
改変型TGF−βスーパーファミリータンパク質
【課題】組換えBMPおよび他のTGF−βファミリータンパク質を、原核生物宿主および真核生物宿主を使用してインビトロで産生するための改善された手段の提供。
【解決手段】N末端短縮化TGF−βファミリータンパク質を含む、適切なリフォールディング条件下でリフォールドさせるための、潜在性TGF−βファミリーメンバー融合タンパク質の成分であって、該短縮化TGF−βファミリータンパク質は、フィンガー1サブドメイン、フィンガー2サブドメイン、およびヒールサブドメインを含む、TGF−βファミリータンパク質C末端7システインドメイン;および該C末端ドメインに作動可能に連結された切断可能な改変型リーダー配列であって、該リーダー配列は、該C末端ドメインと関連する生物学的活性を阻害し、該C末端ドメインは、該リーダー配列の一部または全ての切断の際に活性化する、リーダー配列を含む、融合タンパク質。
【解決手段】N末端短縮化TGF−βファミリータンパク質を含む、適切なリフォールディング条件下でリフォールドさせるための、潜在性TGF−βファミリーメンバー融合タンパク質の成分であって、該短縮化TGF−βファミリータンパク質は、フィンガー1サブドメイン、フィンガー2サブドメイン、およびヒールサブドメインを含む、TGF−βファミリータンパク質C末端7システインドメイン;および該C末端ドメインに作動可能に連結された切断可能な改変型リーダー配列であって、該リーダー配列は、該C末端ドメインと関連する生物学的活性を阻害し、該C末端ドメインは、該リーダー配列の一部または全ての切断の際に活性化する、リーダー配列を含む、融合タンパク質。
【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、改良されたリフォールディング特性、改良された物理的特性(例えば、溶解性および安定性)、改良された生物学的活性(改変されたレセプター結合、改良された標的化能力を含む)を有する組換えタンパク質、潜在性形態のタンパク質、およびこのようなタンパク質を生成する方法に関する。より詳細には、本発明は、TGF−βスーパーファミリーの構造的に関連したタンパク質の生合成メンバーに関する。このような改変型タンパク質構築物は、N末端短縮、「潜在性」タンパク質、融合タンパク質およびヘテロ二量体を有するTGF−βファミリーメンバータンパク質を含む。
【背景技術】
【0002】
(発明の背景)
TGF−βスーパーファミリーは、TGF−βの5個の異なる形態(非特許文献1)、ならびに分化因子Vg−1(非特許文献2)、DPP−Cポリペプチド(非特許文献3)、ホルモンアクチビンおよびインヒビン(inhibin)(非特許文献4;非特許文献5)、Mullerian阻害物質、MIS(非特許文献6)、骨形成タンパク質および形態形成タンパク質OP−1(PCT/US90/05903)、OP−2(PCT/US91/07654)、OP−3(PCT/WO94/10202)、BMP(米国特許第4,877,864号;同第5,141,905号;同第5,013,649号;同第5,116,738号;同第5,108,922号;同第5,106,748号;および同第5,155,058号を参照のこと)、発生的に調節されるタンパク質VGR−1(非特許文献7)、軟骨由来成長因子CDMP−1、CDMP−2およびCDMP−3(またはGDF−5、GDF−6およびGDF−7)、ならびに成長/分化因子GDF−1、GDF−3、GDF−9、およびドーサリン(dorsalin)−1(非特許文献8;非特許文献9)を含む。
【0003】
TGF−βスーパーファミリーのタンパク質は、ジスルフィド結合されたホモ−またはヘテロ二量体である。これらは、疎水性のシグナル配列、長くそして比較的保存されていないN末端の数100アミノ酸のプロ領域配列、切断部位、ならびにファミリーのメンバーの間で変化するN末端領域、および高度に保存されているC末端領域を含む成熟ドメイン、を含む大きな前駆体ポリペプチド鎖として発現される。全ての既知のファミリーのメンバーのプロセシングされた成熟タンパク質中に存在するこのC末端領域は、約100個のアミノ酸を含み、保存されている6個または7個のシステイン骨格を有する特徴的なシステインモチーフを有する。成熟領域とプロ領域との間では切断部位の位置は、ファミリーのメンバーの間で変化するが、全てのタンパク質のC末端のシステインパターンは同一の形式であり、配列Cys−X−Cys−Xで終わる(非特許文献1)。
【0004】
組換えTGF−β1は、クローン化(非特許文献10)、およびチャイニーズハムスター卵巣細胞において発現される(非特許文献11)。さらに、組換えヒトTGF−β2(非特許文献12)、ならびにヒトおよびブタのTGF−β3(非特許文献13;非特許文献14)が、クローン化されている。COS細胞における成熟TGF−β1タンパク質の発現レベルは、セリン残基を有するTGF−β1前駆体のプロ領域に局在したシステイン残基を置換することによって、増加された(非特許文献15)。
【0005】
TGF−βスーパーファミリーのタンパク質の生物学の統一の特徴は、発達のプロセスを調節するそれらの能力である。これらの構造的に関連するタンパク質は、種々の発達的な事象に関与することが同定されている。例えば、TGF−βおよびインヒビン/アクチビンの群のポリペプチドは、細胞の増殖および分化の調節において役割を果たすようである。MISは、哺乳動物の雄性の胚の発達においてミュラー管の退行を生じ、そしてDrosophiaのデカペンタプレジック(decapentaplegic)複合体の遺伝子産物であるdppは、適切な背面−腹面(dorsal−ventral)の特異化に必要である。同様に、Vg−1は、Xenopusにおける中胚葉の誘導に関連し、そしてVgr−1は、発達中のマウスの種々の組織において同定されている。骨形成に関しては、TGF−βスーパーファミリー中の多くのタンパク質(すなわち、OP−1およびBMPのサブセット)が、明らかに主要な役割を果たす。OP−1(BMP−7)および他の骨形成タンパク質は、組換え技術を使用して産生されており(米国特許第5,011,691号およびPCT出願番号第US90/05903号)、そしてインビボで実際に軟骨内の骨の形成を誘導することが可能であることが示されている。BMP−2は、サルCOS−1細胞およびチャイニーズハムスター卵巣細胞において、組換え的に作製される(非特許文献16)。
【0006】
最近、SampathおよびReddi骨形成アッセイによって判定しタンパク質場合に、骨形成活性を有すると教示されたタンパク質のこのファミリーが、形態形成性である(すなわち、成熟した哺乳動物において組織の形態形成の発達のカスケードを誘導し得る)ことが示された(PCT出願番号第US92/01968号を参照のこと)。詳細には、これらのタンパク質は、関連付けられていない前駆細胞の増殖を誘導し得、そして適切な環境条件下で組織特異的様式でこれらの刺激された先祖細胞の分化を誘導し得る。さらに、モルフォゲンは、これらの分化した細胞の増殖および維持を補助し得る。これらの形態形成性の活性は、タンパク質が、適切な形態形成を許された環境において組織の形態形成の発達のカスケードを開始しそして維持すること、幹細胞が組織特異的様式で増殖および分化するように刺激すること、ならびに新しい組織の形成において完了する事象の進行を誘導することを可能にする。これらの形態形成活性はまた、タンパク質がそれらの分化の経路からはぐれるように予め刺激された細胞の「再分化」を誘導することを可能にする。適切な環境条件下では、これらのモルフォゲンもまた関連付けられた細胞の「再分化」を刺激し得ることが予想される。
【0007】
骨形成タンパク質は、一般にTGF−βスーパーファミリーの増殖因子の亜群として、当該分野で分類されている(非特許文献17)、そして「骨形成タンパク質」、「形態形成タンパク質」、「モルフォゲン」、「骨形態形成タンパク質」または「BMP」と様々に呼ばれ、それらのエピトープ性軟骨内性骨形態形成を誘導する能力により同定される。タンパク質のモルフォゲンファミリーのメンバーとしては、哺乳動物の骨形成タンパク質−1(OP−1(BMP−7、およびDrosophilaのホモログである60Aとしてもまた知られている))、骨形成タンパク質−2(OP−2(BMP−8としてもまた知られている))、骨形成タンパク質−3(OP−3)、BMP−2(BMP−2AまたはCBMP−2A、およびDorosphilaのホモログであるDPPとしてもまた知られている)、BMP−3、BMP−4(BMP−2BまたはCMBP−2Bとしてもまた知られている)、BMP−5、BMP−6およびそのマウスのホモログであるVgr−1、BMP−9、BMP−10、BMP−11、BMP−12、GDF3(Vgr2としてもまた知られている)、GDF−8、GDF−9、GDF−10、GDF−11、GDF−12、BMP−13、BMP−14、BMP−15、GDF−5(CDMP−1またはMP52としてもまた知られている)、GDF−6(CDMP−2またはBMP−13としてもまた知られている)、GDF−7(CDMP−3またはBMP−12としてもまた知られている)、XenopusのホモログであるVg1およびNODAL、UNIVIN、SCREW、ADMP、およびNEURALが挙げられる。
【0008】
天然に存在するか、または合成によって調製されたかにはかかわらず、骨形成タンパク質は、前駆細胞の補充および刺激を誘導し得、それによって、軟骨細胞および骨芽細胞へのそれらの分化を誘導し、そしてさらに、中間軟骨の分化、血管新生、骨の形成、再造形、および最終的には骨髄の分化を誘導する。さらに、多数の研究者らが、天然から供給されたマトリックス材料(例えば、コラーゲン)、または合成によって調製されたポリマーマトリックス材料のいずれかと混合した場合に、実際に置換骨が他に生じない条件下で、骨形成(膜性骨形成および軟骨内骨形成含む)を誘導する、骨形成タンパク質の能力を実証した。例えば、マトリックス材料と組み合わせた場合には、これらの骨形成タンパク質は、大きな断片の骨の欠失、脊椎の融合、頭蓋冠の欠損、および骨折において、新しい骨の形成を誘導する。
【0009】
細菌および他の原核生物の発現系は、組換えタンパク質を生成するための好ましい手段として当該分野であてにされている。原核生物(例えば、E.coli)の系は、商業的な量のタンパク質を産生するため、および天然に存在するかまたは生合成変異体およびアナログの生物学的特性および化学的特性を評価するために有用である。代表的には、過剰に発現される真核生物のタンパク質は、原核生物の宿主細胞中で不溶性の細胞内沈殿物(「封入体」)に凝集する。次いで、凝集したタンパク質は封入体から回収され、1つ以上の標準的な変性剤を使用して可溶化され、次いで機能的な状態にリフォールディングされることを可能にされるかまたはそのように誘導される。生物学的に活性なタンパク質構造を形成するために適切なリホールディングは、任意のジスルフィド結合の適切な形成を必要とする。
【0010】
化学合成はまた、タンパク質構築物の産生に用いられ得る。ペプチド鎖の単調な自動化アセンブリを可能にするための技術は広範に利用可能である。合成タンパク質分子にペプチドフラグメントを結合するために酵素法およびキメラ法を利用する技術は、当該分野で公知である。非特許文献18;非特許文献19;非特許文献20;非特許文献21;および非特許文献22を参照のこと。
【0011】
例えば、TGF−β2およびOP−1の両方の三次構造および四次構造が決定されている。TGF−β2およびOP−1は、それらのそれぞれのアミノ酸配列においてわずかに約35%のアミノ酸同一性しか示さないが、両方の分子の三次構造および四次構造は極めて類似している。図1Aに示すように、TGF−β2およびOP−1の両方ともが天然においてダイマーであり、そして7個のC末端のシステイン残基のうちの6個を含む特有のフォールディングパターンを有する。各サブユニットにおいては、4個のシステインが、8個の残基の環を生じるように結合し、そして2個のさらなるシステイン残基が、結び目様の構造を形成するように環を通って通過するジスルフィド結合を形成することが図1Aに示される。番号1に割り当てられた7個の保存性システイン残基の最もN末端側のシステインで開始する番号付けスキームを用いると、2番目と6番目の保存されたシステイン残基は、8個の残基の環の1つの側面を閉じるように結合し、一方、3番目と7番目のシステイン残基は、他の側面を閉じるように結合する。1番目と5番目の保存性システイン残基は、結び目の中心を形成するように、環の中心を通じて結合する。4番目の保存性システインは、他のサブユニット中の対応する残基とともに鎖間ジスルフィド結合を形成する。
【0012】
TGF−β2およびOP−1のモノマーサブユニットは、3つの主要な構造エレメントおよびN末端領域を含む。構造エレメントは、以下の型:(1)ループ、(2)α−へリックス、および(3)β−シートの50%を超える二次構造を保有している、連続しているポリペプチド鎖の領域を構成する。さらに、これらの領域においては、N末端およびC末端の鎖は、7Åを超えては離れていない。保存された1番目のシステインと2番目のシステインとの間の残基(図1A)は、逆平行β−シートフィンガー(本明細書中では、フィンガー1領域(F1)と呼ばれる)によって特徴付けられる構造領域を形成する。フィンガー1ペプチド骨格のリボントレースは、図1Bに示される。同様に、図1Aの保存された5番目のシステインと6番目のシステインとの間の残基もまた、逆平行β−シートフィンガー(本明細書中では、フィンガー2領域(F2)と呼ばれる)を形成する。フィンガーペプチド骨格のリボントレースは、図1Dに示される。β−シートフィンガーは、一本のアミノ酸の鎖であり、これは、1つ以上の逆平行β−シート構造を形成する鎖が入りそして存在するように、β−ターンまたはいくらか大きいループによってそれ自体の上に折れ曲がっているβ鎖を含む。保存された3番目のシステインと4番目のシステインとの間の残基を含む、第3の主要な構造領域は、本明細書中ではヒール領域(H)と呼ばれる3個のターンα−へリックスによって特徴付けられる。ヒールペプチド骨格のリボントレースは、図1Cに示される。
【0013】
このモノマー構造の組織化は、結び目領域が手掌に等しい位置に位置付けされ、フィンガー1がインデックスフィンガーおよびミドルフィンガーに等しく、α−ヘリックスは手のヒールに等しく、そしてフィンガー2が環状フィンガーおよび小フィンガーに等しい、左側の組織化と類似する。N末端領域は(公開された構造において十分規定されていない)は、親指(thumb)に概略的に等しい位置に位置すると予想される。
【0014】
TGF−β2およびOP−1の両方のダイマーの形態においては、サブユニットは、一方のサブユニットのヒール領域が、分子のコアを形成する連結されたサブユニットの結び目の領域を有する他のサブユニットのフィンガー領域と接触するように配向される。4番目のシステインは、第2鎖上のその対応物とジスルフィド結合を形成し、それによって手掌の中心で鎖を等しく連結する。このように形成されたダイマーは、サブユニット間の対称な二倍軸を上から下に見た場合には、楕円形の(葉巻型)の分子である(図2A)。その側から見ると、この分子は湾曲した「葉巻」に似ている。なぜなら、2つのサブユニットが、互いに関してわずかな角度で配向されるからである。
【0015】
しかし、必ずしも全ての可溶化異種タンパク質が、容易にリフォールディングするわけではない。リフォールディングの注意深い操作にもかかわらず、適切にフォールディングされた、生物学的に活性なタンパク質の収量は低いままである。多くのTBF−βファミリーメンバー(BMPを含む)は、少ないリフォールダー(refolder)タンパク質のカテゴリーに入る。TGF−βタンパク質ファミリーのいくつかのメンバーはインビトロで、例えば、E.coliまたは他の原核生物宿主中で産生された場合に比較的効率良くフォールディングされ得るが、他の多く(BMP−5、BMP−6、およびBMP−7を含む)はそうではない。例えば、第EP0433225号、第US5,399,677号、同第5,756,308号、および同第5,804,416号を参照のこと。
【先行技術文献】
【非特許文献】
【0016】
【非特許文献1】SpornおよびRoberts(1990)、Peptide Growth Factors and Their Receptors、SpornおよびRoberts編、Springer−Verlag:Berlin、419−472頁
【非特許文献2】WeeksおよびMelton(1987)、Cell 51:861−867
【非特許文献3】Padgettら(1987)Nature 325:81−84
【非特許文献4】Masonら(1985)、Nature 318:659−663
【非特許文献5】Masonら(1987)、Growth Factors 1:77−88
【非特許文献6】Cateら(1986)、Cell 45:685−698
【非特許文献7】Lyonsら(1989)Proc.Natl.Acad.Sci.USA 86:4554−4558
【非特許文献8】McPherronら(1993)、J.Biol.Chem.268:3444−3449
【非特許文献9】Baslerら(1993)、Cell 73:687−702
【非特許文献10】Derynckら、(1985)Nature 316:701−705
【非特許文献11】Gentryら、(1987)Mol.Cell.Biol.7:3418−3427
【非特許文献12】deMartinら、(1987)EMBO J.6:3673
【非特許文献13】Derynckら、(1988)EMBO J.7:3737−3743
【非特許文献14】Dijkeら、(1988)Proc.Natl.Acad.Sci.USA 85:4715
【非特許文献15】Brunnerら、(1989)J.Biol.Chem.264:13660−13664
【非特許文献16】Wangら、(1990)Proc.Natl.Acad.Sci.USA 87:2220−2224
【非特許文献17】Hogan(1996),Genes&Development,10:1580−1594
【非特許文献18】Hilvert,Chem.Biol.(1994)1(4):201−03
【非特許文献19】Muriら、Proc.Nat’l Acad.Sci.USA(1998)95(12):6705−10
【非特許文献20】Wallace,Curr.Opin.Biotechnol.(1995)6(4):403−10
【非特許文献21】Mirandaら、Proc.Nat’l Acad.Sci.USA(1999)96(4):1181−6
【非特許文献22】Liuら、Proc.Nat’l Acad.Sci.USA(1994)91(14):6584−8
【発明の概要】
【発明が解決しようとする課題】
【0017】
組換えBMPおよび他のTGF−βファミリータンパク質を、原核生物宿主および真核生物宿主を使用してインビトロで産生するための改善された手段の必要性が、残っている。
【課題を解決するための手段】
【0018】
(項目1) 適切なリフォールディング条件下でリフォールドさせる能力のある、生物学的に活性なTGF−βファミリーメンバー融合タンパク質であって、該短縮化TGF−βファミリー融合タンパク質は、以下:
フィンガー1サブドメイン、フィンガー2サブドメイン、およびヒールサブドメインを含む、TGF−βファミリータンパク質C末端7システインドメイン;および
該C末端ドメインに作動可能に連結された異種リーダー配列ドメイン;
を含み、融合タンパク質。
(項目2) 項目1に記載の融合タンパク質であって、ここで前記リーダー配列は、組織標的ドメイン、分子標的ドメイン、金属結合ドメイン、タンパク質結合ドメイン、セラミック結合ドメイン、ヒドロキシアパタイト結合ドメイン、およびコラーゲン結合ドメインからなる群から選択される、融合タンパク質。
(項目3) 前記組織標的ドメインが、骨基質タンパク質に結合する、項目2に記載の融合タンパク質。
(項目4) 前記組織標的ドメインが、細胞表面分子に結合する、項目2に記載の融合タンパク質。
(項目5) 前記細胞表面分子が、前骨芽細胞または軟骨細胞上にある、項目4に記載の融合タンパク質。
(項目6) 適切なリフォールディング条件下でリフォールドさせるための、潜在性TGF−βファミリーメンバー融合タンパク質であって、該短縮化TGF−βファミリータンパク質は、以下:
フィンガー1サブドメイン、フィンガー2サブドメイン、およびヒールサブドメインを含む、TGF−βファミリータンパク質C末端7システインドメイン;および
該C末端ドメインに作動可能に連結された切断可能な改変型リーダー配列であって、該リーダー配列は、該C末端ドメインと関連する生物学的活性を阻害し、そして該C末端ドメインは、該リーダー配列の一部または全ての切断の際に活性化する、リーダー配列;
を含む、融合タンパク質。
(項目7) 項目6に記載の融合タンパク質であって、組織標的ドメインが、前記切断可能なリーダー配列の内部に存在し、それにより該リーダー配列の切断によって、該組織標的ドメインが前記C末端ドメインから切断されない、融合タンパク質。
(項目8) 前記リーダー配列が、前記C末端ドメインから少なくとも7残基は分離されている、項目1または6に記載の融合タンパク質。
(項目9) 前記リーダー配列が、別のTGF−βファミリータンパク質に由来する、項目1に記載の融合タンパク質。
(項目10) 適切なリフォールディング条件下でリフォールドさせる能力のある、生物学的に活性なTGF−βファミリーメンバー融合タンパク質変異体であって、該融合タンパク質は、以下:
フィンガー1サブドメイン、フィンガー2サブドメイン、およびヒールサブドメインを含む、TGF−βファミリーメンバータンパク質C末端7システインドメイン;および
該C末端ドメインに作動可能に連結されるリーダー配列ドメインであって、それによって、該リーダー配列の一部または全てが短縮化される、リーダー配列;
を含み、
ただし、該潜在性TGF−βファミリーメンバー融合タンパク質は、TGF−β1、TGF−β2、またはTGF−β3の該C末端7システインドメインを含まなくても良い、融合タンパク質変異体。
(項目11) 前記短縮が、プロテアーゼ切断によって実施される、項目10に記載のタンパク質変異体。
(項目12) 前記プロテアーゼが、トリプシンである、項目11に記載のタンパク質変異体。
(項目13) 前記短縮が、化学切断によって実施される、項目10に記載の融合タンパク質変異体。
(項目14) 前記化学切断が、酸切断である、項目13に記載の融合タンパク質。
(項目15) 前記リーダー配列の少なくとも1つの塩基性残基が、取り除かれる、項目10に記載のタンパク質変異体。
(項目16) 前記タンパク質変異体が、本質的に、配列番号69のアミノ酸配列からなる、項目10に記載の融合タンパク質。
(項目17) 生物学的に活性なTGF−βファミリーメンバータンパク質のヘテロ二量体であって、以下:
TGF−βファミリーメンバー融合タンパク質である第1のサブユニット;および
第2のサブユニットであって、該第1のサブユニットの融合タンパク質とは異なるTG
F−βファミリーメンバー融合タンパク質および野生型TGF−βファミリタンパク質からなる群より選択される、第2のサブユニット
を含む、ヘテロ二量体。
(項目18) 前記野生型TGF−βファミリータンパク質が、TGF−β1、TGF−β2、TGF−β3、TGF−β4、TGF−β5、dpp、Vg−1、Vgr−1、60A、BMP−2A、BMP−3、BMP−4、BMP−5、BMP−6、ドーサリン、OP−1、OP−2、OP−3、GDF−1、GDF−3、GDMP−1/GDF−5、CDMP−2/GDF−6、GDF−7、GDF−9、インヒビンα、インヒビンβA、およびインヒビンβBからなる群から選択される、項目16に記載のヘテロ二量体。
(項目19) TGF−βファミリータンパク質のヘテロ二量体を精製する方法であって、該方法は、以下:
(a)第1のTGF−βファミリータンパク質サブユニットを提供する工程;
(b)該第1のサブユニットとは異なる第2のTGF−βファミリータンパク質サブユニットを提供する工程;
(c)適切なリフォールディング条件下で、該第1のサブユニットおよび該第2のサブユニットを混合して、以下
(i)2つの該第1のTGF−βファミリータンパク質サブユニットを含む、第1のホモ二量体;
(ii)2つの該第2のTGF−βファミリータンパク質サブユニットを含む、第2のホモ二量体;ならびに
(iii)1つの該第1のTGF−βファミリータンパク質サブユニット、および1つの該第2のTGF−βファミリータンパク質サブユニットを含む、ヘテロ二量体;
を含む混合物を生成する、工程であって、
ここで、該へテロ二量体は、該第1のホモ二量体および該第2のホモ二量体から分離可能である、工程;ならびに
(d)該へテロ二量体を、該第1のホモ二量体および該第2のホモ二量体から、分離する、工程、
を含む、方法。
【0019】
(発明の要旨)
本発明は、C末端活性ドメインのN末端側でのN末端伸長、短縮および他の改変を含む、改変TGF−βファミリータンパク質を提供する。本発明の改変タンパク質は、組換え的に発現される場合に、天然に存在するタンパク質に対して変更されたリフォールディング特性および変更された可溶性を有する。本発明の改変タンパク質はまた、変更された活性プロフィール(増強された特異位的活性を含む)を有し、そして組織特異的標識化または特異的表面結合への影響を受けやすい。
【0020】
これらの発見の結果として、以下を含む変化した生物学的特性を有する新規なBMPおよび他のTGF−βファミリーのメンバーアナログを推測しそして設計するための手段が、ここで利用可能である:インビトロでの改善されたフォールディング能力、改善された可溶性、変化した安定性、変化した等電点、ならびに/または、所望される場合には、変化した生物学的活性。これらの発見はまた、それら自体に、その活性が哺乳動物内の特定の部位に指向され得、そして/またはその活性が調節、阻害および/もしくは誘導され得るタンパク質を作製させる。本発明はまた、フォールディングされたタンパク質のエピトープのマッピングを含む、候補構築物の生物学的特性および/または生化学的特性を容易にかつ迅速に評価するための手段を提供する。
【0021】
本発明は、「少ないリフォールダー(poor refolder)」TGF−βファミリーメンバーのリフォールディング特性を改善する、タンパク質の「変異体」形態を提供する。本明細書中で使用される場合、「少ないリフォールダー」タンパク質とは、適切なリフォールディング条件下でリフォールディングを誘導した場合に、標準的なプロトコール(以下を参照のこと)を使用して測定した場合に、約1%未満の適切にフォールディングされた物質を生じる、任意のタンパク質を意味する。本明細書中で意図される場合は、「適切なリフォールディング条件」とは、その条件下でタンパク質が機能性を付与するために必要とされる程度にリフォールディングされ得る条件である。当業者は、「好ましい実施態様の詳細な説明」の少なくともIC節および実施例3がこのようなリフォールディング条件の非限定的な例であることを認識する。本発明の組成物および方法に関する構造的なパラメーターは、ダイマータンパク質の構造にわたって適切に分布している1つ以上のジスルフィド架橋を含む。そしてこれは、フォールディングされた構造を生じるために、「還元−酸化」(「酸化還元」)反応工程を必要とする。酸化還元反応は、代表的には、天然のpH(すなわち、pH7.0〜8.5の範囲であり、代表的にはpH7.5〜8.5の範囲である)で生じ、そして好ましくは生理学的に適合可能な条件下で生じる。当業者は、成功のための最適な条件を理解しておりそしてそれを認識している。
【0022】
このタンパク質は、好ましくは、ヌクレオチドのアセンブリによって、および/または合成のDNAを生じるようにDNA制限フラグメントを連結することによって、本明細書中で開示されている原理に従って作製される。DNAは、適切なタンパク質発現ビヒクル中に移され、コードされるタンパク質が発現され、必要であればフォールディングされ、そして精製される。特定の構築物は、インビトロでの活性について試験され得る。候補タンパク質の構築物の三次構造は、繰り返し洗練され、そして本明細書中で開示される原理、コンピューターに基づくタンパク質構造のモデリング、および目的の分子の特異的な特性を改善または調節するための最近開発された合理的な薬物設計技術によって補助される、部位特異的変異誘発またはヌクレオチド配列特異的変異誘発によって結合が調節され得る。既知のファージディスプレイまたは他のヌクレオチド発現系は、多数の候補の構築物を同時に産生するために開発され得る。続いて、候補の構築物のプールが、例えば、表面固定化されたレセプターを含むクロマトグラフィーカラム、高結合候補物を選択しそして濃縮するための塩勾配溶出、ならびにインビトロアッセイを使用して、結合特異性についてスクリーニングされ得る。有用な組換えタンパク質の同定には、研究室での使用のため、および究極的には、治療的に有用な分子を産生するための商業的に有用な量のタンパク質を発現する細胞株の産生が続く。今や、適切にフォールディングされる場合に、フィンガー1領域、フィンガー2領域、およびヒール領域を規定する三次構造を想定するアミノ酸配列を含むキメラタンパク質を、設計、作製、試験および使用するための方法が発見された。
【0023】
本発明の全ての構築物は、有用性のために必要とされる領域を定義するアミノ酸配列の領域(すなわち、フィンガー1、フィンガー2、およびヒール領域)、ならびに活性を改変し得るさらなる領域(すなわち、N末端ペプチド配列)を含む。フィンガーおよびヒール領域についての配列は、本明細書中で同定される任意の既知のTGF−βスーパーファミリーのメンバーの、それぞれのフィンガーおよびヒール領域の配列からコピーされ得る。あるいは、フィンガーおよびヒール領域は、本明細書中で下記に開示されている原理を使用して本明細書中で以降に発見されたこのスーパーファミリーの新規のメンバーのアミノ酸配列から選択され得る。
【0024】
フィンガーおよびヒール配列はまた、例えば、Smithら(1990)、Proc.Natl.Acad.Sci.USA 87:118−122(その開示は本明細書中で参考として援用される)に開示されている原理に従って選択される置換アミノ酸残基を利用することによって、アミノ酸置換によって変化され得る。Smithらは、図3に示すアミノ酸の階層の表と同様の、アミノ酸のクラスの階層を開示している。これは、型の全体的な立体構造の変形(これは、さもなくばタンパク質を不活化し得る)を最少にしつつ、別のものについて1つのアミノ酸を合理的に置換するために使用され得る。任意の事象において、天然の領域とわずか70%の相同性、好ましくは80%、そして最も好ましくは少なくとも90%の相同性を有する多くの合成のフィンガー1、フィンガー2、およびヒール領域配列が、活性な形態単位構築物を産生するために使用され得ることが、意図される。本明細書中で開示されているように、構築物の大きさを、鋳型のTGF−βスーパーファミリーのメンバーの天然のフィンガーおよびヒール領域を短縮することによって有意に減少し得ることが意図される。
【0025】
本明細書中で使用される場合は、「酸性」または「負に荷電した残基」は、生理学的条件下で代表的にはそのR基上に負の電荷を保有している、天然に存在するかまたは合成の任意のアミノ酸残基を含むことが理解される。例であり限定的ではないが、アスパラギン酸(「Asp」)、およびグルタミン酸(「Glu」)が挙げられる。同様に、塩基性または正に荷電した残基としては、天然に存在するかまたは合成によって作製された、生理学的条件下で代表的にはそのR基上に正の電荷を保有している、任意のアミノ酸残基が挙げられる。例であり、限定的ではないが、アルギニン(「Arg」)、リジン(「Lys」)、およびヒスチジン(「His」)が挙げられる。本明細書中で使用される場合には、「親水性」残基は、酸性および塩基性の両方のアミノ酸残基、ならびにそれらのR基上にアミド基を保有している荷電していない残基(これらには、限定的ではないが、グルタミン(「Gln」およびアスパラギン(「Asn」)が挙げられる)、ならびに、そのR基上にヒドロキシル基を保有している極性残基(これらには、限定的ではないが、セリン(「Ser」)およびスレオニン(「Thr」)が挙げられる)が挙げられる。当業者は、実際の生理学的pKが変動し、そしてその変化が異なる生理学的環境において変動することを理解する。
【0026】
本明細書中で使用される場合には、「生合成」または「生合成の」は、天然に由来するかまたは合成によって誘導されるフラグメントの連結の結果として生じるか、またはそれに起源することを意味する。例えば、限定的ではないが、本明細書中で開示されている1つ以上のサブドメイン(またはそのフラグメント)に対応する連結ペプチドまたは核酸フラグメントを意味する。「化学合成」または「化学合成の」は、化学的な生成手段の結果として生じるかまたはそれに起源することを意味する。例えば、限定的ではないが、市販の供給業者からの標準的な自動合成装置/シーケンサーを使用するペプチドまたは核酸配列の合成を意味する。天然のアミノ酸および天然ではないアミノ酸の両方が、本明細書中で教示されているような所望される特性を得るように使用され得ることが意図される。「組換え」産生または技術は、遺伝子操作された産生手段の結果として生じるか、またはそれに起源することを意味する。例えば、限定的ではないが、本発明のキメラタンパク質(またはそのフラグメント)をコードする、遺伝子操作されたDMA配列または遺伝子の発現を意味する。少なくともI.B.節;II節、ならびに少なくとも実施例1、および2において以下に示される技術もまた、上記の意味に含まれる。「合成の」は、非天然に存在すること、または非天然に起源すること、すなわち、天然には存在しないことを意味する。
【0027】
本明細書中で使用される場合、「対応する残基位置」は、2つの配列が整列される場合に、OP−1または他の参照TGF−βファミリーメンバーアミノ酸配列中の所定の位置に対応するタンパク質配列中の残基位置をいう。当業者に理解され、そして図1に示すように、BMPファミリーのメンバーの配列は、C末端の活性ドメインにおいて、そして特にフィンガー2サブドメインにおいて高度に保存されている。アミノ酸配列のアラインメントの方法およびプログラムは、当該分野で十分に開発されている。例えば、Alignプログラム(DNAstar,Inc.)のようなコンピュータープログラムによって都合よく実行される、Needlemanら(1970)、J.Mol.Biol.48:443〜453の方法を参照のこと。第2の配列中の内部ギャップおよびアミノ酸の挿入は、アラインメントを計算する目的については無視される。それぞれの記述については、代表的な骨形成性タンパク質として、hOP−1(ヒトOP−1、当該分野において「BMP−7」ともいわれる)が以下に提供される。しかし、OP−1は、単に、タンパク質のTGF−βファミリーの代表的なものにすぎないことが明らかである。
【0028】
本明細書中で使用される「TGF−βファミリーメンバー」または「TGF−βファミリータンパク質」は、TGF−βスーパーファミリーのメンバーとして当業者に公知のタンパク質を意味する。構造的に、そのようなタンパク質は、ジスルフィド結合ホモダイマーまたはヘテロダイマーであり、これは、疎水性シグナル配列、数百アミノ酸のN末端プロ領域、ならびに可変性N末端領域およびより高度に保存されたC末端領域(保存された6または7のシステイン骨格を有する特徴的なシステインモチーフを伴う約100アミノ酸を含む)を含む成熟ドメインを含む大きな前駆体ポリペプチド鎖として発現される。これらの構造的に関連するタンパク質は、種々の発生事象に関与するとして同定されている。TGF−βファミリーメンバーは、TGFβ1およびOP−1によって代表される。本発明の実施において有用な他のTGF−βファミリータンパク質としては、骨形成性タンパク質(以下に定義される)、vg−1、DPP−Cポリペプチド、ホルモンアクチビンおよびインヒビン、MIS、VGR−1および増殖/分化因子GDF−1、GDF−3、GDF−9およびドーサリン−1が挙げられる。
【0029】
TGF−βタンパク質スーパーファミリーの種々のメンバーは、2つの異なる細胞表面レセプター(I型レセプターおよびII型レセプターといわれる)と相互作用し、ヘテロ複合体を形成することによってそれらの活性を媒介することが見出された。I型レセプターおよびII型レセプターはともに、セリン/トレオニンキナーゼであり、そして以下の類似の構造を共有する:本質的にキナーゼからなる細胞内ドメイン、膜を1回貫通するに十分な、短い伸長された疎水性配列、および高濃度の保存されたシステインによって特徴付けられる細胞外リガンド結合ドメイン。種々のI型レセプターおよびII型レセプターは、OP−1および他の形態形成タンパク質、およびそれらのアナログ(本発明の改変されたモルフォゲンを含む)と特異的な結合親和性を有する。
【0030】
「骨形成性タンパク質(osteogenic protein)」または「骨形態形成タンパク質」は、TGF−βスーパーファミリータンパク質を意味する。これらは、軟骨および/または軟骨内での骨の形成を含むがこれらに限定されない、骨格組織の形成を生じる形態形成事象の全体的なカスケードを誘導し得る。本明細書中で有用な骨形成性タンパク質としては、任意の既知の天然に存在するネイティブのタンパク質が挙げられる。これらには、天然に存在するかまたは生合成によって産生されたかにはかかわらず、その対立遺伝子、系統発生的対応物、および他の改変体(例えば、「ムテイン」または「変異タンパク質」を含む)、ならびにタンパク質の一般的な形態形成性のファミリーの新規の骨形成的に活性なメンバーが挙げられる。本明細書中で記載される場合、このクラスのタンパク質は、一般には、ヒトの骨形成性タンパク質1(hOP−1)によって代表される。本発明の実施において有用な他の骨形成性タンパク質としては、以下のリストに含まれる骨形成的に活性な形態のタンパク質が挙げられる:OP−1、OP−2、OP−3、BMP−2、BMP−3、BMP−4、BMP−5、BMP−6、BMP−9、DPP、Vg−1、Vgr、60Aタンパク質、CDMP−1、CDMP−2、CDMP−3、GDF−1、GDF−3、GDF−5、6、7、MP−52、BMP−10、BMP−11、BMP−12、BMP−13、BMP−15、UNIVIN、NODAL、SCREW、ADMP、またはNEURAL(これらのアミノ酸配列改変体、および/またはこれらのヘテロダイマーを含む)。1つの現在好ましい実施態様において、本発明の実施において有用な骨形成性タンパク質は、以下のいずれかのものを含む:OP−1、BMP−2、BMP−4、BMP−12、BMP−13、GDF−5,GDF−6,GDF−7、CDMP−1、CDMP−2、CDMP−3、MP−52、ならびにそれらのアミノ酸配列改変体およびホモログ(これらの種ホモログを含む)。なお別の好ましい実施態様において、有用な骨形成的に活性なタンパク質は、参照の骨形成性配列(例えば、OP−1、OP−2、BMP−2、BMP−4、BMP−5、BMP−6、60A、GDF−5、GDF−6、GDF−7などの保存されている7個のシステインのドメインを規定するC末端配列)をコードするDNAまたはRNAに対して、低い、中程度の、または高いストリンジェンシーのハイブリダイゼーション条件下でハイブリダイズする核酸によってコードされる配列を含むアミノ酸配列を有するポリペプチド鎖を有する。本明細書中で使用される場合、高いストリンジェントなハイブリダイゼーション条件は、公知技術に従って、40%のホルムアミド、5×SSPE、5×デンハルト溶液、および0.1%のSDS中で、37℃にて一晩のハイブリダイゼーション、そして0.1×SSPE、0.1%のSDS中で50℃での洗浄として規定される。標準的なストリンジェンシーの条件は、市販の標準的な分子クローニングのテキストにおいて十分に特徴付けられている。例えば、Molecular Cloning
A Laboratory Manual、第2版、Sambrook、Fritsch、およびManiatis編(Cold Spring Harbor Laboratory Press:1989);DNA Cloning、第1巻および第II巻(D.N.Glover編、1985);Oligonucleotide Synthesis(M.J.Gait編、1984):Nucleic Acid Hybridization(B.D.HamesおよびS.J.Higgins編、1984);ならびにB.Perbal、A
Practical Guide To Molecular Cloning(1984)を参照のこと。上記の開示は、本明細書中で参考として援用される。米国特許第5,750,651号、および同第5,863,758号もまた参照のこと。これらの開示は本明細書中で参考として援用される。
【0031】
本発明の実施において有用性を有する関連するタンパク質のTGF−βスーパーファミリーの他のメンバーとしては、以下のリストの中で、ネイティブの劣ったリフォールディングタンパク質が挙げられる:いくつかを挙げると、TGF−β1、TGF−β2、TGF−β3、TGF−β4、およびTGF−β5、種々のインヒビン、アクチビン、BMP−11、ならびにMIS。図4は、TGF−βスーパーファミリーの種々の既知のメンバーの、フィンガー2サブドメインを規定するC末端の35残基を列挙する。劣ったリフォールディングであるリスト上の任意の1つのタンパク質は、他の既知または発見され得るファミリーのメンバーと同様に、本発明の方法によって改善され得る。本明細書中でさらに記載されているように、本発明での使用に適切な生物学的に活性な骨形成性タンパク質は、当該分野で認識されているような、ReddiおよびSampathによって記載されているバイオアッセイを使用する慣用的な実験によって、同定され得る。有用なタンパク質の詳細な記載が以下に続く。等価物は、慣用的な実験および通常の技術のみを用いて当業者によって同定され得る。
【0032】
「モルフォゲン」または「形態形成タンパク質」は、本明細書中で意図される場合、形態形成性であること、すなわち、成熟した哺乳動物において組織の形態形成の発生カスケード(developmental cascade)を誘導し得ることが認識されている(PCT出願番号第US92/01968号を参照のこと)TGF−βスーパーファミリーのメンバーを含む。詳細には、これらのモルフォゲンは、適切な環境条件下で組織特異的様式で、方向付けられていない前駆細胞の増殖を誘導し得、そしてこれらの刺激された前駆細胞の分化を誘導し得る。さらに、モルフォゲンは、これらの分化した細胞の増殖および維持を支持し得る。これらの形態形成活性は、タンパク質が、適切な形態形成を許容する環境において組織の形態形成の発生カスケードを開始しそして維持することを可能にし、幹細胞が組織特異的様式で増殖および分化するのを刺激し、そして新しい組織の形成を生じる事象の進行を誘導させる。これらの形態形成活性はまた、タンパク質が、それらの分化の経路からはずれるように予め刺激された細胞の「再分化」を誘導することを可能にする。適切な環境条件下では、これらのモルフォゲンはまた方向付けられた細胞の「再分化」を刺激し得ることが予想される。当業者を導くために、種々の組織中で、そして形態形成タンパク質に典型的な種々の特性について形態形成タンパク質を試験するための多数の手段が、本明細書中に記載される。これらの教示が、ネイティブのタンパク質、および本発明の改変されたタンパク質の形態形成特性を評価するために使用され得ることが理解される。
【0033】
本発明の有用なネイティブのタンパク質または親のタンパク質はまた、ヒトOP−1のC末端の7個のシステインのドメイン内で少なくとも70%のアミノ酸配列相同性を共有するものを含む。保存されている7個のシステインのドメインに対する候補のアミノ酸配列のパーセント相同性を決定するために、候補の配列と7個のシステインのドメインとを整列させる。アラインメントを実行するための最初の工程は、Needlemanら、J.Mol.Biol.48:443(1970)(その教示は本明細書中で参考として援用される)に記載されている動的計画法アルゴリズム、およびAlign Program(DNAstar、Inc.によって生産されている市販のソフトウェアパッケージ)のような、アラインメントツールを使用することである。最初のアラインメントをなした後、次いでこれは、関連するタンパク質のファミリーの複数の配列のアラインメントに対する比較によって改善される。一旦、候補配列と7個のシステインのドメインとの間でのアラインメントが行われそして改善されると、パーセント相同性のスコアが計算される。各配列の個々のアミノ酸は、互いに対するそれらの類似性に従って連続して比較される。類似性の因子としては、類似の大きさ、形状、および電荷が挙げられる。アミノ酸の類似性を決定する1つの特に好ましい方法は、Dayhoffら、5 Atlas of Protein Sequence and Structure 345−352(1978および補遣、本明細書中で参考として援用される)に記載されているPAM250マトリックスである。類似性スコアはまず、整列された対となった(pairwise)アミノ酸の類似性スコアの合計として計算される。挿入および欠失は、パーセント相同性および同一性の目的のために無視される。従って、ギャップペナルティーは、この計算においては使用されない。次いで、粗スコアが、候補の化合物および7個のシステインのドメインのスコアの相乗平均でそれを除算することによって正規化される。相乗平均は、これらのスコアの積の平方根である。正規化された粗スコアが、パーセント相同性である。
【0034】
本明細書中で使用される場合は、「保存的置換」は、対応する参照残基に物理的または機能的に類似である残基である。例えば、これは、同様の大きさ、形状、電荷、化学的特性(共有結合または水素結合を形成する能力を含む)などを有する。特に好ましい保存的置換は、Dayhoffら(同書)において、容認された点変異について規定された基準を満たすものである。保存的置換の例として、類似の特徴を有する別のアミノ酸に対する1つのアミノ酸の置換が挙げられる。例えば、以下の群内での置換は周知である:(a)グリシン、アラニン;(b)バリン、イソロイシン、ロイシン;(c)アスパラギン酸、グルタミン酸;(d)アスパラギン、グルタミン;(e)セリン、トレオニン;(f)リジン、アルギニン、ヒスチジン;および(g)フェニルアラニン、チロシン。用語「保存的改変体」または「保存的バリエーション」もまた、結果として生じる置換されたポリペプチド鎖に対して結合特異性を有する抗体がまた、置換されていないかまたは親のポリペプチド鎖についての結合特異性を有する(すなわち、それと「交差反応する」かまたはそれと「免疫反応する」)という条件で、所定のポリペプチド鎖中の置換されていない親のアミノ酸の変わりに、置換されたアミノ酸を使用することを含む。
【0035】
本明細書中で使用される場合、「保存されている残基の位置」は、少なくとも1つの他のメンバーの配列中で、同じアミノ酸またはその保存的な改変体によって占有されている参照アミノ酸配列中の位置をいう。例えば、図4において、参照配列としてOP−1と、BMP−2、BMP−4、BMP−5、およびBMP−6とを比較すると、位置1、5、9、12、14、15、16、17、19、22などは保存されている位置であり、そして残基2、3、4、6、7、8、10、11、13、18、20、21などは保存されていない位置である。
【0036】
本明細書中で使用される場合、フィンガー2サブドメインの「ベース」または「ネック」領域は、OP−1によって例示され、そしてC末端の活性ドメイン中の2つ並んでいるシステインに続く最初の残基から数える場合は、残基1〜10および22〜35によって規定される(図4を参照のこと)。他のTGF−βタンパク質ファミリーメンバーのOP−1との配列アラインメントから容易に明らかであるように、より長いタンパク質(例えば、BMP−9またはDorsalin)についての対応するベースまたはネック領域は、残基1〜10および23〜36によって規定される:より短いタンパク質(例えば、NODAL)については、対応する領域は、残基1〜10および22〜34によって規定される(図4を参照のこと)。配列番号39(ヒトOP−1)において、フィンガー2サブドメインのベースまたはネック領域に対応する残基は、残基397〜406(図4の残基1〜10に対応する)および残基418〜431(図4の残基22〜35に対応する)である。
【0037】
本明細書中で使用される「C末端活性ドメイン」は、成熟TGF−βファミリータンパク質の保存されたC末端領域をいう。C末端活性ドメインは、6または7のシステインの骨格を有する特徴的なシステインモチーフを伴う約100アミノ酸を含む。すべてのタンパク質のC末端のシステインパターンは同一の形式であり、配列Cys−X−Cys−Xで終わる(SpornおよびRoberts(1990)、前出)。
【0038】
本明細書中で使用される場合、「アミノ酸配列の相同性」は、アミノ酸配列の同一性および類似性の両方を含む。相同配列は、同一および/または類似のアミノ酸残基を共有する。ここで、類似の残基は、整列された参照配列中の対応するアミノ酸残基の保存的置換、またはその「許容される点変異」である。
【0039】
本明細書中で使用される場合、用語「キメラタンパク質」、「キメラ」、「キメラポリペプチド鎖」、「キメラ構築物」、および「キメラ変異体」は、任意のBMPまたはTGF−βファミリーのメンバーの合成の構築物をいう。ここで、少なくとも1つの規定された領域、ドメイン、またはサブドメイン(例えば、フィンガー1、フィンガー2、またはヒールサブドメイン)のアミノ酸配列は、全体または一部において、少なくとも1つの他の異なるBMPまたはTGF−βファミリーのメンバーのタンパク質に由来するアミノ酸配列と置き換えられている。その結果、得られる構築物は、異なるタンパク質供給源に由来するとして認識可能なアミノ酸配列を有する。キメラ構築物はまた、1つのモルフォゲンのC末端活性ドメインが、別のモルフォゲンのN末端ドメインに融合されている組換え融合タンパク質を含む。
【0040】
本明細書中で使用される「リーダー配列」は、TGF−βファミリータンパク質のC末端活性ドメイン領域の上流、すなわちC末端からさらに遠くに位置するヌクレオチドの配列に対応するアミノ酸の任意の配列である。リーダー配列における改変は、リフォールディング特性、活性レベル、可溶性および制御活性化を変更し、そして組織標的化および親和性結合能を促進し得る。
【0041】
本明細書中で使用される場合、有用な発現宿主細胞としては、原核生物および真核生物が挙げられる。これには、封入体を産生し得る任意の宿主細胞が挙げられる。特に有用な宿主細胞として、限定的ではないが、E.coliのような細菌宿主、ならびにB.subtilisおよびPseudomonasが挙げられる。他の有用な宿主としては、下等な真核生物(例えば、Saccharomyces cereviceaeまたは他の酵母)および高等真核生物(例えば、Drosophila、CHO細胞、および他の哺乳動物細胞など)が挙げられる。本明細書中において議論されるように、化学合成方法もまた、本発明の改変されたタンパク質を生成するために利用され得る。
【0042】
1つの局面において、本発明は、哺乳動物細胞において容易に発現されない組換えタンパク質(例えば、融合タンパク質など)の構築を提供する。例えば、骨標的化特性を有する融合タンパク質をコードする組換え遺伝子が構築される。ここで、一つの配列は、BMPおよび抗体結合部位(オステオカルシンまたはフィブロネクチンのような骨基質タンパク質に対する特異性を有する)の両方をコードする。同様に、細胞表面レセプター(例えば、前骨芽細胞または軟骨細胞上のレセプター)に結合する融合タンパク質もまた構築され得る。他の組換え遺伝子は、金属または他のタンパク質を特異的に結合する融合タンパク質をコードし得る。結合の特異性は、BMPに付加されたリーダー配列の組成に依存する。これらの遺伝子は、E.coli中で発現され、そしてインビトロでリフォールディングされ得る。
【0043】
別の実施態様において、切断可能な融合タンパク質(プロテアーゼ−例えば、トリプシン、V8、第Xa因子およびその他、または化学的−中程度の酸、ヒドロキシルアミンおよび他の薬剤によって切断可能)が合成され、ここで、TGF−βタンパク質は、活性をブロックするリーダー配列に付着される。なお別の実施態様において、TGF−βファミリーメンバーの活性は、このリーダー配列の一部または全てを切断することによって回復または増強される。活性を阻害する切断可能なリーダー配列を付加することによって、このタンパク質の潜在形態は、実質的に、活性なC末端ドメインを含むタンパク質フラグメントを放出するように切断され得る。
【0044】
さらに別の実施態様において、このリーダー配列はまた、組織標的配列であり、その結果、放出は、インビボにおいてこの標的部位で生じるように制御され得る。切断可能部位の構築はまた、活性なタンパク質の放出を制御することを可能にし得る。例えば、骨組織において、骨の再モデル化に関与する多くのプロテアーゼが代表的に存在し、そして有利に使用され得る。以下に記載される切断可能な「ヘキサ−his」、FBリーダー、またはコラーゲン結合配列は、このタンパク質の潜在形態についての適切なリーダー配列であり得る。例の目的で、この組織標的ドメインは、少なくとも3つの塩基残基の続きを含むリーダー配列によってBMPから分離され得、これは、インビボにおいて切断されることが公知である。
【0045】
なお別の実施態様において、このリーダー配列が構築され得、その結果、特定の活性を阻害するタンパク質の部分は切断され、そして活性は回復されるが、このタンパク質の組織標的部分は、保持される。
【0046】
さらに別の実施態様において、TGF−βファミリータンパク質のリーダー配列は、別のTGF−βメンバーのリーダー配列によって置換される。得られた「キメラ」タンパク質は、変更された可溶性、フォールディングおよび/または組織標的活性、改善された安定性、ならびに/あるいは特定の表面に結合する能力を有し得る。
【0047】
本発明の別の局面において、融合タンパク質は、他のTGF−βファミリータンパク質と組み合わされて、ヘテロダイマーを形成する。ここで、各々のタンパク質の特性が開発される。例えば、組織標的特性を有するが、活性を有さない融合タンパク質は、活性を有するが標的能力を有さない異なるタンパク質とともにヘテロダイマーを形成する。前者のタンパク質は、後者のタンパク質がその機能を実施し得る標的部位にヘテロダイマーを送達する。
【0048】
1つの局面において、本発明は、中性または生理学的条件下で改善されたリフォールディング特性を有する生合成BMPおよびTGF−βファミリーメンバーを提供する。1つの実施態様において、本発明の生合成タンパク質は、約5.0〜10.0の範囲、好ましくは約6.0〜9.0の範囲、より好ましくは約6.0〜8.5の範囲のpH(約pH7.0〜7.5の範囲を含む)で、改善されたリフォールディング特性を有する。
【0049】
別の局面において,本発明は、中性または生理学的条件下で改善された可溶性特性を有する生合成BMPおよびTGF−βファミリーメンバーを提供する。1つの実施態様において、本発明の生合成タンパク質は、約5.0〜10.0の範囲、好ましくは約6.0〜9.0の範囲、より好ましくは約6.0〜8.5の範囲のpH(約pH7.0〜7.5の範囲を含む)で、改善された可溶性を有する。
【0050】
なお別の局面において,本発明は、生理学的条件下でリフォールディングし、そして親配列と比較して変更された等電点を有する、生物学的に活性な生合成BMPおよびTGF−βファミリーメンバー構築物成分を提供する。
【0051】
別の局面においては、本発明は、生理学的条件または中性のpH条件下でほとんどリフォールディングされないホモダイマーおよびヘテロダイマーをフォールディングするための方法を提供する。1つの実施態様においては、この方法は、本発明の1つ以上の可溶化されたTGF−βファミリータンパク質構築物を提供する工程、可溶化されたタンパク質を適切なリフォールディング緩衝液中での酸化還元反応に曝す工程、ならびに所望される場合には、タンパク質のサブユニットがホモダイマーおよび/またはヘテロダイマーにリフォールディングすることを可能にする工程を包含する。別の実施態様においては、本発明の改変TGF−βファミリータンパク質は、それらをこの酸化還元反応に曝す前に変性されない。別の実施態様においては、酸化還元反応系は、グルタチオン、DTT、β−メルカプトメタノール、システイン、およびシスタミンの酸化された形態および還元された形態を利用し得る。別の実施態様においては、酸化還元反応系は、好ましくは金属触媒(例えば、銅)の存在下での、空気の酸化による。なお別の実施態様においては、これらは、約1:10から約10:1の、好ましくは約1:2から2:1の範囲内の酸化剤に対する還元剤の比での酸化還元系として使用され得る。別の好ましい実施態様においては、このタンパク質は、イオン性界面活性剤、非イオン性の界面活性剤(例えば、ジギトニン)、または双性イオン性界面活性剤(例えば、3−[(3−コラミドプロピル(cholamidopropyl))ジメチルアンモニオ]−1−プロパン硫酸(CHAPS)、またはN−オクチルグルコシドを含む、界面活性剤の存在下で可溶化される。なお別の実施態様においては、リフォールディング反応は、約5.0〜10.0の範囲内、好ましくは、約6.0〜9.0の範囲内、より好ましくは、約7.0〜8.5の範囲内の、pHの範囲で生じる。なお別の実施態様においては、リフォールディング反応は、約32〜0℃の範囲内、好ましくは、約25〜4℃の範囲内の温度で生じる。ヘテロダイマーが作製される場合は、2つの異なるサブユニットの添加についての最適な比が、経験的に、そして過度の実験を伴うことなく容易に決定され得る。
【0052】
別の局面において、本発明は、宿主細胞(細菌宿主、または任意の他の宿主細胞を含む)において弱いフォールディングのBMPおよび他のTGF−βファミリーメンバータンパク質を組換え的に生成する方法を提供し、ここで、過剰発現されるタンパク質は、インビトロにおいて可溶化および/またはリフォールディングすることを必要とする形態で凝集する。この方法は、本発明の1つ以上の生合成タンパク質をコードする核酸分子でトランスフェクトされた宿主細胞を提供する工程、生合成タンパク質を発現するために適切な条件下で宿主細胞を培養する工程、凝集したタンパク質を回収する工程、ならびに上記に述べた工程を使用してこのタンパク質を可溶化およびリフォールディングする工程を包含する。別の実施態様において、この方法は、本発明の生合成タンパク質をコードする核酸を用いて宿主細胞をトランスフェクトする、さらなる工程を包含する。
【0053】
本発明の改変モルフォゲンは、生体適合性マトリクス(例えば、コラーゲン、ヒドロキシアパタイト、セラミックスもしくはカルボキシメチルセルロース、および/または他の適切なマトリクス材料(これらに限定されない)と合わせて、骨および/または軟骨を形成するために使用され得る。このような組合せは、再生骨、軟骨および/または他の非鉱化骨格もしくは連結組織(例えば、少数だが、人工軟骨、線維軟骨、関節包、半月板、椎間板(intervertabral disk)、滑膜組織、筋肉、および筋膜(fasica)(これらに限定されない)についての方法において特に有用である。例えば、米国特許第5,674,292号、同第5,840,325号、および米国特許出願第08/253,398号(これらの開示は、本明細書中に参考として援用される)を参照のこと。本発明は、このようなマトリクス材料に対する結合特性および/または接着特性が、タンパク質構築物を生成するために本明細書中に開示される技術を使用して変更され得ることを意図する。本発明の改変タンパク質はまた、腱、靭帯および/または筋肉組織を生成するために利用され得る。
【図面の簡単な説明】
【0054】
【図1A】図1Aは、TGF−βスーパーファミリーメンバーのモノマーサブユニットの構築を記載する際に有用な単純な線図である。説明については、前述の発明の背景を参照のこと。図1B、1Cおよび1Dは、フィンガー1、ヒールおよびフィンガー2領域の代表的な二次構造のそれぞれのペプチド骨格の単眼視野の(monovision)リボン追跡である。
【図1B】図1Aは、TGF−βスーパーファミリーメンバーのモノマーサブユニットの構築を記載する際に有用な単純な線図である。説明については、前述の発明の背景を参照のこと。図1B、1Cおよび1Dは、フィンガー1、ヒールおよびフィンガー2領域の代表的な二次構造のそれぞれのペプチド骨格の単眼視野の(monovision)リボン追跡である。
【図1C】図1Aは、TGF−βスーパーファミリーメンバーのモノマーサブユニットの構築を記載する際に有用な単純な線図である。説明については、前述の発明の背景を参照のこと。図1B、1Cおよび1Dは、フィンガー1、ヒールおよびフィンガー2領域の代表的な二次構造のそれぞれのペプチド骨格の単眼視野の(monovision)リボン追跡である。
【図1D】図1Aは、TGF−βスーパーファミリーメンバーのモノマーサブユニットの構築を記載する際に有用な単純な線図である。説明については、前述の発明の背景を参照のこと。図1B、1Cおよび1Dは、フィンガー1、ヒールおよびフィンガー2領域の代表的な二次構造のそれぞれのペプチド骨格の単眼視野の(monovision)リボン追跡である。
【図2A】図2Aおよび2Bは、以下からの、TGF−βスーパーファミリーメンバータンパク質ダイマーの一般的な三次元形状を示すステレオペプチド骨格リボン追跡図である:A)紙に対して一般に正常なへリックスヒール領域の軸および一般に垂直のフィンガー1およびフィンガー2の各々の軸を有する「トップ」(サブユニット間の対称の2回転軸の下)、ならびにB)紙の代わりにこのサブユニット間の2回転軸、一般に水平のヒールの軸、および一般に垂直の軸を有する「サイド」。このリーダーは、形態単位設計においてよりよい特別な関係を理解するためにウォールアイステレオ(wall eyed stereo)におけるステレオα炭素追跡図を見るために奨励される。
【図2B】図2Aおよび2Bは、以下からの、TGF−βスーパーファミリーメンバータンパク質ダイマーの一般的な三次元形状を示すステレオペプチド骨格リボン追跡図である:A)紙に対して一般に正常なへリックスヒール領域の軸および一般に垂直のフィンガー1およびフィンガー2の各々の軸を有する「トップ」(サブユニット間の対称の2回転軸の下)、ならびにB)紙の代わりにこのサブユニット間の2回転軸、一般に水平のヒールの軸、および一般に垂直の軸を有する「サイド」。このリーダーは、形態単位設計においてよりよい特別な関係を理解するためにウォールアイステレオ(wall eyed stereo)におけるステレオα炭素追跡図を見るために奨励される。
【図3】図3は、SmithおよびSmith(1990)Proc.Natl.Acad.Sci.USA 87:118〜122の教示に従って調製されたパターン規定表である。
【図4】図4は、BMPファミリーの種々の既知のメンバー、およびタンパク質のTGF−βスーパーファミリーについて、2つ並んでいるシステインに続く最初の残基で開始して並べた、フィンガー2サブドメインを規定する、整列したC末端の残基を列挙する。
【図5A】図5A、5B、および5Cは、TGF−βスーパーファミリーの現在既知のメンバーの、それぞれフィンガー1、ヒール、およびフィンガー2領域のアラインメントおよび相同性を示すように並べられた、一文字表記のアミノ酸配列である。以下の各領域を含むそれぞれのアミノ酸が示されている:ヒトのTGF−β1からTGF−β5(TGF−βサブグループ)、dpp、Vg−1、Vgr−1、60A(同時係属中のU.S.S.N.08/271,556号を参照のこと)、BMP−2A(BMP−2としてもまた文献において公知である)、dorsalin、BMP−2B(BMP−4としてもまた文献において公知である)、BMP−3、BMP−5、BMP−6、OP−1(BMP−7としてもまた文献において公知である)、OP−2(PCT/US91/07635号、および米国特許第5,266,683号を参照のこと)、およびOP−3(U.S.S.N.07/971,091号)からなるVg/dppサブグループ、GDF−1、GDF−3、およびGDF−9からなるGDFサブグループ、インヒビンα、インヒビンβA、およびインヒビンβBからなるインヒビンのサブグループ。ダッシュ(−)は、隣接するアミノ酸間のペプチド結合を示す。各サブグループについてのコンセンサス配列パターンを、各サブグループの最下段に示す。
【図5B】図5A、5B、および5Cは、TGF−βスーパーファミリーの現在既知のメンバーの、それぞれフィンガー1、ヒール、およびフィンガー2領域のアラインメントおよび相同性を示すように並べられた、一文字表記のアミノ酸配列である。以下の各領域を含むそれぞれのアミノ酸が示されている:ヒトのTGF−β1からTGF−β5(TGF−βサブグループ)、dpp、Vg−1、Vgr−1、60A(同時係属中のU.S.S.N.08/271,556号を参照のこと)、BMP−2A(BMP−2としてもまた文献において公知である)、dorsalin、BMP−2B(BMP−4としてもまた文献において公知である)、BMP−3、BMP−5、BMP−6、OP−1(BMP−7としてもまた文献において公知である)、OP−2(PCT/US91/07635号、および米国特許第5,266,683号を参照のこと)、およびOP−3(U.S.S.N.07/971,091号)からなるVg/dppサブグループ、GDF−1、GDF−3、およびGDF−9からなるGDFサブグループ、インヒビンα、インヒビンβA、およびインヒビンβBからなるインヒビンのサブグループ。ダッシュ(−)は、隣接するアミノ酸間のペプチド結合を示す。各サブグループについてのコンセンサス配列パターンを、各サブグループの最下段に示す。
【図5C】図5A、5B、および5Cは、TGF−βスーパーファミリーの現在既知のメンバーの、それぞれフィンガー1、ヒール、およびフィンガー2領域のアラインメントおよび相同性を示すように並べられた、一文字表記のアミノ酸配列である。以下の各領域を含むそれぞれのアミノ酸が示されている:ヒトのTGF−β1からTGF−β5(TGF−βサブグループ)、dpp、Vg−1、Vgr−1、60A(同時係属中のU.S.S.N.08/271,556号を参照のこと)、BMP−2A(BMP−2としてもまた文献において公知である)、dorsalin、BMP−2B(BMP−4としてもまた文献において公知である)、BMP−3、BMP−5、BMP−6、OP−1(BMP−7としてもまた文献において公知である)、OP−2(PCT/US91/07635号、および米国特許第5,266,683号を参照のこと)、およびOP−3(U.S.S.N.07/971,091号)からなるVg/dppサブグループ、GDF−1、GDF−3、およびGDF−9からなるGDFサブグループ、インヒビンα、インヒビンβA、およびインヒビンβBからなるインヒビンのサブグループ。ダッシュ(−)は、隣接するアミノ酸間のペプチド結合を示す。各サブグループについてのコンセンサス配列パターンを、各サブグループの最下段に示す。
【図6】図6は、標準的な一文字表記のアミノ酸コードで大文字で示されている、アミノ酸配列の一文字コードのリストであり、そして小文字で、その位置において有用であるアミノ酸の群を同定する。ここで、アミノ酸を表す小文字は、表3に示されているパターン定義の鍵となる表に従って示されている。図6は、本発明の生合成構築物のフィンガー1、ヒール、およびフィンガー2領域を構成するための好ましいパターン配列を同定する。ダッシュ(−)は、隣接するアミノ酸間のペプチド結合を示す。
【図7A】図7(A)は、H2487のヌクレオチドおよび対応するアミノ酸配列を示す。H2487は、7つのシステインのドメインの上流に挿入されたN末端デカペプチドコラーゲン結合部位を含む、改変OP−1である。
【図7B】図7(B)は、H2440のヌクレオチドおよび対応するアミノ酸配列を示す。H2440は、7つのシステインのドメインの第1のシステインの35残基上流に結合されたヘキサ−ヒスチジンドメインを含む、改変OP−1である。
【図7C】図7(C)は、H2521のヌクレオチドおよびアミノ酸配列を示す。H2521は、7つのシステインのドメインの第1のシステインの15残基上流に結合されたプロテインAのFBリーダードメインを含む、改変OP−1である。
【図7D】図7(D)は、H2525のヌクレオチドおよびアミノ酸配列を示す。H2525は、プロテインAのFBリーダードメインおよびヘキサ−ヒスチジンドメインの両方を含む、改変OP−1である。
【図7E】図7(E)は、H2527のヌクレオチドおよびアミノ酸配列を示す。H2527は、FBリーダードメイン、ヘキサ−ヒスチジンドメイン、およびASP−PRO酸切断部位を含む、改変OP−1である。
【図7F】図7(F)は、H2528のヌクレオチドおよびアミノ酸配列を示す。H2528は、FBリーダードメインおよびヘキサ−ヒスチジンドメインを含む、改変CDMP−3である。
【図7G】図7(G)は、H2469のヌクレオチドおよびアミノ酸配列を示す。H2649は、保存された7つのシステインのドメインの第1のシステインの14の元々の残基の上流を含む、改変OP−1(短縮型)である。
【図7H】図7(H)は、H2510のヌクレオチドおよびアミノ酸配列を示す。H2510は、保存された7つのシステインのドメインの第1のシステインの7残基上流に挿入されたコラーゲン結合部位を含む、改変OP−1である。
【図7I】図7(I)は、H2523のヌクレオチドおよびアミノ酸配列を示す。H2523は、コラーゲンペプチドおよび保存された7つのシステインのドメインの第1のシステインから13残基上流に付加されたスペーサーを含む、改変OP−1である。
【図7J】図7(J)は、H2524のヌクレオチドおよびアミノ酸配列を示す。H2524は、ヘキサ−ヒスチジンドメイン、コラーゲンペプチドおよび保存された7つのシステインのドメインの第1のシステインから13残基上流に付加されたスペーサーを含む、改変OP−1である。
【図8】図8は、OP−1 C末端の7つのシステイン活性ドメインをコードする制限マップである。
【図9A】図9Aは、種々の生合成キメラBMP構築物の模式図である。
【図9B】図9Bは、生合成BMP変異体の模式図およびそれらのリフォールディングおよびROS活性である。
【図10】図10は、種々のBMPについてのC末端サブドメインにおける荷電残基の数を示す。
【図11】図11は、濃度(ng/mL)対光学密度(405nmにおける)としてプロットした、OP−1(標準)、変異体H2549タンパク質およびトリプシンで処理したH2549についてのROS活性のグラフである。
【図12】図12は、濃度(ng/mL)対光学密度(405nmにおける)としてプロットした、OP−1(標準)および変異体H2223の種々の画分ならびにこのタンパク質のトリプシン短縮型形態についてのROS活性のグラフである。
【図13】図13(A)は、濃度(ng/mL)対光学密度(405nmにおける)としてプロットした、OP−1ホモダイマー(CHO細胞由来)、BMP−2ホモダイマーおよびヘキサ−his OP−1ヘテロダイマーについてのROS活性のグラフである。
【0055】
図13(B)は、濃度(ng/ml)対光学密度(405nmにおける)としてプロットした、OP−1ホモダイマー(CHO細胞由来)、ヘキサ−his OP−1/BMP−2ヘテロダイマーおよびヘキサ−his OP−1についてのROS活性のグラフである。
【図14】図14は、濃度(ng/mL)対光学密度(405nmにおける)としてプロットした、OP−1(標準)およびBMP−2変異体H2142タンパク質ホモダイマー、変異体H2525タンパク質ホモダイマーおよびH2525/2142へテロダイマーについてのROS活性のグラフである。
【図15】図15は、種々のOP−1変異体のフィンガー2サブドメインのアミノ酸配列、ならびにROS細胞に基づくアルカリホスファターゼアッセイにおけるそれらのフォールディング効率および生物学的活性を示す。
【発明を実施するための形態】
【0056】
(好ましい実施態様の詳細な説明)
本発明は、天然の形態と比較して、変化したリフォールディング特性、および変化した活性プロフィールを有する、改変形態のTGF−βファミリータンパク質を提供する。本発明の改変タンパク質は、天然に存在するTGF−βファミリーメンバー(特に、形態形成タンパク質)のN末端改変を含む。これらの改変は、伸長、短縮、および/または特定の部位でのプロテアーゼ切断または化学切断(例えば、酸またはCNBrによる)による活性化、異なるタンパク質ドメインの結合(融合)ならびに他のTGF−βファミリーメンバー由来のサブユニットとのヘテロダイマーの生成を含む。以下に提供する詳細な説明は、改善した活性および薬学的特性を生じる、多数の例示的な、置換、融合、および伸長を記載する。改変タンパク質を生成する方法もまた、教示する。
【0057】
本発明の1つの局面に従って、ほとんどリフォールディングしないBMPおよびTGF−βスーパーファミリーの他のメンバーのタンパク質(そのヘテロダイマ−およびキメラを含む)のフォールディング能力は、BMPまたはTGF−βファミリーメンバーの既存のN末端ドメイン(次いで、この融合タンパク質内の部位で切断され得る)に、特定の標的領域およびレセプター結合領域を融合させることによって、改善される。この発見の結果として、以下を設計することが可能である:(1)原コア生物細胞または真コア生物細胞中で組換え的に発現されたかまたはポリペプチド合成装置を使用して合成された、BMPおよび他のTGF−βファミリーのタンパク質;(2)変化したフォールディング特性を有する、BMPおよび他のTGF−βファミリーのタンパク質;(3)中性のpH(生理学的条件を含むが、これに限定されない)の下で変化した可溶性を有する、BMPおよび他のTGF−βファミリーのタンパク質;(4)変化した等電点を有する、BMPおよび他のTGF−βファミリーのタンパク質;(5)変化した安定性を有する、BMPおよび他のTGF−βファミリーのタンパク質;(6)固相表面(例えば、生体適合性マトリクスまたは金属)に対する変化した結合特性または付着特性を有する、BMPおよび他のTGF−βファミリーのタンパク質;ならびに/または(7)所望の、変化した生物学的活性(例えば、組織特異性および/またはレセプター特異性)を有する、BMPおよび他のTGF−βファミリーのタンパク質。さらに、本発明は、新規な候補構築物(特に、その候補の生物学的特性または生化学的特性)を迅速に試験するための手段を提供する。本発明はまた、例えば、異なる組み合わせのドメインを有するキメラタンパク質を作製することによって、抗体のエピトープを迅速にマッピングするための手段を提供する。詳細には、本明細書中に開示された発見を使用して、原コア生物宿主(例えば、E.coli)において他の方法では発現され得ないモルフォゲン配列を改変して、E.coliにおける発現およびインビボでのリフォールディングを、今や可能にし得る。
【0058】
従って、本発明は、好ましいキメラタンパク質を含む、速放性、徐放性および/または時限放出性の処方物を設計するための機構を提供し得る。さらに、本発明は、タンパク質構築物の環境的により誘発されて放出するように操作された処方物を設計するための機構を提供する。すなわち、改変タンパク質は、インサイチュでの特定の環境条件下(例えば、pHの変化、特異的プロテアーゼの存在など)で、送達を調節し、そして放出および活性を容易にするように設計され得る。他の利点および特徴は、以下の教示から明らかである。さらに、本明細書中の発見を使用して、変化した表面結合特性/表面付着特性を有する改変タンパク質を、設計および選択し得る。特に重要な表面としては、以下が挙げられるが、これらに限定されない:天然に存在し得る固体表面(例えば、骨);または多孔性粒子表面(例えば、コラーゲンまたは他の生体適合性マトリクス);あるいは人工器官移植片(金属を含む)の加工(fabricated)表面。本明細書中で意図する場合、事実上すべての表面を、構築物の示差的結合についてアッセイし得る。従って、本発明は、その表面結合特性/表面付着特性が変化し、それによってそのような構築物を変化したインビボ適用(徐放性処方物、速放性処方物および/または時限放出性処方物を含む)のために有用にする、多様な機能性分子をを含む。
【0059】
当業者は、1つ以上の任意の上記の特性を混合かつ適合させることによって、カスタマイズされた改変タンパク質(および、それをコードするDNA)の使用を操作するための特定の機会が提供されることを、理解する。例えば、変化した安定性の特性を使用して、インビボでのタンパク質のターンオーバーを操作し得る。さらに、変化したリフォールディングおよび/または機能のような特性を有する改変タンパク質の場合において、フォールディング、機能、および安定性との間に相関関係が存在するようである。例えば、Lipscombら、7、Protein Sci.765−73(1998);およびNikolovaら、95、Proc.Natl.Acad.Sci.USA 14675−80(1998)を参照のこと。本発明の目的のために、安定性の変化を、円偏光二色性、および変性剤の濃度または温度の関数としての安定性の他の指標といった周知の技術を使用して、慣用的にモニターし得る。当業者はまた、慣用的な走査熱分析を使用し得る。同様に、上記の任意の特性と可溶性の特性との間に相関関係が存在するようである。可溶性の場合においては、この特性を操作して、その結果、改変タンパク質が、生理学的適合性状態の下でより可溶性であるかまたはより可溶性ではないかのいずれかであり、そしてその結果として、インビボで投与した場合には、それぞれ、容易に拡散するか、または局在化したままであるようにすることが可能である。
【0060】
適切な生合成タンパク質および本発明の実施において有用な方法、ならびにこれらのタンパク質を使用および試験するための方法の、詳細な説明を、以下に提供する;そして、以下の限定的ではない多数の例を提供する:1)本明細書中に記載されている、生合成タンパク質および方法の適合性を説明する、例;ならびに、2)これらのタンパク質を試験および使用するためのアッセイを提供する、例。
【0061】
(I.タンパク質の考察)
(A.TGF−β2およびOP−1の構造的特徴)
TGF−β2またはOP−1のいずれかの中のサブユニット各々は、特徴的なフォールディングパターン(図1Aに模式的に例示される)を有する。このパターンは、7個のC末端のシステイン残基のうちの6個を含む。簡潔には、各サブユニット中のシステイン残基のうちの4つが2つのジスルフィド結合を形成し、これらは互いに8残基の環を作製する。一方、2つのさらなるシステイン残基が、結び目様の構造を形成するように環を通過する、ジスルフィド結合を形成する。7個の保存されているシステイン残基のうちの最もN末端のシステイン(番号1を割り当てられる)で開始する番号付けスキームを用いると、2番目および6番目のシステイン残基が、8残基の環の1つの側面を閉じるようにジスルフィド結合し、一方、3番目および7番目のシステイン残基が、この環の他の側面を閉じるようにジスルフィド結合する。1番目および5番目の保存されているシステイン残基は、この結び目のコアを形成するように、この環の中心を通ってジスルフィド結合する。アミノ酸配列の整列のパターンは、この構造モチーフが、TGF−βスーパーファミリーのメンバーの間で保存されていることを示唆する。4番目のシステインは半保存されており、そして存在する場合には、代表的に、他のサブユニット中の対応するシステイン残基と鎖間ジスルフィド結合(ICDB)を形成する。
【0062】
TGF−β2およびOP−1中の各サブユニットの構造は、3つの主要な三次構造のエレメントおよびN末端領域を含む。この構造エレメントは、以下の型のうちの50%を超える二次構造を保有する、連続するポリペプチド鎖の領域から構成される:(1)ループ、(2)α−へリックスおよび(3)β−シート。各構造領域について規定する別の特徴は、入口(N末端)および出口(C末端)のペプチド鎖が、約7Åの間隔で、互いにかなり近づいていることである。
【0063】
図1Aに示される、1番目の保存システインと2番目の保存システインとの間のアミノ酸配列は、逆平行β−シートフィンガーによって特徴付けられる構造領域を形成して、本明細書中でフィンガー1領域と呼ばれる。同様に、5番目の保存システインと6番目の保存システインとの間の残基もまた(図1Aに示すように)、逆平行β−シートフィンガーを形成して、フィンガー2領域と本明細書中で呼ばれる。β−シートフィンガーは、一本のアミノ酸鎖であり、β−ターンまたはいくらか長いループによってそれ自身の上にリフォールディングβ−鎖を含む。その結果、この領域に入りかつこの領域から出るポリペプチド鎖は、1つ以上の逆平行β−シート構造を形成する。3番目の保存されているシステインと5番目の保存されているシステインとの間の残基を含む、第3の主要な構造領域(図1Aに示す)は、3個のターンα−へリックスによって特徴付けられ、本明細書中でヒール領域と呼ばれる。その単量体構造の構成は、左手の構成と類似である。この構造においては、その結び目領域が手のひらに相当する位置に位置し、フィンガー1領域が人差し指および中指に相当し、α−へリックス領域またはヒール領域が手首の付け根に相当し、そしてフィンガー2領域が薬指および小指に相当する。N末端領域は、その配列がTGF−βスーパーファミリーを通じて保存されておらず、親指にほぼ相当する位置に位置すると推定される。
【0064】
TGF−β2単量体の3つの主な独立構造エレメントの各々のα炭素骨格の単視的リボントレース(monovision ribbon tracing)を図1B〜1Dに示す。詳細には、第1の逆方向β−シートセグメントを含む例示的フィンガー1領域を図1Bに示し、3つのターンα−へリックスセグメントを含む例示的ヒール領域を図1Cに示し、そして第2および第3の逆方向β−シートセグメントを含む例示的フィンガー2領域を図1Dに示す。
【0065】
図2は、立体構造的に活性なTGF−β2の二量体複合体のペプチド骨格の立体リボントレース図を示す。この二量体複合体中の2つの単量体サブユニットは、2回回転対称性を有して位置しており、その結果、一方のサブユニットのヒール領域が、もう一方のサブユニットのフィンガー領域と接触し、結合したサブユニットの結び目領域が、この分子のコアを形成する。4番目のシステインは、2番目の鎖上の対照物と鎖間ジスルフィド結合を形成し、それによりその掌の中心でこれらの鎖を等しく連結する。このように形成された二量体は、サブユニット間の2回対称軸を上方から見下ろした場合に、楕円形の(葉巻の形状の)分子である(図2A)。側方から見ると、この分子は、まがった「葉巻」に似ている。なぜなら、その2つのサブユニットが、互いに対してわずかな角度で位置しているからである(図2B)。
【0066】
図2に示すように、ともにこの二量体のネイティブな単量体サブユニットを規定する構造エレメントの各々を、22、22’、23、23’、24、24’、25、25’、26、および26’と符号をつける。この図で、エレメント22、23、24、25、および26は、一方のサブユニットによって規定され、そしてエレメント22’、23’、24’、25’および26’はもう一方のサブユニットに所属する。詳細には、22および22’はN末端ドメインを示し;23および23’はフィンガー1領域を示し;24および24’はヒール領域を示し;25および25’はフィンガー2領域を示し;そして26および26’は、各サブユニットの1番目および5番目の保存システインを連結して結び目様の構造を形成する、ジスルフィド結合を示す。図2から、一方のサブユニット由来のヒール領域(例えば、24)ともう一方のサブユニット由来のフィンガー1領域およびフィンガー2領域(例えば、それぞれ、23’および25’)は、互いに相互作用することが理解され得る。これら3つのエレメントは互いに協同して、同族のレセプターの相互作用する表面に結合するリガンドと相互作用しそしてそれらに相補性である、構造を規定する。
【0067】
(フィンガー領域およびヒール領域の選択)
フィンガー領域およびヒール領域を規定するアミノ酸配列が、本明細書中で同定された、TGF−βスーパーファミリーの既知の任意のメンバーの、それぞれのフィンガー領域の配列およびヒール領域の配列に由来し得るか、または、本明細書中で以後に発見される新規のスーパーファミリーのメンバーのアミノ酸配列に由来するものが、利用され得ることが、意図される。
【0068】
図5は、フィンガー1領域(図5A)、ヒール領域(図5B)、およびフィンガー2(図5C)領域へと整列された、現在同定されているTGF−βスーパーファミリーのメンバーのアミノ酸配列を要約する。これらの配列は、コンピューターアルゴリズムによって整列された。このアルゴリズムでは、これらの配列を最適に整列するために、保存的アミノ酸配列または二次構造を有することが既知のアミノ酸配列の領域よりもむしろ、ループ構造を規定することが知られているアミノ酸配列の領域中にギャップが挿入された。例えば、可能である場合には、ギャップは、βシートによって規定されるフィンガー1領域およびフィンガー2領域のアミノ酸配列中か、またはαへリックスによって規定されるヒール領域のアミノ酸配列中には導入されなかった。ダッシュ(−)は、隣接するアミノ酸間でのペプチド結合を示す。各サブグループについてのコンセンサス配列のパターンは、各サブグループの最下段に示される。
【0069】
TGF−βスーパーファミリーのメンバー各々のアミノ酸配列が整列された後、この整列された配列を使用してアミノ酸配列の整列パターンを作製した。このパターンによって、得られた構築物の全体的な三次構造を変更することなく別のアミノ酸またはアミノ酸の群によって置換され得るアミノ酸残基が同定される。このフィンガー領域およびヒール領域中の特定の位置で有用であり得るアミノ酸またはアミノ酸の群は、図3に示されるアミノ酸の階層パターン構造を実行するコンピューターアルゴリズムによって同定された。
【0070】
簡潔には、このアルゴリズムは、4つのレベルの分析を行う。レベルIにおいては、このアルゴリズムは、特定のアミノ酸残基が、そのアミノ酸配列中の特定な位置で75%を超える頻度で生じるかどうかを決定する。例えば、アミノ酸配列中の特定の位置でグリシン残基が10回のうち8回生じる場合には、グリシンがその位置で指定される。試験される位置が全てギャップからなる場合には、ギャップ文字(−)がその位置に割り当てられる。そうでなければ、少なくとも1つのギャップが存在する場合には、「z」(任意の残基またはギャップについての表記)がその位置に割り当てられる。アミノ酸が、特定の位置で候補配列の75%で生じない場合には、アルゴリズムはレベルIIの分析を実行する。
【0071】
レベルIIは、パターンセットa、b、d、l、k、o、n、i、およびhを規定する。ここでは、l、k、およびoは、共通のアミノ酸残基を共有する。次いで、このアルゴリズムは、そのアミノ酸配列中の特定の位置のアミノ酸残基のうちの75%以上が上記のパターンのうちの1つを満たすかどうかを決定する。もしそうであれば、このパターンがその位置に割り当てられる。しかし、パターンlおよびkの両方が同時に満たされ得ることも可能である。なぜなら、これらは、同じアミノ酸(詳細には、アスパラギン酸)を共有するからである。lおよびkの同時の割り当てが生じる場合には、パターンm(レベルIII)が、その位置に割り当てられる。同様に、パターンkおよびoの両方が同時に割り当てられ得ることが可能である。なぜなら、これらは、同じアミノ酸(詳細には、グルタミン酸)を共有するからである。kおよびoの割り当てが同時に起こる場合には、パターンq(レベルIII)がその位置に割り当てられる。レベルIIのパターンおよびレベルIIIのパターン(mおよびq)のいずれもがそのアミノ酸配列中の特定の位置を満たさない場合には、このアルゴリズムはレベルIII分析を実行する。
【0072】
レベルIIIは、パターンセットc、e、m、q、p、およびjを規定する。ここで、m、q、およびpは、共通のアミノ酸残基を共有する。しかし、パターンqは、レベルIIIの分析においては試験されない。パターンmおよびpの両方が同時に満たされ得ることが可能である。なぜなら、これらは同じアミノ酸(詳細には、グルタミン酸)を共有するからである。mおよびpの同時の割り当てが生じる場合には、パターンr(レベルIV)がその位置に割り当てられる。整列されたアミノ酸配列中の予め選択された位置のアミノ酸のうちの75%がレベルIIIのパターンを満たす場合には、レベルIIIのパターンがその位置に割り当てられる。レベルIIIのパターンがその位置に割り当てられ得ない場合には、このアルゴリズムはレベルIVの分析を履行する。
【0073】
レベルIVは、2つの非重複パターンfおよびrを含む。そのアミノ酸配列中の特定の位置のアミノ酸のうちの75%がレベルIVのパターンを満たす場合には、このパターンがその位置に割り当てられる。レベルIVのパターンが割り当てられない場合には、このアルゴリズムは、その位置に任意のアミノ酸を示すX(レベルV)を割り当てる。
【0074】
図3においては、レベルIは、20個の天然に存在するアミノ酸を1文字のアミノ酸コードで大文字で列挙する。レベルII〜Vは、Smithら(前出)において示されているアミノ酸の階層に基づいて、アミノ酸の群を小文字で規定する。図5および6に示されるアミノ酸配列は、上記のコンピューターアルゴリズムを使用して整列された。
【0075】
当業者がTGF−βスーパーファミリーの現在同定されているメンバーに基づいて形態単位構築物を生成することを望む場合には、当業者は、図5に示されるアミノ酸配列を使用して、本発明の形態単位構築物の生成において有用な、フィンガー1領域、フィンガー2領域、およびヒール領域を提供し得る。本明細書以降に発見されたTGF−βスーパーファミリーのメンバーの場合、新規のメンバーのアミノ酸配列は、本発明の実施に有用なヒール領域およびフィンガー領域を規定するために、手動またはコンピューターアルゴリズムのいずれかによって、図5に示されている配列とともに整列され得る。
【0076】
以下の表1は、図5および6に示される配列整列パターンを生成するために使用された、TGF−βスーパーファミリーのメンバーのそれぞれのアミノ酸配列を記載する、刊行物をまとめる。
【0077】
【表1】

【0078】
【0079】
さらに本発明は、周知のタンパク質:GDF−5、GDF−7(米国特許番号第5,801,014号において開示されるように、この全体の開示が、本明細書中に参考として援用される);GDF−6(米国特許番号第5,770,444号において開示されるように、この全体の開示が、本明細書中に参考として援用される);ならびにBMP−12およびBMP−13(米国特許番号第5,658,882号において開示されるように、この全体の開示が、本明細書中に参考として援用される)に由来する対応するフィンガー1サブドメイン配列の使用を意図する。
【0080】
詳細には、本発明の実施において有用なフィンガー1領域を規定するアミノ酸配列が、本明細書中で同定された任意のTGF−βスーパーファミリーのメンバーについてのフィンガー1領域を規定するアミノ酸配列に対応することが、意図される。フィンガー1サブドメインは、ネイティブのタンパク質の特徴である、少なくとも生物学的および/または機能的特性(単数または複数)を付与し得る。有用なインタクトなフィンガー1領域として、以下が挙げられるがこれらに限定されない:
TGF−β1 配列番号40、残基2から29、
TGF−β2 配列番号41、残基2から29、
TGF−β3 配列番号42、残基2から29、
TGF−β4 配列番号43、残基2から29、
TGF−β5 配列番号44、残基2から29、
dpp 配列番号45、残基2から29、
Vg−1 配列番号46、残基2から29、
Vgr−1 配列番号47、残基2から29、
60A 配列番号48、残基2から29、
BMP−2A 配列番号49、残基2から29、
BMP−3 配列番号50、残基2から29、
BMP−4 配列番号51、残基2から29、
BMP−5 配列番号52、残基2から29、
BMP−6 配列番号53、残基2から29、
Dorsalin 配列番号54、残基2から29、
OP−1 配列番号55、残基2から29、
OP−2 配列番号56、残基2から29、
OP−3 配列番号57、残基2から29、
GDF−1 配列番号58、残基2から29、
GDF−3 配列番号59、残基2から29、
GDF−9 配列番号60、残基2から29、
インヒビンα 配列番号61、残基2から29、
インヒビンβA 配列番号62、残基2から29、
インヒビンβB 配列番号63、残基2から29、
CDMP−1/GDF−5 配列番号83、残基2から29、
CDMP−2/GDF−6 配列番号84、残基2から29、
GDF−6(マウス) 配列番号85、残基2から29、
CDMP−2(ウシ) 配列番号86、残基2から29、および
GDF−7(マウス) 配列番号87、残基2から29。
【0081】
本発明はさらに、周知のタンパク質BMP−12およびBMP−13(その全体の開示が本明細書中で参考として援用されている、米国特許第5,658,882号において開示されている)に由来する対応するヒールサブドメイン配列の使用を意図する。
【0082】
本発明の実施において有用なヒール領域を規定するアミノ酸配列が、本明細書中で同定された任意のTGF−βスーパーファミリーのメンバーのインタクトなヒール領域を規定するアミノ酸に対応することもまた、意図される。ヒール領域は、機能的特性および/またはフォールディング特性を含む、ネイティブのタンパク質の特性に少なくとも影響を与え得る。有用なインタクトなヒール領域として、以下が挙げられ得るが、これらに限定されない:
TGF−β1 配列番号40、残基35から62、
TGF−β2 配列番号41、残基35から62、
TGF−β3 配列番号42、残基35から62、
TGF−β4 配列番号43、残基35から62、
TGF−β5 配列番号44、残基35から62、
dpp 配列番号45、残基35から65、
Vg−1 配列番号46、残基35から65、
Vgr−1 配列番号47、残基35から65、
60A 配列番号48、残基35から65、
BMP−2A 配列番号49、残基35から64、
BMP−3 配列番号50、残基35から66、
BMP−4 配列番号51、残基35から64、
BMP−5 配列番号52、残基35から65、
BMP−6 配列番号53、残基35から65、
Dorsalin 配列番号54、残基35から65、
OP−1 配列番号55、残基35から65、
OP−2 配列番号56、残基35から65、
OP−3 配列番号57、残基35から65、
GDF−1 配列番号58、残基35から70、
GDF−3 配列番号59、残基35から64、
GDF−9 配列番号60、残基35から65、
インヒビンα 配列番号61、残基35から65、
インヒビンβA 配列番号62、残基35から69、
インヒビンβB 配列番号63、残基35から68、
CDMP−1/GDF−5 配列番号83、残基35から65、
CDMP−2/GDF−6 配列番号84、残基35から65、
GDF−6(マウス) 配列番号85、残基35から65、
CDMP−2(ウシ) 配列番号86、残基35から65、および
GDF−7(マウス) 配列番号87、残基35から65。
【0083】
本発明はさらに、周知のタンパク質BMP−12およびBMP−13(その全体の開示が本明細書中で参考として援用されている、米国特許第5,658,882号において開示されている)に由来する対応するフィンガー2サブドメイン配列の使用を意図する。
【0084】
本発明の実施において有用なフィンガー2領域を規定するアミノ酸配列が、本明細書中で同定された任意のTGF−βスーパーファミリーのメンバーについてのインタクトなフィンガー2領域を規定するアミノ酸配列に対応することもまた、意図される。フィンガー2サブドメインは、ネイティブのタンパク質の特徴である、少なくともフォールディング特性(単数または複数)を付与し得る。有用なインタクトなフィンガー2領域として、以下が挙げられ得るがこれらに限定されない:
TGF−β1 配列番号40、残基65から94、
TGF−β2 配列番号41、残基65から94、
TGF−β3 配列番号42、残基65から94、
TGF−β4 配列番号43、残基65から94、
TGF−β5 配列番号44、残基65から94、
dpp 配列番号45、残基68から98、
Vg−1 配列番号46、残基68から98、
Vgr−1 配列番号47、残基68から98、
60A 配列番号48、残基68から98、
BMP−2A 配列番号49、残基67から97、
BMP−3 配列番号50、残基69から99、
BMP−4 配列番号51、残基67から97、
BMP−5 配列番号52、残基68から98、
BMP−6 配列番号53、残基68から98、
Dorsalin 配列番号54、残基68から99、
OP−1 配列番号55、残基68から98、
OP−2 配列番号56、残基68から98、
OP−3 配列番号57、残基68から98、
GDF−1 配列番号58、残基73から103、
GDF−3 配列番号59、残基67から97、
GDF−9 配列番号60、残基68から98、
インヒビンα 配列番号61、残基68から101、
インヒビンβA 配列番号62、残基72から102、
インヒビンβB 配列番号63、残基71から101、
CDMP−1/GDF−5 配列番号83、残基68から98、
CDMP−2/GDF−6 配列番号84、残基68から98、
GDF−6(マウス) 配列番号85、残基68から98、
CDMP−2(ウシ) 配列番号86、残基68から98、および
GDF−7(マウス) 配列番号87、残基68から98。
【0085】
さらに、代表的なフィンガー領域およびヒール領域のアミノ酸配列が、アミノ酸置換によって(例えば、本明細書中に開示されているような置換残基を活用することによって)変更され得るか、またはSmithら(1990)、前出において開示されている原理に従って選択され得ることが、意図される。簡潔には、Smithらは、図3にまとめられているアミノ酸のクラスの階層に類似するアミノ酸のクラスの階層を開示する。これらは、タンパク質機能を含み得る型の全体的な立体構造的なひずみを最少にしながら、1つのアミノ酸を別のアミノ酸で合理的に置換するために使用され得る。任意の事象において、天然の領域とわずか70%の相同性、好ましくは80%の、そして最も好ましくは少なくとも90%の相同性を有する、多くの合成の第1のフィンガー領域、第2のフィンガー領域、およびヒール領域の配列が、本発明の構築物を産生するために使用され得ることが意図される。
【0086】
Smithら(1990)前出に記載されている原理に従って推定される、フィンガー領域およびヒール領域の各位置で好ましいアミノ酸を示すアミノ酸配列のパターンもまた、図5および図6に示され、そしてTGF−β;Vg/dpp;GDF;およびインヒビンサブグループのパターンと呼ばれる。各サブグループのフィンガー1、ヒール、およびフィンガー2の配列パターンを規定するアミノ酸配列は、それぞれ、図5A、5B、および5Cに示される。さらに、全体的なTGF−β、Vg/dpp、GDF、およびインヒビンサブグループのパターンを規定するアミノ酸配列は、配列番号64、65、66、および67として、それぞれ配列表に示される。
【0087】
図5A、5B、および5Cに開示され、そして図6にまとめられている、各サブグループについての好ましいアミノ酸配列のパターンは、当業者が、フィンガー1、ヒール、およびフィンガー2のエレメント中の特定の位置で組み込まれ得る代替的アミノ酸を同定することを可能にする。一文字のアミノ酸コードで大文字で示されているアミノ酸は保存されているアミノ酸を示し、これらは共に、フィンガー領域およびヒール領域の構造的および機能的エレメントを規定すると考えられる。図5および6中の大文字の「X」は、任意の天然に存在するアミノ酸がその位置で受容可能であることを示す。図5および6中の小文字の「z」は、ギャップまたは任意の天然に存在するアミノ酸のいずれかがその位置で受容可能であることを示す。小文字は、図5に示されているパターン規定の表に従って示されるアミノ酸を表記し、そしてその位置で有用であるアミノ酸の群を同定する。
【0088】
図5および6に示されているアミノ酸配列のサブグループのパターンに従って、例えば、当業者が、どの適用可能な1つのアミノ酸が、生じるタンパク質構築物中に破壊的な立体化学的変化を誘導することなく別のアミノ酸で置換され得るかを推測し得ることが意図される。例えば、図5Aにおいては、残基番号12でTGF−βサブグループのパターンにおいては、リジン残基(K)またはグルタミン残基(Q)のいずれかが、生じる構築物の構造に影響を与えることなくこの位置に存在し得ることが意図される。従って、位置12での配列パターンは、「n」を含み、これは、図10に従って、リジンまたはグルタミンからなる群から選択されるアミノ酸残基を規定する。従って、天然領域と70%の相同性、好ましくは80%の、そして最も好ましくは少なくとも90%の相同性を有する、多くの合成のフィンガー1領域、フィンガー2領域、およびヒール領域のアミノ酸配列が、本発明の立体構造的に活性なタンパク質を産生するために使用され得ることが意図される。
【0089】
これらの原理に従って、当業者が、図5および6に示される、TGF−β、Vg/dpp、GDF、またはインヒビンのサブグループのパターンに属するアミノ酸配列のパターンを用いて開始することによって合成構築物を設計し得ることが意図される。従って、従来の組換えまたは合成方法論を使用することによって、予め選択されたアミノ酸が、本明細書中の原理によって指示されるように別のアミノ酸で置換され得、そして生じるタンパク質構築物はアゴニスト活性またはアンタゴニスト活性とのいずれかとの組み合わせにおいて結合活性に対して試験される。
【0090】
TGF−βサブグループのパターンである、配列番号64は、TGF−β1、TGF−β2、TGF−β3、TGF−β4、およびTGF−β5を含む、今日までに同定されているTGF−βのサブグループのメンバーの中で共有されている相同性に適応する。以下に示される遺伝子配列は、保存アミノ酸(標準的な3文字コード)ならびに配列内の種々の位置に存在し、そして図3に示される規則によって規定される代替的アミノ酸(Xaa)の両方を含む。
【0091】
【化1】
【0092】
各Xaaは、以下に規定される1つ以上の特定アミノ酸の群から独立的に選択され得、ここで
【0093】
【化2】
【0094】
である。
【0095】
Vg/dppサブグループパターンである配列番号65は、現在までにdpp、vg−1、vgr−1、60A、BMP−2A(BMP−2)、Dorsalin、BMP−2B(BMP−4)、BMP−3、BMP−5、BMP−6、OP−1(BMP−7)、OP−2およびOP−3を含むと同定された、Vg/dppサブグループのメンバー間で共有される相同性を供給する。以下の一般的な配列は、この配列内の可変性位置に存在し、そして図3に示される規則によって規定される、保存されたアミノ酸(標準的な3文字コ
ード)ならびに代替のアミノ酸(Xaa)の両方を含む。
【0096】
【化3】
【0097】
各Xaaは、独立して、以下のように規定される1つ以上の特定のアミノ酸の群から選択され得る。ここで、
【0098】
【化4】
【0099】
【化5】
【0100】
である。
【0101】
GDFサブグループパターンである配列番号66は、現在までにGDF−1、GDF−3およびGDF−9を含むと同定された、GDFサブグループのメンバー間で共有される相同性を供給する。一般的な配列(以下に示される)は、この配列内の可変性位置に存在し、そして図3に示される規則によって規定される、保存されたアミノ酸(標準的な3文字コード)ならびに代替のアミノ酸(Xaa)の両方を含む。
【0102】
【化6】
【0103】
各Xaaは、独立して、以下のように規定される1つ以上の特定のアミノ酸の群から選択され得る。ここで、
【0104】
【化7】
【0105】
【化8】
【0106】
【化9】
【0107】
である。
【0108】
インヒビンサブグループパターンである配列番号67は、現在までにインヒビンα、インヒビンβAおよびインヒビンβBを含むと同定された、インヒビンサブグループのメンバー間で共有される相同性を供給する。一般的な配列(以下に示される)は、この配列内の可変性位置に存在し、そして図3に示される規則によって規定される、保存されたアミノ酸(標準的な3文字コード)ならびに代替のアミノ酸(Xaa)の両方を含む。
【0109】
【化10】
【0110】
各Xaaは、独立して、以下のように規定される1つ以上の特定のアミノ酸の群から選択され得る。ここで、
【0111】
【化11】
【0112】
【化12】
【0113】
【化13】
【0114】
である。
(2)骨形態形成タンパク質の生化学的、構造的、および機能的特性)
その成熟の、ネイティブ形態において、天然に供給される骨形成タンパク質は、グリコシル化されたダイマーであり、代表的には、SDS−PAGEによって決定されるように、約30〜36kDaの見かけの分子量を有する。還元された場合、この30kDaタンパク質は、約16kDaおよび18kDaの見かけの分子量を有するグリコシル化されたペプチドサブユニットを生じる。還元状態において、そのタンパク質は、検出可能な骨形成活性を有さない。グリコシル化されていないタンパク質(これもまた骨形成活性を有する)は、約27kDaの見かけの分子量を有する。還元された場合、この27kDaタンパク質は、約14kDa〜16kDaの分子量を有する2つのグリコシル化されていないペプチド鎖を生じる。代表的には、天然に存在する骨形成タンパク質は、代表的には約30残基未満のN末端シグナルペプチド配列、続いて成熟C末端ドメインを生じるために切断される「プロ」ドメインを有する前駆体として翻訳される。そのシグナルペプチドは、翻訳に際して、Von Heijne(1986)Nucleic Acids Research 14:4683−4691の方法を使用して、所定の配列において予測され得る切断部位で迅速に切断される。本明細書中で有用な骨形成タンパク質は、天然に存在するか、または生合成的に産生される(例えば、「ムテイン」または「変異タンパク質」を含む)かに関わらず、任意の公知の天然に存在するネイティブなタンパク質(その対立遺伝子の系統学的な対応物、および他の改変体を含む)、ならびに、一般的な形態形成ファミリーのタンパク質の骨形成的に活性な新規のメンバーを含む。
【0115】
なお別の好ましい実施態様において、有用な骨形成的に活性なタンパク質は、低ストリンジェンシー、中程度のストリンジェンシー、または高ストリンジェンシーのハイブリダイゼーション条件下で、参照骨形成配列(例えば、OP−1、OP−2、BMP2、4、5、6、60A、GDF5、GDF6、GDF7などの保存性の7つのシステインドメインを規定するC末端配列)をコードするDNAまたはRNAにハイブリダイズする核酸によってコードされる配列を含むアミノ酸配列を有するポリペプチド鎖を有する。本明細書中で使用される場合、高ストリンジェンシーハイブリダイゼーション条件は、40% ホルムアミド、5×SSPE、5×デンハルト溶液、および0.1% SDS中で、一晩37℃での既知の技術に従うハイブリダイゼーション、ならびに0.1×SSPE、0.1% SDS中、50℃での洗浄として規定される。標準的なストリンジェンシー条件は、市販の標準的な分子クローニングテキストにおいて十分に特徴付けられている。例えば、Molecular Cloning A Laboratory Manual、第2版、Sambrook、Fritsch、およびManiatis編(Cold Spring Harbor Laboratory Press:1989);DNA Cloning、第I巻および第II巻(D.N.Glover編、1985);Oligonucleotide Synthesis(M.J.Gait編、1984);Nucleic Acid Hybridization(B.D.Hames & S.J.Higgins編、1984);ならびにB.Perbal、A Practical Guide To Molecular Cloning(1984)を参照のこと。
【0116】
本発明の実施において有用性を有する関連するタンパク質のTGF−βスーパーファミリーの他のメンバーには、以下のリストの中でさほどリフォールディングされていないタンパク質が含まれる:いくつか名前を挙げれば、TGF−β1、TGF−β2、TGF−β3、TGF−β4、およびTGF−β5、種々のインヒビン、アクチビン、BMP−11、ならびにMISである。図5Cは、TGF−βスーパーファミリーの種々の既知のメンバーのフィンガー2サブドメインを規定するC末端残基を列挙する。さほどフォールディングされていない、このリスト上のタンパク質のいずれか1つは、他の公知のファミリーメンバーまたは発見可能なファミリーメンバーであり得るように、本発明の方法によって改善され得る。
【0117】
(B.組換えタンパク質の産生)
上で述べたように、本発明の構築物は、当該分野において周知であり、そして徹底的に実証された従来の組換えDNA方法論の使用によって、ならびに慣用的なペプチド化学またはヌクレオチド化学および自動化したペプチド合成機またはヌクレオチド合成機を用いた周知の生合成の方法論および化学合成の方法論の使用によって、製造され得る。このような慣用的方法論は、例えば以下の出版物に記載され、その教示は本明細書中で参考として援用される:Hilvert,1
Chem.Biol.201−3(1994);Muirら,95 Proc.Natl.Acd.Sci.USA 6705−10(1998);Wallace,6 Curr.Opin.Biotechnol.403−10(1995);Mirandaら,96 Proc.Natl.Acd.Sci.USA 1181−86(1999);Liuら,91 Proc.Natl.Acad.Sci.USA 6584−88(1994)。本発明中の使用に適したものは、天然に存在するアミノ酸およびヌクレオチド;天然には存在しないアミノ酸およびヌクレオチド;修飾されたアミノ酸および異常なアミノ酸;修飾された塩基;翻訳後に修飾されたアミノ酸および/または修飾された連結、架橋およびエンドキャップ、非ペプチジルボンド、などを含むアミノ酸配列であり、そして、付録2中の表1〜6を含むWorld Intellectual Property Organization(WIPO) Handbook on
Industrial Property Information and
Documentation,Standard St.25(1998)に開示された部分がさらに挙げられるが、これらに限定されない(これらは本明細書中で参考として援用される)。これらの等価物は、当該分野の知識と共に慣用的実験法のみに依存して、当業者によって評価され得る。
【0118】
例えば、企図されるDNA構築物は、合成ヌクレオチド配列のアセンブリおよび/またはDNA制限酵素フラグメントを連結して合成DNA分子を産生することによって製造され得る。次いでこのDNA分子は、発現ビヒクル(例えば発現プラスミド)へ連結され、そして適切な宿主細胞(例えば、E.coli)へトランスフェクトされる。このDNA分子によってコードされる企図されたタンパク質構築物が、次いで発現され、精製され、リフォールディングされ、特定の特性(例えば、鋳型TGF−βスーパーファミリーメンバーに対する結合親和性を有するレセプターとの結合活性)に関してインビトロにおいて試験され、そしてこの生合成構築物が、鋳型TGF−βスーパーファミリーメンバーの他の適切な特性を模倣するかどうかを評価するために引き続いて試験された。
【0119】
あるいは、合成DNA構築物のライブラリーが、例えば、前もって選んだ領域内のヌクレオチド組成と異なる合成ヌクレオチド配列のアセンブリによって同時に調製され得る。例えば、特異的TGF−βスーパーファミリーメンバーに基づく構築物の産生中に、当業者が、このようなスーパーファミリーメンバーにとって適切なフィンガーおよびヒールの領域を選び得ることが意図される(例えば、図5〜6より)。一旦、適切なフィンガーおよびヒールの領域が選ばれると、当業者は、次いでこれらの領域をコードする合成DNAを産生し得る。例えば、異なるリンカー配列をコードする複数のDNA分子が、フィンガーおよびヒールの配列をコードするDNA分子を含む連結反応に含まれるならば、適切な制限酵素部位および反応条件の慎重な選択によって、当業者は、各々のDNA構築物がフィンガーおよびヒールの領域をコードするが異なるリンカー配列によって結合されたDNA構築物のライブラリーを産生し得る。その結果生じるDNAは、次いで適切な発現ビヒクル(すなわち、ファージディスプレイライブラリーの調製に有用なプラスミド)へ連結され、宿主細胞へ形質転換され、そして合成DNAによってコードされるポリペプチドは、候補タンパク質のプールを産生するために発現させた。候補タンパク質のプールは、前もって選んだレセプターに対する結合親和性および/または選択性を有する特異的なタンパク質を同定するために引き続いてスクリーニングされ得る。
【0120】
スクリーニングが、候補タンパク質を含む溶液をレセプターが固定化された表面を備えるクロマトグラフィーカラムを通じて通過させることによって実施され得る。次いで、所望の結合特異性を伴うタンパク質が、例えば、塩勾配および/または鋳型TGF−βスーパーファミリーメンバーの濃度勾配によって溶出される。このようなタンパク質をコードするヌクレオチド配列は、引き続き単離され得そして特徴付けられ得る。一旦、適切なヌクレオチド配列が同定されると、リードタンパク質は、従来の組換えDNA方法論またはペプチド合成方法論のいずれかによって、特定の構築物が鋳型TGFβ−スーパーファミリーメンバーの活性を模倣するかどうかを試験するために十分な量が引き続き産生され得る。
【0121】
どちらのアプローチが本発明の構築物をコードするDNA分子を産生するために採用されても、好ましいタンパク質の三次構造が、例えば、本明細書中に記載の原理によって援助されるヌクレオチド変異誘発方法論およびファージディスプレイ方法論の組合せによって、結合活性および/または生物学的活性を最適化するために引き続き調節され得ることが企図される。従って、当業者は、多くのこのようなタンパク質を産生し得、そして同時に試験し得る。
【0122】
(1)遺伝子合成
目的のアミノ酸配列をコードするDNAを製造し、増幅し、そして組換えるためのプロセスは、該して当該分野において周知であり、それゆえ、本明細書中に詳細には記載されない。TGF−βスーパーファミリーメンバーをコードする遺伝子および同族のレセプターを同定しそして単離する方法はまた、十分に理解されており、そして本特許および他の文献に記載される。
【0123】
手短には、本明細書中に開示された生合成構築物をコードするDNAの構築は、配列特異的にDNAを切断し平滑末端または突出末端を産生する種々の制限酵素、DNAリガーゼ、突出末端を平滑末端化されたDNAへの酵素的付加を可能にする技術、短いまたは中程度の長さのオリゴヌクレオチドのアセンブリによる合成DNAの構築、cDNA合成技術、ライブラリーから適切な核酸配列を増幅するためのポリメラーゼ連鎖反応(PCR)技術、ならびにTGF−βスーパーファミリーメンバーおよびそれらの同族のレセプターの遺伝子を単離するための合成プローブの使用を含む周知の技術を用いて実施される。発現の達成に用いられる(少しの例を挙げれば)細菌、哺乳動物、または昆虫由来の種々のプロモーター配列および他の調節DNA配列ならびに種々の型の宿主細胞もまた、公知でありそして入手可能である。従来のトランスフェクションの技術、およびDNAをクローニングおよびサブクローニングするための同等に従来の技術は、本発明の実施に有用であり、そして当業者に周知である。種々の型のベクター(例えばプラスミド、ならびに動物ウイルスおよびバクテリオファージを含むウイルス)が用いられ得る。このベクターは、首尾よくトランスフェクトされた細胞に検出可能な表現型特性(ベクターの組換えDNAを首尾よく組み込まれたクローンのファミリーを同定するために用い得る特性)を与える種々のマーカー遺伝子を利用し得る。
【0124】
本明細書中に開示された生合成構築物をコードするDNAを得るための方法の一つは、従来の、自動化オリゴヌクレオチド合成機において産生される合成オリゴヌクレオチドのアセンブリに引き続いて適切なリガーゼを用いる連結によるものである。例えば、重複した、相補的なDNAフラグメントは、ホスホルアミダイド化学を用いて合成され得、末端セグメントは、連結中に重合を防ぐためにリン酸化されないままである。合成DNAの一端は、特定の制限エンドヌクレアーゼの作用の部位に相当する「突出末端」を伴ったままにし、そして他端は、別の制限エンドヌクレアーゼの作用の部位に相当する末端を伴ったままにする。この相補的DNAフラグメントは、共に連結されて、合成DNA構築物を産生する。
【0125】
あるいはフィンガー1領域、フィンガー2領域およびヒール領域をコードする核酸鎖が、例えば、Sambrookら編(1989)「Molecular Cloning」、Coldspring Harbor Laboratories Press、NYに記載のようなコロニーハイブリダイゼーション手順によって、および/またはInnisら(1990)「PCR Protocols,A guide to methods and applications」、Academic Pressに開示のようなPCR増幅方法論によって、核酸のライブラリーから単離され得る。このフィンガー領域およびヒール領域をコードする核酸は、次いで共に連結され、目的の生合成一本鎖形態単位構築物をコードする合成DNAを産生する。
【0126】
しかし、その複数の形態単位(morphon)をコードするDNA構築物のライブラリーは、例えば、上述したもののような、標準的な組換えDNA方法論によって同時に産生され得る。例えば、カセット変異誘発またはオリゴヌクレオチド特異的変異誘発の使用によって、当業者は、例えば、あらかじめ定義された部位の範囲内(例えば、リンカー配列をコードするDNAカセットの範囲内)に異なるDNA配列を各々が含む一連のDNA構築物を産生し得る。その結果生じるDNA構築物のライブラリーは、例えば、ファージディスプレイライブラリーにおいて発現され得る;そして特異的レセプターに結合する任意のタンパク質構築物は、アフィニティ精製(例えば、レセプターが固定化された表面を備えるクロマトグラフィーカラムを使用して)によって単離され得る(以下のV節を参照のこと)。一旦前もって選択されたレセプターに結合する分子が単離されると、それらの結合特性およびアゴニストの特性は、以下のV節においてまた議論されるように、経験的な精製技術を用いて調節され得る。
【0127】
タンパク質および核酸の変異誘発の方法は、周知であり、当該分野に十分に記載されている。例えば、Sambrookら、(1990) Molecular Cloning:A Laboratory Manual.、第2版(Cold Spring Harbor、N.Y.:Cold Spring Harbor Laboratory
Press)を参照のこと。有用な方法は、PCR(重複伸長、例えば、PCR Primer(DieffenbachおよびDveksler編、Cold Spring
Harbor Press、Cold Spring Harbor、NY、1995、603〜611頁)を参照のこと);クンケル法の後のカセット変異誘発および一本鎖変異誘発を含む。これは、任意の適切な変異誘発方法が利用され得、そしてこの変異誘発法が本発明の重要な局面とみなされないことは当業者によって認識される。アルギニン(Arg)、グルタミン酸(Glu)およびアスパラギン酸(Asp)を含むアミノ酸をコードするのに的確な核酸コドンはまた、周知でありそして当該分野において記載される。例えば、Lehninger、Biochemistry(Worth Publishers、N.Y.、N.Y.)を参照のこと。アルギニン、グルタミン酸およびアスパラギン酸をコードする標準的なコドンは、Arg:CGU、CGC、CGA、CGG,AGA,AGG;Glu:GAA、GAG;およびAsp:GAU、GACである。本発明のキメラの構築物は、切り替えられるタンパク質領域の核酸配列またはドメインを整列化すること、ならびに適合性のスプライス部位を同定することおよび/またはPCR重複伸長を用いた適切な乗り換え配列を構築することによって容易に構築され得る。
【0128】
本発明のTGF−βファミリーメンバーの変異型は、標準的な周知の方法を使用して、細菌中で産生され得る。C末端の7つのシステインドメインのみを規定する全長成熟形態またはより短い配列は、宿主細胞に提供され得る。細菌発現を最適化するために、変異型タンパク質のN末端配列を改変することが好ましくあり得る。例えば、ネイティブなOP−1の細菌発現のために好ましい形態は、C末端の7つのシステインドメイン(例えば、配列番号39の残基330〜431)をコードする、成熟の活性配列(配列番号39の残基293〜431)またはそのフラグメントをコードする配列である。メチオニンは、ネイティブのセリン残基を置き換えて293位において導入され得るか、またはそれは、このセリン残基の前に位置し得る。あるいは、メチオニンは、TGF−βドメインの最初のシステインまでの天然の配列の最初の36残基(残基293〜329)以内のいずれかの場所に導入され得る。そのDNA配列はさらに、精製を改善するために、例えば、IMACカラム上での精製を補助するために「ヘキサhis」テールを付加することによって;またはIgG/カラム上の精製を容易にする、FBリーダー配列を使用することによって、そのN末端において改変され得る。これらおよび他の方法は、十分に記載されており、そして当該分野で周知である。他の細菌種および/またはタンパク質は、そこから得られた変異体BMPの収量を最適化するために、類似の改変からの利点を必要とし得る。このような改変は、十分に当業者のレベル内であり、そして本発明の実質的な局面とはみなされない。
【0129】
合成核酸は、好ましくは、選択した宿主細胞における過剰発現に適切なベクターへ挿入される。任意の発現ベクターは、それが、選択した宿主細胞における異種タンパク質(例えば、BMP)の発現を方向付け得る限り、使用され得る。有用なベクターとしては、少し例を挙げると、プラスミド、ファージミド、ミニクロモソームおよびYACが挙げられるが、これらに限定されない。他のベクター系は周知であり、そして当該分野で特徴付けられている。このベクターは、代表的にはレプリコン、一つ以上の選択マーカー遺伝子配列、および宿主細胞に高コピー数のベクターを維持するための手段を含む。周知の選択マーカー遺伝子は、アンピシリン、テトラサイクリンなどのような抗生物質および重金属耐性を含む。酵母細胞で使用するために有用な選択マーカー遺伝子は、栄養要求性の酵母変異宿主とともに使用するためのURA3遺伝子、LEU2遺伝子、HIS3遺伝子またはTRP1遺伝子を含む。加えて、ベクターもまた、目的の遺伝子を発現するための適切なプロモーター配列(これは誘導性であっても、誘導性でなくてもよい)、ならびに所望であれば、有用な転写開始部位および翻訳開始部位、ターミネーター、ならびに目的の遺伝子の転写および翻訳を最大化し得る他の配列を含む。細菌細胞において特に有用な、十分特徴付けられたプロモーターとしては、少し例を挙げれば、lacプロモーター、tacプロモーター、trpプロモーター、およびtppプロモーターが挙げられる。酵母で有用なプロモーターとしては、例えば、ADHI、ADHII,またはPHO5プロモーターが挙げられる。
【0130】
適切な宿主細胞としては、Bacillus subtilis(B.subtilis)、Pseudomonasの種、Escherichia coli(E.coli)および酵母細胞(例えば、Saccharomyces cerevisiae)のような細菌細胞が挙げられる。他の宿主細胞(例えば、CHOのような哺乳動物細胞)が使用され得る。
【0131】
目的の遺伝子は、標準的な微生物学技術(例えば、エレクトロポレーションまたは塩化カルシウム)を用いて選択した宿主細胞へ形質転換され得、そして適切な条件下でこの細胞を誘導して、増殖させる。細胞培養培地は、多くの周知の基本書を含め、当該分野で十分に記載され、そしてこれらとしては、Sambrookらが挙げられる。有用な培地としては、LB(ルリアブロス)およびダルベッコDMEMが挙げられる。過剰発現されたタンパク質は、標準的な技術(細胞溶解または細胞の機械的破砕(例えば、フレンチプレス、SLM Instruments,Inc,)を含む)による不溶性の屈折封入体(refractile inclusion body)から採取され、次いで、遠心分離および再溶解され得る(以下を参照のこと)。
【0132】
例えば、この遺伝子がE.coliで発現される場合、適切な発現ベクターへクローニングされる。このことは、プロモーター(例えば、TrpまたはTac)の下流に、操作された遺伝子および/またはプロテインAのフラグメントB(FB)のようなリーダーペプチドをコードする遺伝子を配置することにより達成され得る。発現の間に、得られた融合タンパク質は、細胞の細胞質中の屈折体に蓄積し、そしてフレンチプレスまたは超音波処理による細胞の崩壊の後に収集され得る。単離された屈折体は、次いで可溶化され、そして発現したタンパク質は折り畳まれ、そして必要であれば、多くの他の組換えタンパク質に対して既に確立された方法により、リーダーペプチドが切断される。
【0133】
真核生物細胞での操作された遺伝子の発現は、細胞および細胞株を必要とし、そしてこれらの細胞および細胞株はトランスフェクトするのに容易であり、再配列されない配列を伴う外来DNAを安定に維持することが可能であり、これらの細胞および細胞株は、効率的な転写、翻訳、翻訳後修飾、およびタンパク質の分泌に必要な細胞内構成成分を有する。さらに、目的の遺伝子を運ぶ適切なベクターもまた、必要である。哺乳動物細胞へのトランスフェクションのためのDNAベクターの設計は、本明細書中に記載されるような目的の遺伝子の発現を促進するために適切な配列(適切な転写開始配列、終止配列、およびエンハンサー配列ならびに、コザックコンセンサス配列のような翻訳効率を増強する配列を含む)を含むべきである。好ましいDNAベクターはまた、マーカー遺伝子および目的の遺伝子のコピー数を増幅するための手段を含む。哺乳動物細胞での外来タンパク質の産生の技術水準(有用な細胞、タンパク質発現促進配列、マーカー遺伝子、および遺伝子増幅法を含む)の詳細な総説は、Bendig(1988) Genetic Engineering 7:91〜127に開示される。
【0134】
特定の哺乳動物細胞において外来遺伝子を発現するために有用な最も特徴付けられた転写プロモーターは、SV40初期プロモーター、アデノウイルスプロモーター(AdMLP)、マウスメタロチオネインIプロモーター(mMT−I)、ラウス肉腫ウイルス(RSV)長末端反復(LTR),マウス乳腺腫ウイルス長末端反復(MMTV−LTR)、およびヒトサイトメガロウイルス主要即時初期(intermediate−early)プロモーター(hCMV)である。これらのプロモーターのすべてに関するDNA配列は、当該分野において公知であり、そして市販されている。
【0135】
dhfr-細胞株における選択可能なDHFR遺伝子の使用は、哺乳動物細胞系におけ
る遺伝子の増幅に有用な、十分に特徴付けられた方法である。簡単に、このDHFR遺伝子は、目的の遺伝子を運ぶベクター上に提供され、そしてDHFRによって代謝される細胞傷害性の薬剤メトトレキセートの漸増濃度の添加は、目的の関連遺伝子増幅と同様にDHFR遺伝子コピー数の増幅を導く。トランスフェクトされたチャイニーズハムスター卵巣細胞株(CHO細胞)において選択可能で、増幅可能なマーカー遺伝子としてのDHFRは、当該分野において特に十分に特徴付けられている。他の有用な増幅可能なマーカー遺伝子は、アデノシンデアミナーゼ(ADA)遺伝子およびグルタミンシンターゼ(GS)遺伝子を含む。
【0136】
細胞/細胞株の選択はまた、重要であり、そして実験者の要求に依存する。COS細胞は、本発明の生合成構築物を迅速にスクリーニングするために有用な手段を提供する、高いレベルの一過性の遺伝子発現を提供する。COS細胞は、代表的には目的の遺伝子を運ぶシミアンウイルス40(SV40)ベクターと共にトランスフェクトされる。トランスフェクトされたCOS細胞は、最終的に死滅し、それ故に所望のタンパク質産生物の長期間の産生が妨げられる。しかし、一過性の発現は、安定な細胞株の開発のために必要とされる時間のかかる過程を必要とせず、従って結合活性について予備構築物を試験するために有用な技術を提供する。
【0137】
本発明の一本鎖構築物の哺乳動物細胞発現のために使用される種々の細胞、細胞株およびDNA配列は、当該分野で十分に特徴付けられており、そして容易に入手可能である。他のプロモーター、選択マーカー、遺伝子増幅方法および細胞もまた使用して、本発明のタンパク質を発現させ得る。組換えタンパク質のトランスフェクション、発現および精製の具体的な詳細は、当該分野で十分に記載されており、そして当業者により理解される。哺乳動物細胞発現系における外来遺伝子の組換え生成において使用される各工程の種々の技術的局面に対するさらなる詳細は、当該分野における多くの基本書および実験室マニュアルにおいて見出され得る(例えば、F.M.Ausubelら編、Current Protocols in Molecular Biology,John Wiley&Sons,New York(1989))。
【0138】
(c)リフォールディングの考慮事項
一旦封入体から単離されると、タンパク質は、グアニジン塩酸または尿素のような変性剤またはカオトロピック剤によって、好ましくは約4〜9Mの範囲でそして高い温度(例えば25〜37℃)および/または塩基性pH(8〜10)で可溶化される。あるいは、タンパク質は、酸性化によって(例えば、酢酸またはトリフルオロ酢酸を用いて)、一般に1〜4の範囲のpHで可溶化され得る。好ましくは、β−メルカプトエタノールまたはジチオトレイトール(DTT)のような還元剤が、可溶化剤と組合せて用いられる。この可溶化された異種のタンパク質は、透析によっておよび/または公知のカラムクロマトグラフィーの方法によって(例えば、サイズ排除クロマトグラフィー、イオン交換クロマトグラフィー、または逆相高速液体クロマトグラフィー(RP−HPLC)などによって)、可溶化したカオトロープ(caotrope)からさらに精製され得る。
【0139】
可溶化されたタンパク質は、以下のようにリフォールディングされ得る。溶解したタンパク質は、リフォールディング媒体(代表的には、約pH5.0〜10.0の範囲のpH、好ましくは約pH6〜9の範囲のpHを有し、ならびに界面活性剤および/またはカオトロピック剤を含むトリス緩衝化媒体)中に希釈される。有用な市販の入手可能な界面活性剤は、例えば、NP40(Nonidet
40)、CHAPS(3−[(3−コラミドプロピル)ジメチルアンモニオ]−1−プロパン−スルフェートのような)、ジギトニン、デオキシコレート、またはN−オクチルグルコシドのような、イオン性、非イオン性、または双性イオン性であり得る。有用なカオトロピック剤としては、グアニジン、尿素、またはアルギニンが挙げられる。好ましくは、界面活性剤またはカオトロピック剤は、約0.1〜10Mの範囲、好ましくは0.5〜4Mの範囲の濃度で存在する。CHAPSが界面活性剤の場合、これは、好ましくは溶液の約0.5〜5%、さらに好ましくは溶液の約1〜3%含まれる。好ましくは溶液はまた、少し例を挙げると酸化型および還元型のグルタチオン、DTT,β−メルカプトエタノール、β−メルカプトメタノール、システインまたはシスタミンのような適切な酸化還元系を含む。好ましくは、酸化還元系は、還元剤対酸化剤の比が約1:1から約5:1の範囲で存在する。グルタチオン酸化還元系が用いられる場合、還元型グルタチオン対酸化型グルタチオンの比は、還元型対酸化型が好ましくは約0.5対5;さらに好ましくは1対1;そして最も好ましくは2対1の範囲である。好ましくは緩衝液はまた、約0.25〜2.5Mの範囲、好ましくは約0.5〜1.5Mの範囲、最も好ましくは約1Mの範囲にて存在する塩(代表的にはNaCl)を含む。当業者は、上記の条件および媒体が、通常の実験法のみを用いて変化され得ることを認識する。このような変動および改変は、本発明の範囲内である。
【0140】
好ましくは、所定のリフォールディング反応のためのタンパク質濃度は、約0.001〜1.0mg/mlの範囲であり、さらに好ましくはこれは約0.05〜0.25mg/mlの範囲であり、最も好ましくは0.075〜0.125mg/mlの範囲である。当業者によって認識されるように、より高濃度が、凝集物をより多く産生する傾向がある。ヘテロダイマーが産生される場合(例えばOP−1/BMP2またはBMP2/BMP6ヘテロダイマー)、好ましくは個々のタンパク質は、等量のリフォールディング緩衝液に提供される。
【0141】
代表的にリフォールディング反応は、約4℃〜約25℃の温度範囲にて生じる。より好ましくは、リフォールディング反応を4℃にて実行し、そして完了させる。代表的に、リフォールディングは約1日〜7日で完了し、一般的に、タンパク質に依存して、16〜72時間または24〜48時間以内である。当業者に理解されるように、リフォールディング速度はタンパク質により変動し得、そしてより長いリフォールディング時間およびより短いリフォールディング時間が意図され、そして本発明の範囲内である。本明細書中で使用される場合、「良好なリフォルダー(refolder)」タンパク質は、このタンパク質の少なくとも20%が、リフォールディング反応における全タンパク質と比較されるされる場合、本明細書中に記載される任意のリフォールディングアッセイにより測定されるように、そしてさらなる精製を必要とせずに、フォールディング反応後のダイマー形態で存在する場合である。当該分野において「良いリフォルダー」タンパク質と考えられているネイティブなBMPとしては、BMP2、CDMP1、CDMP2およびCDMP3が挙げられる。BMP−3はまた、合理的に十分にリフォールディングされる。対照的に、「乏しいリフォルダー」タンパク質は、1%未満の適切に折り畳まれたタンパク質を産生する。
【0142】
適切にリフォールディングされたダイマータンパク質は、多くの周知かつ十分に特徴付けられたアッセイのいずれかを用いて容易にアッセイされ得る。特に、3つのアッセイ(全てが当該分野で周知かつ十分に記載され、そして以下にさらに記載される)の内の任意の1つ以上を使用して、利益を得ることができる。有用なリフォールディングのアッセイは、1つ以上の以下のものを含む。第1に、ダイマーの存在は、DTTのような還元剤の非存在下で標準的SDS−PAGEによるか、またはHPLC(例えば、C18逆相HPLC)によるかのいずれかで視覚的に検出され得る。BMPダイマータンパク質は、モノマーサブユニット(これは、約14〜18kDaの見かけの分子量を有する)と比較して、約28〜36kDaの範囲の見かけの分子量を有する。ダイマータンパク質は、市販の分子量標準物質との比較により電気泳動ゲル上で容易に視覚化され得る。このダイマータンパク質はまた、約19分で、C18 RP HPLC(45〜50%アセトニトリル:0.1%TFA)から溶出され得る(哺乳動物が生成したhOP−1は、18.95分で溶出する)。
【0143】
第2のアッセイは、ヒドロキシアパタイトに結合する能力によりダイマーの存在を評価する。適切にフォールディングされたダイマーは、モノマーと比較して、0.1〜0.2M NaClの存在下で(ダイマーは0.25M NaClで溶出する)ヒドロキシアパタイトカラムに十分結合する(モノマーは、この濃度では実質的に結合しない)(モノマーは、0.1Mで溶出する)。
【0144】
第3のアッセイは、トリプシンまたはペプシン消化に対するタンパク質の抵抗性によりダイマーの存在を評価する。このフォールディングされたダイマー種は、両酵素(特に、成熟タンパク質のN末端の小部分のみを切断するトリプシン)に対して実質的に抵抗性であり、このことにより、生物学的に活性なダイマー種を未処理ダイマーよりごくわずかにより小さなサイズにする。対照的に、モノマーは、実質的に分解される。このアッセイにおいて、このタンパク質は、標準的な条件(例えば、4M尿素、100mM NaCl、0.3%Tween−80および20mM メチルアミンを含有する、50mM Tris緩衝液(pH8)のような標準的な緩衝液中での消化)を用いて酵素消化に供される。消化は、16時間のオーダーで37℃で生じさせ、そしてこの産物を、任意の適切な手段(好ましくは、SDS−PAGE)により可視化させる。
【0145】
再折りたたみされたTGF−βファミリータンパク質の生物学的活性は、任意の多くの手段によって容易に評価され得る。BMPの軟骨内骨形成を誘導する能力は、十分に特徴付けされたラット皮下骨アッセイを使用して評価され得、当該分野でおよび下記に詳細に記載される。このアッセイにおいて、骨形成は、組織学ならびにアルカリホスファターゼおよび/またはオステオカルシン(osteoclacin)産生によって測定される。さらに、高い特異的骨形成活性を有する骨形成タンパク質(例えば、OP−1、BMP−2、BMP−4、BMP5およびBMP6)はまた、インビトロのラット骨芽細胞または骨肉腫細胞ベースのアッセイにおいてアルカリホスファターゼ活性を誘導する。このようなアッセイは、当該分野においてよく記載されており、本明細書中下記に詳細に記載される。例えば、Sabokdarら、(1994)Bone and Mineral 27:57〜67;Knutsenら、(1993)Biochem.Biophys.Res.Commun.194:1352〜1358;およびMaliakalら、(1994)Growth Factors 1:227〜234)を参照のこと。対照的に、低い特異的骨形成活性を有する骨形成タンパク質(例えば、CDMP−1およびCDMP−2)は、細胞ベースの骨芽細胞アッセイにおけるアルカリホスファターゼ活性の同様なレベルを誘導しない。従って、このアッセイは、BMPの生物学的活性変異体を評価するための容易な方法を提供する。例えば、BMP2、BMP4、BMP5、BMP6またはOP−1よりも低い比活性を有するが、CDMP1、CDMP2およびCDMP3は全て骨形成を誘導する成分である。逆に、CDMP1、CDMP2またはCDMP3よりも低い比活性を有するが、BMP2、BMP4、BMP5、BMP6およびOP−1はすべて関節軟骨形成を誘導し得る。従って、実施例5の細胞ベースのアッセイにおいてアルカリホスファターゼ活性を誘導するコンピテントなCDMP変異体は、ラット動物バイオアッセイにおいてより高い特異的骨形成活性を実証することが予期される。同様に、CDMP1、CDMP2またはCDMP3タンパク質の対応する位置に存在する置換を含み、そしてラットアッセイにおいて骨を誘導するが、細胞ベースのアッセイにおいてアルカリホスファターゼ活性を誘導しないようにコンピテントである、OP−1変異体は、インビボ関節軟骨アッセイにおいて、より高度に特異的な関節性軟骨誘導活性を有することが予期される。本明細書中下記に記載されるように、CDMP活性についての適切なインビトロアッセイは、マウス胚の前骨芽細胞またはマウス癌細胞(例えば、ATDC5細胞)を利用する。下記実施例6を参照のこと。
【0146】
TGF−β活性は、このタンパク質の上皮細胞増殖を阻害する能力によって容易に評価され得る。有用な、十分に特徴付けられたインビトロアッセイは、ミンク肺細胞または黒色腫細胞を利用する。実施例7を参照のこと。TGF−βスーパーファミリーの他のメンバーについての他のアッセイは、文献において十分に記載されており、そして、過度の実験なく実施され得る。
【0147】
(D.処方および生物活性)
生じたキメラタンパク質は、インビボの事象を増強、阻害またはそうでなければ調節をするための治療の一部として個体に提供され得、その事象としては、TGF−βスーパーファミリーメンバーと1つ以上のその同族レセプターとの間の結合相互作用が挙げられるが、これに限定されない。下記のように、この構築物は薬学的組成物に処方され得、そして任意の適切な手段(好ましくは、直接または全身的に、例えば、非経口的または経口的に)よって、形態形成的に有効な量において投与され得る。好ましいキメラタンパク質をコードする、生じたDNA構築物はまた、遺伝子治療の目的のために、レシピエントに直接投与され得、このようなDNAは、キャリア成分を伴うか、もしくは伴わずに、またはマトリクス成分を伴うか、もしくは伴わずに投与され得る。あるいは、このようなDNA構築物を移入された細胞は、レシピエントに移植され得る。このような物質および方法は、当該分野において周知である。
【0148】
本明細書中に開示されるこの任意の構築物が直接(例えば、注射により、所望される組織部位に局所的に)または非経口的に(例えば、静脈内投与、皮下投与、筋肉内投与、眼窩内投与、眼投与、脳室内投与、頭蓋内投与、包内投与、脊髄内投与、槽内投与、腹腔内投与、頬内投与、直腸投与、膣投与、鼻腔内投与によって、またはエアロゾル投与によって)提供されるべきである場合、この治療的組成物は、好ましくは水溶液の部分を含む。この溶液は、好ましくは、患者への所望の構築物の送達に加えて、この溶液が、他の点では患者の電解質バランスおよび体積バランスに不利な影響を及ぼさないように、生理学的に受容可能である。従って、この治療分子のための水性媒体としては、例えば、通常の生理的食塩水(0.9%NaCl、0.15M)、pH7〜7.4またはその他の薬学的に受容可能な塩を含み得る。
【0149】
経口または非経口的投与のための有用な溶液は、薬学的分野において周知の任意の方法によって調製され得る(例えば、Remington’s Pharmaceutical Sciences,(Gennaro,A.編),Mack Pub.,1990に記載される)。例えば、処方物はポリアルキレングリコール(例えば、ポリエチレングリコール)、野菜起源の油、水素化ナフタレンなどを含み得る。特に、直接投与するための処方物は、グリセロールおよび他の高い粘度の組成物を含み得る。生体適合性、好ましくは生体再吸収可能な(bioresorbable)ポリマー(例えば、ヒアルロン酸、コラーゲン、リン酸三カルシウム、ポリ酪酸、ポリラクチド、ポリグリコリド、およびラクチド/グリコリドコポリマーを含む)が、インビボでのモルフォゲンの放出を制御するための有用な賦形剤であり得る。
【0150】
これら治療的分子のための他の潜在的に有用な非経口的送達系は、エチレンビニル酢酸コポリマー粒子、浸透圧ポンプ(osmotic pump)、移植可能な注入系(implantable infusion system)およびリポソームを含む。吸入投与のための処方物は、賦形剤として例えばラクトースを含み得るか、あるいは、例えば、ポリオキシエチレン−9−ラウリルエーテル、グリココール酸およびデオキシコール酸を含む水溶液もしくは点鼻薬の形態における投与のための油性溶液であり得るか、または鼻腔内に適用されるゲルとしてであり得る。
【0151】
最終的に、治療的分子は、単独または他の組織形態形成に影響を及ぼすことが公知である他の分子すなわち、組織を修復ならびに再生し得、そして/または炎症を阻害し得る分子と組み合わせて投与され得る。骨粗鬆症個体における骨組織増殖を刺激するための有用な補因子の例には、例えば、ビタミンD3、カルシトニン、プロスタグランジン、副甲状
腺ホルモン、デキサメサゾン、エストロゲン、およびIGF−IまたはIGF−IIが挙げられるが、これらに限定されない。神経組織の修復および再生のために有用な補因子は、神経増殖因子を含み得る。他の有用な補因子は、症状軽減補因子(symptom alleviating cofactor)が挙げれ、これには防腐剤、抗生物質、抗ウイルス剤および抗真菌剤ならびに鎮痛剤および麻酔薬が含まれる。
【0152】
治療的分子はさらに、薬学的に受容可能な非毒性賦形剤およびキャリアを有する混和剤によって、薬学的組成物に処方され得る。上記のように、そのような組成物は、特に脂質溶液または懸濁液の形態において、非経口的投与のために調製され得;特に錠剤またはカプセルの形態において、経口投与のために調製され得;または鼻腔内では、特に粉末、点鼻薬またはエアロゾルの形態において調製され得る。組織表面への付着が所望される場合、この組成物は、フィブリノゲン−トロンビン組成物または他の生付着性組成物(例えば、PCT US91/09275に開示(その開示は、本明細書中で参考として援用される)されるような)中に分散される生合成構築物を含み得る。次いで、この組成物は、塗布され、噴霧され、または他の方法で所望の組織表面に適用される。この組成物は、治療的に有効量(例えば、その量は、所望される効果を誘導するのに十分な時間、標的組織に適切な濃度の形態単位を提供する)においてヒトまたは他の哺乳動物への非経口的投与または経口的投与のために処方され得る。
【0153】
この治療的分子が、組織保存溶液または器官保存溶液の部分を含む場合、任意の市販の保存溶液が都合よく使用され得る。例えば、当該分野において公知の有用な溶液には、Collins溶液、Wisconsin溶液、Belzer溶液、Eurocollins溶液および乳酸加リンガー溶液が挙げられる。保存溶液および有用な成分の詳細な説明は、例えば、米国特許第5,002,965号において見出され得、その開示は、本明細書中に参考として援用される。
【0154】
いくつかのタンパク質構築物は(例えば、Vg/dppサブグループのメンバーに基づくタンパク質構築物)はまた、マトリックスと組み合わされる場合、インビボで高レベルの活性を示すことが意図される。例えば、米国特許第5,266,683号を参照のこと(この開示は本明細書中に参考として援用される)。現在好ましいマトリックスは、天然における外因性マトリクス、同種異系マトリクスまたは自原性マトリックスである。しかし、ポリ乳酸、ポリグリコール酸、ポリ酪酸、その誘導体およびコポリマーを含む合成材料もまた使用されて適切なマトリックスを産生し得ることが意図される。好ましい合成および天然に由来するマトリックス材料、それらの調製、本発明の形態形成タンパク質とともにそれらを処方するための方法および投与の方法は当該分野において周知であり、そのため本明細書中において詳細に議論されない。例えば、米国特許第5,266,683号を参照のこと(この開示は、本明細書中において参考として援用される)。さらに、プロテーゼデバイスのマトリックスまたは金属表面との結合、接着、または会合は、本明細書中に開示される材料および方法を使用して変えられ得る特性であるということが意図される。例えば、増強されたマトリックス接着特性を有する本発明のマトリックスおよび骨活性構築物を含むデバイスが、徐放デバイスとして使用され得る。当業者は、その変動および操作は、ここで、本明細書中における教示に照らして可能であるということを理解する。
【0155】
当業者によって理解されるように、治療的組成物中の記載される化合物の濃度は、多数の因子(投与されるべき形態形成有効量、使用される化合物の化学的特徴(例えば、疎水性)、および投与経路を含む)に依存して変化する。投与されるべき薬物の好ましい投薬量もまた、おそらく、疾患、組織損失または欠損の型および程度、特定の患者の全体の健康状態、選択された化合物の相対的な生物学的有効性、この化合物の処方、処方物中の賦形剤の存在および型ならびに投与経路のような変数に依存する。概して、本発明の治療的分子は、1日あたり約10ng/kg体重〜約1g/kg体重の範囲の代表的用量で個体に提供され得る(好ましい用量範囲は、約0.1mg/kg体重〜100mg/kg体重である)。
【0156】
(II.特定の改変タンパク質構築物)
一般的に、本発明は、以下の4つの型の改変TGF−βファミリータンパク質構築物に関する:(1)N末端領域で短縮化されたTGF−βファミリータンパク質、(2)切断に際して活性化((例えば、酸切断またはプロテアーゼ処理による)N末端配列の放出を含むが、これに限定されない)され得る「潜伏性」タンパク質、(3)特定の結合能力を有する融合タンパク質、および(4)天然に存在するかまたは改変された、TGF−βファミリーのメンバーのサブユニットからなるヘテロ二量体。これらのモルフォゲン構築物の特定の種を、以下で詳細に記載する。以下に例証される種は、一般的に、改変モルフォゲンまたは骨原性タンパク質構築物に関するが、当業者は、これらの構築物が、TGF−βスーパーファミリーの他のメンバーを用いて生成され得る類似の構築物の代表であることを理解する。
【0157】
本発明に従って、ネイティブなBMPの特性またはTGF−βスーパーファミリーのタンパク質の他のメンバー(そのヘテロダイマーおよびホモダイマーを含む)は、BMPまたはTGF−βスーパーファミリーメンバーの1つ以上の生物学特性を変更するためにネイティブのタンパク質のN末端を改変することによって変えられる。この発見の結果として、TGF−βスーパーファミリータンパク質を、以下のように設計することが可能である;(1)TGF−βスーパーファミリータンパク質が原核生物細胞もしくは真核生物細胞において組換え的に発現されるか、またはポリペプチド合成装置を使用して合成される;(2)TGF−βスーパーファミリータンパク質が、変更されたフォールディング特性を有する;(3)TGF−βスーパーファミリータンパク質が、中性pH(生理学的に適合する条件を含むがこれに限定されない)下で変更された溶解性を有する;(4)TGF−βスーパーファミリータンパク質が、変更された等電点を有する;(5)TGF−βスーパーファミリータンパク質が、変更された安定性を有する;(6)TGF−βスーパーファミリータンパク質が、変更された組織またはレセプター特異性を有する;(7)TGF−βスーパーファミリータンパク質が、再設計されて変更された生物学的活性を有する;および/または(8)TGF−βスーパーファミリータンパク質が、固体表面(例えば、生体適合性マトリックスまたは金属が挙げられるがこれらに限定されない)に対する変更された結合特性または接着特性を有する。従って、本発明は、好ましいタンパク質構築物を含む速放性処方物、徐放性処方物および/または時限放出性処方物を設計するための機構を提供し得る。他の利点および特徴が、以下の教示により明らかである。さらに、本明細書中に開示される発見を利用して、変更された表面結合/表面接着特性を有する改変タンパク質が設計かつ選択され得る。特に重要な表面には、天然に存在し得る固体表面(例えば、骨)、または多孔性粒子表面(例えば、コラーゲン、または他の生体適合性マトリックス)またはプロテーゼインプラントの作製された(flabricate)表面(金属を含む)が挙げられるが、これらに限定されない。本明細書中に意図されるように、実質的に任意の表面が、構築物の差次的結合についてアッセイされ得る。従って、本発明は、その表面結合/表面接着特性における改変を有する多様な機能性分子を含み、それによって、変更されたインビボ適用(徐放性処方物、速放性処方物および/または時限放出性処方物を含む)について有用なこのような構築物を与える。
【0158】
当業者は、1つ以上の上記の任意の特性を混合かつ整合することは、カスタマイズされたタンパク質(およびこのタンパク質をコードするDNA)の用途を操作する特定の機会を提供することを理解する。例えば、変更された安定性の特性が開発され、タンパク質のインビボにおける代謝回転を操作し得る。さらに、タンパク質がまた、変更されたリフォールディングおよび/または機能のような特性を有する場合、おそらく、フォールディングと機能と安定性との間に相互関連が存在する。例えば、Lipscombら、7 Protein Sci.765〜73(1998);およびNikolovaら、95 Proc.Natl.Acad.Sci.USA 14675〜80(1998)を参照のこと。本発明の目的のために、安定性変化は、変性剤濃度または温度の関数として安定性の円偏光二色性の他の指標の周知の技術を使用して慣用的にモニターされ得る。また、慣用的な走査熱分析が使用され得る。同様に、任意の前述の特性と溶解度の特性との間におそらく相互関連が存在する。溶解度の場合、タンパク質構築物が、生理学的に適合する条件下においてより多く可溶性になるか、もしくはあまり可能性でなくなるかのいずれかになるようにこの特性を操作することが可能であり、結果として、インビボにおいて投与される場合、それはそれぞれ容易に拡散するか、または局在化する。
【0159】
改変された特性を有するタンパク質構築物の上記の用途に加えて、改変された安定性を有するタンパク質構築物はまた、有効期間、保存、および/または出荷の検討に関する実用上の利点のために用いられ得る。さらに関連事項について、改変された安定性はまた、投薬量の検討に直接影響し得、それによって、例えば、処置の費用を削減する。
【0160】
構築物の特に重要なクラスは、可溶化されるキャリアまたは賦形剤に対して改変された結合を有するものである。制限されない例として、可溶化キャリア(例えば、ヒアルロン酸)に対する増強された結合を有する、改変されたBMPは、当業者が、拡散剤(diffusion)または体液のいずれかにより、BMPの損失または希釈なしに、欠損部位に注射可能処方物を投与することを可能にする。従って、局在は最大となる。当業者は、本教示により改変が可能であることを理解する。同様に、身体/組織の構成要素に対する改変された結合を有する、構築物の別のクラスが、開発され得る。制限されない例として、インサイチュインヒビターに対する結合の減少を有する、改変されたBMPを用いて、インビトロで特定の組織の修復を増大させ得る。例えば、軟骨組織が、ネイティブのBMPの活性を阻害し得る体液および/または軟骨自体において見出される特定のタンパク質と関連付けられることは、当該分野で周知である。しかし、改変された結合特性を有するキメラ構築物は、これらのインサイチュインヒビターの効果を克服し得、それにより修復などを増強する。当業者は、本教示により改変が可能であることを理解する
(A.短縮化(truncation))
種々の形態のOP−1(例えば、23k、17k、および変動する量の15k)が存在し、それにより、代表的なOP−1調製物はこれらのすべての種を含む。精製された成熟OP−1のN末端配列決定は、異質性を明らかにした。このことは、N末端が多少短縮化され得ることを示す。RP−HPLCからの溶出およびトリプシン切断により回収された種を用いた実験を通して、ROS活性は、15kの種の中で最高である。例えば、短縮化変異体H2469は、CHO由来のOP−1標準と比較して、比較的高い活性を有する。最初の成熟は、プロOP−1においてRXXR部位で生じて17k種を生じるが、異なるプロテアーゼによる二次成熟は、最も活性な15k種を生成する。トリプシン切断は、この二次活性化を模擬し得る。
【0161】
哺乳動物OP−1またはE.coliのリフォールディングしたOP−1のトリプシン処理は、ROS活性の増加を生じる。本明細書中に記載される構築物のN末端(例えば、ヘキサ−his、コラーゲン結合部位、およびBMP−2のN末端)の除去はまた、ROSアッセイにおける活性の増加を生じた。OP−1の短縮化はモルフォゲンの可溶性を増加し得、これはROS活性に影響し得る。従って、構築物は、特異的切断活性を有して作製され得る。すなわち、構築物は、切断の型および切断のタイミングについて選択的である。当業者は、使用される系(哺乳動物または原核生物)に基づいて切断活性が異なり得ることを理解する。例えば、哺乳動物系は、モルフォゲン構築物が、プロ領域(これは、構築物の状況において、フォールディングを破壊し得、そして結果として(哺乳動物系において)最後には全くタンパク質を伴わない完全な細胞内分解を生じる)を含むことを必要とし得る。プロタンパク質形態を含む他の構築物を生成することもまた、所望され得る。このような構築物では、プロドメインは、増加した活性を得るために切断され得る別のN末端エレメントとしてみなされ得る。当業者は、切断されていないプロタンパク質が、その特性(可溶性および活性に関連する)を利用するために使用され得ることを理解する。
【0162】
本発明の変異タンパク質は、改善された生物学的活性ならびに拡大した半減期を示す。さらに、本発明の短縮化タンパク質で観察された活性の増加は、塩基性残基の除去および/またはタンパク質の等電点の低下に起因し得る。生物学的活性および改善されたリフォールディングは、同時係属中の出願[代理人整理番号STK−076、1999年8月16日出願]および[代理人整理番号STK−077、1999年8月16日出願](これらの開示を、本明細書中で参考として援用する)に記載された改変と本発明の改変タンパク質を組み合わせる場合に、増強され得る。
【0163】
(B.特異的特性を有するN末端領域)
本発明のさらなる改変タンパク質は、モルフォゲンの7−システインドメインのN末端に融合された非モルフォゲン起源のペプチドを含む。例えば、図7A〜7Eを参照のこと。生じるN末端融合タンパク質は、この融合物が由来する未改変モルフォゲンにおいては存在しないさらなる生物学的または生化学的な特性を有する。この型の融合物は、そのN末端でタンパク質またはタンパク質フラグメント(例えば、コラーゲン結合ドメイン、プロテインAのFBドメイン、またはヘキサ−ヒスチジン領域)に融合されたモルフォゲンの7−システインドメインを含む。例えば、H2440は、IMAC(固定化金属親和性クロマトグラフィー)樹脂のための結合ドメインとして、そのN末端に付着したヘキサ−hisタグを有するOP−1である(図7B)。このタンパク質は、最初はその非フォールディング状態で、尿素の存在下において、銅IMAC樹脂に対して精製された。IMAC上での非フォールディングタンパク質の精製、次ぐリフォールディングの後、首尾良くリフォールディングされた画分をRP−HPLCにより精製した。このようなN末端融合タンパク質は、ROSアッセイにおいて、ほとんどまたは全く活性を示さないが、N末端非モルフォゲンペプチドの切断に際して活性化されて、活性なC末端モルフォゲンドメインを産生する。
【0164】
特異的部位を標的し得るこれらの操作されたOP−1構築物が特に好ましい。例えば、N末端デカペプチドコラーゲン結合ドメインを有するOP−1、H2487が構築された。ここでこのデカペプチドは、骨マトリックスへのOP−1の特異的かつ堅固な結合を得るために、第1のシステインの上流7残基に配置された(図7Aを参照)。この新規な構築物は、首尾良くリフォールディングされ、そしてROSアッセイにおいて活性であり、それにより特定の骨形成活性を示す。活性を指向するために、他の結合ドメインが同様に用いられ得る。例えば、軟骨修復の場合、OP−1はまた、補綴デバイスに特異的に接着させるため設計され得る。Clostridiumコラゲナーゼから誘導されたペプチドのような他のペプチドはまた、コラーゲン結合特性について探索され得る。
【0165】
本発明の技術が特定の条件下で特定の部位の活性タンパク質の環境誘引放出を可能にする特定の改変タンパク質処方物を生成するために用いられ得ることは、当業者に明白である。例えば、pHの変化または特定のプロテアーゼの存在は、活性タンパク質の送達および誘引放出を調整し得る。
【0166】
BMPまたは他のTGF−βファミリーのメンバーのリーダー配列の改変はまた、このタンパク質の溶解度、活性および発現に影響し得る。例えば、staphylococcus aureusプロテインAのFBサブドメインのリーダー配列で操作されたCDMP−3(腱の修復に有用であると考えられる)を利用する構築物H2528は、骨原性タンパク質の発現が改善されている。
【0167】
当業者は、本発明の構築物が、もし空間的干渉および引き続く生物学的活性の低下が考慮されるならば、TGF−βファミリータンパク質のN末端に結合され得る、種々の特異化された機能的ドメインを含むように操作され得ることを理解する。このような構築物は、活性の阻害またはフォールディングを回避するため7−システインドメインからN末端付加までの少なくとも最小限の間隔を必要とし得る。最小限の間隔の必要条件は、特定されたドメインおよびTGF−βファミリータンパク質の両方がその意図された活性を保持するように、付加された部分の空間特性、および改変された構築物の最終の意図された活性に依存することを、当業者は理解する。
【0168】
(C.潜在性BMP)
本発明はまた、OP−1のN末端の短縮は、タンパク質に負の影響を有さないので、N末端が融合タンパク質の溶解度および活性に影響し得る程度の驚くべき発見を利用する。さらに、OP−1の結晶構造は、N末端に関するどのような位相幾何学的情報も示さなかった。
【0169】
本明細書において記載されるN末端融合タンパク質は、所望の時間および位置で活性タンパク質を生成するために切断され得るタンパク質の潜在性(すなわち、不活性)形態を生成するために有用である。例えば、コラーゲン結合ドメインを含有する改変モルフォゲン(例えば、図7Aに示されるH2487)は、所望の組織位置(例えば、移植されたコラーゲンマトリックスを含む位置)へ不活性形態で送達され得、そしてその位置で切断され活性なモルフォゲンを生成する。切断は、標的位置の内因性条件(例えば、天然に存在するプロテアーゼ)から生じ得るか、または特定のプロテアーゼの投与もしくは他の因子(例えば、位置の酸性化)の結果であり得る。さらに、非常に特異的なプロテアーゼ切断部位が、例えば、骨折部位に見出されるプロテアーゼについて操作され得、移植部位でのOP−1の選択的遅延型活性化および/または漸進的活性化を可能にする。
【0170】
(D.ドメインスワッピング)
リフォールディング、溶解性、活性および発現を変更するためのさらなる構築物が、1つのTGF−βスーパーファミリータンパク質のネイティブリーダー配列を、別のTGF−βファミリーメンバーのネイティブなリーダー配列で置換することにより設計され得る。例えば、構築物H2549は、OP−1上に置き換えられたBMP−2のN末端を有する。
【0171】
(E.ヘテロダイマー)
上記のいくつかのN末端融合タンパク質モノマーは、リーダー配列の切断なしに活性なホモダイマーを形成しないが、活性なヘテロダイマーが、これらのタンパク質とTGF−βファミリータンパク質の未改変モノマーとの間で形成される。従って、このようなヘテロダイマーは、融合タンパク質に付着したN末端非TGF−βファミリータンパク質ドメイン(例えば、コラーゲン結合部位)によってタンパク質を標的部位に提供するために用いられ得る。あるいは、ヘテロダイマーの精製を増強するために設計特徴が用いられ得る。例えば、ヘキサヒスチジンを付加することにより、2種のサブユニットの間の精製の差異を強調することによって精製を容易にし得る。混合されたリフォールディングにより、2つのホモダイマーおよびヘテロダイマーの混合物を提供する。これは3つの分離可能な種を提供する。例えば、ヘキサ−ヒスチジンドメインを含むN末端融合タンパク質(例えば、図7Bに示されるH2440)
(これは、IMACカラムに結合する)は、N末端ドメインの切断により引き続いて活性化され得る融合タンパク質の精製を補助するために有用である。
【0172】
本発明のヘテロダイマーの構築のためにはE.coli発現が好ましい。なぜなら、実施者はヘテロダイマーの至適産生のために各モノマーの比を調節し得るからである。さらに、この方法は非常に迅速である。例えば、荷電したアミノ酸の好ましい改変で改変したヘキサ−ヒスチジンタグ化OP−1のE、D、E、およびR(H2440)(例えば、その全開示が本明細書において参考として援用される、代理人整理番号___を参照のこと)とBMP−2との間でのインビトロにおけるヘテロダイマー形成実験において、ヘテロダイマーの産生は優秀であった。理論的に予期される50%のヘテロダイマーおよび25%の各ホモダイマーよりも、例外的に高い収率のヘテロダイマーが存在する。これは、BMP-2がそれ自体とよりもOP−1とより容易に会合するかまたはそれ自体との再会合より
もOP−1と、より速く会合することによって生じ得る。あるいは、BMP−2は、フォールディングについてシャペロンとして作用し得る。別の実験はまた、BMP−2とH2447変異体であるOP−1(ヘキサ−ヒスチジンタグなし)との間のヘテロダイマー形成を示した。これはまた容易に会合し良好な収率のヘテロダイマーを生じる。ヘテロダイマーはまた、FB−OP−1(H2521)とBMP−2との間で形成され得る。短縮したOP−1であるH2469(第1のシステインの15残基上流を保持している)、およびBMP−5(H2475)のヘテロダイマー;ならびにH2469およびCDMP−2(H2471)のヘテロダイマーがまた構築され得る。
【0173】
リフォールディングにおいて有効であるのと同様、ヘキサ−his−OP−1(H2440)およびBMP−2(H2142)のヘテロダイマーは、ROSアッセイにおいて、ホモダイマーよりも大きい活性を有する。ヘキサ−his−OP−1ホモダイマーは非常に低い活性を有した。BMP−2のホモダイマーは、より良好な活性を有した。しかし、OP−1/BMP−2ヘテロダイマーは、いずれの親ホモダイマーよりもかなり活性であった。このアッセイにおいて、ヘテロダイマーは、CHO誘導型OP−1標準の約1/3の活性しか有さなかった。ヘキサ−his−タグのないOP−1のBMP−2とのヘテロダイマー(H2447)は類似の活性を有した。H2447は、フィンガー−2において改変を有するリフォールディング変異体であり、ホモダイマーと同じ程度の相対的に低い活性を有した。OP−1(H2469)/BMP−5(H2475)およびOP−1(H2469)/CDMP−2(H2471)のヘテロダイマーは、ROSアッセイにおいて良好な結果(2.5−3+)を得た。
【0174】
この同じプロトコールおよび方法論を用いて、E.coliで発現され、そしてインビトロでリフォールディングされたOP−1/BMP−2ヘテロダイマーを構築した。詳細には、H2447/BMP−2ヘテロダイマーおよびH2440/BMP−2ヘテロダイマーは、E.coli発現により生成され、そして生理学的条件下でインビトロでリフォールディングされた。SDS−PAGE分析に基づいて、ほとんどの材料が容易に組み合わされ、ヘテロダイマー種を形成した。非モルフォゲンドメインを含むヘテロダイマーを用いてさらなる種を形成する。このような種の例は、コラーゲン結合ドメイン融合OP−1(H2487)、ヘキサ−ヒスチジン融合OP−1(H2440)およびプロテインAのFBドメイン融合OP−1(H2521)ならびにFB−ドメイン融合ヘキサ−ヒスチジン/OP−1構築物H2440(H2525)のようなN末端融合モルフォゲンである。
【0175】
活性なヘテロダイマーはまた、2つのBMPまたは他のTGF−βファミリータンパク質(異なる系において発現された)から構築され得る。いくつかの構築物が良好に発現され、そして特定の系で他よりも発現された場合、より活性である。当業者は、その発現に最適な環境中で各構築物を発現し得、次いでそれらとの活性なヘテロダイマーを形成し得る。例えば、H2223(OP−1の変異体)は、哺乳動物発現系であるCHO細胞中で発現され、一方、H2525(図7D)(FB−ドメインOP−1)は、細菌発現系であるE.coli中で最も発現される。
【0176】
さらに、ヘテロダイマーの活性は、用いる2つのタンパク質を変化させることにより操作され得る。例えば、デカペプチドコラーゲン結合部位を有するOP−1であるH2487とCDMP3のヘテロダイマーが形成され得る。このヘテロダイマーは、H2487とBMP−2のヘテロダイマーとは異なる活性を有する。
【0177】
(F.構築物の選択および最適化)
本明細書中に教示されるように、本発明は、注文に応じた(customized)キメラタンパク質およびこのキメラタンパク質をコードするDNAを巧妙に作るノウハウを当業者に提供する。キメラタンパク質を特定のインビボでの適用に適切にする、特定の所望される特性を有するキメラタンパク質を設計する手段が、本明細書中にさらに教示および例示される(少なくともI.B.節、II節、およびIII節、前述のキメラタンパク質の例示的な実施態様についての実施例1〜4、8および11を参照のこと)。例えば、変化した溶解度という特性を有するキメラタンパク質は、レシピエントに提供される形態形成的有効量を操作するために、インビボで使用され得る。すなわち、溶解度の増大は、アベイラビリティーの増大を生じ得;溶解度の減少は、アベイラビリティーの減少を生じ得る。従って、このような全身投与されるキメラタンパク質は、即座に利用可能であり得、そして/または即効性の形態形成効果を有し得、一方、局所投与されたキメラタンパク質は、よりゆっくりと利用可能であり得/長期化した形態形成効果を有し得る。当業者は、手近な事実および状況を考慮して、減少した溶解度という特性に対して増大した溶解度という特性が好ましい場合を理解する。このようなパラメーターの最適化は、慣用的な実験法および通常の技能を必要とする。
【0178】
同様に、変化した安定性という特性を有するキメラタンパク質は、レシピエントに提供される形態形成的有効量を操作するために、インビボで使用され得る。すなわち、安定性の増大は、半減期の増大を生じ得る。なぜなら、インビボでの代謝回転は、より小さくなるからであり;安定性の減少は、半減期およびアベイラビリティーの減少を生じ得る。なぜなら、インビボでの代謝回転がより大きくなるからである。従って、このように全身投与されたキメラタンパク質は、迅速に利用可能であり得るか/ボーラス型投薬を達成する即効性の形態形成効果を有し得るか、あるいは長期間にわたってインビボで利用可能であり得るか/徐放型の投薬を達成する長期の形態形成効果を有し得るかのいずれかである。当業者は、手近な事実および状況を考慮して、減少した安定性という特性に対して増大した安定性という特性が好ましい場合を理解する。このようなパラメーターの最適化は、慣用的な実験法および通常の技能を必要とする。
【0179】
さらに、変化した安定性を有するタンパク質がまた、有効期間、貯蔵、および/または運送条件を改善する実質的な利点のために用いられ得る。さらに関連事項に関しては、変化した安定性も投薬条件に直接影響し、それにより例えば、処置のコストを軽減する。
【0180】
さらに変化した特性(例えば、溶解度および安定性の特性であるが、これらに限定されない)の組合せを、レシピエントに提供される形態形成的有効量を操作するためにインビボで用い得る。すなわち、特定の変化した特性の組み合わせを有するキメラタンパク質を設計することにより、形態形成的有効量は、時限放出の様式において投与され得;投薬は、量および持続時間の両方に関して調節され得;処置レジメは、少ないがいくつかの典型を挙げると全身的または局所的に、低用量で開始され、続いて高用量に移され得るか、またはその逆であり得る。当業者は、この事実および手近な状況下で、多い形態形成的有効量に対して少ない形態形成的有効量が適切である場合を理解する。このようなパラメーターの最適化は、慣用的な実験法および通常の技能を必要とする。
【0181】
さらに、1以上の変化した特性を有するキメラタンパク質は、発達における固有の欠損を克服するために有用である。1以上の変化した特性を有するキメラタンパク質は、宿主のネイテイブな形態形成シグナル伝達系における固有の欠損を回避するように設計され得る。非限定的な例として、本発明のキメラタンパク質は、標的組織中のネイティブなレセプターにおける欠損、細胞内シグナル伝達経路における欠損、および/または異なるサブドメインに含まれる機能/生物学的活性と関連する特性と逆であるような、部分自体の認識と関連したサブドメインの特性に依存する他の事象における欠損を迂回するために使用され得る。当業者は、手近な事実および状況を考慮して、このようなキメラタンパク質が適切である場合を理解する。最適化は、慣用的な実験法および通常の技能を必要とする。
【0182】
本発明の実施は、以下の実施例から、さらにより十分に理解され、これらの実施例は、単なる例示のために本明細書中に示され、いかようにも本発明を限定するとは解釈されないべきである。
【実施例】
【0183】
(実施例1.BMP変異体の合成)
図8は、OP−1のC末端の7つのシステインドメインのためのヌクレオチドおよび対応するアミノ酸配列を示す。これらの配列を知ることにより、例えば、カセット変異誘発またはKunkelの周知の方法(ml3から派生した一本鎖の鋳型を使用するプライマー伸長による変異誘発)によって、または周知のPCR方法(重複伸長を含む)によって変異操作のための有用な制限部位の確認が可能になる。OP−1の例示的な変異体は、以下に記載のように構築される、フィンガー−2サブドメインにおいて4つのアミノ酸変化および最後のC末端アミノ酸において1つのアミノ酸変化を有する、H2460である。記載されている変異誘発プロトコルが例示のみであること、および本発明の構築物を作製するための他の手段は周知であり、当該技術分野において十分に記載されていることは当業者によって理解されている。
【0184】
4つのアミノ酸変化が、重複伸長技術を用いる標準的ポリメラーゼ連鎖反応によって、OP−1フィンガー−2のサブドメイン配列に導入され、OP−1変異体H2460を生じた。フィンガー−2の領域の4つの変化は、N6>S、R25>E、N26>DおよびR30>Eであった。この変異体はまた、C末端残基のさらなる変化(H35>R)を含んだ。これらの反応のための鋳型は、野生型OP−1のcDNAクローンの成熟したドメインであった。それは成熟領域の初めのATG開始コドンによって操作されるE.coli発現ベクターに挿入された。ATGは、順プライマーとして以下の配列の合成オリゴヌクレオチドを用いるPCRによって導入された:ATG TCC ACG GGG AGC AAA CAG(配列番号36)、これは、M S T G S K Q(配列番号37)をコードする。PCR反応を、cDNAの3’コード領域に相補的な適当なバック−プライマーと組み合わせて行った。
【0185】
フィンガー−2変異体H2460を構築するために、標準のPCR反応において、市販のPCRキットを用いて、そしてプライマーとして合成オリゴヌクレオチドを用いる製造業者の指示に従って、改変フィンガー−2をコードするPCRフラグメントを作製した。
【0186】
N6>S改変を得るために、配列GCG CCC ACG CAG CTC AGC GCT ATC TCC GTC CTC(配列番号70)の順プライマー(プライマー#1)を用いた。この配列は、アミノ酸配列A P T Q L S A I S V L(配列番号71)をコードする。
【0187】
C末端付近の変化については、バックプライマー(43ヌクレオチド長)(プライマー#2)を用いた。これは、R25>EおよびN26>DおよびR30>EおよびC−末端H35>Rの変化を導入した。このプライマー#2は、以下の配列を含んだ:CTA TCT GCA GCC ACA AGC TTC GAC CAC CAT GTC TTC GTA TTT C(配列番号72)。これは以下のコード配列の相補体である:G AAA TAC GAA GAC ATG GTG GTC GAA GCT TGT GGC TGC AGA TAG(配列番号73)。この配列は、以下のアミノ酸をコードする:K Y E D M V V E A C G C R停止(配列番号74)。
【0188】
次いで、フィンガー2およびC末端変異を有するフラグメントを、N末端、フィンガー−1およびヒールサブドメインを有する成熟OP−1の上流部分をコードする別のPCRフラグメントと組み合わせた。N末端、フィンガー−1およびヒールサブドメインをコードする後者のPCRフラグメントを、鋳型としてE.coliについてのOP−1発現ベクターを用いて再度構築した。このベクターは、T7プロモーターに付着した、成熟OP−1タンパク質をコードするOP−1 cDNAフラグメント、および適切な宿主におけるT7プロモーターか、またはtrpプロモーターの制御下のいずれかの制御下での発現のためのリボソーム結合部位を含んだ。このT7発現ベクターにおいて、成熟したOP−1のXbaI部位のT7プロモーターおよびATGコドン間の配列、Pet 3d(Novagen Inc.(Madison WI))は、以下の通りである:
TCTAGAATAATTTTGTTTAACCTTTAAGAAGGAGATATACG ATG(配列番号75)。
【0189】
この第2のPCR反応を、順プライマー(プライマー#3)TAA TAC GAC TCA CTA TAG G(配列番号76)によってプライムした。これは、T7プロモーター領域およびプライマー#1と重なるバック−プライマー(プライマー#4)においてプライムし、そしてヌクレオチド配列GCT GAG CTG CGT GGG CGC(配列番号77)を有する。この配列は、コード配列GCG CCC ACG CAG CTC AGC(配列番号78)の相補体である。そしてこれは、A P T Q L S(配列番号79)をコードする。
【0190】
第3のPCR反応(実際の重複伸長反応)において、上記の2つのPCRフラグメントの部分は、組み合わされて、PCRによって増幅された。そして、完全な成熟OP−1領域を含む単一のフラグメントを生じた。この反応のために、プライマー#3を順プライマーとして用い、そして新規なプライマー(プライマー#5)を以下の配列GG ATC CTA TCT GCA GCC ACA AGC(配列番号80)を有するバック−プライマーとして用いた。この配列は、コード配列GCT TGT GGC TGC AGA TAGGAT CC(配列番号81)に対する相補体である。この配列は、A C G C R停止(配列番号82)をコードする。このプライマーはまた、遺伝子を発現ベクターに挿入するために便利な3’BamHI部位を付加する。
【0191】
重複伸長PCRから生じる、完全な変異遺伝子を保有する得られたフラグメントを、PCRフラグメントのクローン化のために設計された市販のクローニングベクター(例えば、pCR2.1−topo−TA(Invitrogen Inc.(Carlsbad
CA)))にクローニングした。クローニングしたPCRフラグメントを、XbaIおよびBamHIを用いる制限消化によって回収し、Pet3d(Novagen Inc.(Madison WI))のような市販のT7発現ベクターのXbaIおよびBamHI部位に挿入した。
【0192】
(実施例2.BMPのE.coli発現)
形質転換した細胞を、標準的培養条件下で、標準的SPYE 2YT培地、1:1比(例えば、Sambrookら、を参照のこと)中で、37℃で増殖した。異種タンパク質の過剰発現は、代表的には、8〜48の時間以内に封入体を生じた。封入体を、単離して、次のように可溶化した。1リットルの培養液を、細胞を集めるために遠心分離した。次いで、得られたペレット中の細胞を60mlの25mM Tris、10mM EDTA、pH 8.0(TE緩衝液)+100μg/mlのリゾチーム中に再懸濁し、そして、37℃で2時間インキュベートした。次いで、細胞懸濁液を、氷冷して、超音波処理して細胞を溶解させた。細胞溶解を、検鏡によって確認した。溶解物の体積を、TE緩衝液でほぼ300mlに調整した。次いで、封入体ペレットを得るために遠心分離した。このペレットを、TE緩衝液中の2〜4回の連続再懸濁および遠心分離によって洗浄した。洗浄した封入体ペレットを、40mlの100mM Tris、10mMのEDTA、6MのGuHCl(グアニジウム塩酸塩)、250mMのDTT、pH 8.8中での変性および還元により可溶化した。次いで、タンパク質を、標準的な、市販のC2またはC8カートリッジ(SPICEカートリッジ、400mg、Ananltech、Inc.)を用いて予め精製した。タンパク質溶液を2% TFA(トリフルオロ酢酸)で酸性化し、カートリッジに適用し、0.1% TFA/10%アセトニトリルで洗浄し、そして0.1%TFA/70%アセトニトリルで溶出した。次いで、溶出した物質を、乾燥するかまたは希釈して、C4 RP−HPLCによって分画した。
【0193】
(実施例3.BMPダイマーのリフォールディング)
上記の通りに調製したタンパク質を、リフォールディング前に乾燥するかまたはリフォールディング緩衝液に直接希釈した。用いた好適なリフォールディング緩衝液は、以下であった:100mM Tris、10mM EDTA、1M NaCl、2% CHAPS、5mM GSH(還元グルタチオン)、2.5mM GSSG(酸化グルタチオン)、pH 8.5。リフォールディング(12.5〜200μgタンパク質/ml)を、24〜90時間、代表的には36〜48時間、4℃にて実行し(ただし、いくつかの変異体では、これより長い(数週間まで)と、良好なリフォールディングが提供されると予期される)、続いて、0.1% TFAに対して透析し、次いで0.01% TFA,50%エタノールに対して透析した。次いで、透析した材料のアリコートを種々のアッセイに備えて乾燥した。
【0194】
(実施例4.リフォールディングされたBMPダイマーの精製および試験)
(4A.SDS−PAGE、HPLC)サンプルを乾燥して、Laemmliゲルサンプル緩衝液において再懸濁して、次いで、15%のSDS−ポリアクリルアミドゲルにおいて電気泳動した。全てのアッセイは、比較のための、分子量標準および/または哺乳動物細胞が産生した精製OP−1を含んだ。OP−1ダイマーの解析を、添加した還元剤の非存在下で実行した。一方で、OP−1モノマーをゲルサンプルへの100mMのDTTの添加によって生成した。フォールディングしたダイマーは約30〜36kDaの範囲のみかけの分子量を有する。一方で、モノマー種は、約14〜16kDaの見かけの分子量を有する。
【0195】
あるいは、サンプルを、以下のように、市販のRP−HPLCにてクロマトグラフィーにかけた。サンプルを乾燥して、0.1%のTFA/30%のアセトニトリルの中に再懸濁した。次いで、タンパク質を、0.1%のTFA、30%のアセトニトリル中のC18カラムに適用し、そして、TFA中の30〜60%アセトニトリル勾配を用いて分画した。適切にフォールディングしたダイマーは、孤立したピークとして45〜50%アセトニトリルで溶出する;モノマーは、50〜60%アセトニトリルで溶出する。
【0196】
(4B.ヒドロキシアパタイトクロマトグラフィー)サンプルを10mMリン酸塩、6M 尿素、pH 7.0(カラム緩衝液)のヒドロキシアパタイトにロードした。未結合の物質を、カラム緩衝液を用いる洗浄により除去し、次いでカラム緩衝液+100mM NaClでモノマーを溶出させた。ダイマーを、カラム緩衝液+250mM NaClを用いて溶出した。
【0197】
(4C.トリプシン消化)トリプシン消化は、50mM Tris、4M 尿素、100mM NaCl、0.3% Tween 80、20mM メチルアミン、pH 8.0の消化緩衝液において実行した。酵素対基質の比は、1:50(重量比)であった。37℃での16時間のインキュベーションの後、15μlの消化混合液を、DTTを含まない5μlの4×ゲルサンプル緩衝液と組み合わせて、SDS−PAGEによって分析した。精製した哺乳動物OP−1および未消化のBMPダイマーを、比較のために含んだ。これらの条件の下で、適切にフォールディングされたダイマーを切断して、未切断の標準より僅かに速い移動を有する種を生産する。一方で、モノマー、および誤ってフォールディングしたダイマーは完全に消化され、そして染色したゲルにおいてバンドとしてみられない。
【0198】
(実施例5.骨原性活性のインビトロでの細胞ベースのバイオアッセイ)
本実施例は、本発明に従って骨原性能力または骨形成能力を獲得したモルフォゲン構築物の生物活性を実証する。高い特異的な骨形成活性についての生得の能力または獲得された能力のいずれかを有する骨原性タンパク質は、ラット骨肉腫細胞およびラット頭蓋冠(calveria)細胞を含むラット骨芽細胞においてアルカリホスファターゼ活性を誘導し得る。このアッセイにおいて、ラット骨肉腫または頭蓋冠細胞を、マルチウェルプレート(例えば、48ウェルプレート)の上へ、10%のFBS(ウシ胎仔血清)、L−グルタミンおよびペニシリン/ストレプトマイシンを含有するαMEM(改変イーグル培地、Gibco Inc.、Long Island)中で、1ウェルあたり50,000の骨芽細胞の濃度でプレートした。細胞を、37℃で24時間インキュベートした。この時点で、増殖培地を1%FBS含有αMEMで置換し、そしてこの細胞を、さらに24時間インキュベートし、その結果、この細胞は、実験のこの時点で血清枯渇増殖培地中にあった。
【0199】
次いで、培養した細胞を以下の3つの群に分けた:(1)生合成骨原性(ostegenic)タンパク質の種々の濃度を受けるウェル;(2)哺乳動物が発現したhOP−1のような陽性コントロール;および陰性コントロール(タンパク質、またはTGF−βなし)。試験したタンパク質濃度は、50〜500ng/mlの範囲であった。細胞を、72時間インキュベートした。インキュベーション期間後、細胞層を、0.5mlの1%のTritonX−100を用いて抽出した。得られた細胞抽出物を遠心分離し、100μlの抽出物を90μlのPNPP(パラニトロソフェニルホスフェート)/グリセリン混合液に添加して、37℃の水浴中で30分間インキュベートし、100μlの0.2N NaOHで反応を停止した。次いで、サンプルをプレートリーダー(例えば、Dynatech MR700)を通過させ、そして、標準としてp−ニトロフェノールを用いて400nmで吸光度を測定し、アルカリホスファターゼ活性の存在および量を決定した。タンパク濃度は、標準手段、例えば、Biorad方法、UVスキャンまたは214nmでのRP HPLC面積によって決定した。アルカリホスファターゼ活性を、タンパク質1μgあたりのユニット(単位)で算出した。ここで、1ユニットは、37℃で30分あたりに遊離した1nmolのp−ニトロフェノールに等しい。
【0200】
HOP−1およびBMP2は、100〜200ng/mlで約1.0〜1.4ユニットを生じる。種々のタンパク質構築物についての他の結果を、表1に提供する。
【0201】
(実施例6.CDMP活性のインビトロでの細胞ベースのバイオアッセイ)
本実施例は、本発明に従って増強された組織形態形成能力を獲得した構築物のの生物活性を実証する。ネイティブなCDMPは、実施例5において用いたようなラット骨肉腫細胞において、アルカリホスファターゼ活性を誘導し得ないが、それらは、マウス奇形癌細胞株ATDC−5(軟骨前駆細胞(chondroprogenitor)細胞株(Atsumiら、1990、Cell Differentiation and Development 30:109))においてアルカリホスファターゼ活性を誘導する。ラット骨癌細胞アッセイにおいて陰性であるが、ATDC−5アッセイにおいて陽性であるフォールディングされた変異体は、CDMP様活性を獲得したことが記載されている。ATDC−5アッセイにおいて、細胞を無血清基本培地(BM:Ham’s F−12/DMEM[1:1]以下を含む:ITSTM+培養補充物[Collaborative Biomedical Products、Bedford、MA]、α−ケトグルタレート(1×10-4M)、セルロプラスミン(0.25 U/ml)、コレステロール(5μg/ml)、ホスファチジルエタノールアミン(2μg/ml)、α−トコフェロール酸スクシナート(9×10-7M)、還元グルタチオン(10μg/ml)、タウリン(1.25μg/ml)、トリヨードサイロニン(1.6×10-9M)、副甲状腺ホルモン(5×10-10M)、β−グリセロリン酸(10mM)およびL−アスコルビン酸2−スルフ
ェート(50μg/ml))中、4×104の密度でプレートした。CDMPまたは他の
生合成骨原性タンパク質(0〜300ng/ml)をその翌日、添加し、そして、CDMPまたは生合成骨原性タンパク質を含む培養培地を1日おきに交換した。アルカリホスファターゼ活性を処理の4、6および/または12日後、超音波処理した細胞ホモジネート中で決定した。PBSを用いた大規模な洗浄の後、細胞層を0.05%Triton−X100を含有する500μlのPBS中で超音波処理した。50〜100μlのアリコートを、アッセイ緩衝液(0.1M バルビタールナトリウム緩衝液(pH 9.3))中で、そして基質としてp−ニトロフェニルホスフェートを用いて、酵素活性についてアッセイした。吸光度を、400nmで測定し、そして活性をBradfordタンパク質アッセイ(ウシ血清アルブミン標準)で測定されるタンパク含有量に対して標準化した。
【0202】
CDMP−1およびCDMP−2は、100ng/mlで10日目に約2〜3ユニットの活性を生成した。OP−1は、10日目、100ng/mlで約6〜7ユニットの活性を生成した。
【0203】
(実施例7.TGF−β様活性のインビトロにおける細胞に基づくバイオアッセイ)
本実施例は、本発明に従って生物学的能力を改変された生合成変異体TGF−βタンパク質の生物活性を実証する。TGF−βタンパク質は、上皮細胞増殖を阻害し得る。多数の細胞阻害アッセイが、当該分野において十分に記載されている。例えば、Brownら(1987)J.Immunol.139:2977(ヒト黒色腫A375線維芽細胞を用いる比色アッセイを記載しており、そして、本願明細書において以下に記載される)を参照のこと。別のアッセイは上皮細胞(例えば、ミンク肺上皮細胞)を使用し、そして増殖の効果は3H−チミジン取込みによって決定される。
【0204】
簡潔には、アッセイにおいて、TGF−β生合成構築物を、RPMI−1640培地(Gibco)および5%のウシ胎仔血清を含むマルチウェル組織プレートにおいて階段希釈する。コントロールウェルには、培地のみをいれる。次いで、黒色腫細胞を、ウェルに添加する(1.5×104)。次いで、プレートを5%のCO2中で約72時間37℃でインキュベートする。そして、細胞単層を一度洗って、固定して、そして15分間クリスタルバイオレットで染色する。未結合の染料を洗い流し、次いで染色した細胞を33%の酢酸で溶解し、(細胞核に閉じ込められた)染料を遊離させ、そして試験分子の活性を算出するために標準的な市販の光度計を用いて590nmでODを、測定する。それぞれのウェルにおける染色の強度は、核の数に直接関連する。従って、活性なTGF−β分子は、ウェルを不活性化合物または陰性コントロールウェルより薄く染色すると予期される。
【0205】
別のアッセイにおいては、ミンク肺細胞を用いる。これらの細胞は、標準的培養条件下で生長および増殖するが、ミンク肺上皮細胞株(ATCC番号CCL 64、Rockville、MD)由来の培養細胞を用いる3H−チミジン取込みによって決定されるよう
にTGF−βに曝露された後、停止される。簡潔には、細胞を、10%のFBS、200単位/mlのペニシリンおよび200μg/mlのストレプトマイシンを補充したEMEMを用いて、コンフルエンスになるまで生長させる。これらの細胞を、1ウェルあたり約200,000細胞の細胞密度になるまで培養する。コンフルエンスの時点で、培地を、1%のFBSおよびペニシリン/ストレプトマイシンを含有する0.5mlのEMEMで置換し、そしてこの培養物を、37℃で24時間インキュベートする。次いで、候補タンパク質を各ウェルに添加し、細胞を37℃で18時間インキュベートする。インキュベーションの後、10μl中に1.0μCiの3H−チミジンを、各々のウェルに添加し、そ
して細胞を、37℃で4時間インキュベートした。次いで、培地を、各ウェルから取り除き、細胞を、氷冷リン酸緩衝化生理食塩水を用いて1回洗浄し、そして、各ウェルに0.5mlの10% TCAを添加することによってDNAを、沈殿して、15分間室温でインキュベートした。この細胞を、氷冷蒸留水で3回洗浄し、0.5mlの0.4M NaOHで溶解し、次いで、各ウェルからの溶解物をシンチレーションバイアルに移し、そしてシンチレーションカウンター(Smith−Kline Beckman)を用いて放射能を記録する。生物学的に活性な分子は、細胞増殖を阻害し、その結果、不活性タンパク質および/または陰性コントロールウェル(増殖因子を添加してない)と比べて、より少ないチミジン取込みおよびより少ない計数を生じる。
【0206】
(実施例8.骨原性活性(軟骨内の骨形成および関連特性)のインビボにおけるバイオアッセイ) SampathおよびReddi(Proc.Natl.Acad.Sci.USA(1983)80:6591−6595)、および米国特許第4,968,590号、同第5,266,683号(これらの開示は本明細書において参考として援用される)によって記載される、骨誘導についての当該分野で認識されるバイオアッセイは、所定のタンパク質、デバイスまたは処方物の効力を評価するために用いられ得る。簡潔には、このアッセイは、エーテル麻酔下で試験試料をレシピエントラットの皮下の部位に配置することからなる。垂直の切開(1cm)を、胸の領域の上の皮膚に無菌条件下で作製し、そして、ポケットを、閉鎖(blunt)切開によって準備する。特定の場合において、凍結乾燥のような標準的手順を用いて調製した、所望の量の骨原性タンパク質(10ng〜10μg)を、約25mgのマトリックス材料と混合し、そして、試験試料を、ポケットの内側深くに移植し、そして切開を、金属の皮膚クリップで閉鎖する。異所性部位は、同所性部位の使用から生じる可能なあいまい性なしに骨誘導の研究を可能にする。このインプラントはまた、筋肉内に提供され得る。これはこのデバイスを接近可能な前駆細胞とより近接して設置する。代表的には、筋肉内インプラントを、両方の脚の骨格筋に作製する。
【0207】
異所性部位で生じる連続的な細胞反応は、複雑である。軟骨内骨形成の多段階のカスケードは、以下を包含する:移植したマトリックスに対するフィブリンおよびフィブロネクチンの結合、細胞の走化性、線維芽細胞の増殖、軟骨芽細胞への分化、軟骨形成、血管浸潤、骨形成、再構築、および骨髄分化。
【0208】
首尾良いインプラントは、以下を含むタンパク誘導性軟骨内性骨発生の段階を通じて制御された進行を示す:(1)1日目の多形核白血球による一過性浸潤;(2)2日目および3日目の間葉細胞遊走および増殖;(3)5日目および6日目の軟骨細胞出現;(4)7日目の軟骨基質形成;(5)8日目の軟骨石灰化;(6)9日目および10日目の血管浸潤、骨芽細胞の出現および新しい骨の形成;(7)12〜18日目の骨芽細胞の出現および骨再構築;ならびに(8)21日目の小骨における造血性骨髄分化。
【0209】
組織学的な切片にすることおよび染色は、移植片における骨形成の程度を決定するために好ましい。トルイジンブルーまたはヘマトキシリン(hemotoxylin)/エオシンを用いる染色は、軟骨内性骨の最終的な発達を明らかに実証する。12日目のバイオアッセイは、骨誘導活性が試験サンプルと関連しているか否かを決定するために十分である。
【0210】
さらに、アルカリホスファターゼ活性および/または総カルシウム含量は、骨形成についての生化学的マーカーとして使用され得る。このアルカリホスファターゼ酵素活性は、切り出された試験物質の均質化の後に、分光測光的に決定され得る。この活性は、インビボで9〜10日目でピークとなり、そしてその後にゆっくりと減少する。組織学によって骨の発達を全く示さないサンプルは、これらのアッセイ条件下でアルカリホスファターゼ活性を全く示さないはずである。このアッセイは、定量のために、および試験サンプルがラットから取り出された後に非常に迅速に骨形成の概算を得るために有用である。アルカリホスファターゼ活性レベルおよび組織学的評価によって測定される結果は、「骨形成単位」として表され得る。1骨形成単位は、12日目の最大の骨形成活性の半分に必要とされるタンパク質の量を表す。さらに、用量曲線は、種々の濃度のタンパク質をアッセイすることにより、精製スキームの各工程にて、インビボでの骨誘導活性について作成され得る。従って、当業者は、慣用的な実験のみを使用して、代表的な用量曲線を作成し得る。
【0211】
総カルシウム含量は、例えば、冷たい0.15M NaCl、3mM、NaHCO3、
pH9.0中で均質化し、そして沈渣の酸可溶性画分のカルシウム含量を測定する後に決定され得る。
【0212】
(実施例9:「ドメインスワッピング(Domain Swapping)」変異体の活性)
例えばある型のTGF−βファミリーのメンバーのタンパク質のN末端領域を獲得し、そしてこれを別の型のTGF−βファミリーのメンバーのタンパク質の7つのシステインのドメインに接続する場合、ドメインスワッピングが、生じる。ある変異体構築物は、当業者に一般に公知の慣用的な技術を用いて、OP−1の7つのシステインの活性ドメイン上のBMP−2末端の配列のスプライシングによって作製した。生じた変異体H2549は、MQAKHKQRKRLKSS−CからなるN末端領域を有する。最後のアミノ酸であるシステインは、OP−1の7つのシステインの活性ドメインの最初のシステインである。実施例5に上述したようなROSアッセイを、H2549の活性を試験するために用いた。
【0213】
図11に図示するように、その結果は、H2549がOP−1の活性のレベルに比べて非常に低い活性を有することを示す。しかし、実施例4に記載のダイマーのトリプシン切断に類似の方法を用いたH2549のトリプシン切断に際して、ROS活性が、有意に上昇する。この様式において、TGF−βファミリーのメンバーのタンパク質の活性は、それを不活化するために非天然のN末端配列を接続すること、およびそれを活性化するために非天然の配列を切断することによって選択的に制御され得る。
【0214】
(実施例10:N末端短縮化は活性を上昇させる)
改変されたモルフォゲンタンパク質のN末端領域の、例えば、トリプシン切断による短縮化は、ROS活性を上昇させる。H2223構築物は、CHO細胞において発現される改変されたOP−1変異体である。H2223の2つのHPLC画分である画分13および14が、回収された。各々の画分の量は、実施例4中のダイマーに際して用いたものと類似の様式において、トリプシン分解によって分解された。次いで結果生じた4つのサンプル、すなわちトリプシン未処理の画分13および14およびトリプシン処理した画分13および14は、OP−1活性を標準として用いた上述の実施例5のようにROSアッセイに供した。
【0215】
図12に図示するように、トリプシン処理したおよび未処理の画分14の活性レベルは、相対的に同一である。このことは、部分的に分解されたH2223を含む画分14によって説明され、従って、トリプシンを用いたさらなる消化は、活性を改変しない。対照的に、未処理の画分13は、全長H2223(すなわち、39アミノ酸のN末端全体)から主に構成され、そして画分13のN末端の短縮化は、画分14のROS活性に匹敵するレベルまでROS活性を上昇させない。これらの活性レベルは、OP−1標準のROS活性レベルよりも十分上であり、そして本発明の改変されたタンパク質を用いて得られた活性の改善を実証する。
【0216】
(実施例11:ヘテロダイマー活性)
ヘテロダイマーの活性レベルは、ヘテロダイマーのそれぞれのサブユニット各々より形成されるホモダイマーの活性レベルよりも高い。6−His N末端を伴うOP−1であるH2440構築物、およびBMP−2であるH2142構築物を、上述した実施例3に記載の方法を用いてヘテロダイマーおよびホモダイマーを形成させた。次いでH2440/2142のヘテロダイマーならびにH2440/2440のホモダイマーおよびH2142/2142のホモダイマーを、上述の実施例4および5に記載のようにROSアッセイに供した。
【0217】
図13Aおよび13Bに示すように、N末端に6Hisを伴うOP−1であるH2440のホモダイマーは、非常に低い活性を有する。BMP−2であるH2142のホモダイマーは、より良好な活性を有するが、活性は、まだ相対的には低い。しかし、OP−1 6−HisおよびBMP−2のヘテロダイマーは、これらのホモダイマーのいずれよりもかなり高い活性を有する。このヘテロダイマーは、CHO由来OP−1のたった1/3の活性を有する。
【0218】
類似の実験において、ホモダイマーおよびヘテロダイマーが、FBリーダー配列を伴うOP−1であるH2525とBMP−2であるH2142との間で作製された。これらもまた、OP−1活性のレベルを標準としたROSアッセイに供した。図14に図示するように、FBを伴うOP−1であるH2525のホモダイマーは、実質的に活性がなく、そしてBMP−2であるH2142のホモダイマーは、非常に低い活性を有する。対照的に、H2525/2142の2つのヘテロダイマーは、予期せぬ高い活性レベルを有する。
【技術分野】
【0001】
(発明の分野)
本発明は、改良されたリフォールディング特性、改良された物理的特性(例えば、溶解性および安定性)、改良された生物学的活性(改変されたレセプター結合、改良された標的化能力を含む)を有する組換えタンパク質、潜在性形態のタンパク質、およびこのようなタンパク質を生成する方法に関する。より詳細には、本発明は、TGF−βスーパーファミリーの構造的に関連したタンパク質の生合成メンバーに関する。このような改変型タンパク質構築物は、N末端短縮、「潜在性」タンパク質、融合タンパク質およびヘテロ二量体を有するTGF−βファミリーメンバータンパク質を含む。
【背景技術】
【0002】
(発明の背景)
TGF−βスーパーファミリーは、TGF−βの5個の異なる形態(非特許文献1)、ならびに分化因子Vg−1(非特許文献2)、DPP−Cポリペプチド(非特許文献3)、ホルモンアクチビンおよびインヒビン(inhibin)(非特許文献4;非特許文献5)、Mullerian阻害物質、MIS(非特許文献6)、骨形成タンパク質および形態形成タンパク質OP−1(PCT/US90/05903)、OP−2(PCT/US91/07654)、OP−3(PCT/WO94/10202)、BMP(米国特許第4,877,864号;同第5,141,905号;同第5,013,649号;同第5,116,738号;同第5,108,922号;同第5,106,748号;および同第5,155,058号を参照のこと)、発生的に調節されるタンパク質VGR−1(非特許文献7)、軟骨由来成長因子CDMP−1、CDMP−2およびCDMP−3(またはGDF−5、GDF−6およびGDF−7)、ならびに成長/分化因子GDF−1、GDF−3、GDF−9、およびドーサリン(dorsalin)−1(非特許文献8;非特許文献9)を含む。
【0003】
TGF−βスーパーファミリーのタンパク質は、ジスルフィド結合されたホモ−またはヘテロ二量体である。これらは、疎水性のシグナル配列、長くそして比較的保存されていないN末端の数100アミノ酸のプロ領域配列、切断部位、ならびにファミリーのメンバーの間で変化するN末端領域、および高度に保存されているC末端領域を含む成熟ドメイン、を含む大きな前駆体ポリペプチド鎖として発現される。全ての既知のファミリーのメンバーのプロセシングされた成熟タンパク質中に存在するこのC末端領域は、約100個のアミノ酸を含み、保存されている6個または7個のシステイン骨格を有する特徴的なシステインモチーフを有する。成熟領域とプロ領域との間では切断部位の位置は、ファミリーのメンバーの間で変化するが、全てのタンパク質のC末端のシステインパターンは同一の形式であり、配列Cys−X−Cys−Xで終わる(非特許文献1)。
【0004】
組換えTGF−β1は、クローン化(非特許文献10)、およびチャイニーズハムスター卵巣細胞において発現される(非特許文献11)。さらに、組換えヒトTGF−β2(非特許文献12)、ならびにヒトおよびブタのTGF−β3(非特許文献13;非特許文献14)が、クローン化されている。COS細胞における成熟TGF−β1タンパク質の発現レベルは、セリン残基を有するTGF−β1前駆体のプロ領域に局在したシステイン残基を置換することによって、増加された(非特許文献15)。
【0005】
TGF−βスーパーファミリーのタンパク質の生物学の統一の特徴は、発達のプロセスを調節するそれらの能力である。これらの構造的に関連するタンパク質は、種々の発達的な事象に関与することが同定されている。例えば、TGF−βおよびインヒビン/アクチビンの群のポリペプチドは、細胞の増殖および分化の調節において役割を果たすようである。MISは、哺乳動物の雄性の胚の発達においてミュラー管の退行を生じ、そしてDrosophiaのデカペンタプレジック(decapentaplegic)複合体の遺伝子産物であるdppは、適切な背面−腹面(dorsal−ventral)の特異化に必要である。同様に、Vg−1は、Xenopusにおける中胚葉の誘導に関連し、そしてVgr−1は、発達中のマウスの種々の組織において同定されている。骨形成に関しては、TGF−βスーパーファミリー中の多くのタンパク質(すなわち、OP−1およびBMPのサブセット)が、明らかに主要な役割を果たす。OP−1(BMP−7)および他の骨形成タンパク質は、組換え技術を使用して産生されており(米国特許第5,011,691号およびPCT出願番号第US90/05903号)、そしてインビボで実際に軟骨内の骨の形成を誘導することが可能であることが示されている。BMP−2は、サルCOS−1細胞およびチャイニーズハムスター卵巣細胞において、組換え的に作製される(非特許文献16)。
【0006】
最近、SampathおよびReddi骨形成アッセイによって判定しタンパク質場合に、骨形成活性を有すると教示されたタンパク質のこのファミリーが、形態形成性である(すなわち、成熟した哺乳動物において組織の形態形成の発達のカスケードを誘導し得る)ことが示された(PCT出願番号第US92/01968号を参照のこと)。詳細には、これらのタンパク質は、関連付けられていない前駆細胞の増殖を誘導し得、そして適切な環境条件下で組織特異的様式でこれらの刺激された先祖細胞の分化を誘導し得る。さらに、モルフォゲンは、これらの分化した細胞の増殖および維持を補助し得る。これらの形態形成性の活性は、タンパク質が、適切な形態形成を許された環境において組織の形態形成の発達のカスケードを開始しそして維持すること、幹細胞が組織特異的様式で増殖および分化するように刺激すること、ならびに新しい組織の形成において完了する事象の進行を誘導することを可能にする。これらの形態形成活性はまた、タンパク質がそれらの分化の経路からはぐれるように予め刺激された細胞の「再分化」を誘導することを可能にする。適切な環境条件下では、これらのモルフォゲンもまた関連付けられた細胞の「再分化」を刺激し得ることが予想される。
【0007】
骨形成タンパク質は、一般にTGF−βスーパーファミリーの増殖因子の亜群として、当該分野で分類されている(非特許文献17)、そして「骨形成タンパク質」、「形態形成タンパク質」、「モルフォゲン」、「骨形態形成タンパク質」または「BMP」と様々に呼ばれ、それらのエピトープ性軟骨内性骨形態形成を誘導する能力により同定される。タンパク質のモルフォゲンファミリーのメンバーとしては、哺乳動物の骨形成タンパク質−1(OP−1(BMP−7、およびDrosophilaのホモログである60Aとしてもまた知られている))、骨形成タンパク質−2(OP−2(BMP−8としてもまた知られている))、骨形成タンパク質−3(OP−3)、BMP−2(BMP−2AまたはCBMP−2A、およびDorosphilaのホモログであるDPPとしてもまた知られている)、BMP−3、BMP−4(BMP−2BまたはCMBP−2Bとしてもまた知られている)、BMP−5、BMP−6およびそのマウスのホモログであるVgr−1、BMP−9、BMP−10、BMP−11、BMP−12、GDF3(Vgr2としてもまた知られている)、GDF−8、GDF−9、GDF−10、GDF−11、GDF−12、BMP−13、BMP−14、BMP−15、GDF−5(CDMP−1またはMP52としてもまた知られている)、GDF−6(CDMP−2またはBMP−13としてもまた知られている)、GDF−7(CDMP−3またはBMP−12としてもまた知られている)、XenopusのホモログであるVg1およびNODAL、UNIVIN、SCREW、ADMP、およびNEURALが挙げられる。
【0008】
天然に存在するか、または合成によって調製されたかにはかかわらず、骨形成タンパク質は、前駆細胞の補充および刺激を誘導し得、それによって、軟骨細胞および骨芽細胞へのそれらの分化を誘導し、そしてさらに、中間軟骨の分化、血管新生、骨の形成、再造形、および最終的には骨髄の分化を誘導する。さらに、多数の研究者らが、天然から供給されたマトリックス材料(例えば、コラーゲン)、または合成によって調製されたポリマーマトリックス材料のいずれかと混合した場合に、実際に置換骨が他に生じない条件下で、骨形成(膜性骨形成および軟骨内骨形成含む)を誘導する、骨形成タンパク質の能力を実証した。例えば、マトリックス材料と組み合わせた場合には、これらの骨形成タンパク質は、大きな断片の骨の欠失、脊椎の融合、頭蓋冠の欠損、および骨折において、新しい骨の形成を誘導する。
【0009】
細菌および他の原核生物の発現系は、組換えタンパク質を生成するための好ましい手段として当該分野であてにされている。原核生物(例えば、E.coli)の系は、商業的な量のタンパク質を産生するため、および天然に存在するかまたは生合成変異体およびアナログの生物学的特性および化学的特性を評価するために有用である。代表的には、過剰に発現される真核生物のタンパク質は、原核生物の宿主細胞中で不溶性の細胞内沈殿物(「封入体」)に凝集する。次いで、凝集したタンパク質は封入体から回収され、1つ以上の標準的な変性剤を使用して可溶化され、次いで機能的な状態にリフォールディングされることを可能にされるかまたはそのように誘導される。生物学的に活性なタンパク質構造を形成するために適切なリホールディングは、任意のジスルフィド結合の適切な形成を必要とする。
【0010】
化学合成はまた、タンパク質構築物の産生に用いられ得る。ペプチド鎖の単調な自動化アセンブリを可能にするための技術は広範に利用可能である。合成タンパク質分子にペプチドフラグメントを結合するために酵素法およびキメラ法を利用する技術は、当該分野で公知である。非特許文献18;非特許文献19;非特許文献20;非特許文献21;および非特許文献22を参照のこと。
【0011】
例えば、TGF−β2およびOP−1の両方の三次構造および四次構造が決定されている。TGF−β2およびOP−1は、それらのそれぞれのアミノ酸配列においてわずかに約35%のアミノ酸同一性しか示さないが、両方の分子の三次構造および四次構造は極めて類似している。図1Aに示すように、TGF−β2およびOP−1の両方ともが天然においてダイマーであり、そして7個のC末端のシステイン残基のうちの6個を含む特有のフォールディングパターンを有する。各サブユニットにおいては、4個のシステインが、8個の残基の環を生じるように結合し、そして2個のさらなるシステイン残基が、結び目様の構造を形成するように環を通って通過するジスルフィド結合を形成することが図1Aに示される。番号1に割り当てられた7個の保存性システイン残基の最もN末端側のシステインで開始する番号付けスキームを用いると、2番目と6番目の保存されたシステイン残基は、8個の残基の環の1つの側面を閉じるように結合し、一方、3番目と7番目のシステイン残基は、他の側面を閉じるように結合する。1番目と5番目の保存性システイン残基は、結び目の中心を形成するように、環の中心を通じて結合する。4番目の保存性システインは、他のサブユニット中の対応する残基とともに鎖間ジスルフィド結合を形成する。
【0012】
TGF−β2およびOP−1のモノマーサブユニットは、3つの主要な構造エレメントおよびN末端領域を含む。構造エレメントは、以下の型:(1)ループ、(2)α−へリックス、および(3)β−シートの50%を超える二次構造を保有している、連続しているポリペプチド鎖の領域を構成する。さらに、これらの領域においては、N末端およびC末端の鎖は、7Åを超えては離れていない。保存された1番目のシステインと2番目のシステインとの間の残基(図1A)は、逆平行β−シートフィンガー(本明細書中では、フィンガー1領域(F1)と呼ばれる)によって特徴付けられる構造領域を形成する。フィンガー1ペプチド骨格のリボントレースは、図1Bに示される。同様に、図1Aの保存された5番目のシステインと6番目のシステインとの間の残基もまた、逆平行β−シートフィンガー(本明細書中では、フィンガー2領域(F2)と呼ばれる)を形成する。フィンガーペプチド骨格のリボントレースは、図1Dに示される。β−シートフィンガーは、一本のアミノ酸の鎖であり、これは、1つ以上の逆平行β−シート構造を形成する鎖が入りそして存在するように、β−ターンまたはいくらか大きいループによってそれ自体の上に折れ曲がっているβ鎖を含む。保存された3番目のシステインと4番目のシステインとの間の残基を含む、第3の主要な構造領域は、本明細書中ではヒール領域(H)と呼ばれる3個のターンα−へリックスによって特徴付けられる。ヒールペプチド骨格のリボントレースは、図1Cに示される。
【0013】
このモノマー構造の組織化は、結び目領域が手掌に等しい位置に位置付けされ、フィンガー1がインデックスフィンガーおよびミドルフィンガーに等しく、α−ヘリックスは手のヒールに等しく、そしてフィンガー2が環状フィンガーおよび小フィンガーに等しい、左側の組織化と類似する。N末端領域は(公開された構造において十分規定されていない)は、親指(thumb)に概略的に等しい位置に位置すると予想される。
【0014】
TGF−β2およびOP−1の両方のダイマーの形態においては、サブユニットは、一方のサブユニットのヒール領域が、分子のコアを形成する連結されたサブユニットの結び目の領域を有する他のサブユニットのフィンガー領域と接触するように配向される。4番目のシステインは、第2鎖上のその対応物とジスルフィド結合を形成し、それによって手掌の中心で鎖を等しく連結する。このように形成されたダイマーは、サブユニット間の対称な二倍軸を上から下に見た場合には、楕円形の(葉巻型)の分子である(図2A)。その側から見ると、この分子は湾曲した「葉巻」に似ている。なぜなら、2つのサブユニットが、互いに関してわずかな角度で配向されるからである。
【0015】
しかし、必ずしも全ての可溶化異種タンパク質が、容易にリフォールディングするわけではない。リフォールディングの注意深い操作にもかかわらず、適切にフォールディングされた、生物学的に活性なタンパク質の収量は低いままである。多くのTBF−βファミリーメンバー(BMPを含む)は、少ないリフォールダー(refolder)タンパク質のカテゴリーに入る。TGF−βタンパク質ファミリーのいくつかのメンバーはインビトロで、例えば、E.coliまたは他の原核生物宿主中で産生された場合に比較的効率良くフォールディングされ得るが、他の多く(BMP−5、BMP−6、およびBMP−7を含む)はそうではない。例えば、第EP0433225号、第US5,399,677号、同第5,756,308号、および同第5,804,416号を参照のこと。
【先行技術文献】
【非特許文献】
【0016】
【非特許文献1】SpornおよびRoberts(1990)、Peptide Growth Factors and Their Receptors、SpornおよびRoberts編、Springer−Verlag:Berlin、419−472頁
【非特許文献2】WeeksおよびMelton(1987)、Cell 51:861−867
【非特許文献3】Padgettら(1987)Nature 325:81−84
【非特許文献4】Masonら(1985)、Nature 318:659−663
【非特許文献5】Masonら(1987)、Growth Factors 1:77−88
【非特許文献6】Cateら(1986)、Cell 45:685−698
【非特許文献7】Lyonsら(1989)Proc.Natl.Acad.Sci.USA 86:4554−4558
【非特許文献8】McPherronら(1993)、J.Biol.Chem.268:3444−3449
【非特許文献9】Baslerら(1993)、Cell 73:687−702
【非特許文献10】Derynckら、(1985)Nature 316:701−705
【非特許文献11】Gentryら、(1987)Mol.Cell.Biol.7:3418−3427
【非特許文献12】deMartinら、(1987)EMBO J.6:3673
【非特許文献13】Derynckら、(1988)EMBO J.7:3737−3743
【非特許文献14】Dijkeら、(1988)Proc.Natl.Acad.Sci.USA 85:4715
【非特許文献15】Brunnerら、(1989)J.Biol.Chem.264:13660−13664
【非特許文献16】Wangら、(1990)Proc.Natl.Acad.Sci.USA 87:2220−2224
【非特許文献17】Hogan(1996),Genes&Development,10:1580−1594
【非特許文献18】Hilvert,Chem.Biol.(1994)1(4):201−03
【非特許文献19】Muriら、Proc.Nat’l Acad.Sci.USA(1998)95(12):6705−10
【非特許文献20】Wallace,Curr.Opin.Biotechnol.(1995)6(4):403−10
【非特許文献21】Mirandaら、Proc.Nat’l Acad.Sci.USA(1999)96(4):1181−6
【非特許文献22】Liuら、Proc.Nat’l Acad.Sci.USA(1994)91(14):6584−8
【発明の概要】
【発明が解決しようとする課題】
【0017】
組換えBMPおよび他のTGF−βファミリータンパク質を、原核生物宿主および真核生物宿主を使用してインビトロで産生するための改善された手段の必要性が、残っている。
【課題を解決するための手段】
【0018】
(項目1) 適切なリフォールディング条件下でリフォールドさせる能力のある、生物学的に活性なTGF−βファミリーメンバー融合タンパク質であって、該短縮化TGF−βファミリー融合タンパク質は、以下:
フィンガー1サブドメイン、フィンガー2サブドメイン、およびヒールサブドメインを含む、TGF−βファミリータンパク質C末端7システインドメイン;および
該C末端ドメインに作動可能に連結された異種リーダー配列ドメイン;
を含み、融合タンパク質。
(項目2) 項目1に記載の融合タンパク質であって、ここで前記リーダー配列は、組織標的ドメイン、分子標的ドメイン、金属結合ドメイン、タンパク質結合ドメイン、セラミック結合ドメイン、ヒドロキシアパタイト結合ドメイン、およびコラーゲン結合ドメインからなる群から選択される、融合タンパク質。
(項目3) 前記組織標的ドメインが、骨基質タンパク質に結合する、項目2に記載の融合タンパク質。
(項目4) 前記組織標的ドメインが、細胞表面分子に結合する、項目2に記載の融合タンパク質。
(項目5) 前記細胞表面分子が、前骨芽細胞または軟骨細胞上にある、項目4に記載の融合タンパク質。
(項目6) 適切なリフォールディング条件下でリフォールドさせるための、潜在性TGF−βファミリーメンバー融合タンパク質であって、該短縮化TGF−βファミリータンパク質は、以下:
フィンガー1サブドメイン、フィンガー2サブドメイン、およびヒールサブドメインを含む、TGF−βファミリータンパク質C末端7システインドメイン;および
該C末端ドメインに作動可能に連結された切断可能な改変型リーダー配列であって、該リーダー配列は、該C末端ドメインと関連する生物学的活性を阻害し、そして該C末端ドメインは、該リーダー配列の一部または全ての切断の際に活性化する、リーダー配列;
を含む、融合タンパク質。
(項目7) 項目6に記載の融合タンパク質であって、組織標的ドメインが、前記切断可能なリーダー配列の内部に存在し、それにより該リーダー配列の切断によって、該組織標的ドメインが前記C末端ドメインから切断されない、融合タンパク質。
(項目8) 前記リーダー配列が、前記C末端ドメインから少なくとも7残基は分離されている、項目1または6に記載の融合タンパク質。
(項目9) 前記リーダー配列が、別のTGF−βファミリータンパク質に由来する、項目1に記載の融合タンパク質。
(項目10) 適切なリフォールディング条件下でリフォールドさせる能力のある、生物学的に活性なTGF−βファミリーメンバー融合タンパク質変異体であって、該融合タンパク質は、以下:
フィンガー1サブドメイン、フィンガー2サブドメイン、およびヒールサブドメインを含む、TGF−βファミリーメンバータンパク質C末端7システインドメイン;および
該C末端ドメインに作動可能に連結されるリーダー配列ドメインであって、それによって、該リーダー配列の一部または全てが短縮化される、リーダー配列;
を含み、
ただし、該潜在性TGF−βファミリーメンバー融合タンパク質は、TGF−β1、TGF−β2、またはTGF−β3の該C末端7システインドメインを含まなくても良い、融合タンパク質変異体。
(項目11) 前記短縮が、プロテアーゼ切断によって実施される、項目10に記載のタンパク質変異体。
(項目12) 前記プロテアーゼが、トリプシンである、項目11に記載のタンパク質変異体。
(項目13) 前記短縮が、化学切断によって実施される、項目10に記載の融合タンパク質変異体。
(項目14) 前記化学切断が、酸切断である、項目13に記載の融合タンパク質。
(項目15) 前記リーダー配列の少なくとも1つの塩基性残基が、取り除かれる、項目10に記載のタンパク質変異体。
(項目16) 前記タンパク質変異体が、本質的に、配列番号69のアミノ酸配列からなる、項目10に記載の融合タンパク質。
(項目17) 生物学的に活性なTGF−βファミリーメンバータンパク質のヘテロ二量体であって、以下:
TGF−βファミリーメンバー融合タンパク質である第1のサブユニット;および
第2のサブユニットであって、該第1のサブユニットの融合タンパク質とは異なるTG
F−βファミリーメンバー融合タンパク質および野生型TGF−βファミリタンパク質からなる群より選択される、第2のサブユニット
を含む、ヘテロ二量体。
(項目18) 前記野生型TGF−βファミリータンパク質が、TGF−β1、TGF−β2、TGF−β3、TGF−β4、TGF−β5、dpp、Vg−1、Vgr−1、60A、BMP−2A、BMP−3、BMP−4、BMP−5、BMP−6、ドーサリン、OP−1、OP−2、OP−3、GDF−1、GDF−3、GDMP−1/GDF−5、CDMP−2/GDF−6、GDF−7、GDF−9、インヒビンα、インヒビンβA、およびインヒビンβBからなる群から選択される、項目16に記載のヘテロ二量体。
(項目19) TGF−βファミリータンパク質のヘテロ二量体を精製する方法であって、該方法は、以下:
(a)第1のTGF−βファミリータンパク質サブユニットを提供する工程;
(b)該第1のサブユニットとは異なる第2のTGF−βファミリータンパク質サブユニットを提供する工程;
(c)適切なリフォールディング条件下で、該第1のサブユニットおよび該第2のサブユニットを混合して、以下
(i)2つの該第1のTGF−βファミリータンパク質サブユニットを含む、第1のホモ二量体;
(ii)2つの該第2のTGF−βファミリータンパク質サブユニットを含む、第2のホモ二量体;ならびに
(iii)1つの該第1のTGF−βファミリータンパク質サブユニット、および1つの該第2のTGF−βファミリータンパク質サブユニットを含む、ヘテロ二量体;
を含む混合物を生成する、工程であって、
ここで、該へテロ二量体は、該第1のホモ二量体および該第2のホモ二量体から分離可能である、工程;ならびに
(d)該へテロ二量体を、該第1のホモ二量体および該第2のホモ二量体から、分離する、工程、
を含む、方法。
【0019】
(発明の要旨)
本発明は、C末端活性ドメインのN末端側でのN末端伸長、短縮および他の改変を含む、改変TGF−βファミリータンパク質を提供する。本発明の改変タンパク質は、組換え的に発現される場合に、天然に存在するタンパク質に対して変更されたリフォールディング特性および変更された可溶性を有する。本発明の改変タンパク質はまた、変更された活性プロフィール(増強された特異位的活性を含む)を有し、そして組織特異的標識化または特異的表面結合への影響を受けやすい。
【0020】
これらの発見の結果として、以下を含む変化した生物学的特性を有する新規なBMPおよび他のTGF−βファミリーのメンバーアナログを推測しそして設計するための手段が、ここで利用可能である:インビトロでの改善されたフォールディング能力、改善された可溶性、変化した安定性、変化した等電点、ならびに/または、所望される場合には、変化した生物学的活性。これらの発見はまた、それら自体に、その活性が哺乳動物内の特定の部位に指向され得、そして/またはその活性が調節、阻害および/もしくは誘導され得るタンパク質を作製させる。本発明はまた、フォールディングされたタンパク質のエピトープのマッピングを含む、候補構築物の生物学的特性および/または生化学的特性を容易にかつ迅速に評価するための手段を提供する。
【0021】
本発明は、「少ないリフォールダー(poor refolder)」TGF−βファミリーメンバーのリフォールディング特性を改善する、タンパク質の「変異体」形態を提供する。本明細書中で使用される場合、「少ないリフォールダー」タンパク質とは、適切なリフォールディング条件下でリフォールディングを誘導した場合に、標準的なプロトコール(以下を参照のこと)を使用して測定した場合に、約1%未満の適切にフォールディングされた物質を生じる、任意のタンパク質を意味する。本明細書中で意図される場合は、「適切なリフォールディング条件」とは、その条件下でタンパク質が機能性を付与するために必要とされる程度にリフォールディングされ得る条件である。当業者は、「好ましい実施態様の詳細な説明」の少なくともIC節および実施例3がこのようなリフォールディング条件の非限定的な例であることを認識する。本発明の組成物および方法に関する構造的なパラメーターは、ダイマータンパク質の構造にわたって適切に分布している1つ以上のジスルフィド架橋を含む。そしてこれは、フォールディングされた構造を生じるために、「還元−酸化」(「酸化還元」)反応工程を必要とする。酸化還元反応は、代表的には、天然のpH(すなわち、pH7.0〜8.5の範囲であり、代表的にはpH7.5〜8.5の範囲である)で生じ、そして好ましくは生理学的に適合可能な条件下で生じる。当業者は、成功のための最適な条件を理解しておりそしてそれを認識している。
【0022】
このタンパク質は、好ましくは、ヌクレオチドのアセンブリによって、および/または合成のDNAを生じるようにDNA制限フラグメントを連結することによって、本明細書中で開示されている原理に従って作製される。DNAは、適切なタンパク質発現ビヒクル中に移され、コードされるタンパク質が発現され、必要であればフォールディングされ、そして精製される。特定の構築物は、インビトロでの活性について試験され得る。候補タンパク質の構築物の三次構造は、繰り返し洗練され、そして本明細書中で開示される原理、コンピューターに基づくタンパク質構造のモデリング、および目的の分子の特異的な特性を改善または調節するための最近開発された合理的な薬物設計技術によって補助される、部位特異的変異誘発またはヌクレオチド配列特異的変異誘発によって結合が調節され得る。既知のファージディスプレイまたは他のヌクレオチド発現系は、多数の候補の構築物を同時に産生するために開発され得る。続いて、候補の構築物のプールが、例えば、表面固定化されたレセプターを含むクロマトグラフィーカラム、高結合候補物を選択しそして濃縮するための塩勾配溶出、ならびにインビトロアッセイを使用して、結合特異性についてスクリーニングされ得る。有用な組換えタンパク質の同定には、研究室での使用のため、および究極的には、治療的に有用な分子を産生するための商業的に有用な量のタンパク質を発現する細胞株の産生が続く。今や、適切にフォールディングされる場合に、フィンガー1領域、フィンガー2領域、およびヒール領域を規定する三次構造を想定するアミノ酸配列を含むキメラタンパク質を、設計、作製、試験および使用するための方法が発見された。
【0023】
本発明の全ての構築物は、有用性のために必要とされる領域を定義するアミノ酸配列の領域(すなわち、フィンガー1、フィンガー2、およびヒール領域)、ならびに活性を改変し得るさらなる領域(すなわち、N末端ペプチド配列)を含む。フィンガーおよびヒール領域についての配列は、本明細書中で同定される任意の既知のTGF−βスーパーファミリーのメンバーの、それぞれのフィンガーおよびヒール領域の配列からコピーされ得る。あるいは、フィンガーおよびヒール領域は、本明細書中で下記に開示されている原理を使用して本明細書中で以降に発見されたこのスーパーファミリーの新規のメンバーのアミノ酸配列から選択され得る。
【0024】
フィンガーおよびヒール配列はまた、例えば、Smithら(1990)、Proc.Natl.Acad.Sci.USA 87:118−122(その開示は本明細書中で参考として援用される)に開示されている原理に従って選択される置換アミノ酸残基を利用することによって、アミノ酸置換によって変化され得る。Smithらは、図3に示すアミノ酸の階層の表と同様の、アミノ酸のクラスの階層を開示している。これは、型の全体的な立体構造の変形(これは、さもなくばタンパク質を不活化し得る)を最少にしつつ、別のものについて1つのアミノ酸を合理的に置換するために使用され得る。任意の事象において、天然の領域とわずか70%の相同性、好ましくは80%、そして最も好ましくは少なくとも90%の相同性を有する多くの合成のフィンガー1、フィンガー2、およびヒール領域配列が、活性な形態単位構築物を産生するために使用され得ることが、意図される。本明細書中で開示されているように、構築物の大きさを、鋳型のTGF−βスーパーファミリーのメンバーの天然のフィンガーおよびヒール領域を短縮することによって有意に減少し得ることが意図される。
【0025】
本明細書中で使用される場合は、「酸性」または「負に荷電した残基」は、生理学的条件下で代表的にはそのR基上に負の電荷を保有している、天然に存在するかまたは合成の任意のアミノ酸残基を含むことが理解される。例であり限定的ではないが、アスパラギン酸(「Asp」)、およびグルタミン酸(「Glu」)が挙げられる。同様に、塩基性または正に荷電した残基としては、天然に存在するかまたは合成によって作製された、生理学的条件下で代表的にはそのR基上に正の電荷を保有している、任意のアミノ酸残基が挙げられる。例であり、限定的ではないが、アルギニン(「Arg」)、リジン(「Lys」)、およびヒスチジン(「His」)が挙げられる。本明細書中で使用される場合には、「親水性」残基は、酸性および塩基性の両方のアミノ酸残基、ならびにそれらのR基上にアミド基を保有している荷電していない残基(これらには、限定的ではないが、グルタミン(「Gln」およびアスパラギン(「Asn」)が挙げられる)、ならびに、そのR基上にヒドロキシル基を保有している極性残基(これらには、限定的ではないが、セリン(「Ser」)およびスレオニン(「Thr」)が挙げられる)が挙げられる。当業者は、実際の生理学的pKが変動し、そしてその変化が異なる生理学的環境において変動することを理解する。
【0026】
本明細書中で使用される場合には、「生合成」または「生合成の」は、天然に由来するかまたは合成によって誘導されるフラグメントの連結の結果として生じるか、またはそれに起源することを意味する。例えば、限定的ではないが、本明細書中で開示されている1つ以上のサブドメイン(またはそのフラグメント)に対応する連結ペプチドまたは核酸フラグメントを意味する。「化学合成」または「化学合成の」は、化学的な生成手段の結果として生じるかまたはそれに起源することを意味する。例えば、限定的ではないが、市販の供給業者からの標準的な自動合成装置/シーケンサーを使用するペプチドまたは核酸配列の合成を意味する。天然のアミノ酸および天然ではないアミノ酸の両方が、本明細書中で教示されているような所望される特性を得るように使用され得ることが意図される。「組換え」産生または技術は、遺伝子操作された産生手段の結果として生じるか、またはそれに起源することを意味する。例えば、限定的ではないが、本発明のキメラタンパク質(またはそのフラグメント)をコードする、遺伝子操作されたDMA配列または遺伝子の発現を意味する。少なくともI.B.節;II節、ならびに少なくとも実施例1、および2において以下に示される技術もまた、上記の意味に含まれる。「合成の」は、非天然に存在すること、または非天然に起源すること、すなわち、天然には存在しないことを意味する。
【0027】
本明細書中で使用される場合、「対応する残基位置」は、2つの配列が整列される場合に、OP−1または他の参照TGF−βファミリーメンバーアミノ酸配列中の所定の位置に対応するタンパク質配列中の残基位置をいう。当業者に理解され、そして図1に示すように、BMPファミリーのメンバーの配列は、C末端の活性ドメインにおいて、そして特にフィンガー2サブドメインにおいて高度に保存されている。アミノ酸配列のアラインメントの方法およびプログラムは、当該分野で十分に開発されている。例えば、Alignプログラム(DNAstar,Inc.)のようなコンピュータープログラムによって都合よく実行される、Needlemanら(1970)、J.Mol.Biol.48:443〜453の方法を参照のこと。第2の配列中の内部ギャップおよびアミノ酸の挿入は、アラインメントを計算する目的については無視される。それぞれの記述については、代表的な骨形成性タンパク質として、hOP−1(ヒトOP−1、当該分野において「BMP−7」ともいわれる)が以下に提供される。しかし、OP−1は、単に、タンパク質のTGF−βファミリーの代表的なものにすぎないことが明らかである。
【0028】
本明細書中で使用される「TGF−βファミリーメンバー」または「TGF−βファミリータンパク質」は、TGF−βスーパーファミリーのメンバーとして当業者に公知のタンパク質を意味する。構造的に、そのようなタンパク質は、ジスルフィド結合ホモダイマーまたはヘテロダイマーであり、これは、疎水性シグナル配列、数百アミノ酸のN末端プロ領域、ならびに可変性N末端領域およびより高度に保存されたC末端領域(保存された6または7のシステイン骨格を有する特徴的なシステインモチーフを伴う約100アミノ酸を含む)を含む成熟ドメインを含む大きな前駆体ポリペプチド鎖として発現される。これらの構造的に関連するタンパク質は、種々の発生事象に関与するとして同定されている。TGF−βファミリーメンバーは、TGFβ1およびOP−1によって代表される。本発明の実施において有用な他のTGF−βファミリータンパク質としては、骨形成性タンパク質(以下に定義される)、vg−1、DPP−Cポリペプチド、ホルモンアクチビンおよびインヒビン、MIS、VGR−1および増殖/分化因子GDF−1、GDF−3、GDF−9およびドーサリン−1が挙げられる。
【0029】
TGF−βタンパク質スーパーファミリーの種々のメンバーは、2つの異なる細胞表面レセプター(I型レセプターおよびII型レセプターといわれる)と相互作用し、ヘテロ複合体を形成することによってそれらの活性を媒介することが見出された。I型レセプターおよびII型レセプターはともに、セリン/トレオニンキナーゼであり、そして以下の類似の構造を共有する:本質的にキナーゼからなる細胞内ドメイン、膜を1回貫通するに十分な、短い伸長された疎水性配列、および高濃度の保存されたシステインによって特徴付けられる細胞外リガンド結合ドメイン。種々のI型レセプターおよびII型レセプターは、OP−1および他の形態形成タンパク質、およびそれらのアナログ(本発明の改変されたモルフォゲンを含む)と特異的な結合親和性を有する。
【0030】
「骨形成性タンパク質(osteogenic protein)」または「骨形態形成タンパク質」は、TGF−βスーパーファミリータンパク質を意味する。これらは、軟骨および/または軟骨内での骨の形成を含むがこれらに限定されない、骨格組織の形成を生じる形態形成事象の全体的なカスケードを誘導し得る。本明細書中で有用な骨形成性タンパク質としては、任意の既知の天然に存在するネイティブのタンパク質が挙げられる。これらには、天然に存在するかまたは生合成によって産生されたかにはかかわらず、その対立遺伝子、系統発生的対応物、および他の改変体(例えば、「ムテイン」または「変異タンパク質」を含む)、ならびにタンパク質の一般的な形態形成性のファミリーの新規の骨形成的に活性なメンバーが挙げられる。本明細書中で記載される場合、このクラスのタンパク質は、一般には、ヒトの骨形成性タンパク質1(hOP−1)によって代表される。本発明の実施において有用な他の骨形成性タンパク質としては、以下のリストに含まれる骨形成的に活性な形態のタンパク質が挙げられる:OP−1、OP−2、OP−3、BMP−2、BMP−3、BMP−4、BMP−5、BMP−6、BMP−9、DPP、Vg−1、Vgr、60Aタンパク質、CDMP−1、CDMP−2、CDMP−3、GDF−1、GDF−3、GDF−5、6、7、MP−52、BMP−10、BMP−11、BMP−12、BMP−13、BMP−15、UNIVIN、NODAL、SCREW、ADMP、またはNEURAL(これらのアミノ酸配列改変体、および/またはこれらのヘテロダイマーを含む)。1つの現在好ましい実施態様において、本発明の実施において有用な骨形成性タンパク質は、以下のいずれかのものを含む:OP−1、BMP−2、BMP−4、BMP−12、BMP−13、GDF−5,GDF−6,GDF−7、CDMP−1、CDMP−2、CDMP−3、MP−52、ならびにそれらのアミノ酸配列改変体およびホモログ(これらの種ホモログを含む)。なお別の好ましい実施態様において、有用な骨形成的に活性なタンパク質は、参照の骨形成性配列(例えば、OP−1、OP−2、BMP−2、BMP−4、BMP−5、BMP−6、60A、GDF−5、GDF−6、GDF−7などの保存されている7個のシステインのドメインを規定するC末端配列)をコードするDNAまたはRNAに対して、低い、中程度の、または高いストリンジェンシーのハイブリダイゼーション条件下でハイブリダイズする核酸によってコードされる配列を含むアミノ酸配列を有するポリペプチド鎖を有する。本明細書中で使用される場合、高いストリンジェントなハイブリダイゼーション条件は、公知技術に従って、40%のホルムアミド、5×SSPE、5×デンハルト溶液、および0.1%のSDS中で、37℃にて一晩のハイブリダイゼーション、そして0.1×SSPE、0.1%のSDS中で50℃での洗浄として規定される。標準的なストリンジェンシーの条件は、市販の標準的な分子クローニングのテキストにおいて十分に特徴付けられている。例えば、Molecular Cloning
A Laboratory Manual、第2版、Sambrook、Fritsch、およびManiatis編(Cold Spring Harbor Laboratory Press:1989);DNA Cloning、第1巻および第II巻(D.N.Glover編、1985);Oligonucleotide Synthesis(M.J.Gait編、1984):Nucleic Acid Hybridization(B.D.HamesおよびS.J.Higgins編、1984);ならびにB.Perbal、A
Practical Guide To Molecular Cloning(1984)を参照のこと。上記の開示は、本明細書中で参考として援用される。米国特許第5,750,651号、および同第5,863,758号もまた参照のこと。これらの開示は本明細書中で参考として援用される。
【0031】
本発明の実施において有用性を有する関連するタンパク質のTGF−βスーパーファミリーの他のメンバーとしては、以下のリストの中で、ネイティブの劣ったリフォールディングタンパク質が挙げられる:いくつかを挙げると、TGF−β1、TGF−β2、TGF−β3、TGF−β4、およびTGF−β5、種々のインヒビン、アクチビン、BMP−11、ならびにMIS。図4は、TGF−βスーパーファミリーの種々の既知のメンバーの、フィンガー2サブドメインを規定するC末端の35残基を列挙する。劣ったリフォールディングであるリスト上の任意の1つのタンパク質は、他の既知または発見され得るファミリーのメンバーと同様に、本発明の方法によって改善され得る。本明細書中でさらに記載されているように、本発明での使用に適切な生物学的に活性な骨形成性タンパク質は、当該分野で認識されているような、ReddiおよびSampathによって記載されているバイオアッセイを使用する慣用的な実験によって、同定され得る。有用なタンパク質の詳細な記載が以下に続く。等価物は、慣用的な実験および通常の技術のみを用いて当業者によって同定され得る。
【0032】
「モルフォゲン」または「形態形成タンパク質」は、本明細書中で意図される場合、形態形成性であること、すなわち、成熟した哺乳動物において組織の形態形成の発生カスケード(developmental cascade)を誘導し得ることが認識されている(PCT出願番号第US92/01968号を参照のこと)TGF−βスーパーファミリーのメンバーを含む。詳細には、これらのモルフォゲンは、適切な環境条件下で組織特異的様式で、方向付けられていない前駆細胞の増殖を誘導し得、そしてこれらの刺激された前駆細胞の分化を誘導し得る。さらに、モルフォゲンは、これらの分化した細胞の増殖および維持を支持し得る。これらの形態形成活性は、タンパク質が、適切な形態形成を許容する環境において組織の形態形成の発生カスケードを開始しそして維持することを可能にし、幹細胞が組織特異的様式で増殖および分化するのを刺激し、そして新しい組織の形成を生じる事象の進行を誘導させる。これらの形態形成活性はまた、タンパク質が、それらの分化の経路からはずれるように予め刺激された細胞の「再分化」を誘導することを可能にする。適切な環境条件下では、これらのモルフォゲンはまた方向付けられた細胞の「再分化」を刺激し得ることが予想される。当業者を導くために、種々の組織中で、そして形態形成タンパク質に典型的な種々の特性について形態形成タンパク質を試験するための多数の手段が、本明細書中に記載される。これらの教示が、ネイティブのタンパク質、および本発明の改変されたタンパク質の形態形成特性を評価するために使用され得ることが理解される。
【0033】
本発明の有用なネイティブのタンパク質または親のタンパク質はまた、ヒトOP−1のC末端の7個のシステインのドメイン内で少なくとも70%のアミノ酸配列相同性を共有するものを含む。保存されている7個のシステインのドメインに対する候補のアミノ酸配列のパーセント相同性を決定するために、候補の配列と7個のシステインのドメインとを整列させる。アラインメントを実行するための最初の工程は、Needlemanら、J.Mol.Biol.48:443(1970)(その教示は本明細書中で参考として援用される)に記載されている動的計画法アルゴリズム、およびAlign Program(DNAstar、Inc.によって生産されている市販のソフトウェアパッケージ)のような、アラインメントツールを使用することである。最初のアラインメントをなした後、次いでこれは、関連するタンパク質のファミリーの複数の配列のアラインメントに対する比較によって改善される。一旦、候補配列と7個のシステインのドメインとの間でのアラインメントが行われそして改善されると、パーセント相同性のスコアが計算される。各配列の個々のアミノ酸は、互いに対するそれらの類似性に従って連続して比較される。類似性の因子としては、類似の大きさ、形状、および電荷が挙げられる。アミノ酸の類似性を決定する1つの特に好ましい方法は、Dayhoffら、5 Atlas of Protein Sequence and Structure 345−352(1978および補遣、本明細書中で参考として援用される)に記載されているPAM250マトリックスである。類似性スコアはまず、整列された対となった(pairwise)アミノ酸の類似性スコアの合計として計算される。挿入および欠失は、パーセント相同性および同一性の目的のために無視される。従って、ギャップペナルティーは、この計算においては使用されない。次いで、粗スコアが、候補の化合物および7個のシステインのドメインのスコアの相乗平均でそれを除算することによって正規化される。相乗平均は、これらのスコアの積の平方根である。正規化された粗スコアが、パーセント相同性である。
【0034】
本明細書中で使用される場合は、「保存的置換」は、対応する参照残基に物理的または機能的に類似である残基である。例えば、これは、同様の大きさ、形状、電荷、化学的特性(共有結合または水素結合を形成する能力を含む)などを有する。特に好ましい保存的置換は、Dayhoffら(同書)において、容認された点変異について規定された基準を満たすものである。保存的置換の例として、類似の特徴を有する別のアミノ酸に対する1つのアミノ酸の置換が挙げられる。例えば、以下の群内での置換は周知である:(a)グリシン、アラニン;(b)バリン、イソロイシン、ロイシン;(c)アスパラギン酸、グルタミン酸;(d)アスパラギン、グルタミン;(e)セリン、トレオニン;(f)リジン、アルギニン、ヒスチジン;および(g)フェニルアラニン、チロシン。用語「保存的改変体」または「保存的バリエーション」もまた、結果として生じる置換されたポリペプチド鎖に対して結合特異性を有する抗体がまた、置換されていないかまたは親のポリペプチド鎖についての結合特異性を有する(すなわち、それと「交差反応する」かまたはそれと「免疫反応する」)という条件で、所定のポリペプチド鎖中の置換されていない親のアミノ酸の変わりに、置換されたアミノ酸を使用することを含む。
【0035】
本明細書中で使用される場合、「保存されている残基の位置」は、少なくとも1つの他のメンバーの配列中で、同じアミノ酸またはその保存的な改変体によって占有されている参照アミノ酸配列中の位置をいう。例えば、図4において、参照配列としてOP−1と、BMP−2、BMP−4、BMP−5、およびBMP−6とを比較すると、位置1、5、9、12、14、15、16、17、19、22などは保存されている位置であり、そして残基2、3、4、6、7、8、10、11、13、18、20、21などは保存されていない位置である。
【0036】
本明細書中で使用される場合、フィンガー2サブドメインの「ベース」または「ネック」領域は、OP−1によって例示され、そしてC末端の活性ドメイン中の2つ並んでいるシステインに続く最初の残基から数える場合は、残基1〜10および22〜35によって規定される(図4を参照のこと)。他のTGF−βタンパク質ファミリーメンバーのOP−1との配列アラインメントから容易に明らかであるように、より長いタンパク質(例えば、BMP−9またはDorsalin)についての対応するベースまたはネック領域は、残基1〜10および23〜36によって規定される:より短いタンパク質(例えば、NODAL)については、対応する領域は、残基1〜10および22〜34によって規定される(図4を参照のこと)。配列番号39(ヒトOP−1)において、フィンガー2サブドメインのベースまたはネック領域に対応する残基は、残基397〜406(図4の残基1〜10に対応する)および残基418〜431(図4の残基22〜35に対応する)である。
【0037】
本明細書中で使用される「C末端活性ドメイン」は、成熟TGF−βファミリータンパク質の保存されたC末端領域をいう。C末端活性ドメインは、6または7のシステインの骨格を有する特徴的なシステインモチーフを伴う約100アミノ酸を含む。すべてのタンパク質のC末端のシステインパターンは同一の形式であり、配列Cys−X−Cys−Xで終わる(SpornおよびRoberts(1990)、前出)。
【0038】
本明細書中で使用される場合、「アミノ酸配列の相同性」は、アミノ酸配列の同一性および類似性の両方を含む。相同配列は、同一および/または類似のアミノ酸残基を共有する。ここで、類似の残基は、整列された参照配列中の対応するアミノ酸残基の保存的置換、またはその「許容される点変異」である。
【0039】
本明細書中で使用される場合、用語「キメラタンパク質」、「キメラ」、「キメラポリペプチド鎖」、「キメラ構築物」、および「キメラ変異体」は、任意のBMPまたはTGF−βファミリーのメンバーの合成の構築物をいう。ここで、少なくとも1つの規定された領域、ドメイン、またはサブドメイン(例えば、フィンガー1、フィンガー2、またはヒールサブドメイン)のアミノ酸配列は、全体または一部において、少なくとも1つの他の異なるBMPまたはTGF−βファミリーのメンバーのタンパク質に由来するアミノ酸配列と置き換えられている。その結果、得られる構築物は、異なるタンパク質供給源に由来するとして認識可能なアミノ酸配列を有する。キメラ構築物はまた、1つのモルフォゲンのC末端活性ドメインが、別のモルフォゲンのN末端ドメインに融合されている組換え融合タンパク質を含む。
【0040】
本明細書中で使用される「リーダー配列」は、TGF−βファミリータンパク質のC末端活性ドメイン領域の上流、すなわちC末端からさらに遠くに位置するヌクレオチドの配列に対応するアミノ酸の任意の配列である。リーダー配列における改変は、リフォールディング特性、活性レベル、可溶性および制御活性化を変更し、そして組織標的化および親和性結合能を促進し得る。
【0041】
本明細書中で使用される場合、有用な発現宿主細胞としては、原核生物および真核生物が挙げられる。これには、封入体を産生し得る任意の宿主細胞が挙げられる。特に有用な宿主細胞として、限定的ではないが、E.coliのような細菌宿主、ならびにB.subtilisおよびPseudomonasが挙げられる。他の有用な宿主としては、下等な真核生物(例えば、Saccharomyces cereviceaeまたは他の酵母)および高等真核生物(例えば、Drosophila、CHO細胞、および他の哺乳動物細胞など)が挙げられる。本明細書中において議論されるように、化学合成方法もまた、本発明の改変されたタンパク質を生成するために利用され得る。
【0042】
1つの局面において、本発明は、哺乳動物細胞において容易に発現されない組換えタンパク質(例えば、融合タンパク質など)の構築を提供する。例えば、骨標的化特性を有する融合タンパク質をコードする組換え遺伝子が構築される。ここで、一つの配列は、BMPおよび抗体結合部位(オステオカルシンまたはフィブロネクチンのような骨基質タンパク質に対する特異性を有する)の両方をコードする。同様に、細胞表面レセプター(例えば、前骨芽細胞または軟骨細胞上のレセプター)に結合する融合タンパク質もまた構築され得る。他の組換え遺伝子は、金属または他のタンパク質を特異的に結合する融合タンパク質をコードし得る。結合の特異性は、BMPに付加されたリーダー配列の組成に依存する。これらの遺伝子は、E.coli中で発現され、そしてインビトロでリフォールディングされ得る。
【0043】
別の実施態様において、切断可能な融合タンパク質(プロテアーゼ−例えば、トリプシン、V8、第Xa因子およびその他、または化学的−中程度の酸、ヒドロキシルアミンおよび他の薬剤によって切断可能)が合成され、ここで、TGF−βタンパク質は、活性をブロックするリーダー配列に付着される。なお別の実施態様において、TGF−βファミリーメンバーの活性は、このリーダー配列の一部または全てを切断することによって回復または増強される。活性を阻害する切断可能なリーダー配列を付加することによって、このタンパク質の潜在形態は、実質的に、活性なC末端ドメインを含むタンパク質フラグメントを放出するように切断され得る。
【0044】
さらに別の実施態様において、このリーダー配列はまた、組織標的配列であり、その結果、放出は、インビボにおいてこの標的部位で生じるように制御され得る。切断可能部位の構築はまた、活性なタンパク質の放出を制御することを可能にし得る。例えば、骨組織において、骨の再モデル化に関与する多くのプロテアーゼが代表的に存在し、そして有利に使用され得る。以下に記載される切断可能な「ヘキサ−his」、FBリーダー、またはコラーゲン結合配列は、このタンパク質の潜在形態についての適切なリーダー配列であり得る。例の目的で、この組織標的ドメインは、少なくとも3つの塩基残基の続きを含むリーダー配列によってBMPから分離され得、これは、インビボにおいて切断されることが公知である。
【0045】
なお別の実施態様において、このリーダー配列が構築され得、その結果、特定の活性を阻害するタンパク質の部分は切断され、そして活性は回復されるが、このタンパク質の組織標的部分は、保持される。
【0046】
さらに別の実施態様において、TGF−βファミリータンパク質のリーダー配列は、別のTGF−βメンバーのリーダー配列によって置換される。得られた「キメラ」タンパク質は、変更された可溶性、フォールディングおよび/または組織標的活性、改善された安定性、ならびに/あるいは特定の表面に結合する能力を有し得る。
【0047】
本発明の別の局面において、融合タンパク質は、他のTGF−βファミリータンパク質と組み合わされて、ヘテロダイマーを形成する。ここで、各々のタンパク質の特性が開発される。例えば、組織標的特性を有するが、活性を有さない融合タンパク質は、活性を有するが標的能力を有さない異なるタンパク質とともにヘテロダイマーを形成する。前者のタンパク質は、後者のタンパク質がその機能を実施し得る標的部位にヘテロダイマーを送達する。
【0048】
1つの局面において、本発明は、中性または生理学的条件下で改善されたリフォールディング特性を有する生合成BMPおよびTGF−βファミリーメンバーを提供する。1つの実施態様において、本発明の生合成タンパク質は、約5.0〜10.0の範囲、好ましくは約6.0〜9.0の範囲、より好ましくは約6.0〜8.5の範囲のpH(約pH7.0〜7.5の範囲を含む)で、改善されたリフォールディング特性を有する。
【0049】
別の局面において,本発明は、中性または生理学的条件下で改善された可溶性特性を有する生合成BMPおよびTGF−βファミリーメンバーを提供する。1つの実施態様において、本発明の生合成タンパク質は、約5.0〜10.0の範囲、好ましくは約6.0〜9.0の範囲、より好ましくは約6.0〜8.5の範囲のpH(約pH7.0〜7.5の範囲を含む)で、改善された可溶性を有する。
【0050】
なお別の局面において,本発明は、生理学的条件下でリフォールディングし、そして親配列と比較して変更された等電点を有する、生物学的に活性な生合成BMPおよびTGF−βファミリーメンバー構築物成分を提供する。
【0051】
別の局面においては、本発明は、生理学的条件または中性のpH条件下でほとんどリフォールディングされないホモダイマーおよびヘテロダイマーをフォールディングするための方法を提供する。1つの実施態様においては、この方法は、本発明の1つ以上の可溶化されたTGF−βファミリータンパク質構築物を提供する工程、可溶化されたタンパク質を適切なリフォールディング緩衝液中での酸化還元反応に曝す工程、ならびに所望される場合には、タンパク質のサブユニットがホモダイマーおよび/またはヘテロダイマーにリフォールディングすることを可能にする工程を包含する。別の実施態様においては、本発明の改変TGF−βファミリータンパク質は、それらをこの酸化還元反応に曝す前に変性されない。別の実施態様においては、酸化還元反応系は、グルタチオン、DTT、β−メルカプトメタノール、システイン、およびシスタミンの酸化された形態および還元された形態を利用し得る。別の実施態様においては、酸化還元反応系は、好ましくは金属触媒(例えば、銅)の存在下での、空気の酸化による。なお別の実施態様においては、これらは、約1:10から約10:1の、好ましくは約1:2から2:1の範囲内の酸化剤に対する還元剤の比での酸化還元系として使用され得る。別の好ましい実施態様においては、このタンパク質は、イオン性界面活性剤、非イオン性の界面活性剤(例えば、ジギトニン)、または双性イオン性界面活性剤(例えば、3−[(3−コラミドプロピル(cholamidopropyl))ジメチルアンモニオ]−1−プロパン硫酸(CHAPS)、またはN−オクチルグルコシドを含む、界面活性剤の存在下で可溶化される。なお別の実施態様においては、リフォールディング反応は、約5.0〜10.0の範囲内、好ましくは、約6.0〜9.0の範囲内、より好ましくは、約7.0〜8.5の範囲内の、pHの範囲で生じる。なお別の実施態様においては、リフォールディング反応は、約32〜0℃の範囲内、好ましくは、約25〜4℃の範囲内の温度で生じる。ヘテロダイマーが作製される場合は、2つの異なるサブユニットの添加についての最適な比が、経験的に、そして過度の実験を伴うことなく容易に決定され得る。
【0052】
別の局面において、本発明は、宿主細胞(細菌宿主、または任意の他の宿主細胞を含む)において弱いフォールディングのBMPおよび他のTGF−βファミリーメンバータンパク質を組換え的に生成する方法を提供し、ここで、過剰発現されるタンパク質は、インビトロにおいて可溶化および/またはリフォールディングすることを必要とする形態で凝集する。この方法は、本発明の1つ以上の生合成タンパク質をコードする核酸分子でトランスフェクトされた宿主細胞を提供する工程、生合成タンパク質を発現するために適切な条件下で宿主細胞を培養する工程、凝集したタンパク質を回収する工程、ならびに上記に述べた工程を使用してこのタンパク質を可溶化およびリフォールディングする工程を包含する。別の実施態様において、この方法は、本発明の生合成タンパク質をコードする核酸を用いて宿主細胞をトランスフェクトする、さらなる工程を包含する。
【0053】
本発明の改変モルフォゲンは、生体適合性マトリクス(例えば、コラーゲン、ヒドロキシアパタイト、セラミックスもしくはカルボキシメチルセルロース、および/または他の適切なマトリクス材料(これらに限定されない)と合わせて、骨および/または軟骨を形成するために使用され得る。このような組合せは、再生骨、軟骨および/または他の非鉱化骨格もしくは連結組織(例えば、少数だが、人工軟骨、線維軟骨、関節包、半月板、椎間板(intervertabral disk)、滑膜組織、筋肉、および筋膜(fasica)(これらに限定されない)についての方法において特に有用である。例えば、米国特許第5,674,292号、同第5,840,325号、および米国特許出願第08/253,398号(これらの開示は、本明細書中に参考として援用される)を参照のこと。本発明は、このようなマトリクス材料に対する結合特性および/または接着特性が、タンパク質構築物を生成するために本明細書中に開示される技術を使用して変更され得ることを意図する。本発明の改変タンパク質はまた、腱、靭帯および/または筋肉組織を生成するために利用され得る。
【図面の簡単な説明】
【0054】
【図1A】図1Aは、TGF−βスーパーファミリーメンバーのモノマーサブユニットの構築を記載する際に有用な単純な線図である。説明については、前述の発明の背景を参照のこと。図1B、1Cおよび1Dは、フィンガー1、ヒールおよびフィンガー2領域の代表的な二次構造のそれぞれのペプチド骨格の単眼視野の(monovision)リボン追跡である。
【図1B】図1Aは、TGF−βスーパーファミリーメンバーのモノマーサブユニットの構築を記載する際に有用な単純な線図である。説明については、前述の発明の背景を参照のこと。図1B、1Cおよび1Dは、フィンガー1、ヒールおよびフィンガー2領域の代表的な二次構造のそれぞれのペプチド骨格の単眼視野の(monovision)リボン追跡である。
【図1C】図1Aは、TGF−βスーパーファミリーメンバーのモノマーサブユニットの構築を記載する際に有用な単純な線図である。説明については、前述の発明の背景を参照のこと。図1B、1Cおよび1Dは、フィンガー1、ヒールおよびフィンガー2領域の代表的な二次構造のそれぞれのペプチド骨格の単眼視野の(monovision)リボン追跡である。
【図1D】図1Aは、TGF−βスーパーファミリーメンバーのモノマーサブユニットの構築を記載する際に有用な単純な線図である。説明については、前述の発明の背景を参照のこと。図1B、1Cおよび1Dは、フィンガー1、ヒールおよびフィンガー2領域の代表的な二次構造のそれぞれのペプチド骨格の単眼視野の(monovision)リボン追跡である。
【図2A】図2Aおよび2Bは、以下からの、TGF−βスーパーファミリーメンバータンパク質ダイマーの一般的な三次元形状を示すステレオペプチド骨格リボン追跡図である:A)紙に対して一般に正常なへリックスヒール領域の軸および一般に垂直のフィンガー1およびフィンガー2の各々の軸を有する「トップ」(サブユニット間の対称の2回転軸の下)、ならびにB)紙の代わりにこのサブユニット間の2回転軸、一般に水平のヒールの軸、および一般に垂直の軸を有する「サイド」。このリーダーは、形態単位設計においてよりよい特別な関係を理解するためにウォールアイステレオ(wall eyed stereo)におけるステレオα炭素追跡図を見るために奨励される。
【図2B】図2Aおよび2Bは、以下からの、TGF−βスーパーファミリーメンバータンパク質ダイマーの一般的な三次元形状を示すステレオペプチド骨格リボン追跡図である:A)紙に対して一般に正常なへリックスヒール領域の軸および一般に垂直のフィンガー1およびフィンガー2の各々の軸を有する「トップ」(サブユニット間の対称の2回転軸の下)、ならびにB)紙の代わりにこのサブユニット間の2回転軸、一般に水平のヒールの軸、および一般に垂直の軸を有する「サイド」。このリーダーは、形態単位設計においてよりよい特別な関係を理解するためにウォールアイステレオ(wall eyed stereo)におけるステレオα炭素追跡図を見るために奨励される。
【図3】図3は、SmithおよびSmith(1990)Proc.Natl.Acad.Sci.USA 87:118〜122の教示に従って調製されたパターン規定表である。
【図4】図4は、BMPファミリーの種々の既知のメンバー、およびタンパク質のTGF−βスーパーファミリーについて、2つ並んでいるシステインに続く最初の残基で開始して並べた、フィンガー2サブドメインを規定する、整列したC末端の残基を列挙する。
【図5A】図5A、5B、および5Cは、TGF−βスーパーファミリーの現在既知のメンバーの、それぞれフィンガー1、ヒール、およびフィンガー2領域のアラインメントおよび相同性を示すように並べられた、一文字表記のアミノ酸配列である。以下の各領域を含むそれぞれのアミノ酸が示されている:ヒトのTGF−β1からTGF−β5(TGF−βサブグループ)、dpp、Vg−1、Vgr−1、60A(同時係属中のU.S.S.N.08/271,556号を参照のこと)、BMP−2A(BMP−2としてもまた文献において公知である)、dorsalin、BMP−2B(BMP−4としてもまた文献において公知である)、BMP−3、BMP−5、BMP−6、OP−1(BMP−7としてもまた文献において公知である)、OP−2(PCT/US91/07635号、および米国特許第5,266,683号を参照のこと)、およびOP−3(U.S.S.N.07/971,091号)からなるVg/dppサブグループ、GDF−1、GDF−3、およびGDF−9からなるGDFサブグループ、インヒビンα、インヒビンβA、およびインヒビンβBからなるインヒビンのサブグループ。ダッシュ(−)は、隣接するアミノ酸間のペプチド結合を示す。各サブグループについてのコンセンサス配列パターンを、各サブグループの最下段に示す。
【図5B】図5A、5B、および5Cは、TGF−βスーパーファミリーの現在既知のメンバーの、それぞれフィンガー1、ヒール、およびフィンガー2領域のアラインメントおよび相同性を示すように並べられた、一文字表記のアミノ酸配列である。以下の各領域を含むそれぞれのアミノ酸が示されている:ヒトのTGF−β1からTGF−β5(TGF−βサブグループ)、dpp、Vg−1、Vgr−1、60A(同時係属中のU.S.S.N.08/271,556号を参照のこと)、BMP−2A(BMP−2としてもまた文献において公知である)、dorsalin、BMP−2B(BMP−4としてもまた文献において公知である)、BMP−3、BMP−5、BMP−6、OP−1(BMP−7としてもまた文献において公知である)、OP−2(PCT/US91/07635号、および米国特許第5,266,683号を参照のこと)、およびOP−3(U.S.S.N.07/971,091号)からなるVg/dppサブグループ、GDF−1、GDF−3、およびGDF−9からなるGDFサブグループ、インヒビンα、インヒビンβA、およびインヒビンβBからなるインヒビンのサブグループ。ダッシュ(−)は、隣接するアミノ酸間のペプチド結合を示す。各サブグループについてのコンセンサス配列パターンを、各サブグループの最下段に示す。
【図5C】図5A、5B、および5Cは、TGF−βスーパーファミリーの現在既知のメンバーの、それぞれフィンガー1、ヒール、およびフィンガー2領域のアラインメントおよび相同性を示すように並べられた、一文字表記のアミノ酸配列である。以下の各領域を含むそれぞれのアミノ酸が示されている:ヒトのTGF−β1からTGF−β5(TGF−βサブグループ)、dpp、Vg−1、Vgr−1、60A(同時係属中のU.S.S.N.08/271,556号を参照のこと)、BMP−2A(BMP−2としてもまた文献において公知である)、dorsalin、BMP−2B(BMP−4としてもまた文献において公知である)、BMP−3、BMP−5、BMP−6、OP−1(BMP−7としてもまた文献において公知である)、OP−2(PCT/US91/07635号、および米国特許第5,266,683号を参照のこと)、およびOP−3(U.S.S.N.07/971,091号)からなるVg/dppサブグループ、GDF−1、GDF−3、およびGDF−9からなるGDFサブグループ、インヒビンα、インヒビンβA、およびインヒビンβBからなるインヒビンのサブグループ。ダッシュ(−)は、隣接するアミノ酸間のペプチド結合を示す。各サブグループについてのコンセンサス配列パターンを、各サブグループの最下段に示す。
【図6】図6は、標準的な一文字表記のアミノ酸コードで大文字で示されている、アミノ酸配列の一文字コードのリストであり、そして小文字で、その位置において有用であるアミノ酸の群を同定する。ここで、アミノ酸を表す小文字は、表3に示されているパターン定義の鍵となる表に従って示されている。図6は、本発明の生合成構築物のフィンガー1、ヒール、およびフィンガー2領域を構成するための好ましいパターン配列を同定する。ダッシュ(−)は、隣接するアミノ酸間のペプチド結合を示す。
【図7A】図7(A)は、H2487のヌクレオチドおよび対応するアミノ酸配列を示す。H2487は、7つのシステインのドメインの上流に挿入されたN末端デカペプチドコラーゲン結合部位を含む、改変OP−1である。
【図7B】図7(B)は、H2440のヌクレオチドおよび対応するアミノ酸配列を示す。H2440は、7つのシステインのドメインの第1のシステインの35残基上流に結合されたヘキサ−ヒスチジンドメインを含む、改変OP−1である。
【図7C】図7(C)は、H2521のヌクレオチドおよびアミノ酸配列を示す。H2521は、7つのシステインのドメインの第1のシステインの15残基上流に結合されたプロテインAのFBリーダードメインを含む、改変OP−1である。
【図7D】図7(D)は、H2525のヌクレオチドおよびアミノ酸配列を示す。H2525は、プロテインAのFBリーダードメインおよびヘキサ−ヒスチジンドメインの両方を含む、改変OP−1である。
【図7E】図7(E)は、H2527のヌクレオチドおよびアミノ酸配列を示す。H2527は、FBリーダードメイン、ヘキサ−ヒスチジンドメイン、およびASP−PRO酸切断部位を含む、改変OP−1である。
【図7F】図7(F)は、H2528のヌクレオチドおよびアミノ酸配列を示す。H2528は、FBリーダードメインおよびヘキサ−ヒスチジンドメインを含む、改変CDMP−3である。
【図7G】図7(G)は、H2469のヌクレオチドおよびアミノ酸配列を示す。H2649は、保存された7つのシステインのドメインの第1のシステインの14の元々の残基の上流を含む、改変OP−1(短縮型)である。
【図7H】図7(H)は、H2510のヌクレオチドおよびアミノ酸配列を示す。H2510は、保存された7つのシステインのドメインの第1のシステインの7残基上流に挿入されたコラーゲン結合部位を含む、改変OP−1である。
【図7I】図7(I)は、H2523のヌクレオチドおよびアミノ酸配列を示す。H2523は、コラーゲンペプチドおよび保存された7つのシステインのドメインの第1のシステインから13残基上流に付加されたスペーサーを含む、改変OP−1である。
【図7J】図7(J)は、H2524のヌクレオチドおよびアミノ酸配列を示す。H2524は、ヘキサ−ヒスチジンドメイン、コラーゲンペプチドおよび保存された7つのシステインのドメインの第1のシステインから13残基上流に付加されたスペーサーを含む、改変OP−1である。
【図8】図8は、OP−1 C末端の7つのシステイン活性ドメインをコードする制限マップである。
【図9A】図9Aは、種々の生合成キメラBMP構築物の模式図である。
【図9B】図9Bは、生合成BMP変異体の模式図およびそれらのリフォールディングおよびROS活性である。
【図10】図10は、種々のBMPについてのC末端サブドメインにおける荷電残基の数を示す。
【図11】図11は、濃度(ng/mL)対光学密度(405nmにおける)としてプロットした、OP−1(標準)、変異体H2549タンパク質およびトリプシンで処理したH2549についてのROS活性のグラフである。
【図12】図12は、濃度(ng/mL)対光学密度(405nmにおける)としてプロットした、OP−1(標準)および変異体H2223の種々の画分ならびにこのタンパク質のトリプシン短縮型形態についてのROS活性のグラフである。
【図13】図13(A)は、濃度(ng/mL)対光学密度(405nmにおける)としてプロットした、OP−1ホモダイマー(CHO細胞由来)、BMP−2ホモダイマーおよびヘキサ−his OP−1ヘテロダイマーについてのROS活性のグラフである。
【0055】
図13(B)は、濃度(ng/ml)対光学密度(405nmにおける)としてプロットした、OP−1ホモダイマー(CHO細胞由来)、ヘキサ−his OP−1/BMP−2ヘテロダイマーおよびヘキサ−his OP−1についてのROS活性のグラフである。
【図14】図14は、濃度(ng/mL)対光学密度(405nmにおける)としてプロットした、OP−1(標準)およびBMP−2変異体H2142タンパク質ホモダイマー、変異体H2525タンパク質ホモダイマーおよびH2525/2142へテロダイマーについてのROS活性のグラフである。
【図15】図15は、種々のOP−1変異体のフィンガー2サブドメインのアミノ酸配列、ならびにROS細胞に基づくアルカリホスファターゼアッセイにおけるそれらのフォールディング効率および生物学的活性を示す。
【発明を実施するための形態】
【0056】
(好ましい実施態様の詳細な説明)
本発明は、天然の形態と比較して、変化したリフォールディング特性、および変化した活性プロフィールを有する、改変形態のTGF−βファミリータンパク質を提供する。本発明の改変タンパク質は、天然に存在するTGF−βファミリーメンバー(特に、形態形成タンパク質)のN末端改変を含む。これらの改変は、伸長、短縮、および/または特定の部位でのプロテアーゼ切断または化学切断(例えば、酸またはCNBrによる)による活性化、異なるタンパク質ドメインの結合(融合)ならびに他のTGF−βファミリーメンバー由来のサブユニットとのヘテロダイマーの生成を含む。以下に提供する詳細な説明は、改善した活性および薬学的特性を生じる、多数の例示的な、置換、融合、および伸長を記載する。改変タンパク質を生成する方法もまた、教示する。
【0057】
本発明の1つの局面に従って、ほとんどリフォールディングしないBMPおよびTGF−βスーパーファミリーの他のメンバーのタンパク質(そのヘテロダイマ−およびキメラを含む)のフォールディング能力は、BMPまたはTGF−βファミリーメンバーの既存のN末端ドメイン(次いで、この融合タンパク質内の部位で切断され得る)に、特定の標的領域およびレセプター結合領域を融合させることによって、改善される。この発見の結果として、以下を設計することが可能である:(1)原コア生物細胞または真コア生物細胞中で組換え的に発現されたかまたはポリペプチド合成装置を使用して合成された、BMPおよび他のTGF−βファミリーのタンパク質;(2)変化したフォールディング特性を有する、BMPおよび他のTGF−βファミリーのタンパク質;(3)中性のpH(生理学的条件を含むが、これに限定されない)の下で変化した可溶性を有する、BMPおよび他のTGF−βファミリーのタンパク質;(4)変化した等電点を有する、BMPおよび他のTGF−βファミリーのタンパク質;(5)変化した安定性を有する、BMPおよび他のTGF−βファミリーのタンパク質;(6)固相表面(例えば、生体適合性マトリクスまたは金属)に対する変化した結合特性または付着特性を有する、BMPおよび他のTGF−βファミリーのタンパク質;ならびに/または(7)所望の、変化した生物学的活性(例えば、組織特異性および/またはレセプター特異性)を有する、BMPおよび他のTGF−βファミリーのタンパク質。さらに、本発明は、新規な候補構築物(特に、その候補の生物学的特性または生化学的特性)を迅速に試験するための手段を提供する。本発明はまた、例えば、異なる組み合わせのドメインを有するキメラタンパク質を作製することによって、抗体のエピトープを迅速にマッピングするための手段を提供する。詳細には、本明細書中に開示された発見を使用して、原コア生物宿主(例えば、E.coli)において他の方法では発現され得ないモルフォゲン配列を改変して、E.coliにおける発現およびインビボでのリフォールディングを、今や可能にし得る。
【0058】
従って、本発明は、好ましいキメラタンパク質を含む、速放性、徐放性および/または時限放出性の処方物を設計するための機構を提供し得る。さらに、本発明は、タンパク質構築物の環境的により誘発されて放出するように操作された処方物を設計するための機構を提供する。すなわち、改変タンパク質は、インサイチュでの特定の環境条件下(例えば、pHの変化、特異的プロテアーゼの存在など)で、送達を調節し、そして放出および活性を容易にするように設計され得る。他の利点および特徴は、以下の教示から明らかである。さらに、本明細書中の発見を使用して、変化した表面結合特性/表面付着特性を有する改変タンパク質を、設計および選択し得る。特に重要な表面としては、以下が挙げられるが、これらに限定されない:天然に存在し得る固体表面(例えば、骨);または多孔性粒子表面(例えば、コラーゲンまたは他の生体適合性マトリクス);あるいは人工器官移植片(金属を含む)の加工(fabricated)表面。本明細書中で意図する場合、事実上すべての表面を、構築物の示差的結合についてアッセイし得る。従って、本発明は、その表面結合特性/表面付着特性が変化し、それによってそのような構築物を変化したインビボ適用(徐放性処方物、速放性処方物および/または時限放出性処方物を含む)のために有用にする、多様な機能性分子をを含む。
【0059】
当業者は、1つ以上の任意の上記の特性を混合かつ適合させることによって、カスタマイズされた改変タンパク質(および、それをコードするDNA)の使用を操作するための特定の機会が提供されることを、理解する。例えば、変化した安定性の特性を使用して、インビボでのタンパク質のターンオーバーを操作し得る。さらに、変化したリフォールディングおよび/または機能のような特性を有する改変タンパク質の場合において、フォールディング、機能、および安定性との間に相関関係が存在するようである。例えば、Lipscombら、7、Protein Sci.765−73(1998);およびNikolovaら、95、Proc.Natl.Acad.Sci.USA 14675−80(1998)を参照のこと。本発明の目的のために、安定性の変化を、円偏光二色性、および変性剤の濃度または温度の関数としての安定性の他の指標といった周知の技術を使用して、慣用的にモニターし得る。当業者はまた、慣用的な走査熱分析を使用し得る。同様に、上記の任意の特性と可溶性の特性との間に相関関係が存在するようである。可溶性の場合においては、この特性を操作して、その結果、改変タンパク質が、生理学的適合性状態の下でより可溶性であるかまたはより可溶性ではないかのいずれかであり、そしてその結果として、インビボで投与した場合には、それぞれ、容易に拡散するか、または局在化したままであるようにすることが可能である。
【0060】
適切な生合成タンパク質および本発明の実施において有用な方法、ならびにこれらのタンパク質を使用および試験するための方法の、詳細な説明を、以下に提供する;そして、以下の限定的ではない多数の例を提供する:1)本明細書中に記載されている、生合成タンパク質および方法の適合性を説明する、例;ならびに、2)これらのタンパク質を試験および使用するためのアッセイを提供する、例。
【0061】
(I.タンパク質の考察)
(A.TGF−β2およびOP−1の構造的特徴)
TGF−β2またはOP−1のいずれかの中のサブユニット各々は、特徴的なフォールディングパターン(図1Aに模式的に例示される)を有する。このパターンは、7個のC末端のシステイン残基のうちの6個を含む。簡潔には、各サブユニット中のシステイン残基のうちの4つが2つのジスルフィド結合を形成し、これらは互いに8残基の環を作製する。一方、2つのさらなるシステイン残基が、結び目様の構造を形成するように環を通過する、ジスルフィド結合を形成する。7個の保存されているシステイン残基のうちの最もN末端のシステイン(番号1を割り当てられる)で開始する番号付けスキームを用いると、2番目および6番目のシステイン残基が、8残基の環の1つの側面を閉じるようにジスルフィド結合し、一方、3番目および7番目のシステイン残基が、この環の他の側面を閉じるようにジスルフィド結合する。1番目および5番目の保存されているシステイン残基は、この結び目のコアを形成するように、この環の中心を通ってジスルフィド結合する。アミノ酸配列の整列のパターンは、この構造モチーフが、TGF−βスーパーファミリーのメンバーの間で保存されていることを示唆する。4番目のシステインは半保存されており、そして存在する場合には、代表的に、他のサブユニット中の対応するシステイン残基と鎖間ジスルフィド結合(ICDB)を形成する。
【0062】
TGF−β2およびOP−1中の各サブユニットの構造は、3つの主要な三次構造のエレメントおよびN末端領域を含む。この構造エレメントは、以下の型のうちの50%を超える二次構造を保有する、連続するポリペプチド鎖の領域から構成される:(1)ループ、(2)α−へリックスおよび(3)β−シート。各構造領域について規定する別の特徴は、入口(N末端)および出口(C末端)のペプチド鎖が、約7Åの間隔で、互いにかなり近づいていることである。
【0063】
図1Aに示される、1番目の保存システインと2番目の保存システインとの間のアミノ酸配列は、逆平行β−シートフィンガーによって特徴付けられる構造領域を形成して、本明細書中でフィンガー1領域と呼ばれる。同様に、5番目の保存システインと6番目の保存システインとの間の残基もまた(図1Aに示すように)、逆平行β−シートフィンガーを形成して、フィンガー2領域と本明細書中で呼ばれる。β−シートフィンガーは、一本のアミノ酸鎖であり、β−ターンまたはいくらか長いループによってそれ自身の上にリフォールディングβ−鎖を含む。その結果、この領域に入りかつこの領域から出るポリペプチド鎖は、1つ以上の逆平行β−シート構造を形成する。3番目の保存されているシステインと5番目の保存されているシステインとの間の残基を含む、第3の主要な構造領域(図1Aに示す)は、3個のターンα−へリックスによって特徴付けられ、本明細書中でヒール領域と呼ばれる。その単量体構造の構成は、左手の構成と類似である。この構造においては、その結び目領域が手のひらに相当する位置に位置し、フィンガー1領域が人差し指および中指に相当し、α−へリックス領域またはヒール領域が手首の付け根に相当し、そしてフィンガー2領域が薬指および小指に相当する。N末端領域は、その配列がTGF−βスーパーファミリーを通じて保存されておらず、親指にほぼ相当する位置に位置すると推定される。
【0064】
TGF−β2単量体の3つの主な独立構造エレメントの各々のα炭素骨格の単視的リボントレース(monovision ribbon tracing)を図1B〜1Dに示す。詳細には、第1の逆方向β−シートセグメントを含む例示的フィンガー1領域を図1Bに示し、3つのターンα−へリックスセグメントを含む例示的ヒール領域を図1Cに示し、そして第2および第3の逆方向β−シートセグメントを含む例示的フィンガー2領域を図1Dに示す。
【0065】
図2は、立体構造的に活性なTGF−β2の二量体複合体のペプチド骨格の立体リボントレース図を示す。この二量体複合体中の2つの単量体サブユニットは、2回回転対称性を有して位置しており、その結果、一方のサブユニットのヒール領域が、もう一方のサブユニットのフィンガー領域と接触し、結合したサブユニットの結び目領域が、この分子のコアを形成する。4番目のシステインは、2番目の鎖上の対照物と鎖間ジスルフィド結合を形成し、それによりその掌の中心でこれらの鎖を等しく連結する。このように形成された二量体は、サブユニット間の2回対称軸を上方から見下ろした場合に、楕円形の(葉巻の形状の)分子である(図2A)。側方から見ると、この分子は、まがった「葉巻」に似ている。なぜなら、その2つのサブユニットが、互いに対してわずかな角度で位置しているからである(図2B)。
【0066】
図2に示すように、ともにこの二量体のネイティブな単量体サブユニットを規定する構造エレメントの各々を、22、22’、23、23’、24、24’、25、25’、26、および26’と符号をつける。この図で、エレメント22、23、24、25、および26は、一方のサブユニットによって規定され、そしてエレメント22’、23’、24’、25’および26’はもう一方のサブユニットに所属する。詳細には、22および22’はN末端ドメインを示し;23および23’はフィンガー1領域を示し;24および24’はヒール領域を示し;25および25’はフィンガー2領域を示し;そして26および26’は、各サブユニットの1番目および5番目の保存システインを連結して結び目様の構造を形成する、ジスルフィド結合を示す。図2から、一方のサブユニット由来のヒール領域(例えば、24)ともう一方のサブユニット由来のフィンガー1領域およびフィンガー2領域(例えば、それぞれ、23’および25’)は、互いに相互作用することが理解され得る。これら3つのエレメントは互いに協同して、同族のレセプターの相互作用する表面に結合するリガンドと相互作用しそしてそれらに相補性である、構造を規定する。
【0067】
(フィンガー領域およびヒール領域の選択)
フィンガー領域およびヒール領域を規定するアミノ酸配列が、本明細書中で同定された、TGF−βスーパーファミリーの既知の任意のメンバーの、それぞれのフィンガー領域の配列およびヒール領域の配列に由来し得るか、または、本明細書中で以後に発見される新規のスーパーファミリーのメンバーのアミノ酸配列に由来するものが、利用され得ることが、意図される。
【0068】
図5は、フィンガー1領域(図5A)、ヒール領域(図5B)、およびフィンガー2(図5C)領域へと整列された、現在同定されているTGF−βスーパーファミリーのメンバーのアミノ酸配列を要約する。これらの配列は、コンピューターアルゴリズムによって整列された。このアルゴリズムでは、これらの配列を最適に整列するために、保存的アミノ酸配列または二次構造を有することが既知のアミノ酸配列の領域よりもむしろ、ループ構造を規定することが知られているアミノ酸配列の領域中にギャップが挿入された。例えば、可能である場合には、ギャップは、βシートによって規定されるフィンガー1領域およびフィンガー2領域のアミノ酸配列中か、またはαへリックスによって規定されるヒール領域のアミノ酸配列中には導入されなかった。ダッシュ(−)は、隣接するアミノ酸間でのペプチド結合を示す。各サブグループについてのコンセンサス配列のパターンは、各サブグループの最下段に示される。
【0069】
TGF−βスーパーファミリーのメンバー各々のアミノ酸配列が整列された後、この整列された配列を使用してアミノ酸配列の整列パターンを作製した。このパターンによって、得られた構築物の全体的な三次構造を変更することなく別のアミノ酸またはアミノ酸の群によって置換され得るアミノ酸残基が同定される。このフィンガー領域およびヒール領域中の特定の位置で有用であり得るアミノ酸またはアミノ酸の群は、図3に示されるアミノ酸の階層パターン構造を実行するコンピューターアルゴリズムによって同定された。
【0070】
簡潔には、このアルゴリズムは、4つのレベルの分析を行う。レベルIにおいては、このアルゴリズムは、特定のアミノ酸残基が、そのアミノ酸配列中の特定な位置で75%を超える頻度で生じるかどうかを決定する。例えば、アミノ酸配列中の特定の位置でグリシン残基が10回のうち8回生じる場合には、グリシンがその位置で指定される。試験される位置が全てギャップからなる場合には、ギャップ文字(−)がその位置に割り当てられる。そうでなければ、少なくとも1つのギャップが存在する場合には、「z」(任意の残基またはギャップについての表記)がその位置に割り当てられる。アミノ酸が、特定の位置で候補配列の75%で生じない場合には、アルゴリズムはレベルIIの分析を実行する。
【0071】
レベルIIは、パターンセットa、b、d、l、k、o、n、i、およびhを規定する。ここでは、l、k、およびoは、共通のアミノ酸残基を共有する。次いで、このアルゴリズムは、そのアミノ酸配列中の特定の位置のアミノ酸残基のうちの75%以上が上記のパターンのうちの1つを満たすかどうかを決定する。もしそうであれば、このパターンがその位置に割り当てられる。しかし、パターンlおよびkの両方が同時に満たされ得ることも可能である。なぜなら、これらは、同じアミノ酸(詳細には、アスパラギン酸)を共有するからである。lおよびkの同時の割り当てが生じる場合には、パターンm(レベルIII)が、その位置に割り当てられる。同様に、パターンkおよびoの両方が同時に割り当てられ得ることが可能である。なぜなら、これらは、同じアミノ酸(詳細には、グルタミン酸)を共有するからである。kおよびoの割り当てが同時に起こる場合には、パターンq(レベルIII)がその位置に割り当てられる。レベルIIのパターンおよびレベルIIIのパターン(mおよびq)のいずれもがそのアミノ酸配列中の特定の位置を満たさない場合には、このアルゴリズムはレベルIII分析を実行する。
【0072】
レベルIIIは、パターンセットc、e、m、q、p、およびjを規定する。ここで、m、q、およびpは、共通のアミノ酸残基を共有する。しかし、パターンqは、レベルIIIの分析においては試験されない。パターンmおよびpの両方が同時に満たされ得ることが可能である。なぜなら、これらは同じアミノ酸(詳細には、グルタミン酸)を共有するからである。mおよびpの同時の割り当てが生じる場合には、パターンr(レベルIV)がその位置に割り当てられる。整列されたアミノ酸配列中の予め選択された位置のアミノ酸のうちの75%がレベルIIIのパターンを満たす場合には、レベルIIIのパターンがその位置に割り当てられる。レベルIIIのパターンがその位置に割り当てられ得ない場合には、このアルゴリズムはレベルIVの分析を履行する。
【0073】
レベルIVは、2つの非重複パターンfおよびrを含む。そのアミノ酸配列中の特定の位置のアミノ酸のうちの75%がレベルIVのパターンを満たす場合には、このパターンがその位置に割り当てられる。レベルIVのパターンが割り当てられない場合には、このアルゴリズムは、その位置に任意のアミノ酸を示すX(レベルV)を割り当てる。
【0074】
図3においては、レベルIは、20個の天然に存在するアミノ酸を1文字のアミノ酸コードで大文字で列挙する。レベルII〜Vは、Smithら(前出)において示されているアミノ酸の階層に基づいて、アミノ酸の群を小文字で規定する。図5および6に示されるアミノ酸配列は、上記のコンピューターアルゴリズムを使用して整列された。
【0075】
当業者がTGF−βスーパーファミリーの現在同定されているメンバーに基づいて形態単位構築物を生成することを望む場合には、当業者は、図5に示されるアミノ酸配列を使用して、本発明の形態単位構築物の生成において有用な、フィンガー1領域、フィンガー2領域、およびヒール領域を提供し得る。本明細書以降に発見されたTGF−βスーパーファミリーのメンバーの場合、新規のメンバーのアミノ酸配列は、本発明の実施に有用なヒール領域およびフィンガー領域を規定するために、手動またはコンピューターアルゴリズムのいずれかによって、図5に示されている配列とともに整列され得る。
【0076】
以下の表1は、図5および6に示される配列整列パターンを生成するために使用された、TGF−βスーパーファミリーのメンバーのそれぞれのアミノ酸配列を記載する、刊行物をまとめる。
【0077】
【表1】
【0078】
【0079】
さらに本発明は、周知のタンパク質:GDF−5、GDF−7(米国特許番号第5,801,014号において開示されるように、この全体の開示が、本明細書中に参考として援用される);GDF−6(米国特許番号第5,770,444号において開示されるように、この全体の開示が、本明細書中に参考として援用される);ならびにBMP−12およびBMP−13(米国特許番号第5,658,882号において開示されるように、この全体の開示が、本明細書中に参考として援用される)に由来する対応するフィンガー1サブドメイン配列の使用を意図する。
【0080】
詳細には、本発明の実施において有用なフィンガー1領域を規定するアミノ酸配列が、本明細書中で同定された任意のTGF−βスーパーファミリーのメンバーについてのフィンガー1領域を規定するアミノ酸配列に対応することが、意図される。フィンガー1サブドメインは、ネイティブのタンパク質の特徴である、少なくとも生物学的および/または機能的特性(単数または複数)を付与し得る。有用なインタクトなフィンガー1領域として、以下が挙げられるがこれらに限定されない:
TGF−β1 配列番号40、残基2から29、
TGF−β2 配列番号41、残基2から29、
TGF−β3 配列番号42、残基2から29、
TGF−β4 配列番号43、残基2から29、
TGF−β5 配列番号44、残基2から29、
dpp 配列番号45、残基2から29、
Vg−1 配列番号46、残基2から29、
Vgr−1 配列番号47、残基2から29、
60A 配列番号48、残基2から29、
BMP−2A 配列番号49、残基2から29、
BMP−3 配列番号50、残基2から29、
BMP−4 配列番号51、残基2から29、
BMP−5 配列番号52、残基2から29、
BMP−6 配列番号53、残基2から29、
Dorsalin 配列番号54、残基2から29、
OP−1 配列番号55、残基2から29、
OP−2 配列番号56、残基2から29、
OP−3 配列番号57、残基2から29、
GDF−1 配列番号58、残基2から29、
GDF−3 配列番号59、残基2から29、
GDF−9 配列番号60、残基2から29、
インヒビンα 配列番号61、残基2から29、
インヒビンβA 配列番号62、残基2から29、
インヒビンβB 配列番号63、残基2から29、
CDMP−1/GDF−5 配列番号83、残基2から29、
CDMP−2/GDF−6 配列番号84、残基2から29、
GDF−6(マウス) 配列番号85、残基2から29、
CDMP−2(ウシ) 配列番号86、残基2から29、および
GDF−7(マウス) 配列番号87、残基2から29。
【0081】
本発明はさらに、周知のタンパク質BMP−12およびBMP−13(その全体の開示が本明細書中で参考として援用されている、米国特許第5,658,882号において開示されている)に由来する対応するヒールサブドメイン配列の使用を意図する。
【0082】
本発明の実施において有用なヒール領域を規定するアミノ酸配列が、本明細書中で同定された任意のTGF−βスーパーファミリーのメンバーのインタクトなヒール領域を規定するアミノ酸に対応することもまた、意図される。ヒール領域は、機能的特性および/またはフォールディング特性を含む、ネイティブのタンパク質の特性に少なくとも影響を与え得る。有用なインタクトなヒール領域として、以下が挙げられ得るが、これらに限定されない:
TGF−β1 配列番号40、残基35から62、
TGF−β2 配列番号41、残基35から62、
TGF−β3 配列番号42、残基35から62、
TGF−β4 配列番号43、残基35から62、
TGF−β5 配列番号44、残基35から62、
dpp 配列番号45、残基35から65、
Vg−1 配列番号46、残基35から65、
Vgr−1 配列番号47、残基35から65、
60A 配列番号48、残基35から65、
BMP−2A 配列番号49、残基35から64、
BMP−3 配列番号50、残基35から66、
BMP−4 配列番号51、残基35から64、
BMP−5 配列番号52、残基35から65、
BMP−6 配列番号53、残基35から65、
Dorsalin 配列番号54、残基35から65、
OP−1 配列番号55、残基35から65、
OP−2 配列番号56、残基35から65、
OP−3 配列番号57、残基35から65、
GDF−1 配列番号58、残基35から70、
GDF−3 配列番号59、残基35から64、
GDF−9 配列番号60、残基35から65、
インヒビンα 配列番号61、残基35から65、
インヒビンβA 配列番号62、残基35から69、
インヒビンβB 配列番号63、残基35から68、
CDMP−1/GDF−5 配列番号83、残基35から65、
CDMP−2/GDF−6 配列番号84、残基35から65、
GDF−6(マウス) 配列番号85、残基35から65、
CDMP−2(ウシ) 配列番号86、残基35から65、および
GDF−7(マウス) 配列番号87、残基35から65。
【0083】
本発明はさらに、周知のタンパク質BMP−12およびBMP−13(その全体の開示が本明細書中で参考として援用されている、米国特許第5,658,882号において開示されている)に由来する対応するフィンガー2サブドメイン配列の使用を意図する。
【0084】
本発明の実施において有用なフィンガー2領域を規定するアミノ酸配列が、本明細書中で同定された任意のTGF−βスーパーファミリーのメンバーについてのインタクトなフィンガー2領域を規定するアミノ酸配列に対応することもまた、意図される。フィンガー2サブドメインは、ネイティブのタンパク質の特徴である、少なくともフォールディング特性(単数または複数)を付与し得る。有用なインタクトなフィンガー2領域として、以下が挙げられ得るがこれらに限定されない:
TGF−β1 配列番号40、残基65から94、
TGF−β2 配列番号41、残基65から94、
TGF−β3 配列番号42、残基65から94、
TGF−β4 配列番号43、残基65から94、
TGF−β5 配列番号44、残基65から94、
dpp 配列番号45、残基68から98、
Vg−1 配列番号46、残基68から98、
Vgr−1 配列番号47、残基68から98、
60A 配列番号48、残基68から98、
BMP−2A 配列番号49、残基67から97、
BMP−3 配列番号50、残基69から99、
BMP−4 配列番号51、残基67から97、
BMP−5 配列番号52、残基68から98、
BMP−6 配列番号53、残基68から98、
Dorsalin 配列番号54、残基68から99、
OP−1 配列番号55、残基68から98、
OP−2 配列番号56、残基68から98、
OP−3 配列番号57、残基68から98、
GDF−1 配列番号58、残基73から103、
GDF−3 配列番号59、残基67から97、
GDF−9 配列番号60、残基68から98、
インヒビンα 配列番号61、残基68から101、
インヒビンβA 配列番号62、残基72から102、
インヒビンβB 配列番号63、残基71から101、
CDMP−1/GDF−5 配列番号83、残基68から98、
CDMP−2/GDF−6 配列番号84、残基68から98、
GDF−6(マウス) 配列番号85、残基68から98、
CDMP−2(ウシ) 配列番号86、残基68から98、および
GDF−7(マウス) 配列番号87、残基68から98。
【0085】
さらに、代表的なフィンガー領域およびヒール領域のアミノ酸配列が、アミノ酸置換によって(例えば、本明細書中に開示されているような置換残基を活用することによって)変更され得るか、またはSmithら(1990)、前出において開示されている原理に従って選択され得ることが、意図される。簡潔には、Smithらは、図3にまとめられているアミノ酸のクラスの階層に類似するアミノ酸のクラスの階層を開示する。これらは、タンパク質機能を含み得る型の全体的な立体構造的なひずみを最少にしながら、1つのアミノ酸を別のアミノ酸で合理的に置換するために使用され得る。任意の事象において、天然の領域とわずか70%の相同性、好ましくは80%の、そして最も好ましくは少なくとも90%の相同性を有する、多くの合成の第1のフィンガー領域、第2のフィンガー領域、およびヒール領域の配列が、本発明の構築物を産生するために使用され得ることが意図される。
【0086】
Smithら(1990)前出に記載されている原理に従って推定される、フィンガー領域およびヒール領域の各位置で好ましいアミノ酸を示すアミノ酸配列のパターンもまた、図5および図6に示され、そしてTGF−β;Vg/dpp;GDF;およびインヒビンサブグループのパターンと呼ばれる。各サブグループのフィンガー1、ヒール、およびフィンガー2の配列パターンを規定するアミノ酸配列は、それぞれ、図5A、5B、および5Cに示される。さらに、全体的なTGF−β、Vg/dpp、GDF、およびインヒビンサブグループのパターンを規定するアミノ酸配列は、配列番号64、65、66、および67として、それぞれ配列表に示される。
【0087】
図5A、5B、および5Cに開示され、そして図6にまとめられている、各サブグループについての好ましいアミノ酸配列のパターンは、当業者が、フィンガー1、ヒール、およびフィンガー2のエレメント中の特定の位置で組み込まれ得る代替的アミノ酸を同定することを可能にする。一文字のアミノ酸コードで大文字で示されているアミノ酸は保存されているアミノ酸を示し、これらは共に、フィンガー領域およびヒール領域の構造的および機能的エレメントを規定すると考えられる。図5および6中の大文字の「X」は、任意の天然に存在するアミノ酸がその位置で受容可能であることを示す。図5および6中の小文字の「z」は、ギャップまたは任意の天然に存在するアミノ酸のいずれかがその位置で受容可能であることを示す。小文字は、図5に示されているパターン規定の表に従って示されるアミノ酸を表記し、そしてその位置で有用であるアミノ酸の群を同定する。
【0088】
図5および6に示されているアミノ酸配列のサブグループのパターンに従って、例えば、当業者が、どの適用可能な1つのアミノ酸が、生じるタンパク質構築物中に破壊的な立体化学的変化を誘導することなく別のアミノ酸で置換され得るかを推測し得ることが意図される。例えば、図5Aにおいては、残基番号12でTGF−βサブグループのパターンにおいては、リジン残基(K)またはグルタミン残基(Q)のいずれかが、生じる構築物の構造に影響を与えることなくこの位置に存在し得ることが意図される。従って、位置12での配列パターンは、「n」を含み、これは、図10に従って、リジンまたはグルタミンからなる群から選択されるアミノ酸残基を規定する。従って、天然領域と70%の相同性、好ましくは80%の、そして最も好ましくは少なくとも90%の相同性を有する、多くの合成のフィンガー1領域、フィンガー2領域、およびヒール領域のアミノ酸配列が、本発明の立体構造的に活性なタンパク質を産生するために使用され得ることが意図される。
【0089】
これらの原理に従って、当業者が、図5および6に示される、TGF−β、Vg/dpp、GDF、またはインヒビンのサブグループのパターンに属するアミノ酸配列のパターンを用いて開始することによって合成構築物を設計し得ることが意図される。従って、従来の組換えまたは合成方法論を使用することによって、予め選択されたアミノ酸が、本明細書中の原理によって指示されるように別のアミノ酸で置換され得、そして生じるタンパク質構築物はアゴニスト活性またはアンタゴニスト活性とのいずれかとの組み合わせにおいて結合活性に対して試験される。
【0090】
TGF−βサブグループのパターンである、配列番号64は、TGF−β1、TGF−β2、TGF−β3、TGF−β4、およびTGF−β5を含む、今日までに同定されているTGF−βのサブグループのメンバーの中で共有されている相同性に適応する。以下に示される遺伝子配列は、保存アミノ酸(標準的な3文字コード)ならびに配列内の種々の位置に存在し、そして図3に示される規則によって規定される代替的アミノ酸(Xaa)の両方を含む。
【0091】
【化1】
【0092】
各Xaaは、以下に規定される1つ以上の特定アミノ酸の群から独立的に選択され得、ここで
【0093】
【化2】
【0094】
である。
【0095】
Vg/dppサブグループパターンである配列番号65は、現在までにdpp、vg−1、vgr−1、60A、BMP−2A(BMP−2)、Dorsalin、BMP−2B(BMP−4)、BMP−3、BMP−5、BMP−6、OP−1(BMP−7)、OP−2およびOP−3を含むと同定された、Vg/dppサブグループのメンバー間で共有される相同性を供給する。以下の一般的な配列は、この配列内の可変性位置に存在し、そして図3に示される規則によって規定される、保存されたアミノ酸(標準的な3文字コ
ード)ならびに代替のアミノ酸(Xaa)の両方を含む。
【0096】
【化3】
【0097】
各Xaaは、独立して、以下のように規定される1つ以上の特定のアミノ酸の群から選択され得る。ここで、
【0098】
【化4】
【0099】
【化5】
【0100】
である。
【0101】
GDFサブグループパターンである配列番号66は、現在までにGDF−1、GDF−3およびGDF−9を含むと同定された、GDFサブグループのメンバー間で共有される相同性を供給する。一般的な配列(以下に示される)は、この配列内の可変性位置に存在し、そして図3に示される規則によって規定される、保存されたアミノ酸(標準的な3文字コード)ならびに代替のアミノ酸(Xaa)の両方を含む。
【0102】
【化6】
【0103】
各Xaaは、独立して、以下のように規定される1つ以上の特定のアミノ酸の群から選択され得る。ここで、
【0104】
【化7】
【0105】
【化8】
【0106】
【化9】
【0107】
である。
【0108】
インヒビンサブグループパターンである配列番号67は、現在までにインヒビンα、インヒビンβAおよびインヒビンβBを含むと同定された、インヒビンサブグループのメンバー間で共有される相同性を供給する。一般的な配列(以下に示される)は、この配列内の可変性位置に存在し、そして図3に示される規則によって規定される、保存されたアミノ酸(標準的な3文字コード)ならびに代替のアミノ酸(Xaa)の両方を含む。
【0109】
【化10】
【0110】
各Xaaは、独立して、以下のように規定される1つ以上の特定のアミノ酸の群から選択され得る。ここで、
【0111】
【化11】
【0112】
【化12】
【0113】
【化13】
【0114】
である。
(2)骨形態形成タンパク質の生化学的、構造的、および機能的特性)
その成熟の、ネイティブ形態において、天然に供給される骨形成タンパク質は、グリコシル化されたダイマーであり、代表的には、SDS−PAGEによって決定されるように、約30〜36kDaの見かけの分子量を有する。還元された場合、この30kDaタンパク質は、約16kDaおよび18kDaの見かけの分子量を有するグリコシル化されたペプチドサブユニットを生じる。還元状態において、そのタンパク質は、検出可能な骨形成活性を有さない。グリコシル化されていないタンパク質(これもまた骨形成活性を有する)は、約27kDaの見かけの分子量を有する。還元された場合、この27kDaタンパク質は、約14kDa〜16kDaの分子量を有する2つのグリコシル化されていないペプチド鎖を生じる。代表的には、天然に存在する骨形成タンパク質は、代表的には約30残基未満のN末端シグナルペプチド配列、続いて成熟C末端ドメインを生じるために切断される「プロ」ドメインを有する前駆体として翻訳される。そのシグナルペプチドは、翻訳に際して、Von Heijne(1986)Nucleic Acids Research 14:4683−4691の方法を使用して、所定の配列において予測され得る切断部位で迅速に切断される。本明細書中で有用な骨形成タンパク質は、天然に存在するか、または生合成的に産生される(例えば、「ムテイン」または「変異タンパク質」を含む)かに関わらず、任意の公知の天然に存在するネイティブなタンパク質(その対立遺伝子の系統学的な対応物、および他の改変体を含む)、ならびに、一般的な形態形成ファミリーのタンパク質の骨形成的に活性な新規のメンバーを含む。
【0115】
なお別の好ましい実施態様において、有用な骨形成的に活性なタンパク質は、低ストリンジェンシー、中程度のストリンジェンシー、または高ストリンジェンシーのハイブリダイゼーション条件下で、参照骨形成配列(例えば、OP−1、OP−2、BMP2、4、5、6、60A、GDF5、GDF6、GDF7などの保存性の7つのシステインドメインを規定するC末端配列)をコードするDNAまたはRNAにハイブリダイズする核酸によってコードされる配列を含むアミノ酸配列を有するポリペプチド鎖を有する。本明細書中で使用される場合、高ストリンジェンシーハイブリダイゼーション条件は、40% ホルムアミド、5×SSPE、5×デンハルト溶液、および0.1% SDS中で、一晩37℃での既知の技術に従うハイブリダイゼーション、ならびに0.1×SSPE、0.1% SDS中、50℃での洗浄として規定される。標準的なストリンジェンシー条件は、市販の標準的な分子クローニングテキストにおいて十分に特徴付けられている。例えば、Molecular Cloning A Laboratory Manual、第2版、Sambrook、Fritsch、およびManiatis編(Cold Spring Harbor Laboratory Press:1989);DNA Cloning、第I巻および第II巻(D.N.Glover編、1985);Oligonucleotide Synthesis(M.J.Gait編、1984);Nucleic Acid Hybridization(B.D.Hames & S.J.Higgins編、1984);ならびにB.Perbal、A Practical Guide To Molecular Cloning(1984)を参照のこと。
【0116】
本発明の実施において有用性を有する関連するタンパク質のTGF−βスーパーファミリーの他のメンバーには、以下のリストの中でさほどリフォールディングされていないタンパク質が含まれる:いくつか名前を挙げれば、TGF−β1、TGF−β2、TGF−β3、TGF−β4、およびTGF−β5、種々のインヒビン、アクチビン、BMP−11、ならびにMISである。図5Cは、TGF−βスーパーファミリーの種々の既知のメンバーのフィンガー2サブドメインを規定するC末端残基を列挙する。さほどフォールディングされていない、このリスト上のタンパク質のいずれか1つは、他の公知のファミリーメンバーまたは発見可能なファミリーメンバーであり得るように、本発明の方法によって改善され得る。
【0117】
(B.組換えタンパク質の産生)
上で述べたように、本発明の構築物は、当該分野において周知であり、そして徹底的に実証された従来の組換えDNA方法論の使用によって、ならびに慣用的なペプチド化学またはヌクレオチド化学および自動化したペプチド合成機またはヌクレオチド合成機を用いた周知の生合成の方法論および化学合成の方法論の使用によって、製造され得る。このような慣用的方法論は、例えば以下の出版物に記載され、その教示は本明細書中で参考として援用される:Hilvert,1
Chem.Biol.201−3(1994);Muirら,95 Proc.Natl.Acd.Sci.USA 6705−10(1998);Wallace,6 Curr.Opin.Biotechnol.403−10(1995);Mirandaら,96 Proc.Natl.Acd.Sci.USA 1181−86(1999);Liuら,91 Proc.Natl.Acad.Sci.USA 6584−88(1994)。本発明中の使用に適したものは、天然に存在するアミノ酸およびヌクレオチド;天然には存在しないアミノ酸およびヌクレオチド;修飾されたアミノ酸および異常なアミノ酸;修飾された塩基;翻訳後に修飾されたアミノ酸および/または修飾された連結、架橋およびエンドキャップ、非ペプチジルボンド、などを含むアミノ酸配列であり、そして、付録2中の表1〜6を含むWorld Intellectual Property Organization(WIPO) Handbook on
Industrial Property Information and
Documentation,Standard St.25(1998)に開示された部分がさらに挙げられるが、これらに限定されない(これらは本明細書中で参考として援用される)。これらの等価物は、当該分野の知識と共に慣用的実験法のみに依存して、当業者によって評価され得る。
【0118】
例えば、企図されるDNA構築物は、合成ヌクレオチド配列のアセンブリおよび/またはDNA制限酵素フラグメントを連結して合成DNA分子を産生することによって製造され得る。次いでこのDNA分子は、発現ビヒクル(例えば発現プラスミド)へ連結され、そして適切な宿主細胞(例えば、E.coli)へトランスフェクトされる。このDNA分子によってコードされる企図されたタンパク質構築物が、次いで発現され、精製され、リフォールディングされ、特定の特性(例えば、鋳型TGF−βスーパーファミリーメンバーに対する結合親和性を有するレセプターとの結合活性)に関してインビトロにおいて試験され、そしてこの生合成構築物が、鋳型TGF−βスーパーファミリーメンバーの他の適切な特性を模倣するかどうかを評価するために引き続いて試験された。
【0119】
あるいは、合成DNA構築物のライブラリーが、例えば、前もって選んだ領域内のヌクレオチド組成と異なる合成ヌクレオチド配列のアセンブリによって同時に調製され得る。例えば、特異的TGF−βスーパーファミリーメンバーに基づく構築物の産生中に、当業者が、このようなスーパーファミリーメンバーにとって適切なフィンガーおよびヒールの領域を選び得ることが意図される(例えば、図5〜6より)。一旦、適切なフィンガーおよびヒールの領域が選ばれると、当業者は、次いでこれらの領域をコードする合成DNAを産生し得る。例えば、異なるリンカー配列をコードする複数のDNA分子が、フィンガーおよびヒールの配列をコードするDNA分子を含む連結反応に含まれるならば、適切な制限酵素部位および反応条件の慎重な選択によって、当業者は、各々のDNA構築物がフィンガーおよびヒールの領域をコードするが異なるリンカー配列によって結合されたDNA構築物のライブラリーを産生し得る。その結果生じるDNAは、次いで適切な発現ビヒクル(すなわち、ファージディスプレイライブラリーの調製に有用なプラスミド)へ連結され、宿主細胞へ形質転換され、そして合成DNAによってコードされるポリペプチドは、候補タンパク質のプールを産生するために発現させた。候補タンパク質のプールは、前もって選んだレセプターに対する結合親和性および/または選択性を有する特異的なタンパク質を同定するために引き続いてスクリーニングされ得る。
【0120】
スクリーニングが、候補タンパク質を含む溶液をレセプターが固定化された表面を備えるクロマトグラフィーカラムを通じて通過させることによって実施され得る。次いで、所望の結合特異性を伴うタンパク質が、例えば、塩勾配および/または鋳型TGF−βスーパーファミリーメンバーの濃度勾配によって溶出される。このようなタンパク質をコードするヌクレオチド配列は、引き続き単離され得そして特徴付けられ得る。一旦、適切なヌクレオチド配列が同定されると、リードタンパク質は、従来の組換えDNA方法論またはペプチド合成方法論のいずれかによって、特定の構築物が鋳型TGFβ−スーパーファミリーメンバーの活性を模倣するかどうかを試験するために十分な量が引き続き産生され得る。
【0121】
どちらのアプローチが本発明の構築物をコードするDNA分子を産生するために採用されても、好ましいタンパク質の三次構造が、例えば、本明細書中に記載の原理によって援助されるヌクレオチド変異誘発方法論およびファージディスプレイ方法論の組合せによって、結合活性および/または生物学的活性を最適化するために引き続き調節され得ることが企図される。従って、当業者は、多くのこのようなタンパク質を産生し得、そして同時に試験し得る。
【0122】
(1)遺伝子合成
目的のアミノ酸配列をコードするDNAを製造し、増幅し、そして組換えるためのプロセスは、該して当該分野において周知であり、それゆえ、本明細書中に詳細には記載されない。TGF−βスーパーファミリーメンバーをコードする遺伝子および同族のレセプターを同定しそして単離する方法はまた、十分に理解されており、そして本特許および他の文献に記載される。
【0123】
手短には、本明細書中に開示された生合成構築物をコードするDNAの構築は、配列特異的にDNAを切断し平滑末端または突出末端を産生する種々の制限酵素、DNAリガーゼ、突出末端を平滑末端化されたDNAへの酵素的付加を可能にする技術、短いまたは中程度の長さのオリゴヌクレオチドのアセンブリによる合成DNAの構築、cDNA合成技術、ライブラリーから適切な核酸配列を増幅するためのポリメラーゼ連鎖反応(PCR)技術、ならびにTGF−βスーパーファミリーメンバーおよびそれらの同族のレセプターの遺伝子を単離するための合成プローブの使用を含む周知の技術を用いて実施される。発現の達成に用いられる(少しの例を挙げれば)細菌、哺乳動物、または昆虫由来の種々のプロモーター配列および他の調節DNA配列ならびに種々の型の宿主細胞もまた、公知でありそして入手可能である。従来のトランスフェクションの技術、およびDNAをクローニングおよびサブクローニングするための同等に従来の技術は、本発明の実施に有用であり、そして当業者に周知である。種々の型のベクター(例えばプラスミド、ならびに動物ウイルスおよびバクテリオファージを含むウイルス)が用いられ得る。このベクターは、首尾よくトランスフェクトされた細胞に検出可能な表現型特性(ベクターの組換えDNAを首尾よく組み込まれたクローンのファミリーを同定するために用い得る特性)を与える種々のマーカー遺伝子を利用し得る。
【0124】
本明細書中に開示された生合成構築物をコードするDNAを得るための方法の一つは、従来の、自動化オリゴヌクレオチド合成機において産生される合成オリゴヌクレオチドのアセンブリに引き続いて適切なリガーゼを用いる連結によるものである。例えば、重複した、相補的なDNAフラグメントは、ホスホルアミダイド化学を用いて合成され得、末端セグメントは、連結中に重合を防ぐためにリン酸化されないままである。合成DNAの一端は、特定の制限エンドヌクレアーゼの作用の部位に相当する「突出末端」を伴ったままにし、そして他端は、別の制限エンドヌクレアーゼの作用の部位に相当する末端を伴ったままにする。この相補的DNAフラグメントは、共に連結されて、合成DNA構築物を産生する。
【0125】
あるいはフィンガー1領域、フィンガー2領域およびヒール領域をコードする核酸鎖が、例えば、Sambrookら編(1989)「Molecular Cloning」、Coldspring Harbor Laboratories Press、NYに記載のようなコロニーハイブリダイゼーション手順によって、および/またはInnisら(1990)「PCR Protocols,A guide to methods and applications」、Academic Pressに開示のようなPCR増幅方法論によって、核酸のライブラリーから単離され得る。このフィンガー領域およびヒール領域をコードする核酸は、次いで共に連結され、目的の生合成一本鎖形態単位構築物をコードする合成DNAを産生する。
【0126】
しかし、その複数の形態単位(morphon)をコードするDNA構築物のライブラリーは、例えば、上述したもののような、標準的な組換えDNA方法論によって同時に産生され得る。例えば、カセット変異誘発またはオリゴヌクレオチド特異的変異誘発の使用によって、当業者は、例えば、あらかじめ定義された部位の範囲内(例えば、リンカー配列をコードするDNAカセットの範囲内)に異なるDNA配列を各々が含む一連のDNA構築物を産生し得る。その結果生じるDNA構築物のライブラリーは、例えば、ファージディスプレイライブラリーにおいて発現され得る;そして特異的レセプターに結合する任意のタンパク質構築物は、アフィニティ精製(例えば、レセプターが固定化された表面を備えるクロマトグラフィーカラムを使用して)によって単離され得る(以下のV節を参照のこと)。一旦前もって選択されたレセプターに結合する分子が単離されると、それらの結合特性およびアゴニストの特性は、以下のV節においてまた議論されるように、経験的な精製技術を用いて調節され得る。
【0127】
タンパク質および核酸の変異誘発の方法は、周知であり、当該分野に十分に記載されている。例えば、Sambrookら、(1990) Molecular Cloning:A Laboratory Manual.、第2版(Cold Spring Harbor、N.Y.:Cold Spring Harbor Laboratory
Press)を参照のこと。有用な方法は、PCR(重複伸長、例えば、PCR Primer(DieffenbachおよびDveksler編、Cold Spring
Harbor Press、Cold Spring Harbor、NY、1995、603〜611頁)を参照のこと);クンケル法の後のカセット変異誘発および一本鎖変異誘発を含む。これは、任意の適切な変異誘発方法が利用され得、そしてこの変異誘発法が本発明の重要な局面とみなされないことは当業者によって認識される。アルギニン(Arg)、グルタミン酸(Glu)およびアスパラギン酸(Asp)を含むアミノ酸をコードするのに的確な核酸コドンはまた、周知でありそして当該分野において記載される。例えば、Lehninger、Biochemistry(Worth Publishers、N.Y.、N.Y.)を参照のこと。アルギニン、グルタミン酸およびアスパラギン酸をコードする標準的なコドンは、Arg:CGU、CGC、CGA、CGG,AGA,AGG;Glu:GAA、GAG;およびAsp:GAU、GACである。本発明のキメラの構築物は、切り替えられるタンパク質領域の核酸配列またはドメインを整列化すること、ならびに適合性のスプライス部位を同定することおよび/またはPCR重複伸長を用いた適切な乗り換え配列を構築することによって容易に構築され得る。
【0128】
本発明のTGF−βファミリーメンバーの変異型は、標準的な周知の方法を使用して、細菌中で産生され得る。C末端の7つのシステインドメインのみを規定する全長成熟形態またはより短い配列は、宿主細胞に提供され得る。細菌発現を最適化するために、変異型タンパク質のN末端配列を改変することが好ましくあり得る。例えば、ネイティブなOP−1の細菌発現のために好ましい形態は、C末端の7つのシステインドメイン(例えば、配列番号39の残基330〜431)をコードする、成熟の活性配列(配列番号39の残基293〜431)またはそのフラグメントをコードする配列である。メチオニンは、ネイティブのセリン残基を置き換えて293位において導入され得るか、またはそれは、このセリン残基の前に位置し得る。あるいは、メチオニンは、TGF−βドメインの最初のシステインまでの天然の配列の最初の36残基(残基293〜329)以内のいずれかの場所に導入され得る。そのDNA配列はさらに、精製を改善するために、例えば、IMACカラム上での精製を補助するために「ヘキサhis」テールを付加することによって;またはIgG/カラム上の精製を容易にする、FBリーダー配列を使用することによって、そのN末端において改変され得る。これらおよび他の方法は、十分に記載されており、そして当該分野で周知である。他の細菌種および/またはタンパク質は、そこから得られた変異体BMPの収量を最適化するために、類似の改変からの利点を必要とし得る。このような改変は、十分に当業者のレベル内であり、そして本発明の実質的な局面とはみなされない。
【0129】
合成核酸は、好ましくは、選択した宿主細胞における過剰発現に適切なベクターへ挿入される。任意の発現ベクターは、それが、選択した宿主細胞における異種タンパク質(例えば、BMP)の発現を方向付け得る限り、使用され得る。有用なベクターとしては、少し例を挙げると、プラスミド、ファージミド、ミニクロモソームおよびYACが挙げられるが、これらに限定されない。他のベクター系は周知であり、そして当該分野で特徴付けられている。このベクターは、代表的にはレプリコン、一つ以上の選択マーカー遺伝子配列、および宿主細胞に高コピー数のベクターを維持するための手段を含む。周知の選択マーカー遺伝子は、アンピシリン、テトラサイクリンなどのような抗生物質および重金属耐性を含む。酵母細胞で使用するために有用な選択マーカー遺伝子は、栄養要求性の酵母変異宿主とともに使用するためのURA3遺伝子、LEU2遺伝子、HIS3遺伝子またはTRP1遺伝子を含む。加えて、ベクターもまた、目的の遺伝子を発現するための適切なプロモーター配列(これは誘導性であっても、誘導性でなくてもよい)、ならびに所望であれば、有用な転写開始部位および翻訳開始部位、ターミネーター、ならびに目的の遺伝子の転写および翻訳を最大化し得る他の配列を含む。細菌細胞において特に有用な、十分特徴付けられたプロモーターとしては、少し例を挙げれば、lacプロモーター、tacプロモーター、trpプロモーター、およびtppプロモーターが挙げられる。酵母で有用なプロモーターとしては、例えば、ADHI、ADHII,またはPHO5プロモーターが挙げられる。
【0130】
適切な宿主細胞としては、Bacillus subtilis(B.subtilis)、Pseudomonasの種、Escherichia coli(E.coli)および酵母細胞(例えば、Saccharomyces cerevisiae)のような細菌細胞が挙げられる。他の宿主細胞(例えば、CHOのような哺乳動物細胞)が使用され得る。
【0131】
目的の遺伝子は、標準的な微生物学技術(例えば、エレクトロポレーションまたは塩化カルシウム)を用いて選択した宿主細胞へ形質転換され得、そして適切な条件下でこの細胞を誘導して、増殖させる。細胞培養培地は、多くの周知の基本書を含め、当該分野で十分に記載され、そしてこれらとしては、Sambrookらが挙げられる。有用な培地としては、LB(ルリアブロス)およびダルベッコDMEMが挙げられる。過剰発現されたタンパク質は、標準的な技術(細胞溶解または細胞の機械的破砕(例えば、フレンチプレス、SLM Instruments,Inc,)を含む)による不溶性の屈折封入体(refractile inclusion body)から採取され、次いで、遠心分離および再溶解され得る(以下を参照のこと)。
【0132】
例えば、この遺伝子がE.coliで発現される場合、適切な発現ベクターへクローニングされる。このことは、プロモーター(例えば、TrpまたはTac)の下流に、操作された遺伝子および/またはプロテインAのフラグメントB(FB)のようなリーダーペプチドをコードする遺伝子を配置することにより達成され得る。発現の間に、得られた融合タンパク質は、細胞の細胞質中の屈折体に蓄積し、そしてフレンチプレスまたは超音波処理による細胞の崩壊の後に収集され得る。単離された屈折体は、次いで可溶化され、そして発現したタンパク質は折り畳まれ、そして必要であれば、多くの他の組換えタンパク質に対して既に確立された方法により、リーダーペプチドが切断される。
【0133】
真核生物細胞での操作された遺伝子の発現は、細胞および細胞株を必要とし、そしてこれらの細胞および細胞株はトランスフェクトするのに容易であり、再配列されない配列を伴う外来DNAを安定に維持することが可能であり、これらの細胞および細胞株は、効率的な転写、翻訳、翻訳後修飾、およびタンパク質の分泌に必要な細胞内構成成分を有する。さらに、目的の遺伝子を運ぶ適切なベクターもまた、必要である。哺乳動物細胞へのトランスフェクションのためのDNAベクターの設計は、本明細書中に記載されるような目的の遺伝子の発現を促進するために適切な配列(適切な転写開始配列、終止配列、およびエンハンサー配列ならびに、コザックコンセンサス配列のような翻訳効率を増強する配列を含む)を含むべきである。好ましいDNAベクターはまた、マーカー遺伝子および目的の遺伝子のコピー数を増幅するための手段を含む。哺乳動物細胞での外来タンパク質の産生の技術水準(有用な細胞、タンパク質発現促進配列、マーカー遺伝子、および遺伝子増幅法を含む)の詳細な総説は、Bendig(1988) Genetic Engineering 7:91〜127に開示される。
【0134】
特定の哺乳動物細胞において外来遺伝子を発現するために有用な最も特徴付けられた転写プロモーターは、SV40初期プロモーター、アデノウイルスプロモーター(AdMLP)、マウスメタロチオネインIプロモーター(mMT−I)、ラウス肉腫ウイルス(RSV)長末端反復(LTR),マウス乳腺腫ウイルス長末端反復(MMTV−LTR)、およびヒトサイトメガロウイルス主要即時初期(intermediate−early)プロモーター(hCMV)である。これらのプロモーターのすべてに関するDNA配列は、当該分野において公知であり、そして市販されている。
【0135】
dhfr-細胞株における選択可能なDHFR遺伝子の使用は、哺乳動物細胞系におけ
る遺伝子の増幅に有用な、十分に特徴付けられた方法である。簡単に、このDHFR遺伝子は、目的の遺伝子を運ぶベクター上に提供され、そしてDHFRによって代謝される細胞傷害性の薬剤メトトレキセートの漸増濃度の添加は、目的の関連遺伝子増幅と同様にDHFR遺伝子コピー数の増幅を導く。トランスフェクトされたチャイニーズハムスター卵巣細胞株(CHO細胞)において選択可能で、増幅可能なマーカー遺伝子としてのDHFRは、当該分野において特に十分に特徴付けられている。他の有用な増幅可能なマーカー遺伝子は、アデノシンデアミナーゼ(ADA)遺伝子およびグルタミンシンターゼ(GS)遺伝子を含む。
【0136】
細胞/細胞株の選択はまた、重要であり、そして実験者の要求に依存する。COS細胞は、本発明の生合成構築物を迅速にスクリーニングするために有用な手段を提供する、高いレベルの一過性の遺伝子発現を提供する。COS細胞は、代表的には目的の遺伝子を運ぶシミアンウイルス40(SV40)ベクターと共にトランスフェクトされる。トランスフェクトされたCOS細胞は、最終的に死滅し、それ故に所望のタンパク質産生物の長期間の産生が妨げられる。しかし、一過性の発現は、安定な細胞株の開発のために必要とされる時間のかかる過程を必要とせず、従って結合活性について予備構築物を試験するために有用な技術を提供する。
【0137】
本発明の一本鎖構築物の哺乳動物細胞発現のために使用される種々の細胞、細胞株およびDNA配列は、当該分野で十分に特徴付けられており、そして容易に入手可能である。他のプロモーター、選択マーカー、遺伝子増幅方法および細胞もまた使用して、本発明のタンパク質を発現させ得る。組換えタンパク質のトランスフェクション、発現および精製の具体的な詳細は、当該分野で十分に記載されており、そして当業者により理解される。哺乳動物細胞発現系における外来遺伝子の組換え生成において使用される各工程の種々の技術的局面に対するさらなる詳細は、当該分野における多くの基本書および実験室マニュアルにおいて見出され得る(例えば、F.M.Ausubelら編、Current Protocols in Molecular Biology,John Wiley&Sons,New York(1989))。
【0138】
(c)リフォールディングの考慮事項
一旦封入体から単離されると、タンパク質は、グアニジン塩酸または尿素のような変性剤またはカオトロピック剤によって、好ましくは約4〜9Mの範囲でそして高い温度(例えば25〜37℃)および/または塩基性pH(8〜10)で可溶化される。あるいは、タンパク質は、酸性化によって(例えば、酢酸またはトリフルオロ酢酸を用いて)、一般に1〜4の範囲のpHで可溶化され得る。好ましくは、β−メルカプトエタノールまたはジチオトレイトール(DTT)のような還元剤が、可溶化剤と組合せて用いられる。この可溶化された異種のタンパク質は、透析によっておよび/または公知のカラムクロマトグラフィーの方法によって(例えば、サイズ排除クロマトグラフィー、イオン交換クロマトグラフィー、または逆相高速液体クロマトグラフィー(RP−HPLC)などによって)、可溶化したカオトロープ(caotrope)からさらに精製され得る。
【0139】
可溶化されたタンパク質は、以下のようにリフォールディングされ得る。溶解したタンパク質は、リフォールディング媒体(代表的には、約pH5.0〜10.0の範囲のpH、好ましくは約pH6〜9の範囲のpHを有し、ならびに界面活性剤および/またはカオトロピック剤を含むトリス緩衝化媒体)中に希釈される。有用な市販の入手可能な界面活性剤は、例えば、NP40(Nonidet
40)、CHAPS(3−[(3−コラミドプロピル)ジメチルアンモニオ]−1−プロパン−スルフェートのような)、ジギトニン、デオキシコレート、またはN−オクチルグルコシドのような、イオン性、非イオン性、または双性イオン性であり得る。有用なカオトロピック剤としては、グアニジン、尿素、またはアルギニンが挙げられる。好ましくは、界面活性剤またはカオトロピック剤は、約0.1〜10Mの範囲、好ましくは0.5〜4Mの範囲の濃度で存在する。CHAPSが界面活性剤の場合、これは、好ましくは溶液の約0.5〜5%、さらに好ましくは溶液の約1〜3%含まれる。好ましくは溶液はまた、少し例を挙げると酸化型および還元型のグルタチオン、DTT,β−メルカプトエタノール、β−メルカプトメタノール、システインまたはシスタミンのような適切な酸化還元系を含む。好ましくは、酸化還元系は、還元剤対酸化剤の比が約1:1から約5:1の範囲で存在する。グルタチオン酸化還元系が用いられる場合、還元型グルタチオン対酸化型グルタチオンの比は、還元型対酸化型が好ましくは約0.5対5;さらに好ましくは1対1;そして最も好ましくは2対1の範囲である。好ましくは緩衝液はまた、約0.25〜2.5Mの範囲、好ましくは約0.5〜1.5Mの範囲、最も好ましくは約1Mの範囲にて存在する塩(代表的にはNaCl)を含む。当業者は、上記の条件および媒体が、通常の実験法のみを用いて変化され得ることを認識する。このような変動および改変は、本発明の範囲内である。
【0140】
好ましくは、所定のリフォールディング反応のためのタンパク質濃度は、約0.001〜1.0mg/mlの範囲であり、さらに好ましくはこれは約0.05〜0.25mg/mlの範囲であり、最も好ましくは0.075〜0.125mg/mlの範囲である。当業者によって認識されるように、より高濃度が、凝集物をより多く産生する傾向がある。ヘテロダイマーが産生される場合(例えばOP−1/BMP2またはBMP2/BMP6ヘテロダイマー)、好ましくは個々のタンパク質は、等量のリフォールディング緩衝液に提供される。
【0141】
代表的にリフォールディング反応は、約4℃〜約25℃の温度範囲にて生じる。より好ましくは、リフォールディング反応を4℃にて実行し、そして完了させる。代表的に、リフォールディングは約1日〜7日で完了し、一般的に、タンパク質に依存して、16〜72時間または24〜48時間以内である。当業者に理解されるように、リフォールディング速度はタンパク質により変動し得、そしてより長いリフォールディング時間およびより短いリフォールディング時間が意図され、そして本発明の範囲内である。本明細書中で使用される場合、「良好なリフォルダー(refolder)」タンパク質は、このタンパク質の少なくとも20%が、リフォールディング反応における全タンパク質と比較されるされる場合、本明細書中に記載される任意のリフォールディングアッセイにより測定されるように、そしてさらなる精製を必要とせずに、フォールディング反応後のダイマー形態で存在する場合である。当該分野において「良いリフォルダー」タンパク質と考えられているネイティブなBMPとしては、BMP2、CDMP1、CDMP2およびCDMP3が挙げられる。BMP−3はまた、合理的に十分にリフォールディングされる。対照的に、「乏しいリフォルダー」タンパク質は、1%未満の適切に折り畳まれたタンパク質を産生する。
【0142】
適切にリフォールディングされたダイマータンパク質は、多くの周知かつ十分に特徴付けられたアッセイのいずれかを用いて容易にアッセイされ得る。特に、3つのアッセイ(全てが当該分野で周知かつ十分に記載され、そして以下にさらに記載される)の内の任意の1つ以上を使用して、利益を得ることができる。有用なリフォールディングのアッセイは、1つ以上の以下のものを含む。第1に、ダイマーの存在は、DTTのような還元剤の非存在下で標準的SDS−PAGEによるか、またはHPLC(例えば、C18逆相HPLC)によるかのいずれかで視覚的に検出され得る。BMPダイマータンパク質は、モノマーサブユニット(これは、約14〜18kDaの見かけの分子量を有する)と比較して、約28〜36kDaの範囲の見かけの分子量を有する。ダイマータンパク質は、市販の分子量標準物質との比較により電気泳動ゲル上で容易に視覚化され得る。このダイマータンパク質はまた、約19分で、C18 RP HPLC(45〜50%アセトニトリル:0.1%TFA)から溶出され得る(哺乳動物が生成したhOP−1は、18.95分で溶出する)。
【0143】
第2のアッセイは、ヒドロキシアパタイトに結合する能力によりダイマーの存在を評価する。適切にフォールディングされたダイマーは、モノマーと比較して、0.1〜0.2M NaClの存在下で(ダイマーは0.25M NaClで溶出する)ヒドロキシアパタイトカラムに十分結合する(モノマーは、この濃度では実質的に結合しない)(モノマーは、0.1Mで溶出する)。
【0144】
第3のアッセイは、トリプシンまたはペプシン消化に対するタンパク質の抵抗性によりダイマーの存在を評価する。このフォールディングされたダイマー種は、両酵素(特に、成熟タンパク質のN末端の小部分のみを切断するトリプシン)に対して実質的に抵抗性であり、このことにより、生物学的に活性なダイマー種を未処理ダイマーよりごくわずかにより小さなサイズにする。対照的に、モノマーは、実質的に分解される。このアッセイにおいて、このタンパク質は、標準的な条件(例えば、4M尿素、100mM NaCl、0.3%Tween−80および20mM メチルアミンを含有する、50mM Tris緩衝液(pH8)のような標準的な緩衝液中での消化)を用いて酵素消化に供される。消化は、16時間のオーダーで37℃で生じさせ、そしてこの産物を、任意の適切な手段(好ましくは、SDS−PAGE)により可視化させる。
【0145】
再折りたたみされたTGF−βファミリータンパク質の生物学的活性は、任意の多くの手段によって容易に評価され得る。BMPの軟骨内骨形成を誘導する能力は、十分に特徴付けされたラット皮下骨アッセイを使用して評価され得、当該分野でおよび下記に詳細に記載される。このアッセイにおいて、骨形成は、組織学ならびにアルカリホスファターゼおよび/またはオステオカルシン(osteoclacin)産生によって測定される。さらに、高い特異的骨形成活性を有する骨形成タンパク質(例えば、OP−1、BMP−2、BMP−4、BMP5およびBMP6)はまた、インビトロのラット骨芽細胞または骨肉腫細胞ベースのアッセイにおいてアルカリホスファターゼ活性を誘導する。このようなアッセイは、当該分野においてよく記載されており、本明細書中下記に詳細に記載される。例えば、Sabokdarら、(1994)Bone and Mineral 27:57〜67;Knutsenら、(1993)Biochem.Biophys.Res.Commun.194:1352〜1358;およびMaliakalら、(1994)Growth Factors 1:227〜234)を参照のこと。対照的に、低い特異的骨形成活性を有する骨形成タンパク質(例えば、CDMP−1およびCDMP−2)は、細胞ベースの骨芽細胞アッセイにおけるアルカリホスファターゼ活性の同様なレベルを誘導しない。従って、このアッセイは、BMPの生物学的活性変異体を評価するための容易な方法を提供する。例えば、BMP2、BMP4、BMP5、BMP6またはOP−1よりも低い比活性を有するが、CDMP1、CDMP2およびCDMP3は全て骨形成を誘導する成分である。逆に、CDMP1、CDMP2またはCDMP3よりも低い比活性を有するが、BMP2、BMP4、BMP5、BMP6およびOP−1はすべて関節軟骨形成を誘導し得る。従って、実施例5の細胞ベースのアッセイにおいてアルカリホスファターゼ活性を誘導するコンピテントなCDMP変異体は、ラット動物バイオアッセイにおいてより高い特異的骨形成活性を実証することが予期される。同様に、CDMP1、CDMP2またはCDMP3タンパク質の対応する位置に存在する置換を含み、そしてラットアッセイにおいて骨を誘導するが、細胞ベースのアッセイにおいてアルカリホスファターゼ活性を誘導しないようにコンピテントである、OP−1変異体は、インビボ関節軟骨アッセイにおいて、より高度に特異的な関節性軟骨誘導活性を有することが予期される。本明細書中下記に記載されるように、CDMP活性についての適切なインビトロアッセイは、マウス胚の前骨芽細胞またはマウス癌細胞(例えば、ATDC5細胞)を利用する。下記実施例6を参照のこと。
【0146】
TGF−β活性は、このタンパク質の上皮細胞増殖を阻害する能力によって容易に評価され得る。有用な、十分に特徴付けられたインビトロアッセイは、ミンク肺細胞または黒色腫細胞を利用する。実施例7を参照のこと。TGF−βスーパーファミリーの他のメンバーについての他のアッセイは、文献において十分に記載されており、そして、過度の実験なく実施され得る。
【0147】
(D.処方および生物活性)
生じたキメラタンパク質は、インビボの事象を増強、阻害またはそうでなければ調節をするための治療の一部として個体に提供され得、その事象としては、TGF−βスーパーファミリーメンバーと1つ以上のその同族レセプターとの間の結合相互作用が挙げられるが、これに限定されない。下記のように、この構築物は薬学的組成物に処方され得、そして任意の適切な手段(好ましくは、直接または全身的に、例えば、非経口的または経口的に)よって、形態形成的に有効な量において投与され得る。好ましいキメラタンパク質をコードする、生じたDNA構築物はまた、遺伝子治療の目的のために、レシピエントに直接投与され得、このようなDNAは、キャリア成分を伴うか、もしくは伴わずに、またはマトリクス成分を伴うか、もしくは伴わずに投与され得る。あるいは、このようなDNA構築物を移入された細胞は、レシピエントに移植され得る。このような物質および方法は、当該分野において周知である。
【0148】
本明細書中に開示されるこの任意の構築物が直接(例えば、注射により、所望される組織部位に局所的に)または非経口的に(例えば、静脈内投与、皮下投与、筋肉内投与、眼窩内投与、眼投与、脳室内投与、頭蓋内投与、包内投与、脊髄内投与、槽内投与、腹腔内投与、頬内投与、直腸投与、膣投与、鼻腔内投与によって、またはエアロゾル投与によって)提供されるべきである場合、この治療的組成物は、好ましくは水溶液の部分を含む。この溶液は、好ましくは、患者への所望の構築物の送達に加えて、この溶液が、他の点では患者の電解質バランスおよび体積バランスに不利な影響を及ぼさないように、生理学的に受容可能である。従って、この治療分子のための水性媒体としては、例えば、通常の生理的食塩水(0.9%NaCl、0.15M)、pH7〜7.4またはその他の薬学的に受容可能な塩を含み得る。
【0149】
経口または非経口的投与のための有用な溶液は、薬学的分野において周知の任意の方法によって調製され得る(例えば、Remington’s Pharmaceutical Sciences,(Gennaro,A.編),Mack Pub.,1990に記載される)。例えば、処方物はポリアルキレングリコール(例えば、ポリエチレングリコール)、野菜起源の油、水素化ナフタレンなどを含み得る。特に、直接投与するための処方物は、グリセロールおよび他の高い粘度の組成物を含み得る。生体適合性、好ましくは生体再吸収可能な(bioresorbable)ポリマー(例えば、ヒアルロン酸、コラーゲン、リン酸三カルシウム、ポリ酪酸、ポリラクチド、ポリグリコリド、およびラクチド/グリコリドコポリマーを含む)が、インビボでのモルフォゲンの放出を制御するための有用な賦形剤であり得る。
【0150】
これら治療的分子のための他の潜在的に有用な非経口的送達系は、エチレンビニル酢酸コポリマー粒子、浸透圧ポンプ(osmotic pump)、移植可能な注入系(implantable infusion system)およびリポソームを含む。吸入投与のための処方物は、賦形剤として例えばラクトースを含み得るか、あるいは、例えば、ポリオキシエチレン−9−ラウリルエーテル、グリココール酸およびデオキシコール酸を含む水溶液もしくは点鼻薬の形態における投与のための油性溶液であり得るか、または鼻腔内に適用されるゲルとしてであり得る。
【0151】
最終的に、治療的分子は、単独または他の組織形態形成に影響を及ぼすことが公知である他の分子すなわち、組織を修復ならびに再生し得、そして/または炎症を阻害し得る分子と組み合わせて投与され得る。骨粗鬆症個体における骨組織増殖を刺激するための有用な補因子の例には、例えば、ビタミンD3、カルシトニン、プロスタグランジン、副甲状
腺ホルモン、デキサメサゾン、エストロゲン、およびIGF−IまたはIGF−IIが挙げられるが、これらに限定されない。神経組織の修復および再生のために有用な補因子は、神経増殖因子を含み得る。他の有用な補因子は、症状軽減補因子(symptom alleviating cofactor)が挙げれ、これには防腐剤、抗生物質、抗ウイルス剤および抗真菌剤ならびに鎮痛剤および麻酔薬が含まれる。
【0152】
治療的分子はさらに、薬学的に受容可能な非毒性賦形剤およびキャリアを有する混和剤によって、薬学的組成物に処方され得る。上記のように、そのような組成物は、特に脂質溶液または懸濁液の形態において、非経口的投与のために調製され得;特に錠剤またはカプセルの形態において、経口投与のために調製され得;または鼻腔内では、特に粉末、点鼻薬またはエアロゾルの形態において調製され得る。組織表面への付着が所望される場合、この組成物は、フィブリノゲン−トロンビン組成物または他の生付着性組成物(例えば、PCT US91/09275に開示(その開示は、本明細書中で参考として援用される)されるような)中に分散される生合成構築物を含み得る。次いで、この組成物は、塗布され、噴霧され、または他の方法で所望の組織表面に適用される。この組成物は、治療的に有効量(例えば、その量は、所望される効果を誘導するのに十分な時間、標的組織に適切な濃度の形態単位を提供する)においてヒトまたは他の哺乳動物への非経口的投与または経口的投与のために処方され得る。
【0153】
この治療的分子が、組織保存溶液または器官保存溶液の部分を含む場合、任意の市販の保存溶液が都合よく使用され得る。例えば、当該分野において公知の有用な溶液には、Collins溶液、Wisconsin溶液、Belzer溶液、Eurocollins溶液および乳酸加リンガー溶液が挙げられる。保存溶液および有用な成分の詳細な説明は、例えば、米国特許第5,002,965号において見出され得、その開示は、本明細書中に参考として援用される。
【0154】
いくつかのタンパク質構築物は(例えば、Vg/dppサブグループのメンバーに基づくタンパク質構築物)はまた、マトリックスと組み合わされる場合、インビボで高レベルの活性を示すことが意図される。例えば、米国特許第5,266,683号を参照のこと(この開示は本明細書中に参考として援用される)。現在好ましいマトリックスは、天然における外因性マトリクス、同種異系マトリクスまたは自原性マトリックスである。しかし、ポリ乳酸、ポリグリコール酸、ポリ酪酸、その誘導体およびコポリマーを含む合成材料もまた使用されて適切なマトリックスを産生し得ることが意図される。好ましい合成および天然に由来するマトリックス材料、それらの調製、本発明の形態形成タンパク質とともにそれらを処方するための方法および投与の方法は当該分野において周知であり、そのため本明細書中において詳細に議論されない。例えば、米国特許第5,266,683号を参照のこと(この開示は、本明細書中において参考として援用される)。さらに、プロテーゼデバイスのマトリックスまたは金属表面との結合、接着、または会合は、本明細書中に開示される材料および方法を使用して変えられ得る特性であるということが意図される。例えば、増強されたマトリックス接着特性を有する本発明のマトリックスおよび骨活性構築物を含むデバイスが、徐放デバイスとして使用され得る。当業者は、その変動および操作は、ここで、本明細書中における教示に照らして可能であるということを理解する。
【0155】
当業者によって理解されるように、治療的組成物中の記載される化合物の濃度は、多数の因子(投与されるべき形態形成有効量、使用される化合物の化学的特徴(例えば、疎水性)、および投与経路を含む)に依存して変化する。投与されるべき薬物の好ましい投薬量もまた、おそらく、疾患、組織損失または欠損の型および程度、特定の患者の全体の健康状態、選択された化合物の相対的な生物学的有効性、この化合物の処方、処方物中の賦形剤の存在および型ならびに投与経路のような変数に依存する。概して、本発明の治療的分子は、1日あたり約10ng/kg体重〜約1g/kg体重の範囲の代表的用量で個体に提供され得る(好ましい用量範囲は、約0.1mg/kg体重〜100mg/kg体重である)。
【0156】
(II.特定の改変タンパク質構築物)
一般的に、本発明は、以下の4つの型の改変TGF−βファミリータンパク質構築物に関する:(1)N末端領域で短縮化されたTGF−βファミリータンパク質、(2)切断に際して活性化((例えば、酸切断またはプロテアーゼ処理による)N末端配列の放出を含むが、これに限定されない)され得る「潜伏性」タンパク質、(3)特定の結合能力を有する融合タンパク質、および(4)天然に存在するかまたは改変された、TGF−βファミリーのメンバーのサブユニットからなるヘテロ二量体。これらのモルフォゲン構築物の特定の種を、以下で詳細に記載する。以下に例証される種は、一般的に、改変モルフォゲンまたは骨原性タンパク質構築物に関するが、当業者は、これらの構築物が、TGF−βスーパーファミリーの他のメンバーを用いて生成され得る類似の構築物の代表であることを理解する。
【0157】
本発明に従って、ネイティブなBMPの特性またはTGF−βスーパーファミリーのタンパク質の他のメンバー(そのヘテロダイマーおよびホモダイマーを含む)は、BMPまたはTGF−βスーパーファミリーメンバーの1つ以上の生物学特性を変更するためにネイティブのタンパク質のN末端を改変することによって変えられる。この発見の結果として、TGF−βスーパーファミリータンパク質を、以下のように設計することが可能である;(1)TGF−βスーパーファミリータンパク質が原核生物細胞もしくは真核生物細胞において組換え的に発現されるか、またはポリペプチド合成装置を使用して合成される;(2)TGF−βスーパーファミリータンパク質が、変更されたフォールディング特性を有する;(3)TGF−βスーパーファミリータンパク質が、中性pH(生理学的に適合する条件を含むがこれに限定されない)下で変更された溶解性を有する;(4)TGF−βスーパーファミリータンパク質が、変更された等電点を有する;(5)TGF−βスーパーファミリータンパク質が、変更された安定性を有する;(6)TGF−βスーパーファミリータンパク質が、変更された組織またはレセプター特異性を有する;(7)TGF−βスーパーファミリータンパク質が、再設計されて変更された生物学的活性を有する;および/または(8)TGF−βスーパーファミリータンパク質が、固体表面(例えば、生体適合性マトリックスまたは金属が挙げられるがこれらに限定されない)に対する変更された結合特性または接着特性を有する。従って、本発明は、好ましいタンパク質構築物を含む速放性処方物、徐放性処方物および/または時限放出性処方物を設計するための機構を提供し得る。他の利点および特徴が、以下の教示により明らかである。さらに、本明細書中に開示される発見を利用して、変更された表面結合/表面接着特性を有する改変タンパク質が設計かつ選択され得る。特に重要な表面には、天然に存在し得る固体表面(例えば、骨)、または多孔性粒子表面(例えば、コラーゲン、または他の生体適合性マトリックス)またはプロテーゼインプラントの作製された(flabricate)表面(金属を含む)が挙げられるが、これらに限定されない。本明細書中に意図されるように、実質的に任意の表面が、構築物の差次的結合についてアッセイされ得る。従って、本発明は、その表面結合/表面接着特性における改変を有する多様な機能性分子を含み、それによって、変更されたインビボ適用(徐放性処方物、速放性処方物および/または時限放出性処方物を含む)について有用なこのような構築物を与える。
【0158】
当業者は、1つ以上の上記の任意の特性を混合かつ整合することは、カスタマイズされたタンパク質(およびこのタンパク質をコードするDNA)の用途を操作する特定の機会を提供することを理解する。例えば、変更された安定性の特性が開発され、タンパク質のインビボにおける代謝回転を操作し得る。さらに、タンパク質がまた、変更されたリフォールディングおよび/または機能のような特性を有する場合、おそらく、フォールディングと機能と安定性との間に相互関連が存在する。例えば、Lipscombら、7 Protein Sci.765〜73(1998);およびNikolovaら、95 Proc.Natl.Acad.Sci.USA 14675〜80(1998)を参照のこと。本発明の目的のために、安定性変化は、変性剤濃度または温度の関数として安定性の円偏光二色性の他の指標の周知の技術を使用して慣用的にモニターされ得る。また、慣用的な走査熱分析が使用され得る。同様に、任意の前述の特性と溶解度の特性との間におそらく相互関連が存在する。溶解度の場合、タンパク質構築物が、生理学的に適合する条件下においてより多く可溶性になるか、もしくはあまり可能性でなくなるかのいずれかになるようにこの特性を操作することが可能であり、結果として、インビボにおいて投与される場合、それはそれぞれ容易に拡散するか、または局在化する。
【0159】
改変された特性を有するタンパク質構築物の上記の用途に加えて、改変された安定性を有するタンパク質構築物はまた、有効期間、保存、および/または出荷の検討に関する実用上の利点のために用いられ得る。さらに関連事項について、改変された安定性はまた、投薬量の検討に直接影響し得、それによって、例えば、処置の費用を削減する。
【0160】
構築物の特に重要なクラスは、可溶化されるキャリアまたは賦形剤に対して改変された結合を有するものである。制限されない例として、可溶化キャリア(例えば、ヒアルロン酸)に対する増強された結合を有する、改変されたBMPは、当業者が、拡散剤(diffusion)または体液のいずれかにより、BMPの損失または希釈なしに、欠損部位に注射可能処方物を投与することを可能にする。従って、局在は最大となる。当業者は、本教示により改変が可能であることを理解する。同様に、身体/組織の構成要素に対する改変された結合を有する、構築物の別のクラスが、開発され得る。制限されない例として、インサイチュインヒビターに対する結合の減少を有する、改変されたBMPを用いて、インビトロで特定の組織の修復を増大させ得る。例えば、軟骨組織が、ネイティブのBMPの活性を阻害し得る体液および/または軟骨自体において見出される特定のタンパク質と関連付けられることは、当該分野で周知である。しかし、改変された結合特性を有するキメラ構築物は、これらのインサイチュインヒビターの効果を克服し得、それにより修復などを増強する。当業者は、本教示により改変が可能であることを理解する
(A.短縮化(truncation))
種々の形態のOP−1(例えば、23k、17k、および変動する量の15k)が存在し、それにより、代表的なOP−1調製物はこれらのすべての種を含む。精製された成熟OP−1のN末端配列決定は、異質性を明らかにした。このことは、N末端が多少短縮化され得ることを示す。RP−HPLCからの溶出およびトリプシン切断により回収された種を用いた実験を通して、ROS活性は、15kの種の中で最高である。例えば、短縮化変異体H2469は、CHO由来のOP−1標準と比較して、比較的高い活性を有する。最初の成熟は、プロOP−1においてRXXR部位で生じて17k種を生じるが、異なるプロテアーゼによる二次成熟は、最も活性な15k種を生成する。トリプシン切断は、この二次活性化を模擬し得る。
【0161】
哺乳動物OP−1またはE.coliのリフォールディングしたOP−1のトリプシン処理は、ROS活性の増加を生じる。本明細書中に記載される構築物のN末端(例えば、ヘキサ−his、コラーゲン結合部位、およびBMP−2のN末端)の除去はまた、ROSアッセイにおける活性の増加を生じた。OP−1の短縮化はモルフォゲンの可溶性を増加し得、これはROS活性に影響し得る。従って、構築物は、特異的切断活性を有して作製され得る。すなわち、構築物は、切断の型および切断のタイミングについて選択的である。当業者は、使用される系(哺乳動物または原核生物)に基づいて切断活性が異なり得ることを理解する。例えば、哺乳動物系は、モルフォゲン構築物が、プロ領域(これは、構築物の状況において、フォールディングを破壊し得、そして結果として(哺乳動物系において)最後には全くタンパク質を伴わない完全な細胞内分解を生じる)を含むことを必要とし得る。プロタンパク質形態を含む他の構築物を生成することもまた、所望され得る。このような構築物では、プロドメインは、増加した活性を得るために切断され得る別のN末端エレメントとしてみなされ得る。当業者は、切断されていないプロタンパク質が、その特性(可溶性および活性に関連する)を利用するために使用され得ることを理解する。
【0162】
本発明の変異タンパク質は、改善された生物学的活性ならびに拡大した半減期を示す。さらに、本発明の短縮化タンパク質で観察された活性の増加は、塩基性残基の除去および/またはタンパク質の等電点の低下に起因し得る。生物学的活性および改善されたリフォールディングは、同時係属中の出願[代理人整理番号STK−076、1999年8月16日出願]および[代理人整理番号STK−077、1999年8月16日出願](これらの開示を、本明細書中で参考として援用する)に記載された改変と本発明の改変タンパク質を組み合わせる場合に、増強され得る。
【0163】
(B.特異的特性を有するN末端領域)
本発明のさらなる改変タンパク質は、モルフォゲンの7−システインドメインのN末端に融合された非モルフォゲン起源のペプチドを含む。例えば、図7A〜7Eを参照のこと。生じるN末端融合タンパク質は、この融合物が由来する未改変モルフォゲンにおいては存在しないさらなる生物学的または生化学的な特性を有する。この型の融合物は、そのN末端でタンパク質またはタンパク質フラグメント(例えば、コラーゲン結合ドメイン、プロテインAのFBドメイン、またはヘキサ−ヒスチジン領域)に融合されたモルフォゲンの7−システインドメインを含む。例えば、H2440は、IMAC(固定化金属親和性クロマトグラフィー)樹脂のための結合ドメインとして、そのN末端に付着したヘキサ−hisタグを有するOP−1である(図7B)。このタンパク質は、最初はその非フォールディング状態で、尿素の存在下において、銅IMAC樹脂に対して精製された。IMAC上での非フォールディングタンパク質の精製、次ぐリフォールディングの後、首尾良くリフォールディングされた画分をRP−HPLCにより精製した。このようなN末端融合タンパク質は、ROSアッセイにおいて、ほとんどまたは全く活性を示さないが、N末端非モルフォゲンペプチドの切断に際して活性化されて、活性なC末端モルフォゲンドメインを産生する。
【0164】
特異的部位を標的し得るこれらの操作されたOP−1構築物が特に好ましい。例えば、N末端デカペプチドコラーゲン結合ドメインを有するOP−1、H2487が構築された。ここでこのデカペプチドは、骨マトリックスへのOP−1の特異的かつ堅固な結合を得るために、第1のシステインの上流7残基に配置された(図7Aを参照)。この新規な構築物は、首尾良くリフォールディングされ、そしてROSアッセイにおいて活性であり、それにより特定の骨形成活性を示す。活性を指向するために、他の結合ドメインが同様に用いられ得る。例えば、軟骨修復の場合、OP−1はまた、補綴デバイスに特異的に接着させるため設計され得る。Clostridiumコラゲナーゼから誘導されたペプチドのような他のペプチドはまた、コラーゲン結合特性について探索され得る。
【0165】
本発明の技術が特定の条件下で特定の部位の活性タンパク質の環境誘引放出を可能にする特定の改変タンパク質処方物を生成するために用いられ得ることは、当業者に明白である。例えば、pHの変化または特定のプロテアーゼの存在は、活性タンパク質の送達および誘引放出を調整し得る。
【0166】
BMPまたは他のTGF−βファミリーのメンバーのリーダー配列の改変はまた、このタンパク質の溶解度、活性および発現に影響し得る。例えば、staphylococcus aureusプロテインAのFBサブドメインのリーダー配列で操作されたCDMP−3(腱の修復に有用であると考えられる)を利用する構築物H2528は、骨原性タンパク質の発現が改善されている。
【0167】
当業者は、本発明の構築物が、もし空間的干渉および引き続く生物学的活性の低下が考慮されるならば、TGF−βファミリータンパク質のN末端に結合され得る、種々の特異化された機能的ドメインを含むように操作され得ることを理解する。このような構築物は、活性の阻害またはフォールディングを回避するため7−システインドメインからN末端付加までの少なくとも最小限の間隔を必要とし得る。最小限の間隔の必要条件は、特定されたドメインおよびTGF−βファミリータンパク質の両方がその意図された活性を保持するように、付加された部分の空間特性、および改変された構築物の最終の意図された活性に依存することを、当業者は理解する。
【0168】
(C.潜在性BMP)
本発明はまた、OP−1のN末端の短縮は、タンパク質に負の影響を有さないので、N末端が融合タンパク質の溶解度および活性に影響し得る程度の驚くべき発見を利用する。さらに、OP−1の結晶構造は、N末端に関するどのような位相幾何学的情報も示さなかった。
【0169】
本明細書において記載されるN末端融合タンパク質は、所望の時間および位置で活性タンパク質を生成するために切断され得るタンパク質の潜在性(すなわち、不活性)形態を生成するために有用である。例えば、コラーゲン結合ドメインを含有する改変モルフォゲン(例えば、図7Aに示されるH2487)は、所望の組織位置(例えば、移植されたコラーゲンマトリックスを含む位置)へ不活性形態で送達され得、そしてその位置で切断され活性なモルフォゲンを生成する。切断は、標的位置の内因性条件(例えば、天然に存在するプロテアーゼ)から生じ得るか、または特定のプロテアーゼの投与もしくは他の因子(例えば、位置の酸性化)の結果であり得る。さらに、非常に特異的なプロテアーゼ切断部位が、例えば、骨折部位に見出されるプロテアーゼについて操作され得、移植部位でのOP−1の選択的遅延型活性化および/または漸進的活性化を可能にする。
【0170】
(D.ドメインスワッピング)
リフォールディング、溶解性、活性および発現を変更するためのさらなる構築物が、1つのTGF−βスーパーファミリータンパク質のネイティブリーダー配列を、別のTGF−βファミリーメンバーのネイティブなリーダー配列で置換することにより設計され得る。例えば、構築物H2549は、OP−1上に置き換えられたBMP−2のN末端を有する。
【0171】
(E.ヘテロダイマー)
上記のいくつかのN末端融合タンパク質モノマーは、リーダー配列の切断なしに活性なホモダイマーを形成しないが、活性なヘテロダイマーが、これらのタンパク質とTGF−βファミリータンパク質の未改変モノマーとの間で形成される。従って、このようなヘテロダイマーは、融合タンパク質に付着したN末端非TGF−βファミリータンパク質ドメイン(例えば、コラーゲン結合部位)によってタンパク質を標的部位に提供するために用いられ得る。あるいは、ヘテロダイマーの精製を増強するために設計特徴が用いられ得る。例えば、ヘキサヒスチジンを付加することにより、2種のサブユニットの間の精製の差異を強調することによって精製を容易にし得る。混合されたリフォールディングにより、2つのホモダイマーおよびヘテロダイマーの混合物を提供する。これは3つの分離可能な種を提供する。例えば、ヘキサ−ヒスチジンドメインを含むN末端融合タンパク質(例えば、図7Bに示されるH2440)
(これは、IMACカラムに結合する)は、N末端ドメインの切断により引き続いて活性化され得る融合タンパク質の精製を補助するために有用である。
【0172】
本発明のヘテロダイマーの構築のためにはE.coli発現が好ましい。なぜなら、実施者はヘテロダイマーの至適産生のために各モノマーの比を調節し得るからである。さらに、この方法は非常に迅速である。例えば、荷電したアミノ酸の好ましい改変で改変したヘキサ−ヒスチジンタグ化OP−1のE、D、E、およびR(H2440)(例えば、その全開示が本明細書において参考として援用される、代理人整理番号___を参照のこと)とBMP−2との間でのインビトロにおけるヘテロダイマー形成実験において、ヘテロダイマーの産生は優秀であった。理論的に予期される50%のヘテロダイマーおよび25%の各ホモダイマーよりも、例外的に高い収率のヘテロダイマーが存在する。これは、BMP-2がそれ自体とよりもOP−1とより容易に会合するかまたはそれ自体との再会合より
もOP−1と、より速く会合することによって生じ得る。あるいは、BMP−2は、フォールディングについてシャペロンとして作用し得る。別の実験はまた、BMP−2とH2447変異体であるOP−1(ヘキサ−ヒスチジンタグなし)との間のヘテロダイマー形成を示した。これはまた容易に会合し良好な収率のヘテロダイマーを生じる。ヘテロダイマーはまた、FB−OP−1(H2521)とBMP−2との間で形成され得る。短縮したOP−1であるH2469(第1のシステインの15残基上流を保持している)、およびBMP−5(H2475)のヘテロダイマー;ならびにH2469およびCDMP−2(H2471)のヘテロダイマーがまた構築され得る。
【0173】
リフォールディングにおいて有効であるのと同様、ヘキサ−his−OP−1(H2440)およびBMP−2(H2142)のヘテロダイマーは、ROSアッセイにおいて、ホモダイマーよりも大きい活性を有する。ヘキサ−his−OP−1ホモダイマーは非常に低い活性を有した。BMP−2のホモダイマーは、より良好な活性を有した。しかし、OP−1/BMP−2ヘテロダイマーは、いずれの親ホモダイマーよりもかなり活性であった。このアッセイにおいて、ヘテロダイマーは、CHO誘導型OP−1標準の約1/3の活性しか有さなかった。ヘキサ−his−タグのないOP−1のBMP−2とのヘテロダイマー(H2447)は類似の活性を有した。H2447は、フィンガー−2において改変を有するリフォールディング変異体であり、ホモダイマーと同じ程度の相対的に低い活性を有した。OP−1(H2469)/BMP−5(H2475)およびOP−1(H2469)/CDMP−2(H2471)のヘテロダイマーは、ROSアッセイにおいて良好な結果(2.5−3+)を得た。
【0174】
この同じプロトコールおよび方法論を用いて、E.coliで発現され、そしてインビトロでリフォールディングされたOP−1/BMP−2ヘテロダイマーを構築した。詳細には、H2447/BMP−2ヘテロダイマーおよびH2440/BMP−2ヘテロダイマーは、E.coli発現により生成され、そして生理学的条件下でインビトロでリフォールディングされた。SDS−PAGE分析に基づいて、ほとんどの材料が容易に組み合わされ、ヘテロダイマー種を形成した。非モルフォゲンドメインを含むヘテロダイマーを用いてさらなる種を形成する。このような種の例は、コラーゲン結合ドメイン融合OP−1(H2487)、ヘキサ−ヒスチジン融合OP−1(H2440)およびプロテインAのFBドメイン融合OP−1(H2521)ならびにFB−ドメイン融合ヘキサ−ヒスチジン/OP−1構築物H2440(H2525)のようなN末端融合モルフォゲンである。
【0175】
活性なヘテロダイマーはまた、2つのBMPまたは他のTGF−βファミリータンパク質(異なる系において発現された)から構築され得る。いくつかの構築物が良好に発現され、そして特定の系で他よりも発現された場合、より活性である。当業者は、その発現に最適な環境中で各構築物を発現し得、次いでそれらとの活性なヘテロダイマーを形成し得る。例えば、H2223(OP−1の変異体)は、哺乳動物発現系であるCHO細胞中で発現され、一方、H2525(図7D)(FB−ドメインOP−1)は、細菌発現系であるE.coli中で最も発現される。
【0176】
さらに、ヘテロダイマーの活性は、用いる2つのタンパク質を変化させることにより操作され得る。例えば、デカペプチドコラーゲン結合部位を有するOP−1であるH2487とCDMP3のヘテロダイマーが形成され得る。このヘテロダイマーは、H2487とBMP−2のヘテロダイマーとは異なる活性を有する。
【0177】
(F.構築物の選択および最適化)
本明細書中に教示されるように、本発明は、注文に応じた(customized)キメラタンパク質およびこのキメラタンパク質をコードするDNAを巧妙に作るノウハウを当業者に提供する。キメラタンパク質を特定のインビボでの適用に適切にする、特定の所望される特性を有するキメラタンパク質を設計する手段が、本明細書中にさらに教示および例示される(少なくともI.B.節、II節、およびIII節、前述のキメラタンパク質の例示的な実施態様についての実施例1〜4、8および11を参照のこと)。例えば、変化した溶解度という特性を有するキメラタンパク質は、レシピエントに提供される形態形成的有効量を操作するために、インビボで使用され得る。すなわち、溶解度の増大は、アベイラビリティーの増大を生じ得;溶解度の減少は、アベイラビリティーの減少を生じ得る。従って、このような全身投与されるキメラタンパク質は、即座に利用可能であり得、そして/または即効性の形態形成効果を有し得、一方、局所投与されたキメラタンパク質は、よりゆっくりと利用可能であり得/長期化した形態形成効果を有し得る。当業者は、手近な事実および状況を考慮して、減少した溶解度という特性に対して増大した溶解度という特性が好ましい場合を理解する。このようなパラメーターの最適化は、慣用的な実験法および通常の技能を必要とする。
【0178】
同様に、変化した安定性という特性を有するキメラタンパク質は、レシピエントに提供される形態形成的有効量を操作するために、インビボで使用され得る。すなわち、安定性の増大は、半減期の増大を生じ得る。なぜなら、インビボでの代謝回転は、より小さくなるからであり;安定性の減少は、半減期およびアベイラビリティーの減少を生じ得る。なぜなら、インビボでの代謝回転がより大きくなるからである。従って、このように全身投与されたキメラタンパク質は、迅速に利用可能であり得るか/ボーラス型投薬を達成する即効性の形態形成効果を有し得るか、あるいは長期間にわたってインビボで利用可能であり得るか/徐放型の投薬を達成する長期の形態形成効果を有し得るかのいずれかである。当業者は、手近な事実および状況を考慮して、減少した安定性という特性に対して増大した安定性という特性が好ましい場合を理解する。このようなパラメーターの最適化は、慣用的な実験法および通常の技能を必要とする。
【0179】
さらに、変化した安定性を有するタンパク質がまた、有効期間、貯蔵、および/または運送条件を改善する実質的な利点のために用いられ得る。さらに関連事項に関しては、変化した安定性も投薬条件に直接影響し、それにより例えば、処置のコストを軽減する。
【0180】
さらに変化した特性(例えば、溶解度および安定性の特性であるが、これらに限定されない)の組合せを、レシピエントに提供される形態形成的有効量を操作するためにインビボで用い得る。すなわち、特定の変化した特性の組み合わせを有するキメラタンパク質を設計することにより、形態形成的有効量は、時限放出の様式において投与され得;投薬は、量および持続時間の両方に関して調節され得;処置レジメは、少ないがいくつかの典型を挙げると全身的または局所的に、低用量で開始され、続いて高用量に移され得るか、またはその逆であり得る。当業者は、この事実および手近な状況下で、多い形態形成的有効量に対して少ない形態形成的有効量が適切である場合を理解する。このようなパラメーターの最適化は、慣用的な実験法および通常の技能を必要とする。
【0181】
さらに、1以上の変化した特性を有するキメラタンパク質は、発達における固有の欠損を克服するために有用である。1以上の変化した特性を有するキメラタンパク質は、宿主のネイテイブな形態形成シグナル伝達系における固有の欠損を回避するように設計され得る。非限定的な例として、本発明のキメラタンパク質は、標的組織中のネイティブなレセプターにおける欠損、細胞内シグナル伝達経路における欠損、および/または異なるサブドメインに含まれる機能/生物学的活性と関連する特性と逆であるような、部分自体の認識と関連したサブドメインの特性に依存する他の事象における欠損を迂回するために使用され得る。当業者は、手近な事実および状況を考慮して、このようなキメラタンパク質が適切である場合を理解する。最適化は、慣用的な実験法および通常の技能を必要とする。
【0182】
本発明の実施は、以下の実施例から、さらにより十分に理解され、これらの実施例は、単なる例示のために本明細書中に示され、いかようにも本発明を限定するとは解釈されないべきである。
【実施例】
【0183】
(実施例1.BMP変異体の合成)
図8は、OP−1のC末端の7つのシステインドメインのためのヌクレオチドおよび対応するアミノ酸配列を示す。これらの配列を知ることにより、例えば、カセット変異誘発またはKunkelの周知の方法(ml3から派生した一本鎖の鋳型を使用するプライマー伸長による変異誘発)によって、または周知のPCR方法(重複伸長を含む)によって変異操作のための有用な制限部位の確認が可能になる。OP−1の例示的な変異体は、以下に記載のように構築される、フィンガー−2サブドメインにおいて4つのアミノ酸変化および最後のC末端アミノ酸において1つのアミノ酸変化を有する、H2460である。記載されている変異誘発プロトコルが例示のみであること、および本発明の構築物を作製するための他の手段は周知であり、当該技術分野において十分に記載されていることは当業者によって理解されている。
【0184】
4つのアミノ酸変化が、重複伸長技術を用いる標準的ポリメラーゼ連鎖反応によって、OP−1フィンガー−2のサブドメイン配列に導入され、OP−1変異体H2460を生じた。フィンガー−2の領域の4つの変化は、N6>S、R25>E、N26>DおよびR30>Eであった。この変異体はまた、C末端残基のさらなる変化(H35>R)を含んだ。これらの反応のための鋳型は、野生型OP−1のcDNAクローンの成熟したドメインであった。それは成熟領域の初めのATG開始コドンによって操作されるE.coli発現ベクターに挿入された。ATGは、順プライマーとして以下の配列の合成オリゴヌクレオチドを用いるPCRによって導入された:ATG TCC ACG GGG AGC AAA CAG(配列番号36)、これは、M S T G S K Q(配列番号37)をコードする。PCR反応を、cDNAの3’コード領域に相補的な適当なバック−プライマーと組み合わせて行った。
【0185】
フィンガー−2変異体H2460を構築するために、標準のPCR反応において、市販のPCRキットを用いて、そしてプライマーとして合成オリゴヌクレオチドを用いる製造業者の指示に従って、改変フィンガー−2をコードするPCRフラグメントを作製した。
【0186】
N6>S改変を得るために、配列GCG CCC ACG CAG CTC AGC GCT ATC TCC GTC CTC(配列番号70)の順プライマー(プライマー#1)を用いた。この配列は、アミノ酸配列A P T Q L S A I S V L(配列番号71)をコードする。
【0187】
C末端付近の変化については、バックプライマー(43ヌクレオチド長)(プライマー#2)を用いた。これは、R25>EおよびN26>DおよびR30>EおよびC−末端H35>Rの変化を導入した。このプライマー#2は、以下の配列を含んだ:CTA TCT GCA GCC ACA AGC TTC GAC CAC CAT GTC TTC GTA TTT C(配列番号72)。これは以下のコード配列の相補体である:G AAA TAC GAA GAC ATG GTG GTC GAA GCT TGT GGC TGC AGA TAG(配列番号73)。この配列は、以下のアミノ酸をコードする:K Y E D M V V E A C G C R停止(配列番号74)。
【0188】
次いで、フィンガー2およびC末端変異を有するフラグメントを、N末端、フィンガー−1およびヒールサブドメインを有する成熟OP−1の上流部分をコードする別のPCRフラグメントと組み合わせた。N末端、フィンガー−1およびヒールサブドメインをコードする後者のPCRフラグメントを、鋳型としてE.coliについてのOP−1発現ベクターを用いて再度構築した。このベクターは、T7プロモーターに付着した、成熟OP−1タンパク質をコードするOP−1 cDNAフラグメント、および適切な宿主におけるT7プロモーターか、またはtrpプロモーターの制御下のいずれかの制御下での発現のためのリボソーム結合部位を含んだ。このT7発現ベクターにおいて、成熟したOP−1のXbaI部位のT7プロモーターおよびATGコドン間の配列、Pet 3d(Novagen Inc.(Madison WI))は、以下の通りである:
TCTAGAATAATTTTGTTTAACCTTTAAGAAGGAGATATACG ATG(配列番号75)。
【0189】
この第2のPCR反応を、順プライマー(プライマー#3)TAA TAC GAC TCA CTA TAG G(配列番号76)によってプライムした。これは、T7プロモーター領域およびプライマー#1と重なるバック−プライマー(プライマー#4)においてプライムし、そしてヌクレオチド配列GCT GAG CTG CGT GGG CGC(配列番号77)を有する。この配列は、コード配列GCG CCC ACG CAG CTC AGC(配列番号78)の相補体である。そしてこれは、A P T Q L S(配列番号79)をコードする。
【0190】
第3のPCR反応(実際の重複伸長反応)において、上記の2つのPCRフラグメントの部分は、組み合わされて、PCRによって増幅された。そして、完全な成熟OP−1領域を含む単一のフラグメントを生じた。この反応のために、プライマー#3を順プライマーとして用い、そして新規なプライマー(プライマー#5)を以下の配列GG ATC CTA TCT GCA GCC ACA AGC(配列番号80)を有するバック−プライマーとして用いた。この配列は、コード配列GCT TGT GGC TGC AGA TAGGAT CC(配列番号81)に対する相補体である。この配列は、A C G C R停止(配列番号82)をコードする。このプライマーはまた、遺伝子を発現ベクターに挿入するために便利な3’BamHI部位を付加する。
【0191】
重複伸長PCRから生じる、完全な変異遺伝子を保有する得られたフラグメントを、PCRフラグメントのクローン化のために設計された市販のクローニングベクター(例えば、pCR2.1−topo−TA(Invitrogen Inc.(Carlsbad
CA)))にクローニングした。クローニングしたPCRフラグメントを、XbaIおよびBamHIを用いる制限消化によって回収し、Pet3d(Novagen Inc.(Madison WI))のような市販のT7発現ベクターのXbaIおよびBamHI部位に挿入した。
【0192】
(実施例2.BMPのE.coli発現)
形質転換した細胞を、標準的培養条件下で、標準的SPYE 2YT培地、1:1比(例えば、Sambrookら、を参照のこと)中で、37℃で増殖した。異種タンパク質の過剰発現は、代表的には、8〜48の時間以内に封入体を生じた。封入体を、単離して、次のように可溶化した。1リットルの培養液を、細胞を集めるために遠心分離した。次いで、得られたペレット中の細胞を60mlの25mM Tris、10mM EDTA、pH 8.0(TE緩衝液)+100μg/mlのリゾチーム中に再懸濁し、そして、37℃で2時間インキュベートした。次いで、細胞懸濁液を、氷冷して、超音波処理して細胞を溶解させた。細胞溶解を、検鏡によって確認した。溶解物の体積を、TE緩衝液でほぼ300mlに調整した。次いで、封入体ペレットを得るために遠心分離した。このペレットを、TE緩衝液中の2〜4回の連続再懸濁および遠心分離によって洗浄した。洗浄した封入体ペレットを、40mlの100mM Tris、10mMのEDTA、6MのGuHCl(グアニジウム塩酸塩)、250mMのDTT、pH 8.8中での変性および還元により可溶化した。次いで、タンパク質を、標準的な、市販のC2またはC8カートリッジ(SPICEカートリッジ、400mg、Ananltech、Inc.)を用いて予め精製した。タンパク質溶液を2% TFA(トリフルオロ酢酸)で酸性化し、カートリッジに適用し、0.1% TFA/10%アセトニトリルで洗浄し、そして0.1%TFA/70%アセトニトリルで溶出した。次いで、溶出した物質を、乾燥するかまたは希釈して、C4 RP−HPLCによって分画した。
【0193】
(実施例3.BMPダイマーのリフォールディング)
上記の通りに調製したタンパク質を、リフォールディング前に乾燥するかまたはリフォールディング緩衝液に直接希釈した。用いた好適なリフォールディング緩衝液は、以下であった:100mM Tris、10mM EDTA、1M NaCl、2% CHAPS、5mM GSH(還元グルタチオン)、2.5mM GSSG(酸化グルタチオン)、pH 8.5。リフォールディング(12.5〜200μgタンパク質/ml)を、24〜90時間、代表的には36〜48時間、4℃にて実行し(ただし、いくつかの変異体では、これより長い(数週間まで)と、良好なリフォールディングが提供されると予期される)、続いて、0.1% TFAに対して透析し、次いで0.01% TFA,50%エタノールに対して透析した。次いで、透析した材料のアリコートを種々のアッセイに備えて乾燥した。
【0194】
(実施例4.リフォールディングされたBMPダイマーの精製および試験)
(4A.SDS−PAGE、HPLC)サンプルを乾燥して、Laemmliゲルサンプル緩衝液において再懸濁して、次いで、15%のSDS−ポリアクリルアミドゲルにおいて電気泳動した。全てのアッセイは、比較のための、分子量標準および/または哺乳動物細胞が産生した精製OP−1を含んだ。OP−1ダイマーの解析を、添加した還元剤の非存在下で実行した。一方で、OP−1モノマーをゲルサンプルへの100mMのDTTの添加によって生成した。フォールディングしたダイマーは約30〜36kDaの範囲のみかけの分子量を有する。一方で、モノマー種は、約14〜16kDaの見かけの分子量を有する。
【0195】
あるいは、サンプルを、以下のように、市販のRP−HPLCにてクロマトグラフィーにかけた。サンプルを乾燥して、0.1%のTFA/30%のアセトニトリルの中に再懸濁した。次いで、タンパク質を、0.1%のTFA、30%のアセトニトリル中のC18カラムに適用し、そして、TFA中の30〜60%アセトニトリル勾配を用いて分画した。適切にフォールディングしたダイマーは、孤立したピークとして45〜50%アセトニトリルで溶出する;モノマーは、50〜60%アセトニトリルで溶出する。
【0196】
(4B.ヒドロキシアパタイトクロマトグラフィー)サンプルを10mMリン酸塩、6M 尿素、pH 7.0(カラム緩衝液)のヒドロキシアパタイトにロードした。未結合の物質を、カラム緩衝液を用いる洗浄により除去し、次いでカラム緩衝液+100mM NaClでモノマーを溶出させた。ダイマーを、カラム緩衝液+250mM NaClを用いて溶出した。
【0197】
(4C.トリプシン消化)トリプシン消化は、50mM Tris、4M 尿素、100mM NaCl、0.3% Tween 80、20mM メチルアミン、pH 8.0の消化緩衝液において実行した。酵素対基質の比は、1:50(重量比)であった。37℃での16時間のインキュベーションの後、15μlの消化混合液を、DTTを含まない5μlの4×ゲルサンプル緩衝液と組み合わせて、SDS−PAGEによって分析した。精製した哺乳動物OP−1および未消化のBMPダイマーを、比較のために含んだ。これらの条件の下で、適切にフォールディングされたダイマーを切断して、未切断の標準より僅かに速い移動を有する種を生産する。一方で、モノマー、および誤ってフォールディングしたダイマーは完全に消化され、そして染色したゲルにおいてバンドとしてみられない。
【0198】
(実施例5.骨原性活性のインビトロでの細胞ベースのバイオアッセイ)
本実施例は、本発明に従って骨原性能力または骨形成能力を獲得したモルフォゲン構築物の生物活性を実証する。高い特異的な骨形成活性についての生得の能力または獲得された能力のいずれかを有する骨原性タンパク質は、ラット骨肉腫細胞およびラット頭蓋冠(calveria)細胞を含むラット骨芽細胞においてアルカリホスファターゼ活性を誘導し得る。このアッセイにおいて、ラット骨肉腫または頭蓋冠細胞を、マルチウェルプレート(例えば、48ウェルプレート)の上へ、10%のFBS(ウシ胎仔血清)、L−グルタミンおよびペニシリン/ストレプトマイシンを含有するαMEM(改変イーグル培地、Gibco Inc.、Long Island)中で、1ウェルあたり50,000の骨芽細胞の濃度でプレートした。細胞を、37℃で24時間インキュベートした。この時点で、増殖培地を1%FBS含有αMEMで置換し、そしてこの細胞を、さらに24時間インキュベートし、その結果、この細胞は、実験のこの時点で血清枯渇増殖培地中にあった。
【0199】
次いで、培養した細胞を以下の3つの群に分けた:(1)生合成骨原性(ostegenic)タンパク質の種々の濃度を受けるウェル;(2)哺乳動物が発現したhOP−1のような陽性コントロール;および陰性コントロール(タンパク質、またはTGF−βなし)。試験したタンパク質濃度は、50〜500ng/mlの範囲であった。細胞を、72時間インキュベートした。インキュベーション期間後、細胞層を、0.5mlの1%のTritonX−100を用いて抽出した。得られた細胞抽出物を遠心分離し、100μlの抽出物を90μlのPNPP(パラニトロソフェニルホスフェート)/グリセリン混合液に添加して、37℃の水浴中で30分間インキュベートし、100μlの0.2N NaOHで反応を停止した。次いで、サンプルをプレートリーダー(例えば、Dynatech MR700)を通過させ、そして、標準としてp−ニトロフェノールを用いて400nmで吸光度を測定し、アルカリホスファターゼ活性の存在および量を決定した。タンパク濃度は、標準手段、例えば、Biorad方法、UVスキャンまたは214nmでのRP HPLC面積によって決定した。アルカリホスファターゼ活性を、タンパク質1μgあたりのユニット(単位)で算出した。ここで、1ユニットは、37℃で30分あたりに遊離した1nmolのp−ニトロフェノールに等しい。
【0200】
HOP−1およびBMP2は、100〜200ng/mlで約1.0〜1.4ユニットを生じる。種々のタンパク質構築物についての他の結果を、表1に提供する。
【0201】
(実施例6.CDMP活性のインビトロでの細胞ベースのバイオアッセイ)
本実施例は、本発明に従って増強された組織形態形成能力を獲得した構築物のの生物活性を実証する。ネイティブなCDMPは、実施例5において用いたようなラット骨肉腫細胞において、アルカリホスファターゼ活性を誘導し得ないが、それらは、マウス奇形癌細胞株ATDC−5(軟骨前駆細胞(chondroprogenitor)細胞株(Atsumiら、1990、Cell Differentiation and Development 30:109))においてアルカリホスファターゼ活性を誘導する。ラット骨癌細胞アッセイにおいて陰性であるが、ATDC−5アッセイにおいて陽性であるフォールディングされた変異体は、CDMP様活性を獲得したことが記載されている。ATDC−5アッセイにおいて、細胞を無血清基本培地(BM:Ham’s F−12/DMEM[1:1]以下を含む:ITSTM+培養補充物[Collaborative Biomedical Products、Bedford、MA]、α−ケトグルタレート(1×10-4M)、セルロプラスミン(0.25 U/ml)、コレステロール(5μg/ml)、ホスファチジルエタノールアミン(2μg/ml)、α−トコフェロール酸スクシナート(9×10-7M)、還元グルタチオン(10μg/ml)、タウリン(1.25μg/ml)、トリヨードサイロニン(1.6×10-9M)、副甲状腺ホルモン(5×10-10M)、β−グリセロリン酸(10mM)およびL−アスコルビン酸2−スルフ
ェート(50μg/ml))中、4×104の密度でプレートした。CDMPまたは他の
生合成骨原性タンパク質(0〜300ng/ml)をその翌日、添加し、そして、CDMPまたは生合成骨原性タンパク質を含む培養培地を1日おきに交換した。アルカリホスファターゼ活性を処理の4、6および/または12日後、超音波処理した細胞ホモジネート中で決定した。PBSを用いた大規模な洗浄の後、細胞層を0.05%Triton−X100を含有する500μlのPBS中で超音波処理した。50〜100μlのアリコートを、アッセイ緩衝液(0.1M バルビタールナトリウム緩衝液(pH 9.3))中で、そして基質としてp−ニトロフェニルホスフェートを用いて、酵素活性についてアッセイした。吸光度を、400nmで測定し、そして活性をBradfordタンパク質アッセイ(ウシ血清アルブミン標準)で測定されるタンパク含有量に対して標準化した。
【0202】
CDMP−1およびCDMP−2は、100ng/mlで10日目に約2〜3ユニットの活性を生成した。OP−1は、10日目、100ng/mlで約6〜7ユニットの活性を生成した。
【0203】
(実施例7.TGF−β様活性のインビトロにおける細胞に基づくバイオアッセイ)
本実施例は、本発明に従って生物学的能力を改変された生合成変異体TGF−βタンパク質の生物活性を実証する。TGF−βタンパク質は、上皮細胞増殖を阻害し得る。多数の細胞阻害アッセイが、当該分野において十分に記載されている。例えば、Brownら(1987)J.Immunol.139:2977(ヒト黒色腫A375線維芽細胞を用いる比色アッセイを記載しており、そして、本願明細書において以下に記載される)を参照のこと。別のアッセイは上皮細胞(例えば、ミンク肺上皮細胞)を使用し、そして増殖の効果は3H−チミジン取込みによって決定される。
【0204】
簡潔には、アッセイにおいて、TGF−β生合成構築物を、RPMI−1640培地(Gibco)および5%のウシ胎仔血清を含むマルチウェル組織プレートにおいて階段希釈する。コントロールウェルには、培地のみをいれる。次いで、黒色腫細胞を、ウェルに添加する(1.5×104)。次いで、プレートを5%のCO2中で約72時間37℃でインキュベートする。そして、細胞単層を一度洗って、固定して、そして15分間クリスタルバイオレットで染色する。未結合の染料を洗い流し、次いで染色した細胞を33%の酢酸で溶解し、(細胞核に閉じ込められた)染料を遊離させ、そして試験分子の活性を算出するために標準的な市販の光度計を用いて590nmでODを、測定する。それぞれのウェルにおける染色の強度は、核の数に直接関連する。従って、活性なTGF−β分子は、ウェルを不活性化合物または陰性コントロールウェルより薄く染色すると予期される。
【0205】
別のアッセイにおいては、ミンク肺細胞を用いる。これらの細胞は、標準的培養条件下で生長および増殖するが、ミンク肺上皮細胞株(ATCC番号CCL 64、Rockville、MD)由来の培養細胞を用いる3H−チミジン取込みによって決定されるよう
にTGF−βに曝露された後、停止される。簡潔には、細胞を、10%のFBS、200単位/mlのペニシリンおよび200μg/mlのストレプトマイシンを補充したEMEMを用いて、コンフルエンスになるまで生長させる。これらの細胞を、1ウェルあたり約200,000細胞の細胞密度になるまで培養する。コンフルエンスの時点で、培地を、1%のFBSおよびペニシリン/ストレプトマイシンを含有する0.5mlのEMEMで置換し、そしてこの培養物を、37℃で24時間インキュベートする。次いで、候補タンパク質を各ウェルに添加し、細胞を37℃で18時間インキュベートする。インキュベーションの後、10μl中に1.0μCiの3H−チミジンを、各々のウェルに添加し、そ
して細胞を、37℃で4時間インキュベートした。次いで、培地を、各ウェルから取り除き、細胞を、氷冷リン酸緩衝化生理食塩水を用いて1回洗浄し、そして、各ウェルに0.5mlの10% TCAを添加することによってDNAを、沈殿して、15分間室温でインキュベートした。この細胞を、氷冷蒸留水で3回洗浄し、0.5mlの0.4M NaOHで溶解し、次いで、各ウェルからの溶解物をシンチレーションバイアルに移し、そしてシンチレーションカウンター(Smith−Kline Beckman)を用いて放射能を記録する。生物学的に活性な分子は、細胞増殖を阻害し、その結果、不活性タンパク質および/または陰性コントロールウェル(増殖因子を添加してない)と比べて、より少ないチミジン取込みおよびより少ない計数を生じる。
【0206】
(実施例8.骨原性活性(軟骨内の骨形成および関連特性)のインビボにおけるバイオアッセイ) SampathおよびReddi(Proc.Natl.Acad.Sci.USA(1983)80:6591−6595)、および米国特許第4,968,590号、同第5,266,683号(これらの開示は本明細書において参考として援用される)によって記載される、骨誘導についての当該分野で認識されるバイオアッセイは、所定のタンパク質、デバイスまたは処方物の効力を評価するために用いられ得る。簡潔には、このアッセイは、エーテル麻酔下で試験試料をレシピエントラットの皮下の部位に配置することからなる。垂直の切開(1cm)を、胸の領域の上の皮膚に無菌条件下で作製し、そして、ポケットを、閉鎖(blunt)切開によって準備する。特定の場合において、凍結乾燥のような標準的手順を用いて調製した、所望の量の骨原性タンパク質(10ng〜10μg)を、約25mgのマトリックス材料と混合し、そして、試験試料を、ポケットの内側深くに移植し、そして切開を、金属の皮膚クリップで閉鎖する。異所性部位は、同所性部位の使用から生じる可能なあいまい性なしに骨誘導の研究を可能にする。このインプラントはまた、筋肉内に提供され得る。これはこのデバイスを接近可能な前駆細胞とより近接して設置する。代表的には、筋肉内インプラントを、両方の脚の骨格筋に作製する。
【0207】
異所性部位で生じる連続的な細胞反応は、複雑である。軟骨内骨形成の多段階のカスケードは、以下を包含する:移植したマトリックスに対するフィブリンおよびフィブロネクチンの結合、細胞の走化性、線維芽細胞の増殖、軟骨芽細胞への分化、軟骨形成、血管浸潤、骨形成、再構築、および骨髄分化。
【0208】
首尾良いインプラントは、以下を含むタンパク誘導性軟骨内性骨発生の段階を通じて制御された進行を示す:(1)1日目の多形核白血球による一過性浸潤;(2)2日目および3日目の間葉細胞遊走および増殖;(3)5日目および6日目の軟骨細胞出現;(4)7日目の軟骨基質形成;(5)8日目の軟骨石灰化;(6)9日目および10日目の血管浸潤、骨芽細胞の出現および新しい骨の形成;(7)12〜18日目の骨芽細胞の出現および骨再構築;ならびに(8)21日目の小骨における造血性骨髄分化。
【0209】
組織学的な切片にすることおよび染色は、移植片における骨形成の程度を決定するために好ましい。トルイジンブルーまたはヘマトキシリン(hemotoxylin)/エオシンを用いる染色は、軟骨内性骨の最終的な発達を明らかに実証する。12日目のバイオアッセイは、骨誘導活性が試験サンプルと関連しているか否かを決定するために十分である。
【0210】
さらに、アルカリホスファターゼ活性および/または総カルシウム含量は、骨形成についての生化学的マーカーとして使用され得る。このアルカリホスファターゼ酵素活性は、切り出された試験物質の均質化の後に、分光測光的に決定され得る。この活性は、インビボで9〜10日目でピークとなり、そしてその後にゆっくりと減少する。組織学によって骨の発達を全く示さないサンプルは、これらのアッセイ条件下でアルカリホスファターゼ活性を全く示さないはずである。このアッセイは、定量のために、および試験サンプルがラットから取り出された後に非常に迅速に骨形成の概算を得るために有用である。アルカリホスファターゼ活性レベルおよび組織学的評価によって測定される結果は、「骨形成単位」として表され得る。1骨形成単位は、12日目の最大の骨形成活性の半分に必要とされるタンパク質の量を表す。さらに、用量曲線は、種々の濃度のタンパク質をアッセイすることにより、精製スキームの各工程にて、インビボでの骨誘導活性について作成され得る。従って、当業者は、慣用的な実験のみを使用して、代表的な用量曲線を作成し得る。
【0211】
総カルシウム含量は、例えば、冷たい0.15M NaCl、3mM、NaHCO3、
pH9.0中で均質化し、そして沈渣の酸可溶性画分のカルシウム含量を測定する後に決定され得る。
【0212】
(実施例9:「ドメインスワッピング(Domain Swapping)」変異体の活性)
例えばある型のTGF−βファミリーのメンバーのタンパク質のN末端領域を獲得し、そしてこれを別の型のTGF−βファミリーのメンバーのタンパク質の7つのシステインのドメインに接続する場合、ドメインスワッピングが、生じる。ある変異体構築物は、当業者に一般に公知の慣用的な技術を用いて、OP−1の7つのシステインの活性ドメイン上のBMP−2末端の配列のスプライシングによって作製した。生じた変異体H2549は、MQAKHKQRKRLKSS−CからなるN末端領域を有する。最後のアミノ酸であるシステインは、OP−1の7つのシステインの活性ドメインの最初のシステインである。実施例5に上述したようなROSアッセイを、H2549の活性を試験するために用いた。
【0213】
図11に図示するように、その結果は、H2549がOP−1の活性のレベルに比べて非常に低い活性を有することを示す。しかし、実施例4に記載のダイマーのトリプシン切断に類似の方法を用いたH2549のトリプシン切断に際して、ROS活性が、有意に上昇する。この様式において、TGF−βファミリーのメンバーのタンパク質の活性は、それを不活化するために非天然のN末端配列を接続すること、およびそれを活性化するために非天然の配列を切断することによって選択的に制御され得る。
【0214】
(実施例10:N末端短縮化は活性を上昇させる)
改変されたモルフォゲンタンパク質のN末端領域の、例えば、トリプシン切断による短縮化は、ROS活性を上昇させる。H2223構築物は、CHO細胞において発現される改変されたOP−1変異体である。H2223の2つのHPLC画分である画分13および14が、回収された。各々の画分の量は、実施例4中のダイマーに際して用いたものと類似の様式において、トリプシン分解によって分解された。次いで結果生じた4つのサンプル、すなわちトリプシン未処理の画分13および14およびトリプシン処理した画分13および14は、OP−1活性を標準として用いた上述の実施例5のようにROSアッセイに供した。
【0215】
図12に図示するように、トリプシン処理したおよび未処理の画分14の活性レベルは、相対的に同一である。このことは、部分的に分解されたH2223を含む画分14によって説明され、従って、トリプシンを用いたさらなる消化は、活性を改変しない。対照的に、未処理の画分13は、全長H2223(すなわち、39アミノ酸のN末端全体)から主に構成され、そして画分13のN末端の短縮化は、画分14のROS活性に匹敵するレベルまでROS活性を上昇させない。これらの活性レベルは、OP−1標準のROS活性レベルよりも十分上であり、そして本発明の改変されたタンパク質を用いて得られた活性の改善を実証する。
【0216】
(実施例11:ヘテロダイマー活性)
ヘテロダイマーの活性レベルは、ヘテロダイマーのそれぞれのサブユニット各々より形成されるホモダイマーの活性レベルよりも高い。6−His N末端を伴うOP−1であるH2440構築物、およびBMP−2であるH2142構築物を、上述した実施例3に記載の方法を用いてヘテロダイマーおよびホモダイマーを形成させた。次いでH2440/2142のヘテロダイマーならびにH2440/2440のホモダイマーおよびH2142/2142のホモダイマーを、上述の実施例4および5に記載のようにROSアッセイに供した。
【0217】
図13Aおよび13Bに示すように、N末端に6Hisを伴うOP−1であるH2440のホモダイマーは、非常に低い活性を有する。BMP−2であるH2142のホモダイマーは、より良好な活性を有するが、活性は、まだ相対的には低い。しかし、OP−1 6−HisおよびBMP−2のヘテロダイマーは、これらのホモダイマーのいずれよりもかなり高い活性を有する。このヘテロダイマーは、CHO由来OP−1のたった1/3の活性を有する。
【0218】
類似の実験において、ホモダイマーおよびヘテロダイマーが、FBリーダー配列を伴うOP−1であるH2525とBMP−2であるH2142との間で作製された。これらもまた、OP−1活性のレベルを標準としたROSアッセイに供した。図14に図示するように、FBを伴うOP−1であるH2525のホモダイマーは、実質的に活性がなく、そしてBMP−2であるH2142のホモダイマーは、非常に低い活性を有する。対照的に、H2525/2142の2つのヘテロダイマーは、予期せぬ高い活性レベルを有する。
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1A】

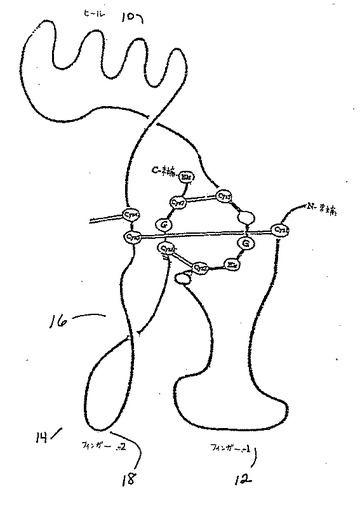
【図1B】
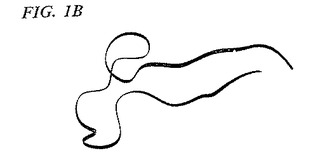
【図1C】
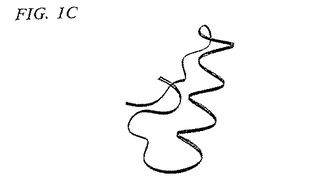
【図1D】

【図2A】
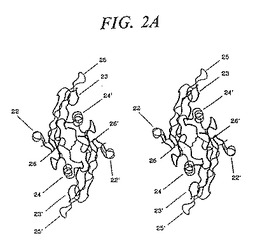
【図2B】
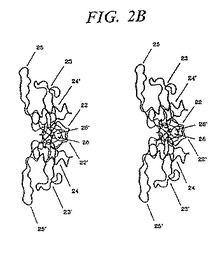
【図3】
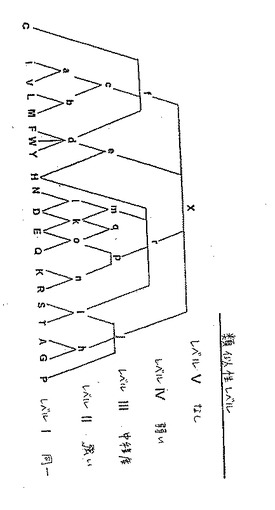
【図4】

【図5A】

【図5B】

【図5C】
【図6】
【図7A】

【図7B】

【図7C】
【図7D】
【図7E】

【図7F】
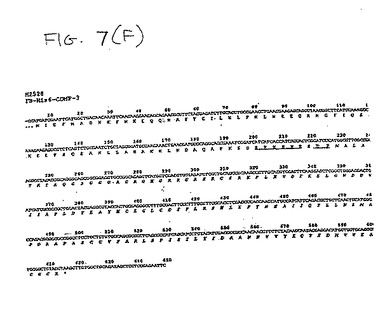
【図7G】

【図7H】

【図7I】
【図7J】

【図8】

【図9A】
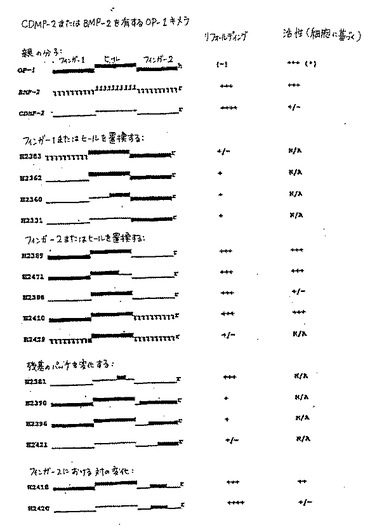
【図9B】
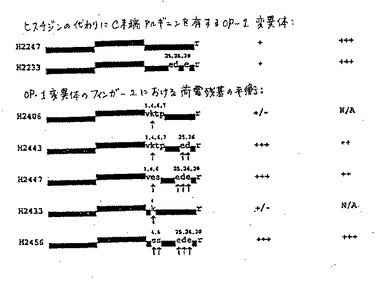
【図10】
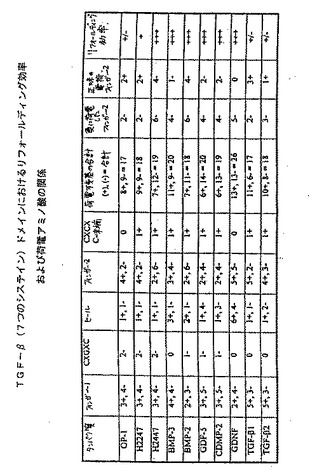
【図11】
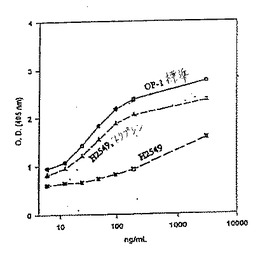
【図12】
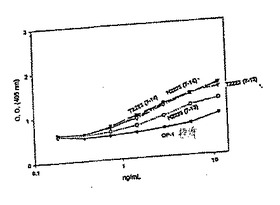
【図13】
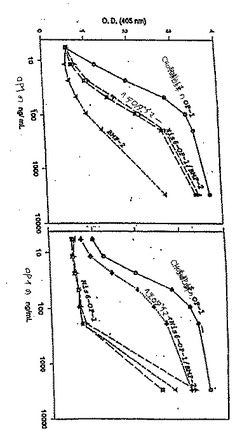
【図14】
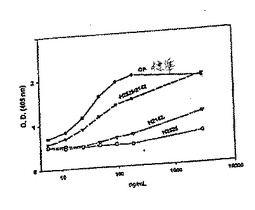
【図15】

【図1B】
【図1C】
【図1D】
【図2A】
【図2B】
【図3】
【図4】
【図5A】
【図5B】
【図5C】
【図6】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図7G】
【図7H】
【図7I】
【図7J】
【図8】
【図9A】
【図9B】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【公開番号】特開2013−14606(P2013−14606A)
【公開日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願番号】特願2012−200388(P2012−200388)
【出願日】平成24年9月12日(2012.9.12)
【分割の表示】特願2010−98044(P2010−98044)の分割
【原出願日】平成11年10月7日(1999.10.7)
【出願人】(595148888)ストライカー コーポレイション (52)
【氏名又は名称原語表記】STRYKER CORPORATION
【Fターム(参考)】
【公開日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願日】平成24年9月12日(2012.9.12)
【分割の表示】特願2010−98044(P2010−98044)の分割
【原出願日】平成11年10月7日(1999.10.7)
【出願人】(595148888)ストライカー コーポレイション (52)
【氏名又は名称原語表記】STRYKER CORPORATION
【Fターム(参考)】
[ Back to top ]