
改変型ホルムアルデヒド脱水素酵素及びその利用
【課題】高温条件下でも熱安定性に優れたホルムアルデヒド脱水素酵素を提供する。
【解決手段】野生型ホルムアルデヒド脱水素酵素のアミノ酸配列中の少なくとも1つの位置のアミノ酸の欠失、置換、付加或いは挿入による改変により得られる改変型ホルムアルデヒド脱水素酵素であって、改変が、野生型ホルムアルデヒド脱水素酵素のアミノ酸配列における第70位−第150位で生じており、野生型ホルムアルデヒド脱水素酵素に比べて安定性が向上している。
【解決手段】野生型ホルムアルデヒド脱水素酵素のアミノ酸配列中の少なくとも1つの位置のアミノ酸の欠失、置換、付加或いは挿入による改変により得られる改変型ホルムアルデヒド脱水素酵素であって、改変が、野生型ホルムアルデヒド脱水素酵素のアミノ酸配列における第70位−第150位で生じており、野生型ホルムアルデヒド脱水素酵素に比べて安定性が向上している。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、改変型ホルムアルデヒド脱水素酵素及びその利用に関する。より詳細には、本発明は、改変型ホルムアルデヒド脱水素酵素、当該酵素をコードする遺伝子、及び当該酵素を含むバイオセンサー、並びに酵素燃料電池に関する。
【背景技術】
【0002】
ホルムアルデヒド脱水素酵素(EC 1.2.1.46)(Formaldehyde dehydrogenase:FDH)は、ホルムアルデヒドを酸化してギ酸を生成する反応を触媒する。ホルアルデヒド脱水素酵素の基質となるホルムアルデヒドは、シックハウス症候群等の慢性的なアレルギー症状を引き起こす原因物質の1つとして知られている。また、稀薄な溶液でも細胞原形質のタンパク質を不可逆的に凝固させ、全ての細胞機能を停止、死滅させる作用があるために、細胞毒としても知られている。そのため、食品中、大気中などのホルムアルデヒドを測定し、これを分解及び除去することが求められており、ホルムアルデヒド脱水素酵素の酵素活性の利用が検討されている。例えば、生体試料中のホルムアルデヒドの定量等のホルムアルデヒド測定技術や、ホルムアルデヒド脱水素酵素を固定化した空気浄化フィルター等のホルムアルデヒドの除去技術等、特に環境技術の分野における応用が期待されている。
【0003】
例えば、ホルムアルデヒドをはじめとするアルデヒド化合物やアルコール化合物を簡易に検出測定できるバイオセンサー用酵素としての利用が報告されている(例えば、特許文献1を参照)。詳細には、気相中や液相中のアルデヒド化合物及びアルコール化合物を簡易に検出測定できる酵素固定化バイオセンサーが開示されており、これは、過酸化水素電極の表面にアルコール酸化酵素が固定化されると共に最表面に高分子被覆が配設され、過酸化水素の酸化電流値、またはその変化よりアルデヒド化合物もしくはアルコール化合物の存在を検出するものである。
【0004】
また、近年、クリーンエネルギー技術として、アルコールなどのバイオマスを燃料とした酵素燃料電池が考案されており、酸化還元酵素が電池の構成部材として利用されている。一例として、メタノールを燃料とした酵素燃料電池の場合、触媒としてメタノールに作用してホルムアルデヒドに酸化するアルコールデヒドロゲナーゼと、ホルムアルデヒドに作用してギ酸に酸化するホルムアルデヒドデヒドロゲナーゼと、ギ酸に作用してCO2に酸化する蟻酸デヒドロゲナーゼとを用いることにより構成することができる。これにより、メタノールはCO2まで分解され、メタノール1分子につき3段階の酸化反応により合計6電子が生成されることとなる。
【0005】
そして、ホルムアルデヒド脱水素酵素は、細菌、酵母から哺乳類に至るまで広く存在していることが知られており、多様な生物由来のホルムアルデヒド脱水素酵素が報告されている。例えば、パン酵母であるサッカロマイセス・セレビシエ(Saccharomyces Cerevisiae)由来のホルムアルデヒド脱水素酵素(例えば、非特許文献1を参照)、パラコッカス・デニトリフィカンス(Paracoccus denitrificans)由来のホルムアルデヒド脱水素酵素(例えば、非特許文献2を参照)、シュードモナス・プチダ(Pseudomonas putida)由来のホルムアルデヒド脱水素酵素(例えば、非特許文献3を参照)等が挙げられる。
【0006】
ホルムアルデヒド脱水素酵素には、反応にグルタチオンを要求するグルタチオン依存性、及びグルタチオンを要求しない非依存性の2つの型がある。一般に広く存在するホルムアルデヒド脱水素酵素はグルタチオン依存性であるのに対して、シュードモナス・プチダ由来のホルムアルデヒド脱水素酵素は、グルタチオン非依存的にホルムアルデヒドの酸化還元反応を触媒する特徴的な酵素であることが知られている。そして、このホルムアルデヒド脱水素酵素は、各サブユニットが376アミノ酸残基からなるホモ4量体酵素であることが報告されている(例えば、非特許文献3を参照)と共に、X線結晶構造解析の結果をもが報告され活性中心をはじめとした高次構造が明らかとなっている(例えば、非特許文献4を参照)。
【0007】
以上の従来文献については、すべて天然に存在する野生型のホルムアルデヒド脱水素酵素に関するものである。一方、分子進化工学的手法を使って酵素の構造を改変し、酵素活性の至適温度を低温化した報告もある(非特許文献5)。これは、Pseudomonas putida由来の野生型のホルムアルデヒド脱水素酵素にアミノ酸変異を導入し、30℃程度の低い温度域において、野生型酵素より酵素活性を1.7倍向上させたものである。
【0008】
ところが、ホルムアルデヒド脱水素酵素に限らず酵素は、触媒作用を選択する性質が非常に強いことが知られている。例えば、酵素は、生体が生存可能な緩和な条件下で最も酵素活性を発揮し、生体内で代謝される機構を有するものであることから、酵素を構成するタンパク質構造に変化を与えるような因子の存在により酵素の活性は影響を受ける。つまり、酵素活性は、温度等の物理的条件、及びpH、塩濃度等の化学的条件の影響を受けることが知られており、これらに対する安定性が低いため用途が限定されるという問題点があった。そして、従来公知のホルムアルデヒド脱水素酵素は、何れも天然に存在する野生型の酵素であることから、熱に対する安定性が低かった。
【0009】
しかしながら、バイオセンサーや燃料電池への適用に際しては、多様な環境において安定して活性を保持できる酵素が不可欠であり、特に、酵素寿命に対して相関があるといわれる熱安定性の向上が要求される。従来公知の天然に存在する野生型ホルムアルデヒド脱水素酵素(非特許文献1〜4)は、何れも熱に対する安定性が低かった。また、野生型のホルムアルデヒド脱水素酵素を改変したホルムアルデヒド脱水素酵素(非特許文献5)についても、活性の至適温度を低温化しているために、高い温度域では野生型酵素より熱安定性が低下しており、酵素の熱失活が生じやすくなっている。
【0010】
そのため、従来公知のホルムアルデヒド脱水素酵素をバイオセンサーや燃料電池への適用した場合は、酵素の保存安定性が低く、酵素の熱失活により酵素の使用量の増大及び精度の低下が生じることから、市場の要望に十分に対応できるものではなかった。従って、高温条件下でも安定的に作用できるホルムアルデヒド脱水素酵素の提供が望まれていた。
【先行技術文献】
【特許文献】
【0011】
【特許文献1】特開2007−139729号公報
【非特許文献】
【0012】
【非特許文献1】Steinman CR. Jakoby WB.著、“Yeast aldehyde dehydrogenase. II. Properties of the homogeneous enzyme preparations.(酵母アルデヒドデヒドロゲナーゼII)” J Biol Chem.、1968年、第243巻、第730〜734頁
【非特許文献2】Ras J. Van Ophem PW. Reijnders WN. Van Spanning RJ. Duine JA. Stouthamer AH. Harms N.著、“Isolation, sequencing, and mutagenesis of the gene encoding NAD- and glutathione-dependent formaldehyde dehydrogenase (GD-FALDH) from Paracoccus denitrificans, in which GD-FALDH is essential for methylotrophicgrowth.” J Bacteriol.、1995年、第177巻、第247〜251頁
【非特許文献3】Ito K. Takahashi M. Yoshimoto T. Tsuru D.著、“Cloning and high-level expression of the glutathione-independent formaldehyde dehydrogenase gene from Pseudomonas putida. ”J Bacteriol. 1994年、第176巻、第2483〜2491頁
【非特許文献4】Tanaka N. Kusakabe Y. Ito K. Yoshimoto T. Nakamura KT.著、“Crystal structure of formaldehyde dehydrogenase from Pseudomonas putida: the structural origin of the tightly bound cofactor in nicotinoprotein dehydrogenases.”J. Mol. Biol.、2002年、第324巻、第519〜533頁
【非特許文献5】Fujii Y, Yamasaki Y, Matsumoto M, Nishida H, Hada M, Ohkubo K. 著、“The artificial evolution of an enzyme by random mutagenesis: the development of formaldehyde dehydrogenase.” Biosci Biotechnol Biochem. 2004, 68(8), 1722-1727.
【発明の概要】
【発明が解決しようとする課題】
【0013】
本発明は、上記事情に鑑みてなされたものであり、高温条件下でも熱安定性に優れたホルムアルデヒド脱水素酵素を提供することにある。
【課題を解決するための手段】
【0014】
本発明者らは、上記課題を解決すべく研究を重ねた結果、野生型ホルムアルデヒド脱水素酵素の特定のアミノ酸配列領域における少なくとも1つの位置のアミノ酸を他のアミノ酸で置換することにより、野生型ホルムアルデヒド脱水素酵素と比較して熱安定性及び保存安定性を向上できることを見出した。更に、かかる酵素を利用することにより、従来の酵素の熱安定性に関する欠点を克服し、より実用面において有利なアルデヒド検出用バイオセンサー及び燃料電池を提供できることをも見出した。本発明者らはこれらの知見に基づき本発明を完成するに至った。
【0015】
即ち、上記目的を達成するため、以下の[1]〜[13]に示す発明を提供する。
[1]野生型ホルムアルデヒド脱水素酵素のアミノ酸配列中の少なくとも1つの位置のアミノ酸の欠失、置換、付加或いは挿入による改変により得られる改変型ホルムアルデヒド脱水素酵素であって、前記改変が、前記野生型ホルムアルデヒド脱水素酵素のアミノ酸配列における第70位−第150位で生じており、前記野生型ホルムアルデヒド脱水素酵素に比べて安定性が向上している改変型ホルムアルデヒド脱水素酵素。
[2]前記改変が、前記アミノ酸配列の第128位に対応する位置で生じている上記[1]に記載の改変型ホルムアルデヒド脱水素酵素。
[3]前記改変が、前記アミノ酸配列の第128位に対応する位置におけるアスパラギン酸のグリシンへの置換である上記[2]に記載の改変型ホルムアルデヒド脱水素酵素。
[4]前記改変が、前記アミノ酸配列の第110位に対応する位置で生じている上記[1]に記載の改変型ホルムアルデヒド脱水素酵素。
[5]前記改変が、前記アミノ酸配列の第110位に対応する位置におけるグリシンのアスパラギン酸への置換である上記[4]に記載の改変型ホルムアルデヒド脱水素酵素。
[6]前記野生型ホルムアルデヒド脱水素酵素が、配列番号2に示すアミノ酸配列を有する上記[1]〜[5]の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素。
[7]上記[1]〜[6]の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素をコードする単離核酸分子。
[8]上記[7]に記載の単離核酸分子を含有する組換えベクター。
[9]上記[8]に記載の組換えベクターを含有する形質転換体。
[10]上記[9]に記載の形質転換体を培養する工程、及び得られた培養物からホルムアルデヒドの脱水素反応を触媒する能力を有するタンパク質を採取する工程を含む、野生型ホルムアルデヒド脱水素酵素に比べて熱安定性が向上している改変型ホルムアルデヒド脱水素酵素の製造方法。
【0016】
上記[1]〜[10]の構成の改変型ホルムアルデヒド脱水素酵素は、野生型ホルムアルデヒド脱水素酵素のアミノ酸配列(具体的には配列番号2に示すアミノ酸配列)に置ける第70位−第150位のアミノ酸の何れかに改変を施している。
この位置は、酵素の補欠分子であるZnの結合部位に近接しており、酵素サブユニット間の界面付近とは離れている。タンパク質の構造安定化には、Znなど補欠分子の結合安定性が大きく影響することが知られていて、一般的にはアミノ酸変異の導入を避けたほうがよいと考えられている。しかし、本願のようにZnの結合部位に近接した位置でアミノ酸に改変を施すことで、熱安定性を向上させた新規なホルムアルデヒド脱水素酵素の提供が可能となる。
好ましくは、前記改変が、前記アミノ酸配列の第128位に対応する位置におけるアスパラギン酸のグリシンへの置換、或いは、第110位に対応する位置におけるグリシンのアスパラギン酸への置換とすれば、野生型ホルムアルデヒド脱水素酵素に比べて顕著な熱安定性および長期安定性を有するようになる。
従って、本発明によって提供されるホルムアルデヒド脱水素酵素は、医療・食品・環境分野等の様々な産業分野におけるホルムアルデヒドの脱水素反応を要する技術に適用できる。更に、熱安定性が向上した実用性の高い本発明の改変型ホルムアルデヒド脱水素酵素のアミノ酸配列、並びに塩基配列が明確になったことから、遺伝子工学的手法を利用して組換え体として当該酵素を低コストかつ工業的に大量生産することが可能となった。
【0017】
[11]上記[1]〜[6]の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素の触媒活性を利用してアルデヒド化合物を検出する、アルデヒド化合物の検出方法。
【0018】
上記[11]の構成によれば、本発明の改変型ホルムアルデヒド脱水素酵素の触媒能力を利用したアルデヒド化合物の検出方法が提供でき、医療、食品、環境分野等、様々な分野に利用することができる。特には、本発明の改変型ホルムアルデヒド脱水素酵素は熱安定性が高いことから、酵素の触媒能力の劣化を招くことなく測定精度の向上を図れる。更には、酵素使用量をも低減できることからコスト削減効果をも奏する。
【0019】
[12]上記[1]〜[6]の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素を電極上に固定化した酵素センサー。
【0020】
上記[12]の構成によれば、本発明の改変型ホルムアルデヒド脱水素酵素の触媒能力を利用したアルデヒド化合物検出用の酵素センサーが提供でき、医療、食品、環境分野等、様々な分野に利用することができる。特には、本発明の改変型ホルムアルデヒド脱水素酵素は熱安定性が高いことから、酵素の触媒能力の劣化を招くことなく測定精度の向上を図れる。更には、酵素使用量をも低減できることからコスト削減効果をも奏する。
【0021】
[13]上記[1]〜[6]の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素を含み、前記改変型ホルムアルデヒド脱水素酵素のアルデヒド化合物の酸化反応に伴って生成する電子を受け取るアノード極、酸素に電子を伝達することのできる触媒および酵素のいずれかを保持するカソード極を備え、前記アノード極と前記カソード極とが電気的に結合されている燃料電池。
【0022】
上記[13]の構成によれば、本発明の改変型ホルムアルデヒド脱水素酵素の触媒能力を利用した燃料電池が提供できる。特に、本発明の改変型ホルムアルデヒド脱水素酵素は熱安定性が高いことから、酵素の触媒能力の劣化を招くことなく持続的に発電を行なうことが可能となり、燃料電池の性能向上を図れる。更には、酵素使用量をも低減できることからコスト削減効果をも奏する。
【図面の簡単な説明】
【0023】
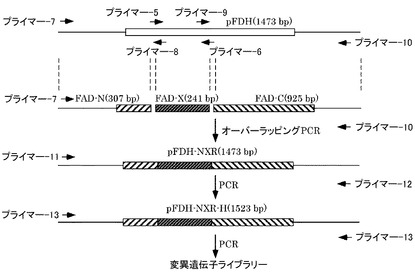
【図1】変異遺伝子ライブラリーの作製概略を示す図である。
【図2】野生型FDH遺伝子とその近傍領域において各プライマーが結合する位置を示す図である。
【図3】野生型FDHの構造解析を行なった結果を示した図である。
【図4】D128Gの構造解析を行なった結果を示した図である((a)野生型FDH、(b)D128G)。
【図5】G110Dの構造解析を行なった結果を示した図である。
【図6】精製したD128Gおよび野生型FDHのアクリルアミドゲル電気泳動の結果を示した図である。
【図7】D128Gおよび野生型FDHのpH依存性を調べた結果を示した図である。
【図8】改変型FDHおよび野生型FDHの温度依存性を調べた結果を示した図である((a)D128G,(b)G110D)。
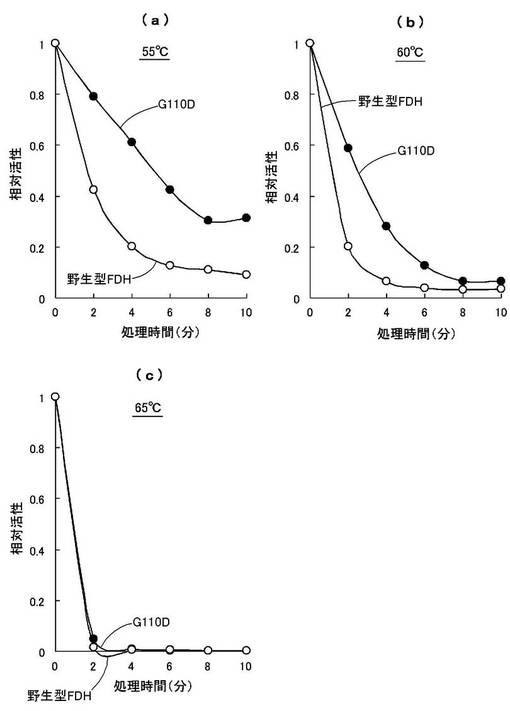
【図9】D128Gおよび野生型FDHの温度安定性を調べた結果を示した図である((a)55℃,(b)60℃,(c)65℃)。
【図10】G110Dおよび野生型FDHの温度安定性を調べた結果を示した図である((a)55℃,(b)60℃,(c)65℃)。
【図11】改変型FDHおよび野生型FDHの安定性を調べた結果を示した図である((a)D128Gおよび野生型FDHの比較、(b)G110Dおよび野生型FDHの比較)。
【図12】D128Gおよび野生型FDHについて常温時(25℃)の安定性を調べた結果を示した図である。
【図13】D128Gおよび野生型FDHについて常温時(30℃)の安定性を調べた結果を示した図である。
【発明を実施するための形態】
【0024】
以下、本発明について詳細に説明する。
近年、クリーンエネルギー技術として、アルコールなどのバイオマスを燃料とした酵素燃料電池が考案されており、そこでは本発明のホルムアルデヒド脱水素酵素のような酸化・還元を触媒する酵素が電池の電極触媒として利用されている。一例として、メタノールを燃料とした酵素燃料電池の場合、触媒としてメタノールに作用してホルムアルデヒドに酸化するアルコールデヒドロゲナーゼと、ホルムアルデヒドに作用して蟻酸に酸化するホルムアルデヒドデヒドロゲナーゼと、蟻酸に作用してCO2に酸化する蟻酸デヒドロゲナーゼとを用いることができる。これにより、メタノールはCO2まで分解され、メタノール1分子につき3段階の酸化反応により合計6電子が生成される。
【0025】
本発明の改変型ホルムアルデヒド脱水素酵素(以下、「改変型FDH」と略する)は、天然に存在する野生型FDHのアミノ酸配列において、少なくとも1つの位置のアミノ酸の欠失、置換、付加或いは挿入により改変されており、そして、野生型FDHに比べて熱安定性が向上している。ここで「少なくとも1つの位置のアミノ酸の欠失、置換、付加或いは挿入により改変」とは、改変の基礎となるタンパク質をコードする遺伝子に対して、公知のDNA組換え技術、及び点変異導入方法等によって、欠失、置換、付加或いは挿入することができる程度の数のアミノ酸が、欠失、置換、付加或いは挿入されることを意味し、これらの組み合わせをも含む。したがって、改変型FDHは、これらの改変は1つ有するものであっても、また2つ以上が組み合わされたものであってもよい。このような改変は、人為的に導入することもできるし、また、自然界において非意図的に生じることもある。したがって、本発明における改変型FDHには、これら双方の改変型が含まれる。
【0026】
野生型FDHとは、自然界より分離されたFDHのアミノ酸配列、及び該酵素をコードする核酸分子の塩基配列が、意図的もしくは非意図的に改変が生じている改変部位を有していないことを意味する。
【0027】
「FDH」とは、ホルムアルデヒドを基質としてギ酸を生成する脱水素反応を触媒する活性を有するタンパク質であり、補酵素としてニコチンアミドアデニンジヌクレオチド(NAD)を要求する。反応式を以下に示す。
ホルムアルデヒド+NAD+(酸化型) → ギ酸+NADH(還元型)+H+
【0028】
FDHには、グルタチオン依存性、非依存性の2つの型が存在する。本発明においては何れもが対象となるが、グルタチオン非依存性が好ましい。
【0029】
改変型の基礎となる野生型FDHは、上記の性質を有する限り何れの生物由来のものであってよい。具体的には、シュードモナス(Pseudomonas)属、パラコッカス(Paracoccus)属、ピロコッカス(Pyrococcus)等の細菌由来、及びサッカロマイセス(Saccharomyces)属等の酵母由来のFDH等が例示される。更には、ウシ(Equus caballus)やラット(Rattus norvergicus)等の哺乳動物由来のFDHをも含む。好ましくは、シュードモナス・プチダ(Pseudomonas putida)、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)、パラコッカス・デニトリフィカンス(Paracoccus denitrificans)由来のFDHである。しかしながら、これらに限定するものではない。
【0030】
「安定性が向上」とは、熱、及び保存に対する当該酵素の安定性が向上することであり、一定期間の熱処理及び保存に対して、当該酵素の触媒機能の失活や活性低下が抑制できることを意味する。具体的には、改変型FDHが、野生型FDHよりも、一定期間の熱処理又は保存に供した後に維持されている酵素の触媒機能の残存率が高いことを意味する。例えば、野生型FDHの活性が維持される25℃での酵素の酵素活性値を100%として、加温処理後の活性値を酵素活性の残存率として算出する。この残存率が、野生型FDHと比較して増大していた場合、当該酵素の熱安定性が向上したと判断できる。また、一定期間の保存に際しての残存率が、野生型FDHと比較して増大していた場合、当該酵素の保存安定性が向上したと判断できる。一例として、50℃にて30分間の加温により60%以上、特には75%以上の残存活性を維持できる熱安定性が好ましい。また、45℃にて10時間の保存で60〜80%、30時間の保存であっても50〜60%程度の残存活性を維持できる保存安定性が好ましい。
【0031】
本発明の改変型FDHはこれに限定されるものではないが、前記改変が、野生型FDHのアミノ酸配列における第70位−第150位で生じたものが、好ましく例示される。
【0032】
野生型FDHのアミノ酸配列において、第70位−第150位におけるアミノ酸の改変は、これらの位置における何れかの改変を有するもの全てが本発明に含まれる。また、かかる位置における1のアミノ酸が改変したものであっても、複数の位置における複数のアミノ酸が改変したものであってもよい。
【0033】
配列番号2に示すアミノ酸配列は、シュードモナス・プチダ由来の野生型FDHのアミノ酸配列であるが、他の生物由来の当該酵素のホモログについても同様の位置が改変されたものは本願発明の改変型FDHに含まれる。
【0034】
ここで、改変としては、特に好ましくは、他のアミノ酸への置換である。そして、他のアミノ酸とは、置換前のアミノ酸以外の何れもが含まれる。しかしながら、タンパク質機能改変の観点から極性、電荷、親水性、若しくは疎水性等の点で置換前のアミノ酸と異なる性質を有するアミノ酸であることが好ましい。
例えば、アラニン、バリン、ロイシン、イソロイシン、プロリン、メチオニン、フェニルアラニン、トリプトファンは、共に非極性アミノ酸に分類されるため、互いに似た性質を有するが、これら以外のアミノ酸への置換はタンパク質機能へ影響を与えることが考えられる。
非荷電性アミノ酸としては、グリシン、セリン、スレオニン、システイン、チロシン、アスパラギン、グルタミンが挙げられる。
酸性アミノ酸としては、アスパラギン酸およびグルタミン酸が挙げられる。
塩基性アミノ酸としては、リシン、アルギニン、ヒスチジンが挙げられる。
これらの各グループ内の以外へのアミノ酸置換は、タンパク質の機能が改変されることが特に予想されるが、これに制限されるものではない。
【0035】
好ましくは、野生型FDHのアミノ酸配列において、当該アミノ酸配列の第128位に対応する位置におけるアスパラギン酸のグリシンへの置換であり、または、配列番号2におけるアミノ酸配列の第110位に対応する位置におけるグリシンのアスパラギン酸への置換が例示される。
【0036】
一例として、野生型FDHとしてシュードモナス・プチダとする場合、上述した第128位の置換を行なったアミノ酸配列を配列番号4に、第110位の置換を行なったアミノ酸配列を配列番号6に示す。
【0037】
更に、前述の改変型FDHの性質を保持している限り、更に特定のアミノ酸に改変が生じている改変部位を有するアミノ酸配列を含むものであってもよい。当業者はアミノ酸配列の改変に際して本発明の改変型FDHの酵素活性を保持する改変を容易に予測することができる。具体的には、例えばアミノ酸置換の場合には、タンパク質構造保持の観点から極性、電荷、親水性、若しくは疎水性等の点で置換前のアミノ酸と類似した性質を有するアミノ酸に置換することができる。このような置換は保守的置換として当業者には周知である。具体例を挙げると、例えば、アラニン、バリン、ロイシン、イソロイシン、プロリン、メチオニン、フェニルアラニン、トリプトファンは、共に非極性アミノ酸に分類されるため、互いに似た性質を有する。また、非荷電性アミノ酸としては、グリシン、セリン、スレオニン、システイン、チロシン、アスパラギン、グルタミンが挙げられる。また、酸性アミノ酸としては、アスパラギン酸およびグルタミン酸が挙げられる。また、塩基性アミノ酸としては、リシン、アルギニン、ヒスチジンが挙げられる。これらの各グループ内のアミノ酸置換は、タンパク質の機能が維持されるとして許容される。
また、その後の精製、固相への固定化等の便宜のため、アミノ酸配列のN、又はC末端にHisタグペプチド、FLAGタグペプチド等を付加したものも好適に例示される。このようなタグペプチドの導入は常法により行なうことができる。また、本発明の酵素活性の喪失を引き起こさない範囲内で、C末端側若しくはN末端側のアミノ酸残基を切断した切断型でもよい。更に、グルコシル化等の化学修飾を付加してもよい。
【0038】
本発明の改変型FDHは公知の方法によって取得することができる。例えば、改変の基礎となる野生型FDHをコードする遺伝子に対して改変を施し、得られた改変型遺伝子を用いて宿主細胞を形質転換し、かかる形質転換体の培養物から上記活性を有するタンパク質を採取することによって取得することができる。
【0039】
改変の基礎となる野生型FDHをコードする遺伝子は、公知の遺伝子クローニング技術を用いて取得することができる。例えば、GenBank等の公知のデータベースを検索することによって取得することができる遺伝子情報を基にしてプライマーを設計し、FDHを産生し得る生物体から抽出したゲノムDNAを鋳型としてPCRを行うことにより取得することができる。また公知の遺伝子情報に基づいて、常法のホスホルアミダイト法等の核酸合成法により合成することによっても取得するができる。ここで、本発明の改変型の基礎として好適なFDHの配列情報として、野生型のシュードモナス・プチダ由来のFDHのアミノ酸配列を配列表の配列番号2に、当該FDHをコードする塩基配列を配列表の配列番号1に示す。
【0040】
野生型FDHをコードする遺伝子に改変を施す方法としては、特に制限はなく、当業者に公知の改変タンパク質作製のための変異導入技術を利用することができる。例えば、部位特異的突然変異誘発法、PCR法等を利用して変異を導入するPCR突然誘発法、あるいは、トランスポゾン挿入突然変異誘発法などの公知の変異導入技術を利用することができる。また、市販の変異導入用キット(例えば、QuikChange(登録商標) Site-directed Mutagenesis Kit(Stratagene社製))を利用してもよい。
【0041】
特には、野生型FDHをコードするDNAを鋳型として、所望の改変(欠失又は置換)を施した配列を含むオリゴヌクレオチドをプライマーとしてPCRを行うことによって取得することが、好ましく例示される。ここで、本発明の改変型FDHの調製において、PCRを利用する場合に用いられるプライマーは、野性型FDHをコードする核酸分子と相補的な配列を含み、かつ所望の改変が生じるように設計されたものであり、常法に基づいて調製することができる。例えば、ホスホアミダイト法等に基づく化学合成法等が利用可能である。化学合成法に基づきプライマーを調製する場合には、合成に先立って標的核酸の配列情報に基づいて設計される。プライマーの設計は、所望の領域を増幅するように、例えばプライマー設計支援ソフト等を利用して設計することができる。プライマーは合成後、HPLC等の手段により精製される。また、化学合成を行う場合には市販の自動合成装置を利用することも可能である。このようなプライマーとしては10以上、好ましくは15以上、更に好ましくは約20〜50の塩基からなるオリゴヌクレオチドが例示される。
【0042】
また、目的とする改変型FDHのアミノ酸配列が定めることにより、それをコードする適当な塩基配列を決定でき、常法のホスホルアミダイト法等の核酸合成技術を利用して本発明の改変型FDHをコードするDNAを化学的に合成することができる。
【0043】
また、このような本発明の改変型FDH改変体は自然又は人工の突然変異により生じた突然変異体の中から前述の理化学的性質を有するタンパク質をスクリーニングすることにより取得できる。
【0044】
(本発明の改変型FDHをコードする核酸分子)
本発明の改変型FDHをコードする核酸分子は、前述の理化学的性質を有するすべての改変型FDHをコードするものを包含する。例えば、配列番号4,6に記載されるアミノ酸配列を含むタンパク質をコードする全てのポリヌクレオチドであり、一具体例としては、配列番号3,5に示す塩基配列を含むポリヌクレオチドが挙げられるが、これに限定するものではない。ここで、本発明におけるポリヌクレオチドにはDNA及びRNAの双方が含まれ、DNAである場合には、1本鎖であると、二本鎖であるとは問わない。
【0045】
本発明の改変型FDHをコードする核酸分子は、本明細書においてその塩基配列が明確になったことから、かかる配列情報に基づいて、常法のホスホルアミダイト法等のDNA合成法を利用して化学的に合成することができる。また、改変の基礎となる野生型FDHをコードするDNAに対して改変を施すことによっても製造することができる。なお、詳細については前述した。
【0046】
(本発明の組換えベクター)
そして、本発明の組換えベクターは、適当なベクターに本発明の改変型FDHをコードする核酸分子を組み込むことによって構築することができる。利用可能なベクターとしては、外来DNAを組み込め、かつ宿主細胞中で自律的に複製可能なものであれば特に制限はない。従って、ベクターは、本発明の改変型FDHをコードする核酸分子を挿入できる少なくとも1つの制限酵素部位の配列を含むものである。例えば、プラスミドベクター(pEX系、pUC系、及びpBR系等)、ファージベクター(λgt10、λgt11、及びλZAP等)、コスミドベクター、ウイルスベクター(ワクシニアウイルス、及びバキュロウイルス等)等が包含される。
【0047】
本発明の組換えベクターは、本発明の改変型FDHをコードする核酸分子がその機能を発現できるように組み込まれている。従って、核酸分子の機能発現に必要な他の既知の塩基配列が含まれていてもよい。例えば、プロモータ配列、リーダー配列、シグナル配列、並びにリボソーム結合配列等が挙げられる。プロモータ配列としては、例えば、宿主が大腸菌の場合にはlacプロモータ、trpプロモータ等が好適に例示される。しかしながら、これに限定するものではなく既知のプロモータ配列を利用できる。
更に、本発明の組換えベクターには、宿主において表現型選択を付与することが可能なマーキング配列等をも含ませることができる。このようなマーキング配列としては、薬剤耐性、栄養要求性などの遺伝子をコードする配列等が例示される。具体的には、カナマイシン耐性遺伝子、クロラムフェニコール耐性遺伝子、アンピシリン耐性遺伝子等が例示される。
【0048】
ベクターへの本発明の改変型FDHをコードする核酸分子等の挿入は、例えば、適当な制限酵素で本発明の遺伝子を切断し、適当なベクターの制限酵素部位、又はマルチクローニング部位に挿入して連結する方法などを用いることができるが、これに限定されない。連結に際しては、DNAリガーゼを用いる方法等、既知の方法を利用できる。また、DNA Ligation Kit(タカラバイオ社)等の市販のライゲーションキットを利用することもできる。
【0049】
(本発明の形質転換体)
本発明の形質転換体は、適当な細胞を本発明の改変型FDHをコードする核酸分子を含む組換えベクターで形質転換することによって構築することができる。ここで、宿主となる細胞としては、本発明の改変型FDHを効率的に発現できる宿主細胞であれば、特に制限はない。原核生物を好適に利用でき、特には大腸菌E.coli(DH5α、BL21、JM109等)を利用することができる。その他、枯草菌、バシラス属細菌、シュードモナス属細菌等をも利用できる。更に、原核生物に限定されず真核生物細胞を利用することが可能である。例えば、サッカロマイセス・セルビシエ(Saccharomyces cerevisiae)等の酵母、Sf9細胞等の昆虫細胞、CHO細胞、COS−7細胞等の動物細胞等を利用することも可能である。形質転換法としては、塩化カルシウム法、エレクトロポレーション法、リポソームフェクション法、マイクロインジェクション法等を既知の方法を利用することができる。
【0050】
(本発明の改変型FDHの製造方法)
本発明の改変型FDHの製造方法は、前述の本発明の形質転換体を培養し、得られた培養物からアルデヒド化合物の脱水素反応を触媒する活性を有するタンパク質を採取することにより行なう。即ち、前述の本発明の形質転換体を培養する培養工程と、前記培養工程で発現した前記タンパク質を回収する回収工程とを備える。このように、適当な宿主で発現させることによって、低コストで本発明の改変型FDHの大量生産が可能となる。
【0051】
培養工程は、本発明の形質転換体を適当な培地に接種し、常法に準じて培養することにより行なわれる。本発明の形質転換体の培養は、宿主細胞の栄養生理学的性質を勘案して、培養条件を選択すればよい。使用される培地としては、宿主細胞が資化し得る栄養素を含み、形質転換体におけるタンパク質の発現を効率的に行えるものであれば特に制限はない。従って、宿主細胞の生育に必要な炭素源、窒素源その他必須の栄養素を含む培地であることが好ましく、天然培地、合成培地の別を問わない。
【0052】
例えば、炭素源として、ホルムアルデヒド、デキストラン、デンプン等が、また、窒素源としては、アンモニウム塩類、硝酸塩類、アミノ酸、ペプトン、カゼイン等が挙げられる。他の栄養素としては、所望により、無機塩類、ビタミン類、抗生物質等とを含ませることができる。宿主細胞が大腸菌の場合には、LB培地、M9培地等が好適利用できる。また、培養形態についても特に制限はないが、大量培養の観点から液体培地が好適に利用できる。
【0053】
本発明の組換えベクターを保持する宿主細胞の選別は、例えば、マーキング配列の発現の有無により行なうことができる。例えば、マーキング配列として薬剤耐性遺伝子を利用する場合には、薬剤耐性遺伝子に対応する薬剤含有培地で培養することによって行うことができる。
【0054】
精製工程は、前述の培養工程において得られた形質転換体の培養物からの本発明の改変型FDHを回収、即ち、単離精製することによって行えばよい。本発明の酵素の存在する画分に応じて、一般的なタンパク質の単離精製方法に準じた手法を適用すればよい。具体的には、本発明のFDHが宿主細胞外に生産される場合には、培養液をそのまま使用するか、遠心分離、濾過等の手段により宿主細胞を除去して培養上清を得る。続いて、培養上清に、既知のタンパク質精製方法を適宜選択することにより、本発明の酵素を単離精製することができる。
【0055】
例えば、硫酸アンモニウム沈殿、透析、SDS−PAGE電気泳動、ゲル濾過、疎水、陰イオン、陽イオン、アフィニティークロマトグラフィ等の各種クロマトグラフィ等の既知の単離精製技術を単独、又は適宜組み合わせて適用することができる。特にアフィニティークロマトグラフィを利用する場合、本発明の酵素をHis Tag等のタグペプチドとの融合タンパク質として発現させて、かかるタグペプチドに対する親和性を利用することが好ましい。また、本発明の改変型FDHが宿主細胞内で産生される場合には、培養物を遠心分離、濾過等の手段により宿主細胞を回収する。
【0056】
続いて、リゾチーム処理などの酵素的破砕方法、又は超音波処理、凍結融解、浸透圧ショック等の物理的破砕方法等により、宿主細胞を破砕する。破砕後、遠心分離、濾過等の手段により可溶化画分を収集する。得られた可溶化画分を、前述の細胞外に生産できる場合と同様に処理することにより単離精製することができる。ここで、本発明で得られる改変型FDHは、熱安定性が高いことから、前述の単離、精製工程において熱処理を併用することが有用かつ便利である。
培養物から得られた宿主細胞及び培養上清には、当該宿主細胞由来の様々なタンパク質を含有する。しかし、熱処理を行なうことにより、宿主細胞由来の夾雑タンパク質は変性し凝縮沈殿する。これに対して、本発明の酵素は、改変型を有するため変性を生じないことから、遠心分離等により宿主由来の夾雑タンパク質と容易に分離できる。また、培養液をそのまま、若しくは粗抽出液を使用する場合においても、熱処理を行なうことにより、他のタンパク質が失活することから、実質的に本発明の改変型FDHのみの酵素液として使用することができる。従って、本発明の改変型FDHを遺伝子工学的手法により製造する場合においても、宿主由来のその他のタンパク質を容易に除去することができる。従って、精製度を向上させることができ、信頼性の高い酵素を製造できるという利点がある。
【0057】
そして、精製されたFDHが所望の改変が生じている改変部位を有する本発明の改変型FDHであるか否かの確認は、その理化学的性質や配列の分析によって行うことができる。理化学的性質の分析による場合、一定期間、熱処理に付した後、酵素の酵素活性を公知の方法により測定し改変部位を有しない野生型FDHと比較して、活性の残存率が高いか否かを確認することによって行うことができる。従って、基質であるアルデヒド化合物に対してNAD+依存的に脱水素反応をする酵素活性を測定することにより行うことができる。
【0058】
酵素の活性は、NAD+依存的にアルデヒド化合物の脱水素反応を触媒する酵素の活性測定法として知られる方法を何れをも利用して行うことができる。例えば、NAD+の存在下で、本発明の酵素をアルデヒド化合物と反応させ、当該酵素の触媒反応で生成するNADH量の変化を340nmの吸光度変化もって検出する。かかる吸光度変化をもって当該酵素の活性とすることができる。従って、FDH活性は、ホルムアルデヒド及びNAD+から、ギ酸及びNADHを生成する触媒反応において、生成したNADHを直接定量することによって測定できる。つまり、酵素反応によりホルムアルデヒドが酸化されギ酸となり、このときNADが還元されNADHが生成する(反応式:HCHO+NAD++H2O → HCOOH+NADH+H+)。このNADからNADHの生成を340nmの吸光度変化を測定して酵素活性とした。この配列分析による場合には、公知のアミノ酸分析法によって行うことができる。例えば、エドマン分解法に基づく自動アミノ酸決定法が利用できる。
【0059】
(アルデヒド化合物の検出方法)
本発明の改変型FDHは、試料中のアルデヒド化合物の検出のために利用することができる。アルデヒド化合物の検出は、例えば、基質であるアルデヒド化合物に対してNAD+依存的に脱水素反応をする酵素活性を測定することにより行うことができる。酵素の活性は、NAD+依存的にアルデヒド化合物の脱水素反応を触媒する酵素の活性測定法として知られる方法をいずれをも利用して行うことができる。例えば、NAD+の存在下で、本発明の酵素を測定対象となる試料と反応させ、当該酵素の触媒反応で生成するNADH量の変化を340nmの吸光度変化もって検出する。かかる吸光度変化をもって当該酵素の活性とすることができ、ひいてはNADH量の変化によりアルデヒド化合物の存在を検出することができる。したがって、FDH活性は、その触媒反応において、生成したNADHを直接定量することによって測定できる。
反応液としては、例えば、ホルムアルデヒド等のアルデヒド基質、2mMのNAD+、25mMのMgCl2、1MのNaCl及び本発明の酵素溶液を、50mMリン酸緩衝液(pH8.8)中にて混合して200μLとすることにより調製することができる。しかしながら、これは標準条件であり、適宜変更することができる。この反応液を、37℃で波長340nmにおける吸光度を測定し、吸光度の増加を求めることにより活性評価試験を行うことができ、吸光度の測定は、既知のマイクロプレートリーダー(モレキュラーデバイス社製)を用いることができる。
そして、適当な試験条件が選択されていれば、この変化は、測定しようとする酵素活性に直線的に比例する。このとき、あらかじめ目的濃度範囲内における標準濃度のアルデヒド化合物溶液により標準曲線を作成することにより、得られた吸光度変化値に基づいてアルデヒド化合物濃度を求めることができる。
【0060】
ここで、試料としては、アルデヒド化合物等、FDHの基質となり得る化合物の存在が予想されるすべての試料を対象とすることができる。例えば、血液、尿、唾液等の生物体由来の生物試料、食品試料、環境試料等が例示されるがこれに限定されるものではない。また、必要に応じて、これらの試料に適当な処理を行った試料をも含み得る。特に、本発明の改変型FDHは優れた熱安定性を示すことから、温度が高い試料の分析に際して好適に利用できる。そして、保存性も高いことから酵素の酵素活性の劣化を招くことなく、測定精度の向上を図れ、ひいては、酵素使用量を軽減できることからコスト削減効果をも奏することができる。
【0061】
(アルデヒド化合物検出用のバイオセンサー)
本発明は、本発明の改変型FDHを利用するアルデヒド化合物検出用のバイオセンサーを提供する。当該検出センサーは、電極材上に本発明の改変型FDHが固定化した作用電極、及びその対極を設けて構成される。必要に応じて、参照電極を設けて三電極方式として構成してもよい。電極としては、カーボン、金、白金等を用いることができる。電極材上への酵素の固定化は既知の方法によって行うことができる。例えば、物理的吸着、イオン結合,共有結合を介して固定化する担体結合法を利用することができる。また、グルタルアルデヒドなどの二価性官能基をもつ試薬で架橋固定する架橋法をも利用でき、更には、アルギン酸,カラギーナン等の多アルデヒド化合物、ポリアクリルアミド等の網目構造をもつゲルや、半透性膜の中に閉じて固定化する包括法等をも利用することができる。そして、本発明のFDHの酵素活性に際して要求される補酵素NAD+、NAD+の還元体であるNADHを酸化する能力を有する酸化酵素(ジアホラーゼ等)、電子メディエーター(フェロセン、ジクロロインドフェノール等)も、必要に応じて電極材上に固定化して構成される。
【0062】
アルデヒド化合物の測定は、例えば、測定対象となる試料を接触させると試料中のアルデヒド化合物が作用極上に固定された本発明の改変型FDHと反応し、続いて電子メディエーターが還元される。そして、電極系に電圧を印加して電子受容体の還元体を酸化し、得られる酸化電流値の変化により試料中のアルデヒド化合物を検出することができる。このとき、あらかじめ目的濃度範囲内における標準濃度のアルデヒド化合物溶液により標準曲線を作成することにより、得られた酸化電流値に基づいてアルデヒド化合物濃度を求めることができる。
【0063】
ここで、試料としては、本発明の改変型FDHの基質となり得るアルデヒド化合物の存在が予想されるすべての試料を対象とすることができる。詳細については前述した。特には、前述の通り、本発明の改変型FDHは、熱安定性が高いことからバイオセンサー製作時における劣化をも防止することができ、また、保存性も高いことから酵素の酵素活性の劣化を招くことなく、測定精度の向上を図れ、ひいては、酵素使用量を軽減できることからコスト削減効果をも奏することができる。
【0064】
(本発明の燃料電池)
本発明は、本発明の改変型FDHを利用する燃料電池を提供する。本発明の燃料電池は、例えば、酸化反応を行うアノード極と、還元反応を行うカソード極から構成され、必要に応じてアソードとカソードを隔離する電解質層を含んで構成される。
アノード電極側では、本発明の改変型FDHがアルデヒド化合物を酸化することによって生じた電子を電極に取り出すと共に、プロトンを発生する。一方、カソード側では、アノード側で発生したプロトンが酸素と反応することによって水を生成するように構成される。電極としては、カーボン、金、白金等を用いることができる。アノード極側には本発明の改変型FDHが供給され、適当な緩衝液中に溶解させた形態で供給してもよいが、電極上に固定化されることが好ましい。このとき、改変型FDHは、好ましくはNADと共に固定化されることが好ましく、特にはNADと結合したホロ酵素の形態で固定化されることが好ましい。また、アポ酵素の形態で固定化し、NADを適当な緩衝液に溶解させた形態で供給してよい。
カソード極側には、酸素に電子を伝達することのできる触媒および酵素を必要に応じて供給してよい。電極材上への酵素の固定化は、公知の方法によって行うことができる。例えば、物理的吸着、イオン結合,共有結合を介して固定化する担体結合法を利用することができる。また、グルタルアルデヒドなどの二価性官能基をもつ試薬で架橋固定する架橋法をも利用でき、更には、アルギン酸,カラギーナン等の多アルデヒド化合物、ポリアクリルアミド等の網目構造をもつゲルや、半透性膜の中に閉じて固定化する包括法等をも利用することができる。
【0065】
以上のように構成することにより、燃料であるアルデヒド化合物を酸化する際に生じた電子がアノード極に電子を受け渡す。そして、アノード極に渡された電子は、外部回路を経てカソード極に到達することで電流が発生する。燃料となるアルデヒド化合物としては、本発明の改変型FDHの基質となり得るアルデヒド化合物であれば制限はない。
【0066】
また、メタノールを燃料とした酵素燃料電池においても本発明の改変型FDHを利用できる。その場合には、触媒として、本発明の改変型FDHに加え、必要に応じてメタノールに作用してホルムアルデヒドに酸化するアルコールデヒドロゲナーゼと、ギ酸に作用してCO2に酸化する蟻酸デヒドロゲナーゼを併用することにより構成することができる。
【0067】
本発明の燃料電池は、熱安定性が高い本発明の改変型FDHの触媒能力を利用することから、持続的に発電を行なうことができ、高性能の燃料電池として構築することができる。また、本発明の改変型FDHは、熱安定性が高いことから燃料電池製作時における劣化をも防止することができ、ひいては、酵素使用量を軽減できることからコスト削減効果をも奏することができる。
【実施例】
【0068】
<実施例1>本発明の改変型FDH遺伝子のクローニング
分子進化工学的手法と称される酵素改変手法を利用して、本発明の改変型FDH遺伝子のクローニングを行った。
【0069】
かかる分子進化工学的手法とは、生物進化の原理をタンパク質に応用する技術である。そのステップとしては、まずランダムに改変の基礎となる遺伝子に変異を導入し、可能な限り多様な改変型遺伝子を作り出す。そして、改変型遺伝子から改変型タンパク質を作製して、その中から所望の性質を保持するタンパク質をコードする遺伝子を選択する。このようにして、設計原理が未知であっても所望の機能や性質を保持する高分子を取得することができ、取得した遺伝子の変異箇所及び変異様式を調べることによって、高分子設計原理の研究においても威力を発揮することができる。
【0070】
Pseudomonas putida由来の野生型FDHの立体構造はX線解析により既に明らかにされており、各サブユニットが398アミノ酸残基からなる4量体構造を有する。野生型FDHの立体構造データは、日本蛋白質構造データバンクで公開されているものを利用した(http://www.rcsb.org/pdb/explore.do?structureId=1KOL)。この野生型FDHのX線結晶構造解析に基づく高次構造を検討すると、4量体酵素のサブユニットの界面付近にループ領域(K76−D79)が存在し、この領域が高次構造におけるサブユニット間相互作に影響を及ぼし、熱安定性を支配していることが示唆される。
熱安定性向上効果が見込める部位を鋭意検討した結果、第70位−第150位のアミノ酸領域(Thr70−Leu150)が4量体酵素のサブユニットの界面のドメイン間の構造安定化に寄与するホットスポットである可能性が高いと考え、このホットスポット周辺へランダム変異を導入したライブラリーを作製し、スクリーニングを行うことより、構造が最適化し熱安定性が向上した変異体を取得できると考えた。なお、これら検討のための一連の酵素の立体構造解析は、アクセリス社製のInsight-II(酵素の分子構造を表示し、かつ構造エネルギー計算が可能なソフトウェア)を用いて行った。分子進化工学的手法を用いた酵素改変の具体的な手順を次に示す。
【0071】
(ステップ1) 酵素遺伝子の取得
バクテリアPseudomonas putida株は、NITE(独立行政法人製品評価技術基盤機構)から入手した(受託番号 NBRC No. NBRC3738)。そのゲノムDNAは、添付資料に記載の培養液を使って菌体を培養し、ゲノムDNA抽出キット(プロメガ社製)を使って抽出と精製を行った。このゲノムDNAを鋳型にして以下のプライマー1,2(配列番号7,8)を用いてPCR反応を行なった。PCR反応は、ポリメラーゼとしてPrimeSTAR GXL DNA Polymerase(タカラバイオ社製)、鋳型として50ngゲノムDNAおよび各0.2μMのプライマーのそれぞれを混合して滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。PCR反応は、98℃で10秒、68℃で60秒のサイクルを30回繰り返した。増幅産物の精製はDNA精製キット(GEヘルスケア社製)を用いた。その結果、予想されるDNA増幅産物(約1.2kbp)を得ることができた。
【0072】
5'-GAGCA TATGT CTGGT AATCG TGGTG T-3'(プライマー1:配列番号7)
5'-CTAAA GCTTA GGCCG CGCTG AGGTC T-3'(プライマー2:配列番号8)
【0073】
(ステップ2) 発現ベクターの構築
PCR増幅で得られたDNA断片(約1.2kbp)を、制限酵素NdeIとHindIIIで切断し、アガロースゲル電気流動で分離しゲルから精製した。DNA断片を、タンパク質発現用プラスミドのpET23bベクター(ノバジェン社製)の制限酵素部位NdeIとHindIIIにライゲーション反応より組み込んで酵素の発現ベクターを構築した。
また無細胞システムを使った酵素合成用の鋳型とする直鎖状DNAを調製するために、酵素遺伝子を組み込んだベクターを鋳型にして、T7プロモーターとT7ターミネーター配列間をPCRにより増幅した。PCR反応は、ポリメラーゼとしてPrimeSTAR GXL DNA Polymerase(タカラバイオ社製)を使用し、鋳型DNA(50ng)、プライマー3,4(配列番号9,10、各0.2μM)を滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。PCR反応は、98℃で10秒、68℃で60秒のサイクルを30回繰り返した。PCR産物の精製には、DNA精製キット(GEヘルスケア社製)を用いた。
【0074】
5'- ATCTCGATCCCGCGAAATTAATAC -3' (プライマー3:配列番号9)
5'- TCCGGATATAGTTCCTCCTTTCAG -3' (プライマー4:配列番号10)
【0075】
(ステップ3) 変異遺伝子ライブラリーの作製
ランダム変異を導入した変異遺伝子ライブラリーの作製は、変異導入のための手法にエラープローンPCRを用いた。ライブラリー作成を行うために、酵素FDH分子内に1〜2箇所の変異導入が得られる条件(エラー率0.5%前後となるエラープローンPCR条件)を検討した。ポリメラーゼの塩基取り込み間違いを促進させるために、マグネシウム濃度とマンガン濃度を[Mg,Mn]=[5,0.5]mMとした。
エラープローンPCR反応は、2.5ユニットのTaq DNAポリメラーゼ(タカラバイオ社製)、各0.5μMのプライマー5,6(配列番号11,12)、10mM Tris-HCl(pH8.3)緩衝液、50mM KCl、5.0mM MgCl2、0.5mM MnCl2のそれぞれを混合して滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。
【0076】
5'-CGGCCTGGTCCTGGGCCACGAAATC-3'(プライマー5:配列番号11)
5'-TGGCCTTGTCGCGCTCAGGCAGCTT-3'(プライマー6:配列番号12)
【0077】
次に、エラープローンPCRによりランダムに変異を導入したFDH遺伝子断片と、変異を導入しないFDH遺伝子の残りの部分をコードするDNA断片を連結するために、オーバーラッピングPCRを行った。
まず変異を導入しないDNA断片を得るために、ステップ2で作製した発現ベクターを鋳型にして、プライマー7,8(配列番号13,14)およびプライマー9,10(配列番号15,16)をそれぞれ用いてPCR増幅を行った。PCR反応は、ポリメラーゼとしてPrimeSTAR GXL DNA Polymerase(タカラバイオ社製)、鋳型DNA(50ng)、プライマー(各0.2μM)のそれぞれを混合して滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。PCR反応は98℃で10秒、68℃で60秒のサイクルを30回繰り返した。増幅産物の精製はDNA精製キット(GEヘルスケア社製)を用いた。
【0078】
5'- ATCTCGATCCCGCGAAATTAATAC -3' (プライマー7:配列番号13)
5'- GATTTCGTGGCCCAGGACCAGGCCG -3' (プライマー8:配列番号14)
5'- TCCGGATATAGTTCCTCCTTTCAG -3' (プライマー9:配列番号15)
5'- AAGCTGCCTGAGCGCGACAAGGCCA -3' (プライマー10:配列番号16)
【0079】
得られた3つのDNA断片を鋳型にして、プライマー7,10を用いてPCR増幅を行った後、さらに反応終了液に含まれるPCR産物を鋳型にしてプライマー11,12(配列番号17,18)を用いてPCR増幅を行った。PCR反応は、ポリメラーゼとしてPrimeSTAR GXL DNA Polymerase(タカラバイオ社製)、鋳型DNA(50ng)、プライマー(各0.2μM)のそれぞれを混合して滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。PCR反応は98℃で10秒、68℃で60秒のサイクルを30回繰り返した。PCR産物の精製には、DNA精製キット(GEヘルスケア社製)を用いた。
次に、得られた酵素遺伝子をコードするDNA断片に、1分子PCR用プライマー配列を付加するために、プライマー13(配列番号19)を用いたPCR増幅を行った。PCR反応は、ポリメラーゼとしてPrimeSTAR GXL DNA Polymerase(タカラバイオ社製)、プライマー(各0.3μM)のそれぞれを混合して滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。PCR反応は94℃で30秒、55℃で30秒、72℃で60秒のサイクルを20回繰り返した。PCR産物の精製には、DNA精製キット(GEヘルスケア社製)を用いた。以上の実施した内容の模式図を図1,2に示す。
【0080】
5'-GGAGCTAGGGTTTACGAGTGGAATG-ATCTCGATCCCGCGAAATTAATAC-3'(プライマー11,配列番号17)
5'-GGAGCTAGGGTTTACGAGTGGAATG-TCCGGATATAGTTCCTCCTTTCAG-3'(プライマー12,配列番号18)
5'-GGAGCTAGGGTTTACGAGTGGAATG-3' (プライマー13,配列番号19)
【0081】
以上により、FDHをコードする遺伝子のコーディング領域中のホットスポット(アミノ酸領域Thr70−Leu150)に、変異を導入した変異遺伝子ライブラリーを作製した。変異遺伝子ライブラリーを、デキストランを含む希釈液で、1.5分子/μLに希釈し、384プレートに分注した。その後、1μL希釈液/1反応ウェルでPCR増幅した。変異遺伝子ライブラリーの希釈倍率算出に使用した計算式を次に示す。
【0082】
DNAの長さ:1400bp
分子量:0.924×106
1μg DNA:1.08×10-6μmol=1.08×10-12mol
1μg DNA:6.02×1023×1.08×10-12=6.5×1011分子に相当
【0083】
この計算から、変異遺伝子(DNA)のゲル切り出し精製したDNA溶液は、9.1×109分子/μLと算出でき、このDNA溶液を3×109希釈したものを、次のPCR反応の鋳型DNAに使用した。PCR反応温度条件は、98℃で10秒、68℃で1分のサイクルを70回繰り返した後、68℃で10分の伸長反応を行なった。
【0084】
(ステップ4) 無細胞タンパク質合成システムを用いた酵素合成とスクリーニング
ラビットトランスレーションシステムRTS100,E.coli HYキット(ロシュ社製)を使って取扱説明書に従い、反応時間は4時間、反応温度は30℃を基本条件として改変FDHを合成した。
変異遺伝子ライブラリーを鋳型にして、大腸菌の無細胞タンパク質合成系でタンパク質を合成し、上述の条件を用いてホルムアルデヒドに対する酵素活性についてスクリーニングし、熱処理後の残存活性が野生型FDHより向上した酵素活性を発現する改変FDHの候補遺伝子を選択した。
【0085】
酵素の活性測定は、8μLの酵素合成終了液に、32μLのリン酸緩衝液を混和して40μLとし、分注装置を使って16μLを新しい384ウェルプレート2枚に移した。2枚のプレートのうち、1枚を熱処理(15分、55℃)した後、氷中に15分間放置し、その後、80μLの活性測定反応液(50mMリン酸緩衝液pH7.4、40mM NAD、0.06%HCHO)を加えて、NAD+からNADH変化に伴う340nmの吸光度変化をプレートリーダで測定した。
【0086】
(ステップ5) スクリーニングで選択したクローンの解析
熱処理後も残存する酵素活性が認められた酵素の合成に使用した変異遺伝子ライブラリーについて、塩基配列を解析してアミノ酸変異を同定し、それらのタンパク質合成と酵素活性の有無も確認した。
タンパク質合成は、無細胞タンパク質合成系ラビットトランスレーションシステムRTS100,E.coli HYキット(ロシュ社製)を用いて、その取り扱い説明書に従って行なった。酵素の有無は、FluoroTect(登録商標) GreenLys in vitro Translation Labeling System(プロメガ社製)を用いて、その取り扱い説明書に従って合成酵素を蛍光標識して確認した。
【0087】
酵素活性の測定は、ホルムアルデヒド脱水素酵素によるホルムアルデヒド分解活性を、NADHの340nmの吸光を指標として行なった。80μLの活性測定反応液(50mMリン酸緩衝液K1K2PO4 pH7.4、1mM NAD+、2mM HCHO)に酵素溶液を加えて、NADからNADH変化に伴う340nmの吸光度変化を5分間(10秒間隔)プレートリーダで測定した。結果を表1に示す。
【0088】
【表1】

【0089】
表中における「アミノ酸変異部位」において、例えば「E76V」との表記は、野生型FDHのアミノ酸配列である配列番号2における第76番目のグルタミン酸(E)がバリン(V)に置換されたことを表す。
本実施例でアミノ酸変異が認められた部位は、クローンP16では第76位、第81位、第219位であり、クローンF12では第127位、第142位であり、クローンK11では第97位、第148位であり、クローンN20では第129位であり、クローンP13では第149位であり、クローンO6,P4では第128位であり、クローンH1では第110位であり、クローンK3では第94位であった。尚、本明細書においては、アミノ酸の位置は、開始コドンであるメチオニンを1として番号付けしている。
【0090】
この中で、タンパク質合成と酵素活性が確認できたものは、クローンO6,P4(第128番目のアスパラギン酸(D)がグリシン(G)に置換されている)と、クローンH1(第110番目のグリシン(G)がアスパラギン酸(D)に置換されている)であった。
【0091】
表1の結果より、アミノ酸変異D128GおよびG110D以外では、変異が1個入っただけの改変で酵素活性が全くなくなるものと認められた。つまり、アミノ酸変異が1個ではタンパク質の構造学的には、10Å程度の距離にしか構造変化の影響を与えないとされているが、このPseudomonas putida由来のFDHは、その微小な構造変化でも酵素の構造的な安定性に影響を与えてしまうデリケートで複雑な構造をしている酵素であるといえる。
その中でも特に、V127DはD128Gの隣のアミノ酸変異であるが、当該V127Dでは酵素活性がなくなるのに対してD128Gでは酵素活性を有していることから、D128GがPseudomonas putida由来のFDHの安定性に重要であると推察されると同時に、従来法での酵素改変は非常に困難であることが推測できる。
【0092】
以下、第128番目のアスパラギン酸がグリシンに改変されたアミノ酸配列を有する改変FDHを「D128G」、第110番目のグリシンがアスパラギン酸に改変されたアミノ酸配列を有する改変FDH「G110D」と称する。
【0093】
<実施例2>改変型FDHの立体構造シミュレーションに基づく高次構造の解析
D128GおよびG110Dの立体構造モデルの構築を行なった。モデリングは、アクセルリス社製Discovery Studio21を用いて、X線結晶解析データ(PDB;1KOL)を参照構造として、ホモロジーモデリングにより実施した。
【0094】
まず、野生型FDH(PDB;1KOL)を用いて変異部位の解析を行なった(図3)。
触媒ドメインとコファクター結合ドメインは、Zn1近傍で上下に分かれる。NADとZn1の相互作用は、両ドメインの位置・構造の安定性に関係していると考えられる。D128(第128位のアスパラギン酸)およびG110(第110位のグリシン)は、両ドメインの界面に位置する。特に、G110はZn2保持ループの根元付近に位置する。Zn2保持ループ周辺は両ドメインの分岐から遠く、パッキングが不十分もしくは安定性が低い可能性が考えられる。
【0095】
D128Gの構造モデルの解析結果は以下の通りである(図4)。
図4(a)には野生型FDH、図4(b)にはD128Gの構造モデルを示した。モデルは分子内の原子に接触する溶媒の接触点によってトレースされるサーフェイス形状で表示した。図4(a)(ii),図4(b)(ii)は、それぞれ図4(a)(i),図4(b)(i)のモデルを90℃回転させたモデルを示した。
野生型FDHではD128とR119(第119位のアルギニン)は静電作用で側鎖が並ぶように位置し、その内側のアミノ酸残基Y126(第126位のチロシン)およびM129(第129位のメチオニン)の間には空間が存在していた。D128G変異体では、D128とR119の側鎖は野生型とは反対方向を向いており、その内側のアミノ酸残基(Y126とM129)の側鎖が内部に向いたことにより、D128とR119も内側にコンパクトにパッキングされたと推測される。余分なキャビティが無くなったことが安定性向上に寄与しているのではないかと推測された。
【0096】
G110Dの構造モデルの解析結果は以下の通りである(図5)。
野生型FDHとG110Dの主鎖の重ね合わせを行った。
G337(第337位のグリシン),Q338(第338位のグルタミン),T339(第339位のトレオニン),P340(第340位のプロリン)は、NADと相互作用するアミノ酸残基、N93(第93位のアスパラギン)はNADと相互作用するアミノ酸残基の隣の残基、C112(第112位のシステイン)はZn2保持アミノ酸残基であったことから、側鎖の向きの変化により、酵素の安定性に何らかの寄与をしたと推測された。
【0097】
<実施例3>組換え大腸菌によるタンパク質の発現と精製
タンパク質発現は、野生型または改変型FDHをコードする遺伝子を、大腸菌用の発現ベクターであるpET22b DNAのマルチクローニングサイトにC末端Hisタグ融合タンパク質として発現するように挿入し、構築されたプラスミドを大腸菌BL21(DE3)に形質転換した。大腸菌の培養は、アンピシリン(50mg/L)入りのLuria−Bertani(LB)培地中を使って37℃で行なった。形質転換した大腸菌は、アンピシリン(50mg/L)を含むLB培地で培養し、培養液の吸光度OD600=0.6で0.5mM IPTG(isopropyl-s-D-thiogalactopyranoside)を添加してタンパク質の合成誘導を行ない、さらに4時間の培養を行なった。
【0098】
発現させたHisタグ融合タンパク質は、アフィニティーカラムを用いて精製した。具体的には、遠心分離により回収した菌体を、リン酸緩衝液(20mM リン酸バッファー、0.5M NaCl、pH7.4)に懸濁した。菌体を超音波で破砕した。破砕は、超音波処理30秒、氷冷30秒のセットを10回繰り返して行なった。遠心分離により得られた上清50mM NaClを加え、酵素液をNi2+カラムクロマトグラフィー通液し、300mMイミダゾールを含む溶液で融合タンパク質を溶出して、活性画分をまとめ、50mMリン酸緩衝液(pH7.4)を透析外液として一晩透析を行った。
得られた酵素溶液を、50mM Tris-HCl緩衝液(pH7.4)にて透析した。タンパク質の確認は、20μLの酵素溶液に20μLの可溶化液を加えて3分間95℃で熱処理したサンプルを、12.5%アクリルアミドゲルで電気泳動し、CBB染色法で酵素を可視化することにより行った。その結果を図6に示す。レーン1は精製した改変型FDH D128G、レーン2は精製した野生型FDHである。
【0099】
この結果、D128Gは、野生型FDHと同様に約50kDaのバンドが認められたため、大腸菌用の発現ベクターで発現することが確認できた。
【0100】
<実施例4>酵素の諸性質の確認(pH依存性)
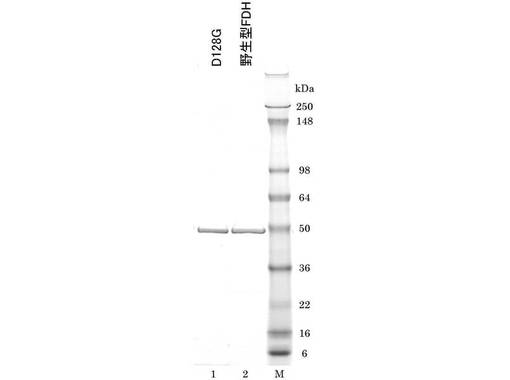
酵素活性をpH5.0〜10.0の範囲で測定した。50mM緩衝液、1mM NAD+、2mM HCHOを含む活性測定液中に等量の酵素液(0.5mg/mL)を加え、25℃でpH8.0の酵素活性の反応速度を1とし、各pHにおける相対活性の変化を比較した結果を図7に示す。
この結果から、D128Gおよび野生型FDHは、pH5.0からpH8.0までは同じpH特性を示し、至適pHはpH8.0付近であることが判明した。一方、pH8.0〜pH10.0では、D128Gより野生型FDHの方が、活性低下が小さいことが判明した。また、G110Dについても、D128Gと同様のpH依存性を示すことが判明した(結果は示さない)。尚、図7においてpH領域A(8以下)についてはリン酸緩衝液を使用し、pH領域B(8以上)についてはTris緩衝液を使用した。
【0101】
<実施例5>酵素の諸性質の確認(温度依存性)
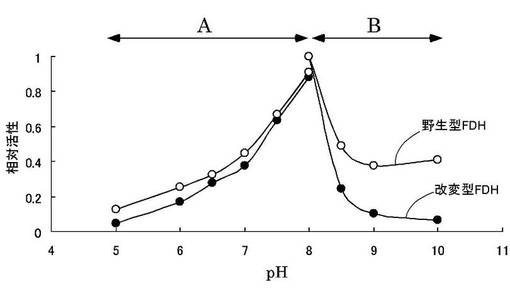
酵素活性を45〜60℃の温度範囲で測定した。50mMリン酸ナトリウム緩衝液(pH7.6)、1mM NAD+、2mM HCHOを含む活性測定液中に等量の酵素液(0.5mg/mL)を加え、45℃での酵素活性の反応速度を1とし、各温度における相対活性の変化を比較した結果を図8に示す。
図8(a)はD128Gおよび野生型FDHの比較を、また、図8(b)はG110Dおよび野生型FDHの比較を示す。この結果から、50〜55℃の温度域において、野生型FDHより改変型FDHの方が2倍程度高い活性を有することが判明した。また、G110DとD128Gは同様の温度依存性を示すことが判明した。
【0102】
<実施例6>酵素の諸性質の確認(温度安定性)
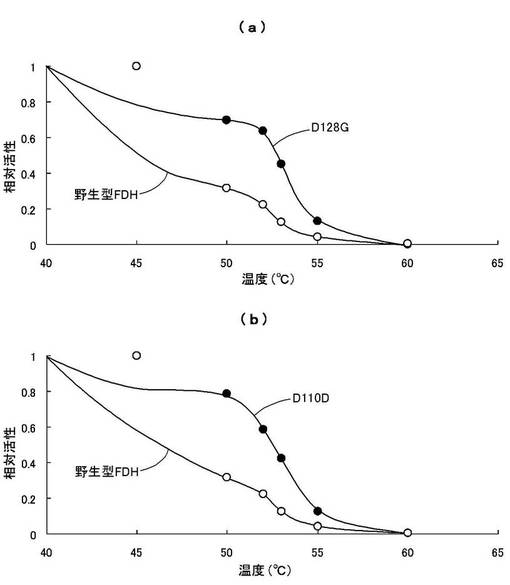
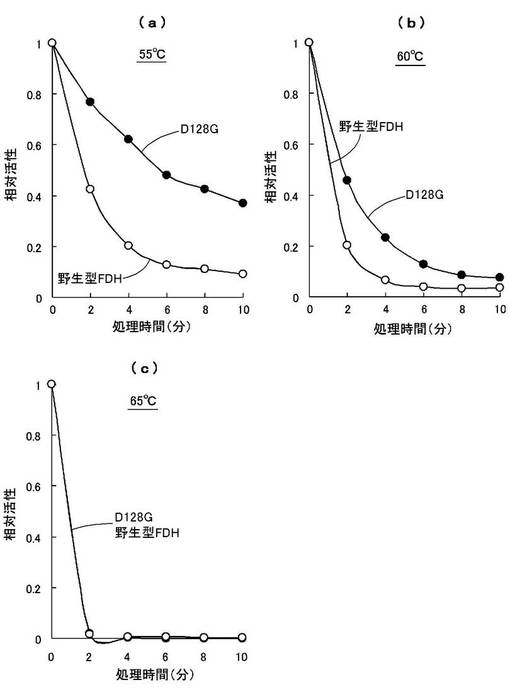
酵素の温度安定性を調べるために、55〜65℃の温度範囲で熱処理後の残存活性を調べた。50mMリン酸ナトリウム緩衝液(pH7.6)中に等量の酵素を加え、55℃、60℃、65℃の各温度で0分〜10分間熱処理し、氷中に30分間置いた後の残存活性を測定した。
50mMリン酸ナトリウム緩衝液(pH7.6)、1mM NAD+、2mM HCHOを含む活性測定液中に熱処理した酵素液(0.5mg/mL)を加え、各熱処理0分時の結果を1とし、各温度と各熱処理時間における相対活性の変化の比較を図9,10に示す。図9はD128Gおよび野生型FDHの比較を、図10はG110Dおよび野生型FDHの比較を示す。
【0103】
この結果、55℃の熱処理では、6分間程度の熱処理において野生型FDHでは相対活性が1/10程度になるのに対し、D128Gでは約50%(図9(a))、G110Dでは約40%(図10(a))の相対活性を有することが判明した。
一方、60℃以上の熱処理では、改変型FDHおよび野生型FDHの何れにおいても残存活性が5分程度で20%以下になり(図9(b),(c)、図10(b),(c))、熱変性により酵素活性を殆ど失うものと認められた。
これより、本発明の改変型FDHは、野生型FDHに比べて熱によって失活し難いものと認められた。
【0104】
<実施例7>酵素の諸性質の確認(安定性)
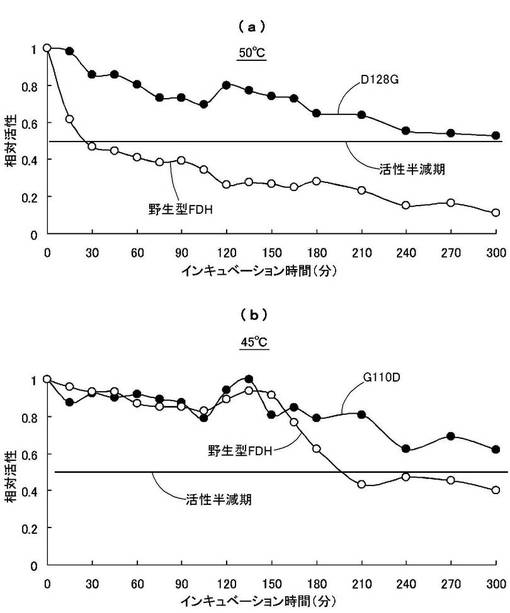
酵素の安定性を調べるために、50℃と45℃の温度下で300分間までの残存活性を調べた。50mMリン酸ナトリウム緩衝液(pH7.6)中に等量の酵素液を加え、0分から300分間置いた後、氷中に30分間置いた後の残存活性を確認した。
50mMリン酸ナトリウム緩衝液(pH7.6)、1mM NAD+、2mM HCHOを含む活性測定液中に熱処理した酵素液(0.5mg/mL)を加え、0分時の酵素活性の反応速度を1としたときの各時間(分)における相対活性の変化の比較を図11に示す。図11(a)は50℃の温度下におけるD128Gおよび野生型FDHの比較を、また、図11(b)は45℃の温度下におけるG110Dおよび野生型FDHの比較を示す。
この結果、50℃での酵素活性の半減期は、野生型FDHでは30分間以内であるのに対し、D128Gでは300分以上であった。また、45℃での酵素活性の半減期は、野生型FDHでは約200分であるのに対し、D128Gでは300分以上であった。
これより、本発明の改変型FDHは、野生型FDHに比べて熱安定性に優れているものと認められた。
【0105】
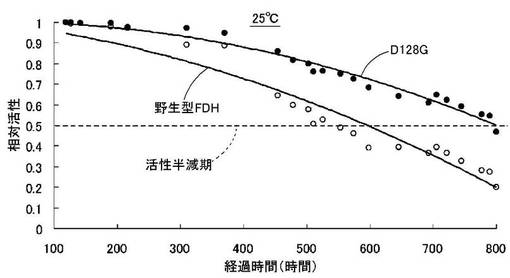
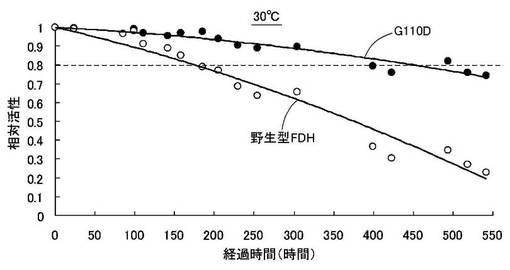
D128Gについて常温時(25℃,30℃)の安定性を調べた。
50mM P1P2PO4、1mM HCHO、0.5mM NADの組成を有する反応液を、D128Gを担持したNi-NTA His・Bind Resinカラム(タカラバイオ社製)、および、野生型FDHを担持したNi-NTA His・Bind Resinカラムにそれぞれ通液した。反応温度は25℃とし、通液速度は0.1mM/分(線流速7.9cm/h)とした。当該カラムを通過した反応済液の340nmの吸光度変化を測定し、通液前の酵素活性の反応速度を1としたときの相対活性の変化の比較を図12に示す。反応温度を30℃とした場合も同様の実験を行なった。
この結果、反応温度を25℃とした場合、野生型FDHの酵素活性の半減期は約500時間程度であるのに対して、D128Gでは約800時間程度であった(図12)。また、反応温度を30℃とした場合、野生型FDHの酵素活性が80%になるのは約175時間程度であるのに対して、D128Gでは約450時間程度であった(図13)。
これより、本発明の改変型FDHは、野生型FDHに比べて長期に亘って安定性に優れているものと認められた。
【0106】
<実施例8>酵素の諸性質の確認(反応速度論解析)
変異型酵素と野生型酵素について酵素反応速度論的な解析を行った。
50mMリン酸ナトリウム緩衝液(pH7.6)、1mM NAD+、0〜4mM HCHOを含む活性測定液中に酵素液(0.5mg/mL)を加え、酵素反応の初期速度のラインウェーバーバークプロットにより、酵素反応の速度定数(Kcat値及びKm値)を求めた。その結果を表2に示す。
【0107】
【表2】
【0108】
この結果、野生型FDHの25℃、pH7.6におけるHCHOに対するKm値は0.35mM、Kcat値は3.2s-1であり、Kcat/Kmは9.1M-1s-1であった。また、D128Gの25℃、pH7.6におけるHCHOに対するKm値は0.35mM、Kcat値は3.2s-1であり、Kcat/Kmは9.1M-1s-1、G110Dの25℃、pH7.6におけるHCHOに対するKm値は0.18mM、Kcat値は2.5s-1であり、Kcat/Kmは13.9M-1s-1であった。
以上より、G110DのKm値が小さく基質親和性が高いこと以外は、本発明の改変型FDHは野生型FDHとほぼ同様であることが判明した。
【0109】
<実施例9>酵素の諸性質の確認(基質選択性)
基質としてアルデヒド類とアルコール類の選択性について調べた。
50mMリン酸ナトリウム緩衝液(pH7.6)、1mM NAD+、2mM各基質を含む活性測定液中に等量の酵素液(0.5mg/mL)を加え、各アルデヒド類とアルコール類を基質としたときの酵素活性の反応速度を1としたときの相対活性について、野生型FDHおよびD128Gを比較したものを表3に示す。
【0110】
【表3】
【0111】
この結果から、D128Gは野生型FDHと同様に、ホルムアルデヒドに対して基質特異性が高いものと認められた。
【産業上の利用可能性】
【0112】
本発明は、改変型ホルムアルデヒド脱水素酵素及びその利用方法に関し、医療、食品、環境分野等の様々な産業分野において利用可能である。
【技術分野】
【0001】
本発明は、改変型ホルムアルデヒド脱水素酵素及びその利用に関する。より詳細には、本発明は、改変型ホルムアルデヒド脱水素酵素、当該酵素をコードする遺伝子、及び当該酵素を含むバイオセンサー、並びに酵素燃料電池に関する。
【背景技術】
【0002】
ホルムアルデヒド脱水素酵素(EC 1.2.1.46)(Formaldehyde dehydrogenase:FDH)は、ホルムアルデヒドを酸化してギ酸を生成する反応を触媒する。ホルアルデヒド脱水素酵素の基質となるホルムアルデヒドは、シックハウス症候群等の慢性的なアレルギー症状を引き起こす原因物質の1つとして知られている。また、稀薄な溶液でも細胞原形質のタンパク質を不可逆的に凝固させ、全ての細胞機能を停止、死滅させる作用があるために、細胞毒としても知られている。そのため、食品中、大気中などのホルムアルデヒドを測定し、これを分解及び除去することが求められており、ホルムアルデヒド脱水素酵素の酵素活性の利用が検討されている。例えば、生体試料中のホルムアルデヒドの定量等のホルムアルデヒド測定技術や、ホルムアルデヒド脱水素酵素を固定化した空気浄化フィルター等のホルムアルデヒドの除去技術等、特に環境技術の分野における応用が期待されている。
【0003】
例えば、ホルムアルデヒドをはじめとするアルデヒド化合物やアルコール化合物を簡易に検出測定できるバイオセンサー用酵素としての利用が報告されている(例えば、特許文献1を参照)。詳細には、気相中や液相中のアルデヒド化合物及びアルコール化合物を簡易に検出測定できる酵素固定化バイオセンサーが開示されており、これは、過酸化水素電極の表面にアルコール酸化酵素が固定化されると共に最表面に高分子被覆が配設され、過酸化水素の酸化電流値、またはその変化よりアルデヒド化合物もしくはアルコール化合物の存在を検出するものである。
【0004】
また、近年、クリーンエネルギー技術として、アルコールなどのバイオマスを燃料とした酵素燃料電池が考案されており、酸化還元酵素が電池の構成部材として利用されている。一例として、メタノールを燃料とした酵素燃料電池の場合、触媒としてメタノールに作用してホルムアルデヒドに酸化するアルコールデヒドロゲナーゼと、ホルムアルデヒドに作用してギ酸に酸化するホルムアルデヒドデヒドロゲナーゼと、ギ酸に作用してCO2に酸化する蟻酸デヒドロゲナーゼとを用いることにより構成することができる。これにより、メタノールはCO2まで分解され、メタノール1分子につき3段階の酸化反応により合計6電子が生成されることとなる。
【0005】
そして、ホルムアルデヒド脱水素酵素は、細菌、酵母から哺乳類に至るまで広く存在していることが知られており、多様な生物由来のホルムアルデヒド脱水素酵素が報告されている。例えば、パン酵母であるサッカロマイセス・セレビシエ(Saccharomyces Cerevisiae)由来のホルムアルデヒド脱水素酵素(例えば、非特許文献1を参照)、パラコッカス・デニトリフィカンス(Paracoccus denitrificans)由来のホルムアルデヒド脱水素酵素(例えば、非特許文献2を参照)、シュードモナス・プチダ(Pseudomonas putida)由来のホルムアルデヒド脱水素酵素(例えば、非特許文献3を参照)等が挙げられる。
【0006】
ホルムアルデヒド脱水素酵素には、反応にグルタチオンを要求するグルタチオン依存性、及びグルタチオンを要求しない非依存性の2つの型がある。一般に広く存在するホルムアルデヒド脱水素酵素はグルタチオン依存性であるのに対して、シュードモナス・プチダ由来のホルムアルデヒド脱水素酵素は、グルタチオン非依存的にホルムアルデヒドの酸化還元反応を触媒する特徴的な酵素であることが知られている。そして、このホルムアルデヒド脱水素酵素は、各サブユニットが376アミノ酸残基からなるホモ4量体酵素であることが報告されている(例えば、非特許文献3を参照)と共に、X線結晶構造解析の結果をもが報告され活性中心をはじめとした高次構造が明らかとなっている(例えば、非特許文献4を参照)。
【0007】
以上の従来文献については、すべて天然に存在する野生型のホルムアルデヒド脱水素酵素に関するものである。一方、分子進化工学的手法を使って酵素の構造を改変し、酵素活性の至適温度を低温化した報告もある(非特許文献5)。これは、Pseudomonas putida由来の野生型のホルムアルデヒド脱水素酵素にアミノ酸変異を導入し、30℃程度の低い温度域において、野生型酵素より酵素活性を1.7倍向上させたものである。
【0008】
ところが、ホルムアルデヒド脱水素酵素に限らず酵素は、触媒作用を選択する性質が非常に強いことが知られている。例えば、酵素は、生体が生存可能な緩和な条件下で最も酵素活性を発揮し、生体内で代謝される機構を有するものであることから、酵素を構成するタンパク質構造に変化を与えるような因子の存在により酵素の活性は影響を受ける。つまり、酵素活性は、温度等の物理的条件、及びpH、塩濃度等の化学的条件の影響を受けることが知られており、これらに対する安定性が低いため用途が限定されるという問題点があった。そして、従来公知のホルムアルデヒド脱水素酵素は、何れも天然に存在する野生型の酵素であることから、熱に対する安定性が低かった。
【0009】
しかしながら、バイオセンサーや燃料電池への適用に際しては、多様な環境において安定して活性を保持できる酵素が不可欠であり、特に、酵素寿命に対して相関があるといわれる熱安定性の向上が要求される。従来公知の天然に存在する野生型ホルムアルデヒド脱水素酵素(非特許文献1〜4)は、何れも熱に対する安定性が低かった。また、野生型のホルムアルデヒド脱水素酵素を改変したホルムアルデヒド脱水素酵素(非特許文献5)についても、活性の至適温度を低温化しているために、高い温度域では野生型酵素より熱安定性が低下しており、酵素の熱失活が生じやすくなっている。
【0010】
そのため、従来公知のホルムアルデヒド脱水素酵素をバイオセンサーや燃料電池への適用した場合は、酵素の保存安定性が低く、酵素の熱失活により酵素の使用量の増大及び精度の低下が生じることから、市場の要望に十分に対応できるものではなかった。従って、高温条件下でも安定的に作用できるホルムアルデヒド脱水素酵素の提供が望まれていた。
【先行技術文献】
【特許文献】
【0011】
【特許文献1】特開2007−139729号公報
【非特許文献】
【0012】
【非特許文献1】Steinman CR. Jakoby WB.著、“Yeast aldehyde dehydrogenase. II. Properties of the homogeneous enzyme preparations.(酵母アルデヒドデヒドロゲナーゼII)” J Biol Chem.、1968年、第243巻、第730〜734頁
【非特許文献2】Ras J. Van Ophem PW. Reijnders WN. Van Spanning RJ. Duine JA. Stouthamer AH. Harms N.著、“Isolation, sequencing, and mutagenesis of the gene encoding NAD- and glutathione-dependent formaldehyde dehydrogenase (GD-FALDH) from Paracoccus denitrificans, in which GD-FALDH is essential for methylotrophicgrowth.” J Bacteriol.、1995年、第177巻、第247〜251頁
【非特許文献3】Ito K. Takahashi M. Yoshimoto T. Tsuru D.著、“Cloning and high-level expression of the glutathione-independent formaldehyde dehydrogenase gene from Pseudomonas putida. ”J Bacteriol. 1994年、第176巻、第2483〜2491頁
【非特許文献4】Tanaka N. Kusakabe Y. Ito K. Yoshimoto T. Nakamura KT.著、“Crystal structure of formaldehyde dehydrogenase from Pseudomonas putida: the structural origin of the tightly bound cofactor in nicotinoprotein dehydrogenases.”J. Mol. Biol.、2002年、第324巻、第519〜533頁
【非特許文献5】Fujii Y, Yamasaki Y, Matsumoto M, Nishida H, Hada M, Ohkubo K. 著、“The artificial evolution of an enzyme by random mutagenesis: the development of formaldehyde dehydrogenase.” Biosci Biotechnol Biochem. 2004, 68(8), 1722-1727.
【発明の概要】
【発明が解決しようとする課題】
【0013】
本発明は、上記事情に鑑みてなされたものであり、高温条件下でも熱安定性に優れたホルムアルデヒド脱水素酵素を提供することにある。
【課題を解決するための手段】
【0014】
本発明者らは、上記課題を解決すべく研究を重ねた結果、野生型ホルムアルデヒド脱水素酵素の特定のアミノ酸配列領域における少なくとも1つの位置のアミノ酸を他のアミノ酸で置換することにより、野生型ホルムアルデヒド脱水素酵素と比較して熱安定性及び保存安定性を向上できることを見出した。更に、かかる酵素を利用することにより、従来の酵素の熱安定性に関する欠点を克服し、より実用面において有利なアルデヒド検出用バイオセンサー及び燃料電池を提供できることをも見出した。本発明者らはこれらの知見に基づき本発明を完成するに至った。
【0015】
即ち、上記目的を達成するため、以下の[1]〜[13]に示す発明を提供する。
[1]野生型ホルムアルデヒド脱水素酵素のアミノ酸配列中の少なくとも1つの位置のアミノ酸の欠失、置換、付加或いは挿入による改変により得られる改変型ホルムアルデヒド脱水素酵素であって、前記改変が、前記野生型ホルムアルデヒド脱水素酵素のアミノ酸配列における第70位−第150位で生じており、前記野生型ホルムアルデヒド脱水素酵素に比べて安定性が向上している改変型ホルムアルデヒド脱水素酵素。
[2]前記改変が、前記アミノ酸配列の第128位に対応する位置で生じている上記[1]に記載の改変型ホルムアルデヒド脱水素酵素。
[3]前記改変が、前記アミノ酸配列の第128位に対応する位置におけるアスパラギン酸のグリシンへの置換である上記[2]に記載の改変型ホルムアルデヒド脱水素酵素。
[4]前記改変が、前記アミノ酸配列の第110位に対応する位置で生じている上記[1]に記載の改変型ホルムアルデヒド脱水素酵素。
[5]前記改変が、前記アミノ酸配列の第110位に対応する位置におけるグリシンのアスパラギン酸への置換である上記[4]に記載の改変型ホルムアルデヒド脱水素酵素。
[6]前記野生型ホルムアルデヒド脱水素酵素が、配列番号2に示すアミノ酸配列を有する上記[1]〜[5]の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素。
[7]上記[1]〜[6]の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素をコードする単離核酸分子。
[8]上記[7]に記載の単離核酸分子を含有する組換えベクター。
[9]上記[8]に記載の組換えベクターを含有する形質転換体。
[10]上記[9]に記載の形質転換体を培養する工程、及び得られた培養物からホルムアルデヒドの脱水素反応を触媒する能力を有するタンパク質を採取する工程を含む、野生型ホルムアルデヒド脱水素酵素に比べて熱安定性が向上している改変型ホルムアルデヒド脱水素酵素の製造方法。
【0016】
上記[1]〜[10]の構成の改変型ホルムアルデヒド脱水素酵素は、野生型ホルムアルデヒド脱水素酵素のアミノ酸配列(具体的には配列番号2に示すアミノ酸配列)に置ける第70位−第150位のアミノ酸の何れかに改変を施している。
この位置は、酵素の補欠分子であるZnの結合部位に近接しており、酵素サブユニット間の界面付近とは離れている。タンパク質の構造安定化には、Znなど補欠分子の結合安定性が大きく影響することが知られていて、一般的にはアミノ酸変異の導入を避けたほうがよいと考えられている。しかし、本願のようにZnの結合部位に近接した位置でアミノ酸に改変を施すことで、熱安定性を向上させた新規なホルムアルデヒド脱水素酵素の提供が可能となる。
好ましくは、前記改変が、前記アミノ酸配列の第128位に対応する位置におけるアスパラギン酸のグリシンへの置換、或いは、第110位に対応する位置におけるグリシンのアスパラギン酸への置換とすれば、野生型ホルムアルデヒド脱水素酵素に比べて顕著な熱安定性および長期安定性を有するようになる。
従って、本発明によって提供されるホルムアルデヒド脱水素酵素は、医療・食品・環境分野等の様々な産業分野におけるホルムアルデヒドの脱水素反応を要する技術に適用できる。更に、熱安定性が向上した実用性の高い本発明の改変型ホルムアルデヒド脱水素酵素のアミノ酸配列、並びに塩基配列が明確になったことから、遺伝子工学的手法を利用して組換え体として当該酵素を低コストかつ工業的に大量生産することが可能となった。
【0017】
[11]上記[1]〜[6]の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素の触媒活性を利用してアルデヒド化合物を検出する、アルデヒド化合物の検出方法。
【0018】
上記[11]の構成によれば、本発明の改変型ホルムアルデヒド脱水素酵素の触媒能力を利用したアルデヒド化合物の検出方法が提供でき、医療、食品、環境分野等、様々な分野に利用することができる。特には、本発明の改変型ホルムアルデヒド脱水素酵素は熱安定性が高いことから、酵素の触媒能力の劣化を招くことなく測定精度の向上を図れる。更には、酵素使用量をも低減できることからコスト削減効果をも奏する。
【0019】
[12]上記[1]〜[6]の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素を電極上に固定化した酵素センサー。
【0020】
上記[12]の構成によれば、本発明の改変型ホルムアルデヒド脱水素酵素の触媒能力を利用したアルデヒド化合物検出用の酵素センサーが提供でき、医療、食品、環境分野等、様々な分野に利用することができる。特には、本発明の改変型ホルムアルデヒド脱水素酵素は熱安定性が高いことから、酵素の触媒能力の劣化を招くことなく測定精度の向上を図れる。更には、酵素使用量をも低減できることからコスト削減効果をも奏する。
【0021】
[13]上記[1]〜[6]の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素を含み、前記改変型ホルムアルデヒド脱水素酵素のアルデヒド化合物の酸化反応に伴って生成する電子を受け取るアノード極、酸素に電子を伝達することのできる触媒および酵素のいずれかを保持するカソード極を備え、前記アノード極と前記カソード極とが電気的に結合されている燃料電池。
【0022】
上記[13]の構成によれば、本発明の改変型ホルムアルデヒド脱水素酵素の触媒能力を利用した燃料電池が提供できる。特に、本発明の改変型ホルムアルデヒド脱水素酵素は熱安定性が高いことから、酵素の触媒能力の劣化を招くことなく持続的に発電を行なうことが可能となり、燃料電池の性能向上を図れる。更には、酵素使用量をも低減できることからコスト削減効果をも奏する。
【図面の簡単な説明】
【0023】
【図1】変異遺伝子ライブラリーの作製概略を示す図である。
【図2】野生型FDH遺伝子とその近傍領域において各プライマーが結合する位置を示す図である。
【図3】野生型FDHの構造解析を行なった結果を示した図である。
【図4】D128Gの構造解析を行なった結果を示した図である((a)野生型FDH、(b)D128G)。
【図5】G110Dの構造解析を行なった結果を示した図である。
【図6】精製したD128Gおよび野生型FDHのアクリルアミドゲル電気泳動の結果を示した図である。
【図7】D128Gおよび野生型FDHのpH依存性を調べた結果を示した図である。
【図8】改変型FDHおよび野生型FDHの温度依存性を調べた結果を示した図である((a)D128G,(b)G110D)。
【図9】D128Gおよび野生型FDHの温度安定性を調べた結果を示した図である((a)55℃,(b)60℃,(c)65℃)。
【図10】G110Dおよび野生型FDHの温度安定性を調べた結果を示した図である((a)55℃,(b)60℃,(c)65℃)。
【図11】改変型FDHおよび野生型FDHの安定性を調べた結果を示した図である((a)D128Gおよび野生型FDHの比較、(b)G110Dおよび野生型FDHの比較)。
【図12】D128Gおよび野生型FDHについて常温時(25℃)の安定性を調べた結果を示した図である。
【図13】D128Gおよび野生型FDHについて常温時(30℃)の安定性を調べた結果を示した図である。
【発明を実施するための形態】
【0024】
以下、本発明について詳細に説明する。
近年、クリーンエネルギー技術として、アルコールなどのバイオマスを燃料とした酵素燃料電池が考案されており、そこでは本発明のホルムアルデヒド脱水素酵素のような酸化・還元を触媒する酵素が電池の電極触媒として利用されている。一例として、メタノールを燃料とした酵素燃料電池の場合、触媒としてメタノールに作用してホルムアルデヒドに酸化するアルコールデヒドロゲナーゼと、ホルムアルデヒドに作用して蟻酸に酸化するホルムアルデヒドデヒドロゲナーゼと、蟻酸に作用してCO2に酸化する蟻酸デヒドロゲナーゼとを用いることができる。これにより、メタノールはCO2まで分解され、メタノール1分子につき3段階の酸化反応により合計6電子が生成される。
【0025】
本発明の改変型ホルムアルデヒド脱水素酵素(以下、「改変型FDH」と略する)は、天然に存在する野生型FDHのアミノ酸配列において、少なくとも1つの位置のアミノ酸の欠失、置換、付加或いは挿入により改変されており、そして、野生型FDHに比べて熱安定性が向上している。ここで「少なくとも1つの位置のアミノ酸の欠失、置換、付加或いは挿入により改変」とは、改変の基礎となるタンパク質をコードする遺伝子に対して、公知のDNA組換え技術、及び点変異導入方法等によって、欠失、置換、付加或いは挿入することができる程度の数のアミノ酸が、欠失、置換、付加或いは挿入されることを意味し、これらの組み合わせをも含む。したがって、改変型FDHは、これらの改変は1つ有するものであっても、また2つ以上が組み合わされたものであってもよい。このような改変は、人為的に導入することもできるし、また、自然界において非意図的に生じることもある。したがって、本発明における改変型FDHには、これら双方の改変型が含まれる。
【0026】
野生型FDHとは、自然界より分離されたFDHのアミノ酸配列、及び該酵素をコードする核酸分子の塩基配列が、意図的もしくは非意図的に改変が生じている改変部位を有していないことを意味する。
【0027】
「FDH」とは、ホルムアルデヒドを基質としてギ酸を生成する脱水素反応を触媒する活性を有するタンパク質であり、補酵素としてニコチンアミドアデニンジヌクレオチド(NAD)を要求する。反応式を以下に示す。
ホルムアルデヒド+NAD+(酸化型) → ギ酸+NADH(還元型)+H+
【0028】
FDHには、グルタチオン依存性、非依存性の2つの型が存在する。本発明においては何れもが対象となるが、グルタチオン非依存性が好ましい。
【0029】
改変型の基礎となる野生型FDHは、上記の性質を有する限り何れの生物由来のものであってよい。具体的には、シュードモナス(Pseudomonas)属、パラコッカス(Paracoccus)属、ピロコッカス(Pyrococcus)等の細菌由来、及びサッカロマイセス(Saccharomyces)属等の酵母由来のFDH等が例示される。更には、ウシ(Equus caballus)やラット(Rattus norvergicus)等の哺乳動物由来のFDHをも含む。好ましくは、シュードモナス・プチダ(Pseudomonas putida)、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)、パラコッカス・デニトリフィカンス(Paracoccus denitrificans)由来のFDHである。しかしながら、これらに限定するものではない。
【0030】
「安定性が向上」とは、熱、及び保存に対する当該酵素の安定性が向上することであり、一定期間の熱処理及び保存に対して、当該酵素の触媒機能の失活や活性低下が抑制できることを意味する。具体的には、改変型FDHが、野生型FDHよりも、一定期間の熱処理又は保存に供した後に維持されている酵素の触媒機能の残存率が高いことを意味する。例えば、野生型FDHの活性が維持される25℃での酵素の酵素活性値を100%として、加温処理後の活性値を酵素活性の残存率として算出する。この残存率が、野生型FDHと比較して増大していた場合、当該酵素の熱安定性が向上したと判断できる。また、一定期間の保存に際しての残存率が、野生型FDHと比較して増大していた場合、当該酵素の保存安定性が向上したと判断できる。一例として、50℃にて30分間の加温により60%以上、特には75%以上の残存活性を維持できる熱安定性が好ましい。また、45℃にて10時間の保存で60〜80%、30時間の保存であっても50〜60%程度の残存活性を維持できる保存安定性が好ましい。
【0031】
本発明の改変型FDHはこれに限定されるものではないが、前記改変が、野生型FDHのアミノ酸配列における第70位−第150位で生じたものが、好ましく例示される。
【0032】
野生型FDHのアミノ酸配列において、第70位−第150位におけるアミノ酸の改変は、これらの位置における何れかの改変を有するもの全てが本発明に含まれる。また、かかる位置における1のアミノ酸が改変したものであっても、複数の位置における複数のアミノ酸が改変したものであってもよい。
【0033】
配列番号2に示すアミノ酸配列は、シュードモナス・プチダ由来の野生型FDHのアミノ酸配列であるが、他の生物由来の当該酵素のホモログについても同様の位置が改変されたものは本願発明の改変型FDHに含まれる。
【0034】
ここで、改変としては、特に好ましくは、他のアミノ酸への置換である。そして、他のアミノ酸とは、置換前のアミノ酸以外の何れもが含まれる。しかしながら、タンパク質機能改変の観点から極性、電荷、親水性、若しくは疎水性等の点で置換前のアミノ酸と異なる性質を有するアミノ酸であることが好ましい。
例えば、アラニン、バリン、ロイシン、イソロイシン、プロリン、メチオニン、フェニルアラニン、トリプトファンは、共に非極性アミノ酸に分類されるため、互いに似た性質を有するが、これら以外のアミノ酸への置換はタンパク質機能へ影響を与えることが考えられる。
非荷電性アミノ酸としては、グリシン、セリン、スレオニン、システイン、チロシン、アスパラギン、グルタミンが挙げられる。
酸性アミノ酸としては、アスパラギン酸およびグルタミン酸が挙げられる。
塩基性アミノ酸としては、リシン、アルギニン、ヒスチジンが挙げられる。
これらの各グループ内の以外へのアミノ酸置換は、タンパク質の機能が改変されることが特に予想されるが、これに制限されるものではない。
【0035】
好ましくは、野生型FDHのアミノ酸配列において、当該アミノ酸配列の第128位に対応する位置におけるアスパラギン酸のグリシンへの置換であり、または、配列番号2におけるアミノ酸配列の第110位に対応する位置におけるグリシンのアスパラギン酸への置換が例示される。
【0036】
一例として、野生型FDHとしてシュードモナス・プチダとする場合、上述した第128位の置換を行なったアミノ酸配列を配列番号4に、第110位の置換を行なったアミノ酸配列を配列番号6に示す。
【0037】
更に、前述の改変型FDHの性質を保持している限り、更に特定のアミノ酸に改変が生じている改変部位を有するアミノ酸配列を含むものであってもよい。当業者はアミノ酸配列の改変に際して本発明の改変型FDHの酵素活性を保持する改変を容易に予測することができる。具体的には、例えばアミノ酸置換の場合には、タンパク質構造保持の観点から極性、電荷、親水性、若しくは疎水性等の点で置換前のアミノ酸と類似した性質を有するアミノ酸に置換することができる。このような置換は保守的置換として当業者には周知である。具体例を挙げると、例えば、アラニン、バリン、ロイシン、イソロイシン、プロリン、メチオニン、フェニルアラニン、トリプトファンは、共に非極性アミノ酸に分類されるため、互いに似た性質を有する。また、非荷電性アミノ酸としては、グリシン、セリン、スレオニン、システイン、チロシン、アスパラギン、グルタミンが挙げられる。また、酸性アミノ酸としては、アスパラギン酸およびグルタミン酸が挙げられる。また、塩基性アミノ酸としては、リシン、アルギニン、ヒスチジンが挙げられる。これらの各グループ内のアミノ酸置換は、タンパク質の機能が維持されるとして許容される。
また、その後の精製、固相への固定化等の便宜のため、アミノ酸配列のN、又はC末端にHisタグペプチド、FLAGタグペプチド等を付加したものも好適に例示される。このようなタグペプチドの導入は常法により行なうことができる。また、本発明の酵素活性の喪失を引き起こさない範囲内で、C末端側若しくはN末端側のアミノ酸残基を切断した切断型でもよい。更に、グルコシル化等の化学修飾を付加してもよい。
【0038】
本発明の改変型FDHは公知の方法によって取得することができる。例えば、改変の基礎となる野生型FDHをコードする遺伝子に対して改変を施し、得られた改変型遺伝子を用いて宿主細胞を形質転換し、かかる形質転換体の培養物から上記活性を有するタンパク質を採取することによって取得することができる。
【0039】
改変の基礎となる野生型FDHをコードする遺伝子は、公知の遺伝子クローニング技術を用いて取得することができる。例えば、GenBank等の公知のデータベースを検索することによって取得することができる遺伝子情報を基にしてプライマーを設計し、FDHを産生し得る生物体から抽出したゲノムDNAを鋳型としてPCRを行うことにより取得することができる。また公知の遺伝子情報に基づいて、常法のホスホルアミダイト法等の核酸合成法により合成することによっても取得するができる。ここで、本発明の改変型の基礎として好適なFDHの配列情報として、野生型のシュードモナス・プチダ由来のFDHのアミノ酸配列を配列表の配列番号2に、当該FDHをコードする塩基配列を配列表の配列番号1に示す。
【0040】
野生型FDHをコードする遺伝子に改変を施す方法としては、特に制限はなく、当業者に公知の改変タンパク質作製のための変異導入技術を利用することができる。例えば、部位特異的突然変異誘発法、PCR法等を利用して変異を導入するPCR突然誘発法、あるいは、トランスポゾン挿入突然変異誘発法などの公知の変異導入技術を利用することができる。また、市販の変異導入用キット(例えば、QuikChange(登録商標) Site-directed Mutagenesis Kit(Stratagene社製))を利用してもよい。
【0041】
特には、野生型FDHをコードするDNAを鋳型として、所望の改変(欠失又は置換)を施した配列を含むオリゴヌクレオチドをプライマーとしてPCRを行うことによって取得することが、好ましく例示される。ここで、本発明の改変型FDHの調製において、PCRを利用する場合に用いられるプライマーは、野性型FDHをコードする核酸分子と相補的な配列を含み、かつ所望の改変が生じるように設計されたものであり、常法に基づいて調製することができる。例えば、ホスホアミダイト法等に基づく化学合成法等が利用可能である。化学合成法に基づきプライマーを調製する場合には、合成に先立って標的核酸の配列情報に基づいて設計される。プライマーの設計は、所望の領域を増幅するように、例えばプライマー設計支援ソフト等を利用して設計することができる。プライマーは合成後、HPLC等の手段により精製される。また、化学合成を行う場合には市販の自動合成装置を利用することも可能である。このようなプライマーとしては10以上、好ましくは15以上、更に好ましくは約20〜50の塩基からなるオリゴヌクレオチドが例示される。
【0042】
また、目的とする改変型FDHのアミノ酸配列が定めることにより、それをコードする適当な塩基配列を決定でき、常法のホスホルアミダイト法等の核酸合成技術を利用して本発明の改変型FDHをコードするDNAを化学的に合成することができる。
【0043】
また、このような本発明の改変型FDH改変体は自然又は人工の突然変異により生じた突然変異体の中から前述の理化学的性質を有するタンパク質をスクリーニングすることにより取得できる。
【0044】
(本発明の改変型FDHをコードする核酸分子)
本発明の改変型FDHをコードする核酸分子は、前述の理化学的性質を有するすべての改変型FDHをコードするものを包含する。例えば、配列番号4,6に記載されるアミノ酸配列を含むタンパク質をコードする全てのポリヌクレオチドであり、一具体例としては、配列番号3,5に示す塩基配列を含むポリヌクレオチドが挙げられるが、これに限定するものではない。ここで、本発明におけるポリヌクレオチドにはDNA及びRNAの双方が含まれ、DNAである場合には、1本鎖であると、二本鎖であるとは問わない。
【0045】
本発明の改変型FDHをコードする核酸分子は、本明細書においてその塩基配列が明確になったことから、かかる配列情報に基づいて、常法のホスホルアミダイト法等のDNA合成法を利用して化学的に合成することができる。また、改変の基礎となる野生型FDHをコードするDNAに対して改変を施すことによっても製造することができる。なお、詳細については前述した。
【0046】
(本発明の組換えベクター)
そして、本発明の組換えベクターは、適当なベクターに本発明の改変型FDHをコードする核酸分子を組み込むことによって構築することができる。利用可能なベクターとしては、外来DNAを組み込め、かつ宿主細胞中で自律的に複製可能なものであれば特に制限はない。従って、ベクターは、本発明の改変型FDHをコードする核酸分子を挿入できる少なくとも1つの制限酵素部位の配列を含むものである。例えば、プラスミドベクター(pEX系、pUC系、及びpBR系等)、ファージベクター(λgt10、λgt11、及びλZAP等)、コスミドベクター、ウイルスベクター(ワクシニアウイルス、及びバキュロウイルス等)等が包含される。
【0047】
本発明の組換えベクターは、本発明の改変型FDHをコードする核酸分子がその機能を発現できるように組み込まれている。従って、核酸分子の機能発現に必要な他の既知の塩基配列が含まれていてもよい。例えば、プロモータ配列、リーダー配列、シグナル配列、並びにリボソーム結合配列等が挙げられる。プロモータ配列としては、例えば、宿主が大腸菌の場合にはlacプロモータ、trpプロモータ等が好適に例示される。しかしながら、これに限定するものではなく既知のプロモータ配列を利用できる。
更に、本発明の組換えベクターには、宿主において表現型選択を付与することが可能なマーキング配列等をも含ませることができる。このようなマーキング配列としては、薬剤耐性、栄養要求性などの遺伝子をコードする配列等が例示される。具体的には、カナマイシン耐性遺伝子、クロラムフェニコール耐性遺伝子、アンピシリン耐性遺伝子等が例示される。
【0048】
ベクターへの本発明の改変型FDHをコードする核酸分子等の挿入は、例えば、適当な制限酵素で本発明の遺伝子を切断し、適当なベクターの制限酵素部位、又はマルチクローニング部位に挿入して連結する方法などを用いることができるが、これに限定されない。連結に際しては、DNAリガーゼを用いる方法等、既知の方法を利用できる。また、DNA Ligation Kit(タカラバイオ社)等の市販のライゲーションキットを利用することもできる。
【0049】
(本発明の形質転換体)
本発明の形質転換体は、適当な細胞を本発明の改変型FDHをコードする核酸分子を含む組換えベクターで形質転換することによって構築することができる。ここで、宿主となる細胞としては、本発明の改変型FDHを効率的に発現できる宿主細胞であれば、特に制限はない。原核生物を好適に利用でき、特には大腸菌E.coli(DH5α、BL21、JM109等)を利用することができる。その他、枯草菌、バシラス属細菌、シュードモナス属細菌等をも利用できる。更に、原核生物に限定されず真核生物細胞を利用することが可能である。例えば、サッカロマイセス・セルビシエ(Saccharomyces cerevisiae)等の酵母、Sf9細胞等の昆虫細胞、CHO細胞、COS−7細胞等の動物細胞等を利用することも可能である。形質転換法としては、塩化カルシウム法、エレクトロポレーション法、リポソームフェクション法、マイクロインジェクション法等を既知の方法を利用することができる。
【0050】
(本発明の改変型FDHの製造方法)
本発明の改変型FDHの製造方法は、前述の本発明の形質転換体を培養し、得られた培養物からアルデヒド化合物の脱水素反応を触媒する活性を有するタンパク質を採取することにより行なう。即ち、前述の本発明の形質転換体を培養する培養工程と、前記培養工程で発現した前記タンパク質を回収する回収工程とを備える。このように、適当な宿主で発現させることによって、低コストで本発明の改変型FDHの大量生産が可能となる。
【0051】
培養工程は、本発明の形質転換体を適当な培地に接種し、常法に準じて培養することにより行なわれる。本発明の形質転換体の培養は、宿主細胞の栄養生理学的性質を勘案して、培養条件を選択すればよい。使用される培地としては、宿主細胞が資化し得る栄養素を含み、形質転換体におけるタンパク質の発現を効率的に行えるものであれば特に制限はない。従って、宿主細胞の生育に必要な炭素源、窒素源その他必須の栄養素を含む培地であることが好ましく、天然培地、合成培地の別を問わない。
【0052】
例えば、炭素源として、ホルムアルデヒド、デキストラン、デンプン等が、また、窒素源としては、アンモニウム塩類、硝酸塩類、アミノ酸、ペプトン、カゼイン等が挙げられる。他の栄養素としては、所望により、無機塩類、ビタミン類、抗生物質等とを含ませることができる。宿主細胞が大腸菌の場合には、LB培地、M9培地等が好適利用できる。また、培養形態についても特に制限はないが、大量培養の観点から液体培地が好適に利用できる。
【0053】
本発明の組換えベクターを保持する宿主細胞の選別は、例えば、マーキング配列の発現の有無により行なうことができる。例えば、マーキング配列として薬剤耐性遺伝子を利用する場合には、薬剤耐性遺伝子に対応する薬剤含有培地で培養することによって行うことができる。
【0054】
精製工程は、前述の培養工程において得られた形質転換体の培養物からの本発明の改変型FDHを回収、即ち、単離精製することによって行えばよい。本発明の酵素の存在する画分に応じて、一般的なタンパク質の単離精製方法に準じた手法を適用すればよい。具体的には、本発明のFDHが宿主細胞外に生産される場合には、培養液をそのまま使用するか、遠心分離、濾過等の手段により宿主細胞を除去して培養上清を得る。続いて、培養上清に、既知のタンパク質精製方法を適宜選択することにより、本発明の酵素を単離精製することができる。
【0055】
例えば、硫酸アンモニウム沈殿、透析、SDS−PAGE電気泳動、ゲル濾過、疎水、陰イオン、陽イオン、アフィニティークロマトグラフィ等の各種クロマトグラフィ等の既知の単離精製技術を単独、又は適宜組み合わせて適用することができる。特にアフィニティークロマトグラフィを利用する場合、本発明の酵素をHis Tag等のタグペプチドとの融合タンパク質として発現させて、かかるタグペプチドに対する親和性を利用することが好ましい。また、本発明の改変型FDHが宿主細胞内で産生される場合には、培養物を遠心分離、濾過等の手段により宿主細胞を回収する。
【0056】
続いて、リゾチーム処理などの酵素的破砕方法、又は超音波処理、凍結融解、浸透圧ショック等の物理的破砕方法等により、宿主細胞を破砕する。破砕後、遠心分離、濾過等の手段により可溶化画分を収集する。得られた可溶化画分を、前述の細胞外に生産できる場合と同様に処理することにより単離精製することができる。ここで、本発明で得られる改変型FDHは、熱安定性が高いことから、前述の単離、精製工程において熱処理を併用することが有用かつ便利である。
培養物から得られた宿主細胞及び培養上清には、当該宿主細胞由来の様々なタンパク質を含有する。しかし、熱処理を行なうことにより、宿主細胞由来の夾雑タンパク質は変性し凝縮沈殿する。これに対して、本発明の酵素は、改変型を有するため変性を生じないことから、遠心分離等により宿主由来の夾雑タンパク質と容易に分離できる。また、培養液をそのまま、若しくは粗抽出液を使用する場合においても、熱処理を行なうことにより、他のタンパク質が失活することから、実質的に本発明の改変型FDHのみの酵素液として使用することができる。従って、本発明の改変型FDHを遺伝子工学的手法により製造する場合においても、宿主由来のその他のタンパク質を容易に除去することができる。従って、精製度を向上させることができ、信頼性の高い酵素を製造できるという利点がある。
【0057】
そして、精製されたFDHが所望の改変が生じている改変部位を有する本発明の改変型FDHであるか否かの確認は、その理化学的性質や配列の分析によって行うことができる。理化学的性質の分析による場合、一定期間、熱処理に付した後、酵素の酵素活性を公知の方法により測定し改変部位を有しない野生型FDHと比較して、活性の残存率が高いか否かを確認することによって行うことができる。従って、基質であるアルデヒド化合物に対してNAD+依存的に脱水素反応をする酵素活性を測定することにより行うことができる。
【0058】
酵素の活性は、NAD+依存的にアルデヒド化合物の脱水素反応を触媒する酵素の活性測定法として知られる方法を何れをも利用して行うことができる。例えば、NAD+の存在下で、本発明の酵素をアルデヒド化合物と反応させ、当該酵素の触媒反応で生成するNADH量の変化を340nmの吸光度変化もって検出する。かかる吸光度変化をもって当該酵素の活性とすることができる。従って、FDH活性は、ホルムアルデヒド及びNAD+から、ギ酸及びNADHを生成する触媒反応において、生成したNADHを直接定量することによって測定できる。つまり、酵素反応によりホルムアルデヒドが酸化されギ酸となり、このときNADが還元されNADHが生成する(反応式:HCHO+NAD++H2O → HCOOH+NADH+H+)。このNADからNADHの生成を340nmの吸光度変化を測定して酵素活性とした。この配列分析による場合には、公知のアミノ酸分析法によって行うことができる。例えば、エドマン分解法に基づく自動アミノ酸決定法が利用できる。
【0059】
(アルデヒド化合物の検出方法)
本発明の改変型FDHは、試料中のアルデヒド化合物の検出のために利用することができる。アルデヒド化合物の検出は、例えば、基質であるアルデヒド化合物に対してNAD+依存的に脱水素反応をする酵素活性を測定することにより行うことができる。酵素の活性は、NAD+依存的にアルデヒド化合物の脱水素反応を触媒する酵素の活性測定法として知られる方法をいずれをも利用して行うことができる。例えば、NAD+の存在下で、本発明の酵素を測定対象となる試料と反応させ、当該酵素の触媒反応で生成するNADH量の変化を340nmの吸光度変化もって検出する。かかる吸光度変化をもって当該酵素の活性とすることができ、ひいてはNADH量の変化によりアルデヒド化合物の存在を検出することができる。したがって、FDH活性は、その触媒反応において、生成したNADHを直接定量することによって測定できる。
反応液としては、例えば、ホルムアルデヒド等のアルデヒド基質、2mMのNAD+、25mMのMgCl2、1MのNaCl及び本発明の酵素溶液を、50mMリン酸緩衝液(pH8.8)中にて混合して200μLとすることにより調製することができる。しかしながら、これは標準条件であり、適宜変更することができる。この反応液を、37℃で波長340nmにおける吸光度を測定し、吸光度の増加を求めることにより活性評価試験を行うことができ、吸光度の測定は、既知のマイクロプレートリーダー(モレキュラーデバイス社製)を用いることができる。
そして、適当な試験条件が選択されていれば、この変化は、測定しようとする酵素活性に直線的に比例する。このとき、あらかじめ目的濃度範囲内における標準濃度のアルデヒド化合物溶液により標準曲線を作成することにより、得られた吸光度変化値に基づいてアルデヒド化合物濃度を求めることができる。
【0060】
ここで、試料としては、アルデヒド化合物等、FDHの基質となり得る化合物の存在が予想されるすべての試料を対象とすることができる。例えば、血液、尿、唾液等の生物体由来の生物試料、食品試料、環境試料等が例示されるがこれに限定されるものではない。また、必要に応じて、これらの試料に適当な処理を行った試料をも含み得る。特に、本発明の改変型FDHは優れた熱安定性を示すことから、温度が高い試料の分析に際して好適に利用できる。そして、保存性も高いことから酵素の酵素活性の劣化を招くことなく、測定精度の向上を図れ、ひいては、酵素使用量を軽減できることからコスト削減効果をも奏することができる。
【0061】
(アルデヒド化合物検出用のバイオセンサー)
本発明は、本発明の改変型FDHを利用するアルデヒド化合物検出用のバイオセンサーを提供する。当該検出センサーは、電極材上に本発明の改変型FDHが固定化した作用電極、及びその対極を設けて構成される。必要に応じて、参照電極を設けて三電極方式として構成してもよい。電極としては、カーボン、金、白金等を用いることができる。電極材上への酵素の固定化は既知の方法によって行うことができる。例えば、物理的吸着、イオン結合,共有結合を介して固定化する担体結合法を利用することができる。また、グルタルアルデヒドなどの二価性官能基をもつ試薬で架橋固定する架橋法をも利用でき、更には、アルギン酸,カラギーナン等の多アルデヒド化合物、ポリアクリルアミド等の網目構造をもつゲルや、半透性膜の中に閉じて固定化する包括法等をも利用することができる。そして、本発明のFDHの酵素活性に際して要求される補酵素NAD+、NAD+の還元体であるNADHを酸化する能力を有する酸化酵素(ジアホラーゼ等)、電子メディエーター(フェロセン、ジクロロインドフェノール等)も、必要に応じて電極材上に固定化して構成される。
【0062】
アルデヒド化合物の測定は、例えば、測定対象となる試料を接触させると試料中のアルデヒド化合物が作用極上に固定された本発明の改変型FDHと反応し、続いて電子メディエーターが還元される。そして、電極系に電圧を印加して電子受容体の還元体を酸化し、得られる酸化電流値の変化により試料中のアルデヒド化合物を検出することができる。このとき、あらかじめ目的濃度範囲内における標準濃度のアルデヒド化合物溶液により標準曲線を作成することにより、得られた酸化電流値に基づいてアルデヒド化合物濃度を求めることができる。
【0063】
ここで、試料としては、本発明の改変型FDHの基質となり得るアルデヒド化合物の存在が予想されるすべての試料を対象とすることができる。詳細については前述した。特には、前述の通り、本発明の改変型FDHは、熱安定性が高いことからバイオセンサー製作時における劣化をも防止することができ、また、保存性も高いことから酵素の酵素活性の劣化を招くことなく、測定精度の向上を図れ、ひいては、酵素使用量を軽減できることからコスト削減効果をも奏することができる。
【0064】
(本発明の燃料電池)
本発明は、本発明の改変型FDHを利用する燃料電池を提供する。本発明の燃料電池は、例えば、酸化反応を行うアノード極と、還元反応を行うカソード極から構成され、必要に応じてアソードとカソードを隔離する電解質層を含んで構成される。
アノード電極側では、本発明の改変型FDHがアルデヒド化合物を酸化することによって生じた電子を電極に取り出すと共に、プロトンを発生する。一方、カソード側では、アノード側で発生したプロトンが酸素と反応することによって水を生成するように構成される。電極としては、カーボン、金、白金等を用いることができる。アノード極側には本発明の改変型FDHが供給され、適当な緩衝液中に溶解させた形態で供給してもよいが、電極上に固定化されることが好ましい。このとき、改変型FDHは、好ましくはNADと共に固定化されることが好ましく、特にはNADと結合したホロ酵素の形態で固定化されることが好ましい。また、アポ酵素の形態で固定化し、NADを適当な緩衝液に溶解させた形態で供給してよい。
カソード極側には、酸素に電子を伝達することのできる触媒および酵素を必要に応じて供給してよい。電極材上への酵素の固定化は、公知の方法によって行うことができる。例えば、物理的吸着、イオン結合,共有結合を介して固定化する担体結合法を利用することができる。また、グルタルアルデヒドなどの二価性官能基をもつ試薬で架橋固定する架橋法をも利用でき、更には、アルギン酸,カラギーナン等の多アルデヒド化合物、ポリアクリルアミド等の網目構造をもつゲルや、半透性膜の中に閉じて固定化する包括法等をも利用することができる。
【0065】
以上のように構成することにより、燃料であるアルデヒド化合物を酸化する際に生じた電子がアノード極に電子を受け渡す。そして、アノード極に渡された電子は、外部回路を経てカソード極に到達することで電流が発生する。燃料となるアルデヒド化合物としては、本発明の改変型FDHの基質となり得るアルデヒド化合物であれば制限はない。
【0066】
また、メタノールを燃料とした酵素燃料電池においても本発明の改変型FDHを利用できる。その場合には、触媒として、本発明の改変型FDHに加え、必要に応じてメタノールに作用してホルムアルデヒドに酸化するアルコールデヒドロゲナーゼと、ギ酸に作用してCO2に酸化する蟻酸デヒドロゲナーゼを併用することにより構成することができる。
【0067】
本発明の燃料電池は、熱安定性が高い本発明の改変型FDHの触媒能力を利用することから、持続的に発電を行なうことができ、高性能の燃料電池として構築することができる。また、本発明の改変型FDHは、熱安定性が高いことから燃料電池製作時における劣化をも防止することができ、ひいては、酵素使用量を軽減できることからコスト削減効果をも奏することができる。
【実施例】
【0068】
<実施例1>本発明の改変型FDH遺伝子のクローニング
分子進化工学的手法と称される酵素改変手法を利用して、本発明の改変型FDH遺伝子のクローニングを行った。
【0069】
かかる分子進化工学的手法とは、生物進化の原理をタンパク質に応用する技術である。そのステップとしては、まずランダムに改変の基礎となる遺伝子に変異を導入し、可能な限り多様な改変型遺伝子を作り出す。そして、改変型遺伝子から改変型タンパク質を作製して、その中から所望の性質を保持するタンパク質をコードする遺伝子を選択する。このようにして、設計原理が未知であっても所望の機能や性質を保持する高分子を取得することができ、取得した遺伝子の変異箇所及び変異様式を調べることによって、高分子設計原理の研究においても威力を発揮することができる。
【0070】
Pseudomonas putida由来の野生型FDHの立体構造はX線解析により既に明らかにされており、各サブユニットが398アミノ酸残基からなる4量体構造を有する。野生型FDHの立体構造データは、日本蛋白質構造データバンクで公開されているものを利用した(http://www.rcsb.org/pdb/explore.do?structureId=1KOL)。この野生型FDHのX線結晶構造解析に基づく高次構造を検討すると、4量体酵素のサブユニットの界面付近にループ領域(K76−D79)が存在し、この領域が高次構造におけるサブユニット間相互作に影響を及ぼし、熱安定性を支配していることが示唆される。
熱安定性向上効果が見込める部位を鋭意検討した結果、第70位−第150位のアミノ酸領域(Thr70−Leu150)が4量体酵素のサブユニットの界面のドメイン間の構造安定化に寄与するホットスポットである可能性が高いと考え、このホットスポット周辺へランダム変異を導入したライブラリーを作製し、スクリーニングを行うことより、構造が最適化し熱安定性が向上した変異体を取得できると考えた。なお、これら検討のための一連の酵素の立体構造解析は、アクセリス社製のInsight-II(酵素の分子構造を表示し、かつ構造エネルギー計算が可能なソフトウェア)を用いて行った。分子進化工学的手法を用いた酵素改変の具体的な手順を次に示す。
【0071】
(ステップ1) 酵素遺伝子の取得
バクテリアPseudomonas putida株は、NITE(独立行政法人製品評価技術基盤機構)から入手した(受託番号 NBRC No. NBRC3738)。そのゲノムDNAは、添付資料に記載の培養液を使って菌体を培養し、ゲノムDNA抽出キット(プロメガ社製)を使って抽出と精製を行った。このゲノムDNAを鋳型にして以下のプライマー1,2(配列番号7,8)を用いてPCR反応を行なった。PCR反応は、ポリメラーゼとしてPrimeSTAR GXL DNA Polymerase(タカラバイオ社製)、鋳型として50ngゲノムDNAおよび各0.2μMのプライマーのそれぞれを混合して滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。PCR反応は、98℃で10秒、68℃で60秒のサイクルを30回繰り返した。増幅産物の精製はDNA精製キット(GEヘルスケア社製)を用いた。その結果、予想されるDNA増幅産物(約1.2kbp)を得ることができた。
【0072】
5'-GAGCA TATGT CTGGT AATCG TGGTG T-3'(プライマー1:配列番号7)
5'-CTAAA GCTTA GGCCG CGCTG AGGTC T-3'(プライマー2:配列番号8)
【0073】
(ステップ2) 発現ベクターの構築
PCR増幅で得られたDNA断片(約1.2kbp)を、制限酵素NdeIとHindIIIで切断し、アガロースゲル電気流動で分離しゲルから精製した。DNA断片を、タンパク質発現用プラスミドのpET23bベクター(ノバジェン社製)の制限酵素部位NdeIとHindIIIにライゲーション反応より組み込んで酵素の発現ベクターを構築した。
また無細胞システムを使った酵素合成用の鋳型とする直鎖状DNAを調製するために、酵素遺伝子を組み込んだベクターを鋳型にして、T7プロモーターとT7ターミネーター配列間をPCRにより増幅した。PCR反応は、ポリメラーゼとしてPrimeSTAR GXL DNA Polymerase(タカラバイオ社製)を使用し、鋳型DNA(50ng)、プライマー3,4(配列番号9,10、各0.2μM)を滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。PCR反応は、98℃で10秒、68℃で60秒のサイクルを30回繰り返した。PCR産物の精製には、DNA精製キット(GEヘルスケア社製)を用いた。
【0074】
5'- ATCTCGATCCCGCGAAATTAATAC -3' (プライマー3:配列番号9)
5'- TCCGGATATAGTTCCTCCTTTCAG -3' (プライマー4:配列番号10)
【0075】
(ステップ3) 変異遺伝子ライブラリーの作製
ランダム変異を導入した変異遺伝子ライブラリーの作製は、変異導入のための手法にエラープローンPCRを用いた。ライブラリー作成を行うために、酵素FDH分子内に1〜2箇所の変異導入が得られる条件(エラー率0.5%前後となるエラープローンPCR条件)を検討した。ポリメラーゼの塩基取り込み間違いを促進させるために、マグネシウム濃度とマンガン濃度を[Mg,Mn]=[5,0.5]mMとした。
エラープローンPCR反応は、2.5ユニットのTaq DNAポリメラーゼ(タカラバイオ社製)、各0.5μMのプライマー5,6(配列番号11,12)、10mM Tris-HCl(pH8.3)緩衝液、50mM KCl、5.0mM MgCl2、0.5mM MnCl2のそれぞれを混合して滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。
【0076】
5'-CGGCCTGGTCCTGGGCCACGAAATC-3'(プライマー5:配列番号11)
5'-TGGCCTTGTCGCGCTCAGGCAGCTT-3'(プライマー6:配列番号12)
【0077】
次に、エラープローンPCRによりランダムに変異を導入したFDH遺伝子断片と、変異を導入しないFDH遺伝子の残りの部分をコードするDNA断片を連結するために、オーバーラッピングPCRを行った。
まず変異を導入しないDNA断片を得るために、ステップ2で作製した発現ベクターを鋳型にして、プライマー7,8(配列番号13,14)およびプライマー9,10(配列番号15,16)をそれぞれ用いてPCR増幅を行った。PCR反応は、ポリメラーゼとしてPrimeSTAR GXL DNA Polymerase(タカラバイオ社製)、鋳型DNA(50ng)、プライマー(各0.2μM)のそれぞれを混合して滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。PCR反応は98℃で10秒、68℃で60秒のサイクルを30回繰り返した。増幅産物の精製はDNA精製キット(GEヘルスケア社製)を用いた。
【0078】
5'- ATCTCGATCCCGCGAAATTAATAC -3' (プライマー7:配列番号13)
5'- GATTTCGTGGCCCAGGACCAGGCCG -3' (プライマー8:配列番号14)
5'- TCCGGATATAGTTCCTCCTTTCAG -3' (プライマー9:配列番号15)
5'- AAGCTGCCTGAGCGCGACAAGGCCA -3' (プライマー10:配列番号16)
【0079】
得られた3つのDNA断片を鋳型にして、プライマー7,10を用いてPCR増幅を行った後、さらに反応終了液に含まれるPCR産物を鋳型にしてプライマー11,12(配列番号17,18)を用いてPCR増幅を行った。PCR反応は、ポリメラーゼとしてPrimeSTAR GXL DNA Polymerase(タカラバイオ社製)、鋳型DNA(50ng)、プライマー(各0.2μM)のそれぞれを混合して滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。PCR反応は98℃で10秒、68℃で60秒のサイクルを30回繰り返した。PCR産物の精製には、DNA精製キット(GEヘルスケア社製)を用いた。
次に、得られた酵素遺伝子をコードするDNA断片に、1分子PCR用プライマー配列を付加するために、プライマー13(配列番号19)を用いたPCR増幅を行った。PCR反応は、ポリメラーゼとしてPrimeSTAR GXL DNA Polymerase(タカラバイオ社製)、プライマー(各0.3μM)のそれぞれを混合して滅菌蒸留水で50μLにメスアップすることにより反応液を調製し、製造業者の指示に従って行った。PCR反応は94℃で30秒、55℃で30秒、72℃で60秒のサイクルを20回繰り返した。PCR産物の精製には、DNA精製キット(GEヘルスケア社製)を用いた。以上の実施した内容の模式図を図1,2に示す。
【0080】
5'-GGAGCTAGGGTTTACGAGTGGAATG-ATCTCGATCCCGCGAAATTAATAC-3'(プライマー11,配列番号17)
5'-GGAGCTAGGGTTTACGAGTGGAATG-TCCGGATATAGTTCCTCCTTTCAG-3'(プライマー12,配列番号18)
5'-GGAGCTAGGGTTTACGAGTGGAATG-3' (プライマー13,配列番号19)
【0081】
以上により、FDHをコードする遺伝子のコーディング領域中のホットスポット(アミノ酸領域Thr70−Leu150)に、変異を導入した変異遺伝子ライブラリーを作製した。変異遺伝子ライブラリーを、デキストランを含む希釈液で、1.5分子/μLに希釈し、384プレートに分注した。その後、1μL希釈液/1反応ウェルでPCR増幅した。変異遺伝子ライブラリーの希釈倍率算出に使用した計算式を次に示す。
【0082】
DNAの長さ:1400bp
分子量:0.924×106
1μg DNA:1.08×10-6μmol=1.08×10-12mol
1μg DNA:6.02×1023×1.08×10-12=6.5×1011分子に相当
【0083】
この計算から、変異遺伝子(DNA)のゲル切り出し精製したDNA溶液は、9.1×109分子/μLと算出でき、このDNA溶液を3×109希釈したものを、次のPCR反応の鋳型DNAに使用した。PCR反応温度条件は、98℃で10秒、68℃で1分のサイクルを70回繰り返した後、68℃で10分の伸長反応を行なった。
【0084】
(ステップ4) 無細胞タンパク質合成システムを用いた酵素合成とスクリーニング
ラビットトランスレーションシステムRTS100,E.coli HYキット(ロシュ社製)を使って取扱説明書に従い、反応時間は4時間、反応温度は30℃を基本条件として改変FDHを合成した。
変異遺伝子ライブラリーを鋳型にして、大腸菌の無細胞タンパク質合成系でタンパク質を合成し、上述の条件を用いてホルムアルデヒドに対する酵素活性についてスクリーニングし、熱処理後の残存活性が野生型FDHより向上した酵素活性を発現する改変FDHの候補遺伝子を選択した。
【0085】
酵素の活性測定は、8μLの酵素合成終了液に、32μLのリン酸緩衝液を混和して40μLとし、分注装置を使って16μLを新しい384ウェルプレート2枚に移した。2枚のプレートのうち、1枚を熱処理(15分、55℃)した後、氷中に15分間放置し、その後、80μLの活性測定反応液(50mMリン酸緩衝液pH7.4、40mM NAD、0.06%HCHO)を加えて、NAD+からNADH変化に伴う340nmの吸光度変化をプレートリーダで測定した。
【0086】
(ステップ5) スクリーニングで選択したクローンの解析
熱処理後も残存する酵素活性が認められた酵素の合成に使用した変異遺伝子ライブラリーについて、塩基配列を解析してアミノ酸変異を同定し、それらのタンパク質合成と酵素活性の有無も確認した。
タンパク質合成は、無細胞タンパク質合成系ラビットトランスレーションシステムRTS100,E.coli HYキット(ロシュ社製)を用いて、その取り扱い説明書に従って行なった。酵素の有無は、FluoroTect(登録商標) GreenLys in vitro Translation Labeling System(プロメガ社製)を用いて、その取り扱い説明書に従って合成酵素を蛍光標識して確認した。
【0087】
酵素活性の測定は、ホルムアルデヒド脱水素酵素によるホルムアルデヒド分解活性を、NADHの340nmの吸光を指標として行なった。80μLの活性測定反応液(50mMリン酸緩衝液K1K2PO4 pH7.4、1mM NAD+、2mM HCHO)に酵素溶液を加えて、NADからNADH変化に伴う340nmの吸光度変化を5分間(10秒間隔)プレートリーダで測定した。結果を表1に示す。
【0088】
【表1】
【0089】
表中における「アミノ酸変異部位」において、例えば「E76V」との表記は、野生型FDHのアミノ酸配列である配列番号2における第76番目のグルタミン酸(E)がバリン(V)に置換されたことを表す。
本実施例でアミノ酸変異が認められた部位は、クローンP16では第76位、第81位、第219位であり、クローンF12では第127位、第142位であり、クローンK11では第97位、第148位であり、クローンN20では第129位であり、クローンP13では第149位であり、クローンO6,P4では第128位であり、クローンH1では第110位であり、クローンK3では第94位であった。尚、本明細書においては、アミノ酸の位置は、開始コドンであるメチオニンを1として番号付けしている。
【0090】
この中で、タンパク質合成と酵素活性が確認できたものは、クローンO6,P4(第128番目のアスパラギン酸(D)がグリシン(G)に置換されている)と、クローンH1(第110番目のグリシン(G)がアスパラギン酸(D)に置換されている)であった。
【0091】
表1の結果より、アミノ酸変異D128GおよびG110D以外では、変異が1個入っただけの改変で酵素活性が全くなくなるものと認められた。つまり、アミノ酸変異が1個ではタンパク質の構造学的には、10Å程度の距離にしか構造変化の影響を与えないとされているが、このPseudomonas putida由来のFDHは、その微小な構造変化でも酵素の構造的な安定性に影響を与えてしまうデリケートで複雑な構造をしている酵素であるといえる。
その中でも特に、V127DはD128Gの隣のアミノ酸変異であるが、当該V127Dでは酵素活性がなくなるのに対してD128Gでは酵素活性を有していることから、D128GがPseudomonas putida由来のFDHの安定性に重要であると推察されると同時に、従来法での酵素改変は非常に困難であることが推測できる。
【0092】
以下、第128番目のアスパラギン酸がグリシンに改変されたアミノ酸配列を有する改変FDHを「D128G」、第110番目のグリシンがアスパラギン酸に改変されたアミノ酸配列を有する改変FDH「G110D」と称する。
【0093】
<実施例2>改変型FDHの立体構造シミュレーションに基づく高次構造の解析
D128GおよびG110Dの立体構造モデルの構築を行なった。モデリングは、アクセルリス社製Discovery Studio21を用いて、X線結晶解析データ(PDB;1KOL)を参照構造として、ホモロジーモデリングにより実施した。
【0094】
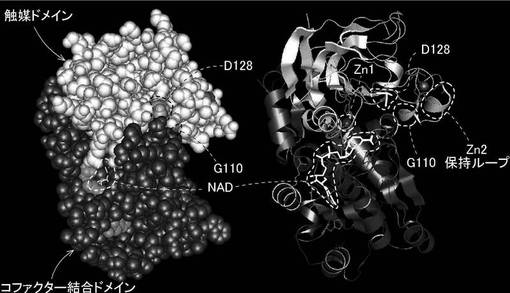
まず、野生型FDH(PDB;1KOL)を用いて変異部位の解析を行なった(図3)。
触媒ドメインとコファクター結合ドメインは、Zn1近傍で上下に分かれる。NADとZn1の相互作用は、両ドメインの位置・構造の安定性に関係していると考えられる。D128(第128位のアスパラギン酸)およびG110(第110位のグリシン)は、両ドメインの界面に位置する。特に、G110はZn2保持ループの根元付近に位置する。Zn2保持ループ周辺は両ドメインの分岐から遠く、パッキングが不十分もしくは安定性が低い可能性が考えられる。
【0095】
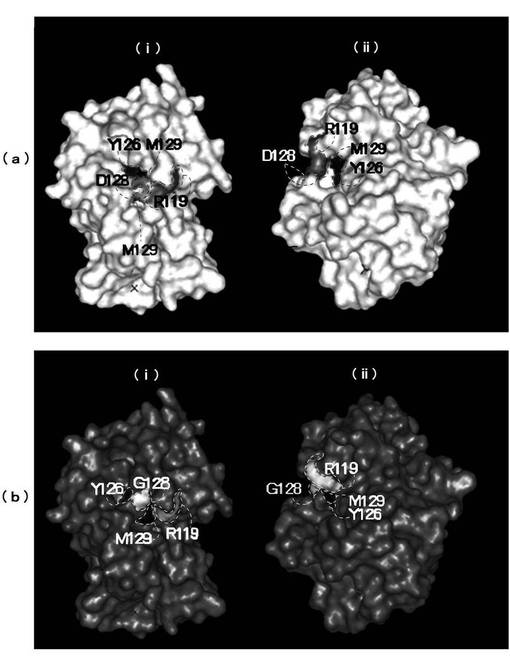
D128Gの構造モデルの解析結果は以下の通りである(図4)。
図4(a)には野生型FDH、図4(b)にはD128Gの構造モデルを示した。モデルは分子内の原子に接触する溶媒の接触点によってトレースされるサーフェイス形状で表示した。図4(a)(ii),図4(b)(ii)は、それぞれ図4(a)(i),図4(b)(i)のモデルを90℃回転させたモデルを示した。
野生型FDHではD128とR119(第119位のアルギニン)は静電作用で側鎖が並ぶように位置し、その内側のアミノ酸残基Y126(第126位のチロシン)およびM129(第129位のメチオニン)の間には空間が存在していた。D128G変異体では、D128とR119の側鎖は野生型とは反対方向を向いており、その内側のアミノ酸残基(Y126とM129)の側鎖が内部に向いたことにより、D128とR119も内側にコンパクトにパッキングされたと推測される。余分なキャビティが無くなったことが安定性向上に寄与しているのではないかと推測された。
【0096】
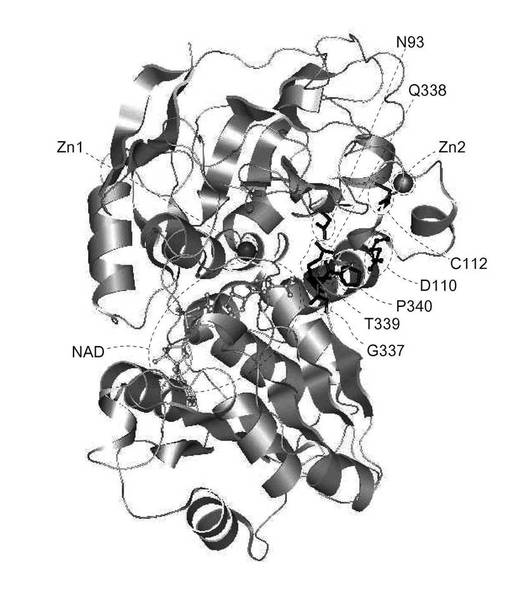
G110Dの構造モデルの解析結果は以下の通りである(図5)。
野生型FDHとG110Dの主鎖の重ね合わせを行った。
G337(第337位のグリシン),Q338(第338位のグルタミン),T339(第339位のトレオニン),P340(第340位のプロリン)は、NADと相互作用するアミノ酸残基、N93(第93位のアスパラギン)はNADと相互作用するアミノ酸残基の隣の残基、C112(第112位のシステイン)はZn2保持アミノ酸残基であったことから、側鎖の向きの変化により、酵素の安定性に何らかの寄与をしたと推測された。
【0097】
<実施例3>組換え大腸菌によるタンパク質の発現と精製
タンパク質発現は、野生型または改変型FDHをコードする遺伝子を、大腸菌用の発現ベクターであるpET22b DNAのマルチクローニングサイトにC末端Hisタグ融合タンパク質として発現するように挿入し、構築されたプラスミドを大腸菌BL21(DE3)に形質転換した。大腸菌の培養は、アンピシリン(50mg/L)入りのLuria−Bertani(LB)培地中を使って37℃で行なった。形質転換した大腸菌は、アンピシリン(50mg/L)を含むLB培地で培養し、培養液の吸光度OD600=0.6で0.5mM IPTG(isopropyl-s-D-thiogalactopyranoside)を添加してタンパク質の合成誘導を行ない、さらに4時間の培養を行なった。
【0098】
発現させたHisタグ融合タンパク質は、アフィニティーカラムを用いて精製した。具体的には、遠心分離により回収した菌体を、リン酸緩衝液(20mM リン酸バッファー、0.5M NaCl、pH7.4)に懸濁した。菌体を超音波で破砕した。破砕は、超音波処理30秒、氷冷30秒のセットを10回繰り返して行なった。遠心分離により得られた上清50mM NaClを加え、酵素液をNi2+カラムクロマトグラフィー通液し、300mMイミダゾールを含む溶液で融合タンパク質を溶出して、活性画分をまとめ、50mMリン酸緩衝液(pH7.4)を透析外液として一晩透析を行った。
得られた酵素溶液を、50mM Tris-HCl緩衝液(pH7.4)にて透析した。タンパク質の確認は、20μLの酵素溶液に20μLの可溶化液を加えて3分間95℃で熱処理したサンプルを、12.5%アクリルアミドゲルで電気泳動し、CBB染色法で酵素を可視化することにより行った。その結果を図6に示す。レーン1は精製した改変型FDH D128G、レーン2は精製した野生型FDHである。
【0099】
この結果、D128Gは、野生型FDHと同様に約50kDaのバンドが認められたため、大腸菌用の発現ベクターで発現することが確認できた。
【0100】
<実施例4>酵素の諸性質の確認(pH依存性)
酵素活性をpH5.0〜10.0の範囲で測定した。50mM緩衝液、1mM NAD+、2mM HCHOを含む活性測定液中に等量の酵素液(0.5mg/mL)を加え、25℃でpH8.0の酵素活性の反応速度を1とし、各pHにおける相対活性の変化を比較した結果を図7に示す。
この結果から、D128Gおよび野生型FDHは、pH5.0からpH8.0までは同じpH特性を示し、至適pHはpH8.0付近であることが判明した。一方、pH8.0〜pH10.0では、D128Gより野生型FDHの方が、活性低下が小さいことが判明した。また、G110Dについても、D128Gと同様のpH依存性を示すことが判明した(結果は示さない)。尚、図7においてpH領域A(8以下)についてはリン酸緩衝液を使用し、pH領域B(8以上)についてはTris緩衝液を使用した。
【0101】
<実施例5>酵素の諸性質の確認(温度依存性)
酵素活性を45〜60℃の温度範囲で測定した。50mMリン酸ナトリウム緩衝液(pH7.6)、1mM NAD+、2mM HCHOを含む活性測定液中に等量の酵素液(0.5mg/mL)を加え、45℃での酵素活性の反応速度を1とし、各温度における相対活性の変化を比較した結果を図8に示す。
図8(a)はD128Gおよび野生型FDHの比較を、また、図8(b)はG110Dおよび野生型FDHの比較を示す。この結果から、50〜55℃の温度域において、野生型FDHより改変型FDHの方が2倍程度高い活性を有することが判明した。また、G110DとD128Gは同様の温度依存性を示すことが判明した。
【0102】
<実施例6>酵素の諸性質の確認(温度安定性)
酵素の温度安定性を調べるために、55〜65℃の温度範囲で熱処理後の残存活性を調べた。50mMリン酸ナトリウム緩衝液(pH7.6)中に等量の酵素を加え、55℃、60℃、65℃の各温度で0分〜10分間熱処理し、氷中に30分間置いた後の残存活性を測定した。
50mMリン酸ナトリウム緩衝液(pH7.6)、1mM NAD+、2mM HCHOを含む活性測定液中に熱処理した酵素液(0.5mg/mL)を加え、各熱処理0分時の結果を1とし、各温度と各熱処理時間における相対活性の変化の比較を図9,10に示す。図9はD128Gおよび野生型FDHの比較を、図10はG110Dおよび野生型FDHの比較を示す。
【0103】
この結果、55℃の熱処理では、6分間程度の熱処理において野生型FDHでは相対活性が1/10程度になるのに対し、D128Gでは約50%(図9(a))、G110Dでは約40%(図10(a))の相対活性を有することが判明した。
一方、60℃以上の熱処理では、改変型FDHおよび野生型FDHの何れにおいても残存活性が5分程度で20%以下になり(図9(b),(c)、図10(b),(c))、熱変性により酵素活性を殆ど失うものと認められた。
これより、本発明の改変型FDHは、野生型FDHに比べて熱によって失活し難いものと認められた。
【0104】
<実施例7>酵素の諸性質の確認(安定性)
酵素の安定性を調べるために、50℃と45℃の温度下で300分間までの残存活性を調べた。50mMリン酸ナトリウム緩衝液(pH7.6)中に等量の酵素液を加え、0分から300分間置いた後、氷中に30分間置いた後の残存活性を確認した。
50mMリン酸ナトリウム緩衝液(pH7.6)、1mM NAD+、2mM HCHOを含む活性測定液中に熱処理した酵素液(0.5mg/mL)を加え、0分時の酵素活性の反応速度を1としたときの各時間(分)における相対活性の変化の比較を図11に示す。図11(a)は50℃の温度下におけるD128Gおよび野生型FDHの比較を、また、図11(b)は45℃の温度下におけるG110Dおよび野生型FDHの比較を示す。
この結果、50℃での酵素活性の半減期は、野生型FDHでは30分間以内であるのに対し、D128Gでは300分以上であった。また、45℃での酵素活性の半減期は、野生型FDHでは約200分であるのに対し、D128Gでは300分以上であった。
これより、本発明の改変型FDHは、野生型FDHに比べて熱安定性に優れているものと認められた。
【0105】
D128Gについて常温時(25℃,30℃)の安定性を調べた。
50mM P1P2PO4、1mM HCHO、0.5mM NADの組成を有する反応液を、D128Gを担持したNi-NTA His・Bind Resinカラム(タカラバイオ社製)、および、野生型FDHを担持したNi-NTA His・Bind Resinカラムにそれぞれ通液した。反応温度は25℃とし、通液速度は0.1mM/分(線流速7.9cm/h)とした。当該カラムを通過した反応済液の340nmの吸光度変化を測定し、通液前の酵素活性の反応速度を1としたときの相対活性の変化の比較を図12に示す。反応温度を30℃とした場合も同様の実験を行なった。
この結果、反応温度を25℃とした場合、野生型FDHの酵素活性の半減期は約500時間程度であるのに対して、D128Gでは約800時間程度であった(図12)。また、反応温度を30℃とした場合、野生型FDHの酵素活性が80%になるのは約175時間程度であるのに対して、D128Gでは約450時間程度であった(図13)。
これより、本発明の改変型FDHは、野生型FDHに比べて長期に亘って安定性に優れているものと認められた。
【0106】
<実施例8>酵素の諸性質の確認(反応速度論解析)
変異型酵素と野生型酵素について酵素反応速度論的な解析を行った。
50mMリン酸ナトリウム緩衝液(pH7.6)、1mM NAD+、0〜4mM HCHOを含む活性測定液中に酵素液(0.5mg/mL)を加え、酵素反応の初期速度のラインウェーバーバークプロットにより、酵素反応の速度定数(Kcat値及びKm値)を求めた。その結果を表2に示す。
【0107】
【表2】
【0108】
この結果、野生型FDHの25℃、pH7.6におけるHCHOに対するKm値は0.35mM、Kcat値は3.2s-1であり、Kcat/Kmは9.1M-1s-1であった。また、D128Gの25℃、pH7.6におけるHCHOに対するKm値は0.35mM、Kcat値は3.2s-1であり、Kcat/Kmは9.1M-1s-1、G110Dの25℃、pH7.6におけるHCHOに対するKm値は0.18mM、Kcat値は2.5s-1であり、Kcat/Kmは13.9M-1s-1であった。
以上より、G110DのKm値が小さく基質親和性が高いこと以外は、本発明の改変型FDHは野生型FDHとほぼ同様であることが判明した。
【0109】
<実施例9>酵素の諸性質の確認(基質選択性)
基質としてアルデヒド類とアルコール類の選択性について調べた。
50mMリン酸ナトリウム緩衝液(pH7.6)、1mM NAD+、2mM各基質を含む活性測定液中に等量の酵素液(0.5mg/mL)を加え、各アルデヒド類とアルコール類を基質としたときの酵素活性の反応速度を1としたときの相対活性について、野生型FDHおよびD128Gを比較したものを表3に示す。
【0110】
【表3】
【0111】
この結果から、D128Gは野生型FDHと同様に、ホルムアルデヒドに対して基質特異性が高いものと認められた。
【産業上の利用可能性】
【0112】
本発明は、改変型ホルムアルデヒド脱水素酵素及びその利用方法に関し、医療、食品、環境分野等の様々な産業分野において利用可能である。
【特許請求の範囲】
【請求項1】
野生型ホルムアルデヒド脱水素酵素のアミノ酸配列中の少なくとも1つの位置のアミノ酸の欠失、置換、付加或いは挿入による改変により得られる改変型ホルムアルデヒド脱水素酵素であって、
前記改変が、前記野生型ホルムアルデヒド脱水素酵素のアミノ酸配列における第70位−第150位で生じており、前記野生型ホルムアルデヒド脱水素酵素に比べて安定性が向上している改変型ホルムアルデヒド脱水素酵素。
【請求項2】
前記改変が、前記アミノ酸配列の第128位に対応する位置で生じている請求項1に記載の改変型ホルムアルデヒド脱水素酵素。
【請求項3】
前記改変が、前記アミノ酸配列の第128位に対応する位置におけるアスパラギン酸のグリシンへの置換である請求項2に記載の改変型ホルムアルデヒド脱水素酵素。
【請求項4】
前記改変が、前記アミノ酸配列の第110位に対応する位置で生じている請求項1に記載の改変型ホルムアルデヒド脱水素酵素。
【請求項5】
前記改変が、前記アミノ酸配列の第110位に対応する位置におけるグリシンのアスパラギン酸への置換である請求項4に記載の改変型ホルムアルデヒド脱水素酵素。
【請求項6】
前記野生型ホルムアルデヒド脱水素酵素が、配列番号2に示すアミノ酸配列を有する請求項1〜5の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素。
【請求項7】
請求項1〜6の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素をコードする単離核酸分子。
【請求項8】
請求項7に記載の単離核酸分子を含有する組換えベクター。
【請求項9】
請求項8に記載の組換えベクターを含有する形質転換体。
【請求項10】
請求項9に記載の形質転換体を培養する工程、及び得られた培養物からホルムアルデヒドの脱水素反応を触媒する能力を有するタンパク質を採取する工程を含む、野生型ホルムアルデヒド脱水素酵素に比べて熱安定性が向上している改変型ホルムアルデヒド脱水素酵素の製造方法。
【請求項11】
請求項1〜6の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素の触媒活性を利用してアルデヒド化合物を検出するアルデヒド化合物の検出方法。
【請求項12】
請求項1〜6の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素を電極上に固定化した酵素センサー。
【請求項13】
請求項1〜6の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素を含み、前記改変型ホルムアルデヒド脱水素酵素のアルデヒド化合物の酸化反応に伴って生成する電子を受け取るアノード極、酸素に電子を伝達することのできる触媒および酵素のいずれかを保持するカソード極を備え、前記アノード極と前記カソード極とが電気的に結合されている燃料電池。
【請求項1】
野生型ホルムアルデヒド脱水素酵素のアミノ酸配列中の少なくとも1つの位置のアミノ酸の欠失、置換、付加或いは挿入による改変により得られる改変型ホルムアルデヒド脱水素酵素であって、
前記改変が、前記野生型ホルムアルデヒド脱水素酵素のアミノ酸配列における第70位−第150位で生じており、前記野生型ホルムアルデヒド脱水素酵素に比べて安定性が向上している改変型ホルムアルデヒド脱水素酵素。
【請求項2】
前記改変が、前記アミノ酸配列の第128位に対応する位置で生じている請求項1に記載の改変型ホルムアルデヒド脱水素酵素。
【請求項3】
前記改変が、前記アミノ酸配列の第128位に対応する位置におけるアスパラギン酸のグリシンへの置換である請求項2に記載の改変型ホルムアルデヒド脱水素酵素。
【請求項4】
前記改変が、前記アミノ酸配列の第110位に対応する位置で生じている請求項1に記載の改変型ホルムアルデヒド脱水素酵素。
【請求項5】
前記改変が、前記アミノ酸配列の第110位に対応する位置におけるグリシンのアスパラギン酸への置換である請求項4に記載の改変型ホルムアルデヒド脱水素酵素。
【請求項6】
前記野生型ホルムアルデヒド脱水素酵素が、配列番号2に示すアミノ酸配列を有する請求項1〜5の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素。
【請求項7】
請求項1〜6の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素をコードする単離核酸分子。
【請求項8】
請求項7に記載の単離核酸分子を含有する組換えベクター。
【請求項9】
請求項8に記載の組換えベクターを含有する形質転換体。
【請求項10】
請求項9に記載の形質転換体を培養する工程、及び得られた培養物からホルムアルデヒドの脱水素反応を触媒する能力を有するタンパク質を採取する工程を含む、野生型ホルムアルデヒド脱水素酵素に比べて熱安定性が向上している改変型ホルムアルデヒド脱水素酵素の製造方法。
【請求項11】
請求項1〜6の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素の触媒活性を利用してアルデヒド化合物を検出するアルデヒド化合物の検出方法。
【請求項12】
請求項1〜6の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素を電極上に固定化した酵素センサー。
【請求項13】
請求項1〜6の何れか一項に記載の改変型ホルムアルデヒド脱水素酵素を含み、前記改変型ホルムアルデヒド脱水素酵素のアルデヒド化合物の酸化反応に伴って生成する電子を受け取るアノード極、酸素に電子を伝達することのできる触媒および酵素のいずれかを保持するカソード極を備え、前記アノード極と前記カソード極とが電気的に結合されている燃料電池。
【図1】

【図2】

【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公開番号】特開2011−67137(P2011−67137A)
【公開日】平成23年4月7日(2011.4.7)
【国際特許分類】
【出願番号】特願2009−220970(P2009−220970)
【出願日】平成21年9月25日(2009.9.25)
【出願人】(000000011)アイシン精機株式会社 (5,421)
【出願人】(000003609)株式会社豊田中央研究所 (4,200)
【Fターム(参考)】
【公開日】平成23年4月7日(2011.4.7)
【国際特許分類】
【出願日】平成21年9月25日(2009.9.25)
【出願人】(000000011)アイシン精機株式会社 (5,421)
【出願人】(000003609)株式会社豊田中央研究所 (4,200)
【Fターム(参考)】
[ Back to top ]