
改良された分泌によるポリペプチドの製造
本発明は、SEC61ポリペプチドの少なくとも1つの活性に対応する活性を有するポリペプチド、これらのポリペプチドをコードするポリヌクレオチド、および目的とするポリペプチドの製造に適切な宿主細胞の調製におけるその使用に関する。かかる宿主細胞は、目的とするポリペプチドを分泌する増大された能力を有しうる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は組換えDNA技術に関する。具体的には、本発明は、酵母SEC61(サッカロミセス(Saccharomyces)ゲノムデータベース−SGD:http://www.yeastgenome.org/)について記載されたタンパク質輸送活性を有するSEC61ポリペプチド、これらのポリペプチドをコードする核酸配列、および目的とするポリペプチドの製造に適切な宿主細胞の調製におけるその使用に関する。
【背景技術】
【0002】
タンパク質の培養培地への分泌は、分泌経路を構成するさまざまな膜に囲まれたコンパートメントを通じたタンパク質の移動を含む。最初にタンパク質は小胞体、ERの内腔へ転位される。そこからタンパク質は膜小胞のゴルジ複合体に輸送され、ゴルジから血漿膜に輸送される。分泌過程は、分泌されたタンパク質を含有する小胞がドナー膜から摘み取られ、受容体膜に標的され、これと融合される幾つかのステップを含む。これらのステップの各々で幾つかの異なるタンパク質の機能が必要である。
【0003】
糸状菌におけるタンパク質分泌を増大させる幾つかの試みが行われている。異種タンパク質の分泌を増大させる一般的な方法が、シグナル配列を使用することである(例えば、EP0215594号明細書を参照)。ランダム突然変異誘発および分泌されたタンパク質のスクリーニング(スミス(Smith)ら、1985年、サカイ(Sakai)ら、1988年、シュスター(Shuster)ら、1989年、スズキ(Suzuki)ら、1989年、スリープ(Sleep)ら、1991年、ラムサ(Lamsa)およびブレバウム(Bloebaum)、1990年、ダン・コールマン(Dunn−Coleman)ら、1991年、および米国特許出願公開第2002/0068325A1号明細書)または異質のタンパク質の有効に分泌された内因性タンパク質との融合(ワード(Ward)ら、1990年、ハルキ(Harkki)ら、1989年、ニーソネン(Nyyssonen)ら、1993年、ニーソネン(Nyyssoenen)ら、1992年)は、異種タンパク質の分泌をより効率的にするために酵母および糸状菌の両方に広く使用されている。これらの方法の両方は使用が限定されている。ランダム突然変異誘発およびスクリーニングによって単離される突然変異はほぼ完全に劣性であり、倍数体である工業上の菌株へ移動されえない。しばしば得られた突然変異はスクリーニングに使用されるタンパク質の改良された分泌能のみを有する。融合タンパク質法は、各々異種または異質のタンパク質の融合構成の適合化を別個に必要とする。融合タンパク質はしばしば機能ではなく、したがって最終生成物はタンパク質分解的切断によって放出される必要があり、これは製造手順を複雑にする。
【0004】
さらに、それらのタンパク質生成因子としての産業上の重要性により、改良された分泌能を有する糸状菌を得る必要がある。
【発明の詳細な説明】
【0005】
本明細書で参照されるすべての特許および刊行物は、かかる特許および刊行物内に開示されたすべての配列および方法を含めて、参照によって明示的に援用される。これらの特許および刊行物としては、EP357127号明細書、EP635574号明細書、国際公開第97/06261号パンフレット、国際公開第98/46772号パンフレットが挙げられる。
【0006】
SEC61活性を表示するポリペプチド
本発明の第1の態様は新規SEC61ポリペプチドに関する。SEC61ポリペプチドは一般にER膜にわたってポリペプチドの輸送を介在することが知られている。本発明によるSEC61ポリペプチドは、SEC61ポリペプチドと関連していることが一般に知られている少なくとも1つの活性を表示するポリペプチドと定義されている。本発明によるSEC61ポリペプチドは、SEC61ポリペプチドと関連していることが一般に知られているすべての活性をも表示しうる。
【0007】
具体的には、本発明は、
(a)配列番号3によるアミノ酸配列を有するポリペプチドと、
(b)配列番号3によるアミノ酸配列と実質的に相同であるアミノ酸配列を有するポリペプチドと
からなる群から選択されるSEC61ポリペプチドを開示する。
【0008】
本発明との関連で、「実質的に相同」という語は、SEC61ポリペプチドの少なくとも1つの活性を表示し、かつ配列番号3に記載されたアミノ酸配列の少なくとも89%、より好ましくは少なくとも90%、さらにより好ましくは少なくとも92%、さらにより好ましくは少なくとも93%、さらにより好ましくは少なくとも94%、さらにより好ましくは少なくとも95%、さらにより好ましくは少なくとも96%、さらにより好ましくは少なくとも97%、さらにより好ましくは少なくとも98%、かつ最も好ましくは少なくとも99%の同一性の程度を有するポリペプチドを包含することが意味される。同一性の程度は、好ましくは、配列番号3の全長にわたって測定される。
【0009】
本発明のために、2つのアミノ酸配列間の同一性の程度は、2つの配列間で同一であるアミノ酸の割合を指す。同一性の程度は、アルトシュール(Altschul)ら、J. Mol.Biol.215:403−410頁(1990年)に記載されているBLASTアルゴリズムを使用して判定される。BLAST分析を実行するためのソフトウェアは、国立バイオテクノロジー情報センター(the National Center for Biotechnology Information)(http://www.ncbi.nlm.nih.gov/)によって公的に入手可能である。BLASTアルゴリズムパラメータW、T、およびXは、アラインメントの感度および速度を決定する。BLASTプログラムではデフォルトとして11の語長(W)、50のBLOSUM62採点マトリックス(ヘニコフ(Henikoff)とヘニコフ(Henikoff)、Proc.Natl. Acad.Sci.USA 89:10915頁(1989年)を参照)アラインメント(B)、10の期待値(E)、M=5、N=−4、および両方の鎖の比較が使用される。
【0010】
好ましい実施形態において、本発明のポリペプチドは配列番号3によるアミノ酸配列を有する。
【0011】
別の実施形態において、配列番号3によるアミノ酸配列と実質的に相同であるポリペプチドのアミノ酸配列は、天然または人工の変異(突然変異誘発)による配列番号3のアミノ酸配列の点で異なりうる。配列番号3によるアミノ酸配列と実質的に相同であるアミノ酸配列を有するポリペプチドは、配列番号3の配列と比べ少なくとも1つの修飾を含有しうる。前記修飾はアミノ酸置換、付加、または欠失でありうる。
【0012】
好ましい実施形態において、本発明のポリペプチドは、5個のアミノ酸が、好ましくは4個のアミノ酸が、より好ましくは3個のアミノ酸が、さらにより好ましくは2個のアミノ酸が、かつ最も好ましくは1個のアミノ酸が、配列番号3に記載されたアミノ酸配列と異なり、かつさらに配列番号3に記載されたアミノ酸配列を有するポリペプチドと同じSEC61活性を有するアミノ酸配列を有する。
【0013】
別の好ましい実施形態において、配列番号3によるアミノ酸配列と実質的に相同であるアミノ酸配列を有するポリペプチドが、配列番号9によるアミノ酸配列を有する。
【0014】
本発明の別の好ましい実施形態において、配列番号3によるアミノ酸配列との少なくとも85%、好ましくは少なくとも87%、より好ましくは少なくとも89%、さらにより好ましくは少なくとも90%、さらにより好ましくは少なくとも92%、さらにより好ましくは少なくとも93%、さらにより好ましくは少なくとも94%、さらにより好ましくは少なくとも95%、さらにより好ましくは少なくとも96%、さらにより好ましくは少なくとも97%、さらにより好ましくは少なくとも98%、かつ最も好ましくは少なくとも99%の同一性の程度を有するSEC61ポリペプチドが提供されるが、ここで少なくとも1個のアミノ酸が、配列番号3の第373〜379位置に対応する、すなわち配列番号3の第373〜379位置に存在するアミノ酸配列ALFSKTWに対応するコンセンサスモチーフで修飾されている。好ましくは、コンセンサスモチーフで修飾されている少なくとも1個のアミノ酸は、配列番号3の第376位置に対応する位置にある。より好ましくは、配列番号3の第376位置に対応するアミノ酸は、トリプトファン、フェニルアラニン、チロシン、またはヒスチジンによって置換される。最も好ましくは、配列番号3の第376位置に対応する位置におけるセリンはトリプトファンによって置換される。かかる修飾SEC61ポリペプチドの例は、配列番号6によるアミノ酸配列を有するポリペプチドである。
【0015】
コンセンサスモチーフにおけるさらなる変異が、配列番号3の第374位置に対応する位置でロイシン、バリン、フェニルアラニン、またはメチノン(methinone)の存在、および/または配列番号3の第377位置に対応する位置におけるリシン、アラニン、トレオニン、またはアルギニンの存在、および/または配列番号3の第378位置に対応する位置におけるトレオニン、またはロイシンの存在でありうる。
【0016】
SEC61ポリペプチドをコードする単離糸状菌核酸配列
本発明の第2の態様は、第1の態様のSEC61ポリペプチドをコードする核酸配列を含んで成る単離ポリヌクレオチドに関する。
【0017】
具体的には、SEC61ポリペプチドをコードするポリヌクレオチドまたは核酸配列は、
(a)配列番号1によるポリヌクレオチドと、
(b)配列番号2による核酸配列と、
(c)配列番号2の核酸配列と少なくとも90%の同一性の程度を有する核酸配列と、
(d)配列番号3のアミノ酸配列と実質的に相同であるアミノ酸配列を有するポリペプチドをコードする核酸配列と
からなる群から選択される。
【0018】
好ましい実施形態において、特徴(c)の核酸配列は、配列番号2に記載された核酸配列と少なくとも91%、より好ましくは少なくとも92%、さらにより好ましくは少なくとも93%、さらにより好ましくは少なくとも94%、さらにより好ましくは少なくとも95%、さらにより好ましくは少なくとも96%、さらにより好ましくは少なくとも97%、さらにより好ましくは少なくとも98%、最も好ましくは少なくとも99%の同一性の程度を有する。
【0019】
本発明のために、2つの核酸配列間の同一性の程度は、同一性表、10のギャップペナルティ、10のギャップ長ペナルティによるClustral法(ヒギンズ(Higgins)、1989年、CABIOS 5:151−153頁)によって判定される。
【0020】
別の好ましい実施形態において、単離核酸配列は、配列番号3のアミノ酸配列を有するポリペプチド、またはSEC61活性を有するその断片をコードする。
【0021】
別の好ましい実施形態において、単離核酸配列は、配列番号9のアミノ酸配列を有するポリペプチド、またはSEC61活性を有するその断片をコードする。
【0022】
別の好ましい実施形態において、SEC61ポリペプチドをコードする核酸配列は配列番号2または配列番号8に記載されている核酸配列を有する。
【0023】
別の好ましい実施形態において、単離核酸配列は、配列番号3によるアミノ酸配列と少なくとも85%の同一性の程度を有するアミノ酸配列を有するSEC61ポリペプチドをコードし、ここで少なくとも1個のアミノ酸が配列番号3の第373〜379位置に対応する、すなわち配列番号3の第373〜379位置に存在するアミノ酸配列ALFSKTWに対応するコンセンサスモチーフで修飾されている。例えば、単離核酸配列は、配列番号6によるアミノ酸配列を有するポリペプチドをコードし、好ましくは配列番号5の核酸配列である。
【0024】
本明細書で使用される「単離ポリヌクレオチドまたは核酸配列」という語は、アガロース電気泳動によって測定されるように、例えば、少なくとも約20%純粋、好ましくは少なくとも約40%純粋、より好ましくは少なくとも約60%純粋、さらにより好ましくは少なくとも約80%純粋、最も好ましくは少なくとも約90%純粋な他の核酸配列を基本的に含んでいないポリヌクレオチドまたは核酸配列を指す。例えば、単離核酸配列が遺伝子工学で使用される標準のクローニング法によって獲得され、核酸配列をその天然位置から再生される異なる部位へ移動させうる。
【0025】
ポリヌクレオチドまたは核酸配列は、ゲノム、cDNA、RNA、半合成、合成起源、またはそのいずれかの組合せでありうる。
【0026】
SEC61ポリペプチドをコードし、かつ配列番号3によるタンパク質と実質的に相同であるアミノ酸配列を有する単離核酸分子が、配列番号2によるコードヌクレオチド配列と比べ、1つもしくはそれ以上のヌクレオチド修飾、すなわち置換、付加、または欠失を含有し、1つもしくはそれ以上のアミノ酸修飾、すなわち置換、欠失、または挿入がコードされたタンパク質に存在するようになりうる。かかる修飾は天然変異によるものであり、または部位特異的突然変異誘発もしくはPCR介在突然変異誘発など標準の突然変異誘発技術によって導入されうる。
【0027】
核酸配列によってコードされるポリペプチドにおける異なるアミノ酸残基をもたらさないsec61コード配列へヌクレオチド置換を導入することも可能である。これは、例えば、コドン使用がSEC61タンパク質の発現を目的とする宿主生物のコドン使用に対応する核酸配列を引き起こすのに有利でありうる。
【0028】
コードされたタンパク質のアミノ酸配列の変更をもたらしうるDNA配列多型が所定の集団内に天然に存在しうることは当業者には明らかであろう。かかる遺伝子多型は、異なる集団からの、または天然対立遺伝子変異による集団内の細胞に存在しうる。
【0029】
本発明によるSEC61ポリペプチドおよびコードする核酸配列は、真核細胞、好ましくは、真菌、より好ましくは、糸状菌から獲得されうる。
【0030】
「糸状菌」としては、真菌類(Eumycota)および卵菌類(Oomycota)の下位分類のすべての糸状形態(ホークスワース(Hawksworth)ら、1995年、上記によって規定)が挙げられる。糸状菌菌株としては、アクレモニウム属(Acremonium)、アスペルギルス属(Aspergillus)、オーレオバシディウム属(Aureobasidium)、クリプトコッカス属(Cryptococcus)、フィリバシジウム属(Filibasidium)、フザリウム属(Fusarium)、フミコーラ属(Humicola)、マグナポルテ属(Magnaporthe)、ムコール属(Mucor)、ミセリオフトラ属(Myceliophthora)、ネオカリマスティックス属(Neocallimastix)、ニューロスポラ属(Neurospora)、パエシロミセス属(Paecilomyces)、ペニシリウム属(Penicillium)、ピロミセス属(Piromyces)、シゾフィラム属(Schizophyllum)、タラロミセス属(Talaromyces)、サーモアスカス属(Thermoascus)、チエラビア属(Thielavia)、トリポクラジウム属(Tolypocladium)、およびトリコデルマ属(Trichoderma)の菌株が挙げられるが、これらに限定されない。
【0031】
より好ましい実施形態において、本発明のSEC61活性を有するポリペプチドをコードする核酸配列は、A.アワモリ(awamori)またはA.ニズランス(nidulans)などアスペルギルス属(Aspergillus)の菌株から得られる。好ましくは、この核酸配列はA.ニゲル(niger)またはA.オリゼー(oryzae)の菌株から得られる。さらにより好ましくは、この核酸配列はA.ニゲル(niger)の菌株の単離株、例えば、配列番号1または配列番号2に記載の核酸配列から得られる。
【0032】
別の実施形態において、本発明のSEC61活性を有するポリペプチドをコードする核酸配列は、F.オキシスポルム(oxysporum)またはF.ヴェネナツム(venenatum)などフザリウム属(Fusarium)の菌株、またはP.クリソゲナム(chrysogenum)などペニシリウム属(Penicillium)の菌株、例えば、配列番号7または配列番号8に記載された核酸配列から得られる。
【0033】
上記の種について、本発明は完全および不完全な状態、および他の分類学的同等物、例えば、それらが知られている種名にかかわらず、アナモルフを包含することが理解されるであろう。当業者は、適切な同等物の同一性を容易に理解するであろう。例えば、ポリペプチドは、それらが知られている種名にかかわらず、レーパー(Raper),K.D.およびフェネル(Fennel),D.I.(1965年。The Genus Aspergillus、ウィルキンズ社(The Wilkins Company)、Baltimore、メリーランド州(MD))によって規定されたアスペルギルス属(Aspergillus)の分類学的同等物である微生物から得られうる。
【0034】
さらに、SEC61活性を有するポリペプチドは、例えば、本発明によるポリヌクレオチドに基づくプローブを使用して自然(例えば、土、堆肥、水等)から単離される微生物を含む、糸状菌以外の供給源から同定され獲得されうる。自然の生息地から微生物を単離するための方法が当業界で公知である。
【0035】
本発明のSEC61ポリペプチドをコードするDNA配列はハイブリダイゼーションによって得られうる。本発明のsec61DNAの変異(例えば、天然対立遺伝子多型)および相同体が、標準のハイブリダイゼーション法によるハイブリダイゼーションプローブとして、好ましくはきわめてストリンジェントなハイブリダイゼーション条件下にこれらの核酸またはその適切な断片を使用して本明細書に開示された核酸との相同性に基づき単離されうる。
【0036】
ハイブリダイゼーション反応の「ストリンジェンシー」は、当業者によって容易に判定される。ハイブリダイゼーション反応のストリンジェンシーの追加の詳細および説明については、オースベル(Ausubel)ら、Current Protocols in Molecular Biology、ワイリー・インターサイエンス・パブリッシャーズ(Wiley Interscience Publishers)、(1995年)を参照。
【0037】
核酸配列は、例えば、問題となっている微生物のゲノムまたはcDNAライブラリーのスクリーニングによって単離されうる。SEC61活性を有するポリペプチドをコードする核酸配列が、例えば、配列番号2由来のプローブで検出されると、配列は当業者に周知である方法を利用して単離またはクローン化されうる(例えば、J.サンブルック(Sambrook)、E.F.フリッチュ(Fritsch)、およびT.マニアチス(Maniatis)、1989年、Molecular Cloning,A Laboratory Manual、第2版、コールド・スプリング・ハーバー(Cold Spring Harbor)、ニューヨーク(New York)を参照)。
【0038】
かかるゲノムDNAからの本発明の核酸配列のクローニングは、例えば、ポリメラーゼ連鎖反応(PCR)または共通の構造的特徴を有するクローン化DNA断片を検出する発現ライブラリーの抗体スクリーニングに基づく方法を使用しても達成されうる(例えば、イニス(Innis)ら、1990年、PCR: A Guide to Methods and Application、アカデミック・プレス(Academic Press)、ニューヨーク(New York)を参照)。
【0039】
本明細書で提供される配列情報は、誤って同定された塩基の包含を必要とするほどに狭く考えられてはならない。本明細書に開示された特定の配列は、糸状菌、具体的にはA.ニゲル(niger)からの完全な遺伝子を単離するために容易に使用され、次いで容易に追加的な配列分析にかけられ、それによって配列決定の誤りを特定することができる。
【0040】
別段の定めがない限り、本明細書のDNA分子の配列決定によって決定されるすべてのヌクレオチド配列は、自動DNAシーケンサーを使用して決定され、本明細書で決定されたDNA分子によってコードされるポリペプチドのすべてのアミノ酸配列は上記の通り決定されたDNA配列の翻訳によって予測された。したがって、この自動的方法によって決定されるDNA配列について当業界で周知のように、本明細書で決定されるヌクレオチド配列は多少の誤りを含有しうる。自動化によって決定されるヌクレオチド配列は、配列決定されたDNA分子の実際のヌクレオチド配列と一般的に少なくとも約90%同一、より一般的には少なくとも約95%〜少なくとも約99.9%同一である。実際の配列は、当業界で公知の手動のDNA配列決定法を含む他の方法によってより正確に決定されうる。当業界で周知であるように、実際の配列と比較して決定されたヌクレオチド配列における単一の挿入または欠失がヌクレオチド配列の翻訳におけるフレームシフトをもたらし、決定されたヌクレオチド配列によってコードされた予測アミノ酸配列が、かかる挿入または欠失の点で開始する、配列決定されたDNA分子によって実際にコードされるアミノ酸配列と完全に異なるようになっている。
【0041】
当業者は、かかる誤って同定された塩基を同定することが可能であり、かかる誤りを修正する方法を知っている。
【0042】
核酸構築物
本発明の別の態様は、適切な発現宿主におけるSEC61活性を有するポリペプチドの発現を指図する1つもしくはそれ以上の制御配列に操作可能に結合されている、本発明によるSEC61ポリペプチドをコードする核酸配列を含んで成る核酸構築物に関する。
【0043】
発現は、ポリペプチドの製造に関与するステップを含むことが理解され、転写、転写後修飾、翻訳、翻訳後修飾、分泌を含みうる。
【0044】
「核酸構築物」は、天然起源遺伝子から単離され、またはそれ以外には天然に存在しないやり方で結合または並列される核酸の部分を含有するように修飾されている一本または二本鎖のいずれかの核酸分子として本明細書では定義される。核酸構築物という語は、核酸構築物が特定の宿主生物におけるコード配列の発現に必要な制御配列をすべて含有する場合に発現ベクターまたはカセットという語と同義である。
【0045】
「制御配列」という語は、本明細書では、ポリペプチドの発現に必要または有利であるすべての要素を含むと定義される。各々の制御配列は天然またはポリペプチドをコードする核酸配列とは異なりうる。かかる制御配列としては、プロモーター、リーダー、最適な翻訳開始配列(コザック(Kozak)、1991年、J.Biol.Chem.266:19867−19870頁)、分泌シグナル配列、プロペプチド配列、ポリアデニル化配列、転写終止が挙げられるが、これらに限定されない。最小限でも、制御配列は、プロモーター、転写および翻訳停止シグナルを含む。
【0046】
「操作可能に結合」という語は、本明細書では、制御配列がDNA配列のコード配列に対する位置で適切に配置され、制御配列がポリペプチドの製造を指図するようになっている構成として定義される。
【0047】
制御配列は、転写制御配列を含有する適切なプロモーター配列でありうる。プロモーターは、突然変異、切断、およびハイブリッドプロモーターを含む細胞における転写調節活性を示す任意の核酸配列であり、かつ細胞外または細胞内ポリペプチドをコードする遺伝子から獲得されうる。プロモーターは、細胞またはポリペプチドと相同または異種のいずれかでありうる。
【0048】
糸状菌の好ましいプロモーターは当業界で周知であり、例えば、グルコース−6−リン酸デヒロゲナーゼgpdAプロモーター、pepA、pepB、pepCなどのプロテアーゼプロモーター、グルコアミラーゼglaAプロモーター、アミラーゼamyA、amyBプロモーター、カタラーゼcatRまたはcatAプロモーター、グルコースオキシダーゼgoxCプロモーター、ベータ−ガラクトシダーゼlacAプロモーター、アルファ−グルコシダーゼaglAプロモーター、翻訳伸長因子tefAプロモーター、xlnA、xlnB、xlnC、xlnDなどのキシラナーゼプロモーター、eglA、eglB、cbhAなどのセルラーゼプロモーター、areA、creA、xlnR、pacC、prfT等などの転写調節因子のプロモーターでありうるとともに、特にNCBIウェブサイト(http://www.ncbi.nlm.nih.gov/entrez/)で確認されうる。
【0049】
好ましい実施形態において、プロモーターは、高く発現される(本明細書では総細胞mRNAの少なくとも0.5%(w/w)のmRNA濃度と定義)遺伝子由来でありうる。別の好ましい実施形態において、プロモーターは、培地発現される(本明細書では総細胞mRNAの少なくとも0.01%〜0.5%(w/w)のmRNA濃度と定義)遺伝子由来でありうる。別の好ましい実施形態において、プロモーターは、低く発現される(本明細書では総細胞mRNAの少なくとも0.01%(w/w)未満のmRNA濃度と定義)遺伝子由来でありうる。
【0050】
制御配列は適切な転写終止配列、転写を終止する糸状菌細胞によって認識される配列でもありうる。終止配列は、ポリペプチドをコードする核酸配列の3’末端と操作可能に結合されている。細胞において機能的であるすべての終止が本発明において使用されうる。
【0051】
糸状菌細胞の好ましい終止は、A.オリゼー(oryzae)TAKAアミラーゼ、A.ニゲル(niger)グルコアミラーゼ、A.ニズランス(nidulans)アントラニル酸シンターゼ、A.ニゲル(niger)アルファ−グルコシダーゼ、trpC遺伝子、およびフザリウム・オキシスポルム(Fusarium oxysporum)トリプシン様プロテアーゼをコードする遺伝子から得られる。
【0052】
制御配列は、糸状菌細胞による翻訳に重要であるmRNAの非翻訳領域の適切なリーダー配列でもありうる。リーダー配列は、ポリペプチドをコードする核酸配列の5’末端と操作可能に結合されている。細胞において機能的であるすべてのリーダー配列が本発明において使用されうる。
【0053】
糸状菌細胞の好ましいリーダーは、A.オリゼー(oryzae)TAKAアミラーゼ、およびA.ニズランス(nidulans)トリオースリン酸イソメラーゼ、およびA.ニゲル(niger)glaAから得られる。
【0054】
制御配列は、核酸配列の3’末端と操作可能に結合されており、転写されると、転写mRNAにポリアデノシン残基を添加するシグナルとしての糸状菌細胞によって認識される配列のポリアデニル化配列でもありうる。細胞において機能的であるすべてのポリアデニル化配列が本発明において使用されうる。
【0055】
糸状菌細胞の好ましいポリアデニル化配列は、A.オリゼー(oryzae)TAKAアミラーゼ、A.ニゲル(niger)グルコアミラーゼ、A.ニズランス(nidulans)アントラニル酸シンターゼ、フザリウム・オキシスポルム(Fusarium oxysporum)トリプシン様プロテアーゼ、およびA.ニゲル(niger)アルファ−グルコシダーゼをコードする遺伝子から得られる。
【0056】
分泌されるポリペプチドには、制御配列は、コードされたポリペプチドを細胞の分泌経路へ指図しうるポリペプチドのアミノ末端と連結されたアミノ酸配列をコードするシグナルペプチドコード領域でもありうる。核酸配列のコード配列の5’末端は、本質的に、分泌されたポリペプチドをコードするコード領域の部分と翻訳リーディングフレームで自然に連結されたシグナルペプチドコード領域を含有しうる。あるいは、コード配列の5’末端は、コード配列とは異なっているシグナルペプチドコード領域を含有しうる。異質のシグナルペプチドコード領域は、コード配列が通常、シグナルペプチドコード領域を含有しない場合に必要とされうる。あるいは、異質のシグナルペプチドコード領域は、ポリペプチドの増強された分泌を得るために天然のシグナルペプチドコード領域を単純に置換しうる。
【0057】
核酸構築物は発現ベクターでありうる。発現ベクターは、組換えDNA法に便利にかけられ、ポリペプチドをコードする核酸配列の発現をもたらしうる任意のベクター(例えば、プラスミドまたはウイルス)でありうる。ベクターの選択は一般的にベクターが導入される細胞とのベクターの適合性に依存する。ベクターは線状または閉環状プラスミドでありうる。
【0058】
ベクターは自己複製ベクター、すなわち、染色体外実体として存在するベクターであり、その複製が染色体複製と無関係であり、例えば、プラスミド、染色体外要素、ミニ染色体、または人工染色体でありうる。糸状菌の自己維持クローニングベクターは、AMA1−配列を含んで成りうる(例えば、アレクセンコ(Aleksenko)とクラッターバック(Clutterbuck)(1997年)、Fungal Genet.Biol.21:373−397頁を参照)。
【0059】
あるいは、ベクターは、細胞へ導入されると、ゲノムへ組込まれ、組込まれている染色体といっしょに複製されるものでありうる。組込みクローニングベクターは、ランダムに、または宿主細胞の染色体における所定の標的遺伝子座で組込まれうる。本発明の好ましい実施形態において、組込みクローニングベクターは、クローニングベクターの組込みをこの所定の遺伝子座にターゲティングするための宿主細胞のゲノムにおける所定の標的遺伝子座のDNA配列と相同であるDNA断片を含んで成る。標的組込みを促進するために、クローニングベクターは、好ましくは、宿主細胞の形質転換前に線状化される。線状化は、好ましくは、少なくとも1つであるが、好ましくは、クローニングベクターの端のいずれかが標的遺伝子座と相同の配列によってフランキングされるように行われる。標的遺伝子座をフランキングする相同配列の長さは、好ましくは、少なくとも0.1kbであり、さらに好ましくは、少なくとも0.2kbであり、より好ましくは、少なくとも0.5kbであり、さらにより好ましくは、少なくとも1kbであり、最も好ましくは、少なくとも2kbである。
【0060】
好ましい実施形態によれば、本発明によるSEC61ポリペプチドをコードする遺伝子を含んで成る発現ベクターの組込みの標的遺伝子座は、sec61遺伝子座である。
【0061】
ベクター系は単一ベクターもしくはプラスミド、または2つもしくはそれ以上のベクターまたはプラスミドでありうるが、これらはいっしょに宿主細胞のゲノム、またはトランスポゾンへ導入されるすべてのDNAを含有する。
【0062】
ベクターは、好ましくは、形質転換細胞の容易な選択を可能にする1つもしくはそれ以上の選択可能なマーカーを含有する。選択可能なマーカーが、その生成物が殺生物性またはウイルス抵抗性、重金属に対する抵抗性、栄養要求体に対する原栄養性(prototrophy)などを提供する遺伝子である。糸状菌細胞において使用するための選択可能なマーカーが、amdS(アセトアミダーゼ)、argB(オルニチンカルバモイルトランスフェラーゼ)、bar(ホスフィノトリシンアセチルトランスフェラーゼ)、hygB(ハイグロマイシンホスホトランスフェラーゼ)、niaD(硝酸還元酵素)、pyrG(オロチジン−5’−リン酸デカルボキシラーゼ)、sC(硫酸アデニルトランスフェラーゼ)、およびtrpC(アントラニル酸シンターゼ)のほか、他の種からの同等物を含むがこれらに限定されない群から選択されうる。アスペルギルス属(Aspergillus)細胞における使用に好ましいのは、A.ニズランス(nidulans)またはA.オリゼー(oryzae)のamdS(EP635574B1号明細書、国際公開第97/06261号パンフレット)およびpyrG遺伝子およびストレプトマイセス・ハイグロスコピクス(Streptomyces hygroscopicus)のbar遺伝子である。より好ましくは、amdS遺伝子が使用され、さらにより好ましくは、A.ニズランス(nidulans)またはA.ニゲル(niger)からのamdS遺伝子である。最も好ましい選択マーカー遺伝子が、A.ニズランス(nidulans)gpdAプロモーターと融合されたA.ニズランス(nidulans)amdSコード配列である(EP635574号明細書を参照)。他の糸状菌からのAmdS遺伝子も使用されうる(国際公開第97/06261号パンフレット)。
【0063】
宿主細胞
別の態様において、本発明は、目的とするポリペプチドの製造に適切な宿主細胞を提供し、前記宿主細胞はSEC61活性を有する発明によるポリペプチドを過剰発現する。前記宿主細胞は、本発明によるSEC61ポリペプチドをコードする核酸構築物または発現ベクターを含んで成る。
【0064】
意外にも、発明者らは、本発明によるSEC61ポリペプチド、具体的には上記の通りコンセンサスモチーフで修飾された突然変異SEC61ポリペプチドが過剰発現している真菌細胞、具体的には糸状菌細胞が、かかるSEC61ポリペプチドを過剰発現することがない真菌細胞と比べ改良されたタンパク質分泌能を有することを見出した。特に、異種タンパク質の分泌が改良されている。場合により、内因性sec61遺伝子は、内因性SEC61ポリペプチドと比べ修飾されているSEC61ポリペプチドを発現する細胞において不活性化されうる。
【0065】
好ましい実施形態においては、本発明によるSEC61ポリペプチドを過剰発現する宿主細胞が、内因性に発現されるSEC61ポリペプチドの量の少なくとも50%以上、好ましくは、少なくとも100%以上、より好ましくは、少なくとも200%以上、最も好ましくは、少なくとも400%以上である。内因性に発現されるSEC61ポリペプチドの量は、本発明によるSEC61ポリペプチドを過剰発現する宿主細胞がそれに由来する親の宿主細胞によって発現されるSEC61ポリペプチドの量である。発現SEC61ポリペプチドの量は、例えば、定量的SDS−PAGE、二次元タンパク質電気泳動、またはウェスタンブロット法によって、当業者に周知の方法によって測定されうる。
【0066】
本発明では、本発明によるSEC61ポリペプチドを過剰発現する宿主細胞のさまざまな実施形態が意図される。
【0067】
1つの実施形態において、天然のSEC61ポリペプチドを過剰発現し、それによって天然のSEC61ポリペプチドが前記宿主によって自然に生成されるSEC61ポリペプチドと同じポリペプチドである宿主細胞が提供される。
【0068】
別の実施形態において、前記宿主によって自然に生成されないSEC61ポリペプチドを発現する宿主細胞が提供される。かかる宿主細胞は天然SEC61ポリペプチドを発現することがない。天然SEC61ポリペプチドの非発現は、例えば、内因性sec61遺伝子の不活性化によって達成されうる。好ましくは、内因性sec61遺伝子の不活性化は、例えば、EP0357127号明細書に記載されている遺伝子置換によって達成されうる。より好ましくは、前記遺伝子置換は、本発明によるSEC61ポリペプチドをコードする核酸配列を含んで成る核酸構築物(発現ベクター)によって内因性sec61遺伝子の置換を含んで成る。
【0069】
本発明の方法における宿主細胞の選択は、目的とするポリペプチドをコードする核酸配列の供給源に大幅に依存する。好ましくは、宿主細胞は真核細胞、より好ましくは、真菌、最も好ましくは、糸状菌である。好ましい実施形態において、糸状菌宿主細胞は、本発明によるSEC61ポリペプチドが獲得されうる種として挙げられている種の細胞である。
【0070】
好ましい実施形態において、糸状菌宿主細胞は、配列番号3のアミノ酸配列と少なくとも89%の同一性を有するアミノ酸配列を有するポリペプチド、好ましくは、配列番号3または配列番号6または配列番号9のアミノ酸配列を有するポリペプチドをコードする核酸配列を有する遺伝子を過剰発現する。
【0071】
得られた宿主細胞は、目的とするポリペプチドを製造するために使用されうる。
【0072】
したがって、別の好ましい実施形態において、宿主細胞は、目的とするポリペプチドをコードする核酸配列を含んで成るポリヌクレオチドの少なくとも1つのコピーをさらに有する。この核酸配列は、sec61遺伝子の発現について以前に記載された発現ベクターへクローン化されうる。本発明の方法は、目的とするポリペプチドを製造する糸状菌細胞を得るための特定の順序に限定されないことが理解されるであろう。場合により内因性sec61遺伝子の修飾と組合される、sec61遺伝子の導入は、目的とするポリペプチドの製造のための細胞の構成における任意のステップで行われうる。
【0073】
ポリペプチドの製造
本発明の別の態様は、以前の態様の宿主細胞、好ましくは、糸状菌宿主細胞における目的とするポリペプチドの製造のための方法に関し、
(a)目的とするポリペプチドの製造のために適切な栄養培地において以前の態様の宿主細胞を培養するステップと、
(b)栄養ブロスから目的とするポリペプチドを回収するステップと
を含んで成る方法に関する。
【0074】
本発明の宿主細胞は、当業界で周知の方法を使用して目的とするポリペプチドの製造に適切な栄養培地中で培養される。例えば、細胞は、目的とするポリペプチドが発現および/または単離されることを可能にする適切な培地および条件下に行われる、実験的または工業的発酵装置における振盪フラスコ培養、小規模または大規模発酵(連続発酵、バッチ発酵、流加発酵、または固相発酵を含む)によって培養されうる。培養は、当業界で周知の方法を使用して少なくとも炭素および窒素および無機塩を含んで成る適切な栄養培地で行われる。目的とするポリペプチドは、当業界で周知の方法によって栄養培地から回収されうる。ポリペプチドが栄養培地へ分泌される場合は、ポリペプチドは培地から直接回収されうる。ポリペプチドが分泌されない場合は、これは細胞溶解物から回収されうる。必要に応じて、回収はさらに従来の手段による単離および精製を含んで成りうる。
【0075】
例えば、ポリペプチドは、遠心分離、ろ過、抽出、噴霧乾燥、蒸発、または沈殿を含むがこれらに限定されない従来の方法によって単離されうる。次いで、単離ポリペプチドは、さらに、クロマトグラフィー(例えば、イオン交換、親和性、疎水性、クロマト分画、およびサイズ排除)、電気泳動法(例えば、予備的等電点電気泳動法)、分画溶解(例えば、硫酸アンモニウム沈殿)、または抽出(例えば、Protein Purification,J.−C.ヤンソン(Janson)とラース ライデン(Lars Ryden)編、VCHパブリッシャーズ(Publischers)、ニューヨーク、1989年を参照)を含むがこれらに限定されない当業者に周知のさまざまな方法によって精製されうる。
【0076】
好都合にも、目的とするポリペプチドは、緩衝剤を含む適切な(固体または液体)担体または希釈剤と混合され、ポリペプチド組成物を生じうる。ポリペプチドは、例えば固体担体上に固定された担体に付着され、またはこれと混合されうる。
【0077】
目的とするポリペプチドは、ポリペプチドに特異的である当業界で周知の方法を使用して検出されうる。これらの検出方法としては、特異的抗体、酵素生成物の形成、酵素基質の消失、またはSDS PAGEの使用を挙げることができる。例えば、酵素アッセイを使用し、ポリペプチドの活性を測定することができる。酵素活性を測定するための方法が、多くの酵素について当業界で周知である。
【0078】
本発明の方法において、宿主細胞、好ましくは、糸状菌宿主細胞は、同一の条件下に培養した場合、SEC61発現に対して本発明による修飾されていないその対応する対応細胞より少なくとも約20%、好ましくは少なくとも約50%、より好ましくは少なくとも約100%、さらにより好ましくは少なくとも約200%、かつ最も好ましくは少なくとも約300%以上の目的とするポリペプチドを産生する。
【0079】
目的とするポリペプチドは、宿主細胞と相同(天然)または異種のポリペプチドでありうる。「ポリペプチド」という語は、本明細書では、コードされた生成物の特定の長さを指すことはなく、したがってペプチド、オリゴペプチド、およびタンパク質を包含する。
【0080】
異種ポリペプチドは、宿主細胞によって自然に産生されないポリペプチドを包含しうる。目的とするポリペプチドは、核酸配列によってコードされる宿主細胞を原産とするポリペプチドをも包含しうるが、その発現はポリペプチドをコードする核酸配列と異質の1つもしくはそれ以上の制御配列によって制御される。
【0081】
目的とするポリペプチドを産生するように遺伝子操作されていないが、かかるポリペプチドを自然に産生する菌株において本発明のSEC61ポリペプチドの過剰発現を達成することも可能である。
【0082】
好ましい実施形態において、目的とするポリペプチドは、抗体もしくはその一部分、抗原、凝固因子、酵素、ホルモン、もしくはホルモン変異体、受容体もしくはその一部分、調節タンパク質、構造的タンパク質、リポーター、または輸送タンパク質である。
【0083】
より好ましい実施形態において、目的とするポリペプチドは酵素であり、最も好ましくは、酵素はオキシドレダクターゼ、トランスフェラーゼ、ヒドラーゼ、リアーゼ、イソメラーゼ、またはリガーゼである。
【0084】
さらにより好ましい実施形態において、酵素はアミノペプチダーゼ、アミラーゼ、カルボヒドラーゼ、カルボキシキペプチダーゼ、カタラーゼ、セルラーゼ、キチナーゼ、クチナーゼ、デオキシリボヌクレアーゼ、デキストラナーゼ、エステラーゼ、アルファ−ガラクトシダーゼ、ベータ−ガラクトシダーゼ、グルコアミラーゼ、アミラーゼ、アルファ−グルコシダーゼ、ベータ−グルコシダーゼ、ハロペルオキシダーゼ、インベルターゼ、ラッカーゼ、リパーゼ、ホスホリパーゼ、マンノシダーゼ、ムタナーゼ、オキシダーゼ、ペクチナーゼ、ペルオキシダーゼ、フィターゼ、ポリフェノールオキシダーゼ、プロテアーゼ、ペプチダーゼ、リボヌクレアーゼ、トランスグルタミナーゼ、またはキシラナーゼである。
【0085】
別のさらにより好ましい実施形態において、ポリペプチドはヒトインスリンもしくはその類似体、ヒト成長ホルモン、エリトロポエチン、またはインスリノトロピンである。
【0086】
目的とするポリペプチドをコードする核酸配列は、原核、真核、または他の供給源から獲得されうる。本発明の目的のために、所定の供給源に関連して本明細書で使用される「から獲得される」という語は、供給源からの遺伝子が挿入されている供給源すなわち細胞によってポリペプチドが製造されることを意味する。
【0087】
本発明は、本発明の範囲の限定として考えるべきではない以下の実施例によってさらに説明される。
【実施例】
【0088】
実験情報
菌株
WT 1:本A.ニゲル(niger)菌株が野生型菌株として使用される。本菌株は寄託番号CBS513.88によりCBS研究所で寄託されている。
【0089】
WT 2:本A.ニゲル(niger)菌株はグルコアミラーゼ(glaA)をコードする遺伝子の欠失を含んで成るWT 1菌株である。WT 2は、CBS 513.88のゲノムにおけるglaA特異的DNA配列を削除するし方が記載されているEP635574号明細書に記載されている「MARKER−GENE FREE」法を使用して構成される。この方法は結果として、異種DNA配列を有さないMARKER−GENE FREE ΔglaA組換えA.ニゲル(niger)CBS513.88菌株をもたらす。
【0090】
WT 3:本A.ニゲル(niger)菌株は主要な細胞外アスパラギン酸プロテアーゼpepAをコードするpepA遺伝子の欠失を含んで成るWT 2菌株である。WT 3はEP0635574号明細書に記載されている「MARKER−GENE FREE」法を使用して構成される。本特許に記載された方法は、ファン・デン・ホンバーグ(van den Hombergh)ら(ファン・デン・ホンバーグ(van den Hombergh)JP、ソレウィン・ゲルプケ(Sollewijn Gelpke)MD、ファン・デ・フォンデルフォルト(Van de Vondervoort)PJ、ブクトン(Buxton)FP、ビセル(Visser)J.(1997年)−アスペルギルス・ニゲル(Aspergillus niger)における3つの酸プロテアーゼの崩壊−−プロテアーゼスペクトル、細胞内タンパク質溶解、および標的タンパク質の分解に対して作用する(Disruption of three acid proteases in Aspergillus niger−−effects on protease spectrum, intracellular proteolysis, and degradation of target proteins)−Eur J Biochem.247(2)605−13頁)によって記載されたCBS513.88のゲノムにおけるpepA特異的DNA配列を削除するために使用される。この方法は結果として、異種DNA配列を有さないMARKER−GENE FREE ΔpepA、ΔglaA組換えA.ニゲル(niger)CBS513.88菌株をもたらす。
【0091】
WT 4:本ペニシリウム・クリゾゲナム(Penicillium chrysogenum)菌株は野生型菌株として使用される。本菌株は寄託番号CBS455.95によりCBS研究所で寄託されている。
【0092】
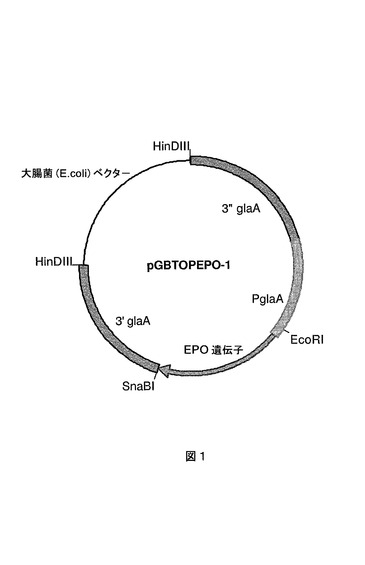
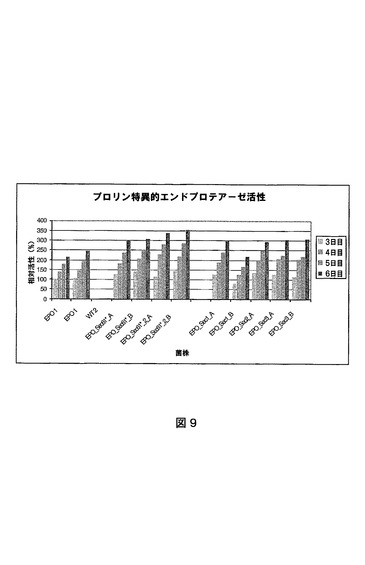
EPO 1:本A.ニゲル(niger)菌株は、他の箇所(国際公開第02/45524号パンフレット)で公開されているプロリン特異的エンドプロテアーゼのA.ニゲル(niger)epo遺伝子の多重コピーを含んで成るWT2菌株である。EPO 1は、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載されているプロリン特異的エンドプロテアーゼの遺伝子コードを含んで成るpGBAAS−1(EP0635574号明細書に記載されている通り構成)およびpGBTOPEPO−1ベクター(図1)で示されているベクターを含有するamdS選択可能なマーカー遺伝子の共形質転換によって構成される。形質転換および対抗選択法は結果としてグルコアミラーゼプロモーターの制御下にプロリン特異的エンドプロテアーゼコード遺伝子の多重コピーを含有するMARKER−GENE FREE EPO 1菌株がもたらされる。
【0093】
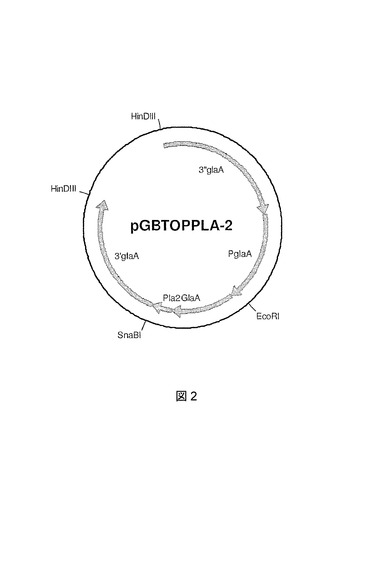
PLA 1:異種ブタホスホリパーゼA2(PLA2)タンパク質は、モデルタンパク質として選択される。本タンパク質は高量でA.ニゲル(niger)において産生するのが困難であることが以前に証明されている(ロベルツ(Roberts)I.N.、ジーンズ(Jeenes)D.J.、マッケンジー(MacKenzie)D.A.、ウィルキンソン(Wilkinson)A.P.、サムナー(Sumner)I.G.およびアーチャー(Archer)D.B.(1992年)−アスペルギルス・ニゲル(Aspergillus niger)における異種遺伝子発現(Heterologous gene expression in Aspergillus niger)。グルコアミラーゼ−ブタ膵ホスホホリパーゼA2融合タンパク質が分泌され加工されて成熟酵素をもたらす。Gene 122:155−161頁)。PLA2の過剰発現の断片は、A.ニゲル(niger)の天然グルコアミラーゼA遺伝子とのプロPLA2の融合として製造され、ロベルツ(Roberts)ら(1992年)によって記載されているように調製される。融合タンパク質は、ゴルジにおいて加工されるためにkex1スプライシング部位を含有する。本glaA−pla2融合遺伝子は、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載されている同じ方法を使用してA.ニゲル(niger)pGBTOP発現ベクターへクローン化され、結果としてpGBTOPPLA−1がもたらされる(図2)。
【0094】
PLA 1 A.ニゲル(niger)菌株は、グルコアミラーゼ−ブタ膵ホスホリパーゼA2融合タンパク質コード遺伝子の多重コピーを含んで成るWT 3菌株である。PLA 1はベクターpGBAAS−1およびpGBTOPPLA−1ベクターを含有するamdS選択可能なマーカー遺伝子の共形質転換によって構成される(図2)。形質転換および対抗選択法の結果として、グルコアミラーゼプロモーターの制御下にグルコアミラーゼ−ブタ膵ホスホリパーゼA2融合タンパク質コード遺伝子の多重コピーを含有するMARKER−GENE FREE PLA 1菌株がもたらされる。
【0095】
A.ニゲル(niger)振盪フラスコ発酵
A.ニゲル(niger)菌株を実施例、すなわち、国際公開第99/32617号パンフレットの「アスペルギルス・ニゲル(Aspergillus niger)振盪フラスコ発酵」の部に記載された前培養培地20ml中で前培養する。一夜増殖後、10mlの本培養液を発酵培地(FM)に移す。発酵培地(FM)は1リットル当り以下を含有する。すなわち、グルコース・1H2O 82.5g、Maldex 15(ブーム・メッペル(Boom Meppel)、オランダ)25g、クエン酸2g、NaH2PO4・1H2O 4.5g、KH2PO4 9g、(NH4)2SO4 15g、ZnCl2 0.02g、MnSO4・1H2O 0.1g、CuSO4・5H2O 0.015g、CoCl2・6H2O 0.015g、MgSO4・7H2O 1g、CaCl2・2H2O 0.1g、FeSO4・7H2O 0.3g、MES(2−[N−モルフォリノ]エタンスルホン酸)30g、pH6。
【0096】
FMにおける発酵は、指示された日数の間、34℃および170rpmで発酵ブロス100mlを含む整流装置を備えた500mlフラスコで実行される。
【0097】
酵素アッセイ
PLA2ホスホリパーゼ活性
アスペルギルス・ニゲル(Aspergillus niger)培養ブロス中のホスホリパーゼPLA2活性(PLA2)を分光光度的に測定するために、人工基質、すなわち、1,2−ジチオジオクタノイルホスファチジルコリン(diC8、基質)を使用する。PLA2はA2位置でのスルフィド結合を加水分解し、チオオクタン酸を解離する。チオオクタン酸は4,4ジチオピリジン(呈色試薬、4−DTDP)と反応し、4−チオピリドンを形成する。4−チオピリドンは、334nmの波長を有する放射線を吸収する4−メルカプトピリジンと互変異性平衡にある。同波長での吸光度の変化を測定する。一単位は、37℃およびpH4.0で1,2−ジチオジオクタノイルホスファチジルコリンからチオ−オクタン酸を毎分1nmol遊離させる酵素の量である。
【0098】
基質溶液は、エタノール66ml当りdiC8結晶1gを溶解し、緩衝酢酸溶液264mlを添加することによって調製される。緩衝酢酸溶液は、0.2%トリトン(Triton)−X100含有0.1M緩衝酢酸溶液pH3.85を含んで成る。呈色試薬は11mM 4,4−ジチオピリジン溶液である。これは2mlエッペンドルフサンプルカップ中に4,4−ジチオピリジン5.0mgを秤量し、エタノール1.00ml中に溶解することによって調製される。1.00mlのmilli−Q水を添加する。
【0099】
プロリン特異的エンドプロテアーゼ活性
プロリン特異的エンドプロテアーゼのタンパク質分解活性は、クエン酸/リン酸2ナトリウム緩衝液pH5中37℃下CBZ−Gly(cine)−Pro(line)−pNAを使用して時間内に410nmで分光光度分析的に測定される。1Uプロリン特異的エンドプロテアーゼを上記の条件下、pH5および37℃でCBZ−Gly(cine)−Pro(line)−pNAを毎分1μmol(マイクロモル)を変換する酵素の量と定義する。
【0100】
グルコアミラーゼ活性アッセイ
グルコアミラーゼ活性は、p−ニトロフェニル α−D−グルコピラノシド(シグマ(Sigma))を使用して国際公開第98/46772号パンフレットに記載されている通りに測定される。
【0101】
実施例1:発現ベクターにおけるsec61遺伝子のクローニング、Sec61転位チャンネルのコード
アスペルギルス・ニゲル(Aspergillus niger)菌株WT 1およびペニシリウム・クリゾゲナム(Penicillium chrysogenum)WT 4のゲノムDNAを配列決定し、分析する。Sec61と相同体と注釈付きの翻訳タンパク質を有する1つの遺伝子が同定され、sec61と命名されている。オープンリーディングフレーム(ORF)(イントロンによる)および約180bpの5’非翻訳領域(UTR)および327bpの3’UTRを含んで成るA.ニゲル(niger)sec61遺伝子座のゲノム配列は、配列番号1に示されている。オープンリーディングフレーム(ORF)(イントロンによる)および約12bpの5’非翻訳領域(UTR)および66bpの3’UTRを含んで成るペニシリウム・クリゾゲナム(Penicillium chrysogenum)sec61遺伝子座のゲノム配列は、配列番号7に示されている。
【0102】
WT 1からのゲノムDNAは、配列番号13および配列番号14のプライマーを使用し、野生型sec61遺伝子を増幅させ、適切な制限部位を付着させるPCR反応におけるテンプレートとして使用される。同時に、同じ実験をcDNAをテンプレートとして使用して実行する。すなわち、PCR反応は配列番号14および15のプライマーを使用して行われ、野生型sec61cDNAを増幅させる。すべてのPCR反応、cDNA合成、ライゲーション、および形質転換は、Molecular Cloning:A Laboratory Manual,サムブルック(Sambrook)ら、ニューヨーク:コールド・スプリング・ハーバー・プレス(Cold Spring Harbour Press)、1989年に記載されている通りに実行される。さまざまに独立して増幅されたPCR断片の配列は配列分析によって決定される。配列番号1は、A.ニゲル(niger)sec61ゲノム配列を含んで成る。A.ニゲル(niger)のSec61タンパク質をコードするcDNA配列のORFは配列番号2として提供されている。配列番号2の翻訳アミノ酸配列は配列番号3で指定され、これはA.ニゲル(niger)WT 1のSec61タンパク質を表す。
【0103】
WT 4からのゲノムDNAは、配列番号10および配列番号11のプライマーを使用し、野生型SEC61遺伝子を増幅させ、適切な制限部位を付着させるPCR反応におけるテンプレートとして使用される。同時に、同じ実験をcDNAをテンプレートとして使用して実行する。すなわち、PCR反応は配列番号11および12のプライマーを使用して行われ、野生型sec61cDNAを増幅させる。さまざまに独立して増幅されたPCR断片の配列は配列分析によって決定される。配列番号7は、ペニシリウム・クリゾゲナム(Penicillium chrysogenum)sec61ゲノム配列を含んで成る。P.クリゾゲナム(chrysogenum)のSec61タンパク質をコードするcDNA配列のORFは配列番号8として提供されている。配列番号8の翻訳アミノ酸配列は配列番号9で指定され、これはP.クリゾゲナム(chrysogenum)WT 4のSec61タンパク質を表す。
【0104】
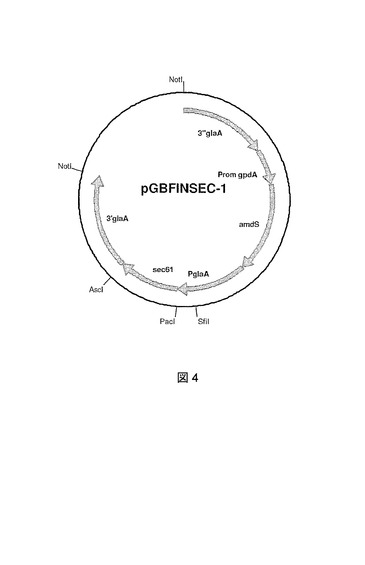
A.ニゲル(niger)sec61ゲノムDNAおよびcDNA配列を含んで成る結果として生じるPCR断片は、メーカーの指示に従ってPaclおよびAsclで切断され、Pacl、Ascl線状化pGBFIN−18A.ニゲル(niger)発現ベクターでライゲートされる(図3)。pGBFIN−18ベクターのプラスミドおよび構成は国際公開第99/32617号パンフレットに記載されている。これは結果として、Sec61がglaAプロモーターの制御下に配置されているpGBFINSEC−1およびpGBFINSEC−2の構築物をもたらした(pGBFINSEC−1のマップについて図4を参照)。
【0105】
アスペルギルス・ニズランス(Aspergillus nidulans)の構成的gpdAプロモーターの制御下、およびpGBFIN−18ベースのA.ニゲル(niger)発現ベクターにも存在するamdS遺伝子は、選択マーカーとして使用され、唯一のN源としてアセトアミドでの増殖が誘発されうる(ケリー(Kelly),J.M.およびハインズ(Hynes),M.J.(1985年)。アスペルギルス・ニズランス(Aspergillus nidulans)のamdS遺伝子によるアスペルギルス・ニゲル(Aspergillus niger)の形質転換。EMBO J.4、475−479頁に記載)。
【0106】
実施例2:遺伝子置換および過剰発現ベクターにおける修飾Sec61転位チャンネルをコードする、sec61*の構成
古典的な分子生物学法をMolecular Cloning:A Laboratory Manual、サンブルック(Sambrook)ら、ニューヨーク:コールド・スプリング・ハーバー・プレス(Cold Spring Harbour Press)、1989年に記載されている通り使用する。A.ニゲル(niger)欠失ベクターの一般的な設計はEP635574号明細書および国際公開第98/46772号パンフレットに記載されている。
【0107】
修飾sec61遺伝子は、セリン376がトリプトファンによって置換されているSec61*タンパク質をコードするsec61*と呼ばれ、配列オーバーラップ伸長PCR(SOE−PCR、Gene.1989年4月15、77(1):51−9頁、ホ(Ho)SN、ハント(Hunt)HD、ホルトン(Horton)RM、プレン(Pullen)JK、ピース(Pease)LR、「ポリメラーゼ連鎖反応を使用するオーバーラップ伸長による部位特異的突然変異誘発(Site−directed mutagenesis by overlap extension using the polymerase chain reaction)」に記載)によって構成される。配列番号18および配列番号17で識別されたオリゴヌクレオチド、およびA.ニゲル(niger)WT 1のゲノムDNAをテンプレートとして使用することにより、断片Aで識別された1.4kbのsec61遺伝子の5’領域がPCRによって増幅される。また、Ascl制限部位が断片Aの5’末端に付着されている。配列番号16および配列番号19で識別されたオリゴヌクレオチドおよびA.ニゲル(niger)WT 1のゲノムDNAをテンプレートとして使用することにより、断片Bで識別された3’−UTRを含む0.6kbのsec61遺伝子の3’領域がPCRによって増幅される。また、NotlおよびEcoRl制限部位が断片Bの3’末端に付着されている。結果として生じる断片AおよびBは、SOE−PCR、配列番号18および配列番号19で識別されるオリゴヌクレオチド、および断片AおよびBによって融合され、2kb断片Cを生成する。本断片CはpCR2.1−TOPO(インビトロジェン(Invitrogen))でクローン化され、pCR_Sec61*を生成する。pCR_Sec61*のPCR生成断片の配列は確認され、配列番号4で提供されている。修飾Sec61*タンパク質のコード配列は配列番号5で提供されている。配列番号5の翻訳アミノ酸配列は配列番号6で指定され、これは修飾Sec61*タンパク質を表す。
【0108】
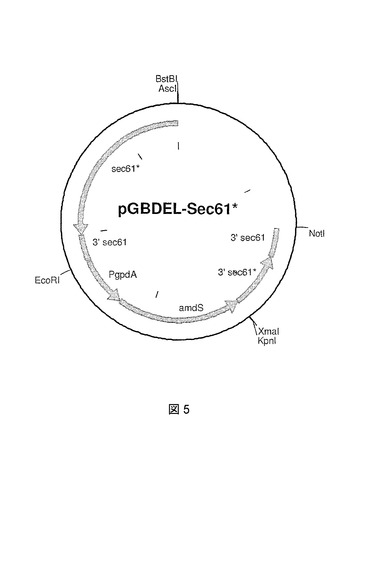
ベクターpCR_Sec61*はAsclおよびEcoRlで消化され、sec61*断片はAsclおよびEcoRl消化ベクターpGBDELへ導入され、pGBDEL−1(データは示さず)を生成する。その後、ベクターpCR_Sec61*はEcoRVおよびNotlで消化され、3’sec61*断片はSmalおよびNotl消化ベクターpGBDEL−1へ導入され、pGBDEL−SEC61*を生成する(図5)。
【0109】
ベクターpCR_Sec61*は配列番号13および配列番号14のプライマーを使用し、修飾sec61遺伝子を増幅させ、適切な制限部位を付着させるPCR反応におけるテンプレートとして使用される。修飾sec61*配列を含んで成る結果として生じるPCR断片は、メーカーの指示に従ってPaclおよびAsclで切断され、Pacl、Ascl線状化pGBFIN−18A.ニゲル(niger)発現ベクターでライゲートされるが、ここでPglaAプロモーターは、pepCプロモーター(フレデリック(Frederick)GD、ロンブーツ(Rombouts)P、ブクストン(Buxton)FP(1993年)−アスペルギルス・ニゲル(Aspergillus niger)からのセリンプロテアーゼをコードする遺伝子、pepCのクローニングおよび特徴づけ(Cloning and characterisation of pepC, a gene encoding a serine protease from Aspergillus niger.)−Gene 125(1):57−64頁)によって置換される。これは結果としてpGBFINSEC−3構築物をもたらしたが、ここで修飾Sec61*遺伝子はpepCプロモーターの制御下に配置される(図6)。
【0110】
実施例3.野生型sec61および修飾sec61*遺伝子を過剰発現し、修飾sec61*転位チャンネルを発現する真菌宿主細胞
構成的gpdAプロモーターの制御下に配置されるアスペルギルス・ニズランス(Aspergillus nidulans)のamdS遺伝子は、EP635574号明細書に記載されている通り選択マーカーとして使用される。原則的には、pGBFINSEC−1、pGBFINSEC−2、pGBFINSEC−3、およびpGBDEL−SEC61*の線状DNAが指示された制限酵素での消化後に単離される。アスペルギルス・ニゲル(Aspergillus niger)PLA1およびEPO 1菌株は形質転換され、その後のアセトアミドでの形質転換体選択は国際公開第98/46772号パンフレット、EP635574号明細書、および国際公開第99/32617号パンフレットに記載されている通りに行われる。その後、コロニーが標準の方法に従って精製される。
【0111】
PLA1 およびEPO 1菌株の増殖コロニーは、それぞれ、形質転換SECプラスミドのコピー数およびpla2およびepo遺伝子コピーの非喪失について診断される。同様に推定されたコピー数、およびpla2およびepo遺伝子コピーの非喪失のpGBFINSEC−1、pGBFINSEC−2、pGBFINSEC−3の形質転換体が選択される。単一コピー(_A、_B、_C、_...)による数個の形質転換体が選択される。これは結果として、野生型のゲノムsec61(pGBFINSEC−1)、cDNAsec61(pGBFINSEC−2)および修飾sec61*遺伝子(pGBFINSEC−3)がもたらした。PLA 1およびEPO 1菌株の形質転換体は、それぞれ、PLA_SEC1、PLA_SEC2、PLA_SEC3、およびEPO_SEC1、EPO_SEC2、EPO_SEC3で指示されている。
【0112】
sec61遺伝子座でゲノムへ組込まれたpGBDEL−SEC61*断片の形質転換体は、図7に示されている通り、sec61*遺伝子およびamdS遺伝子で野生型sec61遺伝子を置換した。標的のために使用された断片および方法により、相同組換え時の置換は100%ではない。したがって、精製pGBDEL−SEC61*形質転換体は、sec61遺伝子座での相同組込みに対する2つのPCR反応およびsec61*遺伝子配列に対するsec61遺伝子の置換によって診断される。sec61*遺伝子によるsec61の本置換は、テンプレートとしてpGBDEL−SEC61*形質転換体のゲノムDNAを使用し、かつ配列番号20および配列番号22のプライマーを使用し、sec61遺伝子座で修飾sec61*遺伝子を増幅するバンドの第1のPCR増幅によって検出可能である。sec61*遺伝子によるsec61の本置換は、テンプレートとしてpGBDEL−SEC61*形質転換体のゲノムDNAを使用し、かつ配列番号20および配列番号21のプライマーを使用し、障害をチェックして野生型sec61遺伝子を増幅させる第2のPCRによって確認される。また、pla2およびepo遺伝子コピーの非喪失が、それぞれ、試験される。その後、胞子がフルオロアセトアミド培地にプレーティングされ、amdSマーカーを喪失する菌株を選択する。さらに、増殖コロニーが、上記の通り、sec61*遺伝子配列およびpla2またはepo遺伝子コピーの非喪失に対するsec61遺伝子の置換についてPCRによって診断され、候補の菌株が、amdSマーカーを喪失する形質転換体および誘導菌株を含有するamdSのサザン分析によって試験される。sec61*遺伝子に対するsec61遺伝子の置換は、両方の菌株のsec61プローブへのハイブリダイゼーション、形質転換体を含有するamdSにおける全遺伝子座をカバーするDNA断片の約2.2kbサイズの増大、およびamdSマーカーを喪失する誘導菌株における野生型断片によって検出可能である。約40%の菌株は、pGBDEL−SEC61*の相同組込みを示したが、そのうち約70%でsec61*遺伝子に対するsec61遺伝子の置換が検出可能である。菌株PLA_SEC61*およびEPO_SEC61*は、修飾sec61*遺伝子によって置換されるsec61遺伝子を有する代表的な菌株として選択される。
【0113】
また、アスペルギルス・ニゲル(Aspergillus niger)PLA_SEC61*およびEPO_SEC61*は、pGBFINSEC−3の線状DNAで形質転換され、その後のアセトアミドでの形質転換体選択およびコロニー単離が上記の通り行われる。菌株PLA_SEC61*_2およびEPO_SEC61*_2は、修飾Sec61*の過剰発現に対する代表的な菌株として選択され、内因性sec61遺伝子が修飾sec61*遺伝子によって置換され、強いglaAプロモーターの制御下に修飾Sec61*遺伝子の追加のコピーを有し、かつpla2またはepo遺伝子のコピーが非喪失である。
【0114】
実施例4.得られたアスペルギルス・ニゲル(Aspergillus niger)菌株によるタンパク質の分泌の改良
pGBTOPEPO−1構築物の同等のコピーを有する一部のA.ニゲル(niger)WT 2およびEPO_SEC1、EPO_SEC2、およびEPO_SEC3、EPO_SEC61*およびEPO_SEC61*_2形質転換体を選択して振盪フラスコ実験を行う。また、pGBTOPPLA−1構築物の同等のコピーを有する一部のA.ニゲル(niger)WT 3およびPLA_SEC1、PLA_SEC2、およびPLA_SEC3、PLA_SEC61*、およびPLA_SEC61* _2形質転換体を選択し、振盪フラスコ実験を行う。振盪フラスコ実験は、実験方法−「A.ニゲル(niger)振盪フラスコ発酵(A. niger shake flask fermentations)」に記載されている通り100mlの発酵培地(FM)中で行われる。培養ブロスは7日間の培養中に採集し、エンドプロテアーゼおよびPLA2活性を上記の通り測定する。
【0115】
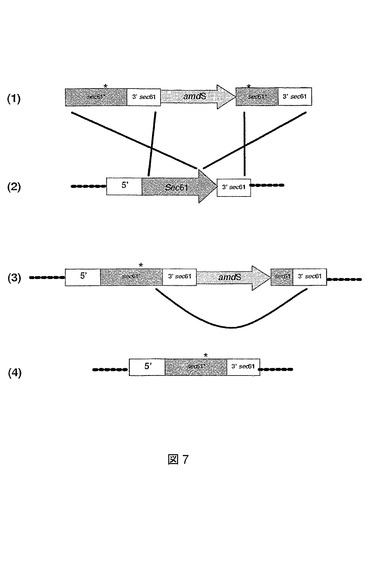
図8は、同じコピー数のA.ニゲル(niger)EPO菌株の形質転換体における相対的プロリン特異的エンドプロテアーゼ活性を示す。発明者らは、Sec61および修飾Sec61*タンパク質の過剰発現が結果としてエンドプロテアーゼ酵素の生産の増大をもたらすと結論づける。過剰発現はグルコアミラーゼglaAおよびプロテアーゼpepCプロモーターを使用して実行されている。また、ゲノムまたはcDNA構築物によるSec61の過剰発現、またはゲノムに依然として存在する内因性sec61遺伝子による修飾Sec61*の過剰発現は結果として、エンドプロテアーゼ酵素の生産の増大をもたらす。
【0116】
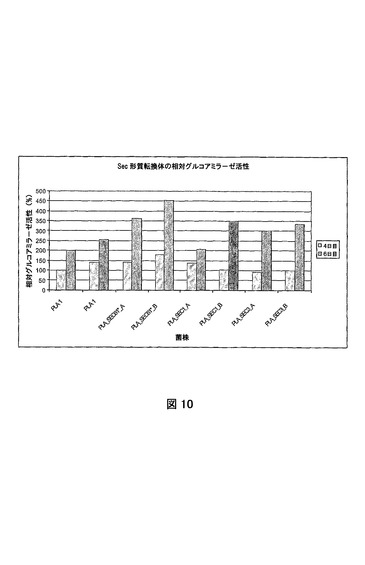
これらの結果は、sec61*およびsec61が、真菌宿主細胞におけるポリペプチドの改良された生産を得る優れた手段であることを示す。図9および10は、それぞれ、A.ニゲル(niger)WT 2、SEC1、SEC2、SEC3、SEC61*、およびSEC61*_2において、およびA.ニゲル(niger)WT 2pGBFIN−PLA2形質転換において測定される際の、PLA2およびグルコアミラーゼ活性を示す。欠失野生型sec61遺伝子を有する菌株において、無傷のA.ニゲル(niger)PLA 1およびPLA_SEC1、PLA_SEC3、およびPLA_SEC61*を有し、同等のコピー数を有する菌株と比べPLA2活性の明らかな増大が確認されることは明らかである。発明者らは、修飾Sec61*タンパク質の発現が結果として、異種ホスホリパーゼA2および相同グルコアミラーゼ酵素の両方の生産の増大をもたらすと結論づける。過剰発現はグルコアミラーゼglaAおよびプロテアーゼpepCプロモーターを使用して実行されている。PLA2および/またはグルコアミラーゼ活性が増大していない菌株は発酵中にコピーを喪失したことが証明されている。したがって、コピー当りの発現は依然としてSec61の過剰発現とともに増大する。これらの結果は、sec61*およびsec61の発現の操作が結果として真菌宿主細胞におけるポリペプチドの生産の改良をもたらすことを示す。
【図面の簡単な説明】
【0117】
【図1】プロリン特異的エンドプロテアーゼ発現ベクターpGBTOPEPO−1のプラスミドマップを示す図である。glaAプロモーターおよびプロリン特異的エンドプロテアーゼepo遺伝子に対するglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素HinDIIIによる消化によって除去されうる。
【図2】PLA2発現ベクターpGBTOPPLA−2のプラスミドマップを示す図である。glaAプロモーター、切断glaA遺伝子、およびプロ−pla2コード配列に対するglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素Notlによる消化によって除去されうる。
【図3】発現ベクターpGBFIN−18のプラスミドマップを示す図である。グルコアミラーゼプロモーターにおける独自のSfilおよびEcoRlクローニング部位の後、PaclおよびAsclクローニング部位によるglaAプロモーターに対するglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素Notlによる消化によって除去されうる。
【図4】発現ベクターpGBFINSEC−1のプラスミドマップを示す図である。glaAプロモーターおよびA.ニゲル(niger)ゲノムsec61遺伝子に対するglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素Notlによる消化によって除去されうる。
【図5】置換ベクターpGBDEL−Sec61*のプラスミドマップを示す図である。amdSマーカーに対するSec61*突然変異遺伝子および3’Sec61*突然変異遺伝子断片が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素AsclおよびNotlによる消化によって除去されうる。
【図6】発現ベクターpGBFINSEC−3のプラスミドマップを示す図である。glaApepCプロモーターおよび修飾sec61*遺伝子に対するglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素Notlによる消化によって除去されうる。
【図7】sec61遺伝子置換を示す概略図である。sec61遺伝子(1)の相同領域によってフランキングされたamdS選択マーカーを含んで成るpGBDEL−Sec61*の線状DNA構築物は、ゲノムsec61遺伝子座(2)で二重相同組換え(X)により結合し、修飾sec61*遺伝子によってゲノムsec61遺伝子コピー(3)を置換する。その後、(修飾)sec61配列(U)の直接の繰返しに及ぶ組換えがamdSマーカーを除去し、結果として修飾sec61*遺伝子(4)によるsec61遺伝子の遺伝子置換イベントが生じる。amdSマーカーが有る場合(3)および無い場合(4)の両方の菌株は機能的修飾sec61*遺伝子を有する。
【図8】さまざまなsec61構築物で形質転換されたEPO 1菌株の相対細胞外プロリン特異的エンドプロテアーゼ活性を示す図である。試料を発酵の数日間に採取する。3日目のEPO 1の発現は100%で設定されている。
【図9】さまざまなsec61構築物で形質転換されたPLA 1菌株の細胞外PLA2活性を示す図である。試料を発酵の数日間に採取する。
【図10】さまざまなsec61構築物で形質転換されたPLA 1菌株の細胞外グルコアミラーゼ活性を示す図である。菌株および試料は図11と同一である。試料を発酵の数日間に採取する。
【技術分野】
【0001】
本発明は組換えDNA技術に関する。具体的には、本発明は、酵母SEC61(サッカロミセス(Saccharomyces)ゲノムデータベース−SGD:http://www.yeastgenome.org/)について記載されたタンパク質輸送活性を有するSEC61ポリペプチド、これらのポリペプチドをコードする核酸配列、および目的とするポリペプチドの製造に適切な宿主細胞の調製におけるその使用に関する。
【背景技術】
【0002】
タンパク質の培養培地への分泌は、分泌経路を構成するさまざまな膜に囲まれたコンパートメントを通じたタンパク質の移動を含む。最初にタンパク質は小胞体、ERの内腔へ転位される。そこからタンパク質は膜小胞のゴルジ複合体に輸送され、ゴルジから血漿膜に輸送される。分泌過程は、分泌されたタンパク質を含有する小胞がドナー膜から摘み取られ、受容体膜に標的され、これと融合される幾つかのステップを含む。これらのステップの各々で幾つかの異なるタンパク質の機能が必要である。
【0003】
糸状菌におけるタンパク質分泌を増大させる幾つかの試みが行われている。異種タンパク質の分泌を増大させる一般的な方法が、シグナル配列を使用することである(例えば、EP0215594号明細書を参照)。ランダム突然変異誘発および分泌されたタンパク質のスクリーニング(スミス(Smith)ら、1985年、サカイ(Sakai)ら、1988年、シュスター(Shuster)ら、1989年、スズキ(Suzuki)ら、1989年、スリープ(Sleep)ら、1991年、ラムサ(Lamsa)およびブレバウム(Bloebaum)、1990年、ダン・コールマン(Dunn−Coleman)ら、1991年、および米国特許出願公開第2002/0068325A1号明細書)または異質のタンパク質の有効に分泌された内因性タンパク質との融合(ワード(Ward)ら、1990年、ハルキ(Harkki)ら、1989年、ニーソネン(Nyyssonen)ら、1993年、ニーソネン(Nyyssoenen)ら、1992年)は、異種タンパク質の分泌をより効率的にするために酵母および糸状菌の両方に広く使用されている。これらの方法の両方は使用が限定されている。ランダム突然変異誘発およびスクリーニングによって単離される突然変異はほぼ完全に劣性であり、倍数体である工業上の菌株へ移動されえない。しばしば得られた突然変異はスクリーニングに使用されるタンパク質の改良された分泌能のみを有する。融合タンパク質法は、各々異種または異質のタンパク質の融合構成の適合化を別個に必要とする。融合タンパク質はしばしば機能ではなく、したがって最終生成物はタンパク質分解的切断によって放出される必要があり、これは製造手順を複雑にする。
【0004】
さらに、それらのタンパク質生成因子としての産業上の重要性により、改良された分泌能を有する糸状菌を得る必要がある。
【発明の詳細な説明】
【0005】
本明細書で参照されるすべての特許および刊行物は、かかる特許および刊行物内に開示されたすべての配列および方法を含めて、参照によって明示的に援用される。これらの特許および刊行物としては、EP357127号明細書、EP635574号明細書、国際公開第97/06261号パンフレット、国際公開第98/46772号パンフレットが挙げられる。
【0006】
SEC61活性を表示するポリペプチド
本発明の第1の態様は新規SEC61ポリペプチドに関する。SEC61ポリペプチドは一般にER膜にわたってポリペプチドの輸送を介在することが知られている。本発明によるSEC61ポリペプチドは、SEC61ポリペプチドと関連していることが一般に知られている少なくとも1つの活性を表示するポリペプチドと定義されている。本発明によるSEC61ポリペプチドは、SEC61ポリペプチドと関連していることが一般に知られているすべての活性をも表示しうる。
【0007】
具体的には、本発明は、
(a)配列番号3によるアミノ酸配列を有するポリペプチドと、
(b)配列番号3によるアミノ酸配列と実質的に相同であるアミノ酸配列を有するポリペプチドと
からなる群から選択されるSEC61ポリペプチドを開示する。
【0008】
本発明との関連で、「実質的に相同」という語は、SEC61ポリペプチドの少なくとも1つの活性を表示し、かつ配列番号3に記載されたアミノ酸配列の少なくとも89%、より好ましくは少なくとも90%、さらにより好ましくは少なくとも92%、さらにより好ましくは少なくとも93%、さらにより好ましくは少なくとも94%、さらにより好ましくは少なくとも95%、さらにより好ましくは少なくとも96%、さらにより好ましくは少なくとも97%、さらにより好ましくは少なくとも98%、かつ最も好ましくは少なくとも99%の同一性の程度を有するポリペプチドを包含することが意味される。同一性の程度は、好ましくは、配列番号3の全長にわたって測定される。
【0009】
本発明のために、2つのアミノ酸配列間の同一性の程度は、2つの配列間で同一であるアミノ酸の割合を指す。同一性の程度は、アルトシュール(Altschul)ら、J. Mol.Biol.215:403−410頁(1990年)に記載されているBLASTアルゴリズムを使用して判定される。BLAST分析を実行するためのソフトウェアは、国立バイオテクノロジー情報センター(the National Center for Biotechnology Information)(http://www.ncbi.nlm.nih.gov/)によって公的に入手可能である。BLASTアルゴリズムパラメータW、T、およびXは、アラインメントの感度および速度を決定する。BLASTプログラムではデフォルトとして11の語長(W)、50のBLOSUM62採点マトリックス(ヘニコフ(Henikoff)とヘニコフ(Henikoff)、Proc.Natl. Acad.Sci.USA 89:10915頁(1989年)を参照)アラインメント(B)、10の期待値(E)、M=5、N=−4、および両方の鎖の比較が使用される。
【0010】
好ましい実施形態において、本発明のポリペプチドは配列番号3によるアミノ酸配列を有する。
【0011】
別の実施形態において、配列番号3によるアミノ酸配列と実質的に相同であるポリペプチドのアミノ酸配列は、天然または人工の変異(突然変異誘発)による配列番号3のアミノ酸配列の点で異なりうる。配列番号3によるアミノ酸配列と実質的に相同であるアミノ酸配列を有するポリペプチドは、配列番号3の配列と比べ少なくとも1つの修飾を含有しうる。前記修飾はアミノ酸置換、付加、または欠失でありうる。
【0012】
好ましい実施形態において、本発明のポリペプチドは、5個のアミノ酸が、好ましくは4個のアミノ酸が、より好ましくは3個のアミノ酸が、さらにより好ましくは2個のアミノ酸が、かつ最も好ましくは1個のアミノ酸が、配列番号3に記載されたアミノ酸配列と異なり、かつさらに配列番号3に記載されたアミノ酸配列を有するポリペプチドと同じSEC61活性を有するアミノ酸配列を有する。
【0013】
別の好ましい実施形態において、配列番号3によるアミノ酸配列と実質的に相同であるアミノ酸配列を有するポリペプチドが、配列番号9によるアミノ酸配列を有する。
【0014】
本発明の別の好ましい実施形態において、配列番号3によるアミノ酸配列との少なくとも85%、好ましくは少なくとも87%、より好ましくは少なくとも89%、さらにより好ましくは少なくとも90%、さらにより好ましくは少なくとも92%、さらにより好ましくは少なくとも93%、さらにより好ましくは少なくとも94%、さらにより好ましくは少なくとも95%、さらにより好ましくは少なくとも96%、さらにより好ましくは少なくとも97%、さらにより好ましくは少なくとも98%、かつ最も好ましくは少なくとも99%の同一性の程度を有するSEC61ポリペプチドが提供されるが、ここで少なくとも1個のアミノ酸が、配列番号3の第373〜379位置に対応する、すなわち配列番号3の第373〜379位置に存在するアミノ酸配列ALFSKTWに対応するコンセンサスモチーフで修飾されている。好ましくは、コンセンサスモチーフで修飾されている少なくとも1個のアミノ酸は、配列番号3の第376位置に対応する位置にある。より好ましくは、配列番号3の第376位置に対応するアミノ酸は、トリプトファン、フェニルアラニン、チロシン、またはヒスチジンによって置換される。最も好ましくは、配列番号3の第376位置に対応する位置におけるセリンはトリプトファンによって置換される。かかる修飾SEC61ポリペプチドの例は、配列番号6によるアミノ酸配列を有するポリペプチドである。
【0015】
コンセンサスモチーフにおけるさらなる変異が、配列番号3の第374位置に対応する位置でロイシン、バリン、フェニルアラニン、またはメチノン(methinone)の存在、および/または配列番号3の第377位置に対応する位置におけるリシン、アラニン、トレオニン、またはアルギニンの存在、および/または配列番号3の第378位置に対応する位置におけるトレオニン、またはロイシンの存在でありうる。
【0016】
SEC61ポリペプチドをコードする単離糸状菌核酸配列
本発明の第2の態様は、第1の態様のSEC61ポリペプチドをコードする核酸配列を含んで成る単離ポリヌクレオチドに関する。
【0017】
具体的には、SEC61ポリペプチドをコードするポリヌクレオチドまたは核酸配列は、
(a)配列番号1によるポリヌクレオチドと、
(b)配列番号2による核酸配列と、
(c)配列番号2の核酸配列と少なくとも90%の同一性の程度を有する核酸配列と、
(d)配列番号3のアミノ酸配列と実質的に相同であるアミノ酸配列を有するポリペプチドをコードする核酸配列と
からなる群から選択される。
【0018】
好ましい実施形態において、特徴(c)の核酸配列は、配列番号2に記載された核酸配列と少なくとも91%、より好ましくは少なくとも92%、さらにより好ましくは少なくとも93%、さらにより好ましくは少なくとも94%、さらにより好ましくは少なくとも95%、さらにより好ましくは少なくとも96%、さらにより好ましくは少なくとも97%、さらにより好ましくは少なくとも98%、最も好ましくは少なくとも99%の同一性の程度を有する。
【0019】
本発明のために、2つの核酸配列間の同一性の程度は、同一性表、10のギャップペナルティ、10のギャップ長ペナルティによるClustral法(ヒギンズ(Higgins)、1989年、CABIOS 5:151−153頁)によって判定される。
【0020】
別の好ましい実施形態において、単離核酸配列は、配列番号3のアミノ酸配列を有するポリペプチド、またはSEC61活性を有するその断片をコードする。
【0021】
別の好ましい実施形態において、単離核酸配列は、配列番号9のアミノ酸配列を有するポリペプチド、またはSEC61活性を有するその断片をコードする。
【0022】
別の好ましい実施形態において、SEC61ポリペプチドをコードする核酸配列は配列番号2または配列番号8に記載されている核酸配列を有する。
【0023】
別の好ましい実施形態において、単離核酸配列は、配列番号3によるアミノ酸配列と少なくとも85%の同一性の程度を有するアミノ酸配列を有するSEC61ポリペプチドをコードし、ここで少なくとも1個のアミノ酸が配列番号3の第373〜379位置に対応する、すなわち配列番号3の第373〜379位置に存在するアミノ酸配列ALFSKTWに対応するコンセンサスモチーフで修飾されている。例えば、単離核酸配列は、配列番号6によるアミノ酸配列を有するポリペプチドをコードし、好ましくは配列番号5の核酸配列である。
【0024】
本明細書で使用される「単離ポリヌクレオチドまたは核酸配列」という語は、アガロース電気泳動によって測定されるように、例えば、少なくとも約20%純粋、好ましくは少なくとも約40%純粋、より好ましくは少なくとも約60%純粋、さらにより好ましくは少なくとも約80%純粋、最も好ましくは少なくとも約90%純粋な他の核酸配列を基本的に含んでいないポリヌクレオチドまたは核酸配列を指す。例えば、単離核酸配列が遺伝子工学で使用される標準のクローニング法によって獲得され、核酸配列をその天然位置から再生される異なる部位へ移動させうる。
【0025】
ポリヌクレオチドまたは核酸配列は、ゲノム、cDNA、RNA、半合成、合成起源、またはそのいずれかの組合せでありうる。
【0026】
SEC61ポリペプチドをコードし、かつ配列番号3によるタンパク質と実質的に相同であるアミノ酸配列を有する単離核酸分子が、配列番号2によるコードヌクレオチド配列と比べ、1つもしくはそれ以上のヌクレオチド修飾、すなわち置換、付加、または欠失を含有し、1つもしくはそれ以上のアミノ酸修飾、すなわち置換、欠失、または挿入がコードされたタンパク質に存在するようになりうる。かかる修飾は天然変異によるものであり、または部位特異的突然変異誘発もしくはPCR介在突然変異誘発など標準の突然変異誘発技術によって導入されうる。
【0027】
核酸配列によってコードされるポリペプチドにおける異なるアミノ酸残基をもたらさないsec61コード配列へヌクレオチド置換を導入することも可能である。これは、例えば、コドン使用がSEC61タンパク質の発現を目的とする宿主生物のコドン使用に対応する核酸配列を引き起こすのに有利でありうる。
【0028】
コードされたタンパク質のアミノ酸配列の変更をもたらしうるDNA配列多型が所定の集団内に天然に存在しうることは当業者には明らかであろう。かかる遺伝子多型は、異なる集団からの、または天然対立遺伝子変異による集団内の細胞に存在しうる。
【0029】
本発明によるSEC61ポリペプチドおよびコードする核酸配列は、真核細胞、好ましくは、真菌、より好ましくは、糸状菌から獲得されうる。
【0030】
「糸状菌」としては、真菌類(Eumycota)および卵菌類(Oomycota)の下位分類のすべての糸状形態(ホークスワース(Hawksworth)ら、1995年、上記によって規定)が挙げられる。糸状菌菌株としては、アクレモニウム属(Acremonium)、アスペルギルス属(Aspergillus)、オーレオバシディウム属(Aureobasidium)、クリプトコッカス属(Cryptococcus)、フィリバシジウム属(Filibasidium)、フザリウム属(Fusarium)、フミコーラ属(Humicola)、マグナポルテ属(Magnaporthe)、ムコール属(Mucor)、ミセリオフトラ属(Myceliophthora)、ネオカリマスティックス属(Neocallimastix)、ニューロスポラ属(Neurospora)、パエシロミセス属(Paecilomyces)、ペニシリウム属(Penicillium)、ピロミセス属(Piromyces)、シゾフィラム属(Schizophyllum)、タラロミセス属(Talaromyces)、サーモアスカス属(Thermoascus)、チエラビア属(Thielavia)、トリポクラジウム属(Tolypocladium)、およびトリコデルマ属(Trichoderma)の菌株が挙げられるが、これらに限定されない。
【0031】
より好ましい実施形態において、本発明のSEC61活性を有するポリペプチドをコードする核酸配列は、A.アワモリ(awamori)またはA.ニズランス(nidulans)などアスペルギルス属(Aspergillus)の菌株から得られる。好ましくは、この核酸配列はA.ニゲル(niger)またはA.オリゼー(oryzae)の菌株から得られる。さらにより好ましくは、この核酸配列はA.ニゲル(niger)の菌株の単離株、例えば、配列番号1または配列番号2に記載の核酸配列から得られる。
【0032】
別の実施形態において、本発明のSEC61活性を有するポリペプチドをコードする核酸配列は、F.オキシスポルム(oxysporum)またはF.ヴェネナツム(venenatum)などフザリウム属(Fusarium)の菌株、またはP.クリソゲナム(chrysogenum)などペニシリウム属(Penicillium)の菌株、例えば、配列番号7または配列番号8に記載された核酸配列から得られる。
【0033】
上記の種について、本発明は完全および不完全な状態、および他の分類学的同等物、例えば、それらが知られている種名にかかわらず、アナモルフを包含することが理解されるであろう。当業者は、適切な同等物の同一性を容易に理解するであろう。例えば、ポリペプチドは、それらが知られている種名にかかわらず、レーパー(Raper),K.D.およびフェネル(Fennel),D.I.(1965年。The Genus Aspergillus、ウィルキンズ社(The Wilkins Company)、Baltimore、メリーランド州(MD))によって規定されたアスペルギルス属(Aspergillus)の分類学的同等物である微生物から得られうる。
【0034】
さらに、SEC61活性を有するポリペプチドは、例えば、本発明によるポリヌクレオチドに基づくプローブを使用して自然(例えば、土、堆肥、水等)から単離される微生物を含む、糸状菌以外の供給源から同定され獲得されうる。自然の生息地から微生物を単離するための方法が当業界で公知である。
【0035】
本発明のSEC61ポリペプチドをコードするDNA配列はハイブリダイゼーションによって得られうる。本発明のsec61DNAの変異(例えば、天然対立遺伝子多型)および相同体が、標準のハイブリダイゼーション法によるハイブリダイゼーションプローブとして、好ましくはきわめてストリンジェントなハイブリダイゼーション条件下にこれらの核酸またはその適切な断片を使用して本明細書に開示された核酸との相同性に基づき単離されうる。
【0036】
ハイブリダイゼーション反応の「ストリンジェンシー」は、当業者によって容易に判定される。ハイブリダイゼーション反応のストリンジェンシーの追加の詳細および説明については、オースベル(Ausubel)ら、Current Protocols in Molecular Biology、ワイリー・インターサイエンス・パブリッシャーズ(Wiley Interscience Publishers)、(1995年)を参照。
【0037】
核酸配列は、例えば、問題となっている微生物のゲノムまたはcDNAライブラリーのスクリーニングによって単離されうる。SEC61活性を有するポリペプチドをコードする核酸配列が、例えば、配列番号2由来のプローブで検出されると、配列は当業者に周知である方法を利用して単離またはクローン化されうる(例えば、J.サンブルック(Sambrook)、E.F.フリッチュ(Fritsch)、およびT.マニアチス(Maniatis)、1989年、Molecular Cloning,A Laboratory Manual、第2版、コールド・スプリング・ハーバー(Cold Spring Harbor)、ニューヨーク(New York)を参照)。
【0038】
かかるゲノムDNAからの本発明の核酸配列のクローニングは、例えば、ポリメラーゼ連鎖反応(PCR)または共通の構造的特徴を有するクローン化DNA断片を検出する発現ライブラリーの抗体スクリーニングに基づく方法を使用しても達成されうる(例えば、イニス(Innis)ら、1990年、PCR: A Guide to Methods and Application、アカデミック・プレス(Academic Press)、ニューヨーク(New York)を参照)。
【0039】
本明細書で提供される配列情報は、誤って同定された塩基の包含を必要とするほどに狭く考えられてはならない。本明細書に開示された特定の配列は、糸状菌、具体的にはA.ニゲル(niger)からの完全な遺伝子を単離するために容易に使用され、次いで容易に追加的な配列分析にかけられ、それによって配列決定の誤りを特定することができる。
【0040】
別段の定めがない限り、本明細書のDNA分子の配列決定によって決定されるすべてのヌクレオチド配列は、自動DNAシーケンサーを使用して決定され、本明細書で決定されたDNA分子によってコードされるポリペプチドのすべてのアミノ酸配列は上記の通り決定されたDNA配列の翻訳によって予測された。したがって、この自動的方法によって決定されるDNA配列について当業界で周知のように、本明細書で決定されるヌクレオチド配列は多少の誤りを含有しうる。自動化によって決定されるヌクレオチド配列は、配列決定されたDNA分子の実際のヌクレオチド配列と一般的に少なくとも約90%同一、より一般的には少なくとも約95%〜少なくとも約99.9%同一である。実際の配列は、当業界で公知の手動のDNA配列決定法を含む他の方法によってより正確に決定されうる。当業界で周知であるように、実際の配列と比較して決定されたヌクレオチド配列における単一の挿入または欠失がヌクレオチド配列の翻訳におけるフレームシフトをもたらし、決定されたヌクレオチド配列によってコードされた予測アミノ酸配列が、かかる挿入または欠失の点で開始する、配列決定されたDNA分子によって実際にコードされるアミノ酸配列と完全に異なるようになっている。
【0041】
当業者は、かかる誤って同定された塩基を同定することが可能であり、かかる誤りを修正する方法を知っている。
【0042】
核酸構築物
本発明の別の態様は、適切な発現宿主におけるSEC61活性を有するポリペプチドの発現を指図する1つもしくはそれ以上の制御配列に操作可能に結合されている、本発明によるSEC61ポリペプチドをコードする核酸配列を含んで成る核酸構築物に関する。
【0043】
発現は、ポリペプチドの製造に関与するステップを含むことが理解され、転写、転写後修飾、翻訳、翻訳後修飾、分泌を含みうる。
【0044】
「核酸構築物」は、天然起源遺伝子から単離され、またはそれ以外には天然に存在しないやり方で結合または並列される核酸の部分を含有するように修飾されている一本または二本鎖のいずれかの核酸分子として本明細書では定義される。核酸構築物という語は、核酸構築物が特定の宿主生物におけるコード配列の発現に必要な制御配列をすべて含有する場合に発現ベクターまたはカセットという語と同義である。
【0045】
「制御配列」という語は、本明細書では、ポリペプチドの発現に必要または有利であるすべての要素を含むと定義される。各々の制御配列は天然またはポリペプチドをコードする核酸配列とは異なりうる。かかる制御配列としては、プロモーター、リーダー、最適な翻訳開始配列(コザック(Kozak)、1991年、J.Biol.Chem.266:19867−19870頁)、分泌シグナル配列、プロペプチド配列、ポリアデニル化配列、転写終止が挙げられるが、これらに限定されない。最小限でも、制御配列は、プロモーター、転写および翻訳停止シグナルを含む。
【0046】
「操作可能に結合」という語は、本明細書では、制御配列がDNA配列のコード配列に対する位置で適切に配置され、制御配列がポリペプチドの製造を指図するようになっている構成として定義される。
【0047】
制御配列は、転写制御配列を含有する適切なプロモーター配列でありうる。プロモーターは、突然変異、切断、およびハイブリッドプロモーターを含む細胞における転写調節活性を示す任意の核酸配列であり、かつ細胞外または細胞内ポリペプチドをコードする遺伝子から獲得されうる。プロモーターは、細胞またはポリペプチドと相同または異種のいずれかでありうる。
【0048】
糸状菌の好ましいプロモーターは当業界で周知であり、例えば、グルコース−6−リン酸デヒロゲナーゼgpdAプロモーター、pepA、pepB、pepCなどのプロテアーゼプロモーター、グルコアミラーゼglaAプロモーター、アミラーゼamyA、amyBプロモーター、カタラーゼcatRまたはcatAプロモーター、グルコースオキシダーゼgoxCプロモーター、ベータ−ガラクトシダーゼlacAプロモーター、アルファ−グルコシダーゼaglAプロモーター、翻訳伸長因子tefAプロモーター、xlnA、xlnB、xlnC、xlnDなどのキシラナーゼプロモーター、eglA、eglB、cbhAなどのセルラーゼプロモーター、areA、creA、xlnR、pacC、prfT等などの転写調節因子のプロモーターでありうるとともに、特にNCBIウェブサイト(http://www.ncbi.nlm.nih.gov/entrez/)で確認されうる。
【0049】
好ましい実施形態において、プロモーターは、高く発現される(本明細書では総細胞mRNAの少なくとも0.5%(w/w)のmRNA濃度と定義)遺伝子由来でありうる。別の好ましい実施形態において、プロモーターは、培地発現される(本明細書では総細胞mRNAの少なくとも0.01%〜0.5%(w/w)のmRNA濃度と定義)遺伝子由来でありうる。別の好ましい実施形態において、プロモーターは、低く発現される(本明細書では総細胞mRNAの少なくとも0.01%(w/w)未満のmRNA濃度と定義)遺伝子由来でありうる。
【0050】
制御配列は適切な転写終止配列、転写を終止する糸状菌細胞によって認識される配列でもありうる。終止配列は、ポリペプチドをコードする核酸配列の3’末端と操作可能に結合されている。細胞において機能的であるすべての終止が本発明において使用されうる。
【0051】
糸状菌細胞の好ましい終止は、A.オリゼー(oryzae)TAKAアミラーゼ、A.ニゲル(niger)グルコアミラーゼ、A.ニズランス(nidulans)アントラニル酸シンターゼ、A.ニゲル(niger)アルファ−グルコシダーゼ、trpC遺伝子、およびフザリウム・オキシスポルム(Fusarium oxysporum)トリプシン様プロテアーゼをコードする遺伝子から得られる。
【0052】
制御配列は、糸状菌細胞による翻訳に重要であるmRNAの非翻訳領域の適切なリーダー配列でもありうる。リーダー配列は、ポリペプチドをコードする核酸配列の5’末端と操作可能に結合されている。細胞において機能的であるすべてのリーダー配列が本発明において使用されうる。
【0053】
糸状菌細胞の好ましいリーダーは、A.オリゼー(oryzae)TAKAアミラーゼ、およびA.ニズランス(nidulans)トリオースリン酸イソメラーゼ、およびA.ニゲル(niger)glaAから得られる。
【0054】
制御配列は、核酸配列の3’末端と操作可能に結合されており、転写されると、転写mRNAにポリアデノシン残基を添加するシグナルとしての糸状菌細胞によって認識される配列のポリアデニル化配列でもありうる。細胞において機能的であるすべてのポリアデニル化配列が本発明において使用されうる。
【0055】
糸状菌細胞の好ましいポリアデニル化配列は、A.オリゼー(oryzae)TAKAアミラーゼ、A.ニゲル(niger)グルコアミラーゼ、A.ニズランス(nidulans)アントラニル酸シンターゼ、フザリウム・オキシスポルム(Fusarium oxysporum)トリプシン様プロテアーゼ、およびA.ニゲル(niger)アルファ−グルコシダーゼをコードする遺伝子から得られる。
【0056】
分泌されるポリペプチドには、制御配列は、コードされたポリペプチドを細胞の分泌経路へ指図しうるポリペプチドのアミノ末端と連結されたアミノ酸配列をコードするシグナルペプチドコード領域でもありうる。核酸配列のコード配列の5’末端は、本質的に、分泌されたポリペプチドをコードするコード領域の部分と翻訳リーディングフレームで自然に連結されたシグナルペプチドコード領域を含有しうる。あるいは、コード配列の5’末端は、コード配列とは異なっているシグナルペプチドコード領域を含有しうる。異質のシグナルペプチドコード領域は、コード配列が通常、シグナルペプチドコード領域を含有しない場合に必要とされうる。あるいは、異質のシグナルペプチドコード領域は、ポリペプチドの増強された分泌を得るために天然のシグナルペプチドコード領域を単純に置換しうる。
【0057】
核酸構築物は発現ベクターでありうる。発現ベクターは、組換えDNA法に便利にかけられ、ポリペプチドをコードする核酸配列の発現をもたらしうる任意のベクター(例えば、プラスミドまたはウイルス)でありうる。ベクターの選択は一般的にベクターが導入される細胞とのベクターの適合性に依存する。ベクターは線状または閉環状プラスミドでありうる。
【0058】
ベクターは自己複製ベクター、すなわち、染色体外実体として存在するベクターであり、その複製が染色体複製と無関係であり、例えば、プラスミド、染色体外要素、ミニ染色体、または人工染色体でありうる。糸状菌の自己維持クローニングベクターは、AMA1−配列を含んで成りうる(例えば、アレクセンコ(Aleksenko)とクラッターバック(Clutterbuck)(1997年)、Fungal Genet.Biol.21:373−397頁を参照)。
【0059】
あるいは、ベクターは、細胞へ導入されると、ゲノムへ組込まれ、組込まれている染色体といっしょに複製されるものでありうる。組込みクローニングベクターは、ランダムに、または宿主細胞の染色体における所定の標的遺伝子座で組込まれうる。本発明の好ましい実施形態において、組込みクローニングベクターは、クローニングベクターの組込みをこの所定の遺伝子座にターゲティングするための宿主細胞のゲノムにおける所定の標的遺伝子座のDNA配列と相同であるDNA断片を含んで成る。標的組込みを促進するために、クローニングベクターは、好ましくは、宿主細胞の形質転換前に線状化される。線状化は、好ましくは、少なくとも1つであるが、好ましくは、クローニングベクターの端のいずれかが標的遺伝子座と相同の配列によってフランキングされるように行われる。標的遺伝子座をフランキングする相同配列の長さは、好ましくは、少なくとも0.1kbであり、さらに好ましくは、少なくとも0.2kbであり、より好ましくは、少なくとも0.5kbであり、さらにより好ましくは、少なくとも1kbであり、最も好ましくは、少なくとも2kbである。
【0060】
好ましい実施形態によれば、本発明によるSEC61ポリペプチドをコードする遺伝子を含んで成る発現ベクターの組込みの標的遺伝子座は、sec61遺伝子座である。
【0061】
ベクター系は単一ベクターもしくはプラスミド、または2つもしくはそれ以上のベクターまたはプラスミドでありうるが、これらはいっしょに宿主細胞のゲノム、またはトランスポゾンへ導入されるすべてのDNAを含有する。
【0062】
ベクターは、好ましくは、形質転換細胞の容易な選択を可能にする1つもしくはそれ以上の選択可能なマーカーを含有する。選択可能なマーカーが、その生成物が殺生物性またはウイルス抵抗性、重金属に対する抵抗性、栄養要求体に対する原栄養性(prototrophy)などを提供する遺伝子である。糸状菌細胞において使用するための選択可能なマーカーが、amdS(アセトアミダーゼ)、argB(オルニチンカルバモイルトランスフェラーゼ)、bar(ホスフィノトリシンアセチルトランスフェラーゼ)、hygB(ハイグロマイシンホスホトランスフェラーゼ)、niaD(硝酸還元酵素)、pyrG(オロチジン−5’−リン酸デカルボキシラーゼ)、sC(硫酸アデニルトランスフェラーゼ)、およびtrpC(アントラニル酸シンターゼ)のほか、他の種からの同等物を含むがこれらに限定されない群から選択されうる。アスペルギルス属(Aspergillus)細胞における使用に好ましいのは、A.ニズランス(nidulans)またはA.オリゼー(oryzae)のamdS(EP635574B1号明細書、国際公開第97/06261号パンフレット)およびpyrG遺伝子およびストレプトマイセス・ハイグロスコピクス(Streptomyces hygroscopicus)のbar遺伝子である。より好ましくは、amdS遺伝子が使用され、さらにより好ましくは、A.ニズランス(nidulans)またはA.ニゲル(niger)からのamdS遺伝子である。最も好ましい選択マーカー遺伝子が、A.ニズランス(nidulans)gpdAプロモーターと融合されたA.ニズランス(nidulans)amdSコード配列である(EP635574号明細書を参照)。他の糸状菌からのAmdS遺伝子も使用されうる(国際公開第97/06261号パンフレット)。
【0063】
宿主細胞
別の態様において、本発明は、目的とするポリペプチドの製造に適切な宿主細胞を提供し、前記宿主細胞はSEC61活性を有する発明によるポリペプチドを過剰発現する。前記宿主細胞は、本発明によるSEC61ポリペプチドをコードする核酸構築物または発現ベクターを含んで成る。
【0064】
意外にも、発明者らは、本発明によるSEC61ポリペプチド、具体的には上記の通りコンセンサスモチーフで修飾された突然変異SEC61ポリペプチドが過剰発現している真菌細胞、具体的には糸状菌細胞が、かかるSEC61ポリペプチドを過剰発現することがない真菌細胞と比べ改良されたタンパク質分泌能を有することを見出した。特に、異種タンパク質の分泌が改良されている。場合により、内因性sec61遺伝子は、内因性SEC61ポリペプチドと比べ修飾されているSEC61ポリペプチドを発現する細胞において不活性化されうる。
【0065】
好ましい実施形態においては、本発明によるSEC61ポリペプチドを過剰発現する宿主細胞が、内因性に発現されるSEC61ポリペプチドの量の少なくとも50%以上、好ましくは、少なくとも100%以上、より好ましくは、少なくとも200%以上、最も好ましくは、少なくとも400%以上である。内因性に発現されるSEC61ポリペプチドの量は、本発明によるSEC61ポリペプチドを過剰発現する宿主細胞がそれに由来する親の宿主細胞によって発現されるSEC61ポリペプチドの量である。発現SEC61ポリペプチドの量は、例えば、定量的SDS−PAGE、二次元タンパク質電気泳動、またはウェスタンブロット法によって、当業者に周知の方法によって測定されうる。
【0066】
本発明では、本発明によるSEC61ポリペプチドを過剰発現する宿主細胞のさまざまな実施形態が意図される。
【0067】
1つの実施形態において、天然のSEC61ポリペプチドを過剰発現し、それによって天然のSEC61ポリペプチドが前記宿主によって自然に生成されるSEC61ポリペプチドと同じポリペプチドである宿主細胞が提供される。
【0068】
別の実施形態において、前記宿主によって自然に生成されないSEC61ポリペプチドを発現する宿主細胞が提供される。かかる宿主細胞は天然SEC61ポリペプチドを発現することがない。天然SEC61ポリペプチドの非発現は、例えば、内因性sec61遺伝子の不活性化によって達成されうる。好ましくは、内因性sec61遺伝子の不活性化は、例えば、EP0357127号明細書に記載されている遺伝子置換によって達成されうる。より好ましくは、前記遺伝子置換は、本発明によるSEC61ポリペプチドをコードする核酸配列を含んで成る核酸構築物(発現ベクター)によって内因性sec61遺伝子の置換を含んで成る。
【0069】
本発明の方法における宿主細胞の選択は、目的とするポリペプチドをコードする核酸配列の供給源に大幅に依存する。好ましくは、宿主細胞は真核細胞、より好ましくは、真菌、最も好ましくは、糸状菌である。好ましい実施形態において、糸状菌宿主細胞は、本発明によるSEC61ポリペプチドが獲得されうる種として挙げられている種の細胞である。
【0070】
好ましい実施形態において、糸状菌宿主細胞は、配列番号3のアミノ酸配列と少なくとも89%の同一性を有するアミノ酸配列を有するポリペプチド、好ましくは、配列番号3または配列番号6または配列番号9のアミノ酸配列を有するポリペプチドをコードする核酸配列を有する遺伝子を過剰発現する。
【0071】
得られた宿主細胞は、目的とするポリペプチドを製造するために使用されうる。
【0072】
したがって、別の好ましい実施形態において、宿主細胞は、目的とするポリペプチドをコードする核酸配列を含んで成るポリヌクレオチドの少なくとも1つのコピーをさらに有する。この核酸配列は、sec61遺伝子の発現について以前に記載された発現ベクターへクローン化されうる。本発明の方法は、目的とするポリペプチドを製造する糸状菌細胞を得るための特定の順序に限定されないことが理解されるであろう。場合により内因性sec61遺伝子の修飾と組合される、sec61遺伝子の導入は、目的とするポリペプチドの製造のための細胞の構成における任意のステップで行われうる。
【0073】
ポリペプチドの製造
本発明の別の態様は、以前の態様の宿主細胞、好ましくは、糸状菌宿主細胞における目的とするポリペプチドの製造のための方法に関し、
(a)目的とするポリペプチドの製造のために適切な栄養培地において以前の態様の宿主細胞を培養するステップと、
(b)栄養ブロスから目的とするポリペプチドを回収するステップと
を含んで成る方法に関する。
【0074】
本発明の宿主細胞は、当業界で周知の方法を使用して目的とするポリペプチドの製造に適切な栄養培地中で培養される。例えば、細胞は、目的とするポリペプチドが発現および/または単離されることを可能にする適切な培地および条件下に行われる、実験的または工業的発酵装置における振盪フラスコ培養、小規模または大規模発酵(連続発酵、バッチ発酵、流加発酵、または固相発酵を含む)によって培養されうる。培養は、当業界で周知の方法を使用して少なくとも炭素および窒素および無機塩を含んで成る適切な栄養培地で行われる。目的とするポリペプチドは、当業界で周知の方法によって栄養培地から回収されうる。ポリペプチドが栄養培地へ分泌される場合は、ポリペプチドは培地から直接回収されうる。ポリペプチドが分泌されない場合は、これは細胞溶解物から回収されうる。必要に応じて、回収はさらに従来の手段による単離および精製を含んで成りうる。
【0075】
例えば、ポリペプチドは、遠心分離、ろ過、抽出、噴霧乾燥、蒸発、または沈殿を含むがこれらに限定されない従来の方法によって単離されうる。次いで、単離ポリペプチドは、さらに、クロマトグラフィー(例えば、イオン交換、親和性、疎水性、クロマト分画、およびサイズ排除)、電気泳動法(例えば、予備的等電点電気泳動法)、分画溶解(例えば、硫酸アンモニウム沈殿)、または抽出(例えば、Protein Purification,J.−C.ヤンソン(Janson)とラース ライデン(Lars Ryden)編、VCHパブリッシャーズ(Publischers)、ニューヨーク、1989年を参照)を含むがこれらに限定されない当業者に周知のさまざまな方法によって精製されうる。
【0076】
好都合にも、目的とするポリペプチドは、緩衝剤を含む適切な(固体または液体)担体または希釈剤と混合され、ポリペプチド組成物を生じうる。ポリペプチドは、例えば固体担体上に固定された担体に付着され、またはこれと混合されうる。
【0077】
目的とするポリペプチドは、ポリペプチドに特異的である当業界で周知の方法を使用して検出されうる。これらの検出方法としては、特異的抗体、酵素生成物の形成、酵素基質の消失、またはSDS PAGEの使用を挙げることができる。例えば、酵素アッセイを使用し、ポリペプチドの活性を測定することができる。酵素活性を測定するための方法が、多くの酵素について当業界で周知である。
【0078】
本発明の方法において、宿主細胞、好ましくは、糸状菌宿主細胞は、同一の条件下に培養した場合、SEC61発現に対して本発明による修飾されていないその対応する対応細胞より少なくとも約20%、好ましくは少なくとも約50%、より好ましくは少なくとも約100%、さらにより好ましくは少なくとも約200%、かつ最も好ましくは少なくとも約300%以上の目的とするポリペプチドを産生する。
【0079】
目的とするポリペプチドは、宿主細胞と相同(天然)または異種のポリペプチドでありうる。「ポリペプチド」という語は、本明細書では、コードされた生成物の特定の長さを指すことはなく、したがってペプチド、オリゴペプチド、およびタンパク質を包含する。
【0080】
異種ポリペプチドは、宿主細胞によって自然に産生されないポリペプチドを包含しうる。目的とするポリペプチドは、核酸配列によってコードされる宿主細胞を原産とするポリペプチドをも包含しうるが、その発現はポリペプチドをコードする核酸配列と異質の1つもしくはそれ以上の制御配列によって制御される。
【0081】
目的とするポリペプチドを産生するように遺伝子操作されていないが、かかるポリペプチドを自然に産生する菌株において本発明のSEC61ポリペプチドの過剰発現を達成することも可能である。
【0082】
好ましい実施形態において、目的とするポリペプチドは、抗体もしくはその一部分、抗原、凝固因子、酵素、ホルモン、もしくはホルモン変異体、受容体もしくはその一部分、調節タンパク質、構造的タンパク質、リポーター、または輸送タンパク質である。
【0083】
より好ましい実施形態において、目的とするポリペプチドは酵素であり、最も好ましくは、酵素はオキシドレダクターゼ、トランスフェラーゼ、ヒドラーゼ、リアーゼ、イソメラーゼ、またはリガーゼである。
【0084】
さらにより好ましい実施形態において、酵素はアミノペプチダーゼ、アミラーゼ、カルボヒドラーゼ、カルボキシキペプチダーゼ、カタラーゼ、セルラーゼ、キチナーゼ、クチナーゼ、デオキシリボヌクレアーゼ、デキストラナーゼ、エステラーゼ、アルファ−ガラクトシダーゼ、ベータ−ガラクトシダーゼ、グルコアミラーゼ、アミラーゼ、アルファ−グルコシダーゼ、ベータ−グルコシダーゼ、ハロペルオキシダーゼ、インベルターゼ、ラッカーゼ、リパーゼ、ホスホリパーゼ、マンノシダーゼ、ムタナーゼ、オキシダーゼ、ペクチナーゼ、ペルオキシダーゼ、フィターゼ、ポリフェノールオキシダーゼ、プロテアーゼ、ペプチダーゼ、リボヌクレアーゼ、トランスグルタミナーゼ、またはキシラナーゼである。
【0085】
別のさらにより好ましい実施形態において、ポリペプチドはヒトインスリンもしくはその類似体、ヒト成長ホルモン、エリトロポエチン、またはインスリノトロピンである。
【0086】
目的とするポリペプチドをコードする核酸配列は、原核、真核、または他の供給源から獲得されうる。本発明の目的のために、所定の供給源に関連して本明細書で使用される「から獲得される」という語は、供給源からの遺伝子が挿入されている供給源すなわち細胞によってポリペプチドが製造されることを意味する。
【0087】
本発明は、本発明の範囲の限定として考えるべきではない以下の実施例によってさらに説明される。
【実施例】
【0088】
実験情報
菌株
WT 1:本A.ニゲル(niger)菌株が野生型菌株として使用される。本菌株は寄託番号CBS513.88によりCBS研究所で寄託されている。
【0089】
WT 2:本A.ニゲル(niger)菌株はグルコアミラーゼ(glaA)をコードする遺伝子の欠失を含んで成るWT 1菌株である。WT 2は、CBS 513.88のゲノムにおけるglaA特異的DNA配列を削除するし方が記載されているEP635574号明細書に記載されている「MARKER−GENE FREE」法を使用して構成される。この方法は結果として、異種DNA配列を有さないMARKER−GENE FREE ΔglaA組換えA.ニゲル(niger)CBS513.88菌株をもたらす。
【0090】
WT 3:本A.ニゲル(niger)菌株は主要な細胞外アスパラギン酸プロテアーゼpepAをコードするpepA遺伝子の欠失を含んで成るWT 2菌株である。WT 3はEP0635574号明細書に記載されている「MARKER−GENE FREE」法を使用して構成される。本特許に記載された方法は、ファン・デン・ホンバーグ(van den Hombergh)ら(ファン・デン・ホンバーグ(van den Hombergh)JP、ソレウィン・ゲルプケ(Sollewijn Gelpke)MD、ファン・デ・フォンデルフォルト(Van de Vondervoort)PJ、ブクトン(Buxton)FP、ビセル(Visser)J.(1997年)−アスペルギルス・ニゲル(Aspergillus niger)における3つの酸プロテアーゼの崩壊−−プロテアーゼスペクトル、細胞内タンパク質溶解、および標的タンパク質の分解に対して作用する(Disruption of three acid proteases in Aspergillus niger−−effects on protease spectrum, intracellular proteolysis, and degradation of target proteins)−Eur J Biochem.247(2)605−13頁)によって記載されたCBS513.88のゲノムにおけるpepA特異的DNA配列を削除するために使用される。この方法は結果として、異種DNA配列を有さないMARKER−GENE FREE ΔpepA、ΔglaA組換えA.ニゲル(niger)CBS513.88菌株をもたらす。
【0091】
WT 4:本ペニシリウム・クリゾゲナム(Penicillium chrysogenum)菌株は野生型菌株として使用される。本菌株は寄託番号CBS455.95によりCBS研究所で寄託されている。
【0092】
EPO 1:本A.ニゲル(niger)菌株は、他の箇所(国際公開第02/45524号パンフレット)で公開されているプロリン特異的エンドプロテアーゼのA.ニゲル(niger)epo遺伝子の多重コピーを含んで成るWT2菌株である。EPO 1は、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載されているプロリン特異的エンドプロテアーゼの遺伝子コードを含んで成るpGBAAS−1(EP0635574号明細書に記載されている通り構成)およびpGBTOPEPO−1ベクター(図1)で示されているベクターを含有するamdS選択可能なマーカー遺伝子の共形質転換によって構成される。形質転換および対抗選択法は結果としてグルコアミラーゼプロモーターの制御下にプロリン特異的エンドプロテアーゼコード遺伝子の多重コピーを含有するMARKER−GENE FREE EPO 1菌株がもたらされる。
【0093】
PLA 1:異種ブタホスホリパーゼA2(PLA2)タンパク質は、モデルタンパク質として選択される。本タンパク質は高量でA.ニゲル(niger)において産生するのが困難であることが以前に証明されている(ロベルツ(Roberts)I.N.、ジーンズ(Jeenes)D.J.、マッケンジー(MacKenzie)D.A.、ウィルキンソン(Wilkinson)A.P.、サムナー(Sumner)I.G.およびアーチャー(Archer)D.B.(1992年)−アスペルギルス・ニゲル(Aspergillus niger)における異種遺伝子発現(Heterologous gene expression in Aspergillus niger)。グルコアミラーゼ−ブタ膵ホスホホリパーゼA2融合タンパク質が分泌され加工されて成熟酵素をもたらす。Gene 122:155−161頁)。PLA2の過剰発現の断片は、A.ニゲル(niger)の天然グルコアミラーゼA遺伝子とのプロPLA2の融合として製造され、ロベルツ(Roberts)ら(1992年)によって記載されているように調製される。融合タンパク質は、ゴルジにおいて加工されるためにkex1スプライシング部位を含有する。本glaA−pla2融合遺伝子は、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載されている同じ方法を使用してA.ニゲル(niger)pGBTOP発現ベクターへクローン化され、結果としてpGBTOPPLA−1がもたらされる(図2)。
【0094】
PLA 1 A.ニゲル(niger)菌株は、グルコアミラーゼ−ブタ膵ホスホリパーゼA2融合タンパク質コード遺伝子の多重コピーを含んで成るWT 3菌株である。PLA 1はベクターpGBAAS−1およびpGBTOPPLA−1ベクターを含有するamdS選択可能なマーカー遺伝子の共形質転換によって構成される(図2)。形質転換および対抗選択法の結果として、グルコアミラーゼプロモーターの制御下にグルコアミラーゼ−ブタ膵ホスホリパーゼA2融合タンパク質コード遺伝子の多重コピーを含有するMARKER−GENE FREE PLA 1菌株がもたらされる。
【0095】
A.ニゲル(niger)振盪フラスコ発酵
A.ニゲル(niger)菌株を実施例、すなわち、国際公開第99/32617号パンフレットの「アスペルギルス・ニゲル(Aspergillus niger)振盪フラスコ発酵」の部に記載された前培養培地20ml中で前培養する。一夜増殖後、10mlの本培養液を発酵培地(FM)に移す。発酵培地(FM)は1リットル当り以下を含有する。すなわち、グルコース・1H2O 82.5g、Maldex 15(ブーム・メッペル(Boom Meppel)、オランダ)25g、クエン酸2g、NaH2PO4・1H2O 4.5g、KH2PO4 9g、(NH4)2SO4 15g、ZnCl2 0.02g、MnSO4・1H2O 0.1g、CuSO4・5H2O 0.015g、CoCl2・6H2O 0.015g、MgSO4・7H2O 1g、CaCl2・2H2O 0.1g、FeSO4・7H2O 0.3g、MES(2−[N−モルフォリノ]エタンスルホン酸)30g、pH6。
【0096】
FMにおける発酵は、指示された日数の間、34℃および170rpmで発酵ブロス100mlを含む整流装置を備えた500mlフラスコで実行される。
【0097】
酵素アッセイ
PLA2ホスホリパーゼ活性
アスペルギルス・ニゲル(Aspergillus niger)培養ブロス中のホスホリパーゼPLA2活性(PLA2)を分光光度的に測定するために、人工基質、すなわち、1,2−ジチオジオクタノイルホスファチジルコリン(diC8、基質)を使用する。PLA2はA2位置でのスルフィド結合を加水分解し、チオオクタン酸を解離する。チオオクタン酸は4,4ジチオピリジン(呈色試薬、4−DTDP)と反応し、4−チオピリドンを形成する。4−チオピリドンは、334nmの波長を有する放射線を吸収する4−メルカプトピリジンと互変異性平衡にある。同波長での吸光度の変化を測定する。一単位は、37℃およびpH4.0で1,2−ジチオジオクタノイルホスファチジルコリンからチオ−オクタン酸を毎分1nmol遊離させる酵素の量である。
【0098】
基質溶液は、エタノール66ml当りdiC8結晶1gを溶解し、緩衝酢酸溶液264mlを添加することによって調製される。緩衝酢酸溶液は、0.2%トリトン(Triton)−X100含有0.1M緩衝酢酸溶液pH3.85を含んで成る。呈色試薬は11mM 4,4−ジチオピリジン溶液である。これは2mlエッペンドルフサンプルカップ中に4,4−ジチオピリジン5.0mgを秤量し、エタノール1.00ml中に溶解することによって調製される。1.00mlのmilli−Q水を添加する。
【0099】
プロリン特異的エンドプロテアーゼ活性
プロリン特異的エンドプロテアーゼのタンパク質分解活性は、クエン酸/リン酸2ナトリウム緩衝液pH5中37℃下CBZ−Gly(cine)−Pro(line)−pNAを使用して時間内に410nmで分光光度分析的に測定される。1Uプロリン特異的エンドプロテアーゼを上記の条件下、pH5および37℃でCBZ−Gly(cine)−Pro(line)−pNAを毎分1μmol(マイクロモル)を変換する酵素の量と定義する。
【0100】
グルコアミラーゼ活性アッセイ
グルコアミラーゼ活性は、p−ニトロフェニル α−D−グルコピラノシド(シグマ(Sigma))を使用して国際公開第98/46772号パンフレットに記載されている通りに測定される。
【0101】
実施例1:発現ベクターにおけるsec61遺伝子のクローニング、Sec61転位チャンネルのコード
アスペルギルス・ニゲル(Aspergillus niger)菌株WT 1およびペニシリウム・クリゾゲナム(Penicillium chrysogenum)WT 4のゲノムDNAを配列決定し、分析する。Sec61と相同体と注釈付きの翻訳タンパク質を有する1つの遺伝子が同定され、sec61と命名されている。オープンリーディングフレーム(ORF)(イントロンによる)および約180bpの5’非翻訳領域(UTR)および327bpの3’UTRを含んで成るA.ニゲル(niger)sec61遺伝子座のゲノム配列は、配列番号1に示されている。オープンリーディングフレーム(ORF)(イントロンによる)および約12bpの5’非翻訳領域(UTR)および66bpの3’UTRを含んで成るペニシリウム・クリゾゲナム(Penicillium chrysogenum)sec61遺伝子座のゲノム配列は、配列番号7に示されている。
【0102】
WT 1からのゲノムDNAは、配列番号13および配列番号14のプライマーを使用し、野生型sec61遺伝子を増幅させ、適切な制限部位を付着させるPCR反応におけるテンプレートとして使用される。同時に、同じ実験をcDNAをテンプレートとして使用して実行する。すなわち、PCR反応は配列番号14および15のプライマーを使用して行われ、野生型sec61cDNAを増幅させる。すべてのPCR反応、cDNA合成、ライゲーション、および形質転換は、Molecular Cloning:A Laboratory Manual,サムブルック(Sambrook)ら、ニューヨーク:コールド・スプリング・ハーバー・プレス(Cold Spring Harbour Press)、1989年に記載されている通りに実行される。さまざまに独立して増幅されたPCR断片の配列は配列分析によって決定される。配列番号1は、A.ニゲル(niger)sec61ゲノム配列を含んで成る。A.ニゲル(niger)のSec61タンパク質をコードするcDNA配列のORFは配列番号2として提供されている。配列番号2の翻訳アミノ酸配列は配列番号3で指定され、これはA.ニゲル(niger)WT 1のSec61タンパク質を表す。
【0103】
WT 4からのゲノムDNAは、配列番号10および配列番号11のプライマーを使用し、野生型SEC61遺伝子を増幅させ、適切な制限部位を付着させるPCR反応におけるテンプレートとして使用される。同時に、同じ実験をcDNAをテンプレートとして使用して実行する。すなわち、PCR反応は配列番号11および12のプライマーを使用して行われ、野生型sec61cDNAを増幅させる。さまざまに独立して増幅されたPCR断片の配列は配列分析によって決定される。配列番号7は、ペニシリウム・クリゾゲナム(Penicillium chrysogenum)sec61ゲノム配列を含んで成る。P.クリゾゲナム(chrysogenum)のSec61タンパク質をコードするcDNA配列のORFは配列番号8として提供されている。配列番号8の翻訳アミノ酸配列は配列番号9で指定され、これはP.クリゾゲナム(chrysogenum)WT 4のSec61タンパク質を表す。
【0104】
A.ニゲル(niger)sec61ゲノムDNAおよびcDNA配列を含んで成る結果として生じるPCR断片は、メーカーの指示に従ってPaclおよびAsclで切断され、Pacl、Ascl線状化pGBFIN−18A.ニゲル(niger)発現ベクターでライゲートされる(図3)。pGBFIN−18ベクターのプラスミドおよび構成は国際公開第99/32617号パンフレットに記載されている。これは結果として、Sec61がglaAプロモーターの制御下に配置されているpGBFINSEC−1およびpGBFINSEC−2の構築物をもたらした(pGBFINSEC−1のマップについて図4を参照)。
【0105】
アスペルギルス・ニズランス(Aspergillus nidulans)の構成的gpdAプロモーターの制御下、およびpGBFIN−18ベースのA.ニゲル(niger)発現ベクターにも存在するamdS遺伝子は、選択マーカーとして使用され、唯一のN源としてアセトアミドでの増殖が誘発されうる(ケリー(Kelly),J.M.およびハインズ(Hynes),M.J.(1985年)。アスペルギルス・ニズランス(Aspergillus nidulans)のamdS遺伝子によるアスペルギルス・ニゲル(Aspergillus niger)の形質転換。EMBO J.4、475−479頁に記載)。
【0106】
実施例2:遺伝子置換および過剰発現ベクターにおける修飾Sec61転位チャンネルをコードする、sec61*の構成
古典的な分子生物学法をMolecular Cloning:A Laboratory Manual、サンブルック(Sambrook)ら、ニューヨーク:コールド・スプリング・ハーバー・プレス(Cold Spring Harbour Press)、1989年に記載されている通り使用する。A.ニゲル(niger)欠失ベクターの一般的な設計はEP635574号明細書および国際公開第98/46772号パンフレットに記載されている。
【0107】
修飾sec61遺伝子は、セリン376がトリプトファンによって置換されているSec61*タンパク質をコードするsec61*と呼ばれ、配列オーバーラップ伸長PCR(SOE−PCR、Gene.1989年4月15、77(1):51−9頁、ホ(Ho)SN、ハント(Hunt)HD、ホルトン(Horton)RM、プレン(Pullen)JK、ピース(Pease)LR、「ポリメラーゼ連鎖反応を使用するオーバーラップ伸長による部位特異的突然変異誘発(Site−directed mutagenesis by overlap extension using the polymerase chain reaction)」に記載)によって構成される。配列番号18および配列番号17で識別されたオリゴヌクレオチド、およびA.ニゲル(niger)WT 1のゲノムDNAをテンプレートとして使用することにより、断片Aで識別された1.4kbのsec61遺伝子の5’領域がPCRによって増幅される。また、Ascl制限部位が断片Aの5’末端に付着されている。配列番号16および配列番号19で識別されたオリゴヌクレオチドおよびA.ニゲル(niger)WT 1のゲノムDNAをテンプレートとして使用することにより、断片Bで識別された3’−UTRを含む0.6kbのsec61遺伝子の3’領域がPCRによって増幅される。また、NotlおよびEcoRl制限部位が断片Bの3’末端に付着されている。結果として生じる断片AおよびBは、SOE−PCR、配列番号18および配列番号19で識別されるオリゴヌクレオチド、および断片AおよびBによって融合され、2kb断片Cを生成する。本断片CはpCR2.1−TOPO(インビトロジェン(Invitrogen))でクローン化され、pCR_Sec61*を生成する。pCR_Sec61*のPCR生成断片の配列は確認され、配列番号4で提供されている。修飾Sec61*タンパク質のコード配列は配列番号5で提供されている。配列番号5の翻訳アミノ酸配列は配列番号6で指定され、これは修飾Sec61*タンパク質を表す。
【0108】
ベクターpCR_Sec61*はAsclおよびEcoRlで消化され、sec61*断片はAsclおよびEcoRl消化ベクターpGBDELへ導入され、pGBDEL−1(データは示さず)を生成する。その後、ベクターpCR_Sec61*はEcoRVおよびNotlで消化され、3’sec61*断片はSmalおよびNotl消化ベクターpGBDEL−1へ導入され、pGBDEL−SEC61*を生成する(図5)。
【0109】
ベクターpCR_Sec61*は配列番号13および配列番号14のプライマーを使用し、修飾sec61遺伝子を増幅させ、適切な制限部位を付着させるPCR反応におけるテンプレートとして使用される。修飾sec61*配列を含んで成る結果として生じるPCR断片は、メーカーの指示に従ってPaclおよびAsclで切断され、Pacl、Ascl線状化pGBFIN−18A.ニゲル(niger)発現ベクターでライゲートされるが、ここでPglaAプロモーターは、pepCプロモーター(フレデリック(Frederick)GD、ロンブーツ(Rombouts)P、ブクストン(Buxton)FP(1993年)−アスペルギルス・ニゲル(Aspergillus niger)からのセリンプロテアーゼをコードする遺伝子、pepCのクローニングおよび特徴づけ(Cloning and characterisation of pepC, a gene encoding a serine protease from Aspergillus niger.)−Gene 125(1):57−64頁)によって置換される。これは結果としてpGBFINSEC−3構築物をもたらしたが、ここで修飾Sec61*遺伝子はpepCプロモーターの制御下に配置される(図6)。
【0110】
実施例3.野生型sec61および修飾sec61*遺伝子を過剰発現し、修飾sec61*転位チャンネルを発現する真菌宿主細胞
構成的gpdAプロモーターの制御下に配置されるアスペルギルス・ニズランス(Aspergillus nidulans)のamdS遺伝子は、EP635574号明細書に記載されている通り選択マーカーとして使用される。原則的には、pGBFINSEC−1、pGBFINSEC−2、pGBFINSEC−3、およびpGBDEL−SEC61*の線状DNAが指示された制限酵素での消化後に単離される。アスペルギルス・ニゲル(Aspergillus niger)PLA1およびEPO 1菌株は形質転換され、その後のアセトアミドでの形質転換体選択は国際公開第98/46772号パンフレット、EP635574号明細書、および国際公開第99/32617号パンフレットに記載されている通りに行われる。その後、コロニーが標準の方法に従って精製される。
【0111】
PLA1 およびEPO 1菌株の増殖コロニーは、それぞれ、形質転換SECプラスミドのコピー数およびpla2およびepo遺伝子コピーの非喪失について診断される。同様に推定されたコピー数、およびpla2およびepo遺伝子コピーの非喪失のpGBFINSEC−1、pGBFINSEC−2、pGBFINSEC−3の形質転換体が選択される。単一コピー(_A、_B、_C、_...)による数個の形質転換体が選択される。これは結果として、野生型のゲノムsec61(pGBFINSEC−1)、cDNAsec61(pGBFINSEC−2)および修飾sec61*遺伝子(pGBFINSEC−3)がもたらした。PLA 1およびEPO 1菌株の形質転換体は、それぞれ、PLA_SEC1、PLA_SEC2、PLA_SEC3、およびEPO_SEC1、EPO_SEC2、EPO_SEC3で指示されている。
【0112】
sec61遺伝子座でゲノムへ組込まれたpGBDEL−SEC61*断片の形質転換体は、図7に示されている通り、sec61*遺伝子およびamdS遺伝子で野生型sec61遺伝子を置換した。標的のために使用された断片および方法により、相同組換え時の置換は100%ではない。したがって、精製pGBDEL−SEC61*形質転換体は、sec61遺伝子座での相同組込みに対する2つのPCR反応およびsec61*遺伝子配列に対するsec61遺伝子の置換によって診断される。sec61*遺伝子によるsec61の本置換は、テンプレートとしてpGBDEL−SEC61*形質転換体のゲノムDNAを使用し、かつ配列番号20および配列番号22のプライマーを使用し、sec61遺伝子座で修飾sec61*遺伝子を増幅するバンドの第1のPCR増幅によって検出可能である。sec61*遺伝子によるsec61の本置換は、テンプレートとしてpGBDEL−SEC61*形質転換体のゲノムDNAを使用し、かつ配列番号20および配列番号21のプライマーを使用し、障害をチェックして野生型sec61遺伝子を増幅させる第2のPCRによって確認される。また、pla2およびepo遺伝子コピーの非喪失が、それぞれ、試験される。その後、胞子がフルオロアセトアミド培地にプレーティングされ、amdSマーカーを喪失する菌株を選択する。さらに、増殖コロニーが、上記の通り、sec61*遺伝子配列およびpla2またはepo遺伝子コピーの非喪失に対するsec61遺伝子の置換についてPCRによって診断され、候補の菌株が、amdSマーカーを喪失する形質転換体および誘導菌株を含有するamdSのサザン分析によって試験される。sec61*遺伝子に対するsec61遺伝子の置換は、両方の菌株のsec61プローブへのハイブリダイゼーション、形質転換体を含有するamdSにおける全遺伝子座をカバーするDNA断片の約2.2kbサイズの増大、およびamdSマーカーを喪失する誘導菌株における野生型断片によって検出可能である。約40%の菌株は、pGBDEL−SEC61*の相同組込みを示したが、そのうち約70%でsec61*遺伝子に対するsec61遺伝子の置換が検出可能である。菌株PLA_SEC61*およびEPO_SEC61*は、修飾sec61*遺伝子によって置換されるsec61遺伝子を有する代表的な菌株として選択される。
【0113】
また、アスペルギルス・ニゲル(Aspergillus niger)PLA_SEC61*およびEPO_SEC61*は、pGBFINSEC−3の線状DNAで形質転換され、その後のアセトアミドでの形質転換体選択およびコロニー単離が上記の通り行われる。菌株PLA_SEC61*_2およびEPO_SEC61*_2は、修飾Sec61*の過剰発現に対する代表的な菌株として選択され、内因性sec61遺伝子が修飾sec61*遺伝子によって置換され、強いglaAプロモーターの制御下に修飾Sec61*遺伝子の追加のコピーを有し、かつpla2またはepo遺伝子のコピーが非喪失である。
【0114】
実施例4.得られたアスペルギルス・ニゲル(Aspergillus niger)菌株によるタンパク質の分泌の改良
pGBTOPEPO−1構築物の同等のコピーを有する一部のA.ニゲル(niger)WT 2およびEPO_SEC1、EPO_SEC2、およびEPO_SEC3、EPO_SEC61*およびEPO_SEC61*_2形質転換体を選択して振盪フラスコ実験を行う。また、pGBTOPPLA−1構築物の同等のコピーを有する一部のA.ニゲル(niger)WT 3およびPLA_SEC1、PLA_SEC2、およびPLA_SEC3、PLA_SEC61*、およびPLA_SEC61* _2形質転換体を選択し、振盪フラスコ実験を行う。振盪フラスコ実験は、実験方法−「A.ニゲル(niger)振盪フラスコ発酵(A. niger shake flask fermentations)」に記載されている通り100mlの発酵培地(FM)中で行われる。培養ブロスは7日間の培養中に採集し、エンドプロテアーゼおよびPLA2活性を上記の通り測定する。
【0115】
図8は、同じコピー数のA.ニゲル(niger)EPO菌株の形質転換体における相対的プロリン特異的エンドプロテアーゼ活性を示す。発明者らは、Sec61および修飾Sec61*タンパク質の過剰発現が結果としてエンドプロテアーゼ酵素の生産の増大をもたらすと結論づける。過剰発現はグルコアミラーゼglaAおよびプロテアーゼpepCプロモーターを使用して実行されている。また、ゲノムまたはcDNA構築物によるSec61の過剰発現、またはゲノムに依然として存在する内因性sec61遺伝子による修飾Sec61*の過剰発現は結果として、エンドプロテアーゼ酵素の生産の増大をもたらす。
【0116】
これらの結果は、sec61*およびsec61が、真菌宿主細胞におけるポリペプチドの改良された生産を得る優れた手段であることを示す。図9および10は、それぞれ、A.ニゲル(niger)WT 2、SEC1、SEC2、SEC3、SEC61*、およびSEC61*_2において、およびA.ニゲル(niger)WT 2pGBFIN−PLA2形質転換において測定される際の、PLA2およびグルコアミラーゼ活性を示す。欠失野生型sec61遺伝子を有する菌株において、無傷のA.ニゲル(niger)PLA 1およびPLA_SEC1、PLA_SEC3、およびPLA_SEC61*を有し、同等のコピー数を有する菌株と比べPLA2活性の明らかな増大が確認されることは明らかである。発明者らは、修飾Sec61*タンパク質の発現が結果として、異種ホスホリパーゼA2および相同グルコアミラーゼ酵素の両方の生産の増大をもたらすと結論づける。過剰発現はグルコアミラーゼglaAおよびプロテアーゼpepCプロモーターを使用して実行されている。PLA2および/またはグルコアミラーゼ活性が増大していない菌株は発酵中にコピーを喪失したことが証明されている。したがって、コピー当りの発現は依然としてSec61の過剰発現とともに増大する。これらの結果は、sec61*およびsec61の発現の操作が結果として真菌宿主細胞におけるポリペプチドの生産の改良をもたらすことを示す。
【図面の簡単な説明】
【0117】
【図1】プロリン特異的エンドプロテアーゼ発現ベクターpGBTOPEPO−1のプラスミドマップを示す図である。glaAプロモーターおよびプロリン特異的エンドプロテアーゼepo遺伝子に対するglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素HinDIIIによる消化によって除去されうる。
【図2】PLA2発現ベクターpGBTOPPLA−2のプラスミドマップを示す図である。glaAプロモーター、切断glaA遺伝子、およびプロ−pla2コード配列に対するglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素Notlによる消化によって除去されうる。
【図3】発現ベクターpGBFIN−18のプラスミドマップを示す図である。グルコアミラーゼプロモーターにおける独自のSfilおよびEcoRlクローニング部位の後、PaclおよびAsclクローニング部位によるglaAプロモーターに対するglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素Notlによる消化によって除去されうる。
【図4】発現ベクターpGBFINSEC−1のプラスミドマップを示す図である。glaAプロモーターおよびA.ニゲル(niger)ゲノムsec61遺伝子に対するglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素Notlによる消化によって除去されうる。
【図5】置換ベクターpGBDEL−Sec61*のプラスミドマップを示す図である。amdSマーカーに対するSec61*突然変異遺伝子および3’Sec61*突然変異遺伝子断片が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素AsclおよびNotlによる消化によって除去されうる。
【図6】発現ベクターpGBFINSEC−3のプラスミドマップを示す図である。glaApepCプロモーターおよび修飾sec61*遺伝子に対するglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、A.ニゲル(niger)菌株への形質転換前に、制限酵素Notlによる消化によって除去されうる。
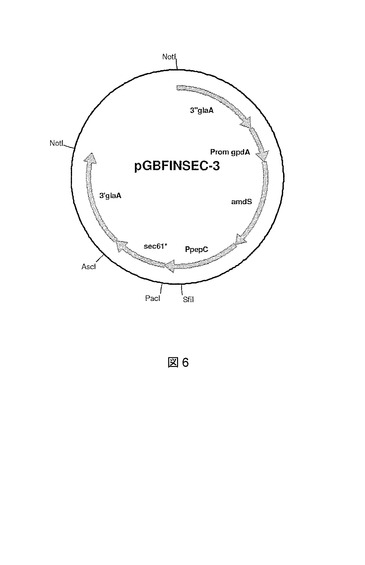
【図7】sec61遺伝子置換を示す概略図である。sec61遺伝子(1)の相同領域によってフランキングされたamdS選択マーカーを含んで成るpGBDEL−Sec61*の線状DNA構築物は、ゲノムsec61遺伝子座(2)で二重相同組換え(X)により結合し、修飾sec61*遺伝子によってゲノムsec61遺伝子コピー(3)を置換する。その後、(修飾)sec61配列(U)の直接の繰返しに及ぶ組換えがamdSマーカーを除去し、結果として修飾sec61*遺伝子(4)によるsec61遺伝子の遺伝子置換イベントが生じる。amdSマーカーが有る場合(3)および無い場合(4)の両方の菌株は機能的修飾sec61*遺伝子を有する。
【図8】さまざまなsec61構築物で形質転換されたEPO 1菌株の相対細胞外プロリン特異的エンドプロテアーゼ活性を示す図である。試料を発酵の数日間に採取する。3日目のEPO 1の発現は100%で設定されている。
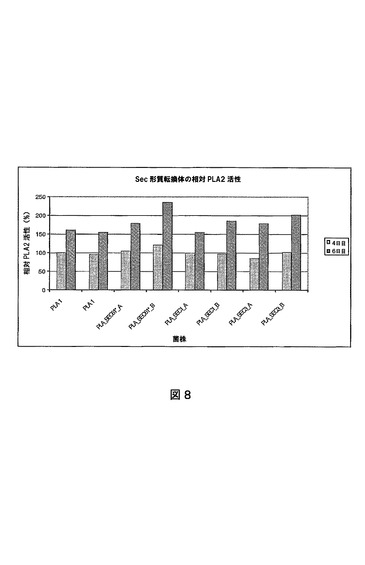
【図9】さまざまなsec61構築物で形質転換されたPLA 1菌株の細胞外PLA2活性を示す図である。試料を発酵の数日間に採取する。
【図10】さまざまなsec61構築物で形質転換されたPLA 1菌株の細胞外グルコアミラーゼ活性を示す図である。菌株および試料は図11と同一である。試料を発酵の数日間に採取する。
【特許請求の範囲】
【請求項1】
(a)配列番号3によるアミノ酸配列を有するポリペプチドと、
(b)配列番号3によるアミノ酸配列との少なくとも89%、より好ましくは少なくとも90%、さらにより好ましくは少なくとも92%、さらにより好ましくは少なくとも93%、さらにより好ましくは少なくとも94%、さらにより好ましくは少なくとも95%、さらにより好ましくは少なくとも96%、さらにより好ましくは少なくとも97%、さらにより好ましくは少なくとも98%、かつ最も好ましくは少なくとも99%の同一性の程度を有するポリペプチドと
からなる群から選択されるSEC61ポリペプチド。
【請求項2】
配列番号9によるアミノ酸配列を有する請求項1に記載のポリペプチド。
【請求項3】
配列番号3によるアミノ酸配列との少なくとも85%の同一性の程度を有し、ここで少なくとも1つのアミノ酸が配列番号第373〜379位置に対応するコンセンサスモチーフで修飾されているポリペプチド。
【請求項4】
修飾されている前記少なくとも1つのアミノ酸が、配列番号3の第376位置に対応する位置にある、請求項2に記載のポリペプチド。
【請求項5】
配列番号3の第376位置に対応する位置における前記アミノ酸が、トリプトファン、フェニルアラニン、チロシン、またはヒスチジン、好ましくは、トリプトファンによって置換される、請求項3に記載のポリペプチド。
【請求項6】
修飾されている前記少なくとも1つのアミノ酸がセリンである、請求項2〜4のいずれか一項に記載のポリペプチド。
【請求項7】
配列番号6に記載のアミノ酸配列を有する請求項1または3〜6のいずれか一項に記載のポリペプチド。
【請求項8】
請求項1〜7のいずれか一項に記載のポリペプチドをコードする核酸配列を含んで成る単離ポリヌクレオチド。
【請求項9】
前記ポリヌクレオチドまたは核酸配列が、
(a)配列番号1に記載のポリヌクレオチドと、
(b)配列番号2に記載の核酸配列と、
(c)配列番号2の核酸配列と少なくとも90%の同一性の程度を有する核酸配列と、
(d)配列番号3のアミノ酸配列と実質的に相同であるアミノ酸配列を有するポリペプチドをコードする核酸配列と
からなる群から選択される、請求項8に記載のポリヌクレオチド。
【請求項10】
請求項8または9の核酸配列をコードするSEC61ポリペプチドを含んで成り、前記核酸配列が、適切な宿主におけるポリペプチドの発現を指図する1つもしくはそれ以上の制御配列に操作可能に結合されている、核酸構築物または発現ベクター。
【請求項11】
前記制御配列が、プロモーターのほか、転写および翻訳停止シグナルを含んで成る、請求項10に記載の核酸構築物または発現ベクター。
【請求項12】
請求項8〜11のいずれか一項に記載のポリヌクレオチド、核酸構築物、または発現ベクターで形質転換された宿主細胞。
【請求項13】
内因性sec61遺伝子が不活性化されている、請求項12に記載の宿主細胞。
【請求項14】
糸状菌細胞である請求項12または13に記載の宿主細胞。
【請求項15】
前記糸状菌細胞が、アスペルギルス(Aspergillus)種、ペニシリウム(Penicillium)種、またはトリコデルマ(Trichoderma)種からなる群から選択される種に属する、請求項14に記載の宿主細胞。
【請求項16】
前記糸状菌細胞が、アスペルギウス・ニゲル(Aspergillus niger)またアスペルギルス・オリゼー(Aspergillus oryzae)の種に属する、請求項15に記載の宿主細胞。
【請求項17】
前記宿主細胞が、目的とするポリペプチドをコードする核酸配列を含んで成るポリヌクレオチドまたは発現ベクターをさらに含んで成る、請求項12〜16のいずれか一項に記載の宿主細胞。
【請求項18】
目的とするポリペプチドの発現をもたらす条件下に請求項16に記載の宿主細胞を培養し、場合により目的とするポリペプチドを回収して培養ブロスを形成するステップを含んで成る、目的とするポリペプチドを製造するための方法。
【請求項1】
(a)配列番号3によるアミノ酸配列を有するポリペプチドと、
(b)配列番号3によるアミノ酸配列との少なくとも89%、より好ましくは少なくとも90%、さらにより好ましくは少なくとも92%、さらにより好ましくは少なくとも93%、さらにより好ましくは少なくとも94%、さらにより好ましくは少なくとも95%、さらにより好ましくは少なくとも96%、さらにより好ましくは少なくとも97%、さらにより好ましくは少なくとも98%、かつ最も好ましくは少なくとも99%の同一性の程度を有するポリペプチドと
からなる群から選択されるSEC61ポリペプチド。
【請求項2】
配列番号9によるアミノ酸配列を有する請求項1に記載のポリペプチド。
【請求項3】
配列番号3によるアミノ酸配列との少なくとも85%の同一性の程度を有し、ここで少なくとも1つのアミノ酸が配列番号第373〜379位置に対応するコンセンサスモチーフで修飾されているポリペプチド。
【請求項4】
修飾されている前記少なくとも1つのアミノ酸が、配列番号3の第376位置に対応する位置にある、請求項2に記載のポリペプチド。
【請求項5】
配列番号3の第376位置に対応する位置における前記アミノ酸が、トリプトファン、フェニルアラニン、チロシン、またはヒスチジン、好ましくは、トリプトファンによって置換される、請求項3に記載のポリペプチド。
【請求項6】
修飾されている前記少なくとも1つのアミノ酸がセリンである、請求項2〜4のいずれか一項に記載のポリペプチド。
【請求項7】
配列番号6に記載のアミノ酸配列を有する請求項1または3〜6のいずれか一項に記載のポリペプチド。
【請求項8】
請求項1〜7のいずれか一項に記載のポリペプチドをコードする核酸配列を含んで成る単離ポリヌクレオチド。
【請求項9】
前記ポリヌクレオチドまたは核酸配列が、
(a)配列番号1に記載のポリヌクレオチドと、
(b)配列番号2に記載の核酸配列と、
(c)配列番号2の核酸配列と少なくとも90%の同一性の程度を有する核酸配列と、
(d)配列番号3のアミノ酸配列と実質的に相同であるアミノ酸配列を有するポリペプチドをコードする核酸配列と
からなる群から選択される、請求項8に記載のポリヌクレオチド。
【請求項10】
請求項8または9の核酸配列をコードするSEC61ポリペプチドを含んで成り、前記核酸配列が、適切な宿主におけるポリペプチドの発現を指図する1つもしくはそれ以上の制御配列に操作可能に結合されている、核酸構築物または発現ベクター。
【請求項11】
前記制御配列が、プロモーターのほか、転写および翻訳停止シグナルを含んで成る、請求項10に記載の核酸構築物または発現ベクター。
【請求項12】
請求項8〜11のいずれか一項に記載のポリヌクレオチド、核酸構築物、または発現ベクターで形質転換された宿主細胞。
【請求項13】
内因性sec61遺伝子が不活性化されている、請求項12に記載の宿主細胞。
【請求項14】
糸状菌細胞である請求項12または13に記載の宿主細胞。
【請求項15】
前記糸状菌細胞が、アスペルギルス(Aspergillus)種、ペニシリウム(Penicillium)種、またはトリコデルマ(Trichoderma)種からなる群から選択される種に属する、請求項14に記載の宿主細胞。
【請求項16】
前記糸状菌細胞が、アスペルギウス・ニゲル(Aspergillus niger)またアスペルギルス・オリゼー(Aspergillus oryzae)の種に属する、請求項15に記載の宿主細胞。
【請求項17】
前記宿主細胞が、目的とするポリペプチドをコードする核酸配列を含んで成るポリヌクレオチドまたは発現ベクターをさらに含んで成る、請求項12〜16のいずれか一項に記載の宿主細胞。
【請求項18】
目的とするポリペプチドの発現をもたらす条件下に請求項16に記載の宿主細胞を培養し、場合により目的とするポリペプチドを回収して培養ブロスを形成するステップを含んで成る、目的とするポリペプチドを製造するための方法。
【図1】


【図2】
【図3】

【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2008−502338(P2008−502338A)
【公表日】平成20年1月31日(2008.1.31)
【国際特許分類】
【出願番号】特願2007−515963(P2007−515963)
【出願日】平成17年6月16日(2005.6.16)
【国際出願番号】PCT/EP2005/052805
【国際公開番号】WO2005/123763
【国際公開日】平成17年12月29日(2005.12.29)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公表日】平成20年1月31日(2008.1.31)
【国際特許分類】
【出願日】平成17年6月16日(2005.6.16)
【国際出願番号】PCT/EP2005/052805
【国際公開番号】WO2005/123763
【国際公開日】平成17年12月29日(2005.12.29)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]