
改変蛍光蛋白質
【課題】蛋白質に加えられる力を感知することができる改変蛋白質の提供。
【解決手段】蛍光蛋白質の円順列変異体とβヘアピン構造を形成できる2つのペプチド(N−β)及び(C−β)とが、N末端側から(N−β)−(円順列変異体)−(C−β)の順で融合されている改変蛋白質であって、該蛋白質に加えられる力を感知するために用いられる改変蛋白質。
【解決手段】蛍光蛋白質の円順列変異体とβヘアピン構造を形成できる2つのペプチド(N−β)及び(C−β)とが、N末端側から(N−β)−(円順列変異体)−(C−β)の順で融合されている改変蛋白質であって、該蛋白質に加えられる力を感知するために用いられる改変蛋白質。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、改変蛍光蛋白質に関する。
【背景技術】
【0002】
蛍光蛋白質、例えば緑色蛍光蛋白質は、基礎研究及び応用研究において、蛋白質を蛍光標識する材料として用いられる。蛍光蛋白質による蛍光標識は、光学顕微鏡下において、蛋白質の局在及び運動の観察を可能とする。しかしながら、蛋白質の局在及び運動は、生体内における機能を、必ずしも反映しているとは限らない。
【0003】
非特許文献1によると、蛍光蛋白質の蛍光共鳴エネルギー転移を利用する事により、蛋白質の多量体化及び立体構造変化を視覚化することができる。蛍光共鳴エネルギー転移には、異なる吸収・蛍光波長を持つ2種の蛍光蛋白質を、連結させて使用する。特許文献1によると、同様の技術で、細胞内のカルシウム濃度を視覚化できる。蛍光共鳴エネルギー転移を用いた発明には、上記の他に、細胞分裂期を視覚化する蛍光蛋白質(非特許文献2)、アデノシン3リン酸の濃度を視覚化する蛍光蛋白質(非特許文献3)、及び、他蛋白質との相互作用を視覚化する蛍光蛋白質(非特許文献4)などがある。しかしながら、これらの方法では、二つの蛍光蛋白質が標識される蛋白質に融合される事が必須となる。そのため、二つの蛍光蛋白質の大きさが標識される蛋白質の機能への阻害効果が懸念される。蛋白質を蛍光標識するための物質は、可能な限り小さい方が好ましい。また、非特許文献5は、蛍光蛋白質の吸収及び蛍光波長が溶媒中のpHによって変化する特性を利用したpH感受性の蛍光蛋白質を開示し、特許文献2は、蛋白質の酵素活性、より具体的には、ヒストンのアセチル化を検出する蛍光蛋白質を開示する。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特開2004−184699号公報
【特許文献2】特開2008−167678号公報
【非特許文献】
【0005】
【非特許文献1】Tsien R.Y. and Miyawaki A. (1998) Science 280, 1954-1955.
【非特許文献2】Sunabori T., Tokunaga A., Nagai T., Sawamoto K., Okabe M., Miyawaki A., Matsuzaki Y., Miyata T. and Okano H. (2008) J Cell Sci. 121, 1204-1212.
【非特許文献3】Imamura H., Nhat K.P.H., Togawa H., Saito K., Iino R., Kato-Yamada Y., Nagai T. and Noji H. (2009) Proc. Natl. Acad. Sci. USA, 106, 15651-15656.
【非特許文献4】Wang H., Nakata E. and Hamachi, I. (2009) ChemBioChem 00, 1-19.
【非特許文献5】Kneen M., Farinas J., Li Y. and Verkman A.S. (1998) Biophys J 74, 1591-1599.
【発明の概要】
【発明が解決しようとする課題】
【0006】
しかしながら、筋肉の収縮を担う蛋白質あるいは細胞環収縮を担う蛋白質あるいは細胞内小胞を輸送する蛋白質に挙げられるような、力を発する事を機能とする蛋白質の力発生を視覚化できる蛍光蛋白質の報告はない。
【0007】
そこで、本発明は、二つの異なる蛍光蛋白質を繋げて得られたものではない改変蛋白質であって、蛋白質に加えられる力を感知することができる改変蛋白質を提供する。
【課題を解決するための手段】
【0008】
本発明は、蛍光蛋白質の円順列変異体とβヘアピン構造を形成できる2つのペプチド(N−β)及び(C−β)とが、N末端側から(N−β)−(円順列変異体)−(C−β)の順で融合されている改変蛋白質であって、該蛋白質に加えられる力を感知するために用いられる改変蛋白質(以下、「本発明の改変蛋白質」ともいう。)に関する。
【発明の効果】
【0009】
本発明の改変蛋白質によれば、二つの異なる蛍光蛋白質を繋げることによる蛍光共鳴エネルギー転移を用いずに、蛋白質に加えられる力を感知することができる。
【図面の簡単な説明】
【0010】
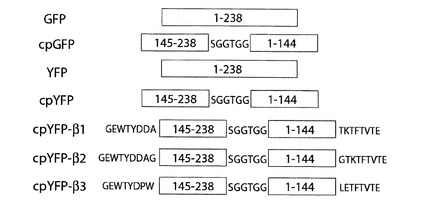
【図1】緑色蛍光蛋白質(GFP)、円順列変異緑色蛍光蛋白質(cpGFP)、黄色蛍光蛋白質(YFP)、円順列変異黄色蛍光蛋白質(cpYFP)、cpYFP−β1、cpYFP−β2、及び、cpYFP−β3の模式図である。
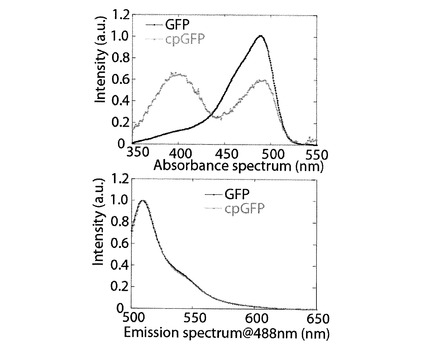
【図2】GFP(黒線)とcpGFP(灰線)のpH8.0の溶媒中における吸光波長特性(上)及び波長488ナノメートルのレーザーで励起した時の蛍光波長特性(下)を示すグラフである。
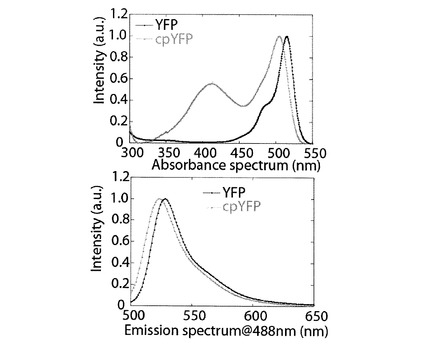
【図3】YFP(黒線)とcpYFP(灰線)のpH8.0の溶媒中における吸光波長特性(左)及び波長488ナノメートルのレーザーで励起した時の蛍光波長特性(右)を示すグラフである。
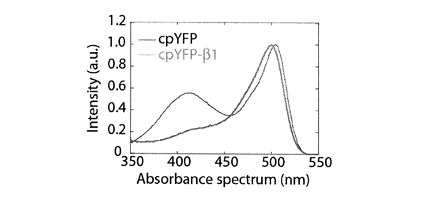
【図4】cpYFP−β1のpH8.0の溶媒中における吸光波長特性を示すグラフである。
【図5】cpYFP−β2のpH8.0の溶媒中における吸光波長特性を示すグラフである。
【図6】cpYFP−β3のpH8.0の溶媒中における吸光波長特性を示すグラフである。
【発明を実施するための形態】
【0011】
蛍光蛋白質は、ベータカン構造と呼ばれる円柱構造を持つ蛋白質である。ベータカン構造の内部に、3つのアミノ酸残基から構成される発光団があり、該発光団は、周囲のベータカン構造によるプロトン配置に対して、敏感に反応する。すなわち、上記ベータカン構造を変化させる事によって、当該蛍光蛋白質の吸収波長特性あるいは蛍光波長特性が変化すると予想される。しかしながら、ベータカン構造は、非常に安定な構造であり、その構造を変化させるためには150ピコニュートン以上の力が必要である。
【0012】
本発明は、まず、リンカー配列の挿入を伴う蛍光蛋白質の円順列変異を行うことで、変異前の蛍光蛋白質と波長特性(例えば、吸収波長及び/又は蛍光波長)が異なる円順列変異体が得られるという知見に基く。この知見に関しては、文献Baird et al.(Proc. Natl. Acad. Sci. USA, vol96, pp11241-11246 1999)を参照できる。
【0013】
次に、本発明は、波長特性が変化した前記円順列変異体の両端に、βヘアピン構造を形成できるペプチドを配置することで、変異により変化した波長特性を変異前の蛍光蛋白質のものと同様なものに戻すことができる、という知見に基く。
【0014】
そして、本発明は、βヘアピン構造を形成できるペプチドが両端に配置された前記円順列変異体の波長特性が該変異体に加えられる力に応じて変化するため、該変異体が力感受性蛍光蛋白質及び力感受性蛍光プローブとして機能しうる、という知見に基く。
【0015】
すなわち、本発明は、蛍光蛋白質の円順列変異体とβヘアピン構造を形成できる2つのペプチド(N−β)及び(C−β)とが、N末端側から(N−β)−(円順列変異体)−(C−β)の順で融合されている改変蛋白質であって、該蛋白質に加えられる力を感知するために用いられる改変蛋白質(以下、「本発明の改変蛋白質」といもいう。)に関する。本発明の改変蛋白質によれば、二つの異なる蛍光蛋白質を繋げることによる蛍光共鳴エネルギー転移を用いずに、蛋白質に加えられる力を感知することができる。
【0016】
[蛍光蛋白質]
本発明において使用されうる蛍光蛋白質は、蛍光を発することができる円順列変異体の作製が可能なものであれば特に制限されず、例えば、クラゲ(オワンクラゲ)由来のGFP及び該GFPに由来するYFP、CFP、EGFP、EYFP、ECFPその他の変異体が挙げられ、市販のものであってもよい。さらに、蛍光を発することができる範囲で欠失、付加、又は置換が行われた変異体の蛍光蛋白質であってもよい。
【0017】
[蛍光蛋白質の円順列変異体]
本明細書において、円順列変異(circular permutation)とは、ある蛋白質の内部に新たにN末とC末を作製し(すなわち、該蛋白質を内部で2分し)、オリジナルのC末とN末とを適当なリンカー配列により繋ぐ変異をいう。蛍光蛋白質における円順列変異は、従来から行われており、例えば、前述の文献Baird et al.(Proc. Natl. Acad. Sci. USA, vol96, pp11241-11246 1999)等を参照できる。本明細書において、円順列変異体とは、円順列変異により得られた変異体をいう。
【0018】
本発明における蛍光蛋白質円順列変異体は、力を感知する観点から、蛍光蛋白質としての性質(励起され蛍光を発光する機能)を維持していることが好ましく、変異前の蛍光蛋白質と波長特性が変化していることがより好ましい。本明細書において、波長特性は、吸光波長及び又は蛍光波長を含む。
【0019】
本発明における蛍光蛋白質円順列変異体の一例として、蛍光蛋白質の(1)N末端からn位までのアミノ酸配列及び(2)n+1位からC末端までのアミノ酸配列、並びに(3)グリシンを含む2〜20残基長のリンカー配列が、N末端側から(2)(3)(1)の順で結合したアミノ酸配列の変異体が挙げられる。円順列変異において、オリジナルの蛍光蛋白質のn位が新たなC末端となり、n+1位が新たなN末端となる。蛍光蛋白質のアミノ酸配列の長さがXとすると、nのとりうる範囲は、1〜X−1である。本発明における蛍光蛋白質円順列変異体は、当業者であれば、前述の文献Baird et al.等を参照して作製することができる。
【0020】
前記リンカー配列は、変異後に蛍光蛋白質としての機能を維持しつつ波長特性を変化させる点から、グリシンを含む2〜20残基長の配列であって、好ましくは4〜18残基長であり、より好ましくは5〜14残基長である。前記リンカー配列の一実施形態としては、(GGX)n、(GXG)n、又は(XGG)nで表されるアミノ酸配列、及びこれらから1又は2残基欠失したアミノ酸配列が挙げられる。前記アミノ酸配列において、Xはそれぞれ独立して任意のアミノ酸残基を表し、nは1〜3の整数を表す。
【0021】
[βヘアピンペプチド(N−β)及び(C−β)]
本発明におけるペプチド(N−β)及び(C−β)は、本発明の改変蛋白質において、それぞれ、本発明における蛍光蛋白質円順列変異体のN末及びC末に結合するペプチドである。ペプチド(N−β)及び(C−β)は、この順で繋がった場合にβヘアピン構造を形成できる配列である。なお、本明細書において、βヘアピン構造は、基本的に、2つのβシートが逆平行に配列し、該βシートが2〜5アミノ酸残基のループで繋がったものをいう。したがって、本発明におけるペプチド(N−β)及び(C−β)は、本発明の改変蛋白質が力を感知する観点から、それぞれ、該2つのβシートの一方及び他方に該当する部分のアミノ酸配列を含むことが好ましい。
【0022】
本発明におけるペプチド(N−β)及び(C−β)は、本発明の改変蛋白質の両端において両者が強く結合する。この強い結合が、蛍光蛋白質円順列変異により構造が変化した蛍光蛋白質のベータカン構造を異なる構造(好ましくは、オリジナル蛍光蛋白質の構造)に近づけ、それとともに波長特性も異なるもの(好ましくは、オリジナル蛍光蛋白質のそれ)に近づけていると考えられる。したがって、本発明の改変蛋白質における(N−β)と(C−β)との結合に外力が加わると再び波長特性が変化する。その外力が大きいほど、好ましくは、波長特性の変化も大きくなる。このようなメカニズムにより、本発明の改変蛋白質は、該蛋白質に加えられる力を感知できるようになると考えられる。但し、本発明はこれらのメカニズムに限定して解釈されなくてもよい。
【0023】
本発明におけるペプチド(N−β)及び(C−β)は、既存のβヘアピン構造を有する蛋白質の配列を参照して決定することができる。βヘアピン構造を有する蛋白質の例としては、ヘパリン、アミロイドβペプチド、グラミシジンSなどが挙げられる。したがって、一実施形態として、これらのβヘアピン構造を有する蛋白質のβヘアピン構造の部位のアミノ酸配列を2分して一方を(N−β)、他方を(C−β)とすることができる。
【0024】
本発明におけるペプチド(N−β)及び(C−β)の合計の長さは、本発明の改変蛋白質が力を感知する観点から、10〜30アミノ酸残基であることが好ましく、12〜26がより好ましく、14〜22がさらに好ましく、16〜20がさらにより好ましい。(N−β)及び(C−β)の長さは、本発明の改変蛋白質が力を感知する観点から、同程度であることが好ましく、両者の差は3アミノ酸残基以内であることがより好ましく、1アミノ酸残基であることがさらに好ましく、同じ長さであることがさらにより好ましい。
【0025】
本発明の改変蛋白質が力を感知する観点からとりわけ好ましい(N−β)の具体例として、Gly Glu Trp Thr Tyr Asp Asp Ala :(配列番号1)が挙げられる。また、本発明の改変蛋白質が力を感知する観点からとりわけ好ましい(C−β)の具体例として、Thr Lys Thr Phe Thr Val Thr Glu :(配列番号2)が挙げられる。したがって、本発明はその他の態様において、円順列変異により波長特性が変化した蛍光蛋白質変異体のN末端に配列表の配列番号1で表されるペプチドが結合し、C末端に配列表の配列番号2で表されるペプチドが結合した改変蛋白質であって、該蛋白質に加えられる力を感知するために用いられる改変蛋白質に関する。なお、本発明の全ての態様において、(N−β)と(C−β)の配列は入れ替えてもよい。
【0026】
本発明の改変蛋白質は、該蛋白質に加えられる力(好ましくは張力)を、波長特性の変化として感知することができる。したがって、本発明はその他の態様において、改変蛋白質の吸収波長及び/又は蛍光波長を検出することを含む、該改変蛋白質に加えられた力を検出する方法に関する。
【0027】
また、本発明の改変蛋白質を、その他の蛋白質の内部に挿入すれば、当該蛋白質の立体構造変化を検出及び視覚化できる。また、本発明の改変蛋白質を、力を発する蛋白質に挿入すれば、当該蛋白質が力を発生した時に、本発明の改変蛋白質の構造に歪みが与えられるため、当該蛋白質の力発生を検出及び視覚化できる。また、本発明の改変蛋白質が内部に挿入された蛋白質に外力を加えると、外力により該蛋白質の構造が歪みを与えられるため、本発明の改変蛋白質の構造にも歪みが与えられ、その外力を検出及び視覚化できる。
【0028】
したがって、本発明はその他の態様において、本発明の改変蛋白質がその他の蛋白質に挿入された組換え蛋白質であって、該その他の蛋白質の立体構造変化の検出及び視覚化、該その他の力発生の検出及び視覚化、並びに該その他の蛋白質に対する外力の検出及び視覚化の少なくとも1つを目的とする組換え蛋白質に関する。本発明はさらにその他の態様において、対象蛋白質の立体構造変化の検出及び視覚化、対象蛋白質の力発生の検出及び視覚化、並びに対象蛋白質に対する外力の検出及び視覚化の少なくとも1つを行う方法であって、該対象蛋白質に本発明の改変蛋白質を挿入する組換え蛋白質を作製すること、及び、組換え蛋白質における本発明の改変蛋白質の波長特性を検出することを含む方法に関する。
【0029】
本発明の蛋白質を挿入する対象となる蛋白質は、特に制限されないが、キネシン及びミオシンのようなモーター蛋白質、カドヘリンのような細胞接着因子が挙げられる。
【0030】
本発明はさらにその他の態様において、本発明の改変蛋白質をコードするベクター(以下、「本発明のベクター」ともいう。)に関する。本発明のベクターは、本発明の改変蛋白質を発現するための発現ベクターであっても良い。該発現ベクターにおいて、発現系は特に制限されず、原核生物、真核生物を問わない。さらに、本発明のベクターは前述の組換え蛋白質を作製するためのベクターであっても良く、本発明の改変蛋白質をコードする塩基配列の上流及び下流にマルチクローニングサイトを備えるベクターであってもよい。さらに、本発明は、前述の組換え蛋白質を作製するためのキットであって、本発明のベクターを含み、任意に必要な試薬、細胞、取扱説明書を含むキットに関しうる。
【0031】
以下、本発明を実施例及び図面を参照しながら説明する。
【実施例】
【0032】
[GFPの円順列変異]
図1に示す、緑色蛍光蛋白質(以下、GFPと表記)の円順列変異を行った。全238位のアミノ酸残基からなる蛍光蛋白質の145位から238位までのアミノ酸残基を新しい1位から93位のアミノ酸残基として、新しい94位から99位のアミノ酸残基をセリン、グリシン、グリシン、スレオニン、グリシン、グリシンとし、蛍光蛋白質の1位から144位までのアミノ酸残基を新しい100位から243位のアミノ酸残基とする(図1;cpGFP)。上記のセリン、グリシン、グリシン、スレオニン、グリシン、グリシンから成るペプチドは、グリシンを含む事が重要であり、それ以外に制限はない。本実施例では、1位から144位及び145位から238位のアミノ酸残基による円順列変異を用いたが、上記の他にも、蛍光発光を保持できるアミノ酸の組み合わせが存在することが、文献Baird et al.(Proc. Natl. Acad. Sci. USA, vol96, pp11241-11246 1999)に記されている。上記の円順列変異を行ったGFPを、以後、cpGFPと表記する。
【0033】
[GFPとcpGFPとの波長特性の比較]
図2に、pH8.0の溶媒中における、GFP及びcpGFPの、吸光波長と(上図)、波長488ナノメートル(以下、nmと記述する)のレーザーで励起した時の蛍光波長(下図)を示す。GFPの吸光波長には、波長485nmの位置に最大吸光が確認された。cpGFPの吸光波長には、波長485nmの位置の最大吸光の他に、波長390nmの位置にも最大吸光が確認された。上記した波長390nmに位置する最大吸光の出現は、円順列変異を行うことで、蛍光蛋白質内にある発光団へのプロトンの配置が変化するためとされている。蛍光波長において、GFPとcpGFPとには、差が見られなかった。すなわち、GFPにおいて、円順列変異による蛍光波長は、変化しない。
【0034】
[YFPの円順列変異]
上記と同様に、黄色蛍光蛋白質(以下、YFPと表記する。アミノ酸配列は、配列表の配列番号3)の円順列変異を行った(図1)。得られた円順列変異体をcpYFPと表記する。cpYFPのアミノ酸配列は、配列表の配列番号4である。図3に、cpYFPの吸光波長(上図)、及び、波長488nmのレーザーで励起した時の蛍光波長を示す(下図)。YFPの吸光波長には、波長513nmの位置に最大吸光が確認された。cpYFPの吸光波長には、513nmの位置の最大吸光は確認できず、波長501nmの位置及び407nmの位置に、最大吸光が確認された。YFPの蛍光波長には、波長527nmの位置に最大蛍光が確認された。cpYFPの蛍光波長には、波長521nmの位置に最大蛍光が確認された。YFPにおいては、円順列変異を行うことにより、吸光波長と蛍光波長の両方ともにおいて、その特性が変化した。
【0035】
上記したように、蛍光蛋白質は円順列変異を行うことにより、波長特性(吸光波長)が、大きく変化する。円順列変異による蛍光蛋白質の構造の歪みが、当該蛍光蛋白質の発光団へのプロトン配置に影響を与えている、と考えられる。
【0036】
[βヘアピンペプチドの導入:cpYFP−β1]
蛍光蛋白質に円順列変異を行うことにより変化した吸光波長特性を、βヘアピン構造を成すペプチドを当該円順列変異蛍光蛋白質に付加することによって、円順列変異を行う前の吸光波長特性に戻した。以下、YFPにおける実施例を用いて説明する。
【0037】
図1に、cpYFPへのペプチド付加を説明する模式図を示す。βヘアピン構造は、2つのベータシートでヘアピン様の構造であり、一次構造上で隣り合った2つのベータシートが逆平行に配列し、2から5アミノ酸残基からなる短いループで繋がったペプチドから構成される。βヘアピン構造を許容するペプチドは、多数存在するが、本実施例では、第1位からグリシン、グルタミン酸、トリプトファン、スレオニン、チロシン、アスパラギン酸、アスパラギン酸、アラニン、スレオニン、リジン、スレオニン、フェニルアラニン、スレオニン、バリン、スレオニン、グルタミン酸の順序で計16残基から構成されるペプチドを用いた。ただし、上記ペプチドのアミノ酸配列は、一実施形態にすぎず、本発明の特許請求の範囲を限定するものではない。
【0038】
cpYFPのN末端に、グリシン、グルタミン酸、トリプトファン、スレオニン、チロシン、アスパラギン酸、アスパラギン酸、アラニンを付加し、当該cpYFPのC末端に、スレオニン、リジン、スレオニン、フェニルアラニン、スレオニン、バリン、スレオニンを、付加した。上記ペプチドを付加したcpYFPを、以下、cpYFP−β1と記載する。cpYFP−β1のアミノ酸配列は、配列表の配列番号5である。
【0039】
図4にcpYFP−β1の吸光波長を示す。cpYFPにおいて確認できた波長407nmに位置する最大吸光は、cpYFP−β1の吸光波長において確認できなかった。cpYFP−β1の吸光波長では、波長500nmの位置にのみ、最大吸光が確認された。従って、上記βヘアピン構造を成すペプチドのcpYFPへの付加は、円順列変異により変化された吸光波長特性を、YFPの光波長特性に近づける効果があることが、証明された。
【0040】
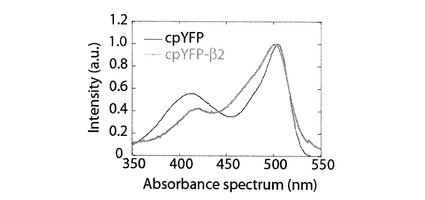
[cpYFP−β2]
上記の証明をさらに強固にするために、以下に記す実験を行った。cpYFP−β1にて、cpYFPのN末端に付加したアミノ酸のうち、cpYFPに最も接近して位置するアラニンの隣にグリシンを挿入し、当該cpYFPのC末端に付加したアミノ酸のうち最もcpYFPに接近して位置するスレオニンの隣にグリシンを挿入した(図1参照)。上記の変異は、グリシン、グルタミン酸、トリプトファン、スレオニン、チロシン、アスパラギン酸、アスパラギン酸、アラニン、スレオニン、リジン、スレオニン、フェニルアラニン、スレオニン、バリン、スレオニン、グルタミン酸から構成されるβヘアピン構造と当該cpYFPの構造の間を柔らかい構造で結合させる。従って、上記の置換は、当該βヘアピンに係る微弱な外力、あるいは、微小な立体構造の変化を模倣する。上記の置換を行ったcpYFP−β1を、以下、cpYFP−β2と記載する。図5にcpYFP−β2の吸光波長を示す。cpYFP−β1において確認できなかった波長407nmに位置する最大吸光が、cpYFP−β2の吸光波長のグラフにおいて確認できた。従って、cpYFP−β1のβヘアピン構造に微弱な外力が係る、あるいは、βヘアピン構造に歪みが与えられたら、cpYFP−β1の吸収波長には、波長407nmに位置する最大吸光が出現すると示唆される。
【0041】
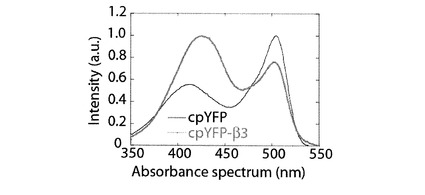
[cpYFP−β3]
cpYFP−β1にてcpYFPのN末端に付加したアミノ酸のうち、cpYFPに最も接近して位置するアスパラギン酸、アラニンを、それぞれ、プロリン、トリプトファンに置換し、当該cpYFPのC末端に付加したアミノ酸のうち最もcpYFPに接近して位置するスレオニン、リジンを、それぞれ、ロイシン、グルタミン酸に置換した(図1参照)。上記の置換を行ったcpYFP−β1を、以下、cpYFP−β3と記載する。アミノ酸のプロリンへの置換は、置換された蛋白質の立体構造を大きく歪ませる効果があり、cpYFP−β3は、cpYFP−β2と比べて、さらに、該βヘアピン構造に歪みを与える。図6にcpYFP−β3の吸光波長を示す。cpYFP−β1において確認できなかった波長407nmに位置する最大吸光が、cpYFP−β3の吸光波長のグラフにおいて確認できた。cpYFP−β3における波長407nmに位置する最大吸光の強度は、cpYFP−β2の波長407nmに位置する最大吸光の強度に比べて、大きい。また、cpYFP−β1において確認された波長501nmに位置する最大吸光の強度が、cpYFP−β3において、減少した。cpYFP−β3のβヘアピン構造とcpYFPとの結合は、cpYFP−β2のβヘアピン構造とcpYFPとの結合よりも、大きく歪みが与えられており、上記の歪みの大きさが、吸収波長の特性に影響を与えている。上記の実験結果は、cpYFP−β1の吸収波長特性は、cpYFPとβヘアピン構造の結合の歪みによって、変化することを示している。さらに詳しくは、cpYFP−β1のβヘアピン構造に微弱な外力が係る、あるいは、βヘアピン構造に歪みが与えられたら、その歪みの大きさに依存して、cpYFP−β1の吸収波長には、波長407nmに位置する最大吸光の強度が増加し、波長501nmに位置する最大吸光の強度が減少することを示している。
【0042】
上述の事から、cpYFP−β1の波長407nmにおける吸光度、かつ、波長501nmにおける吸光度を計測すれば、上記cpYFP−β1に係る微弱な力、あるいは、構造変化を検出することが可能であり、また、光学顕微鏡下で視覚化できる。
【0043】
したがって、cpYFP−β1を蛋白質の内部に挿入すれば、当該蛋白質の立体構造変化を検出及び視覚化できる。また、cpYFP−β1を力を発する蛋白質に挿入すれば、該蛋白質が力を発生した時に、当該cpYFP−β1の構造に歪みが与えられるため、当該蛋白質の力発生を検出及び視覚化できる。また、cpYFP−β1が内部に挿入された蛋白質に、外力を加えると、外力により該蛋白質の構造が歪みを与えられるため、cpYFP−β1の構造にも歪みが与えられ、その外力を検出及び視覚化できる。
【産業上の利用可能性】
【0044】
本発明は、例えば、分子イメージング分野、分子ナノメトリー分野、及び蛋白質の構造解析分野などにおいて有用である。
【配列表フリーテキスト】
【0045】
配列番号1: βヘアピンのN末端側のペプチド。配列番号2のペプチドとともにβヘアピン構造を形成しうる。
配列番号2: βヘアピンのC末端側のペプチド。配列番号1のペプチドとともにβヘアピン構造を形成しうる。
配列番号3: YFP(黄色蛍光蛋白質)
配列番号4: YFPの円順列変異体の一例
配列番号5: 本発明の改変蛋白質の一例
【技術分野】
【0001】
本発明は、改変蛍光蛋白質に関する。
【背景技術】
【0002】
蛍光蛋白質、例えば緑色蛍光蛋白質は、基礎研究及び応用研究において、蛋白質を蛍光標識する材料として用いられる。蛍光蛋白質による蛍光標識は、光学顕微鏡下において、蛋白質の局在及び運動の観察を可能とする。しかしながら、蛋白質の局在及び運動は、生体内における機能を、必ずしも反映しているとは限らない。
【0003】
非特許文献1によると、蛍光蛋白質の蛍光共鳴エネルギー転移を利用する事により、蛋白質の多量体化及び立体構造変化を視覚化することができる。蛍光共鳴エネルギー転移には、異なる吸収・蛍光波長を持つ2種の蛍光蛋白質を、連結させて使用する。特許文献1によると、同様の技術で、細胞内のカルシウム濃度を視覚化できる。蛍光共鳴エネルギー転移を用いた発明には、上記の他に、細胞分裂期を視覚化する蛍光蛋白質(非特許文献2)、アデノシン3リン酸の濃度を視覚化する蛍光蛋白質(非特許文献3)、及び、他蛋白質との相互作用を視覚化する蛍光蛋白質(非特許文献4)などがある。しかしながら、これらの方法では、二つの蛍光蛋白質が標識される蛋白質に融合される事が必須となる。そのため、二つの蛍光蛋白質の大きさが標識される蛋白質の機能への阻害効果が懸念される。蛋白質を蛍光標識するための物質は、可能な限り小さい方が好ましい。また、非特許文献5は、蛍光蛋白質の吸収及び蛍光波長が溶媒中のpHによって変化する特性を利用したpH感受性の蛍光蛋白質を開示し、特許文献2は、蛋白質の酵素活性、より具体的には、ヒストンのアセチル化を検出する蛍光蛋白質を開示する。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】特開2004−184699号公報
【特許文献2】特開2008−167678号公報
【非特許文献】
【0005】
【非特許文献1】Tsien R.Y. and Miyawaki A. (1998) Science 280, 1954-1955.
【非特許文献2】Sunabori T., Tokunaga A., Nagai T., Sawamoto K., Okabe M., Miyawaki A., Matsuzaki Y., Miyata T. and Okano H. (2008) J Cell Sci. 121, 1204-1212.
【非特許文献3】Imamura H., Nhat K.P.H., Togawa H., Saito K., Iino R., Kato-Yamada Y., Nagai T. and Noji H. (2009) Proc. Natl. Acad. Sci. USA, 106, 15651-15656.
【非特許文献4】Wang H., Nakata E. and Hamachi, I. (2009) ChemBioChem 00, 1-19.
【非特許文献5】Kneen M., Farinas J., Li Y. and Verkman A.S. (1998) Biophys J 74, 1591-1599.
【発明の概要】
【発明が解決しようとする課題】
【0006】
しかしながら、筋肉の収縮を担う蛋白質あるいは細胞環収縮を担う蛋白質あるいは細胞内小胞を輸送する蛋白質に挙げられるような、力を発する事を機能とする蛋白質の力発生を視覚化できる蛍光蛋白質の報告はない。
【0007】
そこで、本発明は、二つの異なる蛍光蛋白質を繋げて得られたものではない改変蛋白質であって、蛋白質に加えられる力を感知することができる改変蛋白質を提供する。
【課題を解決するための手段】
【0008】
本発明は、蛍光蛋白質の円順列変異体とβヘアピン構造を形成できる2つのペプチド(N−β)及び(C−β)とが、N末端側から(N−β)−(円順列変異体)−(C−β)の順で融合されている改変蛋白質であって、該蛋白質に加えられる力を感知するために用いられる改変蛋白質(以下、「本発明の改変蛋白質」ともいう。)に関する。
【発明の効果】
【0009】
本発明の改変蛋白質によれば、二つの異なる蛍光蛋白質を繋げることによる蛍光共鳴エネルギー転移を用いずに、蛋白質に加えられる力を感知することができる。
【図面の簡単な説明】
【0010】
【図1】緑色蛍光蛋白質(GFP)、円順列変異緑色蛍光蛋白質(cpGFP)、黄色蛍光蛋白質(YFP)、円順列変異黄色蛍光蛋白質(cpYFP)、cpYFP−β1、cpYFP−β2、及び、cpYFP−β3の模式図である。
【図2】GFP(黒線)とcpGFP(灰線)のpH8.0の溶媒中における吸光波長特性(上)及び波長488ナノメートルのレーザーで励起した時の蛍光波長特性(下)を示すグラフである。
【図3】YFP(黒線)とcpYFP(灰線)のpH8.0の溶媒中における吸光波長特性(左)及び波長488ナノメートルのレーザーで励起した時の蛍光波長特性(右)を示すグラフである。
【図4】cpYFP−β1のpH8.0の溶媒中における吸光波長特性を示すグラフである。
【図5】cpYFP−β2のpH8.0の溶媒中における吸光波長特性を示すグラフである。
【図6】cpYFP−β3のpH8.0の溶媒中における吸光波長特性を示すグラフである。
【発明を実施するための形態】
【0011】
蛍光蛋白質は、ベータカン構造と呼ばれる円柱構造を持つ蛋白質である。ベータカン構造の内部に、3つのアミノ酸残基から構成される発光団があり、該発光団は、周囲のベータカン構造によるプロトン配置に対して、敏感に反応する。すなわち、上記ベータカン構造を変化させる事によって、当該蛍光蛋白質の吸収波長特性あるいは蛍光波長特性が変化すると予想される。しかしながら、ベータカン構造は、非常に安定な構造であり、その構造を変化させるためには150ピコニュートン以上の力が必要である。
【0012】
本発明は、まず、リンカー配列の挿入を伴う蛍光蛋白質の円順列変異を行うことで、変異前の蛍光蛋白質と波長特性(例えば、吸収波長及び/又は蛍光波長)が異なる円順列変異体が得られるという知見に基く。この知見に関しては、文献Baird et al.(Proc. Natl. Acad. Sci. USA, vol96, pp11241-11246 1999)を参照できる。
【0013】
次に、本発明は、波長特性が変化した前記円順列変異体の両端に、βヘアピン構造を形成できるペプチドを配置することで、変異により変化した波長特性を変異前の蛍光蛋白質のものと同様なものに戻すことができる、という知見に基く。
【0014】
そして、本発明は、βヘアピン構造を形成できるペプチドが両端に配置された前記円順列変異体の波長特性が該変異体に加えられる力に応じて変化するため、該変異体が力感受性蛍光蛋白質及び力感受性蛍光プローブとして機能しうる、という知見に基く。
【0015】
すなわち、本発明は、蛍光蛋白質の円順列変異体とβヘアピン構造を形成できる2つのペプチド(N−β)及び(C−β)とが、N末端側から(N−β)−(円順列変異体)−(C−β)の順で融合されている改変蛋白質であって、該蛋白質に加えられる力を感知するために用いられる改変蛋白質(以下、「本発明の改変蛋白質」といもいう。)に関する。本発明の改変蛋白質によれば、二つの異なる蛍光蛋白質を繋げることによる蛍光共鳴エネルギー転移を用いずに、蛋白質に加えられる力を感知することができる。
【0016】
[蛍光蛋白質]
本発明において使用されうる蛍光蛋白質は、蛍光を発することができる円順列変異体の作製が可能なものであれば特に制限されず、例えば、クラゲ(オワンクラゲ)由来のGFP及び該GFPに由来するYFP、CFP、EGFP、EYFP、ECFPその他の変異体が挙げられ、市販のものであってもよい。さらに、蛍光を発することができる範囲で欠失、付加、又は置換が行われた変異体の蛍光蛋白質であってもよい。
【0017】
[蛍光蛋白質の円順列変異体]
本明細書において、円順列変異(circular permutation)とは、ある蛋白質の内部に新たにN末とC末を作製し(すなわち、該蛋白質を内部で2分し)、オリジナルのC末とN末とを適当なリンカー配列により繋ぐ変異をいう。蛍光蛋白質における円順列変異は、従来から行われており、例えば、前述の文献Baird et al.(Proc. Natl. Acad. Sci. USA, vol96, pp11241-11246 1999)等を参照できる。本明細書において、円順列変異体とは、円順列変異により得られた変異体をいう。
【0018】
本発明における蛍光蛋白質円順列変異体は、力を感知する観点から、蛍光蛋白質としての性質(励起され蛍光を発光する機能)を維持していることが好ましく、変異前の蛍光蛋白質と波長特性が変化していることがより好ましい。本明細書において、波長特性は、吸光波長及び又は蛍光波長を含む。
【0019】
本発明における蛍光蛋白質円順列変異体の一例として、蛍光蛋白質の(1)N末端からn位までのアミノ酸配列及び(2)n+1位からC末端までのアミノ酸配列、並びに(3)グリシンを含む2〜20残基長のリンカー配列が、N末端側から(2)(3)(1)の順で結合したアミノ酸配列の変異体が挙げられる。円順列変異において、オリジナルの蛍光蛋白質のn位が新たなC末端となり、n+1位が新たなN末端となる。蛍光蛋白質のアミノ酸配列の長さがXとすると、nのとりうる範囲は、1〜X−1である。本発明における蛍光蛋白質円順列変異体は、当業者であれば、前述の文献Baird et al.等を参照して作製することができる。
【0020】
前記リンカー配列は、変異後に蛍光蛋白質としての機能を維持しつつ波長特性を変化させる点から、グリシンを含む2〜20残基長の配列であって、好ましくは4〜18残基長であり、より好ましくは5〜14残基長である。前記リンカー配列の一実施形態としては、(GGX)n、(GXG)n、又は(XGG)nで表されるアミノ酸配列、及びこれらから1又は2残基欠失したアミノ酸配列が挙げられる。前記アミノ酸配列において、Xはそれぞれ独立して任意のアミノ酸残基を表し、nは1〜3の整数を表す。
【0021】
[βヘアピンペプチド(N−β)及び(C−β)]
本発明におけるペプチド(N−β)及び(C−β)は、本発明の改変蛋白質において、それぞれ、本発明における蛍光蛋白質円順列変異体のN末及びC末に結合するペプチドである。ペプチド(N−β)及び(C−β)は、この順で繋がった場合にβヘアピン構造を形成できる配列である。なお、本明細書において、βヘアピン構造は、基本的に、2つのβシートが逆平行に配列し、該βシートが2〜5アミノ酸残基のループで繋がったものをいう。したがって、本発明におけるペプチド(N−β)及び(C−β)は、本発明の改変蛋白質が力を感知する観点から、それぞれ、該2つのβシートの一方及び他方に該当する部分のアミノ酸配列を含むことが好ましい。
【0022】
本発明におけるペプチド(N−β)及び(C−β)は、本発明の改変蛋白質の両端において両者が強く結合する。この強い結合が、蛍光蛋白質円順列変異により構造が変化した蛍光蛋白質のベータカン構造を異なる構造(好ましくは、オリジナル蛍光蛋白質の構造)に近づけ、それとともに波長特性も異なるもの(好ましくは、オリジナル蛍光蛋白質のそれ)に近づけていると考えられる。したがって、本発明の改変蛋白質における(N−β)と(C−β)との結合に外力が加わると再び波長特性が変化する。その外力が大きいほど、好ましくは、波長特性の変化も大きくなる。このようなメカニズムにより、本発明の改変蛋白質は、該蛋白質に加えられる力を感知できるようになると考えられる。但し、本発明はこれらのメカニズムに限定して解釈されなくてもよい。
【0023】
本発明におけるペプチド(N−β)及び(C−β)は、既存のβヘアピン構造を有する蛋白質の配列を参照して決定することができる。βヘアピン構造を有する蛋白質の例としては、ヘパリン、アミロイドβペプチド、グラミシジンSなどが挙げられる。したがって、一実施形態として、これらのβヘアピン構造を有する蛋白質のβヘアピン構造の部位のアミノ酸配列を2分して一方を(N−β)、他方を(C−β)とすることができる。
【0024】
本発明におけるペプチド(N−β)及び(C−β)の合計の長さは、本発明の改変蛋白質が力を感知する観点から、10〜30アミノ酸残基であることが好ましく、12〜26がより好ましく、14〜22がさらに好ましく、16〜20がさらにより好ましい。(N−β)及び(C−β)の長さは、本発明の改変蛋白質が力を感知する観点から、同程度であることが好ましく、両者の差は3アミノ酸残基以内であることがより好ましく、1アミノ酸残基であることがさらに好ましく、同じ長さであることがさらにより好ましい。
【0025】
本発明の改変蛋白質が力を感知する観点からとりわけ好ましい(N−β)の具体例として、Gly Glu Trp Thr Tyr Asp Asp Ala :(配列番号1)が挙げられる。また、本発明の改変蛋白質が力を感知する観点からとりわけ好ましい(C−β)の具体例として、Thr Lys Thr Phe Thr Val Thr Glu :(配列番号2)が挙げられる。したがって、本発明はその他の態様において、円順列変異により波長特性が変化した蛍光蛋白質変異体のN末端に配列表の配列番号1で表されるペプチドが結合し、C末端に配列表の配列番号2で表されるペプチドが結合した改変蛋白質であって、該蛋白質に加えられる力を感知するために用いられる改変蛋白質に関する。なお、本発明の全ての態様において、(N−β)と(C−β)の配列は入れ替えてもよい。
【0026】
本発明の改変蛋白質は、該蛋白質に加えられる力(好ましくは張力)を、波長特性の変化として感知することができる。したがって、本発明はその他の態様において、改変蛋白質の吸収波長及び/又は蛍光波長を検出することを含む、該改変蛋白質に加えられた力を検出する方法に関する。
【0027】
また、本発明の改変蛋白質を、その他の蛋白質の内部に挿入すれば、当該蛋白質の立体構造変化を検出及び視覚化できる。また、本発明の改変蛋白質を、力を発する蛋白質に挿入すれば、当該蛋白質が力を発生した時に、本発明の改変蛋白質の構造に歪みが与えられるため、当該蛋白質の力発生を検出及び視覚化できる。また、本発明の改変蛋白質が内部に挿入された蛋白質に外力を加えると、外力により該蛋白質の構造が歪みを与えられるため、本発明の改変蛋白質の構造にも歪みが与えられ、その外力を検出及び視覚化できる。
【0028】
したがって、本発明はその他の態様において、本発明の改変蛋白質がその他の蛋白質に挿入された組換え蛋白質であって、該その他の蛋白質の立体構造変化の検出及び視覚化、該その他の力発生の検出及び視覚化、並びに該その他の蛋白質に対する外力の検出及び視覚化の少なくとも1つを目的とする組換え蛋白質に関する。本発明はさらにその他の態様において、対象蛋白質の立体構造変化の検出及び視覚化、対象蛋白質の力発生の検出及び視覚化、並びに対象蛋白質に対する外力の検出及び視覚化の少なくとも1つを行う方法であって、該対象蛋白質に本発明の改変蛋白質を挿入する組換え蛋白質を作製すること、及び、組換え蛋白質における本発明の改変蛋白質の波長特性を検出することを含む方法に関する。
【0029】
本発明の蛋白質を挿入する対象となる蛋白質は、特に制限されないが、キネシン及びミオシンのようなモーター蛋白質、カドヘリンのような細胞接着因子が挙げられる。
【0030】
本発明はさらにその他の態様において、本発明の改変蛋白質をコードするベクター(以下、「本発明のベクター」ともいう。)に関する。本発明のベクターは、本発明の改変蛋白質を発現するための発現ベクターであっても良い。該発現ベクターにおいて、発現系は特に制限されず、原核生物、真核生物を問わない。さらに、本発明のベクターは前述の組換え蛋白質を作製するためのベクターであっても良く、本発明の改変蛋白質をコードする塩基配列の上流及び下流にマルチクローニングサイトを備えるベクターであってもよい。さらに、本発明は、前述の組換え蛋白質を作製するためのキットであって、本発明のベクターを含み、任意に必要な試薬、細胞、取扱説明書を含むキットに関しうる。
【0031】
以下、本発明を実施例及び図面を参照しながら説明する。
【実施例】
【0032】
[GFPの円順列変異]
図1に示す、緑色蛍光蛋白質(以下、GFPと表記)の円順列変異を行った。全238位のアミノ酸残基からなる蛍光蛋白質の145位から238位までのアミノ酸残基を新しい1位から93位のアミノ酸残基として、新しい94位から99位のアミノ酸残基をセリン、グリシン、グリシン、スレオニン、グリシン、グリシンとし、蛍光蛋白質の1位から144位までのアミノ酸残基を新しい100位から243位のアミノ酸残基とする(図1;cpGFP)。上記のセリン、グリシン、グリシン、スレオニン、グリシン、グリシンから成るペプチドは、グリシンを含む事が重要であり、それ以外に制限はない。本実施例では、1位から144位及び145位から238位のアミノ酸残基による円順列変異を用いたが、上記の他にも、蛍光発光を保持できるアミノ酸の組み合わせが存在することが、文献Baird et al.(Proc. Natl. Acad. Sci. USA, vol96, pp11241-11246 1999)に記されている。上記の円順列変異を行ったGFPを、以後、cpGFPと表記する。
【0033】
[GFPとcpGFPとの波長特性の比較]
図2に、pH8.0の溶媒中における、GFP及びcpGFPの、吸光波長と(上図)、波長488ナノメートル(以下、nmと記述する)のレーザーで励起した時の蛍光波長(下図)を示す。GFPの吸光波長には、波長485nmの位置に最大吸光が確認された。cpGFPの吸光波長には、波長485nmの位置の最大吸光の他に、波長390nmの位置にも最大吸光が確認された。上記した波長390nmに位置する最大吸光の出現は、円順列変異を行うことで、蛍光蛋白質内にある発光団へのプロトンの配置が変化するためとされている。蛍光波長において、GFPとcpGFPとには、差が見られなかった。すなわち、GFPにおいて、円順列変異による蛍光波長は、変化しない。
【0034】
[YFPの円順列変異]
上記と同様に、黄色蛍光蛋白質(以下、YFPと表記する。アミノ酸配列は、配列表の配列番号3)の円順列変異を行った(図1)。得られた円順列変異体をcpYFPと表記する。cpYFPのアミノ酸配列は、配列表の配列番号4である。図3に、cpYFPの吸光波長(上図)、及び、波長488nmのレーザーで励起した時の蛍光波長を示す(下図)。YFPの吸光波長には、波長513nmの位置に最大吸光が確認された。cpYFPの吸光波長には、513nmの位置の最大吸光は確認できず、波長501nmの位置及び407nmの位置に、最大吸光が確認された。YFPの蛍光波長には、波長527nmの位置に最大蛍光が確認された。cpYFPの蛍光波長には、波長521nmの位置に最大蛍光が確認された。YFPにおいては、円順列変異を行うことにより、吸光波長と蛍光波長の両方ともにおいて、その特性が変化した。
【0035】
上記したように、蛍光蛋白質は円順列変異を行うことにより、波長特性(吸光波長)が、大きく変化する。円順列変異による蛍光蛋白質の構造の歪みが、当該蛍光蛋白質の発光団へのプロトン配置に影響を与えている、と考えられる。
【0036】
[βヘアピンペプチドの導入:cpYFP−β1]
蛍光蛋白質に円順列変異を行うことにより変化した吸光波長特性を、βヘアピン構造を成すペプチドを当該円順列変異蛍光蛋白質に付加することによって、円順列変異を行う前の吸光波長特性に戻した。以下、YFPにおける実施例を用いて説明する。
【0037】
図1に、cpYFPへのペプチド付加を説明する模式図を示す。βヘアピン構造は、2つのベータシートでヘアピン様の構造であり、一次構造上で隣り合った2つのベータシートが逆平行に配列し、2から5アミノ酸残基からなる短いループで繋がったペプチドから構成される。βヘアピン構造を許容するペプチドは、多数存在するが、本実施例では、第1位からグリシン、グルタミン酸、トリプトファン、スレオニン、チロシン、アスパラギン酸、アスパラギン酸、アラニン、スレオニン、リジン、スレオニン、フェニルアラニン、スレオニン、バリン、スレオニン、グルタミン酸の順序で計16残基から構成されるペプチドを用いた。ただし、上記ペプチドのアミノ酸配列は、一実施形態にすぎず、本発明の特許請求の範囲を限定するものではない。
【0038】
cpYFPのN末端に、グリシン、グルタミン酸、トリプトファン、スレオニン、チロシン、アスパラギン酸、アスパラギン酸、アラニンを付加し、当該cpYFPのC末端に、スレオニン、リジン、スレオニン、フェニルアラニン、スレオニン、バリン、スレオニンを、付加した。上記ペプチドを付加したcpYFPを、以下、cpYFP−β1と記載する。cpYFP−β1のアミノ酸配列は、配列表の配列番号5である。
【0039】
図4にcpYFP−β1の吸光波長を示す。cpYFPにおいて確認できた波長407nmに位置する最大吸光は、cpYFP−β1の吸光波長において確認できなかった。cpYFP−β1の吸光波長では、波長500nmの位置にのみ、最大吸光が確認された。従って、上記βヘアピン構造を成すペプチドのcpYFPへの付加は、円順列変異により変化された吸光波長特性を、YFPの光波長特性に近づける効果があることが、証明された。
【0040】
[cpYFP−β2]
上記の証明をさらに強固にするために、以下に記す実験を行った。cpYFP−β1にて、cpYFPのN末端に付加したアミノ酸のうち、cpYFPに最も接近して位置するアラニンの隣にグリシンを挿入し、当該cpYFPのC末端に付加したアミノ酸のうち最もcpYFPに接近して位置するスレオニンの隣にグリシンを挿入した(図1参照)。上記の変異は、グリシン、グルタミン酸、トリプトファン、スレオニン、チロシン、アスパラギン酸、アスパラギン酸、アラニン、スレオニン、リジン、スレオニン、フェニルアラニン、スレオニン、バリン、スレオニン、グルタミン酸から構成されるβヘアピン構造と当該cpYFPの構造の間を柔らかい構造で結合させる。従って、上記の置換は、当該βヘアピンに係る微弱な外力、あるいは、微小な立体構造の変化を模倣する。上記の置換を行ったcpYFP−β1を、以下、cpYFP−β2と記載する。図5にcpYFP−β2の吸光波長を示す。cpYFP−β1において確認できなかった波長407nmに位置する最大吸光が、cpYFP−β2の吸光波長のグラフにおいて確認できた。従って、cpYFP−β1のβヘアピン構造に微弱な外力が係る、あるいは、βヘアピン構造に歪みが与えられたら、cpYFP−β1の吸収波長には、波長407nmに位置する最大吸光が出現すると示唆される。
【0041】
[cpYFP−β3]
cpYFP−β1にてcpYFPのN末端に付加したアミノ酸のうち、cpYFPに最も接近して位置するアスパラギン酸、アラニンを、それぞれ、プロリン、トリプトファンに置換し、当該cpYFPのC末端に付加したアミノ酸のうち最もcpYFPに接近して位置するスレオニン、リジンを、それぞれ、ロイシン、グルタミン酸に置換した(図1参照)。上記の置換を行ったcpYFP−β1を、以下、cpYFP−β3と記載する。アミノ酸のプロリンへの置換は、置換された蛋白質の立体構造を大きく歪ませる効果があり、cpYFP−β3は、cpYFP−β2と比べて、さらに、該βヘアピン構造に歪みを与える。図6にcpYFP−β3の吸光波長を示す。cpYFP−β1において確認できなかった波長407nmに位置する最大吸光が、cpYFP−β3の吸光波長のグラフにおいて確認できた。cpYFP−β3における波長407nmに位置する最大吸光の強度は、cpYFP−β2の波長407nmに位置する最大吸光の強度に比べて、大きい。また、cpYFP−β1において確認された波長501nmに位置する最大吸光の強度が、cpYFP−β3において、減少した。cpYFP−β3のβヘアピン構造とcpYFPとの結合は、cpYFP−β2のβヘアピン構造とcpYFPとの結合よりも、大きく歪みが与えられており、上記の歪みの大きさが、吸収波長の特性に影響を与えている。上記の実験結果は、cpYFP−β1の吸収波長特性は、cpYFPとβヘアピン構造の結合の歪みによって、変化することを示している。さらに詳しくは、cpYFP−β1のβヘアピン構造に微弱な外力が係る、あるいは、βヘアピン構造に歪みが与えられたら、その歪みの大きさに依存して、cpYFP−β1の吸収波長には、波長407nmに位置する最大吸光の強度が増加し、波長501nmに位置する最大吸光の強度が減少することを示している。
【0042】
上述の事から、cpYFP−β1の波長407nmにおける吸光度、かつ、波長501nmにおける吸光度を計測すれば、上記cpYFP−β1に係る微弱な力、あるいは、構造変化を検出することが可能であり、また、光学顕微鏡下で視覚化できる。
【0043】
したがって、cpYFP−β1を蛋白質の内部に挿入すれば、当該蛋白質の立体構造変化を検出及び視覚化できる。また、cpYFP−β1を力を発する蛋白質に挿入すれば、該蛋白質が力を発生した時に、当該cpYFP−β1の構造に歪みが与えられるため、当該蛋白質の力発生を検出及び視覚化できる。また、cpYFP−β1が内部に挿入された蛋白質に、外力を加えると、外力により該蛋白質の構造が歪みを与えられるため、cpYFP−β1の構造にも歪みが与えられ、その外力を検出及び視覚化できる。
【産業上の利用可能性】
【0044】
本発明は、例えば、分子イメージング分野、分子ナノメトリー分野、及び蛋白質の構造解析分野などにおいて有用である。
【配列表フリーテキスト】
【0045】
配列番号1: βヘアピンのN末端側のペプチド。配列番号2のペプチドとともにβヘアピン構造を形成しうる。
配列番号2: βヘアピンのC末端側のペプチド。配列番号1のペプチドとともにβヘアピン構造を形成しうる。
配列番号3: YFP(黄色蛍光蛋白質)
配列番号4: YFPの円順列変異体の一例
配列番号5: 本発明の改変蛋白質の一例
【特許請求の範囲】
【請求項1】
蛍光蛋白質の円順列変異体とβヘアピン構造を形成できる2つのペプチド(N−β)及び(C−β)とが、N末端側から(N−β)−(円順列変異体)−(C−β)の順で融合されている改変蛋白質であって、該蛋白質に加えられる力を感知するために用いられる、改変蛋白質。
【請求項2】
前記蛍光蛋白質円順列変異体は、前記蛍光蛋白質の(1)N末端からn位までのアミノ酸配列及び(2)n+1位からC末端までのアミノ酸配列、並びに(3)グリシンを含む2〜20残基長のリンカー配列を含み、N末端側から(2)(3)(1)の順で結合したアミノ酸配列の変異体である、請求項1記載の改変蛋白質。
【請求項3】
前記2つのペプチド(N−β)及び(C−β)の合計のアミノ酸配列が、10〜30残基長である、請求項1又は2に記載の改変蛋白質。
【請求項4】
前記(N−β)が、配列表の配列番号1のアミノ酸配列で表されるペプチドであり、前記(C−β)が、配列表の配列番号2のアミノ酸配列で表されるペプチドである、請求項1から3のいずれかに記載の改変蛋白質。
【請求項5】
前記円順列変異体が、前記蛍光蛋白質とは異なる吸収波長及び/又は蛍光波長を示す、請求項1から4のいずれかに記載の改変蛋白質。
【請求項6】
前記円順列変異体が、配列表の配列番号3のアミノ酸配列で表される蛋白質である、請求項1から5のいずれかに記載の改変蛋白質。
【請求項7】
請求項1から6のいずれかに記載の改変蛋白質をコードするベクター。
【請求項8】
請求項1から6のいずれかに記載の改変蛋白質の吸収波長及び/又は蛍光波長を検出することを含む、前記改変蛋白質に加えられた力を検出する方法。
【請求項1】
蛍光蛋白質の円順列変異体とβヘアピン構造を形成できる2つのペプチド(N−β)及び(C−β)とが、N末端側から(N−β)−(円順列変異体)−(C−β)の順で融合されている改変蛋白質であって、該蛋白質に加えられる力を感知するために用いられる、改変蛋白質。
【請求項2】
前記蛍光蛋白質円順列変異体は、前記蛍光蛋白質の(1)N末端からn位までのアミノ酸配列及び(2)n+1位からC末端までのアミノ酸配列、並びに(3)グリシンを含む2〜20残基長のリンカー配列を含み、N末端側から(2)(3)(1)の順で結合したアミノ酸配列の変異体である、請求項1記載の改変蛋白質。
【請求項3】
前記2つのペプチド(N−β)及び(C−β)の合計のアミノ酸配列が、10〜30残基長である、請求項1又は2に記載の改変蛋白質。
【請求項4】
前記(N−β)が、配列表の配列番号1のアミノ酸配列で表されるペプチドであり、前記(C−β)が、配列表の配列番号2のアミノ酸配列で表されるペプチドである、請求項1から3のいずれかに記載の改変蛋白質。
【請求項5】
前記円順列変異体が、前記蛍光蛋白質とは異なる吸収波長及び/又は蛍光波長を示す、請求項1から4のいずれかに記載の改変蛋白質。
【請求項6】
前記円順列変異体が、配列表の配列番号3のアミノ酸配列で表される蛋白質である、請求項1から5のいずれかに記載の改変蛋白質。
【請求項7】
請求項1から6のいずれかに記載の改変蛋白質をコードするベクター。
【請求項8】
請求項1から6のいずれかに記載の改変蛋白質の吸収波長及び/又は蛍光波長を検出することを含む、前記改変蛋白質に加えられた力を検出する方法。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2011−188821(P2011−188821A)
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願番号】特願2010−58120(P2010−58120)
【出願日】平成22年3月15日(2010.3.15)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 日本生物物理学会 「生物物理 vol.49 第47回年会講演予稿集」 平成21年(2009年)9月20日 独立行政法人 科学技術振興機構 「生命現象解明のための計測分析 さきがけ・CREST研究報告会 講演要旨集」 平成22年(2010年)1月12日
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願日】平成22年3月15日(2010.3.15)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 日本生物物理学会 「生物物理 vol.49 第47回年会講演予稿集」 平成21年(2009年)9月20日 独立行政法人 科学技術振興機構 「生命現象解明のための計測分析 さきがけ・CREST研究報告会 講演要旨集」 平成22年(2010年)1月12日
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
[ Back to top ]