
改変型ヒト化抗インターロイキン−18抗体
本発明は、ヒト化抗IL−18抗体、製造方法、及び該抗体を用いた治療方法を開示する。さらに、例えば表面プラズモン共鳴を用いて、治療可能性のある抗体を同定するためのスクリーニング方法を開示する。
【発明の詳細な説明】
【技術分野】
【0001】
1.発明の分野
本発明は、一般的に、ヒトインターロイキン−18により媒介される状態の治療及び診断に有用な、抗体(特にヒト化抗体)などの免疫グロブリンの分野に関する。
【背景技術】
【0002】
2.発明の背景
ヒトインターロイキン−18(hIL−18)は、生物学的に活性を有しない193アミノ酸の前駆体タンパク質として合成されるサイトカインである(Ushioら、J. Immunol. 156:4274, 1996)。前駆体タンパク質の切断、例えばカスパーゼ−1又はカスパーゼ−4による切断によって、156アミノ酸の成熟タンパク質が遊離し(Guら、Science 275:206, 1997;Ghayurら、Nature 386:619, 1997)、これが、T細胞増殖の共刺激、NK細胞の細胞傷害性の増強、T細胞及びNK細胞によるIFN−γ産生の誘導、並びにTヘルパー1型(Th1)分化の増強を含む生物学的活性を示す(Okamuraら、Nature 378:88, 1995;Ushioら、J. Immunol. 156:4274, 1996;Micallefら、Eur. J. Immunol. 26:1647, 1996;Kohnoら、J. Immunol. 158:1541, 1997;Zhangら、Infect. Immunol. 65:3594, 1997;Robinsonら、Immunity 7:571, 1997)。さらに、IL−18は、ヒト単球前炎症性伝達物質、例えばIL−8、腫瘍壊死因子−α(TNF−α)及びプロスタグランジンE2(PGE2)などの効果的な誘導因子である(Ushio, S.ら、J. Immunol. 156:4274-4279, 1996;Puren, A. J.ら、J. Clin. Invest. 10:711-721, 1997;Podolinら、J. Immunol. 提出済, 1999)。
【0003】
以前にクローニングされたIL−1受容体関連タンパク質(IL−1Rrp)(Parnetら、J. Biol. Chem. 271:3967, 1996)は、IL−18受容体(Kd=18nM)のサブユニットとして同定されている(Torigoeら、J. Biol. Chem. 272:25737, 1997)。IL−18受容体の2つ目のサブユニットは、IL−1受容体付属タンパク質に対して相同性を示し、AcPL(付属タンパク質様のため)と称されている。IL−1 Rrp及びAcPLの両方の発現が、IL−18が誘導するNF−κB及びJNKの活性化に必要である(Bornら、J. Biol. Chem. 273:29445, 1998)。NF−κB及びJNKに加えて、IL−1受容体関連キナーゼ(IRAK)、p56lck(LCK)及びマイトジェン活性化タンパク質キナーゼ(MAPK)を介してIL−18はシグナル伝達する(Micallefら、Eur. J. Immunol. 26:1647, 1996;Matsumotoら、Biophys Biochem. Res. Comm. 234:454, 1997;Tsuji-Takayamaら、Biochem. Biophys. Res. Comm. 237:126, 1997)。
【0004】
TH1細胞は、IFN−γ、IL−2及びTNF−βなどの前炎症性サイトカインを産生するが(Mosmannら、J. Immunol. 136:2348, 1986)、これは多くの自己免疫疾患、例えば多発性硬化症(MS)、関節リウマチ(RA)、1型すなわちインスリン依存型糖尿病(IDDM)、炎症性腸疾患(IBD)、及び乾癬などの媒介に関与しているとされている(Mosmann及びSad, Immunol. Today 17:138, 1996)。従って、TH1促進性サイトカイン(IL−18など)のアンタゴニスト作用は、疾患の発症を抑制すると予想される。Il−18特異的mAbは、アンタゴニストとして用いることができるはずである。
【0005】
自己免疫疾患の発症におけるIL−18の役割は証明されている。すなわち、非肥満糖尿病(NOD)マウスの膵臓及び脾臓において、その疾患発症直前にIL−18の発現が有意に増大することが示されている(Rotheら、J. Clin. Invest. 99:469, 1997)。同様に、関節リウマチ患者の滑液においてIL−18レベルが顕著に上昇することが示されている(Kawashimaら、Arthritis and Rheumatism 39:598, 1996)。さらに、IL−18投与によって、Th1媒介自己免疫疾患であるマウス実験的アレルギー性脳脊髄炎(EAE)(多発性硬化症のモデル)の臨床的重篤度が増すことが示されている。加えて、中和抗ラットIL−18抗血清によってLewis雌ラットにおけるEAEの発症が予防されることが示されている(Wildbaumら、J. Immunol. 161:6368, 1998)。従って、IL−18は、自己免疫のための新規な治療薬の開発のための望ましい標的である。
【0006】
Taniguchiら(J. Immunol. Methods 206:107)は、4つの異なる抗原性部位に結合する7種のマウス及び6種のラット抗ヒトIL−18モノクローナル抗体(mAb)を記載している。マウスmAbの1つ(#125−2H)と、6つのラットmAbは、KG−1細胞によるIL−18誘導性IFN−γの産生を阻害し、ラットmAbは#125−2Hよりも10倍低い中和活性を示す。ウエスタンブロット分析によって示されているように、マウスmAbのうち3つ(ただしラットmAbはなし)は、膜に結合したヒトIL−18と強く反応する。さらに、#125−2H及びラットmAbを用いてヒトIL−18を検出するための酵素結合免疫吸着アッセイ(ELISA)が記載されている。このELISAの検出限界は10pg/mlである。
【0007】
欧州特許出願EP0 712 931号は、2種のマウス抗ヒトIL−18 mAb、すなわちH1(IgG1)及びH2(IgM)を開示している。ウエスタンブロット分析によって示されているように、両方のmAbは膜に結合したヒトIL−18と反応するが、膜に結合したヒトIL−12とは反応しない。ヒトIL−18を精製するためにイムノアフィニティークロマトグラフィープロトコールにおいてH1を使用し、そしてヒトIL−18を測定するためにELISAにおいてH1を使用する。ヒトIL−18を測定するためにラジオイムノアッセイにおいてH2を使用する。
【発明の開示】
【発明が解決しようとする課題】
【0008】
中和IL−18抗体は、ヒトにおける自己免疫疾患及び関連する症候の緩和に有用である可能性がある。従って、当該技術分野においては、Th1細胞分化及び増殖を低減し、それゆえ自己免疫疾患及び関連する症候を低減する、高親和性IL−18アンタゴニスト、例えばヒトインターロイキン18に対する中和モノクローナル抗体が必要とされている。
【0009】
本明細書全体で言及する参照文献は全て、明示的かつ全体的に参照により本明細書に組み入れられる。
【課題を解決するための手段】
【0010】
3.発明の概要
本発明により、以下の相補性決定領域(CDR):
CDRH1:配列番号1
CDRH2:配列番号2
CDRH3:配列番号3
CDRL1:配列番号4
CDRL2:配列番号5
CDRL3:配列番号6
を有する重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体が提供される。
【0011】
本発明により、以下のCDR:
CDRH1:配列番号1
CDRH2:配列番号2
CDRH3:配列番号3
CDRL1:配列番号4
CDRL2:配列番号5
CDRL3:配列番号6
を有する重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体であって、該軽鎖の位置71の残基が、前記CDRが由来するドナー抗体中に存在するその対応する残基によって置換されている、ヒト化抗インターロイキン−18抗体が提供される。
【0012】
当業者には明らかなとおり、「由来する」という用語は、それが物質の物理的起源であるという意味で、起源を明らかにするのみならず、その物質と構造的に同一であるもののその基準起源から生じたのではない物質も明示することを意図するものである。したがって、「CDRが由来するドナー抗体フレームワーク中に存在する」対応する残基は、かならずしもドナー抗体フレームワークから精製されたものである必要はない。同様に、「ドナー抗体」由来CDRは、かならずしもドナー抗体から精製されたものである必要はない。
【0013】
CDR及びフレームワーク領域(FR)、並びにアミノ酸の番号付けは、特に指摘しない限り、Kabatら、"Sequences of immunological interest", NIHに記載されているKabat定義に従う。
【0014】
本発明の別の態様において、ヒトアクセプターフレームワークに移植されたドナー抗体由来CDRを含むヒト化抗インターロイキン−18抗体であって、抗インターロイキン−18抗体が配列番号1、2、3、4、5及び6に示される配列を有するCDRを含み、該抗インターロイキン−18抗体の軽鎖の位置71の残基が、該ドナー抗体フレームワーク中のその対応する位置に存在する残基と同一である、ヒト化抗インターロイキン−18抗体が提供される。
【0015】
本発明の別の態様においては、配列番号1、2、3、4、5及び6に示されるCDRを含み、軽鎖の位置71にチロシンを含む、ヒト化抗インターロイキン−18抗体が提供される。
【0016】
本発明の別の態様において、配列番号1、2及び3に示されるCDRを有する重鎖と配列番号4、5及び6に示されるCDRを有する軽鎖を含むヒト化抗インターロイキン−18抗体であって、該軽鎖CDRが、ドナー抗体軽鎖の位置71にチロシンを有するドナー抗体に由来する、ヒト化抗インターロイキン−18抗体が提供される。
【0017】
本発明の別の態様において、ドナー抗体由来CDR及びヒト化抗体軽鎖の位置71にチロシンを含むヒト化抗インターロイキン−18抗体であって、該ドナー抗体が、2C10又はそのフレームワーク変異体である(すなわち、2C10と同じCDRであるが、異なるフレームワークを含む。米国特許第6,706,487号を参照されたい)、ヒト化抗インターロイキン−18抗体が提供される。
【0018】
本発明の別の態様では、
(a)ヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有するCDRを有する重鎖、並びに
(b)ヒト軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有するCDRを有する軽鎖
を含むヒト化抗インターロイキン−18抗体であって、
前記ヒト軽鎖アクセプターフレームワークが配列番号38に由来するフレームワークを含み、かつ配列番号38の位置71がチロシンである、ヒト化抗インターロイキン−18抗体が提供される。
【0019】
本発明の別の態様では、
(a)ヒトIL−18との特異的結合を可能にするCDRを有する重鎖、
(b)アクセプターフレームワークを含み、かつ配列番号4、5及び6に示される配列を有するCDRを有し、そして位置71にチロシン残基を有する軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0020】
軽鎖のCDRは、好ましくは、配列番号35に示される配列内の、配列番号4、5及び6に示される配列のそれぞれの位置に対応するアクセプターフレームワーク内の位置に位置する。軽鎖及び/又は重鎖は、好ましくはヒト患者において非免疫原性のものである。
【0021】
本発明の別の態様において、
(a)配列番号1、2及び3に示される配列を有するCDRを含む重鎖、並びに
(b)ヒト軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有するCDRを含む軽鎖
を含むヒト化抗インターロイキン−18抗体であって、
該ヒト化抗インターロイキン−18抗体の該軽鎖アクセプターフレームワークが、配列番号38に示される配列の変異体に由来するフレームワーク領域を含み、該変異体が位置71にチロシンを含み、かつ該変異体が配列番号38に示される配列を有するフレームワークと75%以上の同一性を有する、ヒト化抗インターロイキン−18抗体が提供される。該変異体は、配列番号38に示されるフレームワークと、好ましくは80%以上、例えば81%、82%、83%、84%、より好ましくは85%以上、例えば86%、87%、88%、89%、よりさらに好ましくは90%以上、例えば91%、92%、93%、94%、最も好ましくは95%以上、例えば96%、97%、98%、99%の同一性を含む。
【0022】
本発明の別の態様において、
(a)ドナー抗体軽鎖の位置71にチロシンを含むドナー抗体に由来する配列番号1、2、3、4、5及び6に示されるCDR、
(b)ヒト軽鎖の位置71にフェニルアラニンを含むヒトアクセプターフレームワーク
を含むヒト化抗インターロイキン−18抗体であって、
該抗インターロイキン−18抗体がその軽鎖の位置71にチロシンを含む、ヒト化抗インターロイキン−18抗体が提供される。
【0023】
本発明の別の態様において、
(a)ドナー抗体軽鎖の位置71に芳香族アミノ酸を含むドナー抗体に由来する配列番号1、2、3、4、5及び6に示されるCDR、
(b)軽鎖アクセプターフレームワークの位置71に、部分(a)の芳香族アミノ酸とは異なる種類の芳香族アミノ酸を含むヒトアクセプターフレームワーク
を含むヒト化抗インターロイキン−18抗体であって、
該抗インターロイキン−18抗体が、位置71に部分(a)の抗体に由来する芳香族アミノ酸を有する軽鎖を含む、ヒト化抗インターロイキン−18抗体が提供される。
【0024】
本発明の別の態様では、37℃において表面プラスモン共鳴(例えば、BiacoreTM、好ましくは以下の第7.4.1節に記載するBiacoreTM3000機器及び条件を用いて)によって測定した場合のヒトIL−18の結合に関して300pM以下の平衡定数(KD)を示すヒト化抗インターロイキン−18抗体が提供される。
【0025】
本発明の別の態様では、配列番号1、2、3、4、5及び6に示されるCDRを含み、かつ37℃において表面プラスモン共鳴(例えば、BiacoreTM、好ましくは以下の第7.4.1節に記載するBiacoreTM3000機器及び条件を用いて)によって測定した場合のヒトIL−18の結合に関して300pM以下の平衡定数(KD)を示す、ヒト化抗インターロイキン−18抗体が提供される。
【0026】
好ましくは、37℃において表面プラスモン共鳴(好ましくは以下の第7.4.2節に記載するBiacoreTMT100機器及び条件を用いて)によって測定した場合のヒトIL−18の結合に関する抗体の平衡定数(KD)は、90pM未満である。平衡定数は、より好ましくは70pM以下、よりさらに好ましくは65pM、60pM、55pM、又は50pM以下である。
【0027】
本発明の別の態様において、37℃において表面プラスモン共鳴(例えば、BiacoreTM、好ましくは以下の第7.4.2節に記載するBiacoreTMT100機器及び条件を用いて)によって測定した場合のヒトIL−18の結合に関して0.0002(1/秒)以上の解離定数又は解離速度(kd)を示すヒト化抗インターロイキン−18抗体が提供される。
【0028】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRが重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、93、39、40、36、71、89、91の一個若しくは複数の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71、及び任意により位置45、83、84、85の一個若しくは複数の(例えば全ての)残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0029】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該抗インターロイキン−18抗体の軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0030】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、39、40、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0031】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、36、39、40、71、89、91、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0032】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0033】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、93、39、40の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0034】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、93、39、40、36、71、89、91の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0035】
本発明の別の態様において、重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体であって、ヒトIL−18と前記抗体との結合の25℃での解離速度(kd)と、ヒトIL−18と前記抗体との結合の37℃での解離速度(kd)との比が、1:5(又は1:5未満)であり、該抗体がドナー抗体由来CDRとヒトアクセプターフレームワークを含み、かつヒトアクセプターフレームワークの軽鎖の位置71の残基が、そのドナー抗体のその対応する残基によって置換されている、ヒト化抗インターロイキン−18抗体が提供される。解離速度は、好ましくは、以下の第7.4.2節に記載するBiacoreTMT100機器及び条件を用いて測定される。
【0036】
本発明の別の態様において、配列番号9、配列番号17、配列番号21からなる群から選択された重鎖と、配列番号13、配列番号29からなる群から選択された軽鎖を含むヒト化抗インターロイキン−18抗体が提供される。
【0037】
特に本発明は、配列番号9の重鎖と配列番号13の軽鎖、又は配列番号9の重鎖と配列番号29の軽鎖を含むヒト化抗インターロイキン−18抗体を提供する。
【0038】
本発明はまた、配列番号17の重鎖と配列番号13の軽鎖、又は配列番号17の重鎖と配列番号29の軽鎖を含むヒト化抗インターロイキン−18抗体を提供する。
【0039】
また本発明は、配列番号21の重鎖と配列番号13の軽鎖、又は配列番号21の重鎖と配列番号29の軽鎖を含むヒト化抗インターロイキン−18抗体を提供する。
【0040】
本発明の別の態様において、本明細書において上述した抗インターロイキン−18抗体、及び薬学的に許容される担体を含む医薬組成物が提供される。
【0041】
本発明の別の態様において、治療に使用する抗体(特に、リガンドと受容体との相互作用を阻害する抗体、抗インターロイキン−18抗体など)を選択する方法であって、
(a)30〜45℃(好ましくは37℃)の温度において前記抗体が特異的に結合する抗原に対する前記抗体の結合親和性を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(b)20〜25℃(好ましくは25℃)の温度において前記抗体が特異的に結合する抗原に対する前記抗体の結合親和性を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(c)(a)の親和性が(b)の親和性を超える場合、好ましくは(a)の親和性がステップ(b)の親和性の2倍以上、より好ましくは4倍以上である場合に、治療に使用する前記抗体を選択するステップ
を含む方法が提供される。
【0042】
本発明の別の態様において、治療に使用する抗体(特に、リガンドと受容体との相互作用を阻害する抗体、抗インターロイキン−18抗体など)を選択する方法であって、
(a)30〜45℃(好ましくは37℃)の温度において前記抗体が特異的に結合する抗原からの前記抗体の解離速度を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(b)20〜25℃(好ましくは25℃)の温度において前記抗体が特異的に結合する抗原からの前記抗体の解離速度を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(c)(a)の解離速度が(b)の解離速度よりも遅い場合、治療に使用する前記抗体を選択するステップ
を含む方法が提供される。
【0043】
「抗インターロイキン−18」という用語は、本発明の抗体を参照する場合、ヒトインターロイキン−18の生物学的活性を中和することが可能な抗体を意味する。しかしながら、そのような抗体が非ヒト霊長類(例えばアカゲザル及び/又はカニクイザル)インターロイキン−18の生物学的活性をさらに中和する可能性があることを排除するものではない。
【発明を実施するための最良の形態】
【0044】
さらに図面を参照しながら本発明を説明する。
【0045】
4.ヒト化抗体
インタクトな非ヒト抗体をヒトの疾患又は障害の治療に使用することは、特に抗体を反復投与した際に、現在十分に立証されている潜在的な免疫原性の問題を引き起こす。すなわち、患者の免疫系はインタクトな非ヒト抗体を非自己と認識して、中和応答を開始する可能性がある。こうした問題を克服するために、完全にヒトの抗体(上記参照)の開発に加えて、長年にわたってさまざまな技法が開発されてきたが、こうした技法は、一般に、免疫した動物、たとえばマウス、ラット又はウサギから非ヒト抗体を得ることの相対的な容易さを維持する一方で、インタクトな治療用抗体における非ヒトアミノ酸配列の構成割合を減らすことを必要とする。概して、これを達成するために2つの手法が用いられてきた。第1はキメラ抗体であって、これは一般に、ヒト定常領域に融合された非ヒト(たとえば、マウスなどの齧歯類)可変領域を含む(Morrison (1984), PNAS, 81, 6851参照)。抗体の抗原結合部位は可変領域内に位置するので、キメラ抗体は、抗原に対するその結合親和性を維持するが、ヒト定常領域のエフェクター機能を獲得しているため、上記のようなエフェクター機能を果たすことができる。キメラ抗体は、典型的には、組換えDNA法を用いて作製される。従来の方法を用いて(たとえば、本発明の抗体のH鎖及びL鎖をコードする遺伝子、例としては上記の配列番号1、2、3、4、5及び6をコードするDNAと、特異的に結合することができるオリゴヌクレオチドプローブを使用することによって)、抗体をコードするDNA(たとえばcDNA)を単離し、配列を決定する。ハイブリドーマ細胞は、こうしたDNAの典型的な供給源となる。キメラ抗体を発現することが望まれる場合には、軽鎖及び重鎖の全長成熟可変領域をコードする単離cDNAを適当な発現ベクターに読み取り枠を合わせて挿入する。該発現ベクターは、特に、適当な免疫グロブリン定常領域(通常はヒト起源のもの)と、シグナル配列、停止コドン、プロモーター、ターミネーター、及び抗体の発現を達成するために必要な他のエレメントを含む。かかるベクターを、次に本来免疫グロブリンを産生しない大腸菌、COS細胞、CHO細胞又は三重ローマ細胞といった宿主細胞にトランスフェクトして、抗体を合成させる。DNAは、ヒトL鎖及びH鎖のコード配列を、対応する非ヒト(たとえばマウス)H及びL定常領域の代わりに用いることによって、改変することができる。たとえば、Morrison; PNAS 81, 6851 (1984)を参照されたい。
【0046】
第2の手法は、可変領域をヒト化することによって抗体の非ヒト含量を減少させる、ヒト化抗体の作製に関する。ヒト化について2つの技術が広く支持されている。第1は、CDRグラフティングによるヒト化である。CDRは抗体のN末端近くにループを作り、そこで、フレームワーク領域によってもたらされるスキャフォールドに嵌め込まれた表面を形成する。抗体の抗原結合特異性は、主として、そのCDR表面のトポグラフィー及び化学的性質によって決まる。次いで、これらの特徴は、個別のCDRのコンフォメーションによって、CDRの相対的配置によって、並びにCDRを構成する残基の側鎖の性質及び配置によって決定される。免疫原性の大幅な低下は、非ヒト(たとえばマウス)抗体(「ドナー」抗体)のCDRのみを、好適なヒトのフレームワーク(「アクセプターフレームワーク」)及び定常領域上にグラフティングすることによって達成することができる(Jonesら、 (1986) Nature 321,522-525、及びVerhoeyen Mら、(1988) Science 239, 1534-1536を参照されたい)。しかしながら、CDRグラフティングは本質的に、抗原結合特性の完全な保持をもたらさない可能性があり、有意な抗原結合親和性を回復したいならば、ドナー抗体のフレームワーク残基(「復帰突然変異」と呼ばれることもある)の一部をヒト化分子内に保持する必要があることが、多くの場合、明らかになっている(Queen C ら、(1989) PNAS 86, 10,029-10,033, Co, Mら、(1991) Nature 351, 501-502を参照されたい)。この場合、ヒトフレームワーク(FR)を用意するために、非ヒトドナー抗体に対して最大の配列相同性(典型的には60%以上)を示すヒトV領域をデータベースから選択する。ヒトFRは、ヒトコンセンサス又は個別のヒト抗体のどちらからも選択することができる。必要ならば、ドナー抗体由来の重要残基を、ヒトアクセプターフレームワーク内に置換して入れて、CDRコンフォメーションを保存する。抗体のコンピューターモデリングを用いることができるが、それは、このような構造的に重要な残基を同定するのに役立つ。国際公開WO99/48523号を参照されたい。
【0047】
別法として、ヒト化は、「veneering」のプロセスによって行うことができる。特異なヒト及びマウス免疫グロブリンの重鎖及び軽鎖可変領域の統計分析によって、露出した残基の正確なパターンがヒトとマウスの抗体で異なること、並びにほとんどの個々の表面位置は少数の異なる残基に強い選択性があることが明らかになった(Padlan E.A.ら、(1991) Mol.Immunol.28, 489-498、及びPedersen J.T.ら、(1994) J.Mol.Biol. 235; 959-973を参照されたい)。したがって、そのフレームワーク領域内の、ヒト抗体に通常存在するものとは異なる、露出した残基を置き換えることによって、非ヒトFvの免疫原性を低下させることができる。タンパク質の抗原性は表面への接近可能性と相関しうるので、表面残基を置き換えることは、マウス可変領域をヒトの免疫系に「見えない」ようにするのに十分であると考えられる(Mark G.E.ら、(1994)、実験薬理学ハンドブック(Handbook of Experimental Pharmacology)第113号より:モノクローナル抗体の薬理学(The pharmacology of モノクローナル 抗体)、Springer-Verlag、105-134ページも参照されたい)。こうしたヒト化の手順は、抗体の表面だけを変更し、支持する残基はそのままになっているので、「veneering」と呼ばれる。さらに別の手法としては、WO04/006955号に記載されている方法、及びHumaneeringTM(Kalobios)のプロセスが挙げられる。この方法は、細菌発現系を使用して、ヒト生殖系と配列が近縁の抗体を産生する(Alfenito-M Advancing Protein Therapeutics, January 2007, San Diego, California)。別の最近のヒト化手法は、ヒトアクセプターフレームワークを、フレームワーク領域などの抗体の他の領域間の相同性よりもむしろ、ドナーマウス抗体CDR領域とのヒトCDR領域の構造類似性に基づいて選択することを含む。このプロセスは、SuperhumanisationTMとしても知られる(Evogenix Inc.; Hwang ら (2005) Methods 36:35-42)。
【0048】
従って本発明は、上の第3節に記載したヒト化抗体に関する。このようなヒト化抗体は、好ましくはIgGアイソタイプ(IgG1又はIgG4など)のヒト定常領域を含む。
【0049】
別の実施形態において、上の第3節で記載したヒト化可変領域は、非ヒト定常領域(「逆キメラ(reverse chimera)」)、例えば非ヒト霊長類、ラット、マウス又はウサギのものなどと融合しうる。
【0050】
当業者には明らかなように、配列番号37及び38に示されるアクセプターフレームワークは、それぞれVH遺伝子及びVκ遺伝子によってコードされる免疫グロブリンのアミノ酸を構成する。これらはそれ自体がアクセプター抗体のフレームワーク領域及びCDRを含む。アクセプター抗体のCDRを配列番号1、2、3、4、5及び6に示されるドナーCDRと置換し、そして得られる配列を好適なフレームワークの4つの配列(配列番号39及び配列番号40に示される配列など)と結合させて、完全な免疫グロブリン可変領域(配列番号11及び配列番号15に示されるものなど)を生成することは当業者の能力の範囲内である。
【0051】
4.1.他の改変
抗体のFc領域とさまざまなFc受容体(FcγR)との相互作用は、抗体のエフェクター機能を仲介すると考えられるが、このエフェクター機能には、抗体依存性細胞傷害(ADCC)、補体結合、食作用、及び抗体の半減期/クリアランスが含まれる。所望のエフェクター特性に応じて、本発明の抗体のFc領域のさまざまな改変を行うことができる。たとえば、本来なら溶解性の抗体を非溶解性にするFc領域内の特異的変異が、欧州特許第0629 240B1号及び同第0307 434B2号に詳述されており、あるいは、抗体にサルベージ(salvage)受容体結合エピトープを組み込んで、血清半減期を延ばすことができる。米国特許第5,739,277号を参照されたい。現在知られているヒトFcγ受容体には5つあり、FcγR(I)、FcRγIIa、FcγRIIb、FcγRIIIa及び新生児FcRnである。Shieldsら、(2001) J. Biol. Chem 276, 6591-6604は、共通のIgG1残基のセットがすべてのFcγRとの結合に関与しているが、FcγRII及びFcγRIIIは、この共通のセットの外側にある別個の部位を利用することを明らかにした。一群のIgG1残基は、Pro−238、Asp−265、Asp−270、Asn−297及びPro−239をアラニンに変えると、すべてのFcγRへの結合を低下させた。これらはいずれもIgG CH2ドメインに存在し、CH1及びCH2をつなぐヒンジ部の近くでクラスターを形成している。FcγRIは、結合のためにIgG1残基の共通セットのみを用いるが、FcγRII及びFcγRIIIは、共通セットに加えて、別個の残基とも相互作用する。一部の残基の変更は、FcγRIIとの結合のみを低下させ(たとえば、Arg−292)、又はFcγRIIIとの結合のみを低下させた(たとえば、Glu−293)。ある変異体は、FcγRII又はFcγRIIIに対する結合の強化を示したが、他の受容体との結合には影響を与えなかった(たとえば、Ser−267Alaは、FcγRIIに対する結合は強化したが、FcγRIIIとの結合には影響しなかった)。また他の変異体は、FcγRII又はFcγRIIIに対する結合の強化を示したが、他の受容体との結合は低下した(たとえば、Ser−298AlaはFcγRIIIに対する結合を強化したが、FcγRIIとの結合は低下した)。FcγRIIIaに関して、もっともよく結合するIgG1変異体は、Ser−298、Glu−333及びLys−334でのアラニン置換の組み合わせを有していた。新生児FcRn受容体は、抗体クリアランス及び組織にわたるトランスサイトーシスのどちらにも関与していると考えられる(Junghans R. P (1997) Immunol. Res 16. 29-57 及びGhetieら、(2000) Annu. Rev. Immunol. 18, 739-766を参照されたい)。ヒトFcRnと直接相互作用すると確定されたヒトIgG1残基には、Ile253、Ser254、Lys288、Thr307、Gln311、Asn434及びHis435がある。本発明は、半減期/クリアランス並びに/又はエフェクター機能(ADCC及び/若しくは補体溶解など)を改変するために、上述した残基の変化のいずれか1(又はそれ以上)を有する本発明の抗体にも関する。
【0052】
他の改変には、本発明の抗体のグリコシル化変異体がある。抗体の定常領域内の保存された位置でのグリコシル化は、抗体機能、特に上記のようなエフェクター機能に重大な影響を及ぼすことが知られている。たとえば、Boydら、(1996), Mol. Immunol. 32, 1311-1318を参照されたい。1又は複数の糖鎖を付加、置換、欠失又は修飾した、本発明の治療用抗体又はその抗原結合フラグメントのグリコシル化変異体が想定される。アスパラギン−X−セリン、又はアスパラギン−X−スレオニンモチーフの導入は、糖鎖を酵素的に連結することができる部位を作り出すので、それを用いて、抗体のグリコシル化を操作することができる。Rajuら、(2001) Biochemistry 40, 8868-8876において、TNFR−IgGイムノアドヘシンの末端シアリル化は、β−1,4−ガラクトシルトランスフェラーゼ及び/又はα−2,3−シアリルトランスフェラーゼを用いる再ガラクトシル化及び/又は再シアリル化により増加した。末端シアリル化の増加は、免疫グロブリンの半減期を長くすると考えられる。抗体は、ほとんどの糖タンパク質と同様に、典型的には複数のグライコフォームの混合物として天然に産生される。こうした混合物は、抗体を真核生物、とりわけ哺乳動物細胞内で産生させたとき、特に明らかである。特定したグライコフォームを製造するために、さまざまな方法が開発されてきた。Zhangら、Science (2004), 303, 371、 Searsら、Science, (2001) 291, 2344、 Wackerら、(2002) Science, 298 1790、Davisら、(2002) Chem. Rev. 102, 579、Hangら、(2001) Acc. Chem. Res 34, 727を参照されたい。したがって、本発明は、前記抗体又はその抗原結合フラグメントの特定された数(たとえば7個以下、例として5個以下、たとえば2又は1個)のグライコフォームを含む、本明細書に記載の複数の治療用抗体(典型的にはモノクローナル抗体)(IgG1アイソタイプ、たとえばIgG1とすることができる)に関する。
【0053】
本発明のさらに他の実施形態には、ポリエチレングリコール(PEG)、ポリプロピレングリコール又はポリオキシアルキレンといった、非タンパク質性ポリマーと結合した本発明の治療用抗体又はその抗原結合フラグメントが含まれる。タンパク質とPEGとの結合は、タンパク質の半減期を長くするのと同時に、タンパク質の抗原性及び免疫原性を低下させるための確立された技術である。分子量及び形状(直鎖若しくは分枝)の異なるPEG付加の使用は、インタクト抗体、及びFab’フラグメントで検討されている。Koumenis I.L.ら、(2000) Int. J. Pharmaceut. 198:83-95を参照されたい。
【0054】
5.作製方法
本発明の抗体は、ヤギ(Pollockら、(1999), J. Immunol. Methods 231:147-157を参照されたい)、ニワトリ(Morrow KJJ (2000) Genet. Eng. News 20:1-55を参照されたい)、マウス(Pollockら、前掲)、又は植物(Doran PM, (2000) Curr. Opinion Biotechnol. 11, 199-204、Ma JK-C (1998), Nat. Med. 4; 601-606、Baez Jら、BioPharm (2000) 13: 50-54、Stoger Eら、(2000) Plant Mol. Biol. 42:583-590を参照されたい)といったトランスジェニック生物内で産生することができる。抗体はまた、化学合成によっても作製することができる。しかしながら、本発明の抗体は、典型的には、当業者に周知の組換え細胞培養技術を用いて作製される。抗体をコードするポリヌクレオチドを単離し、その後の宿主細胞におけるクローニング(増幅)又は発現のために、複製可能なベクター(プラスミドなど)に挿入する。特に宿主細胞がCHO又はNSOである場合(下記を参照されたい)、1つの有用な発現系は、グルタミン酸シンテターゼ系(Lonza Biologicsが販売しているものなど)である。抗体をコードするポリヌクレオチドは、従来の方法(たとえば、オリゴヌクレオチドプローブ)によって、容易に単離され、配列決定される。使用可能なベクターには、プラスミド、ウイルス、ファージ、トランスポゾン、ミニ染色体(プラスミドがその典型的な具体例である)がある。一般にこうしたベクターは、発現を促進するために、軽鎖及び/又は重鎖ポリヌクレオチドに機能的に連結された、シグナル配列、複製起点、1つ若しくは複数のマーカー遺伝子、エンハンサーエレメント、プロモーター、及び転写終結配列をさらに含有する。軽鎖及び重鎖をコードするポリヌクレオチドは、別々のベクター内に挿入し、(例えば形質転換、トランスフェクション、エレクトロポレーション又は形質導入によって)同一の宿主細胞に同時に又は連続的に導入することができるし、あるいは所望であれば、そのような導入の前に重鎖及び軽鎖をともに同一ベクター内に挿入してもよい。
【0055】
遺伝子コードの縮重のため、本発明のポリペプチドをコードする本明細書に記載のポリヌクレオチドの代わりとなるポリヌクレオチドも利用可能であることは、当業者にはすぐにわかることである。
【0056】
5.1.シグナル配列
本発明の抗体は、成熟タンパク質のN末端に特異的な切断部位を有する、異種シグナル配列を含む融合タンパク質として作製することができる。シグナル配列は、宿主細胞によって認識されて、プロセシングされる必要がある。原核生物の宿主細胞については、シグナル配列は、アルカリホスファターゼ、ペニシリナーゼ、又は耐熱性エンテロトキシンIIリーダーとすることができる。酵母分泌のためには、シグナル配列は、酵母インベルターゼリーダー、α因子リーダー、又は酸ホスファターゼリーダーとすることができる。例えばWO90/13646号を参照されたい。哺乳動物細胞系では、ウイルス分泌リーダー、たとえば単純ヘルペスgDシグナル、並びに天然免疫グロブリンシグナル配列(ヒトIg重鎖など)が利用できる。典型的には、シグナル配列は、本発明の抗体をコードするポリヌクレオチドに、リーディングフレーム内で連結される。
【0057】
5.2.複製起点
複製起点は当技術分野で周知であり、pBR322がほとんどのグラム陰性細菌に適合し、2μプラスミドがほとんどの酵母に適合し、さらに、さまざまなウイルス由来のもの、たとえば、SV40、ポリオーマ、アデノウイルス、VSV若しくはBPVはほとんどの哺乳動物細胞に適している。一般に、複製起点の要素は、ベクターの増殖を大腸菌内で行う必要がない限り、組込み型哺乳動物発現ベクターには必要でない。しかしながら、SV40 oriは、初期プロモーターを含有しているので使用することができる。
【0058】
5.3.選択マーカー
典型的な選択遺伝子は、(a)抗生物質若しくは他の毒素(たとえば、アンピシリン、ネオマイシン、メトトレキセート若しくはテトラサイクリン)に対する耐性を付与するタンパク質、又は(b)栄養要求性の欠乏を補完する、若しくは複合培地で利用できない栄養素を補充するタンパク質、あるいは(c)その両方の組み合わせであるタンパク質をコードする。選択方式は、ベクターを含まない又はベクターを含む宿主細胞の増殖の停止を行いうる。本発明の治療用抗体をコードする遺伝子による形質転換が成功した細胞は、たとえば共送達される選択マーカーによって付与された薬剤耐性のために、生き残る。一つの例はDHFR選択系であるが、この場合、形質転換体はDHFR陰性宿主株において作出される(例えばPage and Sydenham 1991 Biotechnology 9: 64-68参照)。この系では、DHFR遺伝子が本発明の抗体ポリヌクレオチド配列と共送達されて、ヌクレオシド除去によりDHFR陽性細胞が選択される。必要に応じて、DHFR阻害剤であるメトトレキサートも使用して、DHFR遺伝子増幅を示す形質転換体を選択する。DHFR遺伝子を本発明の抗体コード配列又はその機能的誘導体に機能的に連結することによって、DHFR遺伝子増幅が目的の所望抗体配列が同時に増幅されるようにする。CHO細胞は、DHFR/メトトレキサート選択に特に有用な細胞株であり、DHFR系を利用した宿主細胞の増幅及び選択方法は当技術分野で十分に確立されている(Kaufman R.J. ら、 J.Mol.Biol. (1982) 159, 601-621を参照のこと。概説については、Werner RG, Noe W, Kopp K,Schluter M,”Appropriate mammalian expression systems for biopharmaceuticals”, Arzneimittel-Forschung. 48(8):870-80, 1998 Augを参照のこと)。さらにもう一つの例は、グルタミン酸シンテターゼ発現系(Lonza Biologics)である。酵母での使用に適した選択遺伝子はtrp1遺伝子である。Stinchcombら、Nature 282, 38, 1979を参照されたい。
【0059】
5.4.プロモーター
本発明の抗体の発現に適したプロモーターは、抗体をコードするDNA/ポリヌクレオチドに、機能するように連結される。原核生物宿主のためのプロモーターには、proAプロモーター、βラクタマーゼ及びラクトースプロモーター系、アルカリホスファターゼ、トリプトファン及びハイブリッドプロモーター、たとえばTacがある。酵母細胞での発現に適したプロモーターには、3−ホスホグリセリン酸キナーゼ又は他の解糖系酵素、たとえばエノラーゼ、グリセルアルデヒド3リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース6リン酸イソメラーゼ、3−ホスホグリセリン酸ムターゼ及びグルコキナーゼがある。誘導性酵母プロモーターには、アルコールデヒドロゲナーゼ2、イソチトクロームC、酸ホスファターゼ、メタロチオネイン、及び窒素代謝又はマルトース/ガラクトース利用を担う酵素が含まれる。
【0060】
哺乳動物細胞系における発現のためのプロモーターには、RNAポリメラーゼIIプロモーター、例えばウイルスプロモーター、たとえば、ポリオーマ、鶏痘及びアデノウイルス(たとえばアデノウイルス2)、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス(具体的には前初期遺伝子プロモーター)、レトロウイルス、B型肝炎ウイルス、アクチン、ラウス肉腫ウイルス(RSV)プロモーター、及び初期若しくは後期シミアンウイルス40、並びに非ウイルスプロモーター、例えばEF−1α(Mizushima and Nagata Nucleic Acids Res 1990 18(17):5322)が挙げられる。プロモーターの選択は、発現に使用される宿主細胞との適切な適合性に基づく。
【0061】
5.5.エンハンサーエレメント
必要に応じて、たとえば、高等真核生物での発現のために、さらなるエンハンサーエレメントを、上述したプロモーターの代わりに、又はそのプロモーター内に位置することが見出されているものと一緒に挿入することができる。適当な哺乳動物エンハンサー配列には、グロビン、エラスターゼ、アルブミン、フェトプロテイン、メタロチオネイン、及びインスリン由来のエンハンサーエレメントが含まれる。あるいはまた、SV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、ポリオーマエンハンサー、バキュロウイルスエンハンサー、又はマウスIgG2a遺伝子座といった、真核細胞ウイルス起源のエンハンサーエレメントを使用してもよい(WO04/009823号を参照されたい)。このようなエンハンサーは典型的に、ベクター上で、プロモーターの上流の部位に位置するが、他の位置、例えば非翻訳領域内又はポリアデニル化シグナルの下流に位置してもよい。エンハンサーの選択及び位置決定は、発現に使用される宿主細胞との適切な適合性に基づく。
【0062】
5.6.ポリアデニル化/終止
真核細胞系では、ポリアデニル化シグナルが、本発明の抗体をコードするポリヌクレオチドに、機能しうるように連結される。こうしたシグナルは典型的には、オープンリーシングフレームの3’側に置かれる。哺乳動物系で、限定的でない例として、シグナルとしては、成長ホルモン、伸長因子1α、並びにウイルス(たとえば、SV40)遺伝子、又はレトロウイルスのロング・ターミナル・リピート由来のものが挙げられる。酵母系では、ポリアデニル化/終結シグナルの限定的でない例として、ホスホグリセリン酸キナーゼ(PGK)及びアルコールデヒドロゲナーゼ1(ADH)遺伝子由来のシグナルが挙げられる。原核細胞系では、ポリアデニル化シグナルは一般的には必要とされず、その代わりにもっと短く明確な転写終結配列を用いるのが通例である。ポリアデニル化/終結配列の選択は、発現に使用される宿主との適切な適合性に基づいて行われる。
【0063】
5.7.収量改善のための他の方法/エレメント
上記に加えて、収量改善のために用いることができる他の特徴としては、クロマチン再構成エレメント、イントロン及び宿主細胞特異的コドン改変が挙げられる。宿主細胞のコドンの偏りに適合させるために本発明の抗体のコドン利用法を改変して、転写及び/又は生成物収率を高めることができる(例えば、Hoekema A ら、Mol Cell Biol 1987 7(8):2914-24)。コドンの選択は、発現に使用される宿主との適切な適合性に基づいて行われる。
【0064】
5.8.宿主細胞
本発明の抗体をコードするベクターをクローニングし、又は発現させるのに適した宿主細胞は、原核細胞、酵母、又は高等真核細胞である。適当な原核細胞には、真正細菌、例を挙げると、腸内細菌科、例えばエシェリキア属(Escherichia)(例として大腸菌(E. coli)(たとえば、ATCC 31,446; 31,537; 27,325))、エンテロバクター属(Enterobacter)、エルウィニア属(Erwinia)、クレブシエラ属(Klebsiella)、プロテウス属(Proteus)、サルモネラ属(Salmonella)(例としてネズミチフス菌(Salmonella typhimurium))、セラチア属(Serratia)(例としてSerratia marcescens)、及び赤痢菌属(Shigella)など、並びにバシラス属(Bacilli)の枯草菌(B. subtilis)及びB. licheniformis(DD 266 710を参照されたい)など、シュードモナス属(Pseudomonas)の緑膿菌(P. aeruginosa)など、さらにストレプトミセス属(Streptomyces)がある。酵母宿主細胞のうち、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(schizosaccharomyces pombe)、クルイベロミセス属(Kluyveromyces)(たとえば、ATCC 16,045; 12,424; 24178; 56,500)、ヤロウィア属(yarrowia)(欧州特許第402,226号)、ピキア・パストリス(Pichia Pastoris)(欧州特許第183,070号、Pengら、J.Biotechnol. 108 (2004) 185-192も参照されたい)、カンジダ属(Candida)、トリコデルマ・リーシア(Trichoderma reesia)(欧州特許第244,234号)、ペニシリン属(Penicillin)、トリポクラジウム属(Tolypocladium)並びにアスペルギルス属(Aspergillus)宿主、たとえばアスペルギルス・ニデュランス(A. nidulans)及びクロコウジカビ(A. niger)も想定される。
【0065】
原核生物及び酵母宿主細胞は、具体的に本発明で想定はされるが、しかしながら、本発明の宿主細胞は、典型的には脊椎動物細胞である。適当な脊椎動物宿主細胞には、哺乳動物細胞、たとえば、COS−1(ATCC番号 CRL 1650)、COS−7(ATCC CRL 1651)、ヒト胚性腎細胞系293、PerC6(Crucell)、仔ハムスター腎細胞(BHK)(ATCC CRL.1632)、BHK570(ATCC番号: CRL 10314)、293(ATCC番号CRL 1573)、チャイニーズハムスター卵巣細胞CHO(たとえば、CHO−K1、ATCC番号:CCL61、DHFR−CHO細胞系、たとえばDG44など(Urlaubら、前掲を参照されたい))、特に懸濁培養に適したCHO細胞系、マウスセルトリ細胞、サル腎細胞、アフリカミドリザル腎細胞(ATCC CRL-1587)、HELA細胞、イヌ腎細胞(ATCC CCL 34)、ヒト肺細胞(ATCC CCL 75)、HepG2及びミエローマ若しくはリンパ腫細胞、たとえばNS0(米国特許第5,807,715号を参照されたい)、Sp2/0、Y0が含まれる。
【0066】
したがって、本発明の一実施形態において、本明細書に記載の治療用抗体の重鎖及び/又は軽鎖をコードするベクターを含む、安定して形質転換された宿主細胞が提供される。こうした宿主細胞は、典型的には、軽鎖をコードする第1のベクター、及び重鎖をコードする第2のベクターを含む。
【0067】
このような宿主細胞はまた、本発明の抗体の質、機能及び/又は収率を改変するためにさらに操作してもよいし、又は適合させてもよい。非限定的な例としては、特定の修飾(例えばグリコシル化)酵素及びタンパク質折りたたみシャペロンの発現が挙げられる。
【0068】
5.9.細胞培養法
本発明の治療用抗体をコードするベクターで形質転換された宿主細胞は、当業者に公知の方法によって培養することができる。宿主細胞は、スピナーフラスコ、振とうフラスコ、ローラーボトル、又は中空糸の系で培養することができるが、大規模生産のためには、特に懸濁培養用に撹拌槽型リアクター又はバッグリアクター(例えば、Wave Biotech, Somerset, New Jersey USA)を使用することが好ましい。撹拌槽は、典型的には、たとえば、スパージャー、バッフル又は低剪断インペラを用いて、通気に適合させる。気泡塔及びエアリフトリアクターには、空気又は酸素気泡による直接通気を用いることができる。宿主細胞を無血清培地中で培養する場合、通気プロセスの結果として細胞が損傷するのを防ぐのに役立つ、プルロニックF−68のような細胞保護物質を添加することが好ましい。宿主細胞の特性に応じて、マイクロキャリアを、足場依存性細胞系の増殖基質として使用することができるし、あるいは細胞を懸濁培養に適応させることもできる(こちらが一般的である)。宿主細胞、特に脊椎動物宿主細胞の培養は、バッチ、流加培養、反復バッチ処理(Drapeauら、(1994) cytotechnology 15: 103-109を参照されたい)、延長バッチ法又は潅流培養といった、さまざまな運転モードを利用することができる。組換えによって形質転換された哺乳動物宿主細胞は、ウシ胎仔血清(FCS)を含む培地のような血清含有培地で培養することができるが、こうした宿主細胞は、Keenら、(1995) Cytotechnology 17:153-163に記載の合成無血清培地、又はProCHO−CDM若しくはUltraCHOTM(Cambrex NJ, USA)といった市販の培地中で、必要ならば、グルコースなどのエネルギー源及び組換えインスリンのような合成増殖因子を添加して、培養することが好ましい。宿主細胞の無血清培養は、その細胞が無血清条件での増殖に適応していることを必要とすると考えられる。1つの適応法は、こうした宿主細胞を血清含有培地で培養し、宿主が無血清条件に適応するように、培地の80%を無血清培地に繰り返し交換することである(たとえば、Scharfenberg Kら、(1995)「動物細胞工学」(Animal Cell technology)より: 21世紀に向けた開発(Developments towards the 21st century)(Beuvery E.C.ら、編)、619-623ページ, Kluwer Academic publishersを参照されたい)。
【0069】
培地中に分泌された本発明の抗体は、さまざまな技法を用いて培地から回収及び精製され、目的の用途に適した精製度を与えることができる。たとえば、本発明の治療用抗体をヒト患者の治療に使用するには、一般に還元SDS−PAGEで測定した場合に少なくとも95%の純度、より典型的には98%、若しくは99%の純度が要求される(治療用抗体を含む粗製の培地と比較した場合)。第1の例では、細胞片を培地から、一般には、遠心によって除去し、次に、たとえば、精密濾過、限外濾過及び/又は深層濾過を用いて、上清の清澄化ステップを行う。あるいは、前もって遠心分離を行わずに、精密濾過、限外濾過又は深層濾過により抗体を回収してもよい。さまざまな他の技法、たとえば、透析及びゲル電気泳動、並びにクロマトグラフ法、例としてヒドロキシアパタイト(HA)、アフィニティクロマトグラフィー(状況に応じて、ポリヒスチジンのようなアフィニティタグ標識システムを含む)及び/又は疎水性相互作用クロマトグラフィー(HIC、米国特許第5,429,746号を参照されたい)が利用できる。一実施形態において、本発明の抗体は、さまざまな清澄化ステップの後、プロテインA若しくはGアフィニティクロマトグラフィーを用いて捕捉し、次いでさらに、イオン交換及び/又はHAクロマトグラフィー、陰イオン若しくは陽イオン交換、サイズ排除クロマトグラフィーといったクロマトグラフィーステップや、硫安沈澱を行う。一般に、さまざまなウイルス除去ステップも行われる(たとえば、DV−20フィルターなどを用いたナノ濾過)。こうしたさまざまなステップの後、少なくとも10mg/ml以上、たとえば100mg/ml以上の本発明の抗体を含む、精製(典型的にはモノクローナル)調製物が得られ、したがってその調製物が本発明の実施形態を構成する。超遠心分離によって100mg/ml以上への濃縮を行うことができる。適切なことに、この調製物は本発明の抗体の凝集体を実質的に含まない。
【0070】
細菌系は、抗体フラグメントの発現に特に適している。こうしたフラグメントは、細胞内又はペリプラズム内に局在する。当業者に公知の方法にしたがって、不溶性ペリプラズムタンパク質を抽出し、リフォールディングして活性タンパク質を形成することができる。Sanchezら、(1999) J. Biotechnol. 72, 13-20及びCupit PMら、(1999) Lett Appl Microbiol, 29, 273-277を参照されたい。
【0071】
6.医薬組成物
上記の本発明の抗体の精製調製物(特にモノクローナル調製物)を、上で概説したヒトの疾患及び障害の治療に使用するための医薬組成物に組み入れることができる。典型的には、こうした組成物は、許容される医薬慣習で知られ、必要とされる、薬学的に許容される(すなわち不活性)担体をさらに含む。たとえば、Remingtons Pharmaceutical Sciences, 第16版、(1980), Mack Publishing Co.を参照されたい。このような担体の例としては、適当なバッファーでpH5〜8の範囲内に緩衝化された、生理食塩水、リンガー溶液又はブドウ糖溶液のような滅菌担体がある。注射用(たとえば、静脈内、腹腔内、皮内、皮下、筋肉内若しくは門脈内投与による)又は持続点滴用の医薬組成物は、好適には目に見える粒子状物質を含まず、0.1ng〜100mgの抗体、典型的には5mg〜25mgの抗体を含有することができる。このような医薬組成物の製造方法は、当業者に周知である。一実施形態において、医薬組成物は、0.1ng〜100mgの本発明の治療用抗体を、状況に応じて使用説明書と共に、単位投与剤形中に含む。本発明の医薬組成物は、当業者に周知の又は明白な方法にしたがって投与前に再構成するために凍結乾燥されていてもよい。本発明の実施形態がIgG1アイソタイプを有する本発明の抗体を含む場合、クエン酸塩(たとえば、クエン酸ナトリウム)又はEDTA又はヒスチジンといった銅のキレート剤を医薬組成物に添加して、このアイソタイプの抗体の、銅を介した分解の度合いを低下させることができる。欧州特許第0612251号を参照されたい。
【0072】
本発明の抗体を投与するための有効量及び治療計画は、一般に、経験的に決定され、患者の年齢、体重及び健康状態、並びに治療すべき疾患又は障害といった要因に左右される。こうした要因は主治医の裁量の範囲内にある。適当な用量を選択する際の指針は、たとえばSmithら、(1977)「ヒトの診断及び治療における抗体」(Antibodies in human diagnosis and therapy)、Raven Press, New Yorkに見出すことができるが、概して1mg〜1000mgである。一実施形態において、RAに罹患したヒト患者の治療用の投与計画は、おおよそ100mg(すなわち50mg〜200mg)の本発明の抗体(又はその抗原結合フラグメント)を毎週又は2週間毎に皮下投与するものである。本発明の組成物はまた予防的に用いることもできる。
【0073】
治療対象の疾患又は障害に応じて、治療上有効な量の本発明の抗体を含む医薬組成物を、有効量の別の薬剤、たとえば、抗炎症薬(たとえば、NSAID、メトトレキセート、、ブシラミン、チオリンゴ酸ナトリウム、又は1以上の抗TNFα治療、例えばEnbrelTM(エタネルセプト)、RemicadeTM(インフリキシマブ)、HumiraTM(アダリムマブ)及び/若しくはCDP870)とともに、同時に、別々に、又は順次、使用することができる。本発明の抗体は、有効量の抗TNF−α受容体抗体と組み合わせて用いることができる。Davis MWら、(2000) Ann Rheum Dis 59(Suppl 1): 41-43を参照されたい。他の実施形態において、本発明の抗体は、以下:IL−1/IL−1R(例えばKineretTM)、CTLA4−Ig、IL−6(Choyら、(2002) Ann.Rheum.Dis 61(suppl 1): 54参照)、IL−8、IL−15、VEGF、IL−17、IL−18(Taylorら、(2001) Curr.Opin.Immunol.13: 611-616参照)、抗ICAM及び/又は抗CD4抗体に対して生起される薬剤、並びにMMPファミリーのメンバー、例えばMMP−1、2、3及び/又は13に対して生起される薬剤の有効量と組み合わせて用いることができる。本発明の抗体はまた、炎症性プロセスに関与することが知られる、細胞を剥離する薬剤と組み合わせて用いることができ、例えばCD20陽性B細胞について例えばMabtheraTM(リツキシマブ)を用いる。本発明の抗体と組み合わせる他の治療法としては、抗血管新生療法、例えばインテグリンαvβ3のアンタゴニスト、クリングル1〜5(Sumariwalla Pら、(2003), Arthritis Res Ther 5:R32-R39参照)、可溶性Flt−1(Miotlaら、(2000) Lab.Invest. 80:1195-1205参照)など;抗COX−2剤又は抗OSM剤、例えば抗OSM抗体(WO2005/095457号参照。その全内容が本明細書中に具体的に組み入れられ、読者はそれを具体的に参照する)などが挙げられる。簡便には、本発明の抗体又はその抗原結合フラグメントの部分の、そのような別の医薬の部分と一緒の、場合によっては使用説明書と一緒の、キット(キットオブパーツ)を含む医薬組成物もまた本発明は意図している。これらの組み合わせは、関節炎疾患/障害(関節リウマチなど)の治療に特に有用である。
【0074】
7.臨床用途
本発明の抗体は、IL−18が媒介する疾患、例えば自己免疫疾患などの治療的処置に用いることができる。具体的には、多発性硬化症、関節炎疾患(関節リウマチなど)、1型糖尿病、炎症性腸疾患(IBD)及び乾癬が挙げられる。従って、本発明はさらに、hIL−18の中和に応答する疾患(多発性硬化症、関節リウマチ、1型糖尿病、IBD、乾癬など)に罹患したヒト患者の治療方法であって、本発明の抗体(特に配列番号9に示される配列を有する重鎖と配列番号13に示される配列を有する軽鎖とを有する抗体)の治療有効量を該患者に投与することを含む方法を包含する。
【0075】
また、上述した疾患/障害のいずれか1つ(又は複数)の治療のための医薬の製造における本発明の抗体の使用も提供される。
【0076】

【0077】
7.例証
以下の実施例により、本発明の様々な態様を説明する。全ての一般的クローニング、ライゲーション、及び他の組換えDNA技術は、Maniatisら, Molecular cloning (A laboratory manual), Cold Spring Harbor Laboratory、又はSambrookら Molecular Cloning (A laboratory manual), Cold Spring Harbor Laboratoryに一般的に教示されているように実施する。本明細書で使用するベクター系及び追加の分子生物学的方法は、その内容全体を参照により本明細書に組み込み、かつ読者が具体的に参照するWO2005/095457号に開示されている。
【0078】
7.1.ハイブリドーマ可変領域のクローニング
親ラット抗体2C10は、米国特許第6,706,487号に記載されている。読者は、具体的にはこの文書を参照されたい。上記公開されている、ヒトIgG1若しくはκC領域に結合しているラットV領域をベースとして、キメラ抗体2C10cを設計した。重鎖及び軽鎖構築体に、一般的免疫グロブリンシグナル配列及び翻訳開始コドンATGを導入した。VLドメインを形成し、既にヒトCκ領域(配列番号36)を含む哺乳動物発現ベクター中にクローニングできるように、HindIII及びBsiWI制限エンドヌクレアーゼ部位を設計した。VHドメインを形成し、既にヒトγ1C領域(配列番号34)を含む哺乳動物発現ベクター中にクローニングできるように、HindIII及びSpeI制限エンドヌクレアーゼ部位を設計した。これにより、配列番号33に示す公開済み配列から得た、2C10のVh領域のフレームワーク4に2個のアミノ酸変化がもたらされた(Kabat残基107及び108)。
【0079】
重複オリゴヌクレオチドを使用して、PCRによってコード配列全体を構築し、上記発現ベクター中にクローニングした。配列を確認後、キメラ抗体をCHO細胞で発現させた。生成された抗体は、rProteinAセファロースでのアフィニティクロマトグラフィーにより、細胞培養上清から精製した。その親ラット2C10に匹敵する効力を実証するために、インビトロ結合アッセイで、2C10キメラ抗体を評価した。これは、ELISAでヒト若しくはアカゲザルIL18との結合のEC50値を測定することによって(図16及び17)、又はKG−1バイオアッセイでIFN−γの放出を阻害することによって(図15を参照されたい)行った。
【0080】
7.2.ヒト化
7.2.1.軽鎖ヒト化手順
ラット2C10可変軽鎖配列について、ラット2C10可変軽鎖配列と64%の同一性(CDR含有)を有するヒト生殖細胞系アクセプターフレームワークを選択した(F_IGKV1D−12−1、配列番号38)。生殖系列V領域は、配列類似性(配列番号40)に基づき、コンピューターで(in silico)好適なFR4、この場合はJ領域κ2ミニ遺伝子(Kabat Vol.II)と組み合わせた。3個のヒト化変異体は、配列比較、及び抗体機能への潜在的影響に基づき生成した。構築体L1は、上で選択したヒトアクセプターフレームワーク中にラットCDR(Kabat定義を使用)を線状に移植(straight graft)したものであった。構築体L2は、L1を基礎にして、残基71に1個の追加の復帰突然変異を有した。構築体L3は、L2を基礎にして、残基45、83、84及び85に4個の追加の復帰突然変異を有した。表1を参照されたい。
【0081】
【0082】
7.2.2.重鎖のヒト化手順
ラット2C10可変重鎖配列について、ラット2C10可変重鎖と59%の同一性(CDR含有)を有する、ヒト生殖細胞系アクセプターフレームワークを選択した(Fp_IGHV1−f_2、配列番号37)。生殖系列V領域は、配列類似性(配列番号39)に基づき、コンピューターで好適なFR4、この場合はJH6ミニ遺伝子(Kabat Vol.II)と組み合わせた。このフレームワークに基づき、3個のヒト化可変重鎖変異体を設計した。H1は、残基27、28、29及び93に4個の追加の復帰突然変異を有するラットCDR(Kabat定義を使用)の移植体である。これにより、(Chothiaが定義するように)CDRの一部を構成しうる親(すなわちドナー)抗体のCDR1の直上流に、極めて珍しいアミノ酸配列を得ることができる。H2は、H1を基礎にして、残基39及び40に2個の追加の復帰突然変異を有した。次いで、H3は、H2を基礎にして、残基36、71、89及び91に別の4個の追加の復帰突然変異を有した。表2を参照されたい。
【0083】
【0084】
7.3.2C10Cのヒト化
重複オリゴヌクレオチドの構築及びPCR増幅によって、ヒト化V領域を新規に合成した。哺乳動物発現ベクター中にクローニングするためにプライマーに制限部位を含め、分泌のためにヒト免疫グロブリンシグナル配列を含めた。ヒト化V領域は、HindIII及びSpeIを使用し、H1、H2及びH3として、ヒトγ1定常領域を含む哺乳動物発現ベクター中にクローン化し、そしてHindIII及びBsiWIを使用し、L1、L2及びL3として、ヒトκ定常領域を含む哺乳動物発現ベクター中にクローン化した。これにより、ヒトIgG1アイソタイプのヒト化重鎖変異体、及びヒトκアイソタイプのヒト化軽鎖変異体が得られた。
【0085】
7.3.1.ヒト化重鎖抗体とヒト化軽鎖抗体の組合せの発現
四回の試験でCHOK1細胞に一過性にトランスフェクトした。上清の抗体濃度をアッセイし、次いで2C10ラット−ヒトキメラと比較することによって、その上清をインビトロ結合アッセイで使用した。
【0086】
全9個の変異体の大規模な一過性発現は、各フラスコについて8mlの培地(OptiMEM/glutamax/5%FBS)中に含まれた、51.4μgの軽鎖プラスミド及び8.6μgの重鎖プラスミドと、240μgのトランスフェクション脂質(この脂質は、WO2006/053783号、実施例13に記載されており、その内容全体を参照により本明細書に組み込む)とを混合し、2個のほぼコンフルエントなCHOK1細胞のT175フラスコに、この混合物を通常の組織培養条件下、72時間添加することによって実施した。シェーカーフラスコを使用して、ポリクローナルCHO細胞系でも抗体をmg量で発現させ、FPLC及びプロテインAを使用して精製した。
【0087】
7.4.インビトロ結合アッセイ
7.4.1.Biacore分析
ヒト化2C10抗体のBiacoreTM動態分析は、BiacoreTM3000機により、HBS−EP緩衝液(BiacoreTM)中でその抗体のプロテインA捕捉を使用し実施した。簡単に説明すると、製造業者が推奨するプロトコルを使用し、一級アミン結合によってCM5チップにプロテインAをおよそ2000〜4000共鳴単位(RU)の密度で固定した。次いで、プロテインA表面にヒト化抗体を流し、安定化期間後およそ200〜500RUのレベルで捕捉させ、捕捉した抗体表面に(ヒト若しくはアカゲザルの)IL18を所定の濃度で流し、結合センサーグラムを得た。酸性溶出条件を使用して再生することにより、捕捉した抗体はプロテインA表面から全て除去され、表面の結合能が著しく低減することはなかった。全ての曲線は、IL18に代わる緩衝液の注入に対して二重に基準を取り、BiaEval 4.1で全体適合パラメーターを使用してデータを1:1結合モデルに適合させた。同じプロテインA捕捉法を使用して解離速度ランキング実験を準備したが、1種類のIL18濃度(10nM)のみを使用した。データは、動態分析の結合モデルと同じ結合モデルを使用し適合させたが、解離速度に1種類の検体濃度のみを使用したことが報告されているので、この値は、正確な動態測定を得るというよりはむしろランキングに有用であり、どの抗体をさらに調査するかの選択法として使用した。
【0088】
最初の25℃での結果では、構築体は全て、ラット2C10親抗体としてヒトIL18と類似する結合親和性を有することが示された。しかし、37℃で解離速度ランキング実験を実施したとき、L1構築体は解離速度に上昇が見られ、L2及びL3構築体と比較して不首尾な結果が示された(表3a及び3b)。
【0089】
【0090】
L1構築体は、アカゲザルIL18との結合親和性の点で、37℃で不首尾な結果を示したのと同様に、25℃でも最低の結果を示した(表4a)。これらの観察に基づき、37℃でのヒトIL18及びアカゲザルIL18との結合について、選択した抗体をより詳細に調査した。表4bに示すヒトIL−18のデータは、6回の別個の測定の平均(及び標準偏差)である。アカゲザルIL−18のデータは、H1L2及びH1L3については2回の実験の平均及び標準偏差を示し、H3L2及びH3L3のデータは1回の実験から得たものである。このデータの比較的高い標準偏差は、おそらく37℃でこの実験を実施した結果である。
【0091】
L1とL2構築体との差が、軽鎖の位置71でのチロシンの復帰突然変異によるフェニルアラニンの置換であることを考慮すると、L1構築体が、相対的に劣る結果を示した事実は驚くべきことである。チロシンとフェニルアラニンはもちろん芳香族アミノ酸であり、従ってフレームワーク構造のそのような些細な変化によって、37℃でBiacoreTM系で観察された(結合親和性の点で)顕著な結果が生じたことは予想外であった(しかし、25℃では観察されなかった)。
【0092】
【0093】
変異体H1L1、H1L2、及びH1L3を選択して、以下第7.4.2節でさらに分析した。
【0094】
7.4.2.Biacore分析T100のデータ
T100 BiacoreTM機を使用してさらにいくつかの変異体抗体の特性決定を実施した。この機器は、高温での緩衝効果を最小化する直列ガス抜き装置(inline degasser)を使用することによって、高温での感受性、温度制御性、及び基線安定性の点で、BiacoreTM3000よりも優れている。また、自動データ分析など、高度なソフトウェアも提供する。
【0095】
方法は、上の第7.4.1節で使用した方法と基本的に同じであり、一級アミン結合によってプロテインAをCM5チップに2000〜6000RUの密度で固定した。HBS−EP(BiacoreTM)で試験を実施した。抗IL18抗体を100〜500RUの密度で捕捉させ、ヒトIL18を16〜0.0625nMの濃度でこの捕捉した表面に流し、0nM濃度(すなわち、緩衝液は捕捉した抗体の注入のみ)を二重基準に使用した。各IL18注入後の再生は、10mMグリシン(pH1.5)の注入を2回を使用する、穏やかな酸性溶出により行なった。この再生ステップにより、プロテインA表面から、捕捉された抗体を除去した(従ってそれに結合するいかなるIL18も除去した)。再生しても、続く抗体パルスと結合するプロテインA表面の能力は大きく変化することなく、次の捕捉事象を行うことができた。得られた結合曲線は、1:1結合モデルを使用するT100機に本来備わる分析ソフトウェアを用いて分析した。示した温度で実験を実施した。
【0096】
15℃、20℃、25℃、32℃及び37℃でのH1L1及びH1L2の結合分析
様々な温度で上記方法を使用して実験を実施した。図1は、温度の結合速度(on-rate)(ka)に対する影響を示し、図2は解離速度(kd)に対する影響、そして図3は平衡定数(KD)に対する影響を示す。表5は、これらの図を作成するために使用した動態値の詳細である。
【0097】
【0098】
データは、試験した2個の抗体の結合速度が、試験した温度範囲にわたり類似することを示し、H1L2の結合速度の方が速い場合は、37℃での最終値まで、H1L2の結合速度の方が一般的に速い。しかし、解離速度に目を向けた場合、より大きな差異が見られ、15℃、20℃、25℃では2個の抗体の解離速度は類似するが、32℃及び37℃で差異を見せ始め、H1L1の解離速度の方が速くなる。これらの変化は、全体的平衡定数(それはkd/kaの関数である)に反映され、H1L1とH1L2との間の差異は、解離速度(kd)によって定義した、主として抗体/IL18複合体の安定性であることを示している。
【0099】
25℃及び37℃でのH1L1、H1L2、H1L3、及びキメラ2C10の結合分析
上記のように実験を実施した。表6は、得られた動態パラメーターの詳細である。データから、25℃及び37℃の平衡定数KDによって定義した結合という点で、H1L2はH1L1よりも良好な抗体であることが示されるが、動態パラメーターは、25℃ではH1L2がH1L1よりも結合速度(ka)が良好であることを示している。37℃では、立場は逆転し、このことは、37℃で見られるH1L2による優れた結合が、むしろ解離速度(kd)によることを示し、L2とL1を区別する変異によって、IL18−抗体複合体の高温での安定性が増大することを示している。
【0100】
【0101】
データは、いくつかの別々のデータセット(n)の平均であり、平均及び標準偏差(括弧内に標準偏差)を示す。5つの異なる温度での分析から得られたH1L1及びH1L2の25℃と37℃での実験の値が、このデータセットに含まれている。
【0102】
7.4.3.IL18結合ELISAにおける2C10cヒト化変異体の評価
精製抗体調製物の様々なバッチを使用して全9個のヒト化変異体のELISAを少なくとも6回実施した。図4A〜4Cには、表7に示すEC50値ランキングを作成した、1回の実験の代表的データを示す。2.5μg/mlの16D10(非中和マウスモノクローナル抗体)を使用し、Nunc Maxisorp96ウェルプレートにヒトIL−18を固定して、5ng/mlの組換えヒトIL−18を捕捉させた。様々な希釈率で抗IL−18ヒト化抗体を加えた。抗ヒトIgG Fc特異的ペルオキシダーゼコンジュゲート(Sigma A0170)を使用して、結合したヒト化抗体を検出した。
【0103】
【0104】
全ての変異体の効力は、2C10キメラに非常に近似しているように見え、このことは、ヒト化による効力の損失はほとんどないことを示している。これらのアッセイを数回反復して得られたEC50値では、変異体ランキングは得られなかったが、ELISA単独ではこれらの変異体を明確に区別することはできなかった(表7及び図4A〜4Cを参照)。BiacoreTMを使用して、変異体をある程度区別することができ(第7.4.1節及び第7.4.2節)、それによってわずか4個の変異体がもたらされ、それらをヒトIL18及びアカゲザルIL18を使用する数回の独立した反復実験でより詳しく試験した(表8、図5[ヒト]及び図6[アカゲザル])。
【0105】
【0106】
7.4.4.IL−18結合ELISAにおける2C10ヒト化H1変異体の評価
H1L1、H1L2及びH1L3という3個のヒト化H1変異体を用いてELISAを実施し、室温及び37℃において、ヒト血清及びブロッキング溶液(1%BSA含有PBS0.05%TWEEN(w/v))中でのヒトIL−18結合を評価した。Nunc Maxisorp96ウェルプレートにヒト化抗体変異体を2.5μg/mlで固定した。5ng/mlの組換えヒトIL−18の捕捉は、室温又は37℃で、ヒト血清又はブロック溶液中で実施した。抗IL−18マウスモノクローナル抗体16D10を加えた。結合したマウス抗体は、抗マウスκペルオキシダーゼコンジュゲート(Serotec MCA 1291P)を使用して検出した。研究データから得られた代表的EC50値を表9に示す。
【0107】
【0108】
このアッセイでは、ヒトIL−18結合ステップを実施する温度を室温から37℃へ変化させても、3個のヒト化H1変異体の効力は影響を受けない。ヒト血清中に抗体が存在すると、低い結合シグナルが観察される。
【0109】
7.4.5.37℃でのヒト化H1変異体抗体の安定性評価
H1L1、H1L2及びH1L3という3個のヒト化H1変異体の保存安定性は、ヒト血清及びリン酸緩衝食塩水中、37℃で14日間にわたって評価した。抗体を50μg/mlに希釈し、安定性は、37℃で0、1、4、6、8及び14日間というインキュベーション時間後に、IL−18結合ELISAによって評価した。IL−18結合ELISAについては、16D10(非中和マウスモノクローナル抗体)をNunc Maxisorpプレートに固定して、5ng/mlの組換えヒトIL−18を捕捉した。37℃での経時インキュベーションの種々の時点でサンプリングした抗IL−18ヒト化抗体を加えた。結合したヒト化抗体は、抗ヒトIgG Fc特異的ペルオキシダーゼコンジュゲートを使用して検出した。
【0110】
このアッセイ型式では、37℃という温度に、0、1、4、6、8及び14日間という長時間曝露を行なっても、ヒトIL−18に結合するその能力に基づく、ヒト化H1抗体変異体の結合効力に影響はなかった。
【0111】
7.5.滑液存在下でのH1L2とヒトIL18との結合
50%ヒト滑液に500ng/ml〜0ng/mlの範囲の組換えヒトIL18を添加しELISAを実施した。次いで、H1L2抗体をコーティングしたMaxisorp96ウェルプレート(Nunc)のウェルに、IL−18を含むこの溶液をアプライした。次いで、ビオチン化した抗IL−18抗体(D045−6, MBL)及びストレプトアビジン−HRPにより、結合したIL−18を検出した。組換えヒトIL18を含む50%滑液(SF)を滴定すると、最大半値のほんのわずかなシフトが見られただけで、ヒト組換えIL−18を含む緩衝液を滴定したものと、ほぼ同じ曲線が生成された。図7を参照されたい。これにより、抗体が治療設定で遭遇する結合環境により類似性が高い50%ヒトSFの存在下でさえ、hIL−18に結合する抗体能力が実証されている。
7.5.1.IL18結合タンパク質(IL18bp)存在下でのIL18に対する結合
BiacoreTM技術(BiacoreTM3000)及びELISAを使用して、ヒトIL18bpの存在下でもH1L2が依然としてヒトIL18と結合できるかどうかを判定した。IL18bpは、IL18に対して高親和性を有し、IL−18機能の天然阻害薬として働く(図8、図9A及び図9B、表10及び表11)。
【0112】
Biacoreによる分析
簡単に説明すると、一級アミン結合によってCM5チップにプロテインAを表面がおよそ4000共鳴単位(RU)の密度になるように固定した。次いで、組換えFc−IL18結合タンパク質(R&D Systems)を濃度3μg/ml、流速10μl/分で1分間流し、これにより、およそ1400RUのIL18結合タンパク質が捕捉された。次いで、捕捉したIL18結合タンパク質表面に、IL18を濃度30nM、流速10μl/分で5分間流した。この後、IL18結合タンパク質/IL18表面に、濃度10nMのラット2C10親抗体を流速30μl/分で3分間流した。2C10抗体によって認識されるIL18上のエピトープが、IL18結合タンパク質との相互作用部位に干渉するならば、いかなる結合シグナルも見られないはずである。図10は、2C10がhIL−18と結合すること、続いてこのhIL−18がhIL−18BPによって捕捉されたことを実証し、このことは、2C10とhIL−18BPの結合部位が重複しないことを示している。
【0113】
【0114】
ELISAによる分析
ヒト化H1L2若しくはラット2C10 MAbと、ヒトIL18が結合した捕捉IL18bpとの結合を、直接結合ELISAで試験したが、その際、Nunc Maxisorpプレートに、組換えヒトIL18bp−Fc融合タンパク質(R&D Systems #119-BP)を0.5μg/mlでコーティングした。組換えヒトIL18(社内試薬)を100ng/mlでブロッキング緩衝液(1%w/vBSA含有PBS)に加えた。精製した抗体を0.5ng/ml〜1μg/mlの濃度範囲で加えた。抗ヒトκ軽鎖特異的HRPコンジュゲート(Sigma)若しくは抗ラットIgG HRPにより結合した抗体を検出した。図18A及び18Bに、得られた結果を示す。
【0115】
7.6.インビトロバイオアッセイ
7.6.1.KG−1細胞系における、IL18の刺激によるIFN−γの放出を中和する、ヒト化構築体の活性
このアッセイでは、IL−18特異的抗体の中和活性を計測するが、アッセイはKG−1細胞におけるIL−18を媒介するIFNγの誘導に基づく。KG−1(ATCC#CCL−246)は、機能性IL−18受容体を構成的に発現し、従って外来性IL−18刺激に応答するヒト骨髄単球細胞系である。
【0116】
全9個のヒト化変異体について、KG−1細胞におおけるヒトIL18に刺激されたIFN−γの放出を阻害するその能力を評価した(表12及び図11)。
【0117】
【0118】
BiacoreTM分析に基づき、組換えヒトIL18及びアカゲザルIL18に最も高い親和性示した、4個の好ましいヒト化変異体で、少なくともさらに6回の反復実験を実施した。図8に、4個の好ましいヒト化変異体及びH1L1の代表的結果を示し、表13に、CHOe1a細胞由来の同じタンパク質バッチ物質によって全て実施した、これらのアッセイの結果の概要を示す。
【0119】
【0120】
H1L1を他のヒト化変異体と比較した事例では、試験し、かつ2C10親MAbとも比較した、その他の4個のヒト化構築体と比べて、H1L1は低い効力を示した。
【0121】
4個の好ましいモノクローナル抗体を用いてさらなる分析を実施したが、KG−1細胞:2C10、並びに2C10から得られたヒト化変異体(H1L1、H1L2及びH1L3)により、IL−18に刺激されたINFγの放出を阻害する能力について、それらのモノクローナル抗体を比較した。96ウェルプレートで、50ng/mlの組換えヒトIL−18、及びIL18に特異的な種々の濃度の抗体(2μg/ml〜7.8ng/mlの範囲、二倍希釈)、又は陰性アイソタイプ対照(Synagis、抗RSV抗体)を37℃及び5%CO2下で1時間インキュベートし、続いてウェル当たり3.105個のKG−1細胞を添加することによって、KG−1バイオアッセイを実施した。最後に、それらのプレートを37℃、5%CO2下で20〜24時間インキュベートした。上清を回収し、市販のヒトIFNγELISAキット(Biosource AHC4432;AHC4539)を使用してIFNγの産生を定量した。
【0122】
3回の実験を実施した。IL−18に刺激されたIFNγの産生の阻害結果を陰性対照に対して正規化した。統計分析は、あらゆるSynagis応答の調整後、各実験について各mAbのIC50推定値を得ることを目的とした。次いで、IC50推定値を統計学的に分析して、95%信頼区間(すなわち統計学的に信頼できる範囲)で各mAbの全体的IC50推定値を生成した。最後に、ダネット検定を使用して2C10に戻って各ヒト化変異体を比較した。
【0123】
図12に、代表的実験を示す。表14及び図13に、95%信頼区間での各モノクローナル抗体の全体的IC50推定値、及びp値と信頼区間と共にラット2C10の変化率(%)を示す。
【0124】
【0125】
H1L1は、2C10よりも統計上有意に効力が低い(p<0.001)。
【0126】
その他のヒト化変異体は、2C10と有意には異ならないが、H1L3対2C10の比較が境界線である。
【0127】
7.6.2.刺激したヒトPBMC中のIFN−γ放出の中和におけるH1L2の活性
3ドナー由来のヒトPBMCは、組換えヒトIL18及び抗CD3抗体で刺激し、4個の選択したヒト化抗体変異体の希釈系列を加えた作用を研究した。比較するために、それぞれにドナー親2C10抗体及びIL18bpを含めた。3ドナーのうち2ドナーでは、IL18及び抗CD3刺激は失敗し、IFN−γは検出されなかった。残りのドナーについては、結果は低濃度では非常に変化したが、IL18が誘発するIFN−γの産生の完全阻害は、ヒト化変異体を含む様々な抗IL18抗体を添加することによって達成することができた。図14を参照されたい。
【0128】
さらにLPSが刺激するヒトPBMCにより実施した実験では、IL18を産生し、付随するIFNγの放出が生じた。濃度依存的にLPSによってIFNγの産生が誘発され、1μg/mlの一定濃度で2C10親モノクローナル抗体を添加すると、この刺激が完全に阻害され、このことは、その効果にはIL18が介在するということ、及び内在性IL18は中和できるということ(データ非掲示)を示している。これは、全血でも実証できたが、2C10による阻害効果は、用量依存性ではあったが、あまり明白ではなかった(データ非掲示)。図9に、3個の独立したドナーによる実験、並びに親ラットモノクローナル2C10、H1L2、及びIL18bpによる阻害の結果を示す。ドナー1及び3は同様の結果を示したが、ドナー2はLPSによる刺激でIFNγを放出しなかった(図示せず)。IL18が媒介するIFNγ放出は、10%若しくは25%のヒト血清の存在下、1μg/mlを超える抗体若しくはIL18bpを加えることによって、完全に阻害することができる。阻害は、10ng/ml以上で既に観察可能であり、10%若しくは25%のヒト血清の存在下でのこの阻害のIC50値を表11に示す。
【0129】
【0130】
7.6.3.IL18オルソログとの結合の概要
ELISA及びBiacore、そしてさらにKG−1細胞バイオアッセイを使用し、抗体と他の種由来のIL18との結合を検査した。これは、当初、アカゲザル/カニクイザルIL18について親2C10及びキメラ2C10cを使用し実施したが、いくつかのヒト化変異体で繰り返した。Biacore及びELISAを使用し、ブタ、マウス及びラットIL18との結合について親2C10を試験し、4個の最も優れたヒト化変異体をアカゲザル/カニクイザル、及びイヌのIL18との結合について試験した。KG−1バイオアッセイは、アカゲザル/カニクイザルのIL18、並びに3個のモノクローナル抗体、2C10キメラ、及びヒト化変異体の代表であるH3L3で実施した。全てのヒト化変異体の類似性を考慮すると、これは、H1L2を含めて生成された他の変異体の代表である可能性が高い(表15、図8を参照)。
【0131】
【0132】
7.7.IL18抗体の円二色性及び熱変性試験
円二色性(CD)試験を使用して、温度、特に25℃〜37℃の関数として、IL18抗体の二次構造変化を試験した。これら同じ抗体の熱変性試験を行って、それらの熱安定性と融解温度(Tm)を測定した。
【0133】
CD方法:CDスペクトルは、0.5nmステップ及びバンド幅1nmで180nm〜280nmの走査を行なうApplied Photophysics Chirascan分光計で取得した。各点ごとの取得時間は5秒であった。サンプルをPBSで約0.2mg/ml希釈し、1mm光路長(pathlength)セルに入れた。各タンパク質のスペクトルは、4℃、25℃及び37℃に設定した恒温槽で取得した。サンプルの実際の温度は、セル内の液体中に置いたプローブによって測定したが、設定温度の約3℃以内であった。
【0134】
Tm方法:Sypro orangeを1:1000で入れたリン酸緩衝食塩水(PBS)溶液で、全てのタンパク質を0.2mg/mlに希釈した。Bioneer Exicycler機器を使用し、0.5℃間隔毎に、620nm(励起490nm)における蛍光発光を測定したが、サンプル温度は各温度点で10秒待機して10℃〜95℃に勾配をつけた。変性曲線は、Grafitを使用して標準的融解等温線に適合させた。
【0135】
予想通り、全4個の抗体のCDスペクトルの形状比較から、それらの構造が高度にβシート状であり、本質的に同じ構造であることが示された。4℃〜37℃の温度範囲では、3個の抗体:
H1L1
H1L2
H1L3
は、CDによって判明した通り、その二次構造において有意な変化は示さなかった。しかし、2C10抗体キメラは、この温度範囲で構造にわずかな減少が見られた。これは、下表16に示す熱安定性傾向に一致する。
【0136】
【0137】
H1L1、H1L2及びH1L3は、体温ではいかなる変性の徴候も見せず、37℃をはるかに超えても明らかに安定している。従って、熱安定性のその差異が、正常な身体及び周囲温度で、何らかの示差的利点を付与する可能性は低い。
【0138】
配列
配列番号1
GYYFH
配列番号2
RIDPEDDSTKYAERFKD
配列番号3
WRIYRDSSGRPFYVMDA
配列番号4
LASEDIYTYLT
配列番号5
GANKLQD
配列番号6
LQGSKFPLT
配列番号7
MAAEPVEDNCINFVAMKFIDNTLYFIAEDDENLESDYFGKLESKLSVIRNLNDQVLFIDQGNRPLFEDMTDSDCRDNAPRTIFIISMYKDSQPRGMAVTISVKCEKISTLSCENKIISFKEMNPPDNIKDTKSDIIFFQRSVPGHDNKMQFESSSYEGYFLACEKERDLFKLILKKEDELGDRSIMFTVQNED
配列番号8
ATGGCTGCTGAACCAGTAGAAGACAATTGCATCAACTTTGTGGCAATGAAATTTATTGACAATACGCTTTACTTTATAGCTGAAGATGATGAAAACCTGGAATCAGATTACTTTGGCAAGCTTGAATCTAAATTATCAGTCATAAGAAATTTGAATGACCAAGTTCTCTTCATTGACCAAGGAAATCGGCCTCTATTTGAAGATATGACTGATTCTGACTGTAGAGATAATGCACCCCGGACCATATTTATTATAAGTATGTATAAAGATAGCCAGCCTAGAGGTATGGCTGTAACTATCTCTGTGAAGTGTGAGAAAATTTCAACTCTCTCCTGTGAGAACAAAATTATTTCCTTTAAGGAAATGAATCCTCCTGATAACATCAAGGATACAAAAAGTGACATCATATTCTTTCAGAGAAGTGTCCCAGGACATGATAATAAGATGCAATTTGAATCTTCATCATACGAAGGATACTTTCTAGCTTGTGAAAAAGAGAGAGACCTTTTTAAACTCATTTTGAAAAAAGAGGATGAATTGGGGGATAGATCTATAATGTTCACTGTTCAAAACGAAGACTAG
配列番号9
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHWVRQAPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTEDTSTDTAYMELSSLRSEDTAVYYCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
配列番号10
CAGGTGCAGCTGGTGCAGAGCGGAGCCGAGGTGAAGAAGCCTGGCGCCAGCGTCAAGGTGTCCTGTAAGGTGTCCGGCGAGATCAGCACCGGCTACTACTTCCACTGGGTGAGGCAGGCCCCTGGCAAGGGCCTGGAGTGGATGGGCAGAATCGACCCCGAGGACGACAGCACCAAGTACGCCGAGCGGTTCAAGGACAGGGTGACCATGACCGAGGACACCAGCACCGATACCGCCTACATGGAGCTGTCCAGCCTGAGAAGCGAGGATACCGCCGTGTACTACTGTACCACCTGGCGGATCTACAGAGACAGCAGCGGCAGACCCTTCTACGTGATGGATGCCTGGGGCCAGGGCACACTAGTGACCGTGTCCAGCGCCAGCACCAAGGGCCCCAGCGTGTTCCCCCTGGCCCCCAGCAGCAAGAGCACCAGCGGCGGCACAGCCGCCCTGGGCTGCCTGGTGAAGGACTACTTCCCCGAACCGGTGACCGTGTCCTGGAACAGCGGAGCCCTGACCAGCGGCGTGCACACCTTCCCCGCCGTGCTGCAGAGCAGCGGCCTGTACAGCCTGAGCAGCGTGGTGACCGTGCCCAGCAGCAGCCTGGGCACCCAGACCTACATCTGTAACGTGAACCACAAGCCCAGCAACACCAAGGTGGACAAGAAGGTGGAGCCCAAGAGCTGTGACAAGACCCACACCTGCCCCCCCTGCCCTGCCCCCGAGCTGCTGGGAGGCCCCAGCGTGTTCCTGTTCCCCCCCAAGCCTAAGGACACCCTGATGATCAGCAGAACCCCCGAGGTGACCTGTGTGGTGGTGGATGTGAGCCACGAGGACCCTGAGGTGAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCACAATGCCAAGACCAAGCCCAGGGAGGAGCAGTACAACAGCACCTACCGGGTGGTGTCCGTGCTGACCGTGCTGCACCAGGATTGGCTGAACGGCAAGGAGTACAAGTGTAAGGTGTCCAACAAGGCCCTGCCTGCCCCTATCGAGAAAACCATCAGCAAGGCCAAGGGCCAGCCCAGAGAGCCCCAGGTGTACACCCTGCCCCCTAGCAGAGATGAGCTGACCAAGAACCAGGTGTCCCTGACCTGCCTGGTGAAGGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAACGGCCAGCCCGAGAACAACTACAAGACCACCCCCCCTGTGCTGGACAGCGATGGCAGCTTCTTCCTGTACAGCAAGCTGACCGTGGACAAGAGCAGATGGCAGCAGGGCAACGTGTTCAGCTGCTCCGTGATGCACGAGGCCCTGCACAATCACTACACCCAGAAGAGCCTGAGCCTGTCCCCTGGCAAGTGA
配列番号11
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHWVRQAPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTEDTSTDTAYMELSSLRSEDTAVYYCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSS
配列番号12
CAGGTGCAGCTGGTGCAGAGCGGAGCCGAGGTGAAGAAGCCTGGCGCCAGCGTCAAGGTGTCCTGTAAGGTGTCCGGCGAGATCAGCACCGGCTACTACTTCCACTGGGTGAGGCAGGCCCCTGGCAAGGGCCTGGAGTGGATGGGCAGAATCGACCCCGAGGACGACAGCACCAAGTACGCCGAGCGGTTCAAGGACAGGGTGACCATGACCGAGGACACCAGCACCGATACCGCCTACATGGAGCTGTCCAGCCTGAGAAGCGAGGATACCGCCGTGTACTACTGTACCACCTGGCGGATCTACAGAGACAGCAGCGGCAGACCCTTCTACGTGATGGATGCCTGGGGCCAGGGCACACTAGTGACCGTGTCCAGC
配列番号13
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPKLLIYGANKLQDGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCLQGSKFPLTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
配列番号14
GATATCCAGATGACCCAGTCCCCCAGCAGCGTGTCCGCCTCTGTGGGCGATAGAGTGACCATCACCTGCCTGGCCAGCGAGGACATCTACACCTACCTGACCTGGTATCAGCAGAAGCCTGGCAAGGCCCCTAAGCTGCTGATCTACGGCGCCAACAAGCTGCAGGACGGCGTGCCCAGCAGATTCAGCGGCAGCGGCTCCGGCACCGACTACACCCTGACCATCAGCAGCCTGCAGCCTGAGGATTTCGCCACCTACTACTGCCTGCAGGGCAGCAAGTTCCCCCTGACCTTCGGCCAGGGCACCAAGCTGGAGATCAAGCGTACGGTGGCCGCCCCCAGCGTGTTCATCTTCCCCCCCAGCGATGAGCAGCTGAAGAGCGGCACCGCCAGCGTGGTGTGTCTGCTGAACAACTTCTACCCCCGGGAGGCCAAGGTGCAGTGGAAGGTGGACAATGCCCTGCAGAGCGGCAACAGCCAGGAGAGCGTGACCGAGCAGGACAGCAAGGACTCCACCTACAGCCTGAGCAGCACCCTGACCCTGAGCAAGGCCGACTACGAGAAGCACAAGGTGTACGCCTGTGAGGTGACCCACCAGGGCCTGTCCAGCCCCGTGACCAAGAGCTTCAACCGGGGCGAGTGC
配列番号15
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPKLLIYGANKLQDGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCLQGSKFPLTFGQGTKLEIK
配列番号16
GATATCCAGATGACCCAGTCCCCCAGCAGCGTGTCCGCCTCTGTGGGCGATAGAGTGACCATCACCTGCCTGGCCAGCGAGGACATCTACACCTACCTGACCTGGTATCAGCAGAAGCCTGGCAAGGCCCCTAAGCTGCTGATCTACGGCGCCAACAAGCTGCAGGACGGCGTGCCCAGCAGATTCAGCGGCAGCGGCTCCGGCACCGACTACACCCTGACCATCAGCAGCCTGCAGCCTGAGGATTTCGCCACCTACTACTGCCTGCAGGGCAGCAAGTTCCCCCTGACCTTCGGCCAGGGCACCAAGCTGGAGATCAAG
配列番号17
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHWVRRRPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTEDTSTDTAYMELSSLRSEDTAVYYCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
配列番号18
CAGGTCCAGCTGGTACAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGTTTCCGGAGAAATAAGTACTGGATACTATTTCCACTGGGTGCGACGAAGGCCTGGAAAAGGGCTTGAGTGGATGGGAAGGATTGATCCTGAGGATGATAGTACTAAATATGCTGAGAGGTTCAAAGACAGAGTCACCATGACCGAGGACACATCTACAGACACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAGGACACGGCCGTGTATTACTGTACCACATGGCGGATATACCGAGATAGTTCTGGCCGCCCCTTCTATGTTATGGATGCCTGGGGCCAAGGGACACTAGTCACAGTCTCCTCAGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAA
配列番号19
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHWVRRRPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTEDTSTDTAYMELSSLRSEDTAVYYCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSS
配列番号20
CAGGTCCAGCTGGTACAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGTTTCCGGAGAAATAAGTACTGGATACTATTTCCACTGGGTGCGACGAAGGCCTGGAAAAGGGCTTGAGTGGATGGGAAGGATTGATCCTGAGGATGATAGTACTAAATATGCTGAGAGGTTCAAAGACAGAGTCACCATGACCGAGGACACATCTACAGACACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAGGACACGGCCGTGTATTACTGTACCACATGGCGGATATACCGAGATAGTTCTGGCCGCCCCTTCTATGTTATGGATGCCTGGGGCCAAGGGACACTAGTCACAGTCTCCTCA
配列番号21
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHFVRRRPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTADTSTDTAYMELSSLRSEDTATYFCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
配列番号22
CAGGTCCAGCTGGTACAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGTTTCCGGAGAAATAAGTACTGGATACTATTTCCACTTTGTGCGACGAAGGCCTGGAAAAGGGCTTGAGTGGATGGGAAGGATTGATCCTGAGGATGATAGTACTAAATATGCTGAGAGGTTCAAAGACAGAGTCACCATGACCGCAGACACATCTACAGACACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAGGACACGGCCACTTATTTTTGTACCACATGGCGGATATACCGAGATAGTTCTGGCCGCCCCTTCTATGTTATGGATGCCTGGGGCCAAGGGACACTAGTCACAGTCTCCTCAGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAA
配列番号23
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHFVRRRPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTADTSTDTAYMELSSLRSEDTATYFCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSS
配列番号24
CAGGTCCAGCTGGTACAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGTTTCCGGAGAAATAAGTACTGGATACTATTTCCACTTTGTGCGACGAAGGCCTGGAAAAGGGCTTGAGTGGATGGGAAGGATTGATCCTGAGGATGATAGTACTAAATATGCTGAGAGGTTCAAAGACAGAGTCACCATGACCGCAGACACATCTACAGACACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAGGACACGGCCACTTATTTTTGTACCACATGGCGGATATACCGAGATAGTTCTGGCCGCCCCTTCTATGTTATGGATGCCTGGGGCCAAGGGACACTAGTCACAGTCTCCTCA
配列番号25
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPKLLIYGANKLQDGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCLQGSKFPLTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
配列番号26
GACATCCAGATGACCCAGTCTCCATCTTCTGTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGTCTGGCAAGTGAGGACATATACACTTATTTAACATGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGGTGCAAATAAGTTGCAAGATGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACTATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCTACAGGGTTCCAAGTTTCCGCTCACGTTTGGCCAGGGGACCAAGCTGGAGATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGACAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGT
配列番号27
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPKLLIYGANKLQDGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCLQGSKFPLTFGQGTKLEIK
配列番号28
GACATCCAGATGACCCAGTCTCCATCTTCTGTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGTCTGGCAAGTGAGGACATATACACTTATTTAACATGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGGTGCAAATAAGTTGCAAGATGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACTATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCTACAGGGTTCCAAGTTTCCGCTCACGTTTGGCCAGGGGACCAAGCTGGAGATCAAA
配列番号29
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPQLLIYGANKLQDGVPSRFSGSGSGTDYTLTISSLQPEDEGDYYCLQGSKFPLTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
配列番号30
GACATCCAGATGACCCAGTCTCCATCTTCTGTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGTCTGGCAAGTGAGGACATATACACTTATTTAACATGGTATCAGCAGAAACCAGGGAAAGCCCCTCAACTCCTGATCTATGGTGCAAATAAGTTGCAAGATGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTATACTCTCACTATCAGCAGCCTGCAGCCTGAAGATGAAGGGGATTACTATTGTCTACAGGGTTCCAAGTTTCCGCTCACGTTTGGCCAGGGGACCAAGCTGGAGATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGACAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAG
配列番号31
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPQLLIYGANKLQDGVPSRFSGSGSGTDYTLTISSLQPEDEGDYYCLQGSKFPLTFGQGTKLEIK
配列番号32
GACATCCAGATGACCCAGTCTCCATCTTCTGTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGTCTGGCAAGTGAGGACATATACACTTATTTAACATGGTATCAGCAGAAACCAGGGAAAGCCCCTCAACTCCTGATCTATGGTGCAAATAAGTTGCAAGATGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTATACTCTCACTATCAGCAGCCTGCAGCCTGAAGATGAAGGGGATTACTATTGTCTACAGGGTTCCAAGTTTCCGCTCACGTTTGGCCAGGGGACCAAGCTGGAGATCAAA
配列番号33
EVQLQQSGAELVRPGTSVKLSCKVSGEISTGYYFHFVRRRPGQGLEWIGRIDPEDDSTKYAERFKDRATLTAQTSSNTAYLNLSSLTSEDTATYFCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
配列番号34
GAGGTCCAGCTACAGCAGTCTGGGGCTGAGCTTGTGAGACCTGGGACCTCTGTGAAGTTATCTTGCAAAGTTTCTGGCGAAATAAGTACAGGATACTATTTCCACTTTGTGAGGCGAAGGCCTGGACAGGGTCTGGAATGGATAGGAAGGATTGATCCTGAGGATGATAGTACTAAATATGCTGAGAGGTTCAAAGACAGGGCGACGCTCACTGCACAAACATCCTCCAACACAGCCTACCTGAACCTCAGCAGCCTGACCTCTGAGGACACTGCAACTTATTTTTGTACCACATGGCGGATATACCGAGATAGTTCTGGCCGCCCCTTCTATGTTATGGATGCCTGGGGTCAAGGAACACTAGTCACAGTCTCCTCAGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAA
配列番号35
DIQMTQSPASLSASLGETVSIECLASEDIYTYLTWYQQKPGKSPQLLIYGANKLQDGVPSRFSGSGSGTQYSLKISGIQPEDEGDYFCLQGSKFPLTFGSGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
配列番号36
GACATTCAAATGACCCAGTCTCCAGCTTCCCTGTCTGCATCTCTGGGAGAAACTGTCTCCATCGAATGTCTGGCAAGTGAGGACATATACACTTATTTAACATGGTATCAGCAGAAACCAGGGAAATCTCCTCAACTCCTGATCTATGGTGCAAATAAGTTGCAAGATGGGGTCCCATCACGGTTCAGTGGCAGTGGATCTGGCACACAGTATTCTCTCAAGATCAGCGGCATACAACCTGAAGATGAAGGGGATTATTTCTGTCTACAGGGTTCCAAGTTTCCGCTCACGTTCGGTTCTGGGACCAAGCTGGAGATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGACAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGT
配列番号37
QVQLVQSGAEVKKPGASVKVSCKVSGYTLTELSMHWVRQAPGKGLEWMGGFDPEDGETIYAQKFQGRVTMTEDTSTDTAYMELSSLRSEDTAVYYCAT
配列番号38
DIQMTQSPSSVSASVGDRVTITCRASQGISSWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQANSFP
配列番号39
WGQGTLVTVSS
配列番号40
FGQGTKLEIK
【図面の簡単な説明】
【0139】
【図1】H1L1及びH1L2の結合速度(ka)に対する温度の影響を示す。
【図2】解離速度(kd)に対する温度の影響を示す。
【図3】平衡定数(KD)に対する温度の影響を示す。
【図4】A〜Cは、表7に示したEC50値を作成した1回の実験の代表的データを示す。
【図5】ヒトIL18に結合する4個の選択したヒト化変異体のEC50値を示す。
【図6】アカゲザルIL18に結合する4個の選択したヒト化変異体のEC50値を示す。
【図7】50%滑液の存在下におけるヒトIL18に対するH1L2の結合を示す。
【図8】KG1アッセイにおけるIL−18が刺激するIFN−γ産生の阻害を示す。
【図9】A及びBはそれぞれヒトPBMCSドナーの10%及び25%自己血清における、LPSが刺激するIFN−γ産生の阻害を示す。
【図10】hIL18BPにより捕捉されたhIL18に対する2C10の結合を示す。
【図11】KG1細胞におけるヒトIL18が刺激するIFN−γ放出を阻害する9個のヒト化変異体の能力を示す。
【図12】KG1細胞におけるH1変異体及び2C10によるIL−18が刺激するIFN−γ産生の阻害を示す。
【図13】H1変異体のIC50データを95%信頼区間と共に示す。
【図14】KG1細胞におけるヒトIL−18が刺激するIFN−γ産生の阻害を示す。
【図15】KG1細胞におけるアカゲザルIL−18が刺激するIFN−γ産生の阻害を示す。
【図16】キメラ2C10を用いたヒトIL−18結合ELISAの結果を示す。
【図17】キメラ2C10を用いたアカゲザルIL−18結合ELISAの結果を示す。
【図18】A及びBはそれぞれヒトIL−18が結合したIL−18BPに対して結合するH1L2及び2C10を用いた結合ELISAの結果を示す。
【技術分野】
【0001】
1.発明の分野
本発明は、一般的に、ヒトインターロイキン−18により媒介される状態の治療及び診断に有用な、抗体(特にヒト化抗体)などの免疫グロブリンの分野に関する。
【背景技術】
【0002】
2.発明の背景
ヒトインターロイキン−18(hIL−18)は、生物学的に活性を有しない193アミノ酸の前駆体タンパク質として合成されるサイトカインである(Ushioら、J. Immunol. 156:4274, 1996)。前駆体タンパク質の切断、例えばカスパーゼ−1又はカスパーゼ−4による切断によって、156アミノ酸の成熟タンパク質が遊離し(Guら、Science 275:206, 1997;Ghayurら、Nature 386:619, 1997)、これが、T細胞増殖の共刺激、NK細胞の細胞傷害性の増強、T細胞及びNK細胞によるIFN−γ産生の誘導、並びにTヘルパー1型(Th1)分化の増強を含む生物学的活性を示す(Okamuraら、Nature 378:88, 1995;Ushioら、J. Immunol. 156:4274, 1996;Micallefら、Eur. J. Immunol. 26:1647, 1996;Kohnoら、J. Immunol. 158:1541, 1997;Zhangら、Infect. Immunol. 65:3594, 1997;Robinsonら、Immunity 7:571, 1997)。さらに、IL−18は、ヒト単球前炎症性伝達物質、例えばIL−8、腫瘍壊死因子−α(TNF−α)及びプロスタグランジンE2(PGE2)などの効果的な誘導因子である(Ushio, S.ら、J. Immunol. 156:4274-4279, 1996;Puren, A. J.ら、J. Clin. Invest. 10:711-721, 1997;Podolinら、J. Immunol. 提出済, 1999)。
【0003】
以前にクローニングされたIL−1受容体関連タンパク質(IL−1Rrp)(Parnetら、J. Biol. Chem. 271:3967, 1996)は、IL−18受容体(Kd=18nM)のサブユニットとして同定されている(Torigoeら、J. Biol. Chem. 272:25737, 1997)。IL−18受容体の2つ目のサブユニットは、IL−1受容体付属タンパク質に対して相同性を示し、AcPL(付属タンパク質様のため)と称されている。IL−1 Rrp及びAcPLの両方の発現が、IL−18が誘導するNF−κB及びJNKの活性化に必要である(Bornら、J. Biol. Chem. 273:29445, 1998)。NF−κB及びJNKに加えて、IL−1受容体関連キナーゼ(IRAK)、p56lck(LCK)及びマイトジェン活性化タンパク質キナーゼ(MAPK)を介してIL−18はシグナル伝達する(Micallefら、Eur. J. Immunol. 26:1647, 1996;Matsumotoら、Biophys Biochem. Res. Comm. 234:454, 1997;Tsuji-Takayamaら、Biochem. Biophys. Res. Comm. 237:126, 1997)。
【0004】
TH1細胞は、IFN−γ、IL−2及びTNF−βなどの前炎症性サイトカインを産生するが(Mosmannら、J. Immunol. 136:2348, 1986)、これは多くの自己免疫疾患、例えば多発性硬化症(MS)、関節リウマチ(RA)、1型すなわちインスリン依存型糖尿病(IDDM)、炎症性腸疾患(IBD)、及び乾癬などの媒介に関与しているとされている(Mosmann及びSad, Immunol. Today 17:138, 1996)。従って、TH1促進性サイトカイン(IL−18など)のアンタゴニスト作用は、疾患の発症を抑制すると予想される。Il−18特異的mAbは、アンタゴニストとして用いることができるはずである。
【0005】
自己免疫疾患の発症におけるIL−18の役割は証明されている。すなわち、非肥満糖尿病(NOD)マウスの膵臓及び脾臓において、その疾患発症直前にIL−18の発現が有意に増大することが示されている(Rotheら、J. Clin. Invest. 99:469, 1997)。同様に、関節リウマチ患者の滑液においてIL−18レベルが顕著に上昇することが示されている(Kawashimaら、Arthritis and Rheumatism 39:598, 1996)。さらに、IL−18投与によって、Th1媒介自己免疫疾患であるマウス実験的アレルギー性脳脊髄炎(EAE)(多発性硬化症のモデル)の臨床的重篤度が増すことが示されている。加えて、中和抗ラットIL−18抗血清によってLewis雌ラットにおけるEAEの発症が予防されることが示されている(Wildbaumら、J. Immunol. 161:6368, 1998)。従って、IL−18は、自己免疫のための新規な治療薬の開発のための望ましい標的である。
【0006】
Taniguchiら(J. Immunol. Methods 206:107)は、4つの異なる抗原性部位に結合する7種のマウス及び6種のラット抗ヒトIL−18モノクローナル抗体(mAb)を記載している。マウスmAbの1つ(#125−2H)と、6つのラットmAbは、KG−1細胞によるIL−18誘導性IFN−γの産生を阻害し、ラットmAbは#125−2Hよりも10倍低い中和活性を示す。ウエスタンブロット分析によって示されているように、マウスmAbのうち3つ(ただしラットmAbはなし)は、膜に結合したヒトIL−18と強く反応する。さらに、#125−2H及びラットmAbを用いてヒトIL−18を検出するための酵素結合免疫吸着アッセイ(ELISA)が記載されている。このELISAの検出限界は10pg/mlである。
【0007】
欧州特許出願EP0 712 931号は、2種のマウス抗ヒトIL−18 mAb、すなわちH1(IgG1)及びH2(IgM)を開示している。ウエスタンブロット分析によって示されているように、両方のmAbは膜に結合したヒトIL−18と反応するが、膜に結合したヒトIL−12とは反応しない。ヒトIL−18を精製するためにイムノアフィニティークロマトグラフィープロトコールにおいてH1を使用し、そしてヒトIL−18を測定するためにELISAにおいてH1を使用する。ヒトIL−18を測定するためにラジオイムノアッセイにおいてH2を使用する。
【発明の開示】
【発明が解決しようとする課題】
【0008】
中和IL−18抗体は、ヒトにおける自己免疫疾患及び関連する症候の緩和に有用である可能性がある。従って、当該技術分野においては、Th1細胞分化及び増殖を低減し、それゆえ自己免疫疾患及び関連する症候を低減する、高親和性IL−18アンタゴニスト、例えばヒトインターロイキン18に対する中和モノクローナル抗体が必要とされている。
【0009】
本明細書全体で言及する参照文献は全て、明示的かつ全体的に参照により本明細書に組み入れられる。
【課題を解決するための手段】
【0010】
3.発明の概要
本発明により、以下の相補性決定領域(CDR):
CDRH1:配列番号1
CDRH2:配列番号2
CDRH3:配列番号3
CDRL1:配列番号4
CDRL2:配列番号5
CDRL3:配列番号6
を有する重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体が提供される。
【0011】
本発明により、以下のCDR:
CDRH1:配列番号1
CDRH2:配列番号2
CDRH3:配列番号3
CDRL1:配列番号4
CDRL2:配列番号5
CDRL3:配列番号6
を有する重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体であって、該軽鎖の位置71の残基が、前記CDRが由来するドナー抗体中に存在するその対応する残基によって置換されている、ヒト化抗インターロイキン−18抗体が提供される。
【0012】
当業者には明らかなとおり、「由来する」という用語は、それが物質の物理的起源であるという意味で、起源を明らかにするのみならず、その物質と構造的に同一であるもののその基準起源から生じたのではない物質も明示することを意図するものである。したがって、「CDRが由来するドナー抗体フレームワーク中に存在する」対応する残基は、かならずしもドナー抗体フレームワークから精製されたものである必要はない。同様に、「ドナー抗体」由来CDRは、かならずしもドナー抗体から精製されたものである必要はない。
【0013】
CDR及びフレームワーク領域(FR)、並びにアミノ酸の番号付けは、特に指摘しない限り、Kabatら、"Sequences of immunological interest", NIHに記載されているKabat定義に従う。
【0014】
本発明の別の態様において、ヒトアクセプターフレームワークに移植されたドナー抗体由来CDRを含むヒト化抗インターロイキン−18抗体であって、抗インターロイキン−18抗体が配列番号1、2、3、4、5及び6に示される配列を有するCDRを含み、該抗インターロイキン−18抗体の軽鎖の位置71の残基が、該ドナー抗体フレームワーク中のその対応する位置に存在する残基と同一である、ヒト化抗インターロイキン−18抗体が提供される。
【0015】
本発明の別の態様においては、配列番号1、2、3、4、5及び6に示されるCDRを含み、軽鎖の位置71にチロシンを含む、ヒト化抗インターロイキン−18抗体が提供される。
【0016】
本発明の別の態様において、配列番号1、2及び3に示されるCDRを有する重鎖と配列番号4、5及び6に示されるCDRを有する軽鎖を含むヒト化抗インターロイキン−18抗体であって、該軽鎖CDRが、ドナー抗体軽鎖の位置71にチロシンを有するドナー抗体に由来する、ヒト化抗インターロイキン−18抗体が提供される。
【0017】
本発明の別の態様において、ドナー抗体由来CDR及びヒト化抗体軽鎖の位置71にチロシンを含むヒト化抗インターロイキン−18抗体であって、該ドナー抗体が、2C10又はそのフレームワーク変異体である(すなわち、2C10と同じCDRであるが、異なるフレームワークを含む。米国特許第6,706,487号を参照されたい)、ヒト化抗インターロイキン−18抗体が提供される。
【0018】
本発明の別の態様では、
(a)ヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有するCDRを有する重鎖、並びに
(b)ヒト軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有するCDRを有する軽鎖
を含むヒト化抗インターロイキン−18抗体であって、
前記ヒト軽鎖アクセプターフレームワークが配列番号38に由来するフレームワークを含み、かつ配列番号38の位置71がチロシンである、ヒト化抗インターロイキン−18抗体が提供される。
【0019】
本発明の別の態様では、
(a)ヒトIL−18との特異的結合を可能にするCDRを有する重鎖、
(b)アクセプターフレームワークを含み、かつ配列番号4、5及び6に示される配列を有するCDRを有し、そして位置71にチロシン残基を有する軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0020】
軽鎖のCDRは、好ましくは、配列番号35に示される配列内の、配列番号4、5及び6に示される配列のそれぞれの位置に対応するアクセプターフレームワーク内の位置に位置する。軽鎖及び/又は重鎖は、好ましくはヒト患者において非免疫原性のものである。
【0021】
本発明の別の態様において、
(a)配列番号1、2及び3に示される配列を有するCDRを含む重鎖、並びに
(b)ヒト軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有するCDRを含む軽鎖
を含むヒト化抗インターロイキン−18抗体であって、
該ヒト化抗インターロイキン−18抗体の該軽鎖アクセプターフレームワークが、配列番号38に示される配列の変異体に由来するフレームワーク領域を含み、該変異体が位置71にチロシンを含み、かつ該変異体が配列番号38に示される配列を有するフレームワークと75%以上の同一性を有する、ヒト化抗インターロイキン−18抗体が提供される。該変異体は、配列番号38に示されるフレームワークと、好ましくは80%以上、例えば81%、82%、83%、84%、より好ましくは85%以上、例えば86%、87%、88%、89%、よりさらに好ましくは90%以上、例えば91%、92%、93%、94%、最も好ましくは95%以上、例えば96%、97%、98%、99%の同一性を含む。
【0022】
本発明の別の態様において、
(a)ドナー抗体軽鎖の位置71にチロシンを含むドナー抗体に由来する配列番号1、2、3、4、5及び6に示されるCDR、
(b)ヒト軽鎖の位置71にフェニルアラニンを含むヒトアクセプターフレームワーク
を含むヒト化抗インターロイキン−18抗体であって、
該抗インターロイキン−18抗体がその軽鎖の位置71にチロシンを含む、ヒト化抗インターロイキン−18抗体が提供される。
【0023】
本発明の別の態様において、
(a)ドナー抗体軽鎖の位置71に芳香族アミノ酸を含むドナー抗体に由来する配列番号1、2、3、4、5及び6に示されるCDR、
(b)軽鎖アクセプターフレームワークの位置71に、部分(a)の芳香族アミノ酸とは異なる種類の芳香族アミノ酸を含むヒトアクセプターフレームワーク
を含むヒト化抗インターロイキン−18抗体であって、
該抗インターロイキン−18抗体が、位置71に部分(a)の抗体に由来する芳香族アミノ酸を有する軽鎖を含む、ヒト化抗インターロイキン−18抗体が提供される。
【0024】
本発明の別の態様では、37℃において表面プラスモン共鳴(例えば、BiacoreTM、好ましくは以下の第7.4.1節に記載するBiacoreTM3000機器及び条件を用いて)によって測定した場合のヒトIL−18の結合に関して300pM以下の平衡定数(KD)を示すヒト化抗インターロイキン−18抗体が提供される。
【0025】
本発明の別の態様では、配列番号1、2、3、4、5及び6に示されるCDRを含み、かつ37℃において表面プラスモン共鳴(例えば、BiacoreTM、好ましくは以下の第7.4.1節に記載するBiacoreTM3000機器及び条件を用いて)によって測定した場合のヒトIL−18の結合に関して300pM以下の平衡定数(KD)を示す、ヒト化抗インターロイキン−18抗体が提供される。
【0026】
好ましくは、37℃において表面プラスモン共鳴(好ましくは以下の第7.4.2節に記載するBiacoreTMT100機器及び条件を用いて)によって測定した場合のヒトIL−18の結合に関する抗体の平衡定数(KD)は、90pM未満である。平衡定数は、より好ましくは70pM以下、よりさらに好ましくは65pM、60pM、55pM、又は50pM以下である。
【0027】
本発明の別の態様において、37℃において表面プラスモン共鳴(例えば、BiacoreTM、好ましくは以下の第7.4.2節に記載するBiacoreTMT100機器及び条件を用いて)によって測定した場合のヒトIL−18の結合に関して0.0002(1/秒)以上の解離定数又は解離速度(kd)を示すヒト化抗インターロイキン−18抗体が提供される。
【0028】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRが重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、93、39、40、36、71、89、91の一個若しくは複数の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71、及び任意により位置45、83、84、85の一個若しくは複数の(例えば全ての)残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0029】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該抗インターロイキン−18抗体の軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0030】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、39、40、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0031】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、36、39、40、71、89、91、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0032】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0033】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、93、39、40の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0034】
本発明の別の態様において、
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、93、39、40、36、71、89、91の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列に由来するフレームワーク領域を含み、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体が提供される。
【0035】
本発明の別の態様において、重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体であって、ヒトIL−18と前記抗体との結合の25℃での解離速度(kd)と、ヒトIL−18と前記抗体との結合の37℃での解離速度(kd)との比が、1:5(又は1:5未満)であり、該抗体がドナー抗体由来CDRとヒトアクセプターフレームワークを含み、かつヒトアクセプターフレームワークの軽鎖の位置71の残基が、そのドナー抗体のその対応する残基によって置換されている、ヒト化抗インターロイキン−18抗体が提供される。解離速度は、好ましくは、以下の第7.4.2節に記載するBiacoreTMT100機器及び条件を用いて測定される。
【0036】
本発明の別の態様において、配列番号9、配列番号17、配列番号21からなる群から選択された重鎖と、配列番号13、配列番号29からなる群から選択された軽鎖を含むヒト化抗インターロイキン−18抗体が提供される。
【0037】
特に本発明は、配列番号9の重鎖と配列番号13の軽鎖、又は配列番号9の重鎖と配列番号29の軽鎖を含むヒト化抗インターロイキン−18抗体を提供する。
【0038】
本発明はまた、配列番号17の重鎖と配列番号13の軽鎖、又は配列番号17の重鎖と配列番号29の軽鎖を含むヒト化抗インターロイキン−18抗体を提供する。
【0039】
また本発明は、配列番号21の重鎖と配列番号13の軽鎖、又は配列番号21の重鎖と配列番号29の軽鎖を含むヒト化抗インターロイキン−18抗体を提供する。
【0040】
本発明の別の態様において、本明細書において上述した抗インターロイキン−18抗体、及び薬学的に許容される担体を含む医薬組成物が提供される。
【0041】
本発明の別の態様において、治療に使用する抗体(特に、リガンドと受容体との相互作用を阻害する抗体、抗インターロイキン−18抗体など)を選択する方法であって、
(a)30〜45℃(好ましくは37℃)の温度において前記抗体が特異的に結合する抗原に対する前記抗体の結合親和性を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(b)20〜25℃(好ましくは25℃)の温度において前記抗体が特異的に結合する抗原に対する前記抗体の結合親和性を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(c)(a)の親和性が(b)の親和性を超える場合、好ましくは(a)の親和性がステップ(b)の親和性の2倍以上、より好ましくは4倍以上である場合に、治療に使用する前記抗体を選択するステップ
を含む方法が提供される。
【0042】
本発明の別の態様において、治療に使用する抗体(特に、リガンドと受容体との相互作用を阻害する抗体、抗インターロイキン−18抗体など)を選択する方法であって、
(a)30〜45℃(好ましくは37℃)の温度において前記抗体が特異的に結合する抗原からの前記抗体の解離速度を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(b)20〜25℃(好ましくは25℃)の温度において前記抗体が特異的に結合する抗原からの前記抗体の解離速度を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(c)(a)の解離速度が(b)の解離速度よりも遅い場合、治療に使用する前記抗体を選択するステップ
を含む方法が提供される。
【0043】
「抗インターロイキン−18」という用語は、本発明の抗体を参照する場合、ヒトインターロイキン−18の生物学的活性を中和することが可能な抗体を意味する。しかしながら、そのような抗体が非ヒト霊長類(例えばアカゲザル及び/又はカニクイザル)インターロイキン−18の生物学的活性をさらに中和する可能性があることを排除するものではない。
【発明を実施するための最良の形態】
【0044】
さらに図面を参照しながら本発明を説明する。
【0045】
4.ヒト化抗体
インタクトな非ヒト抗体をヒトの疾患又は障害の治療に使用することは、特に抗体を反復投与した際に、現在十分に立証されている潜在的な免疫原性の問題を引き起こす。すなわち、患者の免疫系はインタクトな非ヒト抗体を非自己と認識して、中和応答を開始する可能性がある。こうした問題を克服するために、完全にヒトの抗体(上記参照)の開発に加えて、長年にわたってさまざまな技法が開発されてきたが、こうした技法は、一般に、免疫した動物、たとえばマウス、ラット又はウサギから非ヒト抗体を得ることの相対的な容易さを維持する一方で、インタクトな治療用抗体における非ヒトアミノ酸配列の構成割合を減らすことを必要とする。概して、これを達成するために2つの手法が用いられてきた。第1はキメラ抗体であって、これは一般に、ヒト定常領域に融合された非ヒト(たとえば、マウスなどの齧歯類)可変領域を含む(Morrison (1984), PNAS, 81, 6851参照)。抗体の抗原結合部位は可変領域内に位置するので、キメラ抗体は、抗原に対するその結合親和性を維持するが、ヒト定常領域のエフェクター機能を獲得しているため、上記のようなエフェクター機能を果たすことができる。キメラ抗体は、典型的には、組換えDNA法を用いて作製される。従来の方法を用いて(たとえば、本発明の抗体のH鎖及びL鎖をコードする遺伝子、例としては上記の配列番号1、2、3、4、5及び6をコードするDNAと、特異的に結合することができるオリゴヌクレオチドプローブを使用することによって)、抗体をコードするDNA(たとえばcDNA)を単離し、配列を決定する。ハイブリドーマ細胞は、こうしたDNAの典型的な供給源となる。キメラ抗体を発現することが望まれる場合には、軽鎖及び重鎖の全長成熟可変領域をコードする単離cDNAを適当な発現ベクターに読み取り枠を合わせて挿入する。該発現ベクターは、特に、適当な免疫グロブリン定常領域(通常はヒト起源のもの)と、シグナル配列、停止コドン、プロモーター、ターミネーター、及び抗体の発現を達成するために必要な他のエレメントを含む。かかるベクターを、次に本来免疫グロブリンを産生しない大腸菌、COS細胞、CHO細胞又は三重ローマ細胞といった宿主細胞にトランスフェクトして、抗体を合成させる。DNAは、ヒトL鎖及びH鎖のコード配列を、対応する非ヒト(たとえばマウス)H及びL定常領域の代わりに用いることによって、改変することができる。たとえば、Morrison; PNAS 81, 6851 (1984)を参照されたい。
【0046】
第2の手法は、可変領域をヒト化することによって抗体の非ヒト含量を減少させる、ヒト化抗体の作製に関する。ヒト化について2つの技術が広く支持されている。第1は、CDRグラフティングによるヒト化である。CDRは抗体のN末端近くにループを作り、そこで、フレームワーク領域によってもたらされるスキャフォールドに嵌め込まれた表面を形成する。抗体の抗原結合特異性は、主として、そのCDR表面のトポグラフィー及び化学的性質によって決まる。次いで、これらの特徴は、個別のCDRのコンフォメーションによって、CDRの相対的配置によって、並びにCDRを構成する残基の側鎖の性質及び配置によって決定される。免疫原性の大幅な低下は、非ヒト(たとえばマウス)抗体(「ドナー」抗体)のCDRのみを、好適なヒトのフレームワーク(「アクセプターフレームワーク」)及び定常領域上にグラフティングすることによって達成することができる(Jonesら、 (1986) Nature 321,522-525、及びVerhoeyen Mら、(1988) Science 239, 1534-1536を参照されたい)。しかしながら、CDRグラフティングは本質的に、抗原結合特性の完全な保持をもたらさない可能性があり、有意な抗原結合親和性を回復したいならば、ドナー抗体のフレームワーク残基(「復帰突然変異」と呼ばれることもある)の一部をヒト化分子内に保持する必要があることが、多くの場合、明らかになっている(Queen C ら、(1989) PNAS 86, 10,029-10,033, Co, Mら、(1991) Nature 351, 501-502を参照されたい)。この場合、ヒトフレームワーク(FR)を用意するために、非ヒトドナー抗体に対して最大の配列相同性(典型的には60%以上)を示すヒトV領域をデータベースから選択する。ヒトFRは、ヒトコンセンサス又は個別のヒト抗体のどちらからも選択することができる。必要ならば、ドナー抗体由来の重要残基を、ヒトアクセプターフレームワーク内に置換して入れて、CDRコンフォメーションを保存する。抗体のコンピューターモデリングを用いることができるが、それは、このような構造的に重要な残基を同定するのに役立つ。国際公開WO99/48523号を参照されたい。
【0047】
別法として、ヒト化は、「veneering」のプロセスによって行うことができる。特異なヒト及びマウス免疫グロブリンの重鎖及び軽鎖可変領域の統計分析によって、露出した残基の正確なパターンがヒトとマウスの抗体で異なること、並びにほとんどの個々の表面位置は少数の異なる残基に強い選択性があることが明らかになった(Padlan E.A.ら、(1991) Mol.Immunol.28, 489-498、及びPedersen J.T.ら、(1994) J.Mol.Biol. 235; 959-973を参照されたい)。したがって、そのフレームワーク領域内の、ヒト抗体に通常存在するものとは異なる、露出した残基を置き換えることによって、非ヒトFvの免疫原性を低下させることができる。タンパク質の抗原性は表面への接近可能性と相関しうるので、表面残基を置き換えることは、マウス可変領域をヒトの免疫系に「見えない」ようにするのに十分であると考えられる(Mark G.E.ら、(1994)、実験薬理学ハンドブック(Handbook of Experimental Pharmacology)第113号より:モノクローナル抗体の薬理学(The pharmacology of モノクローナル 抗体)、Springer-Verlag、105-134ページも参照されたい)。こうしたヒト化の手順は、抗体の表面だけを変更し、支持する残基はそのままになっているので、「veneering」と呼ばれる。さらに別の手法としては、WO04/006955号に記載されている方法、及びHumaneeringTM(Kalobios)のプロセスが挙げられる。この方法は、細菌発現系を使用して、ヒト生殖系と配列が近縁の抗体を産生する(Alfenito-M Advancing Protein Therapeutics, January 2007, San Diego, California)。別の最近のヒト化手法は、ヒトアクセプターフレームワークを、フレームワーク領域などの抗体の他の領域間の相同性よりもむしろ、ドナーマウス抗体CDR領域とのヒトCDR領域の構造類似性に基づいて選択することを含む。このプロセスは、SuperhumanisationTMとしても知られる(Evogenix Inc.; Hwang ら (2005) Methods 36:35-42)。
【0048】
従って本発明は、上の第3節に記載したヒト化抗体に関する。このようなヒト化抗体は、好ましくはIgGアイソタイプ(IgG1又はIgG4など)のヒト定常領域を含む。
【0049】
別の実施形態において、上の第3節で記載したヒト化可変領域は、非ヒト定常領域(「逆キメラ(reverse chimera)」)、例えば非ヒト霊長類、ラット、マウス又はウサギのものなどと融合しうる。
【0050】
当業者には明らかなように、配列番号37及び38に示されるアクセプターフレームワークは、それぞれVH遺伝子及びVκ遺伝子によってコードされる免疫グロブリンのアミノ酸を構成する。これらはそれ自体がアクセプター抗体のフレームワーク領域及びCDRを含む。アクセプター抗体のCDRを配列番号1、2、3、4、5及び6に示されるドナーCDRと置換し、そして得られる配列を好適なフレームワークの4つの配列(配列番号39及び配列番号40に示される配列など)と結合させて、完全な免疫グロブリン可変領域(配列番号11及び配列番号15に示されるものなど)を生成することは当業者の能力の範囲内である。
【0051】
4.1.他の改変
抗体のFc領域とさまざまなFc受容体(FcγR)との相互作用は、抗体のエフェクター機能を仲介すると考えられるが、このエフェクター機能には、抗体依存性細胞傷害(ADCC)、補体結合、食作用、及び抗体の半減期/クリアランスが含まれる。所望のエフェクター特性に応じて、本発明の抗体のFc領域のさまざまな改変を行うことができる。たとえば、本来なら溶解性の抗体を非溶解性にするFc領域内の特異的変異が、欧州特許第0629 240B1号及び同第0307 434B2号に詳述されており、あるいは、抗体にサルベージ(salvage)受容体結合エピトープを組み込んで、血清半減期を延ばすことができる。米国特許第5,739,277号を参照されたい。現在知られているヒトFcγ受容体には5つあり、FcγR(I)、FcRγIIa、FcγRIIb、FcγRIIIa及び新生児FcRnである。Shieldsら、(2001) J. Biol. Chem 276, 6591-6604は、共通のIgG1残基のセットがすべてのFcγRとの結合に関与しているが、FcγRII及びFcγRIIIは、この共通のセットの外側にある別個の部位を利用することを明らかにした。一群のIgG1残基は、Pro−238、Asp−265、Asp−270、Asn−297及びPro−239をアラニンに変えると、すべてのFcγRへの結合を低下させた。これらはいずれもIgG CH2ドメインに存在し、CH1及びCH2をつなぐヒンジ部の近くでクラスターを形成している。FcγRIは、結合のためにIgG1残基の共通セットのみを用いるが、FcγRII及びFcγRIIIは、共通セットに加えて、別個の残基とも相互作用する。一部の残基の変更は、FcγRIIとの結合のみを低下させ(たとえば、Arg−292)、又はFcγRIIIとの結合のみを低下させた(たとえば、Glu−293)。ある変異体は、FcγRII又はFcγRIIIに対する結合の強化を示したが、他の受容体との結合には影響を与えなかった(たとえば、Ser−267Alaは、FcγRIIに対する結合は強化したが、FcγRIIIとの結合には影響しなかった)。また他の変異体は、FcγRII又はFcγRIIIに対する結合の強化を示したが、他の受容体との結合は低下した(たとえば、Ser−298AlaはFcγRIIIに対する結合を強化したが、FcγRIIとの結合は低下した)。FcγRIIIaに関して、もっともよく結合するIgG1変異体は、Ser−298、Glu−333及びLys−334でのアラニン置換の組み合わせを有していた。新生児FcRn受容体は、抗体クリアランス及び組織にわたるトランスサイトーシスのどちらにも関与していると考えられる(Junghans R. P (1997) Immunol. Res 16. 29-57 及びGhetieら、(2000) Annu. Rev. Immunol. 18, 739-766を参照されたい)。ヒトFcRnと直接相互作用すると確定されたヒトIgG1残基には、Ile253、Ser254、Lys288、Thr307、Gln311、Asn434及びHis435がある。本発明は、半減期/クリアランス並びに/又はエフェクター機能(ADCC及び/若しくは補体溶解など)を改変するために、上述した残基の変化のいずれか1(又はそれ以上)を有する本発明の抗体にも関する。
【0052】
他の改変には、本発明の抗体のグリコシル化変異体がある。抗体の定常領域内の保存された位置でのグリコシル化は、抗体機能、特に上記のようなエフェクター機能に重大な影響を及ぼすことが知られている。たとえば、Boydら、(1996), Mol. Immunol. 32, 1311-1318を参照されたい。1又は複数の糖鎖を付加、置換、欠失又は修飾した、本発明の治療用抗体又はその抗原結合フラグメントのグリコシル化変異体が想定される。アスパラギン−X−セリン、又はアスパラギン−X−スレオニンモチーフの導入は、糖鎖を酵素的に連結することができる部位を作り出すので、それを用いて、抗体のグリコシル化を操作することができる。Rajuら、(2001) Biochemistry 40, 8868-8876において、TNFR−IgGイムノアドヘシンの末端シアリル化は、β−1,4−ガラクトシルトランスフェラーゼ及び/又はα−2,3−シアリルトランスフェラーゼを用いる再ガラクトシル化及び/又は再シアリル化により増加した。末端シアリル化の増加は、免疫グロブリンの半減期を長くすると考えられる。抗体は、ほとんどの糖タンパク質と同様に、典型的には複数のグライコフォームの混合物として天然に産生される。こうした混合物は、抗体を真核生物、とりわけ哺乳動物細胞内で産生させたとき、特に明らかである。特定したグライコフォームを製造するために、さまざまな方法が開発されてきた。Zhangら、Science (2004), 303, 371、 Searsら、Science, (2001) 291, 2344、 Wackerら、(2002) Science, 298 1790、Davisら、(2002) Chem. Rev. 102, 579、Hangら、(2001) Acc. Chem. Res 34, 727を参照されたい。したがって、本発明は、前記抗体又はその抗原結合フラグメントの特定された数(たとえば7個以下、例として5個以下、たとえば2又は1個)のグライコフォームを含む、本明細書に記載の複数の治療用抗体(典型的にはモノクローナル抗体)(IgG1アイソタイプ、たとえばIgG1とすることができる)に関する。
【0053】
本発明のさらに他の実施形態には、ポリエチレングリコール(PEG)、ポリプロピレングリコール又はポリオキシアルキレンといった、非タンパク質性ポリマーと結合した本発明の治療用抗体又はその抗原結合フラグメントが含まれる。タンパク質とPEGとの結合は、タンパク質の半減期を長くするのと同時に、タンパク質の抗原性及び免疫原性を低下させるための確立された技術である。分子量及び形状(直鎖若しくは分枝)の異なるPEG付加の使用は、インタクト抗体、及びFab’フラグメントで検討されている。Koumenis I.L.ら、(2000) Int. J. Pharmaceut. 198:83-95を参照されたい。
【0054】
5.作製方法
本発明の抗体は、ヤギ(Pollockら、(1999), J. Immunol. Methods 231:147-157を参照されたい)、ニワトリ(Morrow KJJ (2000) Genet. Eng. News 20:1-55を参照されたい)、マウス(Pollockら、前掲)、又は植物(Doran PM, (2000) Curr. Opinion Biotechnol. 11, 199-204、Ma JK-C (1998), Nat. Med. 4; 601-606、Baez Jら、BioPharm (2000) 13: 50-54、Stoger Eら、(2000) Plant Mol. Biol. 42:583-590を参照されたい)といったトランスジェニック生物内で産生することができる。抗体はまた、化学合成によっても作製することができる。しかしながら、本発明の抗体は、典型的には、当業者に周知の組換え細胞培養技術を用いて作製される。抗体をコードするポリヌクレオチドを単離し、その後の宿主細胞におけるクローニング(増幅)又は発現のために、複製可能なベクター(プラスミドなど)に挿入する。特に宿主細胞がCHO又はNSOである場合(下記を参照されたい)、1つの有用な発現系は、グルタミン酸シンテターゼ系(Lonza Biologicsが販売しているものなど)である。抗体をコードするポリヌクレオチドは、従来の方法(たとえば、オリゴヌクレオチドプローブ)によって、容易に単離され、配列決定される。使用可能なベクターには、プラスミド、ウイルス、ファージ、トランスポゾン、ミニ染色体(プラスミドがその典型的な具体例である)がある。一般にこうしたベクターは、発現を促進するために、軽鎖及び/又は重鎖ポリヌクレオチドに機能的に連結された、シグナル配列、複製起点、1つ若しくは複数のマーカー遺伝子、エンハンサーエレメント、プロモーター、及び転写終結配列をさらに含有する。軽鎖及び重鎖をコードするポリヌクレオチドは、別々のベクター内に挿入し、(例えば形質転換、トランスフェクション、エレクトロポレーション又は形質導入によって)同一の宿主細胞に同時に又は連続的に導入することができるし、あるいは所望であれば、そのような導入の前に重鎖及び軽鎖をともに同一ベクター内に挿入してもよい。
【0055】
遺伝子コードの縮重のため、本発明のポリペプチドをコードする本明細書に記載のポリヌクレオチドの代わりとなるポリヌクレオチドも利用可能であることは、当業者にはすぐにわかることである。
【0056】
5.1.シグナル配列
本発明の抗体は、成熟タンパク質のN末端に特異的な切断部位を有する、異種シグナル配列を含む融合タンパク質として作製することができる。シグナル配列は、宿主細胞によって認識されて、プロセシングされる必要がある。原核生物の宿主細胞については、シグナル配列は、アルカリホスファターゼ、ペニシリナーゼ、又は耐熱性エンテロトキシンIIリーダーとすることができる。酵母分泌のためには、シグナル配列は、酵母インベルターゼリーダー、α因子リーダー、又は酸ホスファターゼリーダーとすることができる。例えばWO90/13646号を参照されたい。哺乳動物細胞系では、ウイルス分泌リーダー、たとえば単純ヘルペスgDシグナル、並びに天然免疫グロブリンシグナル配列(ヒトIg重鎖など)が利用できる。典型的には、シグナル配列は、本発明の抗体をコードするポリヌクレオチドに、リーディングフレーム内で連結される。
【0057】
5.2.複製起点
複製起点は当技術分野で周知であり、pBR322がほとんどのグラム陰性細菌に適合し、2μプラスミドがほとんどの酵母に適合し、さらに、さまざまなウイルス由来のもの、たとえば、SV40、ポリオーマ、アデノウイルス、VSV若しくはBPVはほとんどの哺乳動物細胞に適している。一般に、複製起点の要素は、ベクターの増殖を大腸菌内で行う必要がない限り、組込み型哺乳動物発現ベクターには必要でない。しかしながら、SV40 oriは、初期プロモーターを含有しているので使用することができる。
【0058】
5.3.選択マーカー
典型的な選択遺伝子は、(a)抗生物質若しくは他の毒素(たとえば、アンピシリン、ネオマイシン、メトトレキセート若しくはテトラサイクリン)に対する耐性を付与するタンパク質、又は(b)栄養要求性の欠乏を補完する、若しくは複合培地で利用できない栄養素を補充するタンパク質、あるいは(c)その両方の組み合わせであるタンパク質をコードする。選択方式は、ベクターを含まない又はベクターを含む宿主細胞の増殖の停止を行いうる。本発明の治療用抗体をコードする遺伝子による形質転換が成功した細胞は、たとえば共送達される選択マーカーによって付与された薬剤耐性のために、生き残る。一つの例はDHFR選択系であるが、この場合、形質転換体はDHFR陰性宿主株において作出される(例えばPage and Sydenham 1991 Biotechnology 9: 64-68参照)。この系では、DHFR遺伝子が本発明の抗体ポリヌクレオチド配列と共送達されて、ヌクレオシド除去によりDHFR陽性細胞が選択される。必要に応じて、DHFR阻害剤であるメトトレキサートも使用して、DHFR遺伝子増幅を示す形質転換体を選択する。DHFR遺伝子を本発明の抗体コード配列又はその機能的誘導体に機能的に連結することによって、DHFR遺伝子増幅が目的の所望抗体配列が同時に増幅されるようにする。CHO細胞は、DHFR/メトトレキサート選択に特に有用な細胞株であり、DHFR系を利用した宿主細胞の増幅及び選択方法は当技術分野で十分に確立されている(Kaufman R.J. ら、 J.Mol.Biol. (1982) 159, 601-621を参照のこと。概説については、Werner RG, Noe W, Kopp K,Schluter M,”Appropriate mammalian expression systems for biopharmaceuticals”, Arzneimittel-Forschung. 48(8):870-80, 1998 Augを参照のこと)。さらにもう一つの例は、グルタミン酸シンテターゼ発現系(Lonza Biologics)である。酵母での使用に適した選択遺伝子はtrp1遺伝子である。Stinchcombら、Nature 282, 38, 1979を参照されたい。
【0059】
5.4.プロモーター
本発明の抗体の発現に適したプロモーターは、抗体をコードするDNA/ポリヌクレオチドに、機能するように連結される。原核生物宿主のためのプロモーターには、proAプロモーター、βラクタマーゼ及びラクトースプロモーター系、アルカリホスファターゼ、トリプトファン及びハイブリッドプロモーター、たとえばTacがある。酵母細胞での発現に適したプロモーターには、3−ホスホグリセリン酸キナーゼ又は他の解糖系酵素、たとえばエノラーゼ、グリセルアルデヒド3リン酸デヒドロゲナーゼ、ヘキソキナーゼ、ピルビン酸デカルボキシラーゼ、ホスホフルクトキナーゼ、グルコース6リン酸イソメラーゼ、3−ホスホグリセリン酸ムターゼ及びグルコキナーゼがある。誘導性酵母プロモーターには、アルコールデヒドロゲナーゼ2、イソチトクロームC、酸ホスファターゼ、メタロチオネイン、及び窒素代謝又はマルトース/ガラクトース利用を担う酵素が含まれる。
【0060】
哺乳動物細胞系における発現のためのプロモーターには、RNAポリメラーゼIIプロモーター、例えばウイルスプロモーター、たとえば、ポリオーマ、鶏痘及びアデノウイルス(たとえばアデノウイルス2)、ウシパピローマウイルス、トリ肉腫ウイルス、サイトメガロウイルス(具体的には前初期遺伝子プロモーター)、レトロウイルス、B型肝炎ウイルス、アクチン、ラウス肉腫ウイルス(RSV)プロモーター、及び初期若しくは後期シミアンウイルス40、並びに非ウイルスプロモーター、例えばEF−1α(Mizushima and Nagata Nucleic Acids Res 1990 18(17):5322)が挙げられる。プロモーターの選択は、発現に使用される宿主細胞との適切な適合性に基づく。
【0061】
5.5.エンハンサーエレメント
必要に応じて、たとえば、高等真核生物での発現のために、さらなるエンハンサーエレメントを、上述したプロモーターの代わりに、又はそのプロモーター内に位置することが見出されているものと一緒に挿入することができる。適当な哺乳動物エンハンサー配列には、グロビン、エラスターゼ、アルブミン、フェトプロテイン、メタロチオネイン、及びインスリン由来のエンハンサーエレメントが含まれる。あるいはまた、SV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、ポリオーマエンハンサー、バキュロウイルスエンハンサー、又はマウスIgG2a遺伝子座といった、真核細胞ウイルス起源のエンハンサーエレメントを使用してもよい(WO04/009823号を参照されたい)。このようなエンハンサーは典型的に、ベクター上で、プロモーターの上流の部位に位置するが、他の位置、例えば非翻訳領域内又はポリアデニル化シグナルの下流に位置してもよい。エンハンサーの選択及び位置決定は、発現に使用される宿主細胞との適切な適合性に基づく。
【0062】
5.6.ポリアデニル化/終止
真核細胞系では、ポリアデニル化シグナルが、本発明の抗体をコードするポリヌクレオチドに、機能しうるように連結される。こうしたシグナルは典型的には、オープンリーシングフレームの3’側に置かれる。哺乳動物系で、限定的でない例として、シグナルとしては、成長ホルモン、伸長因子1α、並びにウイルス(たとえば、SV40)遺伝子、又はレトロウイルスのロング・ターミナル・リピート由来のものが挙げられる。酵母系では、ポリアデニル化/終結シグナルの限定的でない例として、ホスホグリセリン酸キナーゼ(PGK)及びアルコールデヒドロゲナーゼ1(ADH)遺伝子由来のシグナルが挙げられる。原核細胞系では、ポリアデニル化シグナルは一般的には必要とされず、その代わりにもっと短く明確な転写終結配列を用いるのが通例である。ポリアデニル化/終結配列の選択は、発現に使用される宿主との適切な適合性に基づいて行われる。
【0063】
5.7.収量改善のための他の方法/エレメント
上記に加えて、収量改善のために用いることができる他の特徴としては、クロマチン再構成エレメント、イントロン及び宿主細胞特異的コドン改変が挙げられる。宿主細胞のコドンの偏りに適合させるために本発明の抗体のコドン利用法を改変して、転写及び/又は生成物収率を高めることができる(例えば、Hoekema A ら、Mol Cell Biol 1987 7(8):2914-24)。コドンの選択は、発現に使用される宿主との適切な適合性に基づいて行われる。
【0064】
5.8.宿主細胞
本発明の抗体をコードするベクターをクローニングし、又は発現させるのに適した宿主細胞は、原核細胞、酵母、又は高等真核細胞である。適当な原核細胞には、真正細菌、例を挙げると、腸内細菌科、例えばエシェリキア属(Escherichia)(例として大腸菌(E. coli)(たとえば、ATCC 31,446; 31,537; 27,325))、エンテロバクター属(Enterobacter)、エルウィニア属(Erwinia)、クレブシエラ属(Klebsiella)、プロテウス属(Proteus)、サルモネラ属(Salmonella)(例としてネズミチフス菌(Salmonella typhimurium))、セラチア属(Serratia)(例としてSerratia marcescens)、及び赤痢菌属(Shigella)など、並びにバシラス属(Bacilli)の枯草菌(B. subtilis)及びB. licheniformis(DD 266 710を参照されたい)など、シュードモナス属(Pseudomonas)の緑膿菌(P. aeruginosa)など、さらにストレプトミセス属(Streptomyces)がある。酵母宿主細胞のうち、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(schizosaccharomyces pombe)、クルイベロミセス属(Kluyveromyces)(たとえば、ATCC 16,045; 12,424; 24178; 56,500)、ヤロウィア属(yarrowia)(欧州特許第402,226号)、ピキア・パストリス(Pichia Pastoris)(欧州特許第183,070号、Pengら、J.Biotechnol. 108 (2004) 185-192も参照されたい)、カンジダ属(Candida)、トリコデルマ・リーシア(Trichoderma reesia)(欧州特許第244,234号)、ペニシリン属(Penicillin)、トリポクラジウム属(Tolypocladium)並びにアスペルギルス属(Aspergillus)宿主、たとえばアスペルギルス・ニデュランス(A. nidulans)及びクロコウジカビ(A. niger)も想定される。
【0065】
原核生物及び酵母宿主細胞は、具体的に本発明で想定はされるが、しかしながら、本発明の宿主細胞は、典型的には脊椎動物細胞である。適当な脊椎動物宿主細胞には、哺乳動物細胞、たとえば、COS−1(ATCC番号 CRL 1650)、COS−7(ATCC CRL 1651)、ヒト胚性腎細胞系293、PerC6(Crucell)、仔ハムスター腎細胞(BHK)(ATCC CRL.1632)、BHK570(ATCC番号: CRL 10314)、293(ATCC番号CRL 1573)、チャイニーズハムスター卵巣細胞CHO(たとえば、CHO−K1、ATCC番号:CCL61、DHFR−CHO細胞系、たとえばDG44など(Urlaubら、前掲を参照されたい))、特に懸濁培養に適したCHO細胞系、マウスセルトリ細胞、サル腎細胞、アフリカミドリザル腎細胞(ATCC CRL-1587)、HELA細胞、イヌ腎細胞(ATCC CCL 34)、ヒト肺細胞(ATCC CCL 75)、HepG2及びミエローマ若しくはリンパ腫細胞、たとえばNS0(米国特許第5,807,715号を参照されたい)、Sp2/0、Y0が含まれる。
【0066】
したがって、本発明の一実施形態において、本明細書に記載の治療用抗体の重鎖及び/又は軽鎖をコードするベクターを含む、安定して形質転換された宿主細胞が提供される。こうした宿主細胞は、典型的には、軽鎖をコードする第1のベクター、及び重鎖をコードする第2のベクターを含む。
【0067】
このような宿主細胞はまた、本発明の抗体の質、機能及び/又は収率を改変するためにさらに操作してもよいし、又は適合させてもよい。非限定的な例としては、特定の修飾(例えばグリコシル化)酵素及びタンパク質折りたたみシャペロンの発現が挙げられる。
【0068】
5.9.細胞培養法
本発明の治療用抗体をコードするベクターで形質転換された宿主細胞は、当業者に公知の方法によって培養することができる。宿主細胞は、スピナーフラスコ、振とうフラスコ、ローラーボトル、又は中空糸の系で培養することができるが、大規模生産のためには、特に懸濁培養用に撹拌槽型リアクター又はバッグリアクター(例えば、Wave Biotech, Somerset, New Jersey USA)を使用することが好ましい。撹拌槽は、典型的には、たとえば、スパージャー、バッフル又は低剪断インペラを用いて、通気に適合させる。気泡塔及びエアリフトリアクターには、空気又は酸素気泡による直接通気を用いることができる。宿主細胞を無血清培地中で培養する場合、通気プロセスの結果として細胞が損傷するのを防ぐのに役立つ、プルロニックF−68のような細胞保護物質を添加することが好ましい。宿主細胞の特性に応じて、マイクロキャリアを、足場依存性細胞系の増殖基質として使用することができるし、あるいは細胞を懸濁培養に適応させることもできる(こちらが一般的である)。宿主細胞、特に脊椎動物宿主細胞の培養は、バッチ、流加培養、反復バッチ処理(Drapeauら、(1994) cytotechnology 15: 103-109を参照されたい)、延長バッチ法又は潅流培養といった、さまざまな運転モードを利用することができる。組換えによって形質転換された哺乳動物宿主細胞は、ウシ胎仔血清(FCS)を含む培地のような血清含有培地で培養することができるが、こうした宿主細胞は、Keenら、(1995) Cytotechnology 17:153-163に記載の合成無血清培地、又はProCHO−CDM若しくはUltraCHOTM(Cambrex NJ, USA)といった市販の培地中で、必要ならば、グルコースなどのエネルギー源及び組換えインスリンのような合成増殖因子を添加して、培養することが好ましい。宿主細胞の無血清培養は、その細胞が無血清条件での増殖に適応していることを必要とすると考えられる。1つの適応法は、こうした宿主細胞を血清含有培地で培養し、宿主が無血清条件に適応するように、培地の80%を無血清培地に繰り返し交換することである(たとえば、Scharfenberg Kら、(1995)「動物細胞工学」(Animal Cell technology)より: 21世紀に向けた開発(Developments towards the 21st century)(Beuvery E.C.ら、編)、619-623ページ, Kluwer Academic publishersを参照されたい)。
【0069】
培地中に分泌された本発明の抗体は、さまざまな技法を用いて培地から回収及び精製され、目的の用途に適した精製度を与えることができる。たとえば、本発明の治療用抗体をヒト患者の治療に使用するには、一般に還元SDS−PAGEで測定した場合に少なくとも95%の純度、より典型的には98%、若しくは99%の純度が要求される(治療用抗体を含む粗製の培地と比較した場合)。第1の例では、細胞片を培地から、一般には、遠心によって除去し、次に、たとえば、精密濾過、限外濾過及び/又は深層濾過を用いて、上清の清澄化ステップを行う。あるいは、前もって遠心分離を行わずに、精密濾過、限外濾過又は深層濾過により抗体を回収してもよい。さまざまな他の技法、たとえば、透析及びゲル電気泳動、並びにクロマトグラフ法、例としてヒドロキシアパタイト(HA)、アフィニティクロマトグラフィー(状況に応じて、ポリヒスチジンのようなアフィニティタグ標識システムを含む)及び/又は疎水性相互作用クロマトグラフィー(HIC、米国特許第5,429,746号を参照されたい)が利用できる。一実施形態において、本発明の抗体は、さまざまな清澄化ステップの後、プロテインA若しくはGアフィニティクロマトグラフィーを用いて捕捉し、次いでさらに、イオン交換及び/又はHAクロマトグラフィー、陰イオン若しくは陽イオン交換、サイズ排除クロマトグラフィーといったクロマトグラフィーステップや、硫安沈澱を行う。一般に、さまざまなウイルス除去ステップも行われる(たとえば、DV−20フィルターなどを用いたナノ濾過)。こうしたさまざまなステップの後、少なくとも10mg/ml以上、たとえば100mg/ml以上の本発明の抗体を含む、精製(典型的にはモノクローナル)調製物が得られ、したがってその調製物が本発明の実施形態を構成する。超遠心分離によって100mg/ml以上への濃縮を行うことができる。適切なことに、この調製物は本発明の抗体の凝集体を実質的に含まない。
【0070】
細菌系は、抗体フラグメントの発現に特に適している。こうしたフラグメントは、細胞内又はペリプラズム内に局在する。当業者に公知の方法にしたがって、不溶性ペリプラズムタンパク質を抽出し、リフォールディングして活性タンパク質を形成することができる。Sanchezら、(1999) J. Biotechnol. 72, 13-20及びCupit PMら、(1999) Lett Appl Microbiol, 29, 273-277を参照されたい。
【0071】
6.医薬組成物
上記の本発明の抗体の精製調製物(特にモノクローナル調製物)を、上で概説したヒトの疾患及び障害の治療に使用するための医薬組成物に組み入れることができる。典型的には、こうした組成物は、許容される医薬慣習で知られ、必要とされる、薬学的に許容される(すなわち不活性)担体をさらに含む。たとえば、Remingtons Pharmaceutical Sciences, 第16版、(1980), Mack Publishing Co.を参照されたい。このような担体の例としては、適当なバッファーでpH5〜8の範囲内に緩衝化された、生理食塩水、リンガー溶液又はブドウ糖溶液のような滅菌担体がある。注射用(たとえば、静脈内、腹腔内、皮内、皮下、筋肉内若しくは門脈内投与による)又は持続点滴用の医薬組成物は、好適には目に見える粒子状物質を含まず、0.1ng〜100mgの抗体、典型的には5mg〜25mgの抗体を含有することができる。このような医薬組成物の製造方法は、当業者に周知である。一実施形態において、医薬組成物は、0.1ng〜100mgの本発明の治療用抗体を、状況に応じて使用説明書と共に、単位投与剤形中に含む。本発明の医薬組成物は、当業者に周知の又は明白な方法にしたがって投与前に再構成するために凍結乾燥されていてもよい。本発明の実施形態がIgG1アイソタイプを有する本発明の抗体を含む場合、クエン酸塩(たとえば、クエン酸ナトリウム)又はEDTA又はヒスチジンといった銅のキレート剤を医薬組成物に添加して、このアイソタイプの抗体の、銅を介した分解の度合いを低下させることができる。欧州特許第0612251号を参照されたい。
【0072】
本発明の抗体を投与するための有効量及び治療計画は、一般に、経験的に決定され、患者の年齢、体重及び健康状態、並びに治療すべき疾患又は障害といった要因に左右される。こうした要因は主治医の裁量の範囲内にある。適当な用量を選択する際の指針は、たとえばSmithら、(1977)「ヒトの診断及び治療における抗体」(Antibodies in human diagnosis and therapy)、Raven Press, New Yorkに見出すことができるが、概して1mg〜1000mgである。一実施形態において、RAに罹患したヒト患者の治療用の投与計画は、おおよそ100mg(すなわち50mg〜200mg)の本発明の抗体(又はその抗原結合フラグメント)を毎週又は2週間毎に皮下投与するものである。本発明の組成物はまた予防的に用いることもできる。
【0073】
治療対象の疾患又は障害に応じて、治療上有効な量の本発明の抗体を含む医薬組成物を、有効量の別の薬剤、たとえば、抗炎症薬(たとえば、NSAID、メトトレキセート、、ブシラミン、チオリンゴ酸ナトリウム、又は1以上の抗TNFα治療、例えばEnbrelTM(エタネルセプト)、RemicadeTM(インフリキシマブ)、HumiraTM(アダリムマブ)及び/若しくはCDP870)とともに、同時に、別々に、又は順次、使用することができる。本発明の抗体は、有効量の抗TNF−α受容体抗体と組み合わせて用いることができる。Davis MWら、(2000) Ann Rheum Dis 59(Suppl 1): 41-43を参照されたい。他の実施形態において、本発明の抗体は、以下:IL−1/IL−1R(例えばKineretTM)、CTLA4−Ig、IL−6(Choyら、(2002) Ann.Rheum.Dis 61(suppl 1): 54参照)、IL−8、IL−15、VEGF、IL−17、IL−18(Taylorら、(2001) Curr.Opin.Immunol.13: 611-616参照)、抗ICAM及び/又は抗CD4抗体に対して生起される薬剤、並びにMMPファミリーのメンバー、例えばMMP−1、2、3及び/又は13に対して生起される薬剤の有効量と組み合わせて用いることができる。本発明の抗体はまた、炎症性プロセスに関与することが知られる、細胞を剥離する薬剤と組み合わせて用いることができ、例えばCD20陽性B細胞について例えばMabtheraTM(リツキシマブ)を用いる。本発明の抗体と組み合わせる他の治療法としては、抗血管新生療法、例えばインテグリンαvβ3のアンタゴニスト、クリングル1〜5(Sumariwalla Pら、(2003), Arthritis Res Ther 5:R32-R39参照)、可溶性Flt−1(Miotlaら、(2000) Lab.Invest. 80:1195-1205参照)など;抗COX−2剤又は抗OSM剤、例えば抗OSM抗体(WO2005/095457号参照。その全内容が本明細書中に具体的に組み入れられ、読者はそれを具体的に参照する)などが挙げられる。簡便には、本発明の抗体又はその抗原結合フラグメントの部分の、そのような別の医薬の部分と一緒の、場合によっては使用説明書と一緒の、キット(キットオブパーツ)を含む医薬組成物もまた本発明は意図している。これらの組み合わせは、関節炎疾患/障害(関節リウマチなど)の治療に特に有用である。
【0074】
7.臨床用途
本発明の抗体は、IL−18が媒介する疾患、例えば自己免疫疾患などの治療的処置に用いることができる。具体的には、多発性硬化症、関節炎疾患(関節リウマチなど)、1型糖尿病、炎症性腸疾患(IBD)及び乾癬が挙げられる。従って、本発明はさらに、hIL−18の中和に応答する疾患(多発性硬化症、関節リウマチ、1型糖尿病、IBD、乾癬など)に罹患したヒト患者の治療方法であって、本発明の抗体(特に配列番号9に示される配列を有する重鎖と配列番号13に示される配列を有する軽鎖とを有する抗体)の治療有効量を該患者に投与することを含む方法を包含する。
【0075】
また、上述した疾患/障害のいずれか1つ(又は複数)の治療のための医薬の製造における本発明の抗体の使用も提供される。
【0076】
【0077】
7.例証
以下の実施例により、本発明の様々な態様を説明する。全ての一般的クローニング、ライゲーション、及び他の組換えDNA技術は、Maniatisら, Molecular cloning (A laboratory manual), Cold Spring Harbor Laboratory、又はSambrookら Molecular Cloning (A laboratory manual), Cold Spring Harbor Laboratoryに一般的に教示されているように実施する。本明細書で使用するベクター系及び追加の分子生物学的方法は、その内容全体を参照により本明細書に組み込み、かつ読者が具体的に参照するWO2005/095457号に開示されている。
【0078】
7.1.ハイブリドーマ可変領域のクローニング
親ラット抗体2C10は、米国特許第6,706,487号に記載されている。読者は、具体的にはこの文書を参照されたい。上記公開されている、ヒトIgG1若しくはκC領域に結合しているラットV領域をベースとして、キメラ抗体2C10cを設計した。重鎖及び軽鎖構築体に、一般的免疫グロブリンシグナル配列及び翻訳開始コドンATGを導入した。VLドメインを形成し、既にヒトCκ領域(配列番号36)を含む哺乳動物発現ベクター中にクローニングできるように、HindIII及びBsiWI制限エンドヌクレアーゼ部位を設計した。VHドメインを形成し、既にヒトγ1C領域(配列番号34)を含む哺乳動物発現ベクター中にクローニングできるように、HindIII及びSpeI制限エンドヌクレアーゼ部位を設計した。これにより、配列番号33に示す公開済み配列から得た、2C10のVh領域のフレームワーク4に2個のアミノ酸変化がもたらされた(Kabat残基107及び108)。
【0079】
重複オリゴヌクレオチドを使用して、PCRによってコード配列全体を構築し、上記発現ベクター中にクローニングした。配列を確認後、キメラ抗体をCHO細胞で発現させた。生成された抗体は、rProteinAセファロースでのアフィニティクロマトグラフィーにより、細胞培養上清から精製した。その親ラット2C10に匹敵する効力を実証するために、インビトロ結合アッセイで、2C10キメラ抗体を評価した。これは、ELISAでヒト若しくはアカゲザルIL18との結合のEC50値を測定することによって(図16及び17)、又はKG−1バイオアッセイでIFN−γの放出を阻害することによって(図15を参照されたい)行った。
【0080】
7.2.ヒト化
7.2.1.軽鎖ヒト化手順
ラット2C10可変軽鎖配列について、ラット2C10可変軽鎖配列と64%の同一性(CDR含有)を有するヒト生殖細胞系アクセプターフレームワークを選択した(F_IGKV1D−12−1、配列番号38)。生殖系列V領域は、配列類似性(配列番号40)に基づき、コンピューターで(in silico)好適なFR4、この場合はJ領域κ2ミニ遺伝子(Kabat Vol.II)と組み合わせた。3個のヒト化変異体は、配列比較、及び抗体機能への潜在的影響に基づき生成した。構築体L1は、上で選択したヒトアクセプターフレームワーク中にラットCDR(Kabat定義を使用)を線状に移植(straight graft)したものであった。構築体L2は、L1を基礎にして、残基71に1個の追加の復帰突然変異を有した。構築体L3は、L2を基礎にして、残基45、83、84及び85に4個の追加の復帰突然変異を有した。表1を参照されたい。
【0081】
【0082】
7.2.2.重鎖のヒト化手順
ラット2C10可変重鎖配列について、ラット2C10可変重鎖と59%の同一性(CDR含有)を有する、ヒト生殖細胞系アクセプターフレームワークを選択した(Fp_IGHV1−f_2、配列番号37)。生殖系列V領域は、配列類似性(配列番号39)に基づき、コンピューターで好適なFR4、この場合はJH6ミニ遺伝子(Kabat Vol.II)と組み合わせた。このフレームワークに基づき、3個のヒト化可変重鎖変異体を設計した。H1は、残基27、28、29及び93に4個の追加の復帰突然変異を有するラットCDR(Kabat定義を使用)の移植体である。これにより、(Chothiaが定義するように)CDRの一部を構成しうる親(すなわちドナー)抗体のCDR1の直上流に、極めて珍しいアミノ酸配列を得ることができる。H2は、H1を基礎にして、残基39及び40に2個の追加の復帰突然変異を有した。次いで、H3は、H2を基礎にして、残基36、71、89及び91に別の4個の追加の復帰突然変異を有した。表2を参照されたい。
【0083】
【0084】
7.3.2C10Cのヒト化
重複オリゴヌクレオチドの構築及びPCR増幅によって、ヒト化V領域を新規に合成した。哺乳動物発現ベクター中にクローニングするためにプライマーに制限部位を含め、分泌のためにヒト免疫グロブリンシグナル配列を含めた。ヒト化V領域は、HindIII及びSpeIを使用し、H1、H2及びH3として、ヒトγ1定常領域を含む哺乳動物発現ベクター中にクローン化し、そしてHindIII及びBsiWIを使用し、L1、L2及びL3として、ヒトκ定常領域を含む哺乳動物発現ベクター中にクローン化した。これにより、ヒトIgG1アイソタイプのヒト化重鎖変異体、及びヒトκアイソタイプのヒト化軽鎖変異体が得られた。
【0085】
7.3.1.ヒト化重鎖抗体とヒト化軽鎖抗体の組合せの発現
四回の試験でCHOK1細胞に一過性にトランスフェクトした。上清の抗体濃度をアッセイし、次いで2C10ラット−ヒトキメラと比較することによって、その上清をインビトロ結合アッセイで使用した。
【0086】
全9個の変異体の大規模な一過性発現は、各フラスコについて8mlの培地(OptiMEM/glutamax/5%FBS)中に含まれた、51.4μgの軽鎖プラスミド及び8.6μgの重鎖プラスミドと、240μgのトランスフェクション脂質(この脂質は、WO2006/053783号、実施例13に記載されており、その内容全体を参照により本明細書に組み込む)とを混合し、2個のほぼコンフルエントなCHOK1細胞のT175フラスコに、この混合物を通常の組織培養条件下、72時間添加することによって実施した。シェーカーフラスコを使用して、ポリクローナルCHO細胞系でも抗体をmg量で発現させ、FPLC及びプロテインAを使用して精製した。
【0087】
7.4.インビトロ結合アッセイ
7.4.1.Biacore分析
ヒト化2C10抗体のBiacoreTM動態分析は、BiacoreTM3000機により、HBS−EP緩衝液(BiacoreTM)中でその抗体のプロテインA捕捉を使用し実施した。簡単に説明すると、製造業者が推奨するプロトコルを使用し、一級アミン結合によってCM5チップにプロテインAをおよそ2000〜4000共鳴単位(RU)の密度で固定した。次いで、プロテインA表面にヒト化抗体を流し、安定化期間後およそ200〜500RUのレベルで捕捉させ、捕捉した抗体表面に(ヒト若しくはアカゲザルの)IL18を所定の濃度で流し、結合センサーグラムを得た。酸性溶出条件を使用して再生することにより、捕捉した抗体はプロテインA表面から全て除去され、表面の結合能が著しく低減することはなかった。全ての曲線は、IL18に代わる緩衝液の注入に対して二重に基準を取り、BiaEval 4.1で全体適合パラメーターを使用してデータを1:1結合モデルに適合させた。同じプロテインA捕捉法を使用して解離速度ランキング実験を準備したが、1種類のIL18濃度(10nM)のみを使用した。データは、動態分析の結合モデルと同じ結合モデルを使用し適合させたが、解離速度に1種類の検体濃度のみを使用したことが報告されているので、この値は、正確な動態測定を得るというよりはむしろランキングに有用であり、どの抗体をさらに調査するかの選択法として使用した。
【0088】
最初の25℃での結果では、構築体は全て、ラット2C10親抗体としてヒトIL18と類似する結合親和性を有することが示された。しかし、37℃で解離速度ランキング実験を実施したとき、L1構築体は解離速度に上昇が見られ、L2及びL3構築体と比較して不首尾な結果が示された(表3a及び3b)。
【0089】
【0090】
L1構築体は、アカゲザルIL18との結合親和性の点で、37℃で不首尾な結果を示したのと同様に、25℃でも最低の結果を示した(表4a)。これらの観察に基づき、37℃でのヒトIL18及びアカゲザルIL18との結合について、選択した抗体をより詳細に調査した。表4bに示すヒトIL−18のデータは、6回の別個の測定の平均(及び標準偏差)である。アカゲザルIL−18のデータは、H1L2及びH1L3については2回の実験の平均及び標準偏差を示し、H3L2及びH3L3のデータは1回の実験から得たものである。このデータの比較的高い標準偏差は、おそらく37℃でこの実験を実施した結果である。
【0091】
L1とL2構築体との差が、軽鎖の位置71でのチロシンの復帰突然変異によるフェニルアラニンの置換であることを考慮すると、L1構築体が、相対的に劣る結果を示した事実は驚くべきことである。チロシンとフェニルアラニンはもちろん芳香族アミノ酸であり、従ってフレームワーク構造のそのような些細な変化によって、37℃でBiacoreTM系で観察された(結合親和性の点で)顕著な結果が生じたことは予想外であった(しかし、25℃では観察されなかった)。
【0092】
【0093】
変異体H1L1、H1L2、及びH1L3を選択して、以下第7.4.2節でさらに分析した。
【0094】
7.4.2.Biacore分析T100のデータ
T100 BiacoreTM機を使用してさらにいくつかの変異体抗体の特性決定を実施した。この機器は、高温での緩衝効果を最小化する直列ガス抜き装置(inline degasser)を使用することによって、高温での感受性、温度制御性、及び基線安定性の点で、BiacoreTM3000よりも優れている。また、自動データ分析など、高度なソフトウェアも提供する。
【0095】
方法は、上の第7.4.1節で使用した方法と基本的に同じであり、一級アミン結合によってプロテインAをCM5チップに2000〜6000RUの密度で固定した。HBS−EP(BiacoreTM)で試験を実施した。抗IL18抗体を100〜500RUの密度で捕捉させ、ヒトIL18を16〜0.0625nMの濃度でこの捕捉した表面に流し、0nM濃度(すなわち、緩衝液は捕捉した抗体の注入のみ)を二重基準に使用した。各IL18注入後の再生は、10mMグリシン(pH1.5)の注入を2回を使用する、穏やかな酸性溶出により行なった。この再生ステップにより、プロテインA表面から、捕捉された抗体を除去した(従ってそれに結合するいかなるIL18も除去した)。再生しても、続く抗体パルスと結合するプロテインA表面の能力は大きく変化することなく、次の捕捉事象を行うことができた。得られた結合曲線は、1:1結合モデルを使用するT100機に本来備わる分析ソフトウェアを用いて分析した。示した温度で実験を実施した。
【0096】
15℃、20℃、25℃、32℃及び37℃でのH1L1及びH1L2の結合分析
様々な温度で上記方法を使用して実験を実施した。図1は、温度の結合速度(on-rate)(ka)に対する影響を示し、図2は解離速度(kd)に対する影響、そして図3は平衡定数(KD)に対する影響を示す。表5は、これらの図を作成するために使用した動態値の詳細である。
【0097】
【0098】
データは、試験した2個の抗体の結合速度が、試験した温度範囲にわたり類似することを示し、H1L2の結合速度の方が速い場合は、37℃での最終値まで、H1L2の結合速度の方が一般的に速い。しかし、解離速度に目を向けた場合、より大きな差異が見られ、15℃、20℃、25℃では2個の抗体の解離速度は類似するが、32℃及び37℃で差異を見せ始め、H1L1の解離速度の方が速くなる。これらの変化は、全体的平衡定数(それはkd/kaの関数である)に反映され、H1L1とH1L2との間の差異は、解離速度(kd)によって定義した、主として抗体/IL18複合体の安定性であることを示している。
【0099】
25℃及び37℃でのH1L1、H1L2、H1L3、及びキメラ2C10の結合分析
上記のように実験を実施した。表6は、得られた動態パラメーターの詳細である。データから、25℃及び37℃の平衡定数KDによって定義した結合という点で、H1L2はH1L1よりも良好な抗体であることが示されるが、動態パラメーターは、25℃ではH1L2がH1L1よりも結合速度(ka)が良好であることを示している。37℃では、立場は逆転し、このことは、37℃で見られるH1L2による優れた結合が、むしろ解離速度(kd)によることを示し、L2とL1を区別する変異によって、IL18−抗体複合体の高温での安定性が増大することを示している。
【0100】
【0101】
データは、いくつかの別々のデータセット(n)の平均であり、平均及び標準偏差(括弧内に標準偏差)を示す。5つの異なる温度での分析から得られたH1L1及びH1L2の25℃と37℃での実験の値が、このデータセットに含まれている。
【0102】
7.4.3.IL18結合ELISAにおける2C10cヒト化変異体の評価
精製抗体調製物の様々なバッチを使用して全9個のヒト化変異体のELISAを少なくとも6回実施した。図4A〜4Cには、表7に示すEC50値ランキングを作成した、1回の実験の代表的データを示す。2.5μg/mlの16D10(非中和マウスモノクローナル抗体)を使用し、Nunc Maxisorp96ウェルプレートにヒトIL−18を固定して、5ng/mlの組換えヒトIL−18を捕捉させた。様々な希釈率で抗IL−18ヒト化抗体を加えた。抗ヒトIgG Fc特異的ペルオキシダーゼコンジュゲート(Sigma A0170)を使用して、結合したヒト化抗体を検出した。
【0103】
【0104】
全ての変異体の効力は、2C10キメラに非常に近似しているように見え、このことは、ヒト化による効力の損失はほとんどないことを示している。これらのアッセイを数回反復して得られたEC50値では、変異体ランキングは得られなかったが、ELISA単独ではこれらの変異体を明確に区別することはできなかった(表7及び図4A〜4Cを参照)。BiacoreTMを使用して、変異体をある程度区別することができ(第7.4.1節及び第7.4.2節)、それによってわずか4個の変異体がもたらされ、それらをヒトIL18及びアカゲザルIL18を使用する数回の独立した反復実験でより詳しく試験した(表8、図5[ヒト]及び図6[アカゲザル])。
【0105】
【0106】
7.4.4.IL−18結合ELISAにおける2C10ヒト化H1変異体の評価
H1L1、H1L2及びH1L3という3個のヒト化H1変異体を用いてELISAを実施し、室温及び37℃において、ヒト血清及びブロッキング溶液(1%BSA含有PBS0.05%TWEEN(w/v))中でのヒトIL−18結合を評価した。Nunc Maxisorp96ウェルプレートにヒト化抗体変異体を2.5μg/mlで固定した。5ng/mlの組換えヒトIL−18の捕捉は、室温又は37℃で、ヒト血清又はブロック溶液中で実施した。抗IL−18マウスモノクローナル抗体16D10を加えた。結合したマウス抗体は、抗マウスκペルオキシダーゼコンジュゲート(Serotec MCA 1291P)を使用して検出した。研究データから得られた代表的EC50値を表9に示す。
【0107】
【0108】
このアッセイでは、ヒトIL−18結合ステップを実施する温度を室温から37℃へ変化させても、3個のヒト化H1変異体の効力は影響を受けない。ヒト血清中に抗体が存在すると、低い結合シグナルが観察される。
【0109】
7.4.5.37℃でのヒト化H1変異体抗体の安定性評価
H1L1、H1L2及びH1L3という3個のヒト化H1変異体の保存安定性は、ヒト血清及びリン酸緩衝食塩水中、37℃で14日間にわたって評価した。抗体を50μg/mlに希釈し、安定性は、37℃で0、1、4、6、8及び14日間というインキュベーション時間後に、IL−18結合ELISAによって評価した。IL−18結合ELISAについては、16D10(非中和マウスモノクローナル抗体)をNunc Maxisorpプレートに固定して、5ng/mlの組換えヒトIL−18を捕捉した。37℃での経時インキュベーションの種々の時点でサンプリングした抗IL−18ヒト化抗体を加えた。結合したヒト化抗体は、抗ヒトIgG Fc特異的ペルオキシダーゼコンジュゲートを使用して検出した。
【0110】
このアッセイ型式では、37℃という温度に、0、1、4、6、8及び14日間という長時間曝露を行なっても、ヒトIL−18に結合するその能力に基づく、ヒト化H1抗体変異体の結合効力に影響はなかった。
【0111】
7.5.滑液存在下でのH1L2とヒトIL18との結合
50%ヒト滑液に500ng/ml〜0ng/mlの範囲の組換えヒトIL18を添加しELISAを実施した。次いで、H1L2抗体をコーティングしたMaxisorp96ウェルプレート(Nunc)のウェルに、IL−18を含むこの溶液をアプライした。次いで、ビオチン化した抗IL−18抗体(D045−6, MBL)及びストレプトアビジン−HRPにより、結合したIL−18を検出した。組換えヒトIL18を含む50%滑液(SF)を滴定すると、最大半値のほんのわずかなシフトが見られただけで、ヒト組換えIL−18を含む緩衝液を滴定したものと、ほぼ同じ曲線が生成された。図7を参照されたい。これにより、抗体が治療設定で遭遇する結合環境により類似性が高い50%ヒトSFの存在下でさえ、hIL−18に結合する抗体能力が実証されている。
7.5.1.IL18結合タンパク質(IL18bp)存在下でのIL18に対する結合
BiacoreTM技術(BiacoreTM3000)及びELISAを使用して、ヒトIL18bpの存在下でもH1L2が依然としてヒトIL18と結合できるかどうかを判定した。IL18bpは、IL18に対して高親和性を有し、IL−18機能の天然阻害薬として働く(図8、図9A及び図9B、表10及び表11)。
【0112】
Biacoreによる分析
簡単に説明すると、一級アミン結合によってCM5チップにプロテインAを表面がおよそ4000共鳴単位(RU)の密度になるように固定した。次いで、組換えFc−IL18結合タンパク質(R&D Systems)を濃度3μg/ml、流速10μl/分で1分間流し、これにより、およそ1400RUのIL18結合タンパク質が捕捉された。次いで、捕捉したIL18結合タンパク質表面に、IL18を濃度30nM、流速10μl/分で5分間流した。この後、IL18結合タンパク質/IL18表面に、濃度10nMのラット2C10親抗体を流速30μl/分で3分間流した。2C10抗体によって認識されるIL18上のエピトープが、IL18結合タンパク質との相互作用部位に干渉するならば、いかなる結合シグナルも見られないはずである。図10は、2C10がhIL−18と結合すること、続いてこのhIL−18がhIL−18BPによって捕捉されたことを実証し、このことは、2C10とhIL−18BPの結合部位が重複しないことを示している。
【0113】
【0114】
ELISAによる分析
ヒト化H1L2若しくはラット2C10 MAbと、ヒトIL18が結合した捕捉IL18bpとの結合を、直接結合ELISAで試験したが、その際、Nunc Maxisorpプレートに、組換えヒトIL18bp−Fc融合タンパク質(R&D Systems #119-BP)を0.5μg/mlでコーティングした。組換えヒトIL18(社内試薬)を100ng/mlでブロッキング緩衝液(1%w/vBSA含有PBS)に加えた。精製した抗体を0.5ng/ml〜1μg/mlの濃度範囲で加えた。抗ヒトκ軽鎖特異的HRPコンジュゲート(Sigma)若しくは抗ラットIgG HRPにより結合した抗体を検出した。図18A及び18Bに、得られた結果を示す。
【0115】
7.6.インビトロバイオアッセイ
7.6.1.KG−1細胞系における、IL18の刺激によるIFN−γの放出を中和する、ヒト化構築体の活性
このアッセイでは、IL−18特異的抗体の中和活性を計測するが、アッセイはKG−1細胞におけるIL−18を媒介するIFNγの誘導に基づく。KG−1(ATCC#CCL−246)は、機能性IL−18受容体を構成的に発現し、従って外来性IL−18刺激に応答するヒト骨髄単球細胞系である。
【0116】
全9個のヒト化変異体について、KG−1細胞におおけるヒトIL18に刺激されたIFN−γの放出を阻害するその能力を評価した(表12及び図11)。
【0117】
【0118】
BiacoreTM分析に基づき、組換えヒトIL18及びアカゲザルIL18に最も高い親和性示した、4個の好ましいヒト化変異体で、少なくともさらに6回の反復実験を実施した。図8に、4個の好ましいヒト化変異体及びH1L1の代表的結果を示し、表13に、CHOe1a細胞由来の同じタンパク質バッチ物質によって全て実施した、これらのアッセイの結果の概要を示す。
【0119】
【0120】
H1L1を他のヒト化変異体と比較した事例では、試験し、かつ2C10親MAbとも比較した、その他の4個のヒト化構築体と比べて、H1L1は低い効力を示した。
【0121】
4個の好ましいモノクローナル抗体を用いてさらなる分析を実施したが、KG−1細胞:2C10、並びに2C10から得られたヒト化変異体(H1L1、H1L2及びH1L3)により、IL−18に刺激されたINFγの放出を阻害する能力について、それらのモノクローナル抗体を比較した。96ウェルプレートで、50ng/mlの組換えヒトIL−18、及びIL18に特異的な種々の濃度の抗体(2μg/ml〜7.8ng/mlの範囲、二倍希釈)、又は陰性アイソタイプ対照(Synagis、抗RSV抗体)を37℃及び5%CO2下で1時間インキュベートし、続いてウェル当たり3.105個のKG−1細胞を添加することによって、KG−1バイオアッセイを実施した。最後に、それらのプレートを37℃、5%CO2下で20〜24時間インキュベートした。上清を回収し、市販のヒトIFNγELISAキット(Biosource AHC4432;AHC4539)を使用してIFNγの産生を定量した。
【0122】
3回の実験を実施した。IL−18に刺激されたIFNγの産生の阻害結果を陰性対照に対して正規化した。統計分析は、あらゆるSynagis応答の調整後、各実験について各mAbのIC50推定値を得ることを目的とした。次いで、IC50推定値を統計学的に分析して、95%信頼区間(すなわち統計学的に信頼できる範囲)で各mAbの全体的IC50推定値を生成した。最後に、ダネット検定を使用して2C10に戻って各ヒト化変異体を比較した。
【0123】
図12に、代表的実験を示す。表14及び図13に、95%信頼区間での各モノクローナル抗体の全体的IC50推定値、及びp値と信頼区間と共にラット2C10の変化率(%)を示す。
【0124】
【0125】
H1L1は、2C10よりも統計上有意に効力が低い(p<0.001)。
【0126】
その他のヒト化変異体は、2C10と有意には異ならないが、H1L3対2C10の比較が境界線である。
【0127】
7.6.2.刺激したヒトPBMC中のIFN−γ放出の中和におけるH1L2の活性
3ドナー由来のヒトPBMCは、組換えヒトIL18及び抗CD3抗体で刺激し、4個の選択したヒト化抗体変異体の希釈系列を加えた作用を研究した。比較するために、それぞれにドナー親2C10抗体及びIL18bpを含めた。3ドナーのうち2ドナーでは、IL18及び抗CD3刺激は失敗し、IFN−γは検出されなかった。残りのドナーについては、結果は低濃度では非常に変化したが、IL18が誘発するIFN−γの産生の完全阻害は、ヒト化変異体を含む様々な抗IL18抗体を添加することによって達成することができた。図14を参照されたい。
【0128】
さらにLPSが刺激するヒトPBMCにより実施した実験では、IL18を産生し、付随するIFNγの放出が生じた。濃度依存的にLPSによってIFNγの産生が誘発され、1μg/mlの一定濃度で2C10親モノクローナル抗体を添加すると、この刺激が完全に阻害され、このことは、その効果にはIL18が介在するということ、及び内在性IL18は中和できるということ(データ非掲示)を示している。これは、全血でも実証できたが、2C10による阻害効果は、用量依存性ではあったが、あまり明白ではなかった(データ非掲示)。図9に、3個の独立したドナーによる実験、並びに親ラットモノクローナル2C10、H1L2、及びIL18bpによる阻害の結果を示す。ドナー1及び3は同様の結果を示したが、ドナー2はLPSによる刺激でIFNγを放出しなかった(図示せず)。IL18が媒介するIFNγ放出は、10%若しくは25%のヒト血清の存在下、1μg/mlを超える抗体若しくはIL18bpを加えることによって、完全に阻害することができる。阻害は、10ng/ml以上で既に観察可能であり、10%若しくは25%のヒト血清の存在下でのこの阻害のIC50値を表11に示す。
【0129】
【0130】
7.6.3.IL18オルソログとの結合の概要
ELISA及びBiacore、そしてさらにKG−1細胞バイオアッセイを使用し、抗体と他の種由来のIL18との結合を検査した。これは、当初、アカゲザル/カニクイザルIL18について親2C10及びキメラ2C10cを使用し実施したが、いくつかのヒト化変異体で繰り返した。Biacore及びELISAを使用し、ブタ、マウス及びラットIL18との結合について親2C10を試験し、4個の最も優れたヒト化変異体をアカゲザル/カニクイザル、及びイヌのIL18との結合について試験した。KG−1バイオアッセイは、アカゲザル/カニクイザルのIL18、並びに3個のモノクローナル抗体、2C10キメラ、及びヒト化変異体の代表であるH3L3で実施した。全てのヒト化変異体の類似性を考慮すると、これは、H1L2を含めて生成された他の変異体の代表である可能性が高い(表15、図8を参照)。
【0131】
【0132】
7.7.IL18抗体の円二色性及び熱変性試験
円二色性(CD)試験を使用して、温度、特に25℃〜37℃の関数として、IL18抗体の二次構造変化を試験した。これら同じ抗体の熱変性試験を行って、それらの熱安定性と融解温度(Tm)を測定した。
【0133】
CD方法:CDスペクトルは、0.5nmステップ及びバンド幅1nmで180nm〜280nmの走査を行なうApplied Photophysics Chirascan分光計で取得した。各点ごとの取得時間は5秒であった。サンプルをPBSで約0.2mg/ml希釈し、1mm光路長(pathlength)セルに入れた。各タンパク質のスペクトルは、4℃、25℃及び37℃に設定した恒温槽で取得した。サンプルの実際の温度は、セル内の液体中に置いたプローブによって測定したが、設定温度の約3℃以内であった。
【0134】
Tm方法:Sypro orangeを1:1000で入れたリン酸緩衝食塩水(PBS)溶液で、全てのタンパク質を0.2mg/mlに希釈した。Bioneer Exicycler機器を使用し、0.5℃間隔毎に、620nm(励起490nm)における蛍光発光を測定したが、サンプル温度は各温度点で10秒待機して10℃〜95℃に勾配をつけた。変性曲線は、Grafitを使用して標準的融解等温線に適合させた。
【0135】
予想通り、全4個の抗体のCDスペクトルの形状比較から、それらの構造が高度にβシート状であり、本質的に同じ構造であることが示された。4℃〜37℃の温度範囲では、3個の抗体:
H1L1
H1L2
H1L3
は、CDによって判明した通り、その二次構造において有意な変化は示さなかった。しかし、2C10抗体キメラは、この温度範囲で構造にわずかな減少が見られた。これは、下表16に示す熱安定性傾向に一致する。
【0136】
【0137】
H1L1、H1L2及びH1L3は、体温ではいかなる変性の徴候も見せず、37℃をはるかに超えても明らかに安定している。従って、熱安定性のその差異が、正常な身体及び周囲温度で、何らかの示差的利点を付与する可能性は低い。
【0138】
配列
配列番号1
GYYFH
配列番号2
RIDPEDDSTKYAERFKD
配列番号3
WRIYRDSSGRPFYVMDA
配列番号4
LASEDIYTYLT
配列番号5
GANKLQD
配列番号6
LQGSKFPLT
配列番号7
MAAEPVEDNCINFVAMKFIDNTLYFIAEDDENLESDYFGKLESKLSVIRNLNDQVLFIDQGNRPLFEDMTDSDCRDNAPRTIFIISMYKDSQPRGMAVTISVKCEKISTLSCENKIISFKEMNPPDNIKDTKSDIIFFQRSVPGHDNKMQFESSSYEGYFLACEKERDLFKLILKKEDELGDRSIMFTVQNED
配列番号8
ATGGCTGCTGAACCAGTAGAAGACAATTGCATCAACTTTGTGGCAATGAAATTTATTGACAATACGCTTTACTTTATAGCTGAAGATGATGAAAACCTGGAATCAGATTACTTTGGCAAGCTTGAATCTAAATTATCAGTCATAAGAAATTTGAATGACCAAGTTCTCTTCATTGACCAAGGAAATCGGCCTCTATTTGAAGATATGACTGATTCTGACTGTAGAGATAATGCACCCCGGACCATATTTATTATAAGTATGTATAAAGATAGCCAGCCTAGAGGTATGGCTGTAACTATCTCTGTGAAGTGTGAGAAAATTTCAACTCTCTCCTGTGAGAACAAAATTATTTCCTTTAAGGAAATGAATCCTCCTGATAACATCAAGGATACAAAAAGTGACATCATATTCTTTCAGAGAAGTGTCCCAGGACATGATAATAAGATGCAATTTGAATCTTCATCATACGAAGGATACTTTCTAGCTTGTGAAAAAGAGAGAGACCTTTTTAAACTCATTTTGAAAAAAGAGGATGAATTGGGGGATAGATCTATAATGTTCACTGTTCAAAACGAAGACTAG
配列番号9
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHWVRQAPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTEDTSTDTAYMELSSLRSEDTAVYYCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
配列番号10
CAGGTGCAGCTGGTGCAGAGCGGAGCCGAGGTGAAGAAGCCTGGCGCCAGCGTCAAGGTGTCCTGTAAGGTGTCCGGCGAGATCAGCACCGGCTACTACTTCCACTGGGTGAGGCAGGCCCCTGGCAAGGGCCTGGAGTGGATGGGCAGAATCGACCCCGAGGACGACAGCACCAAGTACGCCGAGCGGTTCAAGGACAGGGTGACCATGACCGAGGACACCAGCACCGATACCGCCTACATGGAGCTGTCCAGCCTGAGAAGCGAGGATACCGCCGTGTACTACTGTACCACCTGGCGGATCTACAGAGACAGCAGCGGCAGACCCTTCTACGTGATGGATGCCTGGGGCCAGGGCACACTAGTGACCGTGTCCAGCGCCAGCACCAAGGGCCCCAGCGTGTTCCCCCTGGCCCCCAGCAGCAAGAGCACCAGCGGCGGCACAGCCGCCCTGGGCTGCCTGGTGAAGGACTACTTCCCCGAACCGGTGACCGTGTCCTGGAACAGCGGAGCCCTGACCAGCGGCGTGCACACCTTCCCCGCCGTGCTGCAGAGCAGCGGCCTGTACAGCCTGAGCAGCGTGGTGACCGTGCCCAGCAGCAGCCTGGGCACCCAGACCTACATCTGTAACGTGAACCACAAGCCCAGCAACACCAAGGTGGACAAGAAGGTGGAGCCCAAGAGCTGTGACAAGACCCACACCTGCCCCCCCTGCCCTGCCCCCGAGCTGCTGGGAGGCCCCAGCGTGTTCCTGTTCCCCCCCAAGCCTAAGGACACCCTGATGATCAGCAGAACCCCCGAGGTGACCTGTGTGGTGGTGGATGTGAGCCACGAGGACCCTGAGGTGAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCACAATGCCAAGACCAAGCCCAGGGAGGAGCAGTACAACAGCACCTACCGGGTGGTGTCCGTGCTGACCGTGCTGCACCAGGATTGGCTGAACGGCAAGGAGTACAAGTGTAAGGTGTCCAACAAGGCCCTGCCTGCCCCTATCGAGAAAACCATCAGCAAGGCCAAGGGCCAGCCCAGAGAGCCCCAGGTGTACACCCTGCCCCCTAGCAGAGATGAGCTGACCAAGAACCAGGTGTCCCTGACCTGCCTGGTGAAGGGCTTCTACCCCAGCGACATCGCCGTGGAGTGGGAGAGCAACGGCCAGCCCGAGAACAACTACAAGACCACCCCCCCTGTGCTGGACAGCGATGGCAGCTTCTTCCTGTACAGCAAGCTGACCGTGGACAAGAGCAGATGGCAGCAGGGCAACGTGTTCAGCTGCTCCGTGATGCACGAGGCCCTGCACAATCACTACACCCAGAAGAGCCTGAGCCTGTCCCCTGGCAAGTGA
配列番号11
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHWVRQAPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTEDTSTDTAYMELSSLRSEDTAVYYCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSS
配列番号12
CAGGTGCAGCTGGTGCAGAGCGGAGCCGAGGTGAAGAAGCCTGGCGCCAGCGTCAAGGTGTCCTGTAAGGTGTCCGGCGAGATCAGCACCGGCTACTACTTCCACTGGGTGAGGCAGGCCCCTGGCAAGGGCCTGGAGTGGATGGGCAGAATCGACCCCGAGGACGACAGCACCAAGTACGCCGAGCGGTTCAAGGACAGGGTGACCATGACCGAGGACACCAGCACCGATACCGCCTACATGGAGCTGTCCAGCCTGAGAAGCGAGGATACCGCCGTGTACTACTGTACCACCTGGCGGATCTACAGAGACAGCAGCGGCAGACCCTTCTACGTGATGGATGCCTGGGGCCAGGGCACACTAGTGACCGTGTCCAGC
配列番号13
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPKLLIYGANKLQDGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCLQGSKFPLTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
配列番号14
GATATCCAGATGACCCAGTCCCCCAGCAGCGTGTCCGCCTCTGTGGGCGATAGAGTGACCATCACCTGCCTGGCCAGCGAGGACATCTACACCTACCTGACCTGGTATCAGCAGAAGCCTGGCAAGGCCCCTAAGCTGCTGATCTACGGCGCCAACAAGCTGCAGGACGGCGTGCCCAGCAGATTCAGCGGCAGCGGCTCCGGCACCGACTACACCCTGACCATCAGCAGCCTGCAGCCTGAGGATTTCGCCACCTACTACTGCCTGCAGGGCAGCAAGTTCCCCCTGACCTTCGGCCAGGGCACCAAGCTGGAGATCAAGCGTACGGTGGCCGCCCCCAGCGTGTTCATCTTCCCCCCCAGCGATGAGCAGCTGAAGAGCGGCACCGCCAGCGTGGTGTGTCTGCTGAACAACTTCTACCCCCGGGAGGCCAAGGTGCAGTGGAAGGTGGACAATGCCCTGCAGAGCGGCAACAGCCAGGAGAGCGTGACCGAGCAGGACAGCAAGGACTCCACCTACAGCCTGAGCAGCACCCTGACCCTGAGCAAGGCCGACTACGAGAAGCACAAGGTGTACGCCTGTGAGGTGACCCACCAGGGCCTGTCCAGCCCCGTGACCAAGAGCTTCAACCGGGGCGAGTGC
配列番号15
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPKLLIYGANKLQDGVPSRFSGSGSGTDYTLTISSLQPEDFATYYCLQGSKFPLTFGQGTKLEIK
配列番号16
GATATCCAGATGACCCAGTCCCCCAGCAGCGTGTCCGCCTCTGTGGGCGATAGAGTGACCATCACCTGCCTGGCCAGCGAGGACATCTACACCTACCTGACCTGGTATCAGCAGAAGCCTGGCAAGGCCCCTAAGCTGCTGATCTACGGCGCCAACAAGCTGCAGGACGGCGTGCCCAGCAGATTCAGCGGCAGCGGCTCCGGCACCGACTACACCCTGACCATCAGCAGCCTGCAGCCTGAGGATTTCGCCACCTACTACTGCCTGCAGGGCAGCAAGTTCCCCCTGACCTTCGGCCAGGGCACCAAGCTGGAGATCAAG
配列番号17
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHWVRRRPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTEDTSTDTAYMELSSLRSEDTAVYYCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
配列番号18
CAGGTCCAGCTGGTACAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGTTTCCGGAGAAATAAGTACTGGATACTATTTCCACTGGGTGCGACGAAGGCCTGGAAAAGGGCTTGAGTGGATGGGAAGGATTGATCCTGAGGATGATAGTACTAAATATGCTGAGAGGTTCAAAGACAGAGTCACCATGACCGAGGACACATCTACAGACACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAGGACACGGCCGTGTATTACTGTACCACATGGCGGATATACCGAGATAGTTCTGGCCGCCCCTTCTATGTTATGGATGCCTGGGGCCAAGGGACACTAGTCACAGTCTCCTCAGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAA
配列番号19
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHWVRRRPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTEDTSTDTAYMELSSLRSEDTAVYYCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSS
配列番号20
CAGGTCCAGCTGGTACAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGTTTCCGGAGAAATAAGTACTGGATACTATTTCCACTGGGTGCGACGAAGGCCTGGAAAAGGGCTTGAGTGGATGGGAAGGATTGATCCTGAGGATGATAGTACTAAATATGCTGAGAGGTTCAAAGACAGAGTCACCATGACCGAGGACACATCTACAGACACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAGGACACGGCCGTGTATTACTGTACCACATGGCGGATATACCGAGATAGTTCTGGCCGCCCCTTCTATGTTATGGATGCCTGGGGCCAAGGGACACTAGTCACAGTCTCCTCA
配列番号21
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHFVRRRPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTADTSTDTAYMELSSLRSEDTATYFCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
配列番号22
CAGGTCCAGCTGGTACAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGTTTCCGGAGAAATAAGTACTGGATACTATTTCCACTTTGTGCGACGAAGGCCTGGAAAAGGGCTTGAGTGGATGGGAAGGATTGATCCTGAGGATGATAGTACTAAATATGCTGAGAGGTTCAAAGACAGAGTCACCATGACCGCAGACACATCTACAGACACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAGGACACGGCCACTTATTTTTGTACCACATGGCGGATATACCGAGATAGTTCTGGCCGCCCCTTCTATGTTATGGATGCCTGGGGCCAAGGGACACTAGTCACAGTCTCCTCAGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAA
配列番号23
QVQLVQSGAEVKKPGASVKVSCKVSGEISTGYYFHFVRRRPGKGLEWMGRIDPEDDSTKYAERFKDRVTMTADTSTDTAYMELSSLRSEDTATYFCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSS
配列番号24
CAGGTCCAGCTGGTACAGTCTGGGGCTGAGGTGAAGAAGCCTGGGGCCTCAGTGAAGGTCTCCTGCAAGGTTTCCGGAGAAATAAGTACTGGATACTATTTCCACTTTGTGCGACGAAGGCCTGGAAAAGGGCTTGAGTGGATGGGAAGGATTGATCCTGAGGATGATAGTACTAAATATGCTGAGAGGTTCAAAGACAGAGTCACCATGACCGCAGACACATCTACAGACACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAGGACACGGCCACTTATTTTTGTACCACATGGCGGATATACCGAGATAGTTCTGGCCGCCCCTTCTATGTTATGGATGCCTGGGGCCAAGGGACACTAGTCACAGTCTCCTCA
配列番号25
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPKLLIYGANKLQDGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCLQGSKFPLTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
配列番号26
GACATCCAGATGACCCAGTCTCCATCTTCTGTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGTCTGGCAAGTGAGGACATATACACTTATTTAACATGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGGTGCAAATAAGTTGCAAGATGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACTATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCTACAGGGTTCCAAGTTTCCGCTCACGTTTGGCCAGGGGACCAAGCTGGAGATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGACAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGT
配列番号27
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPKLLIYGANKLQDGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCLQGSKFPLTFGQGTKLEIK
配列番号28
GACATCCAGATGACCCAGTCTCCATCTTCTGTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGTCTGGCAAGTGAGGACATATACACTTATTTAACATGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGGTGCAAATAAGTTGCAAGATGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACTATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTACTATTGTCTACAGGGTTCCAAGTTTCCGCTCACGTTTGGCCAGGGGACCAAGCTGGAGATCAAA
配列番号29
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPQLLIYGANKLQDGVPSRFSGSGSGTDYTLTISSLQPEDEGDYYCLQGSKFPLTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
配列番号30
GACATCCAGATGACCCAGTCTCCATCTTCTGTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGTCTGGCAAGTGAGGACATATACACTTATTTAACATGGTATCAGCAGAAACCAGGGAAAGCCCCTCAACTCCTGATCTATGGTGCAAATAAGTTGCAAGATGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTATACTCTCACTATCAGCAGCCTGCAGCCTGAAGATGAAGGGGATTACTATTGTCTACAGGGTTCCAAGTTTCCGCTCACGTTTGGCCAGGGGACCAAGCTGGAGATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGACAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTAG
配列番号31
DIQMTQSPSSVSASVGDRVTITCLASEDIYTYLTWYQQKPGKAPQLLIYGANKLQDGVPSRFSGSGSGTDYTLTISSLQPEDEGDYYCLQGSKFPLTFGQGTKLEIK
配列番号32
GACATCCAGATGACCCAGTCTCCATCTTCTGTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGTCTGGCAAGTGAGGACATATACACTTATTTAACATGGTATCAGCAGAAACCAGGGAAAGCCCCTCAACTCCTGATCTATGGTGCAAATAAGTTGCAAGATGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTATACTCTCACTATCAGCAGCCTGCAGCCTGAAGATGAAGGGGATTACTATTGTCTACAGGGTTCCAAGTTTCCGCTCACGTTTGGCCAGGGGACCAAGCTGGAGATCAAA
配列番号33
EVQLQQSGAELVRPGTSVKLSCKVSGEISTGYYFHFVRRRPGQGLEWIGRIDPEDDSTKYAERFKDRATLTAQTSSNTAYLNLSSLTSEDTATYFCTTWRIYRDSSGRPFYVMDAWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
配列番号34
GAGGTCCAGCTACAGCAGTCTGGGGCTGAGCTTGTGAGACCTGGGACCTCTGTGAAGTTATCTTGCAAAGTTTCTGGCGAAATAAGTACAGGATACTATTTCCACTTTGTGAGGCGAAGGCCTGGACAGGGTCTGGAATGGATAGGAAGGATTGATCCTGAGGATGATAGTACTAAATATGCTGAGAGGTTCAAAGACAGGGCGACGCTCACTGCACAAACATCCTCCAACACAGCCTACCTGAACCTCAGCAGCCTGACCTCTGAGGACACTGCAACTTATTTTTGTACCACATGGCGGATATACCGAGATAGTTCTGGCCGCCCCTTCTATGTTATGGATGCCTGGGGTCAAGGAACACTAGTCACAGTCTCCTCAGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAA
配列番号35
DIQMTQSPASLSASLGETVSIECLASEDIYTYLTWYQQKPGKSPQLLIYGANKLQDGVPSRFSGSGSGTQYSLKISGIQPEDEGDYFCLQGSKFPLTFGSGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
配列番号36
GACATTCAAATGACCCAGTCTCCAGCTTCCCTGTCTGCATCTCTGGGAGAAACTGTCTCCATCGAATGTCTGGCAAGTGAGGACATATACACTTATTTAACATGGTATCAGCAGAAACCAGGGAAATCTCCTCAACTCCTGATCTATGGTGCAAATAAGTTGCAAGATGGGGTCCCATCACGGTTCAGTGGCAGTGGATCTGGCACACAGTATTCTCTCAAGATCAGCGGCATACAACCTGAAGATGAAGGGGATTATTTCTGTCTACAGGGTTCCAAGTTTCCGCTCACGTTCGGTTCTGGGACCAAGCTGGAGATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGACAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGT
配列番号37
QVQLVQSGAEVKKPGASVKVSCKVSGYTLTELSMHWVRQAPGKGLEWMGGFDPEDGETIYAQKFQGRVTMTEDTSTDTAYMELSSLRSEDTAVYYCAT
配列番号38
DIQMTQSPSSVSASVGDRVTITCRASQGISSWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQANSFP
配列番号39
WGQGTLVTVSS
配列番号40
FGQGTKLEIK
【図面の簡単な説明】
【0139】
【図1】H1L1及びH1L2の結合速度(ka)に対する温度の影響を示す。
【図2】解離速度(kd)に対する温度の影響を示す。
【図3】平衡定数(KD)に対する温度の影響を示す。
【図4】A〜Cは、表7に示したEC50値を作成した1回の実験の代表的データを示す。
【図5】ヒトIL18に結合する4個の選択したヒト化変異体のEC50値を示す。
【図6】アカゲザルIL18に結合する4個の選択したヒト化変異体のEC50値を示す。
【図7】50%滑液の存在下におけるヒトIL18に対するH1L2の結合を示す。
【図8】KG1アッセイにおけるIL−18が刺激するIFN−γ産生の阻害を示す。
【図9】A及びBはそれぞれヒトPBMCSドナーの10%及び25%自己血清における、LPSが刺激するIFN−γ産生の阻害を示す。
【図10】hIL18BPにより捕捉されたhIL18に対する2C10の結合を示す。
【図11】KG1細胞におけるヒトIL18が刺激するIFN−γ放出を阻害する9個のヒト化変異体の能力を示す。
【図12】KG1細胞におけるH1変異体及び2C10によるIL−18が刺激するIFN−γ産生の阻害を示す。
【図13】H1変異体のIC50データを95%信頼区間と共に示す。
【図14】KG1細胞におけるヒトIL−18が刺激するIFN−γ産生の阻害を示す。
【図15】KG1細胞におけるアカゲザルIL−18が刺激するIFN−γ産生の阻害を示す。
【図16】キメラ2C10を用いたヒトIL−18結合ELISAの結果を示す。
【図17】キメラ2C10を用いたアカゲザルIL−18結合ELISAの結果を示す。
【図18】A及びBはそれぞれヒトIL−18が結合したIL−18BPに対して結合するH1L2及び2C10を用いた結合ELISAの結果を示す。
【特許請求の範囲】
【請求項1】
以下の相補性決定領域(CDR):
CDRH1:配列番号1
CDRH2:配列番号2
CDRH3:配列番号3
CDRL1:配列番号4
CDRL2:配列番号5
CDRL3:配列番号6
を有する重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体。
【請求項2】
以下のCDR:
CDRH1:配列番号1
CDRH2:配列番号2
CDRH3:配列番号3
CDRL1:配列番号4
CDRL2:配列番号5
CDRL3:配列番号6
を有する重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体であって、該軽鎖の位置71の残基が、前記CDRが由来するドナー抗体中に存在するその対応する残基によって置換されている、ヒト化抗インターロイキン−18抗体。
【請求項3】
ヒトアクセプターフレームワークに移植されたドナー抗体由来CDRを含むヒト化抗インターロイキン−18抗体であって、抗インターロイキン−18抗体が配列番号1、2、3、4、5及び6に示される配列を有するCDRを含み、該抗インターロイキン−18抗体の軽鎖の位置71の残基が、該ドナー抗体フレームワーク中のその対応する位置に存在する残基と同一である、ヒト化抗インターロイキン−18抗体。
【請求項4】
配列番号1、2、3、4、5及び6に示されるCDRを含み、軽鎖の位置71にチロシンを含む、ヒト化抗インターロイキン−18抗体。
【請求項5】
配列番号1、2及び3に示されるCDRを有する重鎖と配列番号4、5及び6に示されるCDRを有する軽鎖を含むヒト化抗インターロイキン−18抗体であって、該軽鎖CDRが、ドナー抗体軽鎖の位置71にチロシンを有するドナー抗体に由来する、ヒト化抗インターロイキン−18抗体。
【請求項6】
ドナー抗体由来CDR及びヒト化抗体軽鎖の位置71にチロシンを含むヒト化抗インターロイキン−18抗体であって、該ドナー抗体が、2C10又はそのフレームワーク変異体である(すなわち、2C10と同じCDRであるが、異なるフレームワークを含む。米国特許第6,706,487号を参照されたい)、ヒト化抗インターロイキン−18抗体。
【請求項7】
(a)ヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有するCDRを有する重鎖、並びに
(b)ヒト軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有するCDRを有する軽鎖
を含むヒト化抗インターロイキン−18抗体であって、
前記ヒト軽鎖アクセプターフレームワークが配列番号38を含み、かつ軽鎖の位置71がチロシンである、ヒト化抗インターロイキン−18抗体。
【請求項8】
(a)配列番号1、2及び3に示される配列を有するCDRを含む重鎖、並びに
(b)ヒト軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有するCDRを含む軽鎖
を含むヒト化抗インターロイキン−18抗体であって、
該ヒト化抗インターロイキン−18抗体の該軽鎖アクセプターフレームワークが、配列番号38に示される配列の変異体であり、該変異体が位置71にチロシンを含み、かつ該変異体が配列番号38に示される配列を有するフレームワークと75%以上の同一性を有する、ヒト化抗インターロイキン−18抗体。
【請求項9】
(a)ドナー抗体軽鎖の位置71にチロシンを含むドナー抗体に由来する配列番号1、2、3、4、5及び6に示されるCDR、
(b)ヒト軽鎖の位置71にフェニルアラニンを含むヒトアクセプターフレームワーク
を含むヒト化抗インターロイキン−18抗体であって、
該抗インターロイキン−18抗体がその軽鎖の位置71にチロシンを含む、ヒト化抗インターロイキン−18抗体。
【請求項10】
(a)ドナー抗体軽鎖の位置71に芳香族アミノ酸を含むドナー抗体に由来する配列番号1、2、3、4、5及び6に示されるCDR、
(b)軽鎖のアクセプターフレームワークの位置71に、部分(a)の芳香族アミノ酸とは異なる種類の芳香族アミノ酸を含むヒトアクセプターフレームワーク
を含むヒト化抗インターロイキン−18抗体であって、
該抗インターロイキン−18抗体が、位置71に部分(a)の抗体に由来する芳香族アミノ酸を有する軽鎖を含む、ヒト化抗インターロイキン−18抗体。
【請求項11】
37℃で表面プラスモン共鳴(例えば、BiacoreTMT100)によって測定したとき、ヒトIL−18に関して90pM以上の結合親和性(KD)を示すヒト化抗インターロイキン−18抗体。
【請求項12】
配列番号1、2、3、4、5及び6に示されるCDRを含み、かつ37℃で表面プラスモン共鳴(例えば、BiacoreTMT100)によって測定したとき、ヒトIL−18に関して90pM以上の結合親和性(KD)を示す、ヒト化抗インターロイキン−18抗体。
【請求項13】
37℃で表面プラスモン共鳴(例えば、BiacoreTMT100)によって測定したとき、ヒトIL−18の結合に関して0.0002(1/秒)以下の解離速度(kd)を示すヒト化抗インターロイキン−18抗体。
【請求項14】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRが重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、93、39、40、36、71、89、91の一個若しくは複数の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71、及び任意により位置45、83、84、85の一個若しくは複数の(例えば全ての)残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項15】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該抗インターロイキン−18抗体の軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項16】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、39、40、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項17】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、36、39、40、71、89、91、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項18】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項19】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、93、39、40の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項20】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、93、39、40、36、71、89、91の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項21】
配列番号9、配列番号17、配列番号21からなる群から選択された重鎖、及び配列番号13、配列番号29からなる群から選択された軽鎖を含む、ヒト化抗インターロイキン−18抗体。
【請求項22】
配列番号9の重鎖と配列番号13の軽鎖、又は配列番号9の重鎖と配列番号29の軽鎖を含む、ヒト化抗インターロイキン−18抗体。
【請求項23】
配列番号17の重鎖と配列番号13の軽鎖、又は配列番号17の重鎖と配列番号29の軽鎖を含む、ヒト化抗インターロイキン−18抗体。
【請求項24】
配列番号21の重鎖と配列番号13の軽鎖、又は配列番号21の重鎖と配列番号29の軽鎖を含む、ヒト化抗インターロイキン−18抗体。
【請求項25】
(a)ヒトIL−18との特異的結合を可能にするCDRを有する重鎖、
(b)配列番号4、5及び6に示される配列を有するCDRを有し、かつ位置71にチロシン残基を有するアクセプターフレームワークを含む軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項26】
重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体であって、ヒトIL−18と前記抗体との結合の25℃での解離速度(kd)と、ヒトIL−18と前記抗体との結合の37℃での解離速度(kd)との比が、1:5(又は1:5未満)であり、該抗体がドナー抗体由来CDRとヒトアクセプターフレームワークを含み、かつヒトアクセプターフレームワークの軽鎖の位置71の残基が、そのドナー抗体のその対応する残基によって置換されている、ヒト化抗インターロイキン−18抗体。
【請求項27】
前記CDRが配列番号1、2、3、4、5及び6に示される配列を有する、請求項26に記載のヒト化抗体。
【請求項28】
前記ヒトアクセプターフレームワークの軽鎖の位置71の残基が芳香族アミノ酸、好ましくはフェニルアラニンであり、かつ前記ドナー抗体中のその対応する残基が、異なる種類の芳香族アミノ酸、好ましくはチロシンである、請求項26又は27に記載のヒト化抗体。
【請求項29】
請求項1〜28のいずれか一項に記載の抗インターロイキン−18抗体、及び薬学的に許容される担体を含む医薬組成物。
【請求項30】
多発性硬化症、関節リウマチなどの関節炎疾患、1型糖尿病、炎症性腸疾患(IBD)及び乾癬などの自己免疫疾患の治療のための医薬の製造における、請求項1〜28のいずれか一項に記載の抗インターロイキン−18抗体の使用
【請求項31】
関節リウマチの治療のための医薬の製造における、請求項31に記載の使用。
【請求項32】
多発性硬化症、関節リウマチなどの関節炎疾患、1型糖尿病、炎症性腸疾患(IBD)及び乾癬などの自己免疫疾患の治療のための、請求項1〜28のいずれか一項に記載の抗体。
【請求項33】
多発性硬化症、関節リウマチなどの関節炎疾患、1型糖尿病、炎症性腸疾患(IBD)及び乾癬などの自己免疫疾患を患うヒト患者を治療する方法であって、前記患者に請求項1〜28のいずれか一項に記載の抗体の治療有効量を投与することを含む方法。
【請求項34】
請求項1〜28のいずれか一項に記載の抗体を製造する方法であって、前記抗体の発現を可能にする条件下で、前記抗体をコードするポリヌクレオチドを含むベクターで形質転換若しくはトランスフェクトした宿主細胞を培養するステップを含む方法。
【請求項35】
前記条件が、血清不含培地で前記宿主細胞を培養するステップを含む、請求項34に記載の方法。
【請求項36】
前記宿主細胞が哺乳動物細胞である、請求項34又は35に記載の方法。
【請求項37】
前記宿主細胞がCHO又はNS0である、請求項36に記載の方法。
【請求項38】
前記CHO細胞が、前記ポリヌクレオチドでトランスフェクト若しくは形質転換する前の、DHFR−CHO細胞である、請求項37に記載の方法。
【請求項39】
治療に使用する抗体(特に、リガンドと受容体との相互作用を阻害する抗体、本明細書に開示する抗インターロイキン−18抗体など)を選択する方法であって、
(a)30〜45℃(好ましくは37℃)の温度において前記抗体が特異的に結合する抗原に対する前記抗体の結合親和性を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(b)20〜25℃(好ましくは25℃)の温度において前記抗体が特異的に結合する抗原に対する前記抗体の結合親和性を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(c)(a)の親和性が(b)の親和性を超える場合、好ましくは(a)の親和性がステップ(b)の親和性の2倍以上、より好ましくは4倍以上である場合に、治療に使用する前記抗体を選択するステップ
を含む方法。
【請求項40】
治療に使用する抗体(特に、リガンドと受容体との相互作用を阻害する抗体、本明細書に開示する抗インターロイキン−18抗体など)を選択する方法であって、
(a)30〜45℃(好ましくは37℃)の温度において前記抗体が特異的に結合する抗原からの前記抗体の解離速度を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(b)20〜25℃(好ましくは25℃)の温度において前記抗体が特異的に結合する抗原からの前記抗体の解離速度を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(c)(a)の解離速度が(b)の解離速度よりも遅い場合、治療に使用する前記抗体を選択するステップ
を含む方法。
【請求項1】
以下の相補性決定領域(CDR):
CDRH1:配列番号1
CDRH2:配列番号2
CDRH3:配列番号3
CDRL1:配列番号4
CDRL2:配列番号5
CDRL3:配列番号6
を有する重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体。
【請求項2】
以下のCDR:
CDRH1:配列番号1
CDRH2:配列番号2
CDRH3:配列番号3
CDRL1:配列番号4
CDRL2:配列番号5
CDRL3:配列番号6
を有する重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体であって、該軽鎖の位置71の残基が、前記CDRが由来するドナー抗体中に存在するその対応する残基によって置換されている、ヒト化抗インターロイキン−18抗体。
【請求項3】
ヒトアクセプターフレームワークに移植されたドナー抗体由来CDRを含むヒト化抗インターロイキン−18抗体であって、抗インターロイキン−18抗体が配列番号1、2、3、4、5及び6に示される配列を有するCDRを含み、該抗インターロイキン−18抗体の軽鎖の位置71の残基が、該ドナー抗体フレームワーク中のその対応する位置に存在する残基と同一である、ヒト化抗インターロイキン−18抗体。
【請求項4】
配列番号1、2、3、4、5及び6に示されるCDRを含み、軽鎖の位置71にチロシンを含む、ヒト化抗インターロイキン−18抗体。
【請求項5】
配列番号1、2及び3に示されるCDRを有する重鎖と配列番号4、5及び6に示されるCDRを有する軽鎖を含むヒト化抗インターロイキン−18抗体であって、該軽鎖CDRが、ドナー抗体軽鎖の位置71にチロシンを有するドナー抗体に由来する、ヒト化抗インターロイキン−18抗体。
【請求項6】
ドナー抗体由来CDR及びヒト化抗体軽鎖の位置71にチロシンを含むヒト化抗インターロイキン−18抗体であって、該ドナー抗体が、2C10又はそのフレームワーク変異体である(すなわち、2C10と同じCDRであるが、異なるフレームワークを含む。米国特許第6,706,487号を参照されたい)、ヒト化抗インターロイキン−18抗体。
【請求項7】
(a)ヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有するCDRを有する重鎖、並びに
(b)ヒト軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有するCDRを有する軽鎖
を含むヒト化抗インターロイキン−18抗体であって、
前記ヒト軽鎖アクセプターフレームワークが配列番号38を含み、かつ軽鎖の位置71がチロシンである、ヒト化抗インターロイキン−18抗体。
【請求項8】
(a)配列番号1、2及び3に示される配列を有するCDRを含む重鎖、並びに
(b)ヒト軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有するCDRを含む軽鎖
を含むヒト化抗インターロイキン−18抗体であって、
該ヒト化抗インターロイキン−18抗体の該軽鎖アクセプターフレームワークが、配列番号38に示される配列の変異体であり、該変異体が位置71にチロシンを含み、かつ該変異体が配列番号38に示される配列を有するフレームワークと75%以上の同一性を有する、ヒト化抗インターロイキン−18抗体。
【請求項9】
(a)ドナー抗体軽鎖の位置71にチロシンを含むドナー抗体に由来する配列番号1、2、3、4、5及び6に示されるCDR、
(b)ヒト軽鎖の位置71にフェニルアラニンを含むヒトアクセプターフレームワーク
を含むヒト化抗インターロイキン−18抗体であって、
該抗インターロイキン−18抗体がその軽鎖の位置71にチロシンを含む、ヒト化抗インターロイキン−18抗体。
【請求項10】
(a)ドナー抗体軽鎖の位置71に芳香族アミノ酸を含むドナー抗体に由来する配列番号1、2、3、4、5及び6に示されるCDR、
(b)軽鎖のアクセプターフレームワークの位置71に、部分(a)の芳香族アミノ酸とは異なる種類の芳香族アミノ酸を含むヒトアクセプターフレームワーク
を含むヒト化抗インターロイキン−18抗体であって、
該抗インターロイキン−18抗体が、位置71に部分(a)の抗体に由来する芳香族アミノ酸を有する軽鎖を含む、ヒト化抗インターロイキン−18抗体。
【請求項11】
37℃で表面プラスモン共鳴(例えば、BiacoreTMT100)によって測定したとき、ヒトIL−18に関して90pM以上の結合親和性(KD)を示すヒト化抗インターロイキン−18抗体。
【請求項12】
配列番号1、2、3、4、5及び6に示されるCDRを含み、かつ37℃で表面プラスモン共鳴(例えば、BiacoreTMT100)によって測定したとき、ヒトIL−18に関して90pM以上の結合親和性(KD)を示す、ヒト化抗インターロイキン−18抗体。
【請求項13】
37℃で表面プラスモン共鳴(例えば、BiacoreTMT100)によって測定したとき、ヒトIL−18の結合に関して0.0002(1/秒)以下の解離速度(kd)を示すヒト化抗インターロイキン−18抗体。
【請求項14】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRが重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、93、39、40、36、71、89、91の一個若しくは複数の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71、及び任意により位置45、83、84、85の一個若しくは複数の(例えば全ての)残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項15】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該抗インターロイキン−18抗体の軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項16】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、39、40、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項17】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列に由来するフレームワーク領域を含み、該重鎖の位置27、28、29、36、39、40、71、89、91、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項18】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、93の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項19】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、93、39、40の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項20】
(a)ドナー抗体由来CDRを含む重鎖であって、CDRがヒト重鎖アクセプターフレームワークに移植された配列番号1、2及び3に示される配列を有し、該重鎖アクセプターフレームワークが配列番号37に示される配列を有し、該重鎖の位置27、28、29、93、39、40、36、71、89、91の残基が、該ドナー抗体重鎖中のその対応する残基と同一である重鎖、
(b)ドナー抗体由来CDRを含む軽鎖であって、CDRが軽鎖アクセプターフレームワークに移植された配列番号4、5及び6に示される配列を有し、該軽鎖アクセプターフレームワークが配列番号38に示される配列を有し、該軽鎖の位置71、45、83、84、85の残基が、該ドナー抗体軽鎖中のその対応する残基と同一である軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項21】
配列番号9、配列番号17、配列番号21からなる群から選択された重鎖、及び配列番号13、配列番号29からなる群から選択された軽鎖を含む、ヒト化抗インターロイキン−18抗体。
【請求項22】
配列番号9の重鎖と配列番号13の軽鎖、又は配列番号9の重鎖と配列番号29の軽鎖を含む、ヒト化抗インターロイキン−18抗体。
【請求項23】
配列番号17の重鎖と配列番号13の軽鎖、又は配列番号17の重鎖と配列番号29の軽鎖を含む、ヒト化抗インターロイキン−18抗体。
【請求項24】
配列番号21の重鎖と配列番号13の軽鎖、又は配列番号21の重鎖と配列番号29の軽鎖を含む、ヒト化抗インターロイキン−18抗体。
【請求項25】
(a)ヒトIL−18との特異的結合を可能にするCDRを有する重鎖、
(b)配列番号4、5及び6に示される配列を有するCDRを有し、かつ位置71にチロシン残基を有するアクセプターフレームワークを含む軽鎖
を含むヒト化抗インターロイキン−18抗体。
【請求項26】
重鎖と軽鎖を含むヒト化抗インターロイキン−18抗体であって、ヒトIL−18と前記抗体との結合の25℃での解離速度(kd)と、ヒトIL−18と前記抗体との結合の37℃での解離速度(kd)との比が、1:5(又は1:5未満)であり、該抗体がドナー抗体由来CDRとヒトアクセプターフレームワークを含み、かつヒトアクセプターフレームワークの軽鎖の位置71の残基が、そのドナー抗体のその対応する残基によって置換されている、ヒト化抗インターロイキン−18抗体。
【請求項27】
前記CDRが配列番号1、2、3、4、5及び6に示される配列を有する、請求項26に記載のヒト化抗体。
【請求項28】
前記ヒトアクセプターフレームワークの軽鎖の位置71の残基が芳香族アミノ酸、好ましくはフェニルアラニンであり、かつ前記ドナー抗体中のその対応する残基が、異なる種類の芳香族アミノ酸、好ましくはチロシンである、請求項26又は27に記載のヒト化抗体。
【請求項29】
請求項1〜28のいずれか一項に記載の抗インターロイキン−18抗体、及び薬学的に許容される担体を含む医薬組成物。
【請求項30】
多発性硬化症、関節リウマチなどの関節炎疾患、1型糖尿病、炎症性腸疾患(IBD)及び乾癬などの自己免疫疾患の治療のための医薬の製造における、請求項1〜28のいずれか一項に記載の抗インターロイキン−18抗体の使用
【請求項31】
関節リウマチの治療のための医薬の製造における、請求項31に記載の使用。
【請求項32】
多発性硬化症、関節リウマチなどの関節炎疾患、1型糖尿病、炎症性腸疾患(IBD)及び乾癬などの自己免疫疾患の治療のための、請求項1〜28のいずれか一項に記載の抗体。
【請求項33】
多発性硬化症、関節リウマチなどの関節炎疾患、1型糖尿病、炎症性腸疾患(IBD)及び乾癬などの自己免疫疾患を患うヒト患者を治療する方法であって、前記患者に請求項1〜28のいずれか一項に記載の抗体の治療有効量を投与することを含む方法。
【請求項34】
請求項1〜28のいずれか一項に記載の抗体を製造する方法であって、前記抗体の発現を可能にする条件下で、前記抗体をコードするポリヌクレオチドを含むベクターで形質転換若しくはトランスフェクトした宿主細胞を培養するステップを含む方法。
【請求項35】
前記条件が、血清不含培地で前記宿主細胞を培養するステップを含む、請求項34に記載の方法。
【請求項36】
前記宿主細胞が哺乳動物細胞である、請求項34又は35に記載の方法。
【請求項37】
前記宿主細胞がCHO又はNS0である、請求項36に記載の方法。
【請求項38】
前記CHO細胞が、前記ポリヌクレオチドでトランスフェクト若しくは形質転換する前の、DHFR−CHO細胞である、請求項37に記載の方法。
【請求項39】
治療に使用する抗体(特に、リガンドと受容体との相互作用を阻害する抗体、本明細書に開示する抗インターロイキン−18抗体など)を選択する方法であって、
(a)30〜45℃(好ましくは37℃)の温度において前記抗体が特異的に結合する抗原に対する前記抗体の結合親和性を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(b)20〜25℃(好ましくは25℃)の温度において前記抗体が特異的に結合する抗原に対する前記抗体の結合親和性を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(c)(a)の親和性が(b)の親和性を超える場合、好ましくは(a)の親和性がステップ(b)の親和性の2倍以上、より好ましくは4倍以上である場合に、治療に使用する前記抗体を選択するステップ
を含む方法。
【請求項40】
治療に使用する抗体(特に、リガンドと受容体との相互作用を阻害する抗体、本明細書に開示する抗インターロイキン−18抗体など)を選択する方法であって、
(a)30〜45℃(好ましくは37℃)の温度において前記抗体が特異的に結合する抗原からの前記抗体の解離速度を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(b)20〜25℃(好ましくは25℃)の温度において前記抗体が特異的に結合する抗原からの前記抗体の解離速度を測定するステップ(例えば、BiacoreTMなどの表面プラズモン共鳴を使用)、
(c)(a)の解離速度が(b)の解離速度よりも遅い場合、治療に使用する前記抗体を選択するステップ
を含む方法。
【図1】

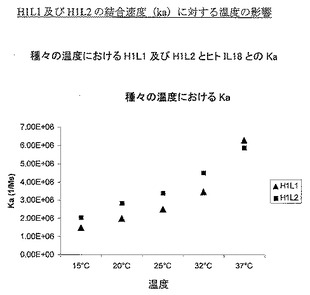
【図2】
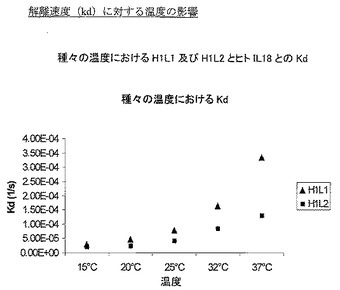
【図3】
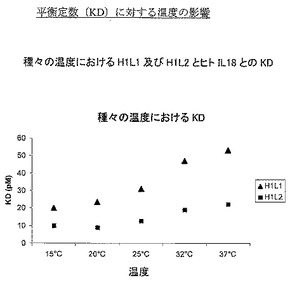
【図4】
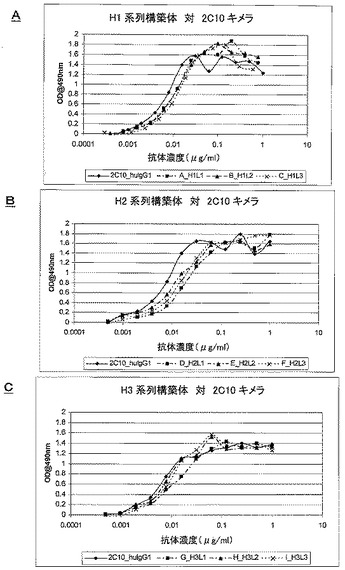
【図5】
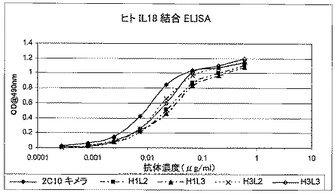
【図6】
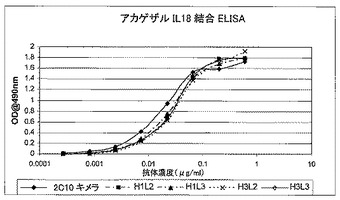
【図7】
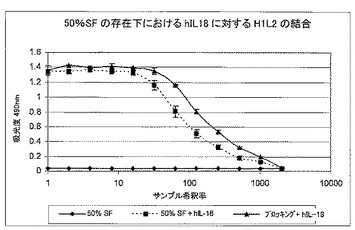
【図8】
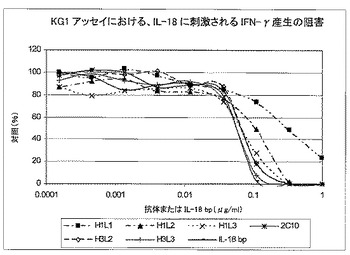
【図9】
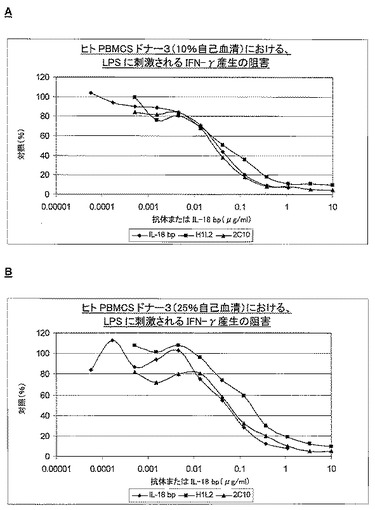
【図10】
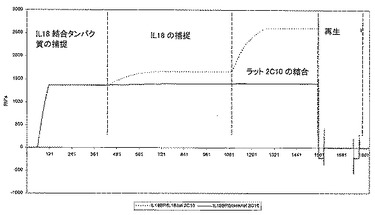
【図11】
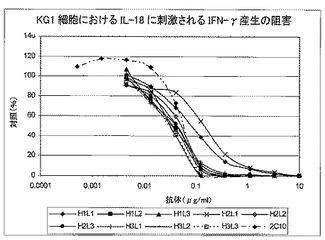
【図12】
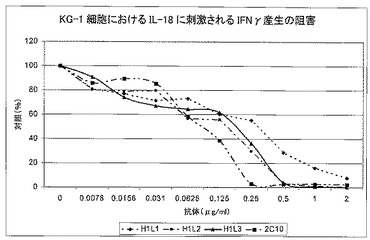
【図13】
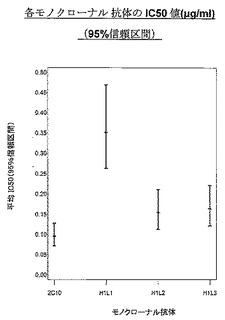
【図14】
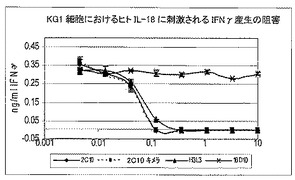
【図15】
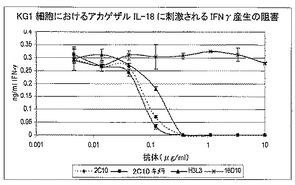
【図16】
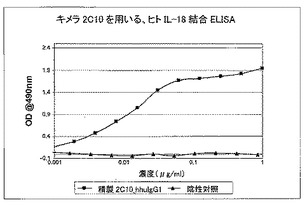
【図17】
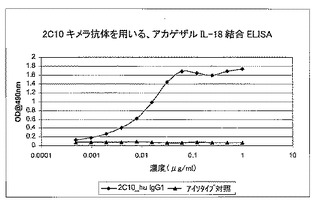
【図18】
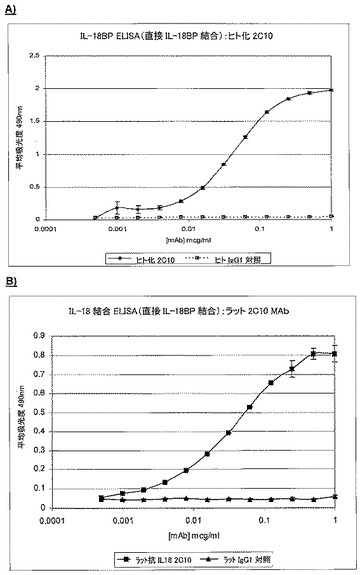
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【公表番号】特表2009−537608(P2009−537608A)
【公表日】平成21年10月29日(2009.10.29)
【国際特許分類】
【出願番号】特願2009−511524(P2009−511524)
【出願日】平成19年5月23日(2007.5.23)
【国際出願番号】PCT/EP2007/055029
【国際公開番号】WO2007/137984
【国際公開日】平成19年12月6日(2007.12.6)
【出願人】(397009934)グラクソ グループ リミテッド (832)
【氏名又は名称原語表記】GLAXO GROUP LIMITED
【住所又は居所原語表記】Glaxo Wellcome House,Berkeley Avenue Greenford,Middlesex UB6 0NN,Great Britain
【Fターム(参考)】
【公表日】平成21年10月29日(2009.10.29)
【国際特許分類】
【出願日】平成19年5月23日(2007.5.23)
【国際出願番号】PCT/EP2007/055029
【国際公開番号】WO2007/137984
【国際公開日】平成19年12月6日(2007.12.6)
【出願人】(397009934)グラクソ グループ リミテッド (832)
【氏名又は名称原語表記】GLAXO GROUP LIMITED
【住所又は居所原語表記】Glaxo Wellcome House,Berkeley Avenue Greenford,Middlesex UB6 0NN,Great Britain
【Fターム(参考)】
[ Back to top ]