
改変候補遺伝子の探索方法
【課題】ランダムな変異導入により取得された微生物変異株において、導入された変異の中から物質生産向上に貢献しうる改変候補遺伝子を探索する方法を提供する。
【解決手段】微生物の遺伝子を改変して特定の機能を向上させる微生物の分子育種における改変候補遺伝子の探索方法であって、野生株と比較して特定の機能が向上している複数の突然変異株間でゲノム比較を行い、複数の突然変異株に変異が導入されている遺伝子を探索することを特徴とする改変候補遺伝子の探索方法である。
【解決手段】微生物の遺伝子を改変して特定の機能を向上させる微生物の分子育種における改変候補遺伝子の探索方法であって、野生株と比較して特定の機能が向上している複数の突然変異株間でゲノム比較を行い、複数の突然変異株に変異が導入されている遺伝子を探索することを特徴とする改変候補遺伝子の探索方法である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、微生物の分子育種を行う際の、改変候補遺伝子の探索方法に関する。
【背景技術】
【0002】
真菌、細菌、古細菌といった微生物は生存、増殖に必要な酵素を自らのために生産するが、そのような微生物が生産する酵素の一部は産業上有用であることが知られている。そして、有用な酵素を生産する微生物を大量に培養し、微生物が分泌した酵素や、微生物内に存在する酵素を分離、精製するといった方法で工業的な酵素生産が行われている。また、近年の遺伝子組み換え技術の進歩により、より効率的に有用物質を生産する微生物の変異株の作製が可能になった。
【0003】
一方、遺伝子組み換え系が確立されていない、もしくは効率的な形質転換が困難な微生物を用いて高生産株を作製しようとする場合、紫外線の照射や化学物質の添加によるランダムな遺伝子の変異の導入により高生産株が作製されている。これらの株が酵素を高生産する機構に関しては不明なものが多い。
【0004】
微生物を利用した酵素の工業的生産の例としては、セルラーゼの生産が挙げられる。セルラーゼは、植物の細胞壁の主成分であるセルロースをグルコース、セロビオースおよびセロオリゴ糖に加水分解する反応を触媒することのできる酵素群の総称である。セルラーゼは、木質バイオマス等の植物由来バイオマスからバイオエタノールを製造する際に有用な酵素である(例えば、特許文献1、2参照)。このことから、酵素の高生産株の開発において、特にセルラーゼの高生産株が過去に多数取得されている。セルラーゼを生産する微生物として、Trichoderma reeseiやAcremonium cellulolyticusなどの糸状菌が用いられており、ランダムな遺伝子変異の導入により多数のセルラーゼを高生産する変異株が取得されている。
【0005】
近年の次世代シークエンサーの進歩により、効率的にゲノム解析が行えるようになり、酵素を高生産する変異株の全ゲノム配列を取得し、野生型株のそれと比較することにより変異が導入された部位を特定できるようになった。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】国際公開1997/33982号パンフレット
【特許文献2】特開2008−271826号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、これらの変異株が野生株と比較してセルラーゼを大量に生産できるメカニズムについては明らかになっていない。また、一般的にランダムに変異を導入した株のゲノムDNAには多数の変異が存在し、どの変異が酵素の高生産に貢献しているのかを特定するのが困難であるという問題点があった。変異株が酵素を大量に生産できるメカニズムの解析や、新たな有用変異株の取得につながることから、酵素の高生産に貢献すると考えられる変異を特定することが望まれている。
【0008】
そこで本発明の目的は、ランダムな変異導入により取得された微生物変異株において、導入された変異の中から物質生産向上に貢献しうる改変候補遺伝子を探索する方法を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題を解決するために鋭意研究した結果、特定の表現形質を呈する複数の突然変異株同志でゲノム比較を行うことにより上記課題を解決しうることを見出し、本発明を完成するに至った。
【0010】
即ち、本発明の改変候補遺伝子の探索方法は、微生物の遺伝子を改変して特定の機能を向上させる微生物の分子育種における改変候補遺伝子の探索方法であって、
野生株と比較して特定の機能が向上している複数の突然変異株間でゲノム比較を行い、複数の突然変異株に変異が導入されている遺伝子を探索することを特徴とするものである。
【0011】
本発明の改変候補遺伝子の探索方法においては、前記特定の機能が、加水分解酵素、DNAおよびRNA合成酵素、制限酵素、酸化酵素、還元酵素、代謝酵素、および、アミノ酸合成酵素からなる群から選ばれる酵素の生産であることが好ましい。
【0012】
また、本発明の改変候補遺伝子の探索方法は、前記特定の機能がセルラーゼまたはヘミセルラーゼの生産であることが好ましい。
【0013】
さらに、本発明の改変候補遺伝子の探索方法は、前記微生物が糸状菌であることが好ましい。
【0014】
本発明の有用微生物の作製方法は前記改変候補遺伝子の探索方法により探索された改変候補遺伝子を改変することを特徴とするものである。また、本発明の有用微生物の作製方法は、糸状菌において、前記探索された遺伝子を改変することが好ましい。
【図面の簡単な説明】
【0015】
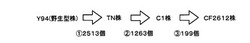
【図1】図1は、本発明の実施例で用いた変異株の関係を表す図である。
【発明を実施するための形態】
【0016】
以下、本発明を実施するための形態について説明する。
【0017】
本発明における微生物としては、酵母、糸状菌等の真菌類、古草菌、放線菌、シアノバクテリア等の細菌類、メタン菌、好熱菌等の古細菌が挙げられる。遺伝子組み換え系が確立されていない、もしくは効率的な形質転換が困難な微生物についても本発明の方法は適用可能である。さらに、セルラーゼ、プロテアーゼ、ペクチナーゼ、ラクターゼ等の有用な酵素を生産する微生物として、工業的に利用され、多数の変異株が存在することから、トリコデルマ(Trichoderma)属、アスペルギルス(Aspergillus)属、ペニシリウム(Penicillium)属、セファロスポリウム(Cephalosporium)属、アクレモニウム(Acremonium)属等の糸状菌が好ましい。
【0018】
本発明においては、特定の機能が向上している複数の突然変異株間でゲノム比較を行う。好ましくは、上記特定の機能とは、加水分解酵素、DNAおよびRNA合成酵素、制限酵素、酸化酵素、還元酵素、代謝酵素、および、アミノ酸合成酵素からなる群から選ばれる酵素の生産である。例えば、セルラーゼ、プロテアーゼ、ペクチナーゼ、ラクターゼ等の有用な酵素の生産量が野生株と比較して高い複数の変異株、好ましくは2〜5系統の変異株それぞれのゲノムDNAの配列をシークエンスし、親株となった野生型株のゲノムDNAも含めて比較する。この場合、酵素の生産量は、ウェスタンブロッティングなど公知の方法を用いて測定・比較することができる。なお、使用する複数の変異株は、同一の野生型株系統を出発株とするものであることが好ましい。同一の野生型株系統由来の変異株同志を比較することで、人為的な突然変異による配列の違いを効率的に比較することができる。複数の変異株は、野生型株から取得された変異株とその変異株にさらに変異を導入して得られた変異株といったように、直列的な関係を有する変異株でもよく、野生型株からそれぞれ独立に得られた変異株であってもよい。上記のうち、直列的な関係を有する変異株が好ましい。つまり、変異が蓄積していった、同一系統由来の株同士を比較することが好ましい。変異が蓄積していくに連れて注目する表現型が顕著になる場合、各変異段階で同じ遺伝子に異なる変異が導入されていれば、その変異が注目する表現型に関与する可能性が高いからである。
【0019】
本発明において、突然変異とは、イオンビーム、紫外線、放射線等の照射、アルキル化剤、化学物質へと曝露等の方法により人為的に誘発された突然変異であり、塩基置換、欠失、挿入といった点突然変異、逆位、転座などいずれの変異であってもよいが、好ましくは、ゲノムDNA上の塩基配列に物理的変化が生じる遺伝子突然変異である。また、点突然変異の場合、同一遺伝子内の異なる塩基に導入された変異であっても、同一の塩基に導入された変異であってもよい。
【0020】
ゲノムDNA配列のシークエンスは公知の方法のいずれも使用できるが、効率やかかる時間の観点から、SOLID system(登録商標 アプライドバイオシステム社)、Genome Sequencer FLX System (GS FLX)(ロシュ社)、Genome Analyzer(イルミナ社)等の高速シークエンスシステムを使用することが好ましい。
【0021】
解析したそれぞれの変異株のゲノムDNA配列を用いて、突然変異が起きた箇所をピックアップする。例えば、得られた配列をアラインメントし、野生型株と比較して、変異が起きている箇所を特定する方法を用いることができる。完全なゲノムDNA配列同志をアラインメントするのみならず、上記の高速シークエンスシステムにより得られた、複数のコンティグ配列を用いてアラインメントを行うこともできる。また、既にゲノムが解読されている近縁種のゲノム配列を参照配列とし、高速シークエンスシステムにより得られた配列をアセンブリして得られたコンティグ配列をマッピングすることで、突然変異の探索を行うこともできる。
【0022】
上記のゲノムDNA配列比較の結果、比較を行った複数の変異株に変異が導入されたことを発見された遺伝子を改変候補遺伝子とする。例えば、同一の野生型株Wから人為的突然変異処理により得られた変異株A、BおよびCのゲノム配列を比較した場合、A株とB株において変異が導入されたことが発見された遺伝子、A株とC株において変異が導入されたことが発見された遺伝子、B株とC株において変異が導入されたことが発見された遺伝子、または、変異株A、B、Cのすべてにおいて変異が導入されたことが発見された遺伝子を改変候補遺伝子とする。
【0023】
セルラーゼは、セルロースをグルコース、セロビオースおよびセロオリゴ糖に加水分解する反応を触媒することのできる酵素群の総称である。分子内部から切断するエンドグルカナーゼ(EC3.2.1.4)と、糖鎖の還元末端と非還元末端のいずれかから分解し、セロビオースを遊離するエキソグルカナーゼ(セロビオヒドロラーゼ)(EC3.2.1.91)に分けられる。例えば、糸状菌Trichoderma reeseiでは、セロビオヒドラーゼとしてCBH1、CBH2、エンドグルカナーゼとしてEG1、EG2、EG3、EG4、EG5、EG6、EG7、EG8等が確認されている。Trichoderma reeseiには、その他に、セロビオースやセロオリゴ糖からグルコースを遊離するβ−グルコシダーゼ(BGL)が存在し、これらの酵素もセルラーゼに含めることができる。ヘミセルラーゼは、ヘミセルロースの分解を触媒することのできる酵素の総称である。ヘミセルロースは、陸上植物細胞の細胞壁を構成する多糖類のうち、セルロースとペクチン以外のものを指す。
【0024】
本発明の方法により、セルラーゼ等の酵素の高生産性変異株など、特定の機能が向上した新たな微生物の変異株を作製するにあたって、改変する遺伝子の候補を効率よく選び出すことができる。特に、ランダムな変異導入により変異株を作製し、所望の形質を発現する株を選択することにより育種を行っていた微生物においては、どの遺伝子の変異が所望の形質発現に寄与しているのか不明なことが多かったことやゲノム比較に用いうる変異株が多数確立されていることから、本発明の方法を好適に使用することができる。
【0025】
本発明の方法により探索された改変候補遺伝子は、ゲノム比較した変異株と同系統の株において改変してもよく、異なる系統の株において改変してもよい。改変候補遺伝子の改変は、相同組み替え、非相同組み替え、プラスミドによる導入など公知の方法を用いることができる。
【実施例】
【0026】
次に実施例を挙げ、本発明をさらに詳しく説明するが、本発明はこれらに限定されるものではない。
【0027】
本実施例においては、セルラーゼ生産菌としてアクレモニウム・セルロリティカス(Acremonium cellulolyticus)Y94株、TN株、C1株、CF−2612株を用いた。図1で示されるように、TN株、C1株、CF−2612株は、Y94株を出発菌株として、紫外線照射や化学変異剤の添加によりランダムな変異が導入されたセルラーゼ高生産変異株である。また、TN株はY94株を、C1株はTN株を、CF−2612株はC1株をそれぞれ親株としてランダムな変異が導入されており、セルラーゼの生産性はCF−2612株、C1株、TN株、Y94株の順に高い。
【0028】
(Y94株のゲノムDNAの配列解析)
野生型株であるY94株のゲノムDNAを物理的切断により約3kbpに断片化した。得られた断片をEcoRIメチラーゼを用いてメチル化した後、末端の平滑化を行った。平滑化したDNA断片の両末端にヘアピンアダプター(ビオチン修飾済)を付加した後、エキソヌクレアーゼ処理を行った。ヘアピンアダプターが付加されたDNAをEcoRIで切断し、ヘアピン部分を除去した後に、ライゲーション反応を行い、環状DNAを調製した。環状DNAを超音波処理によって300〜800bpに断片化した後、アビジンコート磁気ビーズを用いてヘアピンアダプターを含む断片を回収した。回収したDNA断片の両末端を平滑化後、アダプターを付加した。得られたDNAを鋳型にビオチン化したプライマーを用いてPCRを行い増幅した。得られたPCR産物よりアビジンコート磁気ビーズを用いて1本鎖DNAを回収し、シークエンステンプレート用サンプルとした。得られたシークエンステンプレート用サンプルにビーズ(Capture Bead)をアニールさせ、エマルジョンPCRを行った。PCR反応後のビーズを回収、カウントし、適正ビーズ量となるようにプレートに添加し、Genome Sequencer FLX(ロシュ・ダイアグノティクス社)でシークエンスを行った。得られたデータに対し、GS De Novo Assembler(ロシュ・ダイアグノティクス社)を用いてアセンブリを行った。
【0029】
(TN株、C1株、CF−2612株のゲノムDNAの配列解析)
各株のゲノムDNAを物理的に断片化した。得られたDNAの両末端を平滑化・3’-dA突出末端処理を行った後、アダプターを両末端に連結した。アダプターが連結したDNAをアガロース電気泳動により分離した後、300〜400bpのサイズに相当するゲルを切り出し、ゲルからDNAを抽出した。得られたDNAを鋳型とし、PCR法による増幅・濃縮を行い、得られたPCR産物をライブラリーDNAとした。ライブラリーDNAを鋳型として、イルミナ社のGenome analyzer IIxを用い、シークエンスを行った。
【0030】
(各変異株の変異部位の検索)
シークエンスにより得られた配列の先頭35bpを用い、ソフトウェアEdenaを使用してde novoアセンブリを行った。アセンブリにより得られたコンティグ配列をクエリとし、ソフトウェアMUMmerを用いて参照配列へのマッピングを行った。得られたマッピング結果を元に、点変異候補の抽出を行った。その結果、図1に示すように、野生型株であるY94株と比較して、TN株では2513個、C1株では3776個(TN株と比較すると1263個)、CF−2612株では3975個(C1株と比較すると199個)の変異候補箇所を見出した。
【0031】
(変異株間に同時に変異が入った遺伝子の検索)
(1)Y94株からTN株を作製した段階で変異が導入された遺伝子、(2)TN株からC1株を作製した段階で変異が導入された遺伝子、(3)C1株からCF−2612株を作製した段階で変異が導入された遺伝子をそれぞれ検索し、各段階で同時に変異が導入された遺伝子を検索した。その結果、(1)と(2)両方の段階で変異が導入されたP-type proton ATPase 遺伝子(配列番号1)、(2)と(3)両方の段階で変異が導入されたmembrane transporter遺伝子(配列番号2)、Endoplasmic Reticulum Oxidoreductin遺伝子(配列番号3)、Nucleoside hydrolase遺伝子(配列番号4)、および(1)から(3)のすべての段階で変異が導入されたUbiquitin ligase遺伝子(配列番号5)をそれぞれ見出した。
【0032】
図1からも明らかなように、人為的突然変異処理により得られた変異株には、多数の変異が導入されており、どの変異が目的の表現型に貢献しているのか困難である。しかし、本発明の探索方法を用いることにより、糸状菌アクレモニウム・セルロリティカス(Acremonium cellulolyticus)のセルラーゼ生産量の増大に寄与しうる変異遺伝子として、複数の遺伝子をピックアップすることができた。これらの遺伝子を遺伝子組み換え等により改変することで、セルラーゼ、ヘミセルラーゼの生産量が増大した糸状菌アクレモニウム・セルロリティカスの変異株を作製することが期待できる。
【産業上の利用可能性】
【0033】
本発明の方法を利用することにより、微生物の遺伝子を改変して所望の形質を発現する株を作製するにあたって、改変することで所望の形質が得られることが期待される遺伝子を効率良く探索することができる。
【技術分野】
【0001】
本発明は、微生物の分子育種を行う際の、改変候補遺伝子の探索方法に関する。
【背景技術】
【0002】
真菌、細菌、古細菌といった微生物は生存、増殖に必要な酵素を自らのために生産するが、そのような微生物が生産する酵素の一部は産業上有用であることが知られている。そして、有用な酵素を生産する微生物を大量に培養し、微生物が分泌した酵素や、微生物内に存在する酵素を分離、精製するといった方法で工業的な酵素生産が行われている。また、近年の遺伝子組み換え技術の進歩により、より効率的に有用物質を生産する微生物の変異株の作製が可能になった。
【0003】
一方、遺伝子組み換え系が確立されていない、もしくは効率的な形質転換が困難な微生物を用いて高生産株を作製しようとする場合、紫外線の照射や化学物質の添加によるランダムな遺伝子の変異の導入により高生産株が作製されている。これらの株が酵素を高生産する機構に関しては不明なものが多い。
【0004】
微生物を利用した酵素の工業的生産の例としては、セルラーゼの生産が挙げられる。セルラーゼは、植物の細胞壁の主成分であるセルロースをグルコース、セロビオースおよびセロオリゴ糖に加水分解する反応を触媒することのできる酵素群の総称である。セルラーゼは、木質バイオマス等の植物由来バイオマスからバイオエタノールを製造する際に有用な酵素である(例えば、特許文献1、2参照)。このことから、酵素の高生産株の開発において、特にセルラーゼの高生産株が過去に多数取得されている。セルラーゼを生産する微生物として、Trichoderma reeseiやAcremonium cellulolyticusなどの糸状菌が用いられており、ランダムな遺伝子変異の導入により多数のセルラーゼを高生産する変異株が取得されている。
【0005】
近年の次世代シークエンサーの進歩により、効率的にゲノム解析が行えるようになり、酵素を高生産する変異株の全ゲノム配列を取得し、野生型株のそれと比較することにより変異が導入された部位を特定できるようになった。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】国際公開1997/33982号パンフレット
【特許文献2】特開2008−271826号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、これらの変異株が野生株と比較してセルラーゼを大量に生産できるメカニズムについては明らかになっていない。また、一般的にランダムに変異を導入した株のゲノムDNAには多数の変異が存在し、どの変異が酵素の高生産に貢献しているのかを特定するのが困難であるという問題点があった。変異株が酵素を大量に生産できるメカニズムの解析や、新たな有用変異株の取得につながることから、酵素の高生産に貢献すると考えられる変異を特定することが望まれている。
【0008】
そこで本発明の目的は、ランダムな変異導入により取得された微生物変異株において、導入された変異の中から物質生産向上に貢献しうる改変候補遺伝子を探索する方法を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題を解決するために鋭意研究した結果、特定の表現形質を呈する複数の突然変異株同志でゲノム比較を行うことにより上記課題を解決しうることを見出し、本発明を完成するに至った。
【0010】
即ち、本発明の改変候補遺伝子の探索方法は、微生物の遺伝子を改変して特定の機能を向上させる微生物の分子育種における改変候補遺伝子の探索方法であって、
野生株と比較して特定の機能が向上している複数の突然変異株間でゲノム比較を行い、複数の突然変異株に変異が導入されている遺伝子を探索することを特徴とするものである。
【0011】
本発明の改変候補遺伝子の探索方法においては、前記特定の機能が、加水分解酵素、DNAおよびRNA合成酵素、制限酵素、酸化酵素、還元酵素、代謝酵素、および、アミノ酸合成酵素からなる群から選ばれる酵素の生産であることが好ましい。
【0012】
また、本発明の改変候補遺伝子の探索方法は、前記特定の機能がセルラーゼまたはヘミセルラーゼの生産であることが好ましい。
【0013】
さらに、本発明の改変候補遺伝子の探索方法は、前記微生物が糸状菌であることが好ましい。
【0014】
本発明の有用微生物の作製方法は前記改変候補遺伝子の探索方法により探索された改変候補遺伝子を改変することを特徴とするものである。また、本発明の有用微生物の作製方法は、糸状菌において、前記探索された遺伝子を改変することが好ましい。
【図面の簡単な説明】
【0015】
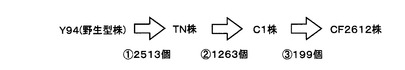
【図1】図1は、本発明の実施例で用いた変異株の関係を表す図である。
【発明を実施するための形態】
【0016】
以下、本発明を実施するための形態について説明する。
【0017】
本発明における微生物としては、酵母、糸状菌等の真菌類、古草菌、放線菌、シアノバクテリア等の細菌類、メタン菌、好熱菌等の古細菌が挙げられる。遺伝子組み換え系が確立されていない、もしくは効率的な形質転換が困難な微生物についても本発明の方法は適用可能である。さらに、セルラーゼ、プロテアーゼ、ペクチナーゼ、ラクターゼ等の有用な酵素を生産する微生物として、工業的に利用され、多数の変異株が存在することから、トリコデルマ(Trichoderma)属、アスペルギルス(Aspergillus)属、ペニシリウム(Penicillium)属、セファロスポリウム(Cephalosporium)属、アクレモニウム(Acremonium)属等の糸状菌が好ましい。
【0018】
本発明においては、特定の機能が向上している複数の突然変異株間でゲノム比較を行う。好ましくは、上記特定の機能とは、加水分解酵素、DNAおよびRNA合成酵素、制限酵素、酸化酵素、還元酵素、代謝酵素、および、アミノ酸合成酵素からなる群から選ばれる酵素の生産である。例えば、セルラーゼ、プロテアーゼ、ペクチナーゼ、ラクターゼ等の有用な酵素の生産量が野生株と比較して高い複数の変異株、好ましくは2〜5系統の変異株それぞれのゲノムDNAの配列をシークエンスし、親株となった野生型株のゲノムDNAも含めて比較する。この場合、酵素の生産量は、ウェスタンブロッティングなど公知の方法を用いて測定・比較することができる。なお、使用する複数の変異株は、同一の野生型株系統を出発株とするものであることが好ましい。同一の野生型株系統由来の変異株同志を比較することで、人為的な突然変異による配列の違いを効率的に比較することができる。複数の変異株は、野生型株から取得された変異株とその変異株にさらに変異を導入して得られた変異株といったように、直列的な関係を有する変異株でもよく、野生型株からそれぞれ独立に得られた変異株であってもよい。上記のうち、直列的な関係を有する変異株が好ましい。つまり、変異が蓄積していった、同一系統由来の株同士を比較することが好ましい。変異が蓄積していくに連れて注目する表現型が顕著になる場合、各変異段階で同じ遺伝子に異なる変異が導入されていれば、その変異が注目する表現型に関与する可能性が高いからである。
【0019】
本発明において、突然変異とは、イオンビーム、紫外線、放射線等の照射、アルキル化剤、化学物質へと曝露等の方法により人為的に誘発された突然変異であり、塩基置換、欠失、挿入といった点突然変異、逆位、転座などいずれの変異であってもよいが、好ましくは、ゲノムDNA上の塩基配列に物理的変化が生じる遺伝子突然変異である。また、点突然変異の場合、同一遺伝子内の異なる塩基に導入された変異であっても、同一の塩基に導入された変異であってもよい。
【0020】
ゲノムDNA配列のシークエンスは公知の方法のいずれも使用できるが、効率やかかる時間の観点から、SOLID system(登録商標 アプライドバイオシステム社)、Genome Sequencer FLX System (GS FLX)(ロシュ社)、Genome Analyzer(イルミナ社)等の高速シークエンスシステムを使用することが好ましい。
【0021】
解析したそれぞれの変異株のゲノムDNA配列を用いて、突然変異が起きた箇所をピックアップする。例えば、得られた配列をアラインメントし、野生型株と比較して、変異が起きている箇所を特定する方法を用いることができる。完全なゲノムDNA配列同志をアラインメントするのみならず、上記の高速シークエンスシステムにより得られた、複数のコンティグ配列を用いてアラインメントを行うこともできる。また、既にゲノムが解読されている近縁種のゲノム配列を参照配列とし、高速シークエンスシステムにより得られた配列をアセンブリして得られたコンティグ配列をマッピングすることで、突然変異の探索を行うこともできる。
【0022】
上記のゲノムDNA配列比較の結果、比較を行った複数の変異株に変異が導入されたことを発見された遺伝子を改変候補遺伝子とする。例えば、同一の野生型株Wから人為的突然変異処理により得られた変異株A、BおよびCのゲノム配列を比較した場合、A株とB株において変異が導入されたことが発見された遺伝子、A株とC株において変異が導入されたことが発見された遺伝子、B株とC株において変異が導入されたことが発見された遺伝子、または、変異株A、B、Cのすべてにおいて変異が導入されたことが発見された遺伝子を改変候補遺伝子とする。
【0023】
セルラーゼは、セルロースをグルコース、セロビオースおよびセロオリゴ糖に加水分解する反応を触媒することのできる酵素群の総称である。分子内部から切断するエンドグルカナーゼ(EC3.2.1.4)と、糖鎖の還元末端と非還元末端のいずれかから分解し、セロビオースを遊離するエキソグルカナーゼ(セロビオヒドロラーゼ)(EC3.2.1.91)に分けられる。例えば、糸状菌Trichoderma reeseiでは、セロビオヒドラーゼとしてCBH1、CBH2、エンドグルカナーゼとしてEG1、EG2、EG3、EG4、EG5、EG6、EG7、EG8等が確認されている。Trichoderma reeseiには、その他に、セロビオースやセロオリゴ糖からグルコースを遊離するβ−グルコシダーゼ(BGL)が存在し、これらの酵素もセルラーゼに含めることができる。ヘミセルラーゼは、ヘミセルロースの分解を触媒することのできる酵素の総称である。ヘミセルロースは、陸上植物細胞の細胞壁を構成する多糖類のうち、セルロースとペクチン以外のものを指す。
【0024】
本発明の方法により、セルラーゼ等の酵素の高生産性変異株など、特定の機能が向上した新たな微生物の変異株を作製するにあたって、改変する遺伝子の候補を効率よく選び出すことができる。特に、ランダムな変異導入により変異株を作製し、所望の形質を発現する株を選択することにより育種を行っていた微生物においては、どの遺伝子の変異が所望の形質発現に寄与しているのか不明なことが多かったことやゲノム比較に用いうる変異株が多数確立されていることから、本発明の方法を好適に使用することができる。
【0025】
本発明の方法により探索された改変候補遺伝子は、ゲノム比較した変異株と同系統の株において改変してもよく、異なる系統の株において改変してもよい。改変候補遺伝子の改変は、相同組み替え、非相同組み替え、プラスミドによる導入など公知の方法を用いることができる。
【実施例】
【0026】
次に実施例を挙げ、本発明をさらに詳しく説明するが、本発明はこれらに限定されるものではない。
【0027】
本実施例においては、セルラーゼ生産菌としてアクレモニウム・セルロリティカス(Acremonium cellulolyticus)Y94株、TN株、C1株、CF−2612株を用いた。図1で示されるように、TN株、C1株、CF−2612株は、Y94株を出発菌株として、紫外線照射や化学変異剤の添加によりランダムな変異が導入されたセルラーゼ高生産変異株である。また、TN株はY94株を、C1株はTN株を、CF−2612株はC1株をそれぞれ親株としてランダムな変異が導入されており、セルラーゼの生産性はCF−2612株、C1株、TN株、Y94株の順に高い。
【0028】
(Y94株のゲノムDNAの配列解析)
野生型株であるY94株のゲノムDNAを物理的切断により約3kbpに断片化した。得られた断片をEcoRIメチラーゼを用いてメチル化した後、末端の平滑化を行った。平滑化したDNA断片の両末端にヘアピンアダプター(ビオチン修飾済)を付加した後、エキソヌクレアーゼ処理を行った。ヘアピンアダプターが付加されたDNAをEcoRIで切断し、ヘアピン部分を除去した後に、ライゲーション反応を行い、環状DNAを調製した。環状DNAを超音波処理によって300〜800bpに断片化した後、アビジンコート磁気ビーズを用いてヘアピンアダプターを含む断片を回収した。回収したDNA断片の両末端を平滑化後、アダプターを付加した。得られたDNAを鋳型にビオチン化したプライマーを用いてPCRを行い増幅した。得られたPCR産物よりアビジンコート磁気ビーズを用いて1本鎖DNAを回収し、シークエンステンプレート用サンプルとした。得られたシークエンステンプレート用サンプルにビーズ(Capture Bead)をアニールさせ、エマルジョンPCRを行った。PCR反応後のビーズを回収、カウントし、適正ビーズ量となるようにプレートに添加し、Genome Sequencer FLX(ロシュ・ダイアグノティクス社)でシークエンスを行った。得られたデータに対し、GS De Novo Assembler(ロシュ・ダイアグノティクス社)を用いてアセンブリを行った。
【0029】
(TN株、C1株、CF−2612株のゲノムDNAの配列解析)
各株のゲノムDNAを物理的に断片化した。得られたDNAの両末端を平滑化・3’-dA突出末端処理を行った後、アダプターを両末端に連結した。アダプターが連結したDNAをアガロース電気泳動により分離した後、300〜400bpのサイズに相当するゲルを切り出し、ゲルからDNAを抽出した。得られたDNAを鋳型とし、PCR法による増幅・濃縮を行い、得られたPCR産物をライブラリーDNAとした。ライブラリーDNAを鋳型として、イルミナ社のGenome analyzer IIxを用い、シークエンスを行った。
【0030】
(各変異株の変異部位の検索)
シークエンスにより得られた配列の先頭35bpを用い、ソフトウェアEdenaを使用してde novoアセンブリを行った。アセンブリにより得られたコンティグ配列をクエリとし、ソフトウェアMUMmerを用いて参照配列へのマッピングを行った。得られたマッピング結果を元に、点変異候補の抽出を行った。その結果、図1に示すように、野生型株であるY94株と比較して、TN株では2513個、C1株では3776個(TN株と比較すると1263個)、CF−2612株では3975個(C1株と比較すると199個)の変異候補箇所を見出した。
【0031】
(変異株間に同時に変異が入った遺伝子の検索)
(1)Y94株からTN株を作製した段階で変異が導入された遺伝子、(2)TN株からC1株を作製した段階で変異が導入された遺伝子、(3)C1株からCF−2612株を作製した段階で変異が導入された遺伝子をそれぞれ検索し、各段階で同時に変異が導入された遺伝子を検索した。その結果、(1)と(2)両方の段階で変異が導入されたP-type proton ATPase 遺伝子(配列番号1)、(2)と(3)両方の段階で変異が導入されたmembrane transporter遺伝子(配列番号2)、Endoplasmic Reticulum Oxidoreductin遺伝子(配列番号3)、Nucleoside hydrolase遺伝子(配列番号4)、および(1)から(3)のすべての段階で変異が導入されたUbiquitin ligase遺伝子(配列番号5)をそれぞれ見出した。
【0032】
図1からも明らかなように、人為的突然変異処理により得られた変異株には、多数の変異が導入されており、どの変異が目的の表現型に貢献しているのか困難である。しかし、本発明の探索方法を用いることにより、糸状菌アクレモニウム・セルロリティカス(Acremonium cellulolyticus)のセルラーゼ生産量の増大に寄与しうる変異遺伝子として、複数の遺伝子をピックアップすることができた。これらの遺伝子を遺伝子組み換え等により改変することで、セルラーゼ、ヘミセルラーゼの生産量が増大した糸状菌アクレモニウム・セルロリティカスの変異株を作製することが期待できる。
【産業上の利用可能性】
【0033】
本発明の方法を利用することにより、微生物の遺伝子を改変して所望の形質を発現する株を作製するにあたって、改変することで所望の形質が得られることが期待される遺伝子を効率良く探索することができる。
【特許請求の範囲】
【請求項1】
微生物の遺伝子を改変して特定の機能を向上させる微生物の分子育種における改変候補遺伝子の探索方法であって、
野生株と比較して特定の機能が向上している複数の突然変異株間でゲノム比較を行い、複数の突然変異株に変異が導入されている遺伝子を探索することを特徴とする改変候補遺伝子の探索方法。
【請求項2】
前記特定の機能が、加水分解酵素、DNAおよびRNA合成酵素、制限酵素、酸化酵素、還元酵素、代謝酵素、および、アミノ酸合成酵素からなる群から選ばれる酵素の生産である請求項1記載の改変候補遺伝子の探索方法。
【請求項3】
前記特定の機能がセルラーゼまたはヘミセルラーゼの生産である請求項1または2記載の改変候補遺伝子の探索方法。
【請求項4】
前記微生物が糸状菌である請求項1〜3のいずれか一項記載の改変候補遺伝子の探索方法。
【請求項5】
請求項1〜4のいずれか一項記載の改変候補遺伝子の探索方法により探索された改変候補遺伝子を改変することを特徴とする有用微生物の作製方法。
【請求項1】
微生物の遺伝子を改変して特定の機能を向上させる微生物の分子育種における改変候補遺伝子の探索方法であって、
野生株と比較して特定の機能が向上している複数の突然変異株間でゲノム比較を行い、複数の突然変異株に変異が導入されている遺伝子を探索することを特徴とする改変候補遺伝子の探索方法。
【請求項2】
前記特定の機能が、加水分解酵素、DNAおよびRNA合成酵素、制限酵素、酸化酵素、還元酵素、代謝酵素、および、アミノ酸合成酵素からなる群から選ばれる酵素の生産である請求項1記載の改変候補遺伝子の探索方法。
【請求項3】
前記特定の機能がセルラーゼまたはヘミセルラーゼの生産である請求項1または2記載の改変候補遺伝子の探索方法。
【請求項4】
前記微生物が糸状菌である請求項1〜3のいずれか一項記載の改変候補遺伝子の探索方法。
【請求項5】
請求項1〜4のいずれか一項記載の改変候補遺伝子の探索方法により探索された改変候補遺伝子を改変することを特徴とする有用微生物の作製方法。
【図1】


【公開番号】特開2011−193773(P2011−193773A)
【公開日】平成23年10月6日(2011.10.6)
【国際特許分類】
【出願番号】特願2010−62959(P2010−62959)
【出願日】平成22年3月18日(2010.3.18)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成23年10月6日(2011.10.6)
【国際特許分類】
【出願日】平成22年3月18日(2010.3.18)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]