
改変rRNAオペロンを有する組換え微生物
【課題】遺伝子特異的に発現を制御することのできる微生物の提供。
【解決手段】改変SD配列を有する標的遺伝子と;当該改変SD配列と相補的な改変アンチSD配列を有する複数個のrrnオペロンと;改変されていないアンチSD配列を有するrrnオペロン、とを有する細菌。
【解決手段】改変SD配列を有する標的遺伝子と;当該改変SD配列と相補的な改変アンチSD配列を有する複数個のrrnオペロンと;改変されていないアンチSD配列を有するrrnオペロン、とを有する細菌。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、改変リボゾームをコードする改変rRNAオペロンを有する微生物と、当該微生物を用いた目的遺伝子産物の生産方法に関する。
【背景技術】
【0002】
リボゾームは遺伝子からタンパク質への翻訳が行われる場所であり、細菌においてはrRNAオペロン(rrn)によってコードされている。rrnオペロンには、16S、23S、5Sという3つのrRNAサブユニットが順にコードされている。細菌ゲノム中でのrrnオペロンのコピー数は種によって様々であり、ある種のマイコプラズマのようにわずか1コピーのものから、大腸菌のように7コピーのものや、枯草菌のように10コピーのものまで存在する。
【0003】
しかし、この細菌ゲノム中でのrrnのコピー数は、当該細菌の成長速度や生理活動の高さを直接反映しているわけではない。例えば、rrnオペロンのコピー数が増えても成長や生理活動量が増加するとは限らず、他方rrnオペロンを一部削除しても成長や生理活動に大きな影響はない。このことから、細菌の生存にとってゲノム中に複数存在するrrnオペロンが同時に全て機能する必要はなく、むしろ細菌はその時々の生育環境の変化にあわせて複数のrrnオペロンを使い分けていると推定される(非特許文献1)。
上記非特許文献1にはまた、10種類のrrnオペロンのうちのいずれか1種を1コピーのみ有する枯草菌株を作製し、それらの成長速度と胞子形成の頻度を調べた結果、各株間で胞子形成能が大きく異なっていたことを報告している。この知見は、複数のrrnオペロンが異種性(heterogeneity)であり、各々が異なる生育局面で異なる機能を担っているという考え方を支持する。
【0004】
このようなrrnオペロンの異種性や機能を鑑みると、細菌におけるrrnオペロンのコピー数とその成長速度や遺伝子発現能との間に相関性を見出すことはできない。
【0005】
シャイン・ダルガノ配列(Shine-Dalgarno sequence:以下、SD配列)は、原核生物のmRNAの開始コドン上流に認められる共通配列(代表的には、−GGAGG−)であり、これが、rrnオペロンの16SrRNAの配列の3’末端領域に存在する相補的なアンチSD配列(代表的には、−CCUCC−)と対合することによって、mRNAとリボゾームが結合し、翻訳が開始される。
佐藤ら(非特許文献2及び3)は、10コピーのrrnオペロンのうちの1つのアンチSD配列を改変した枯草菌株で、改変したSD配列を有するクロラムフェニコール耐性遺伝子(cat)の翻訳効率を調べた結果、改変株では、翻訳効率の低下したSD配列を有するcat遺伝子を持つにもかかわらず、クロラムフェニコール耐性が向上したことを示した。
【0006】
しかし、翻訳効率を向上させるために具体的にどのようにアンチSD配列を改変すればよいかは明らかではなかった。
さらに、非特許文献4及び5によれば、改変rrnオペロンのコピー数を増やしても翻訳効率は増加しないと考えられており、一方で改変rrnオペロンを増やせば、細菌の通常の蛋白合成能が低下して生命活動に影響を及ぼす可能性があるため、アンチSD配列の改変が必ずしも目的蛋白質の生産性向上につながるかどうかは明らかではなかった。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Nanamiya et al. (2010) Microbiology 156:2977-2982
【非特許文献2】佐藤ら、日本農芸化学会2006年度大会 大会講演要旨集、2006年3月5日発行、119頁
【非特許文献3】七宮ら、2006年度グラム陽性細菌のゲノム生物学研究会 プログラム&要旨集、2006年8月31日、pp16
【非特許文献4】Ellwood & Nomura (1980) J. Bacteriol. 143:1077-1080
【非特許文献5】Widom et al. (1988) J. Bacteriol. 170:605-610
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明は、改変リボゾームをコードする改変rrnオペロンを有する細菌、ならびに当該細菌を用いた標的遺伝子の発現制御方法及び目的遺伝子産物の生産方法に関する。
【0009】
SD配列又はアンチSD配列の改変によって、細菌の成長や遺伝子発現を制御できる可能性がある。しかし、各rrnオペロンは上述のように各々異なる機能を担っているため、特定のrrnの改変が細菌の正常な生命活動に影響を及ぼしてその生存率や生理的活動を下げる可能性が考えられた。そのため、SD配列又はアンチSD配列の改変によって細菌の成長や遺伝子発現の制御を満足に行うことができるか否か、あるいは如何なる改変を施せばよいかは不明であった。
【課題を解決するための手段】
【0010】
本発明者らは、遺伝子特異的に発現を制御することのできる微生物株の作製を試みた結果、改変されたSD配列を有する標的遺伝子と、当該改変SD配列に相補的な改変されたアンチSD配列を有するrrnオペロンとに加え、さらに、改変されていないインタクトなアンチSD配列を有するrrnオペロンを有する細菌により、当該標的遺伝子の発現を特異的に制御することが可能であることを見出した。
【0011】
すなわち、本発明は以下を提供する。
(1)改変SD配列を有する標的遺伝子と;当該改変SD配列と相補的な改変アンチSD配列を有する複数個のrrnオペロンと;改変されていないアンチSD配列を有するrrnオペロン、とを有する細菌。
(2)前記改変SD配列が、前記改変されていないアンチSD配列と80%以上の配列同一性を有する(1)記載の細菌。
(3)前記改変SD配列が、CCTCCである(1)又は(2)記載の細菌。
(4)前記改変アンチSD配列が、GGAGGである(1)〜(3)のいずれか1に記載の細菌。
(5)バチルス属細菌である(1)〜(4)のいずれか1に記載の細菌。
(6)前記改変アンチSD配列を有する複数のrrnオペロンが、少なくともrrnIオペロン及びrrnOオペロンである(5)に記載の細菌。
(7)前記改変アンチSD配列を有する複数のrrnオペロンが、rrnIオペロン、rrnOオペロン及びrrnEオペロンである(5)に記載の細菌。
(8)前記改変されていないアンチSD配列を有するrrnオペロンがrrnJオペロンである(6)又は(7)に記載の細菌。
(9)(1)〜(8)のいずれか1に記載の細菌を培養する工程を含む目的遺伝子産物の生産方法。
【発明の効果】
【0012】
本発明によれば、目的遺伝子産物を効率よく生産することができる細菌を提供することができる。また本発明によれば、標的遺伝子の発現を、それ以外の遺伝子の発現に影響を与えることなく制御することができるため、個々の遺伝子やその発現産物の発現や機能の精密な解析を可能にする優れた評価系を提供することができる。
【図面の簡単な説明】
【0013】
【図1】rrnJ+J+株ゲノムにおけるrrnJ遺伝子の概要。
【図2】rrnオペロン改変株の構築手順の概要。
【図3】改変SD配列を有するプラスミドの構築手順の概要。
【発明を実施するための形態】
【0014】
本明細書において、アミノ酸配列及び塩基配列の同一性は、Lipman-Pearson法 (Science, 227, 1435, (1985))によって計算することができる。具体的には、遺伝情報処理ソフトウェアGenetyx-Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0015】
本明細書において、「rrn」又は「rrnオペロン」とは、リボゾームRNA(rRNA)をコードするオペロンを指す。細菌ゲノムに存在するrrnのコピー数は、菌種によって、1コピーのみから10コピー以上まで様々である。各細菌におけるrrnについての情報は、データベース[http://ribosome.mmg.msu.edu/rrndb/]から入手可能である。
枯草菌(Bacillus subtilis)の場合、ゲノム上に10コピーの異なるrrnが存在し、それぞれrrnA、rrnB、rrnD、rrnE、rrnG、rrnH、rrnI、rrnJ、rrnO、rrnWとして知られている。上記10個のrrnの塩基配列及びゲノム上での位置は公知である(Nanamiya et al. Microbiology (2010) 156:2944-2952、あるいはJAFAN(Japan Functional Analysis Network for Bacillus subtilis)により公開されている枯草菌ORFデータベース(BSORF DB)[http://bacillus.genome.ad.jp/])。
上記10個のrrnは、枯草菌等のバチルス属細菌において高度に保存されており、各rrnの配列は、いずれも類似性が高く且つ高度に保存されたアンチSD配列を有する。
【0016】
本明細書において、「SD配列」とは、原核生物のmRNAの開始コドン上流に位置するシャイン・ダルガノ配列(Shine-Dalgarno sequence)、又はこれに対応するゲノム上の配列を指し、文脈によって使い分けられる。mRNAに関して「SD配列」というときは前者を指し、ゲノム上の遺伝子配列に関して「SD配列」というときは後者を指す。
SD配列としては、例えば遺伝子配列上の配列としては、開始コドンのおよそ6〜14bp上流に位置する配列GGAGG等が挙げられる。
【0017】
本明細書において、「アンチSD配列」とは、原核生物の16SrRNAの3’末端付近に位置するアンチ・シャイン・ダルガノ配列(Anti-Shine-Dalgarno sequence)、又はこれに対応するゲノム上の配列を指し、文脈によって使い分けられる。rRNAに関して「アンチSD配列」というときは前者を指し、ゲノム上の遺伝子配列やオペロンに関して「アンチSD配列」というときは後者を指す。
アンチSD配列としては、rRNA上の配列の場合、例えば、16SrRNAの3’末端付近に位置する配列CCUCC等が知られている(McLaughlin et al. (1981) J. Biol. Chem. 256:11283-91、「遺伝子」第8版、B. Lewin 著、東京化学同人、128−130)。したがって、遺伝子配列の場合のアンチSD配列としては、例えば、16SrRNA遺伝子の3’末端付近に位置する配列CCTCC等が挙げられる。
【0018】
本明細書において、SD配列又はアンチSD配列が「改変されていない」又は「インタクト」である場合、上記に挙げたSD配列又はアンチSD配列であり得る。
【0019】
本明細書において、2つの配列が「相補的」である場合、それらの配列は完全に(100%)相補的(互いの塩基の間にミスマッチが全くない)であってもよく、又は2個以下の塩基のミスマッチを含む配列であってもよい。好ましくは、相補的な配列間のミスマッチは1個以下である。さらに好ましくは、相補的な配列は完全に相補的である。
【0020】
本発明の細菌は、改変SD配列を有する標的遺伝子と;当該改変SD配列と相補的な改変アンチSD配列を有する複数個のrrnオペロンと;改変されていないアンチSD配列を有するrrnオペロン、とを有する。
上記改変SD配列は、好ましくは、上記改変されていないアンチSD配列と60%以上の配列同一性を有し、より好ましくは80%以上の配列同一性を有し、さらに好ましくは100%の配列同一性を有する。
したがって、上記改変アンチSD配列は、好ましくは、上記改変されていないアンチSD配列の相補配列と60%以上の配列同一性を有し、より好ましくは80%以上の配列同一性を有し、さらに好ましくは100%の配列同一性を有する。
【0021】
本発明の細菌は、任意の細菌を親株にしてこれを改変することによって得ることができる。親細菌の種及び菌株は特に限定されないが、好ましくはバチルス属細菌が挙げられ、より好ましくは枯草菌(Bacillus subtilis)168株及びその変異株が挙げられる。
【0022】
本発明の細菌がバチルス属細菌である場合、上記改変アンチSD配列を有する複数個のrrnオペロンは、10種のrrnオペロン、すなわちrrnA、rrnB、rrnD、rrnE、rrnG、rrnH、rrnI、rrnJ、rrnO及びrrnWから選ばれるrrnオペロンであればよく、好ましくはrrnIオペロン、rrnOオペロン及びrrnEオペロンから選ばれる。当該複数個のrrnオペロンは、複数個の同じ種類のrrnオペロンであっても、異なる種類のrrnオペロンの組み合わせであってもよい。
当該複数個のrrnオペロンは、好ましくは少なくともrrnIオペロンを含み、より好ましくは少なくともrrnI及びrrnOオペロンを含み、さらに好ましくはrrnI、rrnOオペロン及びrrnEオペロンを含み、なお好ましくはrrnI、rrnOオペロン及びrrnEオペロンからなる。
【0023】
また本発明の細菌がバチルス属細菌である場合、上記改変されていないアンチSD配列を有するrrnオペロンは、上記10種のrrnオペロンのうちのいずれか1以上であればよいが、改変アンチSD配列を有するrrnオペロンとは異なる種類のrrnオペロンであることが好ましく、例えば、rrnJオペロンが好ましい例として挙げられる。また好ましい例として、本発明の細菌は、上記改変されていないアンチSD配列を有するrrnオペロンとしてrrnJオペロンのみを有する。
【0024】
本発明の細菌は、上記改変アンチSD配列を有するrrnオペロン及び改変されていないアンチSD配列を有するrrnオペロンを、各rrnのタイプごとに1コピーずつ有していればよいが、各々1コピー以上有していてもよい。
【0025】
例えば、本発明の細菌は、改変アンチSD配列を有するrrnオペロンとして、好ましくは少なくともrrnI及びrrnOオペロン、より好ましくはrrnI、rrnOオペロン及びrrnEオペロンを有し、且つ記改変されていないアンチSD配列を有するrrnオペロンとしてrrnJオペロンのみを有していればよく、その他のrrnオペロンを有さなくともよい。
また例えば、本発明の細菌は、各1コピーの改変アンチSD配列を有するrrnI及びrrnOオペロンと、2コピーの改変されていないアンチSD配列を有するrrnJオペロンとを有し、他のrrnオペロンを有していない細菌であり得る。
また例えば、本発明の細菌は、各1コピーの改変アンチSD配列を有するrrnI、rrnOオペロン及びrrnEオペロンと、2コピーの改変されていないアンチSD配列を有するrrnJオペロンとを有し、他のrrnオペロンを有していない細菌であり得る。
【0026】
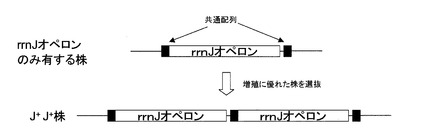
本願実施例では枯草菌168株の10個のrrnオペロンのうちrrnJを除く9個が欠失し、且つ当該rrnJを2個有する株であるrrnJ+J+(ΔrrnHG1 ΔrrnO1 ΔrrnD1 ΔrrnE1 ΔrrnB2 ΔrrnA1 ΔrrnI2 ΔrrnW2)(Nanamiya et al. (2010) Microbiology 156, 2944-2952のSupplementary material中に記載されている。下記参考例2も参照のこと)を元株(RIK530株)として目的の変異株を調製している。rrnJオペロンの5’側先頭付近の配列とrrnJオペロン3’側下流の配列に共通配列(AACAAAATGATCTTTGAAAACTAAACAAGACAAAACGTACCTGTT:配列番号95)が存在する。rrnJ以外の9個のrrnオペロンを欠失した枯草菌168株のゲノムの複製過程における共通配列間での相同組換えにより、rrnJを2個有する株であるrrnJ+J+が出現すると推察される。rrnJ+J+株ゲノムにおけるrrnJ遺伝子の概要を図1に示す。
rrnJ+J+株はrrnJ以外の9個のrrnオペロンを欠失した枯草菌168株において、増殖性が回復した変異体として見出された。rrnオペロンの欠失はrrnオペロン以外の遺伝子の変異では補うことができないため、増殖性はrrnJオペロンの多コピー化でのみ回復されるものと推察される。従って、Nanamiya et al. (2010) Microbiology 156, 2944-2952に記載の方法によりrrnJ以外の9個のrrnオペロンを欠失した枯草菌168株より増殖に優れた株を選抜することにより、rrnJ+J+株を再現良く取得することがでる。
【0027】
本発明の細菌は、細菌内に改変SD配列を有する標的遺伝子と改変アンチSD配列を有する複数個のrrnオペロンと、改変されていないアンチSD配列を有するrrnオペロンとを構築することによって作製することができる。
本発明の細菌において、上記標的遺伝子は、当該細菌が生来有する遺伝子であってもよく、外来遺伝子又は異種遺伝子であってもよい。また本発明の細菌において、上記rrnオペロンは、当該細菌が生来有するオペロンであってもよく、天然に生じた突然変異によって新たに構築されたオペロンであってもよく、又は外部から導入したものでもよい。
したがって、上記改変SD配列及び上記改変アンチSD配列は、当該細菌が生来有するSD配列若しくはアンチSD配列を改変して作製されたものであってもよく、天然に生じた改変SD配列若しくはアンチSD配列であってもよく、又は細胞外から導入された改変されたSD配列若しくはアンチSD配列であってもよい。
【0028】
上記改変SD配列を有する標的遺伝子は、細菌細胞のゲノム内の標的遺伝子のSD配列を改変するか、あるいは改変SD配列を有する標的遺伝子を含むDNA断片を外部から細菌ゲノム内に組み込むか、またあるいは改変SD配列を有する標的遺伝子を含むDNA断片を含有するプラスミドベクター等を細菌内に導入することによって、構築することができる。あるいは、天然に生じた突然変異によって得られた改変SD配列を有する標的遺伝子であってもよい。
【0029】
上記改変アンチSD配列を有するrrnオペロンは、細菌細胞のゲノム内の適当なrrnオペロンのアンチSD配列を改変するか、あるいは改変アンチSD配列を有するrrnオペロンを含むDNA断片を外部から細菌ゲノム内に組み込むか、またあるいは改変アンチSD配列を有するrrnオペロンを含むDNA断片を含有するプラスミドベクター等を細菌内に導入することによって、構築することができる。あるいは、天然に生じた突然変異によって得られた改変アンチSD配列を有するrrnオペロンであってもよい。
【0030】
上記改変されていないアンチSD配列を有するrrnオペロンとしては、細菌が生来有するインタクトなrrnオペロンを利用すればよい。あるいは、改変されていないアンチSD配列を有するrrnオペロンを含むDNA断片を外部から細菌ゲノム内に組み込むか、又は当該DNA断片を含有するプラスミドベクター等を細菌内に導入することによって、構築することができる。またあるいは、天然に生じた突然変異によって得られた改変されていないアンチSD配列を有するrrnオペロンであってもよい。
【0031】
さらに本発明の細菌においては、必要に応じて、細菌が生来有するインタクトなrrnオペロンの一部又は全部が削除されていてもよい。全部を削除する場合は、改変されていないアンチSD配列を有するrrnオペロンを外部から導入するか、又は天然に生じた突然変異によって構築したrrnオペロンを利用することが望ましい。
【0032】
改変SD配列を含むDNA断片、及び改変SD配列を有する標的遺伝子配列を含むDNA断片は、例えば、細菌ゲノムを鋳型とし、改変SD配列を含むプライマーを用いてPCRを行うことによって作製することができる。同様の手順で、改変アンチSD配列を含むDNA断片、及び改変アンチSD配列を有するrrn配列を含むDNA断片を作製することができる。
【0033】
得られたDNA断片を、適当な手順で細菌ゲノム内に組み込むか、あるいは任意の発現ベクターに組み込んだ後当該ベクターを細菌内に導入することにより、細菌内に改変SD配列を有する標的遺伝子や改変アンチSD配列を有するrrnが構築される。
DNA配列の改変、所望の配列を有するDNA断片の構築、DNA断片のベクターへの組み込み、ベクターの細菌細胞への導入、およびDNA断片の細菌ゲノム内への導入の手順は、当業者に周知である。
【0034】
例えば、標的遺伝子又はrrnを含むDNA断片を細菌ゲノム内に組み込むには、相同組換え法を用いることができる。すなわち、組み込み先のゲノム領域の相同配列と導入したい標的遺伝子又はrrnオペロンとを含むDNA断片をPCR等によって作製し、これをプラスミドベクターにクローニングし、得られた組換えプラスミドを細菌内に取り込ませ、プラスミド上の当該相同配列と当該ゲノム領域との間で相同組換えを起こさせることによって、細菌ゲノム上に標的遺伝子又はrrnオペロンを組み込むことができる。
【0035】
同様の手順で、細菌ゲノム上のrrnオペロンを削除することが可能である。すなわち、削除したいrrnの上流及び下流領域のDNA断片をPCR等によって作製し、これらをプラスミドに組み込み、連結させる。得られた組換えプラスミドを細菌内に取り込ませ、当該プラスミド上の領域とゲノム上のrrnの上流及び下流領域との間で相同組換えを起こさせることによって、細菌ゲノム上のrrn領域を削除することができる。
【0036】
同様に相同組換え法を用いて、改変SD配列又はアンチSD配列のみを細菌ゲノム上の標的遺伝子内又はrrnオペロン内の所望の位置に組み込むことができる。
【0037】
本発明はまた、本発明の細菌を培養する工程を含む目的遺伝子産物の生産方法を提供する。当該方法において、本発明の細菌に含まれる改変SD配列を有する標的遺伝子とは、目的遺伝子産物をコードする遺伝子である。目的遺伝子は、細菌が生来有する遺伝子であっても、異種遺伝子であってもよい。当該目的遺伝子としては、タンパク質又はポリペプチドをコードする遺伝子が挙げられる。当該目的遺伝子は、ゲノム上に存在していても細胞内に導入されたプラスミドベクター等のベクター上に存在していてもよいが、好ましくはベクター上にあり、より好ましくは発現ベクター上に存在する。
【0038】
本発明の方法においては、本発明の細菌を、同化性の炭素源、窒素源、その他の必須成分を含む培地に接種し、通常の培養法にて培養する。培養されている細菌内では、改変アンチSD配列を有するrrnによって、改変SD配列を有する目的遺伝子が特異的に翻訳されるため、目的遺伝子産物が効率よく生産される。次いで、当該目的遺伝子産物を培養物から採取・精製すればよい。培養に使用される培地の組成及び培養条件、目的遺伝子産物の採取及び精製等の手順については、使用する細菌、ベクターの種類、目的遺伝子産物の種類等にしたがって、当業者が適宜選択することができる。
【0039】
上記目的遺伝子がコードするタンパク質又はポリペプチドとしては、特に限定されず、洗剤、食品、繊維、飼料、化学品、医療、診断など各種産業用酵素や、生理活性ペプチドなどが含まれる。また、産業用酵素の機能別には、酸化還元酵素(Oxidoreductase)、転移酵素(Transferase)、加水分解酵素(Hydrolase)、脱離酵素(Lyase)、異性化酵素(Isomerase)、合成酵素(Ligase/Synthetase)等が含まれるが、好適にはセルラーゼ、α−アミラーゼ、プロテアーゼ等の加水分解酵素、クロラムフェニコールアセチルトランスフェラーゼ(CAT)等の転移酵素が挙げられる。
【0040】
セルラーゼの具体例としては、多糖加水分解酵素の分類(Bernard H. (1991) Biochem.J. 280:309-316)中でファミリー5に属するセルラーゼが挙げられ、中でも微生物由来、例えば、バチルス属細菌由来のセルラーゼが挙げられる。より具体的な例として、配列番号2及び配列番号4で示されるアミノ酸配列からなる、それぞれバチルス エスピーKSM−S237株(FERM BP−7875)又はバチルス エスピーKSM−64株(FERM BP−2886)由来のアルカリセルラーゼや、当該アミノ酸配列と70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、なお好ましくは98%以上の同一性を有するアミノ酸配列からなるセルラーゼが挙げられる。
【0041】
α−アミラーゼの具体例としては、微生物由来のα−アミラーゼが挙げられ、バチルス属細菌由来の液化型アミラーゼが好ましい。より具体的な例として、配列番号5で示されるアミノ酸配列からなるバチルス エスピーKSM−K38株(FERM BP−6946)由来のアルカリアミラーゼや、当該アミノ酸配列と70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、なお好ましくは98%以上の同一性を有するアミノ酸配列からなるアミラーゼが挙げられる。
【0042】
プロテアーゼの具体例としては、微生物由来、特にバチルス属細菌由来のセリンプロテアーゼや金属プロテアーゼ等が挙げられる。より具体的な例として、配列番号6で示されるアミノ酸配列からなるバチルス クラウジ(Bacillus clausii)KSM−K16株(FERM BP−3376)由来のアルカリプロテアーゼや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるプロテアーゼが挙げられる。
【0043】
クロラムフェニコールアセチルトランスフェラーゼ(CAT)は、アセチル-CoAのアセチル基をクロラムフェニコールの3位に転移する酵素である。具体的には、配列番号7で示されるアミノ酸配列からなるStaphylococcus aureus由来のCATや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有し、クロラムフェニコールアセチルトランスフェラーゼ活性を有するタンパク質が挙げられる。
【0044】
上記タンパク質又はポリペプチドをコードする目的遺伝子は、改変SD配列に加えて、その上流に位置する当該遺伝子の転写、翻訳、分泌に関わる制御領域、即ち、プロモーター及び転写開始点を含む転写開始制御領域、リボソーム結合部位及び開始コドンを含む翻訳開始領域、並びに分泌シグナルペプチド領域から選ばれる1以上の領域と作動可能に連結されていることが望ましい。特に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が連結されていることが好ましく、更に分泌シグナルペプチド領域がバチルス属細菌のセルラーゼ遺伝子由来のものであり、転写開始領域及び翻訳開始領域が当該セルラーゼ遺伝子の開始コドンから始まる長さ0.6〜1kbの上流領域であるものが、目的のタンパク質又はポリペプチド遺伝子と作動可能に連結されていることが望ましい。例えば、特開2000−210081号公報や特開平4−190793号公報等に記載されているバチルス属細菌、すなわちKSM−S237株(FERM BP−7875)、KSM−64株(FERM BP−2886)由来のセルラーゼ遺伝子の転写開始制御領域、翻訳開始領域及び分泌シグナルペプチド領域が目的のタンパク質又はポリペプチドの構造遺伝子と作動可能に連結されていることが望ましい。
【0045】
より具体的には配列番号1で示される塩基配列の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列、また当該塩基配列に対して70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する塩基配列からなるDNA断片、或いは上記いずれかの塩基配列の一部が欠失した塩基配列からなるDNA断片が、改変SD配列に加えて、上記タンパク質又はポリペプチドをコードする目的遺伝子の構造遺伝子と作動可能に連結されていることが望ましい。尚、ここで、上記塩基配列の一部が欠失した塩基配列からなるDNA断片とは、上記塩基配列の一部を欠失しているが、遺伝子の転写、翻訳、分泌に関わる機能を保持しているDNA断片を意味する。
【0046】
本明細書において、プロモーターと遺伝子、又はプロモーターと遺伝子と分泌シグナルとの「作動可能な連結」とは、プロモーターが遺伝子の転写を誘導し得るように、又は遺伝子にコードされたタンパク質が分泌シグナルにより分泌されるように、連結されていることをいう。プロモーターと遺伝子、又はプロモーターと遺伝子と分泌シグナルとの「作動可能な連結」の手順は当業者に周知である。
【実施例】
【0047】
以下、実施例を示し、本発明をより具体的に説明する。
【0048】
実施例1
1.改変rrnを有する変異細菌株の構築
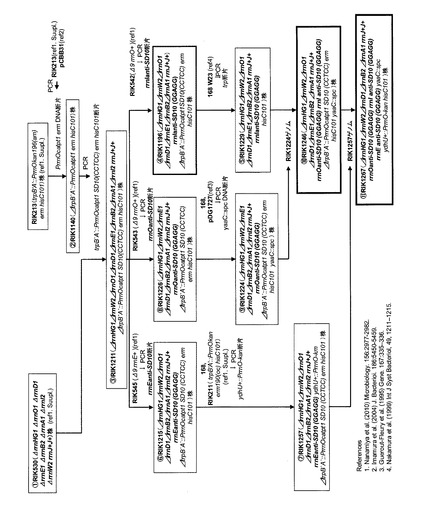
枯草菌168株の10個のrrnオペロンのうちrrnJを除く9個が欠失し、且つ当該rrnJを2個有する株であるrrnJ+J+(ΔrrnHG1 ΔrrnO1 ΔrrnD1 ΔrrnE1 ΔrrnB2 ΔrrnA1 ΔrrnI2 ΔrrnW2)(Nanamiya et al. (2010) Microbiology 156, 2944-2952のSupplementary material中に記載されている。下記参考例2も参照のこと)を元株(RIK530株)として以下の変異株を構築した。変異株構築に用いたプライマーを表1に示し、構築手順の概要を図2に示す。
【0049】
【表1】

【0050】
1)RIK1140(ΔtrpB'A'::PrrnOcatpt1 SD10(CCTCC) erm hisC101)株の構築
RIK213株(trpB'A'::PrrnOkan196(am) erm hisC101)(Nanamiya et al. (2010) Microbiology 156:2944-2952、下記参考例1も参照)のDNAを鋳型としてTRPFプライマーとPROCRプライマー、及びPROCFプライマーとTRPRプライマーをそれぞれ用いてトリプトファン合成遺伝子領域を増幅した。次に、pC194(Horinouchi, S. et al. ( 1982) J. Bacteriol. 150:815-825、Bacillus Genetic Stock Center [http://www.bgsc.org/]より入手可能)を鋳型として、CATFプライマーとCATRプライマーを用いてクロラムフェニコール耐性遺伝子を増幅した。得られた3断片を鋳型としてTRPFプライマーとTRPRプライマーを用いてPrrnOcatpt1 erm DNA断片を作製した。次に、PrrnOcatpt1 erm DNA断片をRIK213株に形質転換し、クロラムフェニコール耐性である形質転換体を選択した。RIK1140株にPrrnOcatpt1 ermが導入されていることをPCRとシークエンシングにより確認した。
【0051】
2)RIK1211(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔrrnI2ΔtrpB'A'::PrrnOcatpt1 SD10(CCTCC) erm hisC101)株の構築
RIK1140株のDNAを鋳型としてTRPFプライマーとCATSDRプライマー、及びCATSDFプライマーとTRPRプライマーを用いてトリプトファン合成遺伝子領域を増幅した。得られた2断片を鋳型としてTRPFプライマーとTRPRプライマーを用いてtrpB'A'::PrrnOcatpt1 SD10(CCTCC) erm hisC101断片を作製した。得られたtrpB'A'::PrrnOcatpt1 SD10(CCTCC) erm hisC101断片をRIK530株に形質転換し、エリスロマイシン耐性である形質転換体を選択した。RIK1211株にtrpB'A'::PrrnOcatpt1 SD10(CCTCC) erm hisC101が導入されていることをPCRとシークエンシングにより確認した。
【0052】
3)RIK1196(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔtrpB'A'::PrrnOcatpt1SD10(CCTCC) erm hisC101 rrnIanti-SD10 (GGAGG))株の構築
rrnI領域内の野生型アンチSD配列CCTCC(rrnIanti-SD)を改変アンチSD配列GGAGG(rrnIanti-SD10)に改変するため、RIK542株(Nanamiya et al. (2010) Microbiology. 156:2977-2982、下記参考例3も参照)のDNAを鋳型としてRRNIFプライマーとRRNISDRプライマー、及びRRNISDFプライマーとRRNIRプライマーをそれぞれ用いてrrnI領域を増幅した。次に、得られた2断片を鋳型としてRRNIFプライマーとRRNIRプライマーを用いて、rrnIanti-SD10を有するDNA断片を作製した。得られたDNA断片をRIK1211株に形質転換し、クロラムフェニコール耐性である形質転換体を選択した。RIK1196株に改変アンチSD(anti-SD10)配列を有するrrnIが導入されていることをシークエンシングにより確認した。
【0053】
4)RIK1229(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1 rrnIanti-SD10 (GGAGG) hisC101)株の構築
枯草菌W168株(Nakamura et al. (1999) Int. J. Syst. Bacteriol. 49:1211-1215、Bacillus Genetic Stock Center [http://www.bgsc.org/]より入手可能)のDNAを鋳型としてTRPF2プライマーとTRPR2プライマーを用いてトリプトファン合成遺伝子領域を増幅した。得られたDNA断片をRIK1196株に形質転換し、トリプトファン非要求性、ヒチジン要求性、および、クロラムフェニコール感受性、エリスロマイシン感受性を示す形質転換体を選択した。RIK1229株に野生型のトリプトファン合成遺伝子が導入されたことをPCRとシークエンシングにより確認した。
【0054】
5)RIK1215(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnB2ΔrrnA1ΔrrnI2 rrnEanti-SD10 (GGAGG)ΔtrpB'A'::PrrnOcatpt1 SD10 (CCTCC) erm hisC101)株の構築
rrnE領域内の野生型アンチSD配列CCTCC(rrnEanti-SD)を改変アンチSD配列GGAGG(rrnEanti-SD10)に改変するため、RIK545株(Nanamiya et al. (2010) Microbiology 156:2977-2982、下記参考例5も参照)のDNAを鋳型としてRRNEFプライマーとRRNESDRプライマー、及びRRNESDFプライマーとRRNERプライマーをそれぞれ用いてrrnE領域を増幅した。次に、得られた2断片を鋳型としてRRNEFプライマーとRRNERプライマーを用いて、rrnEanti-SD10を有するDNA断片を作製した。得られたDNA断片をRIK1211株に形質転換し、クロラムフェニコール耐性である形質転換体を選択した。RIK1215株に改変アンチSD(anti-SD10)配列を有するrrnEが導入されていることをシークエンシングにより確認した。
【0055】
6)RIK1257(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnB2ΔrrnA1ΔrrnI2 rrnEanti-SD10 (GGAGG) ydhU+::PrrnOkanΔtrpB'A'::PrrnOcatpt1 SD10 (CCTCC) erm hisC101)株の構築
ydhU遺伝子の上流領域にrrnOプロモーターを有するカナマイシン耐性遺伝子(PrrnOkan)を導入するため、RIK211株(Nanamiya et al. (2010) Microbiology 156:2977-2982、下記参考例1も参照)を鋳型としてPrrnOkan領域をRRNOPFプライマーとKMRプライマーを用いて増幅した。次に、ydhU遺伝子の上流領域を野生株のDNAを鋳型としてYDHUUPFプライマーとYDHUUPRプライマー、及びYDHUDFプライマーとYDHUDRプライマーをそれぞれ用いて増幅した。得られた2種のDNA断片とPrrnOkan遺伝子を鋳型として、YDHUUPFプライマーとYDHUDRプライマーを用いてydhU遺伝子の上流領域にカナマイシン耐性遺伝子が導入されたDNA断片(ydhU+::PrrnO-kan)を作製した。ydhU+::PrrnO-kan DNA断片をRIK1215株に形質転換し、カナマイシン耐性である形質転換体を選択した。RIK1257株にydhU+::PrrnO-kanが導入されていることをPCRとシークエンシングにより確認した。
【0056】
7)RIK1226(ΔrrnHG1ΔrrnW2ΔrrnE1ΔrrnD1ΔrrnB2ΔrrnA1ΔrrnI2 rrnOanti-SD10 (GGAGG)ΔtrpB'A'::PrrnOcatpt1 SD10 (CCTCC) erm hisC101)株の構築
rrnO領域内の野生型アンチSD配列CCTCC(rrnOanti-SD)を改変アンチSD配列GGAGG(rrnOanti-SD10)に改変するため、RIK543株(Nanamiya et al. (2010) Microbiology 156:2977-2982、下記参考例4も参照)のDNAを鋳型としてRRNOFプライマーとRRNOSDRプライマー、RRNOSDFプライマーとRRNORプライマーをそれぞれ用いてrrnO領域を増幅した。次に、得られた2断片を鋳型としてRRNOFプライマー、RRNORプライマーを用いて、rrnOanti-SD10を有するDNA断片を作製した。得られたDNA断片をRIK1211株に形質転換し、クロラムフェニコール耐性である形質転換体を選択した。RIK1226株に改変アンチSD(anti-SD10)配列を有するrrnOが導入されていることをシークエンシングにより確認した。
【0057】
8)RIK1224(ΔrrnHG1ΔrrnW2ΔrrnE1ΔrrnD1ΔrrnB2ΔrrnA1ΔrrnI2 rrnOanti-SD10 (GGAGG)ΔtrpB'A'::PrrnOcatpt1 SD10 (CCTCC) erm hisC101 yaaC::spc)株の構築
yaaC遺伝子の上流部分をYAAC−F1プライマーとYAAC−R1プライマーを用いて、下流部分をYAAC−F2プライマー、YAAC−R2プライマーを用いて、それぞれ増幅した。次に、プラスミドpDG1727(Guerout-Fleury et al. (1995) Gene. 167:335-336、Bacillus Genetic Stock Center [http://www.bgsc.org/]より入手可能)のスペクチノマイシン耐性遺伝子を鋳型とし、スペクチノマイシン耐性遺伝子をSPC−FプライマーとSPC−Rプライマーを用いて増幅した。得られた3断片を鋳型としてYAAC−F1プライマーとYAAC−R2プライマーを用いて増幅し、yaaC::spc DNA断片を作製した。当該yaaC::spc DNA断片をRIK1226株に形質転換し、スペクチノマイシン耐性である形質転換体を選択した。RIK1224株にyaaC::spc変異が導入されていることをPCRとシークエンシングにより確認した。
【0058】
9)RIK1246(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1 rrnOanti-SD10 (GGAGG) rrnI anti-SD10 (GGAGG)ΔtrpB'A'::PrrnOcatpt1 SD10 (CCTCC) erm hisC101 yaaC::spc)株の構築
RIK1224から抽出した染色体DNAをRIK1229株に形質転換し、スペクチノマイシン耐性である形質転換体を選択した。RIK1246株にrrnO領域が導入されていることをPCRとシークエンシングにより確認した。
【0059】
10)RIK1267(ΔrrnHG1ΔrrnW2ΔrrnD1ΔrrnB2ΔrrnA1 rrnOanti-SD10 (GGAGG) rrnI anti-SD10 (GGAGG)rrnE anti-SD10 (GGAGG) yaaC::spc ydhU+::PrrnO-kan hisC101)株の構築
RIK1257株から抽出した染色体DNAをRIK1246株に形質転換し、カナマイシン耐性である形質転換体を選択した。RIK1267株にrrnE領域が導入されていることをPCRとシークエンシングにより確認した。
【0060】
2.改変SD配列を有する標的遺伝子発現プラスミドpHYS237(pHYS237-SD10)の構築
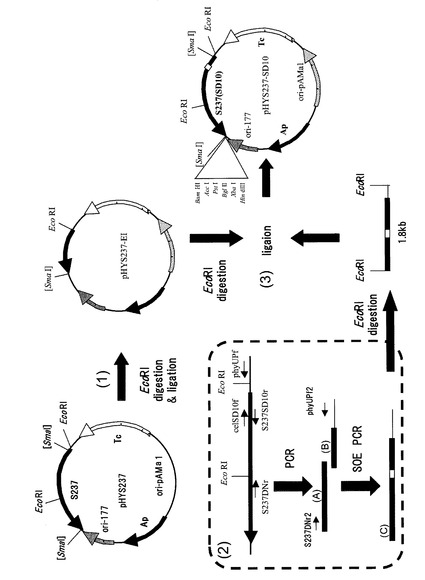
バチルス エスピー(Bacillus sp.)KSM-S237株(FERM BP-7875)由来のアルカリセルラーゼ遺伝子(特開2000−210081号公報)断片(3.1kb)がシャトルベクターpHY300PLKのBamHI制限酵素切断点に挿入された組換えプラスミドpHY-S237のアルカリセルラーゼ遺伝子のSD配列GGAGGを改変SD配列CCTCCに改変したプラスミドpHY237-SD10を、以下のように構築した。プラスミド構築に用いたプライマーを表2に示し、構築手順の概要を図3に示す。
【0061】
【表2】
【0062】
1)pHYS237-EIの構築
バチルス エスピー(Bacillus sp.)KSM−S237株(FERM BP−7875)由来のアルカリセルラーゼ遺伝子(特開2000−210081号公報)の断片(3.1kb)がシャトルベクターpHY300PLKのBamHI制限酵素切断点に挿入された組換えプラスミドであるpHYS237をEcoRI処理した後、アガロースゲル電気泳動を行い、6.2kbのDNA断片をゲルから切り出した。当該DNA断片をプラスミド精製キットHigh Pure PCR Product Purification Kit (Roche)を用いて精製し、この精製したDNAを、LigationHigh(TOYOBO)を用いてライゲーションし、得られたプラスミドベクターをE. coli HB101株に形質転換した。形質転換体から、プラスミドpHYS237-EIを抽出した(図3(1))。
【0063】
2)改変SD配列を含むPCR断片の調製
pHYS237を鋳型にphyUPfとcelSD10fのプライマーセットを用いてPCR断片(A)を調製した。上記DNAを鋳型とし、S237SD10r及びS237DNrのプライマーセットを用いて、断片(B)をPCRにより増幅した。次に、断片(A)と断片(B)を鋳型とし、phyUPf2およびS237DNr2のプライマーセットを用いたPCRにより断片(C)を増幅した(図3(2))。
【0064】
3)pHYS237-SD10の構築
1)で作製したプラスミドpHYS237-EIと2)で作製したDNA断片(C)とを制限酵素EcoRIを用いて消化し、それぞれ線状化したベクター(6.2kb)及びインサート(1.8kb)として精製した。次に、得られた当該ベクターとインサートをLigation High(TOYOBO)を用いて連結した後、得られたベクターを用いてE. coli HB101の形質転換を行った。得られた形質転換体からプラスミドを抽出し、pHYS237-SD10とした(図3(3))。
【0065】
4)組換え細菌株の作製
3)で得られたプラスミドpHYS237-SD10を、プロトプラスト形質転換法によってRIK530株、RIK1196株、RIK1246株、RIK1267株のそれぞれに導入し、組換え細菌株を作製した。
【0066】
試験例1 アルカリセルラーゼ分泌生産評価
上記実施例1で得られたRIK530株、RIK1196株、RIK1246株及びRIK1267株由来の組換え菌株を10mLのLB培地で一夜37℃で振盪培養し、更にこの培養液0.05mLを50mLの2×L−マルトース培地(2%トリプトン、1%酵母エキス、1%NaCl、7.5%マルトース、7.5ppm硫酸マンガン4−5水和物、15ppmテトラサイクリン)に接種し、30℃にて3日間振盪培養を行った。遠心分離によって菌体を除いた培養液上清のアルカリセルラーゼ活性を測定し、培養によって菌体外に分泌生産されたアルカリセルラーゼの量を求めた。
セルラーゼ活性測定については、1/7.5Mリン酸緩衝液(pH7.4、和光純薬)で適宜希釈したサンプル溶液50μLに0.4mM p-nitrophenyl-β-D-cellotrioside(生化学工業)を50μL加えて混和し、30℃にて反応を行った際に遊離するp−ニトロフェノール量を420nmにおける吸光度(OD420nm)変化により定量した。1分間に1μmolのp-ニトロフェノールを遊離させる酵素量を1Uとした。
【0067】
組換え菌株のアルカリセルラーゼの分泌生産能を表3に示す。表3において、セルラーゼの分泌生産能は、RIK1196株にpHYS237-SD10を導入した組換え菌株の生産量を100としたときの相対値で示している。
【0068】
【表3】
【0069】
以下、実施例に用いた菌株の作製方法を参考例として記載する。菌株の作製に使用したプライマーを表4および表5に示す。
【0070】
参考例1 RIK213株(trpB'A'::PrrnOkan erm hisC101)の構築
hisC101は、hisC遺伝子内におけるQ318amber変異を表し、この変異はヒスチジン要求性を示す。またtrpC2変異(トリプトファン要求性)と連鎖しているため、枯草菌の遺伝的マーカーとして使用されている。先ず、trpC2変異とhisC101変異を有し、且つtrpBの+1011位からtrpAの+69位までが欠失した168株のtrpB領域とtrpA領域の間にカナマイシン耐性遺伝子とエリスロマイシン耐性遺伝子をタンデムに導入した。カナマイシン耐性遺伝子はpUB110由来であり、rrnOプロモーターを連結した(PrrnOkan)。エリスロマイシン耐性遺伝子はpMutin由来の遺伝子を用いた。次に、カナマイシン耐性遺伝子にK196amber変異を導入し、カナマイシン感受性であり且つエリスロマイシン耐性を有するRIK213株(trpB'A'::PrrnOkan196(am) erm hisC101)を構築した。また、エリスロマイシン耐性遺伝子内にY190Ocher変異を導入した、カナマイシン耐性、且つエリスロマイシン感受性を示すRIK211(trpB'A'::PrrnOkan erm190(oc) hisC101)株を構築した(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0071】
参考例2 RIK530株の構築
1)rrnHGの欠失
枯草菌168株を鋳型にし、rrnHG領域の上流領域、下流領域をそれぞれrrnHGUFプライマーとrrnHGURプライマー、rrnHGDFプライマーとrrnHGDRプライマーを用いてPCRにて増幅した。得られた断片をpBR322プラスミドに挿入したプラスミド(ptRNA)を調製した。さらにクロラムフェニコール耐性遺伝子(cat)を導入したプラスミドを調製し(ptRNA-Cm)、ptRNA-Cmを用いてゲノム上のrrnHG領域と相同組換えを行なった。次に、ptRNAからrrnHGの上流−下流領域を含むDNA断片を切り出し、カナマイシン耐性遺伝子及び温度感受性変異を有するrepU遺伝子とプラスミド上で連結し、これをゲノム内のptRNA-Cm領域と組換えた。LB培地で30℃で培養し、カナマイシン耐性及びクロラムフェニコール感受性を指標に株を選択し、rrnHG欠失株(ΔrrnHG1)を得た(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0072】
2)rrnOの欠失
rrnO領域の上流領域、下流領域をそれぞれ、rrnOUFプライマーとrrnXURcatプライマー、rrnXDFcatプライマーとrrnODRプライマーを用いて増幅した。次に、CAT−F2プライマー、CAT−R2プライマーを用いてpC194のcat遺伝子を増幅した。得られた3断片をrrnOUFプライマー、rrnODRプライマーを用いてSOE−PCRを行いrrnO領域の上流領域、cat遺伝子、及び下流領域の順に連結されたrrnO::catpt1DNA断片を構築し、枯草菌ゲノムに導入した(ΔrrnO::catpt1)。次に、rrnOを欠失するため、rrnO領域の上流領域、下流領域をそれぞれ、rrnOUFプライマーとrrnXURプライマー、rrnXDFプライマーとrrnODRプライマーにて増幅後、得られた2断片をrrnOUFプライマー、rrnODRプライマーで連結し、ΔrrnO DNA断片を調製した。また、R211株を鋳型として、trpBAUFプライマー及びtrpBADRプライマーを用いてtrpB'A'::PrrnOkan erm190(oc)DNA断片を調製した。得られた、ΔrrnO DNA断片とtrpB'A'::PrrnOkan erm190(oc)DNA断片をΔrrnHG1ΔrrnO::catpt1 trpB'A'::PrrnOkan196(am) erm 株に対して形質転換を行い、ペニシリン濃縮の手法を用いて、カナマイシン耐性を有し、クロラムフェニコール感受性となった株、すなわちcat遺伝子が除かれたΔrrnO株を得た(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0073】
3)rrnD、E、Bの欠失
rrnOと同様にrrnD、E、及びBを欠失した。はじめに、それぞれのrrnにcatpt1変異を導入するため、各rrnの上流、下流領域を以下のプライマーで増幅した。rrnDの上流、下流領域は、rrnDUFプライマーとrrnXURcatプライマー及びrrnXDFcatプライマーとrrnDDRプライマーを、rrnEの上流、下流領域は、rrnEUFプライマーとrrnXURcatプライマー及びrrnXDFcatプライマーとrrnEDRプライマーを、rrnBの上流、下流領域は、rrnBUFプライマーとrrnXURcatプライマー及びrrnXDFcatプライマーとrrnBDRプライマーを用いた。
次に、ΔrrnD1、ΔrrnE1、及びΔrrnB2をそれぞれ有するDNA断片を構築するため、rrnDの上流、下流領域を、rrnDUFプライマーとrrnXURプライマー及びrrnXDFプライマーとrrnDDRプライマー、rrnEの上流、下流領域を、rrnEUFプライマーとrrnXURプライマー及びrrnXDFプライマーとrrnEDRプライマー、rrnBの上流、下流領域を、rrnBUFプライマーとrrnXURプライマー及びrrnXDFプライマーとrrnBDRプライマーを用いて増幅した。得られたDNA断片を用いて、ΔrrnD1、ΔrrnE1、及びΔrrnB2断片を調製した後、rrnOの欠失方法と同様にcatpt1遺伝子を除き、ΔrrnO、ΔrrnD、ΔrrnB2 株を構築した(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0074】
4)rrnAの欠失
rrnAの欠失は、rrnA-16Sの下流に存在するtrnAIleとtrnAAlaを残して構築した。まず、ΔrrnO1::catpt1株と同様にrrnA-16S領域にcatpt1遺伝子を導入した。次に、rrnAプロモーター領域、catpt1遺伝子、tRNA遺伝子を含む領域、及びrrnA−5Sの下流領域を、それぞれrrnAUFプライマーとrrnATRRプライマー、rrnA23SDFプライマーとrrnADRプライマーを用いて増幅した。得られた2種のDNA断片をrrnAUFプライマーとrrnADRプライマーを用いて増幅し、rrnA1::catpt1(trnA-Ile、Ala)DNA断片を構築した。得られたDNA断片をΔrrnHG1、ΔrrnO1、trpB'A'::PrrnOkan196(am) erm hisC101株に形質転換し、クロラムフェニコール耐性である形質転換体を選択した。次に、catpt1遺伝子を除くため、ΔrrnA1::catpt1株のcatpt1遺伝子の上流領域、下流領域、およびrrnA-5Sの下流領域を、それぞれをrrnAUFプライマーとrrnAURプライマー、rrnATRFプライマーとrrnATRRプライマー、rrnA23SDFプライマーとrrnADRプライマーを用いて増幅し、得られた3断片をrrnAUFプライマー、rrnADRプライマーにて連結した。得られたDNA断片をrrnO欠失と同様の手法を用いてcatpt1遺伝子を除いた。ΔrrnA1変異をΔrrnHG1ΔrrnO1ΔrrnD1 ΔrrnE1ΔrrnB2株に導入するため、はじめにΔrrnA1:: catpt1変異を導入し、His+マーカーと同時に形質転換を行なった(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0075】
5)rrnIの欠失
ΔrrnHG1株から抽出したDNAを鋳型にrrnIの上流領域、下流領域をそれぞれrrnIUFプライマーとrrnXURcatプライマー、rrnXDFcatプライマー、rrnXURプライマーを用いて増幅した。ΔrrnO1::catpt1株構築時と同様に、ΔrrnI::catpt1変異を、ΔrrnHG1 ΔrrnO1 ΔrrnD1 ΔrrnE1 trp trpB'A'::PrrnOkan196(am)erm hisC101株に導入した。次に、rrnI欠失株を構築するため、ΔrrnHG1株から抽出したDNAを鋳型に、rrnIUFプライマー、rrnIURプライマー、rrnXDFプライマー、rrnIDRプライマーでDNA断片を増幅し、rrnO欠失株の作成方法と同様にrrnIを欠失した。
ΔrrnHG1ΔrrnO1 ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1株からrrnIを欠失(ΔrrnI2)させるためには、ΔrrnI::catpt1を導入し、その後、ΔrrnI2とTrp+マーカーを同時に形質転換した(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0076】
6)rrnWの欠失
rrnWの上流領域、下流領域をそれぞれrrnWUFプライマーとrrnWURcatプライマー、rrnWDFcatプライマー、rrnWDRプライマーを用いて、また、CAT−F2プライマーとCAT−Rプライマーを用いてcat遺伝子を増幅し、ΔrrnO1::catpt1と同様に、ΔrrnW::catpt1を作製した。次に、rrnWUFプライマー、rrnWURプライマー、rrnWDFプライマー、rrnWDRプライマーを用いて増幅し、rrnO欠失株の作成方法と同様の手法を用いて、rrnWを欠失した(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0077】
7)rrnJ+J+株(ΔrrnHG1ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔrrnI2ΔrrnW2 rrnJ+J+ hisC101)の構築
Δrrn9 rrnJ+株構築の過程で自然突然変異によりrrnJが重複した、rrnJ+J+株が得られ、株をRIK530として取得した(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0078】
参考例3 RIK542株の構築
rrnIオペロンのみを有する株を構築するため、ΔrrnHG1ΔrrnO1ΔrrnE1ΔrrnD1ΔrrnB2ΔrrnA1ΔrrnW2::catpt1 trpB'A'::PrrnOkan196(am) erm and hisC101 株に対し6)の方法と同様にΔrrnW2::catpt1 のcatpt1遺伝子を除去した。次に、trpB'A'::PrrnOkan erm190(oc) 遺伝子を除去した後、ΔrrnJ1::cat、trpC2変異を導入し、RIK542株(ΔrrnHG1 ΔrrnO1ΔrrnE1ΔrrnD1 ΔrrnB2 ΔrrnA1 ΔrrnW2 ΔrrnJ1::cat trpC2)を構築した。(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0079】
参考例4 RIK543株の構築
rrnOオペロンのみを有する株は以下の方法で構築した。はじめに、rrnOプロモーターの下流にcatpt1遺伝子を導入したrrnO2+::catpt1をΔrrnHG1ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔrrnI2ΔrrnW2 rrnJ+J+ hisC101株へ導入した。rrnO2+::catpt1遺伝子断片は、rrnO−catF1プライマーとrrnO−catR1プライマー、rrnO−catF2プライマーとrrnX−catR2プライマー、CAT−F2プライマーとCAT−Rプライマーを用いて増幅し、得られた3断片をrrnO−catF1プライマーとrrnX−catR2プライマーで連結した。次に、rrnOオペロンを有し、かつtrpC2変異を有する染色体DNAを供与DNAに用い、rrnO2+::catpt1 ΔrrnHG1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔrrnI2ΔrrnW2 rrnJ+J+ hisC101株に形質転換を行い、rrnO2+::catpt1とrrnOオペロンを置換した。次に、rrnJオペロンにcat遺伝子を導入するため、rrnJの上流領域、下流領域をそれぞれrrnJUFプライマーとrrnJURcatプライマー、rrnJDFcatプライマーとrrnJDRプライマーを用いて、及び、cat遺伝子をCAT−F2プライマー、CAT−Rプライマーを用いて増幅した。得られた3断片をrrnJUFプライマー、rrnJDRプライマーを用いて連結後、形質転換を行いRIK543株(ΔrrnHG1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔrrnI2ΔrrnW2ΔrrnJ::cat trpC2)を得た(Nanamiya et al. (2010) Microbiology 156:2977-2982、Natori et al. (2009) J. Bacteriol. 191:4555-4561)。
【0080】
参考例5 RIK545株の構築
rrnEオペロンのみを有する株は、参考例4と同様に行なった。rrnE2+::catpt1は、rrnE−catF1プライマーとrrnE−catR1プライマー、rrnE−catF2プライマーとrrnX−catR2プライマーを用いて増幅し、cat遺伝子と連結することで構築した(Nanamiya et al. (2010) Microbiology 156:2977-2982、Natori et al. (2009) J. Bacteriol. 191: 4555-4561)。
【0081】
【表4】
【0082】
【表5】
【技術分野】
【0001】
本発明は、改変リボゾームをコードする改変rRNAオペロンを有する微生物と、当該微生物を用いた目的遺伝子産物の生産方法に関する。
【背景技術】
【0002】
リボゾームは遺伝子からタンパク質への翻訳が行われる場所であり、細菌においてはrRNAオペロン(rrn)によってコードされている。rrnオペロンには、16S、23S、5Sという3つのrRNAサブユニットが順にコードされている。細菌ゲノム中でのrrnオペロンのコピー数は種によって様々であり、ある種のマイコプラズマのようにわずか1コピーのものから、大腸菌のように7コピーのものや、枯草菌のように10コピーのものまで存在する。
【0003】
しかし、この細菌ゲノム中でのrrnのコピー数は、当該細菌の成長速度や生理活動の高さを直接反映しているわけではない。例えば、rrnオペロンのコピー数が増えても成長や生理活動量が増加するとは限らず、他方rrnオペロンを一部削除しても成長や生理活動に大きな影響はない。このことから、細菌の生存にとってゲノム中に複数存在するrrnオペロンが同時に全て機能する必要はなく、むしろ細菌はその時々の生育環境の変化にあわせて複数のrrnオペロンを使い分けていると推定される(非特許文献1)。
上記非特許文献1にはまた、10種類のrrnオペロンのうちのいずれか1種を1コピーのみ有する枯草菌株を作製し、それらの成長速度と胞子形成の頻度を調べた結果、各株間で胞子形成能が大きく異なっていたことを報告している。この知見は、複数のrrnオペロンが異種性(heterogeneity)であり、各々が異なる生育局面で異なる機能を担っているという考え方を支持する。
【0004】
このようなrrnオペロンの異種性や機能を鑑みると、細菌におけるrrnオペロンのコピー数とその成長速度や遺伝子発現能との間に相関性を見出すことはできない。
【0005】
シャイン・ダルガノ配列(Shine-Dalgarno sequence:以下、SD配列)は、原核生物のmRNAの開始コドン上流に認められる共通配列(代表的には、−GGAGG−)であり、これが、rrnオペロンの16SrRNAの配列の3’末端領域に存在する相補的なアンチSD配列(代表的には、−CCUCC−)と対合することによって、mRNAとリボゾームが結合し、翻訳が開始される。
佐藤ら(非特許文献2及び3)は、10コピーのrrnオペロンのうちの1つのアンチSD配列を改変した枯草菌株で、改変したSD配列を有するクロラムフェニコール耐性遺伝子(cat)の翻訳効率を調べた結果、改変株では、翻訳効率の低下したSD配列を有するcat遺伝子を持つにもかかわらず、クロラムフェニコール耐性が向上したことを示した。
【0006】
しかし、翻訳効率を向上させるために具体的にどのようにアンチSD配列を改変すればよいかは明らかではなかった。
さらに、非特許文献4及び5によれば、改変rrnオペロンのコピー数を増やしても翻訳効率は増加しないと考えられており、一方で改変rrnオペロンを増やせば、細菌の通常の蛋白合成能が低下して生命活動に影響を及ぼす可能性があるため、アンチSD配列の改変が必ずしも目的蛋白質の生産性向上につながるかどうかは明らかではなかった。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Nanamiya et al. (2010) Microbiology 156:2977-2982
【非特許文献2】佐藤ら、日本農芸化学会2006年度大会 大会講演要旨集、2006年3月5日発行、119頁
【非特許文献3】七宮ら、2006年度グラム陽性細菌のゲノム生物学研究会 プログラム&要旨集、2006年8月31日、pp16
【非特許文献4】Ellwood & Nomura (1980) J. Bacteriol. 143:1077-1080
【非特許文献5】Widom et al. (1988) J. Bacteriol. 170:605-610
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明は、改変リボゾームをコードする改変rrnオペロンを有する細菌、ならびに当該細菌を用いた標的遺伝子の発現制御方法及び目的遺伝子産物の生産方法に関する。
【0009】
SD配列又はアンチSD配列の改変によって、細菌の成長や遺伝子発現を制御できる可能性がある。しかし、各rrnオペロンは上述のように各々異なる機能を担っているため、特定のrrnの改変が細菌の正常な生命活動に影響を及ぼしてその生存率や生理的活動を下げる可能性が考えられた。そのため、SD配列又はアンチSD配列の改変によって細菌の成長や遺伝子発現の制御を満足に行うことができるか否か、あるいは如何なる改変を施せばよいかは不明であった。
【課題を解決するための手段】
【0010】
本発明者らは、遺伝子特異的に発現を制御することのできる微生物株の作製を試みた結果、改変されたSD配列を有する標的遺伝子と、当該改変SD配列に相補的な改変されたアンチSD配列を有するrrnオペロンとに加え、さらに、改変されていないインタクトなアンチSD配列を有するrrnオペロンを有する細菌により、当該標的遺伝子の発現を特異的に制御することが可能であることを見出した。
【0011】
すなわち、本発明は以下を提供する。
(1)改変SD配列を有する標的遺伝子と;当該改変SD配列と相補的な改変アンチSD配列を有する複数個のrrnオペロンと;改変されていないアンチSD配列を有するrrnオペロン、とを有する細菌。
(2)前記改変SD配列が、前記改変されていないアンチSD配列と80%以上の配列同一性を有する(1)記載の細菌。
(3)前記改変SD配列が、CCTCCである(1)又は(2)記載の細菌。
(4)前記改変アンチSD配列が、GGAGGである(1)〜(3)のいずれか1に記載の細菌。
(5)バチルス属細菌である(1)〜(4)のいずれか1に記載の細菌。
(6)前記改変アンチSD配列を有する複数のrrnオペロンが、少なくともrrnIオペロン及びrrnOオペロンである(5)に記載の細菌。
(7)前記改変アンチSD配列を有する複数のrrnオペロンが、rrnIオペロン、rrnOオペロン及びrrnEオペロンである(5)に記載の細菌。
(8)前記改変されていないアンチSD配列を有するrrnオペロンがrrnJオペロンである(6)又は(7)に記載の細菌。
(9)(1)〜(8)のいずれか1に記載の細菌を培養する工程を含む目的遺伝子産物の生産方法。
【発明の効果】
【0012】
本発明によれば、目的遺伝子産物を効率よく生産することができる細菌を提供することができる。また本発明によれば、標的遺伝子の発現を、それ以外の遺伝子の発現に影響を与えることなく制御することができるため、個々の遺伝子やその発現産物の発現や機能の精密な解析を可能にする優れた評価系を提供することができる。
【図面の簡単な説明】
【0013】
【図1】rrnJ+J+株ゲノムにおけるrrnJ遺伝子の概要。
【図2】rrnオペロン改変株の構築手順の概要。
【図3】改変SD配列を有するプラスミドの構築手順の概要。
【発明を実施するための形態】
【0014】
本明細書において、アミノ酸配列及び塩基配列の同一性は、Lipman-Pearson法 (Science, 227, 1435, (1985))によって計算することができる。具体的には、遺伝情報処理ソフトウェアGenetyx-Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0015】
本明細書において、「rrn」又は「rrnオペロン」とは、リボゾームRNA(rRNA)をコードするオペロンを指す。細菌ゲノムに存在するrrnのコピー数は、菌種によって、1コピーのみから10コピー以上まで様々である。各細菌におけるrrnについての情報は、データベース[http://ribosome.mmg.msu.edu/rrndb/]から入手可能である。
枯草菌(Bacillus subtilis)の場合、ゲノム上に10コピーの異なるrrnが存在し、それぞれrrnA、rrnB、rrnD、rrnE、rrnG、rrnH、rrnI、rrnJ、rrnO、rrnWとして知られている。上記10個のrrnの塩基配列及びゲノム上での位置は公知である(Nanamiya et al. Microbiology (2010) 156:2944-2952、あるいはJAFAN(Japan Functional Analysis Network for Bacillus subtilis)により公開されている枯草菌ORFデータベース(BSORF DB)[http://bacillus.genome.ad.jp/])。
上記10個のrrnは、枯草菌等のバチルス属細菌において高度に保存されており、各rrnの配列は、いずれも類似性が高く且つ高度に保存されたアンチSD配列を有する。
【0016】
本明細書において、「SD配列」とは、原核生物のmRNAの開始コドン上流に位置するシャイン・ダルガノ配列(Shine-Dalgarno sequence)、又はこれに対応するゲノム上の配列を指し、文脈によって使い分けられる。mRNAに関して「SD配列」というときは前者を指し、ゲノム上の遺伝子配列に関して「SD配列」というときは後者を指す。
SD配列としては、例えば遺伝子配列上の配列としては、開始コドンのおよそ6〜14bp上流に位置する配列GGAGG等が挙げられる。
【0017】
本明細書において、「アンチSD配列」とは、原核生物の16SrRNAの3’末端付近に位置するアンチ・シャイン・ダルガノ配列(Anti-Shine-Dalgarno sequence)、又はこれに対応するゲノム上の配列を指し、文脈によって使い分けられる。rRNAに関して「アンチSD配列」というときは前者を指し、ゲノム上の遺伝子配列やオペロンに関して「アンチSD配列」というときは後者を指す。
アンチSD配列としては、rRNA上の配列の場合、例えば、16SrRNAの3’末端付近に位置する配列CCUCC等が知られている(McLaughlin et al. (1981) J. Biol. Chem. 256:11283-91、「遺伝子」第8版、B. Lewin 著、東京化学同人、128−130)。したがって、遺伝子配列の場合のアンチSD配列としては、例えば、16SrRNA遺伝子の3’末端付近に位置する配列CCTCC等が挙げられる。
【0018】
本明細書において、SD配列又はアンチSD配列が「改変されていない」又は「インタクト」である場合、上記に挙げたSD配列又はアンチSD配列であり得る。
【0019】
本明細書において、2つの配列が「相補的」である場合、それらの配列は完全に(100%)相補的(互いの塩基の間にミスマッチが全くない)であってもよく、又は2個以下の塩基のミスマッチを含む配列であってもよい。好ましくは、相補的な配列間のミスマッチは1個以下である。さらに好ましくは、相補的な配列は完全に相補的である。
【0020】
本発明の細菌は、改変SD配列を有する標的遺伝子と;当該改変SD配列と相補的な改変アンチSD配列を有する複数個のrrnオペロンと;改変されていないアンチSD配列を有するrrnオペロン、とを有する。
上記改変SD配列は、好ましくは、上記改変されていないアンチSD配列と60%以上の配列同一性を有し、より好ましくは80%以上の配列同一性を有し、さらに好ましくは100%の配列同一性を有する。
したがって、上記改変アンチSD配列は、好ましくは、上記改変されていないアンチSD配列の相補配列と60%以上の配列同一性を有し、より好ましくは80%以上の配列同一性を有し、さらに好ましくは100%の配列同一性を有する。
【0021】
本発明の細菌は、任意の細菌を親株にしてこれを改変することによって得ることができる。親細菌の種及び菌株は特に限定されないが、好ましくはバチルス属細菌が挙げられ、より好ましくは枯草菌(Bacillus subtilis)168株及びその変異株が挙げられる。
【0022】
本発明の細菌がバチルス属細菌である場合、上記改変アンチSD配列を有する複数個のrrnオペロンは、10種のrrnオペロン、すなわちrrnA、rrnB、rrnD、rrnE、rrnG、rrnH、rrnI、rrnJ、rrnO及びrrnWから選ばれるrrnオペロンであればよく、好ましくはrrnIオペロン、rrnOオペロン及びrrnEオペロンから選ばれる。当該複数個のrrnオペロンは、複数個の同じ種類のrrnオペロンであっても、異なる種類のrrnオペロンの組み合わせであってもよい。
当該複数個のrrnオペロンは、好ましくは少なくともrrnIオペロンを含み、より好ましくは少なくともrrnI及びrrnOオペロンを含み、さらに好ましくはrrnI、rrnOオペロン及びrrnEオペロンを含み、なお好ましくはrrnI、rrnOオペロン及びrrnEオペロンからなる。
【0023】
また本発明の細菌がバチルス属細菌である場合、上記改変されていないアンチSD配列を有するrrnオペロンは、上記10種のrrnオペロンのうちのいずれか1以上であればよいが、改変アンチSD配列を有するrrnオペロンとは異なる種類のrrnオペロンであることが好ましく、例えば、rrnJオペロンが好ましい例として挙げられる。また好ましい例として、本発明の細菌は、上記改変されていないアンチSD配列を有するrrnオペロンとしてrrnJオペロンのみを有する。
【0024】
本発明の細菌は、上記改変アンチSD配列を有するrrnオペロン及び改変されていないアンチSD配列を有するrrnオペロンを、各rrnのタイプごとに1コピーずつ有していればよいが、各々1コピー以上有していてもよい。
【0025】
例えば、本発明の細菌は、改変アンチSD配列を有するrrnオペロンとして、好ましくは少なくともrrnI及びrrnOオペロン、より好ましくはrrnI、rrnOオペロン及びrrnEオペロンを有し、且つ記改変されていないアンチSD配列を有するrrnオペロンとしてrrnJオペロンのみを有していればよく、その他のrrnオペロンを有さなくともよい。
また例えば、本発明の細菌は、各1コピーの改変アンチSD配列を有するrrnI及びrrnOオペロンと、2コピーの改変されていないアンチSD配列を有するrrnJオペロンとを有し、他のrrnオペロンを有していない細菌であり得る。
また例えば、本発明の細菌は、各1コピーの改変アンチSD配列を有するrrnI、rrnOオペロン及びrrnEオペロンと、2コピーの改変されていないアンチSD配列を有するrrnJオペロンとを有し、他のrrnオペロンを有していない細菌であり得る。
【0026】
本願実施例では枯草菌168株の10個のrrnオペロンのうちrrnJを除く9個が欠失し、且つ当該rrnJを2個有する株であるrrnJ+J+(ΔrrnHG1 ΔrrnO1 ΔrrnD1 ΔrrnE1 ΔrrnB2 ΔrrnA1 ΔrrnI2 ΔrrnW2)(Nanamiya et al. (2010) Microbiology 156, 2944-2952のSupplementary material中に記載されている。下記参考例2も参照のこと)を元株(RIK530株)として目的の変異株を調製している。rrnJオペロンの5’側先頭付近の配列とrrnJオペロン3’側下流の配列に共通配列(AACAAAATGATCTTTGAAAACTAAACAAGACAAAACGTACCTGTT:配列番号95)が存在する。rrnJ以外の9個のrrnオペロンを欠失した枯草菌168株のゲノムの複製過程における共通配列間での相同組換えにより、rrnJを2個有する株であるrrnJ+J+が出現すると推察される。rrnJ+J+株ゲノムにおけるrrnJ遺伝子の概要を図1に示す。
rrnJ+J+株はrrnJ以外の9個のrrnオペロンを欠失した枯草菌168株において、増殖性が回復した変異体として見出された。rrnオペロンの欠失はrrnオペロン以外の遺伝子の変異では補うことができないため、増殖性はrrnJオペロンの多コピー化でのみ回復されるものと推察される。従って、Nanamiya et al. (2010) Microbiology 156, 2944-2952に記載の方法によりrrnJ以外の9個のrrnオペロンを欠失した枯草菌168株より増殖に優れた株を選抜することにより、rrnJ+J+株を再現良く取得することがでる。
【0027】
本発明の細菌は、細菌内に改変SD配列を有する標的遺伝子と改変アンチSD配列を有する複数個のrrnオペロンと、改変されていないアンチSD配列を有するrrnオペロンとを構築することによって作製することができる。
本発明の細菌において、上記標的遺伝子は、当該細菌が生来有する遺伝子であってもよく、外来遺伝子又は異種遺伝子であってもよい。また本発明の細菌において、上記rrnオペロンは、当該細菌が生来有するオペロンであってもよく、天然に生じた突然変異によって新たに構築されたオペロンであってもよく、又は外部から導入したものでもよい。
したがって、上記改変SD配列及び上記改変アンチSD配列は、当該細菌が生来有するSD配列若しくはアンチSD配列を改変して作製されたものであってもよく、天然に生じた改変SD配列若しくはアンチSD配列であってもよく、又は細胞外から導入された改変されたSD配列若しくはアンチSD配列であってもよい。
【0028】
上記改変SD配列を有する標的遺伝子は、細菌細胞のゲノム内の標的遺伝子のSD配列を改変するか、あるいは改変SD配列を有する標的遺伝子を含むDNA断片を外部から細菌ゲノム内に組み込むか、またあるいは改変SD配列を有する標的遺伝子を含むDNA断片を含有するプラスミドベクター等を細菌内に導入することによって、構築することができる。あるいは、天然に生じた突然変異によって得られた改変SD配列を有する標的遺伝子であってもよい。
【0029】
上記改変アンチSD配列を有するrrnオペロンは、細菌細胞のゲノム内の適当なrrnオペロンのアンチSD配列を改変するか、あるいは改変アンチSD配列を有するrrnオペロンを含むDNA断片を外部から細菌ゲノム内に組み込むか、またあるいは改変アンチSD配列を有するrrnオペロンを含むDNA断片を含有するプラスミドベクター等を細菌内に導入することによって、構築することができる。あるいは、天然に生じた突然変異によって得られた改変アンチSD配列を有するrrnオペロンであってもよい。
【0030】
上記改変されていないアンチSD配列を有するrrnオペロンとしては、細菌が生来有するインタクトなrrnオペロンを利用すればよい。あるいは、改変されていないアンチSD配列を有するrrnオペロンを含むDNA断片を外部から細菌ゲノム内に組み込むか、又は当該DNA断片を含有するプラスミドベクター等を細菌内に導入することによって、構築することができる。またあるいは、天然に生じた突然変異によって得られた改変されていないアンチSD配列を有するrrnオペロンであってもよい。
【0031】
さらに本発明の細菌においては、必要に応じて、細菌が生来有するインタクトなrrnオペロンの一部又は全部が削除されていてもよい。全部を削除する場合は、改変されていないアンチSD配列を有するrrnオペロンを外部から導入するか、又は天然に生じた突然変異によって構築したrrnオペロンを利用することが望ましい。
【0032】
改変SD配列を含むDNA断片、及び改変SD配列を有する標的遺伝子配列を含むDNA断片は、例えば、細菌ゲノムを鋳型とし、改変SD配列を含むプライマーを用いてPCRを行うことによって作製することができる。同様の手順で、改変アンチSD配列を含むDNA断片、及び改変アンチSD配列を有するrrn配列を含むDNA断片を作製することができる。
【0033】
得られたDNA断片を、適当な手順で細菌ゲノム内に組み込むか、あるいは任意の発現ベクターに組み込んだ後当該ベクターを細菌内に導入することにより、細菌内に改変SD配列を有する標的遺伝子や改変アンチSD配列を有するrrnが構築される。
DNA配列の改変、所望の配列を有するDNA断片の構築、DNA断片のベクターへの組み込み、ベクターの細菌細胞への導入、およびDNA断片の細菌ゲノム内への導入の手順は、当業者に周知である。
【0034】
例えば、標的遺伝子又はrrnを含むDNA断片を細菌ゲノム内に組み込むには、相同組換え法を用いることができる。すなわち、組み込み先のゲノム領域の相同配列と導入したい標的遺伝子又はrrnオペロンとを含むDNA断片をPCR等によって作製し、これをプラスミドベクターにクローニングし、得られた組換えプラスミドを細菌内に取り込ませ、プラスミド上の当該相同配列と当該ゲノム領域との間で相同組換えを起こさせることによって、細菌ゲノム上に標的遺伝子又はrrnオペロンを組み込むことができる。
【0035】
同様の手順で、細菌ゲノム上のrrnオペロンを削除することが可能である。すなわち、削除したいrrnの上流及び下流領域のDNA断片をPCR等によって作製し、これらをプラスミドに組み込み、連結させる。得られた組換えプラスミドを細菌内に取り込ませ、当該プラスミド上の領域とゲノム上のrrnの上流及び下流領域との間で相同組換えを起こさせることによって、細菌ゲノム上のrrn領域を削除することができる。
【0036】
同様に相同組換え法を用いて、改変SD配列又はアンチSD配列のみを細菌ゲノム上の標的遺伝子内又はrrnオペロン内の所望の位置に組み込むことができる。
【0037】
本発明はまた、本発明の細菌を培養する工程を含む目的遺伝子産物の生産方法を提供する。当該方法において、本発明の細菌に含まれる改変SD配列を有する標的遺伝子とは、目的遺伝子産物をコードする遺伝子である。目的遺伝子は、細菌が生来有する遺伝子であっても、異種遺伝子であってもよい。当該目的遺伝子としては、タンパク質又はポリペプチドをコードする遺伝子が挙げられる。当該目的遺伝子は、ゲノム上に存在していても細胞内に導入されたプラスミドベクター等のベクター上に存在していてもよいが、好ましくはベクター上にあり、より好ましくは発現ベクター上に存在する。
【0038】
本発明の方法においては、本発明の細菌を、同化性の炭素源、窒素源、その他の必須成分を含む培地に接種し、通常の培養法にて培養する。培養されている細菌内では、改変アンチSD配列を有するrrnによって、改変SD配列を有する目的遺伝子が特異的に翻訳されるため、目的遺伝子産物が効率よく生産される。次いで、当該目的遺伝子産物を培養物から採取・精製すればよい。培養に使用される培地の組成及び培養条件、目的遺伝子産物の採取及び精製等の手順については、使用する細菌、ベクターの種類、目的遺伝子産物の種類等にしたがって、当業者が適宜選択することができる。
【0039】
上記目的遺伝子がコードするタンパク質又はポリペプチドとしては、特に限定されず、洗剤、食品、繊維、飼料、化学品、医療、診断など各種産業用酵素や、生理活性ペプチドなどが含まれる。また、産業用酵素の機能別には、酸化還元酵素(Oxidoreductase)、転移酵素(Transferase)、加水分解酵素(Hydrolase)、脱離酵素(Lyase)、異性化酵素(Isomerase)、合成酵素(Ligase/Synthetase)等が含まれるが、好適にはセルラーゼ、α−アミラーゼ、プロテアーゼ等の加水分解酵素、クロラムフェニコールアセチルトランスフェラーゼ(CAT)等の転移酵素が挙げられる。
【0040】
セルラーゼの具体例としては、多糖加水分解酵素の分類(Bernard H. (1991) Biochem.J. 280:309-316)中でファミリー5に属するセルラーゼが挙げられ、中でも微生物由来、例えば、バチルス属細菌由来のセルラーゼが挙げられる。より具体的な例として、配列番号2及び配列番号4で示されるアミノ酸配列からなる、それぞれバチルス エスピーKSM−S237株(FERM BP−7875)又はバチルス エスピーKSM−64株(FERM BP−2886)由来のアルカリセルラーゼや、当該アミノ酸配列と70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、なお好ましくは98%以上の同一性を有するアミノ酸配列からなるセルラーゼが挙げられる。
【0041】
α−アミラーゼの具体例としては、微生物由来のα−アミラーゼが挙げられ、バチルス属細菌由来の液化型アミラーゼが好ましい。より具体的な例として、配列番号5で示されるアミノ酸配列からなるバチルス エスピーKSM−K38株(FERM BP−6946)由来のアルカリアミラーゼや、当該アミノ酸配列と70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、なお好ましくは98%以上の同一性を有するアミノ酸配列からなるアミラーゼが挙げられる。
【0042】
プロテアーゼの具体例としては、微生物由来、特にバチルス属細菌由来のセリンプロテアーゼや金属プロテアーゼ等が挙げられる。より具体的な例として、配列番号6で示されるアミノ酸配列からなるバチルス クラウジ(Bacillus clausii)KSM−K16株(FERM BP−3376)由来のアルカリプロテアーゼや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列からなるプロテアーゼが挙げられる。
【0043】
クロラムフェニコールアセチルトランスフェラーゼ(CAT)は、アセチル-CoAのアセチル基をクロラムフェニコールの3位に転移する酵素である。具体的には、配列番号7で示されるアミノ酸配列からなるStaphylococcus aureus由来のCATや、当該アミノ酸配列と70%、好ましくは80%、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有し、クロラムフェニコールアセチルトランスフェラーゼ活性を有するタンパク質が挙げられる。
【0044】
上記タンパク質又はポリペプチドをコードする目的遺伝子は、改変SD配列に加えて、その上流に位置する当該遺伝子の転写、翻訳、分泌に関わる制御領域、即ち、プロモーター及び転写開始点を含む転写開始制御領域、リボソーム結合部位及び開始コドンを含む翻訳開始領域、並びに分泌シグナルペプチド領域から選ばれる1以上の領域と作動可能に連結されていることが望ましい。特に、転写開始制御領域、翻訳開始制御領域及び分泌シグナル領域からなる3領域が連結されていることが好ましく、更に分泌シグナルペプチド領域がバチルス属細菌のセルラーゼ遺伝子由来のものであり、転写開始領域及び翻訳開始領域が当該セルラーゼ遺伝子の開始コドンから始まる長さ0.6〜1kbの上流領域であるものが、目的のタンパク質又はポリペプチド遺伝子と作動可能に連結されていることが望ましい。例えば、特開2000−210081号公報や特開平4−190793号公報等に記載されているバチルス属細菌、すなわちKSM−S237株(FERM BP−7875)、KSM−64株(FERM BP−2886)由来のセルラーゼ遺伝子の転写開始制御領域、翻訳開始領域及び分泌シグナルペプチド領域が目的のタンパク質又はポリペプチドの構造遺伝子と作動可能に連結されていることが望ましい。
【0045】
より具体的には配列番号1で示される塩基配列の塩基番号1〜659の塩基配列、配列番号3で示される塩基配列からなるセルラーゼ遺伝子の塩基番号1〜696の塩基配列、また当該塩基配列に対して70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上の同一性を有する塩基配列からなるDNA断片、或いは上記いずれかの塩基配列の一部が欠失した塩基配列からなるDNA断片が、改変SD配列に加えて、上記タンパク質又はポリペプチドをコードする目的遺伝子の構造遺伝子と作動可能に連結されていることが望ましい。尚、ここで、上記塩基配列の一部が欠失した塩基配列からなるDNA断片とは、上記塩基配列の一部を欠失しているが、遺伝子の転写、翻訳、分泌に関わる機能を保持しているDNA断片を意味する。
【0046】
本明細書において、プロモーターと遺伝子、又はプロモーターと遺伝子と分泌シグナルとの「作動可能な連結」とは、プロモーターが遺伝子の転写を誘導し得るように、又は遺伝子にコードされたタンパク質が分泌シグナルにより分泌されるように、連結されていることをいう。プロモーターと遺伝子、又はプロモーターと遺伝子と分泌シグナルとの「作動可能な連結」の手順は当業者に周知である。
【実施例】
【0047】
以下、実施例を示し、本発明をより具体的に説明する。
【0048】
実施例1
1.改変rrnを有する変異細菌株の構築
枯草菌168株の10個のrrnオペロンのうちrrnJを除く9個が欠失し、且つ当該rrnJを2個有する株であるrrnJ+J+(ΔrrnHG1 ΔrrnO1 ΔrrnD1 ΔrrnE1 ΔrrnB2 ΔrrnA1 ΔrrnI2 ΔrrnW2)(Nanamiya et al. (2010) Microbiology 156, 2944-2952のSupplementary material中に記載されている。下記参考例2も参照のこと)を元株(RIK530株)として以下の変異株を構築した。変異株構築に用いたプライマーを表1に示し、構築手順の概要を図2に示す。
【0049】
【表1】
【0050】
1)RIK1140(ΔtrpB'A'::PrrnOcatpt1 SD10(CCTCC) erm hisC101)株の構築
RIK213株(trpB'A'::PrrnOkan196(am) erm hisC101)(Nanamiya et al. (2010) Microbiology 156:2944-2952、下記参考例1も参照)のDNAを鋳型としてTRPFプライマーとPROCRプライマー、及びPROCFプライマーとTRPRプライマーをそれぞれ用いてトリプトファン合成遺伝子領域を増幅した。次に、pC194(Horinouchi, S. et al. ( 1982) J. Bacteriol. 150:815-825、Bacillus Genetic Stock Center [http://www.bgsc.org/]より入手可能)を鋳型として、CATFプライマーとCATRプライマーを用いてクロラムフェニコール耐性遺伝子を増幅した。得られた3断片を鋳型としてTRPFプライマーとTRPRプライマーを用いてPrrnOcatpt1 erm DNA断片を作製した。次に、PrrnOcatpt1 erm DNA断片をRIK213株に形質転換し、クロラムフェニコール耐性である形質転換体を選択した。RIK1140株にPrrnOcatpt1 ermが導入されていることをPCRとシークエンシングにより確認した。
【0051】
2)RIK1211(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔrrnI2ΔtrpB'A'::PrrnOcatpt1 SD10(CCTCC) erm hisC101)株の構築
RIK1140株のDNAを鋳型としてTRPFプライマーとCATSDRプライマー、及びCATSDFプライマーとTRPRプライマーを用いてトリプトファン合成遺伝子領域を増幅した。得られた2断片を鋳型としてTRPFプライマーとTRPRプライマーを用いてtrpB'A'::PrrnOcatpt1 SD10(CCTCC) erm hisC101断片を作製した。得られたtrpB'A'::PrrnOcatpt1 SD10(CCTCC) erm hisC101断片をRIK530株に形質転換し、エリスロマイシン耐性である形質転換体を選択した。RIK1211株にtrpB'A'::PrrnOcatpt1 SD10(CCTCC) erm hisC101が導入されていることをPCRとシークエンシングにより確認した。
【0052】
3)RIK1196(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔtrpB'A'::PrrnOcatpt1SD10(CCTCC) erm hisC101 rrnIanti-SD10 (GGAGG))株の構築
rrnI領域内の野生型アンチSD配列CCTCC(rrnIanti-SD)を改変アンチSD配列GGAGG(rrnIanti-SD10)に改変するため、RIK542株(Nanamiya et al. (2010) Microbiology. 156:2977-2982、下記参考例3も参照)のDNAを鋳型としてRRNIFプライマーとRRNISDRプライマー、及びRRNISDFプライマーとRRNIRプライマーをそれぞれ用いてrrnI領域を増幅した。次に、得られた2断片を鋳型としてRRNIFプライマーとRRNIRプライマーを用いて、rrnIanti-SD10を有するDNA断片を作製した。得られたDNA断片をRIK1211株に形質転換し、クロラムフェニコール耐性である形質転換体を選択した。RIK1196株に改変アンチSD(anti-SD10)配列を有するrrnIが導入されていることをシークエンシングにより確認した。
【0053】
4)RIK1229(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1 rrnIanti-SD10 (GGAGG) hisC101)株の構築
枯草菌W168株(Nakamura et al. (1999) Int. J. Syst. Bacteriol. 49:1211-1215、Bacillus Genetic Stock Center [http://www.bgsc.org/]より入手可能)のDNAを鋳型としてTRPF2プライマーとTRPR2プライマーを用いてトリプトファン合成遺伝子領域を増幅した。得られたDNA断片をRIK1196株に形質転換し、トリプトファン非要求性、ヒチジン要求性、および、クロラムフェニコール感受性、エリスロマイシン感受性を示す形質転換体を選択した。RIK1229株に野生型のトリプトファン合成遺伝子が導入されたことをPCRとシークエンシングにより確認した。
【0054】
5)RIK1215(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnB2ΔrrnA1ΔrrnI2 rrnEanti-SD10 (GGAGG)ΔtrpB'A'::PrrnOcatpt1 SD10 (CCTCC) erm hisC101)株の構築
rrnE領域内の野生型アンチSD配列CCTCC(rrnEanti-SD)を改変アンチSD配列GGAGG(rrnEanti-SD10)に改変するため、RIK545株(Nanamiya et al. (2010) Microbiology 156:2977-2982、下記参考例5も参照)のDNAを鋳型としてRRNEFプライマーとRRNESDRプライマー、及びRRNESDFプライマーとRRNERプライマーをそれぞれ用いてrrnE領域を増幅した。次に、得られた2断片を鋳型としてRRNEFプライマーとRRNERプライマーを用いて、rrnEanti-SD10を有するDNA断片を作製した。得られたDNA断片をRIK1211株に形質転換し、クロラムフェニコール耐性である形質転換体を選択した。RIK1215株に改変アンチSD(anti-SD10)配列を有するrrnEが導入されていることをシークエンシングにより確認した。
【0055】
6)RIK1257(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnB2ΔrrnA1ΔrrnI2 rrnEanti-SD10 (GGAGG) ydhU+::PrrnOkanΔtrpB'A'::PrrnOcatpt1 SD10 (CCTCC) erm hisC101)株の構築
ydhU遺伝子の上流領域にrrnOプロモーターを有するカナマイシン耐性遺伝子(PrrnOkan)を導入するため、RIK211株(Nanamiya et al. (2010) Microbiology 156:2977-2982、下記参考例1も参照)を鋳型としてPrrnOkan領域をRRNOPFプライマーとKMRプライマーを用いて増幅した。次に、ydhU遺伝子の上流領域を野生株のDNAを鋳型としてYDHUUPFプライマーとYDHUUPRプライマー、及びYDHUDFプライマーとYDHUDRプライマーをそれぞれ用いて増幅した。得られた2種のDNA断片とPrrnOkan遺伝子を鋳型として、YDHUUPFプライマーとYDHUDRプライマーを用いてydhU遺伝子の上流領域にカナマイシン耐性遺伝子が導入されたDNA断片(ydhU+::PrrnO-kan)を作製した。ydhU+::PrrnO-kan DNA断片をRIK1215株に形質転換し、カナマイシン耐性である形質転換体を選択した。RIK1257株にydhU+::PrrnO-kanが導入されていることをPCRとシークエンシングにより確認した。
【0056】
7)RIK1226(ΔrrnHG1ΔrrnW2ΔrrnE1ΔrrnD1ΔrrnB2ΔrrnA1ΔrrnI2 rrnOanti-SD10 (GGAGG)ΔtrpB'A'::PrrnOcatpt1 SD10 (CCTCC) erm hisC101)株の構築
rrnO領域内の野生型アンチSD配列CCTCC(rrnOanti-SD)を改変アンチSD配列GGAGG(rrnOanti-SD10)に改変するため、RIK543株(Nanamiya et al. (2010) Microbiology 156:2977-2982、下記参考例4も参照)のDNAを鋳型としてRRNOFプライマーとRRNOSDRプライマー、RRNOSDFプライマーとRRNORプライマーをそれぞれ用いてrrnO領域を増幅した。次に、得られた2断片を鋳型としてRRNOFプライマー、RRNORプライマーを用いて、rrnOanti-SD10を有するDNA断片を作製した。得られたDNA断片をRIK1211株に形質転換し、クロラムフェニコール耐性である形質転換体を選択した。RIK1226株に改変アンチSD(anti-SD10)配列を有するrrnOが導入されていることをシークエンシングにより確認した。
【0057】
8)RIK1224(ΔrrnHG1ΔrrnW2ΔrrnE1ΔrrnD1ΔrrnB2ΔrrnA1ΔrrnI2 rrnOanti-SD10 (GGAGG)ΔtrpB'A'::PrrnOcatpt1 SD10 (CCTCC) erm hisC101 yaaC::spc)株の構築
yaaC遺伝子の上流部分をYAAC−F1プライマーとYAAC−R1プライマーを用いて、下流部分をYAAC−F2プライマー、YAAC−R2プライマーを用いて、それぞれ増幅した。次に、プラスミドpDG1727(Guerout-Fleury et al. (1995) Gene. 167:335-336、Bacillus Genetic Stock Center [http://www.bgsc.org/]より入手可能)のスペクチノマイシン耐性遺伝子を鋳型とし、スペクチノマイシン耐性遺伝子をSPC−FプライマーとSPC−Rプライマーを用いて増幅した。得られた3断片を鋳型としてYAAC−F1プライマーとYAAC−R2プライマーを用いて増幅し、yaaC::spc DNA断片を作製した。当該yaaC::spc DNA断片をRIK1226株に形質転換し、スペクチノマイシン耐性である形質転換体を選択した。RIK1224株にyaaC::spc変異が導入されていることをPCRとシークエンシングにより確認した。
【0058】
9)RIK1246(ΔrrnHG1ΔrrnW2ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1 rrnOanti-SD10 (GGAGG) rrnI anti-SD10 (GGAGG)ΔtrpB'A'::PrrnOcatpt1 SD10 (CCTCC) erm hisC101 yaaC::spc)株の構築
RIK1224から抽出した染色体DNAをRIK1229株に形質転換し、スペクチノマイシン耐性である形質転換体を選択した。RIK1246株にrrnO領域が導入されていることをPCRとシークエンシングにより確認した。
【0059】
10)RIK1267(ΔrrnHG1ΔrrnW2ΔrrnD1ΔrrnB2ΔrrnA1 rrnOanti-SD10 (GGAGG) rrnI anti-SD10 (GGAGG)rrnE anti-SD10 (GGAGG) yaaC::spc ydhU+::PrrnO-kan hisC101)株の構築
RIK1257株から抽出した染色体DNAをRIK1246株に形質転換し、カナマイシン耐性である形質転換体を選択した。RIK1267株にrrnE領域が導入されていることをPCRとシークエンシングにより確認した。
【0060】
2.改変SD配列を有する標的遺伝子発現プラスミドpHYS237(pHYS237-SD10)の構築
バチルス エスピー(Bacillus sp.)KSM-S237株(FERM BP-7875)由来のアルカリセルラーゼ遺伝子(特開2000−210081号公報)断片(3.1kb)がシャトルベクターpHY300PLKのBamHI制限酵素切断点に挿入された組換えプラスミドpHY-S237のアルカリセルラーゼ遺伝子のSD配列GGAGGを改変SD配列CCTCCに改変したプラスミドpHY237-SD10を、以下のように構築した。プラスミド構築に用いたプライマーを表2に示し、構築手順の概要を図3に示す。
【0061】
【表2】
【0062】
1)pHYS237-EIの構築
バチルス エスピー(Bacillus sp.)KSM−S237株(FERM BP−7875)由来のアルカリセルラーゼ遺伝子(特開2000−210081号公報)の断片(3.1kb)がシャトルベクターpHY300PLKのBamHI制限酵素切断点に挿入された組換えプラスミドであるpHYS237をEcoRI処理した後、アガロースゲル電気泳動を行い、6.2kbのDNA断片をゲルから切り出した。当該DNA断片をプラスミド精製キットHigh Pure PCR Product Purification Kit (Roche)を用いて精製し、この精製したDNAを、LigationHigh(TOYOBO)を用いてライゲーションし、得られたプラスミドベクターをE. coli HB101株に形質転換した。形質転換体から、プラスミドpHYS237-EIを抽出した(図3(1))。
【0063】
2)改変SD配列を含むPCR断片の調製
pHYS237を鋳型にphyUPfとcelSD10fのプライマーセットを用いてPCR断片(A)を調製した。上記DNAを鋳型とし、S237SD10r及びS237DNrのプライマーセットを用いて、断片(B)をPCRにより増幅した。次に、断片(A)と断片(B)を鋳型とし、phyUPf2およびS237DNr2のプライマーセットを用いたPCRにより断片(C)を増幅した(図3(2))。
【0064】
3)pHYS237-SD10の構築
1)で作製したプラスミドpHYS237-EIと2)で作製したDNA断片(C)とを制限酵素EcoRIを用いて消化し、それぞれ線状化したベクター(6.2kb)及びインサート(1.8kb)として精製した。次に、得られた当該ベクターとインサートをLigation High(TOYOBO)を用いて連結した後、得られたベクターを用いてE. coli HB101の形質転換を行った。得られた形質転換体からプラスミドを抽出し、pHYS237-SD10とした(図3(3))。
【0065】
4)組換え細菌株の作製
3)で得られたプラスミドpHYS237-SD10を、プロトプラスト形質転換法によってRIK530株、RIK1196株、RIK1246株、RIK1267株のそれぞれに導入し、組換え細菌株を作製した。
【0066】
試験例1 アルカリセルラーゼ分泌生産評価
上記実施例1で得られたRIK530株、RIK1196株、RIK1246株及びRIK1267株由来の組換え菌株を10mLのLB培地で一夜37℃で振盪培養し、更にこの培養液0.05mLを50mLの2×L−マルトース培地(2%トリプトン、1%酵母エキス、1%NaCl、7.5%マルトース、7.5ppm硫酸マンガン4−5水和物、15ppmテトラサイクリン)に接種し、30℃にて3日間振盪培養を行った。遠心分離によって菌体を除いた培養液上清のアルカリセルラーゼ活性を測定し、培養によって菌体外に分泌生産されたアルカリセルラーゼの量を求めた。
セルラーゼ活性測定については、1/7.5Mリン酸緩衝液(pH7.4、和光純薬)で適宜希釈したサンプル溶液50μLに0.4mM p-nitrophenyl-β-D-cellotrioside(生化学工業)を50μL加えて混和し、30℃にて反応を行った際に遊離するp−ニトロフェノール量を420nmにおける吸光度(OD420nm)変化により定量した。1分間に1μmolのp-ニトロフェノールを遊離させる酵素量を1Uとした。
【0067】
組換え菌株のアルカリセルラーゼの分泌生産能を表3に示す。表3において、セルラーゼの分泌生産能は、RIK1196株にpHYS237-SD10を導入した組換え菌株の生産量を100としたときの相対値で示している。
【0068】
【表3】
【0069】
以下、実施例に用いた菌株の作製方法を参考例として記載する。菌株の作製に使用したプライマーを表4および表5に示す。
【0070】
参考例1 RIK213株(trpB'A'::PrrnOkan erm hisC101)の構築
hisC101は、hisC遺伝子内におけるQ318amber変異を表し、この変異はヒスチジン要求性を示す。またtrpC2変異(トリプトファン要求性)と連鎖しているため、枯草菌の遺伝的マーカーとして使用されている。先ず、trpC2変異とhisC101変異を有し、且つtrpBの+1011位からtrpAの+69位までが欠失した168株のtrpB領域とtrpA領域の間にカナマイシン耐性遺伝子とエリスロマイシン耐性遺伝子をタンデムに導入した。カナマイシン耐性遺伝子はpUB110由来であり、rrnOプロモーターを連結した(PrrnOkan)。エリスロマイシン耐性遺伝子はpMutin由来の遺伝子を用いた。次に、カナマイシン耐性遺伝子にK196amber変異を導入し、カナマイシン感受性であり且つエリスロマイシン耐性を有するRIK213株(trpB'A'::PrrnOkan196(am) erm hisC101)を構築した。また、エリスロマイシン耐性遺伝子内にY190Ocher変異を導入した、カナマイシン耐性、且つエリスロマイシン感受性を示すRIK211(trpB'A'::PrrnOkan erm190(oc) hisC101)株を構築した(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0071】
参考例2 RIK530株の構築
1)rrnHGの欠失
枯草菌168株を鋳型にし、rrnHG領域の上流領域、下流領域をそれぞれrrnHGUFプライマーとrrnHGURプライマー、rrnHGDFプライマーとrrnHGDRプライマーを用いてPCRにて増幅した。得られた断片をpBR322プラスミドに挿入したプラスミド(ptRNA)を調製した。さらにクロラムフェニコール耐性遺伝子(cat)を導入したプラスミドを調製し(ptRNA-Cm)、ptRNA-Cmを用いてゲノム上のrrnHG領域と相同組換えを行なった。次に、ptRNAからrrnHGの上流−下流領域を含むDNA断片を切り出し、カナマイシン耐性遺伝子及び温度感受性変異を有するrepU遺伝子とプラスミド上で連結し、これをゲノム内のptRNA-Cm領域と組換えた。LB培地で30℃で培養し、カナマイシン耐性及びクロラムフェニコール感受性を指標に株を選択し、rrnHG欠失株(ΔrrnHG1)を得た(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0072】
2)rrnOの欠失
rrnO領域の上流領域、下流領域をそれぞれ、rrnOUFプライマーとrrnXURcatプライマー、rrnXDFcatプライマーとrrnODRプライマーを用いて増幅した。次に、CAT−F2プライマー、CAT−R2プライマーを用いてpC194のcat遺伝子を増幅した。得られた3断片をrrnOUFプライマー、rrnODRプライマーを用いてSOE−PCRを行いrrnO領域の上流領域、cat遺伝子、及び下流領域の順に連結されたrrnO::catpt1DNA断片を構築し、枯草菌ゲノムに導入した(ΔrrnO::catpt1)。次に、rrnOを欠失するため、rrnO領域の上流領域、下流領域をそれぞれ、rrnOUFプライマーとrrnXURプライマー、rrnXDFプライマーとrrnODRプライマーにて増幅後、得られた2断片をrrnOUFプライマー、rrnODRプライマーで連結し、ΔrrnO DNA断片を調製した。また、R211株を鋳型として、trpBAUFプライマー及びtrpBADRプライマーを用いてtrpB'A'::PrrnOkan erm190(oc)DNA断片を調製した。得られた、ΔrrnO DNA断片とtrpB'A'::PrrnOkan erm190(oc)DNA断片をΔrrnHG1ΔrrnO::catpt1 trpB'A'::PrrnOkan196(am) erm 株に対して形質転換を行い、ペニシリン濃縮の手法を用いて、カナマイシン耐性を有し、クロラムフェニコール感受性となった株、すなわちcat遺伝子が除かれたΔrrnO株を得た(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0073】
3)rrnD、E、Bの欠失
rrnOと同様にrrnD、E、及びBを欠失した。はじめに、それぞれのrrnにcatpt1変異を導入するため、各rrnの上流、下流領域を以下のプライマーで増幅した。rrnDの上流、下流領域は、rrnDUFプライマーとrrnXURcatプライマー及びrrnXDFcatプライマーとrrnDDRプライマーを、rrnEの上流、下流領域は、rrnEUFプライマーとrrnXURcatプライマー及びrrnXDFcatプライマーとrrnEDRプライマーを、rrnBの上流、下流領域は、rrnBUFプライマーとrrnXURcatプライマー及びrrnXDFcatプライマーとrrnBDRプライマーを用いた。
次に、ΔrrnD1、ΔrrnE1、及びΔrrnB2をそれぞれ有するDNA断片を構築するため、rrnDの上流、下流領域を、rrnDUFプライマーとrrnXURプライマー及びrrnXDFプライマーとrrnDDRプライマー、rrnEの上流、下流領域を、rrnEUFプライマーとrrnXURプライマー及びrrnXDFプライマーとrrnEDRプライマー、rrnBの上流、下流領域を、rrnBUFプライマーとrrnXURプライマー及びrrnXDFプライマーとrrnBDRプライマーを用いて増幅した。得られたDNA断片を用いて、ΔrrnD1、ΔrrnE1、及びΔrrnB2断片を調製した後、rrnOの欠失方法と同様にcatpt1遺伝子を除き、ΔrrnO、ΔrrnD、ΔrrnB2 株を構築した(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0074】
4)rrnAの欠失
rrnAの欠失は、rrnA-16Sの下流に存在するtrnAIleとtrnAAlaを残して構築した。まず、ΔrrnO1::catpt1株と同様にrrnA-16S領域にcatpt1遺伝子を導入した。次に、rrnAプロモーター領域、catpt1遺伝子、tRNA遺伝子を含む領域、及びrrnA−5Sの下流領域を、それぞれrrnAUFプライマーとrrnATRRプライマー、rrnA23SDFプライマーとrrnADRプライマーを用いて増幅した。得られた2種のDNA断片をrrnAUFプライマーとrrnADRプライマーを用いて増幅し、rrnA1::catpt1(trnA-Ile、Ala)DNA断片を構築した。得られたDNA断片をΔrrnHG1、ΔrrnO1、trpB'A'::PrrnOkan196(am) erm hisC101株に形質転換し、クロラムフェニコール耐性である形質転換体を選択した。次に、catpt1遺伝子を除くため、ΔrrnA1::catpt1株のcatpt1遺伝子の上流領域、下流領域、およびrrnA-5Sの下流領域を、それぞれをrrnAUFプライマーとrrnAURプライマー、rrnATRFプライマーとrrnATRRプライマー、rrnA23SDFプライマーとrrnADRプライマーを用いて増幅し、得られた3断片をrrnAUFプライマー、rrnADRプライマーにて連結した。得られたDNA断片をrrnO欠失と同様の手法を用いてcatpt1遺伝子を除いた。ΔrrnA1変異をΔrrnHG1ΔrrnO1ΔrrnD1 ΔrrnE1ΔrrnB2株に導入するため、はじめにΔrrnA1:: catpt1変異を導入し、His+マーカーと同時に形質転換を行なった(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0075】
5)rrnIの欠失
ΔrrnHG1株から抽出したDNAを鋳型にrrnIの上流領域、下流領域をそれぞれrrnIUFプライマーとrrnXURcatプライマー、rrnXDFcatプライマー、rrnXURプライマーを用いて増幅した。ΔrrnO1::catpt1株構築時と同様に、ΔrrnI::catpt1変異を、ΔrrnHG1 ΔrrnO1 ΔrrnD1 ΔrrnE1 trp trpB'A'::PrrnOkan196(am)erm hisC101株に導入した。次に、rrnI欠失株を構築するため、ΔrrnHG1株から抽出したDNAを鋳型に、rrnIUFプライマー、rrnIURプライマー、rrnXDFプライマー、rrnIDRプライマーでDNA断片を増幅し、rrnO欠失株の作成方法と同様にrrnIを欠失した。
ΔrrnHG1ΔrrnO1 ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1株からrrnIを欠失(ΔrrnI2)させるためには、ΔrrnI::catpt1を導入し、その後、ΔrrnI2とTrp+マーカーを同時に形質転換した(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0076】
6)rrnWの欠失
rrnWの上流領域、下流領域をそれぞれrrnWUFプライマーとrrnWURcatプライマー、rrnWDFcatプライマー、rrnWDRプライマーを用いて、また、CAT−F2プライマーとCAT−Rプライマーを用いてcat遺伝子を増幅し、ΔrrnO1::catpt1と同様に、ΔrrnW::catpt1を作製した。次に、rrnWUFプライマー、rrnWURプライマー、rrnWDFプライマー、rrnWDRプライマーを用いて増幅し、rrnO欠失株の作成方法と同様の手法を用いて、rrnWを欠失した(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0077】
7)rrnJ+J+株(ΔrrnHG1ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔrrnI2ΔrrnW2 rrnJ+J+ hisC101)の構築
Δrrn9 rrnJ+株構築の過程で自然突然変異によりrrnJが重複した、rrnJ+J+株が得られ、株をRIK530として取得した(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0078】
参考例3 RIK542株の構築
rrnIオペロンのみを有する株を構築するため、ΔrrnHG1ΔrrnO1ΔrrnE1ΔrrnD1ΔrrnB2ΔrrnA1ΔrrnW2::catpt1 trpB'A'::PrrnOkan196(am) erm and hisC101 株に対し6)の方法と同様にΔrrnW2::catpt1 のcatpt1遺伝子を除去した。次に、trpB'A'::PrrnOkan erm190(oc) 遺伝子を除去した後、ΔrrnJ1::cat、trpC2変異を導入し、RIK542株(ΔrrnHG1 ΔrrnO1ΔrrnE1ΔrrnD1 ΔrrnB2 ΔrrnA1 ΔrrnW2 ΔrrnJ1::cat trpC2)を構築した。(Nanamiya et al. (2010) Microbiology 156:2977-2982)。
【0079】
参考例4 RIK543株の構築
rrnOオペロンのみを有する株は以下の方法で構築した。はじめに、rrnOプロモーターの下流にcatpt1遺伝子を導入したrrnO2+::catpt1をΔrrnHG1ΔrrnO1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔrrnI2ΔrrnW2 rrnJ+J+ hisC101株へ導入した。rrnO2+::catpt1遺伝子断片は、rrnO−catF1プライマーとrrnO−catR1プライマー、rrnO−catF2プライマーとrrnX−catR2プライマー、CAT−F2プライマーとCAT−Rプライマーを用いて増幅し、得られた3断片をrrnO−catF1プライマーとrrnX−catR2プライマーで連結した。次に、rrnOオペロンを有し、かつtrpC2変異を有する染色体DNAを供与DNAに用い、rrnO2+::catpt1 ΔrrnHG1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔrrnI2ΔrrnW2 rrnJ+J+ hisC101株に形質転換を行い、rrnO2+::catpt1とrrnOオペロンを置換した。次に、rrnJオペロンにcat遺伝子を導入するため、rrnJの上流領域、下流領域をそれぞれrrnJUFプライマーとrrnJURcatプライマー、rrnJDFcatプライマーとrrnJDRプライマーを用いて、及び、cat遺伝子をCAT−F2プライマー、CAT−Rプライマーを用いて増幅した。得られた3断片をrrnJUFプライマー、rrnJDRプライマーを用いて連結後、形質転換を行いRIK543株(ΔrrnHG1ΔrrnD1ΔrrnE1ΔrrnB2ΔrrnA1ΔrrnI2ΔrrnW2ΔrrnJ::cat trpC2)を得た(Nanamiya et al. (2010) Microbiology 156:2977-2982、Natori et al. (2009) J. Bacteriol. 191:4555-4561)。
【0080】
参考例5 RIK545株の構築
rrnEオペロンのみを有する株は、参考例4と同様に行なった。rrnE2+::catpt1は、rrnE−catF1プライマーとrrnE−catR1プライマー、rrnE−catF2プライマーとrrnX−catR2プライマーを用いて増幅し、cat遺伝子と連結することで構築した(Nanamiya et al. (2010) Microbiology 156:2977-2982、Natori et al. (2009) J. Bacteriol. 191: 4555-4561)。
【0081】
【表4】
【0082】
【表5】
【特許請求の範囲】
【請求項1】
改変SD配列を有する標的遺伝子と;当該改変SD配列と相補的な改変アンチSD配列を有する複数個のrrnオペロンと;改変されていないアンチSD配列を有するrrnオペロン、とを有する細菌。
【請求項2】
前記改変SD配列が、前記改変されていないアンチSD配列と80%以上の配列同一性を有する請求項1記載の細菌。
【請求項3】
前記改変SD配列が、CCTCCである請求項1又は2記載の細菌。
【請求項4】
前記改変アンチSD配列が、GGAGGである請求項1〜3のいずれか1項に記載の細菌。
【請求項5】
バチルス属細菌である請求項1〜4のいずれか1項に記載の細菌。
【請求項6】
前記改変アンチSD配列を有する複数のrrnオペロンが、少なくともrrnIオペロン及びrrnOオペロンである請求項5に記載の細菌。
【請求項7】
前記改変アンチSD配列を有する複数のrrnオペロンが、rrnIオペロン、rrnOオペロン及びrrnEオペロンである請求項5に記載の細菌。
【請求項8】
前記改変されていないアンチSD配列を有するrrnオペロンがrrnJオペロンである請求項6又は7に記載の細菌。
【請求項9】
請求項1〜8のいずれか1項に記載の細菌を培養する工程を含む目的遺伝子産物の生産方法。
【請求項1】
改変SD配列を有する標的遺伝子と;当該改変SD配列と相補的な改変アンチSD配列を有する複数個のrrnオペロンと;改変されていないアンチSD配列を有するrrnオペロン、とを有する細菌。
【請求項2】
前記改変SD配列が、前記改変されていないアンチSD配列と80%以上の配列同一性を有する請求項1記載の細菌。
【請求項3】
前記改変SD配列が、CCTCCである請求項1又は2記載の細菌。
【請求項4】
前記改変アンチSD配列が、GGAGGである請求項1〜3のいずれか1項に記載の細菌。
【請求項5】
バチルス属細菌である請求項1〜4のいずれか1項に記載の細菌。
【請求項6】
前記改変アンチSD配列を有する複数のrrnオペロンが、少なくともrrnIオペロン及びrrnOオペロンである請求項5に記載の細菌。
【請求項7】
前記改変アンチSD配列を有する複数のrrnオペロンが、rrnIオペロン、rrnOオペロン及びrrnEオペロンである請求項5に記載の細菌。
【請求項8】
前記改変されていないアンチSD配列を有するrrnオペロンがrrnJオペロンである請求項6又は7に記載の細菌。
【請求項9】
請求項1〜8のいずれか1項に記載の細菌を培養する工程を含む目的遺伝子産物の生産方法。
【図1】

【図2】
【図3】
【図2】
【図3】
【公開番号】特開2013−9604(P2013−9604A)
【公開日】平成25年1月17日(2013.1.17)
【国際特許分類】
【出願番号】特願2011−142694(P2011−142694)
【出願日】平成23年6月28日(2011.6.28)
【出願人】(000000918)花王株式会社 (8,290)
【出願人】(300071579)学校法人立教学院 (42)
【Fターム(参考)】
【公開日】平成25年1月17日(2013.1.17)
【国際特許分類】
【出願日】平成23年6月28日(2011.6.28)
【出願人】(000000918)花王株式会社 (8,290)
【出願人】(300071579)学校法人立教学院 (42)
【Fターム(参考)】
[ Back to top ]