
改良された導入遺伝子の発現方法
【課題】欠失の傾向を有する可能性がある配列領域を特定するために、発現ベクターに組み込む前に外来DNA配列をスクリーニングできる方法を提供すること。
【解決手段】外来DNA配列を適切なベクターによる発現に最適化する方法は、(i)前記外来DNAのヌクレオチドコドン使用頻度を最適化して、前記外来DNA配列が発現される宿主細胞のコドン使用頻度に変更する手順と、(ii)コドンが最適化されたDNA配列を改変して、前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得るいずれかの配列部位を変更する手順と、(iii)相同組換えに基づく推定欠失機構に関係する全配列を除去するために、前記外来DNA配列のヌクレオチドコドン使用頻度を変更する手順と、を含む。
【解決手段】外来DNA配列を適切なベクターによる発現に最適化する方法は、(i)前記外来DNAのヌクレオチドコドン使用頻度を最適化して、前記外来DNA配列が発現される宿主細胞のコドン使用頻度に変更する手順と、(ii)コドンが最適化されたDNA配列を改変して、前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得るいずれかの配列部位を変更する手順と、(iii)相同組換えに基づく推定欠失機構に関係する全配列を除去するために、前記外来DNA配列のヌクレオチドコドン使用頻度を変更する手順と、を含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ベクター系、特にレンチウイルスベクターにおいて、改変された導入遺伝子コンストラクトの効果的な転写および翻訳を達成するために改良した方法を提供する。当該方法は、導入遺伝子を含むウイルスベクターゲノムがトランスジェニック鳥類などの動物の生殖細胞系列に安定的に組み込まれることを可能にする無傷の機能導入遺伝子配列を有するウイルスベクターゲノムの生産を容易にする。該導入遺伝子が発現されると、組換え型タンパク質産物が生産され、トランスジェニック鳥類の場合では、目標とするタンパク質をトランスジェニック鳥の卵のなかに生産させることができる。
【背景技術】
【0002】
組換え型タンパク質の伝統的な製造方法には、細菌または哺乳類の細胞を用いた生産方法がある。これに代わる製造方法は、タンパク質の生産にトランスジェニック動物および植物を用いるものである。
【0003】
タンパク質をベースとした多数のバイオ医薬品が、トランスジェニックマウス、ウサギ、ブタ、ヒツジ、ヤギおよびウシなどの種々の哺乳動物の乳汁中に発現されている。当該系は世代時間が長い傾向があり、より大型の哺乳動物では遺伝子導入された初代動物から、乳汁を生産できる段階まで発達するには数年かかる。
【0004】
このほかの問題点は、乳汁の生化学的複雑さと、ヒトと動物との間の進化的保存性に関するものであり、これにより、医薬品を生産する哺乳動物に該医薬品に対する有害反応が起こる可能性がある(Harveyら、2002年)。
【0005】
製薬学的に重要なタンパク質、特に組換え型ヒト抗体のための製造媒体となる可能性があるものとして、ニワトリの卵の使用に関心が高まっている。
【0006】
ニワトリの卵に基づいたタンパク質製造系は、哺乳動物の細胞培養またはトランスジェニック哺乳動物系の使用に比べ利点がいくつかある。ニワトリは世代時間が短く(24週間)、このためトランスジェニックニワトリ群を速やかに確立することができる。さらに、トランスジェニック動物の生産施設のための設備投資は、細胞培養のための設備投資よりはるかに少なくてすむ。細胞培養に比べ、遺伝子導入によるタンパク質生産の促進に必要な特別な処理装置はほんのわずかである。このように設備投資が少なければ、遺伝子導入による治療薬の単位あたりの生産コストは細胞培養で生産される治療薬よりも低くなる。そのうえトランスジェニック系は、精製のバッチサイズおよび頻度に関して多大な柔軟性を提供する。この柔軟性は、バッチサイズを最適化することにより、精製における資本コストおよび操業コストをさらに削減させることにつながる可能性がある。
【0007】
さらに、遺伝子導入タンパク質の生産は市場への導入速度を速めることになる。トランスジェニック哺乳動物は、乳汁1リットルあたり数グラムのタンパク質を生産することができ、大量生産を商業的に実行可能なものとすることができる(Weck、1999年)。またさらに、鳥では世代時間が短いため、生産の迅速なスケールアップが可能である。
【0008】
鳥類の卵、特にニワトリの卵は、タンパク質の生産手段として細胞培養を上回る大きな利点をいくつか提供する。さらに鳥類系は、哺乳動物または植物に基づく他のトランスジェニック生産系を超える重要な利点を提供する。
【0009】
トランスジェニック哺乳動物の生産に使用される方法を鳥の遺伝子操作に直接適用することは、産卵鶏の生殖系の特別な特徴のため、これまで可能ではなかった。
【0010】
卵形成の複雑さにより、ヒヨコ胚発達の初期段階は比較的アクセスしにくい段階である。早期の段階の胚にアクセスするために採用される方法では、通常、ドナーである雌鳥を屠殺して胚を得るか、または輸卵管に直接注射を行なうことが必要である。トランスジェニック哺乳動物の生産方法は、ほぼ例外なく受精卵のマイクロインジェクションに重点が置かれており、この方法ではインビトロで前核にDNAを微量注入し、操作した当該卵を代理母に移して満期まで発育させるが、この方法は雌鳥では実行不可能である。
【0011】
トランスジェニック鳥類の作製には、一般的な方法が4つ開発されている。(i)胚盤の細胞質へのDNAマイクロインジェクションを用いてトランスジェニックニワトリを作製する方法、(ii)インビトロで始原生殖細胞のトランスフェクションを行ない、適切に準備したレシピエントに移植する方法、(iii)発癌性レトロウイルス由来の遺伝子導入ベクターを使用する方法、(iv)ヒヨコ胚細胞をインビトロで培養し、培養した細胞をレシピエント胚に導入することによりキメラ鳥を作製する方法である(Painら、・1996年)。キメラを作製する前に胚細胞をインビトロで遺伝子操作することができ、これによりキメラなトランスジェニック鳥が生じる。
【0012】
レンチウイルスはレトロウイルスの亜群であり、ヒト免疫不全ウイルスHIV−1およびHIV−2、サル免疫不全ウイルス(SIV)などの種々の霊長類ウイルスならびに非霊長類ウイルス(たとえば、マエディビスナウイルス(MW)、ネコ免疫不全ウイルス(FIV)、ウマ伝染性貧血ウイルス(EIAV)、ヤギ関節炎脳炎ウイルス(CAEV)およびウシ免疫不全ウイルス(BIV))が含まれる。これらのウイルスは特に遺伝子治療の開発において関心がもたれており、これは、レンチウイルスが宿主細胞DNAに不可逆的に組み込まれるというレトロウイルスの一般的な特徴を有するだけでなく、非増殖細胞を感染させる能力も有するからである。レンチウイルス感染の生物学はCoffinら(1997年)の総説を参照できる。
【0013】
ウイルスベクターを設計する際に考慮すべき重要事項は、細胞のゲノムに安定的に組み込まれる能力である。これまでの研究から、遺伝子導入媒体として用いるオンコレトロウイルスベクターは、発生過程における遺伝子発現抑制効果により成功率がやや低いことが示されている。マウスに関するPfeiferら(2002年)およびLoisら(2002年)の研究から、HIV−1に基づくレンチウイルスベクターは発生過程で発現が抑制されないことが示された。
【0014】
レンチウイルスベクターに関する開発研究の大半はHIV−1系に焦点を当ててきたが、これは主に、ヒトに病原性を示すためにHIVが最も十分に特徴付けられているレンチウイルスであるという事実によるものである。当該ベクターは、調節遺伝子およびアクセサリー遺伝子を除去して複製を不可能にすることにより、複製能力をなくすように設計される傾向がある。こうしたベクターの最新型は、調節遺伝子のほぼすべて、アクセサリー遺伝子のすべてが除去される程度まで最小化されている。
【0015】
レンチウイルス属のウイルスは、類似したゲノム構成、類似した複製サイクル、成熟マクロファージを感染させる能力など、多数の類似した特徴を有する(ClementsおよびPayne、1994年)。当該レンチウイルスの1つがウマ伝染性貧血ウイルス(EIAV)である。レンチウイルス属の他のウイルスに比べ、EIAVは比較的単純なゲノムを有し、該ゲノムは、レトロウイルスのgag、polおよびenv遺伝子のほかにわずか3つの調節/アクセサリー遺伝子(tat、revおよびS2)からなる。安全かつ効果的なレンチウイルスベクター系の開発は、ベクター自体のデザインに依存することになる。効果的な機能を得るためには、形質導入するベクターとしての機能を保ちながら、ベクターのウイルス構成要素を最小限に抑えることが重要である。
【0016】
オンコレトロウイルスベクター系およびレンチウイルスベクター系は、形質導入可能な細胞の種類および動物種の範囲を広げるために改変することができる。これは、該ウイルスのエンベロープ糖タンパク質を他のウイルスのエンベロープタンパク質に置き換えることによって達成される。EIAVレンチウイルスベクターにパッケージされた導入遺伝子を生殖細胞系列に安定的に発現させることが可能である(McGrewら、2004年)。この方法は、外来DNAの関連断片を合成することと、観察された最適なニワトリの頻度にコドン使用頻度を変えること(口語で「チキナイゼーション(ニワトリ化)」と言われるプロセス)を必要とする。当該プロセスは、特定の外来DNA配列の効果的な転写および翻訳を十分に可能にし、その結果、得られた鳥で目的のタンパク質を発現させると考えられる。しかし一部のタンパク質配列は、安定的に発現できるように改変が必要であることが示されている。
【0017】
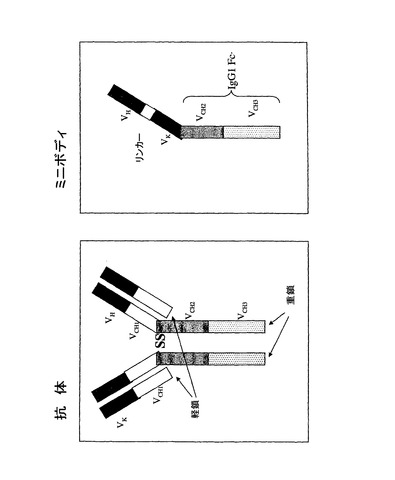
ガングリオシドGD3に特異的であるR24として知られるネズミ抗体を使用して、「ミニボディ」と呼ばれる組換え型抗体様結合分子が作製された。このミニボディの構造は、伝統的な抗体VHおよびVLドメインと、これに結合したリンカーおよびIgG1のFcドメインから構成された。EIAVをベースとしたレンチベクターにこのミニボディのためのコード配列がパッケージされたが、ミニボディタンパク質産物の発現は達成できなかった。
【0018】
その後、R24ミニボディを含有する種々のウイルスゲノムから直接増幅したRT−PCR産物が解析された結果、外来R24ミニボディのコード配列の一部または全部を包含する多数の欠失が起こっていることが確認された。これらの欠失の5’および3’範囲を表す配列が解析された結果、異常スプライシングがこれらの欠失の原因ではないことが示された。当該欠失は小さい(5〜10bp)直列反復配列によって特徴付けられているようであり、このことから、観察された外来DNAコード配列の変化は、これまで知られていない相同的組換えに基づく機構が原因であることが示唆されている。
【0019】
Ch’angらはこれまでに、ネズミ白血病ウイルス(MuLV)の組み込まれたプロウイルスゲノムにおける内部欠失を報告しており、研究中に確認された3つの欠失はいずれも、7つのヌクレオチドの直列反復配列に挟まれたていたと述べている(Ch’angら、1989年)。短い直列反復配列によって挟まれたDNA配列に関する特定の欠失は、他のレトロウイルス遺伝子(Coffinによる総説、1985年)のほか、種々の原核生物および真核生物の遺伝子(Omerら1983年、およびLevyら1985年の論文)においても観察されている。短い直列反復配列に挟まれた欠失は、・鳥肉腫ウイルスsrc遺伝子にも観察されている(Omerら、1983年)。提案された機構は、たとえば、DNAポリメラーゼまたは逆転写酵素などDNA複製機構のずれであることが示唆されている。しかし、R24ミニボディベクター系に観察された欠失は、逆転写されたウイルスRNAゲノムから直接増幅したRT−PCR産物に存在したことから、この機構によって説明することはできない。そうではなく、宿主細胞のRNAポリメラーゼ(Rpol II)が、パッケージング細胞株へのプラスミドのトランスフェクション直後にウイルスゲノムの転写時に欠失を誘導した可能性が高い。この結論を支持するように、一部の宿主DNA依存性RNAポリメラーゼは鋳型スイッチの能力を有することが知られており(Nudlerら、1996年)、さらに、RNA組換えはヘアピンループなどの三次元構造の存在により影響を受けることも知られている(WhiteおよびMorris、1995年)。
【0020】
レンチウイルスベクターのバックボーンに組み込むときに欠失が発生する傾向を検討するために、別の外来遺伝子配列、すなわち、791T/36として知られる組換え型ネズミ抗CD55抗体の遺伝子配列が評価された。欠失に関与することが知られている配列が791T/36に保存されていた。
【0021】
したがって、一部の複合タンパク質をコードする遺伝子内の特定の配列は、レンチウイルスベクターバックボーンに組み込まれると欠失を経験しやすいと考えられる。欠失の程度は遺伝子ごとに劇的に異なる可能性が高く、したがって予測不可能であると考えられる。R24ミニボディの発現に関連して示されているように、導入遺伝子からのタンパク質の発現がもはや不可能である程度まで欠失が起こることがあり、言い換えれば、トランスジェニック系における該タンパク質の発現を妨げることになる。
【0022】
欠失の傾向を有する可能性がある配列の領域を特定するために、発現用ベクターに組み込む前に外来DNA配列をスクリーニングできれば、大いに望ましいことである。
【非特許文献1】Ch’ang LY, Yang WK, Myer FE, Koh CK, Boone LR(1989). Virology 168, 245−255.
【非特許文献2】Clements JE & Payne SL(1994) Virus Res. 32(2),97−109.
【非特許文献3】Coffin J(1985). Genome Structure (R Weiss, N Teich, HE Varmus eds)2,17−74.
【非特許文献4】Graf M, Bojak A, Demi L, Bieler K, Wolf H, Wagner R(2000) J. Virol. 74,10822−826.
【非特許文献5】Harvey AJ, Speksnijder G, Baugh LR, Morris JA, Ivarie R(2002) Poult. Sci .81(2),202−12.
【非特許文献6】Horton RM, Hunt HD, Ho SN, Pullen JK, Pease LR.(1989) Gene77(1),61−8.
【非特許文献7】Levy DE, Lerner RA, Wilson MC(1985). Cell 41,289− 299.
【非特許文献8】Lois c, Hong EJ, Pease S7 Brown EJ, Baltimore D(2002) Science 295(5556),868−72.
【非特許文献9】Martinez−Salas E(1999) Current Opinion Biotechnology 10,458−64.
【非特許文献10】McGrew MJ, Sherman A, Ellard FM, Lillico SG, Gilhooley HJ, Kingsman AJ, Mitrophanous KA & Sang H(2004) EMBO Reports 5(7),728−33.
【非特許文献11】Nudler E, Avetissova E, Markovtsov V, Goldfarb A (1996) Science 273,211−217.
【非特許文献12】Omer CA, Pogue−geile K, Guntaka R, Staskis KA, Faras AJ (1983; J. Virol. 54,889−893.
【非特許文献13】Pain B, Clark ME, Shen M, Nakazawa H, Sakurai M, Samarut J, Etches RJ (1996). Development 122(8),2339−48.
【非特許文献14】Pfeifer A, Ikawa M, dayn Y, Verma IM (2002) PNAS 99(4),2140−45.
【非特許文献15】Schneider R, Campbell M, Nasioulas G, Felber BK, Pavlakis GN (1997) . Journal of Virology 71(7),4892−903.
【非特許文献16】Shaw G, Kamen R(1986). Cell 46(5),659−67.
【非特許文献17】Spies M, Bianco PR, Dillingham MS, Handa N, Baskin RJ, Kowalczykowski SC (2003). Cell 114(5),647−54.
【非特許文献18】Tran DP, Kim SJ, Park NJ, Jew TM, Martinson HG(2001). Molecular and Cellular Biology 21(21),7495−508.
【非特許文献19】Trinh R, Gurbaxani B, Morrison SL, Seyfzadeh M (2004). Molecular Immunology 40,717−722.
【非特許文献20】White KA and Morris TJ (1995)RNA 1,1029−1040. Week, E. 1999 ’Transgenic Animals: ’market opportunities now a reality’ D&MD reports
【発明の開示】
【発明が解決しようとする課題】
【0023】
本発明の発明者らは、驚くべきことに、解析すべき外来DNA配列を欠失または他の配列変異を起こすと予測される配列領域を決定できるスクリーニング方法を開発した。いったん、このような配列領域が特定されれば、これらの配列を改変することができる。さらに、このような改変は、外来DNA配列をベクターバックボーンに組み込む前に、有利に行なうことができる。したがってこの方法は、導入遺伝子を含むウイルスベクターゲノムがトランスジェニック鳥類などの動物の生殖細胞系列に安定的に組み込まれることを可能にする無傷の機能導入遺伝子配列を有するウイルスベクターゲノムの生産を容易にし、これによって、非ヒト動物、特に鳥類などのトランスジェニック系における組換え型タンパク質の生産に使用できる。
【課題を解決するための手段】
【0024】
本発明の第1態様によれば、外来DNA配列を適切なベクターによる発現に最適化する方法であって、(i)前記外来DNAのヌクレオチドコドン使用頻度を最適化して、前記外来DNA配列が発現される宿主細胞のコドン使用頻度に変更する手順と、(ii)コドンが最適化されたDNA配列を改変して、前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得るいずれかの配列部位を変更する手順と、(iii)相同組換えに基づく推定欠失機構に関係する全配列を除去するために、前記外来DNA配列のヌクレオチドコドン使用頻度を変更する手順と、を含む方法が提供される。
【0025】
一実施形態において、この方法は手順(i)及び(iii)を含む。別の実施形態において、この方法は手順(ii)及び(iii)を含む。更に別の実施形態において、この方法は手順(i)、(ii)、及び(iii)を含む。
【0026】
前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得る配列要素としては、陰性要素又は反復配列、スプライシング部位、内部TATAボックス、及び内部リボソーム侵入部位のようなシス活性化モチーフが挙げられる。
【0027】
従って、本発明の実施形態は、前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得る配列要素の存在について外来DNA配列を解析することまで及び、特に前記配列要素としては陰性要素又は反復配列、スプライシング部位、内部TATAボックス、及び内部リボソーム侵入部位のようなシス活性化モチーフが挙げられる。
【0028】
陰性要素は、例えばAT又はGCリッチであったり、重要なRWA二次構造に寄与したり、又は具体的機能が起因するTATAボックス、カイ配列、リボソーム侵入部位、ARE、INS、CRS、スプライシングシグナル、又はポリアデニルかシグナルといったコンセンサス配列と予測されるような一般配列等、2カテゴリの1つに一般に適する。
【0029】
TATAボックスは、転写開始部位(5’ TATAAAA 3’ :この配列は転写開始部位を正確に決定する際に重要となる)の上流25ヌクレオチド付近に位置する真核生物ポリメラーゼIIによって転写される大部分の遺伝子のプロモータ領域において発見されるコンセンサス配列と定義できる。
【0030】
RecBCD酵素は、二重鎖DNAの損傷部位において相同組換えを開始するヘテロ三量体ヘリカーゼ/ヌクレアーゼである。その活性のいくつかは、転移酵素でシスに認識されるDNAカイ配列(5’ GCTGGTGG 3’ )によって調節されている(Spies et al, 2003))。
【0031】
内部リボソーム侵入部位は、機能的基準及び大きな配列相同性を共有することが報告されていないことに基づいて通常定義される。しかし、インシリコ配列解析プログラムは、公知のIRES配列が導入遺伝子内に存在しないことを検証した(Martinez−Salas, 1999で総説される)。アデニンリッチ要素(ARE)は、一過的に発現される遺伝子由来のmRNAの3’UTR内に頻繁に位置するAUリッチ配列と定義される。ARE配列の導入は、mRNAを不安定化するのに充分であり、mRNAのプロセス経路の認識シグナルとして提案されている(Shaw & Kamen, 1986)。
【0032】
阻害要素(INS)及びシス活性リプレッサ配列(CRS)は、いずれもHIVモデル系において当初報告されており、1つの仮説によればmRNAの不安定性に寄与する細胞性因子の結合部位である(Schneider et al, 1997)。かかる配列をHIV転写物から除去すると、その転写物の発現が大きく促進することが実証され、前述したINS又はCRS配列の不存在又は除去を検証することが、導入遺伝子の最適化手順の間望まれる。
【0033】
3タイプのコンセンサススプライシングシグナルが報告されている。第1は、切除される配列(イントロン)の5’末端と定義されるスプライシングドナー(C又はA、A、G/G T、A又はG、A、G、T)である。第2は、切除される配列の3’末端と定義されるスプライシングアダプタ(T又はC, n, N, C又はT, A, G/g)である。第3は、切除される配列内に位置する分岐点配列(TACTAAC)であり、スプライシング反応間のラリアット形成に関与する。
【0034】
RNAポリメラーゼIIによる転写終了は、通常、機能的ポリアデニル化シグナル(ポリA)の存在を要する。脊柱動物のコアポリAシグナルは、ポリA切断部部位に隣接する2つの認識部位からなる。典型的には、ほとんど不変のAAUAAヘキサマーが、U又はGU残基リッチのより可変な要素の20〜50ヌクレオチド上流に位置する。新生転写物の切断は、これら2要素の間で起こり、5’切断産物への250以下のアデノシン(ポリAテイル)の付加につながる(Tran et al, 2001)。
【0035】
前述したいくつか又はすべての配列要素を保持する結果は、保持される配列の性質に応じて変動する。これらは、種々異なる機構によるが異種コード配列の発現を低減することから、陰性要素として記述される。例えば、異種コード配列内に同属スプライシング配列を保持することで、形質転換遺伝子産物の発現を消滅、低減、又は制限するスプライシング及び切除が高効率化し得る。対照的に、INS要素を保持しても、RNA組み込みに影響を及ぼさず、むしろmRNAが所望のコードされる遺伝子産物の大部分の転移が起こる前に高速分解の標的にされる。いずれの機構も、異種タンパク質発現の減少という同じ一般的結果を生み出す。本発明のこの態様の一実施形態において、本発明の発現最適化方法によって解析され任意に改変された外来DNA配列は、トランスジェニック発現系において発現され得るベクターに含まれる。
【0036】
トランスジェニック発現系は、非ヒト哺乳動物であってよい。更に別の実施形態において、トランスジェニック発現系は、鳥類、特にニワトリ又はウズラであってよい。
【0037】
本発明の一実施形態において、外来DNAは、ベクターの内部プロモータの制御下にあり宿主細胞によって発現される異種タンパク質をコードする。
【0038】
本発明の一実施形態において、ベクターはレンチウイルスベクターである。別の実施形態において、ベクターはウマ伝染性貧血ウイルス(EIAV)である。本発明は、レンチウイルスベクターとして、ヒト免疫不全ウイルスHIV−I及びHIV−2、サル免疫不全ウイルス(SIV)、非霊長類ウイルス(例えばマエディ−ビスナウイルス(MW)、ネコ免疫不全ウイルス(FIV)、ウマの伝染性の貧血症ウイルス(EIAV)、ヤギ関節炎脳炎ウイルス(CAEV)、及びウシ免疫不全ウイルス(BIV))も提供する。
【0039】
本発明のこの態様の一実施形態において、外来DNAは、組換え抗体その他の結合フラグメント又は結合物である異種タンパク質をコードしてよい。
【0040】
抗体又は結合物等をコードする外来DNA配列の解析は、抗体又は結合物に組み込まれるためのDNAコード配列から除去された実質的にすべての重複配列を有するリンカー配列を、一次アミノ酸配列中の3重複配列(Gly4Ser1)を保持しながら設計する手順を更に含んでよい。この手順は、本発明のこの態様の方法の一部として実施される場合には手順(iii)の前に行うことが好ましい。
【0041】
具体的には、かかる手順は、手順(ii)の完了後で手順(iii)の実施前に行われるため、以後この手順を本発明のこの態様の方法における手順(iib)と表記する。
【0042】
ここで定義されるように、用語「コドン最適化」は、外来DNA配列のコドン使用頻度が、発現を向上するためにベクターが挿入され発現される非ヒト哺乳類動物細胞型において最も頻繁に使用されるコドンをコードするために意図的にバイアスされるように、コドン使用頻度を変更する手順を指す。例えば、トランスジェニック発現系がニワトリのとき、コドン使用頻度の変更は、ニワトリ種において最も一般に使用されるコドンに発現をバイアスするために特定のコドンを変化させる。ニワトリで実施するとき、ヌクレオチド配列のコドン使用頻度を変更する手順は、外来DNA配列の「ニワトリ化」と口語的に表記され得る。
【0043】
具体的には、ここで定義されるように、用語「ニワトリ化」は、非変更形態においてヌクレオチド配列(コドン)によってコードされるアミノ酸配列をコードするニワトリ種で最も一般的な3ヌクレオチドによってコドンがコードされるように、ヌクレオチド配列内のコドン使用頻度を意図的に変更する手順を指す。トランスジェニックニワトリにおける発現のため、外来DNA配列によって形成されるコドンは、ニワトリにおける最も頻繁なコドン使用パターンに最適化される。しかし、ベクターが発現される任意の鳥類種又は非ヒト哺乳動物の最も頻繁なコドン使用頻度に最適化することもできる。
【0044】
ニワトリ化の方法例として、アミノ酸バリンは4つの異なるコドン、GTG、GTA, GTT、及びGTCによってコードされるが、鳥類ではGTGが最も頻繁に使用される(46% GTG, 11% GTA, 19% GTT、23% GTC)。ヒトIgGFcDNAをニワトリ化するためには、すべてのバリンコドンがAAGに変換される。リジンは2つの異なるコドン、AAG及びAAAによってコードされるが、鳥類ではGTGが最も頻繁に使用される(58%対42%)。配列中のすべてのAAAコドンがAAGに変換される。すべてのコドンの変更が必要とされる訳ではない。例えば、アスパラギン酸としては、2つの異なるコドン、GAT及びGACが略均等に使用されるため、ニワトリ化手順において変更する必要はない。
【0045】
本発明のこの態様の方法の手順(i)及び(ii)で概略されるようなコドン使用頻度及び配列改変は、異種導入遺伝子からの遺伝子発現の最適化として当業者に公知である(例えば、Graf et al . , 2000参照)。
【0046】
本発明のこの態様の方法の手順(i)及び(ii)は、典型的には、同様の配列設計サービスを提供するGeneart GmbH (Germany, www.geneart.com)又は組織と協力して実施される。Geneartによる手順(i)及び(ii)の実施は、配列最適化を行うために配列設計及び解析を可能とするコンピュータ配列設計の実施を含む。この方法は、配列を解析し、コドン使用頻度を交換し、コドン交換による配列変化が陰性要素又は繰返しを導入しないことを確保するために得られる配列を解析する手順を含む。タンパク質発現にヌクレオチド配列を最適化する方法の具体的記載は、国際公開WO2004/059556に見出すことができ、その全容は参照して本明細書に援用される。得られる塩基配列を、手順(iii)に示されるようにして更に改変する。任意に、前述した追加的な手順(iib)を手順(iii)の実施の前に行うことができる。
【0047】
最終的な配列は、外来DNA配列の合成の開始前に、問題配列が再導入されていないかを再確認するために再解析してもよい。
【0048】
この方法は、手順(iib)及び手順(iii)を、発現される外来DNA配列に応じて適切な配列での利用に適合させるだけで、必要に応じてあらゆるタンパク質配列での使用に適合できる。
【0049】
スクリーニング方法のモジュール性は、トランスジェニック動物の作成に使用される場合、レンチウイルスベクター等のベクターに組み込んだ後、欠失の危険があり得る任意の外来DNA配列に適用し得る点で、高度に適合化させる。例えば、サイトカイン又はホルモンのような酵素又は生物活性タンパク質等の標準的導入遺伝子のコード配列を解析してよく、同様に、非ヒト哺乳類トランスジェニック系における発現に適する治療的タンパク質等の他のタンパク質の配列を解析してもよい。
【0050】
更に、スクリーニング方法は、抗体その他の同様の結合フラグメント又は結合物の配列のスクリーニングに使用できる。「抗体」はイムノグロブリンであり、天然、又は部分的もしくは完全に合成的に作成される。この用語は、抗体結合ドメインであり又はそれに相同な結合ドメインを有するいかなるポリペプチド、タンパク質、又はペプチドも包含する。抗体の例は、イムノグロブリン異性体、その異性体サブクラス、Fab, scFv, Fv, dAb, Fdのような抗体結合ドメインを含むフラグメント、組換え型二重特異性抗体である。抗体はヒト化されていてよく、ヒト化抗体には部分的ヒト化(キメラ)又は完全ヒト化された抗体が含まれうる。
【0051】
しかし、本発明のこの態様のスクリーニング方法が組換え抗体系導入遺伝子の発現最適化に使用される場合、改変リンカー配列の使用が推奨される。
【0052】
リンカー配列の発展
広く商業的に入手可能なリンカーの例はRPASマウスscFVモジュール(Amersham Biosciences)であり、リンカー配列は、以下の配列番号1のヌクレオチド配列を有する。
GGT GGA GGC GGT TCA GGC GGA GGT GGC TCT GGC GGT GGC GGA TCG
【0053】
本発明は、設計され、以下の配列番号3のヌクレオチド配列を有する新規リンカーも提供する。
GGG GGA GGG GGC AGC GGC GGA GGG GGA TCC GGC GGT GGG GGA TCT
【0054】
配列番号3のヌクレオチド配列は、配列番号4の配列を有するアミノ酸配列をコードする。
GIy GIy GIy GIy Ser GIy GIy GIy GIy Ser GIy GIy GIy GIy Ser
【0055】
繰返しDNA配列を排除するよう設計するとともに、配列設計及びリンカー配列の解析の間に適用される第2の制限は、隣接コドンとしてGGC及びTCCを回避することであった。例えば、広く商業的に入手可能なリンカーであるRPASマウスscFVモジュール(Amersham Biosciences)(配列番号5)は、隣接配列としてのGGC及びTCCの存在を評価し、以下の配列が確認される。
配列番号5:GGG GGA GGC GGC TCC GGG GGA GGC GGC TCC GGG GGA GGC GGC TCC
【0056】
pRl28(R24ミニボディ発現をドライブするCMVプロモータ)及びpLE38(R24ミニボディ発現をドライブする組織特異的プロモータ)等のいくつかのEIAV系レンチウイルスベクターコンストラクトからの以前のPCRデータから、推定相同組換え系機構における繰返しがR24ミニボディコード配列内の欠失を発生させていると示唆されたことから、再設計を行った。新規リンカーは、それを含む組換えタンパク質の発現レベルの低下を引き起こすことが知られている、コドンのいわゆる「スローペア」であるGGA、GGC(Trinh et al., 2004)の使用も回避する。
【0057】
非推定リンカー配列の使用は公知である。しかし、本発明は、ウイルスベクター導入遺伝子から短い重複配列を除去するために、リンカー内のコドン選択を変更する外来DNA配列の改変を更に提供する。
【0058】
本発明の更なる態様は、異種タンパク質の少なくとも一部をコードする単離DNAであて、本発明のスクリーニング方法で解析されたDNAを提供する。
【0059】
本発明の更なる態様は、組換え抗体系導入遺伝子の発現のためのリンカー配列であって、配列番号3のヌクレオチド配列を有する配列を提供する。
【0060】
本発明の更なる態様は、組換え抗体系導入遺伝子の発現のためのリンカー配列であって、配列番号4のヌクレオチド配列を有する配列を提供する。
【0061】
本発明の更なる態様は、トランジェニック鳥類を作製する方法であって、前記トランジェニック鳥類において発現させる1以上の異種タンパク質をコードする外来DNA配列を準備する手順と、前記外来DNAの異種タンパク質コード領域のヌクレオチドコドン使用頻度を最適化して、前記外来DNA配列が発現されるトランジェニック鳥類のコドン使用頻度に変更する手順と、コドンが最適化されたDNA配列を改変して、前記宿主鳥類内での前記外来DNAの発現を阻害又は下方制御すると予測されるいずれかのコード配列領域を変化させる手順と、相同組換えに基づく推定欠失機構に関係する全配列を除去するために、前記外来DNA配列のヌクレオチドコドン使用頻度を変更する手順と、前記外来DNA配列を含むベクターを鳥類ゲノムに組み込む手順と、前記配列によってコードされる前記異種タンパク質を生産するために前記外来DNA配列を発現させる手順と、を含む方法を提供する。
【0062】
本発明の外来DNA配列を含むベクターを調整する際、外来DNA配列は、関連する制御領域及び発現調節領域とともにパッケージされる。当業者は、ベクターの好ましいパッケージ方法に気づくであろう。
【0063】
本発明は、トランスジェニック鳥類も提供する。トランスジェニック鳥類は、鳥類種のあらゆるメンバーであり、特にニワトリであり、鳥類細胞の1以上は、細胞ゲノム内に組み込まれ、ベクターに含まれる外来遺伝子材料を含む。かかる遺伝子材料の導入に適した遺伝子導入技術は当業者に公知である。
【0064】
本発明の方法は、あらゆるトランスジェニック鳥類の生成に使用でき、特に限定されないが、ニワトリ、シチメンチョウ、カモ、ウズラ、ガチョウ、ダチョウ、キジ、クジャク、ホロホロチョウ、ハト、白鳥、チャボ、及びペンギンが挙げられる。しかしニワトリが好ましい。トランスジェニック鳥類によって発現される異種タンパク質は、特に限定されないが、ヒト及び/又は獣医目的での治療及び診断利用を含む種々の用途を有するタンパク質であってよく、抗体、抗体フラグメント、抗体誘導体、単鎖抗体フラグメント、融合タンパク質、ペプチド、サイトカイン、ケモカイン、ホルモン類、成長因子、またはいかなる組換えタンパク質をコードする配列を含んでよい。
【0065】
本発明は、更にキメラ鳥類又はモザイク鳥類に及び、外来遺伝子材料は鳥類細胞のすべてではないがいくつかにおいて発見される。
【0066】
一実施形態において、トランスジェニック鳥類は、卵管内で外来遺伝子材料を翻訳タンパク質の形で発現し、卵内へ組み込まれる。
【0067】
レンチウイルスベクター発現コンストラクトは、ベクターによってコードされる異種タンパク質の発現を特異的組織に方向付け(組織特異的発現)るために使用できる。一実施形態において、かかる組織特異的発現は、卵内へ異種タンパク質が組み込まれるように方向付けされる。それは卵白又は卵黄内であってよいが、タンパク質が卵白内に存在することが好ましい。次にタンパク質は、当業者に公知の常法によって卵白又は卵黄から単離される。
【0068】
本発明の更なる態様は、鳥類の卵管内で外来タンパク質を発現する方法であって、本発明の方法を用いて解析された外来DNA配列を準備して、前記外来DNA配列によってコードされる異種タンパク質の発現を阻害又は下方制御し得るコード配列のいずれかの部位を除去又は置換する手順と、記外来DNA配列を鳥類ゲノムに組み込む手順と、前記外来DNA配列に操作可能に連結されたプロモータを用いて前記外来DNAコード配列を発現させる手順と、前記トランスジェニック鳥類によって発現される外来タンパク質を得る手順と、を含む方法を提供する。
【0069】
一実施形態において、本発明の第1態様のスクリーニング方法で解析された外来DNAコード配列はウイルスベクターのバックボーンに挿入され、このベクターを鳥類細胞に挿入される。
【0070】
プロモータが、鳥類の輸卵管のマグナム部分の管状腺細胞内で外来DNA配列によってコードされる異種タンパク質の「組織特異的」発現を行うことが好ましい。「組織特異的」発現は、異種タンパク質の特異的組織で発現させ、他の組織では異種タンパク質の発現が排除される。異種タンパク質の組織特異的発現を鳥類の卵管に方向付けると予測されるプロモータの一例は、オバルブミンプロモータである。
【0071】
本発明のこの態様の更なる態様において、プロモータは、外来DNAコード配列によってコードされる異種タンパク質の発現を卵管の他の組織に方向付けるために、必要に応じて改変されてよい。
【0072】
外来タンパク質は、治療的に有用なタンパク質であり得る。特に、発現される異種タンパク質は、抗体又は同様の結合フラグメントもしくは結合物であってよい。
【0073】
本発明の更なる態様は、鳥類において1以上の外来タンパク質を発現する方法であって、発現させる外来タンパク質をコードする外来DNA配列を準備する手順と、本発明のスクリーニング方法を用いて前記外来DNA配列を解析する手順と、前記外来DNA配列を含むベクターを鳥類ゲノム内で発現させる手順と、発現された抗体タンパク質を前記鳥類から得る手順と、を含む方法を提供する。
【0074】
本発明のこの態様の一実施形態において、1以上の異種タンパク質は、組織特異的、最も好ましくは、卵管細胞での組織特異的発現を用いて鳥類の卵管内で発現される。別の実施形態において、外来タンパク質は、鳥類の輸卵管のマグナム部分の管状腺細胞内で発現され、外来タンパク質は卵白に蓄積する。代替的又は追加的に、異種タンパク質は卵白に蓄積し、血液に分泌され得る。
【0075】
更なる実施形態において、鳥類はニワトリである。
【0076】
一実施形態において、卵管内で発現される異種タンパク質は抗体である。更なる実施形態において、抗体は「ヒト化」される。
【0077】
本発明の更なる態様は、本発明の第1態様のスクリーニング方法を用いて解析された外来DNA配列の、外来タンパク質を含む鳥類卵の生産における使用を提供する。一実施形態において、外来タンパク質は卵白内に蓄積される。更なる実施形態において、外来タンパク質は卵黄内に含まれる。
【0078】
本発明の更なる態様は、本発明の第1態様のスクリーニング方法で解析された外来DNA配列の、異種タンパク質製品の生産における使用であって、前記タンパク質製品は前記外来DNA配列の少なくとも一部の転写及び翻訳によって製造される使用を提供する。
【0079】
本発明の更なる態様は、本発明の第1態様のスクリーニング方法で解析された1以上の外来DNA配列を含む発現ベクターを提供する。
【0080】
更なる態様は、前述した発現ベクターが形質転換された宿主細胞を提供する。
【0081】
一実施形態において、発現ベクターはレンチウイルスベクターであり、特にEIAVである。
【0082】
一実施形態において、宿主細胞は非ヒト哺乳類細胞である。更なる実施形態において、宿主細胞は、鳥類細胞であり、特にニワトリ細胞である。
【0083】
本発明の更なる態様において、本発明のいずれかの方法を使用するためのキットであって、使用説明書と、前記方法を使用するプロトコルとを備えるキットが提供される。
【0084】
本発明の各態様の好ましい特徴及び実施形態は、特に要求がない限りにおいて、他の各態様として変更すべき点が変更される。
【0085】
定義
本明細書では、「ベクター」、「ウイルスベクター」および「発現用ベクター」という用語は置き換え可能に使用しており、外来DNA配列の、プロモータにより誘導される発現、すなわち転写とこれに続く翻訳を可能にする任意の核酸、好ましくはDNAを指している。
【0086】
ウイルスベクターゲノムは「複製欠損」であることが好ましく、すなわち、該ベクターのゲノムは、独自に複製して感染性ウイルス粒子が産生されうるほど十分な遺伝情報だけを備えていないことが好ましい。レンチウイルスベクターの場合では、該ゲノムは機能遺伝子gag、envまたはpolを欠いていることになる。
【0087】
「レンチウイルス」という用語は、本発明に特に好ましいレトロウイルス科の亜科を指している。レンチウイルスには、ヒト免疫不全ウイルスHIV−1およびHIV−2、サル免疫不全ウイルス(SIV)などの種々の霊長類ウイルスならびに非霊長類ウイルス(たとえば、マエディビスナウイルス(MW)、ネコ免疫不全ウイルス(FIV)、ウマ伝染性貧血ウイルス(EIAV)、ヤギ関節炎脳炎ウイルス(CAEV)およびウシ免疫不全ウイルス(BIV)・が含まれる。
【0088】
「ウイルスベクターゲノム」とは、ウイルスゲノムからの配列を備えたポリヌクレオチドを指し、該ウイルスゲノムは、かかるポリヌクレオチドのRNA版をウイルス粒子にパッケージすることが十分に可能で、パッケージされたRNAポリヌクレオチドが逆転写されて宿主細胞の染色体に組み込まれるようになるものをいう。プロモータ配列や、異種ペプチドをコードする外来DNA配列などの異種配列も、ウイルスベクターゲノムの一部であってよい。
【0089】
本明細書で核酸分子を説明するために使用している「組換え型」という用語は、ゲノム、cDNA、半合成または合成に由来するポリヌクレオチドを指しており、その由来または操作により、自然界で結び付いているポリヌクレオチドの全部または一部とは結び付いておらず、および/または自然界で連結されているもの以外のポリヌクレオチドと連結されているものをいう。
【0090】
本明細書でタンパク質またはポリペプチドを説明するために使用している「組換え型」という用語は、組換え型ポリヌクレオチドの発現により生産されるポリペプチドを指す。
【0091】
本明細書で使用している「核酸」という用語には、DNA、RNA、mRNA、cDNA、ゲノムDNA、およびそれらの類似体が含まれる。
【0092】
「外来DNA配列」とは、その転写発現を望む核酸配列である。外来DNA配列は一般に、ペプチド、ポリペプチドまたはタンパク質をコードする。
【0093】
「欠失」とは、ウイルスベクターゲノムの元のプラスミドコピーに存在するDNA配列の領域が逆転写の過程で失われる事象をいう。このため欠失した配列は、複製不能なレンチウイルスベクターの粒子が生産されるパッケージング過程で、元のプラスミドから転写された一本鎖RNA分子の一部または全部から失われている。プラスミドDNA配列は常に無傷のまま維持され、欠失は、一本鎖RNAの2つのコピーが逆転写されてタンパク質の外被内で組み立てられるパッケージング過程における転写過程で起こることに注意されたい。
【0094】
さらに、細胞で正常に発現されない未改変の核酸配列またはポリペプチドは異種であるとみなされる。本発明のベクターは、同一のまたは異なる挿入部位に挿入された1つ以上の外来DNA配列を有することが可能で、挿入部位で各外来DNA配列は、該配列の発現を可能にする調節核酸配列に操作可能に連結される。このため、本発明から得られるベクターは、・単量体、二量体および多量体タンパク質などの種々のタイプのタンパク質を発現させるために使用できると考えられる。
【0095】
本発明で説明したベクターは「異種タンパク質」を発現させるために使用できる。
【0096】
本明細書で使用しているように、「異種」という用語は、外来種に由来する核酸配列もしくはポリペプチド、または同一の種から由来する場合はその原形が実質的に改変された核酸配列またはポリペプチドを指す。
【0097】
適切な異種ペプチドは、治療活性または他の商業的関連性のある用途を有する組換え型タンパク質であってもよい。発現されうる異種タンパク質の例として、インターフェロンアルファ、ベータおよび/またはガンマなどのサイトカイン、インターロイキン、第VIII因子などの造血因子が挙げられる。1つの実施形態では、該異種ペプチドは抗体の重鎖または軽鎖をコードするものであってもよく、ネズミ抗体、キメラ抗体、ヒト化抗体およびヒト抗体など任意のタイプの抗体の重鎖または軽鎖であることが可能であり、この2つの鎖は同一の抗体または異なる抗体に由来することができる。
【0098】
別に定義されていない限り、本明細書に使用した技術用語および科学用語はすべて、本発明の技術分野における当業者に広く理解されている意味を有するものである。
【0099】
本明細書の全体を通して、文脈からそうではないことが求められていない限り、「包含する」もしくは「含む」という用語、または「包含している」、「含んでいる」などの変形は、記載した整数または整数群を含めることを意味すると理解されると思われるが、任意の他の整数または整数群を除外することを意味しているのではない。
【実施例】
【0100】
実施例1
R24ミニボディ−RT−PCRデータ
【0101】
EIAVレンチウイルスベクターとともに使用するR24ミニボディの全配列を図1に示す。該組換え型抗体分子は、ヒトIgG1 Fcドメインの上流に挿入された、マウスVH、リンカーおよびマウスVLから構成される標準的scFV断片からなる(図2)。該配列を、2種類のプロモータの下流に導入し、1つは大域的プロモータであるヒトサイトメガロウイルス(hCMV)即時型初期プロモータであった。もう1つは、トランスジェニック鳥類内で空間時間的に限定された様式でR24ミニボディを活発に発現させるように設計した組織特異的プロモータ候補であった。
【0102】
ウイルスゲノムプラスミドpRI28(プラスミドマップは図3、全配列は図4に示す)を作製するために、hCMVプロモータの下流にR24を挿入した。該ゲノムプラスミドをD17イヌ骨肉種細胞に一過性にトランスフェクションし、その後、細胞培地についてELISAを行なった結果、600ng/mlレベルのヒトIgG1が分泌されていることが示された。この結果から、pRI28ゲノムの発現能が確認された。標準的なトランスフェクション手法により、パッケージされたpRI28の複製不能RNAゲノムを得た。次に、D17細胞をpRI28ウイルスで形質導入した。続いて、これらの細胞から収集した培地をELISAにより分析した結果、ヒトIgG1の分泌は検出されなかった。パッケージされたウイルスからウイルスRNAも収集し、pRI28ゲノムの構造をRT−PCRにより解析した。RT−PCRにより、パッケージされたpRI28ウイルスの試料にはゲノム集団が混在していることが示され、そのすべてがpRI28プラスミドの均一な調製物から転写されたものであった。最も著しい違いはゲノムの3’末端に見出され(図5)、そこから、明らかに完全長の産物および不完全長の産物が増幅され得た。明らかに不完全長の多数のRT−PCR産物をクローニングして塩基配列決定を行なった結果、欠失事象はR24コード配列の一部または全部を包含していることが確認された。当該欠失事象の一部の位置を図6に示す(以後は独自の「lt」番号で引用する)。図6に示した欠失事象の性質を考えると、当該ゲノムはR24ミニボディを発現できないと予測されることに注意されたい。
【0103】
上記lt欠失事象を慎重に解析した結果、該欠失は、小さい(5〜10bp)直列反復配列により輪郭が描かれていることが明らかになった。この結果は、当該配列要素がEIAVと適合性がない可能性があることを示している。
【0104】
導入遺伝子欠失事象における短い直列反復配列要素の役割を、同類のウイルスゲノムに関する研究によりさらに確認した。同一のR24ミニボディコード配列を組織特異的プロモータ候補の下流に挿入して、プラスミドpLE38を作製した(ゲノムマップの概略図を図7に示す)。標準的なトランスフェクション手法により、パッケージされたpLE38の複製不能RNAゲノムを得た。pRI28での記述とまったく同様にRT−PCR法を完了したところ、pRI28と同様に、ウイルスゲノムの3’末端から明らかに不完全長のPCR産物が増幅されており、これらはR24コード配列の一部または全部を包含していた。PCR産物のクローニングおよび配列解析により、先にpRI28ウイルスでも検出された1つの特定の欠失産物であるlt1が頻繁に存在することが示された(図6の欠失マップを参照)。lt1欠失産物の全配列を図8に示す。
実施例2−pRI28からのR24ミニボディ配列データの解釈
【0105】
R24ミニボディでは、問題がある可能性のある短い直列反復配列が2種類存在し、1つはscFV領域自体(VH、リンカーおよびVL)のなかにあり、もう1つはIgG1 Fcドメイン内に存在する。R24ミニボディの構造の概略図は図2に示している。
VHドメイン
【0106】
VH内ではR24ミニボディ配列に、問題のある反復配列が4つ確認され、第1は5’末端に位置し(図9のLP、Leu Pro、欠失lt16に関与する)、第2はCDR2内に位置し(KG、欠失lt15に関与する)、第3はFW3にあり(DT、欠失lt11および13に関与する)、第4はリンカー配列より前のVHの3’末端に位置する(LT、欠失lt1に関与する)。
リンカー/VLドメイン
【0107】
リンカーおよびVLドメインには問題のある反復配列が4つ確認された。第1はリンカー内に位置し(図10のGS、欠失lt4および5に関与する)、第2はFW1内に位置し(LS、欠失lt6に関与する)、第3はCDR2にあり(TS、欠失lt3に関与する)、第4はFW3配列にある(YS、欠失lt2に関与する)。
IgG1 Fc
【0108】
上記の項では、R24ミニボディから3’ウイルス由来配列までの欠失を取り上げた。下線を引いた配列はこれらの欠失の5’末端を表している。しかし、R24ミニボディと当該遺伝子の5’末端までの配列との間の組換え事象により生じた可能性のある欠失も検出された。これらの事例では、3’決定因子がR24ミニボディのIgG1 Fcドメイン内に位置していた。現在、該配列内で、これらの欠失に関連または隣接したプロリンリッチ領域が2箇所特定されている。
【0109】
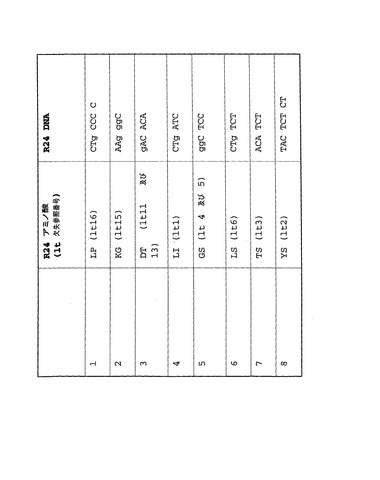
R24ミニボディにおいて欠失(個別のlt番号で引用する)と関連する問題のある可能性のある8つの配列を図11に要約する。先に記述したステップ(iii)の解析により導入遺伝子候補から除去されるのは、該欠失の輪郭を描く短い直列反復配列である。ベクターNtiソフトウェア(Informax Inc., Invitrogen)または同等物を用いて、DNA配列をスクリーニングして該配列の存在を確認できる。導入遺伝子が組換え型抗体でない場合は、上記残基のすべてが保存される可能性は低い。トランスジェニック鳥類発現系は組換え型抗体を発現できると考えられ、この場合は上記残基が保存され、特に一部は、相補性決定領域(CDR)での残基よりも高い保存性を示すことが知られている可変ドメイン下位領域であるフレームワーク領域(FR)内で保存が起こる。
【0110】
これは、市販されている多数の組換え型抗体にとって最適なエフェクタードメインであるために多数の導入遺伝子候補で確実に保存されなければならないIgG1 Fcにも関連していることである。R24ミニボディを用いた研究から、いくつかの欠失決定因子が該ドメイン内に位置している可能性があることが示されており、たとえば、DNAのポリピリミジントラクトによりコードされる2つのプロリンリッチタンパク質領域は、上記欠失に一貫して関連または隣接している。したがって、該ポリピリミジントラクトを除去することが推奨される。ニワトリはほぼ等しい頻度で4つのコドンを使用してPro/Pをコードすることから、得られるタンパク質の複数のプロリン残基をコードしたままの状態でDNA配列のポリピリミジントラクトを除去するために、コドン使用頻度を変えることが可能である。
実施例3−「修復した」R24ミニボディ
【0111】
短い直列反復配列とこれに関連する欠失との関連性を検討して確立するために、R24ミニボディ配列からlt1配列(5’CTG ATC 3’)を除去し、同時に非反復配列を有するリンカーで置き換えることにした。次に、パッケージされたRNAゲノムのかなりの割合にlt1欠失事象が存在することが示されていたpLE38として設計されたベクターを用いて、この修復の効果をテストした。
【0112】
制限酵素BspEIによるpLE38の消化により、5’lt1反復配列および古いリンカーの除去が可能になり、新しいリンカーをコードし、かつlt1配列が除去された新しいDNA断片で置換することができる(図12を参照)。「修復したR24」を作製するためにpLE38に挿入した置換DNA断片の全配列を図13に示す。完成したプラスミドをpLE56と呼んだ。
【0113】
次に、修復および未修復の2つのプラスミドのセットを並行してパッケージし、RNAゲノムの構造のほか、形質導入された細胞のゲノムDNAに組み込まれた導入遺伝子の構造をPCRにより解析した。
実験データ−pLE38およびpLE56
【0114】
修復したR24ミニボディからのウイルスRNAのリアルタイムqPCR法により、明らかに許容レベルの該ゲノムが良好にパッケージされ、lt1修復が力価に悪影響を与えていないことが立証された。ELISA法ではR24ミニボディの発現を検出できなかったが、該ベクターに含まれるプロモータからの発現は原理的には組織特異的でなければならず、該プロモータがインビトロで活性であることを期待しなかったため、これは陽性の結果である。これらのウイルスで形質導入された細胞からのゲノムDNAについて実施したリアルタイムqPCRにより、EIAVパッケージングシグナルにまたがる産物がうまく増幅され、これにより細胞の形質導入状態が確認され、組み込みの不足ではなく、漏出性卵アルブミンプロモータ活性の不足がELISAの陰性結果の原因であることを示す証拠が提供された。
【0115】
さらに、両ウイルスにおけるゲノムの3’末端にまたがるPCR反応を行なった結果、pLE56で形質導入した細胞に限ってゲノムDNAから完全長の産物がうまく増幅された。これは、パッケージされたpLE38(未修復)のRNAゲノムからlt1欠失産物が優勢に増幅された結果とは正反対である。しかし、pLE38テストシステムでは、推定されるより小さい欠失産物の存在を廃するにはlt1の修復のみでは不十分であった。この結果の説明として最も可能性が高いのは、問題の可能性のある他の短い直列反復配列要素の存在であり、5’lt1反復配列のみを除去したため、「修復した」R24内にはほかにも保持されている可能性がある。この可能性は、第1に、図11に列記したEIAVに非適合性の可能性のある配列を他の導入遺伝子に適用できるかどうか評価することと、第2に、EIAVに非適合性の可能性のある配列をすべて除去した導入遺伝子で内部欠失の頻度を評価することによってのみ検討することができる。
細菌における不安定性
【0116】
事例証拠から、R24ミニボディで使用した先のリンカー配列は細菌において不安定であったことが示された。個々の反復配列要素の欠失が検出された。pLE56などの多数の発現用ベクターにうまくクローンニングされた新規のリンカーでは、このような問題に遭遇していない。
実施例4−抗CD55ミニボディ(791T/36)
【0117】
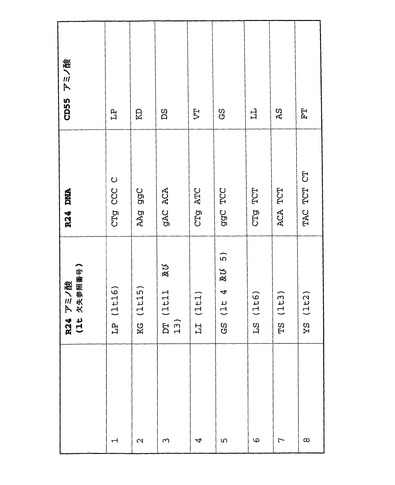
R24ミニボディを用いた研究の成果として、EIAVに非適合の可能性がある多数の配列が特定された。R24をベースとしない導入遺伝子に当該配列が存在するかどうか判断することに関心を向けた。このため、R24で特定したEIAVに非適合の可能性がある配列をEIAVレンチウイルスベクターバックボーンに組み込んだときに該配列に欠失が起こると予測されるとして、該配列を別の導入遺伝子に適用できるかどうか判断するために、抗CD55ミニボディDNA配列を評価した。該ミニボディとR24ミニボディとの間で直接配列比較を行なった。該ミニボディに、問題のある領域が8箇所特定され、これらの領域を図14に要約する。
【0118】
図14の表の行1は、R24ミニボディとCD55ミニボディではlt16欠失事象に関与する残基が完全に一致していることを示している。これは、当該残基が、両コンストラクトが共有する基本的なリゾチームのシグナルペプチドによりコードされているからである。別の導入遺伝子、すなわちサイトカインをベースとした産物を合成する前に、該シグナルペプチドのコドン使用頻度を改変した。これまでに解析したインターフェロンベータ遺伝子に基づいた別の遺伝子コンストラクトでは、改変した該シグナルペプチドには依然としてlt16反復配列が存在しているが、対応するlt16欠失が確認されていない。したがって、少なくともミニボディを含有しないベクターでは、欠失を引き起こすのにlt16反復配列の存在だけでは不十分であるようであり、たとえばリンカードメインなど、別の因子が関与していなければならないと思われる。しかし、シグナルペプチドのコドン使用頻度をさらに改変して、この要素を取り除くことが賢明である。
【0119】
図14の表の行2は、R24ミニボディとCD55ミニボディの間で2つのアミノ酸のうち1つだけがマッチしていることを示している(KG対KD)。ニワトリはLys/Kに対し、ほぼ等しい頻度で2つのコドンを使用するため、アミノ酸特異性を保ったままコドンを変えて、抗CD55からlt15反復配列要素を除去できると考えられる。
【0120】
図14の表の行3は、R24ミニボディとCD55ミニボディとの間で2つのアミノ酸のうち1つだけがマッチしていることを示している(DT対DS)。上記のLys/Kと同じく、ニワトリはAsp/Dに対し、ほぼ等しい頻度で2つのコドンを使用するため、この場合もアミノ酸特異性を保ったままコドンを変えて、抗CD55ミニボディからlt11/13反復要素を除去できると考えられる。
【0121】
この表の行4は、R24ミニボディで最も問題が大きいlt1反復をコードするLI配列に言及している。この欠失は現在、R24ミニボディをベースとしたpRI28およびpLE38の2つのレンチベクターで確認されている。幸い、現時点では、抗CD55ミニボディと配列相同性はない。
【0122】
この表の行5は、R24ミニボディと抗CD55ミニボディではlt4および5欠失事象に関与する残基が完全にマッチしていることを示している。これは、ミニボディのscFV構成要素の構築時にVHドメインとVLドメインを結合するために使用したリンカーがこれらの残基をコードしているためである。いくつかの行の証拠は、該リンカーは発現試験に使用するには最適でない可能性があることを示しており、また、事例証拠から、大腸菌における反復配列の不安定性や、該リンカーに3つの直列反復配列を与える二次構造の可能性が示されており、Geneart社や文献に反復配列とRNAポリメラーゼとの相互作用に関する議論もある。R24ミニボディのリンカーは、図15に示すように、新しいリンカーに取り替えることができる。これは、(GGGS)4アミノ酸パターンを保持しているが、コドン使用頻度を変えて相同性を最小限に抑えている。
【0123】
下線を引いた文字は、元のリンカーで問題のある配列を強調しており、GGC TCCは実際に3回反復されている。新しいリンカーでは、直列反復配列が撤廃されており、GGC TCC配列はまったく存在せず、その交換後のGGA TCTは1回だけ存在する。この新しいリンカーは、EIAVレンチベクター系で使用するための抗CD55もしくは他の任意のscFVまたはミニボディの遺伝子合成時に使用することを推奨する。
【0124】
図14の行6は、lt6反復についてR24ミニボディと抗CD55ミニボディとの間で2つのうち1つがマッチしていることを示している(LS対LL)。ニワトリはLeuに対しCTGコドンを好むため、該配列を変えないことが最良であると考えられる。行7も、lt3反復についてR24と抗CD55との間で2つのうち1つがマッチしていることを示している(TS対AS)。ニワトリはSer/Sに対し6つの異なるコドンを使用するため、lt3反復配列要素を効果的に取り除くために使用できる選択肢はいくつか存在する。最後に行8は、R24ミニボディのlt3欠失に関与する残基YSは抗CD55ミニボディでは保存されないため、この位置では配列の改変が必要ないことを示している(YS対FT)。
IgG1 Fcドメイン
【0125】
Fcドメイン内の2つの多プロリン領域を除去することも推奨される。ニワトリは4つのコドンをほぼ等しい頻度で使用してPro/Pをコードすることから、コドン使用頻度を変えることにより、得られるタンパク質のプロリン残基をコードしたままDNA配列のポリピリミジン領域を除去することが可能である。
【0126】
我々の現在の知見状況を考慮に入れ、上記の推奨事項をすべて使用して、EIAVレンチベクターに使用する最適な抗CD55ミニボディ配列を作製した。最適化した当該配列を図16および17に示す。
【0127】
DNA配列は著しく変わったが、一次アミノ酸配列は単離した元の配列から変化していないことに注目されたい。新しい5’および3’延長配列を追加して、鳥類のトランスジェニック試験系における遺伝子発現を促進し、新しいリンカーを導入して、対応するR24ミニボディ分子に存在する直列反復配列を撤廃した。R24ミニボディと抗CD55ミニボディとの間で保存される位置のほか、コード配列内の他のすべての場所についても、問題の可能性があることが確認された反復配列モチーフをすべて除去した。
【0128】
結論を述べると、抗CD55ミニボディコード配列の上記解析により、R24をベースとしない導入遺伝子に対する当該導入遺伝子最適化方法の妥当性が実際に立証された。
実施例5−抗CD55抗体(791T/36)
【0129】
本文書の実施例4で提示したデータは、EIAVと適合性がない可能性のある短い直列反復配列を除去するという方針が、R24をベースとしない分子、この場合は抗CD55ミニボディに適用できることを示した。この研究の次の段階は、配列最適化のプロセスを本明細書に記載したように正確に適用した後、EIAVレンチウイルスベクターに存在する導入遺伝子配列内の内部欠失の頻度を評価することであった。
【0130】
しかし、実施例4に記載した抗CD55ミニボディをコードする導入遺伝子を作製するのではなく、導入遺伝子最適化の同じ方針を二重鎖マウス/ヒトキメラ抗CD55抗体に適用することとした。図18は、上記両分子の構造を略図で示したものである。
【0131】
当該キメラ抗体は、ヒトIgG1重鎖およびヒトカッパ型軽鎖の上流にそれぞれ挿入したマウス重鎖および軽鎖両方に由来する可変領域からなる。両分子の一次配列は、本明細書に記載した導入遺伝子最適化の段階的プロセスを行う前に、コンピュータ内で組み立てた。図19および20は、キメラ重鎖および軽鎖それぞれの一次アミノ酸配列を示している。両一次アミノ酸配列は、両タンパク質の分泌を可能にするため、内在ニワトリリゾチーム遺伝子からのシグナルペプチドを追加する5’延長配列を含んでいることに注意されたい。
【0132】
本発明の第一の態様において定義したステップに従って最適化のプロセスを実施した。すなわち、望ましい一次アミノ酸配列をGeneart社(ドイツ)に提供し、ニワトリのコドン使用頻度の好みに基づいてDNAコドンが割り当てられ、このプロセスは「チキナイゼーション(ニワトリ化)」と呼ばれる。次に、最適化プロセスのステップ(ii)を完了し、これによりニワトリ化した基本配列を解析して、負の要素すなわち反復配列のほか、スプライス部位、内部TATAボックスまたはリボゾーム進入部位などのシス作用性モチーフなど、遺伝子発現にマイナスの影響を及ぼすと予測される要素を検出した。配列を改変することにより、当該要素をすべて除去した。続いて、ニワトリ化した当該第二世代配列を解析し、図11に示したように問題がある可能性のある配列をすべて取り除いた(最適化プロセスのステップ(iii))。当該第三世代配列をGeneart社に再び送り、上記改変が、負の要素すなわち反復配列のほか、スプライス部位、内部TATAボックスまたはリボゾーム進入部位などのシス作用性モチーフなど、遺伝子発現にマイナスの影響を及ぼすと予測される要素を再導入していないことを確認した。このプロセスを、問題がある可能性があることが確認された反復配列を除去するために設計したすべての改変箇所について繰り返し行ない、コドン使用頻度が依然として最適であり、かつ、負の要素が再導入されていないことを確実にした。遺伝子合成により、キメラ抗CD55重鎖および軽鎖の最終版を作製した。
【0133】
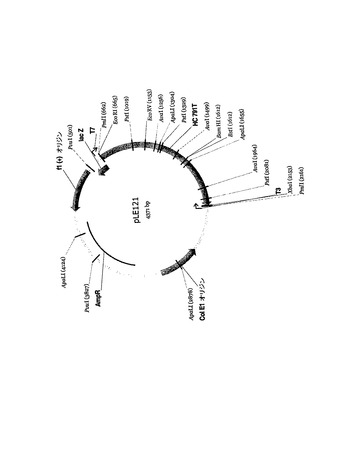
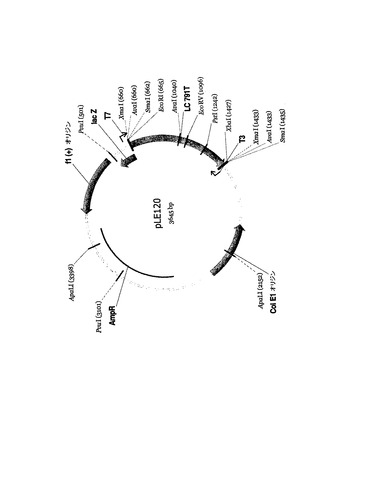
両抗CD55コード配列は、個々のpCRScriptベクターバックボーンに組み込んで供給され、重鎖(図21、pLE121)は制限酵素PmlIにより、軽鎖(図22、pLE120)はSmaIにより切り出すことができた。続いて、組織特異的プロモータ候補の下流に導入遺伝子を導入したベクターゲノムを構築することにより、最適化した導入遺伝子の発現を支援するEIAVレンチウイルスベクター系の能力を分析した。
抗CD55抗体と組織特異的プロモータ候補に基づいた発現コンストラクト
【0134】
重鎖および軽鎖配列を別々に組織特異的プロモータ候補の下流に挿入して、それぞれプラスミドpLE118およびpLE119を作製した。pLE118およびpLE119両方のゲノム構成は、R24が、関連する重鎖または軽鎖配列に置換されていることを除けば、図7のpLE38について示した概略図と同一である。
【0135】
標準的なトランスフェクション手法を用いて、ウイルスゲノムのパッケージングを完了した。ゲノムRNAを収集してRT−PCRにより解析し、さらに、該ウイルス粒子を用いて宿主細胞を形質導入し、続いて該細胞からゲノムDNAを収集した。次に、ゲノム構造のPCR解析を完了した。
【0136】
パッケージされたウイルスゲノムから増幅した産物のRT−PCRと、これに続くクローニングおよびDNA塩基配列決定により、パッケージされたpLE118およびpLE119のゲノム内に無傷の抗CD55のそれぞれ重鎖および軽鎖が存在することが示唆された。興味深いことに、pLE119ゲノムから1つの欠失産物が特定され、この欠失をlt230とした。lt230欠失の範囲を表示したpLE119の3’末端の全配列を図23に示す。当該欠失の5’および3’範囲の輪郭を描いている短い直列反復配列の存在に注目されたい。このデータは、本文書に概略を記載した推定上の相同的組換えに基づく機構により、R24をベースとしないEIAVレンチウイルスベクター導入遺伝子内に内部欠失が発生することを示した最初の証拠である。このため、lt230を挟んでいる反復配列を、導入遺伝子最適化プロセスのステップ(iii)において除去すべき配列のリストに追加した。すべての当該配列を図24に列記する。
【0137】
pLE118およびpLE119を形質導入した細胞のゲノムDNAを解析した結果、主に完全長の増幅産物が得られた。たとえば、組織特異的プロモータ候補内から3’LTRまでまたがり、かつ導入遺伝子コード配列を包含するPCR反応により、pLE118ウイルスで形質導入した細胞のゲノムDNAから、無傷の重鎖配列の存在を示唆する2124bpの産物が生じた(図25、列5)。同一のPCR反応により、pLE119ウイルスで形質導入した細胞のゲノムDNAから、無傷の軽鎖配列の存在を示唆する1398bpの産物が生じた(図25、列13)。両導入遺伝子コード配列は同一のリゾチーム由来リーダーペプチドを共有しており、このため、PCRプライマーを共用できることに注意されたい。pLE119で形質導入した細胞のゲノムDNAからはlt230欠失産物が増幅されなかったことから、該産物は大多数の分子種ではないことが示唆される。
【0138】
この研究から結論がいくつか導き出される。第1は、R24で得られた結果とは対照的に、上記ベクターからは無傷の最適化した抗体コード配列がPCRでうまく増幅されたことである。第2は、CD55配列に新規のlt欠失が発見されたことである。本出願は、R24ミニボディを用いた研究の成果として確認された問題の可能性のある配列をすべて除去する手順を詳しく述べている。抗CD55試験系ではR24でみられた欠失産物がまったく検出できなかったことは、当該配列が欠失機構に直接関与しているという結論を裏付けている。たとえば、抗CD55軽鎖の早期の反復では、lt16反復配列(CTg CCC C)が存在した。これは、問題の可能性のあるこれらの反復配列を除去するためのスクリーニング時に確認されたものであり、後期の反復ではCTg CCT Cに変化していたが、コードされるアミノ酸は未変化のまま維持される。最終的に最適化した抗CD55軽鎖配列にはlt16欠失事象の証拠が決定的にまったく検出されず、先に述べたR24の結果とは対照的であった。
【0139】
しかし、抗CD55抗体の配列に新規のlt欠失が検出されれば、問題のある可能性がある別の配列が提供され、該配列は、本明細書で開示した方法により最適化されるさらなる導入遺伝子では除去されることになる。
実施例6−他の種への移行可能性
【0140】
本明細書に記載した導入遺伝子最適化プロセスは、たとえばウズラ Coturnix coturnixなどの他の種で発現するように設計した異種コード配列に適用できる。図26に示すように、ウズラのコドン使用頻度はニワトリ(Gallus gallus)のコドン使用頻度とほぼ同一である。このため、最適化のプロセスは、本発明の第一の態様で定義したステップに従って実施できると考えられる。すなわち、望ましい一次アミノ酸配列をGeneart社(ドイツ)に提供し、ウズラまたはニワトリのコドン使用頻度に基づいてDNAコドンが割り当てられるが、これはウズラおよびニワトリならびに他の鳥類の間でコドンバイアスに非常に高度の保存性が認められることによるものである。続いて最適化プロセスを完了し、これにより、基本の配列を解析してまず、遺伝子発現にマイナスの影響を及ぼすと予測される配列要素を検出し、次に、図24に示したように問題の可能性のある配列をすべて除去する。
【0141】
本明細書に引用した文書はすべて、参照により本明細書に組み込まれている。本発明の範囲から逸脱することなく、記載した本発明の実施形態に対し種々の変更および変化があることは、当業者には明白である。特定の好ましい実施形態と関連させながら本発明を説明してきたが、特許請求の範囲に示したとおり本発明は、当該実施形態に過度に限定されるべきではないことを理解すべきである。実際に、本発明を実施する上記の態様に種々の変更があることは当業者には明らかであり、種々の変更は本発明によりカバーされるように意図されている。
【図面の簡単な説明】
【0142】
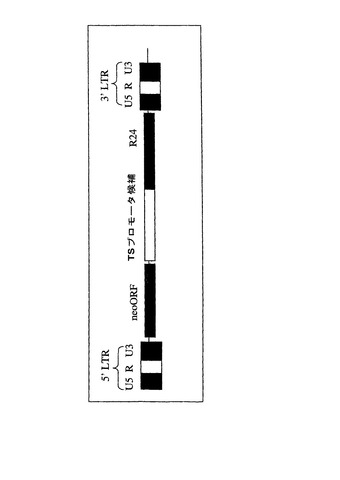
【図1】pRl28及びpLE38の構築に使用したR24ミニボディの全DNA配列を示す。開始コドン及び二重停止コドンは大文字で表す。
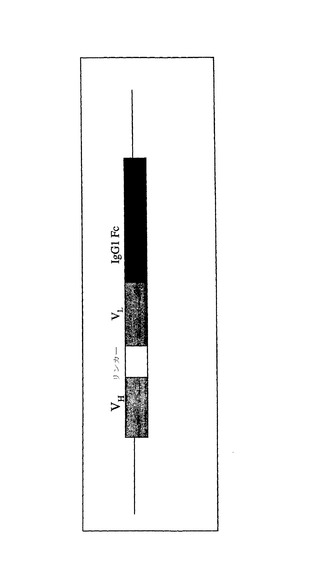
【図2】R24ミニボディの模式的構造を示す。
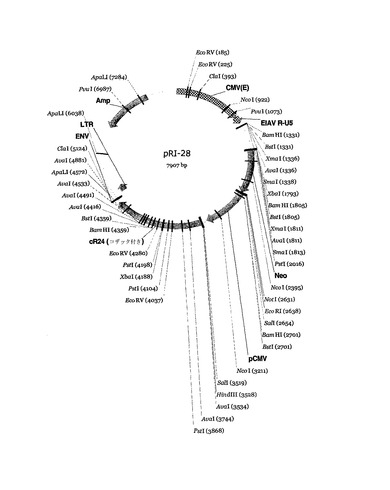
【図3】レンチウイルスベクターゲノムpRl28のプラスミドマップ。
【図4】レンチウイルスベクターゲノムpRl28の完全DNA配列を示す。
【図5】pRl28ウイルスのRNAゲノムの予測構造を示す。
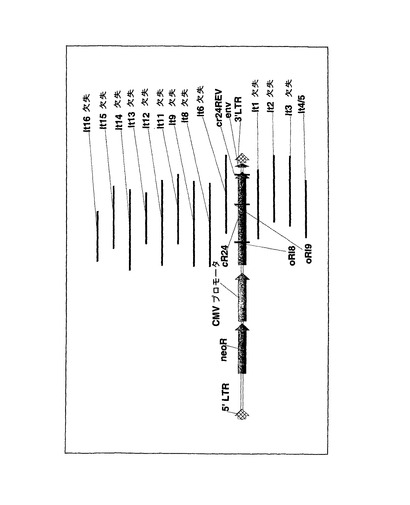
【図6】レンチウイルスベクターpRl28内のR24コード配列内で特定された、欠失の相対位置付きの図を示す(個別の「lt」番号で示す)。
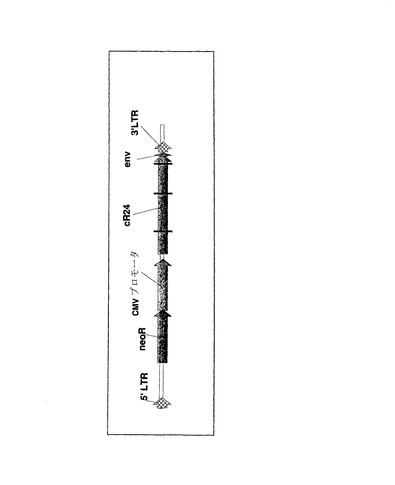
【図7】pLE38のRNAゲノムの予測構造の代表図を示す。
【図8】完全R24コード配列を包含するpLE38ゲノムの3’末端の完全配列を示す(開始コドン及び二重停止コドンを太い大文字で示す)。5’LTR配列も太字で示す。lt1繰返しのコピーはいずれもイタリック体で示し、lt1欠失後に欠損した配列には下線を付す。lt1繰返しの5’コピーが欠失後に保持されているため、下線を付していないことに留意されたい。
【図9】R24ミニボディVHドメインアミノ酸配列を示す。R24ミニボディのアミノ酸配列は1文字コードで示す。イタリック体は、FR及びCDRに位置する、この領域の5’及び3’末端の残基を示す。太字は、3フレームワーク領域を含む残基を示す(図の右側欄のキー)。標準文字はCDRを含む残基を示す。下線文字は、問題のDNA繰返しによってコードされるアミノ酸残基を示す。
【図10】R24ミニボディVLドメインアミノ酸配列を示す。R24ミニボディのアミノ酸配列は1文字コードで示す。イタリック体は、FR及びCDRに位置する、この領域の5’及び3’末端の残基を示す。リンカードメインの残基は、5’末端においてイタリック体にされている。太字は、3フレームワーク領域を含む残基を示す(図の右側欄のキー)。標準文字はCDRを含む残基を示す。下線文字は、問題のDNA繰返しによってコードされるアミノ酸残基を示す。
【図11】R24ミニボディにおける潜在的に問題の8配列、及び関連する欠失(個々のlt番号で示す)を示す。
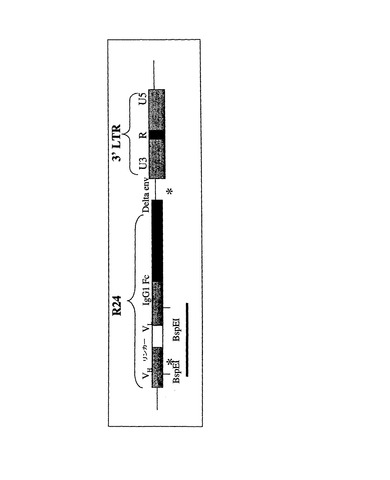
【図12】pLE38における3’末端の図を示す。*は、R24コード配列内の欠失のいくつかに関連し「lt1」で示される短い2つの繰返し配列の位置を示す。「lt1」配列が除去された置換配列である、5’lt1繰返しに隣接する2つのBspEI部位の位置は太い黒線で示す。
【図13】lt1修復手順の間にpLE38に挿入されたBspEIフラグメントの全配列を示し、制限酵素部位は太字で示す。
【図14】R24ミニボディ内の8つの問題配列と、抗CD55ミニボディ内の対応する残基との比較を示す表。
【図15】標準R24内に存在する天然で改変されたリンカーと、修復されたリンカーとによってコードされるDNA及びアミノ酸の配列を示す。
【図16】最適化抗CD55ミニボディの一次アミノ酸配列を示す。
【図17】最適化抗CD55ミニボディのDNA配列を示す。
【図18】抗体及びミニボディの相対的構造を比較する図を示す。
【図19】抗CD55抗体の重鎖の一次アミノ酸配列を示す。
【図20】抗CD55抗体の軽鎖の一次アミノ酸配列を示す。
【図21】pLE121のプラスミドマップを示す。抗CD55抗体重鎖はpCRscriptベクター内でGeneartによって販売される。
【図22】pLE120のプラスミドマップを示す。抗CD55抗体重鎖はpCRscriptベクター内でGeneartによって販売される。
【図23】全抗CD55コード配列を包含するpLE119の3’末端の完全配列を示す(開始コドン及び二重停止コドンを太字及び大文字で示す)。5’LTRも太字で示す。lt230繰返しのコピーはいずれもイタリック体で示し、lt1欠失後に欠損した配列には下線を付す。lt230繰返しの5’コピーが欠失後に保持されているため、下線を付していないことに留意されたい。
【図24】図11に示される表の改訂版であり、R24及び抗CD55を用いて決定された問題の繰返し配列を列挙する。
【図25】pLE118及びpLE119がそれぞれ形質転換された細胞のゲノムDNAから増幅されたPCR産物の、エチジウムブロミド染色1%アガロースゲルを示す。PCRプライマーは、全重鎖及び軽鎖をコードする配列を包含する組織プロモータ候補から3’LTRまで、各ゲノムの3’末端を増幅する。pLE118及びpLE119形質転換細胞から増幅された2124bp及び1398bpの産物は、無傷の抗CD55コード配列の存在を示す。小さい増幅産物が存在しないことに留意されたい。
【図26】ニワトリ(Gallus gallus)及びウズラ(Coturnix coturnix)におけるコドン使用頻度をようやくする2つの表を示す。
【技術分野】
【0001】
本発明は、ベクター系、特にレンチウイルスベクターにおいて、改変された導入遺伝子コンストラクトの効果的な転写および翻訳を達成するために改良した方法を提供する。当該方法は、導入遺伝子を含むウイルスベクターゲノムがトランスジェニック鳥類などの動物の生殖細胞系列に安定的に組み込まれることを可能にする無傷の機能導入遺伝子配列を有するウイルスベクターゲノムの生産を容易にする。該導入遺伝子が発現されると、組換え型タンパク質産物が生産され、トランスジェニック鳥類の場合では、目標とするタンパク質をトランスジェニック鳥の卵のなかに生産させることができる。
【背景技術】
【0002】
組換え型タンパク質の伝統的な製造方法には、細菌または哺乳類の細胞を用いた生産方法がある。これに代わる製造方法は、タンパク質の生産にトランスジェニック動物および植物を用いるものである。
【0003】
タンパク質をベースとした多数のバイオ医薬品が、トランスジェニックマウス、ウサギ、ブタ、ヒツジ、ヤギおよびウシなどの種々の哺乳動物の乳汁中に発現されている。当該系は世代時間が長い傾向があり、より大型の哺乳動物では遺伝子導入された初代動物から、乳汁を生産できる段階まで発達するには数年かかる。
【0004】
このほかの問題点は、乳汁の生化学的複雑さと、ヒトと動物との間の進化的保存性に関するものであり、これにより、医薬品を生産する哺乳動物に該医薬品に対する有害反応が起こる可能性がある(Harveyら、2002年)。
【0005】
製薬学的に重要なタンパク質、特に組換え型ヒト抗体のための製造媒体となる可能性があるものとして、ニワトリの卵の使用に関心が高まっている。
【0006】
ニワトリの卵に基づいたタンパク質製造系は、哺乳動物の細胞培養またはトランスジェニック哺乳動物系の使用に比べ利点がいくつかある。ニワトリは世代時間が短く(24週間)、このためトランスジェニックニワトリ群を速やかに確立することができる。さらに、トランスジェニック動物の生産施設のための設備投資は、細胞培養のための設備投資よりはるかに少なくてすむ。細胞培養に比べ、遺伝子導入によるタンパク質生産の促進に必要な特別な処理装置はほんのわずかである。このように設備投資が少なければ、遺伝子導入による治療薬の単位あたりの生産コストは細胞培養で生産される治療薬よりも低くなる。そのうえトランスジェニック系は、精製のバッチサイズおよび頻度に関して多大な柔軟性を提供する。この柔軟性は、バッチサイズを最適化することにより、精製における資本コストおよび操業コストをさらに削減させることにつながる可能性がある。
【0007】
さらに、遺伝子導入タンパク質の生産は市場への導入速度を速めることになる。トランスジェニック哺乳動物は、乳汁1リットルあたり数グラムのタンパク質を生産することができ、大量生産を商業的に実行可能なものとすることができる(Weck、1999年)。またさらに、鳥では世代時間が短いため、生産の迅速なスケールアップが可能である。
【0008】
鳥類の卵、特にニワトリの卵は、タンパク質の生産手段として細胞培養を上回る大きな利点をいくつか提供する。さらに鳥類系は、哺乳動物または植物に基づく他のトランスジェニック生産系を超える重要な利点を提供する。
【0009】
トランスジェニック哺乳動物の生産に使用される方法を鳥の遺伝子操作に直接適用することは、産卵鶏の生殖系の特別な特徴のため、これまで可能ではなかった。
【0010】
卵形成の複雑さにより、ヒヨコ胚発達の初期段階は比較的アクセスしにくい段階である。早期の段階の胚にアクセスするために採用される方法では、通常、ドナーである雌鳥を屠殺して胚を得るか、または輸卵管に直接注射を行なうことが必要である。トランスジェニック哺乳動物の生産方法は、ほぼ例外なく受精卵のマイクロインジェクションに重点が置かれており、この方法ではインビトロで前核にDNAを微量注入し、操作した当該卵を代理母に移して満期まで発育させるが、この方法は雌鳥では実行不可能である。
【0011】
トランスジェニック鳥類の作製には、一般的な方法が4つ開発されている。(i)胚盤の細胞質へのDNAマイクロインジェクションを用いてトランスジェニックニワトリを作製する方法、(ii)インビトロで始原生殖細胞のトランスフェクションを行ない、適切に準備したレシピエントに移植する方法、(iii)発癌性レトロウイルス由来の遺伝子導入ベクターを使用する方法、(iv)ヒヨコ胚細胞をインビトロで培養し、培養した細胞をレシピエント胚に導入することによりキメラ鳥を作製する方法である(Painら、・1996年)。キメラを作製する前に胚細胞をインビトロで遺伝子操作することができ、これによりキメラなトランスジェニック鳥が生じる。
【0012】
レンチウイルスはレトロウイルスの亜群であり、ヒト免疫不全ウイルスHIV−1およびHIV−2、サル免疫不全ウイルス(SIV)などの種々の霊長類ウイルスならびに非霊長類ウイルス(たとえば、マエディビスナウイルス(MW)、ネコ免疫不全ウイルス(FIV)、ウマ伝染性貧血ウイルス(EIAV)、ヤギ関節炎脳炎ウイルス(CAEV)およびウシ免疫不全ウイルス(BIV))が含まれる。これらのウイルスは特に遺伝子治療の開発において関心がもたれており、これは、レンチウイルスが宿主細胞DNAに不可逆的に組み込まれるというレトロウイルスの一般的な特徴を有するだけでなく、非増殖細胞を感染させる能力も有するからである。レンチウイルス感染の生物学はCoffinら(1997年)の総説を参照できる。
【0013】
ウイルスベクターを設計する際に考慮すべき重要事項は、細胞のゲノムに安定的に組み込まれる能力である。これまでの研究から、遺伝子導入媒体として用いるオンコレトロウイルスベクターは、発生過程における遺伝子発現抑制効果により成功率がやや低いことが示されている。マウスに関するPfeiferら(2002年)およびLoisら(2002年)の研究から、HIV−1に基づくレンチウイルスベクターは発生過程で発現が抑制されないことが示された。
【0014】
レンチウイルスベクターに関する開発研究の大半はHIV−1系に焦点を当ててきたが、これは主に、ヒトに病原性を示すためにHIVが最も十分に特徴付けられているレンチウイルスであるという事実によるものである。当該ベクターは、調節遺伝子およびアクセサリー遺伝子を除去して複製を不可能にすることにより、複製能力をなくすように設計される傾向がある。こうしたベクターの最新型は、調節遺伝子のほぼすべて、アクセサリー遺伝子のすべてが除去される程度まで最小化されている。
【0015】
レンチウイルス属のウイルスは、類似したゲノム構成、類似した複製サイクル、成熟マクロファージを感染させる能力など、多数の類似した特徴を有する(ClementsおよびPayne、1994年)。当該レンチウイルスの1つがウマ伝染性貧血ウイルス(EIAV)である。レンチウイルス属の他のウイルスに比べ、EIAVは比較的単純なゲノムを有し、該ゲノムは、レトロウイルスのgag、polおよびenv遺伝子のほかにわずか3つの調節/アクセサリー遺伝子(tat、revおよびS2)からなる。安全かつ効果的なレンチウイルスベクター系の開発は、ベクター自体のデザインに依存することになる。効果的な機能を得るためには、形質導入するベクターとしての機能を保ちながら、ベクターのウイルス構成要素を最小限に抑えることが重要である。
【0016】
オンコレトロウイルスベクター系およびレンチウイルスベクター系は、形質導入可能な細胞の種類および動物種の範囲を広げるために改変することができる。これは、該ウイルスのエンベロープ糖タンパク質を他のウイルスのエンベロープタンパク質に置き換えることによって達成される。EIAVレンチウイルスベクターにパッケージされた導入遺伝子を生殖細胞系列に安定的に発現させることが可能である(McGrewら、2004年)。この方法は、外来DNAの関連断片を合成することと、観察された最適なニワトリの頻度にコドン使用頻度を変えること(口語で「チキナイゼーション(ニワトリ化)」と言われるプロセス)を必要とする。当該プロセスは、特定の外来DNA配列の効果的な転写および翻訳を十分に可能にし、その結果、得られた鳥で目的のタンパク質を発現させると考えられる。しかし一部のタンパク質配列は、安定的に発現できるように改変が必要であることが示されている。
【0017】
ガングリオシドGD3に特異的であるR24として知られるネズミ抗体を使用して、「ミニボディ」と呼ばれる組換え型抗体様結合分子が作製された。このミニボディの構造は、伝統的な抗体VHおよびVLドメインと、これに結合したリンカーおよびIgG1のFcドメインから構成された。EIAVをベースとしたレンチベクターにこのミニボディのためのコード配列がパッケージされたが、ミニボディタンパク質産物の発現は達成できなかった。
【0018】
その後、R24ミニボディを含有する種々のウイルスゲノムから直接増幅したRT−PCR産物が解析された結果、外来R24ミニボディのコード配列の一部または全部を包含する多数の欠失が起こっていることが確認された。これらの欠失の5’および3’範囲を表す配列が解析された結果、異常スプライシングがこれらの欠失の原因ではないことが示された。当該欠失は小さい(5〜10bp)直列反復配列によって特徴付けられているようであり、このことから、観察された外来DNAコード配列の変化は、これまで知られていない相同的組換えに基づく機構が原因であることが示唆されている。
【0019】
Ch’angらはこれまでに、ネズミ白血病ウイルス(MuLV)の組み込まれたプロウイルスゲノムにおける内部欠失を報告しており、研究中に確認された3つの欠失はいずれも、7つのヌクレオチドの直列反復配列に挟まれたていたと述べている(Ch’angら、1989年)。短い直列反復配列によって挟まれたDNA配列に関する特定の欠失は、他のレトロウイルス遺伝子(Coffinによる総説、1985年)のほか、種々の原核生物および真核生物の遺伝子(Omerら1983年、およびLevyら1985年の論文)においても観察されている。短い直列反復配列に挟まれた欠失は、・鳥肉腫ウイルスsrc遺伝子にも観察されている(Omerら、1983年)。提案された機構は、たとえば、DNAポリメラーゼまたは逆転写酵素などDNA複製機構のずれであることが示唆されている。しかし、R24ミニボディベクター系に観察された欠失は、逆転写されたウイルスRNAゲノムから直接増幅したRT−PCR産物に存在したことから、この機構によって説明することはできない。そうではなく、宿主細胞のRNAポリメラーゼ(Rpol II)が、パッケージング細胞株へのプラスミドのトランスフェクション直後にウイルスゲノムの転写時に欠失を誘導した可能性が高い。この結論を支持するように、一部の宿主DNA依存性RNAポリメラーゼは鋳型スイッチの能力を有することが知られており(Nudlerら、1996年)、さらに、RNA組換えはヘアピンループなどの三次元構造の存在により影響を受けることも知られている(WhiteおよびMorris、1995年)。
【0020】
レンチウイルスベクターのバックボーンに組み込むときに欠失が発生する傾向を検討するために、別の外来遺伝子配列、すなわち、791T/36として知られる組換え型ネズミ抗CD55抗体の遺伝子配列が評価された。欠失に関与することが知られている配列が791T/36に保存されていた。
【0021】
したがって、一部の複合タンパク質をコードする遺伝子内の特定の配列は、レンチウイルスベクターバックボーンに組み込まれると欠失を経験しやすいと考えられる。欠失の程度は遺伝子ごとに劇的に異なる可能性が高く、したがって予測不可能であると考えられる。R24ミニボディの発現に関連して示されているように、導入遺伝子からのタンパク質の発現がもはや不可能である程度まで欠失が起こることがあり、言い換えれば、トランスジェニック系における該タンパク質の発現を妨げることになる。
【0022】
欠失の傾向を有する可能性がある配列の領域を特定するために、発現用ベクターに組み込む前に外来DNA配列をスクリーニングできれば、大いに望ましいことである。
【非特許文献1】Ch’ang LY, Yang WK, Myer FE, Koh CK, Boone LR(1989). Virology 168, 245−255.
【非特許文献2】Clements JE & Payne SL(1994) Virus Res. 32(2),97−109.
【非特許文献3】Coffin J(1985). Genome Structure (R Weiss, N Teich, HE Varmus eds)2,17−74.
【非特許文献4】Graf M, Bojak A, Demi L, Bieler K, Wolf H, Wagner R(2000) J. Virol. 74,10822−826.
【非特許文献5】Harvey AJ, Speksnijder G, Baugh LR, Morris JA, Ivarie R(2002) Poult. Sci .81(2),202−12.
【非特許文献6】Horton RM, Hunt HD, Ho SN, Pullen JK, Pease LR.(1989) Gene77(1),61−8.
【非特許文献7】Levy DE, Lerner RA, Wilson MC(1985). Cell 41,289− 299.
【非特許文献8】Lois c, Hong EJ, Pease S7 Brown EJ, Baltimore D(2002) Science 295(5556),868−72.
【非特許文献9】Martinez−Salas E(1999) Current Opinion Biotechnology 10,458−64.
【非特許文献10】McGrew MJ, Sherman A, Ellard FM, Lillico SG, Gilhooley HJ, Kingsman AJ, Mitrophanous KA & Sang H(2004) EMBO Reports 5(7),728−33.
【非特許文献11】Nudler E, Avetissova E, Markovtsov V, Goldfarb A (1996) Science 273,211−217.
【非特許文献12】Omer CA, Pogue−geile K, Guntaka R, Staskis KA, Faras AJ (1983; J. Virol. 54,889−893.
【非特許文献13】Pain B, Clark ME, Shen M, Nakazawa H, Sakurai M, Samarut J, Etches RJ (1996). Development 122(8),2339−48.
【非特許文献14】Pfeifer A, Ikawa M, dayn Y, Verma IM (2002) PNAS 99(4),2140−45.
【非特許文献15】Schneider R, Campbell M, Nasioulas G, Felber BK, Pavlakis GN (1997) . Journal of Virology 71(7),4892−903.
【非特許文献16】Shaw G, Kamen R(1986). Cell 46(5),659−67.
【非特許文献17】Spies M, Bianco PR, Dillingham MS, Handa N, Baskin RJ, Kowalczykowski SC (2003). Cell 114(5),647−54.
【非特許文献18】Tran DP, Kim SJ, Park NJ, Jew TM, Martinson HG(2001). Molecular and Cellular Biology 21(21),7495−508.
【非特許文献19】Trinh R, Gurbaxani B, Morrison SL, Seyfzadeh M (2004). Molecular Immunology 40,717−722.
【非特許文献20】White KA and Morris TJ (1995)RNA 1,1029−1040. Week, E. 1999 ’Transgenic Animals: ’market opportunities now a reality’ D&MD reports
【発明の開示】
【発明が解決しようとする課題】
【0023】
本発明の発明者らは、驚くべきことに、解析すべき外来DNA配列を欠失または他の配列変異を起こすと予測される配列領域を決定できるスクリーニング方法を開発した。いったん、このような配列領域が特定されれば、これらの配列を改変することができる。さらに、このような改変は、外来DNA配列をベクターバックボーンに組み込む前に、有利に行なうことができる。したがってこの方法は、導入遺伝子を含むウイルスベクターゲノムがトランスジェニック鳥類などの動物の生殖細胞系列に安定的に組み込まれることを可能にする無傷の機能導入遺伝子配列を有するウイルスベクターゲノムの生産を容易にし、これによって、非ヒト動物、特に鳥類などのトランスジェニック系における組換え型タンパク質の生産に使用できる。
【課題を解決するための手段】
【0024】
本発明の第1態様によれば、外来DNA配列を適切なベクターによる発現に最適化する方法であって、(i)前記外来DNAのヌクレオチドコドン使用頻度を最適化して、前記外来DNA配列が発現される宿主細胞のコドン使用頻度に変更する手順と、(ii)コドンが最適化されたDNA配列を改変して、前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得るいずれかの配列部位を変更する手順と、(iii)相同組換えに基づく推定欠失機構に関係する全配列を除去するために、前記外来DNA配列のヌクレオチドコドン使用頻度を変更する手順と、を含む方法が提供される。
【0025】
一実施形態において、この方法は手順(i)及び(iii)を含む。別の実施形態において、この方法は手順(ii)及び(iii)を含む。更に別の実施形態において、この方法は手順(i)、(ii)、及び(iii)を含む。
【0026】
前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得る配列要素としては、陰性要素又は反復配列、スプライシング部位、内部TATAボックス、及び内部リボソーム侵入部位のようなシス活性化モチーフが挙げられる。
【0027】
従って、本発明の実施形態は、前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得る配列要素の存在について外来DNA配列を解析することまで及び、特に前記配列要素としては陰性要素又は反復配列、スプライシング部位、内部TATAボックス、及び内部リボソーム侵入部位のようなシス活性化モチーフが挙げられる。
【0028】
陰性要素は、例えばAT又はGCリッチであったり、重要なRWA二次構造に寄与したり、又は具体的機能が起因するTATAボックス、カイ配列、リボソーム侵入部位、ARE、INS、CRS、スプライシングシグナル、又はポリアデニルかシグナルといったコンセンサス配列と予測されるような一般配列等、2カテゴリの1つに一般に適する。
【0029】
TATAボックスは、転写開始部位(5’ TATAAAA 3’ :この配列は転写開始部位を正確に決定する際に重要となる)の上流25ヌクレオチド付近に位置する真核生物ポリメラーゼIIによって転写される大部分の遺伝子のプロモータ領域において発見されるコンセンサス配列と定義できる。
【0030】
RecBCD酵素は、二重鎖DNAの損傷部位において相同組換えを開始するヘテロ三量体ヘリカーゼ/ヌクレアーゼである。その活性のいくつかは、転移酵素でシスに認識されるDNAカイ配列(5’ GCTGGTGG 3’ )によって調節されている(Spies et al, 2003))。
【0031】
内部リボソーム侵入部位は、機能的基準及び大きな配列相同性を共有することが報告されていないことに基づいて通常定義される。しかし、インシリコ配列解析プログラムは、公知のIRES配列が導入遺伝子内に存在しないことを検証した(Martinez−Salas, 1999で総説される)。アデニンリッチ要素(ARE)は、一過的に発現される遺伝子由来のmRNAの3’UTR内に頻繁に位置するAUリッチ配列と定義される。ARE配列の導入は、mRNAを不安定化するのに充分であり、mRNAのプロセス経路の認識シグナルとして提案されている(Shaw & Kamen, 1986)。
【0032】
阻害要素(INS)及びシス活性リプレッサ配列(CRS)は、いずれもHIVモデル系において当初報告されており、1つの仮説によればmRNAの不安定性に寄与する細胞性因子の結合部位である(Schneider et al, 1997)。かかる配列をHIV転写物から除去すると、その転写物の発現が大きく促進することが実証され、前述したINS又はCRS配列の不存在又は除去を検証することが、導入遺伝子の最適化手順の間望まれる。
【0033】
3タイプのコンセンサススプライシングシグナルが報告されている。第1は、切除される配列(イントロン)の5’末端と定義されるスプライシングドナー(C又はA、A、G/G T、A又はG、A、G、T)である。第2は、切除される配列の3’末端と定義されるスプライシングアダプタ(T又はC, n, N, C又はT, A, G/g)である。第3は、切除される配列内に位置する分岐点配列(TACTAAC)であり、スプライシング反応間のラリアット形成に関与する。
【0034】
RNAポリメラーゼIIによる転写終了は、通常、機能的ポリアデニル化シグナル(ポリA)の存在を要する。脊柱動物のコアポリAシグナルは、ポリA切断部部位に隣接する2つの認識部位からなる。典型的には、ほとんど不変のAAUAAヘキサマーが、U又はGU残基リッチのより可変な要素の20〜50ヌクレオチド上流に位置する。新生転写物の切断は、これら2要素の間で起こり、5’切断産物への250以下のアデノシン(ポリAテイル)の付加につながる(Tran et al, 2001)。
【0035】
前述したいくつか又はすべての配列要素を保持する結果は、保持される配列の性質に応じて変動する。これらは、種々異なる機構によるが異種コード配列の発現を低減することから、陰性要素として記述される。例えば、異種コード配列内に同属スプライシング配列を保持することで、形質転換遺伝子産物の発現を消滅、低減、又は制限するスプライシング及び切除が高効率化し得る。対照的に、INS要素を保持しても、RNA組み込みに影響を及ぼさず、むしろmRNAが所望のコードされる遺伝子産物の大部分の転移が起こる前に高速分解の標的にされる。いずれの機構も、異種タンパク質発現の減少という同じ一般的結果を生み出す。本発明のこの態様の一実施形態において、本発明の発現最適化方法によって解析され任意に改変された外来DNA配列は、トランスジェニック発現系において発現され得るベクターに含まれる。
【0036】
トランスジェニック発現系は、非ヒト哺乳動物であってよい。更に別の実施形態において、トランスジェニック発現系は、鳥類、特にニワトリ又はウズラであってよい。
【0037】
本発明の一実施形態において、外来DNAは、ベクターの内部プロモータの制御下にあり宿主細胞によって発現される異種タンパク質をコードする。
【0038】
本発明の一実施形態において、ベクターはレンチウイルスベクターである。別の実施形態において、ベクターはウマ伝染性貧血ウイルス(EIAV)である。本発明は、レンチウイルスベクターとして、ヒト免疫不全ウイルスHIV−I及びHIV−2、サル免疫不全ウイルス(SIV)、非霊長類ウイルス(例えばマエディ−ビスナウイルス(MW)、ネコ免疫不全ウイルス(FIV)、ウマの伝染性の貧血症ウイルス(EIAV)、ヤギ関節炎脳炎ウイルス(CAEV)、及びウシ免疫不全ウイルス(BIV))も提供する。
【0039】
本発明のこの態様の一実施形態において、外来DNAは、組換え抗体その他の結合フラグメント又は結合物である異種タンパク質をコードしてよい。
【0040】
抗体又は結合物等をコードする外来DNA配列の解析は、抗体又は結合物に組み込まれるためのDNAコード配列から除去された実質的にすべての重複配列を有するリンカー配列を、一次アミノ酸配列中の3重複配列(Gly4Ser1)を保持しながら設計する手順を更に含んでよい。この手順は、本発明のこの態様の方法の一部として実施される場合には手順(iii)の前に行うことが好ましい。
【0041】
具体的には、かかる手順は、手順(ii)の完了後で手順(iii)の実施前に行われるため、以後この手順を本発明のこの態様の方法における手順(iib)と表記する。
【0042】
ここで定義されるように、用語「コドン最適化」は、外来DNA配列のコドン使用頻度が、発現を向上するためにベクターが挿入され発現される非ヒト哺乳類動物細胞型において最も頻繁に使用されるコドンをコードするために意図的にバイアスされるように、コドン使用頻度を変更する手順を指す。例えば、トランスジェニック発現系がニワトリのとき、コドン使用頻度の変更は、ニワトリ種において最も一般に使用されるコドンに発現をバイアスするために特定のコドンを変化させる。ニワトリで実施するとき、ヌクレオチド配列のコドン使用頻度を変更する手順は、外来DNA配列の「ニワトリ化」と口語的に表記され得る。
【0043】
具体的には、ここで定義されるように、用語「ニワトリ化」は、非変更形態においてヌクレオチド配列(コドン)によってコードされるアミノ酸配列をコードするニワトリ種で最も一般的な3ヌクレオチドによってコドンがコードされるように、ヌクレオチド配列内のコドン使用頻度を意図的に変更する手順を指す。トランスジェニックニワトリにおける発現のため、外来DNA配列によって形成されるコドンは、ニワトリにおける最も頻繁なコドン使用パターンに最適化される。しかし、ベクターが発現される任意の鳥類種又は非ヒト哺乳動物の最も頻繁なコドン使用頻度に最適化することもできる。
【0044】
ニワトリ化の方法例として、アミノ酸バリンは4つの異なるコドン、GTG、GTA, GTT、及びGTCによってコードされるが、鳥類ではGTGが最も頻繁に使用される(46% GTG, 11% GTA, 19% GTT、23% GTC)。ヒトIgGFcDNAをニワトリ化するためには、すべてのバリンコドンがAAGに変換される。リジンは2つの異なるコドン、AAG及びAAAによってコードされるが、鳥類ではGTGが最も頻繁に使用される(58%対42%)。配列中のすべてのAAAコドンがAAGに変換される。すべてのコドンの変更が必要とされる訳ではない。例えば、アスパラギン酸としては、2つの異なるコドン、GAT及びGACが略均等に使用されるため、ニワトリ化手順において変更する必要はない。
【0045】
本発明のこの態様の方法の手順(i)及び(ii)で概略されるようなコドン使用頻度及び配列改変は、異種導入遺伝子からの遺伝子発現の最適化として当業者に公知である(例えば、Graf et al . , 2000参照)。
【0046】
本発明のこの態様の方法の手順(i)及び(ii)は、典型的には、同様の配列設計サービスを提供するGeneart GmbH (Germany, www.geneart.com)又は組織と協力して実施される。Geneartによる手順(i)及び(ii)の実施は、配列最適化を行うために配列設計及び解析を可能とするコンピュータ配列設計の実施を含む。この方法は、配列を解析し、コドン使用頻度を交換し、コドン交換による配列変化が陰性要素又は繰返しを導入しないことを確保するために得られる配列を解析する手順を含む。タンパク質発現にヌクレオチド配列を最適化する方法の具体的記載は、国際公開WO2004/059556に見出すことができ、その全容は参照して本明細書に援用される。得られる塩基配列を、手順(iii)に示されるようにして更に改変する。任意に、前述した追加的な手順(iib)を手順(iii)の実施の前に行うことができる。
【0047】
最終的な配列は、外来DNA配列の合成の開始前に、問題配列が再導入されていないかを再確認するために再解析してもよい。
【0048】
この方法は、手順(iib)及び手順(iii)を、発現される外来DNA配列に応じて適切な配列での利用に適合させるだけで、必要に応じてあらゆるタンパク質配列での使用に適合できる。
【0049】
スクリーニング方法のモジュール性は、トランスジェニック動物の作成に使用される場合、レンチウイルスベクター等のベクターに組み込んだ後、欠失の危険があり得る任意の外来DNA配列に適用し得る点で、高度に適合化させる。例えば、サイトカイン又はホルモンのような酵素又は生物活性タンパク質等の標準的導入遺伝子のコード配列を解析してよく、同様に、非ヒト哺乳類トランスジェニック系における発現に適する治療的タンパク質等の他のタンパク質の配列を解析してもよい。
【0050】
更に、スクリーニング方法は、抗体その他の同様の結合フラグメント又は結合物の配列のスクリーニングに使用できる。「抗体」はイムノグロブリンであり、天然、又は部分的もしくは完全に合成的に作成される。この用語は、抗体結合ドメインであり又はそれに相同な結合ドメインを有するいかなるポリペプチド、タンパク質、又はペプチドも包含する。抗体の例は、イムノグロブリン異性体、その異性体サブクラス、Fab, scFv, Fv, dAb, Fdのような抗体結合ドメインを含むフラグメント、組換え型二重特異性抗体である。抗体はヒト化されていてよく、ヒト化抗体には部分的ヒト化(キメラ)又は完全ヒト化された抗体が含まれうる。
【0051】
しかし、本発明のこの態様のスクリーニング方法が組換え抗体系導入遺伝子の発現最適化に使用される場合、改変リンカー配列の使用が推奨される。
【0052】
リンカー配列の発展
広く商業的に入手可能なリンカーの例はRPASマウスscFVモジュール(Amersham Biosciences)であり、リンカー配列は、以下の配列番号1のヌクレオチド配列を有する。
GGT GGA GGC GGT TCA GGC GGA GGT GGC TCT GGC GGT GGC GGA TCG
【0053】
本発明は、設計され、以下の配列番号3のヌクレオチド配列を有する新規リンカーも提供する。
GGG GGA GGG GGC AGC GGC GGA GGG GGA TCC GGC GGT GGG GGA TCT
【0054】
配列番号3のヌクレオチド配列は、配列番号4の配列を有するアミノ酸配列をコードする。
GIy GIy GIy GIy Ser GIy GIy GIy GIy Ser GIy GIy GIy GIy Ser
【0055】
繰返しDNA配列を排除するよう設計するとともに、配列設計及びリンカー配列の解析の間に適用される第2の制限は、隣接コドンとしてGGC及びTCCを回避することであった。例えば、広く商業的に入手可能なリンカーであるRPASマウスscFVモジュール(Amersham Biosciences)(配列番号5)は、隣接配列としてのGGC及びTCCの存在を評価し、以下の配列が確認される。
配列番号5:GGG GGA GGC GGC TCC GGG GGA GGC GGC TCC GGG GGA GGC GGC TCC
【0056】
pRl28(R24ミニボディ発現をドライブするCMVプロモータ)及びpLE38(R24ミニボディ発現をドライブする組織特異的プロモータ)等のいくつかのEIAV系レンチウイルスベクターコンストラクトからの以前のPCRデータから、推定相同組換え系機構における繰返しがR24ミニボディコード配列内の欠失を発生させていると示唆されたことから、再設計を行った。新規リンカーは、それを含む組換えタンパク質の発現レベルの低下を引き起こすことが知られている、コドンのいわゆる「スローペア」であるGGA、GGC(Trinh et al., 2004)の使用も回避する。
【0057】
非推定リンカー配列の使用は公知である。しかし、本発明は、ウイルスベクター導入遺伝子から短い重複配列を除去するために、リンカー内のコドン選択を変更する外来DNA配列の改変を更に提供する。
【0058】
本発明の更なる態様は、異種タンパク質の少なくとも一部をコードする単離DNAであて、本発明のスクリーニング方法で解析されたDNAを提供する。
【0059】
本発明の更なる態様は、組換え抗体系導入遺伝子の発現のためのリンカー配列であって、配列番号3のヌクレオチド配列を有する配列を提供する。
【0060】
本発明の更なる態様は、組換え抗体系導入遺伝子の発現のためのリンカー配列であって、配列番号4のヌクレオチド配列を有する配列を提供する。
【0061】
本発明の更なる態様は、トランジェニック鳥類を作製する方法であって、前記トランジェニック鳥類において発現させる1以上の異種タンパク質をコードする外来DNA配列を準備する手順と、前記外来DNAの異種タンパク質コード領域のヌクレオチドコドン使用頻度を最適化して、前記外来DNA配列が発現されるトランジェニック鳥類のコドン使用頻度に変更する手順と、コドンが最適化されたDNA配列を改変して、前記宿主鳥類内での前記外来DNAの発現を阻害又は下方制御すると予測されるいずれかのコード配列領域を変化させる手順と、相同組換えに基づく推定欠失機構に関係する全配列を除去するために、前記外来DNA配列のヌクレオチドコドン使用頻度を変更する手順と、前記外来DNA配列を含むベクターを鳥類ゲノムに組み込む手順と、前記配列によってコードされる前記異種タンパク質を生産するために前記外来DNA配列を発現させる手順と、を含む方法を提供する。
【0062】
本発明の外来DNA配列を含むベクターを調整する際、外来DNA配列は、関連する制御領域及び発現調節領域とともにパッケージされる。当業者は、ベクターの好ましいパッケージ方法に気づくであろう。
【0063】
本発明は、トランスジェニック鳥類も提供する。トランスジェニック鳥類は、鳥類種のあらゆるメンバーであり、特にニワトリであり、鳥類細胞の1以上は、細胞ゲノム内に組み込まれ、ベクターに含まれる外来遺伝子材料を含む。かかる遺伝子材料の導入に適した遺伝子導入技術は当業者に公知である。
【0064】
本発明の方法は、あらゆるトランスジェニック鳥類の生成に使用でき、特に限定されないが、ニワトリ、シチメンチョウ、カモ、ウズラ、ガチョウ、ダチョウ、キジ、クジャク、ホロホロチョウ、ハト、白鳥、チャボ、及びペンギンが挙げられる。しかしニワトリが好ましい。トランスジェニック鳥類によって発現される異種タンパク質は、特に限定されないが、ヒト及び/又は獣医目的での治療及び診断利用を含む種々の用途を有するタンパク質であってよく、抗体、抗体フラグメント、抗体誘導体、単鎖抗体フラグメント、融合タンパク質、ペプチド、サイトカイン、ケモカイン、ホルモン類、成長因子、またはいかなる組換えタンパク質をコードする配列を含んでよい。
【0065】
本発明は、更にキメラ鳥類又はモザイク鳥類に及び、外来遺伝子材料は鳥類細胞のすべてではないがいくつかにおいて発見される。
【0066】
一実施形態において、トランスジェニック鳥類は、卵管内で外来遺伝子材料を翻訳タンパク質の形で発現し、卵内へ組み込まれる。
【0067】
レンチウイルスベクター発現コンストラクトは、ベクターによってコードされる異種タンパク質の発現を特異的組織に方向付け(組織特異的発現)るために使用できる。一実施形態において、かかる組織特異的発現は、卵内へ異種タンパク質が組み込まれるように方向付けされる。それは卵白又は卵黄内であってよいが、タンパク質が卵白内に存在することが好ましい。次にタンパク質は、当業者に公知の常法によって卵白又は卵黄から単離される。
【0068】
本発明の更なる態様は、鳥類の卵管内で外来タンパク質を発現する方法であって、本発明の方法を用いて解析された外来DNA配列を準備して、前記外来DNA配列によってコードされる異種タンパク質の発現を阻害又は下方制御し得るコード配列のいずれかの部位を除去又は置換する手順と、記外来DNA配列を鳥類ゲノムに組み込む手順と、前記外来DNA配列に操作可能に連結されたプロモータを用いて前記外来DNAコード配列を発現させる手順と、前記トランスジェニック鳥類によって発現される外来タンパク質を得る手順と、を含む方法を提供する。
【0069】
一実施形態において、本発明の第1態様のスクリーニング方法で解析された外来DNAコード配列はウイルスベクターのバックボーンに挿入され、このベクターを鳥類細胞に挿入される。
【0070】
プロモータが、鳥類の輸卵管のマグナム部分の管状腺細胞内で外来DNA配列によってコードされる異種タンパク質の「組織特異的」発現を行うことが好ましい。「組織特異的」発現は、異種タンパク質の特異的組織で発現させ、他の組織では異種タンパク質の発現が排除される。異種タンパク質の組織特異的発現を鳥類の卵管に方向付けると予測されるプロモータの一例は、オバルブミンプロモータである。
【0071】
本発明のこの態様の更なる態様において、プロモータは、外来DNAコード配列によってコードされる異種タンパク質の発現を卵管の他の組織に方向付けるために、必要に応じて改変されてよい。
【0072】
外来タンパク質は、治療的に有用なタンパク質であり得る。特に、発現される異種タンパク質は、抗体又は同様の結合フラグメントもしくは結合物であってよい。
【0073】
本発明の更なる態様は、鳥類において1以上の外来タンパク質を発現する方法であって、発現させる外来タンパク質をコードする外来DNA配列を準備する手順と、本発明のスクリーニング方法を用いて前記外来DNA配列を解析する手順と、前記外来DNA配列を含むベクターを鳥類ゲノム内で発現させる手順と、発現された抗体タンパク質を前記鳥類から得る手順と、を含む方法を提供する。
【0074】
本発明のこの態様の一実施形態において、1以上の異種タンパク質は、組織特異的、最も好ましくは、卵管細胞での組織特異的発現を用いて鳥類の卵管内で発現される。別の実施形態において、外来タンパク質は、鳥類の輸卵管のマグナム部分の管状腺細胞内で発現され、外来タンパク質は卵白に蓄積する。代替的又は追加的に、異種タンパク質は卵白に蓄積し、血液に分泌され得る。
【0075】
更なる実施形態において、鳥類はニワトリである。
【0076】
一実施形態において、卵管内で発現される異種タンパク質は抗体である。更なる実施形態において、抗体は「ヒト化」される。
【0077】
本発明の更なる態様は、本発明の第1態様のスクリーニング方法を用いて解析された外来DNA配列の、外来タンパク質を含む鳥類卵の生産における使用を提供する。一実施形態において、外来タンパク質は卵白内に蓄積される。更なる実施形態において、外来タンパク質は卵黄内に含まれる。
【0078】
本発明の更なる態様は、本発明の第1態様のスクリーニング方法で解析された外来DNA配列の、異種タンパク質製品の生産における使用であって、前記タンパク質製品は前記外来DNA配列の少なくとも一部の転写及び翻訳によって製造される使用を提供する。
【0079】
本発明の更なる態様は、本発明の第1態様のスクリーニング方法で解析された1以上の外来DNA配列を含む発現ベクターを提供する。
【0080】
更なる態様は、前述した発現ベクターが形質転換された宿主細胞を提供する。
【0081】
一実施形態において、発現ベクターはレンチウイルスベクターであり、特にEIAVである。
【0082】
一実施形態において、宿主細胞は非ヒト哺乳類細胞である。更なる実施形態において、宿主細胞は、鳥類細胞であり、特にニワトリ細胞である。
【0083】
本発明の更なる態様において、本発明のいずれかの方法を使用するためのキットであって、使用説明書と、前記方法を使用するプロトコルとを備えるキットが提供される。
【0084】
本発明の各態様の好ましい特徴及び実施形態は、特に要求がない限りにおいて、他の各態様として変更すべき点が変更される。
【0085】
定義
本明細書では、「ベクター」、「ウイルスベクター」および「発現用ベクター」という用語は置き換え可能に使用しており、外来DNA配列の、プロモータにより誘導される発現、すなわち転写とこれに続く翻訳を可能にする任意の核酸、好ましくはDNAを指している。
【0086】
ウイルスベクターゲノムは「複製欠損」であることが好ましく、すなわち、該ベクターのゲノムは、独自に複製して感染性ウイルス粒子が産生されうるほど十分な遺伝情報だけを備えていないことが好ましい。レンチウイルスベクターの場合では、該ゲノムは機能遺伝子gag、envまたはpolを欠いていることになる。
【0087】
「レンチウイルス」という用語は、本発明に特に好ましいレトロウイルス科の亜科を指している。レンチウイルスには、ヒト免疫不全ウイルスHIV−1およびHIV−2、サル免疫不全ウイルス(SIV)などの種々の霊長類ウイルスならびに非霊長類ウイルス(たとえば、マエディビスナウイルス(MW)、ネコ免疫不全ウイルス(FIV)、ウマ伝染性貧血ウイルス(EIAV)、ヤギ関節炎脳炎ウイルス(CAEV)およびウシ免疫不全ウイルス(BIV)・が含まれる。
【0088】
「ウイルスベクターゲノム」とは、ウイルスゲノムからの配列を備えたポリヌクレオチドを指し、該ウイルスゲノムは、かかるポリヌクレオチドのRNA版をウイルス粒子にパッケージすることが十分に可能で、パッケージされたRNAポリヌクレオチドが逆転写されて宿主細胞の染色体に組み込まれるようになるものをいう。プロモータ配列や、異種ペプチドをコードする外来DNA配列などの異種配列も、ウイルスベクターゲノムの一部であってよい。
【0089】
本明細書で核酸分子を説明するために使用している「組換え型」という用語は、ゲノム、cDNA、半合成または合成に由来するポリヌクレオチドを指しており、その由来または操作により、自然界で結び付いているポリヌクレオチドの全部または一部とは結び付いておらず、および/または自然界で連結されているもの以外のポリヌクレオチドと連結されているものをいう。
【0090】
本明細書でタンパク質またはポリペプチドを説明するために使用している「組換え型」という用語は、組換え型ポリヌクレオチドの発現により生産されるポリペプチドを指す。
【0091】
本明細書で使用している「核酸」という用語には、DNA、RNA、mRNA、cDNA、ゲノムDNA、およびそれらの類似体が含まれる。
【0092】
「外来DNA配列」とは、その転写発現を望む核酸配列である。外来DNA配列は一般に、ペプチド、ポリペプチドまたはタンパク質をコードする。
【0093】
「欠失」とは、ウイルスベクターゲノムの元のプラスミドコピーに存在するDNA配列の領域が逆転写の過程で失われる事象をいう。このため欠失した配列は、複製不能なレンチウイルスベクターの粒子が生産されるパッケージング過程で、元のプラスミドから転写された一本鎖RNA分子の一部または全部から失われている。プラスミドDNA配列は常に無傷のまま維持され、欠失は、一本鎖RNAの2つのコピーが逆転写されてタンパク質の外被内で組み立てられるパッケージング過程における転写過程で起こることに注意されたい。
【0094】
さらに、細胞で正常に発現されない未改変の核酸配列またはポリペプチドは異種であるとみなされる。本発明のベクターは、同一のまたは異なる挿入部位に挿入された1つ以上の外来DNA配列を有することが可能で、挿入部位で各外来DNA配列は、該配列の発現を可能にする調節核酸配列に操作可能に連結される。このため、本発明から得られるベクターは、・単量体、二量体および多量体タンパク質などの種々のタイプのタンパク質を発現させるために使用できると考えられる。
【0095】
本発明で説明したベクターは「異種タンパク質」を発現させるために使用できる。
【0096】
本明細書で使用しているように、「異種」という用語は、外来種に由来する核酸配列もしくはポリペプチド、または同一の種から由来する場合はその原形が実質的に改変された核酸配列またはポリペプチドを指す。
【0097】
適切な異種ペプチドは、治療活性または他の商業的関連性のある用途を有する組換え型タンパク質であってもよい。発現されうる異種タンパク質の例として、インターフェロンアルファ、ベータおよび/またはガンマなどのサイトカイン、インターロイキン、第VIII因子などの造血因子が挙げられる。1つの実施形態では、該異種ペプチドは抗体の重鎖または軽鎖をコードするものであってもよく、ネズミ抗体、キメラ抗体、ヒト化抗体およびヒト抗体など任意のタイプの抗体の重鎖または軽鎖であることが可能であり、この2つの鎖は同一の抗体または異なる抗体に由来することができる。
【0098】
別に定義されていない限り、本明細書に使用した技術用語および科学用語はすべて、本発明の技術分野における当業者に広く理解されている意味を有するものである。
【0099】
本明細書の全体を通して、文脈からそうではないことが求められていない限り、「包含する」もしくは「含む」という用語、または「包含している」、「含んでいる」などの変形は、記載した整数または整数群を含めることを意味すると理解されると思われるが、任意の他の整数または整数群を除外することを意味しているのではない。
【実施例】
【0100】
実施例1
R24ミニボディ−RT−PCRデータ
【0101】
EIAVレンチウイルスベクターとともに使用するR24ミニボディの全配列を図1に示す。該組換え型抗体分子は、ヒトIgG1 Fcドメインの上流に挿入された、マウスVH、リンカーおよびマウスVLから構成される標準的scFV断片からなる(図2)。該配列を、2種類のプロモータの下流に導入し、1つは大域的プロモータであるヒトサイトメガロウイルス(hCMV)即時型初期プロモータであった。もう1つは、トランスジェニック鳥類内で空間時間的に限定された様式でR24ミニボディを活発に発現させるように設計した組織特異的プロモータ候補であった。
【0102】
ウイルスゲノムプラスミドpRI28(プラスミドマップは図3、全配列は図4に示す)を作製するために、hCMVプロモータの下流にR24を挿入した。該ゲノムプラスミドをD17イヌ骨肉種細胞に一過性にトランスフェクションし、その後、細胞培地についてELISAを行なった結果、600ng/mlレベルのヒトIgG1が分泌されていることが示された。この結果から、pRI28ゲノムの発現能が確認された。標準的なトランスフェクション手法により、パッケージされたpRI28の複製不能RNAゲノムを得た。次に、D17細胞をpRI28ウイルスで形質導入した。続いて、これらの細胞から収集した培地をELISAにより分析した結果、ヒトIgG1の分泌は検出されなかった。パッケージされたウイルスからウイルスRNAも収集し、pRI28ゲノムの構造をRT−PCRにより解析した。RT−PCRにより、パッケージされたpRI28ウイルスの試料にはゲノム集団が混在していることが示され、そのすべてがpRI28プラスミドの均一な調製物から転写されたものであった。最も著しい違いはゲノムの3’末端に見出され(図5)、そこから、明らかに完全長の産物および不完全長の産物が増幅され得た。明らかに不完全長の多数のRT−PCR産物をクローニングして塩基配列決定を行なった結果、欠失事象はR24コード配列の一部または全部を包含していることが確認された。当該欠失事象の一部の位置を図6に示す(以後は独自の「lt」番号で引用する)。図6に示した欠失事象の性質を考えると、当該ゲノムはR24ミニボディを発現できないと予測されることに注意されたい。
【0103】
上記lt欠失事象を慎重に解析した結果、該欠失は、小さい(5〜10bp)直列反復配列により輪郭が描かれていることが明らかになった。この結果は、当該配列要素がEIAVと適合性がない可能性があることを示している。
【0104】
導入遺伝子欠失事象における短い直列反復配列要素の役割を、同類のウイルスゲノムに関する研究によりさらに確認した。同一のR24ミニボディコード配列を組織特異的プロモータ候補の下流に挿入して、プラスミドpLE38を作製した(ゲノムマップの概略図を図7に示す)。標準的なトランスフェクション手法により、パッケージされたpLE38の複製不能RNAゲノムを得た。pRI28での記述とまったく同様にRT−PCR法を完了したところ、pRI28と同様に、ウイルスゲノムの3’末端から明らかに不完全長のPCR産物が増幅されており、これらはR24コード配列の一部または全部を包含していた。PCR産物のクローニングおよび配列解析により、先にpRI28ウイルスでも検出された1つの特定の欠失産物であるlt1が頻繁に存在することが示された(図6の欠失マップを参照)。lt1欠失産物の全配列を図8に示す。
実施例2−pRI28からのR24ミニボディ配列データの解釈
【0105】
R24ミニボディでは、問題がある可能性のある短い直列反復配列が2種類存在し、1つはscFV領域自体(VH、リンカーおよびVL)のなかにあり、もう1つはIgG1 Fcドメイン内に存在する。R24ミニボディの構造の概略図は図2に示している。
VHドメイン
【0106】
VH内ではR24ミニボディ配列に、問題のある反復配列が4つ確認され、第1は5’末端に位置し(図9のLP、Leu Pro、欠失lt16に関与する)、第2はCDR2内に位置し(KG、欠失lt15に関与する)、第3はFW3にあり(DT、欠失lt11および13に関与する)、第4はリンカー配列より前のVHの3’末端に位置する(LT、欠失lt1に関与する)。
リンカー/VLドメイン
【0107】
リンカーおよびVLドメインには問題のある反復配列が4つ確認された。第1はリンカー内に位置し(図10のGS、欠失lt4および5に関与する)、第2はFW1内に位置し(LS、欠失lt6に関与する)、第3はCDR2にあり(TS、欠失lt3に関与する)、第4はFW3配列にある(YS、欠失lt2に関与する)。
IgG1 Fc
【0108】
上記の項では、R24ミニボディから3’ウイルス由来配列までの欠失を取り上げた。下線を引いた配列はこれらの欠失の5’末端を表している。しかし、R24ミニボディと当該遺伝子の5’末端までの配列との間の組換え事象により生じた可能性のある欠失も検出された。これらの事例では、3’決定因子がR24ミニボディのIgG1 Fcドメイン内に位置していた。現在、該配列内で、これらの欠失に関連または隣接したプロリンリッチ領域が2箇所特定されている。
【0109】
R24ミニボディにおいて欠失(個別のlt番号で引用する)と関連する問題のある可能性のある8つの配列を図11に要約する。先に記述したステップ(iii)の解析により導入遺伝子候補から除去されるのは、該欠失の輪郭を描く短い直列反復配列である。ベクターNtiソフトウェア(Informax Inc., Invitrogen)または同等物を用いて、DNA配列をスクリーニングして該配列の存在を確認できる。導入遺伝子が組換え型抗体でない場合は、上記残基のすべてが保存される可能性は低い。トランスジェニック鳥類発現系は組換え型抗体を発現できると考えられ、この場合は上記残基が保存され、特に一部は、相補性決定領域(CDR)での残基よりも高い保存性を示すことが知られている可変ドメイン下位領域であるフレームワーク領域(FR)内で保存が起こる。
【0110】
これは、市販されている多数の組換え型抗体にとって最適なエフェクタードメインであるために多数の導入遺伝子候補で確実に保存されなければならないIgG1 Fcにも関連していることである。R24ミニボディを用いた研究から、いくつかの欠失決定因子が該ドメイン内に位置している可能性があることが示されており、たとえば、DNAのポリピリミジントラクトによりコードされる2つのプロリンリッチタンパク質領域は、上記欠失に一貫して関連または隣接している。したがって、該ポリピリミジントラクトを除去することが推奨される。ニワトリはほぼ等しい頻度で4つのコドンを使用してPro/Pをコードすることから、得られるタンパク質の複数のプロリン残基をコードしたままの状態でDNA配列のポリピリミジントラクトを除去するために、コドン使用頻度を変えることが可能である。
実施例3−「修復した」R24ミニボディ
【0111】
短い直列反復配列とこれに関連する欠失との関連性を検討して確立するために、R24ミニボディ配列からlt1配列(5’CTG ATC 3’)を除去し、同時に非反復配列を有するリンカーで置き換えることにした。次に、パッケージされたRNAゲノムのかなりの割合にlt1欠失事象が存在することが示されていたpLE38として設計されたベクターを用いて、この修復の効果をテストした。
【0112】
制限酵素BspEIによるpLE38の消化により、5’lt1反復配列および古いリンカーの除去が可能になり、新しいリンカーをコードし、かつlt1配列が除去された新しいDNA断片で置換することができる(図12を参照)。「修復したR24」を作製するためにpLE38に挿入した置換DNA断片の全配列を図13に示す。完成したプラスミドをpLE56と呼んだ。
【0113】
次に、修復および未修復の2つのプラスミドのセットを並行してパッケージし、RNAゲノムの構造のほか、形質導入された細胞のゲノムDNAに組み込まれた導入遺伝子の構造をPCRにより解析した。
実験データ−pLE38およびpLE56
【0114】
修復したR24ミニボディからのウイルスRNAのリアルタイムqPCR法により、明らかに許容レベルの該ゲノムが良好にパッケージされ、lt1修復が力価に悪影響を与えていないことが立証された。ELISA法ではR24ミニボディの発現を検出できなかったが、該ベクターに含まれるプロモータからの発現は原理的には組織特異的でなければならず、該プロモータがインビトロで活性であることを期待しなかったため、これは陽性の結果である。これらのウイルスで形質導入された細胞からのゲノムDNAについて実施したリアルタイムqPCRにより、EIAVパッケージングシグナルにまたがる産物がうまく増幅され、これにより細胞の形質導入状態が確認され、組み込みの不足ではなく、漏出性卵アルブミンプロモータ活性の不足がELISAの陰性結果の原因であることを示す証拠が提供された。
【0115】
さらに、両ウイルスにおけるゲノムの3’末端にまたがるPCR反応を行なった結果、pLE56で形質導入した細胞に限ってゲノムDNAから完全長の産物がうまく増幅された。これは、パッケージされたpLE38(未修復)のRNAゲノムからlt1欠失産物が優勢に増幅された結果とは正反対である。しかし、pLE38テストシステムでは、推定されるより小さい欠失産物の存在を廃するにはlt1の修復のみでは不十分であった。この結果の説明として最も可能性が高いのは、問題の可能性のある他の短い直列反復配列要素の存在であり、5’lt1反復配列のみを除去したため、「修復した」R24内にはほかにも保持されている可能性がある。この可能性は、第1に、図11に列記したEIAVに非適合性の可能性のある配列を他の導入遺伝子に適用できるかどうか評価することと、第2に、EIAVに非適合性の可能性のある配列をすべて除去した導入遺伝子で内部欠失の頻度を評価することによってのみ検討することができる。
細菌における不安定性
【0116】
事例証拠から、R24ミニボディで使用した先のリンカー配列は細菌において不安定であったことが示された。個々の反復配列要素の欠失が検出された。pLE56などの多数の発現用ベクターにうまくクローンニングされた新規のリンカーでは、このような問題に遭遇していない。
実施例4−抗CD55ミニボディ(791T/36)
【0117】
R24ミニボディを用いた研究の成果として、EIAVに非適合の可能性がある多数の配列が特定された。R24をベースとしない導入遺伝子に当該配列が存在するかどうか判断することに関心を向けた。このため、R24で特定したEIAVに非適合の可能性がある配列をEIAVレンチウイルスベクターバックボーンに組み込んだときに該配列に欠失が起こると予測されるとして、該配列を別の導入遺伝子に適用できるかどうか判断するために、抗CD55ミニボディDNA配列を評価した。該ミニボディとR24ミニボディとの間で直接配列比較を行なった。該ミニボディに、問題のある領域が8箇所特定され、これらの領域を図14に要約する。
【0118】
図14の表の行1は、R24ミニボディとCD55ミニボディではlt16欠失事象に関与する残基が完全に一致していることを示している。これは、当該残基が、両コンストラクトが共有する基本的なリゾチームのシグナルペプチドによりコードされているからである。別の導入遺伝子、すなわちサイトカインをベースとした産物を合成する前に、該シグナルペプチドのコドン使用頻度を改変した。これまでに解析したインターフェロンベータ遺伝子に基づいた別の遺伝子コンストラクトでは、改変した該シグナルペプチドには依然としてlt16反復配列が存在しているが、対応するlt16欠失が確認されていない。したがって、少なくともミニボディを含有しないベクターでは、欠失を引き起こすのにlt16反復配列の存在だけでは不十分であるようであり、たとえばリンカードメインなど、別の因子が関与していなければならないと思われる。しかし、シグナルペプチドのコドン使用頻度をさらに改変して、この要素を取り除くことが賢明である。
【0119】
図14の表の行2は、R24ミニボディとCD55ミニボディの間で2つのアミノ酸のうち1つだけがマッチしていることを示している(KG対KD)。ニワトリはLys/Kに対し、ほぼ等しい頻度で2つのコドンを使用するため、アミノ酸特異性を保ったままコドンを変えて、抗CD55からlt15反復配列要素を除去できると考えられる。
【0120】
図14の表の行3は、R24ミニボディとCD55ミニボディとの間で2つのアミノ酸のうち1つだけがマッチしていることを示している(DT対DS)。上記のLys/Kと同じく、ニワトリはAsp/Dに対し、ほぼ等しい頻度で2つのコドンを使用するため、この場合もアミノ酸特異性を保ったままコドンを変えて、抗CD55ミニボディからlt11/13反復要素を除去できると考えられる。
【0121】
この表の行4は、R24ミニボディで最も問題が大きいlt1反復をコードするLI配列に言及している。この欠失は現在、R24ミニボディをベースとしたpRI28およびpLE38の2つのレンチベクターで確認されている。幸い、現時点では、抗CD55ミニボディと配列相同性はない。
【0122】
この表の行5は、R24ミニボディと抗CD55ミニボディではlt4および5欠失事象に関与する残基が完全にマッチしていることを示している。これは、ミニボディのscFV構成要素の構築時にVHドメインとVLドメインを結合するために使用したリンカーがこれらの残基をコードしているためである。いくつかの行の証拠は、該リンカーは発現試験に使用するには最適でない可能性があることを示しており、また、事例証拠から、大腸菌における反復配列の不安定性や、該リンカーに3つの直列反復配列を与える二次構造の可能性が示されており、Geneart社や文献に反復配列とRNAポリメラーゼとの相互作用に関する議論もある。R24ミニボディのリンカーは、図15に示すように、新しいリンカーに取り替えることができる。これは、(GGGS)4アミノ酸パターンを保持しているが、コドン使用頻度を変えて相同性を最小限に抑えている。
【0123】
下線を引いた文字は、元のリンカーで問題のある配列を強調しており、GGC TCCは実際に3回反復されている。新しいリンカーでは、直列反復配列が撤廃されており、GGC TCC配列はまったく存在せず、その交換後のGGA TCTは1回だけ存在する。この新しいリンカーは、EIAVレンチベクター系で使用するための抗CD55もしくは他の任意のscFVまたはミニボディの遺伝子合成時に使用することを推奨する。
【0124】
図14の行6は、lt6反復についてR24ミニボディと抗CD55ミニボディとの間で2つのうち1つがマッチしていることを示している(LS対LL)。ニワトリはLeuに対しCTGコドンを好むため、該配列を変えないことが最良であると考えられる。行7も、lt3反復についてR24と抗CD55との間で2つのうち1つがマッチしていることを示している(TS対AS)。ニワトリはSer/Sに対し6つの異なるコドンを使用するため、lt3反復配列要素を効果的に取り除くために使用できる選択肢はいくつか存在する。最後に行8は、R24ミニボディのlt3欠失に関与する残基YSは抗CD55ミニボディでは保存されないため、この位置では配列の改変が必要ないことを示している(YS対FT)。
IgG1 Fcドメイン
【0125】
Fcドメイン内の2つの多プロリン領域を除去することも推奨される。ニワトリは4つのコドンをほぼ等しい頻度で使用してPro/Pをコードすることから、コドン使用頻度を変えることにより、得られるタンパク質のプロリン残基をコードしたままDNA配列のポリピリミジン領域を除去することが可能である。
【0126】
我々の現在の知見状況を考慮に入れ、上記の推奨事項をすべて使用して、EIAVレンチベクターに使用する最適な抗CD55ミニボディ配列を作製した。最適化した当該配列を図16および17に示す。
【0127】
DNA配列は著しく変わったが、一次アミノ酸配列は単離した元の配列から変化していないことに注目されたい。新しい5’および3’延長配列を追加して、鳥類のトランスジェニック試験系における遺伝子発現を促進し、新しいリンカーを導入して、対応するR24ミニボディ分子に存在する直列反復配列を撤廃した。R24ミニボディと抗CD55ミニボディとの間で保存される位置のほか、コード配列内の他のすべての場所についても、問題の可能性があることが確認された反復配列モチーフをすべて除去した。
【0128】
結論を述べると、抗CD55ミニボディコード配列の上記解析により、R24をベースとしない導入遺伝子に対する当該導入遺伝子最適化方法の妥当性が実際に立証された。
実施例5−抗CD55抗体(791T/36)
【0129】
本文書の実施例4で提示したデータは、EIAVと適合性がない可能性のある短い直列反復配列を除去するという方針が、R24をベースとしない分子、この場合は抗CD55ミニボディに適用できることを示した。この研究の次の段階は、配列最適化のプロセスを本明細書に記載したように正確に適用した後、EIAVレンチウイルスベクターに存在する導入遺伝子配列内の内部欠失の頻度を評価することであった。
【0130】
しかし、実施例4に記載した抗CD55ミニボディをコードする導入遺伝子を作製するのではなく、導入遺伝子最適化の同じ方針を二重鎖マウス/ヒトキメラ抗CD55抗体に適用することとした。図18は、上記両分子の構造を略図で示したものである。
【0131】
当該キメラ抗体は、ヒトIgG1重鎖およびヒトカッパ型軽鎖の上流にそれぞれ挿入したマウス重鎖および軽鎖両方に由来する可変領域からなる。両分子の一次配列は、本明細書に記載した導入遺伝子最適化の段階的プロセスを行う前に、コンピュータ内で組み立てた。図19および20は、キメラ重鎖および軽鎖それぞれの一次アミノ酸配列を示している。両一次アミノ酸配列は、両タンパク質の分泌を可能にするため、内在ニワトリリゾチーム遺伝子からのシグナルペプチドを追加する5’延長配列を含んでいることに注意されたい。
【0132】
本発明の第一の態様において定義したステップに従って最適化のプロセスを実施した。すなわち、望ましい一次アミノ酸配列をGeneart社(ドイツ)に提供し、ニワトリのコドン使用頻度の好みに基づいてDNAコドンが割り当てられ、このプロセスは「チキナイゼーション(ニワトリ化)」と呼ばれる。次に、最適化プロセスのステップ(ii)を完了し、これによりニワトリ化した基本配列を解析して、負の要素すなわち反復配列のほか、スプライス部位、内部TATAボックスまたはリボゾーム進入部位などのシス作用性モチーフなど、遺伝子発現にマイナスの影響を及ぼすと予測される要素を検出した。配列を改変することにより、当該要素をすべて除去した。続いて、ニワトリ化した当該第二世代配列を解析し、図11に示したように問題がある可能性のある配列をすべて取り除いた(最適化プロセスのステップ(iii))。当該第三世代配列をGeneart社に再び送り、上記改変が、負の要素すなわち反復配列のほか、スプライス部位、内部TATAボックスまたはリボゾーム進入部位などのシス作用性モチーフなど、遺伝子発現にマイナスの影響を及ぼすと予測される要素を再導入していないことを確認した。このプロセスを、問題がある可能性があることが確認された反復配列を除去するために設計したすべての改変箇所について繰り返し行ない、コドン使用頻度が依然として最適であり、かつ、負の要素が再導入されていないことを確実にした。遺伝子合成により、キメラ抗CD55重鎖および軽鎖の最終版を作製した。
【0133】
両抗CD55コード配列は、個々のpCRScriptベクターバックボーンに組み込んで供給され、重鎖(図21、pLE121)は制限酵素PmlIにより、軽鎖(図22、pLE120)はSmaIにより切り出すことができた。続いて、組織特異的プロモータ候補の下流に導入遺伝子を導入したベクターゲノムを構築することにより、最適化した導入遺伝子の発現を支援するEIAVレンチウイルスベクター系の能力を分析した。
抗CD55抗体と組織特異的プロモータ候補に基づいた発現コンストラクト
【0134】
重鎖および軽鎖配列を別々に組織特異的プロモータ候補の下流に挿入して、それぞれプラスミドpLE118およびpLE119を作製した。pLE118およびpLE119両方のゲノム構成は、R24が、関連する重鎖または軽鎖配列に置換されていることを除けば、図7のpLE38について示した概略図と同一である。
【0135】
標準的なトランスフェクション手法を用いて、ウイルスゲノムのパッケージングを完了した。ゲノムRNAを収集してRT−PCRにより解析し、さらに、該ウイルス粒子を用いて宿主細胞を形質導入し、続いて該細胞からゲノムDNAを収集した。次に、ゲノム構造のPCR解析を完了した。
【0136】
パッケージされたウイルスゲノムから増幅した産物のRT−PCRと、これに続くクローニングおよびDNA塩基配列決定により、パッケージされたpLE118およびpLE119のゲノム内に無傷の抗CD55のそれぞれ重鎖および軽鎖が存在することが示唆された。興味深いことに、pLE119ゲノムから1つの欠失産物が特定され、この欠失をlt230とした。lt230欠失の範囲を表示したpLE119の3’末端の全配列を図23に示す。当該欠失の5’および3’範囲の輪郭を描いている短い直列反復配列の存在に注目されたい。このデータは、本文書に概略を記載した推定上の相同的組換えに基づく機構により、R24をベースとしないEIAVレンチウイルスベクター導入遺伝子内に内部欠失が発生することを示した最初の証拠である。このため、lt230を挟んでいる反復配列を、導入遺伝子最適化プロセスのステップ(iii)において除去すべき配列のリストに追加した。すべての当該配列を図24に列記する。
【0137】
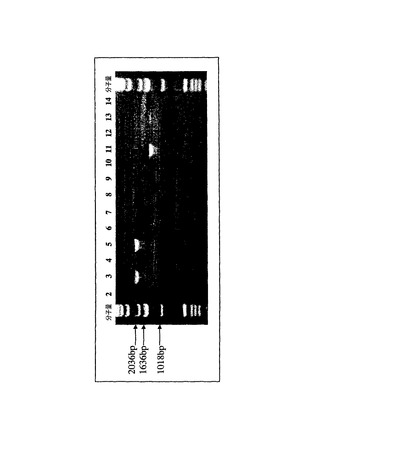
pLE118およびpLE119を形質導入した細胞のゲノムDNAを解析した結果、主に完全長の増幅産物が得られた。たとえば、組織特異的プロモータ候補内から3’LTRまでまたがり、かつ導入遺伝子コード配列を包含するPCR反応により、pLE118ウイルスで形質導入した細胞のゲノムDNAから、無傷の重鎖配列の存在を示唆する2124bpの産物が生じた(図25、列5)。同一のPCR反応により、pLE119ウイルスで形質導入した細胞のゲノムDNAから、無傷の軽鎖配列の存在を示唆する1398bpの産物が生じた(図25、列13)。両導入遺伝子コード配列は同一のリゾチーム由来リーダーペプチドを共有しており、このため、PCRプライマーを共用できることに注意されたい。pLE119で形質導入した細胞のゲノムDNAからはlt230欠失産物が増幅されなかったことから、該産物は大多数の分子種ではないことが示唆される。
【0138】
この研究から結論がいくつか導き出される。第1は、R24で得られた結果とは対照的に、上記ベクターからは無傷の最適化した抗体コード配列がPCRでうまく増幅されたことである。第2は、CD55配列に新規のlt欠失が発見されたことである。本出願は、R24ミニボディを用いた研究の成果として確認された問題の可能性のある配列をすべて除去する手順を詳しく述べている。抗CD55試験系ではR24でみられた欠失産物がまったく検出できなかったことは、当該配列が欠失機構に直接関与しているという結論を裏付けている。たとえば、抗CD55軽鎖の早期の反復では、lt16反復配列(CTg CCC C)が存在した。これは、問題の可能性のあるこれらの反復配列を除去するためのスクリーニング時に確認されたものであり、後期の反復ではCTg CCT Cに変化していたが、コードされるアミノ酸は未変化のまま維持される。最終的に最適化した抗CD55軽鎖配列にはlt16欠失事象の証拠が決定的にまったく検出されず、先に述べたR24の結果とは対照的であった。
【0139】
しかし、抗CD55抗体の配列に新規のlt欠失が検出されれば、問題のある可能性がある別の配列が提供され、該配列は、本明細書で開示した方法により最適化されるさらなる導入遺伝子では除去されることになる。
実施例6−他の種への移行可能性
【0140】
本明細書に記載した導入遺伝子最適化プロセスは、たとえばウズラ Coturnix coturnixなどの他の種で発現するように設計した異種コード配列に適用できる。図26に示すように、ウズラのコドン使用頻度はニワトリ(Gallus gallus)のコドン使用頻度とほぼ同一である。このため、最適化のプロセスは、本発明の第一の態様で定義したステップに従って実施できると考えられる。すなわち、望ましい一次アミノ酸配列をGeneart社(ドイツ)に提供し、ウズラまたはニワトリのコドン使用頻度に基づいてDNAコドンが割り当てられるが、これはウズラおよびニワトリならびに他の鳥類の間でコドンバイアスに非常に高度の保存性が認められることによるものである。続いて最適化プロセスを完了し、これにより、基本の配列を解析してまず、遺伝子発現にマイナスの影響を及ぼすと予測される配列要素を検出し、次に、図24に示したように問題の可能性のある配列をすべて除去する。
【0141】
本明細書に引用した文書はすべて、参照により本明細書に組み込まれている。本発明の範囲から逸脱することなく、記載した本発明の実施形態に対し種々の変更および変化があることは、当業者には明白である。特定の好ましい実施形態と関連させながら本発明を説明してきたが、特許請求の範囲に示したとおり本発明は、当該実施形態に過度に限定されるべきではないことを理解すべきである。実際に、本発明を実施する上記の態様に種々の変更があることは当業者には明らかであり、種々の変更は本発明によりカバーされるように意図されている。
【図面の簡単な説明】
【0142】
【図1】pRl28及びpLE38の構築に使用したR24ミニボディの全DNA配列を示す。開始コドン及び二重停止コドンは大文字で表す。
【図2】R24ミニボディの模式的構造を示す。
【図3】レンチウイルスベクターゲノムpRl28のプラスミドマップ。
【図4】レンチウイルスベクターゲノムpRl28の完全DNA配列を示す。
【図5】pRl28ウイルスのRNAゲノムの予測構造を示す。
【図6】レンチウイルスベクターpRl28内のR24コード配列内で特定された、欠失の相対位置付きの図を示す(個別の「lt」番号で示す)。
【図7】pLE38のRNAゲノムの予測構造の代表図を示す。
【図8】完全R24コード配列を包含するpLE38ゲノムの3’末端の完全配列を示す(開始コドン及び二重停止コドンを太い大文字で示す)。5’LTR配列も太字で示す。lt1繰返しのコピーはいずれもイタリック体で示し、lt1欠失後に欠損した配列には下線を付す。lt1繰返しの5’コピーが欠失後に保持されているため、下線を付していないことに留意されたい。
【図9】R24ミニボディVHドメインアミノ酸配列を示す。R24ミニボディのアミノ酸配列は1文字コードで示す。イタリック体は、FR及びCDRに位置する、この領域の5’及び3’末端の残基を示す。太字は、3フレームワーク領域を含む残基を示す(図の右側欄のキー)。標準文字はCDRを含む残基を示す。下線文字は、問題のDNA繰返しによってコードされるアミノ酸残基を示す。
【図10】R24ミニボディVLドメインアミノ酸配列を示す。R24ミニボディのアミノ酸配列は1文字コードで示す。イタリック体は、FR及びCDRに位置する、この領域の5’及び3’末端の残基を示す。リンカードメインの残基は、5’末端においてイタリック体にされている。太字は、3フレームワーク領域を含む残基を示す(図の右側欄のキー)。標準文字はCDRを含む残基を示す。下線文字は、問題のDNA繰返しによってコードされるアミノ酸残基を示す。
【図11】R24ミニボディにおける潜在的に問題の8配列、及び関連する欠失(個々のlt番号で示す)を示す。
【図12】pLE38における3’末端の図を示す。*は、R24コード配列内の欠失のいくつかに関連し「lt1」で示される短い2つの繰返し配列の位置を示す。「lt1」配列が除去された置換配列である、5’lt1繰返しに隣接する2つのBspEI部位の位置は太い黒線で示す。
【図13】lt1修復手順の間にpLE38に挿入されたBspEIフラグメントの全配列を示し、制限酵素部位は太字で示す。
【図14】R24ミニボディ内の8つの問題配列と、抗CD55ミニボディ内の対応する残基との比較を示す表。
【図15】標準R24内に存在する天然で改変されたリンカーと、修復されたリンカーとによってコードされるDNA及びアミノ酸の配列を示す。
【図16】最適化抗CD55ミニボディの一次アミノ酸配列を示す。
【図17】最適化抗CD55ミニボディのDNA配列を示す。
【図18】抗体及びミニボディの相対的構造を比較する図を示す。
【図19】抗CD55抗体の重鎖の一次アミノ酸配列を示す。
【図20】抗CD55抗体の軽鎖の一次アミノ酸配列を示す。
【図21】pLE121のプラスミドマップを示す。抗CD55抗体重鎖はpCRscriptベクター内でGeneartによって販売される。
【図22】pLE120のプラスミドマップを示す。抗CD55抗体重鎖はpCRscriptベクター内でGeneartによって販売される。
【図23】全抗CD55コード配列を包含するpLE119の3’末端の完全配列を示す(開始コドン及び二重停止コドンを太字及び大文字で示す)。5’LTRも太字で示す。lt230繰返しのコピーはいずれもイタリック体で示し、lt1欠失後に欠損した配列には下線を付す。lt230繰返しの5’コピーが欠失後に保持されているため、下線を付していないことに留意されたい。
【図24】図11に示される表の改訂版であり、R24及び抗CD55を用いて決定された問題の繰返し配列を列挙する。
【図25】pLE118及びpLE119がそれぞれ形質転換された細胞のゲノムDNAから増幅されたPCR産物の、エチジウムブロミド染色1%アガロースゲルを示す。PCRプライマーは、全重鎖及び軽鎖をコードする配列を包含する組織プロモータ候補から3’LTRまで、各ゲノムの3’末端を増幅する。pLE118及びpLE119形質転換細胞から増幅された2124bp及び1398bpの産物は、無傷の抗CD55コード配列の存在を示す。小さい増幅産物が存在しないことに留意されたい。
【図26】ニワトリ(Gallus gallus)及びウズラ(Coturnix coturnix)におけるコドン使用頻度をようやくする2つの表を示す。
【特許請求の範囲】
【請求項1】
外来DNA配列を適切なベクターによる発現に最適化する方法であって、
(i)前記外来DNAのヌクレオチドコドン使用頻度を最適化して、前記外来DNA配列が発現される宿主細胞のコドン使用頻度に変更する手順と、
(ii)コドンが最適化されたDNA配列を改変して、前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得るいずれかの配列部位を変更する手順と、
(iii)相同組換えに基づく推定欠失機構に関係する全配列を除去するために、前記外来DNA配列のヌクレオチドコドン使用頻度を変更する手順と、を含む方法。
【請求項2】
前記外来DNAは異種タンパク質をコードする請求項1記載の方法。
【請求項3】
前記外来DNAは抗体をコードする請求項1記載の方法。
【請求項4】
抗体コード配列に組み込まれるためのリンカー配列を、一次アミノ酸配列中の3重複配列(Gly4Ser1)を保持しつつ設計する設計手順を更に含み、
前記リンカー配列は、DNAコード配列から除去された実質的にすべての重複配列を有する請求項3記載の方法。
【請求項5】
前記抗体又は結合物の設計手順を、手順(iii)の前に行う請求項4記載の方法。
【請求項6】
前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得る配列要素は、陰性要素又は反復配列、スプライシング部位、内部TATAボックス、及び内部リボソーム侵入部位のようなシス活性化モチーフからなる群より選択される請求項1から5いずれか記載の方法。
【請求項7】
前記外来DNA配列の発現に適するベクターにおける、請求項1から6いずれか記載の方法で解析され就職された外来DNA配列の使用。
【請求項8】
前記ベクターはトランスジェニック発現系に導入される請求項7記載の使用。
【請求項9】
前記トランスジェニック発現系はトランスジェニック鳥類である請求項8記載の使用。
【請求項10】
前記トランスジェニック鳥類はニワトリである請求項9記載の使用。
【請求項11】
前記ベクターはレンチウイルスベクターである請求項7から10いずれか記載の使用。
【請求項12】
前記ベクターはウマ伝染性貧血ウイルス(EIAV)である請求項7から11いずれか記載の使用。
【請求項13】
前記リンカー配列は配列番号1の配列を有するリンカー配列。
【請求項14】
組換え抗体のためのリンカー配列であって、そのヌクレオチド配列は、短い重複配列、GGC、及びTCCを、隣接コドンとして有しないリンカー配列。
【請求項15】
組換え抗体系外来遺伝子の発現のためのリンカー配列であって、配列番号3の配列を有するリンカー配列。
【請求項16】
組換え抗体系外来遺伝子の発現のためのリンカー配列であって、配列番号4の配列を有するリンカー配列。
【請求項17】
トランジェニック鳥類を作製する方法であって、
前記トランジェニック鳥類において発現させる1以上の異種タンパク質をコードする外来DNA配列を準備する手順と、
前記外来DNAの異種タンパク質コード領域のヌクレオチドコドン使用頻度を最適化して、前記外来DNA配列が発現されるトランジェニック鳥類のコドン使用頻度に変更する手順と、
コドンが最適化されたDNA配列を改変して、前記宿主鳥類内での前記外来DNAの発現を阻害又は下方制御すると予測されるいずれかのコード配列領域を変化させる手順と、
相同組換えに基づく推定欠失機構に関係する全配列を除去するために、前記外来DNA配列のヌクレオチドコドン使用頻度を変更する手順と、
前記外来DNA配列を含むベクターを鳥類ゲノムに組み込む手順と、
前記配列によってコードされる前記異種タンパク質を生産するために前記外来DNA配列を発現させる手順と、を含む方法。
【請求項18】
前記トランスジェニック鳥類は、ニワトリ、シチメンチョウ、カモ、ウズラ、ガチョウ、ダチョウ、キジ、クジャク、ホロホロチョウ、ハト、白鳥、チャボ、又はペンギンである請求項17記載の方法。
【請求項19】
前記トランスジェニック鳥類は、キメラ鳥類又はモザイク鳥類である請求項17又は18記載の方法。
【請求項20】
前記異種タンパク質の発現を組織特異的に行う請求項17から19いずれか記載の方法。
【請求項21】
前記異種タンパク質の発現を卵管で行う請求項17から20いずれか記載の方法。
【請求項22】
前記異種タンパク質の発現を卵内で行う請求項17から21いずれか記載の方法。
【請求項23】
前記異種タンパク質の発現を卵白で行う請求項17から22いずれか記載の方法。
【請求項24】
鳥類において外来タンパク質を発現する方法であって、
前記鳥類において発現させる1以上の外来タンパク質をコードする外来DNA配列を準備する手順と、
請求項1から6いずれか記載の方法で前記外来DNA配列を解析する手順と、
前記外来DNA配列を含むベクターを鳥類ゲノム内で発現させる手順と、
発現された抗体タンパク質を前記鳥類から得る手順と、を含む方法。
【請求項25】
鳥類の卵管内で外来タンパク質を発現する方法であって、
請求項1から6いずれか記載の方法を用いて解析された外来DNA配列を準備して、前記外来DNA配列によってコードされる異種タンパク質の発現を阻害又は下方制御し得るコード配列のいずれかの部位を除去又は置換する手順と、
記外来DNA配列を鳥類ゲノムに組み込む手順と、
前記外来DNA配列に操作可能に連結されたプロモータを用いて前記外来DNAコード配列を発現させる手順と、
前記トランスジェニック鳥類によって発現される外来タンパク質を得る手順と、を含む方法。
【請求項26】
前記外来DNAコード配列をウイルスベクターのバックボーンに挿入し、このベクターを鳥類細胞に挿入する請求項25記載の方法。
【請求項27】
1以上の外来タンパク質を含む鳥類卵の生産における、請求項1から6いずれか記載の方法を用いて解析された外来DNA配列の使用。
【請求項28】
異種タンパク質製品の生産における、1以上の外来タンパク質を含む鳥類卵の生産における、請求項1から6いずれか記載の方法を用いて解析された外来DNA配列の使用であって、前記タンパク質製品は前記外来DNA配列の少なくとも一部の転写及び翻訳によって製造される使用。
【請求項29】
請求項1から6いずれか記載の方法を用いて解析された1以上の外来DNA配列を含む発現ベクター。
【請求項30】
請求項29記載の発現ベクターが形質転換された宿主細胞。
【請求項31】
請求項1から6、17から26いずれか記載の方法を使用するためのキットであって、使用説明書と、前記方法を使用するプロトコルとを備えるキット。
【請求項1】
外来DNA配列を適切なベクターによる発現に最適化する方法であって、
(i)前記外来DNAのヌクレオチドコドン使用頻度を最適化して、前記外来DNA配列が発現される宿主細胞のコドン使用頻度に変更する手順と、
(ii)コドンが最適化されたDNA配列を改変して、前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得るいずれかの配列部位を変更する手順と、
(iii)相同組換えに基づく推定欠失機構に関係する全配列を除去するために、前記外来DNA配列のヌクレオチドコドン使用頻度を変更する手順と、を含む方法。
【請求項2】
前記外来DNAは異種タンパク質をコードする請求項1記載の方法。
【請求項3】
前記外来DNAは抗体をコードする請求項1記載の方法。
【請求項4】
抗体コード配列に組み込まれるためのリンカー配列を、一次アミノ酸配列中の3重複配列(Gly4Ser1)を保持しつつ設計する設計手順を更に含み、
前記リンカー配列は、DNAコード配列から除去された実質的にすべての重複配列を有する請求項3記載の方法。
【請求項5】
前記抗体又は結合物の設計手順を、手順(iii)の前に行う請求項4記載の方法。
【請求項6】
前記宿主細胞内での前記外来DNAの発現を阻害又は下方制御し得る配列要素は、陰性要素又は反復配列、スプライシング部位、内部TATAボックス、及び内部リボソーム侵入部位のようなシス活性化モチーフからなる群より選択される請求項1から5いずれか記載の方法。
【請求項7】
前記外来DNA配列の発現に適するベクターにおける、請求項1から6いずれか記載の方法で解析され就職された外来DNA配列の使用。
【請求項8】
前記ベクターはトランスジェニック発現系に導入される請求項7記載の使用。
【請求項9】
前記トランスジェニック発現系はトランスジェニック鳥類である請求項8記載の使用。
【請求項10】
前記トランスジェニック鳥類はニワトリである請求項9記載の使用。
【請求項11】
前記ベクターはレンチウイルスベクターである請求項7から10いずれか記載の使用。
【請求項12】
前記ベクターはウマ伝染性貧血ウイルス(EIAV)である請求項7から11いずれか記載の使用。
【請求項13】
前記リンカー配列は配列番号1の配列を有するリンカー配列。
【請求項14】
組換え抗体のためのリンカー配列であって、そのヌクレオチド配列は、短い重複配列、GGC、及びTCCを、隣接コドンとして有しないリンカー配列。
【請求項15】
組換え抗体系外来遺伝子の発現のためのリンカー配列であって、配列番号3の配列を有するリンカー配列。
【請求項16】
組換え抗体系外来遺伝子の発現のためのリンカー配列であって、配列番号4の配列を有するリンカー配列。
【請求項17】
トランジェニック鳥類を作製する方法であって、
前記トランジェニック鳥類において発現させる1以上の異種タンパク質をコードする外来DNA配列を準備する手順と、
前記外来DNAの異種タンパク質コード領域のヌクレオチドコドン使用頻度を最適化して、前記外来DNA配列が発現されるトランジェニック鳥類のコドン使用頻度に変更する手順と、
コドンが最適化されたDNA配列を改変して、前記宿主鳥類内での前記外来DNAの発現を阻害又は下方制御すると予測されるいずれかのコード配列領域を変化させる手順と、
相同組換えに基づく推定欠失機構に関係する全配列を除去するために、前記外来DNA配列のヌクレオチドコドン使用頻度を変更する手順と、
前記外来DNA配列を含むベクターを鳥類ゲノムに組み込む手順と、
前記配列によってコードされる前記異種タンパク質を生産するために前記外来DNA配列を発現させる手順と、を含む方法。
【請求項18】
前記トランスジェニック鳥類は、ニワトリ、シチメンチョウ、カモ、ウズラ、ガチョウ、ダチョウ、キジ、クジャク、ホロホロチョウ、ハト、白鳥、チャボ、又はペンギンである請求項17記載の方法。
【請求項19】
前記トランスジェニック鳥類は、キメラ鳥類又はモザイク鳥類である請求項17又は18記載の方法。
【請求項20】
前記異種タンパク質の発現を組織特異的に行う請求項17から19いずれか記載の方法。
【請求項21】
前記異種タンパク質の発現を卵管で行う請求項17から20いずれか記載の方法。
【請求項22】
前記異種タンパク質の発現を卵内で行う請求項17から21いずれか記載の方法。
【請求項23】
前記異種タンパク質の発現を卵白で行う請求項17から22いずれか記載の方法。
【請求項24】
鳥類において外来タンパク質を発現する方法であって、
前記鳥類において発現させる1以上の外来タンパク質をコードする外来DNA配列を準備する手順と、
請求項1から6いずれか記載の方法で前記外来DNA配列を解析する手順と、
前記外来DNA配列を含むベクターを鳥類ゲノム内で発現させる手順と、
発現された抗体タンパク質を前記鳥類から得る手順と、を含む方法。
【請求項25】
鳥類の卵管内で外来タンパク質を発現する方法であって、
請求項1から6いずれか記載の方法を用いて解析された外来DNA配列を準備して、前記外来DNA配列によってコードされる異種タンパク質の発現を阻害又は下方制御し得るコード配列のいずれかの部位を除去又は置換する手順と、
記外来DNA配列を鳥類ゲノムに組み込む手順と、
前記外来DNA配列に操作可能に連結されたプロモータを用いて前記外来DNAコード配列を発現させる手順と、
前記トランスジェニック鳥類によって発現される外来タンパク質を得る手順と、を含む方法。
【請求項26】
前記外来DNAコード配列をウイルスベクターのバックボーンに挿入し、このベクターを鳥類細胞に挿入する請求項25記載の方法。
【請求項27】
1以上の外来タンパク質を含む鳥類卵の生産における、請求項1から6いずれか記載の方法を用いて解析された外来DNA配列の使用。
【請求項28】
異種タンパク質製品の生産における、1以上の外来タンパク質を含む鳥類卵の生産における、請求項1から6いずれか記載の方法を用いて解析された外来DNA配列の使用であって、前記タンパク質製品は前記外来DNA配列の少なくとも一部の転写及び翻訳によって製造される使用。
【請求項29】
請求項1から6いずれか記載の方法を用いて解析された1以上の外来DNA配列を含む発現ベクター。
【請求項30】
請求項29記載の発現ベクターが形質転換された宿主細胞。
【請求項31】
請求項1から6、17から26いずれか記載の方法を使用するためのキットであって、使用説明書と、前記方法を使用するプロトコルとを備えるキット。
【図1】



【図2】
【図3】
【図4】

【図4】

【図4】

【図5】
【図6】
【図7】
【図8】

【図9】

【図10】

【図11】
【図12】
【図13】

【図14】
【図15】

【図16】

【図17】

【図18】
【図19】

【図20】

【図21】
【図22】
【図23】

【図24】

【図25】
【図26】

【図2】
【図3】
【図4】
【図4】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【公表番号】特表2008−511301(P2008−511301A)
【公表日】平成20年4月17日(2008.4.17)
【国際特許分類】
【出願番号】特願2007−528999(P2007−528999)
【出願日】平成17年9月2日(2005.9.2)
【国際出願番号】PCT/GB2005/003402
【国際公開番号】WO2006/024867
【国際公開日】平成18年3月9日(2006.3.9)
【出願人】(507067180)ヴィラジェン インコーポレイテッド (1)
【Fターム(参考)】
【公表日】平成20年4月17日(2008.4.17)
【国際特許分類】
【出願日】平成17年9月2日(2005.9.2)
【国際出願番号】PCT/GB2005/003402
【国際公開番号】WO2006/024867
【国際公開日】平成18年3月9日(2006.3.9)
【出願人】(507067180)ヴィラジェン インコーポレイテッド (1)
【Fターム(参考)】
[ Back to top ]