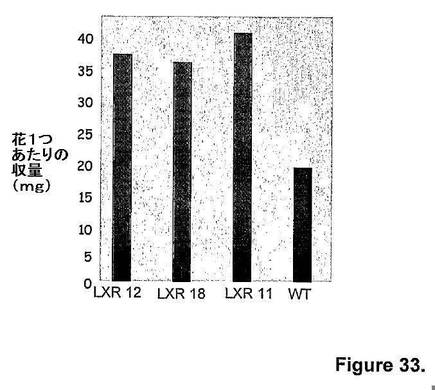
【図34】図34は、隔離圃場条件下での、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12、LXR 18およびLXR 11)と、非トランスジェニック対照シロツメクサ植物(すなわち、WT)の単位面積あたりの種子収量(単位:kg/ヘクタール)を示す。
【図35】図35は、キメラpBMVkATMYB3−900::ipt−nosおよびpBMVkATMYB32xs::ipt−nos遺伝子を含むトランスジェニックアルファルファ植物の生成を示す。A.アルファルファクローンC2−3、C2−4および19−17由来の葉柄外植片をアグロバクテリウム懸濁液の接種に使用し、カナマイシンの存在下での選択後、形質転換胚形成性カルスの発生をもたらす;B〜D.インビトロで培養した体細胞胚からの、pBMVkATMYB3−900::ipt−nosおよびpBMVkATMYB32xs::ipt−nosベクター由来のキメラ遺伝子を有するトランスジェニックアルファルファ小植物の再生。
【図36】図36は、トランスジェニックアブラナ植物(T1 LXRアブラナ植物Line 4)のPCR解析を示す。ゲノムDNAは、T1 LXR04系統の異なるトランスジェニックアブラナ植物から単離し、A.選択可能マーカー(hph)またはB.目的の候補遺伝子(IPT)に特異的なプライマーを用いてPCRに供した。
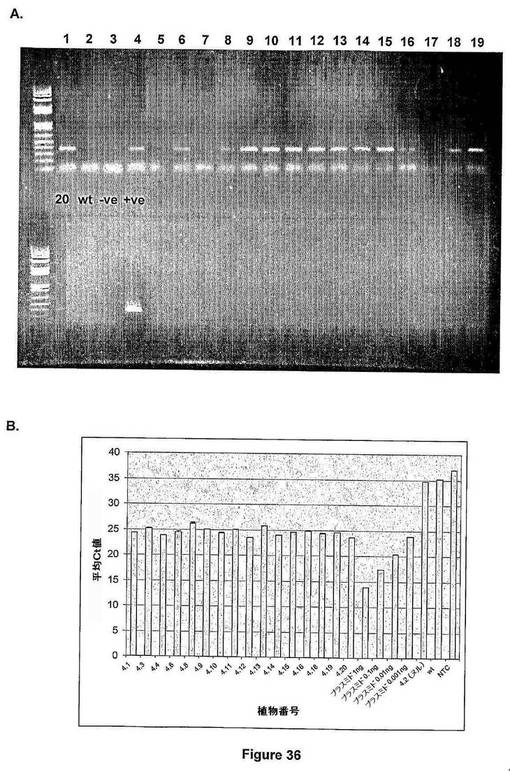
【図37】図37は、T1トランスジェニックアブラナ植物におけるIPT遺伝子の発現解析を示す(T1 LXRアブラナの葉での相対IPT発現)。
【図38】図38は、切り取って7日後、野生型対照子葉と比較したときの、切り取り子葉の老化の遅延を示すトランスジェニックアブラナを示す。
【図39】図39は、切り取って7日後のT1トランスジェニックアブラナ子葉の老化スコアを示す。
【0061】
【化1】

【図40】図40は、切り取って14日後、野生型対照子葉と比較したときの、切り取り若幼葉の老化の遅延を示すトランスジェニックアブラナを示す。
【図41】図41は、切り取って14日後、T1トランスジェニックアブラナの第1の若幼葉の老化スコアを示す。
【0062】
【化2】
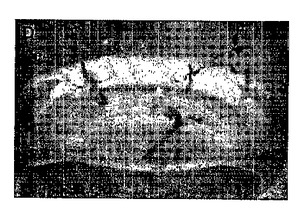
【図42】図42は、トランスジェニック小麦系統のサザンハイブリダイゼーション解析を示す。レーンは、MW−分子量;WT−野生型;トランスジェニック小麦系統LXR3、LXR4、LXR13、LXR16ならびに10pgおよび20pgを含む陽性対照プラスミドを含む。
【図43】図43は、独立したT1トランスジェニック小麦系統の発現解析(小麦におけるIPT定量的発現)を示す。定量的転写物の値は、標的配列を含むプラスミドDNAで得た標準曲線を用いて、フェトモル(fmol)/マイクログラムRNAで測定した。試料は、候補遺伝子IPTに対して高、中および低クラスの発現を表す。
【図44】図44は、温室栽培T1トランスジェニック小麦植物の表現型多様性を示す。A.ヌル対照小麦植物と比較したとき、正常表現型を示すT1 LXR 13小麦植物。B.ヌル対照小麦植物と比較したとき、成長阻害表現型および止め葉数の増加を示すT1 LXR 04小麦植物。
【図45】図45は、切り取って7日後、ヌル対照葉と比較したときの、葉の老化の遅延を示すトランスジェニック小麦を示す。
【発明を実施するための形態】
【0063】
実施例
実施例1
Atmyb32プロモーター配列およびプロモーター配列改変体
Atmybb32プロモーター配列およびその改変体を図1〜4に示す。
【0064】
実施例2
サイトカイニン生合成遺伝子
本発明における使用に適したサイトカイニン生合成遺伝子の例を、図6、8および10に示す。また、好適な遺伝子としては、図7、9および11に示すポリペプチドをコードするものが挙げられる。
【0065】
実施例3
トランスジェニックシロツメクサ植物の生成
トランスジェニックシロツメクサ植物(Trifolium repens cv.HaifaおよびIrrigation)を、キメラatmyb32::ipt遺伝子バイナリーベクター(図12a)を使用し、アグロバクテリウム媒介型形質転換によって生成した。トランスジェニック植物を、iptプライマーおよびnptllプライマー(図12b)を用いてPCRによってスクリーニングした。HindIII消化ゲノムDNA試料をサザンDNAハイブリダイゼーション解析に供すると、iptプローブとnptllプローブの両方によって、4.4kbより大きいDNA断片がすべてのレーンに検出されることが示され、これは、完全長T−DNAの存在およびそのシロツメクサゲノム内への組込みを示す(図12)。トランスジェニック系統Hmi01、Imi06、Imi11およびImi18(それぞれ、レーン1、3、5、8および12)は、ゲノム内に組み込まれた単一コピーの完全長T−DNAを有するようであった。他のトランスジェニック系統は、多コピーのatmyb32::ipt導入遺伝子を有した。
【0066】
実施例4
トランスジェニックシロツメクサ植物におけるIPT遺伝子発現
トランスジェニックシロツメクサ(T.repens)植物におけるatmyb32::ipt導入遺伝子の発現を、RT−PCRによって評価した。ipt mRNAは、調べたatmyb32::iptトランスジェニックシロツメクサ植物すべての葉組織内で検出され、種々のレベルのPCR産物が検出された(図13)。
【0067】
実施例5
トランスジェニックシロツメクサ植物における切り取り葉の老化の遅延
atmyb32::iptトランスジェニック植物の切り取り葉の老化を評価するための実験を行なった。1週間以内に、両方の品種の非形質転換型およびatmyb32::gusAトランスジェニックシロツメクサ植物から切り取られた葉において、急速な黄変が観察された。トランスジェニック系統Hmi01、Hmi08、Imi16およびImi18では、老化の遅延が示されたが、Imi11とImi12では、7日間の最後までに黄変の兆候は示されなかった。2週間後、すべてのatmyb32::iptトランスジェニック植物の葉は、非形質転換型およびatmyb32::gusA対照トランスジェニック植物のものよりもずっと緑色であった(図14)。切除した葉における老化の程度は、cv.Haifaでは、HC、Hmg>Hmi01>Hmi08、ならびにcv.Irrigationでは、ICおよびImg>Imi16>Imi18>Imi11およびImi12の順であった。HCは、Haifa非形質転換対照であり、Hmgは、Haifa atmyb32::gusA対照であり、ICは、Irrigation非形質転換対照であり、Imgは、Irrigation atmyb32::gusA対照である。Hmi01、Hmi08、Imi16、Imi18、Imi11およびImi12は、それぞれ、品種Haifa(H)およびIrrigation(I)に由来する独立したatmyb32::iptトランスジェニックシロツメクサ植物である。
【0068】
実施例6
トランスジェニックシロツメクサ植物における植物の形態と地下部の発育
正常な植物の形態ならびに正常な苗条の発育と正常な地下部の発育が、atmyb32:iptトランスジェニックシロツメクサ植物において観察され(図6)、したがって、atmyb32プロモーターの制御下でのipt遺伝子の発現の調節は、トランスジェニックシロツメクサ植物の地下部発育にも頂芽優性にもマイナスの影響を及ぼさないことが示された(表1)。
【0069】
【表1】
分析した10種類の独立したatmyb32::iptトランスジェニックシロツメクサ系統において、正常な植物の形態および正常な地下部発育が観察された。この10種類の独立したatmyb32::iptトランスジェニックシロツメクサ系統の推定ipt遺伝子コピー数を示す。
【0070】
実施例7
植物形質転換用のベクターの作製
植物のアグロバクテリウム媒介型形質転換のため、4種類のバイナリーベクターを作製した(図16〜19)。各ベクターは、pPZP200ベクターバックボーン(Hajdukiewiczら,1994)を有し、キメラAtmyb32−900::ipt−nosまたはAtmyb32−xs::ipt−nosのいずれかを、キメラ35S::nptll−35stまたは35S::hph−35st選択可能マーカーカセットとともに、またはなしで含む。
【0071】
形質転換ベクターの1つは、微粒子銃形質転換のために構築した(図20および21)。この形質転換ベクターは、キメラAtmyb32−900::ipt−35stをキメラUbi::bar−nos選択可能マーカーカセットとともに含む。
【0072】
Atmyb32プロモーター、プロモーター改変体Atmyb32xs、イソペンチルトランスフェラーゼ遺伝子ならびにターミネーター35stおよびnosを、Gateway(商標)(Invitrogen)適合プライマーを用いてPCRによって増幅し、pDONR221エントリーベクター内にクローニングした。続いて、これらを組換えを用いて、慣用的にクローニングされる選択可能マーカーカセットを含むデスティネーションベクター内にクローニングした。すべてのベクターは、厳密な品質保証プロトコルに従って完全に配列決定した。
【0073】
実施例8
アブラナ(Brassica napus)のアグロバクテリウム媒介型形質転換
キメラipt遺伝子を含むバイナリーベクターpBMVhATMYB3−900::ipt−nos(図20)およびpBMVhATMYB32xs::ipt−nos(図22)を、Atmyb32プロモーター(図1)および地下部特異的モチーフを欠失させたAtmyb32xs改変体プロモーター配列(図2)の制御下で、Brassica napus胚軸セグメントのアグロバクテリウム媒介型形質転換に使用した(図26)。
【0074】
Brassica napusの種子を70%エタノール中で2分間表面消毒し、滅菌水中で3回洗浄し、次いで、1%(w/v)の次亜塩素酸カルシウムおよび0.1%(v/v)のTween 20を含有する溶液中で、30分間さらに表面消毒する。種子を滅菌水中で少なくとも3回洗浄し、1×Murashige and Skoog(Murashige and Skoog Physiol.Plant,15:473−497,1962)多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、2%(w/v)のスクロースを補給、pH5.8)を含有し、4g/LのGelriteを添加した固形発芽培地を入れた120ml容の培養容器内に植え付ける。容器を16時間明/8時間暗条件下、25℃で7日間インキュベートして発芽を促進する。
【0075】
7日後、Brassica napusの苗木(苗木全体)を、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、3%(w/v)のスクロースを補給、pH5.8)からなる液体培地に移す。苗木を集め、地下部と子葉を除去した後、胚軸を7〜10mm切片に切断し、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、3%(w/v)のスクロースを補給、pH5.8)からなり、6.4g/lのBacto−Agarで固化したプレコンディショニング培地を入れた9×1.5cmのペトリ皿の上で平板培養する。
【0076】
胚軸部を24時間培養した後、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、100μMのアセトシリンゴン、3%(w/v)のスクロースを補給、pH5.8)からなるアグロバクテリウム懸濁液OD600=0.2を30分間接種する。
【0077】
接種後、胚軸部を滅菌ペーパータオル上にブロットし、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、100μMのアセトシリンゴン、1mg/Lの2,4−D、3%(w/v)のスクロースを補給、pH5.8、8g/lのBacto−Agarで固化)を入れた9×1.5cmのペトリ皿に移す。外植片を16時間明/8時間暗条件下、25℃で72時間インキュベートして共存培養する。
【0078】
共存培養後、20〜30個の胚軸外植片を1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、1mg/Lの2,4−D、3%(w/v)のスクロースを補給、pH5.8)からなり、8g/lのBacto−Agar(250mg/lのチメンチンおよびハイグロマイシン耐性苗条を選択するための10mg/lのハイグロマイシンを補給)で固化した固形選択培地を入れた9×1.5cmのペトリ皿に移す。プレートを16時間明/8時間暗条件下、25℃でインキュベートする。
【0079】
7日後、胚軸外植片を、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、1mg/Lの2,4−D、3%(w/v)のスクロースを補給、pH5.8)からなり、8g/lのBacto−Agar(4mg/lのBAP、2mg/lのゼアチン、5mg/lの硝酸銀、250mg/lのチメンチンおよび10mg/lのハイグロマイシンを補給)で固化した固形再生培地を入れた9×2.0cmのペトリ皿に移す。プレートを、蛍光灯条件(16時間明/8時間暗の光周期;55μmol m−2秒−1)下の直接照明下、25℃で4週間インキュベートして苗条の発育を促進する。
【0080】
再生を毎週モニタリングし、胚軸外植片を、固形再生培地RM(4mg/lのベンジルアデニン、2mg/lのゼアチン、5mg/lの硝酸銀、250mg/lのチメンチンおよび10mg/lのハイグロマイシンを補給)を入れた新たな9×2.0cmのペトリ皿に6〜8週間移し、苗条の発育を促進する。
【0081】
ハイグロマイシン耐性(Hygr)苗条を、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、1mg/Lの2,4−D、1%(w/v)のスクロースを補給、pH5.8)からなり、8g/lのBacto−Agar(250mg/lのチメンチンを補給)で固化した固形地下部誘導培地RIM1を入れた120ml容器に移す。苗条を、直接蛍光灯(16時間明/8時間暗の光周期;55μmol m−2秒−1)下、25℃でインキュベートして苗条の伸張と地下部の発育を4〜5週間にわたって促進する。苗条と地下部系の発育を有するHygr苗条をすべて土壌に移し、温室条件下で生長させる。
【0082】
実施例9
小麦(Triticum aestivum L.)の微粒子銃形質転換
キメラipt遺伝子を含む形質転換ベクターを、Atmyb32プロモーター(図1)および地下部特異的モチーフを欠失させたAtmyb32xs改変体プロモーター配列(図2)の制御下で、小麦(Triticum aestivum L.MPB Bobwhite 26)の微粒子銃形質転換に使用した。代表的なベクターを図24に示す。小麦の微粒子銃形質転換の手順の概略を図27に示す。形質転換手順は、以下の工程を含む。
【0083】
工程1(ドナー植物生産):
Triticum aestivum(Bobwhite 26)の種子をドナー植物材料の生産に使用する。小麦植物は、堆肥化したマツ樹皮、パーライトおよびバーミキュライトからなる種苗用(nursery)ミックス中で栽培し、鉢1つあたり植物5本とし、最大鉢サイズを20cmとする。植物をほぼ22〜24℃の温室条件下に12〜16週間維持する(図28A)。止め葉から最初の穂状花序(spike)が現れたら植物にタグを付け、開花12〜15日後に最も高い頭部から胚を収集する。
【0084】
工程2(第1日目)
所望の発育段階の穂状花序を採取する(図28B)。穀果を穂状花序から除去し、0.8%(v/v)のNaOCl溶液中で20分間表面消毒し、滅菌蒸留中で少なくとも4回すすぐ。
【0085】
長さ10mmまでの胚を各穀果から、解剖顕微鏡を用いて無菌的に切除し(軸部の取り出し)、軸面を下にして、2×Murashige and Skoog(1962)多量養素、1×微量栄養素および有機ビタミン群、40mg/Lのチアミン、150mg/LのL−アスパラギン(15%(w/v)のマルトース、0.8%(w/v)のSigma製寒天および2.5mg/Lの2,4−Dを補給)からなる浸透圧培地(E3マルトース)上で培養する(図28CおよびD)。胚を、15mLの培地を入れた60mm×15mmの透明なポリプロピレン製ペトリ皿の上で培養する。培養プレートを24℃で、暗所にて4時間インキュベートした後、ボンバードメントを行なう。胚には、900psiで6cmのBioRad PDS1000遺伝子銃で、1μgのベクタープラスミドDNA(0.6μmの金粒子上に沈殿させたもの)を用いてボンバードメントを行なう。ボンバードメント後、胚を浸透圧培地上で、暗所にて一晩インキュベートする。
【0086】
工程3(第2日目):
胚を、2×Murashige and Skoog(1962)多量養素および1×微量栄養素ならびに有機ビタミン群、40mg/Lのチアミン、150mg/LのL−アスパラギン(6%(w/v)のスクロース、0.8%(w/v)のSigma製寒天および2.5mg/Lの2,4−Dを補給)からなるカルス誘導培地(E3カルス)に移す。胚を24℃で暗所にて2週間培養する。
【0087】
工程4(第16日目):
E3カルス上での2週間の培養後、胚から胚誘導性カルスが発生し、2×Murashige and Skoog(1962)多量養素および1×微量栄養素ならびに有機ビタミン群、40mg/Lのチアミン、150mg/LのL−アスパラギン(2%(w/v)のスクロース、0.8%(w/v)のSigma製寒天、5mg/LのD,Lホスフィノトリシン(PPT)を補給、植物生長調節因子なし)からなる選択培地(E3Select)上に継代培養する(図28E〜G)。培養物を、さらに14日間、E3Select上で24℃にて明るい場所で12時間の光周期でインキュベートする。
【0088】
工程5(第30日):
E3Select上での14日間の培養後、胚誘導性カルスを新たなE3Select上で、さらに14日間継代培養する(図28E−G)。
【0089】
工程6(第44日):
E3Select上で約4週間後、生長中の小植物を胚性カルス塊から切除し、地下部誘導培地(RM)を入れた65mm×80mmまたは65mm×150mmのポリカーボネート製組織培養容器内で、さらに3週間生長させる。地下部誘導培地は、1×Murashige and Skoog(1962)多量養素、微量栄養素ならびに有機ビタミン群、40mg/Lのチアミン、150mg/LのL−アスパラギン(2%(w/v)のスクロース、0.8%(w/v)のSigma製寒天、および5mg/LのPPTを補給)からなるものである(図28H)。残りの胚誘導性カルスはE3Select上で、さらに14日間継代培養する。
【0090】
工程7(第65日以降):
RM上で3週間より長く生存し、健常な地下部形成を有する再生小植物を、泥炭と砂(1:1)からなる種苗用ミックス中に植え、種苗用湿度チャンバシステムにて高湿度で22〜24℃に維持する(図28)。2週間後、植物を湿度チャンバから取り出し、手での水やりと、液状肥料Aquasol(商標)の供給を毎週、充分生長するまで与える。T0植物をゲノムDNAと分子解析のために試料採取する。T1種子を採取し、ハイスループットQ−PCR解析のために植える(図28J)。
【0091】
実施例10
トランスジェニックシロツメクサ植物の農業生産力
非トランスジェニック対照シロツメクサ植物のものと比べたatmyb32::iptトランスジェニックシロツメクサ(Trifolium repens)植物の農業生産力を、環境制御生長チャンバ条件下で隔離圃場試験にて評価した(図29)。
【0092】
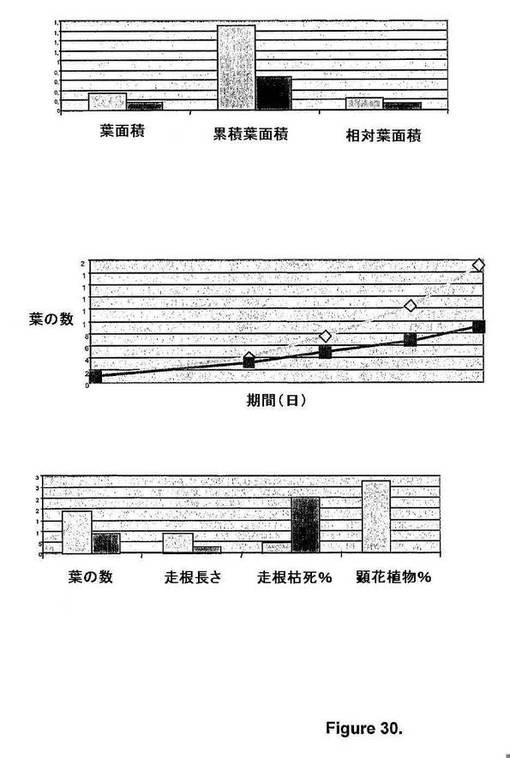
生長制御チャンバ条件下で評価したキメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物は、非トランスジェニック対照シロツメクサ植物と比べて、有意なバイオマス蓄積の向上および老化の兆候の低減を示した(図30)。キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物は、非トランスジェニック対照シロツメクサ植物と比べて、総葉面積の増大、累積葉面積増大、葉の高生長力学(すなわち、経時的な葉の数)、走根長さの増大および顕花植物%の増大ならびに走根の老化および枯死の低減を示した(図30A〜C)。
【0093】
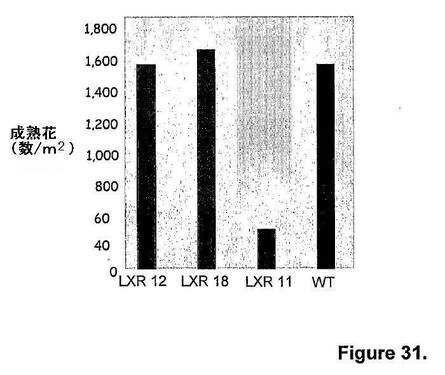
また、トランスジェニックシロツメクサ植物をもたらす3つの独立したatmyb32::ipt(すなわち、LXR 12、LXR 18およびLXR 11)の種子収量の成績を、隔離圃場条件下で、非トランスジェニック対照植物(すなわち、野生型、WT)と比較評価した。非トランスジェニック対照植物(すなわち、WT)と識別不可能な開花強度(すなわち、1m2あたりの成熟花の数)を有するトランスジェニックシロツメクサ植物をもたらした2つの独立したatmyb32::ipt(すなわち、LXR 12およびLXR 18)を、圃場評価に選択した(図31)。
【0094】
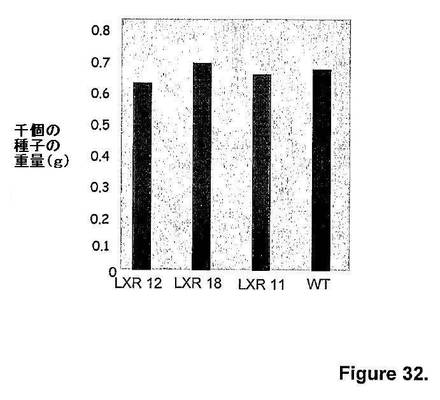
キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12、LXR 18およびLXR 11)の種子重量(すなわち、千個の種子の重量)は、非トランスジェニック対照シロツメクサ植物(すなわち、WT)と識別不可能であったが(図32)、花1つあたり(図33)、および種蒔き面積あたり(図34)の基準で示す総種子収量は、同等の開花強度の非トランスジェニック対照シロツメクサ植物(すなわち、WT)と比べて、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12およびLXR 18)において2倍であった。
【0095】
実施例11
アルファルファ(Medicago sativa)のアグロバクテリウム媒介型形質転換
キメラipt遺伝子を含むバイナリーベクターpBMVkATMYB32xs::ipt−nos(図18)を、地下部特異的モチーフを欠失させたAtmyb32xs改変体プロモーター配列(図2)の制御下で、高度に再生可能なアルファルファ(M.sativa)クローンC2−3、C2−4および19−17由来のMedicago sativaの葉柄外植片のアグロバクテリウム媒介型形質転換に使用した(図35)。
【0096】
バイナリーベクターpBMVkATMYB32xs::ipt−nosを保有するAgrobacterium tumefaciens株LBA 4404との共存培養後、アルファルファ外植片をセフォタキシム含有培地で洗浄し、25mg/lのカナマイシンを含有する選択培地下で、胚誘導性カルスの誘導に使用した。トランスジェニック胚形成性アルファルファカルスを回収し、トランスジェニックアルファルファ苗条を再生させ、これを地下部発育培地に移し、Atmyb32xs改変体プロモーターの制御下で、キメラipt遺伝子発現由来トランスジェニックアルファルファ植物の回収に至るようにした(図35)。
【0097】
本明細書において開示および規定した本発明は、本文または図面に記載し、これらから明白な個々の特徴の2つ以上の択一的なあらゆる組合せに拡張されることは理解されよう。このような種々の組合せはすべて、本発明の種々の択一的態様を構成する。
【0098】
実施例12
トランスジェニックアブラナ植物の生成
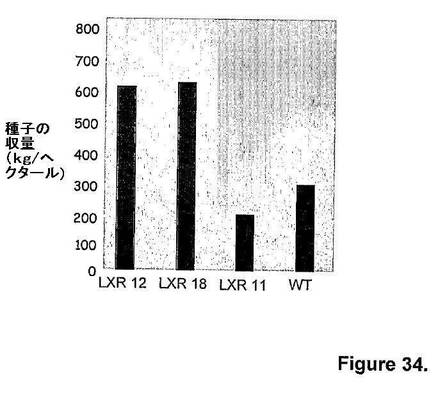
トランスジェニックアブラナ植物(Brassica napus)を、キメラatmyb32::ipt遺伝子を保有するバイナリーベクター(図20および22)を用いて、Agrobacterium媒介型形質転換によって生成した。遺伝子修飾を、候補遺伝子(IPT)または選択可能マーカー(hph)の存在について、T1世代でPCRを用いてキャラクタライズした(図36)。
【0099】
図36は、トランスジェニックアブラナ植物のPCR解析を示す。ゲノムDNAをT1 LXR04系統の種々のトランスジェニックアブラナ植物から単離し、選択可能マーカー(hph遺伝子)または目的の候補遺伝子(IPT)に特異的なプライマーを用いてPCRに供した。図36Aでは、hph特異的プライマーを用いてゲノムDNAから産物を増幅し、アガロースゲル上で可視化した。図36Bは、蛍光PCR法を用いてゲノムDNAを増幅するためのIPT特異的プライマーの使用を示す。これらのプライマーは、PCR産物の蓄積量に反比例する検出可能な蛍光をもたらす標的配列に特異的である。
【0100】
実施例13
トランスジェニックアブラナ植物におけるIPT遺伝子発現
トランスジェニックアブラナにおけるatmyb32::ipt導入遺伝子の発現を、標的配列に特異的な蛍光RT−PCR法を用いて評価した(図37)。組織内のIPT mRNAを検出し、相対発現レベルを系統間およびヌル対照と比較した。ヌル対照は、形質転換プロセスを受けたが、交配後、標的配列を含まない子孫系統である。
【0101】
実施例14
トランスジェニックアブラナ植物における切り取り葉の老化の遅延
atmyb32::iptトランスジェニック植物の切り取り葉の老化を評価するための実験を行なった。図38〜41は、アブラナにおける候補遺伝子の発現に関する切り取り葉の老化(detached senescence)アッセイのデータを示す。切り取り子葉と葉のアッセイは、老化を誘導し、形質転換アブラナの老化表現型を野生型対照と比較して評価するために行なった。切り取り葉の老化アッセイの第7日目および第14日目、老化の進行を、各組織試料について、0−老化の可能性の兆候なし;1−初期の老化の兆候がみられ、緑色が薄くなっている:2−さらに老化が進行、黄変が目立つようになっている;3−組織がほぼ黄変しているが、薄緑色が明白に残っている;4−完全な黄変に進行;5−黄変し、一部脱色および壊死斑を伴う、のいずれかとして定性的にスコアリングした(図39および41)。
【0102】
実施例15
トランスジェニック小麦植物の生成
小麦の遺伝的形質転換は、図27および28に概要を示したTriticum aestivum L Bobwhite 26系統小麦由来の接合子胚の微粒子銃形質転換に基づいたものとした。
【0103】
キメラatmyb32::ipt遺伝子を小麦ゲノム内に、パーティクルボンバードメントによって、ベクターバックボーン配列もまたゲノム内に組み込まれるように完全体プラスミドを用いて挿入した(図24)。
【0104】
形質転換ベクターは充分に配列決定されたものとした(図25)。遺伝子修飾は、候補遺伝子の存在について、T1世代でサザン解析によってキャラクタライズした(図42)。
【0105】
図42は、トランスジェニック小麦植物のサザンハイブリダイゼーション解析を示す。ゲノムDNAを異なるT1系統のトランスジェニック小麦植物から単離し、制限酵素で消化し、候補遺伝子コピー数を測定した。対照は、非形質転換野生型Triticum aestivum「Bobwhite 26」である。消化物を電気泳動させ、ナイロン膜に移し、プローブとしての完全長DIG標識IPT遺伝子とプローブ結合させた。ある範囲のコピー数が観察された。
【0106】
実施例16
トランスジェニック小麦植物におけるIPF遺伝子発現
AtMYB32プロモーターによって駆動させたIPT遺伝子を含む温室栽培トランスジェニックT1小麦植物の若幼葉組織から、RNAを抽出し、第1cDNA鎖を調製した。
【0107】
導入遺伝子の定量的発現は、標的配列のためのプローブ系qRT−PCR法を用いて調べた。各構築物の高、中および低発現系統の代表例を図43に示す。また、内在性スクロースシンターゼ遺伝子に対して設計されたプライマー/プローブセットを対照として使用した。対照遺伝子のすべての増幅プロットは、互いの1サイクル内で始まり、これは、GMOの検出レベル差が発現の多様性によるものであることを示す。
【0108】
PCRプライマーおよびプローブは、ともに、PCR産物の蓄積量に比例する検出可能な蛍光をもたらす標的配列に特異的である。検出対象の標的配列を含む連続希釈プラスミドDNAを用いて、定量のための標準曲線を作成した。
【0109】
実施例17
トランスジェニック小麦植物の植物形態
生長特性の違いを、温室内で、トランスジェニック小麦系統内および系統間で観察した。T1小麦植物間で主に観察された表現型には、植物丈、ひこばえ出現(tillering)強度、葉の数、ならびに植物性バイオマスの抑制が含まれた(図44)。
【0110】
実施例18
トランスジェニック小麦植物における切り取り葉の老化の遅延
切り取り葉アッセイを使用し、ヌル対照と比較したときの形質転換小麦葉の誘導老化および老化表現型を評価(asses)した(図45)。ヌル対照は、形質転換プロセスを受けたが、交配後、標的配列を含まない子孫系統である。
【0111】
また、本明細書で用いる用語「〜を含む/包含する(comprise)」(またはその文法的語尾変化)は、用語「〜を含む(include)」と等価であり、他の要素または特徴の存在を排除するものと解されるべきでないことは理解されよう。
【0112】
本明細書で挙げた文献は、参照目的のためにすぎず、該文献を含めることによって該文献が関連技術分野における周知の一般知識の一部を構成することの承認ではない。
【0113】
最後に、種々の変形、変更および/または付加が、本明細書に概要を示した本発明の精神から逸脱することなく行なわれ得ることを理解されたい。
【技術分野】
【0001】
この出願は、2003年3月5日に出願された表題「Manipulation of plant senescence using an MYB gene promoter and cytokinin biosynthesis gene」の米国特許出願第10/363,723号の一部継続であり、米国特許出願第10/363,723号は、2001年8月30日に出願された表題「Manipulation of Plant Senescence Using An MYB Gene Promoter and Cytokinin Biosynthesis Genes」の国際特許出願PCT/AU01/01092からの優先権を主張し、国際特許出願PCT/AU01/01092は、2000年9月6日に出願された表題「Manipulation of Plant Senescence Using An MYB Gene Promoter and Cytokinin Biosynthesis Genes」の豪国特許出願PQ9946からの優先権を主張する。これらの内容は、それらの全体が参考として本明細書に援用される。
【0002】
本発明は、植物の老化を操作する方法に関する。また、本発明は、かかる方法に有用なベクター、変更された老化特性を有する形質転換植物、ならびに植物細胞、種子およびかかる植物の他の部分に関する。
【背景技術】
【0003】
葉の老化は、細胞死の前に細胞の代謝的および構造的変化を伴う。また、活発に生長している領域への栄養素の再循環も伴う。
【0004】
サイトカイニンによる植物および植物器官の老化の調節は、農業上重要な成果をもたらす。葉内のサイトカイニンレベルの上昇は、老化を遅延させる傾向がある。その産物(イソペンテニルトランスフェラーゼ)がサイトカイニン合成の重要な段階を触媒するipt遺伝子の発現を調節するために、いくつかのプロモーターが使用されている。しかしながら、一般に、ipt遺伝子を過剰発現するトランスジェニック植物は、地下部および苗条の生長の遅延、地下部の無形成、頂芽優性の低下、および葉面積の減少を有することが報告されている。
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明の目的は、先行技術に伴う問題点または欠点の1つ以上を解決または軽減することである。
【課題を解決するための手段】
【0006】
一態様において、本発明は、植物に、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたは機能的に活性な断片またはその改変体を含む遺伝子構築物を導入することを含む、植物の老化を操作する方法を提供する。
【0007】
老化の操作は、植物および/または特定の植物の器官に関連するものである。種々の植物の器官、例えば、葉、地下部、苗条、茎、塊茎、花、走根、および果実などの老化が操作され得る。植物および植物器官の老化の操作は、例えば、園芸製品および切り花における果実、花、葉および塊茎の貯蔵寿命の増大、園芸作物の腐り易さの低減、老化遅延葉内の炭素固定の増大による収穫率の増大、飼料植物におけるバイオマス生産の向上、採種の向上などの農業上重要な成果をもたらし得る。
【0008】
「老化の操作」は、一般的に、非形質転換対照植物と比べて形質転換植物における老化の遅滞に関する。しかしながら、一部の適用用途では、植物の老化を促進あるいは変更することが望ましい場合があり得る。老化は、例えば、アンチセンス遺伝子を利用することにより促進あるいは変更され得る。
【0009】
有効量の前記遺伝子構築物は、任意の適当な手法、例えば、形質導入、トランスフェクションまたは形質転換によって植物に導入され得る。「有効量」とは、前記植物、または該植物に由来する植物、植物種子もしくは他の植物部分において同定可能な表現型形質がもたらされるのに充分な量を意図する。かかる量は、適切な当業者が植物の種類、投与経路および他の関連要素を考慮することによって容易に決定され得る。かかる当業者は、適当な量および投与方法を容易に決定することができよう。例えば、Maniatisら,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory,Cold Spring Harbor(その全開示内容は、引用により本明細書に組み込まれる)を参照のこと。
【0010】
「修飾myb遺伝子プロモーター」とは、通常myb遺伝子と関連しているプロモーターであって、前記プロモーター内の1つ以上の地下部特異的モチーフおよび/または花粉特異的モチーフが欠失もしくは不活化されるように修飾されたプロモーターを意図する。
【0011】
本出願人は理論に拘束されることを望まないが、前記myb遺伝子プロモーター内の1つ以上の地下部特異的モチーフの欠失もしくは不活化により、一部の植物種で地下部の発育に影響し得る植物の成長点におけるサイトカイニン生合成酵素をコードする遺伝子の基底(leaky)発現の問題が軽減または解決され得ると仮定する。また、前記myb遺伝子プロモーター内の1つ以上の花粉特異的モチーフの欠失もしくは不活化により、一部の植物種で花粉の発育に影響し得る花粉におけるサイトカイニン生合成酵素をコードする遺伝子の基底発現の問題が軽減または解決され得ると仮定する。
【0012】
好ましくは、修飾myb遺伝子プロモーターは、修飾myb32遺伝子プロモーターである。好ましくは、修飾myb遺伝子プロモーターはArabidopsis由来のもの、より好ましくはシロイヌナズナ(Arabidopsis thaliana)由来のものである。
【0013】
本発明に従って修飾され得る好適なプロモーターは、Liら,Cloning of three MYB−like genes from Arabidopsis(PGR 99−138)Plant Physiology 121 :313(1999)(その全開示内容は、引用により本明細書に組み込まれる)に記載されたものである。
【0014】
「地下部特異的モチーフ」とは、植物の地下部内の関連遺伝子の発現を指令する3〜7ヌクレオチド、好ましくは4〜6ヌクレオチド、より好ましくは5ヌクレオチドの配列を意図する。
【0015】
好ましくは、地下部特異的モチーフは、コンセンサス配列ATATTまたはAATATを含む。
【0016】
好ましくは、前記myb遺伝子プロモーター内の1〜10個、より好ましくは3〜8個、さらにより好ましくは5〜7個の地下部特異的モチーフを欠失もしくは不活化させ、好ましくは欠失させる。
【0017】
地下部特異的モチーフは、個々のモチーフを取り出すこと、または1つ以上のモチーフを含む該プロモーターの断片を取り出すことにより欠失させ得る。例えば、シロイヌナズナmyb遺伝子プロモーターのヌクレオチド1〜530、好ましくはヌクレオチド110〜530の領域の全部または一部を欠失させ得る。
【0018】
該欠失は、例えば制限エンドヌクレアーゼで核酸を切断し、切断末端をリゲートし、取り出した断片を有するプロモーターを作製することにより行なわれ得る。
【0019】
例えば、修飾シロイヌナズナmyb遺伝子プロモーターは、162〜176位のXcmI部位と520〜525位のSspI部位の間の断片を取り出すことにより作製され得る。これにより、7つの地下部特異的モチーフのうち6つが欠失した修飾myb遺伝子プロモーターが作製される。あるいはまた、例えば、520〜525位のSspI部位の上流の領域を欠失させること、またはヌクレオチド1〜120の領域を、162〜176位のXcmI部位と520〜525位のSspI部位の間の領域とともに欠失させることにより、7つの地下部特異的モチーフすべてを欠失させ得る。
【0020】
地下部特異的モチーフは、該モチーフ内の1つ以上のヌクレオチドを、好ましいコンセンサス配列を有しなくなるように付加、欠失、置換または誘導体化することにより不活化させ得る。
【0021】
好ましくは、修飾myb遺伝子プロモーターは、添付の図2、3および4に示す配列(それぞれ、配列番号2、3および4)からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む。
【0022】
「花粉特異的モチーフ」とは、植物の花粉内の関連遺伝子の発現を指令する3〜7ヌクレオチド、好ましくは4〜6ヌクレオチド、より好ましくは4または5ヌクレオチドの配列を意図する。
【0023】
好ましくは、花粉特異的モチーフは、TTCTおよびAGAAからなる群より選択されるコンセンサス配列を含む。
【0024】
好ましくは、前記myb遺伝子プロモーター内の1〜30個、より好ましくは3〜15個、さらにより好ましくは4〜10個の花粉特異的モチーフを欠失もしくは不活化させ、好ましくは欠失させる。
【0025】
花粉特異的モチーフは、個々のモチーフを取り出すこと、または1つ以上のモチーフを含む該プロモーターの断片を取り出すことにより欠失させ得る。例えば、ヌクレオチド1〜540、好ましくはヌクレオチド390〜540のシロイヌナズナmyb遺伝子プロモーターの領域の全部または一部を欠失させ得る。
【0026】
該欠失は、例えば制限エンドヌクレアーゼで核酸を切断し、切断末端をリゲートし、取り出した断片を有するプロモーターを作製することにより行なわれ得る。
【0027】
例えば、修飾シロイヌナズナmyb遺伝子プロモーターは、162〜176位のXcmI部位と520〜525位のSspI部位の間の断片を取り出すことにより作製され得る。これにより、23個の花粉特異的モチーフのうち4個が欠失した修飾myb遺伝子プロモーターが作製される。あるいはまた、例えば、520〜525位のSspI部位の上流の領域を欠失させることにより10個の花粉特異的モチーフを欠失させ得る。
【0028】
花粉特異的モチーフは、該モチーフ内の1つ以上のヌクレオチドを、好ましいコンセンサス配列がなくなるように付加、欠失、置換または誘導体化することにより不活化させ得る。
【0029】
好ましくは、修飾myb遺伝子プロモーターは、添付の図2、3および4に示す配列(それぞれ、配列番号2、3および4)からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む。
【0030】
本発明のさらなる態様において、植物に、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結されたmyb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む遺伝子構築物を導入することを含む、植物のバイオマスを増大させる方法が提供される。
【0031】
myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体は、完全長のmyb遺伝子プロモーターまたは修飾myb遺伝子プロモーターであり得る。
【0032】
完全長のmyb遺伝子プロモーターは、myb32遺伝子プロモーターであり得る。好ましくは、myb遺伝子プロモーターはArabidopsis由来のもの、より好ましくはシロイヌナズナ由来のものである。最も好ましくはmyb遺伝子プロモーターは、添付の図1に示す配列(配列番号1)からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む。
【0033】
好適なプロモーターは、Liら,Cloning of three MYB−like genes from Arabidopsis(PGR 99−138)Plant Physiology 121 :313(1999)に記載されたものである。
【0034】
修飾myb遺伝子プロモーターは、本明細書において前述の修飾myb遺伝子プロモーターであり得る。
【0035】
「バイオマスの向上」とは、非形質転換対照植物と比べて形質転換植物における、総葉面積、累積葉面積、葉の生長力学(すなわち、経時的な葉の数)、走根長さ、および花1つまたは種蒔き面積あたりの種子収量からなる群より選択される生長特性の向上または増大を意図する。また、「バイオマスの向上」は、非形質転換対照植物と比べて形質転換植物における走根死の割合の低下または減少を包含する。
【0036】
特に、本発明によるトランスジェニック植物の種子重量(すなわち、千個の種子の重量)は非トランスジェニック対照植物と識別不可能であるが、同等の開花強度の非トランスジェニック対照植物と比較すると、花1つまたは種蒔き面積あたりの基準で示す総種子収量は、トランスジェニック植物より有意に高いことを、本出願人らは見い出した。
【0037】
myb遺伝子プロモーターまたは修飾myb遺伝子プロモーターに関連する「機能的に活性な」とは、その断片または改変体(類縁体、誘導体もしくは変異型など)が、本発明の方法による植物の老化の操作能を有することを意図する。かかる改変体には、天然に存在する対立遺伝子改変体および天然に存在しない改変体が包含される。1つ以上のヌクレオチドの付加、欠失、置換および誘導体化が想定されるが、修飾によって該断片または改変体の機能的活性の低下がもたらさないものとする。
【0038】
好ましくは、機能的に活性な断片または改変体は、該断片または改変体に相当する上記配列の関連部分と少なくともほぼ80%の同一性を有し、より好ましくは少なくともほぼ90%の同一性、最も好ましくは少なくともほぼ95%の同一性を有する。好ましくは、該断片は、少なくとも20ヌクレオチド、より好ましくは少なくとも50ヌクレオチド、より好ましくは少なくとも100ヌクレオチド、より好ましくは少なくとも200ヌクレオチド、最も好ましくは少なくとも300ヌクレオチドのサイズを有する。
【0039】
「サイトカイニンの生合成に関与している酵素をコードする遺伝子」とは、カイネチン、ゼアチンおよびベンジルアデニンなどのサイトカインの合成に関与している酵素をコードする遺伝子、例えば、イソペンチルトランスフェラーゼ(ipt)をコードする遺伝子、またはsho遺伝子(例えば、Petunia由来のもの)などのipt様遺伝子を意図する。好ましくは、該遺伝子は、イソペンテニルトランスフェラーゼ(ipt)遺伝子またはsho遺伝子である。好ましい実施形態において、該遺伝子は、Agrobacterium、より好ましくはAgrobacterium tumefaciens;Lotus、より好ましくはミヤコグサ(Lotus japonicus);およびPetunia、より好ましくはツクバネアサガオ(Petunia hybrida)からなる群より選択される種に由来するものである。
【0040】
最も好ましくは、該遺伝子は、添付の図6、8および10に示す配列(配列番号5、7および9)、添付の図7、9および11に示すポリペプチドをコードする配列(配列番号6、8および10)からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む。
【0041】
サイトカイニン生合成酵素をコードする遺伝子に関連する「機能的に活性な」とは、その断片または改変体(類縁体、誘導体または変異型など)が、本発明の方法による植物の老化の操作能を有することを意図する。かかる改変体には、天然に存在する対立遺伝子改変体および天然に存在しない改変体が包含される。ヌクレオチドの1つ以上の付加、欠失、置換および誘導体化が想定されるが、該修飾によって該断片または改変体の機能的活性の低下をもたらさないものとする。好ましくは、機能的に活性な断片または改変体は、該断片または改変体に相当する上記配列の関連部分と少なくともほぼ80%の同一性を有し、より好ましくは少なくともほぼ90%の同一性、最も好ましくは少なくともほぼ95%の同一性を有する。かかる機能的に活性な改変体および断片としては、例えば、同類核酸変化、または対応するアミノ酸配列内の1つ以上の残基の同類アミノ酸置換をもたらす核酸変化を有するものが挙げられる。例えば、機能的に活性な改変体は、図6、8または10に示す配列の1つ以上の同類核酸置換を含むものであり得、得られる機能的に活性な改変体は、それぞれ図7、9または11に示すアミノ酸配列をコードする。好ましくは、該断片は、少なくとも20ヌクレオチド、より好ましくは少なくとも50ヌクレオチド、より好ましくは少なくとも100ヌクレオチド、より好ましくは少なくとも500ヌクレオチドのサイズを有する。
【0042】
遺伝子構築物は植物に、任意の適当な手法によって導入され得る。本発明の遺伝子構築物を植物細胞内に組み込むための手法(例えば、形質導入、トランスフェクションまたは形質転換)は、当業者によく知られたものである。かかる手法としては、アグロバクテリウム媒介型誘導、組織、細胞およびプロトプラストへのエレクトロポレーション、プロトプラスト融合、生殖器官内への注入、未熟胚内への注入ならびに細胞、組織、カルス、未熟および成熟胚に対する高速プロジェクタイル誘導、微粒子銃形質転換ならびにその組合せが挙げられる。手法の選択は、形質転換対象の植物の種類に大きく依存し、適切な当業者によって容易に決定され得る。
【0043】
本発明の遺伝子構築物が組み込まれる細胞は、後述するようにして選択し、次いで、当該技術分野でよく知られた手法を用いて適切な培地中で培養すると、形質転換植物が再生し得る。例えば温度、pHなどの培養条件は当業者には自明であろう。得られた植物は、当該技術分野でよく知られた方法を用い、有性生殖または無性生殖のいずれかによって形質転換植物の後続世代を生成してもよい。
【0044】
本発明の方法は、さまざまな植物、例えば、単子葉[草(例えば、牧草、芝生およびバイオエネルギー草、例えば、ペレニアルライグラス、トールフェスク、イタリアンライグラス、レッドフェスキュー、リードカナリーグラス、ビッグブルーステム、コードグラス、ネピアグラス、野生ライムギ、野生サトウキビ、ススキ属(Miscanthus))、トウモロコシ、オートムギ、小麦および大麦)など]、双子葉[Arabidopsis、タバコ、大豆、クローバ(例えば、シロツメクサ、ムラサキツメクサ、サブタレニアンクローバ)、アルファルファ、アブラナ、アブラナ科野菜、レタス、ホウレンソウなど]および裸子植物に適用され得る。
【0045】
本発明のさらなる態様において、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む植物の老化を操作することができるベクターが提供される。
【0046】
本発明のまたさらなる態様において、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結されたmyb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む、植物のバイオマスを増大させることができるベクターが提供される。
【0047】
myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体は、本明細書において上記の完全長のmyb遺伝子プロモーターまたは修飾myb遺伝子プロモーターであり得る。
【0048】
本発明のこの態様の好ましい実施形態において、ベクターは、さらにターミネーターを含むものであり得、前記プロモーター、遺伝子およびターミネーターは、作動可能に連結されている。
【0049】
「作動可能に連結されている」とは、前記プロモーターが植物細胞において前記遺伝子の発現をもたらし得ること、および前記ターミネーターが植物細胞において前記遺伝子の発現を終了させ得ることを意図する。好ましくは、前記プロモーターは前記遺伝子の上流であり、前記ターミネーターは前記遺伝子の下流である。
【0050】
ベクターは、任意の適当な型のものであり得、ウイルス系または非ウイルス系であり得る。ベクターは、発現ベクターであり得る。かかるベクターとしては、染色体、非染色体および合成の核酸配列、例えば、植物ウイルスの誘導体;細菌系プラスミド;Agrobacterium tumefaciens由来のTiプラスミドの誘導体;Agrobacterium rhizogenes由来のRiプラスミドの誘導体;ファージDNA;酵母人工染色体;細菌系人工染色体;バイナリー細菌系人工染色体;プラスミドとファージDNAの組合せから誘導したベクターが挙げられる。しかしながら、植物細胞内で複製可能または組込み可能またはバイアブルである限り、任意の他のベクターが使用され得る。
【0051】
プロモーター、遺伝子およびターミネーターは、任意の適当な型のものであり得、標的植物細胞に対して内在性であってもよく、外来性であってもよいが、標的植物細胞において機能性であるものとする。
【0052】
また、本発明のベクターに使用され得るさまざまなターミネーターは、当業者に充分知られている。ターミネーターは、プロモーター配列と同じ遺伝子由来のものであってよく、異なる遺伝子由来のものであってもよい。特に好適なターミネーターは、ポリアデニル化シグナル(CaMV 35S ポリAなど)ならびにノパリンシンターゼ(nos)遺伝子およびオクトピンシンターゼ(ocs)遺伝子由来の他のターミネーターである。
【0053】
ベクターには、プロモーター、遺伝子およびターミネーターに加えて、さらに、遺伝子の発現に必要なエレメント、例えば、ベクターバックボーン、複製起点(ori)、マルチクローニング部位、スペーサー配列、エンハンサー、イントロン(トウモロコシユビキチンUbiイントロンなど)、抗生物質耐性遺伝子および他の選択可能マーカー遺伝子[ネオマイシンホスホトランスフェラーゼ(nptll)遺伝子、ハイグロマイシンホスホトランスフェラーゼ(hph)遺伝子、ホスフィノトリシンアセチルトランスフェラーゼ(barまたはpat)遺伝子など]、ならびにレポーター遺伝子(β−グルクロニダーゼ(GUS)遺伝子(gusA)など]が種々の組合せで含まれることがあり得る。また、ベクターには、翻訳開始のためのリボソーム結合部位を含めてもよい。また、ベクターには、発現の増幅のための適切な配列を含めてもよい。
【0054】
形質転換宿主細胞選択のための表現型形質をもたらすのに選択可能マーカー遺伝子を使用する択一法として、形質転換細胞内のベクターの存在を、当該技術分野でよく知られた他の手法、例えば、PCR(ポリメラーゼ連鎖反応)、サザンブロットハイブリダイゼーション解析、組織化学的アッセイ(例えば、GUSアッセイ)、薄層クロマトグラフィー(TLC)、ノーザンおよびウエスタンブロットハイブリダイゼーション解析などによって調べてもよい。
【0055】
当業者には、ベクターの種々の成分は、前記遺伝子の発現がもたらされるように作動可能に連結されていることが認識されよう。本発明のベクターの成分を作動可能に連結させるための手法は、当業者には充分知られている。かかる手法としては、リンカー(例えば、1つ以上の制限酵素部位を含む合成リンカーなど)の使用が挙げられる。
【0056】
本発明のさらなる態様において、変更された老化特性または向上したバイオマスを有するトランスジェニック植物細胞、植物、植物種子、または他の植物部分が提供される。好ましくは、前記植物細胞、植物、植物種子、または他の植物部分は、本発明によるベクターを含む。好ましくは、トランスジェニック植物細胞、植物、植物種子、または他の植物部分は、本発明による方法によって生成されるものである。
【0057】
また、本発明は、本発明の植物細胞由来のトランスジェニック植物、植物種子、または他の植物部分を提供する。
【0058】
また、本発明は、本発明の植物由来のトランスジェニック植物、植物種子、または他の植物部分を提供する。
【0059】
次に、本発明を、以下の実施例および添付の図面に関して、より充分に説明する。しかしながら、以下の説明は例示にすぎず、上記の本発明の一般説明をなんら制限するものと解釈されるべきでないことを理解されたい。
【図面の簡単な説明】
【0060】
【図1】図1は、シロイヌナズナのmyb32遺伝子(atmyb32)由来のプロモーターのヌクレオチド配列であるAtmyb32プロモーター配列を示し(配列番号1)、MYB型、花粉特異的および地下部特異的モチーフを強調表示している。WAACCA(下線/イタリック体)MYB1AT;GTTAGTT(太字/線で囲んだ範囲)MYB1LEPR;CCWACC(線で囲んだ範囲)MYBPZM;GGATA(イタリック体)MYBST1;AGAAA(下線)花粉1LELAT52;ATATT(太字)地下部モチーフAPOX1。
【図2】図2は、XcmI−SspI植物配列欠失型Atmyb32プロモーター配列改変体(Atmyb32xs)を示す。MYB型、花粉特異的および地下部特異的モチーフを強調表示している。WAACCA(下線/イタリック体)MYB1AT;GTTAGTT(太字/線で囲んだ範囲)MYB1LEPR;CCWACC(線で囲んだ範囲)MYBPZM;AGAAA(下線)花粉1LELAT52;ATATT(太字)地下部モチーフAPOX1(配列番号2)。
【図3】図3は、すべての地下部モチーフを欠失させたAtmyb32プロモーター改変体配列を示す。MYB型、花粉特異的および地下部特異的モチーフを強調表示している。WAACCA(下線/イタリック体)MYB1AT;CCWACC(線で囲んだ範囲)MYBPZM;AGAAA(下線)花粉1LELAT52(配列番号3)。
【図4】図4は、SspI部位上流配列を欠失させたAtmyb32プロモーター改変体配列を示す。MYB型、花粉特異的および地下部特異的モチーフを強調表示している。WAACCA(下線/イタリック体)MYB1AT;CCWACC(線で囲んだ範囲)MYBPZM;AGAAA(下線)花粉1LELAT52(配列番号4)。
【図5】図5は、Atmyb32プロモーター配列およびAtmyb32xsプロモーター配列内のモチーフを示す。
【図6】図6は、Agrobacterium tumefaciens由来のイソペンテニルトランスフェラーゼ(ipt)遺伝子のヌクレオチド配列(配列番号5)を示す。
【図7】図7は、Agrobacterium tumefaciens由来のイソペンチルトランスフェラーゼ遺伝子の推定アミノ酸配列(配列番号6)を示す。
【図8】図8は、ミヤコグサ由来のイソペンチルトランスフェラーゼ遺伝子のヌクレオチド配列(配列番号7)を示す。
【図9】図9は、ミヤコグサ由来のイソペンチルトランスフェラーゼ遺伝子の推定アミノ酸配列(配列番号8)を示す。
【図10】図10は、ツクバネアサガオ由来のサイトカイニン生合成Sho遺伝子のヌクレオチド配列(配列番号9)を示す。
【図11】図11は、ツクバネアサガオ由来のサイトカイニン生合成Sho遺伝子の推定アミノ酸配列(配列番号10)を示す。
【図12】図12は、atmyb32::iptトランスジェニックシロツメクサ(Trifolium repens)植物のPCRおよびサザンDNA解析を示す。a)制限酵素部位およびサザンハイブリダイゼーション解析に使用されたプローブの位置を示すpatmyb32:iptのT−DNA領域、b)PCR増幅させた599bp nptll産物および583bp ipt産物の臭化エチジウム染色1%アガロースゲル、c)iptプローブとハイブリダイズさせたPCR陽性シロツメクサ植物から単離したHindIII消化全ゲノムDNAとのサザンブロットハイブリダイゼーション、d)nptllプローブとハイブリダイズさせたPCR陽性シロツメクサ植物から単離したHindIII消化全ゲノムDNAとのサザンブロットハイブリダイゼーション。レーン1〜2:2つの独立したカナマイシン耐性cv.Haifa 再生体(regenerant)、コード:それぞれ、Hmi01、Hmi08;レーン3〜12:12個の独立したカナマイシン耐性cv.Irrigation再生体、コード:それぞれ、Imi06、Imi07、Imi08、Imi09、Imi10、Imi11、Imi12、Imi14、Imi16、Imi18;レーンC:非形質転換シロツメクサ;レーンP:陽性対照プラスミドpatmyb32ipt。
【図13】図13は、atmyb32::iptトランスジェニックシロツメクサ(T.repens)植物におけるipt mRNA発現のRT−PCR解析を示す。レーン1〜11は、図4.8の場合の対応植物コードを有する11種類の独立したトランスジェニック系統由来の試料である;レーンC、対照非形質転換植物;レーンP、陽性対照としてのプラスミド。全RNAを葉組織から単離した。全RNA(13μg)を各逆転写反応に使用し、RT産物の1/5をPCRによって増幅させた。右側のゲル上のDNA産物は、2×30サイクルの集中的(intensive)PCRによって増幅させた。交互のレーンに負荷した対応RT−PCR反応液には、逆転写酵素は添加しなかった。
【図14】図14は、atmyb32::iptトランスジェニックシロツメクサ(T.repens)植物から切除した葉の老化バイオアッセイを示す。各植物系統から少なくとも30枚の葉を、各系統の走根上の類似した位置から収集した。A.切除した葉の総数に対する分数としての黄変葉の数。B.光の下で水上に2週間維持した葉の典型的な外観。植物系統に対するキー:それぞれ、HC、ICおよびHmg、Img、非形質転換およびatmyb32::gusAトランスジェニック植物(cv.HaifaおよびIrrigation);01および08、atmyb32::iptトランスジェニックHaifa系統は、それぞれ、Hmi01およびHmi08;11、12、16および18atmyb32::iptトランスジェニックIrrigation系統は、それぞれ、Imi11、Imi12、Imi16およびImi18。
【図15】図15は、対照植物(左)と比較したatmyb32::iptトランスジェニックシロツメクサ(T.repens)(右)植物における、A)一般的な植物の形態、B)正常な苗条の発育、およびC)正常な地下部の発育を示す。
【図16】図16は、pBMVkAtMYB32−900::ipt[遺伝子:イソペンチルトランスフェラーゼ(IPT);ベクター:pBMVkAtMYB32−900::ipt−nos(バックボーンpPZPRCS2);選択可能マーカー:spec;植物選択可能マーカーカセット:35S::kan::35ST;遺伝子プロモーター:AtMYB32−900;遺伝子ターミネーター:nos.]のベクターの詳細を示す。
【図17】図17は、ベクターpBMVkAtMYB32−900::ipt−nosのヌクレオチド配列(配列番号11)を示す。
【図18】図18は、pBMVkAtMYB32xs::ipt−nos[遺伝子:イソペンチルトランスフェラーゼ(IPT);ベクター:pBMVkAtMYB32XS::ipt−nos(バックボーンpPZPRCS2);選択可能マーカー:spec;植物選択可能マーカーカセット:35S::kan::35ST;遺伝子プロモーター:AtMYB32−xs;遺伝子ターミネーター:nos.]のベクターの詳細を示す。
【図19】図19は、ベクターpBMVkAtMYB32xs::ipt−nosのヌクレオチド配列(配列番号12)を示す。
【図20】図20は、pBMVhAtMYB32−900::ipt−nos[遺伝子:イソペンチルトランスフェラーゼ(IPT);ベクター:pBMVhAtMYB32−900::ipt−nos(バックボーンpPZPRCS2);選択可能マーカー:spec;植物選択可能マーカーカセット:35S::hph::35ST;遺伝子プロモーター:AtMYB32−900;遺伝子ターミネーター:nos.]のベクターの詳細を示す。
【図21】図21は、ベクターpBMVhAtMYB32−900::ipt−nosのヌクレオチド配列(配列番号13)を示す。
【図22】図22は、pBMVhAtMYB32xs::ipt−nos[遺伝子:イソペンチルトランスフェラーゼ(IPT);ベクター:pBMVhAtMYB32XS::ipt−nos(バックボーンpPZPRCS2);選択可能マーカー:spec;植物選択可能マーカーカセット:35S::hph::35ST;遺伝子プロモーター:AtMYB32−xs;遺伝子ターミネーター:nos.]のベクターの詳細を示す。
【図23】図23は、ベクターpBMVhAtMYB32xs::ipt−nosのヌクレオチド配列(配列番号14)を示す。
【図24】図24は、pBSubn−AtMYB32−900::ipt−nos[遺伝子:イソペンチルトランスフェラーゼ(IPT);ベクター:pBSubn−AtMYB32−900::ipt−nos(バックボーンpPZPRCS2);選択可能マーカー:spec;植物選択可能マーカーカセット:Ubi::bar::nos;遺伝子プロモーター:AtMYB32−900;遺伝子ターミネーター:nos.]のベクターの詳細を示す。
【図25】図25は、ベクターpBSAtMYB32900::ipt−nosのヌクレオチド配列(配列番号15)を示す。
【図26】図26は、pBMVhATMYB3−900::ipt−nosおよびpBMVhATMYB32xs::ipt−nosを含むトランスジェニックアブラナの生成を示す。A.アブラナ種子をインビトロで発芽させる;B.胚軸部を7日齢の苗木から切除し、アグロバクテリウム懸濁液を接種する;CおよびD.ハイグロマイシン選択下、接種胚軸部からの再生 ;E〜J.pBMVhATMYB3−900::ipt−nosおよびpBMVhATMYB32xs::ipt−nosベクターを有するトランスジェニックT0アブラナ植物。
【図27】図27は、小麦の微粒子銃形質転換のためのプロセスを示す。
【図28】図28は、小麦(Triticum aestivum L.MPB Bobwhite 26)の微粒子銃形質転換を示す。ドナー植物生産(AおよびB);接合子胚単離(CおよびD);グルホシネート選択下での再生(E〜G);選択下での地下部形成(H);トランスジェニック子孫の回収のための閉鎖温室条件下でのT0植物栽培(I)。
【図29】図29は、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物の隔離圃場試験を示す。
【図30】図30は、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物と、非トランスジェニック対照シロツメクサ植物での生長速度と生長力学の比較評価を示す。A)生長速度 B)生長力学 C)生長特性(45日後) 白抜きの四角 トランスジェニックシロツメクサ 黒塗りの四角 非トランスジェニック対照シロツメクサ (明) (暗)
【図31】図31は、隔離圃場条件下での、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12、LXR 18およびLXR 11)と、非トランスジェニック対照シロツメクサ植物(すなわち、WT)の開花強度(すなわち、1m2あたりの成熟花の数)を示す。
【図32】図32は、隔離圃場条件下での、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12、LXR 18およびLXR 11)と、非トランスジェニック対照シロツメクサ植物(すなわち、WT)の種子重量(すなわち、千個の種子の重量、単位:グラム)を示す。
【図33】図33は、隔離圃場条件下での、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12、LXR 18およびLXR 11)と、非トランスジェニック対照シロツメクサ植物(すなわち、WT)の花1つあたりの種子収量(単位:ミリグラム)を示す。
【図34】図34は、隔離圃場条件下での、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12、LXR 18およびLXR 11)と、非トランスジェニック対照シロツメクサ植物(すなわち、WT)の単位面積あたりの種子収量(単位:kg/ヘクタール)を示す。
【図35】図35は、キメラpBMVkATMYB3−900::ipt−nosおよびpBMVkATMYB32xs::ipt−nos遺伝子を含むトランスジェニックアルファルファ植物の生成を示す。A.アルファルファクローンC2−3、C2−4および19−17由来の葉柄外植片をアグロバクテリウム懸濁液の接種に使用し、カナマイシンの存在下での選択後、形質転換胚形成性カルスの発生をもたらす;B〜D.インビトロで培養した体細胞胚からの、pBMVkATMYB3−900::ipt−nosおよびpBMVkATMYB32xs::ipt−nosベクター由来のキメラ遺伝子を有するトランスジェニックアルファルファ小植物の再生。
【図36】図36は、トランスジェニックアブラナ植物(T1 LXRアブラナ植物Line 4)のPCR解析を示す。ゲノムDNAは、T1 LXR04系統の異なるトランスジェニックアブラナ植物から単離し、A.選択可能マーカー(hph)またはB.目的の候補遺伝子(IPT)に特異的なプライマーを用いてPCRに供した。
【図37】図37は、T1トランスジェニックアブラナ植物におけるIPT遺伝子の発現解析を示す(T1 LXRアブラナの葉での相対IPT発現)。
【図38】図38は、切り取って7日後、野生型対照子葉と比較したときの、切り取り子葉の老化の遅延を示すトランスジェニックアブラナを示す。
【図39】図39は、切り取って7日後のT1トランスジェニックアブラナ子葉の老化スコアを示す。
【0061】
【化1】
【図40】図40は、切り取って14日後、野生型対照子葉と比較したときの、切り取り若幼葉の老化の遅延を示すトランスジェニックアブラナを示す。
【図41】図41は、切り取って14日後、T1トランスジェニックアブラナの第1の若幼葉の老化スコアを示す。
【0062】
【化2】
【図42】図42は、トランスジェニック小麦系統のサザンハイブリダイゼーション解析を示す。レーンは、MW−分子量;WT−野生型;トランスジェニック小麦系統LXR3、LXR4、LXR13、LXR16ならびに10pgおよび20pgを含む陽性対照プラスミドを含む。
【図43】図43は、独立したT1トランスジェニック小麦系統の発現解析(小麦におけるIPT定量的発現)を示す。定量的転写物の値は、標的配列を含むプラスミドDNAで得た標準曲線を用いて、フェトモル(fmol)/マイクログラムRNAで測定した。試料は、候補遺伝子IPTに対して高、中および低クラスの発現を表す。
【図44】図44は、温室栽培T1トランスジェニック小麦植物の表現型多様性を示す。A.ヌル対照小麦植物と比較したとき、正常表現型を示すT1 LXR 13小麦植物。B.ヌル対照小麦植物と比較したとき、成長阻害表現型および止め葉数の増加を示すT1 LXR 04小麦植物。
【図45】図45は、切り取って7日後、ヌル対照葉と比較したときの、葉の老化の遅延を示すトランスジェニック小麦を示す。
【発明を実施するための形態】
【0063】
実施例
実施例1
Atmyb32プロモーター配列およびプロモーター配列改変体
Atmybb32プロモーター配列およびその改変体を図1〜4に示す。
【0064】
実施例2
サイトカイニン生合成遺伝子
本発明における使用に適したサイトカイニン生合成遺伝子の例を、図6、8および10に示す。また、好適な遺伝子としては、図7、9および11に示すポリペプチドをコードするものが挙げられる。
【0065】
実施例3
トランスジェニックシロツメクサ植物の生成
トランスジェニックシロツメクサ植物(Trifolium repens cv.HaifaおよびIrrigation)を、キメラatmyb32::ipt遺伝子バイナリーベクター(図12a)を使用し、アグロバクテリウム媒介型形質転換によって生成した。トランスジェニック植物を、iptプライマーおよびnptllプライマー(図12b)を用いてPCRによってスクリーニングした。HindIII消化ゲノムDNA試料をサザンDNAハイブリダイゼーション解析に供すると、iptプローブとnptllプローブの両方によって、4.4kbより大きいDNA断片がすべてのレーンに検出されることが示され、これは、完全長T−DNAの存在およびそのシロツメクサゲノム内への組込みを示す(図12)。トランスジェニック系統Hmi01、Imi06、Imi11およびImi18(それぞれ、レーン1、3、5、8および12)は、ゲノム内に組み込まれた単一コピーの完全長T−DNAを有するようであった。他のトランスジェニック系統は、多コピーのatmyb32::ipt導入遺伝子を有した。
【0066】
実施例4
トランスジェニックシロツメクサ植物におけるIPT遺伝子発現
トランスジェニックシロツメクサ(T.repens)植物におけるatmyb32::ipt導入遺伝子の発現を、RT−PCRによって評価した。ipt mRNAは、調べたatmyb32::iptトランスジェニックシロツメクサ植物すべての葉組織内で検出され、種々のレベルのPCR産物が検出された(図13)。
【0067】
実施例5
トランスジェニックシロツメクサ植物における切り取り葉の老化の遅延
atmyb32::iptトランスジェニック植物の切り取り葉の老化を評価するための実験を行なった。1週間以内に、両方の品種の非形質転換型およびatmyb32::gusAトランスジェニックシロツメクサ植物から切り取られた葉において、急速な黄変が観察された。トランスジェニック系統Hmi01、Hmi08、Imi16およびImi18では、老化の遅延が示されたが、Imi11とImi12では、7日間の最後までに黄変の兆候は示されなかった。2週間後、すべてのatmyb32::iptトランスジェニック植物の葉は、非形質転換型およびatmyb32::gusA対照トランスジェニック植物のものよりもずっと緑色であった(図14)。切除した葉における老化の程度は、cv.Haifaでは、HC、Hmg>Hmi01>Hmi08、ならびにcv.Irrigationでは、ICおよびImg>Imi16>Imi18>Imi11およびImi12の順であった。HCは、Haifa非形質転換対照であり、Hmgは、Haifa atmyb32::gusA対照であり、ICは、Irrigation非形質転換対照であり、Imgは、Irrigation atmyb32::gusA対照である。Hmi01、Hmi08、Imi16、Imi18、Imi11およびImi12は、それぞれ、品種Haifa(H)およびIrrigation(I)に由来する独立したatmyb32::iptトランスジェニックシロツメクサ植物である。
【0068】
実施例6
トランスジェニックシロツメクサ植物における植物の形態と地下部の発育
正常な植物の形態ならびに正常な苗条の発育と正常な地下部の発育が、atmyb32:iptトランスジェニックシロツメクサ植物において観察され(図6)、したがって、atmyb32プロモーターの制御下でのipt遺伝子の発現の調節は、トランスジェニックシロツメクサ植物の地下部発育にも頂芽優性にもマイナスの影響を及ぼさないことが示された(表1)。
【0069】
【表1】
分析した10種類の独立したatmyb32::iptトランスジェニックシロツメクサ系統において、正常な植物の形態および正常な地下部発育が観察された。この10種類の独立したatmyb32::iptトランスジェニックシロツメクサ系統の推定ipt遺伝子コピー数を示す。
【0070】
実施例7
植物形質転換用のベクターの作製
植物のアグロバクテリウム媒介型形質転換のため、4種類のバイナリーベクターを作製した(図16〜19)。各ベクターは、pPZP200ベクターバックボーン(Hajdukiewiczら,1994)を有し、キメラAtmyb32−900::ipt−nosまたはAtmyb32−xs::ipt−nosのいずれかを、キメラ35S::nptll−35stまたは35S::hph−35st選択可能マーカーカセットとともに、またはなしで含む。
【0071】
形質転換ベクターの1つは、微粒子銃形質転換のために構築した(図20および21)。この形質転換ベクターは、キメラAtmyb32−900::ipt−35stをキメラUbi::bar−nos選択可能マーカーカセットとともに含む。
【0072】
Atmyb32プロモーター、プロモーター改変体Atmyb32xs、イソペンチルトランスフェラーゼ遺伝子ならびにターミネーター35stおよびnosを、Gateway(商標)(Invitrogen)適合プライマーを用いてPCRによって増幅し、pDONR221エントリーベクター内にクローニングした。続いて、これらを組換えを用いて、慣用的にクローニングされる選択可能マーカーカセットを含むデスティネーションベクター内にクローニングした。すべてのベクターは、厳密な品質保証プロトコルに従って完全に配列決定した。
【0073】
実施例8
アブラナ(Brassica napus)のアグロバクテリウム媒介型形質転換
キメラipt遺伝子を含むバイナリーベクターpBMVhATMYB3−900::ipt−nos(図20)およびpBMVhATMYB32xs::ipt−nos(図22)を、Atmyb32プロモーター(図1)および地下部特異的モチーフを欠失させたAtmyb32xs改変体プロモーター配列(図2)の制御下で、Brassica napus胚軸セグメントのアグロバクテリウム媒介型形質転換に使用した(図26)。
【0074】
Brassica napusの種子を70%エタノール中で2分間表面消毒し、滅菌水中で3回洗浄し、次いで、1%(w/v)の次亜塩素酸カルシウムおよび0.1%(v/v)のTween 20を含有する溶液中で、30分間さらに表面消毒する。種子を滅菌水中で少なくとも3回洗浄し、1×Murashige and Skoog(Murashige and Skoog Physiol.Plant,15:473−497,1962)多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、2%(w/v)のスクロースを補給、pH5.8)を含有し、4g/LのGelriteを添加した固形発芽培地を入れた120ml容の培養容器内に植え付ける。容器を16時間明/8時間暗条件下、25℃で7日間インキュベートして発芽を促進する。
【0075】
7日後、Brassica napusの苗木(苗木全体)を、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、3%(w/v)のスクロースを補給、pH5.8)からなる液体培地に移す。苗木を集め、地下部と子葉を除去した後、胚軸を7〜10mm切片に切断し、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、3%(w/v)のスクロースを補給、pH5.8)からなり、6.4g/lのBacto−Agarで固化したプレコンディショニング培地を入れた9×1.5cmのペトリ皿の上で平板培養する。
【0076】
胚軸部を24時間培養した後、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、100μMのアセトシリンゴン、3%(w/v)のスクロースを補給、pH5.8)からなるアグロバクテリウム懸濁液OD600=0.2を30分間接種する。
【0077】
接種後、胚軸部を滅菌ペーパータオル上にブロットし、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、100μMのアセトシリンゴン、1mg/Lの2,4−D、3%(w/v)のスクロースを補給、pH5.8、8g/lのBacto−Agarで固化)を入れた9×1.5cmのペトリ皿に移す。外植片を16時間明/8時間暗条件下、25℃で72時間インキュベートして共存培養する。
【0078】
共存培養後、20〜30個の胚軸外植片を1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、1mg/Lの2,4−D、3%(w/v)のスクロースを補給、pH5.8)からなり、8g/lのBacto−Agar(250mg/lのチメンチンおよびハイグロマイシン耐性苗条を選択するための10mg/lのハイグロマイシンを補給)で固化した固形選択培地を入れた9×1.5cmのペトリ皿に移す。プレートを16時間明/8時間暗条件下、25℃でインキュベートする。
【0079】
7日後、胚軸外植片を、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、1mg/Lの2,4−D、3%(w/v)のスクロースを補給、pH5.8)からなり、8g/lのBacto−Agar(4mg/lのBAP、2mg/lのゼアチン、5mg/lの硝酸銀、250mg/lのチメンチンおよび10mg/lのハイグロマイシンを補給)で固化した固形再生培地を入れた9×2.0cmのペトリ皿に移す。プレートを、蛍光灯条件(16時間明/8時間暗の光周期;55μmol m−2秒−1)下の直接照明下、25℃で4週間インキュベートして苗条の発育を促進する。
【0080】
再生を毎週モニタリングし、胚軸外植片を、固形再生培地RM(4mg/lのベンジルアデニン、2mg/lのゼアチン、5mg/lの硝酸銀、250mg/lのチメンチンおよび10mg/lのハイグロマイシンを補給)を入れた新たな9×2.0cmのペトリ皿に6〜8週間移し、苗条の発育を促進する。
【0081】
ハイグロマイシン耐性(Hygr)苗条を、1×Murashige and Skoog多量養素、1×微量栄養素およびB5有機ビタミン群(500mg/LのMES、1mg/Lの2,4−D、1%(w/v)のスクロースを補給、pH5.8)からなり、8g/lのBacto−Agar(250mg/lのチメンチンを補給)で固化した固形地下部誘導培地RIM1を入れた120ml容器に移す。苗条を、直接蛍光灯(16時間明/8時間暗の光周期;55μmol m−2秒−1)下、25℃でインキュベートして苗条の伸張と地下部の発育を4〜5週間にわたって促進する。苗条と地下部系の発育を有するHygr苗条をすべて土壌に移し、温室条件下で生長させる。
【0082】
実施例9
小麦(Triticum aestivum L.)の微粒子銃形質転換
キメラipt遺伝子を含む形質転換ベクターを、Atmyb32プロモーター(図1)および地下部特異的モチーフを欠失させたAtmyb32xs改変体プロモーター配列(図2)の制御下で、小麦(Triticum aestivum L.MPB Bobwhite 26)の微粒子銃形質転換に使用した。代表的なベクターを図24に示す。小麦の微粒子銃形質転換の手順の概略を図27に示す。形質転換手順は、以下の工程を含む。
【0083】
工程1(ドナー植物生産):
Triticum aestivum(Bobwhite 26)の種子をドナー植物材料の生産に使用する。小麦植物は、堆肥化したマツ樹皮、パーライトおよびバーミキュライトからなる種苗用(nursery)ミックス中で栽培し、鉢1つあたり植物5本とし、最大鉢サイズを20cmとする。植物をほぼ22〜24℃の温室条件下に12〜16週間維持する(図28A)。止め葉から最初の穂状花序(spike)が現れたら植物にタグを付け、開花12〜15日後に最も高い頭部から胚を収集する。
【0084】
工程2(第1日目)
所望の発育段階の穂状花序を採取する(図28B)。穀果を穂状花序から除去し、0.8%(v/v)のNaOCl溶液中で20分間表面消毒し、滅菌蒸留中で少なくとも4回すすぐ。
【0085】
長さ10mmまでの胚を各穀果から、解剖顕微鏡を用いて無菌的に切除し(軸部の取り出し)、軸面を下にして、2×Murashige and Skoog(1962)多量養素、1×微量栄養素および有機ビタミン群、40mg/Lのチアミン、150mg/LのL−アスパラギン(15%(w/v)のマルトース、0.8%(w/v)のSigma製寒天および2.5mg/Lの2,4−Dを補給)からなる浸透圧培地(E3マルトース)上で培養する(図28CおよびD)。胚を、15mLの培地を入れた60mm×15mmの透明なポリプロピレン製ペトリ皿の上で培養する。培養プレートを24℃で、暗所にて4時間インキュベートした後、ボンバードメントを行なう。胚には、900psiで6cmのBioRad PDS1000遺伝子銃で、1μgのベクタープラスミドDNA(0.6μmの金粒子上に沈殿させたもの)を用いてボンバードメントを行なう。ボンバードメント後、胚を浸透圧培地上で、暗所にて一晩インキュベートする。
【0086】
工程3(第2日目):
胚を、2×Murashige and Skoog(1962)多量養素および1×微量栄養素ならびに有機ビタミン群、40mg/Lのチアミン、150mg/LのL−アスパラギン(6%(w/v)のスクロース、0.8%(w/v)のSigma製寒天および2.5mg/Lの2,4−Dを補給)からなるカルス誘導培地(E3カルス)に移す。胚を24℃で暗所にて2週間培養する。
【0087】
工程4(第16日目):
E3カルス上での2週間の培養後、胚から胚誘導性カルスが発生し、2×Murashige and Skoog(1962)多量養素および1×微量栄養素ならびに有機ビタミン群、40mg/Lのチアミン、150mg/LのL−アスパラギン(2%(w/v)のスクロース、0.8%(w/v)のSigma製寒天、5mg/LのD,Lホスフィノトリシン(PPT)を補給、植物生長調節因子なし)からなる選択培地(E3Select)上に継代培養する(図28E〜G)。培養物を、さらに14日間、E3Select上で24℃にて明るい場所で12時間の光周期でインキュベートする。
【0088】
工程5(第30日):
E3Select上での14日間の培養後、胚誘導性カルスを新たなE3Select上で、さらに14日間継代培養する(図28E−G)。
【0089】
工程6(第44日):
E3Select上で約4週間後、生長中の小植物を胚性カルス塊から切除し、地下部誘導培地(RM)を入れた65mm×80mmまたは65mm×150mmのポリカーボネート製組織培養容器内で、さらに3週間生長させる。地下部誘導培地は、1×Murashige and Skoog(1962)多量養素、微量栄養素ならびに有機ビタミン群、40mg/Lのチアミン、150mg/LのL−アスパラギン(2%(w/v)のスクロース、0.8%(w/v)のSigma製寒天、および5mg/LのPPTを補給)からなるものである(図28H)。残りの胚誘導性カルスはE3Select上で、さらに14日間継代培養する。
【0090】
工程7(第65日以降):
RM上で3週間より長く生存し、健常な地下部形成を有する再生小植物を、泥炭と砂(1:1)からなる種苗用ミックス中に植え、種苗用湿度チャンバシステムにて高湿度で22〜24℃に維持する(図28)。2週間後、植物を湿度チャンバから取り出し、手での水やりと、液状肥料Aquasol(商標)の供給を毎週、充分生長するまで与える。T0植物をゲノムDNAと分子解析のために試料採取する。T1種子を採取し、ハイスループットQ−PCR解析のために植える(図28J)。
【0091】
実施例10
トランスジェニックシロツメクサ植物の農業生産力
非トランスジェニック対照シロツメクサ植物のものと比べたatmyb32::iptトランスジェニックシロツメクサ(Trifolium repens)植物の農業生産力を、環境制御生長チャンバ条件下で隔離圃場試験にて評価した(図29)。
【0092】
生長制御チャンバ条件下で評価したキメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物は、非トランスジェニック対照シロツメクサ植物と比べて、有意なバイオマス蓄積の向上および老化の兆候の低減を示した(図30)。キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物は、非トランスジェニック対照シロツメクサ植物と比べて、総葉面積の増大、累積葉面積増大、葉の高生長力学(すなわち、経時的な葉の数)、走根長さの増大および顕花植物%の増大ならびに走根の老化および枯死の低減を示した(図30A〜C)。
【0093】
また、トランスジェニックシロツメクサ植物をもたらす3つの独立したatmyb32::ipt(すなわち、LXR 12、LXR 18およびLXR 11)の種子収量の成績を、隔離圃場条件下で、非トランスジェニック対照植物(すなわち、野生型、WT)と比較評価した。非トランスジェニック対照植物(すなわち、WT)と識別不可能な開花強度(すなわち、1m2あたりの成熟花の数)を有するトランスジェニックシロツメクサ植物をもたらした2つの独立したatmyb32::ipt(すなわち、LXR 12およびLXR 18)を、圃場評価に選択した(図31)。
【0094】
キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12、LXR 18およびLXR 11)の種子重量(すなわち、千個の種子の重量)は、非トランスジェニック対照シロツメクサ植物(すなわち、WT)と識別不可能であったが(図32)、花1つあたり(図33)、および種蒔き面積あたり(図34)の基準で示す総種子収量は、同等の開花強度の非トランスジェニック対照シロツメクサ植物(すなわち、WT)と比べて、キメラAtmyb32::ipt遺伝子発現由来トランスジェニックシロツメクサ植物(すなわち、LXR 12およびLXR 18)において2倍であった。
【0095】
実施例11
アルファルファ(Medicago sativa)のアグロバクテリウム媒介型形質転換
キメラipt遺伝子を含むバイナリーベクターpBMVkATMYB32xs::ipt−nos(図18)を、地下部特異的モチーフを欠失させたAtmyb32xs改変体プロモーター配列(図2)の制御下で、高度に再生可能なアルファルファ(M.sativa)クローンC2−3、C2−4および19−17由来のMedicago sativaの葉柄外植片のアグロバクテリウム媒介型形質転換に使用した(図35)。
【0096】
バイナリーベクターpBMVkATMYB32xs::ipt−nosを保有するAgrobacterium tumefaciens株LBA 4404との共存培養後、アルファルファ外植片をセフォタキシム含有培地で洗浄し、25mg/lのカナマイシンを含有する選択培地下で、胚誘導性カルスの誘導に使用した。トランスジェニック胚形成性アルファルファカルスを回収し、トランスジェニックアルファルファ苗条を再生させ、これを地下部発育培地に移し、Atmyb32xs改変体プロモーターの制御下で、キメラipt遺伝子発現由来トランスジェニックアルファルファ植物の回収に至るようにした(図35)。
【0097】
本明細書において開示および規定した本発明は、本文または図面に記載し、これらから明白な個々の特徴の2つ以上の択一的なあらゆる組合せに拡張されることは理解されよう。このような種々の組合せはすべて、本発明の種々の択一的態様を構成する。
【0098】
実施例12
トランスジェニックアブラナ植物の生成
トランスジェニックアブラナ植物(Brassica napus)を、キメラatmyb32::ipt遺伝子を保有するバイナリーベクター(図20および22)を用いて、Agrobacterium媒介型形質転換によって生成した。遺伝子修飾を、候補遺伝子(IPT)または選択可能マーカー(hph)の存在について、T1世代でPCRを用いてキャラクタライズした(図36)。
【0099】
図36は、トランスジェニックアブラナ植物のPCR解析を示す。ゲノムDNAをT1 LXR04系統の種々のトランスジェニックアブラナ植物から単離し、選択可能マーカー(hph遺伝子)または目的の候補遺伝子(IPT)に特異的なプライマーを用いてPCRに供した。図36Aでは、hph特異的プライマーを用いてゲノムDNAから産物を増幅し、アガロースゲル上で可視化した。図36Bは、蛍光PCR法を用いてゲノムDNAを増幅するためのIPT特異的プライマーの使用を示す。これらのプライマーは、PCR産物の蓄積量に反比例する検出可能な蛍光をもたらす標的配列に特異的である。
【0100】
実施例13
トランスジェニックアブラナ植物におけるIPT遺伝子発現
トランスジェニックアブラナにおけるatmyb32::ipt導入遺伝子の発現を、標的配列に特異的な蛍光RT−PCR法を用いて評価した(図37)。組織内のIPT mRNAを検出し、相対発現レベルを系統間およびヌル対照と比較した。ヌル対照は、形質転換プロセスを受けたが、交配後、標的配列を含まない子孫系統である。
【0101】
実施例14
トランスジェニックアブラナ植物における切り取り葉の老化の遅延
atmyb32::iptトランスジェニック植物の切り取り葉の老化を評価するための実験を行なった。図38〜41は、アブラナにおける候補遺伝子の発現に関する切り取り葉の老化(detached senescence)アッセイのデータを示す。切り取り子葉と葉のアッセイは、老化を誘導し、形質転換アブラナの老化表現型を野生型対照と比較して評価するために行なった。切り取り葉の老化アッセイの第7日目および第14日目、老化の進行を、各組織試料について、0−老化の可能性の兆候なし;1−初期の老化の兆候がみられ、緑色が薄くなっている:2−さらに老化が進行、黄変が目立つようになっている;3−組織がほぼ黄変しているが、薄緑色が明白に残っている;4−完全な黄変に進行;5−黄変し、一部脱色および壊死斑を伴う、のいずれかとして定性的にスコアリングした(図39および41)。
【0102】
実施例15
トランスジェニック小麦植物の生成
小麦の遺伝的形質転換は、図27および28に概要を示したTriticum aestivum L Bobwhite 26系統小麦由来の接合子胚の微粒子銃形質転換に基づいたものとした。
【0103】
キメラatmyb32::ipt遺伝子を小麦ゲノム内に、パーティクルボンバードメントによって、ベクターバックボーン配列もまたゲノム内に組み込まれるように完全体プラスミドを用いて挿入した(図24)。
【0104】
形質転換ベクターは充分に配列決定されたものとした(図25)。遺伝子修飾は、候補遺伝子の存在について、T1世代でサザン解析によってキャラクタライズした(図42)。
【0105】
図42は、トランスジェニック小麦植物のサザンハイブリダイゼーション解析を示す。ゲノムDNAを異なるT1系統のトランスジェニック小麦植物から単離し、制限酵素で消化し、候補遺伝子コピー数を測定した。対照は、非形質転換野生型Triticum aestivum「Bobwhite 26」である。消化物を電気泳動させ、ナイロン膜に移し、プローブとしての完全長DIG標識IPT遺伝子とプローブ結合させた。ある範囲のコピー数が観察された。
【0106】
実施例16
トランスジェニック小麦植物におけるIPF遺伝子発現
AtMYB32プロモーターによって駆動させたIPT遺伝子を含む温室栽培トランスジェニックT1小麦植物の若幼葉組織から、RNAを抽出し、第1cDNA鎖を調製した。
【0107】
導入遺伝子の定量的発現は、標的配列のためのプローブ系qRT−PCR法を用いて調べた。各構築物の高、中および低発現系統の代表例を図43に示す。また、内在性スクロースシンターゼ遺伝子に対して設計されたプライマー/プローブセットを対照として使用した。対照遺伝子のすべての増幅プロットは、互いの1サイクル内で始まり、これは、GMOの検出レベル差が発現の多様性によるものであることを示す。
【0108】
PCRプライマーおよびプローブは、ともに、PCR産物の蓄積量に比例する検出可能な蛍光をもたらす標的配列に特異的である。検出対象の標的配列を含む連続希釈プラスミドDNAを用いて、定量のための標準曲線を作成した。
【0109】
実施例17
トランスジェニック小麦植物の植物形態
生長特性の違いを、温室内で、トランスジェニック小麦系統内および系統間で観察した。T1小麦植物間で主に観察された表現型には、植物丈、ひこばえ出現(tillering)強度、葉の数、ならびに植物性バイオマスの抑制が含まれた(図44)。
【0110】
実施例18
トランスジェニック小麦植物における切り取り葉の老化の遅延
切り取り葉アッセイを使用し、ヌル対照と比較したときの形質転換小麦葉の誘導老化および老化表現型を評価(asses)した(図45)。ヌル対照は、形質転換プロセスを受けたが、交配後、標的配列を含まない子孫系統である。
【0111】
また、本明細書で用いる用語「〜を含む/包含する(comprise)」(またはその文法的語尾変化)は、用語「〜を含む(include)」と等価であり、他の要素または特徴の存在を排除するものと解されるべきでないことは理解されよう。
【0112】
本明細書で挙げた文献は、参照目的のためにすぎず、該文献を含めることによって該文献が関連技術分野における周知の一般知識の一部を構成することの承認ではない。
【0113】
最後に、種々の変形、変更および/または付加が、本明細書に概要を示した本発明の精神から逸脱することなく行なわれ得ることを理解されたい。
【特許請求の範囲】
【請求項1】
植物の老化を操作する方法であって、該植物に、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む遺伝子構築物を導入することを含み、該修飾myb遺伝子プロモーターにおいて、1つ以上の地下部特異的モチーフおよび/または花粉特異的モチーフを欠失もしくは不活化させている、方法。
【請求項2】
各々がコンセンサス配列ATATTまたはAATATを含む1つ以上の地下部特異的モチーフを欠失もしくは不活化させる、請求項1に記載の方法。
【請求項3】
各々がコンセンサス配列TTCTまたはAGAAを含む1つ以上の花粉特異的モチーフを欠失もしくは不活化させる、請求項1に記載の方法。
【請求項4】
前記myb遺伝子プロモーターがArabidopsis由来である、請求項1に記載の方法。
【請求項5】
前記myb遺伝子プロモーターが、配列番号2、3および4からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む、請求項1に記載の方法。
【請求項6】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、イソペンテニルトランスフェラーゼ(ipf)遺伝子またはその機能的に活性な断片もしくは改変体である、請求項1に記載の方法。
【請求項7】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、Agrobacterium、LotusおよびPetuniaからなる群より選択される属由来のものである、請求項6に記載の方法。
【請求項8】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、配列番号5、7および9、配列番号6、8および10の配列を有するポリペプチドをコードする配列からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む、請求項7に記載の方法。
【請求項9】
前記遺伝子構築物が、植物細胞のアグロバクテリウム媒介型または微粒子銃型形質転換(biolistic transformation)によって前記植物に導入される、請求項1に記載の方法。
【請求項10】
前記遺伝子構築物が組み込まれた植物細胞を選択し、次いで培養して形質転換植物を再生させる、請求項9に記載の方法。
【請求項11】
植物の老化を操作することができるベクターであって、該ベクターは、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含み、前記修飾myb遺伝子プロモーターにおいて、1つ以上の地下部特異的モチーフおよび/または花粉特異的モチーフを欠失もしくは不活化させている、ベクター。
【請求項12】
さらにターミネーターを含み、前記プロモーター、遺伝子およびターミネーターが作動可能に連結されている、請求項11に記載のベクター。
【請求項13】
前記修飾myb遺伝子プロモーターにおいて、各々がコンセンサス配列ATATTまたはAATATを含む前記1つ以上の地下部特異的モチーフが欠失もしくは不活化されている、請求項11に記載のベクター。
【請求項14】
前記修飾mybプロモーターにおいて、各々がコンセンサス配列TTCTまたはAGAAを含む1つ以上の花粉特異的モチーフが欠失もしくは不活化されている、請求項11に記載のベクター。
【請求項15】
前記myb遺伝子プロモーターがArabidopsis由来のものである、請求項11に記載のベクター。
【請求項16】
前記myb遺伝子プロモーターが、配列番号2、3および4からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む、請求項11に記載のベクター。
【請求項17】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、イソペンテニルトランスフェラーゼ(ipt)遺伝子またはその機能的に活性な断片もしくは改変体である、請求項11に記載のベクター。
【請求項18】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、Agrobacterium、LotusおよびPetuniaからなる群より選択される属由来のものである、請求項17に記載のベクター。
【請求項19】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、配列番号5、7および9、ならびに配列番号6、8および10の配列を有するポリペプチドをコードする配列からなる群より選択されるヌクレオチド配列を含む、請求項17に記載のベクター。
【請求項20】
変更された老化特性を有するトランスジェニック植物細胞、植物、植物種子、または他の植物部分であって、該植物細胞、植物、植物種子、または他の植物部分は、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む遺伝子構築物を含み、該修飾myb遺伝子プロモーターにおいて、1つ以上の地下部特異的モチーフおよび/または花粉特異的モチーフを欠失もしくは不活化されている、トランスジェニック植物細胞、植物、植物種子、または他の植物部分。
【請求項21】
植物に、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む遺伝子構築物を導入することを含む方法によって生成される、請求項20に記載のトランスジェニック植物細胞、植物、植物種子、または他の植物部分であって、該修飾myb遺伝子プロモーターにおいて、1つ以上の地下部特異的モチーフおよび/または花粉特異的モチーフを欠失もしくは不活化されている、トランスジェニック植物細胞、植物、植物種子、または他の植物部分。
【請求項22】
請求項20に記載の植物細胞由来のトランスジェニック植物、植物種子、または他の植物部分。
【請求項23】
請求項20に記載の植物由来のトランスジェニック植物、植物種子、または他の植物部分。
【請求項24】
植物の老化を操作する方法であって、該方法は、該植物に、コンセンサス配列ATATTまたはAATATを有する1〜10個の地下部特異的モチーフが欠失もしくは不活化されているか、またはコンセンサス配列TTCTまたはAGAAを有する1〜30個の花粉特異的モチーフが欠失もしくは不活化されている修飾myb遺伝子プロモーターを含む遺伝子構築物を導入することを含み、該修飾myb遺伝子プロモーターがiptまたはsho遺伝子に作動可能に連結されている、方法。
【請求項25】
植物のバイオマスを増大させる方法であって、該植物に、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結されたmyb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む遺伝子構築物を導入することを含む、方法。
【請求項26】
サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結されたmyb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む、植物のバイオマスを増大させることができるベクター。
【請求項1】
植物の老化を操作する方法であって、該植物に、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む遺伝子構築物を導入することを含み、該修飾myb遺伝子プロモーターにおいて、1つ以上の地下部特異的モチーフおよび/または花粉特異的モチーフを欠失もしくは不活化させている、方法。
【請求項2】
各々がコンセンサス配列ATATTまたはAATATを含む1つ以上の地下部特異的モチーフを欠失もしくは不活化させる、請求項1に記載の方法。
【請求項3】
各々がコンセンサス配列TTCTまたはAGAAを含む1つ以上の花粉特異的モチーフを欠失もしくは不活化させる、請求項1に記載の方法。
【請求項4】
前記myb遺伝子プロモーターがArabidopsis由来である、請求項1に記載の方法。
【請求項5】
前記myb遺伝子プロモーターが、配列番号2、3および4からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む、請求項1に記載の方法。
【請求項6】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、イソペンテニルトランスフェラーゼ(ipf)遺伝子またはその機能的に活性な断片もしくは改変体である、請求項1に記載の方法。
【請求項7】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、Agrobacterium、LotusおよびPetuniaからなる群より選択される属由来のものである、請求項6に記載の方法。
【請求項8】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、配列番号5、7および9、配列番号6、8および10の配列を有するポリペプチドをコードする配列からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む、請求項7に記載の方法。
【請求項9】
前記遺伝子構築物が、植物細胞のアグロバクテリウム媒介型または微粒子銃型形質転換(biolistic transformation)によって前記植物に導入される、請求項1に記載の方法。
【請求項10】
前記遺伝子構築物が組み込まれた植物細胞を選択し、次いで培養して形質転換植物を再生させる、請求項9に記載の方法。
【請求項11】
植物の老化を操作することができるベクターであって、該ベクターは、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含み、前記修飾myb遺伝子プロモーターにおいて、1つ以上の地下部特異的モチーフおよび/または花粉特異的モチーフを欠失もしくは不活化させている、ベクター。
【請求項12】
さらにターミネーターを含み、前記プロモーター、遺伝子およびターミネーターが作動可能に連結されている、請求項11に記載のベクター。
【請求項13】
前記修飾myb遺伝子プロモーターにおいて、各々がコンセンサス配列ATATTまたはAATATを含む前記1つ以上の地下部特異的モチーフが欠失もしくは不活化されている、請求項11に記載のベクター。
【請求項14】
前記修飾mybプロモーターにおいて、各々がコンセンサス配列TTCTまたはAGAAを含む1つ以上の花粉特異的モチーフが欠失もしくは不活化されている、請求項11に記載のベクター。
【請求項15】
前記myb遺伝子プロモーターがArabidopsis由来のものである、請求項11に記載のベクター。
【請求項16】
前記myb遺伝子プロモーターが、配列番号2、3および4からなる群より選択されるヌクレオチド配列、ならびにその機能的に活性な断片および改変体を含む、請求項11に記載のベクター。
【請求項17】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、イソペンテニルトランスフェラーゼ(ipt)遺伝子またはその機能的に活性な断片もしくは改変体である、請求項11に記載のベクター。
【請求項18】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、Agrobacterium、LotusおよびPetuniaからなる群より選択される属由来のものである、請求項17に記載のベクター。
【請求項19】
サイトカイニンの生合成に関与している酵素をコードする前記遺伝子が、配列番号5、7および9、ならびに配列番号6、8および10の配列を有するポリペプチドをコードする配列からなる群より選択されるヌクレオチド配列を含む、請求項17に記載のベクター。
【請求項20】
変更された老化特性を有するトランスジェニック植物細胞、植物、植物種子、または他の植物部分であって、該植物細胞、植物、植物種子、または他の植物部分は、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む遺伝子構築物を含み、該修飾myb遺伝子プロモーターにおいて、1つ以上の地下部特異的モチーフおよび/または花粉特異的モチーフを欠失もしくは不活化されている、トランスジェニック植物細胞、植物、植物種子、または他の植物部分。
【請求項21】
植物に、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結された修飾myb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む遺伝子構築物を導入することを含む方法によって生成される、請求項20に記載のトランスジェニック植物細胞、植物、植物種子、または他の植物部分であって、該修飾myb遺伝子プロモーターにおいて、1つ以上の地下部特異的モチーフおよび/または花粉特異的モチーフを欠失もしくは不活化されている、トランスジェニック植物細胞、植物、植物種子、または他の植物部分。
【請求項22】
請求項20に記載の植物細胞由来のトランスジェニック植物、植物種子、または他の植物部分。
【請求項23】
請求項20に記載の植物由来のトランスジェニック植物、植物種子、または他の植物部分。
【請求項24】
植物の老化を操作する方法であって、該方法は、該植物に、コンセンサス配列ATATTまたはAATATを有する1〜10個の地下部特異的モチーフが欠失もしくは不活化されているか、またはコンセンサス配列TTCTまたはAGAAを有する1〜30個の花粉特異的モチーフが欠失もしくは不活化されている修飾myb遺伝子プロモーターを含む遺伝子構築物を導入することを含み、該修飾myb遺伝子プロモーターがiptまたはsho遺伝子に作動可能に連結されている、方法。
【請求項25】
植物のバイオマスを増大させる方法であって、該植物に、サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結されたmyb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む遺伝子構築物を導入することを含む、方法。
【請求項26】
サイトカイニンの生合成に関与している酵素をコードする遺伝子またはその機能的に活性な断片もしくは改変体に作動可能に連結されたmyb遺伝子プロモーターまたはその機能的に活性な断片もしくは改変体を含む、植物のバイオマスを増大させることができるベクター。
【図1】


【図2】

【図3】

【図4】

【図6】

【図7】

【図8】

【図9】

【図10】

【図11】

【図13】
【図15】
【図17−1】

【図17−2】

【図17−3】

【図17−4】

【図19−1】

【図19−2】

【図19−3】

【図19−4】

【図21−1】

【図21−2】

【図21−3】

【図21−4】

【図23−1】

【図23−2】

【図23−3】

【図23−4】

【図25−1】

【図25−2】

【図25−3】

【図28】

【図29】

【図35A】

【図35B】

【図35C】

【図35D】
【図42】

【図5】
【図12】
【図14】
【図16】
【図18】

【図20】
【図22】
【図24】

【図26】

【図27】

【図30】
【図31】
【図32】
【図33】
【図34】
【図36】
【図37】
【図38】

【図39】

【図40】

【図41】
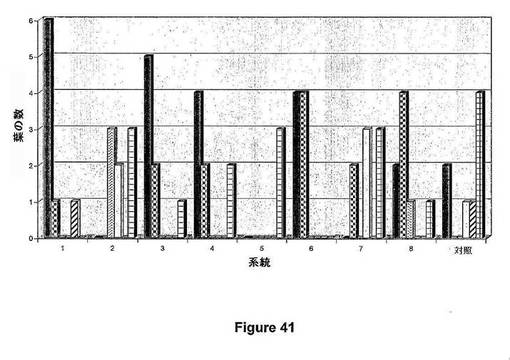
【図43】
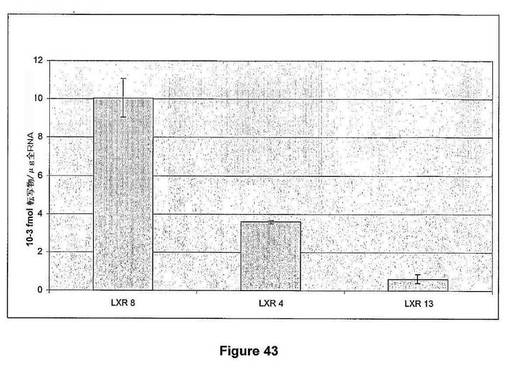
【図44】
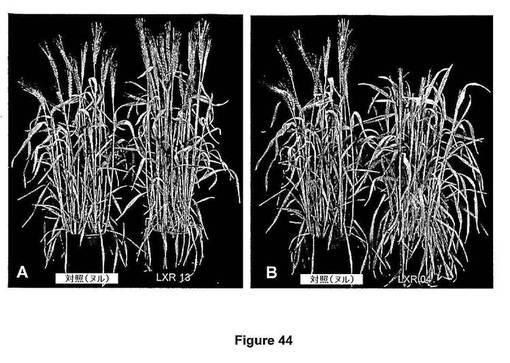
【図45】
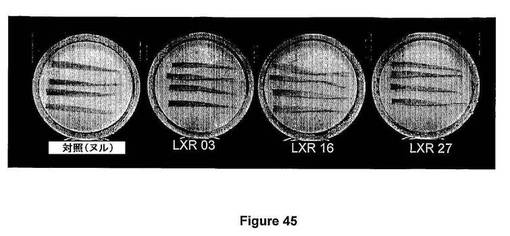
【図2】
【図3】
【図4】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図13】
【図15】
【図17−1】
【図17−2】
【図17−3】
【図17−4】
【図19−1】
【図19−2】
【図19−3】
【図19−4】
【図21−1】
【図21−2】
【図21−3】
【図21−4】
【図23−1】
【図23−2】
【図23−3】
【図23−4】
【図25−1】
【図25−2】
【図25−3】
【図28】
【図29】
【図35A】
【図35B】
【図35C】
【図35D】
【図42】
【図5】
【図12】
【図14】
【図16】
【図18】
【図20】
【図22】
【図24】
【図26】
【図27】
【図30】
【図31】
【図32】
【図33】
【図34】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図43】
【図44】
【図45】
【公表番号】特表2010−524474(P2010−524474A)
【公表日】平成22年7月22日(2010.7.22)
【国際特許分類】
【出願番号】特願2010−504380(P2010−504380)
【出願日】平成20年4月21日(2008.4.21)
【国際出願番号】PCT/AU2008/000556
【国際公開番号】WO2008/128293
【国際公開日】平成20年10月30日(2008.10.30)
【出願人】(503087223)アグリカルチャー ビクトリア サービシーズ プロプライエタリー リミテッド (3)
【出願人】(502155345)ラトローブ ユニバーシティ (2)
【Fターム(参考)】
【公表日】平成22年7月22日(2010.7.22)
【国際特許分類】
【出願日】平成20年4月21日(2008.4.21)
【国際出願番号】PCT/AU2008/000556
【国際公開番号】WO2008/128293
【国際公開日】平成20年10月30日(2008.10.30)
【出願人】(503087223)アグリカルチャー ビクトリア サービシーズ プロプライエタリー リミテッド (3)
【出願人】(502155345)ラトローブ ユニバーシティ (2)
【Fターム(参考)】
[ Back to top ]